Родственные заявки
Настоящая заявка испрашивает приоритет на основании предварительной заявки на патент США № 62/908367, поданной 30 сентября 2019 г., под названием «ВАРИАНТЫ ДОМЕНА CH1, СКОНСТРУИРОВАННЫЕ ДЛЯ ПРЕДПОЧТИТЕЛЬНОГО СПАРИВАНИЯ С ЛЕГКОЙ ЦЕПЬЮ, И СОДЕРЖАЩИЕ ИХ МУЛЬТИСПЕЦИФИЧНЫЕ АНТИТЕЛА», содержание которой включено в настоящий документ посредством ссылки в полном объеме.
Область техники
Настоящее изобретение относится к вариантам домена CH1, которые содержат по меньшей мере одну аминокислотную замену, которая способствует правильному спариванию тяжелых цепей с легкими цепями, и содержащим их тяжелым цепям антител и антителам, в частности, мультиспецифичным антителам. Настоящее изобретение дополнительно относится к композициям, содержащим такие антитела, и их применению, например, в качестве терапевтических или диагностических средств. Настоящее изобретение дополнительно относится к способам получения библиотеки вариантов домена CH1 и способам идентификации одного или более вариантов домена CH1.
Уровень техники
Продолжаются усилия, направленные на разработку терапевтических средств на основе антител, которые характеризуются более чем одной антигенсвязывающей специфичностью, например, биспецифичных антител. Биспецифичные антитела могут применяться для взаимодействия с несколькими поверхностными рецепторами, связанными с раком, воспалительными процессами или другими патологическими состояниями. Биспецифичные антитела также могут применяться для размещения мишеней в непосредственной близости и модулирования образования белкового комплекса или обеспечения контакта между клетками. Получение биспецифичных антител впервые было описано в начале 1960-х годов (Nisonoff et al., Arch Biochem Biophys 1961 93(2): 460-462), и первые моноклональные биспецифичные антитела были получены с применением гибридомной технологии в 1980-х годах (Milstein et al., Nature 1983 305(5934): 537-540). Интерес к биспецифичным антителам значительно возрос за последнее десятилетие из-за их терапевтического потенциала, и теперь биспецифические антитела применяют в клинических целях, например, блинатумомаб и цемиизумаб были одобрены для лечения определенных видов рака (см. Sedykh et al., Drug Des Devel Ther 12:195-208 (2018) и Labrijn et al. Nature Reviews Drug Discovery 18:585-608 (2019), где содержатся недавние обзоры способов получения биспецифичных антител и особенностей биспецифичных антител, одобренных для медицинского применения).
Хотя биспецифичные антитела продемонстрировали значительные преимущества по сравнению с моноспецифичными антителами, широкому коммерческому применению биспецифичных антител препятствует отсутствие эффективных/недорогих способов получения, недостаточная стабильность биспецифичных антител и отсутствие длительного периода полужизни в организме человека. За последние несколько десятилетий было разработано огромное количество способов улучшения получения биспецифичных антител. Они включают рекомбинантную совместную экспрессию двух пар тяжелой и легкой цепей иммуноглобулина с разными специфичностями (см. Milstein and Cuello, Nature 305: 537 (1983)), WO 93/08829 и Traunecker et al., EMBO J. 10: 3655 (1991)); конструирование методом «выступ-во-впадину» (см., например, патент США No 5731168; технологию кроссовера иммуноглобулинов (также известную как обмен Fab-доменами или формат CrossMab) (см., например, WO2009/080253; Schaefer et al., Proc. Natl. Acad. Sci. USA, 108:11187-11192 (2011)); конструирование белков с использованием эффекта электростатического взаимодействия с получением Fc-гетеродимерных молекул антитела (WO 2009/089004A1); перекрестное связывание двух или более антител или фрагментов (см., например, патент США № 4676980 и Brennan et al., Science, 229: 81 (1985)); лейциновые молнии (см., например, Kostelny et al., J. Immunol, 148(5):1547-1553 (1992)); технологию «диатела» (см., например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993)); димеры одноцепочечных Fv (sFv) (см., например, Gruber et al., J. Immunol, 152:5368 (1994)); и триспецифичные антитела, описанные, например, в Tutt et al. J. Immunol 147: 60 (1991).
Несмотря на эти улучшения, получение биспецифичных антител с правильным спариванием тяжелых цепей с легкими цепями остается проблемой. Биспецифичное антитело может быть образовано путем совместной экспрессии двух разных тяжелых цепей и двух разных легких цепей. Надлежащее образование биспецифичных антител в требуемом формате остается проблемой, поскольку тяжелые цепи в ходе эволюции стали характеризоваться относительно беспорядочным связыванием легких цепей. Следовательно, совместная экспрессия двух тяжелых цепей и двух легких цепей может привести к скремблированию пар тяжелой и легкой цепей - сложной смеси из шестнадцати возможных комбинаций, представляющих собой десять различных антител, только одно из которых соответствует требуемому биспецифичному антителу (максимальный выход 12,5% в смеси, если имеет место идеальная беспорядочность). Такое неправильное спаривание (также называемое проблемой ассоциации цепей) остается основной проблемой для получения биспецифичных белков, поскольку однородное спаривание является необходимым условием для обеспечения выполнимости производства и эффективности.
Одна из стратегий, используемых для уменьшения неправильного спаривания, заключается в получении биспецифичных антител, имеющих общую легкую цепь (см., например, Merchant et al., Nat. Biotech. 16:677-681 (1998)). В качестве альтернативы можно использовать одну общую тяжелую цепь и две разные легкие цепи (одну каппа- и одну лямбда-цепь) (см., например, Fischer et al., Nature Commun. 6:6113 (2015)). Однако эта стратегия требует идентификации антитела с общей цепью, что является сложным и склонно нарушать специфичность каждого связывающегося плеча и существенно снижает разнообразие (см., например, Wang et al., MABS 10(8):1226-1235 (2018)).
Другие подходы для улучшения правильного спаривания тяжелых цепей с легкими цепями включают технологию CrossMab (Roche), в которой легкая цепь или один из субдоменов в ней одного плеча антигенсвязывающего фрагмента (Fab) обмениваются соответствующими областями Fd-области тяжелой цепи, и технологию DuetMab (MedImmune), в которой нативную дисульфидную связь в одном плече Fab заменяют сконструированной дисульфидной связью. Однако эти подходы требуют существенных изменений в нативном формате IgG, которые могут привести к тому, что соединения не будут в достаточной степени схожими с природными антителами.
Еще одна стратегия заключается в использовании аминокислотных замен в константных и/или вариабельных областях тяжелых и легких цепей в формате IgG для снижения или устранения неправильного спаривания тяжелых цепей с легкими цепями. Насколько известно авторам настоящего изобретения, до сих пор не было продемонстрировано, что модификация одного лишь домена CH1 решает проблему ассоциации или неправильного спаривания цепей, часто наблюдаемую при экспрессии мультиспецифичных антител. Скорее, мультиспецифичные антитела, сконструированные таким образом, чтобы они содержали варианты домена CH1, дополнительно требуют модификаций и за пределами домена CH1 для решения проблемы ассоциации цепей, например, в домене CL и в некоторых случаях доменах VH, CH2, CH3 и/или VL. Их примеры включают Lewis et al., Nature Biotech. 32(2):191- 198 (2014), которые получали мутантные домены CH1 и CL, CRD1 (с заменами тяжелой цепи D148K, F170T, V185F и заменами легкой цепи K129D, L135F; нумерация EU) и CRD2 (с заменами тяжелой цепи H168A и F170T и заменами легкой цепи L135Y, S176W), в попытке способствовать предпочтительному спариванию измененных тяжелых и легких цепей и не способствовать спариванию доменов тяжелых и легких цепей с константными доменами дикого типа. Однако авторы сообщили, что любая специфичность спаривания, полученная с мутантными доменами CH1 и CL в отсутствие вариабельных доменов, не транслировалась в полноразмерный формат IgG без дополнительных генетических модификаций в пределах области контакта VH-VL, т.е. требовались замены в пределах области контакта VH-VL наряду с заменами домена CL и CH1 для достижения предпочтительного спаривания тяжелых цепей с легкими цепями. Также предполагалось, что конструирование доменов CH1 и CL таким образом, чтобы они содержали заряженные аминокислотные остатки, будет способствовать предпочтительному спариванию тяжелых цепей с легкими цепями (см., например, патент США № 10047163). Также известны биспецифичные антитела, имеющие по меньшей мере два Fab-фрагмента с разными доменами CH1 и CL, в которых один Fab-фрагмент имеет замены в домене CH1 и домене Cκ, чтобы способствовать предпочтительному спариванию (см. US20180022829 и патент США № 9631031, где раскрыты CH1: T187E и Cκ: N137K + S114A; CH1: L145Q + S183V и Cκ: V133T + S176V; CH1: L128A + L145E и Cκ: V133W; CH1: V185A и Cκ: L135W + N137A). Дополнительные примеры конкретных замен домена CH1, которые, как утверждается, способствуют предпочтительному спариванию тяжелых цепей с легкими цепями, или, в некоторых случаях, где CH2, CH3 и/или VH также соответствующим образом заменены для способствования предпочтительному спариванию, включают A141C/L, K147D, G166D, G166K или замену цистеином в положении 128, 129, 162 или 171 (WO2019183406 (Invenra Inc.)); замену цистеина в положении 126 или 220 заменено валином или аланином, или замену отличным от цистеина остатком в положении 128, 141 или 168 цистеином, L145F, K147A, F170V, S183F или V185W/F (патент США № 9527927 (MedImmune)); 172A и 174G (WO2020060924 (Dualogics)); A172R и 174G или замена остатка 190 на M или I (патент США № 10047167 (University of North Carolina Chapel Hill и Eli Lilly)); L128F, A141I/M/T/L, F170S/A/Y/M, S181M/I/T, S183A/E/K/V и V185A/L (US20180177873 (Genentech)); 131C/S, 133R/K, 137E/G, 138S/G 178S/Y, 192N/S и/или 193F/L (патент США № 10487156 (Argenx BVBA)); 145D/E/R/H/K (положение 26 согласно IMGT) (WO2018141894 (Merck)); 124K/E/R/D (патент США № 10392438 (Pfizer)); 133V, 150A, 150D, 152D, 173D или 188W (US20190023810 (MIT)); 133S/W/A, 139W/V/G/I, 143K/E/A, 145E/T/L/Y, 146G, 147T/E, 174V, 175D/R/S, 179K/D/R, 181R, 186R, 188F/L и/или 190S/A/G/Y (US20180179296 и патент США № 9914785 (Zymeworks)); 143A/E/R/K/D и 145T/L (патент США № 10077298 (Zymeworks)); 124A/R/E/W, 145M/T, 143E/R/D/F, 172R/T, 139W/G/C, 179E или 186R (US20170204199 (Zymeworks)); замену цистеином в положении 126, 127, 128, 134, 141, 171 или 173 (Zenyaku Kogyo); L145Q, H168A, F170G, S183V и T187E (WO2020127354 (Alligator Bioscience)); 143D/E, 145T, 190E/D и 124R (WO2017/059551 (Zymeworks)).Также, согласно имеющимся данным в патенте США № 9150639, Kyowa Hakko Kirin получил тяжелые цепи, содержащие A140C, K147C или S183C, с целью введения цистеина для обеспечения химической модуляции. Kirin предполагает, что варианты антител, содержащие эти мутации тяжелой цепи, могут содержать легкие цепи дикого типа, однако, нет никаких доказательств того, что это будет способствовать предпочтительному спариванию тяжелых цепей с легкими цепями.
Еще одной стратегией, используемой для минимизации неправильного спаривания тяжелых цепей с легкими цепями, является использование разных легких цепей, например, легких цепей с разными константными доменами. Например, Loew et al. получили мультиспецифичные антитела, имеющие легкую каппа-цепь и легкую лямбда-цепь, и наблюдали минимальное неправильное спаривание, поскольку некоторые встречающиеся в природе легкие каппа-цепи характеризуются высокой точностью и не спариваются с тяжелыми цепями из антитела с лямбда-цепями, и наоборот (WO2018057955). К сожалению, применимость этой методики ограничена теми легкими цепями, которые характеризуются высокой точностью. Другие авторы получали мультиспецифичные антитела с применением легких каппа-цепей и легких лямбда-цепей, в которых аминокислотные замены используются как в тяжелых, так и в легких цепях для электростатического или стерического обеспечения предпочтительного спаривания (см., например, WO2017059551 (Zymeworks), US20140154254 (Amgen) и патент США № 10047163 (AbbVie Stemcentrx)). Однако введение множества аминокислотных замен как в тяжелые, так и в легкие цепи создает дополнительные технические препятствия, и, кроме того, может оказывать отрицательные эффекты на функцию и/или иммуногенность антител.
Краткое описание изобретения
Целью настоящего изобретения является получение сконструированных биспецифичных антител с надлежащим спариванием тяжелых цепей с легкими цепями. В одном из аспектов в настоящем документе предложены полипептиды варианта домена CH1 (также называемые в настоящем документе вариантами домена CH1), которые способствуют предпочтительному спариванию тяжелой цепи с определенными легкими цепями, и содержащие их полипептиды, такие как антитела. Варианты домена CH1 содержат по меньшей мере одну аминокислотную замену (относительно исходной последовательности, например, последовательности дикого типа).
В некоторых вариантах осуществления варианты домена CH1 содержат по меньшей мере одну аминокислотную замену в положении домена CH1, которое образует область контакта с доменом CL легкой цепи, включая, не ограничиваясь перечисленным, положение 140 и/или 141 или 147 и/или 183 (нумерация EU). Указанная замена способствует предпочтительному спариванию тяжелой цепи, содержащей вариант домена CH1, с определенными легкими цепями, например, вариант 141 домена CH1 предпочтительно спаривается с доменом CL-лямбда, в отличие от домена CL-каппа, тогда как вариант 147F и/или 183R, 183K или 183Y домена CH1 предпочтительно спаривается с доменом CL-каппа, в отличие от домена CL-лямбда.
В некоторых вариантах осуществления варианты домена CH1 содержат по меньшей мере одну аминокислотную замену в положении домена CH1, которое образует область контакта между доменом CH1 и VH, таком как положение 151 CH1 (нумерация EU).
Ожидается, что это предпочтительное спаривание константных доменов будет способствовать спариванию полноразмерной легкой и тяжелой цепи, включая вариабельные домены, таким образом обеспечивая решение проблемы со спариванием цепей для биспецифичных белков. В частности, полипептид варианта домена CH1 содержит аминокислотную замену в одном или более из следующих положений: 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175, 176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216 и 218 согласно нумерации EU. Необязательно такой полипептид варианта домена CH1 предпочтительно спаривается: (i) с доменом константной области легкой каппа-цепи («CL») по сравнению с доменом CL-лямбда и/или с полипептидом легкой каппа-цепи по сравнению с полипептидом легкой лямбда-цепи; (ii) с доменом CL-лямбда по сравнению с доменом CL-каппа и/или с полипептидом легкой лямбда-цепи по сравнению с полипептидом легкой каппа-цепи.
Необязательно в некоторых вариантах осуществления могут быть исключены некоторые варианты домена CH1, и варианты домена CH1 согласно настоящему изобретению могут удовлетворять следующему:
(a) если остаток 141 на CH1 заменен C или L, остаток 166 заменен D или K, остаток 128, 129, 162 или 171 на CH1 заменен C, и/или остаток 147 заменен D, то домен CL, с которым предпочтительно спаривается вариант домена CH1, не содержит аминокислотную замену;
(b) если положение 126 или 220 на CH1 заменено валином или аланином, отличный от цистеина остаток в положении 128, 141 или 168 заменен цистеином, или замены CH1 представляют собой L145F, K147A, F170V, S183F или V185W/F, то домен CL, которым предпочтительно спаривается вариант домена CH1, не содержит аминокислотную замену;
(c) если остаток 172 на CH1 заменен 172R, остаток 174 мутирован в 174G или остаток 190 заменен 190M или 190I, то они не являются единственной заменой (заменами), содержащейся в CH1;
(d) если замены CH1 состоят из L128F, A141I/M/T/L, F170S/A/Y/M, S181M/I/T, S183A/E/K/V и/или V185A/L, то домен CL, с которым предпочтительно спаривается вариант домена CH1, не модифицирован;
(e) если замены CH1 состоят из 131C/S, 133R/K, 137E/G, 138S/G, 178S/Y, 192N/S и/или 193F/L, то они не являются единственными заменами CH1, и/или, в биспецифичном антителе, домены CH1 принадлежат к одному и тому же подтипу или аллотипу человеческого иммуноглобулина;
(f) если замены CH1 состоят из 145D/E/R/H/K (положение 26 согласно IMGT), то отсутствует соответствующая замена LC, 129D/E/R/H/K (положение 18 согласно IMGT);
(g) если замены CH1 состоят из 124K/E/R/D, то в положении 176 LC, с которой предпочтительно спаривается вариант домена CH1, отсутствует соответствующая замена;
(h) если замены CH1 состоят из 133V, 150A, 150D, 152D, 173D и/или 188W, то в LC, с которой предпочтительно спаривается вариант домена CH1, отсутствуют соответствующие замены;
(i) если замены CH1 состоят из 133S/W/A, 139W/V/G/I, 143K/E/A, 145E/T/L/Y, 146G, 147T/E, 174V, 175D/R/S, 179K/D/R, 181R, 186R, 188F/L и/или 190S/A/G/Y, то в LC, с которой предпочтительно спаривается вариант домена CH1, отсутствуют соответствующие замены;
(j) если замены CH1 состоят из 143A/E/R/K/D и 145T/L, то в LC, с которой предпочтительно спаривается вариант домена CH1, отсутствуют соответствующие замены;
(k) если замены CH1 состоят из 124A/R/E/W, 145M/T, 143E/R/D/F, 172R/T и 139W/G/C, 179E и/или 186R, то в LC, с которой предпочтительно спаривается вариант домена CH1, отсутствуют соответствующие замены;
(l) если замены CH1 состоят из замены цистеином в положении 126, 127, 128, 134, 141, 171 или 173, то соответствующие положения LC не модифицированы для образования дисульфидной связи;
(m) если замены CH1 состоят из L145Q, H168A, F170G, S183V и/или T187E, то в LC-каппа или -лямбда, с которой предпочтительно спаривается вариант домена CH1, отсутствуют соответствующие замены;
(n) если замены CH1 состоят из 143D/E, 145T, 190E/D и/или 124R, то в LC, с которой предпочтительно спаривается вариант домена CH1, отсутствуют соответствующие замены; или
(o) если замены CH1 состоят из A140C, K147C и/или S183C, то в LC, с которой предпочтительно спаривается вариант домена CH1, присутствуют замены.
В некоторых вариантах осуществления полипептид варианта домена CH1 содержит аминокислотную замену в одном или более из следующих положений: 118, 124, 126-129, 131, 132, 134, 136, 139, 143, 145, 147-151, 153, 154, 170, 172, 175, 176, 181, 183, 185, 190, 191, 197, 201, 203-206, 210, 212-214 и 218 согласно нумерации EU. Необязательно так, чтобы полипептид варианта домена CH1 предпочтительно спаривался с: (i) доменом CL-каппа (или полипептидом, содержащим CL-каппа), по сравнению с доменом CL-лямбда (или полипептидом, содержащим CL-лямбда); и/или (ii) полипептидом легкой каппа-цепи по сравнению с полипептидом легкой лямбда-цепи.
В отдельных вариантах осуществления такой вариант домена CH1 содержит аминокислотную замену в положении 147, положении 183 или положениях 147 и 183.
В отдельных вариантах осуществления такой вариант домена CH1 содержит одну или более из следующих аминокислотных замен: положение 118 заменено G; положение 124 заменено H, R, E, L или V; положение 126 заменено A, T или L; положение 127 заменено V или L; положение 128 заменено H; положение 129 заменено P; положение 131 заменено A; положение 132 заменено P; положение 134 заменено G; положение 136 заменено E; положение 139 заменено I; положение 143 заменено V или S; положение 145 заменено F, I, N или T; положение 147 заменено F, I, L, R, T, S, M, V, N, E, H, Y, Q, A или G; положение 148 заменено I, Q, Y или G; положение 149 заменено C, S или H; положение 150 заменено L или S; положение 151 заменено A или L; положение 153 заменено S; положение 154 заменено M или G; положение 170 заменено G или L; положение 172 заменено V; положение 175 заменено G, L, E, A; положение 176 заменено P; положение 181 заменено Y, Q или G; положение 183 заменено I, W, F, E, Y, L, K, Q, N, R или H; положение 185 заменено W; положение 190 заменено P; положение 191 заменено I; положение 197 заменено A; положение 201 заменено S; положение 203 заменено S; положение 204 заменено Y; положение 205 заменено Q; положение 206 заменено S; положение 210 заменено R; положение 212 заменено G; положение 213 заменено E или R; положение 214 заменено R; и положение 218 заменено Q.
В отдельных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении каппа-цепи может содержать (i) аминокислотный остаток F, I, L, R, T, S, M, V, N, E, H, Y или Q в положении 147; и/или (ii) аминокислотный остаток I, W, F, E, Y, L, K, Q, N или R в положении 183.
В некоторых предпочтительных вариантах осуществления варианта домена CH1 с предпочтительностью в отношении каппа-цепи полипептид варианта домена CH1 может содержать (i) аминокислотный остаток R, K или Y в положении 183; и/или (ii) аминокислотный остаток F в положении 147.
В дополнительных вариантах осуществления полипептид варианта домена CH1 содержит: (i) аминокислотный остаток F в положении 147 и аминокислотный остаток R в положении 183; (ii) аминокислотный остаток F в положении 147 и аминокислотный остаток K в положении 183; (iii) аминокислотный остаток F в положении 147 и аминокислотный остаток Y в положении 183; (iv) аминокислотный остаток R в положении 183; (v) аминокислотный остаток K в положении 183; или (vi) аминокислотный остаток Y в положении 183. Необязательно такой вариант домена CH1 может содержать аминокислотную последовательность: (i) SEQ ID NO: 137; (ii) SEQ ID NO: 138; (iii) SEQ ID NO: 139; (iv) SEQ ID NO: 60; (v) SEQ ID NO: 41; или (vi) SEQ ID NO: 136.
В некоторых вариантах осуществления полипептид варианта домена CH1 содержит аминокислотную замену в аминокислотном положении CH1 в пределах области контакта между CH1 и VH. Необязательно аминокислотное положение CH1 в пределах такой области контакта представляет собой положение 151. Кроме того, необязательно такой вариант домена CH1 может содержать аминокислотный остаток A или L в положении 151.
В некоторых вариантах осуществления полипептид варианта домена CH1 дополнительно содержит одну или более аминокислотных замен, которые повышают спаривание домена CH1 с: (i) доменом CL-каппа по сравнению с доменом CL-лямбда; и/или (ii) полипептидом легкой каппа-цепи по сравнению с полипептидом легкой лямбда-цепи.
В некоторых вариантах осуществления предложен полипептид варианта домена CH1 по любому из пп. 2-10 формулы изобретения, который обеспечивает повышенное спаривание с: (i) доменом CL-каппа по сравнению с доменом CL-лямбда; и/или (ii) полипептидом легкой каппа-цепи по сравнению с полипептидом легкой лямбда-цепи, на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%. Повышение спаривания с каппа-цепью может необязательно быть измерено с помощью жидкостной хроматографии с масс-спектрометрией (ЖХ-МС).
В некоторых вариантах осуществления предложен полипептид варианта домена CH1 по любому из пп. 2-10 формулы изобретения, который обеспечивает повышенное спаривание с: (i) доменом CL-каппа по сравнению с доменом CL-лямбда; и/или (ii) полипептидом легкой каппа-цепи по сравнению с полипептидом легкой лямбда-цепи, по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз. Повышение спаривания с каппа-цепью можно необязательно быть количественно определено с помощью проточной цитометрии, например, путем сравнения соотношения средней интенсивности флуоресценции (MFI) для окрашивания CL-каппа и окрашивания CL-лямбда.
В некоторых вариантах осуществления полипептид варианта домена CH1 согласно настоящему изобретению содержит аминокислотную замену в одном или более из следующих положений: 119, 124, 126, 127, 130, 131, 133, 134, 138-142, 152, 163, 168, 170, 171, 175, 176, 181, 183-185, 187, 197, 203, 208, 210-214, 216 и 218 согласно нумерации EU. Необязательно вариант домена CH1 предпочтительно спаривается с: (i) доменом CL-лямбда по сравнению с доменом CL-каппа; и/или (ii) полипептидом легкой лямбда-цепи по сравнению с полипептидом легкой каппа-цепи.
В отдельных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи содержит аминокислотную замену в одном или более из положений 141, 170, 171, 175, 181, 184, 185, 187 и 218.
В отдельных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи содержит одну или более из следующих аминокислотных замен: положение 119 заменено R; положение 124 заменено V; положение 126 заменено V; положение 127 заменено G; положение 130 заменено H или S; положение 131 заменено Q, T, N, R, V или D; положение 133 заменено D, T, L, E, S или P; положение 134 заменено A, H, I, P, V, N или L; положение 138 заменено R; положение 139 заменено A; положение 140 заменено I, V, D, Y, K, S, W, R, L или P; положение 141 заменено D, K, E, T, R, Q, V или M; положение 142 заменено M; положение 152 заменено G; положение 163 заменено M; положение 168 заменено F, I или V; положение 170 заменено N, G, E, S или T; положение 171 заменено N, E, G, S, A или D; положение 175 заменено D или M; положение 176 заменено R или M; положение 181 заменено V, L, A, K или T; положение 183 заменено L или V; положение 184 заменено R; положение 185 заменено M, L, S, R или T; положение 187 заменено R, D, E, Y или S; положение 197 заменено S; положение 203 заменено D; положение 208 заменено I; положение 210 заменено T; положение 211 заменено A; положение 212 заменено N; положение 213 заменено E; положение 214 заменено R; положение 216 заменено G; и положение 218 заменено L, E, D, P, A, H, S, Q, N, T, I, M, G, C, K или W.
В некоторых других вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи содержит любой один или более из (i)-(xvii): (i) аминокислотный остаток V в положении 126; (ii) аминокислотный остаток G в положении 127; (iii) аминокислотный остаток V в положении 131; (iv) аминокислотный остаток S в положении 133; (v) аминокислотный остаток R в положении 138; (vi) аминокислотный остаток I или V в положении 140; (vii) аминокислотный остаток D, K, E или T в положении 141; (viii) аминокислотный остаток M в положении 142; (ix) аминокислотный остаток I в положении 168; (x) аминокислотный остаток E, G или S в положении 170; (xi) аминокислотный остаток E, D, G, S или A в положении 171; (xii) аминокислотный остаток M в положении 175; (xiii) аминокислотный остаток R в положении 176; (xiv) аминокислотный остаток K, V, A или L в положении 181; (xv) аминокислотный остаток R в положении 184; (xvi) аминокислотный остаток R в положении 185; (xvii) аминокислотный остаток R в положении 187; и (xviii) аминокислотный остаток L, E, D, P, A, H, S, Q, N, T, I, M, G, C или W в положении 218.
В отдельных предпочтительных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит или состоит из одной или более из следующих замен: 141D, 141E, 171E, 170E, 185R и 187R.
В отдельных предпочтительных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит или состоит из двух или более из следующих замен: 141D, 141E, 171E, 170E, 185R и 187R.
В отдельных предпочтительных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит или состоит из трех или более из следующих замен: 141D, 141E, 171E, 170E, 185R и 187R.
В отдельных предпочтительных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит или состоит из следующих замен: (i) 141E и 185R; (ii) 141E и 187R; (iii) 141E, 170E или 171E и 185R; (iv) 141E, 170E или 171E и 187R; (v) 141D и 185R; (vi) 141D и 187R; (vii) 141D, 170E или 171E и 185R; (viii) 141D, 170E или 171E и 187R; (ix) 141E, 185R и 187R; или (x) 141D, 185R и 187R.
В некоторых других вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит замену в одном или более положениях 141 на D, K или E, необязательно в паре с заменой в положении 181 на K, и дополнительно необязательно в паре с заменой в положении 218 на L, E, D, P, A, H, S, Q, N, T, I, M, G, C или W.
В некоторых других вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит замену в положении 141 на D, K или E в паре с заменой в положении 181 на K, и/или замену в положении 218 на L, E, D, P, A, H, S, Q, N, T, I, M, G, C или W.
В дополнительных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит любой один или более из (i)-(xvii): (i) аминокислотный остаток D, E или K в положении 141; (ii) аминокислотный остаток E в положении 170; (iii) аминокислотный остаток E в положении 171; (iv) аминокислотный остаток M в положении 175; (v) аминокислотный остаток K в положении 181; (vi) аминокислотный остаток R в положении 184; (vii) аминокислотный остаток R в положении 185; (viii) аминокислотный остаток R в положении 187; (ix) аминокислотный остаток P, A или E в положении 218.
В дополнительных вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению содержит: (i) аминокислотный остаток D в положении 141; (ii) аминокислотный остаток D в положении 141 и аминокислотный остаток K в положении 181; (iii) аминокислотный остаток D в положении 141, аминокислотный остаток K в положении 181 и аминокислотный остаток A в положении 218; (iv) аминокислотный остаток D в положении 141, аминокислотный остаток K в положении 181 и аминокислотный остаток P в положении 218; (v) аминокислотный остаток E в положении 141; (vi) аминокислотный остаток E в положении 141 и аминокислотный остаток K в положении 181; (vii) аминокислотный остаток K в положении 141; (viii) аминокислотный остаток K в положении 141 и аминокислотный остаток K в положении 181; (ix) аминокислотный остаток K в положении 141, аминокислотный остаток K в положении 181 и аминокислотный остаток E в положении 218; (x) аминокислотный остаток K в положении 141, аминокислотный остаток K в положении 181 и аминокислотный остаток P в положении 218; (xi) аминокислотный остаток E в положении 141, аминокислотный остаток E в положении 170, аминокислотный остаток V в положении 181 и аминокислотный остаток R в положении 187; (xii) аминокислотный остаток E в положении 141, аминокислотный остаток D в положении 171 и аминокислотный остаток R в положении 185; (xiii) аминокислотный остаток E в положении 141, аминокислотный остаток E в положении 171 и аминокислотный остаток R в положении 185; (xiv) аминокислотный остаток E в положении 141, аминокислотный остаток G в положении 171, аминокислотный остаток R в положении 185 и аминокислотный остаток R в положении 187; (xv) аминокислотный остаток E в положении 141, аминокислотный остаток R в положении 185 и аминокислотный остаток R в положении 187; (xvi) аминокислотный остаток E в положении 141, аминокислотный остаток S в положении 171 и аминокислотный остаток K в положении 181; (xvii) аминокислотный остаток E в положении 141, аминокислотный остаток G в положении 170, аминокислотный остаток M в положении 175, аминокислотный остаток V в положении 181, аминокислотный остаток R в положении 184 и аминокислотный остаток R в положении 187; (xviii) аминокислотный остаток E в положении 141 и аминокислотный остаток R в положении 185;(xix) аминокислотный остаток E в положении 141 и аминокислотный остаток R в положении 187;(xx) аминокислотный остаток E в положении 141, аминокислотный остаток E в положении 170 и аминокислотный остаток R в положении 185; (xxi) аминокислотный остаток E в положении 141, аминокислотный остаток E в положении 170 и аминокислотный остаток R в положении 187; (xxii) аминокислотный остаток D в положении 141 и аминокислотный остаток R в положении 185; (xxiii) аминокислотный остаток D в положении 141 и аминокислотный остаток R в положении 187; (xxiv) аминокислотный остаток D в положении 141, аминокислотный остаток R в положении 185 и аминокислотный остаток R в положении 187; (xxv) аминокислотный остаток D в положении 141, аминокислотный остаток E в положении 170 и аминокислотный остаток R в положении 185; (xxvi) аминокислотный остаток D в положении 141, аминокислотный остаток E в положении 170 и аминокислотный остаток R в положении 187; (xxvii) аминокислотный остаток E в положении 141, аминокислотный остаток E в положении 171 и аминокислотный остаток R в положении 187; (xxiii) аминокислотный остаток D в положении 141, аминокислотный остаток E в положении 171, и аминокислотный остаток R в положении 185; или (xxix) аминокислотный остаток D в положении 141, аминокислотный остаток E в положении 171, и аминокислотный остаток R в положении 187.
Необязательно такой вариант домена CH1 содержит аминокислотную последовательность: (i) SEQ ID NO: 140; (ii) SEQ ID NO: 141; (iii) SEQ ID NO: 142; (iv) SEQ ID NO: 143; (v) SEQ ID NO: 144; (vi) SEQ ID NO: 145; (vii) SEQ ID NO: 146; (viii) SEQ ID NO: 147; (ix) SEQ ID NO: 148; (x) SEQ ID NO: 149; (xi) SEQ ID NO: 155; (xii) SEQ ID NO: 157; (xiii) SEQ ID NO: 159; (xiv) SEQ ID NO: 162; (xv) SEQ ID NO: 163; (xvi) SEQ ID NO: 164; (xvii) SEQ ID NO: 165; (xviii) SEQ ID NO: 178; (xix) SEQ ID NO: 179; (xx) SEQ ID NO: 180; (xxi) SEQ ID NO: 181; (xxii) SEQ ID NO: 182; (xxiii) SEQ ID NO: 183; (xxiv) SEQ ID NO: 184; (xxv) SEQ ID NO: 185; (xxvi) SEQ ID NO: 186; (xxvii) SEQ ID NO: 187; (xxviii) SEQ ID NO: 188; или (xxix) SEQ ID NO: 189.
В некоторых предпочтительных вариантах осуществления вариант домена CH1 с предпочтительностью в отношении лямбда-цепи содержит (i) аминокислотный остаток D в положении 141, аминокислотный остаток E в положении 171 и аминокислотный остаток R в положении 185; или (ii) аминокислотный остаток D в положении 141, аминокислотный остаток E в положении 170 и аминокислотный остаток R в положении 187.
В более предпочтительных вариантах осуществления вариант домена CH1 с предпочтительностью в отношении лямбда-цепи содержит аминокислотные замены, состоящие из: (i) аминокислотного остатка D в положении 141, аминокислотного остатка E в положении 171 и аминокислотного остатка R в положении 185; или (ii) аминокислотного остатка D в положении 141, аминокислотного остатка E в положении 170 и аминокислотного остатка R в положении 187.
В отдельных предпочтительных вариантах осуществления вариант домена CH1 с предпочтительностью в отношении лямбда-цепи содержит аминокислотные замены, состоящие из: (i) SEQ ID NO: 188; или (ii) SEQ ID NO: 186.
В некоторых вариантах осуществления полипептид варианта домена CH1 с предпочтительностью в отношении лямбда-цепи может дополнительно содержать одну или более аминокислотных замен, которые повышают спаривание домена CH1 с: (i) доменом CL-лямбда по сравнению с доменом CL-каппа; и/или (ii) полипептидом легкой лямбда-цепи по сравнению с полипептидом легкой каппа-цепи.
В некоторых вариантах осуществления полипептид варианта домена CH1 может обеспечивать повышенное спаривание с: (i) доменом CL-лямбда по сравнению с доменом CL-каппа; и/или (ii) полипептидом легкой лямбда-цепи по сравнению с полипептидом легкой каппа-цепи, на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%. Повышение спаривания с лямбда-цепью может быть необязательно измерено с помощью жидкостной хроматографии с масс-спектрометрией (ЖХ-МС).
В некоторых вариантах осуществления полипептид варианта домена CH1 может обеспечивать повышенное спаривание с: (i) доменом CL-лямбда по сравнению с доменом CL-каппа; и/или (ii) полипептидом легкой лямбда-цепи по сравнению с полипептидом легкой каппа-цепи, по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, по меньшей мере в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз. Повышение спаривания с лямбда-цепью может быть необязательно измерено с помощью проточной цитометрии, необязательно путем сравнения соотношения значения MFI для окрашивания CL-лямбда и окрашивания CL-каппа.
В еще одном аспекте в настоящем документе дополнительно предложены полипептиды тяжелой цепи антитела, содержащие вариабельную область и константную область, где константная область содержит вариант домена CH1 в соответствии с любым из описанных выше.
В некоторых вариантах осуществления вариант домена CH1 такого полипептида тяжелой цепи антитела содержит аминокислотные замены, состоящие из:
(I) (i) аминокислотного остатка F в положении 147 и аминокислотного остатка R в положении 183; (ii) аминокислотного остатка F в положении 147 и аминокислотного остатка K в положении 183; (iii) аминокислотного остатка F в положении 147 и аминокислотного остатка Y в положении 183; (iv) аминокислотного остатка R в положении 183; (v) аминокислотного остатка K в положении 183; или (vi) аминокислотного остатка Y в положении 183; или
(II) (i) аминокислотного остатка D в положении 141, аминокислотного остатка E в положении 171 и аминокислотного остатка R в положении 185; или (ii) аминокислотного остатка D в положении 141, аминокислотного остатка E в положении 170 и аминокислотного остатка R в положении 187.
В еще одном аспекте в настоящем документе дополнительно предложены антитела или фрагменты антител, содержащие первый полипептид тяжелой цепи и первый полипептид легкой цепи, где (a) первый полипептид тяжелой цепи и первый полипептид легкой цепи образуют первую когнатную пару; и (b) первый полипептид тяжелой цепи содержит первый вариант домена CH1, содержащий аминокислотную замену в одном или более из следующих положений: 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175, 176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216 и 218 согласно нумерации EU, так что первый вариант домена CH1 предпочтительно связывается с первой легкой цепью. Необязательно первый полипептид легкой цепи содержит первый домен CL, который представляет собой домен CL дикого типа. Дополнительно необязательно некоторые варианты домена CH1 могут быть исключены, как описано выше, и варианты домена CH1 согласно настоящему изобретению могут удовлетворять одному или более из пунктов (a)-(o), как описано выше. Также в настоящем документе предложены такие антитела или фрагменты антител, дополнительно содержащие второй полипептид тяжелой цепи и второй полипептид легкой цепи, где: (a) второй полипептид тяжелой цепи и второй полипептид легкой цепи образуют вторую когнатную пару; и (b) второй полипептид тяжелой цепи содержит второй вариант домена CH1, содержащий аминокислотную замену в одном или более из следующих положений: 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175, 176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216 и 218 согласно нумерации EU, так что второй вариант домена CH1 предпочтительно связывается со вторым полипептидом легкой цепи, содержащим второй домен CL. Как и в предыдущем варианте осуществления, необязательно некоторые варианты домена CH1 могут быть исключены, как описано выше, и варианты домена CH1 согласно настоящему изобретению могут удовлетворять одному или более из пунктов (a)-(o), как описано выше. Дополнительно необязательно такое антитело или фрагмент антитела характеризуется одним или более из признаков (i)-(vii): (i) первый домен CL представляет собой домен CL дикого типа; (ii) второй домен CL представляет собой домен CL дикого типа; (iii) первый домен CL представляет собой домен CL-каппа; (iv) первый домен CL представляет собой домен CL-лямбда; (v) второй домен CL представляет собой домен CL-каппа; (vi) второй домен CL представляет собой домен CL-лямбда; (vii) первый вариант домена CH1 представляет собой вариант домена CH1 по любому из пп. 1-20 формулы изобретения; (viii) второй вариант домена CH1 представляет собой вариант домена CH1 по любому из пп. 1-20 формулы изобретения; и/или (ix) аминокислотная замена (замены) в первом варианте домена CH1 отличается от аминокислотной замены (замен) во втором варианте домена CH1.
Дополнительно в настоящем документе предложены антитела или фрагменты антител, содержащие первый полипептид тяжелой цепи и первый полипептид легкой цепи, где: (a) первый полипептид тяжелой цепи и первый полипептид легкой цепи образуют первую когнатную пару; (b) первый полипептид тяжелой цепи содержит первый вариант домена CH1 в соответствии с любым из вариантов домена CH1 с предпочтительностью в отношении каппа-цепи, описанных выше; и (c) первый полипептид легкой цепи содержит домен CL-каппа и необязательно представляет собой полипептид легкой каппа-цепи. Необязательно (i) домен CL-каппа представляет собой домен CL дикого типа; и/или (ii) первый полипептид легкой цепи представляет собой полипептид легкой цепи дикого типа. В отдельных вариантах осуществления первый полипептид тяжелой цепи необязательно содержит одну или более аминокислотных замен за пределами домена CH1, что дополнительно способствует предпочтительному спариванию тяжелой цепи с: (i) доменом CL-каппа по сравнению с доменом CL-лямбда и/или (ii) полипептидом легкой каппа-цепи по сравнению с полипептидом легкой лямбда-цепи. Одна или более аминокислотных замен за пределами домена CH1 могут присутствовать, например, в VH.
Также в настоящем документе предложены антитела или фрагменты антител, содержащие второй полипептид тяжелой цепи и второй полипептид легкой цепи, где: (a) второй полипептид тяжелой цепи и второй полипептид легкой цепи образуют первую когнатную пару; (b) второй полипептид тяжелой цепи содержит второй вариант домена CH1 в соответствии с любым из вариантов домена CH1 с предпочтительностью в отношении лямбда-цепи, описанных выше; и (c) второй полипептид легкой цепи содержит домен CL-лямбда и необязательно представляет собой полипептид легкой лямбда-цепи. Необязательно (i) домен CL-лямбда представляет собой домен CL дикого типа; и/или (ii) второй полипептид легкой цепи представляет собой полипептид легкой цепи дикого типа. В отдельных вариантах осуществления второй полипептид тяжелой цепи необязательно содержит одну или более аминокислотных замен за пределами домена CH1, что дополнительно способствует предпочтительному спариванию тяжелой цепи с: (i) доменом CL-лямбда по сравнению с доменом CL-каппа и/или (ii) полипептидом легкой лямбда-цепи по сравнению с полипептидом легкой каппа-цепи.
Также в настоящем документе предложены антитела или фрагменты антител, содержащие первый полипептид тяжелой цепи, первый полипептид легкой цепи, второй полипептид тяжелой цепи и второй полипептид легкой цепи, где: (a) первый полипептид тяжелой цепи и первый полипептид легкой цепи образуют первую когнатную пару; (b) первый полипептид тяжелой цепи содержит первый домен CH1, содержащий вариант домена CH1 в соответствии с любым из вариантов домена CH1 с предпочтительностью в отношении каппа-цепи, описанных выше; (c) первый полипептид легкой цепи содержит домен CL-каппа и необязательно представляет собой полипептид легкой каппа-цепи;(d) второй полипептид тяжелой цепи и второй полипептид легкой цепи образуют вторую когнатную пару; (e) второй полипептид тяжелой цепи содержит второй домен CH1, содержащий вариант домена CH1 в соответствии с любым из вариантов домена CH1 с предпочтительностью в отношении лямбда-цепи, описанных выше; и (f) второй полипептид легкой цепи содержит домен CL-лямбда и необязательно представляет собой полипептид легкой лямбда-цепи. В отдельных вариантах осуществления первый полипептид тяжелой цепи необязательно содержит одну или более аминокислотных замен за пределами домена CH1, что дополнительно способствует предпочтительному спариванию тяжелой цепи с: (i) доменом CL-каппа по сравнению с доменом CL-лямбда и/или (ii) полипептидом легкой каппа-цепи по сравнению с полипептидом легкой лямбда-цепи. Одна или более аминокислотных замен за пределами домена CH1 могут присутствовать, например, в VH. В отдельных вариантах осуществления второй полипептид тяжелой цепи необязательно содержит одну или более аминокислотных замен за пределами домена CH1, что дополнительно способствует предпочтительному спариванию тяжелой цепи с: (i) доменом CL-лямбда по сравнению с доменом CL-каппа и/или (ii) полипептидом легкой лямбда-цепи по сравнению с полипептидом легкой каппа-цепи.
Любое из антител или фрагментов антител могут быть мультиспецифичными, необязательно биспецифичными. Необязательно структура такого антитела или фрагмента антитела соответствует показанной на любой из фиг. 24-29.
В некоторых вариантах осуществления в мультиспецифичном антителе или фрагменте антитела, как описано выше, первый и второй варианты домена CH1 снижают образование некогтнатных пар тяжелой цепи и легкой цепи на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%. В некоторых вариантах осуществления в мультиспецифичном антителе или фрагменте антитела, как описано выше, первый и второй варианты домена CH1 повышают образование когтнатных пар тяжелой цепи и легкой цепи на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%.
В некоторых вариантах осуществления снижение некогтнатного спаривания тяжелых цепей с легкими и/или повышение когнатного спаривания тяжелых цепей с легкими может быть количественно определено путем трансфекции клеток с помощью HC (или VH плюс CH1), содержащей представляющий интерес CH1, LC-каппа и LC-лямбда с заданным соотношением, таким как HC : LC-каппа : LC-лямбда = 2: 1: 1, и измерения молекул легкой цепи с помощью ЖХ-МС, как показано в примере 7 и на фиг. 23, 30 или 31. В отдельных вариантах осуществления с применением такого или аналогичного способа количественного определения иллюстративный CH1 WT (дикого типа) может обеспечивать образование пар HC-LC, 60% из которых представляют собой когнатные пары и 40% из которых представляют собой некогнатные пары, и с вариантом CH1 согласно настоящему изобретению процент когнатных пар может быть повышен до по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%, и процент некогнатных пар может быть снижен до по меньшей мере 35%, по меньшей мере 30%, по меньшей мере 25%, по меньшей мере 20%, по меньшей мере 15%, по меньшей мере 10%, по меньшей мере 5% или 0%. В частных вариантах осуществления с применением такого или аналогичного способа количественного определения процент когнатной пары может быть повышен до по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%, тогда как процент некогтнатной пары может быть снижен до по меньшей мере 15%, по меньшей мере 10%, по меньшей мере 5% или 0%.
В некоторых вариантах осуществления в мультиспецифичном антителе или фрагменте антитела, как описано выше, первый и второй варианты домена CH1 снижают образование некогтнатных пар тяжелой цепи и легкой цепи по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, по меньшей мере в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз. В некоторых вариантах осуществления в мультиспецифичном антителе или фрагменте антитела, как описано выше, первый и второй варианты домена CH1 повышают образование когтнатных пар тяжелой цепи и легкой цепи по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, по меньшей мере в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз.
В некоторых вариантах осуществления снижение некогтнатной пары тяжелой и легкой цепи и/или повышение когнатной пары тяжелой и легкой цепи может быть количественно определено путем одновременного экспрессирования HC (или VH плюс CH1), содержащей представляющий интерес CH1, LC-каппа и LC-лямбда с заданным соотношением, чтобы обеспечить возможность презентации пар тяжелой и легкой цепи на клетке (например, дрожжевой клетке), окрашивания клеток антителами к каппа- и к лямбда-цепи, и количественного определения присутствия каппа- и лямбда-цепи с помощью FACS (сортировка клеток с активированной флуоресценцией), например, путем сравнения значений MFI, как на фиг. 2-5, 8-13 и 19-22. Для сравнения предпочтительности в отношении каппа-цепи определенного CH1 может быть рассчитано соотношение MFI клеток, окрашенных антителом к каппа-цепи, к MFI клеток, окрашенных антителом к лямбда-цепи, и разделено на такое соотношение для CH1 WT для получения значения кратности относительно исходного (FOP). Для сравнения предпочтительности в отношении лямбда-цепи определенного CH1 может быть рассчитано соотношение MFI клеток, окрашенных антителом к лямбда-цепи, к MFI клеток, окрашенных антителом к каппа-цепи, и разделено на такое соотношение для CH1 WT.
В отдельных вариантах осуществления с применением такого или аналогичного способа количественного определения с вариантом CH1 с предпочтительностью в отношении каппа-цепи согласно настоящему изобретению значение FOP (рассчитанное для предпочтительности в отношении каппа-цепи, т.е. MFI каппа:лямбда) может быть повышено по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз. В отдельных вариантах осуществления с применением такого или аналогичного способа количественного определения с вариантом CH1 с предпочтительностью в отношении лямбда-цепи согласно настоящему изобретению значение FOP (рассчитанное для предпочтительности в отношении лямбда-цепи, т.е. MFI лямбда:каппа) может быть повышено по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз.
В некоторых вариантах осуществления второй вариант домена CH1 содержит замену в положении 141 и снижает образование некогнатных пар тяжелой цепи и легкой цепи на по меньшей мере 50%. В некоторых вариантах осуществления второй вариант домена CH1 содержит замену в положении 141, и первый вариант домена CH1 содержит замену в положении 183 и необязательно в положении 147, или наоборот, и снижает образование некогнатных пар тяжелой цепи и легкой цепи в диапазоне от по меньшей мере 50% до по меньшей мере 75%. В некоторых вариантах осуществления второй вариант домена CH1 содержит 141D или 141E, и второй вариант домена CH1 содержит 183R, 183K или 183Y и необязательно 147F, или наоборот, и снижает образование некогнатных пар тяжелой цепи и легкой цепи в диапазоне от по меньшей мере 50% до по меньшей мере 75%. В некоторых вариантах осуществления второй вариант домена CH1 содержит одну или более из 141D или 141E, 170E, 171E, 181K, 185R, 187R и 218P, и первый вариант домена CH1 содержит 183R, 183K или 183Y и необязательно 147F, или наоборот, и снижает образование некогнатных пар тяжелой цепи и легкой цепи в диапазоне от по меньшей мере 50% до по меньшей мере 75%. В некоторых вариантах осуществления второй вариант домена CH1 содержит комбинацию 141D, 171E и 185R, комбинацию 141D, 171E и 187R или комбинацию 141D, 181K и 218P, и второй вариант домена CH1 содержит 183R, 183K или 183Y и необязательно 147F, или наоборот, и снижает образование некогнатных пар тяжелой цепи и легкой цепи в диапазоне от по меньшей мере 50% до по меньшей мере 75%.
В некоторых вариантах осуществления первый и второй варианты домена CH1 обеспечивают по меньшей мере 75%, по меньшей мере 76%, по меньшей мере 77%, по меньшей мере 78%, по меньшей мере 79%, по меньшей мере 80%, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84%, по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% образование требуемых первой и второй когнатных пар. В некоторых вариантах осуществления первый и второй варианты домена CH1 обеспечивают от приблизительно 85% до приблизительно 95% образования требуемых первой и второй когнатных пар. В некоторых вариантах осуществления второй вариант домена CH1 содержит замену в положении 141, и первый вариант домена CH1 содержит замену в положении 183 и необязательно в положении 147, и обеспечивает от приблизительно 85% до по меньшей мере приблизительно 95% образования требуемых первой и второй когнатных пар. В некоторых вариантах осуществления второй вариант домена CH1 содержит 141D или 141E, и первый вариант домена CH1 содержит 183R, 183K или 183Y и необязательно 147F, или наоборот, и обеспечивает от приблизительно 85% до по меньшей мере приблизительно 95% образования требуемых первой и второй когнатных пар. В некоторых вариантах осуществления первый и второй варианты домена CH1 обеспечивают сниженное образование некогнатных пар тяжелой цепи и легкой цепи, составляющее менее 25%, менее 20%, менее 15%, менее 14%, менее 13%, менее 12%, менее 11% менее 10%, менее 9%, менее 8%, менее 7%, менее 6%, менее 5%, менее 4%, менее 3%, менее 2% или менее 1%. В некоторых вариантах осуществления второй вариант домена CH1 содержит замену в положении 141, 170, 171, 181, 185, 187 и/или 218, и первый вариант домена CH1 содержит замену в положении 183 и необязательно в положении 147, или наоборот, и обеспечивает сниженное образование некогнатных пар тяжелой цепи и легкой цепи, составляющее менее приблизительно 15%, менее приблизительно 10% или менее приблизительно 5%. В некоторых вариантах осуществления второй вариант домена CH1 содержит одну или более из 141D или 141E, 170E, 171E, 181K, 185R, 187R и 218P, и первый вариант домена CH1 содержит 183R, 183K или 183Y и необязательно 147F, или наоборот, и обеспечивает сниженное образование некогнатных пар тяжелой цепи и легкой цепи, составляющее менее приблизительно 15%, менее приблизительно 10% или менее приблизительно 5%.
В еще одном аспекте в настоящем документе дополнительно предложены фармацевтические и диагностические композиции, содержащие: (i) полипептид варианта домена CH1, как описано выше; (ii) полипептид тяжелой цепи антитела, как описано; и/или (iii) антитело или фрагмент антитела, как описано выше.
В еще одном аспекте в настоящем документе дополнительно предложены терапевтические и диагностические применения антител и фармацевтических композиций, содержащих: (i) полипептид варианта домена CH1, как описано выше; (ii) полипептид тяжелой цепи антитела, как описано; и/или (iii) антитело или фрагмент антитела, как описано выше.
В еще одном аспекте в настоящем документе дополнительно предложены нуклеиновые кислоты, кодирующие: (i) полипептид варианта домена CH1, как описано выше; (ii) полипептид тяжелой цепи антитела, как описано; и/или (iii) антитело или фрагмент антитела, как описано выше.
В еще одном аспекте в настоящем документе дополнительно предложены векторы, содержащие клетки, трансфицированные нуклеиновыми кислотами, кодирующими: (i) полипептид варианта домена CH1, как описано выше; (ii) полипептид тяжелой цепи антитела, как описано; и/или (iii) антитело или фрагмент антитела, как описано выше, и их применение для получения вышеуказанного.
В еще одном аспекте настоящего изобретения предложены способы создания библиотеки вариантных доменов CH1, где способ включает стадии (a)-(c): (a) обеспечение (i) одного или более наборов полипептида, содержащего домен CH1 в паре с полипептидом, содержащим домен CL-каппа («набор Cκ»); (ii) одного или более наборов полипептида, содержащего домен CH1 в паре с полипептидом, содержащим домен CL-лямбда («набор Cλ»); и/или (iii) в VH в наборе Cκ и/или в наборе Cλ; (b) выбор одного или более аминокислотных положений домена CH1, которые находятся в контакте с одним или более аминокислотными положениями в домене CL-каппа в наборе Cκ и/или в домене CL-лямбда в наборе Cλ; и (c) получение библиотеки полипептидов вариантов домена CH1 или библиотеки конструкций, кодирующих варианты домена CH1, где одно или более из одного или более аминокислотных положений, выбранных на стадии (b), заменены любой аминокислотой, отличной от дикого типа. Необязательно полипептид, содержащий домен CH1, дополнительно содержит вариабельную область тяжелой цепи (VH), дополнительно где необязательно полипептид, содержащий домен CL-каппа или -лямбда, дополнительно содержит вариабельную область легкой цепи (VL).
Необязательно: (I) на стадии (a) указанный домен CH1, указанный домен CL-каппа и указанный домен CL-лямбда являются доменами дикого типа и/или человеческими; (II) на стадии (a) как (i) указанный полипептид, содержащий домен CH1 в паре с полипептидом, содержащим домен CL-каппа, так и (ii) указанный полипептид, содержащий домен CH1 в паре с полипептидом, содержащим домен CL-лямбда, представляют собой интактное антитело или представляют собой антигенсвязывающий фрагмент («Fab»); (III) на стадии (b) одно или более аминокислотных положений домена CH1 выбирают, если аминокислотный остаток в указанных одном или более аминокислотных положениях домена CH1 имеет атом боковой цепи на расстоянии 5 Å от (i) атома боковой цепи аминокислотного остатка в указанных одном или более аминокислотных положениях в домене CL-каппа, (ii) атома боковой цепи аминокислотного остатка в указанных одном или более аминокислотных положениях в домене CL-лямбда, и/или (iii) атома боковой цепи аминокислотного остатка в указанных одном или более аминокислотных положениях в VH; и/или (IV) указанное получение на стадии (c) осуществляется посредством вырожденного кодона, необязательно вырожденного кодона RMW, представляющего шесть встречающихся в природе аминокислот (D, T, A, E, K и N), или вырожденного кодона NNK, представляющего все 20 встречающихся в природе аминокислотных остатков.
В некоторых вариантах осуществления одно или более аминокислотных положений CH1, выбранных на стадии (b), находятся: (i) в области контакта с доменом CL-каппа в по меньшей мере 10% репрезентативного набора из набора Cκ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора из набора Cκ, (ii) в области контакта с доменом CL-лямбда в по меньшей мере 10% репрезентативного набора из набора Cλ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора из набора Cλ, и/или (iii) в области контакта с VH в по меньшей мере 10% репрезентативного набора из набора Cκ и/или Cλ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора из набора Cκ и/или Cλ.
В некоторых вариантах осуществления аминокислотные положения, выбранные на стадии (b), включают одно или более из положений 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175, 176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216 и 218 согласно нумерации EU. Необязательно некоторые варианты домена CH1 могут быть исключены, как описано выше, и варианты домена CH1 согласно настоящему изобретению могут удовлетворять критериям (a)-(o), как описано выше.
В некоторых вариантах осуществления синтезированные полипептиды, которые кодируют вариантные домены CH1 или библиотеку вариантов домена CH1 на стадии (c), экспрессируются в штамме дрожжей. В некоторых вариантах осуществления штамм дрожжей представляет собой Saccharomyces cerevisiae. В некоторых вариантах осуществления клеточная система, такая как штамм дрожжей, совместно экспрессирует (i) один или более полипептидов, содержащих домен CL-каппа, такой как легкая каппа-цепь, и (ii) один или более полипептидов, содержащих домен CL-лямбда, такой как легкая лямбда-цепь. Необязательно, где домены CL-каппа и/или -лямбда представляют собой домены дикого типа. Кроме того, необязательно домены CL-каппа и/или -лямбда являются человеческими.
В некоторых вариантах осуществления способ согласно настоящему изобретению дополнительно включает подтверждение того, что один или более замененных аминокислотных остатков CH1 способствуют предпочтительному спариванию с легкой каппа-цепью или легкой лямбда-цепью. В некоторых вариантах осуществления для подтверждения того, что один или более замененных аминокислотных остатков CH1 способствуют предпочтительному спариванию с легкой каппа-цепью или легкой лямбда-цепью, применяют сортировку клеток с активированной флуоресценцией.
В некоторых вариантах осуществления один или более константных доменов каппа (Cκ), один или более константных доменов лямбда (Cλ) и один или более доменов CH1 представляют собой домены дикого типа. В некоторых вариантах осуществления один или более константных доменов каппа (Cκ), один или более константных доменов лямбда (Cλ) и один или более доменов CH1 являются человеческими.
В некоторых вариантах осуществления способ создания библиотеки домена CH1 включает стадии (a)-(c): (a) выбор одного или более из следующих аминокислотных положений CH1: 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175, 176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216 и 218 согласно нумерации EU, (b) выбор одного или более аминокислотных положений CH1, представляющих интерес, отличных от положения (положений), выбранных на стадии (a); и (c) получение библиотеки полипептидов вариантов домена CH1 или библиотеки конструкций, кодирующих варианты домена CH1, где одно или более из одного или более аминокислотных положений, выбранных на стадии (a) и (b), заменены любой аминокислотой, отличной от дикого типа. В отдельных вариантах осуществления аминокислотное положение (положения), выбранное на стадии (a), может включать положение 141, 147, 151, 170, 171, 181, 183, 185, 187 или 218 или любую их комбинацию. В отдельных вариантах осуществления указанное получение на стадии (c) осуществляется посредством вырожденного кодона, необязательно вырожденного кодона RMW, представляющего шесть встречающихся в природе аминокислот (D, T, A, E, K и N), или вырожденного кодона NNK, представляющего все 20 встречающихся в природе аминокислотных остатков. В отдельных вариантах осуществления на стадии (c) аминокислотное положение (положения), выбранные на стадии (a), могут быть заменены заданной аминокислотой, и аминокислотное положение (положения), выбранные на стадии (b), могут быть заменены вырожденным кодоном. Необязательно замена на заданную аминокислоту может включать A141D, A141E, K147F, P151A, P151L, F170E, P171E, S181K, S183R, V185R, T187R или K218PP, или любую их комбинацию.
В еще одном аспекте настоящего изобретения предложены способы идентификации одного или более полипептидов варианта домена CH1, которые предпочтительно спариваются с: (A) полипептидом, содержащим домен CL-каппа, по сравнению с полипептидом, содержащим домен CL-лямбда; или (B) полипептидом, содержащим домен CL-лямбда, по сравнению с полипептидом, содержащим домен CL-каппа. Такой способ включает стадии (a)-(c): (a) совместную экспрессию одного или более кандидатных полипептидов вариантов домена CH1 с (i) одним или более полипептидами, содержащими домен CL-каппа, и (ii) одним или более полипептидами, содержащими домен CL-лямбда; (b) сравнение (i) количества кандидатного полипептида варианта домена CH1 в паре с полипептидом, содержащим домен CL-каппа, и (ii) количества кандидатного полипептида варианта домена CH1 в паре с полипептидом, содержащим домен CL-лямбда; (c) на основе сравнения на стадии (b) выбор одного или более вариантов домена CH1, которые обеспечивают предпочтительное спаривание с (A) полипептидом, содержащим домен CL-каппа, по сравнению с полипептидом, содержащим домен CL-лямбда; или (B) полипептидом, содержащим домен CL-лямбда, по сравнению с полипептидом, содержащим домен CL-каппа. На стадии (a) обычно общее количество экспрессируемых кандидатных полипептидов вариантов домена CH1 и общее количество экспрессируемых полипептидов, содержащих домен CL (каппа и лямбда), может быть примерно одинаковым. При этом необязательно на стадии (a) кандидатные полипептиды вариантов домена CH1, полипептиды, содержащие домен CL-каппа, и полипептиды, содержащие домен CL-лямбда, экспрессируются примерно в соотношении 2:1:1.
В некоторых вариантах осуществления на стадии (a) указанные (i) один или более полипептидов, содержащих домен CL-каппа, и (ii) один или более полипептидов, содержащих домен CL-лямбда, представляют собой домены дикого типа и/или человека.
В некоторых вариантах осуществления на стадии (b) количество определяют посредством сортировки клеток с активированной флуоресценцией или посредством жидкостной хроматографии с масс-спектрометрией.
В некоторых вариантах осуществления способ дополнительно включает стадию (d): (d) совместная экспрессия одного или более контрольных вариантов домена CH1 с (i) одним или более полипептидами, содержащими домен CL-каппа, и (ii) одним или более полипептидами, содержащими домен CL-лямбда, где необязательно один или более из указанных одного или более контрольных вариантов домена CH1 соответствуют варианту домена CH1 согласно любому из описанных выше.
Краткое Описание Чертежей
Фиг. 1A-C представляют собой схему связывания варианта домена CH1 с доменом Cλ или доменом Cκ. На фиг. 1A показана гетеродимеризация домена CH1 дикого типа с Cλ и Cκ (домен CH1 дикого типа или немодифицированный домен CH1 обозначен CH1WT). На фиг. 1B показан вариант домена CH1, обладающий предпочтительным спариванием с Cκ (такие варианты домена CH1 с предпочтительным спариванием с Cκ обозначены CH1κ). На фиг. 1C показан вариант домена CH1, обладающий предпочтительным спариванием с Cλ (такие варианты домена CH1 с предпочтительным спариванием с Cλ обозначены CH1λ).
На фиг. 2A и 2B показаны иллюстративные графики FACS для нескольких раундов отбора для идентификации вариантов домена CH1 с предпочтительностью в отношении домена CL-лямбда (фиг. 2A) или домена CL-каппа (фиг. 2B). R1 = первый раунд отбора, R2 = второй раунд отбора, R3 = третий раунд отбора. На оси x показаны легкие лямбда-цепи, меченые PE, а на оси y показаны легкие каппа-цепи, меченые FITC.
На фиг. 3 показаны отдельные уникальные клоны, экспрессирующие вариант домена CH1 с предпочтительностью в отношении домена CL-лямбда или предпочтительностью в отношении домена CL-каппа. Клоны оценивали по соотношению медианной интенсивности флуоресценции (MFI) антитела к каппа-цепи и MFI антитела к лямбда-цепи (соотношение каппа:лямбда). Соотношение каппа:лямбда для любого отдельного клона сравнивали с соответствующим штаммом с последовательностью CH1 дикого типа («исходным»). FOP означает кратность относительно исходного.
На фиг. 4 показаны отдельные уникальные клоны, экспрессирующие вариант домена CH1 с аминокислотной заменой в положении 141, 147 или 183 (нумерация EU). Клоны оценивали по соотношению MFI антитела к каппа-цепи и MFI антитела к лямбда-цепи и сравнивали с исходным для определения FOP. Положения 147 и 183 CH1 были идентифицированы как два положения, обеспечивающие предпочтительность в отношении домена CL-каппа. Положение 141 CH1 было идентифицировано как положение, обеспечивающее предпочтительность в отношении домена CL-лямбда.
На фиг. 5 показаны конкретные аминокислотные замены в положениях 141, 147 и/или 183 (нумерация EU) в домене CH1 с предпочтительностью в отношении домена CL-лямбда (A141T, Q, D или R) или предпочтительностью в отношении домена CL-каппа (K147V, A, F, Y или M; S183K, Y, E, R, W, Q), согласно измерениям соотношения MFI антитела к каппа-цепи и MFI антитела к лямбда-цепи. Аминокислотные замены, показанные в виде белых точек (V134; T141, V147; A151 и K183), были идентифицированы после первоначального отбора из библиотек с диверсификацией в нескольких положениях, а аминокислотные замены, показанные в виде черных точек, были идентифицированы после дополнительных раундов отбора из библиотек с диверсификацией, нацеленной на положения 141, 147 и 183. Исходное соотношение κ:λ (сигнал дикого типа): GAL1 Cκ; GAL10 Cλ: 3,58 и GAL1 Cλ; GAL10 Cκ: 0,3. Исходные соотношения представляют собой средние значения по сравнению с экспериментальными параллельными анализами. Для вариантов CH1 с заменами в обоих положениях 147 и 183 первая указанная аминокислота представляет собой вариант в положении 147, а вторая указанная аминокислота представляет собой вариант в положении 183 (например, Y x F означает вариант CH1 с заменами K147Y и S183F).
На фиг. 6A-E показаны репрезентативные данные по связыванию, демонстрирующие, что вариант домена CH1 не изменял связывание мультиспецифичного антитела (BsAb2-BsAb14) по сравнению с доменом CH1 дикого типа (BsAb1 и BsAb15). На фиг. 6A показаны данные по связыванию IL12B и EGFR для BsAb1-3. На фиг. 6B показаны данные по связыванию IL12B и EGFR для BsAb 5, 7 и 4. На фиг. 6C показаны данные по связыванию IL12B и EGFR для BsAb 9, 10 и 6. На фиг. 6D показаны данные по связыванию IL12B и EGFR для BsAb 11, 12 и 8. На фиг. 6E показаны данные по связыванию IL12B и EGFR для BsAb13-15. Pani = панитумумаб; Uste = устекинумаб.
На фиг. 7 показано повышение правильного спаривания тяжелых цепей с легкими цепями (HC1-LC1 или HC2-LC2) и одновременное снижение неправильного спаривания тяжелых цепей с легкими цепями (HC1-LC2 и HC2-LC1) в биспецифичных антителах, содержащих вариант домена CH1 (BsAb2-BsAb14), по сравнению с биспецифичным антителом, содержащим домен CH1 дикого типа (BsAb1).
На фиг. 8 показаны значения FOP предпочтительности в отношении лямбда-цепи для клона WT, клона A141D и отдельных клонов, имеющих различные аминокислотные замены в положениях 141, 181 и 218 домена CH1, полученных из продукта отбора библиотеки 141x181x218 в примере 5. 13 точек данных, отмеченных в прямоугольнике, соответствуют клонам с наивысшими значениями FOP, и аминокислотные остатки в положениях 141, 181 и 218 CH1, а также значение FOP для каждого клона представлены в таблице 8.
На фиг. 9 показаны значения FOP предпочтительности для клона WT, клона A141D и отдельных клонов, имеющих D в положении 141, K в положении 181 и различные аминокислоты в положении 218 домена CH1 в продукте отбора библиотеки 141x181x218 в примере 5. Незакрашенные точки данных представляют собой FOP отдельных клонов, имеющих одинаковую последовательность CH1, а закрашенные точки данных представляют собой средние значения FOP.
На фиг. 10 показаны значения FOP предпочтительности в отношении лямбда-цепи, измеренные с помощью повторно клонированных клонов и клонов WT и A141D, которые подтверждают сохраняемую предпочтительность в отношении лямбда-цепи.
На фиг. 11 показаны иллюстративные графики рассеяния HEK293, полученные с помощью IgG, характеризующихся CH1 из одного из девяти наилучших результатов 141x181x218, выбранных в примере 5, и WT и A14D, окрашенных на CL-каппа и CL-лямбда. Графики рассеяния отдельных клонов наложены на график WT. На оси x показаны легкие лямбда-цепи, меченые PE, а на оси y показаны легкие каппа-цепи, меченые FITC.
На фиг. 12 показаны значения FOP предпочтительности в отношении лямбда-цепи для девяти наилучших результатов из примера 5 вместе с WT и A141D. Три варианта CH1 с самыми высокими значениями FOP (D_K_WT, D_K_P и D_K_A) были выбраны для последующей трансфекции двух цепей (каппа или лямбда) в HEK293.
На фиг. 13 сравниваются значения FOP предпочтительности в отношении лямбда-цепи для вариантов CH1, имеющих одну и ту же аминокислоту в положении 141. Когда положение 141 представляет собой D, дополнительная аминокислотная замена в положении 181 или в положениях 181 и 218 дополнительно повышает значение FOP.
На фиг. 14 показан % молекул легких цепей (сравнение каппа- и лямбда-) с девятью наилучшими полноразмерными IgG, продуцированными в HEK293, по результатам измерений с помощью жидкостной хроматографии с масс-спектрометрией («ЖХ-МС»). Три варианта CH1 с самыми высокими значениями FOP (D_K_WT, D_K_P и D_K_A) были выбраны для последующей трансфекции в HEK293.
На фиг. 15 показаны иллюстративные выходы процесса для трех наилучших результатов (D_K_WT, D_K_P и D_K_A) и A141D относительно выхода WT, показанные как значения кратности относительно исходного («FOP»).
На фиг. 16 показана Tm (°C) Fab, спаренных с каппа-цепью, и Fab, спаренных с лямбда-цепью, имеющих один из трех наилучших вариантов CH1 (D_K_WT, D_K_P и D_K_A) или A141D или WT.
На фиг. 17 показана относительная Tm (°C) лямбда-цепи, определяемая как: [изменение Tm в варианте Fab, спаренном с лямбда-цепью, относительно Fab WT, спаренного с лямбда-цепью («Δ Tm лямбда»)] - [изменение Tm в варианте Fab, спаренном с каппа-цепью, относительно Fab WT, спаренного с каппа-цепью («Δ Tm каппа»)].
На фиг. 18 представлены результаты секвенирования продуктов повторного клонирования в примере 6, при этом визуализированы частые аминокислотные замены, наблюдаемые среди полученных клонов.
На фиг. 19 показаны значения FOP предпочтительности в отношении лямбда-цепи (MFI лямбда: MFI каппа) для наилучших результатов из продуктов повторного клонирования в примере 6, а также некоторые из наилучших результатов 141x181x218 (DKP, DKA, KKE, KKP и EKK) из примера 5, экспрессируемых в виде IgG в дрожжах. По меньшей мере семь наилучших результатов, помеченных стрелкой, имеют значение FOP, эквивалентное или превышающее значение протестированных наилучших результатов 141x181x218.
На фиг. 20 показаны значения FOP предпочтительности в отношении лямбда-цепи для 14 наилучших результатов из примера 7, а также DKP, идентифицированного в примере 5, A141D и дикого типа. Два наилучших результата, «A414D_P171E_V185R» и «A141D_F170E_T187R», отмеченные стрелкой, продемонстрировали более высокие значения FOP, чем DKP. Все 14 наилучших результатов продемонстрировали более высокое значение FOP, чем у дикого типа.
На фиг. 21 показаны иллюстративные графики FACS, сравнивающие предпочтительность в отношении лямбда-цепи для 14 вариантов домена CH1 в примере 7 и трех контролей (DKP, идентифицированного в примере 5, A141D и дикого типа). На оси x показаны легкие лямбда-цепи, меченые PE, а на оси y показаны легкие каппа-цепи, меченые FITC. Нумерация на каждом графике представляет собой ранжирование, показанное в таблице 14. Например, первые два графика, пронумерованные как «1» и «2», представляют собой графики «A414D_P171E_V185R» и «A141D_F170E_T187R» соответственно.
На фиг. 22 показаны иллюстративные наложения графика FACS для индивидуального графика (отмеченного как «a»), графика дикого типа (отмеченного как «b») и графика DKP (отмеченного как «c») с фиг. 21.
На фиг. 23 показан % молекул легких цепей (сравнение каппа- и лямбда-) с 14 наилучшими результатами и тремя контрольными полноразмерными IgG, продуцированными в HEK293, по результатам измерений с помощью ЖХ-МС в примере 7. Три контроля показаны с помощью незакрашенной стрелки. «A414D_P171E_V185R» и «A141D_F170E_T187R» (закрашенная стрелка) показали более высокий % лямбда-цепей и более низкий % каппа-цепей по сравнению с положительным контролем, «DKP».
На фиг. 24-29 представлены иллюстративные и неограничивающие варианты осуществления различных структур мультиспецифичных антител, с которыми могут быть использованы варианты домена CH1, раскрытые в настоящем документе. На фиг. 24-29 применяется следующее, если не указано иное: (1) каждый домен представлен в виде прямоугольника с текстом, в котором приведено название домена (например, CH1, VH1 и т.д.); (2) закрашенные прямоугольники и прямоугольники с заливкой точками представляют собой варианты домена CH1 с предпочтительностью в отношении каппа- или лямбда-цепи, которые могут представлять собой вариант домена CH1, раскрытый в настоящем документе; (3) «CH1κ» представляет собой вариант домена CH1 с предпочтительностью в отношении CL-каппа, «CH1λ» представляет собой вариант домена CH1 с предпочтительностью в отношении лямбда-цепи, и «CH1» без указания «κ» или «λ» представляет собой любой домен CH1, дикого типа или вариантный, с предпочтительностью в отношении какого-либо изотипа легкой цепи или без нее; (4) «Cκ» представляет собой домен CL-каппа, «Cλ» представляет собой домен CL-лямбда, и «CL» без указания «κ» или «λ», когда показано, что она находится в паре с закрашенным или залитым точками доменом CH1, представляет собой домен CL изотипа (каппа или лямбда), в отношении которого этот спаренный закрашенный или залитый точками домен CH1 обладает предпочтительностью; (5) если в мультиспецифичной структуре присутствует более одного закрашенного и/или залитого точками домена CH1, по меньшей мере один из них представляет собой вариант домена CH1, раскрытый в настоящем документе, а остальные могут представлять или не представлять собой вариант домена CH1, раскрытый в настоящем документе; (6) когда в мультиспецифичной структуре присутствуют как закрашенные, так и залитые точками домены CH1, закрашенные и залитые точками представляют собой домены CH1 с различной предпочтительностью в отношении изотипа легкой цепи (т.е., когда закрашенные представляют собой домен CH1 с предпочтительностью в отношении каппа-цепи, залитые точками представляют собой домен CH1 с предпочтительностью в отношении лямбда-цепи, и наоборот); (7) VH1 и VL1 образуют антигенсвязывающий сайт для первого эпитопа, VH2 и VL2 образуют антигенсвязывающий сайт для второго эпитопа, VH3 и VL3 образуют антигенсвязывающий сайт для третьего эпитопа, VH4 и VL4 образуют антигенсвязывающий сайт для четвертого эпитопа, VH5 и VL5 образуют антигенсвязывающий сайт для пятого эпитопа, и VH6 и VL6 образуют антигенсвязывающий сайт для шестого эпитопа; (8) все эпитопы с первого по шестой могут отличаться друг от друга, или не все эпитопы с первого по шестой могут отличаться друг от друга, при условии, что комбинация специфичности в целом делает представленную структуру мультиспецифичной; (9) набор из нескольких доменов, соединенных друг с другом, представляет собой полипептид (например, полипептид тяжелой цепи, полипептид легкой цепи и т.д.); (10) направление доменов в полипептиде соответствует направлению текста, обозначающего названия доменов, от N-конца к C-концу; (11) между доменами по мере необходимости может быть использован линкер или шарнир, и между полипептидами (и/или внутри домена) может существовать дисульфидная связь (связи), возможно, для обеспечения правильного образования антигенсвязывающего сайта (сайтов), даже если на фигурах в явном виде не показан линкер, шарнир или дисульфидная связь; (12) домен (домены) CH2 и/или CH3, показанный на фигурах, во всех возможных случаях может быть опущен и, при необходимости, заменен шарниром; (13) домены CH1, CH2 и CH3 могут по отдельности быть дикого типа или вариантными и могут по отдельности иметь любой изотип (тяжелой цепи); и (14) когда в структуре присутствует более одного домена CH1, домены CH1 могут иметь или не иметь один и тот же изотип, когда в структуре присутствует более одного домена CH2, домены CH2 могут иметь или не иметь один и тот же изотип, и когда в структуре присутствует более одного домена CH3, домены CH3 могут иметь или не иметь один и тот же изотип.
На фиг. 24A-24C представлены некоторые иллюстративные и неограничивающие варианты осуществления различных структур мультиспецифичных антител, с которыми могут быть использованы варианты домена CH1, раскрытые в настоящем документе. На фиг. 24A в одном из полипептидов использован домен CH1 с предпочтительностью в отношении каппа-цепи («CH1κ»). Другой домен CH1 может иметь или может не иметь предпочтительности в отношении домена CL-лямбда и может представлять или не представлять собой вариант домена CH1, раскрытый в настоящем документе. На фиг. 24B в одном из полипептидов использован домен CH1 с предпочтительностью в отношении лямбда-цепи («CH1λ»). Другой домен CH1 может иметь или может не иметь предпочтительности в отношении домена CL-каппа и может представлять или не представлять собой вариант домена CH1, раскрытый в настоящем документе. На фиг. 24C в одном из полипептидов использован CH1κ, и в одном из полипептидов использован CH1λ. Эта универсальная структура позволяет получать биспецифичное соединение без усилий или с минимальными или меньшими усилиями для удаления неправильно спаренных соединений. По меньшей мере один из доменов CH1κ и CH1λ представляет собой вариант домена CH1, раскрытый в настоящем документе. Как описано выше в (10), направление доменов в полипептиде соответствует направлению текста, обозначающего названия доменов, от N-конца к C-концу. Следовательно, в случае соединения в верхнем левом углу на фиг. 24A первый полипептид содержит VH1-CH1k-CH2-CH3, второй полипептид содержит VL1-Ck, третий полипептид содержит VH2-CH1-CH2-CH3 и четвертый полипептид содержит VL2-CL в направлении от N-конца к C-концу. Треугольник в соединениях вверху в центре и верхнем правом углу на фиг. 24A-24C (и всех других соответствующих фигурах) представляет собой механизм, который способствует гетеродимеризации двух неидентичных полипептидов, такой как конструирование методом «выступ-во-впадину». Соединение внизу в центре на фиг. 24A-24C (и всех других соответствующих фигурах) демонстрирует структуру шарнира, которая соединяет полипептид, содержащий CH1κ, и полипептид, содержащий CH1λ. Несмотря на то, что явным образом показаны две связи (например, дисульфидные связи), соединяющие два полипептида, количество связей и точное расположение/положение связей могут быть варьированы и выбраны нужным образом. В нижнем правом углу на фиг. 24C «+» обозначает смесь двух разных Fab-фрагментов.
На фиг. 25A-25B представлены дополнительные иллюстративные и неограничивающие варианты осуществления различных структур мультиспецифичных антител, с которыми могут быть использованы варианты домена CH1, раскрытые в настоящем документе. Структуры являются схожими с таковыми на фиг. 24A-23C, но порядок доменов отличается. На фиг. 25A CH1κ находится одном и том же полипептиде с VL и CH1λ находится в одном и том же полипептиде с VL. На фиг. 25B Cλ находится в одном и том же полипептиде с CH2 (три верхних и в левом нижнем углу), и Cλ находится в полипептиде, подобном тяжелой цепи (полипептиде, содержащем шарнир) (в правом нижнем углу).
На фиг. 26A-26C представлены дополнительные иллюстративные и неограничивающие варианты осуществления различных структур мультиспецифичных антител, которые содержат два набора из двух антигенсвязывающих сайтов в тандеме и, следовательно, являются четырехвалентными. Структура может быть биспецифичной, триспецифичной или тетраспецифичной, в зависимости от того, что представляют собой первый, второй, третий и четвертый эпитопы. Например, если первый и второй эпитопы отличаются друг от друга, и если четвертый эпитоп является таким же, как первый, второй или третий эпитоп, структура будет представлять собой четырехвалентную триспецифичную структуру.
На фиг. 27A-27C представлены дополнительные иллюстративные и неограничивающие варианты осуществления различных структур мультиспецифичных антител, схожих с таковыми на фиг. 26A-26C, но отличающихся порядком расположения доменов. Как описано выше в (10), направление доменов в полипептиде соответствует направлению текста, обозначающего названия доменов, от N-конца к C-концу. Следовательно, в случае структуры в верхнем левом углу на фиг. 27A первый полипептид содержит VH3-VH1-CH1(закрашенный)-CH2-CH3, второй полипептид содержит VL1-VL3-CL, третий полипептид содержит VH4-VH2-CH1(с заливкой точками)-CH2-CH3, а четвертый полипептид содержит VL2-VL4-CL в направлении от N-конца к C-концу. В любых структурах на фиг. 27A-27C между доменами может использоваться подходящий для обеспечения надлежащего образования антигенсвязывающих сайтов.
На фиг. 28A-28D представлены дополнительные иллюстративные и неограничивающие варианты осуществления различных структур мультиспецифичных антител, которые содержат по меньшей мере один scFv. Любая из структур, представленных на фиг. 24-29, может дополнительно содержать или быть модифицирована так, чтобы она содержала один или более scFv-содержащих фрагментов, например, конъюгированных с любым из константных доменов тяжелой цепи, константных доменов легкой цепи и/или антигенсвязывающих доменов. В качестве примера, на фиг. 28A-28C представлены структуры, в которых структура в верхнем левом углу на фиг. 24A конъюгирована с двумя scFv, что обеспечивает специфичность к максимум четырем эпитопам. На фиг. 28A scFv конъюгированы с доменами CH3. На фиг. 28B scFv конъюгированы с доменами CL. На фиг. 28C scFv конъюгированы с доменами VH. В некоторых случаях может быть конъюгировано более двух scFv. В качестве примера, на фиг. 28A-28C представлены структуры, в которых структура в верхнем левом углу на фиг. 24A конъюгирована с четырьмя scFv, что обеспечивает специфичность к максимум шести эпитопам.
На фиг. 29A-29D представлены дополнительные иллюстративные и неограничивающие варианты осуществления различных структур мультиспецифичных антител, которые содержат два дополнительных Fab-фрагмента. Хотя два Fab-фрагмента конъюгированы с доменами CH3, необходимо отметить, что Fab-фрагменты могут быть конъюгированы с любой другой частью структуры, а также что может быть конъюгирован один (или три или более) вместо двух Fab-фрагментов. На фиг. 29A два домена CH1 находятся в одном и том же полипептиде с доменами CH2 и CH3. В структуре в центре домен CH1 с предпочтительностью в отношении каппа-цепи и домен CH1 с предпочтительностью в отношении лямбда-цепи находятся в одном и том же полипептиде (для обоих из двух CH1-содержащих полипептидов). Когда два CH1-содержащих полипептида являются одинаковыми, эта структура облегчает получение тетравалентных биспецифичных соединений без необходимости в механизме, который способствует гетеродимеризации двух неидентичных полипептидов (таком как метод конструирования «выступ-во-впадину»), например, путем простого применения 3-цепочечной трансфекционной системы, используемой в примерах. На фиг. 29B два полипептида не содержат каких-либо доменов CH1. В структуре в середине, когда два не содержащих CH1 полипептида являются одинаковыми, эта структура облегчает получение тетравалентных биспецифичных соединений без необходимости в механизме, который способствует гетеродимеризации двух неидентичных полипептидов (таком как метод конструирования «выступ-во-впадину»), например, путем простого применения 3-цепочечной трансфекционной системы, используемой в примерах. На фиг. 29C и 29D каждый полипептид содержит домен CH1. В структурах в центре на фиг. 29C и 29D если первый и третий эпитопы являются одним и тем же эпитопом и второй и четвертый эпитопы являются одним и тем же эпитопом, но отличаются от первого и третьего эпитопов, структура является биспецифичной. В такой структуре, если два CH2/CH3-содержащих полипептида являются одинаковыми, эта структура облегчает получение тетравалентных биспецифичных соединений без необходимости в механизме, который способствует гетеродимеризации двух неидентичных полипептидов (таком как метод конструирования «выступ-во-впадину»), например, путем простого применения 3-цепочечной трансфекционной системы, используемой в примерах.
На фиг. 30 показаны иллюстративные выходы процесса для полных IgG, содержащих один из двух наилучших вариантов CH1 с предпочтительностью в отношении лямбда-цепи, идентифицированных в примере 7 («A141D P171E V185R» или «A141D F170E T187R»), или вариант CH1 с предпочтительностью в отношении каппа-цепи, идентифицированный в примере 4 («K147F S183R»), или CH1 WT. Нормированы к выходу процесса для WT. Полосатые столбики (спаренные с каппа) и закрашенные столбики (спаренные с лямбда) представляют собой выход процесса, нормированный к выходу для соответствующего WT. Незакрашенные ромбы (спаренные с каппа) и закрашенные треугольники (спаренные с лямбдой) обозначают непосредственный выход процесса (мг/л).
На фиг. 31 показаны иллюстративные выходы Fab, содержащих один из двух наилучших вариантов CH1 с предпочтительностью в отношении лямбда-цепи, идентифицированных в примере 7 («A141D P171E V185R» или «A141D F170E T187R»), или варианты CH1 с предпочтительностью в отношении лямбда-цепи, идентифицированные в примерах 4 и 5 («A141D» или «A141D S181K K218P»), или вариант CH1 с предпочтительностью в отношении каппа-цепи, идентифицированный в примере 4 («K147F S183R»), или CH1 WT. Нормированы к выходу процесса для WT. Выходы нормированы к выходу для соответствующего WT. Полосатые столбики обозначают Fab, содержащие LC-каппа, а закрашенные столбики обозначают Fab, содержащие LC-лямбда.
На фиг. 32 показана область контакта CH1 дикого типа-Cλ в ее электронной плотности. Иллюстративная электронная плотность в представляющей интерес области для кристаллической структуры Fab вариабельного фрагмента (Fv) панитумумаба и IgG1-CH1 дикого типа в паре с константным доменом лямбда дикого типа (Cλ). Атомы углерода тяжелой цепи (HC) окрашены в светло-серый цвет, атомы углерода легкой лямбда-цепи (λLC) окрашены в белый цвет, атомы азота окрашены в темно-серый цвет, а атомы кислорода окрашены в черный цвет. Белок показан в стержневом виде. Карта электронной плотности с коэффициентами 2Fo-Fc показана в виде серой сетки с контуром на уровне 1σ со срезом 1,6 Å. Данные для этой кристаллической структуры распространяются на атомное разрешение 1,09 Å.
На фиг. 33 показана область контакта CH1 A141D-Cλ в ее электронной плотности. Иллюстративная электронная плотность в представляющей интерес области для кристаллической структуры Fab вариабельного фрагмента (Fv) панитумумаба и IgG1-CH1 с A141D-заменой в паре с константным доменом лямбда дикого типа (Cλ). Атомы углерода тяжелой цепи (HC) окрашены в светло-серый цвет, атомы углерода легкой лямбда-цепи (λLC) окрашены в белый цвет, атомы азота окрашены в темно-серый цвет, а атомы кислорода окрашены в черный цвет. Белок показан в стержневом виде. Карта электронной плотности с коэффициентами 2Fo-Fc показана в виде серой сетки с контуром на уровне 1σ со срезом 2,0 Å. Данные для этой кристаллической структуры распространяются на атомное разрешение 1,2 Å.
На фиг. 34 показана область контакта CH1-Cκ дикого типа в ее электронной плотности. Иллюстративная электронная плотность в представляющей интерес области для кристаллической структуры Fab вариабельного фрагмента (Fv) панитумумаба и IgG1-CH1 дикого типа в паре с константным доменом каппа дикого типа (Cκ). Атомы углерода тяжелой цепи (HC) окрашены в светло-серый цвет, атомы углерода легкой каппа-цепи (κLC) окрашены в белый цвет, атомы азота окрашены в темно-серый цвет, а атомы кислорода окрашены в черный цвет. Белок показан в стержневом виде. Карта электронной плотности с коэффициентами 2Fo-Fc показана в виде серой сетки с контуром на уровне 0,9σ со срезом 1,6 Å. Данные для этой кристаллической структуры распространяются на околоатомное разрешение 2,6 Å.
На фиг. 35 показана область контакта CH1 K147F-S183R-Cκ в ее электронной плотности. Иллюстративная электронная плотность в представляющей интерес области для кристаллической структуры вариабельного фрагмента (Fv) панитумумаба и IgG1-CH1 с заменами K147F-S183R в паре с константным доменом каппа дикого типа (Cκ). Атомы углерода тяжелой цепи (HC) окрашены в светло-серый цвет, атомы углерода легкой каппа-цепи (κLC) окрашены в белый цвет, атомы азота окрашены в темно-серый цвет, а атомы кислорода окрашены в черный цвет. Белок показан в стержневом виде. Карта электронной плотности с коэффициентами 2Fo-Fc показана в виде серой сетки с контуром на уровне 0,9σ со срезом 1,6 Å. Данные для этой кристаллической структуры распространяются на околоатомное разрешение 2,1 Å.
На фиг. 36A-36D показано, что замена HC-A141D обеспечивает возможность водородного связывания с λLC при одновременной дестабилизации спаривания с каппа-цепью посредством стерического столкновения с κLC. На фиг. 36A-36D представлены виды области контакта спаривания, окружающей положение HC-Ala141 между CH1 WT и λLC (фиг. 36A), между CH1 WT и κLC (фиг. 36B), между CH1 A141D и λLC (фиг. 36C) или между CH1 A141D и κLC (фиг. 36D). Область контакта константного домена легкой каппа-цепи (κLC) содержит три гидрофобных остатка Phe116, Phe118 и Leu135, проиллюстрированных на фиг. 36B. Присутствие Thr116 в λLC в структурно эквивалентном положении κLC-Phe116 обеспечивает возможность образования водородной связи с карбоксильной группой HC-Asp141, показанной в виде черной пунктирной линии (фиг. 36C). На фиг. 36D выравнивание HC CH1 A141D-константная лямбда-цепь (Cλ) и CH1 WT-Cκ демонстрирует стерическое столкновение боковой цепи HC-Asp141 с боковой цепью κLC-Phe116. Атомы углерода тяжелой цепи (HC) окрашены в светло-серый цвет, атомы углерода легкой цепи (LC) окрашены в белый цвет, атомы азота окрашены в темно-серый цвет, а атомы кислорода окрашены в черный цвет. Боковые цепи показаны в стержневом виде с прозрачной молекулярной поверхностью, а атомы основной цепи показаны схематично.
На фиг. 37A и 37B показано блокирование HC-Gln175 в последовательности CH1 дикого типа во внутрицепочечной сети водородных связей, которые могут быть нарушены путем замены K147F, обеспечивая свободу взаимодействия HC-Gln175 с κLC-Gln160. На фиг. 37A и 37B представлены виды сети трехчастных водородных связей в HC, включающей Lys147, Asp148 и Gln175, в структуре CH1 панитумумаба дикого типа-константной каппа-цепи (Cκ) (фиг. 37A) и структуре CH1 K147F-S183R панитумумаба-Cκ (фиг. 37B). Атомы углерода тяжелой цепи (HC) окрашены в светло-серый цвет, атомы углерода легкой каппа-цепи (κLC) окрашены в белый цвет, атомы азота окрашены в темно-серый цвет, а атомы кислорода окрашены в черный цвет. Боковые цепи показаны в стержневом виде, а атомы основной цепи показаны схематично. Водородные связи показаны в виде пунктирной линии.
На фиг. 38A-38D показано, что водородная связь между HC-Arg183 и κLC-Thr178 может приводить к спариванию с каппа-цепью, тогда как стерическое столкновение HC-Arg183 с λLC-Tyr178 снижает предпочтительность спаривания с лямбда-цепью. На фиг. 38A-38D представлены виды области, окружающей замену S183R в IgG1-CH1, с водородными связями между HC-Ser183 и λLC-Thr178 в структуре CH1 панитумумаба дикого типа-константной лямбда-цепи (Cλ) (фиг. 38B), а также между HC-Arg183 и κLC-Thr178 в структуре CH1 K147F-S183R панитумумаба-константной каппа-цепи (Cκ) (фиг. 38C). На фиг. 38A показано, что HC-Ser183 и κLC-Thr178 являются слишком удаленными для образования водородной связи. Атомы углерода тяжелой цепи (HC) окрашены в светло-серый цвет, атомы углерода легкой цепи (LC) окрашены в белый цвет, атомы азота окрашены в темно-серый цвет, а атомы кислорода окрашены в черный цвет. Боковые цепи показаны в стержневом виде. Боковая цепь λLC-Tyr178 также показана в виде прозрачной молекулярной поверхности. Водородные связи показаны в виде черных пунктирных линий. На фиг. 38D представлена модель, в которой HC структуры CH1 K147F-S183R панитумумаба-Cκ была наложена на HC структуры CH1 панитумумаба дикого типа-Cλ. Полученная модель демонстрирует явные стерические столкновения между HC-Arg183 и λLC-Tyr178.
Подробное описание изобретения
Если не определено иное, то все технические и научные термины, используемые в настоящем документе, имеют такое же значение, которое обычно понимается специалистом в области техники, к которой относится данное изобретение. Используемый в настоящем документе термин «приблизительно» при использовании в отношении конкретного указанного числового значения означает, что данное значение может отличаться от указанного значения не более чем на 1%. Например, используемое в настоящем документе выражение «приблизительно 100» включает 99 и 101 и все значения между ними (например, 99,1, 99,2, 99,3, 99,4 и т.д.).
Следует понимать, что аспекты и варианты осуществления настоящего изобретения, описанные в настоящем документе, включают «содержащий», «состоящий» и «по существу состоящий из» аспектов и вариантов осуществления.
В настоящем документе предложены сконструированные домены CH1, содержащие по меньшей мере одну аминокислотную замену, которая предотвращает неправильное спаривание тяжелых цепей с легкими цепями, способствуя предпочтительному спариванию содержащей домен CH1 тяжелой цепи с доменом CL-каппа (или легкой каппа-цепью) или доменом CL-лямбда (или легкой лямбда-цепью). Термин «предпочтительное спаривание» относится к спариванию тяжелой цепи (или домена CH1) с легкой цепью (или доменом CL) в полипептиде, например, антителе, например, биспецифичном антителе. Если тяжелая цепь (H1) экспрессируется совместно с двумя различными легкими цепями (L1 и L2), то H1 будет спариваться с каждой из L1 и L2, что приведет к получению смеси H1:L1 и H1:L2. В некоторых случаях H1 может в равной степени хорошо спариваться с L1 и L2, что приводит к получению смеси H1:L1 и H1:L2 примерно в соотношении 50:50. В качестве примера, «предпочтительное спаривание» будет иметь место между H1 и L1, если количество образованного гетеродимера H1:L1 было больше количества образованного гетеродимера H1:L2, когда H1 экспрессируется совместно с L1 и L2. В данном примере H1 предпочтительно спаривается с L1 относительно L2. Если H1 имеет врожденное предпочтение спаривания с L1 по сравнению с L2 (так что соотношение H1:L1 к H1:L2 составляет не 50:50, а, например, 60:40 или 70:30, в этом случае образование H1:L2 по-прежнему нежелательно), то предпочтительное спаривание между требуемой парой, т.е. H1:L1, будет происходить, когда наблюдается улучшение (повышение) величины спаривания между H1:L1 по сравнению с H1:L2. В контексте настоящего документа термин «предпочтительное спаривание» охватывает спаривание тяжелой цепи и легкой цепи (как описано выше), а также спаривание домена CH1 и домена CL. В качестве примера, «предпочтительное спаривание» будет иметь место между доменом CH1 и доменом CL-каппа, если количество образованного CH1:Cκ было больше количества образованного CH1:Cλ, когда CH1 экспрессируется совместно с Cκ и Cλ. Аналогичным образом, «предпочтительное спаривание» будет иметь место между доменом CH1 и доменом CL-лямбда, если количество образованного CH1:Cλ больше количества образованного CH1:Cκ, когда CH1 экспрессируется совместно с Cλ и Cκ.
Было обнаружено, что определенные положения в домене CH1, идентифицированные как часть области контакта CH1-CL (как для Cκ, так и для Cλ), влияют на связывание тяжелой цепи с легкой цепью. Кроме того, было показано, что положения в домене CH1 в области контакта CH1:VH также влияют на связывание тяжелой цепи с легкой цепью. Тяжелая цепь спаривается с легкой цепью через два набора областей контакта доменов: один между доменами VH и VL, а другой между доменами CH1 и CL, и область, где цепи спариваются или встречаются, или приходят в контакт, называется «областью контакта». Кроме того, в пределах тяжелой цепи домен CH1 также приходит в контакт с частью VH, и такое пространство, в котором домен CH1 и VH находятся в непосредственной близости, также охватывается термином «область контакта». Область контакта содержит аминокислотные остатки в тяжелой цепи и аминокислотные остатки в легкой цепи или, в качестве альтернативы, аминокислотные остатки в домене CH1 и аминокислотные остатки в VH, которые контактируют друг с другом в трехмерном пространстве. В некоторых вариантах осуществления область контакта содержит домен CH1 тяжелой цепи и домен CL легкой цепи. В других вариантах осуществления область контакта содержит домен CH1 и домен VH тяжелой цепи. «Область контакта» предпочтительно получена из антитела IgG или его Fab.
Вариантные домены CH1, описанные в настоящем документе, содержат аминокислотную замену в одном или более положениях области контакта CH1:CL (CH1:CL-каппа или CH1:CL-лямбда), например, положениях 141, 147, 170, 171, 175, 181, 183, 184, 185, 187 и/или 218, или одном или более положениях области контакта CH1:VH, например, положении 151, по сравнению с исходным. Термин «исходный» относится к полипептиду (и кодирующей его аминокислотной последовательности), который впоследствии модифицируют для получения варианта. Исходный полипептид может представлять собой полипептид дикого типа или встречающийся в природе полипептид, или его вариант или сконструированную версию. Соответственно, «исходный домен CH1» относится к полипептиду домена CH1 (и аминокислотной последовательности, кодирующей полипептид домена CH1), который впоследствии модифицируют для получения варианта домена CH1. Такой исходный домен CH1 может представлять собой домен CH1 дикого типа или встречающийся в природе домен CH1, или его вариант или сконструированную версию, например, домен CH1 дикого типа, модифицированный для конъюгации токсина или низкомолекулярного лекарственного средства. Такой исходный домен CH1 может быть выделен или быть частью более крупной конструкции, например, Fab, F(ab′)2 или IgG, которая может необязательно содержать дополнительные модификации, например, модификации CH3 для стимуляции гетеродимеризации, модификации CH2 и/или CH3 для изменения связывания Fc-рецептора, продления периода полужизни и/или присоединения дополнительных связывающих доменов.
Полученные в результате вариантные домены CH1 характеризуются предпочтительным спариванием либо с доменом CL-каппа (Cκ), либо с доменом CL-лямбда (Cλ), причем домены Cκ и Cλ могут быть частью легкой цепи. Аминокислотная вариация в одном или в обоих из положений 147 и 183 домена CH1 (нумерация EU) способствует связыванию с Cκ (и одновременно препятствует спариванию с Cλ), тогда как аминокислотная вариация в положении 141 домена CH1 способствует связыванию с Cλ (и одновременно препятствует спариванию с Cκ). Домена CL-каппа и -лямбда могут существовать в любом количестве форматов, включая, не ограничиваясь перечисленным, Fab или IgG, дикого типа или химерные, например, Fab или IgG, содержащие Vκ и Cκ, Vκ и Cλ, Vλ и Cκ или Vλ и Cλ. Такие вариантные домены CH1 могут быть применимы в конструировании мультиспецифичных антител за счет улучшения точности спаривания тяжелых цепей с легкими цепями при сохранении нативной структуры IgG биспецифичного антитела, которое является предпочтительным в связи с его хорошо известными свойствами в качестве терапевтической молекулы, включая длительный период полужизни in vivo и способность вызывать эффекторные функции.
Термин «домен CH1» относится к первому константному домену тяжелой цепи антитела, С-концу вариабельного домена тяжелой цепи и N-концу шарнирной области. В соответствии с IMGT домен CH1 представляет собой аминокислотную последовательность с положений 118-215 (нумерация EU), а шарнирная область представляет собой аминокислотную последовательность с положений 216-230 (нумерация EU). В контексте настоящего документа термин «вариант домена CH1» относится к аминокислотной последовательности, включающей весь домен CH1 (положения 118-215 согласно нумерации EU) или его фрагменты, содержащие по меньшей мере 7 из остатков 118-215 CH1 (согласно нумерации EU), где такие фрагменты включают 1 или более модификаций, раскрытых в настоящем документе, а также часть шарнирной области (положения 216-218). Библиотеки, подвергнутые скринингу для идентификации описанных вариантов домена CH1, включали изменения в шарнирной области, например, положениях 216 и 218.
Домен CH1 спаривается с доменом CL легкой цепи. В некоторых вариантах осуществления легкая цепь представляет собой каппа-цепь. В некоторых вариантах осуществления легкая цепь представляет собой лямбда-цепь. Термин «константный домен каппа», «домен CL-каппа» или «Cκ» относится к константному домену легкой каппа-цепи. Термин «константный домен лямбда», «домен CL-лямбда» или «Cλ» относится к константному домену легкой лямбда-цепи. Одинарная дисульфидная связь ковалентно соединяет CH1 с доменом CL. Домен CH1 в контексте настоящего документа относится ко всем изотипам антител, например, IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD, IgM и IgE.
В настоящем документе термин «антитело» используется в наиболее широком смысле и охватывает различные структуры антитела, включая, не ограничиваясь перечисленным, моноклональные антитела, поликлональные антитела, мультиспецифичные антитела (например, биспецифичные антитела) и/или фрагменты антител (предпочтительно те фрагменты, которые проявляют требуемую антигенсвязывающую активность, которые также называются «антигенсвязывающими фрагментами»).
«Моноклональное антитело» или «mAb» относится к антителу, полученному из популяции по существу однородных антител, т.е. индивидуальные антитела, составляющие популяцию антител, идентичны и/или связывают один и тот же эпитоп, за исключением возможных вариантных антител (например, содержащих встречающуюся в природе или возникающую в ходе получения моноклонального антитела мутацию (мутации) и/или замену (замены)), причем такие варианты обычно присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые, как правило, содержат разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклональных антител направлено на одну детерминанту на антигене.
«Мультиспецифичное антитело», которое также может называться в настоящем документе «мультиспецифичным соединением», относится к антителу, содержащему по меньшей мере два разных антигенсвязывающих домена, которые распознают и специфично связываются с по меньшей мере двумя разными антигенами или по меньшей мере двумя разными эпитопами. В некоторых вариантах осуществления мультиспецифичное антитело содержит (1) первую тяжелую цепь и первую легкую цепь, которые образуют когнатную пару и связываются с первым антигеном, и (2) вторую тяжелую цепь и вторую легкую цепь, которые образуют когнатную пару и связываются со вторым антигеном.
«Биспецифичное антитело», которое также может называться в настоящем документе «биспецифичным соединением», представляет собой тип мультиспецифичного антитела и относится к антителу, содержащему два разных антигенсвязывающих домена, которые распознают и специфично связываются с по меньшей мере двумя разными антигенами или по меньшей мере двумя эпитопами. Указанные по меньшей мере два эпитопа могут находиться или не находиться на одном и том же антигене. Биспецифичное антитело может нацеливаться, например, на два разных поверхностных рецептора на одних и тех же или разных (например, иммунной клетке и раковой клетке) клетках, два разных цитокина/хемокина, рецептор и лиганд. Комбинации антигенов, на которые может нацеливаться биспецифичное антитело, могут включать, не ограничиваясь перечисленным: CD3 и Her2; CD3 и Her3; CD3 и EGFR; CD3 и CD19; CD3 и CD20; CD3 и EpCAM; CD3 и CD33; CD3 и PSMA; CD3 и CEA; CD3 и gp100; CD3 и gpA33; CD3 и B7-H3; CD64 и EGFR; CEA и HSG; TRAIL-R2 и LTbetaR; EGFR и IGFR; VEGFR2 и VEGFR3; VEGFR2 и PDGFR-альфа; PDGFR-альфа и PDGFR-бета; EGFR и MET; EGFR и EDV-miR16; EGFR и CD64; EGFR и Her2; EGFR и Her3; домен ECD2 Her2 и домен ECD4 Her2; Her2 и Her3; IGF-1R и HER3; CD19 и CD22; CD20 и CD22; CD30 и CD16A; FceRI и CD32B; CD32B и CD79B; MP65 и SAP-2; IL-17A и IL-23; IL-1-альфа и IL-1-бета; IL-12 и IL-18; VEGF и остеопонтин; VEGF и Ang-2; VEGF и PDGFR-бета; VEGF и Her2; VEGF и DLL4; FAP и DR5; FcgRII и IgE; CEA и DTPA; CEA и IMP288; и LukS-PV и LukF-PV.
«Отличающийся антиген» может относиться к разным и/или отдельным белкам, полипептидам или молекулам; а также к разным и/или отдельным эпитопам, при этом такие эпитопы могут содержаться в одном белке, полипептиде или другой молекуле. Следовательно, биспецифичное антитело может связываться с двумя эпитопами на одном и том же полипептиде.
Термин «эпитоп» относится к антигенной детерминанте, которая взаимодействует с определенным антигенсвязывающим сайтом в вариабельной области молекулы антитела, известной как паратоп. Один антиген может иметь более одного эпитопа. Таким образом, разные антитела могут связываться с разными областями на антигене и могут оказывать разные биологические эффекты. Термин «эпитоп» также относится к сайту на антигене, на который отвечают B- и/или T-клетки. Он также относится к области антигена, который связывается антителом. Эпитопы могут быть определены как структурные или функциональные. Функциональные эпитопы в целом представляют собой подгруппу структурных эпитопов и содержат остатки, которые напрямую вносят вклад в аффинность взаимодействия. Эпитопы также могут быть конформационными, то есть состоять из нелинейно расположенных аминокислот. В отдельных вариантах осуществления эпитопы могут включать детерминанты, которые представляют собой химически активные поверхностные группы молекул, таких как аминокислоты, боковые цепи сахаров, фосфорильные группы или сульфонатные группы, и в отдельных вариантах осуществления могут обладать специфическими характеристиками трехмерной структуры и/или специфическими характеристиками заряда.
В некоторых случаях антитело содержит четыре полипептидных цепи: две тяжелые (H) цепи и две легкие (L) цепи, соединенные дисульфидными связями. Каждая тяжелая цепь содержит вариабельную область, такую как вариабельная область тяжелой цепи («VH»), и константную область тяжелой цепи («CH»). В случае интактного антитела CH содержит домены CH1, CH2 и CH3. В случае фрагмента антитела CH может содержать домены CH1, CH2 и/или CH3, и в некоторых предпочтительных вариантах осуществления CH содержит по меньшей мере домен CH1. Варианты домена CH1, раскрытые в настоящем документе, могут применяться в комбинации с доменами CH2 и/или CH3 дикого типа или доменами CH2 и/или CH3, содержащими одну или более аминокислотных замен, например, которые изменяют или улучшают стабильность и/или эффекторные функции антител. Каждая легкая цепь состоит из вариабельной области легкой цепи («VL») и константной области легкой цепи («CL»). Области VH и VL можно дополнительно разделить на области гипервариабельности, называемые определяющими комплементарность областями (CDR), чередующиеся с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL содержит три CDR и четыре FR, расположенные от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В отдельных вариантах осуществления настоящего изобретения FR антитела (или его антигенсвязывающего фрагмента) могут быть идентичны последовательностям зародышевой линии человека или могут быть модифицированы естественным или искусственным путем. Аминокислотная консенсусная последовательность может быть определена на основе сравнительного анализа двух или более CDR. Соответственно, CDR в тяжелой цепи обозначаются как «CDRH1», «CDRH2» и «CDRH3», соответственно, а CDR в легкой цепи обозначаются как «CDRL1», «CDRL2» и «CDRL3». В других случаях антитело может вместо этого содержать его мультимеры (например, IgM) или его антигенсвязывающие фрагменты.
В отдельных случаях VH и CL могут существовать в одном полипептиде. В отдельных случаях VL и домен (домены) CH1, CH2 и/или CH3 могут существовать в одном полипептиде. Например, в отдельно взятом антителе или фрагменте антитела, в то время как первый полипептид содержит VH1 и CH1, а второй полипептид содержит VL1 и CL (VH1 и VL образуют антигенсвязывающий сайт для первого эпитопа), третий полипептид содержит VH2 и CL, а четвертый полипептид содержит VL2 и CH1 (VH2 и VL2 образуют антигенсвязывающий сайт для второго эпитопа). В другом отдельно взятом антителе или фрагменте антитела, в то время как первый полипептид содержит VH1 и CH1, а второй полипептид содержит VL1 и CL (VH1 и VL образуют антигенсвязывающий сайт для первого эпитопа), третий полипептид содержит VL2, CL и один или более из доменов CH2 и/или CH3, а четвертый полипептид содержит VH и CH1. Любые антитела или фрагменты антител, которые содержат любой из вариантов CH1, раскрытых в настоящем документе, которые обеспечивают предпочтительное спаривание с CL-каппа или предпочтительное спаривание с CL-лямбда, независимо от того, находится ли домен CH1 в тяжелой цепи или в легкой цепи, охватываются настоящим изобретением.
Термин «когнатная пара» или «когнатное спаривание», используемый в настоящем документе, относится к паре или спариванию двух цепей антитела (например, тяжелой цепи и легкой цепи), каждая из которых содержит вариабельную область (например, VH и VL, соответственно), где комбинация вариабельных областей обеспечивает целевую специфичность связывания с эпитопом или с антигеном. Термин «некогнатная пара» или «некогнатное спаривание», используемый в настоящем документе, относится к паре или спариванию двух цепей антитела (например, тяжелой цепи и легкой цепи), каждая из которых содержит вариабельную область (например, VH и VL, соответственно), где комбинация вариабельных областей не обеспечивает целевую специфичность связывания с эпитопом или с антигеном.
Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно дополнительно разделить на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, соответственно называются α, δ, ε, γ и μ.
Если специально не указано иное, термин «антитело» в контексте настоящего документа охватывает молекулы, содержащие две тяжелые цепи иммуноглобулина и две легкие цепи иммуноглобулина (иногда называемые «полноразмерным антителом», или «интактными антителами», или «целым антителом», и т.п., которые во всех случаях относятся к антителу, имеющему структуру, по существу схожую с нативным антителом), а также их антигенсвязывающие фрагменты. «Антигенсвязывающий фрагмент» или «антигенсвязывающий фрагмент антитела» относится к части интактного антитела или комбинации частей, полученных из интактного антитела или из интактных антител, и связывает антиген (антигены), с которым связывается указанное интактное антитело или антитела.
Антигенсвязывающий фрагмент антитела включает любой встречающийся в природе, получаемый ферментативным путем, синтетический или полученный с помощью генной инженерии полипептид или гликопротеин, который специфично связывает антиген с образованием комплекса. Иллюстративные фрагменты антитела включают, не ограничиваясь перечисленным: Fv; антигенсвязывающий фрагмент («Fab»); Fab'-фрагмент; Fab', содержащий свободную сульфгидрильную группу («Fab'-SH»); F(ab')2-фрагмент; диатела; линейные антитела; молекулы одноцепочечных антител (например, одноцепочечный вариабельный фрагмент («scFv”), нанотело или VHH, или отдельные домены VH или VL); и моноспецифичные или мультиспецифичные соединения, образованные из одного или более фрагментов антител, таких как вышеприведенные. В некоторых вариантах осуществления антигенсвязывающие фрагменты биспецифичных антител, описанных в настоящем документе, представляют собой scFv. В предпочтительных вариантах осуществления антигенсвязывающий фрагмент содержит домен CH1, который предпочтительно спаривается с CL-каппа или CL-лямбда.
Как и в случае с молекулами полных антител, антигенсвязывающие фрагменты могут быть моноспецифичными или мультиспецифичными (например, биспецифичными, триспецифичными, тетраспецифичными и т.д.). Мультиспецифичный антигенсвязывающий фрагмент антитела может содержать по меньшей мере два разных вариабельных домена, где каждый вариабельный домен способен специфично связываться с отдельным антигеном или с отличающимся эпитопом на том же антигене.
В настоящем изобретении предложены варианты домена CH1, которые предпочтительно спариваются с (или связываются) доменом CL легкой каппа-цепи или доменом CL легкой лямбда-цепи. В одном из вариантов осуществления варианты домена CH1 не проявляют или проявляют пониженное связывание с легкой цепью каппа-класса или легкой цепью лямбда-класса и одновременно проявляют исключительную или повышенную предпочтительность в отношении связывания с легкой цепью другого класса (в данном примере соответственно лямбда или каппа). Эти варианты домена CH1 можно применять для полного или частичного решения проблемы неправильного спаривания тяжелой и легкой цепей при получении мультиспецифичных, например, биспецифичных, антител путем стимуляции надлежащего спаривания тяжелой и легкой цепей. В одном из вариантов осуществления варианты домена CH1 могут необязательно применяться в комбинации с другими вариантами за пределами домена CH1 для дополнительного способствования предпочтительному спариванию с доменом CL легкой каппа-цепи или доменом CL легкой лямбда-цепи (например, заменами VH:VL, такими как Q39E/K:Q38K/E (Dillon et al., MAbs 2017 9(2): 213-230); или Q39K + R62E:Q38D + D1R или Q39Y + Q105R: Q38R + K42D (Brinkmann et al., MAbs 2017 9(2): 182-212). Более конкретно, биспецифичные антитела, содержащие эти вариантные домены CH1, будут образовывать меньше нежелательных родственных примесей, т.е. молекул, содержащих неправильно спаренные домены, устранение которых в ходе последующей обработки может представлять сложности. Например, биспецифичное антитело, содержащее (i) тяжелую цепь и легкую цепь из антитела A (где легкая цепь представляет собой легкую каппа-цепь) и (ii) тяжелую цепь и легкую цепь из антитела B (где легкая цепь представляет собой легкую лямбда-цепь), может быть более эффективно получено, т.е. с меньшим количеством родственных примесей, путем конструирования домена CH1 тяжелой цепи антитела A с получением варианта домена CH1 с предпочтительностью в отношении каппа-цепи (такого как, например, 147 Phe и/или 183 Arg, Lys, Tyr) и домена CH1 тяжелой цепи антитела B с получением варианта домена CH1 с предпочтительностью в отношении лямбда-цепи (такого как, например, 141 Asp). В результате тяжелая цепь антитела A будет способствовать связыванию с легкой цепью антитела A (и не будет способствовать связыванию с легкой цепью антитела B), в то время как тяжелая цепь антитела B будет способствовать связыванию с легкой цепью антитела B (и не будет способствовать связыванию с легкой цепью антитела A). См. фиг. 1 и 7 и таблицу 6.
В некоторых вариантах осуществления варианты домена CH1 снижают неправильное спаривание, т.е. образование некогнатных пар HC1-LC2 и/или HC2-LC1, на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75% или по меньшей мере 80%. В некоторых вариантах осуществления варианты домена CH1, содержащие замену в положении 141, например, 141D, отдельно или в комбинации с другими заменами, например, 147F + 183R, 147F + 183K, 147F + 183Y, снижают неправильное спаривание, т.е. образование некогнатных пар HC1-LC2 и/или HC2-LC1, в диапазоне от по меньшей мере 25% до по меньшей мере 80%. В некоторых вариантах осуществления варианты домена CH1, содержащие замену в положении 141, например, 141D, отдельно или в комбинации с другими заменами, например, 183R, 183K, 183Y, 147F + 183R, 147F + 183K, 147F + 183Y, снижают неправильное спаривание, т.е. образование некогнатных пар HC1-LC2 и/или HC2-LC1, на по меньшей мере 50%. В некоторых вариантах осуществления варианты домена CH1, содержащие замену в положении 141, например, 141D, отдельно или в комбинации с другими заменами, например, 183R, 183K, 183Y, 147F + 183R, 147F + 183K, 147F + 183Y, снижают неправильное спаривание, т.е. образование некогнатных пар HC1-LC2 и/или HC2-LC1, на по меньшей мере 75%.
В некоторых вариантах осуществления варианты домена CH1 предпочтительно спариваются с (связываются с) когнатным доменом CL (Cκ или Cλ) или когнатной легкой цепью, содержащей соответствующий домен CL (Cκ или Cλ), что приводит к по меньшей мере 75%, по меньшей мере 76%, по меньшей мере 77%, по меньшей мере 78%, по меньшей мере 79%, по меньшей мере 80%, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84%, по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% образованию требуемых первой и второй когнатных пар, т.е. HC1-LC1 и/или HC2-LC2. В некоторых вариантах осуществления варианты домена CH1 предпочтительно спариваются с (связываются с) когнатным доменом CL (Cκ или Cλ) или когнатной легкой цепью, содержащей соответствующий домен CL (Cκ или Cλ), что приводит к от приблизительно 80% до приблизительно 99%, или, более конкретно, от по меньшей мере приблизительно 85% до по меньшей мере приблизительно 95% образованию требуемых первой и второй когнатных пар, т.е. HC1-LC1 и/или HC2-LC2. В некоторых вариантах осуществления варианты домена CH1, содержащие замену в положении 141, например, 141D, отдельно или в комбинации с другими заменами, например, 183R, 183K, 183Y, 147F + 183R, 147F + 183K, 147F + 183Y, обеспечивают от приблизительно 85% до по меньшей мере приблизительно 95% образования требуемых первой и второй когнатных пар, т.е. HC1-LC1 и/или HC2-LC2.
В некоторых вариантах осуществления варианты домена CH1 обеспечивают сниженное образование гетеродимеров неправильно спаренных тяжелой цепи и легкой цепи, т.е. пар HC1-LC2 и/или HC2-LC1, до менее 25%, менее 20%, менее 15%, менее 14%, менее 13%, менее 12%, менее 11% менее 10%, менее 9%, менее 8%, менее 7%, менее 6%, менее 5%, менее 4%, менее 3%, менее 2% или менее 1%. В некоторых вариантах осуществления варианты домена CH1, содержащие замену в положении 141, например, 141D, отдельно или в комбинации с другими заменами, например, 183R, 183K, 183Y, 147F + 183R, 147F + 183K, 147F + 183Y, обеспечивают сниженное образование гетеродимеров неправильно спаренных тяжелой цепи и легкой цепи до менее приблизительно 15%, менее приблизительно 10% или менее приблизительно 5%.
Несколько положений домена CH1 были идентифицированы как влияющие на предпочтительность связывания легкой цепи, т.е. предпочтительное спаривание с доменом CL-каппа или доменом CL-лямбда, включая положения 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175-176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216 и 218 (нумерация EU). Замена аминокислотного остатка дикого типа в любом из этих положений в домене CH1 вариантным (отличным от дикого типа) аминокислотным остатком приводит к получению тяжелой цепи, которая обладает предпочтительным спариванием с легкой цепью, содержащей либо домен CL-каппа, либо домен CL-лямбда. Например, каждое из положений 147 и 183 было идентифицировано как обладающее предпочтительным спариванием с доменом CL-каппа, и положение 141, 170, 171, 175, 181, 184, 185, 187 и 218 было идентифицировано как обладающее предпочтительным спариванием с доменом CL-лямбда.
Было показано, что замена аминокислотного остатка дикого типа (Ala) в положении 141 домена CH1 на Thr, Asp, Lys, Glu, Arg, Met, Val или Gln повышает предпочтительность тяжелой цепи в отношении связывания с легкой цепью, содержащей домен CL-лямбда. Замена аминокислотного остатка дикого типа (Phe) в положении 170 домена CH1 на Glu, Gly, Ser, Asn или Thr; замена аминокислотного остатка дикого типа (Pro) в положении 171 домена CH1 на Glu, Gly, Ser, Asn, Asp или Ala; замена аминокислотного остатка дикого типа (Met) в положении 175 домена CH1 на Asp или Met; замена аминокислотного остатка дикого типа (Ser) в положении 181 домена CH1 на Val, Leu, Ala, Lys или Thr; замена аминокислотного остатка дикого типа (Ser) в положении 184 домена CH1 на Arg; замена аминокислотного остатка дикого типа (Val) в положении 185 домена CH1 на Met, Leu, Ser, Arg, Thr; замена аминокислотного остатка дикого типа (Thr) в положении 187 домена CH1 на Arg, Asp, Glu, Tyr или Ser; и/или замена аминокислотного остатка дикого типа (Lys) в положении 218 домена CH1 на Leu, Glu, Asp, Pro, Ala, His, Ser, Gln, Asn, Thr, Ile, Met, Gly, Cys, Lys или Trp также способствует повышенному спариванию тяжелой цепи с легкой цепью, содержащей домен CL-лямбда.
Было показано, что замена аминокислотного остатка дикого типа (Lys) в положении 147 домена CH1 на Val, Ala, Phe, Ile, Thr, Ser, Tyr, Leu, Arg, Asn, Glu, His, Met или Gln повышает предпочтительность тяжелой цепи в отношении связывания с легкой цепью, содержащей домен CL-каппа. Было показано, что замена аминокислотного остатка дикого типа (Ser) в положении 183 домена CH1 на Arg, Lys, Tyr, Trp, Glu, Phe, Ile, Leu, Asn или Gln повышает предпочтительность тяжелой цепи в отношении связывания с легкой цепью, содержащей домен CL-каппа (см. фиг. 5). Влияние отдельно взятого вариантного аминокислотного остатка в конкретном положении может варьировать, но все варианты демонстрируют улучшенное предпочтительное спаривание с Cκ или Cλ, в зависимости от аминокислотного положения, содержащего вариантный остаток. Кроме того, учитывая высокую степень сходства в областях CH1 IgG1, IgG2, IgG3 и IgG4, ожидается, что описанные в настоящем документе варианты домена CH1 будут демонстрировать схожие свойства предпочтительного связывания в каждом изотипе.
Первоначальный цикл отбора выявил Thr в положении 141 как способствующий предпочтительному спариванию с Cλ по сравнению с последовательностью домена CH1 дикого типа (Ala в положении 141), но дополнительные раунды отбора выявили Asp, Arg и Gln как обеспечивающие повышенное предпочтительное спаривание по сравнению с Thr (см. фиг. 5). Дополнительная стратегия скрининга выявила Lys и Glu, также обеспечивающие повышенную предпочтительность в отношении лямбда-цепи (см. пример 5, фиг. 10-14). Для Glu в положении 170; Glu в положении 171; Мет в положении 175; Lys в положении 181; Arg в положении 184; Arg в положении 185; Arg в положении 187; и/или Pro, Ala или Glu в положении 218 также было установлено, что они повышают предпочтительность в отношении лямбда-цепи (см. примеры 5-7). Кроме того, конкретные комбинации замен, которые, как показано заявителем, повышают предпочтительность в отношении лямбда-цепи, включают, не ограничиваясь перечисленным, Asp в положении 141 и Lys в положении 181; Asp в положении 141, Lys в положении 181 и Ala в положении 218; Asp в положении 141, Lys в положении 181 и Pro в положении 218; Glu в положении 141, Glu в положении 170, Val в положении 181 и Arg в положении 187; Glu в положении 141, Asp в положении 171 и Arg в положении 185; Glu в положении 141, Glu в положении 171 и Arg в положении 185; Glu в положении 141, Gly в положении 171, Arg в положении 185 и Arg в положении 187; Glu в положении 141, Arg в положении 185 и Arg в положении 187; Glu в положении 141, Ser в положении 171 и Lys в положении 181; Glu в положении 141, Gly в положении 170, Met в положении 175, Val в положении 181, Arg в положении 184 и Arg в положении 187. В дополнительных скрининговых исследованиях выявили, что «Asp в положении 141, Glu в положении 171 и Arg в положении 185» и «Asp в положении 141, Glu в положении 170 и Arg в положении 187» являются комбинациями замен домена CH1, обеспечивающими особую предпочтительность в отношении лямбда-цепи (см. фиг. 20, 23, 30 и 31).
Аналогичным образом, начальный раунд отбора выявил Val или Ala в положении 147 и Lys в положении 183 как способствующие предпочтительному спариванию с Cκ по сравнению с последовательностью домена CH1 дикого типа, но дополнительные раунды отбора выявили Phe, Ile, Thr, Tyr, Leu, Arg, Asn, Glu, His, Met или Gln в положении 147 и/или Arg, Tyr, Trp, Glu, Phe или Gln в положении 183 как обеспечивающие повышенное предпочтительное спаривание по сравнению с 147Val или Ala или 183Lys, соответственно. Эти варианты домена CH1, отдельно или в комбинации с другими аминокислотными заменами, могут улучшать предпочтительное спаривание тяжелой цепи, содержащей такой вариант домена CH1, с легкой цепью, содержащей Cκ или Cλ.
В настоящем документе предложены варианты доменов CH1, которые содержат аминокислотную замену в одном или более из следующих положений и, таким образом, указанные варианты домена CH1 демонстрируют предпочтительное спаривание либо с Cκ, либо с Cλ (или легкой цепью, содержащей такие домены): 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175-176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216, 218 согласно нумерации EU. Как продемонстрировано в настоящем документе, различные замены аминокислотных остатков в одном или более из этих положений могут обеспечивать образование домена CH1, который предпочтительно спаривается либо с Cκ, либо с Cλ (см. таблицу 3 и таблицу 4). В некоторых вариантах осуществления аминокислотная замена в положении 147 (нумерация EU) не представляет собой цистеин. В некоторых вариантах осуществления аминокислотная замена в положении 183 (нумерация EU) не представляет собой цистеин или треонин. В некоторых вариантах осуществления аминокислотная замена в положении 147 (нумерация EU) не представляет собой цистеин, и аминокислотная замена в положении 183 (нумерация EU) не представляет собой цистеин или треонин.
В некоторых вариантах осуществления вариант домена CH1 содержит аминокислотную замену в одном или более из следующих положений для способствования предпочтительному спариванию варианта домена CH1 (или тяжелой цепи, содержащей такой домен) с Cκ (или легкой цепью, содержащей такой домен): 118, 124, 126-129, 131-132, 134, 136, 139, 143, 145, 147-151, 153-154, 170, 172, 175-176, 181, 183, 185, 190-191, 197, 201, 203-206, 210, 212-214 и 218 (нумерация EU). В некоторых вариантах осуществления аминокислотная замена представляет собой одну или более из следующих: положение 118 заменено G; положение 124 заменено H, R, E, L или V; положение 126 заменено A, T или L; положение 127 заменено V или L; положение 128 заменено H; положение 129 заменено P; положение 131 заменено A; положение 132 заменено P; положение 134 заменено G; положение 136 заменено E; положение 139 заменено I; положение 143 заменено V или S; положение 145 заменено F, I, N или T; положение 147 заменено F, I, L, R, T, S, M, V, E, H, Y или Q; положение 148 заменено I, Q, Y или G; положение 149 заменено C, S или H; положение 150 заменено L или S; положение 151 заменено A или L; положение 153 заменено S; положение 154 заменено M или G; положение 170 заменено G или L; положение 172 заменено V; положение 175 заменено G, L, E, A; положение 176 заменено P; положение 181 заменено Y, Q или G; положение 183 заменено I, W, F, E, Y, L, K, Q, N или R; положение 185 заменено W; положение 190 заменено P; положение 191 заменено I; положение 197 заменено A; положение 201 заменено S; положение 203 заменено S; положение 204 заменено Y; положение 205 заменено Q; положение 206 заменено S; положение 210 заменено R; положение 212 заменено G; положение 213 заменено E или R; положение 214 заменено R; и положение 218 заменено Q. В некоторых вариантах осуществления вариант домена CH1 содержит аминокислотную замену в положениях 147 и 183 для способствования предпочтительному спариванию с (связыванию с) легкой каппа-цепью. В некоторых вариантах осуществления аминокислота, замененная в положении 147, выбрана из группы, состоящей из F, I, L, R, T, S, M, V, E, H, Y и Q, и аминокислота, замененная в положении 183, выбрана из группы, состоящей из I, W, F, E, Y, L, K, Q, N и R. В частном варианте осуществления вариант домена CH1 содержит R или K или Y в положении 183, отдельно или в комбинации с F в положении 147. Неограничивающие примеры вариантов домена CH1 с предпочтительностью в отношении каппа-цепи могут содержать аминокислотную последовательность SEQ ID NO: 137, 138, 139, 60, 41 или 136.
В некоторых вариантах осуществления вариант домена CH1 содержит аминокислотную замену в одном или более из следующих положений для способствования предпочтительному спариванию варианта домена CH1 (или тяжелой цепи, содержащей такой домен) в отношении Cλ (или легкой цепи, содержащей такой домен): 119, 124, 126-127, 130-131, 133-134, 138-142, 152, 163, 170-171, 175, 181, 183-185, 187, 197, 203, 208, 210-214, 216 и 218 (нумерация EU). В некоторых вариантах осуществления аминокислотная замена представляет собой одну или более из следующего: положение 119 заменено R; положение 124 заменено V; положение 126 заменено V; положение 127 заменено G; положение 130 заменено H или S; положение 131 заменено Q, T, N, R, V или D; положение 133 заменено D, T, L, E, S или P; положение 134 заменено A, H, I, P, V, N или L; положение 138 заменено R; положение 139 заменено A; положение 140 заменено I, V, D, Y, K, S, W, R, L или P; положение 141 заменено D, T, R, E, K, Q, V или M, предпочтительно D, E или K; положение 142 заменено M; положение 152 заменено G; положение 163 заменено M; положение 168 заменено F, I или V; положение 170 заменено N, G, E, S или T, предпочтительно E или G; положение 171 заменено N, E, G, S, A, D, предпочтительно D, E, G или S; положение 175 заменено D или M, предпочтительно M; положение 181 заменено V, L, A, K или T, предпочтительно K или V; положение 183 заменено L или V; положение 184 заменено R; положение 185 заменено M, L, S, R или T, предпочтительно R; положение 187 заменено R, D, E, Y или S; положение 197 заменено S; положение 203 заменено D; положение 208 заменено I; положение 210 заменено T; положение 211 заменено A; положение 212 заменено N; положение 213 заменено E; положение 214 заменено R; положение 216 заменено G; и положение 218 заменено P, A, L, E, D, H, S, Q, N, T, I, M, G, C, K или W, предпочтительно P или A. В некоторых вариантах осуществления домен CH1 содержит аминокислотную замену при остатке 141 для обеспечения предпочтительного связывания с легкой лямбда-цепью. В некоторых вариантах осуществления аминокислота, замененная при остатке 141, выбрана из группы, состоящей из T, R, E, K, V, D и M. В частном варианте осуществления вариант домена CH1 содержит Asp или Glu в положении 141. В некоторых вариантах осуществления аминокислотная замена в положении 141 может быть скомбинирована с одной или более заменами в CH1, например, Lys в положении 181 или Lys в положении 181 и Ala или Pro в положении 218. Asp или Glu в положении 141 могут быть скомбинированы с одной или более заменами в положениях 170, 171, 175, 181, 184, 185 и/или 187, такими как Glu или Gly в положении 170, Asp, Glu, Gly или Ser в положении 171, Met в положении 175, Val или Lys в положении 181, Arg в положении 184, Arg в положении 185 и/или Arg в положении 187. Неограничивающие примеры предпочтительных вариантов домена CH1 могут содержать аминокислотную последовательность SEQ ID NO: 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 155, 157, 159, 162, 163, 164, 165, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188 или 189.
В частном варианте осуществления вариант домена CH1 содержит комбинацию 141D, 181K и 218P, комбинацию 141D, 171E и 185R или комбинацию 141D, 170E и 187R. В дополнительном варианте осуществления вариант домена CH1 содержит аминокислотную последовательность SEQ ID NO: 188, 186 или 143.
В настоящем изобретении также рассматриваются полипептиды, например, антитела, содержащие варианты домена CH1. Такие полипептиды могут представлять собой мультиспецифичные антитела, содержащие первую тяжелую цепь, содержащую первый вариант домена CH1, и вторую тяжелую цепь, содержащую второй вариант домена CH1. Первая тяжелая цепь и вторая тяжелая цепь могут связываться с разными эпитопами. В некоторых вариантах осуществления антитело содержит первую тяжелую цепь, содержащую первый домен CH1. В некоторых вариантах осуществления антитело дополнительно содержит вторую тяжелую цепь, содержащую второй домен CH1, который содержит аминокислотную последовательность, отличную от аминокислотной последовательности домена CH1 первой тяжелой цепи.
В некоторых вариантах осуществления первый вариант домена CH1 может предпочтительно спариваться с (или связываться с) Cκ, а второй вариант домена CH1 может предпочтительно связываться с Cλ. В этом случае первая легкая цепь содержит домен Cκ, а вторая легкая цепь содержит домен Cλ. В некоторых вариантах осуществления первая легкая цепь представляет собой легкую каппа-цепь (Cκ и Vκ) или химерную легкую цепь (Cκ и Vλ), и вторая легкая цепь представляет собой легкую лямбда-цепь (Cλ и Vλ) или химерную легкую цепь (Cλ и Vκ).
В некоторых вариантах осуществления первый вариант домена CH1 может предпочтительно спариваться с (связываться с) Cλ, а второй домен CH1 может предпочтительно спариваться с (связываться с) Cκ. В этом случае первая легкая цепь содержит домен Cλ, а вторая легкая цепь содержит домен Cκ. В некоторых вариантах осуществления первая легкая цепь представляет собой легкую лямбда-цепь (Cλ и Vλ) или химерную легкую цепь (Cλ и Vκ), и вторая легкая цепь представляет собой легкую каппа-цепь (Cκ и Vκ) или химерную легкую цепь (Cκ и Vλ).
Первая и вторая легкие цепи могут содержать (или могут не содержать) аминокислотную замену, которая способствует предпочтительному спариванию с доменом CH1. В некоторых вариантах осуществления домен CL легкой цепи не модифицирован для изменения связывания с тяжелой цепью, например, с доменом CH1. В некоторых вариантах осуществления первая легкая цепь содержит домен CL дикого типа, например, домен Cκ дикого типа или домен Cλ дикого типа. В некоторых вариантах осуществления вторая легкая цепь содержит домен CL дикого типа, например, домен Cκ дикого типа или домен Cλ дикого типа. Легкая каппа-цепь или домен Cκ дикого типа могут кодироваться IGKC. Легкая лямбда-цепь или домен Cλ дикого типа могут кодироваться IGLC1, IGLC2, IGLC3, IGLC6 или IGLC7.
В некоторых вариантах осуществления антитело представляет собой мультиспецифичное антитело. В некоторых вариантах осуществления антитело представляет собой биспецифичное антитело. Такие мультиспецифичные и биспецифичные антитела могут содержать любой формат, содержащий домен CH1, такой как, не ограничиваясь перечисленным, структуры, показанные на фиг. 24-29. См. также, например, источник Brinkmann and Kontermann, MAbs 9(2):182-212 (2017), таблица 2, включенной в настоящий документ посредством ссылки в полном объеме.
Мультиспецифичное антитело может содержать один или более вариантов домена CH1, имеющих аминокислотную последовательность, перечисленную в таблице 3, 4, 7, 9, 12 или 13. В некоторых вариантах осуществления антитело содержит первую тяжелую цепь, содержащую первый вариант домена CH1, и первую легкую цепь, при этом указанные первая тяжелая цепь и первая легкая цепь образуют первую когнатную пару. Первый вариант домена CH1 может содержать аминокислотную замену в одном или более из следующих положений: 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175-176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216, 218 согласно нумерации EU. Такой первый вариант домена CH1 предпочтительно связывается с первой легкой цепью. Домен CL первой легкой цепи может быть модифицирован или не модифицирован для изменения связывания с первой тяжелой цепью.
В некоторых вариантах осуществления антитело дополнительно содержит вторую тяжелую цепь, содержащую второй вариант домена CH1 и вторую легкую цепь, причем вторая тяжелая цепь и вторая легкая цепь образуют вторую когнатную пару. Второй вариант домена CH1 может содержать аминокислотную замену в одном или более из следующих положений: 118, 119, 124, 126-134, 136, 138-143, 145, 147-154, 163, 168, 170-172, 175-176, 181, 183-185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216, 218 согласно нумерации EU. Такой второй вариант домена CH1 предпочтительно связывается со второй легкой цепью. Домен CL второй легкой цепи может быть модифицирован или не модифицирован для изменения связывания со второй тяжелой цепью.
В отдельных вариантах осуществления мультиспецифичного антитела или фрагмента антитела указанное антитело или фрагмент антитела могут содержать вариант домена CH1 с предпочтительностью в отношении каппа-цепи и вариант домена CH1 с предпочтительностью в отношении лямбда-цепи. В некоторых случаях вариант домена CH1 с предпочтительностью в отношении каппа-цепи может представлять собой вариант домена CH1 с предпочтительностью в отношении каппа-цепи, как раскрыто в настоящем документе, и домен CH1 с предпочтительностью в отношении лямбда-цепи может представлять собой домен CH1 с предпочтительностью в отношении лямбда-цепи, который может быть описан или не описан в настоящем документе. В некоторых случаях вариант домена CH1 с предпочтительностью в отношении лямбда-цепи может представлять собой вариант домена CH1 с предпочтительностью в отношении лямбда-цепи, как раскрыто в настоящем документе, и домен CH1 с предпочтительностью в отношении каппа-цепи может представлять собой домен CH1 с предпочтительностью в отношении каппа-цепи, который может быть описан или не описан в настоящем документе. В некоторых случаях как вариант домена CH1 с предпочтительностью в отношении каппа-цепи, так и вариант домена CH1 с предпочтительностью в отношении лямбда-цепи представляют собой варианты, как раскрыто в настоящем документе.
Любой из вариантов домена CH1, раскрытых в настоящем документе, можно применять для обеспечения предпочтительности спаривания с доменом CL-каппа или с доменом CL-лямбда, и домены CL могут быть дикого типа или отличными от дикого типа. Кроме того, любой из вариантов домена CH1, раскрытых в настоящем документе, может быть использован для обеспечения предпочтительности спаривания с каппа/лямбда в структуре антитела или фрагмента антитела с введением или без введения дополнительного аминокислотного изменения в остальную структуру антитела, например, в домен CH2, CH3, VH, VL или CL. Например, варианты домена CH1, раскрытые в настоящем документе, могут быть использованы с заменой VH, которая может дополнительно усиливать предпочтительность спаривания с легкой цепью (например, заменами VH:VL, такими как Q39E/K:Q38K/E (Dillon et al., MAbs 2017 9(2): 213-230); или Q39K + R62E:Q38D + D1R или Q39Y + Q105R: Q38R + K42D (Brinkmann et al., MAbs 2017 9(2): 182-212).
Без намерения ограничить объем настоящего изобретения авторы подчеркивают, что варианты домена CH1, предложенные в настоящем документе, обеспечивают предпочтительность спаривания с каппа/лямбда в контексте легкой цепи дикого типа (или полипептида, содержащего домен CL дикого типа) без необходимости в другой модификации в CH2, CH3 или вариабельных доменах, хотя такие модификации вне CH1 необязательно могут быть использованы в комбинации с новыми вариантами домена CH1, открытыми авторами настоящего изобретения. Это особенно неожиданно, учитывая большое количество сообщений о неудачах в области получения антител, в частности, мультиспецифичных антител, где модификация только домена CH1 обеспечивает значимую предпочтительность в отношении каппа- или лямбда-цепи.
В некоторых вариантах осуществления антитело является частью фармацевтической композиции. Такая композиция может содержать несколько полипептидов, например, антитела, содержащие варианты домена CH1, описанные в настоящем документе.
Также настоящим изобретением предусмотрены способы получения таких вариантов домена CH1. Варианты домена CH1, описанные в настоящем документе, могут быть идентифицированы с помощью рациональной разработки (in silico) или случайным образом, например, с применением ПЦР с ошибками (ePCR), или других методик мутагенеза, известных из уровня техники. В одном из вариантов осуществления для разработки вариантов домена CH1 применяют рациональный подход к разработке. Для такого подхода набор структур, например, полученных экспериментальным путем белковых структур, например, кристаллических структур Fab, может быть составлен и проанализирован для идентификации положений, доступных для воздействия растворителя, вовлеченных в контакты в области контакта домена CH1-CL (также называемые положениями области контакта домена CH1-CL). Набор может быть подобран путем отбора структур, имеющих определенные свойства, например, высокую процентную идентичность референсному (дикого типа) CH1, Cκ и Cλ. В некоторых вариантах осуществления положения описаны или определены как контактирующие с другим остатком (или как «находящиеся в контакте»), если пара атомов боковой цепи находится на расстоянии отсечки 5 Å. «Остатки области контакта CH1» могут быть определены как остатки в домене CH1, которые контактируют с остатком в домене Cκ или домене Cλ. Термины «остаток» и «положение» могут быть использованы взаимозаменяемо в данном контексте. Кроме того, неожиданное открытие автора изобретения заключается в том, что аминокислотная замена в положении CH1 в области контакта CH1-VH (например, положении 151 CH1) изменяет предпочтительность в отношении изотипа легкой цепи. Таким образом, в некоторых вариантах осуществления положения CH1, контактирующие с остатком VH (например, пара атомов боковой цепи находятся на расстоянии отсечки 5 Å), также могут быть выбраны для рациональной идентификации варианта домена CH1.
Выбор аминокислотных положений, которые могут изменяться, будь то по отдельности или в комбинации (например, синглеты, дуплеты, триплеты и т.д.), может зависеть от множества различных параметров, например, постоянной роли положения в образовании области контакта между CH1 и CL или между CH1 и VH в различных структурах, доступность положения (положений) в структуре в целом, отношение между положением и положениями, которые влияют на связывание антигена, или потенциал влияния остатка на образование области контакта CH1:CL или CH1:VH аллостерическим образом без непосредственного участия в межмолекулярных контактах в указанной области контакта. В некоторых вариантах осуществления аминокислотные остатки в домене CH1 выбирают для варьирования, если: 1) остаток находится в области контакта с константным доменом легкой цепи в по меньшей мере 10% структур в наборе Cκ и имеет относительную доступную для растворителя площадь поверхности (SASA), превышающую 10%, в по меньшей мере 90% структур в наборе Cκ (см. пример 1), ИЛИ 2) остаток находится в области контакта с константным доменом легкой цепи в по меньшей мере 10% структур в наборе Cλ и имеет относительную SASA, превышающую 10%, в по меньшей мере 90% структур в наборе Cλ, ИЛИ 3) остаток находится в области контакта с VH в по меньшей мере 10% репрезентативного набора из набора Cκ и/или Cλ и имеет относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора из набора Cκ и/или Cλ.
Кроме того, для каждой из конкретных аминокислотных замен в домене CH1, которые предусмотрены в настоящем документе для придания предпочтительности в отношении каппа- или лямбда-цепи, аминокислота, включенная в результате замены, может быть дополнительно заменена посредством консервативной аминокислотной замены с получением другого варианта домена CH1, который обеспечивает эквивалентную предпочтительность в отношении каппа- или лямбда-цепи. В качестве альтернативы, для каждого варианта домена CH1 одно или более аминокислотных положений, которые не были затронуты в варианте домена CH1 относительно последовательности дикого типа, могут быть изменены посредством консервативной замены с получением другого варианта домена CH1, который обеспечивает эквивалентную предпочтительность в отношении каппа- или лямбда-цепи.
«Консервативные аминокислотные замены» известны из уровня техники и включают аминокислотные замены, при которых одна аминокислота, обладающая определенными физическими и/или химическими свойствами, заменяется на другую аминокислоту, которая имеет такие же или аналогичные химические или физические свойства. Например, консервативная аминокислотная замена может представлять собой кислую/отрицательно заряженную полярную аминокислоту, замененную другой кислой/отрицательно заряженной полярной аминокислотой (например, Asp или Glu), аминокислоту с неполярной боковой цепью, замененную другой аминокислотой с неполярной боковой цепью (например, Ala, Gly, Val, Ile, Leu, Met, Phe, Pro, Trp, Cys, Val и т.д.), основную/положительно заряженную полярную аминокислоту, замененную другой основной/положительно заряженной полярной аминокислотой (например, Lys, His, Arg и т.д.), незаряженную аминокислоту с полярной боковой цепью, замененную другой незаряженной аминокислотой с полярной боковой цепью (например, Asn, Gln, Ser, Thr, Tyr и т.д.), аминокислоту с β-разветвленной боковой цепью, замененную другой аминокислотой с β-разветвленной боковой цепью (например, Ile, Thr и Val), аминокислоту с ароматической боковой цепью, замещенную другой аминокислотой с ароматической боковой цепью (например, His, Phe, Trp и Tyr), и т.д.
Затем может быть создана библиотека, в которой остатки домена CH1 варьируются. В библиотеке могут быть варьированы один или более остатков домена CH1. В некоторых вариантах осуществления в библиотеке варьируют от одного до шести остатков домена CH1. Аминокислотное разнообразие в отдельных положениях остатков может быть достигнуто посредством вырожденного кодона, например, NNK, для обеспечения представления по меньшей мере всех 20 встречающихся в природе аминокислот в данном положении домена CH1. Выбранные положения домена CH1 могут быть варьированы по отдельности для получения точечных замен (также называемых синглетами), или подмножество комбинаций положений может быть варьировано в комбинации, например, для создания двойных и тройных замен (также называемых дуплетами и триплетами). В некоторых вариантах осуществления создают комбинации вариантов, которые включают положения домена CH1, которые находятся рядом друг с другом в 3D-пространстве, например, положения 147 x [124, 126, 145, 148, 175 и 181].
В некоторых вариантах осуществления способ получения библиотеки вариантов домена CH1 включает: a) обеспечение набора структур, содержащих один или более константных доменов каппа (Cκ), один или более константных доменов лямбда (Cλ) и один или более доменов CH1; b) отбор для замены одного или более положений домена CH1, доступных для воздействия растворителя, в контакте с одним или более положениями домена Cκ и/или одним или более положениями домена Cλ; c) замену одного или более положений домена CH1, идентифицированных на стадии b), любой аминокислотой, отличной от исходной аминокислоты; и d) синтез полипептидов, которые кодируют вариантные домены CH1 на стадии c), для составления библиотеки вариантных доменов CH1.
В некоторых вариантах осуществления один или более доменов Cκ, один или более доменов Cλ и один или более доменов CH1 представляют собой домены дикого типа. В некоторых вариантах осуществления один или более доменов Cκ, один или более доменов Cλ и один или более доменов CH1 являются человеческими (включая все аллельные функциональные варианты). В некоторых вариантах осуществления аминокислотная последовательность Cκ на стадии a) кодируется IGKC. В некоторых вариантах осуществления аминокислотная последовательность Cλ на стадии a) кодируется IGLC1, IGLC2, IGLC3, IGLC6 или IGLC7. В частном варианте осуществления аминокислотная последовательность Cλ на стадии a) кодируется IGLC2. В некоторых вариантах осуществления полученная в результате библиотека домена CH1 разработана таким образом, чтобы требовать взаимодействия во всей области контакта CH1-CL или области контакта CH1-VH.
В некоторых вариантах осуществления один или более аминокислотных остатков CH1, выбранных для замены, находятся: (i) в области контакта с константным доменом легкой цепи в по меньшей мере 10% репрезентативного набора структур CH1:Cκ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора структур CH1:Cκ, (ii) в области контакта с константным доменом легкой цепи в по меньшей мере 10% репрезентативного набора структур CH1:Cλ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора структур CH1:Cλ, и/или (iii) в области контакта с VH в по меньшей мере 10% репрезентативного набора структур Cκ и/или Cλ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора структур Cκ и/или Cλ.
В некоторых вариантах осуществления библиотеку создают путем изменения одного или более положений CH1, которые раскрыты в настоящем документе как изменяющие предпочтительность в отношении изотипа легкой цепи (например, положений 141, 147, 151, 170, 171, 181, 183, 185, 187 или 218, или любой их комбинации), и необязательно одного или более дополнительных положений CH1, представляющих интерес. В отдельных вариантах осуществления библиотека может быть создана путем комбинирования заданной замены в одном или более положениях CH1, которые раскрыты в настоящем документе как изменяющие предпочтительность в отношении изотипа легкой цепи (например, положений 141, 147, 151, 170, 171, 181, 183, 185, 187 или 218, или любой их комбинации), с одним или более дополнительными положениями CH1, представляющими интерес, подвергнутыми изменению. В частных примерах заданная замена может включать A141D, A141E, K147F, P151A, P151L, F170E, P171E, S181K, S183R, V185R, T187R или K218P, или любую их комбинацию.
В некоторых вариантах осуществления библиотеку подвергают скринингу для идентификации вариантов домена CH1, демонстрирующих предпочтительное связывание с легкой каппа-цепью или легкой лямбда-цепью. Такой скрининг может начинаться с экспрессии библиотеки в подходящей клетке-хозяине, например, в эукариотической клетке, например в клетке дрожжей, например, в Saccharomyces cerevisiae. После экспрессии вариантных доменов CH1, включенных в библиотеку в клетке-хозяине, библиотека вариантов может быть подвергнута скринингу для идентификации этих вариантов с требуемыми свойствами связывания, например, с помощью FACS или MACS (магнитно-активированная сортировка клеток).
В некоторых вариантах осуществления способ идентификации варианта домена CH1 с предпочтительным связыванием Cκ- или Cλ-домена включает: a) обеспечение набора структур, содержащих один или более константных доменов каппа (Cκ), один или более константных доменов лямбда (Cλ) и один или более доменов CH1; b) отбор для замены одного или более положений домена CH1, доступных для воздействия растворителя, в контакте с одним или более положениями домена Cκ и/или одним или более положениями домена Cλ; c) замену одного или более положений домена CH1, идентифицированных на стадии b), любой аминокислотой, отличной от исходной аминокислоты; d) синтез полипептидов, которые кодируют вариантные домены CH1 на стадии c), для составления библиотеки вариантных доменов CH1; и e) скрининг библиотеки со стадии d) для идентификации варианта домена CH1 с предпочтительным связыванием Cκ- или Cλ-домена.
В некоторых вариантах осуществления один или более доменов Cκ, один или более доменов Cλ и один или более доменов CH1 представляют собой домены дикого типа. В некоторых вариантах осуществления один или более доменов Cκ, один или более доменов Cλ и один или более доменов CH1 являются человеческими (включая все аллельные функциональные варианты). В некоторых вариантах осуществления аминокислотная последовательность Cκ на стадии a) кодируется IGKC. В некоторых вариантах осуществления аминокислотная последовательность Cλ на стадии a) кодируется IGLC1, IGLC2, IGLC3, IGLC6 или IGLC7. В частном варианте осуществления аминокислотная последовательность Cλ на стадии a) кодируется IGLC2. В некоторых вариантах осуществления полученная в результате библиотека домена CH1 разработана таким образом, чтобы требовать взаимодействия во всей области контакта CH1-CL или в области контакта CH1-VH.
В некоторых вариантах осуществления один или более аминокислотных остатков CH1, выбранных для замены, находятся: (i) в области контакта с константным доменом легкой цепи в по меньшей мере 10% репрезентативного набора структур CH1:Cκ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора структур CH1:Cκ, (ii) в области контакта с константным доменом легкой цепи в по меньшей мере 10% репрезентативного набора структур CH1:Cλ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора структур CH1:Cλ, и/или (iii) в области контакта с VH в по меньшей мере 10% репрезентативного набора структур CH1:Cκ и/или CH1:Cλ и имеют относительную доступную для растворителя площадь поверхности, превышающую 10%, в по меньшей мере 90% репрезентативного набора структур CH1:Cκ и/или CH1:Cλ.
Способы, описанные в настоящем документе, могут дополнительно включать проверку того, что один или более замененных аминокислотных остатков CH1 обеспечивают предпочтительное спаривание тяжелой цепи с доменом CL-каппа (или легкой цепью, содержащей домен CL-каппа) по сравнению с доменом CL-лямбда (или легкой цепью, содержащей домен CL-лямбда), или наоборот. Для оценки предпочтительного спаривания с легкой цепью может быть использован ряд методов, включая, не ограничиваясь перечисленным, сортировку клеток с активированной флуоресценцией (FACS), ЖХ-МС, AlphaLISA и SDS-PAGE (электрофорез белков в полиакриламидном геле). В некоторых вариантах осуществления одно или более положений домена CH1, выбранных для замены на стадии c), находятся в области контакта с легкой цепью с предварительно заданной частотой, например, в любом отдельно взятом наборе структур антитела дикого типа выбранные положения домена CH1 контактируют с доменом CL в по меньшей мере 10% структур. В некоторых вариантах осуществления одно или более положений домена CH1, выбранных для замены на стадии c), имеют относительную доступную для растворителя площадь поверхности, превышающую приблизительно 10%, в по меньшей мере приблизительно 90% или более структур в любом отдельно взятом наборе Cκ или Cλ. В некоторых вариантах осуществления одно или более положений домена CH1, выбранных для замены на стадии c), находятся в области контакта с областью VH с предварительно заданной частотой, например, в любом отдельно взятом наборе структур антитела дикого типа выбранные положения домена CH1 контактируют с VH в по меньшей мере 10% структур.
При использовании способов, описанных в настоящем документе, для идентификации вариантов домена CH1, для замены были выбраны следующие положения домена CH1: 114, 116, 118, 119, 121-124, 124--143, 147-154,160, 162-165, 167, 168, 170-172, 174, 175, 176, 178, 180, 181, 183-185, 187, 190, 191, 197, 201, 203-208, 210-214, 216 и/или 128 (согласно нумерации EU). Замена любого из этих положений домена CH1 или их комбинации может обеспечить домен CH1, обладающий предпочтительным спариванием с определенным доменом CL. В результате, тяжелая цепь, содержащая такой вариант домена CH1, и легкая цепь, содержащая определенный домен CL, с большей вероятностью будут образовывать когнатную пару, т.е. имеет место предпочтительное спаривание между тяжелой цепью и легкой цепью, которые образуют когнатную пару, обусловленное, по меньшей мере частично, одной или более заменами домена CH1.
В одном из вариантов осуществления вариант домена CH1 предпочтительно спаривается с Cκ, что, следовательно, приводит к предпочтительному спариванию легкой цепи, содержащей домен Cκ, и тяжелой цепи, содержащей вариант домена CH1. В еще одном варианте осуществления вариант домена CH1 предпочтительно спаривается с доменом Cλ, что, следовательно, приводит к предпочтительному спариванию легкой цепи, содержащей домен Cλ, и тяжелой цепи, содержащей вариант домена CH1. Некоторые иллюстративные замены домена CH1 были идентифицированы как способствующие предпочтительному спариванию тяжелой цепи с легкой каппа-цепью, например, 147F и/или 183R, 183K или 183Y, тогда как другие замены домена CH1 были идентифицированы как способствующие предпочтительному спариванию тяжелой цепи с легкой лямбда-цепью, например, 141D, 141E, 141K, 170E, 170G, 171E, 171D, 171G, 171S, 175M, 181K, 181B, 184R, 185R, 187R, 218A или 218P. Соответственно, биспецифичные антитела, содержащие такие варианты домена CH1, могут быть получены с улучшенной точностью при спаривании тяжелой цепи с легкой цепью. В некоторых вариантах осуществления биспецифичное антитело содержит первую тяжелую цепь, содержащую CH1λ (такую как 141D, 141E или 141K, в комбинации с 170E, 170G, 171E, 171D, 171G, 171S или 175M, и/или 181K, 181B, 184R, 185R, 187R, 218A, и/или 218P) и вторую тяжелую цепь, содержащую CH1κ (такую как 147F и/или 183R, 183K, или 183Y), каждая из которых предпочтительно спаривается с ее когнатной легкой цепью. В некоторых вариантах осуществления биспецифичное антитело содержит первую тяжелую цепь, содержащую CH1κ (такую как 147F и/или 183R, 183K или 183Y), и вторую тяжелую цепь, содержащую CH1λ (такую как 141D, 141E или 141K, в комбинации с 170E, 170G, 171E, 171D, 171G, 171S или 175M, и/или 181K, 181B, 184R, 185R, 187R, 218A, и/или 218P), например, «141D, 171E и 185R» или «141D, 170E и 187R», каждая из которых предпочтительно спаривается с ее когнатной легкой цепью.
Полипептиды, кодирующие варианты домена CH1, полученные с помощью способов, описанных в настоящем документе, могут быть рекомбинантно экспрессированы в клетке-хозяине, например, в эукариотической клетке. В некоторых вариантах осуществления варианты домена CH1 экспрессируют в дрожжах. В некоторых вариантах осуществления штамм дрожжей представляет собой Saccharomyces cerevisiae. В некоторых вариантах осуществления штамм дрожжей совместно экспрессирует одну или более легких каппа-цепей дикого типа и одну или более легких лямбда-цепей дикого типа.
Ниже представлены примеры для иллюстрации настоящего изобретения. Эти примеры не подразумевают ограничения настоящего изобретения какой-либо конкретной областью применения или принципом действия.
Примеры
Пример 1. Выбор положений домена CH1 для диверсификации в библиотеке in silico
Набор кристаллических структур Fab составляли из Банка данных структур белков (PDB) и использовали для основанного на структуре подхода для идентификации остатков в области контакта CH1-CL для диверсификации.
Исходный набор из 2367 кристаллических структур Fab сужали путем отбора структур с высокой процентной идентичностью референсным последовательностям CH1, Cκ и Cλ (показаны ниже). Референсная последовательность для выравнивания CH1 охватывает собственно CH1 (остатки 118-215 согласно EU) и часть шарнира IgG1 (остатки 216-229 согласно EU). Референсные последовательности Cκ и Cλ охватывают номера остатков 108-214 и 107A-215 согласно EU, соответственно.
Референсная последовательность CH1 (плюс шарнир от верхнего до среднего): ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPC (SEQ ID NO: 1).
Референсная последовательность Cκ: RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 2).
Референсная последовательность Cλ: GQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 3).
Остатки определяли как «находящиеся в контакте», если пара атомов боковой цепи находилась на расстоянии отсечки 5 Å. Остатки в области контакта CH1 определяли как те остатки, которые контактировали с одним или более остатками Cκ или Cλ в отдельных структурах.
Доступную для растворителя площадь поверхности (SASA) отдельных остатков тяжелой и легкой цепей рассчитывали в «свободном состоянии», т.е. без спаривания соответственно с легкой и тяжелой цепями. Относительную SASA определяли как отношение SASA остатка к таковому у модельного выделенного трипептида Gly-X-Gly, содержащего ту же аминокислоту (т.е. X), что и остаток. Доступные для воздействия растворителя остатки определяли как остатки с относительной SASA, превышающей 10%.
Сужение начального набора кристаллических структур с помощью критерия высокой процентной идентичности привело к идентификации набора из 183 структур CH1:Cκ («набор Cκ») и 43 структур CH1:Cλ («набор Cλ»). После учета гэпов в выравнивании из-за отсутствующих в структурах аминокислот все элементы в наборе Cκ были на 100% идентичны референсным последовательностям CH1 и Cκ, тогда как элементы в наборе Cλ были > 99% идентичными референсным последовательностям.
Выравнивание последовательности на основе структуры между Cκ и Cλ показано ниже. CH1 образует стабильную область контакта как с Cκ, так и с Cλ, несмотря на низкую идентичность последовательности между последними доменами. Консервативные и полуконсервативные замены в соответствии с оценками BLOSUM62 показаны с помощью «|» и «:», соответственно. Идентичность последовательности между доменами составляет 38,3% (41 идентичных остатков из 107 остатков Cκ).

Подчеркнутые аминокислоты представляют собой остатки Cκ и Cλ, которые находятся в контакте с доменом CH1. Это определение основано на консенсусных последовательностях по Fab-структурам в наборах Cκ и Cλ. Имеются 25 остатков в области контакта Cκ и 26 остатков в области контакта Cλ. Строили матрицу 2 x 2 с акцентом на положениях, которые находятся в области контакта либо в Cκ, либо в Cλ, (N=28), и в зависимости от (1) того, контактирует ли остаток в данном положении с CH1, и (2) является ли аминокислота в указанном положении идентичной между Cκ и Cλ (см. таблицу 1).
Таблица 1. Аминокислотные положения Cκ и Cλ, находящиеся в области контакта CH1:CL
так и для Cλ
В таблице 1 показано, что имеется набор из 14 положений Cκ и Cλ, которые являются структурно консервативными, т.е. с идентичной нумерацией остатков согласно EU, но с разными аминокислотными остатками, которые контактируют с доменом CH1. В таблице 2 перечислены 14 аминокислотных положений (нумерация согласно EU) и показана аминокислота, присутствующая в легких каппа-цепях и лямбда-цепях. Такие отличия в остатках области контакта Cκ и Cλ могут быть использованы для получения мутантных доменов CH1, которые специфично связываются только с Cκ или Cλ, но не с обоими.
Таблица 2. Структурно консервативные положения, находящиеся в контакте с CH1, с неидентичными аминокислотными остатками для Cκ и Cλ
В качестве начального порогового значения для выбора варьирования в библиотеке отдельные положения домена CH1 должны были соответствовать следующим критериям: 1) положение находится в области контакта с константным доменом легкой цепи в по меньшей мере 10% структур в наборе Cκ, и остаток в этом положении имеет относительную SASA, превышающую 10%, в по меньшей мере 90% структур в наборе Cκ, или 2) положение находится в области контакта с константным доменом легкой цепи в по меньшей мере 10% структур в наборе Cλ, и остаток в этом положении имеет относительную SASA, превышающую 10%, в по меньшей мере 90% структур в наборе Cλ, или 3) положение находится в области контакта с областью VH в по меньшей мере 10% структур в наборе CH1:Cκ (набор Cκ) или наборе CH1: Cλ (набор Cλ), и остаток в этом положении имеет относительную SASA, превышающую 10%, в по меньшей мере 90% структур в наборе Cκ и/или Cλ. Определение области контакта учитывает контакты остатков CH1 с любым остатком домена CL, т.е. включает, не ограничиваясь перечисленным, набор из четырнадцати остатков домена CL, перечисленных в таблице 2, или контакты остатков CH1 с любым остатком VH.
На основании этих пороговых критериев для потенциального включения был определен набор из тридцати аминокислотных положений CH1 (после исключения из рассмотрения Cys220). Из этого более крупного набора для варьирования в библиотеке была выбрана группа из 25 положений CH1. Разнообразие аминокислот в каждом положении получали посредством вырожденного кодона NNK, представляющего все 20 природных аминокислот (Stemmer et al., Proc Natl Acad Sci USA 1994 Oct 25;91(22): 10747-51). Аминокислотные замены получали индивидуально в каждом из 25 положений CH1, и подмножество единичных замен селективно комбинировали, например, с получением двойных и тройных мутантов. Конечная структура библиотеки состояла из 89 олигонуклеотидов CH1, представляющих 25 синглетов (диверсификация кодона NNK в одном положении CH1), 48 двойных мутантов (диверсификация кодона NNK в двух положениях CH1) и 16 тройных мутантов (диверсификация кодона NNK в трех положениях CH1).
Пример 2. Библиотеки варианта домена CH1 в дрожжах, совместно экспрессирующих легкие цепи Cκ и Cλ
Библиотеки вариантов домена CH1 человека создавали и экспрессировали в сконструированных штаммах дрожжей, совместно экспрессирующих легкие цепи Cκ и Cλ человеческого IgG дикого типа (на различных уровнях экспрессии для обеспечения последующего отбора обеспечивающих предпочтительность к Cλ замен CH1 и обеспечивающих предпочтительность к Cκ замен CH1).
Конструировали двунаправленные экспрессионные плазмиды (pAD7064 и pAD4800), каждая из которых содержала промоторную область Saccharomyces cerevisiae Gal1/Gal10, фланкированную константными доменами каппа и лямбда легкой цепи человеческого IgG дикого типа, и ген URA3 S. cerevisiae (маркер отбора). Плазмиды pAD7064 и pAD4800 отличались по ориентации константных доменов каппа и лямбда относительно промоторной области Gal1/10. Уникальные сайты рестриктаз (PME-I и SFI-I) помещали против хода транскрипции относительно константных доменов каппа и лямбда в каждой плазмиде. pAD7064 и pAD4800 индивидуально расщепляли с помощью PME-I и SFI-I, а затем трансформировали в сконструированный штамм дрожжей вместе с ПЦР-амплифицированной ДНК-вставкой (область легкой цепи ADI-26140; промоторная область Gal1/10; и дифференциально кодируемая («вырожденная») вариабельная область легкой цепи ADI-26140 (IDT gblock) с 5’ и 3’-концами для управления сборкой посредством гомологичной рекомбинации в плазмиде). Трансформированные дрожжи высевали на твердые планшеты с агаром без URA3+, выращивали при 30°C в течение 48 часов перед отбором клонов и экстрагировали и очищали ДНК. После секвенирования было идентифицировано две ДНК-конструкции с двумя легкими цепями: (1) Gal10::ADI-26140-VL-Cκ Gal1::ADI-26140-VL-Cλ (Cλ человека под контролем доминантного промотора, что обеспечивает возможность последующего отбора замен CH1, обеспечивающих предпочтительность к Cκ); и (2) Gal10::ADI-26140-VL-Cλ Gal1::ADI-26140-VL-Cκ (Cκ человека под контролем доминантного промотора, что обеспечивает возможность последующего отбора замен CH1, обеспечивающих предпочтительность к Cλ). ADI-26140 представляет собой IgG к лизоциму куриного яйца (HEL).
Для экспрессии тяжелой цепи конструировали ДНК-вектор (pAD4466), содержащий промотор Gal1, сайт рестрикции SFI-I, домены CH2-CH3 тяжелой цепи человеческого IgG (IgG1 (N297A)) и TRP1 (маркер отбора).
Параллельно с этим получали два независимых пула фрагментов ДНК варианта домена CH1 для вставки в pAD4466. Первый пул получали с применением подхода к разработке in silico, описанного в примере 1. Второй пул получали с помощью ПЦР с ошибками (ePCR). Вкратце, мутагенные аналоги нуклеотидов dPTP (0,01 мМ) и 8-оксо-DGTP (0,01 мМ) включали в реакцию ПЦР при разведении (a) 1:100 и 1:100 соответственно или (b) 1:100 и 1:10 соответственно.
pAD4466 расщепляли с помощью SFI-I и вводили в штамм дрожжей, экспрессирующий Cκ и Cλ, вместе с ПЦР-амплифицированной ДНК, кодирующей вариабельную область HC ADI-26140, и ДНК варианта домена CH1, полученной с помощью подхода к рациональной разработке или ePCR. Каждый фрагмент ДНК обладал подходящими последовательностями ДНК на 5’- и 3’-концах для управления сборкой (путем гомологичной рекомбинации) с расщепленной плазмидой или фрагментом ПЦР (вариабельная область тяжелой цепи ADI-26140 или белковый домен CH1).
Сборку отдельных библиотек проводили посредством нативных процессов гомологичной рекомбинации Saccharomyces cerevisiae. Разбавленное количество трансформированных клеток для каждой библиотеки высевали на среду без урацила и триптофана для количественной оценки количества членов каждой библиотеки. Каждая библиотека содержала более 107 элементов. Оставшуюся часть трансформированных клеток культивировали в жидкой среде без урацила и триптофана для отбора по присутствию каждой (HC и двойная LC) плазмиды.
Пример 3. Идентификация положений домена CH1, влияющих на связывание легкой цепи
Библиотеки распространяли, как описано ранее (см., например, WO2009036379; WO2010105256; WO2012009568; Xu et al., Protein Eng Des Sel. 2013 Oct;26(10):663-70). Вкратце, после индукции и презентации IgG клетки дрожжей (~10^7 - 10^8) окрашивали в течение 15 минут при 4°C с помощью козьего античеловеческого F(ab’)2 каппа-FITC, разбавленного 1:100 (Southern Biotech, Бирмингем, Алабама, кат. № 2062-02) и козьего античеловеческого F(ab’)2 лямбда-PE, разбавленного 1:100 (Southern Biotech, Бирмингем, Алабама, кат. № 2072-09) in PBSF. После двукратного промывания ледяным отмывочным буфером клеточные осадки ресуспендировали в 0,4 мл PBSF и переносили в сортировочные пробирки с сетчатыми крышками. Сортировку проводили с применением сортировщика FACS ARIA (BD Biosciences) и определяли гейты для сортировки для того, чтобы либо (1) повысить содержание легкой лямбда-цепи с соразмерной потерей легкой каппа-цепи (фиг. 2A), либо (2) повысить содержание легкой каппа-цепи с соразмерной потерей легкой лямбда-цепи (фиг. 2B). После трех раундов отбора дрожжи высевали на среду без урацила и триптофана для получения отдельных изолятов для идентификации последовательностей.
Отдельные клоны, представляющие уникальные последовательности, культивировали в 96-луночных планшетах. После индукции и презентации IgG ~2x106 клеток дрожжей окрашивали в течение 15 мин при 4°C с помощью козьего античеловеческого F(ab’)2 каппа-FITC, разбавленного 1:100 (Southern Biotech, Бирмингем, Алабама, кат. № 2062-02) и козьего античеловеческого F(ab’)2 лямбда-PE, разбавленного 1:100 (Southern Biotech, Бирмингем, Алабама, кат. № 2072-09) in PBSF. После двукратного промывания ледяным отмывочным буфером клеточные осадки ресуспендировали в 0,1 мл отмывочного буфера и оценивали с помощью прибора BD FACS Canto, присоединенного к 96-луночному манипулятору для планшетов. Отдельные уникальные клоны оценивали в отношении соотношения медианной интенсивности флуоресценции (MFI) антитела к каппа-цепи и MFI антитела к лямбда-цепи (соотношение каппа:лямбда) (фиг. 3), а затем сравнивали с соответствующим штаммом с последовательностью CH1 дикого типа («исходной») для расчета FOP.
Следующие положения домена CH1 (нумерация согласно EU) были идентифицированы как влияющие на предпочтительность связывания легкой цепи, т.е. предпочтительное связывание либо с доменом CL-каппа (или легкой цепью, содержащей домен CL-каппа), либо с доменом CL-лямбда (или легкой цепью, содержащей домен CL-лямбда): 118, 119, 124, 126-134, 136, 139-141, 143, 145, 147-154, 163, 168, 170-172, 175-176, 181, 183, 185, 187, 190, 191, 197, 201, 203-206, 208, 210-214, 216 и 218. В таблице 3 представлен перечень последовательностей CH1, идентифицированных на основе отборов, которые являются предпочтительными в отношении легких каппа-цепей. Аминокислотные остатки, выделенные жирным шрифтом в столбце с последовательностями, обозначают положения с заменами, т.е. аминокислотные замены, которые отличаются от исходного (SEQ ID NO: 1). В таблице 4 представлен перечень последовательностей CH1, идентифицированных на основе отборов, которые являются предпочтительными в отношении легких лямбда-цепей. Аминокислотные остатки, выделенные жирным шрифтом в столбце с последовательностями, обозначают положения с заменами.
Таблица 3. Последовательности домена CH1, которые предпочтительно связывают Cκ
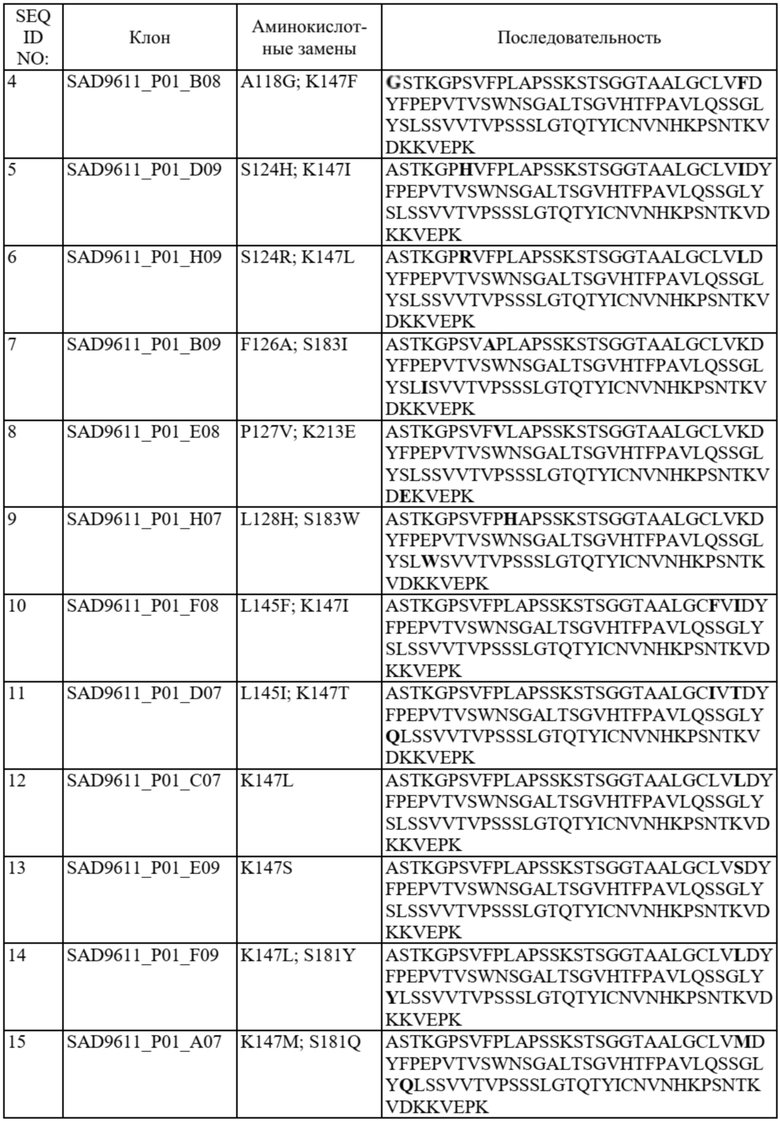
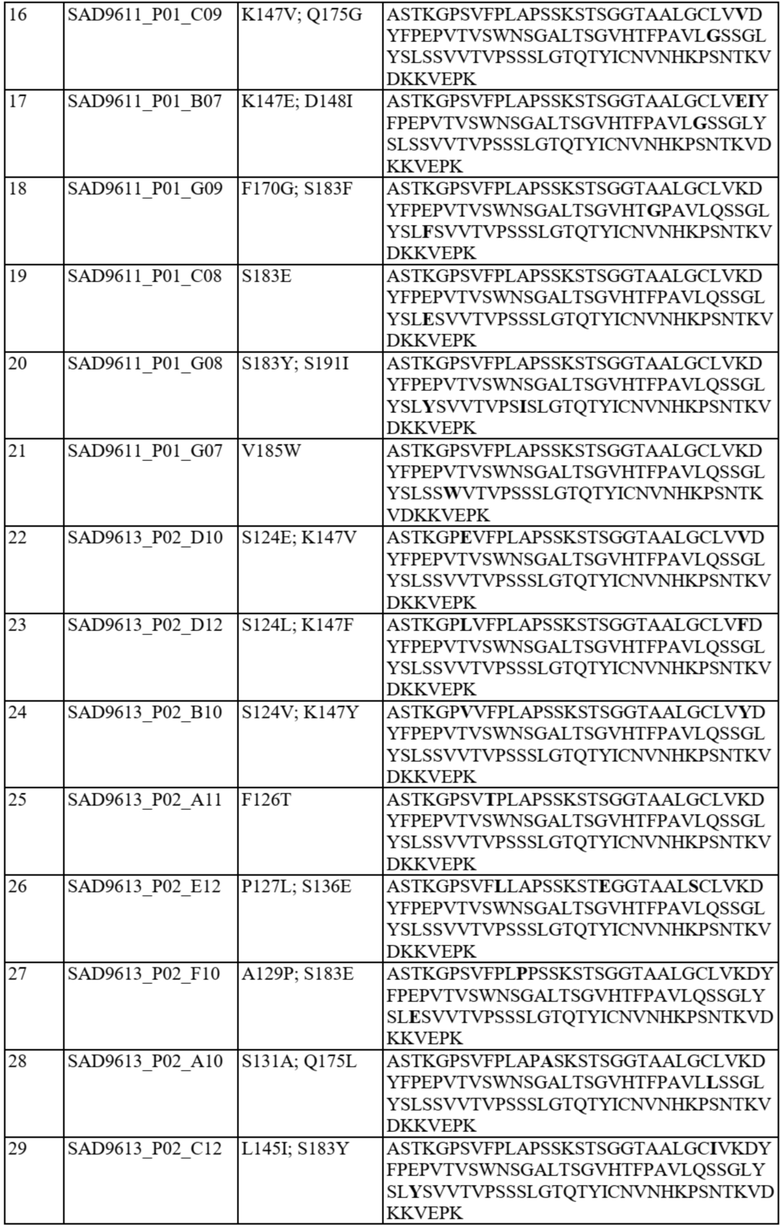
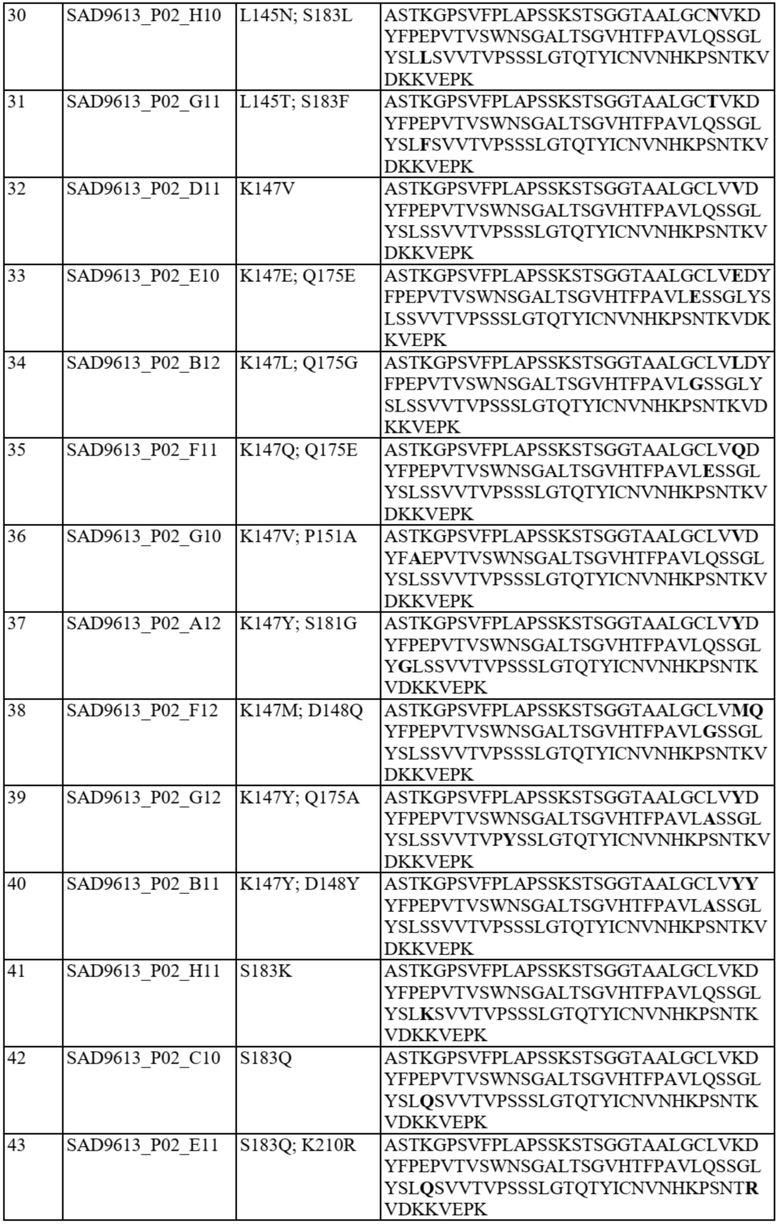
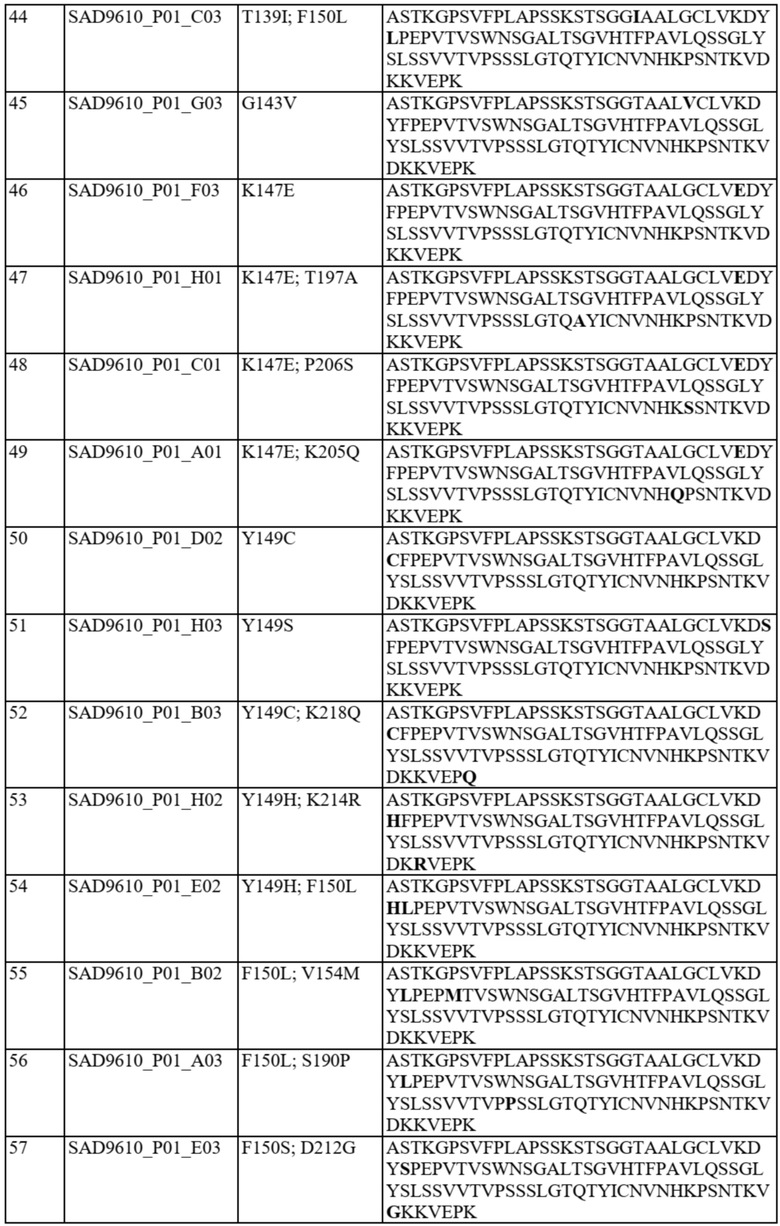
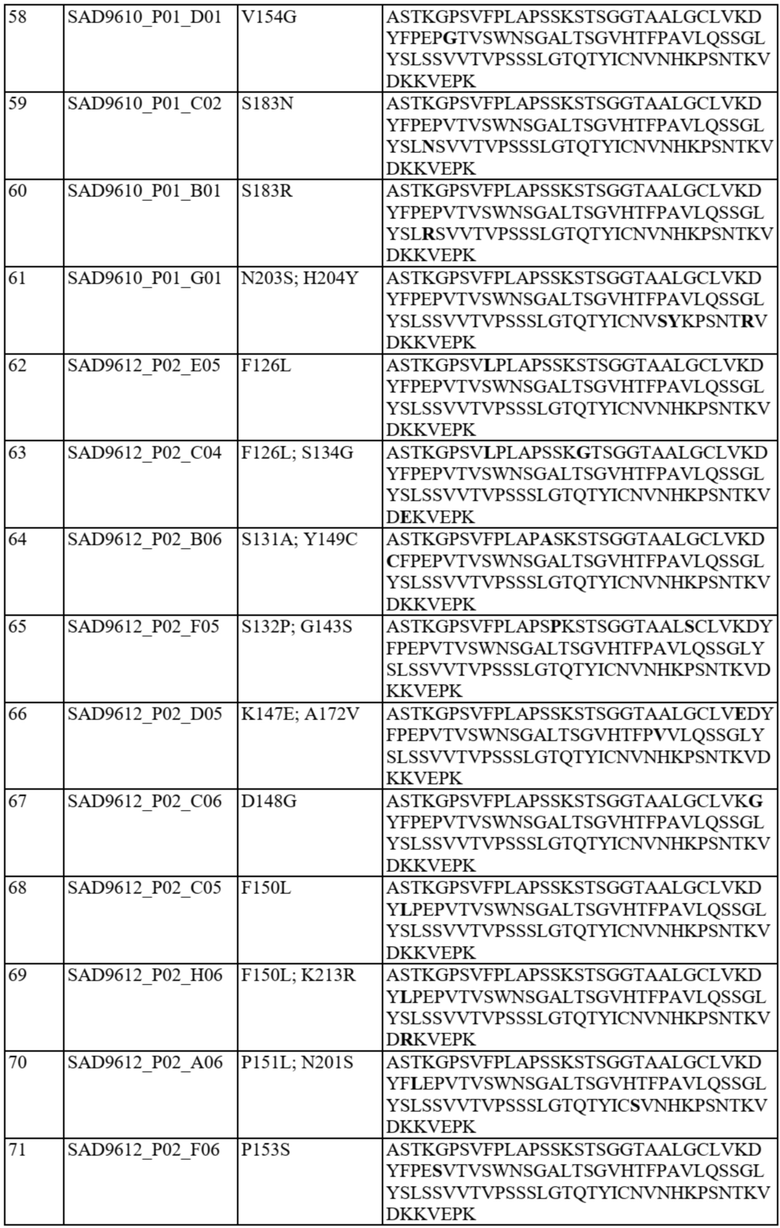
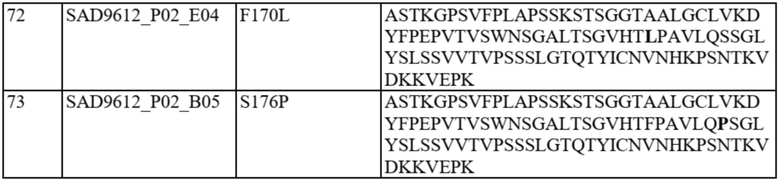
Таблица 4. Последовательности домена CH1, которые предпочтительно связывают Cλ
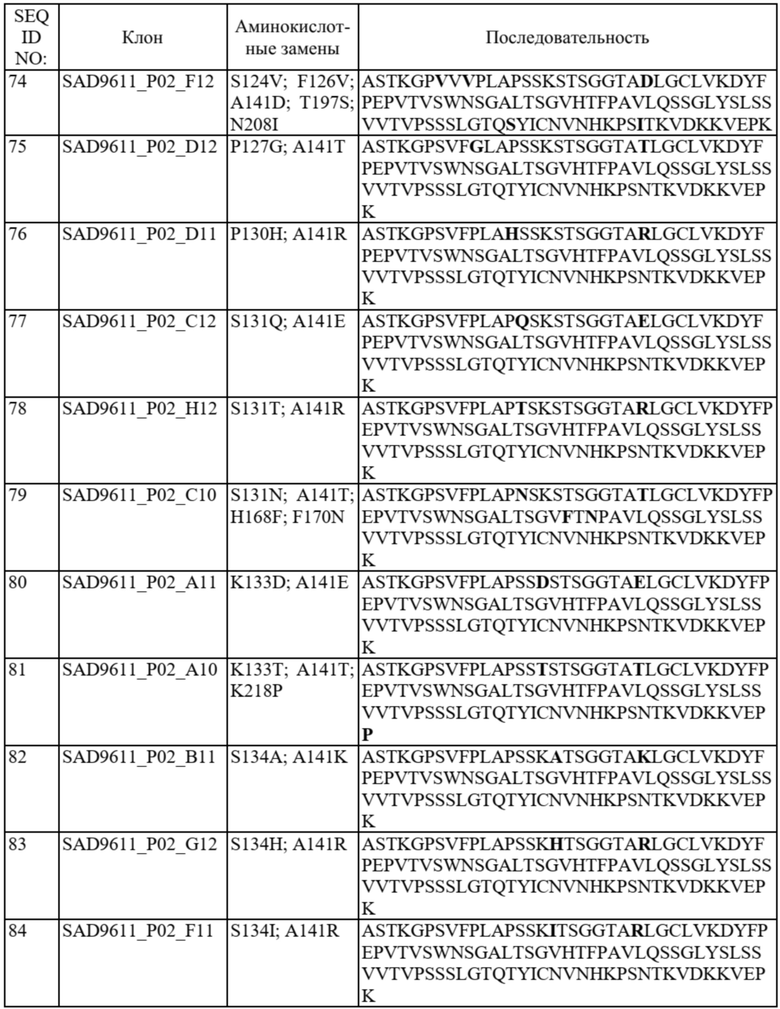
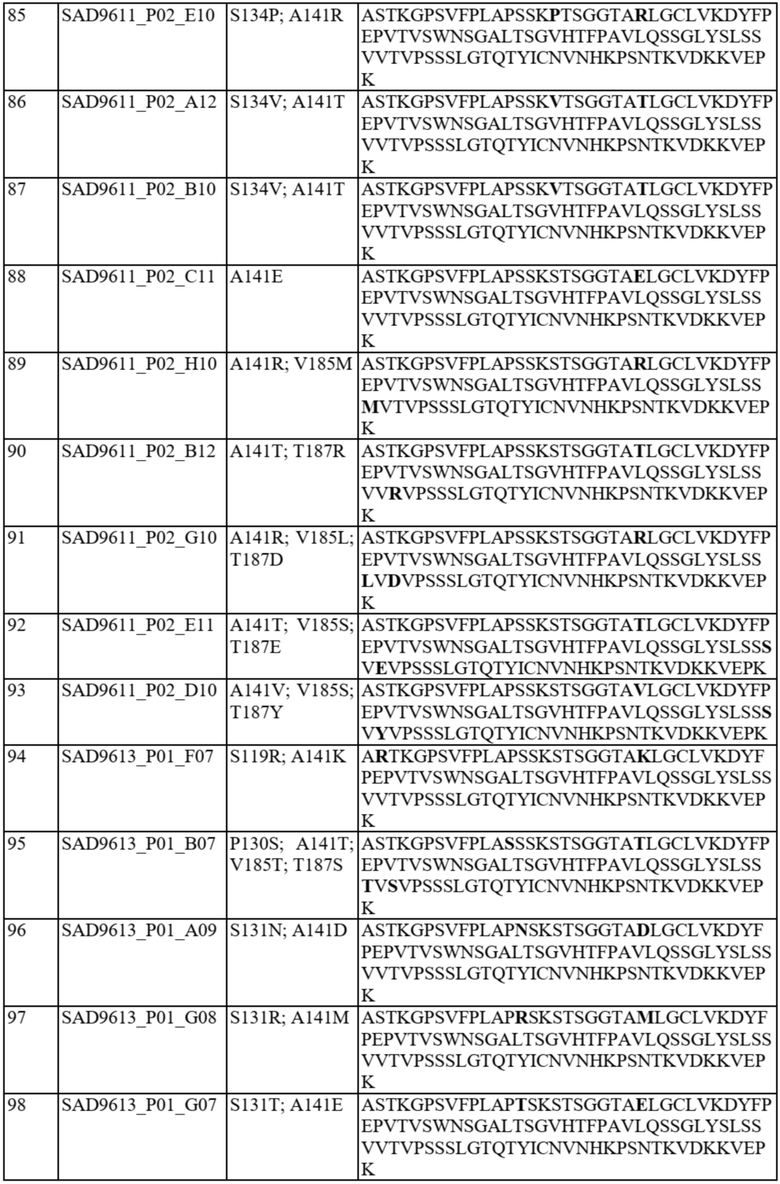
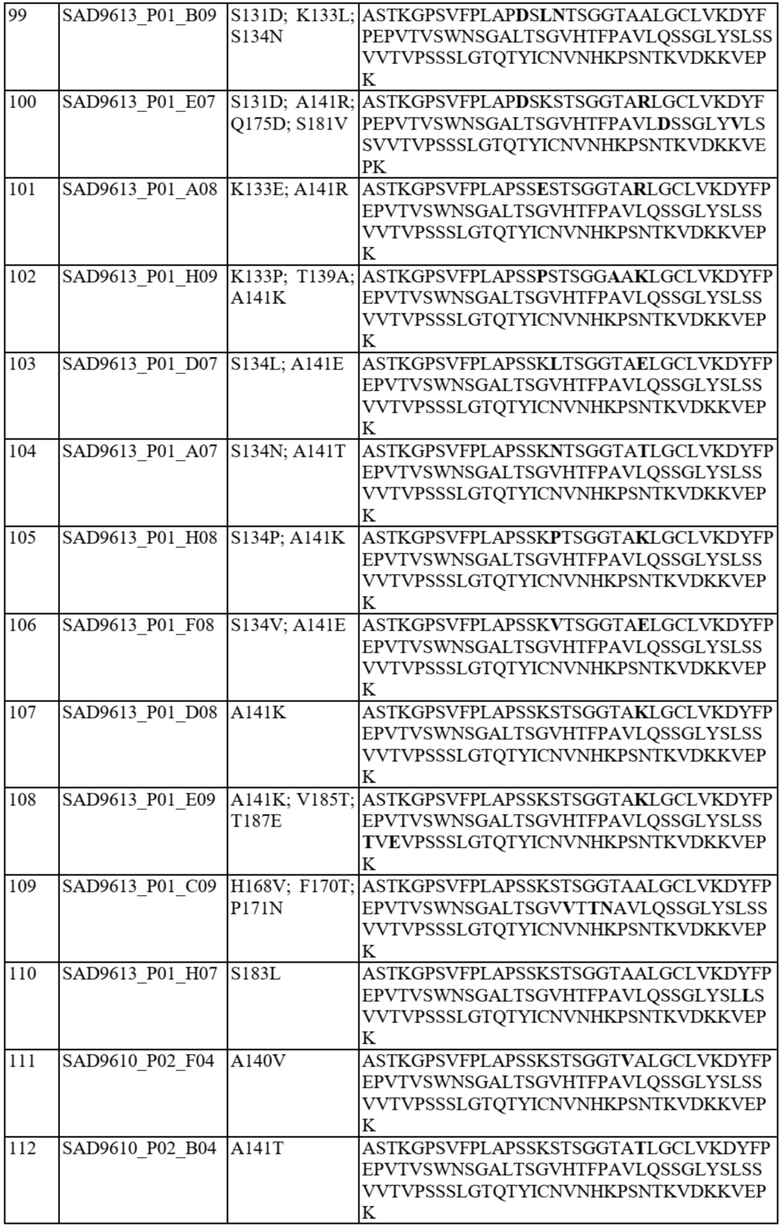
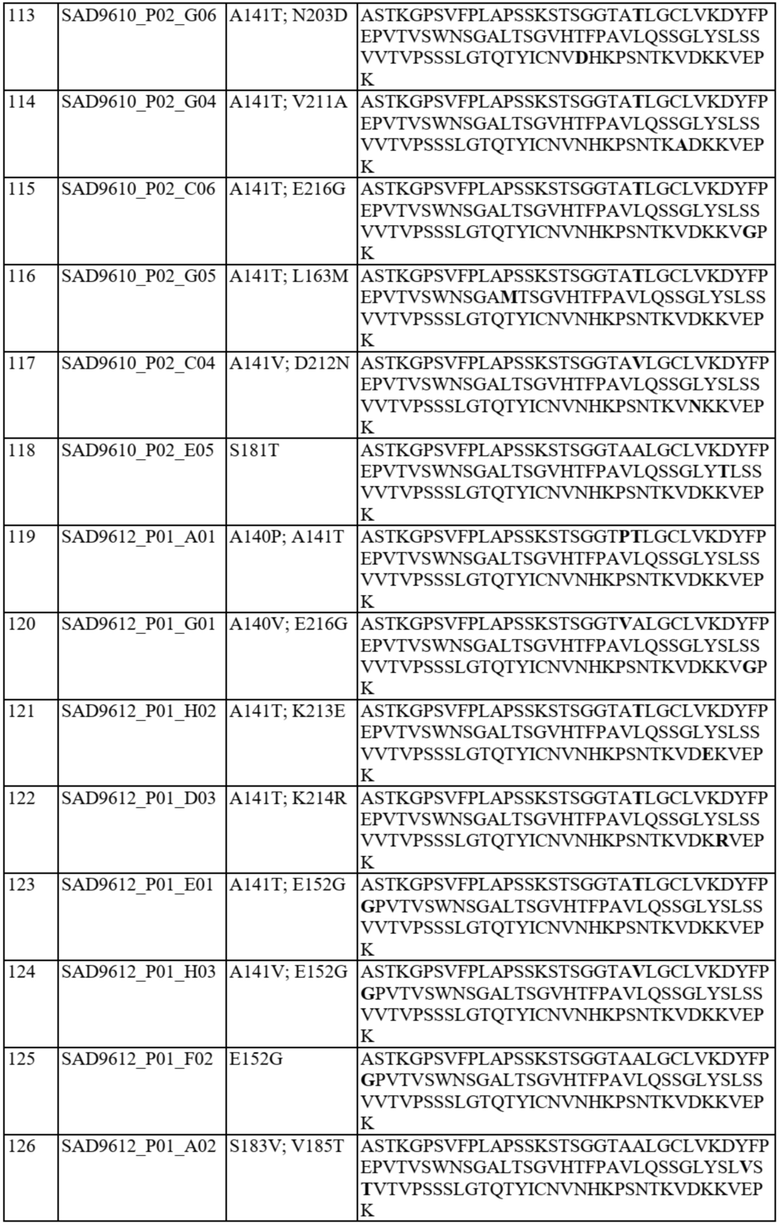
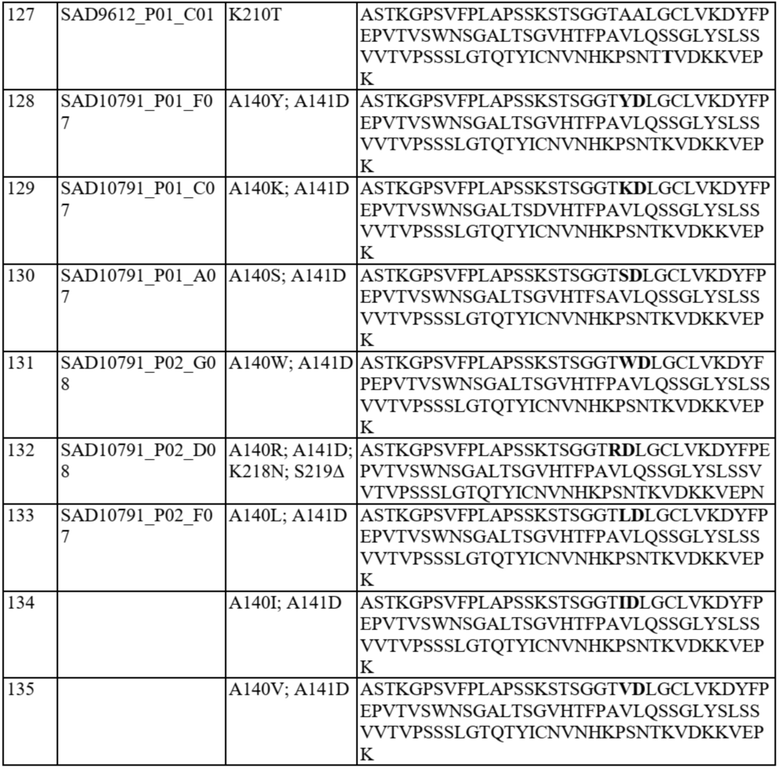
Δ обозначает делецию аминокислоты.
Неожиданно было обнаружено, что некоторые аминокислотные замены CH1, расположенные в области контакта VH:CH1, а не в области контакта CH1:легкая цепь, привели к предпочтительности связывания каппа-цепи в 3-цепочечной системе. В частности, наборы мутаций K147V+P151A и P151L+N201S (SEQ ID NO: 36 и 70, таблица 3) давали значения FOP для каппа-цепи, составляющие соответственно 18,1 и 10,4. Хотя положение CH1:147 находится в области контакта CH1:LC, CH1:201 в ней не находится (оно полностью доступно для воздействия растворителя и не является частью какой-либо междоменной области контакта); следовательно, появление замен P151 в обоих этих клонах с высоким значением FOP указывает на потенциальную роль этого положения при определении предпочтительности каппа по сравнению с лямбда. Не ограничиваясь какой-либо конкретной теорией, полагают, что такие дистальные мутации влияют на спаривание HC:LC по причинам, обсуждаемым ниже, и поэтому можно использовать мутации в области контакта VH:CH1 для предпочтительного спаривания каппа по сравнению с лямбда.
Во-первых, P151 является частью так называемого «соединения по типу шарового шарнира» между доменами VH и CH1 (Lesk A. M. et al., Nature. 1988 Sep 8;335(6186):188-90; Landolfi N. F. et al., J Immunol. 2001 Feb 1;166(3):1748-54). Предполагается, что данное соединение модулирует внутридоменную гибкость посредством его воздействия на «локтевой угол» (Stanfield R. L. et al., J Mol Biol. 2006 Apr 14;357(5):1566-74) между вариабельным и константным доменами антитела. Замены в соединении по типу шарового шарнира могут иметь функциональные последствия, как в случае моноклонального антитела к IFN-гамма со сниженной активностью нейтрализации из-за одной аминокислотной замены в этой области (Landolfi N. F. et al., J Immunol. 2001 Feb 1;166(3):1748-54). Этот эффект был связан с измененной гибкостью и аллостерическим механизмом, а не непосредственными изменениями в области контакта с антигеном. Во-вторых, также известно, что Fab с константными доменами лямбда имеют больший диапазон локтевых углов по сравнению с Fab с доменами каппа (Stanfield R. L. et al., J Mol Biol. 2006 Apr 14;357(5):1566-74. doi: 10.1016/j.jmb.2006.01.023. Epub 2006 Jan 25.). Эту сверхгибкость объясняют вставкой одного остатка в так называемую область переключения между доменами VL и CL. В-третьих, дальнейший анализ кристаллических структур Fab (неопубликованные данные Adimab) выявил различия между каппа- и лямбда-структурами Fab, состоящие в упаковке атомов в области соединения по типу шарового шарнира. Таким образом, модуляция гибкости Fab с помощью соединения по типу шарового шарнира вместе с отличиями, присущими Fab с легкими каппа- и лямбда-цепями, позволяют предположить новый механизм получения дифференциальной предпочтительности в отношении каппа- и лямбда-цепей посредством мутаций в области контакта VH:CH1.
Пример 4. Идентификация и определение характеристик вариантов домена CH1 с предпочтительным в отношении каппа-цепи или предпочтительным в отношении лямбда-цепи спариванием с легкими цепями
Клоны, полученные в результате отбора на повышенную предпочтительность в отношении Cκ и Cλ, отбирали для дальнейшего определения характеристик на основании соотношения MFI между каппа и лямбда (см. фиг. 4). Пул ДНК, подвергнутый единичным мутациям в каждом представляющем интерес положении (141, 147 или 183) в каждую из 20 аминокислот (NNK), выделяли и амплифицировали. С применением подходящего базового штамма легкой цепи эти библиотеки для одного положения конструировали таким образом, как описано ранее. Конструировали четыре библиотеки с вариацией, присутствующей в положении 141, 147, 183 или 147 + 183 соответственно домена CH1. Отбор по предпочтительности в отношении каппа- или лямбда-цепи проводили, как описано выше. Полученные продукты секвенировали, как описано ранее, и проводили количественное определение на основе FACS предпочтительности в отношении каппа- или лямбда-цепи по сравнению с соответствующим исходным белком для определения аминокислотных замен, которые обеспечивали предпочтительное связывание каппа- или лямбда-цепи.
Несколько вариантов домена CH1 с заменами аминокислотных остатков в каждом из положений 141, 147 и 183 идентифицировали как характеризующиеся предпочтительным спариванием либо с доменом CL-каппа (или легкой цепью, содержащей домен CL-каппа), либо с доменом CL-лямбда (или легкой цепью, содержащей домен CL-лямбда). Замена в положении 141 домена CH1 на D, R или Q (по сравнению с A в диком типе) повышает предпочтительное спаривание с доменом CL-лямбда (или легкой цепью, содержащей домен CL-лямбда) (т.е. сниженное соотношение MFI каппа:лямбда) (см. фиг. 5). Замена в положении 147 домена CH1 на F, I, T, Y, L, R, N, E, H, M или Q (по сравнению с K в диком типе) повышает предпочтительное спаривание с доменом CL-каппа (или легкой цепью, содержащей домен CL-каппа) (т.е. повышенное соотношение MFI каппа:лямбда) (см. фиг. 5). Замена в положении 183 домена CH1 на R, K, Y, W, E, F или Q (по сравнению с S в диком типе) повышает предпочтительное спаривание с доменом CL-каппа (или легкой цепью, содержащей домен CL-каппа) (т.е. повышенное соотношение MFI каппа:лямбда) (см. фиг. 5). В таблице 5 показано количество наблюдаемых вариантов домена CH1, характеризующихся конкретными аминокислотными заменами, которые способствуют предпочтительности при спаривании.
Таблица 5. Наблюдаемые аминокислотные замены в вариантах домена CH1 с предпочтительностью в отношении легкой цепи
Затем оценивали влияние идентифицированных вариантов домена CH1 на контрольное стандартное биспецифичное антитело (2 тяжелых цепи x 2 легких цепи) в IgG-подобном формате (2 области Fab, присоединенные N-концом к димерной молекуле Fc). Использовали последовательности VH-CH1, полученные из двух одобренных клинических терапевтических антител, устекинумаба и панитумумаба. Вводили мутации «выступа» (S354C; T366W) и «впадины» (Y349C; T366S; L368A; Y407V) для обеспечения необходимого гетеродимерного спаривания тяжелых цепей. Плазмиды ДНК подтверждали посредством секвенирования по Сэнгеру до трансфекции в клетки HEK293 посредством стандартных протоколов.
Трансфицированные клетки HEK культивировали в среде CD optiCHO (Invitrogen), и на 6 день после трансфекции супернатанты собирали и подвергали аффинной очистке на белке A. Очищенные IgG обрабатывали с помощью GingisKHAN® (Genovis AB) для ферментативного расщепления области Fab из Fc-части.
Проводили ЖХ-МС для очищенных Fab для подтверждения последовательности каждого компонента IgG (2 тяжелых цепи x 2 легких цепи) и для определения относительного процентного содержания каждого компонента (см. фиг. 7). Вкратце, очищенные IgG расщепляли с помощью GingisKHAN для ферментативного расщепления области Fab из Fc-части. Образцы Fab вводили в ВЭЖХ Agilent серии 1100 с колонкой POROS® R2 10 мкм Applied Biosystems (2,1 x 30 мм, 0,1 мл), поддерживаемой при 65°C. После введения образцы элюировали из колонки с применением 0,21-минутного градиента 2-95% ацетонитрила со скоростью потока 2 мл/мин (подвижная фаза A: 0,1% муравьиной кислоты в H2O; подвижная фаза B: 0,1% муравьиной кислоты в ацетонитриле). С помощью отводного клапана 150 мкл/мин общего потока загружали в масс-спектрометр Bruker maXis 4G. Масс-спектрометр работал в режиме положительных ионов с диапазоном масса/заряд 700-2500. Остальные параметры источника задавали следующим образом: капилляр был установлен на 5500 В, распылитель на 4,0 бар, сухой газ на 4,0 л/мин и сухая температура на 200°С. Полученные масс-спектры анализировали с помощью Bruker Compass Data Analysis версии 4.1. Обнаружение интактных молекул Fab подтверждали на основании измерения массы по сравнению с теоретической последовательностью. Относительное количественное определение для каждого вида рассчитывали на основании интенсивности пиков каждого вида по сравнению с суммой всех значений интенсивности пиков.
Когда обе тяжелые цепи относятся к дикому типу, неправильное спаривание происходит примерно в 30% случаев; однако, когда тяжелые цепи содержат вариантный домен CH1, как описано в настоящем документе, наблюдается значительное улучшение правильного спаривания тяжелых цепей с легкими цепями (см. фиг. 7 и таблицу 6). Легкая цепь pani относится к дикому типу. Легкая цепь uste представляет собой лямбда-слияние. HC1 представляет собой pani; LC1 представляет собой каппа pani; HC2 представляет собой uste; LC2 представляет собой лямбда uste. Например, когда первая тяжелая цепь (HC1) содержит K147F и S183R/K/Y, а вторая тяжелая цепь содержит A141D (BsAb 10, 12 и 14 соответственно), неправильное спаривание устраняется по меньшей мере наполовину, т.е. происходит только в 6,8, 10,5 или 11% случаев. Действительно, одиночная замена в положении 141 (141D) приводит к 50% снижению неправильного спаривания, т.е. 6,1% по сравнению с 3,1% HC1-LC2 и 22,8% по сравнению с 9,9% HC2-LC1 (BsAb2). На основе полученных данных заявитель представляет иллюстративные последовательности домена CH1, которые характеризуются предпочтительностью в отношении легкой каппа- или лямбда-цепи/домена CL, в таблице 7.
Таблица 6. Процент образования продукта спаривания тяжелой цепи с легкой цепью
Таблица 7. Домены CH1 с предпочтительностью в отношении каппа- или лямбда-цепи


Экспрессию и качество очищенных антител оценивали с помощью эксклюзионной хроматографии (SEC). Вкратце, для контроля колоночной хроматографии использовали ВЭЖХ Agilent 1 100 (колонка TSCgel Super SW3000). Колонку предварительно подготавливали с помощью высокогликозилированного и агрегированного IgG с целью минимизации потенциала для взаимодействий антитела с колонкой и уравновешивали с помощью промывочного буфера (200 мМ фосфата натрия, 250 мМ хлорида натрия с pH 6,8) перед применением. Примерно 2-5 мкг образца белка вводили в колонку и регулировали скорость потока до 0,400 мл/мин. Миграцию белка отслеживали при длине волны 280 нм. Общее время анализа составляло примерно 11 минут. Данные анализировали с применением программного обеспечения ChemStation. Профили SEC подтверждали, что замены домена CH1 не оказывали влияния на профили вариантов по сравнению с диким типом (данные не показаны).
Измеряли аффинности и кинетику связывания очищенных биспецифичных антител с человеческим IL-12B (Uste) и человеческим EGFR (Pani) для подтверждения того, что вариантный домен CH1 не влиял на связывание с мишенью (см. фиг. 6A-6E). С помощью прибора Octet® QKe (ForteBio) со 100 нМ антигена захватывали биспецифичные образцы IgG на наконечнике сенсора к Fc hIgG и измеряли кинетику связывания с IL12B или EGFR (скорость ассоциации: 180 с, скорость диссоциации: 180 с). Анализ BLI проводили при 29°C с применением 1× буфера для кинетического анализа (ForteBio) в качестве буфера для анализа. Биосенсоры для захвата Fc IgG человека (AHC) (ForteBio) сначала предварительно оставляли в буфере для анализа в течение более пяти минут. Образцы биспецифичных IgG (5 мкг/мл) захватывали на сенсоре в течение 300 секунд. Затем сенсоры помещали в буфер для анализа на 120 секунд для определения исходного уровня перед измерением связывания с белком IL12B или EGFR (концентрация 100 нМ). Диссоциацию IL12B или EGFR измеряли путем перемещения сенсоров в буфер для анализа на 180 секунд. Встряхивание на всех стадиях проводили при 1000 об./мин. Кинетические параметры получали с помощью программного обеспечения для анализа данных Octet® версии 8.2.0.7 с использованием референсного субстрата, межстадийной коррекции на основе диссоциации, модели связывания 1 к 1 и глобальной аппроксимации (Rmax несвязанного с сенсором). Значения константы скорости ассоциации (ka), константы скорости диссоциации (kd) и равновесной константы (KD) были индивидуально назначены для каждого измерения.
Пример 5. Сборка и выбор библиотеки 141x181x218
Также были идентифицированы дополнительные аминокислотные замены CH1, которые обеспечивают предпочтительное спаривание с доменом CL-лямбда. На основании предыдущих данных по отбору, а также структурного анализа, для дополнительного изменения отбирали набор из трех положений CH1 (141, 181 и 218). Аминокислотное разнообразие в положении 141 получали посредством вырожденного кодона RMW, представляющего шесть встречающихся в природе аминокислот (D, T, A, E, K и N). Аминокислотное разнообразие в положениях 181 и 218 получали посредством вырожденного кодона NNK, представляющего все 20 встречающихся в природе аминокислот. Дизайн библиотеки включал все возможные комбинации аминокислот в этих трех положениях с разнообразием 2400. При использовании штамма легкой цепи с легкой лямбда-цепью под контролем промотора GAL10 (GAL1::ADI-26140 VL-Ck x GAL10::ADI-26140 VL - Cl) эту библиотеку конструировали таким образом, как описано ранее. Отбор по предпочтительности к лямбда-цепи проводили путем окрашивания антителами к каппа-FITC человека и к лямбда-PE человека с последующими несколькими раундами сортировки клеток, как описано ранее. Полученные продукты (96 клонов) секвенировали, как описано ранее, и проводили количественное определение на основе FACS предпочтительности в отношении лямбда-цепи по сравнению с исходным штаммом. В анализ включали дикий тип («WT») и ранее идентифицированный наилучший клон A141D. На основании этих данных идентифицировали аминокислотные комбинации, которые обеспечивали наибольшее улучшение в отношении предпочтительного спаривания с лямбда-цепью по сравнению с исходным белком и A141D.
На фиг. 8 показано, что большинство полученных в результате клонов имеют более высокую предпочтительность спаривания с лямбда-цепью, как определено на основании значения FOP. В таблице 8 представлены замены домена CH1 и значения FOP для соотношения MFI лямбда:каппа для 13 лучших клонов, отмеченных на фиг. 8.
Таблица 8. Лучшие 13 значений FOP для полученных в результате клонов
Анализ показал, что замена в положении 141 на D, K или E в паре с заменой в положении 181 на K и заменой в положении 218 на L, E, D, P, A, H, S, Q, N, T, I, M, G, C или W часто встречались среди полученных в результате клонов и увеличивали предпочтительность в отношении легкой лямбда-цепи по сравнению с A141D (повышенное соотношение MFI лямбда:каппа). На фиг. 9 показаны индивидуальные и средние значения FOP, измеренные в клонах, имеющих D в положении 141, K в положении 181 и различные аминокислоты в положении 218 CH1. Основные последовательности CH1 клонировали обратно в штамм LC (этот процесс впоследствии использовали во всех анализах) и подтверждали клоны и предпочтительность в отношении лямбда-цепи путем расчета значений FOP в трех параллельных анализах (фиг. 10).
Дополнительный анализ дал 9 уникальных кандидатных последовательностей CH1 для получения IgG млекопитающих (см. таблицу 9).
Таблица 9. Домены CH1 с предпочтительностью в отношении каппа- или лямбда-цепи
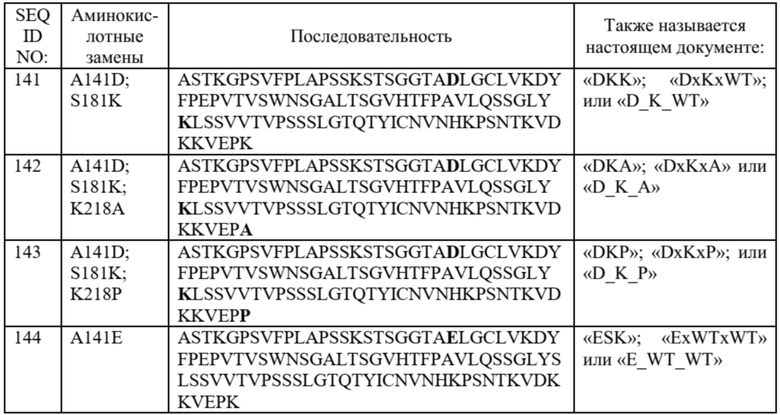
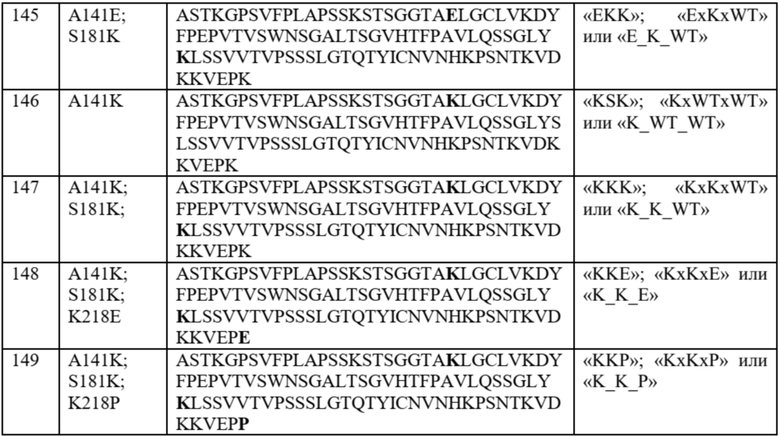
9 кандидатных последовательностей CH1 вместе с WT (т.е. «ASK») и A141D (т.е. «DSK») клонировали в векторы экспрессии млекопитающих стандартными методами. Для определения предпочтительности к лямбда-цепи плазмиды, представляющие требуемую тяжелую цепь, легкую лямбда-цепь и легкую каппа-цепь, трансфицировали в клетки HEK293 при соотношении плазмид 2:1:1. Трансфицированные клетки HEK культивировали и очищали IgG с применением ранее описанных протоколов. Не ограничиваясь какой-либо конкретной теорией, полагают, что экспрессия приблизительно равных количеств всех полипептидов тяжелой цепи и всех полипептидов легкой цепи (в данном случае HC:LC-каппа:LC-каппа = «2:1:1» приводит к общей HC:общей LC = 1:1) (т.е. отсутствие избытка HC и избытка LC), по-видимому, позволила авторам изобретения избежать различных отклонений, что привело к визуализации истинной предпочтительности в отношении каппа- или лямбда-цепи вариантов домена CH1.
Количественное определение предпочтительности в отношении лямбда-цепи на основе FACS проводили для IgG, продуцированных у млекопитающего. На фиг. 11 представлены графики FACS, а на фиг. 12 и в таблице 10 представлены значения FOP (MFI лямбда: каппа) для 9 вариантов CH1 и для WT и A141D (т.е. «DSK»). На фиг. 13 показано, что, когда CH1 имеет D в положении 141, дополнительные замены в положении 181 или в положениях 181 и 218 дополнительно улучшают предпочтительность в отношении лямбда-цепи (на основе соотношения MFI лямбда: каппа).
Таблица 10. Значения FOP для 9 вариантов CH1
Кроме того, данные ЖХ-МС восстановленных полноразмерных IgG использовали для определения относительного количества легкой лямбда-цепи и легкой каппа-цепи в очищенном образце IgG. На фиг. 14 сравнивается % молекул, спаренных с легкой каппа-цепью (LC), и % молекул, спаренных с легкой лямбда-цепью.
Анализ этих данных обеспечил три последовательности CH1 (SEQ ID NO: 143, 142 и 141, имеющие замены DKP, DKA и DKK соответственно) с улучшенной предпочтительностью в отношении лямбда-цепи по сравнению с исходным белком и ранее идентифицированным наилучшим образцом, A141D.
Для определения того, спариваются ли эти последовательности CH1 с легкой каппа-цепью, плазмиды с кандидатными CH1 тяжелой цепи трансфицировали в клетки HE293 с либо 1.) легкой каппа-цепью, либо 2.) легкой лямбда-цепью. K147F S183R в качестве CH1 с предпочтительностью в отношении каппа-цепи, WT, A141D также включали в качестве контролей. Трансфицированные клетки HEK культивировали и очищали стандартными методами. Связанные Fab тяжелой цепи и легкой цепи получали из очищенного IgG с применением описанных ранее способов. Выход процесса определяли с применением стандартных методов и нормировали к выходу процесса для WT для расчета «FOP» выхода процесса. На основании FOP выхода процесса все из A141D, A141D S181K, A141D S181K K218A и A141D S181K K218P все еще связывались с LC-каппа, когда присутствовала только LC-каппа (но не LC-лямбда), но большее связывание происходило с LC-лямбда, чем с LC-каппа (фиг. 15). Tm Fab каппа- и лямбда-Fab измеряли с помощью дифференциальной сканирующей флуориметрии с применением RT-ПЦР BioRad CFX96 (фиг. 16). Для каждого варианта CH1 рассчитывали относительную прибавку Tm Fab, спаренного с лямбда-цепью («относительная прибавка Tm лямбда» или «чистая прибавка Tm лямбда»), определяемую как [изменение Tm в варианте Fab, спаренном с лямбда-цепью, относительно Fab WT, спаренного с лямбда-цепью («Δ Tm лямбда»)] - [изменение Tm в варианте Fab, спаренном с каппа-цепью, относительно Fab WT, спаренного с каппа-цепью («Δ Tm каппа»)] (фиг. 17). Как показано на фиг. 17, относительная прибавка Tm лямбда увеличивался с дополнительной заменой (заменами) при S181 или при S181 и K218. Не ограничиваясь какой-либо конкретной теорией, основываясь на фиг. 16 и 17, дестабилизация спаривания с LC-каппа, по-видимому, способствовала относительной прибавке Tm лямбда-цепи и увеличению спаривания с CL-лямбда.
Пример 6. Построение и выбор библиотеки 141xALL
Дополнительные библиотеки конструировали для тестирования дополнительных остатков в CH1 в отношении способствования предпочтительному связыванию лямбда-цепи в паре с заменой в положении 141. Шесть новых библиотек (LAD11522-LAD11527) были сконструированы таким образом, чтобы они характеризовались максимум тремя заменами в трех областях (DOR1, DOR2 и DOR3) CH1 (таблица 11). Вместе шесть библиотек представляют все возможные наборы замен, которые включают две замены в трех доменах, представляющих интерес, в паре с положением 141. Во всех библиотеках аминокислотное разнообразие в положении 141 получали с помощью вырожденного кодона RMW, а аминокислотное разнообразие в двух других изменяемых положениях получали с помощью вырожденного кодона NNK. Библиотеки конструировали с применением ранее описанных методов. Отбор по предпочтительности в отношении лямбда-цепи проводили, как описано ранее.
Таблица 11. Проектирование и построение библиотеки
Начиная со второго раунда отбора методом FACS продукты отбора разнообразия CH1 выделяли и повторно клонировали в соответствующий штамм легкой цепи с двумя цепями для восстановления сниженной экспрессии легкой каппа-цепи в библиотеке. Разнообразие CH1 выделяли с применением ПЦР-амплификации с соответствующими праймерами и стандартной очистки ДНК. Затем этот пул фрагментов ДНК подвергали электропорации вариабельной областью тяжелой цепи ADI-26140 и расщепляли плазмиду в соответствующий штамм легкой цепи с двумя цепями.
Полученные продукты секвенировали, как описано ранее (фиг. 18), и проводили количественное определение на основе FACS предпочтительности к лямбда-цепи по сравнению с исходным штаммом. В анализ включали ранее идентифицированный наилучший клон A141D S181K K218P. На основании этих данных определяли аминокислотные комбинации с наибольшим улучшением предпочтительного спаривания с лямбда-цепью по сравнению с исходным белком.
Лучшие 46 клонов, содержащие 28 уникальных последовательностей CH1 (таблица 12), экспрессировали в виде IgG в дрожжах. Новые последовательности CH1 вместе с WT, A141D (или «DSK») и некоторыми из лучших результатов из серии 141x181x218 (DKP, DKA, KKE, KKP и EKK) в примере 5 сравнивали по значению FOP, определенному методом проточной цитометрии (MFI лямбда: каппа) (фиг. 19). По меньшей мере семь образцов, имеющих последовательность CH1 согласно SEQ ID NO: 155, 157, 159, 162, 163, 164 или 165, соответствующие точкам данных, помеченным стрелкой на фиг. 19, продемонстрировали значения FOP, эквивалентные или превышающие значения протестированных наилучших результатов 141x181x218.
Таблица 12. 28 уникальных последовательностей CH1 из серий 141xALL с предпочтительностью в отношении лямбда-цепи
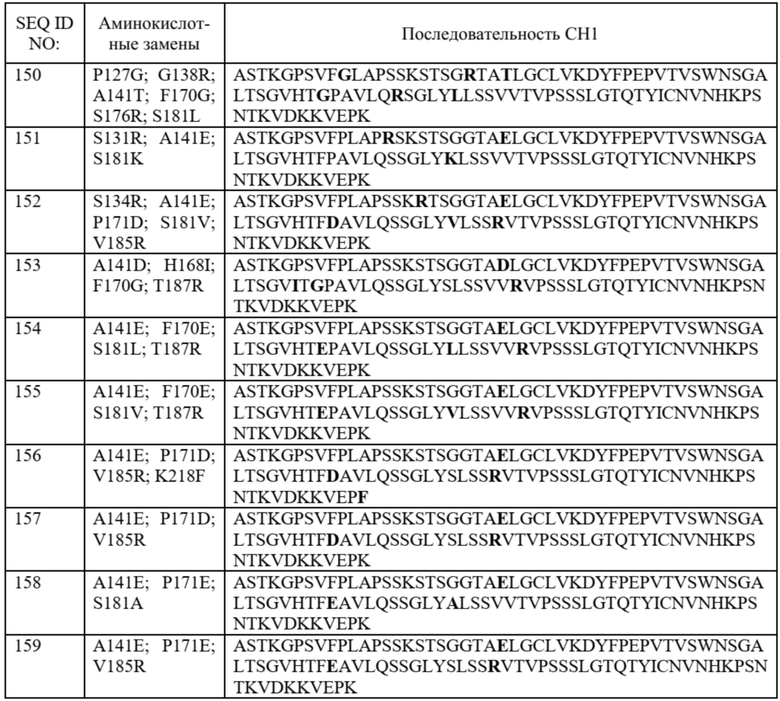
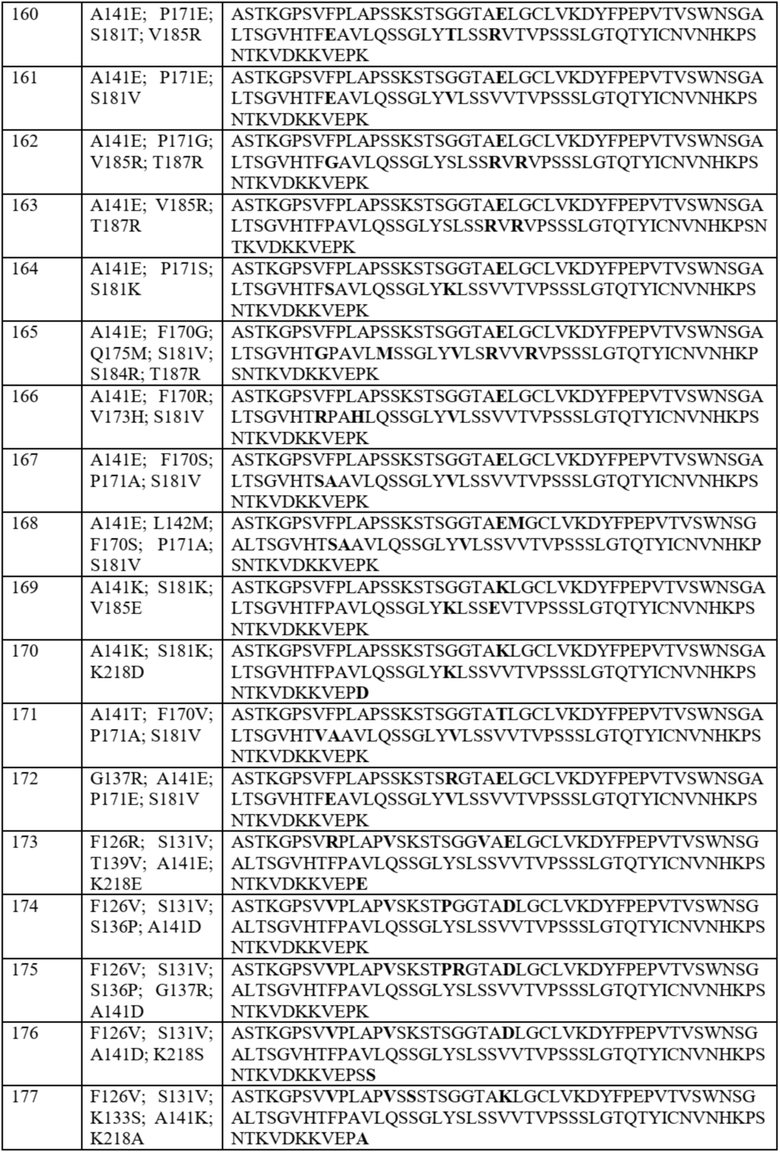
Пример 7. Конструкции и скрининг серии 141x(170/171)x(185/187)
Анализ результатов в примере 6 позволил получить четыре новых положения/остатка, представляющих интерес, включая F170, P171, V185 и T187. На основании аминокислот, часто наблюдаемых в положениях 170, 171, 185, и 187, вместе с 141, которое показало высокие значения FOP в предыдущих исследованиях (например, E и D часто встречаются в положении 141; E часто встречается в положении 170 или 171 в продуктах 141xALL; и R часто встречается в положениях 185 и/или 187, когда положение 141 заменено и независимо с положением 171), были рационально разработаны 14 уникальных вариантов домена CH1, имеющих максимум три аминокислотные замены на домен CH1 (таблица 13), в качестве кандидатов для наборов наилучших замен, обеспечивающих предпочтительность к лямбда-цепи. 14 наилучших результатов в таблице 13 включают «A141E; V185R; T187R» (SEQ ID NO: 163) и «A141E; P171E; V185R (SEQ ID NO: 159)», которые тестировали в примере 6.
Таблица 13. Новые последовательности CH1 из серии 141x(170/171)x(185x187)
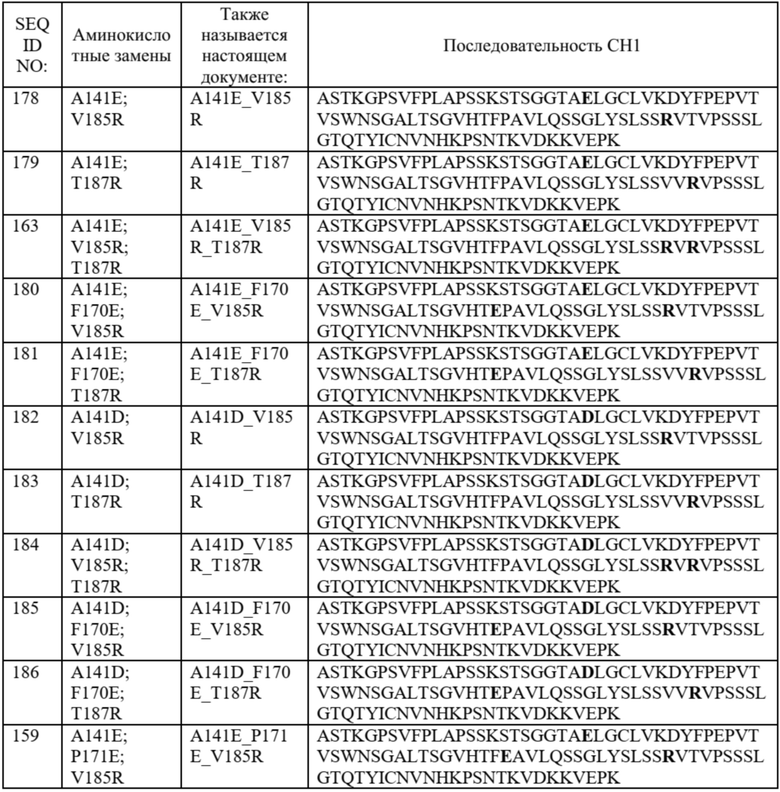

Тяжелые цепи, содержащие одну из 14 последовательностей вариантов домена CH1, клонировали в клетки млекопитающих (HEK), совместно экспрессирующие легкие каппа- и лямбда-цепи (с соотношением тяжелая цепь (HC): легкая лямбда-цепь (LC): LC-каппа = 2:1:1, т.е. соотношение HC:LC всегда составляет 1:1), как описано выше. Также в качестве контролей были включены дикий тип (тяжелая цепь ADI-26140), вариант «A141D» и вариант «A141D_S181K_K218P». Предпочтиость в отношении лямбда-цепи определяли с применением таких же анализов, как описано выше.
Соотношения MFI лямбда к MFI каппа оценивали с помощью проточной цитометрии. Значения FOP для 14 наилучших результатов и индивидуальные графики FACS представлены в таблице 14 и на фиг. 20-22 (нумерация на каждом графике представляет собой ранжирование, показанное в таблице 14). Среди 14 наилучших результатов «A141D_P171E_V185R» и «A141D_F170E_T187R» показали еще более высокие значения FOP, чем «A141D_S181K_K218P» - наилучший результат, идентифицированный в примере 5. Многие другие варианты из 14 наилучших результатов также показали более высокие значения FOP по сравнению с «A141D», и все 14 наилучших результатов показали более высокие значения FOP по сравнению с диким типом.
Таблица 14. Значения FOP для 14 наилучших результатов варианта CH1 и контролей (ранжирования основаны на значении FOP)
Количество LC-каппа и -лямбда на образец количественно определяли с помощью ЖХ-МС (таблица 15 и фиг. 23). Аналогично результатам оценки предпочтительности в отношении лямбда-цепи на основе FACS, «A141D_P171E_V185R» и «A141D_F170E_T187R» продемонстрировали еще более высокий % лямбда-цепей и еще более низкий % каппа-цепей по сравнению с «A141D_S181K_K218P» - наилучшим результатом, идентифицированным в примере 6. Многие другие варианты из 14 наилучших результатов также показали более высокий % лямбда и более низкий % каппа по сравнению с «A141D», и все 14 наилучших результатов показали более высокий % лямбда и более низкий % каппа по сравнению с диким типом.
Таблица 15. % LC-лямбда и % LC-каппа, измеренные с помощью ЖХ-МС (со значениями FOP из таблицы 14)
Для определения того, спариваются ли два обладающих наибольшей предпочтительностью в отношении лямбда-цепи варианта CH1 («A141D_P171E_V185R» и «A141D_F170E_T187R») с легкой каппа-цепью, плазмиды тяжелой цепи с вариантом CH1 трансфицировали в клетки HEK293 с либо 1.) легкой каппа-цепью, либо 2.) легкой лямбда-цепью (с соотношением тяжелая цепь: легкая цепь = 1: 1). K147F S183R в качестве CH1 с предпочтительностью к каппа-цепи, WT также включали в качестве контролей. Трансфицированные клетки HEK культивировали и очищали IgG стандартными методами с применением колонки с белком A. Выход процесса (мг/л) определяли с применением стандартных методов и нормировали к выходу процесса для WT. На основании нормированного выхода процесса как «A141D_P171E_V185R», так и «A141D_F170E_T187R» по-прежнему связывались с LC-каппа, когда присутствовали только LC-каппа (но не LC-лямбда), но большее связывание наблюдалось с LC-лямбда, чем с LC-каппа (фиг. 30).
Также оценивали выходы процесса для Fab-формата. IgG, имеющие тяжелые цепи с вариантом CH1, получали и очищали с применением того же способа. K147F S183R в качестве CH1 с предпочтительностью в отношении каппа-цепи, WT, A141D и A141D S181K K218P также включали в качестве контролей. Связанные Fab тяжелой цепи и легкой цепи получали из очищенного IgG посредством расщепления ферментом папаином и очистки колонки CH1 с применением стандартных методов. Нормированный выход гидролиза Fab рассчитывали как % извлеченного Fab из гидролизата IgG (количество извлеченного Fab/количество IgG в гидролизате), нормированный к % извлечения исходного белка для каждой легкой цепи. Выход процесса определяли с применением стандартных методов и нормировали к выходу процесса для WT. В соответствии с данными на фиг. 15, «A141D» и «A141D S181K K218P» имели более высокие выходы процесса с LC-лямбда по сравнению с LC-каппа, и «K147F S183R» демонстрировало чрезвычайно высокую предпочтительность в отношении каппа-цепи (фиг. 31). Как «A141D_P171E_V185R», так и «A141D_F170E_T187R» по-прежнему связывались с CH1-каппа, когда присутствовали только CH1-каппа (но не CH1-лямбда), но с LC-лямбда был получен заметно более высокий выход, чем с LC-каппа (фиг. 31). Добавление мутации «P171E_V185R» или «F170E_T187R» к «A141D» дополнительно усиливало предпочтительность «A141D» в отношении лямбда-цепи.
Пример 8. Структурный анализ вариантов «A141D» и «K147F S183R»
Методы
Кристаллизация и определение структуры CH1 панитумумаба дикого типа-Cλ
Белок Fab CH1 панитумумаба дикого типа-константной области лямбда (Cλ) в концентрации 6,5 мг/мл центрифугировали при 14000 × g при 4°C в течение 5 минут. 305 нл белка смешивали с каплей резервуарного раствора на 150 нл и 50 нл затравочного раствора и уравновешивали 40 мкл резервуарного раствора при 20°C в 3-луночных планшетах MRC. Затравочные кристаллы, идентифицированные в скрининге BCS (Molecular Dimensions), использовали в экспериментах по матричному скринингу микрозатравки (MMS) (D'Arcy, A., Villard, F., and Marsh, M. (2007) “An automated microseed matrix-screening method for protein crystallization” Acta Crystallogr D Biol Crystallogr 63, 550-554.) с получением кристаллов, выращенных в 0,1 М фосфате/цитрате с pH 5,5 и 36% (об./об.) PEG Smear Low и переносом в 0,1 М фосфат/цитрат с pH 5,5, 38% PEG Smear Low и 4% глицерина, с последующим мгновенным замораживанием в жидком азоте. Дифракционные данные собирали до 1,09 Å при 100 К на станции I03, Diamond Light Source, Дидкот, Англия, оборудованной детектором Eiger2 XE 16M (DECTRIS). Набор данных интегрировали в autoPROC (Vonrhein, C. et al. (2011) “Data processing and analysis with the autoPROC toolbox” Acta Cryst. D67, 293-302.) с использованием XDS (Kabsch W. (2010) “XDS” Acta. Crystallogr. D Biol. Crystallogr. 66, 125-132.) и масштабировали с использованием Aimless (Evans P.R. and Murshudov, G.N. (2013) “How good are my data and what is the resolution” Acta Crystallogr D Biol. Crystallogr. 69, 1204-1214.) пакета программного обеспечения CCP4 (Winn M. D. et al. (2011) “Overview of the CCP4 suite and current developments” Acta Crystallog. D Biol. Crystallogr. 67, 235-242. 235-242.). Кристаллы состояли из 2 молекул на асимметрическую единицу (ASU) в пространственной группе P1211. Структура была решена с помощью автоматизированной системы молекулярной замены MoRDA (Vagin A. and Lebedev A. (2015) “MoRDa, an automatic molecular replacement pipeline” Acta Cryst A. A71, s19.) (включающей MOLREP (Vagin A., Teplyakov A. (1997) “MOLREP: an automated program for molecular replacement” J. Appl. Cryst. 30, 1022-1025.) и Refmac5 (Murshudov, G.N., Skubak, P., Lebedev, A.A., Pannu, N.S., Steiner, R.A., Nicholls, R.A., Winn, M.D. Long, F. and Vagin, A.A. (2011) REFMAC5 for the refinement of macromolecular crystal structures, Acta Crystallogr. D Biol. Crystallogr. 67, 355-367.)), которая выбрала 5N7W и 5SX4 из Банка данных структур белков (Berman H.M. et al. (2000) The “Protein Data Bank” Nucleic Acids Research, 28.) в качестве исходных моделей поиска. Автоматизированное построение модели выполняли с помощью программного обеспечения BUCCANEER (Cowtan K. (2006) “The Buccaneer software for automated model building. 1. Tracing protein chains” Acta Crystallographica D62, 1002-1011.). Модель дополнительно улучшали путем ручного уточнения в Coot (Emsley P., Lohkamp, B., Scott, W.G. and Cowtan K. (2010) “Features and development of Coot” Acta Crystallogr. D Biol. Crystallogr. 66, 486-501.), а также уточнения Refmac5 (Murshudov, G.N., Skubak, P., Lebedev, A.A., Pannu, N.S., Steiner, R.A., Nicholls, R.A., Winn, M.D. Long, F. and Vagin, A.A. (2011) REFMAC5 for the refinement of macromolecular crystal structures, Acta Crystallogr. D Biol. Crystallogr. 67, 355-367.) и Buster (Bricogne G, Blanc E, Brandl M, Flensburg C, Keller P, Paciorek W, Roversi P, Sharff A, Smart O, Vonrhein C, Womack T. (2011). BUSTER version 2.11.7. Global Phasing Ltd, Cambridge, United Kingdom.) до конечных R и Rfree, равных 14,5% и 16,9%, соответственно (фиг. 32).
Кристаллизация и определение структуры CH1 A141D панитумумаба-Cλ, CH1 дикого типа-Cκ и CH1 K147F-S183R-Cκ
Fab CH1 A141D панитумумаба-Cλ, CH1 панитумумаба дикого типа-константный каппа (Cκ) и CH1 K147F-S183R панитумумаба-Cκ центрифугировали при 14000 × g при 4°C в течение 5 минут. Для CH1 A141D панитумумаба-Cλ и CH1 K147F-S183R-Cκ 200 нл 10,0 мг/мл Fab смешивали с 150 нл капли резервуарного раствора и 50 нл затравочного раствора, уравновешенного 40 мкл резервуарного раствора. Затравочные кристаллы, идентифицированные в скрининге BCS, использовали в экспериментах MMS для определения оптимальных условий кристаллизации. 0,1 М фосфатно-цитратный буфер с pH 5,5 и 36% (об./об.) PEG Smear Low применяли для CH1 A141D панитумумаба-Cλ и 0,1 М ацетата натрия с pH 4,5 с 30% об./об. PEG Smear Low для CH1 K147F-S183R панитумумаба-Cκ. 150 нл 19,2 мг/мл CH1 дикого типа-Cκ смешивали с 150 нл капли резервуарного раствора и добавляли к 40 мкл резервуарного раствора и подвергали скринингу с применением PACT Suite (Molecular Dimensions). Конечное условие кристаллизации состояло из 0,1 MES с pH 6,0 с 20 мас./об.% PEG 6000 и 0,2 M дигидрата хлорида кальция. Кристаллы переносили в криорастворы, состоящие из 0,1 М фосфатно-цитратного буфера, pH 5,5, 38% PEG Smear Low, 4% глицерина; 0,07 M MES, pH 6,0, 21% PEG 6000, 0,2 M CaCl2, 23,5% глицерина; и 0,1 M NaAc с pH 4,5, 32,5 % PEG Smear Low, 25% глицерина для CH1 A141D панитумумаба-Cλ, CH1 дикого типа-Cκ и CH1 K147F-S183R-Cκ, соответственно. Все кристаллы мгновенно замораживали в жидком азоте и собирали кристаллографические данные при 100 K на станции I03, Diamond Light Source, Дидкот, Англия, оборудованной детектором Eiger2 XE 16M (DECTRIS) с разрешением 1,2-2,6 Å. Данные индексировали и интегрировали в iMOSFLM (Battye, T. G. G., Kontogiannis, L., Johnson, O., Powell, H. R., & Leslie, A. G. (2011). iMOSFLM: a new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallographica Section D: Biological Crystallography, 67(4), 271-281.), а также масштабировали и объединяли с AIMLESS (Evans P.R. and Murshudov, G.N. (2013) “How good are my data and what is the resolution” Acta Crystallogr D Biol. Crystallogr. 69, 1204-1214.) через CCP4 suite (Winn M. D. et al. (2011) “Overview of the CCP4 suite and current developments” Acta Crystallog. D Biol. Crystallogr. 67, 235-242. 235-242.).
Структура CH1 A141D панитумумаба-Cλ была решена путем молекулярной замены с применением кристаллической структуры CH1 дикого типа-Cλ в качестве модели поиска. Несколько раундов анизотропного B-фактора и простого ограниченного уточнения проводили в Refmac5 (Murshudov, G.N., Skubak, P., Lebedev, A.A., Pannu, N.S., Steiner, R.A., Nicholls, R.A., Winn, M.D. Long, F. and Vagin, A.A. (2011) REFMAC5 for the refinement of macromolecular crystal structures, Acta Crystallogr. D Biol. Crystallogr. 67, 355-367.), с применением фактора размытия в последних раундах уточнения. Позиционные занятости CH1 A141D-Cλ присваивали на основании занятости CH1 дикого типа-Cλ и вручную регулировали в Coot (Emsley P., Lohkamp, B., Scott, W.G. and Cowtan K. (2010) “Features and development of Coot” Acta Crystallogr. D Biol. Crystallogr. 66, 486-501.) во время итеративного уточнения. Конечная структура, решенная в P1211 с 2 молекулами на ASU, характеризовалась значениями R и Rfree 15,2% и 17,0% соответственно (фиг. 33).
Структуры CH1 панитумумаба дикого типа-Cκ и CH1 K147F-S183R-Cκ решали с помощью молекулярной замены с помощью Phaser (McCoy, A. J., Grosse-Kunstleve, R. W., Adams, P. D., Winn, M. D., Storoni, L. C., & Read, R. J. (2007). Phaser crystallographic software. Journal of Applied Crystallography, 40(4), 658-674.) с использованием координат Fab-фрагмента панитумумаба в комплексе с EGFR (код PDB 5SX4) и с решенной структурой CH1 дикого типа-Cκ, соответственно, с последующими итеративными раундами построения модели вручную с использованием Coot (Emsley P., Lohkamp, B., Scott, W.G. and Cowtan K. (2010) “Features and development of Coot” Acta Crystallogr. D Biol. Crystallogr. 66, 486-501.) и автоматическим уточнением в Refmac5 (Murshudov, G.N., Skubak, P., Lebedev, A.A., Pannu, N.S., Steiner, R.A., Nicholls, R.A., Winn, M.D. Long, F. and Vagin, A.A. (2011) REFMAC5 for the refinement of macromolecular crystal structures, Acta Crystallogr. D Biol. Crystallogr. 67, 355-367.). Трансляционную некристаллографическую симметрию наблюдали для структуры CH1 дикого типа-Cκ, поэтому структуру решали в более низкой пространственной группе (P1211) с 6 молекулами Fab в ASU. Структуру уточняли до конечных значений R и Rfree 19,8% и 23,2% соответственно (фиг. 34). Структуру CH1 K147F-S183R-Cκ решали в пространственной группе P31 с 1 молекулой на ASU до конечных значений R и Rfree 19,8% и 23,3% соответственно (фиг. 35).
Анализ структуры и интерпретация
Предпочтиость в отношении легкой лямбда-цепи, опосредованная HC-A141D
Не ограничиваясь какой-либо конкретной теорией, полагают, что повышенная предпочтительность в отношении лямбда-цепи CH1 A141D панитумумаба-Cλ потенциально опосредована межцепочечной водородной связью, образованной между карбоксильной группой боковой цепи HC-Asp141 и гидроксильной группой боковой цепи λLC-Thr116 (фиг. 36C), которая не может образовываться с HC-Ala141 в CH1 панитумумаба дикого типа-Cλ (фиг. 36A). Область κLC, окружающая HC-Ala141, состоит из гидрофобных остатков Phe116, Phe118 и Leu135, тогда как κLC -Phe116 заменен полярным остатком Thr116 в λLC (фиг. 36B). Таким образом, введение заряда посредством мутации A141D может снижать предпочтительность в отношении каппа-цепи за счет нарушения гидрофобности в области контакта CH1-κLC при одновременной стабилизации спаривания CH1-λLC посредством водородной связи с λLC-Thr116. Кроме того, не ограничиваясь какой-либо конкретной теорией, полагают, что предпочтительность в отношении каппа-цепи может быть дополнительно снижена посредством стерического столкновения HC-Asp141 с κLC-Phe116, как показано путем выравнивания CH1 A141D панитумумаба-Cλ и CH1 дикого типа-κLC (фиг. 36D).
В водородной связи между HC-Asp141 и λLC-Thr116 связь образована между атомом акцептора водорода (O) в боковой цепи Asp141 и донорным атомом водорода (H) боковой цепи Thr116. Таким образом, другая аминокислота, которая имеет атом акцептора водорода в боковой цепи, также может образовывать водородную связь с Thr116 в λLC, обеспечивая предпочтительность в отношении лямбда-цепи. Исходя из того, что боковая цепь глутамата также имеет атом акцептора водорода (O), и глутамат по размеру и форме сходен с аспартатом, глутамат, вероятно, образует водородную связь с Thr116 в λLC, вызывая при этом стерическое столкновение с κLC, как показано на фиг. 36D, тем самым обеспечивая предпочтительность в отношении лямбда-цепи. Фактически, замена A141E обеспечивала высокую предпочтительность в отношении лямбда-цепи, как показано в примерах выше, подтверждая структурные анализы, проведенные заявителем.
Предпочтиость в отношении LC-каппа, опосредованная HC-K147F-S183R
Наблюдаемая предпочтительность в отношении каппа-цепи для CH1 K147F-S183R панитумумаба-Cκ может быть опосредована двумя новыми водородными связями в области контакта CH1 и Cκ. В структуре CH1 панитумумаба дикого типа-Cκ сеть водородных связей, координируемая с помощью HC-Lys147 и HC-Asp148, блокирует HC-Gln175, внося вклад в исходную предпочтительность спаривания с каппа-цепью (фиг. 37A). Одним из объяснений является то, что замена CH1 HC-Lys147 фенилаланином в этом положении нарушает эту сеть и освобождает боковую цепь HC-Gln175, которая взаимодействует с κLC посредством водородных связей с формамидным кислородом κLC-Gln160, тем самым увеличивая предпочтительность в отношении каппа-цепи (фиг. 37B). Кроме того, не ограничиваясь какой-либо конкретной теорией, полагают, что замена HC-S183R приводит в результате к дополнительной водородной связи между гуанидиновой группой боковой цепи HC-Arg183 и гидроксильной группой κLC-Thr178 (фиг. 37B, фиг. 38C). И наоборот, не ограничиваясь какой-либо конкретной теорией, полагают, что водородное связывание, наблюдаемое в положении 183 HC между HC-Ser183 и λLC-Tyr178 в CH1 панитумумаба дикого типа-Cλ, устраняется значительным стерическим столкновением боковых цепей HC-Arg183 и λLC-Tyr178 в смоделированном спаривании CH1 K147F-S183R и λLC, дестабилизируя спаривание с лямбда-цепью в пользу κLC (фиг. 38B и 38D).
В водородной связи между HC-Arg183 и κLC-Thr178 связь образуется между донорным атомом водорода (H) в боковой цепи Arg183 и атомом акцептора водорода (O) боковой цепи Thr178. Таким образом, другая аминокислота, которая имеет донорный атом водорода в боковой цепи, также может образовывать водородную связь с Thr178 в κLC, обеспечивая предпочтительность в отношении каппа-цепи. Крупная боковая цепь, такая как Arg, может способствовать генерированию стерического столкновения с Tyr178 в λLC, обеспечивая дополнительную предпочтительность в отношении каппа-цепи. Например, боковая цепь как лизина, так и триптофана имеет крупную боковую цепь, которая содержит донорный атом водорода (H). Таким образом, лизин и триптофан, вероятно, образуют водородную связь с Thr178 в κLC и, вероятно, претерпевают стерическое столкновение с λLC, как показано на фиг. 38D, в целом обеспечивая предпочтительность в отношении каппа-цепи. Боковая цепь треонина также может функционировать в качестве донора водорода посредством атома H в -OH. Таким образом, заявитель дополнительно предполагает, что аминокислота, имеющая относительно крупную. боковую цепь, которая может функционировать в качестве акцептора водорода, также может образовывать водородную связь с Thr178 в κLC, обеспечивая предпочтительность в отношении каппа-цепи. Например, глутамат, глутамин, гистидин или тирозин, которые характеризуются относительно крупной боковой цепью с акцепторным атомом водорода, при размещении в остатке 183 HC также могут обеспечивать предпочтительность в отношении каппа-цепи. На самом деле, большинство из этих новых аминокислотных замен в остатке 183 были фактически идентифицированы как обеспечивающие предпочтительность в отношении каппа-цепи в примере 3 (см. таблицу 3).
Как указано выше, замена Lys147 на Phe нарушила водородную связь между Lys147 и Gln175, тем самым освобождая Gln175 для образования водородной связи с Gln160 в κLC и, таким образом, способствуя предпочтительности в отношении каппа-цепи. Таким образом, замена Lys147 другой аминокислотой, боковая цепь которой не содержит атом донора или акцептора водорода, такой как аланин, глицин, изолейцин, лейцин или валин, также может способствовать предпочтительности в отношении каппа-цепи. На самом деле, большинство из этих новых аминокислотных замен в остатке 147 были фактически идентифицированы как обеспечивающие предпочтительность в отношении каппа-цепи в примере 3 (см. таблицу 3).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Adimab, LLC.
<120> ВАРИАНТЫ ДОМЕНА CH1, СКОНСТРУИРОВАННЫЕ ДЛЯ ИЗБИРАТЕЛЬНОГО
СПАРИВАНИЯ С ЛЕГКОЙ ЦЕПЬЮ, И СОДЕРЖАЩИЕ ИХ
МУЛЬТИСПЕЦИФИЧНЫЕ АНТИТЕЛА
<130> 1160430.002413
<150> 62/908,367
<151> 2019-09-30
<160> 189
<170> PatentIn версии 3.5
<210> 1
<211> 112
<212> Белок
<213> Homo sapiens
<400> 1
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
<210> 2
<211> 107
<212> Белок
<213> Homo sapiens
<400> 2
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 3
<211> 106
<212> Белок
<213> Homo sapiens
<400> 3
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 4
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_B08
<400> 4
Gly Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Phe Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 5
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_D09
<400> 5
Ala Ser Thr Lys Gly Pro His Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Ile Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 6
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_H09
<400> 6
Ala Ser Thr Lys Gly Pro Arg Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Leu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 7
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_B09
<400> 7
Ala Ser Thr Lys Gly Pro Ser Val Ala Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ile Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 8
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_E08
<400> 8
Ala Ser Thr Lys Gly Pro Ser Val Phe Val Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Glu
85 90 95
Lys Val Glu Pro Lys
100
<210> 9
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_H07
<400> 9
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro His Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Trp Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 10
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_F08
<400> 10
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Phe Val Ile Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 11
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_D07
<400> 11
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Ile Val Thr Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Gln
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 12
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_C07
<400> 12
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Leu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 13
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_E09
<400> 13
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Ser Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 14
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_F09
<400> 14
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Leu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Tyr
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 15
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_A07
<400> 15
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Met Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Gln
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 16
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_C09
<400> 16
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Val Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gly Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 17
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_B07
<400> 17
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu Ile Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gly Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 18
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_G09
<400> 18
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Gly Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Phe Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 19
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_C08
<400> 19
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Glu Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 20
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_G08
<400> 20
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Tyr Ser Val Val Thr Val Pro Ser Ile Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 21
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P01_G07
<400> 21
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Trp Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 22
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_D10
<400> 22
Ala Ser Thr Lys Gly Pro Glu Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Val Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 23
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_D12
<400> 23
Ala Ser Thr Lys Gly Pro Leu Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Phe Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 24
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_B10
<400> 24
Ala Ser Thr Lys Gly Pro Val Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Tyr Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 25
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_A11
<400> 25
Ala Ser Thr Lys Gly Pro Ser Val Thr Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 26
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_E12
<400> 26
Ala Ser Thr Lys Gly Pro Ser Val Phe Leu Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Glu Gly Gly Thr Ala Ala Leu Ser Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 27
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_F10
<400> 27
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Pro Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Glu Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 28
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_A10
<400> 28
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ala Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Leu Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 29
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_C12
<400> 29
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Ile Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Tyr Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 30
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_H10
<400> 30
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Asn Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Leu Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 31
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_G11
<400> 31
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Thr Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Phe Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 32
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_D11
<400> 32
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Val Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 33
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_E10
<400> 33
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Glu Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 34
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_B12
<400> 34
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Leu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gly Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 35
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_F11
<400> 35
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Gln Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Glu Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 36
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_G10
<400> 36
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Val Asp Tyr
20 25 30
Phe Ala Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 37
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_A12
<400> 37
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Tyr Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Gly
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 38
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_F12
<400> 38
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Met Gln Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gly Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 39
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_G12
<400> 39
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Tyr Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Ala Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Tyr Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 40
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_B11
<400> 40
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Tyr Tyr Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Ala Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 41
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_H11
<400> 41
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Lys Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 42
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_C10
<400> 42
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Gln Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 43
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P02_E11
<400> 43
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Gln Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Arg Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 44
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_C03
<400> 44
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Ile Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Leu Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 45
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_G03
<400> 45
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Val Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 46
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_F03
<400> 46
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 47
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_H01
<400> 47
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Ala
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 48
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_C01
<400> 48
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Ser Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 49
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_A01
<400> 49
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Gln Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 50
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_D02
<400> 50
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Cys
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 51
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_H03
<400> 51
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Ser
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 52
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_B03
<400> 52
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Cys
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Gln
100
<210> 53
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_H02
<400> 53
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp His
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys
100
<210> 54
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_E02
<400> 54
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp His
20 25 30
Leu Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 55
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_B02
<400> 55
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Leu Pro Glu Pro Met Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 56
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_A03
<400> 56
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Leu Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Pro Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 57
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_E03
<400> 57
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Ser Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Gly Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 58
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_D01
<400> 58
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Gly Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 59
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_C02
<400> 59
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Asn Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 60
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_B01
<400> 60
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Arg Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 61
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P01_G01
<400> 61
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Ser Tyr Lys Pro Ser Asn Thr Arg Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 62
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_E05
<400> 62
Ala Ser Thr Lys Gly Pro Ser Val Leu Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 63
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_C04
<400> 63
Ala Ser Thr Lys Gly Pro Ser Val Leu Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Gly Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Glu
85 90 95
Lys Val Glu Pro Lys
100
<210> 64
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_B06
<400> 64
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ala Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Cys
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 65
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_F05
<400> 65
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Pro Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Ser Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 66
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_D05
<400> 66
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Val Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 67
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_C06
<400> 67
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 68
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_C05
<400> 68
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Leu Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 69
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_H06
<400> 69
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Leu Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Arg
85 90 95
Lys Val Glu Pro Lys
100
<210> 70
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_A06
<400> 70
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Leu Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Ser Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 71
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_F06
<400> 71
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Ser Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 72
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_E04
<400> 72
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Leu Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 73
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P02_B05
<400> 73
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Pro Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 74
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_F12
<400> 74
Ala Ser Thr Lys Gly Pro Val Val Val Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Ser
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Ile Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 75
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_D12
<400> 75
Ala Ser Thr Lys Gly Pro Ser Val Phe Gly Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 76
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_D11
<400> 76
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala His Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 77
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_C12
<400> 77
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Gln Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 78
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_H12
<400> 78
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Thr Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 79
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_C10
<400> 79
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Asn Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val Phe Thr Asn Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 80
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_A11
<400> 80
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Asp
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 81
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_A10
<400> 81
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Thr
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Pro
100
<210> 82
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_B11
<400> 82
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ala Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 83
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_G12
<400> 83
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
His Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 84
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_F11
<400> 84
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ile Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 85
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_E10
<400> 85
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Pro Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 86
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_A12
<400> 86
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Val Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 87
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_B10
<400> 87
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Val Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 88
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_C11
<400> 88
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 89
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_H10
<400> 89
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 90
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_B12
<400> 90
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 91
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_G10
<400> 91
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Leu Val Asp Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 92
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_E11
<400> 92
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Ser Val Glu Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 93
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9611_P02_D10
<400> 93
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Val Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Ser Val Tyr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 94
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_F07
<400> 94
Ala Arg Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 95
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_B07
<400> 95
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Ser Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Thr Val Ser Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 96
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_A09
<400> 96
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Asn Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 97
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_G08
<400> 97
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Arg Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Met Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 98
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_G07
<400> 98
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Thr Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 99
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_B09
<400> 99
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Asp Ser Leu
1 5 10 15
Asn Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 100
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_E07
<400> 100
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Asp Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Asp Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 101
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_A08
<400> 101
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Glu
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Arg Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 102
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_H09
<400> 102
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Pro
1 5 10 15
Ser Thr Ser Gly Gly Ala Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 103
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_D07
<400> 103
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Leu Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 104
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_A07
<400> 104
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Asn Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 105
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_H08
<400> 105
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Pro Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 106
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_F08
<400> 106
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Val Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 107
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_D08
<400> 107
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 108
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_E09
<400> 108
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Thr Val Glu Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 109
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_C09
<400> 109
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val Val Thr Thr Asn Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 110
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9613_P01_H07
<400> 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Leu Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 111
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_F04
<400> 111
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Val Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 112
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_B04
<400> 112
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 113
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_G06
<400> 113
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 114
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_G04
<400> 114
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Ala Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 115
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_C06
<400> 115
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Gly Pro Lys
100
<210> 116
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_G05
<400> 116
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Met Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 117
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_C04
<400> 117
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Val Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asn Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 118
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9610_P02_E05
<400> 118
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Thr
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 119
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_A01
<400> 119
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Pro Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 120
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_G01
<400> 120
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Val Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Gly Pro Lys
100
<210> 121
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_H02
<400> 121
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Glu
85 90 95
Lys Val Glu Pro Lys
100
<210> 122
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_D03
<400> 122
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys
100
<210> 123
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_E01
<400> 123
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Gly Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 124
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_H03
<400> 124
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Val Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Gly Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 125
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_F02
<400> 125
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Gly Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 126
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> S 4
AD9612_P01_A02
<400> 126
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Val Ser Thr Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 127
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD9612_P01_C01
<400> 127
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Thr Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 128
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD10791_P01_F07
<400> 128
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Tyr Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 129
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD10791_P01_C07
<400> 129
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Lys Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Asp Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 130
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD10791_P01_A07
<400> 130
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ser Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Ser Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 131
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD10791_P02_G08
<400> 131
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Trp Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 132
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD10791_P02_D08
<400> 132
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Thr Ser Gly Gly Thr Arg Asp Leu Gly Cys Leu Val Lys Asp Tyr Phe
20 25 30
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
35 40 45
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
50 55 60
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
65 70 75 80
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
85 90 95
Val Glu Pro Asn
100
<210> 133
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> SAD10791_P02_F07
<400> 133
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Leu Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 134
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A140I; A141D
<400> 134
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ile Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 135
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A140V; A141D
<400> 135
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Val Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 136
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> S183Y
<400> 136
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Tyr Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 137
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> K147F; S183R
<400> 137
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Phe Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Arg Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 138
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> K147F; S183K
<400> 138
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Phe Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Lys Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 139
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> K147F S183Y
<400> 139
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Phe Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Tyr Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 140
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D
<400> 140
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 141
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; S181K
<400> 141
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 142
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; S181K; K218A
<400> 142
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Ala
100
<210> 143
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; S181K; K218P
<400> 143
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Pro
100
<210> 144
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E
<400> 144
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 145
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; S181K
<400> 145
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 146
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141K
<400> 146
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 147
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141K; S181K
<400> 147
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 148
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141K; S181K; K218E
<400> 148
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Glu
100
<210> 149
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141K; S181K; K218P
<400> 149
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Pro
100
<210> 150
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> P127G; G138R; A141T; F170G; S176R; S181L
<400> 150
Ala Ser Thr Lys Gly Pro Ser Val Phe Gly Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Arg Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Gly Pro Ala Val Leu Gln Arg Ser Gly Leu Tyr Leu
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 151
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> S131R; A141E; S181K
<400> 151
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Arg Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 152
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> S134R; A141E; P171D; S181V; V185R
<400> 152
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Arg Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Asp Ala Val Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 153
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; H168I; F170G; T187R
<400> 153
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val Ile Thr Gly Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 154
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; F170E; S181L; T187R
<400> 154
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Glu Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Leu
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 155
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; F170E; S181V; T187R
<400> 155
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Glu Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 156
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171D; V185R; K218F
<400> 156
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Asp Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Phe
100
<210> 157
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171D; V185R
<400> 157
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Asp Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 158
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171E; S181A
<400> 158
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Ala
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 159
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171E; V185R
<400> 159
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 160
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171E; S181T; V185R
<400> 160
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Thr
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 161
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171E; S181V
<400> 161
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 162
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171G; V185R; T187R
<400> 162
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Gly Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 163
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; V185R; T187R
<400> 163
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 164
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171S; S181K
<400> 164
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Ser Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 165
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; F170G; Q175M; S181V; S184R; T187R
<400> 165
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Gly Pro Ala Val Leu Met Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Arg Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 166
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; F170R; V173H; S181V
<400> 166
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Arg Pro Ala His Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 167
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; F170S; P171A; S181V
<400> 167
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Ser Ala Ala Val Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 168
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; L142M; F170S; P171A; S181V
<400> 168
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Met Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Ser Ala Ala Val Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 169
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141K; S181K; V185E
<400> 169
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Glu Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 170
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141K; S181K; K218D
<400> 170
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Lys
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Asp
100
<210> 171
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141T; F170V; P171A; S181V
<400> 171
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Thr Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Val Ala Ala Val Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 172
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> G137R; A141E; P171E; S181V
<400> 172
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Arg Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Val
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 173
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> F126R; S131V; T139V; A141E; K218E
<400> 173
Ala Ser Thr Lys Gly Pro Ser Val Arg Pro Leu Ala Pro Val Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Val Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Glu
100
<210> 174
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> F126V; S131V; S136P; A141D
<400> 174
Ala Ser Thr Lys Gly Pro Ser Val Val Pro Leu Ala Pro Val Ser Lys
1 5 10 15
Ser Thr Pro Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 175
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> F126V; S131V; S136P; G137R; A141D
<400> 175
Ala Ser Thr Lys Gly Pro Ser Val Val Pro Leu Ala Pro Val Ser Lys
1 5 10 15
Ser Thr Pro Arg Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 176
<211> 102
<212> Белок
<213> Искусственная последовательность
<220>
<223> F126V; S131V; A141D; K218S
<400> 176
Ala Ser Thr Lys Gly Pro Ser Val Val Pro Leu Ala Pro Val Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Ser Ser
100
<210> 177
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> F126V; S131V; K133S; A141K; K218A
<400> 177
Ala Ser Thr Lys Gly Pro Ser Val Val Pro Leu Ala Pro Val Ser Ser
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Lys Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Ala
100
<210> 178
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; V185R
<400> 178
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 179
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; T187R
<400> 179
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 180
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; F170E; V185R
<400> 180
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Glu Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 181
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; F170E; T187R
<400> 181
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Glu Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 182
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; V185R
<400> 182
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 183
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; T187R
<400> 183
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 184
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; V185R; T187R
<400> 184
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 185
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; F170E; V185R
<400> 185
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Glu Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 186
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; F170E; T187R
<400> 186
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Glu Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 187
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141E; P171E; T187R
<400> 187
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Glu Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 188
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; P171E; V185R
<400> 188
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Arg Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<210> 189
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> A141D; P171E; T187R
<400> 189
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Asp Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Glu Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Arg Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys
100
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ БЕЛКИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2017 |
|
RU2745648C2 |
| АНТИТЕЛО-ПОДОБНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ОБЛАСТЯМИ, ИМЕЮЩИЕ ОРИЕНТАЦИЮ СВЯЗЫВАЮЩИХ ОБЛАСТЕЙ КРЕСТ-НАКРЕСТ | 2012 |
|
RU2823693C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С МУТАНТНЫМИ ДОМЕНАМИ FAB | 2019 |
|
RU2827657C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ, СОДЕРЖАЩИЕ ЛЕГКИЕ ЦЕПИ КАППА И ЛЯМБДА, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2824193C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕСЯ С 5T4 И 4-1BB, И ОТНОСЯЩИЕСЯ К НИМ КОМПОЗИЦИИ И СПОСОБЫ | 2018 |
|
RU2804458C2 |
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕ CD38, CD28 И CD3, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ | 2019 |
|
RU2820351C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ НА ОСНОВЕ ПСЕВДО-FAB | 2019 |
|
RU2820254C2 |
| САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПОСРЕДСТВОМ ГЛИКОИНЖЕНЕРИИ | 2014 |
|
RU2831409C2 |
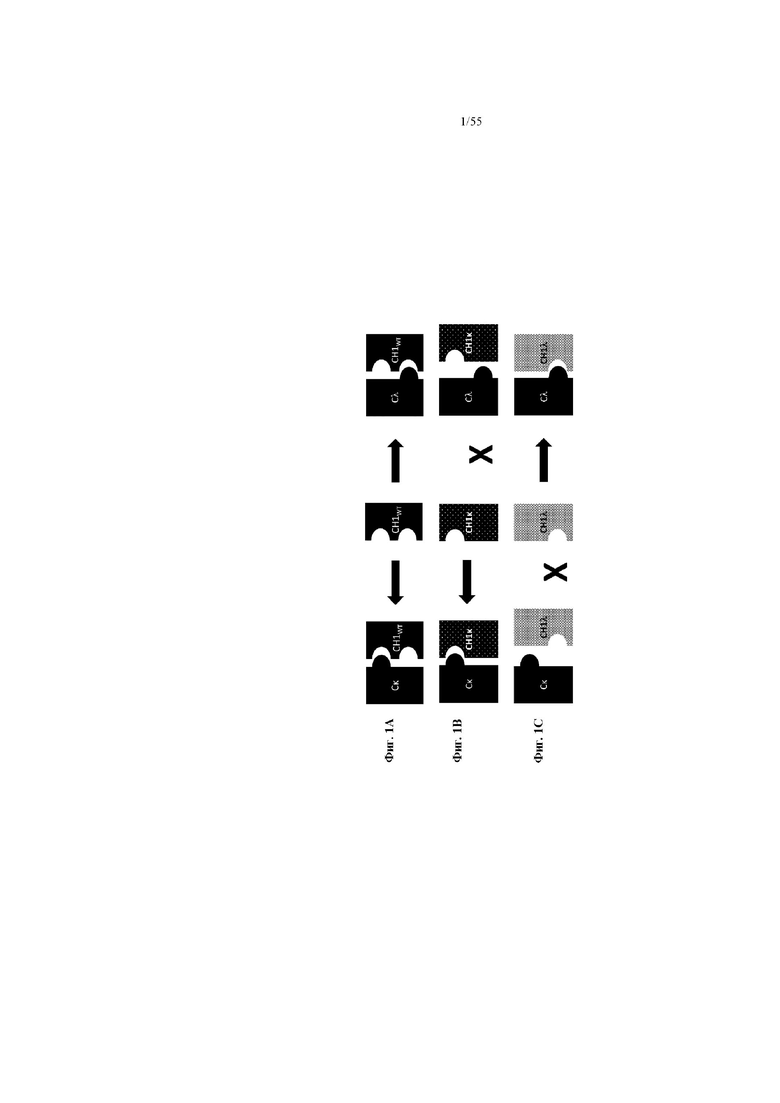
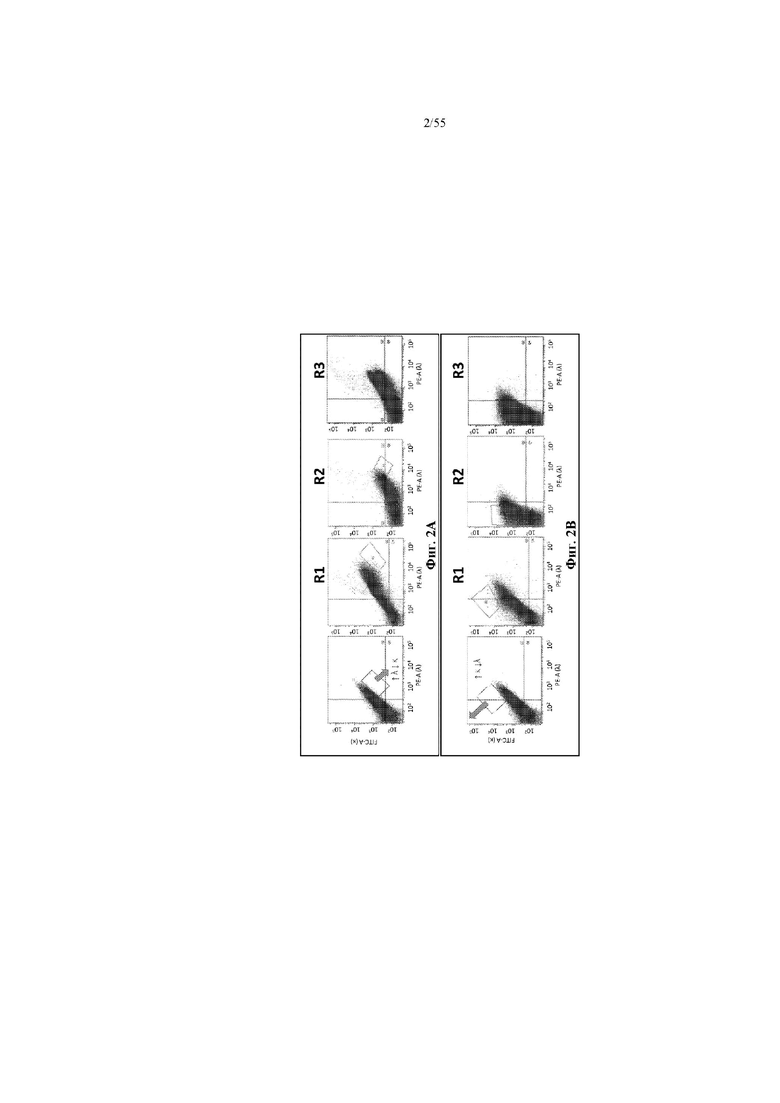



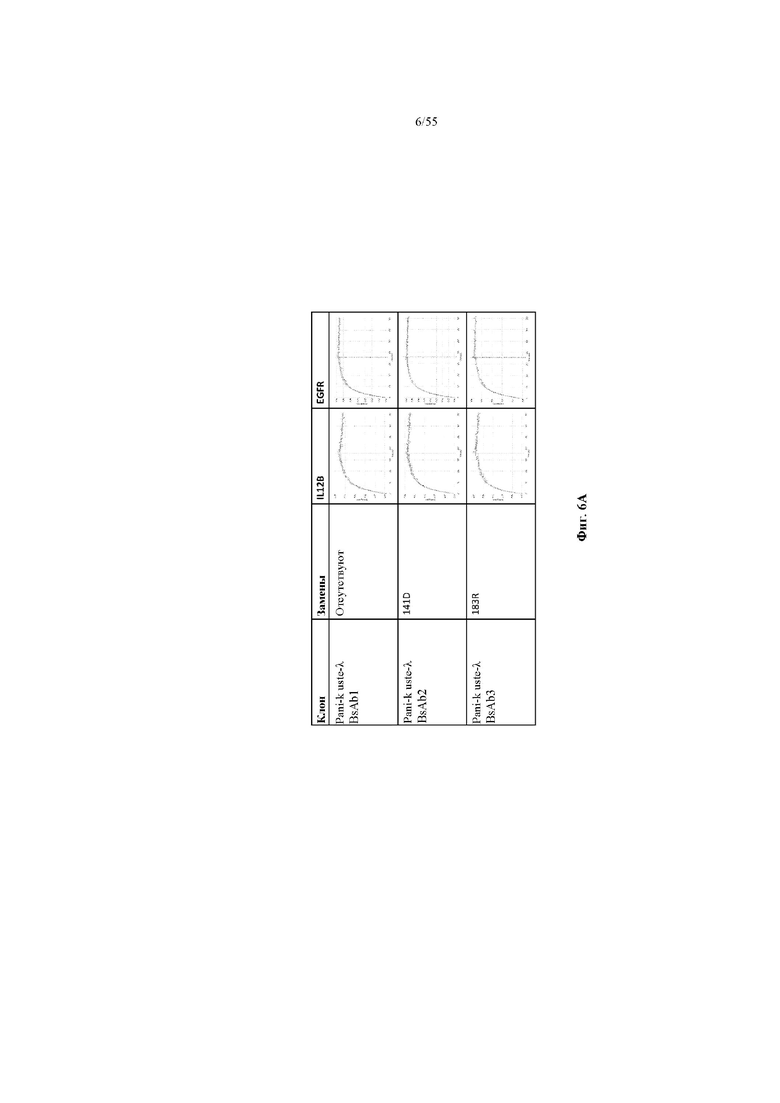
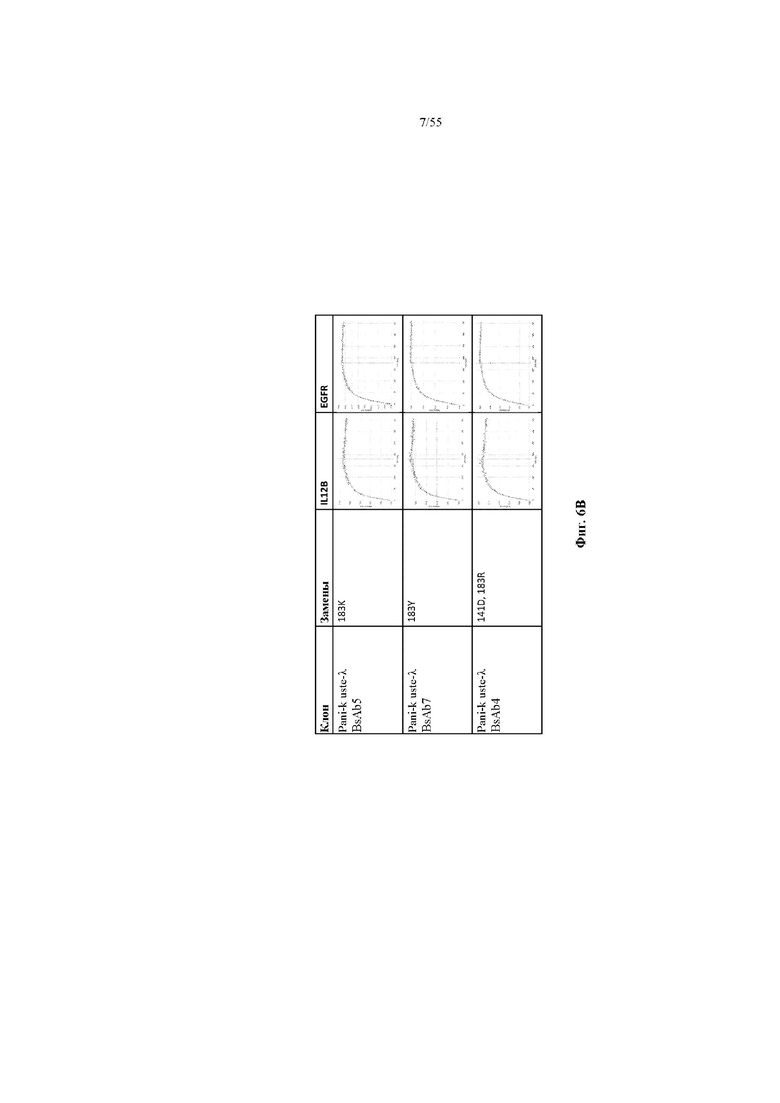
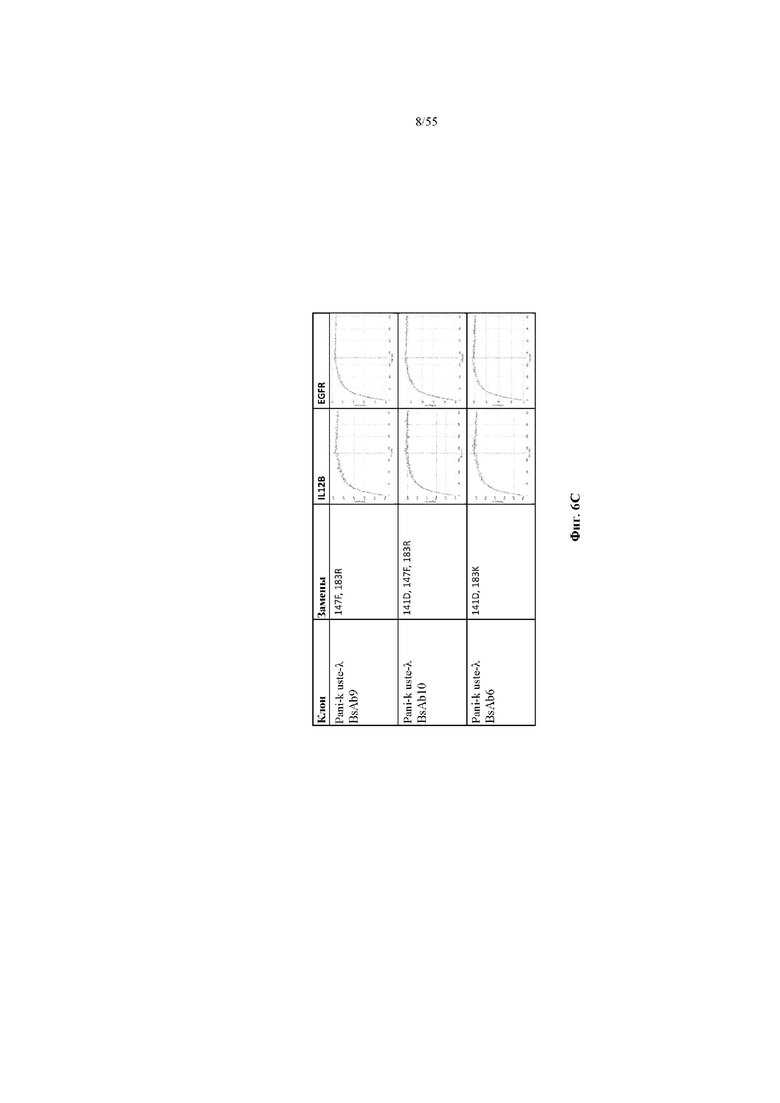
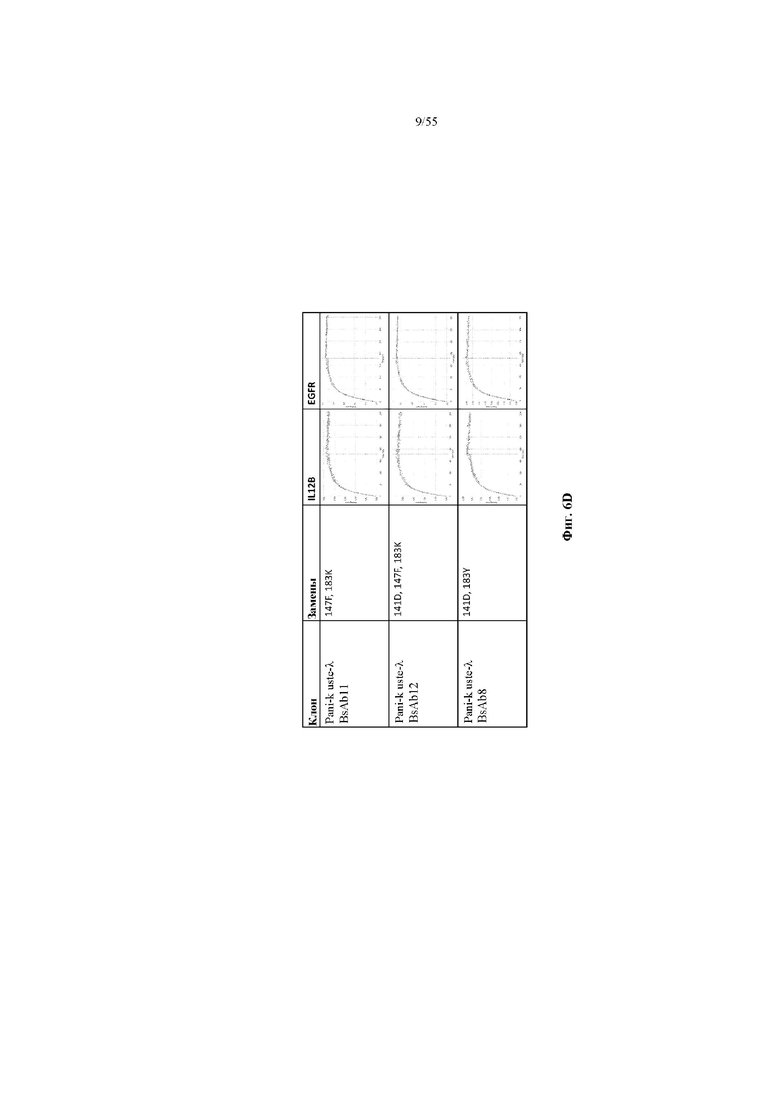
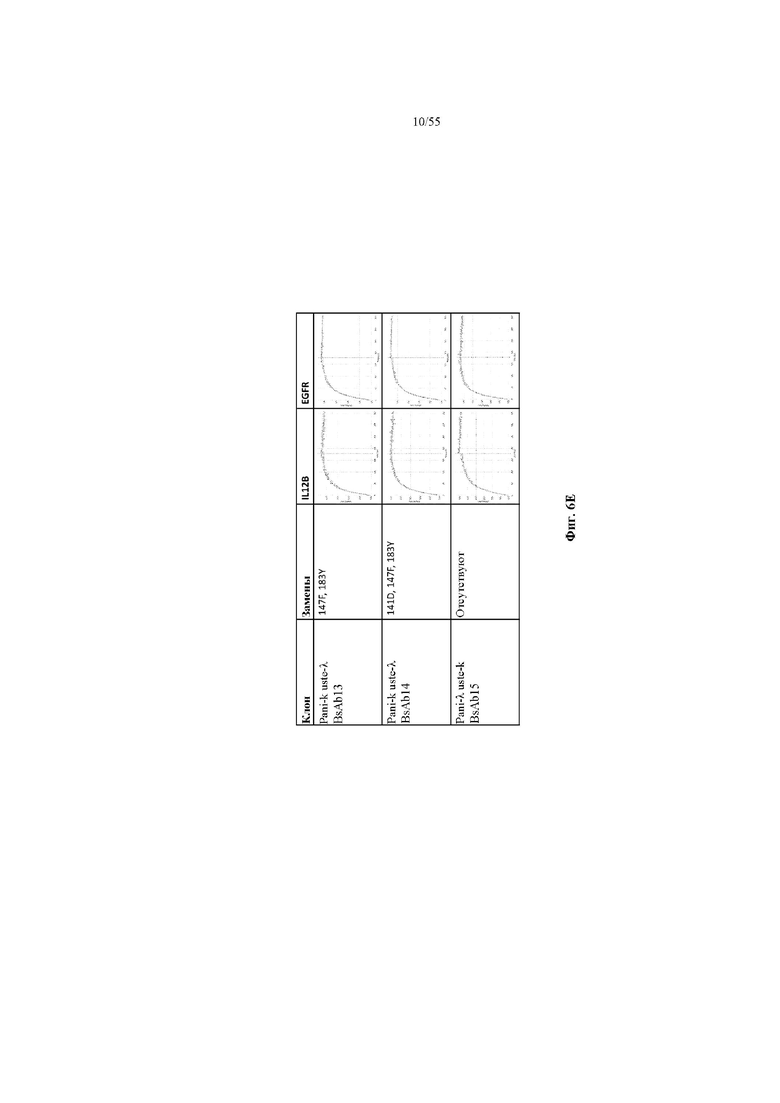

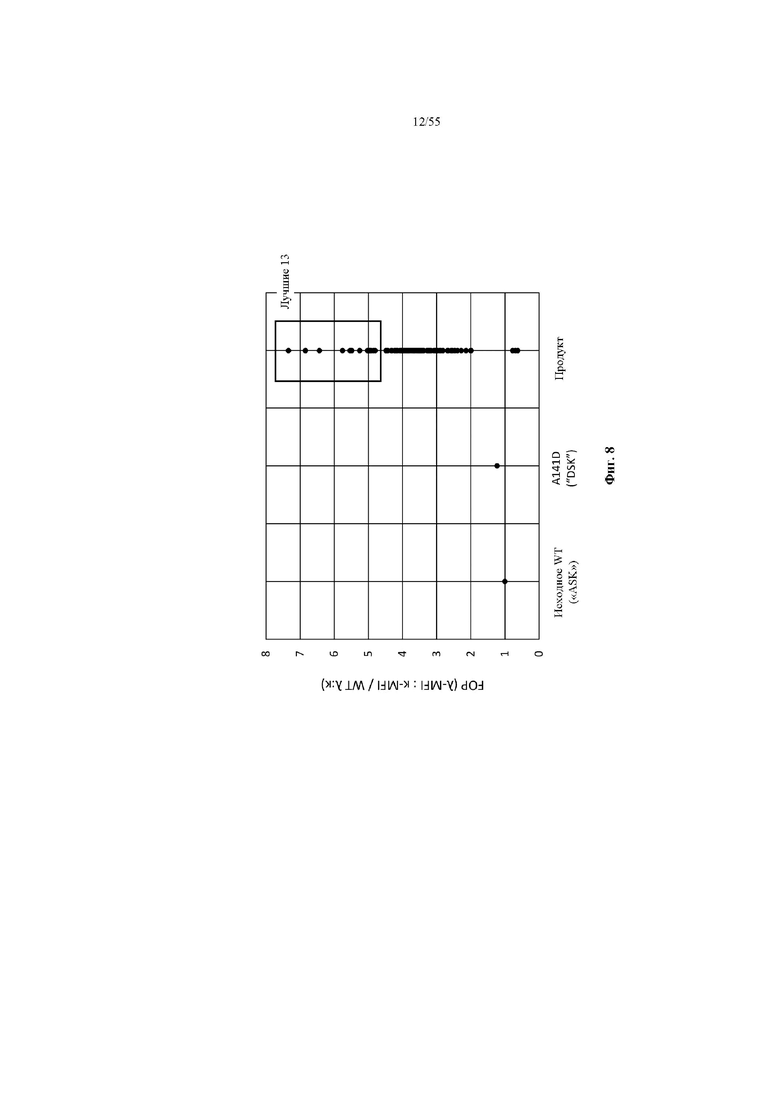

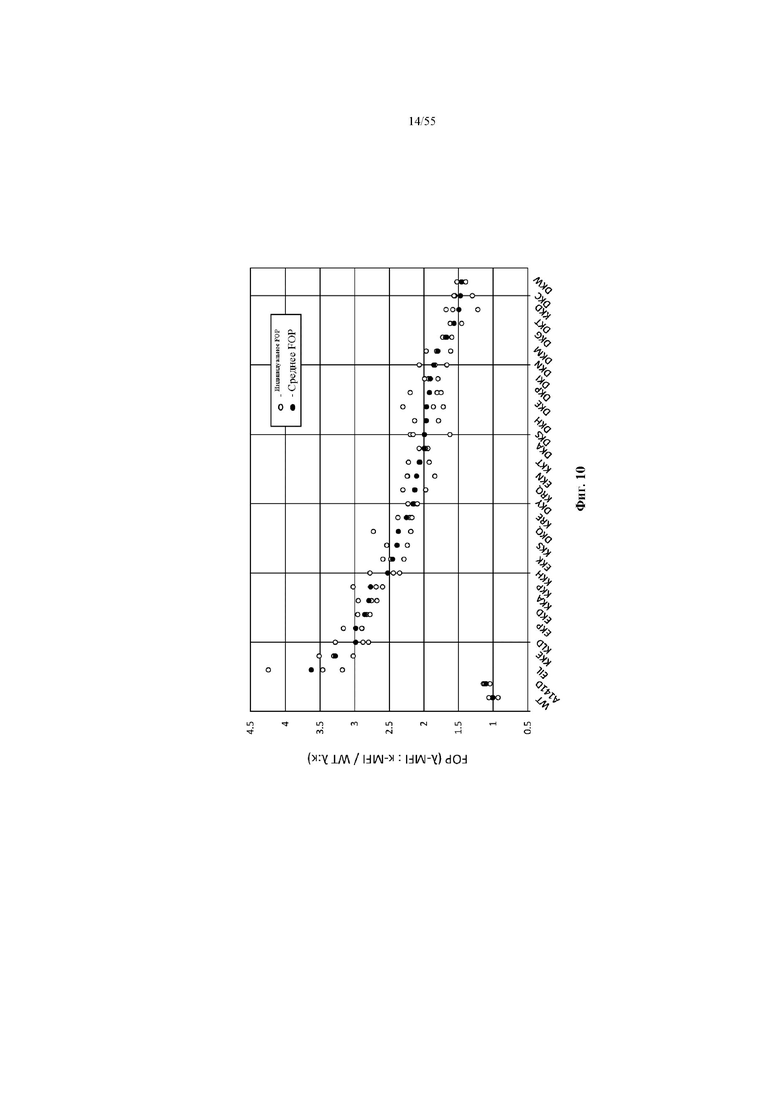
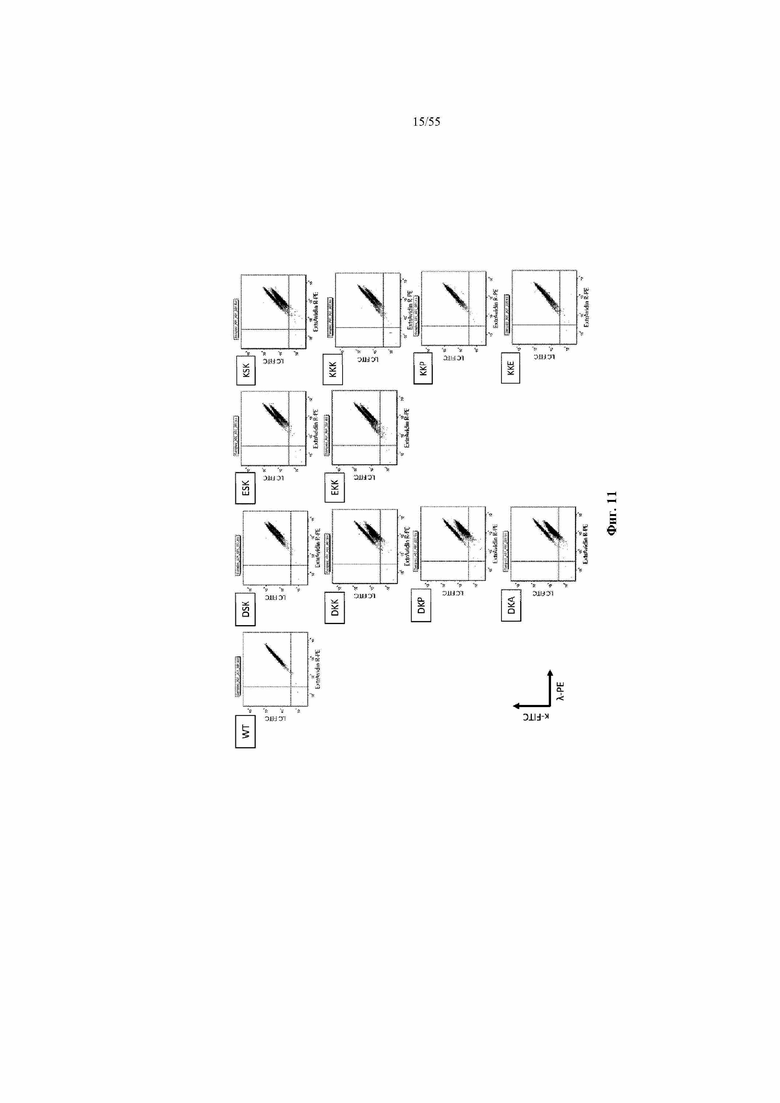
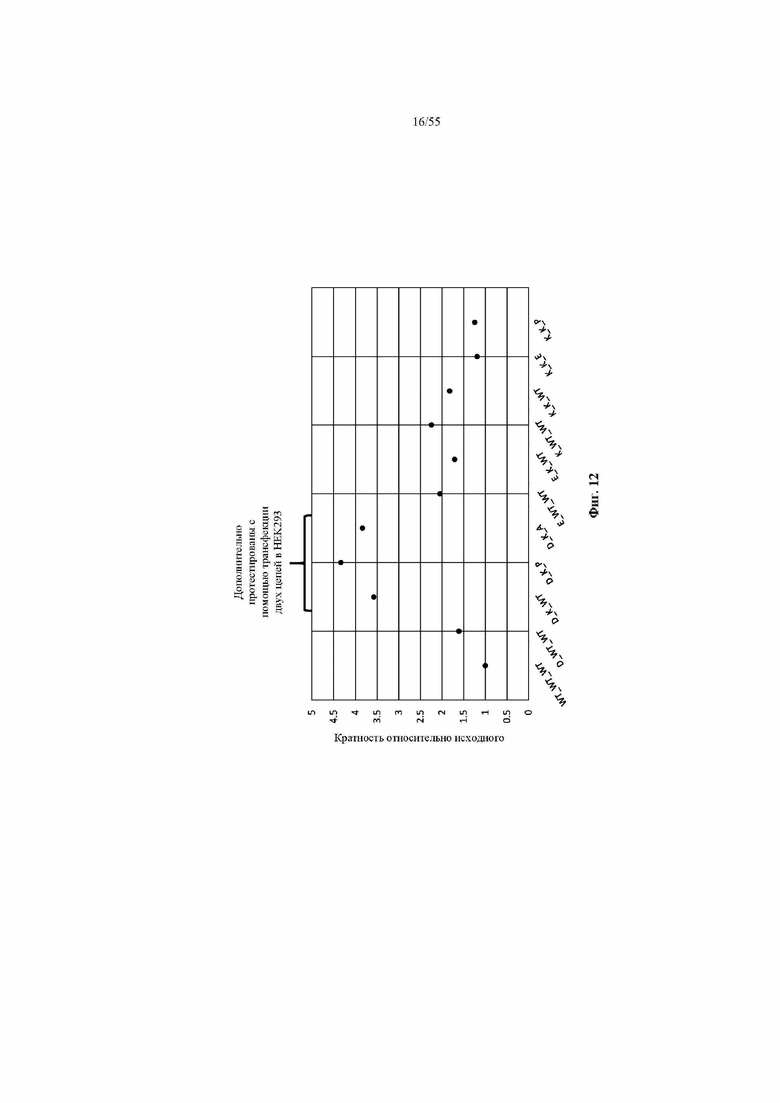
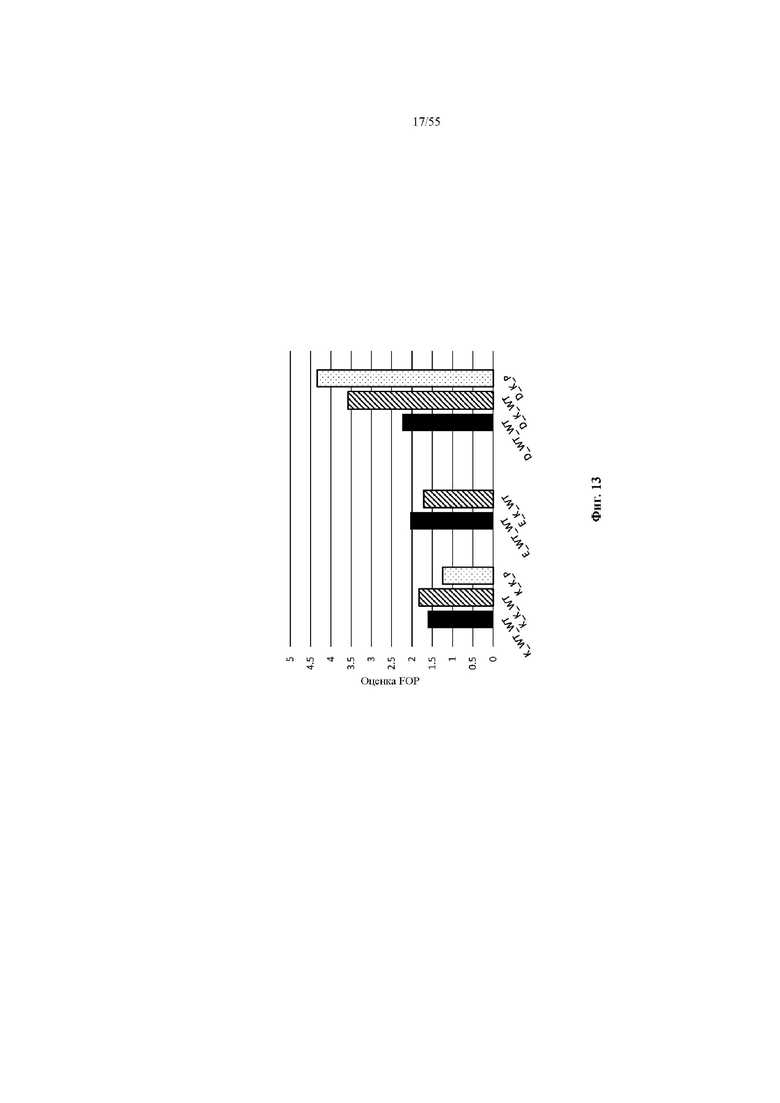


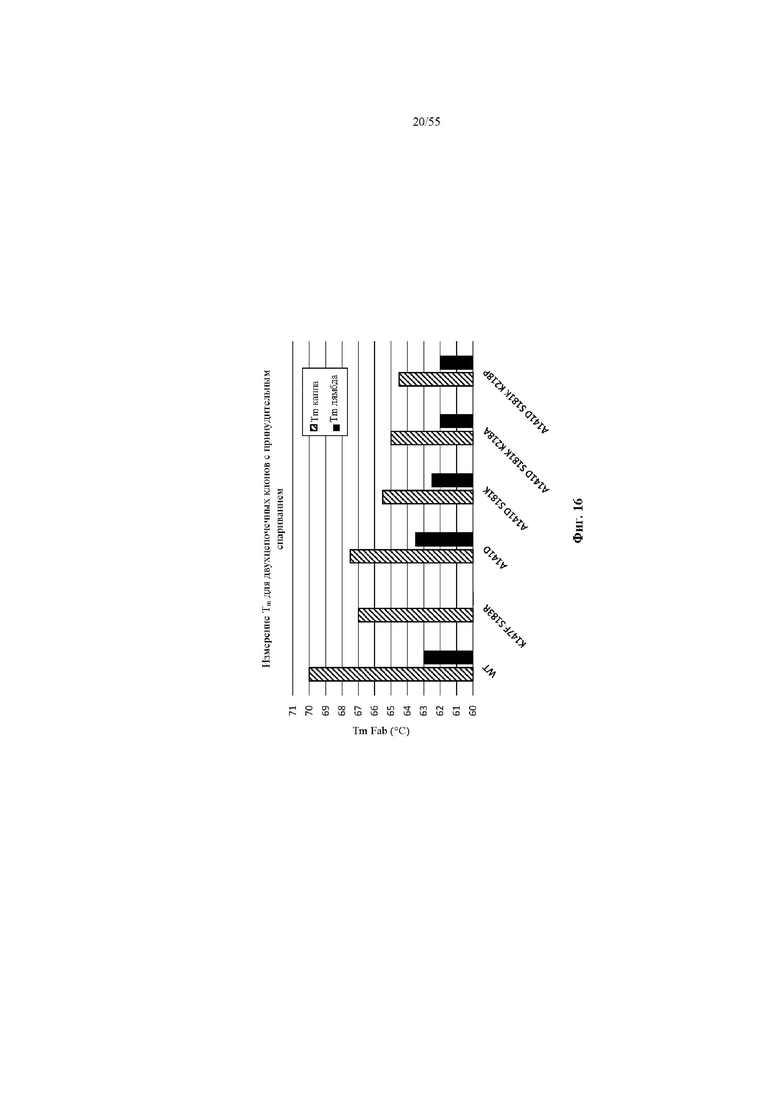
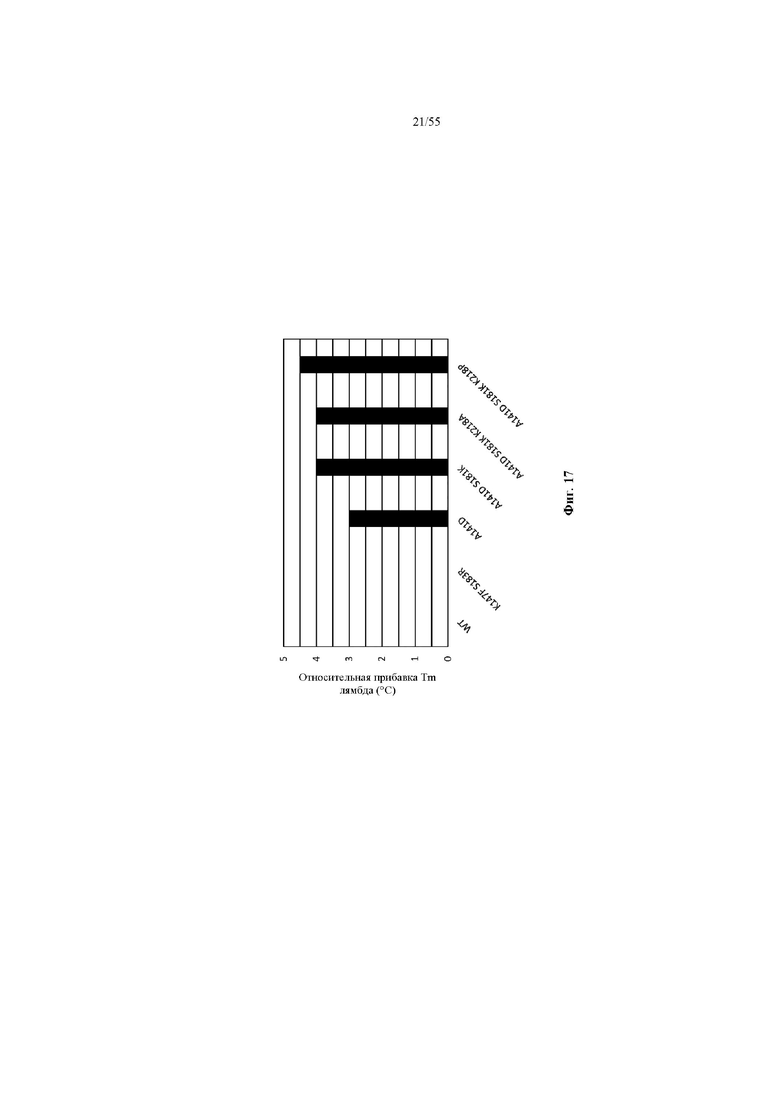

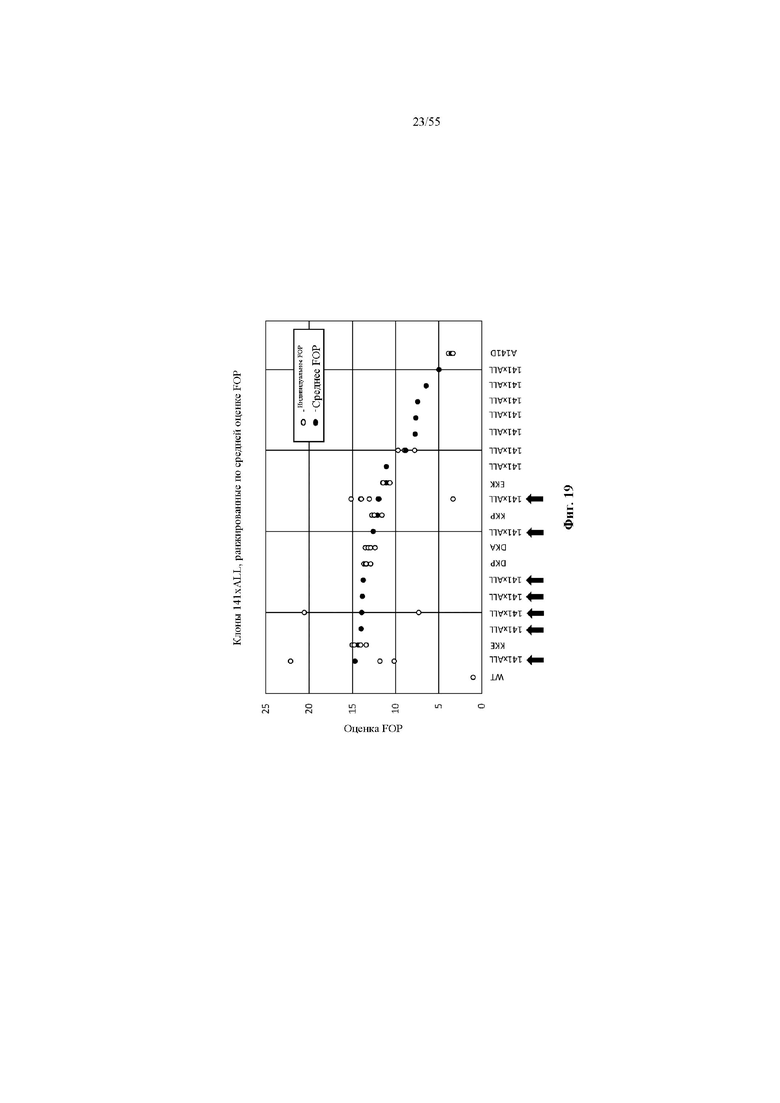

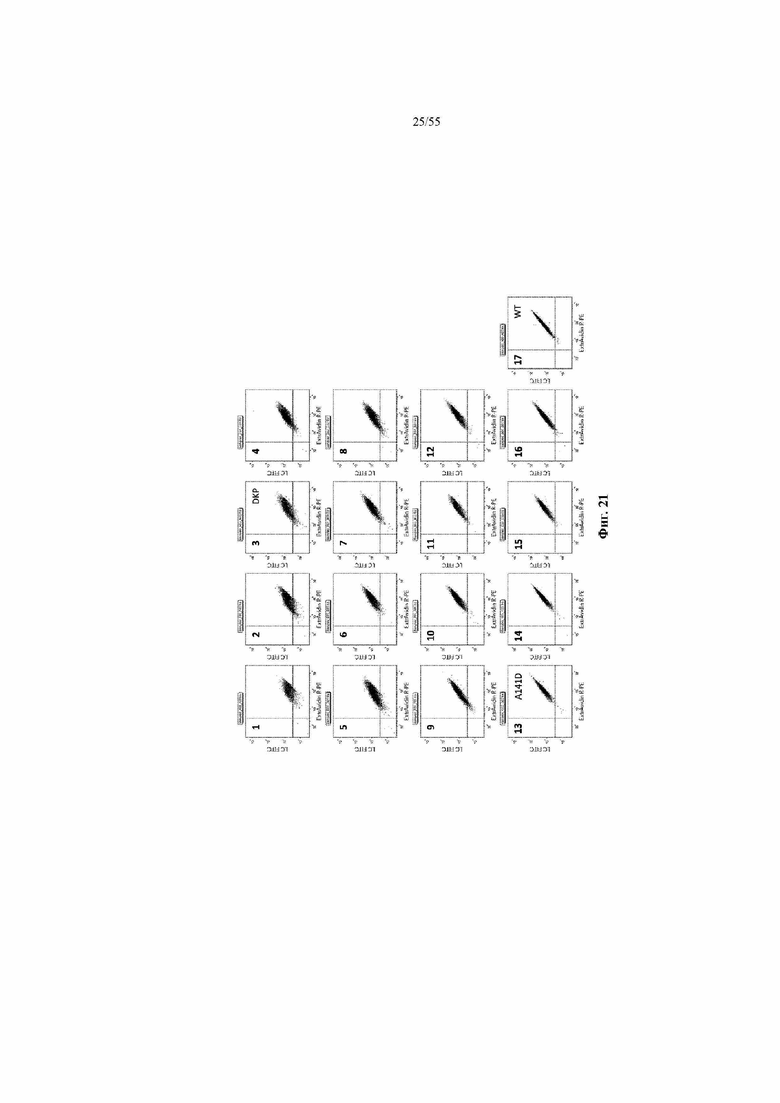
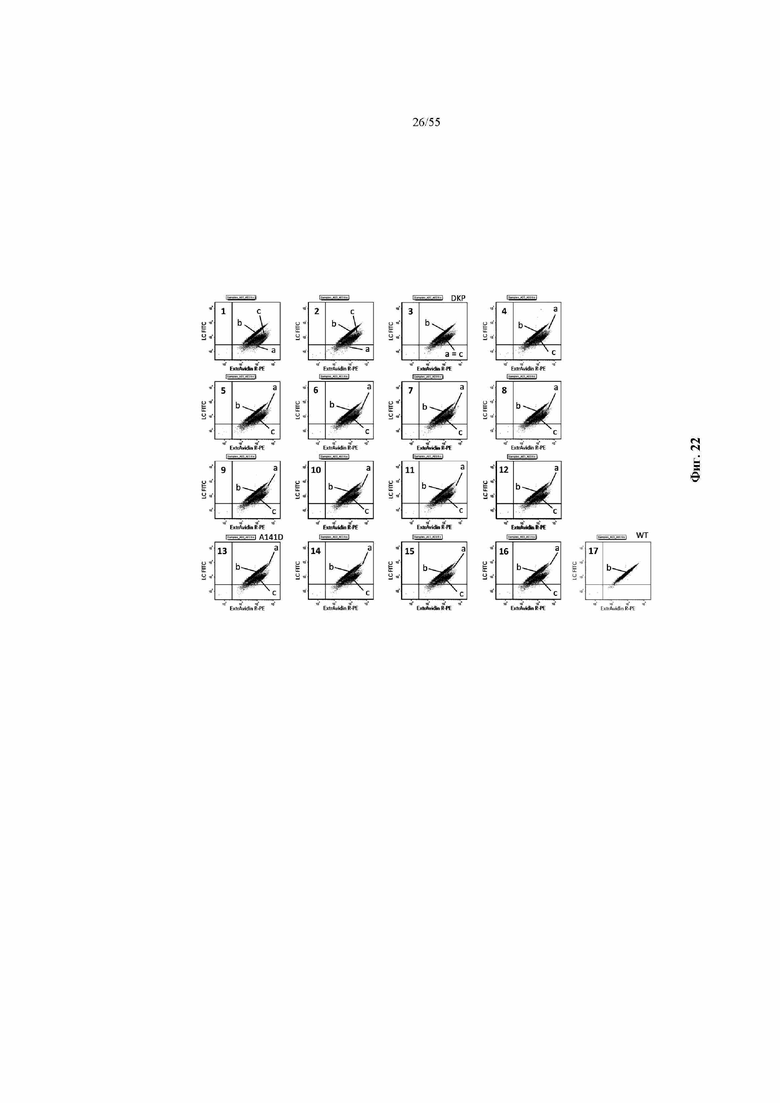
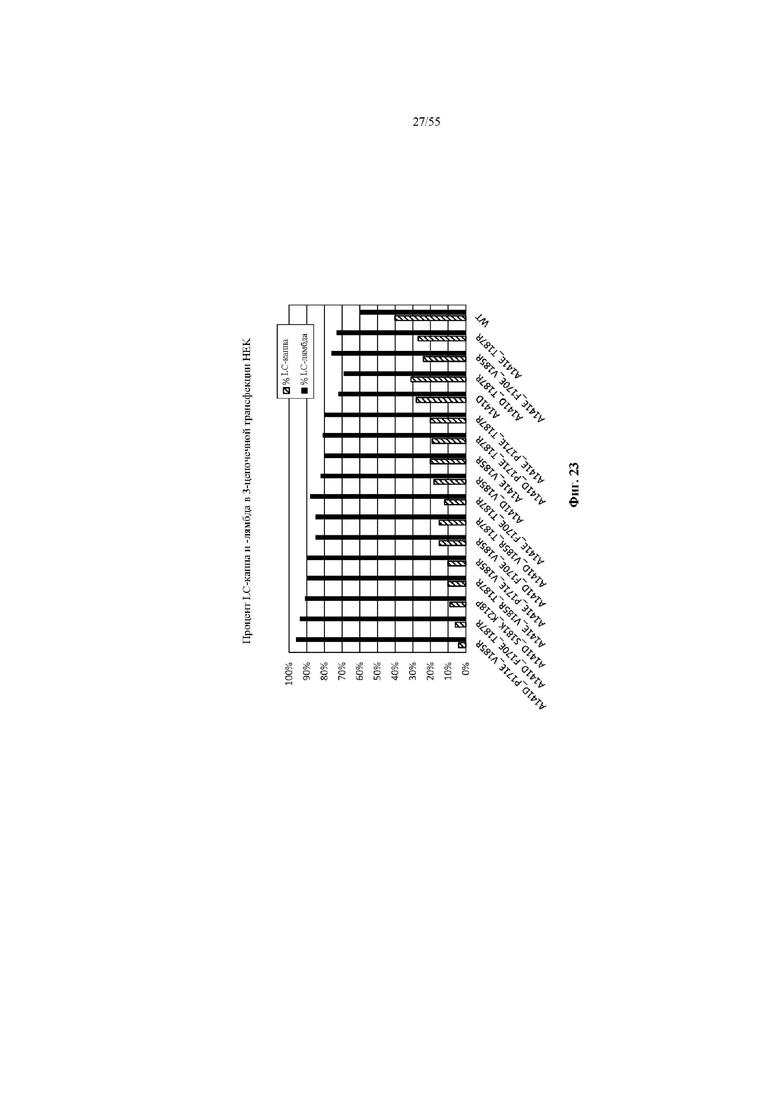
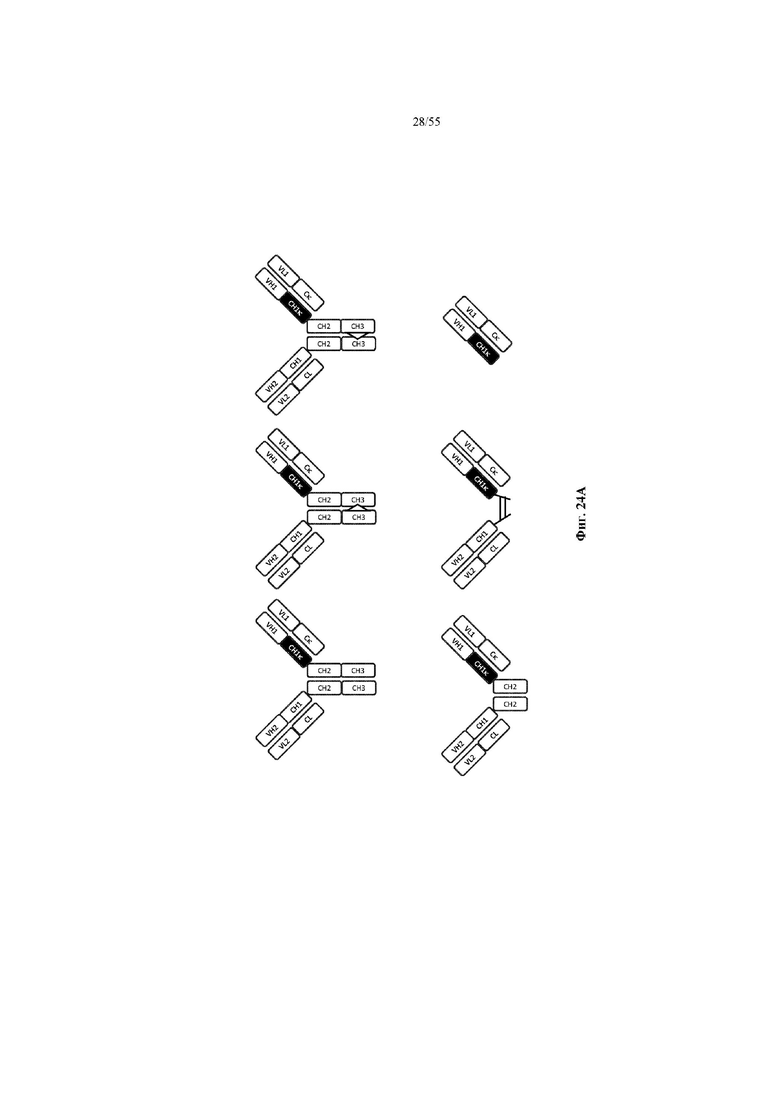
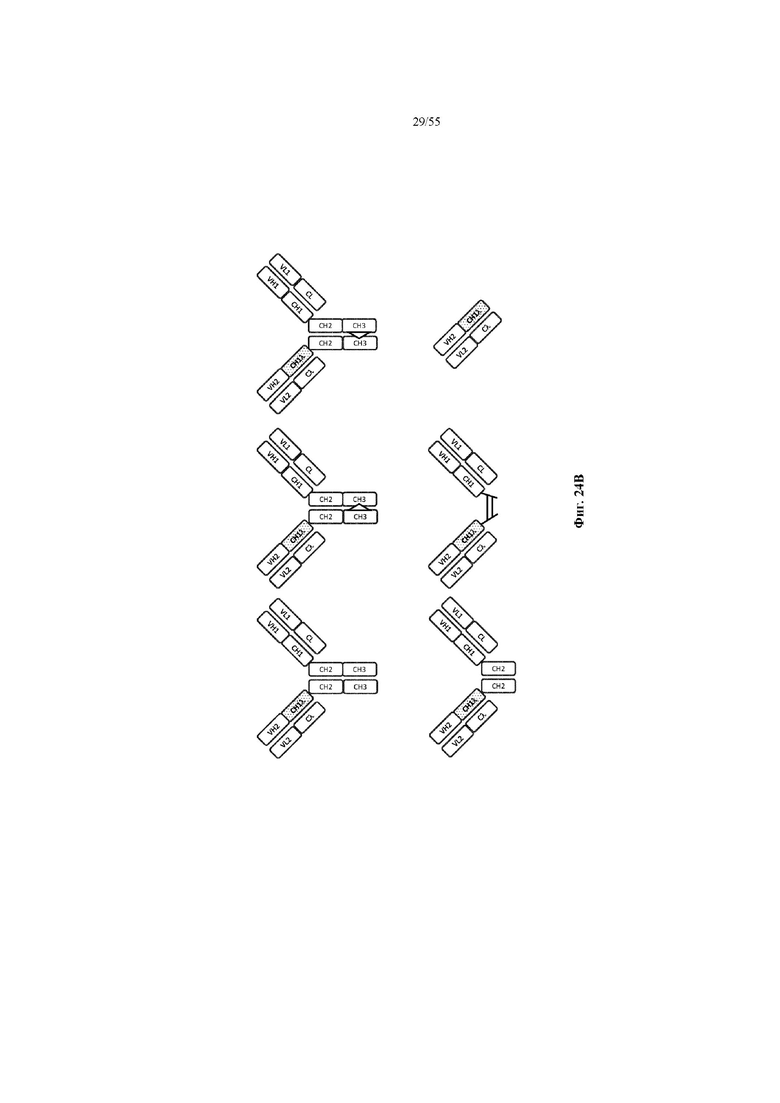
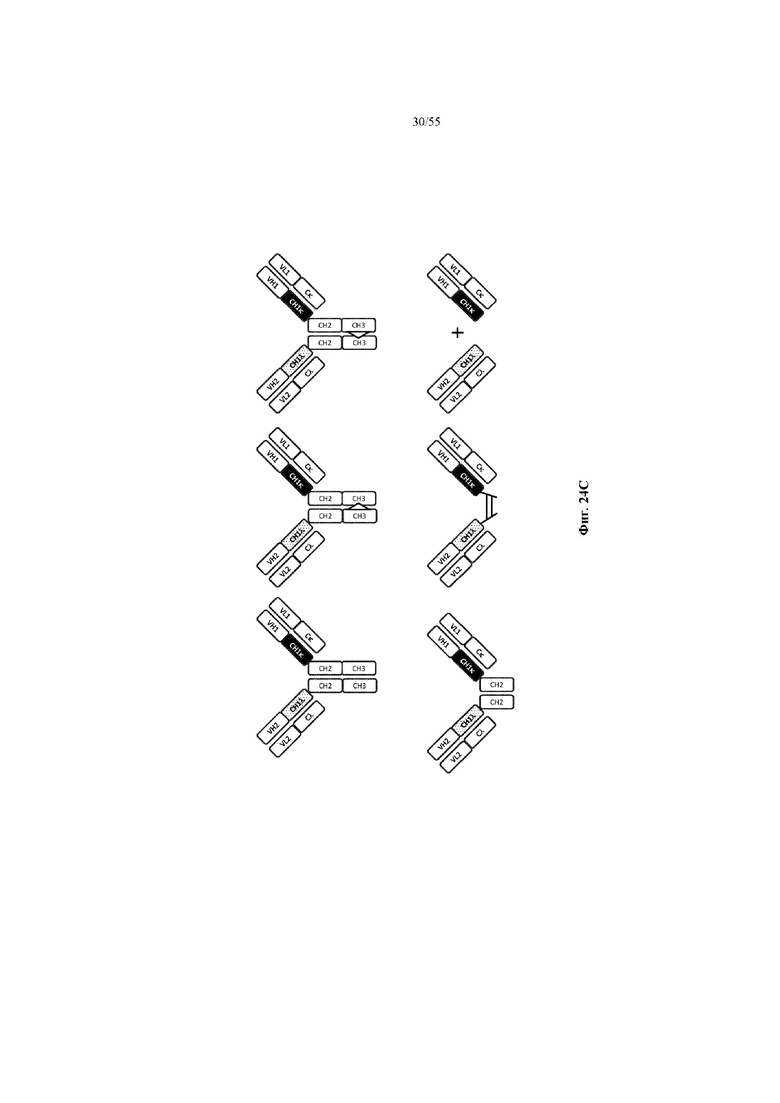
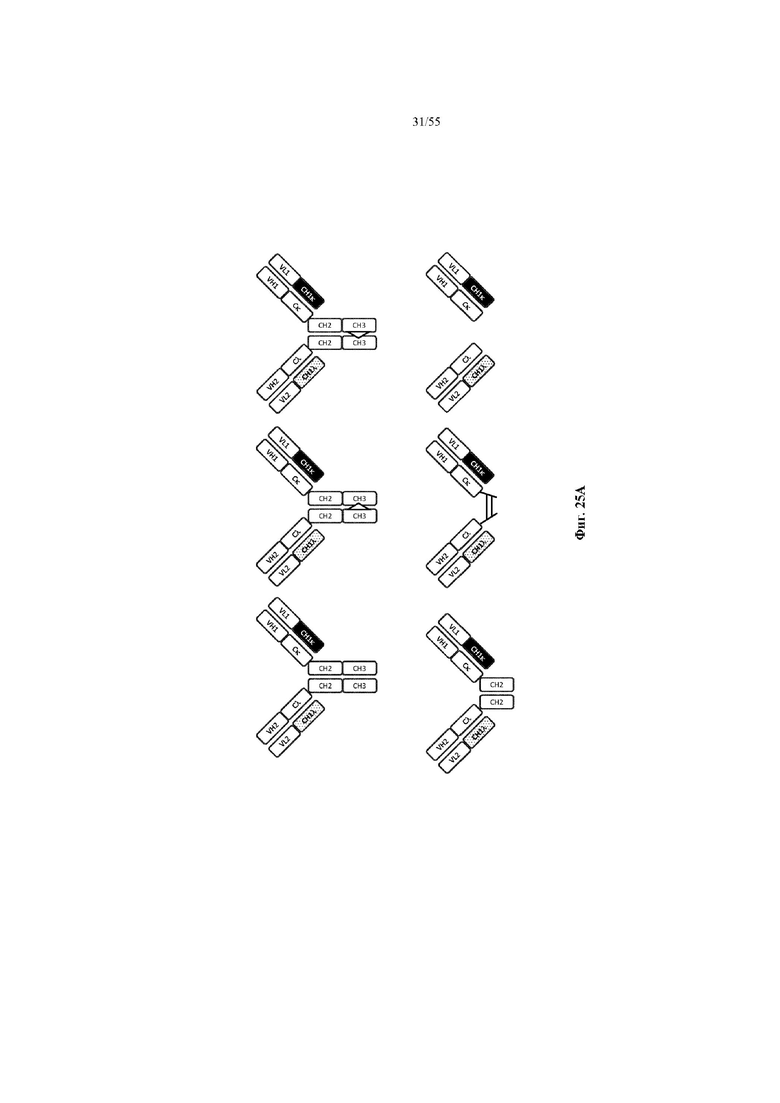
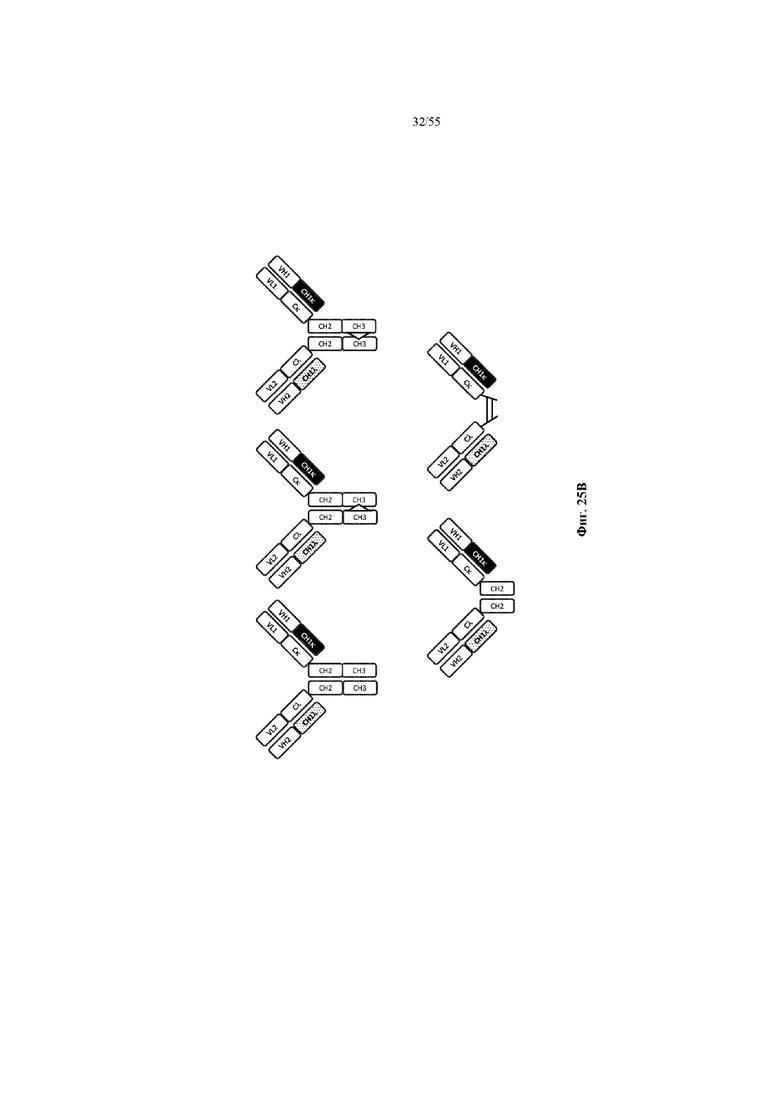
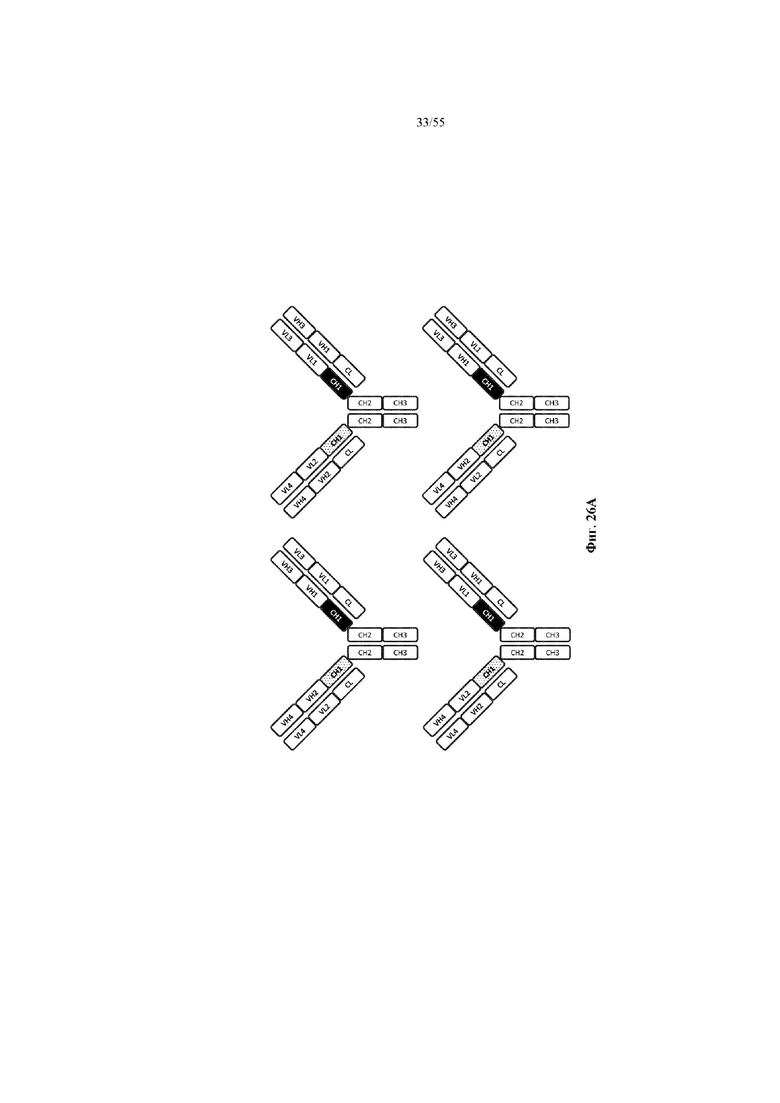
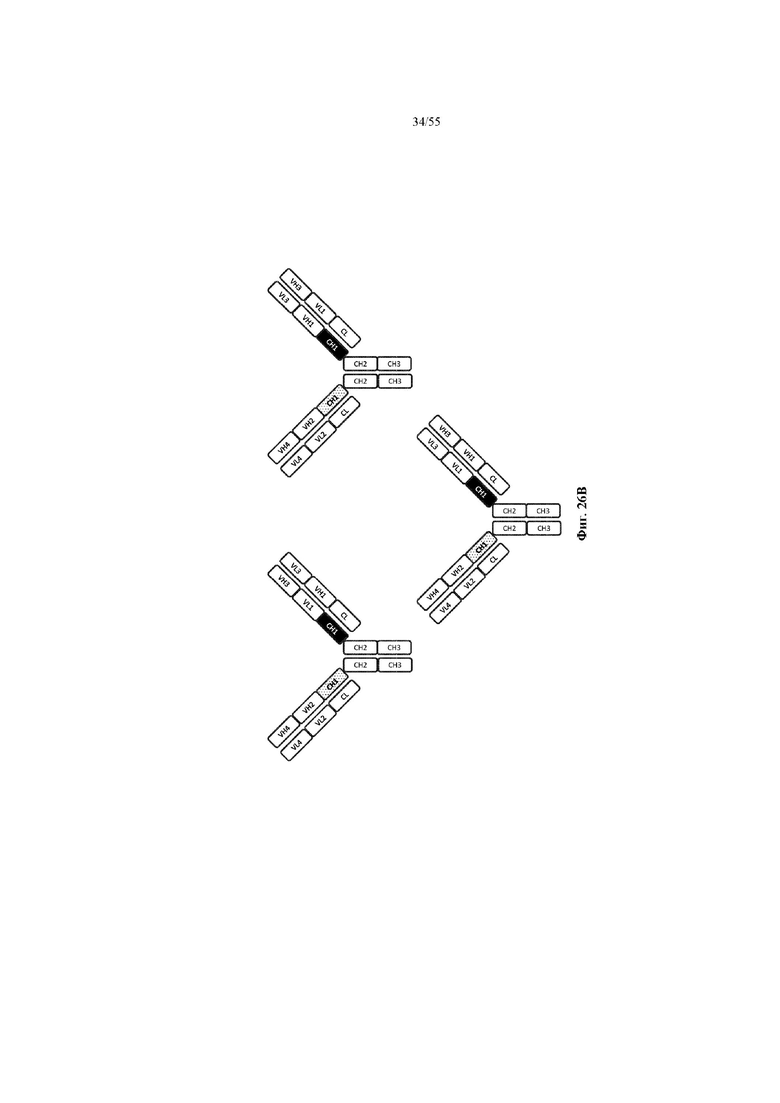
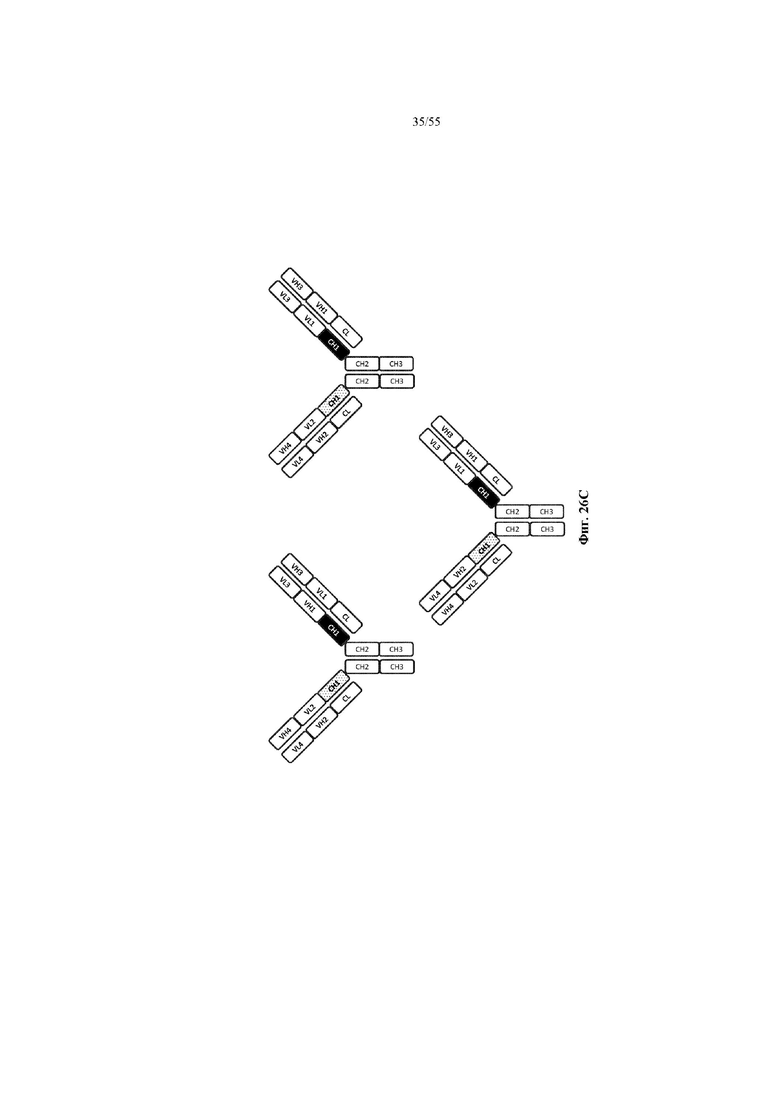
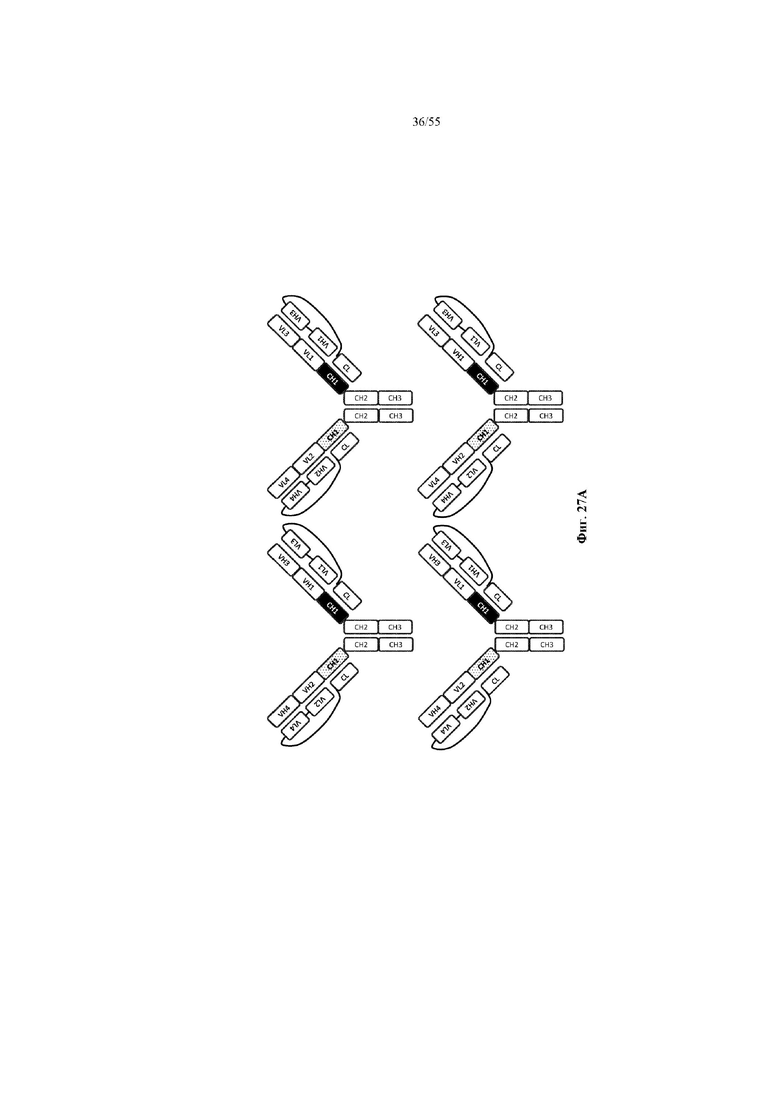
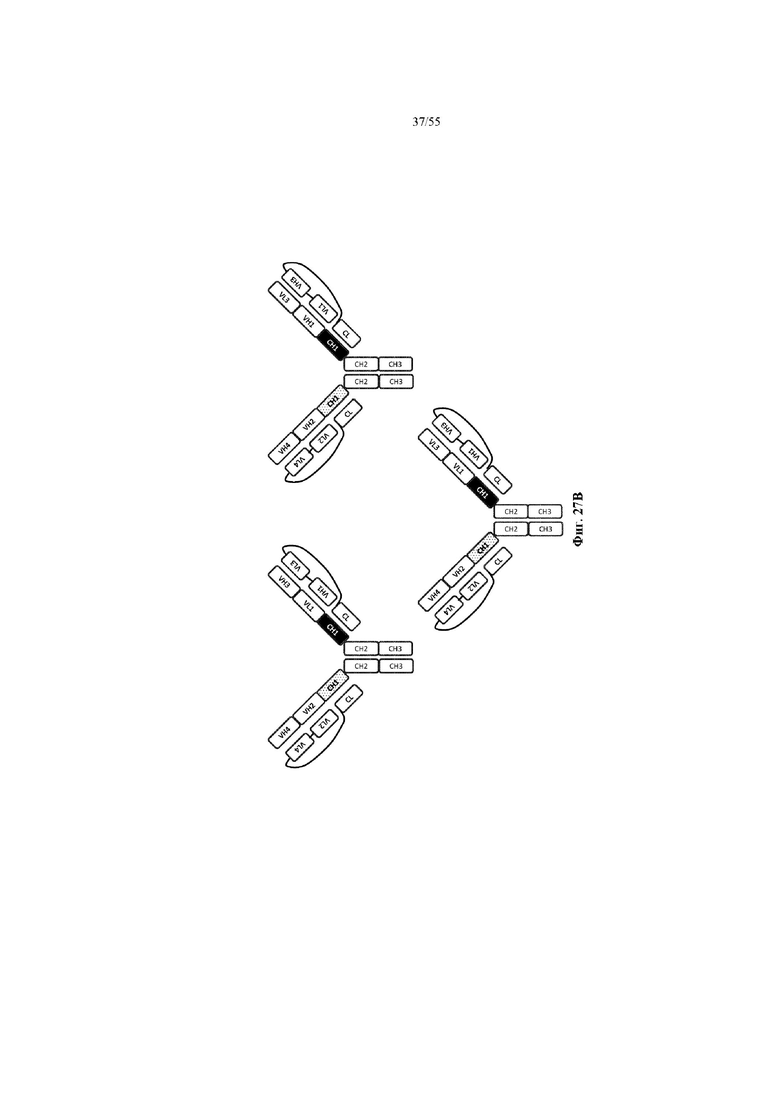
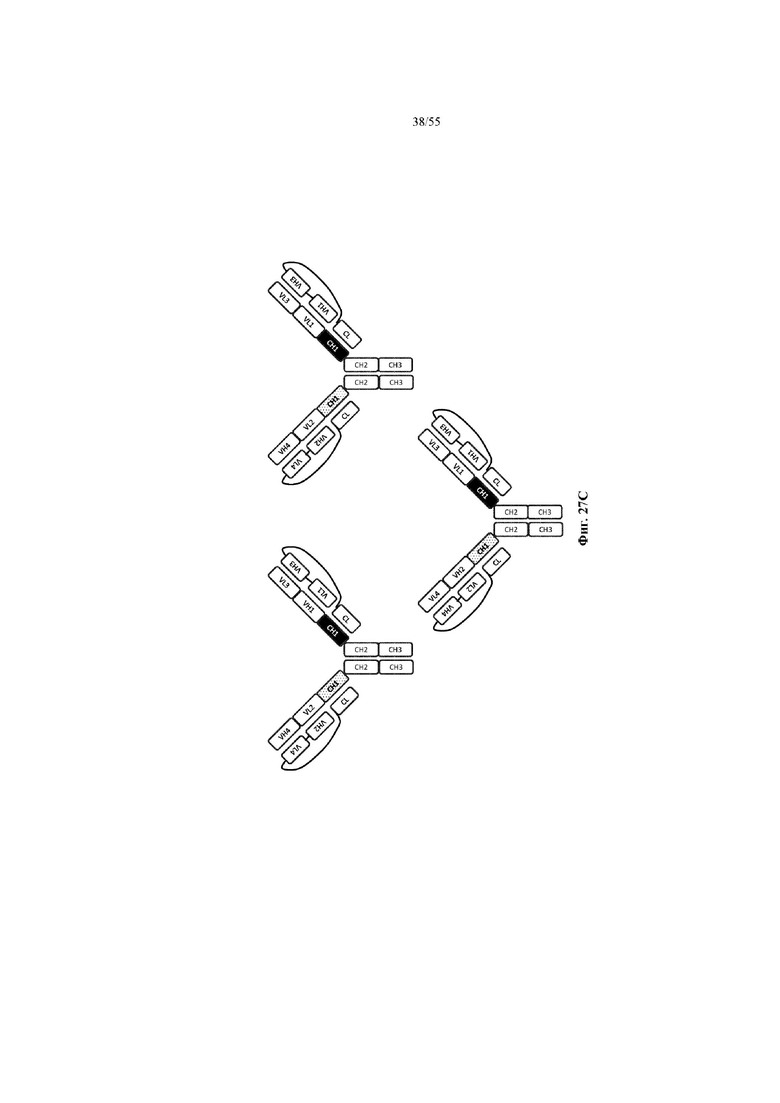
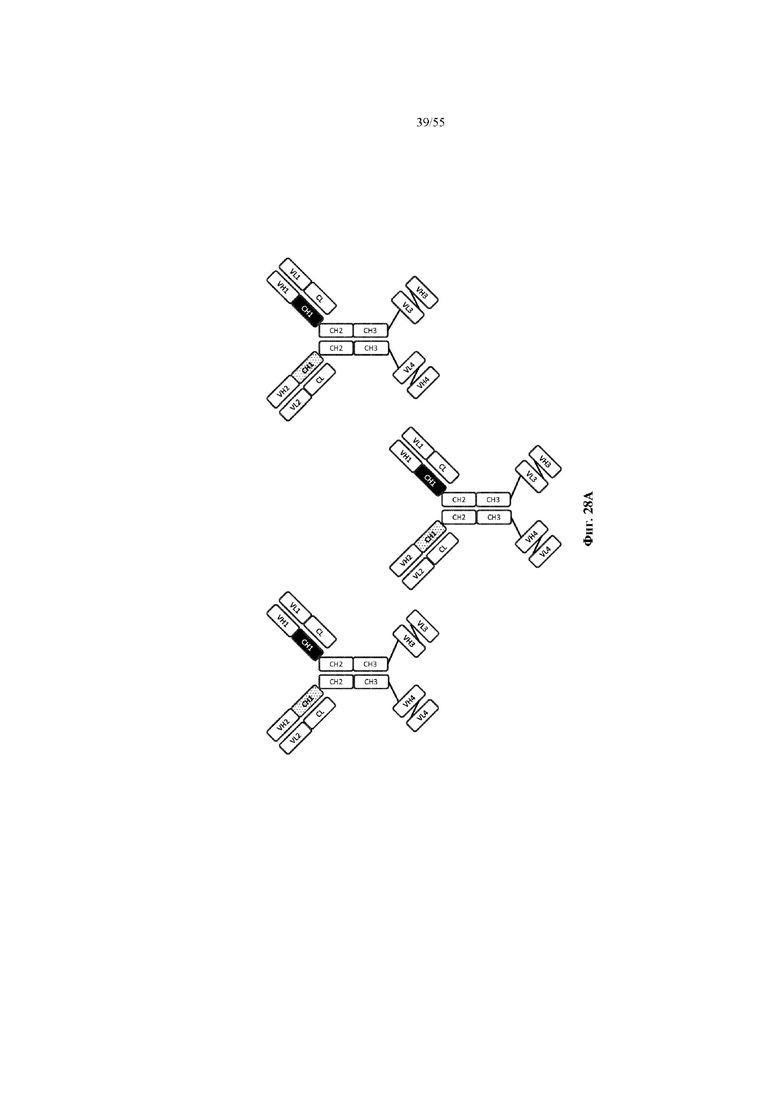
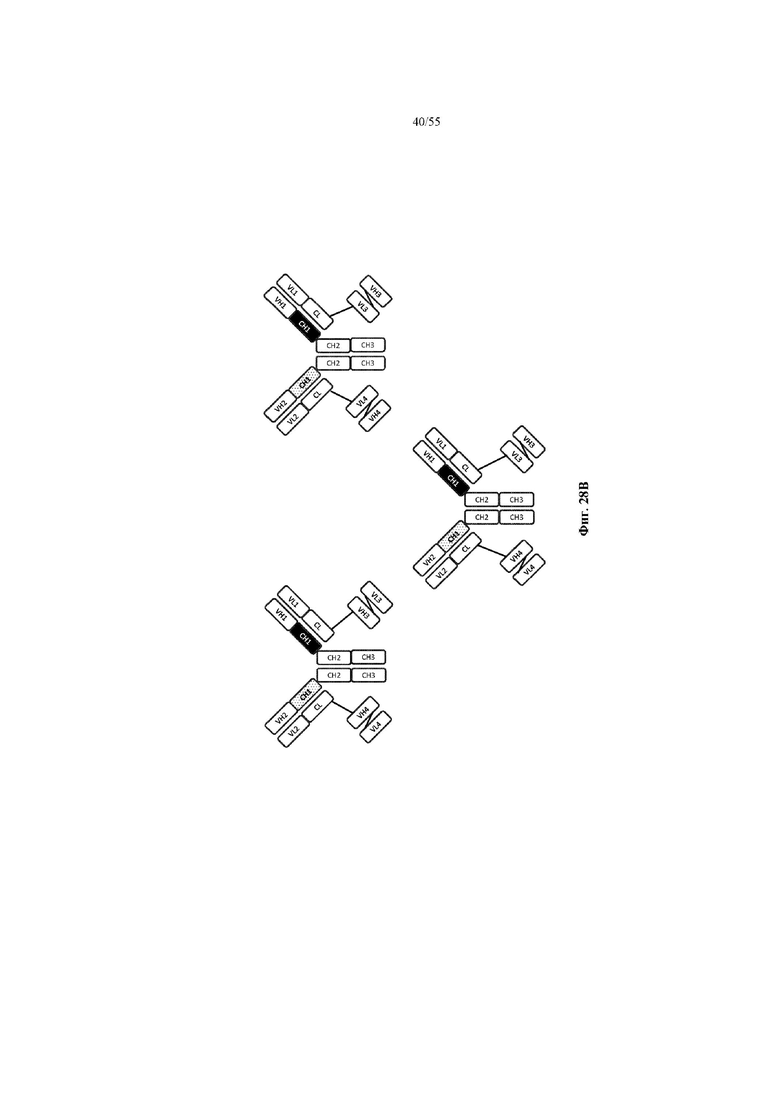
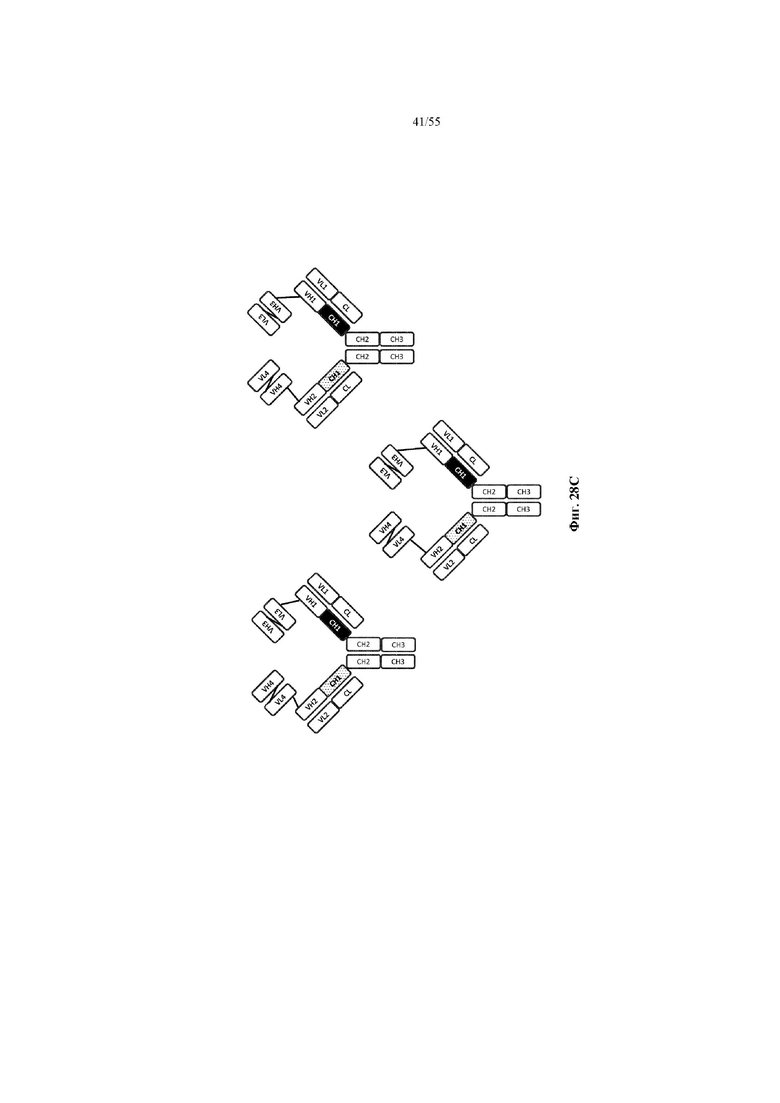
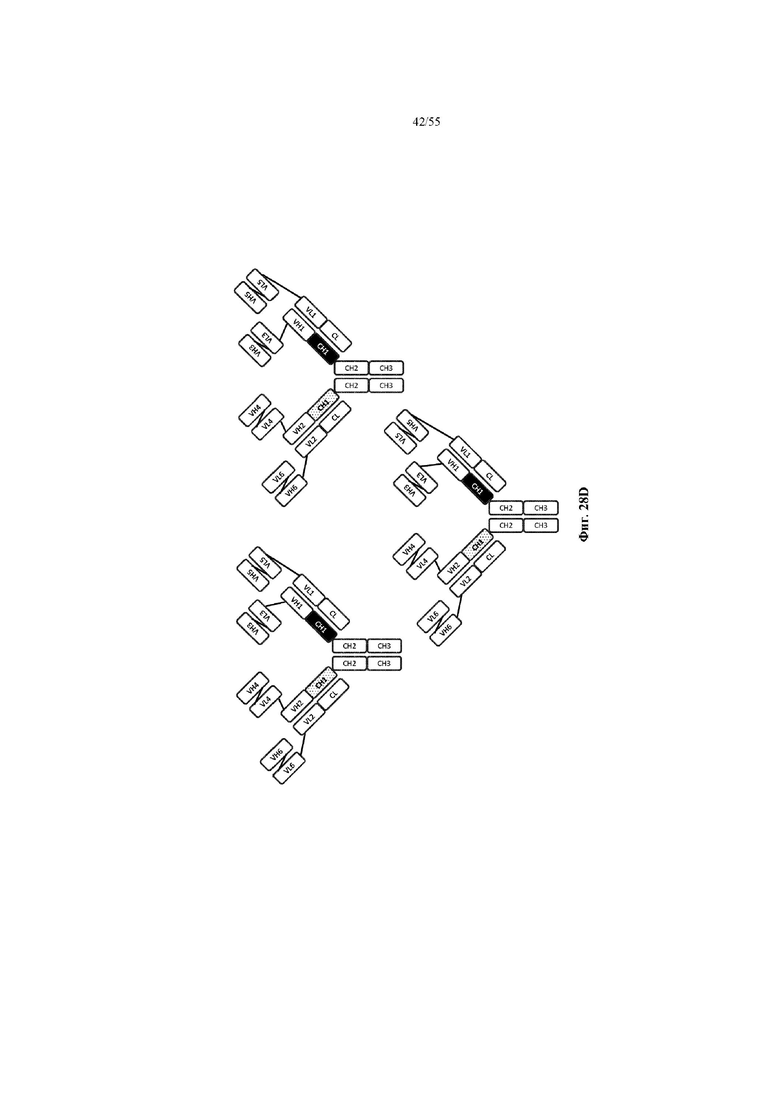
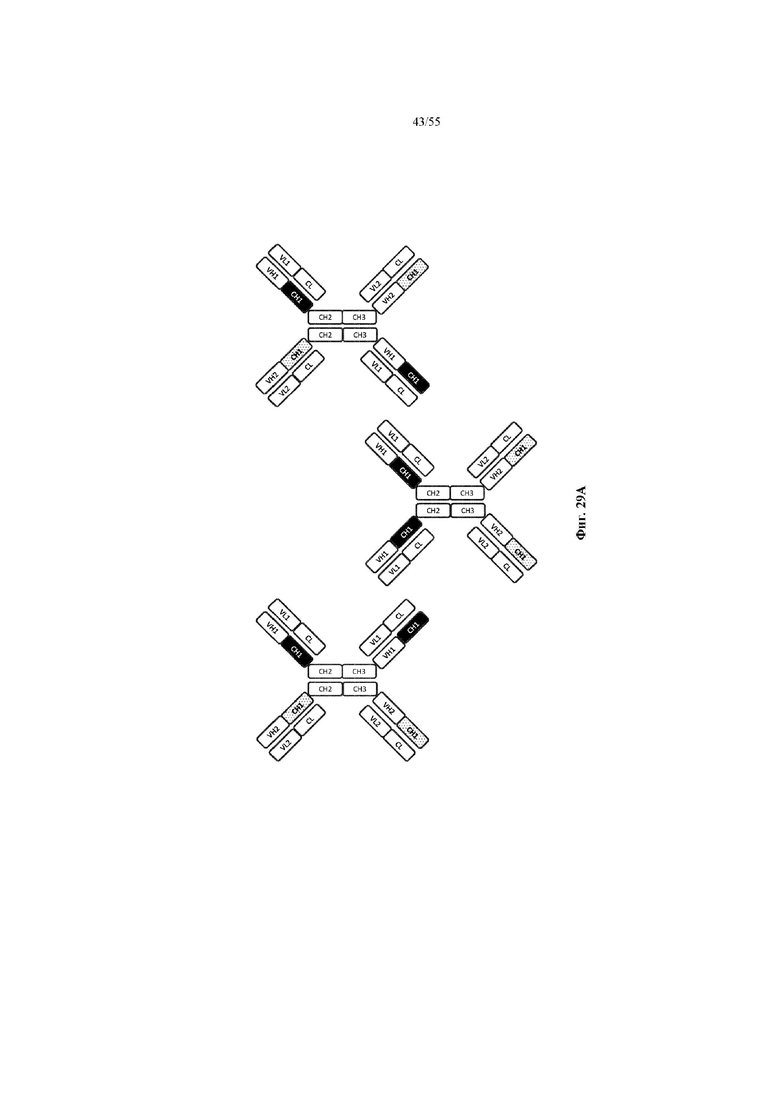
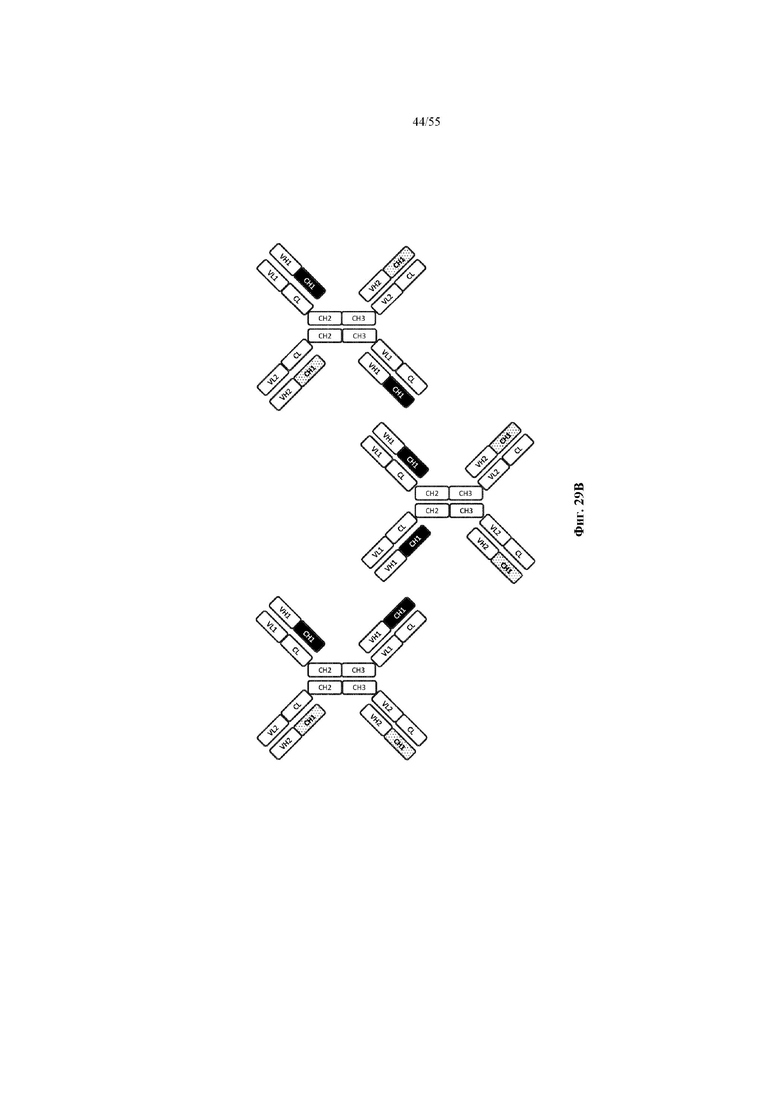
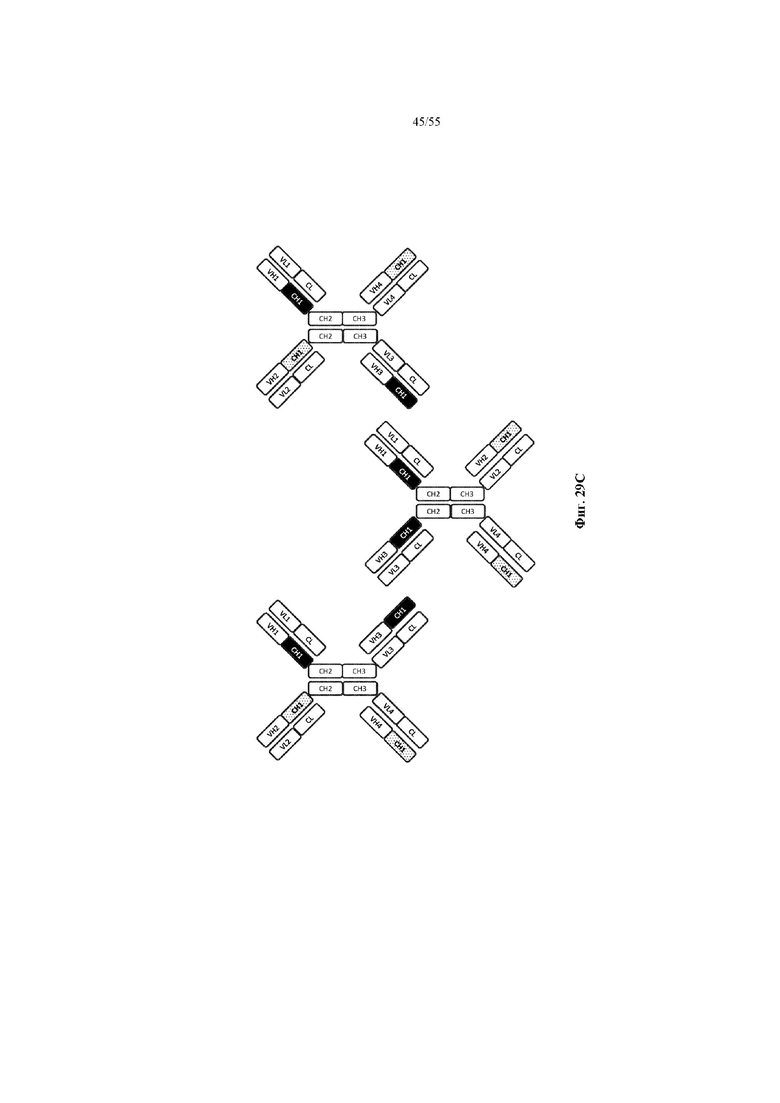
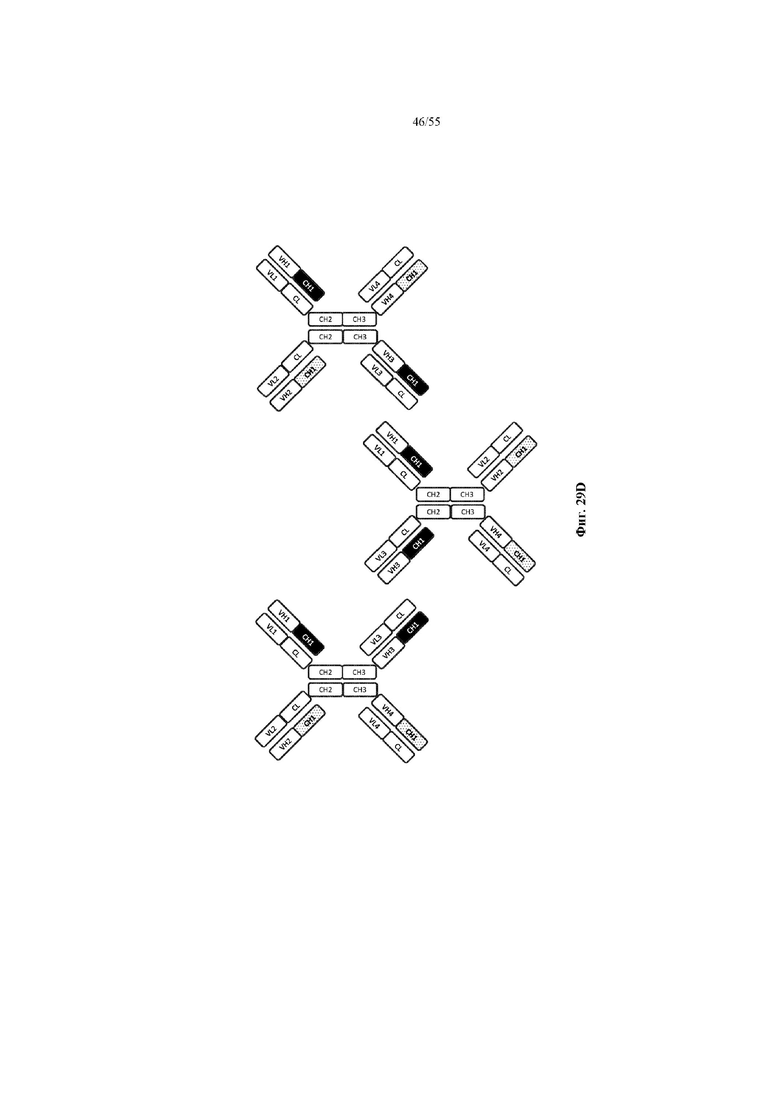

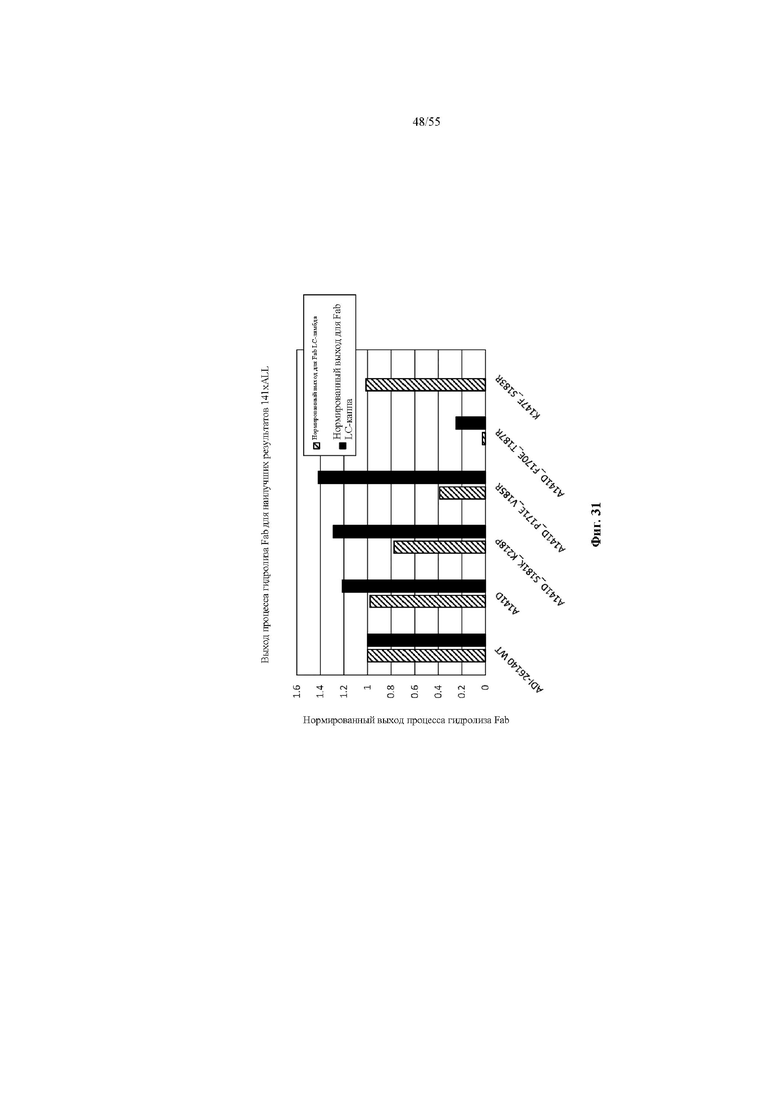


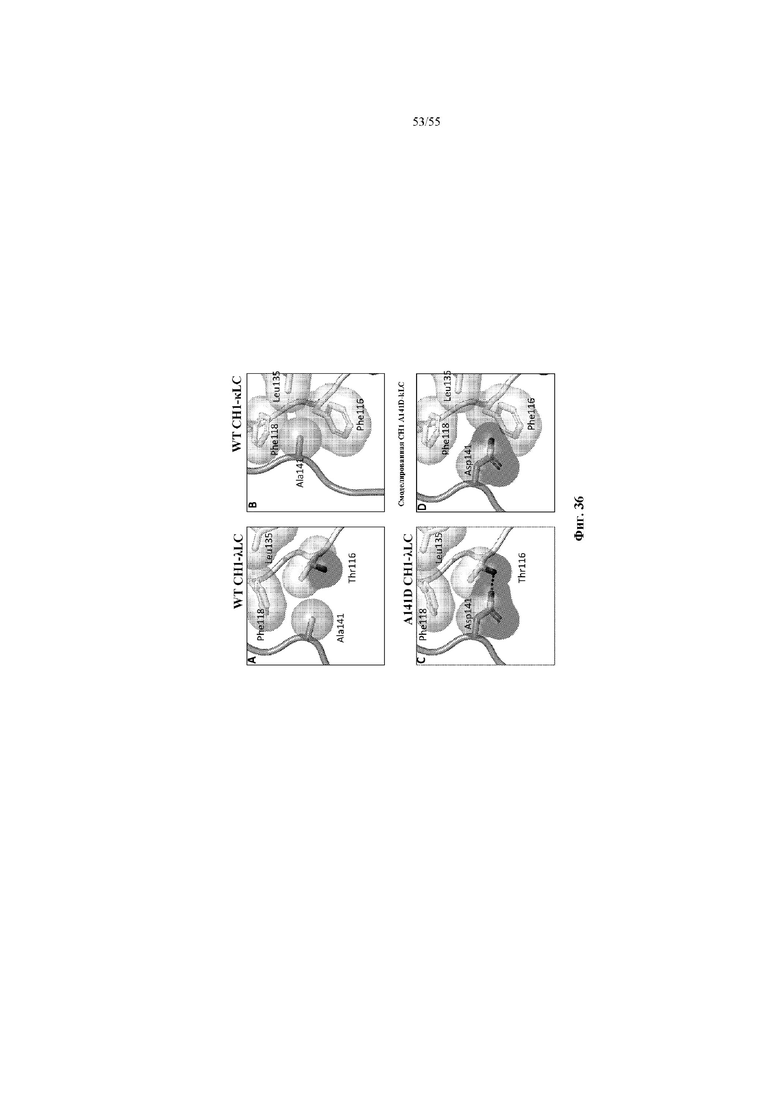
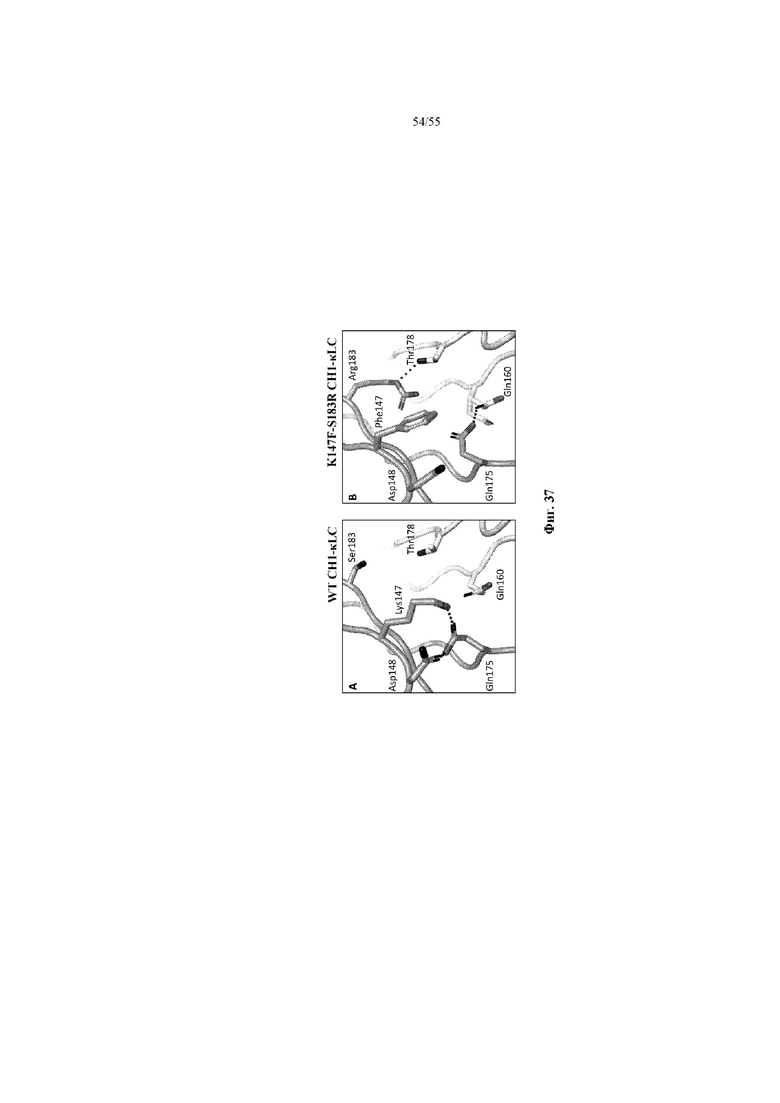

Изобретение относится к области биотехнологии. Предложены варианты домена CH1 иммуноглобулина для избирательного спаривания с полипептидом, содержащим домен CL каппа по сравнению с полипептидом, содержащим домен CL лямбда. Также предложены тяжелые цепи антител и антитела, содержащие указанные варианты домена CH1. Варианты домена CH1 минимизируют неправильное спаривание тяжелых цепей с легкими цепями и способствуют когнатному спариванию тяжелых цепей с легкими цепями, тем самым улучшая получение мультиспецифичных, например, биспецифичных антител. 3 н. и 29 з.п. ф-лы, 58 ил., 15 табл., 8 пр.
1. Полипептид домена константной области 1 тяжелой цепи (CH1) иммуноглобулина для избирательного спаривания с полипептидом, содержащим домен константной области легкой цепи каппа (CL каппа) по сравнению с полипептидом, содержащим домен константной области легкой цепи лямбда иммуноглобулина (CL лямбда), причем этот полипептид домена CH1 представляет собой полипептид домена CH1 из класса IgG человека и содержит последовательность домена CH1, отличного от дикого типа, содержащую:
(i) аминокислотный остаток F в положении 147 и аминокислотный остаток R в положении 183,
(ii) аминокислотный остаток F в положении 147 и аминокислотный остаток K в положении 183 или
(iii) аминокислотный остаток F в положении 147 и аминокислотный остаток Y в положении 183,
где положения указаны согласно нумерации EU.
2. Полипептид домена CH1 по п. 1, который относится к любому из следующих подклассов: IgG1, IgG2, IgG3 или IgG4.
3. Полипептид домена CH1 по п. 1, который представляет собой полипептид домена CH1 подкласса IgG1 человека.
4. Полипептид домена CH1 по п. 1 или 2, где последовательность домена CH1, отличного от дикого типа, содержит следующую(ие) аминокислотную(ые) замену(ы) относительно последовательности домена CH1 дикого типа класса IgG человека:
(i) замену на аминокислоту F в положении 147 и замену на аминокислоту R в положении 183;
(ii) замену на аминокислоту F в положении 147 и замену на аминокислоту K в положении 183;
(iii) замену на аминокислоту F в положении 147 и замену на аминокислоту Y в положении 183,
где положения указаны согласно нумерации EU.
5. Полипептид домена CH1 по п. 4, который является полипептидом домена CH1 подкласса IgG1 человека, и аминокислотная замена(ы) относится(сятся) к последовательности домена CH1 дикого типа подкласса IgG1 человека.
6. Полипептид домена CH1 по п. 4 или 5, где последовательность домена CH1 дикого типа представляет собой или содержит SEQ ID NO: 1.
7. Полипептид домена CH1 по любому из пп. 1-6, где последовательность домена CH1, отличного от дикого типа, содержит аминокислотный остаток A или L в положении 151, где положения указаны согласно нумерации EU.
8. Полипептид домена CH1 по любому из пп. 4-6, где последовательность домена CH1, отличного от дикого типа, дополнительно содержит аминокислотную замену P151A или P151L относительно последовательности домена CH1 дикого типа, где положение указаны согласно нумерации EU.
9. Полипептид домена CH1 по пп. 4-6 или 8, где различия между последовательностью домена CH1, отличного от дикого типа, и последовательностью домена CH1 дикого типа состоят в указанной(ых) аминокислотной(ых) замене(ах).
10. Полипептид домена CH1 по любому из пп. 1-9, в котором последовательность домена CH1, отличного от дикого типа, содержит аминокислотный остаток A в положении 141, где положение указано согласно нумерации EU.
11. Полипептид домена CH1 по любому из пп. 1-10, где последовательность домена CH1, отличного от дикого типа, содержит аминокислотную последовательность:
(i) SEQ ID NO: 137,
(ii) SEQ ID NO: 138 или
(iii) SEQ ID NO: 139.
12. Полипептид домена CH1 по любому из пп. 1-11, который обеспечивает повышенное спаривание с:
(i) доменом CL каппа по сравнению с доменом CL лямбда; и/или
(ii) полипептидом легкой цепи каппа по сравнению с полипептидом легкой цепи лямбда на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%, при измерении с помощью жидкостной хроматографии с масс-спектрометрией (ЖХ-МС), или
по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, по меньшей мере в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз, при измерении с помощью проточной цитометрии, путем сравнения соотношения средней интенсивности флуоресценции (MFI) для окрашивания CL каппа и окрашивания CL лямбда.
13. Полипептид тяжелой цепи антитела для избирательного спаривания с полипептидом, содержащим домен CL каппа, по сравнению с полипептидом, содержащим домен CL лямбда, где указанный полипептид тяжелой цепи антитела содержит вариабельную область и константную область, где константная область содержит полипептид домена CH1 по любому из пп. 1-12.
14. Полипептид тяжелой цепи антитела по п.13, дополнительно содержащий одну или несколько аминокислотных замен вне домена CH1, что дополнительно способствует избирательному спариванию полипептида тяжелой цепи с:
(i) доменом CL каппа по сравнению с доменом CL лямбда, и/или
(ii) полипептидом легкой цепи каппа по сравнению с полипептидом легкой цепи лямбда.
15. Антитело или фрагмент антитела для связывания первого эпитопа, причем указанные антитело или фрагмент антитела содержат первый полипептид тяжелой цепи и первый полипептид легкой цепи, где:
(а) первый полипептид тяжелой цепи и первый полипептид легкой цепи образуют первую когнатную пару;
(b) первый полипептид тяжелой цепи содержит вариабельную область и константную область, где константная область содержит первый полипептид домена CH1 по любому из пп. 1-12,
(c) первый полипептид легкой цепи содержит вариабельную область и константную область, где константная область содержит домен CL каппа, и
(d) вариабельную область полипептида первой тяжелой цепи и вариабельную область полипептида первой легкой цепи из антигенсвязывающего сайта, специфичного к первому эпитопу.
16. Антитело или фрагмент антитела по п. 15, где первый полипептид тяжелой цепи дополнительно содержит одну или несколько аминокислотных замен вне первого домена CH1, что дополнительно способствует избирательному спариванию первого полипептида тяжелой цепи с:
(i) доменом CL каппа по сравнению с доменом CL лямбда и/или
(ii) полипептидом легкой цепи каппа по сравнению с полипептидом легкой цепи лямбда.
17. Антитело или фрагмент антитела по п. 15 или 16, где домен CL каппа представляет собой домен CL каппа дикого типа.
18. Антитело или фрагмент антитела по любому из пп. 15-17, дополнительно содержащие второй полипептид тяжелой цепи и второй полипептид легкой цепи, где:
(a) второй полипептид тяжелой цепи и второй полипептид легкой цепи образуют вторую когнатную пару; и
(b) второй полипептид тяжелой цепи содержит вариабельную область и константную область, где константная область содержит второй полипептид домена CH1;
(c) второй полипептид легкой цепи содержит вариабельную область и константную область, где константная область содержит домен CL лямбда,
где второй домен CH1 содержит последовательность домена CH1, отличного от дикого типа, содержащую следующую(ие) аминокислотный(ые) остаток(тки):
(i) D в положении 141;
(ii) D в положении 141 и K в положении 181;
(iii) D в положении 141, K в положении 181 и A в положении 218;
(iv) D в положении 141, K в положении 181 и P в положении 218;
(v) E в положении 141;
(vi) E в положении 141 и K в положении 181;
(vii) K в положении 141;
(viii) K в положении 141 и K в положении 181;
(ix) K в положении 141, K в положении 181 и E в положении 218;
(x) K в положении 141, K в положении 181 и P в положении 218;
(xi) G в положении 127, R в положении 138, T в положении 141, G в положении 170, R в положении 176 и L в положении 181;
(xii) R в положении 131, E в положении 141 и K в положении 181;
(xiii) R в положении 134, E в положении 141, D в положении 171, V в положении 181 и R в положении 185;
(xiv) D в положении 141, I в положении 168, G в положении 170 и R в положении 187;
(xv) E в положении 141, E в положении 170, L в положении 181 и R в положении 187;
(xvi) E в положении 141, E в положении 170, V в положении 181 и R в положении 187;
(xvii) E в положении 141, D в положении 171, R в положении 185 и F в положении 218;
(xviii) E в положении 141, D в положении 171 и R в положении 185;
(xix) E в положении 141, E в положении 171 и A в положении 181;
(xx) E в положении 141, E в положении 171 и R в положении 185;
(xxi) E в положении 141, E в положении 171, T в положении 181 и R в положении 185;
(xxii) E в положении 141, E в положении 171 и V в положении 181;
(xxiii) E в положении 141, G в положении 171, R в положении 185 и R в положении 187;
(xxiv) E в положении 141, R в положении 185 и R в положении 187;
(xxv) E в положении 141, S в положении 171 и K в положении 181;
(xxvi) E в положении 141, G в положении 170, M в положении 175, V в положении 181, R в положении 184 и R в положении 187;
(xxvii) E в положении 141, R в положении 170, H в положении 173 и V в положении 181;
(xxviii) E в положении 141, S в положении 170, A в положении 171 и V в положении 181;
(xxix) E в положении 141, M в положении 142, S в положении 170, A в положении 171 и V в положении 181;
(xxx) K в положении 141, K в положении 181 и E в положении 185;
(xxxi) K в положении 141, K в положении 181 и D в положении 218;
(xxxii) T в положении 141, V в положении 170, A в положении 171 и V в положении 181;
(xxxiii) R в положении 137, E в положении 141, E в положении 171 и V в положении 181;
(xxxiv) R в положении 126, V в положении 131, V в положении 139, E в положении 141 и E в положении 218;
(xxxv) V в положении 126, V в положении 131, P в положении 136 и D в положении 141;
(xxxvi) V в положении 126, V в положении 131, P в положении 136, R в положении 137 и D в положении 141;
(xxxvii) V в положении 126, V в положении 131, D в положении 141 и S в положении 218;
(xxxviii) V в положении 126, V в положении 131, S в положении 133, K в положении 141 и A в положении 218;
(xxxix) E в положении 141 и R в положении 185;
(xl) E в положении 141 и R в положении 187;
(xli) E в положении 141, R в положении 185 и R в положении 187;
(xlii) E в положении 141, E в положении 170 и R в положении 185;
(vliii) E в положении 141, E в положении 170 и R в положении 187;
(vliv) D в положении 141 и R в положении 185;
(vlv) D в положении 141 и R в положении 187;
(vlvi) D в положении 141, R в положении 185 и R в положении 187;
(vlvii) D в положении 141, E в положении 170 и R в положении 185;
(vlviii) D в положении 141, E в положении 170 и R в положении 187;
(xlix) E в положении 141, E в положении 171 и R в положении 185;
(l) E в положении 141, E в положении 171 и R в положении 187;
(li) D в положении 141, E в положении 171 и R в положении 185;
(lii) D в положении 141, E в положении 171 и R в положении 187;
(liii) V в положении 124, V в положении 126, D в положении 141, S в положении 197 и I в положении 208;
(liv) G в положении 127 и T в положении 141;
(lv) H в положении 130 и R в положении 141;
(lvi) Q в положении 131 и E в положении 141;
(lvii) T в положении 131 и R в положении 141;
(lviii) N в положении 131, T в положении 141, F в положении 168 и N в положении 170;
(lix) D в положении 133 и E в положении 141;
(lx) T в положении 133, T в положении 141 и P в положении 218;
(lxi) A в положении 134 и K в положении 141;
(lxii) H в положении 134 и R в положении 141;
(lxiii) I в положении 134 и R в положении 141;
(lxiv) P в положении 134 и R в положении 141;
(lxv) V в положении 134 и T в положении 141;
(lxvi) V в положении 134 и T в положении 141;
(lxvii) E в положении 141;
(lxviii) R в положении 141 и M в положении 185;
(lxix) T в положении 141 и R в положении 187;
(lxx) R в положении 141, L в положении 185 и D в положении 187;
(lxxi) T в положении 141, S в положении 185 и E в положении 187;
(lxxii) V в положении 141, S в положении 185 и Y в положении 187;
(lxxiii) R в положении 119 и K в положении 141;
(lxxiv) S в положении 130, T в положении 141, T в положении 185 и S в положении 187;
(lxxv) N в положении 131 и D в положении 141;
(lxxvi) R в положении 131 и M в положении 141;
(lxxvii) T в положении 131 и E в положении 141;
(lxxviii) D в положении 131, L в положении 133 и N в положении 134;
(lxxix) D в положении 131, R в положении 141, D в положении 175 и V в положении 181;
(lxxx) E в положении 133 и R в положении 141;
(lxxxi) P в положении 133, A в положении 139 и K в положении 141;
(lxxxii) L в положении 134 и E в положении 141;
(lxxxiii) N в положении 134 и T в положении 141;
(lxxxiv) P в положении 134 и K в положении 141;
(lxxxv) V в положении 134 и E в положении 141;
(lxxxvi) K в положении 141;
(lxxxvii) K в положении 141, T в положении 185 и E в положении 187;
(lxxxviii) V в положении 168, T в положении 170 и N в положении 171;
(lxxxix) L в положении 183;
(xc) V в положении 140;
(xci) T в положении 141;
(xcii) T в положении 141 и D в положении 203;
(xciii) T в положении 141 и A в положении 211;
(xciv) T в положении 141 и G в положении 216;
(xcv) T в положении 141 и M в положении 163;
(xcvi) V в положении 141 и N в положении 212;
(xcvii) T в положении 181;
(xcviii) P в положении 140 и T в положении 141;
(xcix) V в положении 140 и G в положении 216;
(c) T в положении 141 и E в положении 213;
(ci) T в положении 141 и R в положении 214;
(cii) T в положении 141 и G в положении 152;
(ciii) V в положении 141 и G в положении 152;
(civ) G в положении 152;
(cv) V в положении 183 и T в положении 185;
(cvi) T в положении 210;
(cvii) Y в положении 140 и D в положении 141;
(cviii) K в положении 140 и D в положении 141;
(cix) S в положении 140 и D в положении 141;
(cx) W в положении 140 и D в положении 141;
(cxi) R в положении 140, D в положении 141, N в положении 218 и Δ в положении 219;
(cxii) L в положении 140 и D в положении 141;
(cxiii) I в положении 140 и D в положении 141;
(cxiv) V в положении 140 и D в положении 141;
(cxv) E в положении 141, I в положении 181 и L в положении 218;
(cxvi) E в положении 141, K в положении 181 и P в положении 218;
(cxvii) K в положении 141, L в положении 181 и D в положении 218;
(cxviii) K в положении 141, K в положении 181 и A в положении 218;
(cxix) K в положении 141, K в положении 181 и H в положении 218 или
(cxx) E в положении 141, K в положении 181 и D в положении 218,
где положения указаны согласно нумерации EU.
19. Антитело или фрагмент антитела по любому из пп. 15-18, где второй домен CH1 второго полипептида тяжелой цепи относится к любому из следующих классов: IgG, IgA, IgE, IgD или IgM.
20. Антитело или фрагмент антитела по любому из пп. 15-19, где второй домен CH1 второго полипептида тяжелой цепи относится к любому из следующих подклассов: IgG1, IgG2, IgG3, IgG4, IgA1 или IgA2.
21. Антитело или фрагмент антитела по любому из пп. 15-20, где последовательность домена CH1, отличного от дикого типа, второго домена CH1 содержит следующую(ие) аминокислотную(ые) замену(ы) относительно последовательности домена CH1 дикого типа человека:
(i) A141D;
(ii) A141D и S181K;
(iii) A141D, S181K и K218A;
(iv) A141D, S181K и K218P;
(v) A141E;
(vi) A141E и S181K;
(vii) A141K;
(viii) A141K и S181K;
(ix) A141K, S181K и K218E;
(x) A141K, S181K и K218P;
(xi) P127G, G138R, A141T, F170G, S176R и S181L;
(xii) S131R, A141E и S181K;
(xiii) S134R, A141E, P171D, S181V и V185R;
(xiv) A141D, H168I, F170G и T187R;
(xv) A141E, F170E, S181L и T187R;
(xvi) A141E, F170E, S181V и T187R;
(xvii) A141E, P171D, V185R и K218F;
(xviii) A141E, P171D и V185R;
(xix) A141E, P171E и S181A;
(xx) A141E, P171E и V185R;
(xxi) A141E, P171E, S181T и V185R;
(xxii) A141E, P171E и S181V;
(xxiii) A141E, P171G, V185R и T187R;
(xxiv) A141E, V185R и T187R;
(xxv) A141E, P171S и S181K;
(xxvi) A141E, F170G, Q175M, S181V, S184R и T187R;
(xxvii) A141E, F170R, V173H и S181V;
(xxviii) A141E, F170S, P171A и S181V;
(xxix) A141E, L142M, F170S, P171A и S181V;
(xxx) A141K, S181K и V185E;
(xxxi) A141K, S181K и K218D;
(xxxii) A141T, F170V, P171A и S181V;
(xxxiii) G137R, A141E, P171E и S181V;
(xxxiv) F126R, S131V, T139V, A141E и K218E;
(xxxv) F126V, S131V, S136P и A141D;
(xxxvi) F126V, S131V, S136P, G137R и A141D;
(xxxvii) F126V, S131V, A141D и K218S;
(xxxviii) F126V, S131V, K133S, A141K и K218A;
(xxxix) A141E и V185R;
(xl) A141E и T187R;
(xli) A141E, V185R и T187R;
(xlii) A141E, F170E и V185R;
(vliii) A141E, F170E и T187R;
(vliv) A141D и V185R;
(vlv) A141D и T187R;
(vlvi) A141D, V185R и T187R;
(vlvii) A141D, F170E и V185R;
(vlviii) A141D, F170E и T187R;
(xlix) A141E, P171E и V185R;
(l) A141E, P171E и T187R;
(li) A141D, P171E и V185R;
(lii) A141D, P171E и T187R;
(liii) S124V, F126V, A141D, T197S и N208I;
(liv) P127G и A141T;
(lv) P130H и A141R;
(lvi) S131Q и A141E;
(lvii) S131T и A141R;
(lviii) S131N, A141T, H168F и F170N;
(lix) K133D и A141E;
(lx) K133T, A141T и K218P;
(lxi) S134A и A141K;
(lxii) S134H и A141R;
(lxiii) S134I и A141R;
(lxiv) S134P и A141R;
(lxv) S134V и A141T;
(lxvi) S134V и A141T;
(lxvii) A141E;
(lxviii) A141R и V185M;
(lxix) A141T и T187R;
(lxx) A141R, V185L и T187D;
(lxxi) A141T, V185S и T187E;
(lxxii) A141V, V185S и T187Y;
(lxxiii) S119R и A141K;
(lxxiv) P130S, A141T, V185T и T187S;
(lxxv) S131N и A141D;
(lxxvi) S131R и A141M;
(lxxvii) S131T и A141E;
(lxxviii) S131D, K133L и S134N;
(lxxix) S131D, A141R, Q175D и S181V;
(lxxx) K133E и A141R;
(lxxxi) K133P, T139A и A141K;
(lxxxii) S134L и A141E;
(lxxxiii) S134N и A141T;
(lxxxiv) S134P и A141K;
(lxxxv) S134V и A141E;
(lxxxvi) A141K;
(lxxxvii) A141K, V185T и T187E;
(lxxxviii) H168V, F170T и P171N;
(lxxxix) S183L;
(xc) A140V;
(xci) A141T;
(xcii) A141T и N203D;
(xciii) A141T и V211A;
(xciv) A141T и E216G;
(xcv) A141T и L163M;
(xcvi) A141V и D212N;
(xcvii) S181T;
(xcviii) A140P и A141T;
(xcix) A140V и E216G;
(c) A141T и K213E;
(ci) A141T и K214R;
(cii) A141T и E152G;
(ciii) A141V и E152G;
(civ) E152G;
(cv) S183V и V185T;
(cvi) K210T;
(cvii) A140Y и A141D;
(cviii) A140K и A141D;
(cix) A140S и A141D;
(cx) A140W и A141D;
(cxi) A140R, A141D, K218N и S219Δ;
(cxii) A140L и A141D;
(cxiii) A140I и A141D;
(cxiv) A140V и A141D;
(cxv) A141E, S181I и K218L;
(cxvi) A141E, S181K и K218P;
(cxvii) A141K, S181L и K218D;
(cxviii) A141K, S181K и K218A;
(cxix) A141K, S181K и K218H; или
(cxx) A141E, S181K и K218D,
где положения указаны в соответствии с нумерацией EU.
22. Антитело или фрагмент антитела по п. 21, где различия между последовательностью домена CH1, отличного от дикого типа, второго домена CH1 и последовательностью домена CH1 дикого типа состоят из указанных(ой) аминокислотных(ой) замен(ы).
23. Антитело или фрагмент антитела по любому из пп. 15-22, где второй домен CH1 содержит аминокислотную последовательность:
(1) любой из SEQ ID NO: 74-135 и 140-189; или
(2) с первой по 101-ю аминокислоту SEQ ID NO: 1, за исключением аминокислот, включающих:
(cxv) E в положении 141, I в положении 181 и L в положении 218;
(cxvi) E в положении 141, K в положении 181 и P в положении 218;
(cxvii) K в положении 141, L в положении 181 и D в положении 218;
(cxviii) K в положении 141, K в положении 181 и A в положении 218;
(cxix) K в положении 141, K в положении 181 и H в положении 218; или
(cxx) E в положении 141, K в положении 181 и D в положении 218,
где положения указаны в соответствии с нумерацией EU.
24. Антитело или фрагмент антитела по любому из пп. 18-23, где второй полипептид тяжелой цепи приводит к увеличению спаривания с:
(i) доменом CL лямбда по сравнению с доменом CL каппа; и/или
(ii) полипептида легкой цепи лямбда по сравнению с полипептидом легкой цепи каппа на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100%, при измерении с помощью жидкостной хроматографии с масс-спектрометрией (ЖХ-МС), или
по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, по меньшей мере в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз, при измерении с помощью проточной цитометрии, путем сравнения соотношения средней интенсивности флуоресценции (MFI) для окрашивания CL каппа и окрашивания CL лямбда.
25. Антитело или фрагмент антитела по любому из пп. 18-24, в которых второй полипептид тяжелой цепи дополнительно содержит одну или несколько аминокислотных замен за пределами второго домена CH1, что дополнительно способствует избирательному спариванию второго полипептида тяжелой цепи с:
(i) доменом CL лямбда по сравнению с доменом CL каппа, и/или
(ii) полипептидом легкой цепи лямбда по сравнению с полипептидом легкой цепи каппа.
26. Антитело или фрагмент антитела по любому из пп. 18-25, в которых домен CL лямбда представляет собой домен CL лямбда дикого типа.
27. Антитело или фрагмент антитела по любому из пп. 15-26, которые являются мультиспецифичными или биспецифичными.
28. Антитело или фрагмент антитела по п. 27, где структура указанного антитела или фрагмента антитела соответствует показанной на любой из фиг. 24-29.
29. Антитело или фрагмент антитела по любому из пп. 18-26, которые являются мультиспецифичными, где первый и второй домены CH1:
(i) снижают образование некогнатных пар тяжелой цепи и легкой цепи на по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75% или по меньшей мере 80%, или по меньшей мере в 1,2 раза, по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, по меньшей мере в 2,5 раза, по меньшей мере в 3 раза, по меньшей мере в 3,5 раза, по меньшей мере в 4 раза, по меньшей мере в 4,5 раза, по меньшей мере в 5 раз, по меньшей мере в 5,5 раз, по меньшей мере в 6 раз, по меньшей мере в 6,5 раз, по меньшей мере в 7 раз, по меньшей мере в 7,5 раз, по меньшей мере в 8 раз, по меньшей мере в 8,5 раз, по меньшей мере в 9 раз, по меньшей мере в 9,5 раз, по меньшей мере в 10 раз, по меньшей мере в 11 раз, по меньшей мере в 12 раз, по меньшей мере в 13 раз, по меньшей мере в 14 раз, по меньшей мере в 15 раз, по меньшей мере в 16 раз, по меньшей мере в 17 раз, по меньшей мере в 18 раз, по меньшей мере в 19 раз, по меньшей мере в 20 раз, по меньшей мере в 21 раз, по меньшей мере в 22 раза, по меньшей мере в 23 раза, по меньшей мере в 24 раза или по меньшей мере в 25 раз;
(ii) обеспечивают по меньшей мере 75%, по меньшей мере 76%, по меньшей мере 77%, по меньшей мере 78%, по меньшей мере 79%, по меньшей мере 80%, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84%, по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% образование требуемых первой и второй когнатных пар;
(iii) обеспечивают от приблизительно 85% до приблизительно 95% образования требуемых первой и второй когнатных пар; и/или
(iv) обеспечивают сниженное образование некогнатных пар тяжелой цепи и легкой цепи, составляющее менее 25%, менее 20%, менее 15%, менее 14%, менее 13%, менее 12%, менее 11% менее 10%, менее 9%, менее 8%, менее 7%, менее 6%, менее 5%, менее 4%, менее 3%, менее 2% или менее 1%,
где количество когнатных и/или некогнатных пар определяют с помощью ЖХ-МС или проточной цитометрии.
30. Антитело или фрагмент антитела по любому из пп. 18-29, которые являются мультиспецифичными, и где:
(a) последовательность домена CH1, отличного от дикого типа, первого домена CH1 содержит следующие аминокислотные остатки:
(i) F в положении 147 и
(ii) R, K или Y в положении 183, а
(b) последовательность домена CH1, отличного от дикого типа, второго домена CH1 содержит следующие аминокислоты:
(i) D в положении 141, E в положении 171 и R в положении 185;
(ii) D в положении 141, E в положении 170 и R в положении 187; или
(iii) D в положении 141, K в положении 181 и P в положении 218,
где положения указаны в соответствии с нумерацией EU.
31. Антитело или фрагмент антитела по любому из пп. 18-30, где:
(a) первый домен CH1 содержит аминокислотную последовательность:
(i) SEQ ID NO: 137,
(ii) SEQ ID NO: 138 или
(iii) SEQ ID NO: 139, а
(b) второй домен CH1 содержит аминокислотную последовательность:
(i) SEQ ID NO: 188,
(ii) SEQ ID NO: 186 или
(iii) SEQ ID NO: 143.
32. Антитело или фрагмент антитела по любому из пп. 18-31, где первый домен CH1 и второй домен CH1:
(i) снижают образование некогнатных пар тяжелой цепи и легкой цепи в диапазоне от по меньшей мере 50% до по меньшей мере 75%; и/или
(ii) обеспечивает от приблизительно 85% до по меньшей мере приблизительно 95% образования требуемых первой и второй когнатных пар.
| WO 2005092925 A2, 06.10.2005 | |||
| WO 2014138449 A1, 12.09.2014 | |||
| EP 2970469 B1, 03.10.2018 | |||
| EP 1874816 A4, 25.08.2010 | |||
| US 2018177873 A1, 28.06.2018 | |||
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА С РЕГУЛИРУЕМОЙ КОНЪЮГАЦИЕЙ МЕЖДУ ТЯЖЕЛОЙ ЦЕПЬЮ И ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2681885C2 |

