Область техники, к которой относится изобретение
Настоящее изобретение относится к терапевтическим или профилактическим композициям, предназначенным лечения или профилактики связанных с IL-8 заболеваний и т.п.
Предпосылки создания изобретения
Эндометриоз представляет собой не только эстрогензависимое (непатентные документы 1 и 2), но также и воспалительное заболевание (непатентный документ 3).
В одном из объектов изобретения понятие «эндометриоз» относится (но, не ограничиваясь только указанным) к заболеванию, при котором эндометриоидные ткани матки присутствуют и прорастают эктопически за пределы полости матки (как правило, внутрь таза, иногда в брюшную полость и редко в грудную полость). Эндометриоз не является злокачественной опухолью, но болезнь прогрессирует с течением времени и вызывает образование опухоли и адгезию к прилегающим тканям, серьезную боль в брюшной и тазовой области во время менструации или периода между менструациями и бесплодие (нарушенная фертильность). Эндометриоз развивается главным образом у женщин в репродуктивном возрасте 20 лет или старше, он снижает качество жизни (QOL) из-за боли и других симптомов и приводит к хронической боли и бесплодию вследствие усугубления адгезии (образования спаек). В одном из объектов изобретения понятие аденомиоз относится (но, не ограничиваясь только указанным) к заболеванию, напоминающему эндометриоз, но при котором эндометрий встречается внутри миометрия. Аденомиоз вызывает серьезную боль при менструации, гиперменорею, анемию и хроническую боль. (Определение понятий «эндометриоз» и «аденомиоз», применяемых в настоящем описании, приведено ниже в разделе «Варианты осуществления изобретения».)
Принятая терапия эндометриоза или аденомиоза включает применение аналгетиков, гормональной терапии и хирургического вмешательства. Аналгетики оказывают лишь ограниченные эффекты и не могут предупреждать развитие болезненного состояния. Гормональная терапия является эффективной в отношении ослабления боли и замедления развития болезненного состояния; однако с ней связаны проблемы, заключающиеся в неспособности к зачатию во время терапии, поскольку при ее применении отсутствуют менструации, наличии вероятности повторного развития болезненного состояния после прекращения терапии и в неблагоприятных воздействиях гормональных лекарственных средств. Кроме того, отсутствуют сведения о повышении способности к зачатию после применения гормональных лекарственных средств. Даже в случае хирургического лечения шанс рецидива составляет примерно 50% в течение 3-5 лет после хирургического вмешательства (непатентные документы 4 и 5). Для предупреждения послеоперационного рецидива пациенты должны продолжать прием гормональных лекарственных средств и переносить их неблагоприятные воздействия.
IL-8 (интерлейкин 8) является хемокином и он представляет собой белок, для которого установлено, что он имеет формы, состоящие из 72 и 77 аминокислот. IL-8 обозначают также как CXCL8. Мономер IL-8 в случае состоящей из 72 аминокислот формы содержит две дисульфидные перекрестные связи, находящиеся между цистеином 7 и цистеином 34 и между цистеином 9 и цистеином 50. Известно, что IL-8 может присутствовать в форме гомодимера в растворе. Гомодимер не имеет межмолекулярной ковалентной связи и стабилизируется посредством нековалентных связей между β-тяжами двух мономеров.
IL-8 продуцируется различными клетками, такими как моноциты периферической крови, тканевые макрофаги, NK-клетки, фибробласты и сосудистые эндотелиальные клетки, в ответ на стимуляцию провоспалительными цитокинами и т.п. (непатентный документ 6). Считается, что IL-8 обладает, прежде всего, способностью активировать нейтрофилы, усиливает экспрессию ими молекул клеточной адгезии и тем самым повышает их адгезию к сосудистым эндотелиальным клеткам. IL-8 обладает также хемотаксической активностью в отношении нейтрофилов. IL-8, продуцируемый в поврежденных тканях, усиливает миграцию нейтрофилов, прикрепленных к сосудистым эндотелиальным клеткам, в ткани и тем самым индуцирует воспаление, ассоциированное с инфильтрацией нейтрофилов. Кроме того, IL-8 является основным ангиогенным фактором для эндотелиальных клеток, и известно также, что он участвует в ангиогенезе опухолей.
Описано, что, когда бестимусным мышами после овариэктомии, которым внутрибрюшинно трансплантировали человеческую эндометриоидную ткань на фазе пролиферации, имплантировали высвобождающие эстрадиол капсулы, а затем им вводили антитело к IL-8, то был обнаружен регресс повреждений эндометрия (патентный документ 1).
Однако на подопытных животных кроме приматов не удавалось смоделировать менструацию, сходную с менструацией человека. Следовательно, такие животные, как крысы и мыши, не могут служить приемлемой животной моделью эндометриоза человека для выяснения причины эндометриоза человека. Крысы и мыши, прежде всего не имеют IL-8. Кроме того, поскольку к настоящему времени не существует модели in vivo, созданной на приматах кроме человека, с помощью которой можно должным образом изучать эндометриоз человека, то не имеется полученных ранее сведений, подтверждающих воздействие сигнала IL-8 на эндометриоз, которые были бы получены на модели in vivo, созданной на приматах кроме человека.
Кроме того, образование спаек, вызываемое хирургическими операциями, и т.п. является проблемой при различных болезненных состояниях. Применение предупреждающих образование спаек пленок является одним из методов лечения или предупреждения образования спаек. Хотя предупреждающая образование спаек пленка является эффективной в отношения уменьшения послеоперационных спаек, ее действие все-таки является недостаточным, поскольку все еще имеет место образование плотных спаек с частотой 15% (непатентный документ 7).
Перечень ссылок
Патентная литература
Патентный документ 1: WO 2009/026117
Непатентная литература
Непатентный документ 1: Bulun и др., Endometriosis. N Engl J Med 360, 2009, cc. 268-279.
Непатентный документ 2: Giudice и др., Endometriosis. Lancet 364, 2004, сс. 1789-1799.
Непатентный документ 3: Donnez и др., Gynecol Obstet Invest, 54, приложение 1, 2002, сс. 52-58; обсуждение сс. 59-62.
Непатентный документ 4: Vercellini и др., Am J Obstet Gynecol. 198(5), май 2008 г., 504.e1-5.
Непатентный документ 5: Guo и др., Hum Reprod Update. 15(4), июль-август 2009 г., сс. 441-461.
Непатентный документ 6: Remo и др., Expert Rev. Clin. Immunol. 10(5), 2014, сс. 593-619.
Непатентный документ 7: Becker и др., Am Coll Surg. 1 183(4), октябрь 1996 г., сс. 297-306
Краткое изложение сущности изобретения
Задачи, положенные в основе изобретения
Задача, положенная в основу настоящего изобретения, решается с учетом указанных выше обстоятельств. Одним из объектов изобретения, не ограничивающих его объем, являются новые способы и т.п., предназначенные для лечения связанных с IL-8 заболеваний с использованием ингибитора сигнала IL-8.
Более конкретно, одним из объектов изобретения, не ограничивающих его объем, являются новые терапевтические способы и т.п., предназначенные для лечения новых связанных с IL-8 заболеваний с использованием ингибитора сигнала IL-8. Альтернативно этому, другим объектом изобретения, не ограничивающим его объем, являются новые терапевтические способы и т.п., предназначенные для лечения новых или известных связанных с IL-8 болезней с использованием нового антитела к IL-8.
Пути решения задач
Не вдаваясь без необходимости в теорию, при создании настоящего изобретения было сделано концептуальное предположение о том, что истинной патологической основой эндометриоза является хроническое воспалительное пролиферативное заболевание, что было описано ранее (Odagiri, Fertil Steril. 92(5), 2009, сс. 1525-1531). Патологической основой эндометриоза является иммунологический ответ, приводящий к хроническому воспалению и проявляющийся в виде фиброза, метаплазии гладких мышц, нейронной дегенерации, ангиогенеза и т.д. Основываясь на концепции, что IL-8 является важной частью механизма хронического воспаления и что эндометриоз является хроническим воспалительным заболеванием, при создании настоящего изобретения осуществляли оценку патологии и клиническую оценку (исследование с использованием лапароскопа или MP) эффективности лекарственных средств.
Настоящее изобретение основано (но, не ограничиваясь только указанным) на обширных исследованиях, охватывающих диапазон от фундаментальных исследований до клинических испытаний с использованием животных моделей, которые включают создание модели на приматах кроме человека для исследования патологии эндометриоза и т.п., и создание и усовершенствование высоко функциональных антител к IL-8, а также оценку их фармакологических действий. Поскольку надежную оценку человеческих связанных с маткой заболеваний нельзя получить с помощью экспериментов на животных моделях с использованием крыс, мышей или других мелких животных, то при создании настоящего изобретения была создана модель с использованием обезьян циномолгус с индуцированным хирургическим путем эндометриозом, и проведены исследования методом проб и ошибок и специальные исследования, в каждом случае с использованием наилучших технологий, сведений и т.д., например, полученных при проведении основных исследований репродукции на приматах, включая обезьян циномолгус, на основе концепции о патологических состояниях эндометриоза человека и т.д., и их оценке и клинического контроля (лапароскопическая хирургия и обследование, оценка методом ЯМР-томографии (визуализация методом ядерно-магнитного резонанса) и т.д.), что составляет сущность настоящего изобретения
В частности, в одном из вариантов осуществления изобретения (не ограничивающих его объем) был адаптирован подход, касающийся одного из аспектов противовоспалительного действия, сфокусированного на IL-8, который представляет собой основной провоспалительный цитокин при эндометриозе, для поиска терапевтического агента для лечения эндометриоза или аденомиоза, который улучшает их патологические состояния, не оказывая воздействия на половой цикл, что отличает его от существующих гормональных терапий. В результате при создании настоящего изобретения неожиданно было установлено, что патологические состояния облегчались путем введения ингибитора сигнала IL-8.
Кроме того, в другом варианте осуществления изобретения (не ограничивающем его объем) при создании изобретения был адаптирован подход, основанный на противовоспалительном действии, касающийся спаек, вызываемых хирургическими операциями и т.п., и в результате чего неожиданно было установлено, что образование спаек снижалось при введении ингибитора сигнала IL-8.
Кроме того, согласно другому не ограничивающему объем изобретения варианту осуществления изобретения при создании изобретения установлено, что высоко функциональные антитела к IL-8, указанные в настоящем описании, можно применять для лечения и т.д. новых или известных связанных с IL-8 заболеваний. Указанные высоко функциональные антитела к IL-8 включают рН- зависимые антитела к IL-8 (антитела к IL-8, которые связываются с IL-8 в зависимости от рН (рН-зависимым образом) рН-зависимые антитела к IL-8 обладают преимуществом при их введении индивидууму, поскольку по сравнению с референс-антителом они могут обладать по меньшей мере одним или несколькими следующими свойствами: они быстро элиминируют IL-8; они обладают стабильной нейтрализующей IL-8 активностью; их иммуногенность является низкой, и уровни их экспрессии являются высокими. Альтернативно этому, указанные высоко функциональные антитела к IL-8 могут представлять собой антитела, которые обладают по меньшей мере одним или несколькими следующими свойствами: антитела к IL-8 содержат Fc-домен, FcRn-связывающая аффинность которого при кислом рН выше, чем у нативного Fc-домена; антитела к IL-8 содержат Fc-домен, аффинность связывания которых с предсуществующими ADA (антитела к лекарственным средствам) понижена по сравнению с нативным Fc-доменом; антитела к IL-8 содержат Fc-домен, время полужизни в плазме которого повышено по сравнению с нативным Fc-доменом; и рН-зависимые антитела к IL-8 содержат Fc-домен, аффинность связывания которого с эффекторными рецепторами снижена по сравнению с нативным Fc-доменом.
Одним из конкретных вариантов осуществления настоящего изобретения (не ограничивающих его объем) является:
[1] Композиция, предназначенная для лечения или предупреждения связанного с IL-8 заболевания, которая содержит в качестве действующего вещества выделенное антитело к IL-8, которое связывается с человеческим IL-8, где связанное с IL-8 заболевание выбрано из группы, которая состоит из: эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменорее; бесплодия и боли в результате образования спаек, фиброза или воспаления;
где антитело к IL-8 выбрано из группы, которая состоит из:
(1) антитела к IL-8, которое связывается с IL-8 рН-зависимым образом и содержит по меньшей мере одну аминокислотную замену по меньшей мере в одном из следующих участков:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28;
(2) антитела к IL-8, которое связывается с IL-8_рН-зависимым образом и содержит по меньшей мере одну аминокислотную замену по меньшей мере в одном из следующих участков:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28,
где антитело к IL-8 содержит по меньшей мере аминокислотные замены:
тирозина в положении 3 в аминокислотной последовательности SEQ ID NO: 25,
аспарагина в положении 1 и лейцина в положении 5 в аминокислотной последовательности SEQ ID NO: 27 и
глутамина в положении 1 в аминокислотной последовательности SEQ ID NO: 28;
(3) антитела к IL-8, которое связывается с IL-8_pH-зависимым образом и содержит по меньшей мере одну аминокислотную замену по меньшей мере в одном из следующих участков:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28,
где антитело к IL-8 содержит по меньшей мере аминокислотные замены:
тирозина в положении 9 и аргинина в положении 11 в аминокислотной последовательности SEQ ID NO: 24 и
тирозина в положении 3 в аминокислотной последовательности SEQ ID NO:
(4) антитела к IL-8, которое связывается с IL-8 рН-зависимым образом и содержит по меньшей мере одну аминокислотную замену по меньшей мере в одном из следующих участков:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28,
где антитело к IL-8 содержит по меньшей мере аминокислотные замены:
аланина в положении 6, глицина в положении 8, тирозина в положении 9 и аргинина в положении 11 в аминокислотной последовательности SEQ ID NO: 24 и
тирозина в положении 3 в аминокислотной последовательности SEQ ID NO: 25;
(5) антитела к IL-8, которое связывается с IL-8_pH-зависимым образом и содержит по меньшей мере одну аминокислотную замену по меньшей мере в одном из следующих участков:
(a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(6) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28,
где антитело к IL-8 содержит по меньшей мере аминокислотные замены:
аспарагина в положении 1 и лейцина в положении 5 в аминокислотной последовательности SEQ ID NO: 27 и
глутамина в положении 1 в аминокислотной последовательности SEQ ID NO: 28;
(6) антитела к IL-8, которое связывается с IL-8_рН-зависимым образом и содержит по меньшей мере одну аминокислотную замену по меньшей мере в одном из следующих участков:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28,
где антитело к IL-8 содержит по меньшей мере аминокислотные замены:
тирозина в положении 9 и аргинина в положении 11 в аминокислотной последовательности SEQ ID NO: 24,
тирозина в положении 3 в аминокислотной последовательности SEQ ID NO: 25,
аспарагина в положении 1 и лейцина в положении 5 в аминокислотной последовательности SEQ ID NO: 27 и
глутамина в положении 1 в аминокислотной последовательности SEQ ID NO: 28; и
(7) антитела к IL-8, которое связывается с IL-8_рН-зависимым образом и содержит по меньшей мере одну аминокислотную замену по меньшей мере в одном из следующих участков:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28,
где антитело к IL-8 содержит по меньшей мере аминокислотные замены:
аланина в положении 6, глицина в положении 8, тирозина в положении 9 и аргинина в положении 11 в аминокислотной последовательности SEQ ID NO: 24,
тирозина в положении 3 в аминокислотной последовательности SEQ ID NO: 25,
аспарагина в положении 1 и лейцина в положении 5 в аминокислотной последовательности SEQ ID NO: 27 и
глутамина в положении 1 в аминокислотной последовательности SEQ ID NO: 28.
[2] Композиция по п. [1], в которой антитело к IL-8 выбрано из группы, состоящей из:
(8) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит:
(а) аминокислотную последовательность SEQ ID NO: 23 в качестве HVR-H1,
(б) аминокислотную последовательность SEQ ID NO: 29 в качестве HVR-H2 и
(в) аминокислотную последовательность SEQ ID NO: 30 в качестве HVR-Н3;
(9) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит:
(г) аминокислотную последовательность SEQ ID NO: 26 в качестве HVR-L1,
(д) аминокислотную последовательность SEQ ID NO: 31 в качестве HVR-L2 и
(е) аминокислотную последовательность SEQ ID NO: 32 в качестве HVR-L3;
(10) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит:
(а) аминокислотную последовательность SEQ ID NO: 23 в качестве HVR-H1,
(б) аминокислотную последовательность SEQ ID NO: 24 в качестве HVR-H2 и
(в) аминокислотную последовательность SEQ ID NO: 30 в качестве HVR-Н3;
(11) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит:
(г) аминокислотную последовательность SEQ ID NO: 26 в качестве HVR-L1,
(д) аминокислотную последовательность SEQ ID NO: 107 в качестве HVR-L2 и
(е) аминокислотную последовательность SEQ ID NO: 32 в качестве HVR-L3;
(12) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит:
(а) аминокислотную последовательность SEQ ID NO: 23 в качестве HVR-H1,
(б) аминокислотную последовательность SEQ ID NO: 29 в качестве HVR-Н2,
(в) SEQ ID NO: 30 в качестве HVR-H3,
(г) аминокислотную последовательность SEQ ID NO: 26 в качестве HVR-L1,
(д) аминокислотную последовательность SEQ ID NO: 31 в качестве HVR-L2 и
(е) аминокислотную последовательность SEQ ID NO: 32 в качестве HVR-L3;
(13) антитела к IL-8, указанного в подпункте (1) или (2) в п. [1], которое содержит:
(а) аминокислотную последовательность SEQ ID NO: 23 в качестве HVR-Н1,
(б) аминокислотную последовательность SEQ ID NO: 24 в качестве HVR-Н2,
(в) аминокислотную последовательность SEQ ID NO: 30 в качестве HVR-Н3,
(г) аминокислотную последовательность SEQ ID NO: 26 в качестве HVR-L1,
(д) аминокислотную последовательность SEQ ID NO: 107 в качестве HVR-L2 и
(е) аминокислотную последовательность SEQ ID NO: 32 в качестве HVR-L3;
(14) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 34, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 35;
(15) антитела к IL-8, указанного в подпункте (1) или (2) в п. [1], которое содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 108, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 109;
(16) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, последовательности которых идентичны по меньшей мере на 80% последовательности (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23, (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29, (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30, (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26, (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 31, и (е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 32, соответственно;
(17) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3, последовательности которых идентичны по меньшей мере на 80% последовательности (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23, (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24, (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30, (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26, (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 107, и (е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 32, соответственно;
(18) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, последовательности которых идентичны по меньшей мере на 80% последовательности вариабельной области тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 34, и вариабельной области легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 35, соответственно; и
(19) антитела к IL-8, указанного в одном из подпунктов (1)-(7) в п. [1], которое содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, последовательности которых идентичны по меньшей мере на 80% последовательности вариабельной области тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 108, и вариабельной области легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 109, соответственно.
[3] Композиция, предназначенная для лечения или предупреждения связанного с IL-8 заболевания, которая содержит в качестве действующего вещества выделенное антитело к IL-8, которое связывается с человеческим IL-8, где связанное с IL-8 заболевание выбрано из группы, которая состоит из: эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления;
где антитело к IL-8 выбрано из группы, которая состоит из:
(I) антитела к IL-8, которое содержит Fc-область, содержащую аминокислотную замену в одном или нескольких положении(ях), выбранном(ых) из группы, состоящей из положений 235, 236, 239, 327, 330, 331, 428, 434, 436, 438 и 440 согласно EU-нумерации;
(II) антитела к IL-8, которое содержит Fc-область, содержащую аминокислотную замену во всех положениях, выбранных из группы, состоящей из положений 434, 438 и 440 согласно EU-нумерации;
(III) антитела к IL-8, которое содержит Fc-область, содержащую все из следующих аминокислотных замен: аминокислотная замена на Ala в положении 434; аминокислотная замена на Glu, Arg, Ser или Lys в положении 438; и аминокислотная замена на Glu, Asp или Gln в положении 440 согласно EU- нумерации;
(IV) антитела к IL-8, которое содержит Fc-область, содержащую все из следующих аминокислотных замен: аминокислотная замена на Ala в положении 434; аминокислотная замена на Glu, Arg, Ser или Lys в положении 438; и аминокислотная замена на Glu, Asp или Gln в положении 440, и дополнительно содержащую Не или Leu в положении 428, и/или Ile, Leu, Val, Thr или Phe в положении 436 согласно EU-нумерации;
(V) антитела к IL-8, которое содержит Fc-область, содержащую одну или несколько аминокислотную(ых) замену(н), выбранных из группы, состоящей из L235R, G236R, S239K, A327G, A330S, P331S, M428L, N434A, Y436T, Q438R и S440E согласно EU-нумерации;
(VI) антитела к IL-8, которое содержит Fc-область, содержащую комбинацию аминокислотных замен, выбранных из группы, которая состоит из:
N434A/Q438R/S440E;
N434A/Q438R/S440D;
N434A/Q438K/S440E;
N434A/Q438K/S440D;
N434A/Y436T/Q438R/S440E;
N434A/Y436T/Q438R/S440D;
N434A/Y436T/Q438K/S440E;
N434A/Y436T/Q438K/S440D;
N434A/Y436V/Q438R/S440E;
N434A/Y436V/Q438R/S440D;
N434A/Y436V/Q438K/S440E;
N434A/Y436V/Q438K/S440D;
N434A/R435H/F436T/Q438R/S440E; N434A/R435H/F436T/Q438R/S440D;
N434A/R435H/F436T/Q438K/S440E; N434A/R435H/F436T/Q438K/S440D;
N434A/R435H/F436V/Q438R/S440E; N434A/R435H/F436V/Q438R/S440D;
N434A/R435H/F436V/Q438K/S440E; N434A/R435H/F436V/Q438K/S440D;
M428L/N434A/Q438R/S440E; M428L/N434A/Q438R/S440D;
M428L/N434A/Q438K/S440E; M428L/N434A/Q438K/S440D;
M428L/N434A/Y436T/Q438R/S440E; M428L/N434A/Y436T/Q438R/S440D;
M428L/N434A/Y436T/Q438K/S440E; M428L/N434A/Y436T/Q438K/S440D;
M428L/N434A/Y436V/Q438R/S440E; M428L/N434A/Y436V/Q438R/S440D;
M428L/N434A/Y436V/Q438K/S440E и M428L/N434A/Y436V/Q438K/S440D
согласно EU-нумерации;
(VII) антитела к IL-8, которое содержит Fc-область, содержащую комбинацию аминокислотных замен:
L235R/G236R/S239K/M428L/N434A/Y436T/Q438R/S440E или
L235R/G236R/A327G/A330S/P331S/M428L/N434A/Y436T/Q438R/S440E
согласно EU-нумерации;
и
(VIII) антитела к IL-8, которое содержит Fc-область, указанную выше в одном из подпунктов (I)-(VII), где Fc-область обладает по меньшей мере одним из свойств, выбранных из группы, которая состоит из свойств, указанных ниже в подпунктах (а)-(д):
(а) FcRn-связывающая аффинность Fc-области при кислом рН повышена по сравнению с FcRn-связывающей аффинностью нативной Fc-области;
(б) аффинность связывания Fc-области с предсуществующими ADA понижена по сравнению с аффинностью связывания нативной Fc-области с предсуществующими ADA;
(в) время полужизни в плазме Fc-области повышено по сравнению с временем полужизни в плазме нативной Fc-области;
(г) плазменный клиренс Fc-области понижен по сравнению с плазменным клиренсом нативной Fc-области и
(д) аффинность связывания Fc-области с эффекторным рецептором понижена по сравнению с аффинностью связывания нативной Fc-области с эффекторным рецептором.
[4] Композиция, предназначенная для лечения или предупреждения связанного с IL-8 заболевания, которая содержит в качестве действующего вещества выделенное антитело к IL-8, которое связывается с человеческим IL-8, где связанное с IL-8 заболевание выбрано из группы, которая состоит из: эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления;
и где антитело к IL-8 выбрано из группы, которая состоит из:
(A) антитела к IL-8, которое содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 36, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 38;
(Б) антитела к IL-8, которое содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 37, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 38;
(B) антитела к IL-8, которое содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 106, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 44;
(Г) антитела к IL-8, которое содержит тяжелую и легкую цепи, последовательности которых идентичны по меньшей мере на 80% тяжелой цепи, содержащей аминокислотную последовательность SEQ ID NO: 36, и легкой цепи, содержащей аминокислотную последовательность SEQ ID NO: 38, соответственно;
(Д) антитела к IL-8, которое содержит тяжелую и легкую цепи, последовательности которых идентичны по меньшей мере на 80% тяжелой цепи, содержащей аминокислотную последовательность SEQ ID NO: 37, и легкой цепи, содержащей аминокислотную последовательность SEQ ID NO: 38, соответственно; и
(Е) антитела к IL-8, которое содержит тяжелую и легкую цепи, последовательности которых идентичны по меньшей мере на 80% тяжелой цепи, содержащей аминокислотную последовательность SEQ ID NO: 106, и легкой цепи, содержащей аминокислотную последовательность SEQ ID NO: 44, соответственно.
[5] Композиция по одному из п.п. [1]-[4], где фиброзное заболевание выбрано из группы, которая состоит из фиброза при эндометриозе или аденомиозе, хронического обструктивного заболевания легких (COPD), муковисцидоза, псориаза, фиброза печени, фиброза почки и фиброза легких.
[6] Композиция, предназначенная для лечения или предупреждения связанного с IL-8 заболевания, которая содержит в качестве действующего вещества ингибитор сигнала IL-8, где связанное с IL-8 заболевание выбрано из группы, которая состоит из: эндометриоза; аденомиоза; дисменореи; образования спаек; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления.
[7] Композиция по п. [6], предназначенная для подавления инфильтрации иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании.
[8] Композиция по п. [6] или п. [7], предназначенная для ингибирования производства ароматазы или фибротического фактора.
[9] Композиция по одному из п.п. [6]-[8], которая не влияет на половой цикл.
[10] Композиция по одному из п.п. [6]-[9], предназначенная для подавления образования спаек или фиброза при эндометриозе или аденомиозе.
[11] Композиция по одному из п.п. [6]-[10], предназначенная для (1) атрофии эндометриоидных эпителиальных клеток или стромальных клеток у пациенток с эндометриозом или аденомиозом или для (2) уменьшения эндометриоидного интерстиция при эндометриозе.
[12] Композиция по одному из п.п. [6]-[11], где дисменорея представляет собой дисменорею, вызванную эндометриозом или аденомиозом, или представляет собой дисменорею, позволяющую подозревать наличие эндометриоза или аденомиоза.
[13] Композиция по п. [6], где спайки образуются после хирургического вмешательства.
[14] Композиция по одному из п.п. [6]-[13], в которой ингибитор сигнала IL-8 представляет собой ингибитор IL-8, ингибитор CXCR1 или ингибитор CXCR2.
[15] Композиция по п. [14], в которой ингибитор IL-8, ингибитор CXCR1 или ингибитор CXCR2 представляет собой антитело к IL-8, антитело к CXCR1 или антитело к CXCR2 соответственно.
[16] Композиция по одному из п.п. [1]-[5], предназначенная для подавления инфильтрации иммунными клетками повреждения, связанного с IL-8 заболевания или соседней окружающей его среды.
[17] Композиция по одному из п.п. [1]-[5], предназначенная для ингибирования производства ароматазы или фибротического фактора.
[18] Композиция по одному из п.п. [1]-[5], которая не влияет на половой цикл.
[19] Композиция по одному из п.п. [1]-[5], предназначенная для подавления образования спаек или фиброза при эндометриозе или аденомиозе.
[20] Композиция по одному из п.п. [1]-[5], предназначенная для (1) атрофии эндометриоидных эпителиальных клеток или стромальных клеток у пациентов с эндометриозом или аденомиозом или
для (2) уменьшения эндометриоидного интерстиция при эндометриозе.
[21] Композиция по одному из п.п. [1]-[5], где дисменорея представляет собой дисменорею, вызванную эндометриозом или аденомиозом, или представляет собой дисменорею, позволяющую подозревать наличие эндометриоза или аденомиоза.
[22] Композиция по одному из п.п. [1]-[5], где спайки образуются после хирургического вмешательства.
[23] Композиция по одному из п.п. [1]-[22], где связанное с IL-8 заболевание является чувствительным к сигналу IL-8.
[24] Композиция по одному из п.п. [1]-[23], которая дополнительно содержит фармацевтически приемлемый носитель.
[25] Композиция по одному из п.п. [1]-[5] и [16]-[24], где эндометриоз представляет собой эндометриоз человека.
[А1] Антитело к IL-8, указанное в одном из п.п. [1]-[5] и [16]-[25], предназначенное для применения для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из: эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления.
[А2] Ингибитор сигнала IL-8, указанный в одном из п.п. [6]-[15] и [23]-[25], предназначенный для применения для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из: человеческого эндометриоза; аденомиоза; дисменореи; образования спаек; боли при человеческом эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления.
[A3] Способ лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из: эндометриоза; аденомиоза; дисменорея; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления, где способ включает введение индивидууму, который нуждается в этом, антитела к IL-8, указанного в одном из п.п. [1]-[5] и [16]-[25], или композиции по одному из п.п. [1]-[5] и [16]-[25] (где индивидуум, который нуждается в этом, может представлять собой индивидуума, который страдает или предполагается, что будет страдать связанным с IL-8 заболеванием).
[А4] Способ лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из:
человеческого эндометриоза; аденомиоза; дисменореи; образования спаек; боли при человеческом эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления, где способ включает введение индивидууму, который нуждается в этом, ингибитора сигнала IL-8, указанного в одном из п.п. [6]-[15] и [23-[25] или композиции по одному из п.п. [6]-[15] и [23]-[25] (где индивидуум, который нуждается в этом, может представлять собой индивидуума, который страдает или предполагается, что будет страдать связанным с IL-8 заболеванием).
[А5] Применение антитела к IL-8, указанного в одном из п.п. [1]-[5] и [16]-[25], для приготовления лекарственного средства, предназначенного для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из: эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления.
[А6] Применение ингибитора сигнала IL-8, указанного в одном из [6]-[15] и [23]-[25], для приготовления лекарственного средства, предназначенного для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из:
человеческого эндометриоза; аденомиоза; дисменореи; образования спаек; боли при человеческом эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления.
Подразумевается и естественно должно быть понятно обычным специалистам в данной области, что настоящее изобретение включает любые комбинации одного или нескольких элементов, указанных выше, частично или полностью, если это не является технически невозможным согласно общепринятым техническим сведениям, известным обычным специалистам в данной области.
Краткое описание чертежей
На чертежах показано:
на фиг. 1 - уровень связывания Fv4-IgG1, имеющего Fc-домен нативного человеческого IgG1, с ревматоидным фактором в сыворотке страдающих ревматоидным артритом (RA) пациентов;
на фиг. 2 уровень связывания Fv4-YTE с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 3 - уровень связывания Fv4-LS с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 4 - уровень связывания Fv4-N434H с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 5 уровень связывания Fv4-F1847m с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 6 уровень связывания Fv4-F1848m с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 7 - уровень связывания Fv4-F1886m с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 8 - уровень связывания Fv4-F1889m с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 9 уровень связывания Fv4-F1927m с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 10 - уровень связывания Fv4-F1168m с ревматоидным фактором в сыворотке RA-пациентов;
на фиг. 11 - средние уровни связывания с ревматоидным фактором в крови RA-пациентов с каждым из Fv4-IgG1 и с антителами, содержащими различные варианты Fc-области с повышенной FcRn-связывающей активностью;
на фиг. 12 - изменения в зависимости от времени концентрации различных антител к человеческому IgE в плазме обезьян циномолгус при введении ОНВ-IgG1, представляющего собой антитело к человеческому IgE, имеющее Fc-домен нативного человеческого IgG1, и антител, содержащих различные варианты Fc-области с повышенной FcRn-связывающей активностью (OHB-LS, OHB-N434A, OHB-F1847m, OHB-F1848m, OHB-F1886m, OHB-F1889m и OHB-F1927m);
на фиг. 13 - изменения в зависимости от времени концентрации антител в плазме трансгенных по человеческому FcRn мышей при введении Fv4-IgG1, представляющего собой антитело к человеческому IL-6-рецептору, или Fv4-F1718, представляющего собой антитело, полученное в результате повышения FcRn-связывающей активности Fv4-IgG1, в условиях кислого рН;
на фиг. 14 - сенсограммы, характеризующие связывание с IL-8 H998/L63 и Hr9 при рН 7,4 и рН 5,8, полученные с помощью Biacore-анализа;
на фиг. 15 изменения в зависимости от времени концентрации человеческого IL-8 в плазме мышей при введении H998/L63 или H89/L118 мышам в дозе 2 мг/кг в комбинации с человеческим IL-8;
на фиг. 16 - изменения в зависимости от времени концентрации человеческого IL-8 в плазме мышей при введении H89/L118 мышам в дозе 2 мг/кг или 8 мг/кг в комбинации с человеческим IL-8;
на фиг. 17 - изменения в зависимости от времени концентрации человеческого IL-8 в плазме мышей при введении H998/L63 и H89/L118 мышам (каждого в дозе 2 мг/кг или 8 мг/кг) в комбинации с человеческим IL-8;
на фиг. 18-1 - изменения относительной интенсивности химической люминесценции в зависимости от концентраций антител Нг9, H89/L118 и H553/L118 перед хранением в плазме;
на фиг. 18-2 - изменения относительной интенсивности химической люминесценции в зависимости от концентраций антител Hr9, H89/L118 и H553/L118 после хранения в плазме в течение 1 недели;
на фиг. 18-3 - изменения относительной интенсивности химической люминесценции в зависимости от концентраций антител Нг9, H89/L118 и H553/L118 после хранения в плазме в течение 2 недель;
на фиг. 19 - прогнозируемая с использованием EpiMatrix частота встречаемости ADA для каждого из антител к IL-8 (hWS4, Hr9, H89/L118, H496/L118 и H553/L118) и других известных в качестве фармацевтических агентов антител;
на фиг. 20 - прогнозируемая с использованием EpiMatrix частота встречаемости ADA для каждого из антител к IL-8 (H496/L118, H496vl/L118, H496v2/L118, H496v3/L118, H1004/L118 и H1004/L395) и других известных в качестве фармацевтических агентов антител;
на фиг. 21-1 изменения относительной интенсивности химической люминесценции в зависимости от концентраций антител Hr9, H89/L118 и H1009/L395-F1886s перед хранением в плазме;
на фиг. 21-2 - изменения относительной интенсивности химической люминесценции в зависимости от концентраций антител Hr9, H89/L118 и H1009/L395-F1886s после хранения в плазме в течение 1 недели;
на фиг. 21-3 изменения относительной интенсивности химической люминесценции в зависимости от концентраций антител Hr9, H89/L118 и H1009/L395-F1886s после хранения в плазме в течение 2 недель;
на фиг. 22 - изменения в зависимости от времени концентрации человеческого IL-8 в плазме мышей при введении каждого из H1009/L395, H553/L118 и H998/L63H89/L118 мышам в комбинации с человеческим IL-8;
на фиг. 23 уровень связывания с внеклеточным матриксом при добавлении Hr9, H89/L118 или H1009/L395 к клеточному матриксу индивидуально или в комбинации с человеческим IL-8;
на фиг. 24 - изменения в зависимости от времени концентрации антител в плазме трансгенных по человеческому FcRn мышей при введении антитела, имеющего вариабельную область H1009/L395 и Fc-домен (F1942m), который не связывается с FcRn, индивидуально или в комбинации с человеческим IL-8;
на фиг. 25 - прогнозируемая с использованием EpiMatrix частота встречаемости ADA для H1009/L395 и H1004/L395 и других известных в качестве фармацевтических агентов антител;
на фиг. 26 - изменения в зависимости от времени концентрации в плазме обезьян циномолгус каждого из антител к человеческому IL-8 при введении H89/L118-IgG1, содержащего вариабельную область H89/L118 и Fc-домен нативного человеческого IgG1, и антител, содержащих различные варианты Fc-области с повышенной FcRn-связывающей активностью (H89/L118-F1168m, H89/L118-F1847m, H89/L118-F1848m, H89/L118-F1886m, H89/L118-F1889m и H89/L118-F1927m);
на фиг. 27 связывающая активность антител с различными FcγR, где антитела содержат вариабельную область H1009/L395 и Fc-домены которых представляют собой варианты (F1886m, F1886s и F1974m);
на фиг 28 - изменения в зависимости от времени концентрации человеческого IL-8 в плазме трансгенных по человеческому FcRn мышей при введении мышам антител к IL-8 в комбинации с человеческим IL-8. Антитела к IL-8 представляли собой: H1009/L395-IgG1 (2 мг/кг), которое содержит вариабельную область H1009/L395 и Fc-домен нативного человеческого IgG1; и H1009/L395-F1886s (2, 5 или 10 мг/кг), которое содержит вариабельную область H1009/L395 и модифицированный Fc-домен;
на фиг. 29 - изменения в зависимости от времени концентрации антител в плазме обезьян циномолгус при введении Hr9-IgG1 или H89/L118-IgG1, которые содержат Fc-область нативного человеческого IgG1, или H1009/L395-F1886s или H1009/L395-F1974m, которые содержат модифицированную Fc-область;
на фиг. 30 - график, демонстрирующий концентрацию IL-8 в кистозной жидкости страдающих эндометриозом пациентов;
на фиг. 31 таблица данных, полученных путем лапароскопического обследования, в которой представлены характеристики повреждений, образовавшихся в результате высевания эндометриоидных тканей, разрезанных на маленькие кусочки, и повреждений, образовавшихся в результате спаек и шва;
на фиг. 32 - фотографии, демонстрирующие образование узловых повреждений и спаек через 4 месяца после индукции и 12 месяцев после индукции (6 месяцев после введения);
на фиг. 33 фотография, демонстрирующая образование подобного эндометриозу пролиферирующего эндометриоидного эпителия и стромы через 12 месяцев после индукции (6 месяцев после введения);
на фиг. 34-1 - график, демонстрирующий высокую корреляцию между баллом спаек согласно шкале r-AFS (балльная оценка тяжести эндометриоза, разработанная Американским обществом репродукции (AFS)) и концентрацией IL-8 в кистозной жидкости на модели эндометриоза, созданной на обезьянах;
на фиг. 34-2 график, демонстрирующий относительные объемы эндометриоидных узловых повреждений после введения наполнителя или антитела H1009/L395-F1974m;
на фиг. 35-1 - серия графиков, демонстрирующих изменения общего r-AFS-балла, r-AFS-балла спаек и r-AFS-балла размера повреждений до и после введения наполнителя или антитела H1009/L395-F1974m;
на фиг. 35 график, демонстрирующий изменения общего r-AFS-балла до и после введения наполнителя или антитела H1009/L395-F1974m;
на фиг. 36 - гистопатологические изображения области трансплантации в обработанной наполнителем группе и в группе, которой вводили антитело H1009/L395 F1974m, через 12 месяцев после индукции (6 месяцев после введения). Атрофия пролиферирующего эпителия и стромальных клеток и уменьшение интерстиция обнаружены в группе, которой вводили антитело H1009/L395-F1974m, по сравнению с обработанной наполнителем группой;
на фиг. 37 - серия фотографий, демонстрирующих эндометриоидные ткани обработанной наполнителем и антителом H1009/L395-F1974m групп обезьян с сопутствующим аденомиозом. Атрофия эндометриоидного эпителия и атрофия и уменьшение стромальных клеток обнаружены в обработанной H1009/L395-F1974m группе по сравнению с обработанной наполнителем группой;
на фиг. 38 набор фотографий, касающихся послеоперационного образования спаек на модели эндометриоза, созданной на обезьянах, на которых продемонстрированы изображения брюшных полостей в обработанной наполнителем группе и группе, которой вводили антитело H1009/L395-F1974m;
на фиг. 39 - результаты оценки относительной способности к миграции нейтрофилов в соответствующие лунки, в которые дополнительно вносили различные реагенты, относительно способности нейтрофилов к миграции в лунку без IL-8;
на фиг. 40 - результаты анализа уровня экспрессии ароматазы при добавлении супернатанта культуры нейтрофилов к эндометриоидным клеткам;
на фиг. 41 - результаты анализа концентрации МСР-1 в культуральной жидкости нейтрофилов при добавлении IL-8 и антитела к IL-8;
на фиг. 42 - результаты анализа экспрессии CTGF в макрофагах при добавлении IL-8 и антитела к IL-8.
Описание вариантов осуществления изобретения
Ниже представлены варианты осуществления изобретения, не ограничивающие его объем.
Подразумевается, что все элементы, указанные ниже в примерах, естественно, соответствуют представленным в разделе «Описание вариантов осуществления изобретения» без ограничений, обусловленных практикой патентования, указами, регламентами или иными правилами, которые могли бы приводить к более узкой интерпретации представленного в примерах содержания в странах, в которых предполагается получать патентное право на основе настоящей заявки на патент.
Подразумевается и это, естественно, должно быть очевидно обычным специалистам в данной области, что настоящее описание включает любые комбинации некоторых или всех из одного или нескольких указанных в настоящем описании элементов, если это не является технически противоречивым с точки зрения обычных компетентных специалистов в данной области техники.
В контексте настоящего описания понятие «антитело» используется в его наиболее широком смысле и включает (но, не ограничиваясь только этим) моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они обладают требуемой антигенсвязывающей активностью.
В настоящем описании понятие «антитело, которое связывается с тем же эпитопом», что и описанное референс-антитело (например, референс-антитело к IL-8, референс антитело к CXCR1 или референс-антитело к CXCR2), относится в одном из вариантов осуществления изобретения к антителу, ингибирующему связывание с антигеном референс-антитела, которое связывается с требуемым антигеном ((например, IL-8, CXCR1 или CXCR2), например, на 50%, 60%, 70% или 80% или более. И, наоборот, референс-антитело ингибирует связывание антитела с антигеном, например, на 50%, 60%, 70%, 80%, 90%, 95% или более. Примеры типичных анализов в условиях конкуренции (конкурентных анализов), которые можно применять, представлены в настоящем описании (но они не ограничены только указанными).
В настоящем описании понятие «моноклональное антитело» относится к антителу, полученному из популяции практически гомогенных антител, В частности, например, индивидуальные антитела, входящие в популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантов антител, например, содержащих мутации, встречающиеся в естественных условиях или возникающие в процессе производства препарата моноклонального антитела, указанные варианты, как правило, присутствуют в небольших количествах. В отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминаты антигена. Таким образом, прилагательное «моноклональный» определяет особенность антитела, характеризуя его как полученное из практически гомогенной популяции антител, а не сконструированное в соответствии с требованиями к получению антитела с помощью какого-либо конкретного метода. Например, моноклональные антитела, предлагаемые в настоящем изобретении, можно создавать с помощью различных технологий, включая (но, не ограничиваясь только ими) метод гибридом, методы рекомбинантной ДНК, методы фагового дисплея и методы, основанные на использовании трансгенных животных, которые содержат все или часть локусов человеческого иммуноглобулина. В одном из вариантов осуществления изобретения антитела, указанные в настоящем описании, могут представлять собой моноклональные антитела. В контексте настоящего описания понятие «нативное антитело» относится к встречающимся в естественных условиях молекулам иммуноглобулинов с различными структурами. В одном из объектов изобретения нативные антитела в виде IgG представляют собой, например (но, не ограничиваясь только ими), гетеротетрамерные гликопротеины с молекулярной массой примерно 150000 Да, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидными мостиками. В направлении от N-конца к С-концу, каждая тяжелая цепь имеет вариабельную область (VH), за которой следует три константных области (CH1, СН2 и СН3). Аналогично этому, в направлении от N- конца к С-концу, каждая легкая цепь имеет вариабельную область (VL), за которой следует константная область (CL). Легкая цепь антитела в зависимости от аминокислотной последовательности ее константного домена может принадлежать к одному из двух типов, которые обозначают каппа (κ) и лямбда (λ). Константные области, которые применяют в контексте настоящего описания, включают области любого из указанных аллотипов (аллелей) или любого подкласса/изотипа. Например, константная область тяжелой цепи, которую можно применять согласно настоящему описанию, включает (но, не ограничиваясь только ими) константную область нативного антитела IgG-типа (IgG1, IgG2, IgG3 и IgG4). Например, известные аллели IgG1 представляют собой IGHG1*01-05 (см. http://www.imgt.org/), и любой из них можно применять в качестве последовательности нативного человеческого IgG1. При этом, последовательность константного домена можно получать из одного аллеля или подкласса/изотипа или из нескольких аллелей или подклассов/изотипов. В частности, указанные антитела включают (но, не ограничиваясь только ими) антитела, СН1 которых происходит из IGHG1*01, СН2 происходит из IGHG1*02, а СН3 происходит из IGHG1*01. Константные области тяжелых цепей нативных человеческих антител в виде IgG включают, например, константную область человеческого IgG1 (SEQ ID NO: 100), константную область человеческого IgG2 (SEQ ID NO: 101), константную область человеческого IgG3 (SEQ ID NO: 102) и константную область человеческого IgG4 (SEQ ID NO: 103). А константные области легкой цепи нативных человеческих антител к виде IgG, включают например, константную область человеческой κ-цепи (SEQ ID NO: 104) и константную область человеческой λ-цепи (SEQ ID NO: 105).
В контексте настоящего описания понятие=«каркасный участок» или «FR» означает остатки вариабельного домена, отличные от остатков гипервариабельного участка (HVR). FR вариабельной области, как правило, состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Таким образом, последовательности HVR и FR, как правило, имеют следующее расположение в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
В контексте настоящего описания понятие «человеческий консенсусный каркасный участок» представляет собой каркасный участок, в который входят наиболее часто встречающиеся аминокислотные остатки в выбранных последовательностях каркасных участков VL или VH человеческого иммуноглобулина. Как правило, последовательности VL или VH человеческого иммуноглобулина выбирают из подгруппы последовательностей вариабельных областей. Как правило, подгруппа последовательностей представляет собой подгруппу, описанную у Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-е изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991. В одном из вариантов осуществления изобретения касательно VL подгруппа может представлять собой подгруппу κI согласно Kabat и др., выше. В одном из вариантов осуществления изобретения касательно VH подгруппа может представлять собой подгруппу III согласно Kabat и др., выше.
В контексте настоящего описания=«акцепторный человеческий каркасный участок» означает каркасный участок, который содержит аминокислотную последовательность каркасного участка VL или VH, происходящую из каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка. Акцепторный человеческий каркасный участок, «происходящий из» каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка, может иметь такую же аминокислотную последовательность или может содержать уже существующие замены в аминокислотной последовательности. В одном из вариантов осуществления изобретения количество уже существующих аминокислотных замен составляет 10, 9, 8, 7, 6, 5, 4, 3 2 или менее. В одном из вариантов осуществления изобретения последовательность человеческого акцепторного каркасного участка VL идентична последовательности каркасного участка VL человеческого иммуноглобулина или последовательности человеческого консенсусного каркасного участка.
В контексте настоящего описания понятие «вариабельная область» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с требуемым антигеном. Вариабельные области тяжелой цепи и легкой цепи (VH и VL соответственно) нативного антитела, как правило, имеют сходные структуры, при этом каждый домен содержит четыре консервативных каркасных участка (FR) и три гипервариабельных участка (HVR) (Kindt и др., Kuby Immunology, 6-е изд., изд-во W.H. Freeman and Co., 2007, с. 91). Согласно одному из вариантов осуществления изобретения одного (но, не ограничиваясь только им) VH-или VL-домена может быть достаточно для обеспечения специфичности связывания антигена. Кроме того, антитела, которые связываются с конкретным антигеном, можно выделять, используя VH- или VL-домен из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных VL- или VH-доменов соответственно (см., например, Portolano и др., J. Immunol. 150, 1993, сс. 880-887; Clarkson и др., Nature 352, 1991, сс. 624-628).
В контексте настоящего описания понятие гипервариабельный участок» или «HVR» относится к каждому из участков вариабельного домена антитела, последовательности которых являются гипервариабельными («определяющие комплементарность участки» или «CDR») и/или образуют петли определенной структуры («гипервариабельные петли»), и/или содержат контактирующие с антигеном остатки («контакты с антигеном»). Как правило, антитела содержат шесть HVR; три в VH (H1, Н2 и Н3) и три в VL (L1, L2 и L3).
Примеры HVR включают (но, не ограничиваясь только ими):
(а) гипервариабельные петли, включающие аминокислотные остатки 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia и Lesk, J. Mol. Biol. 196, 1987, cc. 901-917);
(б) CDR, включающие аминокислотные остатки 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat и др., Sequences of Proteins of Immunological Interest, 5-е изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991);
(в) области контакта с антигеном, включающие аминокислотные остатки 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum и др., J. Mol. Biol. 262, 1996, cc. 732-745); и
(г) комбинации остатков, указанных в подпунктах (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Если не указано иное, то HVR-остатки и другие остатки в вариабельных областях (например, остатки в FR) нумеруют в настоящем описании согласно Kabat и др., выше.
В контексте настоящего описания «индивидуум» представляет собой млекопитающее. К млекопитающим относятся (но, не ограничиваясь только ими) одомашненные животные (например, коровы, овцы, кошки, собаки и лошади), приматы (например, человек и приматы кроме человека, например, обезьяны), кролики и грызуны (например, мыши и крысы). В одном из вариантов осуществления изобретения «индивидуум» предпочтительно представляет собой млекопитающие, в естественных условиях содержащее в организме IL-8, более предпочтительно животное с менструацией, сходной с менструацией людей например, примата кроме человека и еще более предпочтительно человека. В контексте настоящего описания понятие «индивидуум» применяют взаимозаменяемо с понятием «субъект», если из контекста не следует иное. В контексте настоящего описания «выделенное» антитело представляет собой антитело, которое отделено от компонента его естественного окружения. В одном из вариантов осуществления изобретения антитело можно очищать до чистоты 95% или выше или 99% или выше, например, с помощью хроматографических методов (таких, например, как ионообменная хроматография или ЖХВР с обращенной фазой) или=электрофоретических анализов (таких, например, как ДСН-ПААГ, изоэлектрическое фокусирование (ИЭФ) или капиллярный электрофорез). Обзор методов оценки чистоты антител см., например, у Flatman и др., J. Chrom. В 848, 2007, сс. 79-87.
В одном из объектов изобретения в контексте настоящего описания понятие «выделенное» антитело может относиться к «очищенному» антителу.
В контексте настоящего описания «выделенная» нуклеиновая кислота представляет собой молекулу нуклеиновой кислоты, которая отделена от компонента ее естественного окружения. Нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетке, которая в норме включает молекулу нуклеиновой кислоты, но в которой молекула нуклеиновой кислоты присутствует вне хромосомы или в которой ее локализация на хромосоме отличается от ее встречающейся в естественных условиях локализации на хромосоме.
В контексте настоящего описания понятие «аффинность», как правило, может относиться к суммарной силе всех нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела или химического соединения) и ее партнером по связыванию (например, антигеном). Если не указано иное, то понятие «аффинность связывания» в контексте настоящего описания относится к присущей молекуле аффинности связывания, отражающей взаимодействие по типу 1:1 между компонентами связывающейся пары (например, антителом или химическим соединением и антигеном). Аффинность молекулы X к ее партнеру Y, как правило, можно характеризовать с помощью константы диссоциации (KD). Аффинность связывания можно оценивать с помощью методов, известных в данной области, включая те, которые представлены в настоящем описании.
В одном из вариантов осуществления изобретения антитело, которое связывается с антигеном, таким как IL-8, CXCR1 или CXCR2, может иметь константу диссоциации (KD), составляющую например, ≤1000 нМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1нМ, ≤0,01 нM или ≤0,001нМ (например, 10-8М или менее, от 10-8М до 10-13М или от 10-9М до 10-13М).
В контексте настоящего описания понятия «клетка-хозяин» и «линия клеток-хозяев» используют взаимозаменяемо, и они относятся к клеткам, в которые интродуцирована экзогенная нуклеиновая кислота (включая потомство указанных клеток). К клеткам-хозяевам относятся «трансформанты» и «трансформированные клетки», которые включают первично трансформированную клетку и полученное из нее потомство безотносительно к количеству пересевов. Потомство может не быть полностью идентичным по составу нуклеиновых кислот родительской клетке, но может содержать мутации. Настоящее описание относится также к мутантному потомству, обладающему такой же функцией или биологической активностью, которая обнаружена в результате скрининга или селекции у исходной трансформированной клетки.
В контексте настоящего описания понятие «вектор» относится к молекуле нуклеиновой кислоты, которая обладает способностью к размножению другой нуклеиновой кислоты, с которой она связана. Понятие включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он интродуцирован. Некоторые векторы обладают способностью контролировать экспрессию нуклеиновых кислот, с которыми они функционально связаны. Указанные векторы в контексте настоящего описания также обозначают как «экспрессионные векторы».
В одном из вариантов осуществления изобретения антитело, указанное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) может представлять собой фрагмент антитела. Фрагменты антител могут включать, например, Fab, Fab', Fab'-SH, F(ab')2, Fv, scFv-фрагменты, димерные антитела (диабоди) и однодоменные антитела. Обзор фрагментов антител см., у Hudson и др., Nat. Med. 9, 2003, сс. 129-134. Обзор scFv-фрагментов см., например, у Pluckthun в: The Pharmacology of Monoclonal Antibodies, под ред. Rosenburg и Moore, изд-во Springer, New York, т. 113, 1994, cc. 269-315; см. также WO 93/16185; и US №5571894 и US №5587458.
В контексте настоящего описания «димерные антитела» представляют собой фрагменты антител с двумя антигенсвязывающими центрами, которые могут быть двухвалентными или биспецифическими (см., например, ЕР 0404097; WO 1993/01161; Hudson и др., Nat. Med. 9, 2003, сс. 129-134; и Holliger и др., Proc. Natl. Acad. Sci. USA 90, 1993, сс. 6444-6448). Тримерные и тетрамерные антитела описаны также у Hudson и др., Nat. Med. 9, 2003, сс. 129-134).
В контексте настоящего описания «однодоменное антитело» представляет собой фрагмент антитела, содержащий весь вариабельный домен тяжелой цепи или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В одном из вариантов осуществления изобретения, когда антитело, указанное в настоящем описании, представляет собой однодоменное антитело, то оно может представлять собой человеческое однодоменное антитело (фирма Domantis, Inc., Уолтхэм, шт. Массачусетс; см., например, US №6248516 В1).Фрагменты антител можно создавать различными методиками, включая (но, не ограничиваясь только ими) протеолитическое расщепление интактного антитела, а также получать с помощью рекомбинантных клеток-хозяев, как указано в настоящем описании.
В контексте настоящего описания понятие «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи происходит из конкретного источника или вида, а остальная часть тяжелой и/или легкой цепи происходит из другого источника или вида.
В контексте настоящего описания понятие «гуманизированное» антитело относится к химерному антителу, которое содержит аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В одном из вариантов осуществления изобретения гуманизированное антитело состоит практически по меньшей мере из одной и, как правило, двух вариабельных областей, в которых все (или практически все) HVR соответствуют HVR (например, CDR) нечеловеческого антитела, а все (или практически все) FR соответствуют участкам человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела, происходящей из человеческого антитела.
В одном из вариантов осуществления изобретения антитело, указанное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) может представлять собой химерное антитело. Химерные антитела описаны, например, в US №4816567; и у Morrison и др., Proc. Nat1. Acad. Sci. USA 81, 1984, сс. 6851-6855. Химерное антитело может содержать нечеловеческую вариабельную область (например, вариабельную область из антитела приматов кроме человека, таких как обезьяны, или мышей, крыс, хомяков или кроликов) и человеческую константную область.
В одном из вариантов осуществления изобретения антитело, указанное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) может представлять собой гуманизированное антитело. Как правило, нечеловеческое антитело гуманизируют для снижения иммуногенности для людей, сохраняя при этом специфичность и аффинность родительского нечеловеческого антитела. Как правило, гуманизированное антитело содержит один или несколько вариабельных доменов, в которых HVR, например, CDR (или их участки), получают из нечеловеческого антитела, a FR (или их участки) получают из последовательностей человеческого антитела. Гуманизированное антитело необязательно может содержать также по меньшей мере часть человеческой константной области. В одном из вариантов осуществления изобретения некоторые аминокислотные остатки FR в гуманизированном антителе заменены на соответствующие аминокислотные остатки из нечеловеческого антитела (например, антитела, из которого происходят остатки HVR), например, для сохранения или улучшения специфичности или аффинности антитела.
Сведения о гуманизированных антителах и методах их создания обобщены, например, у Almagro и Fransson, Front. Biosci. 13, 2008, cc. 1619-1633), и описаны также у Riechmann и др., Nature 332, 1988, cc. 323-329; Queen и др., Proc. Nat1. Acad. Sci. USA 86, 1989, cc. 10029-10033; US №№5821337, 7527791, 6982321 и 7087409; Kashmiri и др., Methods 36, 2005, cc. 25-34 (описание трансплантации SDR (a-CDR)); Padlan, Mol. Immunol. 28, 1991, cc. 489-498 (описание «нанесения нового покрытия»); Dall' Acqua и др., Methods 36, 2005, cc. 43-60 (описание «перестановки FR»); и Osbourn и др., Methods 36, 2005, cc. 61-68 и Klimka и др., Br. J. Cancer 83, 2000, cc. 252-260 (описание подхода «направленной селекции» для перестановки FR).
В одном из вариантов осуществления изобретения человеческий каркасный участок, который можно применять для гуманизации, может включать, например, каркасные участки, выбранные с использованием методов «наилучшего подбора» (Sims и др., J. Immunol. 151, 1993, cc. 2296-2308); каркасные участки, происходящие из консенсусной последовательности конкретной подгруппы вариабельных областей легких и тяжелых цепей человеческих антител (Carter и др., Proc. Natl. Acad. Sci. USA 89, 1992, сс. 4285-4289 и Presta и др., J. Immunol. 151, 1993, сс. 2623-2632); и каркасные участки, полученные в результате скрининга FR-библиотек (см., например, Васа и др., J. Biol. Chem. 272, 1997, сс. 10678-10684 и Rosok и др., J. Biol. Chem. 271, 1996, сс. 22611-22618).
В одном из вариантов осуществления изобретения антитело, указанное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) может представлять собой человеческое антитело. Человеческие антитела можно получать различные методами. Человеческие антитела описаны в целом у van Dijk и van de Winkel, Curr. Op in. Pharmacol. 5, 2001, cc. 368-374 и Lonberg, Curr. Opin. Immunol. 20, 2008, cc. 450-459. Человеческие антитела можно получать путем введения антигена (например, IL- 8, CXCR1 или CXCR2) трансгенному животному, модифицированному таким образом, чтобы оно продуцировало интактные человеческие антитела или интактные антитела с человеческими вариабельными областями в ответ на антиген. Указанные животные, как правило, содержат все или часть локусов человеческого иммуноглобулина вместо эндогенных иммуноглобулиновых локусов, которые либо присутствуют вне хромосом, либо интегрированы произвольно в хромосомы животного. У таких трангенных мышей эндогенные иммуноглобулиновые локусы, как правило, инактивированы. Обзор методов получения человеческих антител в трансгенных животных см. у Lonberg, Nat. Biotech. 23, 2005, сс. 1117-1125 (см., например, также в US №6075181 и US №6150584 описание технологии XENOMOUSE™; в US №5770429 описание технологии HUMAB™; в US №7041870 описание технологии K-М MOUSE™ и в публикации заявки на патент США 2007/0061900 описание технологии VELOCIMOUSE™).
Человеческие вариабельные области из интактных антител, полученных с помощью указанных животных, можно дополнительно модифицировать, например, путем их объединения с другой человеческой константной областью.
В другом варианте осуществления изобретения, человеческие антитела можно создавать с помощью методов на основе гибридом. Ниже описаны клеточные линии человеческой миеломы и мышиной-человеческой гетеромиеломы, предназначенные для получения человеческих моноклональных антител (см., например, Kozbor, J. Immunol. 133, 1984, сс. 3001-3005; Brodeur и др., Monoclonal Antibody Production Techniques and Applications, изд-во Marcel Dekker, Inc., New York, 1987, cc. 51-63 и Boerner и др., J. Immunol. 147, 1991, cc. 86-95). Человеческие антитела, созданные с использованием технологии на основе человеческих В-клеточных гибридом, описаны у Li и др., Proc. Natl. Acad. Sci. USA 103, 2006, cc. 3557-3562. Дополнительные методы включают методы, описанные, например, в US №7189826 (описание получения моноклональных человеческих антител типа IgM из клеточных линий гибридом), и у Ni, Xiandai Mianyixue 26, 2006, cc. 265-268 (описание человеческих-человеческих гибридом). Технология человеческих гибридом (технология Trioma) описана также у Vollmers и Brandlein, Histology and Histopathology 20, 2005, cc. 927-937 и Vollmers и Brandlein, Methods and Findings in Experimental and Clinical Pharmacology 27, 2005, cc. 185-191.
В альтернативном варианте осуществления изобретения человеческие антитела можно создавать также путем выделения последовательностей вариабельных доменов Fv-клонов, выбранных из фаговых дисплейных библиотек, для создания которых использовали человеческие антитела. Указанные последовательности вариабельных областей затем можно объединять с требуемой человеческой константной областью. Методики отбора человеческих антител из библиотек антител описаны ниже.
В одном из вариантов осуществления изобретения антитело, указанное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) можно выделять путем скрининга комбинаторных библиотек антител с требуемой активностью или видами активности. Например, в данной области известны разнообразные методы для создания фаговых дисплейных библиотек и скрининга указанных библиотек в отношении антител, обладающих требуемыми характеристиками связывания. Указанные методы обобщены, например, у Hoogenboom и др., Methods in Molecular Biology 178, 2001, cc. 1-37 (под. ред.  и др., изд-во Human Press, Totowa, NJ, 2001), и описаны также, например, у McCafferty и др., Nature 348, 1990; Clackson и др., Nature 352, 1991, сс. 624-628; Marks и др., J. Mol. Biol. 222, 1992, сс. 581-597; Marks и Bradbury, Molecular Biology 248, 2003, cc. 161-175 (под ред. Lo и др., изд-во Human Press, Totowa, NJ, 2003); Sidhu и др., J. Mol. Biol. 338(2), 2004, cc. 299-310; Lee и др., J. Mol. Biol. 340(5), 2004, cc. 1073-1093; Fellouse, Proc. Natl. Acad. Sci. USA 101(34), 2004, cc. 12467-12472; и Lee и др., J. Immunol. Methods 284(1-2), 2004, cc. 119-132.
и др., изд-во Human Press, Totowa, NJ, 2001), и описаны также, например, у McCafferty и др., Nature 348, 1990; Clackson и др., Nature 352, 1991, сс. 624-628; Marks и др., J. Mol. Biol. 222, 1992, сс. 581-597; Marks и Bradbury, Molecular Biology 248, 2003, cc. 161-175 (под ред. Lo и др., изд-во Human Press, Totowa, NJ, 2003); Sidhu и др., J. Mol. Biol. 338(2), 2004, cc. 299-310; Lee и др., J. Mol. Biol. 340(5), 2004, cc. 1073-1093; Fellouse, Proc. Natl. Acad. Sci. USA 101(34), 2004, cc. 12467-12472; и Lee и др., J. Immunol. Methods 284(1-2), 2004, cc. 119-132.
Согласно одному из вариантов осуществления изобретения при осуществлении некоторых методов фагового дисплея популяции VH и VL можно клонировать по отдельности с помощью полимеразной цепной реакции (ПЦР) и рекомбинируют произвольно в фаговых библиотеках, которые затем подвергают скринингу в отношении антигенсвязывающего фага согласно методу, описанному у Winter и др., Ann. Rev. Immunol. 12, 1994, cc. 433-455. Фаги, как правило, экспонируют фрагменты антител, например scFv и Fab. Библиотеки из иммунизированных источников могут обеспечивать получение высокоаффинных антител к иммуногену без необходимости в создании гибридом. В альтернативном варианте осуществления изобретения можно клонировать также необработанную («наивную») популяцию (например, из организма человека), получая один источник антител к широкому спектру чужих, а также своих антигенов без какой-либо иммунизации, что описано у Griffiths и др., EMBO J, 12, 1993, сс. 725-734. В следующем варианте осуществления изобретения наивные библиотеки можно получать также методами синтеза путем клонирования неперегруппированных сегментов V-гена из стволовых клеток и с помощью ПЦР-праймеров, содержащих случайную последовательность, для кодирования гипервариабельных CDR3-участков и для осуществления перегруппировки in vitro согласно методу, описанному у Hoogenboom и Winter, J. Mol. Biol., 227, 1992, cc. 381-388. Фаговые библиотеки человеческих антител описаны, например, в следующих патентных публикациях: US №5750373; US 2005/0079574; US 2005/0119455; US 2005/0266000; US 2007/0117126; US 2007/0160598; US 2007/0237764; US 2007/0292936 и US 2009/0002360.
Антитела или фрагменты антител, выделенные из человеческих библиотек антител, в контексте настоящего описания рассматриваются как человеческие антитела или фрагменты человеческих антител.
В одном из вариантов осуществления изобретения антитело, представленное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) представляет собой мультиспецифическое антитело (например, биспецифическое антитело). Мультиспецифические антитела представляют собой антитела (например, моноклональные антитела), которые имеют специфичности, связывающиеся по меньшей мере с двумя различными сайтами.
В одном из вариантов осуществления изобретения одна из связывающих специфичностей обеспечивает связывание с антигеном (например, IL-8, CXCR1 или CXCR2), а другая с любым другим антигеном. В другом варианте осуществления изобретения биспецифическое антитело может связываться с различными двумя эпитопами антигена (например, IL-8, CXCR1 или CXCR2). Биспецифические антитела можно применять для локализации цитотоксических агентов в клетках, которые экспрессируют антиген (например, IL-8, CXCR1 или CXCR2). Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
Методики создания мультиспецифических антител включают (но, не ограничиваясь только ими) рекомбинантную совместную экспрессию двух пар тяжелых цепей-легких цепей иммуноглобулинов, обладающих различными специфичностями (см., например, Milstein и Cuello, Nature 305, 1983, сс. 537-540, WO 93/08829 и Traunecker и др., EMBO J. 10, 1991, сс.3655-3659), и технологию «knob-in-hole» (обеспечение взаимодействия по типу «выступ-во впадину», см., например, US №5731168). Мультиспецифические антитела можно получать также путем создания регулируемых электростатических воздействий для получения молекул антитела с гетеродимерными Fc (см., например, WO 2009/089004); перекрестного связывания двух или большего количества антител или фрагментов (см., например, US №4676980 и Brennan и др., Science 229, 1985, сс. 81-83); применения лейциновых молний для получения биспецифических антител (см., например, Kostelny и др., J. Immunol. 148, 1992, сс. 1547-1553); применения технологии «димерных антител (диабоди)» для создания фрагментов биспецифических антител (см., например, Hollinger и др., Proc. Natl. Acad. Sci. USA 90, 1993, сс. 6444-6448); и применения димеров scFv (см., например, Gruber и др., J. Immunol. 152, 1994, сс. 5368-5374); и получения триспецифических антител согласно методу, описанному, например, у Tutt и др., J. Immunol. 147, 1991, сс. 60-69).
Сконструированные антитела с тремя или большим количеством функциональных антигенсвязывающих сайтов, включая «антитела-осьминоги» (см., например, US 2006/0025576), также включены в контекст настоящего описания.
Согласно одному из вариантов осуществления изобретения антитело, представленное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) или фрагмент антитела может представлять собой «Fab двойного действия» или «DAF», содержащий антигенсвязывающий сайт, который связывается с антиген (например, IL-8, CXCR1 или CXCR2), а также с другим, отличным от него антигеном (см, например, US 2008/0069820).
Согласно одному из вариантов осуществления изобретения можно получать варианты аминокислотной последовательности (мутанты) антитела, представленного в настоящем описании (например, антитела к IL-8, антитела к CXCR1 или антитела к CXCR2) путем интродукции соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем синтеза пептидов. Для достижения указанных модификаций можно применять интродукцию одной или нескольких произвольных делеций, инсерций, замен произвольных аминокислот (остатков) или их комбинации в аминокислотные последовательности. Можно применять любую комбинацию делеций, инсерций и замен, при условии, что конечная конструкция обладает требуемыми характеристиками (например, связыванием антигена).
Согласно одному из вариантов осуществления изобретения, если варианты антител (мутанты) имеют одну или несколько аминокислотных замен, то сайты, представляющие интерес для замещающего мутагенеза, могут включать HVR и FR.
Консервативные замены, являющиеся одним из вариантов осуществления изобретения, представлены в таблице 1 под заголовком «предпочтительные замены», а более значимые замены представлены в таблице 1 под заголовком «приведенные в качестве примера замены», и они описаны ниже со ссылкой на классы боковых цепей аминокислот.
Аминокислотные замены можно интродуцировать в представляющее интерес антитело и продукты можно подвергать скринингу в отношении требуемой активности, например, сохранения/улучшения связывания антигена, пониженной иммуногенности или улучшенной ADCC или CDC.

Аминокислоты можно группировать на основе общих свойств боковых цепей следующим образом: (1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile; (2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln; (3) кислотные: Asp, Glu; (4) основные: His, Lys, Arg; (5) остатки, которые влияют на ориентацию цепи: Gly, Pro; (6) ароматические: Trp, Tyr, Phe.
Под неконсервативными заменами подразумевают замену представителя одного из указанных классов на представителя из другого класса.
Согласно одному из вариантов осуществления изобретения аминокислотные инсерций включают амино- и/или карбоксиконцевые слияния, варьирующие по длине от одного остатка до полипептидов, содержащих сто или большее количество остатков, а также инсерций одного или нескольких аминокислотных остатков внутри последовательности. Примером результата осуществления концевых инсерций является антитело с N-концевым метионильным остатком. Другие примеры инсерционных вариантов (мутантов) включают слияние N- или С-концов антитела с ферментом (например, для ADEPT (антитело-направляемый фермент пролекарственной терапии) или полипептидом, который удлиняет время полужизни антитела в сыворотке.
В контексте настоящего описания «процент (%) идентичности аминокислотной последовательности» относительно полипептидной референс-последовательности определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в полипептидной референс-последовательности, после выравнивая последовательностей и при необходимости интродукции брешей для достижения максимального процента идентичности последовательностей, и не рассматривая какие-либо консервативные замены в качестве компонента при оценке идентичности последовательностей. Сравнительный анализ для целей определения процента идентичности аминокислотных последовательностей можно осуществлять различными путями, известными в данной области, используя, например, публично доступные компьютерные программы, такие как программа BLAST, BLAST-2, ALIGN, Megalign (DNASTAR) или GENETYX®. Специалисты в данной области могут определять соответствующие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
Авторство компьютерной программы для сравнения последовательностей ALIGN-2 принадлежит фирме Genentech, Inc., и исходный код был представлен в комплекте с документацией для пользователей в U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован как U.S. Copyright Registration № TXU510087. Программа ALIGN-2 публично доступна от фирмы Genentech, Inc., Южный Сан-Франциско, шт. Калифорния, или ее можно компилировать на основе исходного кода. Программу ALIGN-2 можно компилировать для применения в операционной системе UNIX, включая цифровую систему UNIX V4.0D. Все параметры, требуемые для сравнения последовательностей, устанавливаются программой ALIGN-2 и не должны изменяться.
В ситуациях, в которых ALIGN-2 применяют для сравнения аминокислотных последовательностей, % идентичности аминокислотных последовательностей данной аминокислотной последовательности А по сравнению, с или относительно данной аминокислотной последовательности Б (что в альтернативном варианте можно обозначать как данная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности по сравнению, с или относительно данной аминокислотной последовательности Б), рассчитывают следующим образом: 100 × дробь X/Y, где X обозначает количество аминокислотных остатков, определенных как идентичные совпадения при оценке с помощью программы для сравнительного анализа последовательностей ALIGN-2, где с помощью программы осуществляли сравнение А и Б, и где Y обозначает общее количество аминокислотных остатков в Б. Должно быть очевидно, что, если длина аминокислотной последовательности А не равна длине аминокислотной последовательности Б, то % идентичности аминокислотных последовательностей А относительно Б не может быть равен % идентичности аминокислотной последовательности Б относительно А. Если специально не указано иное, то все величины % идентичности аминокислотных последовательностей, указанные в настоящем описании, получали с использованием описанной в предыдущем параграфе компьютерной программы ALIGN-2.
В одном из вариантов осуществления изобретения аминокислотную последовательность, имеющую соответствующую замену, делецию или инсерцию одной или нескольких аминокислот в аминокислотной последовательности CDR-участка, вариабельной области тяжелой цепи, вариабельной области легкой цепи, константной области тяжелой цепи, константной области легкой цепи, полной тяжелой цепи, полной легкой цепи или любой конкретной области антитела, представленного в настоящем описании (например, антитела к IL-8, антитела к CXCR1, антитела к CXCR2), и обладающего связывающей активностью с антигеном (например, IL-8, CXCR1 или CXCR2), можно получать путем гибридизации нуклеиновых кислот, которые гибридизуются в строгих условиях с нуклеиновыми кислотами, которые содержат нуклеотидную последовательность, кодирующую аминокислотную последовательность указанной области. Примеры строгих условий гибридизации для выделений нуклеиновой кислоты, которая гибридизуется в строгих условиях, включают такие условия, как 6М мочевина, 0,4% ДСН, 0,1 × SSC и 42°С, или условия гибридизации, эквивалентные им по строгости. Выделение нуклеиновых кислот с более высокой гомологией можно ожидать при использовании более строгих условий, например, таких как 6М мочевина, 0,4% ДСН, 0,1 × SSC и 42°С. Условия промывки после гибридизации представляют собой, например, промывку с использованием 0,5 × SSC (где 1 × SSC включает, например, 0,15М NaCl и 0,015М цитрат натрия, рН 7,0) и 0,1% ДСН при 60°С, более предпочтительно 0,2 × SSC и 0,1% ДСН при 60°С, более предпочтительно 0,2 × SSC и 0,1% ДСН при 62°С, еще более предпочтительно 0,2 × SSC и 0,1% ДСН при 65°С и еще более предпочтительно 0,1 × SSC и 0,1% ДСН при 65°С. Продолжительность промывки и количество промывок можно соответствующим образом регулировать, например, 20-минутную промывку можно осуществлять три раза. Выделенные нуклеиновые кислоты можно секвенировать с помощью известных методов.
В альтернативном варианте осуществления изобретения вместо вышеописанных методов гибридизации можно применять также методы амплификации генов, например, ПЦР с использованием праймеров, синтезированных на основе информации о нуклеотидной последовательности, которая кодирует аминокислотную последовательность CDR-участка, вариабельной области тяжелой цепи, вариабельной области легкой цепи, константной области тяжелой цепи, константной области легкой цепи, полной области тяжелой цепи, полной области легкой цепи или любой ее конкретной области, для выделения нуклеиновой кислоты, которая гибридизуется в строгих условиях с нуклеиновой кислотой, которая содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность указанной области.
Выделенная таким образом нуклеиновая кислота имеет по меньшей мере 50% или более, предпочтительно 70% или более, 75% или более, 80% или более, 85% или более и более предпочтительно 90% или более (например, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% или более) идентичность всей последовательности с нуклеотидной последовательностью представляющей интерес области. Антитело, имеющее аминокислотную последовательность, кодируемую указанной выделенной нуклеиновой кислотой, является предпочтительным, если его функциональность (практически) эквивалентна (например, (практически) эквивалентными являются один или несколько показателей или характеристик, такие как связывание антигена или сохранение в крови) функциональности антитела, которое имеет аминокислотную последовательность представляющей интерес области; однако не ограничено указанным. В контексте настоящего описания подразумевается, что понятие «практически» означает сохранение функции предпочтительно на 50% или более, предпочтительно 70% или более, 75% или более, 80% или более 85% или более и более предпочтительно 90% или более (например, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% или более) (например, можно рассматривать любой один или несколько показателей или характеристик, таких как связывание антигена и сохранение в крови), по сравнению с функцией, которой обладает антитело, имеющее аминокислотную последовательность представляющей интерес области, при оценке методами, известными специалистам в данной области.
В одном из вариантов осуществления изобретения антитело, представленное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2), может быть гликозилированным. Добавление или делецию сайтов гликозилирования в антителе можно легко осуществлять путем такого изменения аминокислотной последовательности, которое позволяет создавать или удалять сайты гликозилирования.
В одном из вариантов осуществления изобретения, когда антитело, представленное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2), содержит Fc-область, то сахарные цепи, которые можно присоединять к Fc-домену, могут варьироваться. Нативные антитела, которые продуцируются клетками животных, как правило, содержат разветвленный биантенный олигосахарид, который, как правило, присоединен с помощью N-связи к Asn297 СН2-домена Fc-области (Wright и др., TIBTECH 15, 1997, сс. 26-32). Олигосахарид может включать, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стебле» биантенной олигосахаридной структуры. В одном из вариантов осуществления изобретения олигосахарид в антителе, представленном в настоящем описании, можно модифицировать для создания вариантов антитела с определенными улучшенными свойствами.
В контексте настоящего описания понятие «эффекторные функции» относится к тем видам биологической активности, которые связаны с Fc-областью антитела (например, антитела к IL-8, антитела к CXCR1 или антитела к CXCR2), которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают (но, не ограничиваясь только ими) связывание C1q и комплементзависимую цитотоксичность (CDC); связывание Fc-рецептора; антитело-обусловленную клеточнозависимую цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора) и активацию В-клеток.
Понятие «Fc-область» в контексте настоящего описания применяют для определения С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Понятие включает нативные Fc-области и варианты Fc-области. Нативная Fc-область относится к Fc-области нативного антитела. Примеры Fc-области нативного человеческого IgG, включают, например, Fc-область, входящую в константную область человеческого IgG1 (SEQ ID NO: 100), константную область человеческого IgG2 (SEQ ID NO: 101), константную область человеческого IgG3 (SEQ ID NO: 102) константную область человеческого IgG4 (SEQ ID NO: 103), описанные выше. В одном из вариантов осуществления изобретения Fc-область тяжелой цепи человеческого IgG простирается с Cys226 или с Pro230 до С-конца тяжелой цепи. Однако С-концевой лизин (Lys447) Fc-области или С-концевой глицин (Gly446) и лизин (Lys447) Fc-домена могут присутствовать или отсутствовать. Если специально не указано иное, то в контексте настоящего описания нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, которую обозначают также как EU-индекс, описанной у Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
В отличие от FcγR, который принадлежит к суперсемейству иммуноглобулинов, «FcRn», указанный в настоящем описании, в частности человеческий FcRn, обладает структурным сходством с полипептидами молекул главного комплекса гистосовместимости (ГКГС) класса I и его последовательность идентична на 22-29% последовательности молекул ГКГС класса I (Ghetie и др., Immunol. Today 18(12), 1997, сс. 592-598). FcRn экспрессируется в виде гетеродимера, состоящего из растворимой (β- или легкой цепи (β2-микроглобулин), образующей комплекс с трансмембранной α - цепью или тяжелой цепью. Аналогично ГКГС α-цепь FcRn содержит три внеклеточных домена (α1, α2 и α3) и короткий цитоплазматический домен, связывающий белки с клеточной поверхностью, α1- и α2-домены взаимодействуют с FcRn-связывающим доменом Fc-области антитела (Raghavan и др., Immunity 1, 1994, cc. 303-315). FcRn экспрессируется в материнской плаценте и желточной мешке млекопитающих и участвует в переносе IgG от матери к плоду. Кроме того, в тонком кишечнике новорожденных грызунов при экспрессии FcRn он участвует в переносе материнских IgG через щеточную каемку эпителия из потребленного молозива или молока. FcRn экспрессируется в широком разнообразии других тканей и эндотелиальных клеточных систем различных видов. У взрослых людей FcRn экспрессируется также в сосудистом эндотелии, мышечной сосудистой системе и синусоидальных капиллярах печени. FcRn, вероятно, играет роль в поддержании концентрации IgG в плазме путем связывания с IgG и рециклинга IgG в сыворотке. Как правило, связывание FcRn с молекулами IgG строго зависит от рН. Оптимальное связывание обнаружено в кислом диапазоне рН ниже 7,0. Полинуклеотидные и аминокислотные последовательности человеческого FcRn можно получать, например, из предшественников, которые представлены в NM_004107.4 и NP_004098.1 (содержащих сигнальную последовательность) соответственно (в скобках представлены коды доступа в RefSeq). Предшественники образуют комплексы с человеческим β2-микроглубулином in vivo. Таким образом, с использованием известных методов рекомбинантной экспрессии можно получать растворимый человеческий FcRn, обладающий способностью образовывать комплекс с человеческим β2-микроглубулином, для соответствующего применения в различных экспериментальных системах. Указанный растворимый человеческий FcRn можно применять для оценки антител или вариантов Fc-области в отношении их FcRn-связывающей активности. FcRn не ограничен конкретным FcRn, если он находится в форме, в которой может связываться с FcRn-связывающим доменом и является человеческий FcRn.
В одном из вариантов осуществления изобретения, если антитело, представленное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2), или вариант их Fc-области, обладают FcRn-связывающей активностью, то антитело или вариант их Fc-области предпочтительно имеют «FcRn-связывающий домен», более предпочтительно человеческий FcRn-связывающий домен. FcRn-связывающий домен не ограничен конкретным доменом, если антитело (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) обладает связывающей активностью или аффинностью к FcRn при кислом рН и/или нейтральном рН. Альтернативно этому, FcRn-связывающий домен может обладать способностью прямо или косвенно связываться с FcRn. Такие домены включают, например (но, не ограничиваясь только ими), Fc-области иммуноглобулинов IgG-типа, альбумина, домена 3 альбумина, антител к FcRn, анти-FcRn пептидов и анти-FcRn скаффолд-молекул, которые обладают способностью непосредственно связываться с FcRn, и молекулы, которые связываются с IgG или альбумином, которые обладают способностью опосредованно связываться с FcRn. Альтернативно этому, можно также применять, например, домены, которые обладают FcRn-связывающей активностью при кислом рН и/или нейтральном рН. Если домен исходно обладает FcRn-связывающей активностью при кислом рН и/или нейтральном рН, то их можно применять без дополнительной модификации. Если домен обладает лишь слабой или не обладают вообще FcRn-связывающей активностью при кислом рН и/или нейтральном рН, то аминокислотные остатки в FcRn-связывающем домене антитела или варианте Fc-области можно модифицировать так, чтобы они приобретали FcRn-связывающую активность при кислом рН и/или нейтральном рН. Альтернативно этому, аминокислоты домена, который исходно обладает FcRn-связывающей активностью при кислом рН и/или нейтральном рН, можно модифицировать для дополнительного повышения их FcRn-связывающей активности. Для выявления представляющих интерес аминокислотных модификаций для FcRn-связывающего домена можно сравнивать FcRn-связывающую активность при кислом рН и/или нейтральном рН до и после осуществления аминокислотных модификаций.
FcRn-связывающий домен предпочтительно представляет собой область, которая непосредственно связывается с FcRn. Предпочтительные примеры FcRn-связывающего домена включают константные области и Fc-области антител. Однако области, способные связываться с полипептидом, обладающим FcRn-связывающей активностью, таким как альбумин или IgG, могут связываться с FcRn косвенным образом посредством альбумина или IgG. Таким образом, FcRn-связывающие области могут представлять собой области, которые связываются с полипептидом, обладающим связывающей альбумин или IgG активностью. Известно (но это ограничивает объем изобретения), что для повышения элиминации антигена из плазмы FcRn-связывающая активность FcRn-связывающего домена предпочтительно должна быть более высокой при нейтральном рН, в то время как для повышения удерживания антитела в плазме FcRn-связывающая активность FcRn-связывающего домена предпочтительно должна быть более высокой при кислом рН. Например, можно отбирать и применять FcRn-связывающий домен, исходная FcRn-связывающая активность которого является более высокой при нейтральном рН или при кислом рН. Альтернативно этому можно модифицировать аминокислоты антитела или Fc-области для придания FcRn-связывающей активности при нейтральном рН или при кислом рН. Альтернативно этому можно повышать уже существующую FcRn-связывающую активность при нейтральном рН или при кислом рН.
Для решения вопроса о том, повышается ли, (практически) сохраняется или снижается FcRn-связывающая активность антитела (например, антитела к IL-8, антитела к CXCR1 или антитела к CXCR2) или Fc-области (варианта)) по сравнению с ними до модификации, можно применять любые методы, известные специалистам в данной области (но, не ограничиваясь только ими), а также можно применять методы, описанные в разделе «Примеры» настоящего описания. Например, можно применять BIACORE, график Скэтчарда, проточную цитометрию и т.п. (WO 2013/046722). Внеклеточный домен человеческого FcRn можно применять в качестве растворимого антигена в этих анализах. Специалист в данной области легко может выбирать условия помимо рН для измерения FcRn-связывающей активности антитела или Fc-области (варианта) (но, не ограничиваясь только ими). Такие анализы можно осуществлять, например, в условиях MES-буфера и 37°С, как описано в WO 2009/125825. FcRn-связывающую активность антитела или Fc-области (варианта) можно оценивать, например, путем пропускания FcRn в качестве аналита по поверхности чипа с иммобилизованным на нем антителом.
Кроме того, FcRn-связывающую активность антитела или Fc-области (варианта) можно оценивать на основе константы диссоциации (KD), кажущейся константы диссоциации (кажущаяся KD), константы скорости реакции диссоциации (kd), кажущейся константы скорости реакции диссоциации (кажущаяся kd).
В качестве условий рН для измерения связывающей активности между FcRn и FcRn-связывающим доменом, входящим в антитело (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2) или Fc-область (вариант), можно использовать кислые условия рН или нейтральные условия рН. В качестве температурных условий для измерения FcRn-связывающей активности (аффинность связывания) FcRn-связывающего домена можно использовать любую температуру от 10°С до 50°С. Предпочтительно для определения FcRn-связывающей активности (аффинность связывания) человеческого FcRn-связывающего домена можно использовать температуру от 15°С до 40°С. Более предпочтительно температура, используемая для измерения FcRn-связывающей активности (аффинность связывания) FcRn-связывающего домена может представлять собой любую температуру от 20°С до 35°С, такую как любая из следующих температур: 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35°С. В качестве примера указанной температуры можно использовать (но, не ограничиваясь только ею) 25°С.
В одном из вариантов осуществления изобретения аминокислоты, входящие в аминокислотную последовательность антитела, представленного в настоящем описании (например, антитела к IL-8, антитела к CXCR1 или антитела к CXCR2), можно подвергать пост-трансляционным модификациям (например, модификация N-концевого глутамина в пироглутамат путем пироглутамилирования хорошо известна специалистам в данной области). Очевидно, что указанные аминокислотные последовательности с подвергнутыми пост-трансляционной модификации аминокислотами включены также в качестве эквивалентов аминокислотных последовательностей, указанных в настоящем описании.
В одном из вариантов осуществления изобретения антитело, представленное в настоящем описании (например, антитело к IL-8, антитело к CXCR1 или антитело к CXCR2), можно дериватизировать таким образом, чтобы оно содержало дополнительные небелковые фрагменты. Фрагменты, пригодные для дериватизации антител, включают, например, водорастворимые полимеры.
Примерами водорастворимых полимеров являются полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (либо гомополимеры, либо статистические сополимеры), декстран- или поли(н-винилпиролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры пропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликольпропиональдегид может иметь преимущество при производстве благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество полимеров, присоединенных к антителу, может варьироваться и, если присоединено более одного полимера, то они могут представлять собой одинаковые или различные молекулы. В целом, количество и/или тип полимеров, применяемых для дериватизации, можно определять с учетом таких особенностей (но, не ограничиваясь только ими), как конкретные свойства или функции антитела, подлежащие усовершенствованию, предполагается ли применение производного антитела в терапии в определенных условиях, и т.д.
В качестве примера указанных фрагментов полиэтиленгликольпропиональдегид может иметь преимущество при производстве благодаря его стабильности в воде. Полимеры могут иметь любую молекулярную массу и могут быть разветвленными или неразветвленными. Количество полимеров, присоединенных к антителу, может варьироваться и, если присоединен один или несколько полимеров, то они могут представлять собой одинаковые или различные молекулы. В целом, количество и/или тип полимеров, применяемых для дериватизации, можно определять с учетом, например, таких особенностей как конкретные свойства или функции антитела, подлежащие усовершенствованию, предполагается ли применение производного антитела в терапии в определенных условиях, и т.д.
Одним из вариантов осуществления изобретения являются конъюгаты антитела, представленного в настоящем описании (например, антитела к IL-8, антитела к CXCR1 или антитела к CXCR2), и небелкового фрагмента, которые можно избирательно нагревать, воздействуя на них излучением. Небелковый фрагмент может представлять собой, например, углеродную нанотрубку (Kam и др., Рrос. Natl. Acad. Sci. USA 102, 2005, cc. 11600-11605). Излучение может иметь любую длину волны, если оно позволяет достигать требуемой цели, и может иметь длины волн, которые не являются опасными для обычных клеток, но которые нагревают небелковый фрагмент для температуры, при которой уничтожаются клетки, ближайшие к конъюгату: антитело-небелковый фрагмент.
Понятие «IL-8» в контексте настоящего описания относится к любому нативному IL-8, полученному из любых позвоночных животных, приматов (например, человека, обезьян циномолгус, макак резус) и других млекопитающих (например, собак и кроликов; следует отметить, что у мышей и крыс отсутствует эндогенный IL-8), если специально не указано иное.
IL-8 включает интактный IL-8, любую модифицированную in vivo форму IL-8 и встречающиеся в естественных условиях производные IL-8, такие как сплайсинговые варианты и аллельные варианты. Типичный пример аминокислотной последовательности человеческого IL-8 представлен в SEQ ID NO: 22. Кроме того, примером аминокислотной последовательности человеческого IL-8 является (но, не ограничиваясь только ею) последовательность предшественника человеческого интерлейкина-8, зарегистрированная как NP_000575.1 в NCBI. Кроме того, примером аминокислотной последовательности IL-8 обезьяны циномолгус является (но, не ограничиваясь только ею) последовательность, зарегистрированная как ХР_005555144.1 в NCBI.
CXCR1 обозначают также как альфа-рецептор интерлейкина 8, IL8RA или CD181, a CXCR2 обозначают также как бета-рецептор интерлейкина 8 или IL8RB, они оба представляют собой хемокиновые рецепторы. Известно, что IL-8 проявляет свои физиологические действия посредством связывания с его рецепторами CXCR1 и CXCR2 (Science 253, 1991, cc. 1278-1280; Science 253, 1991, cc. 1280-1283). Установлено, что CXCR1 активируется только IL-8 и гранулоцитарным хемотаксическим белком-2, a CXCR2 активируется посредством связывания с IL-8 и различными молекулами, такими как GROα, β и γ, нейтрофилы-активирующим пептидом и гранулоцитарным хемотаксическим белком-2 (Neuro-oncol. 7, 2005, cc. 122-133). К настоящему времени не описаны никакие рецепторы IL-8 кроме CXCR1 и CXCR2. Как будет описано в разделе «Примеры», при создании изобретения продемонстрировано, что способность нейтрофилов мигрировать по направлению к IL-8 блокируется нейтрализующим IL-8 антителом, а также ингибитором CXCR2, поэтому можно легко прогнозировать, что не только антитела к IL-8, но также и ингибиторы сигнала IL-8, включая ингибиторы CXCR1 и ингибиторы CXCR2, могут оказывать благоприятное действие при эндометриозе, спайках и аденомиозе, что указано в разделе «Примеры» настоящего описания.
Понятие «CXCR1» в контексте настоящего описания относится, если специально не указано иное, к нативному CXCR1, который присутствует у любых позвоночных животных, приматов (например, у человека, обезьян циномолгус, макак резус) и других млекопитающих (например, кроликов и собак). CXCR1 включает интактный CXCR1, любую модифицированную in vivo форму CXCR1 и встречающиеся в естественных условиях производные CXCR1, такие как сплайсинговые варианты и аллельные варианты. Примером аминокислотной последовательности человеческого CXCR1 является (но, не ограничиваясь только ею) последовательность, зарегистрированная как NP_000625.1 в NCBI.
Понятие «CXCR2» в контексте настоящего описания относится, если специально не указано иное, к нативному CXCR2, который присутствует у любых позвоночных животных, приматов (например, у человека, обезьян циномолгус, макак резус) и других млекопитающих (например, кроликов и собак). CXCR2 включает интактный CXCR2, любую модифицированную in vivo форму CXCR2 и встречающиеся в естественных условиях производные CXCR2, такие как сплайсинговые варианты и аллельные варианты. Примерами аминокислотных последовательностей человеческого CXCR2 являются (но, не ограничиваясь только ими) последовательности, зарегистрированные как NP_001161770.1 и NP_001548.1 в NCBI.
IL-8 продуцируется иммунными клетками, такими как моноциты, макрофаги, нейтрофилы и лимфоциты, кожными фибробластами, кератиноцитами, сосудистыми эндотелиальными клетками, меланоцитами, гепатоцитами и различными линиями опухолевых клеток. IL-8 является эффективными хемокином для нейтрофилов и участвует в миграции нейтрофилов к областям воспаления. Установлено, что при связывании IL-8 с его высокоаффинными рецепторами (CXCR1 и CXCR2) на поверхности нейтрофилов, он активирует нейтрофилы путем усиления дегрануляции и повышения концентрации цитоплазматического свободного Са2+ и индуцирует их миграцию, тем самым разрушая инфильтрованные ткани (WO 2004/058797).
Как указано в разделе «Примеры» настоящего описания, при создании изобретения установлено, что CXCR1 и CXCR2 экспрессировались в иммунных клетках, таких как нейтрофилы, мононуклеарные клеток и макрофаги, эндометриоидные эпителиальные клетки и сосудистые эндотелиальные клетки, но не экспрессировались в стромальных клетках и мышечных фибробластах. Кроме того, добавление IL-8 или антитела к IL-8 к эндометриоидным стромальным клеткам и мышечным фибробластам, которые получали из организма страдающих эндометриозом женщин и культивировали, не приводило к изменениям, такими как пролиферация или атрофия (данные не представлены).
Эти результаты не согласуются с данными, полученными Ulukus с соавторами (Human Reproduction 20(3), 2005, сс. 794-780), о том, что для CXCR1 характерен высокой уровень экспрессии в эпителии, строме и областях фиброза эндометриоидных тканей. В статье Ulukus общая интенсивность окрашивания была высокой. CXCR2 также окрашивался, хотя окрашивание было слабо позитивным, на стромальных клетках, а также в сосудистых эндотелиальных клетках. Кроме того, вышеуказанные результаты не согласовались с данными, полученными Iwabe и др., Fertility and Sterility 69(5), 1998, сс. 924-930, которые в экспериментах in vitro продемонстрировали, что пролиферация стромальных клеток, полученных из страдающих эндометриозом пациентов, повышалась под действием IL-8, но подавлялась добавлением антитела к IL-8. В экспериментах, описанных в статье Iwabe, из эндометриоидных повреждений, полученных из организма страдающих эндометриозом женщин, удаляли эпителиальные клетки для получения стромальных клеток и затем добавляли IL-8 для оценки клеточной пролиферации. Ответ был слабым и составлял примерно 20%.
Однако, как указано в разделе «Примеры» настоящего описания, при создании изобретения на модели эндометриоза приматов кроме человека продемонстрировано, что в пролиферирующих эпителиальных и стромальных клетках происходили атрофические изменения после введения ингибитора сигнала IL-8, такого как антитело H1009/L395-F1974m.
Не вдаваясь в конкретную теорию, одним из предположений является то, что в экспериментах, описанных у Iwabe, могли одновременно присутствовать иммунные клетки, поскольку их проводили вскоре после пересева клеток. Поскольку CXCR1 и CXCR2 не экспрессируются в стромальных клетках, считается, что опосредуемый CXCR1 или CXCR2 сигнал IL-8 не действует непосредственно на стромальные клетки в качестве его мишени, а действует на эндометриоидный эпителий и иммунные клетки в качестве непосредственных мишеней и опосредованно действует на стромальные клетки через сигнал из эндометриоидного эпителия и иммунных клеток.
Понятие «ингибитор сигнала IL-8» в контексте настоящего описания не ограничен конкретным ингибитором, если он может ингибировать сигнал IL-8 непосредственно или косвенно и частично или полностью. Например, ингибитор сигнала IL-8 может представлять собой нуклеиновую кислоту (например, двухцепочечную нуклеиновую кислоту, такую как siPHK), белок (включая антитела и фрагменты антител), пептид или другое соединение (например, низкомолекулярное соединение). В одном из объектов изобретения ингибитор сигнала IL-8 может представлять собой ингибитор IL-8, ингибитор CXCR1 или ингибитор CXCR2, и в этом случае они включают, например, нуклеиновые кислоты (например, двухцепочечные нуклеиновые кислоты, такие как siPHK), белки (включая антитела и фрагменты антител, более конкретно антитела к IL-8, антитела к CXCR1, антитела к CXCR2 и фрагменты антител, которые полностью или частично сохраняют свои функции), пептиды или другие соединения (например, низкомолекулярные соединения).
Если ингибитор сигнала IL-8 полностью или частично ингибирует связывание IL-8 с его рецептором CXCR1 и/или CXCR2, то ингибирование IL-8 предпочтительно снижает или изменяет нормальный уровень или тип активности, который имеет место при связывании IL-8 с его рецептором без ингибирования. Указанное снижение или изменение может проявляться, например, как ингибирование индуцированного IL-8 высвобождения эластазы или потока кальция, ингибирование индуцированного IL-8 повышения экспрессии CD11b (Мас-1) или ингибирование уменьшения экспрессии L-селектина. Указанное ингибирование включает снижение или изменение вышеуказанного уровня или типа активности, например, по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99% или 100% по сравнению с отсутствием ингибитора сигнала IL-8.
В одном из вариантов осуществления изобретения ингибитор CXCR1, представленный в настоящем описании, может связываться (обладать специфичностью) с CXCR1, но не с CXCR2, или может связываться (обладать специфичностью) и с CXCR1, и с CXCR2 (который обозначают также как CXCR1/ CXCR2-ингибитор).
В одном из вариантов осуществления изобретения ингибитор CXCR2, представленный в настоящем описании, может связываться (обладать специфичность) с CXCR2, но не с CXCR1, или может связываться (обладать специфичность) и с CXCR1, и с CXCR2 (который обозначают также как CXCR1/ CXCR2-ингибитор).
В одном из вариантов осуществления изобретения конкретные примеры ингибитора CXCR1 и/или ингибитора CXCR2, указанные в настоящем описании, включают, например, антитела к CXCR1, репартаксин и производные репартаксина, описанные в WO 2010/056753; ингибиторы CXCR1 и/или CXCR2, описанные в WO 2011/042466; антагонисты хемокиновых рецепторов CXCR1 и CXCR2, описанные в WO 2005/113534 и WO 2003/057676; CXCR2-связывающие полипептиды, описанные в WO 2012/062713 и WO 2013/168108; ингибиторы CXCR2, описанные в WO 2008/061740, WO 2008/061741, WO 2008/062026, WO 2009/106539, WO 2010/015613 и WO 2013/030803.
В одном из вариантов осуществления изобретения конкретные примеры ингибитора IL-8, указанные в настоящем описании, включают, например, миметики IL-8, описанные в JP 2006-117633А; ингибиторы экспрессии IL-8, описанные в JP 2013-180992 A (или JP 5496232B); ингибиторы производства IL-8, описанные в JP H9-2954A (или JP3008010B); комплексы фрагмент антитела-полимер, описанные в WO 1998/037200; антитела к IL-8, мультиспецифические антитела, которые связываются с антигенами, включая IL-8 и его варианты, описанные в WO 1995/023865, WO 2009/026117, WO 2013/166099, WO 2006/113643, WO 2004/058797, WO 2008/130969, WO 2011/100271, WO 98/58671, WO 2014/149733, US 2003/0077283A1 и т.д. В другом варианте осуществления изобретения указанные варианты антител к IL-8 могут дополнительно обладать одним или несколькими свойствами, указанными ниже в контексте настоящего описания для высокофункциональных антител к IL-8.
В одном из вариантов осуществления изобретения понятие «антитело к IL-8» или «антитело, которое связывается с IL-8» в контексте настоящего описания относится к антителу, которое обладает способностью связываться с IL-8 с достаточной аффинностью, что позволяет применять его в качестве композиции, предназначенной для обнаружения, диагностирования, терапии и/или предупреждения путем таргетинга IL-8.
В одном из вариантов осуществления изобретения уровень неспецифического связывания антитела к IL-8 с нерелевантными не представляющими собой IL-8 белками составляет менее чем примерно 10% от связывания антитела с IL-8.
В одном из вариантов осуществления изобретения антитело к IL-8, представленное в настоящем описании, обладает нейтрализующей IL-8 активностью. IL-8-нейтрализующая активность может представлять собой активность ингибирования биологической активности, характерной для IL-8, или может представлять собой активность ингибирования связывания IL-8 с его рецептором (CXCR1 или CXCR2).
В одном из вариантов осуществления изобретения антитело к IL-8, представленное в настоящем описании, может представлять собой химерное антитело, гуманизированное антитело или человеческое антитело.
В одном из вариантов осуществления изобретения антитело к IL-8, представленное в настоящем описании, предпочтительно обладает перекрестной реактивностью с эндогенным IL-8 животных (например, кроликов) или в некоторых случаях предпочтительно представляет собой антитело к IL-8 приматов (предпочтительно антитело против IL-8 примата кроме человека с менструацией, сходной с человеческой менструацией, таких как обезьяны циномолгус или бабуины, или антитело к человеческому IL-8 (например, античеловеческое IL-8-нейтрализующее антитело)).
Как указано в приведенных в настоящем описании примерах, при создании настоящего изобретения разработана (но, не ограничиваясь только ею) технология аминокислотных модификаций, которая придает высокую функциональность антителу (например, антителу к IL-8, антителу к CXCR1 или антителу к CXCR2), и на основе указанного открытия созданы обладающие высокой функциональностью антитела к IL-8. Не ограничивающие объем изобретения примеры указанных антител описаны ниже.
В одном из вариантов осуществления изобретения антитело к IL-8, указанное в настоящем описании, может представлять собой антитело к IL-8, которое может обладать зависящей от рН аффинностью к IL-8. Указанное антитело к IL-8 содержит, например, последовательности, содержащие замену по меньшей мере одной, двух, трех, четырех, пяти, шести, семи, восьми или большего количества аминокислот в следующих аминокислотных последовательностях:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28,
Если конкретно не указано иное, то аминокислоты можно заменять на любые другие аминокислоты. В следующем варианте осуществления изобретения антитело к IL-8, указанное в настоящем описании, может дополнительно по меньшей мере содержать замену на другую аминокислоту или на аминокислоту, указанную ниже в положении(ях), выбранном(ых) из группы, которая состоит из указанных ниже положений:
(1) серии в положении 8 в последовательности SEQ ID NO: 26; аспарагин в положении 1 и лейцин в положении 5 в последовательности SEQ ID NO: 27; и глутамин в положении 1 в последовательности SEQ ID NO: 28;
(2) замену аланина в положении 6 на аспарагиновую кислоту и замену аргинина в положении 11 на пролин в последовательности SEQ ID NO: 24; и замену тирозина в положении 3 на гистидин в последовательности SEQ ID NO: 25;
(3) замену глицина в положении 8 на тирозин и замену тирозина в положении 9 на гистидин в последовательности SEQ ID NO: 24;
(4) замену аланина в положении 6 на аспарагиновую кислоту, замену глицина в положении 8 на тирозин, замену тирозина в положении 9 на гистидин и замену аргинина в положении 11 на пролин в последовательности SEQ ID NO: 24; и замену тирозина в положении 3 на гистидин в последовательность SEQ ID NO: 25;
(5) замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в последовательности SEQ ID NO: 28;
(6) замену серина в положении 8 на глутаминовую кислоту в последовательности SEQ ID NO: 26; замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в последовательности SEQ ID NO: 28;
(7) замену тирозина в положении 9 на гистидин и замену аргинина в положении 11 на пролин в последовательности SEQ ID NO: 24; замену тирозина в положении 3 на гистидин в последовательности SEQ ID NO: 25; замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в последовательности SEQ ID NO: 28; и
(8) замену аланина в положении 6 на аспарагиновую кислоту, замену глицина в положении 8 на тирозин, замену тирозина в положении 9 на гистидин и замену аргинина в положении 11 на пролин в последовательности SEQ ID NO: 24; замену тирозина в положении 3 на гистидин в последовательности SEQ ID NO: 25; замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в последовательности SEQ ID NO: 28.
В дополнительном или альтернативном варианте осуществления изобретения антитело к IL-8, которое может обладать зависящей от рН аффинностью к IL-8, содержит одну или несколько аминокислотных последовательностей, выбранных из группы, которая состоит из:
(а) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 23, или HVR-H1, который имеет последовательность, идентичную ей по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более;
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 24 или 29, или HVR-H2, который имеет последовательность, идентичную им по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более;
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 25 или 30, или HVR-H3, который имеет последовательность, идентичную им по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более;
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 26, или HVR- L1, который имеет последовательность, идентичную ей по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более;
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 27 или 107, или HVR- L2, который имеет последовательность, идентичную им по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более;
и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 28 или 32, или HVR-L3, который имеет последовательность, идентичную им по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более.
В данном контексте HVR, последовательность которого идентична на конкретный процент аминокислотной референс-последовательности, может являться функциональным эквивалентом HVR, который содержит аминокислотную референс-последовательность. Кроме того, антитело к IL-8 может содержать, например, HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29, и HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30, и/или HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 31, и HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 32. Кроме того, антитело к IL-8 может дополнительно по меньшей мере содержать замену на другую аминокислоту или на аминокислоту, указанную ниже в положении(ях), выбранном(ых) из группы, которая состоит из указанных ниже положений:
(1) серии в положении 8 в аминокислотной последовательности SEQ ID NO: 26; аспарагин в положении 1 и лейцин в положении 5 в аминокислотной последовательности SEQ ID NO: 27; и глутамин в положении 1 в аминокислотной последовательности SEQ ID NO: 28;
(2) замену аланина в положении 6 на аспарагиновую кислоту и замену аргинина в положении 11 на пролин в аминокислотной последовательности SEQ ID NO: 24; и замену тирозина в положении 3 на гистидин в аминокислотной последовательности в аминокислотной последовательности SEQ ID NO: 25;
(3) замену глицина в положении 8 на тирозин и замену тирозина в положении 9 на гистидин в аминокислотной последовательности SEQ ID NO: 24;
(4) замену аланина в положении 6 на аспарагиновую кислоту, замену глицина в положении 8 на тирозин, замену тирозина в положении 9 на гистидин и замену аргинина в положении 11 на пролин в аминокислотной последовательности SEQ ID NO: 24; и замену тирозина в положении 3 на гистидин в аминокислотной последовательность SEQ ID NO: 25;
(5) замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в аминокислотной последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в аминокислотной последовательности SEQ ID NO: 28;
(6) замену серина в положении 8 на глутаминовую кислоту в аминокислотной последовательности SEQ ID NO: 26; замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в аминокислотной последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в аминокислотной последовательности SEQ ID NO: 28;
(7) замену тирозина в положении 9 на гистидин и замену аргинина в положении 11 на пролин в аминокислотной последовательности SEQ ID NO: 24; замену тирозина в положении 3 на гистидин в аминокислотной последовательности SEQ ID NO: 25; замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в аминокислотной последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в аминокислотной последовательности SEQ ID NO: 28; и
(8) замену аланина в положении 6 на аспарагиновую кислоту, замену глицина в положении 8 на тирозин, замену тирозина в положении 9 на гистидин и замену аргинина в положении 11 на пролин в аминокислотной последовательности SEQ ID NO: 24; замену тирозина в положении 3 на гистидин в аминокислотной последовательности SEQ ID NO: 25; замену аспарагина в положении 1 на лизин и замену лейцина в положении 5 на гистидин в аминокислотной последовательности SEQ ID NO: 27; и замену глутамина в положении 1 на лизин в аминокислотной последовательности SEQ ID NO: 28.
Специалистам в данной области, естественно, должны быть очевидны все возможные комбинации аминокислотных последовательностей, указанных в контексте настоящего описания.
В одном из вариантов осуществления изобретения подразумевается, что понятие «функциональный эквивалент» в контексте настоящего описания, если конкретно не указано иное, относится к сохранению по меньшей мере на 50% или более, предпочтительно на 70% или более, 75% или более, 80% или более, 85% или более и более предпочтительно на 90% или более (например, на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% или более) при оценке методами, известными специалистам в данной области, функции, характерной для антитела, которое содержит аминокислотную референс-последовательность, представляющей собой физико-химические свойства и/или биологическую активность и т.д. (например, при рассмотрении одного из показателей или одной из характеристик, таких как связывание антигена и сохранение в крови),
В дополнительном или альтернативном варианте осуществления изобретения антитело к IL-8, указанное в настоящем описании, которое может обладать зависящей от рН аффинностью к IL-8, содержит одну или несколько аминокислотных последовательностей, выбранных из группы, которая состоит из:
(А) вариабельной области тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 33, 34 или 108, или вариабельной области тяжелой цепи, которая имеет последовательность, идентичную им по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более, и
(Б) вариабельной области легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 35 или 109, или вариабельной области легкой цепи, которая имеет последовательность, идентичную им по меньшей мере на 65%, 70%, 75% или более, предпочтительно на 80%, 85%, 90%, или более, и более предпочтительно на 95% или более.
В данном контексте вариабельная область, последовательность которой идентична на конкретный процент аминокислотной референс-последовательности, может являться функциональным эквивалентом вариабельной области, которая содержит аминокислотную референс-последовательность. Кроме того, антитело к IL-8 может содержать вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 34, и вариабельную область легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 35. Указанное антитело может обладать преимуществом благодаря тому, что его IL-8-нейтрализующая активность является стабильной in vivo (например, в плазме) или тому, что его иммуногенность является низкой. Кроме того, когда антитело к IL-8 содержит вариант аминокислотной последовательности SEQ ID NO: 33, оно может дополнительно содержать по меньшей мере замену на другую аминокислоту или на аминокислоту, указанную ниже в положении(ях), выбранном(ых) из группы, которая состоит из указанных ниже положений:
(I) замену аланина в положении 55 на другую аминокислоту, например, аспарагиновую кислоту, в аминокислотной последовательности SEQ ID NO: 33;
(II) замену глицина в положении 57 на другую аминокислоту, например, тирозин, в аминокислотной последовательности SEQ ID NO: 33;
(III) замену тирозина в положении 58 на другую аминокислоту, например, гистидин, в аминокислотной последовательности SEQ ID NO: 33;
(IV) замену аргинина в положении 60 на другую аминокислоту, например, пролин, в аминокислотной последовательности SEQ ID NO: 33;
(V) замену глутамина в положении 84 на другую аминокислоту, например, треонин, в аминокислотной последовательности SEQ ID NO: 33;
(VI) замену серина в положении 87 на другую аминокислоту, например, аспарагиновую кислоту, в аминокислотной последовательности SEQ ID NO: 33; и
(VII) замену тирозина в положении 103 на другую аминокислоту, например, гистидин, в аминокислотной последовательности SEQ ID NO: 33.
Специалистам в данной области, естественно, должны быть очевидны все возможные комбинации аминокислотных последовательностей, указанных в настоящем описании.
В другом варианте осуществления изобретения антитело к IL-8, указанное в настоящем описании, может представлять собой антитело, которое содержит по меньшей мере 1-6 из аминокислотных последовательностей, указанных ниже в подпунктах (а)-(е) в каждом из пунктов (1)-(7), или антитело, которое содержит по меньшей мере одну аминокислотную замену по меньшей мере в одной или нескольких из аминокислотных последовательностей, указанных ниже в каждом из подпунктов (а)-(е). Антитело может представлять собой антитело к IL-8 с зависящей от рН аффинностью к IL-8. Если конкретно не указано иное, то аминокислоты могут быть заменены на любые другие аминокислоты:
(1) (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 58,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 59,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 60,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 61,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 62, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 63.
(2) (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 64,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 65,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 66,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 67,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 68, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 69.
(3) (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 70,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 71,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 72,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 73,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 74, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 75.
(4) (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 76,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 77,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 78,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 79,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 80, и
(e) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 81.
(5) (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 82,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 83,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 84,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 85,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 86, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 87.
(6) (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 88,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 89,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 90,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 91,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 92, и
(е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 93.
(7) (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 94,
(б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 95,
(в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 96,
(г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 97,
(д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 98, и
(е) HVR-L3 который содержит аминокислотную последовательность SEQ ID NO: 99.
В контексте настоящего описания понятие кислый рН» означает рН, которое можно выбирать, например, из рН от 4,0 до 6,5. В одном из вариантов осуществления изобретения кислое рН означает (но, не ограничиваясь только ими) рН 4,0, рН 4,1, рН 4,2, рН 4,3, рН 4,4, рН 4,5, рН 4,6, рН 4,7, рН 4,8, рН 4,9, рН 5,0, рН 5,1, рН 5,2, рН 5,3, рН 5,4, рН 5,5, рН 5,6, рН 5,7, рН 5,8, рН 5,9, рН 6,0, рН 6,1, рН 6,2, рН 6,3, рН 6,4 или рН 6,5.
В контексте настоящего описания понятие = «нейтральное рН» означает рН, который можно выбирать, например, из рН от 6,7 до 10,0. В одном из вариантов осуществления изобретения нейтральное рН означает (но, не ограничиваясь только ими) рН 6,7, рН 6,8, рН 6,9, рН 7,0, рН 7,1, рН 7,2, рН 7,3, рН 7,4, рН 7,5, рН 7,6, рН 7,7, рН 7,8, рН 7,9, рН 8,0, рН 8,1, рН 8,2, рН 8,3, рН 8,4, рН 8,5, рН 8,6, рН 8,7, рН 8,8, рН 8,9, рН 9,0, рН 9,5 или рН 10,0.
В одном из вариантов осуществления изобретения антитело к IL-8, представленное в настоящем описании, означает антитело к IL-8, которое связывается с IL-8 рН-зависимым образом. В одном из объектов настоящего изобретения антитело к IL-8, которое связывается с IL-8 рН-зависимым образом, означает антитело, аффинность связывания которого с IL-8 при кислом рН ниже по сравнению с аффинностью связывания с IL-8 при нейтральном рН. Указанные рН-зависимые антитела к IL-8 включают, например, антитела, которые характеризуются более высокой аффинностью к IL-8 при нейтральном рН, чем при кислом рН. В одном из вариантов осуществления изобретения антитела к IL-8, представленные в настоящем описании, обладают аффинностью к IL-8 при нейтральном рН, которая по меньшей мере в 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 раз или выше, чем аффинность при кислом рН. Аффинность связывания можно измерять методами поверхностного плазмонного резонанса (например, BIACORE и т.д.) (но, не ограничиваясь только ими). Константу скорости реакции ассоциации (kon) и константу скорости реакции диссоциации (koff) можно рассчитывать с помощью программного обеспечения BIACORE Т200 Evaluation (фирма GE Healthcare) с использованием простой модели связывания Ленгмюра 1:1 путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Константу равновесия реакции диссоциации (KD) рассчитывают как отношение koff/kon. Для скрининга в отношении антител, аффинность связывания которых зависит от рН, можно применять (но, не ограничиваясь только ими), ELISA, анализ кинетического исключения (KinExA™), а также методы поверхностного плазмонного резонанса (такие как BIACORE и т.д.). рН-зависимая IL-8-связывающая способность означает способность связываться с IL-8 рН-зависимым образом. Кроме того, обладает ли антитело способностью многократно связываться с IL-8, можно оценивать методами, описанными в WO 2009/125825.
В одном из вариантов осуществления изобретения предпочтительно, чтобы антитело к IL-8, представленное в настоящем описании, имело низкую константу диссоциации (KD) для IL-8 при нейтральном рН (например, при рН 7,4). Константа диссоциации антитела, характеризующая связывание антитела с IL-8 при нейтральном рН (например, при рН 7,4), может составлять, например, 0,ЗнМ или менее, 0,1нМ или менее или 0,03нМ или менее.
В одном из вариантов осуществления изобретения предпочтительно, чтобы антитело к IL-8, представленное в настоящем описании, имело высокую константу диссоциации (KD) для IL-8 при кислом рН (например, при рН 5,8). Константа диссоциации антитела, характеризующая связывание антитела с IL-8 при кислом рН (например, при рН 5,8), может составлять, например, 3 нМ или более,10 нМ или более или 30 нМ или более.
В одном из вариантов осуществления изобретения отношение величины константы диссоциации при кислом рН (например, при рН 5,8) к величине константы диссоциации при нейтральном рН (например, при рН 7,4) [KD (кислое pH)/KD (нейтральное рН)] (например, KD(pH 5,8)/KD(pH 7,4)) антитела к IL-8, представленного в настоящем описании, составляет, например, 30 или более 50 или более или, например, 100 или более, например, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000, 5500, 6000, 6500, 7000, 7500, 8000, 8500, 9000 или 9500 или более.
В одном из вариантов осуществления изобретения предпочтительно, чтобы антитело к IL-8, представленное в настоящем описании, имело высокую константу скорости реакции диссоциации (koff) при кислом рН (например, при рН 5,8). Константа скорости реакции диссоциации антитела при кислом рН (например, при рН 5,8) может составлять, например, 0,003 (1/с) или более, 0,005 (1/с) или более или 0,01 (1/с) или более.
В WO 2013/046704 описано, что вариант Fc-области, в который интродуцирована специфическая мутация (репрезентативным примером которой является мутация двух остатков Q438R/S440E согласно EU-нумерации) в комбинации с мутацией, которая повышает FcRn-связывание при кислом рН, характеризовался в значительной степени пониженным связыванием с ревматоидным фактором. Однако в WO 2013/046704 не описано, что вариант Fc-области, связывание которого с ревматоидным фактором снижено благодаря модификации Q438R/S440E, обладает удлиненным временем удерживания в плазме по сравнению с антителом с нативной Fc-областью. Таким образом, существует необходимость в безопасных и более ценных вариантах Fc-области с улучшенным временем удерживания в плазме, но которые не связываются с предсуществующими ADA. При создании изобретения установлено, что варианты Fc-области, которые содержат замену на Ala (А) аминокислоты в положении 434 согласно EU-нумерации, и специфическую мутацию двух остатков (репрезентативным примером является Q438R/S440E), в качестве комбинации мутаций аминокислотных остатков, являются предпочтительными в качестве безопасных и представляют собой более предпочтительные варианты Fc-области, характеризующиеся улучшенным временем удерживания в плазме, но не связывающиеся с антителами к лекарственным средствам (предсуществующие ADA и т.д.).
Таким образом, согласно дополнительному или альтернативному варианту осуществления изобретения антитело к IL-8, представленное в настоящем описании, содержит вариант Fc-области, который содержит FcRn-связывающий домен, и FcRn-связывающий домен может содержать, например, Ala в положении 434; Glu, Arg, Ser или Lys в положении 438; и Glu, Asp или Gln в положении 440 согласно EU-нумерации, в отличие от нативной Fc-области В антителе к IL-8 FcRn-связывающий домен может представлять собой FcRn связывающий домен, обладающий повышенной FcRn-связывающей активностью при кислом рН и нейтральном рН, в частности, при кислом рН. Альтернативно этому, в антителе его FcRn-связывающий домен предпочтительно содержит вариант Fc-области, содержащий Ala в положении 434; Glu, Arg, Ser или Lys в положении 438; и Glu, Asp или Gin в положении 440, более предпочтительно содержащий Ala в положении 434; Arg или Lys в положении 438; и Glu или Asp в положении 440 согласно EU-нумерации. Альтернативно этому, вариант Fc-области предпочтительно дополнительно содержит Не или Leu в положении 428; и/или Ile, Leu, Val, Thr или Phe в положении 436, более предпочтительно дополнительно содержит Leu в положении 428; и/или Val или Thr в положении 436 согласно EU-нумерации.
В одном из вариантов осуществления изобретения вариант Fc-области антитела к IL-8, представленного в настоящем описании, предпочтительно представляет собой вариант Fc-области нативного антитела в виде Ig и более предпочтительно вариант Fc-области нативного антитела IgG-типа (IgG1-, IgG2-, IgG3- или IgG4-типа), и еще более предпочтительно полученный из нативного человеческого IgG1, IgG2, IgG3 или IgG4. Вариант Fc-области может, например, иметь происхождение из человеческого IgG1.
Касательно Fc-областей нативных человеческих антител IgG1-, IgG2-, IgG3- и IgG4-типа, за исключением положения 436, положения 428, 434, 438 и 440 согласно EU-нумерации, которые представляют собой описанные выше сайты модификации, являются общими для Fc-областей всех нативных человеческих антител IgG1-, IgG2-, IgG3- и IgG4-типа. С другой стороны, в положении 436 в Fc-области нативные человеческие антитела IgG1-, IgG2- и IgG4-типа несут Tyr (Y), а нативное человеческое антитело IgG3-типа имеет Phe (F) Однако Stapleton и др., (Competition for FcRn-mediated transport gives rise to short half-life of human IgG3 and offers therapeutic potential, Nature Communications, декабрь 2011 г. №599) описали, что человеческие IgG3-аллотипы, которые содержат аминокислотную замену R435H согласно EU-нумерации, обладают временем полужизни в плазме человека, сопоставимым с IgG1. Отсюда высказано предположение о том, что время удерживания в плазме можно улучшать посредством повышения связывания FcRn в кислых условиях путем интродукции аминокислотной замены R435H в дополнение к аминокислотной замене в положении 436.
В WO 2013/046704 конкретно описаны замены двух аминокислотных остатков Q438R/S440E, Q438R/S440D, Q438K/S440E и Q438K/S440D согласно EU нумерации, которые могут приводить к существенному снижению связывания с ревматоидным фактором при объединении с аминокислотной заменой, которая повышает связывание FcRn при кислом рН.
Таким образом, в одном из вариантов осуществления изобретения аминокислотная последовательность варианта Fc-области антитела к IL-8, представленного в настоящем описании, по сравнению с нативной Fc-областью может содержать в ее FcRn-связывающем домене комбинацию замененных аминокислот, выбранную из группы, которая состоит из:
N434A/Q438R/S440E;
N434A/Q438R/S440D;
N434A/Q438K/S440E;
N434A/Q438K/S440D;
N434A/Y436T/Q438R/S440E;
N434A/Y436T/Q438R/S440D;
N434A/Y436T/Q438K/S440E;
N434A/Y436T/Q438K/S440D;
N434A/Y436V/Q438R/S440E;
N434A/Y436V/Q438R/S440D;
N434A/Y436V/Q438K/S440E;
N434A/Y436V/Q438K/S440D;
N434A/R435H/F436T/Q438R/S440E;
N434A/R435H/F436T/Q438R/S440D;
N434A/R435H/F436T/Q438K/S440E;
N434A/R435H/F436T/Q438K/S440D;
N434A/R435H/F436V/Q438R/S440E;
N434A/R435H/F436V/Q438R/S440D;
N434A/R435H/F436V/Q438K/S440E;
N434A/R435H/F436V/Q438K/S440D;
M428L/N434A/Q438R/S440E;
M428L/N434A/Q438R/S440D;
M428L/N434A/Q438K/S440E;
M428L/N434A/Q438K/S440D;
M428L/N434A/Y436T/Q438R/S440E;
M428L/N434A/Y436T/Q438R/S440D;
M428L/N434A/Y436T/Q438K/S440E;
M428L/N434A/Y436T/Q438K/S440D;
M428L/N434A/Y436V/Q438R/S440E;
M428L/N434A/Y436V/Q438R/S440D;
M428L/N434A/Y436V/Q438K/S440E; и
M428L/N434A/Y436V/Q438K/S440D;
L235R/G236R/S239K/M428L/N434A/Y436T/Q438R/S440E; и
L235R/G236R/A327G/A330S/P331S/M428L/N434A/Y436T/Q438R/S440E
согласно EU нумерации.
Альтернативно этому, в другом варианте осуществления изобретения вариант Fc-области антитела к IL-8, представленного в настоящем описании, может содержать замены на другие аминокислоты в одном или нескольких положениях (например, в шести, семи или во всех положениях), выбранных из группы, которая состоит из положений 235, 236, 239, 327, 330, 331, 428, 434, 436, 438 и 440 согласно EU-нумерации, по сравнению с нативной Fc-областью, например, одну или несколько аминокислотных замен (например, шесть, семь или все замены), выбранных из группы, которая состоит из L235R, G236R, S239K, A327G, A330S, P331S, M428L, N434A, Y436T, Q438R и S440E.
В одном из вариантов осуществления изобретения предпочтительно, чтобы FcRn-связывающая активность варианта Fc-области антитела к IL-8, представленного в настоящем описании, повышалась при кислом рН по сравнению с Fc-областью нативного IgG.
Повышение аффинности связывания FcRn FcRn-связывающего домена при определенном рН может соответствовать повышению измеренной аффинности связывания FcRn по сравнению с измеренной аффинностью связывания FcRn нативного FcRn-связывающего домена. В этом случае величина отношения KD (нативная Fc-область)/KD (вариант Fc-области антитела к IL-8, представленного в настоящем описании), которое характеризует различие аффинности связывания, может составлять по меньшей мере 1, 5, 2, 3, 4, 5, 10, 15, 20, 50, 70, 80, 100, 500 или 1000. Указанное увеличение аффинности связывания FcRn FcRn-связывающего домена может иметь место при кислом рН и/или при нейтральном рН; однако, в частности, предпочтительно, если FcRn-связывающая активность возрастает при кислом рН.
Кроме того, аффинность связывания FcRn варианта Fc-области антитела к IL-8, представленного в настоящем описании, у которого аффинность связывания FcRn повышалась при кислом рН, может быть выше по сравнению с Fc-областью нативного IgG, например, при рН 6,0 и 25°С в 1, 5, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 50, 75, 100, 200, 500, 1000 раз или более. В контексте настоящего описания понятие «антитело к лекарственному средству» или «ADA» относится к эндогенному антителу, которое обладает способностью связываться с эпитопом, локализованном на терапевтическом антителе, и в результате может связываться с терапевтическим антителом. В контексте настоящего описания понятие «предсуществующее антитело к лекарственному средству» или «предсуществующее ADA» относится к антителу к лекарственному средству, которое присутствует и поддается обнаружению в крови пациента до введения пациенту терапевтического антитела. В некоторых вариантах осуществления изобретения предсуществующее ADA представляет собой ревматоидный фактор, поликлональное или моноклональное аутоантитело против Fc-области человеческого антитела в виде IgG. Эпитоп ревматоидного фактора локализован в области раздела СН2/СН3, а также в СН3-домене, и может изменяться в зависимости от клона. Для «варианта Fc-области, аффинность связывания которого с предсуществующим ADA является низкой» аффинность может быть понижена, например, до 1/10 или менее, 1/50 или менее или 1/100 или менее по сравнению с аффинносью связывания ADA антитела, содержащего Fc-область нативного IgG.
В одном из вариантов осуществления изобретения аффинность связывания Fc-области (варианта) антитела в отношении предсуществующего ADA можно, например, характеризовать с помощью электрохемилюминесцентной (ECL) реакции при кислом рН и/или при нейтральном рН; однако для определения аффинности связывания предсуществующего ADA можно применять также другие соответствующие методы, известные специалистам в данной области. ECL-анализ описан, например, у Moxness и др., Clin Chem. 51, 2005, сс. 1983-1985 и в примерах, представленных в настоящем описании. Анализы можно осуществлять, например, в условиях применения MES-буфера и 37°С. Антигенсвязывающую активность антитела можно определять, например, с помощью BIACORE-анализа. В одном из вариантов осуществления изобретения аффинность связывания предсуществующего ADA можно оценивать при любой температуре от 10°С до 50°С. Активность связывания между человеческой Fc-областью и предсуществующим человеческим ADA предпочтительно определяют при температуре от 15°С до 40°С. Температура предпочтительно составляет 20°С-25°С, более предпочтительно 25°С. Активность можно измерять при рН 7,4 (или рН 7,0) и 25°С.
В одном из вариантов осуществления изобретения предпочтительно, чтобы вариант Fc-области антитела к IL-8, представленного в настоящем описании, не обладал существенно повышенной аффинностью связывания с предсуществующим ADA, предпочтительно, ревматоидным фактором (RF), при нейтральном рН и/или обладал повышенной FcRn-связывающей активностью при кислом рН по сравнению с Fc-областью нативного IgG; в результате предпочтительно, чтобы он характеризовался пониженным плазменным клиренсом (CL), пролонгированным временем удерживания в плазме или пролонгированным временем полужизни в плазме (t1/2). Корреляция между плазменным клиренсом антитела (CL), временем удерживания в плазме и временем полужизни в плазме (t1/2) известна специалистам в данной области. В одном из вариантов осуществления изобретения вариант Fc-области антитела к IL-8, представленного в настоящем описании, обладает улучшенным временем удерживания в плазме по сравнению с референс-вариантом Fc-области, который содержит комбинацию аминокислотных замен N434Y/Y436V/Q438R/S440E согласно EU-нумерации.
В примерах 1-3, приведенных в настоящего описания, представлено сравнение времени удерживания в плазме двух вариантов Fc-области: варианта Fc-области, обозначенного F1718 (Fc-область с мутациями, интродуцированными в четыре сайта: N434Y/Y436V/Q438R/S440E), который описан в WO 2013/046704, и варианта Fc-области F1848m (Fc-область с мутациями, интродуцированными в четыре сайта: N434A/Y436V/Q438R/S440E). Аминокислотные мутации для двух вариантов Fc-области отличаются только тем, что в положении 434 согласно EU-нумерации интродуцированная аминокислотная мутация представляет собой Y (тирозин) в случае F1718 и А (аланин) в случае F 1848m. Однако по сравнению с нативным IgG1 F 1848m характеризуется улучшенным временем удерживания в плазме, в то время как F1718 не обладает таким улучшенным временем удерживания в плазме (см. в частности пример (3-2) в настоящем описании). Кроме того, результаты экспериментов, описанных в примерах (1-2) и (3-3) в настоящем описании, продемонстрировали, что среди различных вариантов Fc-области время удерживания в плазме F1847m, F1886m, F1889m и F1927m оказалось в большей степени улучшенным по сравнению с F1848m. Таким образом, обычным специалистам в данной области должно быть очевидно, что варианты Fc-области, включая F1847m, F1886m, F1889m и F1927m, а также F1848m, обладают улучшенным временем удерживания в плазме по сравнению с референс-вариантом Fc-области, содержащим комбинацию аминокислотных замен N434Y/Y436V/Q438R/S440E согласно EU-нумерации.
Антитела с удлиненным временем полужизни в крови и улучшенной FcRn-связывающей активностью при кислом рН описаны также в US 2005/0014934 (Hinton и др.). Антитела могут содержать вариант Fc-области, который содержит одну или несколько аминокислотных замен, которые повышают связывание Fc-области с FcRn, и могут содержать одну или несколько аминокислотную(ых) замену(н) в положении(ях), выбранном(ых) из положений: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 и 434 (EU- нумерация), например, аминокислотную замену в положении 434 в Fc-области. Другие примеры, касающиеся других вариантов Fc-области, описаны у Duncan, Nature 322, 1988, сс. 738-740; в US №5648260; US №5624821 и WO 94/29351. Соответствующую модификацию указанных аминокислот можно интродуцировать в Fc-область (вариант) антитела к IL-8, представленного в настоящем описании.
Связывание с FcγR или белком комплемента может также оказывать неблагоприятное действие (например, несоответствующую активацию тромбоцитов). Варианты Fc-области, которые не связываются с эффекторными рецепторами, такими как FcγRIIa-рецептор, могут быть безопасными и/или более предпочтительными.
В одном из вариантов осуществления изобретения антитело к IL-8, которое содержит вариант Fc-области, представленный в настоящем описании, обладает лишь слабой способностью связывать белок комплемента или не связывается с белками комплемента. Предпочтительный белок комплемента представляет собой C1q. Низкая способность связываться с белком комплемента может означать способность связываться с белком комплемента, сниженную например, до 1/10 или менее, 1/50 или менее или 1/100 или менее по сравнению со способностью связываться с белком комплемента нативного IgG или антитела, содержащего Fc-область нативного IgG. Способность связываться с белком комплемента Fc-области (варианта) можно уменьшать с помощью модификации аминокислотной последовательности, такой как аминокислотная замена, инсерция и делеция.
В дополнительном или альтернативном варианте осуществления изобретения антитело к IL-8, содержащее вариант Fc-области, представленный в настоящем описании, предпочтительно обладает лишь слабой связывающей активностью в отношении эффекторного рецептора или не связывается с эффекторными рецепторами. Эффекторные рецепторы включают, например, активирующий FcγR, в частности FcγRI, FcγRII и FcγRIII. FcγRI включает, например, FcγRIa, FcγRIb и FcγRIc, а также их подтипы. FcγRII включает, например, FcγRIIa (который имеет два аллотипа: R131 и Н131) и FcγRIIb. FcγRIII включает, например, FcγRIIIa (который имеет два аллотипа: V158 и F158) и FcγRIIIb (который имеет два аллотипа: FcγRIIIb-NA1 и FcγRIIIb-NA2). Антитела, которые обладают лишь слабой аффинностью связывания с эффекторными рецепторами или не связываются с рецепторами, включают антитела, содержащие вариант Fc-области, аффинность связывания которых с эффекторными рецепторами снижена, например, до 1/10 или менее, 1/50 или менее или 1/100 или менее по сравнению с аффинностью связывания антитела, которое содержит Fc-область нативного IgG. Конкретный пример включат антитела, содержащие молчащую Fc-область, и антитела, которые не имеют Fc-области (например, Fab, F(ab)'2, scFv, sc(Fv)2 и диабоди).
Примеры вариантов Fc-областей, которые обладают лишь слабой связывающей активностью в отношении эффекторных рецепторов или не связываются с эффекторными рецепторами, описаны, например, у Strohl и др., Current Opinion in Biotechnology 20(6), 2009, cc. 685-691, и они включают дегликозилированные Fc-области (N297A и N297Q) и молчащие Fc-области, полученные в результате манипуляции с Fc-областями для придания им молчащих эффекторных функцией (или для подавления иммунитета) (IgG1-L234A/L235A, IgG1-H268Q/A330S/P331S, IgG1-C226S/C229S, IgG1-C226S/C229S/E233P/L234V/L235A, IgG1-L234F/L235E/P331S, IgG2-V234A/G237A, IgG2-H268Q/V309L/A330S/A33IS, IgG4-L235A/G237A/E318A и IgG4-L236E). В WO 2008/092117 описаны антитела, содержащие молчащую Fc-область, которая включает замены G236R/L328R, L235G/G236R, N325A/L328R или N325L/L328R (согласно EU-нумерации). В WO 2000/042072 описаны антитела, содержащие молчащую Fc-область, которая включает замены в одном или нескольких положениях EU233, EU234, EU235 и EU237. В WO 2009/011941 описаны антитела, содержащие молчащую Fc-область, в которой отсутствуют остатки EU231-EU238. У Davis и др., Journal of Rheumatology 34(11), 2007, cc. 2204-2210 описаны антитела с молчащей Fc-областью, содержащей замены C220S/C226S/C229S/P238S. У Shields и др., Journal of Biological Chemistry 276(9), 2001, cc. 6591-6604 описаны антитела, содержащие молчащую Fc-область, которая включает замену D265A. В US №. 6737056 описаны антитела с пониженной эффекторной функцией, которые содержат одну или несколько аминокислотных замен в EU238, EU265, EU269, EU270, EU297, EU327 и EU329. В US №7332581 описаны варианты Fc-области, которые содержат две или большее количество аминокислотных замен в EU265, EU269, EU270, EU297 и EU327, а также варианты Fc-области, несущие так называемую мутацию «DANA», которая приводит к замене на аланин в положениях EU265 и EU297. Модификацию указанных аминокислотных остатков можно также соответственно интродуцировать в варианты Fc-областей антител к IL-8, представленных в настоящем описании.
Понятие «слабое связывание с эффекторными рецепторами» означает, что связывающая активность в отношении эффекторных рецепторов составляет, например, 95% или менее, предпочтительно 90% или менее, 85% или менее, 80% или менее, 75% или менее, более предпочтительно 70% или менее, 65% или менее, 60% или менее, 55% или менее, 50% или менее, 45% или менее, 40% или менее, 35% или менее, 30% или менее, 25% или менее, 20% или менее, 15% или менее, 10% или менее, 9% или менее, 8% или менее, 7% или менее, 6% или менее, 5% или менее, 4% или менее, 3% или менее, 2% или менее или 1% или менее относительно связывающей активности в отношении эффекторных рецепторов нативного IgG (или антитела, содержащего Fc-область нативного IgG). FcγR-связывающая активность может снижаться до 1/10 или менее, 1/50 или менее или 1/100 или менее по сравнению со связывающей активностью в отношении эффекторных рецепторов нативного IgG (или антитела, содержащего Fc-область нативного IgG).
В контексте настоящего описания «молчащая Fc-область» означает вариант Fc-области, содержащий одну или большее количество аминокислотных замен, инсерций, делеций и других модификаций, которые снижают связывание с эффекторными рецепторами по сравнению с нативной Fc-областью. Поскольку связывающая активность в отношении эффекторных рецепторов может быть существенно снижена, такие молчащие Fc-области более (практически) не могут связываться с эффекторными рецепторами. Молчащие Fc-области включают, например, Fc-области, содержащие аминокислотные замены в одном или нескольких положениях, выбранных из группы, которая состоит из: EU234, EU235, EU236, EU237, EU238, EU239, EU265, EU266, EU267, EU269, EU270, EU271, EU295, EU296, EU297, EU298, EU300, EU324, EU325, EU327, EU328, EU329, EU331 и EU332. В одном из вариантов осуществления изобретения модификации указанных аминокислотных остатков можно также соответственно интродуцировать в Fc-область (вариант) антитела к IL-8, представленного в настоящем описании.
В одном из вариантов осуществления изобретения способ модификации константных областей антител с получением варианта Fc-области антитела к IL-8, представленного в настоящем описании, может базироваться, например, на оценке нескольких изотипов константных областей (IgG1, IgG2, IgG3 и IgG4) с последующей интродукцией, например, соответствующих аминокислотных замен в их последовательности для выбора изотипов, которые обладают пониженной антигенсвязывающей активностью при кислом рН и/или обладают повышенной скоростью реакции диссоциации при кислом рН.
В дополнительном или альтернативном варианте осуществления изобретения антитело к IL-8, представленное в настоящем описании, может содержать:
(А) тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 36, 37 или 106, или тяжелую цепь, которая имеет последовательность, идентичную ей по меньшей мере на 65%, 70% или 75%, или более, предпочтительно на 80%, 85% или 90% или более, и более предпочтительно на 95% или более; и/или
(Б) легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 38 или 44, или легкую цепь, которая имеет последовательность, идентичную ей по меньшей мере на 65%, 70% или 75%, или более, предпочтительно на 80%, 85% или 90% или более, и более предпочтительно на 95% или более.
В данном контексте тяжелая или легкая цепь, последовательность которой идентична на конкретный процент (%) аминокислотной референс-последовательности, может являться функциональным эквивалентом тяжелой или легкой цепи, которая содержит аминокислотную референс-последовательность. Кроме того, антитело к IL-8 может содержать вариант Fc-области, имеющий по меньшей мере одно из пяти указанных ниже свойств:
(а) аффинность связывания FcRn варианта Fc-области повышена относительно аффинности связывания с FcRn нативной Fc-области при кислом рН;
(б) аффинность связывания варианта Fc-области с предсуществующим ADA снижена относительно аффинности связывания нативной Fc-области с предсуществующими ADA;
(в) время полужизни в плазме варианта Fc-области удлинено относительно времени полужизни в плазме нативной Fc-области;
(г) клиренс из плазмы варианта Fc-области понижен относительно клиренса из плазмы нативной Fc-области;
(д) аффинность связывания варианта Fc-области с эффекторным рецептором снижена относительно аффинности связывания нативной Fc-области с эффекторным рецептором; и
(е) способность связываться с IL-8 рН-зависимым образом.
Специалистам в данной области должны быть очевидны все возможные комбинации аминокислотных последовательностей, указанных в контексте настоящего описания.
В одном из вариантов осуществления изобретения антитело к IL-8, представленное в настоящем описании, можно получать, например, с использованием методов, описанных в US №4816567. Одним из вариантов осуществления изобретения являются выделенные нуклеиновые кислоты, кодирующие антитело к IL-8, представленное в настоящем описании. Указанная нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL и/или аминокислотную последовательность, содержащую VH антитела (например, легкую цепь и/или тяжелую цепь антитела). Выделенную нуклеиновую кислоту, кодирующую антитело к IL-8, можно встраивать в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетках-хозяевах. Указанную нуклеиновую кислоту можно легко выделять и секвенировать с использованием общепринятых процедур (например, с использованием олигонуклеотидных зондов, которые специфически связываются с генами, кодирующими тяжелые и/или легкие цепи антитела).
Другим вариантом осуществления изобретения является/являются один или несколько векторов (содержащих указанную нуклеиновую кислоту (например, экспрессионные векторы). Другим вариантом осуществления изобретения являются клетки-хозяева, содержащие указанную нуклеиновую кислоту. Клетки-хозяева содержат: (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и/или аминокислотную последовательность, содержащую VH антитела; или (2) первый вектор, который содержит нуклеиновую кислоту, кодирующую аминокислотную последовательность, которая содержит VL антитела, и второй вектор, который содержит нуклеиновую кислоту, кодирующую аминокислотную последовательность, которая содержит VH антитела (например, клетки, трансформированные векторами).
В одном из вариантов осуществления изобретения хозяин представляет собой эукариотическую клетку (например, (СНО-клетки) или лимфоциты (например, Y0-, NS0- или=SP20-клетки)).
Одним из вариантов осуществления изобретения является способ получения антитела к IL-8, представленного в настоящем описании. Способ включает, например, культивирование клеток-хозяев, содержащих вышеуказанную нуклеиновую кислоту, в условиях, пригодных для экспрессии антитела, и необязательно сбор антитела из клеток-хозяев (или среды для культивирования клетки-хозяина).
Приемлемые клетки-хозяева для клонирования или экспрессии кодирующих антитело векторов включают прокариотические или эукариотические клетки. Клетки, полученные из многоклеточных организмов (беспозвоночных и позвоночных), можно применять для экспрессии гликозилированных антител. Примерами клеток беспозвоночных являются клетки насекомых, а также можно применять клетки растений.
В одном из вариантов осуществления изобретения антитело к IL-8, полученное культивированием, в условиях, пригодных для экспрессии антитела, при котором клетки-хозяева, содержащие нуклеиновую кислоту, которая кодирует антитело, можно выделять изнутри или снаружи клетки-хозяина (среды, молоко и т.д.) для очистки его в виде практически чистого гомогеннного антитела. Методы выделения/очистки, которые, как правило, применяют для очистки полипептидов (но, не ограничиваясь только ими), можно соответственно применять для отделения и очистки антитела. Антитело можно соответственно отделять и очищать, например, с помощью соответствующего отбора и объединения хроматографии на колонках, фильтрации, ультрафильтрации, обессоливания, осаждения растворителем, экстракции растворителем, дистилляции, иммунопреципитации, электрофореза в полиакриламидном геле в присутствии ДСН, изоэлектрического фокусирования, диализа, перекристаллизации. Хроматография включает, например, аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гель-фильтрацию, хроматографию с обращенной фазой и адсорбционную хроматографию. Указанную хроматографию можно осуществлять, например, с использованием жидкостной хроматографии, такой как ЖХВР и ЖХБР. Колонки для применения в аффинной хроматографии включают, например, колонку с белком А или колонку с белком G. Колонка с белком А включает, например, Hyper D, POROS и Sepharose F.F. (фирма Pharmacia).
В одном из вариантов осуществления изобретения в настоящем описании представлены прежде всего характеристики антител к IL-8, такие как повышенная активность связывания с внеклеточным матриксом и усиленное поглощение клетками комплекса, образованного между IL-8 и антителом к IL-8, и, таким образом, настоящее описание относится к способам отбора антител с повышенной активностью связывания внеклеточного матрикса и антител с усиленным поглощением клетками. В одном из вариантов осуществления изобретения в настоящем описании представлены способы получения антитела к IL-8, содержащего вариабельную область, связывающая активность которой с IL-8 зависит от рН, включающие стадии: (I) оценки связывания между антителом к IL-8 и внеклеточным матриксом; (II) отбора антитела к IL-8, отличающегося сильным связыванием с внеклеточным матриксом; (III) культивирования хозяина, который содержит вектор, содержащий нуклеиновую кислоту, которая кодирует антитело; и (IV) выделения антитела из клеточной среды (супернатанта культуры и т.д.). Связывание с внеклеточным матриксом можно оценивать, например, используя систему ELISA для обнаружения связывания между антителом и внеклеточным матриксом, для чего антитело добавляют к планшету с иммобилизованным внеклеточным матриксом и к нему добавляют меченое антитело против антитела. Альтернативно этому, связывание можно оценивать, например, следующим образом: смесь антитела и меченного рутением антитела добавляют к планшету с иммобилизованным внеклеточным матриксом и связывание между антителом и внеклеточным матриксом оценивают методом электрохемилюминесценции (ECL) на основе электрохемилюминесценции рутения.
В одном из вариантов осуществления изобретения предпочтительно, чтобы антитело к IL-8, представленное в настоящем описании, сохраняло стабильную нейтрализующую IL-8 активность в растворе (например, в ЗФР). Сохраняется ли активность в стабильном состоянии в растворе, можно оценивать, определяя, изменяется ли нейтрализующая IL-8 активность антитела к IL-8, представленного в настоящем описании, добавленного в раствор, после хранения в течение определенного периода времени при определенной температуре, относительно активности до добавления в раствор. В одном из вариантов осуществления изобретения период хранения составляет, например, 1-4 недели, а температура хранения составляет, например, 25°С, 30°С, 35°С, 40°С или 50°С.
В одном из вариантов осуществления изобретения предпочтительно, чтобы антитело к IL-8, представленное в настоящем описании, сохраняло стабильную нейтрализующую IL-8 активность in vivo (например, в плазме). Сохраняется ли активность в стабильном состоянии in vivo, можно оценивать, определяя, изменяется ли нейтрализующая IL-8 активность антитела к IL-8, представленного в настоящем описании, добавленного в плазму животного кроме человека (например, мыши) или человека, после хранения в течение определенного периода времени при определенной температуре, относительно активности до добавления в плазму. В одном из вариантов осуществления изобретения период хранения составляет, например, 1-4 недели, а температура хранения составляет, например, 25°С, 30°С, 35°С или 40°С.
В одном из вариантов осуществления изобретения скорость клеточного поглощения антитела к IL-8, представленного в настоящем описании, выше, когда антитело образует комплекс с IL-8, чем не входящего в комплекс антитела.
В одном из вариантов осуществления изобретения предпочтительно, чтобы прогнозируемая иммуногенность антитела к IL-8, представленного в настоящем описании, ожидаемая для человека-хозяина, снижалась. «Низкая иммуногенность» может означать, например, что после введения антитело к IL-8 не индуцирует иммунный ответ in vivo, по меньшей мере у большинства индивидуумов, которым антитело вводят в достаточном количестве в течение периода времени, достаточного для достижения терапевтического действия. Индукция иммунного ответа может включать производство антител против лекарственного средства. Понятие «низкий уровень производства антител против лекарственного средства» применяют взаимозаменяемо с понятием «низкая иммуногенность». Уровень иммуногенности в организме человека можно определять с использованием программы предсказания Т-клеточных эпитопов. Указанные программы предсказания Т-клеточных эпитопов включают Epibase (фирма Lonza), iTope/TCED (фирма Antitope) и EpiMatrix (фирма EpiVax). Последовательности с пониженной иммуногенностью можно создавать например, путем анализов с использованием программ предсказания Т-клеточных эпитопов. Примеры сайтов аминокислотных модификаций включают (но, не ограничиваясь только ими) положение 81 и/или положение 82b согласно нумерации Кэбота в последовательности тяжелой цепи антитела к IL-8, представленной в SEQ ID NO: 34.
В одном из вариантов осуществления изобретения референс-антитело, применяемое для функционального сравнения с антителом к IL-8, указанным в контексте настоящего описания, может представлять собой антитело, содержащее аминокислотные последовательности SEQ ID NO: 39 и 40. В конкретном варианте осуществления изобретения референс-антитело, применяемое для ФК-теста, может представлять собой антитело, содержащее аминокислотные последовательности SEQ ID NO: 43 и 45.
В некоторых вариантах осуществления изобретения ингибиторы сигнала IL-8 (например, антитела к IL-8), представленные в настоящем описании, идентифицируют, осуществляют их скрининг или характеризуют их физические/химические свойства и/или виды биологической активности с помощью различных известных методов.
Измерения связывания и другие измерения
Согласно одному из объектов изобретения антитела, представленные в настоящем описании, можно оценивать в отношении их антигенсвязывающей активности с помощью любых методов, таких, например, как ELISA, Вестерн-блоттинг, анализ кинетического исключения (KinExA™) и поверхностный плазмонный резонанс с использованием такого устройства, как BIACORE (фирма GE Healthcare).
Согласно одному из вариантов осуществления изобретения аффинность связывания можно измерять с использованием Biacore Т200 (фирма GE Healthcare) следующим образом. Взятый в соответствующем количестве «захватывающий» белок (например, белок A/G (фирма PIERCE)) иммобилизуют на сенсорном чипе СМ4 (фирма GE Healthcare) методом аминного сочетания и дают ему «захватить» представляющее интерес антитело. Затем разведенный раствор антигена и подвижный буфер (в качестве референс-раствора: например, 0,05% Твин 20, 20 мМ ACES, 150 мМ NaCl, рН 7,4) инъецируют для взаимодействия молекулы антигена с антителом, «захваченным» на сенсорном чипе. Сенсорный чип регенерируют, используя 10 мМ раствор глицина × HCl (рН 1,5). Измерения осуществляют при предварительной определенной температуре (например, 37°С, 25°С или 20°С). KD (М) каждого антитела в отношении антигена рассчитывают на основе константы скорости реакции ассоциации kon (1/Мс) и константы скорости реакции диссоциации koff (1/с), которые обе представляют собой кинетические параметры, рассчитанные на основе сенсограмм, полученных при измерении. Каждый параметр рассчитывают с помощью программного обеспечения BIACORE Т200 Evaluation (фирма GE Healthcare).
В одном из вариантов осуществления изобретения IL-8 можно количественно определять согласно описанному ниже методу. Антитело к человеческому IL-8, которое содержит константную область мышиного IgG, иммобилизуют на планшете. На иммобилизованный планшет добавляют в виде аликвот раствор, содержащий IL-8, связанный с гуманизированным антителом к IL-8, которое не конкурирует с вышеуказанным антителом к человеческому IL-8. После перемешивания добавляют биотинилированное антитело к человеческой легкой цепи Igκ и дают прореагировать в течение определенного периода времени. Затем добавляют также меченый с помощью сульфо-метки стрептавидин и дают прореагировать в течение определенного периода времени. Затем добавляют буфер для анализа и немедленно осуществляют измерения с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery).
Анализы для оценки активности
Одним из объектов изобретения являются анализы для идентификации антитела, обладающего биологической активностью. Биологическая активность включает, например, нейтрализующую антиген (например, IL-8) активность и активность в отношении блокады сигналов от антигена (например, IL-8).
В одном из вариантов осуществления изобретения можно определять уровень IL-8-нейтрализующей активности, например, с помощью описанных ниже методов. Клеточная линия PathHunter™ СНО-К1 CXCR2 β-аррестин (фирма DiscoveRx, каталожный №93-0202 С2) представляет собой искусственную клеточную линию, созданную для экспрессии человеческого CXCR2, известного как человеческий IL-8-рецептор, и испускающую хемилюминесценцию после трансдукции сигналов от человеческого IL-8. После добавления человеческого IL-8 в культуральную среду клеток клетки испускают хемилюминесценцию в зависимости от концентрации добавленного человеческого IL-8. При добавлении человеческого IL-8 в сочетании с антителом к человеческому IL-8 в культуральную среду хемилюминесценция клеток снижается или не поддается обнаружению по сравнению с вариантом без добавления антитела, поскольку антитело к человеческому IL-8 может блокировать трансдукцию сигналов через IL-8. В частности, более сильной нейтрализующей активности антитела в отношении человеческого IL-8 соответствует более низкий уровень хемилюминесценции; а более слабой нейтрализующей активности антитела в отношении человеческого IL-8 соответствует более высокий уровень хемилюминесценции. Таким образом, нейтрализующую активность в отношении человеческого IL-8 антитела к человеческому IL-8 можно определять путем оценки указанного выше различия.
В контексте настоящего описания понятия «диагностическая композиция», «терапевтическая композиция» и «профилактическая композиция» используют взаимозаменяемо с понятиями «диагностический агент», «терапевтический агент» и «профилактический агент» соответственно. В одном из вариантов осуществления изобретения, когда «терапевтическая композиция» или «профилактическая композиция» предназначена для лечения или предупреждения заболевания или симптома у рассматриваемого индивидуума, то композицию обозначают также как «фармацевтическая композиция»; как правило, фармацевтическая композиция относится к агенту, предназначенному для лечения или предупреждения заболевания или симптома.
В контексте настоящего описания понятие «фармацевтически приемлемый носитель» относится к компоненту в фармацевтической композиции, отличному от действующих веществ, входящих в композицию. Фармацевтически приемлемые носители, нетоксичные для индивидуумов, как правило, выбирают с учетом пользы/риска, при этом выбор зависит от поставленной цели. Фармацевтически приемлемые носители включают, например, буферы, эксципиенты, стабилизаторы и консерванты.
В одном из вариантов осуществления изобретения терапевтическая или профилактическая композиция, представленная в настоящем описании, содержит фармацевтически приемлемый носитель и ее можно получать в форме высушенного сушкой вымораживанием агента или в жидкой форме. В одном из вариантов осуществления изобретения, фармацевтически приемлемые носители, как правило, являются нетоксичными для реципиентов в используемых дозах и концентрациях, и они включают, например: буферы, такие как фосфат, цитрат и гистидин; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (например, хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и мета-крезол); полипептиды с низкой молекулярной массой (содержащие менее приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин и иммуноглобулины; гидрофильные полимеры, такие как поли(винилпирролидон); аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу и декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза и сорбит; солеобразующие противоионы, такие как натрий; содержащие металл комплексы (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ).
Альтернативно этому, фармацевтически приемлемые носители могут включать также диспергирующие агенты интерстициальных лекарственных средств, такие как растворимые гликопротеины гиалуронидаз (sHASEGP) (US 2005/0260186; US 2006/0104968). Можно также объединять sHASEGP с одним или несколькими гликозаминогликаназами, такими как хондроитиназы.
В контексте настоящего описания понятие «эффективное количество» действующего вещества (ингибитора сигнала IL-8, такого как ингибитор IL-8 (например, антитело к IL-8 антитело), ингибитора CXCR1 или ингибитора CXCR2) в фармацевтической композиции, означает количество, эффективное при применении в дозах и в течение периода времени, необходимых для достижения требуемого терапевтического или профилактического результата.
Основой одного из объектов изобретения, представленных в настоящем описании, является применимость ингибиторов сигнала IL-8 (например, ингибиторов IL-8 (например, антител к IL-8), ингибиторов CXCR1 и ингибиторов CXCR2) в качестве фармацевтических композиций. Ингибиторы сигнала IL-8 (например, ингибиторы IL-8 (например, антитела к IL-8), ингибиторы CXCR1 и ингибиторы CXCR2), представленные в настоящем описании, можно применять, например, для диагностирования, лечения или предупреждения заболеваний, при которых IL-8 присутствует в избыточном количестве.
В одном из вариантов осуществления изобретения ингибиторы сигнала IL-8 (например, ингибитор IL-8 (например, антитело к IL-8), ингибитор CXCR1 и ингибитор CXCR2), представленные в настоящем описании, вводят любыми приемлемыми путями, включая парентеральный, внутрилегочный и интраназальный и при необходимости применяют для местной обработки, введения в повреждение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное и подкожное введение. Дозирование можно осуществлять любым приемлемым путем, например, путем инъекций, таких как внутривенные или подкожные инъекции, в зависимости, в том числе, от того, является ли лечение кратковременным или хроническим. Можно применять различные схемы дозирования
В одном из вариантов осуществления изобретения ингибитор сигнала IL-8, представленный в настоящем описании, можно включать в состав композиций, дозировать и вводить в соответствии с надлежащей клинической практикой.
Один из вариантов осуществления изобретения, представленных в настоящем описании, относится к продуктам, содержащим ингибиторы IL-8, которые можно применять для диагностирования, лечения и/или предупреждения заболеваний, указанных в контексте настоящего описания. Указанный продукт включает контейнер и этикетку или сопровождающий его документ, присоединенный к контейнеру. Приемлемыми контейнерами являются, например банки, пузырьки и пакеты для внутривенного раствора.
Приемлемую дозу и интервалы между введениями ингибитора сигнала IL-8 (например, ингибитора IL-8 (например, антитела к IL-8), ингибитора CXCR1 или ингибитора CXCR2), представленного в настоящем описании, для предупреждения или лечения заболевания можно определять с учетом типа заболевания, подлежащего лечению, типа ингибитора, серьезности и течения болезни, цели введения, предшествующей терапии, истории болезни пациента и ответа на ингибитор и предписания лечащего врача.
Согласно одному из вариантов осуществления изобретения терапевтические воздействия включают, например, предупреждение развития или рецидива, ремиссию симптомов, облегчение любого прямого или косвенного патологического последствия заболевания, предупреждение метастазов, уменьшение скорости прогрессирования заболевания, излечивание, облегчение и ремиссию патологического состояния, и улучшение прогноза.
В одном из вариантов осуществления изобретения, представленных в настоящем описании, заболевания или симптомы, при которых IL-8 присутствует в избыточном количестве, и заболевания, при которых сигналы IL-8 участвуют или могут участвовать в возникновении, прогрессировании, обострении, рецидиве и т.п. патологических состояний, в целом обозначают как «связанные с IL-8 заболевания».
В этом контексте одним из вариантов осуществления изобретения, представленных в настоящем описании, являются терапевтические или профилактические композиции, предназначенные для лечения связанных с IL-8 заболеваний, которые содержат ингибитор сигнала IL-8 (например, ингибитор IL-8 (например, антитело к IL-8), ингибитор CXCR1 или ингибитор CXCR2), в качестве действующего вещества. В дополнительном варианте осуществления изобретения композиции могут дополнительно содержать фармацевтически приемлемый носитель.
В этом контексте одним из вариантов осуществления изобретения, представленных в настоящем описании, являются ингибиторы сигнала IL-8 (например, ингибиторы IL-8 (например, антитела к IL-8), ингибиторы CXCR1 и ингибиторы CXCR2), предназначенные для применения для лечения или предупреждения связанных с IL-8 заболеваний (например, эндометриоза, аденомиоза, дисменореи, спаек и фиброзных заболеваний).
В этом контексте одним из вариантов осуществления изобретения, представленных в настоящем описании, является применение ингибиторов сигнала IL-8 (например, ингибиторов IL-8 (например, антител к IL-8), ингибитора CXCR1 и ингибиторов CXCR2), в качестве диагностических, терапевтических или профилактических композиций для связанных с IL-8 заболеваний (например, эндометриоза, аденомиоза, дисменорея, спаек и фиброзных заболеваний).
В этом контексте одним из вариантов осуществления изобретения, представленных в настоящем описании, являются способы лечения или предупреждения связанных с IL-8 заболеваний (например, эндометриоза, аденомиоза, дисменореи, спаек и фиброзных заболеваний), которые включают введение (в эффективном количестве) ингибитора сигнала IL-8 (например, ингибитора IL-8 (например, антитела к IL-8), ингибитора CXCR1 или ингибитора CXCR2), или содержащей его терапевтической или профилактической композиции индивидууму, который нуждается в этом. В контексте настоящего описания индивидуум, который нуждается в этом, может представлять собой индивидуума, который страдает или который предположительно может страдать связанным с IL-8 заболеванием. В другом варианте осуществления изобретения композиции могут дополнительно содержать фармацевтически приемлемый носитель.
В этом контексте одним из вариантов осуществления изобретения, представленных в настоящем описании, является применение ингибитора сигнала IL-8 (например, ингибитора IL-8 (например, антитела к IL-8), ингибитора CXCR1 или ингибитора CXCR2), представленного в настоящем описании, или содержащей его терапевтической или профилактической композиции для приготовления фармацевтических агентов, предназначенных для лечения или предупреждения связанных с IL-8 заболеваний (например, эндометриоза, аденомиоза, дисменореи, спаек и фиброзных заболеваний). В другом варианте осуществления изобретения композиции могут дополнительно содержать фармацевтически приемлемый носитель.
В одном из вариантов осуществления изобретения связанные с IL-8 заболевания, которые являются мишенью лечения или предупреждения с использованием ингибитора сигнала IL-8 (например, ингибитора IL-8 (например, антитела к IL-8), ингибитора CXCR1 или ингибитора CXCR2), представленного в настоящем описании, включают, например: эндометриоз; аденомиоз; дисменорею; спайки (синехии), такие как синдром Ашермана; бесплодие; боль при эндометриозе, аденомиозе или дисменореи; боль при спайках, фиброзе или воспалении; воспалительные заболевания кожи, такие как воспалительный кератоз, атопический дерматит, контактный дерматит, ладонно-подошвенный пустулез и псориаз; хронические воспалительные заболевания, такие как хронический ревматоидный артрит, системная красная волчанка (SLE) и болезнь Бехчета, представляющие собой аутоиммунные заболевания: воспалительные заболевания кишечника, такие как болезнь Крона и неспецифический язвенный колит; воспалительные заболевания печени, такие как гепатит В, гепатит С, алкогольный гепатит и индуцированный лекарственным средством аллергический гепатит; воспалительные заболевания почек, такие как гломерулонефрит и ассоциированный с ANCA гепатит; воспалительные респираторные заболевания, такие как бронхит и астма; воспалительные хронические сосудистые заболевания, такие как, атеросклероз; рассеянный склероз; стоматит; хордит, синдром пост-трансплантационной ишемии-реперфузии; воспаление, ассоциированное с применением искусственных органов и/или искусственных кровеносных сосудов; злокачественные опухоли, такие как рак яичника, рак легкого, рак предстательной железы, рак желудка, рак молочной железы, меланома (злокачественная меланома), раки головы и шеи, рак почки, рак мочевого пузыря и глиобластома; сепсис, связанный с инфекцией или т.п. (включая септический шок); острое легочное повреждение; фиброзные заболевания, такие как фиброз при эндометриозе или аденомиозе, муковисцидоз, фиброз печени, фиброз почки и легочный фиброз, такой как хроническое обструктивное заболевание легких (COPD); нарушения головного мозга, выбранные из мозгового удара, инфаркта головного мозга (например, ишемического мозгового инфаркта), церебрального тромбоза, церебральной эмболии, геморрагического поражения сосудов головного мозга, внутримозгового кровоизлияния и субарахноидального кровоизлияния; инфаркта сердца (включая острый инфаркт сердца, нестабильную стенокардию и ишемическое/реперфузное повреждение сердца) (см., например, раздел «Примеры» в контексте настоящего описания; документы, представленные в контексте настоящего описания; Remo и др., Expert Rev. Clin. Immunol. 10(5), 2014, cc. 593-619; Mian и др., Clin Cancer Res. 9(8), 1 августа 2003 г., cc.3167-3175; Huang и др., Am J Pathol. 161(1), июль, 2002 г., cc. 125-134; WO 97/49426; WO 97/39775; JP 10-045622; JP 10-182488; JP 10-053536). Связанные с IL-8 заболевания могут представлять собой заболевания млекопитающих и предпочтительно заболевания приматов и более предпочтительно заболевания человека (например, человеческий эндометриоз в случае эндометриоза, человеческий аденомиоз в случае аденомиоза; и это далее используется в контексте настоящего описания).
В одном из вариантов осуществления изобретения пациенты (индивидуумы) со связанным с IL-8 заболеванием, которые являются мишенью терапии или профилактики с использованием ингибитора сигнала IL-8, представленного в настоящем описании, могут представлять собой индивидуумов, которые страдают или предположительно могут страдать связанным с IL-8 заболеванием. В контексте настоящего описания индивидуумы, которые предположительно могут страдать связанным с IL-8 заболеванием, включают (но, не ограничиваясь только ими) индивидуумов, которые уже один раз страдали связанным с IL-8 заболеванием и у них существует возможность рецидива заболевания, и индивидуумов, которые предположительно могут страдать связанным с IL-8 заболеванием до постановки подтвержденного диагноза или т.п. врачом или другими специалистами на основе патологических проявлений, свидетельствующих о связанном с IL-8 заболевании. В одном из вариантов осуществления изобретения предупреждение и терапию связанного с IL-8 заболевания можно в некоторых случаях интерпретировать в качестве синонимов. Альтернативно этому, предупреждение связанного с IL-8 заболевания можно, если это допустимо, можно интерпретировать на основе общей концепции, хорошо известной специалистам в данной области. В одном из вариантов осуществления изобретения, представленных в настоящем описании, связанные с IL-8 заболевания чувствительны к сигналам IL-8.
В контексте настоящего описания понятие «эндометриоз» относится (но, не ограничиваясь только указанным) к заболеванию, симптомы которого проявляются в том, что ткани, имеющие морфологию и функцию такую же или сходную с эндометриоидными тканями матки, развиваются и прорастают эктопически за пределы полости матки (как правило, внутрь таза, иногда в брюшную полость и редко в грудную полость) (Japan Society of Obstetrics and Gynecology, 1993). Эндометриоз не является злокачественной опухолью, однако эта болезнь прогрессирует с течением времени и вызывает образование опухоли и спайки в окружающих тканях, серьезную боль в брюшной и тазовой области во время менструации или между периодами, боль при половом контакте, боль при дефекации и бесплодие (нарушенная фертильность). Эндометриоз развивается главным образом у женщин в репродуктивном возрасте 20 лет или старше, снижает качество жизни (QOL) из-за боли и других симптомов и может приводить к хронической боли и бесплодию вследствие серьезных спаек. Фиброз является результатом прогрессирования патологического состояния в области спаек в эндометриоидных повреждениях (в пораженных областях). В областях фиброза образуется коллаген, и это приводит к фиброзу. Прогрессирование фиброза при эндометриозе, вероятно, как правило, является необратимым. Так, отсутствуют данные о методах прекращения развития фиброза или облегчения фиброза при эндометриозе. Как описано выше, эндометриоз представляет собой заболевание, которое усиливается с течением времени, имеющее различные стадии, характеризуясь при этом различными патологическими особенностями в процессе прогрессирования относительно начала болезни; однако заболевание в целом называют эндометриозом. Однако в настоящее время создана модель in vivo эндометриоза с использованием приматов кроме человека с менструацией, сходной с менструацией человека, что в конечном итоге облегчает понимание патологических состояний заболевания. При создании настоящего изобретения путем хирургической индукции эндометриоза создана модель in vivo эндометриоза на обезьянах циномолгус с менструацией, сходной с менструацией человека. Используя указанную надежную модель in vivo при создании настоящего изобретения впервые в мире было успешно доказано, что разрастание эндометриоидных повреждений можно подавлять путем введения ингибиторов сигнала IL-8 в живой организм, достигая тем самым атрофических изменений эпителиальных клеток и стромальных клеток в эндометриоидных повреждениях или уменьшения интерстиция при эндометриозе, что приводит к уменьшению количества гладкомышечных клеток. Из-за атрофии эндометриоидных тканей, в частности, интерстиция, можно ожидать также уменьшения количества эктопических нейронов. Кроме того, при создании настоящего изобретения обнаружена регенерация мышечной ткани в группе, которой вводили ингибиторы сигнала IL-8, что не выявлено в обработанной наполнителем группе. Таким образом, при создании настоящего изобретения обнаружен регресс повреждений, внедрившихся в миометрий, и замещение их мышечными тканями для восстановления. Присутствие глубоких повреждений оценивали по тому, имело ли место внедрение протоков эндометриоидной железы в мышечный слой брюшной стенки, и было высказано предположение о том, что ингибиторы сигнала IL-8 могут подавлять образование глубоких повреждений. Кроме того, при создании настоящего изобретения также впервые было обнаружено, что ингибиторы сигнала IL-8 обладают способностью подавлять даже спайки и фиброз при эндометриозе. Таким образом, согласно одному из вариантов осуществления изобретения ингибиторы сигнала IL-8, представленные в настоящем описании, можно применять для лечения или предупреждения симптома, такого как фиброз, на конкретной стадии эндометриоза, который рассматривается в качестве фиброзной болезни. Кроме того, на модели in vivo на обезьянах циномолгус установлено, что одно животное имело нерегулярный половой цикл, вероятно, вследствие влияния процесса создания модели; однако за исключением этого животного, половой цикл был регулярным, поэтому можно предположить, что ингибиторы сигнала IL-8 не оказывали влияния на половой цикл (см. раздел «Примеры» в настоящем описании). Эти результаты свидетельствуют о том, что ингибиторы сигнала IL-8 являются перспективными терапевтическими или профилактическими агентами по сравнению с общепринятыми терапевтическими агентами, применяемыми при эндометриозе. Кроме того, неожиданно было установлено, что эндометриоз можно лечить или предупреждать, используя ингибитор сигнала IL-8 в качестве единственного лекарственного средства без его объединения с гормональным лекарственным средством. В одном из вариантов осуществления изобретения эндометриоз, указанный в настоящем описании, является чувствительным к сигналам IL-8.
В контексте настоящего описания понятие «аденомиоз» относится (но, не ограничиваясь только указанным) к состоянию, при котором эндометриоидные ткани, в частности, эндометриоидные железы и окружающая матку строма, присутствуют в мускулатуре матки, и при этом заболевании имеет место гипертрофия и гиперплазия окружающих гладких мышц, и это вызывает серьезную боль при менструации, гиперменорею, анемию и хроническую боль. Хотя 6-20% страдающих аденомиозом пациентов имеют также эндометриоз (Taketani и др., Study report on mental and physical disorders sponsored by the Ministry of Health and Welfare. 1998, cc. 99-104), аденомиоз представляет собой болезнь, отличную от эндометриоза, согласно классификации заболеваний. В настоящее время специалисты в данной области обычно рассматривают их в качестве двух отдельных отличных друг от друга заболеваний (Frankl и др., 10, 1995, сс. 680-684; Brosens и др., Lancet., 341(8838), 16 января 1993 г., сс. 181-182; Benagiano и др., Best Pract Res Clin Obstet Gynaecol. 20(4), август 2006 г., сс. 449-463). Хотя аденомиоз напоминает эндометриоз по развитию эндометрия и эктопическому разрастанию и тем изменениям, которые происходят зависимым от эстрогена образом, и боли при менструации, что является основным симптомом, аденомиоз часто развивается у женщин относительно более старшего возраста по сравнению с эндометриозом, что является его эпидемиологическим отличием от эндометриоза. Невозможно точно определить причину, приводящую к разрушению пограничного слоя между миометрием и базальным слоем эндометрия; однако для разработки предложена следующая гипотеза: повреждения матки (чрезмерное количество выскабливаний или повреждения при родах) приводят к непосредственному включению эндометрия в миометрий (Pappas и др., Obset Gynecol. 13, 1959, с. 714; Benagiano и др., Fertil Steril. 98(3), сентябрь 2012 г., сс. 572-579). Состояние, при котором эндометриоидные ткани отсутствуют на вплоть до 90% миометрия на обращенной в полость стороне и они распределены только на серозной стороне миометрия, называют эндометриозом тазовой брюшины (Bergeron и др., Best Pract Res Clin Obset Gynaecol 2006; 20(4):511-521).
При создании изобретения было обнаружено и подтверждено спонтанное начало аденомиоза на вышеописанной модели in vivo на обезьянах циномолгус и впервые в мире успешно продемонстрировано, что разрастание связанных с аденомиозом повреждений можно подавлять путем введения ингибиторов сигнала IL-8 в живой организм. Ингибиторы сигнала IL-8 не влияют на половой цикл на модели in vivo на обезьянах циномолгус (см. раздел «Примеры» в настоящем описании). Таким образом, ингибиторы сигнала IL-8 являются перспективными терапевтическими или профилактическими агентами по сравнению с общепринятыми терапевтическими агентами, применяемыми при аденомиозе. Кроме того, неожиданно было установлено, что аденомиоз можно лечить или предупреждать, используя ингибитор сигнала IL-8 в качестве единственного лекарственного средства без его объединения с гормональным лекарственным средством. Введение ингибитора сигнала IL-8 может приводить к истончению гипертрофированной стенки матки при аденомиозе и поэтому можно ожидать, что он будет оказывать терапевтические или профилактические воздействия на фиброз при аденомиозе. В одном из вариантов осуществления изобретения аденомиоз, указанный в настоящем описании, является чувствительным в сигналам IL-8.
Понятие «дисменорея» не ограничено в контексте настоящего описания, но может прежде всего относиться к обозначенному таким образом симптому, который нарушает повседневную жизнь. Известно, что эндометриоз, аденомиоз, гиперменорея и т.п. подпадают под понятие дисменорея, и ее типичные симптомы включают, например, поясничную боль, головную боль, тошноту, анемию и утомляемость, а также боль в области живота в процессе менструации, в части, боль в нижней части живота. На основе ее причин дисменорею грубо классифицируют как относящуюся к двум типам: функциональная (первичная) дисменорея и органическая (вторичная) дисменорея. Причины функциональной дисменореи включают, например, избыточную секрецию простагландинов, а также причины, связанные с прогрессированием заболевания, такого как фиброид матки и эндометриоз. Причины органической дисменореи включают, например, причины, связанные с прогрессированием заболевания, такого как фиброид матки и эндометриоз. Кроме того, известно, что дисменорея вызывается воспалением внутри или снаружи матки. При этом известно, что IL-8 представляет собой провоспалительный цитокин. Таким образом, согласно одному из вариантов осуществления изобретения ингибитор сигнала IL-8, предлагаемый в настоящем описании, может лечить или предупреждать дисменорею. Согласно предпочтительному не ограничивающему объем изобретения варианту осуществления изобретения ингибитор сигнала IL-8, представленный в настоящем описании, может лечить или предупреждать дисменорею при эндометриозе или аденомиозе, дисменорею, при которой предполагается наличие эндометриоза или аденомиоза, дисменорею, сопровождающую воспаление, или дисменорею с болью, вызываемой спайками. Альтернативно этому, в другом варианте осуществления изобретения дисменорея, указанная в настоящем описании, является чувствительной к сигналу IL-8.
В контексте настоящего описания понятие «дисменорея, при которой предполагается наличие эндометриоза или аденомиоза» включает (но, не ограничиваясь только ими) дисменорею у индивидуума, который уже страдал один раз эндометриозом или аденомиозом и имеет вероятность реактивации заболевания, и дисменорею у индивидуума, у которого предполагается эндометриоз или аденомиоз до постановки диагноза или т.п. лечащим врачом или другим специалистом, например, на основе патологических признаков, свидетельствующих о эндометриозе или аденомиозе.
В одном из вариантов осуществления изобретения, в котором ингибитор сигнала IL-8, представленный в настоящем описании, применяют для лечения или предупреждения дисменореи, сопровождающей эндометриоз или аденомиоз, или дисменорею, при которой предполагается наличие эндометриоза или аденомиоза, ингибитор сигнала IL-8 обладает преимуществом, поскольку не влияет на половой цикл и его можно применять для лечения или предупреждения эндометриоза или аденомиоза в качестве единственного лекарственного средства без его объединения с гормональным лекарственным средством, как описано выше.
Кроме того, при создании настоящего изобретения было сделано предположение и продемонстрировано, что ингибиторы сигнала IL-8 помимо эндометриоза и аденомиоза могут лечить или предупреждать также спайки, что описано в разделе «Примеры» в настоящем описании. Дисменорея также характеризуется болью в области живота. Поэтому естественно было предположить, что не только спайки (спаечный процесс) при эндометриозе, но также спайки, связанные с другими симптомами или нарушениями, вызывают боль у пациентов вследствие адгезивного контакта тканей или органов. Например, известно, что при спайках в тазовой области боль коррелирует с образованием спайек в дугласовом кармане, и при этом не коррелирует со спайками между придатками и репродуктивным органом (Porpora и др., The Journal of the American Association of Gynecologic Laparoscopists 6, 1999, cc. 429-434), а спайки между органами вызывают судороги, приводящие к боли. Кроме того, известно, что эктопические нервные волокна и производство NGF вызывают боль и обострение боли при эндометриозе (Anaf и др., Hum Reprod 15, 2000, сс. 1744-1750; Berkley и др., Proc Natl Acad Sci USA 101, 2004, cc. 11094-11098; Odagiri и др., Fertil Steril 92, 2009, cc. 1525-1531). Кроме того, известна более ранняя публикация (Odagiri и др., Fertil Steril 92, 2009, cc. 1525-1531), в которой высказано предположение о том, что интерстициальная ткань с увеличенным гладкомышечным окружением эндометриоидных повреждений усиливает боль в процессе сокращения. Установлено, что введение антител к IL-8, указанных в разделе «Примеры» в контексте настоящего описания, уменьшает интерстициальную ткань эндометриоидных тканей и, как ожидается, должно уменьшать количество эктопических нейронов и таким образом уменьшать боль.
Согласно одному из вариантов осуществления изобретения ингибитор сигнала IL-8, представленный в настоящем описании, может лечить или предупреждать боль при эндометриозе или аденомиозе (хроническую боль, боль в процессе менструации и т.д.), боль при дисменорее или боль, связанную со спайками, фиброзом или воспалением. В данном контексте хроническая боль включает (но, не ограничиваясь только ею) хроническую боль в нижней части живота и хроническую боль в области таза.
В одном из вариантов осуществления изобретения в контексте настоящего описания понятие «не влияет на половой цикл» может означать, что практически не обнаружено воздействия на периодический менструальный цикл и овуляцию у индивидуума (женщина или самка животного кроме человека). Периодичность цикла можно оценивать по наличию кровотечения в процессе менструации, что описано в разделе «Примеры» в настоящем описании, или на основе изменений уровней эстрогена или прогестерона в крови или моче. Известно, что гормональная (например, лекарственное средство на основе комбинации эстрогена/прогестерона, препарат прогестерона, агонист GnRH и даназол) терапия влияет на менструальный цикл и ингибирует овуляцию. Наличие значительного воздействия можно оценивать (но, не ограничиваясь только указанным), например, путем изучения тенденции или путем статистической оценки воздействия на менструальный цикл в группе, которую обрабатывали ингибитором сигнала IL-8, по сравнению с референс-группой, которую подвергали гормональной терапии, которая, как известно, влияет на половой цикл. Согласно одному из вариантов осуществления изобретения, поскольку ингибитор сигнала IL-8, представленный в настоящем описании, не влияет на половой цикл у индивидуумов с эндометриозом, аденомиозом или дисменореей, которым его вводят, сделано предположение о том, что он не будет ингибировать овуляцию и дает шанс забеременеть в отличие от общепринятой гормональной терапии. Кроме того, ингибитор обладает преимуществом, состоящим в том, что позволяет исключить применение гормональных лекарственных средств, и поэтому позволяет индивидуумам избегать неблагоприятных воздействий, таких как менопауза, связанных с низким уровнем эстрогена.
Организм обладает способностью заживлять раны, исцеляя место повреждения или ранения при повреждении или нарушении клеток, тканей, органов или внутренних органов. Однако в процессе заживления раны могут образовываться в норме нежелательные слипания, которые могут иметь место между клетками, тканями, органами или внутренними органами. Как правило (но, не ограничиваясь только указанным) указанное состояние слипания обозначают как образование спаек (адгезия). Для некоторых пациентов не характерен симптом образования спаек; однако незначительное количество пациентов нуждается в лечении из-за сопутствующих симптомов, таких как боль, бесплодие и чувство тяжести в брюшной области. Спайки включают, например, спайки, образовавшиеся (образующиеся) после хирургического вмешательства (например, спайки, которые прямо или косвенно сформировались (формируются) на поверхности хирургического разреза, в областях вокруг места наложения шва, местах потертостей, например, марлей; внутриматочные спайки (синехии) (синдром Ашермана), образовавшиеся (образующиеся) после выскабливания матки; спайки, вызываемые лекарственными средствами (например, местной обработкой органов лекарственными средствами); спайки, вызываемые заболеваниями (например, эндометриоз, аденомиоз, инфильтрующие раковые клетки или ткани и воспалительные заболевания, связанные с инфекцией (сальпингит, оофорит, тазовый перитонит и т.д.); атрезию кишечника в результате образования спаек; и спонтанное образование спаек. При создании настоящего изобретения случайно на модели эндометриоза in vivo на обезьянах циномолгус было обнаружено, что введение in vivo ингибитора сигнала IL-8 снижало образование спаек в области лапаротомии, отличных от спаек при пост-хирургическом эндометриозе. В одном из вариантов осуществления настоящего изобретения спайки чувствительны к сигналам IL-8.
В контексте настоящего описания бесплодие означает (но, не ограничиваясь только указанным), что, несмотря на половой контакт без контрацепции, у здоровой пары, в которой супруги хотят наступления беременности, беременность не достигается в течение определенного периода времени (как правило, в течение 1 года) (Japan Society of Obstetrics and Gynecology, 2016). В этом контексте бесплодие включает (но, не ограничиваясь только указанным) бесплодие вследствие образования спаек (в частности, вызываемое адгезией) и бесплодие, вызываемое воспалением внутри брюшной области. В одном из вариантов осуществления настоящего изобретения бесплодие является чувствительным к сигналам IL-8.
В контексте настоящего описания ладонно-подошвенный пустулез означает (но, не ограничиваясь только указанным) заболевание, при котором образуются многочисленные высыпания (наполненные гноем пузыри) на ладонях или подошвах. Наполненные гное пузыри являются стерильными и не содержат бактерий или грибов, в них нейтрофилы накапливаются в кератине кожи и IL-8 присутствует в высокой концентрации (Skov и др., J Immunol 181, 2008, cc. 669-679). Симптом является хроническим с частыми рецидивами в течение длительного периода времени. В клинических испытаниях установлено, что при ладонно-подошвенном пустулезе введение HuMablOFS, представляющего собой антитело к IL-8, патологическое состояние облегчалось (Skov и др., J Immunol 181, 2008, cc. 669-679). Таким образом, в одном из вариантов осуществления настоящего изобретения ладонно-подошвенный пустулез является чувствительным к сигналам IL-8.
В контексте настоящего описания ANCA-ассоциированный нефрит означает нефрит, позитивный по ANCA (антитело к цитоплазме нейтрофилов). ANCA-ассоциированный нефрит представляет собой заболевание, при котором имеет место воспаление в почечных кровеносных сосудах, таких как клубочки, и типичным патологическим состоянием является некротизирующий серповидный гломерулонефрит. По мере того, как нефрит становится хроническим, фиброз прогрессирует и клубочки затвердевают, что приводит к почечной недостаточности («Evidence-based Clinical Guideline for rapidly progressive glomerulonephritis (PRGN) 2014» в Japanese; Nature Review Rheumatology 10, 2014, c. 463). При ANCA-ассоциированном нефрите в повреждениях обнаружен высокий уровень экспрессии IL-8, и способность к миграции нейтрофилов пациентов дополнительно повышалась при обработке ANCA и подавлялась при обработке антителом к IL-8 (Cockwell и др., Kidney Int. 55(3), март 1999 г., сс. 852-863). В одном из вариантов осуществления изобретения, представленных в настоящем описании, ANCA-ассоциированный нефрит является чувствительным к сигналам IL-8.
В контексте настоящего описания муковисцидоз означает системное аутосомно-рецессивное наследственное нарушение, вызываемое генетической мутацией в регуляторе трансмембранной проводимости при муковисцидозе (CFTR). Секреторная жидкость и мукозная жидкость, такая как жидкость дыхательных путей, жидкость брюшной полости и жидкость поджелудочной железы, становятся слишком вязкими во всем организме, что приводит к окклюзии полостей, и обусловливает предрасположенность к инфекции и проявляется в виде таких симптомов, как мекониевая непроходимость кишечника, расстройство пищеварения и нарушение всасывания, из-за атрофии поджелудочных желез и экзокринной недостаточности, и респираторная недостаточность из-за рецидивирующей инфекции дыхательных путей. Повреждения и окружающая их область, например, жидкость дыхательных путей или жидкость бронхоальвеолярного лаважа, содержит в высокой концентрации IL-8 (Marcos и др., Nat Med. 16(9), сентябрь 2010 г., сс. 1018-1023; Khan и др., Am J Respir Crit Care Med. 151(4), апреля 1995 г., сс. 1075-1082) и в них присутствует большое количество нейтрофилов, мигрирующих под действием IL-8. Предполагается, что возникает избыточное и персестирующее воспаление и нейтрофилы продуцируют ферменты, такие как эластаза, и подвергаются нетозу, и в результате вязкость общей воды организма повышается, приводя к появлению указанных симптомов (Cheng и др., Front Immunol. 4, 24 января 2013 г., с. 1). Обнаружено, что местная миграция нейтрофилов подавлялась в клинических условиях ингибитором CXCR2 элубриксином, средством против муковисцидоза (J Cyst Fibros. 12(3), май 2013 г., сс. 241-248). Можно ожидать, что, когда производство ферментов и нетоз подавляются путем ингибирования миграции нейтрофилов посредством блокады пути IL-8, то местная вязкость снижается и симптомы облегчаются. Таким образом, в одном из вариантов осуществления изобретения, представленных в настоящем в настоящем описании, муковисцидоз является чувствительным к сигналам IL-8.
В контексте настоящего описания, не ограничивая его объем, псориаз также относят к воспалительному заболеванию. Его типичным симптомом являются четко ограниченные красные папулы или выпуклые образования, покрытые серебристой перхотью. Обычные провоцирующие факторы включают наружное повреждение, инфекцию и некоторые лекарственные средства. Как правило, симптомы практически отсутствуют, но неожиданно появляется слабый зуд. Некоторые основные жалобы могут быть обусловлены косметическими недостатками. Некоторые люди в качестве симптома имеют артрит, сопровождающийся болью, что повышает серьезность заболевания. В US 2003/0077283 А1 описана местная терапия при псориазе с использованием нейтрализующего IL-8 антитела. Кроме того, в продажу подступает терапевтический агент в виде нейтрализующего антитела к IL-8 под названием Abcream TM для лечения инфекции. Таким образом, в одном из вариантов осуществления изобретения, представленных в настоящем в настоящем описании, псориаз является чувствительным к сигналам IL-8.
В основе фиброза лежит следующий механизм: воспаление после повреждения, вызванного кровотечением или заболеванием, служит в качестве провоцирующего фактора, и хемокины и цитокины, включая IL-8, приводят к рекрутингу иммунных клеток, таких как нейтрофилы и моноциты, из окружающей ткани; продуцируются цитокины и хемокины, и это индуцирует активацию и пролиферацию мышечных фибробластов (или фиброзных клеток), а также избыточное производство или пониженное расщепление внеклеточного матрикса, такого как коллаген, что приводит к фиброзу. Это является общим для фиброза в различных органах (N Engl J Med 372, 19 марта 2015 г., cc. 1138-1149).
В контексте настоящего описания заболевания и состояния с таким фиброзом обозначают в целом как «фиброзные заболевания», фокусируя внимание на фиброзном состоянии.
Поскольку ингибиторы сигнала IL-8 оказывают ингибирующее действие на миграцию нейтрофилов, которые продуцируют МСР-1 (известный как фактор миграции макрофагов и усиливающий фиброз фактор) и т.п., при оценке в анализе in vitro, в дополнение к тому факту, что ингибиторы сигнала IL-8 подавляют инфильтрацию иммунных клеток и уменьшают богатый коллагеном интерстиций на модели in vivo на обезьянах циномолгус (см. раздел «Примеры» в настоящем описании), ингибиторы сигнала IL-8 являются перспективными терапевтическими или профилактическими агентами при описанном выше фиброзе.
Кроме того, различные антитела к IL-8 подавляли повышение экспрессии фактора роста соединительной ткани (СТОР)являющегося усиливающим фиброз фактором, в анализе in vitro, который представлен в разделе «Примеры» в настоящем описании. Этот результат подтвердил тот факт, что ингибиторы сигнала IL-8, такие как антитела к IL-8, являются перспективными терапевтическими или профилактическими агентами при фиброзных заболеваниях.
В одном из вариантов осуществления изобретения, представленных в настоящем описании, спайки и фиброз при эндометриозе могут возникать во время процесса, при котором ткань матки развивается и прорастает наружу от матки (например, в органы таза, в брюшную полость или в грудную полость).
В контексте настоящего описания печеночный фиброз означает (но, не ограничиваясь только указанным) заболевание, при котором хроническое воспаление, вызываемое вирусной инфекцией, отложением жира или т.п., провоцирует избыточное производство или пониженное расщепление внеклеточного матрикса (ЕСМ), такого как коллаген, в печени, что приводит к фиброзу. Печеночный фиброз включает, например, цирроз, вызванный хроническим воспалением, таким как вирусный гепатит, алкогольный гепатит, неалкогольная жировая болезнь печени (NAFLD) или первичный билиарный цирроз (РВС).
Известны следующие данные: уровень IL-8 повышается при хронических заболеваниях печени, таких как вирусный гепатит А и В, алкогольный гепатит и первичный билиарный цирроз (Mediators Inflamm. 2015, 2015, с. 276850); и уровень IL-8 в крови является высоким при хронических заболеваниях печени и особенно высоким у пациентов с запущенными патологическими состояниями; и уровень экспрессии рецептора CXCR1 является высоким в моноцитах; и IL-8 сильно повышает воспаление в печени; и указанные результаты позволяют предположить участие IL-8 в прогрессировании фиброза (PLoS One. 2011; 6(6):е21381).
Кроме того, концентрация IL-8 в плазме существенно увеличена у пациентов с NASH, имеющих фиброз (Gastroenterology, 149(3), сентябрь 2015 г., сс. 635-648.е14), что позволяет предположить наличие корреляции между IL-8 и фиброзом. Таким образом, в одном из вариантов осуществления изобретения, представленных в настоящем в настоящем описании, печеночный фиброз является чувствительным к сигналам IL-8.
В контексте настоящего описания почечный фиброз означает (но, не ограничиваясь указанным) заболевание, при котором воспаление провоцирует избыточное производство или пониженное расщепление внеклеточного матрикса, такого как коллаген, в почке, что приводит к фиброзу. Почечный фиброз включает, например, хронические болезни почек, вызванные хроническим воспалением, таким как хронический нефрит и диабет. По мере прогрессирования хронических заболеваний почек фиброз развивается в почках вне зависимости от являющегося причиной заболевания. Начавшись, фиброз становится необратимым, и в случае фиброза почек требуется диализ или трансплантация почки.
Опубликованы следующие данные: в почечных фибробластах повышается производство IL-8 посредством IL-1 (Kidney Int. 47(3), март 1995 г., сс. 845-854); почечный фиброз уменьшается при подавлении сигнала МСР-1 (Kitagawa и др., Am J Pathol. 165(1), июль 2004 г., сс. 237-246); активацию и механизм фиброза при диабетическом состоянии изучали с использованием человеческих фиброзных клеток, и результаты позволили предположить, что человеческие фиброзные клетки участвуют в прогрессировании диабетической нефропатии благодаря повышенным уровням глюкозы и MCP-1/CCR2 (Clin Exp Nephrol. 17(6), декабрь 2013 г., сс. 793-804). Поскольку ингибиторы сигнала IL-8 ингибируют миграцию продуцирующих МСР-1 нейтрофилов к повреждениям (см. раздел «Примеры» в настоящем описании), можно ожидать, что ингибиторы будут подавлять развитие или облегчать почечный фиброз посредством МСР-1. В одном из вариантов осуществления изобретения, представленных в настоящем в настоящем описании, почечный фиброз является чувствительным к сигналам IL-8.
В контексте настоящего описания легочный фиброз означает (но, не ограничиваясь указанным) заболевание, при котором фиброз вызывается избыточным производством или пониженным расщеплением внеклеточного матрикса, такого как коллаген, в легких или бронхах, что провоцируется воспалением. Легочный фиброз включает, например, хроническое обструктивное заболевание легких (COPD), комбинированный фиброз легких и эмфизему легких (CPFE), идиопатические интерсцитиальные пневмонии (IIP) и идиопатический легочный фиброз (IPF).
Хроническое обструктивное заболевание легких (COPD) означает (но, не ограничиваясь только указанным) воспалительное заболевание легких, вызванное продолжительным ингаляционным воздействием вредных субстанций, прежде всего сигаретного дыма (Guideline for diagnosis and therapy of chronic obstructive pulmonary disease (COPD), 4-ое изд.). В дыхательных путях и в легких хроническое воспаление снижает эластичность альвеол и приводит к уменьшению просвета бронхов, в результате чего воздушный поток к легким становится слабым, а препятствие воздушному потоку вызывает затрудненность дыхания, хронический кашель, образование мокроты и т.д. Известно, что в мокроте страдающих COPD пациентов повышен уровень IL-8 и присутствует большое количество нейтрофилов, и хемотаксический индекс, полученный при анализе миграции нейтрофилов пациентов, коррелирует с легочной функцией FEV1/FVC (Yamamoto и др., Chest. 112(2), август 1997 г., сс. 505-510; Wu и др., PLoS One. 10(5), 11 мая 2015 г.). Кроме того, описано, что в клинических испытаниях ингибитор CXCR2 МК7123 улучшал легочную функцию (FEV1) у страдающих COPD пациентов в группе, которая получала активное лекарственное средство, в то время как их функция ухудшалась в обработанной плацебо группе (Am J Respir Crit Care Med. 191(9), 1 мая 2015 г., сс. 1001-1011).
Комбинированный фиброз легких и эмфизема легких (CPFE) представляет собой (но, не ограничиваясь только указанным) плохо прогнозируемое заболевание, при котором патологическое изменение, называемое эмфиземой, которое разрушает и расширяет легкие, осложняется фиброзом легких (Cottin и др., Eur Respir J. 26(4), октябрь 2005 г., сс. 586-593). Известны исследования, в которых установлено, что при CPFE увеличивается концентрация IL-8 в жидкости бронхоальвеолярного лаважа пациентов, и концентрация IL-8 в жидкости бронхоальвеолярного лаважа коррелирует с областью малого затухания (LAA), что свидетельствует о связанном с эмфиземой повреждении, выявляемом при КТ грудной клетки (Respirology. 17(5), июль 2012 г., сс. 814-820); и LAA, свидетельствующая о связанных с эмфиземой повреждениях, выявляемых при КТ грудной клетки, коррелирует с прогнозом (Johannessen А. и др., Am J Respir Crit Care Med 187, 2013, cc. 602-608).
IIP является общим названием интерстициальных пневмоний, причина которых не известна, они представляют собой заболевания, при которых воспаление и поражения затрагивают стенки альвеол, и это приводит к фиброзу, в результате чего стенки утолщаются и затвердевают, что приводит к нарушению газообменной функции (Guideline for diagnosis and therapy of idiopathic interstitial pneumonias, 2-е пересмотренное изд.; Travis и др., Am J Respir Crit Care Med. 88(6), 15 сентября 2013 г., cc. 733-748). Среди IIP IPF является частым и наиболее трудно поддающимся лечению и плохо прогнозируемым заболеванием, для которого характерно хроническое и прогрессирующее течение и которое приводит к развитию серьезного фиброза, вызывая необратимое образование сотового (решетчатого) легкого (Guideline for diagnosis and therapy of idiopathic interstitial interstitial pneumonias, 2-oe пересмотренное изд. Raghu и др., Am J Respir Crit Care Med. 2011 Mar 15;183(6):788-824). Уровень IL-8 является высоким в сыворотке и жидкости бронхоальвеолярного лаважа у страдающих IPF пациентов (Car и др., Am J Respir Crit Care Med 149, 1994, cc. 655-659; Ziegenhagen и др., Am J Respir Crit Care Med 157, 1998, cc. 762-768); а количество нейтрофилов, мигрирующих в ответ на IL-8, увеличивается в жидкости бронхоальвеолярного лаважа, и прогноз является тем хуже, чем выше их количество (Haslam и др., Thorax 35, 1980, сс. 328-339; Turner-Warwick М. и др., Am Rev Respir Dis 135, 1987, cc. 26-34). Это позволяет предположить, что IL-8 и нейтрофилы принимают большое участие в патологическом состоянии, связанном с IPF. Предполагается также, что IL-8 является причиной заболевания, основано на данных о том, что страдающие IPF пациенты имеют мутацию SNIP (одиночный нуклеотидный полиморфизм) IL-8 и повышенную местную концентрацию IL-8 (Ahn и др., Respir Res. 12, 8 июня 2011 г., с. 73). В одном из вариантов осуществления изобретения, представленных в настоящем в настоящем описании, фиброз легких, COPD, CPFE, IIP и IPF являются чувствительными к сигналам IL-8.
Согласно дополнительному или альтернативному варианту осуществления изобретения в настоящем описании представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для ингибирования производства фибротического фактора. Альтернативно этому, в настоящем описании представлены способы ингибирования производства фибротического фактора у индивидуума, которые включают введение в эффективном количестве ингибитора сигнала IL-8 (например, антитела к IL-8) индивидууму, или представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для применения в способах. Альтернативно этому, в настоящем описании представлены фармацевтические композиции, предназначенные для ингибирования производства фибротического фактора, которые содержат в эффективном количестве ингибитор сигнала IL-8 (например, антитело к IL-8). Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для приготовления фармацевтического агента, предназначенного для ингибирования производства фибротического фактора. Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для ингибирования производства фибротического фактора. Альтернативно этому, в настоящем описании представлены способы приготовления фармацевтической композиции, предназначенной для ингибирования производства фибротического фактора, которые включают стадию смешения ингибитора сигнала IL-8 (например, антитела к IL-8) с фармацевтически приемлемым носителем. В одном из вариантов осуществления изобретения фибротический фактор включает, например, МСР-1 (моноцитарный хемотаксический белок-1). Ингибирование производства указанного фибротического фактора может происходить, например, в иммунных клетках, таких как нейтрофилы.
Согласно дополнительному или альтернативному варианту осуществления изобретения в настоящем описании представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для применения для ингибирования ангиогенеза. Альтернативно этому, в настоящем описании представлены способы ингибирования ангиогенеза у индивидуума, которые включают введение индивидууму в эффективном количестве ингибитора сигнала IL-8 (например, антитела к IL-8), или представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для применения в способах. Альтернативно этому, в настоящем описании представлены фармацевтические композиции, предназначенные для ингибирования ангиогенеза, которые включают в эффективном количестве ингибитор сигнала IL-8 (например, антитело к IL-8). Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для приготовления фармацевтического агента, предназначенного для ингибирования ангиогенеза. Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для ингибирования ангиогенеза. Альтернативно этому, в настоящем описании представлены способы приготовления фармацевтической композиции, предназначенной для ингибирования ангиогенеза, которые включают стадию смешения ингибитора сигнала IL-8 (например, антитела к IL-8) с фармацевтически приемлемым носителем.
Как указано в разделе «Примеры» в контексте настоящего описания с помощью анализа in vitro было продемонстрировано, что миграцию нейтрофилов можно ингибировать не только с помощью поступающих в продажу антител к IL-8, но также с помощью ингибиторов IL-8-рецептора, а также антител к IL-8, которые применяли в экспериментах in vivo. Таким образом, специалистам в данной области должно быть очевидно, что данные, полученные с помощью модели in vivo, которые представлены в разделе «Примеры» в настоящем описании, можно обобщать и распространять на применение ингибиторов сигнала IL-8 в целом.
Таким образом, согласно дополнительному или альтернативному варианту осуществления изобретения в настоящем описании представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для применения для подавления инфильтрации (которую согласно одному из аспектов можно обозначать как миграция) иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании. Альтернативно этому, в настоящем описании представлены способы подавления инфильтрации иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании, которые включают введение индивидууму в эффективном количестве ингибитора сигнала IL-8 (например, антитела к IL-8), или представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для применения в способах. Альтернативно этому, в настоящем описании представлены фармацевтические композиции, предназначенные для подавления инфильтрации иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании, которые содержат в эффективном количестве ингибитор сигнала IL-8 (например, антитело к IL-8). Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для приготовления фармацевтического агента, предназначенного для подавления инфильтрации иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании. Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для подавления инфильтрации иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании. Альтернативно этому, в настоящем описании представлены способы приготовления фармацевтической композиции для подавления инфильтрации иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании, которые включают стадию смешения ингибитора сигнала IL-8 (например, антитела к IL-8) с фармацевтически приемлемым носителем.
В одном из вариантов осуществления изобретения иммунные клетки предпочтительно включают, например, нейтрофилы, макрофаги, моноциты, Т-клетки-хелперы, Т-клетки-киллеры, дендритные клетки и NK-клетки, и предпочтительно включают (но, не ограничиваясь только ими) нейтрофилы, макрофаги и моноциты. Касательно «инфильтрации иммунными клетками повреждения при связанном с IL-8 заболевании» связанное с IL-8 заболевание может представлять собой любое из описанных выше заболеваний, например, эндометриоз, аденомиоз, дисменорею, образование спаек или фиброзное заболевание. Подразумевается, что иммунные клетки поступают или мигрируют к повреждению (области болезни) или его окружению при связанном с IL-8 заболевании. Кроме того, при ссылке на «инфильтрацию иммунными клетками повреждения или окружающей его среды при связанном с IL-8 заболевании» специалисту в данной области должно быть очевидно, что, например, в случае эндометриоза, подразумевается инфильтрация или миграция в брюшную полость или асциты, или например, в случае фиброза легких, подразумевается инфильтрация или миграция в грудную полость или плевральную эффузию.
Согласно дополнительному или альтернативному варианту осуществления изобретения в настоящем описании представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для применения для ингибирования производства ароматазы в повреждении, связанном с эндометриозом или аденомиозом. Альтернативно этому, в настоящем описании представлены способы ингибирования производства ароматазы у индивидуума, которые включают введение в эффективном количестве ингибитора сигнала IL-8 (например, антитела к IL-8) индивидууму, или представлены ингибиторы сигнала IL-8 (например, антитела к IL-8), предназначенные для применения в способах. Альтернативно этому, в настоящем описании представлены фармацевтические композиции, предназначенные для ингибирования производства ароматазы, которые содержат в эффективном количестве ингибитор сигнала IL-8 (например, антитело к IL-8). Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для приготовления фармацевтического агента, предназначенного для ингибирования производства ароматазы. Альтернативно этому, в настоящем описании представлено применение ингибитора сигнала IL-8 (например, антитела к IL-8) для ингибирования производства ароматазы в области повреждения, связанного с эндометриозом или аденомиозом. Альтернативно этому, в настоящем описании представлены способы приготовления фармацевтической композиции, предназначенной для ингибирования производства ароматазы в области повреждения, связанного с эндометриозом или аденомиозом, которые включают стадию смешения ингибитора сигнала IL-8 (например, антитела к IL-8) с фармацевтически приемлемым носителем.
Специалистам в данной области должно быть очевидно на основе общеизвестных в данной области технических знаний, что описание включает любые комбинации указанных в настоящем описании полных вариантов осуществления изобретения или их частей, за исключением тех случаев, когда это является технически неприемлемым.
Все процитированные документы, описывающие данный уровень техники, включены в настоящее описание в качестве ссылок, которые характеризуют известные специалистам в данной области технические знания.
В контексте настоящего описания подразумевается, что понятие «и/или» включает комбинации всех понятий, указанных до и после «и/или», включая при необходимости комбинацию понятий «и» и «или».
Хотя различные элементы обозначены в настоящем описании с помощью таких понятий как первый, второй, третий, четвертый и т.д., следует иметь в виду, что такие понятия не накладывают ограничения на элементы. Эти понятия используются только для того, чтобы отличать один элемент от других элементов, и следует иметь в виду, что, например, первый элемент может быть обозначен как второй элемент, и аналогично этому, второй элемент может быть обозначен как первый элемент, без отклонения от объема настоящего описания.
Если специально не указано иное, или, если это не является неприемлемым в контексте описания, подразумевается, что все понятия в единственном числе в контексте настоящего описания означают «по меньшей мере один».
В контексте настоящего описания применяемая терминология используется для целей описания конкретных вариантов осуществления изобретения и не направлена на ограничение изобретения. Если не указано иное, то все понятия (включая технические и научные понятия), применяемые в настоящем описании, следует интерпретировать как имеющие то же самое значение, которое, как правило, подразумевают обычные специалисты в данной области, и их не следует интерпретировать в идеализированном или излишне формальном смысле.
В контексте настоящего описания подразумевается, что понятие «содержит» определяет присутствие описанных элементов (представителей, стадий, элементов, чисел и т.д.), если из контекста ясно не следует иное; и понятие не исключает присутствия других элементов (представителей, стадий, элементов, чисел и т.д.).
Варианты осуществления изобретения, представленные в настоящем описании, даны со ссылкой на схематические иллюстрации, которые для ясности могут быть увеличены.
Если из контекста настоящего описания не следует иное, то численные значения, используемые в контексте настоящего описания, следует понимать как значения в определенном диапазоне, что находится в технической компетенции обычного специалиста в данной области. Например, выражение «1 мг» следует понимать как «примерно 1 мг» с определенными вариациями. Кроме того, например, выражение «1-5 элементов», применяемое в контексте настоящего описания, следует понимать так, как если бы было специально написано «1 элемент, 2 элемента, 3 элемента, 4 элемента, 5 элементов», если это не противоречит контексту.
Примеры
Пример 1. Получение новых вариантов Fc с повышенным связыванием с FcRn в условиях кислого рН для увеличения времени удерживания в плазме
Известно, что в условиях кислого рН в эндосомах антитела IgG-типа, поглощенные клетками, возвращаются в плазму посредством связывания с FcRn. Таким образом, антитела IgG-типа, как правило, обладают продолжительным временем полужизни в плазме по сравнению с белками, которые не связываются с FcRn. Уже известны методы, в которых это свойство увеличения времени удерживания антител в плазме путем повышения их способности к связыванию с FcRn (аффинности к связыванию) в условиях кислого рН реализуют посредством интродукции аминокислотных модификаций в Fc-область антитела. Более конкретно, известны методы увеличения времени удерживания антител в плазме путем повышения их способности к связыванию с FcRn в условиях кислого рН посредством аминокислотных модификаций, таких как модификация M252Y/S254T/T256E (YTE) (J. Biol. Chem. 281, 2006, сс. 23514-23524), модификация M428L/N434S (LS) (Nat. Biotechnol. 28, 2010, сс. 157-159) и модификация N434H.
С другой стороны, как указано выше, известно также, что варианты Fc-области с повышенной способностью к связыванию с FcRn в условиях кислого рН характеризуются связыванием с ревматоидным фактором (RF) (WO 2013/046704). Поэтому осуществляли представленные ниже исследования с целью получения вариантов Fc-области, которые позволяют повышать время удерживания в плазме и не обладают способностью к связыванию с ревматоидным фактором.
(1-1) Получение антител, содержащих новый вариант Fc-области
Согласно описанному ниже методу получали варианты Fc-области с повышенной способностью к связыванию с FcRn в условиях кислого рН, содержащие известные модификации: YTE, LS или N434H, и несколько новых обнаруженных вариантов Fc-области (F1847m, F1848m, F1886m, F1889m, F1927m и F1168m).
Гены тяжелых цепей, в которые интродуцировали аминокислотные модификации в Fc-область тяжелой цепи (VH3-IgG1m) Fv4-IgG1, представляющего собой антитело к человеческому рецептору IL-6, получали с помощью метода, описанного в референс-примере 1. Эти тяжелые цепи использовали для получения указанных ниже антител с помощью метода, описанного в референс-примере 2:
Fv4-IgG1, состоящего из VH3-IgG1m (SEQ ID NO: 2) в качестве тяжелой цепи и VL3-CK (SEQ ID NO: 110) в качестве легкой цепи;
Fv4-YTE, состоящего из VH3-YTE (SEQ ID NO: 3) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи;
Fv4-LS, состоящего из VH3-LS (SEQ ID NO: 4) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи;
Fv4-N434H, состоящего из VH3-N434H (SEQ ID NO: 5) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи;
Fv4-F1847m, состоящего из VH3-F1847m (SEQ ID NO: 6) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи;
Fv4-F1848m, состоящего из VH3-F1848m (SEQ ID NO: 7) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи;
Fv4-F1886m, состоящего из VH3-F1886m (SEQ ID NO: 8) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи;
Fv4-F1889m, состоящего из VH3-F1889m (SEQ ID NO: 9) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи;
Fv4-F1927m, состоящего из VH3-F1927m (SEQ ID NO: 10) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи; и
Fv4-F1168m, состоящего из VH3-F1168m (SEQ ID NO: 11) в качестве тяжелой цепи и VL3-CK в качестве легкой цепи.
(1-2) Кинетический анализ связывания с человеческим FcRn
Антитела, содержащие VH3-IgG1m или указанный выше вариант в качестве тяжелой цепи и L(WT) (SEQ ID NO: 1) в качестве легкой цепи, получали с помощью метода, описанного в референс-примере 2, и оценивали связывающую активность в отношении человеческого FcRn следующим образом.
Кинетические анализы для человеческого FcRn и каждого из антител осуществляли с помощью устройства BIACORE T100 (фирма GE Healthcare). Белок L (фирма ACTIGEN) в соответствующем количестве фиксировали на сенсорном чипе СМ4 (фирма GE Healthcare) с помощью метода аминного сочетания для захвата представляющих интерес антител. Затем осуществляли взаимодействие человеческого FcRn с антителами, захваченными на сенсорном чипе, путем инъекции разведенного раствора FcRn и подвижного буфера (применяемого в качестве референс-раствора). В качестве подвижного буфера использовали 50 мМ фосфат натрия, 150 мМ NaCl и 0,05% (мас./об.) Твин 20 с рН 6,0, и соответствующий буфер использовали также для разведения FcRn. Для регенерации сенсорного чипа использовали 10 мМ глицин-HCl с рН 1,5. Все измерения осуществляли при 25°С. KD (М) для человеческого IgE рассчитывали для каждого антитела на основе константы скорости реакции ассоциации ka (1/Мс) и константы скорости реакции диссоциации kd (1/с), которые представляют собой кинетические параметры, рассчитанные на основе сенсограмм, полученных при измерениях. Для расчета каждого параметра применяли программное обеспечение BIACORE T100 Evaluation (фирма GE Healthcare).
Результаты представлены в таблице 2.

Пример 2. Оценка связывания антител к ревматоидному фактору, содержащих новый вариант Fc-области, характеризующийся повышенным связыванием с FcRn в условиях кислого рН
Антитела к лекарственным средствам (ADA) влияют на эффективность и фармакокинетические характеристики терапевтических антител и иногда приводят к серьезным побочным действиям; таким образом, клиническая применимость и эффективность терапевтических антител могут ограничиваться в результате производства ADA. Многие факторы влияют на иммуногенность терапевтических антител, и присутствие эпитопов эффекторных Т-клеток является одним из таких факторов. Кроме того, присутствие ADA у пациента перед введением терапевтического антитела (которое обозначают также как «предсуществующее антитело») также может приводить к сходным проблемам. Специфически, в случае терапевтических антител для пациентов с аутоиммунными заболеваниями, такими как ревматоидный артрит (RA), ревматоидный фактор (RF), который является аутоантителом к человеческому IgG, может становиться представляющим проблему «предсуществующим ADA». В настоящее время описано гуманизированное анти-CD4 антитело IgG1-типа с мутацией N434H, обеспечивающее значительное связывание с ревматоидным фактором (Clin Pharm Ther.; 89(2), февраль 2011 г., сс. =283-290). Подробное исследование подтвердило, что мутация Asn434His в человеческом IgG1 увеличивает связывание ревматоидного фактора с Fc-областью антител по сравнению с родительским человеческим IgG1.
Ревматоидный фактор представляет собой поликлональное аутоантитело против человеческого IgG и его эпитопы в человеческом IgG варьируются в зависимости от клона, и они, по-видимому, локализованы в области раздела СН2/СН3, а также в СН3-домене, который может перекрываться с FcRn-связывающим эпитопом. Таким образом, мутации, приводящие к повышению связывающей активности (связывающей аффинности) в отношении FcRn, могут повышать также связывающую активность (связывающую аффинность) в отношении конкретных клонов ревматоидного фактора.
Фактически, касательно Fc с повышенной способностью к связыванию с FcRn при кислом рН или нейтральном рН, известно также, что не только модификация N434H, но также и многие другие аминокислотные модификации сходным образом повышают связывание с ревматоидным фактором (WO 2013/046704).
С другой стороны, в WO 2013/046704 в качестве примеров представлены также несколько аминокислотных модификаций, которые избирательно подавляют связывание с ревматоидным фактором, не оказывая влияния на связывание с FcRn, и среди них указаны комбинации двух аминокислотных мутаций, а именно Q438R/S440E, Q438R/S440D, Q438K/S440E и Q438K/S440D. Поэтому проводили исследование того, можно ли снижать связывание новой созданной в настоящее время Fc с повышенной способностью к связыванию при кислом значении рН с ревматоидными факторами путем интродукции Q438R/S440E.
(2-1) Анализ связывания ревматоидного фактора антителами, содержащими новый вариант Fc-области
Анализ связывания с ревматоидным фактором осуществляли с помощью электрохемилюминесценции (ECL) при рН 7,4 с использованием индивидуальных образцов сыворотки (фирма Proteogenex), полученных от 30 страдающих RA пациентов. Смешивали образец 50-кратно разведенной сыворотки, каждое биотинилированное тестируемое антитело (1 мкг/мл) и каждое меченное сложным NHS-эфиром, содержащим сульфо-метку (фирма Meso Scale Discovery), тестируемое антитело (1 мкг/мл) и инкубировали при комнатной температуре в течение 3 ч. После этого смесь добавляли в покрытый стрептавидином 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Discovery) и планшет инкубировали при комнатной температуре в течение 2 ч, после чего промывали. После добавления в каждую лунку буфера для считывания Т(×4) (фирма Meso Scale Discovery) планшет сразу помещали на ридер SECTOR imager 2400 (фирма Meso Scale Discovery) и измеряли хемилюминесценцию.
Результаты анализа представлены на фиг. 8-10. Fv4-IgG1 (фиг. 1), представляющее собой нативный человеческий IgG1, характеризовался лишь слабым связыванием с ревматоидным фактором, в то время как существующие Fc-варианты с повышенным FcRn-связыванием, Fv4-YTE (фиг. 2), Fv4-LS (фиг. 3) и Fv4-N434H (фиг. 4), все характеризовались существенно повышенным связыванием с ревматоидным фактором многих доноров. С другой стороны, все новые варианты Fc-области с повышенным связыванием с FcRn, Fv4-F1847m (фиг. 5), Fv4-F1848m (фиг. 6), Fv4-F1886m (фиг. 7), Fv4-F1889m (фиг. 8), Fv4-F1927m (фиг. 9) и Fv4-F1168m (фиг. 10), характеризовались лишь слабым связыванием с ревматоидным фактором, и это свидетельствовало о том, что связывание с ревматоидным фактором существенно ингибировалось в результате модификаций, повышающих связывание с FcRn.
Кроме того, на фиг. 11 для каждого из вариантов представлены средние величины связывания с ревматоидным фактором в крови 30 страдающих RA пациентов. Все шесть новых вариантов характеризовались более низким связыванием, чем три известных ранее варианта (YTE, LS и N434H), и они характеризовались также более низким связыванием с ревматоидным фактором по сравнению с нативным человеческим IgG1. Таким образом, с точки зрения создания для клинических целей терапевтических антител с более высокой способностью к связыванию с FcRn для лечения аутоиммунных заболеваний и т.п., таких как ревматоидный артрит и т.п., риск, ассоциированный с ревматоидным фактором, который является проблемой при использовании существующих вариантов Fc-области, был уменьшен для новых представленных в настоящем описании вариантов Fc-области, и, следовательно, они могут быть более безопасными для применения.
Пример 3. Оценка ФК новых вариантов Fc-области с повышенным связыванием с FcRn в условиях кислого рН на обезьянах циномолгус
В примере 3 оценивали эффект увеличения времени удерживания в плазме обезьян циномолгус с помощью представленного ниже метода с использованием содержащих новый вариант Fc-области антител, для которых было подтверждено подавление связывания с ревматоидным фактором.
(3-1) Получение содержащих новый вариант Fc-области антител
Получали следующие антитела к человеческому IgE:
OHB-IgG1, состоящее из OHBH-IgG1 (SEQ ID NO: 12) в качестве тяжелой цепи и OHBL-CK (SEQ ID NO: 13) в качестве легкой цепи;
OHB-LS, состоящее из OHBH-LS (SEQ ID NO: 14) в качестве тяжелой цепи и OHBL-CK в качестве легкой цепи;
OHB-N434A, состоящее из OHBH-N434A (SEQ ID NO: 15) в качестве тяжелой цепи и OHBL-CK в качестве легкой цепи;
OHB-F1847m, состоящее из OHBH-F1847m (SEQ ID NO: 16) в качестве тяжелой цепи и OHBL-CK в качестве легкой цепи;
OHB-F1848m, состоящее из OHBH-F1848m (SEQ ID NO: 17) в качестве тяжелой цепи и OHBL-CK в качестве легкой цепи;
OHB-F1886m, состоящее из OHBH-F1886m (SEQ ID NO: 18) в качестве тяжелой цепи и OHBL-CK в качестве легкой цепи;
OHB-F1889m, состоящее из OHBH-F1889m (SEQ ID NO: 19) в качестве тяжелой цепи и OHBL-CK в качестве легкой цепи и
OHB-F1927m, состоящее из OHBH-F1927m (SEQ ID NO: 20) в качестве тяжелой цепи и OHBL-CK в качестве легкой цепи.
(3-2) Анализ ФК антител, содержащих новый вариант Ес-области, в организме обезьян
Оценивали кинетику in vivo антител к человеческому IgE в плазме после введения антител к человеческому IgE обезьянам циномолгус. Раствор антитела к человеческому IgE вводили однократно внутривенно в дозе 2 мг/кг. Сбор крови осуществляли через 5 мин, (два часа), семь часов, один день, два дня, три дня, (четыре дня), семь дней, 14 дней, 21 день, 28 дней, 35 дней, 42 дня, 49 дней и 56 дней после введения. Собранную кровь сразу же подвергали центрифугированию при 4°С и 15000 об/мин в течение 5 мин для получения плазмы. Отделенную плазму хранили в морозильной камере при -80°С или ниже до осуществления измерений. Применяли восемь типов антител к человеческому IgE, а именно OHB-IgG1, OHB-LS, OHB-N434A, OHB-F1847m, OHB-F1848m, OHB-F1886m, OHB-F1889m и OHB-F1927m.
(3-3) Измерение концентрации антител к человеческому IgE в плазме с помощью ELISA
Концентрацию антител к человеческому IgE в плазме обезьян циномолгус измеряли с помощью ELISA. Сначала вносили антитело к каппа-цепи человеческого IgG (фирма Antibody Solution) в планшет Nunc-Immuno Plate, MaxiSorp (фирма Nalge Nunc International) и давали выстаиваться в течение ночи при 4°C с получением планшета с иммобилизованным антителом к каппа-цепи человеческого IgG. Приготавливали образцы для калибровочной кривой с концентрацией в плазме 640, 320, 160, 80, 40, 20 или 10 нг/мл и образцы для измерений в плазме обезьян циномолгус, разведенные в 100 раз или более. Эти образцы для калибровочной кривой и образцы для измерений в плазме, получали путем добавления IgE обезьян циномолгус (продукт, полученный компанией заявителя) в концентрации 1 мкг/мл. После этого образцы вносили в планшет с иммобилизованных антителом к каппа-цепи человеческого IgG и давали выстаиваться при комнатной температуре в течение 2 ч. Затем вносили конъюгированное с HRP антитело к гамма-цепи человеческого IgG (фирма Southern Biotech) и давали выстаиваться при комнатной температуре в течение 1 ч. После этого осуществляли хромогенную реакцию с использованием хромогенного раствора ТМВ (фирма Life Technologies) в качестве субстрата и после прекращения реакции путем добавления 1н. серной кислоты (фирма Wako) измеряли абсорбцию при 450 нм с помощью ридера для микропланшетов. Концентрацию антитела к человеческому IgE в плазме обезьян рассчитывали на основе абсорции с использованием калибровочной кривой с помощью аналитического программного обеспечения SOFTmax PRO (фирма Molecular Devices). Результаты измерений изменения концентрации антитела к человеческому IgE в плазме обезьян представлены на фиг. 12. На основе измеренного изменения концентрации антитела к человеческому IgE в плазме обезьян рассчитывали клиренс в результате элиминации с помощью анализа в определенные моменты времени с использованием программы Phoenix WinNonlin, версия 6.2 (фирма Pharsight Corporation). Рассчитанные фармакокинетические параметры представлены в таблице 3. Образцы, полученные от индивидуумов, которые были позитивные по антителам против введенного образца в плазме, исключали при расчете изменения концентрации антитела к человеческому IgE и клиренса в плазме обезьян.
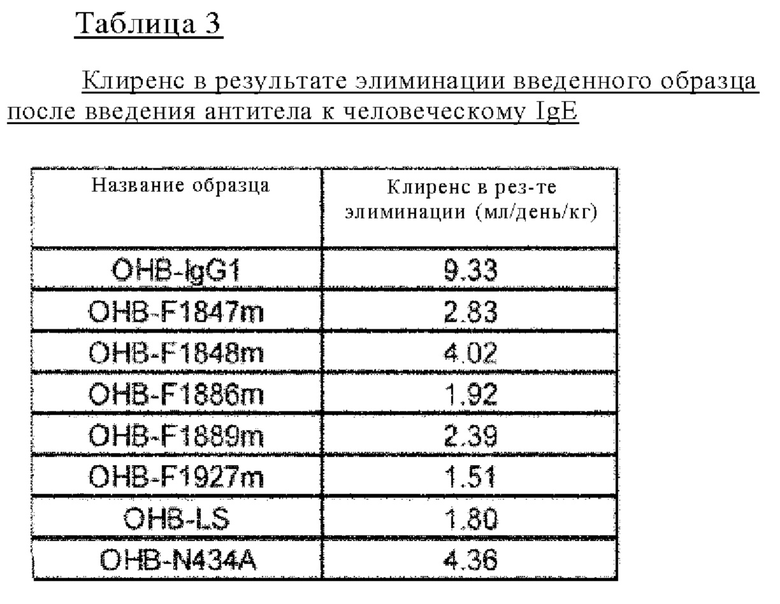
(3-4) Оценка антител к введенным образцам в плазме с помощью электрохемилюминесцентного метода
Антитела в плазме обезьян к введенным образцам измеряли электрохемилюминесцентным методом. Вводимый образец, который был мечен рутением с использованием сложного несущего сульфо-метку NHS-эфира (фирма Meso Scale Discovery), вводимый образец, который был биотинилирован с использованием набора EZ-Link Micro Sulfo-NHS-Biotinylation (фирма Pierce) и образец для измерений в плазме обезьян циномолгус, смешивали в равных количествах и давали выстаиваться в течение ночи при 4°С. Образцы добавляли в 96-луночный покрытый стрептавидином планшет Gold MULTI-ARRAY (фирма Meso Scale Discovery), затем давали осуществляться реакции при комнатной температуре в течение 2 ч и промывали. Затем, сразу после внесения на планшет буфера для считывания Т (×4) (фирма Meso Scale Discovery) осуществляли измерения с использованием устройства SECTOR Imager 2400 (фирма Meso Scale Discovery).
В результате было подтверждено, что все новые варианты Fc-области характеризовались значительно увеличенным временем удерживания в плазме по сравнению со случаем нативного IgG1.
(3-5) Анализ ФК вариантов Fc в организме мышей
Описанный ниже эксперимент осуществляли для сравнения F1718, представляющего собой вариант Fc-области, описанный в WO 2013/046704, и F 1848m, который представляет собой новый вариант Fc-области, полученный при создании настоящего изобретения, в качестве вариантов Fc, предназначенных для повышения связывания FcRn при кислом рН.
Гены тяжелых цепей, в которые интродуцировали аминокислотные модификации в Fc-область тяжелой цепи (VH3-IgG1) Fv4-IgG1, антитела к человеческому рецептору IL-6, получали с помощью метода, описанного в референс-примере 1. С использованием этих тяжелых цепей получали представленные ниже антитела с помощью метода, описанного в референс-примере 2:
Fv4-IgG1, состоящее из VH3-IgG1 в качестве тяжелой цепи и VL3-CK в качестве легкой цепи; и
Fv4-F1718, состоящее из VH3-F1718 (SEQ ID NO: 21) в качестве тяжелой цепи и VL3-CK качестве легкой цепи.
Указанные выше антитела к человеческому рецептору IL-6 вводили однократно в дозе 1 мг/кг в хвостовую вену трансгенных по человеческому FcRn мышей (B6.mFcRn-/-.hFcRn Tg-мыши линии 32+/+; фирма Jackson Laboratories, Methods Mol. Biol. 602, 2010, сс. 93-104). Кровь собирали через 15 мин, 7 ч, 1 день, 2 дня, 3 дня, 7 дней, 14 дней, 21 день и 28 дней после введения антител к человеческому рецептору IL-6. Собранную кровь сразу же центрифугировали при 15000 об/мин и 4°С в течение 15 мин для получения плазмы. Отделенную плазму хранили в морозильной камере при температуре, установленной на -20°С или ниже, до осуществления измерений.
(3-6) Измерение концентрации антител к человеческому рецептору IL-6 в плазме с помощью ELISA
Концентрацию антител к человеческому рецептору IL-6 в мышиной плазме измеряли с помощью ELISA. Сначала антитело к F(ab')2-фрагменту человеческого IgG (специфическое в отношении гамма-цепи) (фирма SIGMA) вносили в планшет Nunc-Immuno Plate, MaxiSorp (фирма Nalge nunc International) и давали выстаиваться в течение ночи при 4°С для получения планшета с иммобилизованным антителом к человеческому IgG. Приготавливали образцы для калибровочной кривой, содержащие антитело к человеческому рецептору IL-6, при концентрации в плазме 0,8, 0,4, 0,2, 0,1, 0,05, 0,025 или 0,0125 мкг/мл и образцы для измерений в мышиной плазме, разведенные в 100 раз или более. Добавляли 200 мкл растворимого человеческого рецептора IL-6 в концентрации 20 нг/мл к 100 мкл образцов для калибровочной кривой или образцов для измерений в плазме и затем содержащим смесь растворам давали выстаиваться в течение 1 ч при комнатной температуре. После этого смешанные растворы вносили в каждую лунку планшета с иммобилизованным антителом к человеческому IgG и планшету давали выстаиваться в течение 1 ч при комнатной температуре. Затем добавляли биотинилированное антитело к человеческому IL-6R (фирма R&D) и давали осуществляться реакции в течение 1 ч при комнатной температуре. После этого добавляли конъюгат стрептавидин-PolyHRP80 (фирма Stereospecific Detection Technologies) и давали осуществляться реакции в течение 1 ч при комнатной температуре и осуществляли хромогенную реакцию этого реакционного раствора с использованием в качестве субстрата ТМВ One Component HRP Microwell Substrate (фирма BioFX Laboratories). После прекращения реакции путем добавления 1н. серной кислоты (фирма Showa Chemical) измеряли абсорбцию в реакционном растворе в каждой лунке при 450 нм с помощью ридера для микропланшетов. Концентрацию антитела в мышиной плазме рассчитывали на основе абсорбции с использованием калибровочной кривой с помощью аналитического программного обеспечения SOFTmax PRO (фирма Molecular Devices).
Результаты представлены на фиг. 13. Конструкция F1718, которая представляла собой вариант Fc-области для повышения связывания FcRn при кислом рН, описанный в WO 2013/046704, не оказывала никакого пролонгирующего эффекта на PK антитела, но обладало временем удерживания в плазме, эквивалентным времени удерживания нативного IgG1.
Представленная в настоящем описании конструкция F1718, которая описана в WO 2013/046704, имеет четыре мутации, а именно N434Y/Y436V/Q438R/S440E интродуцированные в Fc-область. В отличие от этого, в F1848m, впервые полученную при создании настоящего изобретения, были интродуцированы четыре мутации, а именно N434A/Y436V/Q438R/S440E. Единственная разница между аминокислотными мутациями, интродуцированными в указанные два типа Fc-областей, состояла в том, что аминокислотная мутация, интродуцированная в положение 434 согласно EU-нумерации, представляла собой Y (тирозин) в F1718, и А (аланин) в F1848m. В примере (3-2) продемонстрировано, что вариант F1848m характеризовался увеличенным временем удерживания в плазме по сравнению с нативным IgG1, в то время как конструкция F1718 не характеризовалась никаким увеличением времени удерживания в плазме. Следовательно, можно считать, что А (аланин) является предпочтительным (но это не накладывает конкретных ограничений на объем изобретения) для осуществления аминокислотной мутации, которую следует интродуцировать в положение 434 для увеличения времени удерживания в плазме.
Пример 4. Получение гуманизированных антител к человеческому IL-8
(4-1) Получение гуманизированного антитела hWS-4 к человеческому IL-8
Гуманизированные антитела к IL-8, описанное в US №6245894 (WO 1996/002576), связываются с человеческим IL-8 (в контексте настоящего описания обозначено как hIL-8) и блокируют его физиологическую функцию. Можно получать путем объединения последовательностей вариабельных областей тяжелой и легкой цепей, описанных в US №6245894, с любой из известных последовательностей константной области человеческого антитела. На последовательности константной области человеческого антитела не накладываются конкретные ограничения, но в качестве последовательностей константных областей тяжелой цепи можно использовать последовательности нативного человеческого IgG1 или последовательности нативного человеческого IgG4, а в качестве последовательностей константных областей легкой цепи можно использовать последовательности нативной человеческой каппа-цепи.
Так, из числа гуманизированных антител к IL-8, описанных в US №6245894, ген hWS4H-IgG1 (SEQ ID NO: 39), в котором были объединены последовательность вариабельной области тяжелой цепи RVHg и нативной последовательность константной области тяжелой цепи человеческого антитела к IgG1, получали методом, описанным в референс-примере 1. Кроме того, ген hWS4L-k0MT (SEQ ID NO: 40), в котором были объединены последовательность вариабельной области легкой цепи RVLa и последовательность нативной константной области человеческой легкой канна-цепи, получали согласно методу, описанному в референс-примере 1. Получали антитело, в котором указанные выше тяжелая цепь и легкая цепь были объединены, и его обозначили как гуманизированное антитело WS-4 (ниже в настоящем описании обозначено как hWS-4)
(4-2) Получение гуманизированного антитела Hr9 к человеческому IL-8
Новое гуманизированное антитело получали с использованием человеческих консенсусных последовательностей каркасного участка, отличных от FR, которые использовали в hWS-4.
Более конкретно, гибридную последовательность VH3-23 и VH3-64 использовали в качестве FR1 тяжелой цепи, последовательность, представленную в VH3-15 и VH3-49, использовали в качестве FR2, последовательность, представленную в VH3-72, использовали в качестве FR3 (при условии, что исключали 82а согласно нумерации Кэбота), а последовательность, представленную в JH1 и т.д., использовали в качестве FR4. Эти последовательности связывали с последовательностями CDR тяжелой цепи hWS-4 с получением Hr9-IgG1 (SEQ ID NO: 41), тяжелой цепи нового гуманизированного антитела.
Затем получали два типа антител, а именно, hWS-4, имеющее hWS4H-IgG1 в качестве тяжелой цепи и hWS4L-k0MT в качестве легкой цепи, и Hr9, имеющее Hr9-IgG1 в качестве тяжелой цепи и hWS4L-k0MT в качестве легкой цепи. В контексте настоящего описания при ссылке на легкую цепь, в частности на Hr9 (последовательность вариабельной области его тяжелой цепи представлена в SEQ ID NO: 33), ее обозначают как Hr9/hWS4L. Экспрессию антител осуществляли с использованием клеток FreeStyle 293F (фирма Invitrogen) согласно протоколу, приложенному к продукту. Антитела очищали из супернатанта культуры с помощью метода, описанного в референс-примере 2. В результате получали антитела в количествах, указанных в таблице 4. Неожиданно было установлено, что уровень экспрессии Нг9 был примерно в 8 раз выше, чем уровень экспрессии hWS-4.

(4-3) Активности hWS-4 и Hr9 в отношении связывания человеческого IL-8
Аффинности, характеризующие связывание hWS-4 и Hr9 с человеческим IL-8, определяли согласно описанному ниже методу с использованием устройства BIACORE T200 (фирма GE Healthcare).
Применяли подвижный буфер, имеющий следующий состав: 0,05% Твин 20, 20 мМ ACES и 150 мМ NaCl (pH 7,4). Белок A/G (фирма PIERCE) в соответствующем количестве иммобилизовали на сенсорном чипе СМ4 (фирма GE Healthcare) с помощью метода аминного сочетания и осуществляли захват представляющего интерес антитела. Затем осуществляли взаимодействие человеческого IL-8 с антителом, захваченным на сенсорном чипе, путем инъекции разведенного раствора человеческого IL-8 и подвижного буфера (который применяли в качестве референс-раствора). В качестве подвижного буфера использовали раствор описанного выше состава, и этот буфер использовали также для разведения человеческого IL-8. Для регенерации сенсорного чипа использовали 10 мМ глицин-HCl с pH 1,5. Все измерения осуществляли при 37°С. KD (М) для человеческого IgE рассчитывали для каждого антитела на основе константы скорости реакции ассоциации kon (1/Mc) и константы скорости реакции диссоциации koff (1/с), которые представляют собой кинетические параметры, рассчитанные на основе сенсограмм, полученных при измерениях. Для расчета каждого параметра применяли программное обеспечение BIACORE Т100 Evaluation (фирма GE Healthcare).
Результаты представлены в таблице 5. Было подтверждено, что hWS-4 и Hr9 обладали эквивалентными аффинностями связывания с человеческим IL-8

Для создания содержащих антитело фармацевтических средств важным фактором является уровень производства молекул антитела, и, как правило, желательно, чтобы уровень производства был высоким. Следует особо отметить, что на основе описанных выше исследований была выбрана более пригодная полученная из человеческого консенсусного каркасного участка последовательность для объединения с HVR-последовательностью hWS-4 и получено Hr9, характеризующееся более высоким уровнем производства при сохранении аффинности связывания с человеческим IL-8.
Пример 5. Получение антител с рН-зависимой способностью к связыванию с IL-8
(5-1) Получение антител на основе модификации Hr9 для придания зависимости от рН
Были проведены исследования с целью придания рН-зависимой способностью к связыванию с IL-8 Hr9, полученному в примере 4.
Не вдаваясь в какую-либо конкретную теорию, можно считать, что антитела, обладающие рН-зависимой способностью к связыванию с IL-8, могут иметь следующие характеристики in vivo. Антитела, введенные в живой организм, могут более сильно связываться с IL-8 в условиях, в которых поддерживается нейтральное значение рН (например, в плазме), и блокировать его функцию. Часть таких комплексов IL-8/антитело поглощается клетками в результате неспецифического взаимодействия с клеточной мембраной (пиноцитоз) (ниже в настоящем описании обозначен как неспецифическое поглощение). В условиях кислого рН в эндосомах аффинности связывания указанных выше антител с IL-8 становятся слабыми и поэтому антитело освобождает IL-8. Затем антитела, освободившие IL-8 в результате диссоциации, могут возвращаться в окружающую клетки среду посредством FcRn. Указанные выше антитела, которые возвратились наружу из клетки (в плазму) таким путем, могут снова связываться с другим IL-8 и блокировать его функцию. Предполагается, что антитела, обладающие рН-зависимой способностью к связыванию с IL-8, должны быть также способны связываться с IL-8 несколько раз посредством указанного выше механизма.
В отличие от этого, антитело, которое не обладает свойством, присущим указанному выше антителу, представляет собой молекулу антитела, которая способна нейтрализовать антиген только один раз, но не может нейтрализовать антиген несколько раз. В целом, поскольку антитело IgG-типа имеет два Fab, то одна молекула антитела может нейтрализовать две молекулы IL-8. С другой стороны, антитела, которые могут связываться с IL-8 несколько раз, могут связываться с IL-8 любое количество раз в течение того периода времени, пока они остаются в живом организме. Например, одна молекула обладающего рН-зависимым связыванием с IL-8 антитела, которая поглощается клетками десять раз с момента ее введения до элиминации, может нейтрализовать как максимум 20 молекул IL-8. Следовательно, антитело, которое может связываться несколько раз с IL-8, обладает преимуществом, заключающимся в способности нейтрализовать несколько молекул IL-8 даже в случае небольшого количества антитела. С другой точки зрения, антитело, которое может связываться несколько раз с IL-8, обладает преимуществом, заключающимся в способности поддерживать состояние, в котором оно может нейтрализовать IL-8, в течение более длинного периода времени по сравнению с тем случаем, когда вводят в том же самом количестве антитело. С другой точки зрения, антитело, которое может связываться несколько раз с IL-8, обладает преимуществом, заключающимся в том, что оно способно более сильно блокировать биологическую активность IL-8 по сравнению с тем случаем, когда вводят в том же самом количестве антитело, которое не обладает этим свойством.
Для достижения этих преимуществ интродуцировали аминокислотные модификации, главным образом гистидин, в вариабельные области Hr9-IgG1 и WS4L-k0MT с целью получения антител, которые могут связываться с IL-8 несколько раз. Более конкретно, получали варианты, представленные в таблице 6, с помощью методов, описанных в референс-примерах 1 и 2.
В обозначениях, таких как «Y97H», приведенных в таблице 6, указано положение, в которое интродуцируют мутацию, согласно нумерации Кэбота, аминокислота до интродукции мутации и аминокислота после интродукции мутации. Конкретно, обозначение «Y97H» означает, что аминокислотный остаток в положении 97 согласно нумерации Кэбота заменен с Y (тирозин) на Н (гистидин). Кроме того, когда интродуцируют комбинацию нескольких аминокислотных замен, то ее обозначают следующим образом «N50H/L54H».
(5-2) рН-зависимая активность связывания с IL-8
Аффинность связывания с человеческим IL-8 антител, полученных в примере 5-1, определяли согласно описанному ниже методу с использованием устройства BIACORE T200 (фирма GE Healthcare). Применяли два следующих подвижных буфера: -
0,05% Твин 20, 20 мМ ACES, 150 мМ NaCl, pH 7,4;
0,05% Твин 20, 20 мМ ACES, 150 мМ NaCl, pH 5,8.
Белок А/G (фирма PIERCE) в соответствующем количестве иммобилизовали на сенсорном чипе СМ4 (фирма GE Healthcare) с помощью метода аминного сочетания и осуществляли захват представляющего интерес антитела. Затем осуществляли взаимодействие человеческого IL-8 с антителом, захваченным на сенсорном чипе, путем инъекции разведенного раствора человеческого IL-8 и подвижного буфера (который применяли в качестве референс-раствора). В качестве подвижного буфера использовали любой из растворов описанного выше состава, и этот буфер использовали также для разведения человеческого IL-8. Для регенерации сенсорного чипа использовали 10 мМ глицин-HCl с pH 1,5. Все измерения осуществляли при 37°С. KD (М) для каждого из антител к человеческому IgE рассчитывали на основе константы скорости реакции ассоциации kon (1/Mc) и константы скорости реакции диссоциации koff (1/c), которые представляют собой кинетические параметры, рассчитанные на основе сенсограмм, полученных при измерениях. Для расчета каждого параметра применяли программное обеспечение BIACORE Т100 Evaluation (фирма GE Healthcare).
Результаты представлены в таблице 7. Во-первых, по сравнению с Hr9, Hr9/L16, которое содержит модификацию L54H в легкой цепи, обладало немного более высокой аффинностью связывания с человеческим IL-8 при нейтральном рН (рН 7,4), но более низкой аффинностью связывания с человеческим IL-8 при кислом рН (рН 5,8). С другой стороны, антитела к IL-8 (H89/WS4L, H89/L12 и H89/L16), полученные путем объединения различных легких цепей с Н89, содержащим модификацию Y97H в тяжелой цепи, все характеризовались сниженной аффинностью связывания с человеческим IL-8 при кислом рН, а также сниженной аффинностью связывания с человеческим IL-8 при нейтральном рН.
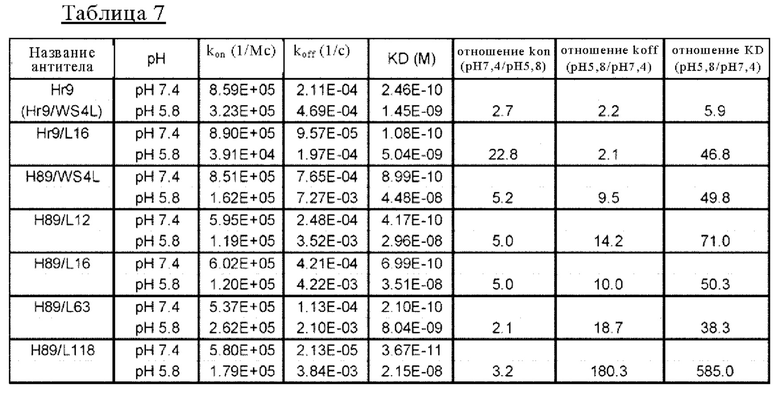
(5-3) Получение и оценка антител, модифицированных для придания рН-зависимости
Далее оценивали комбинации перспективных модификаций, выявленных в примере 5-2, и новых аминокислотных мутаций и в результате были выявлены следующие комбинации.
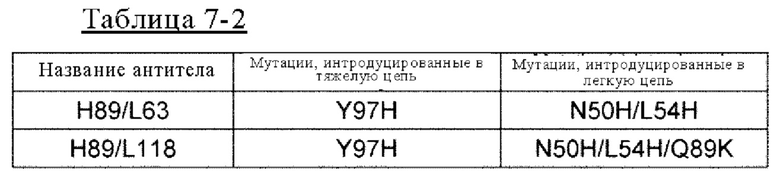
Указанные варианты получали с помощью методов, описанных в референс-примерах 1 и 2, а аффинность связывания с человеческим IL-8 оценивали с помощью метода, сходного с методом, описанным в примере 5-2. Результаты представлены также в таблице 7. H89/L63, которое имело H89-IgG1 (SEQ ID NO: 42) в качестве тяжелой цепи и L63-k0MT (SEQ ID NO: 43) в качестве легкой цепи, характеризовались аффинностью связывания с человеческим IL-8 при нейтральном рН (рН 7,4), эквивалентной аффинности Hr9, и сниженной при кислом рН (рН 5,8). Конкретно, обе величины, и koff (константа скорости реакции диссоциации) и KD (константа диссоциации) H89/L63, при рН 5,8 были выше, чем соответствующие величины для Hr9. Это означает, что в условиях кислого рН в эндосомах, H89/L63 обладало способностью быстро высвобождать человеческий IL-8.
Кроме того, неожиданно было установлено, что H89/L118, которое имеет H89-IgG1 в качестве тяжелой цепи и L118-k0MT (SEQ ID NO: 44) в качестве легкой цепи, обладало более высокой аффинностью связывания (KD) с человеческим IL-8 в условиях нейтрального рН по сравнению с аффинностью Hr9, но более слабой аффинностью связывания (KD) с человеческим IL-8 в условиях кислого рН по сравнению с аффинностью Hr9. Как правило (но это не является ограничением), когда антитела, которые могут несколько раз связываться с антигенами, применяют в качестве фармацевтического продукта, то обладающие рН-зависимым связыванием с антигеном антитела предпочтительно обладают сильной аффинностью связывания (малая величина KD), так что они могут эффективно нейтрализовать антигены в условиях нейтрального рН (например, в плазме). С другой стороны, антитела предпочтительно имеют большую константу скорости реакции диссоциации (koff) и/или низкую аффинностью связывания (большая величина KD), так что они могут быстрее высвобождать антигены в условиях кислого рН (например, в эндосомах). По сравнению с Hr9, H89/L118 приобрело более предпочтительные свойства как при нейтральном рН, так и при кислом рН.
Таким образом, для Hr9 были идентифицированы пригодные аминокислотные модификации, такие как Y97H для его тяжелой цепи и N50H/L54H/Q89K для его легкой цепи. При этом было установлено, что можно создавать характеризующиеся рН-зависимым связыванием с IL-8 антитела, обладающие преимуществами в качестве фармацевтических средств, путем интродукция одной или комбинации нескольких аминокислотных модификаций, выбранных из указанных модификаций (но, не ограничиваясь только ими). Не вдаваясь ни в какую-либо конкретную теорию, можно считать, что при использовании обладающего рН-зависимым связыванием с антигеном антитела в качестве фармацевтического средства важным показателем является то, может или нет введенное в организм антитело высвобождать антиген в эндосоме. В этой связи, по-видимому, важно, чтобы имело место достаточно слабое связывание (большая величина константы диссоциации (KD)) в условиях кислого рН или достаточно быстрая скорость реакции диссоциации (большая величина константы скорости диссоциации (koff)). Поэтому в представленном ниже эксперименте изучали, достаточна ли величина KD или koff H89/L118, измеренная методом BIACORE, для отделения антигена путем диссоциации в эндосоме in vivo.
Пример 6
Получение обладающих высокой аффинностью антител для анализа ФК в организме мышей
Не накладывается конкретных ограничений на методы для подтверждения воздействия антитела на скорость элиминации человеческого IL-8 из организма мышей. Например, метод включает введение антитела в виде смеси с человеческим IL-8 мышам и последующее сравнение скорости элиминации человеческого IL-8 из мышиной плазмы.
В рассматриваемом случае желательно, чтобы референс антитело, предназначенное для применения в анализе ФК в организме мышей, обладало достаточно сильной аффинностью связывания как в условиях нейтрального рН, так и в условиях кислого рН. Затем осуществляли поиск модификаций, которые придают Hr9 высокую аффинность, и в результате было создано антитело H998/L63, имеющее H998-IgG1 (SEQ ID NO: 45) в качестве тяжелой цепи и L63-k0MT в качестве легкой цепи.
H998/L63 использовали для оценки аффинности связывания с человеческим IL-8 с помощью метода, сходного с методом, описанным в примере 5-2. Полученные сенсограммы представлены на фиг. 14.
H998/L63 характеризовалось весьма низкой скоростью реакции диссоциации как в условиях нейтрального рН, так и в условиях кислого рН, и было установлено, что оно обладало более сильной аффинности связывания с IL-8, чем Hr9. Однако, известно, что вследствие механических ограничений метода BIACORE аналитические величины, такие как константа скорости реакции диссоциации (koff) и константа диссоциации (KD) не могут быть точно рассчитаны в тех случаях, когда белок-белковые взаимодействия характеризуются низкой скоростью реакции диссоциации. Поскольку точные аналитические величины не удалось получить для H998/L63, то аналитические величины для него не представлены в настоящем описании. Однако полученные в эксперименте результаты свидетельствовали о том, что H998/L63 обладало очень сильной аффинностью связывания как при нейтральном рН, так и при кислом рН, и его можно применять в качестве антитела, используемого для сравнения в анализе ФК в организме мышей.
Пример 7
Анализ ФК в организме мышей с использованием обладающего рН-зависимым связыванием с IL-8 антитела H89/L118
(7-1) Анализ ФК в организме мышей с использованием H89/L118
Скорость элиминации человеческого IL-8 in vivo оценивали с использованием H89/L118, полученного в примере 5, и H998/L63, полученного в примере 6.
Фармакокинетические характеристики человеческого IL-8 оценивали после одновременного введения человеческого IL-8 и антител к человеческому IL-8 мышам (линия C57BL/6J, фирма Charles river). Раствор, содержащий смесь человеческого IL-8 и антитела к человеческому IL-8 (10 мкг/мл и 200 мкг/мл соответственно), вводили в виде однократной дозы 10 мл/кг в хвостовую вену. Поскольку антитело к человеческому IL-8 присутствовало в достаточном избытке, то считалось, что в этот момент времени, почти весь человеческий IL-8 должен быть связан с антителом. Кровь собирали через пять минут, 2 ч, 4 ч, 7 ч, 1 день, 2 дня, 3 дня, 7 дней, 14 дней, 21 день и 28 дней после введения. Собранную кровь сразу же центрифугировали при 15000 об/мин и 4°С в течение 15 мин для получения плазмы. Отделенную плазму хранили в морозильной камере при -20°С или ниже до осуществления измерений.
(7-2) Измерение концентрации человеческого IL-8 в плазме
Концентрацию человеческого IL-8 в мышиной плазме определяли электрохемилюминесцентным методом. Сначала антитело к человеческому IL-8 (полученное в лаборатории заявителей), содержащее константную область мышиного IgG, вносили в 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Discovery) и давали выстаиваться при комнатной температуре в течение 1 ч. Затем для получения планшета с иммобилизованным антителом к человеческому IL-8 применяли раствор ЗФР-Твин, содержащий 5% БСА (мас./об.), для блокирования при комнатной температуре в течение 2 ч. Приготавливали образцы для построения калибровочной кривой, содержащие человеческий IL-8, с концентрацией в плазме 275, 91,7, 30,6, 10,2, 3,40, 1,13 или 0,377 нг/мл и образцы для измерений в мышиной плазме, разведенные в 25 раз или более. Образцы смешивали с hWS-4 и давали прореагировать в течение ночи при 37°С. После этого в каждую лунку планшета с иммобилизованным антителом к человеческому IL-8 вносили по 50 мкл содержащих смесь растворов и раствор перемешивали при комнатной температуре в течение 1 ч. Конечную концентрацию hWS-4 доводили до 25 мкг/мл. Затем после взаимодействия в течение 1 ч с биотинилированным мышиным антителом к легкой каппа-цепи человеческого Ig (фирма BD Pharmingen) при комнатной температуре и затем взаимодействия в течение 1 ч с меченным сульфа-меткой стрептавидином (фирма Meso Scale Discovery) при комнатной температуре вносили буфер Т для считывания (1×) (фирма Meso Scale Discovery) и сразу же осуществляли измерения с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию человеческого IL-8 рассчитывали на основе ответа с использованием калибровочной кривой с помощью аналитического программного обеспечения SOFT Max PRO (фирма Molecular Devices).
Полученные данные о концентрации человеческого IL-8 в плазме представлены на фиг. 15, а величины клиренса (CL) человеческого IL-8 из мышиной плазмы представлены в таблице 8.
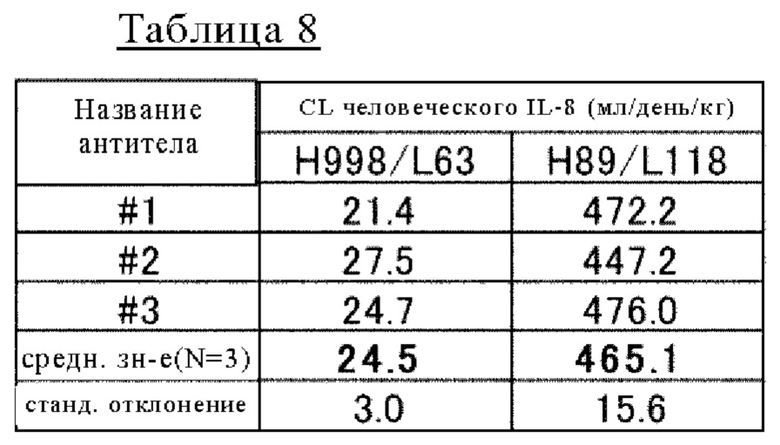
Как следует из фиг. 15, оказалось, что по сравнению с человеческим IL-8, введенным одновременно с H998/L63, человеческий IL-8, введенный одновременно с H89/L118, весьма быстро элиминировался из мышиной плазмы. Кроме того, величины CL, которые качественно отражают скорость элиминации человеческого IL-8 из мышиной плазмы, свидетельствуют о том, что скорость элиминации человеческого IL-8 была примерно в 19 раз более высокой в случае H89/L118 по сравнению с H998/L63.
Не вдаваясь в какую-либо конкретную теорию, на основе полученных данных можно сделать следующий вывод. Большая часть человеческого IL-8, введенного одновременно с антителом, связывается с антителом в плазме и находится в составе комплексов. Человеческий IL-8, связанный с H998/L63, существует в связанном с антителом состоянии даже в условиях кислого рН в эндосоме благодаря присущей антителу сильной аффинности. Следовательно, H998/L63 возвращается в плазму посредством FcRn, находясь все еще в составе комплекса с человеческим IL-8; следовательно, когда это происходит, то человеческий IL-8 также возвращается в плазму в то же самое время. Следовательно, большая часть человеческого IL-8, поглощенного клетками, снова возвращается в плазму. Это означает, что уровень элиминации человеческого IL-8 из плазмы существенно снижается, если одновременно вводят H998/L63. С другой стороны, как указано ранее, человеческий IL-8, поглощенный клетками в составе комплекса с H89/L118, рН-зависимым IL-8-связывающим антителом, отделяется в результате диссоциации от антитела в условиях кислого рН в эндосоме. Человеческий IL-8, отделенный в результате диссоциации от антитела, будет расщепляться после переноса в лизосому. Следовательно, рН-зависимое IL-8-связывающие антитела могут значительно ускорять элиминацию человеческого IL-8 по сравнению с IL-8-связывающим антителом, таким как H998/L63, которое обладает сильной аффинностью связывания как при кислом рН, так и при нейтральном рН.
(7-3) Анализ ФК в организме мышей при увеличенной дозе H89/L118
Затем проводили эксперимент по проверке воздействия варьирования дозы H89/L118, который заключался в следующем. После одновременного введения человеческого IL-8 и H89/L118 (2 мг/кг или 8 мг/кг) мышам (линия C57BL/6J, фирма Charles river) оценивали фармакокинетические характеристики человеческого IL-8. Раствор, содержащий смесь человеческого IL-8 (2,5 мкг/мл) и антитела к человеческому IL-8 (200 мкг/мл или 800 мкг/мл), вводили однократно в хвостовую вену в дозе 10 мл/кг. Считалось, что, поскольку антитело к человеческому IL-8 присутствует в достаточном избытке по сравнению с человеческим IL-8, то в это время почти весь человеческий IL-8 должен быть связан с антителом. Кровь собирали через 5 мин, 7 ч, один день, два дня, три дня, семь дней, 14 дней, 21 день и 28 дней после введения. Собранную кровь сразу же центрифугировали при 15000 об/мин и 4°С в течение 15 мин для получения плазмы. Отделенную плазму хранили в морозильной камере при -20°С или ниже до осуществления измерений.
Измерение концентрации человеческого IL-8 в мышиной плазме осуществляли методом, сходным с методом, описанным в примере 7-2. Полученные данные о концентрации человеческого IL-8 в плазме представлены на фиг. 16, а величины клиренса (CL) человеческого IL-8 из мышиной плазмы представлены в таблице 9.
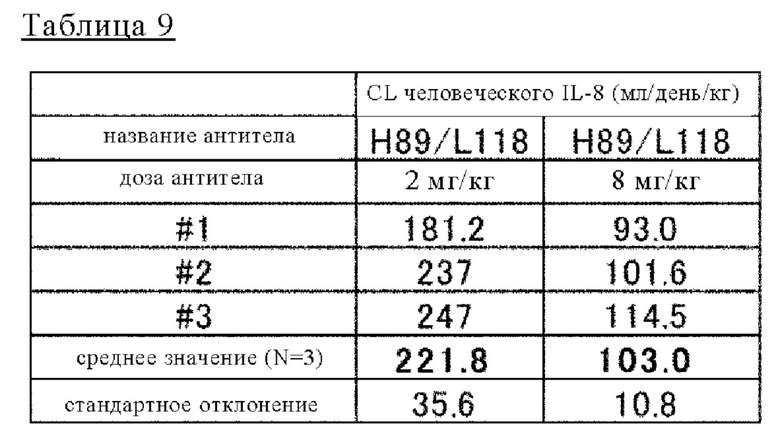
В результате было подтверждено, что по сравнению с группой, которой вводили H89/L118 в дозе 2 мг/кг, в группе, которой вводили антитело в дозе 8 мг/кг, скорость элиминации человеческого IL-8 была примерно в 2 раза медленнее.
Ниже в настоящем описании, не вдаваясь в конкретную теорию, описан один из возможных факторов, обусловливающих указанные выше результаты.
Предпочтительно, чтобы среди антител, которые возвращаются из эндосомы в плазму посредством FcRn, доля связанных с человеческим IL-8 антител была низкой. Касательно человеческого IL-8, присутствующего в эндосоме, желательно, чтобы его большая доля находилась в свободной форме, не связанной с антителом. Когда человеческий IL-8 вводят вместе с антителом, которое не обладает рН-зависимой способностью связывать IL-8, то считается, что большая часть (почти 100%) человеческого IL-8 в эндосоме присутствует в составе комплексов с антителом, а в свободной форме находится небольшое количество (близкое к 0%). С другой стороны, при введении вместе с обладающим рН-зависимым связыванием с IL-8 антителом (например, H89/L118), определенная доля человеческого IL-8 должна присутствовать в эндосоме в свободной форме. Гипотетически долю свободной формы в этом случае можно определять следующим образом: [доля свободного человеческого IL-8 в эндосоме (%)] = [концентрация свободного человеческого IL-8 в эндосоме]/[общая концентрация человеческого IL-8 в эндосоме] × 100.
Желательно, чтобы доля свободного человеческого IL-8 в эндосоме, которую рассчитывают согласно приведенному выше уравнению, была более высокой и, например, 20% является более предпочтительной величиной, чем 0%, 40% более предпочтительно, чем 20%, 60% более предпочтительно, чем 40%, 80% более предпочтительно, чем 60% и 100% более предпочтительно, чем 80%.
Так, можно с большой доли вероятности предположить, что существует корреляция между долей свободного человеческого IL-8 в эндосоме, указанной выше, и аффинностью связывания (KD) и/или константой скорости реакции диссоциации (koff) для человеческого IL-8 при кислом рН. Она заключается в следующем: чем слабее аффинность связывания и/или чем больше скорость реакции диссоциации для человеческого IL-8 при кислом рН, тем должна быть выше доля свободного человеческого IL-8 в эндосоме. Однако, в случае обладающих рН-зависимым связыванием с IL-8 антител, которые приводят к тому, что доля свободного человеческого IL-8 в эндосоме уже близка к 100%, дополнительное ослабление аффинности связывания и/или повышение скорости реакции диссоциации при кислом рН не обязательно приводит к эффективному увеличению доли свободного человеческого IL-8. Легко можно понять, например, что если доля свободного человеческого IL-8 повышается с 99,9% даже до 99,99%, то такая степень повышения может не являться существенной.
Кроме того, согласно общей теории химического равновесия, когда антитело к IL-8 и человеческий IL-8 присутствуют вместе и реакция их связывания и реакция диссоциации достигли равновесия, то доля свободного человеческого IL-8 однозначно определяется тремя параметрами: концентрацией антитела, концентрацией антигена и константой диссоциации (KD). Следовательно, когда концентрация антитела высока, когда концентрация антигена является высокой или когда константа диссоциации (KD) мала, комплексы образуются легко и доля свободного человеческого IL-8 снижается. С другой стороны, когда концентрация антитела низка, когда концентрация антигена низка или когда константа диссоциации (KD) велика, образование комплексов затрудняется и доля свободного человеческого IL-8 увеличивается.
Между тем в рассматриваемом эксперименте скорость элиминации человеческого IL-8 при введении H89/L118 в дозе 8 мг/кг была ниже, чем в случае, когда антитело вводили в дозе 2 мг/кг. Следовательно, это позволяет предположить, что в эндосоме доля свободного человеческого IL-8 снижалась, когда антитело вводили в дозе 8 мг/кг, по сравнению с тем случаем, когда антитело вводили в дозе 2 мг/кг. Причина этого могла заключаться в том, что увеличение дозы антитела в четыре раза повышало концентрацию антитела в эндосоме и тем самым облегчало формирование комплекса IL-8-антитело в эндосоме. Это означает, что в группе, которой вводили повышенную дозу антитела, вероятно доля свободного человеческого IL-8 в эндосоме снижалась и, следовательно, снижалась скорость элиминации человеческого IL-8. Это позволяет также предположить, что когда антитело вводили в дозе 8 мг/кг, величина константы диссоциации (KD) H89/L118 в условиях кислого рН являлась недостаточной для доведения содержания свободного человеческого IL-8 почти до 100%. Более конкретно, если антитело, которое характеризуется большей величиной константы диссоциации (KD) (более слабое связывание) в условиях кислого рН, то оно может достигать состояния, в котором доля свободного IL-8 составляет почти 100%, даже тогда, когда антитело вводят в дозе 8 мг/кг и скорость элиминации человеческого IL-8 эквивалентна той, которая имеет место при введении антитела в дозе 2 мг/кг.
На основе представленных выше данных, для того, чтобы проверить, может ли представляющее интерес обладающее рН-зависимым связыванием с IL-8 антитело обеспечивать долю свободного человеческого IL-8 в эндосоме, составляющую, как указано выше, почти 100%, можно проверить (но, не ограничиваясь только этим), имеется ли возможность для увеличения степени, обусловливающего элиминацию антигена действия in vivo или нет. Например, в одном из методов сравнивают скорость элиминации человеческого IL-8 при использовании нового обладающего рН-зависимым связыванием с IL-8 антитела со скоростью при использовании H89/L118, где новое антитело обладает более слабой аффинностью связывания при кислом рН и/или повышенной скоростью реакции диссоциации при кислом рН по сравнению с H89/L118. В том случае, когда указанное выше новое рН-зависимое антитело к IL-8 обусловливает скорость элиминации человеческого IL-8, эквивалентную той, которую обусловливает H89/L118, это позволяет предположить, что аффинность связывания и/или скорость реакции диссоциации H89/L118 при кислом рН уже находится на уровне, достаточном для достижения доли свободного человеческого IL-8 в эндосоме, составляющей почти 100%. С другой стороны, в тех случаях, когда вышеуказанное новое о рН-зависимое антитело к IL-8 обусловливает более высокую скорость элиминации человеческого IL-8, это позволяет предположить, что имеется возможность повышения аффинности связывания и/или скорости реакции диссоциации H89/L118 при кислом рН.
Пример 8
Получение и оценка обладающего рН-зависимым связыванием с IL-8 антитела H553/L118
(8-1) Получение нового H553/L118, обладающего рН-зависимой IL-8-связывающей способностью
В данном случае цель авторов изобретения состояла в том, чтобы создать антитела, обладающие еще более слабой аффинностью связывания с человеческим IL-8 в условиях кислого рН и/или более высокой скоростью реакции диссоциации, чем H89/L118.
Для получения модифицированных антител, представленных в таблице 10, интродуцировали аминокислотные модификации, главным образом с использованием гистидина, с помощью метода, сходного с методом, описанным в примере 5, применяя в качестве основы H89/L118. Кроме того, для этих антител определяли аффинность связывания с человеческим IL-8 с помощью метода, сходного с методом, описанным в примере 5-2.
Часть этих результатов представлена в таблице 10. Продемонстрировано, что антитело H553/L118, содержащее H553-IgG1 (SEQ ID NO: 46) в качестве тяжелой цепи и L118-k0MT в качестве легкой цепи, и антитело H496/L118, содержащее H496-IgG1 (SEQ ID NO: 57) в качестве тяжелой цепи и L118-k0MT в качестве легкой цепи, обладали более сильной зависимостью от рН, чем H89/L118.
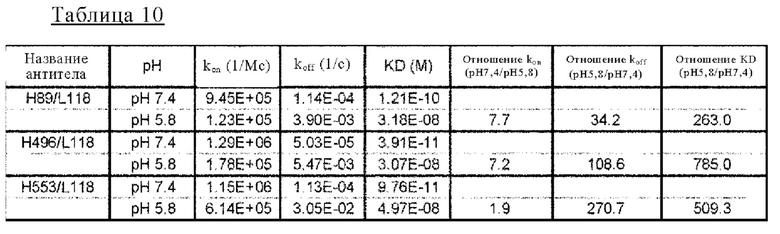
Полученное H553/L118 содержало две аминокислотные модификации, а именно Y55H и R57P, интродуцированные в тяжелую цепь H89/L118. С другой стороны, H496/L118, полученное путем интродукции только R57P в тяжелую цепь H89/L118, обладало повышенной аффинностью связывания с человеческим IL-8 при нейтральный рН, но слабо измененной аффинностью связывания с человеческим IL-8 при кислом рН, по сравнению с H89/L118. Более конкретно, модификация R57P, интродуцированная в H89/L118 представляет собой модификацию, которая повышает аффинность связывания с человеческим IL-8 только при нейтральном рН, не изменяя аффинность связывания при кислом рН. Кроме того, H553/L118, полученное путем интродукции модификации Y55H в тяжелую цепь H496/L118, обладало такой же или слегка повышенной аффинностью связывания при нейтральном рН, но, с другой стороны, пониженной аффинностью связывания при кислом рН по сравнению с H89/L118. Таким образом, интродукция комбинации двух аминокислотных модификаций, Y55H и R57P, в H89/L118 позволяла еще больше повышать способность к снижению аффинности связывания при кислом рН при сохранении или небольшом повышении аффинности связывания при нейтральном рН.
(8-2) Анализ ФК в организме мышей с использованием H553/L118
Оценку скорости элиминации человеческого IL-8 из организма мышей с использованием H553/L118 осуществляли с помощью метода, сходного с методом, описанным в примере 7-2. Полученные данные о концентрации человеческого IL-8 в плазме представлены на фиг. 17, а величины клиренса (CL) человеческого IL-8 из мышиной плазмы представлены в таблице 11.
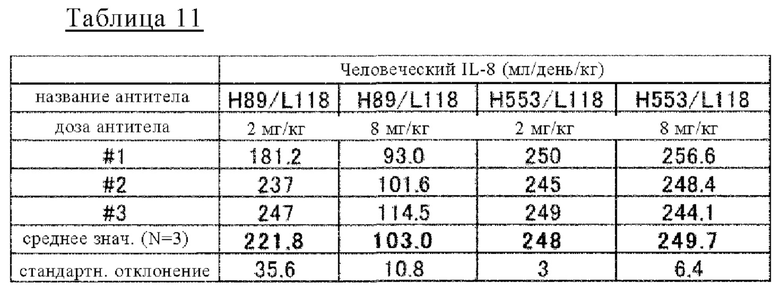
В результате не было выявлено больших различий между H553/L118 и H89/L118 при сравнении данных, полученных в группах, которым вводили антитело в дозе 2 мг/кг; однако, при сравнении данных, полученных в группах, которым вводили антитело в дозе 8 мг/кг, было подтверждено, что H553/L118 ускоряет элиминацию человеческого IL-8 в 2,5 раза или около этого по сравнению с H89/L118. С другой точки зрения, для H553/L118 не было выявлено различия в скорости элиминации человеческого IL-8 при его введении в дозах 2 мг/кг и 8 мг/кг, а для H89/L118 не было обнаружено снижения скорости элиминации антигена, обусловленного увеличением дозы антитела.
Одной из причин (но, не ограничиваясь только ей), по которой были получены такие результаты, может быть следующее. H533/L118 обеспечивало эквивалентную скорость элиминации человеческого IL-8 при введении в дозах 2 мг/кг и 8 мг/кг. Это свидетельствует о том, что доля свободного IL-8 в эндосоме достигает уровня, близкого к 100%, поскольку связывание IL-8 H553/L118 при кислом рН даже в условиях введения в дозе 8 мг/кг является достаточно слабым. Иными словами, это позволяет предположить, что хотя H89/L118 может обеспечивать максимальное действие в отношении элиминации человеческого IL-8 при его введении в дозе 2 мг/кг, его действия могут ослабевать при введении в высокой дозе, составляющей около 8 мг/кг. С другой стороны, H553/L118 может обеспечивать максимальное действие в отношении элиминации человеческого IL-8 даже при введении в высокой дозе, составляющей примерно 8 мг/кг.
(8-3) Оценка стабильности с использованием H553/L118
Было продемонстрировано, что H553/L118 представляет собой антитело, которое может ускорять элиминацию человеческого IL-8 из организма мышей в большей степени, чем H89/L118. Однако для того, чтобы это антитело сохраняло ингибирующее действие на человеческий IL-8 в течение длительного периода времени in vivo, важно также, чтобы нейтрализующая IL-8 активность оставалась стабильной (стабильность нейтрализующей IL-8 активности этого антитела) в течение периода времени, пока введенное антитело присутствует in vivo (например, в плазме). Поэтому проводили оценку стабильности указанных антител в мышиной плазме с помощью описанного ниже метода.
Мышиную плазму собирали из крови мышей линии C57BL/6J (фирма Charles River) с помощью метода, известного в данной области. 200 мкл 200 мМ ЗФР (фирма Sigma, P4417) добавляли к 800 мкл мышиной плазмы с получением объема 1 мл. Кроме того, добавляли азид натрия до конечной концентрации 0,1% в качестве антисептика. Затем добавляли к указанной выше плазме каждое из антител (Hr9, H89/L118 и H553/L118) до конечной концентрации 0,2 мг/мл. В этот момент времени отбирали часть образца в качестве исходного образца. Оставшуюся часть образца хранили при 40°С.После хранения в течение одной недели и двух недель отбирали часть каждого образца и использовали в качестве образца, хранившегося в течение одной недели, и образца, хранившегося в течение двух недель. Все образцы замораживали при -80°С и хранили до осуществления анализа.
Затем анализировали антитела к IL-8, содержащиеся в мышиной плазме в отношении их нейтрализующей человеческий IL-8 активности следующим образом: CXCR1 и CXCR2 представляют собой известные рецепторы для человеческого IL-8. PathHunter®, клеточная линия СНО-K1 CXCR2 β-аррестин (фирма DiscoveRx Co., каталожный номер 93-0202С2) экспрессирует человеческий CXCR2 и представляет собой клеточную линию, искусственно созданную таким образом, чтобы испускать электрохемилюминесценцию при передаче опосредованных человеческим IL-8 сигналов. Нейтрализующую человеческий IL-8 активность, которой обладает антитело к человеческому IL-8, можно оценивать (но, не ограничиваясь только этим) с использованием указанной клетки. Когда человеческий IL-8 добавляют к раствору культуры клеток, происходит испускание определенного уровня хемилюминесценции, который зависит от концентрации добавленного человеческого IL-8. Во-первых, когда человеческий IL-8 и антитело к человеческому IL-8 добавляют вместе к раствору культуры, то трансдукция сигнала человеческого IL-8 может блокироваться при связывании антитела к человеческому IL-8 с человеческим IL-8. В результате хемилюминесценция, вызываемая добавлением человеческого IL-8, будет ингибироваться антителом к человеческому IL-8 и хемилюминесценция должна быть слабее, чем в случае, когда антитело не добавляют, или хемилюминесценции может не быть совсем. Следовательно, когда нейтрализующая человеческий IL-8 активность, которой обладает антитело, становится сильнее, то степень хемилюминесценции становится слабее; а когда нейтрализующая человеческий IL-8 активность, которой обладает антитело, становится слабее, то степень хемилюминесценции становится сильнее.
То же самое имеет место и в случае антитела, которое добавляли к мышиной плазме и хранили в течение определенного периода времени. Если нейтрализующая активность антитела в мышиной плазме не изменяется в процессе хранения, то степень указанной выше хемилюминесценции до и после хранения не должна изменяться. С другой стороны, в случае антитела, нейтрализующая активность которого снижается при хранении в мышиной плазме, степень хемилюминесценции при использовании находившегося на хранении антитела должна возрастать по сравнению со степенью до хранения.
Затем указанную выше клеточную линию использовали для изучения вопроса о том, сохраняется ли нейтрализующая активность антитела при хранении в мышиной плазме. Сначала клеточную линию суспендировали в реагенте Cell Plating 0 AssayComplete™ и затем высевали в 384-луночный планшет из расчета 5000 клеток/лунку. Через 1 день после начала культивирования клеток, проводили описанный ниже эксперимент для определения концентрации предназначенного для добавления человеческого IL-8. Серийно разведенные растворы человеческого IL-8, которые содержали конечные концентрации человеческого IL-8 от 45 нМ (400 нг/мл) до 0,098 нМ (0,1 нг/мл), добавляли к раствору клеточной культуры. Затем добавляли реагент для обнаружения согласно протоколу, прилагаемому к продукту, и определяли относительный уровень хемилюминесценции с использованием хемилюминесцентного детектора. На основе полученных данных подтверждали реактивность клеток в отношении человеческого IL-8 и определяли концентрацию человеческого IL-8, пригодную для подтверждения нейтрализующей активности антител к человеческому IL-8. В данном случае концентрацию человеческого IL-8 доводили до 2 нМ.
Затем мышиную плазму с добавленным вышеуказанным антителом к человеческому IL-8 использовали для оценки нейтрализующей активности содержащихся в ней антител. Человеческий IL-8 в определенной выше концентрации и содержащую вышеуказанное антитело к человеческому IL-8 мышиную плазму добавляли к клеточной культуре. Подлежащее добавлению количество мышиной плазмы определяли таким образом, чтобы она содержала ступенчатые концентрации антитела к человеческому IL-8 в диапазоне от 2 мкг/мл (13,3 нМ) до 0,016 мкг/мл (0,1 нМ). Затем добавляли реагенты для обнаружения согласно протоколу, прилагаемому к продукту, и проводили определение относительных уровней хемилюминесценции с использованием хемилюминесцентного детектора.
В данном случае рассчитывали относительные величины, представляющие собой относительные уровни хемилюминесценции для каждой концентрации антитела, принимая средний относительный уровень хемилюминесценции в лунках без добавления человеческого IL-8 и антитела за 0%, и принимая средний относительный уровень хемилюминесценции в лунках, в которые добавляли только человеческий IL-8, но не добавляли антитело, за 100%.
Результаты анализа ингибирования человеческого IL-8 с использованием экспрессирующих человеческий CXCR2 клеток представлены на фиг. 18-1, где приведены результаты, полученные для исходного образца (без консервирующей обработки в мышиной плазме), на фиг. 18-2, где приведены результаты, полученные для образцов, хранившихся при 40°С в течение одной недели, и на фиг. 18-3, где приведены результаты, полученные для образцов, хранившихся при 40°С в течение двух недель.
Результаты свидетельствуют о том, что не было обнаружено различий между Hr9 и H89/L118 в нейтрализующей человеческий IL-8 активности до и после хранения в мышиной плазме. С другой стороны, для H553/L118 продемонстрировано снижение нейтрализующей человеческий IL-8 активности после хранения в течение двух недель. Следовательно, нейтрализующая человеческий IL-8 активность H553/L118 быстро снижается в мышиной плазме по сравнению с активностью Hr9 и H89/L118, и было установлено, что H553/L118 представляет собой антитело, обладающее нестабильными свойствами с точки зрения нейтрализующей IL-8 активности.
Пример 9
Получение антител с пониженным баллом предсказанной иммуногенности, рассчитанным с использованием системы in silico
(9-1) Балл предсказанной иммуногенности различных связывающих IL-8 антител
Образование антител к лекарственным средствам (ADA) влияет на эффективность и фармакокинетические характеристики терапевтических антител и приводит в некоторых случаях к серьезным побочным действиям; и следовательно, клиническая применимость и лекарственная эффективность терапевтических антител могут ограничиваться образованием ADA. Известно, что на иммуногенность терапевтических антител влияют многие факторы и, в частности, много раз отмечалась важность эпитопов эффекторных Т-клеток, которые несут терапевтические антитела.
Разработаны «инструменты» in silico для предсказания Т-клеточных эпитопов, такие как Epibase (фирма Lonza), iTope/TCED (фирма Antitope) и EpiMatrix (фирма EpiVax). С использованием этих «инструментов» in silico можно предсказывать Т-клеточные эпитопы в любой аминокислотной последовательности (Expert Opin. Biol. Ther. 7(3), март 2007 г., сс. 405-418) и можно оценивать потенциальную иммуногенность терапевтических антител.
Для расчета баллов иммуногенности каждого из антител к IL-8 применяли EpiMatrix. EpiMatrix представляет собой систему предсказания иммуногенности представляющего интерес белка на основе автоматического конструирования последовательностей пептидных фрагментов путем секционирования аминокислотной последовательности белка, для которого требуется предсказать его иммуногенность, на девять аминокислот и затем расчета их способности связываться с восемью основными аллелями ГКС класса II (DRB1*0101, DRB1*0301, DRB1*0401, DRB1*0701, DRB1*0801, DRB1*1101, DRB1*1301 и DRB1*1501) (Clin. Immunol. 131(2), май 2009 г. cc. 189-201).
Баллы иммуногенности тяжелых цепей и легких цепей каждого антитела к IL-8, которые были рассчитаны согласно описанному выше методу, представлены в таблице 12 в колонке, озаглавленной «EpiMatrix Score». Кроме того, в колонке, озаглавленной «tReg Adjusted Ерх Score», представлены баллы иммуногенности, указанные в EpiMatrix Score, которые были скорректированы на содержание Tregitope. Tregitope представляет собой последовательность пептидного фрагмента, присутствующую в больших количествах главным образом в последовательностях встречающихся в естественных условиях антител, и представляет собой последовательность, которая, как считается, ингибирует иммуногенность путем активации супрессорных Т-клеток (Treg).
Кроме того, для этих баллов в колонке, озаглавленной «Сумма», представлены сумма баллов для тяжелой и легкой цепей.
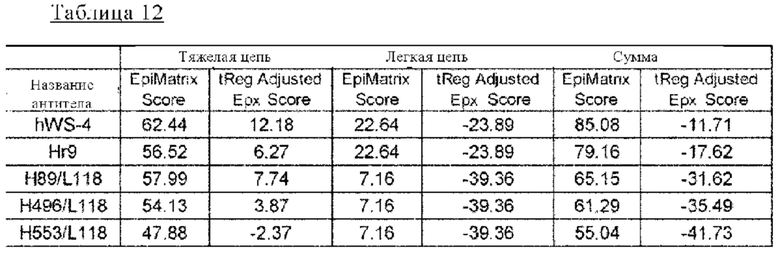
Согласно этим результатам данные, представленные как в «EpiMatrix Score», так и в «tReg Adjusted Ерх Score», свидетельствуют о том, что баллы иммуногенности для H89/L118, H496/L118 и H553/L118 являются более низкими по сравнению баллами для hWS-4, которое представляет собой известное гуманизированное антитело к человеческому IL-8.
Кроме того, с использованием EpiMatrix, можно сравнивать предсказанную частоту образования ADA для молекулы антитела как целого путем сравнения баллов для тяжелой цепи и легкой цепи с фактической частотой образования ADA, обусловленного различными поступающими в продажу антителами. Результаты осуществления такого анализа представлены на фиг. 19. Вследствие ограничений, накладываемых системой, на фиг. 19 использованы следующие обозначения: «WS4» для hWS-4, «HR9» для Hr9, «H89L118» для H89/L118, «H496L118» для H496/L118 и «H553L118» для H553/L118.
Известно, что частота образования ADA в организме человека для различных поступающих в продажу антител составляет: 45% для кампата (алемтузумаб), 27% для ритуксана (ритуксимаб) и 14% для зенапакса (даклизумаб), что представлено на фиг. 19. С другой стороны, в то время как частота образования ADA, предсказанная на основе аминокислотных последовательностей, составляла 10,42% для hWS-4, которое представляет собой известное гуманизированное антитело к человеческому IL-8, частоты для новых идентифицированных при создании изобретения антител H89/L118 (5,52%), H496/L118 (4,67%) или H553/L118 (3,45%) были существенно меньше по сравнению с частотой для hWS-4.
(9-2) Получение модифицированных антител с пониженными баллами предсказанной иммуногенности
Как указано выше, баллы иммуногенности для H89/L118, H496/L118 и H553/L118 были меньше, чем для hWS-4; однако, как следует из таблицы 12, баллы иммуногенности для тяжелой цепи больше, чем баллы для легких цепей, это позволяет предположить, что в данном случае еще есть возможность улучшения аминокислотных последовательностей тяжелой цепи, прежде всего с точки зрения иммуногенности. Поэтому осуществляли поиск аминокислотных модификаций вариабельной области тяжелой цепи Н496, которые могут снижать балл иммуногенности. В результате тщательного поиска были найдены три варианта, H496v1, в котором аланин в положении 52 с согласно нумерации Кэбота был заменен на аспарагиновую кислоту, H496v2, в котором глутамин в положении 81 был заменен на треонин, и H496v3, в котором серии в положении 82b был заменен на аспарагиновую кислоту. Кроме того, был получен вариант Н1004, в который были интродуцированы все три указанные модификации.
Результаты расчета баллов иммуногенности с помощью метода, сходного с методом, описанным в примере 9-1, представлены в таблице 13.
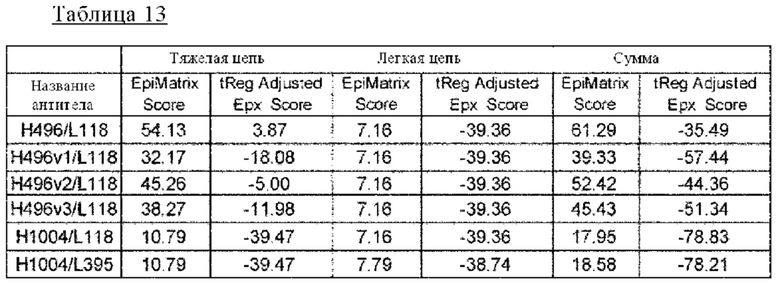
Три тяжелые цепи, H496v1, H496v2 и H496v3, каждая из которых содержит одну модификацию, характеризовались более низкими баллами иммуногенности по сравнению с Н496. Кроме того, в случае H1004, в которую интродуцирована комбинацию трех модификаций, было достигнуто существенное улучшение балла иммуногенности.
В данном случае было установлено, что помимо L118, L395 представляет собой легкую цепь, пригодную для использования в комбинации с H1004. Поэтому при расчете баллов иммуногенности использовали как комбинацию с L118, так и комбинацию с L395. Как продемонстрировано в таблице 13, H1004/L118 и H1004/L395, представляющие собой комбинации тяжелой цепи и легкой цепи, также характеризовались очень низкими баллами иммуногенности.
Затем для них предсказывали частоту образования ADA с помощью метода, сходного с методом, описанным в примере 9-1. Результаты представлены на фиг. 20. На фиг. 20 использованы следующие обозначения: «V1» для H496v1/L118, «V2» для H496v2/L118, «V3» для H496v3/L118, «H1004L118» для H1004/L118 и «H1004L395» для H1004/L395.
Неожиданно было установлено, что H1004/L118 и H1004/L395, которые имели существенно более низкие баллы иммуногенности, характеризовались также улучшенными величинами частоты образования ADA, для них предсказанная величина равна 0%.
(9-3) Измерение IL-8-связывающей аффинности H1004/L395
Получали H1004/L395, которое представляет собой антитело, имеющее H1004-IgG1m (SEQ ID NO: 47) в качестве тяжелой цепи и L395-k0MT (SEQ ID NO: 82) в качестве легкой цепи. Аффинность связывания H1004/L395 с человеческим IL-8 измеряли согласно описанному ниже методу с использованием устройства BIACORE Т200 (фирма GE Healthcare).
Применяли два указанных ниже подвижных буфера, измерения проводили при соответствующих температурах:
0,05% Твин20, 40 мМ ACES, 150 мМ NaCl, рН 7,4, 40°С и
0,05% Твин20, 40 мМ ACES, 150 мМ NaCl, рН 5,8, 37°С.
Белок A/G (фирма PIERCE) в соответствующем количестве иммобилизовали на сенсорном чипе СМ4 (фирма GE Healthcare) с помощью метода аминного сочетания и захватывали представляющие интерес антитела. Затем инъецировали разведенный раствор человеческого IL-8 или подвижный буфер (который применяли в качестве референс-раствора) для осуществления взаимодействия антител, захваченных на сенсорном чипе, с человеческим IL-8. В качестве подвижного буфера использовали один из указанных выше растворов и соответствующие буферы использовали также для разведения человеческого IL-8. Для регенерации сенсорного чипа применяли 25 мМ NaOH и 10 мМ глицин-HCl (рН 1,5). KD (М) для каждого антитела, характеризующую связывание с человеческим IL-8, рассчитывали на основе константы скорости реакции ассоциации kon (1/Мс) и константы скорости реакции диссоциации koff (1/с), которые представляют собой кинетические параметры, рассчитанные на основе сенсограмм, полученных при измерениях. Для расчета каждого параметра применяли программное обеспечение BIACORE Т200 Evaluation (фирма GE Healthcare).
Результаты измерений представлены в таблице 14. По сравнению с H89/L118, H1004/L395, имеющее более низкий балл иммуногенности, имело эквивалентную величину KD для человеческого IL-8 при нейтральном рН, но более высокие величины KD и koff при кислом рН; и было установлено, что оно обладало способностью быстро отделяться в результате диссоциации от IL-8 в эндосоме.
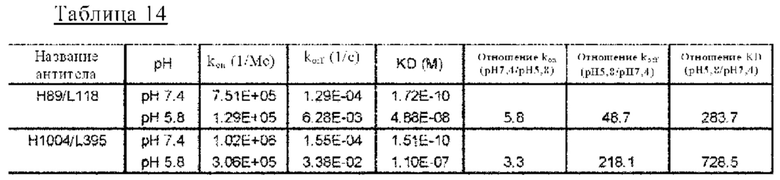
Пример 10
Получение и оценка обладающего рН-зависимым связыванием с IL-8 антитела H1009/L395
(10-1) Получение различных обладающих рН-зависимым связыванием с IL-8 антител
Получали H1004/L395, которое обладает рН-зависимой IL-8-связывающей способностью, а также пониженным баллом иммуногенности, осуществляя оценку согласно методу, описанному в примере 9. После этого проводили специальное исследование для получения вариантов, которые обладали указанными предпочтительными свойствами, а также стабильностью в мышиной плазме.
На основе H1004/L395 получали представленные ниже модифицированные антитела путем интродукции различных модификаций.


Кроме того, было получено в общей сложности 36 типов антител путем объединения 18 типов тяжелых цепей и двух типов легких цепей, описанных выше. Для этих антител проводили различные оценки согласно описанному ниже методу.
Аффинности связывания с человеческим IL-8 в условиях нейтрального и кислого рН измеряли методом, сходным с методом, описанном в примере 9-3. Из числа полученных результатов в таблице 15 представлены KD при рН 7,4 и KD и koff при рН 5,8.
Затем с помощью описанного ниже метода оценивали стабильность в отношении связывания IL-8 при хранении антител в ЗФР.
Соответствующие антитела подвергали в течение ночи диализу в противотоке D-ЗФР (фирма Sigma-Aldrich) и затем концентрацию каждого из антител доводили до 0,1 мг/мл. В это момент времени некоторые образцы антител собирали в качестве исходных образцов. Оставшиеся образцы хранили при 50°С в течение одной недели и затем собирали в качестве образцов для ускоренного теста при повышенной температуре.
Затем осуществляли измерение методом BIACORE аффинности связывания с IL-8 с использованием исходных образцов и образцов для ускоренного теста при повышенной температуре.
Уровни связывания человеческого IL-8 с модифицированными антителами анализировали с использованием устройства BIACORE Т200 (фирма GE Healthcare). Измерения осуществляли при 40°С с использованием 0,05% Твин20, 40 мМ ACES и 150 мМ NaCl с рН 7,4 в качестве подвижного буфера.
Белок A/G (фирма PIERCE) в соответствующем количестве иммобилизовали на сенсорном чипе СМ4 (фирма GE Healthcare) с помощью метода аминного сочетания и захватывали представляющие интерес антитела. Затем инъецировали разведенный раствор человеческого IL-8 или подвижный буфер (который применяли в качестве референс-раствора) для осуществления взаимодействия антител, захваченных на сенсорном чипе, с человеческим IL-8.
Подвижный буфер использовали также для разведения человеческого IL-8. Для регенерации сенсорного чипа применяли 25 мМ NaOH и 10 мМ глицин-HCl (рН 1,5). Измеренный уровень связывания человеческого IL-8 и количество антител, захваченных при этом уровне связывания, рассчитывали с помощью программного обеспечения BIACORE Т200 Evaluation (фирма GE Healthcare).
Рассчитывали количество связываний человеческого IL-8 на соответствующее 1000 RU количество захваченного антитела для исходных образцов и образцов для ускоренного теста при повышенной температуре. Кроме того, рассчитывали отношение уровня связывания человеческого IL-8 для исходных образцов к уровню связывания для образцов для ускоренного теста при повышенной температуре.
Полученные отношения уровня связывания человеческого IL-8 для исходных образцов к уровню связывания для образцов для ускоренного теста при повышенной температуре также представлены в таблице 15.
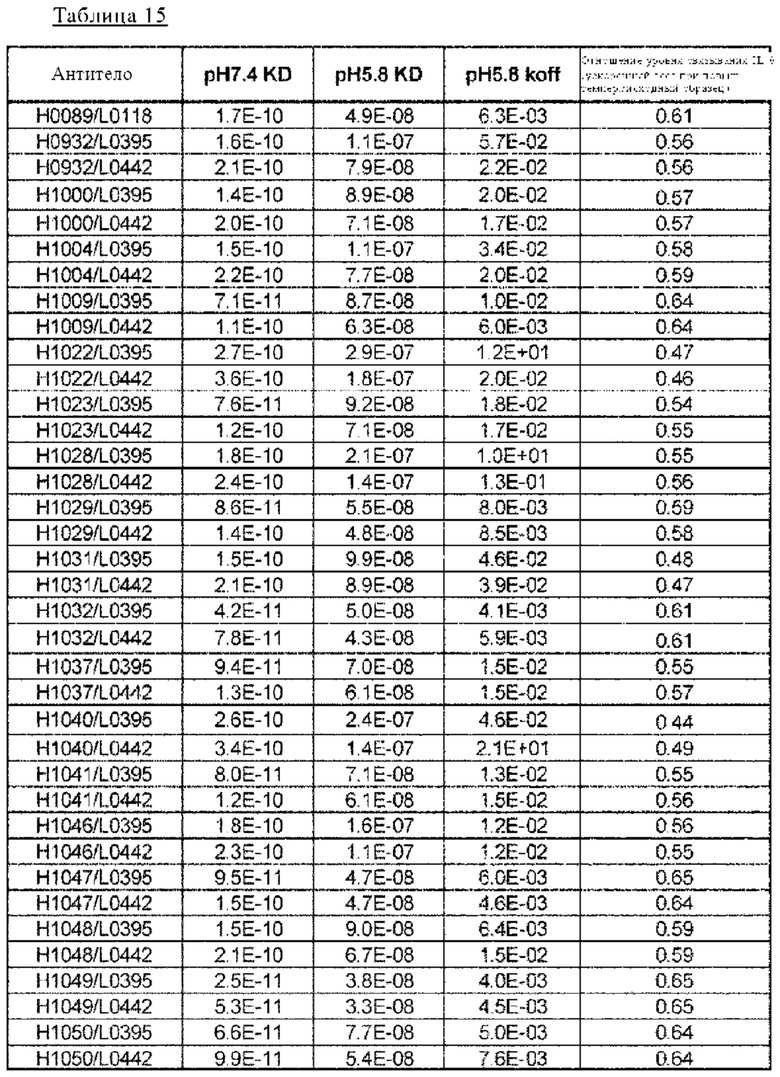
На основе описанного выше исследования было получено антитело H1009/L395, которое представляло собой антитело, имеющее H1009-IgG1m (SEQ ID NO: 48) в качестве тяжелой цепи и L395-k0MT в качестве легкой цепи.
Как продемонстрировано в таблице 15, по сравнению с H89/L118 H1009/L395 обладало немного более высокой аффинностью связывания с человеческим IL-8 при нейтральном рН, но, с другой стороны, пониженной аффинностью связывания при кислый рН, это означает, что его рН-зависимость была еще более сильной. Кроме того, при его нахождении в строгих условиях, таких как 50°С в ЗФР, H1009/L395 обладало несколько улучшенной стабильностью касательно связывания с IL-8 по сравнению со стабильностью H89/L118.
Поэтому H1009/L395 было выбрано в качестве антитела, чья нейтрализующая активность в мышиной плазме может поддерживаться на стабильном уровне, при сохранении его рН-зависимой способности к связыванию с IL-8.
(10-2) Оценка стабильности H1009/L395
Затем, методом, аналогичным методу, описанному в примере 8-3, оценивали, сохраняется ли нейтрализующая IL-8 активность H1009/L395 в стабильном состоянии в мышиной плазме. В этом опыте использовали H1009/L395-F1886s, которое более подробно будет описано в примере 15. Это антитело имело такую же вариабельную область, что и H1009/L395, а его константная область имела модификации, повышающие связывание FcRn в условиях кислого рН, и модификации, снижающие его связывание с FcγR, по сравнению с нативным человеческим IgG1. Вариабельная область H1009/L395, прежде всего область вокруг HVR, ответственна за связывание с человеческим IL-8 и нейтрализующую IL-8 активность этого антитела, и считается, что модификации, интродуцированные в константную область, не влияют на указанные свойства.
Оценку стабильности в мышиной плазме осуществляли следующим образом. 150 мкл 200 мМ фосфатного буфера (рН 6,7) добавляли к 585 мкл мышиной плазмы. Далее добавляли азид натрия в качестве антисептика в конечной концентрации 0,1%. Каждое антитело (Hr9, H89/L118 или H1009/L395-F1886s) добавляли к указанной выше мышиной плазме в конечной концентрации 0,4 мг/мл. В этот момент времени часть образца отбирали в качестве исходного образца. Оставшийся образец хранили при 40°С. Через 1 неделю и две недели после начала хранения часть каждого образца отбирали и их использовали в качестве образца, хранившегося в течение 1 недели, и образца, хранившегося в течение 2 недель. Все образцы замораживали при -80°С и хранили вплоть до осуществления каждого анализа.
Измерение нейтрализующей человеческий IL-8 активности осуществляли с использованием экспрессирующих человеческий CXCR2 клеток с помощью метода, аналогичного описанному в примере 8-3. Однако концентрация человеческого IL-8, которую применяли для подтверждения нейтрализующей активности антитела к человеческому IL-8, в этом опыте составляла 1,2 нМ.
Результаты анализа ингибирования человеческого IL-8, полученные с использованием указанных выше антител и экспрессирующих человеческий CXCR2 клеток, приведены на фиг. 21-1, где представлены результаты для исходного образца (который не подвергали хранению в мышиной плазме), фиг. 21-2, где представлены результаты для образцов, которые хранили при 40°С в течение 1 недели, и на фиг. 21-3, где представлены результаты для образцов, которые хранили при 40°С в течение 2 недель.
В результате неожиданно было установлено, что, нейтрализующая активность H1009/L395-F1886s в отношении человеческого IL-8 сохранялась даже после его хранения в мышиной плазме при 40°С в течение 2 недель и нейтрализующая человеческий IL-8 активность оказалась более стабильной, чем в случае H553/L118.
(10-3) Анализ ФК в организме мышей с использованием H1009/L395
Скорость элиминации человеческого IL-8 антителом H1009/L395 в организме мышей оценивали с использованием следующего метода. В качестве антител применяли H1009/L395, H553/L118 и H998/L63. Введение мышам и сбор крови, а также измерение концентрации человеческого IL-8 в плазме мышей осуществляли с помощью метода, описанного в примере 7.
Полученные в результате данные о концентрации человеческого IL-8 в плазме, представлены на фиг. 22, а данные о величине клиренса человеческого IL-8 (CL) из плазмы мышей представлены в таблице 16.
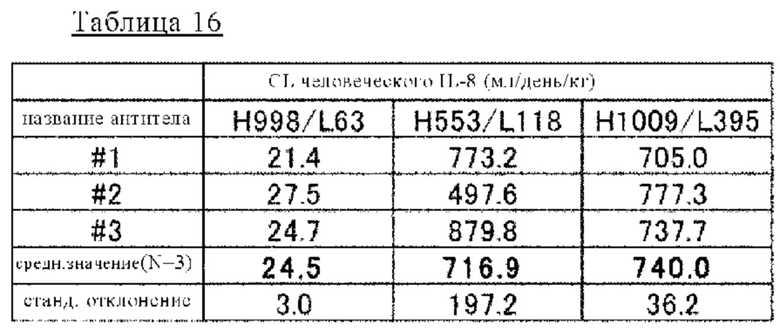
В результате установлено, что скорость элиминации человеческого IL-8 из организма мышей при введении H1009/L395 в дозе 2 мг/кг, была эквивалентна скорости элиминации при введении H553/L118, и установлено, что при применении H1009/L395 достигался практически 100%-ный уровень свободного IL-8 в эндосоме. Установлено, что величина клиренса (CL), которая является количественным показателем скорости элиминации человеческого IL-8 из плазмы мышей, примерно в 30 раз выше, чем при применении H998/L63.
Воздействие повышение скорости элиминации человеческого IL-8 можно объяснять следующим образом (но, не ограничиваясь только им). Как правило, в живом организме, в котором концентрации антигенов поддерживаются примерно на постоянном уровне, скорости образования и скорости элиминации антигенов должны также поддерживаться примерно на постоянных уровнях. Когда в таких условиях вводят антитела, даже в тех случаях, когда не оказывается воздействия на скорость образования антигенов, скорости элиминации антигенов могут изменяться из-за образования комплексов антигена с антителом. Как правило, поскольку скорость элиминации антигена выше, чем скорость элиминации антитела, то в таких случаях скорость элиминации антигенов, образующих комплексы с антителом, снижается. Когда скорость элиминации антигена снижается, концентрация антигена в плазме возрастает, но уровень повышения в этом случае можно определят также как соотношение скорости элиминации, когда антиген присутствует в несвязанном состоянии, и скорости элиминации, когда антиген образует комплекс. Это означает, что по сравнению со скоростью элиминации, когда антиген присутствует в несвязанном состоянии, если скорость элиминации при образовании комплекса снижается на 1/10, то концентрация антигена в плазме организма, которому вводят антитело, может повышаться примерно вплоть до 10 раз по сравнению с концентрацией до введения антитела. В контексте настоящего описания клиренс (CL) можно использовать в качестве скорости элиминации. Более конкретно, увеличение концентрации антигена (накопление антигена), которое имеет место после введения антитела в организм, можно определять по величине CL антигена в каждом варианте условий, т.е. перед введением антитела и после введения антитела.
В настоящем описания наличие примерно 30-кратного различия в CL человеческого IL-8 при введении H998/L63 и H1009/L395 позволяет предположить, что может быть примерно 30-кратное различие между уровнями повышения концентрации человеческого IL-8 в плазме при введении этих антител людям. Кроме того, создание 30-кратного различия в концентрации человеческого IL-8 в плазме свидетельствует о том, что может иметь место также примерно 30-кратное различие в количестве антител, необходимых для полной блокады биологической активности человеческого IL-8 в соответствующих условиях. Это означает, что по сравнению с H998/L63 H1009/L395 может блокировать биологическую активность IL-8 в плазме при использовании примерно 1/30 количества, что представляет собой очень небольшое количество антитела. Кроме того, когда H1009/L395 и H998/L63 вводят индивидуально человеку в одинаковой дозе, то H1009/L395 может блокировать биологическую активность IL-8 в течение более длительного периода времени с большей силой. Для блокады биологической активности IL-8 в течение длительного периода времени необходимо, чтобы IL-8-нейтрализующая активность стабильно сохранялась. Как продемонстрировано в примере 10, эксперименты с использованием мышиной плазмы свидетельствуют о том, что H1009/L395 может сохранять нейтрализующую человеческий IL-8 активность в течение длительного периода времени. Установлено также, что H1009/L395, которое обладает такими важными отличительными свойствами, является антителом с предпочтительными воздействиями с точки зрения эффективности в отношении нейтрализации IL-8 in vivo.
Пример 11
Оценка связывания с внеклеточным матриксом с использованием рН-зависимого IL-8-связывающего антитела H1009/L395
Очень высокое (увеличенное в 30 раз) воздействие H1009/L395 в отношении элиминации человеческого IL-8, описанное в примере 10, являлось неожиданным. Известно, что скорость элиминации антигена при введении рН-зависимого антигенсвязывающего антитела зависит от скорости поглощения комплекса антитело-антиген клетками. Это означает, что, если скорость поглощения клетками рН-зависимого антитела повышается при образовании комплекса антиген-антитело комплекс по сравнению с вариантом без образования комплекса, то элиминация антигена рН-зависимым антителом может увеличиваться. Известные методы повышения скорости поглощения антитела клетками включают метод придания антителу FcRn-связывающей способности в условиях нейтрального рН (WO 2011/122011), метод повышения связывающей способности антитела в отношении FcγR (WO 2013/047752) и метод, в котором применяют ускорение формирования комплексов, содержащих многовалентное антитело и многовалентный антиген (WO 2013/081143).
Однако указанные выше методики не использовали для константных областей H1009/L395. Кроме того, хотя известно, что IL-8 образует гомодимер, установлено, что человеческий IL-8, связанный H1009/L395, присутствует в форме мономера, поскольку H1009/L395 распознает образующую гомодимер поверхность человеческого IL-8. Следовательно, эти антитела не могут образовывать многовалентные комплексы.
Более конкретно, хотя вышеописанные методики не применялись для H1009/L395, H1009/L395 обладает повышенным в 30 раз элиминирующим человеческий IL-8 действием.
Человеческий IL-8 представляет собой белок с высокой изоэлектрической точкой (pI), при этом теоретическая изоэлектрическая точка, рассчитанная с помощью известного метода, составляет примерно 10. Это означает, что в условиях нейтрального рН человеческий IL-8 представляет собой белок, заряд которого сдвинут в сторону положительных значений. рН-зависимые IL-8-связывающие антитела, представителем которых является H1009/L395, также представляют собой белки, заряд которых сдвинут в сторону положительных значений, и теоретическая изоэлектрическая точка H1009/L395 составляет примерно 9. Это означает, что изоэлектрическая точка комплекса, образовавшегося в результате связывания H1009/L395, т.е. белка, который имеет высокую изоэлектрическую точку и исходно богат положительными зарядами, с человеческим IL-8, который имеет высокую изоэлектрическую точку, должна быть выше, чем у H1009/L395 в несвязанном состоянии.
Повышение изоэлектрической точки антитела, что включает увеличение количества положительных зарядов и/или уменьшение количества отрицательных зарядов на антителе, возможно может повышать неспецифическое поглощение клетками комплекса антитело-антиген. Существует также возможность того, что изоэлектрическая точка комплекса, образованного между антителом к IL-8 и человеческим IL-8, который имеет высокую изоэлектрическую точку, выше, чем изоэлектрическая точка антитела к IL-8 в несвязанном состоянии, и комплекс может быстрее поглощаться клетками.
Как описано ранее, связывание с внеклеточным матриксом также является фактором, который может влиять на поглощение клетками. Поэтому при создании изобретения определяли, существует ли различие в способности связывать внеклеточный матрикс между антителом в несвязанном состоянии и входящим в комплекс антителом к человеческому IL-8.
(11-1), Оценка количества антитела, связывающегося с внеклеточным матриксом, с помощью метода ECL (электрохемилюминесценция)
Внеклеточный матрикс (BD Matrigel Basement Membrane Matrix; производство фирмы BD) разводили до концентрации 2 мг/мл с использованием TBS (фирма Takara). Разведенный внеклеточный матрикс вносили на 96-луночный планшет MULTI-ARRAY, High bind, Bare (производство фирмы Meso Scale Discovery:MSD) из расчета 5 мкл на лунку и иммобилизовали в течение ночи при 4°С. Затем осуществляли блокирование с помощью 20 мМ ACES-буфера, рН 7,4, содержащего 150 мМ NaCl, 0,05% Твин 20, 0,5% БСА и 0,01% NaN3.
Предназначенные для оценки антитела получали следующим образом. Образцы антител, предназначенные для индивидуального добавления, получали путем разведения каждого антитела до концентрации 9 мкг/мл, используя буфер 1, указанный ниже, а затем дополнительно разводили их буфером 2 до конечной концентрации 3 мкг/мл.
Буфер-1: 20 мМ ACES-буфер содержащий 150 мМ NaCl, 0,05% Твин20 и 0,01% NaN3, рН 7,4.
Буфер-2: 20 мМ ACES-буфер, содержащий 150 мМ NaCl, 0,05% Твин20, 0,1% БСА и 0,01% NaN3, рН 7,4.
С другой стороны, образцы антител, предназначенные для добавления в виде комплекса с человеческим IL-8, получали путем добавления человеческого IL-8 в 10-кратной молярной концентрации относительно антитела к образцу антитела, затем разводили каждое антитело с помощью буфера-1 до получения концентрации антитела 9 мкг/мл соответственно, а затем дополнительно разводили каждое из них с помощью буфера-2 до конечной концентрации антитела 3 мкг/мл. При этом концентрация человеческого IL-8 составляла примерно 0,6 мкг/мл. Смесь встряхивали при комнатной температуре в течение 1 ч для образования комплекса.
Затем разведенные растворы только антитела или комплекса добавляли на планшет, из которого был удален блокирующий раствор, и планшет встряхивали при комнатной температуре в течение 1 ч. Затем, после удаления раствора, содержащего несвязанное антитело, или раствора, содержащего комплекс, добавляли буфер-1, содержащий 0,25% глутарового альдегида. Затем после выстаивания планшета в течение 10 мин его промывали D-ЗФР (производство фирмы Wako Pure Chemical Industries), содержащим 0,05% Твин 20. Антитела для ECL-обнаружения получали путем конъюгирования с сульфо-меткой козьего античеловеческого IgG (гамма) (производство фирмы Zymed Laboratories) с использованием сложного NHS-эфира с сульфо-меткой (производство фирмы MSD). Антитела для ECL-обнаружения разводили в буфере-2 до концентрации 1 мкг/мл, добавляли на планшет и затем встряхивали в темноте при комнатной температуре в течение 1 ч. Антитела для ECL-обнаружения удаляли и добавляли 2-кратно разведенный раствор, полученный путем разведения буфера для считывания MSD Read Buffer Т (4×) (производство фирмы MSD) ультрачистой водой; и затем измеряли уровень люминесценции с помощью устройства SECTOR Imager 2400 (производство фирмы MSD).
Результаты представлены на фиг. 23. Важно отметить, что было установлено, что все антитела к IL-8, такие как H1009/L395, характеризовались слабой способностью связываться с внеклеточным матриксом в виде несвязанного антитела (-IL8), но связывались с внеклеточным матриксом после образования комплекса с человеческим IL-8 (+hIL8).
Как описано выше, способность антител к IL-8 к приобретению способности связываться с внеклеточным матриксом путем связывания с человеческим IL-8 не была объяснена на основе известной из прототипов информации. Кроме того, объединение указанных свойств с рН-зависимым связыванием у IL-8-связывающих антител (но, не ограничиваясь только указанным) позволяет повышать скорость элиминации IL-8 более эффективно.
Пример 12
Анализ ФК в организме мышей с использованием не связывающих FcRn антител
Анализ ФК в организме мышей с использованием не связывающих FcRn антител
Приведенный ниже метод применяли для подтверждения образования комплекса между человеческим IL-8 и рН-зависимым IL-8-связывающим антителом и повышения поглощения указанного комплекса клетками in vivo.
Сначала получали мутант, содержащий вариабельную область H1009/L395 и Fc-область с дефицитом аффинности связывания с различными Fc-рецепторами. В частности, в качестве модификаций, предназначенных для устранения способности связывания с человеческим FcRn в условиях кислого рН, тяжелую цепь H1009-IgG1 подвергали замене на аланин изолейцина в положении 253 и на аспарагиновую кислоту серина в положении 254 согласно EU-нумерации. Кроме того, в качестве модификаций, предназначенных для устранения связывания с мышиными FcγR, лейцин в положении 235 заменяли на аргинин, глицин в положении 236 заменяли на аргинин и серии в положении 239 заменяли на лизин. Получали 1009-F1942m (SEQ ID NO: 49) в качестве тяжелой цепи, содержащей четыре указанные модификации. Кроме того, получали антитело H1009/L395-F1942m, имеющее H1009-F1942m в качестве тяжелой цепь и L395-k0MT в качестве легкой цепи.
Поскольку антитело, которое имеет указанную Fc-область, обладает дефицитом аффинности связывания с FcRn в условиях кислого рН, оно не переносится из эндосомы в плазму. Следовательно, указанное антитело быстро элиминируется из плазмы в живом организме относительно антитела, которое содержит нативную Fc-область. В этом случае после включения в клетки антител, которые имеют нативную Fc-область, только их часть, которая не спасается с помощью FcRn, расщепляется после переноса в лизосому, но в случае антител, имеющих Fc-область, которая не обладает аффинностью связывания с FcRn, все антитела, включенные в клетки, расщепляются в лизосомах. Более конкретно, в случае антител, которые имеют указанную модифицированную Fc-область, скорость элиминации введенного антитела из плазмы может быть эквивалентной скорости включения в клетки. Это означает, что скорость внутриклеточного поглощения антитела, аффинность связывания с FcRn которого устранена, можно определять также путем измерения скорости элиминации этих антител из плазмы.
Далее определяли, повышается ли внутриклеточное поглощение комплекса, образовавшегося между H1009/L395-F1942m и человеческим IL-8, по сравнению с поглощением H1009/L395-F1942m. В частности, определяли, будет ли изменяться скорость элиминации антитела из плазмы, когда антитело вводят индивидуально и когда антитело вводят после образования комплекса с человеческим IL-8.
Соответствующую биокинетику антитела к человеческому IL-8 оценивали в случаях, когда антитело к человеческому IL-8 вводили индивидуально трансгенным по человеческому FcRn мышам (B6.mFcRn-/-.hFcRn Tg-мыши линии 32 +/+; фирма Jackson Laboratories; Methods Mol. Biol. 602, 2010, cc. 93-104) и когда человеческий IL-8 и антитело к человеческому IL-8 вводили одновременно трансгенным по человеческому FcRn мышам. Раствор антитела к человеческому IL-8 (200 мкг/мл) и раствор, содержащий смесь человеческого IL-8 (10 мкг/мл) и антитела к человеческому IL-8 (200 мкг/мл), вводили индивидуально однократно в дозе 10 мл/кг в хвостовую вену. В этом случае, поскольку антитело к человеческому IL-8 присутствовало в существенном избытке относительно человеческого IL-8, почти весь человеческий IL-8 должен связываться с антителом. Образцы крови получали через 5 мин, 2 ч, 7 ч, 1 день и 2 дня после введения. Собранную кровь сразу же центрифугировали при 4°С и 15000 об/мин в течение 15 мин для получения плазмы. Отделенную плазму хранили в морозильной камере при -20°С или ниже, до осуществления измерений.
Концентрацию антитела к человеческому IL-8 в плазме мышей измеряли электрохемилюминесцентным методом. Сначала давали прореагировать биотинилированному козьему антителу к легкой каппа-цепи человеческого Ig (фирма IBL) с покрытым стрептавидином планшетом Gold Multi-ARRAY (фирма Meso Scale Discovery), который предварительно блокировали в течение ночи при комнатной температуре с использованием раствора ЗФР-Твин, содержащего 5% (мас./об.) БСА, при комнатной температуре в течение 1 ч с получением планшета, с иммобилизованным античеловеческим антителом. Приготавливали образцы для калибровочной кривой, содержащие антитело к человеческому IL-8 в концентрациях 3,20, 1,60, 0,800, 0,400, 0,200, 0,100 и 0,0500 мкг/мл в плазме, и образцы для измерения в мышиной плазме, разведенные в 100 раз или более. Каждый образец смешивали с человеческим IL-8 и затем вносили в планшет с иммобилизованный античеловеческим антителом из расчета 50 мкл на лунку и затем перемешивали при комнатной температуре в течение 1 ч. Конечную концентрацию человеческого IL-8 доводили до 333 нг/мл.
Затем добавляли в планшет антитело к человеческому IL-8 (полученное в лаборатории заявителей), содержащее константную область мышиного IgG, и давали прореагировать при комнатной температуре в течение 1 ч. Кроме того, в планшет добавляли антимышиный IgG (фирма BECKMAN COULTER), меченный рутением, с использованием сложного NHS-эфира с сульфо-меткой (фирма Meso Scale Discovery) и давали прореагировать в течение 1 ч. Затем сразу после добавления в планшет буфера для считывания Т(×1) (фирма Meso Scale Discovery) осуществляли измерение с использованием SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию антитела к человеческому IL-8 рассчитывали на основе ответа с использованием калибровочной кривой с помощью аналитического программного обеспечения SOFTmax PRO (фирма Molecular Devices).
Полученные в результате концентрации антитела в мышиной плазме, представлены на фиг. 24, а данные о клиренсе антитела в соответствующих условиях представлены в таблице 17.
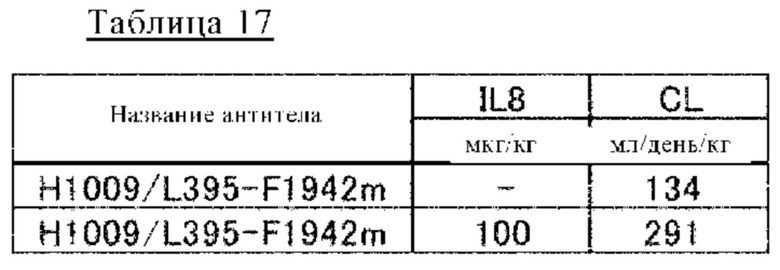
Установлено, что скорость внутриклеточного поглощения комплекса H1009/L395-F1942m и человеческого IL-8 увеличивалась по меньшей мере в 2,2 раза по сравнению со скоростью поглощения H1009/L395-F1942m. Следует отметить, что в контексте настоящего описания указано «по меньшей мере в 2,2 раза», поскольку по указанной ниже причине не исключается возможность того, что значение может фактически составлять 5, 10 или 30 раз. Поскольку скорость элиминации человеческого IL-8 из мышиной плазмы является очень быстрой по сравнению со скоростью элиминации H1009/L395-F1942m, то доля H1009/L395-F1942m, связанного с человеческим IL-8 в плазме, быстро снижается после введения. Более конкретно, даже при введении одновременно с человеческим IL-8, не все H1009/L395-F1942m, присутствующие в плазме, находятся в связанной с человеческим IL-8 форме, и фактически примерно через 7 ч после введения большая часть антител уже находится в свободной форме. Поскольку скорость поглощения исследовали в указанных условиях, то даже, если скорость внутриклеточного поглощения комплекса H1009/L395-F1942m и человеческого IL-8 фактически повышалась в 5, 10 или 30 раз по сравнению со скоростью поглощения H1009/L395-F1942m, результаты, полученные с использованием этой экспериментальной системы, отражают это только частично; следовательно, существует вероятность того, что эффект может повышаться более чем в 2,2 раза или т.д. Таким образом, исходя из полученных результатов, согласно которым скорость внутриклеточного поглощения комплекса H1009/L395 и IL-8, как установлено, должна превышать фактическую скорость внутриклеточного поглощения H1009/L395 in vivo, этот эффект не ограничен полученным повышением в 2,2 раза.
Полученные данные можно интерпретировать следующим образом (но, не ограничиваясь только указанным). Когда H1009/L395, которое представляет собой рН-зависимое IL-8-связывающее антитело, образует комплекс с человеческим IL-8, то комплекс имеет более высокую изоэлектрическую точку и больший сдвиг в сторону положительного заряда, чем антитело, находящееся в несвязанном состоянии. Одновременно связывание комплекса с внеклеточным матриксом является более высокой, чем связывание антитела в несвязанном состоянии. Такие свойства, как повышение изоэлектрической точки и увеличение связывания с внеклеточным матриксом, можно рассматривать в качестве факторов, которые способствуют поглощению антитела клетками in vivo. Кроме того, в проведенных на мышах экспериментах установлено, что внутриклеточное поглощение комплекса H1009/L395 и человеческого IL-8 также возрастало в 2,2 раза или более по сравнению со скоростью поглощения H1009/L395. Исходя из вышесказанного, теоретическое объяснение, а также свойства in vitro и явления, обнаруженные in vivo, убедительно подтверждают гипотезу о том, что H1009/L395 и человеческий IL-8 образуют комплекс, что способствует поглощению комплекса клетками и приводит к существенному увеличению элиминации человеческого IL-8.
К настоящему времени известно несколько антител к IL-8, однако отсутствуют данные о повышении связывания с внеклеточным матриксом после образования комплекса с IL-8 и увеличении поглощения комплексов клетками.
Кроме того, на основе полученных данных об увеличении внутриклеточного поглощения антител к IL-8, когда антитела образуют комплексы с IL-8, можно предположить, что антитела к IL-8, которые могут образовывать комплексы с IL-8 в плазме, быстро поглощаются клетками, а антитела в свободном состоянии, которые не могут образовывать комплексы с IL-8, имеют тенденцию сохраняться в плазме без поглощения клетками. В том случае, когда антитело к IL-8 является рН-зависимым, антитело к IL-8, поглощенное клетками, высвобождает молекулу IL-8 в клетках и затем возвращается в среду вне клеток и после этого может связываться с другой молекулой IL-8; и следовательно увеличение внутриклеточного поглощения после образования комплекса может обладать дополнительным эффектом более сильной элиминации IL-8. Это означает, что отбор антител к IL-8 с повышенной способностью к связыванию с внеклеточным матриксом или антител к IL-8 с повышенным поглощением клетками также является еще одним аспектом, указанным в настоящем описании. Пример 13
Предсказание иммуногенности рН-зависимого IL-8-связывающего антитела H1009/L395 с использование системы in silico
Далее балл иммуногенности и частоту образования ADA предсказывали для H1009/L395 с помощью метода, аналогичного описанному в примере 9-1. Результаты представлены в таблице 18 и на фиг. 25. На фиг. 25 H1009/L395 обозначен как «H1009L395».
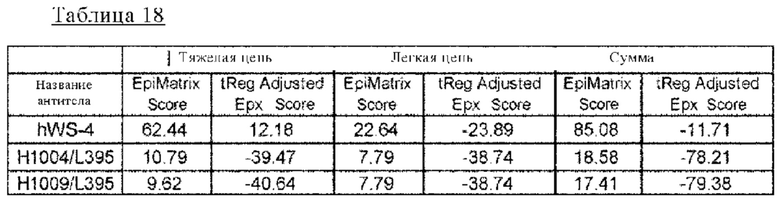
Результаты, представленные в таблице 18, демонстрируют, что H1009/L395 имеет такой же уровень баллов иммуногенности, что и H1004/L395. Кроме того, частота образования ADA, предсказанная для H1009/L395, исходя из результатов, представленных на фиг. 25, составляла 0%, и это также сходно с величиной этого параметра, указанной для H1004/L395.
Таким образом, предсказанная иммуногенность существенно снижалась у H1009/L395 по сравнению с известным антителом к человеческому IL-8 hWS-4. Следовательно, можно предположить, что H1009/L395 должно обладать очень низкой иммуногенностью для людей и должно обладать способностью сохранять стабильную нейтрализующую активность в отношении IL-8 в течение длительного периода времени.
Пример 14
Анализ ФК в организме обезьян циномолгус с использованием варианта H89/L118 с повышенной FcRn-связывающей способностью в условиях кислого рН
Как описано в представленных выше примерах, среди антител, которые имеют константную область нативного IgG1 в качестве их константной области, рН-зависимое IL-8-связывающее антитело H1009/L395 представляло собой антитело с улучшенными свойствами. Однако указанные антитела можно применять также в качестве антител, содержащих аминокислотные замены в константной области, например, содержащих Fc-область с повышенной способностью связываться с FcRn при кислом рН, примеры которых представлены в примере 5. Таким образом, H89/L118 применяли для подтверждения того, что Fc-область с повышенной способностью связываться с FcRn при кислом рН может функционировать также в рН-зависимом IL-8-связывающем антителе.
(14-1) Получение H89/L118 с модифицированной Fc-областью с повышенной способностью связывать FcRn при кислом рН
Различные модификации, повышающие FcRn-связывание, описанные в примере 5-1, интродуцировали в Fc-область H89/L118. В частности, путем интродукции модификаций, применяемых в F1847m, F1848m, F1886m, F1889m, F1927m и F1168m, в Fc-область H89-IgG1 получали следующие варианты конструкций:
H89/L118-IgG1, имеющую H89-IgG1m (SEQ ID NO: 50) в качестве тяжелой цепи и L118-K0MT в качестве легкой цепи;
H89/L118-F1168m, имеющую H89-F1168m (SEQ ID NO: 51) в качестве тяжелой цепи L118-K0MT в качестве легкой цепи;
H89/L118-F1847m, имеющую H89-F1847m (SEQ ID NO: 52) в качестве тяжелой цепи и L118-K0MT в качестве легкой цепи;
H89/L118-F1848m, имеющую H89-F1848m (SEQ ID NO: 53) в качестве тяжелой цепи и L118-K0MT в качестве легкой цепи;
H89/L118-F1886m, имеющую H89-F1886m (SEQ ID NO: 54) в качестве тяжелой цепи и L118-K0MT в качестве легкой цепи;
H89/L118-F1889m, имеющую H89-F1889m (SEQ ID NO: 55) в качестве тяжелой цепи и L118-K0MT в качестве легкой цепи;
H89/L118-F1927m, имеющую H89-F1927m (SEQ ID NO: 56) в качестве тяжелой цепи и L118-K0MT в качестве легкой цепи.
Анализы ФК в организме обезьян циномолгус с использованием этих антител осуществляли с применением описанного ниже метода.
Описанную ниже в примерах конструкцию H89/L118-F22 (имеющую Н89-F22 (SEQ ID NO: 106) в качестве тяжелой цепи и L118-K0MT в качестве легкой цепи) получали также аналогичным путем.
(14-2) Анализы ФК в организме обезьян циномолгус с использованием содержащих новый вариант Fc-области антител
После введения антител к человеческому IL-8 обезьянам циномолгус оценивали биокинетику антител к человеческому IL-8. Раствор антитела к человеческому IL-8 вводили внутривенно однократно в дозе 2 мг/кг. Образцы крови получали через 5 мин, 4 ч, 1 день, 2 дня, 3 дня, 7 дней, 10 дней, 14 дней, 21 день, 28 дней, 35 дней, 42 дня, 49 дней и 56 дней после введения. Собранную кровь сразу же центрифугировали при 4°С и 15000 об/мин в течение 10 мин для получения плазмы. Отделенную плазму хранили в морозильной камере при -60°С или ниже, до осуществления измерений.
Концентрацию антитела к человеческому IL-8 в плазме обезьян циномолгус измеряли электрохемилюминесцентным методом. Сначала вносили захватывающее Ат к человеческой каппа-цепи (h-каппа) (фирма Antibody Solutions) в 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Discovery) и перемешивали при комнатной температуре в течение 1 ч. Затем раствор ЗФР-Твин, содержащий 5% (мас./об.) БСА, применяли для блокады в течение 2 ч с получением планшета с иммобилизованным античеловеческим антителом. Приготавливали образцы для калибровочной кривой, содержащие антитело к человеческому IL-8 в концентрациях 40,0, 13,3, 4,44, 1,48, 0,494, 0,165 и 0,0549 мкг/мл в плазме, и приготавливали образцы для измерения в плазме обезьян циномолгус, разведенные в 500 раз или более. Вносили по 50 мкл растворов в каждую лунку планшета с иммобилизованным античеловеческим антителом и растворы перемешивали при комнатной температуре в течение 1 ч. Затем добавляли конъюгированное с биотином репортерное Ат к h-каппа (фирма Antibody Solutions) в вышеуказанный планшет и давали прореагировать при комнатной температуре в течение 1 ч. После дополнительного добавления меченного сульфо-меткой стрептавидина (фирма Meso Scale Discovery) и выдерживания в течение 1 ч комнатной температуре для обеспечения прохождения реакции добавляли в планшет буфер для считывания Т (×1) (фирма Meso Scale Discovery) и сразу же проводили измерение с использованием SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию антитела к человеческому IL-8 рассчитывали на основе ответа с использованием калибровочной кривой с помощью аналитического программного обеспечения SOFTmax PRO (фирма Molecular Devices).
Полученные результаты, характеризующие время полужизни (t1/2) и клиренс (CL) каждого из антител, представлены в таблице 19, и изменения в концентрации антител в плазме обезьян циномолгус представлены на фиг. 26.
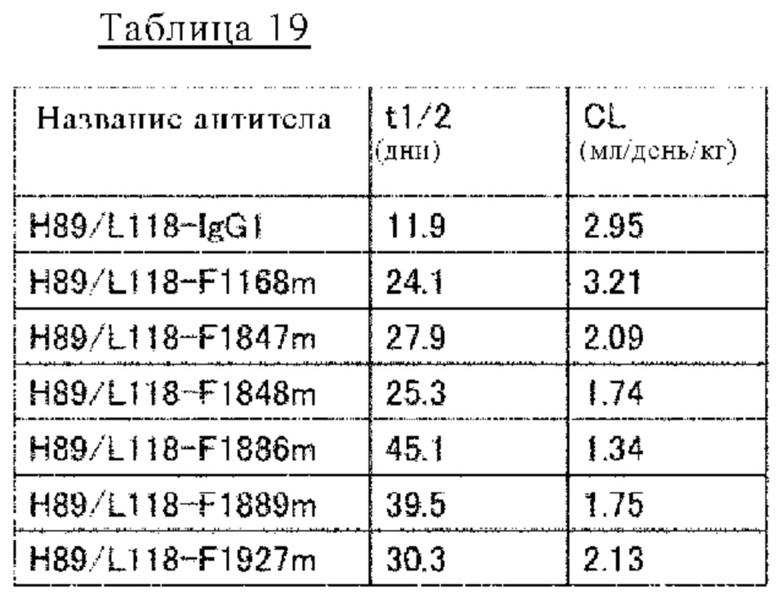
Приведенные выше результаты подтвердили, что все варианты Fc-области характеризовались улучшенным временем удерживания в плазме по сравнению с антителом, имеющим Fc-область нативного IgG1. В частности, для H89/L118-F1886m обнаружена наиболее предпочтительная кинетика в крови.
Пример 15
Fc-область с пониженной связывающей способностью в отношении FcγR
Известно, что Fc-область нативного человеческого IgG1 связывается с Fcγ-рецептором(ами) (обозначенные далее как FcγR) на различных клетках иммунной системы и обладает эффекторными функциями, такими как ADCC и ADCP, в отношении клеток-мишеней.
С другой стороны, IL-8 представляет собой растворимый цитокин, и применяемые в качестве фармацевтических препаратов антитела к IL-8, как ожидается, главным образом должны обладать фармакологическими действиями посредством нейтрализации функций IL-8 в областях, в которых IL-8 присутствует в избыточном количестве. Указанные области, в которых IL-8 присутствует в избыточном количестве, не ограничены конкретными областями, и, например, могут представлять собой области воспаления. Как хорошо известно в указанных областях воспаления собираются и активируются иммунные клетки. Передача нежелательных сигналов активации к этим иммунным клеткам через Fc-рецепторы и индукция активности, такой как ADCC и ADCP, в нежелательных клетках не всегда являются нужными. Следовательно, с точки зрения безопасности (но, не ограничиваясь только этим) может оказаться предпочтительным, чтобы антитела к IL-8, вводимые in vivo, обладали низкой аффинностью к FcγR.
(15-1) Получение модифицированных антител с пониженной способностью связываться с FcγR
Аминокислотные модификации дополнительно интродуцировали в Fc-область H1009/L395-F1886m для снижения способности связываться с различными FcγR человека и обезьян циномолгус. В частности, H1009-F1886s (SEQ ID NO: 37) получали путем осуществления в тяжелой цепи H1009-F1886m каждой из следующих замен: замена L на R в положении 235, замена G на R в положении 236 и замена S на K в положении 239 согласно EU-нумерации. Аналогично этому, H1009-F1974m (SEQ ID NO: 36) получали путем осуществления в H1009-F1886m замены L на R в положении 235 и замены G на R в положении 236 согласно EU-нумерации и замены области, простирающейся от положения 327 до положения 331 согласно EU-нумерации в последовательности нативного человеческого IgG4. H1009/L395-F1886s и H1009/L395-F1974m получали в виде антител, имеющих указанные тяжелые цепи и L395-k0MT в качестве легкой цепи.
(15-2) Подтверждение связывания с различными человеческими FcγR
Затем связывание полученных H1009/L395-F1886s или H1009/L395-F1974m к растворимым формам FcγRIa или FcγRIIIa человека или обезьян циномолгус подтверждали следующим методом.
Анализы для оценки связывания полученных H1009/L395-F1886s или H1009/L395-F1974m с растворимыми формами FcγRIa или FcγRIIIa человека или обезьян циномолгус осуществляли с использованием устройства BIACORE Т200 (фирма GE Healthcare). Растворимые FcγRIa и FcγRIIIa как человека, так и обезьян циномолгус получали в форме меченных His молекул с помощью методов, известных обычным специалистам в данной области. Взятый в соответствующем количестве рекомбинантный белок (r-белок) L (фирма BioVision) иммобилизовывали на сенсорном чипе СМ4 (фирма GE Healthcare) с помощью метода аминного сочетания и захватывали антитело. Затем растворимый FcγRIa или FcγRIIIa инъецировали с использованием подвижного буфера (применяемого в качестве референс-раствора) и давали взаимодействовать с антителами, захваченными на сенсорном чипе. HBS-EP+ (фирма GE Healthcare) применяли в качестве подвижного буфера, a HBS-EP + применяли для разведения растворимого FcγRIa или FcγRIIIa. Для регенерации сенсорного чипа применяли 10 мМ глицин-HCl, рН 1,5. Все измерения проводили при 20°С.
Результаты представлены на фиг. 27. На указанном чертеже для человеческого FcγRIa, человеческого FcγRIIIa, FcγRIa обезьян циномолгус и FcγRIIIa обезьян циномолгус применяли следующие обозначения: hFcγRIa, hFcγRIIIa, cynoFcγRIa и cynoFcγRIIIa соответственно. Установлено, что конструкция H1009/L395-F1886m связывалась со всеми FcγR, но, с другой стороны, подтверждено, что вновь созданные H1009/L395-F1886s и H1009/L395-F1974m не связывались ни с одним из FcγR.
(15-3) Анализ элиминации мышиного IL-8 при применении вариантов Fc
Далее для полученных H1009/L395-F1886s и H1009/L395-F1974m изучали скорость элиминации человеческого IL-8 и удерживание антител в плазме мышей с использованием следующего эксперимента. В нем использовали три дозы H1009/L395-F1886s, составляющие 2, 5 и 10 мг/кг, для того, чтобы оценить воздействия возрастающей дозы антитела H1009/L395-F1886s.
После одновременного введения человеческого IL-8 и антитела к человеческому IL-8 трансгенным по человеческому FcRn мышам (мыши B6.mFcRn-/-.hFcRn Tg линии 32+/+; фирма Jackson Laboratories; Methods Mol. Biol. 602, 2010, cc. 93-104) оценивали биокинетические характеристики человеческого IL-8. Смешанный раствор человеческого IL-8 (10 мкг/мл) и антитела к человеческому IL-8 (200, 500 или 1000 мкг/мл) вводили однократно в дозе 10 мл/кг в хвостовую вену. В этом случае, поскольку антитело к человеческому IL-8 присутствовало в достаточном избытке относительно человеческого IL-8, считалось, что практически весь человеческий IL-8 связывался с антителом. Образцы крови собирали через 5 мин, 1 ч, 4 ч, 7 ч, 1 день, 2 дня, 3 дня, 7 дней, 14 дней, 21 день и 28 дней после введения. Собранную кровь немедленно центрифугировали при 4°С и 15000 об/мин в течение 15 мин для получения плазмы. Отделенную плазму хранили в холодильнике, установленном на -20°С или ниже, до осуществления измерений.
Концентрацию человеческого IL-8 в плазме мышей измеряли методом, аналогичным описанному в примере 7. Полученные данные, касающиеся концентрации человеческого IL-8 в плазме, представлены на фиг. 28, а величины клиренса (CL) человеческого IL-8 из плазмы мышей представлены в таблице 20.
Прежде всего, было продемонстрировано, что антитело H1009/L395, имеющее Fc-область нативного IgG1, и H1009/L395-F1886s, имеющее модифицированную Fc-область, обладали эквивалентной способностью элиминировать человеческий IL-8 при сравнении групп, которых обрабатывали дозой 2 мг/кг.
Затем при изменении дозы антитела H1009/L395-F1886s существенного различия в величинах клиренса человеческого IL-8 не обнаружено при применении доз 2 мг/кг и 10 мг/кг, хотя выявлено небольшое различие концентрации IL-8 в плазме на следующий день после введения. Эти данные с большой доли уверенности позволяют предположить, что антитела, содержащие вариабельную область H1009/L395, обладают достаточно сильной способностью элиминировать IL-8, даже, если антитела вводят в высоких дозах.
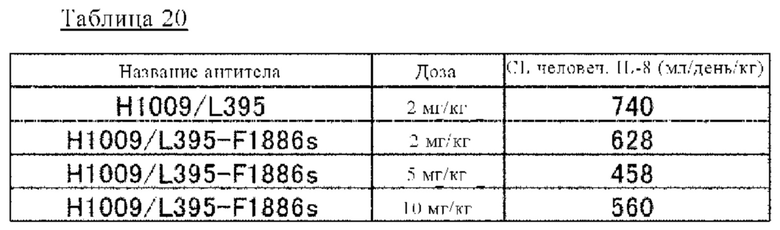
15-4) Анализ ФК-характеристик вариантов Fc-области на обезьянах циномолгус
Затем оценивали удерживание антител в плазме обезьян циномолгус с помощью описанного ниже метода с использованием антитела H1009/L395-F1886s или H1009/L395-F1974m.
Биокинетические характеристики антитела к человеческому IL-8 оценивали после введения антитела к человеческому IL-8 индивидуально или после одновременного введения человеческого IL-8 и антитела к человеческому IL-8 обезьянам циномолгус. Раствор, содержащий антитело к человеческому IL-8 (2 мг/мл), или раствор, содержащий смесь человеческого IL-8 (100 мкг/кг) и антитела к человеческому IL-8 (2 мг/кг), вводили внутривенно в дозе 1 мл/кг. Образцы крови собирали через 5 минут, 4 часа, 1 день, 2 дня, 3 дня, 7 дней, 10 дней, 14 дней, 21 день, 28 дней, 35 дней, 42 дня, 49 дней и 56 дней после введения. Собранную кровь сразу же центрифугировали при 4°С и 15000 об/мин в течение десяти минут с получением плазмы. Отделенную плазму хранили в морозильной камере при -60°С или ниже до осуществления измерений.
Концентрацию антитела к человеческому IL-8 в плазме обезьян циномолгус измеряли с помощью метода, описанного в примере 14. Полученные данные о концентрации антитела к человеческому IL-8 представлены на фиг. 29, а величины времени полужизни (t1/2) и клиренса (CL) антитела к человеческому IL-8 из плазмы обезьян циномолгус представлены в таблице 21.
Прежде всего, было продемонстрировано, что по сравнению с Hr9 и H89/L118, которые имеют Fc-область нативного человеческого IgG1, H1009/L395-F1886s, которое имеет Fc-область с улучшенными функциями, характеризовалось значительно более пролонгированным удерживанием в плазме.
Кроме того, когда H1009/L395-F1886s вводили одновременно с человеческим IL-8, изменение концентрации в плазме было эквивалентным изменению, которое имело место при введении только одного антитела. Не вдаваясь в конкретную теорию, на основе этих данных можно высказать следующие соображения. Как описано выше, было продемонстрировано, что внутриклеточное поглощение комплекса H1009/L395 и человеческого IL-8 являлось более высоким по сравнению с поглощением только одного H1009/L395. Как правило, считается, что высокомолекулярные белки должны проникать в клетки неспецифически или зависимым от рецептора путем, затем переноситься в лизосому и расщепляться различными расщепляющими ферментами, присутствующими в лизосоме. Следовательно, если скорость (уровень!!!) поглощения белка клетками возрастает, то, по-видимому, при этом (также!!!) ухудшается удерживание белка в плазме. Однако, в случае антитела, оно обладает свойством возвращаться в плазму посредством FcRn в эндосоме; и следовательно, пока «спасение» посредством функций FcRn является достаточным, увеличение скорости внутриклеточного поглощения может не влиять на удерживание в плазме. В рассматриваемом случае влияние на удерживание в плазме не имело место даже в том случае, когда H1009/L395-F1886s вводили обезьянам циномолгус одновременно с человеческим IL-8. Это указывает на возможность того, что, хотя скорость поглощения антитела H1009/L395-F1886s клетками возрастает, антитело в достаточной степени «спасается» посредством FcRn, поскольку оно может возвращаться в плазму.
Кроме того, продемонстрировано, что имеющее другой вариант Fc H1009/L395-F1974m также характеризовалось удерживанием в плазме, эквивалентным удерживанию H1009/L395-F1886s. Хотя в указанные варианты Fc были интродуцированы различные модификации, которые снижали способность к связыванию с различными FcγR, как описано выше, было продемонстрировано, что они не влияли на удерживание самих антител в плазме. Полученные выше данные продемонстрировали, что удерживание в плазме обезьян циномолгус как H1009/L395-F1886s, так и H1009/L395-F1974m, существенно пролонгировалось и оказалось в высшей степени удовлетворительным по сравнению с удерживанием антител, которые имеют Fc-область нативного IgG1.
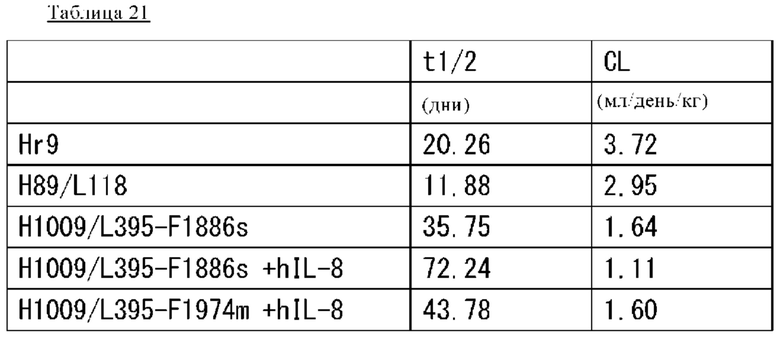
Как указано в описанных выше примерах, H1009/L395 представляет собой антитело, которое впервые создано как антитело, которое может существенно повышать скорость элиминации человеческого IL-8 in vivo благодаря тому, что оно обладает способностью к рН-зависимому связыванию IL-8, а также способностью к быстрому встраиванию в клетки после образования комплекса с IL-8. Кроме того, поскольку аффинность H1009/L395 к связыванию с IL-8 в условиях нейтрального рН также повышена по сравнению с аффинностью известного антитела hWS-4, то H1009/L395 может в более значительной степени нейтрализовать человеческий IL-8 в условиях нейтрального рН, таких как в плазме. Кроме того, оно представляет собой антитело, которое обладает более высокой стабильностью в условиях плазмы и его нейтрализующая IL-8 активность не снижается даже после введения in vivo. Кроме того, H1009/L395, сконструированное на основе Hr9, уровень производства которого был значительно повышен по сравнению с hWS-4, представляет собой антитело, которое пригодно для производства с точки зрения объема производства. Кроме того, согласно in silico-прогнозу иммунногенности H1009/L395 имел очень низкий балл иммуногенности, который был значительно меньше по сравнению с баллом известного антитела hWS-4 и некоторых существующих антител, поступающих в продажу. Более конкретно, можно ожидать, что H1009/L395 представляет антитело, которое можно безопасно применять в течение продолжительного периода времени, поскольку для него с меньшей вероятностью могут образовываться ADA в человеческом организме. Таким образом, H1009/L395 представляет собой антитело, которое с различных точек зрения обладает улучшенными характеристиками по сравнению с известными антителами к человеческому IL-8 и которое является очень полезным в качестве фармацевтического агента.
Как указано выше, H1009/L395, имеющее Fc-область нативного IgG, является достаточно ценным; однако, можно применять также варианты H1009/L395, содержащие Fc-область с улучшенной функцией, в качестве антител, обладающих повышенной пользой. Более конкретно, его действие может сохраняться в течение более продолжительного периода времени за счет усиления связывания с FcRn в условиях кислого рН и улучшения удерживания в плазме. Кроме того, варианты, содержащие Fc-область, в которую были интродуцированы модификации для снижения способности к связыванию с FcγR, позволяют избегать нежелательной активации клеток иммунной системы, развития цитотоксической активности и т.п. в организме, в который их вводят, и их можно применять в качестве обладающих высокой безопасностью фармацевтических антител. Такие Fc наиболее предпочтительно включают (но, не ограничиваясь только ими) F1886s и F1974m, представленные в настоящем описании. В качестве вариантов осуществления настоящего изобретения можно применять также фармацевтические антитела, содержащие другие, модифицированные Fc-области, если они обладают сходными функциями.
Таким образом, антитела, представленные в настоящем описании, включая H1009/L395-F1886s и H1009/L395-F1974m, могут сохранять состояние, позволяющее осуществлять сильное ингибирование биологической активности человеческого IL-8 безопасно и в течение продолжительного периода времени. Антитела позволяют достигать уровня, недостижимого для существующих антител к IL-8, и можно ожидать, что их можно будет применять в качестве обладающих исключительно высоким качеством фармацевтических средств на основе антитела к IL-8.
Пример 16. Измерение концентрации IL-8 в кистозной жидкости пациентов с эндометриозом
Концентрацию IL-8 в кистозной жидкости пациентов с эндометриозом определяли с целью оценки участия IL-8 в эндометриозе. В качестве образца использовали кистозную жидкость, собранную из подвергнутых хирургическому вмешательству тканей пациентов с эндометриозом, которым осуществляли операцию в Медицинском Университете Джичи, и после первой анонимизации образцы подвергали второй анонимизации в компании Chugai Pharmaceutical Co., Ltd. Концентрацию IL-8 в кистозной жидкости измеряли методом электрохемилюминесценции. Сначала, после мечения биотином вносили аликвоты антитела к человеческому IL-8 (фирма Hycult Biotech) на сенсибилизированный стрептавидином 96-луночный планшет MSD GOLD 96-Well Streptavidin SECTOR (фирма Meso Scale Discovery), который блокировали в течение 2 ч с использованием раствора ЗФР-Твин, содержащего 5 мас./об. % БСА, и встряхивали в течение 1 ч при комнатной температуре в темноте с получением планшета с иммобилизованным антителом к человеческому IL-8. Для построения стандартной кривой приготавливали образцы, содержащие человеческий IL-8 (полученный в лаборатории заявителя согласно NP_000575.1) в кистозной жидкости в концентрации 900, 300, 100, 33,3, 11,1, 3,70 или 1,23 пг/мл, и предназначенные для измерений образцы человеческой кистозной жидкости, разведенные в 5 раз или более, и вносили в виде аликвот объемом по 25 мкл в каждую лунку планшета с иммобилизованным антителом к человеческому IL-8, после чего осуществляли перемешивание в течение 1 ч при комнатной температуре в темноте. Затем после взаимодействия с меченным SULFO-TAG антителом к человеческому IL-8 (приготовленным в лаборатории заявителя) в течение 1 ч при комнатной температуре в темноте, вносили аликвоты буфера для считывания Read Buffer Т (×1) (фирма Meso Scale Discovery) и сразу же осуществляли измерение с использованием визуализирующего устройства SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию человеческого IL-8 рассчитывали на основе стандартной кривой ответов с использованием программного обеспечения для анализа SOFT Max PRO (фирма Molecular Devices).
Как представлено на фиг. 30, результаты свидетельствуют о том, что концентрация IL-8 в кистозной жидкости пациентов с эндометриозом составляла 10000 пг/мл или более, и, таким образом, была весьма высокой. Это свидетельствует о важной роли IL-8 в эндометриозе.
Пример 17 Создание модели индуцированного хирургическим путем эндометриоза на обезьянах и оценка патологических состояний
Для изучения эффективности антитела к IL-8 в качестве лекарственного средства в отношении эндометриоза создавали модель индуцированного хирургическим путем эндометриоза и проводили исследование. Модель патологии создавали следующим образом.
Самок обезьян циномолгус возрастом 8-14 лет с регулярным половым циклом (полученных от исследовательского центра приматов Цукуба (Tsukuba Primate Research Center) при Национальных институтах биомедицинских инноваций, здоровья и питания (National Institutes of Biomedical Innovation, Health and Nutrition)) подвергали лапаротомии под анестезией в период времени, соответствующий лютеиновой фазе. Тело матки удаляли путем V-образного рассечения. Затем тело матки разрезали на кубики размером 5-10 мм, сохраняя при этом гладкомышечный слой матки, и трансплантировали один и два кусочка на правую абдоминальную стенку и левую абдоминальную стенку (краниальная и каудальная стороны) соответственно таким образом, чтобы содержащая эндометрий часть находилась в контакте с брюшиной, осуществляя сшивание с использованием рассасывающегося шовного материала и формируя тем самым трансплантат. После иссечения матку зашивали с использованием рассасывающегося шовного материала. Затем оставшиеся эндометриоидные ткани разрезали на мелкие кусочки и суспендировали в 5 мл раствора фактора роста гепатоцитов (фирма R&D systems) (2 нг/мл), полученным продуктом засевали перитонеальную полость с целью трансплантации, после чего зашивали брюшную полость. Анестезию осуществляли с использованием сбалансированной смеси кетамин/ксилазин (смешанные в соотношении, соответствующем примерно 2:1); при этом анестезию поддерживали с использованием изофлурана в зависимости от периода, в течение которого требовалось поддерживать анестезию. Температуру тела поддерживали и контролировали в процессе осуществления хирургического вмешательства с использованием грелки-матраца и осуществляли мониторинг состояния с помощью электрокардиографа. В момент пробуждения после хирургического вмешательства вводили антагонист антиседан (примерно в таком же количестве, что и ксилазин) и антибиотик цефрадин. Обезболивающее средство залбан вводили в течение двух дней после хирургического вмешательства, обезьян регулярно обследовали каждый день для подтверждения отсутствия послеоперационных патологий. При проведении лапароскопического обследования анестезию и послеоперационный мониторинг осуществляли таким же образом.
Лапароскопическое обследование осуществляли через 4 месяца, 9 месяцев (через 3 месяца после введения) и 12 месяцев (через 6 месяцев после введения) после описанной выше индукции эндометриоза хирургическим путем и осуществляли оценку обусловленных эндометриозом патологических состояний следующим образом.
Под анестезией осуществляли медианный разрез брюшной полости и проводили лапароскопическое обследование путем введения троакара и последующего введения лапароскопа. Лапароскоп был присоединен к видеосистеме (фирма KARL STORZ) и монитору. Монитор использовали для внутрибрюшинного обследования, а видеосистему применяли для регистрации. Затем брюшную полость заполняли газообразным диоксидом углерода, используя для этого внутрибрюшинный инсуффлятор (фирма OLYMPUS) для расширения брюшной полости и из вентрального участка вводили градуированные линейки и пинцет. Размер повреждения измеряли с помощью градуированной линейки или градуированного пинцета.
Измеряли размеры (ширину, высоту и глубину) узловых повреждений, образовавшихся при сшивании эндометриоидных тканей, и рассчитывали их объемы по формуле: ширина (мм) × высота (мм) × глубина (мм). Повреждения и спайки, образовавшиеся в результате высевания эндометриоидных тканей после их измельчения, оценивали с точки зрения области, площади и глубины спайки и размера повреждения путем лапароскопического обследования на основе модифицированной шкалы r-AFS, модифицированной для обезьян шкалы r-AFS, которую применяют в клинических исследованиях. Как продемонстрировано на фиг. 31, модифицированная шкала r-AFS включает две следующие модификации для применения на обезьянах:
1. Добавлен пункт, касающийся спайки с пузырно-маточным углублением, поскольку на обезьяньей модели частота встречаемости спайки с мочевым пузырем больше, чем у человека. В методе оценки спайки с мочевым пузырем использовали ту же самую шкалу, которую применяют для оценки спаек с Дугласовым карманом.
2. Поскольку размеры тела и органов у обезьян меньше, чем у человека, то критерии оценки размера повреждений изменяли на следующие: <3 мм, 3-10 мм и >10 мм.
Окончательные лапароскопические оценки делали позже с использованием для их подтверждения видеозаписей, сделанных при лапароскопических обследованиях с использованием видеосистемы.
На основе результатов лапароскопического обследования, проведенного через четыре месяца после индукции, отбирали особей, которые удовлетворяли критериям, и группировали их. Критерии отбора были следующими:
1. Приживление эндометриоидного трансплантата, подтвержденное путем лапароскопического обследования через четыре месяца после индукции.
2. Наличие менструальных кровотечений, подтвержденное три или более раз в течение периода времени с момента индукции до момента введения.
Разделение на группы осуществляли в два отдельных этапа. После рандомизации на основе среднего объема узловых повреждений и количества поддающихся измерению повреждений у каждой особи проводили окончательное разделение: шесть особей включали в группу, которую обрабатывали наполнителем, и семь особей включали в группу, которой вводили антитело H1009/L395-F1974m. Для подтверждения того, что между группами не было различий, проводили оценку на основе критерия Крускала-Уоллиса с использованием данных для всех окончательно сгруппированных особей. Оцениваемыми параметрами были: объем узлового повреждения (для каждого повреждения), вес тела, балл размера по шкале r-AFS и балл спаек по шкале r-AFS.
Введение осуществляли, начиная с шестого месяца после индукции, каждые 4 недели в течение 6 месяцев, в общей сложности шесть раз. H1009/L395-F1974m вводили животным в группе, которую обрабатывали антителом, путем внутривенной инъекции в дозе 10 мг/кг, а His-буфер (20 мМ His-Asp, 150 мМ Arg-Asp (рН 6,0)) вводили подкожно животным в группе, которую обрабатывали наполнителем. Для оценки обусловленного эндометриозом патологического состояния осуществляли лапароскопическое обследование каждые три месяца после начала введения, а именно, через 3 и 6 месяцев после первого введения.
После конечного лапароскопического обследования, которое проводили через шесть месяцев после начала введения, обезьян подвергали эвтаназии путем обескровливания после дополнительной анестезии. Осуществляли аутопсию для сбора органов полости таза (правый и левый яичники, правый и левый яйцеводы, правый и левый мезосальпингсы, правая и левая фимбрии фаллопиевых труб, правая и левая широкие связки матки, матка, влагалище, Дугласов карман и пузырно-маточное углубление), включая участки, в которые осуществляли трансплантацию. Собранные ткани фиксировали в 10%-ном нейтральном забуференном формалине и затем тонкие срезы образцов залитых в парафин тканей окрашивали гематоксилином и эозином (НЕ). Разрезали также ткани и повреждения, индуцированные при наложении швов, для сбора скопившейся в них кистозной ткани, если она присутствовала.
Результаты лапароскопического обследования обработанной наполнителем контрольной группы продемонстрировали образование узловых повреждений, возникших при наложении швов на эндометриоидные ткани, через четыре месяца после индукции, и дополнительное образование узловых повреждений и примыкающих к ним черных (шоколадных) кист через 12 месяцев после индукции (через шесть месяцев после введения). Кроме того, было выявлено образование спайки в дугласовом кармане и мочевом пузыре, которое было обусловлено засеванием тонко измельченных эндометриоидных тканей (фиг. 32). Кроме того, как продемонстрировано на фиг. 33, НЕ-окрашивание образцов ткани из обработанной наполнителем группы, собранных в момент аутопсии через 12 месяцев после индукции, продемонстрировало, что имело место образование компактного пролиферирующего эндометриоидного эпителия, примыкающего к трансплантированным эндометриоидным тканям (исходный трансплантат), вплоть до мышечного слоя брюшной стенки. Области, окружающие эндометриоидный эпителий, состояли из нескольких слоев стромальных клеток и интерстиция с высоким содержанием коллагеновых волокон, и, таким образом, наблюдалось образование структуры, сходной со структурой человеческих эндометриозных тканей. Эти результаты продемонстрировали, что рассматриваемая модель является пригодной моделью эндометриоза (12-й Всемирный конгресс по эндометриозу, 2014, Р-221). Аналогично этому, опубликованы данные о методе, в котором использовали бабуинов для создания индуцированной хирургическим путем модели эндометриоза (Fertil Steril.; 99(3), 2013, сс. 783-789, Fertil Steril.; 100(4), 2013, сс. 1144-1150).
Кроме того, осуществляли аутопсию после конечного лапароскопического обследования через 12 месяцев после индукции (через шесть месяцев после введения). Собирали скопившуюся кистозную жидкость и измеряли методом электрохемилюминесценции концентрацию IL-8 в кистозной жидкости у животных в группе, обработанной наполнителем. Сначала вносили аликвоты раствора антитела H1009/L395-F1974m на 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Discovery) и планшет выдерживали при 4°С в течение ночи для подготовки планшета с иммобилизованным антителом H1009/L395-F1974m. Приготавливали предназначенные для построения стандартной кривой образцы раствора обезьяньего IL-8 (полученного в лаборатории согласно ХМ_005555087.1) в буфере с концентрацией 269, 108, 43,0, 17,2, 6,89, 2,75 или 1,10 пг/мл и предназначенные для измерения образцы кистозной жидкости обезьян, разведенные в 50 раз или более, и вносили в виде аликвот в каждую лунку планшета с иммобилизованным антителом H1009/L395-F1974m, после чего перемешивали в течение двух часов при комнатной температуре в темноте. Затем проводили взаимодействие с кроличьим антителом к IL-8 (фирма Hycult Biotech) в течение 1 ч при комнатной температуре в темноте. Затем после взаимодействия с меченным SULFO-TAG антикроличьим антителом (фирма Meso Scale Discovery) в течение 1 ч при комнатной температуре в темноте вносили аликвоты буфера для считывания Read Buffer Т (×1) (фирма Meso Scale Discovery) и сразу же проводили измерение с использованием визуализирующего устройства SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию обезьяньего IL-8 рассчитывали на основе стандартной кривой ответов с использованием программного обеспечения для анализа SOFT Max PRO (фирма Molecular Devices). Как продемонстрировано в таблице 23, результаты свидетельствуют о том, что концентрация IL-8 оказалась выраженно больше в кистозной жидкости обезьян, на которых моделировали эндометриоз посредством его индукции хирургическим путем, что сходно с тем, что имеет место в кистозной жидкости больных эндометриозом женщин. Эти данные подтверждают то, что результаты, полученные на обезьяньей модели, очень хорошо могут быть экстраполированы на человеческий эндометриоз. Кроме того, оценивали корреляцию концентрации IL-8 в кистозной жидкости каждой особи с баллом спаек по шкале r-AFS. Важно отметить, что, как продемонстрировано на фиг. 34-1, имеется тенденция к корреляции концентрации IL-8 в кистозной жидкости с баллом спаек по шкале r-AFS. Если у особи было определено несколько концентраций IL-8 в кистозной жидкости, то в качестве репрезентативной величины использовали наивысшее значение. Этот результат позволяет предположить, что IL-8 участвует в спайкообразовании при эндометриозе.
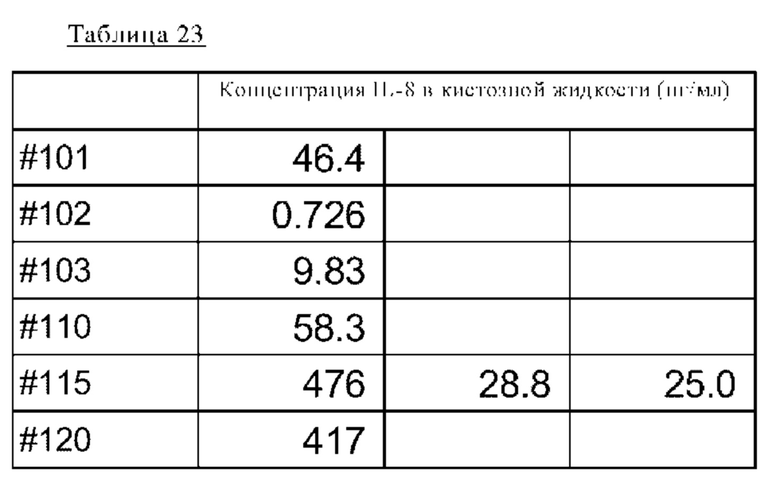
Оценку эффективности лекарственного средства проводили путем лапароскопического обследования через шесть месяцев после начала введения антитела H1009/L395-F1974m животным в группе, которой вводили антитело, и в группе, обработанной наполнителем. При проведении анализа исключали из анализа особей, у которых было установлено, что количество менструальных кровотечений в течение шестимесячного периода введения было меньше трех. Среди особей, на которых проводили данное исследование, особь №117 из группы, которой вводили антитело H1009/L395-F1974m (группа, которой вводили дозу 10 мг/кг), подпадала под это ограничение и, таким образом, она была исключена из анализа. В группе, которой вводили антитело H1009/L395-F1974m, осуществляли обнаружение анти-антитело с помощью метода, известного специалистам в данной области. Анти-антитела были обнаружены и было установлено, что минимальная концентрация API в крови оказалась сниженной у двух особей из шести. Описанный ниже анализ проводили для позитивных по анти-антителам особей.
Сначала рассчитывали объемы каждого из узловых повреждений, образовавшихся в результате наложения швов на эндометриоидные ткани, и классифицировали на основе их размеров и изменений с течением времени, как представлено ниже в таблице 24. Повреждения, зависимость от времени которых нельзя было оценить, такие как обусловленные спайками, не включали в анализ.

Как продемонстрировано в таблице 25, результаты свидетельствуют о том, что в обработанной наполнителем группе частота встречаемости типов А и Б, которые характеризуются разрастанием (увеличением размера), составляла 54% (7/13 повреждений), а типа Г, который характеризовался уменьшением, составляла 15% (2/13 повреждений), в то время как в группе, которой вводили антитело H1009/L395-F1974m, частота встречаемости типа Г, который характеризовался уменьшением, составляла 58% (7/12 повреждений). Таким образом, продемонстрировано, что введение антитела H1009/L395-F1974m оказывало уменьшающее узловые повреждения действие.

Кроме того, для каждой особи рассчитывали относительный объем узловых повреждений с использованием описанной ниже процедуры расчета для оценки эффективности антитела H1009/L395-F1974m в качестве лекарственного средства в отношении узловых повреждений (относительный объем узловых повреждений (%) = сумма объемов выбранных повреждений, измеренных через шесть месяцев после начала введения/сумма объемов выбранных повреждений, измеренных за два месяца до начала введения) × 100). Результат представлен на фиг. 34-2. Относительный объем повреждений составил 141% (67-331) (медианное значение (минимальное значение - максимальное значение)) для группы, обработанной наполнителем, и 49% (15-156) для группы, которой вводили антитело H1009/L395-F1974m. Узловые повреждения значимо уменьшались при введении антитела H1009/L395-F1974m по сравнению с повреждениями в обработанной наполнителем группе (Р=0,0492). В статистическом анализе использовали параметрический t-критерий (уровень значимости: Р<0,05). Результат свидетельствует о том, что воздействие антитела к IL-8 подавляет увеличение объема узловых повреждений.
Затем для изучения эффективности лекарственного средства оценивали с использованием «модифицированного r-AFS-балла» повреждения, образовавшиеся в результате тонкого измельчения и высевания эндометриоидных тканей, и спаек в группе, которой вводили антитело H1009/L395-F1974m (N=6), и обработанной наполнителем группе (N=6). «Изменение модифицированного r-AFS-балла» рассчитывали согласно описанной ниже процедуре для выявления изменения между повреждениями до и после введения антитела у каждой особи. Результат представлен на фиг. 35-1.
Изменение общего r-AFS-балла = общий r-AFS-балл (после введения) - общий r-AFS-балл (до введения)
Изменение r-AFS-балла спаек = r-AFS-балл спаек (после введения) - r-AFS-балл спаек (до введения)
Изменение r-AFS-балла размера = r-AFS-балл размера (после введения) - r-AFS-балл (до введения)
«Изменение общего r-AFS-балла» и «изменение балла спаек» для группы, которой вводили антитело H1009/L395-F1974m, выраженно уменьшались по сравнению с обработанной наполнителем группой (р=0,0070, р=0,0096). «Изменение r-AFS-балла размера» также имело тенденцию к уменьшению для группы, которой вводили антитело H1009/L395-F1974m по сравнению с обработанной наполнителем группой. Статистический анализ осуществляли с использованием непараметрического критерия Стила (уровень значимости: 5% для двустороннего критерия) с использованием программного обеспечения для статистического анализа (компания SAS). Результат, заключающийся в том, что «изменение балла спаек» значимо снижалось при введении антитела H1009/L395-F1974m, и в том, что, как продемонстрировано выше на фиг. 34-1 имелась тенденция к корреляции между концентрацией IL-8 в кистозной жидкости и «r-AFS-баллом спаек», свидетельствует о том, что IL-8 играет значительную роль в образовании спаек при эндометриозе и что антитела к IL-8 можно применять для уменьшения спаек при эндометриозе.
На фиг. 35-1 представлено «изменение модифицированного r-AFS-балла», которое оценивали на основе изображений, зарегистрированных при лапароскопическом хирургическом вмешательстве. На фиг. 35-2 представлен «общий модифицированный r-AFS-балл», полученный на основе баллов, зарегистрированных практикующим врачом при лапароскопическом хирургическом вмешательстве. Баллы, полученные за два месяца до первого введения, использовали в качестве баллов до введения. Статистический анализ изменения «общего модифицированного r-AFS-балла» до и после введения осуществляли для каждой группы с использованием знакового рангового критерия Уилкоксона, который применяли для соответствующих пар. Результат (уровень значимости: Р<0,05) продемонстрировал, что балл после введения значимо возрастал в группе, обработанной наполнителем (Р=0,0313), но он не изменялся в группе, которой вводили антитело H1009/L395-F1974m (Р=0,875). Поскольку «общий модифицированный r-AFS-балл» представляет собой оценку степени патологической злокачественности эндометриоза, то данный результат демонстрирует действие антитела к IL-8 в отношении подавления патологического обострения эндометриоза.
После конечного лапароскопического обследования трансплантаты и области приживления трансплантатов, собранные при проведении аутопсии, окрашивали НЕ и проводили их гистопатологическое обследование с помощью светового микроскопа. В результате этого, как продемонстрировано на фиг. 36, были обнаружены атрофические изменения пролиферирующего эпителия и стромальных клеток, а также уменьшение интерстиция, основным компонентом которого являются коллагеновые волокна, в группе, которой вводили антитело H1009/L395-F1974m, по сравнению с группой, обработанной наполнителем, и этот эффект был выражен наиболее сильно у негативных по анти-антителам особей, у которых сохранялась концентрация API в крови (таблица 26). Хотя в группе, обработанной наполнителем, были выявлены повреждения типа Г (№102-L2 и №115-R), которые имели тенденцию к уменьшению объема узловых повреждений, в результате обследования патологических узловых повреждений не было обнаружено атрофии или уменьшения интерстиция, так что повреждения сохранялись. Хотя группа, которой вводили антитело H1009/L395-F1974m, включала особей с повреждениями типа Б, которые имели тенденцию к увеличению объема узловых повреждений, в результате обследования патологических узловых повреждений было обнаружено атрофическое изменение стромы и уменьшение интерстиция у №106-L1, что свидетельствовало об улучшении патологического состояния. Кроме того, гемосидерин был обнаружен у 56% (9/16) особей в группе, обработанной наполнителем, и только у 24% (4/17) особей в группе, которой вводили антитело, это позволяет предположить, что введение антитела подавляла инфильтрацию макрофагов или моноцитов. Кроме того, в группе, которой вводили антитело, наблюдалась регенерация мышечной ткани, которая не была обнаружена в группе, обработанной наполнителем, и эти данные позволяют предположить, что имел место регресс повреждений, которые распространились вплоть до мышечного слоя, и их замещение мышечными тканями. При оценке наличия глубоких повреждений на основе присутствия протоков эндометриоидных желез, которые проникали вплоть до мышечного слоя стенки брюшины, не было обнаружено значительных различий между группой, обработанной наполнителем и группой, которой вводили антитело H1009/L395-F1974m, включающей позитивных по анти-антителам особей; однако, когда были исключены позитивные по анти-антителам особи, то частота встречаемости в группе, которой вводили антитело H1009/L395-F1974m, составляла 1/11 повреждений по сравнению с 5/18 повреждениями в группе, обработанной наполнителем. Таким образом, можно предположить, что антитело может оказывать также влияние на образование глубоких повреждений.

Кроме того, проводили также тем же самым методом оценку антитела H89/L118-F22, которое представляет собой антитело до модификации, в результате которой получают антитело H1009/L395-F1974m, в отношении его эффективности в качестве лекарственного средства с использованием обезьяньей модели индуцированного хирургическим путем эндометриоза. В группе, которой вводили антитело H89/L118-F22, также была обнаружена атрофия пролиферирующего эпителия и стромальных клеток, а также уменьшение интерстиция, основным компонентом которого являются коллагеновые волокна.
Эти результаты свидетельствуют о том, что антитела к IL-8 антитела обладают эффективностью в качестве лекарственного средства в отношении уменьшения объема эндометриоидных повреждений, уменьшения спаек, индукции атрофических изменений эпителия и стромальных клеток, снижения инфильтрации иммунных клеток и ослабления фиброза, это демонстрирует, что антитела к IL-8 можно применять в качестве терапевтических средств против эндометриоза. На основе описанных выше результатов для специалистов в данной области, естественно, должно быть понятно, что ингибиторы сигнала IL-8 можно применять в качестве терапевтических или профилактических агентов против эндометриоза.
Пример 18. Лекарственная эффективность антитела к IL-8 в отношении аденомиоза
После конечного лапароскопического обследования через шесть месяцев после начала введения антитела обезьянам, на которых моделировали эндометриоз посредством его индукции хирургическим путем, собранные при аутопсии матки фиксировали с помощью 10%-ного нейтрального забуференного формалина и окрашивали НЕ- для обследования. Явно выраженные эндометриоидные ткани, аналогичные тканям при аденомиозе, были обнаружены в матке в одном случае в группе, обработанной наполнителем, а также в одном случае в группе, которой вводили антитело. Эндометриоидные ткани, обнаруженные у этих двух обезьян циномолгус, имели патологические признаки аденомиоза, которые наблюдаются в человеческих матках, в том числе эндометриоидные железы и окружающую эндометриоидную строму в миометрии. Таким образом, обезьян рассматривали в качестве модели для изучения человеческого аденомиозного осложнения и осуществляли сравнение эндометриоидных тканей в миометрии для этих отдельных случаев. Как представлено на фиг. 37, в группе, которой вводили антитело H1009/L395-F1974m, были выявлены атрофия эндометриоидного эпителия в аденомиозных повреждениях, а также уменьшение и атрофия стромальных клеток по сравнению с обработанной наполнителем группой. Эти результаты свидетельствуют о том, что антитела к IL-8 оказывают также подавляющее аденомиоз действие. На основе описанных выше результатов для специалистов в данной области, естественно, должно быть очевидно, что ингибиторы сигнала IL-8 можно применять в качестве терапевтических или профилактических агентов против эндометриоза.
Пример 19. Анализ экспрессии CXCR1 и CXCR2 в эндометриоидных тканях
CXCR1 и CXCR2 известны в качестве человеческих рецепторов IL-8. Для изучения клеток, на которые действует IL-8 в эндометриоидных повреждениях, залитые в парафин блоки вырезанных хирургическим путем человеческих эндометриоидных тканей подвергали иммуногистохимическому окрашиванию с использованием мышиного моноклонального антитела к человеческому CXCR1 (фирма R&D Systems, каталожный номер: 42705) и мышиного моноклонального антитела к человеческому CXCR2 (фирма Abcam, каталожный номер: 19). Из этих парафиновых блоков приготавливали тонкие срезы и инкубировали с антителами к CXCR1 и к CXCR2 в качестве первичных антител. Затем осуществляли взаимодействие с козьим антителом к мышиному IgG (фирма Dako), конъюгированным с полимером-HRP, в качестве вторичного антитела, и осуществляли визуализацию с использованием диаминобензидина (фирма Wako Pure Chemical Industries). Результаты, представленные в таблице 27, свидетельствовал о том, что стромальные клетки были негативными по CXCR1 и CXCR2. В то же время, нейтрофилы, инфильтрованные в ткани, а также макрофаги и мононуклеарные клетки с отложениями гемосидерина, были позитивными как по CXCR1, так и по CXCR2, и было установлено, что в эпителиальных клетках имела место экспрессия CXCR1 и CXCR2 от слабой до умеренной. Кроме того, сосудистые эндотелиальные клетки были слабо позитивными по CXCR1 и CXCR2. Эти данные позволяют предположить, что IL-8 может оказывать непосредственное фармакологическое действие на эндометриоидные эпителиальные клетки, а также фармакологическое действие посредством инфильтрующих иммунных клеток, таких как нейтрофилы и макрофаги.
Пример 20. Эффективность антитела к IL-8 в качестве лекарственного средства в отношении спаек после хирургического вмешательства
При использовании описанных выше моделей индуцированного хирургическим путем эндометриоза на обезьянах циномолгус у всех особей при лапароскопическом обследовании через четыре месяца после индукции (за два месяца до введения антитела) были выявлены спайки с областью, окружающей область иссечения в стенке брюшины, которые отличались от спаек при эндометриозе, возникшем после применяемой для его индукции лапаротомии. Для изучения эффективности антитела к IL-8 к качестве лекарственного средства в отношении спаек после хирургического вмешательства анонимизировали несколько особей и оценивали спайки с областью, окружающей область иссечения на стенке брюшины путем сравнения результатов лапароскопических обследований, проведенных за два месяца до введения, с фотографиями, сделанными при конечных лапароскопических обследованиях, проведенных через шесть месяцев после начала введения. Более конкретно, пять осуществлявших обследование специалистов сравнивали данные о спайке с областью, окружающей область иссечения на стенке брюшины, полученные до и после введения, и оценивали по трехбалльной шкале: (1) «Улучшенная», (2) «Стабильная» и (3) «Прогрессирующая». Оценки принимали при наличии консенсуса между тремя или большим количеством из пяти проводивших обследование специалистов; в противном случае использовали оценку (2) «Стабильная". Оценки проводили на шести обезьянах в группе, обработанной наполнителем, а также на семи обезьянах в группе, которой вводили антитело H1009/L395-F1974m. На фиг. 38 представлены репрезентативные фотографии брюшных полостей у животных в группе, обработанной наполнителем, и в группе, которой вводили антитело H1009/L395-F1974m. Как представлено в таблице 28, из шести обезьян в группе, обработанной наполнителем, пять получили оценку «Стабильная» и одна оценку «Прогрессирующая», в то время как в группе, которой вводили антитело H1009/L395-F1974m, из семи обезьян шесть получили оценку «Улучшенная», а одна оценку «Прогрессирующая». Обезьяна с оценкой «Прогрессирующая» представляла собой животное, у которого концентрация антитела H1009/L395-F1974m в крови снизилась вследствие производства анти-антител. Описанные выше данные позволяют предположить, что антитела к IL-8 можно применять в качестве терапевтических агентов или профилактических агентов для улучшения послеоперационных спаек. На основе описанных выше результатов для специалистов в данной области, естественно, должно быть понятно, что ингибиторы сигнала IL-8 можно применять в качестве терапевтических или профилактических агентов против спаек.
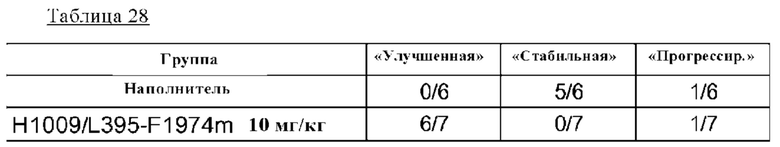
Пример 21. Выделение нейтрофилов и оценка миграции нейтрофилов, обусловленной IL-8
IL-8 обладает хемотаксической активностью в отношении нейтрофилов. Считается, что IL-8, продуцируемый поврежденными тканями, стимулирует миграцию нейтрофилов, присоединенных к сосудистым эндотелиальным клеткам, к тканям и индуцирует воспаление, ассоциируемое с инфильтрацией нейтрофилов. Как представлено в таблице 27, инфильтрация нейтрофилов, макрофагов и мононуклеарных клеток наблюдалась также в областях эндометриоидного повреждения и указанные иммунные клетки были позитивными по экспрессии как CXCR1, так и CXCR2. Кроме того, эпителиальные клетки также были от слабо позитивных до позитивных по экспрессии, это позволяет предположить, что IL-8 оказывает непосредственное фармакологическое действие на эндометриоидные эпителиальные клетки, а также фармакологическое действие посредством инфильтрующих иммунных клеток, таких как нейтрофилы и макрофаги. С учетом этого при создании изобретения в качестве первой стадии оценивали способность нейтрофилов мигрировать к IL-8. Нейтрофилы собирали из периферической крови здоровых особей и осуществляли выделение и очистку согласно описанной ниже процедуре.
Обработанную гепарином цельную кровь объединяли с взятым в 1/5 объема HetaSep (фирма STEMCELL Technologies). Супернатант, содержащий ядерные клетки, собирали согласно протоколу получения продукта и затем к нему добавляли раствор ACK (фирма GIBCO) для индукции гемолиза. После гемолиза клетки суспендировали в 0,1% смеси БСА/ЗФР для определения количества клеток. После измерения количества клеток количество клеток стандартизовали и выделяли нейтрофилы с использованием набора для обогащения человеческих нейтрофилов EasySep™ (фирма STEMCELL technologies) и магнита EasySep™ Magnet (фирма STEMCELL technologies) согласно протоколу получения продукта. После выделения нейтрофилы оценивали методом FACS с использованием антитела к CD66b и антитела к CD45RA для подтверждения их чистоты.
Выделенные нейтрофилы оценивали в отношении их способности к миграции с использованием 96-луночного планшета для оценки миграции с помощью анализа клеточной миграции CytoSelect™ (фирма Cell Biolabs, каталожный номер: СВА-104). Нижний слой, находящийся под мембраной, представлял собой лоток для питательной среды, в который добавляли 1% ЗФР или 1% ЗФР+IL-8 (100 нг/мл) и 150 мкл среды RPMI-1640 (фирма SIGMA), дополненной различными антителами, химическими соединениями или контрольными растворителями. Объектами для оценки были следующие: в качестве антител (каждое в концентрации 10 мкг/мл): H1009/L395-F1974m, нейтрализующие антитела к IL-8 антитело к IL-8 [807] (фирма abcam, каталожный номер: ab18672) и антитело к IL-8 [6217] (фирма abcam, каталожный номер: ab10768), и IgG1 (фирма Sigma, каталожный номер: 15154) в качестве отрицательного контроля; в качестве химических соединений - ингибитор CXCR2 SRT3109 (10 мкг/мл) (фирма Shanghai Haoyuan Chemexpress, каталожный номер: HY-15462); и в качестве контрольных сред - ЗФР и ДМСО. Верхний слой, находящийся над мембраной, представлял собой мембранную камеру, и нейтрофилы, суспендированные в бессывороточной среде RPMI-1640, высевали в камеру с плотностью 50000 клеток/лунку. После инкубации в течение 2 ч определяли количество жизнеспособных клеток (нейтрофилов), которые мигрировали в нижний слой в лоток для питательной среды, на основе уровня хемолюминесценции с помощью люминесцентного анализа оценки жизнеспособности клеток CellTiter-Glo™ (фирма Promega). Средний относительный уровень хемолюминесценции в лунке, в которую не добавляли IL-8, принимали за 1 при расчете относительных уровней относительной хемолюминесценции после добавления различных реагентов. Результат представлен на фиг. 39.
Результат свидетельствует о том, что миграция нейтрофилов стимулируется IL-8 и различные нейтрализующие антитела к IL-8 и ингибиторы CXCR2 ингибируют миграцию нейтрофилов. Описанный выше результат убедительно подтверждает ту точку зрения, что не только ингибиторы IL-8, но также и широкий спектр ингибиторов сигнала IL-8, таких как ингибиторы CXCR1 и ингибиторы CXCR2, которые, как известно, участвуют в ингибировании сигнала IL-8, подавляют миграцию нейтрофилов. Так, данный результат убедительно свидетельствует о том, что ингибиторы сигнала IL-8 подавляют инфильтрацию нейтрофилов при эндометриозе, а также в повреждения, где концентрация IL-8 является высокой. Таким образом, обобщая и расширяя сферу приложения этих результатов, специалисты в данной области имеют достаточно оснований считать, что ингибиторы сигнала IL-8 обладают способностью предупреждать эндометриоз, а также воспалительные заболевания, в отношении которых известно, что они сопровождаются высокой концентрацией IL-8.
Пример 22. Действие нейтрофилов на эндометриоидные клетки
Далее, проводили изучение эндометриоидных клеток для выяснения того, каким образом действуют нейтрофилы, которые инфильтруют эндометриоидные повреждения. Эндометриоидные клетки получали с помощью следующей процедуры. Человеческие эндометриоидные ткани, вырезанные путем хирургической операции, измельчали в среде DMEM/F-12, HEPES (фирма GIBCO), содержащей 0,5 мг/мл коллагеназы и 0,1 мг/мл ДНКазы 1, и после инкубации при 37°С собирали клетки. Клеточную суспензию пропускали через 100-микрометровый клеточный фильтр и затем к ней добавляли буфер для лизиса (фирма BD Biosciences) для индукции гемолиза. После гемолиза клетки высевали и после пересева клетки замораживали в жидком азоте и помещали на хранение в качестве эндометриоидных клеток. Культивирование осуществляли с использованием среды DMEM/F-12, HEPES, дополненной 10% ЗФР, 2,5 мкг/мл амфотерицина В, 100 ед./мл пенициллина и 100 мкг/мл стрептомицина. Хранившиеся в замороженном состоянии клетки инициировали перед их использованием в экспериментах.
Супернатант культуры нейтрофилов добавляли к эндометриоидным клеткам, полученным с помощью описанного выше метода, и оценивали реактивность эндометриоидных клеток. Для получения супернатанта культуры нейтрофилов выделенные нейтрофилы суспендировали в среде RPMI-1640, содержащей 1% ЗФР, и высевали из расчета 3,0×106 клеток/лунку в 6-луночный планшет. Супернатант культуры собирали после культивирования в течение одного дня. Эндометриоидные клетки высевали из расчета 10000 клеток/лунку в 96-луночный планшет и к ним добавляли собранный супернатант культуры нейтрофилов в объеме, составляющем половину объема среды. Аналогично этому, в случае контрольной группы (в примерах, приведенных в настоящем описании обозначена также как «Ctrl») добавляли среду RPMI-1640, содержащую 1% FBS, в объеме, составляющем половину объема среды. Через 8 ч среду удаляли и очищали РНКазы с использованием набора RNeasy 96 (фирма QIAGEN) согласно протоколу получения продукта. кДНК синтезировали на основе очищенных РНК с использованием мастер-смеси SuperScript™ VILO™ (фирма Invitrogen) и сравнивали уровень экспрессии мРНК ароматазы с помощью ПЦР в реальном времени. В качестве зондов применяли зонд Taqman Hs00903411_m1 (фирма Thermo Fisher) для ароматазы и в качестве эндогенного контроля человеческий GAPD (GAPDH) (фирма Applied Biosystems, каталожный номер: 4326317Е) для GAPDH-анализа.
Относительный уровень экспрессии ароматазы (в группе Ctrl принимали за 1 после стандартизации уровня экспрессии ароматазы относительно уровня GAPDH) представлен на фиг. 40. Продемонстрировано, что уровень экспрессии ароматазы был повышен в эндометриоидных клетках, к которым добавляли супернатант культуры нейтрофилов.
Ароматаза представляет собой эстроген-синтетазу, а эстроген, как известно, стимулирует пролиферацию эндометриоидных эпителиальных клеток. Экспрессия ароматазы повышалась в эндометриоидных клетках при добавлении супернатанта культуры нейтрофилов, и этот результат позволяет предположить, что инфильтрация нейтрофилов в эндометриоидные повреждения повышает уровень экспрессии ароматазы в эндометриоидных клетках, что усиливает пролиферацию эндометриоидных эпителиальных клеток. Таким образом, можно предположить, что в результате подавления инфильтрации нейтрофилов в эндометриоидные повреждения с помощью ингибитора сигнала IL-8 происходит подавление пролиферации эндометриоидных эпителиальных клеток. Описанное выше предположение подтверждается тем установленным фактом, что пролиферирующие эпителиальные клетки были атрофированы у обезьян, на которых моделировали индуцированный хирургическим путем эндометриоз, в группе, которой вводили антитело H1009/L395-F1974m.
Пример 23. Анализ хемокинов и цитокинов, продуцируемых нейтрофилами
Далее анализировали, какие типы хемокинов, цитокинов и факторов роста продуцируются нейтрофилами в присутствии или отсутствии IL-8 или в присутствии антитела к IL-8. Выделенные нейтрофилы суспендировали в среде RPMI-1640, содержащей 1% FBS, и высевали из расчета 1,5×106 клеток/лунку в 6-луночный планшет. В лунки добавляли или не добавляли IL-8, или добавляли IL-8 и различные антитела, и нейтрофилы культивировали в течение одного дня. Добавляли антитела H1009/L395-F1974m, антитело к IL-8 [807] (ab18672) и IgG1, каждое в конечной концентрации 10 мкг/мл, и добавляли IL-8 в конечной концентрации 100 нг/мл. Через один день собирали раствор клеточной культуры и определяли концентрации различных цитокинов, хемокинов и факторов роста в растворе клеточной культуры с использованием магнитной 30-плексной панели для определения человеческих цитокинов (Cytokine Human Magnetic 30-Plex Panel) для платформы Luminex™ (фирма Thermo Fisher) согласно протоколу получения продукта.
Результаты, представленные на фиг. 41, демонстрируют, что концентрация МСР-1 (моноцитарный хемоаттрактантный белок 1) в растворе клеточной культуры сильно повышалась при добавлении IL-8 и что повышение подавлялось нейтрализующими антителами к IL-8.
МСР-1 обозначают также как CCL2 и он обладает хемотаксической активностью в отношении моноцитов. Считается, что МСР-1 стимулирует миграцию и инфильтрацию моноцитов в области локального воспаления. Как представлено в таблице 26, уровень гемосидерина был снижен в узловых повреждениях у обезьян, на которых моделировали индуцированный хирургическим путем эндометриоз, в группе, которой вводили антитело H1009/L395-F1974m, и этот результат позволяет предположить, что введение антитела приводит к подавлению инфильтрации макрофагов или моноцитов. Поскольку МСР-1, который обладает хемотаксической активностью в отношении моноцитов, продуцировался при стимулировании IL-8 нейтрофилов, то описанный выше результат позволяет предположить, что в ответ на IL-8, происходит инфильтрация нейтрофилов в эндометриоидные повреждения и они продуцируют МСР-1, что приводит к миграции и инфильтрации моноцитов и макрофагов в области повреждения.
Кроме того, известно, что МСР-1 воздействует на фибробласты и стимулирует производство коллагена (J Biol Chem.; 271(30), 26 июля 1996 г., сс. 17779-17784), и имеется много публикаций, в которых описано, что МСР-1 усиливает фиброз ткани in vivo (J Immunol., 153(10), 15 ноября 1994 г., сс. 4733-4441; Am J Physiol Lung Cell Mol Physiol., 286(5), май 2004 г., сс. L1038-L1044; J Invest Dermatol.; 121(3), сентябрь 2003 г., сс. 510-516). Это позволяет предположить возможность того, что в ответ на IL-8, происходит инфильтрация нейтрофилов в области эндометриоидных повреждений и дополнительное производство ими МСР-1, что приводит к усилению фиброза в областях эндометриоидных повреждений в дополнение к усилению миграции и инфильтрации моноцитов и макрофагов. Описанный выше результат позволяет предположить, что фиброз в областях эндометриоидных повреждений можно ослаблять путем подавления инфильтрации нейтрофилов с использованием ингибиторов сигнала IL-8. Действительно, как продемонстрировано на фиг. 36 и в таблице 26, было выявлено уменьшение интерстиция, основным компонентом которого являются коллагеновые волокна, в областях узловых повреждений у обезьян, на которых моделировали индуцированный хирургическим путем эндометриоз, в группе, которой вводили антитело H1009/L395-F1974m, что демонстрирует эффективность лекарственного агента в отношении ослабления фиброза. Описанный выше результат убедительно подтверждает, что ингибиторы сигнала IL-8 можно применять против фиброза при эндометриозе. Таким образом, обобщая и расширяя сферу приложения этих результатов, специалисты в данной области имеют достаточно оснований считать, что ингибиторы сигнала IL-8 можно применять для лечения или предупреждения фиброза при эндометриозе, а также при различных фиброзных заболеваниях.
Пример 24. Создание обезьяньей модели индуцированных послеоперационных спаек и оценка патологических состояний
(24-1) Создание обезьяньей модели индуцированных послеоперационных спаек
Для создания обезьяньей модели индуцированных послеоперационных спаек самок обезьян циномолгус возрастом 9-16 лет (полученных от исследовательского центра приматов Цукуба при Национальных институтах биомедицинской инновации, здоровья и питания обрабатывали под анестезией следующим образом. Анестезию осуществляли с использованием сбалансированной смеси для анестезии кетамин/ксилазин (смешанные в адекватном соотношении примерно 2:1) и изофлурана. Делали разрез длиной 5-6 см по медианной линии брюшной стенки. После открытия брюшины делали разрез длиной 1 см на теле матки и зашивали с помощью 4 стежков с использованием одной шовной нити. Затем всю матку вычищали пять раз с помощью марли. Затем делали разрез длиной 1 см на перитонеальном участке правой брюшной стенки на расстоянии 2 см от медианного разреза брюшной стенки и зашивали, накладывая шов с 4 стежками с использованием рассасывающегося шовного материала. После наложения шва зашитую часть вычищали пять раз с помощью марли. Аналогичным образом разрезали, зашивали и вычищали с помощью марли брюшину на левой брюшной стенке. И наконец, разрез по медианной линии брюшной стенки зашивали, накладывая шов с 10-11 стежками. Все стадии осуществляли в течение одного часа или менее. В момент пробуждения после хирургического вмешательства вводили антагонист антиседан (примерно в таком же количестве, что и ксилазин) и антибиотик цефрадин. Затем в течение двух дней после хирургического вмешательства вводили анальгетик залбан, и обезьян регулярно обследовали каждый день для подтверждения отсутствия послеоперационных патологий. При проведении лапароскопического обследования анестезию и послеоперационный мониторинг осуществляли таким же образом.
(24-2) Оценка эффективности антитела к IL-8 в качестве лекарственного средства на моделях индуцированных послеоперационных спаек
Животным в группе, которой вводили антитело к IL-8, вводили внутривенно антитело к IL-8 H1009/L395-F1974m в дозе 10 мг/кг через 1 ч или менее после хирургического вмешательства для индукции послеоперационных спаек, как описано выше. Контрольную группу не обрабатывали. Через 24-35 дней после введения антитела к IL-8 осуществляли лапароскопию для обследования спаек, образовавшихся в результате разреза, наложения швов и вычищения марлей. Под анестезией осуществляли медианный разрез брюшной полости и проводили лапароскопическое обследование путем введения троакара и последующего введения лапароскопа. Лапароскоп был присоединен к видеосистеме (фирма KARL STORZ) и монитору. Видеосистему применяли для регистрации, а монитор использовали для внутрибрюшинного обследования. Размер спайки измеряли с помощью градуированной линейки или градуированного пинцета. В процессе лапароскопического обследования оценивали наличие, размер и положение спайки. Результат представлен в таблице 29. Результат лапароскопического обследования продемонстрировал, что у двух обезьян из необработанной группы была обнаружена спайка на всей области разреза. Хотя у особи №206 в группе, которой вводили антитело к IL-8, была обнаружена спайка, у особи №201 не было выявлено спайки в брюшине правой и левой брюшной стенки. Таким образом, продемонстрировано предупреждающее образование спаек действие антител к IL-8. Этот результат свидетельствует о том, что антитела к IL-8 можно применять в качестве предупреждающих образование спаек агентов. Специалистам в данной области естественно должно быть очевидно, что антитела к IL-8 могут оказывать более сильное препятствующее образованию спаек действие при соответствующем изменении графика введения или т.п. Кроме того, на основе описанных выше результатов, специалистам в данной области естественно должно быть очевидно, что в качестве предупреждающих образование спаек агентов можно применять ингибиторы сигнала IL-8.
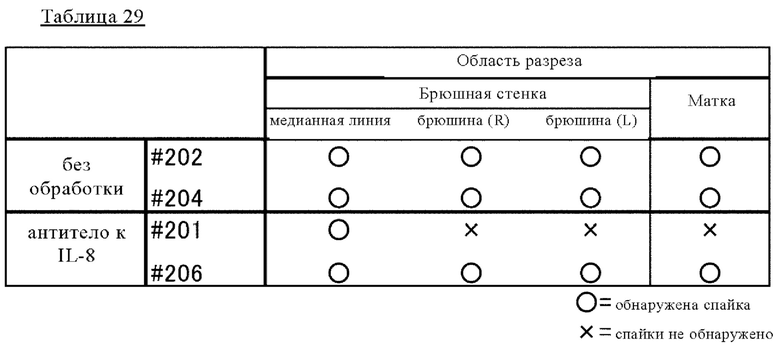
Пример 25. Функциональный анализ IL-8 в макрофагах, образовавшихся в результате дифференцировки из моноцитов, и оценка антител к IL-8 в отношении их эффективности в качестве лекарственных средств
Получали моноциты (фирма All Cells, каталожный номер: РВ011-P-F-2), выделенные в виде CD14-позитивной фракции из периферической крови здоровых индивидуумов, и помещали на хранение при -80°С. При применении в экспериментах клетки подвергали оттаиванию и использовали, как описано ниже. Клетки подвергали оттаиванию при 37°С в водяной бане и к ним добавляли среду, содержащую ДНКазу I (фирма STEMCELL Technologies, каталожный номер: 07900) и 10% FBS. После центрифугирования в высокоскоростной центрифуге удаляли супернатант. После повторения этой стадии клетки суспендировали в соответствующем объеме среды для определения количества клеток. Моноциты культивировали для дифференцировки in vitro в макрофаги и анализировали в отношении изменений экспрессии фактора роста соединительной ткани (CTGF) в присутствии и в отсутствии IL-8. CTGF представляет собой фактор, который стимулирует фиброз. Моноциты суспендировали в среде RPMI-1640 (фирма SIGMA), содержащей 10% FBS и 25 нг/мл GM-CSF (фирма SIGMA), и высевали на культуральный планшет. Клетки культивировали в течение семи дней, заменяя половину супернатанта культуры каждые два или три дня. Через семь дней супернатант культуры удаляли и добавляли 100 нг/мл IL-8 и различные антитела к среде RPMI-1640, содержащей 10% FBS, 20 нг/мл IFN-γ (фирма SIGMA) и 10 пг/мл LPS (фирма Wako Pure Chemical Industries), и клетки культивировали в течение трех дней.
Оценивали следующие антитела: антитело H1009/L395-F1974m, антитело к IL-8 [807] (фирма Abcam, каталожный номер: ab18672), антитело к IL-8 [6217] (Abcam, каталожный номер: ab10768), антитело к IL-8 (фирма Becton Dickinson, каталожный номер: BD554726) и антитело к IL-8 (фирма Becton Dickinson, каталожный номер: BD555717); указанные антитела применяли в концентрации 10 мкг/мл. В качестве отрицательного контроля применяли IgG1 (фирма SIGMA, каталожный номер: 15154) в концентрации 10 мкг/мл.
После культивирования в течение трех дней клетки собирали и очищали РНК с использованием мининабора RNeasy (фирма QIAGEN) согласно протоколу получения продукта. кДНК синтезировали с очищенных РНК с использованием мастер-смеси SuperScript VILO (фирма Thermo Fisher) и определяли уровень экспрессии мРНК CTGF с помощью ПЦР в реальном времени. В качестве зондов применяли Taqman-зонд Hs00170014_m1 (фирма Thermo Fisher) для CTGF и в качестве эндогенного контроля эукариотическую 18S rРНК (фирма Thermo Fisher, каталожный номер: Е4319413Е) для анализа 18S rРНК.
Уровень экспрессии CTGF стандартизовали относительно 18S rРНК. Стандартизованная величина представлена на фиг. 42. В том случае, когда IL-8 не добавляли, экспрессия CTGF не была обнаружена в макрофагах, образовавшихся в результате дифференцировки из моноцитов. В то же время в том случае, когда добавляли IL-8, была выявлена повышенная экспрессия CTGF в образовавшихся в результате дифференцировки макрофагах. Этот результат свидетельствует о том, что IL-8 обладает функцией, заключающейся в индукции повышения экспрессии CTGF в макрофагах, образовавшихся в результате дифференцировки из моноцитов. Кроме того, когда антитело к IL-8 добавляли одновременно с IL-8, то повышения экспрессии CTGF не было обнаружено. Продемонстрировано, что повышение экспрессии CTGF, обусловленное IL-8, подавлялось различными антителами к IL-8. На основе описанного выше результата специалистам в данной области, естественно, должно быть очевидно, что IL-8 представляет собой фактор, участвующий в прогрессировании фиброза, и что ингибиторы сигнала IL-8, такие как антитела к IL-8 антитела, можно применять в качестве терапевтических или профилактических агентов против фиброзных заболеваний.
Референс-пример 1
Конструирование экспрессионных векторов для антител IgG-типа с заменой аминокислот
Представляющие интерес экспрессионные векторы для Н-цепи и L-цепи конструировали путем встраивания в экспрессионный вектор для клеток животных фрагмента плазмиды, содержащего вариант, полученный с использованием системы для сайт-направленного мутагеназа QuickChange (фирма Stratagene) с помощью метода, описанного в прилагаемой инструкции по эксплуатации. Нуклеотидные последовательности полученных экспрессионных векторов определяли методами, известными специалистам в данной области.
Референс-пример 2
Экспрессия и очистка антител IgG-типа
Экспрессию антител осуществляли с помощью следующего метода Полученную из клеток рака почки человеческого эмбриона клеточную линию HEK293H (фирма Invitrogen) суспендировали в среде DMEM (фирма Invitrogen), дополненной 10% фетальной бычьей сыворотки (фирма Invitrogen), и высевали в объеме 10 мл на чашку в чашки для прикрепленных клеток (диаметром 10 см; фирма CORNING) с плотностью клеток от 5×105 до 6×105 клеток/мл. После культивирования в СО2-инкубаторе (37°С, 5% СО2) в течение одного дня и одной ночи среду удаляли аспирацией и добавляли 6,9 мл среды CHO-S-SFM-II (фирма Invitrogen). Полученные плазмиды интродуцировали в клетки методом липофекции. Образовавшийся супернатант культуры собирали, центрифугировали (примерно при 2000 g в течение 5 мин при комнатной температуре) для удаления клеток и стерилизовали фильтрацией через 0,22-мкм фильтр MILLEX®-GV (фирма Millipore) с получением супернатанта культуры. Полученный супернатант культуры очищали с помощью известного в данной области метода с использованием rProtein A Sepharose™ Fast Flow (фирма Amersham Biosciences). Концентрацию очищенного антитела определяли на основе абсорбции, измеренной при 280 нм с помощью спектрофотометра. Концентрации антител рассчитывали на основе установленных величин, используя коэффициент экстинкции, рассчитанный методом, который описан в Protein Science, 4, 1995, сс. 2411-2423.
Промышленная применимость
В настоящем изобретении продемонстрировано, что антитела к IL-8 и т.п. можно применять в качестве терапевтических и/или профилактических агентов против эндометриоза и т.п. Настоящее изобретение можно применять для уменьшения объема эндометриоидных повреждений, уменьшения спаек, индуцирования атрофических изменений в эпителии и стромальных клетках, снижения инфильтрации иммунных клеток, ослабления фиброза и т.п.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЧУГАИ СЕЙЯКУ КАБУСИКИ КАЙСЯ
<120> Композиция для профилактики или лечения связанных
с IL-8 заболеваний
<130> C1-A1608P
<150> JP 2016-154174
<151> 2016-08-05
<160> 110
<170> PatentIn version 3.5
<210> 1
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 1
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 2
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 2
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 3
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 3
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr
245 250 255
Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 4
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 4
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 5
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 5
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His His His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 6
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 6
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 7
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 7
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Ala His Val Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 8
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 8
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 9
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 9
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Val Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 10
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 10
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 11
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 11
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 12
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 12
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 13
<211> 218
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 13
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Asn Ile Tyr Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Ser Tyr Tyr Phe Ile Ser Ser
85 90 95
Thr Asp Phe Asn Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 14
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 14
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 15
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 15
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 16
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 16
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 17
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 17
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Ala His Val Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 18
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 18
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 19
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 19
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Val Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 20
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 20
Gln Val Thr Leu Lys Glu Ser Gly Gly Arg Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Phe
20 25 30
Asn Met Asp Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Val Ser Thr Gly Gly Ser Ala Tyr Tyr Ala Lys Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Ile Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Val Asp Ser Ser Gly Trp Gly Tyr Phe Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 21
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 21
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Tyr His Val Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 22
<211> 77
<212> PRT
<213> Homo sapiens
<400> 22
Ala Val Leu Pro Arg Ser Ala Lys Glu Leu Arg Cys Gln Cys Ile Lys
1 5 10 15
Thr Tyr Ser Lys Pro Phe His Pro Lys Phe Ile Lys Glu Leu Arg Val
20 25 30
Ile Glu Ser Gly Pro His Cys Ala Asn Thr Glu Ile Ile Val Lys Leu
35 40 45
Ser Asp Gly Arg Glu Leu Cys Leu Asp Pro Lys Glu Asn Trp Val Gln
50 55 60
Arg Val Val Glu Lys Phe Leu Lys Arg Ala Glu Asn Ser
65 70 75
<210> 23
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 23
Asp Tyr Tyr Leu Ser
1 5
<210> 24
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 24
Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala Ser
1 5 10 15
Val Lys Gly
<210> 25
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 25
Glu Asn Tyr Arg Tyr Asp Val Glu Leu Ala Tyr
1 5 10
<210> 26
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 26
Arg Ala Ser Glu Ile Ile Tyr Ser Tyr Leu Ala
1 5 10
<210> 27
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 27
Asn Ala Lys Thr Leu Ala Asp
1 5
<210> 28
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 28
Gln His His Phe Gly Phe Pro Arg Thr
1 5
<210> 29
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 29
Leu Ile Arg Asn Lys Asp Asn Tyr His Thr Pro Glu Tyr Ser Ala Ser
1 5 10 15
Val Lys Gly
<210> 30
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 30
Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr
1 5 10
<210> 31
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 31
Lys Ala Lys Thr His Ala Asp
1 5
<210> 32
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 32
Lys His His Phe Gly Phe Pro Arg Thr
1 5
<210> 33
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 33
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn Tyr Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 34
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 34
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Asp Asn Tyr His Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Thr Met Ser Asp Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 35
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 35
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ile Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Lys Thr His Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Lys His His Phe Gly Phe Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 36
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 36
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Asp Asn Tyr His Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Thr Met Ser Asp Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 37
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 37
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Asp Asn Tyr His Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Thr Met Ser Asp Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Lys Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 38
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ile Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Lys Thr His Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Lys His His Phe Gly Phe Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 39
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 39
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Glu Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Leu Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn Tyr Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 40
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ile Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Ala Lys Thr Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln His His Phe Gly Phe Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 41
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 41
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn Tyr Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 42
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 42
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 43
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 43
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ile Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Lys Thr His Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln His His Phe Gly Phe Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 44
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 44
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ile Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Lys Thr His Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Lys His His Phe Gly Phe Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 45
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 45
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Leu Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Thr Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn Tyr Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 46
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 46
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly His Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 47
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 47
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Asp Asn Gly Tyr Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Thr Met Ser Asp Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 48
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 48
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Asp Asn Tyr His Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Thr Met Ser Asp Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 49
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 49
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Asp Asn Tyr His Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Thr Met Ser Asp Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Arg
225 230 235 240
Arg Gly Pro Lys Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ala Asp Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 50
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 50
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 51
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 51
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 52
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 52
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 53
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 53
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Ala His Val Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 54
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 54
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Thr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 55
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 55
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Val Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 56
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 56
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu
435 440 445
Ser Pro
450
<210> 57
<211> 450
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 57
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Pro Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 58
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 58
Ser Tyr Gly Met Leu
1 5
<210> 59
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 59
Asp Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 60
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 60
Asp Arg Ile Ala Val Ala Asp Tyr
1 5
<210> 61
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 61
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 62
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 62
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 63
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 63
Gln Gln Tyr Gly Ser Ser Phe Thr
1 5
<210> 64
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 64
Asn Tyr Gly Met His
1 5
<210> 65
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 65
Val Ile Tyr Phe Glu Gly Ser Asn Lys Tyr Asn Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 66
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 66
Ser Pro Tyr Gly Asp Tyr Leu Asp Tyr
1 5
<210> 67
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 67
Arg Ala Ser Gln Thr Ile Asp Tyr Asn Tyr Leu His
1 5 10
<210> 68
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 68
Gly Thr Phe Ile Arg Ala Thr
1 5
<210> 69
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 69
Gln Gln Phe Gly Arg Ser Pro Leu Thr
1 5
<210> 70
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 70
Ser Tyr Gly Met Leu
1 5
<210> 71
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 71
Asp Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 72
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 72
Asp Arg Ile Ala Val Ala Asp Tyr
1 5
<210> 73
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 73
Arg Ala Ser Gln Ser Val Ser Ser Ser Phe Leu Ala
1 5 10
<210> 74
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 74
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 75
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 75
Gln Gln Tyr Asp Ser Ser Phe Thr
1 5
<210> 76
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 76
Gly Tyr Tyr Trp Thr
1 5
<210> 77
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 77
Glu Val Ile His His Gly Ser Thr Asn Tyr Ser Pro Ser Leu Lys Ser
1 5 10 15
<210> 78
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 78
Gly Gly Ala Ala Ala Ala Leu Asp Ser
1 5
<210> 79
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 79
Lys Ser Ser Gln Ser Val Leu Phe Ser Ser Asn Asn Arg Lys Tyr Leu
1 5 10 15
Ala
<210> 80
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 80
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 81
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 81
Gln Gln Tyr Tyr Ser Thr Pro Phe Thr
1 5
<210> 82
<211> 5
<212> PRT
<213> Mus musculus
<400> 82
Ser Tyr Trp Met His
1 5
<210> 83
<211> 17
<212> PRT
<213> Mus musculus
<400> 83
Glu Ile Asp Pro Ser Asp Ser Asn Thr Asn Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 84
<211> 7
<212> PRT
<213> Mus musculus
<400> 84
Glu Leu Leu His Ala Val Tyr
1 5
<210> 85
<211> 11
<212> PRT
<213> Mus musculus
<400> 85
Thr Ala Ser Gln Asp Ile His Lys Tyr Ile Ser
1 5 10
<210> 86
<211> 6
<212> PRT
<213> Mus musculus
<400> 86
Thr Ser Thr Leu Gln Pro
1 5
<210> 87
<211> 8
<212> PRT
<213> Mus musculus
<400> 87
Leu Gln Tyr Asp Asn Leu Trp Thr
1 5
<210> 88
<211> 5
<212> PRT
<213> Mus musculus
<400> 88
Asn Tyr Trp Ile Val
1 5
<210> 89
<211> 17
<212> PRT
<213> Mus musculus
<400> 89
Asp Leu Tyr Ser Gly Gly Gly Tyr Thr Phe Tyr Ser Glu Asn Phe Lys
1 5 10 15
Gly
<210> 90
<211> 10
<212> PRT
<213> Mus musculus
<400> 90
Ser Gly Tyr Asp Arg Thr Trp Phe Ala His
1 5 10
<210> 91
<211> 11
<212> PRT
<213> Mus musculus
<400> 91
Gln Ala Ser Gln Asp Ile Glu Ser Tyr Leu Ser
1 5 10
<210> 92
<211> 7
<212> PRT
<213> Mus musculus
<400> 92
Tyr Ala Thr Arg Leu Ala Asp
1 5
<210> 93
<211> 9
<212> PRT
<213> Mus musculus
<400> 93
Leu Gln His Gly Glu Ser Pro Pro Thr
1 5
<210> 94
<211> 5
<212> PRT
<213> Homo sapiens
<400> 94
His Tyr Gly Met Tyr
1 5
<210> 95
<211> 17
<212> PRT
<213> Homo sapiens
<400> 95
Val Ile Trp Tyr Asp Gly Ser Tyr Glu Tyr Asn Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 96
<211> 8
<212> PRT
<213> Homo sapiens
<400> 96
Asp Arg Val Gly Leu Phe Asp Tyr
1 5
<210> 97
<211> 12
<212> PRT
<213> Homo sapiens
<400> 97
Arg Ala Ser Gln Ser Ile Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 98
<211> 7
<212> PRT
<213> Homo sapiens
<400> 98
Gly Pro Ser Ser Arg Ala Thr
1 5
<210> 99
<211> 8
<212> PRT
<213> Homo sapiens
<400> 99
Gln Gln Tyr Ala Gly Ser Leu Thr
1 5
<210> 100
<211> 330
<212> PRT
<213> Homo sapiens
<400> 100
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 101
<211> 326
<212> PRT
<213> Homo sapiens
<400> 101
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 102
<211> 377
<212> PRT
<213> Homo sapiens
<400> 102
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 103
<211> 327
<212> PRT
<213> Homo sapiens
<400> 103
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 104
<211> 107
<212> PRT
<213> Homo sapiens
<400> 104
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 105
<211> 105
<212> PRT
<213> Homo sapiens
<400> 105
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 106
<211> 450
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 106
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Leu His Glu Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro
450
<210> 107
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 107
His Ala Lys Thr His Ala Asp
1 5
<210> 108
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 108
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Arg Asn Lys Ala Asn Gly Tyr Thr Arg Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Asn His Arg Tyr Asp Val Glu Leu Ala Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 109
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 109
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ile Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr His Ala Lys Thr His Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Lys His His Phe Gly Phe Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 110
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 110
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |
| Новые варианты интерлейкина-2 для лечения рака | 2020 |
|
RU2833537C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2728861C1 |
| АНТИ-IL-5 АНТИТЕЛА | 2017 |
|
RU2758008C2 |
| КОМПОЗИЦИЯ ДЛЯ КУЛЬТИВИРОВАНИЯ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ И СПОСОБ ПОЛУЧЕНИЯ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2020 |
|
RU2824216C1 |
| CD131-СВЯЗЫВАЮЩИЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773927C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2698655C2 |
| ТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО, СОДЕРЖАЩЕЕ АНТАГОНИСТ IL-33, ДЛЯ ЛЕЧЕНИЯ ЭНДОМЕТРИОЗА | 2018 |
|
RU2791705C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2746550C2 |
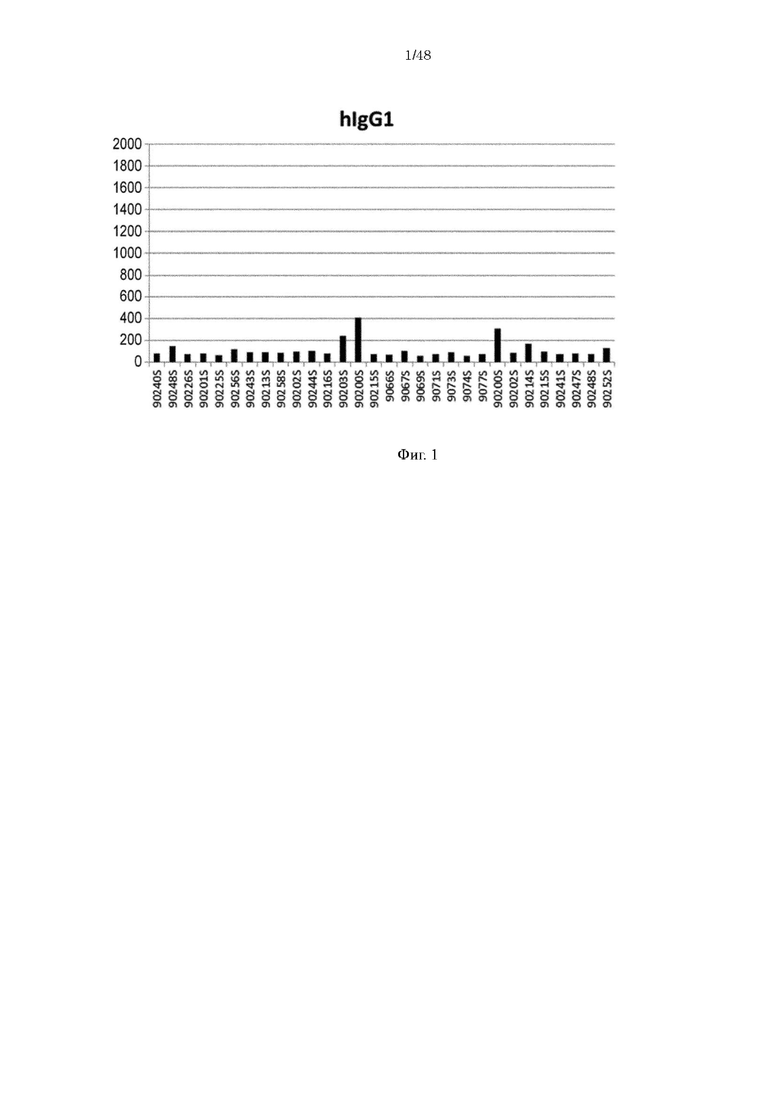
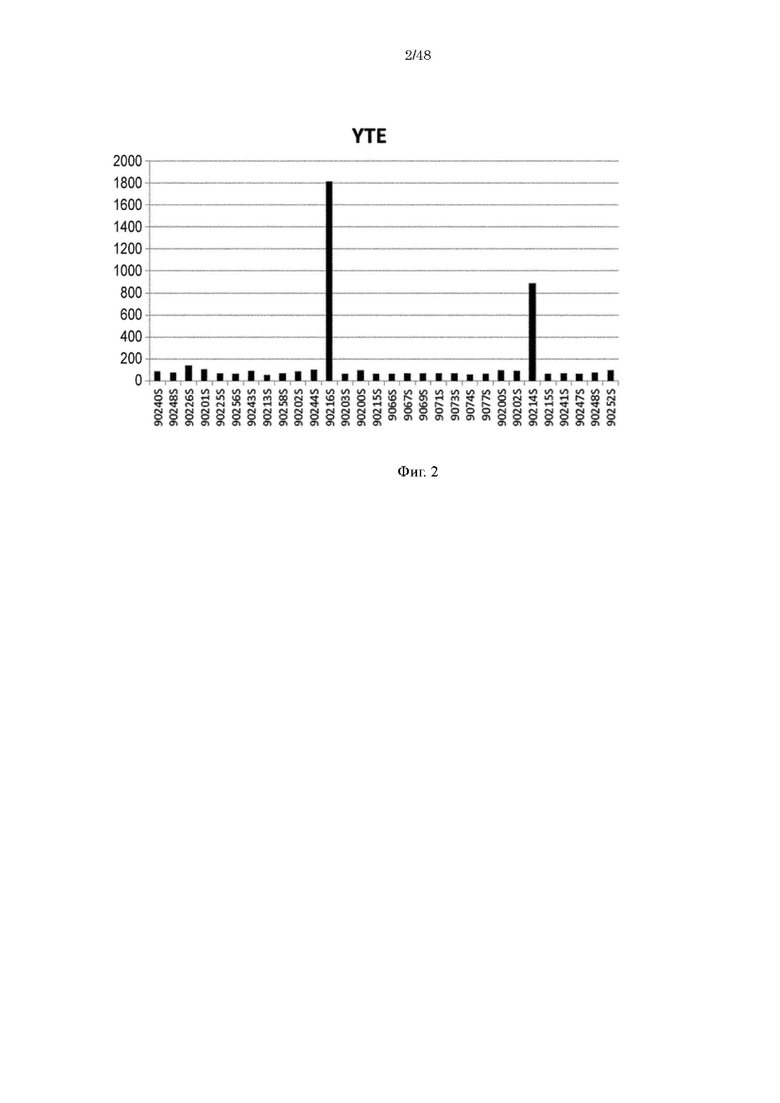
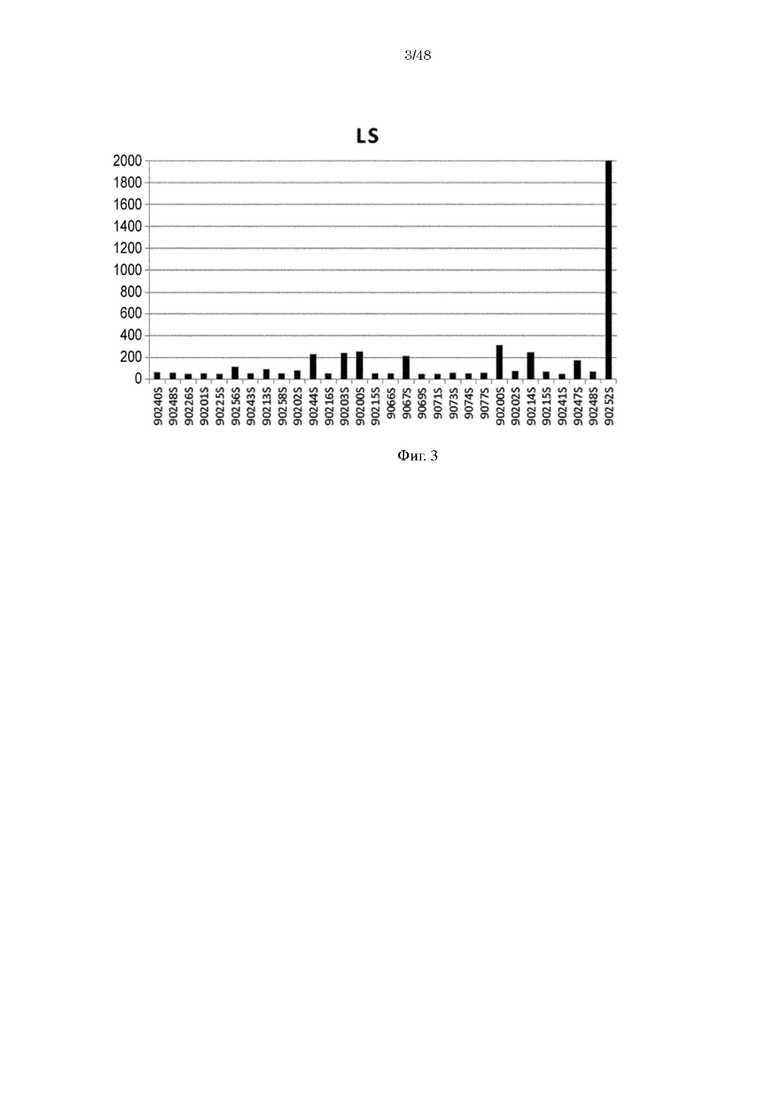
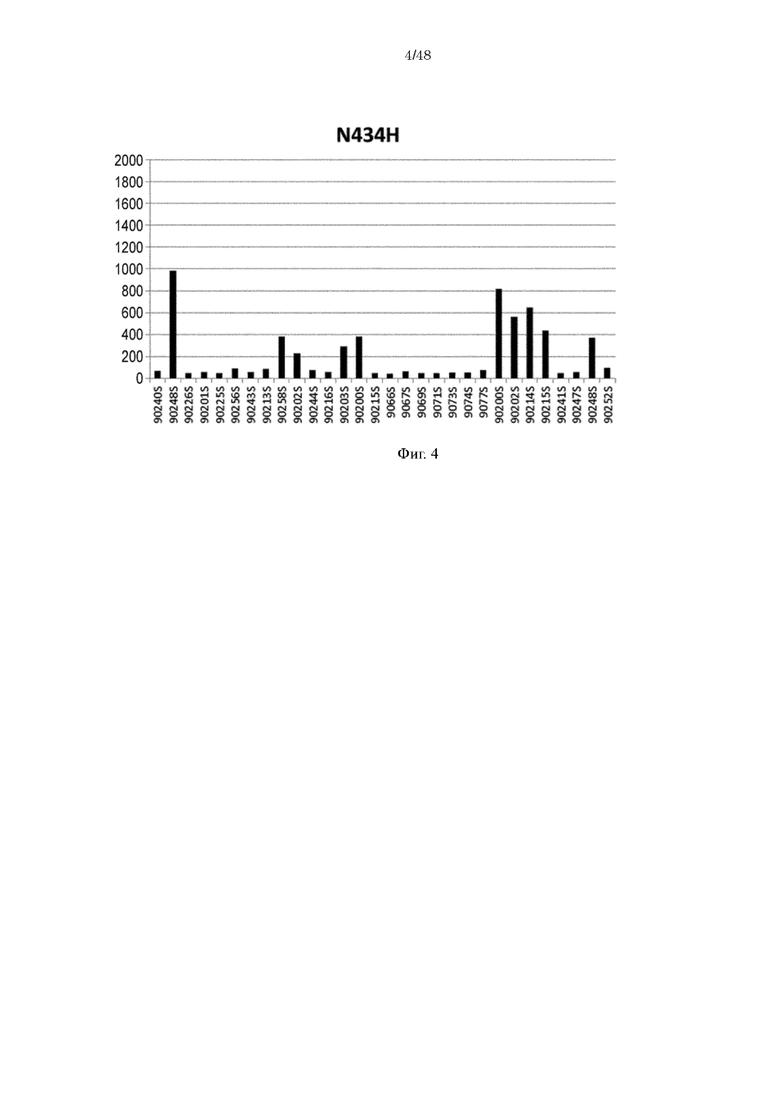


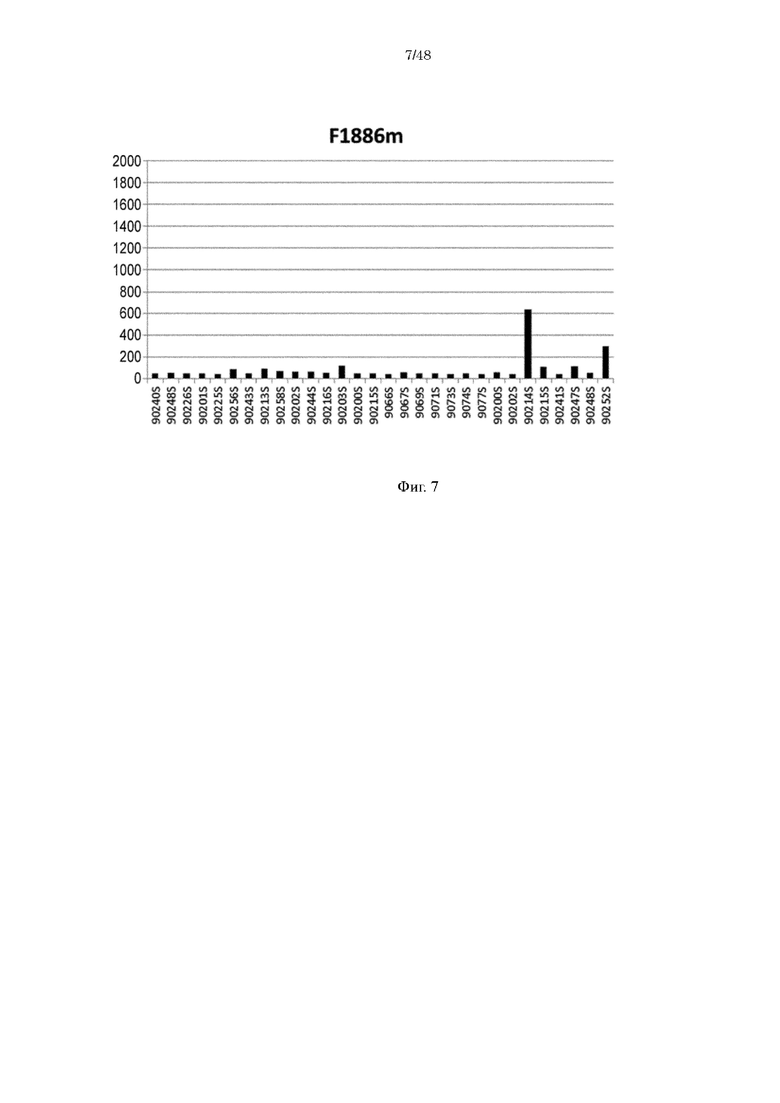



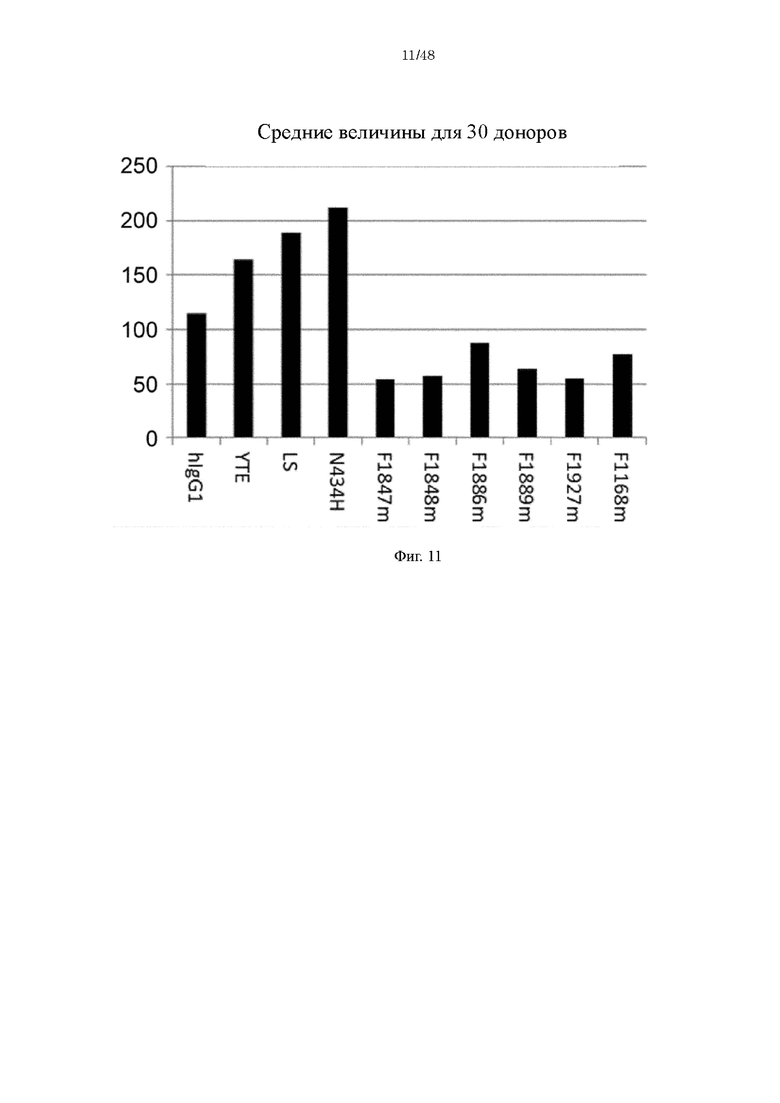
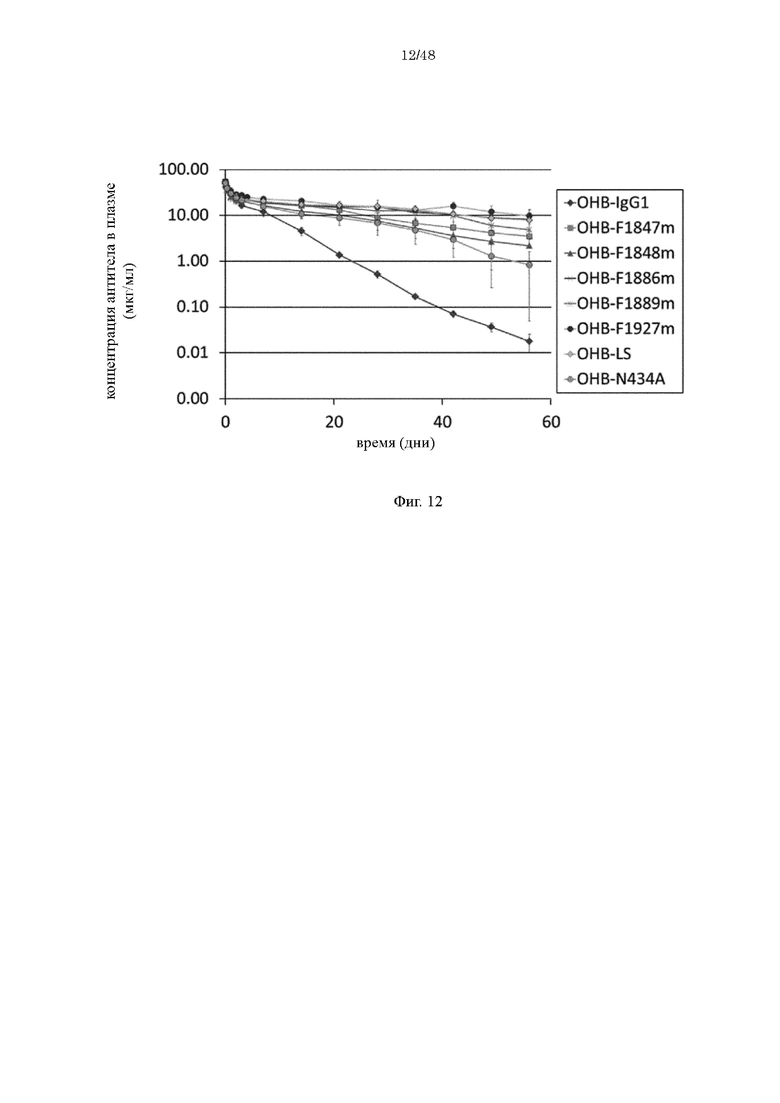

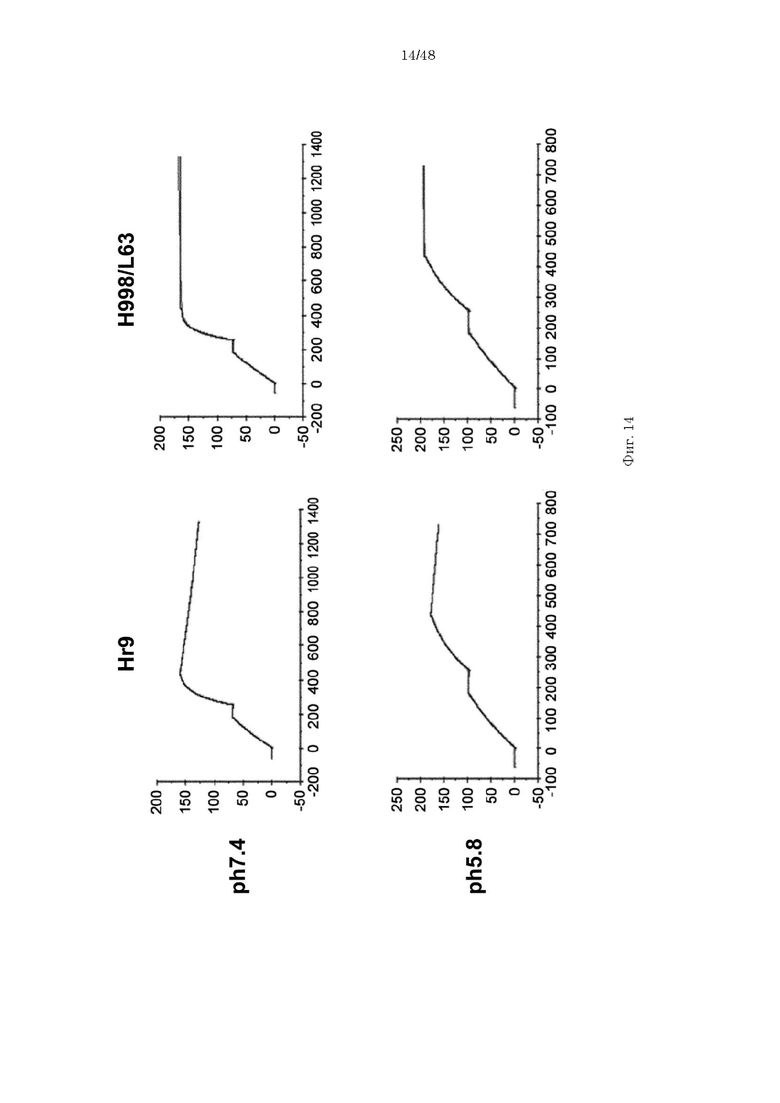
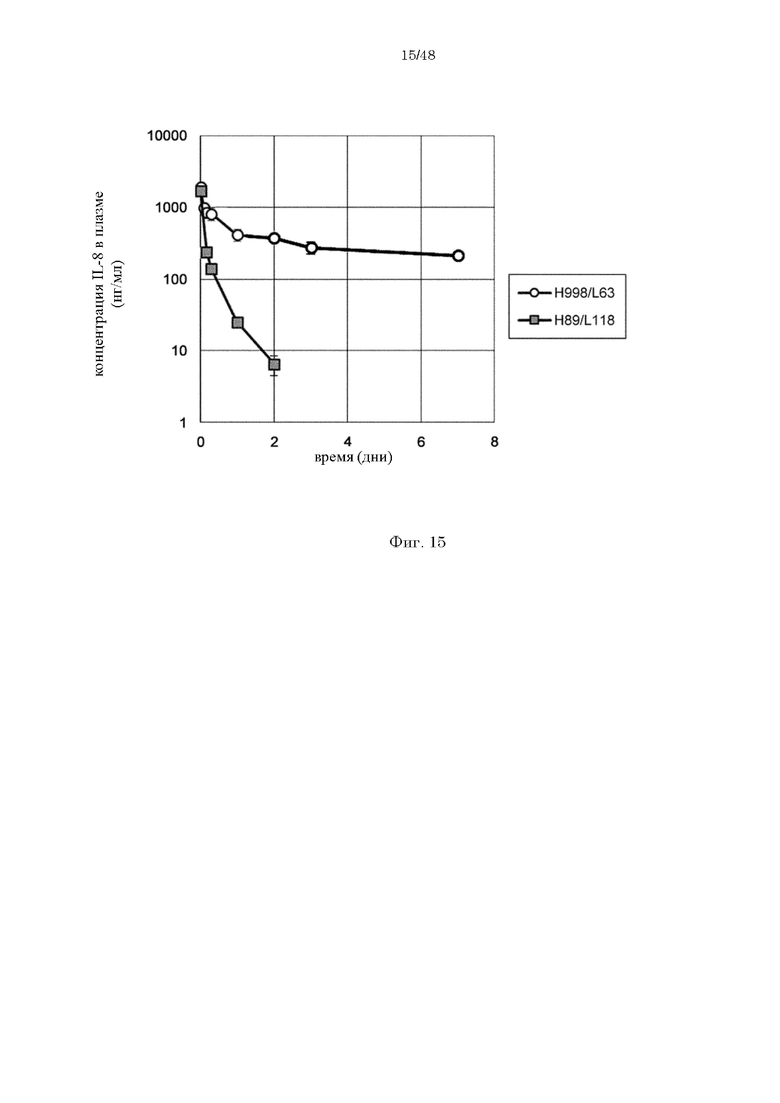
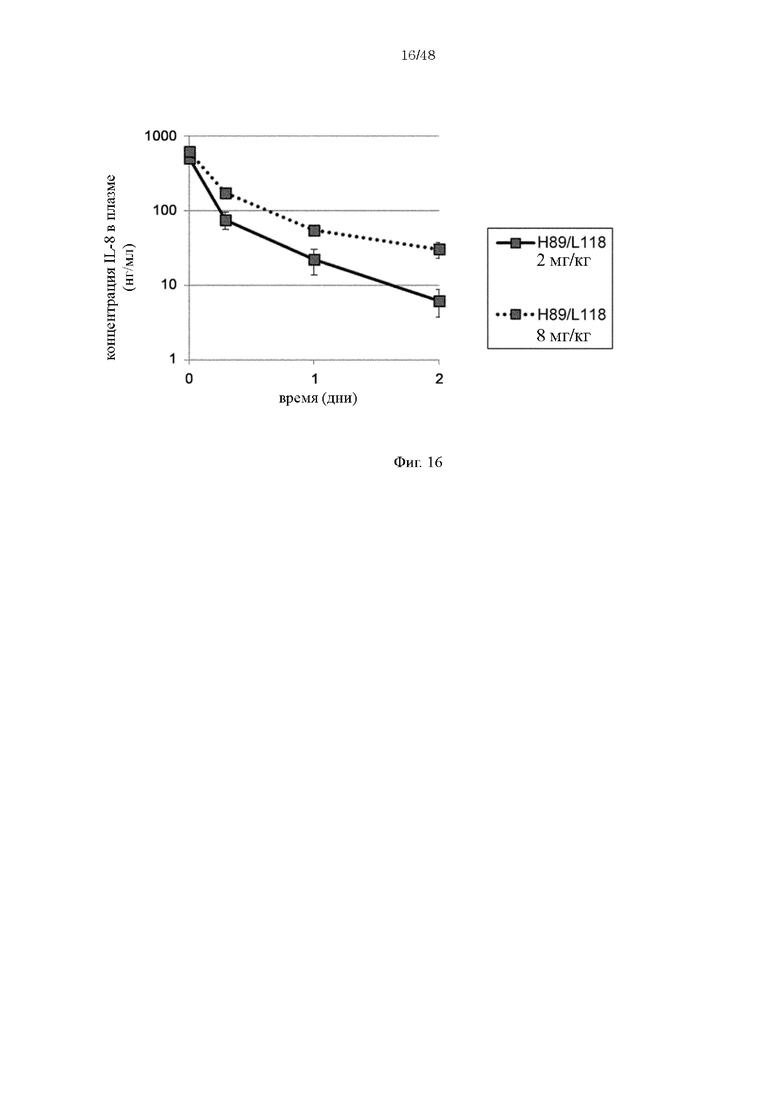
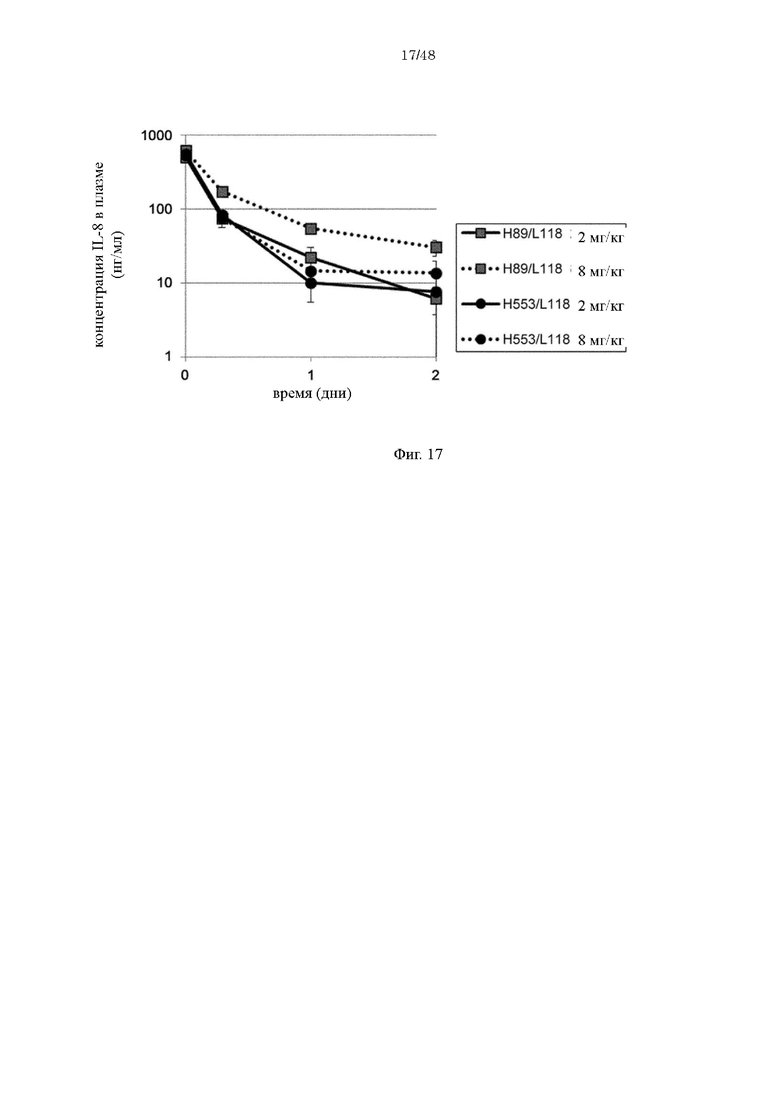
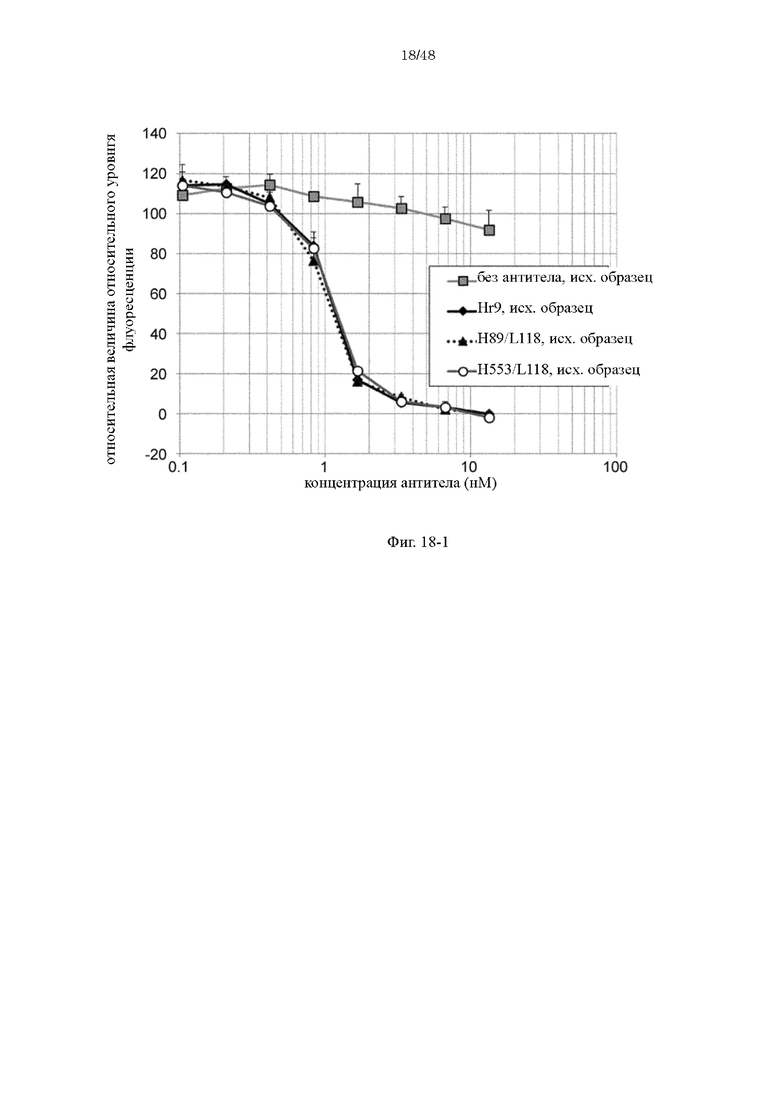
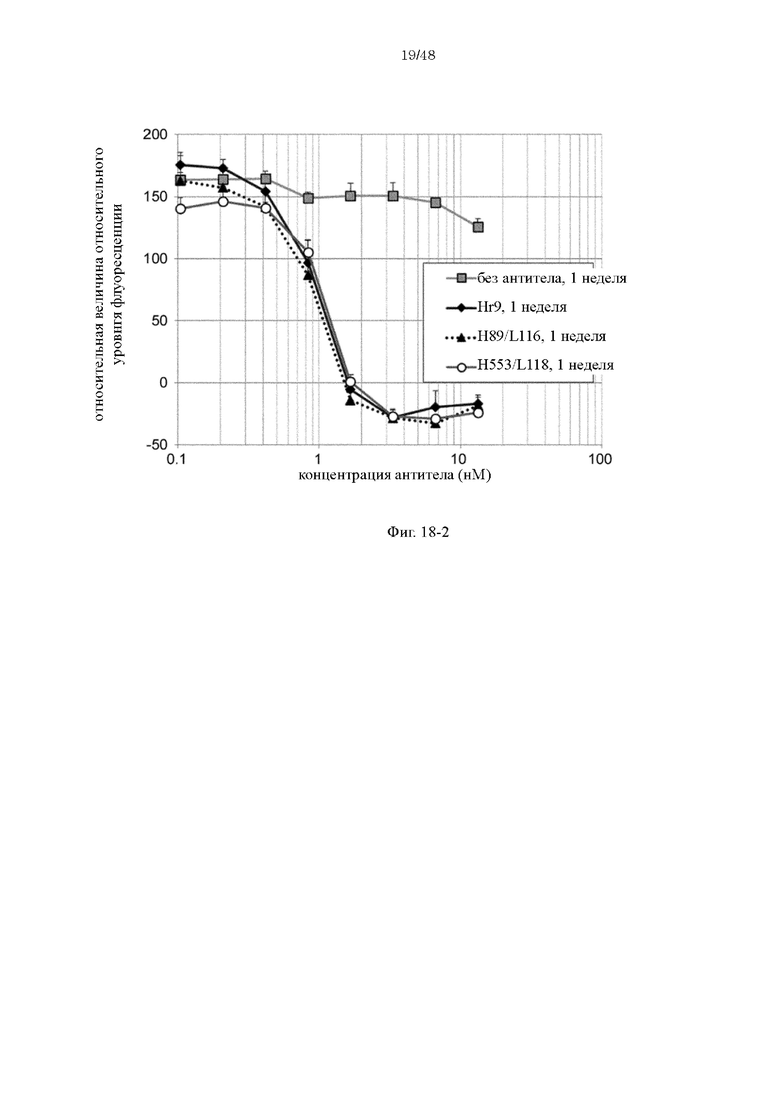
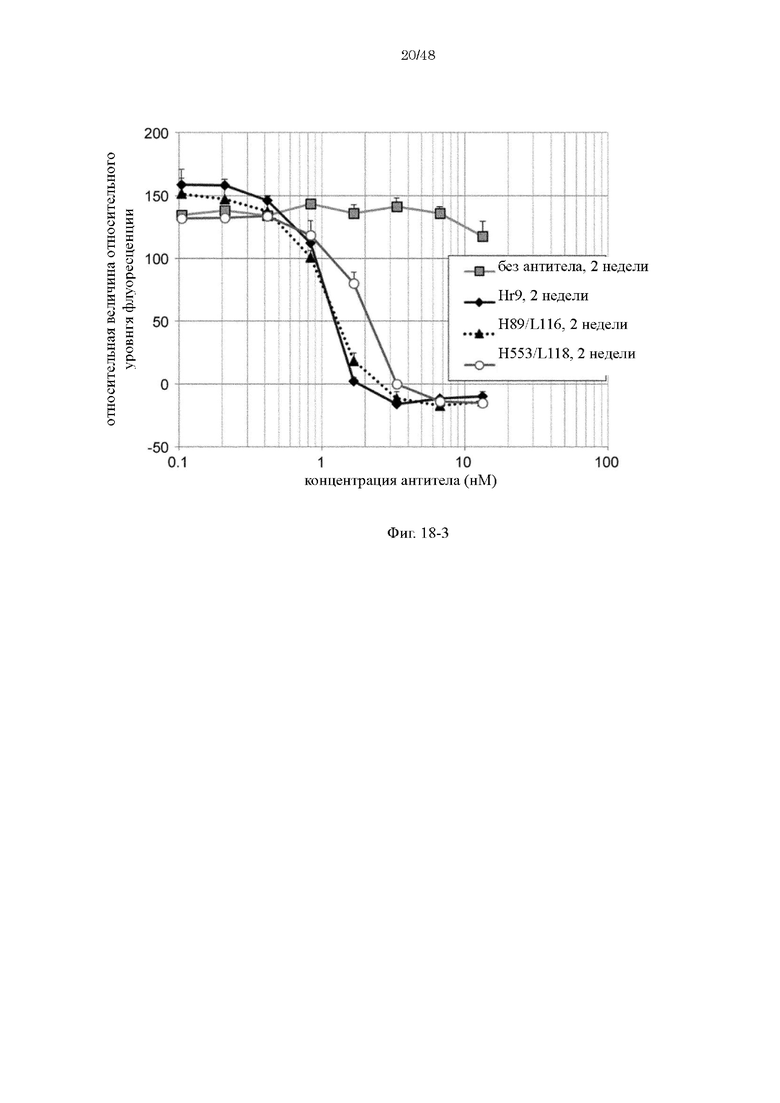
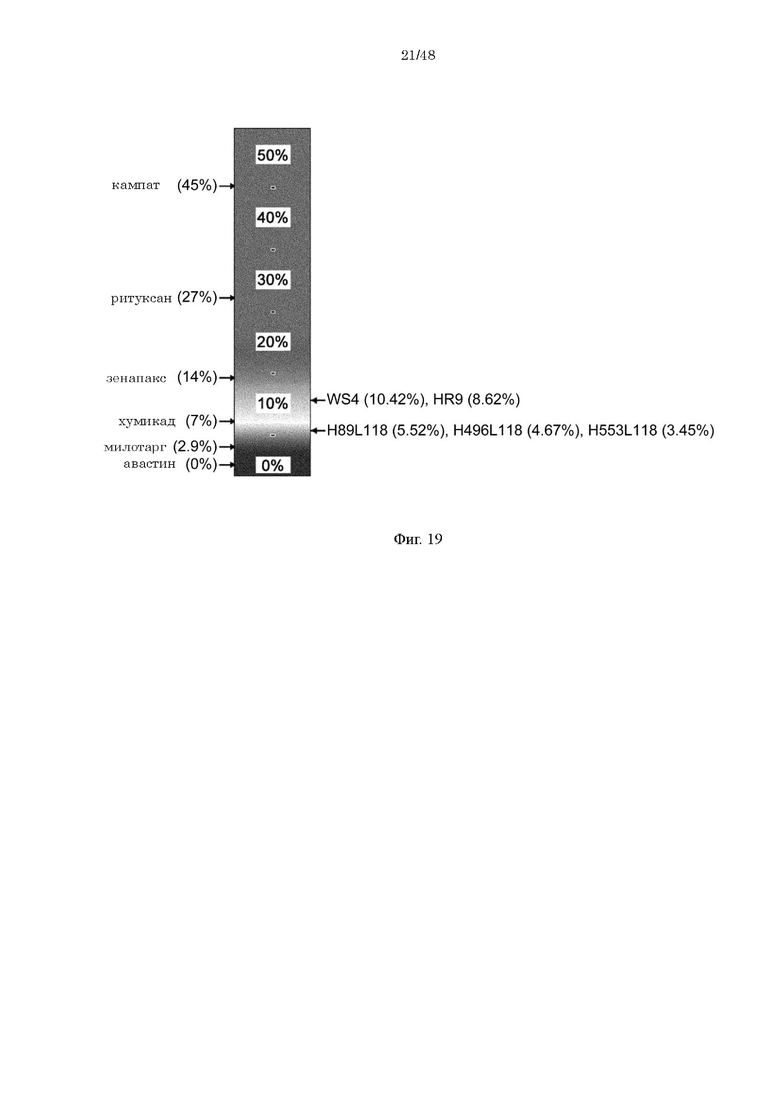
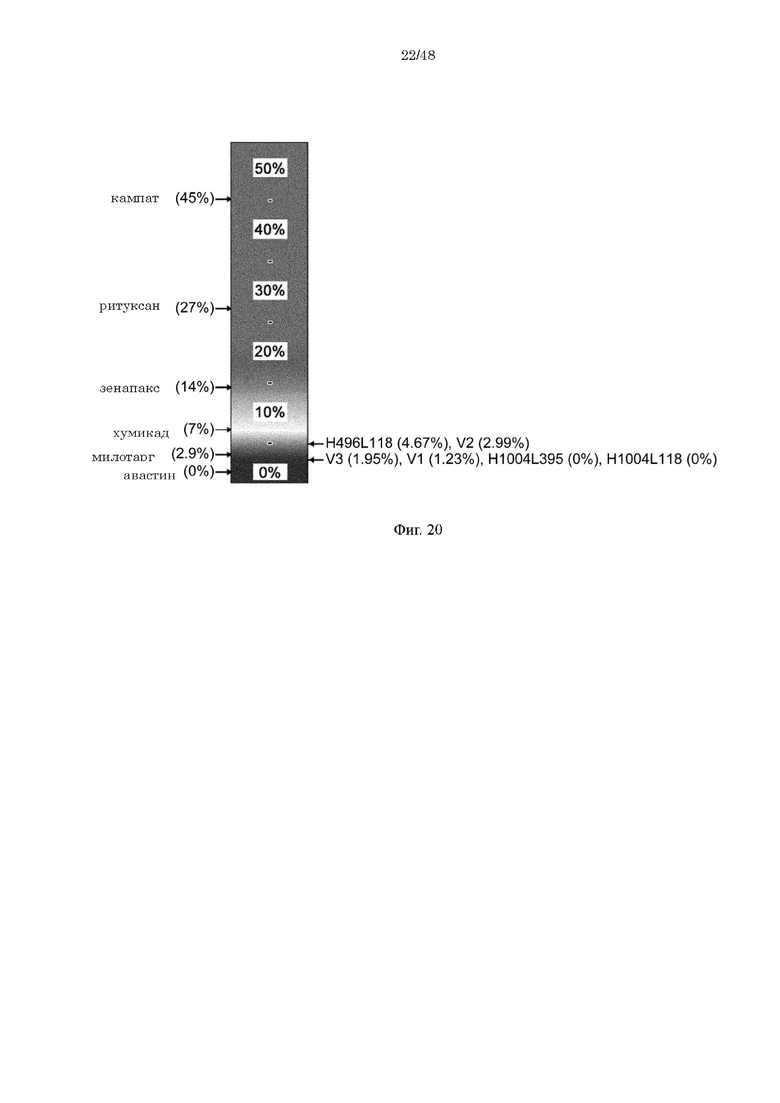
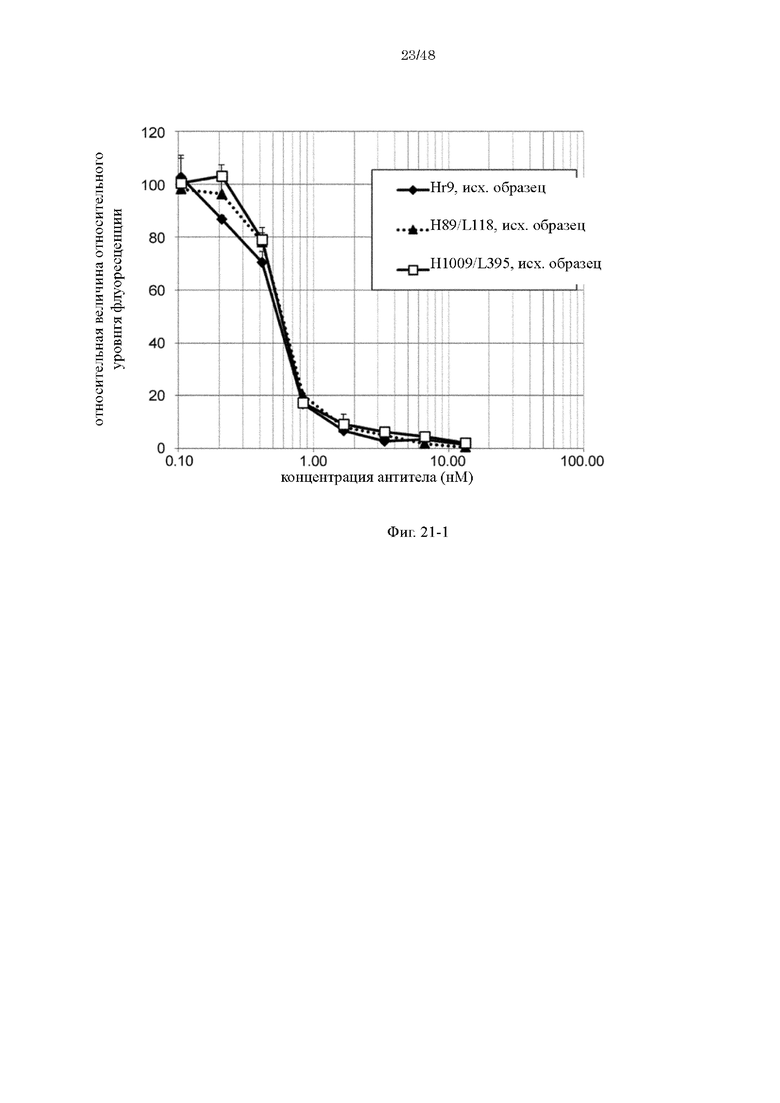
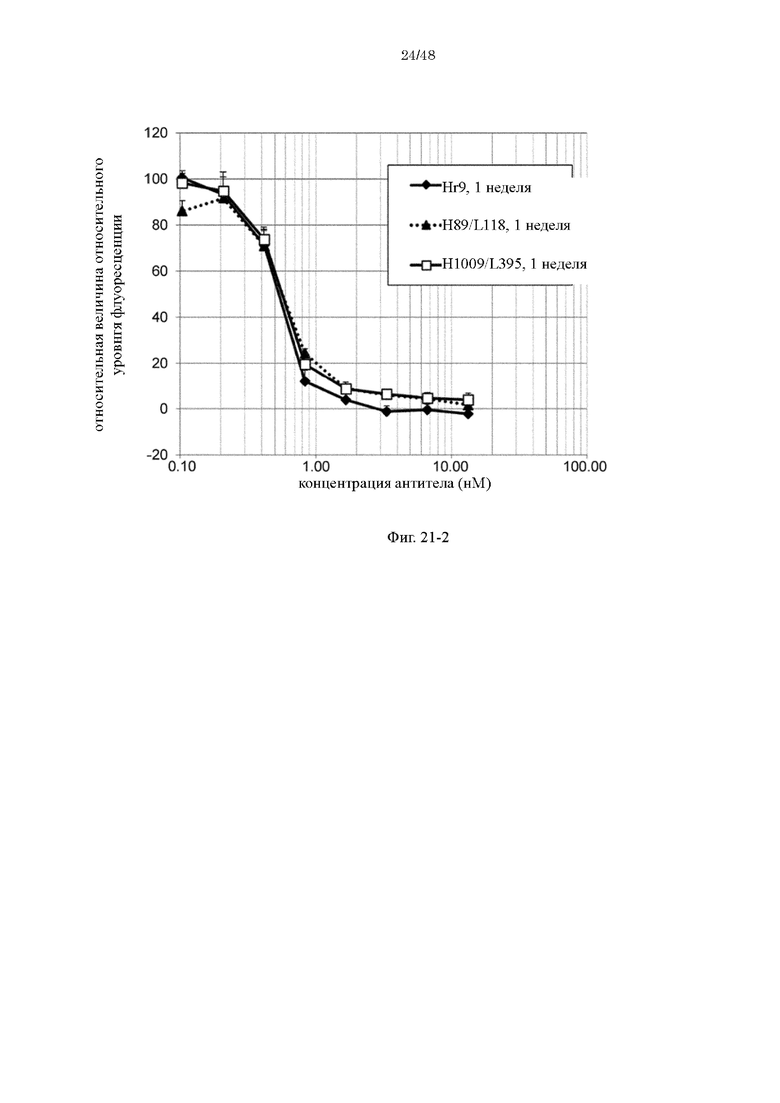
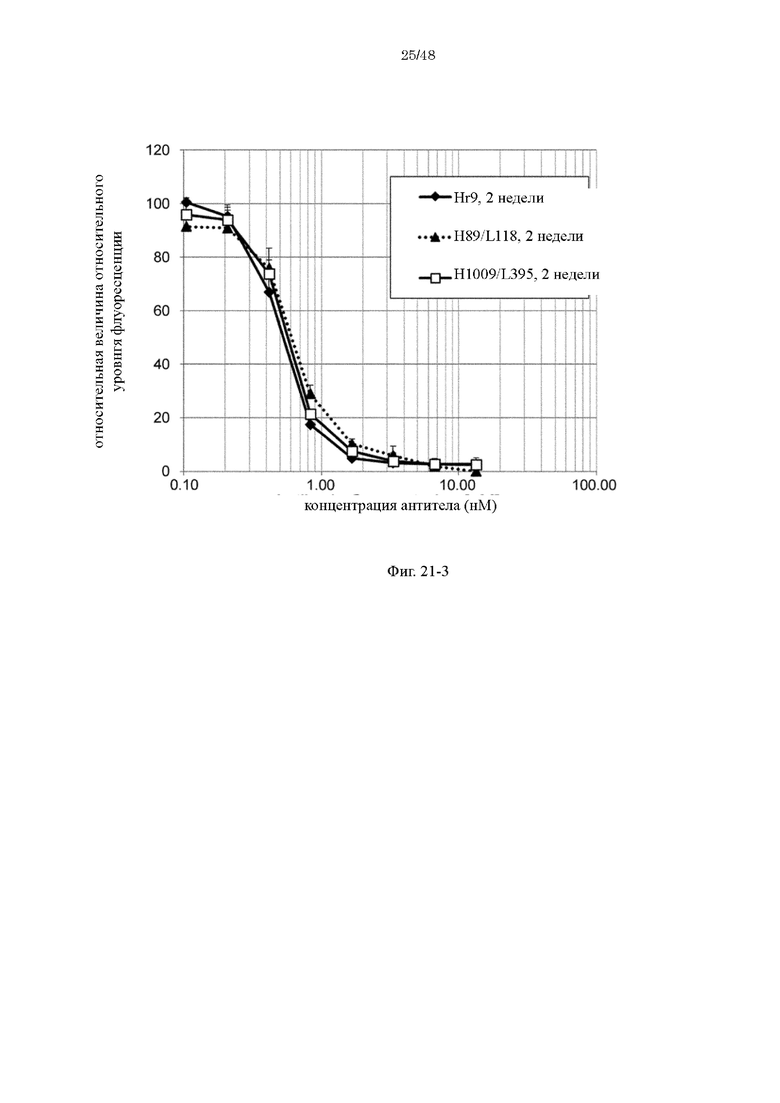

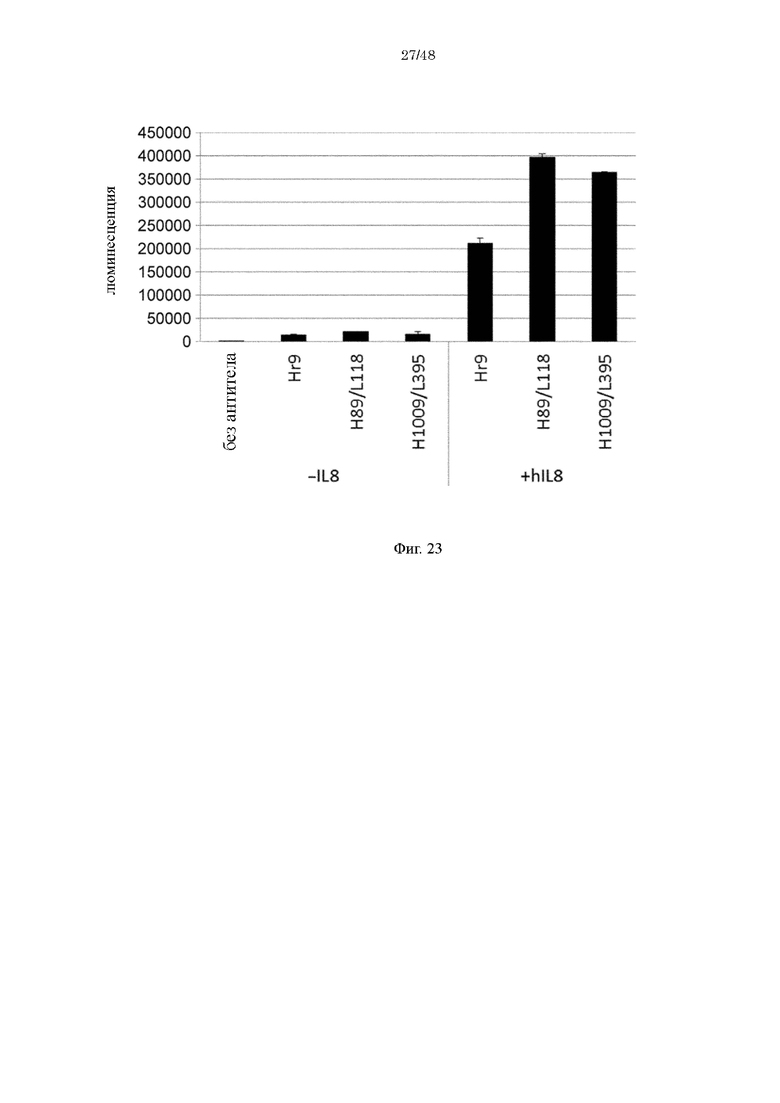
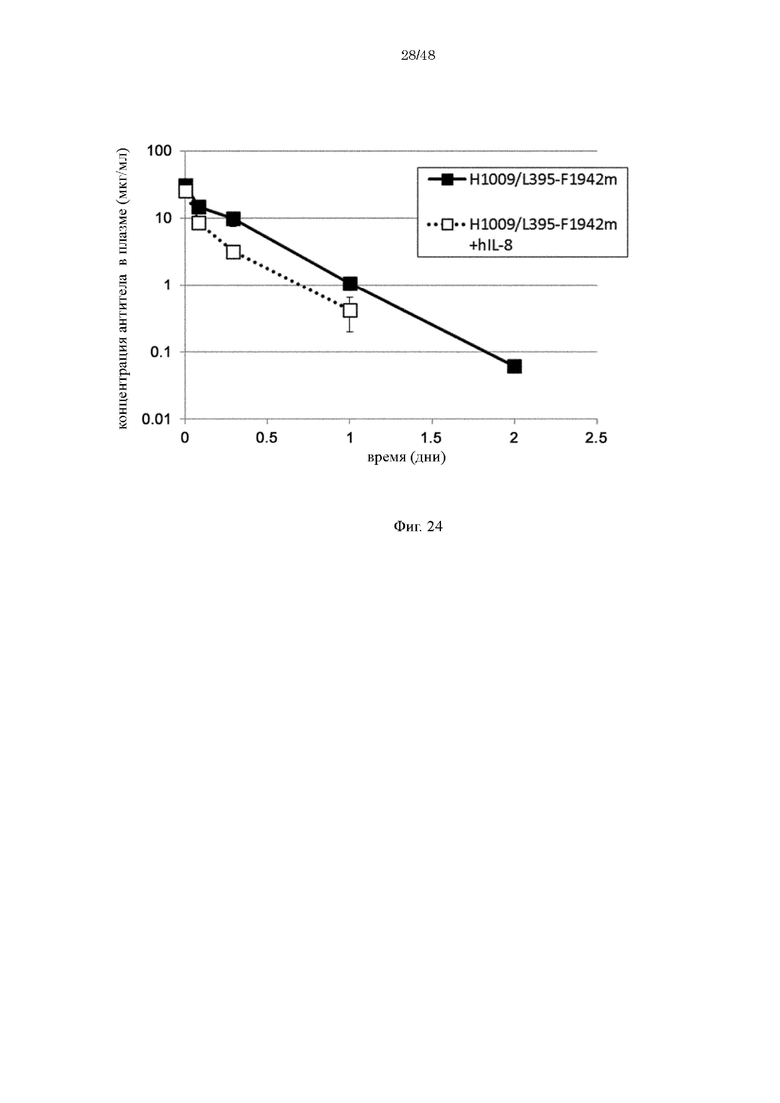
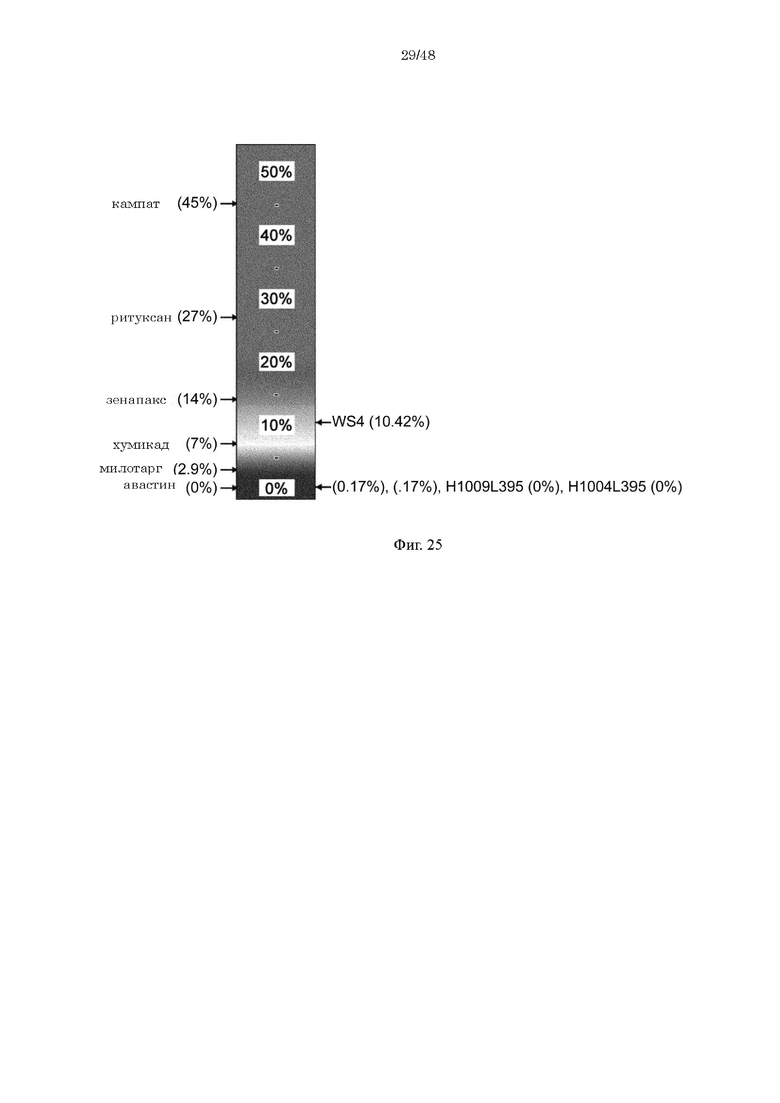
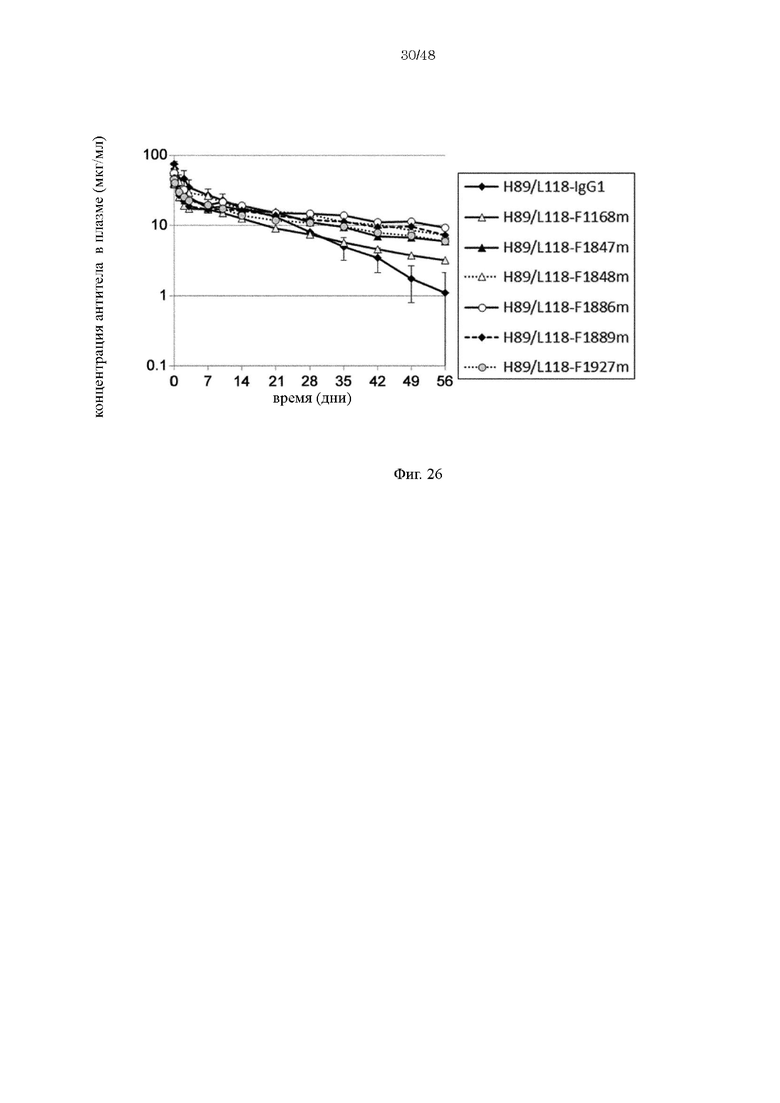
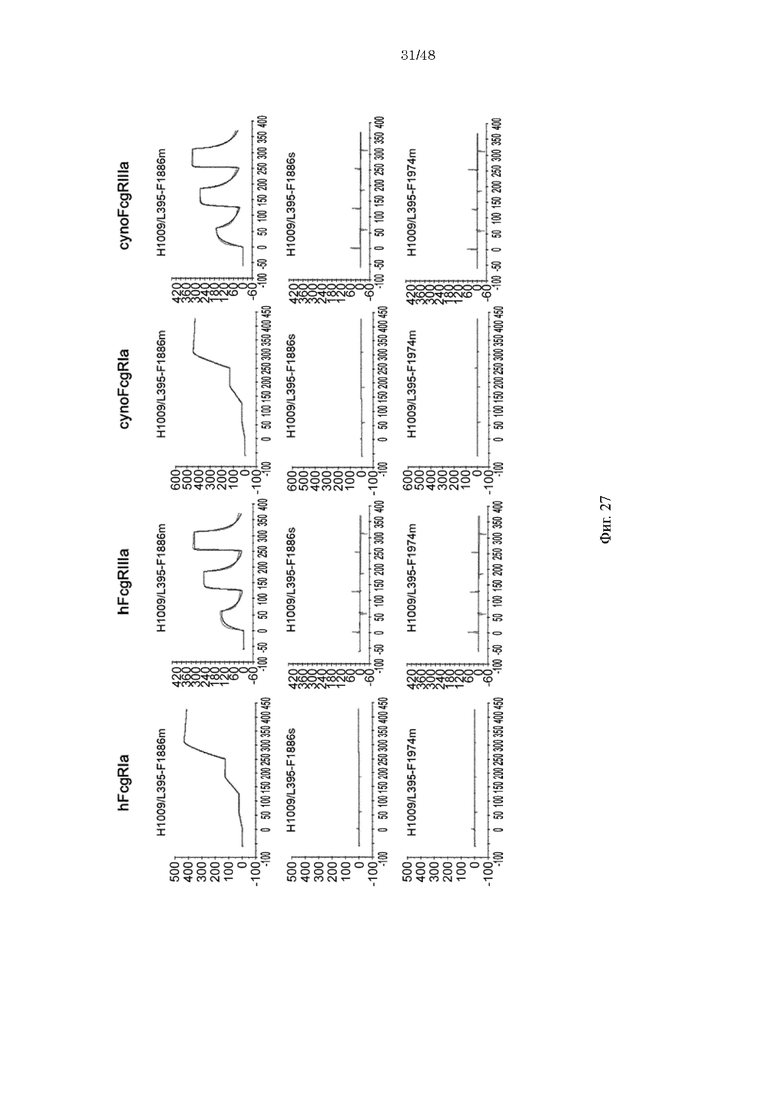
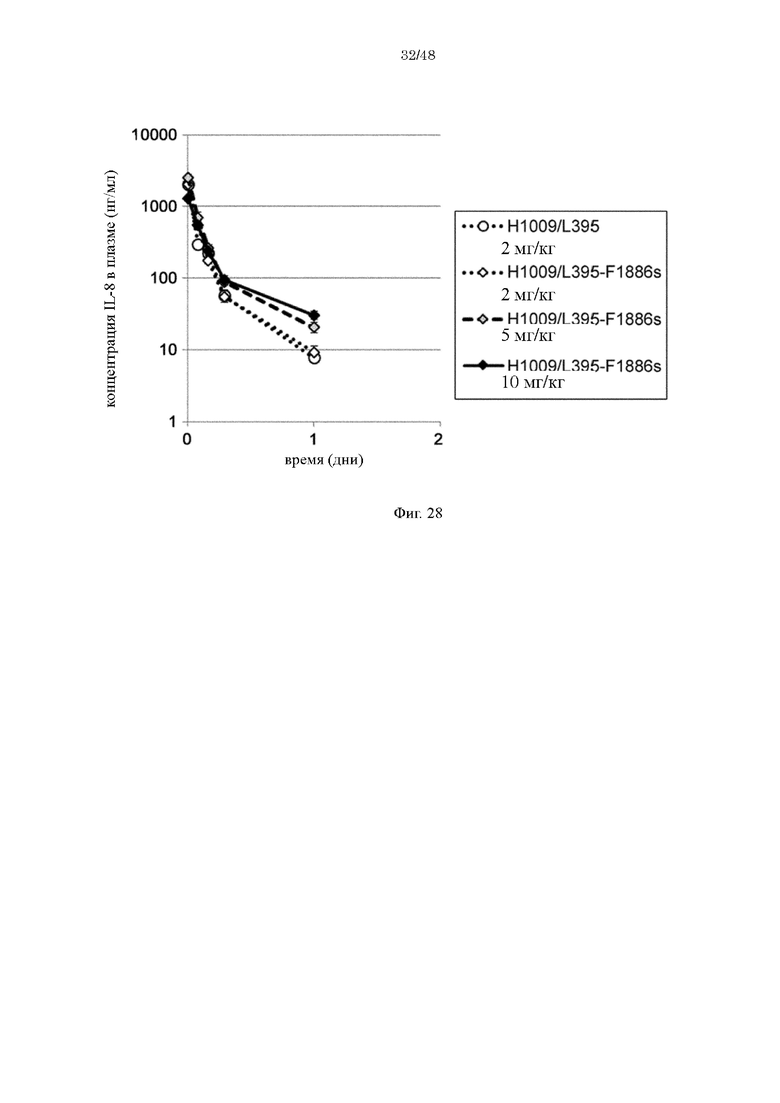

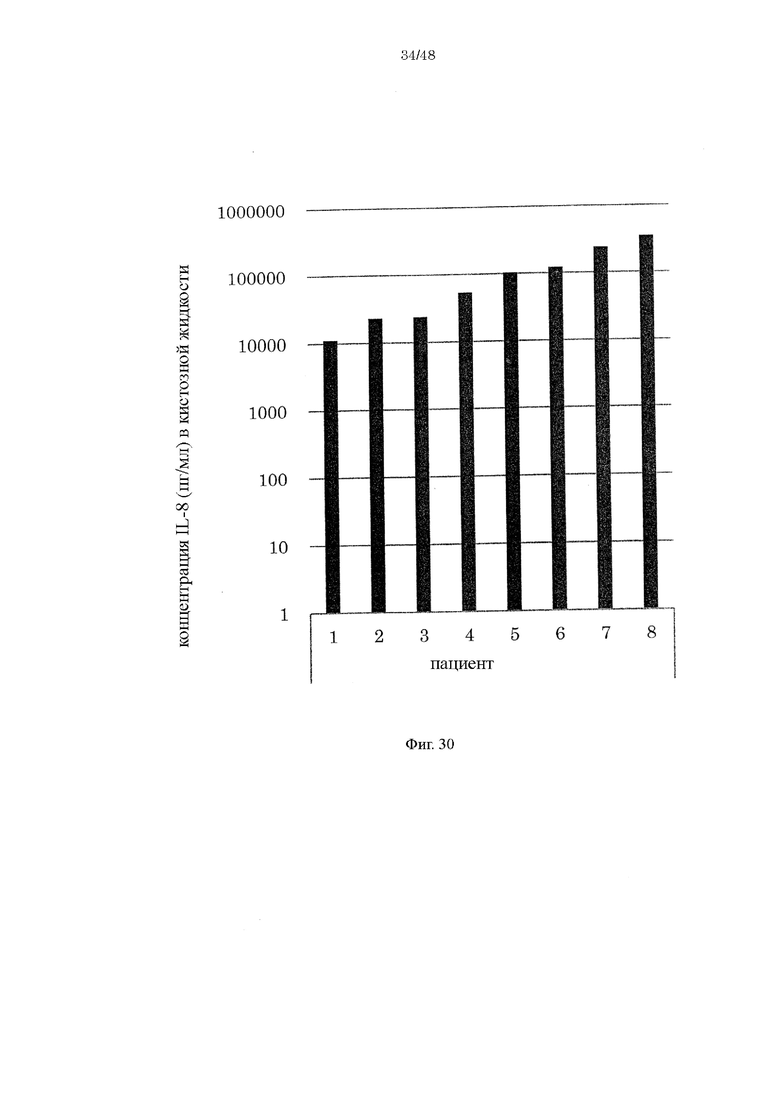
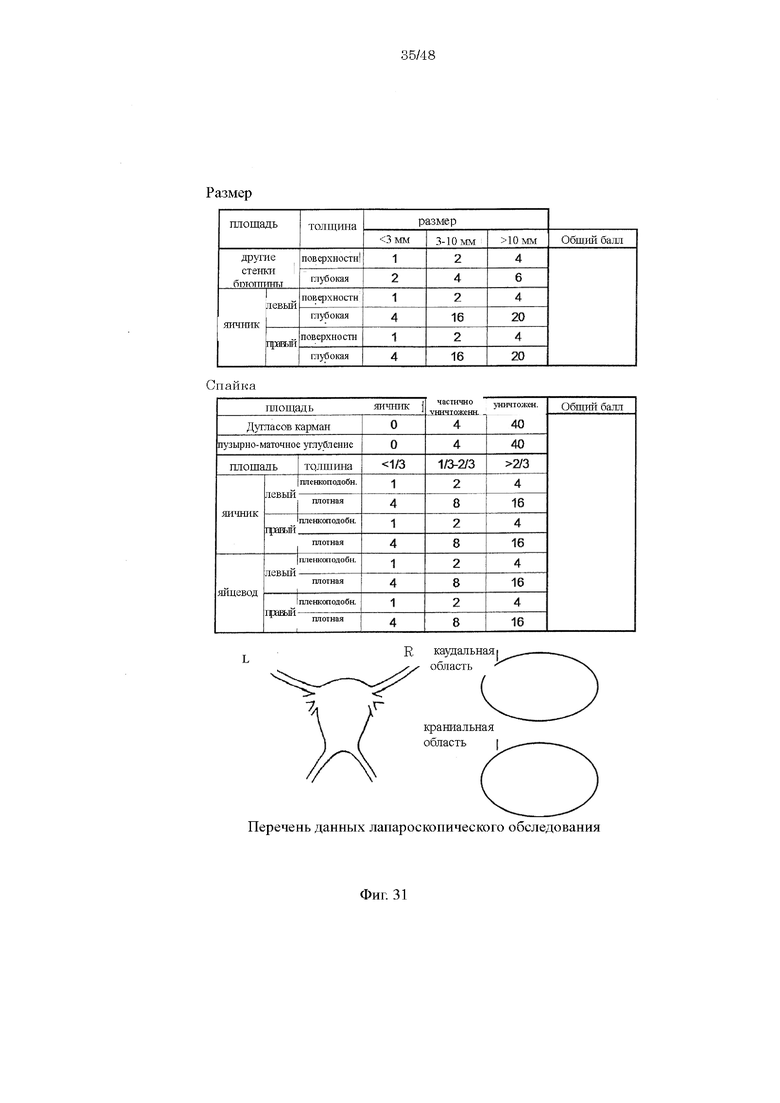
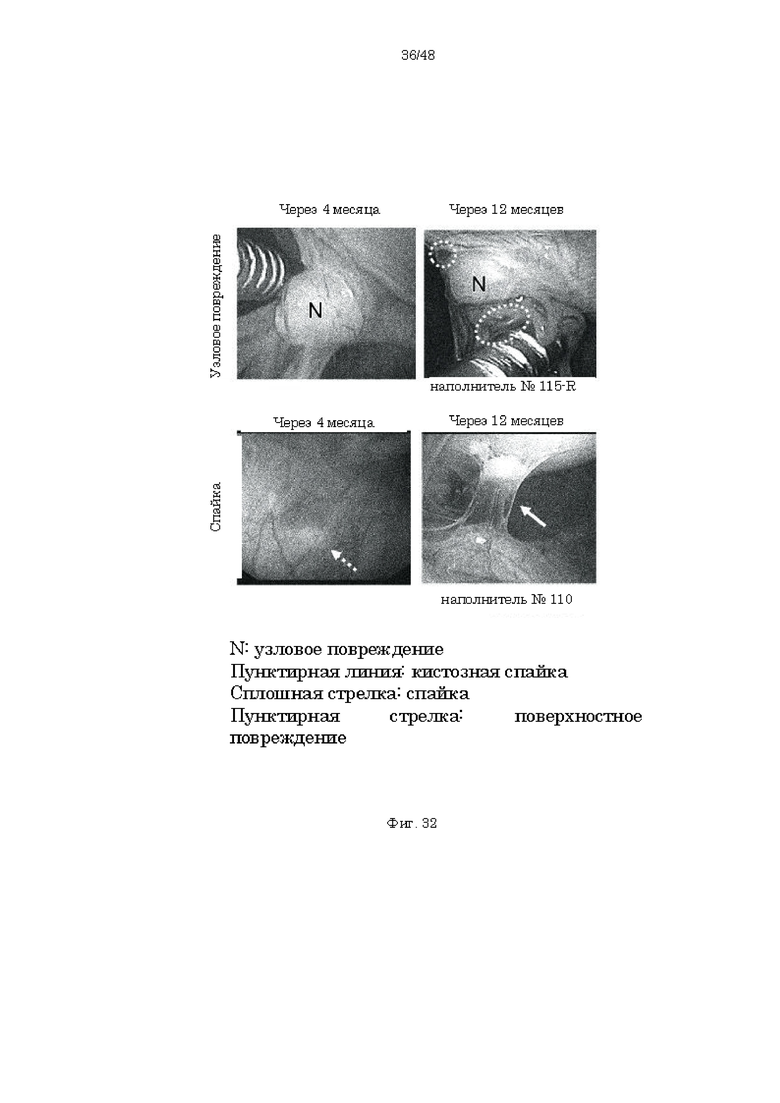

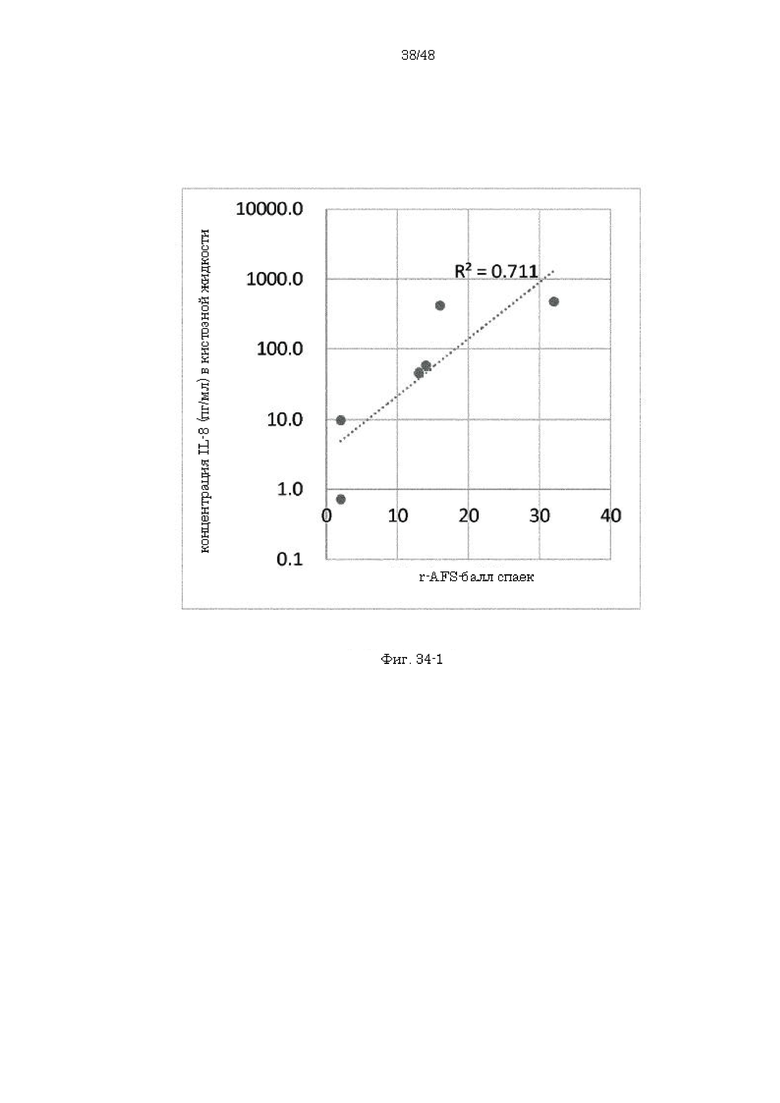
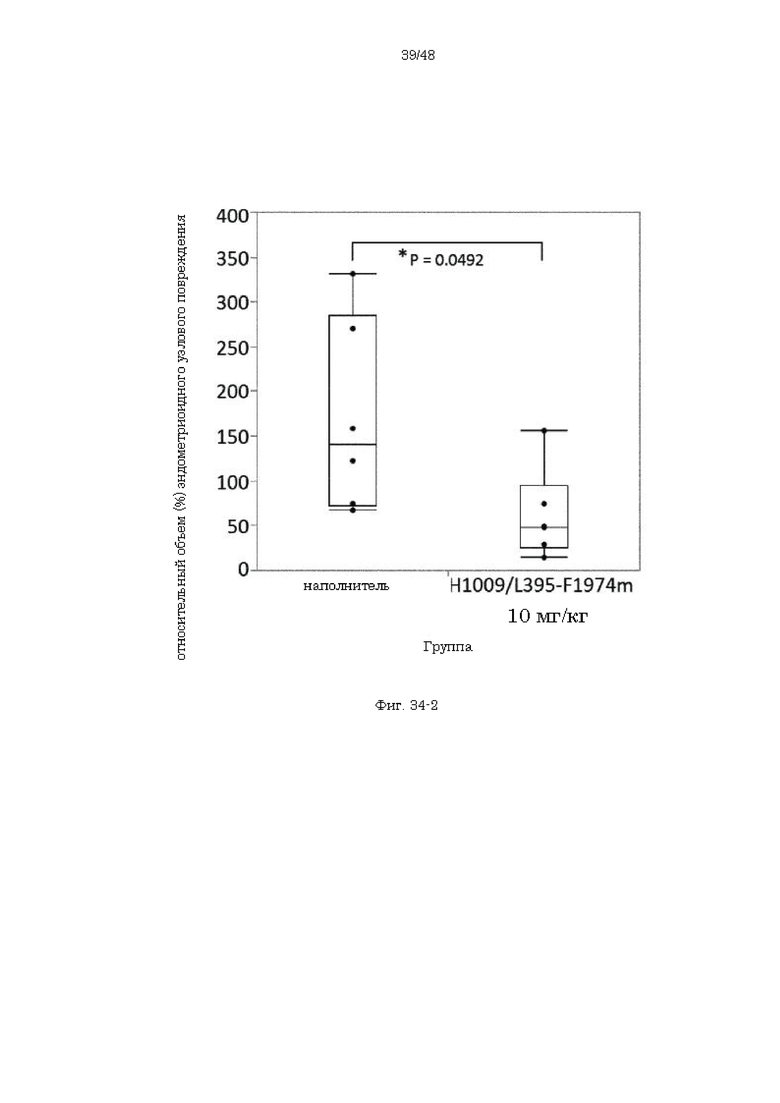
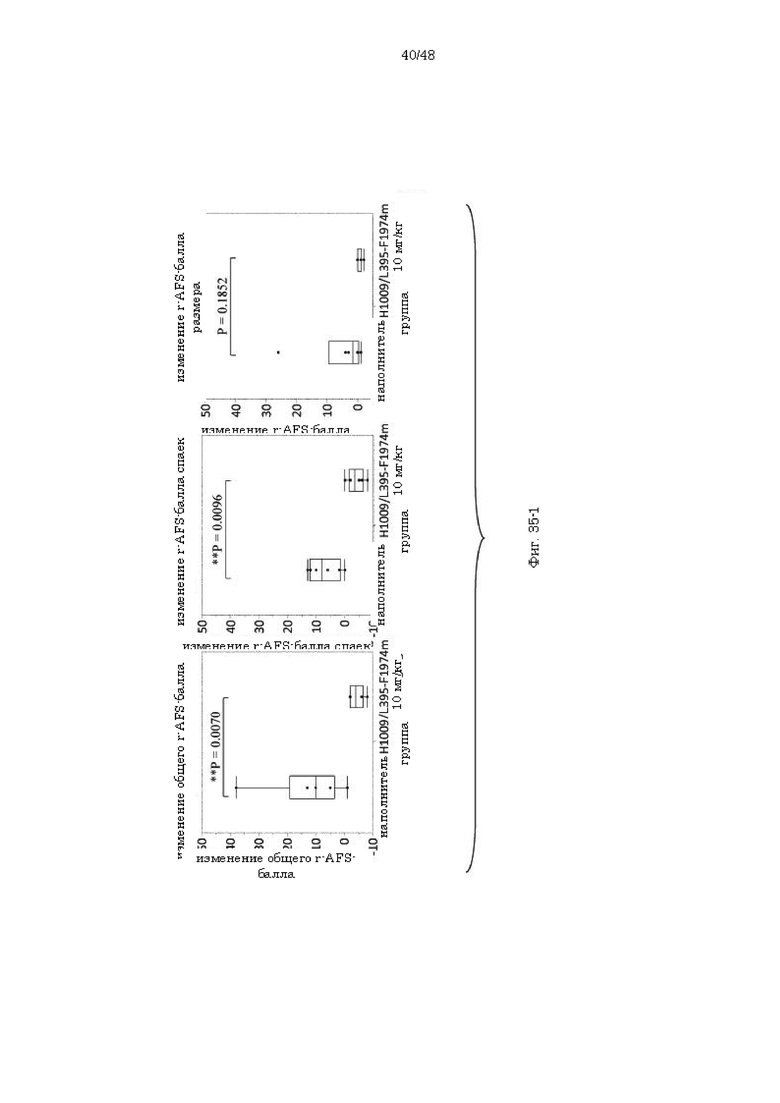
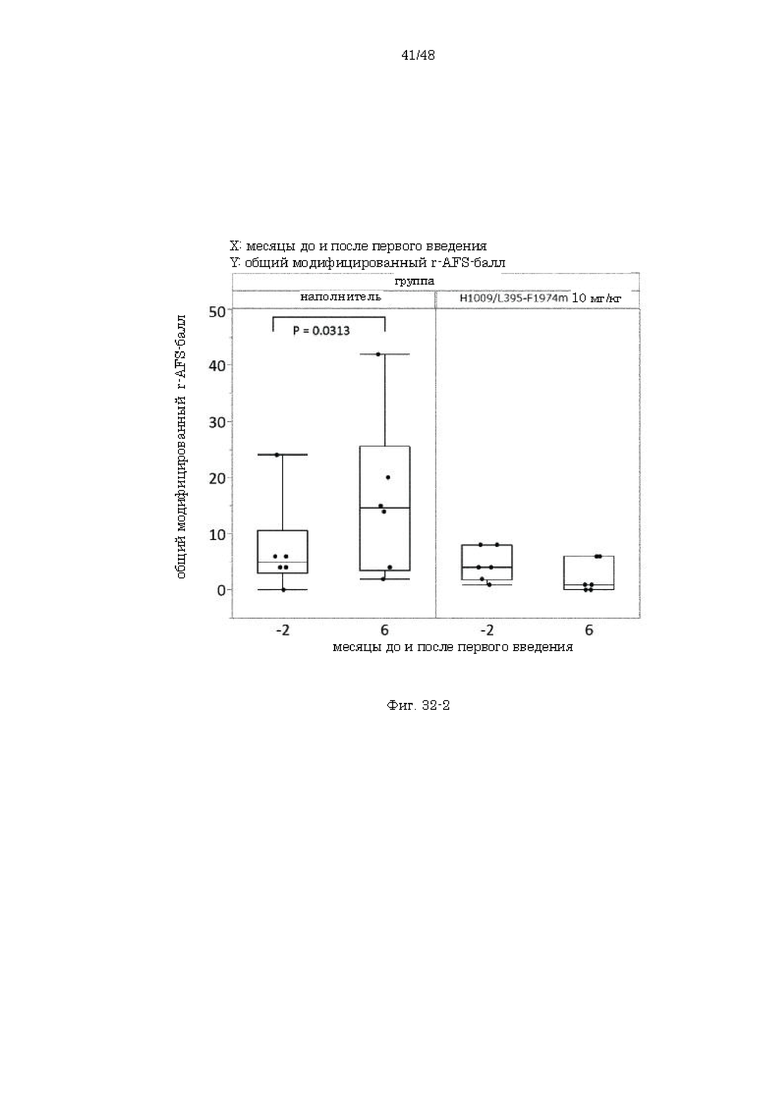
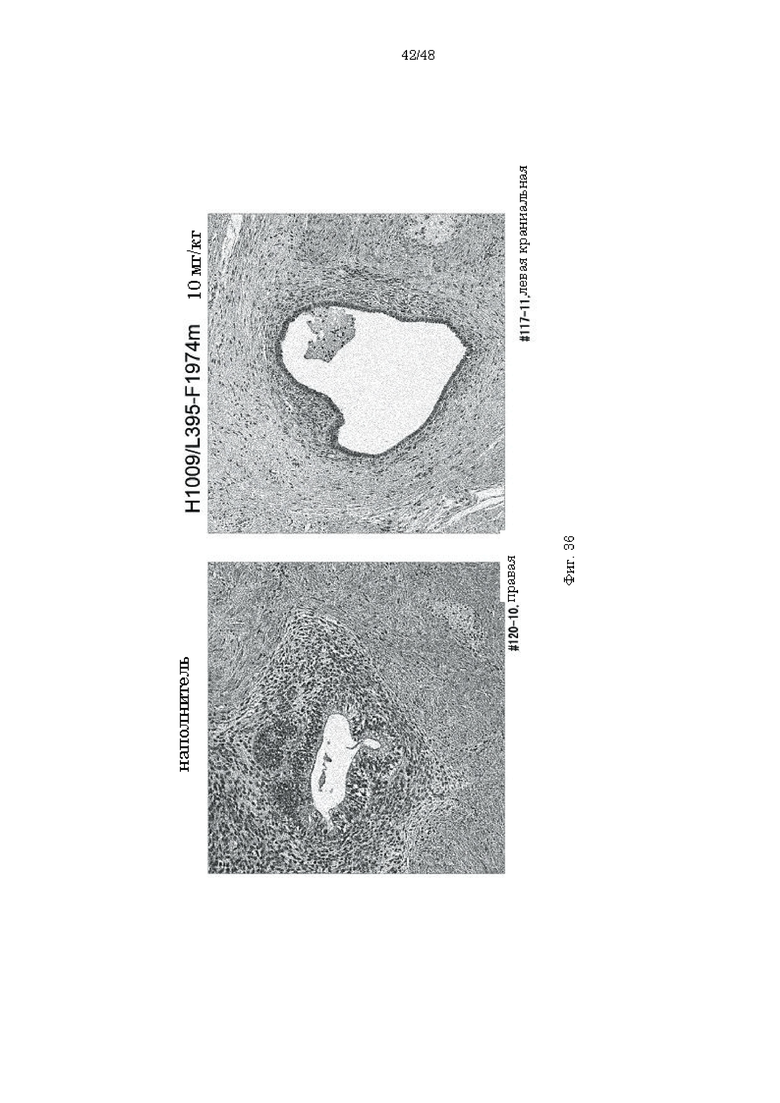
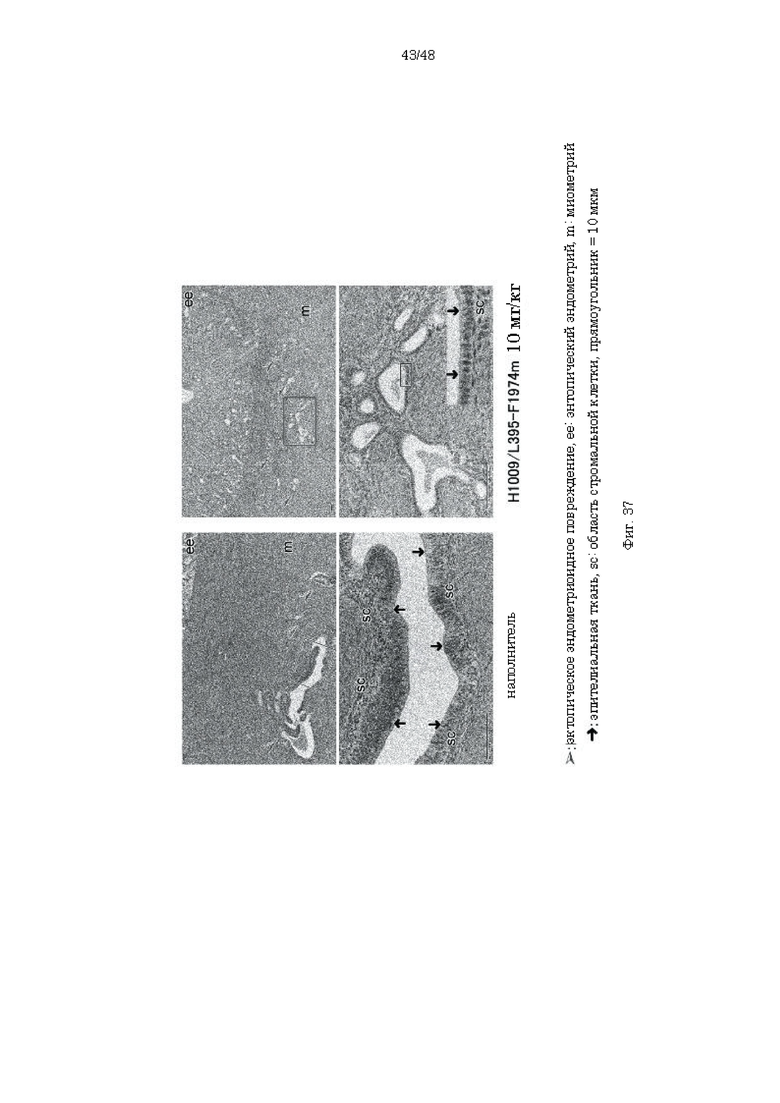
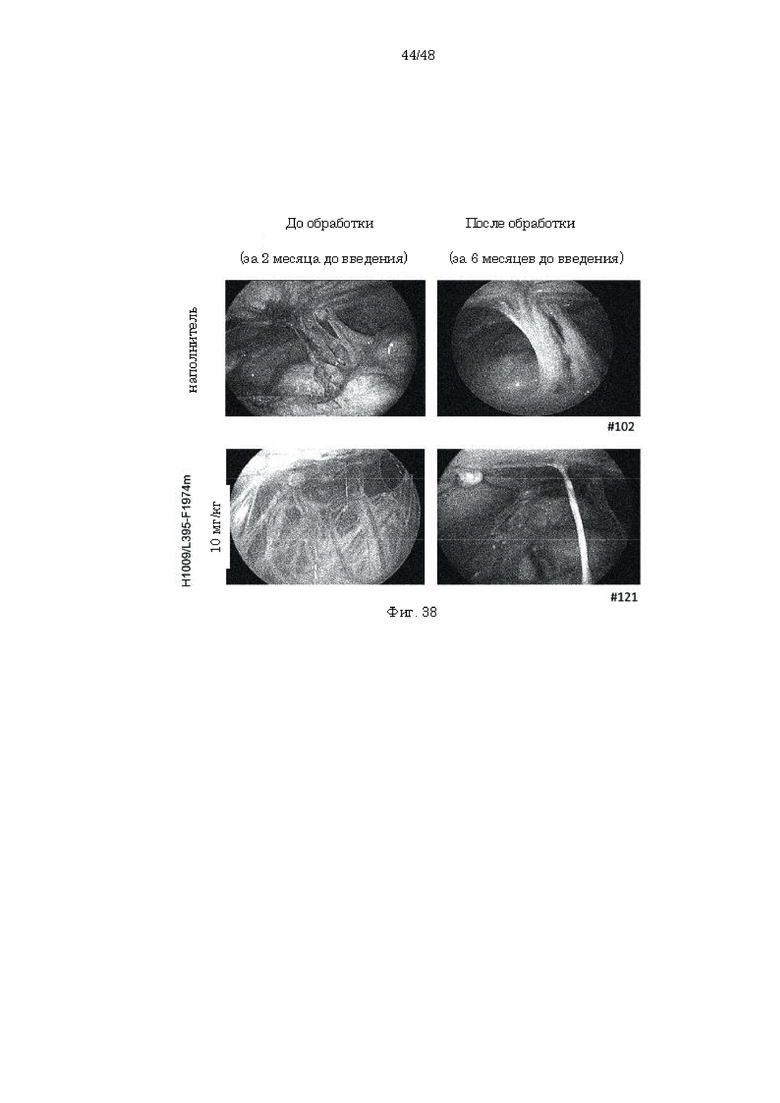
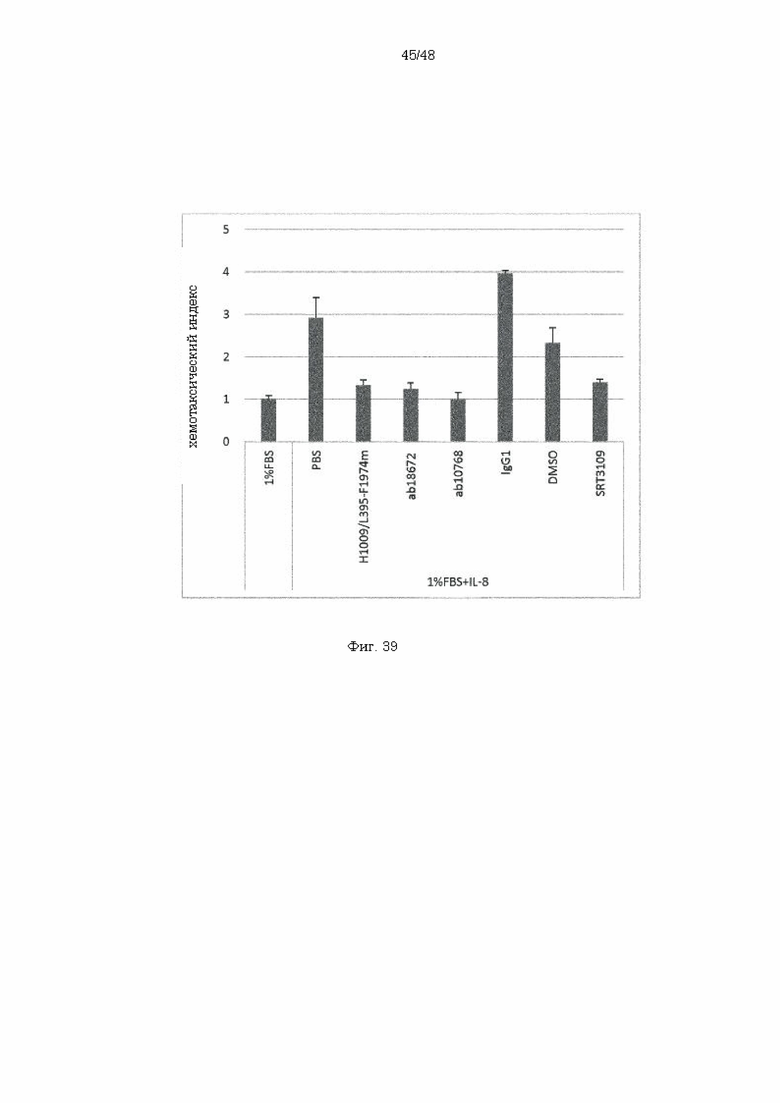
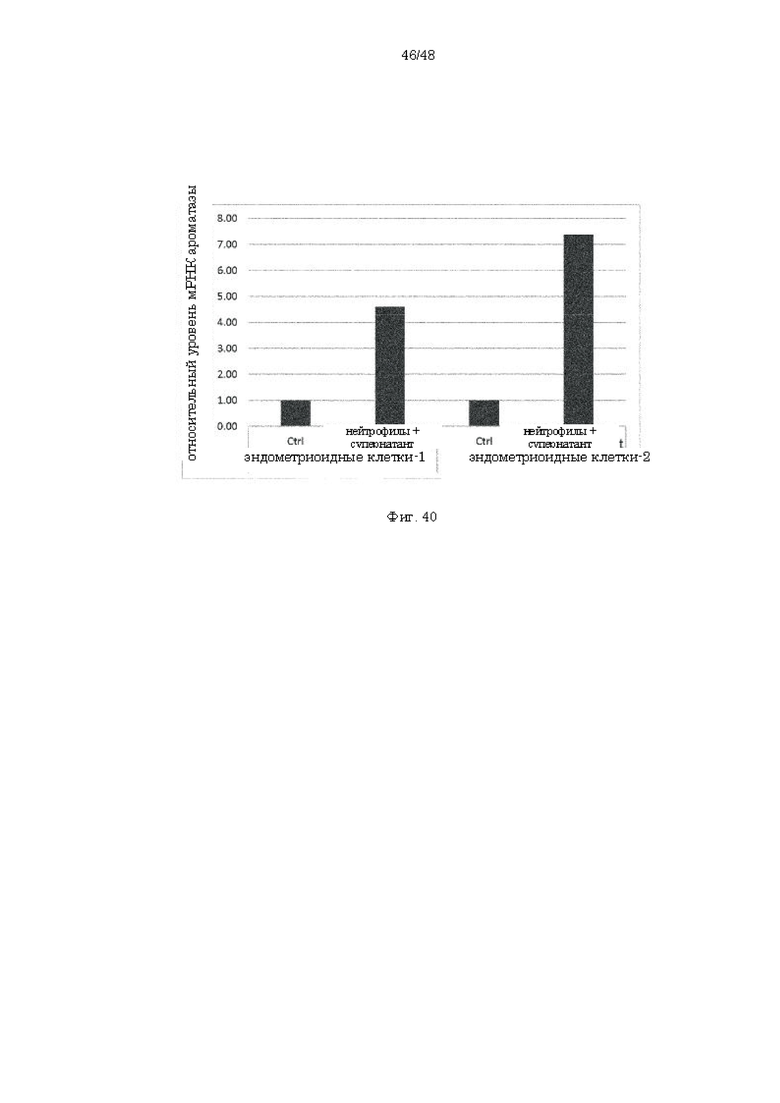
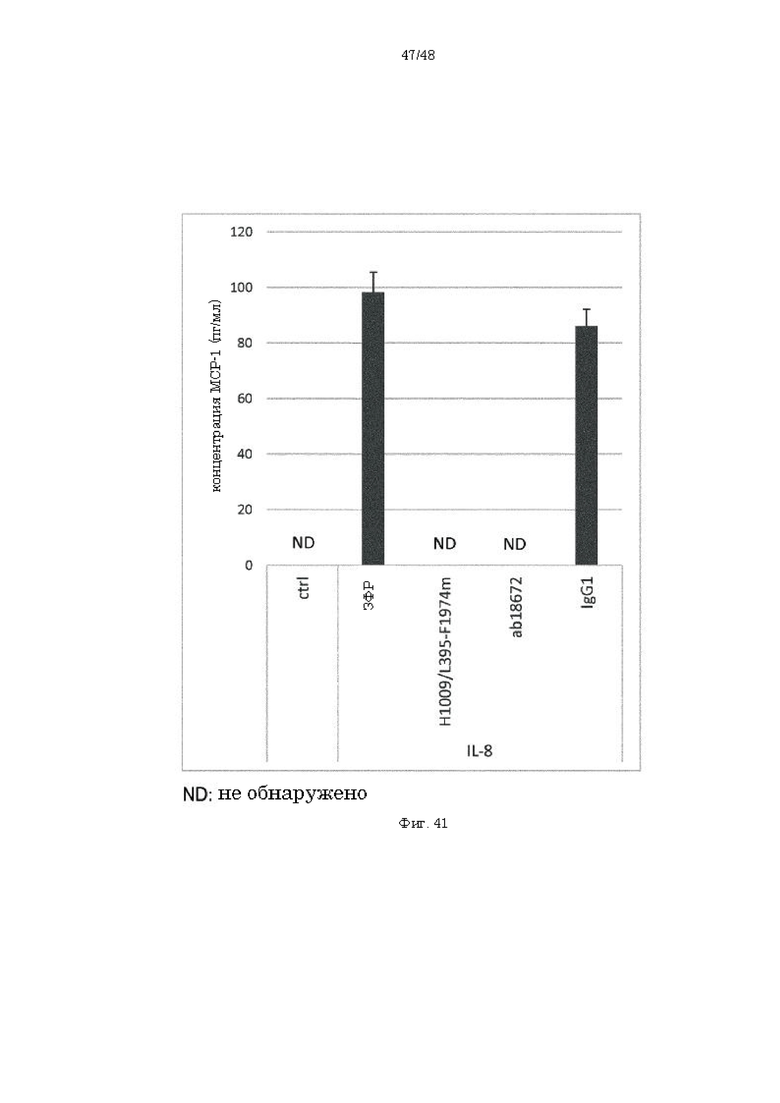
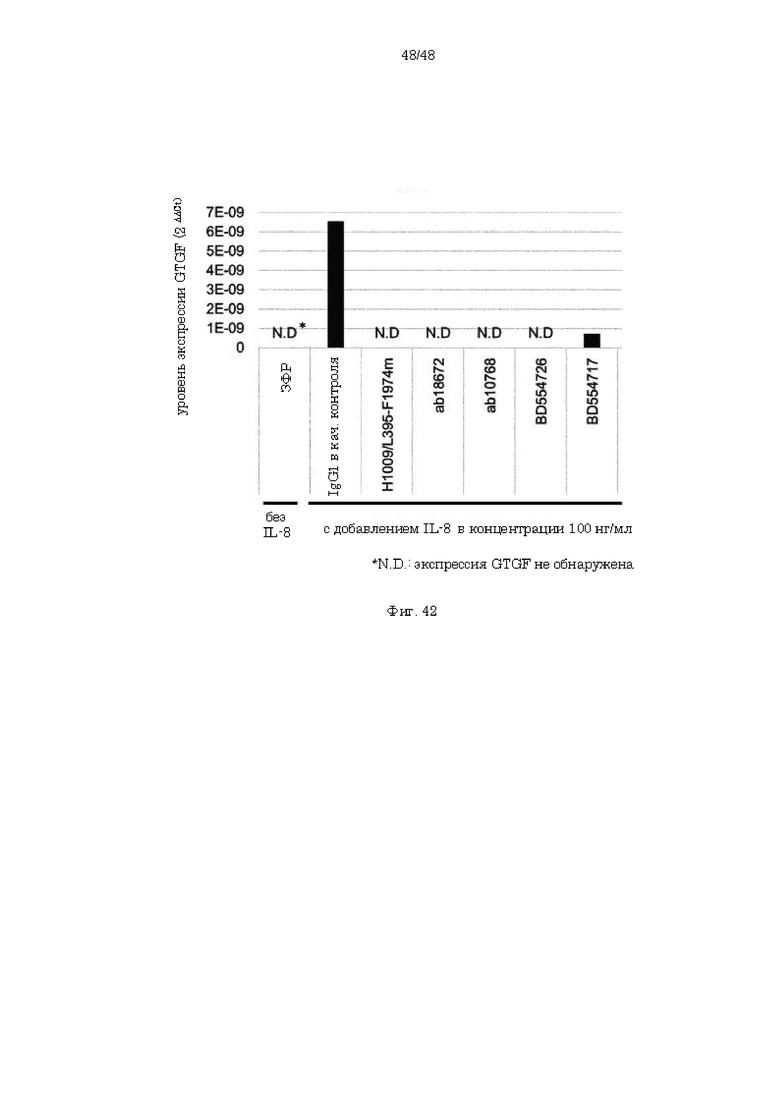
Изобретение относится к применение композиции, которая содержит в качестве действующего
вещества выделенное антитело к IL-8, которое связывается с человеческим IL-8, для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из:
эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления. 3 н. и 1 з.п. ф-лы, 42 ил., 29 табл., 25 пр.
1. Применение композиции, которая содержит в качестве действующего вещества выделенное антитело к IL-8, которое связывается с человеческим IL-8,
где антитело к IL-8 содержит:
(а) аминокислотную последовательность SEQ ID NO: 23 в качестве HVR-H1,
(b) аминокислотную последовательность SEQ ID NO: 29 в качестве HVR-H2,
(c) аминокислотную последовательность SEQ ID NO: 30 в качестве HVR-H3,
(d) аминокислотную последовательность SEQ ID NO: 26 в качестве HVR-L1,
(e) аминокислотную последовательность SEQ ID NO: 31 в качестве HVR-L2, и
(f) аминокислотную последовательность SEQ ID NO: 32 в качестве HVR-L3,
для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из:
эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления.
2. Применение композиции, которая содержит в качестве действующего вещества выделенное антитело к IL-8, которое связывается с человеческим IL-8,
где антитело к IL-8 содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 34, и вариабельную область легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 35,
для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, состоящей из:
эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия; и боли в результате спаек, фиброза или воспаления.
3. Применение композиции, которая содержит в качестве действующего вещества выделенное антитело к IL-8, которое связывается с человеческим IL-8, где антитело к IL-8 выбрано из группы, которая состоит из:
(A) антитела к IL-8, которое содержит тяжелую цепь, включающую аминокислотную последовательность SEQ ID NO: 36, и легкую цепь, включающую аминокислотную последовательность SEQ ID NO: 38; и
(Б) антитела к IL-8, которое содержит тяжелую цепь, включающую аминокислотную последовательность SEQ ID NO: 37, и легкую цепь, включающую аминокислотную последовательность SEQ ID NO: 38;
для лечения или предупреждения связанного с IL-8 заболевания, выбранного из группы, которая состоит из:
эндометриоза; аденомиоза; дисменореи; образования спаек; фиброзных заболеваний; боли при эндометриозе, аденомиозе или дисменореи; бесплодия и боли в результате образования спаек, фиброза или воспаления.
4. Применение по одному из пп. 1-3, где фиброзное заболевание выбрано из группы, которая состоит из фиброза при эндометриозе или аденомиозе, хронического обструктивного заболевания легких (COPD), муковисцидоза, псориаза, фиброза печени, фиброза почки и фиброза легких.
| WO 2005023193 A2, 17.03.2005 | |||
| WO 2009026117 A2, 26.02.2009 | |||
| РЕКОНСТРУИРОВАННЫЕ ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-8 | 1995 |
|
RU2159776C2 |

