Область техники
Настоящее изобретение относится к новым биспецифическим антителам (contorsbodies - конторстелам), состоящим из двух гибридизированных полипептидов, содержащих два антигенсвязывающих домена, способных к специфическому связыванию с первой мишенью, и один антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью, к способам получения этих молекул и к способам их применения.
Предшествующий уровень техники
Со времени разработки в 1974 году Koehler и Milstein первых моноклональных антител, много усилий было направлено на разработку антител, пригодных для лечения людей. Первые доступные моноклональные антитела были разработаны в организмах мышей и крыс. Эти антитела при лечении людей вызывают нежелательные побочные эффекты из-за выработки антител против антител грызунов. Много усилий было предпринято для снижения проявления таких нежелательных побочных эффектов или даже их устранения. В последние годы на рынке появились моноклональные антитела человека или гуманизированные моноклональные антитела, количество которых постоянно возрастает.
Биспецифические антитела представляют все больший интерес для диагностических и терапевтических целей. Хотя природные антитела являются моноспецифическими, биспецифические антитела распознают два разных эпитопа либо на одном, либо на разных антигенах. За последние годы было разработано множество форматов новых антител. Применение сложного молекулярного дизайна и генной инженерии решило многие технические проблемы, связанные с созданием биспецифических антител, например, такие как стабильность, растворимость и другие свойства, которые необходимы лекарственным препаратам и которые в целом обобщают под понятием «необходимые свойства для разработки». Кроме того, для придания различных необходимых свойств разрабатываемому биспецифическому антителу (то есть профили целевого продукта) необходим доступ к разнообразным панелям форматов антител. Эти форматы могут варьировать по размеру, геометрии связывающих модулей, валентности, гибкости, а также по фармакокинетическим свойствам (Brinkmann U., Kontermann R.E., MABS, 9(2), 2017, 182-212).
Тем не менее, предполагают, что не существует единого наилучшего формата, соответствующего всем потребностям, и что по-прежнему имеется потенциал для оптимизации форматов антител, полученных из четырех-цепочечного формата Y-образных антител дикого типа. Для применения в качестве фармацевтического продукта биспецифические антитела необходимо получать в больших количествах воспроизводимым способом, предпочтительно с высоким выходом. Более сложный состав (например, 3-4 цепи в отличие от 2-цепочечных IgG) часто требует более экстенсивной оптимизации систем экспрессии. Кроме того, наличие или отсутствие нежелательных побочных продуктов может иметь большое значение.
Краткое описание изобретения
Настоящее изобретение относится к биспецифическим антителам, состоящим из двух цепей, хотя они содержат три антигенсвязывающих домена. Антитела по настоящему изобретению отличаются по пространственной ориентации антигенсвязывающего домена от классических антител в формате IgG.
Настоящее изобретение относится к биспецифическому антителу, состоящему из двух гибридизированных полипептидов и содержащему два антигенсвязывающих домена, способных специфически связываться с первой мишенью, и один антигенсвязывающий домен, способный специфически связываться со второй мишенью, в котором
(а) первый гибридизированный полипептид содержит первую часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, спейсерный домен, вторую часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и первую часть антигенсвязывающего домена, способную к специфическому связыванию со второй мишенью, в котором
- спейсерный домен является полипептидом и содержит по меньшей мере 25 аминокислотных остатков,
- первая часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована или непосредственно, или через первый пептидный линкер, с N-концом спейсерного домена,
- вторая часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована или непосредственно, или через второй пептидный линкер, с С-концом спейсерного домена, и
- первая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована или непосредственно, или через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, или гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и
(б) второй гибридизированный полипептид, содержащий первую часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, спейсерный домен, вторую часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, и вторую часть антигенсвязывающего домена, способную к специфическому связыванию со второй мишенью, в котором
спейсерный домен является полипептидом и содержит по меньшей мере 25 аминокислотных остатков,
- первая часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через первый пептидный линкер с N-концом спейсерного домена,
- вторая часть второго антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, гибридизирована либо непосредственно, либо через второй пептидный линкер с С-концом спейсерного домена, и
- вторая часть антигенсвязывающего домена, способного к специфическому связыванию со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с С-концом второй части второго антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, или гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части второго антигенсвязывающего домена, способного специфически связываться с первой мишенью,
где первая часть и вторая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, связаны друг с другом с формированием антигенсвязывающего домена, способного специфически связываться со второй мишенью, причем первая часть и вторая часть первого и второго антигенсвязывающих доменов, способных специфически связываться с первой мишенью, связаны друг с другом с образованием кольцевого гибридного полипептида, и
где спейсерный домен первого гибридизированного полипептида и спейсерный домен второго гибридизированного полипептида ковалентно связаны друг с другом дисульфидной связью и содержат модификации, способствующие ассоциации первого и второго гибридизированных полипептидов.
В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, описанное выше в настоящем описании, в котором в первом гибридизированном полипептиде первая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и в котором во втором гибридизированном полипептиде вторая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью.
В другом варианте осуществления настоящего изобретения предусматривают биспецифическое антитело, описанное выше в настоящем описании, в котором в первом гибридизированном полипептиде первая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и в котором во втором гибридизированном полипептиде вторая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью.
В еще одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, описанное выше в настоящем описании, в котором третий пептидный линкер, соединяющий первую часть или вторую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, включает, по меньшей мере, 15 аминокислот. В другом варианте осуществления настоящего изобретения третий пептидный линкер, соединяющий первую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, и третий пептидный линкер, соединяющий вторую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, являются идентичными. В еще одном из вариантов осуществления настоящего изобретения предусматривают третий пептидный линкер, включающий от 15 до 25 аминокислот. В одном из вариантов осуществления настоящего изобретения третий пептидный линкер содержит аминокислотную последовательность SEQ ID NO:84.
В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, описанное выше в настоящем описании, в котором в первый гибридизированный полипептид содержит вариабельный домен тяжелой цепи антигенсвязывающего домена, способного специфически связываться со второй мишенью, и второй гибридизированный полипептид содержит вариабельный домен легкой цепи антигенсвязывающего домена, способного специфически связываться со второй мишенью, или наоборот. В другом варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена является фрагментом Fab тяжелой цепи антитела, и вторая часть антигенсвязывающего домена является фрагментом Fab легкой цепи антитела, или наоборот. В еще одном варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена и вторая часть антигенсвязывающего домена ковалентно связаны друг с другом дисульфидной связью.
Таким образом, в одном из вариантов осуществления настоящего изобретения первая часть антигенсвязывающего домена является фрагментом Fab тяжелой цепи антитела (VH-CH1), и вторая часть антигенсвязывающего домена является фрагментом Fab легкой цепи антитела (VL-Ckappa). В другом варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена является фрагментом Fab легкой цепи антитела, и вторая часть антигенсвязывающего домена является фрагментом Fab тяжелой цепи антитела. В еще одном из вариантов осуществления настоящего изобретения первая часть антигенсвязывающего домена является фрагментом Fab перекрестного антитела, содержащего VH-Ckappa, и вторая часть антигенсвязывающего домена является кросс-Fab фрагментом антитела, содержащим VL-CH1. В другом варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена является кросс-Fab фрагментом антитела, содержащим VL-CH1, и вторая часть антигенсвязывающего домена является кросс-Fab фрагментом антитела, содержащим VH-Ckappa.
Выше описано, что биспецифическое антитело состоит из первого и второго гибридизированных полипептидов, которые оба содержат спейсерный домен, причем спейсерный домен первого гибридизированного полипептида и спейсерный домен второго гибридизированного полипептида ковалентно связаны друг с другом посредством дисульфидной связи и содержат модификации, способствующие ассоциации первого и второго гибридизированных полипептидов. Спейсерный домен содержит, по меньшей мере, 25 аминокислот.
В одном из вариантов осуществления настоящего изобретения спейсерный домен содержит шарнирную область антитела или его (С-концевой) фрагмент и домен СН2 антитела или его (N-концевой) фрагмент. В еще одном из вариантов осуществления настоящего изобретения спейсерный домен содержит шарнирную область антитела или ее фрагмент, домен СН2 антитела и домен СН3 антитела или их фрагмент. Кроме того, спейсерный домен первого гибридизированного полипептида и спейсерный домен второго гибридизированного полипептида содержат модификации, способствующие ассоциации первого и второго гибридизированных полипептидов. В одном из вариантов настоящего изобретения спейсерный домен первого гибридизированного полипептида содержит впадины, а спейсерный домен второго гибридизированного полипептида содержит выступы в соответствии с принципом «выступ-во-впадину». В еще одном варианте осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором спейсерный домен содержит шарнирную область антитела или его фрагмент и домен Fc IgG1. В частности, домен Fc IgG1 включает одно или несколько аминокислотных замещений, которые уменьшают связывание с рецептором Fc, в частности, с рецептором Fcγ. Точнее, домен Fc IgG1 содержит аминокислотные замещения L234A, L235A и P329G (нумерация по EU индексу Kabat).
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором один антигенсвязывающий домен, способный специфически связываться со второй мишенью, является антигенсвязывающий доменом, способным специфически связываться с опухолеассоциированным антигеном (tumor associated antigen - ТАА). В частности, опухолеассоциированный антиген является белком активации фибробластов (Fibroblast Activation Protein - FAP). В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный специфически связываться со второй мишенью, является антигенсвязывающим доменом, способным специфически связываться с белком активации фибробластов (FAP).
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с FAP, содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:1, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:2, и (iii) CDR-Н3, содержащий аминокислотную последовательность SEQ ID NO:3 и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:4, (v) CDR-L2, включающую аминокислотная последовательность SEQ ID NO:5 и (vi) CDR-L3, содержащая аминокислотную последовательность SEQ ID NO:6, или
(б) вариабельную область тяжелой цепи (VHFAP), содержащую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:9, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:10, и (iii) CDR-Н3, содержащий аминокислотную последовательность SEQ ID NO:11, и вариабельную область легкой цепи (VLFAP), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:12, (v) CDR-L2, включающую аминокислотная последовательность SEQ ID NO:13 и (vi) CDR-L3, содержащая аминокислотную последовательность SEQ ID NO:14.
Точнее, антигенсвязывающий домен, способный специфически связываться с FAP, содержит
(а) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:7, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:8, или
(б) вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:16.
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный специфически связываться с первой мишенью, является антигенсвязывающим доменом, способным специфически связываться с рецептором TNF, в частности костимулирующим рецептором TNF. В частности, костимулирующим рецептором TNF является ОХ40. В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный специфически связываться с первой мишенью, является антигенсвязывающим доменом, способным специфически связываться с ОХ40. В частности, биспецифическое антитело по настоящему изобретению содержит два антигенсвязывающих домена, способных специфически связываться с ОХ40.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с ОХ40, включает
(а) вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:22, и вариабельную область легкой цепи (VLOX40), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:35, или
(б) вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:21, и вариабельную область легкой цепи (VLOX40), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:34, или
(в) вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-Н2, содержащую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:23, и вариабельную область легкой цепи (VLOX40), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:36, или
(г) вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:24, и вариабельную область легкой цепи (VLOX40), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:37, или
(д) вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:18, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:25, и вариабельную область легкой цепи (VLOX40), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:29, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:32, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:38, или
(е) вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:18, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:26, и вариабельную область легкой цепи (VLOX40), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:29, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:32, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:38, или
(ж) вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:18, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:27, и вариабельную область легкой цепи (VLOX40), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:30, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:33, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:39.
В частности, антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит вариабельную область тяжелой цепи (VHOX40), включающую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:19 и (iii) CDR-H3, содержащая аминокислотную последовательность SEQ ID NO:22, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, содержащий аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, содержащий аминокислотную последовательность SEQ ID NO:35.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит
(а) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:40, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:41, или
(б) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:42, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:43, или
(в) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:44, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:45, или
(г) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:46, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:47, или
(а) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:48, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:49, или
(а) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:50, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:51, или
(а) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:52, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:53.
В другом предпочтительном варианте осуществления настоящего изобретения, антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит
(а) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:40, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:41.
В частности, настоящее изобретение предусматривает биспецифическое антитело, которое содержит
(а) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:54, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:55,
(б) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:56, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:57,
(в) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:58, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:59,
(г) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:60, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:61,
(д) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:62, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:63,
(е) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:64, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:65, или
(ж) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:66, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:67.
Кроме того, настоящее изобретение предусматривает биспецифическое антитело, которое содержит
(а) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:116, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:117,
(б) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:118, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:119,
(в) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:120, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:121,
(г) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:122, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:123,
(д) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:124, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:125,
(е) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:126, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:128,
(ж) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:129, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95, 96, 97, 98, 99 или 100% идентична аминокислотной последовательности SEQ ID NO:130,
(з) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:131, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:132, или
(и) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:133, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:134.
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором костимулирующим рецептором TNF является 4-1ВВ. В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный к специфическому связыванию с первой мишенью, представляет собой антигенсвязывающий домен, способный к специфическому связыванию с 4-1ВВ. В частности, биспецифическое антитело по настоящему изобретению содержит два антигенсвязывающих домена, способных специфически связываться с 4-1ВВ.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с 4-1ВВ, вариабельной области тяжелой цепи (VH4-1BB), включает (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:135, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:136, и (iii) CDR-Н3, содержащую аминокислотную последовательность SEQ ID NO:137, и вариабельную область легкой цепи (Vl4-1BB), включающую (iv) CDR-L1, содержащую аминокислотную SEQ ID NO:138, (v) CDR-L2, содержащую аминокислотную SEQ ID NO:139, и (vi) CDR-L3, содержащую аминокислотную SEQ ID NO:140. В одном из вариантов осуществления настоящего изобретения, антигенсвязывающий домен, способный специфически связываться с 4-1ВВ, содержит вариабельную область тяжелой цепи (VH4-1BB), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:141, и вариабельную область легкой цепи (VL4-1ВВ), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:142.
В частности, настоящее изобретение предусматривает биспецифическое антитело, которое содержит
(а) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:143, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:144, или
(б) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:145, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:146.
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором костимулирующим рецептором TNF является CD40. В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный специфически связываться с первой мишенью, является антигенсвязывающий доменом, способным специфически связываться с CD40. В частности, биспецифическое антитело по настоящему изобретению содержит два антигенсвязывающих домена, способных специфически связываться с CD40.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с CD40, содержит вариабельную область тяжелой цепи (VHCD40), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:147, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:148, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:149, и вариабельную область легкой цепи (VLCD40), включающую (iv) CDR-L1, содержащую аминокислотную SEQ ID NO:150, (v) CDR-L2, содержащую аминокислотную SEQ ID NO:151, и (vi) CDR-L3, содержащую аминокислотную SEQ ID NO:152. В одном из вариантов осуществления настоящего изобретения, антигенсвязывающий домен, способный специфически связываться с CD40, содержит вариабельную область тяжелой цепи (VHCD40), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:153, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:154.
В другом варианте осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с первой мишенью, является антигенсвязывающим доменом, способным специфически связываться с CD40, и содержит
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO:167, SEQ ID NO:168, SEQ ID NO:169 и SEQ ID NO:170, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO:171, SEQ ID NO:172, SEQ ID NO:173 и SEQ ID NO:174, или
(ii) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO:175, SEQ ID NO:176, SEQ ID NO:177, SEQ ID NO:178, SEQ ID NO:179 и SEQ ID NO:180, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO:181, SEQ ID NO:182, SEQ ID NO:183 и SEQ ID NO:184.
В частности, настоящее изобретение предусматривает биспецифическое антитело, которое содержит
(а) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:155, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:156,
(б) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:157, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:158,
(в) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:159, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:160, или
(г) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:161, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:162, или
(д) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:163, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:164, или
(е) гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:165, и гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:166.
Настоящее изобретение также предусматривает выделенную нуклеиновую кислоту, кодирующую биспецифическое антитело по настоящему изобретению. Также предусматривают вектор экспрессии, содержащий нуклеиновую кислоту по настоящему изобретению, и, кроме того, клетку-хозяина, содержащую выделенную нуклеиновую кислоту или вектор экспрессии по настоящему изобретению. Также предусмотрен метод получения биспецифического антитела, включающий культивирование клетки-хозяина по настоящему изобретению в условиях, пригодных для экспрессии биспецифического антитела. Этот метод также может включать стадию выделения биспецифического антитела.
Настоящее изобретение также предусматривает фармацевтическую композицию, содержащую биспецифическое антитело по настоящему изобретению и, по меньшей мере, один фармацевтически приемлемый эксципиент.
Настоящее изобретение также предусматривает биспецифическое антитело по настоящему изобретению или фармацевтическую композицию по настоящему изобретению для применения в качестве лекарственного средства. Более конкретно, также в настоящем изобретении предусматривают биспецифическое антитело по настоящему изобретению для лечения рака или инфекционных заболеваний. В частности, предложено биспецифическое антитело по настоящему изобретению для применения при лечении рака.
В другом варианте осуществления настоящего изобретения предусматривают применение биспецифического антитела по настоящему изобретению или фармацевической композиции по настоящему изобретению для получения лекарственного средства, предназначенного для
(i) стимулирования ответа Т-клеток,
(ii) поддержки выживаемости активированных Т-клеток,
(iii) лечения инфекций,
(iv) лечения рака,
(v) задержки прогрессирования рака, или
(vi) продления выживаемости пациента, больного раком.
Настоящее изобретение также предусматривает метод лечения индивидуума, больного раком или инфекционным заболеванием, включающий введение индивидууму эффективного количества биспецифического антитела по настоящему изобретению или фармацевтической композиции по настоящему изобретению.
Настоящее изобретение также предусматривает применение биспецифического антитела по настоящему изобретению или фармацевтической композиции по настоящему изобретению для получения лекарственного средства для усиления или продления цитотоксической активности Т-клеток. Также предусмотрен метод усиления или продления цитотоксической активности Т-клеток у больного раком индивидуума, включающий введение индивидууму эффективного количества биспецифического антитела по настоящему изобретению или фармацевтической композиции по настоящему изобретению. В некоторых вариантах осуществления настоящего изобретения в соответствии с различными аспектами настоящего изобретения индивидуумом является млекопитающее, в частности человек.
Краткое описание фигур
На фигурах 1А, 1Б, 1В и 1Г показывают примеры того, как могут быть составлены конторстела настоящего изобретения. На фиг. 1А конторстело состоит из первого гибридизированного полипептида, содержащего первую половину Fab, способного к специфическому связыванию с первой мишенью (Fab тяжелой цепи), связанной через пептидный линкер (черная линия) со второй половиной Fab, способной специфически связываться с первой мишенью (Fab легкой цепи), которая дополнительно связана с первой половиной перекрестного Fab, способного специфически связываться со второй мишенью (VH-Ckappa) (в направлении от N к С), и второго гибридизированного полипептида, содержащего первую половину Fab, способного к специфическому связыванию с первой мишенью (Fab тяжелой цепи), связанной через пептидный линкер (черная линия) со второй половиной Fab, способной специфически связываться с первой мишенью (Fab легкой цепи), которая дополнительно связана с первой половиной перекрестного Fab, способного специфически связываться со второй мишенью (VL-CH1) (в направлении от N к С). Два спейсерных домена отличаются друг от друга и содержат модификации, способствующие ассоциации первого и второго гибридизируемых полипептидов. Примером конторстела такого типа является CD134-0093 (см. пример 2.1). На фиг. 1Б конторстело состоит из первого гибридизированного полипептида, содержащего первую половину перекрестного Fab, способного к специфическому связыванию с первой мишенью (VH-Ckappa), связанного через пептидный линкер (черная линия) со спейсерным доменом, связанным через пептидный линкер (черная линия) со второй половиной перекрестного Fab, способного специфически связываться с первой мишенью (VL-CH1), которая дополнительно связана с первой половиной Fab, способной специфически связываться со второй мишенью (Fab легкой цепи) (в направлении от N к С), и второго гибридизированного полипептида, содержащего первую половину перекрестного Fab, способного к специфическому связыванию с первой мишенью (VH-Ckappa), связанной через пептидный линкер (черная линия) со спейсерным доменом, связанным через пептидный линкер (черная линия) со второй половиной перекрестного Fab, способного специфически связываться с первой мишенью (VL-CH1), которая дополнительно связана со второй половиной Fab, способной специфически связываться со второй мишенью (Fab тяжелой цепи) (в направлении от N к С). Два спейсерных домена отличаются друг от друга и содержат модификации, способствующие ассоциации первого и второго гибридизируемых полипептидов. Примером конторстела такого типа является CD134-0094 (см. пример 2.2). На фиг. 1В конторстело состоит из первого гибридизированного полипептида, содержащего первую половину Fab, способную к специфическому связыванию с первой мишенью (VL-Ckappa), связанного через пептидный линкер (черная линия) со спейсерным доменом, связанным через пептидный линкер (черная линия) со второй половиной Fab, способной специфически связываться с первой мишенью (VH-CH1), которая дополнительно связана с первой половиной перекрестного Fab, способной специфически связываться со второй мишенью (VH-Ckappa) (в направлении от N к С), и второго гибридизированного полипептида, содержащего первую половину Fab, способную к специфическому связыванию с первой мишенью (VL-Ckappa), связанную через пептидный линкер (черная линия) со спейсерным доменом, связанным через пептидный линкер (черная линия) со второй половиной Fab, способной специфически связываться с первой мишенью (VH-СН1), которая дополнительно связана со второй половиной перекрестного Fab, способной специфически связываться со второй мишенью (VL-CH1) (в направлении от N к С). Два спейсерных домена отличаются друг от друга и содержат модификации, способствующие ассоциации первого и второго гибридизируемых полипептидов. Примером конторстела такого типа является Р1АЕ0821 (см. пример 2.7). На фиг. 1Г конторстело состоит из первого гибридизированного полипептида, содержащего первую половину перекрестного Fab, способную специфически связываться со второй мишенью (VH-Ckappa), связанную через пептидный линкер (черная линия) с первой половиной перекрестного Fab, способного специфически связываться с первой мишенью (VH- Скарра), которая дополнительно связана через пептидный линкер (черная линия) со спейсерным доменом, связанным через пептидный линкер (черная линия) со второй половиной перекрестного Fab, способной специфически связываться с первой мишенью (VH-CH1) (в направлении от N к С), из второго гибридизированного полипептида, содержащего первую половину перекрестного Fab, способную специфически связываться со второй мишенью (VH-CH1), связанной через пептидный линкер (черная линия) с первой половиной перекрестного Fab, способной специфически связываться с первой мишенью (VH-Ckappa), которая дополнительно связана через пептидный линкер (черная линия) со спейсерным доменом, связанным через пептидный линкер (черная линия) со второй половиной перекрестного Fab, способной специфически связываться с первой мишенью (VL-CH1) (в направлении от N к С). Два спейсерных домена отличаются друг от друга и содержат модификации, способствующие ассоциации первого и второго гибридизируемых полипептидов. Примером конторстела такого типа является Р1АЕ2735 (см. пример 2.14).
Фиг. 1Д представляет схему собранной структуры конторстела CD134-0093 (пример 2.1).
Фиг. 1Е представляет схему собранной структуры конторстела CD134-0094 (пример 2.2).
Фиг. 1Ж представляет схему собранной структуры конторстел Р1АЕ0085 и Р1АЕ0086 (примеры 2.3 и 2.4). Антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью (перекрестный Fab), связан через удлиненный пептидный линкер, который изменяет геометрию молекулы.
Фиг. 13 представляет схему собранной структуры конторстела Р1АЕ0087 (пример 2.5) и конторстела Р1АЕ0839 (пример 2.6). В этом случае антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью, является Fab, и оба антигенсвязывающих домена, способных специфически связываться с первой мишенью, являются перекрестными Fab.
Фиг. 1И представляет схему собранной структуры конторстела Р1АЕ0821 (конторстело 11, пример 2.7).
Фиг. 1К представляет схему собранной структуры конторстела Р1АЕ1122 (конторстело 1, пример 2.8).
Фиг. 1Л представляет схему собранной структуры конторстела Р1АЕ1887 (конторстело 3, пример 2.10).
Фиг. 1М представляет схему собранной структуры конторстела Р1АЕ2254 (конторстело 5, пример 2.12). В СН и Ckappa, гибридизированных с VL и VH в ОХ40, соответственно, аминокислотные мутации (так называемые заряженные остатки) интродуцируют для предупреждения образования белков Бен-Джонса и для дополнительного облегчения и корректировки спаривания.
Фиг. 1Н представляет схему собранной структуры конторстела Р1АЕ2340 (конторстело 6, пример 2.13). В этом примере молекула состоит из двух гибридизированных белков и легкой цепи.
Фиг. 1O представляет схему собранной структуры конторстела Р1АЕ2735 (конторстело 8, пример 2.14).
Фиг. 1П представляет схему биспецифического антитела 2+1 ОХ40 х FAP с бивалентным связыванием с ОХ40 и моновалентным связыванием с FAP (молекула положительного контроля). Была получена молекула отрицательного контроля с такой же структурой, в которой FAP-связывающий домен заменен зародышевой линией DP47 (антитело 2+1 ОХ40 х DP47).
Фиг. 1Р представляет схему биспецифического антитела 4+1 ОХ40 х FAP с тетравалентным связыванием ОХ40 и моновалентным связыванием FAP. Эти контрольные молекулы подробно описаны в примере 2.18.
Фиг. 2А, 2Б, 2В и 2Г показывают связывание биспецифических антител ОХ40 х FAP с активированными CD4+ и CD8+ Т-клетками (фиг. 2А и 2В, соответственно) и покоящимися CD4+ и CD8+ Т-клетками (фиг. 2Б и 2Г, соответственно). Тетравалентное биспецифическое антитело 4+1 ОХ40 х FAP (4В9) показало наиболее сильное связывание с CD4+ и CD8+ Т-клетками. 2+1 форматы 2+1 ОХ40 (49В4) х FAP (28Н1) и 2+1 ОХ40 (49В4) х DP47 показали промежуточное связывание. Конторстело CD134-0093 показало более сильное связывание, чем форматы 2+1, тогда как конторстело CD 134-0094 связывалось слабее. Связывание с CD4+ Т-клетками было гораздо сильнее, чем с CD8+ Т-клетками. Отрицательный контроль DP47 hu IgG1 P329G LALA не связывался с CD4+ или CD8+ Т-клетками. Ни одна из молекул не связана с покоящимися CD4+ или CD8+ Т-клетками (фиг. 2Б и 2Г).
Фиг. 2Д, 2Е, 2Ж и 2З представляют результаты, полученные с конторстелами Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087. Все 2+1 форматы ОХ40 (49В4) х FAP (28Н1), ОХ40 (49В4) х FAP (4В9) и ОХ40 (49В4) х DP47 продемонстрировали схожее хорошее связывание с активированными CD4+ и CD8+ Т-клетками (фиг. 2Д и 2Ж, соответственно), тогда как связывание с ОХ40 было частично ослаблено для молекул конторстел, особенно для конторстела Р1АЕ0087. DP47 hu IgG1 P329G LALA не связывалось с CD4+ или CD8+ Т-клетками, как ожидалось. Не обнаружено связывания с покоящимися CD4+ или CD8+ Т-клетками (фиг. 2Е и 2З, соответственно) для каких-либо исследованных молекул.
Связывание с опухолевыми клетками, экспрессирующими FAP человека, показано на фиг. 3А, 3Б, 3В и 3Г. В первом эксперименте конторстела CD134-0093 и CD134-0094 сравнивают с контрольными молекулами. Связывание с WM266-4 (FAP+ положительными) и A549NLR (FAP- отрицательными) опухолевыми клетками показано на фиг. 3А и 3Б, соответственно. Все биспецифические антитела ОХ40 х FAP связывались эффективно с клетками-мишенями, экспрессирующими FAP человека. Тетравалентное 4+1 ОХ40 х FAP (4 В9, высокое сродство с FAP) биспецифическое антитело связывается наиболее сильно с FAP+ клетками, затем конторстело CD134-0093, конторстело CD1334-0094 и 2+1 ОХ40 (49В4) х FAP (28Н1) биспецифическое антитело. Не нацеленное 2+1 ОХ40 (49 В4) х DP47 и отрицательный контроль (DP47 hu IgG1 P329G LALA) не связываются с какими-либо клетками FAP+. Фиг. 3В и 3Г демонстрируют связывание конторстел Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087 с NIH/3T3huFAP клоном 19 (FAP) (фиг. 3В) и опухолевыми клетками A549NLR (FAP-) (фиг. 3Г). Все FAP-нацеленные анти-ОХ40 антитела связываются эффективно с клетками-мишенями, экспрессирующими FAP человека. Связывание молекул конторстел несколько ослаблено по сравнению с соответствующими контролями. К тому же не нацеленное 2+1 ОХ40 (49В4) х DP47 и отрицательный контроль (DP47 hu IgG1 P329G LALA) не проявляют связывания с клетками FAP+.
На фиг. 4А, 4Б, 4В, 4Г, 4Д и 4Е показывают NFκB активацию с разными типами перекрестного связывания. Используя FAP-экспрессирующие клетки (NIH/3T3 huFAP клон 19) в качестве перекрестных связующих, FAP-нацеленная 2+1 ОХ40 (49 В4) х FAP (28Н1) конструкция индуцирует самую сильную активацию NFκB, за которой следуют конторстела CD134-0093 и CD134-0094, хотя CD134-0094 немного более сильную активацию, чем CD134-0093. Не нацеленные и, следовательно, не перекрестно сшивающие 2 не нацеленных 2+1 ОХ40 (49В4) х DP47 индуцируют слабую активацию NFκB (фиг. 4А). При использовании вторичного анти-IgG1 человека Fcγ-специфического антитела в качестве перекрестного связующего, две 2+1 конструкции ОХ40 (49В4) х FAP (28Н1) и ОХ40 (49В4) х DP47 действуют схожим образом. Конторстела CD134-0093 и CD134-0094 также действует аналогично, но на более низком уровне, чем конструкции 2+1 (фиг. 4Б). Наименьшую активацию NFκB наблюдают без применения перекрестных сшивающих агентов. 2+1 конструкции ОХ40 (49В4) х FAP (28Н1) и ОХ40 (49В4) х DP47 показывают умеренную активацию NFκB, за ними следуют еще менее активные конторстела CD134-0093 и CD134-0094. DP47 hu IgG1 P329G LALA не индуцирует какой-либо активации NFκB (фиг. 4В). Способность конторстел Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087 индуцировать активацию NFκB с разными типами перекрестного связывания показана на фиг. 4Г-4Е. В отсутствие перекрестного связывания может быть обнаружен только самый слабый сигнал при самой высокой концентрации антител (фиг. 4Г). Три молекулы конторстела индуцируют весьма схожую активацию NFκB относительно FAP-нацеленных 2+1 форматов ОХ40 (49В4) х FAP (28Н1) и ОХ40 (49В4) х FAP (4В9) при перекрестном сшивании FAP-экспрессирующими клетками человека (фиг. 4Д). При перекрестном сшивании вторичного антитела все три молекулы конторстел проявляют несколько заниженную активацию NFκB по сравнению с 2+1 контрольными молекулами (фиг. 4Е). DP47 hu IgG1 P329G LALA не индуцирует какой-либо активации NFκB.
ОХ40-опосредованная костимуляция субоптимально TCR-стимулированных покоящихся МКПК человека и гипер-перекрестное сшивание FAP на поверхности клеток показаны на фиг. 5А, 5Б, 5В, 5Г, 5Д и 5Е. Фиг. 5А и 5Б показывают FSC-A ("area" of a Forward Side Scatter область прямого светорассеяния; (FSC) импульс), соответственно, размер CD4 и CD8 Т-клеток, соответственно, после субоптимальной CD3 стимуляции измеренный как интенсивность света, рассеянного под малыми углами, с помощью анализа FACS. Нацеленная на FAP 2+1 конструкция ОХ40 (49В4) х FAP (28Н1) показывает промежуточное повышение размера, тогда как конторстело CD134-0093 показывает более сильное повышение. Не нацеленные 2+1 ОХ40 (49В4) х DP47, конторстело CD134-0094 и отрицательный контроль (DP47 hu IgG1 P329G LALA) не меняют размер ни CD4, ни CD8 Т-клеток. Фиг. 5В и 5Г показывают активацию CD4 и CD8 Т-клеток, используя поверхностный маркер CD25. В отношении CD4 Т-клеток (фиг. 5В) конторстела CD134-0093 и CD134-0094 проявляют наивысшую активацию, за ними следует несколько слабее нацеливающееся 2+1 ОХ40 (49 В4) х FAP (28Н1). Не нацеленное ОХ40 (49 В4) х DP47 и отрицательный контроль не показывают какой-либо активации после поправки концентрации по нулевому уровню. В отношении CD8 Т-клеток (фиг. 5Г) конторстело CD134-0094 проявляет повышенную активацию по сравнению с 2+1 ОХ40 (49В4) х FAP (28Н1) и CD134-0093. Фиг. 5Д и 5Е показывают при активации пониженную регуляцию IL-7Rα (CD127). Конторстело CD134-0093 проявляет самое сильное снижение регуляции CD127 для CD4 и CD8 Т-клеток, за ним следует гацеленная конструкция 2+1 ОХ40 (49В4) х FAP (28Н1) и конторстело CD134-0094.
Фиг. 6 показывает величины нормализованной площади под кривой (area under the curve, AUC) для FSA-A и CD25 на CD4 и CD8 Т-клетках. Заштрихованные обозначения относятся к CD4 Т-клеткам, незаштрихованные к CD8 Т-клеткам. Все значения нормализованы по верхним значениям AUC для конторстела CD134-0093 (= 100%). Не нацеленная конструкция 2+1 ОХ40 (49В4) x DP47 проявляет только минимальную активацию на CD4 и CD8 Т-клетках, а FAP нацеленная конструкция 2+1 ОХ40 (49В4) х FAP (28Н1) проявляет повышенную активность (нормализованные значения между 60 и 100 относительно FSC-A и CD25 на CD4 и CD8 Т-клетках, но меньше по сравнению с CD134-0093. Для конторстела CD134-0094 только значения среднекратного прироста (mean fold increase, MFI) CD25 AUC для CD4 и CD8 Т-клеток показывают сходную активацию по сравнению с CD134-0093.
Влияние конторстел Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087 на ОХ40-опосредованную костимуляцию субоптимально запускаемых TCR МКПК и гипер-перекрестное сшивание FAP на клеточной поверхности показаны на фиг. 7А, 7Б, 7В и 7Г. Фиг. 7А и 7Б показывают FSC-A, соответственно, размер CD4 и CD8 Т-клеток после субоптимальной стимуляции CD3. Фиг. 7В и 7Г показывают активацию CD4 и CD8 Т-клеток, соответственно, с использованием экспрессии на клеточной поверхности маркера CD25. Все молекулы, нацеленные на FAP (контроля и все три молекулы конторстел), индуцируют дозозависимое увеличение зоны прямого светорассеяния и экспрессию CD25 как на CD4, так и на CD8 Т-клетках. Не нацеленная 2+1 конструкция ОХ40 (49В4) х DP47 и отрицательный контроль (DP47 hu IgG1 P329G LALA) не проявляют какой-либо активации после базовой коррекции.
Фиг. 8А, 8Б, 8В и 8Г показывают связывание различных ОХ40 х FAP конторстел с активированными CD4+ Т-клетками. Не нацеленную 2+1 конструкцию ОХ40 (49В4) х DP47 применяют в качестве положительного контроля и DP47 hu IgG1 P329G LALA в качестве отрицательного контроля. Фиг. 8А показывает связывание конторстела 1 (Р1АЕ1122), конторстела 2 (Р1АЕ1942) и конторстела 3 (Р1АЕ1887), а фиг. 8Б показывает связывание конторстела 4 (Р1АЕ1888), конторстела 5 (Р1АЕ2254) и конторстела 6 (Р1АЕ2340). Связывание конторстела 7 (Р1АЕ0086) и конторстела 8 (Р1АЕ2735) показано на фиг. 8В, и связывание конторстела 9 (Р1АЕ2743) и конторстела 10 (Р1АЕ2762) показаны на фиг. 8Г. На фиг. 9А, 9Б, 9В и 9Г, соответственно, показывают, что ни одно из исследованных конторстел не связывается с покоящимися CD4 Т-клетками. На фигурах 10А, 10Б, 10В и 10Г показывают связывание тех же ОХ40 х FAP конторстел с активированными CD8+ Т-клетками. Не нацеленную 2+1 конструкцию ОХ40 (49В4) х DP47 применяют в качестве положительного контроля и DP47 hu IgG1 P329G LALA в качестве отрицательного контроля. На фиг. фиг. 11А, 11Б, 11В и 11Г, соответственно, показывают, что ни одно из исследованных конторстел не связывается с покоящимися CD8 Т-клетками. Сумма значений площадей под кривыми как показатель связывания с активированными или покоящимися CD4+ Т-клетками показана на фиг. 12А, а связывания с активированными или покоящимися CD8+ Т-клетками показана на фиг. 12Б.
Связывание с опухолевыми клетками, экспрессирующими FAP человека, конторстел с 1 по 10 показано на фиг. 13А, 13Б, 13В и 13Г и фиг. 14А, 14Б, 14В и 14Г, соответственно. Связывание с NIH/3T3-huFAP клон 19 (FAP+) опухолевыми клетками (FAP+ положительными) показано на фиг. с 12А по 12Г, соответственно. На фиг. 13А показано связывание конторстела 1 (Р1АЕ1122), конторстела 2 (Р1АЕ1942) и конторстела 3 (Р1АЕ1887) и фиг. 13Б демонстрирует связывание конторстела 4 (Р1АЕ1888), конторстела 5 (Р1АЕ2254) и конторстела 6 (Р1АЕ2340). Связывание конторстела 7 (Р1АЕ0086) и конторстела 8 (Р1АЕ2735) показано на фиг. 13В, а связывание конторстела 9 (Р1АЕ2743) и конторстела 10 (Р1АЕ2762) показано на фиг. 13Г. Все ОХ40 х FAP биспецифические антитела эффективно связывают целевые клетки, экспрессирующие FAP человека. Контортело 8 (фиг. 12В) связывается наиболее крепко с FAP+ клетками, затем следует конторстело 10 (фиг. 12Г) и контортело 6 (фиг. 12Б), намного сильнее, чем 2+1 биспецифическое антитело ОХ40 (49В4) х FAP (4В9). Не нацеленную 2+1 конструкция ОХ40 (49В4) х DP47 и отрицательный контроль (DP47 hu IgG1 P329G LALA) не связываются с какими-либо FAP+ клетками. Соответствующее связывание контортел с 1 по 10 с A549NLR (FAP" отрицательными) опухолевыми клетками показано на фигурах с 14А по 14Г. Ни одна из FAP-нацеленных молекул (конторстел с 1 по 10 или 2+1 биспецифическое антитело ОХ40 (49В4) х FAP (4В9)) не может связываться с FAP- целевыми клетками.
На фигурах 15А, 15Б, 15В и 15Г показана NFκB активация конторстел с 1 по 11 с перекрестным связыванием. При использовании FAP-экспрессирующих клеток (NIH/3T3 huFAP клон 19) в качестве перекрестных связующих, активация NFκB сопоставима со всеми конторстелами, причем с контортелом 11 (Р1АЕ0821) активация самая слабая. Не нацеленное и следовательно не перекрестно связанное 2 не нацеленное 2+1 ОХ40 (49В4) х DP47 антитело (7718) индуцируют слабую NFκB активацию, тогда как биспецифическое ОХ40 (49В4) х FAP (4В9) антитело (7719) вызывает наивысшую активацию NFκB. DP47 hu IgG1 P329G LALA (8105) не индуцирует какой-либо активации NFκB. NFκB активация без перекрестного связывания FAP показана на фигурах 16А, 16Б, 16В и 16Г. В отсутствие перекрестного связывания только слабый сигнал может быть обнаружен при повышенных концентрациях антитела. Не нацеленное 2+1 ОХ40 (49 В4) х DP47 антитело (7718) показывает умеренную активацию NFκB, за ним следует конторстело 7 (фиг. 16В). Другие конторстела индуцируют минорные или нулевые уровни. Отрицательный контроль (8105) не индуцирует какой-либо активации NFκB. Значения нормализованных площадей под кривой для активации NFκB в клетках HeLa с/без перекрестного сшивания с FAP+ клетками суммированы на фиг. 17. Отрицательный контроль (8105) не индуцирует какой-либо NFκB активации в обоих случаях, хотя не нацеленное 2+1 ОХ40 (49В4) х DP47 антитело (7718) показывает только минимальную активацию в обоих случаях. Биспецифическое ОХ40 (49В4) х FAP (4В9) антитело (7719) проявляет повышенные уровни активации, за которым следуют конторстела 8, 9 и 10. Пониженные уровни наблюдают для конторстела 11 (Р1АЕ0821).
ОХ40 опосредует ко-стимуляцию суб-оптимально TCR-стимулированных покоящихся МКПК человека и гипер-перекрестное сшивание поверхностным FAP контортел 7, 8, 9 и 10, что показано на фиг. 18А, 18Б, 18В и 18Г. Фигуры 18А и 18Б показывают активацию CD4 Т-клеток, а фигуры 18В и 18Г показывают активацию CD8 Т-клеток, используя поверхностный маркер CD25 после субоптимальной CD3 стимуляции. Для CD4 Т-клеток (фигуры 18А и 18Б) биспецифическое ОХ40 (49В4) х FAP (4В9) антитело (7719) проявляет наивысшую активацию, за ним следует конторстело 8 с минимально меньшей активностью. 8. Не нацеленные 2+1 ОХ40 (49В4) х DP47 антитела (7718) и отрицательный контроль (8105) не показывают какой-либо активации после поправки концентрации по нулевому уровню. Для CD8 Т-клеток (фигуры 18В и 18Г), конторстела 7, 8 и 9, а также биспецифическое ОХ40 (49В4) х FAP (4В9) антитело (7719) показывает более сильную активацию по сравнению с конторстелом 10. Фигуры 19А, 19Б, 19В и 19Г показывают FSC-A, соответственно, размер CD4 (фигуры 19А и 19Б) и CD8 Т-клеток (фигуры 19В и 19Г) после субоптимальной CD3 стимуляции. Все конторстела показывают сопоставимые средние показатели увеличения. Добавление не нацеленной 2+1 анти-ОХ40 молекулы (7718) и отрицательного контроля (8105) не изменяет размера ни CD4, ни CD8 Т-клеток. Фигуры 20А, 20Б, 20В и 20Г показывают уровни eFluor 670, соответственно, пролиферацию CD4 (фигуры 20А и 20Б) и CD8 Т-клеток (фигуры 20В и 20Г) после субоптимальной CD3 стимуляции. В CD4 субпопуляциях конторстела 8 и 10 показывают значительное снижение уровней eFluor 670, что свидетельствует об усиленной пролиферации. В CD8 субпопуляциях конторстела 7 и 9 показывает большее снижение уровней eFluor 670. Не нацеленная 2+1 анти-ОХ40 молекула (7718) и отрицательный контроль (8105) показывают лишь незначительное снижение во всех субпопуляциях. Фигуры 21А, 21Б, 21В и 21Г показывают после активации подавление IL-7Rα (CD127). Биспецифическое ОХ40 (49В4) х FAP (4В9) антитело (7719) показывает самое сильное подавление CD4 и CD8 Т-клеток, за ним следует конторстело 10. Большее снижение регуляции CD127 наблюдают в субпопуляциях клеток CD4, а не CD8. Величины нормализованных областей под кривой для CD25 (фиг. 22А), FSC-A (фиг. 22Б), eFluor 670 (фиг. 22В) и CD127 (фиг. 22Г) на CD4 и CD8 Т-клетках суммированы на фигурах 22А, 22Б, 22В и 22Г. Не нацеленная 2+1 анти-ОХ40 молекула (7718) проявляет лишь минимальную активацию на CD4 и CD8 Т-клетках, тогда как биспецифическое ОХ40 (49В4) х FAP (4В9) антитело (7719) показывает повышенную активность, касающуюся eFluor 679 и CD127 на CD4 и CD8 Т-клетках по сравнению с конторстелами. CD25 и FSC-A указывают на одинаковые уровни активации как в молекуле 7719, так и в конторстелах в субпопуляциях CD4 и CD8.
Фигуры 23А и 23Б показывают NFκB-опосредованную активность по экспрессии люциферазы в 4-1ВВ экспрессирующих репортерных клетках линии Jurkat-hu4-1BB-NFκB-luc2. Для определения функциональности инкубируют молекулы 2+1 анти-4-1ВВ (20Н4.9) х анти-РАР(4В9) конторстел в сравнении с 2+1 анти-4-1ВВ (20Н4.9) х анти-FAP (4В9) антигенсвязывающими молекулами и с контролями. Концентрации антигенсвязывающих молекул или их контролей для сравнения наносят против единиц высвобожденного света (released light units, RLU), измеренных через 6 ч инкубации. Для всех показателей вносят поправки концентраций по нулевому уровню (например, без добавления антител). На фиг. 23А показана FAP-мишень-независимая 4-1ВВ активация, посредством которой 4-1ВВ-связывание индуцирует NFκB-контролируемую экспрессию люциферазы в линии репортерных клеток без какого-либо FAP-опосредованного перекрестного связывания. На фигуре 23Б добавлена FAP-экспрессирующая на высоком уровне линия клеток NIH/3T3-huFAP клон 19 (линия клеток FAP-человека-трансгенных фибробластов мыши). FAP-экспрессирующие опухолевые клетки приводят к перекрестному скрещиванию биспецифических 4-1ВВ (20Н4.9) х FAP (4В9) антигенсвязывающих молекул и сильно повышают их потенциал по индукции NFκB-индуцированной /люциферазной активации в 4-1 ВВ-экспрессирующей репортерной клеточной линии. Биспецифические 2+1 анти-4-1ВВ (20Н4.9) х анти-FAP(4B9) антигенсвязывающие молекулы (черные звездочка и линия) показывают немного лучшую активацию (пониженные значения ЕС50) по сравнению с конторстелами. Тем не менее, активация, вызванная контортелами, намного выше, чем активация, проявляемая не нацеленными 4-1 ВВ антителами.
На фиг. 24А, 24Б, 24В, 24Г, 24Д, 24Е, 24Ж и 243 показывают активацию in vitro В-клеток человека бивалентными анти-CD40 человека х FAP конторстелами в присутствии гранул Dynabeads® с покрытием FAP (фиг. 24А, 24В, 24Д и 24Ж) или без покрытия (фиг. 24Б, 24Г, 24Е и 24 и 243) после двух суток инкубирования. По сравнению с FAP-независимым повышением активации CD69, CD80, CD86 и HLA-DR, индуцированной перекрестно связанным SGN40, повышение активации таких маркеров активации, индуцированных FAP-зависимыми биспецифическими антигенсвязывающими молекулами в присутствии гранул, покрытых FAP, было несколько ниже. В отсутствие FAP (гранулы без покрытия) не наблюдают увеличения экспрессии CD69, CD80, CD86 или HLA-DR с нацеленными на FAP анти-CD-40 конторстелами, хотя перекрестно связанное антитело SGN40 положительного контроля индуцирует повышенную регуляцию таких маркеров активации. Показан процент CD69, CD80, CD86 или HLA-DR-положительных живых В-клеток после двухдневной инкубации с указанными оттитрованными конторстелами или контрольным антителом. XL обозначает перекрестное связывание с F (ab')2 фрагментом козы анти-IgG FGγ-фрагмента человека. На оси абсцисс показана концентрация конструкций конторстел или контрольного антитела.
Подробное описание изобретения
Определения
Если не указано иное, технические и научные термины, используемые в настоящем изобретении, имеют то же значение, которое обычно используют в области техники, к которой относится настоящее изобретение. Для целей понимания данного описания будут применяться следующие определения, и при необходимости термины, используемые в единственном числе, будут также подразумевать множественное число и наоборот.
В контексте настоящего изобретения понятие «антигенсвязывающая молекула» относится к в самом широком смысле к молекуле, которая специфически связывает антигенную детерминанту. Примерами антигенсвязывающих молекул являются антитела, фрагменты антител и каркасные антигенсвязывающие белки.
Понятие «антигенсвязывающий домен» относится к части антигенсвязывающей молекулы, которая специфически связывается с антигенной детерминантой. В одном из вариантов осуществления настоящего изобретения антигенсвязывающий домен способен активировать передачу сигнала через соответствующий антиген клетки-мишени. Антигенсвязывающие домены включают область или фрагмент антитела, который специфически связывается и является комплементарным части антигена или целому антигену. Кроме того, антигенсвязывающие домены содержат каркасные антигенсвязывающие белки, как дополнительно определено в настоящем изобретении, например, связывающие домены, которые основаны на сконструированных повторяющихся белках или сконструированных повторяющихся доменах (см., например, WO 2002/020565). В частности, антигенсвязывающий домен состоит из первой части и второй части, где первая часть содержит вариабельную область легкой цепи антитела (VL), а вторая часть содержит вариабельную область тяжелой цепи антитела (VH), или наоборот.
Понятие «антитело» в описании настоящего изобретения используют в самом широком смысле, и оно охватывает различные структуры антител, включая, но ими не ограничиваясь, моноклональные антитела, поликлональные антитела, моноспецифические и мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они проявляют требуемую антигенсвязывающая активность.
Понятие «моноклональное антитело» в описании настоящего изобретения относится к антителу, полученному из популяции по существу гомогенных антител, то есть отдельные антитела, составляющие популяцию, являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантов антител, например, содержащих природные мутации или возникающие во время получения препарата моноклонального антитела, такие варианты обычно содержатся в популяции в небольших количествах. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело препарата моноклональных антител направлено против одной детерминанты на антигене.
Понятие «моноспецифическое» антитело в контексте настоящего изобретения обозначает антитело, которое имеет один или несколько сайтов связывания, каждый из которых связывается с одним и тем же эпитопом одного и того же антигена. Понятие «биспецифический» означает, что антигенсвязывающая молекула способна специфически связываться, по меньшей мере, с двумя разными антигенными детерминантами. Биспецифическая антигенсвязывающая молекула содержит, по меньшей мере, два антигенсвязывающих сайта, каждый из которых специфичен для определенного антигенного детерминанта. В некоторых вариантах осуществления настоящего изобретения биспецифическая антигенсвязывающая молекула способна одновременно связывать два антигенных детерминанта, в частности два антигенных детерминанта, экспрессированных на двух разных клетках. Например, антигенсвязывающие молекулы по настоящему изобретению являются биспецифическими, содержащими антигенсвязывающий домен, способный специфически связываться с первой мишенью, и антигенсвязывающий домен, способный специфически связываться со второй мишенью. В одном из вариантов осуществления настоящего изобретения антитело по настоящему изобретению содержит антигенсвязывающий домен, способный специфически связываться с ОХ40, и антигенсвязывающий домен, способный специфически связываться с FAP. В другом варианте осуществления настоящего изобретения антитело по настоящему изобретению содержит антигенсвязывающий домен, способный специфически связываться с 4-1ВВ, и антигенсвязывающий домен, способный специфически связываться с FAP. В еще одном варианте осуществления настоящего изобретения антитело по настоящему изобретению содержит антигенсвязывающий домен, способный специфически связываться с CD40, и антигенсвязывающий домен, способный специфически связываться с FAP.
Понятие «валентность», используемое в настоящем изобретении, означает наличие определенного числа сайтов связывания в антигенсвязывающей молекуле. Таким образом, понятия «двухвалентная», «четырехвалентная» и «шестивалентная» означают наличие двух антигенсвязывающих сайтов, четырех антигенсвязывающих сайтов, шести антигенсвязывающих сайтов, соответственно, в молекуле. Валентность антигенсвязывающей молекулы также может быть выражена по отношению к числу сайтов связывания для определенного антигенного детерминанта. Например, биспецифические антитела по настоящему изобретению являются двухвалентными по отношению к первой мишени и одновалентными по отношению ко второй мишени.
Понятия «антитело полной длины», «интактное антитело» и «цельное антитело» в настоящем изобретении применяют взаимозаменяемо для обозначения антитела, имеющего структуру, которая весьма сходна со структурой нативного антитела. «Нативные антитела» относятся к природным молекулам иммуноглобулина разной структуры. Например, нативными антителами класса IgG являются гетеротетрамерные гликопротеины размером, примерно, 150000 Да, состоящие из двух легких цепей и двух тяжелых цепей, которые связаны дисульфидной связью. От N- до С-конца каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным доменом тяжелой цепи, за которой следуют три константных домена (CH1, СН2 и СН3), также называемые константной областью тяжелой цепи. Сходным образом, от N- до С-конца каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным доменом легкой цепи, за которой следует константный домен легкой цепи (CL), также называемый константной областью легкой цепью. Тяжелая цепь антитела может быть отнесена к одному из пяти типов, называемых α (IgA), δ (IgD), ε (IgE), γ (IgG) или μ (IgM), некоторые из которых могут быть дополнительно разделены на подтипы, например γ1 (IgG1), γ2 (IgG2), γ3 (IgG3), γ4 (IgG4), α1 (IgA1) и α2 (IgA2). Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основе аминокислотной последовательности ее константного домена.
Понятие «фрагмент антитела» относится к молекуле, отличающейся от интактного антитела, которая содержит часть интактного антитела, связывающуюся с антигеном, с которым связывается интактное антитело. К примерам фрагментов антител относятся, но ими перечень не ограничивается, Fv, Fab, Fab', Fab'-SH, F(ab')2; диатела, триатела, тетратела, кросс-Fab-фрагменты; линейные антитела; молекулы одноцепочечных антител (например, scFv); и однодоменные антитела. См. обзор по некоторым фрагментам антител в публикации Hudson с соавт., Nat Med 9, 2003, 129-134. По фрагментам scFv см. обзор  в кн. «The Pharmacology of Monoclonal Antibodies» под ред. Rosenburg и Moore, изд. Springer-Verlag, том 113, 1994, 269-315; также см. WO 93/16185 и US 5571894 и US 5587458. Обсуждение фрагментов Fab и F(ab')2, включающих остатки эпитопа связывания рецептора реутилизации и обладающих повышенной полужизнью in vivo, см. в US 5869046. Диатела и фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть бивалентными или биспецифичными, описаны, например, в ЕР 404097, WO 1993/01161, Hudson с соавт., Nat Med 9, 2003, 129-134, и Hollinger с соавт., Proc Natl Acad Sci USA 90, 1993, 6444-6448. Триатела и тетратела также описаны в публикации Hudson с соавт., Nat Med 9, 2003, 129-134. Однодоменные антитела являются фрагментами антител, включающими целиком или частично вариабельный домен тяжелой цепи, или целиком или частично вариабельный домен легкой цепи антитела. В некоторых вариантах осуществления настоящего изобретения однодоменным антителом является однодоменное антитело человека (Domantis, Inc., Уолтем, Массачусетс; см., например, US 6248516 В1). Фрагменты антитела могут быть получены разными методами, в том числе, например, ограничением протеолитического расщепления интактного антитела, а также выработкой с применением рекомбинантных клеток-хозяев (например, Е. coli или фагов), согласно описанному в настоящем изобретении.
в кн. «The Pharmacology of Monoclonal Antibodies» под ред. Rosenburg и Moore, изд. Springer-Verlag, том 113, 1994, 269-315; также см. WO 93/16185 и US 5571894 и US 5587458. Обсуждение фрагментов Fab и F(ab')2, включающих остатки эпитопа связывания рецептора реутилизации и обладающих повышенной полужизнью in vivo, см. в US 5869046. Диатела и фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть бивалентными или биспецифичными, описаны, например, в ЕР 404097, WO 1993/01161, Hudson с соавт., Nat Med 9, 2003, 129-134, и Hollinger с соавт., Proc Natl Acad Sci USA 90, 1993, 6444-6448. Триатела и тетратела также описаны в публикации Hudson с соавт., Nat Med 9, 2003, 129-134. Однодоменные антитела являются фрагментами антител, включающими целиком или частично вариабельный домен тяжелой цепи, или целиком или частично вариабельный домен легкой цепи антитела. В некоторых вариантах осуществления настоящего изобретения однодоменным антителом является однодоменное антитело человека (Domantis, Inc., Уолтем, Массачусетс; см., например, US 6248516 В1). Фрагменты антитела могут быть получены разными методами, в том числе, например, ограничением протеолитического расщепления интактного антитела, а также выработкой с применением рекомбинантных клеток-хозяев (например, Е. coli или фагов), согласно описанному в настоящем изобретении.
Расщепление папаином интактных антител приводит к формированию двух идентичных антигенсвязывающих фрагментов, называемых «Fab-фрагментами», каждый из которых содержит вариабельные домены тяжелой и легкой цепей, а также константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. В настоящем изобретении понятие «Fab-фрагмент» относится к фрагменту антитела, содержащему фрагмент легкой цепи, содержащий VL и константный домен легкой цепи (CL), а также VH и первый константный домен (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на С-конце домена СН1 тяжелой цепи, включая один или несколько цистеинов из шарнирной области антитела. Fab'-SH-фрагменты относятся к Fab'-фрагментам, в которых остаток (остатки) цистеина константного домена несут свободную тиольную группу. В результате обработки пепсином образуется фрагмент F(ab')2, который содержит два антигенсвязывающих сайта (два фрагмента Fab) и часть области Fc. Согласно настоящему изобретению, понятие «Fab-фрагмент» также включает «кросс-Fab-фрагменты» или «перекрестные Fab-фрагменты», что подробно представлено ниже.
Понятие «Kpocc-Fab-фрагмент», или «xFab-фрагмент», или «перекрестный Fab-фрагмент» относится к фрагменту Fab, в котором заменены либо вариабельные области, либо константные области тяжелой и легкой цепи. Возможны две разные композиции цепей молекулы кросс-Fab, которые входят в состав биспецифических антител по настоящему изобретению. С одной стороны, обмениваются вариабельные области тяжелой и легкой цепей Fab, т.е. перекрестная молекула Fab содержит пептидную цепь, состоящую из вариабельной области легкой цепи (VL) и константной области тяжелой цепи (СН1), и пептидную цепь, состоящую из вариабельной области тяжелой цепи (VH) и константной области легкой цепи (CL). Такая перекрестная молекула Fab также называется кросс-Fab (VLVH). С другой стороны, когда константные области тяжелой и легкой цепи Fab обмениваются, перекрестная молекула Fab содержит пептидную цепь, состоящую из вариабельной области тяжелой цепи (VH) и константной области легкой цепи (CL), и пептидную цепь, состоящую из вариабельной области легкой цепи (VL) и константной области тяжелой цепи (СН1). Такая перекрестная молекула Fab также называется CrossFab (CLCH1).
«Одноцепочечный Fab-фрагмент» или «scFab (single chain Fab fragment)» является полипептидом, состоящим из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 антитела (СН1), вариабельного домена легкой цепи антитела (VL), константного домена легкой цепи антитела (CL) и линкера, где указанные домены антитела и указанный линкер расположены одним из следующих образов в направлении от N-конца к С-концу: a) VH-CH1-линкер-VL-CL, б) VL-CL-линкер -VH-CH1, в) VH-CL-линкер-VL-СН1 или г) VL-CH1-линкер-VH-CL; причем указанный линкер является полипептидом, состоящим, по меньшей мере, из 30 аминокислот, предпочтительно из 32-50 аминокислот. Указанные одноцепочечные Fab-фрагменты стабилизируются за счет естественной дисульфидной связи между доменом CL и доменом СН1. Кроме того, эти одноцепочечные молекулы Fab могут быть дополнительно стабилизированы путем формирования межцепочечных дисульфидных связей вставки из остатков цистеина (например, в положении 44 в вариабельной тяжелой цепи и в положении 100 в вариабельной легкой цепи согласно нумерации Kabat).
«Перекрестный одноцепочечный Fab-фрагмент» или «х-scFab» является полипептидом, состоящим из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 антитела (СН1), вариабельного домена легкой цепи антитела (VL), константного домена легкой цепи (CL) антитела и линкера, где указанные домены антитела и указанный линкер расположены одним из следующих образов в направлении от N-конца к С-концу: а) VH-CL-линкер-VL-СН1 и б) VL-CH1-линкер-VH-CL; где VH и VL вместе образуют антигенсвязывающий сайт, который специфически связывается с антигеном, и где указанный линкер является полипептидом, состоящим, по меньшей мере, из 30 аминокислот. Кроме того, молекулы x-scFab могут быть дополнительно стабилизированы путем формирования межцепочечных дисульфидных связей за счет вставки из остатков цистеина (например, в положении 44 в вариабельной тяжелой цепи и в положении 100 в вариабельной легкой цепи согласно нумерации Kabat).
«Одноцепочечный вариабельный фрагмент (scFv)» является гибридизированным белком, сформированным за счет слияния вариабельных областей тяжелой (VH) и легкой (VL) цепей антитела, связанных коротким пептидным линкером из 10-25 аминокислот. Линкер обычно обогащен глицином для гибкости, а также серином или треонином для растворимости, и может либо соединять N-конец VH с С-концом VL, либо наоборот. Этот белок сохраняет специфичность исходного антитела, несмотря на удаление константных областей и введение линкера. Антитела scFv описаны, например, в публикации Houston J.S., Methods in Enzymol. 203, 1991, 46-96. Кроме того, фрагменты антител содержат одноцепочечные полипептиды, имеющие свойства VH, а именно, способность соединяться с VL, или VL, а именно, способность соединяться с VH, с функциональным антигенсвязывающим сайтом, тем самым обеспечивая антигенсвязывающую способность полноразмерных антител.
«Каркасные антигенсвязывающие белки» известны в данной области, например, фибронектин и разработанные белки с анкириновым повтором (DARPin designed ankyrin repeat protein), используют в качестве альтернативных каркасов для антигенсвязывающих доменов, см., например, публикацию Gebauer и Skerra, Curr Opin Chem Biol 13, 2009, 245-255, а также Stumpp с соавт., Drug Discovery Today 13, 2008, 695-701. В одном из вариантов осуществления настоящего изобретения каркасный антигенсвязывающий белок выбран из группы, включающей CTLA-4 (Evibody), липокалины (Anticalin), белок А-производные молекулы, например, Z-домен белка A (Affibody), А-домен (Avimer/Maxibody), трансферин сыворотки (trans-тело); разработанный белок с анкириновым повтором (DARPin), вариабельный домен легкой цепи или тяжелой цепи антитела (однодоменное антитело, single-domain antibody - sdAb), вариабельный домен тяжелой цепи антитела (нанотело, aVH), VNAR-фрагменты, фибронектин (AdNectin), лектиновый домен С-типа (тетранектин); вариабельный домен нового рецептора антигена бета-лактамазы (VNAR-фрагменты), гамма-кристаллин человека или убиквитин (молекулы Affilin); домен типа Куница ингибиторов протеазы человека, микротела, например, белки из семейства ноттина, пептидные аптамеры и фибронектин (аднектин). CTLA-4 (Cytotoxic Т Lymphocyte-associated Antigen 4 - антиген 4, ассоциированный с цитотоксическими Т-лимфоцитами) относится к семейству рецепторов CD28 и экспрессируется в основном на CD4+ Т-клетках. Его внеклеточный домен имеет подобную вариабельному домену Ig складчатость. Петли, соответствующие CDR антител, могут быть замещены гетерологичной последовательностью для придания различных связывающих свойств. Молекулы CTLA-4, сконструированные так, чтобы иметь различную специфичность связывания, также известны как эвитела (Evibodies), например, описанные в US 7166697 B1. Эвитела имеют примерно тот же размер, что и выделенная вариабельная область антитела (например, доменного антитела). Для получения дополнительной подробной информации см. публикацию Journal of Immunological Methods 248(1-2), 2001, 31-45. Липокалины составляют семейство внеклеточных белков, которые транспортируют небольшие гидрофобные молекулы, такие как стероиды, билины, ретиноиды и липиды. Они имеют жесткую бета-складчатую вторичную структуру с несколькими петлями на открытом конце конической структуры и могут быть сконструированы для связывания с различными целевыми антигенами. Антикалины состоят из 160-180 аминокислот и являются производными липокалинов. Подробнее информацию можно почерпнуть из Biochim Biophys Acta 1482, 2000, 337-350, US 7250297 B1 и US 20070224633. Аффитело - это каркас, производный от белка A Staphylococcus aureus, который может быть сконструирован для связывания с антигеном. Домен состоит из трехспирального пучка приблизительно из 58 аминокислот. Библиотеки были получены путем рандомизации поверхностных остатков. Для получения дополнительной подробной информации см. Protein Eng. Des. Sel. 17, 2004, 455-462 и ЕР 1641818 А1. Авимеры это мультидоменные белки, производные от семейства А-доменных каркасов. Нативные домены, состоящие приблизительно из 35 аминокислот, принимают определенную структуру, связанную дисульфидной связью. Разнообразие создается за счет перетасовки естественных вариаций, свойственных представителям семейства А-доменов. Для получения дополнительной подробной информации см. Nature Biotechnology 23(12), 2005, 1556 - 1561 и Expert Opinion on Investigational Drugs 16(6), 2007, 909-917. Трансферин является мономерным сывороточным транспортным гликопротеином. Трансферины могут быть сконструированы для связывания разных целевых антигенов путем инсерции пептидных последовательностей в пермиссивной петле на поверхности. К примерам сконструированных каркасов относят транс-тело. Для получения дополнительной подробной информации см. J. Biol. Chem 274, 1999, 24066-24073. Разработанные белки с анкириновым повтором (DARPin) получены из анкирина, относящегося к семейству белков, которые обеспечивают прикрепление интегральных мембранных белков к цитоскелету. Один анкириновый повтор представляет собой мотив из 33 остатков, состоящий из двух альфа-спиралей и бета-изгиба. Они могут быть сконструированы так, чтобы связывать различные целевые антигены путем рандомизации остатков в первой альфа-спирали и бета-изгибе каждого повтора. Их интерфейс связывания может быть увеличен путем увеличения числа модулей (метод созревания аффинности). Для получения дополнительной подробной информации см. J. Mol. Biol. 332, 2003, 489-503; PNAS 100(4), 2003, 1700-1705; J. Mol. Biol. 369, 2007, 1015-1028; US 20040132028 A1. Однодоменное антитело является фрагментом антитела, состоящий из одного мономерного вариабельного домена антитела. Первые единичные домены были получены из вариабельного домена тяжелой цепи антитела верблюдов (нанотела или фрагменты VHH). Кроме того, термин однодоменное антитело включает автономный вариабельный домен тяжелой цепи человека (aVH) или фрагменты VNAR, полученные из акул. Фибронектин представляет собой каркас, который может быть сконструирован для связывания с антигеном. Аднектины в своей основе состоят из природной аминокислотной последовательности 10-го домена из 15 повторяющихся звеньев человеческого фибронектина типа III (FN3). Три петли на одном конце бета-сэндвича могут быть сконструированы так, чтобы позволить аднектину специфически распознавать терапевтическую мишень, представляющую интерес. Для получения дополнительной подробной информации см. Protein Eng. Des. Sel. 18, 2005, 435- 444; US 20080139791; WO 2005056764 и US 6818418 B1. Пептидные аптамеры являются молекулами комбинаторного распознавания, которые состоят из белка с постоянным каркасом, обычно тиоредоксина (TrxA), содержащего ограниченную вариабельную пептидную петлю, инсертированную в активный сайт. Для получения дополнительной подробной информации см. Expert Opin. Biol. Ther. 5, 2005, 783-797. Микротела производны от встречающихся в природе микропротеинов размером 25-50 аминокислот, которые содержат 3-4 цистеиновых мостика. К примерам микропротеинов относят Kalata BI, конотоксин и ноттины. Микропротеины имеют петлю, которая может содержать до 25 аминокислот, в целом не затрагивая складку микропротеина. Для получения дополнительной подробной информации см. конструирование доменов ноттина в WO 2008098796.
Понятие «антигенсвязывающая молекула, которая связывается с тем же эпитопом», что и эталонная молекула, относится к антигенсвязывающей молекуле, которая блокирует связывание эталонной молекулы с ее антигеном в конкурентном анализе на 50% или более, и наоборот, эталонная молекула блокирует связывание антигенсвязывающей молекулы с ее антигеном в конкурентном анализе на 50% или более.
В контексте настоящего изобретения понятие «антигенный детерминант» является синонимом понятий «антиген» и «эпитоп» и относится к сайту (например, непрерывному участку аминокислот или конформационной конфигурации, состоящей из различных областей несмежных аминокислот) на макромолекуле полипептида, с которой связывается антигенсвязывающий фрагмент, образуя комплекс антигенсвязывающий фрагмент-антиген. Полезные антигенные детерминанты могут быть обнаружены, например, на поверхностях опухолевых клеток, на поверхностях инфицированных вирусом клеток, на поверхностях других клеток с признаками болезни, на поверхности иммунных клеток, в свободном состоянии в сыворотке крови и/или на внеклеточном матриксе (extracellular matrix ЕСМ). В настоящем изобретении белки, используемые в качестве антигенов, могут представлять собой какую-либо нативную форму белков из какого-либо позвоночного, включая млекопитающих, например, приматов (например, людей) и грызунов (например, мышей и крыс), если не указано иное. В одном из вариантов осуществления настоящего изобретения антигеном является белок человека. Если имеют ввиду конкретный белок, понятие охватывает «полноразмерный» нативный белок, а также какую-либо форму белка, которая возникает в результате процессинга в клетке. Это понятие также охватывает встречающиеся в природе варианты белка, например сплайс-варианты или аллельные варианты.
Понятие «паратоп» относится к той части молекулы данного антитела, которая необходима для специфического связывания мишени и сайта связывания. Паратоп может быть непрерывным, то есть образованным соседними аминокислотными остатками, присутствующими в сайте связывания, или прерывистым, то есть сформированным аминокислотными остатками, которые находятся в разных положениях в первичной последовательности аминокислотных остатков, например в аминокислотной последовательности CDR, но в непосредственной близости в трехмерной структуре, которую принимает сайт связывания.
Понятие «специфическое связывание» означает, что связывание является селективным для антигена и его следует отличать от нежелательных или неспецифических взаимодействий. Способность антигенсвязывающей молекулы связываться с определенным антигеном может быть измерена либо с помощью иммуноферментного анализа (enzyme-linked immunosorbent assay, ELISA), либо с помощью других методов, известных специалистам в данной области, например, методом поверхностного плазмонного резонанса (Surface Plasmon Resonance, SPR; измерение на приборе BIAcore) (Liljeblad с соавт., Glyco J 17, 2000, 323-329), а также традиционными методами определения связывания (Heeley, Endocr Res 28, 2002, 217-229). В одном из вариантов осуществления настоящего изобретения степень связывания антигенсвязывающей молекулы с неродственным белком составляет менее чем примерно 10% от связывания антигенсвязывающей молекулы с антигеном, что может быть измерено, например, с помощью SPR. В различных вариантах осуществления настоящего изобретения константа диссоциации (Kd) молекулы, которая связывается с антигеном, составляет ≤ 1 мкМ, ≤ 100 нМ, ≤ 10 нМ, ≤ 1 нМ, ≤ 0.1 нМ, ≤ 0.01 нМ или ≤ 0.001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
«Аффинность» или «связывающее сродство» относится к совокупной силе нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его связывающим партнером (например, антигеном). Если не указано иначе, в настоящем изобретении «связывающее сродство» относится к внутренней аффинности, которая отражает взаимодействие 1:1 между составляющими связывающейся пары (например, между антителом и антигеном). Аффинность молекулы X с ее партнером Y обычно может быть представлено константой диссоциации (Kd), которая является соотношением констант диссоциации и ассоциации (соответственно, koff и kon). Таким образом, эквивалентные аффинности могут содержать разные константы до тех пор, пока соотношение констант остается неизменным. Сродство может быть измерено общепринятыми методами, известными в данной области, в том числе описанными в настоящем изобретении. Одним из признанных методов определения аффинности является поверхностный плазмонный резонанс (SPR).
Антитело с «созревшей аффинностью» относится к антителу с одним или несколькими изменениями в одной или нескольких гипервариабельных областях (hypervariable region, HVR) по сравнению с исходным антителом, которое не обладает такими изменениями, такие изменения приводят к улучшению аффинности антитела по отношению к антигену.
В настоящем изобретении понятие «опухолеассоциированный антиген» относится к антигенному детерминанту на поверхности клетки-мишени, которая является клеткой опухоли, например, раковой клеткой, клеткой стромы опухоли или В-клеткой. В некоторых вариантах осуществления настоящего изобретения опухолеассоциированный антиген является белком активации фибробластов (Fibroblast Activation Protein, FAP).
Понятие «способная к специфическому связыванию с белком активации фибробластов (FAP)» относится к антигенсвязывающей молекуле, которая способна связывать FAP с достаточной степенью аффинности таким образом, что антигенсвязывающая молекула может быть применима в качестве диагностического и/или терапевтического агента при нацеливании на FAP. К антигенсвязывающий молекулам относят, но ими перечень не ограничивается, антитела, молекулы Fab, перекрестные молекулы Fab, одноцепочечные молекулы Fab, молекулы Fv, молекулы scFv, однодоменные антитела, а также VH и каркасный антигенсвязывающий белок. В одном из вариантов осуществления настоящего изобретения степень связывания анти-FAP антигенсвязывающей молекулы с неродственным, не являющимся FAP белком, составляет примерно менее 10% от связывания антигенсвязывающей молекулы с FAP, согласно измерению, например, методом поверхностного плазмонного резонанса (SPR). В частности, антигенсвязывающая молекула, которая способна специфически связываться с FAP, имеет константу диссоциации (Kd) ≤ 1 мкМ, ≤ 100 нМ, ≤ 10 нМ, ≤ 1 нМ, ≤ 0,1 нМ, ≤ 0,01 нМ или ≤ 0,001. нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В некоторых вариантах осуществления настоящего изобретения анти-FAP антигенсвязывающая молекула связывается с FAP из разных видов. В частности, анти-FAP антигенсвязывающая молекула связывается с FAP человека, макака крабоеда и мыши.
Понятие «белок активации фибробластов (Fibroblast activation protein, FAP)», также известный как пролилэндопептидаза FAP или сепраза (ЕС 3.4.21), относится к какому-либо нативному FAP из какого-либо позвоночного, включая млекопитающих, таких как приматы (например, люди), приматы, отличные от человека (например, макаки-крабоеды) и грызуны (например, мыши и крысы), если не указано иное. Понятие охватывает «полноразмерный» не подвергавшийся обработке FAP, а также какую-либо форму FAP, образующуюся в клетке. Это понятие также относится к встречающимся в природе вариантам FAP, например, сплайс-вариантам или аллельным вариантам. В одном варианте осуществления настоящего изобретения антигенсвязывающая молекула по настоящему изобретению способна специфически связываться с FAP человека, мыши и/или макаки-крабоеда. Аминокислотная последовательность FAP человека показана в UniProt (www.uniprot.org) номер доступа Q12884 (версия 149, SEQ ID NO:97) или NCBI (www.ncbi.nlm.nih.gov/) RefSeq NP_004451.2. Внеклеточный домен (ECD) FAP человека расположен от аминокислоты в положении 26 до аминокислоты в положении 761. Предпочтительно, анти-FAP-связывающая молекула по настоящему изобретению связывается с внеклеточным доменом FAP. Примеры анти-FAP-связывающих молекул описаны в WO 2012/020006 А2.
Понятие «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антигенсвязывающей молекулы с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно имеют сходные структуры, причем каждый домен содержит четыре консервативные каркасные области (framework region, FR) и три гипервариабельные области (hypervariable region, HVR). См., например, Kindt с соавт., Kuby Immunology, 6е изд., 2007, W.H. Freeman and Co., стр. 91. Одного VH или VL может быть достаточно для придания антигенсвязывающей специфичности.
Понятие «гипервариабельные области (hypervariable region, HVR)» в настоящем изобретении относится к каждой области вариабельного домена антитела, которая гипервариабельна по своей последовательности («область, определяющая комплементарность; complementarity determining region, CDR»), и/или формирует петли определенной структуры («гипервариабельные петли») и/или содержит антиген-контактирующие остатки («антигенные контакты»). Как правило, антитела содержат шесть HVR: три в VH (H1, Н2, Н3) и три в VL (L1, L2, L3). Примеры HVR в настоящем изобретении включают:
(а) гипервариабельные петли, имеющиеся в аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia и Lesk, J. Mol. Biol. 196, 1987, 901-917);
(б) CDR имеющиеся в аминокислотных остатках 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (Н2) и 95-102 (Н3) (Kabat с соавт., Sequences of Proteins of Immunological Interest, 5e изд., 1991, Public Health Service, National Institutes of Health, Бетесда, Мэриленд);
(в) антигенные контакты, имеющиеся в аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum с соавт. J. Mol. Biol. 262, 1996, 732-745); и
(г) комбинации (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Если не указано иначе, нумерацию остатков HVR (например, CDR) и других остатков в вариабельном домене (например, остатков FR) в настоящем изобретении осуществляют по Kabat с соавт. (см. ссылку выше).
Kabat с соавт. также разработали систему нумерации для последовательностей вариабельных областей, которая применима к какому-либо антителу. Специалист в данной области техники может применить систему «нумерации по Kabat» к любой последовательности вариабельной области, не полагаясь на какие-либо экспериментальные данные, достаточно самой последовательности. Применяемая в настоящем изобретении «нумарация Kabat) является системой нумерации, установленной Kabat с соавт., U.S. Dept. of Health and Human Services, "Sequence of Proteins of Immunological Interest" (1983). Если не указано иначе, ссылки на нумерацию определенных положений аминокислотных остатков в вариабельной области антитела соответствуют системе нумерации Kabat.
В настоящем изобретении понятие «зрелая аффинность» в контексте антигенсвязывающих молекул (например, антител) относится к антигенсвязывающей молекуле, которая получена из эталонной антигенсвязывающей молекулы, например, путем мутации, связывается с тем же антигеном, предпочтительно связывается с тем же эпитопом, что и контрольное антитело; имеет более высокое сродство к антигену, чем у эталонной антигенсвязывающей молекулы. Созревание аффинности обычно включает модификацию одного или нескольких аминокислотных остатков в одной или нескольких CDR антигенсвязывающей молекулы. Обычно антигенсвязывающая молекула со зрелой аффинностью связывается с тем же эпитопом, что и исходная контрольная антигенсвязывающая молекула.
«Каркас (Framework, FR)» относится к остаткам вариабельного домена, отличающимся от остатков гипервариабельной области (hypervariable region, HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно появляются в следующей последовательности в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
В контексте настоящего изобретения понятие «акцепторный каркас человека» означает каркас, включающий аминокислотную последовательность каркаса вариабельного домена легкой цепи (VL) или каркаса вариабельного домена тяжелой цепи (VH), которые являются производными каркаса иммуноглобулина человека или консенсусной последовательности каркаса человека, как показано ниже. Акцепторный каркас человека, «производный от» каркаса иммуноглобулина человека или консенсусного каркаса человека, может содержать ту же самую аминокислотную последовательность или может содержать изменения в аминокислотной последовательности. В некоторых вариантах осуществления настоящего изобретения число замещений аминокислот составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления настоящего изобретения последовательность VL акцепторного каркаса человека идентична последовательности VL каркаса иммуноглобулина человека или конценсусной последовательности каркаса человека
Понятие «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи является производной от определенного источника или вида, тогда как остальная часть тяжелой и/или легкой цепи происходит от другого источника или вида.
«Класс» антитела определяют по типу константного домена или константной области его тяжелой цепи. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть далее разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам иммуноглобулинов, называют α, δ, ε, γ и μ, соответственно.
Понятие «гуманизированное» антитело означает химерное антитело, содержащее аминокислотные остатки HVR, происходящие не от человека, и аминокислотные остатки от FR человека. В некоторых вариантах осуществления настоящего изобретения гуманизированное антитело может включать, по существу, все из, по меньшей мере, одного, обычно двух вариабельных доменов, в которых все или по существу все HVR (например, CDR) соответствуют доменам антитела, не являющегося антителом человека, и все или практически все FR соответствуют FR антитела человека. Гуманизированное антитело необязательно может содержать, по меньшей мере, часть константной области антитела, производную от антитела человека. «Гуманизированная форма» антитела, например, антитела, не являющегося антителом человека и подвергшегося гуманизации. Другими формами «гуманизированных антител», охватываемых настоящим изобретением, являются те, в которых константная область дополнительно модифицирована или изменена по сравнению с формой исходного антитела для придания свойств, описанных в настоящем изобретении, особенно касающихся связывания с C1q и/или Fc рецептором (FcR).
«Антитело человека» является антителом, которое обладает аминокислотной последовательностью, соответствующей последовательности антитела, продуцируемого человеком или клеткой человека или полученного из другого источника, не от человека, в котором используют популяционный спектр антител человека или другие последовательности, кодирующие антитела человека. Это определение антитела человека конкретно исключает гуманизированные антитела, содержащие антигенсвязывающие остатки, происходящие не от человека.
Понятие «домен СН1» означает часть полипептида тяжелой цепи антитела, которая расположена примерно от положения EU 118 до положения EU 215 (нумерация по Kabat, индекс EU). В одном из вариантов осуществления настоящего изобретения домен СН1 имеет аминокислотную последовательность ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALT SGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKKV(SEQ ID NO:94). Обычно сегмент, имеющий аминокислотную последовательность EPKSC (SEQ ID NO:99), следует для связывания домена СН1 с шарнирной областью.
Понятие «шарнирная область» означает часть полипептида тяжелой цепи антитела, которая соединяет в тяжелой цепи антитела дикого типа домен СН1 и домен СН2, т.е. примерно от положения 216 примерно до положения 230 по системе нумерации EU по Kabat, или примерно от положения 226 до примерно положения 230 по системе нумерации EU по Kabat. Шарнирные области других подклассов IgG могут быть определены путем выравнивания с остатками цистеина шарнирной области последовательности подкласса IgG1. Шарнирная область обычно является димерной молекулой, состоящей из двух полипептидов с идентичной аминокислотной последовательностью. Шарнирная область обычно содержит до 25 аминокислотных остатков и является гибкой, позволяя связанным с мишенью сайтам связывания перемещаться независимо. Шарнирная область может быть подразделена на три домена: верхний, средний и нижний шарнирный домен (см., например Roux, с соавт., J. Immunol. 161, 1998, 4083).
В одном из вариантов осуществления настоящего изобретения шарнирная область имеет аминокислотную последовательность DKTHTCPXCP (SEQ ID NO:100), в которой X означает S или Р. В другом варианте осуществления настоящего изобретения шарнирная область имеет аминокислотную последовательность НТСРХСР (SEQ ID NO:101), в которой X означает S или Р. В еще одном из вариантов осуществления настоящего изобретения шарнирная область имеет аминокислотную последовательность СРХСР (SEQ ID NO:102), в которой X означает S или Р.
Понятие «домен Fc» или «область Fc» в настоящем изобретении используют для определения С-концевой области тяжелой цепи антитела, которая содержит, по меньшей мере, часть константной области. Понятие включает области Fc нативной последовательностью и варианты областей Fc. В одном из вариантов осуществления настоящего изобретения Fc-домен тяжелой цепи IgG человека расположен от Cys226, или от Pro230, или от Ala231 до С конца тяжелой цепи. Однако С-концевой лизин (Lys447) Fc-области может быть или не быть. Область Fc IgG содержит домен СН2 IgG и домен СН3 IgG.
«Домен СН2» Fc-области IgG человека обычно располагается от аминокислотного остатка примерно в положении EU 231 до аминокислотного остатка примерно в положении EU 340 (нумерация по Kabat, индекс EU). В одном из вариантов осуществления настоящего изобретения домен СН2 имеет аминокислотную последовательность APELLGGPSV FLFPPKPKDT LMISRTPEVT CVWDVSHEDP EVKFNWYVDG VEVHNAKTKP REEQESTYRW SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTI SKAK (SEQ ID NO:95). Домен СН2 уникален тем, что он не тесно спарен с другим доменом. Скорее, две N-связанные разветвленные углеводные цепи расположены между двумя доменами СН2 интактной нативной Fc-области. Предполагают, что углевод может быть заменой парного сочленения домен-домен и стабилизировать домен СН2. Burton, Mol. Immunol. 22, 1985, 161-206. В одном варианте осуществления настоящего изобретения углеводная цепь присоединена к домену СН2. В настоящем изобретении домен СН2 может быть нативной последовательностью домена СН2 с или вариантом домена СН2.
«Домен СН3» включает участок остатков с С-конца к домену СН2 в области Fc, обозначает часть полипептида тяжелой цепи антитела, которая локализована приблизительно от положения EU 341 до положения EU 446 (нумерация по Kabat, индекс EU). В одном из вариантов осуществления настоящего изобретения домен СН3 имеет аминокислотную последовательность GQPRE PQVYT LPPSRDELTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPG (SEQ ID NO:96). Область СН3 в настоящем изобретении может быть нативной последовательностью домена СН3 или вариантом домена СН3 (например, доменом СН3 с индуцированной «выпуклостью» («выступом») в одной его цепи и соответствующей «полостью» («впадиной») в другой его цепи; см. патент US 5821333, включенный в настоящее описание в виде ссылки). Такой вариант доменов СН3 может применяться для индукции гетеродимеризации двух идентичных тяжелых цепей антитела, согласно описанному в настоящем изобретении. В одном из вариантов осуществления настоящего изобретения область Fc тяжелой цепи IgG имеет протяженность от Cys226 или Pro230 до карбокси-конца тяжелой цепи. Однако С-концевой лизин (Lys447) в области Fc может быть, или может отсутствовать. Если в настоящем изобретении не указано иначе, нумерация аминокислотных остатков в области Fc или в константной области дана в соответствии с системой нумерации EU, также называемой индексом EU и описанной в публикации Kabat с соавт. в кн.: «Sequences of Proteins of Immunological Interest», 5e изд., 1991, Public Health Service, National Institutes of Health, Бетесда, Мэриленд.
Метод «выступ-во-впадину» описан, например, в US 5731168, US 7695936, Ridgway с соавт., Prot Eng 9, 1996, 617-621, Carter, J Immunol Meth 248, 2001, 7-15. Обычно метод включает введение выпуклости («выступа») на границе раздела первого полипептида и соответствующей полости («впадины») на границе раздела второго полипептида так, что выпуклость может быть расположена в полости таким образом, чтобы способствовать образованию гетеродимера и препятствовать образованию гомодимера. Выпуклости конструируют путем замены небольших аминокислотных боковых цепей на границе первого полипептида более крупными боковыми цепями (например, тирозином или триптофаном). Компенсаторные полости идентичного или сходного размера с выпуклостями создаются на границе раздела второго полипептида путем замены больших боковых цепей аминокислот более мелкими (например, аланином или треонином). Выпуклость и полость могут быть получены путем изменения нуклеиновой кислоты, кодирующей полипептиды, например, путем сайт-специфического мутагенеза или пептидного синтеза. В одном из вариантов осуществления настоящего изобретения модификация выступа включает аминокислотное замещение T366W в одной из двух субъединиц домена Fc, и модификация впадины включает аминокислотные замещения T366S, L368A и Y407V в другой из двух субъединиц домена Fc. В еще одном из вариантов осуществления настоящего изобретения субъединица домена Fc, содержащая модификацию выступа, дополнительно включает аминокислотное замещение S354C, а субъединица домена Fc, содержащая модификацию впадины, дополнительно включает аминокислотное замещение Y349C. Интродукция этих двух остатков цистеина приводит к формированию дисульфидного мостика между двумя субъединицами области Fc, таким образом дополнительно стабилизируя димер (Carter, J Immunol Methods 248, 2001, 7-15). Нумерация соответствует индексу EU по Kabat с соавт. в кн.: «Sequences of Proteins of Immunological Interest», 5e изд., 1991, Public Health Service, National Institutes of Health, Бетесда, Мэриленд).
«Область, эквивалентную области Fc иммуноглобулина» предназначена для того, чтобы включить природные аллельные варианты области Fc иммуноглобулина, а также варианты, имеющие изменения, которые вызывают замещения, инсерции или делеции, но которые существенно не снижают способность иммуноглобулин для опосредования эффекторных функций (таких как антитело-зависимая клеточная цитотоксичность). Например, одна или несколько аминокислот могут быть делетированы с N-конца или С-конца Fc-области иммуноглобулина без существенной потери биологической функции. Такие варианты могут быть выбраны в соответствии с общими правилами, известными в данной области, таким образом, чтобы оказывать минимальное влияние на активность (см., например, Bowie J.U. с соавт., Science 247, 1990, 1306-1310).
Понятие «домен Fc дикого типа» означает аминокислотную последовательность, идентичную аминокислотной последовательности природного Fc-домена. К Fc-доменам человека дикого типа относят нативную Fc-область IgG1 человека (не аллотипы не-А и А), нативную Fc-область IgG2 человека, нативную Fc-область IgG3 человека и нативную Fc-область IgG4 человека, а также встречающиеся в природе их варианты. Fc-области дикого типа обозначены как SEQ ID NO:102 (IgG1, кавказский аллотип), SEQ ID NO:103 (IgG1, афроамериканский аллотип), SEQ ID NO:104 (IgG2), SEQ ID NO:105 (IgG3) и SEQ ID NO:106 (IgG4).
Понятие «вариант Fc-домена (человека)» означает аминокислотную последовательность, которая отличается от аминокислотной последовательности Fc-домена «дикого типа» (человека) домена из-за, по меньшей мере, одной «аминокислотной мутации». В одном из вариантов осуществления настоящего изобретения вариант Fc-области имеет, по меньшей мере, одну аминокислотную мутацию по сравнению с нативной Fc-областью, например, от одной до десяти аминокислотных мутаций, и в одном варианте осуществления настоящего изобретения примерно от одной до примерно пяти аминокислотных мутаций в нативной Fc-области. В другом варианте осуществления настоящего изобретения Fc-область (вариант) имеет, по меньшей мере, примерно 95% гомологии с Fc-областью дикого типа.
Понятие «эффекторные функции» относится к тем биологическим активностям, которые вязаны с областью Fc антитела, которые варьируют в зависимости от изотипа антитела. Примерами эффекторных функций антител являются: связывание C1q и комплемент-зависимая цитотоксичность (CDC), связывание с Fc-рецептором, антителозависимая клеточно-опосредованную цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP), секреция цитокинов, опосредованное иммунным комплексом поглощение антигена антигенпрезентирующими клетками, снижение регуляции рецепторов на поверхности клеток (например, рецептора В-клеток) и активация В-клеток.
Зависящие от рецепторов Fc эффекторные функции могут быть опосредованы взаимодействием Fc-области антитела с рецепторами Fc (FcR), которые являются специализированными рецепторами клеточной поверхности клеток кроветворения. Fc-рецепторы принадлежат к суперсемейству иммуноглобулинов и, как было показано, опосредуют как удаление покрытых антителами патогенов путем фагоцитоза иммунных комплексов, так и лизис эритроцитов и различных других клеточных мишеней (например, опухолевых клеток), покрытых соответствующим антителом, через антитело-зависимую клеточно-опосредованную цитотоксичность (antibody dependent cell mediated cytotoxicity, ADCC) (см., например Van de Winkel J.G., Anderson C.L., J. Leukoc. Biol. 49, 1991, 511-524). FcR определяются по специфичностью в отношении изотипов иммуноглобулина: Fc-рецепторы для антител IgG обозначаются как FcγR. Связывание с Fc-рецептором описано, например, в публикациях Ravetch J.V., Kinet J.P., Annu. Rev. Immunol. 9, 1991, 457-492; Capel P.J. с соавт., Immunomethods 4, 1994, 25-34; de Haas M. с соавт., J. Lab. Clin. Med. 126, 1995, 330-341; Gessner J.E. с соавт., Ann. Hematol. 76, 1998, 231-248.
Перекрестное связывание рецепторов для Fc-области антител IgG (FcγR) запускает широкий спектр эффекторных функций, включая фагоцитоз, антителозависимую клеточную цитотоксичность и высвобождение медиаторов воспаления, а также клиренс иммунных комплексов и регуляцию выработки антител. У людей описаны три класса FcγR, а именно:
- FcγRI (CD64) связывает мономерный IgG с высокой аффинностью и экспрессируется на макрофагах, моноцитах, нейтрофилах и эозинофилах. Модификация в Fc-области IgG по меньшей мере по одному из аминокислотных остатков E233-G236, Р238, D265, N297, А327 и Р329 (нумерация по Kabat, индекс EU) уменьшает связывание с FcγRI. Остатки IgG2 в положениях 233-236, замещенные на IgG1 и IgG4, снижают связывание с FcγRI в 103 раз и устраняют ответ моноцитов человека на сенсибилизированные антителами эритроциты (Armour K.L. с соавт., Eur. J. Immunol. 29, 1999, 2613 2624).
- FcγRII (CD32) связывает комплексный IgG со средним или низким сродством и широко экспрессируется. Этот рецептор можно разделить на два подтипа, FcγRIIA и FcγRIIB. FcγRIIA выявляют во многих клетках, участвующих в уничтожении (например, макрофагах, моноцитов, нейтрофилов) и, по-видимому, способен активировать процесс уничтожения. FcγRIIB, по-видимому, играет роль в процессах ингибирования и обнаруживается на В-клетках, макрофагах, тучных клетках и эозинофилах. На В-клетках он, по-видимому, подавляет дальнейшую выработку иммуноглобулина и переключение изотипа, например, на класс IgE. На макрофагах FcγRIIB действует, ингибируя фагоцитоз, что опосредуется через FcγRIIA. На эозинофилах и тучных клетках В-форма может помочь подавить активацию этих клеток через связывание IgE с соответствующим рецептором. Пониженное связывание с FcγRIIA установлено, например, для антител, содержащих IgG Fc-область с мутациями по меньшей мере одного аминокислотного остатка из E233-G236, Р238, D265, N297, А327, Р329, D270, Q295, А327, R292 и К414 (нумерация по Kabat, индекс EU).
- FcγRIII (CD16) связывает IgG со средним или низким сродством и существует в виде двух типов. FcγRIIIA обнаруживается на клетках NK, макрофагах, эозинофилах и некоторых моноцитах и Т-клетках и опосредует ADCC. FcγRIIIB на высоком уровне экспрессируется на нейтрофилах. Пониженное связывание с FcγRIIIA обнаружено, например, для антител, содержащих Fc-область IgG с мутацией по меньшей мере в одном из аминокислотных остатков E233-G236, Р238, D265, N297, А327, Р329, D270, Q295, А327, S239, Е269, Е293, Y296, V303, А327, K338 и D376 (нумерация по Kabat, индекс EU).
Картирование сайтов связывания на IgG1 человека для Fc-рецепторов, указанные выше сайты мутаций и методы измерения связывания с FcγRI и FcγRIIA описаны в публикации Shields R.L., с соавт. J. Biol. Chem. 276, 2001, 6591-6604.
Понятие «антитело-зависимая клеточная цитотоксичность (antibody-dependent cellular cytotoxicity, ADCC)» означает функцию, опосредованную связыванием Fc-рецептора, и относится к лизису клеток-мишеней антителом, согласно описанному в настоящем изобретении, в присутствии эффекторных клеток. Способность антитела индуцировать начальные стадии, опосредующие ADCC, исследуют путем измерения их связывания с Fcγ-рецепторами, экспрессируемыми клетками, например, клетками, рекомбинантно экспрессирующими FcγRI, и/или FcγRIIA, или NK (экспрессирующими по существу FcγRIIIA). В частности, измеряют связывание FcγR с клетками NK.
«Активирующий Fc-рецептор» является Fc-рецептором, который после захвата антитела Fc-областью вызывает сигнальные события, которые стимулируют несущую рецептор клетку к выполнению эффекторных функций. Активирующие Fc-рецепторы включают FcγRIIIa (CD 16а), FcγRI (CD64), FcγRIIa (CD32) и FcaRI (CD89). Примером активирующего Fc-рецептора является FcγRIIIa человека (см. UniProt no. Р08637, версия 141).
«Суперсемейство рецепторов фактора некроза опухолей» или «суперсемейство TNF (Necrosis factor receptor, TNF)» в настоящее время состоит из 27 рецепторов. Это группа цитокиновых рецепторов, характеризующаяся способностью связывать факторы некроза опухоли (TNF) через внеклеточный домен, богатый цистеином (extracellular cysteine-rich domain, CRD). Эти псевдоповторы определяются внутрицепочечными дисульфидами, образованными высококонсервативными остатками цистеина в цепях рецепторов. За исключением фактора роста нервов (nerve growth factor, NGF), все TNF гомологичны архетипическому TNF-альфа. В активной форме большинство рецепторов TNF образуют тримерные комплексы в плазматической мембране. Соответственно, большинство рецепторов TNF содержит трансмембранные домены (transmembrane domain, TMD). Некоторые из этих рецепторов также содержат внутриклеточные домены смерти (death domain, DD), которые рекрутируют взаимодействующие с каспазой белки с последующим связывание лиганда для инициации внешнего метаболического пути активации каспазы. Другие рецепторы суперсемейства TNF, у которых отсутствуют домены смерти, связывают ассоциированные с TNF-рецептором факторы и активируют внутриклеточные сигнальные метаболические пути, которые могут привести к пролиферации или дифференциации. Эти рецепторы также могут инициировать апоптоз, но через непрямые механизмы. Помимо регуляции апоптоза, некоторые рецепторы суперсемейства TNF участвуют в регуляции функций иммунных клеток, таких как гомеостаз и активация В-клеток, активация природных клеток-киллеров и костимуляция Т-клеток. Некоторые другие регулируют ответы, специфичные для клеток определенного типа, например, развитие развитие волосяного фолликула и развитие остеокластов. Представителями суперсемейства TNF-рецепторов являются: рецептор фактора некроза опухоли 1 (1А) (TNFRSF1A, CD120a), рецептор фактора некроза опухоли 2 (1В) (TNFRSF1B, CD120b), рецептор бета-лимфотоксина (LTBR, CD18), ОХ40 (TNFRSF4, CD134), CD40 (Вр50), Fas-рецептор (Аро-1, CD95, FAS), рецептор-ловушка 3 (TR6, М68, TNFRSF6B), CD27 (S152, Тр55), CD30 (Ki-1, TNFRSF8), 4-1 ВВ (CD137, TNFRSF9), DR4 (TRAILR1, Аро-2, CD261, TNFRSF10A), DR5 (TRAILR2, CD262, TNFRSF10B), рецептор-ловушка 1 (TRAILR3, CD263, TNFRSF10C), рецептор-ловушка 2 (TRAILR4, CD264, TNFRSF10D), RANK (CD265, TNFRSF11A), остеопротегерин (OCIF, TR1, TNFRSF11B), рецептор TWEAK (Fn14, CD266, TNFRSF12A), TACI (CD267, TNFRSF13B), рецептор BAFF (CD268, TNFRSF13C), медиатор проникновения вируса герпеса (HVEM, TR2, CD270, TNFRSF14), рецептор фактора роста нервов (p75NTR, CD271, NGFR), антиген созревания В-клеток (CD269, TNFRSF17), глюкокортикоид-индуцированный TNFR-связанный (GITR, AITR, CD357, TNFRSF18), TROY (TNFRSF19), DR6 (CD358, TNFRSF21), DR3 (Аро-3, TRAMP, WS-1, TNFRSF25) и рецептор эктодисплазина А2 (XEDAR, EDA2R).
Несколько представителей семейства рецепторов фактора некроза опухоли (tumor necrosis factor receptor, TNFR) функционируют после начальной активации Т-клеток, чтобы поддерживать Т-клеточные ответы. Понятие «представитель семейства костимулирующих рецепторов TNF» или «рецептор семейства костимулирующих TNF» относится к подгруппе представителей семейства рецепторов TNF, которые способны костимулировать пролиферацию и выработку цитокинов Т-клетками. Понятие относится к какому-либо природному рецептору семейства TNF от какого-либо позвоночного животного, включая млекопитающих, например, приматов (например, людей), приматов, но не от людей (например, от макак крабоедов) и грызунов (например, от мышей и крыс), если не указано иное. В некоторых вариантах осуществления настоящего изобретения представители семейства костимулирующих рецепторов TNF выбраны из группы, состоящей из ОХ40 (CD134), 4-1ВВ (CD 137), CD40, CD27, HVEM (CD270), CD30 и GITR, которые все могут иметь костимулирующее воздействие на Т-клетки. Точнее, антигенсвязывающая молекула по настоящему изобретению содержит, по меньшей мере, фрагмент, способный специфически связываться с членом семейства костимуляторных рецепторов ОХ40.
Дополнительная информация, в частности последовательности, касающаяся представителей семейства рецепторов TNF, может быть получена из общедоступных баз данных, таких как Uniprot (www.uniprot.org). Например, костимулирующие рецепторы TNF человека имеют следующие аминокислотные последовательности: ОХ40 человека (UniProt номер в каталоге Р43489, SEQ ID NO:108), 4-1 ВВ человека (UniProt номер в каталоге Q07011, SEQ ID NO:109), CD27 человека (UniProt номер в каталоге Р26842, SEQ ID NO:110), HVEM человека (UniProt номер в каталоге Q92956, SEQ ID NO:111), CD30 человека (UniProt номер в каталоге Р28908, SEQ ID NO:112), GITR человека (UniProt accession no. Q9Y5U5, SEQ ID NO:113) и CD40 человека (UniProt номер в каталоге Р25942, SEQ ID NO:115).
В настоящем изобретении понятие «ОХ40» относится к какому-либо нативному ОХ40 от какого-либо позвоночного животного, включая млекопитающих, например приматов (например, людей) и грызунов (например, мышей и крыс), если не указано иное. Понятие охватывает не подвергшийся изменениям ОХ40 «полной длины», а также какую-либо форму ОХ40, полученную в результате протекающих в клетке процессов. Понятие также охватывает встречающиеся в природе варианты ОХ40, например, сплайс-варианты или аллельные варианты. Аминокислотная последовательность примера ОХ40 человека представлена последовательностью SEQ ID NO:107 (Uniprot Р43489, версия 112), а аминокислотная последовательность примера ОХ40 мыши представлена последовательностью SEQ ID NO:113 (Uniprot Р47741, версия 101).
Среди нескольких костимулирующих молекул представитель семейства рецепторов фактора некроза опухолей (TNF) ОХ40 (CD134) играет ключевую роль в выживании и гомеостазе эффекторных Т-клеток и Т-клеток памяти (Croft М. с соавт., Immunological Reviews 229, 2009, 173-191). ОХ40 (CD134) экспрессируется в клетках нескольких типов и регулирует иммунные ответы против инфекций, опухолей и аутоантигенов, причем его экспрессия продемонстрирована на поверхности Т-клеток, NKT-клеток и NK-клеток, а также нейтрофилов (Baumann R. с соавт., Eur. J. Immunol. 34, 2004, 2268-2275), и показано, что он строго индуцибелен или сильно активирован в ответ на различные стимулирующие сигналы. Функциональная активность молекулы продемонстрирована на клетках каждого типа, экспрессирующих ОХ40, что предполагает сложную регуляцию активности, опосредованную ОХ40, in vivo. В сочетании с запуском рецептора Т-клеток, зацепление ОХ40 на Т-клетках с помощью его природного лиганда или агонистических антител приводит к синергетической активации сигнальных путей PI3K и NFkB (Song J. с соавт., J. Immunology 180(11), 2008, 7240-7248). В свою очередь, это приводит к повышенной пролиферации, увеличению цитокиновых рецепторов и выработки цитокинов, а также лучшей выживаемости активированных Т-клеток. Недавно было показано, что помимо костостимулирующей активности в эффекторных CD4+ или CD8+ Т-клетках, запуск ОХ40 ингибирует развитие и иммуносупрессорную функцию регуляторных Т-клеток. Этот эффект, вероятно, ответственен, по крайней мере частично, за усиление активности ОХ40 в отношении противоопухолевых или антимикробных иммунных ответов. Учитывая, что вовлечение ОХ40 может приводить к возрастанию популяции Т-клеток, стимуляции секреции цитокинов и поддержке Т-клеток памяти, агонисты, включая антитела и растворимые формы лиганда OX40L, были успешно использованы на различных моделях доклинических опухолей (Weinberg с соавт., J. Immunol. 164, 2000, 2160-2169).
Понятия «анти-ОХ40 антитело», «анти-ОХ40», «антитело ОХ40» и «антитело, которое специфически связывается с ОХ40» относятся к антителу, которое способно связывать ОХ40 с достаточной аффинностью таким образом, что антитело может быть использовано в качестве диагностического средства и/или терапевтического агента для нацеливания на ОХ40. В одном варианте осуществления настоящего изобретения степень связывания анти-ОХ40-антитела с неродственным белком, не являющимся ОХ40, составляет приблизительно менее чем 10% от связывания антитела с ОХ40, что измерено, например, с помощью жидкостной цитометрии (FACS). В некоторых вариантах осуществления настоящего изобретения антитело, связывающееся с ОХ40, имеет константу диссоциации (KD) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-6 М или менее, например, от 10-68 М до 10-13 М, например, от 10-8 М до 10-10 М).
Понятие «4-1 ВВ» или «CD137», используемое в настоящем изобретении, относится к какому-либо нативному 4-1 ВВ от какого-либо позвоночного животного, включая млекопитающих, например, приматов (например, людей), и грызунов (например, мышей и крыс), если не указано иное. Понятие охватывает «полной длины» не подвергшийся обработке рецептор 4-1 ВВ, а также любую форму 4-1 ВВ, полученную в результате процессинга в клетке. Понятие также охватывает встречающиеся в природе варианты 4-1 ВВ, например, сплайс-варианты или аллельные варианты. Аминокислотная последовательность примера 4-1 ВВ человека представлена последовательностью SEQ ID NO:109 (Uniprot номер в каталоге Q07011).
Понятия «анти-4-1 ВВ-антитело», «анти-4-1 ВВ», «антитело 4-1 ВВ» и «антитело, которое специфически связывается с 4-1 ВВ» относятся к антителу, которое способно связывать 4-1 ВВ с достаточной аффинностью таким образом, что антитело может быть использовано в качестве диагностического средства и/или терапевтического агента для нацеливания на 4-1 ВВ. В одном варианте осуществления настоящего изобретения степень связывания анти-4-1 ВВ-антитела с неродственным белком, не являющимся 4-1 ВВ, составляет приблизительно менее чем 10% от связывания антитела с 4-1 ВВ, что измерено, например, методом радиоиммунного анализа (RIA) или с помощью жидкостной цитометрии (FACS). В некоторых вариантах осуществления настоящего изобретения антитело, связывающееся с 4-1 ВВ, имеет константу диссоциации (KD)≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-6 М или менее, например, от 10-68 М до 10-13 М, например, от 10-8 М до 10-10 М). В частности, анти-4-1 ВВ антитела является клон 20Н4.9, описанный в US 7288638.
В настоящем изобретении понятие «CD40» относится к какому-либо нативному CD40 от какого-либо позвоночного животного, включая млекопитающих, например, приматов (например, людей), и грызунов (например, мышей и крыс), если не указано иное. Понятие охватывает «полной длины» не подвергшийся обработке рецептор CD40, а также какую-либо форму CD40, полученную в результате процессинга в клетке. Понятие также охватывает встречающиеся в природе варианты CD40, например, сплайс-варианты или аллельные варианты. Аминокислотная последовательность примера CD40 человека представлена последовательностью SEQ ID NO:115 (Uniprot номер в каталоге Р25942, version 200). Антиген CD40 представляет собой гликопротеин массой 50 кДа на поверхности клеток, который принадлежит к семейству рецепторов фактора некроза опухоли (TNF-R). (Stamenkovic с соавт., EMBO J. 8, 1989, 1403-1410). CD40 экспрессируется во многих здоровых и опухолевых типах клеток, включая В-лимфоциты, дендритные клетки, моноциты, макрофаги, эпителий тимуса, эндотелиальные клетки, фибробласты и клетки гладких мышц. CD40 экспрессируется во всех В-лимфомах и в 70% всех солидных опухолей и активируется в антигенпрезентирующих клетках (antigen presenting cell, АРС) сигналами созревания, такими как IFN-гамма и GM-CSF. Активация CD40 также индуцирует дифференциацию моноцитов в функциональные дендритные клетки (DC) и усиливает цитолитическую активность клеток NK через APC-CD40 индуцированные цитокины. Таким образом, CD40 играет существенную роль в инициации и усилении иммунных ответов, индуцируя созревание АРС, секрецию цитокинов-хэлперов, активацию костимулирующих молекул и усиление эффекторных функций.
В настоящем изобретении понятие «агонист CD40» означает какой-либо фрагмент, который агонизирует взаимодействие CD40/CD40L. Используемый в данном контексте CD40 относится предпочтительно к CD40 человека, таким образом, агонист CD40 предпочтительно является агонистом CD40 человека. Как правило, часть молекулы может быть агонистическим CD40 антителом или фрагментом антитела.
Понятия «анти-CD40 антитело», «анти-CD40», «CD40 антитело» и «антитело, которое специфически связывается с CD40» относятся к антителу, которое способно связывать CD40 с достаточной аффинностью таким образом, что антитело может быть использовано в качестве диагностического и/или терапевтического агента для нацеливания на CD40. В одном из вариантов осуществления настоящего изобретения степень связывания анти-CD40-антитела с неродственным, не-CD40-белком, составляет менее чем примерно 10% от связывания антитела с CD40 по данным измерений, например, методом радиоиммуноанализа (radioimmunoassay, RIA) или жидкостной цитометрии (FACS). В определенных вариантах осуществления настоящего изобретения антитело, которое связывается с CD40, имеет константу диссоциации (KD)<1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0.1 нМ, ≤0.01 нМ или<0.001 нМ (например 10-6 М или менее, например, от 10-68 М до 10-13 М, например, от 10-8 М до 10-10 М).
Понятие «пептидный линкер» относится к пептиду, содержащему один или несколько аминокислот, обычно от 2 до 20 аминокислот. Пептидные линкеры известны в данной области, а также описаны в настоящем изобретении. Пригодными не иммуногенными пептидными линкерами являются, например, пептидные линкеры (G4S)n, (SG4)n или G4(SG4)n, в которых «n» обычно означают число от 1 до 10, обычно от 1 до 4, особенно 2, то есть пептиды, выбранные из группы, включающей GGGGS (SEQ ID NO:77), GGGGSGGGGS (SEQ ID NO:78), SGGGGSGGGG (SEQ ID NO:79), GGGGGSGGGGSSGGGGS (SEQ ID NO:80), (G4S)3 or GGGGSGGGGSGGGGS (SEQ ID NO:81), GGGGSGGGGSGGGG or G4(SG4)2 (SEQ ID NO:82), (G4S)4 or GGGGSGGGGSGGGGSGGGGS (SEQ ID NO:83) и GGGGSGGGGSGGGSGGGGS (SEQ ID NO:84), а также последовательности GSPGSSSSGS (SEQ ID NO:85), GSGSGSGS (SEQ ID NO:86), GSGSGNGS (SEQ ID NO:87), GGSGSGSG (SEQ ID NO:88), GGSGSG (SEQ ID NO:89), GGSG (SEQ ID NO:90), GGSGNGSG (SEQ ID NO:91), GGNGSGSG (SEQ ID NO:92) и GGNGSG (SEQ ID NO:93). Пептидными линкерами, представляющими особый интерес, являются (G4S)2 или GGGGS GGGGS (SEQ ID NO:78) и GGGGSGGGGSGGGSGGGGS (SEQ ID NO:84).
В настоящем изобретении понятие «спейсерный домен» означает полипептид, образующий структурный домен после укладки. Таким образом, спейсерный домен может быть меньше, чем 100 аминокислотных остатков, но должен быть структурно ограничен, чтобы фиксировать мотивы связывания. Примерами спейсерных доменов являются пентамерные суперспирали, шарнирные области антител, или области Fc антитела, или их фрагменты. Спейсерный домен является доменом димеризации, т.е. спейсерный домен содержит аминокислоты, которые способны обеспечивать функциональность димеризации.
Понятие «аминокислота», используемое в настоящем изобретении, означает группу природных карбокси-α-аминокислот, включающую аланин (трехбуквенный код: ala, однобуквенный код: А), аргинин (arg, R), аспарагин (asn, N), аспарагиновую кислоту (asp, D), цистеин (cys, С), глутамин (gin, Q), глутаминовую кислоту (glu, Е), глицин (gly, G), гистидин (his, Н), изолейцин (Ile, I), лейцин (leu, L), лизин (lys, К), метионин (met, М), фенилаланин (phe, F), пролин (pro, Р), серии (ser, S), треонин (thr, Т), триптофан (trp, W), тирозин (tyr, Y) и валин (val, V).
Понятие «гибридизированный» или «присоединенный» означает, что компоненты (например, тяжелая цепь антитела и фрагмент Fab) связаны пептидными связями либо непосредственно, либо через один или несколько пептидных линкеров.
В настоящем изобретении понятие «гибридизированный полипептид» или «полипептид слияния» относится к одноцепочечному полипептиду, состоящему из различных компонентов, таких как эктодомен представителя семейства лигандов TNF, которые гибридизированы либо непосредственно, либо через пептидный линкер. Понятие «гибридизированный» или «присоединенный» означает, что компоненты (например, полипептид и эктодомен указанного представителя семейства лигандов TNF) связаны пептидными связями либо напрямую, либо через один или несколько пептидных линкеров.
«Процент (%) идентичности аминокислотной последовательности» в отношении контрольной полипептидной (белковой) последовательности определяют в виде процента аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в контрольной полипептидной последовательности после выравнивания последовательностей и введения гэпов в случае необходимости для достижения максимального процента идентичности последовательности, и не рассматривая какие-либо консервативные замещения как часть идентичности последовательности. Выравнивание в целях определения процента идентичности аминокислотной последовательности может быть достигнуто различными способами, которые известны специалисту в данной области, например, с использованием общедоступного компьютерного программного обеспечения, такого как BLAST, BLAST-2, ALIGN. SAWI или Megalign (DNASTAR). Специалист в данной области может определить подходящие параметры для выравнивания последовательностей, включая какие-либо алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Однако для целей настоящего изобретения процент идентичности аминокислотной последовательности получают с использованием компьютерной программы сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 разработана фирмой Genentech, Inc., а программный код был подан вместе с документацией пользователя в Бюро по охране авторских прав США, Вашингтон, округ Колумбия, 20559, где она зарегистрирована под номером регистрации авторского права США № TXU510087. Программа ALIGN-2 общедоступна от фирмы Genentech, Inc., Южный Сан-Франциско, Калифорния, или может быть скомпилирована из программного кода. Программа ALIGN-2 должна быть скомпилирована для использования в операционной системе UNIX, включая цифровую UNIX V4.0D. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и не меняются. В ситуациях, когда ALIGN-2 используют для сравнений аминокислотных последовательностей, процент идентичности аминокислотной последовательности данной аминокислотной последовательности А с или против данной аминокислотной последовательности Б (которая в другом варианте может быть определена как заданная аминокислотная последовательность А, которая имеет или включает определенный процент идентичности аминокислотной последовательности с или против данной аминокислотной последовательности Б), рассчитывают следующим образом:
100 раз фракция X/Y,
где X - количество аминокислотных остатков, которые оценены программой выравнивания последовательностей ALIGN-2 как идентичные совпадения при выравнивании А и Б, и где Y - общее количество аминокислотных остатков в Б. Учитывая, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности Б, процент идентичности аминокислотной последовательности А относительно Б не будет равен проценту идентичности аминокислотной последовательности от Б относительно А. Если специально не указано иное, все проценты аминокислотной идентичности в настоящем описании получают согласно описанию, приведенному непосредственно в предшествующем абзаце с использованием компьютерной программы ALIGN-2.
В некоторых вариантах осуществления настоящего изобретения предусматривают варианты аминокислотных последовательностей антигенсвязывающих молекул. Например, может потребоваться улучшение аффинности и/или других биологических свойств антигенсвязывающих молекул. Варианты аминокислотных последовательностей антигенсвязывающих молекул могут быть получены путем введения соответствующих модификаций в нуклеотидную последовательность, кодирующую молекулы, или путем пептидного синтеза. Такие модификации включают, например, делеции, и/или инсерции, и или замещения остатков в аминокислотных последовательностях антитела. Какая-либо комбинация делеции, вставки и замещения может быть сделана для достижения итоговой конструкции, при условии, что итоговая конструкция обладает желаемыми характеристиками, например, антигенсвязывающим свойством. Сайты, представляющие интерес для замещающего мутагенеза, включают HVR и каркас (FR). Консервативные замещения представлены в таблице Б под заголовком «Предпочтительные замещения» и дополнительно описаны ниже со ссылкой на классы боковых цепей аминокислот (1)-(6). Аминокислотные замещения могут быть интродуцированы в интересующую молекулу, после чего проводят скрининг на требуемую активность, например, сохраненное/улучшенное связывание антигена, пониженную иммуногенность или улучшенные ADCC или CDC.
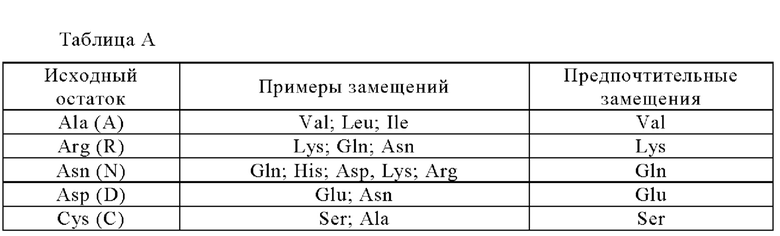
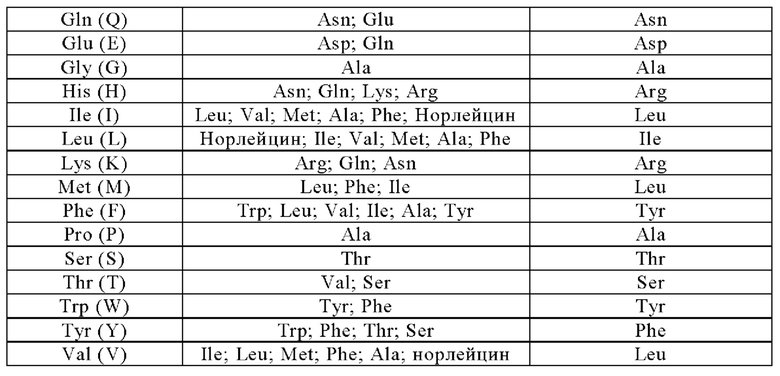
Аминокислоты могут быть сгруппированы в соответствии с общими свойствами боковой цепи:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, влияющие на ориентацию цепи: Gly, Pro;
(6) ароматические: Тгр, Tyr, Phe.
Неконсервативные замены вызовут замену члена одного из этих классов на представителя другого класса.
Понятие «варианты аминокислотной последовательности» включает существенные варианты, в которых имеются аминокислотные замещения в одном или нескольких остатках гипервариабельной области исходной антигенсвязывающей молекулы (например, гуманизированного антитела или антитела человека). В целом, итоговый вариант (варианты), выбранные для дальнейшего изучения, могут иметь модификации (например, улучшения) в определенных биологических свойствах (например, повышенную аффинность, пониженную иммуногенность) по отношению к родительской антигенсвязывающей молекуле и/или будут по существу сохранять определенные биологические свойства исходной антигенсвязывающей молекулы. Примером вариантом замещения является антитело со зрелой аффинностью, которое может быть обычным образом получено, например, с использованием метода созревания аффинности на основе фагового дисплея, например, по описания в настоящем изобретении. Вкратце, один или несколько остатков HVR мутируют, и вариантные антигенсвязывающие молекулы представляют на фаге и подвергают скринингу для выявления определенной биологической активности (например, аффинности). В некоторых вариантах осуществления настоящего изобретения замещения, инсерции или делеции могут происходить в одной или нескольких гипервариабельных областях (HVR) до тех пор, пока такие изменения существенно не снижают способности антигенсвязывающей молекулы связывать антиген. Например, консервативные изменения (например, консервативные замены, представленные в настоящем изобретении), которые существенно не снижают аффинность, могут быть сделаны в HVR. Полезный метод идентификации остатков или областей антитела, на которые может быть нацелен мутагенез, называется «аланин-сканирующим мутагенезом», как описано Cunningham and Wells (1989) Science, 244: 1081-1085. В этом методе остаток или группа целевых остатков (например, заряженные остатки, такие как Arg, Asp, His, Lys и Glu) идентифицируют и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином) для того, чтобы определить затрагивается ли взаимодействие антитела с антигеном. Дополнительные замещения могут быть интродуцированы в положениях аминокислот, демонстрирующих функциональную чувствительность к исходным замещениям. В другом варианте или дополнительно рассматривают кристаллическую структуру комплекса антиген-антигенсвязывающая молекула для идентификации точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки могут быть предметом нацеливания или могут быть исключены из числа кандидатов на замещение. Варианты могут быть просканированы, чтобы определить, содержат ли они требуемые свойства.
Инсерции аминокислотных последовательностей включают гибридизацию с амино- и/или карбоксильного конца с последовательностями разной длины, от одного остатка до полипептидов, содержащих сто или более остатков, а также инсерции внутри последовательности одного или многих аминокислотных остатков. Примеры концевых инсерции включают биспецифические антигенсвязывающие молекулы по настоящему изобретению с N-концевым метионильным остатком. Другие инсерционные варианты молекулы включают гибридизацию с N- или С-конца с полипептидом, что увеличивает период полужизни в сыворотке биспецифических антигенсвязывающих молекул.
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифические антигенсвязывающие молекулы, которые изменены для повышения или понижения степени гликозилирования антитела. Варианты гликозилирования молекул несложно получить путем изменения аминокислотной последовательности таким образом, что один или несколько сайтов гликозилирования создают или удаляют. Если биспецифическая антигенсвязывающая молекула содержит Fc-область, присоединенный к ней углевод может быть изменен. Нативные антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленный биантенарный олигосахарид, который обычно присоединен через N-связь к Asn297 домена СН2 области Fc. См., например, Wright с соавт. TIBTECH 15, 1997, 26-32. К олигосахаридам могут относить различные углеводы, например, маннозу, N-ацетилглюкозамин (N-acetyl glucosamine, GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стволовой» части биантенарной олигосахаридной структуры. В некоторых вариантах осуществления настоящего изобретения модификации олигосахарида в антигенсвязывающей молекуле могут быть разработаны для создания вариантов с определенными улучшенными свойствами. В одном варианте осуществления настоящего изобретения предлагают варианты биспецифических антигенсвязывающих молекул или антител по настоящему изобретению, которые имеют углеводную структуру без фукозы, присоединенную (прямо или опосредованно) к области Fc. Такие варианты фукозилирования могут иметь улучшенную функцию ADCC, см., например, US 2003/0157108 или US 2004/0093621. В другом варианте осуществления настоящего изобретения предусматривают варианты биспецифических антигенсвязывающих молекул или антител по настоящему изобретению обеспечивают двурассеченными олигосахаридами, например, в которых биантенарные олигосахариды, присоединенные к области Fc, разделены на две части GlcNAc. Такие варианты могут иметь пониженное фукозилирование и/или улучшенную функцию ADCC, см. например, WO 2003/011878; US 6602684; US 2005/0123546. Также предусматривают варианты по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к области Fc. Такие варианты антител могут иметь улучшенную функцию CDC и описаны, например, в WO 1997/30087; WO 1998/58964; WO 1999/22764.
В некоторых вариантах осуществления настоящего изобретения может быть желательным создание цистеин-сконструированных вариантов биспецифических антигенсвязывающих молекул по настоящему изобретению, например, молекул «thioMAb», в которых один или несколько остатков молекулы замещены остатками цистеина. В конкретных вариантах осуществления настоящего изобретения замещенные остатки встречаются в доступных сайтах молекулы. При замещении этих остатков цистеином реакционноспособные тиоловые группы позиционируются в доступных сайтах антитела и могут использоваться для конъюгирования антитела с другими фрагментами, такими как фрагменты лекарственного средства или фрагменты лекарственного средства-линкера, для создания иммуноконъюгата. В других вариантах осуществления настоящего изобретения какой-либо один или несколько из следующих остатков могут быть замещены цистеином: V205 (нумерация по Kabat) в легкой цепи; А118 (EU нумерация) в тяжелой цепи; S400 (EU нумерация) в Fc области тяжелой цепи. Цистеин-сконструированные антигенсвязывающие молекулы могут быть получены согласно описанию, например, в US 7521541.
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, которое может быть дополнительно модифицировано для содержания дополнительных небелковых частей молекулы, которые известны в данной области и доступны. К фрагментам, пригодным для дериватизации антитела, относятся водорастворимые полимеры, но не только они. Примерами водорастворимых полимеров являются, но ими перечень не ограничивается, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или статистические сополимеры) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры пропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликоль-пропионовый альдегид может иметь преимущества при получении из-за его стабильности в воде. Полимер может иметь какую-либо молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединенных к антителу, может варьировать, и если присоединено более одного полимера, то они могут быть одинаковыми или разными молекулами. В общем, число и/или тип полимеров, используемых для дериватизации, могут быть определены на основании свойств и функций улучшаемого антитела, но ими анализ не ограничивается, независимо от того, будет ли производное биспецифического антитела использоваться в терапии при определенных условиях и т.д. В другом варианте осуществления настоящего изобретения предусматривают конъюгаты антитела и небелкового фрагмента, которые можно избирательно нагревать при облучении. В еще одном варианте осуществления настоящего изобретения небелковым фрагментом является углеродная нанотрубка (Kam N.W. с соавт., Proc. Natl. Acad. Sci. USA 102, 2005, 11600-11605). Облучение может иметь какую-либо длины волны и включает длины волн (но ими не ограничиваются), которые не наносят вреда обычным клеткам, но нагревают небелковый фрагмент до температуры, при которой клетки, проксимальные по отношению к небелковому фрагменту антитела, уничтожаются.
В другом варианте осуществления настоящего изобретения предусматривают иммуноконъюгаты биспецифических антител. Понятие «иммуноконъюгат» означает антитело, конъюгированное с одной или несколькими гетерологичными молекулами, включая, но не ограничиваясь им, цитотоксический агент.
Понятие «нуклеиновая кислота» относится к выделенной молекуле или конструкции нуклеиновой кислоты, например, мессенджер-РНК (мРНК), вирусной РНК или плазмидной ДНК (пДНК). Полинуклеотид может содержать обычную фосфодиэфирную связь или необычную связь (например, амидную связь, такую как найденная в пептидных нуклеиновых кислотах (peptide nucleic acid, PNA). Понятие «молекула нуклеиновой кислоты» относится к какому-либо одному или нескольким сегментам нуклеиновой кислоты, например ДНК или фрагментов РНК, присутствующих в полинуклеотиде.
Под «выделенной» молекулой нуклеиновой кислоты или полинуклеотидом подразумевают молекулу нуклеиновой кислоты, ДНК или РНК, которая была удалена из естественной для нее среды. Например, рекомбинантный полинуклеотид, кодирующий полипептид, содержащийся в векторе, является выделенным для целей настоящего изобретения. Дополнительные примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, поддерживаемые в гетерологичных клетках-хозяевах, или очищенные (частично или в высокой степени) полинуклеотиды в растворе. Выделенный полинуклеотид включает полинуклеотидную молекулу, содержащуюся в клетках, которые обычно содержат полинуклеотидную молекулу, но полинуклеотидная молекула присутствует внехромосомно или при хромосомной локализации отличаются от ее естественного хромосомного расположения. Выделенные молекулы РНК включают in vivo или in vitro транскрипты РНК по настоящему изобретению, а также формы положительных и отрицательных цепей и двухцепочечные формы. Выделенные полинуклеотиды или нуклеиновые кислоты по настоящему изобретению дополнительно включают такие молекулы, полученные синтетически. Кроме того, полинуклеотид или нуклеиновая кислота могут быть или могут включать регуляторный элемент, например промотор, сайт связывания рибосомы или терминатор транскрипции.
Под нуклеиновой кислотой или полинуклеотидом, имеющим нуклеотидную последовательность, по меньшей мере, например, на 95% «идентичную» контрольной нуклеотидной последовательности по настоящему изобретению, подразумевают, что нуклеотидная последовательность полинуклеотида идентична контрольной последовательности, при условии, что полинуклеотидная последовательность может включать до пяти точечных мутаций на каждые 100 нуклеотидов контрольной нуклеотидной последовательности. Иначе говоря, для получения полинуклеотида, имеющего нуклеотидную последовательность, по меньшей мере, на 95% идентичную контрольной нуклеотидной последовательности, до 5% нуклеотидов в контрольной последовательности может быть делетировано или замещено другими нуклеотидами или числом нуклеотидов до 5% от общего количества нуклеотидов в контрольной последовательности может быть инсертировано в контрольную последовательность. Эти изменения контрольной последовательности могут происходить в 5'- или 3'-концевых положениях контрольной нуклеотидной последовательности или в каком-либо положении между этими концевыми положениями, перемежаясь либо индивидуально среди остатков в контрольной последовательности, либо в виде одной или нескольких смежных групп внутри контрольной последовательности. На практике, является ли какая-либо конкретная полинуклеотидная последовательность, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной нуклеотидной последовательности по настоящему изобретению, можно определить обычным путем, используя известные компьютерные программы, такие как обсуждаемые выше для полипептидов (например, ALIGN-2).
Понятие «кассета экспрессии» относится к полинуклеотиду, получаемому рекомбинантно или синтетически, с серией определенных элементов нуклеиновой кислоты, которые допускают транскрипцию конкретной нуклеиновой кислоты в клетке-мишени. Рекомбинантная кассета экспрессии может быть включена в плазмиду, хромосому, митохондриальную ДНК, пластидную ДНК, вирус или фрагмент нуклеиновой кислоты. Как правило, рекомбинантная часть кассеты экспрессии вектора экспрессии включает, помимо других последовательностей, последовательности нуклеиновой кислоты для транскрибирования и промотора. В некоторых вариантах осуществления настоящего изобретения кассета экспрессии содержит полинуклеотидные последовательности, которые кодируют биспецифические антигенсвязывающие молекулы по настоящему изобретению или их фрагменты.
Понятие «вектор» или «вектор экспрессии» является синонимом «конструкции экспрессии» и относится к молекуле ДНК, которую используют для введения и направления экспрессии конкретного гена, с которым он функционально связан в клетке-мишени. Это понятие включает вектор как самореплицирующуюся структуру нуклеиновой кислоты, а также вектор, встроенный в геном клетки-хозяина, в которую он был интродуцирован. Вектор экспрессии по настоящему изобретению содержит кассету экспрессии. Векторы экспрессии допускают транскрипцию больших количеств стабильных мРНК. Когда вектор экспрессии находится внутри клетки-мишени, молекула рибонуклеиновой кислоты или белок, который кодируется этим геном, продуцируется в результате клеточной транскрипции и/или трансляции. В некоторых вариантах осуществления настоящего изобретения вектор экспрессии по настоящему изобретению содержит кассету экспрессии, которая содержит полинуклеотидные последовательности, кодирующие биспецифические антигенсвязывающие молекулы по настоящему изобретению или их фрагменты.
Понятия «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используют и их относят к клеткам, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформантов» и «трансформированные клетки», которые включают первичные трансформированные клетку и их потомство, независимо от количества пассажей. Потомство может не быть полностью идентичным по содержанию нуклеиновой кислоты родительской клетке и может содержать мутации. Мутантное потомство, которое имеет ту же функцию или биологическую активность, что и скрининг или отбор для первоначально трансформированной клетки, включено в рамки охвата настоящего изобретения. Клетка-хозяин является клеточной системой какого-либо типа, которая может быть применена для выработки биспецифических антигенсвязывающих молекул по настоящему изобретению. Клетки-хозяева включают культивируемые клетки, например культивируемые клетки млекопитающих, такие как клетки СНО, клетки ВНК, клетки NS0, клетки SP2/0, клетки миеломы YO, клетки миеломы мыши РЗХ63, клетки PER, клетки PER.C6 или клетки гибридомы, клетки дрожжей, клетки насекомых и растительные клетки, если перечислить только несколько клеток, но также клетки, содержащиеся в трансгенном животном, трансгенном растении или в культивируемых растительных или животных тканях.
«Эффективное количество» агента означает такое количество, которое необходимо для того, чтобы привести к физиологическим изменениям в клетке или ткани, которым этот агент вводят.
«Терапевтически эффективное количество» агента, например фармацевтической композиции, относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для достижения желаемого терапевтического или профилактического результата. Терапевтически эффективное количество средства, например, устраняет, уменьшает, задерживает, минимизирует или предотвращает неблагоприятные воздействия заболевания.
Понятия «индивидуум» или «субъект» относится к млекопитающему. Млекопитающие включают, но ими перечень не ограничивают, одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и других приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В особенности, индивидуумом или субъектом является человек.
Понятие «фармацевтическая композиция» относится к препарату, который находится в такой форме, которая позволяет проявить эффективность биологической активности активного ингредиента, содержащегося в нем, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому предназначено введение композиции.
«Фармацевтически приемлемый эксципиент» относится к ингредиенту в фармацевтической композиции, отличному от активного ингредиента, который нетоксичен для субъекта. Фармацевтически приемлемый эксципиент включает, но ими перечень не ограничивают, буфер, стабилизатор или консервант.
Понятие «вкладыш в упаковку» означает инструкции, обычно включаемые в коммерческие упаковки терапевтических продуктов и содержащие информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях относительно использования таких терапевтических продуктов.
В настоящем изобретении понятие «лечение» (и его грамматические производные, например «лечить» или «лечиться») относится к клиническому вмешательству и попытке изменить естественное состояние подвергаемого лечению индивидуума, и может осуществляться либо для профилактики, либо во время курса клинической патологии. Желательные эффекты лечения включают, но ими перечень не ограничивают, предотвращение возникновения или рецидива заболевания, ослабление симптомов, уменьшение любых прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, уменьшение скорости прогрессирования заболевания, улучшение или смягчение болезненного состояния, ремиссию или улучшение прогноза. В некоторых вариантах осуществления молекулы по настоящему изобретению используют для задержки развития заболевания или для замедления прогрессирования заболевания.
В настоящем изобретении понятие «рак» означает пролиферативные заболевания, например лимфомы, карциномы, лимфомы, бластомы, саркомы, лейкозы, лимфолейкозы, рак легких, немелкоклеточный рак легких (non-small cell lung, NSCL), бронхиолоальвиолярный клеточный рак легких, рак костей, рак поджелудочной железы, рак кожи, рак головы или шеи, кожную или интраокулярную меланому, рак матки, рак яичников, рак прямой кишки, рак анальной области, рак желудка, колоректальный рак (colorectal cancer, CRC), рак поджелудочной железы, рак молочной железы, рак молочной железы с тройным негативным фенотипом, рак матки, карциному маточных труб, карциному эндометрия, карциному шейки матки, рак влагалища, карциному вульвы, болезнь Ходжкина, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркому мягких тканей, рак мочеиспускательного канала, рак полового члена, рак предстательной железы, рак мочевого пузыря, рак почки или мочеточника, почечноклеточный рак, рак почечной лоханки, мезотелиому, гепатоцеллюлярный рак, рак желчных путей, новообразования центральной нервной системы (ЦНС), опухоли спинного мозга, глиому ствола головного мозга, мультиформную глиобластому, астроцитому, шваномы, эпендимоны, медуллобластомы, менингиомы, плоскоклеточный рак, аденому гипофиза и саркому Юинга, меланому, множественную миелому, В-клеточный рак (лимфому), хронический лимфолейкоз (ХЛЛ), острый лимфобластный лейкоз (ОЛЛ), волосатоклеточный лейкоз, хронический миелобластный лейкоз, включая невосприимчивые версии какого-либо из вышеуказанных видов рака или комбинацию одного или нескольких из указанных видов рака.
Биспецифические антигенсвязывающие молекулы по настоящему изобретению
Настоящее изобретение предусматривает новые биспецифические антитела с особенно полезными свойствами, такими как продуктивность, стабильность, аффинность, биологическая активность, эффективность нацеливания и пониженная токсичность. Новые биспецифические антитела состоят из двух гибридизированных полипептидов, содержащих два антигенсвязывающих домена, способных к специфическому связыванию с первой мишенью, и один антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью. Удивительно, но эти два гибридизированных полипептида сконструированы таким образом, что три антигенсвязывающих домена могут собираться должным образом и что биспецифическое связывание является полностью функциональным.
Для молекулы, предназначенной для клинического применения, следует избегать агрегатов функционально активных молекул, что означает, что чистота и стабильность сборки различных гибридизированных полипептидов, образующих антигенсвязывающую молекулу, критически важны. Важно отметить, что в биспецифических антителах по настоящему изобретению все три антигенсвязывающих домена гибридизированы таким образом, чтобы обеспечить правильную сборку трех антигенсвязывающих доменов. Все три антигенсвязывающих домена расположены таким образом, что каждый антигенсвязывающий домен может связываться с их соответствующими мишенями. Новые биспецифичные антигенсвязывающие молекулы по настоящему изобретению, кроме того, состоят только из двух гибридизированных полипептидов и не содержат каких-либо легких цепей. Таким образом, можно избежать проблемы неправильного спаривания между тяжелой и легкой цепями. Также важно отметить, что конструкции могут экспрессироваться с достаточно хорошими титрами и вырабатывают хорошую долю требуемого продукта. Антитело-подобная архитектура, содержащая спейсерный домен для димеризации, является стабильной по сравнению с другими белками; их экспрессия также в высокой степени устойчива при использовании различных линий клеток.
Новые биспецифические антигенсвязывающие молекулы по настоящему изобретению называются 2+1 конторстелами.
Таким образом, 2+1 конторстело является биспецифическим антителом, состоящим из двух гибридизированных полипептидов и содержащим два антигенсвязывающих домена, способных специфически связываться с первой мишенью, и один антигенсвязывающий домен, способный специфически связываться со второй мишенью, где
(а) первый гибридизированный полипептид содержит первую часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, спейсерный домен, вторую часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и первую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, где
- спейсерный домен является полипептидом и содержит, по меньшей мере, 25 аминокислотных остатков,
- первая часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через первый пептидный линкер с N-концом спейсерного домена,
- вторая часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через второй пептидный линкер с С-концом спейсерного домена, и
- первая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, или гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и
(б) второй гибридизированный полипептид содержит первую часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, спейсерный домен, вторую часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, и вторую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, где
- спейсерный домен является полипептидом и содержит, по меньшей мере, 25 аминокислотных остатков,
- первая часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через первый пептидный линкер с N-концом спейсерного домена,
- вторая часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через второй пептидный линкер с С-концом спейсерного домена, и
- вторая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с С-концом второй части второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, или гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части второго антигенсвязывающего домена, способного специфически связываться с первой мишенью,
где первая часть и вторая часть антигенсвязывающего домена, способного к специфическому связыванию со второй мишенью, связаны друг с другом с образованием антигенсвязывающего домена, способного к специфическому связыванию со второй мишенью, причем первая часть и вторая часть первого и второго антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, связаны друг с другом с образованием циклического гибридизированного полипептида, причем спейсерный домен первого гибридизированного полипептида и спейсерный домен второго гибридизированного полипептида ковалентно связаны друг с другом дисульфидной связью и содержат модификации, способствующие ассоциации первого и второго гибридизированного полипептида.
В одном варианте осуществления настоящего изобретения предусматривают биспецифическое антитело, согласно описанному выше, в котором в первом гибридизированном полипептиде первая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, и во втором гибридизированном полипептиде вторая часть антигенсвязывающего домена, способного к специфическому связыванию со второй мишенью, гибридизирована либо непосредственно, либо через третью пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного к специфически связываться с первой мишенью.
В другом варианте осуществления настоящего изобретения предусматривают биспецифическое антитело, согласно описанному выше, в котором в первом гибридизированном полипептиде первая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части первого антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, и во втором гибридизированном полипептиде вторая часть антигенсвязывающего домена, способного к специфическому связыванию со второй мишенью, гибридизирована либо непосредственно, либо через третью пептидный линкер с N-концом первой части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью.
Таким образом, в биспецифическом антителе, согласно описанному выше, исходные домены антитела гибридизированы гибкими пептидными линкерами. Эти линкеры обеспечивают правильную ассоциацию домена в молекуле Конторстела, а также правильную складчатость антитела. Эта новая топология цепей приводит к пространственной ориентации плечей Fab и части Fc, которая отличается от классического формата IgG1. Из-за параллельной ориентации своих антигенсвязывающих сайтов, Конторстело является в высокой степени пригодным форматом антител для агонистических механизмов.
В одном из вариантов осуществления настоящего изобретения предусматривают описанное выше антитело, в котором третий пептидный линкер, соединяющий первую часть или вторую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, содержащей, по меньшей мере, 15 аминокислот. В одном из вариантов осуществления настоящего изобретения третий пептидный линкер, соединяющий первую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, и третий пептидный линкер, соединяющий вторую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, являются идентичными. В одном из вариантов осуществления настоящего изобретения третий пептидный линкер содержит от 15 до 25 аминокислот. В одном из предпочтительных вариантов осуществления настоящего изобретения третий пептидный линкер содержит аминокислотную последовательность SEQ ID NO:83 или SEQ ID NO:84. Точнее, третий пептидный линкер содержит аминокислотную последовательность SEQ ID NO:84. В еще одном варианте осуществления настоящего изобретения третий пептидный линкер (в обоих гибридизированных полипептидах) содержит аминокислотную последовательность SEQ ID NO:83 или SEQ ID NO:84, a первый и второй пептидный линкер содержит аминокислотную последовательность SEQ ID NO:78.
В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, согласно описанному выше, в котором первый гибридизированный полипептид содержит вариабельный домен тяжелой цепи антгенсвязывающего домена, способного специфически связываться со второй мишенью, и второй гибридизированный полипептид содержит вариабельный домен легкой цепи антитела антигенвязывающего домена, способного специфически связываться со второй мишенью, или наоборот.
В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, согласно описанному выше, в котором первый гибридизированный полипептид содержит вариабельный домен тяжелой цепи антигенсвязывающего домена, способного специфически связываться со второй мишенью, и второй гибридизированный полипептид содержит вариабельный домен легкой цепи антитела антигенсвязывающего домена, способный специфически связываться со второй мишенью, или наоборот. В одном из вариантов осуществления настоящего изобретения первая часть антигенсвязывающего домена является Fab-фрагментом тяжелой цепи антитела, и вторая часть антигенсвязывающего домена является Fab-фрагментом легкой цепи антитела, или наоборот. В одном из вариантов осуществления настоящего изобретения первая часть антигенсвязывающего домена и вторая часть антигенсвязывающего домена ковалентно связаны друг с другом дисульфидной связью.
В другом варианте осуществления настоящего изобретения биспецифическое антитело, состоящее из двух гибридизированных полипептидов и содержащее два антигенсвязывающих домена, способных к специфическому связыванию с первой мишенью, и один антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью, где
(а) первый гибридизированный полипептид содержит первую часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, спейсерный домен, вторую часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и первую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, где
- спейсерный домен является полипептидом и содержит, по меньшей мере, 25 аминокислотных остатков,
- первая часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через первый пептидный линкер с N-концом спейсерного домена,
- вторая часть первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через второй пептидный линкер с С-концом спейсерного домена, и
- первая часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, гибридизирована либо непосредственно, либо через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, или гибридизирована либо непосредственно, либо через третий пептидный линкер с N-концом первой части первого антигенсвязывающего домена, способного специфически связываться с первой мишенью, и
(б) второй гибридизированный полипептид содержит первую часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, спейсерный домен, вторую часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, и вторую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью, где
- спейсерный домен является полипептидом и содержит, по меньшей мере, 25 аминокислотных остатков,
- первая часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через первый пептидный линкер с N-концом спейсерного домена,
- вторая часть второго антигенсвязывающего домена, способного специфически связываться с первой мишенью, гибридизирована либо непосредственно, либо через второй пептидный линкер с С-концом спейсерного домена, и
(в) легкая цепь, содержащая вторую часть антигенсвязывающего домена, способного специфически связываться со второй мишенью,
где первая часть и вторая часть антигенсвязывающего домена, способного к специфическому связыванию со второй мишенью, связаны друг с другом с образованием антигенсвязывающего домена, способного к специфическому связыванию со второй мишенью, причем первая часть и вторая часть первого и второго антигенсвязывающего домена, способного к специфическому связыванию с первой мишенью, связаны друг с другом с образованием гибридного циклического полипептида, и
где спейсерный домен первого гибридизированного полипептида и спейсерный домен второго гибридизированного полипептида ковалентно связаны друг с другом дисульфидной связью и содержат модификации, способствующие ассоциации первого и второго гибридизированных полипептидов.
В некоторых вариантах осуществления настоящего изобретения первая часть антигенсвязывающего домена является Fab-фрагментом тяжелой цепи антитела, а вторая часть антигенсвязывающего домена является Fab-фрагментом легкой цепи антитела, или наоборот. В одном из вариантов осуществления настоящего изобретения первая часть антигенсвязывающего домена и вторая часть антигенсвязывающего домена ковалентно связаны друг с другом дисульфидной связью.
В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, описанное выше, в котором как в первом гибридизированном полипептиде, так и во втором гибридизированном полипептиде, первая часть антигенсвязывающего домена, способного специфически связываться с первой мишенью, является фрагментом Fab тяжелой цепи антитела и вторая часть антигенсвязывающего домена, способного специфически связываться с первой мишенью, является фрагментом Fab легкой цепи антитела.
Если антигенсвязывающий домен является Fab-фрагментом, то Fab может быть обычным Fab, кросс-Fab или DutaFab.
В случае обычного Fab первая часть антигенсвязывающего домена содержит вариабельный домен тяжелой цепи антитела (VH) и, по меньшей мере, N-концевой фрагмент константного домена (или полный) тяжелой цепи первого антитела (СН1) и соответствующая вторая часть антигенсвязывающего домена содержит вариабельный домен легкой цепи (VL) антитела и, по меньшей мере, N-концевой фрагмент константного домена (или полный) легкой цепи антитела (CL или Скарра). Порядок этих доменов может быть каким-либо до тех пор, пока их ассоциация и образование (функционального) антигенсвязывающего домена возможно (т.е. не предотвращено). В одном из вариантов осуществления настоящего изобретения одна часть антигенсвязывающего домена содержит в направлении от N- к С-концу VH-CH1, а другая часть антигенсвязывающего домена содержит в направлении от N- к С-концу VL-CL (Скарра).
В случае кросс-Fab каждая из двух частей антигенсвязывающего домена содержит вариабельный домен антитела и, по меньшей мере, N-концевой фрагмент константного домена (или полный) антитела, причем пары вариабельного домена и константного домена не являются природными связанными друг с другом и их получают путем кроссовера/обмена домена тяжелой цепи и домена легкой цепи. Это может быть обмен VH с VL или СН1 с CL. Порядок этих доменов может быть каким-либо до тех пор, пока их ассоциация и образование (функционального) сайта связывания возможны (то есть не предотвращены). В одном из вариантов осуществления настоящего изобретения первая часть антигенсвязывающего домена содержит в направлении от N- к С-концуа VL-CH1, а вторая часть связывающего домена содержит в направлении от N- к С-концу VH-CL (Скарра). В другом предпочтительном варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена содержит в направлении от N- к С-концу VH-CL, а вторая часть связывающего домена содержит в направлении от N- к С-концу VL-СН1.
В случае DutaFab первая часть антигенсвязывающего домена содержит вариабельный домен тяжелой цепи антитела (VH) и, по меньшей мере, N-концевой фрагмент константного домена (или полный) тяжелой цепи первого антитела (СН1), и соответствующий второй антигенсвязывающий домен содержит вариабельный домен легкой цепи антитела (VL) и, по меньшей мере, N-концевой фрагмент константного домена (или полный) легкой цепи антитела (CL), где указанный антигенсвязывающий домен содержит два непересекающихся паратопа в комплементарной паре вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VL), где первый паратоп содержит остатки из CDR1 и CDR3 домена VL и CDR2 домена VH, а второй паратоп содержит остатки из CDR1 и CDR3 домена VH и CDR2 домена VL.
Таким образом, в одном варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена является Fab-фрагментом тяжелой цепи антитела (VH-CH1), а вторая часть антигенсвязывающего домена является Fab-фрагментом легкой цепи антитела (VL-Ckappa). В другом варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена является Fab-фрагментом легкой цепи антитела, а вторая часть антигенсвязывающего домена является Fab-фрагментом тяжелой цепи антитела. В другом варианте осуществления настоящего изобретения первая часть антигенсвязывающего домена является Kpocc-Fab-фрагментом антитела, содержащим VH-Ckappa, а вторая часть антигенсвязывающего домена является Kpocc-Fab-фрагментом антитела, содержащим VL-CH1. В еще одном из вариантов осуществления настоящего изобретения первая часть антигенсвязывающего домена является Kpocc-Fab-фрагментом антитела, содержащим VL-CH1, а вторая часть антигенсвязывающего домена является Kpocc-Fab-фрагментом антитела, содержащим VH-Ckappa.
В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, в котором антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью, является Kpocc-Fab-фрагментом, и где оба антигенсвязывающих домена, способные к специфическому связыванию с первой мишенью, являются обычными Fab.
В другом варианте осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью, является обычным Fab, и в котором оба антигенсвязывающих домена, способные к специфическому связыванию с первой мишенью, являются кросс-Fab-фрагментами.
Выше было описано, что биспецифическое антитело состоит из первого и второго гибридизированных полипептидов, которые оба содержат спейсерный домен, спейсерный домен первого гибридизированного полипептида и спейсерный домен второго гибридизированного полипептида ковалентно связаны друг с другом посредством дисульфидной связи и содержат модификации, способствующие ассоциации первого и второго гибридизированных полипептидов. Спейсерный домен содержит по меньшей мере 25 аминокислот.
В одном варианте осуществления настоящего изобретения спейсерный домен содержит шарнирную область антитела или ее (С-концевой) фрагмент и СН2-домен антитела или его (N-концевой) фрагмент. В другом варианте осуществления настоящего изобретения спейсерный домен содержит шарнирную область антитела или ее фрагмент, домен СН2 антитела и домен СН3 антитела или его фрагмент. В другом варианте осуществления настоящего изобретения спейсерным доменом гибридизированного полипептида, согласно описанному выше, является Fc-домен антитела, в частности подкласса IgG1, IgG2 или IgG4, более конкретно подкласса IgG1.
В одном из вариантов осуществления настоящего изобретения спейсерный домен содержит домен Fc с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO:103, Seq ID NO:104, SEQ ID NO:105, SEQ ID NO:106 и SEQ ID NO:107, или ее вариантом с 95% гомологии.
Кроме того, спейсерный домен первого гибридизированного полипептида и спейсерный домен второго гибридизированного полипептида содержат модификации, способствующие ассоциации первого и второго гибридизированных полипептидов. В одном из вариантов осуществления настоящего изобретения спейсерный домен первого гибридизированного полипептида содержит впадины, а спейсерный домен второго гибридизированного полипептида содержит выступы в соответствии с методом «выступ-во впадину». В другом варианте осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором спейсерный домен содержит шарнирную область антитела или ее фрагмент и Fc-домен IgG1. В частности, Fc-домен IgG1 включает одну или несколько аминокислотных замен, которые уменьшают связывание с Fc-рецептором, в частности, с Fcγ-рецептором. В одном из вариантов осуществления настоящего изобретения Fc-домен IgG1 включает аминокислотные замещения L234A и L235A. В другом варианте осуществления настоящего изобретения Fc-домен IgG1 содержит мутацию P329G. Более того, Fc-домен IgG1 содержит аминокислотные замещения L234A, L235A и P329G (нумерация по Kabat, индекс EU нумерация по Kabat, индекс EU).
В другом варианте осуществления настоящего изобретения домен Fc относится к подклассу IgG1 или IgG2 и содержит мутации PVA236, GLPSS331, и/или L234A/L235A/P329G (нумерация по Kabat, индекс EU). В еще одном варианте осуществления настоящего изобретения описанный домен Fc относится к подклассу IgG4 и включает мутацию L235E. В одном варианте осуществления настоящего изобретения описанный домен Fc дополнительно содержит мутацию S228P. В другом варианте осуществления настоящего изобретения домен Fc подкласса IgG4 включает мутацию P329G. В еще одном варианте осуществления настоящего изобретения описанный домен Fc подкласса IgG4 включает мутации S228P/L235E/P329G (нумерация по Kabat, индекс EU).
В некоторых вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором одним антигенсвязывающим доменом, способным к специфическому связыванию со второй мишенью, является антигенсвязывающий домен, способный специфически связываться с опухолеассоциированным антигеном (tumor associated antigen, ТАА). В частности, опухолеассоциированный антиген является белком активации фибробластов (Fibroblast Activation Protein, FAP). В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, специфически связываться со второй мишенью, является антигенсвязывающий доменом, способным к специфическому связыванию с белком активации фибробластов (FAP).
Модификации домена Fc, способствующие гетеродимеризации
одном варианте осуществления настоящего изобретения предусматривают биспецифические антитела, которые могут содержать (а) первый гибридизированный полипептид, описанный в настоящем изобретении выше, и второй гибридизированный слитый полипептид, описанный в настоящем изобретении выше, причем первый и второй гибридизированные полипептиды содержат модификации, способствующие ассоциации первого и второго гибридизированных полипептида. Обычно такие модификации интродуцируют в домены Fc. Рекомбинантная совместная экспрессия двух структурно различных гибридизированных полипептидов и последующая димеризация привели бы к нескольким возможным комбинациям двух полипептидов. Таким образом, для повышения выхода и чистоты биспецифических антител при рекомбинантной продукции будет целесообразно интродуцировать в Fc-домен биспецифических антигенсвязывающих молекул по настоящему изобретению модификации, способствующие ассоциации требуемых полипептидов.
Сайт наиболее выраженного взаимодействия белок-белок между двумя субъединицами домена IgG Fc человека находятся в домене СН3 домена Fc. Таким образом, указанная модификация находится, в частности, в домене СН3 домена Fc.
В настоящем изобретении указанная модификация является модификацией «выступ-во-впадину», содержащей модификацию «выступа» в одной из двух субъединиц домена Fc и модификацию «впадины» во второй из двух субъединиц домена Fc. Таким образом, в одном из вариантов осуществления настоящего изобретения предусматривают биспецифическую антигенсвязывающую молекулу, согласно описанному выше, которая включает молекулу IgG, в которой часть Fc первой тяжелой цепи содержит первый модуль димеризации, и Fc-часть второй тяжелой цепи содержит второй модуль димеризации, обеспечивающий гетеродимеризацию двух тяжелых цепей молекулы IgG, и первый модуль димеризации содержит выступы, а второй модуль димеризации содержит впадины в соответствии с технологией «выступ-во-впадину».
Метод «выступ-во-впадину» описан, например, в US 5731168; US 7695936; Ridgway с соавт., Prot Eng 9, 1996, 617-621; Carter, J Immunol Meth 248, 2001, 7-15. В общем, метод включает введение выпуклости («выступа») на границе раздела первого полипептида и соответствующей полости («впадины») на границе раздела второго полипептида так, что выпуклость может быть расположена в полости таким образом, чтобы способствовать образованию гетеродимера и препятствовать образованию гомодимера. Выпуклости конструируют путем замены небольших аминокислотных боковых цепей на границе первого полипептида более крупными боковыми цепями (например, тирозином или триптофаном). Компенсаторные полости идентичного или сходного размера с выпуклостями создаются на границе раздела второго полипептида путем замены больших боковых цепей аминокислот более мелкими (например, аланином или треонином).
Домены СН3 в первом и втором гибридизированном полипептиде, которые описаны в настоящем изобретении, могут быть изменены методом «выступ-во-впадину», который подробно описан в нескольких примерах, например в WO 96/027011; Ridgway J.B., с соавт., Protein Eng. 9, 1996, 617-621; Merchant A.M., с соавт., Nat. Biotechnol. 16, 1998, 677-681. По этому методу взаимодействующие поверхности двух доменов СН3 изменяют для повышения гетеродимеризации обеих тяжелых цепей, содержащих эти два домена СН3. Каждый из двух доменов СН3 (двух тяжелых цепей) может быть «выступом», а другой «впадиной». Введение дисульфидного мостика дополнительно стабилизирует гетеродимеры (Merchant A.M., с соавт., Nature Biotech. 16, 1998, 677-681; Atwell S., с соавт., J. Mol. Biol. 270, 1997, 26-35) и повышает выход.
Соответственно, в одном из вариантов осуществления настоящего изобретения в домене СН3 первой субъединицы домена Fc биспецифических антигенсвязывающих молекул по настоящему изобретению аминокислотный остаток заменяют аминокислотным остатком, имеющим больший объем боковой цепи, создавая тем самым выступ в домене СН3 первой субъединицы, которая расположена в полости внутри домена СН3 второй субъединицы, а в домене СН3 второй субъединицы домена Fc аминокислотный остаток заменяют аминокислотным остатком, имеющим меньший объем боковой цепи, тем самым создавая полость в домене СН3 второй субъединицы, внутри которой расположен выступ в домене СН3 первой субъединицы.
В еще одном из вариантов осуществления настоящего изобретения в домене СН3 первой субъединицы домена Fc («цепь с выступом») остаток треонина в положении 366 заменен остатком триптофана (T366W), и в домене СН3 второй субъединицы Fc-домен остатка тирозина в положении 407 заменен остатком валина (Y407V). Точнее, во второй субъединице домена Fc («цепь с впадиной») дополнительно остаток треонина в положении 366 заменен остатком серина (T366S), а остаток лейцина в положении 368 заменен остатком аланина (L368A). Точнее, в первой субъединице домена Fc дополнительно сериновый остаток в положении 354 заменен остатком цистеина (S354C), и во второй субъединице домена Fc дополнительно остаток тирозина в положении 349 заменен остатком цистеина (Y349C). Введение этих двух остатков цистеина приводит к образованию дисульфидного мостика между двумя субъединицами Fc-домена. Дисульфидный мостик дополнительно стабилизирует димер (Carter, J Immunol Methods 248, 2001, 7-15).
Другие методы «выступ-во-впадину» также описаны в ЕР 1870459 А1 и могут применяться в другом варианте осуществления настоящего изобретения или дополнительно. В одном из вариантов осуществления настоящего изобретения кратно-круговой гибридизированный полипептид, согласно указанному в настоящем изобретении, включает мутации R409D и K370E в домене СН3 «цепи с выступом» и мутации D399K и E357K в домене СН3 «цепи со впадиной» (нумерация по Kabat, индекс EU нумерация по Kabat, индекс EU).
В еще одном из вариантов осуществления настоящего изобретения молекула, связывающая биспецифический антиген, может содержать мутации Y349C и T366W в одном из двух доменов СН3 и мутации S354C, T366S, L368A и Y407V в другом из двух доменов СН3, или молекула, связывающая биспецифический антиген по настоящему изобретению, содержит мутации Y349C и T366W в одном из двух доменов СН3 и мутации S354C, T366S, L368A и Y407V в другом из двух доменов СН3, и дополнительно мутации R409D и K370E в домене СН3 в «цепи с впадиной» и мутации D399K и E357K в домене СН3 в «цепи с впадиной» (нумерация по Kabat, индекс EU).
В альтернативном варианте осуществления настоящего изобретения модификация, стимулирующая ассоциацию первой и второй субъединиц Fc-домена, содержит модификацию, опосредующую эффекты электростатического управления, например, как описано в публикации РСТ WO 2009/089004. В целом этот метод включает замену одного или нескольких аминокислотных остатков на границе раздела двух субъединиц Fc-домена заряженными аминокислотными остатками таким образом, что образование гомодимера становится электростатически неблагоприятным, а гетеродимеризация электростатически выгодной.
Помимо метода «выступ-во-впадину» в данной области техники известны другие методы модификации доменов СН3 тяжелых цепей для обеспечения гетеродимеризации. Эти методы, особенно те, которые описаны в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954 и WO 2013/096291, рассматриваются здесь в качестве альтернативы метода «выступ-во-впадину» в сочетании с биспецифической антигенсвязывающей молекулой по настоящему изобретению.
В одном из вариантов осуществления настоящего изобретения заряженные аминокислоты с противоположными зарядами в определенных положениях аминокислот на границе раздела СН3/СН3-доменов первой и второй тяжелых цепей интродуцируют для дальнейшего стимулирования ассоциации требуемых полипептидов. Соответственно, этот вариант осуществления настоящего изобретения относится к биспецифическим антигенсвязывающим молекулам по настоящему изобретению, причем в третичной структуре антитела домен СН3 первой тяжелой цепи и домен СН3 второй тяжелой цепи образуют поверхность контакта, которая расположена между соответствующими доменами СН3 антитела, где соответствующие аминокислотные последовательности домена СН3 первой тяжелой цепи и домена СН3 второй тяжелой цепи содержат набор аминокислот, который расположен внутри указанной поверхности контакта в третичной структуре кругового гибридизированного полипептида, и где из набора аминокислот, который расположен на поверхности контакта в домене СН3 одной тяжелой цепи, первая аминокислота заменена положительно заряженной аминокислотой и из набора аминокислот, который расположен на поверхности контакта в домене СН3 другой тяжелой цепи вторая аминокислота замещена отрицательно заряженной аминокислотой. Биспецифическую антигенсвязывающую молекула по настоящему изобретению также обозначают как «СН3(+/-)-сконструированную из семейства TNF лигандный тример-содержащую антигенсвязывающую молекулу» («+/-» означает противоположно заряженные аминокислоты, которые вводят в соответствующие домены СН3). В еще одном варианте осуществления настоящего изобретения в указанной СН3(+/-)- сконструированной биспецифической антигенсвязывающей молекуле по настоящему изобретению положительно заряженная аминокислота выбрана из К, R и Н, а отрицательно заряженная аминокислота выбрана из Е или D. В другом варианте осуществления настоящего изобретения в указанной СН3(+/-)-сконструированной биспецифической антигенсвязывающей молекуле по настоящему изобретению положительно заряженная аминокислота выбрана из К и R, и отрицательно заряженная аминокислота выбрана из Е или D. В еще одном из вариантов осуществления настоящего изобретения в указанной СН3(+/-)-сконструированной биспецифической антигенсвязывающей молекуле по настоящему изобретению положительно заряженной аминокислотой является K, а отрицательно заряженной аминокислотой - Е. В одном варианте осуществления настоящего изобретения в указанной СН3(+/-)-сконструированной биспецифической антигенсвязывающей молекуле по настоящему изобретению в домене СН3 одной тяжелой цепи аминокислота R в положении 409 замещена на D, аминокислота К в положении замещена на Е, и в домене СН3 другой тяжелой цепи аминокислота D в положении 399 замещена на K и аминокислота Е а позиция 357 замещена на K (нумерация по Kabat, индекс EU).
В еще одном из вариантов осуществления настоящего изобретения домен Fc IgG1 включает одно или несколько аминокислотных замещений, которые уменьшают связывание с Fc-рецептором, в частности, с рецептором Fey.
Модификации домена Fc. снижающие связывание рецептора Fc и/или эффекторную функцию
Биспецифические антитела по настоящему изобретению могут содержать в качестве спейсерного домена домены тяжелой цепи молекулы иммуноглобулина. Например, домен Fc молекулы иммуноглобулина G (IgG) является димером, каждая субъединица которого содержит константные домены СН2 и СН3 тяжелых цепей IgG. Две субъединицы домена Fc способны стабильно ассоциироваться друг с другом. Область Fc придает благоприятные фармакокинетические свойства биспецифическим антителам по настоящему изобретению, включая длительное время полужизни в сыворотке, которое способствует хорошему накоплению в целевой ткани и благоприятное соотношение распределения между тканью и кровью. В то же время это может, однако, привести к нежелательному нацеливанию биспецифических антител по настоящему изобретению на клетки, экспрессирующие рецепторы Fc, а не на предпочтительные несущие антиген клетки. Соответственно, в определенных вариантах осуществления настоящего изобретения область Fc биспецифических антител по настоящему изобретению проявляет пониженное связывающее сродство с рецептором Fc и/или пониженную эффекторную функцию по сравнению с нативной областью Fc IgG, в частности областью Fc IgG1 или областью Fc IgG4. Более конкретно, областью Fc является область Fc IgG1.
В одном таком варианте осуществления настоящего изобретения область Fc (или биспецифическая антигенсвязывающая молекула по настоящему изобретению, содержащая указанную область Fc) проявляет менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и наиболее предпочтительно менее 5% аффинности к Fc-рецептору по сравнению с нативной Fc-областью IgG1 (или биспецифической антигенсвязывающей молекулой по настоящему изобретению, содержащей нативную Fc-область IgG1), и/или менее 50%, предпочтительно менее 20%, более предпочтительно менее чем 10% и наиболее предпочтительно менее 5% эффекторной функции по сравнению с нативной Fc-областью IgG1 (или биспецифической антигенсвязывающей молекулой по настоящему изобретению, содержащей нативную Fc-область IgG1). В одном варианте осуществления настоящего изобретения область Fc (или биспецифическая антигенсвязывающая молекула по настоящему изобретению, содержащая указанную область Fc) существенным образом не связывается с рецептором Fc и/или индуцирует эффекторную функцию. В одном из вариантов осуществления настоящего изобретения рецептором Fc является рецептор Fey. В другом варианте осуществления настоящего изобретения рецептором Fc является рецептор Fc человека. В еще одном варианте осуществления настоящего изобретения рецептором Fc является активирующий рецептор Fc. В другом варианте осуществления настоящего изобретения Fc-рецептором является активирующий Fcγ-рецептор человека, точнее FcγRIIIa, FcγRI или FcγRIIa человека, наиболее конкретно -FcγRIIIa человека. В другом варианте осуществления настоящего изобретения рецептором Fc является ингибирующий рецептор Fc. В конкретном варианте осуществления настоящего изобретения Fc-рецептором является ингибирующий Fcγ-рецептор человека, более конкретно FcγRIIB человека. В одном варианте осуществления настоящего изобретения эффекторная функция представляет собой одну или несколько из числа CDC, ADCC, ADCP и секреции цитокинов. В конкретном варианте осуществления настоящего изобретения эффекторной функцией является ADCC. В одном варианте осуществления настоящего изобретения домен области Fc проявляет существенно сходную аффинность с неонатальным рецептором Fc (FcRn) по сравнению с нативной областью Fc IgG1. Существенно сходное связывание с FcRn достигают, если область Fc (или биспецифическая антигенсвязывающая молекула по настоящему изобретению, содержащая указанную область Fc) демонстрирует примерно более чем 70%, особенно примерно более чем 80%, точнее примерно более 90% аффинности нативной области Fc IgG1 (или биспецифической антигенсвязывающей молекулы по настоящему изобретению, содержащей нативную область Fc IgG1) с FcRn.
В одном из вариантов осуществления настоящего изобретения область Fc сконструирована таким образом, чтобы иметь пониженную аффинность связывания с рецептором Fc и/или пониженную эффекторную функцию по сравнению с областью Fc, не подвергавшейся конструированию. В еще одном варианте осуществления настоящего изобретения область Fc биспецифической антигенсвязывающей молекулы по настоящему изобретению содержит одну или несколько аминокислотных мутаций, которые снижают аффинность связывания области Fc с рецепторной и/или эффекторной функцией Fc. Обычно одни и те же одна или несколько аминокислотных мутаций присутствуют в каждой из двух субъединиц области Fc. В одном варианте осуществления настоящего изобретения аминокислотная мутация снижает аффинность связывания области Fc с рецептором Fc. В другом варианте осуществления настоящего изобретения аминокислотная мутация снижает аффинность области Fc с рецептором Fc, по меньшей мере, в 2 раза, по меньшей мере, в 2,5 раза, по меньшей мере, в 5 раз, или по меньшей мере, в 10 раз. В еще одном из вариантов осуществления настоящего изобретения биспецифическая антигенсвязывающая молекула по настоящему изобретению, включающая сконструированную область Fc, проявляет менее 20%, в частности, менее 10%, более конкретно, менее 5% аффинности с рецептором Fc по сравнению с биспецифическими антителами по настоящему изобретению, содержащими не сконструированную область Fc. В одном из вариантов осуществления настоящего изобретения рецептор Fc является рецептором Fey. В других вариантах осуществления настоящего изобретения рецептор Fc является рецептором Fc человека. В еще одном из вариантов осуществления настоящего изобретения рецептор Fc является ингибирующим рецептором Fc. В другом варианте осуществления настоящего изобретения рецептор Fc является ингибирующим рецептором Fey человека, точнее рецептором FcγRIIB человека. В некоторых вариантах осуществления настоящего изобретения рецептор Fc является активирующим рецептором Fc. В еще одном из вариантов осуществления настоящего изобретения рецептор Fc является активирующим рецептором Fey человека, точнее активирующим рецептором FcγRIIIa, FcγRI или FcγRIIa человека, и наиболее точно FcγRIIIa человека. Предпочтительно, связывание с каждым из этих рецепторов понижено. В некоторых вариантах осуществления настоящего изобретения аффинность с компонентом комплемента, специфическая аффинность с C1q, также понижена. В одном из вариантов осуществления настоящего изобретения аффинность с неонатальным рецептором Fc (neonatal Fc receptor, FcRn) не понижена. По существу аналогичное связывание с FcRn, то есть сохранение аффинности области Fc с указанным рецептором, достигается, если область Fc (или биспецифическая антигенсвязывающая молекула по настоящему изобретению, включающая указанную Fc область) проявляет более чем примерно 70%-ную аффинность не сконструированной формы области Fc (или биспецифическая антигенсвязывающая молекула по настоящему изобретению, включающая указанную не сконструированную форму Fc области) относительно FcRn. Область Fc, или биспецифическая антигенсвязывающая молекула по настоящему изобретению, содержащая указанную область Fc, могут проявлять более примерно 80% и даже более примерно 90% такой аффинности. В некоторых вариантах осуществления настоящего изобретения область Fc биспецифической антигенсвязывающей молекулы по настоящему изобретению конструируют таким образом, чтобы они имели пониженную эффекторную функцию по сравнению с не измененной областью Fc. Пониженная эффекторная функция может включать одну или несколько из следующих функций, хотя ими перечень не ограничивается: пониженную комплемент-зависимую цитотоксичность (complement dependent cytotoxicity - CDC), пониженную антитело-зависимую клеточно-опосредованную цитотоксичность (antibody-dependent cell-mediated cytotoxicity - ADCC), пониженный антитело-зависимый клеточный фагоцитоз (antibody-dependent cellular phagocytosis ADCP), пониженную секрецию цитокинов, пониженное опосредованное иммунным комплексом поглощение антигена антиген-презентирующими клетками, пониженное связывание с клетками NK, пониженное связывание с макрофагами, пониженное связывание с моноцитами, пониженное связывание с полиморфно-ядерными клетками, пониженную индуцированную апоптозом прямую передачу сигнала, уменьшенное созревание дендритных клеток или пониженное примирование Т-клеток.
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, которое обладает некоторыми, но не всеми эффекторными функциями, что делает его желательным кандидатом для применения в тех случаях, когда период полужизни in vivo важен, но некоторые эффекторные функции (такие как комплемент и ADCC) не нужны или вредны. In vitro и/или in vivo анализ цитотоксичности может проводиться для подтверждения снижения/истощения активности CDC и/или ADCC. Например, анализы связывания с рецептором Fc (FcR) могут проводиться для подтверждения, что циклический гибридизированный полипептид не связывается с FcγR (следовательно, вероятно, не обладает активностью ADCC), но сохраняет способность связываться с FcRn.
Соответственно, в конкретных вариантах осуществления настоящего изобретения домен Fc биспецифического антитела по настоящему изобретению проявляет пониженную аффинность связывания с рецептором Fc и/или пониженную эффекторную функцию по сравнению с нативным Fc-доменом IgG1. В одном варианте осуществления настоящего изобретения Fc, по существу, не связывается с рецептором Fc и/или не индуцирует эффекторную функцию. В одном из вариантов осуществления настоящего изобретения рецептор Fc является рецептором Fcγ. В одном из вариантов осуществления настоящего изобретения рецептор Fc является рецептором Fc человека. В другом варианте осуществления настоящего изобретения Fc-рецептор является активирующий рецептор Fcγ человека, точнее FcγRIIIa, FcγRI или FcγRIIa человека, наиболее конкретно FcγRIIIa человека. В другом варианте осуществления настоящего изобретения домен Fc не индуцирует эффекторной функции. Пониженная эффекторная функция может включать, но ими перечень не ограничивается, одну или несколько из следующих функций: пониженная комплемент-зависимая цитотоксичность (CDC), пониженная антителозависимая клеточно-опосредованная цитотоксичность (ADCC), пониженный антителозависимый клеточный фагоцитоз (ADCP), пониженная секреция цитокинов, пониженное опосредованное иммунным комплексом поглощение антигена антигенпрезентирующими клетками, пониженное связывание с NK-клетками, пониженное связывание с макрофагами, пониженное связывание с моноцитами, пониженное связывание с полиморфно-ядерными клетками, пониженная прямая передача сигнала, вызывающая апоптоз, пониженное созревание дендритных клеток, пониженное примирование Т-клеток.
В некоторых вариантах осуществления настоящего изобретения одна или несколько аминокислотных модификаций могут быть интродуцированы в область в Fc биспецифической антигенсвязывающей молекулы, предусмотренной в настоящем изобретении, тем самым образуя вариант области Fc. Вариант области Fc может содержать последовательность области Fc человека (например, область Fc IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замещение) по одному или нескольким аминокислотным положениям.
В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическую антигенсвязывающую молекулу, в которой спейсерный домен включает домен Fc, который содержит одно или несколько аминокислотных замещений, которые уменьшают связывание с рецептором Fc, в частности, с рецептором Fcγ.
В некоторых вариантах осуществления настоящего изобретения домен Fc биспецифической антигенсвязывающей молекулы по настоящему изобретению содержит одну или несколько аминокислотных мутаций, которые снижают связывающее сродство (аффинность) домена Fc с рецептором Fc и/или эффекторную функцию. Обычно одна и та же одна или несколько аминокислотных мутаций присутствует в каждой из двух субъединиц домена Fc. В частности, домен Fc содержит аминокислотную замену в положении Е233, L234, L235, N297, Р331 и Р329 (нумерация EU). В частности, домен Fc содержит аминокислотные замещения в положениях 234 и 235 (нумерация EU) и/или 329 (нумерация EU) тяжелых цепей IgG. В частности, предложена биспецифическая антигенсвязывающая молекула по настоящему изобретению, которая содержит домен Fc с аминокислотными заменами L234A, L235A и P329G («P329G LALA», нумерация по Kabat, индекс EU) в тяжелых цепях IgG. Аминокислотные замещения L234A и L235A относятся к так называемым мутациям LALA. Комбинация «P329G LALA» аминокислотных замещений почти полностью устраняет связывание Fcγ-рецептора Fc-домена IgG1 человека и описана в патентной заявке WO 2012/130831 А1, которая также описывает методы получения таких мутантных доменов Fc и методы определения их свойств, например, связывания с Fc-рецептором или эффекторные функции. «Нумерация EU» означает нумерацию по индексу EU в публикации Kabat с соавт. в кн.: «Sequences of Proteins of Immunological Interest)), 5e изд., 1991, Public Health Service, National Institutes of Health, Бетесда, Мэриленд).
Домены Fc с пониженным связыванием рецептора Fc и/или пониженной эффекторной функцией также включают домены с заменой одного или нескольких остатков в домене Fc 238, 265, 269, 270, 297, 327 и 329 (US 6737056). Такие мутанты Fc включают те мутанты Fc, в которых есть заменами в двух или нескольких положениях аминокислот 265, 269, 270, 297 и 327, включая так называемый мутант Fc «DANA» с заменой остатков 265 и 297 на аланин (US 7332581).
В другом варианте осуществления настоящего изобретения доменом Fc является домен Fc IgG4. Антитела IgG4 проявляют пониженную аффинность связывания с Fc-рецепторами и пониженные эффекторные функции по сравнению с антителами IgG1. В более конкретном варианте доменом Fc является домен Fc IgG4, содержащий аминокислотную замену в положении S228 (нумерация по Kabat), в частности аминокислотную замену S228P. В более конкретном варианте осуществления настоящего изобретения домен Fc является домен Fc IgG4, содержащий аминокислотные замены L235E, S228P и P329G (нумерация EU). Такие мутанты Fc-домена IgG4 и их способность связывания с рецептором Fcγ также описаны в WO 2012/130831.
Мутантные Fc-домены могут быть получены путем делеции, замещения, вставки или модификации аминокислот с использованием генетических или химических методов, хорошо известных в данной области. Генетические методы могут включать сайт-специфический мутагенез кодирующей последовательности ДНК, ПЦР, синтез генов и другие. Правильность изменения нуклеотидов можно проверить, например, секвенированием.
Связывание с Fc-рецепторами может быть легко определено, например, с помощью ELISA или поверхностного плазмонного резонанса (SPR) с использованием стандартного инструментария, например, BIAcore (фирма GE Healthcare), и Fc-рецепторов, которые могут быть получены рекомбинантной экспрессией. Применимый анализ связывания описан в настоящем изобретении. В другом варианте осуществления настоящего изобретения аффинность Fc-доменов или активирующих клетки биспецифических антигенсвязывающих молекул, содержащих Fc-домен для Fc-рецепторов, может быть оценена с использованием клеточных линий, о которых известно, что они экспрессируют конкретные Fc-рецепторы, например, NK-клетки человека, экспрессирующие FcγIIIa-рецептор.
Эффекторная функция Fc-домена или биспецифических антител по настоящему изобретению, содержащих Fc-домен, может быть измерена способами, известными в данной области. В настоящем изобретении описан анализ для измерения ADCC, который может быть применим. Другие примеры исследований in vitro для оценки активности ADCC молекул, представляющих интерес, описаны в US 5500362; Hellstrom с соавт. Proc Natl Acad Sci USA 83, 1986, 7059-7063; Hellstrom с соавт., Proc Natl Acad Sci USA 82, 1985, 1499-1502; US 5821337; Bruggemann с соавт., J Exp Med 166, 1987, 1351-1361. В другом варианте, могут быть применены не радиоактивные методы исследования (см., например, ACTI™ не радиоактивный метод исследования для жидкостной цитометрии (фирма CellTechnology, Inc., Mountain View, Калифорния); и CytoTox 96® не радиоактивное исследование цитотоксичности (фирма Promega, Мэдисон, Висконсин)). К эффекторным клеткам, которые могут применяться в таком исследовании, относят мононуклеарные клетки переферической крови (МКПК) и природные клетки-киллеры (Natural Killer, NK). В другом варианте, или дополнительно, активность ADCC молекул, представляющих интерес, может быть оценена in vivo, например, в модельных животных, например, как описано в публикации Clynes с соавт., Proc Natl Acad Sci USA 95, 1998, 652-656.
В некоторых вариантах осуществления настоящего изобретения понижено связывание домена Fc с компонентом комплемента, специфичным в отношении C1q. Соответственно, в определенных вариантах осуществления настоящего изобретения, в которых домен Fc сконструирован для понижения эффекторной функции, указанная пониженная эффекторная функция включает пониженную комплемент-зависимую цитотоксичность (complement dependent cytotoxicity, CDC). Исследования связывания C1q могут быть выполнены, чтобы определить, способны ли биспецифические антитела по настоящему изобретению связывать C1q и, следовательно, обладают ли они активностью CDC. См., например, связывание методом ELISA C1q и С3с в WO 2006/029879 и WO 2005/100402. Для оценки активирования комплемента может быть проведен анализ CDC (см., например, Gazzano-Santoro с соавт., J Immunol Methods 202, 1996, 163; Cragg с соавт., Blood 101, 2003, 1045-1052; Cragg и Glennie, Blood 103, 2004, 2738-2743).
В некоторых вариантах осуществления настоящего изобретения биспецифические антигенсвязывающие молекулы содержат все положения в соответствии с нумерацией по Kabat, индекс EU:
i) гомодимерную Fc-область подкласса IgG1 человека необязательно с мутациями P329G, L234A и L235A, или
ii) гомодимерную Fc-область подкласса IgG4 человека необязательно с мутациями P329G, S228P и L235E, или
iii) гомодимерную Fc-область подкласса IgG1 человека необязательно с мутациями P329G, L234A, L235A, I253A, Н310А и Н435А, или необязательно с мутациями P329G, L234A, L235A, Н310А, Н433А и Y436A, или
iv) гомодимерную Fc-область, в которой
а) один полипептид Fc-области включает мутацию T366W, и другой полипептид Fc-области включает мутации T366S, L368A и Y407V, или
б) один полипептид Fc-области включает мутации T366W и Y349C, и другой полипептид Fc-области включает мутации T366S, L368A, Y407V и S354C, или
в) один полипептид Fc-области включает мутации T366W и S354C, и другой полипептид Fc-области включает мутации T366S, L368A, Y407V и Y349C, или
v) гетеродимерную Fc-область подкласса IgG1 человека, в которой оба полипептида Fc-области включают мутации P329G, L234A и L235A, и
а) один полипептид Fc-области включает мутацию T366W, и другой полипептид Fc-области включает мутации T366S, L368A и Y407V, или
б) один полипептид Fc-области включает мутации T366W и Y349C, и другой полипептид Fc-области включает мутации T366S, L368A, Y407V и S354C, или
в) один полипептид Fc-области включает мутации T366W и S354C, и другой полипептид Fc-области включает мутации T366S, L368A, Y407V и Y349C, или
vi) гетеродимерную Fc-область подкласса IgG4 человека, в которой оба полипептида Fc-области включают мутации P329G, S228P и L235E, и
а) один полипептид Fc-области включает мутацию T366W, и другой полипептид Fc-области включает мутации T366S, L368A и Y407V, или
б) один полипептид Fc-области включает мутации T366W и Y349C, и другой полипептид Fc-области включает мутации T366S, L368A, Y407V и S354C, или
в) один полипептид Fc-области включает мутации T366W и S354C, и другой полипептид Fc-области включает мутации T366S, L368A, Y407V и Y349C,
или
vii) комбинация i), ii) и iii) с одним из vi), v) и vi).
С-конец гибридизированных полипептидов, содержащихся в биспецифических антителах, как указано в настоящем изобретении, может быть полным С-концом, заканчивающимся аминокислотными остатками PGK. С-конец может быть укороченным С-концом, в котором один или два С-концевых аминокислотных остатка удалены. В одном предпочтительном варианте осуществления настоящего изобретения С-конец представляет собой укороченный С-конец, заканчивающийся аминокислотными остатками PG.
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором один антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью, является антигенсвязывающим доменом, способным к специфическому связыванию с опухолеассоциированным антигеном (ТАА). В частности, опухолеассоциированный антиген является белком активации фибробластов (Fibroblast Activation Protein, FAP). В одном варианте осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный к специфическому связыванию со второй мишенью, является антигенсвязывающим доменом, способным к специфическому связыванию с белком активации фибробластов (Fibroblast Activation Protein, FAP).
Биспецифические антитела, связывающиеся с рецептором TNF и FAP
В некоторых вариантах осуществления настоящего изобретения
антигенсвязывающий домен, способный специфически связываться с FAP, содержит
(а) вариабельную область тяжелой цепи (VHFAP), включающую (i) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:1, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:2, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:3, и вариабельную область легкой цепи (VLFAP), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:4, (v) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:5, и (vi) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:6, или
(б) вариабельную область тяжелой цепи (VHFAP), включающую (i) CDR-Н1, содержащую аминокислотную последовательность SEQ ID NO:9, (ii) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:10, и (iii) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:11, и вариабельную область легкой цепи (VLFAP), включающую (iv) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:12, (v) CDR-L2, включающую SEQ ID NO:13, и (vi) CDR-L3, включающую SEQ ID NO:14.
Точнее, антигенсвязывающий домен, способный специфически связываться с FAP, содержит
(а) вариабельную область тяжелой цепи (VHFAP), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:7, и вариабельную область легкой цепи (VLFAP), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:8, или
(б) вариабельную область тяжелой цепи (VHFAP), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:7, и вариабельную область легкой цепи (VLFAP), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:16.
В одном варианте осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO:7, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO:8. В другом варианте осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с FAP, состоит из вариабельной области тяжелой цепи (VHFAP), содержащей аминокислотную последовательность SEQ ID NO:7, и вариабельной области легкой цепи (VLFAP), содержащей аминокислотную последовательность SEQ ID NO:8.
В еще одном варианте осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO:15, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO:16. В другом варианте осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с FAP, состоит из вариабельной области тяжелой цепи (VHFAP), содержащей аминокислотную последовательность SEQ ID NO:15, и вариабельной области легкой цепи (VLFAP), содержащей аминокислотную последовательность SEQ ID NO:16.
В другом варианте осуществления настоящего изобретения биспецифические антитела, предусмотренные в настоящем изобретении, связываются моновалентно с FAP.
Биспецифические антитела, связывающиеся с ОХ40 и FAP
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифичное антитело, в котором антигенсвязывающий домен, способный специфически связываться с первой мишенью, является антигенсвязывающим доменом, способным специфически связываться с рецептором TNF, в частности костимулирующим рецептором TNF. В частности, костимулирующим рецептором TNF является ОХ40. В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный к специфическому связыванию с первой мишенью, является антигенсвязывающим доменом, способным к специфическому связыванию с ОХ40. В частности, биспецифическое антитело по настоящему изобретению содержит два антигенсвязывающих домена, способных специфически связываться с ОХ40.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит
(а) вариабельную область тяжелой цепи (VHOX40), содержащую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:22, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:35, или
(б) вариабельную область тяжелой цепи (VHOX40), содержащую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:21, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:34, или
(в) вариабельную область тяжелой цепи (VHOX40), содержащую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:23, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:36, или
(г) вариабельную область тяжелой цепи (VHOX40), включающую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:24, и вариабельную область легкой цепи (VlOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:37, или
(д) вариабельную область тяжелой цепи (VHOX40), содержащую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:18, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:20, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:25, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:29, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:32, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:38, или
(е) вариабельную область тяжелой цепи (VHOX40), содержащую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:18, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:20, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:26, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:29, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:32, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:38, или
(ж) вариабельную область тяжелой цепи (VHOX40), содержащую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:18, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:20, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:27, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:30, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:33, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:39.
В частности, антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит вариабельную область тяжелой цепи (VHOX40), содержащую (i) CDR-H1, включающую аминокислотную последовательность SEQ ID NO:17, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:19, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:22, и вариабельную область легкой цепи (VLOX40), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:28, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:31, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:35.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит
(а) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:40, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:41, или
(б) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:42, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:43, или
(в) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:44, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:45, или
(г) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:46, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:47, или
(д) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:48, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:49, или
(е) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:50, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:51, или
(ж) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:52, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:53.
В одном из вариантов осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит (а) вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:40, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:41.
В одном из вариантов осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с ОХ40, содержит вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:40, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:41.
Точнее, настоящее изобретение предусматривает биспецифическое антитело, которое содержит
(а) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:54, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:55,
(б) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:56, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:57,
(в) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:58, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:59,
(г) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:60, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:61,
(д) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:62, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:63,
(е) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:64, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:65, или
(ж) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:66, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:67.
В одном из вариантов осуществления настоящего изобретения биспецифическое антитело содержит
(а) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:54, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:55,
(б) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:56, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:57,
(в) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:58, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:59,
(г) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:60, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:61,
(д) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:62, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:63,
(е) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:64, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:65,
(ж) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:66, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:67.
Кроме того, в настоящем изобретении предусматривают биспецифическое антитело, которое содержит
(а) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:116, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:117,
(б) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:118, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:119,
(в) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:120, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:121,
(г) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:122, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:123,
(д) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:124, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:125,
(е) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:126, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:128,
(ж) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:129, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:130,
(з) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:131, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:132, или
(и) первый гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:133, и второй гибридизированный полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:134.
В одном из вариантов осуществления настоящего изобретения биспецифическое антитело содержит
(а) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:116, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:117,
(б) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:118, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:119,
(в) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:120, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:121,
(г) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:122, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:123,
(д) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:1246, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:125,
(е) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:126, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:127,
(ж) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:129, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:130,
(з) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:131, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:132, или
(и) первый гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:133, и второй гибридизированный полипептид, содержащий аминокислотную последовательность SEQ ID NO:134.
Биспецифические антитела, связывающиеся с 4-1ВВ и FAP
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный к специфическому связыванию с первой мишенью, является антигенсвязывающим доменом, способным к специфическому связыванию с рецептором TNF, где костимулирующим рецептором TNF является 4-1ВВ. В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный к специфическому связыванию с первой мишенью, является антигенсвязывающим доменом, способным к специфическому связыванию с 4-1ВВ. В частности, биспецифическое антитело по настоящему изобретению содержит два антигенсвязывающих домена, способных специфически связываться с 4-1ВВ.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий агент, способный специфически связываться с 4-1ВВ, содержит вариабельную область тяжелой цепи (VH4-1BB), содержащую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:135, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:136, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:137, и вариабельную область легкой цепи (VH4-1BB), содержащую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:138, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:139, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:140. В одном из вариантов осуществления настоящего изобретения антигенсвязывающий агент, способный специфически связываться с 4-1ВВ, содержит вариабельный домен тяжелой цепи (VH4-1BB), содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:141, и вариабельный домен легкой цепи (VL4-1ВВ), содержащий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:142.
В одном из вариантов осуществления настоящего изобретения антигенсвязывающий агент, способный специфически связываться с 4-1ВВ, содержит вариабельную область тяжелой цепи (VH4-1BB), содержащую аминокислотную последовательность SEQ ID NO:141, и вариабельную область легкой цепи (VL4-1BB), включающую аминокислотную последовательность SEQ ID NO:142. В еще одном из вариантов осуществления настоящего изобретения антигенсвязывающий агент, способный специфически связываться с 4-1ВВ, состоит из вариабельной области тяжелой цепи (VH4-1BB), включающей аминокислотную последовательность SEQ ID NO:141, и вариабельной области легкой цепи (VL4-1BB), включающей аминокислотную последовательность SEQ ID NO:142.
В частности, настоящее изобретение предусматривает биспецифическое антитело, которое содержит
(а) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:143, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:144, или
(б) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:145, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:146.
В одном из вариантов осуществления настоящего изобретения биспецифическое антитело содержит
(а) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:143, и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:144, или
(б) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:145, и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:146.
Биспецифические антитела, связывающиеся с CD40 и FAP
В некоторых вариантах осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный специфически связываться с первой мишенью, является антигенсвязывающим доменом, способным специфически связываться с рецептором TNF, причем костимулирующим рецептором TNF является CD40. В одном из вариантов осуществления настоящего изобретения предусматривают биспецифическое антитело, в котором антигенсвязывающий домен, способный специфически связываться с первой мишенью, является антигенсвязывающий домен, способный специфически связываться с CD40. В частности, биспецифическое антитело по настоящему изобретению содержит два антигенсвязывающих домена, способных специфически связываться с CD40.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с CD40, содержит вариабельную область тяжелой цепи (VHCD40), включающую (i) CDR-Н1, включающую аминокислотную последовательность SEQ ID NO:147, (ii) CDR-H2, включающую аминокислотную последовательность SEQ ID NO:148, и (iii) CDR-H3, включающую аминокислотную последовательность SEQ ID NO:149, и вариабельную область легкой цепи (VLCD40), включающую (iv) CDR-L1, включающую аминокислотную последовательность SEQ ID NO:150, (v) CDR-L2, включающую аминокислотную последовательность SEQ ID NO:151, и (vi) CDR-L3, включающую аминокислотную последовательность SEQ ID NO:152. В одном из вариантов осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с CD40, содержит вариабельную область тяжелой цепи (VHCD40), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:153, и вариабельную область легкой цепи (VLCD40), включающую аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:154.
В другом варианте осуществления настоящего изобретения антигенсвязывающий домен, способный к специфическому связыванию с первой мишенью, является антигенсвязывающим доменом, способным к специфическому связыванию с CD40, который включает
(i) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:167, SEQ ID NO:168, SEQ ID NO:169 и SEQ ID NO:170, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:171, SEQ ID NO:172, SEQ ID NO:173 и SEQ ID NO:174, или
(ii) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:175, SEQ ID NO:176, SEQ ID NO:177, SEQ ID NO:178, SEQ ID NO:179 и SEQ ID NO:180, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:181, SEQ ID NO:182, SEQ ID NO:183 и SEQ ID NO:184.
В одном варианте осуществления настоящего изобретения антигенсвязывающий домен, способный к специфическому связыванию с первой мишенью, является антигенсвязывающим доменом, способным к специфическому связыванию с CD40, который включает
(а) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO:153, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO:154, или
(б) вариабельную область тяжелой цепи (VHCD40), содержащую аминокислотную последовательность SEQ ID NO:167, и вариабельную область легкой цепи (VLCD40), содержащую аминокислотную последовательность SEQ ID NO:171.
В частности, настоящее изобретение предусматривает биспецифическое антитело, которое содержит
(а) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:155, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:156,
(б) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:157, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:158,
(в) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:159, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:160,
(г) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:161, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:162,
(д) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:163, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:164, или
(е) первый гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:165, и второй гибридизированный полипептид, включающий аминокислотную последовательность, которая, по меньшей мере, примерно на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO:166.
В одном из вариантов осуществления настоящего изобретения биспецифическое антитело содержит
(а) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:155 и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:156,
(б) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:157 и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:158,
(в) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:159 и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:160,
(г) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:161 и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:162,
(д) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:163 и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:164, или
(е) первый гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:165 и второй гибридизированный полипептид, включающий аминокислотную последовательность SEQ ID NO:166.
Модификации в доменах Fab
В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, содержащему (а) два фрагмента Fab, способных специфически связываться с ОХ40, и (б) один фрагмент Fab, способный специфически связываться с FAP, где в одном из (а) и (б) во фрагментах Fab либо вариабельные домены VH и VL, либо константные домены СН1 и CL заменяются. Биспецифичные антитела получают по технологии Crossmab. Биспецифические антитела получают методом Crossmab.
Мультиспецифические антитела с перемещением/заменой домена в одном связывающем плече (CrossMabVH-VL или CrossMabCH-CL) подробно описаны в WO 2009/080252 и публикации Schaefer W. с соавт., PNAS, 108, 2011, 11187-1191. Они явно уменьшают побочные продукты, вызванные несоответствием легкой цепи против первого антигена с неправильной тяжелой цепью против второго антигена (по сравнению с подходами без такого обмена доменов).
В еще одном варианте осуществления настоящее изобретение относится к биспецифическому антителу, содержащему (а) два Fab-фрагмента, способных специфически связываться с ОХ40, и (б) Kpocc-Fab-фрагмент, способный специфически связываться с FAP, в котором константные домены CL (Ckappa) и СН1 заменены друг на друга таким образом, что домен СН1 гибридизирован с доменом VH.
В другом варианте осуществления настоящее изобретение относится к биспецифическому антителу, содержащему (а) два Kpocc-Fab-фрагмента, способных специфически связываться с ОХ40, в котором домен VH гибридизирован с доменом CL (Ckappa) и домен VL гибридизирован с доменом СН1, и (б) Fab-фрагмент, способный специфически связываться с FAP.
В другом варианте осуществления настоящего изобретения и для дополнительного улучшения правильного спаривания биспецифическое антитело может содержать замены различными заряженными аминокислотами (так называемыми «заряженными остатками»). Эти модификации интродуцируют в скрещенные или нескрещенные домены СН1 и CL. В одном из вариантов осуществления настоящее изобретение относится к биспецифическому антителу, в котором в одном из доменов CL аминокислота в положении 123 (нумерация EU) заменена на аргинин (R), а аминокислота в положении 124 (нумерация EU) заменена на лизин (К) (положительные заряды), и в котором в одном из доменов СН1 аминокислоты в положении 147 (нумерация EU) и в положении 213 (нумерация EU) были замещены на глутаминовую кислоту (Е) (отрицательные заряды).
Более конкретно, настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, содержащей Fab, причем в домене CL аминокислота в положении 123 (нумерация EU) заменена на аргинин (R) и аминокислота в положении 124 (нумерация EU) была замещена на лизин (K), и где в домене СН1 аминокислоты в положении 147 (нумерация EU) и в положении 213 (нумерация EU) замещены на глутаминовую кислоту (Е).
Соответственно, в некоторых вариантах осуществления настоящего изобретения один или несколько фрагментов Fab (например, фрагментов Fab, способных специфически связываться с ОХ40) биспецифической антигенсвязывающей молекулы по настоящему изобретению содержат домен CL, содержащий аргинин (R) в положении 123 (Нумерация EU) и лизин (К) в положении 124 (нумерация EU), а также домен СН1, содержащий глутаминовую кислоту (Е) в положении 147 (нумерация EU) и глутаминовую кислоту (Е) в положении 213 (нумерация в EU).
Полинуклеотиды
Настоящее изобретение также предусматривает выделенную нуклеиновую кислоту, кодирующую биспецифическое антитело, описанное в настоящем изобретении, или его фрагмент.
Выделенная нуклеиновая кислота, кодирующая биспецифическое антитело по настоящему изобретению, может быть выражена в виде одного полинуклеотида, который кодирует всю антигенсвязывающую молекулу, или в виде нескольких (например, двух или более) полинуклеотидов, которые совместно экспрессируются. Полипептиды, кодируемые полинуклеотидами, которые совместно экспрессируются, могут связываться, например, через дисульфидные связи или другими способами, с образованием функциональной антигенсвязывающей молекулы. При совместной экспрессии гибридизированные полипептиды могут связываться с образованием антигенсвязывающего домена, способного специфически связываться со второй мишенью (например, FAP). Антигенсвязывающие домены, способные специфически связываться с первой мишенью (например, ОХ40), могут кодироваться одним полинуклеотидом. При совместной экспрессии гибридизированные полипептиды объединяются с образованием биспецифического антитела.
В некоторых вариантах осуществления настоящего изобретения полинуклеотидом или нуклеиновой кислотой является ДНК. В других вариантах осуществления настоящего изобретения полинуклеотидом является РНК, например, информационная (или матричная) РНК (мРНК). РНК по настоящему изобретению может быть одноцепочечной или двухцепочечной.
В другом варианте осуществления настоящего изобретения предусматривают выделенный полинуклеотид, кодирующий гибридизированный полипептид, описанный выше. Настоящее изобретение также предусматривает вектор, в частности экспрессирующий вектор, содержащий выделенный полинуклеотид по изобретению, и клетку-хозяина, содержащую выделенный полинуклеотид или вектор по настоящему изобретению. В некоторых вариантах осуществления настоящего изобретения клетка-хозяин представляет эукариотическую клетку, в частности клетку млекопитающего.
В другом варианте осуществления настоящего изобретения предусматривают метод получения биспецифического антитела по настоящему изобретению, включающий стадии (i) культивирования клетки-хозяина по настоящему изобретению в условиях, пригодных для экспрессии указанного антитела, и (ii) выделения указанного биспецифического антитела. Изобретение также охватывает биспецифическое антитело, полученное указанным методом.
Методы рекомбинации
Биспецифические антигенсвязывающие молекулы по настоящему изобретению могут быть получены, например, твердофазным пептидным синтезом (например, твердофазным синтезом Меррифильда) или рекомбинантным получением. Для рекомбинантного получения один или несколько полинуклеотидов, кодирующих антигенсвязывающую молекулу или ее полипептидные фрагменты, например, описанные выше, выделяют и инсертируют в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такой полинуклеотид может быть легко выделен и секвенирован с использованием обычных процедур. В одном варианте осуществления настоящего изобретения предусматривают вектор, предпочтительно вектор экспрессии, содержащий один или несколько полинуклеотидов по настоящему изобретению. Методы, которые хорошо известны специалистам в данной области, могут быть использованы для конструирования векторов экспрессии, содержащих кодирующую последовательность биспецифической антигенсвязывающей молекулы (фрагмента) вместе с соответствующими сигналами контроля транскрипции/трансляции. Эти методы включают методы рекомбинации ДНК in vitro, методы синтеза и рекомбинации/генетической рекомбинации in vivo. См., например, методы в работах Maniatis с соавт., MOLECULAR CLONING: А LABORATORY MANUAL, изд. Cold Spring Harbor Laboratory, Нью-Йорк (1989); and Ausubel с соавт., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, изд. Greene Publishing Associates and Wiley Interscience, Нью-Йорк (1989). Вектор экспрессии может быть частью плазмиды, вируса или может быть фрагментом нуклеиновой кислоты. Вектор экспрессии включает кассету экспрессии, в которой полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу или ее полипептидные фрагменты (то есть кодирующую область), клонируют в функциональной ассоциации с промотором и/или другими элементами контроля транскрипции или трансляции. В настоящем изобретении понятие «кодирующая область» означает часть нуклеиновой кислоты, которая состоит из кодонов, транслируемых в аминокислоты. Хотя «стоп-кодон» (TAG, TGA или ТАА) не транслируется в аминокислоту, его можно рассматривать как часть кодирующей области, если она присутствует, но какие-либо фланкирующие последовательности, например промоторы, сайты связывания рибосом, терминаторы транскрипции, интроны, 5' и 3' нетранслируемые области и т.п. не являются частью кодирующей области. Две или более кодирующих областей могут быть в одной полинуклеотидной конструкции, например, на в векторе или в отдельных полинуклеотидных конструкциях, например на отдельных (разных) векторах. Кроме того, какой-либо вектор может содержать одну кодирующую область или может содержать две или более кодирующих областей, например, вектор по настоящему изобретению может кодировать один или несколько полипептидов, которые после или одновременно с трансляцией разделяются на итоговые белки посредством протеолитического расщепления. Кроме того, вектор, полинуклеотид или нуклеиновая кислота по настоящему изобретению могут кодировать гетерологичные кодирующие области, либо гибридизированные, либо не гибридизированные с полинуклеотидом, кодирующим биспецифическую антигенсвязывающую молекулу по настоящему изобретению или ее полипептидные фрагменты, или их варианты, или производные. Гетерологичные кодирующие области включают без ограничения специализированные элементы или мотивы, например секреторный сигнальный пептид или гетерологичный функциональный домен. Функциональную ассоциацию наблюдают в тех случаях, когда область кодирования генного продукта, например, полипептида, связана с одной или несколькими регуляторными последовательностями таким образом, чтобы поставить экспрессию продукта гена под воздействие или контроль регуляторной последовательности (последовательностей). Два фрагмента ДНК (например область, кодирующая полипептид и связанный с ним промотор) являются «функционально связанными», если индукция функции промотора приводит к транскрипции мРНК, кодирующей требуемый генный продукт, и если природа связи между двумя фрагментами ДНК не мешает способности последовательностей регуляции экспрессии направлять экспрессию генного продукта или мешать способности транскрибировать матрицу ДНК. Таким образом, область промотора была бы функционально связана с нуклеиновой кислотой, кодирующей полипептид, если бы промотор был способен осуществлять транскрипцию этой нуклеиновой кислоты. Промотор может быть специфичным для клеток промотором, который направляет существенную транскрипцию ДНК только в заранее определенные клетки. Другие элементы контроля транскрипции, помимо промотора, например энхансеры, операторы, репрессоры и сигналы терминации транскрипции, могут быть функционально связаны с полинуклеотидом для направления специфичной для клетки транскрипции.
Подходящие промоторы и другие области контроля транскрипции предусмотрены в настоящем изобретении. Специалистам в данной области известно множество областей контроля транскрипции. К ним относят, но ими перечень не ограничивается, области контроля транскрипции, которые функционируют в клетках позвоночных, например, но ими перечень не ограничивается, сегменты промотора и энхансера цитомегаловирусов (например, немедленно-ранний промотор в сочетании с интроном-А), вируса обезьяны 40 (например, ранний промотор) и ретровирусов (например, вируса саркомы Рауса). Другие области контроля транскрипции включают области, полученные из генов позвоночных, например, генов актина, белка теплового шока, бычьего гормона роста и α-глобина кролика, а также другие последовательности, способные контролировать экспрессию генов в эукариотических клетках. Дополнительные подходящие области контроля транскрипции включают тканеспецифичные промоторы и энхансеры, а также индуцируемые промоторы (например, промоторы, индуцируемые тетрациклинами). Аналогично, специалистам в данной области техники известны различные элементы контроля трансляции. К ним относят, но ими перечень не ограничивается, сайты связывания рибосомы, кодоны инициации и терминации трансляции, а также элементы, производные от вирусных систем (в частности, внутренний участок посадки рибосомы (internal ribosome entry site, IRES), также называемый последовательностью CITE). Кассета экспрессии может также включать другие признаки, например, точку начала репликации и/или элементы интеграции хромосом, например, ретровирусные длинные концевые повторы (long terminal repeat, LTR) или аденоассоциированные вирусные (adeno-associated viral, AAV) инвертированные концевые повторы (inverted terminal repeat, ITR).
Полинуклеотидные и нуклеиновые кислоты, кодирующие области по настоящему изобретению, могут быть связаны с дополнительными кодирующими областями, которые кодируют секреторные или сигнальные пептиды, которые направляют секрецию полипептида, кодируемого полинуклеотидом по настоящему изобретению. Например, если желательна секреция биспецифической антигенсвязывающей молекулы или ее полипептидных фрагментов, ДНК, кодирующая сигнальную последовательность, может быть размещена выше последовательности нуклеиновой кислоты, кодирующей биспецифическую антигенсвязывающую молекулу по настоящему изобретению или ее полипептидные фрагменты. Согласно сигнальной гипотезе, белки, секретируемые клетками млекопитающих, имеют сигнальный пептид или секреторную лидерную последовательность, которая отщепляется от зрелого белка после начала экспорта растущей белковой цепи через шероховатую эндоплазматическую сеть. Специалистам в данной области известно, что полипептиды, секретируемые клетками позвоночных, обычно имеют сигнальный пептид, слитый с N-концом полипептида, который отщепляется от транслированного полипептида с образованием секретированной или «зрелой» формы полипептида. В некоторых вариантах осуществления настоящего изобретения используют нативный сигнальный пептид, например, сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина, или функциональное производное этой последовательности, которое сохраняет способность направлять секрецию полипептида, который функционально с ним связан. В другом варианте осуществления настоящего изобретения может быть использован гетерологичный сигнальный пептид млекопитающего или его функциональное производное. Например, лидерная последовательность дикого типа может быть заменена лидерной последовательностью тканевого активатора плазминогена человека (tissue plasminogen activator, ТРА) или β-глюкуронидазы мыши.
ДНК, кодирующая короткую белковую последовательность, которая может быть использована для облегчения последующей очистки (например, гистидиновая метка) или для нанесения метки на гибридизированный белок, может быть включена в состав полинуклеотида, например, на его концах, который кодирует биспецифическую антигенсвязывающую молекулу по настоящему изобретению или его полипептидные фрагменты.
В другом варианте осуществления настоящего изобретения предусматривают клетку-хозяина, содержащую один или несколько полинуклеотидов по настоящему изобретению. В некоторых вариантах осуществления настоящего изобретения предусматривают клетку-хозяина, содержащую один или несколько векторов по настоящему изобретению. Полинуклеотиды и векторы могут включать какие-либо признаки, по отдельности или в комбинации, описанные в настоящем изобретении в отношении полинуклеотидов и векторов, соответственно. В одном варианте осуществления настоящего изобретения предусматривают клетку-хозяина, которая содержит (например, была трансформирована или трансфицирована) вектор, содержащий полинуклеотид, который кодирует (частично) биспецифическую антигенсвязывающую молекулу по настоящему изобретению. Используемое в настоящем изобретении понятие «клетка-хозяин» относится к какому-либо типу клеточной системы, которая может быть сконструирована для выработки гибридизированных белков по настоящему изобретению или их фрагментов. Клетки-хозяева, подходящие для репликации и поддержания экспрессии антигенсвязывающих молекул, известны в данной области. Такие клетки могут быть трансфицированы или трансдуцированы, в зависимости от ситуации, определенным вектором экспрессии, и большие количества клеток, содержащих вектор, могут быть выращены для высева в ферментеры большого объема для получения достаточных количеств антигенсвязывающей молекулы для клинических применений. Подходящие клетки-хозяева включают прокариотические микроорганизмы, например, Е. coli, или различные эукариотические клетки, например, клетки яичника китайского хомячка (Chinese hamster ovary cell, CHO), клетки эмбриональной почки человека (human embryonic kidney, НЕК), клетки насекомых или другие. Например, полипептиды могут продуцироваться в бактериях, в частности, когда не требуется гликозилирование. После экспрессии полипептид может быть выделен из биомассы бактериальных клеток в виде растворимой фракции и может быть очищен дополнительно. Помимо прокариотов могут быть применены эукариотические микроорганизмы, например, мицелиальные грибы или дрожжи, в качестве хозяев для клонирования или для экспрессии векторов, кодирующих полипептиды, включая такие грибы и штаммы дрожжей, у которых пути гликозилирования были «гуманизированы», что приводит к получению полипептида человека с частичным или полным гликозилированием. См. Gerngross, Nat Biotech 22, 2004, 1409-1414, и Li с соавт., Nat Biotech 24, 2006, 210-215).
Подходящие клетки-хозяева для экспрессии (гликозилированных) полипептидов также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток-хозяев также являются клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать вместе с клетками насекомых, особенно для трансфекции клеток Spodoptera frugiperda. Клетки растений также могут применяться в качестве клеток-хозяев. См., например, US 5959177, 6040498, 6420548, 7125978 и 6417429 (описание технологии PLANTIBODIES™ для получения антител в трансгенных растениях). Клетки позвоночных также могут применяться в качестве клеток-хозяев. Например, могут быть полезны линии клеток млекопитающих, которые адаптированы для роста в виде суспензии. Другими примерами полезных линий клеток-хозяев млекопитающих являются линия CV1 почек обезьян, трансформированная SV40 (COS-7); линия эмбриональной почки человека (клетки 293 или 293Т описаны, например, в публикации Graham с соавт., J Gen Virol 36, 1977, 59), клетки почки новорожденного хомяка (baby hamster kidney cell, BHK), клетки Сертоли мыши (клетки ТМ4, описанные, например, Mather, Biol Reprod 23, 1980, 243-251), клетки почки обезьяны (CV1), клетки почки африканской зеленой мартышки (VERO-76), клетки карциномы шейки матки человека (human cervical carcinoma cells, HELA), клетки почки собаки (MDCK), клетки печени серой крысы (buffalo rat liver, BRL 3А), клетки легких человека (W138), клетки печени человека (Hep G2), клетки опухоли молочной железы мыши (mouse mammary tumor, ММТ 060562), клетки TRI (описанные, например, Mather с соавт., Annals N.Y. Acad Sci 383, 1982, 44-68), клетки MRC 5 и клетки FS4. К другим линиям клеток млекопитающих, которые могут быть применены, относят клетки яичника китайского хомячка (СНО), включая клетки dhfr- СНО (Urlaub с соавт., Proc Natl Acad Sci USA 77, 1980, 4216); и линии клеток миеломы, например, YO, NS0, Р3Х63 и Sp2/0. Обзор некоторых линий клеток-хозяев млекопитающих, которые могут быть применены для выработки белка, см., например, Yazaki и Wu в кн.: Methods in Molecular Biology, том 248, 2003, 255-268, под ред. В.K.С. Lo, изд-во Humana Press, Тотова, Нью-Джерси. К клеткам-хозяевам относят культивируемые клетки, например культивируемые клетки млекопитающих, дрожжевые клетки, клетки насекомых, бактериальные клетки и клетки растений, и это лишь некоторые из них, а также клетки от трансгенных животных, трансгенных растении или культивируемых ткани растений или животных. В одном из вариантов осуществления настоящего изобретения клетка-хозяин является эукариотической клеткой, предпочтительно клеткой млекопитающих, например, клетки яичника китайского хомячка (СНО), клетки эмбриональной почки человека (human embryonic kidney, НЕК) или лимфоидные клетки (например, клетки Y0, NS0, Sp20). В данной области известны стандартные методы для экспрессии чужеродных генов в таких системах. Клетки, экспрессирующие полипептид, содержащий либо тяжелую, либо легкую цепь антигенсвязывающего домена, могут быть сконструированы так, чтобы также экспрессировать и вторую из цепей иммуноглобулина таким образом, что экспрессируемый продукт является антигенсвязывающим доменом, который содержит и тяжелую, и легкую цепь.
В другом варианте осуществления настоящего изобретения предусматривают метод получения биспецифического антитела по настоящему изобретению, включающий стадии (i) культивирования клеток-хозяев по настоящему изобретению в условиях, подходящих для экспрессии указанного биспецифического антитела, и (ii) выделения указанного биспецифического антитела из клеток-хозяев или из культуральной среды клеток-хозяев.
Компоненты биспецифического антитела генетически соединены друг с другом. Биспецифические антигенсвязывающие молекулы могут быть сконструированы таким образом, чтобы их компоненты были гибридизированы непосредственно друг с другом или опосредованно через линкерную последовательность. Состав и длина линкера могут быть определены в соответствии с методами, известными в данной области, и могут быть проверены на эффективность. Примеры линкерных последовательностей между различными компонентами биспецифических антигенсвязывающих молекул можно найти в настоящем описании. Дополнительные последовательности также при необходимости могут быть включены в сайт для расщепления и разделения на отдельные компоненты продукта гибридизации, если желательно, например, последовательности распознавания эндопептидазы.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий домен, способный специфически связываться с FAP (например, фрагментами Fab или scFv), образующими часть антитела, содержит, по меньшей мере, вариабельную область иммуноглобулина, способную связываться с FAP. Аналогично, в определенных вариантах осуществления настоящего изобретения фрагменты, способные специфически связываться с ОХ40 (например, фрагментами Fab или scFv), образующими часть биспецифического антитела, содержат, по меньшей мере, вариабельную область иммуноглобулина, способную связываться с ОХ40. Вариабельные области могут образовывать часть природных или не встречающихся в природе антител и их фрагментов и быть полученными из них. Способы получения поликлональных антител и моноклональных антител известны в данной области (см., например, Harlow и Lane, «Antibodies, a laboratory manual», Cold Spring Harbor Laboratory, 1988). Не встречающиеся в природе антитела могут быть сконструированы с использованием твердофазного пептидного синтеза, могут быть получены рекомбинантно (например, согласно описанному в US 4186567) или могут быть получены, например, путем скрининга комбинаторных библиотек, содержащих вариабельные тяжелые цепи и вариабельные легкие цепи (см., например, US 5969108).
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающие домены, способные специфически связываться с соответствующей мишенью (например, Fab-фрагменты или scFv), содержащейся в антигенсвязывающих молекулах по настоящему изобретению, сконструированы так, чтобы иметь повышенную аффинность, например, в соответствии с методами, описанными в публикации РСТ WO 2012/020006 (см. примеры, касающиеся созревания аффинности) или в Appl. Публ. №2004/0132066. Способность антигенсвязывающих молекул по настоящему изобретению связываться со специфическим антигенным детерминантом может быть измерена либо с помощью иммуносорбентного анализа (ELIS А), либо с помощью других методов, известных специалисту в данной области, например, метода поверхностного плазмонного резонанса (Liljeblad с соавт., Glyco J 17, 2000, 323-329) и традиционные методы анализа связывания (Heeley, Endocr Res 28, 2002, 217-229). Конкурентные анализы могут быть использованы для идентификации антигенсвязывающей молекулы, которая конкурирует с контрольным антителом за связывание с конкретным антигеном. В некоторых вариантах осуществления настоящего изобретения такая конкурирующая антигенсвязывающая молекула связывается с тем же эпитопом (например, линейным или конформационным эпитопом), который связан с контрольной антигенсвязывающей молекулой. Подробные варианты методов картирования эпитопа, с которым связывается антигенсвязывающая молекула, представлены в публикации Morris «Epitope Mapping Protocols)) в Methods in Molecular Biology, т. 66, 1996, изд. Humana Press, Тотова, Нью-Джерси. В одном из примеров конкурентного анализа иммобилизованный антиген инкубируют в растворе, содержащем первую меченую антигенсвязывающую молекулу, которая связывается с антигеном, и вторую немеченую антигенсвязывающую молекулу, у которой тестируют способность конкурировать с первой антигенсвязывающей молекулой за связывание с антигеном. Вторая антигенсвязывающая молекула может присутствовать в супернатанте гибридомы. В качестве контроля иммобилизованный антиген инкубируют в растворе, содержащем первую меченую антигенсвязывающую молекулу, но не содержащим вторую немеченую антигенсвязывающую молекулу. После инкубации в условиях, разрешающих связывание первого антитела с антигеном, избыток несвязанного антитела удаляют и измеряют количество метки, связанной с иммобилизованным антигеном. Если количество метки, связанной с иммобилизованным антигеном, существенно снижается в тестируемом образце по сравнению с контрольным образцом, значит вторая антигенсвязывающая молекула конкурирует с первой антигенсвязывающей молекулой за связывание с антигеном. См. Harlow и Lane в кн.: «Antibodies: A Laboratory Manual» глава 14, 1988, изд. Cold Spring Harbor Laboratory, Cold Spring Harbor, Нью-Йорк.
Биспецифические антитела по настоящему изобретению, полученные, как описано в настоящем изобретении, могут быть очищены известными в данной области методами, например, высокоэффективной жидкостной хроматографией, ионообменной хроматографией, гель-электрофорезом, аффинной хроматографией, эксклюзионной хроматографией и другими. Определенные условия, используемые для очистки конкретного белка, частично зависят от таких факторов, как суммарный заряд, гидрофобность, гидрофильность и т.д., и известны специалистам в данной области. Для очистки аффинной хроматографией могут быть использованы антитело, лиганд, рецептор или антиген, с которыми связывается биспецифическая антигенсвязывающая молекула. Например, для очистки аффинной хроматографией гибридизированных белков по настоящему изобретению можно использовать матрицу с белком А или белком G. Последовательная аффинная хроматография с белком А или G и эксклюзионная хроматография могут быть использованы для выделения антигенсвязывающей молекулы, по существу, как это описано в примерах. Чистота молекулы биспецифического антигенсвязывающего вещества или ее фрагментов может быть определена любым из множества известных аналитических методов, включая гель-электрофорез, жидкостную хроматографию высокого давления и другие. Например, было показано, что биспецифичные антигенсвязывающие молекулы, экспрессируемые, как описано в примерах, являются интактными и правильно собраны, что установлено методами восстанавливающего и невосстанавливающего SDS-PAGE (электрофорез в полиакриламидном геле (ПААГ) в присутствии додецилсульфата натрия (ДСН).
Изобретение также охватывает биспецифичные антитела, продуцируемые методами настоящего изобретения.
Исследования
В настоящем изобретении биспецифические антигенсвязывающие молекулы могут быть идентифицированы, подвергнуты скринингу или охарактеризованы по их физическим / химическим свойствам и/или биологическим активностям с помощью различных анализов, известных в данной области.
1. Исследования сродства
Аффинность биспецифической антигенсвязывающей молекулы, предусмотренной в настоящем изобретении, с ОХ40 или FAP может быть определена методами, описанными далее в примерах, например, методом поверхностного плазмонного резонанса (surface plasmon resonance - SPR) с использованием стандартных приборов, например, BIAcore (фирма GE Healthcare), и рецепторов белков-мишеней, например тех, которые могут быть получены рекомбинантной экспрессией. Пример 3 является конкретной иллюстрацией и вариантом осуществления измерения связывающего сродства (аффинности). В одном из вариантов осуществления настоящего изобретения измеряют методом поверхностного плазмонного резонанса с использованием прибора BIACORE® Т200 (фирма GE Healthcare) при 25°С.
2. Исследования связывания и другие
Связывание биспецифической антигенсвязывающей молекулы, предусмотренной в настоящем изобретении, с соответствующими экспрессирующими ОХ40 и/или FAP клетками, может быть оценено с использованием линий клеток, экспрессирующих конкретный рецептор или целевой антиген, например, с помощью проточной цитометрии (FACS). В одном из вариантов осуществления настоящего изобретения свежие мононуклеарные клетки периферической крови (МКПК), экспрессирующие ОХ40, используют для анализа связывания. Эти клетки используются непосредственно после выделения (наивные МКПК) или после стимуляции (активированные МКПК). Пример 4.1 является конкретной иллюстрацией и примером варианта осуществления измерения связывания с ОХ40.
В другом варианте осуществления настоящего изобретения для демонстрации связывания биспецифических антител с FAP используют линии раковых клеток, экспрессирующих FAP (см. пример 4.2).
В еще одном из вариантов осуществления настоящего изобретения конкурентные анализы могут быть применены для идентификации антигенсвязывающей молекулы, которая конкурирует со специфическим антителом или антигенсвязывающей молекулой за связывание с FAP или ОХ40, соответственно. В некоторых вариантах осуществления настоящего изобретения такое конкурирующее антитело связывается с одним и тем же эпитопом (например, линейным или конформационным эпитопом), который связывается с определенным aHTH-FAP-антителом или определенным анти-ОХ40-антителом. Подробные иллюстративные способы картирования эпитопа, с которым связывается антитело, представлены в работе Morris в кн.: Methods in Molecular Biology 1996, т. 66 (изд. Humana Press, Totowa, Нью-Джерси, США).
3. Исследования активности
В одном из вариантов осуществления настоящего изобретения предусматривают методы идентификации биспецифических антигенсвязывающих молекул, которые связываются с FAP и с ОХ40, обладающими биологической активностью. Биологическая активность может включать, например, агонистическую передачу сигналов через ОХ40 на клетки, экспрессирующие ОХ40. Также предусматривают идентифицированные с помощью таких методов биспецифические антигенсвязывающие молекулы, обладающие такой биологической активностью in vitro. В частности, предусматривают метод анализа репортерных клеток, детектирующий активацию NFκB в клетках Hela, экспрессирующих ОХ40 человека, и совместно культивируемых с FAP-экспрессирующими опухолевыми клетками (см., например, пример 5.1).
В другом варианте осуществления настоящего изобретения предусматривают анализы для идентификации биспецифических антигенсвязывающих молекул, которые связываются с FAP и с 4-1ВВ, обладающими биологической активностью. В частности, предусмотрен анализ репортерных клеток, обнаруживающий активацию NFκB в линии репортерных клеток Jurkat-hu4-1BB-NFκB-luc2, экспрессирующих 4-1ВВ человека и репортерный ген NFκB-люциферазы (см., например, пример 7.2).
В другом варианте осуществления настоящего изобретения предусматривают анализы для идентификации биспецифических антигенсвязывающих молекул, которые связываются с FAP и CD40, обладающими биологической активностью. В частности, предусмотрен метод измерения активации В-клеток человека FAP-нацеленными анти-CD40 человека связывающими молекулами, используя гранулы с FAP-покрытием Dynabeads® в качестве источника антигена (см., например, пример 10.1).
В некоторых вариантах осуществления настоящего изобретения биспецифические антигенсвязывающие молекулы по настоящему изобретению тестируют на такую биологическую активность. Анализы для определения биологической активности молекул по настоящему изобретению описаны в примере 5. Кроме того, анализы для определения лизиса клеток (например, путем измерения высвобождения LDH), индуцированной кинетики апоптоза (например, путем измерения активности каспазы 3/7) или апоптоза (например, с использованием анализа TUNEL) хорошо известен в данной области. Кроме того, биологическая активность таких комплексов может быть определена путем оценки их влияния на выживание, пролиферацию и секрецию лимфокинов различными подгруппами лимфоцитов, например, NK-клетками, NKT-клетками или γδ Т-клетками, или путем оценки их способности модулировать фенотип и функцию антигенпрезентирующих клеток, например, дендритных клеток, моноцитов/макрофагов или В-клеток.
Фармацевтические композиции, составы и способы введения
В еще одном варианте осуществления настоящее изобретение относится к фармацевтическим композициям, содержащим какое-либо из биспецифических антител, предусмотренных в настоящем изобретении, например, для применения в каком-либо из приведенных ниже терапевтических методов. В одном варианте осуществления настоящего изобретения фармацевтическая композиция содержит какое-либо из биспецифических антител, предусмотренных в настоящем изобретении, и, по меньшей мере, один фармацевтически приемлемый наполнитель. В другом варианте осуществления настоящего изобретения фармацевтическая композиция содержит какое-либо из биспецифических антител, предусмотренных в настоящем изобретении, и, по меньшей мере, один дополнительный терапевтический агент, например, согласно приведенному ниже описанию.
Фармацевтические композиции по настоящему изобретению содержат терапевтически эффективное количество одного или нескольких биспецифических антител, растворенных или диспергированных в фармацевтически приемлемом наполнителе. Фразы «фармацевтический или фармакологически приемлемый» относятся к молекулам веществ и композициям, которые обычно нетоксичны для реципиентов в используемых дозировках и концентрациях, т.е. не вызывают неблагоприятных, аллергических или других неблагоприятных реакций при введении животному, например, человеку, при назначении. Приготовление фармацевтической композиции, которая содержит, по меньшей мере, одно биспецифическое антитело и, необязательно, дополнительный активный ингредиент, будет понятно специалистам в данной области при ознакомлении с настоящим описанием, которое согласуется с примерами в кн.: «Remington's Pharmaceutical Sciences», 18-е изд., 1990, изд-во Ed. Mack Printing Company, включенной в настоящее описание в виде ссылки. В частности, композиции представляют собой лиофилизированные составы или водные растворы. Используемое в настоящем изобретении понятие «фармацевтически приемлемый эксципиент» включает какие-либо и все растворители, буферы, дисперсионные среды, покрытия, поверхностно-активные вещества, антиоксиданты, консерванты (например, антибактериальные агенты, противогрибковые агенты), изотонические агенты, соли, стабилизаторы и их комбинации, применяемые по решению специалиста в данной области.
Парентеральные композиции разработаны для введения путем инъекции, например, подкожной, внутрикожной, внутрираневой, внутривенной, внутриартериальной, внутримышечной, интратекальной или внутрибрюшинной. Для инъекции биспецифические антитела по настоящему изобретению могут быть приготовлены в водных растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хенкса, раствор Рингера или физиологический солевой буфер. Раствор может содержать рецептурные агенты, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. В другом варианте осуществления настоящего изобретения гибридизированные белки могут находиться в форме порошка для разведения перед использованием подходящим носителем, например стерильной апирогенной водой. Стерильные растворы для инъекций получают путем включения гибридизированных белков по настоящему изобретению в необходимом количестве в соответствующем растворителе с различными другими ингредиентами, перечисленными ниже, по мере необходимости. Стерильность может быть легко достигнута, например, фильтрованием через стерильные фильтрующие мембраны. Обычно дисперсии готовят путем включения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и/или другие ингредиенты. В случае стерильных порошков для приготовления стерильных растворов, суспензий или эмульсий для инъекций предпочтительными способами приготовления являются методы вакуумной сушки или сушки вымораживанием, которые дают порошок активного ингредиента плюс какой-либо дополнительный желаемый жидкий ингредиент, ранее простерилизованный фильтрацией. Жидкая среда должна быть надлежащим образом забуферена, если необходимо, и жидкий разбавитель должен быть изотоническим перед инъекцией с достаточным количеством физиологического раствора или глюкозы. Композиция должна быть стабильной в условиях получения и хранения, и должна быть защищена от загрязнения микроорганизмами, например, бактериями и грибами. Очевидно, что загрязнение эндотоксином должно быть минимально на безопасном уровне, например, менее 0,5 нг/мг белка. Подходящие фармацевтически приемлемые наполнители включают, но не ограничиваются ими: буферы, например, фосфатный, цитратный и с другими органическими кислотами; антиоксид анты, включая аскорбиновую кислоту и метионин; консерванты (например, хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутил или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, например, сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатообразующие агенты, например, ЭДТА; сахара, например, сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, например, натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, например, полиэтиленгликоль (ПЭГ). Водные инъекционные суспензии могут содержать соединения, которые увеличивают вязкость суспензии, например, натрий-карбоксиметилцеллюлоза, сорбит, декстран или другие подобные. Необязательно, суспензия также может содержать подходящие стабилизаторы или агенты, которые увеличивают растворимость соединений, что позволяет получать высококонцентрированные растворы. Кроме того, суспензии активных соединений могут быть приготовлены в виде подходящих масляных инъекционных суспензий. Подходящие липофильные растворители или носители включают жирные масла, например, кунжутное масло, или синтетические сложные эфиры жирных кислот, например, этиловые эфиры олеиновой кислоты или триглицериды, или липосомы.
Активные ингредиенты могут быть приготовлены в микрокапсулах, приготовленных, например, методами коацервации или межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметацилатные) микрокапсулы, соответственно, в коллоидных системах доставки лекарственных средств (например, в липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие методы раскрыты в кн.: «Remington's Pharmaceutical Sciences», 18-е изд., 1990, изд-во Ed. Mack Printing Company. Могут быть приготовлены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих полипептид, причем эти матрицы имеют определенную форму, например, в виде пленок или микрокапсул. В конкретных вариантах осуществления настоящего изобретения пролонгированное всасывание инъецируемой композиции может быть обусловлено включением в композиции агентов, замедляющих абсорбцию, например, моностеарат алюминия, желатин или их комбинации.
К примерам фармацевтически приемлемых эксципиентов в настоящем изобретении дополнительно относят агенты для диспергирования лекарственных средств, например, растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, растворимые гликопротеины гиалуронидазы человека РН-20, например, rHuPH20 (HYLENEX®, фирма Baxter International, Inc.). Некоторые типичные sHASEGP и методы их использования, включая rHuPH20, описаны в патентных публикациях US 2005/0260186 и 2006/0104968. В одном варианте осуществления настоящего изобретения sHASEGP комбинируют с одной или несколькими дополнительными гликозаминогликаназами, например, хондроитиназами.
Примеры лиофилизированных составов антител описаны в US 6267958. Водные составы антител включают препараты, описанные в патенте US 6171586 и WO 2006/044908, причем последние составы включают гистидин-ацетатный буфер.
В дополнение к композициям, описанным ранее, гибридизированные белки также могут быть составлены в виде депо-препаратов. Такие препараты длительного действия могут вводиться путем имплантации (например, подкожно или внутримышечно) или внутримышечной инъекцией. Таким образом, например, гибридизированные белки могут быть составлены из подходящих полимерных или гидрофобных материалов (например, в виде эмульсии в приемлемом масле), или ионообменных смол, или в виде труднорастворимых производных, например, в виде труднорастворимой соли.
Фармацевтические композиции, содержащие гибридизированные белки по настоящему изобретению, могут быть изготовлены с помощью обычных процессов смешивания, растворения, эмульгирования, инкапсуляции, захвата или лиофилизации. Фармацевтические композиции могут быть составлены общепринятым способом с использованием одного или нескольких физиологически приемлемых носителей, разбавителей, эксципиентов или вспомогательных веществ, которые облегчают переработку белков в препараты, которые могут быть использованы фармацевтически. Правильный состав зависит от выбранного способа введения.
Биспецифичные антитела могут быть переработаны в композицию в форме свободной кислоты или основания, нейтральной или солевой формы. Фармацевтически приемлемые соли являются солями, которые, которые по существу сохраняют биологическую активность свободной кислоты или основания. Они включают кислые аддитивные соли, например, соли, образованные со свободными аминогруппами белковой композиции, или которые образованы неорганическими кислотами, например, соляной или фосфорной кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная или миндальная кислота. Соли, образованные за счет свободных карбоксильных групп, также могут быть получены из неорганических оснований, например, гидроксиды натрия, калия, аммония, кальция или железа; или такие органические основания, как изопропиламин, триметиламин, гистидин или прокаин. Фармацевтические соли имеют тенденцию быть более растворимыми в водных и других протонных растворителях, чем соответствующие формы свободного основания.
Композиция в настоящем изобретении также может содержать более одного активного ингредиента, если это необходимо для определенного состояния, подвергаемого лечению, предпочтительно те, которые обладают дополнительными активностями, но которые не оказывают вредного влияния друг на друга. Такие активные ингредиенты образом присутствуют в комбинациях в количествах, которые эффективны для назначенной цели.
Составы, используемые для введения in vivo, обычно являются стерильными. Стерильность может быть легко достигнута, например, фильтрованием через стерильные фильтрующие мембраны.
Терапевтические методы и композиции
Какое-либо из биспецифических антител, предусмотренных в настоящем изобретении, может быть использовано в терапевтических целях. Для применения в терапевтических методах антигенсвязывающие молекулы по настоящему изобретению могут быть составлены, дозированы и введены способом, соответствующим медицинской практике. В этом контексте оцениваемые факторы включают конкретное расстройство, подвергаемое лечению, конкретное млекопитающее, нуждающееся в лечении, клиническое состояние конкретного пациента, причину расстройства, место доставки агента, способ введения, график введения и другие факторы, известные врачам.
В одном варианте осуществления настоящего изобретения биспецифические антитела по настоящему изобретению предназначены для применения в качестве лекарственного средства. В других вариантах осуществления настоящего изобретения биспецифические антигенсвязывающие молекулы по настоящему изобретению предназначены для применения в лечении заболевания, в частности, для применения в лечении рака. В определенных вариантах осуществления настоящего изобретения биспецифические антитела по настоящему изобретению предусмотрены для применения в способе лечения. В одном варианте осуществления настоящее изобретение относится к биспецифическому антителу, описанному в настоящем изобретении, для применения при лечении заболевания у индивидуума, нуждающегося в этом. В определенных вариантах осуществления настоящее изобретение относится к биспецифическому антителу для применения в методе лечения больного индивидуума, включающем введение индивидууму терапевтически эффективного количества молекул биспецифического антигенсвязывающего вещества. В некоторых вариантах осуществления настоящего изобретения подлежащим лечению заболеванием является рак. В определенных вариантах осуществления настоящего изобретения подлежащим лечению заболеванием является пролиферативное расстройство, в частности рак. Примеры раковых заболеваний включают рак мочевого пузыря, рак головного мозга, рак головы и шеи, рак поджелудочной железы, рак легких, рак молочной железы, рак яичников, рак матки, рак шейки матки, рак эндометрия, рак пищевода, рак толстой кишки, рак прямой кишки, рак желудка, рак простаты, рак крови, рак кожи, плоскоклеточный рак, рак костей и рак почек. Другие нарушения пролиферации клеток, которые можно лечить с использованием биспецифической антигенсвязывающей молекулы по настоящему изобретению, включают, но не ограничиваются ими, новообразования, расположенные в: брюшной полости, кости, молочной железе, пищеварительной системе, печени, поджелудочной железе, брюшине, эндокринных железах (надпочечниках, паращитовидной железе, гипофизе, яичках, яичниках, тимусе, щитовидной железе), глазах, голове и шее, нервной системе (центральной и периферической), лимфатической системе, в области таза, коже, мягких тканях, селезенке, области груди и мочеполовой системе. Также включены предраковые состояния или поражения и метастазы. В некоторых вариантах осуществления настоящего изобретения рак выбирают из группы, состоящей из почечно-клеточного рака, рака кожи, рака легких, колоректального рака, рака молочной железы, рака головного мозга, рака головы и шеи. Субъект, пациент или «индивидуум», нуждающийся в лечении, обычно является млекопитающим, более конкретно - человеком.
Настоящее изобретение также охватывает биспецифическое антитело по настоящему изобретению, или фармацевтическую композицию по настоящему изобретению для применения при повышающей регуляции или продлении цитотоксической активности Т-клеток.
В другом варианте осуществления настоящего изобретения предусматривают применение биспецифического антитела по настоящему изобретению при получении лекарственного средства для лечения заболевания у индивидуума, нуждающегося в этом. В одном варианте осуществления настоящего изобретения лекарственное средство предназначено для применения в лечении заболевания, включающем введение больному индивидууму терапевтически эффективного количества лекарственного средства. В некоторых вариантах осуществления настоящего изобретения заболевание, подлежащее лечению, является пролиферативным расстройством, в частности раком. Примерами раковых заболеваний являются рак мочевого пузыря, рак головного мозга, рак головы и шеи, рак поджелудочной железы, рак легких, рак молочной железы, рак яичников, рак матки, рак шейки матки, рак эндометрия, рак пищевода, рак толстой кишки, рак прямой кишки, рак желудка, рак простаты, рак крови, рак кожи, плоскоклеточный рак, рак кости и рак почки. Другие нарушения пролиферации клеток, которые можно лечить с использованием биспецифической антигенсвязывающей молекулы по настоящему изобретению, включают, но ими перечень не ограничивают, новообразования, расположенные в: брюшной полости, кости, молочной железе, пищеварительной системе, печени, поджелудочной железе, брюшине, эндокринных железах (надпочечниках, паращитовидной железе, гипофизе, яичке, яичнике, тимусе, щитовидной железе), глазах, голове и шее, нервной системе (центральной и периферической), лимфатической системе, в области таза, коже, мягких тканях, селезенке, области груди и мочеполовой системе. Также включены предраковые состояния или поражения и метастазы. В некоторых вариантах осуществления настоящего изобретения рак выбирают из группы, состоящей из почечно-клеточного рака, рака кожи, рака легких, колоректального рака, рака молочной железы, рака головного мозга, рака головы и шеи. Специалист в данной области может определить, что во многих случаях биспецифическая антигенсвязывающая молекула может не обеспечить излечение и может принести только частичную пользу. В некоторых вариантах осуществления настоящего изобретения физиологическое изменение, имеющее некоторую пользу, также считают терапевтически полезным. Таким образом, в некоторых вариантах осуществления количество биспецифического антитела, которое обеспечивает физиологическое изменение, считается «эффективным количеством» или «терапевтически эффективным количеством». В каком-либо из вышеуказанных вариантов осуществления настоящего изобретения индивидуумом предпочтительно является млекопитающее, особенно человек.
В другом варианте осуществления настоящее изобретение относится к применению биспецифического антитела по настоящему изобретению для получения лекарственного средства для лечения инфекционных заболеваний, в частности, для лечения вирусных инфекций или для лечения аутоиммунных заболеваний, например, волчанки.
В еще одном варианте осуществления настоящее изобретение относится к методу лечения заболевания у индивидуума, включающему введение указанному индивидууму терапевтически эффективного количества биспецифической антигенсвязывающей молекулы по настоящему изобретению. В одном варианте осуществления настоящего изобретения указанному индивидууму вводят композицию, содержащую гибридизированный белок по настоящему изобретению в фармацевтически приемлемой форме. В некоторых вариантах осуществления настоящего изобретения заболевание, подлежащее лечению, является пролиферативным расстройством. В конкретном варианте осуществления настоящего изобретения заболевание является раком. В других вариантах осуществления настоящего изобретения метод лечения дополнительно включает введение индивидууму терапевтически эффективного количества, по меньшей мере, одного дополнительного терапевтического агента, например противоракового агента, если подлежащее лечению заболевание является раком. «Индивидуум» в соответствии с любым из вышеуказанных вариантов осуществления может быть млекопитающим, предпочтительно человеком.
Для предотвращения или лечения заболевания необходимая доза биспецифического антитела по настоящему изобретению (при использовании отдельно или в сочетании с одним или несколькими другими дополнительными терапевтическими агентами) будет зависеть от типа заболевания, которое нужно лечить, пути введения, массы тела пациента, типа гибридизированного белка, тяжести и течения заболевания, вводится ли слитый белок в профилактических или терапевтических целях, предыдущих или одновременных терапевтических вмешательства, истории болезни пациента и реакции на гибридизированный белок, а также от мнения лечащего врача. В любом случае специалист, ответственный за введение, будет определять концентрацию активного ингредиента(ов) в композиции и соответствующую дозу(ы) для конкретного субъекта. В настоящем изобретении рассматривают различные схемы дозирования, включая, но ими не ограничиваясь, однократное или множественное введение в различные моменты времени, болюсное введение и пульс-инфузию.
Биспецифическое антитело вводят пациенту однократно или в виде серии введений. В зависимости от типа и тяжести заболевания начальная доза кандидата для введения пациенту может составлять примерно от 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг 10 мг/кг) антигенсвязывающей молекулы, будь то однократное введение, несколько отдельных введений или непрерывная инфузия. Одна типичная суточная доза может варьировать примерно от 1 мкг/кг до 100 мг/кг или более, в зависимости от выше приведенных факторов. Для повторных введений в течение нескольких дней или дольше, в зависимости от состояния, лечение обычно будет продолжаться до тех пор, пока не произойдет желаемое подавление симптомов заболевания. Одна примерная доза гибридизированного белка может быть в диапазоне примерно от 0,005 мг/кг до примерно 10 мг/кг. В других примерах доза также может включать примерно от 1 мкг/кг массы тела, примерно 5 мкг/кг массы тела, примерно 10 мкг/кг массы тела, примерно 50 мкг/кг массы тела, примерно 100 мкг/кг массы тела, примерно 200 мкг/кг массы тела, примерно 350 мкг/кг массы тела, примерно 500 мкг/кг массы тела, примерно 1 мг/кг массы тела, примерно 5 мг/кг массы тела, примерно 10 мг/кг массы тела, примерно 50 мг/кг массы тела, примерно 100 мг/кг массы тела, примерно 200 мг/кг массы тела, примерно 350 мг/кг массы тела, примерно 500 мг/кг массы тела, до примерно 1000 мг/кг массы тела, или более чем за одно введение, и в каком-либо диапазоне указанных доз. В примерах диапазона, исходя из указанных выше чисел, диапазон от около 5 мг/кг массы тела до около 100 мг/кг массы тела, от около 5 мкг/кг массы тела до около 500 мг/кг массы тела и т.д., может вводить, основываясь на числах, описанных выше. Таким образом, одна или несколько доз в количестве примерно 0,5 мг/кг, 2,0 мг/кг, 5,0 мг/кг или 10 мг/кг (или какая-либо их комбинация) могут быть введены пациенту. Такие дозы можно вводить периодически, например, каждую неделю или каждые три недели (например, так, чтобы пациент получал примерно от двух до примерно двадцати или, например, примерно шесть доз биспецифической антигенсвязывающей молекулы). Может быть назначена начальная повышенная нагрузочная доза, за которой следует одна или несколько более низких доз. Однако другие схемы дозировки также могут быть применимы. Прогресс такой терапии легко контролировать с помощью традиционных методов и анализов.
Биспецифическое антитело по настоящему изобретению обычно применяют в количестве, эффективном для достижения намеченной цели. Для лечения или предотвращения болезненного состояния биспецифические антигенсвязывающие молекулы по настоящему изобретению или их фармацевтические композиции вводят или применяют в терапевтически эффективном количестве. Терапевтически эффективного количество определяют специалисты в данной области, особенно с учетом подробного описания настоящего изобретения.
Для системного введения терапевтически эффективную дозу можно первоначально определить с помощью анализов in vitro, таких как анализы с применением культур клеток. Затем на животных моделях можно составить дозу для достижения диапазона концентраций в кровяном русле, который включает IC50 по результатам исследований в культуре клеток. Такая информация может быть использована для более точного определения полезных доз для людей. Начальные дозы также могут быть оценены по данным in vivo, например, на животных моделях, с использованием методик, которые известны в данной области. Специалист в данной области может оптимизировать введение людям на основании данных, полученных на животных. Количество и интервал дозировки можно регулировать индивидуально для обеспечения в плазме уровней биспецифических антигенсвязывающих молекул, которые достаточны для поддержания терапевтического эффекта. Обычные дозировки для пациентов для введения путем инъекции составляют примерно от 0,1 до 50 мг/кг/день, обычно от примерно 0,5 до 1 мг/кг день. Терапевтически эффективные уровни в плазме могут быть достигнуты путем введения нескольких доз каждый день. Уровни в плазме могут быть измерены, например, с помощью ВЭЖХ.
В случаях местного введения или селективного поглощения эффективная локальная концентрация биспецифических антигенсвязывающих молекул может не быть связана с концентрацией в плазме. Специалист в данной области сможет оптимизировать терапевтически эффективные дозы для местного введения местные без чрезмерных экспериментов.
Терапевтически эффективная доза биспецифического антитела, описанная в настоящем изобретении, обычно обеспечивает положительный терапевтический ответ без существенной токсичности. Токсичность и терапевтическая эффективность гибридизированного белка могут быть определены стандартными фармацевтическими процедурами в культуре клеток или на экспериментальных животных. Анализы с применением клеточных культур и исследования на животных можно использовать для определения LD50 (доза, летальная для 50% популяции) и ED50 (доза, терапевтически эффективная в 50% популяции). Соотношение доз между токсическим и терапевтическим эффектами называют терапевтическим индексом, который может быть выражен как соотношение LD50/ED50. Предпочтительными являются биспецифические антигенсвязывающие молекулы, которые проявляют большие терапевтические показатели. В одном варианте осуществления биспецифическая антигенсвязывающая молекула по настоящему изобретению проявляет высокий терапевтический индекс. Данные, полученные из анализов клеточных культур и исследований на животных, могут быть использованы при разработке диапазона доз, подходящих для применения людьми. Дозировка предпочтительно находится в диапазоне циркулирующих в кровяном русле концентраций, которые включают ED50 с минимальной токсичностью или без нее. Дозировка может варьировать в пределах этого диапазона в зависимости от множества факторов, например, используемой лекарственной формы, используемого способа введения, состояния субъекта и других обстоятельств. Точный состав, способ введения и дозировка могут быть выбраны лечащим врачом с учетом состояния пациента (см., например, публикацию Fingl с соавт., 1975, в кн.: «The Pharmacological Basis of Therapeutics», глава 1, часть 1, сущность которой включена в настоящее описание в виде ссылки).
Лечащий врач пациентов, которых лечат биспецифическими антигенсвязывающими молекулами по настоящему изобретению, будет знать, как и когда прекратить, прервать или скорректировать введение из-за токсичности, дисфункции органов и других подобных обстоятельств. И наоборот, лечащий врач также должен знать, как скорректировать лечение, доведя его до более высоких уровней дозирования, если клинический ответ не был адекватным (исключая токсичность). Величина введенной дозы при лечении определенного расстройства будет варьировать в зависимости от тяжести состояния, подлежащего лечению, пути введения и других подобных обстоятельств. Тяжесть состояния может, например, оцениваться частично стандартными методами прогностической оценки. Кроме того, доза и, возможно, частота введения дозы, также будут варьировать в зависимости от возраста, массы тела и реакции конкретного пациента.
Другие агенты и методы лечения
Биспецифические антитела по настоящему изобретению можно вводить в комбинации с одним или несколькими другими терапевтическими агентами. Например, биспецифическое антитело по настоящему изобретению можно вводить, по меньшей мере, с одним дополнительным терапевтическим агентом. Понятие «терапевтический агент» означает какой-либо агент, который можно вводить для лечения симптома или заболевания у индивидуума, нуждающегося в таком лечении. Такой дополнительный терапевтический агент может содержать какие-либо активные ингредиенты, подходящие для лечения конкретного показания, предпочтительно те, которые обладают комплементарными активностями, которые не оказывают вредного воздействия друг на друга. В определенных вариантах осуществления настоящего изобретения дополнительный терапевтический агент представляет собой другой противораковый агент.
Присутствие таких других агентов целесообразно в комбинации в количествах, которые эффективны для предполагаемой цели. Эффективное количество таких других агентов зависит от количества используемого гибридизированного белка, типа расстройства или лечения и других факторов, обсуждаемых выше. Биспецифические антигенсвязывающие молекулы обычно используются в тех же дозировках и способами введения, описанными в настоящем изобретении, или примерно от 1 до 99% дозировок, описанных в настоящем изобретении, или в какой-либо дозировке и каким-либо способом, который эмпирически/клинически определен как пригодный.
Такие комбинированные терапии, отмеченные выше, включают комбинированное введение (т.е. два или более терапевтических агента включены в одну и ту же или отдельные композиции) и раздельное введение, и в этом случае введение биспецифической антигенсвязывающей молекулы по изобретению может происходить до, одновременно и/или после введения дополнительного терапевтического агента и/или адъюванта.
Получаемые изделия
В другом варианте осуществления настоящего изобретения предусматривают изделие, которое содержит материалы для лечения, профилактики и/или диагностики описанных выше расстройств. Получаемое изделие содержит контейнер и на нем этикетку или в нем вкладыш. Таким контейнерами, например, могут быть бутыли, флаконы, шприцы, пакеты для внутривенного введения и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит композицию, которая, сама по себе или в комбинации с другой композицией, эффективна для лечения, предотвращения и/или диагностики расстройства, и может иметь стерильный порт доступа (например, контейнер может быть пакетом для внутривенного введения раствора или флаконом с пробкой, которая прокалывается иглой для подкожных инъекций). По меньшей мере, один активный агент в композиции представляет собой биспецифическое антитело, описанное в настоящем изобретении.
Этикетка или вкладыш в упаковку указывает, что композицию используют для лечения определенного состояния. Кроме того, изделие может содержать (а) первый контейнер с содержащейся в нем композицией, где композиция содержит биспецифическое антитело по настоящему изобретению; и (б) второй контейнер с содержащейся в нем композицией, которая содержит дополнительный цитотоксический или иной терапевтический агент. Получаемое изделие в этом варианте осуществления настоящего изобретения может дополнительно содержать вкладыш в упаковку, указывающий, что композиции можно использовать для лечения определенного состояния.
В другом варианте осуществления настоящего изобретения или дополнительно, изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, например, бактериостатическую воду для инъекций (БВДИ), фосфатно-солевой буфер (ФСБ), раствор Рингера и раствор декстрозы. Кроме того, оно может содержать другие материалы, необходимые с коммерческой точки зрения или полезные для потребителей, например, другие буферы, разбавители, фильтры, иглы и шприцы.
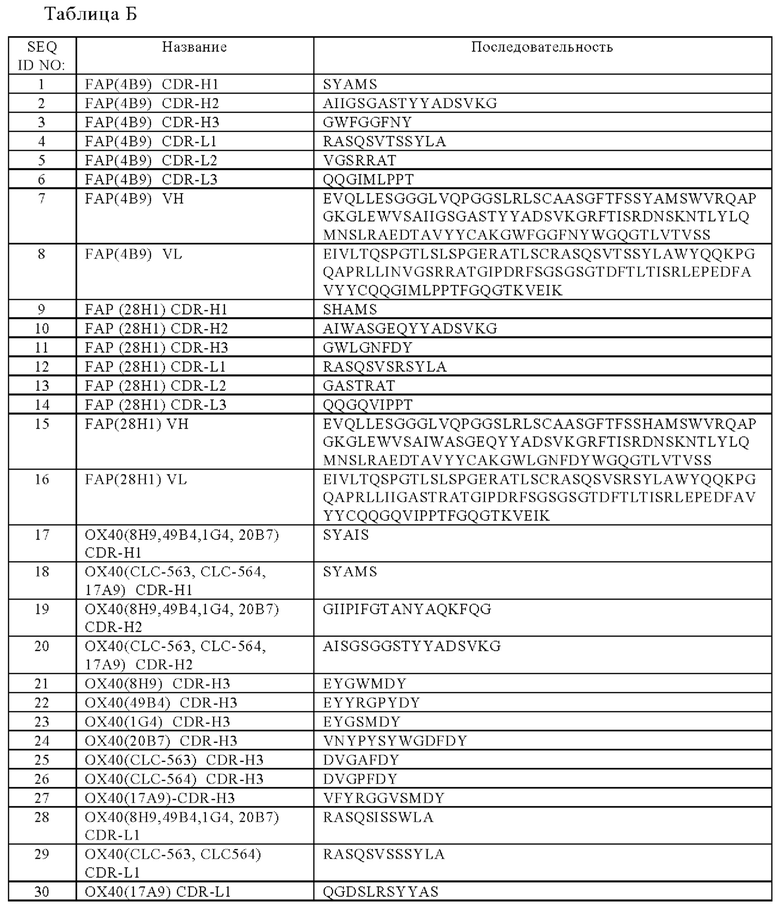

Общая информация, касающаяся нуклеотидных последовательностей легкой и тяжелой цепей иммуноглобулинов человека, приводится в публикации Kabat с соавт. в кн.: «Sequences of Proteins of Immunological Interest», 5e изд., 1991, Public Health Service, National Institutes of Health, Бетесда, Мэриленд. Аминокислоты цепей антител пронумерованы и обозначены в соответствии с системой нумерации EU по Kabat (Kabat с соавт. в кн.: «Sequences of Proteins of Immunological Interest)), 5e изд., 1991, Public Health Service, National Institutes of Health, Бетесда, Мэриленд) согласно описанному выше.
Примеры
Ниже приведены примеры методов и композиций, предусматриваемых в настоящем изобретении. Очевидно, что различные другие варианты настоящего изобретения могут быть осуществлены на практике, учитывая общее описание, приведенное выше.
Методы рекомбинантной ДНК
Стандартные методы применяют для манипулирования ДНК, как описано в публикации Sambrook с соавт., Molecular cloning: A laboratory manual; изд. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк, 1989. Реагенты для молекулярной биологии применяют в соответствии с инструкциями производителя. Общая информация о нуклеотидных последовательностях легкой и тяжелой цепей иммуноглобулина человека приведена Kabat с соавт.в кн.: «Sequences of Proteins of Immunological Interest)), 5e изд., 1991, Public Health Service, National Institutes of Health, Бетесда, Мэриленд.
Секвенирование ДНК
Последовательности ДНК определяют методом двухцепочечного секвенирования.
Синтез генов
Необходимые генные сегменты получают химическим синтезом в фирме Geneart AG (Регенсбург, Германия) из синтетических олигонуклеотидов путем автоматического синтеза генов. Синтезированные фрагменты гена клонируют в плазмиде Е. coli для размножения/амплификации. Последовательности ДНК субклонированных фрагментов гена проверяют секвенированием ДНК. В другом варианте короткие синтетические фрагменты ДНК собирают путем отжига химически синтезированных олигонуклеотидов или с помощью ПЦР. Соответствующие олигонуклеотиды получают в фирме metabion GmbH (Planegg-Martinsried, Германия).
Методы культивирования клеток
Применяют стандартные методы культивирования клеток, описанные в кн.: Current Protocols in Cell Biology, под ред. Bonifacino J.S., Dasso M., Harford J.B., Lippincott-Schwartz J. и Yamada K.M., 2000, изд. John Wiley & Sons, Inc.
Реагенты
Все коммерческие реагенты, антитела и наборы применяют в соответствии с протоколом производителя, если не указано иначе.
Пример 1
Получение биспецифических конторстел
1.1. Конструирование и экспрессия плазмид для получения биспецифических конторстел
Для экспрессии биспецифических конторстел, описанных в настоящем изобретении, используют единицу транскрипции, содержащую следующие функциональные элементы:
- немедленно-ранний энхансер и промотор от цитомегаловируса человека (cytomegalovirus, P-CMV) включая интрон А,
- 5'-нетранслируемая область тяжелой цепи иммуноглобулина человека (5'-untranslatedregion, 5'UTR),
- сигнальная последовательность тяжелой цепи иммуноглобулина грызунов,
- нуклеиновая кислота, кодирующая соответствующий циклический гибридизированный полипептид, и
- последовательность полиаденилирования бычьего гормона роста (bovine growth hormone polyadenylation sequence, BGH pA).
Помимо экспрессирующей единицы/кассеты, включающей требуемый экспрессируемый ген, основная/стандартная плазмида экспрессии млекопитающих содержит
- точку начала репликации из вектора pUC18, который допускает репликацию этой плазмиды в Е. coli, и
- ген бета-лактамазы, который придает кишечной палочке устойчивость к ампициллину.
1.2. Экспрессия биспецифических конторстел
Временную экспрессию биспецифических антигенсвязывающих молекул проводят в адаптированных к суспензии клетках HEK293F (клетки FreeStyle 293-F; фирма Invitrogen) с реагентом для трансфекции без клеток 293 (Transfection Reagent 293-free; фирма Novagen).
Клетки пересевают путем разбавления, по меньшей мере, четыре раза (объем 30 мл) после оттаивания в колбе объемом 125 мл при встряхивании (инкубируют/встряхивают при 37°С, в атмосфере 7% СО2, влажности 85%, 135 об/мин). Клетки размножают до титра 3×105 клеток/мл в объеме 250 мл. Через трое суток клетки разделяют и новые высевают с плотностью 7×105 клеток/мл в объеме 250 мл в качалочную колбу объемом 1 литр. Трансфекцию осуществляют через 24 ч при плотности клеток примерно 1,4-2,0×106 клеток/мл.
Перед трансфекцией 250 мкг плазмидной ДНК разводят в конечном объеме 10 мл предварительно нагретой (на водяной бане при 37°) среде Opti-MEM (фирма Gibco). Раствор осторожно перемешивают и инкубируют при комнатной температуре не более 5 минут. Затем 333,3 мкл реагента для трансфекции без клеток 293 добавляют к раствору ДНК-OptiMEM. После этого раствор осторожно перемешивают и инкубируют при комнатной температуре в течение 15-20 минут. Весь объем смеси добавляют в качалочную колбу объемом 1 литр с 250 мл НЕК-клеточной культуры.
Инкубируют/встряхивают при 37°, в атмосфере 7% СО2, влажности 85%, 135 об/мин в течение 6 или 7 суток.
Супернатант собирают на первой стадии центрифугирования при 2000 об/мин, 4°, в течение 10 мин. Затем супернатант переносят в новую центрифужную колбу для второго центрифугирования при 4000 об/мин, 4°, в течение 20 мин. После этого бесклеточный супернатант фильтруют через фильтр с порами 0,22 мкм на крышке бутыли и хранят в морозильной камере (-20°).
1.3. Очистка биспецифических конторстел
Антигенсвязывающие молекулы, содержащие культуральные супернатанты, фильтруют и очищают за два этапа хроматографии. Антитела улавливают с помощью аффинной хроматографии с использованием колонки HiTrap MabSelectSuRe (фирма GE Healthcare), уравновешенной ФСБ (1 мМ KH2PO4, 10 мМ Na2HPO4, 137 мМ NaCl, 2,7 мМ KCl), рН 7,4. Несвязанные белки удаляют промыванием уравновешивающим буфером, и антигенсвязывающую молекулу извлекают 50 мМ цитратным буфером, рН 2,8, и сразу после элюирования нейтрализуют до рН 6,0 с помощью 1 М трис-основания, рН 9,0. Эксклюзионную хроматографию по размеру на сорбенте Superdex 200ТМ (фирма GE Healthcare) используют на второй стадии очистки. Эксклюзионную хроматографию по размеру проводят в 20 мМ гистидиновом буфере, 0,14 М NaCl, рН 6,0. Растворы, содержащие биспецифические антигенсвязывающие молекулы, концентрируют с помощью центробежного фильтра Ultrafree-CL, снабженного мембраной Biomax-SK (фирма Millipore, Биллерика, Массачусетс) и хранят при -80°С.
1.4. Анализ методом масс-спектрометрии биспецифических конторстел
PNGase F получают от фирмы Roche Diagnostics GmbH (14.3 Ед/мкл; в растворе фосфата натрия, EDTA и глицерина). Протеазу, специфически расщепляющую в шарнирной области антитела IgG, восстанавливают из лиофилизата перед применением.
Ферментативное дегликозилирование с помощью PNGase F
50 мкг антигенсвязывающей молекулы разводят до конечной концентрации 0,6 мг/мл буфером 10 мМ фосфата натрия, рН 7,1, и дегликозилируют с применением 1 мкл PNGase F при 37°С в течение 16 ч.
Ферментативное расщепление
Дегликозилированный образец разводят до конечной концентрации 0,5 мг/мл в 200 мМ Трис-буфере, рН 8,0, и затем расщепляют IgG-специфической протеазой при 37°С в течение 1 ч.
ESI-OTOF масс-спектрометрия
Образец обессоливают методом ВЭЖХ в колонке Sephadex G25 (фирма Kronlab, 5×250 мм, TAC05/250G0-SR) используя 40% ацетонитрилл с 2% муравьиной кислоты (объем/объем). Общую массу определяют методом ESI-QTOF MS в системе maXis 4G UHR-QTOF MS (фирма Bruker Daltonik), оборудованной TriVersa NanoMate источником (фирма Advion). Калибровку проводят раствором иодида натрия (Waters ToF G2-Sample Kit 2 Part: 700008892-1). Для расщепленной антигенсвязывающей молекулы сбор данных осуществляют при 1000-4000 m/z (ISCID: 30 eV). Необработанные масс-спектры оценивают и преобразуют в отдельные относительные мольные массы. Для визуализации результатов использовалось проприетарное программное обеспечение для генерации деконволютивных масс-спектров.
Пример 2. Получение биспецифических антител с двумя антигенсвязывающими доменами, связывающимися с ОХ40, и одним антигенсвязывающим доменом, связывающимся с FAP (FAP-OX40 конторстела)
Получение и подготовку связующих FAP описывают в WO 2012/020006 А2, который включен в настоящее описание в виде ссылки. Связующее ОХ40 описано в WO 2017/055398 А2.
2.1 Получение FAP (4В9)-ОХ40 (49В4) конторстела CD134-0093
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1А:
- первый гибридизированный полипептид (ot N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор (G4S)2, VH(FAP)-Ckappa
- второй гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор (G4S)2, VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
Таблица 1 представляет аминокислотные последовательности биспецифического антитела CD134-0093.

Схема сборки структуры показана на фиг. 1Д.
2.2. Получение FAP (4B9)-OX40 (49В4) конторстела CD134-0094
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1Б:
- первый гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-Скарра, коннектор (G4S)2, шарнир IgG1, выступ Fc, коннектор (G4S)2, VL(OX40)-CH1, коннектор (G4S)2, VL(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-Скарра, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-CH1, коннектор (G4S)2, VH(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 2 показаны аминокислотные последовательности биспецифического антитела CD134-0094.

Схема сборки структуры показана на фиг. 1Е.
2.3 Получение FAP Г28Н1УОХ40 (49В4) конторстела Р1АЕ0085
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1 А:
- первый гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 3 показаны аминокислотные последовательности биспецифического антитела Р1АЕ0085.
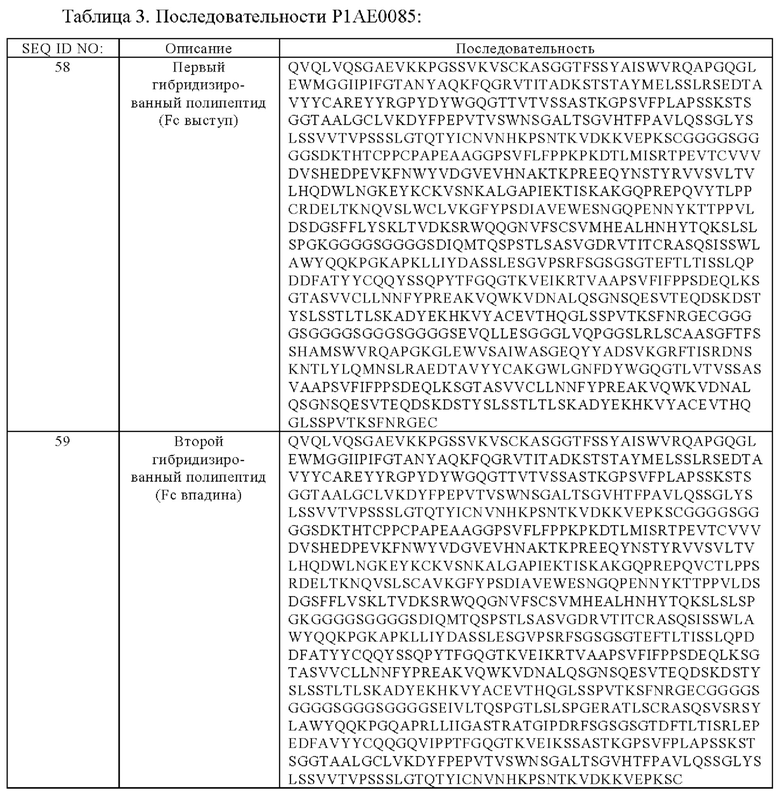
Схема сборки структуры показана на фиг. 1Ж.
2.4 Получение FAP (4В9)-ОХ40 (49В4) конторстела Р1АЕ0086
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1А:
- первый гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO:84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 4 показаны аминокислотные последовательности биспецифического антитела (Конторстело 7).
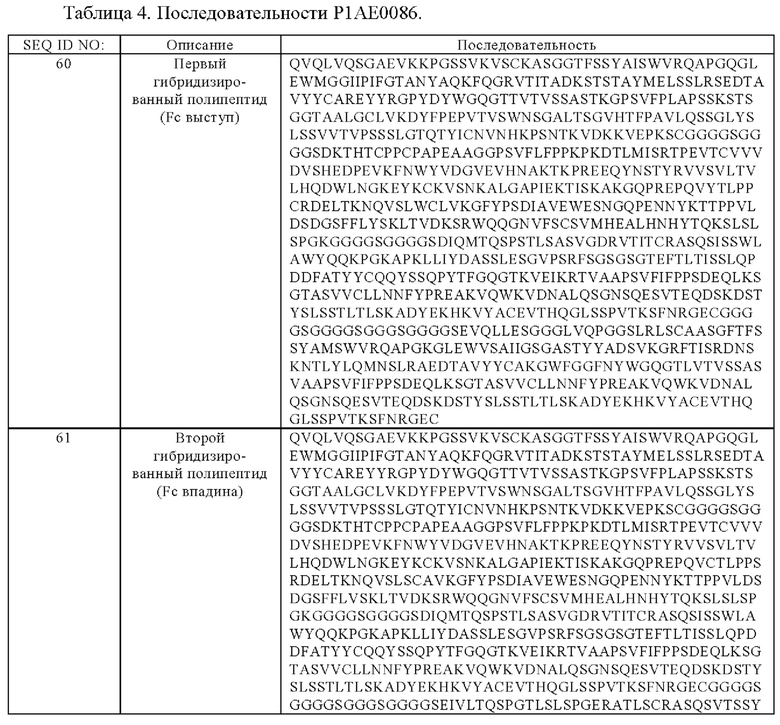

Схема сборки структуры показана на фиг. 1Ж.
2.5 Получение FAP (28Н1)-ОХ40 (49В4) конторстела Р1АЕ0087
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1Б:
- первый гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VH (OX40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-CH1, коннектор GGGGS GGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 5 показаны аминокислотные последовательности биспецифического антитела Р1АЕ0087.

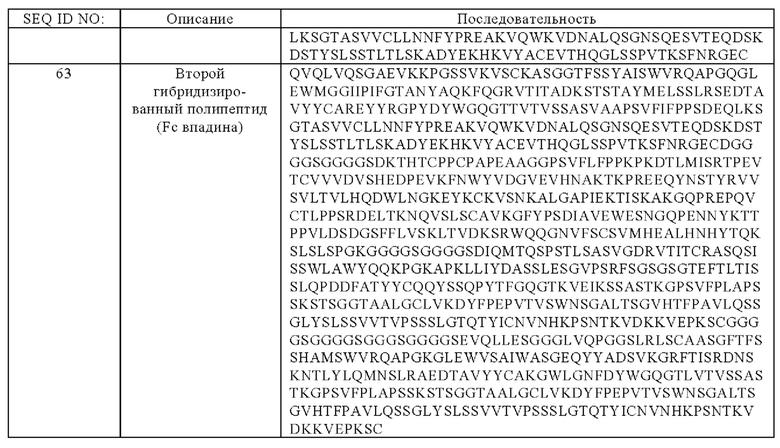
Схема сборки структуры показана на фиг. 1З.
2.6 Получение FAP (4В9)-ОХ40 (49В4) конторстела P1AE0839
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1Б:
- первый гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 6 показаны аминокислотные последовательности биспецифического антитела Р1АЕ0839.
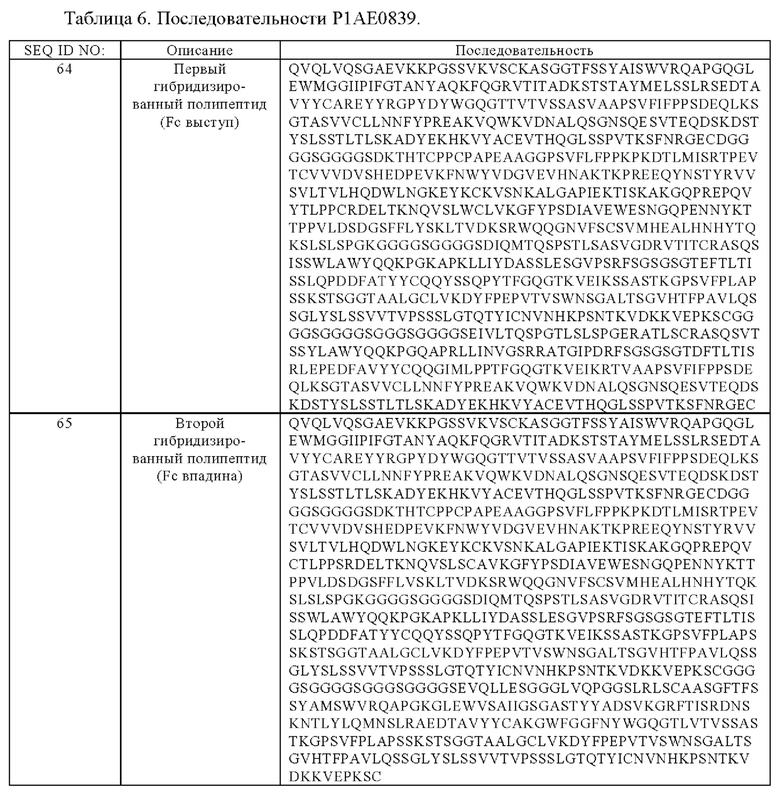
Схема сборки структуры показана на фиг. 1З.
2.7 Получение FAP (4В9)-ОХ40 (49В4) конторстела P1AE0821
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1В:
- первый гибридизированный полипептид (от N- к С-концу): VL (ОХ40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VH(OX40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
- второй гибридизированный полипептид (от N- к С-концу): VL (ОХ40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VH(OX40)-CH1, коннектор GGGGSGGGGSGGG SGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 7 показаны аминокислотные последовательности биспецифического антитела Р1АЕ0821.
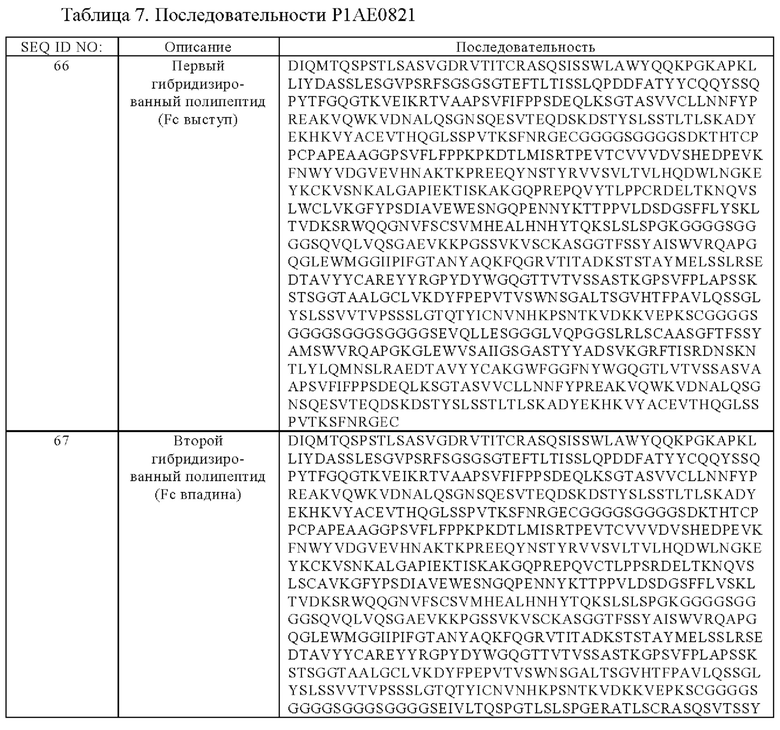

Схема сборки структуры показана на фиг. 1И.
2.8 Получение FAP (4В9)-ОХ40 (49В4) конторстела Р1АЕ1122 (конторстело 1)
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1Г:
- первый гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
- второй гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 8 показаны аминокислотные последовательности биспецифического антитела Р1АЕ1122.

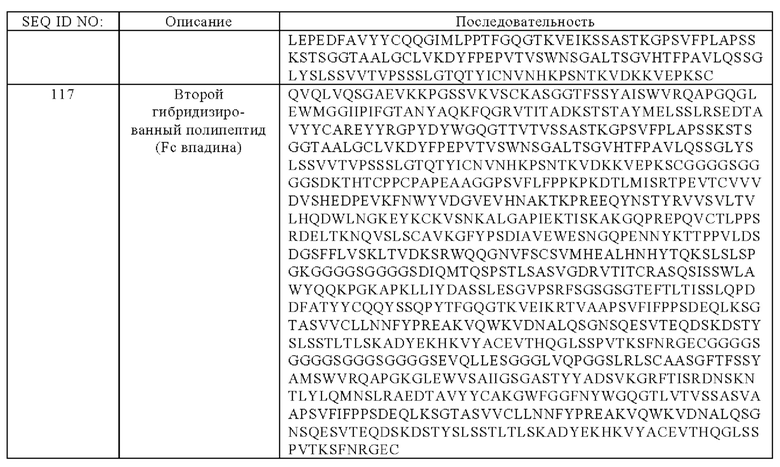
Схема сборки структуры показана на фиг. 1К.
2.9 Получение FAP (4В9)-ОХ40 (49В4) конторстела P1AE1942 (конторстело 2)
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VL (ОХ40)-Скарра, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VH(OX40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1;
- второй гибридизированный полипептид (от N- к С-концу): VL (OX40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VH(OX40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 9 показаны аминокислотные последовательности биспецифического антитела Р1АЕ1942.
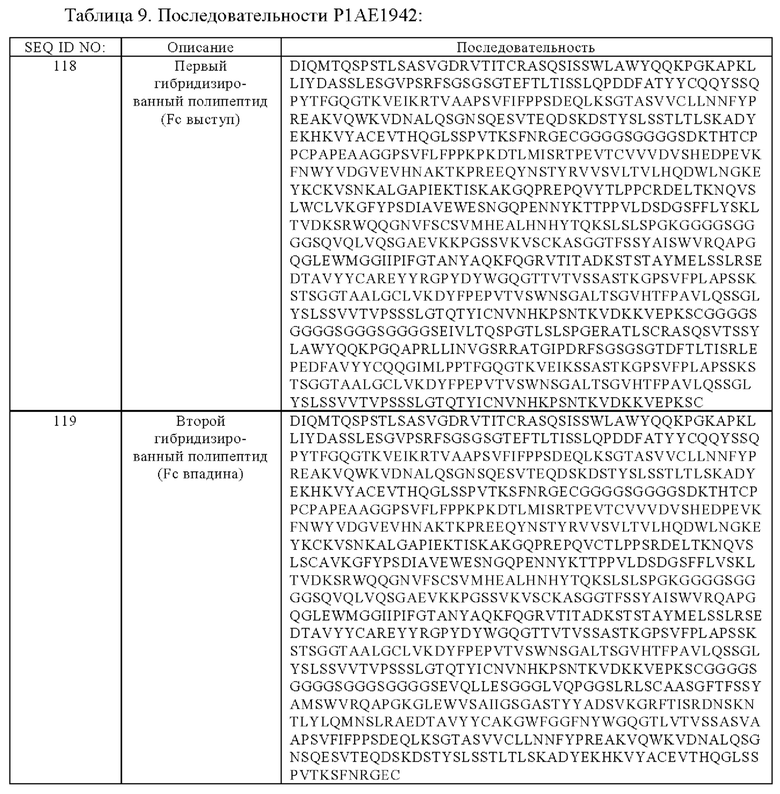
2.10 Получение FAP (4В9)-ОХ40 (49В4) конторстела Р1АЕ1887 (конторстело 3)
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VL (ОХ40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VH(OX40)-CH1, коннектор GGGGS GGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VH (ОХ40)-СН1, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGG SGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 10 показаны аминокислотные последовательности биспецифического антитела Р1АЕ1887.
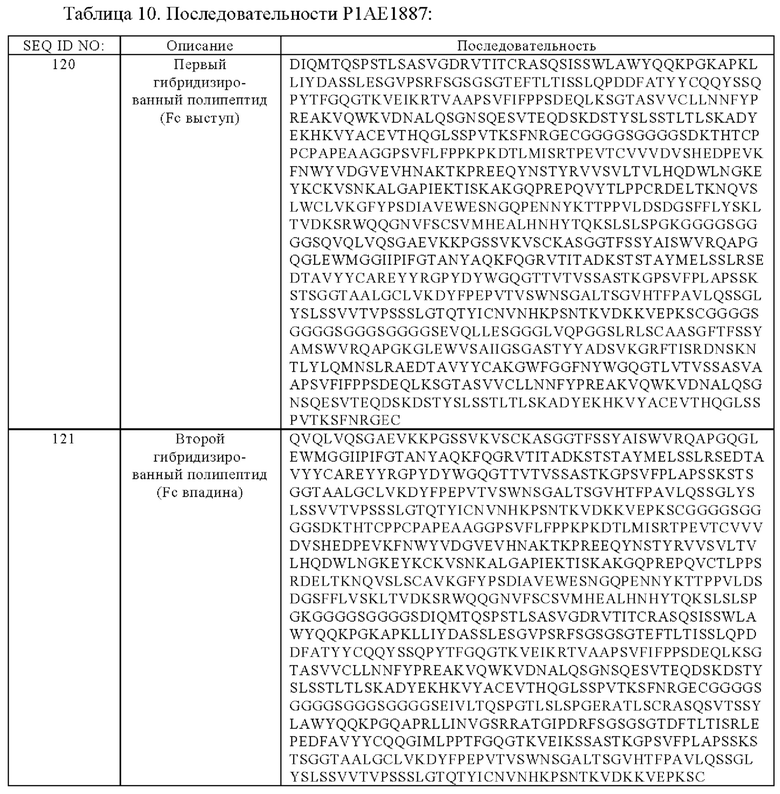
Схема сборки структуры показана на фиг. 1Л.
2.11 Получение FAP (4В9)-ОХ40 (49В4) конторстела Р1АЕ1888 (аонторстело 4)
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(OX40)-CH1, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VL(OX40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VH(OX40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235A1a интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 11 показаны аминокислотные последовательности биспецифического антитела Р1АЕ1888.


2.12 Получение FAP (4B9)-OX40 (49B4) конторстела P1AE2254 (конторстело 5)
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(OX40)-CH1 ЕЕ (K147Е, K213Е), коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa RK (E123R, Q124K), коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
- второй гибридизированный полипептид (от N- к С-концу): VH(OX40)-CH1_EE (K147Е, K213Е), коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa RK (E123R, Q124K), коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235A1a интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15). Кроме того, СН и Ckappa, гибридизированные с VL и VH в ОХ40, соответственно, аминокислотные мутации (так называемые заряженные остатки) интродуцируют для предупреждения образования белков Бен-Джонса и для дополнительного облегчения корректного спаривания, т.е. отрицательные заряды в домене CHI (K147Е, K213Е, нумерация по Kabat, индекс EU) и положительные заряды в домене CL анти-ОХ40 связующего 49 В4 (E123R и Q124K, нумерация по Kabat, индекс EU).
В таблице 12 показаны аминокислотные последовательности биспецифического антитела Р1АЕ2254.
Схема сборки структуры показана на фиг. 1М.
2.13 Получение FAP f4B9VOX40 (49В4) конторстела Р1АЕ2340 (конторстело 6)
Биспецифическое антитело, содержащее два гибридизиро ванных полипептида и легкую цепь, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(OX40)-CH1_EE (K147Е, K213Е), коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa RK (E123R, Q124K), коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VH(OX40)-CH1_ЕЕ (К147Е, K213Е), коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa_RK (E123R, Q124K).
- легкая цепь: VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15). Кроме того, СН и Ckappa, гибридизированные с VL и VH в ОХ40, соответственно, аминокислотные мутации (так называемые заряженные остатки) интродуцируют для предупреждения образования белков Бен-Джонса и для дополнительного облегчения корректного спаривания, т.е. отрицательные заряды в домене CHI (K147Е, К213Е, нумерация по Kabat, индекс EU) и положительные заряды в домене CL анти-ОХ40 связующего 49 В4 (E123R и Q124K, нумерация по Kabat, индекс EU).
В таблице 13 показаны аминокислотные последовательности биспецифического антитела Р1АЕ2340.

Схема сборки структуры показана на фиг. 1Н.
2.14 Получение FAP (4В9)-ОХ40 (49В4) конторстела PIAE2735 (конторстело 8)
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(FAP)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(OX40)-CH1, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-Ckappa;
- второй гибридизированный полипептид (от N- к С-концу): VL(FAP)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(OX40)-CH1, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-Ckappa.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 14 показаны аминокислотные последовательности биспецифического антитела Р1АЕ2735.
Схема сборки структуры показана на фиг. 1O.
2.15 Получение FAP (4В9)-ОХ40 (49В4) конторстела Р1АЕ2743 (конторстело 9) Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(FAP)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(OX40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VH(OX40)-CH1.
- второй гибридизированный полипептид (от N- к С-концу): VL(FAP)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(OX40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VH(OX40)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 15 показаны аминокислотные последовательности биспецифического антитела Р1АЕ2743.

2.16 Получение FAP (4 В9УОХ40 (49B4) конторстела P1AE2762 (конторстело 10)
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(FAP)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(OX40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc выступ, коннектор (G4S)2, VL(OX40)-CH1.
- второй гибридизированный полипептид (от N- к С-концу): VH(FAP)-Ckappa, коннектор GGGGSGGGGSGGG SGGGGS (SEQ ID NO: 84), VH(OX40)-Ckappa, коннектор (G4S)2, IgG1 шарнир, Fc впадина, коннектор (G4S)2, VL(OX40)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
В таблице 16 показаны аминокислотные последовательности биспецифического антитела Р1АЕ2762.

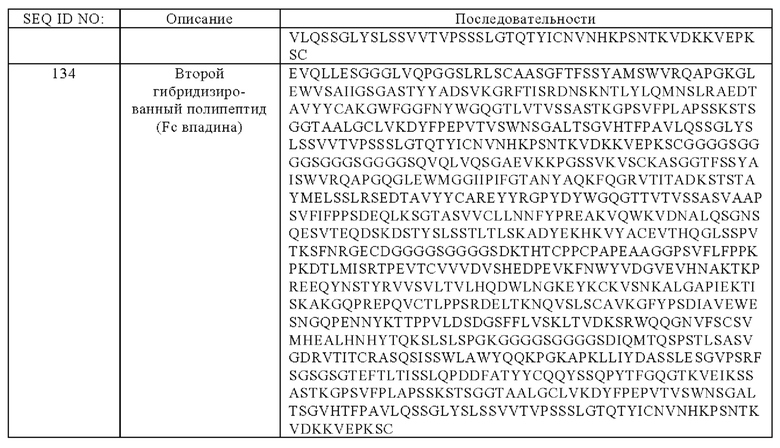
2.17 Биохимический анализ молекул после очистки
Табл. 17 суммирует выход и итоговое содержание мономера FAP ОХ40 конторстел.
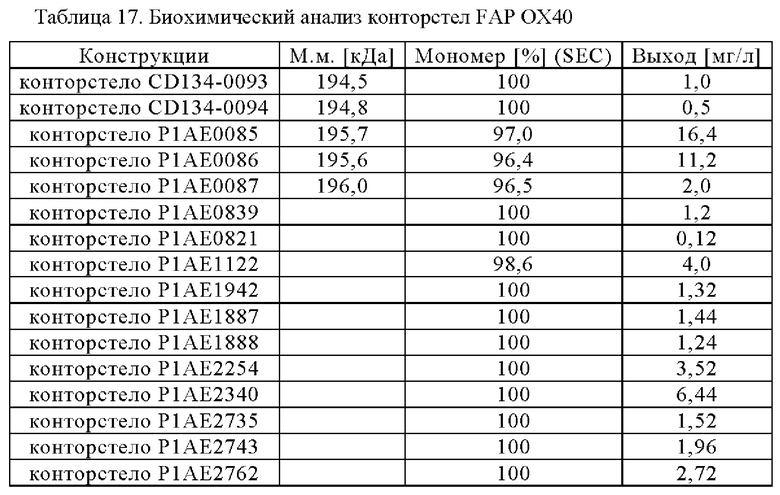
2.18 Получение биспецифических ОХ40 антител как молекул контроля
Для применения в качестве контролей получают следующие биспецифические анти-ОХ40-антитела:
а) биспецифическое антитело с двухвалентным связыванием ОХ40 и моновалентным связыванием FAP получают по аналогии с примером 4.4 в WO 2017/055398 А2 (формат 2+1). В этой молекуле первая тяжелая цепь (НС 1) состоит из одной единицы Fab (VHCH1) анти-ОХ40 49В4 связующего, за которой следует цепь Fc с выступом, гибридизированная через линкер (G4S) с доменом VH анти-FAP связующего 28Н1 или 4 В9. Вторая тяжелая цепь (НС 2) конструкции состоит из одной единицы Fab (VHCH1) анти-ОХ40 связующего 49В4, за которой следует цепь Fc с впадиной, гибридизированная через линкер (G4S) с доменом VL анти-FAP связующего 28Н1 или 4В9. Схема молекул показана на фиг. 1П.
б) антитело с двухвалентным связыванием ОХ40, согласно указанному выше, в котором домены VH и VL связующего анти-FAP заменены контролем зародышевой линии, называемой DP47, не связывающейся с антигеном. Эту молекулу применяют в качестве отрицательного, «не нацеливающегося» контроля.
в) биспецифическое антитело с двухвалентным связыванием ОХ40 и моновалентным связыванием FAP получают по аналогии с примером 4.4 в WO 2017/060144 А1 (формат 4+1). В этой молекуле первая тяжелая цепь (НС 1) состоит из двух единиц Fab (VHCH1 и VHCH1) анти-ОХ40 связующего 49 В4, за которым следует цепь Fc с выступом, гибридизированная через линкер (G4S) с доменом VH анти-FAP связующего 4В9. Вторая тяжелая цепь (НС 2) конструкции состоит из двух единиц Fab (VHCH1 и VHCH1) анти-ОХ40 связующего 49В4, за которыми следует цепь Fc с впадиной, гибридизированная через линкер (G4S) с доменом VL связующего вещества против анти-FAP связующего 4 В9. Схема молекулы показана на фиг. 1Р.
Биспецифические агонистические ОХ40 антитела с четырехвалентным связыванием ОХ40 и моновалентным связыванием FAP получают методом «выступ-во-впадину», позволяющим собирать две разные тяжелые цепи. Мутации Pro329Gly, Leu234Ala и Leu235Ala вводят в константную область тяжелых цепей для прекращения связывания с Fc-гамма-рецепторами в соответствии с методом, описанным в патентной заявке WO 2012/130831 А1.
Пример 3. Свойства антител FAP OX
3.1 Связывание OX40 человека (кинетическая аффинность)
Связывание биспецифических антител FAP-OX40 с ОХ40 человека исследуют методом поверхностного плазмонного резонанса с использованием прибора BIACORE Т100 (фирма GE Healthcare). Около 8000 резонансных единиц (resonance unit, RU) системы захвата (20 мкг/мл анти-IgG человека (Fc); код заказа: BR100839; фирма GE Healthcare Bio-Sciences АВ, Швеция) связаны на чипе СМ5 (фирма GE Healthcare BR-1005). -30) при рН 5,0 с использованием набора для захвата аминов, поставляемого фирмой GE Healthcare. Подвижным буфером является PBS-P, рН 7,4 (20 мМ фосфатный буфер, 2,7 мМ KCl, 137 мМ NaCl, 0,05% поверхностно-активное вещество Р20). В проточной кювете устанавливают температуру 25°С, а в блоке пробы 12°С, и дважды заполняют подвижным буфером. Биспецифическое антитело захватывают путем впрыскивания раствора 2 мкг/мл в течение 60 сек при скорости потока 5 мкл/мин. Ассоциацию измеряют путем введения ОХ40 человека в течение 120 сек при скорости потока 30 мкл/мин, начиная с 600 нМ в разведении 1:3. Фазу диссоциации подвергают мониторингу в течение вплоть до 720 сек и запускают переключением с раствора образца на подвижный буфер. Поверхность регенерируют промывкой 3 М MgCl2 в течение 60 сек при скорости потока 10 мкл/мин. Различия в индексе объемного преломления корректируют путем вычитания ответа, полученного от поверхности анти-IgG человека (Fc). Пустые инъекции также вычитают (= двойное сопоставление). Для расчета KD и кинетических параметров используют модель Ленгмюра 1:1.
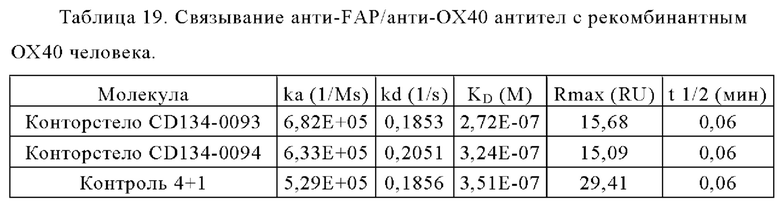
Оба конторстела имеют сходные значения KD по сравнению с «4+1» IgG-подобным форматом для четырехвалентного анти-ОХ40 антитела; аффинность сопоставима между молекулами, и Rmax указывает на валентность различных протестированных молекул.
Дополнительные FAP-OX40 конторстела тестируют и полученные значения KD указаны в табл. 20 ниже.
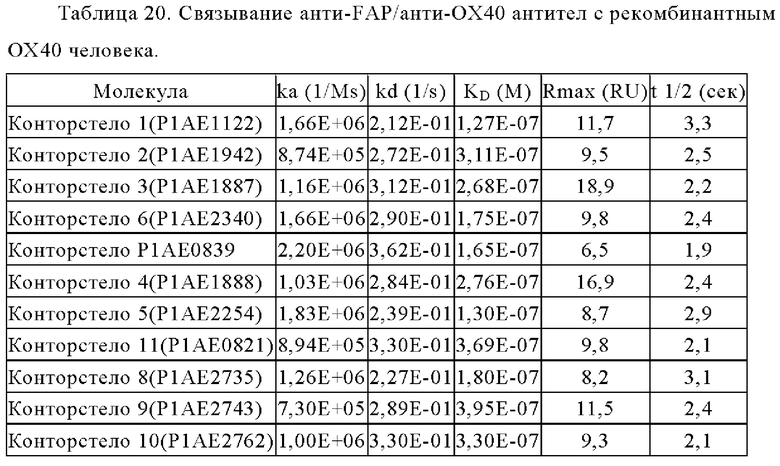
3.2 Связывание с FAP человека (кинетическая аффинность)
Связывание биспецифических антител FAP-OX40 с FAP человека исследуют методом поверхностного плазмонного резонанса с использованием прибора BIACORE Т100 (фирма GE Healthcare). Около 12000 резонансных единиц (RU) системы захвата (15 мкг/мл антигистидинового антитела; код заказа: 28995056; фирма GE Healthcare Bio-Sciences АВ, Швеция) связаны на чипе СМ5 (фирма GE Healthcare BR-1005-30) при рН 4,5 с использованием набора для присоединения амина, поставляемого фирмой GE Healthcare. Подвижный буфер для иммобилизации представляет HBS-N, рН 7,4 (10 мМ HEPES, 150 мМ NaCl, рН 7,4, фирма GE Healthcare). Для следующей кинетической характеристики подвижным буфером является PBS-P, рН 7,4 (20 мМ фосфатный буфер, 2,7 мМ KCl, 137 мМ NaCl, 0,05% сурфактанта Р20). В проточной кювете устанавливают температуру 25°С, а в блоке пробы 12°С, и ее дважды заполняют подвижным буфером. Рекомбинантный FAP человека захватывают путем впрыскивания раствора 25 мкг/мл в течение 60 сек при скорости потока 5 мкл/мин. Ассоциацию измеряют путем инъекции биспецифического антитела в течение 120 сек при скорости потока 30 мкл/мин, начиная с 300 нМ в разведении 1:2. Фазу диссоциации подвергают мониторингу в течение вплоть до 720 сек и запускают переключением с раствора образца на подвижный буфер. Поверхность регенерируют промыванием 10 мМ глицином, рН 1,5, в течение 60 сек при скорости потока 30 мкл/мин. Различия в индексе объемного преломления корректируют путем вычитания ответа, полученного от поверхности анти-гистидина. Пустые инъекции также вычитают (= двойное сопоставление). Для расчета KD и кинетических параметров используют модель Ленгмюра 1:1.
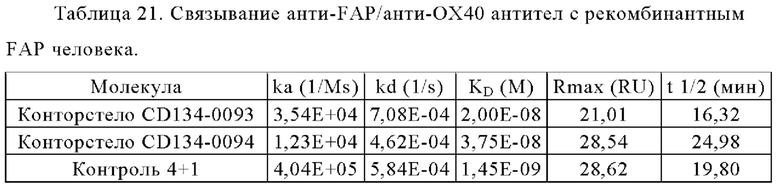
Обе молекулы имеют одинаковые значения KD. Ассоциация с внеклеточным доменом (ECD) FAP менее оптимальна в формате конторстело по сравнению со стандартной контрольной молекулой. Однако в контрольной молекуле два пептидных линкера 4xG4S применяют для связывания С-концевой анти-БАР-фрагмент с Fc-частью, тогда как в конторстелах два пептидных линкера 2xG4S применяют для связывания анти-FAP-фрагмента с конторстелом. Другие FAP-OX40 конторстела тестируют и их значения KD перечислены в табл. 22 ниже.
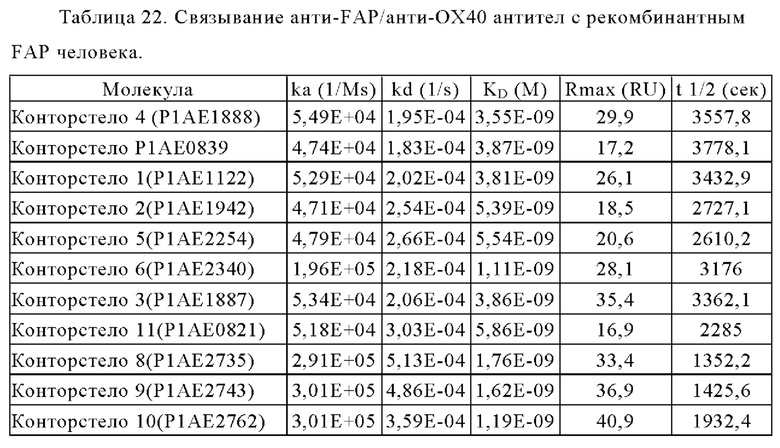
3.3 Одновременное связывание ОХ40 человека и FAP человека (кинетическая аффинность)
Способность одновременного связывания ОХ40 человека и FAP человека также оценивают методом поверхностного плазмонного резонанса (SPR), используя прибор BIACORE Т100 (фирма GE Healthcare).
Около 8000 резонансных единиц (resonance unit, RU) системы захвата (20 мкг/мл анти-IgG человека (Fc); код заказа: BR100839; фирма GE Healthcare Bio-Sciences АВ, Швеция) связаны на чипе СМ5 (фирма GE Healthcare BR-1005-30) при рН 5,0 с использованием набора для захвата аминов, поставляемого фирмой GE Healthcare. Подвижным буфером является PBS-P, рН 7,4 (20 мМ фосфатный буфер, 2,7 мМ KCl, 137 мМ NaCl, 0,05% поверхностно-активное вещество Р20). В проточной кювете устанавливают температуру 25°, а в блоке пробы 12°, и дважды заполняют подвижным буфером. Биспецифическое антитело захватывают путем впрыскивания раствора 2 мкг/мл в течение 60 сек при скорости потока 5 мкл/мин. Ассоциацию измеряют путем введения первого исследуемого вещества (ОХ40 человека или FAP человека, соответственно) в течение 120 сек при скорости потока 30 мкл/мин. Затем впрыскивают второе исследуемое вещество (FAP человека или ОХ40 человека, соответственно) в течение 120 сек при скорости потока 30 мкл/мин. Фазу диссоциации подвергают мониторингу в течение вплоть до 720 сек и запускают переключением с раствора образца на подвижный буфер. Поверхность регенерируют промывкой 3 М MgCl2 в течение 60 сек при скорости потока 10 мкл/мин. Различия в индексе объемного преломления корректируют путем вычитания ответа, полученного от поверхности анти-IgG человека (Fc). Пустые инъекции также вычитают (=двойное сопоставление). Для расчета KD и кинетических параметров используют модель Ленгмюра 1:1. Все конторстела способны одновременно и независимо связываться с обоими антигенами.
Пример 4. Связывание на клетках
4.1 Связывание с нативными МКПК человека и сравнение связывания с активированными МКПК человека
Лейкоцитарные пленки получают от Цюрихского фонда донорства крови SRK (Stiftung Zurcher Blutspendedienst SRK). Мононуклеарные клетки периферической крови (МКПК) человека выделяют центрифугированием в градиенте плотности фиколла. Для выделения свежих МКПК лейкоцитарные пленки разводят равным объемом DPBS (фирма Gibco by Life Technologies, номер в каталоге 14190 326). В полипропиленовые центрифужные пробирки объемом 50 мл (фирма ТРР, номер в каталоге 91050) вносят по 15 мл раствора Histopaque 1077 (фирма SIGMA Life Science, номер в каталоге 10771, полиахароза и диатриозат натрия, плотность скорректирована до 1,077 г/мл) и раствор лейкоцитарных пленок наслаивают поверх раствора Histopaque 1077. Пробирки центрифугируют в режиме 400 × g в течение 30 мин при комнатной температуре с низким ускорением и без торможения. После этого МКПК собирают с области контакта, трижды промывают ФСБ, ресуспендируют в среде для Т-клеток, состоящей из RPMI 1640 (фирма Gibco by Life Technology, номер в каталоге 42401-042) с добавлением 10% фетальной сыворотки теленка (ФСТ, фирма Gibco by Life Technology, номер в каталоге 16000-044, партия 941273, стерилизованной радиацией, не содержащей микоплазм и инактивированной нагреванием при 56°С в течение 35 мин), 1% (объем/объем) GlutaMAX I (фирма Gibco by Life Technology, номер в каталоге 35050 038), 1 мМ пирувата натрия (фирма SIGMA, номер в каталоге S8636), 1% (объем/объем) минимальной питательной среды с аминокислотами, не являющимися незаменимыми (MEM non-essential amino acids; фирма SIGMA, номер в каталоге М7145) и 50 мкМ β-меркаптоэтанола (фирма SIGMA, М3148). МКПК замораживают в ФСБ, содержащем 10% (объем/объем) диметилсульфоксида.
Замороженные МКПК оттаивают в среде для Т-клеток и МКПК применяют сразу после выделения (связывание с покоящимися МКПК человека) или их стимулируют для получения сильной экспрессии ОХ40 человека на поверхности Т-клеток (связывание с активированными МКПК человека). Поэтому наивные МКПК культивируют в течение двух дней в среде для Т-клеток, обогащенной 200 ед./мл пролейкина и 2 мкг/мл PHA-L в 6-луночном планшете для культуры ткани, а затем 1 день 6-луночных планшетах для культуры ткани с предварительно нанесенном покрытием [4 мкг/мл анти-CD3 человека (клон ОКТЗ) и 2 мкг/мл анти-С028 (клон CD28.2)] в среде для Т-клеток.
Для обнаружения ОХ40 наивные МКПК человека и активированные ч МКПК человека смешивают. Чтобы различать наивные и активированные МКПК человека, перед анализом связывания наивные клетки метят красителем для детекции пролиферации клеток eFluor670 (фирма eBioscience, номер в каталоге 65-0840-85). Смесь 1:1 клеток в количестве 1×105, наивных МКПК меченых eFluor670 и немеченых активированных МКПК человека, затем вносят в каждую лунку круглодонного 96-луночного планшета (фирма greiner bio-one; cellstar, номер в каталоге 650185) и исследуют связывание.
Клетки окрашивают в течение 120 мин при 4° в темноте в 50 мкл/лунку холодного буфера FACS (температура 4°), содержащего оттитрованные конструкции анти-ОХ40 антител. После трехкратной промывки избытком буфера FACS клетки окрашивают в течение 45 мин при 4° в темноте в 25 мкл/лунку холодного буфера FACS (температура 4°), содержащего смесь флуоресцентно меченного анти-С04 человека (клон RPA-T4, IgG1k мыши, фирма BioLegend, номер в каталоге 300532), анти-С08 человека (клон RPA-T8, IgG1k мыши, фирма BioLegend, номер в каталоге 3010441) и флуоресцеин-изотиоцианат (FITC)-конъюгированный AffiniPure анти-IgG человека Fcγ-фрагмент-специфичный козий IgG F(ab')2 фрагмент (фирма Jackson ImmunoResearch, номер в каталоге 109-096-098). В итоге планшеты ресуспендируют в 85 мкл/лунку FACS-буфера, содержащего 0,2 мкг/мл DAPI (фирма Santa Cruz Biotec, номер в каталоге Sc-3598) и исследуют в тот же день, применяя флуориметр 5-лазерный LSR-Fortessa (BD Bioscience с программным обеспечением DIVA).
На фиг. 2Б и 2Г показано, что ни одна антигенсвязывающая молекула, специфичная в отношении ОХ40, не связана с покоящимися CD4 Т-клетками или CD8 Т-клетками человека. Напротив, все антигенсвязывающие молекулы (ОХ40 (49 В4) FAP (28Н1) 2+1 биспецифическое антитело, ОХ40 (49 В4) DP47 2+1 биспецифическое антитело, ОХ40 (49 В4) FAP (4 В9) 4+1 биспецифическое антитело, конторстела CD134-0093 и CD134-0094) связаны с активированными CD8 + или CD4 + Т-клетками (фигуры 2А и 2В). Связывание с CD4 + Т-клетками намного сильнее, чем с CD8 + Т-клетками. Все форматы конструкции 2+1 связаны со сходной силой с ОХ40-экспрессирующими (положительными) клетками, независимо от связывающего фрагмента второй специфичности. Кроме того, конструкция 4+1 показывает самое сильное связывание. Конторстело CD134-0093 показывает промежуточное связывание между форматами 2+1 и 4+1, второе конторстело CD134-0094 связывается с CD4 + и CD8 + Т-клетками слабее, чем форматы 2+1. Антитело отрицательного контроля DP47 hu IgG1 (P329G LALA) не связывается ни с активированными, ни с покоящимися Т-клетками. Поскольку ОХ40 не активируется на покоящихся CD4 или CD8 Т-клетках, ни одна из протестированных молекул не связывается с покоящимися клетками. Более того, связывание всех конструкций (кроме отрицательного контроля) сильнее на Т-клетках CD4, поскольку экспрессия ОХ40 на этих клетках выше, чем на CD8 Т-клетках.
Результаты второго эксперимента показаны на фиг. 2Д-2З. Как показано на этих фигурах, ни одна из протестированных молекул (ОХ40 (49 В4) FAP (28Н1) 2+1 биспецифическое антитело, ОХ40 (49В4) FAP (4В9) 2+1 биспецифическое антитело, ОХ40 (49В4) DP47 2+1 биспецифичное антитело, конторстела Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087), связанных с покоящимися CD4 + Т-клетками или CD8 + Т-клетками человека, как и ожидалось, поскольку ОХ40 не экспрессируется в покоящихся клетках (фиг. 2Е и 2З).
Напротив, все антигенсвязывающие молекулы демонстрируют связывание с активированными CD8 + и CD4 + Т-клетками (фигуры 2Д и 2Ж). Амплитуда сигнала была ниже на CD8 + Т-клетках, чем на CD4 Т-клетках, что коррелирует с уровнем экспрессии ОХ40 (высокий на CD4 + Т-клетках). Однако характер связывания каждой тестируемой молекулы сопоставим у CD4+ и CD8+ Т-клеток. Молекулы всех трех антител не связываются с ОХ40-экспрессирующими клетками, однако они показывают пониженную связывающую способность с ОХ40 по сравнению с контрольными молекулами 2+1 независимо от клона, связывающего FAP. В частности, связывание конторстела Р1АЕ0087 несколько хуже, чем связывание конторстела Р1АЕ0085.
Связывание дополнительных конторстел (конторстела 1-11) с активированными CD4+ Т-клетками показано на фиг. 8А-8Г, а связывание с активированными CD8+ Т-клетками показано на фиг. 10А-10Г. На фиг. 9А-9Г и фиг. 11А-11Г, соответственно, показано, что ни одно из конторстел 1-11 не связывается с покоящимися CD4 Т-клетками или CD8 Т-клетками человека, что ожидалось. Напротив, все антигенсвязывающие молекулы связаны с активированными CD8+ или CD4+ Т-клетками. Связывание с CD4+ Т-клетками намного сильнее, чем с CD8+ Т-клетками. Все форматы конструкции 2+1 связываются с силой, сходной с ОХ40-положительными клетками, независимо от связывающего фрагмента второй специфичности. Отрицательный контроль (DP47 hu IgG1 P329G LALA) не связывается с активированными или покоящимися Т-клетками. Поскольку ОХ40 не активируется на покоящихся CD4 или CD8 Т-клетках, ни одна из тестируемых молекул не связывается с покоящимися клетками. Более того, связывание всех конструкций (кроме отрицательного контроля) сильнее на CD4+ Т-клетках, поскольку экспрессия ОХ40 выше в этих клетках, чем на CD8+ Т-клетках.
4.2 Связывание с опухолевыми клетками, экспрессирующими FAP человека
Связывание с FAP клеточной поверхности тестируют с использованием белка, активирующего фибробласты человека (huFAP), экспрессируемого клетками WM266-4 (АТСС CRL-1676). Отсутствие связывания с ОХ40-отрицательными FAP-отрицательными опухолевыми клетками тестируют с использованием А549 NucLight™ Red Cells (фирма Essen Bioscience, номер в каталоге 4491), экспрессирующих флуоресцентный белок NucLight Red, ограниченный ядром, чтобы осуществить отделение от немеченных FAP человека положительных клеток WM266-4. Исходный штамм А549 (АТСС CCL-185) трансдуцируют CellPlayer NucLight Red Lentivirus (фирма Essen Bioscience, номер в каталоге 4476; EF1α, пуромицин) при MOI 3 (TU/клетку) в присутствии 8 мкг/мл полибрена по стандартному протоколу Essen. Эффективность трансдукции составляет ≥70%.
Смесь 5×104 немеченных клеток WM266-4 и немеченных А549 NucLight™ Red Cells в буфере FACS добавляют в каждую лунку 96-луночных круглодонных планшетов для суспензий клеток (фирма greiner bio-one; cellstar, номер в каталоге 650185) и исследуют связывание. Планшеты центрифугируют в течение 4 мин в режиме 400×g при 4° и супернатанты удаляют. Клетки промывают однократно 200 мкл DPBS, и осадки ресуспендируют с помощью короткого и аккуратного встряхивания. Все образцы ресуспендируют в 50 мкл/лунку холодного буфера FACS (температура 4°), содержащего биспецифические антигенсвязывающие молекулы (первичного антитела) в указанном диапазоне концентраций (оттитрованного антитела), и инкубируют в течение 120 мин при 4°. После этого клетки промывают четыре раза 200 мкл холодного буфера FACS (температура 4°) и ресуспендируют быстрым встряхиванием. Затем клетки окрашивают 25 мкл/лунку раствора холодного вторичного антител (температура 4°), содержащего флуоресцеин-изотиоцианат (Р1ТС)-конъюгированный AffiniPure анти-IgG человека Fcγ-фрагмент-специфичный козий IgG F(ab')2 фрагмент (фирма Jackson ImmunoResearch, номер в каталоге 109-096-098) и инкубируют в течение 60 мин при 4° в темноте.
В итоге планшеты ресуспендируют в 90 мкл/лунку FACS-буфера, содержащего 0,2 мкг/мл DAPI (фирма Santa Cruz Biotec, номер в каталоге Sc-3598) и исследуют в тот же день, применяя флуориметр 5-лазерный LSR-Fortessa (BD Bioscience с программным обеспечением DIVA).
На фиг. 3А и 3Б показано, что FAP-нацеленные анти-ОХ40-биспецифичные антитела эффективно связываются с клетками-мишенями, экспрессирующими FAP человека. Следовательно, только анти-ОХ40-связывающие молекулы, нацеленные на FAP, проявляют свойство прямого нацеливания на опухоль. FAP4B9 имеет высокую аффинность к FAP человека, тогда как 28Н1 имеет высокую аффинность к FAP человека. Нацеленное на FAP (ОХ40 (49В4) FAP (4В9) 4+1 биспецифическое антитело показывает самое сильное связывание с клетками FAP+, за которым следует 2+1 конторстело CD134-0093, конторстело CD134-0094 и затем ОХ40 (49В4) FAP (28Н1) 2+1 биспецифичное антитело. Следует отметить, что FAP 4В9 обладает повышенной аффинностью к FAP человека по сравнению с 28Н1. Связывание с FAP+ клетками обоих конторстел было немного улучшено по сравнению с VHVL-содержащей нацеливающейся конструкцией антитела 2+1. Ненацеливающееся биспецифическое антитело ОХ40 (49В4) DP47 2+1 и отрицательный контроль (антитело DP47 hu IgG1 (P329G LALA)) не связываются с какими-либо FAP+ клетками. Значения связывания ЕС50 с активированными CD4 Т-клетками человека и клетками FAP-положительной опухоли приведены в табл. 23.
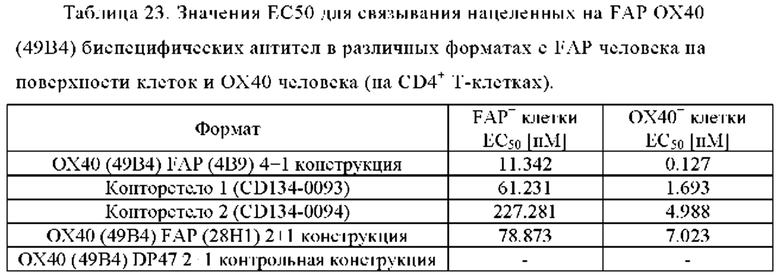
В другом эксперименте конторстела Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087 тестируют и сравнивают с ОХ40 (49В4) FAP (28Н1) 2+1 биспецифическим антителом, ОХ40 (49В4) FAP (4В9) 2+1 биспецифическим антителом и ОХ40 (49В4) DP47 2+1 биспецифическим антителом (отрицательный контроль). Результаты показаны на фиг. 3В и 3Г. Все FAP-нацеленные анти-ОХ40-антигенсвязывающие молекулы связаны с FAP-человека экспрессирующими клетками. FAP 4В9 клон имеет повышенную аффинность к FAP человека, чем 28Н1. Следовательно, наилучшими связывающими свойствами обладает биспецифическое антитело ОХ40 (49В4) FAP (4В9) 2+1, а биспецифическое антитело ОХ40 (49В4) FAP (28Н1) 2+1 показало пониженное связывание, что и ожидалось. Все три контортела показывают связывание с FAP, хотя связывание с клетками FAP+ было слегка нарушено по сравнению с биспецифическими антителами ОХ40 (49В4) FAP 2+1 для обоих FAP клонов (4В9 и 28Н1). Длина линкера, по-видимому, не влияет на связывание с huFAP, поскольку конторстело Р1АЕ0085 и Р1АЕ0087 демонстрируют очень похожие свойства связывания. Ненацеленное биспецифическое антитело 2 ОХ40 (49В4) DP47 2+1 и отрицательный контроль (антитело DP47 hu IgG1 (P329G LALA)) не связываются с какими-либо клетками FAP+. Кроме того, ни одна из молекул не показывает связывания с FAP- A549NLR клетками, что свидетельствует о специфичности связывания с FAP человека. Значения связывания ЕС50 с активированными CD4 и CD8 Т-клетками и FAP-положительными клетками человека суммированы в табл. 24.
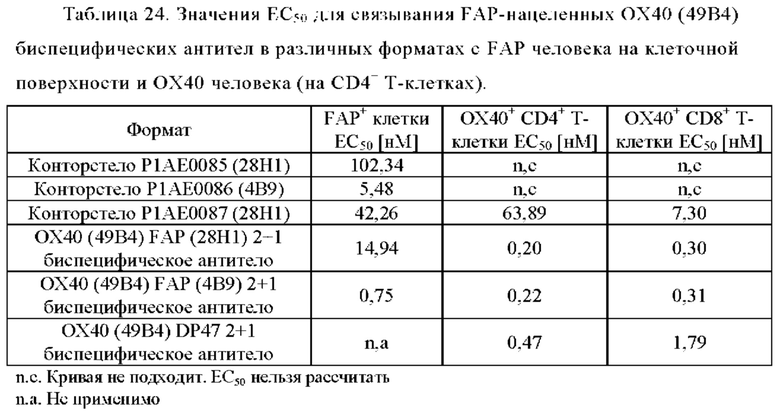
Результаты связывания конторстел 1-10 с клетками FAP+ (опухолевые клетки NIH/3T3-huFAP) показаны на фиг. 13А-13Г. Все FAP-нацеленные анти-ОХ40-антигенсвязывающие молекулы связываются с клетками, экспрессирующими FAP человека. Как показано на фиг. 14А-14Г, ни одна из FAP-нацеленных антигенсвязывающих анти-ОХ40 молекул не способна связываться с опухолевыми клетками A549NLR (FAP-отрицательными). Конторстело 8 сильнее всего связывается с клетками FAP+, за которым следуют конторстело 10 и конторстело 6. Связывание с FAP+ клетками обоих конторстел немного улучшено по сравнению с VHVL-содержащей нацеливающейся конструкцией антител 2+1. Не способная к нацеливанию 2+1 анти-ОХ40 конструкция (7718) и отрицательный контроль (8105) не связываются с какими-либо FAP+ клетками. Значения связывания ЕС50 с активированными CD4+ Т-клетками человека и FAP-положительными опухолевыми клетками суммированы в табл. 25.
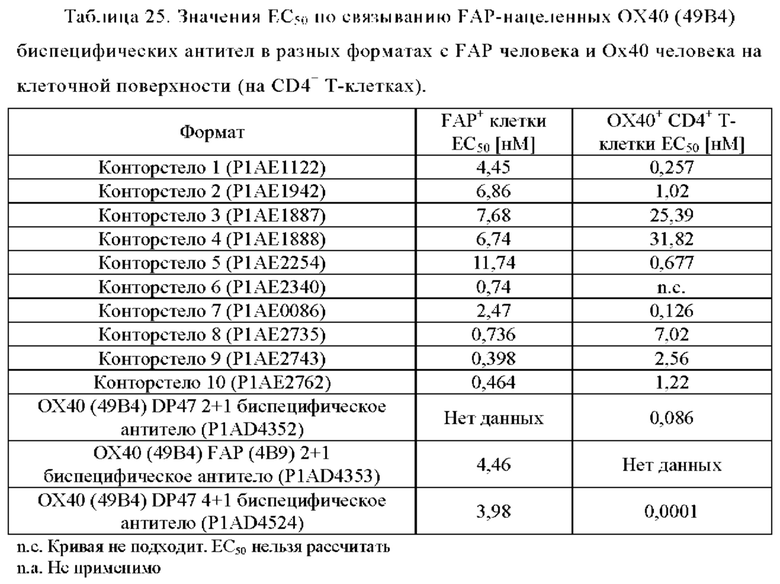
Пример 5. Функциональные свойства биспецифических анти-ОХ40 человека связывающих молекул
5.1 Клетки HeLa, экспрессирующие ОХ40 человека и репортерный ген NFκB-люциферазы
Агонистическое связывание ОХ40 с соответствующим лигандом индуцирует передачу сигналов вниз по цепи посредством активации ядерного фактора каппа В (NFkB) (A.D. Weinberg с соавт., J. Leukoc. Biol. 75(6), 2004, 962-972). Линия рекомбинантных репортерных клеток HeLa_hOx40_NFκB_Luc1 была создана для экспрессии Ох40 человека на их поверхности. Кроме того, эти клетки несут репортерную плазмиду, содержащую ген люциферазы под контролем NFκB-чувствительного сегмента энхансера. В качестве триггера ОХ40 индуцирует дозозависимую активацию NFκB, который транслоцируется в ядре, где связывается с чувствительным к NFκB энхансером репортерной плазмиды для увеличения экспрессии белка люциферазы. Люцифераза катализирует окисление люциферина, в результате чего получают оксилюциферин, который излучает свет. Эмиссию света можно определить количественно с помощью люминометра.
Таким образом, способность различных молекул анти-ОХ40 индуцировать активацию NFκB в репортерных клетках HeLa_hOx40_NFκB_Luc1 рассматривают в качестве меры биоактивности.
Тестируют NFκB активирующую способность отобранных биспецифических ОХ40 (49В4) антител в двухвалентном FAP-нацеленном VHVL или конторстела формате, отдельно и с гипер-перекрестным связыванием конструкций либо вторичным антителом, либо линией FAP+ опухолевых клеток. Перекрестное связывание FAP-связывающих антител FAP на клеточной поверхности исследуют с помощью активирующего фибробласты человека белка (huFAP), экспрессируемым NIH/3T3-huFAP клоном 19. Эту линию клеток получают путем трансфекции клеточной линии эмбриональных фибробластов мыши NIH/3T3 (АТСС CRL-1658) вектором экспрессии pETR4921 для экспрессии huFAP на фоне отбора с помощью 1,5 мкг/мл пуромицина.
Адгезивные клетки HeLa_hOX40_NFκB_Luc1 культивируют в течение ночи при плотности клеток 0,2×105 клеток на лунку и стимулируют в течение 5 ч средой для анализа, содержащей оттитрованные биспецифические анти-ОХ40 (49В4) антитела (ОХ40 (49В4) FAP (28Н1) 2+1 биспецифическое антитело, ОХ40 (49В4) DP47 2+1 биспецифическое антитело, конторстела CD134-0093 и CD134-0094). Для тестирования эффекта гипер-перекрестного сшивания вторичными антителами, 25 мкл/лунку среды, содержащей вторичное антитело анти-IgG человека Fcγ-фрагмент-специфичный козьего IgG F(at')2 фрагмент (фирма Jackson ImmunoResearch, номер в каталоге 109-006-098), добавляют в пропорции 1:2 (первичное ко вторичному антителу). Для тестирования эффекта гиперперекрестного сшивания связыванием FAP на клеточной поверхности, 25 мкл/лунку среды с опухолевыми клетками FAP+ (NIH/3T3-huFAP клон 19) совместно культивируют в соотношении 4:1 (в четыре раза больше опухолевых клеток FAP+ относительно репортерных клеток в лунке).
После инкубирования надосадочную жидкость для анализа отсасывают и планшеты дважды промывают буфером DPBS. Эмиссию света определяют количественно с помощью системы анализа люциферазы 100 и репортерного лизирующего буфера (оба реактива от фирмы Promega, номера в каталоге Е4550 и Е3971, соответственно) по инструкциями производителя. Вкратце, клетки лизируют в течение 10 мин при -20°С за счет добавления 30 мкл на лунку 1х лизирующего буфера. Клетки оттаивают в течение 20 мин при 37°С до внесения в лунки по 90 мкл предусмотренного реагента для исследования с люциферазой. Эмиссию света определяют сразу же с помощью ридера для микропланшетов SpectraMax М5/М5е (фирма Molecular Devices, США), используя время интеграции 500 мсек, без какого-либо фильтра для сбора всех длин волн. Относительные единицы испускаемого света (relative light units, URL) корректируют с помощью базальной люминесценции клеток HeLa_hOx40_NFNB_Luc1 и наносят против логарифма концентрации первичного антитела, используя программное обеспечение Prism4 (фирма GraphPad Software, США). Кривые подбирают, используя встроенный сигмоидальный дозовый ответ.
На фиг. 4А-4В представлены все конструкции анти-ОХ40 индуцировало активацию NFκB. На фиг. 4А-4В показано, что присутствие анти-ОХ40 конструкций индуцирует NFκB активацию. Гипер-перекрестное сшивание через вторичное анти-huIgG Fcγ-специфическое антитело повышает активацию NFκB для всех связующих FAP-независимым образом. Два биспецифических антитела, 2+1 конструкции ОХ40 (49В4) FAP (28Н1) и ОХ40 (49В4) DP47, действуют сходным образом, потому что нацеливание FAP, по-видимому, не влияет на индукцию NFκB. Конторстела CD134-0093 и CD134-0094 показывают сопоставимую, но не такую сильную активацию NFκB при перекрестном сшивании (их влияние, по-видимому, является внутренним и не зависит от перекрестного сшивания). FAP-экспрессирующие опухолевые клетки сильно увеличивают индукцию NFκB-опосредованной зависимым от концентрации образом при использовании FAP-нацеленных молекул (закрашенный треугольник, полу-закрашенный круг, закрашенный круг). Такой эффект не наблюдают с биспецифическим антителом ОХ40 (49В4) DP47 2+1 (незакрашенный кружок), поскольку конструкция не может быть дополнительно гипер-перекрестно сшита опухолевыми клетками FAP+. Кроме того, FAP-нацеленное биспецифическое антитела ОХ40 (49В4) FAP (28Н1) 2+1 индуцирует более сильную NFκB активацию по сравнению с конторстелом CD134-0093 и конторстелом CD134-0094, тогда как CD134-0094 показывает более сильную активацию по сравнению с CD134-0093 (фиг. 4А). В целом, конструкции конторстел способны индуцировать ОХ40-опосредованную активацию NFκB, однако они менее активными, чем FAP-нацеленное биспецифическое антитело ОХ40 (49В4) FAP (28Н1) 2+1, либо перекрестно сшитое вторичным антителом, либо FAP, экспрессируемым на клеточной поверхности. Но также наблюдают пониженную активность при добавлении без вторичной олигомеризации.
Результаты второго эксперимента показаны на фиг. 4Г-4Е, из которых следует, что присутствие всех анти-ОХ40 протестированных молекул (биспецифическое антитело ОХ40 (49 В4) FAP (28Н1) 2+1, биспецифическое антитело ОХ40 (49В4) FAP (4В9) 2+1, биспецифичное антитело ОХ40 (49В4) DP47 2+1, конторстела Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087) индуцирует очень низкую активацию NFκB в отсутствие перекрестного связывания (фигура 4Г). Все три контрольных молекулы 2+1 (биспецифическое антитело ОХ40 (49В4) FAP (28Н1) 2+1, биспецифическое антитело ОХ40 (49В4) FAP (4В9) 2+1 и биспецифическое антитело ОХ40 (49В4) DP47 2+1) проявляют очень близкую NFκB активацию, если гипер-перекрестное сшивание обеспечивает вторичное антитело (фиг. 4Е). Три молекулы конторстел показывают немного пониженную активность по сравнению с контролями. Это различие в NFkB активации между тестируемыми конторстелами и контрольными молекулами не наблюдают в присутствии FAP-экспрессирующих клеток. Кривые, соответствующие молекулам конторстел и соответствующие им FAP-нацеленным контрольным молекулам, анализируют с помощью программного обеспечения GraphPad Prism как разделяющую одну глобальную подборку кривых, показывают, что нет значительного различия в эффективности. Несмотря на то, что между двумя протестированными клонами (4В9 и 28Н1) существует разная аффинность с FAP, в этом анализе это не привело к заметной разнице в эффективности. DP47-нацеленная 2+1 контрольная молекула проявляет только минимальную активность, поскольку она не имеет FAP-связывающего фрагмента. Отрицательный контроль (DP47 hu IgG1 антитело (P329G LALA)) не проявляет активности, независимо от поперечного сшивания.
Дополнительно проверяют NFkB активирующую способность конторстел 1-11 и гипер-перекрестного сшивания конструкций биспецифических антител с помощью линии FAP+ опухолевых клеток. На фиг. 15А-15Г показывают NFκB активацию в присутствии FAP-экспрессирующих опухолевых клеток. Активация NFκB без перекрестного сшивания с помощью FAP показана на фиг. 16А-16Г.
Сумма площади под кривой значений по NFκB активации в клетках HeLa с/без перекрестного сшивания с FAP+ клетками показана на фиг. 17. Наличие всех анти-ОХ40 конструкций индуцирует NFκB активацию. Гипер-перекрестное сшивание через FAP+ опухолевые клетки увеличивает NFκB активацию для всех протестированных биспецифических антител. FAP-экспрессирующая опухолевая клетка сильно увеличивает индукцию NFκB-опосредованную активацию люциферазы зависимым от концентрации образом при использовании FAP-нацеливающихся молекул. Такого эффекта не наблюдают, если в формате 2+1 FAP-связывающий фрагмент заменен не связывающей единицей DP47 (незаштрихованный треугольник), поскольку конструкция не может быть дополнительно гипер-перекрестно сшита опухолевыми клетками FAP+. Все конторстела выполнены сопоставимо с 2+1 биспецифическим антителом ОХ40 (49В4) FAP (4В9).
5.2. ОХ40-опосредованная костимуляция субоптимально TCR-стимулированных покоящихся МКПК человека и гипер-перекрестное сшивание FAP на клеточной поверхности
В разделе 5.1 показано, что добавление FAP+ опухолевых клеток может сильно увеличить активность NFκB, индуцированную FAP-нацеленным двухвалентными анти-ОХ40-антителами в линиях ОХ40-положительных репортерных клеток человека, обеспечивая сильную олигомеризацию рецепторов ОХ40. Аналогичным образом, тестируют FAP-нацеленные двухвалентные анти-ОХ40-антитела в присутствии клеток NIH/3T3-huFAP клона 19 на способность восстанавливать субоптимальную TCR-стимуляцию покоящихся клеток МКПК человека.
Препараты МКПК человека содержат (1) покоящиеся ОХ40-отрицательные CD4+ и CD8+ Т-клетки и (2) антиген-презентирующие клетки с различными молекулами рецептора Fcγ на поверхности, например, В-клетки и моноциты. Анти-CD3 человека антитело изотипа IgG1 человека может связываться своей Fc-частью с присутствующими молекулами рецептора Fey и опосредовать длительную TCR активацию на покоящихся ОХ40-отрицательных CD4+ и CD8+ Т-клетках. Эти клетки затем начинают экспрессировать ОХ40 в течение нескольких часов. Функциональные агонистические соединения против ОХ40 могут передавать сигналы через рецептор ОХ40, присутствующий на активированных CD8+ и CD4+ Т-клетках, и поддерживать TCR-опосредованную стимуляцию.
Покоящиеся МКПК человека стимулируют в течение пяти дней субоптимальной концентрацией анти-CD3-антитела в присутствии облученных клеток FAP+ NIH/3T3-huFAP клона 19 и оттитрованных анти-ОХ40 конструкций. Воздействие на выживание и пролиферацию Т-клеток исследуют путем мониторинга общего количества клеток и совместного окрашивания флуоресцентно-меченными антителами против маркеров активации и созревания Т-клеток (CD25/CD127) методом жидкостной цитометрии.
Эмбриональные фибробласты мыши NIH/3T3-huFAP клон 19 собирают, используя буфер для диссоциации клеток (фирма Invitrogen, номер по каталогу 13151-014) в течение 10 мин при 37°С. Клетки однократно промывают буфером DPBS. Клетки NIH/3T3-huFAP клона 19 культивируют при плотности 0,2×105 клеток на лунку в среде для Т-клеток в стерильном 96-луночном круглодонном планшете для культивирования адгезивной ткани (фирма ТРР, номер в каталоге 92097) в течение ночи при 37°С в атмосфере 5% CO2 в инкубаторе (Hera Cell 150). На следующий день их облучают рентгеном из облучателя, используя дозу 4500 RAD, чтобы предотвратить последующее чрезмерное размножение МКПК человека линией опухолевых клеток.
МКПК человека выделяют центрифугированием в плотности фиколла. Клетки добавляли в каждую лунку в количестве 0,6×105 клеток на лунку. Анти-CD3 человека антитело (клон V9, IgG1 человека) в конечной концентрации [10 нМ] и FAP-нацеленные двухвалентные анти-ОХ40 антигенсвязывающие молекулы и конторстела добавляют в номинальных концентрациях. Клетки активируют в течение четырех дней при 37°С и в атмосфере 5% СО2 в инкубаторе (Hera Cell 150).
Затем поверхность клеток окрашивают флуоресцентными конъюгированными с красителем анти-CD4 человека антителами (клон RPA-T4, фирма BioLegend, номер по каталогу 300532), CD8 (клон RPa-T8, фирма BioLegend, номер по каталогу 3010441), CD25 (клон М-А251, фирма BioLegend, номер по каталогу 356112) и CD127 (клон A019D5, фирма BioLegend, номер по каталогу 351324) в течение 20 минут при 4°С. Осадки клеток однократно промывают буфером FACS. В итоге планшеты ресуспендируют в 85 мкл/лунку FACS-буфера, содержащего 0,2 мкг/мл DAPI (фирма Santa Cruz Biotec, номер по каталогу Sc-3598), и исследуют в тот же день, применяя флуориметр 5-лазерный LSR-Fortessa (фирма BD Bioscience с программным обеспечением DIVA).
Как показано на фиг. 5А-5Е, костимуляция не нацеленным ОХ40 (49В4) DP47 2+1 биспецифическим антителом (незаштрихованный треугольник) не восстанавливает субоптимально TCR-стимулируемые CD4 и CD8 Т-клетки. Гипер-перекрестное сшивание FAP-нацеленных ОХ40 (49В4) FAP (28Н1) 2+1 биспецифического антитела (заштрихованный треугольник) и 2+1 конторстел CD134-0093 и CD134-0094 в присутствии клеток NIH/3T3-huFAP клона 19 сильно способствует выживанию и индуцирует повышено активированный фенотип в CD4 и CD8 Т-клетках человека. Кроме того, особенно конторстело CD134-0093 (заштрихованный кружок), по-видимому, демонстрирует наиболее сходные свойства по сравнению с нацеленным ОХ40 (49В4) FAP (28Н1) 2+1 биспецифическим антителом в отношении активации. Кроме того, конторстело CD134-0094 (наполовину заштрихованные кружки) показывает повышенную активацию CD8 Т-клеток по сравнению с активацией CD4 Т-клеток (фиг. 5Г), но оно проявляет сопоставимую активацию с конторстелом CD134-0093 и FAP-нацеленным ОХ40 (49В4) FAP (28Н1) 2+1 биспецифическим антителом на CD4 Т-клетках. В целом, результаты по биоактивности и активации Т-клеток нормализуют и суммируют на фиг. 6, где агонистическая способность каждой конструкции определена количественно для анализируемых маркеров как площадь под кривой и нанесены на график для сопоставления. Как показано на рисунке 6, конторстело CD134-0093 показывает наиболее сильную активацию Т-клеток CD4 и CD8 в отношении FSC-A и CD25. Эта молекула, по-видимому, более эффективна, чем FAP-нацеленное ОХ40 (49В4) FAP (28Н1) 2+1 биспецифическое антитело, которое по эффективности примерно на 75% равно CD134-0093. Конторстело CD134-0094 представляется немного менее мощным, поскольку только CD25 получает усиленную регуляцию, но размер (FSC-A) CD4 и CD8 Т-клеток не увеличивается. Кроме того, не нацеленная молекула 2+1 и отрицательный контроль не показывают какой-либо активации (очень низкие нормализованные значения AUC).
В следующем эксперименте костимуляция не нацеленным ОХ40 (49В4) DP47 2+1 биспецифическим антителом не восстанавливает субоптимально TCR простимулированные CD4+ и CD8+ Т-клетки. Гипер-перекрестное сшивание FAP-нацеленных двухвалентных анти-ОХ40-антител ОХ40 (49В4) FAP (28Н1) 2+1 биспецифического антитела и ОХ40 (49В4) FAP (4В9) биспецифического антитела, а также молекул трех конторстел (Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087), благодаря наличию клеток NIH/3T3-huFAP клона 19 значительно усиливает активацию первичных CD4 и CD8 Т-клеток человека. Р1АЕ0086 индуцирует очень сходную активацию Т-клеток, в отличие от соответствующего 2+1 4В9 контрольного ОХ40 (49В4) FAP (4В9) биспецифического антитела. Они приводят к более активированному фенотипу, чем молекулы с 28Н1 FAP-связующим, которые, как известно, имеют пониженную аффинность к FAP человека. Конторстела Р1АЕ0085 и Р1АЕ0087 проявляют сходную экспрессию CD25 на CD4 и CD8 Т-клетках и немного более высокий показатель FFI-A MFI по сравнению с 2+1 контрольной молекулой ОХ40 (49В4) FAP (28Н1) 2+1 биспецифического антитела. Этот анализ с первичными МКПК показывает, что активность FAP-нацеленных молекул, содержащих связующее с высокой аффинностью (4В9), выше, чем у молекул с связующим веществом с низкой аффинностью FAP (28Н1). Результаты биологической активности и активации Т-клеток показаны на фиг. 7А-7Г.
Также исследуют способность конторстел 7, 8, 9 и 10 восстанавливать субоптимально TCR-простимулированные CD4+ и CD8+ Т-клетки. Фиг. 18А и 18Б показывают активацию CD4 Т-клеток, отмеченных поверхностным маркером CD25, после субоптимальной CD3 стимуляции. На фиг. 18В и 18Г показана активация CD8 Т-клеток, обозначенных поверхностным маркером CD25. На фигурах 19А и 19Б показана FSC-A MFI CD4 Т-клеток, а на фиг. 19В и 19Г показана FSC-A MFI CD8 Т-клеток. eFluor 670 является красителем, применяемым для выявления пролиферации и измерения деления клеток. Уровни eFluor 670, соответственно, пролиферация CD4 Т-клеток, показаны на фиг. 20А и 20Б, а пролиферация CD8 Т-клеток показана на фиг. 20В и 20Г. Пониженная регуляция IL-7Ra (CD127) показана для CD4 Т-клеток на фиг. 21А и 21Б и для CD8 Т-клеток на фиг. 21В и 21Г. Все результаты демонстрируют, что гипер-перекрестное сшивание контортел и ОХ40 (49В4) FAP (4В9) биспецифического антитела (заштрихованный треугольник) в присутствии клеток NIH/3Т3-huFAP клон 19 в высокой степени способствует выживанию и индуцирует усиленный активированный фенотип в CD4 и, в меньшей степени, в CD8 Т-клетках человека. На фиг. 22А-22Г результаты различных анализов биологической активности суммированы как площадь под кривой.
5.3 Суммирование результатов
В целом, 2+1 конторстело CD134-0093 действует активнее, чем 2+1 конторстело CD134-0094. Связывание CD134-0093 с активированными CD4 Т-клетками сильнее, чем у FAP-нацеленного ОХ40 (49В4) FAP (28Н1) 2+1 биспецифического антитела и CD 134-0094, которые оба проявляют сходные связывающие свойства (фиг. 2А). Оба конторстела проявляют хорошее связывание с FAP человека на поверхности клеток WM266-4 (фиг. 3А), сила связывания занимает промежуточное положение между ОХ40 (49В4) FAP (4В9) 4+1 биспецифическим антителом (содержащим высоко аффинное связующее 4В9) и ОХ40 (49В4) FAP (28Н1) 2+1 биспецифическим антителом (содержащим связующее с низкой аффинностью 28Н1). Даже если формат конторстел отличается от ОХ40 (49В4) FAP (28Н1) 2+1 биспецифического антитела, согласно описанию в примере 2.9, связывающие свойства представляются хорошими или даже лучшими. Тем не менее, они не достигают величин MFI ОХ40 (49В4) FAP (4В9) 4+1 биспецифического антитела (пример 2.9). Исходя из результатов активации NFκB с помощью перекрестного сшивания клеток NIH 3Т3 huFAP, следует что конторстела продемонстрировали промежуточную активацию NFκB по сравнению с (49В4) FAP (28Н1) 2+1 биспецифическим антителом, но все же более сильную активацию, чем у ненацеленного ОХ40 (49В4) DP47 2+1 биспецифического антитела (фиг. 4А). Домены FAP обоих конторстел, по-видимому, способны действовать в качестве перекрестных связующих и активировать передачу сигнала через ОХ40. Используя вторичное Fcγ-специфическое антитело в качестве перекрестного связующего, оба конторстела проявляют сходную активацию NFκB, но менее эффективны, чем нацеленные контрольные молекулы (фиг. 4Б). Исследование биоактивности конторстел показывает, что CD134-0093, по-видимому, индуцирует более выраженный активированный фенотип на CD4 и CD8 Т-клетках, чем CD134-0094 (фиг. 5А-Г). Конторстело CD134-0093 даже проявляет повышенную активацию Т-клеток, чем двухвалентный формат 2+1 (фиг. 6).
Конторстела Р1АЕ0085, Р1АЕ0086 и Р1АЕ 0087 проявляют несколько сниженную способность связываться с ОХ40, экспрессированным на поверхности активированных CD4+ и CD8+ Т-клеток, по сравнению с двухвалентными анти-ОХ40 контрольными молекулами (фиг. 2Д-2З). Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087 также показывают частично нарушенное связывание с FAP человека, экспрессируемого на поверхности клеток NIH-3T3 huFAP клон 19 (фиг. 3В и 3Г) при сравнении с соответствующими контролями (2+1 4В9 и 28Н1 FAP связующими). Молекулы, содержащие высоко аффинное FAP связующее 4В9, показывают превосходную связывающую способность с FAP человека, по сравнению с молекулами со связующим 28Н1 с пониженной аффинностью. Анализ активации NFκB показывает, что три молекулы конторстел Р1АЕ0085, Р1АЕ0086 и Р1АЕ 0087 индуцируют сходную активацию NFκB при сравнении с контрольными FAP-нацеленными молекулами, если перекрестное связывание происходит через клетки NIH-3T3 huFAP клона 19 (фиг. 4Е). Не обнаружено различий в активности FAP-нацеленных анти-ОХ40 молекул с низкой и высокой активностью при проведении HeLa NFκB репортерного анализа. Биоактивность молекул конторстел также тестируют, используя первичные МКПК человека и клетки NIH-3T3 huFAP клона 19 в качестве перекрестно связывающим клеток, в пятисуточном анализе активации. Молекулы с 4В9 FAP связующим индуцируют более активированный фенотип в CD4+ и CD8+ Т-клетках по сравнению с 28Н1 связующим. Молекулы конторстел проявили себя так же, как соответствующие им контрольные молекулы. Взятые вместе, эти данные показывают, что молекулы конторстел Р1АЕ0085, Р1АЕ0086 и Р1АЕ0087 имеют близкую активность, отличающуюся от активности контрольных молекул, несмотря на слегка сниженную способность к связыванию с ОХ40 и huFAP.
В целом, несмотря на разный формат, конторстела (и особенно CD134-0093), по-видимому, обладают сопоставимыми, если не лучшими свойствами в отношении связывания и активации Т-клеток, чем FAP-нацеленные 2+1 анти-ОХ40-антитела, описанные в примере 2.18.
Пример 6. Получение биспецифических антител с двумя антигенсвязывающими доменами, связывающимися с 4-1ВВ, и одним антигенсвязывающим доменом, связывающимся с FAP (FAP-4-1BB конторстела)
Выработка и приготовление связующих FAP описано в патентной заявке WO 2012/020006 А2, которая включена в настоящее изобретение в виде ссылки. Для связующего 4-1ВВ последовательности VH и VL клона 20Н4.9 получают в соответствии с описанием US 7288638 В2 или US 7659384 В2.
6.1 Получение FAP (4В9)-4-1ВВ (20Н4.9) конторстела Р1АЕ1899
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1А:
- первый гибридизированный полипептид (от N- к С-концу): VH (4-1ВВ)-СН1, коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VL(4-1BB)-Ckappa, коннектор (GGGGSGGGGSGGGSGGGGS (SEQ ID NO:84), VH(FAP)-Ckappa
- второй гибридизированный полипептид (от N- к С-концу): VH (4-1ВВ)-СН1, коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VL(4-1BB)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
Таблица 26 представляет аминокислотные последовательности биспецифического антитела Р1АЕ1899.

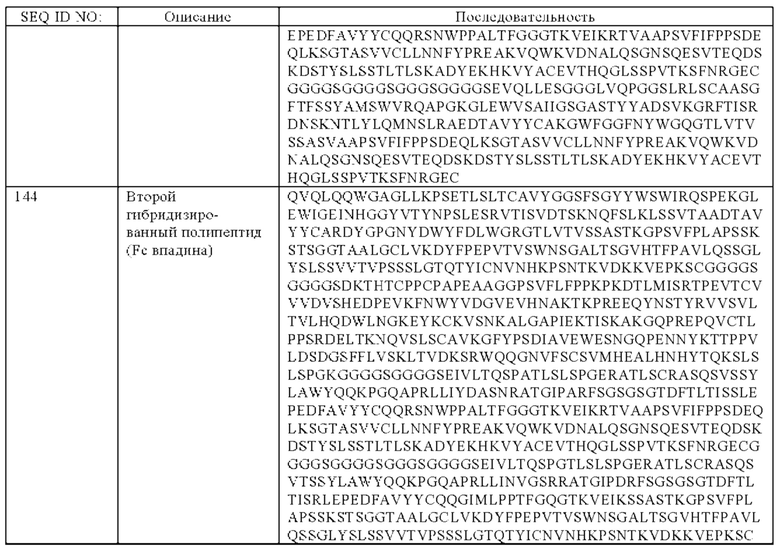
Схема собранной структуры показана на фиг. 1Д (ОХ40 заменен на CD40).
6.2 Получение FAP (4В9)-4-1ВВ (20Н4.9) конторстела Р1АЕ2051
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(4-1BB)-CH1_ЕЕ (K147E, K213E), коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VL(4-1BB)-Ckappa_RK (E123R, Q124K), коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO:84), VH(FAP)-Ckappa.
- второй гибридизированный полипептид (от N- к С-концу): VH(4-1BB)-CH1EE (K147E, K213E), коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VL(4-1BB)-Ckappa_RK (E123R, Q124K), коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO:84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15). Кроме того, в СН и Ckappa, гибридизированных с VL и VH в ОХ40, соответственно, аминокислотные мутации (так называемые заряженные остатки) интродуцируют для предупреждения выработки белков Бен-Джонса и для дальнейшего облегчения правильного спаривания, то есть отрицательных зарядов в домене СН1 (K147E, K213E, нумерация по Kabat, индекс EU) и положительных зарядов в домене CL анти-ОХ40 связующего 49В4 (E123R и Q124K, нумерация по Kabat, индекс EU).
Таблица 26 представляет аминокислотные последовательности биспецифического антитела Р1АЕ2051.
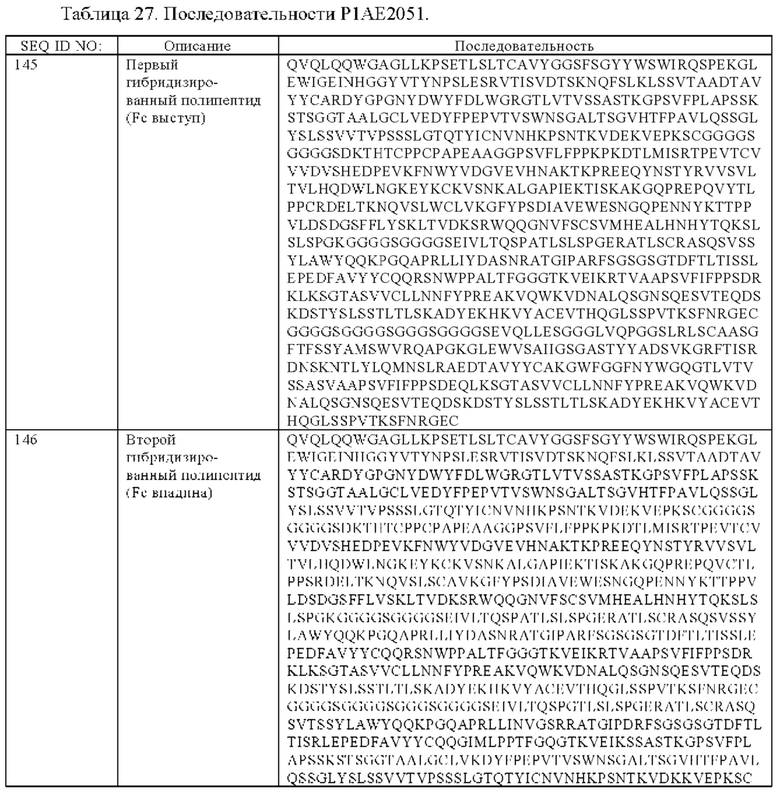
Схема собранной структуры показана на фиг. 1М (ОХ40 заменен на 4-1ВВ).
6.3 Биохимический анализ молекул после очистки
Табл. 28 суммирует выход и итоговое содержание мономера конторстел FAP-4-1ВВ.

Пример 7. Свойства конторстел FAP-4-BB
7.1 Связывание FAP человека (кинетическая аффинность)
Связывание биспецифических антител FAP-4-1BB с FAP человека исследуют методом плазмонного резонанса, используя прибор BIACORE Т100 (фирма GE Healthcare), согласно описанию в примере 3.2. Для расчета величины KD и кинетических параметров применяют модель связывания Ленгмюра.

Обе молекулы имеют близкие значения KD.
7.2 Активация NF-κВ в линии репортерных клеток Jurkat-hu4-1BB-NFκB-luc2, экспрессирующих 4-1ВВ человека и NFκB-люцифераза репортерный ген
Агонистическое связывание рецептора 4-1ВВ (CD137) с его лигандом (4-1BBL) вызывает 4-1ВВ-нисходящую передачу сигнала через активацию ядерного фактора каппа В (NFκB) и стимулирует выживание и активность CD8 Т-клеток (Lee H.W. с соавт., J Immunol, 169, 2002, 4882-4888). Для мониторинга такой NFκB-активации, опосредованной биспецифическим антителом 2+1 Н2Н анти-4-1ВВ, анти-FAP huIgGI PGLALA, получают линию репортерных клеток Jurkat-hu4-1BB-NFκB-luc2 от фирмы Promega (Германия). Клетки культивируют в виде суспензии в среде RPMI 1640 (фирма GIBCO by Life Technologies, номер в каталоге 42401-042), обогащенной 10% (объем/объем) фетальной сыворотки теленка (fetal bovine serum, FBS; фирма GIBCO by Life Technologies, номер в каталоге 16000-044, Lot 941273, обработана радиацией и не содержит микоплазмы, инактивирована нагреванием), 2 мМ дипептида L-аланил-L-глутамина (Glutqa-MAX-I; фирма GIBCO by Life Technologies, номер в каталоге 35050-038), 1 мМ пирувата натрия (SIGMA-Aldrich, номер в каталоге S8636), 1% (объем/объем) MEM-раствор аминокислот, не являющихся незаменимыми 100х (фирма SIGMA-Aldrich, номер в каталоге М7145), 600 мкг/мл G-418 (фирма Roche, номер в каталоге 04727894001), 400 мкг/мл гигромицина В (фирма Roche, номер в каталоге 10843555001) и 25 мМ HEPES (фирма Sigma Life Sience, номер в каталоге Н0887-100 мл. Для анализа клетки собирают и ресуспендируют в среде для анализа RPMI 1640, обогащенной 10% ФСБ (объем/объем) и 1% (объем/объем) GlutaMAX-I. 10 мкл с 2×103 репортерными клетками Jurkat-hu4-1BB-NFκB-luc2 вносят в лунки стерильного белого 384-луночного плоскодонного планшета для культуры ткани с крышкой (фирма Corning, номер в каталоге 3826). Добавляют по 10 мкл среды для анализа, содержащей оттитрованные концентрации конторстел, 2+1 биспецифическое агонистическое анти-4-1ВВ (20Н4.9) х анти-FAP (4В9) huIgG1 P329GLALA антитело, анти-4-1ВВ (20Н4.9) huIgG1 P329GLALA антитело, анти-4-1ВВ (20Н4.9) huIgG4 и контроль изотипа (антитело DP47 hu IgG1 P329GLALA). В итоге, по 10 мкл среды для анализа, без добавок или содержащей 1×104 FAP-экспрессирующих клеток, NIH/3T3-huFAP клон 19 (согласно описанному выше), вносят и планшеты инкубируют в течение 6 ч при 37°С в атмосфере 5% СО2 в инкубаторе клеток. В лунки добавляют по 6 мкл только что оттаянного раствора для обнаружения One-Glo Luciferase (фирма Promega, номер в каталоге Е6110) и сразу измеряют эмиссию света от люциферазы, используя ридер для микропланшет Tecan (время интеграции 500 мсек, без фильтра, анализ в расширенном диапазоне волн).
На фигуре 23А показано, что в отсутствие FAP-экспрессирующих клеток ни одна из молекул не может индуцировать сильную активацию рецептора 4-1ВВ человека в линии репортерных клеток Jurkat-hu4-1BB-NFκB-luc2, приводящую к активации NFkB и, следовательно, к экспрессии люциферазы. В присутствии FAP-экспрессирующих клеток, таких как NIH/3T3-huFAP клон 19 (клеточная линия фибробластов трансгенной мыши с FAP-человека) (см. фиг. 23Б), перекрестное сшивание биспецифических 2+1 FAP 4-1ВВ конторстел, а также 2+1 биспецифического агонистического анти-4-1ВВ (20Н4.9) х анти-FAP (4В9) huIgG1 P329GLALA антитела (черная звездочка) приводит к сильному увеличению NFκB-активированной активности люциферазы в линии репортерных клеток Jurkat-hu4-1BB-NFκB-luc2, которая была выше активации, опосредованной ненацеленными контрольными 4-1ВВ антителами. Значения ЕС50 и площадь под кривой (AUC) кривых активации приведены в табл. 30 и табл. 31.


Пример 8. Получение биспецифических антител с двумя антигенсвязывающими доменами, связывающимися с CD40, и одним антигенсвязывающим доменом, связывающимся с FAP (конторстела FAP-CD40)
Разработка и получение связующих FAP описаны в патенте WO 2012/020006 А2, который включен в настоящее изобретение в виде ссылки. Для связующего CD40, последовательности VH и VL клона 20Н4.9 получают в соответствии с SEQ ID NO: 10 и SEQ ID NO: 16 из WO 2006/128103.
8.1 Получение FAP (4 В9УСР40 конторстела P1AE1799
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют как показано на фиг. 1А:
- первый гибридизированный полипептид (от N- к С-концу): VH (CD40)-CH1, коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VL(CD40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa
- второй гибридизированный полипептид (от N- к С-концу): VH (CD40)-CH1, коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VL(CD40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
Таблица 32 представляет аминокислотные последовательности биспецифического антитела Р1АЕ1799.

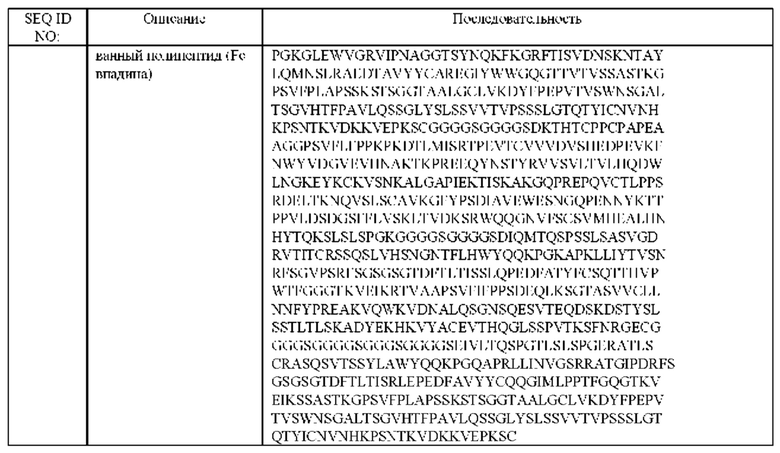
Схема собранной структуры показана на фиг. 1Д (ОХ40 заменен на CD40).
8.2 Получение FAP (4В9)-CD40 конторстела Р1АЕ1902
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH (CD40)-CH1, коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VL(CD40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
- второй гибридизированный полипептид (от N- к С-концу): VH (CD40)-CH1, коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VL(CD40)-Ckappa, коннектор GGGGS GGGGS GGG SGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
Таблица 33 представляет аминокислотные последовательности биспецифического антитела Р1АЕ1902.
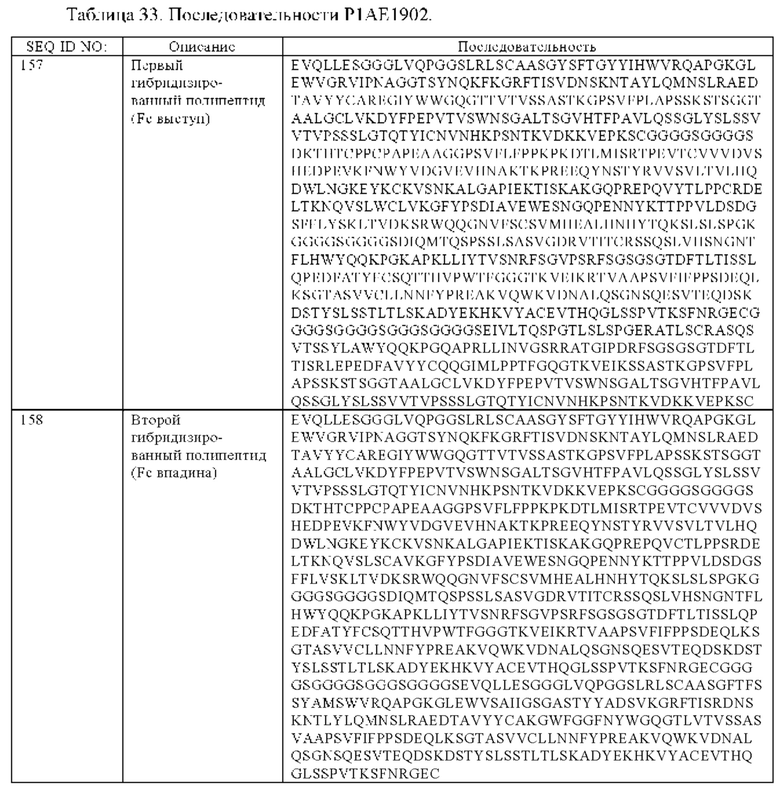
8.3 Получение FAP (4B9VCD40 конторстела P1AE1800
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH (CD40)-CH1, коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VL(CD40)-Ckappa, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
- второй гибридизированный полипептид (от N- к С-концу): VL(CD40)-Ckappa, коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VH(CD40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
Таблица 34 представляет аминокислотные последовательности биспецифического антитела Р1АЕ1800.
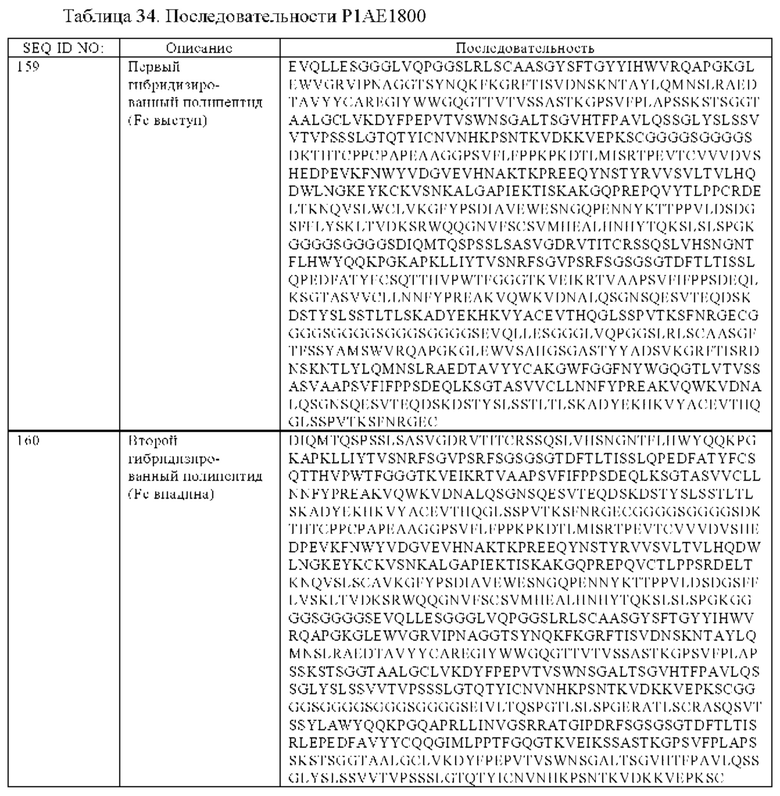
8.4 Получение FAP (4B9)-CD40 конторстела P1AE2052
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VL(CD40)-Ckappa, коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VH(CD40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
- второй гибридизированный полипептид (от N- к С-концу): VL(CD40)-Ckappa, коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VH(CD40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
Таблица 35 представляет аминокислотные последовательности биспецифического антитела Р1АЕ2052.


8.5 Получение FAP (4B9) CD40 конторстела Р1АЕ1901
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VL(CD40)-Ckappa, коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VH(CD40)-CH1,
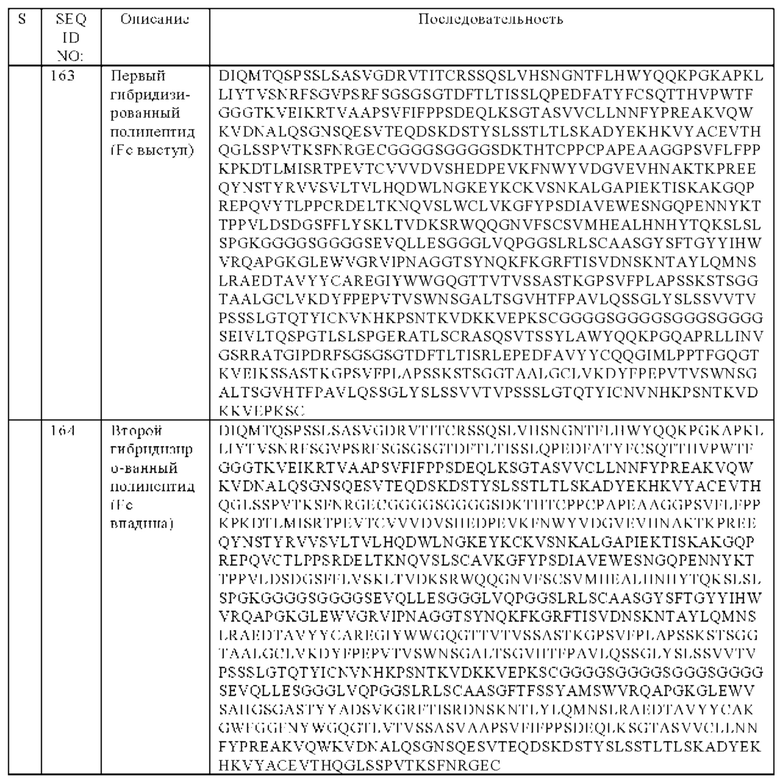
коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VL(FAP)-CH1.
- второй гибридизированный полипептид (от N- к С-концу): VL(CD40)-Ckappa, коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VH(CD40)-CH1, коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15).
Таблица 36 представляет аминокислотные последовательности биспецифического антитела Р1АЕ1901.
Таблица 36. Последовательности Р1АЕ1901
8.6 Получение FAP (4B9VCD40 конторстела Р1АЕ2255
Биспецифическое антитело, содержащее два гибридизированных полипептида, клонируют следующим образом:
- первый гибридизированный полипептид (от N- к С-концу): VH(CD40)-CH1 ЕЕ (K147E, K213E), коннектор (G4S)2, шарнир IgG1, Fc выступ, коннектор (G4S)2, VL(CD40)-Ckappa RK (E123R, Q124K), коннектор GGGGSGGGGSGGGSGGGGS (SEQ ID NO: 84), VH(FAP)-Ckappa.
- второй гибридизированный полипептид (от N- к С-концу): VH(CD40)-CH1_EE (K147E, K213E), коннектор (G4S)2, шарнир IgG1, Fc впадина, коннектор (G4S)2, VL(CD40)-Ckappa_RK (E123R, Q124K), коннектор GGGGS GGGGSGGGSGGGGS (SEQ ID NO:84), VL(FAP)-CH1.
Мутации Pro329Gly, Leu234Ala и Leu235Ala интродуцируют в константную область с выступом и с впадиной тяжелых цепей, чтобы отменить связывание с рецепторами Fc гамма, согласно методу, описанному в International Patent Appl. Publ. No. WO 2012/130831. Метод гетеродимеризации выступ-во-впадину применяют, используя мутации S354C/T366W в домене СН3 цепи с выступом и, соответственно, мутации Y349C/T366S/L368A/Y407V в домене СН3 цепи с впадиной (Carter, J Immunol Methods 248, 2001, 7-15). Кроме того, в СН и Скарра, гибридизированных с VL и VH в ОХ40, соответственно, аминокислотные мутации (так называемые заряженные остатки) интродуцируют для предупреждения выработки белков Бен-Джонса и для дальнейшего облегчения правильного спаривания, то есть отрицательных зарядов в домене СН1 (K147E, K213E, нумерация по Kabat, индекс EU) и положительных зарядов в домене CL анти-ОХ40 связующего 49В4 (E123R и Q124K, нумерация по Kabat, индекс EU).
Таблица 37 представляет аминокислотные последовательности биспецифического антитела Р1АЕ2255.
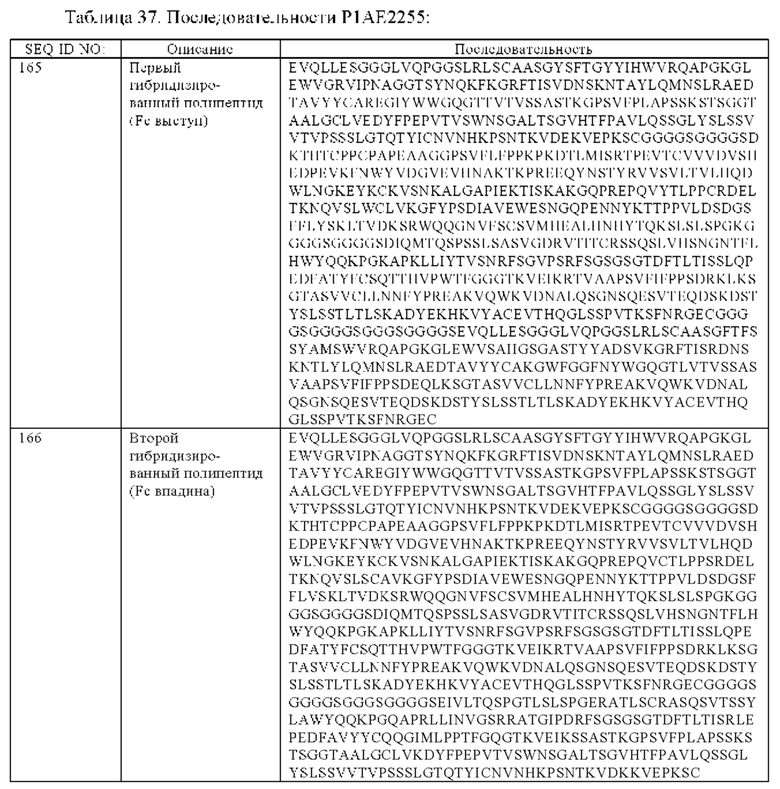
Схема собранной структуры показана на фиг. 1М (ОХ40 заменен на CD40).
8.7 Биохимический анализ молекул после очистки
В табл. 38 суммирован выход и содержание мономера конторстел FAP-4-1BB.
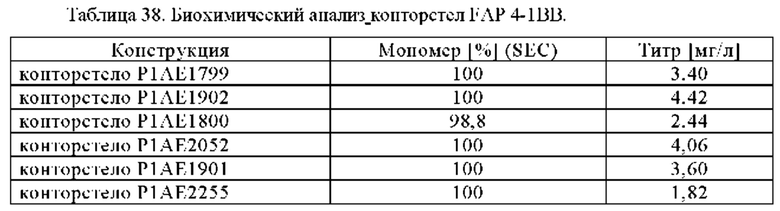
Пример 9
Характеристика антител FAP CD40
9.1 Связывание с CD40 человека
Способность биспецифических конструкций связываться с CD40 человека оценивают методом поверхностного плазмонного резонанса (SPR). Все эксперименты SPR проводят на приборе Biacore Т200 (фирма Biacore) при 25°С с HBS-EP в качестве подвижного буфера (0,01 М HEPES рН 7,4, 0,15 М NaCl, 3 мМ EDTA, 0,005% сурфактанта Р20, (фирма Biacore). Ассоциацию измеряют путем впрыскивания внеклеточного домена CD40 человека в различных концентрациях в растворе в течение 300 сек при скорости потока 30 мкл/мин, начиная с 300 нМ в разведении 1:3. Фазу диссоциации подвергают мониторингу в течение вплоть до 1200 сек и запускают переключением с раствора образца на подвижный буфер. Поверхность регенерируют путем в течение 60 сек раствором глицина с рН 2,1 при скорости потока 30 мкл/мин. Значительные различия в показателях преломления корректируют путем вычитания отклика, полученного от поверхности козьего анти-Е(ab’)2 человека. Пустые инъекции также вычитают (=двойная ссылка). Для расчета кажущейся Kd и других кинетических параметров используют модель Ленгмюра 1:1. Кажущуюся Kd рассчитывают с использованием оценочного программного обеспечения BiacoreTM В4000 (версия 1.1).
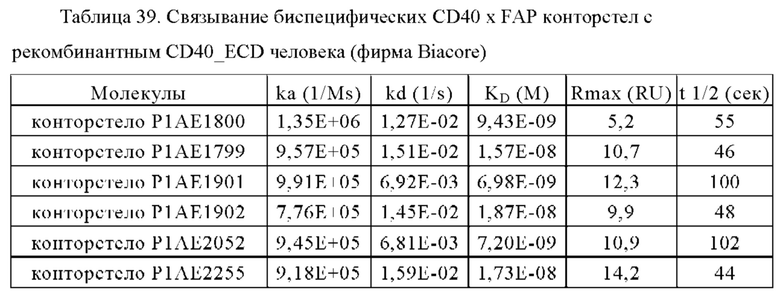
9.2. Связывание FAP человека (кинетическая аффинность)
Связывание биспецифических антител FAP-OX40 с FAP человека исследуют методом поверхностного плазмонного резонанса с использованием прибора BIACORE Т100 (фирма GE Healthcare). Около 12000 резонансных единиц (RU) системы захвата (15 мкг/мл антигистидинового антитела; код заказа: 28995056; фирма GE Healthcare Bio-Sciences АВ, Швеция) связаны с чипом СМ5 (фирма GE Healthcare BR-1005-30) при рН 4,5 с использованием набора для присоединения амина, поставляемого фирмой GE Healthcare. Подвижным буфером для иммобилизации является HBS-N, рН 7,4 (10 мМ HEPES, 150 мМ NaCl, рН 7,4, фирма GE Healthcare). Для следующей кинетической характеристики подвижным буфером является PBS-P, рН 7,4 (20 мМ фосфатный буфер, 2,7 мМ KCl, 137 мМ NaCl, 0,05% сурфактанта Р20). В проточной кювете устанавливают температуру 25°, а в блоке пробы 12°, и дважды заполняют подвижным буфером. Рекомбинантный FAP человека захватывают путем впрыскивания раствора 25 мкг/мл в течение 60 сек при скорости потока 5 мкл/мин. Ассоциацию измеряют путем введения биспецифического антитела течение 120 сек при скорости потока 30 мкл/мин, начиная с 300 нМ в разведении 1:2. Фазу диссоциации подвергают мониторингу в течение вплоть до 720 сек и запускают переключением с раствора образца на подвижный буфер. Поверхность регенерируют промывкой 10 мМ глицином, рН 1,5, в течение 60 сек при скорости потока 30 мкл/мин. Различия в индексе объемного преломления корректируют путем вычитания ответа, полученного от поверхности антигистидина. Пустые инъекции также вычитают (=двойное сопоставление). Для расчета KD и кинетических параметров используют модель Ленгмюра 1:1.
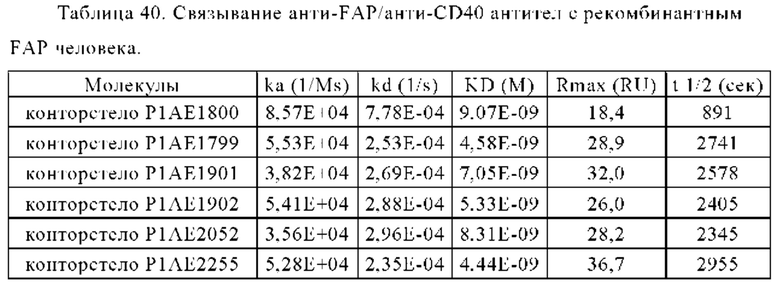
9.3 Одновременное связывание CD40 человека и FAP человека (кинетическая аффинность)
Способность связывать одновременно CD40 человека и FAP человека также оценивают с помощью поверхностного плазмонного резонанса (SPR) с использованием прибора BIACORE Т100 (фирма GE Healthcare). Около 8000 резонансных единиц (RU) системы захвата (20 мкг/мл анти-IgG человека (Fc); код заказа: BR100839; фирма GE Healthcare Bio-Sciences АВ, Швеция) связаны с чипом СМ5 (фирма GE Healthcare BR-1005-30) при рН 5,0 с использованием набора для присоединения амина, поставляемого фирмой GE Healthcare. Подвижным буфером для иммобилизации является HBS-N, рН 7,4 (20 мМ фосфатный буфер, 2,7 мМ KCl, 137 мМ NaCl, 0,05% сурфактанта Р20). В проточной кювете устанавливают температуру 25°, а в блоке пробы 12°, и дважды заполняют подвижным буфером. Биспецифическое антитело захватывают путем впрыскивания раствора 2 мкг/мл в течение 60 сек при скорости потока 5 мкл/мин. Ассоциацию измеряют путем введения первого анализируемого вещества (CD40 человека или FAP человека, соответственно) в течение 120 сек при скорости потока 30 мкл/мин. Затем вводят второе анализируемое вещество (FAP человека или CD40 человека, соответственно) в течение 120 сек при скорости потока 30 мкл/мин. Фазу диссоциации подвергают мониторингу в течение вплоть до 720 сек и запускают переключением с раствора образца на подвижный буфер. Поверхность регенерируют промывкой 3 М MgCl2 в течение 60 сек при скорости потока 10 мкл/мин. Различия в индексе объемного преломления корректируют путем вычитания ответа, полученного от поверхности анти-IgG человека (Fc). Пустые инъекции также вычитают (=двойное сопоставление). Для расчета Kd и кинетических параметров используют модель Ленгмюра 1:1. Все конторстела FAP-CD40 способны связывать одновременно или независимо с двум антигенами.
Пример 10. Функциональные свойства молекул, связывающих FAP-нацеленные анти-CD40 человека
10.1 Активация В-клеток человека с помощью молекул, связывающих FAP-нацеленные анти-CD40 человека, с использованием в качестве источника антигена гранул Dynabeads® с FAP-покрытием
В-клетки выделяют из лейкоцитарных пленок, полученных от Цюрихский фонд донорства крови SRK (Stiftung Zurcher Blutspendedienst SRK). Чтобы выделить мононуклеарные клетки периферической крови (МКПК), 50 мл лейкоцитарных пленок разводят равным объемом ФСБ (фирма gibco, номер в каталоге 10010023). В полипропиленовые центрифужные пробирки объемом 50 мл (фирма ТРР, номер в каталоге 91050) вносят по 15 мл среды Lymphoprep™ (фирма STEMCELL Technologies, номер в каталоге 07851) и по 25 мл раствора для лейкоцитарных пленок аккуратно наслаивают поверх среды Lymphorep™. Пробирки центрифугируют в режиме 2000 об/мин в течение 24 мин при комнатной температуре с низким ускорением и без торможения. После этого МКПК собирают с области контакта, трижды промывают ФСБ, ресуспендируют в 10 мл ФСБ и клетки исследуют для определения их типа и количества с применением счетчика клеток Beckman Coulter cell counter Ac⋅T™ 5diff OV (фирма Beckman Coulter, номер в каталоге 6605580). Перед выделением В-клеток из МКПК CD14-положительную фракцию удаляют с помощью нанесения магнитной метки на CD14-положительные клетки с помощью CD14 микрогранул (фирма Miltenyi, номер в каталоге 130-050-201) с последующим выделением на сепараторе autoMACS® Pro Separator (фирма Miltenyi, номер в каталоге 130-092-545). CD14-отрицательную фракцию применяют для последующего выделения В-клеток с применением набора II для выделения клеток Miltenyi (номер в каталоге 130-091-151) и сепарированием в autoMACS® 1×105 В-клеток в 100 мкл среды R10, состоящей из среды RPMI 1640 (Roswell Park Memorial Institute) (фирма gibco, номер в каталоге 31870-025) с добавлением 10% (объем/объем) фетальной сыворотки теленка (ФСТ) (фирма life technologies, номер в каталоге 16140, партия 1797306А), 1% (объем/объем) пенициллина/стрептомицина (фирма gibco, номер в каталоге 15070-063), 1% (объем/объем) L-глутамина (фирма gibco, номер в каталоге 25030-024), 1% (объем/объем) пирувата натрия (фирма gibco, номер в каталоге 11360-039), 1% (объем/объем) минимальная питательная среда с аминокислотами, не являющимися незаменимыми (MEM non-essential amino acids, MEM; фирма gibco, номер в каталоге 11140-035) и 50 мкМ β-меркаптоэтанола (фирма gibco, номер в каталоге 31350-010) вносят в лунки 96-луночного плоскодонного планшета. Гранулы Dynabeads® со стрептовидином (фирма ThermoFisher Scientific, номер в каталоге 11205D) покрывают биотилинированным FAP человека (получают самостоятельно, связывающая способность 6,5×104 гранул: 0,01 мкг белка) по протоколу производителя и добавляют к В-клеткам при соотношении гранул к клеткам 2:1 в 50 мкл среды R10. В качестве контроля к В-клеткам добавляют гранулы без покрытия.
FAP-нацеленные анти-CD40 человека конторстела добавляют в 50 мкл среды R10 к В-клеткам в концентрациях, варьирующих от 6,7 до 0,003 нМ (серии 3-кратных разведений). В качестве FAP-независимого контроля применяют агонистическое анти-CD40 человека антитело SGN40 (IgG1, INN: дацетузумаб). Поскольку в литературе описано, что антитело SGN40 требует перекрестного сшивания Fc-рецептора для биологической активности (С. Law с соавт., Cancer Res, 65, 2005, 8331-8338), антитело инкубируют с поперечно сшивающим козьим анти-IgG Fey фрагмент-специфичным Р(ab’)2-фрагментом (фирма Jackson ImmunoResearch, номер в каталоге 109-006-008) в течение 30 мин перед добавлением к В-клеткам. Через 48 ч клетки переносят в 96-луночный круглодонный планшет, один раз промывают ФСБ и инкубируют с 50 мкл 3 мкг/мл Fc-рецептора, блокирующего контроль изотип IgG мыши, (фирма ThermoFisher Scientific, номер в каталоге 10400С) в ФСБ. Через 15 мин инкубирования при 4°С клетки промывают ФСБ и 50 мкл смеси флуоресцентно меченых антител в ФСБ добавляют к клеткам. Применяют следующие флуоресцентно меченые антитела: анти-CD80 человека BV605 (фирма BD Biosciences, клон L307.4, номер в каталоге 563315), анти-CD69 человека Alexa Fluor® 488 (фирма Biolegend, клон FN50, номер в каталоге 310916), анти-CDH человека PerCP-Су5.5 (фирма Biolegend, клон HCD14, номер в каталоге 325622), анти-CD3 человека PerCP-Су5.5 (фирма Biolegend, клон UCHT1, номер в каталоге 300430), анти-CD86 человека PE-CF594 (фирма BD Biosciences, клон FUN-1, номер в каталоге 562390), анти-HLA-DR BUV395 (фирма BD Biosciences, клон G46-6, номер в каталоге 564040) и анти-CD19 человека АРС-Н7 (фирма BD Biosciences, клон SJ25C1, номер в каталоге 560177). Чтобы отличить живые клетки от мертвых, прижизненный краситель Zombie Aqua™ (фирма Biolegend, номер в каталоге 423102) добавляют к смеси антител. Через 30 мин инкубирования при 4°С клетки дважды промывают ФСБ и ресуспендируют в 200 мкл ФСБ. Клетки анализируют в тот же день, используя 5-laser LSR-Fortessa (фирма BD Bioscience с программным обеспечением DIVA). Анализ данных проводят с использованием программного обеспечения Flow Jo версии 10 (FlowJo LLC). Живые (аква-негативные) клетки, негативные по CD14 и CD3 и позитивные по CD19, анализируют на экспрессию CD69, CD80, CD86 и HLA-DR.
В-клетки, проанализированные через 2 дня инкубации с агонистическими анти-CD40 конторстелами или перекрестно-связанным антителом SGN40, показывают увеличение экспрессии CD69, CD80, CD86 и HLA-DR для всех исследуемых конструкций (см. фиг. 24А-24З). Повышенная регуляция этих маркеров экспрессии зависит от FAP в случае различных FAP-нацеленных конторстел, и увеличение экспрессии, индуцированной этими FAP-зависимыми антителами, сопоставимо или немного ниже, чем увеличение, индуцированное перекрестно-связанным антителом SGN40.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ф.ХОФФМАНН-ЛЯ РОШ АГ
<120> БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА
<130> P34510-WO
<150> EP17199537.6
<151> 2017-11-01
<160> 184
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) CDR-H1
<400> 1
Ser Tyr Ala Met Ser
1 5
<210> 2
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) CDR-H2
<400> 2
Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 3
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) CDR-H3
<400> 3
Gly Trp Phe Gly Gly Phe Asn Tyr
1 5
<210> 4
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) CDR-L1
<400> 4
Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr Leu Ala
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) CDR-L2
<400> 5
Val Gly Ser Arg Arg Ala Thr
1 5
<210> 6
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) CDR-L3
<400> 6
Gln Gln Gly Ile Met Leu Pro Pro Thr
1 5
<210> 7
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) VH
<400> 7
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 8
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(4B9) VL
<400> 8
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 9
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) CDR-H1
<400> 9
Ser His Ala Met Ser
1 5
<210> 10
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) CDR-H2
<400> 10
Ala Ile Trp Ala Ser Gly Glu Gln Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 11
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) CDR-H3
<400> 11
Gly Trp Leu Gly Asn Phe Asp Tyr
1 5
<210> 12
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) CDR-L1
<400> 12
Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr Leu Ala
1 5 10
<210> 13
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) CDR-L2
<400> 13
Gly Ala Ser Thr Arg Ala Thr
1 5
<210> 14
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) CDR-L3
<400> 14
Gln Gln Gly Gln Val Ile Pro Pro Thr
1 5
<210> 15
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) VH
<400> 15
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Trp Ala Ser Gly Glu Gln Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Lys Gly Trp Leu Gly Asn Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 16
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> FAP(28H1) VL
<400> 16
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln Val Ile Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 17
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9,49B4,1G4, 20B7) CDR-H1
<400> 17
Ser Tyr Ala Ile Ser
1 5
<210> 18
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563, CLC-564, 17A9) CDR-H1
<400> 18
Ser Tyr Ala Met Ser
1 5
<210> 19
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9,49B4,1G4, 20B7) CDR-H2
<400> 19
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 20
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563, CLC-564, 17A9) CDR-H2
<400> 20
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 21
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9) CDR-H3
<400> 21
Glu Tyr Gly Trp Met Asp Tyr
1 5
<210> 22
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(49B4) CDR-H3
<400> 22
Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr
1 5
<210> 23
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(1G4) CDR-H3
<400> 23
Glu Tyr Gly Ser Met Asp Tyr
1 5
<210> 24
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(20B7) CDR-H3
<400> 24
Val Asn Tyr Pro Tyr Ser Tyr Trp Gly Asp Phe Asp Tyr
1 5 10
<210> 25
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563) CDR-H3
<400> 25
Asp Val Gly Ala Phe Asp Tyr
1 5
<210> 26
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-564) CDR-H3
<400> 26
Asp Val Gly Pro Phe Asp Tyr
1 5
<210> 27
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(17A9)-CDR-H3
<400> 27
Val Phe Tyr Arg Gly Gly Val Ser Met Asp Tyr
1 5 10
<210> 28
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9,49B4,1G4, 20B7) CDR-L1
<400> 28
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 29
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563, CLC564) CDR-L1
<400> 29
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 30
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(17A9) CDR-L1
<400> 30
Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala Ser
1 5 10
<210> 31
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9,49B4,1G4, 20B7) CDR-L2
<400> 31
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 32
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563, CLC564) CDR-L2
<400> 32
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 33
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(17A9) CDR-L2
<400> 33
Gly Lys Asn Asn Arg Pro Ser
1 5
<210> 34
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9) CDR-L3
<400> 34
Gln Gln Tyr Leu Thr Tyr Ser Arg Phe Thr
1 5 10
<210> 35
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(49B4) CDR-L3
<400> 35
Gln Gln Tyr Ser Ser Gln Pro Tyr Thr
1 5
<210> 36
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(1G4) CDR-L3
<400> 36
Gln Gln Tyr Ile Ser Tyr Ser Met Leu Thr
1 5 10
<210> 37
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(20B7) CDR-L3
<400> 37
Gln Gln Tyr Gln Ala Phe Ser Leu Thr
1 5
<210> 38
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563, CLC-164) CDR-L3
<400> 38
Gln Gln Tyr Gly Ser Ser Pro Leu Thr
1 5
<210> 39
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(17A9) CDR-L3
<400> 39
Asn Ser Arg Val Met Pro His Asn Arg Val
1 5 10
<210> 40
<211> 116
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9) VH
<400> 40
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Gly Trp Met Asp Tyr Trp Gly Gln Gly Thr Thr Val
100 105 110
Thr Val Ser Ser
115
<210> 41
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(8H9) VL
<400> 41
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Thr Tyr Ser Arg
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 42
<211> 118
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(49B4) VH
<400> 42
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 43
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(49B4) VL
<400> 43
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 44
<211> 116
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(1G4) VH
<400> 44
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Gly Ser Met Asp Tyr Trp Gly Gln Gly Thr Thr Val
100 105 110
Thr Val Ser Ser
115
<210> 45
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(1G4) VL
<400> 45
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ile Ser Tyr Ser Met
85 90 95
Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 46
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(20B7) VH
<400> 46
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Asn Tyr Pro Tyr Ser Tyr Trp Gly Asp Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 47
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(20B7) VL
<400> 47
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gln Ala Phe Ser Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 48
<211> 116
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563) VH
<400> 48
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Asp Val Gly Ala Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 49
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-563) VL
<400> 49
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 50
<211> 116
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-564) VH
<400> 50
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Asp Val Gly Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 51
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(CLC-564) VL
<400> 51
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 52
<211> 120
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(17A9) VH
<400> 52
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Phe Tyr Arg Gly Gly Val Ser Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 53
<211> 106
<212> PRT
<213> Artificial sequence
<220>
<223> OX40(17A9) VL
<400> 53
Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Val Arg Ile Thr Cys Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Asn Ser Arg Val Met Pro His Asn Arg
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val
100 105
<210> 54
<211> 916
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) CD134-0093
<400> 54
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val
690 695 700
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
705 710 715 720
Phe Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly
725 730 735
Leu Glu Trp Val Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr
740 745 750
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
755 760 765
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
770 775 780
Val Tyr Tyr Cys Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly
785 790 795 800
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser
805 810 815
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
820 825 830
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
835 840 845
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
850 855 860
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
865 870 875 880
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
885 890 895
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
900 905 910
Arg Gly Glu Cys
915
<210> 55
<211> 905
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) CD134-0093
<400> 55
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser
690 695 700
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser
705 710 715 720
Val Thr Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala
725 730 735
Pro Arg Leu Leu Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro
740 745 750
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
755 760 765
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly
770 775 780
Ile Met Leu Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
785 790 795 800
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
805 810 815
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
820 825 830
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
835 840 845
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
850 855 860
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
865 870 875 880
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
885 890 895
Asp Lys Lys Val Glu Pro Lys Ser Cys
900 905
<210> 56
<211> 910
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) CD134-0094
<400> 56
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His
225 230 235 240
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly
450 455 460
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
465 470 475 480
Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
485 490 495
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu
515 520 525
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe
530 535 540
Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys
565 570 575
Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
580 585 590
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
595 600 605
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
610 615 620
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
625 630 635 640
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
645 650 655
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
660 665 670
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
675 680 685
Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly
690 695 700
Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
705 710 715 720
Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro
725 730 735
Gly Gln Ala Pro Arg Leu Leu Ile Asn Val Gly Ser Arg Arg Ala Thr
740 745 750
Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
755 760 765
Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys
770 775 780
Gln Gln Gly Ile Met Leu Pro Pro Thr Phe Gly Gln Gly Thr Lys Val
785 790 795 800
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
805 810 815
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
820 825 830
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
835 840 845
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
850 855 860
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
865 870 875 880
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
885 890 895
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
900 905 910
<210> 57
<211> 915
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) CD134-0094
<400> 57
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His
225 230 235 240
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro
355 360 365
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly
450 455 460
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
465 470 475 480
Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
485 490 495
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu
515 520 525
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe
530 535 540
Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys
565 570 575
Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
580 585 590
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
595 600 605
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
610 615 620
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
625 630 635 640
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
645 650 655
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
660 665 670
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
675 680 685
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly
690 695 700
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
705 710 715 720
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro
725 730 735
Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ile Gly Ser Gly Ala Ser
740 745 750
Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
755 760 765
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
770 775 780
Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly Trp Phe Gly Gly Phe Asn
785 790 795 800
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
805 810 815
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
820 825 830
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
835 840 845
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
850 855 860
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
865 870 875 880
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
885 890 895
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
900 905 910
Lys Ser Cys
915
<210> 58
<211> 924
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE0085
<400> 58
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Trp Ala Ser Gly Glu Gln Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe
755 760 765
Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn
770 775 780
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly Trp
785 790 795 800
Leu Gly Asn Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
805 810 815
Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
820 825 830
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
835 840 845
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
850 855 860
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
865 870 875 880
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
885 890 895
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
900 905 910
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920
<210> 59
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE0085
<400> 59
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Ile Gly
740 745 750
Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln Val Ile Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 60
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE0086
<400> 60
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 61
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE0086
<400> 61
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val
740 745 750
Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 62
<211> 919
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE0087
<400> 62
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His
225 230 235 240
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly
450 455 460
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
465 470 475 480
Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
485 490 495
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu
515 520 525
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe
530 535 540
Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys
565 570 575
Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
580 585 590
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
595 600 605
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
610 615 620
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
625 630 635 640
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
645 650 655
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
660 665 670
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
675 680 685
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser
690 695 700
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
705 710 715 720
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser
725 730 735
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
740 745 750
Ile Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
755 760 765
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
770 775 780
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln Val Ile Pro
785 790 795 800
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
805 810 815
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
820 825 830
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
835 840 845
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
850 855 860
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
865 870 875 880
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
885 890 895
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
900 905 910
Ser Phe Asn Arg Gly Glu Cys
915
<210> 63
<211> 923
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE0087
<400> 63
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His
225 230 235 240
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro
355 360 365
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly
450 455 460
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
465 470 475 480
Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
485 490 495
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu
515 520 525
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe
530 535 540
Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys
565 570 575
Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
580 585 590
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
595 600 605
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
610 615 620
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
625 630 635 640
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
645 650 655
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
660 665 670
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
675 680 685
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser
690 695 700
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
705 710 715 720
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
725 730 735
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
740 745 750
Ser Ala Ile Trp Ala Ser Gly Glu Gln Tyr Tyr Ala Asp Ser Val Lys
755 760 765
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
770 775 780
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
785 790 795 800
Lys Gly Trp Leu Gly Asn Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
805 810 815
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
820 825 830
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
835 840 845
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
850 855 860
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
865 870 875 880
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
885 890 895
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
900 905 910
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
915 920
<210> 64
<211> 919
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE0839
<400> 64
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His
225 230 235 240
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly
450 455 460
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
465 470 475 480
Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
485 490 495
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu
515 520 525
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe
530 535 540
Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys
565 570 575
Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
580 585 590
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
595 600 605
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
610 615 620
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
625 630 635 640
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
645 650 655
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
660 665 670
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
675 680 685
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser
690 695 700
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
705 710 715 720
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
725 730 735
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
740 745 750
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
755 760 765
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
770 775 780
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
785 790 795 800
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
805 810 815
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
820 825 830
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
835 840 845
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
850 855 860
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
865 870 875 880
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
885 890 895
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
900 905 910
Ser Phe Asn Arg Gly Glu Cys
915
<210> 65
<211> 924
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE0839
<400> 65
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
130 135 140
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
145 150 155 160
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
165 170 175
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
180 185 190
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
195 200 205
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
210 215 220
Cys Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His
225 230 235 240
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro
355 360 365
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly
450 455 460
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
465 470 475 480
Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
485 490 495
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys
500 505 510
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu
515 520 525
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe
530 535 540
Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr
545 550 555 560
Cys Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys
565 570 575
Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
580 585 590
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
595 600 605
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
610 615 620
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
625 630 635 640
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
645 650 655
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
660 665 670
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
675 680 685
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser
690 695 700
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
705 710 715 720
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
725 730 735
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
740 745 750
Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val
755 760 765
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
770 775 780
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
785 790 795 800
Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu
805 810 815
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
820 825 830
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
835 840 845
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
850 855 860
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
865 870 875 880
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
885 890 895
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
900 905 910
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
915 920
<210> 66
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE0821
<400> 66
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
450 455 460
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys
465 470 475 480
Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser
485 490 495
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile
500 505 510
Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg
515 520 525
Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu
530 535 540
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu
545 550 555 560
Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr
565 570 575
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
580 585 590
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
595 600 605
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
610 615 620
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
625 630 635 640
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
645 650 655
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
660 665 670
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 67
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE0821
<400> 67
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
450 455 460
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys
465 470 475 480
Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser
485 490 495
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile
500 505 510
Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg
515 520 525
Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu
530 535 540
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu
545 550 555 560
Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr
565 570 575
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
580 585 590
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
595 600 605
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
610 615 620
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
625 630 635 640
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
645 650 655
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
660 665 670
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val
740 745 750
Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 68
<211> 583
<212> PRT
<213> Artificial Sequence
<220>
<223> (49B4) VHCH1 Fc knob VH (28H1) (heavy chain 1)
<400> 68
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
465 470 475 480
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
485 490 495
Ser Ser His Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
500 505 510
Glu Trp Val Ser Ala Ile Trp Ala Ser Gly Glu Gln Tyr Tyr Ala Asp
515 520 525
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
530 535 540
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
545 550 555 560
Tyr Cys Ala Lys Gly Trp Leu Gly Asn Phe Asp Tyr Trp Gly Gln Gly
565 570 575
Thr Leu Val Thr Val Ser Ser
580
<210> 69
<211> 575
<212> PRT
<213> Artificial Sequence
<220>
<223> (49B4) VHCH1 Fc hole VL (28H1) (heavy chain 2)
<400> 69
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
355 360 365
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu
465 470 475 480
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val
485 490 495
Ser Arg Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
500 505 510
Arg Leu Leu Ile Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
530 535 540
Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln
545 550 555 560
Val Ile Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
565 570 575
<210> 70
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> (49B4) VLCL-light chain
<400> 70
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 71
<211> 584
<212> PRT
<213> Artificial Sequence
<220>
<223> (49B4) VHCH1 Fc knob VH (4B9) (heavy chain 1)
<400> 71
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
465 470 475 480
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
485 490 495
Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
500 505 510
Glu Trp Val Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala
515 520 525
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
530 535 540
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
545 550 555 560
Tyr Tyr Cys Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln
565 570 575
Gly Thr Leu Val Thr Val Ser Ser
580
<210> 72
<211> 575
<212> PRT
<213> Artificial Sequence
<220>
<223> (49B4) VHCH1 Fc hole VL (4B9) (heavy chain 2)
<400> 72
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
355 360 365
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu
465 470 475 480
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val
485 490 495
Thr Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
500 505 510
Arg Leu Leu Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
530 535 540
Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile
545 550 555 560
Met Leu Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
565 570 575
<210> 73
<211> 582
<212> PRT
<213> Artificial Sequence
<220>
<223> (49B4) VHCH1 Fc knob VH (DP47) (heavy chain 1)
<400> 73
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
465 470 475 480
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
485 490 495
Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
500 505 510
Glu Trp Val Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala
515 520 525
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
530 535 540
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
545 550 555 560
Tyr Tyr Cys Ala Lys Gly Ser Gly Phe Asp Tyr Trp Gly Gln Gly Thr
565 570 575
Leu Val Thr Val Ser Ser
580
<210> 74
<211> 575
<212> PRT
<213> Artificial Sequence
<220>
<223> (49B4) VHCH1 Fc hole VL (DP47) (heavy chain 2)
<400> 74
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
355 360 365
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
435 440 445
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu
465 470 475 480
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val
485 490 495
Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
500 505 510
Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
530 535 540
Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly
545 550 555 560
Ser Ser Pro Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
565 570 575
<210> 75
<211> 816
<212> PRT
<213> Artificial Sequence
<220>
<223> HC 1(49B4) VHCH1_VHCH1 Fc knob VH (4B9)
<400> 75
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly
225 230 235 240
Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ala
245 250 255
Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser Trp Val Arg Gln Ala
260 265 270
Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile Ile Pro Ile Phe Gly
275 280 285
Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala
290 295 300
Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser
305 310 315 320
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Tyr Tyr Arg Gly Pro
325 330 335
Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser
340 345 350
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
355 360 365
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
370 375 380
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
385 390 395 400
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
405 410 415
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
420 425 430
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
435 440 445
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
450 455 460
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
465 470 475 480
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
485 490 495
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
500 505 510
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
515 520 525
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
530 535 540
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
545 550 555 560
Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
565 570 575
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr
580 585 590
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
595 600 605
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
610 615 620
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
625 630 635 640
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
645 650 655
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
660 665 670
Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly
675 680 685
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Leu
690 695 700
Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
705 710 715 720
Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser Trp Val
725 730 735
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ile Gly
740 745 750
Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
755 760 765
Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
770 775 780
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly Trp Phe
785 790 795 800
Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
805 810 815
<210> 76
<211> 807
<212> PRT
<213> Artificial Sequence
<220>
<223> HC 2 (49B4) VHCH1_VHCH1 Fc hole VL (4B9)
<400> 76
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly
225 230 235 240
Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ala
245 250 255
Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser Trp Val Arg Gln Ala
260 265 270
Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile Ile Pro Ile Phe Gly
275 280 285
Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala
290 295 300
Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser
305 310 315 320
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Tyr Tyr Arg Gly Pro
325 330 335
Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser
340 345 350
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
355 360 365
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
370 375 380
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
385 390 395 400
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
405 410 415
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
420 425 430
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
435 440 445
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
450 455 460
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
465 470 475 480
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
485 490 495
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
500 505 510
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
515 520 525
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
530 535 540
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
545 550 555 560
Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
565 570 575
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
580 585 590
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
595 600 605
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
610 615 620
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
625 630 635 640
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
645 650 655
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
660 665 670
Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly
675 680 685
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr
690 695 700
Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu
705 710 715 720
Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala Trp Tyr
725 730 735
Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val Gly Ser
740 745 750
Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly
755 760 765
Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala
770 775 780
Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe Gly Gln
785 790 795 800
Gly Thr Lys Val Glu Ile Lys
805
<210> 77
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker G4S
<400> 77
Gly Gly Gly Gly Ser
1 5
<210> 78
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker (G4S)2
<400> 78
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 79
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker (SG4)2
<400> 79
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10
<210> 80
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker particular
<400> 80
Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly
1 5 10 15
Ser
<210> 81
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker (G4S)3
<400> 81
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 82
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker G4(SG4)2
<400> 82
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10
<210> 83
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker (G4S)4
<400> 83
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 84
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GGGGSGGGGSGGGSGGGGS
<400> 84
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly
1 5 10 15
Gly Gly Ser
<210> 85
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GSPGSSSSGS
<400> 85
Gly Ser Pro Gly Ser Ser Ser Ser Gly Ser
1 5 10
<210> 86
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GSGSGSGS
<400> 86
Gly Ser Gly Ser Gly Ser Gly Ser
1 5
<210> 87
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GSGSGNGS
<400> 87
Gly Ser Gly Ser Gly Asn Gly Ser
1 5
<210> 88
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GGSGSGSG
<400> 88
Gly Gly Ser Gly Ser Gly Ser Gly
1 5
<210> 89
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GGSGSG
<400> 89
Gly Gly Ser Gly Ser Gly
1 5
<210> 90
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GGSG
<400> 90
Gly Gly Ser Gly
1
<210> 91
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> GGSGNGSG
<400> 91
Gly Gly Ser Gly Asn Gly Ser Gly
1 5
<210> 92
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide inker GGNGSGSG
<400> 92
Gly Gly Asn Gly Ser Gly Ser Gly
1 5
<210> 93
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide linker GGNGSG
<400> 93
Gly Gly Asn Gly Ser Gly
1 5
<210> 94
<211> 98
<212> PRT
<213> Homo sapiens
<400> 94
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 95
<211> 107
<212> PRT
<213> Homo sapiens
<400> 95
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Trp Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
35 40 45
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
50 55 60
Glu Ser Thr Tyr Arg Trp Ser Val Leu Thr Val Leu His Gln Asp Trp
65 70 75 80
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
85 90 95
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105
<210> 96
<211> 106
<212> PRT
<213> Homo sapiens
<400> 96
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 97
<211> 760
<212> PRT
<213> Homo sapiens
<400> 97
Met Lys Thr Trp Val Lys Ile Val Phe Gly Val Ala Thr Ser Ala Val
1 5 10 15
Leu Ala Leu Leu Val Met Cys Ile Val Leu Arg Pro Ser Arg Val His
20 25 30
Asn Ser Glu Glu Asn Thr Met Arg Ala Leu Thr Leu Lys Asp Ile Leu
35 40 45
Asn Gly Thr Phe Ser Tyr Lys Thr Phe Phe Pro Asn Trp Ile Ser Gly
50 55 60
Gln Glu Tyr Leu His Gln Ser Ala Asp Asn Asn Ile Val Leu Tyr Asn
65 70 75 80
Ile Glu Thr Gly Gln Ser Tyr Thr Ile Leu Ser Asn Arg Thr Met Lys
85 90 95
Ser Val Asn Ala Ser Asn Tyr Gly Leu Ser Pro Asp Arg Gln Phe Val
100 105 110
Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp Arg Tyr Ser Tyr Thr Ala
115 120 125
Thr Tyr Tyr Ile Tyr Asp Leu Ser Asn Gly Glu Phe Val Arg Gly Asn
130 135 140
Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys Trp Ser Pro Val Gly Ser
145 150 155 160
Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile Tyr Leu Lys Gln Arg Pro
165 170 175
Gly Asp Pro Pro Phe Gln Ile Thr Phe Asn Gly Arg Glu Asn Lys Ile
180 185 190
Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu Glu Glu Met Leu Ala Thr
195 200 205
Lys Tyr Ala Leu Trp Trp Ser Pro Asn Gly Lys Phe Leu Ala Tyr Ala
210 215 220
Glu Phe Asn Asp Thr Asp Ile Pro Val Ile Ala Tyr Ser Tyr Tyr Gly
225 230 235 240
Asp Glu Gln Tyr Pro Arg Thr Ile Asn Ile Pro Tyr Pro Lys Ala Gly
245 250 255
Ala Lys Asn Pro Val Val Arg Ile Phe Ile Ile Asp Thr Thr Tyr Pro
260 265 270
Ala Tyr Val Gly Pro Gln Glu Val Pro Val Pro Ala Met Ile Ala Ser
275 280 285
Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp Val Thr Asp Glu Arg Val
290 295 300
Cys Leu Gln Trp Leu Lys Arg Val Gln Asn Val Ser Val Leu Ser Ile
305 310 315 320
Cys Asp Phe Arg Glu Asp Trp Gln Thr Trp Asp Cys Pro Lys Thr Gln
325 330 335
Glu His Ile Glu Glu Ser Arg Thr Gly Trp Ala Gly Gly Phe Phe Val
340 345 350
Ser Thr Pro Val Phe Ser Tyr Asp Ala Ile Ser Tyr Tyr Lys Ile Phe
355 360 365
Ser Asp Lys Asp Gly Tyr Lys His Ile His Tyr Ile Lys Asp Thr Val
370 375 380
Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys Trp Glu Ala Ile Asn Ile
385 390 395 400
Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr Ser Ser Asn Glu Phe Glu
405 410 415
Glu Tyr Pro Gly Arg Arg Asn Ile Tyr Arg Ile Ser Ile Gly Ser Tyr
420 425 430
Pro Pro Ser Lys Lys Cys Val Thr Cys His Leu Arg Lys Glu Arg Cys
435 440 445
Gln Tyr Tyr Thr Ala Ser Phe Ser Asp Tyr Ala Lys Tyr Tyr Ala Leu
450 455 460
Val Cys Tyr Gly Pro Gly Ile Pro Ile Ser Thr Leu His Asp Gly Arg
465 470 475 480
Thr Asp Gln Glu Ile Lys Ile Leu Glu Glu Asn Lys Glu Leu Glu Asn
485 490 495
Ala Leu Lys Asn Ile Gln Leu Pro Lys Glu Glu Ile Lys Lys Leu Glu
500 505 510
Val Asp Glu Ile Thr Leu Trp Tyr Lys Met Ile Leu Pro Pro Gln Phe
515 520 525
Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile Gln Val Tyr Gly Gly Pro
530 535 540
Cys Ser Gln Ser Val Arg Ser Val Phe Ala Val Asn Trp Ile Ser Tyr
545 550 555 560
Leu Ala Ser Lys Glu Gly Met Val Ile Ala Leu Val Asp Gly Arg Gly
565 570 575
Thr Ala Phe Gln Gly Asp Lys Leu Leu Tyr Ala Val Tyr Arg Lys Leu
580 585 590
Gly Val Tyr Glu Val Glu Asp Gln Ile Thr Ala Val Arg Lys Phe Ile
595 600 605
Glu Met Gly Phe Ile Asp Glu Lys Arg Ile Ala Ile Trp Gly Trp Ser
610 615 620
Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu Ala Ser Gly Thr Gly Leu
625 630 635 640
Phe Lys Cys Gly Ile Ala Val Ala Pro Val Ser Ser Trp Glu Tyr Tyr
645 650 655
Ala Ser Val Tyr Thr Glu Arg Phe Met Gly Leu Pro Thr Lys Asp Asp
660 665 670
Asn Leu Glu His Tyr Lys Asn Ser Thr Val Met Ala Arg Ala Glu Tyr
675 680 685
Phe Arg Asn Val Asp Tyr Leu Leu Ile His Gly Thr Ala Asp Asp Asn
690 695 700
Val His Phe Gln Asn Ser Ala Gln Ile Ala Lys Ala Leu Val Asn Ala
705 710 715 720
Gln Val Asp Phe Gln Ala Met Trp Tyr Ser Asp Gln Asn His Gly Leu
725 730 735
Ser Gly Leu Ser Thr Asn His Leu Tyr Thr His Met Thr His Phe Leu
740 745 750
Lys Gln Cys Phe Ser Leu Ser Asp
755 760
<210> 98
<211> 761
<212> PRT
<213> Mus musculus
<400> 98
Met Lys Thr Trp Leu Lys Thr Val Phe Gly Val Thr Thr Leu Ala Ala
1 5 10 15
Leu Ala Leu Val Val Ile Cys Ile Val Leu Arg Pro Ser Arg Val Tyr
20 25 30
Lys Pro Glu Gly Asn Thr Lys Arg Ala Leu Thr Leu Lys Asp Ile Leu
35 40 45
Asn Gly Thr Phe Ser Tyr Lys Thr Tyr Phe Pro Asn Trp Ile Ser Glu
50 55 60
Gln Glu Tyr Leu His Gln Ser Glu Asp Asp Asn Ile Val Phe Tyr Asn
65 70 75 80
Ile Glu Thr Arg Glu Ser Tyr Ile Ile Leu Ser Asn Ser Thr Met Lys
85 90 95
Ser Val Asn Ala Thr Asp Tyr Gly Leu Ser Pro Asp Arg Gln Phe Val
100 105 110
Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp Arg Tyr Ser Tyr Thr Ala
115 120 125
Thr Tyr Tyr Ile Tyr Asp Leu Gln Asn Gly Glu Phe Val Arg Gly Tyr
130 135 140
Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys Trp Ser Pro Val Gly Ser
145 150 155 160
Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile Tyr Leu Lys Gln Arg Pro
165 170 175
Gly Asp Pro Pro Phe Gln Ile Thr Tyr Thr Gly Arg Glu Asn Arg Ile
180 185 190
Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu Glu Glu Met Leu Ala Thr
195 200 205
Lys Tyr Ala Leu Trp Trp Ser Pro Asp Gly Lys Phe Leu Ala Tyr Val
210 215 220
Glu Phe Asn Asp Ser Asp Ile Pro Ile Ile Ala Tyr Ser Tyr Tyr Gly
225 230 235 240
Asp Gly Gln Tyr Pro Arg Thr Ile Asn Ile Pro Tyr Pro Lys Ala Gly
245 250 255
Ala Lys Asn Pro Val Val Arg Val Phe Ile Val Asp Thr Thr Tyr Pro
260 265 270
His His Val Gly Pro Met Glu Val Pro Val Pro Glu Met Ile Ala Ser
275 280 285
Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp Val Ser Ser Glu Arg Val
290 295 300
Cys Leu Gln Trp Leu Lys Arg Val Gln Asn Val Ser Val Leu Ser Ile
305 310 315 320
Cys Asp Phe Arg Glu Asp Trp His Ala Trp Glu Cys Pro Lys Asn Gln
325 330 335
Glu His Val Glu Glu Ser Arg Thr Gly Trp Ala Gly Gly Phe Phe Val
340 345 350
Ser Thr Pro Ala Phe Ser Gln Asp Ala Thr Ser Tyr Tyr Lys Ile Phe
355 360 365
Ser Asp Lys Asp Gly Tyr Lys His Ile His Tyr Ile Lys Asp Thr Val
370 375 380
Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys Trp Glu Ala Ile Tyr Ile
385 390 395 400
Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr Ser Ser Asn Glu Phe Glu
405 410 415
Gly Tyr Pro Gly Arg Arg Asn Ile Tyr Arg Ile Ser Ile Gly Asn Ser
420 425 430
Pro Pro Ser Lys Lys Cys Val Thr Cys His Leu Arg Lys Glu Arg Cys
435 440 445
Gln Tyr Tyr Thr Ala Ser Phe Ser Tyr Lys Ala Lys Tyr Tyr Ala Leu
450 455 460
Val Cys Tyr Gly Pro Gly Leu Pro Ile Ser Thr Leu His Asp Gly Arg
465 470 475 480
Thr Asp Gln Glu Ile Gln Val Leu Glu Glu Asn Lys Glu Leu Glu Asn
485 490 495
Ser Leu Arg Asn Ile Gln Leu Pro Lys Val Glu Ile Lys Lys Leu Lys
500 505 510
Asp Gly Gly Leu Thr Phe Trp Tyr Lys Met Ile Leu Pro Pro Gln Phe
515 520 525
Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile Gln Val Tyr Gly Gly Pro
530 535 540
Cys Ser Gln Ser Val Lys Ser Val Phe Ala Val Asn Trp Ile Thr Tyr
545 550 555 560
Leu Ala Ser Lys Glu Gly Ile Val Ile Ala Leu Val Asp Gly Arg Gly
565 570 575
Thr Ala Phe Gln Gly Asp Lys Phe Leu His Ala Val Tyr Arg Lys Leu
580 585 590
Gly Val Tyr Glu Val Glu Asp Gln Leu Thr Ala Val Arg Lys Phe Ile
595 600 605
Glu Met Gly Phe Ile Asp Glu Glu Arg Ile Ala Ile Trp Gly Trp Ser
610 615 620
Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu Ala Ser Gly Thr Gly Leu
625 630 635 640
Phe Lys Cys Gly Ile Ala Val Ala Pro Val Ser Ser Trp Glu Tyr Tyr
645 650 655
Ala Ser Ile Tyr Ser Glu Arg Phe Met Gly Leu Pro Thr Lys Asp Asp
660 665 670
Asn Leu Glu His Tyr Lys Asn Ser Thr Val Met Ala Arg Ala Glu Tyr
675 680 685
Phe Arg Asn Val Asp Tyr Leu Leu Ile His Gly Thr Ala Asp Asp Asn
690 695 700
Val His Phe Gln Asn Ser Ala Gln Ile Ala Lys Ala Leu Val Asn Ala
705 710 715 720
Gln Val Asp Phe Gln Ala Met Trp Tyr Ser Asp Gln Asn His Gly Ile
725 730 735
Ser Ser Gly Arg Ser Gln Asn His Leu Tyr Thr His Met Thr His Phe
740 745 750
Leu Lys Gln Cys Phe Ser Leu Ser Asp
755 760
<210> 99
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> CH1 connector
<400> 99
Glu Pro Lys Ser Cys
1 5
<210> 100
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> hinge with X being S or P
<220>
<221> X
<222> (8)..(8)
<223> X is S or P
<400> 100
Asp Lys Thr His Thr Cys Pro Xaa Cys Pro
1 5 10
<210> 101
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> hinge with X being S or P
<220>
<221> X
<222> (5)..(5)
<223> X is S or P
<400> 101
His Thr Cys Pro Xaa Cys Pro
1 5
<210> 102
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hinge with X being S or P
<220>
<221> X
<222> (3)..(3)
<223> X is S or P
<400> 102
Cys Pro Xaa Cys Pro
1 5
<210> 103
<211> 330
<212> PRT
<213> Homo sapiens
<400> 103
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 104
<211> 330
<212> PRT
<213> Homo sapiens
<400> 104
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 105
<211> 326
<212> PRT
<213> Homo sapiens
<400> 105
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 106
<211> 377
<212> PRT
<213> Homo sapiens
<400> 106
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 107
<211> 327
<212> PRT
<213> Homo sapiens
<400> 107
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 108
<211> 277
<212> PRT
<213> Homo sapiens
<400> 108
Met Cys Val Gly Ala Arg Arg Leu Gly Arg Gly Pro Cys Ala Ala Leu
1 5 10 15
Leu Leu Leu Gly Leu Gly Leu Ser Thr Val Thr Gly Leu His Cys Val
20 25 30
Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys His Glu Cys Arg Pro
35 40 45
Gly Asn Gly Met Val Ser Arg Cys Ser Arg Ser Gln Asn Thr Val Cys
50 55 60
Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val Val Ser Ser Lys Pro
65 70 75 80
Cys Lys Pro Cys Thr Trp Cys Asn Leu Arg Ser Gly Ser Glu Arg Lys
85 90 95
Gln Leu Cys Thr Ala Thr Gln Asp Thr Val Cys Arg Cys Arg Ala Gly
100 105 110
Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val Asp Cys Ala Pro Cys
115 120 125
Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln Ala Cys Lys Pro Trp
130 135 140
Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu Gln Pro Ala Ser Asn
145 150 155 160
Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro Pro Ala Thr Gln Pro
165 170 175
Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Ile Thr Val Gln Pro Thr
180 185 190
Glu Ala Trp Pro Arg Thr Ser Gln Gly Pro Ser Thr Arg Pro Val Glu
195 200 205
Val Pro Gly Gly Arg Ala Val Ala Ala Ile Leu Gly Leu Gly Leu Val
210 215 220
Leu Gly Leu Leu Gly Pro Leu Ala Ile Leu Leu Ala Leu Tyr Leu Leu
225 230 235 240
Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly Gly
245 250 255
Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His Ser
260 265 270
Thr Leu Ala Lys Ile
275
<210> 109
<211> 255
<212> PRT
<213> Homo sapiens
<400> 109
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His Cys Leu Gly
85 90 95
Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Ile Ile Ser Phe Phe Leu
180 185 190
Ala Leu Thr Ser Thr Ala Leu Leu Phe Leu Leu Phe Phe Leu Thr Leu
195 200 205
Arg Phe Ser Val Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
210 215 220
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
225 230 235 240
Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
245 250 255
<210> 110
<211> 260
<212> PRT
<213> Homo sapiens
<400> 110
Met Ala Arg Pro His Pro Trp Trp Leu Cys Val Leu Gly Thr Leu Val
1 5 10 15
Gly Leu Ser Ala Thr Pro Ala Pro Lys Ser Cys Pro Glu Arg His Tyr
20 25 30
Trp Ala Gln Gly Lys Leu Cys Cys Gln Met Cys Glu Pro Gly Thr Phe
35 40 45
Leu Val Lys Asp Cys Asp Gln His Arg Lys Ala Ala Gln Cys Asp Pro
50 55 60
Cys Ile Pro Gly Val Ser Phe Ser Pro Asp His His Thr Arg Pro His
65 70 75 80
Cys Glu Ser Cys Arg His Cys Asn Ser Gly Leu Leu Val Arg Asn Cys
85 90 95
Thr Ile Thr Ala Asn Ala Glu Cys Ala Cys Arg Asn Gly Trp Gln Cys
100 105 110
Arg Asp Lys Glu Cys Thr Glu Cys Asp Pro Leu Pro Asn Pro Ser Leu
115 120 125
Thr Ala Arg Ser Ser Gln Ala Leu Ser Pro His Pro Gln Pro Thr His
130 135 140
Leu Pro Tyr Val Ser Glu Met Leu Glu Ala Arg Thr Ala Gly His Met
145 150 155 160
Gln Thr Leu Ala Asp Phe Arg Gln Leu Pro Ala Arg Thr Leu Ser Thr
165 170 175
His Trp Pro Pro Gln Arg Ser Leu Cys Ser Ser Asp Phe Ile Arg Ile
180 185 190
Leu Val Ile Phe Ser Gly Met Phe Leu Val Phe Thr Leu Ala Gly Ala
195 200 205
Leu Phe Leu His Gln Arg Arg Lys Tyr Arg Ser Asn Lys Gly Glu Ser
210 215 220
Pro Val Glu Pro Ala Glu Pro Cys His Tyr Ser Cys Pro Arg Glu Glu
225 230 235 240
Glu Gly Ser Thr Ile Pro Ile Gln Glu Asp Tyr Arg Lys Pro Glu Pro
245 250 255
Ala Cys Ser Pro
260
<210> 111
<211> 283
<212> PRT
<213> Homo sapiens
<400> 111
Met Glu Pro Pro Gly Asp Trp Gly Pro Pro Pro Trp Arg Ser Thr Pro
1 5 10 15
Lys Thr Asp Val Leu Arg Leu Val Leu Tyr Leu Thr Phe Leu Gly Ala
20 25 30
Pro Cys Tyr Ala Pro Ala Leu Pro Ser Cys Lys Glu Asp Glu Tyr Pro
35 40 45
Val Gly Ser Glu Cys Cys Pro Lys Cys Ser Pro Gly Tyr Arg Val Lys
50 55 60
Glu Ala Cys Gly Glu Leu Thr Gly Thr Val Cys Glu Pro Cys Pro Pro
65 70 75 80
Gly Thr Tyr Ile Ala His Leu Asn Gly Leu Ser Lys Cys Leu Gln Cys
85 90 95
Gln Met Cys Asp Pro Ala Met Gly Leu Arg Ala Ser Arg Asn Cys Ser
100 105 110
Arg Thr Glu Asn Ala Val Cys Gly Cys Ser Pro Gly His Phe Cys Ile
115 120 125
Val Gln Asp Gly Asp His Cys Ala Ala Cys Arg Ala Tyr Ala Thr Ser
130 135 140
Ser Pro Gly Gln Arg Val Gln Lys Gly Gly Thr Glu Ser Gln Asp Thr
145 150 155 160
Leu Cys Gln Asn Cys Pro Pro Gly Thr Phe Ser Pro Asn Gly Thr Leu
165 170 175
Glu Glu Cys Gln His Gln Thr Lys Cys Ser Trp Leu Val Thr Lys Ala
180 185 190
Gly Ala Gly Thr Ser Ser Ser His Trp Val Trp Trp Phe Leu Ser Gly
195 200 205
Ser Leu Val Ile Val Ile Val Cys Ser Thr Val Gly Leu Ile Ile Cys
210 215 220
Val Lys Arg Arg Lys Pro Arg Gly Asp Val Val Lys Val Ile Val Ser
225 230 235 240
Val Gln Arg Lys Arg Gln Glu Ala Glu Gly Glu Ala Thr Val Ile Glu
245 250 255
Ala Leu Gln Ala Pro Pro Asp Val Thr Thr Val Ala Val Glu Glu Thr
260 265 270
Ile Pro Ser Phe Thr Gly Arg Ser Pro Asn His
275 280
<210> 112
<211> 595
<212> PRT
<213> Homo sapiens
<400> 112
Met Arg Val Leu Leu Ala Ala Leu Gly Leu Leu Phe Leu Gly Ala Leu
1 5 10 15
Arg Ala Phe Pro Gln Asp Arg Pro Phe Glu Asp Thr Cys His Gly Asn
20 25 30
Pro Ser His Tyr Tyr Asp Lys Ala Val Arg Arg Cys Cys Tyr Arg Cys
35 40 45
Pro Met Gly Leu Phe Pro Thr Gln Gln Cys Pro Gln Arg Pro Thr Asp
50 55 60
Cys Arg Lys Gln Cys Glu Pro Asp Tyr Tyr Leu Asp Glu Ala Asp Arg
65 70 75 80
Cys Thr Ala Cys Val Thr Cys Ser Arg Asp Asp Leu Val Glu Lys Thr
85 90 95
Pro Cys Ala Trp Asn Ser Ser Arg Val Cys Glu Cys Arg Pro Gly Met
100 105 110
Phe Cys Ser Thr Ser Ala Val Asn Ser Cys Ala Arg Cys Phe Phe His
115 120 125
Ser Val Cys Pro Ala Gly Met Ile Val Lys Phe Pro Gly Thr Ala Gln
130 135 140
Lys Asn Thr Val Cys Glu Pro Ala Ser Pro Gly Val Ser Pro Ala Cys
145 150 155 160
Ala Ser Pro Glu Asn Cys Lys Glu Pro Ser Ser Gly Thr Ile Pro Gln
165 170 175
Ala Lys Pro Thr Pro Val Ser Pro Ala Thr Ser Ser Ala Ser Thr Met
180 185 190
Pro Val Arg Gly Gly Thr Arg Leu Ala Gln Glu Ala Ala Ser Lys Leu
195 200 205
Thr Arg Ala Pro Asp Ser Pro Ser Ser Val Gly Arg Pro Ser Ser Asp
210 215 220
Pro Gly Leu Ser Pro Thr Gln Pro Cys Pro Glu Gly Ser Gly Asp Cys
225 230 235 240
Arg Lys Gln Cys Glu Pro Asp Tyr Tyr Leu Asp Glu Ala Gly Arg Cys
245 250 255
Thr Ala Cys Val Ser Cys Ser Arg Asp Asp Leu Val Glu Lys Thr Pro
260 265 270
Cys Ala Trp Asn Ser Ser Arg Thr Cys Glu Cys Arg Pro Gly Met Ile
275 280 285
Cys Ala Thr Ser Ala Thr Asn Ser Cys Ala Arg Cys Val Pro Tyr Pro
290 295 300
Ile Cys Ala Ala Glu Thr Val Thr Lys Pro Gln Asp Met Ala Glu Lys
305 310 315 320
Asp Thr Thr Phe Glu Ala Pro Pro Leu Gly Thr Gln Pro Asp Cys Asn
325 330 335
Pro Thr Pro Glu Asn Gly Glu Ala Pro Ala Ser Thr Ser Pro Thr Gln
340 345 350
Ser Leu Leu Val Asp Ser Gln Ala Ser Lys Thr Leu Pro Ile Pro Thr
355 360 365
Ser Ala Pro Val Ala Leu Ser Ser Thr Gly Lys Pro Val Leu Asp Ala
370 375 380
Gly Pro Val Leu Phe Trp Val Ile Leu Val Leu Val Val Val Val Gly
385 390 395 400
Ser Ser Ala Phe Leu Leu Cys His Arg Arg Ala Cys Arg Lys Arg Ile
405 410 415
Arg Gln Lys Leu His Leu Cys Tyr Pro Val Gln Thr Ser Gln Pro Lys
420 425 430
Leu Glu Leu Val Asp Ser Arg Pro Arg Arg Ser Ser Thr Gln Leu Arg
435 440 445
Ser Gly Ala Ser Val Thr Glu Pro Val Ala Glu Glu Arg Gly Leu Met
450 455 460
Ser Gln Pro Leu Met Glu Thr Cys His Ser Val Gly Ala Ala Tyr Leu
465 470 475 480
Glu Ser Leu Pro Leu Gln Asp Ala Ser Pro Ala Gly Gly Pro Ser Ser
485 490 495
Pro Arg Asp Leu Pro Glu Pro Arg Val Ser Thr Glu His Thr Asn Asn
500 505 510
Lys Ile Glu Lys Ile Tyr Ile Met Lys Ala Asp Thr Val Ile Val Gly
515 520 525
Thr Val Lys Ala Glu Leu Pro Glu Gly Arg Gly Leu Ala Gly Pro Ala
530 535 540
Glu Pro Glu Leu Glu Glu Glu Leu Glu Ala Asp His Thr Pro His Tyr
545 550 555 560
Pro Glu Gln Glu Thr Glu Pro Pro Leu Gly Ser Cys Ser Asp Val Met
565 570 575
Leu Ser Val Glu Glu Glu Gly Lys Glu Asp Pro Leu Pro Thr Ala Ala
580 585 590
Ser Gly Lys
595
<210> 113
<211> 241
<212> PRT
<213> Homo sapiens
<400> 113
Met Ala Gln His Gly Ala Met Gly Ala Phe Arg Ala Leu Cys Gly Leu
1 5 10 15
Ala Leu Leu Cys Ala Leu Ser Leu Gly Gln Arg Pro Thr Gly Gly Pro
20 25 30
Gly Cys Gly Pro Gly Arg Leu Leu Leu Gly Thr Gly Thr Asp Ala Arg
35 40 45
Cys Cys Arg Val His Thr Thr Arg Cys Cys Arg Asp Tyr Pro Gly Glu
50 55 60
Glu Cys Cys Ser Glu Trp Asp Cys Met Cys Val Gln Pro Glu Phe His
65 70 75 80
Cys Gly Asp Pro Cys Cys Thr Thr Cys Arg His His Pro Cys Pro Pro
85 90 95
Gly Gln Gly Val Gln Ser Gln Gly Lys Phe Ser Phe Gly Phe Gln Cys
100 105 110
Ile Asp Cys Ala Ser Gly Thr Phe Ser Gly Gly His Glu Gly His Cys
115 120 125
Lys Pro Trp Thr Asp Cys Thr Gln Phe Gly Phe Leu Thr Val Phe Pro
130 135 140
Gly Asn Lys Thr His Asn Ala Val Cys Val Pro Gly Ser Pro Pro Ala
145 150 155 160
Glu Pro Leu Gly Trp Leu Thr Val Val Leu Leu Ala Val Ala Ala Cys
165 170 175
Val Leu Leu Leu Thr Ser Ala Gln Leu Gly Leu His Ile Trp Gln Leu
180 185 190
Arg Ser Gln Cys Met Trp Pro Arg Glu Thr Gln Leu Leu Leu Glu Val
195 200 205
Pro Pro Ser Thr Glu Asp Ala Arg Ser Cys Gln Phe Pro Glu Glu Glu
210 215 220
Arg Gly Glu Arg Ser Ala Glu Glu Lys Gly Arg Leu Gly Asp Leu Trp
225 230 235 240
Val
<210> 114
<211> 272
<212> PRT
<213> Mus musculus
<400> 114
Met Tyr Val Trp Val Gln Gln Pro Thr Ala Leu Leu Leu Leu Ala Leu
1 5 10 15
Thr Leu Gly Val Thr Ala Arg Arg Leu Asn Cys Val Lys His Thr Tyr
20 25 30
Pro Ser Gly His Lys Cys Cys Arg Glu Cys Gln Pro Gly His Gly Met
35 40 45
Val Ser Arg Cys Asp His Thr Arg Asp Thr Leu Cys His Pro Cys Glu
50 55 60
Thr Gly Phe Tyr Asn Glu Ala Val Asn Tyr Asp Thr Cys Lys Gln Cys
65 70 75 80
Thr Gln Cys Asn His Arg Ser Gly Ser Glu Leu Lys Gln Asn Cys Thr
85 90 95
Pro Thr Gln Asp Thr Val Cys Arg Cys Arg Pro Gly Thr Gln Pro Arg
100 105 110
Gln Asp Ser Gly Tyr Lys Leu Gly Val Asp Cys Val Pro Cys Pro Pro
115 120 125
Gly His Phe Ser Pro Gly Asn Asn Gln Ala Cys Lys Pro Trp Thr Asn
130 135 140
Cys Thr Leu Ser Gly Lys Gln Thr Arg His Pro Ala Ser Asp Ser Leu
145 150 155 160
Asp Ala Val Cys Glu Asp Arg Ser Leu Leu Ala Thr Leu Leu Trp Glu
165 170 175
Thr Gln Arg Pro Thr Phe Arg Pro Thr Thr Val Gln Ser Thr Thr Val
180 185 190
Trp Pro Arg Thr Ser Glu Leu Pro Ser Pro Pro Thr Leu Val Thr Pro
195 200 205
Glu Gly Pro Ala Phe Ala Val Leu Leu Gly Leu Gly Leu Gly Leu Leu
210 215 220
Ala Pro Leu Thr Val Leu Leu Ala Leu Tyr Leu Leu Arg Lys Ala Trp
225 230 235 240
Arg Leu Pro Asn Thr Pro Lys Pro Cys Trp Gly Asn Ser Phe Arg Thr
245 250 255
Pro Ile Gln Glu Glu His Thr Asp Ala His Phe Thr Leu Ala Lys Ile
260 265 270
<210> 115
<211> 277
<212> PRT
<213> Homo sapiens
<400> 115
Met Val Arg Leu Pro Leu Gln Cys Val Leu Trp Gly Cys Leu Leu Thr
1 5 10 15
Ala Val His Pro Glu Pro Pro Thr Ala Cys Arg Glu Lys Gln Tyr Leu
20 25 30
Ile Asn Ser Gln Cys Cys Ser Leu Cys Gln Pro Gly Gln Lys Leu Val
35 40 45
Ser Asp Cys Thr Glu Phe Thr Glu Thr Glu Cys Leu Pro Cys Gly Glu
50 55 60
Ser Glu Phe Leu Asp Thr Trp Asn Arg Glu Thr His Cys His Gln His
65 70 75 80
Lys Tyr Cys Asp Pro Asn Leu Gly Leu Arg Val Gln Gln Lys Gly Thr
85 90 95
Ser Glu Thr Asp Thr Ile Cys Thr Cys Glu Glu Gly Trp His Cys Thr
100 105 110
Ser Glu Ala Cys Glu Ser Cys Val Leu His Arg Ser Cys Ser Pro Gly
115 120 125
Phe Gly Val Lys Gln Ile Ala Thr Gly Val Ser Asp Thr Ile Cys Glu
130 135 140
Pro Cys Pro Val Gly Phe Phe Ser Asn Val Ser Ser Ala Phe Glu Lys
145 150 155 160
Cys His Pro Trp Thr Ser Cys Glu Thr Lys Asp Leu Val Val Gln Gln
165 170 175
Ala Gly Thr Asn Lys Thr Asp Val Val Cys Gly Pro Gln Asp Arg Leu
180 185 190
Arg Ala Leu Val Val Ile Pro Ile Ile Phe Gly Ile Leu Phe Ala Ile
195 200 205
Leu Leu Val Leu Val Phe Ile Lys Lys Val Ala Lys Lys Pro Thr Asn
210 215 220
Lys Ala Pro His Pro Lys Gln Glu Pro Gln Glu Ile Asn Phe Pro Asp
225 230 235 240
Asp Leu Pro Gly Ser Asn Thr Ala Ala Pro Val Gln Glu Thr Leu His
245 250 255
Gly Cys Gln Pro Val Thr Gln Glu Asp Gly Lys Glu Ser Arg Ile Ser
260 265 270
Val Gln Glu Arg Gln
275
<210> 116
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1122
<400> 116
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val
740 745 750
Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 117
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1122
<400> 117
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 118
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1942
<400> 118
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
450 455 460
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys
465 470 475 480
Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser
485 490 495
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile
500 505 510
Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg
515 520 525
Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu
530 535 540
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu
545 550 555 560
Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr
565 570 575
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
580 585 590
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
595 600 605
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
610 615 620
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
625 630 635 640
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
645 650 655
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
660 665 670
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val
740 745 750
Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 119
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1942
<400> 119
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
450 455 460
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys
465 470 475 480
Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser
485 490 495
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile
500 505 510
Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg
515 520 525
Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu
530 535 540
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu
545 550 555 560
Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr
565 570 575
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
580 585 590
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
595 600 605
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
610 615 620
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
625 630 635 640
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
645 650 655
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
660 665 670
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 120
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1887
<400> 120
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
450 455 460
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys
465 470 475 480
Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser
485 490 495
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile
500 505 510
Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg
515 520 525
Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu
530 535 540
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu
545 550 555 560
Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr
565 570 575
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
580 585 590
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
595 600 605
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
610 615 620
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
625 630 635 640
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
645 650 655
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
660 665 670
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 121
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1887
<400> 121
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val
740 745 750
Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 122
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1888
<400> 122
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 123
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1888
<400> 123
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
450 455 460
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys
465 470 475 480
Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser
485 490 495
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile
500 505 510
Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg
515 520 525
Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu
530 535 540
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu
545 550 555 560
Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr
565 570 575
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
580 585 590
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
595 600 605
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
610 615 620
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
625 630 635 640
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
645 650 655
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
660 665 670
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val
740 745 750
Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 124
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2254
<400> 124
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 125
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2254
<400> 125
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val
690 695 700
Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala
705 710 715 720
Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu Ala
725 730 735
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn Val
740 745 750
Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly
755 760 765
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp
770 775 780
Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr Phe
785 790 795 800
Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 126
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2340
<400> 126
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
625 630 635 640
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
645 650 655
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
660 665 670
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln
690 695 700
Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg
705 710 715 720
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
725 730 735
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile
740 745 750
Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
755 760 765
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
770 775 780
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gly
785 790 795 800
Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
805 810 815
Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 127
<211> 688
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2340
<400> 127
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Gly Gly Gly
210 215 220
Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
225 230 235 240
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
275 280 285
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
290 295 300
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
305 310 315 320
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
325 330 335
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
340 345 350
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu
355 360 365
Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
485 490 495
Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
500 505 510
Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser
515 520 525
Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
530 535 540
Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser
545 550 555 560
Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
565 570 575
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys
580 585 590
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
595 600 605
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
610 615 620
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Val Thr Glu Gln
625 630 635 640
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
645 650 655
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
660 665 670
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
675 680 685
<210> 128
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain P1AE2340
<400> 128
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser
100 105 110
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
115 120 125
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
130 135 140
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
145 150 155 160
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
165 170 175
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
180 185 190
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
195 200 205
Glu Pro Lys Ser Cys
210
<210> 129
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2735
<400> 129
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe
115 120 125
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
130 135 140
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
145 150 155 160
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
165 170 175
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
180 185 190
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
195 200 205
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly
225 230 235 240
Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
245 250 255
Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe
260 265 270
Ser Ser Tyr Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
275 280 285
Glu Trp Met Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala
290 295 300
Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser
305 310 315 320
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
325 330 335
Tyr Tyr Cys Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly
340 345 350
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
355 360 365
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
370 375 380
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
385 390 395 400
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
405 410 415
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
420 425 430
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
435 440 445
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys
465 470 475 480
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
485 490 495
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
500 505 510
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
515 520 525
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
530 535 540
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
545 550 555 560
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
565 570 575
Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys
580 585 590
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys
595 600 605
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
610 615 620
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
625 630 635 640
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
645 650 655
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
660 665 670
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
675 680 685
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly
690 695 700
Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser
705 710 715 720
Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala
725 730 735
Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly
740 745 750
Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly
755 760 765
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu
770 775 780
Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln
785 790 795 800
Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu
805 810 815
Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
820 825 830
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
835 840 845
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
850 855 860
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
865 870 875 880
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
885 890 895
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
900 905 910
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 130
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2735
<400> 130
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser
100 105 110
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
115 120 125
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
130 135 140
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
145 150 155 160
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
165 170 175
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
180 185 190
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
195 200 205
Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly
225 230 235 240
Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ala
245 250 255
Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser Trp Val Arg Gln Ala
260 265 270
Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile Ile Pro Ile Phe Gly
275 280 285
Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala
290 295 300
Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser
305 310 315 320
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Tyr Tyr Arg Gly Pro
325 330 335
Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser
340 345 350
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
355 360 365
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
370 375 380
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
385 390 395 400
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
405 410 415
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
420 425 430
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
435 440 445
Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp
450 455 460
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
465 470 475 480
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
485 490 495
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
500 505 510
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
515 520 525
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
530 535 540
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
545 550 555 560
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu
565 570 575
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
580 585 590
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
595 600 605
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
610 615 620
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
625 630 635 640
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
645 650 655
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
660 665 670
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
675 680 685
Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met
690 695 700
Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr
705 710 715 720
Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr
725 730 735
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser
740 745 750
Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
755 760 765
Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala
770 775 780
Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln
785 790 795 800
Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe
805 810 815
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
820 825 830
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
835 840 845
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
850 855 860
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
865 870 875 880
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
885 890 895
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
900 905 910
Glu Cys
<210> 131
<211> 925
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2743
<400> 131
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe
115 120 125
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
130 135 140
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
145 150 155 160
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
165 170 175
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
180 185 190
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
195 200 205
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly
225 230 235 240
Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala
245 250 255
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile
260 265 270
Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
275 280 285
Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg
290 295 300
Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser
305 310 315 320
Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser
325 330 335
Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr
340 345 350
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
355 360 365
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
370 375 380
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
385 390 395 400
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
405 410 415
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
420 425 430
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
435 440 445
Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly
450 455 460
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
465 470 475 480
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
485 490 495
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
500 505 510
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
515 520 525
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
530 535 540
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
545 550 555 560
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
565 570 575
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
580 585 590
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn
595 600 605
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
610 615 620
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
625 630 635 640
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
645 650 655
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
660 665 670
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
675 680 685
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
690 695 700
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
705 710 715 720
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
725 730 735
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
740 745 750
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
755 760 765
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
770 775 780
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
785 790 795 800
Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr
805 810 815
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
820 825 830
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
835 840 845
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
850 855 860
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
865 870 875 880
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
885 890 895
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
900 905 910
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
915 920 925
<210> 132
<211> 914
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2743
<400> 132
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser
100 105 110
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
115 120 125
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
130 135 140
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
145 150 155 160
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
165 170 175
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
180 185 190
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
195 200 205
Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro
225 230 235 240
Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
245 250 255
Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro
260 265 270
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser
275 280 285
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr
290 295 300
Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys
305 310 315 320
Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val
325 330 335
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
340 345 350
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
355 360 365
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
370 375 380
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
385 390 395 400
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
405 410 415
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
420 425 430
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly
435 440 445
Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro
450 455 460
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
465 470 475 480
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
485 490 495
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
500 505 510
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
515 520 525
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
530 535 540
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
545 550 555 560
Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
565 570 575
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp
580 585 590
Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
595 600 605
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
610 615 620
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
625 630 635 640
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
645 650 655
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
660 665 670
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val
690 695 700
Lys Lys Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly
705 710 715 720
Thr Phe Ser Ser Tyr Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln
725 730 735
Gly Leu Glu Trp Met Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn
740 745 750
Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser
755 760 765
Thr Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr
770 775 780
Ala Val Tyr Tyr Cys Ala Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr
785 790 795 800
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
805 810 815
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
820 825 830
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
835 840 845
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
850 855 860
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
865 870 875 880
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
885 890 895
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
900 905 910
Ser Cys
<210> 133
<211> 919
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2762
<400> 133
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln
225 230 235 240
Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser Val Lys Val Ser Cys
245 250 255
Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala Ile Ser Trp Val Arg
260 265 270
Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Gly Ile Ile Pro Ile
275 280 285
Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Ile
290 295 300
Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu
305 310 315 320
Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Tyr Tyr Arg
325 330 335
Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
340 345 350
Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
355 360 365
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
370 375 380
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
385 390 395 400
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
405 410 415
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
420 425 430
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
435 440 445
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Asp Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro
465 470 475 480
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
485 490 495
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
500 505 510
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
515 520 525
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
530 535 540
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
545 550 555 560
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
565 570 575
Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
580 585 590
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu
595 600 605
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
610 615 620
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
625 630 635 640
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
645 650 655
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
660 665 670
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
675 680 685
Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly
690 695 700
Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala
705 710 715 720
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile
725 730 735
Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
740 745 750
Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg
755 760 765
Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser
770 775 780
Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser
785 790 795 800
Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser
805 810 815
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
820 825 830
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
835 840 845
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
850 855 860
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
865 870 875 880
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
885 890 895
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
900 905 910
Lys Val Glu Pro Lys Ser Cys
915
<210> 134
<211> 924
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2762
<400> 134
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly
210 215 220
Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
225 230 235 240
Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser Ser
245 250 255
Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr Ala
260 265 270
Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly
275 280 285
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
290 295 300
Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr Met
305 310 315 320
Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala
325 330 335
Arg Glu Tyr Tyr Arg Gly Pro Tyr Asp Tyr Trp Gly Gln Gly Thr Thr
340 345 350
Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe
355 360 365
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
370 375 380
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
385 390 395 400
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
405 410 415
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
420 425 430
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
435 440 445
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
450 455 460
Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr
465 470 475 480
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
485 490 495
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
500 505 510
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
515 520 525
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
530 535 540
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
545 550 555 560
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
565 570 575
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
580 585 590
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro
595 600 605
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val
610 615 620
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
625 630 635 640
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
645 650 655
Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
660 665 670
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
675 680 685
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly
690 695 700
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro
705 710 715 720
Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
725 730 735
Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro
740 745 750
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Ser Leu Glu Ser
755 760 765
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr
770 775 780
Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys
785 790 795 800
Gln Gln Tyr Ser Ser Gln Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val
805 810 815
Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
820 825 830
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
835 840 845
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
850 855 860
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
865 870 875 880
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
885 890 895
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
900 905 910
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
915 920
<210> 135
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) CDR-H1
<400> 135
Gly Tyr Tyr Trp Ser
1 5
<210> 136
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) CDR-H2
<400> 136
Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu Ser
1 5 10 15
<210> 137
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) CDR-H3
<400> 137
Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu
1 5 10
<210> 138
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) CDR-L1
<400> 138
Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala
1 5 10
<210> 139
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) CDR-L2
<400> 139
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 140
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) CDR-L3
<400> 140
Gln Gln Arg Ser Asn Trp Pro Pro Ala Leu Thr
1 5 10
<210> 141
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) VH
<400> 141
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 142
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> 4-1BB (20H4.9) VL
<400> 142
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 143
<211> 930
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1899
<400> 143
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys
225 230 235 240
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
245 250 255
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
260 265 270
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
275 280 285
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
290 295 300
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
305 310 315 320
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
325 330 335
Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys
340 345 350
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys
355 360 365
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
370 375 380
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
385 390 395 400
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
405 410 415
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
420 425 430
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
435 440 445
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Ala
465 470 475 480
Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
485 490 495
Ser Gln Ser Val Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
500 505 510
Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly
515 520 525
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
530 535 540
Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
545 550 555 560
Gln Arg Ser Asn Trp Pro Pro Ala Leu Thr Phe Gly Gly Gly Thr Lys
565 570 575
Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
580 585 590
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
595 600 605
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
610 615 620
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
625 630 635 640
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
645 650 655
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
660 665 670
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly
690 695 700
Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
705 710 715 720
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
725 730 735
Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
740 745 750
Trp Val Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp
755 760 765
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
770 775 780
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
785 790 795 800
Tyr Cys Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly
805 810 815
Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe
820 825 830
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
835 840 845
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
850 855 860
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
865 870 875 880
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
885 890 895
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
900 905 910
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
915 920 925
Glu Cys
930
<210> 144
<211> 919
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1899
<400> 144
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys
225 230 235 240
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
245 250 255
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
260 265 270
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
275 280 285
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
290 295 300
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
305 310 315 320
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
325 330 335
Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys
340 345 350
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser
355 360 365
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys
370 375 380
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
385 390 395 400
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
405 410 415
Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
420 425 430
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
435 440 445
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Ala
465 470 475 480
Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
485 490 495
Ser Gln Ser Val Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
500 505 510
Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly
515 520 525
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
530 535 540
Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
545 550 555 560
Gln Arg Ser Asn Trp Pro Pro Ala Leu Thr Phe Gly Gly Gly Thr Lys
565 570 575
Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
580 585 590
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
595 600 605
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
610 615 620
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
625 630 635 640
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
645 650 655
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
660 665 670
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly
690 695 700
Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser
705 710 715 720
Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr
725 730 735
Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg
740 745 750
Leu Leu Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg
755 760 765
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg
770 775 780
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met
785 790 795 800
Leu Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser
805 810 815
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
820 825 830
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
835 840 845
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
850 855 860
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
865 870 875 880
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
885 890 895
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
900 905 910
Lys Val Glu Pro Lys Ser Cys
915
<210> 145
<211> 930
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2051
<400> 145
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys
225 230 235 240
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
245 250 255
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
260 265 270
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
275 280 285
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
290 295 300
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
305 310 315 320
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
325 330 335
Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys
340 345 350
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys
355 360 365
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
370 375 380
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
385 390 395 400
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
405 410 415
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
420 425 430
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
435 440 445
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Ala
465 470 475 480
Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
485 490 495
Ser Gln Ser Val Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
500 505 510
Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly
515 520 525
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
530 535 540
Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
545 550 555 560
Gln Arg Ser Asn Trp Pro Pro Ala Leu Thr Phe Gly Gly Gly Thr Lys
565 570 575
Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
580 585 590
Pro Ser Asp Arg Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
595 600 605
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
610 615 620
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
625 630 635 640
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
645 650 655
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
660 665 670
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly
690 695 700
Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
705 710 715 720
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
725 730 735
Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
740 745 750
Trp Val Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp
755 760 765
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
770 775 780
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
785 790 795 800
Tyr Cys Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly
805 810 815
Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe
820 825 830
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
835 840 845
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
850 855 860
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
865 870 875 880
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
885 890 895
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
900 905 910
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
915 920 925
Glu Cys
930
<210> 146
<211> 919
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2051
<400> 146
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys
225 230 235 240
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
245 250 255
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
260 265 270
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
275 280 285
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
290 295 300
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
305 310 315 320
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
325 330 335
Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys
340 345 350
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser
355 360 365
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys
370 375 380
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
385 390 395 400
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
405 410 415
Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
420 425 430
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
435 440 445
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Ala
465 470 475 480
Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
485 490 495
Ser Gln Ser Val Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly
500 505 510
Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly
515 520 525
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
530 535 540
Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln
545 550 555 560
Gln Arg Ser Asn Trp Pro Pro Ala Leu Thr Phe Gly Gly Gly Thr Lys
565 570 575
Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
580 585 590
Pro Ser Asp Arg Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
595 600 605
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
610 615 620
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
625 630 635 640
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
645 650 655
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
660 665 670
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly
675 680 685
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly
690 695 700
Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser
705 710 715 720
Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr
725 730 735
Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg
740 745 750
Leu Leu Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg
755 760 765
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg
770 775 780
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met
785 790 795 800
Leu Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser
805 810 815
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
820 825 830
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
835 840 845
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
850 855 860
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
865 870 875 880
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
885 890 895
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
900 905 910
Lys Val Glu Pro Lys Ser Cys
915
<210> 147
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-H1
<400> 147
Gly Tyr Tyr Ile His
1 5
<210> 148
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-H2
<400> 148
Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 149
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-H3
<400> 149
Glu Gly Ile Tyr Trp
1 5
<210> 150
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-L1
<400> 150
Arg Ser Ser Gln Ser Leu Val His Ser Asn Gly Asn Thr Phe Leu His
1 5 10 15
<210> 151
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-L2
<400> 151
Thr Val Ser Asn Arg Phe Ser
1 5
<210> 152
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 CDR-L3
<400> 152
Ser Gln Thr Thr His Val Pro Trp Thr
1 5
<210> 153
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 VH
<400> 153
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 154
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> hu CD40 VL
<400> 154
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 155
<211> 926
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1799
<400> 155
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly
210 215 220
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
465 470 475 480
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
485 490 495
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
545 550 555 560
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
565 570 575
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
580 585 590
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
595 600 605
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
610 615 620
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
625 630 635 640
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
645 650 655
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
660 665 670
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
690 695 700
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
705 710 715 720
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met
725 730 735
Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala
740 745 750
Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
755 760 765
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
770 775 780
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
785 790 795 800
Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr
805 810 815
Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
820 825 830
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
835 840 845
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
850 855 860
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
865 870 875 880
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
885 890 895
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
900 905 910
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 156
<211> 915
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1799
<400> 156
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly
210 215 220
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
465 470 475 480
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
485 490 495
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
545 550 555 560
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
565 570 575
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
580 585 590
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
595 600 605
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
610 615 620
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
625 630 635 640
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
645 650 655
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
660 665 670
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile
690 695 700
Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg
705 710 715 720
Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu
725 730 735
Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn
740 745 750
Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser
755 760 765
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
770 775 780
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr
785 790 795 800
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys
805 810 815
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
820 825 830
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
835 840 845
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
850 855 860
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
865 870 875 880
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
885 890 895
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
900 905 910
Lys Ser Cys
915
<210> 157
<211> 915
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1902
<400> 157
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly
210 215 220
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
465 470 475 480
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
485 490 495
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
545 550 555 560
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
565 570 575
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
580 585 590
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
595 600 605
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
610 615 620
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
625 630 635 640
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
645 650 655
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
660 665 670
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile
690 695 700
Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg
705 710 715 720
Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu
725 730 735
Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn
740 745 750
Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser
755 760 765
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
770 775 780
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr
785 790 795 800
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys
805 810 815
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
820 825 830
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
835 840 845
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
850 855 860
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
865 870 875 880
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
885 890 895
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
900 905 910
Lys Ser Cys
915
<210> 158
<211> 926
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1902
<400> 158
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly
210 215 220
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
465 470 475 480
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
485 490 495
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
545 550 555 560
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
565 570 575
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
580 585 590
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
595 600 605
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
610 615 620
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
625 630 635 640
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
645 650 655
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
660 665 670
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
690 695 700
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
705 710 715 720
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met
725 730 735
Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala
740 745 750
Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
755 760 765
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
770 775 780
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
785 790 795 800
Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr
805 810 815
Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
820 825 830
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
835 840 845
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
850 855 860
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
865 870 875 880
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
885 890 895
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
900 905 910
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 159
<211> 926
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1800
<400> 159
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly
210 215 220
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
465 470 475 480
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
485 490 495
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
545 550 555 560
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
565 570 575
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
580 585 590
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
595 600 605
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
610 615 620
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
625 630 635 640
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
645 650 655
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
660 665 670
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
690 695 700
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
705 710 715 720
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met
725 730 735
Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala
740 745 750
Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
755 760 765
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
770 775 780
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
785 790 795 800
Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr
805 810 815
Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
820 825 830
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
835 840 845
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
850 855 860
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
865 870 875 880
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
885 890 895
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
900 905 910
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 160
<211> 915
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1800
<400> 160
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
465 470 475 480
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr
485 490 495
Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
500 505 510
Trp Val Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln
515 520 525
Lys Phe Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr
530 535 540
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
545 550 555 560
Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val
565 570 575
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
580 585 590
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
595 600 605
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
610 615 620
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
625 630 635 640
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
645 650 655
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
660 665 670
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile
690 695 700
Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg
705 710 715 720
Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu
725 730 735
Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn
740 745 750
Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser
755 760 765
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
770 775 780
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr
785 790 795 800
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys
805 810 815
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
820 825 830
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
835 840 845
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
850 855 860
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
865 870 875 880
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
885 890 895
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
900 905 910
Lys Ser Cys
915
<210> 161
<211> 926
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2052
<400> 161
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
465 470 475 480
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr
485 490 495
Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
500 505 510
Trp Val Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln
515 520 525
Lys Phe Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr
530 535 540
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
545 550 555 560
Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val
565 570 575
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
580 585 590
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
595 600 605
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
610 615 620
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
625 630 635 640
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
645 650 655
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
660 665 670
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
690 695 700
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
705 710 715 720
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met
725 730 735
Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala
740 745 750
Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
755 760 765
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
770 775 780
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
785 790 795 800
Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr
805 810 815
Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
820 825 830
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
835 840 845
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
850 855 860
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
865 870 875 880
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
885 890 895
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
900 905 910
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 162
<211> 915
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2052
<400> 162
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
465 470 475 480
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr
485 490 495
Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
500 505 510
Trp Val Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln
515 520 525
Lys Phe Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr
530 535 540
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
545 550 555 560
Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val
565 570 575
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
580 585 590
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
595 600 605
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
610 615 620
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
625 630 635 640
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
645 650 655
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
660 665 670
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile
690 695 700
Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg
705 710 715 720
Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu
725 730 735
Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn
740 745 750
Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser
755 760 765
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
770 775 780
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr
785 790 795 800
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys
805 810 815
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
820 825 830
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
835 840 845
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
850 855 860
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
865 870 875 880
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
885 890 895
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
900 905 910
Lys Ser Cys
915
<210> 163
<211> 915
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE1901
<400> 163
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
465 470 475 480
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr
485 490 495
Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
500 505 510
Trp Val Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln
515 520 525
Lys Phe Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr
530 535 540
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
545 550 555 560
Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val
565 570 575
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
580 585 590
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
595 600 605
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
610 615 620
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
625 630 635 640
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
645 650 655
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
660 665 670
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile
690 695 700
Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg
705 710 715 720
Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu
725 730 735
Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn
740 745 750
Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser
755 760 765
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
770 775 780
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr
785 790 795 800
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys
805 810 815
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
820 825 830
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
835 840 845
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
850 855 860
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
865 870 875 880
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
885 890 895
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
900 905 910
Lys Ser Cys
915
<210> 164
<211> 926
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE1901
<400> 164
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
465 470 475 480
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr
485 490 495
Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
500 505 510
Trp Val Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln
515 520 525
Lys Phe Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr
530 535 540
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
545 550 555 560
Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val
565 570 575
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
580 585 590
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
595 600 605
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
610 615 620
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
625 630 635 640
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
645 650 655
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
660 665 670
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
690 695 700
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
705 710 715 720
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met
725 730 735
Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala
740 745 750
Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
755 760 765
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
770 775 780
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
785 790 795 800
Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr
805 810 815
Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
820 825 830
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
835 840 845
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
850 855 860
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
865 870 875 880
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
885 890 895
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
900 905 910
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 165
<211> 926
<212> PRT
<213> Artificial Sequence
<220>
<223> first fusion polypeptide (Fc knob) P1AE2255
<400> 165
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly
210 215 220
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
465 470 475 480
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
485 490 495
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
545 550 555 560
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
565 570 575
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg
580 585 590
Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
595 600 605
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
610 615 620
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
625 630 635 640
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
645 650 655
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
660 665 670
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
690 695 700
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
705 710 715 720
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met
725 730 735
Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala
740 745 750
Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
755 760 765
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
770 775 780
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
785 790 795 800
Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr
805 810 815
Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
820 825 830
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
835 840 845
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
850 855 860
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
865 870 875 880
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
885 890 895
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
900 905 910
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
915 920 925
<210> 166
<211> 915
<212> PRT
<213> Artificial Sequence
<220>
<223> second fusion polypeptide (Fc hole) P1AE2255
<400> 166
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Gly Gly Gly Gly Ser Gly Gly
210 215 220
Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
465 470 475 480
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
485 490 495
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
545 550 555 560
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
565 570 575
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg
580 585 590
Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
595 600 605
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
610 615 620
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
625 630 635 640
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
645 650 655
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
660 665 670
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser
675 680 685
Gly Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile
690 695 700
Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg
705 710 715 720
Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser Tyr Leu
725 730 735
Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Asn
740 745 750
Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser
755 760 765
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
770 775 780
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro Pro Thr
785 790 795 800
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys
805 810 815
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
820 825 830
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
835 840 845
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
850 855 860
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
865 870 875 880
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
885 890 895
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
900 905 910
Lys Ser Cys
915
<210> 167
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1a (CD40)
<400> 167
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 168
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1b (CD40)
<400> 168
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 169
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1c (CD40)
<400> 169
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 170
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH1d (CD40)
<400> 170
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Ser Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 171
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1a (CD40)
<400> 171
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 172
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1b (CD40)
<400> 172
Asp Ile Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 173
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1c (CD40)
<400> 173
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 174
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL1d (CD40)
<400> 174
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 175
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2a (CD40)
<400> 175
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 176
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2b (CD40)
<400> 176
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 177
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2c (CD40)
<400> 177
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 178
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2d (CD40)
<400> 178
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 179
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2ab (CD40)
<400> 179
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 180
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> VH2ac (CD40)
<400> 180
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 181
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2a (CD40)
<400> 181
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 182
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2b (CD40)
<400> 182
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 183
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2ab (CD40)
<400> 183
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 184
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> VL2ac (CD40)
<400> 184
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ КОСТИМУЛЯТОРНОГО TNF-РЕЦЕПТОРА | 2016 |
|
RU2761115C1 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РАКОВЫМИ КЛЕТКАМИ И ОБЕСПЕЧИВАЮТ НАПРАВЛЕННЫЙ ПЕРЕНОС РАДИОНУКЛИДОВ К УКАЗАННЫМ КЛЕТКАМ | 2020 |
|
RU2833191C2 |
| УЛУЧШЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ РЕЦЕПТОРЫ | 2018 |
|
RU2825816C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
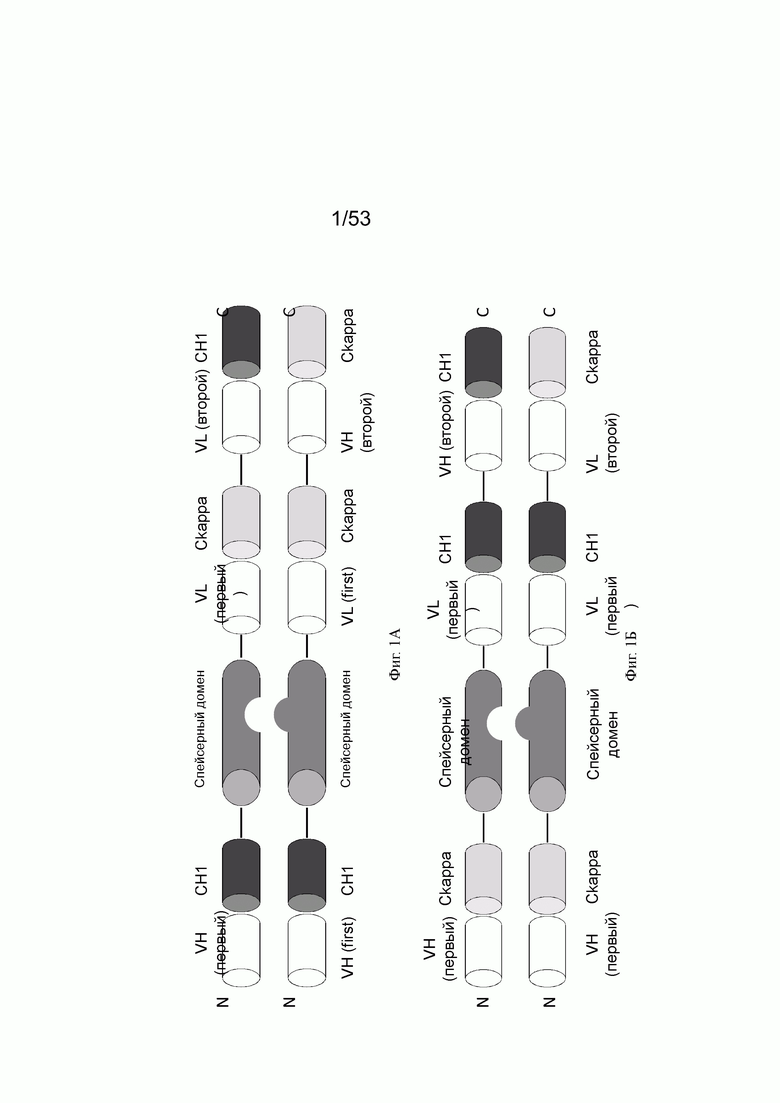
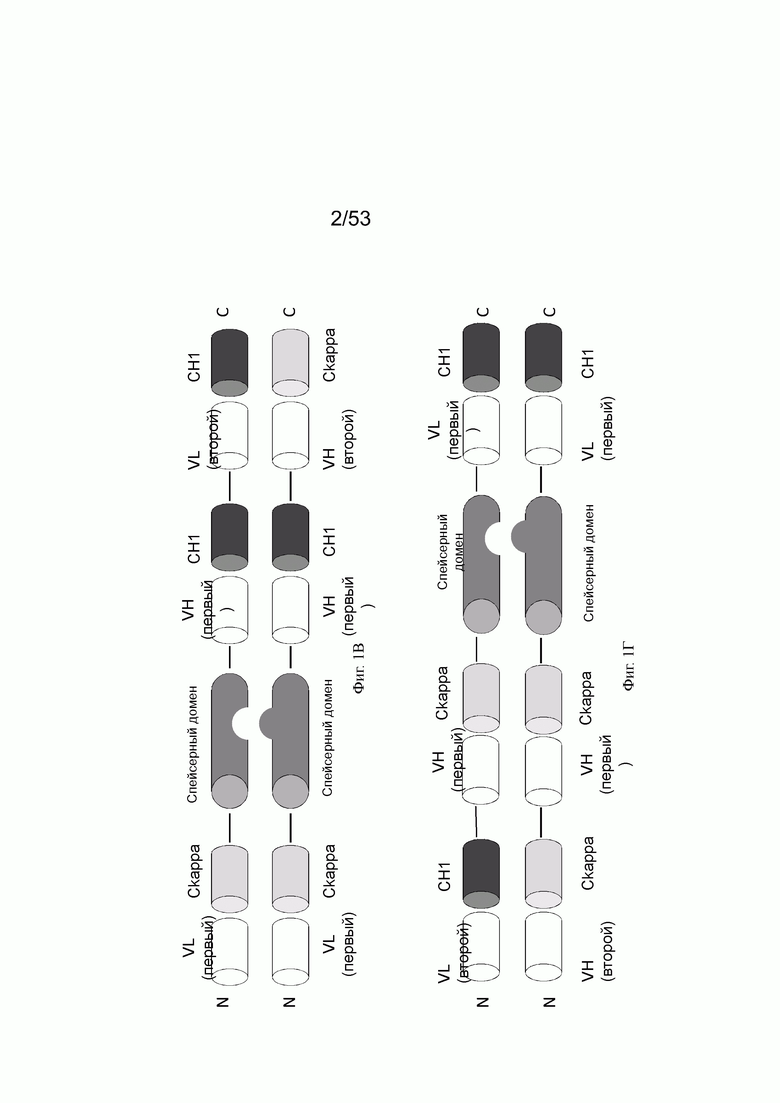
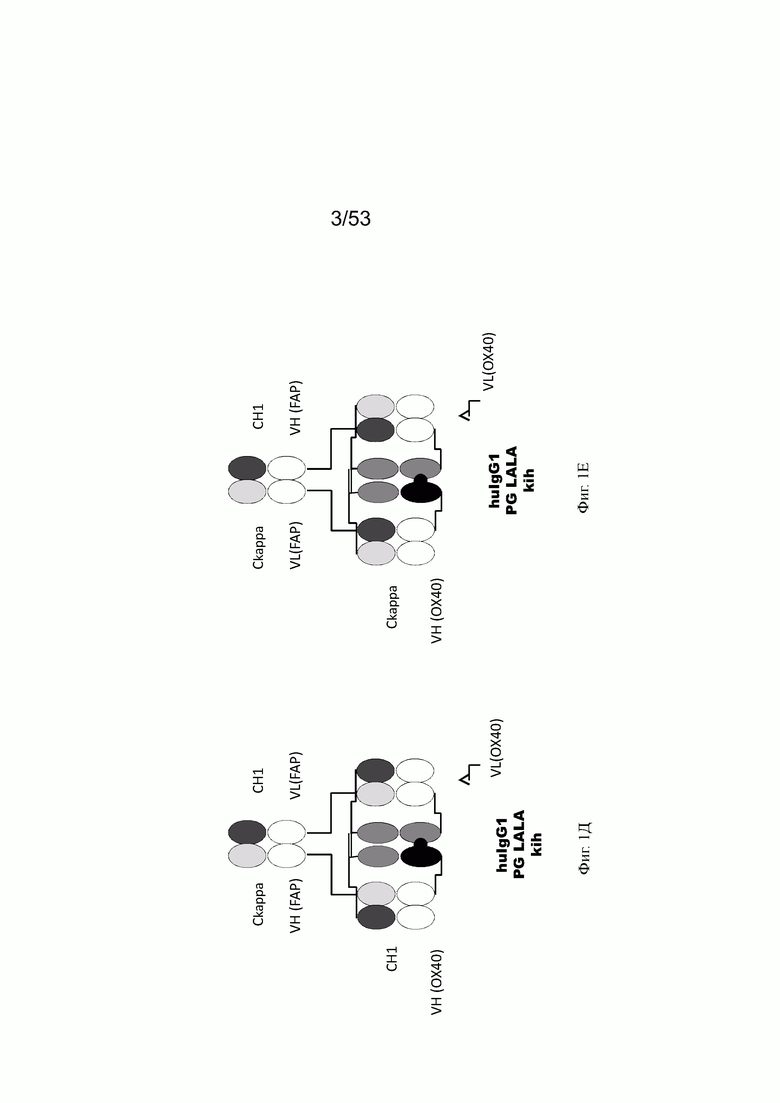
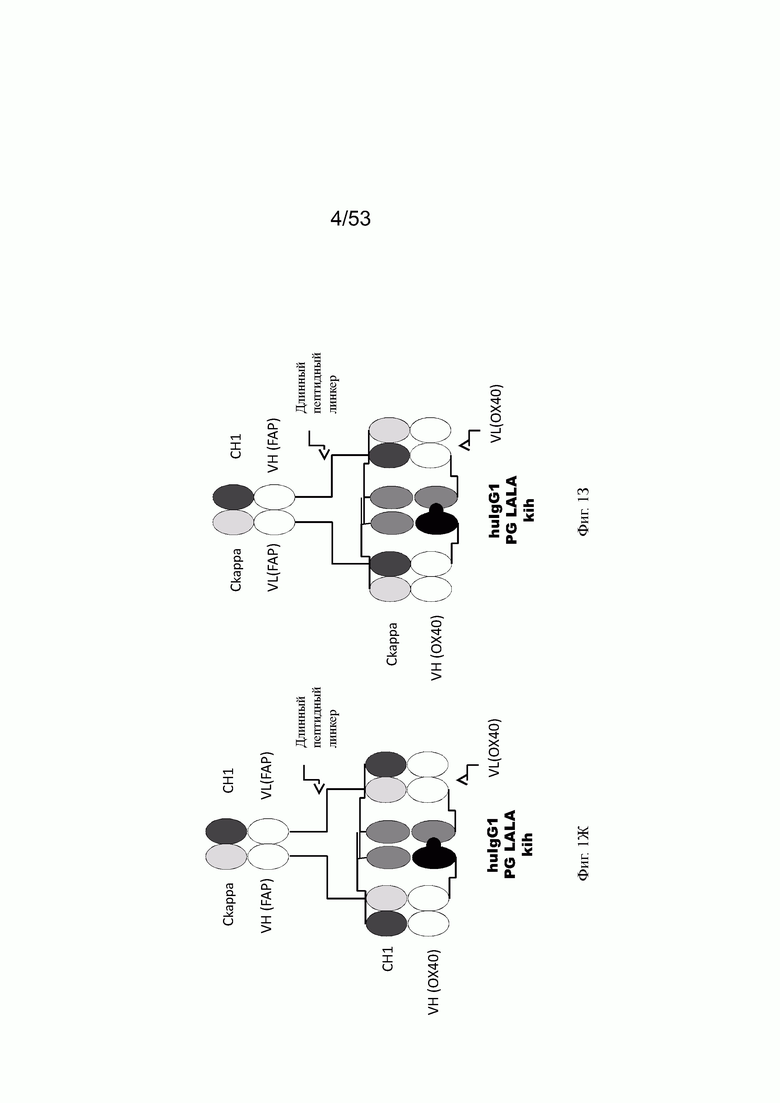
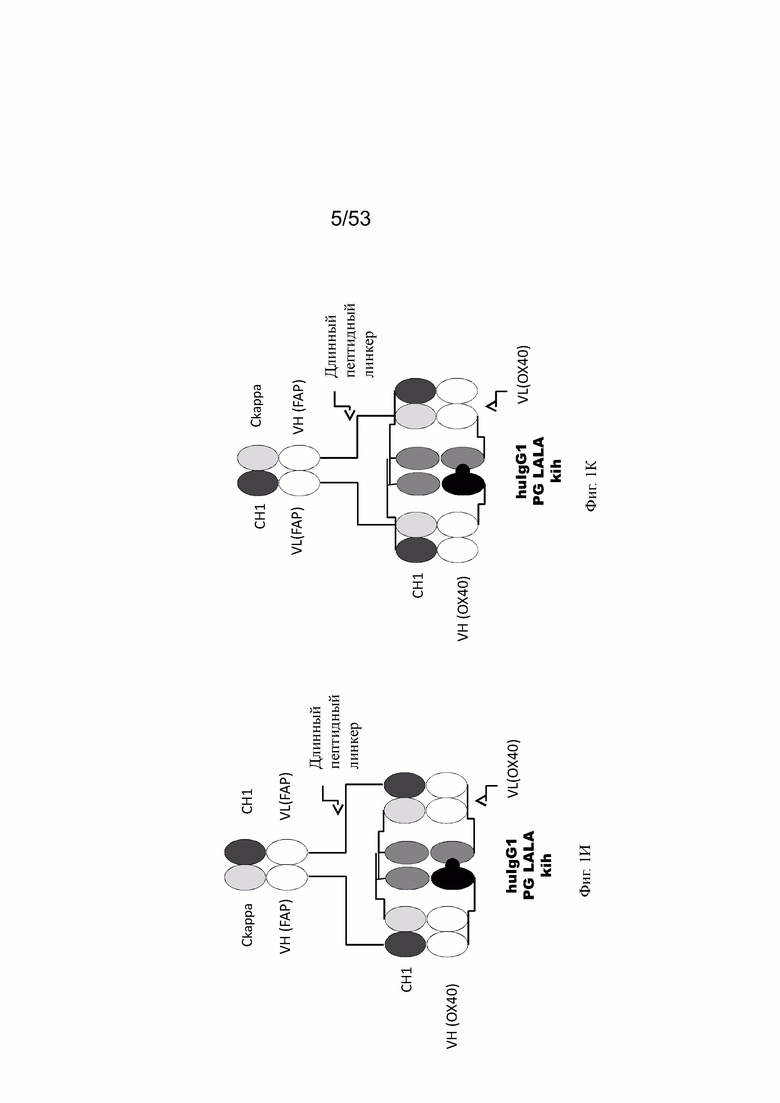
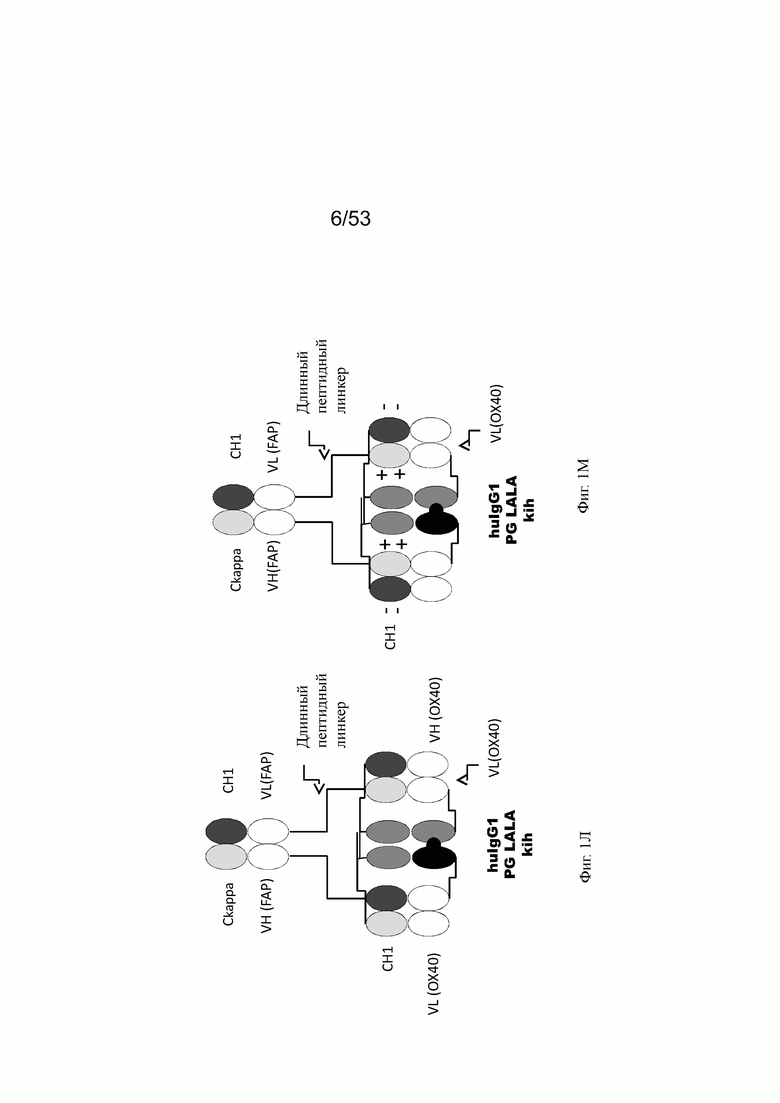
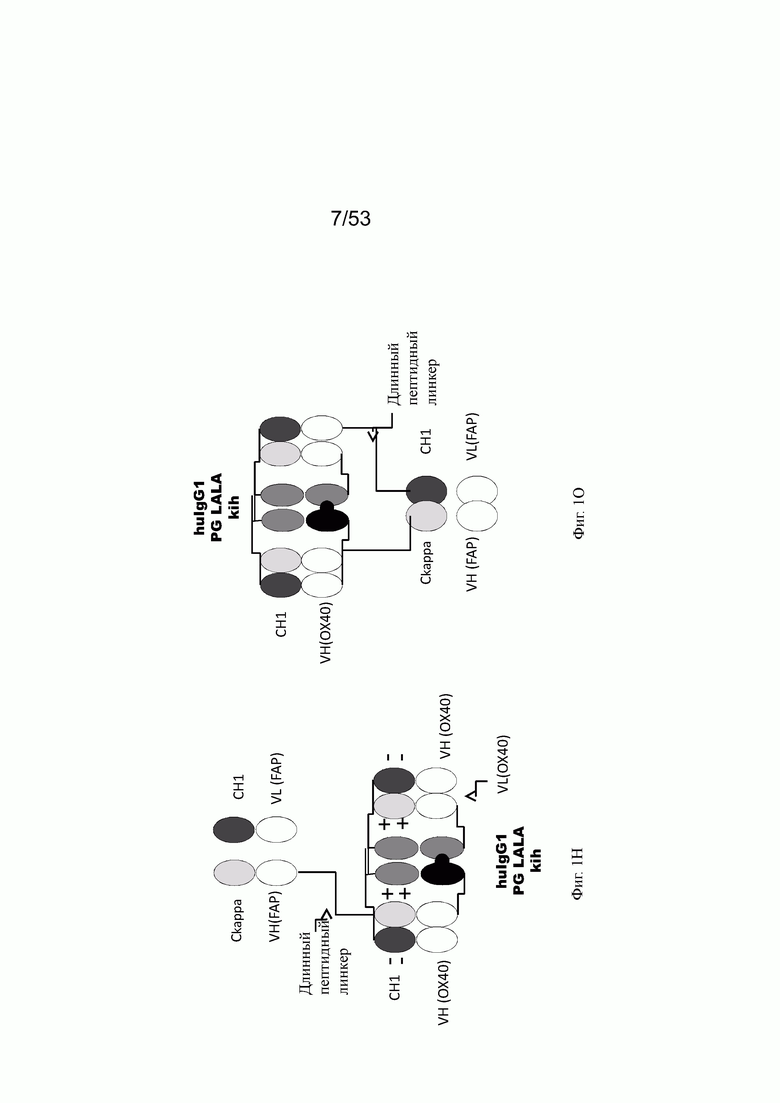
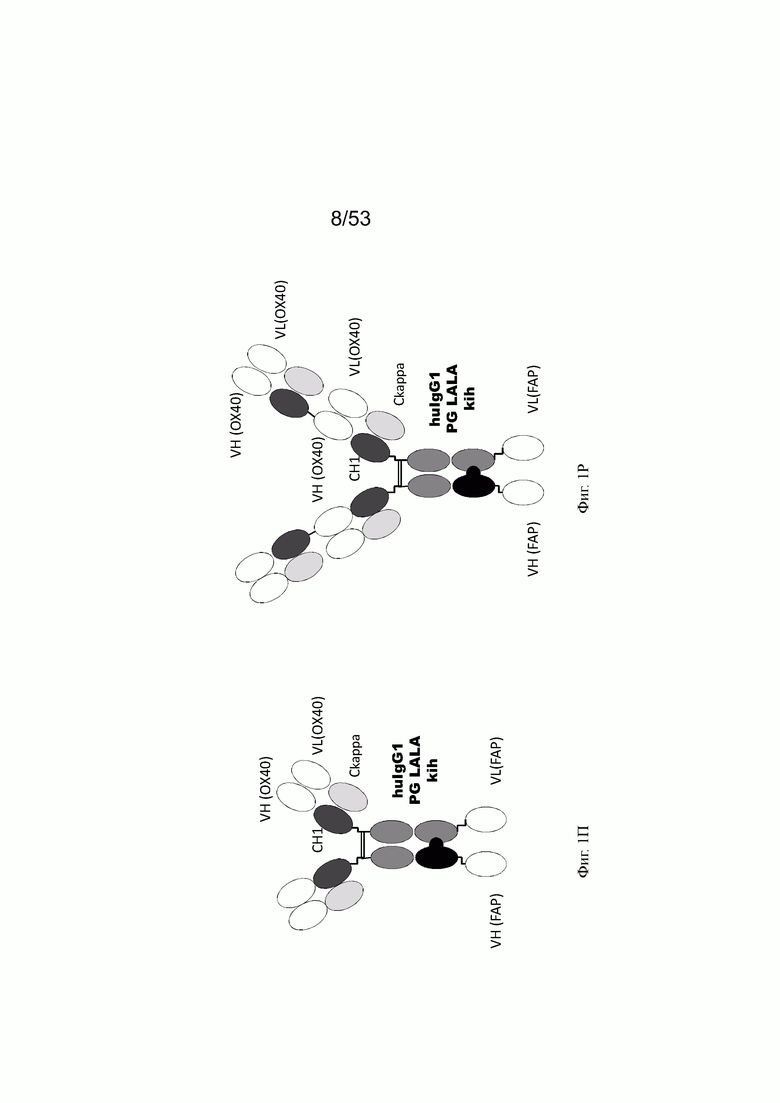
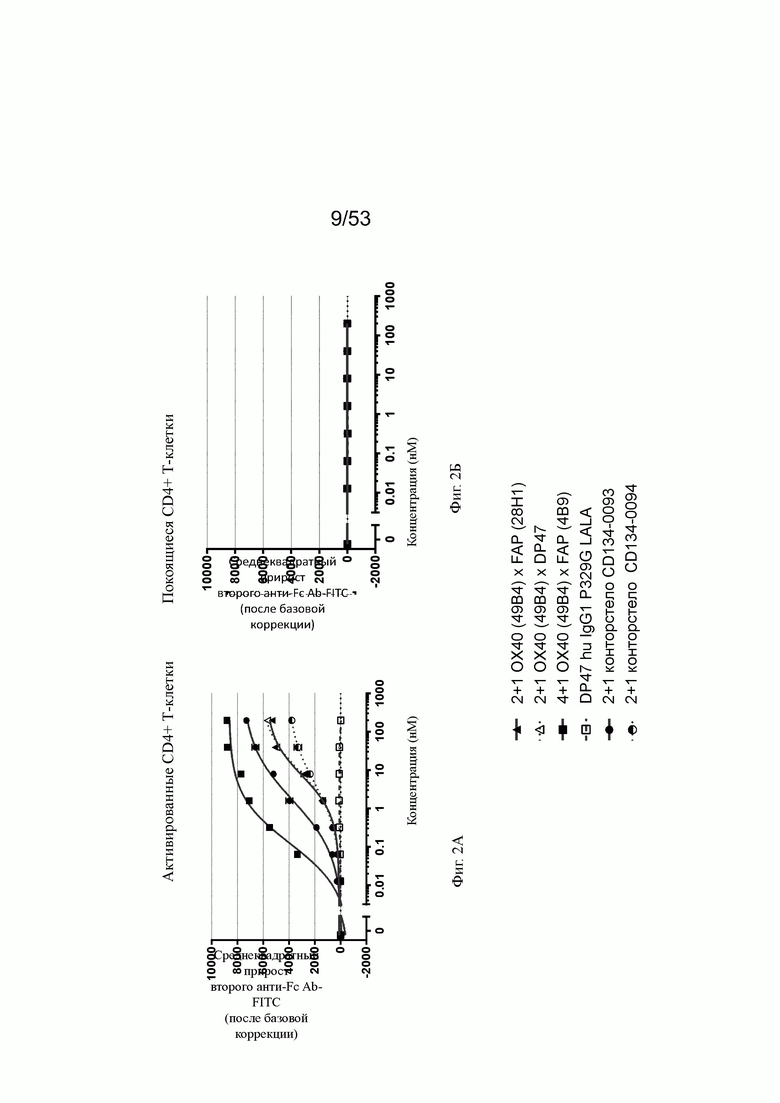
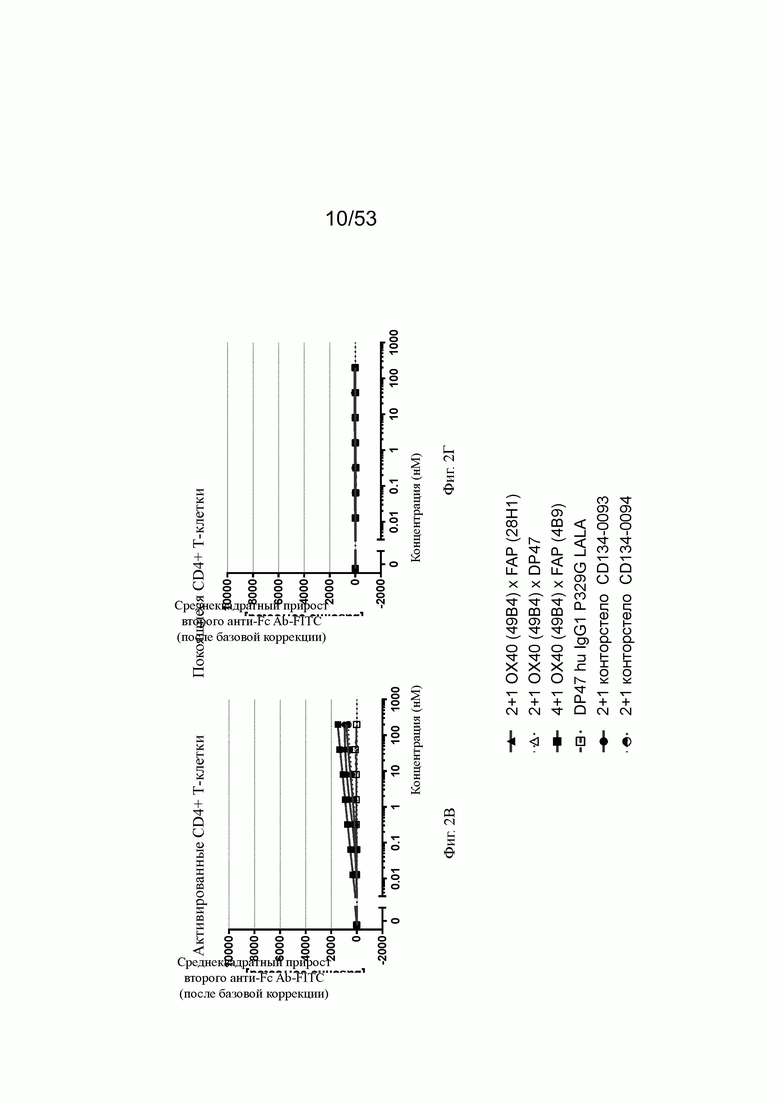
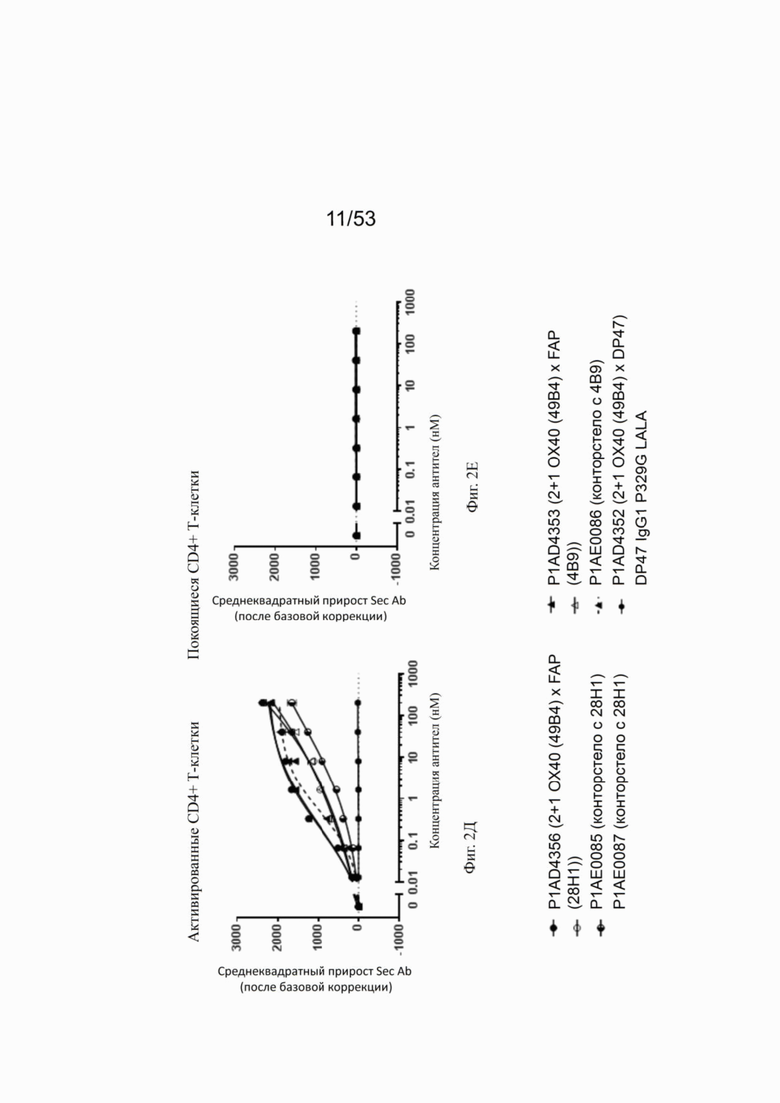
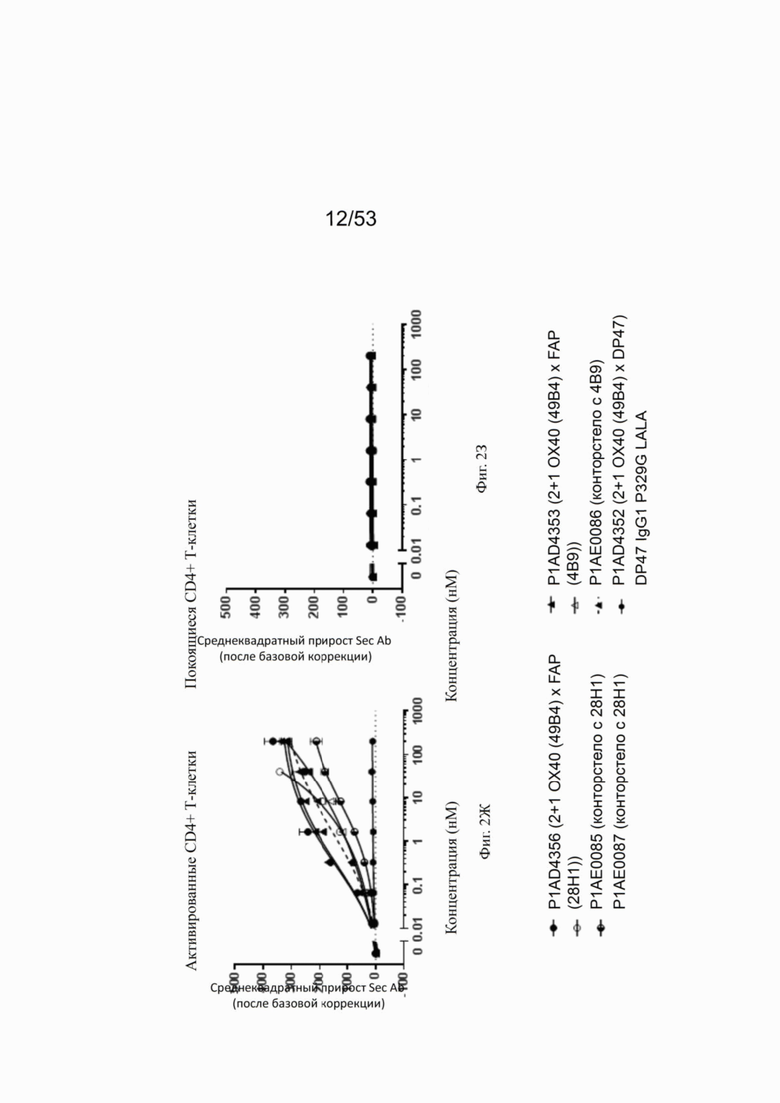

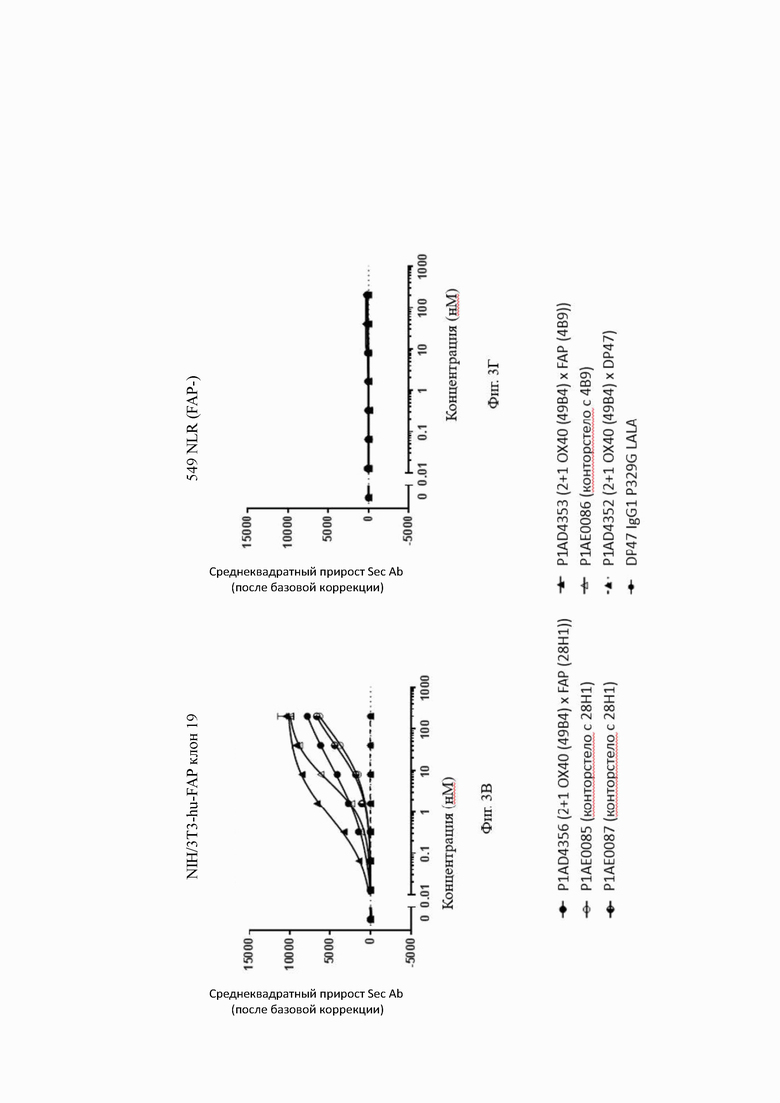
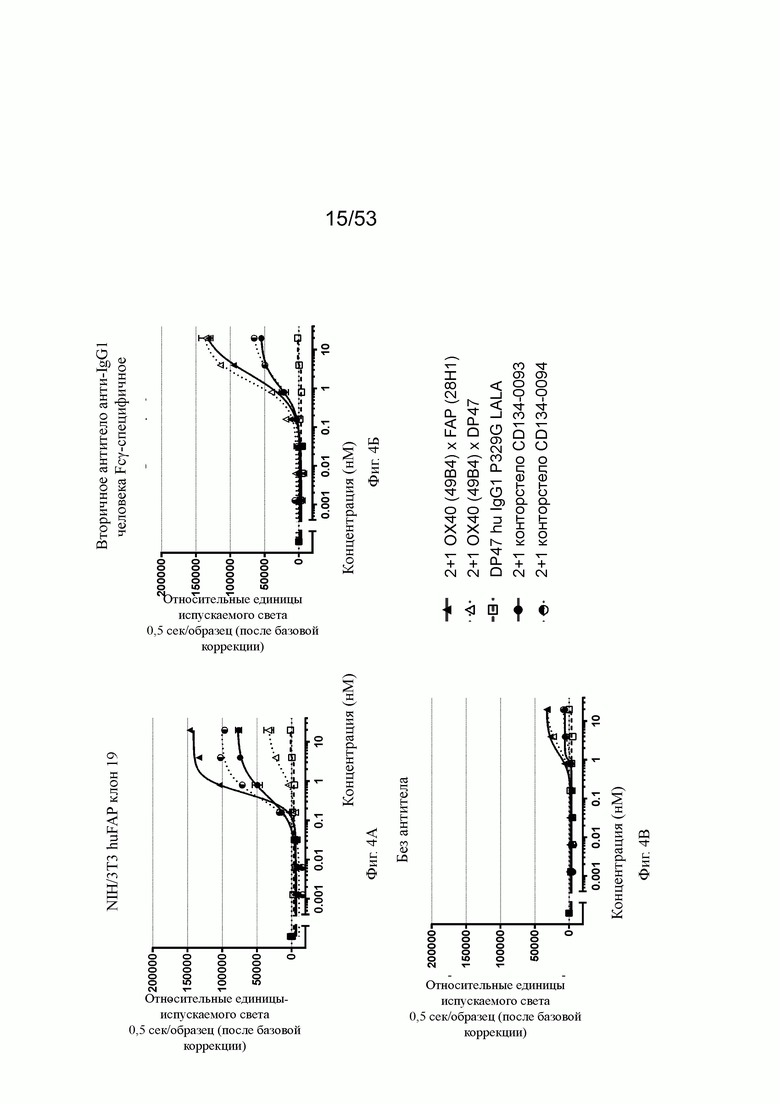
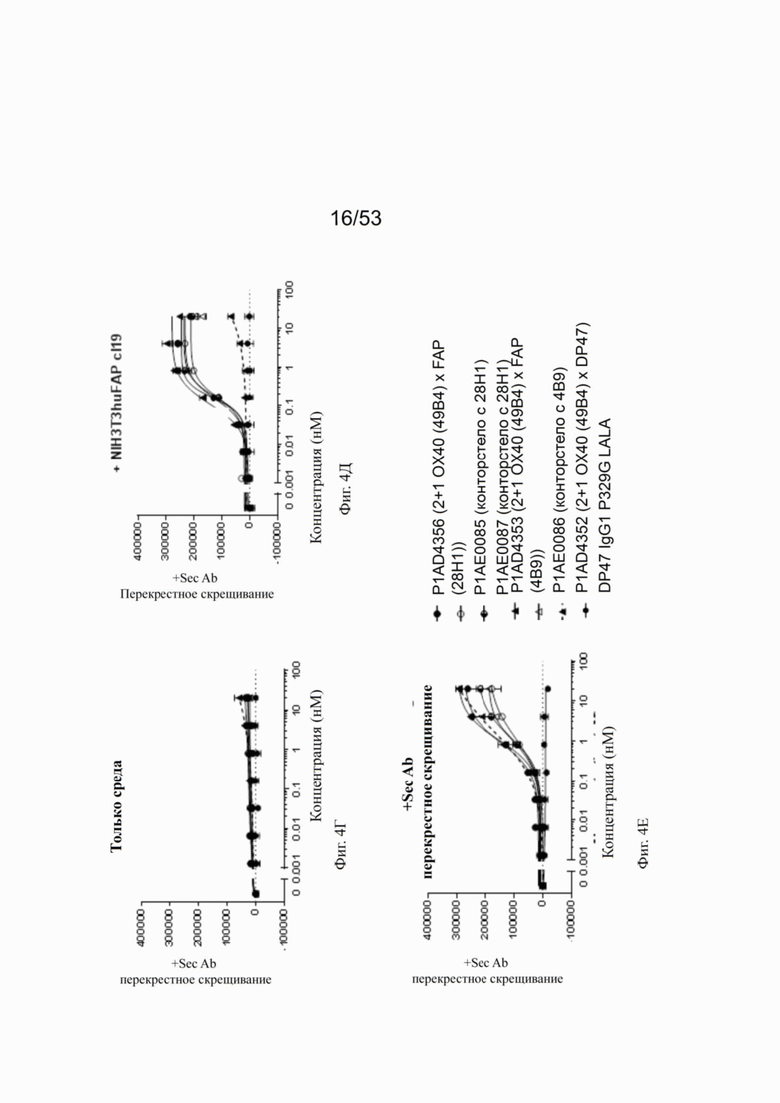
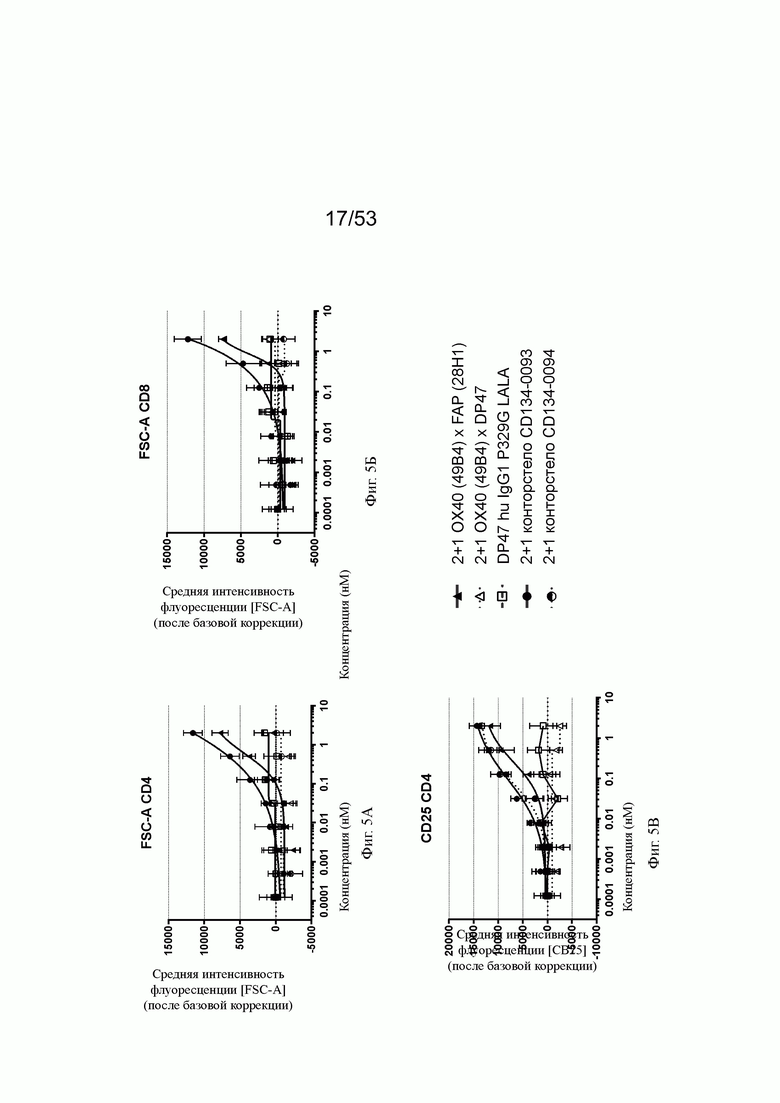
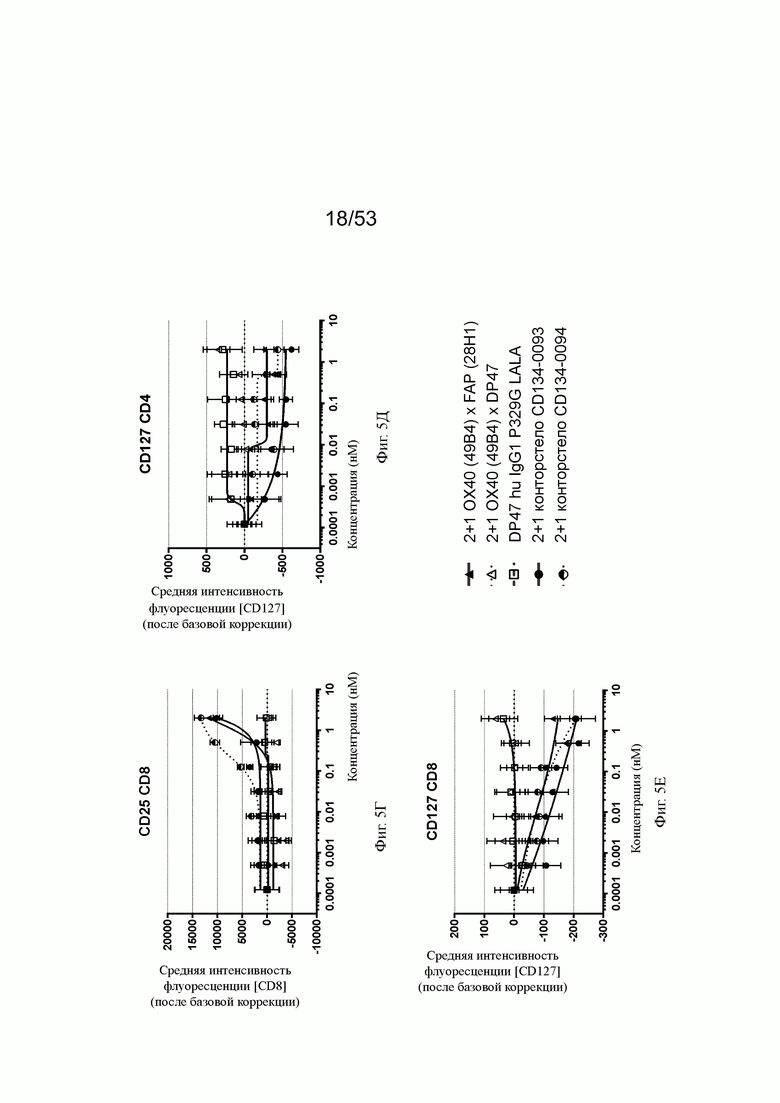
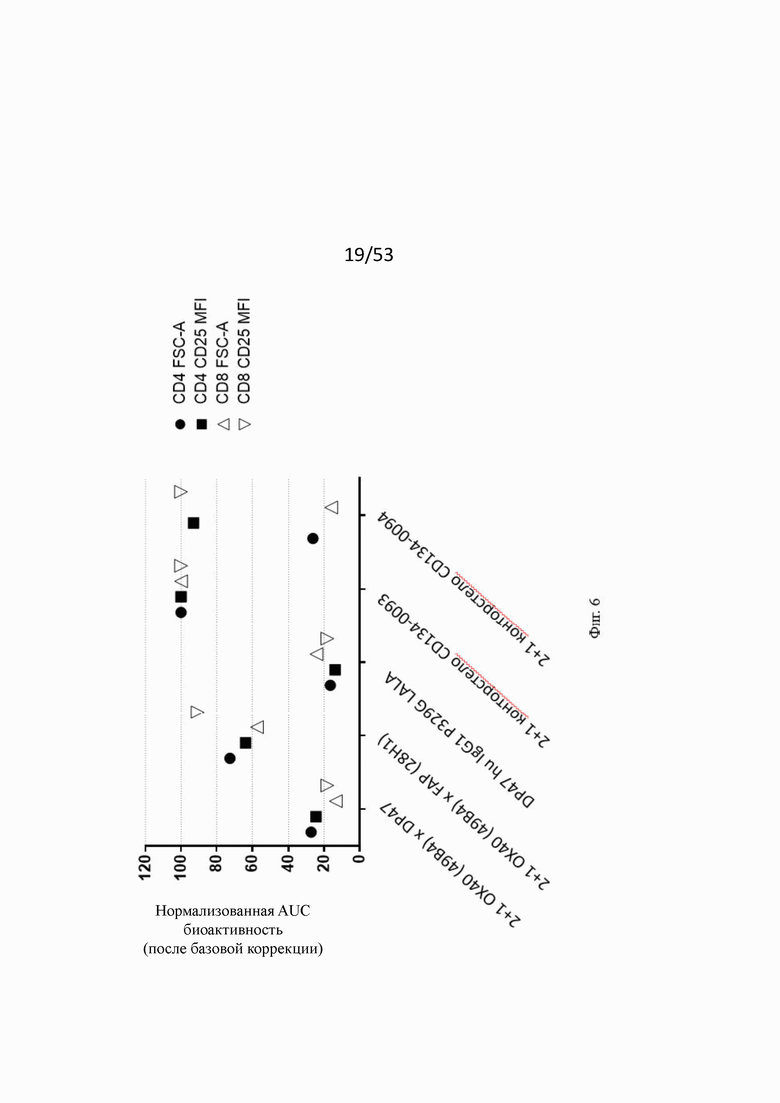
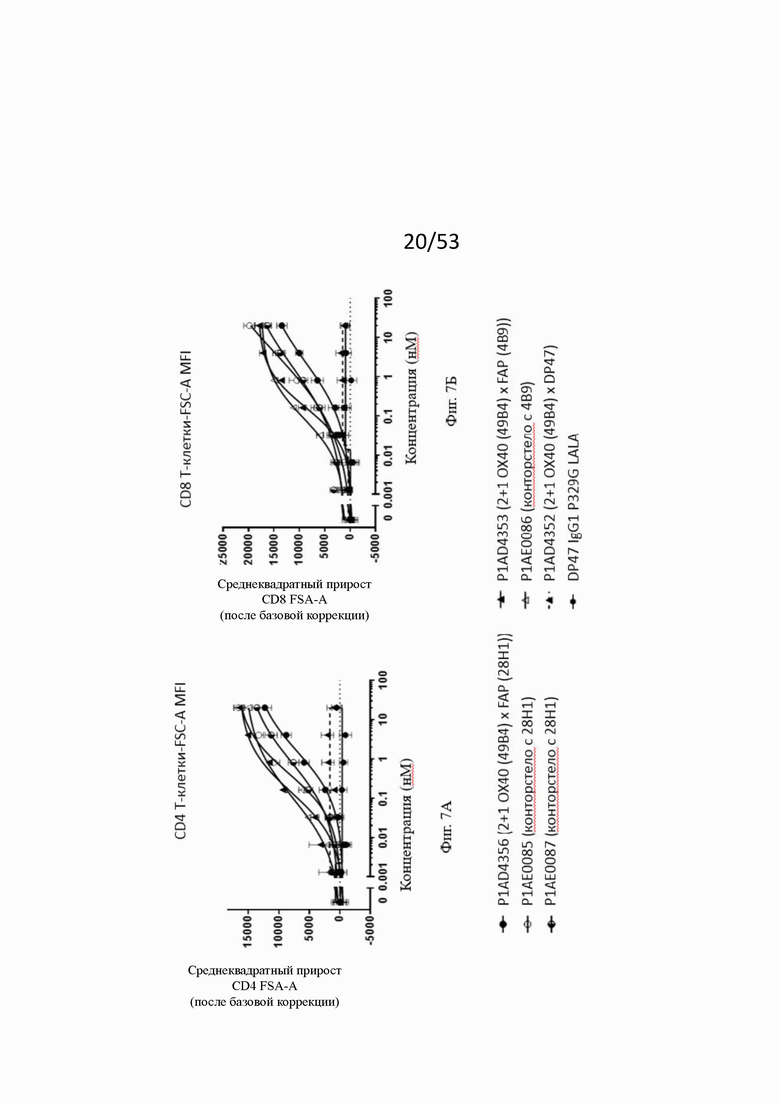
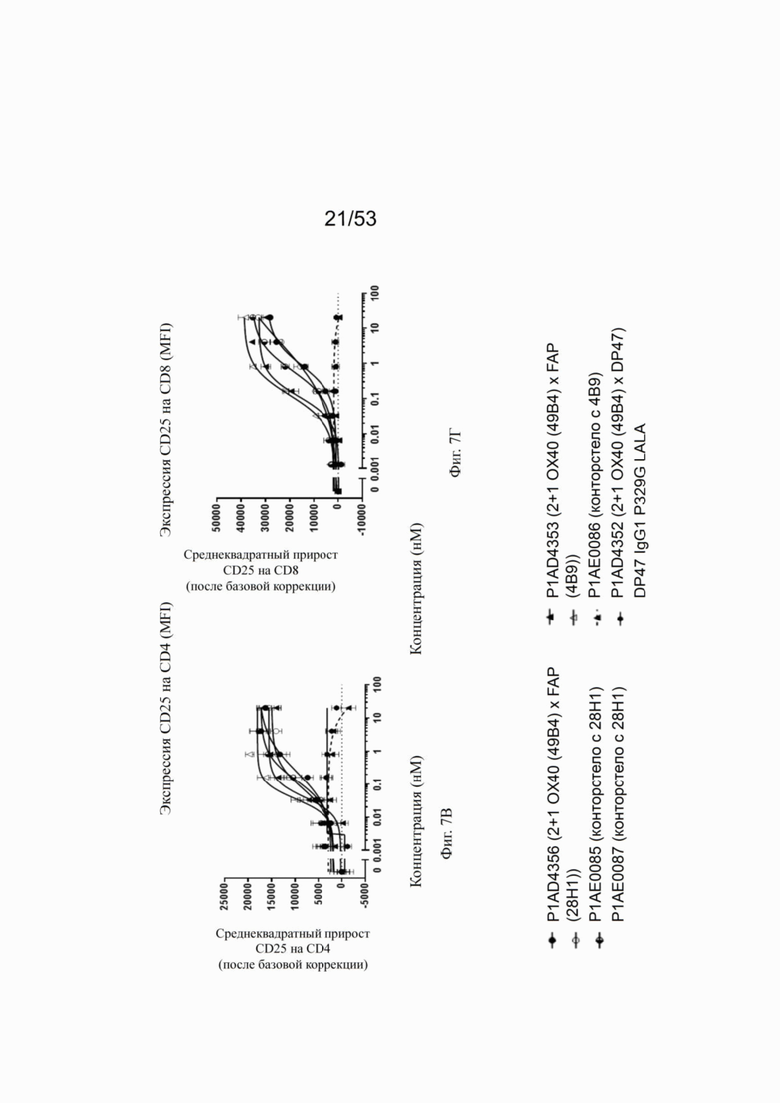
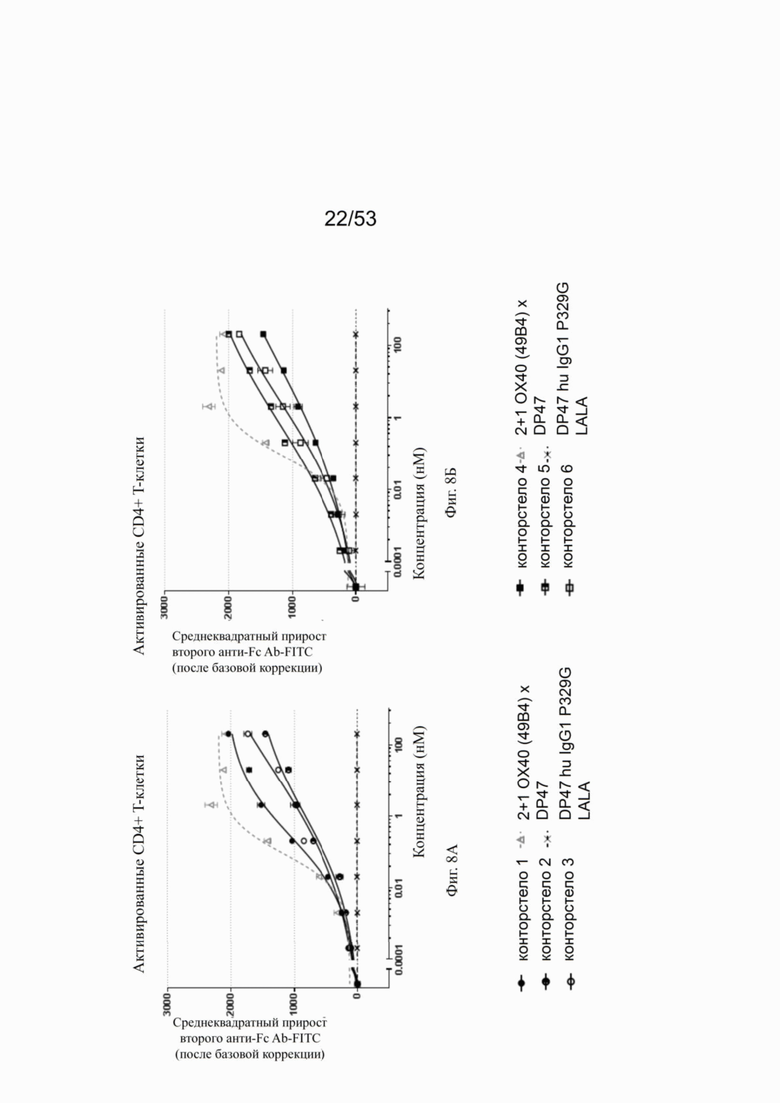
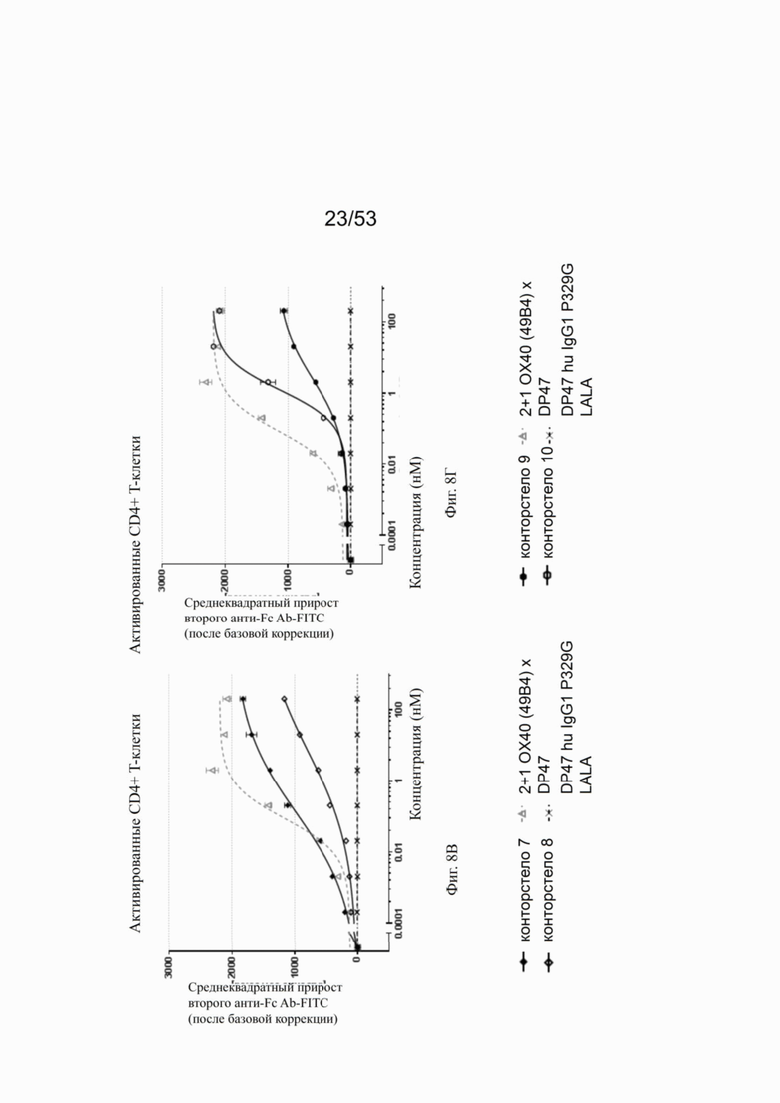
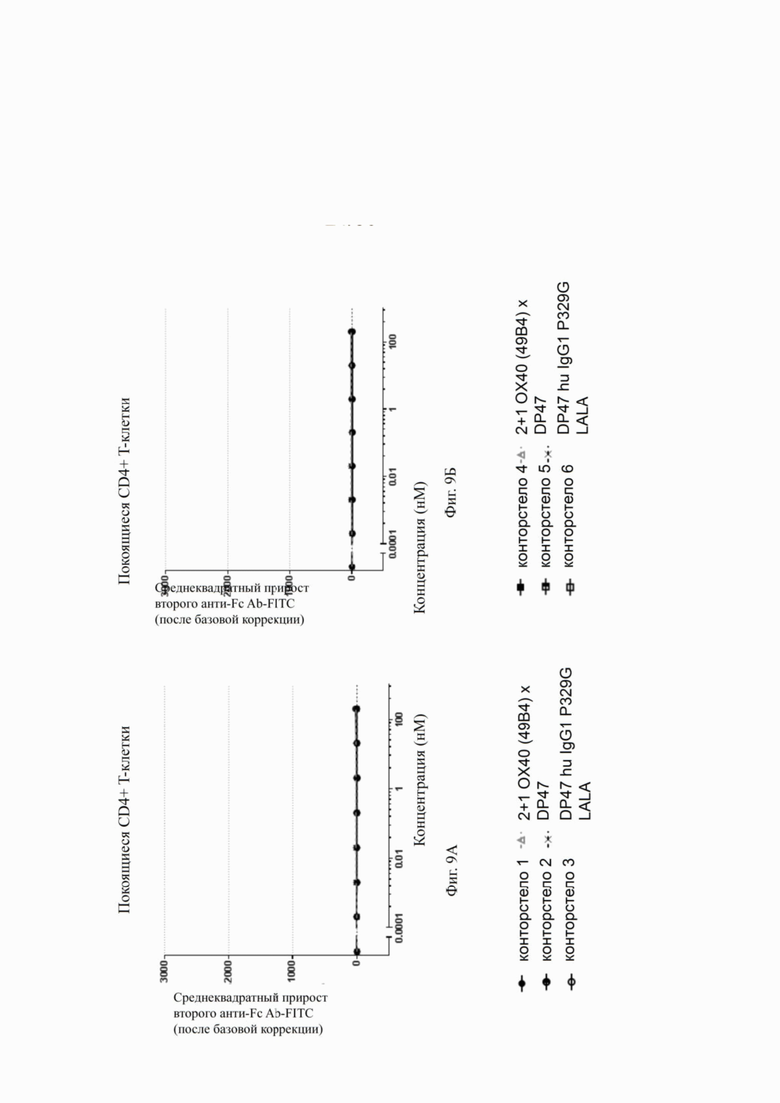
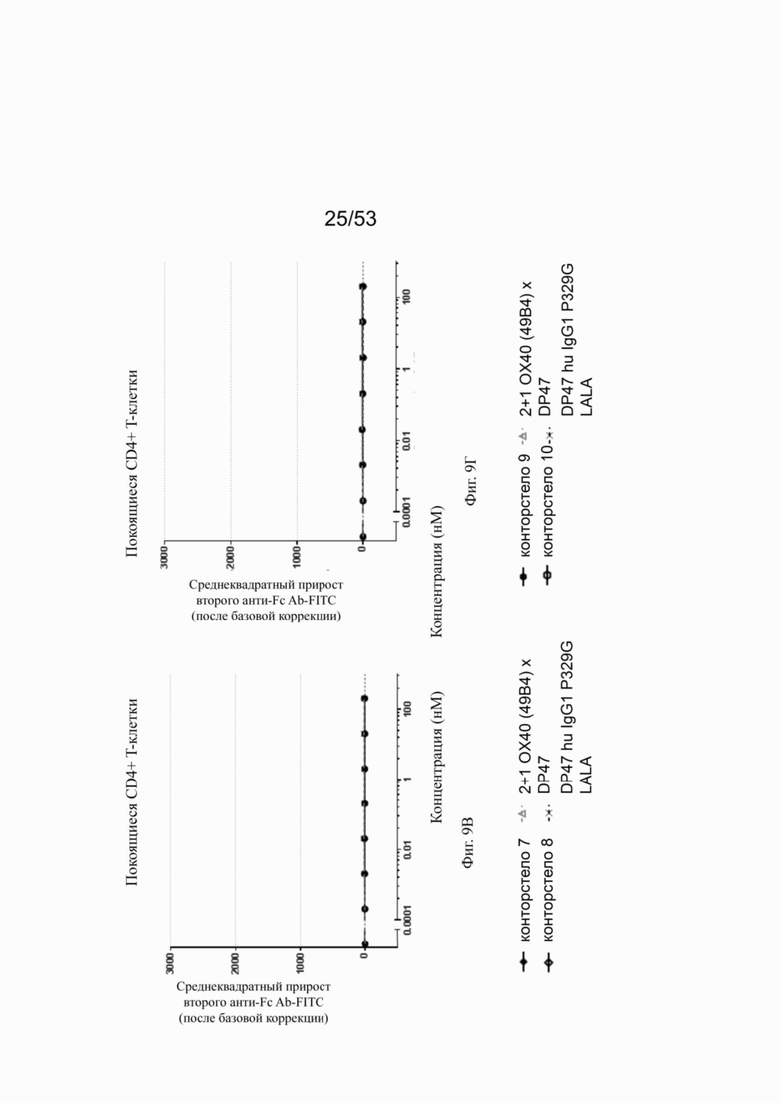
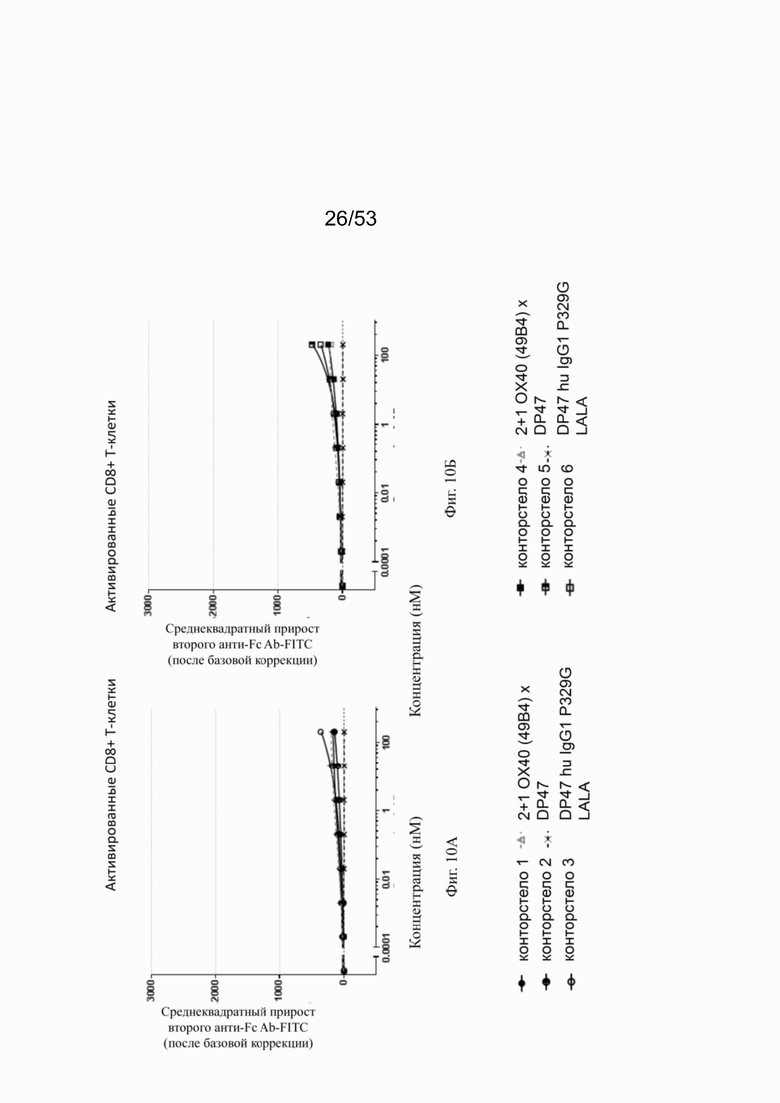

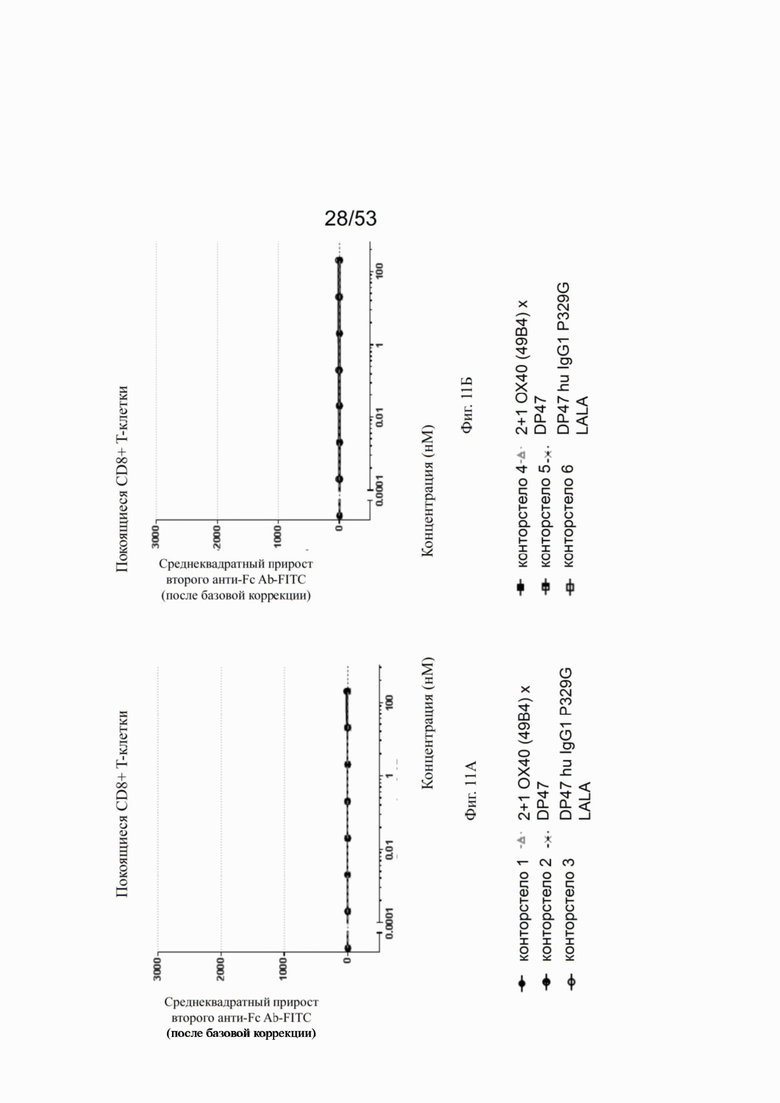
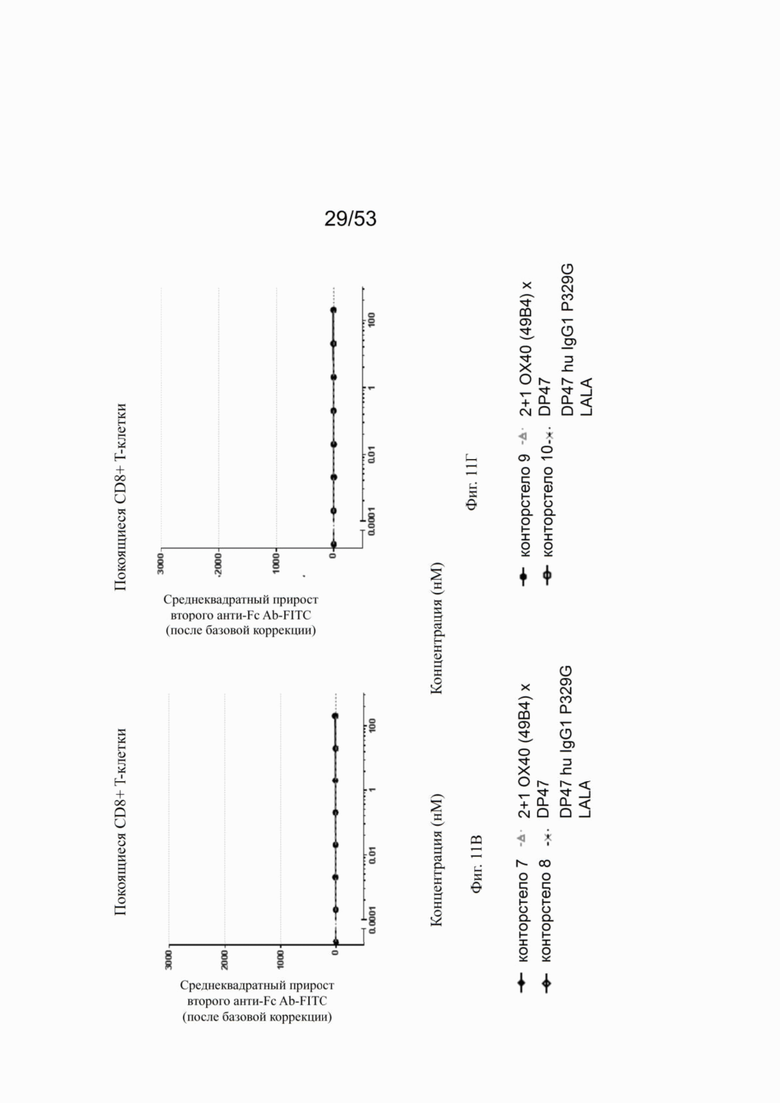
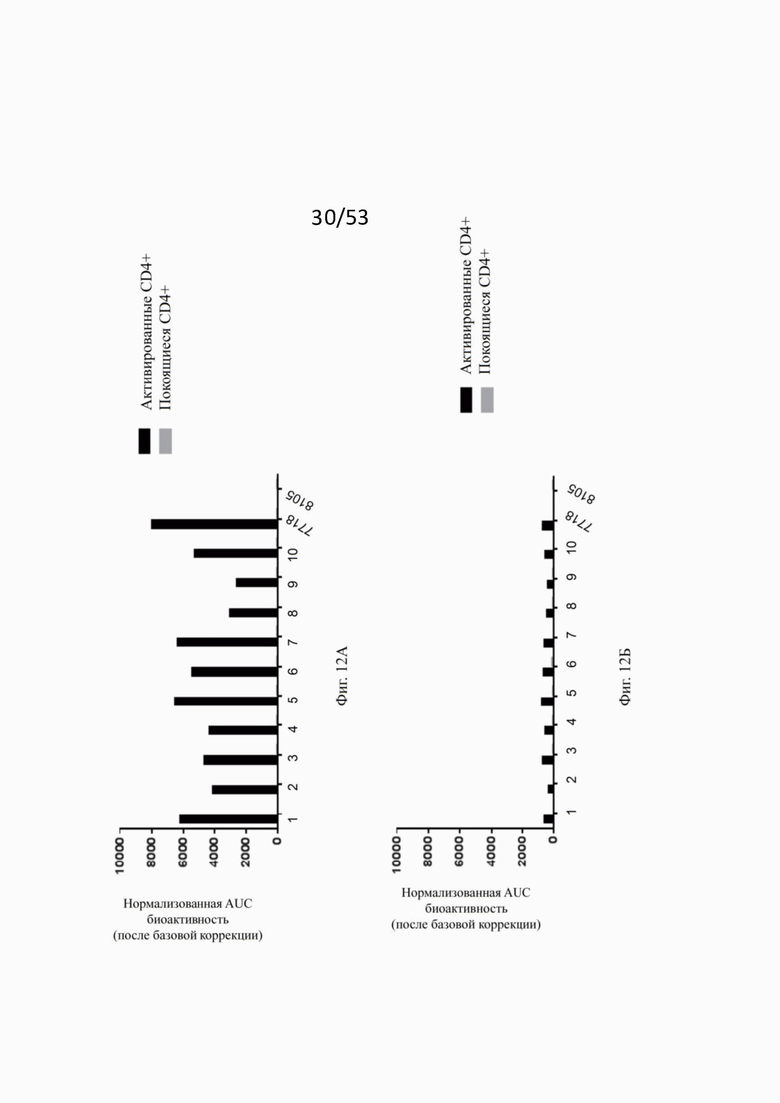
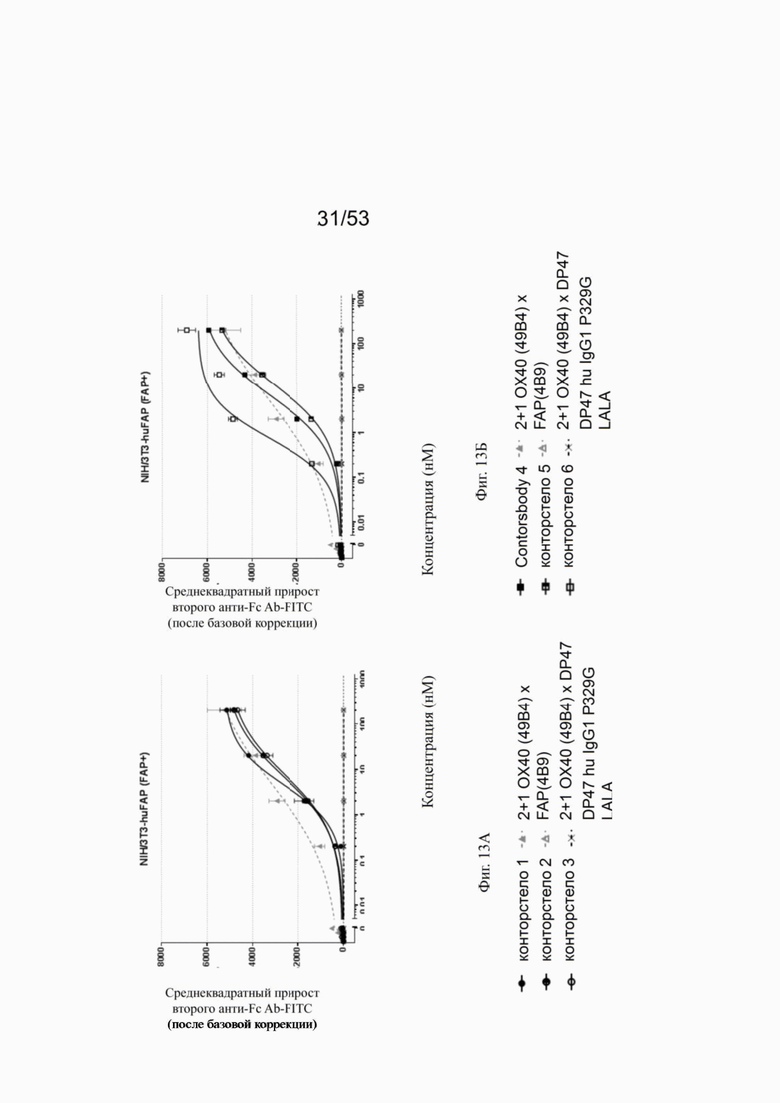
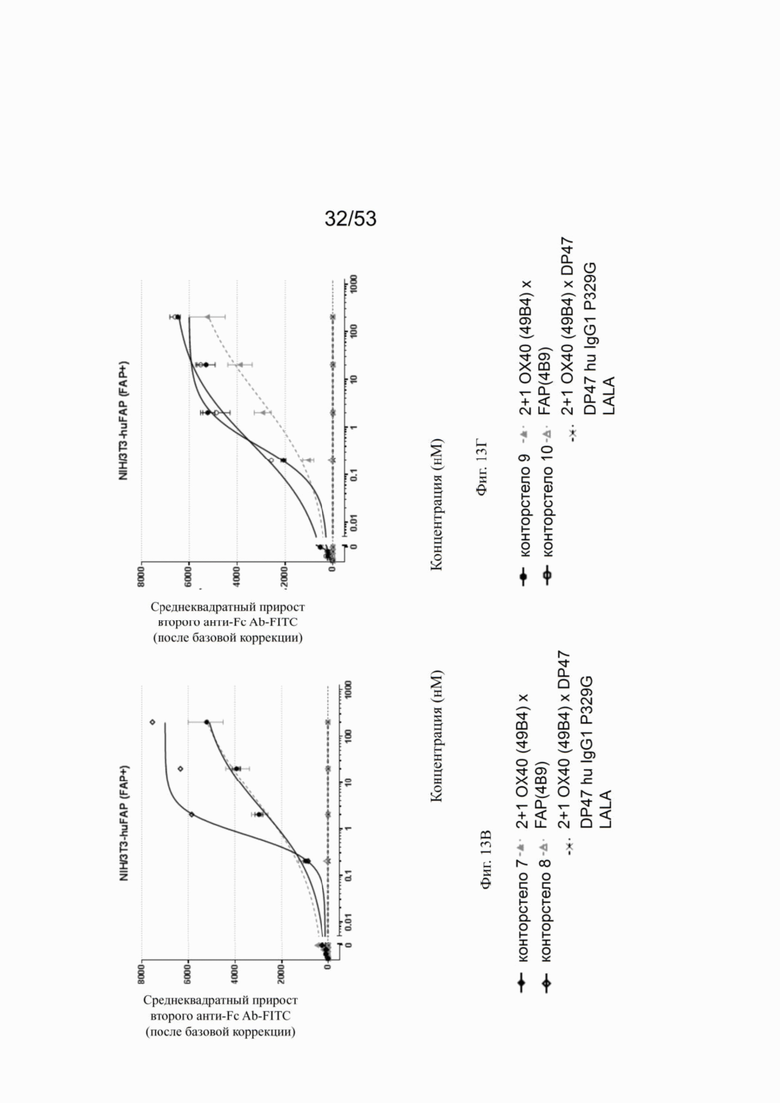

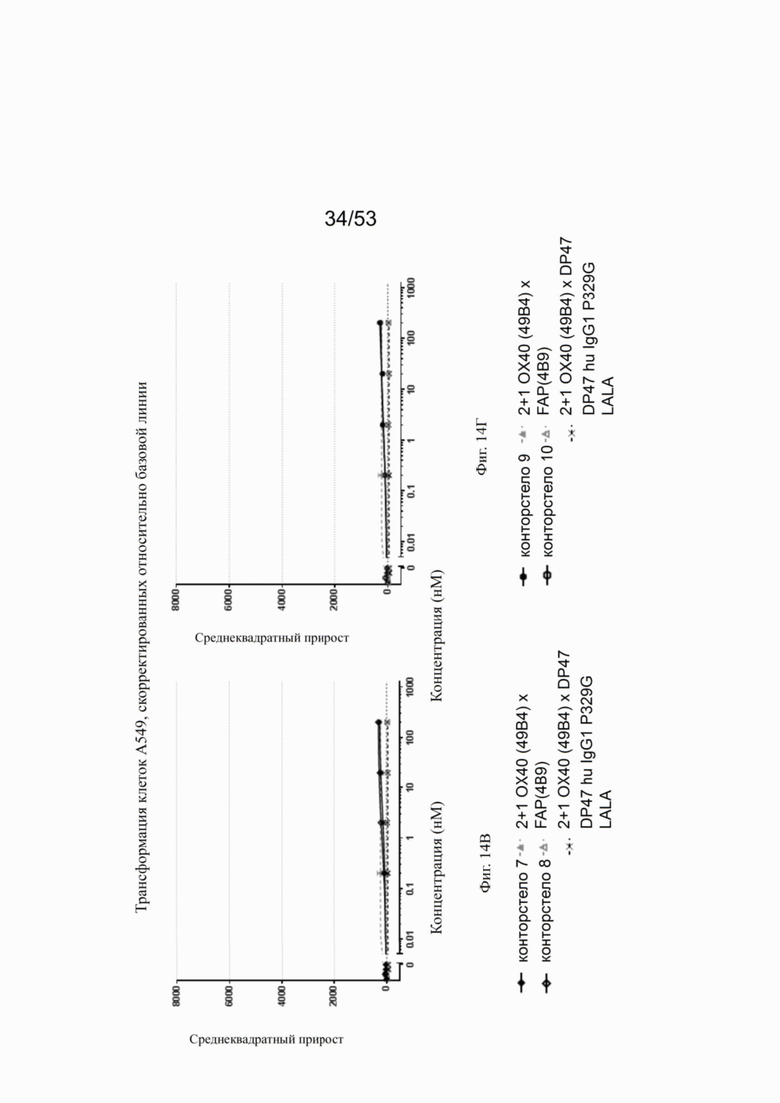
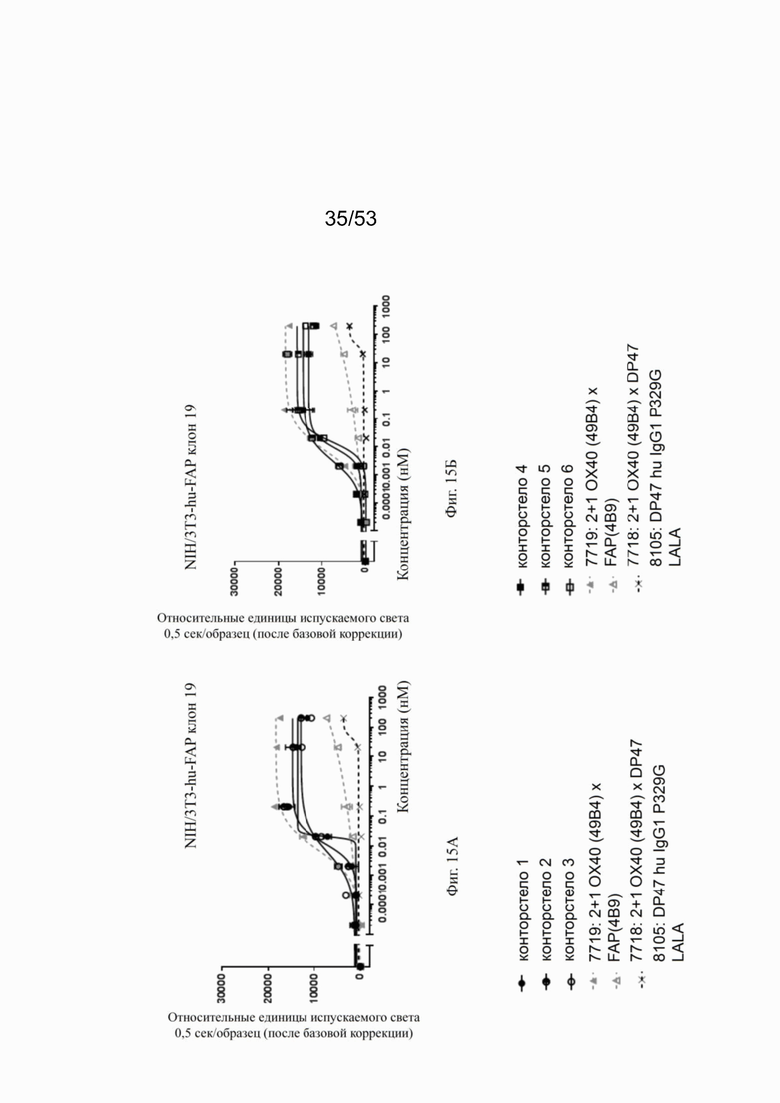
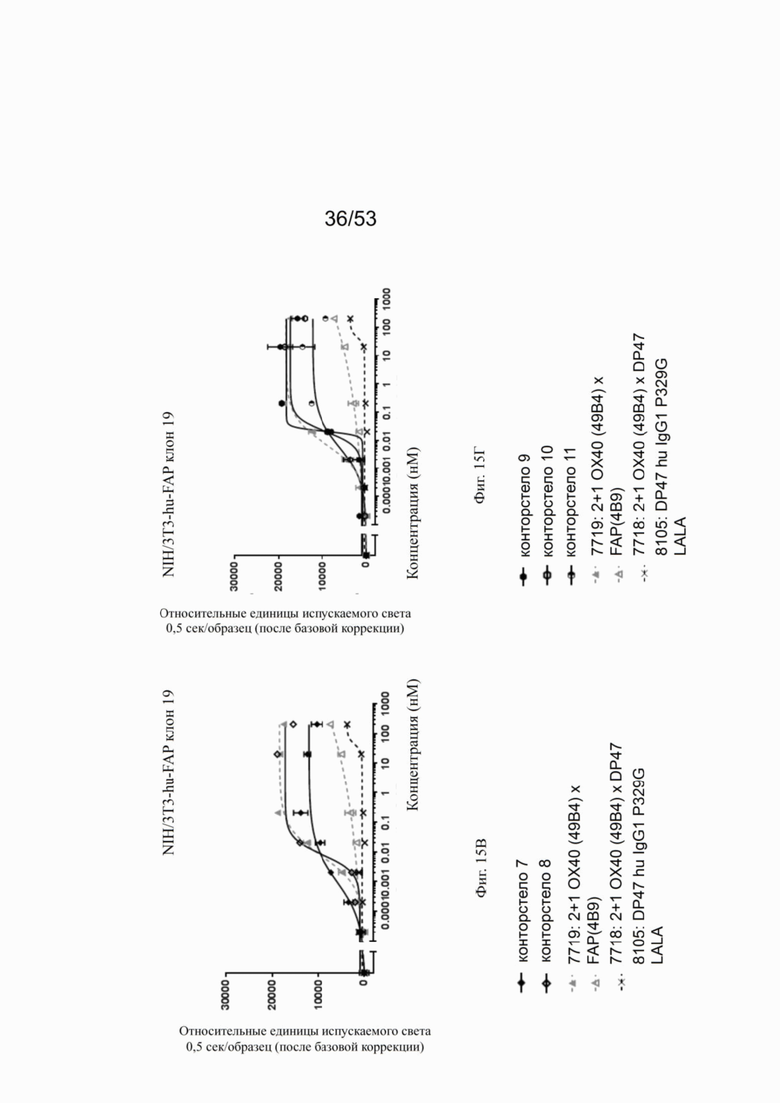
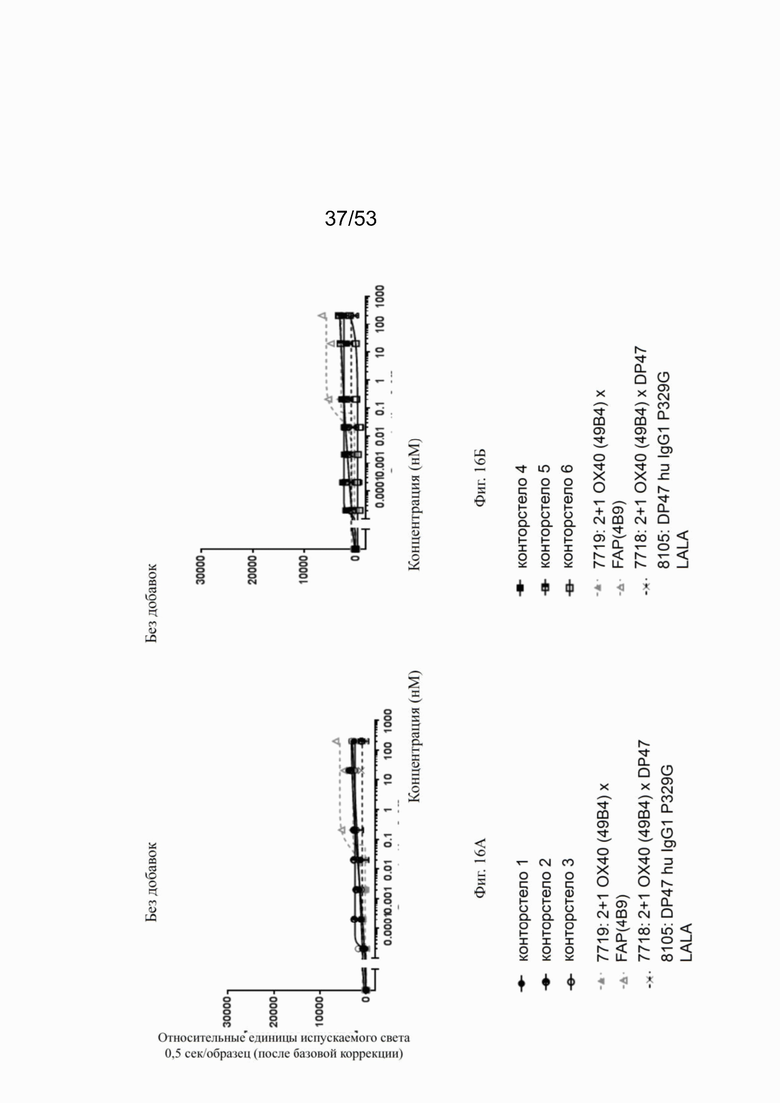
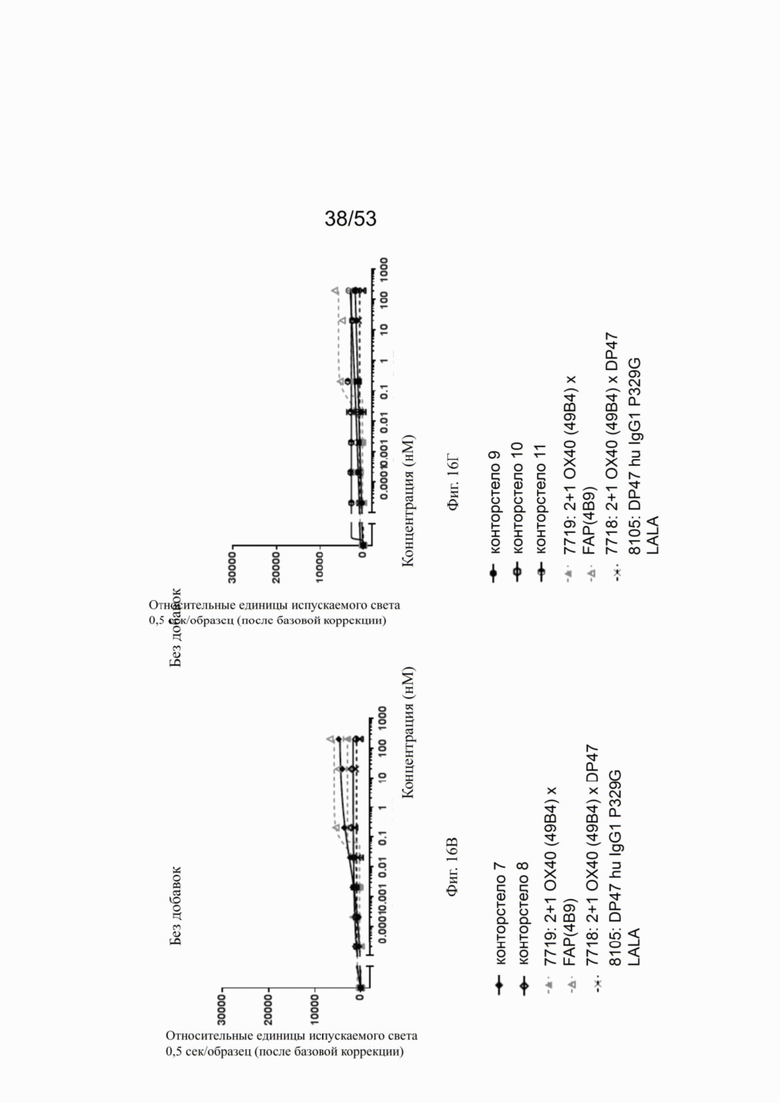

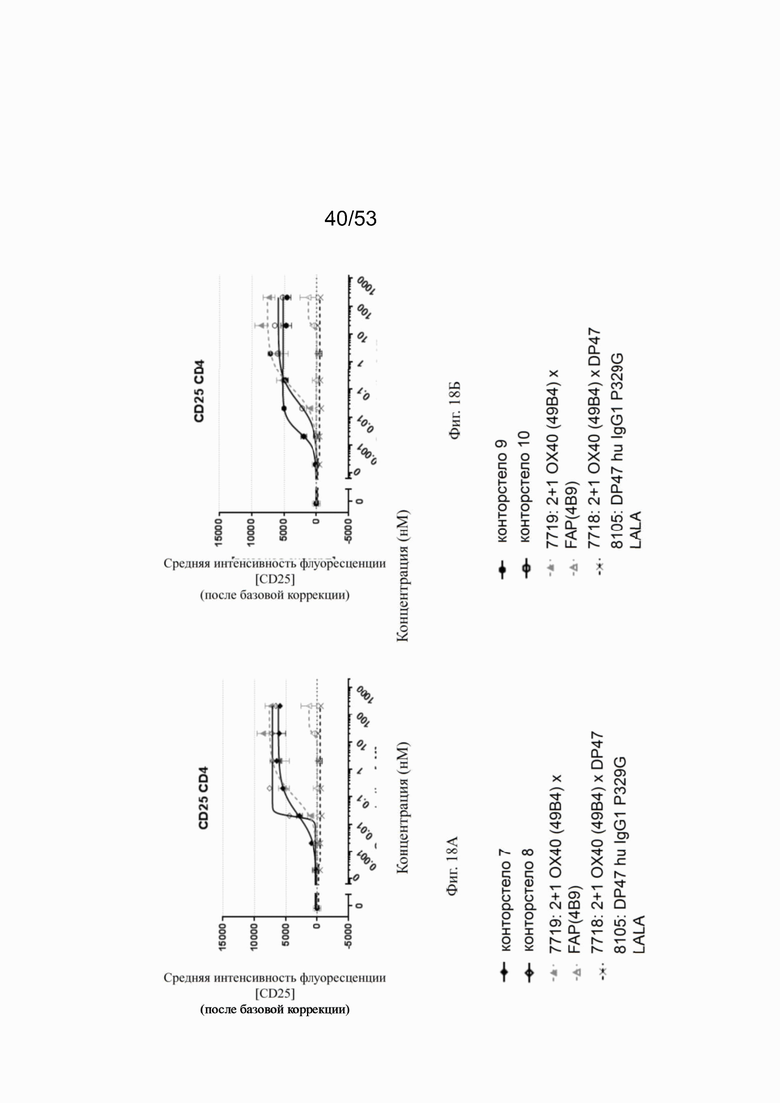
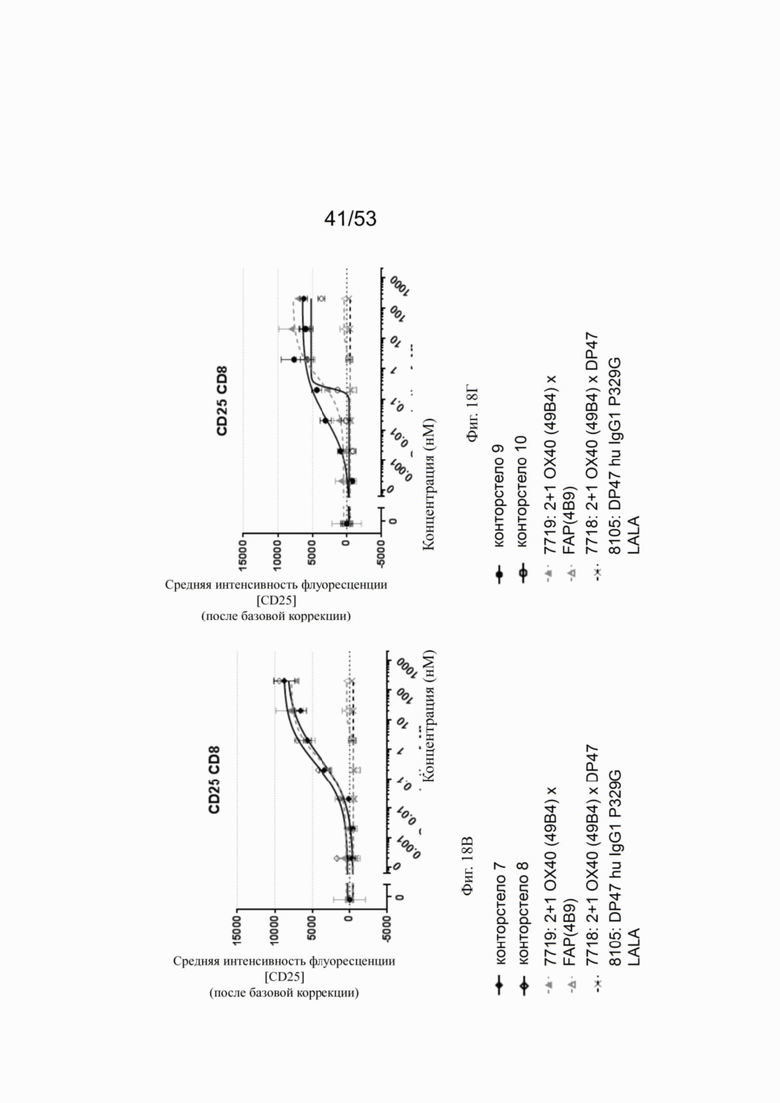
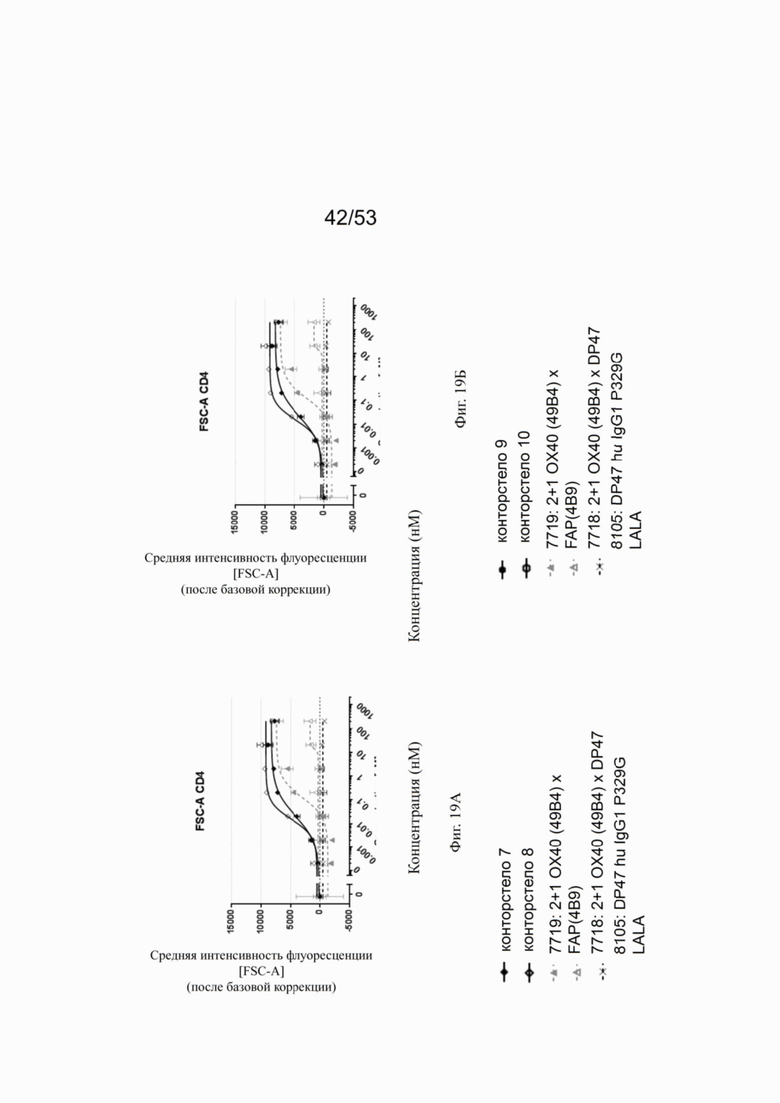
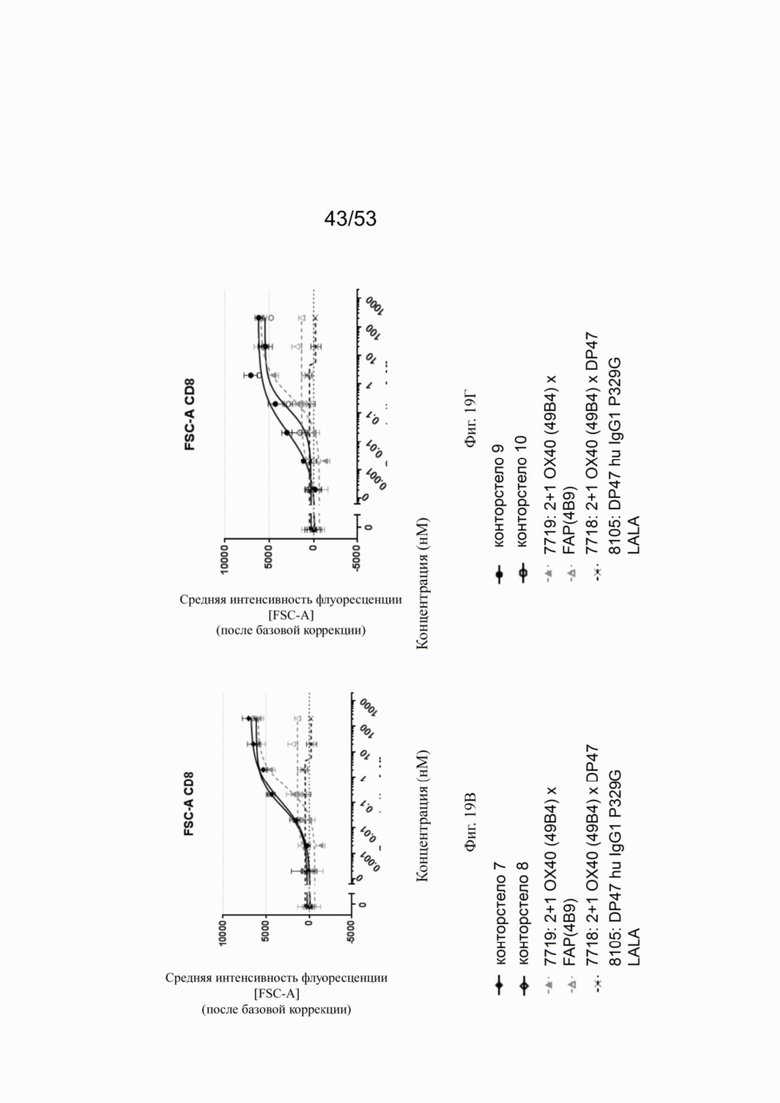
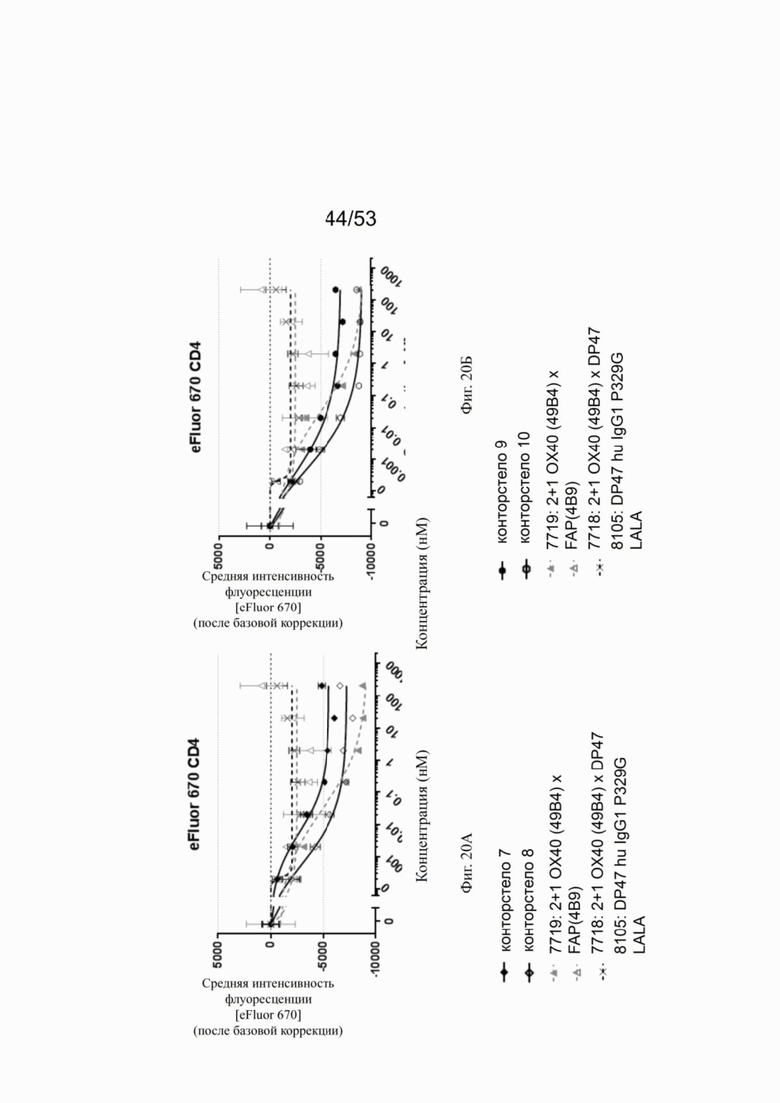
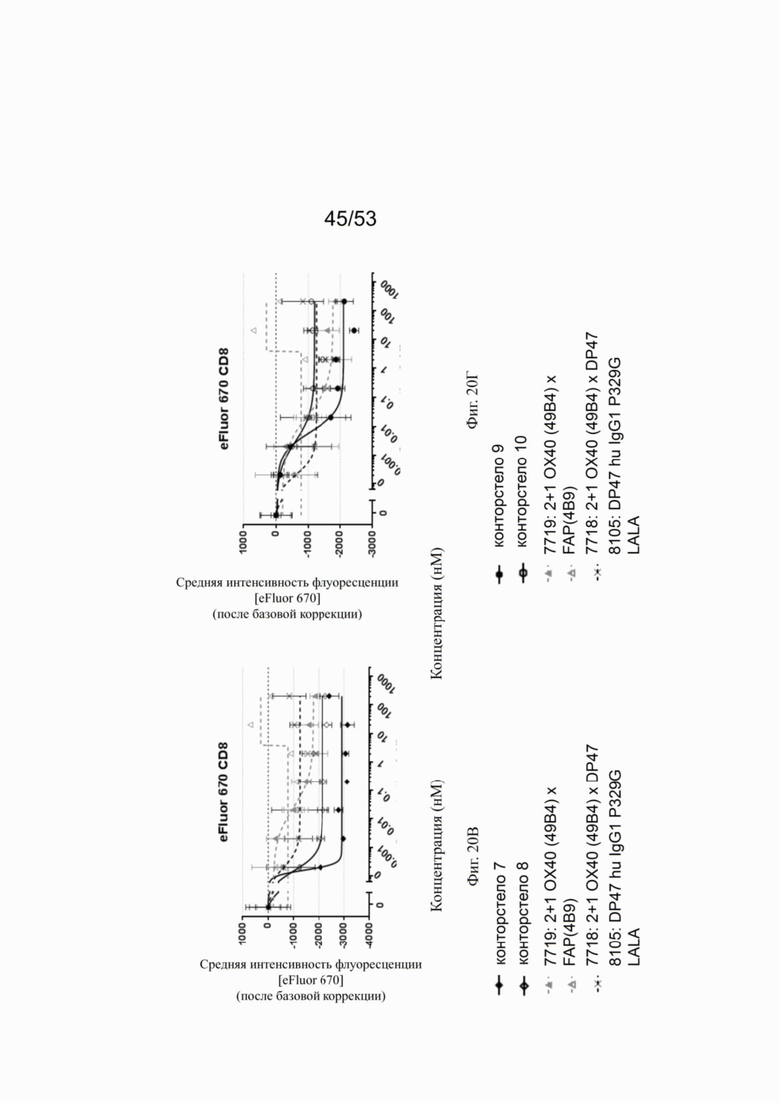
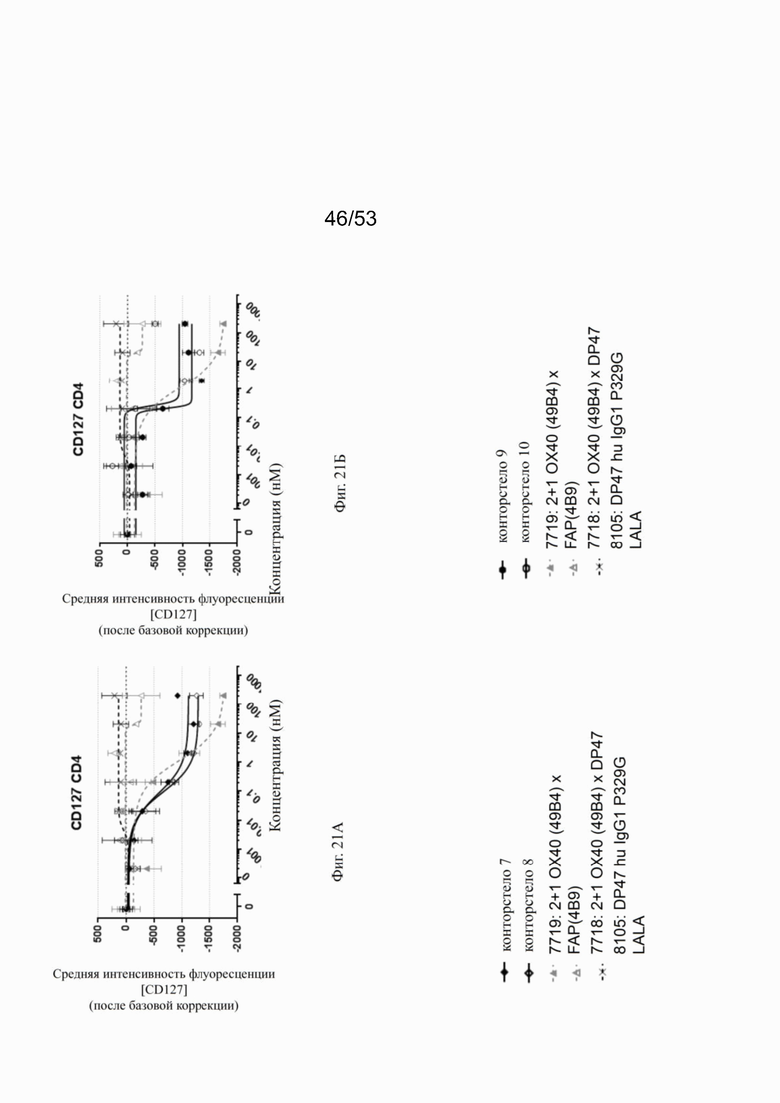
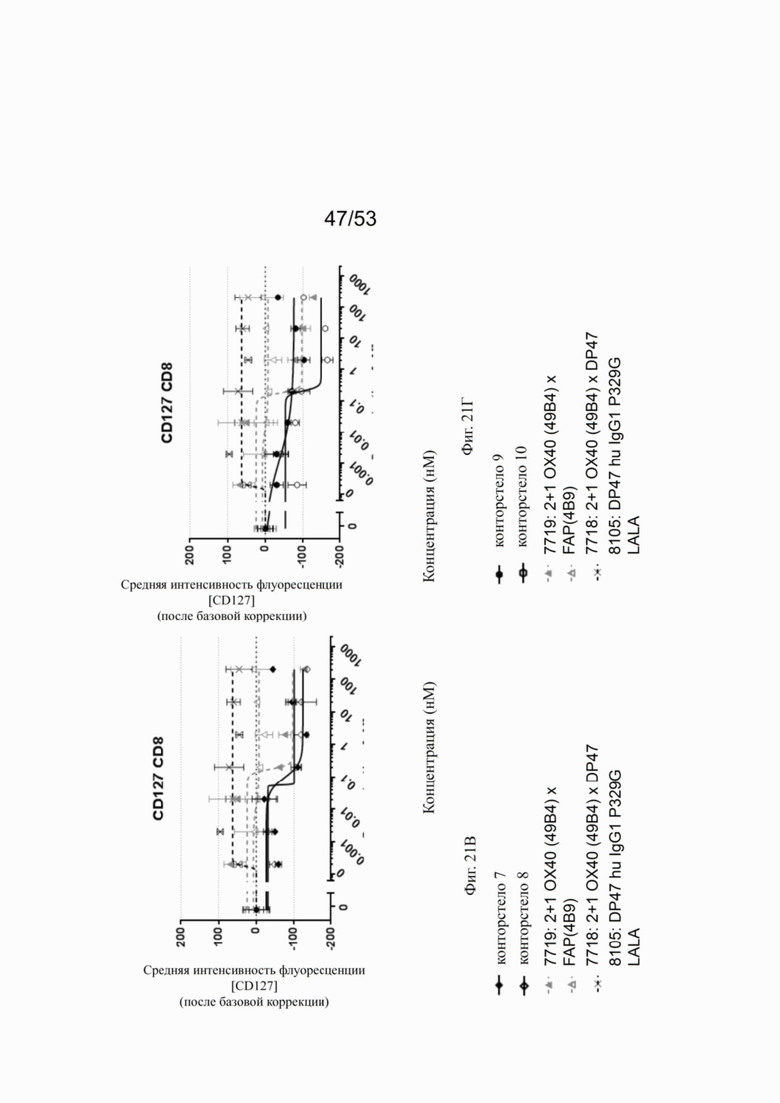
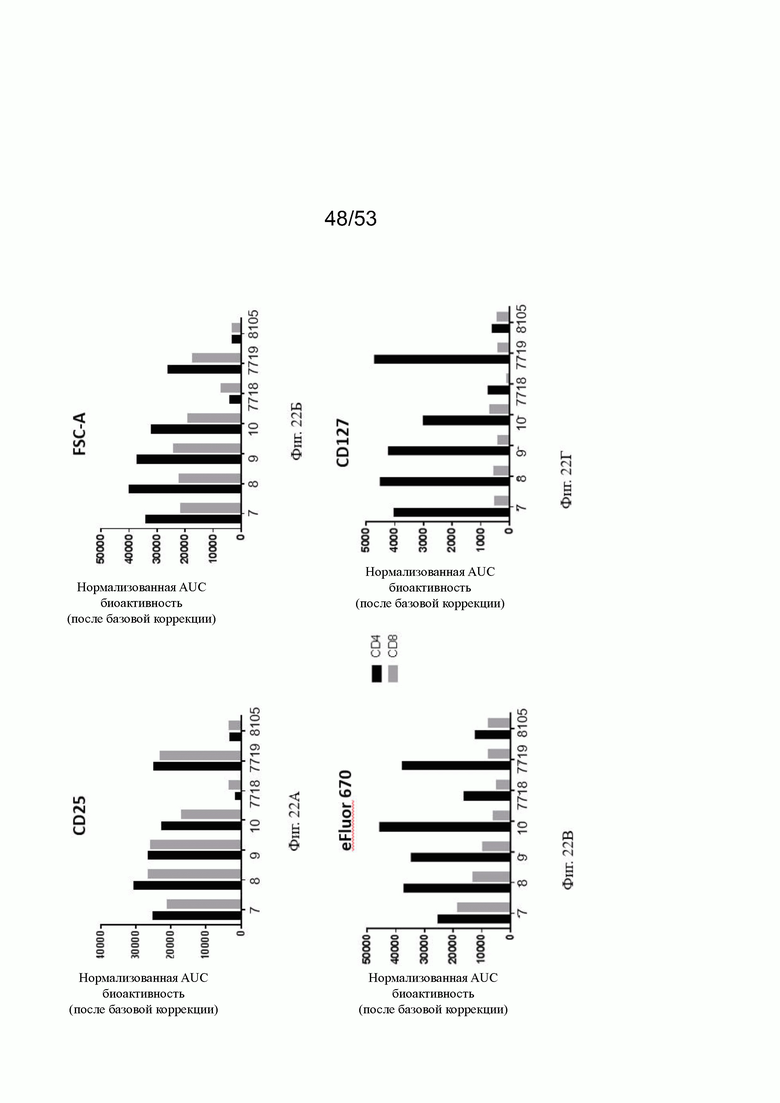
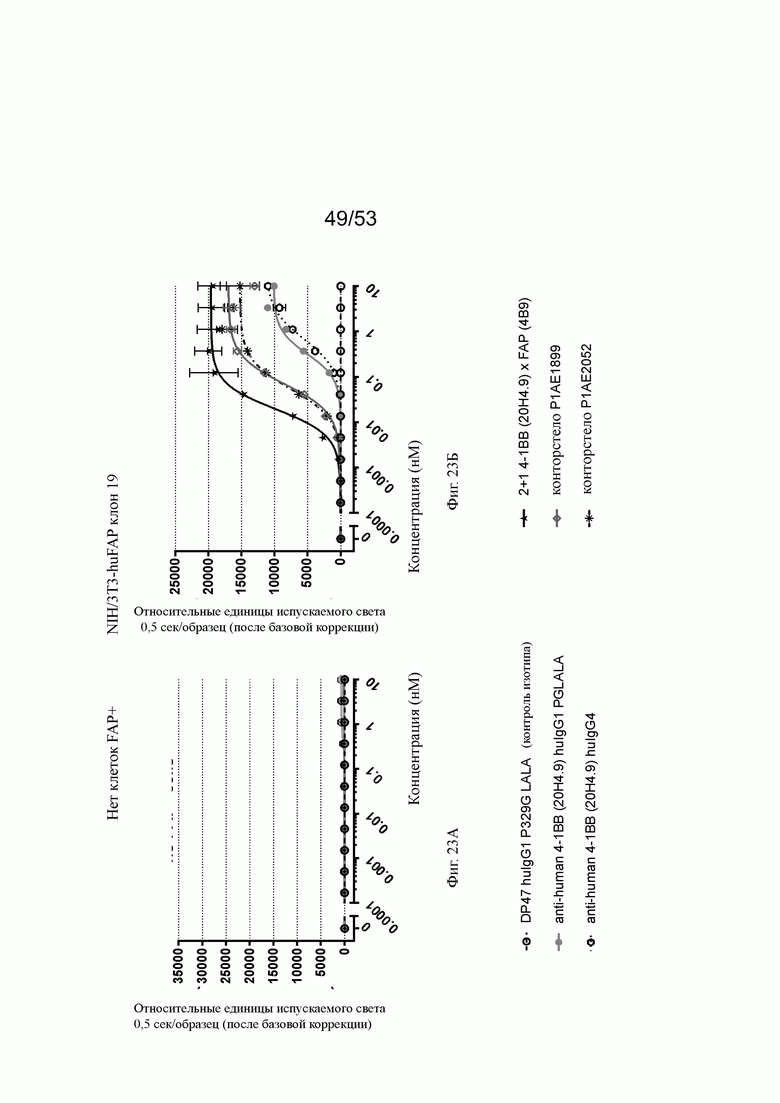
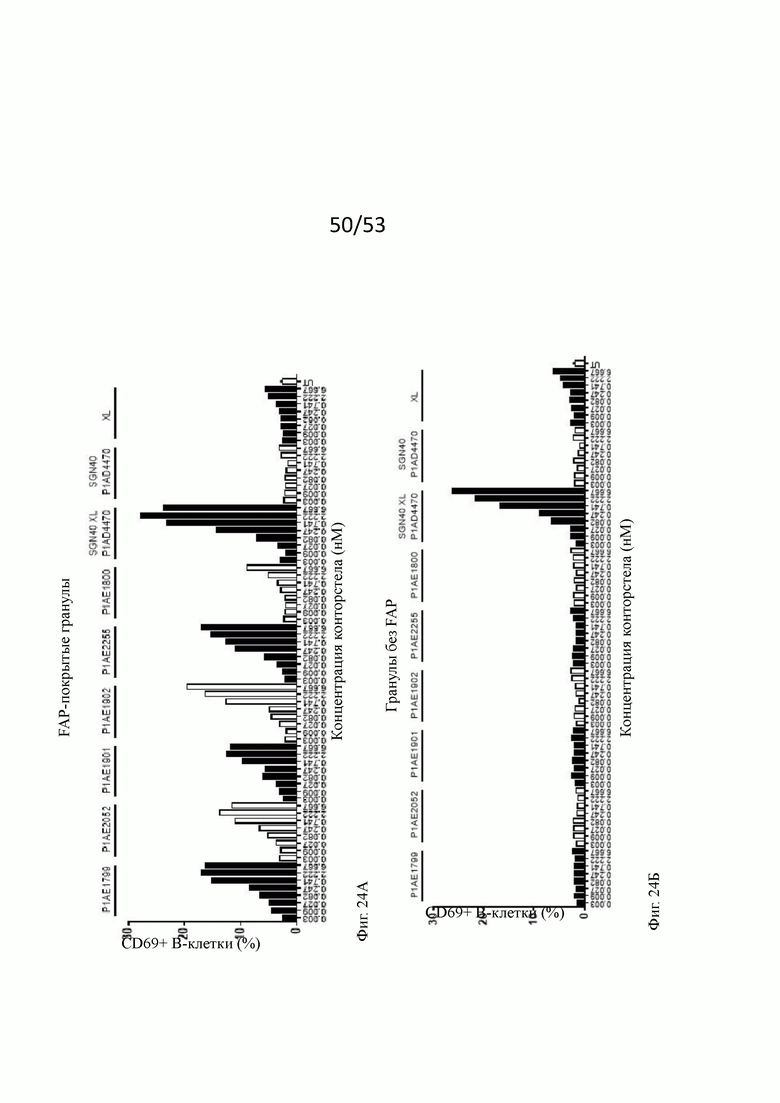
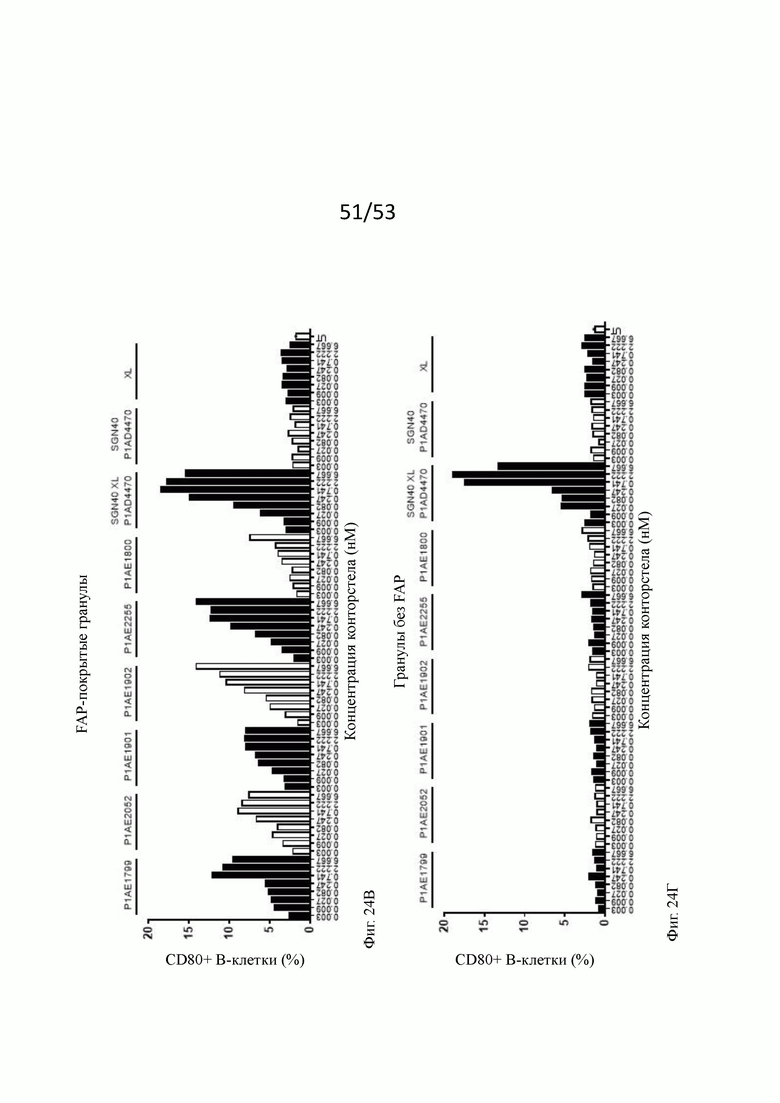
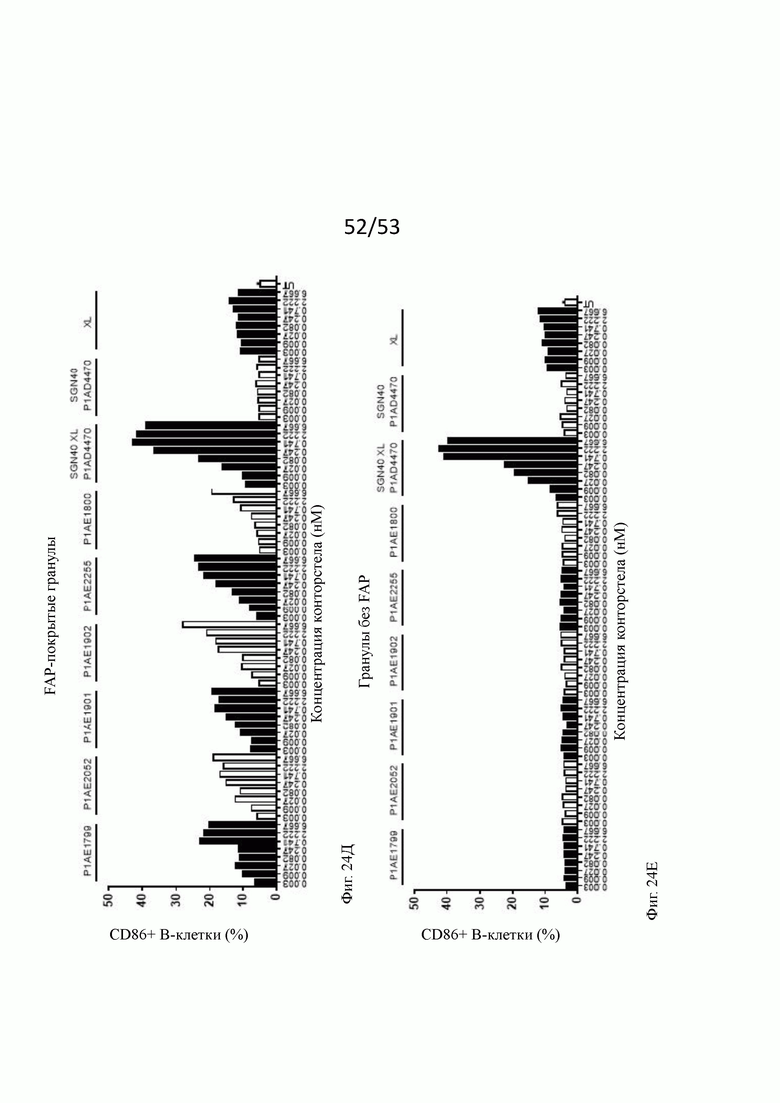
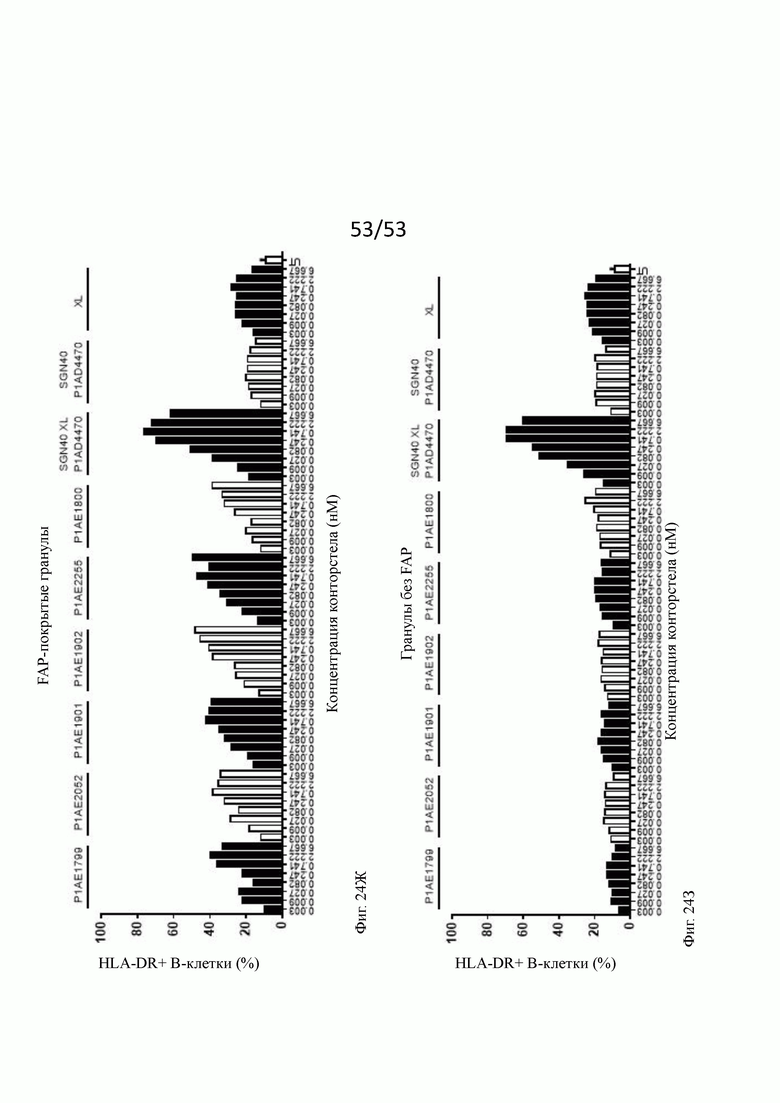
Изобретение относится к области биотехнологии. Предложено биспецифическое антитело, которое специфически связывается с белком активации фибробластов (FAP) и с OX40. Также изобретение относится к вектору экспрессии, кодирующему указанное антитело, клетке-хозяину, способу получения антитела, фармацевтической композиции. Биспецифические антитела по изобретению могут применяться для лечения инфекций и/или для лечения рака. 6 н. и 5 з.п. ф-лы, 110 ил., 42 табл., 10 пр.
1. Биспецифическое антитело, которое специфически связывается с белком активации фибробластов (FAP) и с OX40, состоящее из двух слитых полипептидов и содержащее два антигенсвязывающих домена, способных к специфическому связыванию с OX40, и один антигенсвязывающий домен, способный к специфическому связыванию с белком активации фибробластов(FAP), включающее
(a) первый слитый полипептид, содержащий первую часть первого антигенсвязывающего домена, способного к специфическому связыванию с OX40, спейсерный домен, вторую часть первого антигенсвязывающего домена, способного к специфическому связыванию с OX40, и первую часть антигенсвязывающего домена, способного к специфическому связыванию с FAP, в котором
- спейсерный домен содержит шарнирную область антитела или ее фрагмент, домен CH2 антитела и домен CH3 антитела или его фрагмент,
- первая часть первого антигенсвязывающего домена, способного специфически связываться с OX40, слита либо непосредственно, либо через первый пептидный линкер с N-концом спейсерного домена,
- вторая часть первого антигенсвязывающего домена, способного к специфическому связыванию с OX40, слита либо непосредственно, либо через второй пептидный линкер с C-концом спейсерного домена, и
- первая часть антигенсвязывающего домена, способного специфически связываться с FAP, слита либо непосредственно, либо через третий пептидный линкер с C-концом второй части первого антигенсвязывающего домена, способного специфически связываться с OX40, или слита либо непосредственно, либо через третий пептидный линкер, с N-концом первой части первого антигенсвязывающего домена, способного специфически связываться с OX40, и
(б) второй слитый полипептид, содержащий первую часть второго антигенсвязывающего домена, способного специфически связываться с OX40, спейсерный домен, вторую часть второго антигенсвязывающего домена, способного специфически связываться с OX40, и вторую часть антигенсвязывающего домена, способного специфически связываться с FAP, в котором
- спейсерный домен содержит шарнирную область антитела или ее фрагмент, домен CH2 антитела и домен CH3 антитела или его фрагмент,
- первая часть второго антигенсвязывающего домена, способного специфически связываться с OX40, слита либо непосредственно, либо через первый пептидный линкер с N-концом спейсерного домена,
- вторая часть второго антигенсвязывающего домена, способного к специфическому связыванию с OX40, слита либо непосредственно, либо через второй пептидный линкер с C-концом спейсерного домена, и
- вторая часть антигенсвязывающего домена, способная специфически связываться с FAP, слита либо непосредственно, либо через третий пептидный линкер, с С-концом второй части второго антигенсвязывающего домена, способного специфически связываться с OX40, или слита либо непосредственно, либо через третий пептидный линкер с N-концом первой части второго антигенсвязывающего домена, способного специфически связываться с OX40,
и где как в первом слитом полипептиде, так и во втором слитом полипептиде первая часть антигенсвязывающего домена, способного к специфическому связыванию с OX40, представляет собой фрагмент Fab тяжелой цепи антитела, содержащий VH-CH1 или VH-Ckappa, а вторая часть антигенсвязывающего домена, способного к специфическому связыванию с OX40, представляет собой фрагмент Fab легкой цепи антитела, содержащий VL-Ckappa или VL-CH1 соответственно,
и где первая часть антигенсвязывающего домена, способного к специфическому связыванию с FAP, содержит вариабельный домен тяжелой цепи антитела (VH), а вторая часть антигенсвязывающего домена, способного к специфическому связыванию с FAP, содержит вариабельный домен легкой цепи антитела (VL), и первая часть и вторая части антигенсвязывающего домена, способного к специфическому связыванию с FAP, соединены друг с другом, образуя антигенсвязывающий домен, способный специфически связываться с FAP, причем первая и вторая части первого и второго антигенсвязывающих доменов, способных специфически связываться с OX40, соединены друг с другом с образованием кольцевого слитого полипептида, и
где спейсерный домен первого слитого полипептида и спейсерный домен второго слитого полипептида ковалентно соединены друг с другом дисульфидной связью и содержат модификации, способствующие соединению первого и второго слитых полипептидов,
и при этом антигенсвязывающий домен, способный к специфическому связыванию с FAP, содержит вариабельную область тяжелой цепи (VHFAP), содержащую аминокислотную последовательность SEQ ID NO:7, и вариабельную область легкой цепи (VLFAP), содержащую аминокислотную последовательность SEQ ID NO:8,
и при этом антигенсвязывающий домен, способный к специфическому связыванию с OX40, содержит вариабельную область тяжелой цепи (VHOX40), содержащую аминокислотную последовательность SEQ ID NO:40, и вариабельную область легкой цепи (VLOX40), содержащую аминокислотную последовательность SEQ ID NO:41.
2. Биспецифическое антитело по п. 1, в котором в первом слитом полипептиде первая часть антигенсвязывающего домена, способного специфически связываться с FAP, слита или непосредственно, или через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного специфически связываться с OX40, и в котором во втором слитом полипептиде вторая часть антигенсвязывающего домена, способного специфически связываться с FAP, слита или непосредственно, или через третий пептидный линкер с С-концом второй части первого антигенсвязывающего домена, способного специфически связываться с OX40.
3. Биспецифическое антитело по п. 1 или 2, в котором третий пептидный линкер, соединяющий первую часть или вторую часть антигенсвязывающего домена, способного специфически связываться с FAP, содержит по меньшей мере 15 аминокислот.
4. Биспецифическое антитело по любому из пп. 1-3, в котором спейсерный домен, содержащий шарнирную область антитела или ее фрагмент, и Fc домен IgG1.
5. Биспецифическое антитело по любому из пп. 1-4, в котором спейсерный домен первого слитого полипептида содержит впадины, а спейсерный домен второго слитого полипептида содержит выступы согласно методу выступ-во-впадину.
6. Биспецифическое антитело по любому из пп. 1-5, в котором домен FcIgG1 содержит аминокислотные замены L234A, L235A и P329G (нумерация по Kabat, EU).
7. Вектор экспрессии, который содержит нуклеиновую кислоту, кодирующую биспецифическое антитело по любому из пп. 1-6.
8. Клетка-хозяин для получения биспецифического антитела по любому из пп. 1-6, содержащая вектор по п. 7.
9. Способ получения биспецифического антитела, включающий стадию (i) культивирования клетки-хозяина по п. 8 в условиях, пригодных для экспрессии биспецифического антитела и выделения указанного биспецифического антитела из клетки-хозяина или среды для культивирования клеток-хозяев.
10. Фармацевтическая композиция для лечения инфекций и/или рака, содержащая биспецифическое антитело по любому из пп. 1-6 и фармацевтически приемлемый эксципиент.
11. Применение биспецифического антитела по любому из пп. 1-6 или фармацевтической композиции по п. 10 для получения лекарственного средства для лечения инфекций и/или лечения рака.
| WO 2017180913 A2, 19.10.2017 | |||
| WO 2014116846 A2, 31.07.2014 | |||
| WO 2017060144 A1, 13.04.2017 | |||
| WO 2012020006 A2, 16.02.2012 | |||
| HORNIG N | |||
| et al | |||
| Evaluating combinations of costimulatory antibody-ligand fusion proteins for targeted cancer immunotherapy, Cancer Immunol Immunother, vol.62(8), pp.1369-1380; | |||
| АНТИТЕЛА ПРОТИВ ОХ40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2562874C1 |
