Настоящее изобретение относится к мультиспецифическим антителам с улучшенными свойствами, полезным в области медицины.
Предшествующий уровень техники
Терапевтические гибридные белки стали важным средством разработки лекарственных средств. Зачастую пептидные линкеры используются для конструирования мультидоменных белков из различных функциональных белковых модулей. Получающиеся в результате мультидоменные белки предназначены для связывания с мишенью, узнаваемой индивидуальными модулями (или одновременно для выполнения биологической функции отдельных модулей), чтобы либо усиливать биологические эффекты, связанные с отдельными одиночными доменами, либо создавать новые биологические активности, недостижимые посредством отдельных одиночных доменов. Существует множество примеров молекул, в которых используются пептидные линкеры: одноцепочечные вариабельные домены антител (scFv), иммуноцитокины (гибриды цитокинов и антител), биспецифичные антитела (BsAb) и т.д. Выбор линкера(ов) для конкретного гибридного белка продиктовано такими соображениями, как: 1) требуется ли линкеру(ам) гибкость, чтобы позволить сворачивать различные домены в конкретную третичную структуру (например, антитела на основе scFv), 2) требуется ли линкеру(ам) жесткость чтобы обеспечить необходимое разделение между белковыми доменами, или 3) требуется ли линкеру(ам) быть расщепляемым, чтобы позволить разделение доменов in vivo для получения искомой активности (Xiaoying Chen, et al., Adv Drug Deliv Rev. 2013. 65(10): 1357-1369). Выбор линкеров может быть критическим, поскольку неподходящие линкеры могут снижать или устранять искомую активность гибридного белка (Yumi Maeda, et al., Anal. Biochem. 1997. 249(2): 147-152).
Для применения при конструировании гибридных белков были идентифицированы различные линкерные последовательности (Richard George and Jaap Heringa, Protein Engineering. 2003. 15(11): 871-879; Xiaoying Chen et al, Adv Drug Deliv Rev. 2013. 65(10): 1357-1369). Существуют также различные доступные базы данных, в которых собраны линкерные последовательности, используемые для конструирования гибридных белков: 1) SynLinker, скомпилированный Национальным университетом Сингапура (http://synlinker.syncti.org), и 2) Международный конкурс по генно-инженерным машинам (http://parts.igem.org/Protein_domains/Linker); Центр интегративной биоинформатики при Университете Врие в Амстердаме (http://www.ibi.vu.nl/programs/linkerdbwww).
Международная патентная заявка WO2013/005194 раскрывает мультиспецифичные антитела, сконструированные с линкерами, разработанными из шарнирной последовательности IgG1, за которой следует N-конец последовательности домена CH2 из IgG1, за которым следует полужесткая линкерная последовательность из 8 аминокислот из центральной части шарнира IgA1. Этот полужесткий линкер, составляющий центральную часть шарнира IgA1, важен для обеспечения достаточного разделения обоих Fab-доменов антител, чтобы избежать стерических помех от С-конца внешнего Fab1, на который воздействует антигенсвязывающий паратоп внутреннего Fc-проксимального Fab2.
Однако авторы изобретения определили, что присутствие большого количества гликоформ делает проблематичным получение единообразных препаратов таких мультиспецифических антител, необходимых для разработки терапевтических средств. Кроме того, характеризация таких препаратов также будет довольно сложной, что сделает трудным сравнение различных произведенных партий.
Сущность изобретения
По этой причине изобретатели пришли к выводу, что можно провести улучшение, изменив конструкцию линкера, в частности, удалив из линкера сайты гликозилирования.
Таким образом, изобретение обеспечивает линкерный полипептид, который включает или состоит из аминокислотной последовательности EPKX1CDKX2НХ3Х4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 1),
где X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, одинаковые или разные, представляют собой любую аминокислоту, как определено в данном документе; при условии, что полипептид не включает и не состоит из последовательностей EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 5) или EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
Такой полипептид полезен в качестве линкера в гибридных белках, в частности, в мультиспецифических, в частности, в биспецифических антителах.
Таким образом, объектом изобретения является мультиспецифический антигенсвязывающий фрагмент, содержащий, по меньшей мере, два Fab-фрагмента с разными доменами CH1 и CL, причем каждый Fab-фрагмент распознает иной представляющий интерес эпитоп, и указанные Fab-фрагменты расположены тандемно в любом порядке, так что C-конец домена CH1 первого фрагмента Fab связан с N-концом домена VH следующего фрагмента Fab через полипептидный линкер,
отличающийся тем, что последовательность полипептидного линкера включает или состоит из аминокислотной последовательности EPKX1CDKX2HX3X4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 1), где X1, Х2, Х3, Х4, Х5, Х6, Х7, Х8, Х9, Х10, одинаковые или разные, представляют собой любую аминокислоту; при условии, что линкерная последовательность не включает и не состоит из последовательностей EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 5) или EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
Кроме того, предложено мультиспецифическое антитело, имеющее два идентичных антигенсвязывающих плеча, каждое из которых состоит из мультиспецифического антигенсвязывающего фрагмента, как определено в настоящем документе.
В предпочтительном воплощении предлагается мультиспецифическое антитело, которое имеет иммуноглобулиноподобную структуру, включающее:
- два идентичных антигенсвязывающих плеча, каждое из которых состоит из мультиспецифического антигенсвязывающего фрагмента, как определено в настоящем документе;
- димеризованные домены СН2 и СН3 иммуноглобулина;
- шарнирную область IgA, IgG или IgD, связывающую C-концы доменов CH1 антигенсвязывающих плеч с N-концами доменов CH2.
Таким образом, настоящее изобретение более конкретно относится к мультиспецифическому, предпочтительно биспецифическому антителу, содержащему две тяжелые цепи и четыре легкие цепи,
где каждая тяжелая цепь включает
а. область Fc иммуноглобулина, содержащую домены Шарнира-CH2-CH3,
b. где область Fc связана с CH1-VH тяжелой цепи Fab антитела 1 (Ab1) указанным шарнирным доменом,
с. который, в свою очередь, связан с CH1-VH тяжелой цепи Fab антитела 2 (Ab2) посредством полипептидной линкерной последовательности, где полипептидная линкерная последовательность связывает N-конец указанного VH-домена тяжелой цепи Fab Ab1 с C-концом указанного домена CH1 из Ab2,
а четыре легкие цепи включают легкие цепи Ab1 и легкие цепи Ab2, связанные с их когнатными доменами тяжелых цепей;
которое отличается тем, что последовательность полипептидного линкера включает или состоит из аминокислотной последовательности EPKX1CDKX2НХ3Х4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 1),
где X1, X2, X3, X4, X5, X6, X7, X8, X9, X10,одинаковые или разные, представляют собой любые аминокислоты; при условии, что линкерная последовательность не включает и не состоит из последовательностей EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 5) или EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
В конкретном воплощении полипептидная линкерная последовательность включает или состоит из последовательности, выбранной из группы, состоящей из
EPKSCDKTHTSPPAPAPELLGGPGGPPGPGPGGG (SEQ ID NO: 2);
EPKSCDKTHTSPPAPAPELLGGPAAPPAPAPAGG (SEQ ID NO: 3); и
EPKSCDKTHTSPPAPAPELLGGPAAPPGPAPGGG (SEQ ID NO: 4).
Другим объектом изобретения является полипептид, который включает, предпочтительно состоит из тяжелой цепи мультиспецифического антигенсвязывающего фрагмента, или мультиспецифического, предпочтительно биспецифического антитела, как определено в настоящем документе.
Изобретение дополнительно относится к полинуклеотиду, содержащему последовательность, кодирующую такой полипептид.
Клетка-хозяин, трансфицированная экспрессирующим вектором, содержащим указанный полинуклеотид, также является частью изобретения.
Следующим объектом изобретения является способ получения мультиспецифического антитела, предпочтительно биспецифического антитела, как описано в данном документе, причем указанный способ включает следующие стадии: а) культивирование в подходящей среде и условиях культивирования клетки-хозяина, экспрессирующей тяжелую цепь антитела, как определено в данном документе, и экспрессирующей легкую цепь антитела, как определено в данном документе; и b) извлечение указанных продуцируемых антител из культуральной среды или из указанных культивируемых клеток.
Подписи к чертежам
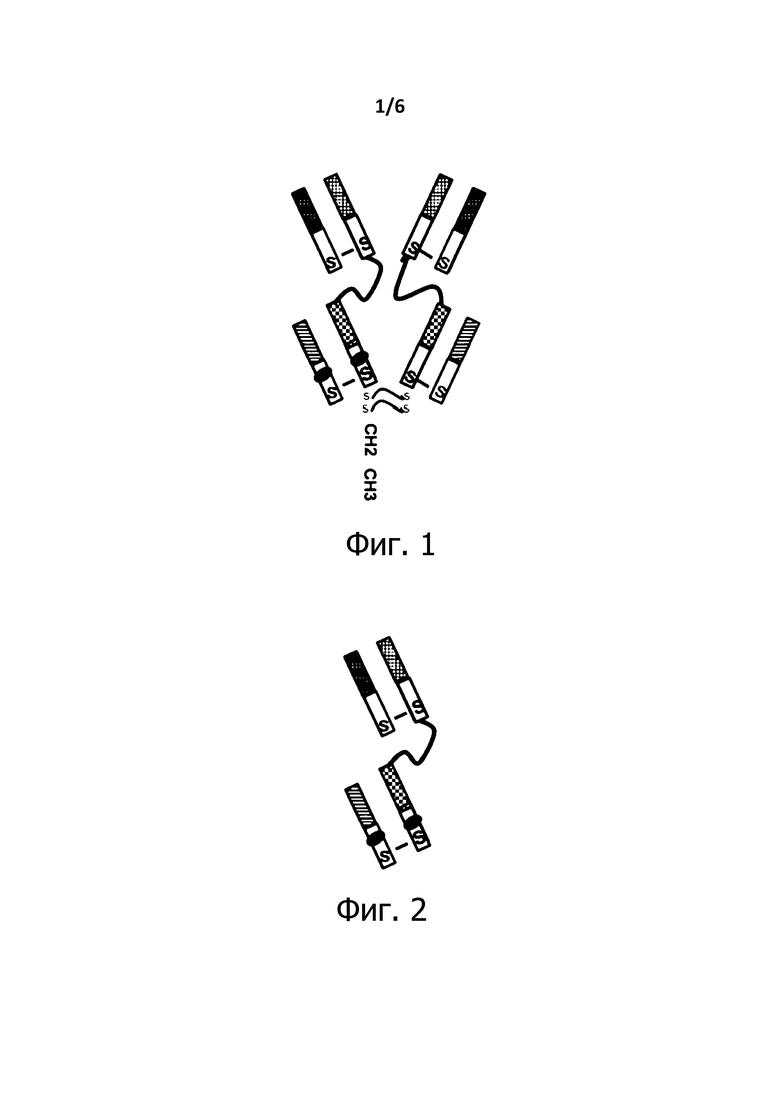
Фигура 1 представляет собой схематическое изображение полноразмерного биспецифического антитела BiXAb по изобретению.
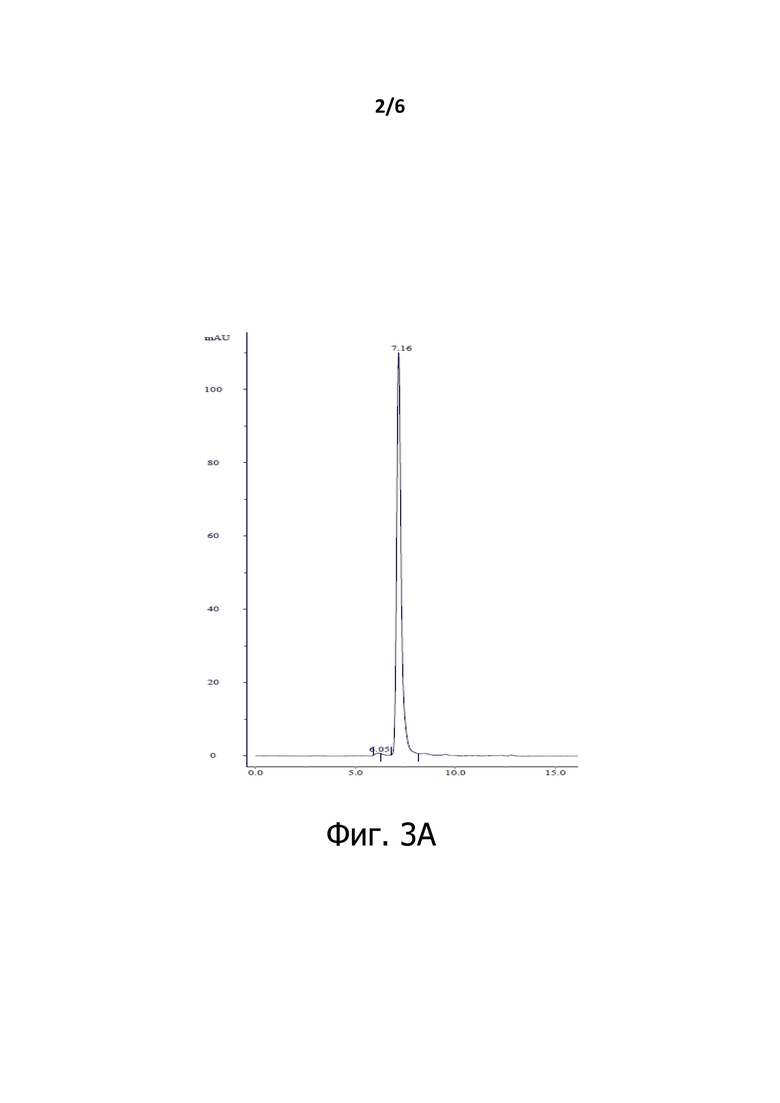
Фигура 2 представляет собой схематическое изображение биспецифической конструкции, которая включает только два Fab-домена, соединенных линкером без Fc-домена (Fab-Fab).
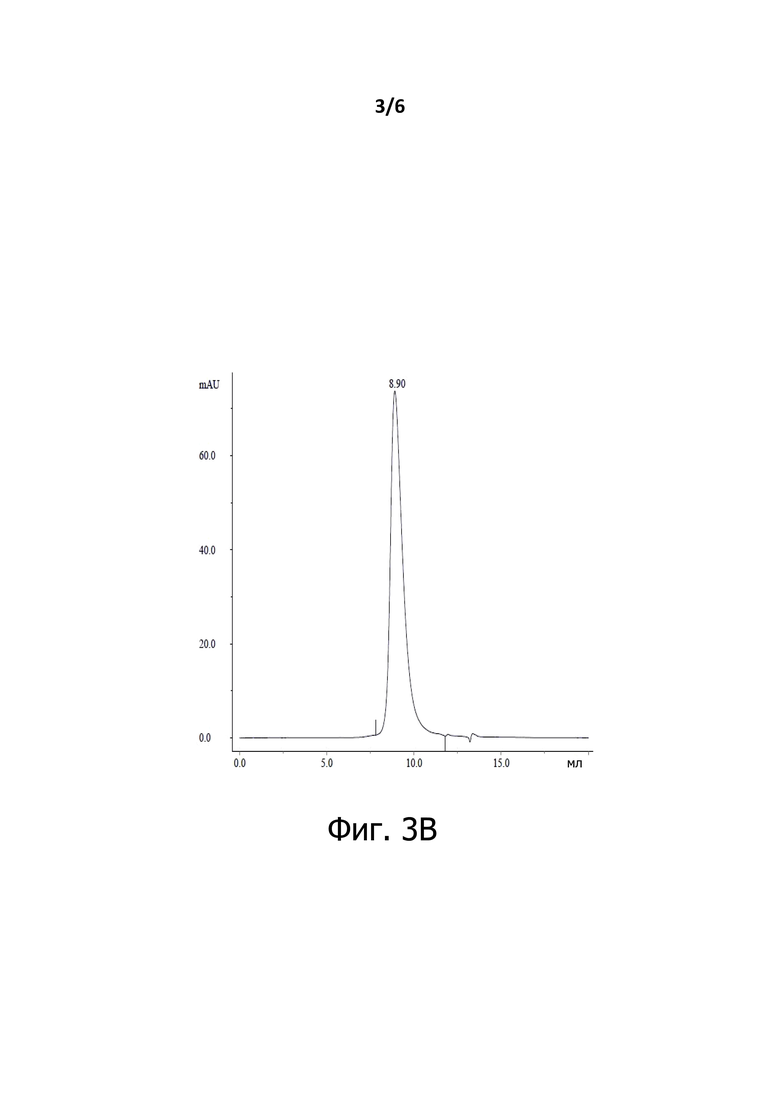
На Фигуре 3А показан анализ BiXAb2b эксклюзионной хроматографией.
На Фигуре 3B показан анализ BiXAb3b эксклюзионной хроматографией.
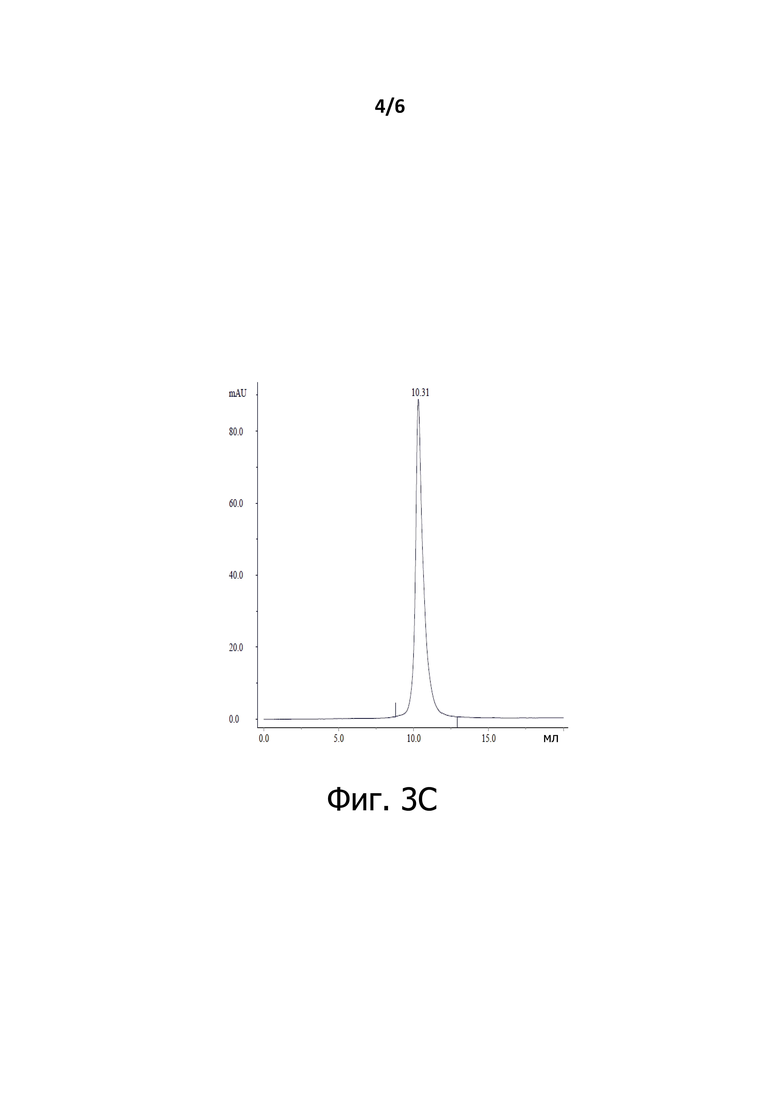
На Фигуре 3C показан анализ Fab-Fab3b эксклюзионной хроматографией.
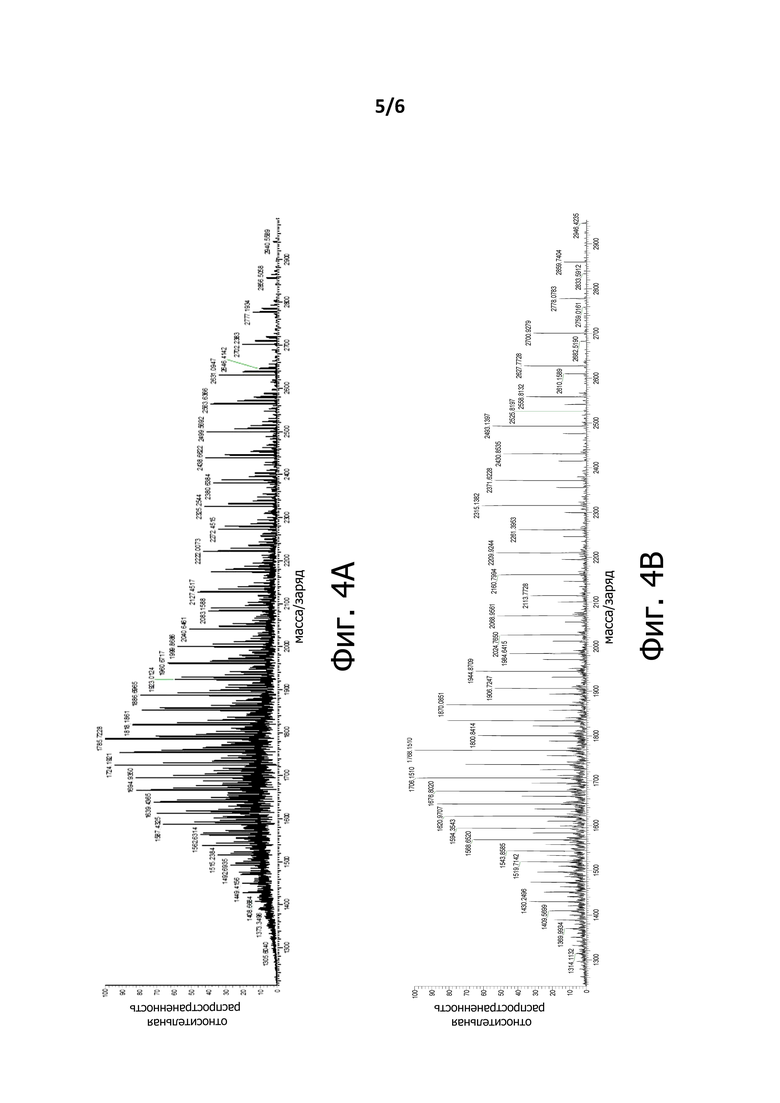
На Фигуре 4А показан MS-спектр LC-MS-анализа Fab-Fab3a.
На Фигуре 4В показан MS-спектр LC-MS-анализа Fab-Fab3b.
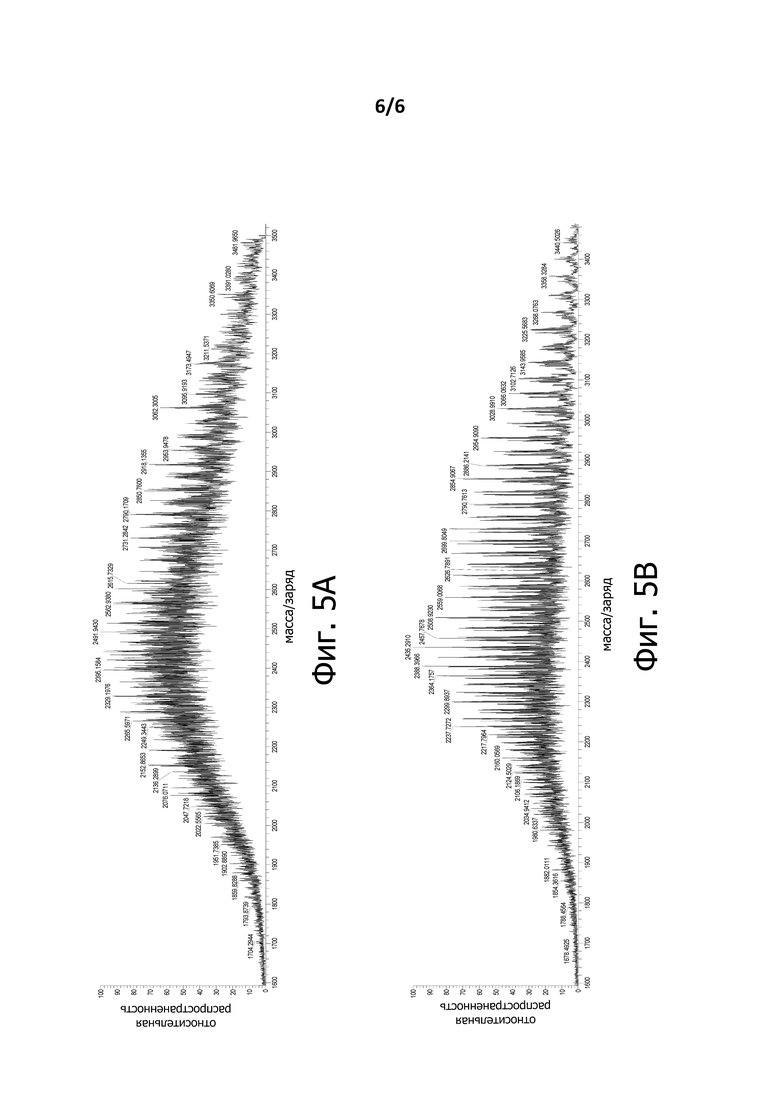
На Фигуре 5A показан MS-спектр LC-MS-анализа BiXAb3a.
На Фигуре 5В показан MS-спектр LC-MS-анализа BiXAb3b.
Подробное описание изобретения
Определения
Базовая структура молекулы антитела природного происхождения представляет собой тетрамерную четвертичную структуру Y-образной формы, состоящую из двух идентичных тяжелых цепей и двух идентичных легких цепей, удерживаемых вместе нековалентными взаимодействиями и межцепочечными дисульфидными связями.
У видов млекопитающих существует пять типов тяжелых цепей: α, δ, ε, γ и μ, которые определяют класс (изотип) иммуноглобулина: IgA, IgD, IgE, IgG и IgM, соответственно. За N-концевым вариабельным доменом тяжелой цепи (VH) следует константная область, содержащая три домена (пронумерованных CHI, СН2 и СН3 от N-конца к С-концу) в γ, α и δ тяжелых цепях, в то время как константные области тяжелых цепей μ и ε состоят из четырех доменов (пронумерованных CHI, СН2, СН3 и СН4 от N-конца к С-концу). Домены СН1 и СН2 из IgA, IgG и IgD разделены гибким шарниром, длина которого варьирует между различными классами, а в случае IgA и IgG - между различными подтипами: lgGl, lgG2, IgG3 и IgG4 имеют, соответственно, шарниры из 15, 12, 62 (или 77) и 12 аминокислот, a IgAl и IgA2 имеют, соответственно, шарниры из 20 и 7 аминокислот.
Существует два типа легких цепей: λ и κ, которые могут ассоциироваться с любым изотипом тяжелой цепи, но обе легкие цепи принадлежат к одному типу в данной молекуле антитела. Обе легкие цепи по всей видимости являются функционально идентичными. За их N-концевым вариабельным доменом (VL) следует константная область, состоящая из одного домена, называемого CL.
Тяжелые и легкие цепи спариваются белок/белковым взаимодействием между доменами СН1 и CL и между доменами VH и VL, а две тяжелые цепи связываются белок/белковым взаимодействием между их доменами СН3.
Антигенсвязывающие области соответствуют плечам Y-образной структуры, каждое из которых состоит из полной легкой цепи в паре с доменами VH и СН1 тяжелой цепи и называются Fab-фрагментами (Fragment antigen binding - фрагмент, связывающий антиген). Fab-фрагменты сначала были получены из молекул нативного иммуноглобулина путем расщепления папаином, который расщепляет молекулу антитела в шарнирной области на аминоконцевой стороне межцепочечных дисульфидных связей, освобождая, таким образом, два идентичных антигенсвязывающих плеча. Другие протеазы, такие как пепсин, также расщепляют молекулу антитела в шарнирной области, но на карбоксиконцевой стороне межцепочечных дисульфидных связей, высвобождая фрагменты, состоящие из двух идентичных фрагментов Fab, и оставаясь связанными через дисульфидные связи; восстановление дисульфидных связей в фрагментах F(ab')2 приводит к образованию фрагментов Fab'.
Часть антигенсвязывающей области, соответствующая доменам VH и VL, называется фрагментом Fv (Fragment variable - вариабельный фрагмент); она содержит CDR (определяющие комплементарность области), которые образуют антигенсвязывающий сайт (также называемый паратопом).
Эффекторная область антитела, которая отвечает за его связывание с эффекторными молекулами на иммунных клетках, соответствует стволу Y-образной структуры и содержит спаренные домены CH2 и CH3 тяжелой цепи (или CH2, CH3 и CH4 доменов, в зависимости от класса антитела), и называется Fc-область (Fragment crystallisable - кристаллизуемый фрагмент).
Из-за идентичности двух тяжелых цепей и двух легких цепей природные молекулы антител имеют два идентичных антигенсвязывающих сайта и, таким образом, связываются одновременно с двумя идентичными эпитопами.
В контексте настоящего изобретения «мультиспецифический антигенсвязывающий фрагмент» определяется в данном документе как молекула, имеющая две или более антигенсвязывающих области, каждая из которых распознает отличающиеся эпитоп. Различные эпитопы могут переноситься одной и той же антигенной молекулой или разными антигенными молекулами. Термин «распознающий» или «распознает» означает, что фрагмент специфически связывает целевой антиген.
Антитело «специфически связывается» с антигеном-мишенью, если оно связывается с большей аффинностью, авидностью, более легко и/или с большей продолжительностью, чем оно связывается с другими веществами. «Специфическое связывание» или «преференциальное связывание» не обязательно требует (хотя может включать) исключительное связывание. Как правило, но необязательно, ссылка на связывание означает преференциальное связывание.
Термины «объект», «индивидуум» и «пациент» используются в данном документе взаимозаменяемо и относятся к млекопитающему, которого оценивают на предмет лечения и/или подвергают лечению. Объектами могут быть люди, но также могут быть и другие млекопитающие, в частности те млекопитающие, которые могут использоваться в качестве лабораторных моделей заболеваний человека, например, мышь, крыса, кролик, собака и т.д.
Термин «лечение» или «лечить» относится к действию, применению или терапии, в которых объект, включая человека, подвергается медицинской помощи с целью улучшения состояния объекта, прямо или косвенно. В частности, этот термин относится к уменьшению заболеваемости или облегчению симптомов, устранению рецидивов, предотвращению рецидивов, предупреждению заболеваемости, улучшению симптомов, улучшению прогноза или их комбинации в некоторых воплощениях. Специалисту в данной области понятно, что лечение не обязательно приводит к полному отсутствию или устранению симптомов. Например, в отношении онкологического заболевания «лечение» или «лечить» может относиться к замедлению роста, пролиферации или метастазирования опухолевых или злокачественных клеток, предотвращению или задержке развития роста, пролиферации или метастазирования опухолевых или злокачественных клеток, или некоторых их комбинаций.
Конструирование мультиспецифичных антител
В данном документе предоставлены конструкции мультиспецифического антигенсвязывающего(их) фрагмента(ов) и мультиспецифических антител, содержащих указанные фрагменты, где каждый мультиспецифический антигенсвязывающий фрагмент состоит по существу из тандемно расположенных Fab-фрагментов, разделенных линкером по изобретению.
Такие фрагменты и конструкции предпочтительно содержат цепи из человеческих иммуноглобулинов, предпочтительно из IgG, а еще более предпочтительно из IgG1.
В случае мультиспецифического антигенсвязывающего фрагмента, содержащего более двух разных фрагментов Fab, полипептидные линкеры, разделяющие фрагменты Fab, могут быть одинаковыми или разными.
В соответствии с предпочтительным воплощением мультиспецифического антитела по изобретению, оно имеет два идентичных антигенсвязывающих плеча, каждое из которых состоит из мультиспецифического антигенсвязывающего фрагмента, как определено выше. Антигенсвязывающие ветви могут быть связаны друг с другом различными способами, в зависимости от предполагаемого применения антитела.
Если желательно получить антитело без Fc-опосредованных эффектов, антитело не будет содержать Fc-области. В этом случае две антигенсвязывающие ветви могут быть связаны вместе, например:
- гомодимеризацией антигенсвязывающих плеч через межцепочечные дисульфидные связи, обеспечиваемые полипептидным(и) линкером(ами), разделяющим(и) фрагменты Fab; и/или
- посредством добавления на С-конце каждого антигенсвязывающего плеча полипептидного удлинения, содержащего остатки цистеина, позволяющего образование межцепочечных дисульфидных связей, и гомодимеризации указанного полипептидного удлинения, приводящего к шарнирообразной структуре; в качестве неограничивающих примеров указанное удлинение полипептида может быть, например, шарнирной последовательностью IgG1, IgG2 или IgG3;
- через полужесткий линкер, соединяющий С-концы тяжелых цепей двух антигенсвязывающих плеч с образованием единой полипептидной цепи и поддержание указанных антигенсвязывающих плеч на достаточном расстоянии друг от друга.
Альтернативно, если желательны эффекторные функции, такие как CDC, ADCC или ADP, мультиспецифическое антитело по изобретению может дополнительно содержать домен Fc, обеспечивающий эти эффекторные функции. Выбор домена Fc будет зависеть от типа требуемых эффекторных функций.
В этом случае мультиспецифическое антитело по изобретению имеет иммуноглобулиноподобную структуру, включающую:
- два идентичных мультиспецифических антигенсвязывающих плеча, как определено выше;
- димеризованные домены СН2 и СН3 иммуноглобулина;
- либо шарнирную область IgA, IgG или IgD, связывающую C-концы доменов CH1 антигенсвязывающих плеч с N-концами доменов CH2, или, альтернативно, когда домены CH4, следующие за доменами CH3, которые происходят из IgM или IgE, причем C-концы доменов CH1 антигенсвязывающих плеч в этом случае могут быть связаны непосредственно с N-концами доменов CH2.
Предпочтительно домены СН2 и СН3, шарнирная область и/или домены СН4 происходят из того же иммуноглобулина или из иммуноглобулинов того же изотипа и подкласса, что и домены СН1 антигенсвязывающего плеча.
Могут быть использованы домены СН2, СН3 и, необязательно, СН4, и шарнирные области из нативных иммуноглобулинов. При желании их также можно мутировать, например, для модуляции эффекторной функции антитела. В некоторых случаях весь или часть домена CH2 или CH3 могут быть отброшены.
Более конкретно, изобретение относится к биспецифическим тетравалентным антителам, содержащим два сайта связывания с каждой из их мишеней и функциональный домен Fc, позволяющий активировать эффекторные функции, такие как антитело-зависимая клеточная цитотоксичность (ADCC) и фагоцитоз.
Такие предпочтительные антитела по изобретению представляют собой полноразмерные антитела. Они предпочтительно содержат тяжелые цепи и легкие цепи из иммуноглобулинов человека, предпочтительно IgG, еще более предпочтительно IgG1.
Легкие цепи могут быть лямбда- или каппа-легкими цепями; предпочтительно они представляют собой каппа-легкие цепи.
В предпочтительном воплощении линкер по изобретению связывает Fab-домены IgG в формате биспецифического антитела тетра-Fab, аминокислотная последовательность которого содержит последовательности тяжелой цепи, по меньшей мере, двух Fab, соединенных указанным полипептидным линкером, за которым следует последовательность нативного шарнира, за которой следует последовательность Fc из IgG, коэкспрессируемая с соответствующими последовательностями легкой цепи IgG.
Пример антител по изобретению, названных антителами BiXAb, которые имеют IgG-подобную структуру, показан на Фигуре 1. Антитела, обозначенные BiXAb2a, BiXAb2b, BiXAb2c и BiXAb3b, описанные ниже, являются конкретными примерами.
Биспецифичные антитела по изобретению обычно включают
- непрерывную тяжелую цепь, сконструированную из Fc (шарнир-CH2-CH3)
- затем следует тяжелая цепь Fab (CH1-VH) антитела 1 и последующая тяжелая цепь Fab (CH1-VH) антитела 2, последняя присоединена полипептидной линкерной последовательностью по изобретению,
- и во время экспрессии белка полученная тяжелая цепь собирается в димеры, в то время как коэкспрессированные легкие цепи антитела 1 и антитела 2 (VL-CL) связываются с когнатными тяжелыми цепями с образованием конечной тандемной молекулы F(ab)'2-Fc,
при этом антитело 1 (Ab1) и антитело 2 (Ab2) являются разными.
В предпочтительном воплощении описаны биспецифичные антитела, которые включают
• два Fab-фрагмента с разными доменами CH1 и CL, состоящие из
a) Fab-фрагмента, имеющего домены CH1 и C-Каппа, полученные из человеческого IgG1/Kappa, и домены VH и VL из Ab1,
b) Fab-фрагмента, имеющего домены CH1 и C-каппа, полученные из IgG1/Kappa человека и доменов VH и VL из Ab2,
с) мутированного константного домена CL легкой цепи, полученного из константного домена Каппа человека,
d) мутированного константного домена CH1 тяжелой цепи
при этом Fab-фрагменты располагаются тандемно в следующем порядке
- С-конец домена СН1 фрагмента Fab из Ab1 связан с N-концом домена VH фрагмента Fab из Ab2 через полипептидный линкер,
- шарнирная область человеческого IgG1, связывающая C-концы домена CH1 фрагмента Ab2 с N-концом домена CH2,
- димеризованные домены CH2 и CH3 человеческого IgG1.
Ab1 и Ab2 могут представлять собой любое антитело, представляющее интерес, особенно любое антитело, представляющее терапевтический интерес.
В конкретном воплощении Ab1 и Ab2, будучи различными, независимо выбираются из группы, состоящей из антитела против EGFR и антитела против HER2/neu. В предпочтительном воплощении Ab1 и Ab2, будучи различными, независимо выбираются из группы, состоящей из цетуксимаба или его мутированного производного, с одной стороны, и трастузумаба или его мутированного производного, с другой стороны.
В другом конкретном воплощении Ab1 и Ab2, отличающиеся друг от друга, независимо выбраны из группы, состоящей из антитела против CD38 и антитела против PD-L1.
Такие антитела полезны в качестве лекарственного средства, в частности, при лечении онкологических заболеваний.
На всем протяжении настоящего описания аминокислотные последовательности определены согласно Kabat et al, Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991).
Другой пример конструкций по изобретению, который представляет собой мультиспецифический антигенсвязывающий фрагмент Fab-Fab, который не содержит Fc-домен, проиллюстрирован на Фигуре 2. Конкретный пример, обозначенный Fab-Fab3a, описан ниже.
Такие конструкции Fab-Fab обычно содержат два разных домена Fab. Такие антитела обладают только одним Fab-доменом, каждый из которых связывается с антигеном 1 и антигеном 2. Они обладают теми же легкими цепями, что и в соответствующих антителах BiXAb; однако тяжелая цепь Fab-Fab укорачивается таким образом, что их последний С-концевой остаток представляет собой цистеин-220 (в нумерации EU).
Дизайн линкеров
Полипептидная последовательность линкера согласно изобретению включает или состоит из аминокислотной последовательности EPKX1CDKX2НХ3Х4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 1),
где X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, одинаковые или разные, представляют собой любую аминокислоту; при условии, что линкерная последовательность не включает и не состоит из последовательностей EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 5) или EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
Последовательность полипептидного линкера состоит из менее чем 80 аминокислот, предпочтительно менее чем 60 аминокислот, еще более предпочтительно менее чем 40 аминокислот.
В конкретном воплощении X1, X2 и X3, одинаковые или разные, представляют собой треонин (T) или серин (S).
В другом конкретном воплощении X1, X2 и X3, одинаковые или разные, представляют собой любые аминокислоты, отличные от треонина (T) или серина (S), предпочтительно, где X1, X2 и X3, одинаковые или разные, представляют собой выбран из группы, состоящей из Ala (A), Gly (G), Val (V), Asn (N), Asp (D) и Ile (I), все еще предпочтительно X1, X2 и X3, одинаковых или разных, может быть Ala (A) или Gly (G).
Альтернативно, X1, X2 и X3, одинаковые или разные, могут быть Leu (L), Glu (E), Gln (Q), Met (M), Lys (K), Arg (R), Phe (F), Tyr (T), His (H), Trp (W), предпочтительно Leu (L), Glu (E) или Gln (Q).
В конкретном воплощении X4 и X5, одинаковые или разные, представляют собой любую аминокислоту, выбранную из группы, состоящей из серина (S), цистеина (C), аланина (A) и глицина (G).
В предпочтительном воплощении X4 представляет собой серин (S) или цистеин (C).
В предпочтительном аспекте X5 представляет собой аланин (A) или цистеин (C).
В конкретном воплощении X6, X 7, X8, X 9, X10, одинаковые или разные, представляют собой любую аминокислоту, отличную от треонина (T) или серина (S). Предпочтительно Х 6, Х 7, X8, X9, X10, одинаковые или разные, выбраны из группы, состоящей из Ala (A), Gly (G), Val (V), Asn (N), Asp (D) и Ile (I).
Альтернативно, X6, X7, Х8, Х9, Х10, одинаковые или разные, могут быть Leu (L), Glu (E), Gln (Q), Met (M), Lys (K), Arg (R), Phe (F), Tyr (T), His (H) Trp (W), предпочтительно Leu (L), Glu (E) или Gln (Q).
В предпочтительном воплощении X6, X7, X8, X9, X10, одинаковые или разные, выбраны из группы, состоящей из Ala (A) и Gly (G).
В еще одном предпочтительном воплощении X6 и X7 являются одинаковыми и предпочтительно выбраны из группы, состоящей из Ala (A) и Gly (G).
В предпочтительном воплощении полипептидная линкерная последовательность включает или состоит из последовательности SEQ ID NO: 1, где
X1, X2 и X3, одинаковые или разные, представляют собой треонин (T), серин (S);
X4 представляет собой серин (S) или цистеин (C);
X5 представляет собой аланин (A) или цистеин (C);
Х6, Х7, X8, X9, X10, одинаковые или разные, выбраны из группы, состоящей из Ala (A) и Gly (G).
В конкретном воплощении полипептидная линкерная последовательность включает или состоит из последовательности, выбранной из группы, состоящей из
EPKSCDKTHTSPPAPAPELLGGPGGPPGPGPGGG (SEQ ID NO: 2);
EPKSCDKTHTSPPAPAPELLGGPAAPPAPAPAGG (SEQ ID NO: 3); и
EPKSCDKTHTSPPAPAPELLGGPAAPPGPAPGGG (SEQ ID NO: 4).
В другом предпочтительном воплощении полипептидная линкерная последовательность включает или состоит из последовательности SEQ ID NO: 1, где
X1, X2 и X3, одинаковые или разные, представляют собой Ala (A) или Gly (G);
X4 представляет собой серин (S) или цистеин (C);
X5 представляет собой аланин (A) или цистеин (C);
Х6, Х7, X8, X9, X10, одинаковые или разные, выбраны из группы, состоящей из Ala (A) и Gly (G).
Получение антител
Нуклеиновые кислоты, кодирующие тяжелые и легкие цепи антител по изобретению, встраивали в экспрессирующие векторы. Легкие и тяжелые цепи могут быть клонированы в одном или разных экспрессирующих векторах. Сегменты ДНК, кодирующие цепи иммуноглобулина, функционально связаны с контрольными последовательностями в экспрессирующем(их) векторе(ах), которые обеспечивают экспрессию полипептидов иммуноглобулина. Такие контрольные последовательности включают сигнальную последовательность, промотор, энхансер и последовательность терминации транскрипции. Векторы экспрессии обычно реплицируются в организмах-хозяевах либо в виде эписом, либо в качестве составной части хромосомной ДНК хозяина. Обычно экспрессирующие векторы содержат маркеры отбора, например, тетрациклин или неомицин, чтобы позволить обнаруживать те клетки, которые трансформированы искомыми последовательностями ДНК.
В одном примере кодирующие последовательности как тяжелой, так и легкой цепи (например, последовательности, кодирующие VH и VL, VH-CH1 или VL-CL) включены в один экспрессирующий вектор. В другом примере каждая из тяжелых и легких цепей антитела клонируется в отдельный вектор. В последнем случае экспрессирующие векторы, кодирующие тяжелые и легкие цепи, могут быть совместно трансфицированы в одну клетку-хозяина для экспрессии обеих цепей, которые могут быть собраны с образованием интактных антител либо in vivo, либо in vitro.
В конкретном воплощении клетка-хозяин котрансфицировали с тремя независимыми экспрессирующими векторами, такими как плазмиды, что приводит к совместному продуцированию всех трех цепей (а именно тяжелой цепи HC и двух легких цепей LC1 и LC2, соответственно) и к секреции мультиспецифического антитела.
Более конкретно, три вектора могут быть преимущественно использованы при следующем молекулярном соотношении 3: 2: 2 (HC: LC1: LC2).
Рекомбинантные векторы для экспрессии антител, описанных в данном документе, обычно содержат нуклеиновую кислоту, кодирующую аминокислотные последовательности антител, функционально связанные с промотором, конститутивным или индуцибельным. Векторы могут подходить для репликации и интеграции у прокариот, эукариот или обоих. Типичные векторы содержат терминаторы транскрипции и трансляции, последовательности инициации и промоторы, полезные для регуляции экспрессии нуклеиновой кислоты, кодирующей антитело. Векторы необязательно содержат общие экспрессирующие кассеты, содержащие, по меньшей мере, одну независимую терминаторную последовательность, последовательности, позволяющие реплицировать кассету как у эукариот, так и у прокариот, т.е. челночные векторы, и маркеры отбора как для прокариотической, так и для эукариотической систем.
Мультиспецифичные антитела, как описано в настоящем документе, могут продуцироваться в прокариотических или эукариотических системах экспрессии, таких как бактерии, дрожжи, мицелиальные грибы, клетки насекомых и млекопитающих. Нет необходимости, чтобы рекомбинантные антитела по изобретению были гликозилированы или экспрессированы в эукариотических клетках; однако экспрессия в клетках млекопитающих обычно является предпочтительной. Примерами полезных линий клеток-хозяев млекопитающих являются эмбриональная почечная линия человека (клетки 293), клетки почки новорождённого хомяка (клетки BHK), клетки яичника китайского хомяка/- или + DHFR (клетки CHO, CHO-S, CHO-DG44, Flp-in CHO), клетки почки африканской зеленой мартышки (клетки VERO) и клетки печени человека (клетки Hep G2).
Культура клеток тканей млекопитающих является предпочтительной для экспрессии и продуцирования полипептидов, поскольку в данной области техники было разработано несколько подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, и они включают линии клеток СНО, различные линии клеток Cos, клетки HeLa, предпочтительно клеточные линии миеломы или трансформированные В-клетки или гибридомы.
В наиболее предпочтительном воплощении мультиспецифические, предпочтительно биспецифичные антитела по изобретению получают с помощью клеточной линии CHO, наиболее предпочтительно клеточной линии CHO-S.
Экспрессирующие векторы для этих клеток могут включать последовательности контроля экспрессии, такие как точка начала репликации, промотор и энхансер, и необходимые сайты обработки информации, такие как сайты связывания рибосом, сайты сплайсинга РНК, сайты полиаденилирования и последовательности терминатора транскрипции. Предпочтительными последовательностями, контролирующими экспрессию, являются промоторы, полученные из генов иммуноглобулина, SV40, аденовируса, вируса бычьей папилломы, цитомегаловируса и тому подобного.
Векторы, включающие представляющие интерес полинуклеотидные последовательности (например, последовательности, кодирующие тяжелую и легкую цепь и последовательности, контролирующие экспрессию), могут быть перенесены в клетку-хозяина хорошо известными способами, которые варьируются в зависимости от типа клеточного хозяина. Например, обработка фосфатом кальция или электропорация могут быть использованы для других клеточных хозяев (См. в целом Sambrook et al., Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press, 2nd ed., 1989). Когда тяжелые и легкие цепи клонируют на отдельных экспрессирующих векторах, векторы совместно трансфицируют для получения экспрессии и сборки интактных иммуноглобулинов.
Клетки-хозяева трансформируют или трансфицируют векторами (например, способами химической трансфекции или электропорации) и культивируют в обычных питательных средах (или модифицируют соответствующим образом) для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих искомые последовательности.
После экспрессии целые антитела, их димеры, отдельные легкие и тяжелые цепи или другие формы иммуноглобулинов по настоящему изобретению могут быть дополнительно выделены или очищены для получения препаратов, которые являются по существу гомогенными для дальнейших анализов и применений. Могут быть использованы стандартные способы очистки белка, известные в данной области. Например, подходящие процедуры очистки могут включать фракционирование на иммуноаффинных или ионообменных колонках, осаждение этанолом, высокоэффективную жидкостную хроматографию (ВЭЖХ), электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), осаждение сульфатом аммония и гель-фильтрацию (см. в целом Scopes, Protein Purification (Springer-Verlag, N.Y., 1982). По существу чистые иммуноглобулины с гомогенностью, по меньшей мере, примерно от 90 до 95% являются предпочтительными, а гомогенность от 98 до 99% или более наиболее предпочтительна для фармацевтического применения.
Производство in vitro позволяет увеличить масштаб для получения больших количеств желаемых мультиспецифических, предпочтительно биспецифичных, антител по изобретению. Такие способы могут использовать гомогенную суспензионную культуру, например, в аэролифтном реакторе или в реакторе с непрерывным перемешиванием, или культуру иммобилизованных или захваченных клеток, например, в полых волокнах, микрокапсулах, на агарозных микрогранулах или керамических картриджах.
Терапевтические применения
Еще одним аспектом изобретения является фармацевтическая композиция, содержащая антитело по изобретению. Другим аспектом изобретения является применение антитела по изобретению для изготовления фармацевтической композиции. Еще одним аспектом изобретения является способ изготовления фармацевтической композиции включающий антитело по изобретению.
В другом аспекте настоящее изобретение относится к композиции, например, фармацевтической композиции, содержащей антитело, как определено в данном документе, которое включено в композицию вместе с фармацевтическим носителем.
Композицию по настоящему изобретению можно вводить различными способами, известными в данной области.
Настоящее изобретение, таким образом, в целом описанное выше, будет понятнее со ссылкой на следующие примеры, которые представлены в качестве иллюстрации и не предназначены для ограничения настоящего изобретения.
ПРИМЕРЫ
Дизайн
Первым биспецифичным антителом по изобретению является антитело, обозначенное BiXAb2a, имеющее следующую структуру:
i) непрерывная тяжелая цепь, которая включает
- вариабельную область тяжелой цепи (VH) трастузумаба, соответствующую SEQ ID NO: 7
- Константный домен CH1 дикого типа (остаток в положении Kabat 192 представляет собой треонин) из человеческого IgG1, соответствующий SEQ ID NO: 8
Полипептидный линкер, соединяющий 2 тяжелые цепи Fab, состоящий из EPKSCDKTHTSPPAPAPELLGGPGGPPGPGPGGG (SEQ ID NO: 2);
- Вариабельная область тяжелой цепи цетуксимаба (VH), соответствующая SEQ ID NO: 9
- Мутированный константный домен CH1 (остаток в положении Kabat 192 был мутирован из треонина в глутаминовую кислоту) из человеческого IgG1, соответствующего SEQ ID NO: 10
- Шарнирный участок дикого типа из человеческого IgG1, соответствующий SEQ ID NO: 11
- Домен CH2 дикого типа человеческого IgG1, соответствующий SEQ ID NO: 12
- Домен CH3 дикого типа человеческого IgG1, соответствующий SEQ ID NO: 13
Так, биспецифическое антитело по изобретению имеет непрерывную тяжелую цепь (701 остаток) SEQ ID NO: 14
ii) легкая цепь трастузумаба дикого типа, состоящая из SEQ ID NO: 15
iii) легкая цепь цетуксимаба с мутированным константным доменом (остатки в положениях Kabat Ser 114 и Asn 137 были мутированы в Ala и Lys, соответственно) из Каппа человека, соответствующего SEQ ID NO: 16.
Вторым биспецифичным антителом по изобретению является антитело, обозначенное BiXAb2b, которое состоит из тех же последовательностей, за исключением линкера, который представляет собой
EPKSCDKTHTSPPAPAPELLGGPAAPPAPAPAGG (SEQ ID NO: 3).
Третьим биспецифичным антителом по изобретению является антитело, обозначенное BiXAb2c, которое состоит из тех же последовательностей, за исключением линкера, который представляет собой
EPKSCDKTHTSPPAPAPELLGGPAAPPGPAPGGG (SEQ ID NO: 4).
Четвертым биспецифичным антителом по изобретению является антитело, обозначенное BiXAb3b, имеющее следующую структуру:
i) непрерывная тяжелая цепь, которая включает
- Вариабельную область тяжелой цепи (VH) атезолизумаба, соответствующую SEQ ID NO: 23
(EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISP YGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQ GTLVTVSS)
- Мутированный константный домен CH1 (остаток в положении 192 Kabat был мутирован из треонина в глутаминовую кислоту) из человеческого IgG1, соответствующего SEQ ID NO: 10
Полипептидный линкер, соединяющий 2 тяжелые цепи Fab, состоит из SEQ ID NO: 3;
Вариабельную область тяжелой цепи даратумумаба (VH), соответствующую SEQ ID NO: 17
(EVQLLESGGGLVQPGGSLRLSCAVSGFTFNSFAMSWVRQAPGKGLEWVSAISGS GGGTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYFCAKDKILWFGEPVFDYW GQGTLVTVSS)
- Константный домен CH1 дикого типа (остаток в положении Kabat 192 представляет собой треонин) из человеческого IgG1, соответствующий SEQ ID NO: 8
- Шарнирный участок дикого типа из человеческого IgG1, соответствующий SEQ ID NO: 11
- Домен CH2 дикого типа человеческого IgG1, соответствующий SEQ ID NO: 12
- Домен CH3 дикого типа человеческого IgG1, соответствующий SEQ ID NO: 13
[Таким образом, тяжелая цепь по изобретению имеет непрерывную тяжелую цепь SEQ ID NO: 18
ii) легкая цепь атезолизумаба с мутированным константным доменом (остатки в положениях Kabat Ser 114 и Asn 137 были мутированы в Ala и Lys, соответственно) из Каппа человека, соответствующая SEQ ID NO: 19
DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKRTVAAPAVFIFPPSDEQLKSGTASVVCLLKNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
iii) легкая цепь даратумумаба дикого типа, состоящая из SEQ ID NO: 20
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPPTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Для сравнения было также получено антитело, обозначенное BiXAb3a, которое отличается от антитела BiXAb3b линкером, который состоит из
EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
Другая конструкция изобретения обозначена Fab-Fab3b; он состоит из той же последовательности, что и BiXAb3b, за исключением того, что в тяжелой цепи отсутствуют домены шарнира, CH2 и CH3. Итак, Fab-Fab3b имеет непрерывную тяжелую цепь SEQ ID NO: 21
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVEVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTSPPAPAPELLGGPAAPPAPAPAGGEVQLLESGGGLVQPGGSLRLSCAVSGFTFNSFAMSWVRQAPGKGLEWVSAISGSGGGTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYFCAKDKILWFGEPVFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
Для целей сравнения также была получена конструкция, обозначенная Fab-Fab3a, которая состоит из той же последовательности, что и BiXAb3a, за исключением того, что в тяжелой цепи отсутствуют домены петли, CH2 и CH3.
SEQ ID NO: 7-16 показаны ниже.
- SEQ ID NO: 7
EVQLVESGGGLVQPGGSLRLSCAASGFNIKDTYIHWVRQAPGKGLEWVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDGFYAMDYWGQGTLVTVSS
- SEQ ID NO: 8
ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKV
- SEQ ID NO: 9
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSNDTAIYYCARALTYYDYEFAYWGQGTLVTVSA
- SEQ ID NO: 10
ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVEVPSSSLGTQTYICNVNHKPSNTKVDKKV
- SEQ ID NO: 11
EPKSCDKTHTCPPCP
- SEQ ID NO: 12
APELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAK
- SEQ ID NO: 13
GQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
- SEQ ID NO: 14
EVQLVESGGGLVQPGGSLRLSCAASGFNIKDTYIHWVRQAPGKGLEWVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDGFYAMDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTSPPAPAPELLGGPGGPPGPGPGGGQVQLKQSGPGLVQPSQSLSITCTVSGFSLTNYGVHWVRQSPGKGLEWLGVIWSGGNTDYNTPFTSRLSINKDNSKSQVFFKMNSLQSNDTAIYYCARALTYYDYEFAYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVEVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
- SEQ ID NO: 15
DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
- SEQ ID NO: 16
DILLTQSPVILSVSPGERVSFSCRASQSIGTNIHWYQQRTNGSPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQNNNWPTTFGAGTKLELKRTVAAPAVFIFPPSDEQLKSGTASVVCLLKNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Синтез генов
Аминокислотные последовательности анти-HER2 (трастузумаб, клон humAb4D5-8) и анти-EGFR (цетуксимаб) использовали для конструирования последовательностей ДНК после оптимизации кодонов для экспрессии в млекопитающих с использованием программы GeneScript. Для тяжелой цепи за ДНК, кодирующими сигнальные пептиды, вариабельную область и константный домен CH1 из Fab1, следовали псевдо-шарнирный линкер, а вариабельную область и константный домен CH1 из Fab2 с фланкирующими последовательностями для расщепления рестриктазой синтезировали с помощью GeneScript. Для легкой цепи GeneScript синтезировал ДНК, кодирующие сигнальные пептиды и вариабельные и константные каппа-области.
Реакции ПЦР с использованием PfuTurbo Hot Start проводили для амплификации вставок, которые затем расщепляли NotI + ApaI и NotI + HindIII для тяжелых и легких цепей, соответственно. Фрагменты тяжелой цепи, подвергнутые двойному расщеплению, лигировали с обработанным NotI + ApaI экспрессирующим вектором pcDNA3.1 (Invitrogen), в который уже были встроены домены CH1 + шарнир + CH2 + CH3 из IgG1 человека. Фрагменты легкой цепи с двойным расщеплением лигировали с обработанным NotI + HindIII экспрессирующим вектором pcDNA3.1 (Invitrogen). Плазмидные ДНК были проверены путем секвенирования двухцепочечной ДНК.
Экспрессия и очистка
Биспецифичные антитела по изобретению получали транзиторной экспрессией генов путем котрансфекции 3 генов, кодируемых отдельными векторами в молярном соотношении 2: 3: 3 = HC: LC1: LC2 (1 непрерывная тяжелая цепь (HC) и 2 легкие цепи (LC)) в клетках CHO-S, адаптированных к бессывороточной среде в суспензии (среда CHO SFM-II от Life Technologies™). Как правило, для тестирования экспрессии в среднем объеме 50 мл 50 мг плазмидной ДНК (25 мкг тяжелой цепи 1, 12,5 мкг легкой цепи тратузумаба и 12,5 мкг легкой цепи цетуксимаба) смешивали в пробирке Eppendorf объемом 1,5 мл, добавляли 1 мл среды СНО SFM, содержащей 25 мкл 3 мг/мл реагента для трансфекции PEI (Polyplus) pH 7,0, инкубировали при комнатной температуре в течение 20 минут. Смесь ДНК-PEI загружали в 49 мл клеток Invitrogen FreeStyle ™ CHO-S от Life Technologies при 1 ~ 2 × 10 6/мл во встряхиваемой колбе объемом 125 мл. Клетки встряхивали еще 6 дней. Надосадочную жидкость собирали центрифугированием клеток при 3000 об/мин в течение 15 минут. Титр экспрессии BiXAb в надосадочной жидкости определяли с использованием биосенсоров FortéBio с протеином А (Octet® Systems). Биспецифическое моноклональное антитело (BiXAb) затем очищали в аффинной среде с протеином А с использованием MabSelect SuRe (GE Healthcare Life Sciences). Антитело элюировали из протеина А, с помощью 0,1 М глицина, рН 3,5, с нейтрализацией в 1 М TRIS. Очищенное антитело в PBS Дульбекко (Lonza BE17-512Q) подвергали стерильной фильтрации (0,2 мкМ стерильные фильтры от Techno Plastic Products AG) и определяли конечную концентрацию путем считывания OD при 280 нм с использованием спектрофотометра Eppendorf BioSpectrometer®.
Анализ ДСН-ПААГ
Электрофорез проводили в восстанавливающих и невосстанавливающих условиях, используя гели Biorad Stain-Free 4-15% и соответствующий рабочий буфер. Образцы готовили объединением очищенных антител BiXAb или Fab-Fab с буфером для образцов 2X SDS и нагреванием в течение 5 минут при 95 °C. Подготовка восстановленных образцов включала добавление восстановителя NuPAGE перед нагреванием. Кажущуюся MW определяли с использованием стандартов неокрашенных белков Ladder Precision Plus (Biorad).
Анализ эксклюзионной хроматографией
В сконструированных белковых молекулах часто наблюдается агрегация белка. Мы выполнили аналитическую эксклюзионную хроматографию (SEC) для анализа содержания высокомолекулярных частиц в наших антителах. Мы использовали колонку SEC-s3000 (300x7,8 мм) (BioSep) и систему Aktapurifier 10 (GE Healthcare); анализ проводили при скорости потока 1 мл/мин с использованием буфера PBS pH 7,4.
Хроматограммы SEC BiXAb2b (Фигура 3А), BiXAb3a, BiXAb3b (Фигура 3В), Fab-Fab3a и Fab-Fab3b (Фигура 3С) показали, что основной пик соответствует ожидаемым размерам мономерных антител BiXAb и Fab-Fab; Эти пики составляли 99,9-100,0 % от общего образца. Таким образом, мы пришли к выводу, что антитела, содержащие новый линкер, не имеют высокомолекулярных разновидностей. Узкая и симметричная форма мономерных пиков позволяет предположить, что все BiXAb и Fab-Fabs были правильно собраны и представлены одной разновидностью.
Характеризация BiXAb с помощью дифференциальной сканирующей калориметрии
Дифференциальная сканирующая калориметрия (DSC) была использована для проверки термостабильности BiXAb2b. Для проведения экспериментов по дифференциальной сканирующей калориметрии использовали систему Microcal™ VP-Capillary DSC (Malvern Instruments).
Образцы центрифугировали (20000 × g, 5 мин, 4 °C), и содержание белка в них определяли количественно до анализа DSC с использованием спектрофотометра Nanodrop ND-1000 (Thermo Scientific) с использованием программы анализа IgG. Для анализа образцы разводили в PBS до конечной концентрации 1 мг/мл.
Время предварительного уравновешивания составляло 3 мин, и полученные термограммы были получены при температуре от 20 до 110 °С при скорости сканирования 60 °С/ч, периоде фильтрации 25 с и обратной связи со средой. Перед анализом образца было проведено 5 сканирований буфера/буфера для стабилизации прибора, и между каждым сканированием белка/буфера проводилось сканирование буфера/буфера. Данные были сопоставлены с моделью развертывания, не состоящей из двух состояний, с предварительным и последующим переходом, скорректированным путем вычитания исходного уровня.
Результаты DSC продемонстрировали, что профиль DSC BiXAb2b показал два перехода. Меньший пик имел Cp max 96 ккал/моль/°C и Tm1 71,5°C, что соответствует разворачиванию обоих доменов CH2 и Fab, а более крупный пик имел Cp max 190 ккал/моль/° C и Tm2 80,5°C, что соответствует разворачиванию домена CH3.
Анализ методом жидкостной хроматографии/масс-спектроскопии (LC-MS)
Данные LC-MS/MS были получены с использованием системы Dionex Ultimate 3000, соединенной с масс-спектрометром Q-Exactive (Thermo Fisher Scientific, Бремен, Германия) и колонкой с обращенной фазой Proswift RP-4H (250 мм × 1 мм; Thermo Fisher). Температура в колоночном термостате была установлена на 65 °С. Десять микролитров инъецировали для разделения LC. Градиент подвижных фаз, состоящих из воды категории LC-MS с 0,1% муравьиной кислоты (фаза A) и ацетонитрила с 0,1% муравьиной кислотой (фаза B), подавали со скоростью потока 0,2 мл/мин (общее время прогона - 20 минут). Элюированные разновидности антител вводили в прибор Q-Exactive с помощью электрораспылительной ионизации (ESI), которая работала в режиме положительных ионов с использованием полного сканирования с разрешением 15000. Программное обеспечение Xcalibur 2.2 (Thermo Fisher Scientific, Бремен, Германия) использовали для инструментального контроля и обработки файлов данных.
На Фигурах 4A и 4B представлен анализ LC-MS Fab-Fab3a (содержащего линкер с SEQ ID NO: 6) и Fab-Fab3a (содержащего линкер с SEQ ID NO: 3), соответственно. Фигура 4А демонстрирует, что спектр LC-MS Fab-Fab3a с линкером, соответствующим SEQ ID NO: 6, является значительно более гетерогенным, чем у родственного антитела Fab-Fab3b, которое отличалось только по составу линкера (SEQ ID NO : 3). Линкерная последовательность в антителе Fab-Fab3a содержит последовательность PSTPPTPSPS (SEQ ID NO: 22), которая обнаружена в петле IgA1 человека и, как известно, подвергается О-сцепленному гликозилированию по 2 треониновым и 2 сериновым остаткам. Поскольку гликозилирование этих сайтов является гетерогенным, продукт обычно обладает множественными гликоформами, чьи популяции сильно зависят от условий экспрессии рекомбинантных белков. Общее число N-связанных и О-связанных сайтов гликозилирования в Fab-Fab3a составляет, по меньшей мере, 4, что объясняет сложный спектр MS, наблюдаемый на Фигуре 4А. Последовательность линкера, соответствующая SEQ ID NO: 3, была разработана для того, чтобы уменьшить гетерогенность вследствие негомогенного О-связанного гликозилирования. Как таковые, несколько остатков серина и треонина в части линкера, чья последовательность совпадает с последовательностью петли человеческого IgA1, о которой известно, что она подвергается О-связанному гликозилированию, были заменены остатками глицина; Кроме того, в линкере некоторые другие остатки серина и треонина были заменены глицином. На Фигуре 4В показано, что спектр MS для Fab-Fab3b, содержащего линкер с SEQ ID NO: 3, существенно упрощен; это связано с тем, что аналит Fab-Fab3b является менее сложным, чем аналит Fab-Fab3a, спектр которого представлен на Фигуре 4А. Таким образом, мы пришли к выводу, что удаление 4 O-связанных сайтов гликозилирования из антитела Fab-Fab3b путем замены линкера SEQ ID NO: 6 на линкер SEQ ID NO: 3 привело к существенно более гомогенному препарату BiXAb.
Разница в гомогенности биспецифичных антител, содержащих линкеры SEQ ID NO: 6 и SEQ ID NO: 3, была особенно очевидна из анализа полноразмерных антител BiXAb, которые представляют собой симметричные молекулы, которые имеют две линкерные последовательности и, таким образом, обладают увеличенным числом гликоформ в препарате BiXAb. Кроме того, BiXAb обладают двумя N-связанными сайтами гликозилирования, по одному на каждую тяжелую цепь Fc-домена. Спектры LC-MS BiXAb3a (созданного с помощью линкера SEQ ID NO: 6) и BiXAb3b (созданного с помощью линкера SEQ ID NO: 3) представлены на Фигурах 5A и 5B, соответственно. Ожидается, что BiXAb3a будет содержать 8 дополнительных O-связанных сайтов гликозилирования относительно BiXAb3b, основываясь на различиях в последовательностях линкера. Большое количество O-связанных сайтов гликозилирования в BiXAb3a объясняет очень сложный спектр MS на Фигуре 5А. Оба антитела BiXAb обладают 2 сайтами N-гликозилирования в Fc-домене, что приводит к дополнительным гликоформам, способствующим гетерогенности обоих BiXAb, что объясняет остаточное количество гетерогенности в BiXAb3b, наблюдаемое на Фигуре 5B.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> BIOMUNEX pharmaceuticals
<120> A polypeptide linker for preparing multispecific antibodies
<130> B2412PC
<160> 23
<170> PatentIn version 3.5
<210> 1
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (10)..(11)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (14)..(14)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (24)..(25)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (28)..(28)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (30)..(30)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (32)..(32)
<223> Xaa can be any naturally occurring amino acid
<400> 1
Glu Pro Lys Xaa Cys Asp Lys Xaa His Xaa Xaa Pro Pro Xaa Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Xaa Xaa Pro Pro Xaa Pro Xaa Pro Xaa
20 25 30
Gly Gly
<210> 2
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 2
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ala Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Gly Gly Pro Pro Gly Pro Gly Pro Gly
20 25 30
Gly Gly
<210> 3
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 3
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ala Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ala Ala Pro Pro Ala Pro Ala Pro Ala
20 25 30
Gly Gly
<210> 4
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 4
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ala Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ala Ala Pro Pro Gly Pro Ala Pro Gly
20 25 30
Gly Gly
<210> 5
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 5
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
20 25 30
Gly Gly
<210> 6
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 6
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ser Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
20 25 30
Gly Gly
<210> 7
<211> 120
<212> PRT
<213> Homo sapiens
<400> 7
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 8
<211> 98
<212> PRT
<213> Homo sapiens
<400> 8
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 9
<211> 119
<212> PRT
<213> Homo sapiens
<400> 9
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala
115
<210> 10
<211> 98
<212> PRT
<213> Homo sapiens
<400> 10
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Glu Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 11
<211> 15
<212> PRT
<213> Homo sapiens
<400> 11
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 12
<211> 110
<212> PRT
<213> Homo sapiens
<400> 12
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 13
<211> 107
<212> PRT
<213> Homo sapiens
<400> 13
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
100 105
<210> 14
<211> 701
<212> PRT
<213> Homo sapiens
<400> 14
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Ser Pro Pro Ala Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Gly Gly Pro Pro Gly Pro Gly Pro Gly Gly Gly Gln Val Gln Leu
245 250 255
Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln Ser Leu Ser Ile
260 265 270
Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr Gly Val His Trp
275 280 285
Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu Gly Val Ile Trp
290 295 300
Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr Ser Arg Leu Ser
305 310 315 320
Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe Lys Met Asn Ser
325 330 335
Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala Arg Ala Leu Thr
340 345 350
Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
355 360 365
Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
370 375 380
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
385 390 395 400
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
405 410 415
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
420 425 430
Leu Tyr Ser Leu Ser Ser Val Val Glu Val Pro Ser Ser Ser Leu Gly
435 440 445
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
450 455 460
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
465 470 475 480
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
485 490 495
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
500 505 510
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
515 520 525
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
530 535 540
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
545 550 555 560
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
565 570 575
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
580 585 590
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
595 600 605
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
610 615 620
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
625 630 635 640
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
645 650 655
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
660 665 670
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
675 680 685
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
690 695 700
<210> 15
<211> 214
<212> PRT
<213> Homo sapiens
<400> 15
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 16
<211> 214
<212> PRT
<213> Homo sapiens
<400> 16
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Lys Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 17
<211> 122
<212> PRT
<213> Homo sapiens
<400> 17
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Phe Asn Ser Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Lys Ile Leu Trp Phe Gly Glu Pro Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 18
<211> 702
<212> PRT
<213> Homo sapiens
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Glu Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Ser Pro Pro Ala Pro Ala Pro Glu Leu Leu Gly Gly Pro Ala
225 230 235 240
Ala Pro Pro Ala Pro Ala Pro Ala Gly Gly Glu Val Gln Leu Leu Glu
245 250 255
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
260 265 270
Ala Val Ser Gly Phe Thr Phe Asn Ser Phe Ala Met Ser Trp Val Arg
275 280 285
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser Gly Ser
290 295 300
Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
305 310 315 320
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
325 330 335
Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys Ala Lys Asp Lys Ile Leu
340 345 350
Trp Phe Gly Glu Pro Val Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
355 360 365
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
370 375 380
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
385 390 395 400
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
405 410 415
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
420 425 430
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
435 440 445
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
450 455 460
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
465 470 475 480
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
485 490 495
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
500 505 510
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
515 520 525
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
530 535 540
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
545 550 555 560
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
565 570 575
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
580 585 590
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
595 600 605
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
610 615 620
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
625 630 635 640
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
645 650 655
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
660 665 670
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
675 680 685
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
690 695 700
<210> 19
<211> 214
<212> PRT
<213> Homo sapiens
<400> 19
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Lys Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 20
<211> 214
<212> PRT
<213> Homo sapiens
<400> 20
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 21
<211> 475
<212> PRT
<213> Homo sapiens
<400> 21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Glu Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Ser Pro Pro Ala Pro Ala Pro Glu Leu Leu Gly Gly Pro Ala
225 230 235 240
Ala Pro Pro Ala Pro Ala Pro Ala Gly Gly Glu Val Gln Leu Leu Glu
245 250 255
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
260 265 270
Ala Val Ser Gly Phe Thr Phe Asn Ser Phe Ala Met Ser Trp Val Arg
275 280 285
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser Gly Ser
290 295 300
Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
305 310 315 320
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
325 330 335
Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys Ala Lys Asp Lys Ile Leu
340 345 350
Trp Phe Gly Glu Pro Val Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
355 360 365
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
370 375 380
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
385 390 395 400
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
405 410 415
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
420 425 430
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
435 440 445
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
450 455 460
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
465 470 475
<210> 22
<211> 10
<212> PRT
<213> Homo sapiens
<400> 22
Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
1 5 10
<210> 23
<211> 118
<212> PRT
<213> Homo sapiens
<400> 23
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| СКОНСТРУИРОВАННЫЕ МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И ДРУГИЕ МУЛЬТИМЕРНЫЕ БЕЛКИ С АСИММЕТРИЧНЫМИ МУТАЦИЯМИ В ОБЛАСТИ CH2-CH3 | 2018 |
|
RU2804031C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ ПРОТИВ GITR И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2795625C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ НА ОСНОВЕ ПСЕВДО-FAB | 2019 |
|
RU2820254C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2811477C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
Настоящее изобретение относится к области биотехнологии. Изобретение позволяет получить мутантный полипептидный линкер, разработанный из шарнирной последовательности IgG1 с удаленными сайтами гликозилирования. Указанный линкер может быть использован в получении разнообразных мультиспецифических антител. В сравнении с известными аналогами использование линкера согласно настоящему изобретению позволяет получить более единообразные препараты мультиспецифических антител. 4 н. и 16 з.п. ф-лы, 5 ил., 1 пр.
1. Мультиспецифический антигенсвязывающий фрагмент, содержащий по меньшей мере два Fab-фрагмента, где каждый Fab-фрагмент распознает отличный представляющий интерес эпитоп, а указанные Fab-фрагменты расположены тандемно в любом порядке, где C-конец домена CH1 первого фрагмента Fab связан с N-концом домена VH следующего фрагмента Fab через полипептидный линкер, отличающийся тем, что последовательность полипептидного линкера включает или состоит из аминокислотной последовательности
EPKX1CDKX2HX3X4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 1),
где X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, одинаковые или разные, представляют собой любую аминокислоту; при условии, что линкерная последовательность не включает и не состоит из последовательностей
EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 5) или
EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
2. Мультиспецифический антигенсвязывающий фрагмент по п. 1, где указанный мультиспецифический антигенсвязывающий фрагмент содержит по меньшей мере два Fab фрагмента с различными доменами CH1 и CL .
3. Мультиспецифическое антитело, имеющее два идентичных антигенсвязывающих плеча, каждое из которых состоит из мультиспецифического антигенсвязывающего фрагмента по п. 1 или 2.
4. Мультиспецифическое антитело по п. 3, которое имеет иммуноглобулиноподобную структуру, включающее:
- два идентичных антигенсвязывающих плеча, каждое из которых состоит из мультиспецифического антигенсвязывающего фрагмента, как определено в п. 1;
- димеризованные домены СН2 и СН3 иммуноглобулина;
- шарнирную область IgA, IgG или IgD, связывающую C-концы доменов CH1 антигенсвязывающих плеч с N-концами доменов CH2.
5. Мультиспецифическое антитело, предпочтительно биспецифическое антитело, включающее две тяжелые цепи и четыре легкие цепи,
отличающееся тем, что каждая тяжелая цепь включает
- область Fc иммуноглобулина, содержащую домены Шарнир-CH2-CH3,
- где область Fc связана с CH1-VH из тяжелой цепи Fab антитела 1 (Ab1) указанным шарнирным доменом,
- который, в свою очередь, связан с CH1-VH из тяжелой цепи Fab антитела 2 (Ab2) посредством полипептидной линкерной последовательности, где полипептидная линкерная последовательность связывает N-конец указанного домена VH из тяжелой цепи Fab Ab1 с C-концом указанного домена CH1 из Ab2,
а четыре легкие цепи включают легкие цепи Ab1 и легкие цепи Ab2, связанные с их когнатными доменами тяжелых цепей;
где последовательность полипептидного линкера включает или состоит из аминокислотной последовательности
EPKX1CDKX2HX3X4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 1),
где X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, одинаковые или разные, представляют собой любую аминокислоту; при условии, что линкерная последовательность не включает и не состоит из последовательностей EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 5) или EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
6. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-5, где X1, X2 и X3, одинаковые или разные, представляют собой треонин (T) или серин (S).
7. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-5, отличающиеся тем, что X1, X2 и X3, одинаковые или разные, представляют собой любую аминокислоту, отличную от треонина (T) или серина (S), предпочтительно, где X1, X2 и X3, одинаковые или разные, выбраны из группы, состоящей из Ala (A), Gly (G), Val (V), Asn (N), Asp (D) и Ile (I).
8. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-7, отличающиеся тем, что X4 и X5, одинаковые или разные, представляют собой любую аминокислоту, выбранную из группы, состоящей из серина (S), цистеина (C), аланина (А) и глицина (G).
9. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-8, отличающиеся тем, что X6, X7, X8, X9, X10, одинаковые или разные, представляют собой любую аминокислоту, отличную от треонина (T) или серина (S).
10. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-9, отличающиеся тем, что X4 представляет собой серин (S) или цистеин (C).
11. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-10, отличающиеся тем, что X5 представляет собой аланин (A) или цистеин (C).
12. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-9, отличающиеся тем, что X6, X7, X8, X9, X10, одинаковые или разные, выбраны из группы, состоящей из Ala (A), Gly (G), Val (V), Asn (N), Asp (D) и Ile (I).
13. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по п. 12, отличающиеся тем, что X6, X7, X8, X9, X10, одинаковые или разные, выбраны из группы, состоящей из Ala (A) и Gly (G).
14. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по п. 13, отличающиеся тем, что X6 и X7 являются идентичными и предпочтительно выбраны из группы, состоящей из Ala (A) и Gly (G).
15. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по п. 13, отличающиеся тем, что полипептидная линкерная последовательность включает или состоит из последовательности, выбранной из группы, состоящей из
EPKSCDKTHTSPPAPAPELLGGPAAPPAPAPAGG (SEQ ID NO: 3);
EPKSCDKTHTSPPAPAPELLGGPGGPPGPGPGGG (SEQ ID NO: 2); и
EPKSCDKTHTSPPAPAPELLGGPAAPPGPAPGGG (SEQ ID NO: 4).
16. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-15, которое распознает как EGFR, так и HER2/neu.
17. Мультиспецифическое антитело по п. 5, отличающееся тем, что Ab1 и Ab2, отличающиеся друг от друга, независимо выбраны из группы, состоящей из цетуксимаба или его мутированного производного, с одной стороны, и трастузумаба или его мутированного производного, с другой стороны.
18. Мультиспецифический антигенсвязывающий фрагмент или мультиспецифическое антитело по любому из пп. 1-15, которое распознает как CD38, так и PD-L1.
19. Линкерный полипептид, который включает или состоит из аминокислотной последовательности
EPKX1CDKX2HX3X4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 1),
где Х1, Х2, Х3, Х4, Х5, Х6, Х7, X8, X9, X10, одинаковые или разные, представляют собой любую аминокислоту, как определено в любом из пп. 1-11; при условии, что полипептид не включает и не состоит из последовательностей
EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 5) или
EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 6).
20. Линкерный полипептид по п. 19, который включает или состоит из последовательности, выбранной из группы, включающей
EPKSCDKTHTSPPAPAPELLGGPAAPPAPAPAGG (SEQ ID NO: 3);
EPKSCDKTHTSPPAPAPELLGGPGGPPGPGPGGG (SEQ ID NO: 2); и
EPKSCDKTHTSPPAPAPELLGGPAAPPGPAPGGG (SEQ ID NO: 4).
| WO 2013005194 A2, 10.01.2013 | |||
| GOLAY J | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| РОЙТ А | |||
| и др | |||
| Иммунология, Москва, "Мир", 2000, стр | |||
| Прибор, автоматически записывающий пройденный путь | 1920 |
|
SU110A1 |
| СИНГЕР М | |||
| и др | |||
| Гены и геномы, Москва, "Мир", 1998, том 1, стр | |||
| Способ приготовления сернистого красителя защитного цвета | 1915 |
|
SU63A1 |
| MARIUZZA | |||

