Родственная заявка
[0001] Это заявка заявляет интересы Предварительной Заявки США № 61/374159, зарегистрированной 16 августа 2010, Предварительной Заявки США № 61/443008, зарегистрированной 15 февраля 2011, Предварительной Заявки США № 61/509260, зарегистрированной 19 июля 2011, содержание которых включено в данный документ ссылкой в полном объеме.
Область техники, к которой относится изобретение
[0002] Изобретение относится к получению новых биспецифичных моноклональных антител, имеющих различную специфичность для каждого сайта связывания молекулы иммуноглобулина. Антитела по изобретению состоят из одной тяжелой цепи и двух различных легких цепей, одна из которых содержит константный домен Каппа, а другая - константный домен Лямбда. Изобретение в частности относится к выделению антител различной специфичности, но имеющих общую тяжелую цепь. Изобретение дополнительно относится к контролируемой ко-экспрессии двух легких цепей и одной тяжелой цепи, что приводит к сборке моноспецифичных и биспецифичных антител. В изобретении предлагается способ получения полностью человеческого биспецифичного и двухвалентного антитела, который не изменяет последовательность и не включает применения линкеров или других последовательностей с происхождением, отличным от человеческого, а также предлагается способ получения смеси антител из двух моноспецифических антител и одного биспецифичного антитела. В изобретении также предложены способы эффективной очистки биспецифичного антитела.
Предшествующий уровень техники
[0003] Антитело состоит из четырех полипептидов: две тяжелые цепи и две легкие цепи. Антиген-связывающий участок антитела образуется с помощью вариабельного домена легкой цепи (VL) и вариабельного домена тяжелой цепи (VH). На одном конце этих доменов шесть петель образуют антиген-связывающий сайт, также обозначаемый как гипервариабельный участок (CDR). Три CDR локализованы на VH-домене (H1, H2 и H3) и три других располагаются на VL-домене (LI, L2 и L3). Во время развития В-клетки уникальный участок иммуноглобулина образуется с помощью соматической рекомбинации, известной как V(D)J-рекомбинация. Вариабельный участок тяжелой или легкой цепи иммуноглобулина кодируется различными генными сегментами. Тяжелая цепь кодируется тремя сегментами, называемыми вариабельным сегментом (V), D-сегментом (определяющим разнообразие) (D) и соединительным (J) сегментом, тогда как вариабельность легкой цепи формируется с помощью рекомбинации только двух сегментов, V и J. Большое количество паратопов антител может быть получено с помощью рекомбинации между одной из множества копий V, D и J-сегментов, которые присутствуют в геноме. V-сегмент кодирует CDR1 и CDR2, тогда как CDR3 получается в процессе рекомбинационных событий. В процессе иммунного ответа дополнительное разнообразие вводится в антиген-связывающий сайт с помощью процесса, называемого соматической сверхмутацией (SHM). Во время этого процесса точечные мутации вводятся в вариабельные гены тяжелой и легкой цепей и, конкретно, в участки, кодирующие CDR. Эта дополнительная вариабельность дает возможность селекции и размножения В-клеток, экспрессирующих варианты антител с повышенной аффинностью к распознаваемому ими антигену.
[0004] Огромное число иммуноглобулинов представляет собой двухвалентные и моноспецифичные молекулы, несущие одинаковую специфичность на обоих плечах, поскольку они состоят из двух идентичных полипептидов тяжелой цепи и двух идентичных полипептидов легкой цепи. Однако практически сразу еще в процессе развития гибридомной технологии было выяснено, что гибридные гибридомы могут быть созданы с помощью слияния двух гибридом (Suresh MR et al., Methods Enzymol 1986; 121: 210- 228). Эти 'квадромы (quadromas)' экспрессируют две различные тяжелые и две различные легкие цепи и, таким образом, вырабатывают множество различных видов антител, получаемых в результате случайного спаривания тяжелой и легкой цепей. Среди этих различных видов получаются биспецифичные антитела (bsAb), несущие различные специфичности на каждом плече. Другое существующее в природе исключение представляет собой иммуноглобулин IgG4-изотипа, который способен подвергаться обмену тяжелых цепей благодаря менее стабильной димеризации, опосредованной шарнирным участком этого изотипа (van der Neut Kolfschoten M et al., Science. 2007 317(5844): 1554-7). Хотя этот обмен, по-видимому, происходит in vivo, его биологическое значение остается невыясненным.
[0005] Моноклональные антитела появились как успешный и привлекательный класс молекул для терапевтического введения, применяемых при некоторых заболеваниях человека. Однако направленного воздействия или нейтрализации одного белка не всегда достаточно для достижения эффективности при некоторых заболеваниях, которые ограничивают терапевтическое применение моноклональных антител. Еще более очевидно, что в ряде случаев нейтрализации одного компонента биологической системы недостаточно для достижения эффективности. Одно из решений проблемы это совместное введение нескольких моноклональных антител. Однако этот способ затрудняется регуляторными аспектами, если объединяемые антитела не были ранее улучшены в индивидуальном порядке. Кроме того, комбинаторные способы также дороги с точки зрения производственной перспективы. Соответственно, существует необходимость создания антител и лекарственных средств, которые дают возможность направленного воздействия на множество антигенов с помощью одной молекулы.
Сущность изобретения
[0006] Изобретение обеспечивает возможность идентифицировать, получать и очищать биспецифичные антитела, которые не отличимы по последовательности от стандартных антител. Изобретение также обеспечивает возможность получения и очистки простой смеси антител, состоящей из трех и более антител, несущих одинаковые тяжелые цепи. Немодифицированный характер антител по изобретению обеспечивает их предпочтительными производственными характеристиками, аналогичными как для стандартных моноклональных антител.
[0007] Биспецифичные антитела по изобретению получают с использованием следующих стадий:
- выделяли два антитела с различной специфичностью и общим одинаковым вариабельным доменом тяжелой цепи, но с различными вариабельными доменами легких цепей. Эту стадию облегчали использованием библиотек антител, содержащих фиксированные тяжелые цепи, или использованием трансгенных животных, содержащих одинаковые VH-гены.
- вариабельный домен тяжелой цепи сшивали с константным участком тяжелой цепи, вариабельный домен одной легкой цепи сшивали с константным доменом Каппа, и вариабельный домен другой легкой цепи сшивали с константным доменом Лямбда. Предпочтительно, вариабельный домен легкой цепи, сшитый с константным доменом цепи Каппа, относится к Каппа-типу, а вариабельный домен, сшитый с константным доменом Лямбда, относится к Лямбда-типу. Однако изобретение также дает возможность получения гибридных легких цепей так, чтобы два вариабельных домена легких цепей одного типа можно было использовать для получения биспецифичных антител по изобретению.
- три цепи ко-экспрессировали в клетках млекопитающего, что приводило к сборке и секреции в надосадочную жидкость смеси из трех антител: двух моноспецифичных и одного биспецифичного антитела, несущего две различные легкие цепи. Соотношение различных антител зависит от относительной экспрессии цепей и их сборки в IgG. В изобретении предлагаются способы регуляции этих соотношений и максимизации получения биспецифичного антитела.
- смесь антител очищали, используя стандартные хроматографические методы, используемые для очистки антител. Смесь антител может быть охарактеризована и использована в качестве агента с множественным направленным воздействием.
- биспецифичное антитело очищали с использованием ступенчатого аффинного хроматографического материала, который специфично связывается с константными участками Каппа человека и Лямбда человека. Этот процесс очистки не зависит от последовательности вариабельных доменов легких цепей и, таким образом, является характерным для всех биспецифичных антител по изобретению.
- выделенное биспецифичное антитело, несущее легкую цепь, содержащую константный домен Каппа, и легкую цепь, содержащую константный домен Лямбда, охарактеризовывали с использованием различных биохимических и иммунологических методов.
- биспецифичное антитело по изобретению может использоваться для терапевтического воздействия или в качестве исследовательского или диагностического реагента.
[0008] Изобретение относится к моноклональному антителу, имеющему различную специфичность в каждом объединенном сайте и включающее две копии полипептида одной тяжелой цепи и первую легкую цепь и вторую легкую цепь, где первая и вторая легкие цепи различны.
[0009] В некоторых антителах, по меньшей мере, первая часть первой легкой цепи относится к Каппа-типу и, по меньшей мере, часть второй легкой цепи относится к Лямбда-типу. В некоторых антителах, первая легкая цепь включает, по меньшей мере, константный участок Каппа. В некоторых антителах, первая легкая цепь дополнительно включает вариабельный участок Каппа. В некоторых антителах, первая легкая цепь дополнительно включает вариабельный участок Лямбда. В некоторых антителах, вторая легкая цепь включает, по меньшей мере, константный участок Лямбда. В некоторых антителах, вторая легкая цепь дополнительно включает вариабельный участок Лямбда. В некоторых антителах, вторая легкая цепь дополнительно включает вариабельный участок Каппа. В некоторых антителах, первая легкая цепь включает константный участок Каппа и вариабельный участок Каппа, а вторая легкая цепь включает константный участок Лямбда и вариабельный участок Лямбда.
[0010] В некоторых вариантах осуществления последовательности константного и вариабельного каркасного участка являются человеческими.
[0011] Изобретение относится к способам получения биспецифичного антитела с помощью a) выделения антитела или участка фрагмента антитела, имеющего специфичность, определяемую вариабельным доменом тяжелой цепи вместе с вариабельным доменом первой легкой цепи; b) выделения антитела или участка фрагмента антитела, имеющего другую специфичность, определяемую тем же вариабельным доменом тяжелой цепи как у антитела стадии a) вместе с вариабельным доменом второй легкой цепи; c) ко-экспрессии в клетке: (i) вариабельного домена общей тяжелой цепи, сшитого с константным участком тяжелой цепи иммуноглобулина; (ii) вариабельного домена первой легкой цепи, сшитого или с константным доменом легкой цепи Каппа-типа, или с константным доменом легкой цепи Лямбда-типа; и (iii) вариабельного домена второй легкой цепи, сшитого с константным доменом легкой цепи другого типа, отличного от типа цепи первого вариабельного константного домена.
[0012] Некоторые способы также включают дополнительную стадию d) выделения биспецифичных антител, получаемых из полученных моноспецифичных антител. Например, в некоторых способах выделение выполняется с использованием стадии очистки с помощью аффинной хроматографии. В некоторых способах, стадию очистки осуществляют с использованием аффинного хроматографического материала, специфичного константному домену Каппа, к константному домену Лямбда, специфичному к ним обоим - константному домену Каппа и константному домену Лямбда.
[0013] В некоторых способах вариабельный домен легкой цепи Каппа сшит с константным участком Каппа-типа. В некоторых способах, вариабельный домен легкой цепи Каппа сшит с константным участком Лямбда-типа. В некоторых способах, вариабельный домен легкой цепи Лямбда сшит с константным участком Каппа-типа. В некоторых способах, вариабельный домен легкой цепи Лямбда сшит с константным участком Лямбда-типа.
[0014] В некоторых способах стадия a) и b) облегчаются применением библиотек антител, имеющих общую тяжелую цепь, и разнообразие ограничивается вариабельными доменами легкой цепи. В основе вариабельного домена тяжелой цепи, который имеется в одной из таких библиотек, могут быть различные вариабельные зародышевые гены, и они могут иметь различные последовательности в обоих участках, в CDR и каркасном участке. В некоторых способах, такие библиотеки конструируются с использованием различных типов вариабельных доменов тяжелой цепи и могут использоваться для получения антител по изобретению.
[0015] В некоторых способах библиотека антител представлена на нитевидном бактериофаге, на поверхности дрожжей, бактерий или клеток млекопитающих или используется для рибосом или другого типа in vitro дисплея.
[0016] Изобретение также относится к способам получения биспецифичного антитела, которое специфично связывается с первым антигеном и со вторым антигеном, где первый и второй антигены являются различными, с помощью a) предоставления первой молекулы нуклеиновой кислоты, кодирующей первый полипептид, включающий вариабельный участок тяжелой цепи полипептида иммуноглобулина или его фрагмент, который связывается с первым антигеном совместно с константным участком иммуноглобулина; b) предоставление второй молекулы нуклеиновой кислоты, кодирующей второй полипептид, включающий вариабельный участок легкой цепи полипептида иммуноглобулина или его фрагмент, который связывается с первым антигеном совместно с константным участком первой легкой цепи Каппа-типа или Лямбда-типа; c) предоставление третьей молекулы нуклеиновой кислоты, кодирующей третий полипептид, включающий вариабельный участок легкой цепи полипептида иммуноглобулина или его фрагмент, который связывается с вторым антигеном совместно с константным участком второй легкой цепи Каппа-типа или Лямбда-типа, где константные домены первой легкой цепи и второй легкой цепи относятся к различным типам; и (d) культивирование клетки-хозяина, включающей молекулы первой, второй и третьей нуклеиновых кислот при условиях, которые дают возможность экспрессии первого, второго и третьего полипептидов.
[0017] Некоторые способы также включают дополнительную стадию e) извлечения биспецифичного антитела. Например, в некоторых способах, биспецифичное антитело извлекают на стадии e) с использованием стадии очистки с помощью аффинной хроматографии. В некоторых способах, стадию очистки осуществляют с использованием аффинного хроматографического материала, специфичного константному домену Каппа, к константному домену Лямбда, специфичному к ним обоим - константному домену Каппа и константному домену Лямбда.
[0018] В некоторых способах вторая нуклеиновая кислота кодирует вариабельный домен легкой цепи Каппа-типа. В некоторых способах, вторая нуклеиновая кислота кодирует константный участок Каппа-типа. В некоторых способах, вторая нуклеиновая кислота кодирует константный участок Лямбда-типа. В некоторых способах, вторая нуклеиновая кислота кодирует вариабельный домен легкой цепи Лямбда-типа. В некоторых способах, вторая нуклеиновая кислота кодирует константный участок Каппа-типа. В некоторых способах, вторая нуклеиновая кислота кодирует константный участок Лямбда-типа. В некоторых способах, третья нуклеиновая кислота кодирует вариабельный домен легкой цепи Каппа-типа. В некоторых способах, третья нуклеиновая кислота кодирует константный участок Каппа-типа. В некоторых способах, третья нуклеиновая кислота кодирует константный участок Лямбда-типа. В некоторых способах, третья нуклеиновая кислота кодирует вариабельный домен легкой цепи Лямбда-типа. В некоторых способах, третья нуклеиновая кислота кодирует константный участок Каппа-типа. В некоторых способах, третья нуклеиновая кислота кодирует константный участок Лямбда-типа.
[0019] Изобретение также относится к смеси антител, которая включает два моноспецифичных антитела и одно биспецифичное антитело, которые имеют общую тяжелую цепь. Например, биспецифичное антитело представляет собой любое из биспецифичных антител, описанных в данном документе или полученных с использованием способов, описанных в данном документе. Изобретение также относится к способам получения такой смеси антител с помощью a) выделения антитела или участка фрагмента антитела, имеющего специфичность, определяемую вариабельным доменом тяжелой цепи вместе с вариабельным доменом первой легкой цепи; b) выделения антитела или участка фрагмента антитела, имеющего другую специфичность, определяемую тем же вариабельным доменом тяжелой цепи как у антитела стадии a) вместе с вариабельным доменом второй легкой цепи; c) ко-экспрессии в клетке: (i) общего вариабельного домена тяжелой цепи, сшитого с константным участком тяжелой цепи иммуноглобулина; (ii) вариабельного домена первой легкой цепи, сшитого или с константным доменом легкой цепи Каппа-типа, или с константным доменом легкой цепи Лямбда-типа; и (iii) вариабельного домена второй легкой цепи, сшитого с константным доменом легкой цепи Каппа-типа, или сшитого с константнм доменом легкой цепи Лямбда-типа. Некоторые способы также включают дополнительную стадию d) выделения смеси антител, полученной в стадии c) из надосадочной жидкости клеточной культуры.
[0020] Изобретение также относится к способам получения двух или более, например, трех или более неидентичных антител в одной рекомбинантной клетке-хозяине с помощью a) экспрессии в одной рекомбинантной клетке-хозяине одной или нескольких последовательностей нуклеиновых кислот, кодирующих общую тяжелую цепь иммуноглобулина и, по меньшей мере, две, например, по меньшей мере, три различные легкие цепи иммуноглобулина, которые способны спариваться с общей тяжелой цепью иммуноглобулина с образованием функциональных антиген-связывающих доменов и с получением двух или более, например, трех или более неидентичных антител, которые включают общую тяжелую цепь. Некоторые способы также включают стадию сбора клеток или другую очистку двух или более, например, трех или более неидентичных антител из рекомбинатной клетки-хозяина или из культуры клеток-хозяев. Клетка-хозяин может быть, например, клеткой млекопитающего. В некоторых способах, неидентичные антитела включают моноспецифичные и биспецифичные антитела.
[0021] В некоторых способах неидентичные антитела направленно воздействуют на различные эпитопы одного антигена-мишени. В некоторых способах неидентичные антитела обладают различными аффинностями по отношению к одному эпитопу-мишени. В некоторых способах, неидентичные антитела связываются с различными антигенами.
[0022] В некоторых способах два или более, например, три или более неидентичных антитела независимо выбирают из группы, состоящей из: IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD, IgE и IgM.
[0023] В некоторых вариантах осуществления два или более, например, три или более неидентичных антитела содержат модифицированный участок Fc, который модифицирует эффекторные функции антитела, такие как антиген-зависимая клеточно-опосредованная цитотоксичность (ADCC), комплемент-зависимая цитотоксичность (CDC), антиген-зависимый клеточный фагоцитоз (ADCP), или модифицирует их фармакокинетические свойства путем изменения их связывания с неонатальными Fc-рецепторами.
[0024] В некоторых способах два или более, например, три или более различных иммуноглобулина представлены в формате F(ab')2.
[0025] В некоторых способах одна или более последовательностей нуклеиновой кислоты стабильно экспрессируются в клетке-хозяине.
[0026] В некоторых способах два или более, например, три или более неидентичных антитела вырабатываются клеткой-хозяином in vitro.
[0027] Некоторые способы также включают дополнительные стадии селекции, по меньшей мере, одной клетки-хозяина путем оценки двух или более, например, трех или более неидентичных антител, вырабатываемых рекомбинантной клеткой-хозяином, на предмет их способности связываться с антигеном-мишенью; стадию культивирования рекомбинантной клетки-хозяина; и выделения трех или более неидентичных антител. Антитела могут быть выделены с использованием любого из методов, описанных в данном документе, или с помощью любых других известных в данной области методов.
[0028] В некоторых способах различные легкие цепи иммуноглобулинов содержат идентичные константные участки. В некоторых способах, различные легкие цепи иммуноглобулинов содержат различные константные участки.
Краткое описание чертежей
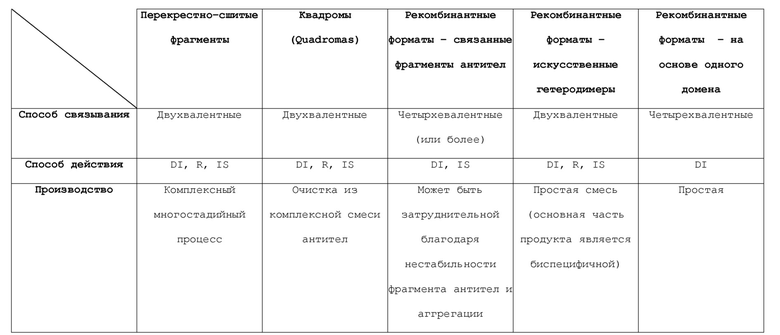
[0029] Фигура 1 представляет собой схематичное изображение различных форматов биспецифичного антитела. A. Форматы на основе фрагментов антител: X-связаннй Fab, перекрестно-связанные Fab-фрагменты; tascFv/BiTE, двойной-scFv/биспецифичный T-клеточный захватчик; Db, диатело; taDb, двойное диатело. B. Форматы на основе Fc-гибридов: Db-Fc, гибрид диатело-Fc; гибрид taDb-Fc, гибрид двойное диатело-Fc; taDb-CH3, гибрид двойное диатело-CH3; (scFv)4-Fc, гибрид тетра-scFv-Fc; DVD-Ig, иммуноглобулин с двойным вариабельным доменом. C. форматы IgG: "knob-hole" и SEED, домен, сконструированный путем обмена цепей; CrossMab, "knob-hole" вместе с обменом доменов тяжелой и легкой цепи; bsAb, биспецифичное антитело, выделенное из квадромы (quadroma); sdAb, антитело на основе одного домена.
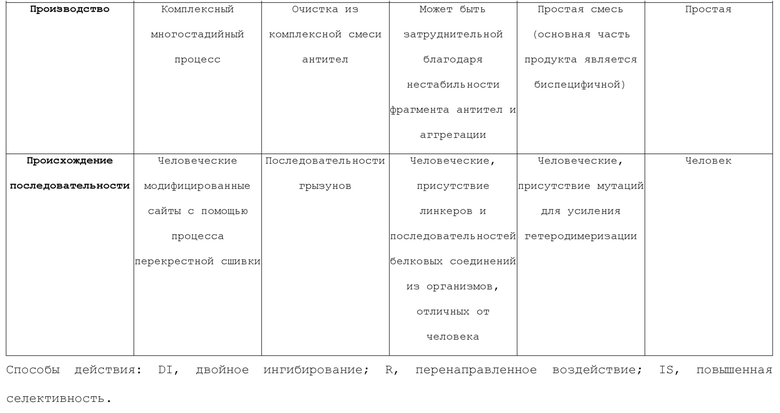
[0030] Фигура 2 представляет собой схематичное изображение возможных способов действия с поддержкой биспецифичных антител. 2A. Направленное воздействие на два антигена. 2B. Повторное направленное воздействие токсического компонента или активности на клетку-мишень. 2C. Повышение селективности, опосредованной авидностью.
[0031] Фигуры 3A-3C представляют собой схематическое изображение структуры различных биспецифичных антител по изобретению, состоящих из двух копий уникального полипептида тяжелой цепи и двух различных полипептидов легких цепей. Подразумевается, что локализации и/или расположения легкой цепи Каппа и легкой цепи Лямбда (или их частей), представленные на этих фигурах, не являются ограничениями. Специалистам в данной области понятно, что легкая цепь Каппа и легкая цепь Лямбда (или их части) также могут располагаться так, чтобы получилось зеркальное изображение биспецифичных антител, представленных на Фигурах 3A-3C. Специалистам в данной области также понятно, что биспецифичные антитела, которые представлены в формате полноразмерного IgG на Фигурах 3A-3C, также могут быть получены с использованием других иммуноглобулиновых изотипов или в других иммуноглобулиновых форматах, таких как F(ab')2. 3А. Вариабельный домен Каппа, сшитый с константным доменом Каппа, и вариабельный домен Лямбда, сшитый с константным доменом Лямбда. 3B. Вариабельные домены Каппа, сшитые с константным доменом Каппа и с константным доменом Лямбда. 3С. Вариабельные домены Лямбда, сшитые с константным доменом Каппа и с константным доменом Лямбда.
[0032] Фигура 4 представляет собой иллюстрацию анализа ELISA, тестирующего клоны, специфичные к hCXCL10-NusA или hIL6RC и несущие одинаковый вариабельный домен тяжелой цепи. Каждый клон тестировали против обеих мишеней для демонстрации специфичности.
[0033] Фигуры 5A-C представляют собой ряд иллюстраций, изображающих три типа библиотек, используемых в Примерах, для каждого типа библиотеки, Vκ и Vλ, библиотеки поддерживали отдельно. Фигуры 5A и C: Два набора библиотек, которые содержат фиксированный вариабельный домен VH3-23, который отличается только их последовательностью CDR H3, которая указана ниже H3 (CDR определение согласно IMGT). Комбинацию легкой цепи изменяли либо путем вставки случайных последовательностей в CDRL3 выбранных вариабельных генов легкой цепи (Фигуры 5A и 5C), либо путем захвата естественно перестроенных вариабельных доменов легких цепей, выделенных из человеческих доноров, и которые могут включать все человеческие вариабельные гены и содержат расхождения во всех 3 CDR (Фигура 5B). Различные стратегии получения разнообразия проиллюстрированы с помощью горизонтальных линий ниже измененных участков комбинаций легкой цепи.
[0034] Фигуры 6A-6B представляют собой графики, изображающие результаты ELISA с использованием моноспецифичных IgGλ, и IgGκ, селектированных против hIFNγ и IL6RC, соответственно, и несущих общую тяжелую цепь. Форматы ELISA схематично представлены после каждого графика. На Фигуре 6A, INFγ иммобилизовали на планшете, инкубировали с анти-IFNγ IgGλ или с анти-IL6RC (т.e., растворимый комплекс рецептор IL-6R/IL-6) IgGκ, и оба детектировали с помощью антител, связывающих человеческий Gκ, или антител, связывающих человеческий Cλ, связанных с пероксидазой хрена. Сигнал обнаруживали с помощью колориметрии и оценивали количественно с использованием ридера для микропланшетов.
[0035] Фигура 7 представляет собой схематичное изображение векторов, используемых для ко-экспрессии одной тяжелой цепи и двух легких цепей в клетках млекопитающего. Оба вектора содержат три промотора для управления генной экспрессией, ген глутамин синтетазы для селекции стабильной клеточной линии. Во втором векторе, pNovI kHλk, экспрессия дополнительной легкой цепи Каппа управляется с помощью сайта внутренней посадки рибосомы (IRES). Указаны различные гены и генетические контрольные элементы. hCMV, человеческий цитомегаловирусный промотор; SV40, V40 промотор; pA сигнал полиаденилирования; VH, вариабельный домен тяжелой цепи; VK, вариабельный домен легкой цепи Каппа; CK константный домен легкой цепи Каппа; VL, вариабельный домен легкой цепи Лямбда; CL2, константный домен 2 легкой цепи Лямбда; GS кДНК, кДНК глутамин синтетазы; AmpR; маркер селекции на устойчивость к ампициллину. Указано выбранное количество сайтов рестрикции.
[0036] Фигура 8A представляет собой схематическое изображение процесса очистки для биспецифичных антител по изобретению. Фигура 8B представляет собой иллюстрацию, изображающую ко-экспрессию, очистку и анализ SDS-PAGE биспецифичных антител по изобретению. Гель окрашивали с использованием красителя simply blue. PA: Протеин A; K: селекция Каппа; λ: селекция Лямбда; FT: проточная колонка; E: элюированная фракция.
[0037] Фигура 9 представляет собой иллюстрацию анализа SDS-PAGE суммарного IgG, очищенного из клеток млекопитающего, трансфицированных векторами, дающими возможность получения различного уровня экспрессии легкой цепи Каппа с использованием различных элементов IRES внутри вектора pNovI kHλk (дорожка 1-5), и сравнивали с вектором pNovI kHλ. Относительные интенсивности легкой цепи Каппа и Лямбда выявляют, что уровень экспресии может модулироваться.
[0038] Фигура 10A представляет собой иллюстрацию, изображающую анализ на IEF-геле очищенного моноспецифиченого IgG (κκ и λλ) и биспецифичного IgG (κλ). Фигура 10B представляет собой иллюстрацию анализа IEX-HPLC моноспецифичных и биспецифичных антител. Три антитела инъецировали независимо и их профиль элюции наносили на график. Представлен градиент, используемый в эксперименте.
[0039] Фигура 11 изображает анализ ELISA, используемый для определения способности биспецифичного антитела связываться как с мишенью, так и с легкой цепью Каппа и Лямбда в молекуле. Фигура 11A представляет собой схематичное изображение формата ELISA. Фигура 11B представляет собой график, изображающий результаты ELISA с INFγ, иммобилизованным на планшете. Фигура 11С представляет собой график, изображающий результаты ELISA с IL-6RC, иммобилизованным на планшете. IgGк, моноспецифичное антитело к IL6RC; IgGλ, моноспецифичное антитело к IFNγ; IgGкλ, биспецифичное антитело к IL6RC/INFγ. Указаны антитела вторичного детектирования, антитело к человеческому белку Лямбда HRP и антитело к человеческому белку Каппа HRP.
[0040] Фигура 12 представляет собой иллюстрацию SPR-аализа биспецифичных антител IgGκλ. На Фигуре 12A, INFγ иммобилизовали на поверхности чипа Biacore и инъецировали на поверхность моноспецифичное антитело к IL6RC (IgGκ), моноспецифичное антитело к INFγ (IgGλ) и биспецифичное антитело к IL6RC/INFγ (IgGκλ) с последующей инъекцией IL6RC. На Фигуре 12B, биспецифичное антитело к IL6RC/INFγ (IgGκλ) иммобилизовали на поверхности чипа и инъецировали антитело к человеческому белку Каппа и антитело к человеческому белку Лямбда в такой же концентрации. Эксперимент повторяли, инвертируя порядок инъекции антител с получением идентичных результатов.
[0041] Фигура 13 представляет собой схематичное изображение краткого описания одного способа получения биспецифичных и мультиспецифичных антител, описанных в данном документе, в клетках CHO.
[0042] Фигура 14A представляет собой график, изображающий профили роста и выработки антител пулами клеток CHO на уровне лабораторного производства в конической колбе Эрленмейера. Уровень выработки антител определяли с помощью анализа Протеин A-ВЭЖХ. VCC означает концентрацию жизнеспособных клеток и Ab означает антитело. Фигура 14B представляет собой график, изображающий сравнение профиля роста и профиля продуцирования антитела между лабораторной и полупромышленной ферментацией. Уровень продуцирования антител определяли с помощью анализа Протеин A-ВЭЖХ. VCC означает концентрацию жизнеспособных клеток и Ab означает антитело.
[0043] Фигура 15A и 15В представляют собой ряд графиков, изображающих продуцирование антител в 96-луночном планшете (96wpl) клеточными линиями, экспрессирующими моноспецифичное антитело к Каппа (KK), моноспецифичное антитело к Лямбда (LL) и биспецифичное антитело к Каппа и к Лямбда (KL) через пять недель после трансфекции в двух независимых экспериментах. Уровень продуцирования антитела определяли с помощью ELISA, где m Ab означает моноклональное антитело. Фигуры 15C и 15D представляют собой ряд графиков, изображающих продуцирование атитела во встряхиваемом 24-луночном планшете (24wpl) ночными периодическими культурами моноспецифичного антитела к Каппа (KK), мноспецифичного антитела к Лямбда (LL) и биспецифичного антитела к Каппа.
[0044] Фигура 16A представляет собой иллюстрацию, изображающую результаты анализа с помощью восстанавливающего SDS-PAGE моноспецифичных молекул κ IgG (т.e. моноспецифичные молекулы, содержащие легкие цепи Каппа, также обозначаемые в данном документе как "моно κ" молекулы), моноспецифичные λ IgG молекулы (т.e. моноспецифичные молекулы, содержащие легкие цепи Лямбда, также обозначаемые как "моно λ" молекулы), и κλ антитела (т.e. антитела, содержащие обе легкие цепи, Каппа и Лямбда), посредством стадий очистки, описанных выше. Фигура 16B представляет собой иллюстрацию, изображающую результаты анализа с помощью восстанавливающего SDS-PAGE антител моно κ, моно λ и κλ, полученных после стадий элюции, описанных выше. На Фиг.16A и 16B, гель окрашивали с помощью красителя "simply blue", и с помощью E обозначали фракцию элюции, с помощью FT обозначали проточную фракцию и с помощью MM обозначали маркер молекулярной массы. Фигура 16C представляет собой иллюстрацию, изображающую анализ на геле изоэлектрического фокусирования (IEF) очищенных моноспецифичных молекул IgG (κκ и λλ) и биспецифичной молекулы IgG (kλ).
[0045] Фигуры 17A-D представляют собой ряд графиков и иллюстраций, изображающих, что с помощью способов получения биспецифичных антител по изобретению получают антитела, которые включают обе легкие цепи, Каппа и Лямбда, и что очищенные антитела демонстрируют биспецифичность. Графики изображают результаты ELISA с использованием антитела к κλ против hIFNγ и IL6RC. ELISA осуществляли с использованием указанных детектирующих антител к Каппа и к Лямбда. Фигуры 17A-D иллюстрируют, что легкая цепь Лямбда связывается с hIFNγ, в то время как легкая цепь Каппа связывается с IL6RC. NI-0501 представляет собой контрольное антитело для легкой цепи Лямбда, антитело к hIFNγ, и NI-1201 представляет собой контрольное антитело для легко цепи Каппа, антитело к IL6RC.
[0046] Фигура 18 представляет иллюстрацию анализа с использованием IEF-геля для различных моноспецифичных и биспецифичных антител, выявляющего, что различие в pI может варьироваться в зависимости от вариабельной последовательности легкой цепи антитела. Дорожка 1, анти-NusA IgGκ; Дорожка 2, анти-NusA/анти-IFNγ IgGκλ; Дорожка 3, анти-IFNγ IgGλ; Дорожка 4, анти-IL6RC IgGκ; Дорожка 5, анти-IL6RC/анти-IL6RC IgGκλ; Дорожка 6, анти-IL6RC IgGλ.
[0047] Фигура 19 представляет собой схематичное изображение трех различных гибридных белков, получаемых путем объединения вариабельного гена Лямбда и константного гена Каппа. Точки сшивки отличаются между различными гибридами: на 19A, VLambda, сшитый с CKappa; на 19B, VLambda до CDR3, сшитый с VKappa FR4 и CKappa; и на 19C, VLambda и первые четыре аминокислоты CLambda и CKappa за исключением первых четырех аминокислот. CDR, гипервариабельный участок; FR, каркасный участок.
[0048] Фигура 20 представляет собой иллюстрацию анализа конструктов двух легких цепей на системе Bionalyzer 2100 с использованием чипа Protein 80 (Agilent Technologies). Показана электроферограмма, соответствующая изображению геля.
[0049] Фигура 21 представляет собой ряд графиков, изображающих результаты эффекта дозы ELISA с использованием scFv, специфичных для IFNγ (A) или IL6RC (B), в которых домен VH был или общим исходно выбранным VH (верхние кривые), или он представлял собой другие домены VH, которые дают возможность экспрессии и очистки scFv (нижние кривые).
[0050] Фигура 22 представляет собой график, изображающий результаты, получаемые для IgGκλ, количественная оценка биспецифичного антитела с использованием формата сэндвич-ELISA. Эффекта дозы добивались с использованием или индивидуального очищенного биспецифичного антитела, или смешанного с моноспецифичными антителами Каппа и Лямбда в различных указанных соотношениях, с целью оценки этих молекул в анализе.
Подробное описание
[0051] С целью преодоления ограничений терапевтических средств, содержащих моноклональные и одновалентные антитела, которые могут направленно воздействовать только на один антиген, или с целью преодоления ограничений комбинаций терапевтических средств, содержащих одновалентные антитела, были предприняты интенсивные усилия, направленные на получения форматов биспецифичных антител с направленным воздействием на множество антигенов. Такие антитела, несущие более чем одну специфичность, представляют интерес с точки зрения биотехнологии и обладают большим потенциалом в качестве терапевтических агентов, дающих возможность осуществления новых терапевтических приемов (Fischer and Léger, Pathobiology 2007; 74:3-14; Morrison SL Nature Biotechnol 2007; 25:1233-1234). Биспецифичные антитела являются предпочтительными, поскольку они дают возможность множественного направленного воздействия, они повышают терапевтический потенциал, они имеют отношение к созданию альтернативных возможностей биологических систем, и они обеспечивают новые механизмы действия посредством таких возможностей, как повторное направленное воздействие и/или повышенная специфичность. Как установлено, одиночные терапевтические мишени становятся более и более исчерпанными, поэтому комбинации, представленные биспецифичными антителами, обеспечивают новые и обширные множества мишеней для терапевтических агентов и применений.
[0052] Для получения таких биспецифичных молекул использовались несколько стратегий, таких как химическая перекрестная сшивка фрагментов антител, принудительная гетеродимеризация, квадромная технология, сшивка фрагментов антител посредством полипептидных линкеров и использование антител с одним доменом. Доступность технологий рекомбинантной ДНК приводит к получению множества антител биспецифичного формата (см., например, Ridgway JB et al. (1996) Protein Eng 9: 617-621). Линкеры и мутации часто вводятся в различные участки антитела для усиления образования гетеродимера или для соединения различных связывающих компонентов в единую молекулу. Однако эти сконструированные молекулы часто обладают плохими производственными характеристиками, а также повышенным риском иммуногенности, которые ограничивают или предотвращают их продвижение к клиническим применениям. Кроме того, прежние попытки разработок биспецифичных форматов были ограничены благодаря таким факторам, как плохая стабильность и/или экспрессия. Эти способы дополнительно описаны ниже и обсуждаемые форматы проиллюстрированы на Фигуре 1.
[0053] Химическая перекрестная сшивка. Использование реагентов химической перекрестной сшивки для ковалентного связывания двух антител является по существу эффективным способом. Фрагменты антител, полученные из их соответствующих антител с помощью ферментативного гидролиза или полученные с помощью рекомбинантных технологий, конъюгируются с использованием бифункциональных реагентов (Glennie MJ et al., J Exp Med 1992; 175:217-225). Гомогенность продукта является главным ограничением данного способа, поскольку биспецифичные образцы следует очищать из гомодимеров, и стадии модификаций могут изменять целостность и стабильность белков. Множество включенных стадий делает данный способ затруднительным в смысле производства и гомогенности продукта.
[0054] Квадромы. Квадромы и триомы могут быть получены посредством слияния либо двух гибридом, либо одной гибридомы с В лимфоцитом, соответственно (Suresh MR et al., Methods Enzymol 1986; 121: 210-228). В данном случае одновременная экспрессия двух тяжелых и двух легких цепей приводит к случайной сборке 10 комбинаций антител, и целевое bsAb представляет собой только небольшую фракцию секретированных антител. bsAb следует очищать с использованием комбинации хроматографических методов и при этом существенно снижается выход продукта. Основное ограничение состоит в том, что квадромы вырабатывают bsAb с происхождением из грызунов, что ограничивает их терапевтический потенциал благодаря проблемам иммуногенности.
[0055] Рекомбинатные биспецифичные антитела. Большинство антител биспецифичного формата получали с помощью методов генетической инженерии с использованием фрагментов антител, таких как scFv- или Fab-фрагменты, в качестве структурных элементов, соединенных посредством полипептидных линкеров. Форматы на основе связанных фрагментов антител включают тандем scFv (BiTE), диатела и тандем-диатела (Kipriyanov SM. Methods Mol Biol 2003; 207:323-333; Korn T et al., Int J Cancer 2002; 100:690-697). Эти структурные элементы могут дополнительно связываться с Fc-участком иммуноглобулина с получением 'IgG-подобных' молекул. Эти форматы включают диатело-Fc, тандем диатело-Fc, тандем диатело-CH3, (scFv)4-Fc и DVD-Ig (Lu D et al., J Immunol Methods 2003; 279: 219-232; Lu D et al., J Biol Chem 2005; 280: 19665-19672; Lu D et al., J Biol Chem 2004; 279: 2856-2865; Wu C et al., Nat Biotechnol 2007 25:1290-7). Потенциальное ограничение использования линкеров состоит в том, что гибкая природа этих пептидов делает их более чувствительными к протеолитическому расщеплению, потенциально приводя к слабой стабильности антител, к агрегации и к повышенной иммуногенности. Кроме того, эти чужеродные пептиды могут вызывать иммунный ответ против соединительной части между белком и линкером или против самого линкера. Вообще, bsAb на основе связанных структурных элементов представляют собой трудоемкие компоненты в смысле производства, что ограничивает их терапевтическое применение.
[0056] Идеальная биспецифичная молекула для терапии человека должна быть неотличима от нормального IgG. Изучали стратегии на основе принудительной гетеродимеризации двух тяжелых цепей. Первый разработанный способ 'knob into hole' имеет целью форсировать спаривание двух различных тяжелых цепей IgG путем введения мутаций в CH3-домены для модификации контактной поверхности (Ridgway JB et al., Protein Eng 1996; 9: 617-621). На одну цепь вводили аминокислоты с большими боковыми цепями для создания 'knob'. С другой стороны, большие аминокислоты заменяли на аминокислоты с короткими боковыми цепями для создания 'hole' в другом CH3-домене. С помощью ко-экспрессии этих двух тяжелых цепей наблюдали более чем 90%-образование гетеродимера ('knob-hole') по сравнению с образованием гомодимеров ('hole-hole' или 'knob-knob'). Аналогичную концепцию разрабатывали с использованием человеческих CH3-доменов, полученных с помощью технологии образования доменов путем обмена цепей (SEED), на основе человеческих последовательностей IgG и IgA (Davis JH et al., 2010, PEDS 23:195-202). Эти сконструированные домены приводят к образованию гетеродимерных молекул, которые могут нести две различные специфичности. Эти два способа являются привлекательными, поскольку они благоприятствуют выработке представляющих интерес гетеродимеров (до 95%), но при этом полностью не предотвращается образование гомодимеров. Таким образом, все еще существует необходимость процедуры последующей очистки, способной удалить гомодимеры от гетеродимеров. Другое потенциальное свойство этих способов состоит в том, что мутированные домены не являются полностью человеческими и могут приводить к иммуногенности, а также могут воздействовать на стабильность домена и склонность молекулы к агрегации. Поскольку эти стратегии дают возможность форсированного спаривания тяжелых цепей, то различные легкие цепи могут образовывать случайные пары с любой из тяжелых цепей и приводить к получению различных антител, которые необходимо очистить друг от друга. Недавно было описано улучшение, касающееся способа 'knob into hole' для решения проблемы спаривания легких цепей (WO 2009/080253 A1). Способ включает обмен некоторых из доменов легкой цепи и тяжелой цепи дополнительно к мутациям 'knob into hole'. Основное преимущество данного метода состоит в том, что биспецифичное двухвалентное антитело, имеющее два различных вариабельных домена тяжелой цепи и два различных вариабельных домена легкой цепи, может быть получено и обозначается "CrossMab." Однако последовательности данного биспецифичного антитела не являются полностью человеческими, поскольку оно содержит как мутации в Fc для принудительной гетеродимеризации, так и искусственные точки соединения между различными доменами иммуноглобулина. Кроме того, эти модификации приводят к пониженному уровню экспрессии антител биспецифичного формата по сравнению с монокональным антителом (Schaefer et al., PNAS 2011; 108:11187-11192).
[0057] Антитела на основе одного домена. Иммунные системы кам (camelid) (лама и верблюд) и хрящевой рыбы (акула-нянька) используют одиночные V-домены, сшитые с Fc, демонстрируя, что одиночный домен может придавать антигену высокую аффинность связывания. V-домены кам, акул и даже человека представляют собой альтернативы антителам, но они также могут использоваться для получения bsAb. Они могут переформатироваться в классические IgG, в которых каждое плечо имеет потенциал связывания с двумя мишенями либо посредством VH-домена, либо посредством VL-домена. Этот однодоменный IgG будет обладать биохимическим свойствами, аналогичными свойствам IgG, и потенциально решает проблемы, связанные с другими форматами bsAb в смысле продуцирования и гетерогенности. Однако вероятно, что стерические затруднения будут часто предотвращать одновременное связывание обоих антигенов на обоих плечах антитела.
[0058] Характеристики форматов биспецифичных антител, описанных выше, представлены на Фигуре 1. Некоторые из этих характеристик форматов взяты из Fischer and Léger, Pathobiology 2007; 74:3-14; and Morrison SL Nature Biotechnol 2007; 25:1233-1234.
[0059] В отличие от этих предыдущих форматов, биспецифичные антитела, мультиспецифичные антитела, композиции и способы, представленные в данном документе, преодолевают такие препятствия при разработках. Биспецифичные антитела, представленные в данном документе, имеют общую тяжелую цепь, две легких цепи - одна Каппа (к), одна Лямбда (λ) - каждая из которых имеет отличную специфичность (т.е. две легких цепи, две специфичности). Предпочтительно, чтобы биспецифичные антитела не содержали никаких линкеров или других модификаций, включая аминокислотные мутации. С помощью способов, представленных в данном документе, получают молекулы, обладающие специфичностью связывания, где разнообразие ограничивается VL-участком. С помощью этих способов получают биспецифичные антитела посредством контролируемой ко-экспрессии трех цепей (одной VH-цепи, двух VL-цепей), и посредством очистки биспецифичного антитела. Биспецифичные и/или мультиспецифичные антитела, описанные в данном документе, демонстрируют аналогичные аффинности для данной мишени по сравнению с аффинностями моноспецифичных антител для той же мишени. Предпочтительно, биспецифичные и/или мультиспецифичные антитела, описанные в данном документе, фактически неотличимы от стандартных молекул IgG.
[0060] Способы, представленные в данном документе, также представляют средства получения простых смесей антител из двух моноспецифичных антител и одного биспецифичого антитела, которые применяются, например, для множественного направленного воздействия без очистки биспецифичного антитела из смеси.
[0061] Возможные способы действия биспецифичных антител
[0062] Одновременное ингибирование двух мишеней. По определению биспецифичные антитела несут две специфичности и, таким образом, могут ингибировать более чем одну мишень. Эти мишени могут представлять собой растворимые факторы или локализованные на поверхности клетки. Успешно получали ряд форматов множественного направленного воздействия на цитокины (Wu C et al., Nat Biotechnol 2007 25:1290-7).
[0063] Перенаправленное воздействие. Поскольку большинство форматов биспецифичных антител способны связываться с двумя молекулами одновременно, они, таким образом, могут использоваться в качестве связующих молекул для перенаправленного воздействия на цитотоксические эффекторные клетки или на цитотоксические агенты по отношению к клеткам, вовлеченным в заболевание. Данное применение было изучено в отношении онкологии. В некоторых случаях, одна специфичность антитела направлена против опухолевых клеточных маркеров, таких как CD 19, CD20, HER2, карциноэмбриональный антиген (CEA). Второе плечо антитела несет на себе в непосредственной близости токсический компонент или активность, такую как лекарственные средства, токсины, цитокины или эффекторную клетку иммунной системы (T-клетки, NK-клетки, моноциты и нейтрофилы с направленным воздействием на CD3, CD16, CD64 и CD89, соответственно) (Thielemans K, Blood 1996; 87: 4390-4398; Goldstein J et al., J Immunol 1997; 158: 872-879).
[0064] Повышенная специфичность через авидность. В классическом формате IgG, связывание антитела направляется как с помощью аффинности каждого объединенного сайта для его антигена, так и с помощью эффектов авидности, обеспечиваемых двухвалентным связыванием. Эффект авидности существенно повышает явную аффинность антитела к маркерам клеточной поверхности, поскольку происходят два события диссоциации для высвобождаемого антитела. Некоторые из биспецифичных форматов, описанных выше, являются двухвалентными (т.е. один сайт связывания для каждой мишени), тогда как другие являются четырехвалентными. Последние имеют четыре сайта связывания или более и имеют потенциал для связывания каждой мишени двухвалентным образом. Двухвалентные биспецифичные антитела применяются в качестве терапевтических агентов, селективно воздействующих на клеточные популяции, которые экспрессируют комбинацию клеточных поверхностных маркеров. Данный уникальный способ действия в основном ограничен молекулами, которые могут извлекать пользу из компонента авидности, чтобы проводить различия между клетками, экспрессирующими оба антигена, и теми клетками, которые экспрессируют только один маркер.
[0065] Характеристики возможных способов действия, опосредованных биспецифичными антителами, представлены на Фигуре 2. Данную характеристику взяли из Fischer and Léger, Pathobiology 2007; 74:3-14.
[0066] Характеристики и ограничения форматов биспецифичных антител
[0067] Ключевые характеристики данных форматов биспецифичных антител суммированы в Таблице I. Все форматы за исключением тех, что основаны на человеческих доменах, содержат последовательности, которые не имеют человеческого происхождения или содержат белковые последовательности организма, отличного от человека, полученные путем сшивки различных белковых доменов. Большинство форматов, использующих линкеры, приводит к потенциальным проблемам производства благодаря доменной агрегации. Присутствие чужеродных последовательностей и неблагоприятные характеристики стабильности потенциально могут значительно повышать риск иммуногенности. Ключевое отличие между форматами состоит в валентности их сайтов связывания, которые непосредственно связаны со способностью опосредовать перенаправленное воздействие или селективное связывание, опосредованное авидностью. Таким образом, все форматы не могут обладать возможностью всех способов действия. Конкретно, единственный формат, который не отличим от полностью человеческого иммуноглобулина, не может опосредовать активности перенаправленного воздействия или повышенной селективности. Таким образом, существует необходимость получения новых биспецифичных антител с благоприятными свойствами для разработки терапевтических средств, т.е. таких, которые неотличимы от полностью человеческой иммуноглобулиновой молекулы, с хорошими свойствами возможности производства и с возможностью широкого спектра возможных способов действия.
[0068] Улучшенные способы получения биспецифичных и двухвалентных антител.
[0069] В настоящем изобретении предлагаются способы получения биспецифичных антител, которые идентичны по структуре с человеческим иммуноглобулином. Данный тип молекул состоит из двух копий уникального полипептида тяжелой цепи, вариабельный участок первой легкой цепи сшит с константным доменом Каппа, и вариабельный участок второй легкой цепи сшит с константным доменом Лямбда. Каждый объединенный сайт демонстрирует различную антигенную специфичность, которой способствуют как тяжелая цепь, так и легкая. Вариабельные участки легкой цепи могут относиться к семейству Лямбда или Каппа и предпочтительно сшиты с константными доменами Лямбда и Каппа, соответственно. Это является предпочтительным для того, чтобы избежать получения неестественных полипептидных соединений. Однако также возможно получение биспецифичных антител по изобретению путем сшивки вариабельного домена легкой цепи Каппа с константным доменом Лямбда для получения первой специфичности, а также сшивка вриабельного домена легкой цепи Лямбда с константным доменом Каппа для второй специфичности (Фигура 3). Биспецифичные антитела, описанные в данном документе, также обозначаются как антитела IgGκλ или "κλ-антитела", новые полноразмерные человеческие биспецифичные антитела формата IgG. Этот формат κλ-антитела дает возможность аффинной очистки биспецифичного антитела, которое неотличимо от стандартной молекулы IgG с характеристиками, которые неотличимы от стандартного моноклонального антитела и, таким образом, является предпочтительным по сравнению с предыдущими форматами.
[0070] Существенной стадией способа является идентификация двух Fv-участков антитела (каждый состоит из вариабельного участка легкой цепи и вариабельного домена тяжелой цепи), обладающих различными антигенными специфичностями, которые имеют общий вариабельный домен тяжелой цепи. Было описано множество способов для получения моноклональных антител и их фрагментов. (Смотри, например, Antibodies: A Laboratory Manual, Harlow E, and Lane D, 1988, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, публикация введена в данный документ ссылкой). Полностью человеческие антитела и молекулы антител, в которых последовательность обеих цепей, легкой и тяжелой, включая CDR 1 и 2, происходят из человеческих генов. CDR3-участок может иметь человеческое происхождение или может быть сконструирован синтетическими средствами. Такие антитела обозначаются в данном документе как "человеческие антитела", или "полностью человеческие антитела". Человеческие моноклональные антитела могут быть получены путем использования триомного метода (trioma); гибридомного метода человеческих B-клеток (см. Kozbor, et al., 1983 Immunol Today 4: 72); и EBV-гибридомного метода с получением моноклональных антител человека (см. Cole, et al., 1985 In: Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96). Человеческие моноклональные антитела могут применяться и могут быть получены с использованием человеческих гибридом (с. Cote, et al., 1983. Proc Natl Acad Sei USA 80: 2026-2030) или с помощью трансформации человеческих B-клеток вирусом Эпштейна Барра in vitro (см. Cole, et al., 1985 In: monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96).
[0071] Моноклональные антитела получают, например, путем иммунизации животного антигеном-мишенью или его иммуногенным фрагментом, производным или вариантом. Альтернативно, животное иммунизируют клетками, трансфицированными вектором, содержащим молекулу нуклеиновой кислоты, кодирующую антиген-мишень, так что он экспрессируется и ассоциируется с поверхностью трансфицированной клетки. Множество методов хорошо известно в данной области для получения ксеногенных животных, отличных от человека. Смотри, например, патент США № 6075181 и № 6150584, которые введены в данный документ ссылкой в полном объеме.
[0072] Альтернативно, антитела получают путем скрининга библиотеки, которая содержит последовательности антител или антиген-связывающих доменов для связывания с антигеном-мишенью. Данную библиотеку получают, например, в бактериофаге в виде белковых или пептидных гибридов с белком оболочки бактериофага, который экспрессируется на поверхности собранных фаговых частиц, и которые кодируются ДНК-последовательностями, содержащимися в фаговых частицах (т.e. "библиотека фагового дисплея").
[0073] Гибридомы, полученные в результате слияния миеломы/B-клетки, затем скринируют на предмет их реактивности с атигеном-мшенью. Моноклональные антитела получают, например, с использованием гибридомных методов, таких как описанные у Kohler and Milstein, Nature, 256:495 (1975). В гибридомном методе мышь, хомяка или другое подходящее животное-хозяин, как правило, иммунизируют с помощью иммунизирующего агента для того, чтобы стимулировать лимфоциты, которые продуцируют или способны продуцировать антитела, которые будут специфично связываться с иммунизирующим агентом. Альтернативно, лимфоциты могут быть иммунизированы in vitro.
[0074] Хотя и не строго невозможна, но маловероятна идентификация различных антител, обладающих одинаковыми вариабельными доменами тяжелой цепи, но направленных против различных антигенов. Действительно, в большинстве случаев тяжелая цепь в большей степени вносит вклад в антиген-связывающую поверхность, а также является наиболее вариабельной по последовательности. Конкретно, CDR3 на тяжелой цепи является наиболее отличным CDR по последовательности, длине и структуре. Таким образом, два антитела, специфичные к различным антигенам, будут почти неизменно нести различные вариабельные домены тяжелых цепей.
[0075] Способ по изобретению преодолевает это ограничение и существенно облегчает выделение антител, имеющих одинаковый вариабельный домен тяжелой цепи, путем использования библиотек антител, в которых вариабельный домен тяжелой цепи является одинаковым для всех компонентов библиотеки и, таким образом, многообразие ограничивается вариабельными доменами легкой цепи. Такие библиотеки описаны, например, в приложенной заявке PCT/US2010/035619, от 20.5.2010 и опубликованной 25.11.2010 в виде PCT публикации № WO 2010/135558, а также в приложенной заявке PCT/US2010/057780, от 23.11.2010, каждая из которых включена в данный документ ссылкой в полном объеме. Однако поскольку вариабельный домен легкой цепи экспрессируется вместе с вариабельным доменом тяжелой цепи, оба домена могут вносить вклад в связывание антигена. Для дополнительного облегчения процесса, библиотеки антител, содержащие одинаковые вариабельные домены тяжелой цепи и различающиеся вариабельным доменом легкой цепи Лямбда или вариабельным доменом легкой цепи Каппа, могут использоваться параллельно для in vitro селекции антител против различных антигенов. Данный способ дает возможность идентификации двух антител, имеющих общую тяжелую цепь, но одно из антител несет вариабельный домен легкой цепи Лямбда, а другое вариабельный домен легкой цепи Каппа, которые могут использоваться в качестве структурных элементов для получения биспецифичного антитела в формате полноразмерного иммуноглобулина по изобретению. Биспецифичные антитела по изобретению могут относиться к различным изотипам и их Fc-участок может быть модифицирован с целью изменения свойств связывания с различными рецепторами Fc, и таким образом модифицируют эффекторные функции антитела, а также его фармакокинетические свойства. Множество способов модификации Fc-участка описано и применяется к антителам по изобретению, (см., например, Strohl, WR Curr Opin Biotechnol 2009 (6):685-91; патент США № 6528624; PCT/US2009/0191199 от 9.01.2009). Способы по изобретению также могут использоваться для получения биспецифичных антител и смесей антител в формате F(ab')2, который лишен Fc-участка.
[0076] Другая ключевая стадия изобретения это оптимизация ко-экспрессии общей тяжелой цепи и двух различных легких цепей в одной клетке для возможности сборки биспецифичного антитела п изобретению. Если все полипептиды экспрессируются на одинаковом уровне и одинаково собираются с образованием иммуноглобулиновой молекулы, то соотношение моноспецифичных антител (одинаковые легкие цепи) и биспецифичных антител (две различные легкие цепи) должно составить 50%. Однако вероятно, что различные легкие цепи экспрессируются на различном уровне и/или не собираются с одинаковой эффективностью. Таким образом, способы по изобретению также предлагают средства для модулирования относительной экспрессии различных полипептидов для компенсации характеристик их внутренней экспрессии или различной склонности к сборке с использованием общей тяжелой цепи. Это модулирование может быть достигнуто посредством силы промотора, использования сайтов внутренней посадки рибосомы (IRES), характеризующих различные эффективности, или посредством других типов регуляторных элементов, которые могут действовать на транскрипционном или трансляционном уровне, а также действуя на стабильность мРНК. Различные промоторы различной силы могут включать CMV (предранний промотор цитомегаловируса); EF1-1α (промотор субъединицы человеческого фактора элонгации 1α); Ubc (промотор человеческого убиквитина С); SV40 (промотор обезьяньего вируса 40). Также были описаны различные IRES из млекопитающего и имеющие вирусное происхождение. (Смотри, например, Hellen CU and Sarnow P. Genes Dev 2001 15: 1593-612). Эти IRES могут по большей части отличаться по их длине и эффективности привлечения рибосомы. Кроме того, возможна дополнительная регуляция активности путем введения множества копий IRES (Stephen et al. 2000 Proc Natl Acad Sei USA 97: 1536-1541). Модулирование экспрессии также может быть достигнуто с помощью множества последовательных трансфекций клеток для повышения количества копий индивидуальных генов, экспрессирующих одну или другую легкую цепь и, таким образом, модифицирующих их относительную экспрессию. Примеры, представленные в данном документе, демонстрируют, что контроль относительной экспрессии различных цепей является критическим для максимизации сборки и общего выхода биспецифичного антитела.
[0077] В результате ко-экспрессии тяжелой цепи и двух легких цепей получают смесь из трех различных антител в надосадочной жидкости клеточной культуры: два моноспецифичных двухвалентных антитела и одно биспецифичное двухвалентное антитело. Последнее следует очистить из смеси для получения молекулы, представляющей интерес. Способ, описанный в данном документе, существенно облегчает данную процедуру очистки путем применения аффинного хроматографического материала, который специфично взаимодействует с константными доменами легкой цепи Каппа или Лямбда, такого как аффинные матрицы CaptureSelect Fab Kappa и CaptureSelect Fab Lambda (BAC BV, Holland). Данный способ многостадийной аффинной хроматографической очистки эффективен и, как правило, применим к антителам по изобретению. Это по форме контрастирует со способами специфичной очистки, которые разрабатывали и оптимизировали для каждого биспецифичного антитела, полученного из квадром или из других клеточных линий, экспрессирующих смеси антител. Действительно, если биохимические характеристики различных антител в смеси похожи, то их разделение с использованием стандартного хроматографического метода, такого как ион-обменная хроматография, затруднительно или совсем невозможно.
[0078] Изобретение также относится к новым средствам получения простых смесей антител из двух или более моноспецифичных антител и одного или более биспецифичных антител, которые имеют одинаковую тяжелую цепь и могут быть очищены с использованием стандартных хроматографических методов, используемых для очистки моноклональных антител. (Смотри, например, Lowy, I et al. N Engl J Med 2010; 362:197-205; Goudsmit, J. et al. J Infect Dis. 2006. 193, 796-801). Такие простые смеси могут использоваться в качестве агентов с множественным направленным воздействием для терапевтического применения.
[0079] Успешная ко-экспрессия, очистка и характеризация тяжелой цепи и двух легких цепей и очистка биспецифичных антител представлены в Примерах. Гены, кодирующие общую тяжелую цепь и две легкие цепи, клонировали в вектор, содержащий три промотора. После транзиторной трансфекции собирали надосадочную жидкость клеток PEAK.
[0080] Ко-экспрессия трех цепей приводила к сборке трех различных антител: двух моноспецифичных и одного биспецифичного антитела. Их теоретические относительные соотношения должны составлять 1:1:2 при условии, что уровень экспрессии и скорость сборки одинаковы для обеих легких цепей. Биспецифичные антитела очищали с использованием трехстадийной процедуры аффинной хроматографии: (1) Протеин A: захват IgG (моно- и би-), (2) Kappaselect: захват IgG, содержащего легкие цепи Каппа, и (3) Lambdaselect: захват IgG, содержащего легкую цепь Лямбда. Kappaselect и Lambdaselect представляют собой аффинный хроматографический материал, разработанный BAC, BV и GE Healthcare.
[0081] Очищенные биспецифичные антитела характеризовали, как описано ниже. Анализировали проточную и элюированную фракции из каждой стадии аффинной очистки с помощью SDS-PAGE. Результаты выявили, что на каждой стадии биспецифичные антитела обогащаются (Фигура 8). κλ-антитело содержало эквивалентные количества легких цепей Каппа и Лямбда. κλ-антитело демонстрировало образец с промежуточной миграцией на геле с изоэлектрическим фокусированием и в ион-обменной хроматографии по сравнению с моноспецифичными антителами (Фигура 10). Специфичность и аффинность κλ-антител определяли с помощью ELISA и поверхностного плазмонного резонанса. Способы по изобретению дают возможность идентификации антител с аффинностями в интервале от субнаномолярного до наномолярного без оптимизации. Это не очевидно, поскольку разнообразие библиотек антител, описанных в данном документе, ограничено легкой цепью, которая вносит меньший вклад в энергию связывания в стандартных антителах.
[0082] Чтобы избежать требования наличия доступа к двум антителам, имеющим вариабельные домены легкой цепи Каппа-типа и Лямбда-типа, которое казалось ограничением настоящего изобретения, способы, описанные в данном документе, дают возможность получения гибридной легкой цепи, в которой вариабельный домен Лямбда может быть сшит с константным доменом Каппа и, наоборот, вариабельный домен Каппа может быть сшит с константным доменом Лямбда, как описано на Фигуре 3. Это расширяет применения изобретения по отношению к парам антител, которые имеют общую легкую цепь одного типа. Как описано в Примерах, представленных в данном документе, точка сшивки между вариабельным и константным доменами является важной и может влиять на процесс очистки биспецифичного антитела.
[0083] Обзор одного способа получения биспецифичных и/или мультиспецифичных антител по изобретению представлен на Фигуре 13. В некоторых вариантах осуществления в способах получения биспецифичных и/или мультиспецифичных антител используют полностью бессывороточный химически обусловленный процесс. Эти способы включают наиболее широко используемую в фармацевтической промышленности клеточную линию, клетки яичника китайского хомяка (CHO). Способы, описанные в данном документе, используются для получения как полустабильных, так и стабильных клеточных линий. Способы могут использоваться для лабораторного производства биспецифичных и/или мультиспецифичных антител по изобретению (например, в колбе Эрленмейера) и в среднем масштабе (например, в 25-л баллонах "Wave bag"). Способы также легко адаптируются для более крупномасштабного производства бисецифичных и/или мультиспецифичных антител, а также смесей антител по изобретению.
[0084] Способы получения биспецифичных и/или мультиспецифичных антител по изобретению являются предпочтительными, так как они применяют универсальные процессы очистки, как показано на Фигуре 8A. Фигура 16 демонстрирует очистку и тестирование целостности продукта биспецифичных антител, очищенных из полустабильной клеточной линии. Биспецифичные антитела очищали с использованием трехстадийной процедуры аффинной хроматографии: (i) очистка с использованием Протеина A с захватом молекул IgG, включающих как моноспецифичные, так и биспецифичные антитела; (ii) очистка на KappaSelect с захватом IgG, содержащих легкие цепи Каппа; (iii) очистка на LambdaSelect с захватом IgG, содержащих легкие цепи Лямбда. Анализировали проточную и элюированную фракции из каждой стадии аффинной очистки с помощью SDS-PAGE. Результаты демонстрировали удаление каждой моноспецифичной формы (т.е. моноспецифичных молекул IgG, имеющих легкие цепи Каппа, и моноспецифичных молекул IgG, имеющих легкие цепи Лямбда) во время процесса очистки (Фигура 16A). Очищенные κλ-содержащие антитела (т.e. антитела, имеющие легкие цепи как Каппа, так и Лямбда) содержали эквивалентное количество легких цепей Каппа и Лямбда (Фигура 16B). Очищенные κλ-содержащие антитела представляли собой образец промежуточной миграции на геле с изоэлектрическим фокусированием по сравнению с двумя моноспецифичными антителами (Фигура 16C).
[0085] Химически обусловленные процессы для производства биспецифичных и/или моноспецифичных антител по изобретению могут использоваться или с пулами клеток CHO, или с созданными клеточными линиями. Результаты, полученные с использованием химически обусловленных процессов с использованием либо пулов, либо созданных клеточных линий, демонстрируют характеристики продуктивности и роста, сравнимые с клетками, экспрессирующими соответствующие Каппа- или Лямбда-содержащие моноспецифичные антитела. Таким образом, κλ-антитело сохраняет как структуру, так и характеристики производительности классических человеческих IgG.
[0086] Предыдущие способы получения биспецифичного формата антител, направленные на форсирование выработки гомогенной биспецифичной молекулы с использованием различных способов конструирования, описанных выше, осуществлялись за счет продуктивности, масштабируемости и стабильности продукта. Настоящее изобретение представляет собой отличный способ, который дает возможность получения простой смеси антител, которые обладают стандартными характеристиками продуктивности и масштабируемости моноклональных антител, и обеспечивает универсальные средства очистки биспецифичного антитела из смеси или очистки смеси антител.
ПРИМЕРЫ
ПРИМЕР 1: Получение библиотек антител, имеющих фиксированные тяжелые цепи
[0087] Библиотеки антител, в которых вариабельный домен тяжелой цепи идентичен для всех компонентов библиотеки, получали, как описано далее. Во-первых, вариабельный домен VH3-23 тяжелой цепи, содержащий определенную последовательность CDR3 AKSYGAFDY (SEQ ID NO: 1) (номенклатура CDR согласно IMGT) и определенную последовательность FR4, клонировали в вектор pNDS с использованием сайтов рестрикции SfiI и XhoI с получением фагмидного вектора pNDS_VHfixed. Аминокислотная последовательность VK FR4 соответствует участку FR4, кодируемому зародышевыми J генами JK1. Аминокислотная последовательность Vλ, FR4 соответствует участку FR4, кодируемому зародышевыми J генами JL2 и JL3. Два варианта последовательности Vk FR4 получали с заменой одной аминокислоты в положении 106 (Аргинин или Глицин). Всего клонировали 6 генов вариабельных доменов Каппа (VK1-33, VK1-39, VK3-11, VK3-15, VK3-20, VK4-1) и 5 Лямбда (Vλ1-44, Vλ1-51, Vλ6-57, Vλ2-14, Vλ1-40), содержащих дополнительный фрагмент вместо CDR3-кодирующей последовательности, в вектор pNDS_VHfixed с целью получения 17 акцепторных векторов, в которые можно было клонировать множество генов синтетического или естественного происхождения и можно было получать библиотеки с высокой степенью разнообразия согласно способам, описанным в прилагаемой заявке PCT/US2010/035619, от 20.05.2010, опубликованной как W02010/135558, и согласно способам, описанным в Ravn et al. (2010) Nucl Acid Res 38(21):el93, каждая из которых введена в данный документ ссылкой в полном объеме. Процесс, приводящий в результате к получению 17 библиотек, содержащих общий домен VH3-23 и различные домены VKappa или VLambda, располагающиеся в гипервариабельном участке легкой цепи 3 (CDRL3-участок), содержащих в целом 6,9×109 вариантов (Фигура 5A). Секвенирование 180 случайно отобранных трансформантов выявило, что >90% клонов интегрировали CDRL3-последовательности, которые были в рамке, и, таким образом, являлись потенциально функциональными.
ПРИМЕР 2: Восстановление фаговых библиотек
[0088] Каждую библиотеку восстанавливали независимо согласно стандартным процедурам фагового дисплея, которые вкратце суммированы ниже. Объем клеток и замороженных аликвот библиотеки, достаточных для перекрывания, по меньшей мере, в 10 раз теоретического разнообразия библиотеки, добавляли к 500 мл 2xTYAG (100 мкг/мл ампициллина; 2% глюкозы) и наращивали при 37°C с перемешиванием (240 об/мин) до достижения OD600, составляющего 0,3-0,5. Затем культуру суперинфицировали с использованием фага-помощника MK13KO7 в течение одного часа при 37°C (150 об/мин). Затем среду меняли путем центрифугирования клеток при 2000 об/мин в течение 10 минут, удаляли среду и ресуспендировали осадок в 500 мл 2xTY-AK (100 мкг/мл ампициллина; 50 мкг/мл канамицина). Затем наращивали культуру в течение ночи при 30°C (240 об/мин). Культуру центрифугировали при 4000 об/мин в течение 20 минут для осаждения клеток. Надосадочную жидкость собирали и добавляли 30% (об./об.) PEG 8000 (20%)/2,5 M NaCl для осаждения фаговых частиц путем инкубации смеси 1 час на льду. Фаговые частицы собирали путем центрифугирования при 10000 об/мин в течение 30 минут и ресуспендировали в 10 мл TE буфера (10 мM tris-HCl pH 8,0; 1 мM EDTA). Ресуспендированный раствор центрифугировали при 10000 об/мин для очистки от бактериального дебриса и повторяли процедуру осаждения. После конечного ресуспендирования, фаг титровали путем инфекции E. coli и измерении поглощения при 260 нм. Представленный уровень scFv на поверхности фага также оценивали с помощью Вестерн-блот-анализа с использованием моноклонального антитела к c-myc. Очищенный фаг из различных библиотек сохраняли замороженным при -80°C после добавления глицерина до конечной концентрации 15% (масс./об.).
ПРИМЕР 3: Селекция фагового дисплея с использованием библиотек с фиксированной тяжелой цепью
[0089] Жидкофазные селекции против биотинилированного гибридного белка hCXCL10-NusA (hCXCL10-NusA) и биотинилированного рецепторного комплекса hIL6 (hIL6RC): Аликвоты VH-Vk и VH-Vλ фаговых библиотек (1011-1012 БОЕ) поддерживали отдельно и блокировали с использованием PBS, содержащего 3% (масс./об.) обезжиренного молока, в течение одного часа при комнатной температуре на роторном миксере. Блокированный фаг затем отбирали на покрытые стрептавидином магнитные микросферы (Dynal M-280) в течение одного часа при комнатной температуре на роторном миксере. Отобранный фаг затем инкубировали вместе с in vivo биотинилированным hCXCL10-NusA или hIL6RC (100 нM) в течение двух часов при комнатной температуре на роторном миксере. Микросферы добавляли к мишени и захватывались с использованием магнитного штатива с последующими четырьмя промывками с помощью PBS/0,1% Tween 20 и 3 промывками с помощью PBS. Микросферы затем добавляли непосредственно к 10 мл экспоненциальной культуры TG1-клеток и инкубировали в течение одного часа при 37°C с медленным встряхиванием (100 об/мин). Аликвоту инфицированных TG1 последовательно разводили для титрования результатов селекции. Оставшиеся инфицированные TG1 центрифугировали при 3000 об/мин в течение 15 минут и ресуспендировали в 0,5 мл 2xTYAG (2xTY среда, содержащая 100 мкг/мл ампициллина и 2% глюкозы) и высевали на чашки с 2xTYAG агаром Bioassay. После ночной инкубации при 30°C, 10 мл 2xTYAG добавляли к чашкам и клетки соскребали с поверхности и переносили в 50-мл полипропиленовые пробирки. К клеточной суспензии добавляли 2xTYAG, содержащую 50% глицерина с получением конечной концентрации глицерина 17%. Аликвоты раунда селекции поддерживали при -80°C.
[0090] Восстановление фага: 100 мкл клеточной суспензии, полученной в предыдущих раундах селекции, добавляли к 20 мл 2xTYAG и наращивали при 37°C с перемешиванием (240 об/мин) до достижения значения OD600 составляющего 0,3-0,5. Культуру затем суперинфицировали с помощью 3,3×1010 фага-помощника MK13KO7 и инкубировали в течение одного часа при 37°C (150 об/мин). Затем среду меняли путем центрифугирования клеток при 3800 об/мин в течение 10 минут, удаляли среду и ресуспендировали осадок в 20 мл 2xTY-AK (100 мкг/мл ампициллина; 50 мкг/мл канамицина). Затем наращивали культуру в течение ночи при 30°C (240 об/мин). На следующий день аликвоту центрифугированной надосадочной жидкости использовали в качестве исходного количества следующего раунда селекции.
[0091] Восстановление моноклонального фага для ELISA: Одиночные клоны отбирали в микропланшет, содержащий 150 мкл среды 2xTYAG (2% глюкозы) на лунку, и наращивали при 37°C (100- 120 об/мин) в течение 5-6 ч. Фаг-помощник M13KO7 добавляли к каждой лунке с получением множественности заражения (MOI) 10 (т.e. 10 фагов на каждую клетку в культуре) и инкубировали при 37°C (100 об/мин) в течение 1 ч. После наращивания планшеты центрифугировали при 3200 об/мин в течение 10 мин. Надосадочную жидкость тщательно удаляли, клетки ресуспендировали в 150 мкл среды 2xTYAK и наращивали в течение ночи при 30°C (120 об/мин). Для ELISA, фаг блокировали путем добавления 150 мкл 2× концентрации PBS, содержащего 5% сухого обезжиренного молока с последующей инкубацией в течение часа при комнатной температуре. Планшеты затем центрифугировали 10 минут при 3000 об/мин, и надосадочную жидкость, содержащую фаг, использовали для ELISA.
[0092] ELISA фага: на ELISA-планшеты (Maxisorp, NUNC) наносили покрытие в течение ночи с помощью 2 мкг/мл hCXCL10-NusA в PBS или 2 мкг/мл hIL6RC в PBS. Планшеты блокировали с помощью 3% обезжиренного молока/PBS при комнатной температуре в течение 1 ч. Планшеты промывали 3 раза с помощью PBS 0,05% Tween 20 перед переносом предварительно блокированных фаг-содержащих надосадочных жидкостей и инкубацией в течение одного часа при комнатной температуре. Каждый клон тестировали против обеих мишеней для демонстрации специфичности. Планшеты промывали 3 раза с помощью PBS 0,05% Tween 20. Добавляли к каждой лунке 50 мкл 3% обезжиренного молока в PBS, содержащего (HRP)-конъюгированное антитело к M13 (Amersham, разведенное 1:10000). После инкубации при комнатной температуре в течение 1 ч, планшеты промывали 5 раз с помощью PBS 0,05% Tween 20. Затем осуществляли ELISA путем добавления 50 мкл TMB (Sigma) и 50 мкл 2N H2SO4 для остановки реакции. Интенсивность поглощения считывали при 450 нм. Можно было идентифицировать клоны, специфичные к hCXCL10-NusA или hIL6RC, несущим одинаковый вариабельный домен тяжелой цепи. (Фигура 4).
[0093] Секвенирование фаговых клонов: одиночные клоны наращивали в 5 мл среды 2xTYAG (2% глюкозы) на лунку и при 37°C (120 об/мин) в течение ночи. На следующий день фагмидную ДНК очищали и использовали для ДНК-секвенирования с использованием праймера, специфичного для pNDS1: mycseq, 5'-CTCTTCTGAGATGAGTTTTTG. (SEQ ID NO:2).
[0094] Крупномасштабная очистка scFv: исходную культуру в количестве 1 мл 2xTYAG инокулировали одиночной колонией со свежей чашки с агаром 2xTYAG и инкубировали со встряхиванием (240 об/мин) при 37°C в течение 5 часов. 0,9 мл данной культуры использовали для инокуляции 400 мл культуры той же среды и наращивали в течение ночи при 30°C с интенсивным встряхиванием (300 об/мин). На следующий день культуру индуцировали путем добавления 400 мкл 1M IPTG и продолжали инкубацию в течение дополнительных 3 часов. Клетки собирали с помощью центрифугирования при 5000 об/мин в течение 10 минут при 4°C. Осажденные клетки ресуспендировали в 10 мл ледяного буфера TES с добавлением протеазных ингибиторов, как описано выше. Осмотического шока достигали путем добавления 15 мл разведенного 1:5 буфера TES и инкубации в течение 1 часа на льду. Культуру центрифугировали при 10000 об/мин в течение 20 минут при 4°C для осаждения клеток. Надосадочную жидкость тщательно переносили в свежую пробирку. К надосадочной жидкости добавляли имидазол до конечной концентрации 10 мM. 1 мл смолы Ni-NTA (Qiagen), уравновешенной в PBS, добавляли в каждую пробирку и инкубировали на роторном миксере при 4°C (20 об/мин) в течение 1 часа. Пробирки центрифугировали при 2000 об/мин в течение 5 минут и надосадочную жидкость тщательно удаляли. Осажденную смолу ресуспендировали в 10 мл холодного (4°C) промывочного буфера 1 (50 мM NaH2PO4 300 мM NaCl, 10 мM имидазол, pH до 8). Суспензию добавляли к колонке "polyprep" (Biorad). 8 мл холодного промывочного буфера 2 (50 мM NaH2PO4, 300 мM NaCl, 20 мM имидазол, pH до 8) использовали для промывки колонки путем самотека. scFv элюировали из колонки с помощью 2 мл элюирующего буфера (50 мM NaH2PO4 300 мM NaCl, 250 мM имидазол, pH до 8). Фракции анализировали путем измерения поглощения при 280 нм и белок-содержащие фракции собирали перед заменой буфера на обессоливающей колонке NAP5 (Amersham), уравновешенной с помощью PBS. scFv в PBS анализировали с помощью SDS-PAGE и оценивали количественно с помощью поглощения при 280 нм. Очищенные scFv разделяли на аликвоты и сохраняли при -20°C и при 4°C. Очищенные scFv использовали в ELISA для подтверждения специфичного связывания с мишенью, против которой они были отобраны.
ПРИМЕР 4: Дополнительные селекции с использованием различных библиотек с фиксированными VH
[0095] Библиотеки антител также получали путем захвата естественно перестроенных вариантов легких цепей и путем их клонирования применительно к VH-домену, описанному в Примере 1. В данном случае цельный участок вариабельного гена легкой цепи амплифицировали из человеческой кДНК с использованием праймеров, которые соответствуют 5' и 3' участку человеческих перестроенных вариабельных участков, и клонировали в вектор pNDS_VHfixed, описанный в Примере 1. Другой набор библиотек получали, как описано в Примере 1, но с использованием фиксированного домена VH3-23, содержащего другую CDRH3-последовательность ARGDDVS (SEQ ID NO:3). Библиотеки, описанные выше, схематично представлены на Фигуре 5. Эти библиотеки с фиксированными VH использовали против панели белков-мишеней с использованием методологии селекции и скрининга, описанных в Примере 2 и 3. Селекцию осуществляли с использованием следующих мишеней: hCXCL10-NusA, IL-6RC, CD47, CD16, CD8 и hIFNγ. Идентифицированные кандидаты, которые, как было показано, являются специфичными для их мишеней, перечислены в Таблице II. Эти результаты демонстрируют, что могу быть получены антитела, связывающиеся с различными мишенями и обладающие общей тяжелой цепью, и что разнообразия, ограниченного легкой цепью, достаточно для придания антигенной специфичности. Кандидаты могут быть получены из библиотек, содержащих домен VH3-23 с различными последовательностями CDRH3 или имеющих варианты VL, отличающиеся в результате использования различных стратегий. Таким образом, результаты демонстрируют, что способ не ограничивается конкретной последовательностью VH или конкретной стратегией получения различий вариабельных доменов легкой цепи.
Ряд независимых клонов, идентифицированных против панели мишеней с использованием библиотек, содержащих фиксированную последовательность VH с двумя различными последовательностями CDRH3. NA: селекцию не осуществляли.
ПРИМЕР 5: Кандидаты с фиксированными VH, преобразующиеся в IgG, и транзиторная экспрессия в клетках млекопитающего
[0096] После скрининга scFv-кандидаты преобразовывались в IgG и экспрессировались при транзиторной трансфекции клеток PEAK. Получали несколько IgGλ (n=5) и IgGκ (n=9), имеющих общую тяжелую цепь, и специфичные к IFNγ и IL6RC, соответственно, и имеющие различные последовательности легкой цепи, как указано ниже. Последовательности VH и VL селектированных scFv амплифицировали с помощью специфичных олигонуклеотидов и клонировали в экспрессирующий вектор, содержащий константные участки тяжелой и легкой цепи, и полученные конструкты были подтверждены с помощью секвенирования. Экспрессирующие векторы трансфицировали в клетки млекопитающего с использованием реагента для трансфекции Fugene 6 (Roche, Базель, Швейцария). Вкратце, клетки Peak культивировали в 6-луночных планшетах в концентрации 6×105 клеток на лунку в 2 мл культуральной среды, содержащей фетальную бычью сыворотку. Экспрессирующие векторы, кодирующие кандидатные последовательности VH и VL, ко-трансфицировали в клетки с использованием реагента для трансфекции Fugene 6 согласно инструкциям производителя. Через день после трансфекции, культуральную среду удаляли и добавляли к клеткам 3 мл свежей бессывороточной среды и культивировали в течение трех дней при 37°C. Через три дня периода культивирования надосадочную жидкость собирали для получения IgG, очищенного на быстропроточных колонках с протеин G-Сефарозой 4B (Sigma, Сент-Луис, Миссури) согласно инструкциям производителя. Вкратце, надосадочные жидкости из трансфицированных клеток инкубировали при 4°C в присутствии IgG-связывающего буфера ImmunoPure (G) (Pierce, Рокфорд, Иллинойс). Образцы затем пропускали через быстропроточные колонки с Протеин G-Сефарозой 4B, и затем очищали IgG с использованием элюирующего буфера. Элюированная фракцию IgG затем диализовали против PBS, и содержание IgG оценивали количественно с помощью поглощения при 280 нм. Чистоту и целостность IgG подтверждали с помощью SDS-PAGE. IgG затем тестировали на предмет связывания с IFNγ с комплексом рецепторов IL-6/IL-6R, обозначенных в данном документе как IL6RC, с помощью ELISA. Биотинилированные IFNγ и IL6RC иммобилизовали на покрытых стрептавидином микропланшетах (Streptawell, Roche), и планшеты затем блокировали с помощью PBS с добавлением 2% BSA при комнатной температуре в течение 1 ч. Планшеты промывали 3 раза с помощью PBS 0,05% Tween 20 перед добавлением очищенных анти-IFNγ IgGλ или анти-IL6RC IgGк к лункам, покрытым с помощью каждой мишени. Через один час инкубации при комнатной температуре и промывки связанных планшетов, антитела детектировали с использованием связанных с пероксидазой хрена антител к человеческому Сk или к человеческому Cλ. Затем осуществляли ELISA путем добавления 50 мкл TMB (Sigma) и 50 мкл 2N H2SО4 для остановки реакции. Интенсивность поглощения считывали при 450 нм. Результаты выявляют, что специфичность связывания scFv, выделенных из библиотек с фиксированными VH, поддерживали в формате IgG и продемонстрировали, что два IgG, имеющих одинаковую тяжелую цепь, но различные легкие цепи, могут быть специфичны к различным мишеням (Фигура 6).
[0097] Аминокислотные последовательности тяжелой и легкой цепи анти-IFNγ IgGλ или анти-IL6RC IgGк указаны ниже:
Анти-IL6RC VKappa легкая цепь
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSG SGSGTDFTLTISSLEPEDFAVYYCQQWLPTTPATFGQGTKVEIKRTVAAPSVFIFPPSDEQLKS GTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKV YACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 4)
Анти-IFNγ VLambda легкая цепь
NFMLTQPHSVSESPGKTVTISCTRSSGSIASNYVQWYQQRPGSSPTTVIYEDNQRPSGVPDRFS GSIDSSSNSASLTISGLKTEDEADYYCQSQSWDGNHIVFGGGTKLTVLGQPKAAPSVTLFPPSS EELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWK SHRSYSCQVTHEGSTVEKTVAPTECS(SEQ ID NO: 5)
Общая тяжелая цепь
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSV KGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKSYGAFDYWGQGTLVTVSSASTKGPSVFPLA PSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSL GTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTP EVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKC KVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG (SEQ ID NO: 6).
ПРИМЕР 6: Ко-экспрессия одной тяжелой цепи и двух легких цепей в клетках млекопитающего
[0098] Одновременная экспрессия одной тяжелой цепи и двух легких цепей в одной клетке может приводить к сборке трех различных антител (Фигура 8A). Одновременная экспрессия может достигаться различными путями, как например, при трансфекции множества векторов, экспрессирующих одну из цепей для их ко-экспрессии, или путем использования векторов, которые управляют экспрессией множества генов. Вектор pNovi kНλ конструировали для возможности ко-экспрессии одной тяжелой цепи, одной легкой цепи Каппа и одной легкой цепи Лямбда (Фигура 7A). Экспрессия трех генов управляется промоторами человеческого цитомегаловируса (hCMV), и векторы также сдержат ген глутамин синтетазы (GS) который дает возможность селекции и создания стабильных клеточных линий. Также конструировали другой вектор, pNovi kНλk, в котором вторую копию легкой цепи Каппа вставляли после легкой цепи Лямбда и их экспрессией управляли с помощью внутреннего сайта посадки рибосомы (IRES). С использованием данной бицистронной конструкции можно было увеличить относительную экспрессию легкой цепи Каппа. Два вектора схематично представлены на Фигуре 7.
ПРИМЕР 7: Транзиторная ко-экспрессия одной тяжелой цепи и двух легких цепей в клетках млекопитающего и очистка суммарного IgG.
[0099] Ген VH и VL анти-IFNγ IgGλ, или анти-IL6RC IgGк клонировали в вектор pNovi kНλ, описанный в Примере 5, для транзиторной экспрессии в клетках млекопитающего. Клетки Peak культивировали в 6-луночных планшетах в концентрации 6×105 клеток на лунку в 2 мл культуральной среды, содержащей фетальную бычью сыворотку. 2 мкг плазмидной ДНК трансфицировали в клетки с использованием реагента для трансфекции TransIT-LT1 (Mirus) согласно инструкциям производителя. Через день после трансфекции, культуральную среду удаляли и добавляли к клеткам 3 мл свежей бессывороточной среды и культивировали в течение пяти дней при 37°C. Через пять дней периода культивирования надосадочную жидкость собирали, центрифугировали при 4000 об/мин и применяли на смоле MabSelect Sure PA (GE Healthcare) согласно инструкциям производителя. Суммарный IgG эллюировали при кислых условиях с последующей нейтрализацией и заменой буфера на PBS. Содержание IgG оценивали количественно с помощью поглощения при 280 нм и анализировали на SDS-PAGE. Присутствие двух полос, соответствующих двум легким цепям, и одной полосы, соответствующей тяжелой цепи, выявило, что три цепи могут экспрессироваться одновременно в одной клетке (Фигура 8B).
ПРИМЕР 8: Очистка биспецифичных антител, несущих легкую цепь Лямбда и Каппа
[00100] Коэкспрессия тяжелой цепи и двух легких цепей в одних и тех же клетках может привести к сборке трех различных антител: два моноспецифичных антитела, несущих одну и ту же легкую цепь на обоих плечах, и биспецифичное антитело, несущее различные легкие цепи, ассоциированные с тяжелой цепью на каждом плече. Последнее может быть отделено от моноспецифичных антител методами хроматографии, в которых используются преимущества различных биохимических свойств моноспецифичных и биспецифичных антител. Для разработки общего подхода очистки, который применим к получению всех биспецифичных антител, мы применили подход аффинной хроматографии, схематически изображенный на Фигуре 8А. Культуральные надосадочные жидкости из трансфицированных клеток наносили на колонку со смолой MabSelect Sure PA (GE Healthcare) и очищали общий IgG как описано в Примере 7. Общие IgG затем наносили на колонку, содержащую аффинную матрицу CaptureSelect Fab Kappa (BAC BV, Голландия), уравновешенную 10 объемами PBS. Колонку затем промывали PBS, 5-10 объемами колонки. Иммуноглобулиновые молекулы, несущие легкую цепь Каппа, элюировали из колонки путем нанесения 5 объемов колонки 0,1 M глицина pH 2,0 и фракции собирали и нейтрализовали. Фракции, содержащие антитело, объединяли перед заменой буфера на обессоливающей колонке PD10 (Amersham), уравновешенной PBS. Антитело затем наносили на вторую колонку, содержащую аффинную матрицу CaptureSelect Fab Lambda, уравновешенную 10 объемами PBS. Колонку затем промывали PBS, 5-10 объемами колонки. Иммуноглобулиновые молекулы, несущие только легкую цепь Каппа, связывались с колонкой, и обнаруживались в элюате. Антитела, несущие легкую цепь Лямбда, элюировали из колонки нанесением 5 объемов колонки 0,1 M глицина pH 3,0 и собирали фракции. Фракции, содержащие биспецифичное антитело, объединяли перед заменой буфера на обессоливающей колонке PD10 (Amersham), уравновешенной PBS. Иммуноглобулиновые молекулы, несущие легкую цепь Каппа, элюировали из колонки путем нанесения 5 объемов колонки 0,1 M глицина pH 2,0, и фракции собирали и нейтрализовали. Фракции, содержащие антитело, объединяли перед заменой буфера на обессоливающей колонке PD10 (Amersham), уравновешенной PBS. Антитело затем наносили на вторую колонку, содержащую аффинную матрицу CaptureSelect Fab Lambda, уравновешенную 10 объемами PBS. Колонку затем промывали PBS, 5-10 объемов колонки. Иммуноглобулиновые молекулы, несущие только легкую цепь Каппа, связывались с колонкой, и обнаруживались в элюате. Антитела, несущие легкую цепь Лямбда, элюировали из колонки нанесением 5 объемов колонки 0,1 M глицина pH 3,0 и собирали фракции. Фракции, содержащие биспецифичное антитело, объединяли перед заменой буфера на обессоливающей колонке PD10 (Amersham), уравновешенной PBS. Проточную фракцию и элюированную фракцию каждой стадии очистки анализировали с помощью SDS-PAGE, и было показано, что антитела, несущие легкие цепи Лямбда, были обнаружены в проточной фракции аффинной матрицы CaptureSelect Fab Kappa и, наоборот, что антитела, несущие легкие цепи Каппа, были обнаружены в проточной фракции аффинной матрицы CaptureSelect Fab Lambda (Фигура 8B). Как ожидалось, в конечной элюированной фракции интенсивность полос, соответствующих двум легким цепям, была эквивалентна. Таким образом, этот трехстадийный способ дает возможность очистки биспецифичных антител, несущих легкие цепи Каппа и Лямбда. Конечное извлечение биспецифичного антитела, несущего легкие цепи Каппа и Лямбда, составляло приблизительно 10-21% в различных экспериментах. Фракция, элюированная с использованием Протеина A, также выявила, что большее количество легкой цепи Лямбда, чем Каппа участвовало в сборке до очищенного IgG, предполагая, что повышенная экспрессия легкой цепи Каппа повышает сборку и извлечение биспецифичного антитела.
ПРИМЕР 9: Модуляция экспрессии легкой цепи для оптимизации сборки биспецифичного антитела
[00101] Для повышения экспрессии легкой цепи Каппа, общий VH ген, ген VLambda анти-IFNγ антитела и две копии VKappa анти-IL6RC клонировали в вектор pNovi kНλk описанный в Примере 6. Экспрессия второй копии легкой цепи VKappa управляется посредством IRES. Различные элементы IRES тестировали для достижения различных уровней экспрессии легкой цепи VKappa. Это привело к созданию пяти независимых векторов: pNovi kНλκ 1-5. Эти векторы были использованы для временных трансфекций в клетки млекопитающих как описано в Примере 7, и общий IgG очищали из надосадочной жидкости с помощью аффинной очистки на протеине А. SDS-PAGE - анализ показал, что различные векторы pNovi kНλk повышают экспрессию и сборку легкой цепи Каппа в IgG по сравнению с экспрессией с помощью вектора pNovi kНλ, (Фигура 9). Фракции общего IgG, полученные после очистки на протеине А для каждого из этих конструктов, дополнительно очищали в двух последовательных стадиях с помощью CaptureSelect Fab Kappa и CaptureSelect Fab Lambda, как описано в Примере 8. Выход биспецифичных антител составил 20% для pNovi kНλ, и находился в диапазоне от 33 до 41% для конструктов pNovi kНλk, что указывает на то, что повышенная экспрессия легкой цепи Каппа приводит к улучшенной сборке биспецифичного антитела. Для дополнительного подтверждения этих результатов, проводили котрансфекции с pNovi kНλ, и вектором, экспрессирующим моноспецифичное анти-IL6RC IgGк. Таким образом, относительные уровни легкой цепи Каппа, а также выход биспецифичного антитела также были повышены, что указывает на то, что несколько подходов могут быть предприняты для коррекции относительной экспрессии легкой цепи.
ПРИМЕР 10: Описание очищенного биспецифичного антитела
Очищенные биспецифичные антитела, выделенные, как описано в Примерах выше, описывали с помощью различных методов.
[00102] Изоэлектрическое фокусирование (IEF). Очищенные биспецифичные антитела, выделенные как описано в Примере 9, анализировали с помощью пластин "IsoGel Agarose IEF" с диапазоном pH 3-10 (Lonza) и сравнивали с моноспецифичными антителами анти-IFNγ IgGλ и анти-IL6RC IgGк. После фокусирования, гель был помещен в фиксирующий раствор на 30 мин, отмыт водой в течение 5 мин и затем высушен при КТ. Гель окрашивали Кумасси в течение 15 мин, вкратце, споласкивали водой и обесцвечивающим раствором 2 раза по 15 мин. В конце гель сушили при КТ перед получением изображения. Результаты, показанные на Фигуре 10А, указывают на то, что IgGкλ, биспецифичное антитело, имеет отличный и промежуточный pI по сравнению с pI двух моноспецифичных антител, что прогнозируется форматом антитела и теоретическими расчетами. Окрашивание также продемонстрировало, что биспецифичное антитело является в высокой степени чистым после трехстадийного процесса очистки, описанного в Примере 8.
[00103] Ионообменная хроматография (IEX). 50 мкг очищенных моноспецифичных и биспецифичных антител анализировали ионообменной высокоэффективной жидкостной хроматографией (IEX-ВЭЖХ) (HPLC Waters e2695/детектор 2489) на колонке Agilent Bio Mab, NP5, (Agilent): мобильными фазами являлись: A: Na2HPO4/NaH2PO4 10 мМ, pH 6,5; B: Na2HPO4/NaH2PO4 10 мM, NaCl 100 мМ, pH 6,5; при потоке 0,8 мл/мин и 20%-60% градиенте в течение 133 минут. Три различных антитела имеют различные периоды удержания с пиком, соответствующими биспецифичному антителу, имеющему промежуточный профиль по сравнению с моноспецифичными антителами (Фигура 10В).
[00104] ELISA. Моно специфичное анти-IFNγ IgGλ, анти-IL6RC IgGк и биспецифичное IgGкλ, антитела были протестированы ELISA на предмет связывания с IFNγ и IL6RC. Результаты, показанные на Фигуре 11, указывают на то, что биспецифичное антитело IgGкλ способно связывать обе мишени и может быть обнаружено как вторичными антителами, связывающими человеческий Ск, так и вторичными антителами, связывающими человеческий Сλ.
[00105] Поверхностный плазмонный резонанс (SPR). Аффинность и кинетика связывания моноспецифичного анти-IFNγ IgGλ, анти-IL6RC IgGк и биспецифичного IgGкλ антитела были описаны на приборе Biacore 2000 (Biacore AB, Упсала, Швеция). 200RU козьих поликлональных антител против IgG человека (ahIgG; Biacore) иммобилизовали с помощью химического состава EDC/NHS на чипе CM5 Biacore. Эту поверхность использовали для захвата моноспецифичного или биспецифичного человеческого IgG. Поверхность обновляли после каждого цикла инъекцией 10 мМ глицина pH 1,5 при 30 мкл/мин, в течение 30 сек, с последующей 1 мин стабилизацией в буфере HBS-EP. Данные согласовывали согласно 1:1 модели Ленгмюра и определяли значения Kon, Koff и KD (Таблица III). Похожие значения аффинности были получены для моноспецифичных антител и биспецифичных антител IgGкλ. Данные указывают на то, что два антиген-связыващих активных центра антитела связывают hIFNγ и IL6RC похожим образом в моноспецифичном и биспецифичном формате.
[00106] Способность биспецифичного IgGкλ взаимодействовать с обоими мишенями одновременно тестировали с помощью SPR. Биотинилированный IFNγ иммобилизовали на покрытом стрептавидином чипе CM5 Biacore. Моноспецифичное и биспецифичное антитела были инъецированы на эту поверхность с последующей инъекцией IL6RC. Сенсограмма, показанная на Фигуре 11A, демонстрирует, что биспецифичное антитело IgGкλ было способно связывать иммобилизованный IFNγ, и было способно одновременно захватывать IL6RC. SPR также использовали для оценки относительных количеств легких цепей Каппа и Лямбда в очищенном биспецифичном антителе. IgGкλ напрямую иммобилизовали присоединением через амин на поверхности чипа CM5 Biacore и инъецировали антитело, связывающее человеческий CKappa, за которым следовало антитело, связывающее человеческий CLambda, в такой же концентрации. Эквивалентные ответы были получены как антителами против легкой цепи, что указывает на то, что как спрогнозировано форматом, эквивалентные количества Каппа и Лямбда легких цепей присутствуют в биспецифическом антителе IgGкλ (Фигура 12В).
Анализ кинетики связывания с IFNγ и IL6RC для моноспецифичного и биспецифичного IgGкλ антител измеряли на системе Biacore 2000.
ПРИМЕР 11: Производство биспецифичных антител IgGкλ
[00107] Экспрессия IgGкλ, биспецифичного антитела, также была осуществлена в клетках Яичников Китайского Хомяка (CHO), которые широко используются для производства моноклональных антител. В примере, представленном в данном случае, для выработки биспецифичных антител IgGкλ были созданы как полустабильные пулы трансфицированных клеток CHO, так и стабильные линии клеток CHO. В исследовании, представленном в данном случае, стабильные линии CHO были получены и выращены с помощью процесса производства без животных компонентов и с определенным химическим составом (CDACF). Полный процесс изображен на Фигуре13.
[00108] Пулы CHO. Клетки CHO электропорировали линеаризованным вектором pNovi kНλk, кодирующим IgGкλ, анти-IFNγ/анти-IL6RC биспецифичное антитело, описанное в примерах выше, а также плазмидами, экспрессирующими моноспецифичные анти-IFNγ IgGλ и анти-IL6RC IgGк. После электропорации, пулы трансфицированных клеток выращивали в 10 дневных условиях перероста с отсутствием подкормки. Клетки, трансфицированые биспецифичным конструктом, представляют похожие профили роста по сравнению с клетками, трансфицированными векторами для экспрессии моноспецифичных антител. Кроме того, продуктивности также были сравнимы и достигали типичного диапазона продуктивности антител: между 100-200 мг/л для культур, перерощенных пулов с отсутствием подкормки (Фигура 14А). Масштабирование от мелкомасшатабной выработки в колбе Эрленмейера (100 мл) до среднемасштабной выработки в 25 л Wave Bag было успешно осуществлено (Фигура 14В). Эти результаты указывают на то, что похожие кривые роста и продуктивности получаются в ходе экспрессии биспецифичного антитела IgGкλ в клеточной линии CHO и соответствующих моноспецифичных антител.
[00109] Стабильные линии CHO. Рекомбинантные клеточные линии, вырабатывающие биспицефичное антитело, были получены электропорацией клеток CHO вектором pNovi кНкλ. После трансфекции, рекомбинантные клеточные линии отбирали разведением клеточной культуры в присутствии конечной концентрации 50 мкM метионин сульфоксимина (MSX). После 6 недель инкубации, колонии рекомбинантных клеточных линии подвергали скринингу по общей продуктивности IgG c помощью FastELISA® (R&D Biotech) (Фигура 15A-B). Выбранные клеточные линии наращивали в клеточной культуральной среде, содержащей 25 мкМ MSX, переносили в 24-луночные микропланшеты и подвергали скринингу на продуктивность и особенность роста в суспензионной культуре (Фигура 15C-D). Результаты выявили, что IgGкλ, биспецифичное антитело, может быть выработано в перемешиваемых периодических условиях перероста на уровне, сравнимом с клеточной линией, экспрессирующей стандартные моноспецифичные антитела. Клеточные линии с наивысшей выработкой были отобраны и обработаны в 50 мл периодических переросших культурах в перемешиваемых колбах в течение максимум 10 дней. Протеин А ВЭЖХ использовали для количественной оценки суммарного IgG в надосадочной жидкости. Суммарный IgG из надосадочной жидкости 10 клеточных линий с наивысшей выработкой были очищены хроматографией "MabSelect SuRE" с помощью 1 мл предварительно наполненных колонок HiTrap (GE Healthcare). Относительные количества моноспецифичных и биспецифичных антител в суммарном очищенном IgG оценивали с помощью IEX-ВЭЖХ, как описано в Примере 10. Для основной части клеточных линий, фракция IgGкλ, биспецифичного антитела варьирует между 37-42% от общего IgG, и две клеточные линии экспрессируют меньшие количества IgGкλ (22 и 25%). Результаты 10 клеточных линий CHO суммированы в Таблице IV.
SuRr (мг)
[00110] Очистка и описание биспецифичного антитела IgGкλ, экспрессированного в CHO. Надосадочную жидкость пула клеток CHO, трансфицированных биспецифичными и моноспецифичными конструктами, использовали для очистки. Моноспецифические анти-IFNγ IgGλ и анти-IL6RC IgGк очищали с помощью афинной хроматографии с протеином А и обессоливали в PBS, тогда как биспецифичные антитела IgGкλ очищали с помощью трехстадийного процесса афинной хроматографии, описанного в Примере 8. Элюированные фракции и проточную фракцию различных стадий, а также конечные очищенные образцы анализировали с помощью SDS-PAGE и IEF (Фигура 16). Специфичность IgGкλ отслеживали после каждой стадии очистки с помощью ELISA (Фигура 17). Результаты говорят о том, что процесс очистки был надежным и сравнимым с экспрессией в CHO и давал биспецифичное антитело IgGкλ высокой чистоты.
ПРИМЕР 12: Дополнительные примеры биспецифичных антител IgGкλ
[00111] Другие примеры биспецифичных антител IgGкλ были выделены, экспрессированы и очищены, как описано в Примерах выше. Они включают биспецифичные антитела анти-NusA/анти-IFNγ IgGкλ и анти-IL6RC/анти-IL6RC IgGкλ, в которых оба объединенных сайта связывают IL6RC, но несут легкую цепь Каппа и Лямбда. Эти очищенные биспецифичные антитела анализировали с помощью IEF вместе с их соответствующими моноспецифичными эквивалентами (Фигура 18). Результаты говорят о том, что pI биспецифичного антитела всегда находится посередине между pI моноспецифичных антител, но что данные различия могут варьировать значительно в зависимости от последовательности вариабельного домена легкой цепи. Данный пример является иллюстрацией того, что очистка биспецифичного антитела, основанная на различиях заряда, может быть затруднительной, если две легкие цепи имеют одинаковые биохимические свойства, и подчеркивает преимущество подхода аффинной очистки изобретения.
ПРИМЕР 13: Получение биспецифических антител IgGкλ с помощью двух Каппа или двух Лямбда вариабельных доменов
[00112] Биспецифичные антитела могут быть получены с помощью двух вариабельных доменов одного и того же типа (Лямбда и Каппа). Поскольку стадии аффинной очистки требуют наличия константного Каппа и константного Лямбда доменов легких цепей, любой вариабельный домен легкой цепи может в принципе быть объединен с этими константными доменами для получения гибридных легких цепей, как показано на Фигуре 3. Это было продемонстрировано с помощью двух антител, направленных против IFNγ и IL6RC, выделенных из библиотек антител с фиксированным VH, описанных в Примере 1. В этом случае обе библиотеки совместно используют одинаковые VH и имеют вариабельный домен легкой цепи Лямбда. Домен VLamda антитела анти-IFNγ объединяли с константным доменом Лямбда, тогда как домен VLamda анти-IL6RC антитела объединяли с константным доменом Каппа. В случае последнего получили три различных конструкта, включая различные точки объединения между доменами VLamda и доменом CKappa. В одном конструкте (гибрид 1) точка объединения была на конце каркасного участка 4 (FR4) домена VLambda и включала целый константный домен Каппа (Фигура 19A). В другом конструкте (гибрид 2), участок FR4 лямбда был заменена на участок FR4 Каппа (Фигура 19B). В третьем конструкте (гибрид 3), первые 4 аминокислоты константного домена Каппа были заменены 4 аминокислотами константного домена Лямбда (Фигура 19C). Эти различные Лямбда-Каппа гибриды легких цепей были клонированы вместе с обычной тяжелой цепью и легкой цепью Лямбда в вектор pNovi kНλ. Трансфекцию клеток млекопитающих, экспрессию белков и три стадии аффинной очистки IgGкλ осуществляли, как описано в Примерах выше. Анализ элюированных фракций указывает на то, что легкая цепь гибрида 3 не связывается со смолой, селективной для Каппа и следовательно не позволяет осуществить эффективную очистку биспецифичного антитела. Гибрид 1 и гибрид 2 делает возможной эффективную биспецифичную очистку IgGкλ. Эти очищенные IgGкλ, несущие гибридную легкую цепь, анализировали на Agilent Bionalyzer 2100 с помощью чипа Protein 80 (Agilent Technologies) и пики на электроферограмме, соответствующие легким цепям, были показаны как эквивалентные (Фигура 20). Результаты говорят о том, что биспецифичные антитела по изобретению могут быть получены с помощью двух антител, имеющих общую тяжелую цепь и две легкие цепи, имеющие вариабельные домены одного типа (Каппа или Лямбда).
[00113]
ПРИМЕР 14: VH и VL описанных антител вовлечены в связывание антигена
[00114] Для проверки вклада общего VH в связывание антигена, различные легкие цепи scFv, несущие общий VH, выбранные в Примерах 1 и 4, были объединены с двумя различными VH, которые отличаются от исходного общего VH. Различные scFv могут быть экспрессированы и очищены, как описано в Примере 3 и протестированы на предмет связывания с мишенью, против которой они были получены. Примеры, представленные на Фигуре 21, демонстрируют, что scFv, специфичное для IFNγ, и scFv, специфичное для IL6RC, могут только связывать соответствующие мишени, при объединении с VH доменом, с которым они изначально были отобраны, и не наблюдалось связывания при объединении с двумя другими VH доменами, которые делают возможными их экспрессию и очистку. Результаты указывают на то, что несмотря на тот факт, что вариабельность относится исключительно к VL домену, и общий VH и VL вносят вклад в связывание антигена.
ПРИМЕР 15: Разработка ELISA для IgGкλ, количественная оценка с биспецифичным антителом
[00115] Разработка ELISA для IgGкλ, количественная оценка с биспецифичным антителом были разработаны биспецифичные антитела для сендвич ELISA для определения количества IgGкλ. 96-луночные планшеты Maxisorp (Nunc) покрывали 10 мкг/мл мышиного антитела против человеческой Лямдба (Southern Biotech) и инкубировали при 4°C в течение ночи. После отмывки (PBS Tween 20 при 0,05%, 3 отмывки), планшет блокировали PBS-BSA 3% (Sigma) в течение 2 часов при комнатной температуре. Очищенный IgGкλ стандарт серийно разводили в PBS-BSA 1% от 500 нг/мл до 1 нг/мл для получения хорошего диапазона линейности для количественной оценки образцов. В зависимости от их происхождения образцы разводили для вхождения в диапазон количественной оценки следующим образом: необработанную надосадочную жидкость CHO 1/1500; Протеин A, очищенный 1/15000. Затем образцы разводили серийно 1:2. После отмывки планшетов, 50 мкл каждого приготовленного разведения добавляли в двух повторах и инкубировали в течение 1 часа при комнатной температуре. Планшеты повторно отмывали и инкубировали в течение 1 часа при комнатной температуре с 50 мкл 1:2000 антитела к человеческой Каппа (Southern Biotech). После последней отмывки (PBST 0,05%, 5 отмывок), реакцию проявляли с помощью 50 мкл субстрата TMB (Sigma) и останавливали через 15 мин добавлением 50 мкл H2SO4 (Sodium hydroxide 2N). Поглощение при 450 нм записывали с помощью точного ридера микропланшетов (Epoch, Witec). Поскольку моноспецифичные антитела могут воздействовать на анализ путем связывания, используемого для покрытия захватывающего антитела, осуществляли эксперименты с добавлением возрастающих количеств моноспецифичного IgGк и моноспецифичного IgGλ антитела к IgGкλ, биспецифичному стандарту. Были протестированы различные соотношения: (50% IgGкλ биспецифичный, 25% моноспецифичный IgGк, 25% моноспецифичный IgGλ); (67% IgGкλ, биспецифичный; 33% моноспецифичный IgGλ); (50% IgGкλ биспецифичный, 50% моноспецифичный IgGλ); (25% IgGкλ, биспецифичный, 75% моноспецифичный IgGλ); (50% IgGкλ биспецифичный, 50% моноспецифичный IgGк).
[00116] Результаты, представленные на Фигуре 22, указывают на то, что на анализ не оказывают значимого воздействия моноспецифичные антитела и что, следовательно, он может быть использован для количественной оценки биспецифичного антитела IgGкλ в комплексных образцах, таких как надосадочные жидкости клеточных культур. ELISA использовали для количественной оценки биспецифичного антитела IgGкλ в надосадочной жидкости из стабильных клеточных линий CHO, и затем суммарный IgG очищали из тех же самых надосадочных жидкостей с помощью аффинной хроматографии с протеином А. Результаты количественной оценки с помощью ELISA сравнивали с суммарным содержанием IgG, определенным с помощью ВЭЖХ с протеином А или по поглощению при 280 нм и суммировали в таблице V. Концентрации, полученные с помощью ELISA, соответствовали 30-40% от суммарного IgG, доли ожидаемых биспецифичных антител. Данные говорят о том, что данный анализ может быть использован для определения количества IgGкλ, биспецифичного антитела в надосадочной жидкости клеточной культуры, и он облегчает скрининг стабильных клеточных линий по их продуктивности специфичного антитела IgGкλ.
Количественная оценка биспецифичного антитела IgGкλ различных стабильных клеточных линий CHO с помощью необработанных надосадочных жидкостей или суммарного IgG с помощью ELISA, и сравнение с ВЭЖХ с протеином А и результатами количественной оценки суммарного IgG по A280.
Другие варианты осуществления
[00117] Хотя изобретение приведено вместе с подробным описанием, представленное выше описание предназначено для иллюстрации и не ограничивает объем притязаний изобретения, который определен объемом прилагаемой формулы изобретения. Другие аспекты, преимущества и модификации находятся в рамках представленной ниже формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Fischer, Nicolas
Magistrelli, Giovanni
Gueneau, Franck
Ravn, Ulla
Elson, Greg
<120> СПОСОБЫ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧНЫХ И МУЛЬТИВАЛЕНТНЫХ
АНТИТЕЛ
<130> 23135-423001US
<140> PCT/IB2011/002664
<141> 2011-08-16
<150> US 61/374159
<151> 2010-08-16
<150> US 61/443008
<151> 2011-02-15
<150> US 61/509260
<151> 2011-07-19
<160> 6
<170> PatentIn version 3.5
<210> 1
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> химически синтезированная
<400> 1
Ala Lys Ser Tyr Gly Ala Phe Asp Tyr
1 5
<210> 2
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> химически синтезированная
<400> 2
ctcttctgag atgagttttt g 21
<210> 3
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> химически синтезированная
<400> 3
Ala Arg Gly Asp Asp Val Ser
1 5
<210> 4
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> химически синтезированная
<400> 4
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Leu Pro Thr Thr Pro
85 90 95
Ala Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 5
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> химически синтезированная
<400> 5
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Ile Ala Ser Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ser Pro Thr Thr Val
35 40 45
Ile Tyr Glu Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Gln Ser Trp
85 90 95
Asp Gly Asn His Ile Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 6
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> химически синтезированная
<400> 6
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ser Tyr Gly Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ДОМЕНА CH1, СКОНСТРУИРОВАННЫЕ ДЛЯ ИЗБИРАТЕЛЬНОГО СПАРИВАНИЯ С ЛЕГКОЙ ЦЕПЬЮ, И СОДЕРЖАЩИЕ ИХ МУЛЬТИСПЕЦИФИЧНЫЕ АНТИТЕЛА | 2020 |
|
RU2839961C1 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛЫЕ ЦЕПИ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С CD19 | 2019 |
|
RU2824170C2 |
| ПОЛУЧЕНИЕ МУЛЬТИСПЕЦИФИЧЕСКИХ СМЕСЕЙ АНТИТЕЛ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2815826C2 |
| НОВЫЕ БИСПЕЦИФИЧНЫЕ ПОЛИПЕПТИДНЫЕ КОМПЛЕКСЫ ПРОТИВ CD3/CD19 | 2018 |
|
RU2788127C2 |
| АНТИТЕЛО-ПОДОБНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ОБЛАСТЯМИ, ИМЕЮЩИЕ ОРИЕНТАЦИЮ СВЯЗЫВАЮЩИХ ОБЛАСТЕЙ КРЕСТ-НАКРЕСТ | 2012 |
|
RU2823693C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СВОЙСТВАМИ СВЯЗЫВАНИЯ FcRn И БЕЛКА А | 2015 |
|
RU2713131C1 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ FcRn И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714116C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CD19 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2731156C1 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2831724C2 |
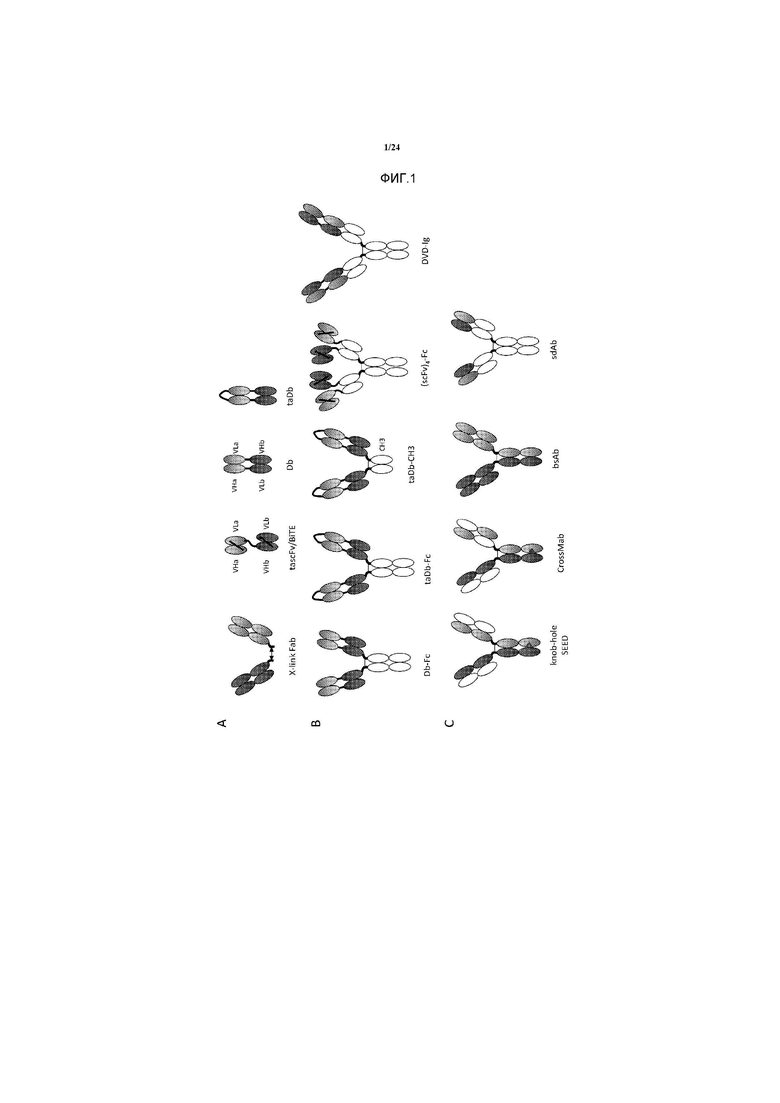
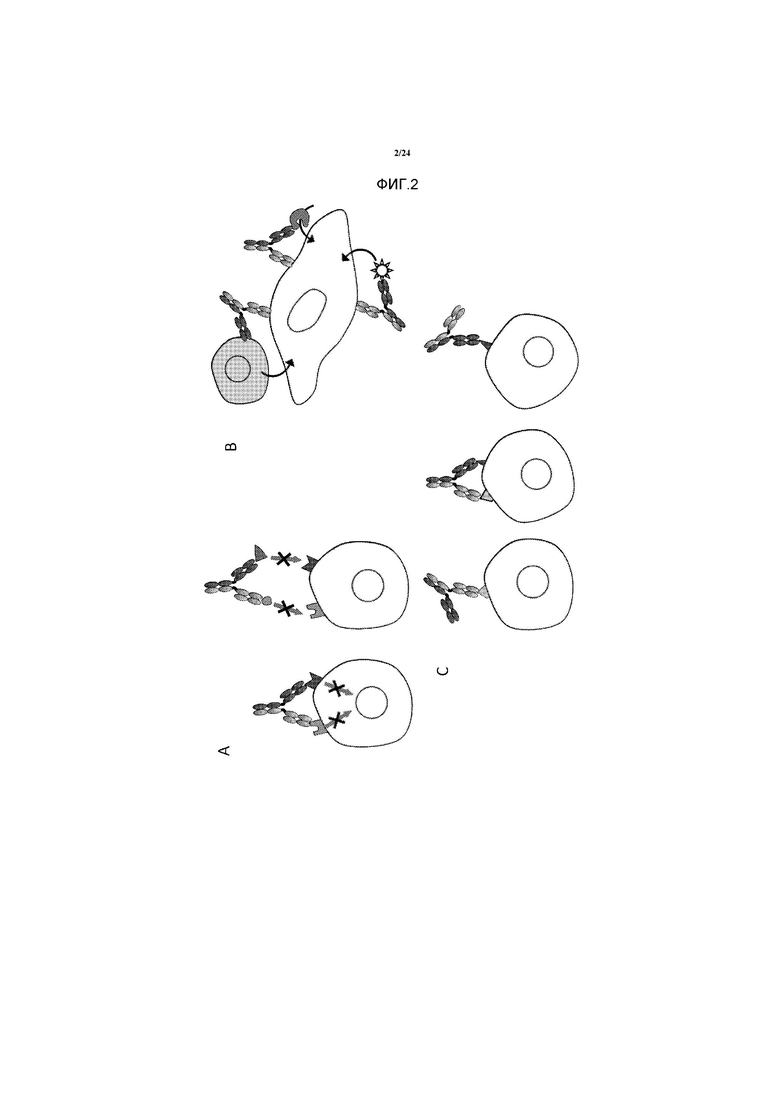
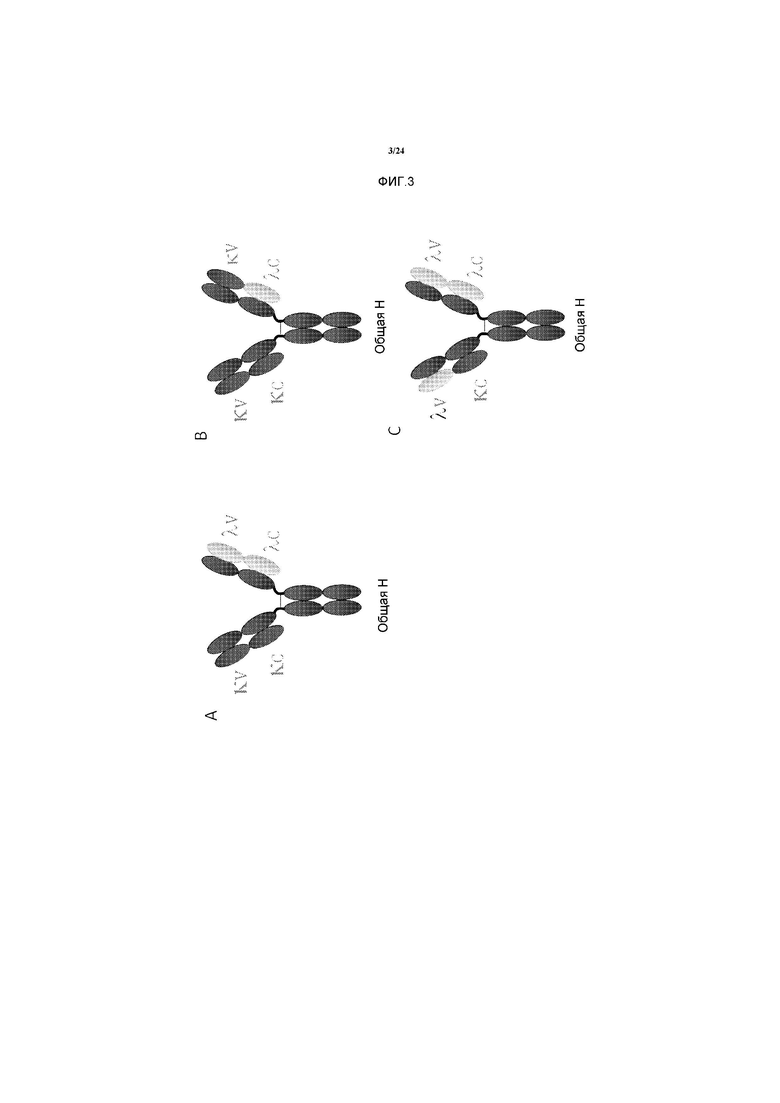
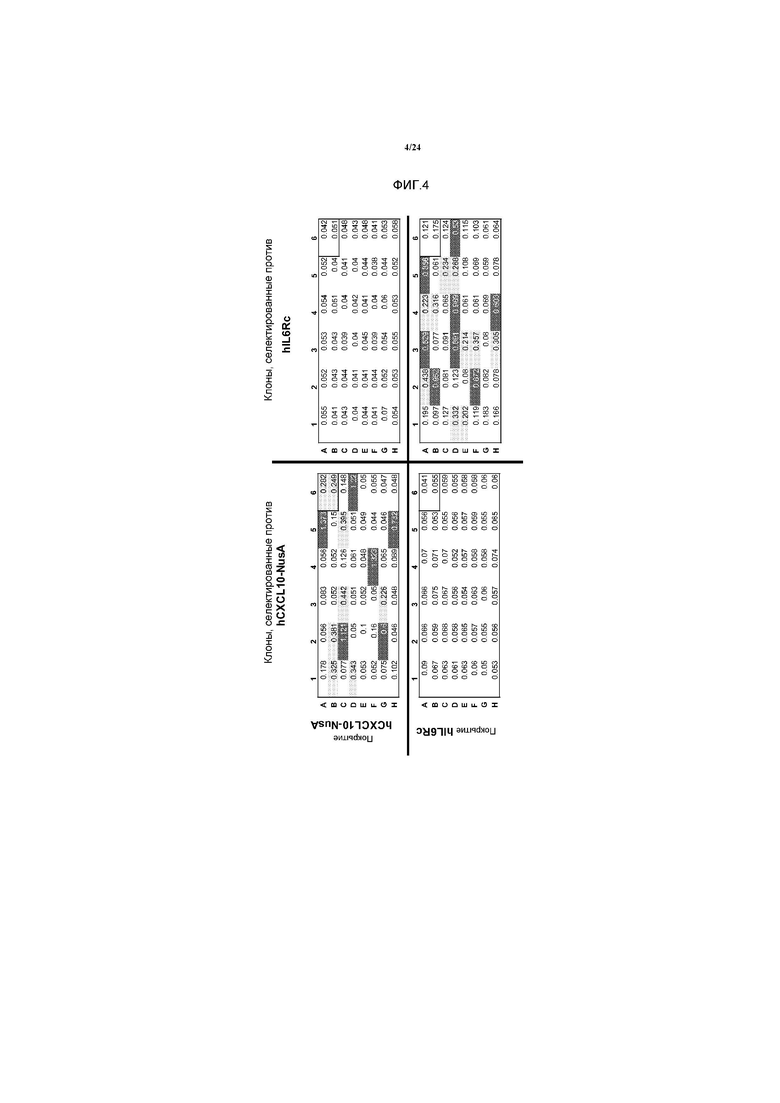



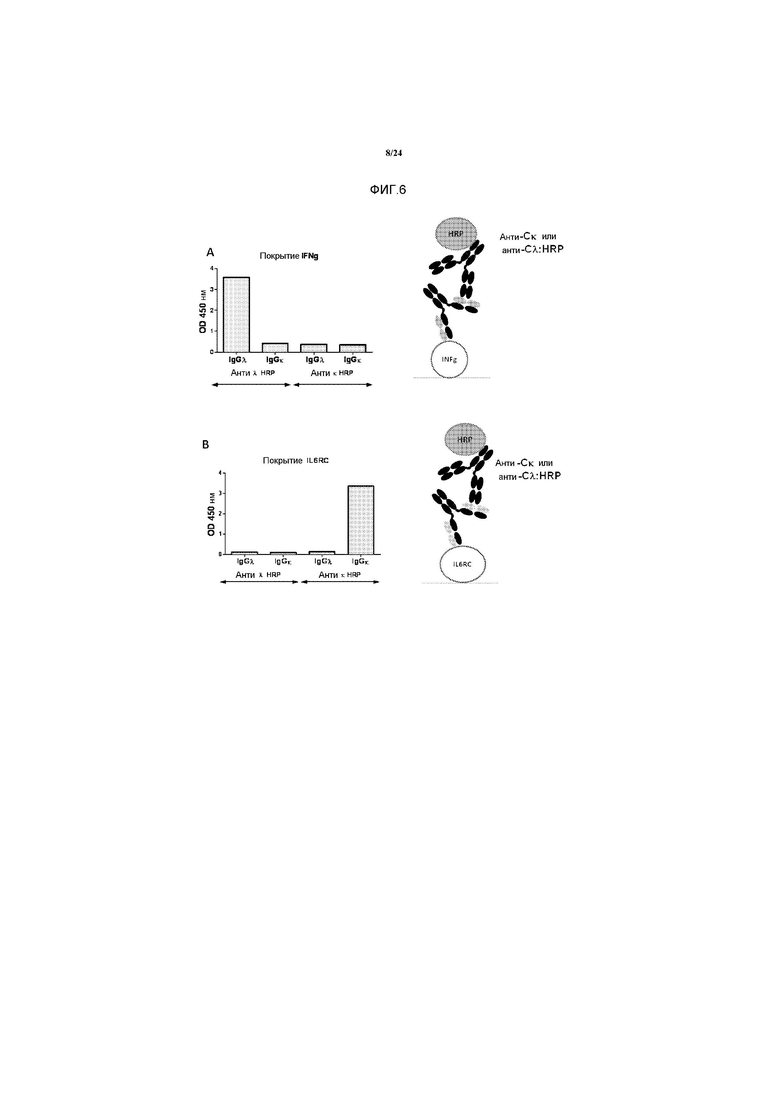
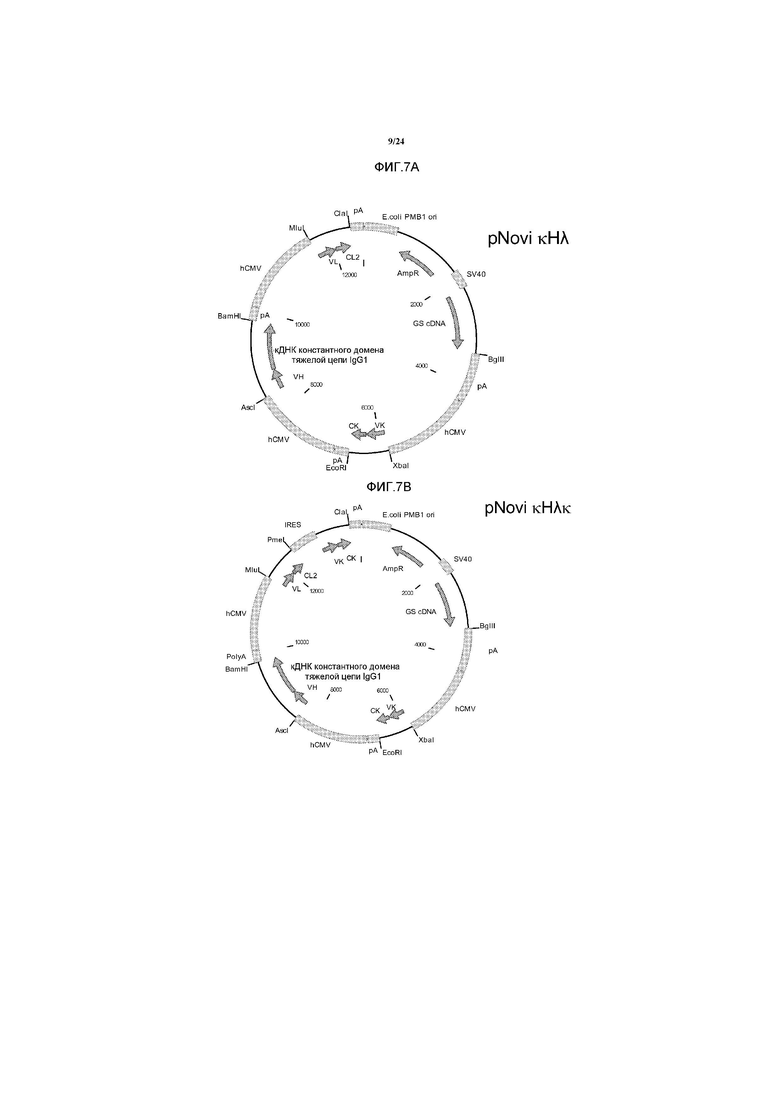

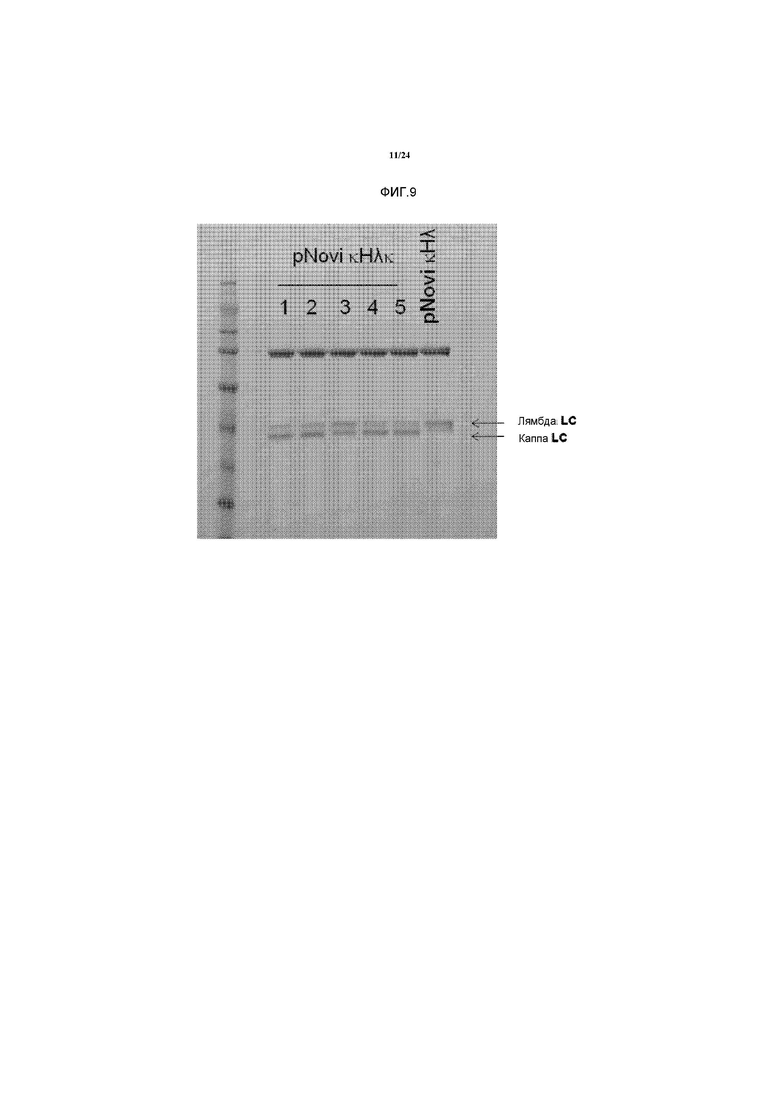
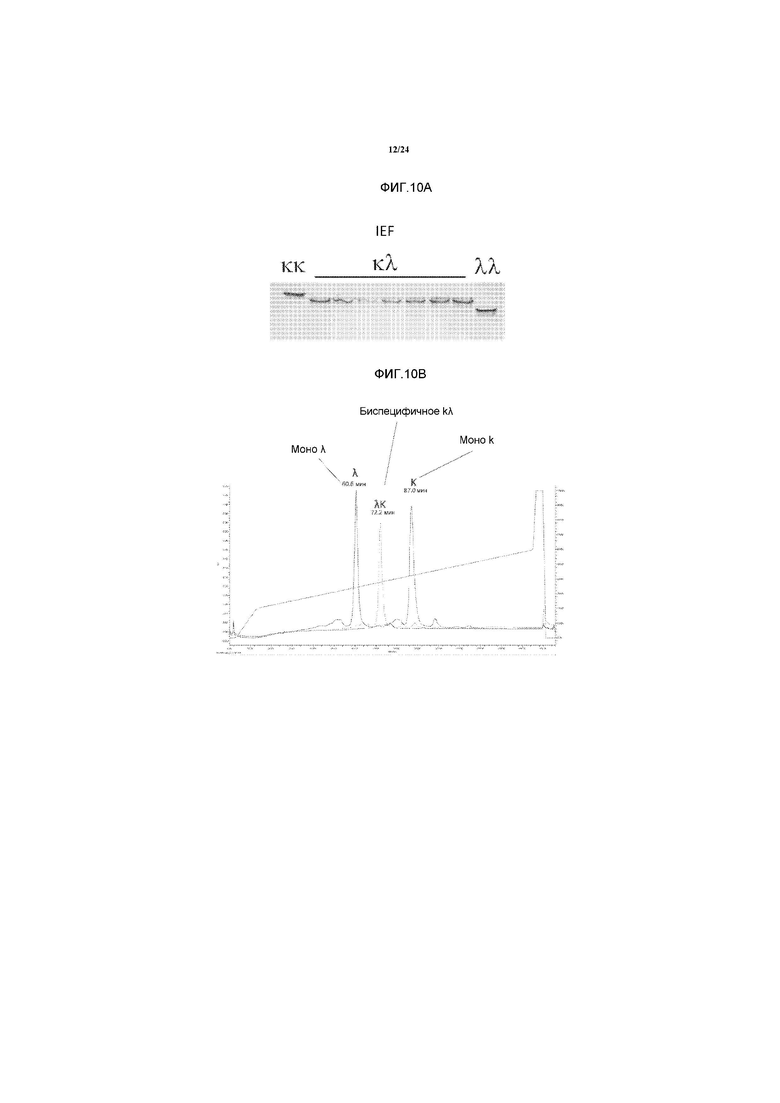
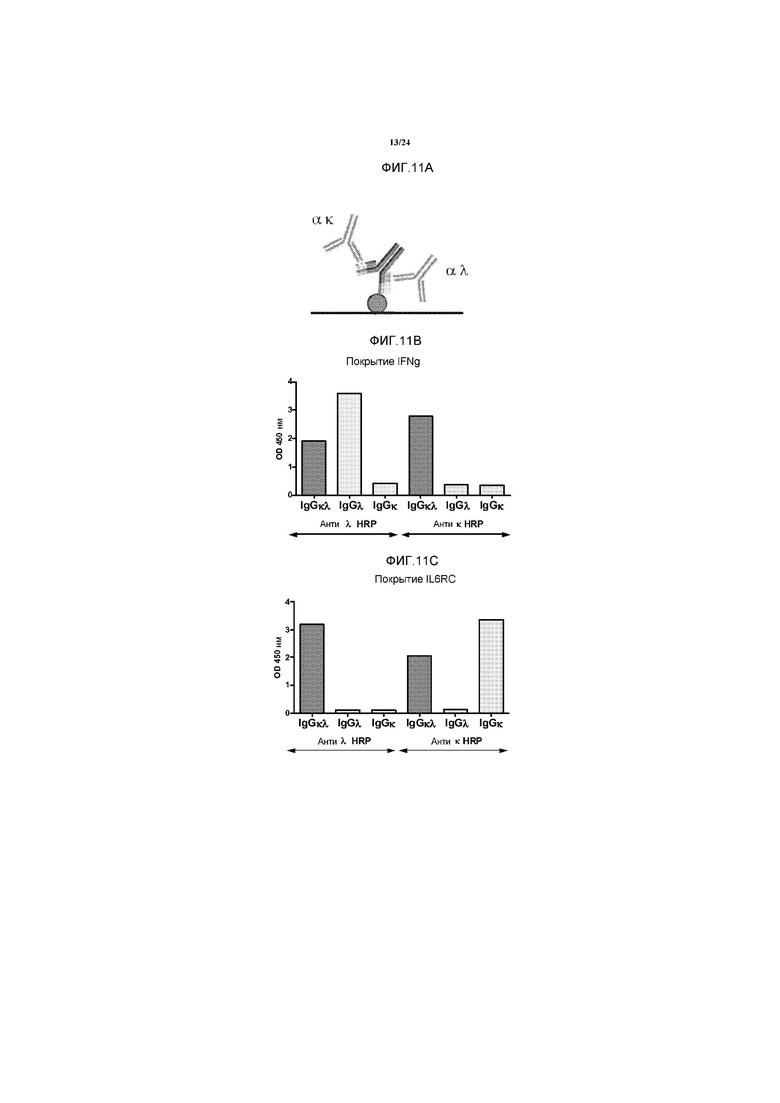

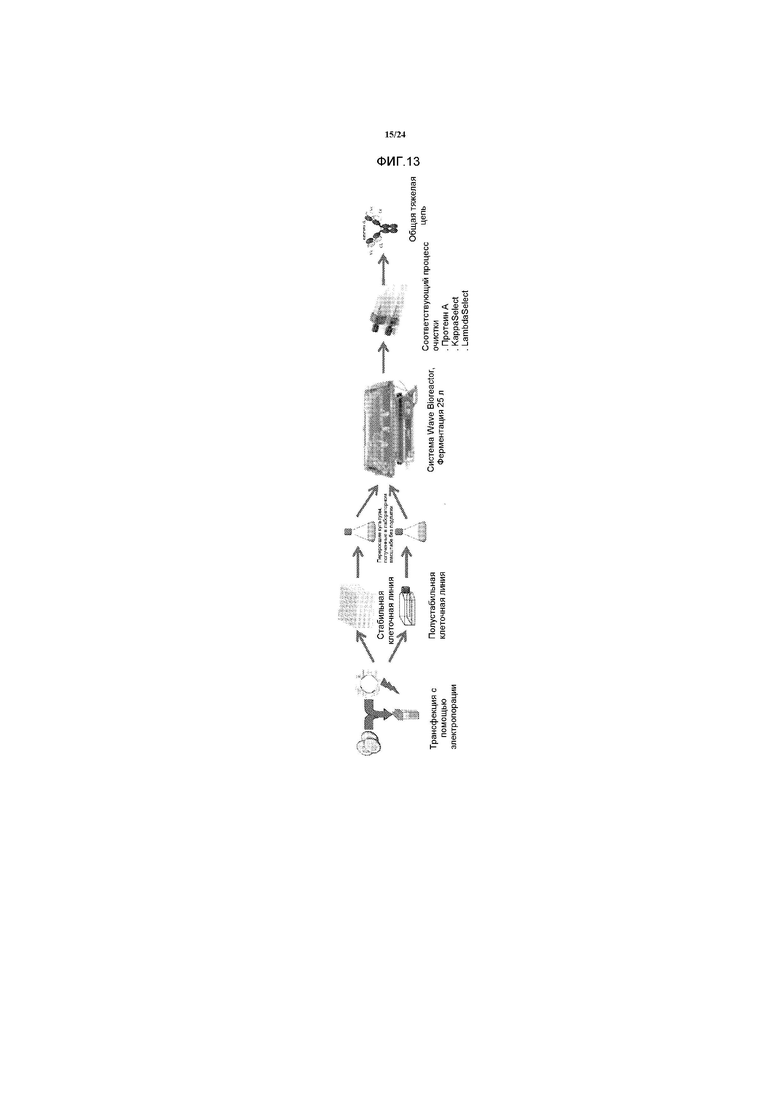
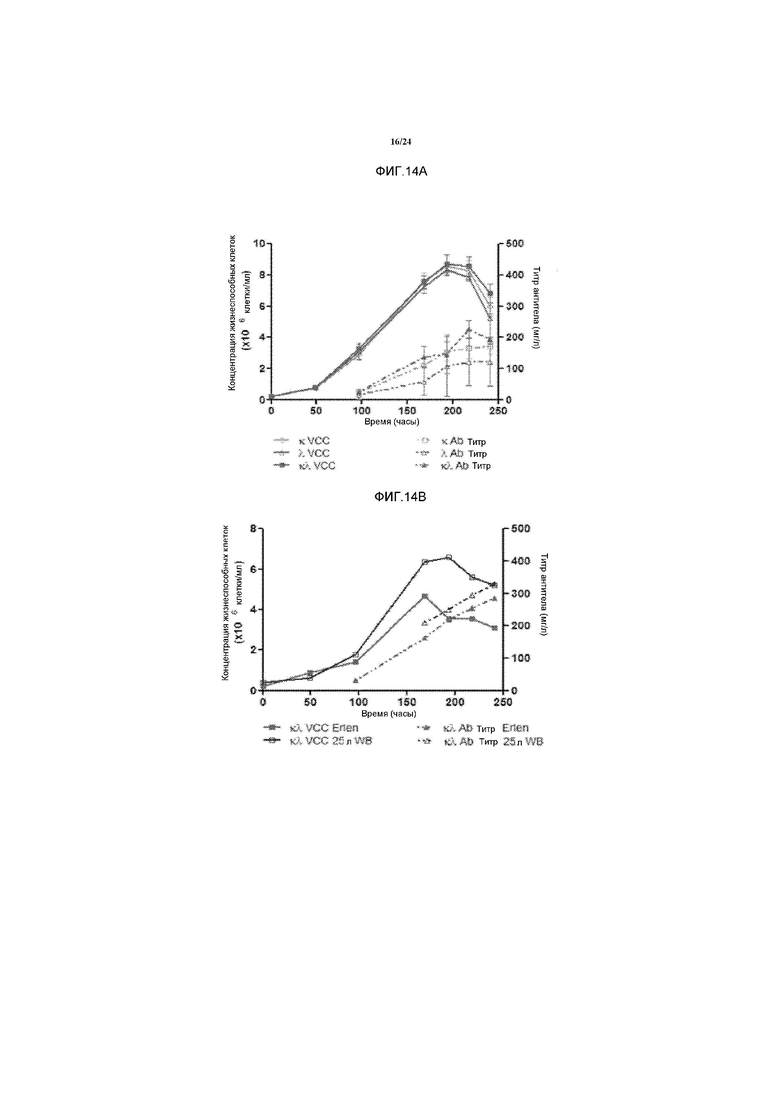
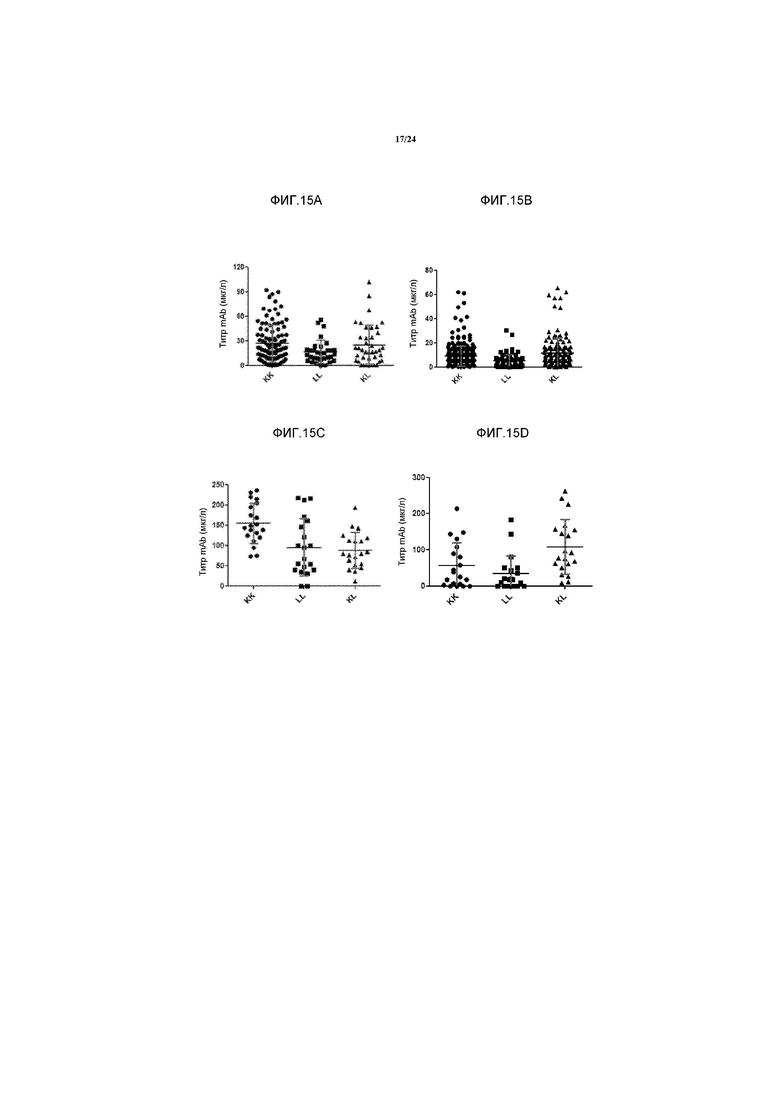
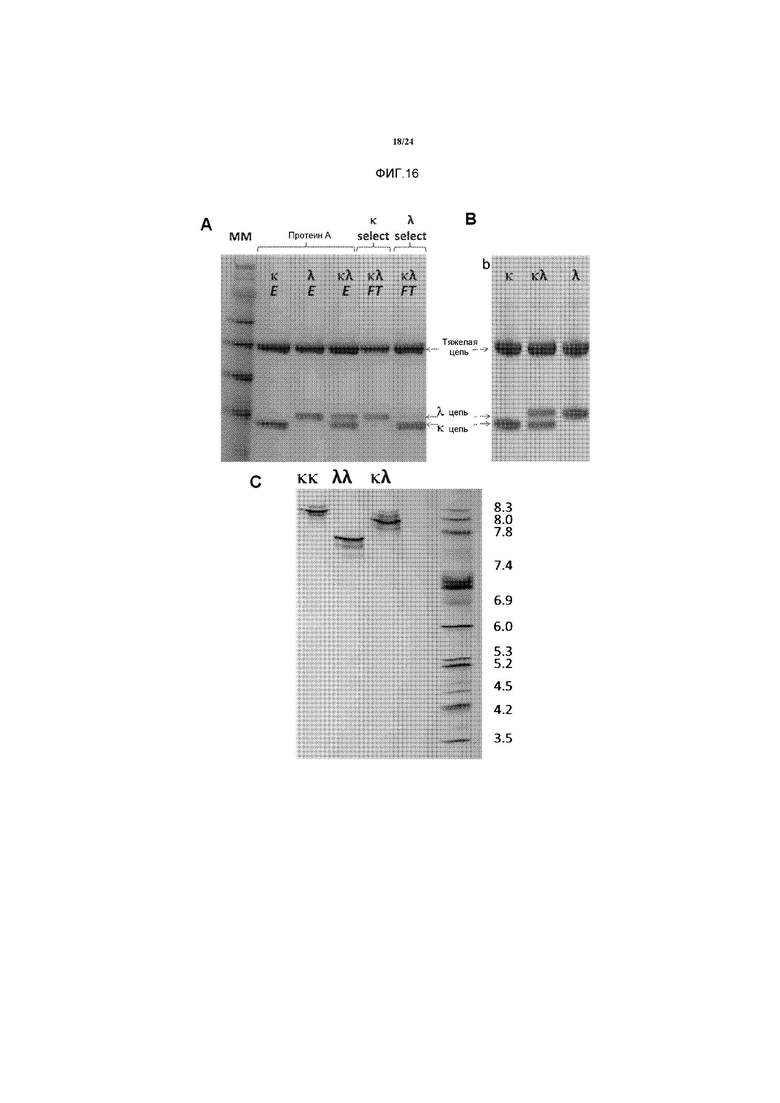
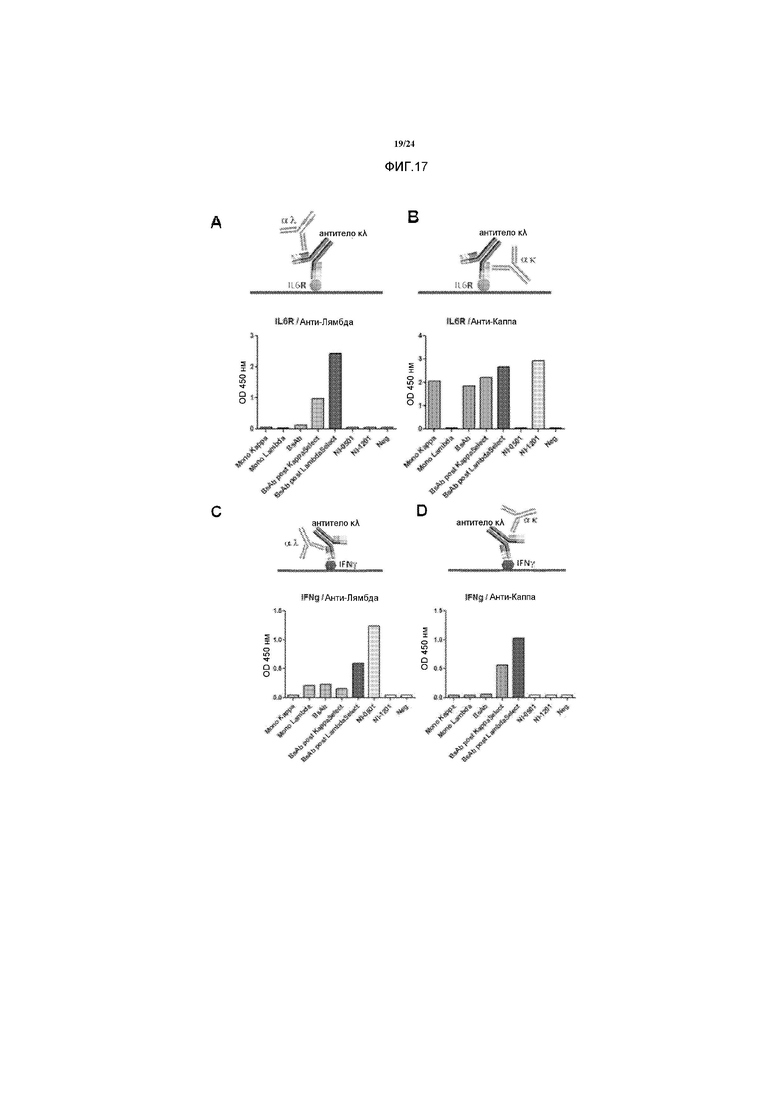



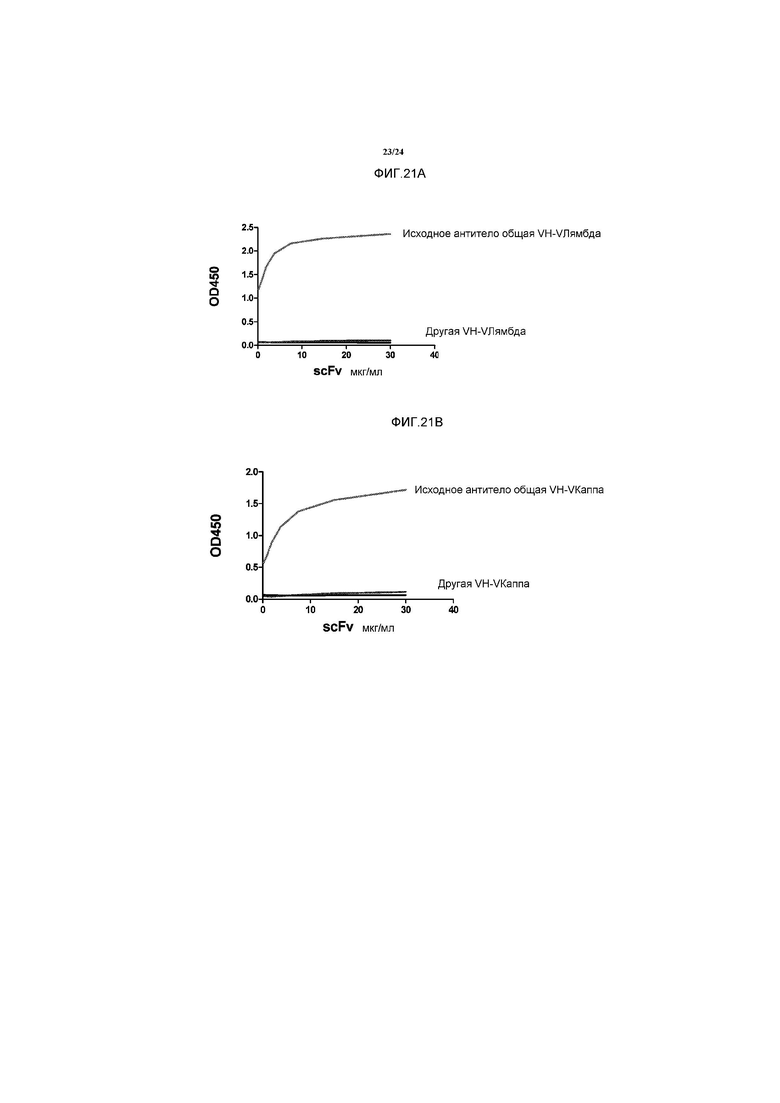

Изобретение относится к области биотехнологии. Предложен способ получения смеси антител, содержащей два моноспецифических антитела и одно биспецифическое антитело, где все антитела имеют общую тяжелую цепь. Выделяют два антитела с различной специфичностью, определяемой одним и тем же вариабельным доменом тяжелой цепи. Затем коэкспрессируют в клетке две различных легких цепи и одну тяжелую цепь указанных антител, что приводит к сборке моноспецифических и биспецифических антител. Одна из указанных легких цепей содержит константный домен Каппа, а другая - константный домен Лямбда. Также предложена смесь антител для выделения двух моноспецифических антител и одного биспецифического антитела, полученная указанным способом. Изобретение обеспечивает регуляцию соотношения различных антител в получаемой смеси для максимизации получения биспецифического антитела. 2 н. и 6 з.п. ф-лы, 37 ил., 5 табл., 15 пр.
1. Способ получения смеси антител, содержащей два моноспецифических антитела и одно биспецифическое антитело, где все эти антитела имеют общую тяжелую цепь, где способ включает:
а. выделение антитела или участка фрагмента антитела, специфичность которого определяется вариабельным доменом тяжелой цепи, объединенным с первым вариабельным доменом легкой цепи;
b. выделение антитела или участка фрагмента антитела, имеющего другую специфичность, определяемую тем же самым вариабельным доменом тяжелой цепи, что и антитело стадии а), в сочетании со вторым вариабельным доменом легкой цепи;
c. коэкспрессию в клетке:
i. вариабельного домена общей тяжелой цепи, слитого с константной областью тяжелой цепи иммуноглобулина;
ii. вариабельного домена первой легкой цепи, слитого с константным доменом легкой цепи каппа; и
iii. вариабельного домена второй легкой цепи, слитого с константным доменом легкой цепи лямбда,
тем самым получая смесь антител, содержащую два моноспецифических антитела и одно биспецифическое антитело,
где все эти антитела имеют общую тяжелую цепь,
где биспецифическое антитело имеет различную специфичность в первом сайте связывания антигена и втором сайте связывания антигена и состоит из двух копий полипептида общей тяжелой цепи и первой легкой цепи и второй легкой цепи,
где первая и вторая легкие цепи отличаются друг от друга, и
где первый сайт связывания специфически связывается с первым антигеном и второй сайт связывания специфически связывается со вторым антигеном.
2. Способ по п. 1, где:
(a) на стадии (c) (ii) вариабельный домен легкой цепи каппа сливается с константным доменом легкой цепи каппа;
(b) на стадии (c) (ii) вариабельный домен легкой цепи лямбда сливается с константным доменом легкой цепи каппа;
(с) на стадии (c) (iii) вариабельный домен легкой цепи каппа сливается с константным доменом легкой цепи лямбда; или
(d) на стадии (c) (iii) вариабельный домен легкой цепи лямбда сливается с константным доменом легкой цепи лямбда.
3. Способ по п. 1, дополнительно включающий стадию (d) очистки смеси антител, полученной на стадии (c), от супернатанта клеточной культуры.
4. Способ по п. 3, где стадия (d) очистки смеси антител представляет собой стадию аффинной хроматографии.
5. Способ по п. 4, где стадия очистки представляет собой стадию очистки аффинной хроматографией, проводимой с помощью среды для аффинной хроматографии, специфичной для константного домена каппа, специфичной для константного домена лямбда, или среды, специфичной как для константного домена каппа, так и для константного домена лямбда.
6. Смесь антител для выделения двух моноспецифических антител и одного биспецифического антитела, содержащая два моноспецифических антитела и одно биспецифическое антитело, где все эти антитела имеют общую тяжелую цепь, полученная способом по п. 1.
7. Способ по п. 1, где проведение стадий (а) и (b) упрощают благодаря использованию библиотек антител, имеющих общую тяжелую цепь и разнообразие в вариабельных доменах легкой цепи.
8. Способ по п. 7, где для библиотеки антител используется дисплей нитчатого бактериофага, поверхность клеток дрожжей, бактерий или млекопитающих, или используются библиотеки рибосом или другие in vitro библиотеки.
| US 2007098712 A1, 03.05.2007 | |||
| FISCHER N., LEGER O., Bispecific Antibodies: Molecules That Enable Novel Therapeutic Strategies, PATHOBIOLOGY, 2007, vol.74, no.1, pp.3-14 | |||
| ДЕЕВ С.М., ЛЕБЕДЕНКО Е.Н., Современные технологии создания неприродных антител для клинического применения, Acta Naturae (русскоязычная версия), 2009, N 1, с.32-50 | |||
| US 2008108794 |

