ПЕРЕКРЕСТНАЯ ССЫЛКА
По настоящей заявке испрашивается приоритет в соответствии с международной заявкой на патент № PCT/CN2017/103032, поданной 22 сентября 2017 года.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение в целом относится к биспецифичным полипептидным комплексам против CD3×CD19, содержащим вариабельные области антитела, конденсированные с константными областями TCR.
УРОВЕНЬ ТЕХНИКИ
Биспецифичные антитела становятся новым классом терапевтических антител. Они могут связывать две разные мишени или два разных эпитопа на мишени, создавая аддитивный или синергетический эффект, превосходящий эффект отдельных антител. Для конструирования новых биспецифичных форматов, таких как DVD-Ig, CrossMab, BiTE и т.д., было направлено много усилий (Spiess et al., Molecular Immunology, 67(2), pp.95-106 (2015)). Однако эти форматы могут потенциально иметь различные ограничения по стабильности, растворимости, коротком периоде полураспада и иммуногенности.
В числе указанных форматов биспецифичных антител, IgG-подобное биспецифичное антитело является распространенным форматом: одно плечо связывается с мишенью А, а другое плечо связывается с мишенью В. Структурно оно состоит из половины антитела А и половины антитела В с размером и формой, аналогичными природному IgG. Для облегчения последующей модификации, желательно, чтобы такие биспецифичные молекулы можно было легко изготавливать, подобно нормальным IgG, из одной клетки-хозяина с высоким уровнем экспрессии и правильно собранной формой. К сожалению, спаривание когнатных легких и тяжелых цепей, а также сборка двух разных полуантител не могут контролироваться автоматически. Все виды ошибочных спариваний могут привести к значительной гетерогенности продукта.
Предпочтительная гетеродимерная сборка двух разных тяжелых цепей осуществлялась посредством мутаций в Fc-области, таких как “выступ-в-углубление” (Ridgway et al., Protein Engineering, 9(7), pp.617-21(1996); Merchant et al., Nature Biotechnology, 16(7), pp.677-681(1998)), электростатического управления (Gunasekaran et al., Journal of Biological Chemistry, 285(25), pp.19637-19646 (2010)) или негативного дизайна (Kreudenstein et al., mAbs, 5(5), pp.646-654 (2013); Leaver-Fay et al., Structure, 24(4), pp.641-651 (2016)). Однако селективное спаривание легких и тяжелых цепей каждого отдельного антитела остается все еще сложной задачей. Область контакта между легкими и тяжелыми цепями включает вариабельный домен (VH-VL) и константный домен (CH1-CL). В разработке ортогональных областей контакта, для облегчения когнатного спаривания, применяли различные методы. В компании “Roche” меняли домены CH1 и CL и создали платформу CrossMab (Schaefer et al., Proceedings of the National Academy of Sciences of the United States of America, 108(27), pp.11187-11192 (2011)), в компании “Medimmmune” вводили альтернативно дисульфидную связь (Mazor et al., mAbs, 7(2), pp.377-389 (2015)), в компании “Amgen” осуществляли дополнительное электростатическое управление в области CH1-CL (Liu et al., Journal of Biological Chemistry, 290(12), pp.7535-7562 (2015)), а в компаниях “Lilly” (Lewis et al., Nature Biotechnology, 32(2), pp.191-198 (2014)) и “Genentech” (Dillon et al., mAbs, 9(2), pp.213-230 (2017)) осуществляли мутации как в вариабельных, так и в константных доменах.
CD19 человека представляет собой трансмембранный белок типа I, принадлежащий суперсемейству иммуноглобулинов (Carter et al., Curr Dir Autoimmun, 7: 4-32 (2004)). Он экспрессируется на большинстве В-клеток, но не обнаруживается на плазматических клетках, стволовых клетках или нормальных клетках миелоидной линии (Tedder, Nat Rev Rheumatol, 5 (10): 572-577 (2009)). CD19 принимают немаловажное участие в установлении эндогенных порогов передачи сигналов B-клеток посредством модуляции как зависимых от B-клеточных рецепторов (BCR), так и независимых сигналов (Wang et al., Experimental Hematology & Oncology, 1:36 (2012)). CD19 более широко экспрессируется, чем CD20. Характер экспрессии CD19 сохраняется при В-клеточных злокачественных опухолях, включая все подтипы В-клеточной лимфомы, от медленно растущих до агрессивных форм, а также В-клеточную хроническую лимфоцитарную лейкемию и не Т-клеточный острый лимфобластный лейкоз, а также обеспечивает нацеливание на опухоли из недифференцированных В-клеток, такие как острый лимфобластный лейкоз (ALL), которые не являются мишенью для ритуксимаба. Для терапии лимфомы, исследованию подвергали несколько моноклональных антител к CD19 (публикация заявки на патент США № 20140072587 A1, патент США № 8242252 B2 и патент США № 8097703 B2).
Т-клеточный ко-рецептор CD3 представляет собой белковый комплекс, состоящий из четырех различных цепей: CD3 гамма-цепи, CD3 дельта-цепи и двух CD3 эпсилон-цепей. Четыре цепи ассоциируются с молекулой, известной как Т-клеточный рецептор (TCR) и дзета-цепью с генерированием сигнала активации в Т-лимфоцитах. TCR, зета-цепь и молекулы CD3 образуют комплекс TCR, в котором TCR как субъединица распознает и связывается с антигеном, а CD3, как субъединица, переносит и передает антигенную стимуляцию по сигнальному пути и, в конечном итоге, регулирует T-клеточную активность. Белок CD3 присутствует практически во всех Т-клетках. Комплекс CD3-TCR модулирует функции Т-клеток как врожденного, так и адаптивного иммунного ответа, а также клеточные и гуморальные иммунные функции. К ним относятся уничтожение патогенных организмов и контроль роста опухоли с помощью широкого спектра цитотоксических эффектов. Моноклональные антитела мыши, специфичные к CD3 человека, такие как OKT3 (Kung et al., Science, 206: 347-9 (1979)), были антителами CD3 первого поколения, разработанными для лечения. Хотя OKT3 обладает сильным иммуносупрессивным действием, его клиническое применение было затруднено вследствие серьезных побочных эффектов, связанных с его иммуногенным и митогенным потенциалами. (Chatenoud, Nature Reviews, 3:123-132 (2003)). ОКТ3 индуцирует антиглобулиновый ответ, способствуя своему быстрому выведению и нейтрализации (Chatenoud et al., Eur. J. Immunol., 137:830-8 (1982)). Кроме того, OKT3 индуцирует пролиферацию Т-клеток и продукцию цитокинов in vitro и приводит к крупномасштабному высвобождению цитокинов in vivo. (Hirsch et al., J. Immunol, 142: 737-43 (1989)). Такие серьезные побочные эффекты ограничивают более широкое применение OKT3 при трансплантации, а также распространение его применения на другие клинические области, такие как аутоиммунитет.
Биспецифичное антитело, нацеленное на CD3 и CD19, может связываться с Т-клетками и В-клетками одновременно. Как только биспецифичное антитело связывается с CD3-позитивной Т-клеткой и CD19-позитивной В-клеткой, образуется цитолитический синапс. Затем цитотоксичность индуцируется высвобождением перфорина и гранзимов из гранул в цитотоксической Т-клетке, причем последняя вызывает апоптоз и лизис злокачественной В-клетки.
Существует большая потребность в конструировании биспецифичных молекул с желаемым уровнем экспрессии и временем полужизни in vivo как для CD3, так и для CD19. Такие биспецифичные полипептидные комплексы против CD3×CD19 применяются для лечения состояний, связанных с CD19, включая злокачественную опухоль.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В одном из аспектов, настоящее изобретение относится к биспецифичному полипептидному комплексу, содержащему первый антигенсвязывающий фрагмент, ассоциированный со вторым антигенсвязывающим фрагментом, где:
первый антигенсвязывающий фрагмент содержит:
первый полипептид, содержащий, от N-конца до C-конца, первый вариабельный домен тяжелой цепи (VH1) первого антитела, функционально связанный с первой константной областью (C1) рецептора Т-клеток (TCR), и
второй полипептид, содержащий, от N-конца до C-конца, первый вариабельный домен легкой цепи (VL1) первого антитела, функционально связанный со второй константной областью TCR (C2),
где:
C1 содержит сконструированную СБета, содержащую SEQ ID NO: 1, и C2 содержит сконструированную САльфа, содержащую SEQ ID NO: 2,
аминокислота C48 в SEQ ID NO: 1 и аминокислота C41 в SEQ ID NO: 2 способны образовывать ненативную межцепочечную дисульфидную связь,
С1 и С2 способны образовывать димер, причем ненативная межцепочечная дисульфидная связь способна стабилизировать димер,
и
второй антигенсвязывающий фрагмент содержит:
второй вариабельный домен тяжелой цепи (VH2) второго антитела, функционально связанный с доменом CH1 тяжелой цепи антитела, и
второй вариабельный домен легкой цепи (VL2) второго антитела, функционально связанный с константным доменом легкой цепи антитела (CL),
где:
один из первого и второго антигенсвязывающих фрагментов представляет собой анти-CD3-связывающий фрагмент, а другой представляет собой анти-CD19-связывающий фрагмент,
анти-CD3-связывающий фрагмент получен из анти-CD3-антитела, содержащего:
а) CDR1 тяжелой цепи, содержащую SEQ ID NO: 3, b) CDR2 тяжелой цепи, содержащую SEQ ID NO: 4, c) CDR3 тяжелой цепи, содержащую SEQ ID NO: 5, d) CDR1 каппа-легкой цепи, содержащую SEQ ID NO: 6, e) CDR2 каппа-легкой цепи, содержащую SEQ ID NO: 7, и f) CDR3 каппа-легкой цепи, содержащую SEQ ID NO: 8,
анти-CD19 связывающий фрагмент получен из анти-CD19 антитела, содержащего:
а) CDR1 тяжелой цепи, содержащую SEQ ID NO: 9, b) CDR2 тяжелой цепи, содержащую SEQ ID NO: 10, c) CDR3 тяжелой цепи, содержащую SEQ ID NO: 11, d) CDR1 каппа-легкой цепи, содержащую SEQ ID NO: 12, e) CDR2 каппа-легкой цепи, содержащую SEQ ID NO: 13, и f) CDR3 каппа-легкой цепи, содержащую SEQ ID NO: 14,
и
первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент менее подвержены ошибочному спариванию, чем, если бы и первый и второй антигенсвязывающие фрагменты были аналогами природного Fab.
В некоторых вариантах осуществления, анти-CD3 связывающий фрагмент биспецифичного полипептидного комплекса получают из анти-CD3 антитела, содержащего последовательность вариабельного домена тяжелой цепи, содержащую SEQ ID NO: 15, и последовательность вариабельного домена легкой цепи, содержащую SEQ ID NO: 17.
В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент биспецифичного полипептидного комплекса получают из антитела против CD19, содержащего последовательность вариабельного домена тяжелой цепи, содержащую SEQ ID NO: 19, и последовательность вариабельного домена легкой цепи, содержащую SEQ ID NO: 21.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс содержит сочетание четырех полипептидных последовательностей: SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 29.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс содержит сочетание четырех полипептидных последовательностей: SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 30.
В одном из аспектов, настоящее изобретение относится к конъюгату, содержащему биспецифичный полипептидный комплекс, предусмотренный в настоящем документе, конъюгированный с фрагментом.
В одном из аспектов, настоящее изобретение относится к выделенному полинуклеотиду, кодирующему биспецифичный полипептидный комплекс, предусмотренный в настоящем документе.
В одном из аспектов, настоящее изобретение относится к выделенному вектору, содержащему полинуклеотид, представленный в настоящем документе.
В одном из аспектов, настоящее изобретение относится к клетке-хозяину, содержащей выделенный полинуклеотид, представленный в настоящем документе, или выделенный вектор, представленный в настоящем документе.
В одном из аспектов, настоящее изобретение относится к способу экспрессии биспецифичного полипептидного комплекса, представленного в настоящем документе, включающему культивирование клетки-хозяина, предоставленной в настоящем документе, в условиях, при которых экспрессируется биспецифичный полипептидный комплекс.
В одном из аспектов, настоящее изобретение относится к композиции, содержащей биспецифичный полипептидный комплекс, представленный в настоящем документе.
В одном из аспектов, настоящее изобретение относится к фармацевтической композиции, содержащей биспецифичный полипептидный комплекс, представленный в настоящем документе, и фармацевтически приемлемый носитель.
В одном из аспектов, настоящее изобретение относится к способу лечения заболевания или состояния, связанного с CD19, у субъекта, нуждающегося в таком лечении, включающему введение субъекту терапевтически эффективного количества биспецифичного полипептидного комплекса, представленного в настоящем документе. В некоторых вариантах осуществления, заболевание или состояние можно облегчать, устранять, лечить или предотвращать в случае, когда первый антиген и второй антиген, оба, модулированы.
В некоторых вариантах осуществления, первый VH функционально связан с СБета на первом домене конъюгирования, который содержит C-концевой фрагмент V/C конъюгирования антитела и имеет аминокислотную последовательность SEQ ID NO: 23 (LEDLKNVFPP), и первый VL функционально связан с САльфа на втором домене конъюгирования, который содержит N-концевой фрагмент V/C конъюгирования TCR и имеет аминокислотную последовательность SEQ ID NO: 24 (KPDIQNPDP).
В некоторых вариантах осуществления, первый антигенсвязывающий фрагмент связан с первым доменом димеризации, а второй антигенсвязывающий фрагмент связан со вторым доменом димеризации, где первый и второй домены димеризации ассоциированы. В некоторых вариантах осуществления, ассоциация происходит через линкер, дисульфидную связь, водородную связь, электростатическое взаимодействие, солевой мостик или гидрофобно-гидрофильное взаимодействие или их сочетание.
В некоторых вариантах осуществления, первый и/или второй димеризационный домен содержит, по меньшей мере, часть шарнирной области антитела, необязательно полученную из IgG1, IgG2 или IgG4.
В некоторых вариантах осуществления, C1 содержит сконструированную СБета, и первый димеризационный домен функционально связан с сконструированной СБета на третьем домене конъюгирования, который содержит SEQ ID NO: 25 (YGPPCPPCPAPEFLGGP).
В некоторых вариантах осуществления, второй димеризационный домен функционально связан с вариабельным доменом тяжелой цепи второго антигенсвязывающего фрагмента.
В некоторых вариантах осуществления, первый и второй домены димеризации различны и ассоциируются таким образом, чтобы препятствовать гомодимеризации и/или способствовать гетеродимеризации.
В некоторых вариантах осуществления, первый и второй димеризационные домены способны ассоциироваться в гетеродимеры методом “выступ-в-углубление”, гидрофобного взаимодействия, электростатического взаимодействия, гидрофильного взаимодействия или повышенной пластичности.
В другом аспекте, настоящее изобретение относится к набору, содержащему полипептидный комплекс, предусмотренный в настоящем документе, для выявления, диагностики, прогноза или лечения заболевания или состояния.
Вышеописанные и другие признаки и преимущества изобретения станут более очевидными из следующего подробного описания нескольких вариантов осуществления со ссылкой на прилагаемые чертежи.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 представлены схематические представления изученных форматов антител. Были разработаны как антитело против CD3 T3, так и антитело против CD19 U4. Для создания уникальной области контакта легкой и тяжелой цепей, которая ортогональна регулярному антителу, константная область (CL и CH1) T3 была заменена константными доменами TCR. Для конструирования биспецифичных антител в форматах E17 и F16, использовали TCR-модифицированное T3 и нативное U4 в сочетании с мутациями “выступ-в-углубление” в Fc-домене.
На фигурах 2A-2D представлены перекрывающие положения модели Fv антитела и структуры TCR, обеспечивающие контроль по слиянию Fv антитела и константной области TCR. На фигуре 2А представлена модель структуры Fv антитела, которая была построена на основе последовательности антитела T3 против CD3, разработанного в рамках лаборатории. На рисунке 2В представлена структура TCR от PDB 4L4T. На фигуре 2C представлена структурная модель Fv антитела, перекрывающая вариабельную область TCR в различных направлениях. Шероховатые химерные белки были созданы путем удаления вариабельного домена TCR в перекрывающих положениях, как показано на фиг.2D. Перекрытые остатки в области конъюгирования способствовали конструированию области конъюгирования. Цепь VL антитела и альфа-цепь TCR окрашены в белый цвет. VH и бета цепи окрашены в черный цвет.
На фиг.3А показана последовательность нативной альфа-цепи TCR и соответствующая ей последовательность с мутированными остатками цистеина. TRAC_Human соответствует природной последовательности константной области альфа-цепи. 4L4T_Alpha_Crystal соответствует последовательности кристаллической структуры (PDB код 4L4T) с мутациями S55C, которые могут образовывать межцепную дисульфидную связь. Область серого цвета соответствует константной области, используемой в качестве скелета химерного белка в настоящем изобретении.
На фиг.3В показана последовательность нативной бета-цепи TCR и соответствующая ей последовательность с мутированными цистеиновыми остатками.
На фиг.4А-4В показаны последовательности и нумерация константных областей TCR САльфа и СБета с удаленным N-гликозилированием. На фиг.4А показаны последовательности и нумерация константной области TCR альфа. На фиг.4B показаны последовательности и нумерация константной области TCR бета.
На фиг.5 показаны гистограммы проточной цитометрии линии клеток, трансфицированных CD19 яванского макака, WBP701.CHO-K1.cpro1.FL.C9 и линии исходных клеток CHO-K1.
На фиг.6 показан ДНС-ПААГ W3438-T3U4.F16-1.uIgG4.SP. М: белковый маркер; дорожка 1: W3438-T3U4.F16-1.uIgG4.SP, в невосстанавливающих условиях; Дорожка 3: W3438-T3U4.F16-1.uIgG4.SP, в восстанавливающих условиях.
На фиг.7 показана хроматограмма эксклюзионной ВЭЖХ W3438-T3U4.F16-1.uIgG4.
На фиг.8 показан ДНС-ПААГ W3438-T3U4.E17-1.uIgG4.SP. M: белковый маркер; дорожка 1: W3438-T3U4.E17-1.uIgG4.SP, в невосстанавливающих условиях; дорожка 2: W3438-T3U4.E17-1.uIgG4.SP, в восстанавливающих условиях.
На фиг.9 показана хроматограмма эксклюзионной ВЭЖХ W3438-T3U4.E17-1.uIgG4.SP.
На фиг.10A-10B показано связывание W3438-T3U4.E17-1.uIgG4.SP с клетками линии Ramos (фиг.10A) и клетками линии Jurkat (фиг.10B) посредством флуоресцентного сортинга (FACS).
На фиг.11A-11B показано связывание W3438-T3U4.F16-1.uIgG4.SP с клетками линии Ramos (фигура 11A) и клетками Jurkat (фигура 11B) посредством флуоресцентного сортинга (FACS).
На фиг.12 показано связывание W3438-T3U4.E17-1.uIgG4.SP с клеткой, экспрессирующей CD19 яванского макака, посредством флуоресцентного сортинга (FACS).
На фиг.13 показано связывание W3438-T3U4.E17-1.uIgG4.SP с CD3 яванского макака посредством ИФА (ELISA).
На фиг.14A-14B показано сродство W3438-T3U4.E17-1.uIgG4.SP к CD19 и CD3 человека, измеренное по связыванию с клетками Ramos (фиг.14A) и Jurkat (фиг.14B).
На фиг.15А-15В показано двойное связывание W3438-T3U4.E17-1.uIgG4.SP с CD19 и CD3 (фиг.15А); и отрицательный контроль (фиг.15B).
На фиг.16A-16B показана цитотоксическая активность W3438-T3U4.E17-1.uIgG4.SP на клетках Raji (фиг.16A) и цитотоксическая активность W3438-T3U4.F16-1.uIgG4.SP на клетках Raji (фиг.16B).
На фиг.17A-17D показана экспрессия CD69 и CD25 на Т-клетках в присутствии или в отсутствие CD19+ клеток-мишеней. Выраженная в процентах экспрессия CD69 на Т-клетках в субпопуляции CD4+ Т-клеток (фиг.17А); выраженная в процентах экспрессия CD69 на Т-клетках в CD8+ Т-клеточной субпопуляции (фиг.17B); выраженная в процентах экспрессия CD25 на Т-клетках в CD4+ Т-клеточной субпопуляции (фиг.17C); выраженная в процентах экспрессия CD25 на Т-клетках в CD8+ Т-клеточной субпопуляции (фиг.17D).
На фиг.18A-18D показано высвобождение цитокинов IFN-γ и TNF-α Т-клетками в присутствии или в отсутствие клеток-мишеней CD19+. Высвобождение IFN-γ в CD4+ T клеточной субпопуляции (фиг.18A); Высвобождение TNF-α в CD4+ T клеточнй субпопуляции (фиг.18B); Высвобождение IFN-γ в CD8+ T клеточной субпопуляция (фиг.18C); Высвобождение TNF-α в CD8+ T клеточной субпопуляции (фиг.18D).
На фиг.19А-19В показана стабильность W3438-T3U4.E17-1.uIgG4.SP в сыворотке человека. Связывание инкубированных в сыворотке образцов W3438-T3U4.E17-1.uIgG4.SP с клетками Ramos в указанные дни (фиг.19A); связывание инкубированных в сыворотке образцов W3438-T3U4.E17-1.uIgG4.SP с клетками Jurkat в указанные дни (фиг.19B).
На фиг.20 показано связывание W3438-T3U4.E17-1.uIgG4.SP с C1Q с помощью метода ИФА (ELISA). Антитело IgG1 использовали в качестве аналитического контроля.
На фиг.21 показана кривая объема опухоли после введения W3438-T3U4.E17-1.uIgG4.SP при различных дозах гуманизированным PBMC мышам с ксенотрансплантатами опухоли из клеток Raji. Эспериментальные точки представляют среднее по группе, “усы” представляют стандартную ошибку среднего (SEM). Нерелевантное антитело IgG4 использовали в качестве отрицательного контроля.
На фиг.22 показаны концентрации W3438-T3U4.E17-1.uIgG4.SP в сыворотке яванского макака после однократного введения 1 мг/кг. Образцы сыворотки от двух обезьян анализировали методом ИФА (ELISA).
На фиг.23А-23В показано антитело к лекарственному препарату (ADA) в образцах сыворотки от обезьяны № 1 (фиг.23А) и обезьяны № 2 (фиг.23В), включая как до введения, так и после введения W3438-T3U4.E17-1.uIgG4.SP.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следующее описание изобретения предназначено только для иллюстрации различных вариантов осуществления изобретения.
Определения
В настоящем документе, существительные в единственном числе используются для обозначения одного или более (то есть, по меньшей мере, одного) объекта. Например, “полипептидный комплекс” означает один полипептидный комплекс или более полипептидных комплексов.
Используемый в настоящем документе термин “около” или “приблизительно” относится к количеству, уровню, значению, числу, частоте, проценту, степени, размеру, объему, массе или длине, которые изменяются на целых 30, 25, 20, 25 , 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1% относительно эталонного количества, уровня, значения, числа, частоты, процента, степени, размера, объема, массы или длины. В конкретных вариантах осуществления, термины “около” или “приблизительно”, стоящий перед числовом значением, означают отклонения значений в сторону уменьшения или увеличения на 15%, 10%, 5% или 1%.
По всему объему настоящего изобретения, если контекст не требует иного, слова “содержат”, “содержит” и “содержащий” будут пониматься как подразумевающие включение указанной стадии или элемента или группы стадий или элементов, но не исключение любой другой стадии или элемента или группы стадий или элементов. Под “состоящий из” подразумевается включение и ограничение всего, что следует за фразой “состоящий из”. Таким образом, фраза “состоящий из” указывает, что перечисленные элементы являются необходимыми или обязательными, и что никакие другие элементы не могут присутствовать. Термин “состоящий по существу из” означает включение любых элементов, перечисленных после фразы, и ограничивается другими элементами, которые не препятствуют или не способствуют активности или действию, указанному в изобретении в отношении перечисленных элементов. Таким образом, фраза “состоящий по существу из” указывает, что перечисленные элементы являются необходимыми или обязательными, однако другие элементы являются необязательными и могут присутствовать или не присутствовать в зависимости от того, оказывают ли они влияние на активность или действие перечисленных элементов.
Ссылка по всему настоящему изобретению на “один из вариантов осуществления”, “вариант осуществления”, “конкретный вариант осуществления”, “связанный вариант осуществления”, “определенный вариант осуществления”, “дополнительный вариант осуществления” или “следующий вариант осуществления” или их сочетания означает, что конкретный признак, структура или характеристика, описанные в связи с вариантом осуществления, включены, по меньшей мере, в один вариант осуществления настоящего изобретения. Таким образом, появления вышеуказанных фраз в различных местах настоящего описания не обязательно все относятся к одному и тому же варианту осуществления. Кроме того, конкретные признаки, структуры или характеристики могут быть объединены любым подходящим способом в одном или более вариантах осуществления.
Термины “полипептид”, “пептид” и “белок” используются в настоящем документе взаимозаменяемо для обозначения полимера из аминокислотных остатков или совокупности нескольких полимеров из аминокислотных остатков. Термины применяются к аминокислотным полимерам, в которых один или более аминокислотных остатков представляют собой синтетический химический миметик соответствующей природной аминокислоты, а также к природным аминокислотным полимерам и неприродному аминокислотному полимеру. Термин “аминокислота” относится к природным и синтетическим аминокислотам, а также к аминокислотным аналогам и аминокислотным миметикам, которые функционируют аналогично природным аминокислотам. Природные аминокислоты представляют собой аминокислоты, кодируемые генетическим кодом, а также аминокислоты, которые в дальнейшем подвергаются модификации, например, гидроксипролин, гамма-карбоксиглутамат и O-фосфосерин. Аминокислотные аналоги относятся к соединениям, которые имеют основную химическую структуру, аналогичную встречающейся в природе аминокислоте, то есть альфа-углерод, который связан с водородом, карбоксильной группой, аминогруппой и группой R, например, гомосерин, норлейцин, метионинсульфоксид, метионин метилсульфоний. Такие аналоги имеют модифицированные группы R (например, норлейцин) или модифицированные пептидные остовы, но сохраняют аналогичную основную химическую структуру, что и встречающаяся в природе аминокислота. Альфа-углерод относится к первому атому углерода, который присоединяется к функциональной группе, такой как карбонил. Бета-углерод относится ко второму атому углерода, связанному с альфа-углеродом, и система продолжает обозначать атомы углерода в алфавитном порядке с греческими буквами. Аминокислотные миметики относятся к химическим соединениям, структура которых отличается от общей химической структуры аминокислоты, но которая функционирует аналогично природной аминокислоте. Термин “белок” обычно относится к длинноцепочечным полипептидам. Термин “пептид” обычно относится к коротким полипептидам. Полипептидные последовательности обычно характеризуются тем, что левым концом полипептидной последовательности является аминоконец (N-конец); правым концом полипептидной последовательности является карбоксильный конец (С-конец). “Полипептидный комплекс” в контексте настоящего описания относится к комплексу, содержащему один или более полипептидов, которые связаны для выполнения определенных функций. В некоторых вариантах осуществления, полипептиды являются иммунозависимыми.
Используемый в настоящем документе термин “антитело” включает любой иммуноглобулин, моноклональное антитело, поликлональное антитело, полиспецифическое антитело или биспецифичное (двухвалентное) антитело, которое связывается со специфическим антигеном. Нативное интактное антитело содержит две тяжелые цепи и две легкие цепи. Каждая тяжелая цепь состоит из вариабельной области (“HCVR”) и первой, второй и третьей константной области (CH1, CH2 и CH3), при этом каждая легкая цепь состоит из вариабельной области (“LCVR”) и константной области (CL). Тяжелые цепи млекопитающих классифицируются как α, δ, ε, γ и μ, причем легкие цепи млекопитающих классифицируются как λ или κ. Антитело имеет форму “Y”, причем “ствол” Y состоит из второй и третьей константных областей двух тяжелых цепей, связанных вместе посредством дисульфидной связи. Каждое “плечо” Y включает вариабельную область и первую константную область одной тяжелой цепи, связанную с вариабельной и константной областями одной легкой цепи. Вариабельные области легкой и тяжелой цепей ответственны за связывание антигена. Вариабельные области в обеих цепях обычно содержат три высоковариабельные петли, называемые областями, определяющими комплементарность (CDR) (CDR легкой (L) цепи, включая LCDR1, LCDR2 и LCDR3, CDR тяжелой (H) цепи, включая HCDR1, HCDR2, HCDR3). Границы CDR для антител могут быть определены или идентифицированы в соответствии с определениями Kabat, Chothia или Al-Lazikani (Al-Lazikani, B., Chothia, C., Lesk, A. M., J. Mol. Biol., 273(4), 927 (1997); Chothia, C. et al., J Mol Biol. Dec 5;186(3):651-63 (1985); Chothia, C. and Lesk, A.M., J.Mol.Biol., 196,901 (1987); Chothia, C. et al., Nature. Dec 21-28; 342(6252):877-83 (1989); Kabat E.A. et al., National Institutes of Health, Bethesda, Md. (1991)). Три CDR вставлены между фланкирующими участками, известными как каркасные области (FR), которые являются более консервативными, чем CDR, и образуют каркас для поддержки гипервариабельных петель. Каждая HCVR и LCVR содержит четыре FR, и CDR и FR располагаются от аминоконца до карбоксиконца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Константные области тяжелой и легкой цепей не участвуют в связывании антигена, но демонстрируют различные эффекторные функции. Антитела относятся к классам на основе аминокислотной последовательности константной области их тяжелой цепи. Пятью основными классами или изотипами антител являются IgA, IgD, IgE, IgG и IgM, которые характеризуются присутствием тяжелых цепей α, δ, ε, γ и μ соответственно. Некоторые из основных классов антител подразделяются на подклассы, такие как IgG1 (тяжелая цепь γ1), IgG2 (тяжелая цепь γ2), IgG3 (тяжелая цепь γ3), IgG4 (тяжелая цепь γ4), IgA1 (тяжелая цепь α1) или IgA2 (α2 тяжелая цепь).
Используемый в настоящем документе термин “вариабельный домен”, в отношении антитела, относится к вариабельной области антитела или его фрагменту, содержащему одну или более CDR. Хотя вариабельный домен может содержать интактную вариабельную область (такую как HCVR или LCVR), возможно также включение меньшей, чем интактная вариабельная область, но с сохранением способности связывания с антигеном или формирования антигенсвязывающего сайта.
Используемый в настоящем документе термин “антигенсвязывающий фрагмент” относится к фрагменту антитела, сформированному из части антитела, содержащей одну или более CDR, или любого другого фрагмента антитела, который связывается с антигеном, но не содержит интактную нативную структуру антитела. Примеры антигенсвязывающего фрагмента включают, без ограничения, вариабельный домен, вариабельную область, диатело, Fab, Fab', F(ab')2, фрагмент Fv, стабилизированный дисульфидом фрагмент Fv (dsFv), (dsFv)2, биспецифичный dsFv (dsFv-dsFv'), стабилизированное дисульфидом диатело (ds диатело), мультиспецифическое антитело, верблюжье однодоменное антитело, нанотело, доменное антитело и двухвалентное доменное антитело. Антигенсвязывающий фрагмент способен связываться с тем же антигеном, с которым связывается исходное антитело. В некоторых вариантах осуществления, антигенсвязывающая часть может содержать одну или более CDR от конкретного человеческого антитела, привитого на каркасную область от одного или более различных антител человека. Более подробные форматы антигенсвязывающего фрагмента описаны в Spiess et al., 2015 (Выше) и Brinkman et al., MAbs, 9 (2), pp.182-212 (2017), которые включены в настоящий документ посредством ссылки полностью.
Термин “Fab”, в отношении антитела, относится к той части антитела, которая состоит из одной легкой цепи (как вариабельной, так и константной области), ассоциированной с вариабельной областью и первой константной областью одной тяжелой цепи посредством дисульфидной связи. В некоторых вариантах осуществления, константные области, как легкой цепи, так и тяжелой цепи, заменены TCR константными областями.
“Fab” относится к Fab-фрагменту, который включает часть шарнирной области.
“F(ab')2” относится к димеру Fab’.
Термин “Fd-область (Fd)”, в отношении антитела, относится к аминоконцевой половине фрагмента тяжелой цепи, которая может быть объединена с легкой цепью с образованием Fab.
Термин “Fc”, в отношении антитела, относится к той части антитела, которая состоит из второй (СН2) и третьей (СН3) константных областей первой тяжелой цепи, связанных со второй и третьей константными областями второй тяжелой цепи посредством дисульфидной связи. Fc-часть антитела отвечает за различные эффекторные функции, такие как ADCC и CDC, но не учавствует в связывании антигена.
Термин “шарнирная область”, в отношении антитела, включает часть молекулы тяжелой цепи, которая присоединяет домен CH1 к домену CH2. Эта шарнирная область содержит приблизительно 25 аминокислотных остатков и является гибкой, что позволяет двум N-концевым антигенсвязывающим областям перемещаться независимо.
Используемый в настоящем документе термин “домен CH2” относится к части молекулы тяжелой цепи, которая простирается, например, от примерно 244 аминокислоты до 360 аминокислоты антитела IgG с применением традиционных схем нумерации (аминокислоты с 244 по 360, система нумерации Кэйбота; и аминокислоты 231-340, система нумерации ЕС; см. Kabat, E., et al., U.S. Department of Health and Human Services, (1983)).
“Домен СН3” простирается от домена СН2 до С-конца молекулы IgG и содержит приблизительно 108 аминокислот. Некоторые классы иммуноглобулинов, например, IgM, дополнительно включают область CH4.
“Fv”, в отношении антитела, относится к наименьшему фрагменту антитела, который содержит полный участок связывания антигена. Фрагмент Fv состоит из вариабельного домена одной легкой цепи, связанного с вариабельным доменом одной тяжелой цепи. Был предложен ряд конструкций Fv, включая dsFv, в которых связь между двумя доменами усиливается введенной дисульфидной связью; и scFv могут быть получены с использованием пептидного линкера для связывания двух доменов вместе в виде одного полипептида. Также получали конструкции Fv, содержащие вариабельный домен тяжелой или легкой цепи иммуноглобулина, ассоциированный с вариабельным и константным доменом соответствующей тяжелой или легкой цепи иммуноглобулина. Fv также подвергались мультимеризации с образованием диател и триател (Maynard et al., Annu Rev Biomed Eng 2 339-376 (2000)).
“ScFab” относится к слитому полипептиду с Fd, связанному с легкой цепью через полипептидный линкер, что приводит к образованию одноцепочечного фрагмента Fab (scFab).
“TriFab” относится к трехвалентному биспецифичному слитому белку, состоящему из трех единиц с Fab функциональными группами. TriFab содержат два регулярных Fab, слитых с асимметричным Fab-подобным фрагментом.
“Fab-Fab” относится к слитому белку, образованному путем слияния цепи Fd первого плеча Fab с N-концом цепи Fd второго плеча Fab.
“Fab-Fv” относится к слитому белку, образованному путем слияния HCVR с C-концом цепи Fd и LCVR с C-концом легкой цепи. Молекула “Fab-dsFv” может быть образована путем введения междоменной дисульфидной связи между доменом HCVR и доменом LCVR.
“MAb-Fv” или “IgG-Fv” относится к слитому белку, образованному слиянием домена HCVR с C-концом одной цепи Fc, и доменом LCVR, либо экспрессированным отдельно, либо слитым с C-концом другой с получением биспецифичного трехвалентного слитого белка IgG-Fv (mAb-Fv), причем Fv стабилизируется междоменной дисульфидной связью.
Термины “ScFab-Fc-scFv2” и “ScFab-Fc-scFv” относятся к слитому белку, образованному слиянием одноцепочечного Fab с Fc и стабилизированным дисульфидом Fv доменами.
Термин “присоединенный IgG” относится к слитому белку с плечом Fab, слитым с IgG с формированием формата биспецифичного (Fab)2-Fc. Он может образовывать “IgG-Fab” или “Fab-IgG”, причем Fab, слит с C-концом или N-концом молекулы IgG с или без линкера. В некоторых вариантах осуществления, присоединенный IgG может быть дополнительно модифицирован до формата IgG-Fab4 (см. Brinkman et al., 2017, Выше).
“DVD-Ig” относится к антителу с двумя вариабельными доменами, которое образуется путем слияния дополнительного домена HCVR и домена LCVR второй специфичности с тяжелой цепью и легкой цепью IgG. “CODV-Ig” относится к связанному формату, в котором два домена HCVR и два домена LCVR связаны таким образом, чтобы обеспечить перекрестное спаривание вариабельных доменов HCVR-LCVR, которые расположены либо (от N- до C-конца) в порядке HCVRA-HCVRB и LCVRB-LCVRA, или в порядке HCVRB-HCVRA и LCVRA-LCVRB.
“CrossMab” относится к технологии спаривания немодифицированной легкой цепи с соответствующей немодифицированной тяжелой цепью и спаривания модифицированной легкой цепи с соответствующей модифицированной тяжелой цепью, в результате чего образуется антитело с уменьшенным ошибочным спаривание в легкой цепи.
“BiTE” представляет собой биспецифичную молекулу-рекрутер Т-клеток, содержащую первый scFv с первой специфичностью к антигену в ориентации LCVR-HCVR, связанный со вторым scFv со второй специфичностью в ориентации HCVR-LCVR.
“WuXiBody” представляет собой биспецифичное антитело, содержащее растворимый химерный белок с вариабельными доменами антитела и константными доменами TCR, где субъединицы (такие как альфа- и бета-домены) константных доменов TCR связаны посредством сконструированной дисульфидной связи.
“Процент (%) идентичности последовательностей”, в отношении аминокислотной последовательности (или последовательности нуклеиновой кислоты), определяется как процентное содержание аминокислотных (или нуклеиновой кислоты) остатков в последовательности-кандидате, которые идентичны аминокислотным (или нуклеиновой кислоте) остаткам в контрольной последовательности, после выравнивания последовательностей и, при необходимости, введения пробелов, для достижения максимального количества идентичных аминокислот (или нуклеиновых кислот). Консервативная замена аминокислотных остатков может рассматриваться или не рассматриваться в качестве идентичных остатков. Выравнивание с целью определения процента идентичности аминокислотной (или нуклеиновой кислоты) последовательности может быть достигнуто, например, посредством общедоступных инструментов, таких как BLASTN, BLASTp (доступны на веб-сайте Национального центра биотехнологической информации США (NCBI), см. также Altschul S.F. et al., J. Mol. Biol., 215:403-410 (1990); Stephen F. et al., Nucleic Acids Res., 25:3389-3402 (1997)), ClustalW2 (доступно на сайте Европейского института биоинформатики, см. также Higgins D.G. et al., Methods in Enzymology, 266:383-402 (1996); Larkin M.A. et al., Bioinformatics (Oxford, England), 23(21): 2947-8 (2007)) и программного обеспечения ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут использовать стандартные параметры, предоставляемые инструментом, или могут настроить параметры в соответствии с настройкой, например, путем выбора подходящего алгоритма.
Термин “антиген” или “Ag”, в контексте настоящего описания, относится к соединению, композиции, пептиду, полипептиду, белку или веществу, которые могут стимулировать выработку антител или Т-клеточный ответ в клеточной культуре или у животного, включая композиции (например, такие, которые содержат опухоль-специфичный белок), которые добавляют в клеточную культуру (такую как гибридома), или вводят или абсорбируют животному. Антиген взаимодействует с продуктами специфического, гуморального или клеточного, иммунитета (например антитело), включая индуцированные гетерологичными антигенами.
“Эпитоп” или “антигенная детерминанта” относится к участку антигена, с которым связывается связывающий агент (такой как антитело). Эпитопы могут быть образованы как из смежных аминокислот (также называемые линейным или последовательным эпитопом), так и из несмежных аминокислот, совмещенных фолдингом третичной белковой структуры (также называемые конфигурационным или конформационным эпитопом). Эпитопы, образованные из смежных аминокислот, обычно, располагаются линейно вдоль первичных аминокислотных остатков на белке, причем небольшие сегменты смежных аминокислот могут отщепляться от антигена, связывающегося с молекулами основного комплекса гистосовместимости (МНС), или сохраняются при воздействии денатурирующих растворителей, тогда как эпитопы, образованные фолдингом третичной структуры, обычно теряются при обработке денатурирующими растворителями. Эпитоп обычно включает, по меньшей мере, 3 и более, обычно, по меньшей мере, 5, около 7 или около 8-10 аминокислот в уникальной пространственной конформации.
Используемый в настоящем документе термин “специфическое связывание” или “специфически связывается” относится к неслучайной реакции связывания между двумя молекулами, например, между антителом и антигеном. В некоторых вариантах осуществления, полипептидный комплекс и биспецифичный полипептидный комплекс, представленный в настоящем документе, специфически связывают антиген с аффинностью связывания (KD) ≤ 10-6 М (например, ≤ 5×10-7 М, ≤ 2×10-7 М, ≤ 10-7 M, ≤ 5×10-8 M, ≤ 2×10-8 M, ≤ 10-8 M, ≤ 5×10-9 M, ≤ 2×10-9 M, ≤ 10-9 M или ≤ 10-10 M). Используемый в настоящем документе KD относится к отношению скорости диссоциации к скорости ассоциации (koff/kon) и может быть определен с использованием методов поверхностного плазмонного резонанса, например, с использованием прибора, такого как Biacore.
Термин “функциональная связь” или “функционально связанный” относится к совмещению, с или без спейсера или линкера, двух или более представляющих интерес биологических последовательностей таким образом, что они находятся в взаимоотношениях, позволяющих им функционировать определенным образом. При использовании по отношению к полипептидам, подразумевается, что полипептидные последовательности связаны таким образом, что позволяет связанному продукту выполнять предполагаемую биологическую функцию. Например, вариабельная область антитела может быть функционально связана с константной областью с обеспечением стабильного продукта с антигенсвязывающей активностью. Термин также может быть использован в отношении полинуклеотидов. Например, когда полинуклеотид, кодирующий полипептид, функционально связан с регуляторной последовательностью (например, последовательностью промотора, энхансера, сайленсера и т.д.), это означает, что полинуклеотидные последовательности связаны таким образом, что допускается контролируемая экспрессия полипептида из полинуклеотида.
Термин “слияние” или “слитый”, при использовании в отношении аминокислотных последовательностей (например, пептида, полипептида или белка), относится к объединению двух или более аминокислотных последовательностей, например, посредством химической связи или рекомбинантных средств, в отдельную аминокислотную последовательность, которая не является природной. Слитая аминокислотная последовательность может быть получена путем генетической рекомбинации двух кодирующих полинуклеотидных последовательностей и может быть экспрессирована методом введения конструкции, содержащей рекомбинантные полинуклеотиды, в клетку-хозяина.
Используемый в настоящем документе термин “спейсер” относится к искусственной аминокислотной последовательности, имеющей 1, 2, 3, 4 или 5 аминокислотных остатков или длину от 5 до 15, 20, 30, 50 или более аминокислотных остатков, связанных пептидными связями и используемых для связывания одного или более полипептидов. Спейсер может иметь или не иметь вторичную структуру. Спейсерные последовательности известны в данной области техники, см., например, Holliger et al., Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993); Poljak et al., Structure 2:1121-1123 (1994). Могут быть использованы любые подходящие спейсеры, известные в данной области.
Термин “антигенная специфичность” относится к конкретному антигену или его эпитопу, который избирательно распознается антигенсвязывающей молекулой.
Используемый в настоящем документе термин “замещение”, в отношении аминокислотного остатка, относится к природной или индуцированной замене одной или более аминокислот на другую в пептиде, полипептиде или белке. Замена в полипептиде может привести к снижению, усилению или устранению полипептидной функции.
Замена также может быть “консервативной заменой”, применительно к аминокислотной последовательности, которая относится к замене аминокислотного остатка другим аминокислотным остатком, имеющим боковую цепь со сходными физико-химическими свойствами, или замене тех аминокислот, которые не являются критическими для активности полипептида. Например, консервативные замены могут быть сделаны среди аминокислотных остатков с неполярными боковыми цепями (например, Met, Ala, Val, Leu и Ile, Pro, Phe, Trp), среди остатков с незаряженными полярными боковыми цепями (например, Cys, Ser Thr, Asn, Gly и Gln), среди остатков с кислотными боковыми цепями (например, Asp, Glu), среди аминокислот с основными боковыми цепями (например, His, Lys и Arg), среди аминокислот с бета-разветвленными боковыми цепями (например, Thr, Val и Ile), среди аминокислот с серосодержащими боковыми цепями (например, Cys и Met) или среди остатков с ароматическими боковыми цепями (например, Trp, Tyr, His и Phe). В некоторых вариантах осуществления, замены, делеции или добавления также могут рассматриваться как “консервативные замены”. Количество аминокислот, которые встраиваются или удаляются, может находиться в диапазоне от 1 до 5. Консервативное замещение обычно не вызывает значительного изменения конформационной структуры белка и, следовательно, может сохранять биологическую активность белка.
Используемый в настоящем документе термин “мутация” или “мутированный”, в отношении аминокислотного остатка, относится к замене, вставке или добавлению аминокислотного остатка.
Используемый в настоящем документе термин “Т-клетка” относится к типу лимфоцитов, которые играют основную роль в клеточном иммунитете, включая Т-хелперы (например, CD4+ Т-клетки, Т-хелперы 1 типа, Т-хелперы 2 типа, Т-хелперы 3 типа, T-хелперы 17 типа), цитотоксические T-клетки (например, CD8+ T-клетки), T-клетки памяти (например, T-клетки центральной памяти (клетки TCM), эффекторные T-клетки памяти (TEM-клетки и TEMRA-клетки) и резидентные T-клетки памяти (TRM), которые являются либо CD8+, либо CD4+), естественные киллерные T-клетки (NKT) и ингибирующие T-клетки.
Нативный “Т клеточный рецептор” или нативный “TCR” представляет собой гетеродимерный белок на поверхности Т-клеток, который связан с инвариантными цепями CD3 с образованием комплекса, способного опосредовать передачу сигнала. TCR принадлежит к суперсемейству иммуноглобулинов и аналогичен полуантителу с одной тяжелой цепью и одной легкой цепью. Нативный TCR имеет внеклеточную часть, трансмембранную часть и внутриклеточную часть. Внеклеточный домен TCR имеет мембранно-проксимальную константную область и мембранно-дистальную вариабельную область.
Используемый в настоящем документе термин “субъект” или “индивидуум” или “животное” или “пациент” относится к животному или отличному от человека животному, включая млекопитающее или примата, нуждающемуся в диагностике, прогнозе, уменьшении интенсивности, профилактике и/или лечении заболевания или расстройства. Субъекты-млекопитающие включают людей, домашних животных, сельскохозяйственных животных и зоопарковых животных, спортивных или домашних животных, таких как собаки, кошки, морские свинки, кролики, крысы, мыши, лошади, свиньи, коровы, медведи и так далее.
Биспецифичный полипептидный комплекс
В одном из аспектов, настоящее изобретение относится к биспецифичному полипептидному комплексу. Используемый в настоящем документе термин “биспецифичный” означает, что существуют два антигенсвязывающих фрагмента, каждый из которых способен специфически связываться с другим антигеном или другим эпитопом на одном и том же антигене. Биспецифичный полипептидный комплекс, представленный в настоящем документе, содержит первый антигенсвязывающий фрагмент, связанный со вторым антигенсвязывающим фрагментом, и один из них специфически связывается с CD3, а другой специфически связывается с CD19. Другими словами, первый антигенсвязывающий фрагмент может специфически связываться с CD3, а второй антигенсвязывающий фрагмент может специфически связываться с CD19. Альтернативно, первый антигенсвязывающий фрагмент может специфически связываться с CD19, а второй антигенсвязывающий фрагмент может специфически связываться с CD3.
В некоторых вариантах осуществления, настоящее изобретение относится к биспецифичному полипептидному комплексу, содержащему первый антигенсвязывающий фрагмент, связанный со вторым антигенсвязывающим фрагментом, где:
первый антигенсвязывающий фрагмент включает:
первый полипептид, содержащий, от N-конца до C-конца, первый вариабельный домен тяжелой цепи (VH1) первого антитела, функционально связанный с константной областью первого Т-клеточного рецептора (TCR) (C1), и
второй полипептид, содержащий, от N-конца до C-конца, первый вариабельный домен легкой цепи (VL1) первого антитела, функционально связанный со второй константной областью TCR (C2),
где:
C1 содержит сконструированную СБета, содержащую SEQ ID NO: 1, и C2 содержит сконструированную САльфа, содержащую SEQ ID NO: 2,
аминокислота C48 в SEQ ID NO: 1 и аминокислота C41 в SEQ ID NO: 2 способны образовывать ненативную межцепочечную дисульфидную связь,
С1 и С2 способны образовывать димер, и ненативная межцепочечная дисульфидная связь способна стабилизировать димер,
и
второй антигенсвязывающий фрагмент включает:
второй вариабельный домен тяжелой цепи (VH2) второго антитела, функционально связанный с доменом CH1 тяжелой цепи антитела, и
вариабельный домен второй легкой цепи (VL2) второго антитела, функционально связанный с константным доменом легкой цепи антитела (CL),
где:
один из первого и второго антигенсвязывающих фрагментов представляет собой анти-CD3 связывающий фрагмент, а другой - анти-CD19 связывающий фрагмент,
анти-CD3 связывающий фрагмент получен из анти-CD3 антитела, содержащего:
а) CDR1 тяжелой цепи, содержащую SEQ ID NO: 3, b) CDR2 тяжелой цепи, содержащую SEQ ID NO: 4, c) CDR3 тяжелой цепи, содержащую SEQ ID NO: 5, d) CDR1 легкой цепи каппа, содержащую SEQ ID NO: 6, e) CDR2 каппа-легкой цепи, содержащую SEQ ID NO: 7, и f) CDR3 каппа-легкой цепи, содержащую SEQ ID NO: 8,
анти-CD19 связывающий фрагмент получен из анти-CD19 антитела, содержащего:
а) CDR1 тяжелой цепи, содержащую SEQ ID NO: 9, b) CDR2 тяжелой цепи, содержащую SEQ ID NO: 10, c) CDR3 тяжелой цепи, содержащую SEQ ID NO: 11, d) CDR1 каппа-легкой цепи, содержащую SEQ ID NO: 12, e) CDR2 каппа-легкой цепи, содержащую SEQ ID NO: 13 и f) CDR3 каппа-легкой цепи, содержащую SEQ ID NO: 14,
и
первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент меньше подвержены ошибочному спариванию, чем, если бы и первый и второй антигенсвязывающие фрагменты были бы аналогами природного Fab.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс, представленный в настоящем документе, содержит первый антигенсвязывающий фрагмент, содержащий последовательность, полученную из константной области TCR, но при этом второй антигенсвязывающий фрагмент не содержит последовательность, полученную из константной области TCR.
Биспецифичный полипептидный комплекс, представленный в настоящем документе, значительно в меньшей степени подвержен ошибочному спариванию вариабельных доменов тяжелой цепи и легкой цепи. Не желая ограничиваться какой-либо теорией, полагают, что стабилизированные константные области TCR в первом антигенсвязывающем фрагменте могут специфически ассоциироваться друг с другом и, следовательно, вносить вклад в высокоспецифичное спаривание предполагаемых VH1 и VL1, препятствуя нежелательным ошибочным спариваниям VH1 или VL1 с другими вариабельными областями, которые не предусматривают предполагаемые антигенсвязывающие участки.
В некоторых вариантах осуществления, второй антигенсвязывающий фрагмент дополнительно содержит константный домен СН1 антитела, функционально связанный с VH2, и константный домен легкой цепи антитела, функционально связанный с VL2. Таким образом, второй антигенсвязывающий фрагмент содержит Fab.
Когда первый, второй, третий и четвертый вариабельные домены (например, VH1, VH2, VL1 и VL2) экспрессируются в одной клетке, крайне желательно, чтобы VH1 специфически связывался с VL1, и VH2 специфически связывался с VL2 с тем, чтобы, в результате, биспецифичный белковый продукт имел правильную антигенсвязывающую специфичность. Однако в существующих технологиях, таких как межвидовая гибридома (или квадрома), происходит случайное спаривание VH1, VH2, VL1 и VL2, что, соответственно, приводит к образованию до десяти различных видов, из которых только один является функциональной биспецифичной антигенсвязывающей молекулой. Это не только снижает выход продукта, но и усложняет очистку целевого продукта.
Представленные в настоящем документе биспецифичные полипептидные комплексы являются исключительными в том, что вариабельные домены менее подверженными ошибочному спариванию, чем первый и второй антигенсвязывающие фрагменты, являющиеся аналогами природного Fab. В иллюстративном примере первый антигенсвязывающий домен содержит VH1-C1 в паре с VL1-C2, а второй антигенсвязывающий домен содержит VH2-CH1 в паре с VL2-CL. Неожиданно было обнаружено, что C1 и C2 предпочтительно ассоциируются друг с другом и менее склонны ассоциироваться с CL или CH1, вследствие чего образование нежелательных пар, таких как C1-CH, C1-CL, C2-CH и C2-CL, не желательно и значительно сокращено. В результате специфической ассоциации C1-C2, VH1 специфически связывается с VL1, тем самым предоставляя первый антигенсвязывающий участок, и CH1 специфически связывается с CL, тем самым обеспечивая специфическое спаривание VH2-VL2, что обеспечивает второй антигенсвязывающий участок. Соответственно, первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент менее подвержены ошибочному спариванию, и ошибочное спаривание между, например, VH1-VL2, VH2-VL1, VH1-VH2, VL1-VL2 значительно снижено по сравнению с тем, что могло бы быть, если бы и первый и второй антигенсвязывающие фрагменты были аналогами природных Fabs, например, в форме VH1-CH1, VL1-CL, VH2-CH1 и VL2-CL.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс, представленный в настоящем документе, при экспрессии из клетки имеет значительно меньше продуктов ошибочного спаривания (например, меньше, по крайней мере, на 1, 2, 3, 4, 5 или более продуктов ошибочного спаривания) и/или значительно выше выход продукта (например, по крайней мере, на 10%, 20%, 30%, 40%, 50%, 60% или более высокий выход), чем у эталонной молекулы, экспрессируемой в сопоставимых условиях, где эталонная молекула в остальном идентична биспецифичному полипептидному комплексу за исключением наличия нативного CH1 вместо С1 и нативного CL вместо С2.
Антигенсвязывающий фрагмент, включающий сконструированные САльфа и СБета
Первый антигенсвязывающий фрагмент, представленный в настоящем документе, содержит первый вариабельный домен тяжелой цепи антитела, функционально связанный с константной областью первого T-клеточного рецептора (TCR), и первый вариабельный домен легкой цепи антитела, функционально связанный со второй константной областью TCR, где первая константная область TCR и вторая константная область TCR связаны через, по крайней мере, одну ненативную межцепочечную дисульфидную связь. Первый антигенсвязывающий фрагмент содержит, по меньшей мере, две полипептидные цепи, каждая из которых содержит вариабельный домен, полученный из антитела, и константный участок, полученный из TCR. Таким образом, первый антигенсвязывающий фрагмент содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, которые функционально связаны с парой константных областей TCR соответственно. В некоторых вариантах осуществления, изобретения пара константных областей TCR в первом антигенсвязывающем фрагменте представляет собой константные области альфа/бета TCR. Константные области TCR в полипептидных комплексах, представленных в настоящем документе, способны связываться друг с другом с образованием димера через, по крайней мере, одну ненативную дисульфидную связь.
Неожиданно было обнаружено, что первый антигенсвязывающий фрагмент, представленный в настоящем документе, по крайней мере с одной ненативной дисульфидной связью может быть рекомбинантно экспрессирован и собран в желаемую конформацию, которая стабилизирует димер константной области TCR, обеспечивая при этом хорошую антигенсвязывающую активность вариабельных областей антитела. Кроме того, обнаружено, что первый антигенсвязывающий фрагмент допускает рутинную инженерию антител, например, модификацию сайтов гликозилирования и удаление некоторых природных последовательностей. Кроме того, полипептидные комплексы, представленные в настоящем документе, могут быть включены в биспецифичный формат, который может быть легко экспрессирован и собран с минимальным ошибочным спариванием антигенсвязывающих последовательностей, или практически без него, вследствие присутствия константных областей TCR в первом антигенсвязывающем фрагменте. Дополнительные преимущества первого антигенсвязывающего фрагмента и конструкций, представленных в настоящем документе, станут более очевидными в следующем описании ниже.
Таким образом, первый антигенсвязывающий фрагмент, представленный в настоящем документе, содержит первый полипептид, содержащий, от N-конца до C-конца, первый вариабельный домен тяжелой цепи (VH) первого антитела, функционально связанный с первой константной областью (C1) Т-клеточного рецептора (TCR), и второй полипептид, содержащий, от N-конца до C-конца, первый вариабельный домен легкой цепи (VL) первого антитела, функционально связанный со второй константной областью TCR (C2), где: C1 содержит сконструированную СБета, содержащую SEQ ID NO: 1, и C2 содержит сконструированную САльфа, содержащую SEQ ID NO: 2, аминокислота C48 в SEQ ID NO: 1 и аминокислота C41 в SEQ ID NO: 2 способны образовывать ненативную межцепочечной дисульфидную связь, C1 и C2 способны образовывать димер, и ненативная межцепочечная дисульфидная связь между C1 и C2 способна стабилизировать димер.
Константная область TCR
Первый антигенсвязывающий фрагмент, представленный в настоящем документе, содержит константную область альфа или бета, полученную из TCR.
На фиг.3А и 3В представлены аминокислотные последовательности константных областей нативного TCR альфа- и бета-цепей TCR. Для ясности и последовательности, каждый из аминокислотных остатков в этих последовательностях пронумерован на фиг.4А и 4В, и такая нумерация используется в настоящем описании для обозначения конкретного аминокислотного остатка в конкретной константной области TCR.
Константная область альфа-цепи TCR человека известна как TRAC с NCBI-номером доступа P01848 или аминокислотной последовательностью SEQ ID NO: 31.
Константная область бета-цепи TCR человека имеет два различных варианта, известных как TRBC1 и TRBC2 (номенклатура IMGT) (см. также Toyonaga B, et al., PNAs, Vol. 82, pp.8624-8628, Immunology (1985)). Последовательность TRBC1 (SEQ ID NO: 33) была выбрана в качестве основной цепи для конструирования полипептидных комплексов, описанных в настоящем документе.
В частности, бета-цепь нативного TCR содержит нативный остаток цистеина в положении 74 (см. Фиг. 4B), который является неспаренным и, следовательно, не образует дисульфидную связь в нативном альфа/бета TCR. В полипептидных комплексах, представленных в настоящем документе, этот нативный остаток цистеина в положении 74 бета-цепи TCR мутирован в аланиновый остаток. Это может использоваться для устранения неправильного внутрицепочечного или межцепочечного спаривания. В некоторых вариантах осуществления, замена, в некоторых вариантах осуществления, может улучшить эффективность рефолдинга TCR in vitro.
В настоящем описании, первая и вторая константные области TCR первого антигенсвязывающего фрагмента, представленного в настоящем документе, способны образовывать димер, содержащий между константными областями TCR, по меньшей мере, одну ненативную межцепочечную дисульфидную связь, которая способна стабилизировать димер.
Используемый в нстоящем документе термин “димер” относится к связанной структуре, образованной двумя молекулами, такими как полипептиды или белки, посредством ковалентных или нековалентных взаимодействий. Гомодимер или гомодимеризация возникает посредством двух одинаковых молекул, а гетеродимер или гетеродимеризация возникает посредством двух разных молекул. Димер, образованный первой и второй константными областями TCR, представляет собой гетеродимер.
“Мутированный” аминокислотный остаток относится к остатку, который замещен, вставлен или добавлен и он отличается от своего нативного остатка-аналога в соответствующей нативной константной области TCR. Например, если аминокислотный остаток в определенном положении в константной области TCR дикого типа указывается как “нативный” остаток, то его мутированным аналогом является любой остаток, который отличается от нативного остатка, но находится в том же положении в константной области TCR. Мутированный остаток может представлять собой другой остаток, который замещает нативный остаток в том же положении, или который вставлен перед нативным остатком и, соответственно, занимает свое исходное положение.
В полипептидных комплексах, представленных в настоящем документе, первая и/или вторая константные области TCR были сконструированы так, чтобы содержать один или более мутированных аминокислотных остатков, которые отвечают за образование ненативной межцепочечной дисульфидной связи. Чтобы ввести такой мутированный остаток в константную область TCR, можно манипулировать кодирующей последовательностью области TCR, например, для замены кодона, кодирующего нативный остаток, на кодон, кодирующий мутированный остаток, или для вставки кодона, кодирующего мутированный остаток, перед кодоном нативного остатка.
В полипептидных комплексах, представленных в настоящем документе, первая и/или вторая константные области TCR были сконструированы так, чтобы они содержали один или более мутированных цистеиновых остатков вследствие чего после замены на цистеиновые остатки могла образовываться ненативная межцепочечная дисульфидная связь между двумя константными областями TCR.
Ненативная межцепочечная дисульфидная связь способна стабилизировать первый антигенсвязывающий фрагмент. Такие эффекты в стабилизации могут быть реализованы различными способами. Например, присутствие мутированного аминокислотного остатка или ненативной межцепочечной дисульфидной связи может позволить полипептидному комплексу стабильно экспрессироваться и/или экспрессироваться на высоком уровне и/или объединяться в стабильный комплекс, имеющий желаемую биологическую активность (например, антигенсвязывающую активность) и/или экспрессироваться и собираться в высокоэффективный желаемый стабильный комплекс, имеющий желаемую биологическую активность. Способность межцепной дисульфидной связи стабилизировать первую и вторую константные области TCR можно оценить с использованием соответствующих методов, известных в данной области техники, таких как определение молекулярной массы посредством ДНС-ПААГ, или определение термостабильности дифференциальной сканирующей калориметрией (DSC) или дифференциальной сканирующей флуориметрией (DSF). В иллюстративном примере, образование стабильного первого антигенсвязывающего фрагмента, представленного в настоящем документе, может быть подтверждено с помощью ДНС-ПААГ, если продукт демонстрирует молекулярную массу, сопоставимую с общей молекулярной массой первого и второго полипептидов. В некоторых вариантах осуществления, первый антигенсвязывающий фрагмент, представленный в настоящем документе, является стабильным в том смысле, что его термическая стабильность составляет не менее 50, 60, 70, 80 или 90% стабильности природного Fab. В некоторых вариантах осуществления, первый антигенсвязывающий фрагмент, представленный в настоящем документе, стабилен в том смысле, что его термическая стабильность сопоставима со стабильностью природного Fab.
Не желая быть связанными какой-либо теорией, полагают, что ненативная межцепочечная дисульфидная связь, образованная между первой и второй константными областями TCR в первом антигенсвязывающем фрагменте, способна стабилизировать гетеродимер константных областей TCR, тем самым повышая уровень корректного фолдинга, структурную стабильность и/или уровень экспрессии гетеродимера и первого антигенсвязывающего фрагмента. В отличие от нативного TCR, “заякоренного” на мембране Т-клеточной поверхности, гетеродимеры внеклеточных доменов нативного TCR оказались значительно менее стабильными, несмотря на свое сходство с антителом Fab в трехмерной структуре. К тому же нестабильность нативного TCR в растворимом состоянии была существенным препятствием, мешающим определению его кристаллической структуры. (см. Wang, Protein Cell, 5(9), pp.649-652 (2014)). Путем введения пары цистеиновых (Cys) мутаций в константные области TCR и, тем самым, возможности образования межцепочечной ненативной дисульфидной связи, первый антигенсвязывающий фрагмент может стабильно экспрессироваться, хотя при этом антигенсвязывающие способности вариабельной области антитела сохраняются.
Константная область TCR, содержащая мутированный остаток, также называется в настоящем документе “сконструированной” константной областью TCR. В некоторых вариантах осуществления, первая константная область TCR (C1) полипептидного комплекса содержит сконструированную альфа-цепь TCR (САльфа), а вторая константная область TCR (C2) содержит сконструированную бета-цепь TCR (СБета). В полипептидных комплексах, представленных в настоящем документе, С1 включает сконструированную СБета, а С2 содержит сконструированную САльфа.
В представленных в настоящем документе полипептидных комплексах сконструированная константная область TCR содержит один или более мутированных остатков цистеина в пределах контактной области первой и/или второй сконструированных константных областей TCR.
Используемый в настоящем документе термин “область контакта” относится к конкретной области(областям) на полипептидах, где полипептиды взаимодействуют/связываются друг с другом. Область контакта включает один или более аминокислотных остатков, которые способны взаимодействовать с соответствующим аминокислотным остатком(остатками), который вступает в контакт или ассоциацию, когда происходит взаимодействие. Аминокислотные остатки в контактной области могут быть или не быть в непрерывной последовательности. Например, когда область контакта является трехмерной, аминокислотные остатки внутри области контакта могут быть разделены, находясь в разных положениях линейной последовательности.
В некоторых вариантах осуществления, одна или более дисульфидных связей могут быть образованы между сконструированной САльфа и сконструированной СБета. В некоторых вариантах осуществления, мутированный цистеиновый остаток в СБета представляет собой S56C (соответствует аминокислоте C48 в SEQ ID NO: 1), а мутированный остаток цистеина в САльфа представляет собой T49C (соответствующий аминокислоте C41 в SEQ ID NO: 2), и где пара цистеиновых остатков способна образовывать ненативную межцепочечную дисульфидную связь.
Используемый по свему настоящему документу термин “XnY”, в отношении константной области TCR, предназначен для обозначения того, что n-й аминокислотный остаток X в константной области TCR (на основе нумерации на фиг.4A и 4B, представленной в данном документе) заменяется аминокислотным остатком Y, где X и Y представляют собой соответственно однобуквенную аббревиатуру конкретного аминокислотного остатка. Следует отметить, что число n основано только на нумерации, представленной на фиг.4А и 4В, и может отличаться от своего фактического положения. В качестве иллюстрации, последовательность СБета (S56C) (N69Q), показанная в SEQ ID NO: 1, используется в качестве примера. Хотя замена S на C происходит на 48-м остатке в SEQ ID NO: 1, сам остаток обозначен как 56-й остаток на основе системы нумерации на фиг. 4B, и, следовательно, эта замена S на C обозначена как S56C, а не S48C. Аналогично, замена N на Q также обозначается как N69Q на основе системы нумерации на фиг.4А и 4В. Это правило обозначения замены аминокислотных остатков применимо ко всем константным областям TCR в настоящем описании, если не указано иное.
В полипептидных комплексах, представленных в настоящем документе, сконструированная СБета содержит или представляет собой SEQ ID NO: 1, а сконструированная САльфа содержит или представляет собой SEQ ID NO: 2.
Аминокислотные последовательности SEQ ID NO: 1 и 2 представлены ниже.
SEQ ID NO: 1
EVAVFEPSEAEISHTQKATLVCLATGFYPDHVELSWWVNGKEVHSGVCTDPQPLKEQPALQDSRYALSSRLRVSATFWQNPRNHFRCQVQFYGLSENDEWTQDRAKPVTQIVSAEA
SEQ ID NO: 2
AVYQLRDSKSSDKSVCLFTDFDSQTQVSQSKDSDVYITDKCVLDMRSMDFKSNSAVAWSQKSDFACANAFQNSIIPEDTFFPSPESS
В представленных в настоящем документе пептидных комплексах один или более нативных участков гликозилирования, присутствующих в нативных константных областях TCR, были модифицированы (например, удалены) в первом антигенсвязывающем фрагменте, представленном в настоящем описании. Используемый в настоящем документе термин “участок гликозилирования”, в отношении полипептидной последовательности, относится к аминокислотному остатку с боковой цепью, к которой может быть присоединен углеводный фрагмент (например, структура олигосахарида). Гликозилирование полипептидов, подобных антителам, обычно является либо N-связанным, либо O-связанным. N-связанное относится к присоединению углеводной части к боковой цепи аспарагинового остатка, например аспарагинового остатка в трипептидной последовательности, такой как аспарагин-Х-серин и аспарагин-Х-треонин, где Х представляет собой любую аминокислоту кроме пролина. О-связанное гликозилирование относится к присоединению одного из сахаров (N-ацетилгалактозамина, галактозы или ксилозы) к гидроксиаминокислоте, обычно к серину или треонину. Удаление нативных участков гликозилирования может быть удобно осуществлено путем изменения аминокислотной последовательности, таким образом, что одна или более из описанных выше трипептидных последовательностей (для N-связанных участков гликозилирования) или один или более сериновых или треониновых остатков (для O-связанных участков гликозилирования) заменяются.
В первом антигенсвязывающем фрагменте, представленном в настоящем документе, по меньшей мере, один нативный участок гликозилирования отсутствует в сконструированных константных областях TCR, например, в первой и/или второй константных областях TCR. Не желая быть связанными какой-либо теорией, полагают, что первый антигенсвязывающий фрагмент, представленный в настоящем документе, может допускает удаление всех или части участков гликозилирования, не оказывая влияние на экспрессию и стабильность белка, в отличие от существующих представлений, что присутствие участков N-связанного гликозилирования в константной области TCR, например САльфа (т.е. N34, N68 и N79) и СБета (т.е. N69), необходимо для экспрессии и стабильности белка (см. Wu et al., Mabs, 7:2, 364-376, 2015).
В первом антигенсвязывающем фрагменте, представленном в настоящем документе, участки N-гликозилирования в сконструированной САльфа на N34, N68 и N79 отсутствуют. Сконструированная последовательность САльфа, в которой отсутствут участок гликозилирования, содержит или соответствует SEQ ID NO: 2. В первом антигенсвязывающем фрагменте, представленном в настоящем документе, участок N-гликозилирования в сконструированной СБета на N69 отсутствует. Сконструированная последовательность СБета (TRBC1), в которой отсутствут участок гликозилирования, содержит или соответствует SEQ ID NO: 1.
В первом антигенсвязывающем фрагменте, представленном в настоящем документе, константные области, полученные из TCR, функционально связаны с вариабельными областями, полученными из антитела.
В некоторых вариантах осуществления, первый вариабельный домен антитела (VH) слит с первой константной областью TCR (C1) на первом домене конъюгирования, а первый вариабельный домен антитела (VL) слит со второй константной областью TCR (C2) на втором домене конъюгирования.
Термин “домен конъюгирования”, используемый в настоящем документе, относится к границе или граничной области, где две аминокислотные последовательности слиты или объединены. В некоторых вариантах осуществления, первый домен конъюгирования содержит, по меньшей мере, часть C-концевого фрагмента V/C конъюгирования антитела, а второй домен конъюгирования содержит, по меньшей мере, часть N-концевого фрагмента V/C конъюгирования TCR.
Используемый в настоящем документе термин “V/C конъюгирование антител” относится к границе вариабельного домена и константного домена антитела, например, границы между вариабельным доменом тяжелой цепи и доменом CH1 или между вариабельным доменом легкой цепи и константным доменом легкой цепи. Аналогично, термин “V/C конъюгирование TCR” относится к границе вариабельного домена TCR и константного домена, например, границы между вариабельнымом доменом TCR альфа и константным доменом или между вариабельным доменом TCR бета и константным доменом.
В некоторых вариантах осуществления, первый полипептид содержит последовательность, содержащую домены, функционально связанные так, как например в формуле (I): VH-HCJ-C1, и второй полипептид содержит последовательность, содержащую домены, функционально связанные так, как например в формуле (II): VL-LCJ-C2, где:
VH представляет собой вариабельный домен тяжелой цепи антитела;
HCJ является первым доменом конъюгирования, как определено выше;
C1 является первым константным доменом TCR, как определено выше;
VL представляет собой вариабельный домен легкой цепи антитела;
LCJ является вторым доменом конъюгирования, как определено выше;
C2 является вторым константным доменом TCR, как определено выше.
В таких вариантах осуществления, C1 представляет собой сконструированную СБета, которая содержит или соответствует SEQ ID NO: 1, и C2 представляет собой сконструированную САльфа, которая содержит или соответствует SEQ ID NO: 2, HCJ содержит или соответствует SEQ ID NO: 23, и LCJ содержит или соответствует SEQ ID NO: 24.
Вариабельная область антитела
Биспецифичный полипептидный комплекс, представленный в настоящем документе, содержит первый антигенсвязывающий фрагмент, связанный со вторым антигенсвязывающим фрагментом, и один из них специфически связывается с CD3, тогда как другой специфически связывается с CD19. В представленном в настоящем документе полипептидном комплексе, первый антигенсвязывающий фрагмент содержит первый вариабельный домен тяжелой цепи (VH1) и первый вариабельный домен легкой цепи (VL1) первого антитела, а второй антигенсвязывающий фрагмент содержит второй вариабельный домен тяжелой цепи (VH2) и второй вариабельный домен легкой цепи (VL2) второго антитела, где первое антитело и второе антитело отличаются и выбраны из группы, состоящей из антитела против CD3 и антитела против CD19. В некоторых вариантах осуществления, первое антитело представляет собой антитело против CD3, а второе антитело представляет собой антитело против CD19. В некоторых других вариантах осуществления, первое антитело представляет собой антитело против CD19, а второе антитело представляет собой антитело против CD3.
В обычном нативном антителе вариабельная область содержит три области CDR, расположенные между фланкирующими каркасными областями (FR), например, как указано в следующей формуле: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, от N-конца к C-концу.
a) Анти-CD3 связывающий фрагмент
В полипептидном комплексе, представленном в настоящем документе, первый антигенсвязывающий фрагмент или второй антигенсвязывающий фрагмент представляет собой анти-CD3 связывающий фрагмент. В некоторых вариантах осуществления, анти-CD3 связывающий фрагмент получен из анти-CD3 антитела WBP3311_2.306.4-z1, показанного в таблице A ниже. Последовательности CDR антитела WBP3311_2.306.4-z1 представлены ниже.
Последовательности вариабельной области тяжелой и каппа-легкой цепи антитела WBP3311_2.306.4-z1 представлены ниже.
WBP3311_2.306.4-z1-VH
Аминокислотная последовательность (SEQ ID NO: 15):
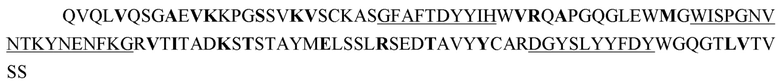 Последовательность нуклеиновой кислоты (SEQ ID NO: 16):
Последовательность нуклеиновой кислоты (SEQ ID NO: 16):
CAGGTGCAGCTTGTGCAGTCTGGGGCAGAAGTGAAGAAGCCTGGGTCTAGTGTCAAGGTGTCATGCAAGGCTAGCGGGTTCGCCTTTACTGACTACTACATCCACTGGGTGCGGCAGGCTCCCGGACAAGGGTTGGAGTGGATGGGATGGATCTCCCCAGGCAATGTCAACACAAAGTACAACGAGAACTTCAAAGGCCGCGTCACCATTACCGCCGACAAGAGCACCTCCACAGCCTACATGGAGCTGTCCAGCCTCAGAAGCGAGGACACTGCCGTCTACTACTGTGCCAGGGATGGGTACTCCCTGTATTACTTTGATTACTGGGGCCAGGGCACACTGGTGACAGTGAGCTCC
WBP3311_2.306.4-z1-VK
Аминокислотная последовательность (SEQ ID NO: 17):
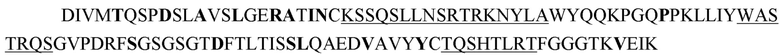
Последовательность нуклеиновой кислоты (SEQ ID NO: 18):
GATATCGTGATGACCCAGAGCCCAGACTCCCTTGCTGTCTCCCTCGGCGAAAGAGCAACCATCAACTGCAAGAGCTCCCAAAGCCTGCTGAACTCCAGGACCAGGAAGAATTACCTGGCCTGGTATCAGCAGAAGCCCGGCCAGCCTCCTAAGCTGCTCATCTACTGGGCCTCCACCCGGCAGTCTGGGGTGCCCGATCGGTTTAGTGGATCTGGGAGCGGGACAGACTTCACATTGACAATTAGCTCACTGCAGGCCGAGGACGTGGCCGTCTACTACTGTACTCAGAGCCACACTCTCCGCACATTCGGCGGAGGGACTAAAGTGGAGATTAAG
Известно, что CDR ответственны за связывание антигена.
В некоторых вариантах осуществления, анти-CD3 связывающий фрагмент, представленный в настоящем документе, содержит последовательность CDR3 тяжелой цепи WBP3311_2.306.4-z1. В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, содержит CDR3 тяжелой цепи, содержащую SEQ ID NO: 5. Области CDR3 тяжелой цепи расположены в центре антигенсвязывающего участка, и, соответственно, полагают, что они в наибольшей степени контактируют с антигеном и обеспечивают наибольшую свободную энергию для сродства антитела к антигену. Также полагают, что CDR3 тяжелой цепи является определенно самым разнообразным CDR антигенсвязывающего участка с точки зрения длины, аминокислотного состава и конформации посредством различных механизмов диверсификации. (Tonegawa S., Nature. 302:575-81 (1983)). Разнообразие в CDR3 тяжелой цепи является достаточным для получения большей специфичности антител (Xu JL, Davis MM., Immunity. 13: 37-45 (2000)), а также желаемой антигенсвязывающей аффинности. (Schier R, et al., J Mol Biol. 263:551-67 (1996)).
Анти-CD3-связывающий фрагмент, представленный в настоящем документе, дополнительно содержит подходящие последовательности каркасной области (FR), при условии, что анти-CD3-связывающий фрагмент может специфически связываться с CD3.
Гуманизированное антитело против CD3 WBP3311_2.306.4-z1 обладает специфической аффинностью связывания с CD3-экспрессирующей клеткой (например, CD4 T-клеткой) и может активировать человеческие T-клетки и запускать высвобождение цитокинов TNF-альфа и IFN-гамма.
В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, содержит последовательность вариабельного домена тяжелой цепи, представляющую SEQ ID NO: 15, и последовательность вариабельного домена легкой цепи, представляющую SEQ ID NO: 17.
Аффинность связывания анти-CD3-связывающего фрагмента, представленного в настоящем документе, может быть представлена значением KD, которое представляет собой отношение скорости диссоциации к скорости ассоциации (koff/kon), когда связывание между антигеном и антигенсвязывающей молекулой достигает равновесия. Антигенсвязывающую аффинность (например, KD) можно соответствующим образом определить, используя подходящие способы, известные в данной области техники, включая, например, исследование методом проточной цитометрии. В некоторых вариантах осуществления, связывание антитела с антигеном в различных концентрациях может быть определено методом проточной цитометрии, определенная средняя интенсивность флуоресценции (MFI) может сначала быть нанесена на график в зависимости от концентрации антитела, затем значение KD может быть рассчитано путем подгонки зависимости интенсивности специфической связывающей флуоресценции (Y) и концентрации антител (X) в уравнении насыщенности одного участка: Y=Bmax*X/(KD+X) с использованием программного обеспечения Prism, версия 5 (GraphPad Software, San Diego, CA), где Bmax относится к максимальному специфическому связыванию исследуемого антитела с антигеном.
В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, способен специфически связываться с CD3 человека, экспрессированным на поверхности клетки, или с рекомбинантным CD3 человека. CD3 является рецептором, экспрессируемым на клетке. Рекомбинантный CD3 представляет собой растворимый CD3, который экспрессируется рекомбинантно и не связан с клеточной мембраной. Рекомбинантный CD3 может быть получен посредством различных рекомбинантных технологий. В одном из примеров, последовательность ДНК CD3, кодирующая внеклеточный домен CD3 человека (NP_000724.1) (Met1-Asp126), может быть слита с полигистидиновой меткой на С-конце в векторе экспрессии, и затем трансфицирована и экспрессирована в клетках 293E и очищена с помощью Ni-аффинной хроматографии.
В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, способен специфически связываться с CD3 человека, экспрессируемым на поверхности клеток, со сродством связывания (KD) не более 5×10-9 М, не более 4×10-9 М, не более 3×10-9 М, не более 2×10-9 М, не более 10-9 М, не более 5×10-10 М, не более 4×10-10 М, не более 3×10-10 М, не более 2×10-10 M, не более 10-10 M, не более 5×10-11 M или не более 4×10-11 M, не более 3×10-11 M или не более 2×10-11 M или не более 10-11 М, что определено методом проточной цитометрии.
В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, вступает в перекрестную реакцию с CD3 яванского макака, например, CD3 яванского макака, экспрессированном на поверхности клетки, или с растворимым рекомбинантным CD3 яванского макака.
Связывание анти-CD3-связывающего фрагмента с рекомбинантным CD3 или CD3, экспрессируемым на поверхности клеток, можно измерить способами, известными в данной области техники, например, посредством сэндвич-анализа, такого как ELISA, вестерн-блоттинга, анализа проточной цитометрией и другого анализа связывания. В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, специфически связывается с рекомбинантным CD3 человека при значении EC50 (т.е. концентрации связывания 50%) не более 0,01 нМ, не более 0,02 нМ, не более 0,03 нМ, не более 0,04 нМ, не более 0,05 нМ, не более 0,06 нМ, не более 0,07 нМ и не более 0,08 нМ, определяемом методом ELISA. В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, специфически связывается с CD3 человека, экспрессируемым на поверхности клеток при значении EC50 не более 0,5 нМ, не более 0,6 нМ, не более 0,7 нМ, не более 0,8 нМ, не более 0,9 нМ, не более 1 нМ, не более 2 нМ, не более 3 нМ, не более 4 нМ, не более 5 нМ, не более 6 нМ, не более 7 нМ не более 8 нМ, не более 9 нМ или не более 10 нМ, определяемом методом проточной цитометрии.
В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент связывается с CD3 яванского макака со сродством связывания, аналогичным сродству связывания CD3 человека. Например, связывание типичного антитела WBP3311_2.306.4 с CD3 яванского макака происходит при сродстве или значении ЕС50, аналогичном для CD3 человека.
В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, специфически связывается с рекомбинантным CD3 яванского макака при значении EC50 не более 0,001 нМ, не более 0,005 нМ, не более 0,01 нМ, не более 0,02 нМ, не более 0,03 нМ, не более 0,04 нМ или не более 0,05 нМ, определяемом методом ELISA.
В некоторых вариантах осуществления, анти-CD3-связывающий фрагмент, представленный в настоящем документе, обладает специфическим сродством связывания с человеческим CD3, которое является достаточным для обеспечения диагностического и/или терапевтического применения. Ряд терапевтических стратегий модулирует Т-клеточный иммунитет путем нацеливания на передачу сигналов TCR, в частности, на моноклональные антитела против CD3 человека, которые клинически используются.
b) Анти-CD19 антитело
В полипептидном комплексе, представленном в настоящем документе, первый антигенсвязывающий фрагмент или второй антигенсвязывающий фрагмент представляет собой анти-CD19-связывающий фрагмент. В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент получен из анти-CD19-антитела W7011-4.155.8-z1-P15, показанного в таблице B ниже. Последовательности CDR анти-CD19-антитела представлены ниже.
Последовательности вариабельной области тяжелой и каппа-легкой цепи антитела WBP7011_4.155.8-z1-P15 представлены ниже.
W7011-4.155.8-z1-P15-VH
Аминокислотная последовательность (SEQ ID NO: 19):
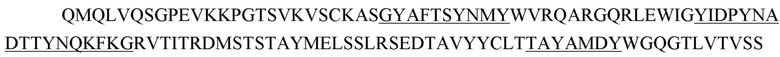
Последовательность нуклеиновой кислоты (SEQ ID NO: 20):
CAAATGCAGCTCGTCCAGTCTGGACCTGAAGTGAAGAAGCCCGGGACATCCGTCAAGGTCTCATGTAAGGCTAGCGGGTACGCATTCACTTCCTACAACATGTACTGGGTGCGCCAGGCCAGAGGACAGAGGTTGGAGTGGATCGGCTACATCGACCCATACAACGCCGATACTACCTACAATCAGAAGTTTAAAGGGCGGGTGACCATTACCCGGGATATGTCCACCTCCACCGCCTACATGGAGCTGAGCAGCCTGAGGAGCGAGGACACAGCCGTGTACTACTGCCTGACAACAGCCTATGCCATGGACTATTGGGGCCAGGGCACACTTGTGACTGTGAGCAGT
W7011-4.155.8-z1-P15-VK
Аминокислотная последовательность (SEQ ID NO: 21):
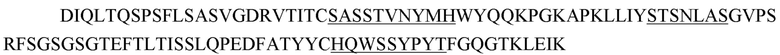
Последовательность нуклеиновой кислоты (SEQ ID NO: 22):
GACATCCAGCTCACCCAATCCCCTTCTTTCCTCTCCGCAAGTGTCGGAGATAGGGTGACTATCACCTGCTCAGCTTCTTCAACCGTGAACTACATGCATTGGTACCAGCAGAAGCCCGGGAAAGCCCCAAAGCTGCTGATCTACAGCACCTCCAATCTGGCCAGTGGAGTGCCAAGCCGGTTTAGCGGGAGCGGCTCCGGCACTGAATTCACTTTGACAATTAGCAGCCTTCAGCCTGAGGACTTTGCCACATATTACTGTCACCAGTGGTCCAGCTACCCCTACACATTCGGGCAGGGCACAAAGCTGGAGATTAAG
Известно, что CDR ответственны за связывание антигена.
В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, описанный в настоящем документе, содержит последовательность CDR3 тяжелой цепи антитела против CD19 W7011-4.155.8-z1-P15. В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, представленный в настоящем документе, содержит последовательность CDR3 тяжелой цепи, содержащую SEQ ID NO: 11. Области CDR3 тяжелой цепи расположены в центре антигенсвязывающего участка, и, соответственно, полагают, что они в наибольшей степени контактируют с антигеном и обеспечивают наибольшую свободную энергию для сродства антитела к антигену. Также полагают, что CDR3 тяжелой цепи является определенно самым разнообразным CDR антигенсвязывающего участка с точки зрения длины, аминокислотного состава и конформации посредством различных механизмов диверсификации. (Tonegawa S., Nature. 302:575-81 (1983)). Разнообразие в CDR3 тяжелой цепи является достаточным для получения большей специфичности антител (Xu JL, Davis MM. Immunity. 13: 37-45 (2000)), а также желаемой антигенсвязывающей аффинности (Schier R, et al., J Mol Biol. 263:551-67 (1996)).
Анти-CD19-связывающий фрагмент, представленный в настоящем документе, дополнительно содержит подходящие последовательности каркасной области (FR), при условии, что анти-CD19-связывающий фрагмент может специфически связываться с CD19.
В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, представленный в настоящем документе, содержит последовательность вариабельного домена тяжелой цепи, представляющую SEQ ID NO: 19, и последовательность вариабельного домена легкой цепи, представляющую SEQ ID NO: 21.
В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, представленный в настоящем документе, способен специфически связываться с CD19 человека, экспрессируемым на поверхности клеток со сродством связывания (KD) не более 5×10-9 M, не более 1×10-9 M, нет. более 9×10-10 M, не более 8×10-10 M, не более 7×10-10 M, не более 6×10-10 M, не более 5×10-10 M, не более 4×10-10 M, не более 3×10-10 М, не более 2×10-10 М или не более 1×10-10 М, определенным методом проточной цитометрии.
В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, представленный в настоящем документе, вступает в перекрестную реакцию с CD19 яванского макака, например, с CD19 яванского макака, экспрессированной на поверхности клетки, или с растворимой рекомбинантной CD19 яванского макака.
Связывание анти-CD19-связывающего фрагмента с CD19, экспрессированного в клетке, может быть измерено способами, известными в данной области техники, например, посредством сэндвич-анализа, такого как ELISA, вестерн-блоттинга, анализа методом проточной цитометрии и другого анализа связывания. В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, представленный в настоящем документе, специфически связывается с CD19 человека, экспрессированным в клетке при значении EC50 не более 0,01 нМ, не более 0,02 нМ, не более 0,03 нМ, не более 0,04 нМ, не более 0,05 нМ, не более 0,1 нМ, не более 0,2 нМ, не более 0,3 нМ, не более 0,4 нМ, не более 0,5 нМ, не более 0,5 нМ, не более 0,6 нМ, не более 0,7 нМ, не более 0,8 нМ, не более 0,9 нМ или не более 1 нМ, определенном методом проточной цитометрии.
В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент связывается с CD19 яванского макака со сродством связывания, аналогичным сродству связывания CD19 человека. В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, представленный в настоящем документе, специфически связывается с CD19 яванского макака, экспрессируемой на клетке, при значении EC50 не более 0,2 нМ, не более 0,5 нМ, не более 0,8 нМ, не более 1 нМ, не более 2 нМ или не более 3 нМ, определенном методом проточной цитометрии.
В некоторых вариантах осуществления, анти-CD19-связывающий фрагмент, представленный в настоящем документе, интернализуется CD19-экспрессирующей клеткой при значении EC50 не более 1 пМ, не более 2 пМ, не более 3 пМ, не более 4 пМ. не более 5 пм, не более 6 пм, не более 7 пм, не более 8 пм, не более 9 пм, не более 10 пм, не более 11 пм, не более 12 пм, не более 13 пм, не более 14 пм, не более 15 пм, не более 16 пм, не более 17 пм, не более 18 пм, не более 19 пм, не более 20 пм, не более 21 пм, не более 22 пм, не более 23 пм, не более 24 пм, не более 25 пм, не более 30 пм, не более 35 пм, не более 40 пм, не более 45 пм или не более 50 пМ, определенном методом анализа Fab-Zap.
Биспецифичный полипептидный комплекс
В некоторых вариантах осуществления, первый и/или второй антигенсвязывающий фрагмент является многовалентным, например двухвалентным, трехвалентным, четырехвалентным. Используемый в настоящем документе термин “валентность” относится к наличию определенного количества участков связывания антигена в данной молекуле. Как таковые, термины “двухвалентный”, “четырехвалентный” и “шестивалентный” обозначают наличие двух антигенсвязывающих молекул, соответственно, двух участков связывания, четырех участков связывания и шести участков связывания, соответственно. Двухвалентная молекула может быть моноспецифичной, если оба участка связывания предназначены для специфического связывания одного и того же антигена или одного и того же эпитопа. Аналогично, трехвалентная молекула может быть биспецифичной, например, когда два участка связывания являются моноспецифичными для первого антигена (или эпитопа), а третий сайт связывания является специфичным для второго антигена (или эпитопа). В некоторых вариантах осуществления, первый и/или второй антигенсвязывающий фрагмент в биспецифичном полипептидном комплексе, представленном в настоящем документе, может быть двухвалентным, трехвалентным или четырехвалентным, по меньшей мере с двумя участками связывания, специфичными для одного и того же антигена или эпитопа. Это, в некоторых вариантах осуществления, обеспечивает более сильное связывание с антигеном или эпитопом, чем моновалентный аналог. В некоторых вариантах осуществления, в двухвалентном антигенсвязывающем фрагменте, первый участок связывания и второй участок связывания структурно идентичны (т.е. имеют одинаковые последовательности) или структурно различны (т.е. имеют разные последовательности, хотя и с одинаковой специфичностью).
В некоторых вариантах осуществления первый и/или второй антигенсвязывающий фрагмент является многовалентным и содержит два или более антигенсвязывающих участка, функционально связанных вместе, со спейсером или без него.
В некоторых вариантах осуществления, второй антигенсвязывающий фрагмент содержит два или более Fab второго антитела. Два Fab могут быть функционально связаны друг с другом, например, первый Fab может быть ковалентно присоединен ко второму Fab посредством тяжелой цепи со спейсером между ними или без него.
В некоторых вариантах осуществления, первый антигенсвязывающий фрагмент связан с первым доменом димеризации, а второй антигенсвязывающий фрагмент связан со вторым доменом димеризации. Используемый в настоящем документе термин “димеризационный домен” относится к пептидному домену, который способен ассоциироваться другим пептидным доменом с образованием димера или, в некоторых примерах, обеспечивает возможность спонтанной димеризации двух пептидов.
В некоторых вариантах осуществления, первый димеризационный домен может быть связан со вторым доменом димеризации. Ассоциация может происходить посредством любого подходящего взаимодействия или связи или соединения, например, через линкер, дисульфидную связь, водородную связь, электростатическое взаимодействие, солевой мостик или гидрофобно-гидрофильное взаимодействие или их сочетание. Типичные димеризационные домены включают, без ограничения, шарнирный участок антитела, домен CH2 антитела, домен CH3 антитела и другие подходящие белковые мономеры, способные димеризоваться и связываться друг с другом. Шарнирная область, домен CH2 и/или CH3 могут быть получены из любых изотипов антител, таких как IgG1, IgG2 и IgG4.
Термин “дисульфидная связь” относится к ковалентной связи со структурой R-S-S-R’. Аминокислота цистеин содержит тиоловую группу, которая может образовывать дисульфидную связь со второй тиольной группой, например, из другого цистеинового остатка. Дисульфидная связь может образовываться между тиольными группами двух цистеиновых остатков, находящихся соответственно на двух полипептидных цепях, образуя тем самым межцепочечный мостик или межцепочечную связь.
Водородная связь образуется в результате электростатического притяжения между двумя полярными группами, когда атом водорода ковалентно связан атомом с высокой электроотрицательностью, таким как азот, кислород или фтор. Водородная связь может образовываться в полипептиде между атомами кислорода основной цепи (например, группами халькогенов) и амидными атомами водорода (группой азота) двух остатков, соответственно, таких как группа азота в Asn и группа кислорода в His, или группа кислорода в Asn и группа азота в Lys. Водородная связь сильнее, чем ван-дер-ваальсово взаимодействие, но слабее, чем ковалентные или ионные связи, и имеет решающее значение для поддержания вторичной структуры и третичной структуры. Например, альфа-спираль образуется при регулярном расстоянии между аминокислотными остатками в положениях i и i+4, и бета-лист представляет собой участок пептидной цепи длиной 3-10 аминокислот, образуемый, когда два пептида соединены, по меньшей мере, двумя или тремя остовными водородными связями с образованием закрученного складчатого листа.
Электростатическое взаимодействие является нековалентным взаимодействием и является важным для фолдинга белка, стабильности, пластичности и функции, включая ионные взаимодействия, водородную связь и галогенную связь. Электростатические взаимодействия могут образовываться в полипептиде, например, между Lys и Asp, между Lys и Glu, между Glu и Arg или между Glu, Trp в первой цепи и Arg, Val или Thr во второй цепи.
Солевой мостик представляет собой электростатические взаимодействия ближнего порядка, которые, в основном, возникают из анионного карбоксилата Asp или Glu, и катионного аммония из Lys или гуанидиния Arg, которые являются пространственно близкими парами противоположно заряженных остатков в нативных белковых структурах. Заряженные и полярные остатки в основном в гидрофобных поверхностях раздела могут действовать как “горячие точки” для связывания. Среди прочего, остатки с ионизируемыми боковыми цепями, например His, Tyr и Ser, также могут участвовать в образовании солевого мостика.
Гидрофобное взаимодействие может возникать между одним или более Val, Tyr и Ala в первой цепи и одним или более Val, Leu и Trp во второй цепи или His и Ala в первой цепи и Thr и Phe во второй цепи (см. Brinkmann, et al, 2017, Выше).
В некоторых вариантах осуществления, первый и/или второй димеризационный домен содержит, по меньшей мере, часть шарнирной области антитела. В некоторых вариантах осуществления, первый и/или второй димеризационный домен может дополнительно содержать домен СН2 антитела и/или домен СН3 антитела. В некоторых вариантах осуществления, первый и/или второй димеризационный домен содержит, по меньшей мере, часть области Hinge-Fc, т.е. домен Hinge-CH2-CH3. В некоторых вариантах осуществления, первый димеризационный домен может быть функционально связан с C-концом первой константной области TCR. В некоторых вариантах осуществления, второй димеризационный домен может быть функционально связан с C-концом константной области CH1 антитела второго антигенсвязывающего фрагмента.
В полипептидном комплексе, представленном в настоящем документе, С1 содержит сконструированную СБета, и первый димеризационный домен функционально связан с сконструированной СБета на третьем домене конъюгирования, который содержит SEQ ID NO: 25.
В представленном в настоящем документе полипептидном комплексе первый димеризационный домен функционально связан с С-концом сконструированной константной области TCR и вместе образует химерную константную область. Другими словами, химерная константная область содержит первый димеризационный домен, функционально связанный с сконструированной константной областью TCR.
В некоторых вариантах осуществления, химерная константная область содержит сконструированную СБета, присоединенную к первой шарнирной-Fc области, полученной из IgG1, IgG2 или IgG4. Примерные последовательности такой химерной константной области приведены в примере 2.
В некоторых вариантах осуществления, химерная константная область дополнительно содержит первый домен СН2 антитела и/или первый домен СН3 антитела. Например, химерная константная область дополнительно содержит первый домен СН2-СН3 антитела, присоединенный к С-концу третьего домена конъюгирования. Иллюстративные последовательности такой химерной константной области приведены в примере 2.
Эти пары химерных константных областей и вторых константных доменов TCR являются практичными в том смысле, что ими можно манипулировать для слияния с желаемой вариабельной областью антитела с целью обеспечения полипептидного комплекса, как описано в настоящем документе. Например, вариабельная область тяжелой цепи антитела может быть слита с химерной константной областью (содержащей С1), предоставляя, таким образом, первую полипептидную цепь полипептидного комплекса, предусмотренного в настоящем документе; и аналогично, вариабельная область легкой цепи антитела может быть слита со вторым константным доменом TCR (содержащим C2), предоставляя, таким образом, вторую полипептидную цепь полипептидного комплекса, предусмотренного в настоящем документе.
В некоторых вариантах осуществления, второй димеризационный домен содержит шарнирную область. Шарнирная область может быть получена от антитела, такого как IgG1, IgG2 или IgG4. В некоторых вариантах осуществления, второй димеризационный домен может необязательно дополнительно содержать домен CH2 антитела и/или домен CH3 антитела, например, такой как шарнирная-Fc область. Шарнирная область может быть присоединена к тяжелой цепи антитела второго участка связывания антигена (например, Fab).
В биспецифичном полипептидном комплексе первый и второй димеризационные домен способны ассоциироваться в димер. В некоторых вариантах осуществления, первый и второй димеризационные домены являются различными и ассоциируются т ассоциируются таким образом, чтобы препятствовать гомодимеризации и/или способствовать гетеродимеризации. Например, первый и второй димеризационные домены могут быть выбраны так, чтобы они не были идентичными и чтобы они предпочтительно образовывали гетеродимеры между собой, а не образовывали гомодимеры в пределах своего типа. В некоторых вариантах осуществления, первый и второй димеризационные домены способны ассоциироваться в гетеродимеры путем образования “выступ-в-углубление”, гидрофобного взаимодействия, электростатического взаимодействия, гидрофильного взаимодействия или повышенной пластичности.
В некоторых вариантах осуществления, первый и второй димеризационные домены содержат СН2- и/или СН3-домены, которые соответственно мутированы с целью возможности образования “выступ-в-углубление”. Выступ может быть получен путем замены небольшого аминокислотного остатка более крупным в первом СН2/СН3 полипептиде, а углубление может быть получено путем замены большого остатка меньшим. Подробнее об участках мутаций для выступов в углублениях смотри Ridgway et al., 1996 выше, Spiess et al., 2015 выше, и Brinkmann et al., 2017 выше.
Биспецифичный формат
В полипептидном комплексе, представленном в настоящем документе, первый антигенсвязывающий фрагмент и второй связывающий фрагмент ассоциированы в Ig-подобную структуру. Ig-подобная структура аналогична природному антителу, имеющему Y-образную конструкцию, с двумя “плечами” для связывания антигена и одним “стволом” для ассоциации и стабилизации. Сходство с природным антителом может обеспечивать различные преимущества, такие как хорошая фармакокинетика in vivo, желаемый иммунологический ответ, стабильность и т.д. Было обнаружено, что Ig-подобная структура, содержащая первый антигенсвязывающий фрагмент, представленный в настоящем документе, связана со вторым антигенсвязывающим фрагментом, представленным в настоящем документе, обладает термостабильностью, сопоставимой с термостабильностью Ig (например, IgG). В некоторых вариантах осуществления, представленная в настоящем документе Ig-подобная структура составляет, по меньшей мере, 70%, 80%, 90%, 95% или 100% структуры природного IgG.
Представленный в настоящем документе биспецифичный полипептидный комплекс включает четыре полипептидные цепи: i) VH1-C1-шарнир-CH2-CH3; ii) VL1-C2; iii) VH2-CH1-шарнир-CH2-CH3 и iv) VL2-CL, где C1 и C2 способны образовывать димер, содержащий, по меньшей мере, одну ненативную межцепную связь, и две шарнирные области и/или два СН3-домена способны образовывать одну или более межцепных связей, которые могут способствовать димеризации.
Описанные в настоящем документе биспецифичные полипептидные комплексы имеют более длительный период полужизни in vivo и относительно проще в производстве при их сравнинии с биспецифичными полипептидными комплексами в других форматах.
Последовательности биспецифичных комплексов
В некоторых вариантах осуществления, первый антигенсвязывающий фрагмент биспецифичного комплекса способен специфически связываться с CD3, а второй антигенсвязывающий фрагмент способен специфически связываться с CD19. В других вариантах осуществления, первый антигенсвязывающий фрагмент биспецифичного комплекса способен специфически связываться с CD19, а второй антигенсвязывающий фрагмент способен специфически связываться с CD3.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс содержит сочетание четырех полипептидных последовательностей: SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 29 (E17), как показано в примере 2. В некоторых вариантах осуществления, биспецифичный полипептидный комплекс содержит сочетание четырех полипептидных последовательностей: SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 30 (F16), как показано в примере 2. В таких вариантах осуществления, первый антигенсвязывающий фрагмент связывается с CD3, а второй антигенсвязывающий фрагмент связывается с CD19. Конструкция E17 представляет собой биспецифичное двухвалентное антитело, а конструкция F16 представляет собой биспецифичный и трехвалентный антигенсвязывающий комплекс с двумя повторами Fab анти-CD19 антитела.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс содержит четыре полипептидные цепи, включающие: i) VH1, функционально связанный с первой химерной константной областью; ii) VL1, функционально связанный со второй химерной константной областью; iii) VH2, функционально связанный с константной областью легкой цепи обычного антитела, и iv) VL2, функционально связанный с константной областью легкой цепи стандартного антитела. В некоторых вариантах осуществления, первая химерная константная область может содержать C1-шарнир-CH2-CH3, где каждый является таким, как определено выше. В некоторых вариантах осуществления, вторая химерная константная область может содержать С2, как определено выше. В некоторых вариантах осуществления, константная область тяжелой цепи стандартного антитела может содержать CH1-шарнир-CH2-CH3, где каждый является таким, как определено выше. В некоторых вариантах осуществления, константная область легкой цепи стандартного антитела может содержать CL, как определено выше.
Способ получения
Настоящее изобретение относится к выделенным нуклеиновым кислотам или полинуклеотидам, которые кодируют полипептидный комплекс, и к биспецифичному анти-CD3 × CD19 полипептидному комплексу, предусмотренному в настоящем документе.
Используемый в настоящем документе термин “нуклеиновая кислота” или “полинуклеотид” относится к дезоксирибонуклеиновой кислоте (ДНК) или рибонуклеиновой кислоте (РНК) и их полимерам в одно- или двухцепочечной форме. Если конкретно не ограничено, термин охватывает полинуклеотиды, включая известные аналоги природных нуклеотидов, которые обладают аналогичными свойствами связывания, как эталонная нуклеиновая кислота, и метаболизируются аналогично природным нуклеотидам. Если не указано иное, конкретная полинуклеотидная последовательность также в неявной форме включает свои консервативно модифицированные варианты (например, замены вырожденных кодонов), аллели, ортологи, SNP и комплементарные последовательности, а также последовательность, указанную в явной форме. В частности, замены вырожденных кодонов могут быть получены в результате создания последовательностей, в которых в третьем положении в одном или в нескольких выбранных (или во всех) кодонах произведена замена смешанным основанием и/или остатками дезоксиинозина (см. Batzer et al., Nucleic Acid Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); и Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)).
Нуклеиновые кислоты или полинуклеотиды, кодирующие полипептидный комплекс и биспецифичный полипептидный комплекс, представленные в настоящем документе, могут быть сконструированы с использованием методов рекомбинации. С этой целью, ДНК, кодирующая антигенсвязывающий фрагмент исходного антитела (такого как CDR или вариабельная область), может быть выделена и секвенирована с использованием обычных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Аналогичным образом может быть получена ДНК, кодирующая константную область TCR. В качестве примера, полинуклеотидная последовательность, кодирующая вариабельный домен (VH), и полинуклеотидная последовательность, кодирующая первую константную область TCR (C1), получены и функционально связаны с обеспечением транскрипции и экспрессии в клетке-хозяине для продукции первого полипептида. Аналогичным образом, полинуклеотидная последовательность, кодирующая VL, функционально связана с полинуклеотидной последовательностью, кодирующей С1, с обеспечением экспрессии второго полипептида в клетке-хозяине. При необходимости, кодирующие полинуклеотидные последовательности для одного или более спейсеров также функционально связаны с другими кодирующими последовательностями с обеспечением экспрессии желаемого продукта.
Кодирующие полинуклеотидные последовательности могут быть дополнительно функционально связаны с одной или более регуляторными последовательностями, необязательно, в экспрессирующем векторе, благодаря чему экспрессия или продуцирование первого и второго полипептидов осуществимы и находятся под соответствующим контролем.
Кодирующая полинуклеотидная последовательность(и) может быть вставлена в вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии с использованием методов рекомбинации, известных в данной области техники. В другом варианте осуществления, полипептидный комплекс и биспецифичный полипептидный комплекс, представленные в настоящем документе, могут быть получены посредством гомологичной рекомбинацией, известной в данной области техники. Многие векторы являются доступными. Компоненты вектора обычно включают, но ими не ограничиваются, один или более из следующих: сигнальная последовательность, источник репликации, один или более маркерных генов, энхансерный элемент, промотор (например, SV40, CMV, EF-1α) и последовательность терминации транскрипции.
Используемый в настоящем документе термин “вектор” относится к носителю, в который может быть функционально встроен полинуклеотид, кодирующий белок, чтобы вызвать экспрессию этого белка. Обычно конструкция также включает соответствующие регуляторные последовательности. Например, полинуклеотидная молекула может включать регуляторные последовательности, расположенные в 5'-фланкирующей области нуклеотидной последовательности, кодирующей направляющую РНК, и/или нуклеотидную последовательность, кодирующую сайт-направленный модифицирующий полипептид, функционально связанную с кодирующими последовательностями способом, допускающим экспрессию желаемого транскрипта/гена в клетке-хозяине. Вектор может быть использован для трансформации, трансдукции или трансфекции клетки-хозяина, чтобы вызвать экспрессию генетического элемента, который он несет в клетке-хозяине. Примеры векторов включают плазмиды, фагемиды, космиды, искусственные хромосомы, такие как искусственная хромосома дрожжей (YAC), искусственная бактериальная хромосома (BAC) или искусственная хромосома на основе бактериофага P1 (PAC), бактериофаги, такие как фаг лямбда или фаг M13, и вирусы животных. Категории животных вирусов, используемых в качестве векторов, включают ретровирус (включая лентивирус), аденовирус, аденоассоциированный вирус, герпесвирус (например, вирус простого герпеса), поксвирус, бакуловирус, вирус папилломы и паповавирус (например, SV40). Вектор может содержать множество элементов для контроля экспрессии, включая последовательности промотора, последовательности инициации транскрипции, последовательности энхансера, селектируемые элементы и репортерные гены. Кроме того, вектор может содержать точку начала репликации. Вектор может также включать вещества, способствующие его проникновению в клетку, включая, но ими не ограничиваясь, вирусную частицу, липосому или белковую оболочку.
В некоторых вариантах осуществления, векторная система включает системы млекопитающих, бактериальные системы, дрожжевые системы и т.д., и содержит плазмиды, такие как, но ими не ограничиваясь, pALTER, pBAD, pcDNA, pCal, pL, pET, pGEMEX, pGEX, pCI, pCMV, pEGFP, pEGFT, pSV2, pFUSE, pVITRO, pVIVO, pMAL, pMONO, pSELECT, pUNO, pDUO, Psg5L, pBABE, pWPXL, pBI, p15TV-L, pPro18, pTD, pRS420, pLexA, pACT2.2 и т.д. и коммерчески доступные векторы. Подходящие векторы могут включать плазмидные или вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы).
Векторы, содержащие представленную в настоящем документе полинуклеотидную последовательность(и), могут быть введены в клетку-хозяина для клонирования или экспрессии гена. Используемая в настоящем документе фраза “клетка-хозяин” относится к клетке, в которую был введен экзогенный полинуклеотид и/или вектор.
Подходящими клетками-хозяевами для клонирования или экспрессии ДНК в векторах по настоящему изобретению являются клетки прокариот, дрожжей или высших эукариот, описанные выше. Подходящие прокариоты для этой цели включают эубактерии, такие как грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как Escherichia, например, E.coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans и Shigella, а также Bacilli, такие как B. subtilis и B. licheniformis, Pseudomonas, такие как P. aeruginosa и Streptomyces.
Помимо прокариот, эукариотические микробы, такие как нитчатые грибы или дрожжи, являются подходящими хозяевами клонирования или экспрессии для векторов, кодирующих полипептидный комплекс и биспецифичный полипептидный комплекс. Saccharomyces cerevisiae, или обычные пекарские дрожжи, наиболее часто используются среди низших эукариотических микроорганизмов-хозяев. Тем не менее, ряд других родов, видов и штаммов являются общедоступными и применимыми в данном документе, например, организмы-хозяева Schizosaccharomyces pombe; Kluyveromyces, такие как, например, K. lactis, K. fragilis (ATCC 12,424), K. bulgaricus (ATCC 16,045), K. wickeramii (ATCC 24,178), K. waltii (ATCC 56,500), K. drosophilarum (ATCC 36,906), K. Thermotolerans и K. marxianus; yarrowia (EP 402,226); Pichia pastoris (EP 183,070); Candida; Trichoderma reesia (EP 244,234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и filamentous fungi, такие как, например, Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A. nidulans и A. niger.
Подходящие клетки-хозяева для экспрессии гликозилированного полипептидного комплекса и биспецифичного полипептидного комплекса, представленного в настоящем документе, получены из многоклеточных организмов. Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы и варианты и соответствующие пермиссивные клетки-хозяева насекомых из таких хозяев, как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Разнообразные штаммы вирусов для трансфекции являются общедоступными, например, вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV, и такие вирусы могут быть использованы в качестве вируса в соответствии с настоящим изобретением, в частности для трансфекции клеток Spodoptera frugiperda. Культуры растительных клеток хлопка, кукурузы, картофеля, сои, петунии, томатов и табака также могут быть использованы в качестве хозяев.
Однако наибольший интерес вызывают клетки позвоночных, и размножение клеток позвоночных в культуре (культура ткани) стало обычной процедурой. Примерами используемых линий клеток-хозяев млекопитающих являются линия CV1 почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); клеточная линия эмбриональных почек человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36:59 (1977)), например Expi293; клетки почек детенышей хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 4216 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почек африканских зеленых мартышек (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почек собак (MDCK, ATCC CCL 34); клетки печени крыс линии Buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383: 44-68 (1982)); клетки MRC 5; клетки FS4; и линия клеток гепатомы человека (Hep G2).
Клетки-хозяева, трансформированные описанными выше векторами экспрессии или клонирования, можно культивировать в обычных питательных средах, модифицированных, при необходимости, для индукции промоторов, выбора трансформантов или амплификации клонирующих векторов.
Для продуцирования полипептидного комплекса и биспецифичного полипептидного комплекса, представленного в настоящем документе, клетки-хозяева, трансформированные экспрессирующим вектором, можно культивировать в различных средах. Коммерчески доступные среды, такие как Ham's F10 (Sigma), минимальная питательная среда (MEM), (Sigma), RPMI-1640 (Sigma) и модифицированная по Дульбекко среда Игла (DMEM), Sigma), подходят для культивирования клеток-хозяев. Кроме того, любой из носителей, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102: 255 (1980), патент США № 4767704; 4657866; 4927762; 4560655; или 5122469; WO 90/03430; WO 87/00195; или патент США RE 30985, можно использовать в качестве культуральной среды для клеток-хозяев. Любая из этих сред может, при необходимости, дополняться гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальция, магния и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как препарат GENTAMYCINTM), микроэлементами (определяемые как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Любые другие необходимые добавки также могут быть включены в соответствующих концентрациях, которые известны специалистам в данной области техники. Условия культивирования, такие как температура, рН и тому подобное, являются такими, которые ранее использовались с клеткой-хозяина, выбранной для экспрессии, и будут очевидны для специалиста в данной области техники.
В одном из аспектов, настоящее изобретение относится к способу экспрессии полипептидного комплекса и биспецифичного полипептидного комплекса, предусмотренного в настоящем документе, включающему культивирование клетки-хозяина, предоставленной в настоящем документе, в условиях, при которых экспрессируется полипептидный комплекс или биспецифичный полипептидный комплекс.
В некоторых вариантах осуществления, настоящее изобретение обеспечивает способ получения биспецифичного полипептидного комплекса, представленного в настоящем документе, включающий а) введение в клетку-хозяина: одного или более полинуклеотидов, кодирующих первый антигенсвязывающий фрагмент, включающий первый полинуклеотид, кодирующий первый полипептид, содержащий от N-конца до C-конца первый вариабельный домен тяжелой цепи (VH) первого антитела, функционально связанный с первой константной областью TCR (C1), второй полинуклеотид, кодирующий второй полипептид, содержащий от N-конца до C-конца первый вариабельный домен легкой цепи (VL) первого антитела, функционально связанный со второй константной областью TCR (C2), и один или более дополнительных полинуклеотидов, кодирующих второй антигенсвязывающий фрагмент, где: C1 содержит сконструированную СБета, содержащую SEQ ID NO: 1, и C2 содержит сконструированную САльфа, содержащую SEQ ID NO: 2, аминокислота C48 в SEQ ID NO: 1 и аминокислота C41 в SEQ ID NO: 2 способны образовывать ненативную межцепочечную дисульфидную связь, С1 и С2 способны образовывать димер, и ненативная межцепочечная дисульфидная связь способна стабилизировать димер С1 и С2, первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент меньше подвержены ошибочному спариванию, чем, если бы и первый и второй антигенсвязывающие фрагменты были бы аналогами природного Fab, и первое антитело обладает первой антигенной специфичностью, и второе антитело обладает второй антигенной специфичностью, b) предоставление возможности клетке-хозяину экспрессировать биспецифичный полипептидный комплекс.
В некоторых вариантах осуществления, способ дополнительно включает выделение биспецифичного полипептидного комплекса.
При использовании методов рекомбинации, биспецифичный полипептидный комплекс, представленный в настоящем документе, может продуцироваться внутриклеточно, в периплазматическом пространстве, или непосредственно секретироваться в среду. Если продукт продуцируется внутриклеточно, на первой стадии, частицы твердых частиц, либо клетки-хозяева, либо лизированные фрагменты, удаляются, например, центрифугированием или ультрафильтрацией. В Carter et al., Bio/Technology 10: 163-167 (1992) описана процедура выделения антител, которые секретируются в периплазматическое пространство E. coli. Вкратце, клеточную пасту размораживают в присутствии ацетата натрия (pH 3,5), EDTA и фенилметилсульфонилфторида (PMSF) в течение примерно 30 минут. Клеточный дебрис может быть удален центрифугированием. Когда продукт секретируется в среду, супернатанты из таких систем экспрессии обычно сначала концентрируют с использованием коммерчески доступного фильтра для концентрации белка, например, ультрафильтрационной установки Amicon или Millipore Pellicon. Ингибитор протеазы, такой как PMSF, может быть включен в любую из вышеуказанных стадий для ингибирования протеолиза, и антибиотики могут быть включены для предотвращения роста случайных загрязнителей.
Биспецифичный полипептидный комплекс, предоставленный в настоящем документе, полученный из клеток, может быть очищен с использованием, например, хроматографии с гидроксиапатитом, гель-электрофореза, диализа, ионообменной хроматографии на DEAE-целлюлозе, осаждения сульфатом аммония, высаливания и аффинной хроматографии, при этом аффинная хроматография является предпочтительным методом очистки.
Когда биспецифичный полипептидный комплекс, представленный в настоящем документе, содержит Fc-домен иммуноглобулина, тогда белок A можно использовать в качестве аффинного лиганда в зависимости от вида и изотипа Fc-домена, который присутствует в полипептидном комплексе. Белок A можно использовать для очистки полипептидных комплексов на основе тяжелых цепей γ1, γ2 или γ4 человека (Lindmark et al., J. Immunol. Meth. 62: 1-13 (1983)). Белок G рекомендуется для всех мышиных изотипов и для γ3 человека (Guss et al., EMBO J. 5: 1567 1575 (1986)). Матрица, к которой присоединен аффинный лиганд, чаще всего является агарозной, но доступны и другие матрицы. Механически стабильные матрицы, такие как стекло с заданным размером пор или полистиролдивинилбензол, обеспечивают более высокие скорости потока и более короткое время обработки, чем это может быть достигнуто с агарозой.
Когда биспецифичный полипептидный комплекс, представленный в настоящем документе, содержит домен СН3, для очистки можно использовать смолу Bakerbond ABX (J. T. Baker, Phillipsburg, N.J.). Другие методы очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на силикагеле, хроматография на гепарине, хроматография на анионообменной или катионообменной смоле SEPHAROSETM (например колонка с полиаспарагиновой кислотой), хроматофокусировка, ДНС-ПААГ и осаждение сульфатом аммония, также доступны в зависимости от антитела, которое будет извлекаться.
После любой стадии(стадий) предварительной очистки, смесь, содержащую представляющий интерес полипептидный комплекс и загрязняющие вещества, может быть подвергнута хроматографии гидрофобного взаимодействия с низким pH с использованием элюирующего буфера при значении pH, составляющем около 2,5-4,5, предпочтительно выполняемого при низких концентрациях соли (например, от 0 до 0,25 М соли).
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс, представленный в настоящем документе, может быть легко очищен с высокими выходами при использовании обычных способов. Одним из преимуществ биспецифичного полипептидного комплекса является значительное снижение ошибочного спаривания между последовательностями вариабельного домена тяжелой цепи и легкой цепи. Это снижает продукцию нежелательных побочных продуктов и позволяет получать продукт высокой чистоты с высоким выходом, используя относительно простые способы очистки.
Производные
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс может быть использован в качестве основы для конъюгации с желаемыми конъюгатами.
Предполагается, что различные конъюгаты могут быть связаны с полипептидным комплексом или биспецифичным полипептидным комплексом, представленным в настоящем документе (см., например, “Conjugate Vaccines”, Contributions to Microbiology and Immunology, J. M. Cruse and R. E. Lewis, Jr. (eds.), Carger Press, New York, (1989)). Эти конъюгаты могут быть связаны с полипептидным комплексом или биспецифичным полипептидным комплексом, в числе других способов, путем ковалентного связывания, аффинного связывания, интеркаляции, координатного связывания, комплексообразования, ассоциации, смешивания или добавления.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс, представленный в настоящем документе, может быть сконструирован таким образом, чтобы он содержал специфические сайты за пределами эпитоп-связывающей части, которые можно использовать для связывания с одним или более конъюгатами. Например, такой участок может включать один или более реакционноспособных аминокислотных остатков, таких как, например, цистеиновые остатки или гистидиновые остатки, для облегчения ковалентной связи с конъюгатом.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс может быть связан с конъюгатом непосредственно или опосредованно, например, через другой конъюгат или через линкер.
Например, биспецифичный полипептидный комплекс, имеющий реактивный остаток, такой как цистеин, может быть связан с тиол-реактивным агентом, в котором реактивной группой является, например, малеимид, иодацетамид, пиридилдисульфид или другой тиол-реактивный партнер по конъюгации (Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Hermanson, G. in Bioconjugate Techniques (1996) Academic Press, San Diego, pp. 40-55, 643-671).
В другом примере, биспецифичный полипептидный комплекс может быть конъюгирован с биотином, затем опосредованно конъюгирован со вторым конъюгатом, который конъюгирован с авидином. Еще в одном примере, полипептидный комплекс или биспецифичный полипептидный комплекс может быть связан с линкером, который дополнительно связывается с конъюгатом. Примеры линкеров включают бифункциональные связующие агенты, такие как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидо соединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и гис-активные фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). В частности, предпочтительные связующие агенты включают N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173: 723-737 (1978)) и N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP) для обеспечения дисульфидной связи.
Конъюгат может представлять собой детектируемую метку, фармакокинетически модифицирующий фрагмент, очищающий фрагмент или цитотоксический фрагмент. Примеры детектируемой метки могут включать флуоресцентные метки (например, флуоресцеин, родамин, дансил, фикоэритрин или техасский красный), фермент-субстратные метки (например, пероксидаза хрена, щелочная фосфатаза, люцерифераза, глюкоамилаза, лизоцим, сахаридоксидаза или β-D-галактозидаза), радиоизотопы (например, 123I, 124I, 125I, 131I, 35S, 3H, 111In, 112In, 14C, 64Cu, 67Cu, 86Y, 88Y, 90Y, 177Lu, 211At, 186Re, 188Re, 153Sm, 212Bi и 32P, другие лантаноиды, люминесцентные метки), хромофорный фрагмент, дигоксигенин, биотин/авидин, молекулу ДНК или золото для детекции. В некоторых вариантах осуществления, конъюгат может представлять собой фармакокинетически модифицирующий фрагмент, такой как PEG, который способствует увеличению времени полужизни антитела. Другие подходящие полимеры включают, например, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, сополимеры этиленгликоля и пропиленгликоля и тому подобное. В некоторых вариантах осуществления, конъюгат может представлять собой очищающий фрагмент, такой как магнитный микроноситель. “Цитотоксический фрагмент” может быть любым агентом, который является пагубным для клеток или может повредить клетки или привести к их гибели. Примеры цитотоксического фрагмента включают, без ограничения, таксол, цитохалазин B, грамицидин D, этидий бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин и его аналоги, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил декарбазин), алкилирующие агенты (например, мехлоретамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, диброманнит, стрептозотоцин, митомицин C и цис-дихлордиаминплатина (II) (DDP), цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)) и антимитотические агенты (например, винкристин и винбластин).
Способы конъюгирования конъюгатов с белками, такими как антитела, иммуноглобулины или их фрагменты, описаны, например, в патенте США № 5208020; патенте США 6441163; WO2005037992; WO2005081711; и WO2006/034488, которые включены в настоящем докумене посредством ссылки полностью.
Фармацевтическая композиция
Настоящее изобретение также относится к фармацевтической композиции, содержащей биспецифичный полипептидный комплекс, представленный в настоящем документе, и фармацевтически приемлемый носитель.
Термин “фармацевтически приемлемый” указывает на то, что указанный носитель, наполнитель, разбавитель, вспомогательное(ые) вещество(а) и/или соль обычно химически и/или физически совместимы с другими ингредиентами, составляющими композицию, и физиологически совместимы с его реципиентом.
Термин “фармацевтически приемлемый носитель” относится к ингредиенту в фармацевтической композиции, отличному от активного ингредиента, который является биологически активным и нетоксичным для субъекта. Фармацевтически приемлемые носители для использования в описанных в настоящем документе фармацевтических композициях могут включать, например, фармацевтически приемлемые жидкие, гелеобразные или твердые носители, водные носители, неводные носители, антимикробные агенты, изотонические агенты, буферы, антиоксиданты, анестетики, суспендирующие/диспергирующие агенты, изолирующие или хелатирующие агенты, разбавители, адъюванты, вспомогательные вещества или нетоксичные вспомогательные вещества, другие компоненты, известные в данной области техники, или их различные сочетания.
Подходящие компоненты могут включать, например, антиоксиданты, наполнители, связующие, разрыхлители, буферы, консерванты, смазывающие вещества, ароматизаторы, загустители, красители, эмульгаторы или стабилизаторы, такие как сахара и циклодекстрины. Подходящие антиоксиданты могут включать, например, метионин, аскорбиновую кислоту, EDTA, тиосульфат натрия, платину, каталазу, лимонную кислоту, цистеин, тиоглицерин, тиогликолевую кислоту, тиосорбит, бутилгидроксанизол, бутилгидрокситолуол и/или пропилгаллат. Как описано в настоящем документе, включение одного или более антиоксидантов, таких как метионин, в фармацевтическую композицию, предусмотренную в настоящем документе, уменьшает окисление полипептидного комплекса или биспецифичного полипептидного комплекса. Это уменьшение окисления предотвращает или уменьшает потерю аффинности связывания, тем самым улучшая стабильность белка и максимизируя срок годности. Следовательно, в определенных вариантах осуществления, предусмотрены композиции, которые включают полипептидный комплекс или биспецифичный полипептидный комплекс, описанный в настоящем документе, и один или более антиоксидантов, таких как метионин.
Для дополнительной иллюстрации, фармацевтически приемлемые носители могут включать, например, водные носители, такие как физиологический раствор для инъекций, раствор Рингера для инъекций, изотонической раствор декстрозы для инъекций, стерильная вода для инъекций или декстрозный и лактатный раствор Рингера для инъекций, неводные носители, такие как нелетучие масла растительного происхождения, хлопковое масло, кукурузное масло, кунжутное масло или арахисовое масло, антимикробные агенты в бактериостатических или фунгистатических концентрациях, изотонические агенты, такие как хлорид натрия или декстроза, буферы, такие как фосфатные или цитратные буферы, антиоксиданты, такие как бисульфат натрия, местные анестетики, такие как прокаин-гидрохлорид, суспендирующие и диспергирующие агенты, такие как карбоксиметилцеллюлоза натрия, гидроксипропилметилцеллюлоза или поливинилпирролидон, эмульгирующие агенты, такие как полисорбат 80 (TWEEN-80), секвестрирующие или хелатообразующие агенты, такие как EDTA (этилендиаминтетрауксусная кислота) или EGTA (этиленгликольтетрауксусная кислота), этиловый спирт, полиэтиленгликоль, пропиленгликоль, гидроксид натрия, соляная кислота, лимонная кислота или молочная кислота. Противомикробные агенты, используемые в качестве носителей, могут добавляться к фармацевтическим композициям в упаковках лекарственных средств для многократного приема, которые включают фенолы или крезолы, ртутесодержащие вещества, бензиловый спирт, хлорбутанол, метиловые и пропиловые сложные эфиры п-гидроксибензойной кислоты, тимеросал, хлорид бензалкония и хлорид бензетония. Подходящие вспомогательные вещества могут включать, например, воду, физиологический раствор, декстрозу, глицерин или этанол. Подходящие нетоксичные вспомогательные вещества могут включать, например, смачивающие или эмульгирующие агенты, рН-буферные агенты, стабилизаторы, усилители растворимости или агенты, такие как ацетат натрия, сорбитанмонолаурат, триэтаноламинолеат или циклодекстрин.
Фармацевтические композиции могут представлять собой жидкий раствор, суспензию, эмульсию, пилюлю, капсулу, таблетку, состав с замедленным высвобождением или порошок. Пероральные препараты могут включать стандартные носители, такие как фармацевтической степени чистоты маннит, лактозу, крахмал, стеарат магния, поливинилпирролидон, сахарин натрия, целлюлозу, карбонат магния и т.д.
В некоторых вариантах осуществления, фармацевтические композиции получены в виде инъецируемой композиции. Инъецируемые фармацевтические композиции могут быть получены в любой обычной форме, такой как, например, жидкий раствор, суспензия, эмульсия или твердые формы, подходящие для получения жидкого раствора, суспензии или эмульсии. Препараты для инъекций могут включать стерильные и/или апирогенные растворы, готовые для инъекции, стерильные сухие растворимые продукты, такие как лиофилизированные порошки, готовые к объединению с растворителем непосредственно перед применением, включая таблетки для подкожных инъекций, стерильные суспензии, готовые для инъекции, стерильные сухие нерастворимые продукты, готовые к объединению с носителем непосредственно перед применением, и стерильные и/или апирогенные эмульсии. Растворы могут быть водными или неводными.
В некоторых вариантах осуществления, парентеральные препараты, содержащие стандартную дозу, упаковывают в ампулу, флакон или шприц с иглой. Все препараты для парентерального введения должны быть стерильными и апирогенными, как это известно и применяется на практике в данной области.
В некоторых вариантах осуществления, стерильный лиофилизированный порошок получают растворением полипептидного комплекса или биспецифичного полипептидного комплекса, как описано в настоящем документе, в подходящем растворителе. Растворитель может содержать вспомогательное вещество, которое улучшает стабильность или другие фармакологические компоненты порошка или восстановленного раствора, приготовленного из порошка. Вспомогательные вещества, которые могут использоваться, включают, но ими не ограничиваются, воду, декстрозу, сорбит, фруктозу, кукурузный сироп, ксилит, глицерин, глюкозу, сахарозу или другой подходящий агент. Растворитель может содержать буфер, такой как цитрат, фосфат натрия или калия, или другой подобный буфер, известный специалистам в данной области техники, в одном из вариантов осуществления, с приблизительно нейтральным pH. Последующая стерилизующая фильтрация раствора с дальнейшей лиофилизацией в стандартных условиях, известных специалистам в данной области, обеспечивает желаемую композицию. В одном из вариантов осуществления, полученный раствор будет распределен в сосуды для лиофилизации. Каждый сосуд может содержать однократную дозу или более доз полипептидного комплекса, биспецифичного полипептидного комплекса, представленного в настоящем документе, или его композиции. Переполнение сосудов незначительным количеством, превышающим необходимое для одной дозы или более доз (например, около 10%), является приемлемым для обеспечения точного отбора пробы и точного дозирования. Лиофилизированный порошок можно хранить в подходящих условиях, например при температуре от 4°C до значения комнатной температуры.
Восстановление лиофилизированного порошка водой для инъекций обеспечивает лекарственную форму для применения при парентеральном введении. В одном из вариантов осуществления, стерильную и/или апирогенную воду или другой жидкий подходящий носитель добавляют к лиофилизированному порошку для восстановления. Точное количество зависит от выбранной терапии и может быть определено эмпирически.
Способ лечения
Также предложены способы терапии, включающие: введение терапевтически эффективного количества полипептидного комплекса или биспецифичного полипептидного комплекса, представленного в настоящем документе, субъекту, нуждающемуся в таком лечении, для лечения или профилактики состояния или расстройства. В некоторых вариантах осуществления, субъект идентифицирован как имеющий расстройство или состояние, которое может отвечать на полипептидный комплекс или биспецифичный полипептидный комплекс, представленный в настоящем документе.
Используемый в настоящем документе термин “лечить” или “лечение” состояния включает предотвращение или облегчение состояния, замедление возникновения или скорости развития состояния, уменьшение риска развития состояния, предотвращение или задержку развития симптомов, ассоциированных с состоянием, ослабление или устранение симптомов, связанных с состоянием, инициирование полного или частичного регресса состояния, исцеление состояния или сочетание некоторых из указанных эффектов.
Терапевтически эффективное количество биспецифичного полипептидного комплекса, представленного в настоящем документе, будет зависеть от различных факторов, известных в данной области, таких как, например, масса тела, возраст, анамнез, принимаемые в настоящее время лекарственные средства, состояние здоровья субъекта и возможность перекрестной реакции, аллергия, чувствительность и побочные эффекты, а также способ введения и степень развития заболевания. Рядовой специалист в данной области (например, врач или ветеринар) может пропорционально уменьшать или увеличивать дозу в зависимости от указанных и других факторов или требований.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс, представленный в настоящем документе, можно вводить в терапевтически эффективной дозе от около 0,01 до около 100 мг/кг (например, около 0,01 мг/кг, около 0,5 мг/кг, около 1 мг/кг, около 2 мг/кг, около 5 мг/кг, около 10 мг/кг, около 15 мг/кг, около 20 мг/кг, около 25 мг/кг, около 30 мг/кг, около 35 мг/кг, около 40 мг/кг, около 45 мг/кг, около 50 мг/кг, около 55 мг/кг, около 60 мг/кг, около 65 мг/кг, около 70 мг/кг, около 75 мг/кг, около 80 мг/кг, около 85 мг/кг, около 90 мг/кг, около 95 мг/кг или около 100 мг/кг). В некоторых из этих вариантов осуществления, полипептидный комплекс или биспецифичный полипептидный комплекс, представленный в настоящем документе, вводят в дозе около 50 мг/кг или меньше, и в некоторых из этих вариантов осуществления доза составляет 10 мг/кг или меньше, 5 мг/кг или меньше, 1 мг/кг или меньше, 0,5 мг/кг или меньше или 0,1 мг/кг или меньше. В некоторых вариантах осуществления, доза введения может изменяться в течение курса лечения. Например, в некоторых вариантах осуществления, начальная доза введения может быть выше, чем последующая доза введения. В некоторых вариантах осуществления, доза введения может изменяться в течение курса лечения в зависимости от реакции субъекта.
Режимы дозирования могут быть скорректированы с обеспечением оптимального желаемого ответа (например, терапевтического ответа). Например, может быть введена одна доза или более разделенных доз в течение времени.
Биспецифичный полипептидный комплекс, представленный в настоящем документе, может вводиться любым способом, известным в данной области, таким как, например, парентеральный (например, подкожный, внутрибрюшинный, внутривенный, включая внутривенную инфузию, внутримышечную или внутрикожную инъекцию) или непарентеральный (например, пероральный, интраназальный, внутриглазный, сублингвальный, ректальный или местный).
В некоторых вариантах осуществления, состояние или расстройство, которое лечат с помощью биспецифичного полипептидного комплекса, представленного в настоящем документе, представляет собой злокачественную опухоль или раковое состояние, аутоиммунные заболевания, инфекционные и паразитарные заболевания, сердечно-сосудистые заболевания, невропатии, психоневрологические состояния, травмы, воспаления или нарушение свертываемости крови.
Используемый в настоящем документе термин “злокачественная опухоль” или “раковое состояние” относится к любому медицинскому состоянию, опосредованному ростом, пролиферацией или метастазированием опухолевых или злокачественных клеток, и включает как солидную злокачественную опухоль, так и несолидную злокачественную опухоль, такую как лейкоз. Используемый в настоящем документе термин “опухоль” относится к твердой массе опухолевых и/или злокачественных клеток.
Применительно к злокачественной опухоли, термин “лечить” или “лечение” может относиться к ингибированию или замедлению роста, пролиферации или метастазирования опухолевых или злокачественных клеток, предотвращению или задержке развития роста, пролиферации или метастазирования опухолевых или злокачественных клеток, или любому сочетанию этих эффектов. Применительно к опухоли, термин “лечить” или “лечение” включает устранение всей или части опухоли, ингибирование или замедление роста опухоли и метастазирования, предотвращение или задержку развития опухоли, или к любому сочетанию этих эффектов.
Например, применительно к использованию описанного в настоящем документе биспецифичного полипептидного комплекса для лечения злокачественной опухоли, терапевтически эффективным количеством является доза или концентрация полипептидного комплекса, способная устранять всю или часть опухоли, ингибировать или замедлять рост опухоли, ингибировать рост или пролиферацию клеток, опосредующих раковое состояние, ингибировать метастазирования опухолевых клеток, уменьшать интенсивность любого симптома или признака, ассоциированного с опухолью или раковым состоянием, предотвращать или задерживать развития опухоли или ракового состояния, или способная оказывать эти эффекты в любом их сочетании.
В некоторых вариантах осуществления, состояния и расстройства включают опухоли и злокачественные опухоли, например немелкоклеточный рак легких, мелкоклеточный рак легких, почечно-клеточный рак, колоректальный рак, рак яичников, рак молочной железы, рак поджелудочной железы, рак желудка, рак мочевого пузыря, рак пищевода, мезотелиома, меланома, рак головы и шеи, рак щитовидной железы, саркома, рак предстательной железы, глиобластома, рак шейки матки, рак тимуса, лейкоз, лимфомы, миеломы, фунгоидная гранулема, рак клеток Меркеля и другие гематологические злокачественные новообразования, такие как классическая лимфома Ходжкина (CHL), первичная медиастинальная В-крупноклеточная лимфома, Т-клеточная/богатая гистиоцитами В-клеточная лимфома, ВЭБ-положительный и ВЭБ-отрицательный посттрансплантационный лимфопролиферативный синдром, и ВЭБ-ассоциированная диффузная В-клеточная крупноклеточная лимфома (DLBCL), плазмобластическая лимфома, экстранодальная NK/Т-клеточная лимфома, рак носоглотки и HHV8-ассоциировнная первичная эффузионная лимфома, лимфома Ходжкина, новообразования центральной нервной системы (ЦНС), такие как первичная лимфома ЦНС, опухоль спинного мозга, глиома ствола головного мозга.
В некоторых вариантах осуществления, состояния и расстройства включают CD19-связанное состояние, такое как В-клеточная лимфома, необязательно лимфома Ходжкина или неходжкинская лимфома, где неходжкинская лимфома включает: диффузную крупноклеточную В-клеточную лимфому (DLBCL), фолликулярную лимфому, В-клеточную лимфому маргинальной зоны (MZL), лимфому лимфоидной ткани слизистых оболочек (MALT), мелкоклеточную лимфоцитарную лимфому (хронический лимфоцитарный лейкоз, CLL) или лимфому из клеток мантийной зоны (MCL), острый лимфобластный лейкоз или макроглобулинемию Вальденстрема (WM).
Биспецифичный полипептидный комплекс можно вводить отдельно или в сочетании с одним или более дополнительными терапевтическими средствами или агентами.
В некоторых вариантах осуществления, при применении для лечения злокачественной опухоли или опухоли или пролиферативного заболевания, биспецифичный полипептидный комплекс, представленный в настоящем документе, может вводиться в сочетании с химиотерапией, лучевой терапией, хирургическим вмешательством для лечения злокачественной опухоли (например, туморэктомия), одним или более противорвотными средствами или другими средствами для лечения осложнений, возникающих в результате химиотерапии, или любого другого терапевтического средства для применения при лечении злокачественной опухоли или любого связанного медицинского расстройства. Используемый в настоящем документе термин “вводимый в сочетании”, подразумевает одновременное введение в виде части одной и той же фармацевтической композиции, одновременное введение в виде отдельных композиций, или в разные моменты времени в виде отдельных композиций. Полагают, что композицию, вводимую до или после другого агента, вводят “в сочетании” с этим агентом, поскольку фраза используется в настоящем документе, даже если композицию и второй агент вводят разными способами. В тех случаях, когда это возможно, дополнительные терапевтические средства, вводимые в сочетании с полипептидным комплексом или биспецифичным полипептидным комплексом, представленным в настоящем документе, вводят в соответствии с графиком, приведенным в информационном листке для дополнительного терапевтического средства, или в соответствии со справочником лекарственных средств (Physicians' Desk Reference, 70th Ed (2016)) или в соответствии с протоколами, хорошо известными в данной области.
В некоторых вариантах осуществления, терапевтические агенты могут индуцировать или усиливать иммунный ответ против злокачественной опухоли. Например, противоопухолевая вакцина может применяться для вызывания иммунного ответа на определенную опухоль или злокачественную опухоль. Также может быть использована цитокиновая терапия для усиления презентации опухолевого антигена иммунной системе. Примеры цитокиновой терапии включают, без ограничения, интерфероны, такие как интерферон-α, -β и -γ, колониестимулирующие факторы, такие как макрофаг-CSF, гранулоцитарный макрофаг CSF и гранулоцит-CSF, интерлейкины, такие как IL-1, IL-1α IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11 и IL-12, факторы некроза опухоли, такие как TNF-α и TNF-β. Также могут быть применяться агенты, которые инактивируют иммуносупрессивные мишени, например, ингибиторы TGF-бета, ингибиторы IL-10 и ингибиторы Fas-лиганда. Другая группа агентов включает агенты, которые активируют иммунную восприимчивость к опухолевым клеткам или клеткам злокачественной опухоли, например, агенты, которые усиливают активацию Т-клеток (например, агонист костимулирующих молекул Т-клеток, такие как CTLA-4, ICOS и OX-40), и агенты, которые усиливают функцию дендритных клеток и презентацию антигена.
Наборы
Настоящее изобретение также относится к наборам, содержащим биспецифичный полипептидный комплекс, представленный в настоящем документе. В некоторых вариантах осуществления, наборы используются для обнаружения присутствия или уровня, или для захвата или обогащения, одной или более представляющих интерес целей в биологическом образце. Биологический образец может содержать клетку или ткань.
В некоторых вариантах осуществления, набор содержит представленный в настоящем документе биспецифичный полипептидный комплекс, который конъюгирован с детектируемой меткой. В некоторых других вариантах осуществления, набор содержит немеченый биспецифичный полипептидный комплекс, представленный в настоящем документе, и также содержит вторичное меченое антитело, которое способно связываться с немеченым биспецифичным полипептидным комплексом, представленным в настоящем документе. Набор может дополнительно содержать инструкцию по применению и упаковку, которая разделяет каждый из компонентов в наборе.
В некоторых вариантах осуществления, биспецифичный полипептидный комплекс, представленный в настоящем документе, связан с подложкой или устройством. Используемой подложкой или устройством могут быть, например, магнитные микроносители, панель микротитратора или тест-полоска. Они могут быть использованы для анализа связывания (такого как ELISA), иммунографического анализа, захвата или обогащения молекулы-мишени в биологическом образце.
Следующие примеры приведены для лучшей иллюстрации заявленного изобретения и не должны интерпретироваться как ограничивающие объем изобретения. Все конкретные композиции, вещества и способы, описанные ниже, полностью или частично, включены в объем настоящего изобретения. Эти конкретные композиции, вещества и способы предназначены не с целью ограничения изобретения, а только лишь для иллюстрации конкретных вариантов осуществления, включенных в объем изобретения. Специалист в данной области может разработать эквивалентные композиции, вещества и способы без применения изобретательских способностей и без выхода за рамки объема настоящего изобретения. Понятно, что в описанных в настоящем документе процедурах могут быть осуществлены различные изменения, при этом они все еще остаются в рамках настоящего изобретения. Авторы изобретения предполагают, что такие варианты включены в объем настоящего изобретения.
ПРИМЕРЫ
ПРИМЕР 1: Дизайн и конструирование химерных белков антител и TCR
Последовательности TCR
TCR представляют собой гетеродимерные белки, состоящие из двух цепей. Около 95% Т-клеток человека имеют TCR, состоящие из альфа- и бета-цепей. Принимая во внимание, что для TRBC1 бета-цепи доступно больше кристаллических структур, последовательности TRBC1 были выбраны в качестве основного остова для конструирования полипептидного комплекса, описанного в настоящем документе (“WuXiBody”). Типичная аминокислотная последовательность TRBC1 может быть найдена в базе данных белковых структур (PDB), структура 4L4T.
Межцепочечная дисульфидная связь TCR
Для формирования описываемого дизайна WuXiBody использовали кристаллические структуры TCR. В отличие от нативного TCR, заякоренного в мембране T-клеточной поверхности, растворимые молекулы TCR менее стабильны, хотя их трехмерная структура в значительной степени аналогична структуре антитела Fab. Фактически, нестабильность TCR в растворимом состоянии была значительным препятствием, мешающим выяснить его кристаллическую структуру (Wang 2014, выше). Авторы настоящего изобретения приняли стратегию введения пары мутаций Cys в константную область TCR и обнаружили, что она может значительно улучшить сборку цепи и повысить экспрессию.
Для создания стабильного и функционального WuXiBody были тщательно подобраны связи, связывающие вариабельные домены антитела и константные домены TCR, их относительные ориентации слияния, а также Fc-связывающие связи. Поскольку структура TCR очень похожа на структуру антитела Fab, авторы настоящего изобретения наложили модель гомологии Fv антитела на вариабельную область TCR (PDB 4L4T, фиг.2). Наложенная структура указывает на то, что антитело Fv является структурно совместимым с константным доменом TCR. На основе этого структурного выравнивания и соответствующих последовательностей были разработаны все соответствующие технические параметры. Подходящие области конъюгирования описаны в примере 2.
ПРИМЕР 2: Биспецифичное анти-CD13×CD19 WuXiBody
Генерация CD19-экспрессирующей линии клеток яванского макака
Ген полноразмерного CD19 человека или яванского макака клонировали в вектор pcDNA3.3. Затем каждый вектор экспрессии трансфицировали в клетки CHO-K1, соответственно, с использованием Lipofectamine 2000. Клетки культивировали в F12-K с 10% ФБС. После трансфекции через 24-48 часов добавляли бластидин. После двух-трех пассирований отбора, клетки обогащали PE-конъюгированным анти-CD19-антителом и анти-PE-микрогранулами (Miltenyi-013-048-801). Стабильные моноклеточные клоны выделяли путем предельного разведения и скринировали с помощью FACS с использованием анти-CD19 антитела.
Мишень-экспрессирующие опухолевые клеточные линии
Клетки линии Raji и Jurkat получали из Американской коллекции типовых культур (АТСС). Клетки Ramos получали из Европейской коллекции клеточных культур (ECACC). Все опухолевые клетки культивировали в RPMI1640/10% ФБС.
Генерация WuXiBody W3438-T3U4.E17-1.uIgG4.SP и W3438-T3U4.F16-1.uIgG4.SP
Гены VL, VH, Ck, CH1 амплифицировали посредством ПЦР из существующих внутрилабораторных ДНК-матриц. Гены САльфа и СБета были синтезированы компанией Genewiz Inc. Гены легкой цепи анти-CD19-нативного или анти-CD3-химерного антитела встраивали в линеаризованный вектор, содержащий промотор CMV и сигнальный пептид каппа. Фрагменты ДНК VH-СБета анти-CD3 встраивали в линеаризованный вектор, содержащий константную область IgG4S228P CH2-CH3 человека с мутацией “выступ”. Фрагменты ДНК анти-CD19 VH-CH1 встраивали в линеаризованный вектор, содержащий константную область IgG4S228P CH2-CH3 человека с мутацией “углубление”. Вектор содержит промотор CMV и сигнальный пептид тяжелой цепи антитела человека.
Экспрессия и очистка W3438-T3U4.E17-1.uIgG4.SP и W3438-T3U4.F16-1.uIgG4.SP
Плазмиды экспрессии тяжелой цепи и легкой цепи совместно трансфицировали в клетки Expi293 с использованием набора системы экспрессии Expi293 (ThermoFisher-A14635) в соответствии с инструкциями производителя. Через пять дней после трансфекции, супернатанты собирали и белок очищали с использованием колонки Protein A (GE Healthcare-17543802) и дополнительной эксклюзионной колонки (GE Healthcare-17104301). Концентрацию антител измеряли с помощью прибора Nano Drop. Чистоту белков оценивали с помощью ДНС-ПААГ и ВЭЖХ-ЭХ.
Целевое связывание в соответствии с FACS
Связывание би-специфических антител с CD3- и CD19-экспрессирующими клетками оценивали с использованием клеток линии Jurkat и Ramos соответственно. Нерелевантное антитело использовали в качестве изотипического контроля. Клетки распределяли в 96-луночные планшеты (Corning-3799) с плотностью 105 клеток/лунку и промывали ФСБ/1% БСА. Антитела серийно разводили и инкубировали с клетками при 4°С в течение 1 часа. Для детекции использовали PE-конъюгированное козье антитело против IgG Fc человека (Jackson-109-115-098). После промывания и ресуспендирования, клетки анализировали проточной цитометрией (Canto II, BD Biosciences). Данные анализировали с использованием программного обеспечения FlowJo. Четырех-параметрический нелинейный регрессионный анализ использовали для вычисления значений EC50 с использованием программного обеспечения Prism GraphPad.
Связывания CD3 яванского макака
Связывание биспецифичного антитела CD3×CD19 с CD3 яванского макака исследовали посредством анализа связывания белка ELISA. 96-луночные планшеты для анализа ELISA с высоким связыванием белка (Nunc MaxiSorp, ThermoFisher, Thermo-442404) покрывали в течение ночи при 4°С 100 мкл CD3-эпсилон белка яванского макака в концентрации 1 мкг/мл (Acro, # CDE-C5226) в карбонатно-бикарбонатном буфере (20 мМ Na2CO3, 180 мМ NaHCO3, PH9,2). Все лунки промывали один раз 300 мкл на лунку ФСБ/0,5% Твин-20 (об./об.). Затем в течение одного часа при комнатной температуре лунки блокировали 200 мкл на лунку ФСБ/2% БСА (BOVOGEN, #BSAS) и трижды промывали 300 мкл на лунку ФСБ/0,5% Твин-20 (об./об.). Для первичного связывания антител в соответствующие лунки добавляли биспецифичное антитело CD3×CD19, серийно разбавленное в ФСБ/2% БСА, и инкубировали при комнатной температуре в течение двух часов. Планшеты промывали три раза, как ранее, перед добавлением 100 мкл 100 нг/мл вторичного антитела козы к Fc-HRP IgG человека (Bethyl, # A80-304P). Планшеты инкубировали при комнатной температуре в течение одного часа, а затем шесть раз промывали, как описано выше. Для определения связывания, 100 мкл раствора субстрата тетраметилбензидина (ТМВ) (Sigma-860336) добавляли во все лунки в течение 10 минут при комнатной температуре в темноте до остановки реакции с помощью 100 мкл 2М HCl. Степень связывания биспецифичного антитела с CD3 яванского макака определяли путем измерения оптической плотности OD450 с использованием микропланшета-ридера SpectraMax® M5e. Везде, где это уместно, значения связывания EC50 получали с помощью четырехпараметрического нелинейного регрессионного анализа с использованием программного обеспечения GraphPad Prism5.
Связывание CD19 яванского макака
Связывание биспецифичного антитела CD3×CD19 с белком-мишенью CD19 яванского макака, экспрессируемым на клетках CHOK1, определяли методом проточной цитометрии. Вкратце, сверхэкспрессированную стабильную клеточную линию CD19 яванского макака (WBP701.CHOK1.cPro1.C9, WuXi Biologics) собирали с трипсином и разбавляли до концентрации 1×106 клеток/мл в 1% БСА/1×ФСБ. В каждую лунку 96-луночного U-образного планшета (Corning, # 3799) добавляли 1×105 клеток/лунку (100 мкл) и центрифугировали при 1500 об/мин (Eppendorf, # 5810R) в течение 5 минут перед удалением супернатанта. Антитела, серийно разведенные в 1% БСА/1×ФСБ, добавляли при 100 мкл/лунку к осажденным клеткам и инкубировали при 4°С в течение 1 часа. Несвязанное антитело hIgG4 использовали в качестве изотипического контроля. Клетки дважды отмывали 180 мкл/лунку 1% БСА/1×ФСБ центрифугированием при 1500 об/мин в течение 5 минут при 4°С. Гранулированные клетки ресуспендировали в 100 мкл/лунку флуоресцентно-меченного антитела к Fc IgG человека (Jackson, № 109-115-098) 1:150, разведенного в 1% БСА/1×ФСБ в течение 30 минут при 4°С в темноте. Затем клетки промывали два раза способом, описанным выше. После заключительной промывки, клетки ресуспендировали в 80 мкл 1% БСА/1×ФСБ и значения флуоресценции измеряли с помощью цитометра FACS Canto II (BD Biosciences). Количество анти-CD19 и CD3 биспецифичного антитела, связанного с клеточной поверхностью, оценивали путем измерения средней флуоресценции (MFI). Исходные данные FACS анализировали с помощью программного обеспечения FlowJo, лунки, не содержащие антитело или только вторичное антитело, использовали для установления фоновой флуоресценции. Значения EC50 для связывания получали с помощью четырехпараметрического нелинейного регрессионного анализа с использованием программного обеспечения GraphPad Prism 5.
Аффинность в соответствии с FACS
Аффинность связывания с CD3 и CD19 определяли проточной цитометрией с использованием клеток линии Jurkat и Ramos, соответственно. Клетки переносили в 96-луночные планшеты с U-образным дном (BD) при плотности 5×104 клеток/лунку. Антитела, подлежащие исследованию, серийно 1:2-кратно разводили в 1% 1×ФСБ/1% БСА и инкубировали с клетками при температуре 4°С в течение 1 часа. Затем планшеты центрифугировали при 1500 об/мин в течение 4 минут и супернатант отбраковывали. Второе конъюгированное с Alexa647 антитело козы к Fc IgG человека, (Jackson, Cat # 109-605-098) или конъюгированное с FITC антитело к His козы (Bethyl, Cat # A190-113F) добавляли к ресуспендированным клеткам и инкубировали при 4°С в темноте в течение 30 мин. Клетки промывали один раз и ресуспендировали в 100 мкл 1×ФСБ/1% БСА. Интенсивность флуоресценции измеряли с помощью проточной цитометрии (BD Canto II) и анализировали с помощью FlowJo. Интенсивность флуоресценции преобразовывали в связанные молекулы/клетку на основе количественных частиц (наборы QuantumTM MESF, Bangs Laboratories). KD вычисляли с помощью программы Graphpad Prism5.
Двойное связывание на клетках-мишенях
Способность биспецифичных антител связываться с CD3 T-клетками и CD19 B-клетками исследовали методом FACS. Клетки линии Jurkat и клетки Raji предварительно метили отдельно 20 нМ CellTrace Far Red (Invitrogen-C34564) и 50 нМ Calcein-AM (Invitrogen-C3099) при 37°C в течение 30 мин при плотности 1*106 клеток/мл. Предварительно меченную клеточную массу дважды промывали ФСБ/1% БСА, затем смешивали 1:1 до конечной плотности 1*106 клеток/мл. Клеточную смесь центрифугировали и ресуспендировали с 10 нМ антителом с последующей инкубацией в течение 1 часа. Клеточную смесь анализировали проточной цитометрией непосредственно после инкубации. Выраженное в процентах мостиковое конъюгировние определяли как процент событий, которые одновременно мечены Far-Red и Calcein.
Исследование цитотоксичности
Мононуклеарные клетки периферической крови (РВМС) были свежеизолированы путем центрифугирования в градиенте плотности Ficoll-Paque PLUS (GE Healthcare-17-1440-03) из гепаринизированной венозной крови. Затем полученные PBMC пропускали через колонки EasySep (Stemcell-19053) для обогащения CD8+ T-клеток, которые использовали в качестве эффекторных клеток. Эффективность антител для опосредования лизиса опухолевых клеток CD8+ Т-клетками определяли методом проточной цитометрии. В анализе циотоксичности, CD19 В-клетки Raji в качестве клеток-мишеней предварительно метили 20 нМ CellTrace Far Red (Invitrogen-C34564) при 37°C в течение 30 минут с последующим двойным промыванием клеточной массы средой RPMI 1640 без фенола (Invitrogen-11835030) с добавлением 1% ФБС. Far Red-окрашенные клетки Raji (20000 клеток на лунку) инкубировали в 96-луночном круглодонном планшете (Corning-3799) с изолированными CD8+ Т-клетками (соотношение эффектор и клеток-мишеней 5:1) и серийно разведенными антителами при 37°С в течение 4 ч. После инкубации 3 мкМ иодида пропидия (PI, Invitrogen-P3566) тщательно перемешивали для идентификации мертвых клеток. Через 15 мин, клетки анализировали проточной цитометрией с использованием цитометра FACSCanto II. Ab-опосредованная цитотоксичность может быть определена как процент PI-позитивных клеток-мишеней в Far Red-позитивных клетках-мишенях. Значение EC50 цитотоксичности определяли с использованием программы Prism.
Исследование активации Т-клеток - секретируемые цитокины TNFα и IFNɣ
Активацию Т-клеток определяли по количеству TNFα и IFNɣ, секретируемых в супернатант. Процедура выделения CD4 и CD8-позитивных Т-клеток описана в разделе “T Cell Activation (Intracellular Cytokine TNFα & IFNɣ Staining)”. Смесь В-клеток Raji человека (2 × 104 клеток/лунка), CD4 или CD8 Т-клеток (1 × 105 клеток/лунка) и антител совместно инкубировали при 37°С в течение 24 часов. Супернатант собирали с последующим центрифугированием реакционной смеси при 1500 об/мин в течение 5 минут. Количество TNFα и IFNɣ в супернатанте определяли с помощью набора ELISA для TNF человека (R&D-DY210) и набора ELISA для IFNɣ человека (Capture Ab: Thermo Fisher-M700A, Detection Ab: Thermo Fisher-M701B, Standard substance: PEROTECH-300-02) соответственно.
Процедура сэндвич-ELISA была следующей. В течение ночи при 4°С или комнатной температуре, 96-луночные планшеты для ELISA с высоким связыванием белка (ThermoFisher-442404) покрывали 50 мкл/лунку иммобилизованного антитела в карбонат-бикарбонатном буфере (20 мМ Na2CO3, 180 мМ NaHCO3, pH 9,2) в соответствии с инструкцией для набора. Все лунки трижды промывали 300 мкл на лунку ФСБ/0,5% Твин-20 (об./об.) и все последующие стадии промывки в анализе проводили аналогичным образом. Потом лунки блокировали в течение одного часа с помощью ФСБ/2% БСА (BovoGen Biologicals-BSAS) для TNFα и 100% казеина (Pierce-37528) для IFNɣ, затем трижды промывали с последующим связыванием собранного супернатанта или стандартного вещества (50 мкл/лунку) в течение 1 часа при комнатной температуре, а также три раза после этого. Для детекции связывания антител, соответствующие антитела, разведенные в ФСБ/2% БСА для TNFα и 50% казеина для IFNɣ, добавляли в соответствующие лунки и инкубировали при комнатной температуре в течение двух часов. Планшеты трижды промывали перед добавлением 50 мкл второго антитела SA-HRP. Планшеты инкубировали при комнатной температуре в течение одного часа, а затем шесть раз промывали способом, описанным выше. Для обнаружения связывания, 50 мкл раствора субстрата тетраметилбензидина (ТМВ) (Sigma-860336) добавляли во все лунки в течение 10 минут перед тем, как остановить реакцию с помощью 50 мкл 2М HCl. Количество TNFα и IFNɣ определяли путем измерения оптической плотности OD450 с использованием считывающего устройства для микропланшетов SpectraMax® M5e.
Исследование активации Т-клеток - экспрессия поверхностных маркеров CD25 и CD69
Активацию Т-клеток определяли по окрашиванию сигналов поверхностных рецепторов CD25 и CD69. Процедура выделения CD4 и CD8-положительных Т-клеток была описана в разделе “T Cell Activation (Intracellular Cytokine TNFα & IFNɣ Staining)”. Смесь В-клеток Raji человека (2 × 104 клеток/лунка), CD4 или CD8 Т-клеток (1 × 105 клеток/лунка) и антител совместно инкубировали при 37°С в течение 24 часов. После однократного промывания 1% БСА, клеточный осадок ресуспендировали с помощью окрашивающего буфера, FITC-меченное мышиное актитело к CD4 человека (BD-550628) или PerCpCy5.5-меченное мышиное актитело к CD8 человека (BD-560662), PE-меченное мышиное актитело к CD69 человека (BD-560968) и APC-меченное мышиное актитело к CD25 человека (BD-555434) с последующей 30-минутной инкубацией при 4°С. После двухкратного промывания клеток, процентное содержание PE- и APC-позитивных клеток в FITC- или PerCpCy5,5-позитивных клетках определяли проточной цитометрией.
Термостабильность (DSF)
Температуру плавления (Tm) антител исследовали с использованием системы ПЦР в реальном времени QuantStudio® Flex (Applied Biosystems). 19 мкл раствора антител смешивали с 1 мкл раствора 62.5 Х SYPRO Orange (Invitrogen) и переносили в 96-луночный планшет (Biosystems). Планшет нагревали от 26 до 95°С со скоростью 0,9°С/мин, и полученные собирали данные флуоресценции. Определяли отрицательные производные изменений флуоресценции относительно различных температур, и максимальное значение определяли как температуру плавления Tm. Если белок имел множественные конформационные переходы с разворачиванием, сообщалось о первых двух Tm, названных Tm1 и Tm2. Сбор данных и определение Tm проводились автоматически с помощью программного обеспечения.
Стабильность в сыворотке крови
Свежевзятую кровь человека у отобранных доноров помещали в полистирольные пробирки без антикоагулянта. После выдерживания в течение 30 минут при комнатной температуре, кровь человека центрифугировали при 4000 об/мин в течение 10 минут для сбора сывороточного слоя. Стадию центрифугирования повторяли до осветления сыворотки. В процессе обнаружения антитела смешивали с собранной сывороткой в соотношении 1:9, и аликвоты отбирали при 37°С в течение указанного времени: 0 дней, 1 день, 4 дня, 7 дней и 14 дней. Образцы быстро замораживали в разные моменты времени в жидком азоте и хранили при -80°С перед использованием. Образцы анализировали с помощью FACS для оценки способности к связыванию CD3 T-клеток Jurkat и CD19 B-клеток Ramos по сравнению с соответствующими антителами без сывороточной обработки.
Аффинность связывания Fcγ-рецептора в соответствии с ППР (SPR)
Аффинность связывания антитела с FcγR определяли с использованием Biacore T200 (или Biacore 8K). Каждый рецептор фиксировали на сенсорном чипе CM5 с иммобилизованном антителом к гистидину (GE). Антитела в различных концентрациях инъецировали на сенсорный чип со скоростью подачи 30 мкл/мин в течение фазы ассоциации 60 сек с последующей диссоциацией в течение 60 сек. Затем чип регенерировали 10 мМ глицином (рН 1,5) после каждого цикла связывания.
Сенсограммы пустой поверхности и буферного канала вычитали из аналитических сенсограмм. Экспериментальные данные аппроксимировали по 1:1 модели с использованием анализа Ленгмюра (для FcγRI) или равновесной модели (для других рецепторов). Для вычисления молярной концентрации антител использовали молекулярную массу, составляющую 150 кДа.
Связывание C1q в соответствии с ИФА (ELISA)
Планшеты для ELISA (Nunc) покрывали образцами антител при концентрации 3 мкг/мл в течение ночи при 4°C. После блокирования и промывания, C1q градиентно разбавляли, начиная с 600 мкг/мл, и инкубировали при комнатной температуре в течение 2 часов. Затем планшеты промывали и далее инкубировали с овечьим антителом к C1q Ab-HRP человека в течение 1 часа. После промывки, добавляли субстрат TMB, и взаимодействие останавливали 2М HCl. Оптическую плотность при 450 нм считывали с использованием микропланшета-ридера (Molecular Device).
Аффинность связывания FcRn в соответствии с ППР (SPR)
Аффинность связывания антитела с FcRn определяли посредством Biacore T200 (или Biacore 8K). Каждое антитело иммобилизовали на сенсорном чипе CM5 (GE). FcRn при различных концентрациях в подвижном буфере (50 мМ Na2HPO4/NaH2PO4, 150 мМ NaCl, 0,05% Tween 20, pH 6,0) инъецировали на сенсорный чип при скорости подачи 30 мкл/мин в течение фазы ассоциации 60 сек с последующей диссоциацией в течение 60 сек. Затем чип регенерировали 1×ФСБ (рН 7,4) после каждого цикла связывания.
Сенсограммы пустой поверхности и буферного канала вычитали из аналитических сенсограмм. Экспериментальные данные аппроксимировали по равновесной модели. Для вычисления молярной концентрации FcRn использовали молекулярную массу 45 кДа.
Исследование эффективности на Raji/PBMC мышиной модели
Опухолевые клетки Raji (ATCC® CCL-86 ™) сохраняли in vitro в виде монослойной культуры в среде RPMI-1640, дополненной 10% эмбриональной бычьей сывороткой, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина при 37ºC в атмосфера 5% CO2 в воздухе. Опухолевые клетки пассировали дважды в неделю в установленном порядке. Клетки, растущие в фазе экспоненциального роста, собирали и подсчитывали для инокуляции опухоли.
Мононуклеарные клетки периферической крови (РВМС) человека выделяли из гепаринизированной цельной крови с использованием Ficoll-Paque Plus в соответствии с инструкциями производителя.
Каждую мышь подкожно в правый бок инокулировали смесью опухолевых клеток Raji с матригелем и свежими РВМС в 0,2 мл ФСБ в день 0. Инъекцию антител проводили начиная со дня 3 (внутривенно раз в две недели (BIW) × 4 раза).
мг/мл
Измерения опухоли и ожидаемые результаты
Основной ожидаемый результат заключался в том, чтобы увидеть, можно ли задержать рост опухоли или вылечить мышей. Размер опухоли измеряли два раза в неделю в двух измерениях, используя штангенциркуль, и объем выражали в мм3, используя формулу: V=0,5 a × b2, где a и b представляют собой больший и меньший диаметры опухоли соответственно. Значение T/C (в процентах) является показателем противоопухолевой эффективности.
Ингибирование роста опухоли (TGI) вычисляли для каждой группы по формуле: TGI (%) = [1-(Ti-T0)/(Vi-V0)]×100; Ti - средний объем опухоли в группе лечения в данный день, T0 - средний объем опухоли группы лечения в день начала лечения, Vi - средний объем опухоли контрольной группы с носителем в тот же день, что и Ti, V0 - средний объем опухоли в группе носителя в день начала лечения.
Анализ фармакокинетики, токсичности и иммуногенности препарата на яванском макаке
Одному самцу и одной самке яванской макаки однократно вводили WBP3438 в дозе 1 мг/кг путем внутривенного болюсного введения. Препарат изготавливали в 20 мМ NaAc-HAc, 7,0% (масса/масса) сахарозе, 0,02% (масса/масса) PS80, рН 5,0. ФК образцы крови собирали до введения препарата (день -1), через 0,25 ч, 0,5 ч, 1 ч, 4 ч, 8 ч, 24 ч, в день 3, в день 7, в день 14, в день 21 и в день 28. Образцы противолекарственных антител (ADA) собирали в день 3, день 14 (через 312 ч) и день 28 (через 480 ч).
Концентрации WBP3438 и ADA в образцах сыворотке крови определяли методом ИФА (ELISA). Концентрацию WBP3438 в сыворотке у обезьян подвергали некомпартментному фармакокинетическому анализу с использованием программного обеспечения Phoenix WinNonlin (версия 6.3, Pharsight, Mountain View, CA). При получении ФК-параметров применяли линейно-логарифмическое правило трапеций.
Осуществляли наблюдение у клетки за общим состоянием здоровья и внешнего вида, в частности за раздражением кожи. Анализ образцов цельной крови для гематологии (CBC) и анализ сыворотки для химического детектирования осуществляли посредством гематологического анализатора (ADVIA2120) и анализатора биохимии (HITACHI 7180) соответственно.
Результаты
Генерация линии клеток, экспрессирующих CD19, яванского макака
Экспрессия линии клеток WBP701.CHO-K1.cpro1.FL.C9, экспрессирующей CD19, яванского макака была обнаружена с использованием антитела к CD19 методом проточной цитометрии. WBP701.CHO-K1.cpro1.FL.C9 показала высокую экспрессию CD19 обезьяны (фиг.5).
Генерация и оптимизация WuXiBody
На фиг.1 представлены схематические изображения исследуемых антител и их форматы. Были разработаны как антитело против CD3 T3, так и антитело против CD19 U4. Константная область (CL и CH1) T3 была заменена константной областью TCR для конструирования уникальной области легкой-тяжелой цепи, которая ортогональна регулярному антителу. TCR-модифицированное T3 и нативное U4 в сочетании с мутациями “выступ-в-углубление” в домене Fc были использованы для конструирования биспецифичных антител формата E17 и F16.
Последовательности вариабельных областей тяжелой цепи и легкой цепи анти-CD3 и анти-CD19-связывающих фрагментов из W3438-T3U4.E17-1.uIgG4.SP и W3438-T3U4.F16-1.uIgG4.SP представлены ниже:
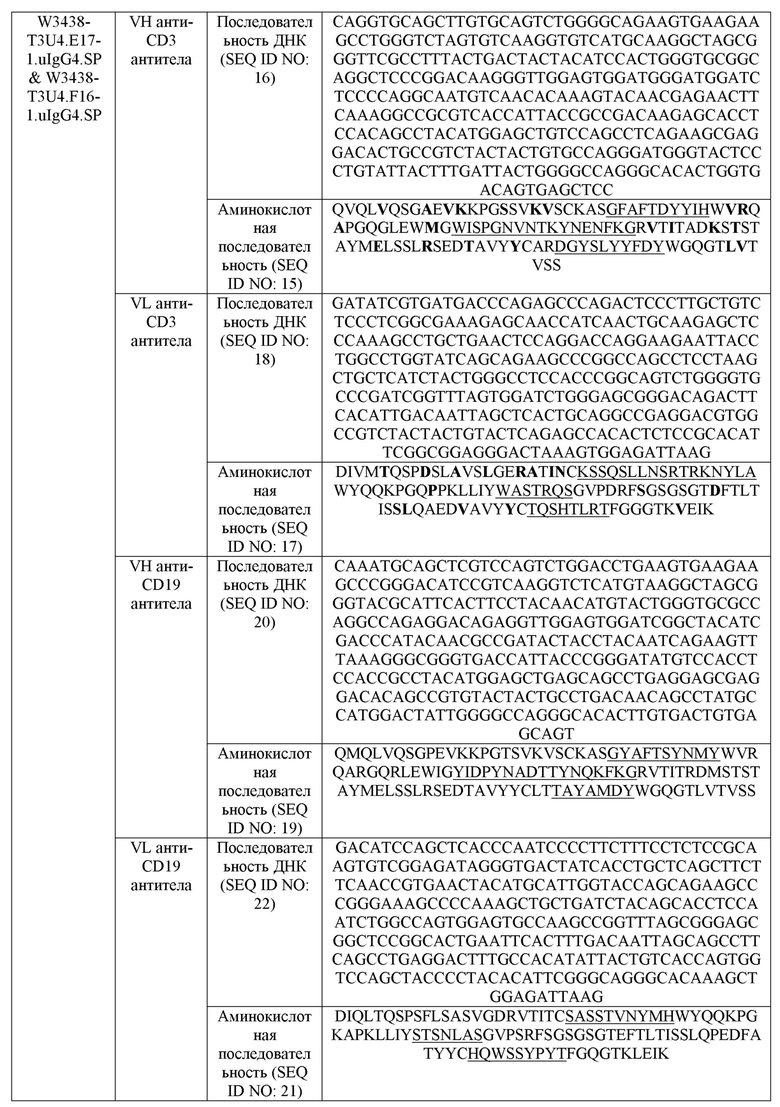
Полноразмерные последовательности W3438-T3U4.E17-1.uIgG4.SP и W3438-T3U4.F16-1.uIgG4.SP представлены ниже:


Продукция W3438-T3U4.F16-1.uIgG4.SP
В результате временной экспрессии, титр экспрессии антитела W3438-T3U4.F16-1.uIgG4.SP выше 90 мг/л. После двухступенчатой очистки, чистота W3438-T3U4.F16-1.uIgG4.SP достигает 97,5% (эксклюзионная ВЭЖХ, фиг.7). W3438-T3U4.F16-1.uIgG4.SP мигрирует со средней молекулярной массой 75 кДа, 55 кДа и 25 кДа в ДНС-ПААГ в восстанавливающих условиях, соответствуя двум тяжелым цепям и двум легким цепям. Две легкие цепи могут перекрываться из-за сходных молекулярных масс. Антитело мигрирует со средней молекулярной массой 200 кДа в невосстанавливающих условиях, что указывает на интактную биспецифичную молекулу (фиг.6).
Продукция W3438-T3U4.E17-1.uIgG4.SP
В результате временной экспрессии, титр экспрессии антитела W3438-T3U4.E17-1.uIgG4.SP выше, чем 100 мг/л. После двухступенчатой очистки, чистота W3438-T3U4.E17-1.uIgG4.SP достигает 95% (эксклюзионная ВЭЖХ, фиг.9). W3438-T3U4.E17-1.uIgG4.SP мигрирует со средней молекулярной массой 54 кДа, 56 кДа и 25 кДа в ДНС-ПААГ в восстанавливающих условиях, соответствуя двум тяжелым цепям и двум легким цепям. Две легкие цепи могут перекрываться из-за аналогичных молекулярных масс. Антитело мигрирует со средней молекулярной массой 150 кДа в невосстанавливающих условиях, что указывает на интактную биспецифичную молекулу (фиг.8).
Связывание с мишенью
Связывание W3438-T3U4.E17-1.uIgG4.SP с CD19 и CD3 исследовали на клетках Ramos и Jurkat посредством проточной цитометрии (фигуры 10A-10B). Антитело W3438-T3U4.E17-1.uIgG4.SP продемонстрировало сильно выраженную активность связывания с клетками Ramos и Jurkat со значениями EC50 15,6 нМ и 47 нМ соответственно.
Связывание W3438-T3U4.F16-1.uIgG4.SP с CD19 и CD3 исследовали на клетках Ramos и Jurkat посредством проточной цитометрии (фигуры 11A-11B). Антитело W3438-T3U4.F16-1.uIgG4.SP показало сильно выраженную активность связывания с клетками Ramos и Jurkat, со значениями EC50 1,8 нМ и 19,3 нМ соответственно.
Межвидовое связывание
Связывание W3438-T3U4.E17-1.uIgG4.SP с CD19 яванского макака исследовали на клетке WBP701.CHO-K1.cpro1.FL.C9 (клетка, экспрессирующая CD19) посредством проточной цитометрии (фиг.12). Значение EC50 для связывания составляло 26 нМ. Связывание W3438-T3U4.E17-1.uIgG4.SP с CD3 яванского макака исследовали с использованием W331-cynoPro1.ECD.His (CD3-эпсилон-белок яванского макака) посредством ИФА (ELISA) (фиг.13). Значение EC50 для связывания составляло 0,04 нМ.
Сродство к клеткам-мишеням
Аффинность связывания W3438-T3U4.E17-1.uIgG4.SP с CD19 и CD3 человека исследовали на клетках Ramos и Jurkat посредством проточной цитометрии. Соотношение связанный IgG/свободный IgG относительно связанного IgG наносили на график, как показано на фиг. 14A-14B. Подходящие значения KD связывания с CD19 и CD3 составляли 23 нМ и 9,0 нМ соответственно.
Двойное связывание на клетках-мишенях
Активность W3438-T3U4.E17-1.uIgG4.SP для связывания CD3 T-клеток и CD19 B-клеток исследовали с использованием предварительно меченых клеток Jurkat и Raji посредством проточной цитометрии (фиг. 15A-15B). Q2 показывает популяцию мостиковых клеток Jurkat и Raji. По сравнению с отрицательным контролем, примерно 18% клеток были связаны через биспецифическое антитело W3438-T3U4.E17-1.uIgG4.SP.
Анализ цитотоксичности
Цитотоксическую активность W3438-T3U4.E17-1.uIgG4.SP оценивали с использованием CD8+ Т-клеток и клеток линии Raji. W3438-T3U4.E17-1.uIgG4.SP вызывал быстрый и эффективный лизис клеток после 4-часовой инкубации (фиг.16А) со значением ЕС50 15 нМ. Максимальный процент гибели клеток составил 90%.
Цитотоксическую активность W3438-T3U4.F16-1.uIgG4.SP оценивали с использованием CD8+ Т-клеток и клеток линии Raji. W3438-T3U4.F16-1.uIgG4.SP вызывал быстрый и эффективный лизис клеток после 4-часовой инкубации (фиг.16B) со значением EC50 3,2 нМ. Максимальный процент гибели клеток составил 90%.
Мишень-специфическая активация Т-клеток
W3438-T3U4.E17-1.uIgG4.SP исследовали в анализах, которые указывают на активацию Т-клеток через маркеры активации CD69 и CD25 в присутствии или отсутствии CD19+ клеток-мишеней. Результаты показали, что W3438-T3U4.E17-1.uIgG4.SP индуцирует экспрессию маркеров активации Т-клеток CD25 и CD69 дозозависимым образом только в присутствии клеток-мишеней CD19+ (фигуры 17A-17D). Когда B-клетка отсутствует, экспрессия CD25 и CD69 не наблюдалась как в CD4+, так и в CD8+ T-клеточной субпопуляции.
W3438-T3U4.E17-1.uIgG4.SP также исследовали в анализах активации T-клеток на высвобождение цитокинов в присутствии или в отсутствие клеток-мишеней CD19+. Результаты показали, что W3438-T3U4.E17-1.uIgG4.SP индуцирует высвобождение IFN-γ и TNF-α дозозависимым образом только в присутствии клеток-мишеней CD19+ (фиг. 18A-18D). Когда B-клетка отсутствует, IFN-γ и TNF-α не обнаруживались как в CD4+, так и в CD8+ T-клеточной субпопуляции.
Термостабильность
Термостабильность W3438-T3U4.E17-1.uIgG4.SP исследовали с использованием ПЦР в реальном времени. Tm1 и Tm2 для W3438-T3U4.E17-1.uIgG4.SP составляют 60,2°С и 72,7°С.
Стабильность в сыворотке
W3438-T3U4.E17-1.uIgG4.SP инкубировали в сыворотке при 37°С в течение 14 дней. Активность связывания антитела, инкубированного в течение 0, 1, 4, 7 и 14 дней, определяли посредством проточной цитометрии. Результаты показали, что активность связывания W3438-T3U4.E17-1.uIgG4.SP как с клетками CD3 так и CD19 не изменялась после инкубации в сыворотке человека в течение 14 дней (фигуры 19А и 19В).
Связывание с рецептором Fcγ
Активность связывания W3438-T3U4.E17-1.uIgG4.SP с FcγRI, FcγRIIa (H167), FcγRIIa (R167), FcγRIIb, FcγRIIIa (F176), FcγRIIIa (V176) и FcγRIIIb исследовали посредством ППР (SPR). Аффинности приведены в таблице 1. W3438-T3U4.E17-1.uIgG4.SP показал типичную аффинность связывания IgG4 человека со всеми Fcγ-рецепторами.
Активность связывания антител с C1Q исследовали методом ИФА (ELISA). W3438-T3U4.E17-1.uIgG4.SP показал отсутствие сигнала связывания в анализе ИФА (ELISA) (фиг.20), причем контрольное антитело IgG1 человека показало нормальный сигнал связывания.
Связывание FcRn
Связывание W3438-T3U4.E17-1.uIgG4.SP с FcRn исследовали посредством ППР (SPR) при pH 6,0. Аффинность составляла 2,58 мкМ, что является типичным сродством IgG4 человека к FcRn.
Характеристика in vivo
Исследование эффективности на модели ксенотрансплантата клеток МКПК(PBMC)/Raji
В этом анализе исследовали противоопухолевую эффективность W3438-T3U4.E17-1.uIgG4.SP в смешанной гуманизированной модели МКПК, несущей клетку Raji, у мышей NOG. Кривая роста опухоли показана на фиг.21.
В день 14, средний размер опухоли в группе обработки изотипическим контролем достигал 342 мм3. Обработка 1,5 мг/кг и 0,5 мг/кг W3438-T3U4.E17-1.uIG4.SP обеспечивал значительную противоопухолевую активность. Средний размер опухоли составлял соответственно 78 мм3 (T/C=23,0%, TGI=93,9%, p=0,016) и 75 мм3 (T/C=22,0%, TGI=95,3%, p=0,014), причем опухоль у одного животного в группе с высокой дозировкой была полностью устранена. W3438-T3U4.E17-1.uIgG4.SP в очень низкой дозе (0,06 мг/кг) не показал противоопухолевой активности.
Фармакокинетика WuXiBody у яванского макака
Концентрацию W3438-T3U4.E17-1.uIgG4.SP в сыворотке яванского макака исследовали методом ИФА (ELISA) (фиг.22). Вычисленные фармакокинетические (ФК) параметры приведены в таблице 2. Период полураспада W3438-T3U4.E17-1.uIgG4.SP для однократной внутривенной инъекции в дозе 1 мг/кг составил 152 часа. W3438-T3U4.E17-1.uIgG4.SP показал намного более длительный период полураспада у обезьян, чем блинатумомаб с очень коротким периодом полураспада (1,5-2,6 часа) у шимпанзе (экспертный отчет Европейского агентства лекарственных средств EMA/CHMP/469312/2015).
Токсичность
Все обезьяны хорошо переносили препарат в течение всего курса исследования. Никаких побочных эффектов не наблюдали в течение жизненной стадии исследования. Каких-либо очевидных изменений в потреблении корма и массе не наблюдали. Параметры гематологии и клинической химии, включая AST (аспарагинаминотрансфераза), ALT (аланинаминотрансфераза), WBC (лейкоциты), HGB (гемоглобин) и HCT (гематокрит), в целом находились в пределах референтного диапазона.
Иммуногенность
Результаты противолекарственного антитела (ADA) W3438-T3U4.E17-1.uIgG4.SP показаны на фиг.23A-23B. Титры ADA против W3438-T3U4.E17-1.uIgG4.SP в сыворотке обезьяны через 3, 14 и 28 дней после введения не показали значимого отличия от значений до введения препарата. Соответственно, однократная внутривенная инъекция W3438-T3U4.E17-1.uIgG4.SP в дозе 1 мг/кг оказалась не иммуногенной у обезьян.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> WuXi Biologics (Shanghai) CO., LTD.
WUXI BIOLOGICS (CAYMAN) INC.
<120> НОВЫЕ БИСПЕЦИФИЧНЫЕ ПОЛИПЕПТИДНЫЕ КОМПЛЕКСЫ ПРОТИВ CD3/CD19
<130> IEC180126PCT
<160> 40
<170> PatentIn version 3.5
<210> 1
<211> 116
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 1
Glu Val Ala Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln
1 5 10 15
Lys Ala Thr Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val
20 25 30
Glu Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys
35 40 45
Thr Asp Pro Gln Pro Leu Lys Glu Gln Pro Ala Leu Gln Asp Ser Arg
50 55 60
Tyr Ala Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asn
65 70 75 80
Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu
85 90 95
Asn Asp Glu Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val
100 105 110
Ser Ala Glu Ala
115
<210> 2
<211> 87
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 2
Ala Val Tyr Gln Leu Arg Asp Ser Lys Ser Ser Asp Lys Ser Val Cys
1 5 10 15
Leu Phe Thr Asp Phe Asp Ser Gln Thr Gln Val Ser Gln Ser Lys Asp
20 25 30
Ser Asp Val Tyr Ile Thr Asp Lys Cys Val Leu Asp Met Arg Ser Met
35 40 45
Asp Phe Lys Ser Asn Ser Ala Val Ala Trp Ser Gln Lys Ser Asp Phe
50 55 60
Ala Cys Ala Asn Ala Phe Gln Asn Ser Ile Ile Pro Glu Asp Thr Phe
65 70 75 80
Phe Pro Ser Pro Glu Ser Ser
85
<210> 3
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 3
Gly Phe Ala Phe Thr Asp Tyr Tyr Ile His
1 5 10
<210> 4
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 4
Trp Ile Ser Pro Gly Asn Val Asn Thr Lys Tyr Asn Glu Asn Phe Lys
1 5 10 15
Gly
<210> 5
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 5
Asp Gly Tyr Ser Leu Tyr Tyr Phe Asp Tyr
1 5 10
<210> 6
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 6
Lys Ser Ser Gln Ser Leu Leu Asn Ser Arg Thr Arg Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 7
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 7
Trp Ala Ser Thr Arg Gln Ser
1 5
<210> 8
<211> 8
<212> БЕЛОК
<213> Mus musculus
<400> 8
Thr Gln Ser His Thr Leu Arg Thr
1 5
<210> 9
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 9
Gly Tyr Ala Phe Thr Ser Tyr Asn Met Tyr
1 5 10
<210> 10
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 10
Tyr Ile Asp Pro Tyr Asn Ala Asp Thr Thr Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 11
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 11
Thr Ala Tyr Ala Met Asp Tyr
1 5
<210> 12
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 12
Ser Ala Ser Ser Thr Val Asn Tyr Met His
1 5 10
<210> 13
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 13
Ser Thr Ser Asn Leu Ala Ser
1 5
<210> 14
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 14
His Gln Trp Ser Ser Tyr Pro Tyr Thr
1 5
<210> 15
<211> 119
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Ala Phe Thr Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Pro Gly Asn Val Asn Thr Lys Tyr Asn Glu Asn Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Leu Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 16
<211> 357
<212> ДНК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 16
caggtgcagc ttgtgcagtc tggggcagaa gtgaagaagc ctgggtctag tgtcaaggtg 60
tcatgcaagg ctagcgggtt cgcctttact gactactaca tccactgggt gcggcaggct 120
cccggacaag ggttggagtg gatgggatgg atctccccag gcaatgtcaa cacaaagtac 180
aacgagaact tcaaaggccg cgtcaccatt accgccgaca agagcacctc cacagcctac 240
atggagctgt ccagcctcag aagcgaggac actgccgtct actactgtgc cagggatggg 300
tactccctgt attactttga ttactggggc cagggcacac tggtgacagt gagctcc 357
<210> 17
<211> 112
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 17
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Gln Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser His Thr Leu Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 18
<211> 336
<212> ДНК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 18
gatatcgtga tgacccagag cccagactcc cttgctgtct ccctcggcga aagagcaacc 60
atcaactgca agagctccca aagcctgctg aactccagga ccaggaagaa ttacctggcc 120
tggtatcagc agaagcccgg ccagcctcct aagctgctca tctactgggc ctccacccgg 180
cagtctgggg tgcccgatcg gtttagtgga tctgggagcg ggacagactt cacattgaca 240
attagctcac tgcaggccga ggacgtggcc gtctactact gtactcagag ccacactctc 300
cgcacattcg gcggagggac taaagtggag attaag 336
<210> 19
<211> 116
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 19
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Ser Tyr
20 25 30
Asn Met Tyr Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asp Pro Tyr Asn Ala Asp Thr Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Leu Thr Thr Ala Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 20
<211> 348
<212> ДНК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 20
caaatgcagc tcgtccagtc tggacctgaa gtgaagaagc ccgggacatc cgtcaaggtc 60
tcatgtaagg ctagcgggta cgcattcact tcctacaaca tgtactgggt gcgccaggcc 120
agaggacaga ggttggagtg gatcggctac atcgacccat acaacgccga tactacctac 180
aatcagaagt ttaaagggcg ggtgaccatt acccgggata tgtccacctc caccgcctac 240
atggagctga gcagcctgag gagcgaggac acagccgtgt actactgcct gacaacagcc 300
tatgccatgg actattgggg ccagggcaca cttgtgactg tgagcagt 348
<210> 21
<211> 106
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 21
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Thr Val Asn Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys His Gln Trp Ser Ser Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 22
<211> 318
<212> ДНК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 22
gacatccagc tcacccaatc cccttctttc ctctccgcaa gtgtcggaga tagggtgact 60
atcacctgct cagcttcttc aaccgtgaac tacatgcatt ggtaccagca gaagcccggg 120
aaagccccaa agctgctgat ctacagcacc tccaatctgg ccagtggagt gccaagccgg 180
tttagcggga gcggctccgg cactgaattc actttgacaa ttagcagcct tcagcctgag 240
gactttgcca catattactg tcaccagtgg tccagctacc cctacacatt cgggcagggc 300
acaaagctgg agattaag 318
<210> 23
<211> 10
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 23
Leu Glu Asp Leu Lys Asn Val Phe Pro Pro
1 5 10
<210> 24
<211> 9
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 24
Lys Pro Asp Ile Gln Asn Pro Asp Pro
1 5
<210> 25
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 25
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly
1 5 10 15
Pro
<210> 26
<211> 207
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 26
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Gln Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Ser His Thr Leu Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Pro Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Gln Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Gln Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Gln Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
195 200 205
<210> 27
<211> 472
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 27
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Ala Phe Thr Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Pro Gly Asn Val Asn Thr Lys Tyr Asn Glu Asn Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Ser Leu Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Leu Glu Asp Leu Lys Asn Val Phe Pro Pro Glu
115 120 125
Val Ala Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys
130 135 140
Ala Thr Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu
145 150 155 160
Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr
165 170 175
Asp Pro Gln Pro Leu Lys Glu Gln Pro Ala Leu Gln Asp Ser Arg Tyr
180 185 190
Ala Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro
195 200 205
Arg Asn His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn
210 215 220
Asp Glu Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser
225 230 235 240
Ala Glu Ala Trp Gly Arg Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala
245 250 255
Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
260 265 270
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
275 280 285
Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
290 295 300
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
305 310 315 320
Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
325 330 335
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
340 345 350
Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
355 360 365
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu Glu Met Thr
370 375 380
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
385 390 395 400
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
405 410 415
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
420 425 430
Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe
435 440 445
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
450 455 460
Ser Leu Ser Leu Ser Leu Gly Lys
465 470
<210> 28
<211> 213
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 28
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Thr Val Asn Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys His Gln Trp Ser Ser Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 29
<211> 443
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 29
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Ser Tyr
20 25 30
Asn Met Tyr Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asp Pro Tyr Asn Gly Asp Thr Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Leu Thr Thr Ala Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Val Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 30
<211> 667
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 30
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Ser Tyr
20 25 30
Asn Met Tyr Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asp Pro Tyr Asn Gly Asp Thr Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Leu Thr Thr Ala Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
225 230 235 240
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Ser Tyr
245 250 255
Asn Met Tyr Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
260 265 270
Gly Tyr Ile Asp Pro Tyr Asn Gly Asp Thr Thr Tyr Asn Gln Lys Phe
275 280 285
Lys Gly Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
290 295 300
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
305 310 315 320
Leu Thr Thr Ala Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val
325 330 335
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
340 345 350
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
355 360 365
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
370 375 380
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
385 390 395 400
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
405 410 415
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
420 425 430
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
435 440 445
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
450 455 460
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
465 470 475 480
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
485 490 495
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
500 505 510
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
515 520 525
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
530 535 540
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
545 550 555 560
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
565 570 575
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
580 585 590
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
595 600 605
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
610 615 620
Phe Leu Val Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
625 630 635 640
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
645 650 655
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
660 665
<210> 31
<211> 142
<212> БЕЛОК
<213> Homo sapiens
<400> 31
Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
1 5 10 15
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
35 40 45
Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
50 55 60
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
65 70 75 80
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys
85 90 95
Asp Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn
100 105 110
Phe Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val
115 120 125
Ala Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 32
<211> 95
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 32
Pro Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
1 5 10 15
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
35 40 45
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
50 55 60
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
65 70 75 80
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
85 90 95
<210> 33
<211> 177
<212> БЕЛОК
<213> Homo sapiens
<400> 33
Glu Asp Leu Asn Lys Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala Asp Cys Gly Phe Thr Ser Val Ser Tyr Gln Gln Gly Val Leu Ser
130 135 140
Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala
145 150 155 160
Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
Phe
<210> 34
<211> 130
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 34
Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala Asp
130
<210> 35
<211> 95
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 35
Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
1 5 10 15
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
35 40 45
Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
50 55 60
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
65 70 75 80
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
85 90 95
<210> 36
<211> 95
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 36
Pro Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
1 5 10 15
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
35 40 45
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
50 55 60
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
65 70 75 80
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
85 90 95
<210> 37
<211> 95
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 47
Pro Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
1 5 10 15
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Thr Gln Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
35 40 45
Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
50 55 60
Ala Trp Ser Gln Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Gln Asn
65 70 75 80
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
85 90 95
<210> 38
<211> 129
<212> БЕЛОК
<213> Homo sapiens
<400> 38
Glu Asp Leu Asn Lys Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala
<210> 39
<211> 130
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 39
Leu Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu
1 5 10 15
Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys
20 25 30
Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val
35 40 45
Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu
50 55 60
Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu Ser Ser Arg
65 70 75 80
Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg
85 90 95
Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln
100 105 110
Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly
115 120 125
Arg Ala
130
<210> 40
<211> 130
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> синтетическая
<400> 40
Leu Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu
1 5 10 15
Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys
20 25 30
Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val
35 40 45
Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu
50 55 60
Lys Glu Gln Pro Ala Leu Gln Asp Ser Arg Tyr Ala Leu Ser Ser Arg
65 70 75 80
Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg
85 90 95
Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln
100 105 110
Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly
115 120 125
Arg Ala
130
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| ХИМЕРНЫЕ КОНСТРУКЦИИ АНТИТЕЛО/Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2767209C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛЫЕ ЦЕПИ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С CD19 | 2019 |
|
RU2824170C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| МОЛЕКУЛЫ IMMTAC, СВЯЗЫВАЮЩИЕ РЕСТРИКТИРОВАННЫЙ ПО HLA-A*02 ПЕПТИД, С УВЕЛИЧЕННЫМ ПЕРИОДОМ ПОЛУВЫВЕДЕНИЯ | 2020 |
|
RU2833530C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ ДОМЕНЫ ПРОТИВ TRBC1 | 2018 |
|
RU2791327C2 |
| CD3-СВЯЗЫВАЮЩИЙ ДОМЕН | 2015 |
|
RU2742691C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2826991C2 |
| АНТИТЕЛЬНЫЕ АГЕНТЫ, СПЕЦИФИЧНЫЕ К CD19 ЧЕЛОВЕКА, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773317C2 |
| ТРИФУНКЦИОНАЛЬНАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2753882C2 |
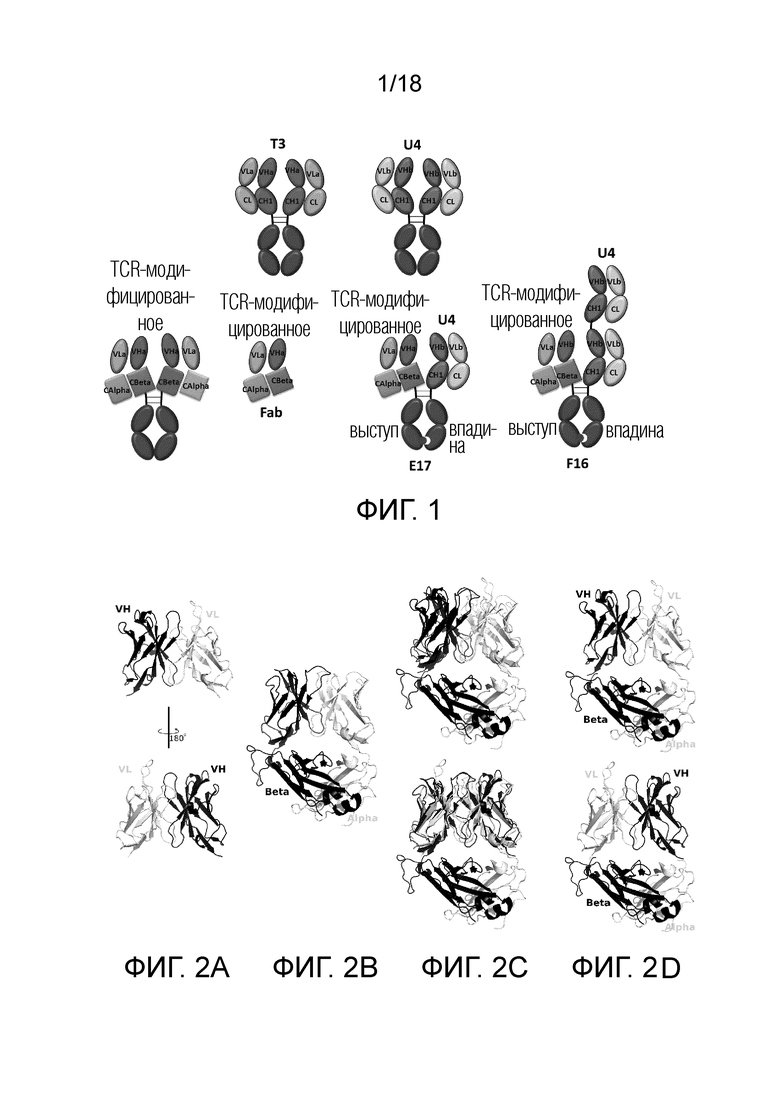


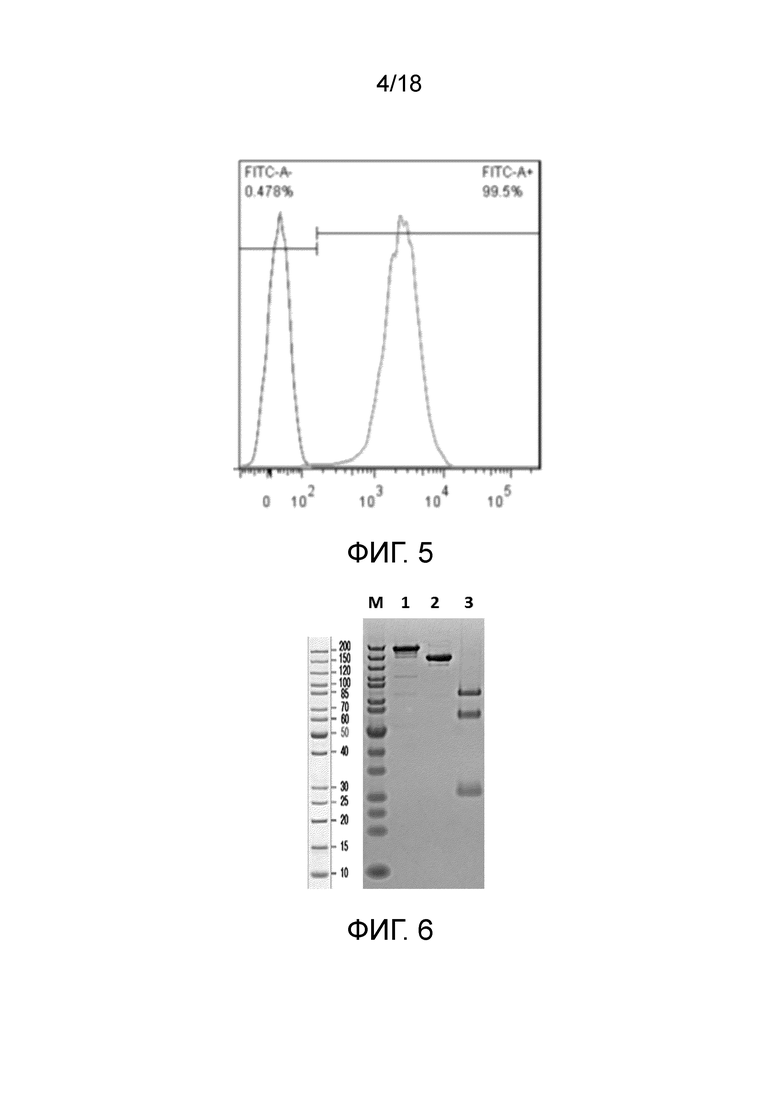
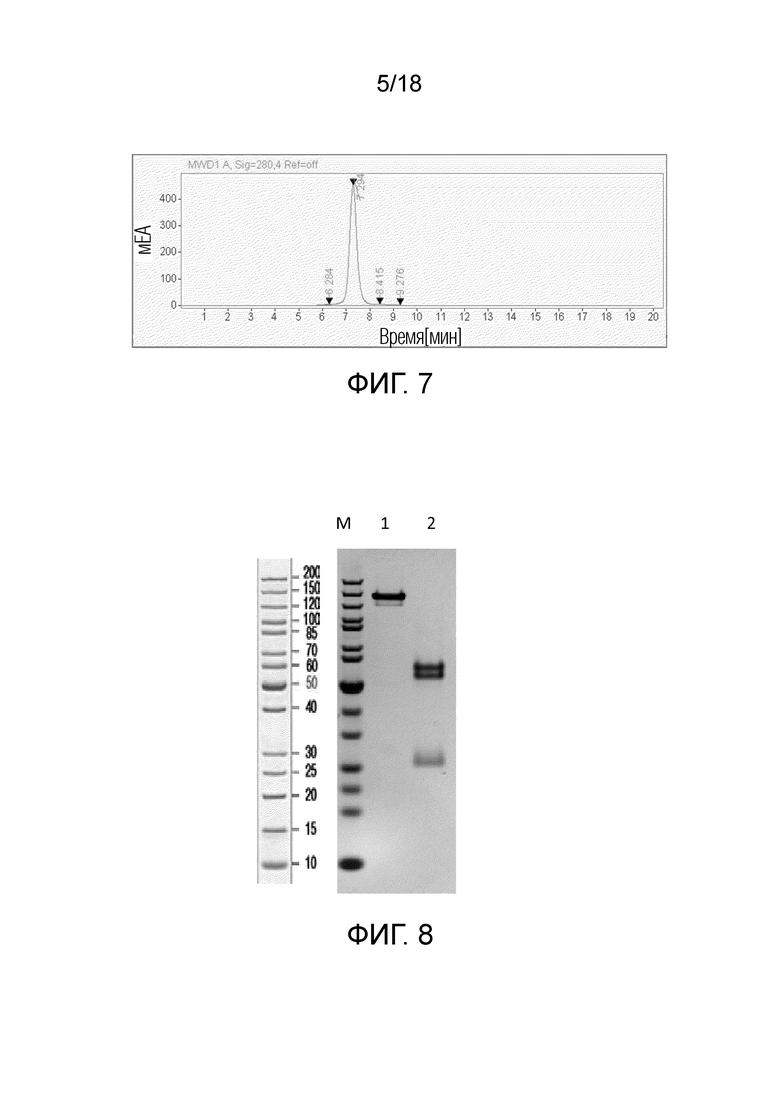
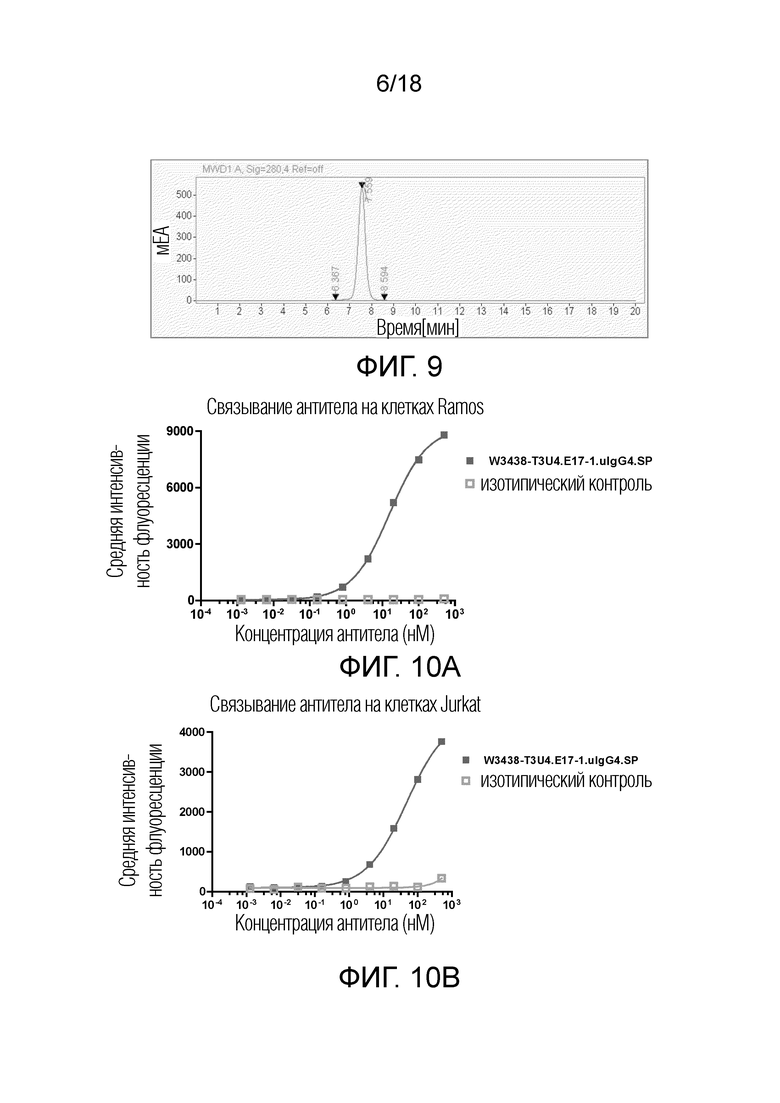
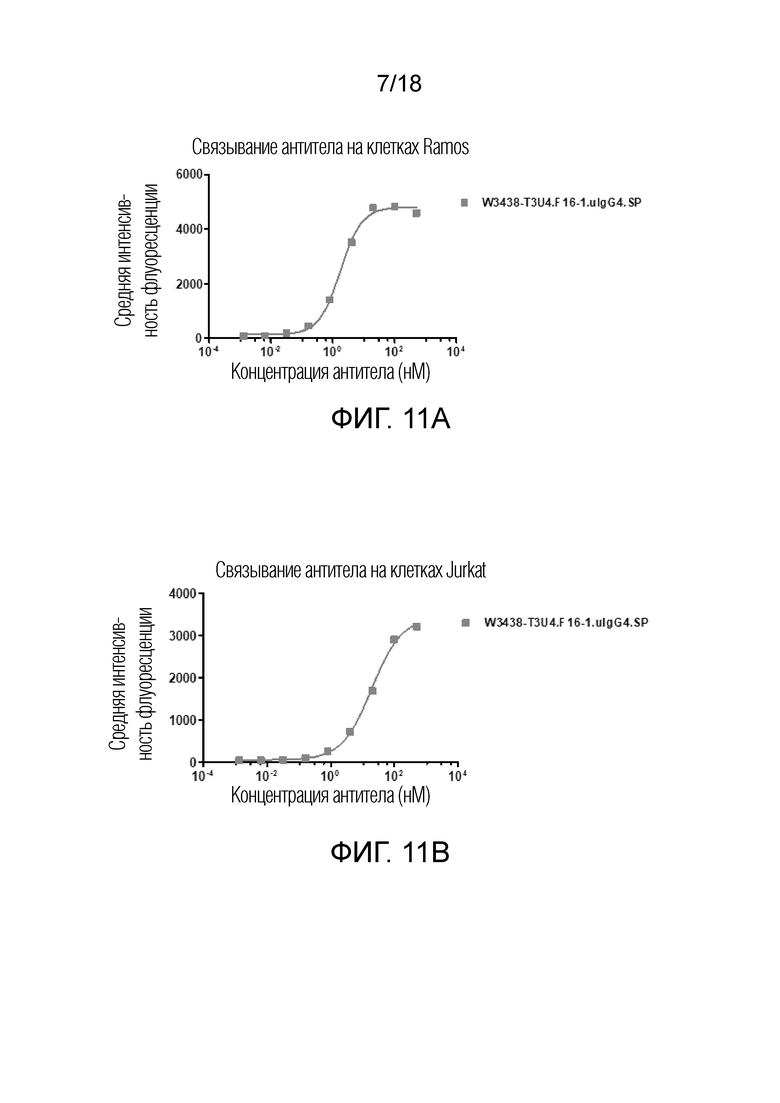
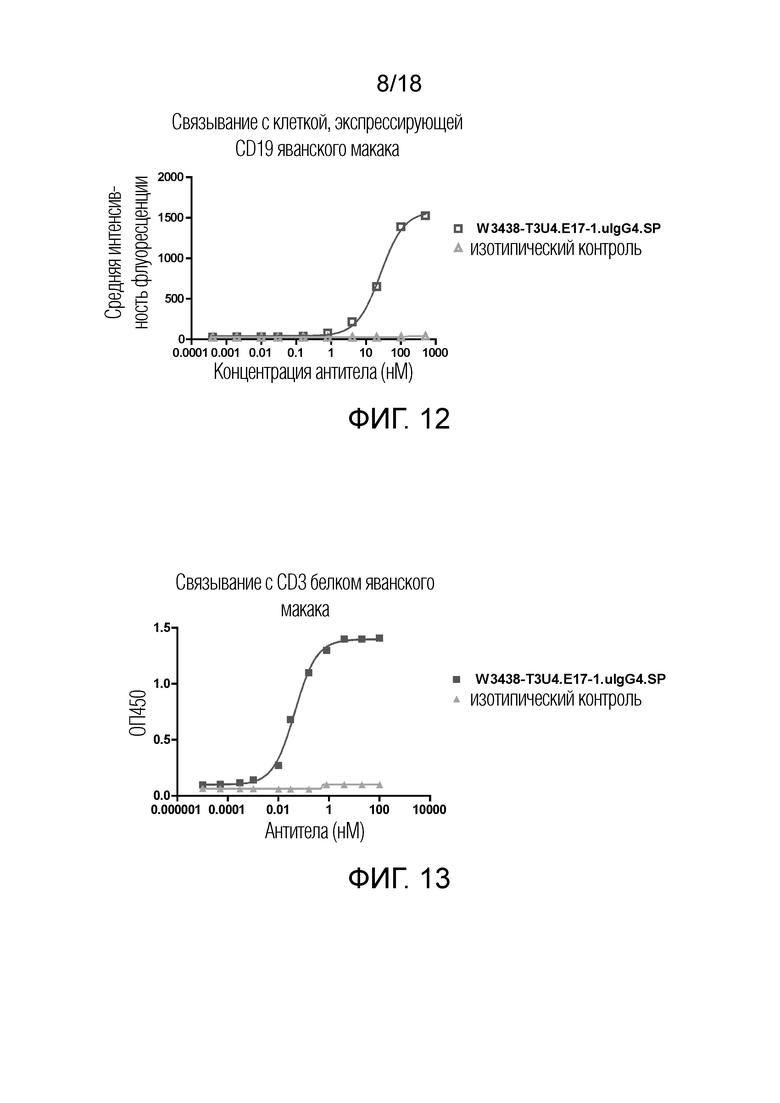
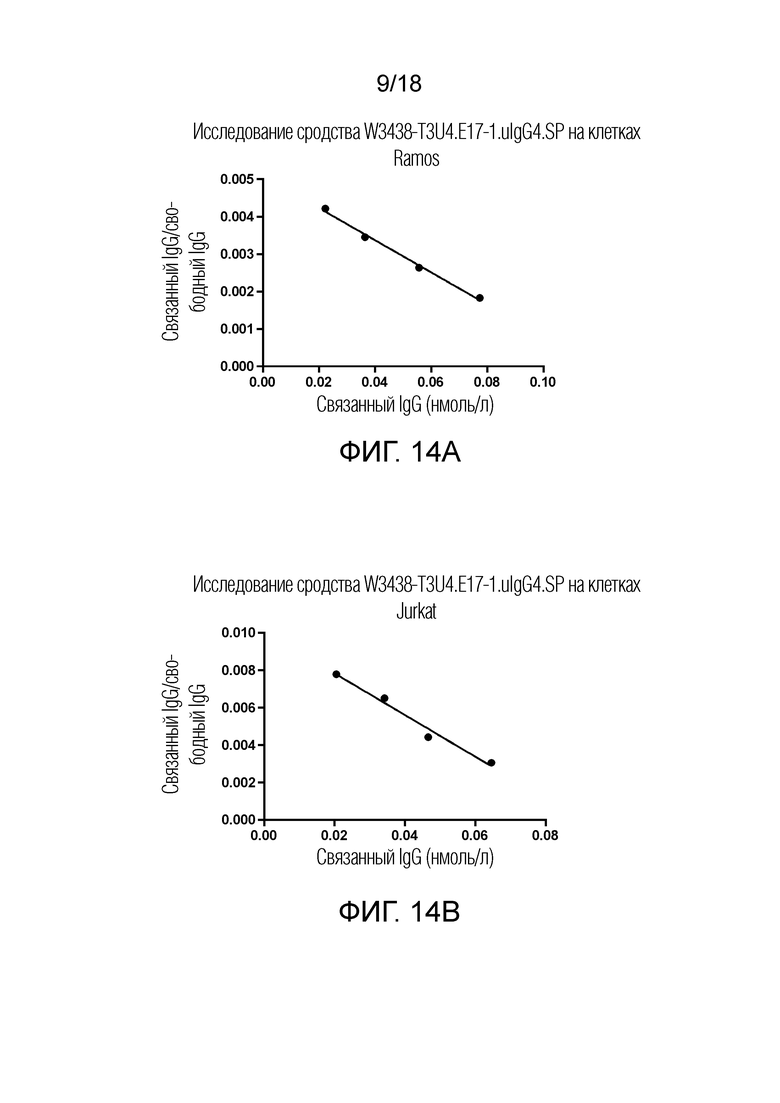
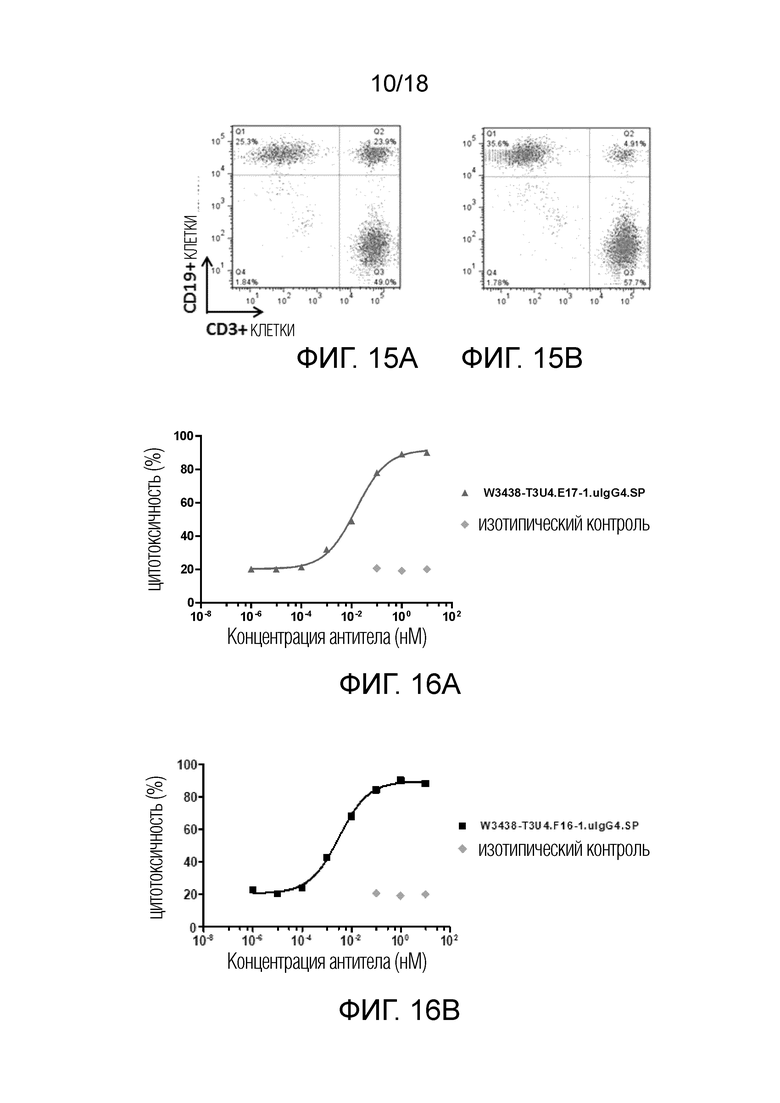
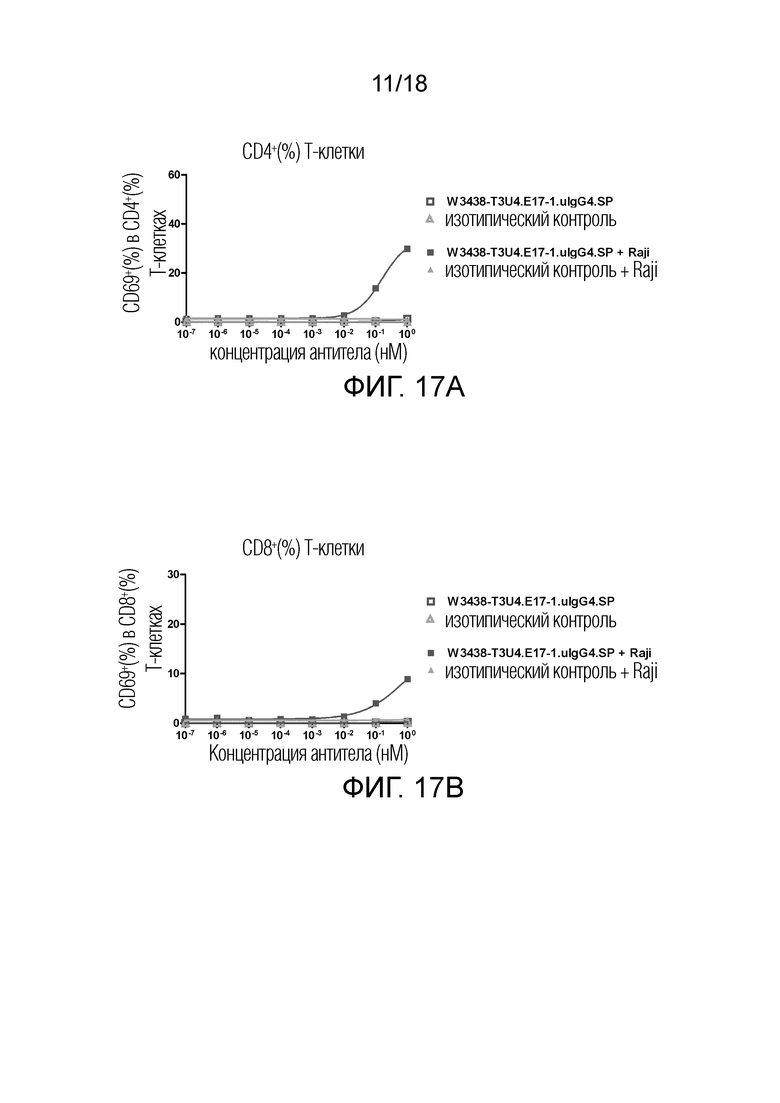
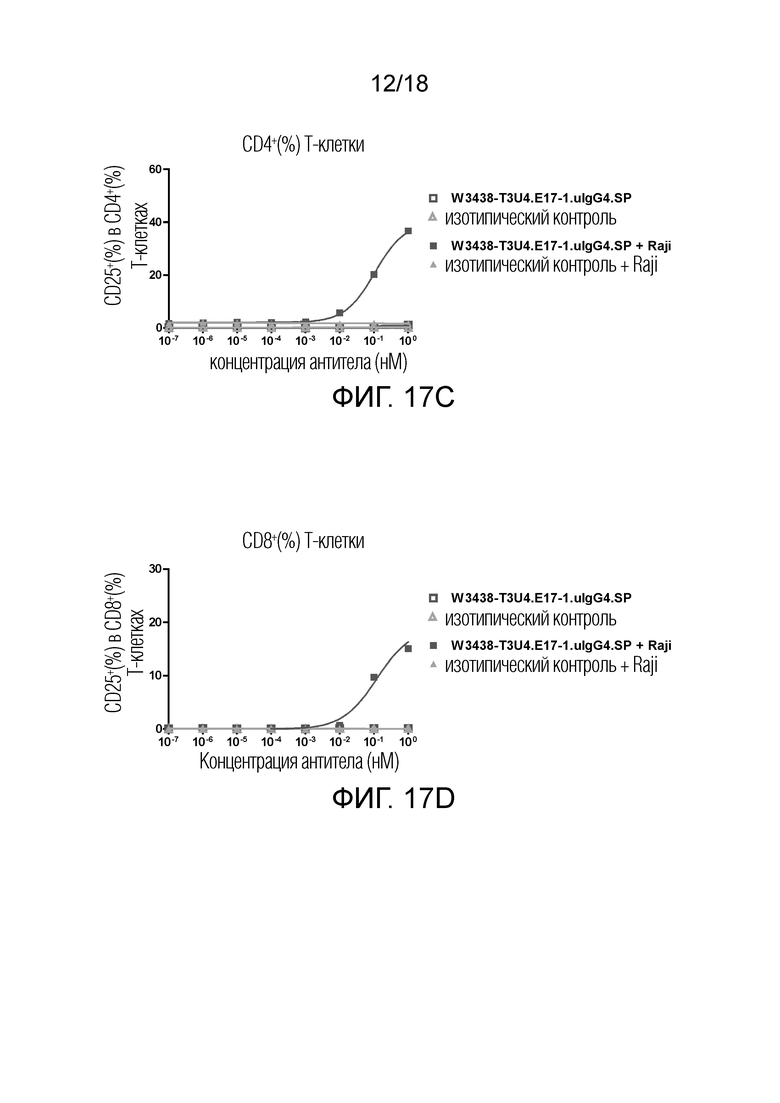
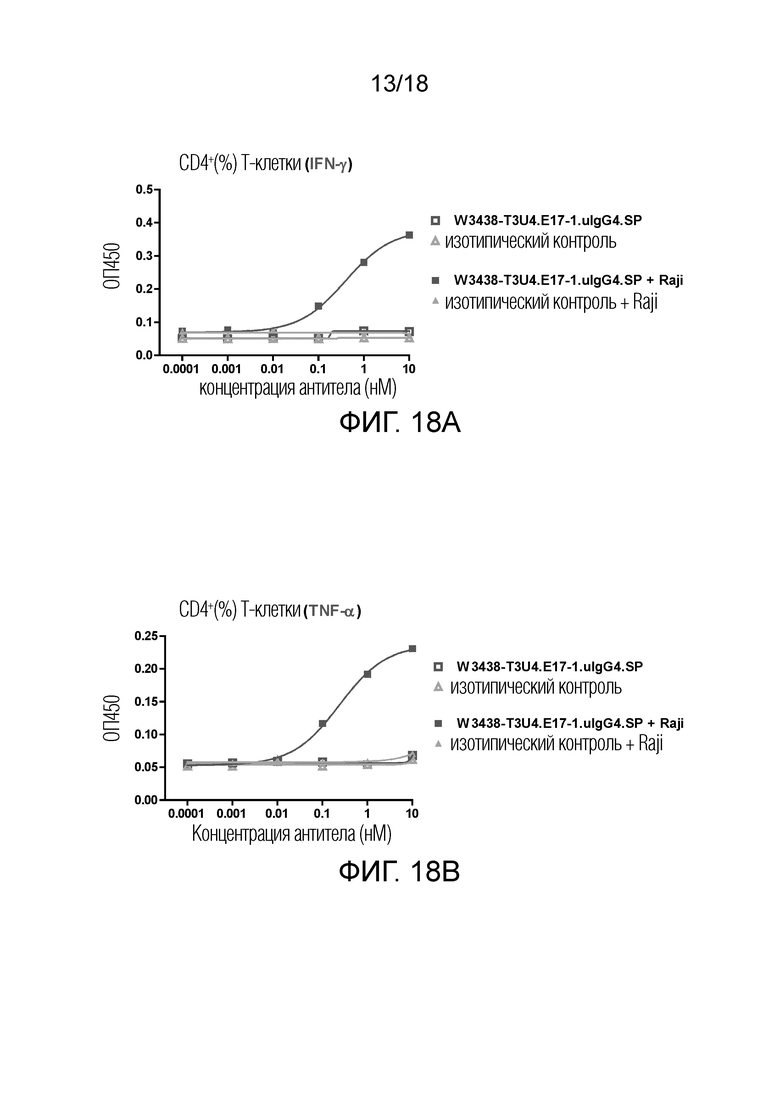
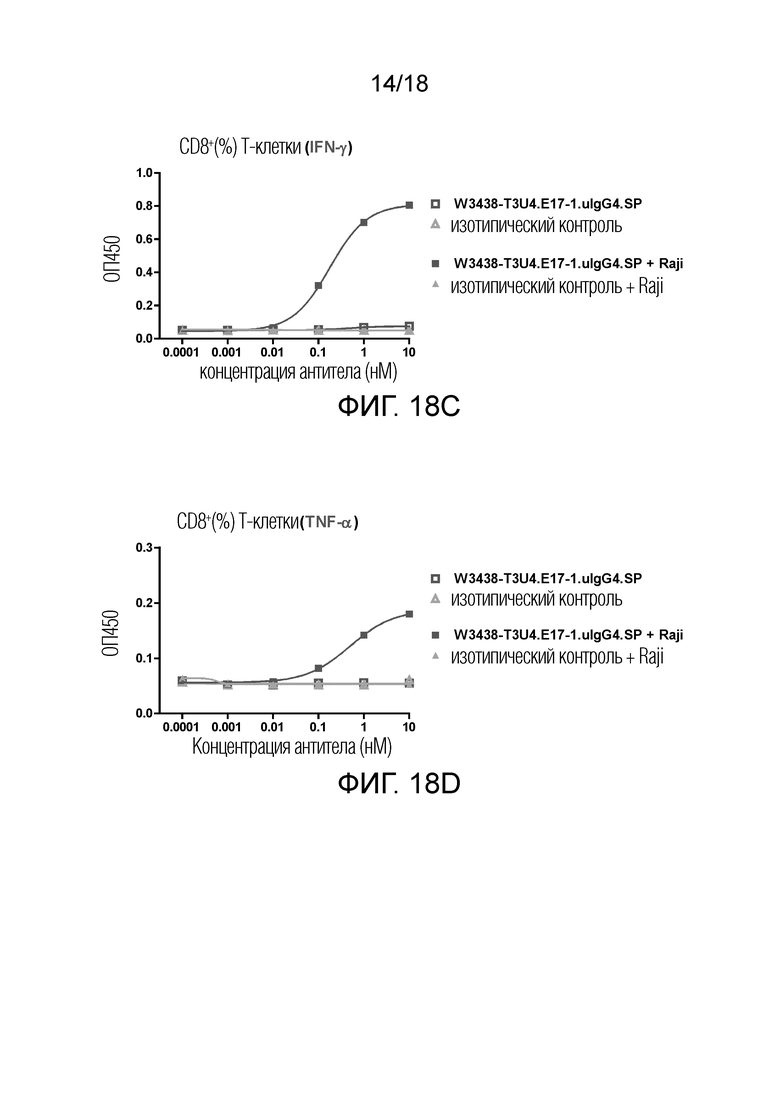
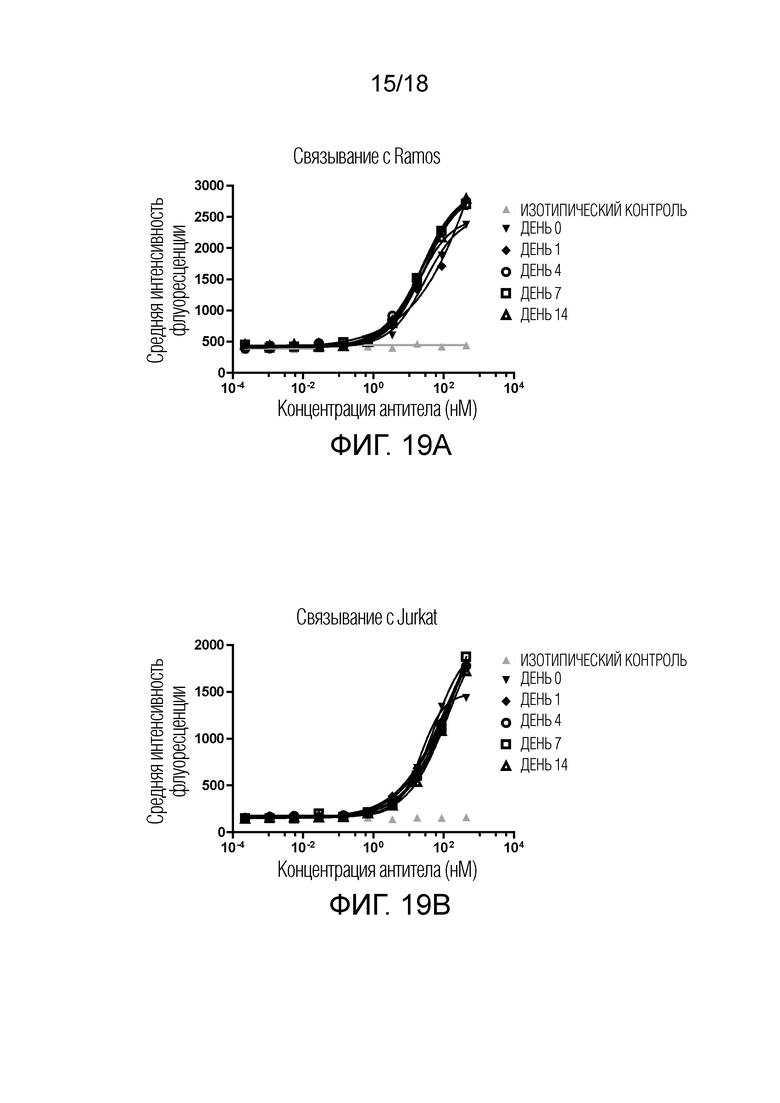
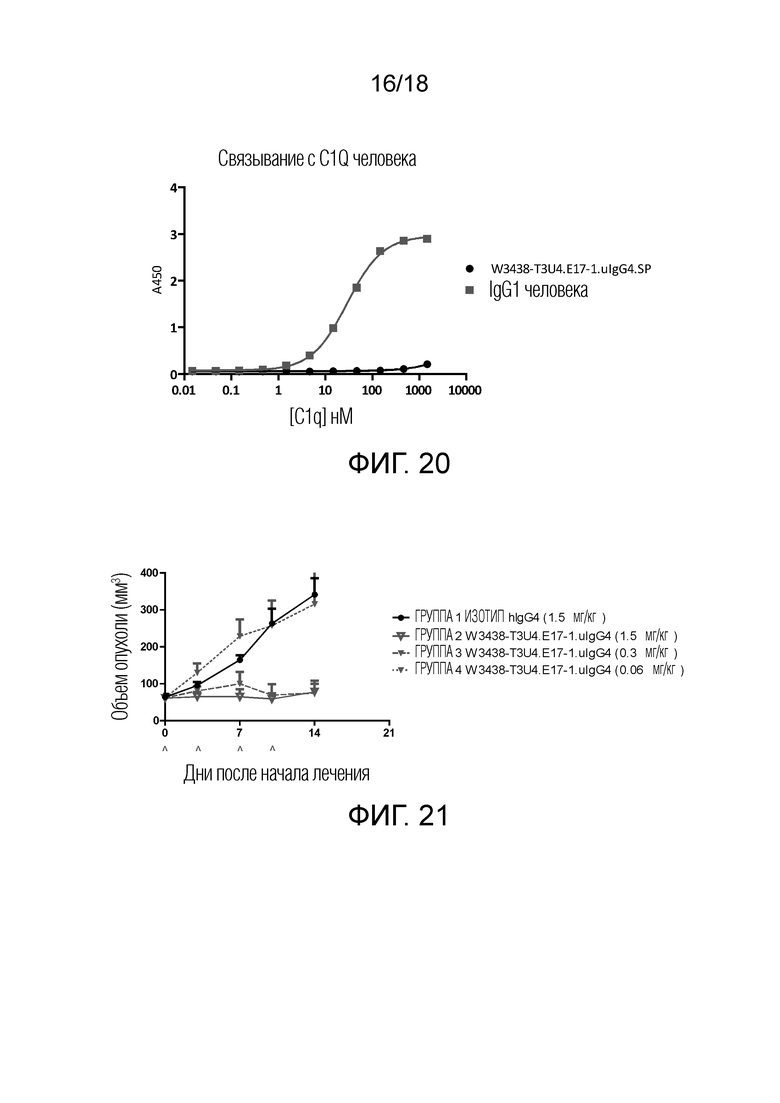
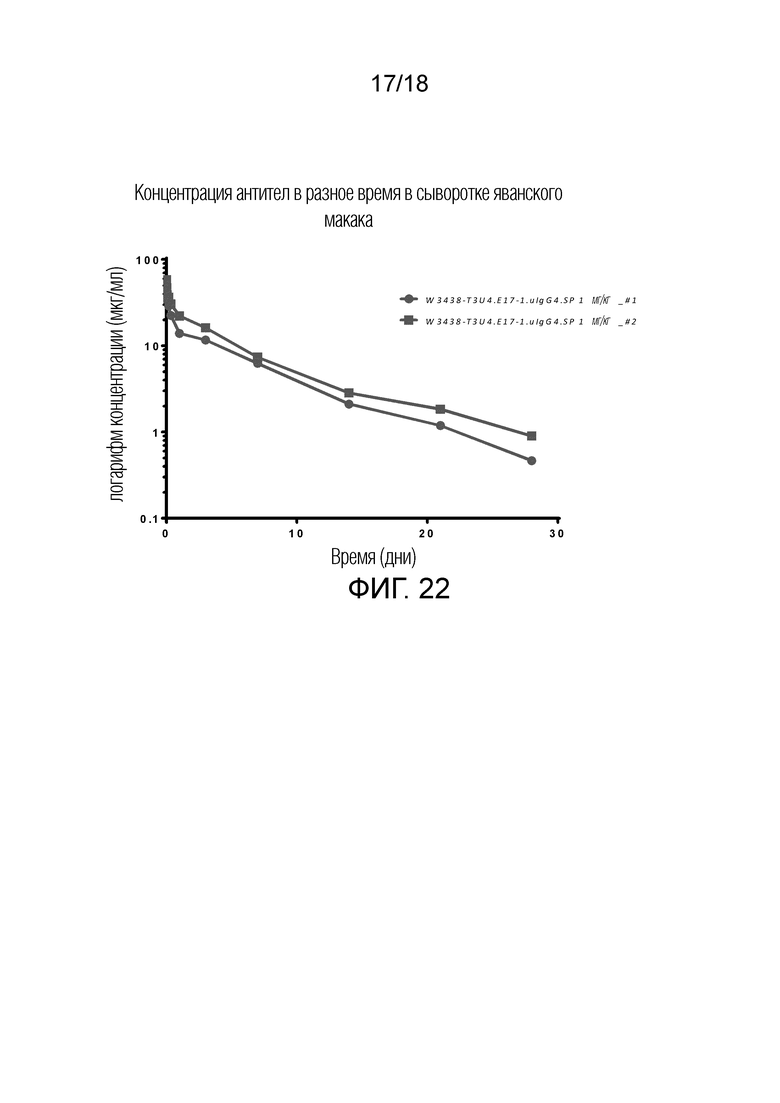
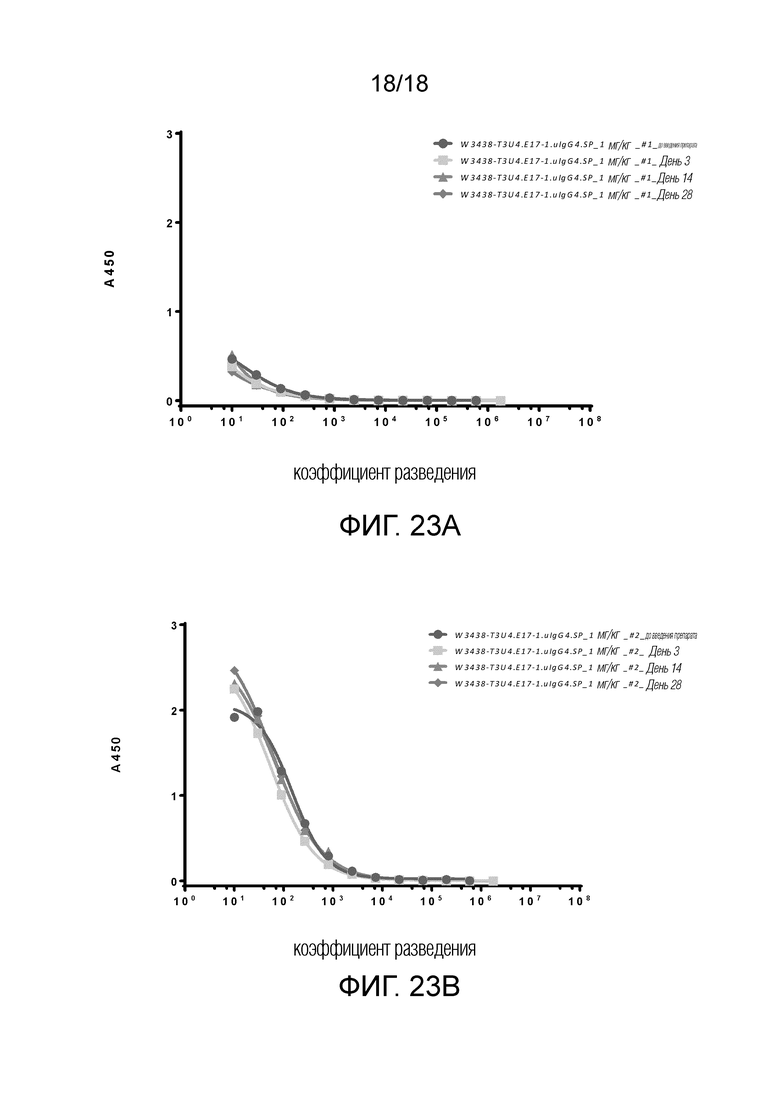
Изобретение относится к области биотехнологии, конкретно к биспецифическим полипептидным комплексам, специфически связывающимся с CD3 и CD19, и может быть использовано в медицине для лечения В-клеточной лимфомы. Рекомбинантным путем получают СD3/CD19 биспецифические полипептидные комплексы, которые содержат сочетание четырех полипептидных последовательностей: SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 29 или SEQ ID NO: 30. Изобретение позволяет получить биспецифические полипептидные комплексы, которые способны одновременно связываться с CD3 и CD19, с Т-клетками и В-клетками и вызывать последующий лизис злокачественной CD19-позитивной В-клетки. 11 н. и 2 з. п. ф-лы, 23 ил., 2 табл., 2 пр.
1. Биспецифичный полипептидный комплекс, который специфически связывается с CD3 и CD19, отличающийся тем, что биспецифичный полипептидный комплекс содержит сочетание четырех полипептидных последовательностей: SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 29.
2. Биспецифичный полипептидный комплекс, который специфически связывается с CD3 и CD19, отличающийся тем, что биспецифичный полипептидный комплекс содержит сочетание четырех полипептидных последовательностей: SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 30.
3. Набор выделенных полинуклеотидных молекул для экспрессии биспецифичного полипептидного комплекса, который специфически связывается с CD3 и CD19, по п.1 или 2, где каждая из указанных выделенных полинуклеотидных молекул кодирует каждую полипептидную цепь биспецифичного полипептидного комплекса по п.1 или 2.
4. Набор выделенных векторов для экспрессии биспецифичного полипептидного комплекса, который специфически связывается с CD3 и CD19, по п.1 или 2, где каждый из указанных выделенных векторов содержит каждую из полинуклеотидных молекул по п.3.
5. Клетка-хозяин для экспрессии биспецифичного полипептидного комплекса, который специфически связывается с CD3 и CD19, по п.1 или 2, содержащая набор выделенных полинуклеотидных молекул по п.3 или набор выделенных векторов по п.4.
6. Способ получения биспецифичного полипептидного комплекса, который специфически связывается с CD3 и CD19, по п.1 или 2, включающий культивирование клетки-хозяина по п.5 в условиях, при которых каждая полипептидная цепь указанного биспецифичного полипептидного комплекса экспрессируется и экспрессированные полипептидные цепи собираются в биспецифичный полипептидный комплекс.
7. Способ получения биспецифичного полипептидного комплекса, который специфически связывается с CD3 и CD19, по п.1 или 2, включающий:
a) введение в клетку-хозяина выделенных полинуклеотидных молекул, каждая из которых кодирует каждую полипептидную цепь биспецифичного полипептидного комплекса по п.1 или 2; и
b) предоставление возможности клетке-хозяину экспрессировать каждую полипептидную цепь биспецифичного полипептидного комплекса и собрать экспрессированные полипептидные цепи в биспецифичный полипептидный комплекс.
8. Способ по п.6 или 7, дополнительно включающий выделение биспецифичного полипептидного комплекса.
9. Композиция для лечения заболевания или состояния, связанного с CD19, у субъекта, содержащая эффективное количество биспецифичного полипептидного комплекса по п.1 или 2, где указанное заболевание или состояние, связанное с CD19, представляет собой В-клеточную лимфому.
10. Фармацевтическая композиция для лечения заболевания или состояния, связанного с CD19, у субъекта, содержащая эффективное количество биспецифичного полипептидного комплекса по п.1 или 2 и фармацевтически приемлемый носитель, где указанное заболевание или состояние, связанное с CD19, представляет собой В-клеточную лимфому.
11. Способ лечения заболевания или состояния, связанного с CD19, у субъекта, нуждающегося в таком лечении, включающий введение субъекту терапевтически эффективного количества биспецифичного полипептидного комплекса по п.1 или 2 или терапевтически эффективного количества фармацевтической композиции по п.10, где указанное заболевание или состояние, связанное с CD19, представляет собой В-клеточную лимфому.
12. Способ по п. 11, отличающийся тем, что В-клеточная лимфома представляет собой лимфому Ходжкина или неходжкинскую лимфому и где неходжкинская лимфома включает: диффузную крупноклеточную В-клеточную лимфому (DLBCL), фолликулярную лимфому, В-клеточную лимфому маргинальной зоны (MZL), лимфому лимфоидной ткани слизистых оболочек (MALT), мелкоклеточную лимфоцитарную лимфому, хронический лимфоцитарный лейкоз (CLL), лимфому из клеток мантийной зоны (MCL), острый лимфобластный лейкоз или макроглобулинемию Вальденстрема (WM).
13. Набор для лечения заболевания или состояния, связанного с CD19, у субъекта, содержащий биспецифичный полипептидный комплекс по п.1 или 2, где указанное заболевание или состояние, связанное с CD19, представляет собой В-клеточную лимфому.
| WO 2017055314 A1, 06.04.2017 | |||
| WU X | |||
| et al., Protein design of IgG/TCR chimeras for the co-expression of Fab-like moieties within bispecific antibodies, MAbs, 2015, V | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| KIPRIYANOV S | |||
| M | |||
| et al., Bispecific CD3 X CD19 diabody for T cell‐mediated lysis of malignant human B cells, International journal of cancer, 1998, V | |||
| Спускная труба при плотине | 0 |
|
SU77A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |

