Перекрестные ссылки на родственные заявки
В заявке заявлен приоритет по предварительной заявке на патент США с порядковым номером 62/560574, от 19 сентября 2017 года, и предварительной заявке на патент США с порядковым номером 62/692386, от 29 июня 2018 года, которые непосредственно включены в данный документ ссылкой в полном объеме.
Область техники, к которой относится изобретение
Настоящее изобретение относится в некоторых аспектах к молекулам, связывающим HLA-A2, в частности к гуманизированным анти-HLA-A2 антителам. Настоящее изобретение также относится к рекомбинантным рецепторам, содержащим такие антитела, включая химерные антигенные рецепторы (CAR), которые включают такие антитела. Раскрытие дополнительно относится к генно-инженерным клеткам, экспрессирующим такие рецепторы и антитела, и к их применению в клеточной терапии.
Предшествующий уровень техники
HLA-антигены класса I представляют собой полиморфные белки, экспрессируемые на всех ядросодержащих клетках, и являются критическими мишенями для иммунного распознавания в контексте трансплантации. В действительности развитие Т-клеток и/или антител, специфических по HLA класса I, является основным фактором риска острого и хронического отторжения аллотрансплантата, а присутствие предварительно сформированных антител против донорных HLA класса I может привести к гиперострому отторжению (Konvalinka et al., 2015). Таким образом, поиск путей контроля иммунного ответа на белки HLA класса I был бы главным прорывом в трансплантации.
Классические молекулы HLA класса I полиморфны и кодируются многими различными аллелями, которые эволюционировали в ответ на эволюционное давление от инфекций. Есть три локуса, которые кодируют классические белки HLA класса I, которые называются локусы A, B и C. В пределах локуса HLA-A семейство аллелей HLA-A2 является самым большим и самым разнообразным семейством, по меньшей мере, с 31 различными аллелями HLA-A2, которые, как известно, существуют у людей. Интересно, что в отличие от многих других семейств аллелей HLA, HLA-A2 часто встречается во всех этнических группах, и встречается у 50% европеоидов и 35% афроамериканцев (Ellis et al., 2000). Многие аллели HLA-A2 отличаются только от 1 до 9 аминокислот, причем большая часть полиморфизма сосредоточена вокруг пептидсвязывающей канавки (Hilton et al., 2013). Аллели HLA-A2 разбиты на две основные ветви: те, которые получены посредством событий интераллельной конверсии генов из A*0201 или A*0205 (Ellis et al., 2000).
Адоптивная иммунотерапия T-регуляторными (Treg) клетками как способ контроля нежелательного иммунитета к белкам HLA и другим антигенам, которые управляют отторжением трансплантата, является многообещающим методом лечения отторжения аллотрансплантата и болезни «трансплантат против хозяина» (GVHD). Сообщалось о применении поликлонального переноса Treg-клеток в профилактике болезни «трансплантат против хозяина» (GVHD) после аллогенной трансплантации гемопоэтических стволовых клеток (HSCT) (Brunstein et al., 2011; Di Ianni et al., 2011; Trzonkowski et al., 2009). Также сообщалось о применении переноса Treg-клеток для поддержания уровня c-пептида при диабете 1 типа (Bluestone et al., 2015; Marek-Trzonkowska et al., 2012). Примечательно, что сообщалось, что может быть временный риск генерализованной иммуносупрессии, связанный с применением поликлональных Treg-клеток для такой клеточной терапии (Brunstein et al., 2013).
Данные исследований на животных показывают, что эффективность и специфичность клеточной терапии Treg-клетками могут быть значительно повышены при применении антигенспецифических клеток. Например, в моделях аутоиммунитета антиген-специфичные Treg-клетки превосходят поликлональные Treg-клетки по снижению заболеваемости: Treg-клетки, выделенные из лимфатических узлов поджелудочной железы или активированные островковым антигеном, значительно лучше предотвращают или излечивают диабет 1 типа, чем поликлональные Treg-клетки (Green et al., 2002; Masteller et al., 2005; Tang et al., 2004; Tarbell et al., 2007; Tarbell et al., 2004), и Treg-клетки, экспрессирующие аутоантигенспецифический трансгенный Т-клеточный рецептор (TCR), превосходят поликлональные Treg-клетки по подавлению воспаления центральной нервной системы в модели экспериментального аутоиммунного энцефаломиелита (EAE) (Stephens et al., 2009). Аналогично, аллоантиген-специфичные Treg-клетки, обогащенные стимулированным аллоантигеном экспансией in vitro или сконструированные для экспрессии TCR-трансгена, более эффективны, чем поликлональные Treg-клетки, в предотвращении отторжения трансплантатов органов и тканей (Golshayan et al., 2007; Joffre et al., 2008; Nishimura et al., 2004; Sanchez-Fueyo et al., 2006; Tsang et al., 2008). Существуют некоторые доказательства того, что Treg-клетки, размноженные с аллоантигенами, эффективно предотвращают GVHD (Trenado et al., 2006) и что индукция антиген-специфических Treg-клеток in vivo способствует принятию гемопоэтических аллотрансплантатов без GVHD (Verginis et al., 2008). Модели гуманизированных мышей показали сходные результаты: размноженные с аллоантигеном человеческие Treg-клетки являются более сильными супрессорами отторжения кожного трансплантата, чем поликлональные Treg-клетки (Putnam et al., 2013; Sagoo et al., 2011).
Альтернативным подходом к сверхэкспрессии трансгенных TCR или стимулированной антигеном экспансии для обогащения антиген-специфическими Т-клетками является применение химерных антигенных рецепторов (CAR). В адоптивной иммунотерапии на основе клеток иммунные клетки, выделенные у пациента, могут быть модифицированы для экспрессии синтетических белков, которые позволяют клеткам выполнять новые терапевтические функции после того, как они впоследствии передаются обратно пациенту. Примером такого синтетического белка является CAR. Примером используемого в настоящее время CAR является слияние внеклеточного домена распознавания (например, антигенсвязывающего домена), трансмембранного домена и одного или нескольких внутриклеточных сигнальных доменов. После включения антигена внутриклеточная сигнальная часть CAR может инициировать связанный с активацией ответ в иммунной клетке. Например, Т-клетки могут быть генетически сконструированы для экспрессии внеклеточных антигенсвязывающих доменов одноцепочечных антител (scFv), слитых с внутриклеточными сигнальными доменами (Gill and June, 2015; June et al., 2015). В частности, сообщалось о Treg-клетках, экспрессирующих CAR, специфичных для модельных антигенов (Blat et al., 2014; Elinav et al., 2009; Elinav et al., 2008; Fransson et al., 2012; Hombach et al., 2009, Boardman et al., 2016; MacDonald et al., 2016; Noyan et al., 2016).
Сущность изобретения
Аспекты настоящего раскрытия включают анти-HLA-A2 антитела. Также предлагаются химерные антигенные рецепторы (CAR), включающие внеклеточный домен, включающий любое из анти-HLA-A2 антител по настоящему изобретению. Также предлагаются нуклеиновые кислоты, кодирующие анти-HLA-A2 антитела и CAR по настоящему изобретению, экспрессирующие векторы, содержащие их, и клетки-хозяева, содержащие такие экспрессирующие векторы. Аспекты настоящего раскрытия также включают гуманизированные анти-HLA-A2 антитела. Также предлагаются химерные антигенные рецепторы (CAR), включающие внеклеточный домен, включающий любое из гуманизированных анти-HLA-A2 антител по настоящему изобретению. Также предлагаются нуклеиновые кислоты, кодирующие гуманизированные анти-HLA-A2 антитела и CAR по настоящему изобретению, экспрессирующие векторы, содержащие их, и клетки-хозяева, включающие такие экспрессирующие векторы. Также предлагаются иммунные клетки, например, иммунорегуляторные клетки, которые включают CAR и/или экспрессирующие векторы по настоящему изобретению, композиции и фармацевтические композиции, включающие такие иммунные клетки, наборы частей, включающие такие иммунные клетки и/или реагенты (например, нуклеиновую кислоту или вектор, кодирующий антитело против HLA-A или CAR по настоящему изобретению) для получения таких иммунных клеток, и способы получения таких иммунных клеток. Также предлагаются способы применения анти-HLA-A2 антител, CAR, иммунных клеток и фармацевтических композиций по настоящему изобретению. Например, рассматриваемые анти-HLA-A2 антитела, CAR, иммунные клетки (например, иммунорегуляторные клетки) и фармацевтические композиции находят применение, например, для стимулирования иммунной толерантности у объекта, предотвращения или лечения болезни трансплантат против хозяина (GVHD) у объекта, предотвращение или лечение отторжения трансплантата органа или ткани у объекта и т.п.
В некоторых воплощениях предлагается гуманизированное анти-HLA-A2 антитело, где антитело способно составлять антигенсвязывающий домен химерного антигенного рецептора (CAR), где CAR способен экспрессироваться в клетке человека (например, иммунной клетке человека, такой как иммунная регуляторная клетка человека), так, что CAR специфически связывается с HLA-A2. В определенных аспектах антитело по настоящему изобретению конкурирует за связывание с HLA-A2 с антителом, включающим: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190.
В определенных аспектах предлагается гуманизированное анти-HLA-A2 антитело, где антитело конкурирует за связывание с HLA-A2 с антителом, включающим: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190.
В определенных аспектах гуманизированное анти-HLA-A2 антитело, изложенное выше, связывается с тем же эпитопом HLA-A2, где антитело включает: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190.
В некоторых воплощениях гуманизированное анти-HLA-A2 антитело по настоящему изобретению имеет меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 и любых их комбинаций по сравнению с антителом BB7.2. Например, в некоторых воплощениях гуманизированное анти-HLA-A2 антитело по настоящему изобретению имеет меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*25, HLA-A*29, HLA-A*30 и любой их комбинации по сравнению с антителом BB7.2.
В определенных аспектах гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, выбранную из группы, состоящей из: SYHIQ (SEQ ID NO: 1) и GYTFTSY (SEQ ID NO: 2).
Согласно определенным воплощениям осуществления гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, выбранную из группы, состоящей из: YPGDGS (SEQ ID NO: 4) и WIYPGDGSTX10YX12X13KFX16G (SEQ ID NO: 10), где X10 представляет собой Q или K, X12 представляет собой N или S, X13 представляет собой E или Q и X16 представляет собой K или Q. Такое антитело может включать, например, вариабельную область тяжелой цепи, включающую аминокислотную последовательность, выбранную из группы, состоящей из: WIYPGDGSTQYNEKFKG (SEQ ID NO: 3) и YPGDGS (SEQ ID NO: 4). Также в качестве примера такое антитело может включать, например, вариабельную область тяжелой цепи, включающую аминокислотную последовательность WIYPGDGSTKYSQKFQG (SEQ ID NO: 5). В определенных аспектах гуманизированное антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность EGTYYAMDY (SEQ ID NO: 6).
В некоторых воплощениях гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую аминокислотную последовательность RSSQSIVHSNGNTYLE (SEQ ID NO: 7). В определенных аспектах гуманизированное антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую аминокислотную последовательность KVSNRFS (SEQ ID NO: 8). Согласно некоторым воплощениям гуманизированное антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую аминокислотную последовательность FQGSHVPRT (SEQ ID NO: 9).
В определенных аспектах гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую каркасную область 1 (VH FR1), включающую аминокислотную последовательность, выбранную из группы, состоящей из: QVQLVQSGAEVKKPGASVKVSCKAS (SEQ ID NO: 11) и QVQLVQSGAEVKKPGASVKVSCKASGYTFT (SEQ ID NO: 12).
Согласно определенным воплощениям гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую каркасную область 2 (VH FR2), включающую аминокислотную последовательность, выбранную из группы, состоящей из:
WVRQAPGQX9LEWMGX15 (SEQ ID NO: 13),
WVRQAPGQX9LEWMGX15WI (SEQ ID NO: 17),
HIQWVRQAPGQX12LEWMGX18WI (SEQ ID NO: 21), и
HIQWVRQAPGQX12LEWMGX18 (SEQ ID NO: 25), где:
X9 представляет собой R или G и X15 представляет собой I или отсутствует в SEQ ID NO: 13;
X9 представляет собой R или G и X15 представляет собой I или отсутствует в SEQ ID NO: 17;
X12 представляет собой R или G и X18 представляет собой I или отсутствует в SEQ ID NO: 21; и
X12 представляет собой R или G, а X18 представляет собой I или отсутствует в SEQ ID NO: 25.
В некоторых воплощениях гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую каркасную область 3 (VH FR3), включающую аминокислотную последовательность, выбранную из группы, состоящей из:
X1VTX4TX6DTSX10STAYMX16LSX19LRSX23DX25AVYYCAR (SEQ ID NO: 29),
TX2YX4X5KFX8GX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34
AVYYCAR (SEQ ID NO: 35),
TQYNEKFKGX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34
AVYYCAR (SEQ ID NO: 36) и
TKYSQKFQGX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34AVYYCAR (SEQ ID NO: 37), где:
X1 представляет собой R или отсутствует, X4 представляет собой I или M, X6 представляет собой R или A, X10 представляет собой A, T или I, X16 представляет собой E или L, X19 представляет собой S или R, X23 представляет собой E или D и X25 представляет собой T или M в SEQ ID NO: 29;
X2 представляет собой Q или K, X4 представляет собой N или S, X5 представляет собой E или Q, X8 представляет собой K или Q, X10 представляет собой R или отсутствует, X13 представляет собой I или M, X15 представляет собой R или A, X19 представляет собой A, T или I, X25 представляет собой E или L, X28 представляет собой S или R, X32 представляет собой E или D и X34 представляет собой T или M в SEQ ID NO: 35;
X10 представляет собой R или отсутствует, X13 представляет собой I или M, X15 представляет собой R или A, X19 представляет собой A, T или I, X25 представляет собой E или L, X28 представляет собой S или R, X32 представляет собой E или D и X34 представляет собой T или M в SEQ ID NO: 36; и
X10 представляет собой R или отсутствует, X13 представляет собой I или M, X15 представляет собой R или A, X19 представляет собой A, T или I, X25 представляет собой E или L, X28 представляет собой S или R, X32 представляет собой E или D и X34 представляет собой T или M в SEQ ID NO: 37.
В определенных аспектах гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую каркасную область 4 (VH FR4), включающую аминокислотную последовательность WGQGTTVTVSS (SEQ ID NO: 44). Согласно некоторым воплощениям гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 61-66.
Согласно определенным воплощениям гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую каркасную область 1 (VL FR1), включающую аминокислотную последовательность DX2VMTQX7PLSX11X12VTX15GQPASISX23 (SEQ ID NO: 46), где X2 представляет собой V или I, X7 представляет собой S или T, X11 представляет собой L или S, X12 представляет собой P или S, X15 представляет собой L или P и X23 представляет собой C или F.
В некоторых воплощениях гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую каркасную область 2 (VL FR2), включающую аминокислотную последовательность WX2X3QX5PGQX9PX11X12LIY (SEQ ID NO: 51), где X2 представляет собой F или Y, X3 означает Q или L, X5 означает R или K, X9 означает S или P, X11 означает R или Q, а X12 означает R или L.
В определенных аспектах гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую каркасную область 3 (VL FR3), включающую аминокислотную последовательность GVPDRFSGSGX11GTDFTLKISRVEAEDVGVYYC (SEQ ID NO: 56), где X11 представляет собой S или А. Согласно некоторым воплощениям гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую каркасную область 4 (VL FR4), включающую аминокислотную последовательность FGGGTKVEIK (SEQ ID NO: 59). В некоторых воплощениях гуманизированное анти-HLA-A2 антитело по настоящему изобретению включает вариабельную область легкой цепи, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 67-71. В некоторых воплощениях гуманизированное анти-HLA-A2 антитело настоящего изобретения представляет собой полноразмерное антитело, одноцепочечное антитело, димерное одноцепочечное антитело, Fv, scFv, Fab, F(ab)'2, дефукозилированное антитело, биспецифическое антитело, диатело, триатело, тетратело, фрагмент антитела, выбранный из группы, состоящей из унитела, доменного антитела, и нанотела или миметика антитела, выбранного из группы, состоящей из аффитела, альфа-тела, каркаса на основе белка с повтором armadillo, кноттина, пептида домена Куница, аффилина, аффитина, аднектина, атримера, эвазина, DARPin, антикалина, авимера, финомера, версатела или дуокалина.
Согласно определенным воплощениям гуманизированное анти-HLA-A2 антитело по настоящему изобретению представляет собой scFv. Например, гуманизированное анти-HLA-A2 антитело по настоящему изобретению может представлять собой scFv, включающее аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 72-91.
В некоторых воплощениях предлагается гуманизированное анти-HLA-A2 антитело, как указано выше, где антитело способно составлять антигенсвязывающий домен химерного антигенного рецептора (CAR), где CAR способен экспрессироваться в иммунной клетке (например, регуляторной T-клетке (Treg)), так что CAR специфически связывается с HLA-A2. В определенных аспектах предлагается гуманизированное анти-HLA-A2 антитело, как указано выше, где антитело способно составлять антигенсвязывающий домен химерного антигенного рецептора (CAR), где CAR способен экспрессироваться в иммунной клетке (например, регуляторной T-клетке (Treg)), так что иммунная клетка активируется HLA-A2.
В определенных аспектах предлагается нуклеиновая кислота, кодирующая любое из гуманизированных анти-HLA-A2 антител, указанных выше. Также предлагаются экспрессирующий вектор и векторы для генной терапии, которые включают такую нуклеиновую кислоту. Также предлагается клетка-хозяин, включающая такой экспрессирующий вектор или вектор для генной терапии.
Аспекты настоящего раскрытия дополнительно включают химерные антигенные рецепторы (CAR). Например, предлагается CAR, включающий: (i) внеклеточный домен, включающий любое из гуманизированных анти-HLA-A2 антител, указанных выше; (ii) трансмембранный домен; и (iii) цитоплазматический домен, включающий внутриклеточный сигнальный домен; где CAR способен экспрессироваться в иммунной клетке, так что CAR специфически связывается с HLA-A2. Такой CAR может иметь меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 и любой их комбинации по сравнению с CAR, включающим антитело BB7.2. Например, в некоторых воплощениях такой CAR имеет меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из HLA-A*25, HLA-A*29, HLA-A*30 и любой их комбинации, по сравнению с CAR, включающим антитело BB7.2. CAR по настоящему изобретению может быть способен экспрессироваться в иммунной клетке (например, регуляторной T-клетке (Treg)), так что иммунная клетка активируется HLA-A2. CAR по настоящему изобретению может включать шарнирную область. В определенных аспектах шарнирная область включает область стебля CD8α.
CAR по настоящему изобретению может включать трансмембранный домен, который включает трансмембранный домен белка, выбранного из группы, состоящей из: CD3 гамма, CD3 дельта, CD3 эпсилон, CD3 дзета, альфа-цепи Т-клеточного рецептора, бета-цепи Т-клеточного рецептора, гамма-цепи Т-клеточного рецептора, дельта-цепи Т-клеточного рецептора, CD28, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154 и любой их комбинации. В некоторых воплощениях трансмембранный домен включает трансмембранный домен CD28.
В соответствии с некоторыми воплощениями, CAR по настоящему изобретению включает внутриклеточный сигнальный домен, который включает функциональный сигнальный домен белка, выбранного из группы, состоящей из: CD3 гамма, CD3 дельта, CD3 эпсилон, CD3 дзета, FcR гамма, FcR альфа, FcR эпсилон, CD5, CD22, CD79a, CD79b и CD66d и любой их комбинации. В некоторых воплощениях внутриклеточный сигнальный домен включает функциональный сигнальный домен CD3 дзета. В определенных аспектах внутриклеточный сигнальный домен дополнительно включает костимулирующий домен. Такой костимулирующий домен может включать функциональный сигнальный домен белка, выбранного из группы, состоящей из OX40, CD27, CD28, антигена-1, связанного с функцией лимфоцитов (LFA-1) (CD11a/CD18), TNFR1 (CD120a/TNFRSF1A), TNFR2 (CD120b/TNFRSF1B), CTLA-4 (CD152), CD95, ICOS (CD278), 4-1BB (CD137), CD2, CD30, CD40, PD-1, CD7, LIGHT, NKG2C, B7-H3, ICAM- 1, лиганд, который специфически связывается с CD83, IL2ra (CD25), IL6Ra (CD126), IL-7Ra (CD127), IL-13RA1, IL-13RA2, IL-33R (IL1RL1), IL-10RA, IL-10RB, IL-4R, IL-5R (CSF2RB), ARHR, рецептором BAFF, IL-21R, TGFbR1, TGFbR2, TGFbR3, общей гамма-цепи и любой их комбинации. Согласно некоторым воплощениям костимулирующий домен включает функциональный сигнальный домен белка, выбранного из CD28 и 4-1BB. Например, костимулирующий домен может включать функциональный сигнальный домен CD28.
Также предлагаются модифицированные иммунные клетки, включающие любой из CAR настоящего изобретения. В некоторых воплощениях модифицированная иммунная клетка представляет собой регуляторную T-клетку (Treg).
Настоящее раскрытие относится к нуклеиновым кислотам, кодирующим любой из CAR настоящего раскрытия. Также предлагаются экспрессирующие векторы, включающие такие нуклеиновые кислоты, а также иммунные клетки (например, регуляторные T-клетки (Treg)), включающие такие экспрессирующие векторы.
Также предлагаются композиции (например, фармацевтические композиции). В определенных аспектах предлагается фармацевтическая композиция, включающая множество модифицированных иммунных клеток или иммунных клеток по настоящему изобретению. Также предлагаются наборы. В некоторых воплощениях указанные наборы содержат в первой части иммунные клетки по настоящему изобретению и во второй части другой терапевтический агент, такой как, например, иммуносупрессирующий агент. В некоторых воплощениях указанные наборы включают один или несколько реагентов (например, нуклеиновую кислоту или экспрессирующий вектор, кодирующий антитело против HLA-A или CAR по настоящему изобретению) для получения клеток по настоящему изобретению. Также предлагаются способы получения модифицированных иммунных клеток по настоящему изобретению. В некоторых воплощениях такие способы включают трансдукцию иммунной клетки экспрессирующим вектором по настоящему изобретению, в результате чего получают модифицированную иммунную клетку.
Также предлагаются способы применения антител, CAR, иммунных клеток, модифицированных иммунных клеток и фармацевтических композиций по настоящему изобретению. В определенных аспектах предлагаются способы стимулирования иммунной толерантности у объекта, причем способы включают введение объекту фармацевтической композиции по настоящему изобретению, например, фармацевтической композиции, включающей множество модифицированных иммунных клеток или иммунных клеток по настоящему изобретению. В некоторых воплощениях иммунная толерантность представляет собой толерантность к пересаженному органу или ткани. В соответствии с определенными воплощениями предлагаются способы профилактики или лечения заболевания «трансплантат против хозяина» (GVHD) у объекта, причем способы включают введение объекту фармацевтической композиции по настоящему изобретению, например, фармацевтической композиции, включающей множество модифицированных иммунных клетки или иммунные клетки по настоящему изобретению. В определенных аспектах объект подвергается или перенес трансплантацию гемопоэтических стволовых клеток. Также предлагаются способы предотвращения или лечения отторжения органа или ткани у объекта, причем способы включают введение объекту фармацевтической композиции по настоящему изобретению, например, фармацевтической композиции, включающей множество модифицированных иммунных клеток или иммунных клеток по настоящему раскрытию. В некоторых воплощениях объект дополнительно получает иммуносупрессирующий агент. Согласно определенным воплощениям предлагаются способы предотвращения или лечения отторжения трансплантата органа или ткани, или заболевания «трансплантат против хозяина» (GVHD) у объекта, причем способы включают введение объекту комбинации иммунной клетки по настоящему изобретению, по меньшей мере, с одним иммуносупрессирующим агентом для индукции иммунной толерантности. В любом из способов применения антител, CAR, иммунных клеток, модифицированных иммунных клеток и фармацевтических композиций по настоящему изобретению объект может быть человеком.
Краткое описание чертежей
Фигура 1. Конструирование гуманизированных анти-HLA-A2 CAR. Схематическое представление лентивирусных конструкций. Вверху: укороченная контрольная конструкция NGFR (без CAR); Внизу: конструкция гуманизированного анти-HLA-A2 CAR. «SP»: сигнальный пептид; «GS»: глицин-сериновый линкер; «ТМ»: трансмембранный регион; «Hs»: гуманизированный.
Фигура 2. Экспрессия на клеточной поверхности и специфичность гуманизированных анти-HLA-A2 CAR. Клетки 293T транзиторно трансфицировали указанной конструкцией, и через 48 часов измеряли экспрессию и специфичность антигена окрашиванием и проточной цитометрией с помощью anti-ΔNGFR mAb и тетрамеров HLA-A2. A & B показывают точечные графики для конструкций, которые сохраняют или не сохраняют свою способность связываться с HLA-A2, соответственно. Данные являются характерными для двух независимых экспериментов.
Фигура 3. Сравнение силы связывания гуманизированных анти-HLA-A2 CAR. Клетки 293T трансфицировали указанными гуманизированными конструкциями анти-HLA-A2 CAR и окрашивали указанными разведениями тетрамера HLA-A2. На Фигурах A, B и C представлены графики, изображающие среднюю геометрическую интенсивность флуоресценции связывания тетрамера HLA-A2 в гейтированных клетках ΔNGFR+, с конструкциями, сгруппированными в соответствии с использованием легкой цепи.
Фигура 4. Экспрессия и специфичность гуманизированных анти-HLA-A2 CAR на Treg. CD4+CD25hiCD127lo Treg активировали, через день трансдуцировали указанным лентивирусом, а затем давали возможность размножаться. Через семь дней после активации экспрессирующие ΔNGFR клетки отбирали путем разделения на магнитных гранулах. Эффективность трансдукции и связывание HLA-A2 определяли проточной цитометрией до и после разделения клеток ΔNGFR+ (A, B, C и D). Числа представляют долю ΔNGFR+ тетрамер+ клеток. Данные являются репрезентативными для независимых экспериментов. (E) обобщенные данные процента или средней интенсивности флуоресценции связывания A*02:01-тетрамера.
Фигура 5. HLA-A2 CAR-опосредованная активация Treg. CD4+CD25hiCD127lo Treg активировали, трансдуцировали указанным лентивирусом и давали возможность размножаться. Через 7 дней Treg оставляли с 100 Ед./мл IL-2 в течение ночи, затем оставляли нестимулированными или стимулированными совместным культивированием с соотношением 2: 1 (Treg-клетки: K562) анти-CD3/28-нагруженных клеток CD64-K562 (TCR) или клеток HLA-A2-K562 (CAR). Через 24 часа экспрессию CD69, CD154, CTLA-4 и LAP измеряли проточной цитометрией на живых клетках CD4+. (A и B) показывают репрезентативные гистограммы и (C и D) показывают усредненные данные из двух независимых экспериментов (E). Контрольные ΔNGFR/CAR Treg совместно культивировали в соотношении 2:1 (Treg: K562) с HLA-A2-экспрессирующими клетками К562. Через 16 часов экспрессию CD69, CD71, CTLA-4 и LAP измеряли проточной цитометрией. Процент положительного и кратного увеличения по сравнению с исходным уровнем (без K562) экспрессии CD69 и CD71. (F) Процент положительного и кратного увеличения по сравнению с исходным уровнем (без K562) экспрессии CTLA-4 и LAP. Данные представляют собой n = 2-4 для каждой конструкции, по меньшей мере, из двух независимых экспериментов. Односторонний дисперсионный анализ и апостериорный тест Holm-Sidak, сравнивающие все конструкции с mA2-CAR Treg. Среднее ± SEM. * р <0,05, ** р <0,01, *** р <0,001, **** р <0,0001.
Фигура 6. Определение перекрестной реактивности гуманизированных анти-HLA-A2 CAR с распространенными аллельными вариантами HLA-A и HLA-B. (A) показывает схематическую диаграмму экспериментальных установок и стратегии гейтирования для анализа клеток FlowPRT. ΔNGFR+ Treg, экспрессирующие указанные гуманизированные CAR, инкубировали с гранулами Flow Panel Reactive Single Antigen и фиксируемым красителем для оценки жизнеспособности в течение 30 минут при комнатной температуре. Затем образцы промывали, фиксировали и анализировали с помощью проточной цитометрии. (B) Связывание с гранулами, покрытыми HLA-A*02:01, для каждого m/hA2-CAR Treg относительно связывания контроля ΔNGFR Treg. (C и D) Корреляция между средним значением связывания HLA-A*02:01, измеренным с помощью анализа клеток FlowPRT, и либо (C) MFI HLA-A*02:01-тетрамера, оцененным с помощью проточной цитометрии, либо (D) увеличением доли клеток CD69+ через 16 ч после совместного культивирования с HLA-A*02:01 по сравнению с отрицательным контролем HLA-A*24:01 клеток K562. Данные в E, F, G и H показывают процент связывания относительно контрольных Treg, экспрессирующих только укороченный NGFR, нормализованный по количеству HLA-негативных гранул, собранных цитометром. Точки данных над заштрихованной серой областью (E, F и G) или горизонтальной пунктирной линией (H) представляют значения, которые отличались более чем на два стандартных отклонения от среднего значения контроля только с гранулами и, таким образом, статистически значимы (р <0,05). Данные являются усреднением трех независимых экспериментов. (I) ΔNGFR или m/hA2-CAR Treg совместно культивировали с указанными клетками K562, трансдуцированными для экспрессии отобранных аллелей HLA-A. Через 16 часов экспрессию CD69, CD71, LAP и CTLA-4 измеряли на живых CD4+ Т-клетках. n = 2-6, по меньшей мере, из 2 независимых экспериментов. Статистическая значимость определяется односторонним дисперсионным анализом и апостериорным тестом Holm-Sidak по сравнению с mA2-CAR. среднее ± SEM, ** р <0,01.
Фигура 7. Treg, экспрессирующие гуманизированный CAR HLA-A2, мощно подавляют пролиферацию Т-клеток, стимулируемую дендритными клетками HLA-A2+. (А) Схематическое представление установок эксперимента. Зрелые дендритные клетки HLA-A2+ использовали для стимуляции HLA-A2neg CD4+ «респондерных» Т-клеток, меченных красителем клеточной пролиферации (CPD)-e450. Меченные CPD-e660 Treg, которые либо не трансдуцировали, либо трансдуцировали контрольным лентивирусом, кодирующим ΔNGFR, или гуманизированными A2 CAR-экспрессирующими Treg. (B, C, D и E) Указанные соотношения клеток совместно культивировали в течение шести дней, затем величину пролиферации CD4+ Т-клеток-респондеров, меченных CPD-e450, измеряли проточной цитометрией. B показывает репрезентативные точечные графики, а C, D и E показывают графические данные для нескольких соотношений клеток. C, D и E демонстрируют средние данные для n = 3-7, по меньшей мере, из 3 независимых экспериментов. Статистические данные были выполнены с использованием двухстороннего дисперсионного анализа с апостериорным тестом Holm-Sidak в сравнении с контролем ΔNGFR Treg. * р <0,05, среднее значение ± стандартная ошибка среднего.
Фигура 8. Treg, экспрессирующие гуманизированный HLA-A2 CAR, мощно подавляют ксеногенную болезнь «трансплантат против хозяина». Облученным мышам NSG инъецировали PBS (n = 3), 8 × 106 HLA-A2+ PBMC (n = 5) или 4 × 106 H1k2-CAR-экспрессирующих Treg (n = 6). Приживление клеток человека в крови контролировали каждые 7 дней. (A) Кривая выживания и (B) процент изменения массы относительно начала эксперимента. (C) Доля общих мононуклеарных клеток (живых синглетов), экспрессирующих CD45 человека в крови. (D) стратегия гейтирования для различения общего приживления CD45+ и CAR Treg (hCD45+hCD4+HLA-A2-) клеток человека. (E) Приживление клеток in vivo после адоптивного переноса на мышиной модели ксеногенной GVHD, что показано абсолютным количеством приживления PBMC и CAR Treg на мкл крови с течением времени. Количество PBMC рассчитывали как hCD45+ минус общее количество CAR Treg, как указано в D.
Фигура 9. Экспрессия m/hA2 CAR обеспечивает Treg быстрым и стойким хомингом к аллотрансплантатам кожи HLA-A2:01+. Treg совместно трансдуцировали лентивирусом, кодирующим люциферазу, и либо контрольным HER2-CAR, mA2-CAR либо hA2-CAR (H1k2). Двойные трансдуцированные клетки сортировали в FACS, размножали в течение 5 дней, а затем инъецировали мышам NSG, которым ранее трансплантировали смежные кожные трансплантаты как от трансгенных мышей NSG, так и NSG-HLA-A*02:01. (А) Схематическое изображение экспериментальных условий. (B) Репрезентативная люциферазная визуализация трансплантатов кожи (слева) через 72 часа или (справа) через 21 день после инъекции Treg. Количество люциферазного излучения определяли с использованием среднего количества фотонов/сек/см2/стерадиан и наносили на график как соотношение между (C) кожными трансплантатами HLA-A*02:01-NSG и NSG через 72 часа после инъекции Treg или (D) в зависимости от времени. n = 2-3 на группу из трех независимых экспериментов, среднее значение ± SEM. Повторные измерения дисперсионного анализа с коррекцией Бонферрони.
Фигура 10. Отслеживание с помощью проточной цитометрии m/hA2 CAR Treg с быстрым и стойким хомингом к кожным аллотрансплантатам HLA-A2:01+. Treg совместно трансдуцировали лентивирусом, содержащим люциферазу и конструкции HER2-CAR, mA2-CAR или hA2-CAR, размножали и инъецировали трансплантированным мышам NSG, как показано на Фигуре 9. (A) Предварительное гейтирование для графиков проточной цитометрии было основано на клетках селезенки контрольных HER2-CAR. (B) профиль проточной цитометрии hCD4/hCD8 для указанных конструкций. На графиках проводили предварительно гейтирование на FvD-hCD45+ как в (A). (C) Графики проточной цитометрии, показывающие окрашивание для m/hA2-CAR Treg в селезенке и дренирующем лимфатическом узле по окончании эксперимента n = 1 на группу из одного независимого эксперимента. * p < 0,05.
Фигура 11. hA2-CAR-Treg уменьшают отторжение аллотрансплантата кожи человека. Мышам NSG трансплантировали человеческую кожу HLA-A*02:01+ и спустя три недели инъецировали: PBS (n = 3); HLA-A*02: 01neg PBMC отдельно (n = 4) или с соотношением 2: 1 аутологичных H1k2 CAR Treg (n = 6). PBMC/hA2-CAR Treg были от двух отдельных доноров, протестированных в одном эксперименте. (A) Массу тела контролировали трижды в неделю, и (B) измеряли долю CD45+ клеток человека в крови (слева) и селезенке (справа) по экспериментальной конечной точке. (C) Кумулятивный гистологический балл трансплантированных участков кожи, определенный по окраске гематоксилином-эозином. (D) Трансплантированные кожные трансплантаты иммуноокрашивали в конце эксперимента для количественной оценки степени экспрессии инволюкрина и доли клеток Ki-67+ в эпидермисе. (E) экспрессию мРНК указанных генов в трансплантированных участках кожи определяли с помощью qRT-PCR. (F) Трансплантированные кожные трансплантаты иммуноокрашивали в конце эксперимента для количественного определения доли клеток FOXP3+ в клетках CD45+ человека. (G) Трансплантированные кожные трансплантаты, срезы кишечника, легких и печени иммуноокрашивали в конце эксперимента, чтобы показать долю клеток FOXP3+ в клетках CD45+ человека в каждой ткани. Каждая точка данных представляет одну мышь. Диаграммы типа «ящик с усами» демонстрируют среднее значение ± диапазон. Статистическую значимость определяли с помощью двустороннего критерия Манна-Уитни, сравнивающего РВМС с H1k2. * p < 0,05.
Фигура 12: Отслеживание проточной цитометрией hA2 CAR Treg в крови на модели трансплантата кожи человека. Мышам NSG трансплантировали кожу человека HLA-A*02+ и инъецировали клетки, как описано на Фигуре 11. (A) Стратегия гейтирования для различения общего приживления CD45+ (PBMC) человека и CAR Treg (hCD45+ hCD4+ NGFR+). (B) Абсолютное количество приживления РВМС и CAR Treg на мкл крови с течением времени. Количество РВМС было рассчитано как hCD45 + минус общее количество Treg CAR, как указано в (A).
Фигура 13: Активация hA2 CAR Treg с использованием искусственных антигенпрезентирующих клеток. Контроль ΔNGFR/CAR Treg совместно культивировали в течение 16 часов без стимуляции или с соотношением 2: 1 (Treg : K562) CD64-экспрессирующих клеток K562, нагруженных моноклональными антителами против CD3 и против CD28 (стимуляция TCR). (A) Пример стратегии гейтирования. (B) Экспрессию CD69, CD71, CTLA-4 и LAP измеряли с помощью проточной цитометрии (слева). Было рассчитано кратное увеличение каждого маркера активации по сравнению с исходным уровнем (без стимуляции) (справа). Данные представляют собой n = 2-4 для каждой конструкции, по меньшей мере, из двух независимых экспериментов. Односторонний дисперсионный анализ и тест множественных сравнений Holm-Sidak, сравнивающие все конструкции с mA2-CAR Treg. * p < 0.05, ** p < 0.01, **** p < 0.0001.
ПОДРОБНОЕ ОПИСАНИЕ
I. Общие Методы
При практическом применении настоящего изобретения будут применяться до тех пор, пока не указано иное, стандартные методы молекулярной биологии (включая рекомбинантные методы), микробиологии, клеточной биологии, биохимии и иммунологии, которые известны специалисту в данной области. Такие методы полностью описаны в литературе, такие как «Molecular Cloning: A Laboratory Manual», second edition (Sambrook et al., 1989); «Oligonucleotide Synthesis» (M. J. Gait, ed., 1984); «Animal Cell Culture» (R. I. Freshney, ed., 1987); «Methods in Enzymology» (Academic Press, Inc.); «Current Protocols in Molecular Biology» (F. M. Ausubel et al., eds., 1987, and periodic updates); «PCR: The Polymerase Chain Reaction», (Mullis et al., ed., 1994); «A Practical Guide to Molecular Cloning» (Perbal Bernard V., 1988); «Phage Display: A Laboratory Manual» (Barbas et al., 2001).
Перед тем как настоящее изобретение будет дополнительно описано, следует понять, что это изобретение не ограничено описанными конкретными воплощениями, и в силу этого может, конечно, варьировать. Также следует понимать, что используемая в данном документе терминология предназначена только для описания конкретных воплощений и не предназначена для ограничения, поскольку объем настоящего изобретения будет ограничен только прилагаемой формулой изобретения.
Если иное не определено, все технические и научные термины, использованные в данном документе, имеют те же смыслы, которые вкладываются в них обычным специалистом области, к которой принадлежит данное изобретение. Хотя любые способы и материалы, подобные или эквивалентные тем, что описаны в данном документе, могут также быть применены на практике или при тестировании настоящего изобретения, далее будут описаны предпочтительные способы и материалы. Специалисту в данной области техники известно множество способов и материалов, аналогичных или эквивалентных описанным в настоящем документе, которые могут быть использованы при практическом осуществлении настоящего изобретения. Действительно, настоящее изобретение никоим образом не ограничивается описанными способами и материалами. Для целей настоящего изобретения далее определены нижеследующие термины. Все публикации, упомянутые в данном документе, включены в данный документ посредством ссылки для раскрытия и описания способов и/или материалов, в связи с которыми публикации цитируются. В случае если любое из приведенных определений вступает в противоречие с любым документом, включенным в настоящий документ посредством ссылки, определение, изложенное ниже, имеет преимущественную силу.
11. ОПРЕДЕЛЕНИЯ
Термины, приведенные в единственном числе, также охватывают и свое множественное число. Например, «элементt» означает один элемент или более одного элемента.
Термин «около», когда он относится к измеряемому значению, такому как величина, временная длительность и т.п., подразумевает, что он охватывает колебания ±20% или в некоторых случаях ±10%, а в некоторых случаях ±5%, или в некоторых случаях ±1% или в некоторых случаях ±0,1% от указанного значения, поскольку такие вариации являются подходящими для выполнения раскрытых способов.
Там, где предлагается диапазон значений, следует понимать, что каждое промежуточное значение, до десятой значения нижнего предела, если контекст ясно не указывает на иное, между верхним и нижним лимитом этого диапазона и любое другое заданное или промежуточное значение в этом заданном диапазоне, охватывается изобретением. Верхний и нижний пределы этих более мелких диапазонов могут быть независимо включены в более мелкие диапазоны, и также охвачены изобретением, подчинены любому конкретному исключенному пределу в указанном диапазоне. Если заданный диапазон включает один или оба предела, диапазоны, исключающие либо один, либо оба этих включенных предела, также включены в изобретение.
Используемый в данном документе термин «HLA-A2» и «A2» относится к белкам человеческого лейкоцитарного антигена (HLA), включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*02 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные термином «HLA-A2» и «A2», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*02 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*02 включают «HLA-A2», HLA-A02» и «HLA-A*2». Были разработаны различные системы присваивания названий, которые идентифицируют белки HLA, кодируемые этим семейством аллелей, включая систему присваивания названий HLA, разработанную в 2010 году Комитетом ВОЗ по факторам системы HLA. Термин «HLA-A2» и «A2» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, которые начинаются с «HLA-A*02», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*02:01», «HLA-A*02:02», «HLA-A*02:03», «HLA-A*02:04», «HLA-A*02:05», «HLA-A*02:06», «HLA-A*02:07», «HLA-A*02:08», «HLA-A*02:09», «HLA-A*02:10» и «HLA-A*02:11». В дополнение к числам, которые следуют за «HLA-A*02:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*02:01P или HLA-A*02:01:01G). Обозначения аллелей, начинающиеся с «HLA-A*02:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термины «HLA-A2» и «A2» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*02» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*02:01», «HLA-A*02:02», «HLA-A*02:03», «HLA-A*02:04», «HLA-A*02:05», «HLA-A*02:06», «HLA-A*02:07», «HLA-A*02:08», «HLA-A*02:09», «HLA-A*02:10», и «HLA-A*02:11».
Используемый в данном документе термин «подтип HLA-A» относится к белку, кодируемому аллелем гена HLA-A.
Используемый в данном документе термин «HLA-A*03» относится к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*03 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные термином «HLA-A*03», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*03 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*03 включают «HLA-A03» и «HLA-A3». Термин «HLA-A*03» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*03», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*03:01», «HLA-A*03:02», «HLA-A*03:04», «HLA-A*03:05», «HLA-A*03:06», «HLA-A*03:07», «HLA-A*03:08», «HLA-A*03:09», «HLA-A*03:10» и «HLA-A*03:12». В дополнение к числам, которые следуют за «HLA-A*03:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*03:01P или HLA-A*03:01:01G). Обозначения аллелей, начинающиеся с «HLA-A*03:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*03» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*03:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*03:01», «HLA-A*03:02», «HLA-A*03:04», «HLA-A*03:05», «HLA-A*03:06», «HLA-A*03:07», «HLA-A*03:08», «HLA-A*03:09», «HLA-A*03:10» и «HLA-A*03:12».
Термины «HLA-A*25», «HLA-A25» и «A25», используемые в данном документе, относятся к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*25 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные терминами «HLA-A*25», «HLA-A25» и «A25», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*25 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*25 включают «HLA-A25». Термин «HLA-A*25», «HLA-A25» и «A25» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*25:», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*25:01», «HLA-A*25:02», «HLA-A*25:03», «HLA-A*25:04», «HLA-A*25:05», «HLA-A*25:06», «HLA-A*25:07», «HLA-A*25:08», «HLA-A*25:09», «HLA-A*25:10» и «HLA-A*25:11». В дополнение к числам, которые следуют за «HLA-A*25:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*25:01P или HLA-A*25:01:01G). Обозначения аллелей, начинающиеся с «HLA-A*25:», за которыми следуют 2, 3 или 4 дополнительные цифры, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*25», «HLA-A25» и «A25» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*25:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*25:01», «HLA-A*25:02», «HLA-A*25:03», «HLA-A*25:04», «HLA-A*25:05», «HLA-A*25:06», «HLA-A*25:07», «HLA-A*25:08», «HLA-A*25:09», «HLA-A*25:10» и «HLA-A*25:11».
Термины «HLA-A*29», «HLA-A29» и «A29», используемые в данном документе, относятся к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*29 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные терминами «HLA-A*29», «HLA-A29» и «A29», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*29 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*29 включают «HLA-A29». Термин «HLA-A*29», «HLA-A29» и «A29» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*29:», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*29:01», «HLA-A*29:02», «HLA-A*29:03», «HLA-A*29:04», «HLA-A*29:05», «HLA-A*29:06», «HLA-A*29:07», HLA-A*29:09», «HLA-A*29:10» и «HLA-A*29:11». В дополнение к числам, которые следуют за «HLA-A*29:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*29:01P или HLA-A*29:02:01G). Обозначения аллелей, начинающиеся с «HLA-A*29:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*29», «HLA-A29» и «A29» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*25:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*29:01», «HLA-A*29:02», «HLA-A*29:03», «HLA-A*29:04», «HLA-A*29:05», «HLA-A*29:06», «HLA-A*29:07», «HLA-A*29:09», «HLA-A*29:10» и «HLA-A*29:11».
Термины «HLA-A*30», «HLA-A30» и «A30», используемые в данном документе, относятся к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*30 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные терминами «HLA-A*30», «HLA-A30» и «A30», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*30 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*30 включают «HLA-A30». Термин «HLA-A*30», «HLA-A30» и «A30» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*30:», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*30:01», «HLA-A*30:02», «HLA-A*30:03», «HLA-A*30:04», «HLA-A*30:06», «HLA-A*30:07», «HLA-A*30:08», HLA-A*30:09», «HLA-A*30:10» и «HLA-A*30:11». В дополнение к числам, которые следуют за «HLA-A*30:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*30:01P, HLA-A*30:02P, HLA-A*30:04P, HLA-A*30:01:01G, HLA-A*30:02:01G или HLA-A*30:04:01G). Обозначения аллелей, начинающиеся с «HLA-A*30:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*30», «HLA-A30» и «A30» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*25:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*30:01», «HLA-A*30:02», «HLA-A*30:03», «HLA-A*30:04», «HLA-A*30:05», «HLA-A*30:06», «HLA-A*30:07», «HLA-A*30:09», «HLA-A*30:10» и «HLA-A*30:11».
Термины «HLA-A*31», «HLA-A31» и «A31», используемые в данном документе, относятся к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*31 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные терминами «HLA-A*31», «HLA-A31» и «A31», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*31 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*31 включают «HLA-A31». Термин «HLA-A*31», «HLA-A31» и «A31» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*31:», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*31:01», «HLA-A*31:02», «HLA-A*31:03», «HLA-A*31:04», «HLA-A*31:05», «HLA-A*31:06», «HLA-A*31:07», «HLA-A*31:08», «HLA-A*31:09», «HLA-A*31:10» и «HLA-A*31:11». В дополнение к числам, которые следуют за «HLA-A*31:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*31:01P или HLA-A*31:01:01G). Обозначения аллелей, начинающиеся с «HLA-A*31:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*31», «HLA-A31» и «A31» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*31:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*31:01», «HLA-A*31:02», «HLA-A*31:03», «HLA-A*31:04», «HLA-A*31:05», «HLA-A*31:06», «HLA-A*31:07», «HLA-A*31:08», «HLA-A*31:09», «HLA-A*31:10» и «HLA-A*31:11».
Термины «HLA-A*33», «HLA-A33» и «A33», используемые в данном документе, относятся к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*33 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные терминами «HLA-A*33», «HLA-A33» и «A33», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*33 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*33 включают «HLA-A33». Термины «HLA-A*33», «HLA-A30» и «A30» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*33:», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*33:01», «HLA-A*33:03», «HLA-A*33:04», «HLA-A*33:05», «HLA-A*33:06», «HLA-A*33:07», «HLA-A*33:08», HLA-A*33:09», «HLA-A*33:10» и «HLA-A*33:11». В дополнение к числам, которые следуют за «HLA-A*33:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*33:01P или HLA-A*33:01:01G). Обозначения аллелей, начинающиеся с «HLA-A*33:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*33», «HLA-A33» и «A33» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*25:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*33:01», «HLA-A*33:02», «HLA-A*33:03», «HLA-A*33:04», «HLA-A*33:05», «HLA-A*33:06», «HLA-A*33:07», «HLA-A*33:09», «HLA-A*33:10» и «HLA-A*33:11».
Термины «HLA-A*36», «HLA-A36» и «A36», используемые в данном документе, относятся к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*36 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные терминами «HLA-A*36», «HLA-A36» и «A36», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*36 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*36 включают «HLA-A36». Термин «HLA-A*36», «HLA-A36» и «A36» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*36:», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*36:01», «HLA-A*36:02», «HLA-A*36:03», «HLA-A*36:04» и «HLA-A*36:05». В дополнение к числам, которые следуют за «HLA-A*36:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G». Обозначения аллелей, начинающиеся с «HLA-A*36:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*36», «HLA-A36» и «A36» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*25:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*36:01», «HLA-A*36:02», «HLA-A*36:03», «HLA-A*36:04», «HLA-A*36:05» и «HLA-A*36:06».
Термины «HLA-A*68», «HLA-A68» и «A68», используемые в данном документе, относятся к белкам HLA, включая белки клеточной поверхности, кодируемые семейством аллелей HLA-A*68 в локусе HLA-A генного комплекса HLA. Белки HLA, охваченные терминами «HLA-A*68», «HLA-A68» и «A68», включают белки HLA, идентифицированные как принадлежащие к типу антигена HLA-A*68 с помощью серологического тестирования или генотипирования. Дополнительные названия для типа антигена HLA-A*68 включают «HLA-A68». Термин «HLA-A*68», «HLA-A68» и «A68» относится к белкам HLA, кодируемым аллелями, имеющими обозначения в соответствии с этой системой присваивания названий, разработанной в 2010 году комитетом ВОЗ по Факторам системы HLA, которые начинаются с «HLA-A*68:», включая, без ограничения указанным, обозначения, начинающиеся с «HLA-A*68:01», «HLA-A*68:02», «HLA-A*68:03», «HLA-A*68:04», «HLA-A*68:05», «HLA-A*68:06», «HLA-A*68:07», «HLA-A*68:08», «HLA-A*68:09» и «HLA-A*68:10». В дополнение к числам, которые следуют за «HLA-A*68:», обозначения аллелей могут также содержать заглавные буквы, включая, без ограничения указанным, заглавные буквы «P» и «G» (например, HLA-A*68:01P, HLA-A*68:01:01G или HLA-A*68:01:02G). Обозначения аллелей, начинающиеся с «HLA-A*68:», за которыми следуют 2, 3 или 4 дополнительных числа, могут составлять полное обозначение или начальную часть обозначения. Обозначения аллелей могут быть выделены курсивом. Термин «HLA-A*68», «HLA-A68» и «A68» также относится к белкам HLA, идентифицированным с обозначениями, которые начинаются с «HLA-A*25:» в соответствии с этой системой присваивания названий, включая, без ограничения указанным, обозначения «HLA-A*68:01», «HLA-A*68:02», «HLA-A*68:03», «HLA-A*68:04», «HLA-A*68:05», «HLA-A*68:06», «HLA-A*68:07», «HLA-A*68:08», «HLA-A*68:09» и «HLA-A*68:10».
Конкретные белки HLA могут упоминаться в настоящем документе с использованием обозначений белков в соответствии с системой именования HLA, разработанной в 2010 году Комитетом ВОЗ по факторам системы HLA. Например, термин «HLA-A*02: 01», используемый в данном документе, относится к белку HLA с обозначением «HLA-A*02: 01» в соответствии с этой системой именования. Аналогично, термины «HLA-A*03:01», «HLA-A*25:01», «HLA-A*29:02», «HLA-A*30:01», «HLA-A*31:01», «HLA-A*33:01», «HLA-A*36:01» и «HLA-A*68:01» относятся к белкам HLA с обозначением «HLA-A*03:01», «HLA-A*25:01», «HLA-A*29:02», «HLA-A*30:01», «HLA-A*31:01», «HLA-A*33:01», «HLA-A*36:01» и «HLA-A*68:01», соответственно.
Используемый в данном документе термин «анти-HLA-A2 антитело» относится к антителу, которое предпочтительно или специфически связывается с HLA-A2.
Используемый в данном документе термин «BB7.2» относится к мышиной гибридоме, идентифицированной как депозит ATCC HB-82. Клетки гибридомы BB7.2 секретируют мышиное моноклональное антитело изотипа IgG2b каппа, которое было охарактеризовано Parham P. et al. и Hilton et al. (Parham, P. et al., 1981; Hilton et al., 2013). Аминокислотные последовательности шести определяющих комплементарность областей (CDR) моноклонального антитела, секретируемого BB7.2, являются следующими:
CDR1 тяжелой цепи (HCDR1): SYHIQ (SEQ ID NO: 183);
CDR2 тяжелой цепи (HCDR2): WIYPGDGSTQYNEKFKG (SEQ ID NO: 185);
CDR3 тяжелой цепи (HCDR3): EGTYYAMDY (SEQ ID NO: 187);
CDR1 легкой цепи (LCDR1): RSSQSIVHSNGNTYLE (SEQ ID NO: 188)
CDR2 легкой цепи (LCDR2): KVSNRFS (SEQ ID NO: 189);
CDR3 легкой цепи (LCDR3): FQGSHVPRT (SEQ ID NO: 190).
Используемый в данном документе термин «антитело BB7.2» представляет собой антитело, имеющее VH (SEQ ID NO: 191) и VL (SEQ ID NO: 192) моноклонального антитела, секретируемого BB7.2. Антитело BB7.2 может быть полноразмерным антителом или его фрагментом, имеющим VH и VL моноклонального антитела, секретируемого BB7.2, таким как scFv, имеющее моноклонального антитела VH и VL, секретируемого BB7.2.
Термины «антитела» и «иммуноглобулин» включают антитела или иммуноглобулины любого изотипа, фрагменты антител, которые сохраняют специфическое связывание с антигеном, включая, без ограничения указанным, Fab, Fab', F(ab')2, Fv, scFv, Fd, диатела, однодоменные антитела (sdAb), линейные антитела, молекулы одноцепочечных антител и мультиспецифичные антитела, образованные из фрагментов антител, химерные антитела, гуманизированные антитела, одноцепочечные антитела и гибридные белки, содержащие антигенсвязывающую часть антитела, и белок, не являющийся антителом.
Антитела могут быть поликлональными или моноклональными, многоцепочечными или одноцепочечными или интактными иммуноглобулинами и могут быть получены из природных источников или из рекомбинантных источников. Антитела могут быть тетрамерами молекул иммуноглобулина. Основное 4-цепочечное антитело представляет собой гетеротетрамерный гликопротеин, состоящий из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. В случае IgG 4-цепочечная единица обычно имеет массу около 150000 дальтон. Каждая L-цепь связана с H-цепью одной ковалентной дисульфидной связью, тогда как две H-цепи связаны друг с другом одной или несколькими дисульфидными связями в зависимости от изотипа H-цепи. Каждая H- и L-цепь также имеет регулярно расположенные внутрицепочечные дисульфидные мостики. Каждая Н-цепь имеет на N-конце вариабельный домен (VH), за которым следуют три константных домена (СН) для каждой из цепей α и γ и четыре домена СН для изотипов μ и ε. Каждая L-цепь имеет на N-конце вариабельный домен (VL), за которым следует константный домен (CL) на другом конце. VL выровнен с VH, а CL выровнен с первым константным доменом тяжелой цепи (CH1). Считается, что конкретные аминокислотные остатки образуют поверхность раздела между вариабельными доменами легкой цепи и тяжелой цепи. Спаривание VH и VL вместе образует один антигенсвязывающий сайт. О структуре и свойствах различных классов антител см., например, Basic and Clinical Immunology, 8th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, CT, 1994, стр. 71 и главу 6. L-цепь из любого вида позвоночных может быть отнесена к одному из двух четко различимых типов, называемых каппа и лямбда, на основе аминокислотных последовательностей их константных доменов. В зависимости от аминокислотной последовательности константного домена их тяжелых цепей (СН) иммуноглобулины могут быть отнесены к разным классам или изотипам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, имеющих тяжелые цепи, обозначаемые как α, δ, ε, γ и μ, соответственно. Классы γ и α далее подразделяются на подклассы на основании относительно незначительных различий в последовательности и функции СН, например, люди экспрессируют следующие подклассы: IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2.
«Вариабельный участок» или «вариабельный домен» антитела относится к N-концевым доменам тяжелой или легкой цепи антитела. Вариабельный домен тяжелой цепи может обозначаться как «VH», «VH» или «H». Вариабельный домен легкой цепи может называться «VL», «VL» или «L». Эти домены обычно являются наиболее вариабельными частями антитела и включают антигенсвязывающие сайты.
Термин «вариабельный» относится к тому факту, что определенные сегменты V-доменов сильно различаются по последовательности среди антител. V домен опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельность неравномерно распределена по диапазону от 110 до 130 аминокислот вариабельных доменов. Между тем V-области состоят из относительно инвариантных участков, называемых каркасными областями (FR) из 15-30 аминокислот, разделенных более короткими областями черезмерной вариабельности, называемыми «гипервариабельными областями», каждая из которых имеет длину 9-12 аминокислот. Каждый из вариабельных доменов нативных тяжелых и легких цепей включает четыре FR, в значительной степени принимающих конфигурацию β-листа, соединенных тремя гипервариабельными областями, которые образуют петли, соединяющие и в некоторых случаях являющиеся частью структуры β-листа. Гипервариабельные области в каждой цепи удерживаются вместе в непосредственной близости FR и, вместе с гипервариабельными областями из другой цепи, способствуют образованию антигенсвязывающего сайта антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)).
«Интактным» антителом является антитело, которое включает антигенсвязывающий сайт, а также CL и, по меньшей мере, константные домены тяжелой цепи, CH1, CH2 и CH3. Константные домены могут представлять собой константные домены с нативной последовательностью (например, константные домены с нативной последовательностью человека) или их варианты с аминокислотной последовательностью. В одном воплощении интактное антитело может иметь одну или несколько эффекторных функций.
Термин «фрагмент антитела» относится, по меньшей мере, к одной части интактного антитела или его рекомбинантным вариантам и относится к антигенсвязывающему домену, например, к области, определяющей антигенную вариабельность интактного антитела, которая достаточна для придания узнаваемости и специфичности связывания фрагмента антитела с мишенью, такой как антиген. Примеры фрагментов антител включают Fab, Fab', F(ab')2, Fv-фрагменты, scFv-фрагменты; димеры; однодоменные антитела (sdAb); линейные антитела (см. пат. США № 5641870, Пример 2; Zapata et al., Protein Eng. 8 (10): 1057-1062 [1995]); молекулы одноцепочечных антител; и мультиспецифичные антитела, образованные из фрагментов антител. В одном воплощении фрагмент антитела содержит антигенсвязывающий сайт интактного антитела и, таким образом, сохраняют способность связывать антиген. Среди фрагментов анти-HLA-A2 антител также включены части анти-HLA-A2 антител (и комбинации частей анти-HLA-A2 антител, например, scFv), которые можно использовать в качестве нацеливающих плечей, направленных на антиген HLA-A2, в химерных антигенных рецепторах CAR-модифицированных иммунных клеток. Такие фрагменты не обязательно являются протеолитическими фрагментами, а скорее являются частями полипептидных последовательностей, которые могут придавать аффинность к мишени. Кроме того, среди фрагментов анти-HLA-A2 антител включены однодоменные антитела (sdAb) (см., Например, Li et al. (2017); Jamnani et al. (2014)). Такие однодоменные антитела могут быть использованы в качестве нацеливающих плечей в CAR-модифицированных иммунных клетках по настоящему изобретению.
При расщеплении антител папаином образуются два идентичных антигенсвязывающих фрагмента, называемых фрагментами «Fab», и остаточный фрагмент «Fc», обозначение, отражающее способность легко кристаллизоваться. Фрагмент Fab состоит из всей L-цепи вместе с доменом вариабельной области H-цепи (VH) и первым константным доменом одной тяжелой цепи (CH1). Каждый фрагмент Fab является одновалентным в отношении связывания антигена, то есть он имеет один антигенсвязывающий сайт. Обработка антителом пепсином дает один большой фрагмент F(ab')2, который грубо соответствует двум дисульфидно-связанным фрагментам Fab, обладающим двухвалентной антигенсвязывающей активностью, с сохранением способности к перекрестному связыванию антигена. Фрагменты Fab' отличаются от фрагментов Fab наличием нескольких дополнительных остатков на карбоксиконце домена СН1, включая один или несколько цистеинов из шарнирной области антитела. Fab'-SH обозначен в данном документе как Fab', в котором остаток(ки) цистеина константных доменов несут свободную тиоловую группу. Антительные фрагменты F(ab')2 исходно были получены в виде пар фрагментов Fab', которые содержат шарнирные цистеины между ними. Другие химические сочетания фрагментов антител также известны.
Фрагмент Fc антитела включает карбоксиконцевые части обеих Н-цепей, удерживаемые вместе дисульфидами. Эффекторные функции антител определялись последовательностями в области Fc, причем эта область также является частью, распознаваемой рецепторами Fc (FcR), обнаруженными в определенных типах клеток.
«Fv» представляет собой минимальный фрагмент антитела, который включает полный сайт узнавания антигена и связывания антигена. Этот фрагмент состоит из димера одного домена вариабельной области тяжелой цепи и одного домена вариабельной области легкой цепи в тесной, нековалентной ассоциации. В одноцепочечном типе Fv (scFv) один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи может быть ковалентно связан с гибким пептидным линкером, так что легкая и тяжелая цепи могут ассоциировать в «димерную» структуру, аналогичную структуре двухцепочечного типа Fv. В результате свертывания этих двух доменов образуются шесть гипервариабельных петель (по 3 петли каждой из цепей H и L), которые вносят аминокислотные остатки для связывания антигена и придают антителу специфичность связывания антигена. Однако даже один вариабельный домен (или половина Fv, содержащего только три CDR, специфичных для антигена) обладает способностью распознавать и связывать антиген, хотя и с более низкой аффинностью, чем весь сайт связывания.
«Одноцепочечный Fv», также сокращенно обозначаемый как «sFv» или «scFv», представляет собой фрагменты антител, которые содержат домены антител VH и VL, связанные в одну полипептидную цепь. Полипептид scFv может дополнительно содержать полипептидный линкер между доменами VH и VL, который позволяет scFv образовывать искомую структуру для связывания антигена. Если не указано иное, используемый в данном документе scFv может иметь вариабельные области VL и VH в любом порядке, например, относительно N-концевых и С-концевых концов полипептида, scFv может содержать VL-линкер-VH или может содержать VH-линкер-VL. Для обзора scFv, см. Pluckthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); Antibody Engineering, ed. Borrebaeck, Oxford University Press, New York (1995). В одном воплощении scFv, полученный из анти-HLA-A2 антитела, может быть использован в качестве нацеливающего плеча CAR-модифицированной иммунной клетки, раскрытой в данном документе.
Термин «Аднектин», также известный как монотело, хорошо известен в данной области и относится к белкам, предназначенным для связывания с высокой аффинностью и специфичностью с антигенами. Они принадлежат к классу молекул, которые в совокупности называются «миметиками антител».
Термин «альфатело», который также может упоминаться как проникающие в клетки альфатела, относится к типу миметиков антител, состоящих из небольших белков с молекулярной массой 10 кДа, сконструированных для связывания с различными антигенами. Альфатела способны достигать и связываться с внутриклеточными белковыми мишенями.
Термин «аффитело» хорошо известен в данной области и относится к аффинным белкам на основе белкового домена из 58 аминокислотных остатков, полученного из одного из IgG-связывающих доменов стафилококкового протеина А.
Термин «Антикалин» хорошо известен в данной области и относятся к технологии миметиков антител, в которой специфичность связывания получена из липокалина. Антикалин также может быть отформатирован как белок двойного нацеливания, называемый дуокалином.
Термин «каркас на основе белка с повторами armadillo» относится к типу миметиков антител, соответствующих искусственным пептидсвязывающим каркасам на основе белков с повторами armadillo. Белки с повторами armadillo характеризуются доменом armadillo, состоящим из тандемных повторов armadillo около из 42 аминокислот, который обеспечивает взаимодействие с пептидами или белками.
«Авимеры» хорошо известны в данной области и относятся к технологии миметиков антител. Термин «DARPins» (сконструированные белки с анкириновыми повторами) хорошо известны в данной области и относятся к технологии миметика антитела DRP (сконструированный белок с повторами), разработанной для применения способности связывания полипептидов, не являющихся антителами.
Термин «диатела» относится к небольшим фрагментам антител, полученным путем конструирования фрагментов scFv с короткими линкерами (около 5-10 остатков) между доменами VH и VL, так что достигается межцепочечное, но не внутрицепочечное спаривание доменов V, что приводит к двухвалентному фрагменту, то есть фрагменту, имеющем два антигенсвязывающих сайта. Биспецифичные диатела представляют собой гетеродимеры двух «перекрестных» scFv-фрагментов, в которых домены VH и VL двух антител присутствуют в разных полипептидных цепях. Диатела более полно описаны, например, в ЕР 0404097; WO 93/11161; и Holliger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993).
Термин «эвазин» хорошо известен в данной области и относится к классу хемокин-связывающих белков.
Термин «феномер» хорошо известен в данной области и относится к белкам, которые относятся к классу миметиков антител. Они являются привлекательными связывающими молекулами благодаря их высокой термостабильности и пониженной иммуногенности.
Термин «кноттин» (который также может называться ингибиторным цистиновым узлом) относится к миметику антитела, содержащему структурный мотив белка, содержащий три дисульфидных мостика.
Термин «пептид домена Куница» относится к типу миметиков антител и основан на активных доменах белков, ингибирующих функцию протеаз.
Термин «нанотело» хорошо известен в данной области техники и относится к полученному из антител терапевтическому белку, который включает уникальные структурные и функциональные свойства природных антител, состоящих только из тяжёлых цепей. Эти антитела, состоящие только из тяжёлых цепей, содержат один вариабельный домен (VHH) и два константных домена (CH2 и CH3).
Термин «унитело» хорошо известен в данной области и относится к фрагменту антитела, в котором отсутствует шарнирная область антител IgG4. Удаление области шарнира приводит к молекуле, которая по существу в два раза меньше традиционных антител IgG4 и имеет унивалентную область связывания, в отличие от двухвалентной области связывания антител IgG4.
Термин «версатело» хорошо известен в данной области и относится к другой технологии миметиков антител. Это небольшие белки 3-5 кДа с >15% цистеинов, которые образуют каркас с высокой дисульфидной плотностью, заменяющий гидрофобное ядро, которое имеют типичные белки.
Термин «гибкий полипептидный линкер» или «линкер», используемый в контексте scFv, относится к пептидному линкеру, который состоит из аминокислот, таких как остатки глицина и/или серина, используемых отдельно или в комбинации, для связывания вместе вариабельных областей тяжелой и легкой цепей. В одном воплощении гибкий полипептидный линкер представляет собой линкер Gly/Ser и содержит аминокислотную последовательность (Gly-Gly-Gly-Ser)n, где n представляет собой положительное целое число, равное или большее 1. Например, n=1, n=2, n=3, n=4, n=5, n=6, n=7, n=8, n=9 и n=10. В другом воплощении гибкий полипептидный линкер представляет собой линкер Gly/Ser и содержит аминокислотную последовательность (Gly-Gly-Gly-Gly-Ser)n, где n представляет собой положительное целое число, равное или большее 1. Например, n=1, n=2, n=3, n=4, n=5, n=6, n=7, n=8, n=9 и n=10. В одном воплощении гибкие полипептидные линкеры включают, без ограничения указанным, (Gly4 Ser)4 или (Gly4 Ser)3. В другом воплощении линкеры включают несколько повторов (Gly2Ser), (GlySer) или (Gly3Ser). Также в объем изобретения включены линкеры, описанные в WO2012/138475, включенной в настоящий документ ссылкой).
Термин «тяжелая цепь» относится к большему из двух типов полипептидных цепей, присутствующих в молекулах антитела в их естественных конформациях, и который обычно определяет класс, к которому относится антитело.
Термин «легкая цепь» относится к меньшему из двух типов полипептидных цепей, присутствующих в молекулах антитела в их естественных конформациях. Легкие цепи каппа (K) и лямбда (λ) относятся к двум основным изотипам легкой цепи антитела.
При использовании в данном документе термин «гипервариабельный участок», «HVR» или «HV» относится к участкам вариабельного домена антитела, которые гипервариабельны по последовательности и/или образуют структурно определенные петли. Как правило, антитела содержат шесть гипервариабельных участков; три в VH (H1, H2, H3), и три в VL (L1, L2, L3). Ряд трансдифференцировок гипервариабельных участков применяются и охвачены настоящим изобретением. Участки, определяющие комплементарность по Kabat (CDR), основываются на вариабельности последовательности и являются наиболее часто используемыми (Kabat et al., 1991). Напротив, Chothia ссылается на локализацию структурных петель (Chothia et al., 1987). Конец петли CDR-H1 по Chothia при нумерации с использованием соглашения о нумерации Kabat варьирует между H32 и H34 в зависимости от длины петли (это связано с тем, что схема нумерации по Kabat размещает вставки в H35A и H35B; если 35A или 35B отсутствуют, петля заканчивается в 32, если присутствует только 35А, петля заканчивается в 33; если присутствуют обе 35А и 35В, петля заканчивается в 34). Гипервариабельные участки AbM представляют собой компромисс между CDR по Kabat и структурными петлями Chothia и используются Оксфордской программой молекулярного моделирования антител Oxford Molecular's AbM. «Контактные» гипервариабельные участки основаны на анализе доступных комплексных кристаллических структур. Остатки из каждого из этих гипервариабельных участков отмечены ниже. Остатки из каждого из этих гипервариабельных участков отмечены ниже.
Петля Kabat AbM Chothia Контакт
------- -------------- -------------- -------------- -------------
L1 L24-L34 L24-L34 L24-L34 L30-L36
L2 L50-L56 L50-L56 L50-L56 L46-L55
L3 L89-L97 L89-L97 L89-L97 L89-L96
H1 H31-H35B H26-H35B H26-H32.34 H30-H35B
(Нумерация по Kabat)
H1 H31-H35 H26-H35 H26-H32 H30-H35
(Нумерация по Chothia)
H2 H50-H65 H50-H58 H52-H56 H47-H58
H3 H95-H102 H95-H102 H95-H102 H93-H101
Термины «гипервариабельный участок» и «область, определяющая комплементарность» и их соответствующие сокращения (HVR, HV, CDR) используются в данном документе взаимозаменяемо. Кроме того, следующие пары терминов также используются в данном документе взаимозаменяемо:
«VH CDR1» и «HCDR1»;
«VH CDR2» и «HCDR2»;
«VH CDR3» и «HCDR3»;
«VL CDR1» и «LCDR1»;
«VL CDR2» и «LCDR2»; и
«VL CDR3» и «LCDR3».
Гипервариабельные участки могут содержать «протяженные гипервариабельные участки», как указанные далее: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 (L3) в VL и 26-35B (HI). 50-65, 47-65 или 49-65 (H2) и 93-102, 94-102 или 95-102 (H3) в VH. Остатки вариабельного домена пронумерованы согласно Kabat et al. (Kabat et al., 1991) для каждого из этих определений.
Остатки «каркаса» или «FR» представляют собой остатки вариабельного домена, отличные от остатков гипервариабельной области, определенных в настоящем документе.
Термин «нумерация остатков вариабельного домена как у Kabat» или «нумерция аминокислотного положения как у Kabat» и его вариации относится к системе нумерации, используемой для вариабельных доменов тяжелой цепи или для вариабельных доменов легкой цепи сборки антител в (Kabat et al., 1991). С использованием данной системы нумерации, фактическая линейная аминокислотная последовательность может содержать меньше или дополнительные аминокислоты, соответствующие укорачиванию или вставке в FR или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может включать единичную аминокислотную вставку (остаток 52a согласно Kabat) после остатка 52 из H2 и вставленные остатки (например, остатки 82a, 82b, и 82c, и т.д. согласно Kabat) после остатка 82 в FR тяжелой цепи. Нумерация остатков по Kabat может быть определена для данного антитела путем выравнивания по областям гомологии последовательности антитела со «стандартной» последовательностью, пронумерованной по Kabat.
Система нумерации Kabat, как правило, используется при ссылке на остаток в вариабельном домене (приблизительно остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., 1991). «Система нумерации EU» или «индекс EU», как правило, используется при ссылке на остаток в константном участке тяжелой цепи иммуноглобулина (например, индекс EU, сообщенный в Kabat et al., выше). «Индекс EU как у Kabat» относится к нумерации остатков человеческого антитела IgG1 EU. Если в данном документе не указано иное, ссылки на номера остатков в вариабельном домене антител означают нумерацию остатков с помощью системы нумерации Kabat.
Термин «специфически связывается» относится к лиганду (например, гуманизированному антителу против HLA-A2), который распознает и связывается с белком когнатного партнера связывания (например, HLA-A2), присутствующим в образце, но этот лиганд существенно не распознает или не связывает молекулы в образце. Неспецифическое связывание будет относиться к связыванию с аффинностью менее 10-7 М, например, связыванию с аффинностью 10-6 М, 10-5 М, 10-4 М и т.д.
Антитело, которое «специфически связывает» интересующий антиген или эпитоп, представляет собой антитело, которое связывает антиген или эпитоп с достаточной аффинностью, которая заметно отличается от неспецифического взаимодействия. Специфическое связывание может быть измерено, например, путем определения связывания молекулы по сравнению со связыванием контрольной молекулы.
Термин «рекомбинантное антитело» относится к антителу, которое получают с использованием технологии рекомбинантной ДНК, такому как, например, антитело, экспрессируемое системой экспрессии на основе бактериофага или дрожжей. Термин также следует истолковывать как означающий антитело, которое было получено в результате синтеза молекулы ДНК, кодирующей антитело, и каковая молекула ДНК экспрессирует белок антитела, или аминокислотную последовательность, определяющую антитело, где последовательность ДНК или аминокислотная последовательность были получены с использованием технологии рекомбинантных ДНК или технологии аминокислотных последовательностей, которые доступны и хорошо известны в данной области.
Термин «моноклональное антитело», используемый в данном документе, относится к антителу, полученному из популяции частично гомогенных антител, т.е. индивидуальных антител составляющих популяцию, которые являются идентичными за исключением возможных естественных мутаций, которые могут присутствовать в минорных количествах. Моноклональные антитела являются высокоспецифичными и направлены против одного антигенного сайта. Кроме того, в отличие от препаратов поликлональных антител, которые включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. В дополнение к их специфичности, моноклональные антитела имеют то преимущество, что они могут быть синтезированы без загрязнения другими антителами. Модификатор «моноклональный» не следует истолковывать как требующий получения антитела каким-либо конкретным способом. Например, моноклональные антитела могут быть получено по методике гибридом, впервые описанной Kohler et al., Nature, 256: 495 (1975), или могут быть получены с использованием способов рекомбинантной ДНК в клетках бактерий, эукариотических животных или растений (см., например, пат. США No. 4816567). «Моноклональное антитело» также может быть выделено из библиотек фаговых антител с использованием методик, описанных, например, в Clackson et al., Nature, 352: 624-628 (1991) и Marks et al., J. Mol. Biol., 222: 581-597 (1991).
Термин «антиген» относится к молекуле, которая вызывает иммунный ответ. Этот иммунный ответ может включать либо выработку антител, либо активацию специфических иммунологически компетентных клеток, или то и другое. Специалист в данной области поймет, что любая макромолекула, включая практически все белки или пептиды, может служить антигеном. Кроме того, антигены могут быть получены из рекомбинантной или геномной ДНК. Специалист в данной области поймет, что любая ДНК, которая включает нуклеотидную последовательность или частичную нуклеотидную последовательность, кодирующую белок, который вызывает иммунный ответ, следовательно, кодирует «антиген», как этот термин используется в данном документе. Кроме того, специалист в данной области поймет, что антиген не должен кодироваться исключительно полноразмерной нуклеотидной последовательностью гена. Очевидно, что настоящее изобретение включает, без ограничения указанным, применение частичных нуклеотидных последовательностей более чем одного гена и что эти нуклеотидные последовательности расположены в различных комбинациях для кодирования полипептидов, которые вызывают желаемый иммунный ответ. Кроме того, специалист в данной области поймет, что антиген вовсе не должен кодироваться «геном». Очевидно, что антиген может быть создан, синтезирован или может быть получен из биологического образца или может быть макромолекулой помимо полипептида. Такой биологический образец может включать, без ограничения указанным, образец ткани, образец опухоли, клетку или жидкость с другими биологическими компонентами.
Используемый в данном документе термин «аффинность» относится к константе равновесия для обратимого связывания двух агентов и выражается в виде константы диссоциации (Kd). Аффинность может быть, по меньшей мере, в 1 раз больше, по меньшей мере, в 2 раза больше, по меньшей мере, в 3 раза больше, по меньшей мере, в 4 раза больше, по меньшей мере, в 5 раз больше, по меньшей мере, в 6 раз больше, по меньшей мере, в 7 раз больше, по меньшей мере, в 8 раз больше, по меньшей мере, в 9 раз больше, по меньшей мере, в 10 раз больше, по меньшей мере, в 20 раз больше, по меньшей мере, в 30 раз больше, по меньшей мере, в 40 раз больше, по меньшей мере, в 50 раз больше, по меньшей мере, в 60 раз больше, по меньшей мере, в 70 раз больше, по меньшей мере, в 80 раз больше, по меньшей мере, в 90 раз больше, по меньшей мере, в 100 раз или, по меньшей мере, в 1000 раз больше, или более, чем аффинность антитела для неродственных аминокислотных последовательностей. Аффинность антитела к белку-мишени может составлять, например, от около 100 нМ до около 0,1 нМ, от около 100 нМ до около 1 пикомоля (пМ) или от около 100 нМ до около 1 фемтомоля (фМ) или больше. Используемый в данном документе термин «авидность» относится к устойчивости комплекса из двух или более агентов к диссоциации после разбавления. Термины «иммунореактивный», «предпочтительно связывает» и «специфически связывает» используются в данном документе взаимозаменяемо в отношении антител и их фрагментов. Анти-HLA-A2-антитела по изобретению, включая гуманизированные анти-HLA-A2-антитела, а также их фрагменты в том смысле, в каком в данном документе используется этот термин, специфически связываются с HLA-A2.
Термин «связывание» относится к прямой ассоциации между двумя молекулами, например, вследствие ковалентных, электростатических, гидрофобных и ионных и/или водородных связей, включая такие взаимодействия, как солевые и водяные мостики. Неспецифическое связывание будет относиться к связыванию с аффинностью менее 10-7 М, например, связыванию с аффинностью 10-6 М, 10-5 М, 10-4 М и т.д.
Используемый в данном документе термин «реактивность» относится к способности антитела реагировать (то есть связываться) с молекулой (например, специфически связываться с молекулой). Первое антитело обладает «меньшей реактивностью» к молекуле (например, молекуле HLA), чем второе антитело, когда первое антитело проявляет пониженное связывание с молекулой по сравнению со вторым антителом. Известны подходы для быстрого сравнения реактивности первого и второго антител с одной или несколькими конкретными молекулами HLA. Один примерный подход представлен в разделе «Примеры» в данном документе, где гранулы FlowPRA® Single Antigen (One Lambda) были использованы для исследования антител на способность реагировать (или связываться) с конкретными молекулами HLA. Такие подходы на основе проточной цитометрии позволяют осуществить высокопроизводительный анализ реактивности антител.
Используемый в данном документе термин «шарнирная область» относится к гибкой полипептидной соединительной области (также называемой в данном документе «шарниром» или «спейсером»), обеспечивающей структурную гибкость и расстояние от фланкирующих полипептидных областей, и может состоять из природных или синтетических полипептидов. Шарнирная область может влиять на активность иммунной клетки, экспрессирующей CAR (см., Например, Watanabe et al. (2016)). «Шарнирная область», полученная из иммуноглобулина (например, IgG1), обычно определяется как участок от Glu216 до Pro230 человеческого IgG1 (Burton (1985) Molec. Immunol., 22:161-206). Шарнирные области других изотипов IgG могут быть выровнены с последовательностью IgG1 путем размещения первого и последнего остатков цистеина, образующих дисульфидные (S-S) связи между тяжелыми цепями, в тех же положениях. Шарнирная область может быть естественной или неестественной, включая, без ограничения указанным, измененную шарнирную область, описанную в Пат. США No. 5677425. Шарнирная область может включать полную шарнирную область, полученную из антитела другого класса или подкласса, чем у домена CH1. Термин «шарнирная область» может также включать области, происходящие из CD8 и других рецепторов, которые обеспечивают аналогичную функцию в обеспечении гибкости и расстояния до фланкирующих областей.
Используемый в данном документе термин «иммунные клетки» обычно включает белые кровяные клетки (лейкоциты), которые получены из гемопоэтических стволовых клеток (HSC), продуцируемых в костном мозге. «Иммунные клетки» включают, например, лимфоциты (Т-клетки, В-клетки, клетки-натуральные киллеры (NK)) и миелоидные клетки (нейтрофилы, эозинофилы, базофилы, моноциты, макрофаги, дендритные клетки).
«Т-клетки» включают все типы иммунных клеток, экспрессирующих CD3, включая T-хелперные клетки (CD4+ клетки), CD8+ T-клетки (например, цитотоксические CD8+ T-клетки, регуляторные CD8+ T-клетки), регуляторные T-клетки (Treg), гамма-дельта Т-клетки и двойные отрицательные Т-клетки.
«Цитотоксическая клетка» включает цитотоксические CD8+ T-клетки, натуральные киллеры (NK) и нейтрофилы, которые способны опосредовать цитотоксические реакции.
Используемый в данном документе термин «регуляторная иммунная клетка» относится к иммунной клетке, которая действует «регуляторным» способом, подавляя активацию иммунной системы и, таким образом, поддерживает гомеостаз иммунной системы и толерантность к аутоантигенам. «Регуляторные иммунные клетки» также могут оказывать влияние на неиммунные клетки, что приводит к улучшению клинического состояния, например, способствует восстановлению или регенерации тканей. Регуляторные иммунные клетки могут включать, регуляторные T-клетки, CD4+ регуляторные T-клетки, CD8+ регуляторные T-клетки, регуляторные γδ T-клетки, регуляторные DN T-клетки, регуляторные B-клетки, регуляторные NK-клетки, регуляторные макрофаги и регуляторные дендритные клетки.
«Регуляторный T-лимфоцит», «регуляторная T-клетка», «регуляторная T-клетка», «Treg- клетка» или «Treg», используемые в настоящем описании, являются синонимами и имеют стандартное определение, используемое в данной области. Treg-клетки представляют собой специализированную субпопуляцию Т-клеток, которые действуют «регуляторным» способом, подавляя активацию иммунной системы и тем самым поддерживая гомеостаз иммунной системы и толерантность к аутоантигенам. Treg иногда называют супрессорными Т-клетками. Treg-клетки часто, но не всегда, характеризуются экспрессией фактора транскрипции семейства forkhead, Foxp3 (forkhead бокс p3). Они также могут экспрессировать поверхностные белки CD4 или CD8. Они обычно также экспрессируют CD25. При использовании в настоящем описании и формуле изобретения, и если не указано иное, Treg включают «естественные» Treg, которые развиваются в тимусе, индуцированные/адаптивные/периферические Treg, которые возникают в результате процесса дифференцировки, который происходит вне тимуса (например, в тканях или вторичном лимфоидном органе, или в лабораторных условиях при определенных условиях культивирования), и Treg, которые были созданы с использованием технологии рекомбинантных ДНК, например, сконструированные для экспрессии FOXP3. Встречающиеся в природе Treg-клетки (CD4+ CD25+ Foxp3+) возникают, как и все другие Т-клетки в тимусе. Напротив, индуцированные/адаптивные/периферические Treg-клетки (которые включают CD4+ CD25+ Foxp3+ Treg, Tr1-клетки, Th3-клетки и другие) возникают вне тимуса. Одним из способов индукции Treg является воздействие IL-10 или TGF-β на эффекторные T-клетки. Т-клетки также могут быть превращены в Treg-клетки путем трансфекции или трансдукции гена Foxp3 в смешанную популяцию Т-клеток. Т-клетка, в которой вызвана экспрессия Foxp3, принимает фенотип Treg, и такие рекомбинантные Treg также определены в данном документе как «Treg».
Используемый в данном документе термин «иммунная эффекторная клетка» относится к клетке иммунной системы, которая находится в форме, способной вызывать специфический иммунный ответ.
Используемый в данном документе термин «иммунный ответ» включает иммунные ответы, опосредованные Т-клетками и/или В-клетками. Типичные иммунные ответы включают Т-клеточные ответы, например, выработку цитокинов и клеточную цитотоксичность. Кроме того, термин «иммунный ответ» включает иммунные ответы, на которые косвенно влияют активация Т-клеток, например, выработка антител (гуморальные ответы) и активация чувствительных к цитокинам клеток, например, макрофагов. Иммунные клетки, участвующие в иммунном ответе, включают лимфоциты, такие как В-клетки и Т-клетки (CD4+, CD8+, клетки Th1 и Th2); антигенпрезентирующие клетки (например, профессиональные антигенпрезентирующие клетки, такие как дендритные клетки, макрофаги, В-лимфоциты, клетки Лангерганса, и непрофессиональные антигенпрезентирующие клетки, такие как кератиноциты, эндотелиальные клетки, астроциты, фибробласты, олигодендроциты); естественные клетки-киллеры; миелоидные клетки, такие как макрофаги, эозинофилы, тучные клетки, базофилы и гранулоциты.
Термин «отторжение» относится к состоянию, при котором пересаженный орган или ткань не воспринимается организмом реципиента. Отторжение происходит в результате воздействия иммунной системы реципиента на пересаженный орган или ткань. Отторжение может происходить от нескольких дней до нескольких недель после трансплантации (острое) или от нескольких месяцев до нескольких лет после трансплантации (хроническое).
Термин «заболевание «трансплантат против хозяина» или «GVHD» в контексте настоящего описания относится к медицинскому осложнению после получения трансплантированной ткани от генетически отличного человека. Иммунные клетки в донорской ткани (трансплантат) распознают реципиента (хозяина) как чужеродные. Трансплантированные иммунные клетки затем атакуют клетки организма хозяина. GVHD обычно ассоциируется с трансплантацией стволовых клеток; однако этот термин включает GVHD, возникающие из-за других форм тканевого трансплантата. GVHD может также возникнуть после переливания крови.
Используемый в данном документе термин «иммунологическая толерантность» или «иммунная толерантность» относится к методам, выполняемым на части подвергнутых лечению объектов по сравнению с необработанными объектами, где: а) сниженный уровень специфического иммунологического ответа (который считается опосредованным, по меньшей мере, частично антиген-специфическими эффекторными Т-лимфоцитами, В-лимфоцитами, антителами или их эквивалентами); b) задержка начала или прогрессирования специфического иммунологического ответа; или c) сниженный риск возникновения или прогрессирования специфического иммунологического ответа. «Специфическая» иммунологическая или иммунная толерантность возникает, когда иммунологическая или иммунная толерантность вызывается преимущественно против определенных антигенов по сравнению с другими.
Используемый в данном документе термин «функциональная толерантность» относится к клинической ситуации, когда имеется стабильная функция трансплантата, при которой отсутствуют гистологические признаки отторжения, включая острое или хроническое отторжение, при отсутствии какой-либо иммуносупрессивной лекарственной терапии в течение, по меньшей мере, 1 года, в иммунокомпетентном хозяине способном реагировать на другие вызовы, включая инфекции.
Используемый в данном документе термин «иммунная аккомодация» относится к состоянию реципиента трансплантата, при котором трансплантат органа или ткани функционирует нормально, несмотря на наличие у реципиента антител, специфичных для трансплантата органа или ткани.
Используемый в данном документе термин «стволовая клетка» обычно включает плюрипотентные или мультипотентные стволовые клетки. «Стволовые клетки» включают, без ограничения указанным, эмбриональные стволовые клетки (ES); мезенхимальные стволовые клетки (MSC); индуцированные плюрипотентные стволовые клетки (iPS); и коммитированные клетки-предшественники (гематопоэтические стволовые клетки (HSC); клетки, полученные из костного мозга, и т.д.).
Термин «лечение», «лечить» и т.п. относятся к получению требуемого фармакологического и/или физиологического эффекта. Эффект может быть терапевтическим в плане частичного или полного лечения заболевания и/или неблагоприятного эффекта, присущего заболеванию. Термин «лечение», при использовании в данном документе, охватывает любое лечение заболевания у млекопитающего, например, у человека, и включает облегчение заболевания, то есть вызывание регрессии заболевания и/или ослабление одного или нескольких симптомов заболевания.
Используемые в данном документе термины «предотвращение», «предотвращающий» и «предотвращает» означают предоставление профилактического или защитного лечения заболевания или болезненного состояния. Эффект может быть профилактическим в плане полного или частичного предотвращения заболевания или его симптома. «Предотвращение» в контексте настоящего описания охватывает любое профилактическое воздействие на заболевание у млекопитающего, например, у человека, и включает: (а) предотвращение возникновения заболевания у объекта, который может быть предрасположен к заболеванию, но еще не был диагностирован как имеющий его; и (b) подавление заболевания, то есть прекращение его развития.
Термины «пациент», «объект», «индивидуум», «хозяин» и тому подобное используются в данном документе взаимозаменяемо и относятся к любому животному или его клеткам, будь то in vitro или in situ, поддающимся описанным в данном документе способам. Термины «пациент», «объект», «индивидуум», «хозяин» и тому подобное подразумевают включение живых организмов, у которых может быть вызван иммунный ответ (например, млекопитающие). Примеры «пациента», «объекта», «индивидуума», «хозяина» включают мышей (например, крыс, мышей), зайцеобразных (например, кроликов), приматов, не являющихся людьми, людей, собак, кошек, копытных (например, лошадей, крупного рогатого скота, овец, свиней, коз и т.д. и их трансгенных разновидностей. В некоторых неограничивающих воплощениях пациентом, объектом, хозяином или индивидуумом является человек.
Термин «эффективное количество» или «терапевтически эффективное количество» используется в данном документе взаимозаменяемо и относится к количеству терапевтического агента или объединенным количествам более чем одного терапевтического агента, которые будут вызывать биологический или медицинский ответ ткани, системы или объекта, который изыскивается исследователем, ветеринаром, лечащим врачом или другим клиническим врачом. Термин «терапевтически эффективное количество» включает такое количество терапевтического средства, которое при введении является достаточным для предотвращения развития или до некоторой степени ослабления одного или нескольких признаков, или симптомов расстройства или заболевания, подвергаемого лечению. Терапевтически эффективное количество будет варьировать в зависимости от терапевтического агента, заболевания и его тяжести, а также возраста, массы и т.д. объекта, подлежащего лечению.
Используемый в данном документе термин «активация» относится к состоянию Т-клетки (например, регуляторной Т-клетки), которая была достаточно стимулирована для индукции обнаруживаемого клеточного ответа. Активация также может быть связана с обнаруживаемой(ыми) эффекторной(ыми) функцией(ями), такой как продуцирование цитокинов или супрессирующая активность. Термин «активированные» регуляторные T-клетки относится, среди прочего, к регуляторным T-клеткам, которые способны подавлять иммунный ответ.
Термин «химерный антигенный рецептор» или, альтернативно, «CAR» относится к рекомбинантной полипептидной конструкции, содержащей внеклеточный домен, содержащий антигенсвязывающий домен; трансмембранный домен; и цитоплазматический домен, содержащий внутриклеточный сигнальный домен. В одном воплощении CAR необязательно включает шарнир. Термины «химерный рецептор» или «химерный антигенный рецептор» или «CAR» могут, в частности, относится к одному полипептиду или к набору полипептидов, обычно двух в простейших воплощениях, которые, когда они находятся в иммунной клетке, придают клетке специфичность к лиганду-мишени и генерируют внутриклеточный сигнал. В некоторых воплощениях набор полипептидов является смежным друг с другом. В некоторых воплощениях химерный рецептор представляет собой химерный слитый белок, содержащий набор полипептидов. В некоторых воплощениях набор полипептидов включает переключатель димеризации, который при наличии молекулы димеризации может связывать полипептиды друг с другом, например, может связывать лигандсвязывающий домен с внутриклеточным сигнальным доменом. В одном воплощении химерный рецептор включает необязательную лидерную последовательность на амино-конце (N-ter) химерного рецепторного гибридного белка. В одном воплощении химерный рецептор дополнительно включает лидерную последовательность на N-конце внеклеточного лиганд-связывающего домена, где лидерная последовательность необязательно отщепляется от лиганд-связывающего домена во время клеточного процессинга и локализации химерного рецептора на клеточной мембране.
Термин «сигнальный домен» относится к функциональной части белка, которая действует путем передачи информации в клетке, чтобы регулировать клеточную активность через определенные сигнальные пути, генерируя вторичные мессенджеры или функционируя как эффекторы, реагирующие на такие мессенджеры.
Термин «аутологичный» относится к любому материалу, полученному от того же объекта, которому он позднее будет повторно введен.
Термин «аллогенный» относится к любому материалу, полученному от другого объекта того же вида, что и объект, которому этот материал вводится. Говорят, что два или более объекта аллогенны друг другу, когда гены в одном или нескольких локусах не идентичны. В некоторых аспектах аллогенный материал от объектов одного и того же вида может быть достаточно непохож генетически, чтобы взаимодействовать антигенно. Термин «аллотрансплантат» относится к трансплантату, полученному от другого объекта того же вида.
Термин «ксеногенный» относится к любому материалу, полученному от объекта другого вида. Термин «ксенотрансплантат» относится к трансплантату, полученному от объекта другого вида.
Используемый в настоящем документе термин «инструкции» включает публикацию, запись, диаграмму или любой другой носитель выражения, который можно использоваться для сообщения о применимости композиций и способов по изобретению. Инструкции набора по изобретению может, например, быть прикреплен к контейнеру, который включает нуклеиновую кислоту, пептид, клетку и/или композицию по изобретению, или поставляться вместе с контейнером, который включает нуклеиновую кислоту, пептид, клетку и/или композицию. В качестве альтернативы, инструкции могут быть доставлены отдельно от контейнера с целью совместного применения реципиентом инструкций и нуклеиновой кислоты, пептида, клетки и/или композиции.
«Модификация» аминокислотного остатка/положения, при использовании в данном документе, относится к изменению первичной аминокислотной последовательности по сравнению с исходной аминокислотной последовательностью, где изменение является результатом изменения последовательности, включающего указанные аминокислотный остаток/положения. Например, типичные модификации включают замену остатка (или в указанном положении) другой аминокислотой (например, консервативная или неконсервативная замена), вставка одной или нескольких (обычно менее 5 или 3) аминокислот рядом с указанным остатком/положением и удаление указанного остатка/положения. «Замещение аминокислоты» или его разновидность относится к замене существующего аминокислотного остатка в предварительно определенной (исходной) аминокислотной последовательности другим аминокислотным остатком. Обычно и предпочтительно модификация приводит к изменению, по меньшей мере, одной физико-биохимической активности вариантного полипептида по сравнению с полипептидом, содержащим исходную (или «дикого типа») аминокислотную последовательность. Например, в случае антитела физико-биохимическая активность, которая изменяется, может представлять собой аффинность связывания, способность связывания и/или эффект связывания с молекулой-мишенью.
Термин «модификация консервативной последовательности» относится к модификации аминокислоты, которая не оказывает существенного влияния или не изменяет характеристики связывания антитела или фрагмента антитела, содержащего аминокислотную последовательность. Такие консервативные модификации включают аминокислотные замены, добавления и делеции. Модификации могут быть введены в антитело или фрагмент антитела по изобретению стандартными способами, известными в данной области, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Консервативные аминокислотные замены представляют собой такие, в которых аминокислотный остаток заменяют на аминокислотный остаток, имеющий сходную боковую цепь. Семейства аминокислотных остатков, имеющих похожие боковые цепи, известны с данной области. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотные боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или несколько аминокислотных остатков в CAR по изобретению могут быть заменены другими аминокислотными остатками из того же семейства боковых цепей, а измененный CAR может быть протестирован с использованием функциональных анализов, описанных в данном документе.
Термин «стимуляция» относится к первичному ответу, индуцированному связыванием стимулирующей молекулы (например, комплекса TCR/CD3) с его когнатным лигандом, тем самым опосредуя событие передачи сигнала, такое как, без ограничения указанным, передачу сигнала через комплекс TCR/CD3. Стимуляция может опосредовать измененную экспрессию определенных молекул, такую как повышенная регуляция или пониженная регуляция цитокинов и белков клеточной поверхности и/или реорганизация структур цитоскелета и т.п.
Термин «стимулирующая молекула» относится к молекуле, экспрессируемой Т-клеткой, которая обеспечивает первичную цитоплазматическую сигнальную последовательность (последовательности), которая регулирует первичную активацию комплекса TCR стимулирующим образом, по меньшей мере, для некоторого аспекта пути передачи сигнала Т-клетки. В одном аспекте сигнал является первичным сигналом, который инициируется, например, связыванием комплекса TCR/CD3 с молекулой МНС, загруженной пептидом, и который приводит к опосредованию ответа Т-клеток, включая, без ограничения указанным, пролиферацию, активацию, дифференцировку и т.п. Первичная цитоплазматическая сигнальная последовательность (также называемая «первичным сигнальным доменом»), которая действует стимулирующим образом, может содержать сигнальный мотив, известный как иммунорецепторный мотив активации на основе тирозина (ITAM).
Термин «антигенпрезентирующая клетка» или «APC» относится к клетке иммунной системы, такой как добавочная клетка (например, B-клетка, дендритная клетка, макрофаги, клетки Лангерганса и т.п.), которая презентирует чужеродный антиген, образующий комплекс с основными комплексами гистосовместимости (MHC) на его поверхности для распознавания определенными лимфоцитами, такими как Т-клетки. Т-клетки могут распознавать эти комплексы, используя свои T-клеточные рецепторы (TCR). Антигенпрезентирующие клетки могут обрабатывать антигены для презентации в сочетании с МНС. Используемый в данном документе термин «антигенпрезентирующая клетка» или «APC» включает состояния, в которых APC презентируют антиген, и состояния, в которых APC не презентируют антиген. В некоторых случаях АРС процессируют антигены и презентируют их Т-клеткам. В других случаях Т-клетки могут распознавать APC в отсутствие презентации антигена, когда TCR напрямую связывается с белком MHC. Например, в контексте трансплантации APC могут напрямую стимулировать T-клетки посредством экспрессии чужеродных белков MHC.
«Внутриклеточный сигнальный домен», при использовании в данном документе, относится к внутриклеточной части CAR. Внутриклеточный сигнальный домен генерирует сигнал, который способствует иммунной эффекторной функции CAR-содержащей клетки, например, Treg-клетки с CAR. Примеры иммунной эффекторной функции, например, в Treg-клетке c CAR, могут включать супрессию или отрицательную регуляцию эффекторной функции других иммунных клеток. Другие иммунные клетки включают лейкоциты любого типа, например, (без ограничения указанным), Т-клетки, В-клетки, NK-клетки. Кроме того, иммунная эффекторная функция Treg может включать воздействие на неиммунные клетки, которое приводит к улучшению клинического состояния, например, способствует восстановлению или регенерации тканей.
Термин «дзета» или, альтернативно, «дзета-цепь» или «CD3-дзета» определяется как белок, предоставленный как GenBank Acc. No. BAG36664.1, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, человекообразной обезьяны и т.п., и «дзета-стимулирующий домен» или «CD3-дзета-стимулирующий домен» определяется как аминокислотные остатки цитоплазматического домена дзета-цепи, которые достаточны для функциональной передачи исходного сигнала, необходимого для активации Т-клеток. В одном воплощении цитоплазматический домен дзета включает остатки с 52 по 164 из GenBank Acc. No. BAG36664.1 или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, человекообразной обезьяны и т.п., которые являются их функциональными ортологами.
«Костимулирующий лиганд», используемый в данном документе, включает молекулу антигенпрезентирующей клетки (например, АРС, дендритную клетку, В-клетку и тому подобное), которая специфически связывает когнатную костимулирующую молекулу на Т-клетке, обеспечивая тем самым сигнал, который, в дополнение к первичному сигналу, обеспечиваемому, например, связыванием комплекса TCR/CD3 с молекулой МНС, загруженной пептидом, опосредует ответ Т-клеток, включая, без ограничения указанным, пролиферацию, активацию, дифференцировку и т.п. Костимулирующий лиганд может включать, без ограничения указанным, CD7, B7-1 (CD80), B7-2 (CD86), PD-L1, PD-L2, 4-1BBL, OX40L, индуцибельный костимулирующий лиганд (ICOS- L), молекулу межклеточной адгезии (ICAM), CD30L, CD40, CD70, CD83, HLA-G, MICA, MICB, HVEM, бета-рецептор лимфотоксина, 3/TR6, ILT3, ILT4, HVEM, агонист или антитело, которое связывает рецептор Toll-лиганда и лиганд, который специфически связывается с B7-H3. Костимулирующий лиганд также включает, помимо прочего, лиганд, включая антитело, которое специфически связывается с костимулирующей молекулой, присутствующей на Т-клетке, такой как, без ограничения указанным, молекула МНС класса I, BTLA, рецептор Toll-лиганда, OX40, CD27, CD28, антиген-1, связанный с функцией лимфоцитов (LFA-1) (CD11a/CD18), TNFR1 (CD120a/TNFRSF1A), TNFR2 (CD120b/TNFRSF1B), CTLA-4 (CD152), CD95, ICOS (CD278), 4-1BB (CD137), CD2, CD30, CD40, PD-1, CD7, LIGHT, NKG2C, B7-H3, ICAM-1, GITR, HVEM, SLAMF7, NKp80, CD160, IL2ra, IL6Ra, IL-7Ra, IL-13RA1/RA2, IL-33R(IL1RL1), IL-10RA/RB, IL-4R, IL-5R (CSF2RB), ARHR, рецептор BAFF, IL-21R, TGFbR1/2/3, общая гамма-цепь, лиганд, который специфически связывается с CD83, и любую их комбинацию.
Термин «костимулирующая молекула» относится к родственному связывающему партнеру на Т-клетке, который специфически связывается с костимулирующим лигандом, тем самым опосредуя костимулирующий ответ Т-клетки, такой как, без ограничения указанным, пролиферация. Костимулирующие молекулы представляют собой молекулы клеточной поверхности, отличные от антигенных рецепторов или их лигандов, которые способствуют эффективному иммунному ответу. Костимулирующая молекула может быть представлена в следующих семействах белков: белки рецептора TNF, иммуноглобулиноподобные белки, рецепторы цитокинов, интегрины, сигнальные молекулы активации лимфоцитов (белки SLAM) и активирующие рецепторы NK-клеток. Костимулирующие молекулы включают, без ограничения указанным, молекулу МНС класса I, BTLA, рецептор Toll-лиганда, OX40, CD27, CD28, антиген-1, связанный с функцией лимфоцитов (LFA-1) (CD11a/CD18), TNFR1 (CD120a/TNFRSF1A), TNFR2 (CD120b/TNFRSF1B), CTLA-4 (CD152), CD95, ICOS (CD278), 4-1BB (CD137), CD2, CD30, CD40, PD-1, CD7, LIGHT, NKG2C, B7-H3, ICAM-1, GITR, HVEM, SLAMF7, NKp80, CD160, IL2ra, IL6Ra, IL-7Ra, IL-13RA1/RA2, IL-33R(IL1RL1), IL-10RA/RB, IL-4R, IL-5R (CSF2RB), ARHR, рецептор BAFF, IL-21R, TGFbR1/2/3, общую гамма-цепь, лиганд, который специфически связывается с CD83, и любую их комбинацию.
«Костимулирующий внутриклеточный сигнальный домен» или «костимулирующий домен» может быть внутриклеточной частью костимулирующей молекулы. Примеры таких молекул включают молекулу МНС класса I, BTLA, рецептор Toll-лиганда, OX40, CD27, CD28, антиген-1, связанный с функцией лимфоцитов (LFA-1) (CD11a/CD18), TNFR1 (CD120a/TNFRSF1A), TNFR2 (CD120b/TNFRSF1B), CTLA-4 (CD152), CD95, ICOS (CD278), 4-1BB (CD137), CD2, CD30, CD40, PD-1, CD7, LIGHT, NKG2C, B7-H3, ICAM-1, GITR, HVEM, SLAMF7, NKp80, CD160, IL2ra, IL6Ra, IL-7Ra, IL-13RA1/RA2, IL-33R(IL1RL1), IL-10RA/RB, IL-4R, IL-5R (CSF2RB), ARHR, рецептор BAFF, IL-21R, TGFbR1/2/3, общую гамма-цепь, лиганд, который специфически связывается с CD83, и т.п.
Используемый в данный момент термин «костимулирующий сигнал» относится к сигналу, который в сочетании с первичным сигналом, таким как лигирование TCR/CD3, приводит к пролиферации Т-клеток и/или повышающей или понижающей регуляции ключевых молекул.
Внутриклеточный сигнальный домен может включать всю внутриклеточную часть или весь нативный внутриклеточный сигнальный домен молекулы, из которой он получен, или его функциональный фрагмент.
«Заболевание» - это состояние здоровья животного, при котором животное не может поддерживать гомеостаз, и если состояние при заболевании не улучшается, то здоровье животного продолжает ухудшаться. Напротив, «расстройство» у животного - это состояние здоровья, при котором животное способно поддерживать гомеостаз, но состояние здоровья животного менее благоприятное, чем оно было бы в отсутствие расстройства. При отсутствии лечения расстройство не обязательно вызывает дальнейшее снижение состояния здоровья животного.
Термин «кодирование» относится к наследуемому свойству специфических последовательностей нуклеотидов в полинуклеотиде, таких как, например, ген, кДНК или мРНК, которые служат в качестве матриц для синтеза других полимеров и макромолекул в биологических процессах, имеющих либо определенную последовательность нуклеотидов (например, рРНК, тРНК и мРНК), либо определенную последовательность аминокислот и биологические свойства, полученные из них. Таким образом, ген, кДНК или РНК кодирует белок, если транскрипция и трансляция мРНК, соответствующие этому гену, продуцируют белок в клетке или другой биологической системе. Как кодирующая цепь, нуклеотидная последовательность которой идентична последовательности мРНК и обычно указывается в перечнях последовательностей, так и некодирующая цепь, используемая в качестве матрицы для транскрипции гена или кДНК, могут упоминаться как кодирующие белок или другой продукт этого гена или кДНК.
Если не указано иное, «нуклеотидная последовательность или последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность» включает все нуклеотиды или нуклеотидные последовательности, которые являются вырожденными версиями друг друга и которые кодируют одну и ту же аминокислотную последовательность. Фраза нуклеотидная последовательность, кодирующая белок или РНК может также включать интроны в той степени, в которой нуклеотидная последовательность, кодирующая белок, может в некоторых версиях содержать интрон(ы).
Термин «трансплантат», используемый в настоящем документе, относится к клеткам, тканям или органам, которые вводятся объекту. Источником трансплантированного материала могут быть культивируемые клетки, клетки из другого объекта или клетки от того же объекта (например, после того, как клетки культивируются in vitro). Примерами трансплантации органов являются почка, печень, сердце, легкое и поджелудочная железа. Примером трансплантации ткани являются островковые клетки. Примером трансплантации клеток является аллогенная трансплантация гемопоэтических стволовых клеток.
Термин «экзогенный» относится к любому материалу, введенному или произведенному вне организма, клетки, ткани или системы.
Термин «экспрессия» относится к транскрипции и/или трансляции конкретной нуклеотидной последовательности, управляемой промотором.
Термин «экспрессирующий вектор» относится к вектору, содержащему рекомбинантный полинуклеотид, содержащий последовательности контроля экспрессии, функционально связанные с нуклеотидной последовательностью, которая должна быть экспрессирована. Экспрессирущий вектор включает достаточное количество цис-действующих элементов для экспрессии; другие элементы для экспрессии могут поставляться клеткой-хозяином или в системе экспрессии in vitro. Экспрессирующие векторы включают все известные в данной области техники, включая космиды, плазмиды (например, «голые» или содержащиеся в липосомах) и вирусы (например, лентивирусы, ретровирусы, аденовирусы и аденоассоциированные вирусы), которые включают рекомбинантный полинуклеотид.
Термин «лентивирус» относится к роду семейства Retroviridae. Лентивирусы уникальны среди ретровирусов по способности инфицировать неделящиеся клетки; они могут доставлять значительное количество генетической информации в ДНК клетки-хозяина, поэтому они являются одним из наиболее эффективных способов для вектора доставки генов. HIV, SIV и FIV являются примерами лентивирусов.
Термин «лентивирусный вектор» относится к вектору, полученному, по меньшей мере, из части генома лентивируса, включая, в частности, самоинактивирующийся лентивирусный вектор, представленный в Milone et al., Mol. Ther. 17 (8): 1453-1464 (2009). Другие примеры лентивирусных векторов, которые могут использоваться в клинике, включают, без ограничения указанным, например, технологию доставки генов LENTIVECTOR® от Oxford BioMedica, векторную систему LENTIMAX™ от Lentigen и т.п. Неклинические типы лентивирусных векторов также доступны и будут известны специалисту в данной области.
Термин «гомологичный» или «идентичность» относится к идентичности последовательности субъединицы между двумя полимерными молекулами, например, между двумя молекулами нуклеиновой кислоты, такими как две молекулы ДНК или две молекулы РНК, или между двумя полипептидными молекулами. Когда положение субъединицы в обеих из двух молекул занимает одна и та же мономерная субъединица; например, если положение в каждой из двух молекул ДНК занято аденином, то они гомологичны или идентичны в этом положении. Гомология между двумя последовательностями является прямой функцией числа совпадающих или гомологичных положений; например, если половина (например, пять положений в полимере из десяти субъединиц длиной) из положений в двух последовательностях гомологична, эти две последовательности являются гомологичными на 50%; если 90% положений (например, 9 из 10) совпадают или гомологичны, две последовательности гомологичны на 90%.
«Гуманизированные» формы нечеловеческих (например, мышиных) антител представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как, например, Fv, scFv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), которые содержат минимальную последовательность, полученную из нечеловеческого иммуноглобулина. По большей части гуманизированные антитела и их фрагменты антител представляют собой человеческие иммуноглобулины (реципиентное антитело или фрагмент антитела), в которых остатки из области, определяющей комплементарность (CDR) реципиента, заменены остатками из CDR нечеловеческого вида (донорное антитело), такое как мышь, крыса или кролик, имеющие искомую специфичность, аффинность и потенциал. В некоторых случаях остатки каркасной области Fv (FR) человеческого иммуноглобулина заменены соответствующими нечеловеческими остатками. Кроме того, фрагмент гуманизированного антитела/антитела может содержать остатки, которые не обнаружены ни в реципиентном антителе, ни в импортированных CDR или каркасных последовательностях. Эти модификации могут дополнительно улучшить и оптимизировать характеристики антитела или фрагмента антитела. Как правило, гуманизированное антитело или его фрагмент антитела будут включать, по существу, все, по меньшей мере, один, и, как правило, два вариабельных домена, в которых все или по существу все области CDR соответствуют таковым для нечеловеческого иммуноглобулина и все или значительных часть FR-областей представляет собой последовательности иммуноглобулина человека. Гуманизированное антитело или фрагмент антитела также может содержать, по меньшей мере, часть константной области иммуноглобулина (Fc), обычно таковой иммуноглобулина человека. Для получения дополнительной информации см. Jones et al., Nature, 321: 522-525, 1986; Reichmann et al., Nature, 332: 323-329, 1988; Presta, Curr. Op. Struct. Biol., 2: 593-596, 1992.
«Человеческий» иммуноглобулин, антитело или фрагмент антитела относится к иммуноглобулину, такому как антитело или фрагмент антитела, где целая молекула имеет человеческое происхождение или состоит из аминокислотной последовательности, идентичной человеческой форме антитела или иммуноглобулина.
Термин «выделенный» означает измененный или удаленный из естественного состояния. Например, нуклеиновая кислота или пептид, естественно присутствующие в живом животном, не являются «выделенными», но та же самая нуклеиновая кислота или пептид, частично или полностью отделенные от сосуществующих материалов их естественного состояния, являются «выделенными». Выделенная нуклеиновая кислота или пептид могут существовать по существу в очищенной форме или могут существовать в ненативной среде, такой как, например, клетка-хозяин. «Выделенное» антитело представляет собой антитело, которое было идентифицировано и отделено и/или извлечено из компонента его естественной среды. Загрязняющие компоненты его естественной среды представляют собой вещества, которые будут препятствовать терапевтическому применению антитела и могут включать ферменты, гормоны и другие белковые или небелковые растворы.
В контексте настоящего изобретения используются следующие сокращения наиболее часто встречающихся оснований нуклеиновых кислот. «А» относится к аденину, «С» относится к цитозину, «G» относится к гуанину, «Т» относится к тимину и «U» относится к урацилу.
Термин «функционально связанный» относится к функциональной связи между регуляторной последовательностью и последовательностью гетерологичной нуклеиновой кислоты, приводящей к экспрессии последней. Например, первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты находится в функциональной связи со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с кодирующей последовательностью, когда промотор воздействует на транскрипцию или экспрессию кодирующей последовательности. Функционально связанные последовательности ДНК могут быть смежными друг с другом и, например, при необходимости соединять две белок-кодирующие области, которые находятся в одной рамке считывания.
Термин «парентеральное» введение иммуногенной композиции включает, например, подкожные (п.к.), внутрикожные, интранодальные, интрамедуллярные, внутрибрюшинные, внутривенные (в.в.), внутримышечные (в.м.) или надчревные инъекции, внутриопухолевые или инфузионные методы.
Термины «нуклеиновая кислота» или «полинуклеотид» используются взаимозаменяемо и относятся к полимерной форме нуклеотидов любой длины, либо дезоксирибонуклеотидов, либо рибонуклеотидов. Таким образом, этот термин включает, без ограничения указанным, одно-, двух- или многоцепочечные ДНК или РНК, геномные ДНК, кДНК, ДНК-РНК-гибриды или полимер, содержащий пуриновые и пиримидиновые основания или другие природные, химические или биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания. За исключением случаев конкретного ограничения, термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые обладают свойствами связывания, сходными с эталонной нуклеиновой кислотой, и метаболизируются способом, подобным встречающимся в природе нуклеотидам. Если не указано иное, конкретная последовательность нуклеиновой кислоты также неявно включает ее консервативно модифицированные варианты (например, замены вырожденных кодонов), аллели, ортологи, SNP и комплементарные последовательности, а также явно указанная последовательность. В частности, вырожденные замены кодонов могут быть достигнуты путем генерирования последовательностей, в которых третье положение одного или нескольких выбранных (или всех) кодонов замещено остатками смешанного основания и/или дезоксиинозина (Batzer et al., Nucleic Acid Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); и Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)).
Термины «пептид», «полипептид» и «белок» используются взаимозаменяемо и относятся к соединению, состоящему из аминокислотных остатков, ковалентно связанных пептидными связями. Белок или пептид должен содержать, по меньшей мере, две аминокислоты, и никакие ограничения не налагаются на максимальное количество аминокислот, которое может включать последовательность белка или пептида. Полипептиды включают любой пептид или белок, содержащий две или более аминокислоты, соединенные друг с другом пептидными связями. Используемый в данном документе термин относится как к коротким цепям, которые также обычно упоминаются в данной области, например, как пептиды, олигопептиды и олигомеры, так и к более длинным цепям, которые обычно упоминаются в данной области как белки, и которых существует множество типов. «Полипептиды» включают, например, биологически активные фрагменты, по существу гомологичные полипептиды, олигопептиды, гомодимеры, гетеродимеры, варианты полипептидов, модифицированные полипептиды, производные, аналоги, гибридные белки наряду с прочим. Полипептид включает природный пептид, рекомбинантный пептид или их комбинацию.
Термин «промотор/регуляторная последовательность» относится к последовательности нуклеиновой кислоты, которая необходима для экспрессии генного продукта, функционально связанного с промотором/регуляторной последовательностью. В некоторых случаях эта последовательность может быть последовательностью основного промотора, а в других случаях эта последовательность может также включать последовательность энхансера и другие регуляторные элементы, которые необходимы для экспрессии продукта гена. Промотор/регуляторная последовательность может, например, представлять собой последовательность, которая экспрессирует генный продукт тканеспецифическим образом.
Термин «конститутивный промотор» относится к нуклеотидной последовательности, которая, когда функционально связана с полинуклеотидом, который кодирует или специфицирует продукт гена, вызывает образование генного продукта в клетке в большинстве или во всех физиологических условиях клетки.
Термин «индуцибельный промотор» относится к нуклеотидной последовательности, которая, когда функционально связана с полинуклеотидом, который кодирует или специфицирует генный продукт, заставляет генный продукт продуцироваться в клетке, по существу, только когда присутствует индуктор, который соответствует промотору, присутствующему в клетке.
Используемый в данном документе термин «транзиторная» относится к экспрессии неинтегрированного трансгена в течение периода часов, дней или недель, где период времени экспрессии меньше, чем период времени для экспрессии гена, если он интегрирован в геном или содержится в стабильном плазмидном репликоне в клетке-хозяине.
Термин «по существу очищенная» клетка относится к клетке, которая по существу не включает других типов клеток. По существу, очищенная клетка также относится к клетке, которая была отделена от клеток других типов, с которыми она обычно связана в своем естественном состоянии. В некоторых случаях популяция по существу очищенных клеток относится к гомогенной популяции клеток. В других случаях этот термин относится просто к клеткам, которые были отделены от клеток, с которыми они естественным образом связаны в своем естественном состоянии. В некоторых аспектах клетки культивируют in vitro. В других аспектах клетки не культивируются in vitro.
Термин «трансфицированный» или «трансформированный» или «трансдуцированный» относится к процессу, посредством которого экзогенная нуклеиновая кислота переносится или вводится в клетку-хозяина. «Трансфицированная» или «трансформированная» или «трансдуцированная» клетка представляет собой клетку, которая была трансфицирована, трансформирована или трансдуцирована экзогенной нуклеиновой кислотой. Клетка включает первичную клетку объекта и ее потомство.
Различные аспекты изобретения могут быть представлены во всем этом раскрытии в формате диапазона. Следует понимать, что описание в формате диапазона дано просто для удобства и краткости и не должно рассматриваться как негибкое ограничение объема изобретения. Соответственно, следует считать, что описание диапазона конкретно раскрывает все возможные поддиапазоны, а также отдельные числовые значения в этом диапазоне. Например, описание диапазона, такого как от 1 до 6, следует считать специально раскрытыми поддиапазонами, такими как от 1 до 3, от 1 до 4, от 1 до 5, от 2 до 4, от 2 до 6, от 3 до 6 и т.д., а также отдельные числа в этом диапазоне, например, 1, 2, 2,7, 3, 4, 5, 5,3 и 6. В качестве другого примера, диапазон, такой как идентичность 95-99%, включает что-то с идентичностью 95%, 96%, 97%, 98% или 99% и включает поддиапазоны, такие как 96-99%, 96-98%, 96-97%, 97-99%, 97-98% и 98-99% идентичности. Это применимо независимо от широты диапазона.
III. АНТИ-HLA-A2 АНТИТЕЛА
В одном аспекте настоящее изобретение относится к анти-HLA-A2 антителам. Примерные антитела включают моноклональные антитела, поликлональные антитела, рекомбинантные антитела, химерные антитела, антитела человека, гуманизированные антитела и их антигенсвязывающие фрагменты.
В одном воплощении изобретение относится к гуманизированным анти-HLA-A2 антителам. Представленные в данном документе гуманизированные анти-HLA-A2-антитела специфически связываются с HLA-A2. В одном воплощении гуманизированные анти-HLA-A2 антитела, представленные в настоящем документе, специфически связываются с HLA-A*02:01. Как будет понятно специалисту в данной области техники, способность антитела связываться с HLA-A2 может быть обнаружена с использованием методов, известных в данной области. Например, связывание антитела с HLA-A2 может быть обнаружено с использованием тетрамера HLA-A2, как проиллюстрировано в данном документе. В одном воплощении гуманизированные анти-HLA-A2 антитела, представленные в настоящем документе, конкурируют за связывание с HLA-A2 с антителом, включающим: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении гуманизированные анти-HLA-A2 антитела, представленные в настоящем документе, связываются с тем же эпитопом HLA-A2, что и антитело, включающее: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении гуманизированные анти-HLA-A2 антитела, представленные в настоящем документе, конкурируют за связывание с HLA-A2 с антителом BB7.2.
В одном воплощении гуманизированные анти-HLA-A2 антитела, представленные в настоящем документе, связываются с тем же эпитопом HLA-A2, что и антитело BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из двух или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33,HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из трех или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из четырех или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из пяти или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из шести или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из семи или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из каждого из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*25, HLA-A*29, HLA-A*30 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из двух или нескольких из числа HLA-A*25, HLA-A*29, HLA-A*30 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из каждого из числа HLA-A*25, HLA-A*29, HLA-A*30 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к двум из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к трем из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к четырем из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к пяти из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к шести из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к семи из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к двум из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01, по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*25:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*29:02 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*30:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*03:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*31:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*33:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*36:01 по сравнению с антителом BB7.2. В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*68:01 по сравнению с антителом BB7.2. Антитело BB7.2 может быть выделено из гибридомы BB7.2 (депозитный номер ATCC HB-82).
Методы определения реактивности гуманизированных анти-HLA-A2 антител к подтипам HLA-A будут известны специалистам в данной области. Например, реактивность гуманизированных анти-HLA-A2 антител к подтипам HLA-A может быть определена с помощью анализа с использованием анализа на гранулах с индивидуальными антигенами. Такие анализы на гранулах с индивидуальными антигенами коммерчески доступны (например, FlowPRA Single Antigen Antibody; ONE LAMBDA).
В одном воплощении гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, и любой их комбинации по сравнению с антителом BB7.2, например, по сравнению с антителом BB7.2 scFv при измерении в условиях теста А. Например, в некоторых воплощениях гуманизированные анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из HLA-A*25, HLA-A*29, HLA-A*30, и любой их комбинации по сравнению с антителом BB7.2, например, по сравнению с scFv BB7.2 при измерении в условиях теста А.
Тест А:
0.25⋅106 Т-клеток, экспрессирующих CAR, содержащего гуманизированное анти-HLA-A2 антитело или антитело BB7.2, например, BB7.2 scFv (mA2 CAR)), инкубировали с панелью гранул с индивидуальными антигенами FlowPRA (FL1HD01, FL1HD02, FL1HD03). FL1HD04, FL1HD06 и FL1HD08, One Lambda) и фиксируемым красителем для оценки жизнеспособности (FVD, ThermoFisher, 65-0865-14, eBioscience) в течение 30 минут при комнатной температуре. Образцы промывали, фиксировали 0,5% формальдегидом и анализировали с помощью проточной цитометрии. Двести гранул отрицательного контроля использовали на образец. В качестве отрицательного контроля использовали гранулы в чистом виде. Для анализа сначала удаляли мертвые клетки с помощью фиксируемого красителя для оценки жизнеспособности. Гранулы с индивидуальным антигеном затем отбирали по сигналу выше порогового значения после исключения мертвых клеток и дублетов. Затем количество гранул на HLA определяли их соответствующим пиком интенсивности PE. Данные нормализовали путем умножения количества представляющих интерес гранул в каждом пике HLA на 200, деленного на количество отрицательных гранул в образце. Для каждого пика HLA процент относительного связывания CAR Treg по сравнению с контролем (клетками, не экспрессирующими CAR) определяли путем вычитания количества гранул в CAR-Treg из числа гранул в контрольном образце с последующим делением среднего числа гранул в не-CAR-экспрессирующем контроле на 100.
В одном воплощении гуманизированное анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 статистически уступающей реактивности антитела BB7.2, например, при измерении в условиях теста А.
В одном воплощении анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, статистически уступающей реактивности антитела BB7.2, например, при измерении в условиях теста А.
В одном воплощении термин «статистически уступающий» означает, что реактивность (то есть, например, относительное связывание в условиях теста А), измеренная для анти-HLA-A2 антитела по изобретению, ниже реактивности, измеренной для антитела BB7.2, с p-значением, по большей мере, около 0,05, предпочтительно, по большей мере, около 0,01, более предпочтительно, по большей мере, около 0,005, и даже более предпочтительно, по большей мере, около 0,001, в частности, при анализе с помощью двухфакторного дисперсионного анализа, апостериорного критерия Даннетта.
В одном воплощении анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 уступающей реактивности антитела BB7.2. В некоторых воплощениях такое анти-HLA-A2 антитело имеет относительное связывание, по меньшей мере, с одним подтипом HLA-A, выбранным из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 уступающей связыванию антитела BB7.2 при измерении в условиях теста А. В определенных аспектах относительное связывание, измеренное для такого анти-HLA-A2 антитела, составляет, по большей мере, около 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или менее относительного связывания, измеренного для антитела BB7.2.
В одном воплощении анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, уступающей реактивности антитела BB7.2. В некоторых воплощениях такое анти-HLA-A2 антитело имеет относительное связывание, по меньшей мере, с одним подтипом HLA-A, выбранным из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, уступающей связыванию антитела BB7.2 при измерении в условиях теста А. В определенных аспектах относительное связывание, измеренное для такого анти-HLA-A2 антитела, составляет, по большей мере, около 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или менее относительного связывания, измеренного для антитела BB7.2.
Кроме того, гуманизированные анти-HLA-A2 антитела, представленные в настоящем документе, обладающие антигенсвязывающей активностью, способны образовывать антигенсвязывающие домены химерных антигенных рецепторов (CAR), где такие CAR способны экспрессироваться в клетках человека, так что CAR специфически связываются с HLA-A2. В одном воплощении CAR специфически связываются с HLA-A*02:01. Как будет понятно специалисту в данной области техники, способность антитела связываться с HLA-A2 может быть обнаружена с использованием методов, известных в данной области. Например, связывание CAR с HLA-A2 может быть обнаружено с использованием тетрамера HLA-A2, как проиллюстрировано в данном документе. В одном воплощении клетка человека представляет собой иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg. Кроме того, гуманизированные анти-HLA-A2 антитела, представленные в настоящем документе, обладающие антигенсвязывающей активностью, способны образовывать антигенсвязывающие домены химерных антигенных рецепторов (CAR), где такие CAR способны экспрессироваться в регуляторных Т-клетках, так что CAR специфически связываются с HLA-A2. В одном воплощении CAR специфически связываются с HLA-A*02:01. В одном воплощении Treg представляет собой Treg человека.
В одном воплощении гуманизированное анти-HLA-A2 антитело способно составлять антигенсвязывающий домен CAR, где такой CAR способен экспрессироваться в иммунной клетке, так что иммунная клетка активируется HLA-A2. В одном воплощении иммунная клетка активируется HLA-A*02:01. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении Т-клетка представляет собой человеческую Т-клетку. В одном воплощении Treg представляет собой Treg человека.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: SYHIQ (SEQ ID NO: 1) и GYTFTSY (SEQ ID NO: 2).
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 1, определяющую комплементарность (VH CDR1), выбранную из SEQ ID NO: 1-2. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую VH CDR1, представленную SEQ ID NO: 1. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую VH CDR1, представленную SEQ ID NO: 2.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: YPGDGS (SEQ ID NO: 4) и WIYPGDGSTX10YX12X13KFX16G (SEQ ID NO: 10), где в SEQ ID NO: 10, аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, и аминокислота в положении 16 (X16) представляет собой K или Q.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность WIYPGDGSTX10YX12X13KFX16G (SEQ ID NO: 10), где аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, а аминокислота в положении 16 (X16) представляет собой K или Q.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: YPGDGS (SEQ ID NO: 4) и WIYPGDGSTX10YX12X13KFX16G (SEQ ID NO: 10), где в SEQ ID NO: 10, аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, и аминокислота в положении 16 (X16) представляет собой K или Q.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), содержащую аминокислотную последовательность WIYPGDGSTX10YX12X13KFX16G (SEQ ID NO: 10), где аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, а аминокислота в положении 16 (X16) представляет собой K или Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой Q или K, аминокислота в положении 12 представляет собой S, аминокислота в положении 13 представляет собой Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой K, аминокислота в положении 12 представляет собой N или S, аминокислота в положении 13 представляет собой Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой K, аминокислота в положении 12 представляет собой S, аминокислота в положение 13 представляет собой Е или Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой K, аминокислота в положении 12 представляет собой S, аминокислота в положение 13 представляет собой Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой Q или K, аминокислота в положении 12 представляет собой N или S, аминокислота в положении 13 представляет собой Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой Q или K, аминокислота в положении 12 представляет собой S, аминокислота в положение 13 представляет собой Е или Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой Q или K, аминокислота в положении 12 представляет собой S, аминокислота в положение 13 представляет собой Q, а аминокислота в положении 16 представляет собой K или Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой K, аминокислота в положении 12 представляет собой N или S, аминокислота в положение 13 представляет собой Е или Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой K, аминокислота в положении 12 представляет собой N или S, аминокислота в положении 13 представляет собой Q, а аминокислота в положении 16 представляет собой K или Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой K, аминокислота в положении 12 представляет собой S, аминокислота в положение 13 представляет собой Е или Q, а аминокислота в положении 16 представляет собой K или Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой Q или K, аминокислота в положении 12 представляет собой N или S, аминокислота в положение 13 представляет собой Е или Q, а аминокислота в положении 16 представляет собой Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой Q или K, аминокислота в положении 12 представляет собой N или S, аминокислота в положении 13 представляет собой Q, а аминокислота в положении 16 представляет собой K или Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой Q или K, аминокислота в положении 12 представляет собой S, аминокислота в положение 13 представляет собой Е или Q, а аминокислота в положении 16 представляет собой K или Q.
В одном воплощении вариабельная область тяжелой цепи включает VH CDR2, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 10, где аминокислота в положении 10 представляет собой K, аминокислота в положении 12 представляет собой N или S, аминокислота в положение 13 представляет собой Е или Q, а аминокислота в положении 16 представляет собой K или Q.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: WIYPGDGSTQYNEKFKG (SEQ ID NO: 3), YPGDGS (SEQ ID NO: 4) и WIYPGDGSTKYSQKFQG (SEQ ID NO: 5). В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: WIYPGDGSTQYNEKFKG (SEQ ID NO: 3) и YPGDGS (SEQ ID NO: 4). В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: YPGDGS (SEQ ID NO: 4) и WIYPGDGSTKYSQKFQG (SEQ ID NO: 5).
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 3. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 4. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 5.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), выбранную из SEQ ID NO: 3-5. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), выбранную из SEQ ID NO: 3-4. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), выбранную из SEQ ID NO: 4-5. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), представленную SEQ ID NO: 5. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую VH CDR2, представленную SEQ ID NO: 3. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую VH CDR2, представленную SEQ ID NO: 4.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность EGTYYAMDY (SEQ ID NO: 6).
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 3, определяющую комплементарность (VH CDR3), представленную SEQ ID NO: 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую, по меньшей мере, одну из следующих CDR:
VH CDR1, представленную в SEQ ID NO: 1; или
VH CDR2, представленную в SEQ ID NO: 10, где аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, и аминокислота в положении 16 (X16) представляет собой K или Q; или
VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую, по меньшей мере, одну из следующих CDR:
VH CDR1, представленную в SEQ ID NO: 1; или
VH CDR2, представленную в SEQ ID NO: 3; или
VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую, по меньшей мере, одну из следующих CDR:
VH CDR1, представленную в SEQ ID NO: 1; или
VH CDR2, представленную в SEQ ID NO: 5; или
VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR1, представленную в SEQ ID NO: 1; одну VH CDR2, представленную в SEQ ID NO: 10, где аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, и аминокислота в положении 16 (X16) представляет собой K или Q; и одну VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR1, представленную в SEQ ID NO: 1; одну VH CDR2, представленную в SEQ ID NO: 3; и одну VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR1, представленную в SEQ ID NO: 1; одну VH CDR2, представленную в SEQ ID NO: 5; и одну VH CDR3, представленную в SEQ ID NO: 6.
Согласно настоящему изобретению любая из CDR 1, 2 или 3 тяжелой цепи может быть охарактеризована как имеющая аминокислотную последовательность, которая имеет, по меньшей мере, около 60%, 70%, 75%, 80%, 85%, 90%. 95%, 96%, 97%, 98% или 99% идентичности с конкретными наборами CDR, перечисленными в соответствующих SEQ ID NO: 1, 3, 5, 6 и 10.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую, по меньшей мере, одну из следующих CDR:
VH CDR1, представленную в SEQ ID NO: 2; или
VH CDR2, представленную в SEQ ID NO: 4; или
VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR1, представленную в SEQ ID NO: 2; одну VH CDR2, представленную в SEQ ID NO: 4; и одну VH CDR3, представленную в SEQ ID NO: 6.
Согласно настоящему изобретению любая из CDR 1, 2 или 3 тяжелой цепи может быть охарактеризована как имеющая аминокислотную последовательность, которая имеет, по меньшей мере, около 60%, 70%, 75%, 80%, 85%, 90%. 95%, 96%, 97%, 98% или 99% идентичности с конкретными наборами CDR, перечисленными в соответствующих SEQ ID NO: 2, 4 и 6.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность RSSQSIVHSNGNTYLE (SEQ ID NO: 7).
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 1, определяющую комплементарность (VH CDR1), представленную в SEQ ID NO: 7.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность KVSNRFS (SEQ ID NO: 8).
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), представленную в SEQ ID NO: 8.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность FQGSHVPRT (SEQ ID NO: 9).
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 3, определяющую комплементарность (VH CDR3), представленную в SEQ ID NO: 9.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую, по меньшей мере, одну из следующих CDR:
VL CDR1, представленную в SEQ ID NO: 7; или
VL CDR2, представленную в SEQ ID NO: 8; или
VL CDR3, представленную в SEQ ID NO: 9.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую одну VL CDR1, представленную в SEQ ID NO: 7; одну VL CDR2, представленную в SEQ ID NO: 8; и одну VL CDR3, представленную в SEQ ID NO: 9.
Согласно настоящему изобретению любая из CDR 1, 2 или 3 легкой цепи может быть охарактеризована как имеющая аминокислотную последовательность, которая имеет, по меньшей мере, около 60%, 70%, 75%, 80%, 85%, 90%. 95%, 96%, 97%, 98% или 99% идентичности с конкретными наборами CDR, перечисленными в соответствующих SEQ ID NO: 7, 8 и 9.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность, выбранную из группы, состоящей из: QVQLVQSGAEVKKPGASVKVSCKAS (SEQ ID NO: 11) и QVQLVQSGAEVKKPGASVKVSCKASGYTFT (SEQ ID NO: 12).
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность QVQLVQSGAEVKKPGASVKVSCKAS (SEQ ID NO: 11). В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85 %, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 11.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность QVQLVQSGAEVKKPGASVKVSCKASGYTFT (SEQ ID NO: 12). В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85 %, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 12.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, выбранную из группы, состоящей из:
WVRQAPGQX9LEWMGX15 (SEQ ID NO: 13),
WVRQAPGQX9LEWMGX15WI (SEQ ID NO: 17),
HIQWVRQAPGQX12LEWMGX18WI (SEQ ID NO: 21), и
HIQWVRQAPGQX12LEWMGX18 (SEQ ID NO: 25), где:
X9 представляет собой R или G и X15 представляет собой I или отсутствует в SEQ ID NO: 13;
X9 представляет собой R или G и X15 представляет собой I или отсутствует в SEQ ID NO: 17;
X12 представляет собой R или G и X18 представляет собой I или отсутствует в SEQ ID NO: 21; и
X12 представляет собой R или G, а X18 представляет собой I или отсутствует в SEQ ID NO: 25.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность WVRQAPGQX9LEWMGX15 (SEQ ID NO: 13), где аминокислота в положении 9 (X9) обозначает R или G, а аминокислота в положении 15 (X15) обозначает I или отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 13, где аминокислота в положении 9 представляет собой R или G, а аминокислота в положении 15 отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 14, 15 или 16. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88 %, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 14, 15 или 16.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность HIQWVRQAPGQX12LEWMGX18 (SEQ ID NO: 25), где аминокислота в положении 12 (X12) представляет собой R или G, а аминокислота в положении 18 (X18) представляет собой I или отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 25, где аминокислота в положении 12 представляет собой R или G, а аминокислота в положении 18 отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 26, 27 или 28. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 26, 27 или 28.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность WVRQAPGQX9LEWMGX15WI (SEQ ID NO: 17), где аминокислота в положении 9 (X9) обозначает R или G, а аминокислота в положении 15 (X15) обозначает I или отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 17, где аминокислота в положении 9 представляет собой R или G, а аминокислота в положении 15 отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 18, 19 или 20. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 18, 19 или 20.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность HIQWVRQAPGQX12LEWMGX18WI (SEQ ID NO: 21), где аминокислота в положении 12 (X12) представляет собой R или G, а аминокислота в положении 18 (X18) представляет собой I или отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 25, где аминокислота в положении 12 представляет собой R или G, а аминокислота в положении 18 отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 22, 23 или 24. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 22, 23 или 24.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 3 (VH FR2), содержащую аминокислотную последовательность, выбранную из группы, состоящей из:
X1VTX4TX6DTSX10STAYMX16LSX19LRSX23DX25AVYYCAR (SEQ ID NO: 29),
TX2YX4X5KFX8GX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34
AVYYCAR (SEQ ID NO: 35),
TQYNEKFKGX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34
AVYYCAR (SEQ ID NO: 36) и
TKYSQKFQGX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34AVYYCAR (SEQ ID NO: 37), где:
X1 представляет собой R или отсутствует, X4 представляет собой I или M, X6 представляет собой R или A, X10 представляет собой A, T или I, X16 представляет собой E или L, X19 представляет собой S или R, X23 представляет собой E или D и X25 представляет собой T или M в SEQ ID NO: 29;
X2 представляет собой Q или K, X4 представляет собой N или S, X5 представляет собой E или Q, X8 представляет собой K или Q, X10 представляет собой R или отсутствует, X13 представляет собой I или M, X15 представляет собой R или A, X19 представляет собой A, T или I, X25 представляет собой E или L, X28 представляет собой S или R, X32 представляет собой E или D и X34 представляет собой T или M в SEQ ID NO: 35;
X10 представляет собой R или отсутствует, X13 представляет собой I или M, X15 представляет собой R или A, X19 представляет собой A, T или I, X25 представляет собой E или L, X28 представляет собой S или R, X32 представляет собой E или D и X34 представляет собой T или M в SEQ ID NO: 36; и
X10 представляет собой R или отсутствует, X13 представляет собой I или M, X15 представляет собой R или A, X19 представляет собой A, T или I, X25 представляет собой E или L, X28 представляет собой S или R, X32 представляет собой E или D и X34 представляет собой T или M в SEQ ID NO: 37.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 3 (VH FR3), содержащую аминокислотную последовательность X1VTX4TX6DTSX10STAYMX16LSX19LRSX23DX25AVYYCAR (SEQ ID NO: 29), где аминокислота в положении 1 (X1) представляет собой R или отсутствует, аминокислота в положении 4 (X4) представляет собой I или M, аминокислота в положении 6 (X6) представляет собой R или A, аминокислота в положении 10 (X10) представляет собой A, T или I, аминокислота в положении 16 (X16) представляет собой E или L, аминокислота в положении 19 (X19) представляет собой S или R, аминокислота в положении 23 (X23) представляет собой E или D, и аминокислота в положении 25 (Х25) представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 29, где аминокислота в положении 1 представляет собой R, аминокислота в положении 4 представляет собой I или M, аминокислота в положении 6 представляет собой R, аминокислота в положении 10 представляет собой A или I, аминокислота в положении 16 представляет собой E, аминокислота в положении 19 представляет собой S или R, аминокислота в положении 23 представляет собой E или D, и аминокислота в положении 25 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 29, где аминокислота в положении 1 представляет собой R, аминокислота в положении 4 представляет собой I или M, аминокислота в положении 6 представляет собой R, аминокислота в положении 10 представляет собой A или I, аминокислота в положении 16 представляет собой E, аминокислота в положении 19 представляет собой S или R, аминокислота в положении 23 представляет собой E или D, и аминокислота в положении 25 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 29, где аминокислота в положении 1 представляет собой R, аминокислота в положении 4 представляет собой I, аминокислота в положении 6 представляет собой R, аминокислота в положении 10 представляет собой A, аминокислота в положении 16 представляет собой E, аминокислота в положении 19 представляет собой S, аминокислота в положении 23 представляет собой E, и аминокислота в положении 25 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 30, 31, 32, 33 или 34. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 30, 31, 32, 33 или 34.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 3 (VH FR3), содержащую аминокислотную последовательность TX2YX4X5KFX8GX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34AVYYCAR (SEQ ID NO: 35), где аминокислота в положении 2 (X2) представляет собой Q или K, аминокислота в положении 4 (X4) представляет собой N или S, аминокислота в положении 5 (X5) представляет собой E или Q, аминокислота в положении 8 (X8) представляет собой K или Q, аминокислота в положении 10 (X10) представляет собой R или отсутствует, аминокислота в положении 13 (X13) представляет собой I или M, аминокислота в положении 15 (X15) представляет собой R или A, аминокислота в положении 19 (X19) представляет собой A, T или I, аминокислота в положении 25 (X25) представляет собой E или L, аминокислота в положении 28 (X28) представляет собой S или R, аминокислота в положении 32 (X32) представляет собой E или D, и аминокислота в положении 34 (X34) представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S, аминокислота в положении 32 представляет собой E, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q, аминокислота в положении 4 представляет собой N, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q, аминокислота в положении 4 представляет собой N, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q, аминокислота в положении 4 представляет собой N, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q или K, аминокислота в положении 4 представляет собой N, аминокислота в положении 5 представляет собой E, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 35, где аминокислотой в положении 2 представляет собой Q, аминокислота в положении 4 представляет собой N или S, аминокислота в положении 5 представляет собой E или Q, аминокислота в положении 8 представляет собой K или Q, аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 3 (VH FR3), содержащую аминокислотную последовательность TQYNEKFKGX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34AVYYCAR (SEQ ID NO: 36), где аминокислота в положении 10 (X10) представляет собой R или отсутствует, аминокислота в положении 13 (X13) представляет собой I или M, аминокислота в положении 15 (X15) представляет собой R или A, аминокислота в положении 19 (X19) представляет собой A, T или I, аминокислота в положении 25 (X25) представляет собой E или L, аминокислота в положении 28 (X28) представляет собой S или R, аминокислота в положении 32 (X32) представляет собой E или D, и аминокислота в положении 34 (Х34) представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 36, где аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 36, где аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 36, где аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S, аминокислота в положении 32 представляет собой E, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 3 (VH FR3), содержащую аминокислотную последовательность TKYSQKFQGX10VTX13TX15DTSX19STAYMX25LSX28LRSX32DX34AVYYCAR (SEQ ID NO: 37), где аминокислота в положении 10 (X10) представляет собой R или отсутствует, аминокислота в положении 13 (X13) представляет собой I или M, аминокислота в положении 15 (X15) представляет собой R или A, аминокислота в положении 19 (X10) представляет собой A, T или I, аминокислота в положении 25 (X25) представляет собой E или L, аминокислота в положении 28 (X28) представляет собой S или R, аминокислота в положении 32 (X32) представляет собой E или D, и аминокислота в положении 34 (Х34) представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 37, где аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 37, где аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I или M, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A или I, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S или R, аминокислота в положении 32 представляет собой E или D, и аминокислота в положении 34 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 37, где аминокислота в положении 10 представляет собой R, аминокислота в положении 13 представляет собой I, аминокислота в положении 15 представляет собой R, аминокислота в положении 19 представляет собой A, аминокислота в положении 25 представляет собой E, аминокислота в положении 28 представляет собой S, аминокислота в положении 32 представляет собой E, и аминокислота в положении 34 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 38, 39, 40, 41, 42 или 43. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 38, 39, 40, 41, 42 или 43.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 4 (VH FR1), содержащую аминокислотную последовательность WGQGTTVTVSS (SEQ ID NO: 44). В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR4), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 44.
В одном воплощении вариабельная область тяжелой цепи включает:
VH CDR1, представленную в SEQ ID NO: 1;
VH CDR2, представленную в SEQ ID NO: 10, где аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, и аминокислота в положении 16 (X16) представляет собой K или Q;
VH CDR3, представленную в SEQ ID NO: 6;
VH FR1, представленную в SEQ ID NO: 12;
VH FR2, представленную в SEQ ID NO: 13, где аминокислота в положении 9 (X9) представляет собой R или G, а аминокислота в положении 15 (X15) представляет собой I или отсутствует;
VH FR3, представленную в SEQ ID NO: 29, где аминокислота в положении 1 (X1) представляет собой R или отсутствует, аминокислота в положении 4 (X4) представляет собой I или M, аминокислота в положении 6 (X6) представляет собой R или A, аминокислота в положении 10 (X10) представляет собой A, T или I, аминокислота в положении 16 (X16) представляет собой E или L, аминокислота в положении 19 (X19) представляет собой S или R, аминокислота в положении 23 (X23) представляет собой E или D, а аминокислота в положении 25 (X25) представляет собой T или M; и
VH FR4, представленную в SEQ ID NO: 44.
В одном воплощении вариабельная область тяжелой цепи включает:
VH CDR1, представленную SEQ ID NO: 2;
VH CDR2, представленную в SEQ ID NO: 10, где аминокислота в положении 10 (X10) представляет собой Q или K, аминокислота в положении 12 (X12) представляет собой N или S, аминокислота в положении 13 (X13) представляет собой E или Q, и аминокислота в положении 16 (X16) представляет собой K или Q;
VH CDR3, представленную в SEQ ID NO: 6;
VH FR1, представленную в SEQ ID NO: 11;
VH FR2, представленную в SEQ ID NO: 25, где аминокислота в положении 12 (X12) представляет собой R или G, а аминокислота в положении 18 (X18) представляет собой I или отсутствует;
VH FR3, представленную в SEQ ID NO: 29, где аминокислота в положении 1 (X1) представляет собой R или отсутствует, аминокислота в положении 4 (X4) представляет собой I или M, аминокислота в положении 6 (X6) представляет собой R или A, аминокислота в положении 10 (X10) представляет собой A, T или I, аминокислота в положении 16 (X16) представляет собой E или L, аминокислота в положении 19 (X19) представляет собой S или R, аминокислота в положении 23 (X23) представляет собой E или D, а аминокислота в положении 25 (X25) представляет собой T или M; и
VH FR4, представленную в SEQ ID NO: 44.
В одном воплощении вариабельная область тяжелой цепи включает:
VH CDR1, представленную SEQ ID NO: 2;
VH CDR2, представленную в SEQ ID NO: 4;
VH CDR3, представленную в SEQ ID NO: 6;
VH FR1, представленную в SEQ ID NO: 11;
VH FR2, представленную в SEQ ID NO: 21, где аминокислота в положении 12 (X12) представляет собой R или G, а аминокислота в положении 18 (X18) представляет собой I или отсутствует;
VH FR3, представленную в SEQ ID NO: 35, где аминокислота в положении 2 (X2) представляет собой Q или K, аминокислота в положении 4 (X4) представляет собой N или S, аминокислота в положении 5 (X5) представляет собой E или Q, аминокислота в положении 8 (X8) представляет собой K или Q, аминокислота в положении 10 (X10) представляет собой R или отсутствует, аминокислота в положении 13 (X13) представляет собой I или M, аминокислота в положении 15 (X15) представляет собой R или A, аминокислота в положении 19 (X19) представляет собой A, T или I, аминокислота в положении 25 (X25) представляет собой E или L, аминокислота в положении 28 (X28) представляет собой S или R, аминокислота в положении 32 (X32) представляет собой E или D, а аминокислота в положении 34 (X34) представляет собой T или M; и
VH FR4, представленную в SEQ ID NO: 44.
В одном воплощении вариабельная область тяжелой цепи включает:
VH CDR1, представленную в SEQ ID NO: 1;
VH CDR2, представленную в SEQ ID NO: 4;
VH CDR3, представленную в SEQ ID NO: 6;
VH FR1, представленную в SEQ ID NO: 12;
VH FR2, представленную в SEQ ID NO: 17, где аминокислота в положении 9 (X9) представляет собой R или G, а аминокислота в положении 15 (X15) представляет собой I или отсутствует;
VH FR3, представленную в SEQ ID NO: 35, где аминокислота в положении 2 (X2) представляет собой Q или K, аминокислота в положении 4 (X4) представляет собой N или S, аминокислота в положении 5 (X5) представляет собой E или Q, аминокислота в положении 8 (X8) представляет собой K или Q, аминокислота в положении 10 (X10) представляет собой R или отсутствует, аминокислота в положении 13 (X13) представляет собой I или M, аминокислота в положении 15 (X15) представляет собой R или A, аминокислота в положении 19 (X19) представляет собой A, T или I, аминокислота в положении 25 (X25) представляет собой E или L, аминокислота в положении 28 (X28) представляет собой S или R, аминокислота в положении 32 (X32) представляет собой E или D, а аминокислота в положении 34 (X34) представляет собой T или M; и
VH FR4, представленную в SEQ ID NO: 44.
В одном воплощении изобретения гуманизированным анти-HLA-A2-антитело содержит вариабельный участок тяжелой цепи, содержащий аминокислотную последовательность QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYHIQWVRQAPGQX44LEWMGX50WIYPG DGSTX60YX62X63KFX66GX68VTX71TX73DTSX77STAYMX83LSX86LRSX90DX92AVYYCARE GTYYAMDYWGQGTTVTVSS (SEQ ID NO: 45), в котором аминокислота в положении 44 (X44) является R или G, аминокислота в положении 50 (X50) представляет собой I или отсутствует, аминокислота в положении 60 (X60) представляет собой Q или K, аминокислота в положении 62 (X62)представляет собой N или S, аминокислота в положении 63 (X63) представляет собой E или Q, аминокислота в положении 66 (X66) представляет собой K или Q, аминокислота в положении 68 (X68) представляет собой R или отсутствует, аминокислота в положении 71 (X71) представляет собой I или M, аминокислота в положении 73 (X73) представляет собой R или A, аминокислота в положении 77 (X77) представляет собой A, T или I, аминокислота в положении 83 (X83) представляет собой E или L, аминокислота в положении 86 (X86) представляет собой S или R, аминокислота в положении 90 (X90) представляет собой E или D, а аминокислота в положении 92 (X92) представляет собой T или M.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G; аминокислота в положении 50 отсутствует; аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R; аминокислота в положении 71 представляет собой I или М; аминокислота в положении 73 представляет собой R; аминокислота в положении 77 представляет собой А или I; аминокислота в положении 83 представляет собой E; аминокислота в положении 86 представляет собой S или R; аминокислота в положении 90 представляет собой E или D; и аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G; аминокислота в положении 50 отсутствует; аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R; аминокислота в положении 71 представляет собой I или М; аминокислота в положении 73 представляет собой R; аминокислота в положении 77 представляет собой А или I; аминокислота в положении 83 представляет собой E; аминокислота в положении 86 представляет собой S или R; аминокислота в положении 90 представляет собой E или D; и аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S, аминокислота в положении 90 представляет собой Е, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q; аминокислота в положении 62 представляет собой N; аминокислота в положении 63 представляет собой Е; аминокислота в положении 66 представляет собой K; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q; аминокислота в положении 62 представляет собой N; аминокислота в положении 63 представляет собой Е; аминокислота в положении 66 представляет собой K; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q; аминокислота в положении 62 представляет собой N; аминокислота в положении 63 представляет собой Е; аминокислота в положении 66 представляет собой K; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S, аминокислота в положении 90 представляет собой Е, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 является S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 это K; аминокислота в положении 62 представляет собой S; аминокислота в положении 63 является Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 представляет собой S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S, аминокислота в положении 90 представляет собой Е, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 является Q или K; аминокислота в положении 62 является S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 является Q; аминокислота в положении 66 является Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 является S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 это K; аминокислота в положении 62 представляет собой S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 является S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 является Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 представляет собой S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 представляет собой S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой Q или K; аминокислота в положении 62 является S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 45, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 60 представляет собой K; аминокислота в положении 62 представляет собой N или S; аминокислота в положении 63 представляет собой Е или Q; аминокислота в положении 66 представляет собой K или Q; аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 61, 62, 63, 64, 65 или 66. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88 %, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 61, 62, 63, 64, 65 или 66.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую каркасную область 1 (VL FR1), содержащую аминокислотную последовательность DX2VMTQX7PLSX11X12VTX15GQPASISX23 (SEQ ID NO: 46), где аминокислота в положении 2 (X2) представляет собой V или I, аминокислота в положении 7 (X7) представляет собой S или T, аминокислота в положении 11 (X11) представляет собой L или S; аминокислота в положении 12 (X12) представляет собой P или S; аминокислота в положении 15 (X15) представляет собой L или P, а аминокислота в положении 23 (X23) представляет собой C или F.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 1 (VL FR1), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 46, где аминокислота в положении 2 представляет собой V или I; аминокислота в положении 7 представляет собой S или T; аминокислота в положении 11 представляет собой L или S; аминокислота в положении 12 представляет собой P или S; аминокислота в положении 15 представляет собой L или P; и аминокислота в положении 23 представляет собой С.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 1 (VL FR1), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 46, где аминокислота в положении 2 представляет собой I; аминокислота в положении 7 представляет собой Т; аминокислота в положении 11 представляет собой L или S; аминокислота в положении 12 представляет собой P или S; аминокислота в положении 15 представляет собой L или P; и аминокислота в положении 23 представляет собой С.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 1 (VL FR1), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 46, где аминокислота в положении 2 представляет собой V или I; аминокислота в положении 7 представляет собой S или T; аминокислота в положении 11 представляет собой L; аминокислота в положении 12 представляет собой P или S; аминокислота в положении 15 представляет собой L или P; и аминокислота в положении 23 представляет собой С.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 1 (VL FR1), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 46, где аминокислота в положении 2 представляет собой V или I; аминокислота в положении 7 представляет собой S или T; аминокислота в положении 11 представляет собой L или S; аминокислота в положении 12 представляет собой P; аминокислота в положении 15 представляет собой L; и аминокислота в положении 23 представляет собой С.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 1 (VL FR1), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 47, 48, 49 или 50. В одном воплощении вариабельная область легкой цепи содержит каркасную область 1 (VL FR1), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88 %, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 47, 48, 49 или 50.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую каркасную область 2 (VL FR2), содержащую аминокислотную последовательность WX2X3QX5PGQX9PX11X12LIY (SEQ ID NO: 51), где аминокислота в положении 2 (X2) представляет собой F или Y, аминокислота в положении 3 (X3) представляет собой Q или L, аминокислота в положении 5 (X5) представляет собой R или K, аминокислота в положении 9 (X9) представляет собой S или P, аминокислота в положении 11 (X11) представляет собой R или Q, а аминокислота в положении 12 (X12) представляет собой R или L.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 2 (VL FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 51, где аминокислотой в положении 2 является Y, аминокислотой в положении 3 является Q или L, аминокислота в положении 5 представляет собой R или K, аминокислота в положении 9 представляет собой S или P, аминокислота в положении 11 представляет собой R или Q, а аминокислота в положении 12 представляет собой L.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 2 (VL FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 51, где аминокислотой в положении 2 является Y, аминокислотой в положении 3 является Q или L, аминокислота в положении 5 представляет собой R или K, аминокислота в положении 9 представляет собой S, аминокислота в положении 11 представляет собой R или Q, а аминокислота в положении 12 представляет собой L.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 2 (VL FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 51, где аминокислотой в положении 2 является Y, аминокислотой в положении 3 является Q, аминокислота в положении 5 представляет собой R, аминокислота в положении 9 представляет собой S или P, аминокислота в положении 11 представляет собой R, а аминокислота в положении 12 представляет собой L.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 2 (VL FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 51, где аминокислотой в положении 2 является F или Y, аминокислотой в положении 3 является Q или L, аминокислота в положении 5 представляет собой R или K, аминокислота в положении 9 представляет собой S, аминокислота в положении 11 представляет собой R или Q, а аминокислота в положении 12 представляет собой R или L.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 2 (VL FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 51, где аминокислотой в положении 2 представляет собой F или Y, аминокислотой в положении 3 представляет собой Q, аминокислота в положении 5 представляет собой R, аминокислота в положении 9 представляет собой S или P, аминокислота в положении 11 представляет собой R, а аминокислота в положении 12 представляет собой R или L.
В одном воплощении вариабельная область легкой цепи содержит каркасную область 2 (VL FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 52, 53, 54 или 55. В одном воплощении вариабельная область легкой цепи содержит каркасную область 1 (VL FR2), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88 , 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 52, 53, 54 или 55.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую каркасную область 3 (VL FR3), содержащую аминокислотную последовательность GVPDRFSGSGX11GTDFTLKISRVEAEDVGVYYC (SEQ ID NO: 56), где аминокислота в положении 11 (X11) представляет собой S или A.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую каркасную область 3 (VL FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 57 или 58. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 3 (VL FR3), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85 %, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 57 или 58.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую каркасную область 4 (VL FR4), содержащую аминокислотную последовательность FGGGTKVEIK (SEQ ID NO: 59). В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 4 (VL FR4), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85 %, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 59.
В одном воплощении изобретения гуманизированным анти-HLA-A2-антитело содержит вариабельный участок легкой цепи, содержащий аминокислотной последовательности DX2VMTQX7PLSX11X12VTX15GQPASISX23RSSQSIVHSNGNTYLEWX41X42QX44PGQX48P X50X51LIYKVSNRFSGVPDRFSGSGX72GTDFTLKISRVEAEDVGVYYCFQGSHVPRTFGG GTKVEIK (SEQ ID NO: 60), в котором аминокислота в положении 2 (X2) представляет собой В или I, аминокислота в положении 7 (X7) представляет собой S или T, аминокислота в положении 11 (X11) представляет собой L или S, аминокислота в положении 12 (X12) представляет собой P или S, аминокислота в положении 15 (X15) представляет собой L или P, аминокислота в положении 23 (X23) представляет собой C или F, аминокислота в положении 41 (X41) представляет собой F или Y, аминокислота в положении 42 (X42) представляет собой Q или L, аминокислота в положении 44 (X44) представляет собой R или K, аминокислота в положении 48 (X48) представляет собой S или P, аминокислота в положении 50 (X50) представляет собой R или Q, аминокислота в положении 51 (X51) представляет собой R или L и аминокислота в положении 72 (X72) представляет собой S или A.
В одном воплощении вариабельная область легкой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 60, где аминокислота в положении 2 представляет собой V или I, аминокислота в положении 7 представляет собой S или T, аминокислота в положение 11 представляет собой L или S, аминокислота в положении 12 представляет собой P или S, аминокислота в положении 15 представляет собой L или P, аминокислота в положении 23 представляет собой C, аминокислота в положении 41 представляет собой Y, аминокислота в положении 42 представляет собой Q или L, аминокислота в положении 44 представляет собой R или K, аминокислота в положении 48 представляет собой S или P, аминокислота в положении 50 представляет собой R или Q, аминокислота в положении 51 представляет собой L, и аминокислота в положении 72 представляет собой S или A.
В одном воплощении вариабельная область легкой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 60, где аминокислота в положении 2 представляет собой I, аминокислота в положении 7 представляет собой T, аминокислота в положение 11 представляет собой L или S, аминокислота в положении 12 представляет собой P или S, аминокислота в положении 15 представляет собой L или P, аминокислота в положении 23 представляет собой C, аминокислота в положении 41 представляет собой Y, аминокислота в положении 42 представляет собой Q или L, аминокислота в положении 44 представляет собой R или K, аминокислота в положении 48 представляет собой S или P, аминокислота в положении 50 представляет собой R или Q, аминокислота в положении 51 представляет собой L, и аминокислота в положении 72 представляет собой S или A.
В одном воплощении вариабельная область легкой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 60, где аминокислота в положении 2 представляет собой V или I, аминокислота в положении 7 представляет собой S или T, аминокислота в положение 11 представляет собой L, аминокислота в положении 12 представляет собой P или S, аминокислота в положении 15 представляет собой L или P, аминокислота в положении 23 представляет собой C, аминокислота в положении 41 представляет собой Y, аминокислота в положении 42 представляет собой Q или L, аминокислота в положении 44 представляет собой R или K, аминокислота в положении 48 представляет собой S, аминокислота в положении 50 представляет собой R или Q, аминокислота в положении 51 представляет собой L, и аминокислота в положении 72 представляет собой S.
В одном воплощении вариабельная область легкой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 60, где аминокислота в положении 2 представляет собой V или I, аминокислота в положении 7 представляет собой S или T, аминокислота в положение 11 представляет собой L или S, аминокислота в положении 12 представляет собой P или S, аминокислота в положении 15 представляет собой L, аминокислота в положении 23 представляет собой C, аминокислота в положении 41 представляет собой Y, аминокислота в положении 42 представляет собой Q, аминокислота в положении 44 представляет собой R, аминокислота в положении 48 представляет собой S или P, аминокислота в положении 50 представляет собой R, аминокислота в положении 51 представляет собой L, и аминокислота в положении 72 представляет собой S или A.
В одном воплощении вариабельная область легкой цепи содержит аминокислотную последовательность, представленную в SEQ ID NO: 60, где аминокислота в положении 2 представляет собой V или I, аминокислота в положении 7 представляет собой S или T, аминокислота в положение 11 представляет собой L или S, аминокислота в положении 12 представляет собой P или S, аминокислота в положении 15 представляет собой L или P, аминокислота в положении 23 представляет собой C, аминокислота в положении 41 представляет собой F или Y, аминокислота в положении 42 представляет собой Q или L, аминокислота в положении 44 представляет собой R или K, аминокислота в положении 48 представляет собой S или P, аминокислота в положении 50 представляет собой R или Q, аминокислота в положении 51 представляет собой R или L, и аминокислота в положении 72 представляет собой S или A.
В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 67, 68, 69, 70 или 71. В одном воплощении гуманизированное анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88 %, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO:67, 68, 69, 70 или 71.
В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFv, scFab или sdAb. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFv или scFab. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой sdAb. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFab. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFv. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFv, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90 или 91. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFv, содержащий аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89 %, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90 или 91.
В одном воплощении гуманизированное анти-HLA-A2 антитело содержит аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90 %, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO, указанными выше. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFv или scFab, содержащее аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO, указанными выше.
В одном аспекте изобретение относится к антителу против HLA, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность WIYPGDGSTKYSQKFQG (SEQ ID NO: 5).
В одном воплощении указанное антитело представляет собой миметик антитела, выбранный из группы, состоящей из аффитела, альфа-тела, каркаса на основе белка с повтором armadillo, кноттина, пептида домена Кунитца, аффилина, аффитина, аднектина, атримера, эвазина, DARPin, антикалина, авимера, финомера, версатела и дуокалина.
Анти-HLA-A2 антитела, представленные в настоящем документе, специфически связываются с HLA-A2. В одном воплощении анти-HLA-A2 антитела, представленные в настоящем документе, специфически связываются с HLA-A*02:01. Как будет понятно специалисту в данной области техники, способность антитела связываться с HLA-A2 может быть обнаружена с использованием методов, известных в данной области. Например, связывание антитела с HLA-A2 может быть обнаружено с использованием тетрамера HLA-A2, как проиллюстрировано в данном документе. В одном воплощении анти-HLA-A2 антитела, представленные в настоящем документе, конкурируют за связывание с HLA-A2 с антителом, включающим: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении анти-HLA-A2 антитела, представленные в настоящем документе, связываются с тем же эпитопом HLA-A2, что и антитело, включающее: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении анти-HLA-A2 антитела, представленные в настоящем документе, конкурируют за связывание с HLA-A2 с антителом BB7.2. В одном воплощении анти-HLA-A2 антитела, представленные в настоящем документе, связываются с тем же эпитопом HLA-A2, что и антитело BB7.2. В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), представленную SEQ ID NO: 5. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из двух или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из трех или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из четырех или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из пяти или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из шести или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из семи или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из каждого из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*25, HLA-A*29, HLA-A*30 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из двух или нескольких из числа HLA-A*25, HLA-A*29, HLA-A*30 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из каждого из числа HLA-A*25, HLA-A*29, HLA-A*30 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к двум из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к трем из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к четырем из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к пяти из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к шести из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к семи из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к двум из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01, по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*25:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*29:02 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*30:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*03:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*31:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*33:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*36:01 по сравнению с антителом BB7.2. В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*68:01 по сравнению с антителом BB7.2. Антитело BB7.2 может быть выделено из гибридомы BB7.2 (депозитный номер ATCC HB-82). Методы определения реактивности анти-HLA-A2 антител к подтипам HLA-A будут известны специалистам в данной области. Например, реактивность анти-HLA-A2 антител к подтипам HLA-A может быть определена с помощью анализа с использованием анализа на гранулах с индивидуальными антигенами. Такие анализы на гранулах с индивидуальными антигенами коммерчески доступны (например, FlowPRA Single Antigen Antibody; ONE LAMBDA).
В одном воплощении анти-HLA-A2-антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 по сравнению с антителом BB7.2 при измерении в условиях теста А.
Например, в некоторых воплощениях анти-HLA-A2 антитела, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A* 25, HLA-A* 29, HLA-A* 30 и любой их комбинации по сравнению с антителом BB7.2 при измерении в условиях теста А.
Тест А:
0.25⋅106 Т-клеток, экспрессирующих CAR, содержащего анти-HLA-A2 антитело или антитело BB7.2, например, BB7.2 scFv (mA2 CAR)), инкубировали с панелью гранул с индивидуальными антигенами FlowPRA (FL1HD01, FL1HD02, FL1HD03). FL1HD04, FL1HD06 и FL1HD08, One Lambda) и фиксируемым красителем для оценки жизнеспособности (FVD, ThermoFisher, 65-0865-14, eBioscience) в течение 30 минут при комнатной температуре. Образцы промывали, фиксировали 0,5% формальдегидом и анализировали с помощью проточной цитометрии. Двести гранул отрицательного контроля использовали на образец. В качестве отрицательного контроля использовали гранулы в чистом виде. Для анализа сначала удаляли мертвые клетки с помощью фиксируемого красителя для оценки жизнеспособности. Гранулы с индивидуальными антигенами затем отбирали по сигналу выше порогового значения после исключения мертвых клеток и дублетов. Затем количество гранул на HLA определяли по их соответствующим пикам интенсивности PE. Данные нормализовали путем умножения количества представляющих интерес гранул в каждом пике HLA на 200, деленного на количество отрицательных гранул в образце. Для каждого пика HLA процент относительного связывания CAR Treg по сравнению с контролем (клетками, не экспрессирующими CAR) определяли путем вычитания количества гранул в CAR-Treg из числа гранул в контрольном образце с последующим делением среднего числа гранул в не-CAR-экспрессирующем контроле на 100. В одном воплощении анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, статистически уступающей реактивности антитела BB7.2, например, при измерении в условиях теста А.
В одном воплощении анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, статистически уступающей реактивности антитела BB7.2, например, при измерении в условиях теста А.
В одном воплощении термин «статистически уступающий» означает, что реактивность (то есть, например, относительное связывание в условиях теста А), измеренная для анти-HLA-A2 антитела по изобретению, ниже реактивности, измеренной для антитела BB7.2, с p-значением, по большей мере, около 0,05, предпочтительно, по большей мере, около 0,01, более предпочтительно, по большей мере, около 0,005, и даже более предпочтительно, по большей мере, около 0,001, в частности, при анализе с помощью двухфакторного дисперсионного анализа, апостериорного критерия Даннетта.
В одном воплощении анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, уступающей антителу BB7.2. В некоторых воплощениях такое анти-HLA-A2 антитело имеет относительное связывание, по меньшей мере, с одним подтипом HLA-A, выбранным из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 уступающее антителу BB7.2 при измерении в условиях теста А. В определенных аспектах измеряется относительное связывание для такого анти-HLA-A2 антитела, по большей мере, составляет около 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или менее относительного связывания, измеренного для антитела BB7.2.
В одном воплощении анти-HLA-A2 антитело по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, уступающей реактивности антитела BB7.2. В некоторых воплощениях такое анти-HLA-A2 антитело имеет относительное связывание, по меньшей мере, с одним подтипом HLA-A, выбранным из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, уступающей связыванию антитела BB7.2 при измерении в условиях теста А. В определенных аспектах относительное связывание, измеренное для такого анти-HLA-A2 антитела, составляет, по большей мере, около 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или менее относительного связывания, измеренного для антитела BB7.2.
Кроме того, анти-HLA-A2-антитела, представленные в данном документе, с антигенсвязывающей активностью, способны составлять антигенсвязывающие домены CAR, где такие CAR способны экспрессироваться в клетках человека, так что CAR специфически связываются с HLA-A2. В одном воплощении CAR специфически связываются с HLA-A*02:01. Как будет понятно специалисту в данной области техники, способность антитела связываться с HLA-A2 может быть обнаружена с использованием методов, известных в данной области. Например, связывание CAR с HLA-A2 может быть обнаружено с использованием тетрамера HLA-A2, как проиллюстрировано в данном документе. В одном воплощении клетка человека представляет собой иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg.
Кроме того, анти-HLA-A2 антитела, представленные в настоящем документе, обладающие антигенсвязывающей активностью, способны образовывать антигенсвязывающие домены химерных антигенных рецепторов (CAR), где такие CAR способны экспрессироваться в регуляторных Т-клетках (Treg), так что CAR специфически связываются с HLA-A2. В одном воплощении CAR специфически связываются с HLA-A*02:01. В одном воплощении Treg представляет собой Treg человека.
В одном воплощении анти-HLA-A2 антитело способно составлять антигенсвязывающий домен CAR, где такой CAR способен экспрессироваться в иммунной клетке, так что иммунная клетка активируется HLA-A2. В одном воплощении иммунная клетка активируется HLA-A*02:01. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении Т-клетка представляет собой человеческую Т-клетку. В одном воплощении Treg представляет собой Treg человека.
В одном воплощении такое анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из: SYHIQ (SEQ ID NO: 1) и GYTFTSY (SEQ ID NO: 2).
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 1, определяющую комплементарность (VH CDR1), выбранную из SEQ ID NO: 1-2. В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую VH CDR1, представленную SEQ ID NO: 1. В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую VH CDR1, представленную SEQ ID NO: 2.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность EGTYYAMDY (SEQ ID NO: 6).
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 3, определяющую комплементарность (VH CDR3), представленную SEQ ID NO: 6.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR2, представленную в SEQ ID NO: 5, и, по меньшей мере, одну из следующих CDR:
VH CDR1, представленную в SEQ ID NO: 1; или
VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR1, представленную в SEQ ID NO: 1; одну VH CDR2, представленную в SEQ ID NO: 5; и одну VH CDR3, представленную в SEQ ID NO: 6.
Согласно настоящему изобретению любая из CDR 1, 2 или 3 тяжелой цепи может быть охарактеризована как имеющая аминокислотную последовательность, которая имеет, по меньшей мере, около 60%, 70%, 75%, 80%, 85%, 90%. 95%, 96%, 97%, 98% или 99% идентичности с конкретными наборами CDR, перечисленными в соответствующих SEQ ID NO: 1, 5 и 6.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR2, представленную в SEQ ID NO: 5, и, по меньшей мере, одну из следующих CDR:
VH CDR1, представленную в SEQ ID NO: 2; или
VH CDR3, представленную в SEQ ID NO: 6.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую одну VH CDR1, представленную в SEQ ID NO: 2; одну VH CDR2, представленную в SEQ ID NO: 5; и одну VH CDR3, представленную в SEQ ID NO: 6.
Согласно настоящему изобретению любая из CDR 1, 2 или 3 тяжелой цепи может быть охарактеризована как имеющая аминокислотную последовательность, которая имеет, по меньшей мере, около 60%, 70%, 75%, 80%, 85%, 90%. 95%, 96%, 97%, 98% или 99% идентичности с конкретными наборами CDR, перечисленными в соответствующих SEQ ID NO: 2, 5 и 6.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность RSSQSIVHSNGNTYLE (SEQ ID NO: 7).
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 1, определяющую комплементарность (VH CDR1), представленную в SEQ ID NO: 7.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность KVSNRFS (SEQ ID NO: 8).
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 2, определяющую комплементарность (VH CDR2), представленную в SEQ ID NO: 8.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую аминокислотную последовательность FQGSHVPRT (SEQ ID NO: 9).
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую область 3, определяющую комплементарность (VH CDR3), представленную в SEQ ID NO: 9.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую, по меньшей мере, одну из следующих CDR:
VL CDR1, представленную в SEQ ID NO: 7; или
VL CDR2, представленную в SEQ ID NO: 8; или
VL CDR3, представленную в SEQ ID NO: 9.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область легкой цепи, содержащую одну VL CDR1, представленную в SEQ ID NO: 7; одну VL CDR2, представленную в SEQ ID NO: 8; и одну VL CDR3, представленную в SEQ ID NO: 9.
Согласно настоящему изобретению любая из CDR 1, 2 или 3 легкой цепи может быть охарактеризована как имеющая аминокислотную последовательность, которая имеет, по меньшей мере, около 60%, 70%, 75%, 80%, 85%, 90%. 95%, 96%, 97%, 98% или 99% идентичности с конкретными наборами CDR, перечисленными в соответствующих SEQ ID NO: 7, 8 и 9.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность, выбранную из группы, состоящей из: QVQLVQSGAEVKKPGASVKVSCKAS (SEQ ID NO: 11) и QVQLVQSGAEVKKPGASVKVSCKASGYTFT (SEQ ID NO: 12).
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность QVQLVQSGAEVKKPGASVKVSCKAS (SEQ ID NO: 11). В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85 %, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 11.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность QVQLVQSGAEVKKPGASVKVSCKASGYTFT (SEQ ID NO: 12). В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 1 (VH FR1), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85 %, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 12.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, выбранную из группы, состоящей из: WVRQAPGQX9LEWMGX15 (SEQ ID NO: 13) и HIQWVRQAPGQX12LEWMGX18 (SEQ ID NO: 25), где: X9 представляет собой R или G и X15 представляет собой I или отсутствует в SEQ ID NO: 13; и X12 представляет собой R или G, и X18 представляет собой I или отсутствует в SEQ ID NO: 25.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность WVRQAPGQX9LEWMGX15 (SEQ ID NO: 13), где аминокислота в положении 9 (X9) обозначает R или G, а аминокислота в положении 15 (X15) обозначает I или отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 13, где аминокислота в положении 9 представляет собой R или G, а аминокислота в положении 15 отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 14, 15 или 16. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 14, 15 или 16.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 2 (VH FR2), содержащую аминокислотную последовательность HIQWVRQAPGQX12LEWMGX18 (SEQ ID NO: 25), где аминокислота в положении 12 (X12) представляет собой R или G, а аминокислота в положении 18 (X18) представляет собой I или отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 25, где аминокислота в положении 12 представляет собой R или G, а аминокислота в положении 18 отсутствует.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 26, 27 или 28. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 2 (VH FR2), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 26, 27 или 28.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 3 (VH FR3), содержащую аминокислотную последовательность X1VTX4TX6DTSX10STAYMX16LSX19LRSX23DX25AVYYCAR (SEQ ID NO: 29), где аминокислота в положении 1 (X1) представляет собой R или отсутствует, аминокислота в положении 4 (X4) представляет собой I или M, аминокислота в положении 6 (X6) представляет собой R или A, аминокислота в положении 10 (X10) представляет собой A, T или I, аминокислота в положении 16 (X16) представляет собой E или L, аминокислота в положении 19 (X19) представляет собой S или R, аминокислота в положении 23 (X23) представляет собой E или D, и аминокислота в положении 25 (Х25) представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 29, где аминокислота в положении 1 представляет собой R, аминокислота в положении 4 представляет собой I или M, аминокислота в положении 6 представляет собой R, аминокислота в положении 10 представляет собой A или I, аминокислота в положении 16 представляет собой E, аминокислота в положении 19 представляет собой S или R, аминокислота в положении 23 представляет собой E или D, и аминокислота в положении 25 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 29, где аминокислота в положении 1 представляет собой R, аминокислота в положении 4 представляет собой I или M, аминокислота в положении 6 представляет собой R, аминокислота в положении 10 представляет собой A или I, аминокислота в положении 16 представляет собой E, аминокислота в положении 19 представляет собой S или R, аминокислота в положении 23 представляет собой E или D, и аминокислота в положении 25 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 29, где аминокислота в положении 1 представляет собой R, аминокислота в положении 4 представляет собой I, аминокислота в положении 6 представляет собой R, аминокислота в положении 10 представляет собой A, аминокислота в положении 16 представляет собой E, аминокислота в положении 19 представляет собой S, аминокислота в положении 23 представляет собой E, и аминокислота в положении 25 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 30, 31, 32, 33 или 34. В одном воплощении вариабельная область тяжелой цепи включает каркасную область 3 (VH FR3), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 30, 31, 32, 33 или 34.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 4 (VH FR4), содержащую аминокислотную последовательность WGQGTTVTVSS (SEQ ID NO: 44). В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую каркасную область 4 (VH FR4), содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85 %, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 44.
В одном воплощении вариабельная область тяжелой цепи включает:
VH CDR1, представленную в SEQ ID NO: 1;
VH CDR2, представленную в SEQ ID NO: 5;
VH CDR3, представленную в SEQ ID NO: 6;
VH FR1, представленную в SEQ ID NO: 12;
VH FR2, представленную в SEQ ID NO: 13, где аминокислота в положении 9 (X9) представляет собой R или G, а аминокислота в положении 15 (X15) представляет собой I или отсутствует;
VH FR3, представленную в SEQ ID NO: 29, где аминокислота в положении 1 (X1) представляет собой R или отсутствует, аминокислота в положении 4 (X4) представляет собой I или M, аминокислота в положении 6 (X6) представляет собой R или A, аминокислота в положении 10 (X10) представляет собой A, T или I, аминокислота в положении 16 (X16) представляет собой E или L, аминокислота в положении 19 (X19) представляет собой S или R, аминокислота в положении 23 (X23) представляет собой E или D, а аминокислота в положении 25 (X25) представляет собой T или M; и
VH FR4, представленную в SEQ ID NO: 44.
В одном воплощении вариабельная область тяжелой цепи включает:
VH CDR1, представленную SEQ ID NO: 2;
VH CDR2, представленную в SEQ ID NO: 5;
VH CDR3, представленную в SEQ ID NO: 6;
VH FR1, представленную в SEQ ID NO: 11;
VH FR2, представленную в SEQ ID NO: 25, где аминокислота в положении 12 (X12) представляет собой R или G, а аминокислота в положении 18 (X18) представляет собой I или отсутствует;
VH FR3, представленную в SEQ ID NO: 29, где аминокислота в положении 1 (X1) представляет собой R или отсутствует, аминокислота в положении 4 (X4) представляет собой I или M, аминокислота в положении 6 (X6) представляет собой R или A, аминокислота в положении 10 (X10) представляет собой A, T или I, аминокислота в положении 16 (X16) представляет собой E или L, аминокислота в положении 19 (X19) представляет собой S или R, аминокислота в положении 23 (X23) представляет собой E или D, а аминокислота в положении 25 (X25) представляет собой T или M; и
VH FR4, представленную в SEQ ID NO: 44.
В одном воплощении изобретения антитело анти-HLA-A2 включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYHIQWVRQAPGQX44LEWMGX50WIYPGDGSTKYSQKFQGX68VTX71TX73DTSX77STAYMX83LSX86LRSX90DX92AVYYCAREGTYYAMDYWGQGTTVTVSS (SEQ ID NO: 92), в которой аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 представляет собой I или отсутствует, аминокислота в положении 68 представляет собой R или отсутствует, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R или A, аминокислота в положении 77 представляет собой A, T или I, аминокислота в положении 83 представляет собой Е или L, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой Е или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 92, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой Е или D, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 92, где аминокислота в положении 44 представляет собой R или G, аминокислота в положении 50 отсутствует, аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I или M, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A или I, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S или R, аминокислота в положении 90 представляет собой E или D, а аминокислота в положении 92 представляет собой Т.
В одном воплощении вариабельная область тяжелой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 92, где аминокислота в положении 44 представляет собой R, аминокислота в положении 50 отсутствует, аминокислота в положении 68 представляет собой R, аминокислота в положении 71 представляет собой I, аминокислота в положении 73 представляет собой R, аминокислота в положении 77 представляет собой A, аминокислота в положении 83 представляет собой E, аминокислота в положении 86 представляет собой S, аминокислота в положение 90 представляет собой Е, а аминокислота в положении 92 представляет собой Т или М.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 66. В одном воплощении анти-HLA-A2 антитело включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 66.
В одном воплощении анти-HLA-A2 антитело включает вариабельную область легкой цепи, как определено в данном документе. Например, в одном воплощении анти-HLA-A2 антитело может содержать вариабельную область легкой цепи, как определено в данном документе для гуманизированного анти-HLA-A2 антитела.
В одном воплощении анти-HLA-A2 антитело представляет собой scFv, scFab или sdAb. В одном воплощении анти-HLA-A2 антитело представляет собой scFv или scFab. В одном воплощении анти-HLA-A2 антитело представляет собой sdAb. В одном воплощении анти-HLA-A2 антитело представляет собой scFab. В одном воплощении анти-HLA-A2 антитело представляет собой scFv. В одном воплощении гуманизированное анти-HLA-A2 антитело представляет собой scFv, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 91. В одном воплощении анти-HLA-A2 антитело представляет собой scFv, содержащий аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89 %, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 91.
В одном воплощении анти-HLA-A2 антитело представляет собой гуманизированное анти-HLA-A2 антитело. В одном воплощении анти-HLA-A2 антитело представляет собой антитело человека. В одном воплощении анти-HLA-A2 антитело представляет собой негуманизированное антитело. В одном воплощении анти-HLA-A2 антитело представляет собой нечеловеческое антитело.
В одном воплощении анти-HLA-A2 антитело содержит аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90 %, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO, указанными выше. В одном воплощении анти-HLA-A2 антитело представляет собой scFv или scFab, содержащее аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO, указанными выше.
Также предлагается композиция, содержащая, по существу состоящая или состоящая из, по меньшей мере, одного анти-HLA-A2 антитела по изобретению.
Используемый в данном документе термин «состоящий по существу из» со ссылкой на композицию означает, что, по меньшей мере, одно анти-HLA-A2 антитело по изобретению, как описано в данном документе выше, является единственным терапевтическим агентом или агентом с биологической активностью в указанной композиции.
В другом воплощении предлагается фармацевтическая композиция, содержащая, по меньшей мере, одно анти-HLA-A2 антитело по изобретению и фармацевтически приемлемый носитель.
В одном воплощении анти-HLA-A2 антитело представляет собой гуманизированное анти-HLA-A2 антитело.
Примеры фармацевтически приемлемых носителей включают, без ограничения указанным, среды, растворители, покрытия, изотонические и замедляющие абсорбцию агенты, добавки, стабилизаторы, консерванты, поверхностно-активные вещества, вещества, которые ингибируют ферментативную деградацию, спирты, агенты, регулирующие рН, антиоксиданты; хелатирующие агенты, такие как ЭДТА или глутатион; адъюванты (например, гидроксид алюминия); консерванты и пропелленты.
Примеры фармацевтически приемлемых сред включают, без ограничения указанным, воду, нейтральный солевой буферный раствор, фосфатно-солевой буферный раствор, физиологический раствор или другой физиологически забуференный солевой раствор или другой растворитель, такой как гликоль, глицерин, и масло, такое как оливковое масло или органический эфир для инъекций. Фармацевтически приемлемая среда также может содержать липосомы или мицеллы.
Примеры материалов для покрытия включают, без ограничения указанным, лецитин.
Примеры изотонических агентов включают, без ограничения указанным, сахара, хлорид натрия и тому подобное.
Примеры агентов, которые замедляют абсорбцию, включают, без ограничения указанным, моностеарат алюминия и желатин.
Примеры добавок включают, без ограничения указанным, маннит, декстран, углеводы (такие как, например, глюкоза, манноза, сахароза или декстран); глицин, лактоза или поливинилпирролидон или другие добавки, такие как антиоксиданты или инертный газ, стабилизаторы или рекомбинантные белки (например, г. сывороточный альбумин человека), подходящий для введения in vivo.
Примеры подходящих стабилизаторов включают, без ограничения указанным, сахарозу, желатин, пептон, расщепленные белковые экстракты, такие как NZ-амин или NZ-амин AS.
Фармацевтически приемлемые носители, которые могут быть использованы в этих композициях, дополнительно включают, без ограничения указанным, ионообменники, оксид алюминия, стеарат алюминия, лецитин, белки сыворотки, такие как человеческий сывороточный альбумин, буферные вещества, такие как фосфаты, глицин, сорбиновая кислота, калий сорбат, частичные глицеридные смеси насыщенных растительных жирных кислот, воду, соли или электролиты, таких как протаминсульфат, динатрийгидрофосфат, гидрофосфат калия, хлорид натрия, соли цинка, коллоидный диоксид кремния, трисиликат магния, поливинилпирролидон, вещества на основе целлюлозы, полиэтиленгликоль, натрий карбоксиметилцеллюлозу, полиакрилаты, воски, полиэтиленполиоксипропилен-блок полимеры, полиэтиленгликоль и ланолин.
Также предлагается лекарственное средство, содержащее, состоящее или состоящее по существу, по меньшей мере, из одного анти-HLA-A2 антитела по изобретению, как описано выше. В одном воплощении анти-HLA-A2 антитело представляет собой гуманизированное анти-HLA-A2 антитело.
IV. Химерные антигенные рецепторы (CAR)
В одном аспекте настоящее изобретение относится к химерным антигенным рецепторам (CAR). CAR являются химерными белковыми молекулами, которые сочетают антительную специфичность к целевому антигену с рецептор-активированным внутриклеточным доменом иммунной клетки.
CAR по изобретению содержат внеклеточный домен, который специфически связывается с HLA-A2. Внеклеточный домен включает анти-HLA-A2 антитело по изобретению. В одном воплощении анти-HLA-A2 антитело представляет собой гуманизированное анти-HLA-A2 антитело. CAR по изобретению дополнительно содержат трансмембранный домен и цитоплазматический домен, включающий внутриклеточный сигнальный домен. В одном воплощении CAR по изобретению способны экспрессироваться в клетке человека, так что CAR специфически связываются с HLA-A2. В других воплощениях CAR по изобретению способны экспрессироваться в иммунной клетке, так что CAR специфически связываются с HLA-A2. В одном воплощении CAR специфически связываются с HLA-A*02:01. Как будет понятно специалисту в данной области техники, способность антитела связываться с HLA-A2 может быть обнаружена с использованием методов, известных в данной области. Например, связывание CAR с HLA-A2 может быть обнаружено с использованием тетрамера HLA-A2, как проиллюстрировано в данном документе. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из двух или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из трех или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из четырех или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из пяти или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из шести или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из семи или нескольких из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из каждого из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в данном документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из одного или нескольких из числа HLA-A*25, HLA-A*29, HLA-A*30, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в данном документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из двух или нескольких из числа HLA-A*25, HLA-A*29, HLA-A*30, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в данном документе, имеют меньшую реактивность, по меньшей мере, к одному подтипу HLA-A, выбранному из каждого из числа HLA-A*25, HLA-A*29, HLA-A*30, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к двум из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к трем из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к четырем из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к пяти из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к шести из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к семи из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к одному из HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность, по меньшей мере, к двум из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01, по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*25:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*29:02 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*30:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*03:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*31:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*33:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*36:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении CAR, представленные в настоящем документе, имеют меньшую реактивность к HLA-A*68:01 по сравнению с CAR, содержащим антитело BB7.2. В одном воплощении клетка человека представляет собой иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg.
Методы определения реактивности CAR по изобретению к подтипам HLA-A будут известны специалистам в данной области. Например, реактивность CAR по изобретению к подтипам HLA-A может быть определена с помощью анализа с использованием анализа на гранулах с индивидуальными антигенами. Такие анализы на гранулах с индивидуальными антигенами коммерчески доступны (например, FlowPRA Single Antigen Antibody; ONE LAMBDA).
В одном воплощении CAR по изобретению имеет меньшую реактивность к подтипу HLA-A, выбранному из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68 и любую их комбинацию, по сравнению с CAR, содержащими антитело BB7.2, при измерении в условиях Теста A, как описано выше.
В одном воплощении CAR по изобретению имеет меньшую реактивность к подтипу HLA-A, выбранному из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30 и любую их комбинацию, по сравнению с CAR, содержащими антитело BB7.2, при измерении в условиях Теста A, как описано выше.
В одном воплощении CAR по изобретению по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, статистически уступающей реактивности антитела BB7.2, например, при измерении в условиях теста А.
В одном воплощении CAR по изобретению по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A*25, HLA-A*29, HLA-A*30, статистически уступающей реактивности антитела BB7.2, например, при измерении в условиях теста А.
В одном воплощении термин «статистически уступающий» означает, что реактивность (то есть, например, относительное связывание в условиях теста А), измеренная для CAR по изобретению, ниже реактивности, измеренной для CAR, включающего антитело BB7.2, с p-значением, по большей мере, около 0,05, предпочтительно, по большей мере, около 0,01, более предпочтительно, по большей мере, около 0,005 и более предпочтительно, по большей мере, около 0,001 в частности, при анализе с помощью двухфакторного дисперсионного анализа, апостериорного критерия Даннетта.
В одном воплощении CAR по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, уступающей реактивности антитела BB7.2. В некоторых воплощениях такой CAR имеет относительное связывание, по меньшей мере, с одним подтипом HLA-A, выбранным из группы, включающей HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68, уступающее связыванию CAR, включающего антитело BB7.2 при измерении в условиях теста А. В определенных аспектах относительное связывание, измеренное для такого CAR, составляет, по большей мере, около 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или менее относительного связывания, измеренного для CAR, включающего антитело BB7.2.
В одном воплощении CAR по изобретению обладает реактивностью, по меньшей мере, к одному подтипу HLA-A, выбранному из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, уступающей реактивности антитела BB7.2. В некоторых воплощениях такой CAR имеет относительное связывание, по меньшей мере, с одним подтипом HLA-A, выбранным из группы, включающей HLA-A*25, HLA-A*29, HLA-A*30, уступающее связыванию CAR, включающего антитело BB7.2 при измерении в условиях теста А. В определенных аспектах относительное связывание, измеренное для такого CAR, составляет, по большей мере, около 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или менее относительного связывания, измеренного для CAR, включающего антитело BB7.2.
В одном воплощении CAR способен экспрессироваться в иммунной клетке так, что иммунная клетка активируется HLA-A2. В одном воплощении иммунная клетка активируется HLA-A*02:01. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении Т-клетка представляет собой человеческую Т-клетку. В одном воплощении Treg представляет собой Treg человека.
В одном воплощении CAR конкурирует за связывание с HLA-A2 с антителом, включающим: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении CAR связывается с тем же эпитопом HLA-A2, что и антитело, включающее: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении CAR конкурирует за связывание с HLA-A2 с антителом BB7.2. В одном воплощении CAR связывается с тем же эпитопом HLA-A2, что и антитело BB7.2. Антитело BB7.2 может быть выделено из гибридомы BB7.2 (депозитный номер ATCC HB-82).
В одном воплощении CAR по изобретению включает: внеклеточный домен, содержащий анти-HLA-A2 антитело; трансмембранный домен; и цитоплазматический домен, содержащий внутриклеточный сигнальный домен. В одном воплощении анти-HLA-A2 антитело представляет собой гуманизированное анти-HLA-A2 антитело.
В одном воплощении химерный рецептор дополнительно включает метку и/или лидерную последовательность.
В одном воплощении химерный рецептор дополнительно включает метку, такую как, например, метка для контроля качества, обогащения, отслеживания in vivo и тому подобное. Указанная метка может быть локализована на N-конце, C-конце и/или внутри. Примеры меток, которые можно использовать в химерном рецепторе по изобретению, хорошо известны специалисту в данной области. Например, без ограничения указанным, метка, используемая в изобретении, может быть меткой, выбранной из группы, включающей или состоящей из метки гемагглютинина, метки полиаргинина, метки полигистидина, Myc-метки, Strep-метки, S-метки, HAT-метки, 3x Flag-метки, метки кальмодуллин-связывающего пептида, SBP-метки, метки хитин-связывающего домена, GST-метки, метки мальтозосвязывающего белка, метки флуоресцентного белка (например, eGFP), T7-метки, V5-метки и Xpress-метки.
Внеклеточный домен представляет собой специфический для мишени связывающий элемент, который также иногда называют нацеливающим плечом CAR. Внеклеточный домен выбирается для распознавания лиганда, который действует как маркер клеточной поверхности на клетках-мишенях, связанных с конкретным болезненным состоянием. CAR по настоящему изобретению разработан для нацеливания на клетку, презентирующую HLA-A2, путем создания соответствующего внеклеточного домена, который специфически связывается с эпитопом HLA-A2. Специфичный для мишени связывающий элемент или антигенсвязывающий домен CAR по настоящему изобретению может упоминаться в данном документе как домен, связывающий анти-HLA-A2. В некоторых воплощениях домен, связывающий анти-HLA-A2, может представлять собой гуманизированный домен, связывающий анти-HLA-A2.
Трансмембранный домен присоединен к внеклеточному домену и цитоплазматическому домену CAR. Трансмембранный домен способен передавать сигналы во внутриклеточный сигнальный домен(домены) цитоплазматического домена всякий раз, когда внеклеточный домен CAR связывается с мишенью.
Цитоплазматический домен, который включает внутриклеточный сигнальный домен CAR, отвечает за активацию, по меньшей мере, одной из физиологических эффекторных функций иммунной клетки (например, регуляторной T-клетки), в которую был помещен CAR. Термин «эффекторная функция» относится к специализированной функции иммунной клетки. Например, эффекторная функция регуляторной Т-клетки может включать супрессию или отрицательное регулирование индукции и/или пролиферации других иммунных клеток. Кроме того, эффекторная функция Treg может включать воздействие на неиммунные клетки, которое приводит к улучшению клинического состояния, например, способствует восстановлению или регенерации тканей. Таким образом, термин «внутриклеточный сигнальный домен» относится к части белка, которая преобразовывает сигнал эффекторной функции и направляет иммунную клетку для выполнения своей специализированной функции. Хотя обычно можно использовать весь внутриклеточный сигнальный домен, во многих случаях нет необходимости использовать всю цепь. В той степени, в которой используется укороченная часть внутриклеточного сигнального домена, такая укороченная часть может использоваться вместо интактной цепи, если она передает сигнал эффекторной функции. Термин «внутриклеточный сигнальный домен», таким образом, подразумевает включение любой усеченной части внутриклеточного сигнального домена, достаточной для преобразования сигнала эффекторной функции.
В одном воплощении может иметь место спейсерный домен (или линкер или шарнир), включенный между антигенсвязывающим доменом и трансмембранным доменом CAR или между внутриклеточным сигнальным доменом и трансмембранным доменом CAR. Как определено выше, спейсерный домен, линкер и шарнир представляют собой олиго- или полипептиды, которые функционируют для связывания трансмембранного домена либо с антигенсвязывающим доменом, либо с внутриклеточным сигнальным доменом в полипептидной цепи. Спейсерный домен может включать, например, вплоть до 300 аминокислот, от 10 до 100 аминокислот, от 25 до 75 аминокислот или от 25 до 50 аминокислот, или аминокислоты любых поддиапазонов или отдельных числовых значений в этих диапазонах.
Внеклеточный домен
Внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению. В одном воплощении внеклеточный домен включает гуманизированное анти-HLA-A2 антитело по настоящему изобретению.
В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой scFv, scFab или sdAb. В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой scFv или scFab. В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой sdAb. В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой scFv. В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой scFab. В одном воплощении внеклеточный домен содержит любое гуманизированное анти-HLA-A2 антитело по настоящему изобретению, где гуманизированное анти-HLA-A2 антитело представляет собой scFv, scFab или sdAb. В одном воплощении внеклеточный домен содержит любое гуманизированное анти-HLA-A2 антитело по настоящему изобретению, где гуманизированное анти-HLA-A2 антитело представляет собой scFv или sdAb. В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой sdAb. В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой scFv. В одном воплощении внеклеточный домен содержит анти-HLA-A2 антитело по настоящему изобретению, которое представляет собой scFab.
В некоторых воплощениях внеклеточный домен может содержать шарнир, где трансмембранный домен прикреплен к внеклеточной области CAR, например, антигенсвязывающему домену CAR, через шарнир. В одном воплощении шарнир может быть из человеческого белка. Например, в одном воплощении шарнир может быть шарниром человеческого Ig (иммуноглобулина), например, шарниром IgG4 или шарниром CD8α. В некоторых случаях внеклеточный домен CAR по изобретению может содержать шарнир CD8α. В одном воплощении шарнирная область содержит область стебля CD8α. В одном воплощении шарнир CD8 может кодироваться последовательностью нуклеиновой кислоты SEQ ID NO: 15 в патенте США № 9102760. В одном воплощении шарнир CD8 может содержать аминокислотную последовательность SEQ ID NO: 21 в патенте США № 9102760. В другом воплощении шарнир CD8 может содержать аминокислотную последовательность SEQ ID NO: 21 в патенте США № 9102760. В одном воплощении шарнир или спейсер может включать аминокислотную последовательность SEQ ID NO: 115 или 219 в таблице 3. В одном воплощении шарнир или спейсер могут кодироваться последовательностью нуклеиновой кислоты SEQ ID NO: 159 или 220 в таблице 4.
Трансмембранный домен
Трансмембранный домен может быть получен либо из природного источника, либо из синтетического источника. В одном воплощении трансмембранный домен может быть получен из природного источника, например, из любого мембраносвязанного или трансмембранного белка.
В одном воплощении трансмембранный домен CAR может быть получен из трансмембранного домена, который естественным образом связан с одним из доменов CAR. В других воплощениях трансмембранный домен может быть выбран или модифицирован аминокислотной заменой, чтобы избежать связывания таких доменов с трансмембранными доменами одного и того же или разных белков поверхностной мембраны, чтобы минимизировать взаимодействия с другими членами рецепторного комплекса.
В одном воплощении трансмембранный домен может включать одну или несколько дополнительных аминокислот, смежных с трансмембранной областью, например, одну или несколько аминокислот, связанных с внеклеточной областью белка, из которого получают трансмембранный домен (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислот внеклеточной области) и/или одну или несколько дополнительных аминокислот, связанных с цитоплазматической областью белка? из которого получают трансмембранный домен (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислот цитоплазматической области).
В одном воплощении трансмембранный домен может содержать трансмембранный домен белка, выбранного из группы, состоящей из CD3-гамма, CD3-дельта, CD3-эпсилон, CD3-дзета, альфа-цепи Т-клеточного рецептора, бета-цепи Т-клеточного рецептора, гамма-цепь Т-клеточного рецептора, дельта-цепь Т-клеточного рецептора, CD28, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137, KIRDS2, OX40, CD2, CD27, LFA-1 (CD1a, CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD40, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRFl), CD160, CD19, IL2R бета, IL2R гамма, IL7Ra, ITGA1, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, PD1, ITGAX, CDl1c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, TNFR2, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (тактильный), CEACAM1, CRT AM, Ly9 (CD229), CD160 (BY55), PSGL1, CDIOO (SEMA4D), SLAMF6 (NTB-A, Lyl08), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LT BR, PAG/Cbp, NKp44, NKp30, NKp46, NKG2D и/или NKG2C и CD154 и любой их комбинации. В одном воплощении трансмембранный домен может содержать трансмембранный домен CD28. В одном воплощении трансмембранный домен CD28 кодируется последовательностью нуклеиновой кислоты SEQ ID NO: 160 в таблице 4. В одном воплощении трансмембранный домен CD28 содержит аминокислотную последовательность SEQ ID NO: 116 в таблице 3. В другом воплощении трансмембранный домен CD28 содержит аминокислотную последовательность SEQ ID NO: 116 в таблице 3.
В одном воплощении трансмембранный домен может содержать трансмембранный домен CD8. В одном воплощении трансмембранный домен CD8 кодируется последовательностью нуклеиновой кислоты SEQ ID NO: 16 в патенте США № 9102760. В одном воплощении трансмембранный домен CD8 содержит аминокислотную последовательность SEQ ID NO: 22 в патенте США № 9102760. В другом воплощении трансмембранный домен CD8 содержит аминокислотную последовательность SEQ ID NO: 22 в патенте США № 9102760.
В одном воплощении трансмембранный домен CD8 кодируется последовательностью нуклеиновой кислоты SEQ ID NO: 224 в таблице 4. В одном воплощении трансмембранный домен CD8 содержит аминокислотную последовательность SEQ ID NO: 223 в таблице 3.
В других воплощениях трансмембранный домен может быть синтетическим, и в этом случае он может содержать преимущественно гидрофобные остатки, включая лейцин и валин. В одном воплощении триплет фенилаланина, триптофана и валина может быть обнаружен на каждом конце синтетического трансмембранного домена.
В одном воплощении короткий олиго- или полипептидный линкер может образовывать связь между трансмембранным доменом и цитоплазматическим доменом CAR. В одном воплощении линкер может иметь длину в диапазоне 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот. В одном воплощении линкер может содержать глицин-сериновый дублет.
Цитоплазматический домен
В одном воплощении внутриклеточные сигнальные домены для применения в CAR по изобретению могут включать цитоплазматические последовательности Т-клеточного рецептора (TCR) и корецепторов, которые действуют совместно, чтобы инициировать трансдукцию сигнала после включения антигенного рецептора, а также любое производное или вариант этих последовательностей и любой синтетической последовательности, которая имеет одинаковые функциональные возможности.
Первичные цитоплазматические сигнальные последовательности регулируют первичную активацию комплекса TCR либо стимулирующим, либо ингибирующим образом. Первичные цитоплазматические сигнальные последовательности, которые действуют стимулирующим образом, могут включать сигнальные мотивы, которые известны как иммунорецепторные мотивы активации на основе тирозина или ITAM. Примеры ITAM, содержащих первичные цитоплазматические сигнальные последовательности, которые особенно применимы в изобретении, включают последовательности, полученные из CD3-дзета, FcR-гамма, CD3-гамма, CD3-дельта, CD3-эпсилон, CD5, CD22, CD79a, CD79b и CD66d. В одном воплощении первичный сигнальный домен содержит модифицированный домен ITAM, например, мутированный домен ITAM, который изменил (например, увеличил или уменьшил) активность по сравнению с собственным доменом ITΑΜ. В одном воплощении первичный сигнальный домен включает модифицированный ITAM-содержащий первичный внутриклеточный сигнальный домен, например, оптимизированный и/или укороченным ITAM-содержащий первичный внутриклеточный сигнальный домен. В воплощении первичный сигнальный домен включает один, два, три, четыре или более мотивов ITAM.
В одном воплощении внутриклеточный сигнальный домен включает функциональный сигнальный домен белка, выбранного из группы, состоящей из CD3-гамма, CD3-дельта, CD3-эпсилон, CD3-дзета, FcR-гамма (например, FCγRI, RCγRIIA, FcγRIIB1, FcγRIIB2, FcγRIIIA или FcγRIIIB), FcR альфа (например, FcαRI), FcR эпсилон (например, FcεRI или FcεRII), CD5, CD22, CD79a, CD79b, DAP10, DAP12 и CD66d и любой их комбинации. В одном воплощении внутриклеточный сигнальный домен содержит или состоит из первичного сигнального домена CD3-дзета.
Известно, что сигналов, генерируемых только через TCR, может быть недостаточно для полной активации Т-клеток и что также может потребоваться вторичный или костимулирующий сигнал. Таким образом, в некоторых воплощениях активация Т-клеток может быть опосредована двумя классами цитоплазматической сигнальной последовательности: теми, которые инициируют антиген-зависимую первичную активацию через TCR (первичные цитоплазматические сигнальные последовательности), и теми, которые действуют антиген-независимым образом, чтобы обеспечить вторичный или костимулирующий сигнал (вторичные цитоплазматические сигнальные последовательности). В одном воплощении внутриклеточный сигнальный домен может дополнительно содержать костимулирующий домен.
В одном воплощении внутриклеточный сигнальный домен CAR может содержать CD3-дзета-сигнальный домен сам по себе или он может быть объединен с любым другим искомым внутриклеточным сигнальным доменом (доменами), полезным в контексте CAR по изобретению. Например, внутриклеточный сигнальный домен CAR может содержать участок дзета-цепи CD3 и костимулирующий сигнальный домен. Костимулирующий сигнальный домен относится к части CAR, содержащей внутриклеточный домен костимулирующей молекулы. Костимулирующая молекула представляет собой молекулу клеточной поверхности, отличную от антигенного рецептора или его лигандов, которая необходима для эффективного ответа лимфоцитов на антиген. Примеры таких молекул включают OX40, CD27, CD28, антиген-1, связанный с функцией лимфоцитов (LFA-1) (CD11a/CD18), TNFR1 (CD120a/TNFRSF1A), TNFR2 (CD120b/TNFRSF1B), CTLA-4 (CD152), CD95, ICOS (CD278), 4-1BB (CD137), CD2, CD30, CD40, PD-1, CD7, LIGHT, NKG2C, B7-H3, ICAM-1, лиганд, который специфически связывается с CD83, IL2ra (CD25), IL6Ra (CD126), IL-7Ra (CD127), IL-13RA1, IL-13RA2, IL-33R (IL1RL1), IL-10RA, IL-10RB, IL-4R, IL-5R (CSF2RB), ARHR, рецептор BAFF, IL-21R, TGFbR1, TGFbR2, TGFbR3, общая гамма-цепь, молекула MHC класса I, BTLA и рецептор Toll-лиганда, лиганд, который специфически связывается с CD83, CDS, ICAM-1, GITR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), NKp44, NKp30, NKp46, CD160 (BY55), CD19, CD19a, CD4, CD8-альфа, CD8-бета, IL2R-бета, IL2R-гамма, IL7R-альфа, ITGA4, VLA1, CD49a, IT4, CD49D, IT4, CD49 V, IT4, CD49 V, IT4, CD49A, IT4, CD49 -6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CitgbD18, ITGB7, TRANCE/RANKL, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Тактильный), CEACAM1, CRTAM, Ly9 (CD229), PSGL1, CD100 (SEMA4D), CD69, SLAMF6 (NTB-A, Lyl08), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, LAT, GADS, SLP-76, PAG/Cbp, NKG2D и т.п.
Внутриклеточные сигнальные последовательности в цитоплазматической части CAR по изобретению могут быть связаны друг с другом в случайном или заданном порядке. Необязательно, короткий олиго- или полипептидный линкер, например, длиной от 2 до 10 аминокислот (например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот) может образовывать связь между внутриклеточными сигнальными последовательностями. В одном воплощении глицин-сериновый дублет может быть использован в качестве подходящего линкера. В одном воплощении в качестве подходящего линкера может быть использована одна аминокислота, например, аланин, глицин.
В одном воплощении внутриклеточный сигнальный домен предназначен для включения двух или более, например, 2, 3, 4, 5 или более, костимулирующих сигнальных доменов. В одном воплощении два или более, например, 2, 3, 4, 5 или более, костимулирующих сигнальных доменов разделены молекулой линкера, например, молекулой линкера, описанной в данном документе. В одном воплощении внутриклеточный сигнальный домен содержит два костимулирующих сигнальных домена. В некоторых воплощениях молекула линкера представляет собой глициновый остаток. В некоторых воплощениях линкер представляет собой остаток аланина.
В одном воплощении цитоплазматический домен может включать сигнальный домен CD3-дзета и сигнальный домен CD28. В другом воплощении цитоплазматический домен может включать сигнальный домен CD3-дзета и сигнальный домен 4-IBB. В еще одном воплощении цитоплазматический домен может включать сигнальный домен CD3-дзета и сигнальные домены CD28 и 4-1BB.
В одном воплощении цитоплазматический домен может включать сигнальный домен CD28 и сигнальный домен CD3-дзета, где сигнальный домен CD28 кодируется последовательностью нуклеиновой кислоты, представленной в SEQ ID NO: 161 в таблице 4, и передачей сигналов домен CD3-дзета кодируется последовательностью нуклеиновой кислоты, представленной в SEQ ID NO: 162 в таблице 4.
В одном воплощении цитоплазматический домен может включать сигнальный домен CD28 и сигнальный домен CD3-дзета, где сигнальный домен CD28 содержит аминокислотную последовательность SEQ ID NO: 117 в таблице 3 и сигнальный домен CD3- дзета содержит аминокислотную последовательность SEQ ID NO: 118 в таблице 3.
В одном воплощении цитоплазматический домен может включать сигнальный домен CD28 и сигнальный домен CD3-дзета, где сигнальный домен CD28 содержит аминокислотную последовательность, представленную в SEQ ID NO: 117 в таблице 3, и сигнальный домен CD3-дзета содержит аминокислотную последовательность, представленную в SEQ ID NO: 118 в таблице 3.
В одном воплощении цитоплазматический домен может включать сигнальный домен 4-1BB и сигнальный домен CD3-дзета, где сигнальный домен 4-1BB кодируется последовательностью нуклеиновой кислоты, представленной в SEQ ID NO: 17 из пат. США № 9102760 и сигнальный домен CD3-дзета кодируется последовательностью нуклеиновой кислоты, представленной в SEQ ID NO: 162 в таблице 4.
В одном воплощении цитоплазматический домен может включать сигнальный домен 4-1BB и сигнальный домен CD3-дзета, где сигнальный домен 4-1BB содержит аминокислотную последовательность SEQ ID NO: 23 в патенте США № 9102760 и сигнальный домен CD3-дзета содержит аминокислотную последовательность SEQ ID NO: 118 в таблице 3.
В одном воплощении цитоплазматический домен может включать сигнальный домен 4-1BB и сигнальный домен CD3-дзета, где сигнальный домен 4-1BB содержит аминокислотную последовательность, представленную в SEQ ID NO: 23 в патенте США № 9102760 и сигнальный домен CD3-дзета содержит аминокислотную последовательность, представленную в SEQ ID NO: 118 в таблице 3.
В одном воплощении CAR включает аминокислотную последовательность, представленную в SEQ ID NO: 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138 или 213. В одном воплощении CAR содержит аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92 %, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138 или 213.
V. НУКЛЕИНОВЫЕ КИСЛОТЫ И ВЕКТОРЫ
В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей анти-HLA-A2 антитело по настоящему изобретению. В некоторых воплощениях в изобретении предлагается нуклеиновая кислота, кодирующая анти-HLA-A2 антитело по настоящему изобретению. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90 или 91. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111 или 112. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90 или 91. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111 или 112. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, содержащей последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157 или 158. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, содержащей последовательность нуклеиновой кислоты, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157 или 158. В некоторых воплощениях изобретение относится к белку, кодируемому нуклеиновой кислотой, содержащей последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157 или 158. В некоторых воплощениях изобретение относится к белку, кодируемому нуклеиновой кислотой, содержащей последовательность нуклеиновой кислоты, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157 или 158. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 61, 62, 63, 64, 65, 66, 67, 68, 69, 70 или 71. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 61, 62, 63, 64, 65, 66, 67, 68, 69, 70 или 71.
В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей химерный антигенный рецептор (CAR) по настоящему изобретению. Последовательности нуклеиновой кислоты, кодирующие CAR, по изобретению могут кодировать CAR, которые включают внеклеточный домен, описанный где-либо в данном документе.
Внеклеточный домен может содержать анти-HLA-A2 антитело по настоящему изобретению. Внеклеточный домен может содержать гуманизированное анти-HLA-A2 антитело по настоящему изобретению. В одном воплощении внеклеточный домен может дополнительно включать лидерную последовательность. В одном воплощении лидерная последовательность включает аминокислотную последовательность, представленную в SEQ ID NO: 113. В одном воплощении внеклеточный домен содержит шарнирную область, где анти-HLA-A2-связывающий домен соединен с трансмембранным доменом посредством шарнирной области. В одном воплощении шарнирная область содержит область стебля CD8α.
Последовательности нуклеиновых кислот по изобретению могут кодировать CAR, которые содержат трансмембранный домен, описанный где-либо в данном документе. Например, в одном воплощении трансмембранный домен включает трансмембранный домен белка, выбранного из группы, состоящей из CD3-гамма, CD3-дельта, CD3-эпсилон, CD3-дзета, альфа-цепи Т-клеточного рецептора, бета-цепи Т-клеточного рецептора, гамма-цепь Т-клеточного рецептора, дельта-цепь Т-клеточного рецептора, CD28, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137, KIRDS2, OX40, CD2, CD27, LFA-1 (CD1a, CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD40, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRFl), CD160, CD19, IL2R бета, IL2R гамма, IL7Ra, ITGA1, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, PD1, ITGAX, CDl1c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, TNFR2, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (tactile), CEACAM1, CRT AM, Ly9 (CD229), CD160 (BY55), PSGL1, CDIOO (SEMA4D), SLAMF6 (NTB-A, Lyl08), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LT BR, PAG/Cbp, NKp44, NKp30, NKp46, NKG2D и/или NKG2C и CD154 и любой их комбинации. В одном воплощении трансмембранный домен содержит трансмембранный домен CD28.
Последовательности нуклеиновых кислот по изобретению могут кодировать CAR, которые содержат цитоплазматический домен, как описано в данном документе в любом месте. Кроме того, цитоплазматический домен может включать внутриклеточный сигнальный домен, как описано в данном документе в любом месте. Например, в одном воплощении внутриклеточный сигнальный домен включает функциональный сигнальный домен белка, выбранного из группы, состоящей из CD3-гамма, CD3-дельта, CD3-эпсилон, CD3-дзета, FcR-гамма (например, FCγRI, RCγRIIA, FcγRIIB1, FcγRIIB2, FcγRIIIA или FcγRIIIB), FcR альфа (например, FcαRI), FcR эпсилон (например, FcεRI или FcεRII), CD5, CD22, CD79a, CD79b, DAP10, DAP12 и CD66d и любой их комбинации. В одном воплощении внутриклеточный сигнальный домен включает функциональный сигнальный домен CD3-дзета. В одном воплощении внутриклеточный сигнальный домен дополнительно включает костимулирующий домен. Костимулирующий домен CAR, кодируемых последовательностями нуклеиновых кислот по изобретению, может представлять собой костимулирующий домен, как описано в данном документе в любом месте. Например, в одном воплощении костимулирующий домен содержит функциональный сигнальный домен белка, выбранного из группы, которая включает OX40, CD27, CD28, антиген-1, связанный с функцией лимфоцитов (LFA-1) (CD11a/CD18), TNFR1 (CD120a/TNFRSF1A), TNFR2 (CD120b/TNFRSF1B), CTLA-4 (CD152), CD95, ICOS (CD278), 4-1BB (CD137), CD2, CD30, CD40, PD-1, CD7, LIGHT, NKG2C, B7-H3, ICAM-1, лиганд, который специфически связывается с CD83, IL2ra (CD25), IL6Ra (CD126), IL-7Ra (CD127), IL-13RA1, IL-13RA2, IL-33R (IL1RL1), IL-10RA, IL-10RB, IL-4R, IL-5R (CSF2RB), ARHR, рецептор BAFF, IL-21R, TGFbR1, TGFbR2, TGFbR3, общая гамма-цепь, молекулу MHC класса I, BTLA и рецептор Toll-лиганда, лиганд, который специфически связывается с CD83, CDS, ICAM-1, GITR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), NKp44, NKp30, NKp46, CD160 (BY55), CD19, CD19a, CD4, CD8-альфа, CD8-бета, IL2R-бета, IL2R-гамма, IL7R-альфа, ITGA4, VLA1, CD49a, IT4, CD49D, IT4, CD49 V, IT4, CD49 V, IT4, CD49A, IT4, CD49 -6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CitgbD18, ITGB7, TRANCE/RANKL, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Тактильный), CEACAM1, CRTAM, Ly9 (CD229), PSGL1, CD100 (SEMA4D), CD69, SLAMF6 (NTB-A, Lyl08), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, LAT, GADS, SLP-76, PAG/Cbp, NKG2D и любую их комбинацию. В одном воплощении костимулирующий домен содержит функциональный сигнальный домен белка, выбранного из группы, состоящей из CD28, 4-1BB и их комбинации. В одном воплощении костимулирующий домен содержит функциональный сигнальный домен CD28. В одном воплощении костимулирующий домен содержит функциональный сигнальный домен 4-1BB. В одном воплощении последовательности, содержащие внутриклеточный сигнальный домен, экспрессируются в одном и том же каркасе и в виде одной полипептидной цепи.
Последовательности нуклеиновых кислот по настоящему изобретению могут быть выделенными последовательностями нуклеиновых кислот.
В одном воплощении нуклеиновая кислота предлагается в качестве транскрипта информационной РНК. В одном воплощении нуклеиновая кислота представлена в виде ДНК-конструкции.
В одном воплощении предложена конструкция рекомбинантной ДНК, содержащая последовательности, кодирующие CAR, где CAR включает: (i) внеклеточный домен, содержащий анти-HLA-A2 антитело; (ii) трансмембранный домен; и (iii) цитоплазматический домен, содержащий внутриклеточный сигнальный домен, где кодированный CAR способен экспрессироваться в клетке человека, так что CAR способен специфически связываться с HLA-A2. В одном воплощении CAR способен специфически связываться с HLA-A*02:01. В одном воплощении анти-HLA-A2 антитело представляет собой гуманизированное анти-HLA-A2 антитело. В одном воплощении клетка человека представляет собой иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg.
В одном воплощении предложена конструкция рекомбинантной ДНК, содержащая последовательности, кодирующие CAR, где CAR включает: (i) внеклеточный домен, содержащий анти-HLA-A2 антитело; (ii) трансмембранный домен; и (iii) цитоплазматический домен, содержащий внутриклеточный сигнальный домен, где кодированный CAR способен экспрессироваться в клетке человека, так что CAR способен специфически связываться с HLA-A2. В одном воплощении CAR способен специфически связываться с HLA-A*02:01. В одном воплощении связывание CAR с HLA-A2 может быть обнаружено посредством применения тетрамера HLA-A2, как проиллюстрировано в данном документе. В одном воплощении анти-HLA-A2 антитело представляет собой гуманизированное анти-HLA-A2 антитело. В одном воплощении Treg представляет собой Treg человека.
В одном воплощении кодируемый CAR способен экспрессироваться в иммунной клетке так, что иммунная клетка активируется HLA-A2. В одном воплощении иммунная клетка активируется HLA-A*02:01. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка представляет собой Treg. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении Т-клетка представляет собой человеческую Т-клетку. В одном воплощении Treg представляет собой Treg человека.
В одном воплощении кодированный CAR конкурирует за связывание с HLA-A2 с антителом, включающим: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении CAR связывается с тем же эпитопом HLA-A2, что и антитело, включающее: область 1, определяющую комплементарность тяжелой цепи (HCDR1), имеющую аминокислотную последовательность SEQ ID NO: 183; область 2, определяющую комплементарность тяжелой цепи (HCDR2), имеющую аминокислотную последовательность SEQ ID NO: 185; область 3, определяющую комплементарность тяжелой цепи (HCDR3), имеющую аминокислотную последовательность SEQ ID NO: 187; область 1, определяющую комплементарность легкой цепи (LCDR1), имеющую аминокислотную последовательность SEQ ID NO: 188; область 2, определяющую комплементарность легкой цепи (LCDR2), имеющую аминокислотную последовательность SEQ ID NO: 189; и область 3, определяющую комплементарность легкой цепи (LCDR3), имеющую аминокислотную последовательность SEQ ID NO: 190. В одном воплощении кодируемый CAR конкурирует за связывание с HLA-A2 антителом BB7.2. В одном воплощении кодируемый CAR связывается с тем же эпитопом HLA-A2, что и антитело BB7.2.
В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138 или 213. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, кодирующей белок, содержащий аминокислотную последовательность, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138 или 213. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, содержащей последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 или 214. В некоторых воплощениях изобретение относится к нуклеиновой кислоте, содержащей последовательность нуклеиновой кислоты, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 или 214. В некоторых воплощениях изобретение относится к белку, кодируемому нуклеиновой кислотой, содержащей последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 или 214. В некоторых воплощениях изобретение относится к белку, кодируемому нуклеиновой кислотой, содержащей последовательность нуклеиновой кислоты, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 или 214.
Последовательности нуклеиновых кислот, кодирующие искомые молекулы, могут быть получены с использованием рекомбинантных способов, известных в данной области, таких как, например, скрининг библиотек из клеток, экспрессирующих ген, путем получения гена из вектора, о котором известно, что он его включает, или путем выделения непосредственно из клеток и тканей, содержащих его, используя стандартные методы. Альтернативно, представляющая интерес нуклеиновая кислота может быть получена синтетическим путем, а не клонирована.
Настоящее изобретение также относится к вектору, содержащему молекулу нуклеиновой кислоты, кодирующую анти-HLA-A2 антитело, или молекулу нуклеиновой кислоты, кодирующую CAR. В одном воплощении изобретение относится к вектору, содержащему молекулу нуклеиновой кислоты, кодирующую гуманизированное анти-HLA-A2 антитело. В одном воплощении изобретение относится к векторам, в которые вставлена такая молекула нуклеиновой кислоты.
В одном воплощении настоящее изобретение относится к вектору, включающему последовательность нуклеиновой кислоты, описанной где-либо в данном документе.
В некоторых воплощениях настоящее изобретение относится к вектору, включающему нуклеиновую кислоту, которая кодирует любое из анти-HLA-A2 антител, описанных в данном документе. В других воплощениях настоящее изобретение относится к вектору, включающему нуклеиновую кислоту, которая кодирует любое из гуманизированных анти-HLA-A2 антител, описанных в данном документе. В некоторых воплощениях настоящее изобретение относится к вектору, включающему нуклеиновую кислоту, содержащую последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157 или 158. В некоторых воплощениях изобретение относится к вектору, включающему нуклеиновую кислоту, содержащую последовательность нуклеиновой кислоты, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157 или 158. В определенных аспектах предлагается клетка-хозяин, которая включает такой вектор.
В одном воплощении настоящее изобретение относится к вектору, содержащему нуклеиновую кислоту, кодирующую CAR, по изобретению. В некоторых воплощениях настоящее изобретение относится к вектору, содержащему последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 или 214. В некоторых воплощениях изобретение относится к вектору, включающему нуклеиновую кислоту, содержащую последовательность нуклеиновой кислоты, имеющую около 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182 или 214.
В одном воплощении векторы по изобретению могут быть трансдуцированы в клетку. В одном воплощении векторы по изобретению могут быть трансдуцированы или сконструированы посредством способов на основе невирусных векторов в клетку человека. В одном воплощении векторы по изобретению могут быть трансдуцированы в иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении иммунная клетка представляет собой регуляторную T-клетку (Treg). В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка может представлять собой Treg. В одном воплощении вектор способен экспрессировать CAR в иммунных клетках млекопитающих. В одном воплощении иммунная клетка млекопитающего представляет собой иммунную клетку человека. В одном воплощении вектор способен экспрессировать CAR в регуляторных иммунных клетках млекопитающих. В одном воплощении регуляторная иммунная клетка млекопитающего представляет собой регуляторную иммунную клетку человека. В одном воплощении вектор способен экспрессировать CAR в Т-клетках млекопитающих. В одном воплощении T-клетки млекопитающих представляют собой регуляторные T-клетки млекопитающих. В одном воплощении Т-клетка млекопитающего представляет собой Т-клетку человека. В одном воплощении Т-клетка млекопитающего представляет собой регуляторную Т-клетку человека.
В одном воплощении вектор представляет собой клонирующий или экспрессирующий вектор, например, вектор, включающий, без ограничения указанным, одну или несколько плазмид (например, экспрессирующие плазмиды, клонирующие векторы, миникольца, минивекторы, двойные микрохромосомы), ретровирусные и лентивирусные векторные конструкты.
Настоящее изобретение включает ретровирусные и лентивирусные векторные конструкции, экспрессирующие CAR, которые могут быть непосредственно трансдуцированы в клетку. Векторы, полученные из ретровирусов, таких как лентивирус, являются подходящими инструментами для достижения долгосрочного переноса гена, поскольку они обеспечивают долговременную стабильную интеграцию трансгена и его размножение в дочерних клетках. Лентивирусные векторы имеют дополнительное преимущество перед векторами, полученными из онкоретровирусов, таких как вирусы лейкоза мыши, в том, что такие векторы могут трансдуцировать непролиферирующие клетки, такие как гепатоциты. Они также обладают дополнительным преимуществом низкой иммуногенности.
Вкратце, экспрессия природных или синтетических нуклеиновых кислот, кодирующих CAR, обычно достигается путем функционального связывания нуклеиновой кислоты, кодирующей полипептид CAR или его частей, с промотором и включения конструкции в экспрессирующий вектор. Векторы могут подходить для репликации и интеграции в эукариот. Типичные клонирующие векторы включают терминаторы транскрипции и трансляции, последовательности инициации и промоторы, полезные для регуляции экспрессии искомой последовательности нуклеиновой кислоты.
В дополнение к способам, описанным выше, могут использоваться следующие способы.
Экспрессирующие конструкции по настоящему изобретению также можно использовать для иммунизации нуклеиновыми кислотами и генной терапии с использованием стандартных протоколов доставки генов. Способы доставки генов известны в данной области. См., например, Пат. США No. 5399346, 5580859, 5589466, полностью включенные в настоящий документ ссылкой. В другом воплощении изобретение обеспечивает генно-терапевтический вектор. В другом воплощении способы редактирования генома, такие как методы CRISPR-Cas9 или TALEN, могут использоваться для введения нуклеиновых кислот, кодирующих CAR по настоящему изобретению, в геном иммунных клеток-реципиентов (Delhove, JMKM и др., 2017; Eyquem, J. et al., 2017).
Нуклеиновая кислота может быть клонирована во множество типов векторов. Например, нуклеиновая кислота может быть клонирована в вектор, включающий, без ограничения указанным, плазмиду, фагмиду, производное фага, вирус животных и космиду. Векторы, представляющие особый интерес, включают экспрессирующие векторы, репликационные векторы, векторы для получения зондов и векторы для секвенирования.
Кроме того, экспрессирующий вектор может быть предоставлен клетке в форме вирусного вектора. Технология вирусных векторов хорошо известна в данной области и описана, например, в Sambrook et al. (2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York), и в других руководствах по вирусологии и молекулярной биологии. Вирусы, которые полезны в качестве векторов, включают, без ограничения указанным, ретровирусы, аденовирусы, аденоассоциированные вирусы, вирусы герпеса, поксвирусы и лентивирусы (см., например, Пат. США No. 5350674 и 5585362). В общем, подходящий вектор содержит точку начала репликации, функционирующую, по меньшей мере, в одном организме, промоторную последовательность, удобные сайты эндонуклеаз рестрикции и один или несколько селектируемых маркеров (например, WO 01/96584; WO 01/29058; и Пат. США № 6326193).
Был разработан ряд вирусных систем для переноса генов в клетки млекопитающих. Например, ретровирусы предоставляют удобную платформу для систем доставки генов. Выбранный ген может быть вставлен в вектор и упакован в ретровирусные частицы с использованием методов, известных в данной области. Затем рекомбинантный вирус может быть выделен и доставлен в клетки объекта либо in vivo, либо ex vivo. Ряд ретровирусных систем известен в данной области. В некоторых воплощениях используются аденовирусные векторы. Ряд аденовирусных векторов известен в данной области. В одном воплощении используются лентивирусные векторы.
Дополнительные транскрипционно активные элементы, например, промоторы и энхансеры, могут регулировать частоту инициации транскрипции. Как правило, в случае основного промотора, они расположены в области 30-110 п.н. выше по последовательности от стартового сайта, хотя недавно было показано, что ряд промоторов также содержат функциональные элементы ниже по последовательности от стартового сайта, а энхансерные элементы обычно расположены -2000 п.о. выше по последовательности от стартового сайта. Интервал между элементами промотора часто является гибким, так что функция промотора сохраняется, когда элементы инвертированы или перемещены относительно друг друга. В промоторе тимидинкиназы (tk) расстояние между промоторными элементами может быть увеличено до 50 п.н., прежде чем активность начнет снижаться. В зависимости от промотора оказывается, что отдельные элементы могут функционировать совместно или независимо для активации транскрипции.
Одним примером подходящего промотора является последовательность немедленно-раннего промотора цитомегаловируса (CMV). Эта промоторная последовательность является сильной конститутивной промоторной последовательностью, способной стимулировать высокие уровни экспрессии любой полинуклеотидной последовательности, функционально связанной с ней. Другим примером подходящего промотора является фактор элонгации-1α (EF-1α). Другим примером подходящего промотора является промотор фосфоглицераткиназы (PGK). Однако могут также использоваться другие последовательности конститутивного промотора, включая, без ограничения указанным, ранний промотор обезьяньего вируса 40 (SV40), вирус опухоли молочной железы мыши (MMTV), промотор длинного концевого повтора (LTR) вируса иммунодефицита человека (HIV), промотор MoMuLV, промотор вируса птичьего лейкоза, немедленно-ранний промотор вируса Эпштейна-Барра, промотор вируса саркомы Рауса, а также промоторы генов человека, такие как, без ограничения указанным, промотор актина, промотор миозина, промотор гемоглобина и промотор креатинкиназы. Кроме того, изобретение не должно быть ограничено применением конститутивных промоторов. Индуцибельные промоторы также рассматриваются как часть изобретения. Использование индуцибельного промотора обеспечивает молекулярный переключатель, способный включать экспрессию полинуклеотидной последовательности, с которой он функционально связан, когда такая экспрессия желательна, или отключать экспрессию, когда экспрессия нежелательна. Примеры индуцибельных промоторов включают, без ограничения указанным, металлотиониновый промотор, глюкокортикоидный промотор, прогестероновый промотор и тетрациклиновый промотор.
Чтобы оценить экспрессию полипептида CAR или его частей, экспрессирующий вектор, который должен быть введен в клетку, может также содержать ген селектируемого маркера или ген-репортер, или и то, и другое для облегчения идентификации и отбора экспрессирующих клеток из популяции клеток, которые пытались трансфицировать или инфицировать через вирусные векторы. В других аспектах селектируемый маркер может быть нанесен на отдельный фрагмент ДНК и использован в процедуре котрансфекции. Как селектируемые маркеры, так и репортерные гены могут быть фланкированы соответствующими регуляторными последовательностями для обеспечения экспрессии в клетках-хозяевах. Полезные селектируемые маркеры включают, например, гены устойчивости к антибиотикам, такие как neo и т.п.
Репортерные гены используются для идентификации потенциально трансфицированных клеток и для оценки функциональности регуляторных последовательностей. Как правило, репортерный ген представляет собой ген, который не присутствует или не экспрессируется организмом или тканью реципиента и который кодирует полипептид, экспрессия которого проявляется в некотором легко обнаруживаемом свойстве, например, ферментативной активности. Экспрессия репортерного гена анализируется в подходящее время после введения ДНК в клетки реципиента. Подходящие репортерные гены могут включать гены, кодирующие люциферазу, бета-галактозидазу, хлорамфениколацетилтрансферазу, секретируемую щелочную фосфатазу или ген зеленого флуоресцентного белка (например, Ui-Tei et al., 2000 FEBS Letters 479: 79-82). Подходящие системы экспрессии хорошо известны и могут быть получены с использованием известных методов или получены коммерчески. В целом, конструкция с минимальной 5'-фланкирующей областью, показывающая самый высокий уровень экспрессии репортерного гена, идентифицируется как промотор. Такие промоторные области могут быть связаны с репортерным геном и использоваться для оценки агентов на способность модулировать управляемую промотором транскрипцию.
Способы введения и экспрессии генов в клетку известны в данной области. В контексте экспрессирующего вектора вектор может быть легко введен в клетку-хозяина, например, в клетку млекопитающего, бактерии, дрожжей или насекомого, любым способом, известным в данной области. Например, экспрессирующий вектор может быть перенесен в клетку-хозяина физическим, химическим или биологическим способом.
Физические способы введения полинуклеотида в клетку-хозяина включают осаждение с фосфатом кальция, липофекцию, бомбардировку частицами, микроинъекцию, электропорацию и т.п. Способы получения клеток, содержащих векторы и/или экзогенные нуклеиновые кислоты, хорошо известны в данной области (см., например, Sambrook et al., 2001). Например, полинуклеотид может быть введен в клетку-хозяина путем трансфекции с фосфатом кальция. Альтернативно, клетка-хозяин может быть трансфицирована полинуклеотидом, таким как РНК, путем электропорации (см., например, WO 2007/065957).
Химические средства для введения полинуклеотида в клетку-хозяина включают коллоидные дисперсионные системы, такие как макромолекулярные комплексы, нанокапсулы, микросферы, гранулы и системы на основе липидов, включая эмульсии масло-в-воде, мицеллы, смешанные мицеллы и липосомы. Типичной коллоидной системой для применения в качестве средства доставки in vitro и in vivo является липосома (например, везикула с искусственной мембраной). Доступны другие способы современной направленной доставки нуклеиновых кислот, такие как доставка полинуклеотидов с таргетными наночастицами или другой подходящей системой доставки субмикронного размера.
В случае, когда используется невирусная система доставки, примерным средством доставки является липосома. Предполагается применение липидных составов для введения нуклеиновых кислот в клетку-хозяина (in vitro, ex vivo или in vivo). В другом аспекте нуклеиновая кислота может быть связана с липидом. Нуклеиновая кислота, связанная с липидом, может быть инкапсулирована в водной внутренней части липосомы, помещена в липидный бислой липосомы, присоединена к липосоме через связывающую молекулу, которая связана как с липосомой, так и с олигонуклеотидом, захваченным в липосому, образует комплекс с липосомой, диспергируется в растворе, содержащем липид, смешивается с липидом, объединяется с липидом, содержится в виде суспензии в липиде, содержится или образует комплекс с мицеллой, или иным образом связан с липидом. Композиции, связанные с липидом, липидом/ДНК или липидным/экспрессирующим вектором, не ограничиваются какой-либо конкретной структурой в растворе. Например, они могут присутствовать в двухслойной структуре, в виде мицелл, или в «коллапсированной» структуре. Они также могут просто помещаться в раствор, возможно, образуя агрегаты, которые не являются однородными по размеру или форме. Липиды представляет собой жирные вещества, которые могут встречаться в природе или синтетические липиды. Например, липиды включают жировые капли, которые естественным образом встречаются в цитоплазме, а также класс соединений, которые содержат длинноцепочечные алифатические углеводороды и их производные, такие как жирные кислоты, спирты, амины, аминоспирты и альдегиды.
Липиды, пригодные для применения, могут быть получены из коммерческих источников. Например, димиристилфосфатидилхолин («DMPC») может быть получен в Sigma, Сент-Луис, Миссури; дицетилфосфат («DCP») можно получить в K&K Laboratories (Плейн-вью, Нью-Йорк); холестерин («Choi») может быть получен в Calbiochem-Behring; димиристилфосфатидилглицерин («DMPG») и другие липиды могут быть получены в Avanti Polar Lipids, Inc. (Бирмингем, Алабама). Исходные растворы липидов в хлороформе или хлороформе/метаноле можно хранить при температуре около -20°C. Хлороформ используется в качестве единственного растворителя, поскольку он легче испаряется, чем метанол. «Липосома» представляет собой общий термин, охватывающий множество одно- и многослойных липидных носителей, образованных в результате образования закрытых липидных бислоев или агрегатов. Липосомы могут быть охарактеризованы как имеющие везикулярные структуры с фосфолипидной двухслойной мембраной и внутренней водной средой. Мультиламеллярные липосомы имеют несколько липидных слоев, разделенных водной средой. Они образуются спонтанно, когда фосфолипиды суспендируют в избытке водного раствора. Липидные компоненты подвергались самоорганизации перед образованием закрытых структур и захватывают воду и растворенные растворы между липидными бислоями (Ghosh et al., 1991). Однако также охватываются композиции, которые имеют другие структуры в растворе, чем нормальная везикулярная структура. Например, липиды могут иметь мицеллярную структуру или просто существовать в виде неоднородных агрегатов молекул липидов. Также рассматриваются комплексы липофектамин-нуклеиновая кислота.
Независимо от способа, используемого для введения экзогенных нуклеиновых кислот в клетку-хозяина, для подтверждения присутствия последовательности рекомбинантной ДНК в клетке-хозяине могут быть выполнены различные анализы. Такие анализы включают, например, «молекулярно-биологические» анализы, такие как Саузерн- и Нозерн-блотинг, ОТ-ПЦР и ПЦР; «биохимические» анализы, такие как обнаружение присутствия или отсутствия конкретного пептида, например, иммунологическими средствами (ELISA и вестерн-блоттингом) или анализами, описанными в настоящем документе, для идентификации агентов, попадающих в объем изобретения.
VI. ИММУННЫЕ КЛЕТКИ, МОДИФИЦИРОВАННЫЕ С ПОМОЩЬЮ CAR
Нуклеиновые кислоты, кодирующие CAR, по изобретению могут быть введены в иммунные клетки для получения CAR-модифицированных иммунных клеток, экспрессирующих CAR по настоящему изобретению, где модифицированные иммунные клетки находят применение, как описано в данном документе. CAR-модифицированные иммунные клетки могут также упоминаться в данном документе как «CAR-сконструированные иммунные клетки». В воплощении предлагается модифицированная иммунная клетка, содержащая CAR, как описано в данном документе в любом месте. В другом воплощении предлагается иммунная клетка, содержащая экспрессирующий вектор, описанный где-либо в данном документе. Иммунная клетка может представлять собой любую иммунную клетку, подходящую для применения в клеточной терапии. В одном воплощении иммунная клетка может представлять собой иммунную клетку человека.
В одном воплощении иммунная клетка выбрана из группы, включающей лимфоцит, миелоидную клетку и любую их комбинацию. В одном воплощении лимфоцит выбран из группы, состоящей из Т-клеток, В-клеток, клеток-натуральных киллеров (NK) и любой их комбинации. В одном воплощении иммунная клетка представляет собой Т-клетку. В одном воплощении Т-клетка выбрана из группы, состоящей из CD4+ Т-клетки, CD8+ Т-клетки, γδ Т-клетки, двойной отрицательной (DN) Т-клетки и любой их комбинации. В одном воплощении CD4+ T-клетка выбрана из группы, состоящей из T-хелперной клетки, регуляторной T-клетки и любой их комбинации. В одном воплощении иммунная клетка представляет собой регуляторную Т-клетку. В одном воплощении Treg представляет собой Treg, полученный из тимуса, или адаптивный или индуцированный Treg. В одном воплощении Treg представляет собой регуляторную CD4+FOXP3+ T-клетку или регуляторную T-клетку CD4+FOXP3- T-клетку (TR1-клетку). В одном воплощении Treg представляет собой регуляторную CD4+FOXP3+ T-клетку. В одном воплощении Treg представляет собой D4+FOXP3- регуляторную T-клетку (TR1-клетку). В одном воплощении CD8+ T-клетка представляет собой цитотоксическую CD8+ T-клетку или CD8+ регуляторную T-клетку. В одном воплощении CD8+ T-клетка представляет собой CD8+ регуляторную T-клетку. В одном воплощении CD8+ регуляторную T-клетку выбирают из группы, состоящей из CD8+ CD28- регуляторной T-клетки, CD8+CD103+ регуляторной T-клетки, CD8+ FoxP3+ регуляторной T-клетки, CD8+CD122+ регуляторной T-клетки и любой их комбинации. В одном воплощении CD8+ регуляторная T-клетка представляет собой INFγ+IL10+IL34+CD8+CD45RClow регуляторную T-клетку. В одном воплощении γδ T-клетка представляет собой регуляторную γδ T-клетку. В одном воплощении DN T-клетка представляет собой регуляторную DN T-клетку. В одном воплощении иммунная клетка представляет собой B-клетку. В одном воплощении B-клетка представляет собой регуляторную B-клетку. В одном воплощении регуляторная B-клетка представляет собой B-клетку CD19+ CD24hiCD38hi. В одном воплощении NK-клетка представляет собой регуляторную NK-клетку. В одном воплощении, миелоидная клетка выбрана из группы, состоящей из нейтрофилов, эозинофилов, базофилов, моноцитов, макрофагов, дендритных клеток или любой их комбинации. В одном воплощении макрофаг является регуляторным макрофагом. В одном воплощении дендритная клетка представляет собой регуляторную дендритную клетку.
В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. Иммунная клетка может представлять собой любую регуляторную иммунную клетку, подходящую для применения в клеточной терапии (см., например, Wood, K.J. et al., 2012; Papp, G. et al., 2017). В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В одном воплощении регуляторная иммунная клетка представляет собой CD8+ регуляторную Т-клетку (см., например, Guillonneau, C. et al., 2010). В одном воплощении CD8+ регуляторную T-клетку выбирают из группы, состоящей из CD8+ CD28- регуляторной T-клетки, CD8+ CD103+ регуляторной T-клетки, CD8+ FoxP3+ регуляторной T-клетки, CD8+ CD122+ регуляторной T-клетки и любой их комбинации. В одном воплощении CD8+ регуляторная T-клетка представляет собой INFγ+IL10+IL34+ CD8+CD45RClow регуляторную T-клетку. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную γδ T-клетку (см., например, Wesch, D., et al., 2014). В одном воплощении иммунная клетка представляет собой DN T-клетку (см., например, Juvet, S.C., et al., 2012). В одном воплощении регуляторная иммунная клетка представляет собой регуляторную B-клетку (см., например, Chong, A.S., et al., 2017). В одном воплощении регуляторная B-клетка представляет собой B-клетку CD19+ CD24hiCD38hi. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную NK-клетку (см., например, Fu, B. et al., 2013; Crome, S. Q., et al., 2013; Crome, S. Q., et al., 2017). В одном воплощении регуляторная иммунная клетка представляет собой регуляторный макрофаг (см., например, Hutchinson, J.A. et al., 2017). В одном воплощении регуляторная иммунная клетка представляет собой регуляторную дендритную клетку. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную Т-клетку. В одном воплощении Treg представляет собой Treg, полученный из тимуса, или адаптивный или индуцированный Treg. В одном воплощении Treg представляет собой регуляторную CD4+ FOXP3+ T-клетку или регуляторную T-клетку CD4+FOXP3- T-клетку (TR1-клетку). В одном воплощении Treg представляет собой регуляторную CD4+FOXP3+ T-клетку. В одном воплощении Treg представляет собой D4+FOXP3- регуляторную T-клетку (TR1-клетку).
В одном воплощении регуляторная иммунная клетка имеет следующий фенотип: CD4+ CD25+, такой как, например, CD4+ CD25+ CD127-, такой как, например, CD4+ CD25+ CD127-CD45RA+. В дополнительных воплощениях регуляторная иммунная клетка имеет следующий фенотип: FoxP3+CD4+CD25+, такой как, например, FoxP3+CD4+CD25+CD127-, такой как, например, FoxP3+CD4+CD25+CD127-CD45RA+.
В одном воплощении иммунная регуляторная клетка имеет, по меньшей мере, один из следующих фенотипов: CD4+CD25+, FoxP3+, CD127lo/-, CTLA-4+, CD39+, Helios+, CD62L+/hi, VLA4+, LFA1+, CD49bint, ITGb7int, PSGL-1+, ICOS+, GITR+, PD-1int, Perflo/-, CCR7+. В одном воплощении иммунная регуляторная клетка не экспрессирует гранзим A и/или гранзим B.
В одном воплощении определение уровня экспрессии молекул проводят с помощью проточной цитометрии, иммунофлуоресценции или анализа изображений, например, анализа высокого содержания. Предпочтительно определение уровня экспрессии молекул проводят проточной цитометрией. В одном воплощении перед проведением анализа проточной цитометрией клетки фиксировали и проницаемы, что позволяет обнаруживать внутриклеточные белки.
В одном воплощении определение уровня экспрессии молекулы в клеточной популяции включает определение процента клеток клеточной популяции, экспрессирующих молекулу (то есть клетки «+» для молекулы). Предпочтительно указанный процент клеток, экспрессирующих молекулу, измеряют с помощью FACS.
Термины «экспрессирующий (или +)» и «неэкспрессирующий (или -)» хорошо известны в данной области и относятся к уровню экспрессии представляющего интерес клеточного маркера, в котором уровень экспрессии клеточного маркера соответствующий «+» представляет собой высокий или средний, также называемый «+/-», а уровень экспрессии маркера клетки, соответствующего «-», равен нулю.
Термины «low» или «lo» или «lo/-» хорошо известны в данной области и относятся к уровню экспрессии представляющего интерес клеточного маркера, в котором уровень экспрессии клеточного маркера является низким по сравнению с уровнем экспрессии этого клеточного маркера в популяции анализируемых клеток в целом. Более конкретно, термин «lo» относится к отдельной популяции клеток, которые экспрессируют клеточный маркер на более низком уровне, чем одна или несколько других различных популяций клеток.
Термин «high» или «hi» или «bright» хорошо известен в данной области техники и относится к уровню экспрессии представляющего интерес клеточного маркера в том смысле, что уровень экспрессии клеточного маркера является высоким по сравнению с уровнем экспрессии этого клеточного маркера в популяции клеток, которые анализировались в целом.
Обычно клетки в верхних 2, 3, 4 или 5% интенсивности окрашивания обозначаются как «hi», а клетки, попадающие в верхнюю половину популяции, классифицируются как «+».Клетки, демонстрирующие ниже 50% интенсивности флуоресценции обозначаются как «lo», а клетки ниже 5% - как »-» клетки.
Уровень экспрессии представляющего интерес клеточного маркера определяли путем сравнения средней интенсивности флуоресценции (MFI) клеток из популяции клеток, окрашенных флуоресцентномеченнм антителом, специфичным для этого маркера, с интенсивностью флуоресценции (FI) клеток из той же популяции клеток, окрашенных флуоресцентномеченым антителом с нерелевантной специфичностью, но с тем же изотипом, тем же флуоресцентным зондом и происходящим из одного и того же вида (обозначается как изотипный контроль). Клетки из популяции, окрашенные флуоресцентномечеными антителами, специфичными для этого маркера и демонстрирующие эквивалентную MFI или более низкую MFI, чем клетки, окрашенные изотипическими контролями, не экспрессируют этот маркер, и поэтому обозначаются (-) или отрицательно. Клетки из популяции, окрашенные флуоресцентномеченым антителом, специфичным для этого маркера и демонстрирующие значение MFI, превосходящее значение у клеток, окрашенных изотипическими контролями, экспрессируют этот маркер и поэтому обозначаются (+) или положительно.
В одном воплощении иммунная клетка по изобретению экспрессирует на своей клеточной поверхности CAR по изобретению и другой рецептор (в настоящем документе называемый «вторым рецептором»), который связывается с другим лигандом, отличным от HLA-A2. Согласно изобретению этот второй рецептор включает внеклеточный лиганд-связывающий домен, необязательно шарнир, необязательно трансмембранный домен и внутриклеточный сигнальный домен, как описано ранее.
В одном воплощении второй рецептор является эндогенным (таким как, например, эндогенный TCR). В другом воплощении второй рецептор является экзогенным, и его экспрессия индуцируется в иммунной клетке по изобретению путем трансформации или трансдукции кодирующей его нуклеиновой кислоты. Указанный экзогенный рецептор может представлять собой экзогенный TCR или химерный антигенный рецептор. Следовательно, в одном воплощении иммунная клетка по изобретению экспрессирует два химерных рецептора антигена, где первый распознает HLA-A2, а второй распознает отдельный лиганд.
В другом воплощении иммунная клетка по изобретению экспрессирует на своей клеточной поверхности CAR по изобретению и другой рецептор (называемый в данном документе «вторым рецептором»), который связывается с другим эпитопом в HLA-A2. Согласно изобретению этот второй рецептор включает внеклеточный лиганд-связывающий домен, необязательно шарнир, необязательно трансмембранный домен и внутриклеточный сигнальный домен, как описано ранее.
В другом воплощении иммунная клетка по изобретению экспрессирует два CAR, причем первый распознает первый эпитоп HLA-A2, а второй распознает эпитоп, отличный от HLA-A2.
В одном воплощении CAR по изобретению включает первый внутриклеточный сигнальный домен, а второй рецептор включает отдельный второй внутриклеточный сигнальный домен. В первом воплощении CAR по изобретению включает первичный сигнальный домен T-клеток (такой как, например, CD3дзета), а второй рецептор включает костимулирующий сигнальный домен (такой как, например, 4-1BB, CD28 или комбинация костимулирующего сигнального домена 4-1BB и CD28). Во втором воплощении CAR по изобретению включает костимулирующий сигнальный домен (такой как, например, 4-1BB, CD28 или комбинацию костимуляторного сигнального домена 4-1BB и CD28), и второго рецептора включающего первичный сигнальный домен T-клетки (такой как, например, CD3дзета).
Следовательно, в соответствии с этими воплощениями полная активация иммунной клетки по изобретению требует как связывания CAR по изобретению с HLA-A2, так и связывания второго рецептора с лигандом, на который он нацелен.
В одном воплощении лиганд, распознаваемый вторым рецептором, экспрессируется или присутствует в пораженной ткани или органе или в месте аутоиммунного ответа.
Примеры лигандов, которые могут распознаваться вторым рецептором, включают, без ограничения указанным, пищевые антигены из обычной диеты человека, аутоантигены, вдыхаемые аллергены, проглоченные аллергены или контактные аллергены.
Термин «пищевой антиген из обычной диеты человека» относится к иммуногенному пептиду, который поступает из пищевых продуктов, общих для человека, таких как пищевые антигены из следующего неограничивающего списка: бычьи антигены, такие как липокалин, Ca-связывающий S100, альфа-лактальбумин, лактоглобулины, такие как бета-лактоглобулин, бычий сывороточный альбумин, казеины. Пищевые антигены также могут представлять собой антигены атлантического лосося, такие как парвальбумин; куриные антигены, такие как, например, овомукоид, овальбумин, Ag22, кональбумин, лизоцим или куриный сывороточный альбумин; антигены арахиса; антигены креветок, такие как тропомиозин; пшеничные антигены, такие как агглютинин или глиадин; антигены сельдерея, такие как профилин сельдерея; морковные антигены, такие как морковный профилин; яблочные антигены, такие как тауматин, белок, переносящий липид яблока, или яблочный профилин; грушевые антигены, такие как грушевый профилин или изофлавоновая редуктаза; антигены авокадо, такие как эндохитиназа; абрикосовые антигены, такие как белок переноса абрикосовых липидов; персиковые антигены, такие как персиковый белок-переносчик липидов или персиковый профилин; соевые антигены, такие как HPS, соевый профилин или (SAM22) PR-10 pro; фрагменты, их варианты и смеси.
В одном воплощении указанный аутоантиген представляет собой антиген, ассоциированный с рассеянным склерозом, антиген, ассоциированный с суставом, антиген, ассоциированный с глазами, антиген HSP человека, антиген, связанный с кожей или антиген, участвующий в отторжении трансплантата или GVHD.
Термин «антиген, ассоциированный с рассеянным склерозом» относится к основному белку миелина (MBP), ассоциированному с миелином гликопротеину (MAG), миелин-олигодендроцитарноу гликопротеину (MOG), протеолипидному белку (PLP), олигодендроцитному миелиновому олигопротеину (OMGP), миелин-ассоциированному олигодендроцитарному основному белку (MOBP), олигодендроцит-специфическому белку (OSP/Клаудин 1), белкам теплового шока, олигодендроцит-специфическим белкам (OSP), NOGO A, гликопротеину Po, периферическому миелиновому белку 22 (PMP22), 3’-фосфодиэстеразе 2,3-циклических нуклеотидов (CNPase), их фрагментам, вариантам и смесям.
Термин «сустав-ассоциированный антиген» относится к цитруллинзамещенным циклическим и линейным пептидам филагрина, пептидам коллагена II типа, пептидам гликопротеина хряща 39 человека (HCgp39), HSP, пептидам гетерогенного ядерного рибонуклеопротеина (hnRNP) A2, hnRNP B1, hnRNP D, Ro60/52, HSP60, HSP65, HSP70 и HSP90, BiP, кератину, виментину, фибриногену, пептиду коллагена типа I, III, IV и V, аннексину V, глюкозо-6-фосфат-изомеразе (GPI), ацетил-кальпастатину, пируватдегидрогеназе (PDH), альдолазе, топоизомеразе I, snRNP, PARP, Scl-70, Scl-100, фосфолипидным антигенам, включая анионный кардиолипин и фосфатидилсерин, нейтрально заряженный фосфатидилэтаноламин и фосфатидилхолин, матриксную металлопротеиназу, фибриллину, аггрекану, их фрагментам, вариантам и смесям.
Термин «глазной антиген» относится к коллагену типа II, аррестину сетчатки, S-аррестину, интерфорецепторным ретиноид-связывающим белкам (IRBP1), бета-кристаллину В1, белкам сетчатки, белкам хориоида и их фрагментам, вариантам и смесям.
Термин «антиген HSP человека» относится к HSP60, HSP70, HSP90 человека, их фрагментам, вариантам и смесям.
Примеры ассоциированных с кожей антигенов включают, без ограничения указанным, антигены кератиноцитов, антиген, присутствующий в дерме или эпидермисе, антиген меланоцитов (такой как, например, меланин или тирозиназа), десмоглеин (например, десмоглеин 1 или 3, которые также могут упоминаться как Dsg1/3), BP180, BP230, плектин, интегрины (например, интегрин α4β6), коллагены (например, коллаген типа VII), ламинины (например, ламинин 332 или ламинин γ1), плакины (например, энвоплакин, периплакин или десмоплакин), кератины (например, KRT5, KRT8, KRT15, KRT17 и KRT31), белки, связанные с кератиновыми филаментами, филагрин, корнеодесмосин и эластин.
В одном воплощении лиганд представляет собой антиген, участвующий в отторжении трансплантата или GVHD. Примеры таких антигенов включают, без ограничения указанным, MHC, специфичные для трансплантированной ткани или к хозяину, β2-микроглобулин, антигены из системы ABO, антигены системы резус (в частности, антигены из системы C, c, E et e и D) и изогемагглютинины. Другие примеры антигенов, которые могут быть вовлечены в отторжение трансплантата или GVHD, включают, без ограничения указанным, HLA-DR (в частности, в течение первых шести месяцев после трансплантации), HLA-B (в частности, в течение первых двух лет после трансплантации), второстепенные антигены гистосовместимости (miHA, например, HLA-E, HLA-F и HLA-G), HLA, соответствующие MHC класса I (B и C), HLA, соответствующие MHC класса II (DP, DM, DOA, DOB, DQ и DR) и HLA, соответствующие классу III MHC (например, компоненты системы комплемента).
Другие примеры аутоантигенов включают, без ограничения указанным, водные каналы аквапоринов (такие как, например, водный канал аквапорина-4 (AQP4)), Hu, Ma2, белок-медиатор ответа на коллапсин 5 (CRMP5) и амфифизин, потенциалозависимый калиевый канал (VGKC), N-метил-d-аспартатный рецептор (NMDAR), α-амино-3-гидрокси-5-метил-4-изоксазолепропионовую кислоту (AMPAR), тиреоидную пероксидазу, тиреоглобулин, анти-N-метил-D-аспартатный рецептор (субъединица NR1), антигены резус-группы крови, антиген I, десмоглеин 1 или 3 (Dsg1/3), BP180, BP230, никотиновые постсинаптические рецепторы ацетилхолина, интегрин тромбоцитов, GpIIb: IIIa, коллаген (например, цепь альфа-3 (IV) коллагена), ревматоидный фактор, кальпастатин, цитруллинированные белки, миелиновый основной белок (MBP), пептиды миелинового олигодендроцитного гликопротеина (MOG), альфа-бета-кристаллин, ДНК, гистон, рибосомы, RNP, тканевую трансглутаминазу (TG2), внутренний фактор, антиген 65 кДа, фосфатидилсерин, рибосомные фосфобелки, антинейтрофильное цитоплазматическое антитело, Scl-70, U1-RNP, ANA, SSA, анти-SSB, антиядерные антитела (ANA), антинейтрофильные цитоплазматические антитела (ANCA), Jo-1, антимитохондриальные антитела, gp210, p62, sp100, антифосфолипидные антитела, U1-70 kd snRNP, ганглиозид GQ1b, GM1, асиало-GM1, GD1b, антитела против гладких мышц (ASMA), антитела к микросомам печени и почек-1 (ALKM-1), антитела против цитозоля печени-1 (ALC-1), IgA антиэндомизальные антитела, белки гранул нейтрофилов, антиген клеточной стенки стрептококка, внутренний фактор париетальных клеток желудка, инсулин (IAA), декарбоксилаза глутаминовой кислоты (GAA или GAD) и протеинтирозинфосфатаза (такая как, например, IA2 или ICA512) PLA2R1 и THSD7A1.
В одном воплощении указанный лиганд выбран из группы, включающей фрагменты овальбумина, MOG, коллагена II типа, их варианты и смеси.
В одном воплощении указанный лиганд представляет собой овальбумин, его фрагменты, варианты и смеси.
В другом воплощении указанный лиганд представляет собой MOG, его фрагменты, варианты и смеси.
В другом воплощении указанный лиганд представляет собой коллаген II типа, его фрагменты, варианты и смеси.
В другом воплощении указанный лиганд представляет собой IL23R, его фрагменты, варианты и смеси.
В одном воплощении CAR по изобретению дополнительно включает внеклеточный лиганд-связывающий домен, распознающий лиганд, отличный от HLA-A2. В одном воплощении указанный лиганд-связывающий домен представляет собой антитело или его антигенсвязывающий фрагмент.
В одном воплощении лиганд-связывающий домен CAR по изобретению представляет собой многофункциональное антитело, распознающее множество различных эпитопов на HLA-A2. В одном воплощении лиганд-связывающий домен CAR по изобретению представляет собой бифункциональное антитело, распознающее два разных эпитопа на HLA-A2.
В одном воплощении изобретения CAR по изобретению включает внеклеточный лиганд-связывающий домен, содержащий HLA-A2-связывающий домен и другой лиганд-связывающий домен, распознающий лиганд, отличный от HLA-A2. В одном воплощении указанный лиганд-связывающий домен представляет собой бифункциональное антитело, распознающее как HLA-A2, так и отдельный лиганд.
Примеры лигандов, отличных от HLA-A2, которые могут быть распознаны с помощью CAR по изобретению, перечислены выше.
В одном воплощении CAR-модифицированные иммунные клетки по изобретению могут быть получены введением в клетки лентивирусного вектора, содержащего искомый CAR. В одном воплощении предлагается способ получения иммунной клетки, модифицированной для экспрессии CAR, при этом способ включает трансдукцию иммунной клетки вектором, описанным где-либо в данном документе, с образованием, таким образом, указанной модифицированной иммунной клетки.
В одном воплощении генетически модифицированные иммунные клетки по изобретению модифицируют путем введения РНК. В одном воплощении транскрибированная РНК-CAR in vitro может быть введена в клетку в форме транзиторной трансфекции. РНК может быть получена путем транскрипции in vitro с использованием матрицы, полученной с помощью полимеразной цепной реакции (ПЦР). Интересующая ДНК из любого источника может быть непосредственно преобразована с помощью ПЦР в матрицу для синтеза мРНК in vitro с использованием соответствующих праймеров и РНК-полимеразы. Источником ДНК может быть, например, геномная ДНК, плазмидная ДНК, фаговая ДНК, кДНК, синтетическая последовательность ДНК или любой другой подходящий источник ДНК. Желаемым шаблоном для транскрипции in vitro является CAR по настоящему изобретению.
В одном воплощении ДНК, предназначенная для ПЦР, включает открытую рамку считывания. ДНК может происходить из встречающейся в природе последовательности ДНК из генома организма. В одном воплощении ДНК представляет собой представляющий интерес полноразмерный ген или часть гена. Ген может включать некоторые или все 5' и/или 3' нетранслируемые области (UTR). Ген может включать экзоны и интроны. В одном воплощении ДНК, используемая для ПЦР, представляет собой ген человека. В другом воплощении ДНК, используемая для ПЦР, представляет собой ген человека, включающий 5' и 3' UTR. Альтернативно, ДНК может представлять собой искусственную последовательность ДНК, которая обычно не экспрессируется в естественном организме. Примером последовательности искусственной ДНК является та, которая включает части генов, которые лигированы вместе, чтобы образовать открытую рамку считывания, которая кодирует слитый белок. Части ДНК, которые лигированы вместе, могут быть из одного организма или из более чем одного организма.
ПЦР может использоваться для создания матрицы для транскрипции мРНК in vitro, которая используется для трансфекции. Способы проведения ПЦР хорошо известны в данной области. Праймеры для применения в ПЦР предназначены для того, чтобы иметь области, которые по существу комплементарны областям ДНК, которые будут использоваться в качестве матрицы для ПЦР. «По существу комплементарный», при использовании в настоящем документе, относится к последовательностям нуклеотидов, где большинство или все основания в последовательности праймера являются комплементарными или одно или несколько оснований не являются комплементарными или не совпадают. Существенно комплементарные последовательности способны отжигать или гибридизоваться с предполагаемой ДНК-мишенью в условиях отжига, используемых для ПЦР. Праймеры могут быть сконструированы так, чтобы они были существенно комплементарны любой части матрицы ДНК. Например, праймеры могут быть разработаны для амплификации части гена, которая обычно транскрибируется в клетках (открытая рамка считывания), включая 5' и 3' UTR. Праймеры также могут быть сконструированы для амплификации части гена, которая кодирует определенный интересующий домен. В одном воплощении праймеры предназначены для амплификации кодирующей области кДНК человека, включая все или части 5' и 3' UTR. Праймеры, пригодные для ПЦР, получают синтетическими способами, которые хорошо известны в данной области.
«Прямые праймеры» представляют собой праймеры, которые содержат область нуклеотидов, которые по существу комплементарны нуклеотидам на матрице ДНК, которые находятся выше последовательности ДНК, которая должна быть амплифицирована. «Вверх по поверхности» используется в данном документе для обозначения местоположения 5' последовательности ДНК, которая должна быть амплифицирована относительно кодирующей цепи. «Обратные праймеры» представляют собой праймеры, которые содержат область нуклеотидов, которые по существу комплементарны матрице двухцепочечной ДНК, которая находится ниже последовательности ДНК, которая должна быть амплифицирована. Термин «ниже по последовательности» используется в данном документе для обозначения местоположения 3' последовательности ДНК, подлежащей амплификации, относительно кодирующей цепи.
Любая ДНК-полимераза, пригодная для ПЦР, может быть использована в способах, раскрытых в данном документе. Реагенты и полимераза коммерчески доступны из ряда источников.
Также могут быть использованы химические структуры, способные обеспечивать стабильность и/или эффективность трансляции. РНК предпочтительно имеет 5' и 3' UTR. В одном воплощении 5' -UTR имеет длину от нуля до 3000 нуклеотидов. Длина 5'- и 3'-последовательностей UTR, которые должны быть добавлены в область кодирования, может быть изменена различными способами, включая, без ограничения указанным, конструирование праймеров для ПЦР, которые отжигаются в различных областях UTR. Используя этот подход, специалист в данной области техники может модифицировать 5' и 3' длины UTR, необходимые для достижения оптимальной эффективности трансляции после трансфекции транскрибированной РНК. 5' и 3' UTR могут представлять собой встречающиеся в природе эндогенные 5' и 3' UTR для представляющего интерес гена. Альтернативно, последовательности UTR, которые не являются эндогенными для интересующего гена, могут быть добавлены путем включения последовательностей UTR в прямой и обратный праймеры или с помощью любых других модификаций матрицы. Использование последовательностей UTR, которые не являются эндогенными для интересующего гена, может быть полезным для изменения стабильности и/или эффективности трансляции РНК. Например, известно, что богатые AU элементы в 3'-последовательностях UTR могут снижать стабильность мРНК. Следовательно, 3'-UTR могут быть выбраны или разработаны для увеличения стабильности транскрибированной РНК на основе свойств UTR, которые хорошо известны в данной области.
В одном воплощении 5'-UTR может содержать последовательность Kozak эндогенного гена. Альтернативно, когда 5' -UTR, который не является эндогенным для интересующего гена, добавляется с помощью ПЦР, как описано выше, консенсусная последовательность Kozak может быть изменена путем добавления 5'-последовательности UTR. Последовательности Kozak могут повысить эффективность трансляции некоторых РНК-транскриптов, но, по-видимому, не требуются, чтобы все РНК обеспечивали эффективную трансляцию. Требование последовательностей Kozak для многих мРНК известно в данной области. В других воплощениях 5'-UTR может быть получен из РНК-вируса, чей РНК-геном стабилен в клетках. В других воплощениях различные нуклеотидные аналоги можно использовать в 3' или 5' UTR, чтобы препятствовать деградации мРНК экзонуклеазой.
Чтобы сделать возможным синтез РНК с матрицы ДНК без необходимости клонирования гена, промотор транскрипции должен быть присоединен к матрице ДНК перед транскрибируемой последовательностью. Когда последовательность, которая функционирует в качестве промотора для РНК-полимеразы, добавляется к 5' концу прямого праймера, промотор РНК-полимеразы становится включенным в продукт ПЦР перед открытой рамкой считывания, которая должна быть транскрибирована. В одном предпочтительном воплощении промотор представляет собой T7-полимеразный промотор, описанный где-либо в данном документе. Другие полезные промоторы включают, без ограничения указанным, промоторы РНК-полимеразы Т3 и SP6. Консенсусные нуклеотидные последовательности для промоторов T7, T3 и SP6 известны в данной области.
В предпочтительном воплощении мРНК имеет как кэп на 5' конце, так и 3' поли (А) хвост, которые определяли связывание рибосомы, инициацию трансляции и стабильность мРНК в клетке. На кольцевой матрице ДНК, например, плазмидной ДНК, РНК-полимераза продуцирует длинный конкатемерный продукт, который не подходит для экспрессии в эукариотических клетках. Транскрипция плазмидной ДНК, линеаризованной в конце 3 'UTR, приводит к мРНК нормального размера, которая не эффективна для эукариотической трансфекции, даже если она полиаденилируется после транскрипции.
На линейной ДНК-матрице РНК-полимераза фага Т7 может удлинять 3'-конец транскрипта за пределы последнего основания матрицы (Schenborn and Mierendorf, Nuc Acids Res., 13:6223-36 (1985); Nacheva and Berzal-Herranz, Eur. J. Biochem., 270: 1485-65 (2003).
Общепринятым способом интеграции полиА/Т-участков в матрицу ДНК является молекулярное клонирование. Однако последовательность polyA/T, интегрированная в плазмидную ДНК, может вызывать нестабильность плазмиды, поэтому матрицы плазмидной ДНК, полученные из бактериальных клеток, часто сильно загрязнены делециями и другими аберрациями. Это делает процедуры клонирования не только трудоемкими и длительными, но зачастую и ненадежными. Вот почему способ, который позволяет конструировать ДНК-матрицы с полиА/Т 3'-участком без клонирования, крайне желателен.
Сегмент полиA/T транскрипционной ДНК-матрицы может быть получен во время ПЦР с использованием обратного праймера, содержащего хвост полиT, такой как хвост 100T (размер может быть 50-5000 Т) или после ПЦР любым другим способом, включая, без ограничения указанным, лигирование ДНК или рекомбинацию in vitro. Поли(А)-хвосты также обеспечивают стабильность РНК и уменьшают их деградацию. Как правило, длина поли (А)-хвоста положительно коррелирует со стабильностью транскрибированной РНК. В одном воплощении поли (А) хвост составляет от 100 до 5000 аденозинов.
Поли(А)-хвосты РНК могут быть дополнительно удлинены после транскрипции in vitro с использованием поли(А)-полимеразы, такой как полиА-полимераза Е.coli (E-PAP). В одном воплощении увеличение длины поли(А)-хвоста от 100 нуклеотидов до 300-400 нуклеотидов приводит к около двукратному увеличению эффективности трансляции РНК. Кроме того, присоединение различных химических групп к 3'-концу может повысить стабильность мРНК. Такое присоединение может содержать модифицированные/искусственные нуклеотиды, аптамеры и другие соединения. Например, в поли (А) хвост могут быть включены аналоги АТФ с использованием поли(А)-полимеразы. Аналоги АТФ могут еще больше повысить стабильность РНК.
5'-кэпы на РНК также обеспечивают стабильность молекул РНК. В предпочтительном воплощении РНК, полученные способами, раскрытыми в данном документе, включают 5'-кэп. 5'-кэп предлагается с использованием методов, известных в данной области техники и описанных в данном документе (Cougot et al., Trends in Biochem. Sci., 29:436-444 (2001); Stepinski, et al., RNA, 7: 1468-95 (2001); Elango, et al., Biochim. Biophys. Res. Commun., 330:958-966 (2005)).
РНК, полученные способами, раскрытыми в данном описании, также могут содержать последовательность участка внутренней посадки рибосомы (IRES). Последовательность IRES может быть любой вирусной, хромосомной или искусственно сконструированной последовательностью, которая инициирует кэп-независимое связывание рибосомы с мРНК и облегчает инициацию трансляции. Могут быть включены любые растворы, подходящие для электропорации клеток, которые могут содержать факторы, способствующие проницаемости и жизнеспособности клеток, такие как сахара, пептиды, липиды, белки, антиоксиданты и поверхностно-активные вещества.
РНК может быть введена в клетки-мишени с использованием любого из ряда различных способов, например, коммерчески доступных способов, которые включают, без ограничения указанным, электропорацию (Amaxa Nucleofector-II (Amaxa Biosystems, Кельн, Германия)) (ECM 830 (BTX) (Harvard Instruments, Бостон, штат Массачусетс) или Gene Pulser II (BioRad, Denver, Colo.), Multiporator (Eppendort, Гамбург, Германия), трансфекцию, опосредованную катионными липосомами, с использованием липофекции, инкапсуляцию полимером, трансфекцию, опосредованную пептидами, или баллистические системы доставки частиц, такие как «генные пушки» (см., например, Nishikawa, et al. Hum Gene Ther., 12(8):861-70 (2001).
В одном воплощении последовательности CAR доставляются в клетки с использованием ретровирусного или лентивирусного вектора. CAR-экспрессирующие ретровирусные и лентивирусные векторы могут доставляться в различные типы эукариотических клеток, а также в ткани и целые организмы с использованием трансдуцированных клеток в качестве носителей или бесклеточной локальной или системной доставки инкапсулированных, связанных или «голых» векторов. Используемый способ может использоваться для любых целей, где требуется или достаточна стабильная экспрессия.
В другом воплощении последовательности CAR доставляются в клетки с использованием транскрибированной мРНК in vitro. Транскрибированная мРНК CAR in vitro может быть доставлена в различные типы эукариотических клеток, а также в ткани и целые организмы, используя трансфицированные клетки в качестве носителей или бесклеточную локальную или системную доставку инкапсулированной, связанной или «голой» мРНК. Используемый способ может использоваться для любых целей, где требуется или достаточна транзиторная экспрессия.
В другом воплощении искомый CAR может быть экспрессирован в клетках посредством транспозонов.
В одном воплощении иммунная клетка по изобретению представляет собой Т-клетку. До размножения и генетической модификации Т-клеток по изобретению источник Т-клеток получают из объекта. Т-клетки могут быть получены из ряда источников, включая мононуклеарные клетки периферической крови, костный мозг, ткань лимфатического узла, пуповинную кровь, ткань тимуса, ткань из места инфекции, асцит, плевральный выпот, ткань селезенки и опухоли. В определенных воплощениях настоящего изобретения может использоваться любое количество линий Т-клеток, доступных в данной области. В некоторых воплощениях настоящего изобретения Т-клетки могут быть получены из единицы крови, взятой у объекта, с использованием любого количества методик, известных специалисту в данной области, таких как разделение на Ficoll™ или система разделения Sepax. В одном предпочтительном воплощении клетки из циркулирующей крови индивидуума получают аферезом. Продукт афереза обычно включает лимфоциты, включая Т-клетки, моноциты, гранулоциты, В-клетки, другие ядросодержащие лейкоциты, эритроциты и тромбоциты. В одном воплощении клетки, собранные с помощью афереза, могут быть промыты для удаления фракции плазмы и помещения клеток в соответствующий буфер или среду для последующих стадий обработки. В одном воплощении изобретения клетки промывали фосфатно-солевым буфером (PBS). В альтернативном воплощении моющему раствору не хватает кальция и может не хватать магния или может не хватать многих, если не всех двухвалентных катионов. После промывания клетки могут быть ресуспендированы в различных биосовместимых буферах, таких как, например, не содержащий Ca2+, не содержащий Mg 2+ PBS, PlasmaLyte A или другой физиологический раствор с буфером или без него. Альтернативно, нежелательные компоненты образца афереза могут быть удалены, и клетки непосредственно ресуспендируются в культуральной среде.
Источники, активация и размножение Т-клеток
В одном воплощении иммунная клетка может представлять собой Т-клетку. До размножения и генетической модификации Т-клеток по изобретению источник Т-клеток или клеток-предшественников, из которых могут быть получены Т-клетки, получают из объекта. Т-клетки могут быть получены из любого источника, в котором находятся эти клетки, включая мононуклеарные клетки периферической крови, костный мозг, ткань лимфатического узла, пуповинную кровь, ткань тимуса, зародышевые центры, ткани из места инфекции, асциты, плевральный выпот, селезенку, опухоли и пересаженные органы/ткани. В определенных воплощениях настоящего изобретения Т-клетки могут быть получены из единицы крови, взятой у объекта, с использованием любого количества методов, известных специалисту в данной области, таких как разделение на Ficoll™. В одном воплощении клетки из циркулирующей крови объекта получают аферезом. Продукт для афереза обычно включает лимфоциты, включая Т-клетки, моноциты, гранулоциты, В-клетки, другие ядросодержащие лейкоциты, эритроциты и тромбоциты. В одном воплощении клетки, собранные с помощью афереза, могут быть промыты для удаления фракции плазмы и помещения клеток в соответствующий буфер или среду для последующих стадий обработки. В одном воплощении изобретения клетки могут быть промыты промывочным раствором (например, фосфатно-солевым буферным раствором (PBS)). Специалистам в данной области техники легко понять, что стадия промывки может быть выполнена способами, известными специалистам в данной области, такими как применение полуавтоматической «проточной» центрифуги (например, клеточный процессор Cobe 2991, Baxter CytoMate или Haemonetics Cell Saver 5) в соответствии с инструкциями производителя. После промывания клетки могут быть ресуспендированы в различных биосовместимых буферах, таких как, например, не содержащий Ca2+, не содержащий Mg 2+ PBS, PlasmaLyte A или другой физиологический раствор с буфером или без него. Альтернативно, нежелательные компоненты образца афереза могут быть удалены, и клетки непосредственно ресуспендируются в культуральной среде.
В другом воплощении Т-клетки выделяют из лимфоцитов периферической крови путем лизиса эритроцитов и истощения моноцитов, например, путем центрифугирования в градиенте PERCOLL™ или путем противоточного центрифугирования. Специфическая субпопуляция Т-клеток, таких как CD3+, CD25+, CD127neg, HLA-DR+, CD28+, CD4+, CD8+, CD45RA+ и/или CD45RO+ T-клетки, могут быть дополнительно выделены методами положительной или отрицательной селекции. Специалист в данной области техники признает, что множественные раунды селекции также могут быть использованы в контексте этого изобретения. В определенных воплощениях может быть желательно выполнить процедуру селекции и использовать «неотобранные» клетки в процессе активации и размножения. «Неотобранные» клетки также могут подвергаться дополнительным раундам отбора.
Обогащение популяции Т-клеток путем отрицательного отбора может быть достигнуто с помощью комбинации антител, направленных на поверхностные маркеры, уникальные для отрицательно отобранных клеток. Одним из способов является сортировка и/или селекция клеток с помощью отрицательной магнитной иммуноадгезии или проточной цитометрии, в которой используется набор моноклональных антител, направленных на маркеры клеточной поверхности, присутствующие на отрицательно отобранных клетках. Например, для обогащения клеток CD4+ путем отрицательного отбора коктейль из моноклональных антител обычно включает антитела к CD14, CD20, CD11b, CD16, HLA-DR и CD8. В некоторых воплощениях может быть желательным обогащение или положительный отбор регуляторных Т-клеток, как правило экспрессирующих CD4+, CD25+, CD62Lhi, GITR+ и FoxP3+.
Для выделения искомой популяции клеток с помощью положительного или отрицательного отбора можно варьировать концентрацию клеток и поверхности (например, частиц, таких как гранулы). В определенных воплощениях может быть желательным значительно уменьшить объем, в котором гранулы и клетки смешиваются друг с другом (то есть увеличить концентрацию клеток), чтобы обеспечить максимальный контакт клеток и гранул. Например, в одном воплощении используется концентрация 2 миллиарда клеток/мл. В одном воплощении используется концентрация 1 миллиард клеток/мл. В дополнительном воплощении используют более 100 миллионов клеток/мл. В следующем воплощении используется концентрация клеток 10, 15, 20, 25, 30, 35, 40, 45 или 50 миллионов клеток/мл. В еще одном воплощении используется концентрация клеток от 75, 80, 85, 90, 95 или 100 миллионов клеток/мл. В других воплощениях могут использоваться концентрации 125 или 150 миллионов клеток/мл. Использование высоких концентраций может привести к увеличению выхода клеток, активации клеток и размножения клеток. Кроме того, применение высоких концентраций клеток позволяет более эффективно захватывать клетки, которые могут слабо экспрессировать представляющие интерес антигены-мишени, или из образцов, где присутствует много опухолевых клеток (то есть лейкозная кровь, опухолевая ткань и т.д.). Такие популяции клеток могут иметь терапевтическую ценность и их было бы желательно получить.
В связанном воплощении может быть желательно использовать более низкие концентрации клеток. Благодаря значительному разбавлению смеси Т-клеток и поверхности (например, частиц, таких как гранулы), взаимодействие между частицами и клетками сводится к минимуму. Таким образом, выбираются клетки, которые экспрессируют большое количество искомых антигенов, которые будут связаны с частицами. Например, CD4+ T-клетки экспрессируют более высокие уровни CD28 и захватываются более эффективно, чем CD8+ T-клетки в разбавленных концентрациях. В одном воплощении используемая концентрация клеток составляет 5×106/мл. В других воплощениях используемая концентрация может составлять от около 1×105/мл до 1×106/мл, и любое целочисленное значение между ними.
В других воплощениях клетки можно инкубировать на вращающемся устройстве в течение различных промежутков времени с разными скоростями либо при 2-10°С, либо при комнатной температуре.
Т-клетки для стимуляции также могут быть заморожены после промывки. Не желая быть связанными теорией предполагаем, что стадия замораживания и последующего оттаивания обеспечивает более однородный продукт за счет удаления гранулоцитов и в некоторой степени моноцитов в клеточной популяции. После стадии промывания, которая удаляет плазму и тромбоциты, клетки могут быть суспендированы в замораживающем растворе. Хотя многие растворы и параметры для замораживания известны в данной области техники и будут полезны в этом контексте, один способ включает применение PBS, содержащего 20% DMSO и 8% человеческого сывороточного альбумина, или культуральную среду, содержащую 10% декстрана 40 и 5% декстрозы, 20% человеческого сывороточного альбумина и 7,5% DMSO или 31,25% плазмалита-А, 31,25% декстрозы 5%, 0,45% NaCl, 10% декстрана 40 и 5% декстрозы, 20% человеческого сывороточного альбумина и 7,5% DMSO или других подходящих сред для замораживания клеток, содержащих, например, Hespan и PlasmaLyte A, клетки затем замораживали до -80°C со скоростью 1°C в минуту и хранили в паровой фазе резервуара для хранения жидкого азота. Могут быть использованы другие методы контролируемого замораживания, а также неконтролируемое замораживание сразу при -20 °C или в жидком азоте.
В определенных воплощениях криоконсервированные клетки оттаивают и промывают, как описано в данном документе, и оставляют на один час при комнатной температуре перед активацией с использованием способов по настоящему изобретению.
В контексте изобретения также рассматривается отбор образцов крови или продукта для афереза у объекта в период времени, предшествующий тому, когда могут потребоваться размноженные клетки, как описано в данном документе. Как таковой, источник клеток, подлежащих размножению, может быть собран в любой момент времени, когда необходимо, и искомые клетки, такие как Т-клетки, могут быть выделены и заморожены для последующего применения в Т-клеточной терапии для любого числа заболеваний или состояний, которые будут полезны Т-клеточной терапии, такой как описанная в данном документе. В некоторых воплощениях образец крови или аферез забирают у объекта, у которого есть риск развития заболевания, но у которого еще не развилось заболевание, и представляющие интерес клетки выделяют и замораживают для последующего применения. В определенных воплощениях Т-клетки могут быть размножены, заморожены и использованы позднее. В определенных воплощениях образцы отбирают у объекта вскоре после диагностики конкретного заболевания, как описано в настоящем документе, но до начала любого лечения. В дополнительном воплощении клетки выделяют из образца крови или афереза из объекта до любого количества соответствующих способов лечения, включая, без ограничения указанным, лечение такими агентами, как натализумаб, эфализумаб, противовирусные агенты, химиотерапия, облучение, иммуносупрессивные агенты, такие как циклоспорин, азатиоприн, метотрексат, микофенолат и FK506, антитела или другие иммуноабляционные агенты, такие как CAMPATH, анти-CD3 антитела, цитоксан, флударабин, циклоспорин, FK506, рапамицин, микофеноловая кислота, стероиды и облучение. Эти лекарственные средства ингибируют кальций-зависимую фосфатазу кальциневрин (циклоспорин и FK506) или ингибируют киназу p70S6, которая важна для передачи сигналов, индуцированных фактором роста (рапамицин) (Liu et al., Cell 66: 807-815, 1991; Henderson et al., Immun 73: 316-321, 1991; Bierer et al., Curr. Opin. Immun 5:763-773, 1993). В дополнительном воплощении клетки выделяют для пациента и замораживают для последующего применения в сочетании (например, до, одновременно или после) трансплантации костного мозга или стволовых клеток, абляционной терапии Т-клеток с использованием любых химиотерапевтических агентов, таких как флударабин, наружная дистанционная лучевая терапия (XRT), циклофосфамид или антитела, такие как OKT3 или CAMPATH. В другом воплощении клетки выделяют до и могут быть заморожены для последующего применения для лечения после B-клеточной абляционной терапии, например, агентами, которые реагируют с CD20, например, ритуксимабом.
В дополнительном воплощении настоящего изобретения Т-клетки получают от пациента непосредственно после лечения. В связи с этим было отмечено, что после определенных иммуносупрессивных процедур, в частности лечения лекарственными средствами, которые повреждают иммунную систему, вскоре после лечения в течение периода, когда пациенты, как правило, оправляются от лечения, качество полученных Т-клеток может быть оптимальным или улучшенным по их способности размножаться ex vivo. Аналогично, после манипуляций ex vivo с использованием способов, описанных в данном документе, эти клетки могут находиться в предпочтительном состоянии для усиленного приживления и размножения in vivo. Таким образом, в контексте настоящего изобретения предполагается собирать иммунные клетки во время этой фазы извлечения. Кроме того, в определенных воплощениях схемы мобилизации (например, мобилизация с помощью GM-CSF) и кондиционирования могут использоваться для создания состояния у объекта, в котором предпочтительны репопуляция, рециркуляция, регенерация и/или размножение определенных типов клеток, особенно во время определенного интервала времени после терапии.
До или после генетической модификации Т-клеток для экспрессии искомого CAR, Т-клетки можно активировать и размножать, как правило, с использованием способов, как описано, например, в пат. США No. 6352694; 6534055; 6905680; 6692964; 5858358; 6887466; 6905681; 7144575; 7067318; 7172869; 7232566; 7175843; 5883223; 6905874; 6797514; 6867041; и публикации патентной заявки США № 20060121005.
Как правило, Т-клетки по изобретению размножаются при контакте с поверхностью, на которой прикреплен агент, стимулирующий сигнал, связанный с комплексом CD3/TCR, и лиганд, который стимулирует костимулирующую молекулу на поверхности Т-клеток. В частности, популяции T-клеток может быть стимулирована, как описано в настоящем документе, например, путем контакта с анти-CD3-антителом или его антигенсвязывающим фрагментом или анти-CD2-антителом, иммобилизованным на поверхности, или путем контакта с активатором протеинкиназы С (например, бриостатином) в сочетании с ионофором кальция. Для костимуляции вспомогательной молекулы на поверхности Т-клеток используется лиганд, который связывает вспомогательную молекулу. Например, популяция Т-клеток может контактировать с анти-CD3 антителом и анти-CD28 антителом в условиях, подходящих для стимуляции пролиферации Т-клеток. Для стимуляции пролиферации CD4+ T-клеток, анти-CD3 антитела и анти-CD28 антитела. Примеры анти-CD28 антитела включают 9.3, B-T3, XR-CD28 (Diaclone, Безансон, Франция) могут быть использованы, как и другие способы, обычно известные в данной области (Berg et al., Transplant Proc. 30(8):3975-3977, 1998; Haanen et al., J. Exp. Med. 190(9):13191328, 1999; Garland et al., J. Immunol Meth. 227(1-2):53-63, 1999).
В некоторых воплощениях первичный стимулирующий сигнал и костимулирующий сигнал для Т-клеток могут предоставляться различными протоколами. Например, агенты, обеспечивающие каждый сигнал, могут быть в растворе или связаны с поверхностью. Когда они связаны с поверхностью, агенты могут быть связаны с одной и той же поверхностью (то есть в «цис»-образование) или с другими поверхностями (то есть в «транс»-образование). Альтернативно, один агент может быть связан с поверхностью, а другой агент может находиться в растворе. В одном воплощении агент, обеспечивающий костимулирующий сигнал, связан с поверхностью клетки, а агент, обеспечивающий первичный сигнал активации, находится в растворе или связан с поверхностью. В определенных воплощениях оба агента могут находиться в растворе. В другом воплощении агенты могут быть в растворимой форме и затем сшиты с поверхностью, такой как клетка, экспрессирующая Fc-рецепторы, или антитело или другой связывающий агент, который будет связываться с агентами. В связи с этим см., например, публикации патентных заявок США No. 20040101519 и 20060034810 об искусственных антигенпрезентирующих клетках (aAPC), которые предполагаются для применения в активации и размножении Т-клеток в настоящем изобретении.
В одном воплощении два агента иммобилизованы на гранулах, либо на одной и той же грануле, то есть «цис», либо на отдельных гранулах, то есть «транс». Например, агент, обеспечивающий первичный сигнал активации, представляет собой анти-CD3 антитело или его антигенсвязывающий фрагмент, а агент, обеспечивающий сигнал костимуляции, представляет собой анти-CD28 антитело или его антигенсвязывающий фрагмент; и оба агента совместно иммобилизованы на одной и той же грануле в эквивалентных молекулярных количествах. В одном воплощении используется соотношение 1: 1 каждого антитела, связанного с гранулами, для размножения CD4+ T-клеток и роста T-клеток. В некоторых аспектах настоящего изобретения используется соотношение анти-CD3:CD28-антител, связанных с гранулами, так что наблюдается увеличение размножения Т-клеток по сравнению с размножением, наблюдаемым с использованием соотношения 1:1. В одном конкретном воплощении наблюдается увеличение от около 1 до около 3 раз по сравнению с размножением, наблюдаемым с использованием соотношения 1:1. В одном воплощении отношение антитела CD3:CD28, связанного с гранулами, находится в диапазоне от 100:1 до 1:100 и всех целочисленных значений между ними. В одном аспекте настоящего изобретения большее количество анти-CD28-антитела связано с частицами, чем анти-CD3-антитела, то есть соотношение CD3:CD28 меньше единицы. В некоторых воплощениях изобретения отношение анти-CD28-антитела к анти-CD3-антителу, связанному с гранулами, составляет более 2:1. В одном конкретном воплощении используется соотношение 1:100 CD3:CD28 антител, связанных с гранулами. В другом варианте используют соотношение 1:75 CD3:CD28 антител, связанных с гранулами. В дополнительном воплощении используется соотношение 1:50 CD3:CD28 антител, связанных с гранулами. В другом воплощении используют соотношение 1:30 CD3:CD28 антител, связанных с гранулами. В одном предпочтительном воплощении используется соотношение 1:10 CD3:CD28 антител, связанных с гранулами. В другом воплощении используют соотношение 1: 3 CD3:CD28 антител, связанных с гранулами. В еще одном воплощении используется соотношение 3:1 CD3:CD28 антител, связанных с гранулами.
Соотношения частиц к клеткам от 1:500 до 500:1 и любые целочисленные значения между ними могут использоваться для стимуляции Т-клеток или других клеток-мишеней. Как легко могут оценить специалисты в данной области, соотношение частиц к клеткам может зависеть от размера частиц относительно клетки-мишени. Например, мелкие гранулы могут связывать только несколько клеток, в то время как большие гранулы могут связывать много клеток. В некоторых воплощениях отношение клеток к частицам находится в диапазоне от 1: 100 до 100: 1, и любые целочисленные значения между ними, а в других воплощениях отношение составляет от 1:9 до 9:1, и любые целочисленные значения между ними также могут быть использованы для стимуляции Т-клеток. Соотношение анти-CD3- и анти-CD28-связанных частиц к Т-клеткам, которые приводят к стимуляции Т-клеток, может варьировать, как отмечено выше. Например, такие значения могут включать 1:100, 1:50, 1:40, 1:30, 1:20, 1:10, 1:9, 1:8, 1:7, 1:6, 1:5, 1:4, 1:3, 1:2, 1:1, 2:1, 3:1, 4:1, 5:1, 6:1, 7:1, 8:1, 9:1, 10:1, 15:1 и 1:1 частиц на Т-клетку. В одном воплощении используется соотношение частиц к клеткам 1:1 или менее. В одном конкретном воплощении соотношение частиц:клеток может составлять 1:5. В других воплощениях отношение частиц к клеткам может варьировать в зависимости от дня стимуляции. Например, в одном воплощении отношение частиц к клеткам составляет от 1:1 до 10:1 в первый день, и дополнительные частицы добавляют к клеткам каждый день или через день после этого в течение до 10 дней в конечных соотношениях. от 1:1 до 1:10 (на основе количества клеток в день добавления). В одном конкретном воплощении соотношение частиц к клеткам составляет 1:1 в первый день стимуляции и доводят до 1:5 в третий и пятый дни стимуляции. В другом воплощении изобретения частицы добавляют ежедневно или через день до конечного соотношения 1: 1 в первый день и 1: 5 в третий и пятый дни стимуляции. В другом воплощении отношение частиц к клеткам составляет 2:1 в первый день стимуляции и доводят до 1:10 в третий и пятый дни стимуляции. В другом воплощении изобретения частицы добавляют ежедневно или через день до конечного соотношения 1: 1 в первый день и 1:10 в третий и пятый дни стимуляции. Специалист в данной области поймет, что множество других соотношений могут быть подходящими для применения в настоящем изобретении. В частности, соотношения будут варьировать в зависимости от размера частиц, а также от размера и типа клеток.
В других воплощениях настоящего изобретения клетки, такие как Т-клетки (например, Treg-клетки) объединяют с гранулами, покрытыми агентом, гранулы и клетки затем разделяют, а клетки затем культивируют. В альтернативном воплощении перед культивированием покрытые агентом гранулы и клетки не разделяют, а культивируют вместе. В дополнительном воплощении гранулы и клетки сначала концентрируют путем приложения силы, такой как магнитная сила, что приводит к усилению лигирования маркеров клеточной поверхности, вызывая тем самым стимуляцию клеток.
Например, белки клеточной поверхности можно лигировать, позволяя парамагнитным гранулам, к которым присоединены анти-CD3 и анти-CD28 (3 × 28 гранулы), контактировать с T-клетками. В одном воплощении клетки (например, 104-109 Т-клеток) и гранулы (например, парамагнитные гранулы DYNABEADS® M-450 CD3/CD28 T в соотношении 1: 1) объединяют в буфере, предпочтительно PBS (без двухвалентных катионов, таких как кальций и магний). Опять же специалисты в данной области техники могут легко понять, что можно использовать любую концентрацию клеток.
Например, клетка-мишень может быть очень редкой в образце и содержать только 0,01% образца, или весь образец (т.е. 100%) может содержать интересующую ячейку-мишень. Соответственно, любое количество клеток находится в контексте настоящего изобретения. В определенных воплощениях может быть желательным значительно уменьшить объем, в котором частицы и клетки смешиваются (то есть увеличить концентрацию клеток), чтобы обеспечить максимальный контакт клеток и частиц. Например, в одном воплощении используется концентрация около 2 миллиардов клеток/мл. В другом воплощении используют более 100 миллионов клеток/мл. В следующем воплощении используется концентрация клеток 10, 15, 20, 25, 30, 35, 40, 45 или 50 миллионов клеток/мл. В еще одном воплощении используется концентрация клеток от 75, 80, 85, 90, 95 или 100 миллионов клеток/мл. В других воплощениях могут использоваться концентрации 125 или 150 миллионов клеток/мл. Использование высоких концентраций может привести к увеличению выхода клеток, активации клеток и размножения клеток. Кроме того, применение высоких концентраций клеток позволяет более эффективно захватывать клетки, которые могут слабо экспрессировать представляющие интерес антигены-мишени. Такие популяции клеток могут иметь терапевтическую ценность и их было бы желательно получить в определенных воплощениях.
В одном воплощении настоящего изобретения смесь может быть культивирована в течение от нескольких часов (около 3 часов) до около 14 дней или любого часового целочисленного значения между ними. В другом воплощении смесь можно культивировать в течение 21 дня. В одном воплощении изобретения гранулы и Т-клетки культивируют вместе в течение около восьми дней. В другом воплощении гранулы и Т-клетки культивируют вместе в течение 2-3 дней. Также могут быть желательными несколько циклов стимуляции, так что время культивирования Т-клеток может составлять 60 дней или более. Условия, подходящие для культуры Т-клеток, включают подходящие среды (например, минимальная питательная среда или среды RPMI 1640 или X-vivo 15 (Lonza)), которые могут содержать факторы, необходимые для пролиферации и жизнеспособности, включая сыворотку (например, эмбриональная бычья или человеческая сыворотка), интерлейкин-2 (IL-2), инсулин, IFN-γ, IL-4, IL-7, GM-CSF, IL-10, IL-12, IL-15, TGFβ и TNF-α или любые другие добавки для роста клеток, известные специалисту в данной области. Другие добавки для роста клеток включают, без ограничения указанным, поверхностно-активное вещество, плазманат и восстановители, такие как N-ацетилцистеин и 2-меркаптоэтанол. Среда может включать RPMI 1640, Immunocult (StemCell), AIM-V, DMEM, MEM, α-MEM, F-12, X-Vivo 15 и X-Vivo 20, Optimizer, с добавлением аминокислот, пирувата натрия и витаминов, либо без сыворотки или дополненный соответствующим количеством сыворотки (или плазмы) или определенного набора гормонов и/или количеством цитокинов, достаточным для роста и размножения Т-клеток. Антибиотики, например, пенициллин и стрептомицин, включены только в экспериментальные культуры, но не в культуры клеток, которые должны быть введены объекту. Клетки-мишени поддерживают в условиях, необходимых для поддержания роста, например, при подходящей температуре (например, 37°C) и атмосфере (например, воздух плюс 5% CO2).
Т-клетки, которые подвергались различным временам стимуляции, могут иметь различные характеристики. Например, типичные продукты мононуклеарных клеток крови или периферической крови с периферической кровью имеют популяцию вспомогательных Т-клеток (Th, CD4+), которая больше, чем популяция цитотоксических или супрессорных Т-клеток (Tc, CD8+). Экспрессия Т-клеток ex vivo путем стимуляции рецепторов CD3 и CD28 приводит к образованию популяции Т-клеток, которая до около 8-9 дней состоит преимущественно из Th-клеток, в то время как через около 8-9 дней популяция Т-клеток включает все большую популяцию Тс-клеток.
Кроме того, помимо маркеров CD4 и CD8, другие фенотипические маркеры значительно варьируют, но в значительной степени воспроизводимы в ходе процесса размножения клеток. Таким образом, такая воспроизводимость дает возможность адаптировать активированный Т-клеточный продукт для конкретных целей.
В одном воплощении регуляторные Т-клетки могут быть получены путем выделения CD4+ Т-клеток от доноров HLA-A2- через RosetteSep (Stemcell) и обогащены по CD25+-клеткам (Miltenyi) перед сортировкой живых CD4+CD127loCD25hi Treg с помощью FACSAria II (BD Biosciences). Сортированные Treg могут быть стимулированы с помощью L-клеток и αCD3 mAb (ОКТ3; 200 нг/мл) в 1000 Ед/мл IL-2.
В одном воплощении изобретения Т-клетки можно культивировать в присутствии рапамицина для получения регуляторных Т-клеток, как описано, например, в WO2007110785, включенном в настоящий документ ссылкой. Другой способ получения регуляторных Т-клеток описан в US2016024470, включенном в настоящий документ ссылкой, где Т-клетки культивируют с активатором Т-клеточного рецептора (TCR)/CD3, таким как, например, антитела TCR/CD3, активатор костимулятора TCR, такой как, например, антитела CD28, CD137 (4-1 BB), GITR, B7-1/2, CD5, ICOS, OX40, CD40 или CD137 и рапамицин.
В одном воплощении изобретения Т-клетки, генетически модифицированные путем экспрессии CAR, также могут быть генетически модифицированы путем экспрессии, по меньшей мере, одного внутриклеточного фактора, такого как ROR-C, Foxo1, T-bet или Gata 3, c-Maf, AhR. В одном воплощении генетически модифицированная иммунная клетка по изобретению экспрессирует Foxo1.
В одном воплощении генетически модифицированные клетки по настоящему изобретению могут представлять собой аллогенные иммунные клетки, такие как, например, аллогенные T- или Treg-клетки. Например, аллогенная иммунная клетка может представлять собой иммунную клетку, в которой отсутствует экспрессия функционального человеческого лейкоцитарного антигена (HLA), например, HLA класса I и/или HLA класса II, или отсутствует экспрессия функционального рецептора Т-клеток (TCR).
В одном воплощении T-клетка, не имеющая функционального TCR, может быть сконструирована таким образом, чтобы она не экспрессировала какой-либо функциональный TCR на своей поверхности, сконструирована таким образом, чтобы она не экспрессировала одну или несколько субъединиц, которые содержат функциональный TCR, или сконструирована таким образом, что она продуцировала очень маленький функциональный TCR на своей поверхности. Альтернативно, Т-клетка может экспрессировать существенно ослабленный TCR, например, путем экспрессии мутированных или укороченной форм одной или нескольких субъединиц TCR. Термин «существенно ослабленный TCR» означает, что этот TCR не будет вызывать неблагоприятную иммунную реакцию у хозяина.
В другом воплощении генетически модифицированные клетки, описанные в данном документе, могут быть сконструированы таким образом, чтобы они не экспрессировали функциональный HLA на своей поверхности. Например, иммунная клетка, описанная в данном документе, может быть сконструирована так, чтобы снижалась экспрессия HLA на клеточной поверхности, например, HLA класса 1 и/или HLA класса II, или неклассических молекул HLA.
В другом воплощении Т-клетка может не иметь функционального TCR и функционального HLA, такого как HLA класса I и/или HLA класса II.
Модифицированные иммунные клетки (такие как, например, модифицированные T- или Treg-клетки), у которых отсутствует экспрессия функционального TCR и/HLA, могут быть получены любым подходящим способом, включая нокаут или нокдаун одной или нескольких субъединиц TCR и/или HLA. Например, иммунная клетка может включать нокдаун TCR и/или HLA с использованием siRNA, shRNA, кластерных коротких палиндромных повторов, разделенных регулярными промежутками (CRISPR), таких как эффекторная нуклеаза, подобная активаторам транскрипции (TALEN), эндонуклеаза цинкового пальца (ZFN), мегануклеаза (mn, также известная как хоминг-эндонуклеаза) или megaTAL (объединяющий эффектор TAL с доменом расщепления mn) (Osborn et al, «Evaluation of TCR Gene Editing Achieved by TALENs, CRISPR/Cas9, and megaTAL Nucleases» Mol Ther. 2016 Mar;24(3):570-81).
В одном воплощении экспрессия TCR и/или экспрессия HLA могут быть ингибированы с использованием siРНК или shRNA, которая нацелена на нуклеиновую кислоту, кодирующую TCR и/или HLA в Т-клетке. Экспрессия siRNA и shRNA в Т-клетках может быть достигнута с использованием любой обычной системы экспрессии, например, такой как система экспрессии лентивируса. Типичные siRNA и shRNA, которые подавляют экспрессию генов HLA класса I и/или HLA класса II, описаны, например, в US 2007/0036773. Типичные shRNA, которые подавляют экспрессию компонентов TCR, описаны, например, в US2012/0321667.
Используемый в данном документе термин «CRISPR» или «CRISPR для TCR и/или HLA» или «CRISPR для ингибирования TCR и/или HLA» относится к набору кластерных коротких палиндромных повторов, разделенные регулярными промежутками, или к системе, содержащей такой набор повторов. «Cas», при использовании в данном документе, относится к CRISPR-ассоциированному белку. A Система «CRISPR/Cas» относится к системе, полученной из CRISPR и Cas, которая может использоваться для подавления или мутации гена TCR и/или HLA.
Встречающиеся в природе системы CRISPR/Cas обнаруживаются в около 40% секвенированных геномов эубактерий и в 90% секвенированных архей. Grissa et al. (2007) BMC Bioinformatics 8: 172. Эта система представляет собой тип прокариотической иммунной системы, которая придает устойчивость к чужеродным генетическим элементам, таким как плазмиды и фаги, и обеспечивает форму приобретенного иммунитета. Barrangou et al. (2007) Science 315: 1709-1712; Marragini et al. (2008) Science 322: 1843-1845. Система CRISPR/Cas была модифицирована для применения при редактировании генов (сайленсинг, усиление или изменение определенных генов) у эукариот, таких как мыши или приматы. Wiedenheft et al. (2012) Nature 482: 331-8. Это достигается путем введения в эукариотическую клетку плазмиды, содержащей специально сконструированный CRISPR и один или несколько подходящих Cas. Последовательность CRISPR, иногда называемая локусом CRISPR, включает чередующиеся повторы и спейсеры. Встречающиеся в природе спейсеры CRISPR обычно содержат последовательности, чужеродные для бактерии, такие как последовательность плазмиды или фага; в системе TCR и/или HLA CRISPR/Cas спейсеры получены из последовательности гена TCR или HLA. РНК из локуса CRISPR конститутивно экспрессируется и перерабатывается белками Cas в небольшие РНК. Они содержат спейсер, фланкированный повторяющейся последовательностью. РНК направляют другие белки Cas, чтобы заставить отключить экзогенные генетические элементы на уровне РНК или ДНК. Horvath et al. (2010) Science 327: 167-170; Makarova et al. (2006) Biology Direct 1: 7. Спейсеры, таким образом, служат матрицами для молекул РНК, аналогично миРНК. Pennisi (2013) Science 341: 833-836. Таким образом, система CRISPR/Cas может использоваться для редактирования гена TCR и/или HLA (добавления или удаления пары оснований) или для введения преждевременного стоп-кодона, который, таким образом, уменьшает экспрессию TCR и/или HLA. В качестве альтернативы можно использовать систему CRISPR/Cas, подобно РНК-интерференции, обратимым образом отключая ген HLA. Например, в клетке млекопитающего РНК может направлять белок Cas к промотору TCR и/или HLA, стерически блокируя РНК-полимеразы.
Могут быть созданы искусственные системы CRISPR/Cas, которые ингибируют TCR и/или HLA, используя технологию, известную в данной области, например, описанную в US 20140068797 и Cong (2013) Science 339: 819-823. Также могут быть получены другие искусственные системы CRISPR/Cas, известные в данной области техники, которые ингибируют TCR и/или HLA, например, которые описаны в Tsai (2014) Nature Biotechnol., 32: 6 569-576, US 8,871,445; 8865406; 8795965; 8771945; и 8 697 359.
«TALEN» или «TALEN для TCR и/или HLA» или «TALEN для ингибирования TCR и/или HLA» относится к эффекторной нуклеазе, подобной активатору транскрипции, искусственной нуклеазе, которую можно использовать для редактирования TCR и/или HLA. TALEN получают искусственно путем слияния TAL-эффекторного ДНК-связывающего домена с доменом расщепления ДНК. Активаторы, подобные активаторам транскрипции (TALE), могут быть сконструированы так, чтобы связывать любую искомую последовательность ДНК, включая часть гена TCR и/или HLA. Комбинируя сконструированный TALE с доменом расщепления ДНК, можно получить рестриктазу, которая специфична для любой искомой последовательности ДНК, включая последовательность TCR и/или HLA. Затем они могут быть введены в клетку, где они могут быть использованы для редактирования генома. Boch (2011) Nature Biotech. 29: 135-6; и Boch et al. (2009) Science 326: 1509-12; Moscou et al. (2009) Science 326: 3501.
TALE представляет собой белки, секретируемые бактериями Xanthomonas. ДНК-связывающий домен включает повторяющуюся высококонсервативную 33-34 аминокислотную последовательность, за исключением 12-й и 13-й аминокислот. Эти две позиции сильно варьируют, показывая сильную корреляцию с распознаванием специфических нуклеотидов. Таким образом, они могут быть сконструированы так, чтобы связываться с искомой последовательностью ДНК. Чтобы получить TALEN, белок TALE сливают с нуклеазой (N), которая является эндонуклеазой Fokl дикого типа или мутированной. Было получено несколько мутаций Fokl для ее использования в TALEN; например, они улучшают специфичность или активность расщепления. Cermak et al. (2011) Nucl. Acids Res. 39: e82; Miller et al. (2011) Nature Biotech. 29: 143-8; Hockemeyer et al. (2011) Nature Biotech. 29: 731-734; Wood et al. (2011) Science 333: 307; Doyon et al. (2010) Nature Methods 8: 74-79; Szczepek et al. (2007) Nature Biotech. 25: 786-793; и Guo et al. (2010)/. Mol. Biol. 200: 96. Домен Fokl функционирует как димер, требуя двух конструкций с уникальными ДНК-связывающими доменами для сайтов в геноме-мишени с правильной ориентацией и расстоянием. Количество аминокислотных остатков между ДНК-связывающим доменом TALE и доменом расщепления Fokl, а также количество оснований между двумя отдельными сайтами связывания TALEN, по-видимому, являются важными параметрами для достижения высоких уровней активности. Miller et al. (2011) Nature Biotech. 29: 143-8. TCR и/или HLA TALEN могут использоваться внутри ячейки для создания двухцепочечного разрыва (DSB). Мутация может быть введена в месте разрыва, если ремонтные механизмы неправильно восстанавливают разрыв через негомологичное соединение конца. Например, неправильное восстановление может привести к мутации со сдвигом рамки. Альтернативно, чужеродная ДНК может быть введена в клетку вместе с TALEN; в зависимости от последовательностей чужеродной ДНК и хромосомной последовательности этот процесс можно использовать для исправления дефекта в гене TCR и/или HLA или для введения такого дефекта в ген wt TCR и/или HLA, таким образом уменьшая экспрессию TCR и/или HLA. TALEN, специфичные для последовательностей в TCR и/или HLA, могут быть созданы с использованием любого способа, известного в данной области техники, включая различные схемы с использованием модульных компонентов. Zhang et al. (2011) Nature Biotech. 29: 149-53; Geibler et al. (2011) PLoS ONE 6: el9509.
«ZFN» или «нуклеаза с доменом цинковые пальцы» или «ZFN для TCR и/или HLA» или «ZFN для ингибирования TCR и/или HLA» относятся к нуклеазе с доменом цинковые пальцы, искусственной нуклеазе, которую можно использовать для редактирования гена TCR и/или HLA. Как и TALEN, ZFN включает нуклеазный домен Fokl (или его производное), слитые с ДНК-связывающим доменом. В случае ZFN ДНК-связывающий домен включает один или несколько цинковых пальцев. Carroll et al. (2011) Genetics Society of America 188: 773-782; и Kim et al. (1996) Proc. Natl. Acad. Sci. USA 93: 1156-1160. Цинковый палец представляет собой небольшой структурный мотив белка, стабилизированный одним или несколькими ионами цинка. Цинковый палец может содержать, например, Cys2His2 и может распознавать последовательность длиной около 3 п.н. Различные цинковые пальцы известной специфичности могут быть объединены для получения полипептидов с несколькими пальцами, которые распознают последовательности длиной 6, 9, 12, 15 или 18 п.н. Доступны различные методы отбора и модульной сборки для создания цинковых пальцев (и их комбинаций), распознающих специфические последовательности, которые включают фаговый дисплей, дрожжевые одногибридные системы, бактериальные одногибридные и двухгибридные системы и клетки млекопитающих.
Как и TALEN, ZFN должен димеризоваться, чтобы расщеплять ДНК. Таким образом, для нацеливания на непалиндромные сайты ДНК необходима пара ZFN. Два отдельных ZFN должны связывать противоположные цепи ДНК с их нуклеазами, правильно расположенными друг относительно от друга. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10570-5. Также как TALEN, ZFN может создать двухцепочечный разрыв в ДНК, который может создавать мутацию со смещением рамки, при неправильной репарации, что приводит к снижению экспрессии и количества TCR и/или HLA в клетке. ZFN также можно использовать с гомологичной рекомбинацией для мутации в гене TCR и/или HLA. ZFN, специфичные для последовательностей в TCR и/или HLA, могут быть сконструированы с использованием любого способа, известного в данной области. См., Например, Provasi (2011) Nature Med. 18: 807-815; Torikai (2013) Blood 122: 1341-1349; Cathomen et al. (2008) Mol. Ther. 16: 1200-7; Quo et al. (2010) /. Mol. Biol. 400: 96; US2011/0158957; и US2012/0060230.
«Мегануклеаза» или «мегануклеаза для TCR и/или HLA» или «мегануклеаза для ингибирования TCR и/или HLA» относится к мономерной эндонуклеазе с большими (> 14 пар оснований) сайтами распознавания, которые можно использовать для редактирования гена TCR и/или HLA. Мегануклеазы (mn) представляют собой мономерные белки с врожденной нуклеазной активностью, которые получены из бактериальных самонаводящихся эндонуклеаз и сконструированы для уникального сайта-мишени. Хоминг-эндонуклеазы являются ДНК-расщепляющими ферментами, которые могут генерировать двухцепочечные разрывы в отдельных локусах в своих геномах-хозяевах и, таким образом, управлять сайт-специфическими событиями конверсии генов. (Stoddard, Structure. 2011 Jan 12;19(1):7-15). Несмотря на их небольшой размер, хоминг-эндонуклеазы распознают длинные последовательности ДНК (обычно от 20 до 30 пар оснований). Хоминг-эндонуклеазы чрезвычайно распространены и обнаружены в микробах, а также в фагах и вирусах. Бокс-ферменты LAGLIDADG и His-Cys (которые являются наиболее специфичными для последовательности из этих ферментов) полагаются на антипараллельные β-листы, которые закрепляются в основных бороздках ДНК их сайтов-мишеней (Flick et al., 1998; Jurica et al., 1998). Там они устанавливают коллекцию специфичных для последовательности и неспецифических контактов, которые распределены неравномерно по нескольким последовательным базовым парам (Chevalier et al., 2003; Scalley-Kim et al., 2007).
Семейство хоминг-эндонуклеаз LAGLIDADG (LHE) является основным источником сконструированных ферментов, используемых в приложениях для направленного воздействия на гены. Семейство LHE в основном кодируется в археях и в хлоропластных и митохондриальных геномах водорослей и грибов (Chevalier et al., 2005; Dalgaard et al., 1997; Sethuraman et al., 2009). Мегануклеазы, которые обладают одним консервативным мотивом LAGLIDADG (SEQ ID NO: 85) на цепь белка, образуют гомодимерные белки, которые расщепляют последовательности-мишени палиндромной и почти палиндромной ДНК, тогда как те, которые содержат два таких мотива на цепь белка, образуют более крупные псевдосимметричные мономеры, которые могут направленно воздействовать на полностью асимметричные последовательности ДНК.
Мегануклеазы могут быть сконструированы для направленного воздействия на TCR и/или HLA и, таким образом, создавать двухцепочечный разрыв в ДНК, который может создать мутацию со сдвигом рамки при неправильной репарации, что приводит к снижению экспрессии и количеству TCR и/или HLA в клетке.
«MegaTAL» или «megaTAL для TCR и/или HLA» или «megaTAL для ингибирования TCR и/или HLA» относится к искусственной нуклеазе, которую можно использовать для редактирования гена TCR и/или HLA. MegaTAL представляют собой гибридные мономерные нуклеазы, полученные путем слияния минимальных TAL (подобных активатору транскрипции) эффекторных доменов с N-концом мегануклеаз, происходящих из семейства хоминг-эндонуклеаз LAGLIDADG (Nucleic Acids Res. 2014 Feb;42(4):2591-601; Takeuchi et al, Methods Mol Biol. 2015;1239:105-32. doi: 10.1007/978-1-4939-1862-1_6). MegaTAL, таким образом, состоят из сайт-специфической головки расщепления мегануклеазой с дополнительной аффинностью и специфичностью, обеспечиваемой связывающим доменом эффекторной ДНК TAL. MegaTAL могут быть сконструированы так, чтобы нацеливаться на TCR и/или HLA и, таким образом, создавать двухцепочечный разрыв в ДНК, который может создать мутацию со сдвигом рамки при неправильной репарации, что приведет к снижению экспрессии и количества HLA в клетке. Вариант мегануклеазы I-OnuI (mn) использовали для конструирования TCRα-megaTAL для нокаута гена T-клеточного рецептора альфа (TCRα). TCRα mn слили с массивом из 10,5-повторов TALE, предназначенным для связывания последовательности ДНК перед сайтом связывания TCRα mn. Было обнаружено, что megaTAL, нацеленный на TCRα, достигал чрезвычайно высокого разрушения гена без заметного расщепления вне цели в первичных Т-клетках человека. (Boissel et al., Nucleic Acids Res. 2014 Feb;42(4):2591-601).
Не желая быть связанными какой-либо конкретной теорией, полагаем, что в некоторых воплощениях терапевтическая Т-клетка имеет кратковременное персистирование у пациента вследствие укороченных теломер в Т-клетке; соответственно, трансфекция геном теломеразы может удлинить теломеры Т-клетки и улучшить устойчивость Т-клетки у пациента. См. Carl June, «Adoptive T cell therapy for cancer in the clinic», Journal of Clinical Investigation, 117: 1466-1476 (2007). Таким образом, в одном воплощении генетически модифицированная иммунная клетка по изобретению эктопически экспрессирует субъединицу теломеразы, например, каталитическую субъединицу теломеразы, например, TERT, например, hTERT. В некоторых аспектах это раскрытие относится к способу получения CAR-экспрессирующей иммунной клетки, включающему контактирование иммунной клетки с нуклеиновой кислотой, кодирующей субъединицу теломеразы, например, каталитическую субъединицу теломеразы, например, TERT, например, hTERT. Клетка может контактировать с нуклеиновой кислотой до, одновременно или после контакта с конструкцией, кодирующей CAR.
VII. ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ
В изобретении предлагаются иммунные клетки, модифицированные для экспрессии CAR, нацеленного на антиген HLA-A2, в качестве средства для привлечения указанных иммунных клеток к участкам специфического иммунного или воспалительного ответа и для активации иммунных клеток для подавления иммунологической активности иммунных эффекторных клеток в этих участки. Таким образом, CAR-опосредованный рекрутинг и активация иммунных клеток обеспечивает способ предотвращения или лечения ряда иммунологических заболеваний или нарушений, для которых такая иммуносупрессивная активность является полезной.
В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. Иммунная клетка может представлять собой любую регуляторную иммунную клетку, подходящую для применения в клеточной терапии (см., например, Wood, K.J. et al., 2012; Papp, G. et al., 2017). В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации.
В одном воплощении иммунная клетка может представлять собой регуляторную Т-клетку. В другом воплощении регуляторная T-клетка может представлять собой регуляторную T-клетку человека. Хотя активация регуляторных Т-клеток является антиген-зависимой, подавляющее действие этих клеток не зависит от антигена, TCR и MHC. Соответственно, экспрессия CAR в регуляторных T-клетках перенаправляет эти клетки и их активацию в соответствующую ткань-мишень, так что они активируются антигенспецифическим образом; однако их супрессирующие эффекты происходят без необходимости дальнейшего распознавания антигенов, связанных с заболеванием. Следовательно, до тех пор, пока регуляторные Т-клетки находятся в правильной близости от того места, где находятся иммунные эффекторные клетки и опосредуют свои нежелательные эффекты, перенаправленные регуляторные Т-клетки могут запускаться или активироваться в этом месте для обеспечения своих супрессирующих эффектов.
В одном воплощении целевой антиген HLA-A2 может присутствовать или экспрессироваться в сайте или ткани-мишени нежелательного иммунного или воспалительного ответа, опосредованного иммунными эффекторными клетками.
В одном воплощении CAR-модифицированные иммунные клетки по изобретению можно использовать для лечения одного или нескольких заболеваний, расстройств, симптомов или состояний, связанных с трансплантацией органа или ткани (например, отторжение/дисфункция органа или ткани, GVHD и/или связанные с этим условия). Отторжение трансплантата включает разрушение трансплантированной ткани донора иммунными клетками реципиента посредством иммунного ответа. Иммунный ответ также участвует в GVHD; однако в этом случае ткани реципиента разрушаются иммунными клетками донора, переданными реципиенту посредством трансплантации. Соответственно, CAR-обусловленное перенаправление и активация иммунных клеток обеспечивают способ подавления отторжения несоответствующих клеток и/или тканей иммунными эффекторными клетками у реципиентов трансплантата или ингибирования патогенного действия трансплантированных иммунокомпетентных клеток в случае GVHD. В одном воплощении несоответствующие клетки и/или ткани содержат клетки и/или ткани несоответствующие по HLA-A2.
Таким образом, другим воплощением настоящего изобретения является способ лечения одного или нескольких заболеваний, расстройств, симптомов или состояний, связанных с трансплантацией органа или ткани (например, отторжения/дисфункции органа или ткани, GVHD и/или состояний, связанных с ними) объекту, где указанный способ включает введение объекту сконструированной иммунной клетки с CAR или композиции, как описано в настоящем документе.
В одном воплощении способ представляет собой способ клеточной терапии. В одном воплощении клеточная терапия является аутологичной. В одном воплощении клеточная терапия является гетерологичной. В одном воплощении клеточная терапия является аллогенной. В одном воплощении способ представляет собой способ генной терапии.
Таким образом, другим воплощением настоящего изобретения является иммунная клетка, сконструированная с помощью CAR, или композиция, описанная в настоящем документе, для применения при лечении одного или нескольких заболеваний, расстройств, симптомов или состояний, связанных с трансплантацией органа или ткани (например, отторжение/дисфункция органа или ткани, GVHD и/или связанные с ним состояния) у объекта.
CAR-модифицированные иммунные клетки по изобретению можно использовать для стимуляции иммунной толерантности, функциональной толерантности и/или иммунной аккомодации у объекта. CAR-модифицированные иммунные клетки по настоящему изобретению могут использоваться для стимуляции иммунной толерантности, функциональной толерантности и/или иммунной аккомодации у объекта после трансплантации органа или ткани. В одном воплощении предлагается способ стимуляции иммунной толерантности, функциональной толерантности и/или иммунной аккомодации у объекта, причем способ включает введение объекту CAR-модифицированной иммунной клетки, описанной где-либо в данном документе. В другом воплощении предлагается способ стимуляции иммунной толерантности, функиональной толерантности и/или иммунной аккомодации у объекта, причем способ включает введение объекту фармацевтической композиции, описанной где-либо в данном документе. В одном воплощении применение может быть для стимуляции иммунной толерантности, функциональной толерантности и/или иммунной аккомодации к трансплантированному органу или ткани у объекта. В одном воплощении CAR-модифицированную иммунную клетку вводят одновременно с, до или после трансплантации органа или ткани. В одном воплощении CAR-модифицированную иммунную клетку вводят одновременно с трансплантацией органа или ткани. В другом воплощении CAR-модифицированную иммунную клетку вводят перед трансплантацией органа или ткани. В одном воплощении CAR-модифицированную иммунную клетку вводят после трансплантации органа или ткани. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В одном воплощении иммунная клетка представляет собой регуляторную Т-клетку. В одном воплощении иммунная клетка представляет собой регуляторную CD4+ Т-клетку. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В другом воплощении иммунная клетка представляет собой регуляторную Т-клетку человека. В одном воплощении иммунная клетка представляет собой CD4+ регуляторную Т-клетку человека.
CAR-модифицированные иммунные клетки по настоящему изобретению можно использовать для предотвращения или лечения отторжения трансплантированного органа или ткани. В одном воплощении CAR-модифицированные иммунные (например, CAR-сконструированные Treg-клетки) по настоящему изобретению можно использовать для предотвращения или лечения чрезмерного отторжения пересаженного органа или ткани. В одном воплощении CAR-модифицированные иммунные клетки (например, CAR-сконструированные Treg-клетки) по настоящему изобретению можно использовать для предотвращения или лечения опосредованного антителами отторжения трансплантированного органа или ткани. В одном воплощении способ включает введение CAR-сконструированных иммунных клеток (например, CAR-сконструированных Treg-клеток) по настоящему изобретению объекту, подвергающемуся воздействию трансплантированного органа или ткани. В одном воплощении трансплантированные орган или ткань может включать трансплантацию костного мозга, трансплантацию органа, переливание крови или любую другую инородную ткань или клетку, которые целенаправленно вводятся объекту. В одном воплощении CAR-модифицированные иммунные клетки (например, CAR-сконструированные Treg-клетки) по настоящему изобретению могут использоваться в качестве терапии для ингибирования отторжения трансплантата после трансплантации. В одном воплощении отторжение трансплантата может представлять собой отторжение аллотрансплантата. В одном воплощении отторжение трансплантата может представлять собой отторжение ксенотрансплантата. В одном воплощении предоставлен способ предотвращения или лечения отторжения трансплантата органа или ткани у объекта, причем способ включает введение объекту CAR-модифицированной иммунной клетки, описанной где-либо в данном документе. В одном воплощении предложен способ предотвращения или лечения отторжения трансплантата органа или ткани у объекта, причем способ включает введение объекту фармацевтической композиции, описанной где-либо в данном документе. Таким образом, другим воплощением настоящего изобретения является CAR-сконструированная иммунная клетка (CAR-сконструированная Treg-клетка) по изобретению, или фармацевтическая композиция, содержащая указанные иммунные клетки, для применения в профилактике или лечении отторжения трансплантата органа или ткани у объекта. В одном воплощении настоящее изобретение относится к способу увеличения периода выживания трансплантата у объекта, включающему введение объекту CAR-модифицированной иммунной клетки (например, CAR-сконструированной Treg-клетки), описанной где-либо в данном документе. В одном воплощении настоящее изобретение относится к способу увеличения периода выживаемости трансплантата у объекта, включающему введение объекту фармацевтической композиции, описанной где-либо в данном документе. В одном воплощении способ обеспечивает период выживания трансплантата, составляющий 1 неделю, 2 недели, 3 недели, 1 месяц, 2 месяца, 3 месяца, 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев, 1 год, 2 года, 3 года, 4 года, 5 лет, 6 лет, 7 лет, 8 лет, 9 лет, 10 лет, 20 лет, 30 лет, 40 лет, 50 лет, 60 лет, 70 лет, 80 лет, 90 лет, 100 лет или все время жизни объекта. В одном воплощении объект не подвергается какой-либо терапии иммуносупрессирующими агентами. В одном воплощении введение иммунной клетки или композиции по изобретению позволяет снизить количество терапии иммуносупрессантом, получаемого объектом. В одном воплощении трансплантат может быть аллотрансплантатом. В одном воплощении трансплантат может подвергаться воздействию CAR-модифицированных иммунных клеток по настоящему изобретению в то же время, до, или после трансплантации трансплантата реципиенту. В одном воплощении трансплантат органа или ткани может представлять собой сердце, сердечный клапан, легкое, почку, печень, поджелудочную железу, кишечник, кожу, кровеносные сосуды, костный мозг, стволовые клетки, кости или островковые клетки. Однако изобретение не ограничено конкретным типом трансплантации. В одном воплощении донорский трансплантат может быть «предварительно кондиционирован» или «предварительно обработан» путем обработки органа или тканевого трансплантата перед трансплантацией реципиенту CAR-модифицированных иммунных клеток по изобретению, чтобы снизить иммуногенность трансплантата против реципиента, тем самым уменьшая или предотвращая отторжение трансплантата. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного подтипа HLA-A, выбранного из одного из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из двух из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного подтипа HLA-A, выбранного из одного из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из двух из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*03. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*25. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*29. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*31. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*33. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*36. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*25:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*29:02. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*03:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*31:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*33:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*36:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*68:01. В одном воплощении трансплантат является HLA-A2-положительным.
В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В одном воплощении иммунная клетка представляет собой регуляторную Т-клетку. В одном воплощении иммунная клетка представляет собой регуляторную CD4+ Т-клетку. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В другом воплощении иммунная клетка представляет собой регуляторную Т-клетку человека. В одном воплощении иммунная клетка представляет собой CD4+ регуляторную Т-клетку человека.
CAR-модифицированные иммунные клетки (например, CAR-сконструированные Treg-клетки) по настоящему изобретению могут быть использованы для предотвращения или лечения болезни трансплантат против хозяина (GVHD). В одном воплощении способ включает введение CAR-модифицированной иммунной клетки (например, CAR-сконструированных Treg-клеток) по настоящему изобретению объекту, подвергающемуся воздействию трансплантированного органа или ткани. В одном воплощении трансплантированный орган или ткань может включать трансплантацию костного мозга, трансплантацию органа, переливание крови или любую другую инородную ткань или клетку, которые целенаправленно вводятся объекту. Например, GVHD может возникать после трансплантации сердца, сердечного клапана, легких, почек, печени, поджелудочной железы, кишечника, кожи, кровеносных сосудов, костного мозга, стволовых клеток, костей или островковых клеток. Однако изобретение не ограничено конкретным типом трансплантации. В одном воплощении GVHD может возникать после трансплантации гемопоэтических стволовых клеток. В одном воплощении предоставлен способ предотвращения или лечения болезни «трансплантат против хозяина» (GVHD) у объекта, причем способ включает введение объекту CAR-модифицированной иммунной клетки, описанной где-либо в данном документе. В одном воплощении предлагается способ предотвращения или лечения болезни «трансплантат против хозяина» (GVHD) у объекта, включающий введение объекту фармацевтической композиции, описанной где-либо в данном документе. В одном воплощении изобретение относится к способу контакта донорного трансплантата, например, биосовместимой решетки или донорской ткани, органа или клетки, с CAR-модифицированными иммунными клетками по настоящему изобретению одновременно, до или после пересадки трансплантата реципиенту. В одном воплощении CAR-модифицированные иммунные клетки по настоящему изобретению могут быть использованы для ослабления, ингибирования или снижения неблагоприятного ответа донорного трансплантата против реципиента, тем самым осуществляя предотвращение или лечение GVHD. В одном воплощении настоящее изобретение относится к способу предотвращения или отсрочки возникновения GVHD у объекта, включающему введение объекту CAR-модифицированной иммунной клетки, описанной где-либо в данном документе. В одном воплощении настоящее изобретение относится к способу предотвращения или отсрочки возникновения GVHD у объекта, включающему введение объекту фармацевтической композиции, описанной где-либо в данном документе. В одном воплощении начало GVHD задерживается на 1 неделю, 2 недели, 3 недели, 1 месяц, 2 месяца, 3 месяца, 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев, 1 год, 2 года, 3 года, 4 года, 5 лет, 6 лет, 7 лет, 8 лет, 9 лет, 10 лет, 20 лет, 30 лет, 40 лет, 50 лет, 60 лет, 70 лет, 80 лет, 90 лет, 100 лет или на все время жизни объекта. В одном воплощении объект не подвергается какой-либо терапии иммуносупрессирующими агентами. В одном воплощении объект подвергается иммуносупрессирующей терапии. В одном воплощении иммунные клетки по изобретению вводят объекту с целью уменьшения терапевтически эффективного количества указанной иммуносупрессирующей терапии. В одном воплощении GVHD может быть острой GVHD или хронической GVHD. В одном воплощении донорский трансплантат может быть «предварительно кондиционирован» или «предварительно обработан» путем обработки трансплантата перед трансплантацией реципиенту CAR-модифицированными иммунными клетками (например, CAR-сконструированными Treg-клетками) по изобретению для снижения иммуногенности трансплантата против реципиента, тем самым уменьшая или предотвращая GVHD. В одном воплощении трансплантат может контактировать с клетками или тканью реципиента до трансплантации, чтобы активировать Т-клетки, которые могут быть связаны с трансплантатом. После обработки трансплантата клетками или тканью реципиента клетки или ткани могут быть удалены из трансплантата. Затем обработанный трансплантат может быть далее приведен в контакт с CAR-модифицированными иммунными клетками (например, CAR-сконструированными Treg-клетками) по настоящему изобретению, чтобы снизить, ингибировать или устранить активность иммунных эффекторных клеток, которые были активированы при обработке клеток или ткани из реципиента. После этой обработки трансплантата с помощью CAR-модифицированных иммунных клеток по настоящему изобретению CAR-модифицированные иммунные клетки могут быть удалены из трансплантата до трансплантации реципиенту. Однако некоторые CAR-модифицированные иммунные клетки могут прилипать к трансплантату и, следовательно, могут быть введены реципиенту с помощью трансплантата. В этой ситуации CAR-модифицированные иммунные клетки, введенные реципиенту, могут подавлять иммунный ответ против реципиента, вызванный клеткой, связанной с трансплантатом. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного подтипа HLA-A, выбранного из одного из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из двух из числа HLA-A*03, HLA-A*25, HLA-A*29, HLA-A*30, HLA-A*31, HLA-A*33, HLA-A*36, HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного подтипа HLA-A, выбранного из одного из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного или двух из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из одного из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух различных подтипов HLA-A, выбранных из двух из числа HLA-A*25, HLA-A*29, HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*03. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*25. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*29. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*30. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*31. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*33. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*36. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для HLA-A-подтипа HLA-A*68. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух из числа HLA-A*03:01, HLA-A*25:01, HLA-A*29:02, HLA-A*30:01, HLA-A*31:01, HLA-A*33:01, HLA-A*36:01 и HLA-A*68:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного или двух из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для одного из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным для двух из числа HLA-A*25:01, HLA-A*29:02 и HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*25:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*29:02. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*30:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*03:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*31:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*33:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*36:01. В одном воплощении трансплантат хозяина или реципиента является HLA-A2-отрицательным и является положительным по HLA-A*68:01. В одном воплощении трансплантат является HLA-A2-положительным. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В одном воплощении иммунная клетка представляет собой регуляторную Т-клетку. В одном воплощении иммунная клетка представляет собой регуляторную CD4+ Т-клетку. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В другом воплощении иммунная клетка представляет собой регуляторную Т-клетку человека. В одном воплощении иммунная клетка представляет собой CD4+ регуляторную Т-клетку человека.
В одном воплощении настоящее изобретение относится к способу размножения популяции иммунных клеток у объекта, где иммунные клетки модифицированы для экспрессии химерного антигенного рецептора (CAR), причем способ включает введение объекту иммунной клетки, описанной где-либо в данном документе, где введенные модифицированные иммунные клетки продуцируют популяцию потомственных иммунных клеток у объекта. В одном воплощении иммунная клетка представляет собой регуляторную иммунную клетку. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В одном воплощении иммунные клетки могут представлять собой регуляторные Т-клетки. В одном воплощении иммунные клетки могут быть CD4+ регуляторными Т-клетками. В одном воплощении иммунная клетка представляет собой иммунную клетку человека. В одном воплощении регуляторная иммунная клетка представляет собой регуляторную иммунную клетку человека. В одном воплощении регуляторная иммунная клетка выбрана из группы, состоящей из регуляторной T-клетки, CD4+ регуляторной T-клетки, CD8+ регуляторной T-клетки, регуляторной γδ T-клетки, регуляторной DN T-клетки, регуляторной B-клетки, регуляторной NK-клетки, регуляторного макрофага, регуляторной дендритной клетки и любой их комбинации. В другом воплощении иммунная клетка представляет собой регуляторную Т-клетку человека. В одном воплощении иммунная клетка представляет собой CD4+ регуляторную Т-клетку человека. В одном воплощении CAR-модифицированные Treg-клетки по настоящему изобретению способны реплицироваться in vivo, что приводит к длительной персистенции, которая может приводить к длительному подавлению иммунного ответа клетки-мишени и иммунной толерантности. В одном воплощении предлагается способ создания персистирующей популяции регуляторных Т-клеток у объекта, где регуляторные Т-клетки модифицированы для экспрессии химерного антигенного рецептора (CAR), причем способ включает введение объекту регуляторных Т-клеток, описанных где-либо в данном документе, где персистирующая популяция модифицированных регуляторных Т-клеток сохраняется у объекта в течение, по меньшей мере, 1 дня, 2 дней, 3 дней, 4 дней, 5 дней, 6 дней, 7 дней, 8 дней, 9 дней, 10 дней, 11 дней, 12 дней, 13 дней, 14 дней, 15 дней, 16 дней, 17 дней, 18 дней, 19 дней, 20 дней, 30 дней, 40 дней, 50 дней, 60 дней, 70 дней, 80 дни, 90 дней, 100 дней, 200 дней, 300 дней, 400 дней, 500 дней, 600 дней, 700 дней, 800 дней, 900 дней или 1000 дней после введения. В одном воплощении CAR-Treg-клетки могут быть способны к самообновлению и повторной активации in vivo для подавления иммунного ответа клетки-мишени. В одном воплощении CAR-Treg-клетки могут представлять собой CAR-Treg-клетки памяти, которые можно повторно активировать для подавления иммунного ответа клетки-мишени.
Иммунные клетки могут быть получены из любого источника. Например, в одном воплощении иммунные клетки могут быть получены от донора ткани, реципиента трансплантата или иного неродственного источника (другого объекта или вида в целом) для получения CAR-модифицированных иммунных клеток по настоящему изобретению. Соответственно, модифицированные с помощью CAR иммунные клетки по настоящему изобретению могут быть аутологичными, аллогенными или ксеногенными для реципиента трансплантата или иным образом не связанным источником. В одном воплощении CAR-Treg-клетки по настоящему изобретению могут быть аутологичными, аллогенными или ксеногенными для реципиента трансплантата. В одном воплощении CAR-Treg-клетки по настоящему изобретению могут быть аутологичными для реципиента трансплантата.
В одном воплощении объект может быть млекопитающим. В одном воплощении объектом может быть человеком.
В одном воплощении может быть желательным вводить активированные Т-клетки объекту, а затем впоследствии повторно отбирать кровь (или проводить аферез), активировать Т-клетки из нее согласно настоящему изобретению и повторно вводить объекту эти активированные и размноженные Т-клетки. Этот процесс может выполняться несколько раз каждые несколько недель. В некоторых воплощениях Т-клетки могут быть активированы при заборе крови от 10 до 400 см3. В некоторых воплощениях Т-клетки активируются при заборе крови объемом 20 см3, 30 см3, 40 см3, 50 см3, 60 см3, 70 см3, 80 см3, 90 см3 или 100 см3. Не желая быть связанными теорией, полагаем, что применение этого протокола множественного забора крови/множественной реинфузии может служить для отбора определенных популяций Т-клеток.
CAR-модифицированные иммунные клетки по настоящему изобретению можно вводить отдельно или в виде фармацевтической композиции в комбинации с разбавителями и/или с другими компонентами, такими как IL-2 или другие цитокины или клеточные популяции. В одном воплощении фармацевтическая композиция может содержать множество модифицированных иммунных клеток, описанных где-либо в данном документе. В одном воплощении фармацевтические композиции по настоящему изобретению могут содержать CAR-модифицированную иммунную клетку или клеточную популяцию, описанные где-либо в данном документе, в комбинации с одним или несколькими фармацевтически или физиологически приемлемыми носителями, разбавителями или наполнителями. Такие композиции могут включать буферы, например, нейтральный буферный физиологический раствор, фосфатно-солевой буферный раствор и т.п.; углеводы, такие как глюкоза, манноза, сахароза или декстраны, маннит; белки; полипептиды или аминокислоты, такие как глицин; антиоксиданты; хелатирующие агенты, такие как EDTA или глутатион; адъюванты (например, гидроксид алюминия); и консерванты. Фармацевтические композиции по настоящему изобретению можно вводить объекту любым подходящим способом, включая вдыхание аэрозоля, инъекцию, прием внутрь, переливание, имплантацию или трансплантацию. Фармацевтические композиции, описанные в данном документе, могут быть введены объекту путем парентерального введения. Фармацевтические композиции, описанные в данном документе, могут быть введены объекту подкожно, внутрикожно, внутриопухолево, интранодально, интрамедуллярно, внутримышечно, интрастернально, внутривенно (в/в) инъекцией, методами инфузии или внутрибрюшинно. В одном воплощении композиции модифицированных с помощью CAR иммунных клеток по настоящему изобретению можно вводить объекту путем внутрикожной или подкожной инъекции. В другом воплощении композиции CAR-модифицированных иммунных клеток по настоящему изобретению можно вводить внутривенной инъекцией. В одном воплощении композиции модифицированных с помощью CAR иммунных клеток могут быть инъецированы непосредственно в лимфатический узел, сайт инфекции, сайт воспаления или сайт отторжения ткани или органа.
Фармацевтические композиции по настоящему изобретению можно вводить способом, подходящим для предотвращения или лечения заболевания. Количество и частота введения будут определяться такими факторами, как состояние объекта, а также тип и тяжесть заболевания объекта, хотя соответствующие дозы могут быть определены клиническими испытаниями.
Когда указано «эффективное количество» или «терапевтическое количество», количество композиций по настоящему изобретению для введения может быть определено с учетом индивидуальных различий в возрасте, массе, титре антител и состоянии объекта. Как правило, можно утверждать, что фармацевтическую композицию, содержащую CAR-модифицированные иммунные клетки, описанные в данном документе, можно вводить в дозировке от 1×104 до 1×109 клеток/кг массы тела. В одном воплощении CAR-модифицированные иммунные клетки можно вводить в дозировке от 1×105 до 100×105 клеток/кг массы тела, включая все целочисленные значения в этих диапазонах. Композиции CAR-модифицированных иммунных клеток также могут вводиться многократно в этих дозировках. CAR- модифицированные иммунные клетки можно вводить с использованием способов инфузии, которые широко известны в иммунотерапии (см., например, Rosenberg et al, 1988). Оптимальную дозировку и схему лечения для конкретного объекта можно легко определить путем мониторинга объекта на наличие признаков заболевания и соответствующей корректировки лечения.
В одном воплощении, по меньшей мере, одну CAR- сконструированную иммунную клетку по изобретению вводят объекту, нуждающемуся в этом, в сочетании с другим активным агентом. Согласно одному воплощению, по меньшей мере, одну популяцию иммунных клеток вводят до, одновременно или после введения другого активного агента.
В одном воплощении CAR-модифицированные иммунные клетки по настоящему изобретению могут вводиться объекту в сочетании (например, до, одновременно или после) с любым количеством подходящих способов лечения, включая, без ограничения указанным, лечение средствами, такими как противовирусные терапия, химиотерапия, облучение, иммуносупрессанты, антитела, иммуноаблативные агенты, цитокины и облучение. В одном воплощении CAR-модифицированные иммунные клетки по настоящему изобретению можно вводить в сочетании с иммуносупрессирующим агентом. Может быть использован любой иммуносупрессирующий агент, известный в данной области. Например, иммуносупрессирующий агент может представлять собой ингибиторы кальциневрина, такие как циклоспорин, такролимус, азатиоприн, метотрексат, метоксален, рапамицин, микофенолат мофетил, микофеноловая кислота, микофенолат натрия, 6-меркаптопурин, 6-тиогуанин, ритуксимаб, ингибиторы mTOR, такие как сиролимус, эверолимус, базиликсимаб, даклизумаб, белатацепт, алемтузумаб, муромонаб-CD3, анти-тимоцитарный глобулин, глюкокортикостероиды или адренокортикальные стероиды, такие как преднизон и преднизолон, или любую их комбинацию. CAR-модифицированные иммунные клетки по настоящему изобретению можно вводить объекту до, после или одновременно с иммуносупрессирующим агентом. Например, CAR-модифицированные иммунные клетки по настоящему изобретению можно вводить после того, как объекту вводят иммуносупрессирующий агент, или CAR-модифицированные иммунные клетки по изобретению можно вводить до того, как объекту вводят иммуносупрессирующий агент. Альтернативно или в дополнение, CAR-модифицированные иммунные клетки по изобретению могут вводиться одновременно с иммуносупрессирующим агентом, вводимым объекту. CAR-модифицированные иммунные клетки по настоящему изобретению и/или иммуносупрессирующий агент могут быть введены объекту после трансплантации. Альтернативно или в дополнение, CAR-модифицированные иммунные клетки по настоящему изобретению и/или иммуносупрессирующий агент могут быть введены объекту перед трансплантацией. CAR-модифицированные иммунные клетки по настоящему изобретению и/или иммуносупрессирующий агент могут быть введены объекту во время трансплантационной хирургии. В одном воплощении способ по изобретению введения CAR-модифицированных иммунных клеток объекту осуществляют после начала иммуносупрессивной терапии. В некоторых воплощениях способ осуществляют более одного раза, например, для мониторинга реципиента трансплантата во времени и, если это применимо, в различных режимах иммуносупрессивной терапии. В некоторых воплощениях иммуносупрессивная терапия снижается, если спрогнозировано, что реципиент трансплантата является толерантным к трансплантату. В некоторых воплощениях иммуносупрессивную терапию не назначают, например, иммуносупрессивную терапию прекращают, если спрогнозировано, что реципиент трансплантата будет толерантным к трансплантату.
Также предлагается способ предотвращения или лечения отторжения органа или ткани у объекта, включающий введение объекту (i), по меньшей мере, одного иммуносупрессивного агента и (ii) CAR-сконструированных иммунных клеток (например, CAR-сконструированных Treg-клеток) по изобретению или фармацевтическую композицию, содержащую указанные иммунные клетки.
Таким образом, другим воплощением настоящего изобретения является комбинация CAR-сконструированной иммунной клетки (предпочтительно CAR-сконструированной Treg-клетки) по изобретению или фармацевтической композиции, содержащей указанные иммунные клетки, и, по меньшей мере, одного иммуносупрессивного агента для применения в предотвращении или лечении отторжения трансплантата органа или ткани у объекта.
Также предлагается способ увеличения периода выживания трансплантата у объекта, включающий введение объекту, по меньшей мере, одного иммуносупрессирующего агента и CAR-сконструированных иммунных клеток (предпочтительно CAR-сконструированных Treg-клеток) по настоящему изобретению, или фармацевтической композиции, содержащей их.
В одном воплощении комбинация, по меньшей мере, одного иммуносупрессивного агента с CAR-сконструированными иммунными клетками (CAR-сконструированными Treg-клетками) по настоящему изобретению используется для предотвращения или лечения болезни трансплантат против хозяина (GVHD). В одном воплощении GVHD может возникать после трансплантации гемопоэтических стволовых клеток.
Таким образом, другим воплощением изобретения является способ предотвращения или лечения болезни трансплантат против хозяина (GVHD) у объекта, включающий введение объекту, по меньшей мере, одного иммуносупрессивного агента и CAR-сконструированных иммунных клеток (CAR-сконструированных Treg-клеток) или фармацевтической композиции, как описано в данном документе.
В одном воплощении изобретение относится к способу контакта донорного трансплантата, например, биосовместимой сетки или донорской ткани, органа или клетки, по меньшей мере, с одним иммуносупрессивным агентом и с CAR-сконструированными иммунными клетками (CAR-сконструированными Treg-клетками) по настоящему изобретению, одновременно, до или после пересадки трансплантата реципиенту.
В одном воплощении комбинация, по меньшей мере, одного иммуносупрессивного агента с CAR-сконструированными иммунными клетками (CAR-сконструированными Treg-клетками) по настоящему изобретению можно использовать для ослабления, ингибирования или снижения неблагоприятного ответа донорного трансплантата против реципиента, таким образом, предотвращая или подвергая лечению GVHD.
Таким образом, другим воплощением настоящего изобретения является способ предотвращения или отсрочки возникновения GVHD у объекта, включающий введение объекту, по меньшей мере, одного иммуносупрессирующего агента и CAR-сконструированных иммунных клеток (предпочтительно CAR-сконструированных Treg-клеток), или фармацевтической композиции, как описано в данном документе.
В одном воплощении иммунные клетки, сконструированные с помощью CAR, по изобретению и, по меньшей мере, один иммуносупрессивный агент вводят одновременно или последовательно.
В одном воплощении CAR-сконструированные иммунные клетки по настоящему изобретению и, необязательно, по меньшей мере, один другой активный агент, предпочтительно иммуносупрессивный агент, вводят в сочетании с (например, до, одновременно или после) трансплантатом.
CAR-сконструированные иммунные клетки по настоящему изобретению можно вводить после диагностики отторжения органа или ткани трансплантата с последующими дозами как сконструированных с помощью CAR-сконструированных иммунных клеток по изобретению, так и иммуносупрессирующего агента до тех пор, пока симптомы отторжения органа или ткани не исчезнут. В дополнительном воплощении композиции CAR-сконструированных иммунных клеток по настоящему изобретению могут вводиться объекту в сочетании (например, до, одновременно или после) трансплантации костного мозга. В другом воплощении CAR-сконструированные иммунные клетки по настоящему изобретению можно вводить после абляционной терапии B-клетками, например, агентами, которые реагируют с CD20, например, ритуксимабом. Например, в одном воплощении объекты могут проходить стандартное лечение химиотерапией с высокой дозой с последующей трансплантацией стволовых клеток периферической крови. В определенных воплощениях после трансплантации объекты могут получать инфузию размноженных CAR-сконструированных иммунных клеток по настоящему изобретению. В дополнительном воплощении размноженные CAR-сконструированные иммунные клетки можно вводить до или после операции.
В одном воплощении объект (например, человек) получает начальное введение, по меньшей мере, одной иммунной клетки или популяции по изобретению и одного или несколько последующих введений, при этом одно или несколько последующих введений вводят менее чем за 15 дней, например, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3 или 2 дня после предыдущего введения. В одном воплощении терапевтически эффективное количество иммунных клеток по изобретению вводят или подлежат введению объекту. В одном воплощении количество иммунных клеток, по меньшей мере, одной популяции иммунных клеток по изобретению, вводимых объекту, составляет, по меньшей мере, 102, 103, 104, 105, 106, 107, 108 или 109 клеток. В одном воплощении количество иммунных клеток по изобретению, вводимых объекту, составляет от около 102 до около 109, от около 103 до около 108, от около 104 до около 107 или от около 105 до около 106 клеток. В другом воплощении количество иммунных клеток по изобретению, вводимых объекту, составляет от около 106 до около 109, от около 106 до 107, от около 106 до 108, от около 107 до 109, от около 107 до 108, от около 108 до 109. В другом воплощении количество иммунных клеток по изобретению, вводимых объекту, составляет около 106, около 107, около 108 или около 109. В одном воплощении количество иммунных клеток, по меньшей мере, одной популяции иммунных клеток по изобретению, вводимых объекту, составляет, по меньшей мере, 102, 103, 104, 105, 106, 107, 108 или 109 клеток/кг тела. В одном воплощении количество иммунных клеток по изобретению, вводимых объекту, составляет от около 104 до 109 клеток/кг массы тела или от 105 до 108 клеток/кг массы тела, включая все целочисленные значения в этих диапазонах. В одном воплощении объекту (например, человеку) вводят более одного введения, по меньшей мере, одной иммунной клетки или популяции по изобретению в неделю, например, проводят 2, 3 или 4 введения генетически модифицированной иммунной клетки или популяции по изобретению в неделю.
В некоторых воплощениях может быть желательным включение гена «самоубийства» в CAR-модифицированные иммунные клетки по настоящему изобретению, чтобы сделать возможным избирательное разрушение CAR-модифицированных иммунных клеток in vivo. В одном воплощении CAR-модифицированные иммунные клетки по настоящему изобретению могут дополнительно содержать систему гена «самоубийства». В одном воплощении CAR-модифицированные иммунные клетки по настоящему изобретению могут дополнительно содержать нуклеиновую кислоту, кодирующую систему гена «самоубийства». Термины «терапия на основе гена «самоубийства»», «ген «самоубийства»» и «система гена «самоубийства»», как описано в настоящем документе, относятся к способам селективного разрушения клетки посредством апоптоза. Такие способы используют ген «самоубийства», который заставит клетку убить себя апоптозом. Ген «самоубийства» кодирует продукт, который вызывает гибель клеток сам по себе или в присутствии других соединений. Ген «самоубийства» может происходить из любой системы генов «самоубийства», подходящих для применения в клеточной терапии (см., Например, Jones et al., 2014; Wang et al., 2017; Zhang et al., 2017; Casucci et al., 2011; Gargett et al., 2014; Khaleghi, 2012; Philip et al., 2014).
В одном воплощении система гена «самоубийства» может включать генно-направленную ферментную пролекарственную терапию (GDEPT), где нетоксичное лекарственное средство (то есть пролекарство гена «самоубийства») превращается в токсичное соединение в клетках, модифицированных геном «самоубийства». Например, в одном воплощении система гена «самоубийства» может представлять собой систему гена «самоубийства» на основе тимидинкиназы вируса простого герпеса (HSV-TK)/ганцикловира (GCV), которая использует ген тимидинкиназы из вируса простого герпеса в сочетании с пролекарственным ганцикловиром. В других воплощениях система гена «самоубийства» может представлять собой систему гена «самоубийства» на основе цитозин-деаминазы/5-фторцитозина. В некоторых альтернативах способов, представленных в настоящем документе, где CAR-модифицированные иммунные клетки содержат систему гена «самоубийства», способы дополнительно включают введение объекту пролекарства гена «самоубийства».
В других воплощениях система гена «самоубийства» может включать механизм индукции димеризации, при котором апоптотические гены, такие как каспазы, разрушают клетки, индуцируя апоптоз. Системы гена «самоубийства» на основе индукции димеризации могут включать химерные белки, состоящие из домена, связывающего лекарственное средство, соединенного с компонентами пути апоптоза, позволяющего условную димеризацию и апоптоз модифицированных иммунных клеток после введения химического индуктора димеризации (CID). Например, в одном воплощении система гена «самоубийства» может представлять собой систему гена «самоубийства» на основе каспазы. Неограничивающие примеры индуцибельной системы гена «самоубийства» на основе каспазы могут включать индуцибельную систему гена «самоубийства» на основе каспазы-9 (iCasp9) и систему гена «самоубийства» на основе каспазы-8 (iCasp8), где каспазу активируют с помощью CID, FK506 или их аналога. В некоторых альтернативах способов, представленных в настоящем документе, где CAR-модифицированные иммунные клетки содержат систему гена «самоубийства», способы дополнительно включают введение объекту химического индуктора димеризации.
В других воплощениях система гена «самоубийства» может быть опосредована терапевтическим моноклональным антителом. Терапевтические mAb-опосредованные системы гена «самоубийства» включают экспрессию белков на клеточной поверхности, которые делают модифицированные иммунные клетки чувствительными к терапевтическим mAb. Модифицированные иммунные клетки могут быть избирательно разрушены после введения специфического терапевтического mAb. Неограничивающие примеры терапевтических mAb-опосредованных систем гена «самоубийства» могут включать систему гена «самоубийства» на основе CD20 и систему гена «самоубийства» на основе RQR8, где терапевтическое mAb представляет собой mAb против CD20. В некоторых воплощениях mAb против CD20 может представлять собой ритуксимаб. В некоторых альтернативах способов, представленных в настоящем документе, где CAR-модифицированные иммунные клетки содержат систему гена «самоубийцы», способы дополнительно включают введение объекту терапевтического моноклонального антитела.
VIII. ГОТОВЫЕ ИЗДЕЛИЯ И НАБОРЫ
В другом воплощении изобретение предоставляет изделие, содержащее материалы, полезные для профилактики и/или лечения отторжения трансплантата или GVHD. Изделие может содержать контейнер и этикетку или вкладыш в контейнер или ассоциированный с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер включает композицию, которая эффективна для предотвращения и/или лечения иммунологического состояния, такого как отторжение трансплантата или GVHD, и может иметь стерильное отверстие для доступа (например, контейнер может представлять собой мешок для внутривенного раствора или флакон с пробкой, пригодной для прокалывания иглой для подкожных инъекций). По меньшей мере, один активный агент в композиции представляет собой CAR-модифицированную иммунную клетку по настоящему изобретению. В одном воплощении CAR-модифицированная иммунная клетка представляет собой CAR-модифицированную Treg. В одном воплощении CAR-модифицированный Treg представляет собой CAR-модифицированную Treg человека. Этикетка или вкладыш в упаковку могут указывать, что композиция используется для предотвращения или лечения отторжения трансплантата или GVHD. Изделие, этикетка или вкладыш в упаковку могут дополнительно содержать инструкции для введения композиции CAR-сконструированных иммунных клеток, пациенту. Кроме того изделие может дополнительно содержать второй контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатно-солевой раствор, раствор Рингера и раствор декстрозы. Кроме того, он может включать другие материалы, желательные с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Также предлагаются наборы, которые полезны для различных целей (например, для предотвращения или лечения отторжения трансплантата или GVHD). Могут быть предлагаются наборы, которые содержат CAR-сконструированные иммунные клетки. Как и в случае с изделием, набор может содержать контейнер и этикетку или вкладыш в контейнер или ассоциированный с контейнером. Контейнер содержит композицию, содержащую, по меньшей мере, одну CAR-модифицированную иммунную клетку по настоящему изобретению. Могут быть включены дополнительные контейнеры, которые содержат, например, разбавители и буферы. Этикетка или вкладыш в упаковку могут содержать описание композиции, а также инструкции по предполагаемому применению. В одном воплощении CAR-модифицированные иммунные клетки представляет собой CAR-модифицированные Treg. В одном воплощении CAR-модифицированные Treg представляет собой CAR-модифицированные Treg человека.
Таким образом, настоящее изобретение, кроме того, относится к набору частей, включающему, в первую очередь, иммунную клетку или популяцию иммунных клеток по настоящему изобретению, а во второй части - другой активный агент, включая, без ограничений указанным, противовирусную терапию, химиотерапию, лучевую терапию, иммуносупрессанты, антитела, иммуноаблативные агенты, цитокины и облучение. В некоторых воплощениях набор из части настоящего изобретения включает в первой части иммунную клетку или популяцию иммунных клеток по настоящему изобретению и во второй части один или несколько иммуносупрессивных агентов. Примеры иммуносупрессивных агентов включают, без ограничения указанным, ингибиторы кальциневрина, такие как циклоспорин, такролимус, азатиоприн, метотрексат, метоксален, рапамицин, микофенолат мофетил, микофеноловая кислота, микофенолат натрия, 6-меркаптопурин, 6-тиогуанин, ритуксимаб, ингибиторы mTOR, такие как сиролимус, эверолимус, базиликсимаб, даклизумаб, белатацепт, алемтузумаб, муромонаб-CD3, анти-тимоцитарный глобулин, глюкокортикостероиды или адренокортикальные стероиды, такие как пре-предолидон, или любую их комбинацию.
Таблица 1
Таблица 2
Таблица 3
(без лидера)
(без лидера)
(без лидера)
(без лидера)
(без лидера)
(без лидера)
(с лидером)
(с лидером)
(с лидером)
(с лидером)
(без лидера)
(без лидера)
(без лидера)
(без лидера)
(SEQ ID NO: 23 из US 9,102,760)
Таблица 4
(SEQ ID NO: 17 или US9,102,760)
Таблица 5
VH CDR1
VH CDR2
VH CDR3
Таблица 6
VL CDR1
VL CDR2
VL CDR3
Таблица 7
VH
VL
scFv
(без лидера)
(без лидера)
(с лидером)
(с лидером)
ПРИМЕРЫ
Способы
Получение гуманизированных HLA-A*02-специфических scFv. Гуманизированные гены были оптимизированы по кодонам с использованием оптимизатора кодонов сервиса Invitrogen GeneArt Gene Synthesis с использованием настроек для Homo sapiens. Генные фрагменты gBlocks® были заказаны у компании Integrated DNA Technologies (Коралвилл, Айова) таким образом, чтобы 5'-область химерного антигеннного рецептора содержала последовательность Kozak и перекрытие из 38 нуклеотидов с плазмидой pcDNA3. 3'-конец содержит сайт BamHI и перекрытие с шарнирной последовательностью CD8 для облегчения сборки Гибсона в плазмиду в рамке с шарниром CD8 и внутриклеточными сигнальными доменами химерного антигенного рецептора.
Получение hA2-CAR. Варианты scFv были слиты с областью стебля из CD8α человека, трансмембранного и внутриклеточного доменов CD28 человека и CD3ζ человека, как описано (Sadelain et al., 2013). Полученные кДНК были клонированы в лентивирусный вектор, который кодирует экспрессию на клеточной поверхности укороченного рецептора фактора роста нервов (TrkA) (ΔNGFR) в качестве маркера (Фигура 1). Поверхностную экспрессию определяли проточной цитометрией транзиторно трансфицированных клеток HEK 293T (jetPRIME®, Polyplus Transfection). Вирусные частицы получали, как описано (Allan et al., 2008).
Получение HLA-экспрессирующих клеточных линий K562. Клетки K562, экспрессирующие CD64 (K562.64), были подарены Джеймсом Райли, Университет Пенсильвании, Филадельфия, США. кДНК для HLA-A*02:01 и A*24:02 выделяли из мРНК мононуклеарных клеток периферической крови от донора, гомозиготного по A*02:01 или A*24:02, соответственно, на локусе HLA-A, используя последовательности праймеров: 5'-TTTTCTAGACGCGTGCCACCATGGCCGTCATGGCGCC-3 '(прямой) (SEQ ID NO: 197) и 5'-AAGTCGACGCTAGCTCACACTTTACAAGCTGTGAGAGACA-3' (обратный) (SEQ ID NO: 198). Полученная последовательность была подтверждена секвенированием по Сангеру, выровнена с ожидаемыми последовательностями из базы данных IPD и IMGT/HLA (Robinson et al., 2015) и трансдуцирована в клетки K562, соответственно, с использованием экспрессирующего лентивирусного вектора. Для получения клеток A25- и A68-K562 последовательности HLA для A*25:01, A*68:01 извлекали из базы данных IPD и IMGT/HLA (Robinson et al., 2015), оптимизировали по кодонам с использованием инструмента оптимизации кодонов (для Homo sapiens) сервиса генного синтеза ThermoFisher Invitrogen GeneArt, затем клонировали в лентивирусный экспрессирующий вектор и трансдуцировали в клетки K562. Полученные клеточные линии K562 затем сортировали на FACSAria II (BD Biosciences) с использованием анти-HLA-ABC (ThermoFisher, 12-9983-41) для обеспечения эквивалентной поверхностной экспрессии трансдуцированного HLA, и анти-HLA-A2 (BB7.2) (ThermoFisher, 17-9876-42) для обеспечения чистоты.
Сортировка, трансдукция и размножение Treg. CD4+ Т-клетки выделяли из доноров HLA-A2- через RosetteSep (Stemcell) и обогащали для CD25+ клеток (Miltenyi) перед сортировкой живых CD4+CD25hiCD127lo Treg (для экспериментов in vitro и люциферазы) или CD4+CD127loCD25hi CD45RA+ Treg (для ксеногенной GvHD и экспериментов по пересадке кожи) с использованием MoFlo® Astrios (Beckman Coulter) или FACSAria II (BD Biosciences), соответственно. Сортированные Treg стимулировали L-клетками и αCD3 mAb (OKT3; 100 нг/мл) в среде для размножения Т-клеток immunocult-XF (STEMCELL Technologies) с 1000 Ед./мл IL-2 (Пролейкин). Один день спустя клетки трансдуцировали лентивирусом при множественной инфекции 10 вирусных частиц:1 клетку. На 7-й день клетки NGFR+ очищали магнитной селекцией (Miltenyi), затем использовали в анализах или повторно стимулировали L-клетками, как описано выше, и размножали в течение 5 дней для экспериментов in vivo. Чтобы проверить эффекты HLA-A2-опосредованной стимуляции, Treg культивировали с ограничением IL-2 (100 Ед/мл) в течение 24 часов, а затем повторно стимулировали облученными анти-CD3/анти-CD28 нагруженными клетками K562.64 или клетками K562.64, экспрессирующими HLA-A*02:01, A*24:02, A*25:01 или A*68:01, в соотношении 1:2 (К562:Т-клетки) в течение 24 часов.
Проточная цитометрия. Для фенотипического анализа клетки окрашивали фиксируемым красителем для оценки жизнеспособности (FVD, 65-0865-14, ThermoFisher; 423102, Biolegend) и по поверхностным маркерам перед фиксацией/перманеабилизацией с помощью набора FOXP3/Transcription Factor Staining Buffer (eBioscience) и окрашиванием внутриклеточных белков. Образцы считывали на Cytoflex (Beckman-Coulter), а результаты анализировали с использованием программного обеспечения FlowJo версий 9.9.4 и 10.3 (Tree Star).
Окрашивание поверхности проводили для ΔNGFR (130-091-885, Miltenyi), CD3 (564465, BD Biosciences), CD4 (317410, Biolegend), CD25 (130-091-024, Miltenyi), LAP (25-9829-42, ThermoFisher), CD69 (310946, Biolegend), CD154 (555702, BD Biosciences), CD71 (BD Biosciences, 563768) и CD127 (48-1278-42, ThermoFisher). Окрашивание тетрамера проводили мономерами HLA-A*02:01, превращенными в тетрамеры со стрептавидин-аллофикоцианином (PJ27S, Prozyme). Внутриклеточное окрашивание проводили по CTLA-4 (369606, Biolegend).
Для экспериментов in vivo 50 мкл крови собирали еженедельно и в конечных точках. Хлорид аммония использовали для лизиса эритроцитов. Клетки ресуспендировали в PBS с анти-CD16/32 мыши (ThermoFisher, 14-0161-82) и окрашивали на внеклеточные маркеры с использованием фиксируемого красителя для оценки жизнеспособности (FVD; ThermoFisher, 65-0865-14), с анти-CD45 мыши (ThermoFisher, 25 -0451-82) и анти-CD45 человека (BD Biosciences, 560777), анти-CD4 (Biolegend, 300554, 317434), анти-CD8 (ThermoFisher, 48-0087-42), анти-CD271 человека (NGFR; BD Biosciences, 557196) анти-HLA-A2 (BD Biosciences, 551285). Внутриклеточное окрашивание по FOXP3 (ThermoFisher, 12-4777-42) выполняли с помощью набора FOXP3/Transcription Factor Staining Buffer (ThermoFisher, 00-5523-00). 10000 отсчитанных гранул добавляли к каждому образцу (ThermoFisher, 01-1234-42). Стратегии гейтирования в экспериментах с ксеногенной GvHD, люциферазой и трансплантатом кожи показаны на Фигурах 8D, 10A и 12A, соответственно.
Анализ перекрестной реактивности аллелей HLA. 0,25×106 CAR Treg (приготовленных, как описано выше, после 7 дней культивирования) инкубировали с панелями гранул с индивидуальными антигенами для определения антител FlowPRA (FL1HD01, FL1HD02, FL1HD03, FL1HD04, FL1HD06 и FL1HD08, One Lambda) и с фиксируемым красителем для оценки жизнеспособности (FVD, 65 -0865-14, ThermoFisher) в течение 30 минут при комнатной температуре, затем промывали, фиксировали 0,5% формальдегидом и анализировали с помощью проточной цитометрии. На образец использовали двести гранул отрицательного контроля. Для анализа мертвые клетки сначала удаляли, используя фиксируемый краситель для определения жизнеспособности. Одиночные гранулы затем подвергали гейтированию после исключения мертвых клеток и дублетов. Затем количество гранул для каждого антигена HLA определяли по их соответствующей интенсивности PE. Данные нормализовали путем деления числа отрицательных гранул в образце на количество отрицательных гранул в образце Treg-NGFR, умноженного на количество отрицательных гранул в образце Treg-NGFR. Относительное процентное связывание представляет собой количество гранул в образце NGFR для гранул специфичных по HLA минус нормализованное количество гранул в образце для этих HLA, деленное на количество гранул в образце NGFR, и умноженное на 100.
Супрессия реакций смешанных лимфоцитов. Адгерентные клетки из РВМС от здоровых доноров HLA-A2 + дифференцировали в моноцитарные дендритные клетки, как описано (Dijke et al., 2015). Для реакций смешанных лимфоцитов клетки-респондеры HLA-A2-PBMC метили красителем клеточной пролиферации eF450 (65-0842-85, ThermoFisher), затем рассевали с 5x104 HLA-A2+ моноцитарных дендритных клеток и увеличивали соотношение размноженных ΔNGFR- или hA2-CAR-экспрессирующих Treg, меченных красителем клеточной пролиферации e670 (65-0840-90, ThermoFisher). Через 6 дней деление HLA-A2-CD4+ Т-клеток-респондеров измеряли с помощью проточной цитометрии. Процент супрессии рассчитывали на основе индекса пролиферации данной комбинации клеток и соотношения в сравнении с положительным контролем (только HLA-A2+ моноцитарные дендритные клетки с HLA-A2- CD4+ T-клетками-респондерами), как описано (McMurchy and Levings, 2012). Данные были нормализованы, вначале путем рассчета процента супрессии следующим образом: 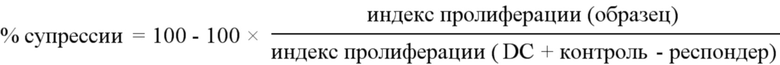 , затем нормализовали полученные значения от 0 до 100%, согласно формуле для каждого независимого эксперимента:
, затем нормализовали полученные значения от 0 до 100%, согласно формуле для каждого независимого эксперимента: 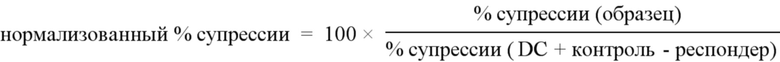
Супрессия ксеногенной болезни «трансплантат против хозяина» in vivo. Самки мышей NSG в возрасте от 8 до 12 недель (лаборатория Джексона, размноженные самостоятельно) получали облучение всего тела (150 сГр, RS-2000 Pro Biological System) за 1 день до введения 8×106 HLA-A2+ РВМС с или без 4 х 106 hA2-CAR Treg внутривенно в хвостовую вену. Мыши с инъекцией физиологического раствора служили в качестве контроля. hA2-CAR Treg получали от четырех разных здоровых доноров. GVHD оценивали на основе массы, текстуры меха, позы, уровня активности и целостности кожи, с 0 до 2 баллов на категорию, как описано (Cooke, et al., 1996; Hill et al., 1997). Оценка GVHD проводилась двумя исследователями, не имеющими конкретной информации об эксперименте. Периферическую кровь из подкожной вены центрифугировали; затем эритроциты лизировали и лейкоциты измеряли проточной цитометрией.
Одобрение исследования. В отношении РВМС человека здоровые добровольцы дали письменное информированное согласие в соответствии с протоколами, утвержденными Советом по этике клинических исследований Университета Британской Колумбии и Канадскими службами крови. Образцы кожи человека, выброшенные после пластической хирургии, были получены из Гарвардского центра кожных ресурсов, Skin Works или хирургической клиники Cambie в соответствии с протоколами, утвержденными Советом по этике клинических исследований Университета Британской Колумбии. Протоколы экспериментов на животных были одобрены Комитетом по уходу за животными UBC.
Трансплантация кожи. Для оценки хоминга A2-CAR Treg и способности ингибировать отторжение кожи самкам мышей NSG в возрасте от 8 до 12 недель (лаборатория Джексона, размноженные самостоятельно) трансплантировали кожу от трансгенных мышей HLA-A2+ NSG (лаборатория Джексона, размноженные самостоятельно) и кожу NSG (HLA-A2-отрицательные). Для мышиных трансплантатов кожи кожу разрезали на круглые кусочки с использованием 8-миллиметрового биопсийного штампа и кожу помещали на свежие чашки с PBS и хранили при низкой температуре (4-8°C) до пересадки (~1–4 часа). Экспрессию HLA-A2 кожи человека оценивали с помощью проточной цитометрии и кПЦР. Расщеплённые экспланты получали путем обрезки жира из образца. Экспланты промывали стерильным PBS и хранили в RPMI. Качественные кожные экспланты разрезали на кусочки по 1 см2, помещали на свежие чашки с PBS и хранили при низкой температуре (4-8°C) до трансплантации (~1-4 часа). Для трансплантации кожи мыши и человека предварительно выбритых мышей анестезировали, кожу на спине разрезали возле плеча, и удаляли кожу мыши аналогичного размера, затем трансплантаты помещали на открытую область и стабилизировали стерильными полосками (3M, Nexcare). Трансплантаты покрывали вазелиновой марлей и оборачивали бинтом CoFlex шириной 2 см (3M, Nexcare) для закрепления трансплантата в течение 14 дней до инъекции клеток.
Гистология
Человеческий кожный трансплантат и окружающую мышиную кожу собирали через 27 дней после введения клеток, фиксировали в течение ночи при 4°С в 10% формалине (соотношение ткани и формалина 1:10 по объему) и хранили в 70% этаноле перед заливкой парафином. Получали парафиновые срезы (толщиной 5 мкм) и проводили окрашивание гематоксилином-эозином в Службе гистологических исследований Детского госпиталя BC (Ванкувер, Британская Колумбия, Канада). Для иммуноокрашивания, срезы депарафинизировали и повторно гидратировали с помощью серий промывок ксилолом (×3), набором спиртовых растворов изменяющейся концентрации (2×100% этанол, 1× 95% этанол и 1×70% этанол) и 1×PBS. Высокотемпературную демаскировку антигена (HIER) проводили на предметных стеклах с использованием микроволновой печи для достижения 93-95°C (5 минут высокая мощность, затем 20 минут низкая мощность) в 10 мМ буфере цитрата натрия (0,5% Tween-20, pH 6,0). После HIER предметные стекла промывали проточной водопроводной водой, деионизированной водой и PBS. Срезы инкубировали с белковым блокатором DAKO®, не содержащим сыворотки (Dako, Берлингтон, Канада, X0909), для ограничения окрашивания неспецифических антител. Затем срезы инкубировали при 4°С в течение ночи со следующими первичными антителами: FOXP3 (Invitrogen, клон PCH101, 14-4776-82), CD45 (eBioscience, клон H130, 17-0459), Ki67 (eBioscience, клон 20Raji, 17- 5699), Инволукрин (Abcam, ab53112). На следующий день срезы аккуратно промывали PBS несколько раз, затем окрашивали в течение 1 часа при комнатной температуре следующими вторичными антителами: ослиное антитело против антител крысы 488 (Life Tech, A11006), козлиное антитело против мышиных антител APC (Invitrogen, 1834696), ослиное антитело против антител кролика 488 (Life Tech, A21206). Наконец, срезы контрастировали 4',6-диамидино-2-фенилиндолом (DAPI) для идентификации ядер клеток и заключали в среду для заливки VECTASHIELD® с DAPI (Vector Laboratories, Берлингейм, Калифорния, США, H-1200). Все антитела разводили в разбавителе антител (Dako, Берлингтон, Канада, S3022). Изображения были сняты с помощью флуоресцентного микроскопа Olympus BX61 и автоматического прямого микроскопа в светлом поле с камерой QImaging Retiga Exi и цветной камерой Olympus DP71. Количественный анализ флуоресцентных изображений, выполняли с использованием Fiji, с помощью плагина Olympus Viewer (Schneider et al., 2012; Eliceiri et al., 2012).
Предметные стекла, окрашенные гематоксилином-эозином, оценивались клиническим патологом, не имеющим конкретной информации об эксперименте, с использованием системы оценок, определяемой 8 факторами, каждый из которых был оценен от 0 до 3-4; Степень Лернера (0, 1 - очаговая или диффузная вакуолярная дегенерация, 2 - дискератоз, 3 - трещины в базальном или поверхностном слоях, 4 - явная потеря эпидермиса), спонгиоз (0, 1 - только базальный слой, 2 - до половины пути, 3 - полная толщина), некротические кератиноциты (0, 1 - редко (1/hpf), 2 - случайно (2-3/hpf), 3 - много (> 4/hpf)), расположение некротических кератиноцитов (0, 1 - только базальное, 2 - до верхней половины, 3 - полная толщина), сатертоз (только 0, 1 - 1, 2 - 2-3/hpf, 3 -> 4/hpf), экзоцитоз (0, 1 - очаговый, 2 - <50% биопсия, 3 -> 50% биопсия), вовлечение прилежащих органов (0, 1 - незначительное вовлечение любых прилежащих органов, 2 - заметное вовлечение <50% прилежащих органов, 3 - заметное вовлечение> 50% прилежащих органов) и лимфоидные манжеты в дерме (0, 1 - незначительные, 2 - обильные, 3-полосные) (Massi et al., 1999; Fischer et al., 2015, Kanitakis et al., 2008).
кПЦР. РНК собирали из образцов кожи человека в соответствии с инструкциями производителя (RNeasy Plus Mini Kit; Qiagen) и превращали в комплементарную ДНК (кДНК). кПЦР проводили с использованием SYBR-зеленого (Biorad) и праймеров для IL17, IL6, IL1B, DEFB4, IFNg, TNFa, рибосомальной 18S РНК (Таблица 8). Собирали данные кривой плавления и данные эмиссии SYBR-зеленого. Относительные концентрации рассчитывали с использованием стандартной кривой, и значения нормализовали относительно продуктов амплификации рибосомальной 18S РНК. Значения Log2 (RQ) для каждого образца получали с использованием способа двойной дельты Ct (∆∆Ct) (Schmittgen et al., 2008). Значение ∆Ct каждого образца получали путем расчета усредненного Ct (гена, представляющего интерес) - Ct (гена домашнего хозяйства). Чтобы получить ∆∆Ct, ∆Ct контрольного образца вычитали из ∆Ct обработанного образца. Затем кратную экспрессию гена рассчитывали по 2- (∆∆Ct).
Таблица 8: Праймеры, использованные в анализе кПЦР
олигонуклеотида
Прямой (FWD) или Обратный (RVS)
Люцифераза. Чтобы оценить хоминг Treg к HLA-A2-экспрессирующим трансплантатам кожи мыши in vivo, отсортированные Treg (CD4+CD25hiCD127lo) стимулировали L-клетками, как описано выше. На следующий день клетки трансдуцировали лентивирусом HER2-CAR, mA2-CAR или h1k2 hA2-CAR при MOI 10 и через 8 ч с помощью лентивирусом с люциферазой и GFP при MOI 5. Лентивирусную плазмиду, кодирующую гибридный белок люциферазы жука и GFP (pELNS.CBG-T2A-GFP (CBR)), использовали, как описано ранее (Barrett et al., 2014). После 7 дней культивирования, дважды трансдуцированные GFP+ΔNGFR+ Treg, экспрессирующие CAR и люциферазу, сортировали перед рестимуляцией L-клетками, как описано выше. На 12 день культивирования 1-3×106 люцифераза-CAR Treg и 6×106 человеческих аллогенных HLA-A2-РВМС инъецировали внутривенно мышам NSG с трансплантированной кожей. Для получения биолюминесцентных изображений интраперитонеально вводили калиевую соль D-люциферина (150 мг/кг, Gold Bio) непосредственно перед анестезией изофлураном и получали изображения в течение 15-20 минут на Ami-X (Spectral Instruments Imaging). Данные были проанализированы с помощью программного обеспечения AmiView (Spectral Instruments Imaging, версия 1.7.06), и люминесцентный сигнал был количественно определен как отношение фотонов/сек/см2/стерадиан в HLA-A2+ по сравнением с HLA-A2- кожным трансплантом. В конечной точке эксперимента собирали подмышечные лимфатические узлы, дренирующие кожу и селезенку, помещали в 70 мкм клеточное сито (BD Falcon), затем фрагментировали и фильтровали с помощью поршня шприца объемом 1 см3. Затем клетки окрашивали для проточной цитометрии.
Статистический анализ Вся статистика была сделана с использованием Prism 7.0b. Для Фигуры 9 использовали IBM*SPSS Statistics версии 24.0.0.0.
Результаты
Получение гуманизированных HLA-A2-специфических CAR
Аминокислотная последовательность вариабельных областей тяжелой и легкой цепей mAb мыши BB7.2 была приведена в соответствие с последовательностями иммуноглобулина человека, полученными из информационной системы международной базы данных ImMunoGeneTics® (IMGT®) с использованием инструмента IgBLAST, доступного в Национальном центре биотехнологической информации (NCBI). Система разграничения V-генов была установлена на последовательности Kabat для получения CDR, определенных Kabat (Kabat et al., 1991). Кроме того, было идентифицировано определение по Chothia (Chothia et al., 1987).
Ряд различных генов зародышевой линии человека тестировали в качестве каркасных последовательностей для прививки CDR. В дополнение к идентификации последовательностей зародышевой линии человека, которые были наиболее сопоставимы с последовательностью мыши, длины CDR также рассматривались как поддерживающие структуру в максимально возможной степени. Человеческие CDR (нумерация по Kabat) были заменены мышиными аналогами CDR из антитела BB7.2. Кроме того, дополнительные аминокислоты CDR тяжелой цепи были заменены путем объединения нумерации Chothia и Kabat. Были получены 6 различных гуманизированных тяжелых цепей и 5 различных гуманизированных легких цепей, и в общей сложности было получено 20 различных рецепторов химерных антигенов путем объединения гуманизированных тяжелых и легких цепей.
Экспрессия и антигенная специфичность гуманизированных HLA-A2-специфических CAR
Для проверки специфичности антигена различные конструкции hA2-CAR транзиторно трансфицировали в клетки 293T, окрашивали тетрамерами HLA-A*02:01 и анализировали проточной цитометрией. Одиннадцать hA2-CAR конструкций экспрессировали и связывали с тетрамером A*02:01 (Фигура 2).
Затем мы сравнили силу связывания различных конструкций hA2-CAR. Транзиторно трансфицированные клетки 293Т окрашивали увеличивающимися разведениями тетрамера A*02:01, затем анализировали проточной цитометрией. Как показано на Фигуре 3 (A, B и C), одиннадцать конструкций hA2-CAR в высокой степени связывают тетрамер A*02:01 в зависимости от концентрации тетрамера.
hA2-CAR-опосредованная стимуляция активирует Treg
Чтобы проверить функцию A2-CAR в Treg, клетки CD4+CD25hiCD127lo были отсортированы из периферической крови. Как показано на Фигуре 4 (A, B и C), Treg стимулировали, трансдуцировали и культивировали в течение дополнительных 6-7 дней. В конце 7-8 дней культивирования клетки очищали как клетки NGFR+ и экспрессию на клеточной поверхности варианта hA2-CAR тестировали проточной цитометрией. Как показано на Фигуре 4, Treg, трансдуцированные конструкциями hA2-CAR, показали положительное окрашивание тетрамером HLA-A2 по сравнению с Treg, трансдуцированными контрольной конструкцией ΔNGFR, что указывает на сохранение антигенной специфичности для Treg, экспрессирующих hA2-CAR. При получении лентивируса с высоким титром для (H1k3) возникли технические трудности, и этот CAR не был протестирован в Treg в этом эксперименте. Экспрессию и специфичность клеточной поверхности CAR тестировали путем окрашивания тетрамером A*02:01, показав, что H2k2, H4k4 и H5k4 имели бимодальный паттерн экспрессии в Treg с более низкой средней интенсивностью флуоресценции (MFI) клеток A2-тетрамера+ (Фигура 4D и 4E).
Далее мы исследовали, стимулирует ли стимуляция с помощью hA2-CAR-активированных Treg (hA2-CAR-Treg) по сравнению с Treg, экспрессирующими только укороченный маркер трансдукции NGFR (NGFR Treg). NGFR Treg и hA2-CAR Treg оставляли нестимулированными, стимулировали через CAR с помощью клеток K562, экспрессирующих HLA-A*02:01, или стимулировали через TCR с помощью клеток K562, экспрессирующих CD64, Fc-рецептор с высокой аффинностью, который был предварительно загружен анти-CD3/анти-CD28 антителами. Сравнение между NGFR Treg и hA2-CAR Treg показало значительно более высокую CAR-стимулированную экспрессию CD69 и CD71 в большинстве hA2-CAR Treg в течение нескольких временных точек. Напротив, при стимуляции через TCR, как NGFR Treg, так и hA2-CAR Treg были способны положительно регулировать CD69, что указывает на то, что они сохраняют свою способность передавать сигналы через TCR после трансдукции ДНК CAR (как показано на Фигуре 13). Опосредованная hA2-CAR стимуляция Treg также вызывала значительно более сильную положительную регуляцию белков, связанных с функцией Treg, демонстрируя, что стимуляция через hA2-CAR сохраняет ожидаемую высокую экспрессию CTLA-4 и LAP (Фигуры 5A-F).
Перекрестная реактивность конструкций hA2-CAR на других аллелях HLA класса
Существует множество разных аллелей HLA, которые эволюционировали со временем из меньшего числа аллелей предков. Следовательно, существуют семейства аллелей, которые могут различаться только несколькими аминокислотами, и одно антитело против HLA может распознавать несколько аллелей в эволюционно-связанном семействе. Известно, что мышиное моноклональное антитело (BB7.2) обладает перекрестной реактивностью к дополнительным аллелям HLA-A (Hilton & Parham, 2013.). В частности, при тестировании в твердофазном анализе было обнаружено, что BB7.2 распознает пять подтипов HLA-A2 (*02:01, *02:02, *02:03, *02:05, *02:06) и явлется перекрестно реактивным с HLA-A*69: 01, а при испытаниях в высоких концентрациях также с HLA-A*23:01, A*24:02, A*24:03, A*68:01 и A*68:02 (Hilton & Parham, 2013). В контексте трансплантации знание специфичности alloAg необходимо для обеспечения специфического нацеливания на аллогенные клетки, ткани и/или органы. Традиционный способ измерения аллореактивности Т-клеток является неточным и не количественным, поскольку включает функциональные MLR с большими банками гаплотипированных PBMC. Для оценки перекрестной реактивности гуманизированных анти-HLA-A2-антител по изобретению и сравнения с антителом BB7.2, мы адаптировали твердофазный анализ ONE Lambda, который предназначен для измерения анти-HLA-антител в сыворотке, путем измерения связывания гранул, покрытых HLA, с интересующими гуманизированными CAR. BB7.2 CAR создали, как описано в MacDonald, K.G. (2016). NGFR+ Treg, экспрессирующие указанные гуманизированные CAR, инкубировали с гранулами Flow Panel Reactive Single Antigen с количественным определением связывания с одним классом гранул в виде потери сигнала в «воротах» гранул на графике прямого светорассеяния/бокового светорассеяния (подробности см. в методах). Данные были нормализованы к количеству отрицательных контрольных гранул в образце, и количество относительного связывания с hCAR-экспрессирующими Treg рассчитывали относительно количества связывания NGFR-контрольными Treg по количеству отрицательных гранул в образце NGFR, умноженном на количество отрицательных гранул в образце NGFR. Чтобы подтвердить методологию, относительное связывание каждой конструкции m/hA2-CAR с HLA-A*02:01, определенное с помощью анализа клеток FlowPRT (Фигура 6B), сравнивали с MFI связывания тетрамера (Фигура 6C). Этот анализ выявил сильную прямую корреляцию между двумя способами обнаружения связывания A*02:01. Мы также поинтересовались, коррелирует ли количество связывания A*02:01, определенное в анализе клеток FlowPRT, с биологическим эффектом воздействия A*02:01. В действительности мы обнаружили, что существует прямая корреляция между количеством связывания A*02:01, количественно определенным с помощью анализа клеток FlowPRT, и стимуляцией активации Treg, что определили по положительной регуляции CD69 после воздействия A*02:01-экспрессирующих APC (Фигура 6D). Эти данные демонстрируют полезность клеточного метода FlowPRT для измерения способности alloAg-специфических CAR связываться с различными аллелями HLA.
Как показано на Фигуре 6 и в Таблице 9, все конструкции hA2-CAR значительно связывают HLA-A*02:01, подтверждая анализы по связыванию тетрамера. Когда мы проверили связывание антитела BB7.2 в одном анализе одиночных антигенов FlowPRA, мы подтвердили высокое связывание с A*69:01, но не смогли подтвердить перекрестную реактивность с A*23:01 или A*24:02 (Фигура 6). При тестировании относительной способности m/hA2-CAR Treg связываться с различными аллелями HLA-A, мы обнаружили, что mA2-CAR-Treg значительно связаны с HLA-A*03:01, A*25:01, A*29:02, A*30:01. A*31:01, A*33:01, A*36:01, A*68:01 и A*69:01 (Фигура 6, Таблица 9). Напротив, все варианты hA2-CAR Treg неожиданно продемонстрировали пониженную перекрестную реактивность по сравнению с mA2-CAR Treg (Фигура 6, таблица 9). Как и ожидалось, все конструкции CAR связывались с HLA-A*69:01, вариантом A*02:01, отличающимся только 6 аминокислотами. Ни одна из CAR-конструкций не показала какого-либо значительного связывания с HLA-B (Фигура 6).
Связь между степенью связывания CAR-Treg Ag и биологической активностью неизвестна. Для определения биологической значимости перекрестной реактивности HLA, мы создали APC, экспрессирующие HLA-A*24:02, A*25:01 или A*68:01; все клетки имели сравнимые уровни экспрессии HLA-A (данные не показаны). Мы обнаружили, что только совместное культивирование с HLA-A*02:01-экспрессирующими клетками приводило к значительной активации m/hCAR-Treg, о чем свидетельствует повышенная экспрессия CD69, CD71, LAP, CTLA-4 (Фигура 6I) или CD40L (данные не показаны). Эти данные свидетельствуют о том, что эффективная CAR-опосредованная активация Treg требует высокой аффинности и/или авидности взаимодействий. Соответственно, хотя некоторые hA2-CAR демонстрируют связывание A*25:01 и A*68:01 в анализе FlowPRT, сила связывания недостаточна для клеточной активации.
Таблица 9. Связывающие взаимодействия между m/hA2-CAR и белком HLA-A/B оказались статистически значимыми (двухфакторний дисперсионный анализ, апостериорный критерий Даннетта).
Мы разработали новый способ систематического тестирования специфичности CAR для аллоантигенов, создав новую платформу для комплексной идентификации конструкций с определенной аллельной специфичностью. Удивительно, но мы обнаружили, что по сравнению с mA2-CAR гуманизация CAR снизила перекрестную реактивность к нескольким аллельным вариантам HLA-A.
hA2-CAR Treg опосредуют HLA-A2-специфическую супрессию in vitro
Чтобы проверить способность hA2-CAR стимулировать антигенспецифическую супрессивную активность в Treg, мы использовали реакции смешанных лимфоцитов (MLR) в соответствии с экспериментальным планом, показанным на Фигуре 7A. В частности, пролиферацию HLA-A*02:01-отрицательных Т-клеток стимулировали совместной культурой со зрелыми HLA-A*02:01-положительными дендритными клетками в отсутствие или в присутствии возрастающих соотношений Treg, которые либо не трансдуцированы, либо трансдуцированы контрольным NGFR-лентивирусом или лентивирусом, кодирующим H1k2 hA2-CAR. Как показано на Фигурах 7B-E, hA2-CAR-экспрессирующие Treg были значительно лучше способны супрессировать аллоантиген-стимулированную пролиферацию CD4+ T-клеток по сравнению с контрольными Treg, трансдуцированными контрольной лентивирусной конструкцией с NGFR.
hA2-CAR Treg опосредуют HLA-A2-специфическую супрессию ксеногенной GVHD in vivo
Чтобы протестировать функциональную способность hA2-CAR Treg in vivo, мы использовали модель мыши, в которой человеческие PBMC, вживленные в иммунодефицитных мышей NSG, вызывали ксеногенную GVHD. 8×106 PBMC от HLA-A2+ донора инъецировали облученным мышам NSG с или без Treg, экспрессирующих h12-CAR H1k2. Соотношение протестированных PBMC и hA2-CAR Treg составляло 1: 2 (то есть 8×106 PBMC к 4×106 Treg). Мышей отслеживали в течение 7 недель по клинической оценке, описанной в методах. В соответствии с данными in vitro на Фигуре 7, мыши, получавшие hA2-CAR-экспрессирующие Treg, имели улучшенную выживаемость, уменьшенную потерю массы и отсроченное начало xenoGVHD по сравнению с мышами, которые не получали hA2-CAR Treg (сплошная серая линия) (Фигура 8А, В и С). Несмотря на то, что наблюдался биологический эффект Treg, мы не обнаружили циркулирующих m/hA2-CAR Treg, измеренных через 14 дней после введения (Фигура 8D и 8E).
Движение hA2-CAR Treg к HLA-A2+ трансплантатам кожи in vivo
Чтобы проверить, как CAR-направленная специфичность влияет на перемещение Treg, мы провели параллельную пересадку кожи от NSG- или NSG-A*02:01-трансгенных мышей мышам NSG (Фигура 9А). После извлечения трансплантата РВМС инъецировали в отсутствие или в присутствие m/hA2-CAR Treg или с HER2-CAR Treg в качесте неспецифических поликлональных контрольных Treg (MacDonald et al., 2016). В дополнение к CAR, Treg ко-трансдуцировали лентивирусом, кодирующим гибридный белок люцифераза-GFP. Биолюминесцентную визуализацию проводили после инъекции D-люциферина в течение 21 дня после инъекции Treg (Barrett et al., 2014). Поликлональные HER2-CAR Treg продемонстрировали тенденцию направленного движения к аллотрансплантатам, но были в равной степени распределены между A2-положительными и A2-отрицательными трансплантатами. Эти поликлональные Treg могут мигрировать в ответ на воспалительные сигналы, исходящие от послеоперационной кожи, потому что у необработанных иммунодефицитных мышей Т-клетки человека обычно попадают в легкие (Nervi et al., 2007). В отличие от поликлональных HER2-CAR Treg, которые перемещались в равной степени к А2-отрицательной и А2-положительной коже, оба m/hA2-CAR Treg быстро перемещаются на А2-экспрессирующую кожу. Кроме того, m/hA2-CAR Treg сохранялись более длительно, чем неспецифичные HER2-CAR Treg. В то время как HER2-CAR Treg не обнаруживались к 21 дню, оставался сильный сигнал m/hA2-CAR Treg (Фигура 9B). Количественная оценка соотношения люминесценции в A2-положительном и A2-отрицательном транспланте выявила значительный Ag-управляемое перемещение как H1k2-, так и mA2-CAR Treg в A2-экспрессирующий трансплантат (Фигура 9C и 9D).
В дополнение к локализованным в трансплантате m/hA2-CAR Treg, на 21-й день мы отметили смежный сигнал, согласующийся с расположением локального дренирующего лимфатического узла. После умерщвления собрали лимфатические узлы, дренирующие трансплантат, и анализ с помощью проточной цитометрии показал значительную долю hCD4+FOXP3+ΔNGFR+A2-тетрамер+ CAR Treg. Напротив, очень мало этих клеток было обнаружено в селезенке (Фигура 10B и C). Ранее сообщалось, что этот двухстадийный миграционный процесс от трансплантата к лимфатическому узлу необходим для индукции толерантности (Zhang et al., 2009). Важно, что CAR Treg, находящиеся в лимфатических узлах, сохраняли экспрессию FOXP3 и CAR, иллюстрируя их стабильный фенотип.
hA2-CAR Treg предотвращают отторжение аллотрансплантата кожи человека
Чтобы оценить иммунорегуляторный потенциал hA2-CAR Treg в модели трансплантации твердого органа, мы использовали гуманизированную модель трансплантации кожи, в которой мышам NSG трансплантировали кожные трансплантаты HLA-A2pos человека. Через 6 недель мышам вводили РВМС HLA-A2neg с или без аутологичных H1k2 hA2-CAR Treg. Через четыре недели после введения клеток мышей умерщвляли и собирали кожные трансплантаты для оценки патологии и экспрессии воспалительных цитокинов. Все мыши сохраняли стабильную массу тела, что указывает на отсутствие ксеногенной GvHD (Фигура 11А), с аналогичными уровнями приживления лейкоцитов человека в крови и селезенке (Фигура 11В). Срезы, окрашенные гематоксилином-эозином, оценивали с использованием 25-балльной шкалы, выявляя значительное снижение кумулятивного показателя патологического отторжения у мышей, которые получали H1k2 hA2-CAR Treg по сравнению с РВМС (Фигура 11C). Иммуноокрашивание показало, что по сравнению с мышами, получавшими только PBMC, мыши, получавшие PBMC и H1k2 hA2-CAR Treg, имели значительное снижение Ki67+ кератиноцитов и имели пониженное разрушение инволюкрина (Фигура 11D). Количественный анализ кПЦР также показал общее снижение воспалительных цитокинов в трансплантате H1k2 hA2-CAR Treg-обработанных мышей (Фигура 11E).
Что касается ксеногенной модели GVHD, в то время как PBMC обнаруживались в крови, CAR Treg не обнаруживались (Фигура 12). Однако иммуноокрашивание показало, что по сравнению с мышами, получавшими только PBMC, у мышей, получавших PBMC и H1k2 hA2-CAR Treg, была значительно более высокая доля клеток FOXP3+ в трансплантате (Фигура 11F). Присутствие FOXP3+ клеток было уникальным для трансплантированного кожного трансплантата, поскольку они не были обнаружены в кишечнике, печени или легких (Фигура 11G). Эти данные показывают, что, как и в случае модели с A2-трансгенной NSG-кожей, H1k2 hA2-CAR Treg специфически связываются с A2+ аллотрансплантатом кожи человека, где они сохраняются в течение неопределенного времени.
Эта заявка включает список последовательностей в электронной форме в виде текстового файла «CDRD-003WO_SEQ LISTING_ST25», созданного 18 сентября 2018 года и имеющего размер 320 КБ. Содержимое текстового файла включено в данный документ ссылкой полностью.
Список литературы
Allan, S.E., A.N. Alstad, N. Merindol, N.K. Crellin, M. Amendola, R. Bacchetta, L. Naldini, M.G. Roncarolo, H. Soudeyns, and M.K. Levings. 2008. Generation of potent and stable human CD4+ T regulatory cells by activation-independent expression of FOXP3. Molecular therapy: the journal of the American Society of Gene Therapy 16:194-202.
Barrett, D. M., Singh, N., Liu, X., Jiang, S., June, C.H., Grupp, S.A., and Zhao, Y. 2014. Relation of clinical culture method to T-cell memory status and efficacy in xenograft models of adoptive immunotherapy. Cytotherapy 16: 619-630.
Blat, D., E. Zigmond, Z. Alteber, T. Waks, and Z. Eshhar. 2014. Suppression of murine colitis and its associated cancer by carcinoembryonic antigen-specific regulatory T cells. Molecular therapy: the journal of the American Society of Gene Therapy 22:1018-1028.
Bluestone, J.A., J.H. Buckner, M. Fitch, S.E. Gitelman, S. Gupta, M.K. Hellerstein, K.C. Herold, A. Lares, M.R. Lee, K. Li, W. Liu, S.A. Long, L.M. Masiello, V. Nguyen, A.L. Putnam, M. Rieck, P.H. Sayre, and Q. Tang. 2015. Type 1 diabetes immunotherapy using polyclonal regulatory T cells. Science translational medicine 7:315ra189.
Boardman, D.A., C. Philippeos, G.O. Fruhwirth, M.A. Ibrahim, R.F. Hannen, D. Cooper, F.M. Marelli-Berg, F.M. Watt, R.I. Lechler, J. Maher, L.A. Smyth, and G. Lombardi. 2016. Expression of a Chimeric Antigen Receptor Specific for Donor HLA Class I Enhances the Potency of Human Regulatory T Cells in Preventing Human Skin Transplant Rejection. American journal of transplantation: official journal of the American Society of Transplantation and the American Society of Transplant Surgeons
Brunstein, C.G., B.R. Blazar, J.S. Miller, Q. Cao, K.L. Hippen, D.H. McKenna, J. Curtsinger, P.B. McGlave, and J.E. Wagner. 2013. Adoptive transfer of umbilical cord blood-derived regulatory T cells and early viral reactivation. Biology of blood and marrow transplantation: journal of the American Society for Blood and Marrow Transplantation 19:1271-1273.
Brunstein, C.G., J.S. Miller, Q. Cao, D.H. McKenna, K.L. Hippen, J. Curtsinger, T. Defor, B.L. Levine, C.H. June, P. Rubinstein, P.B. McGlave, B.R. Blazar, and J.E. Wagner. 2011. Infusion of ex vivo expanded T regulatory cells in adults transplanted with umbilical cord blood: safety profile and detection kinetics. Blood 117:1061-1070.
Casucci M, Bondanza A. 2011. Suicide Gene Therapy to Increase the Safety of Chimeric Antigen Receptor-Redirected T Lymphocytes. Journal of Cancer, 2:378-382.
Chong, A.S. and S.H. Khiew. Transplantation tolerance: don't forget about the B cells. Clin Exp Immunol. 2017, 189(2):171-180.
Chothia and Lesk J. Mol. Biol. 196:901-917 (1987).
Cooke KR, et al. An experimental model of idiopathic pneumonia syndrome after bone marrow transplantation: I. The roles of minor H antigens and endotoxin. Blood. 1996;88(8):3230–3239.
Crome, S.Q., Lang, P.A., Lang, K.S., and P.S. Ohashi. Natural killer cells regulate diverse T cell responses. Trends Immunol. 2013 Jul;34(7):342-9.
Crome, S.Q., Nguyen, L.T., Lopez-Verges, S., Yang, S.Y., Martin, B., Yam, J.Y., Johnson, D.J., Nie, J., Pniak, M., Yen, P.H., Milea, A., Sowamber, R., Katz, S.R., Bernardini, M.Q., Clarke, B.A., Shaw, P.A., Lang, P.A., Berman, H.K., Pugh, T.J., Lanier, L.L., and P.S. Ohashi. A distinct innate lymphoid cell population regulates tumor-associated T cells. Nat Med. 2017 Mar;23(3):368-375.
Delhove, J. M. K. M. et al., Genome-Edited T Cell Therapies, Curr Stem Cell Rep. 2017; 3(2): 124–136.
Di Ianni, M., F. Falzetti, A. Carotti, A. Terenzi, F. Castellino, E. Bonifacio, B. Del Papa, T. Zei, R.I. Ostini, D. Cecchini, T. Aloisi, K. Perruccio, L. Ruggeri, C. Balucani, A. Pierini, P. Sportoletti, C. Aristei, B. Falini, Y. Reisner, A. Velardi, F. Aversa, and M.F. Martelli. 2011. Tregs prevent GVHD and promote immune reconstitution in HLA-haploidentical transplantation. Blood 117:3921-3928.
Dijke, I.E., R.E. Hoeppli, T. Ellis, J. Pearcey, Q. Huang, A.N. McMurchy, K. Boer, A.M. Peeters, G. Aubert, I. Larsen, D.B. Ross, I. Rebeyka, A. Campbell, C.C. Baan, M.K. Levings, and L.J. West. 2016. Discarded Human Thymus Is a Novel Source of Stable and Long-Lived Therapeutic Regulatory T Cells. American journal of transplantation. 16(1):58-71.
Eliceiri, K. W., Berthold, M.R., Goldberg, I.G., Ibanez, L. Manjunath, B.S., Martone, M.E., Murphy, R.F., Peng, H., Plant, A.L., Roysam, B., Stuurman, N., Swedlow, J.R., Tomancak, P., Carpenter, A.E. 2012. Biological imaging software tools. Nature Methods 9: 697-710.
Elinav, E., N. Adam, T. Waks, and Z. Eshhar. 2009. Amelioration of colitis by genetically engineered murine regulatory T cells redirected by antigen-specific chimeric receptor. Gastroenterology 136:1721-1731.
Elinav, E., T. Waks, and Z. Eshhar. 2008. Redirection of regulatory T cells with predetermined specificity for the treatment of experimental colitis in mice. Gastroenterology 134:2014-2024.
Ellis, J. M., Henson, V., Slack, R., Ng, J., Hartzman, R. J., and C. Katovich Hurley. Frequencies of HLA-A2 alleles in five U.S. population groups. Predominance Of A*02011 and identification of HLA-A*0231. Hum Immunol. 61(3):334-40 (2000).
Eyquem, J. et al. Targeting a CAR to the TRAC locus with CRISPR/Cas9 enhances tumour rejection, Nature 543, 113–117 (2017).
Fischer, A., Jakubowski, A.A., Lacouture, M.E., Hollman, T.J., Drucker, A.M., Maloy, M., Prockop, S., Querfeld, C., Busam, K.J., Pulitzer, M.P2015. Histopathologic Features of Cutaneous Acute Graft-Versus-Host Disease in T-Cell-Depleted Peripheral Blood Stem Cell Transplant Recipients. The American Journal ofDdermatopathology 37: 523-529.
Fransson, M., E. Piras, J. Burman, B. Nilsson, M. Essand, B. Lu, R.A. Harris, P.U. Magnusson, E. Brittebo, and A.S. Loskog. 2012. CAR/FoxP3-engineered T regulatory cells target the CNS and suppress EAE upon intranasal delivery. Journal of neuroinflammation 9:112.
Fu, B., Tian, Z., and H. Wei. Subsets of human natural killer cells and their regulatory effects. Immunology. 2014 Apr; 141(4):483-9.
Gargett T, Brown MP. 2014. The inducible caspase-9 suicide gene system as a «safety switch» to limit on-target, off-tumor toxicities of chimeric antigen receptor T cells. Frontiers in Pharmacology, 5:235.
Ghosh et al., 1991 Glycobiology 5: 505-10.
Gill, S., and C.H. June. 2015. Going viral: chimeric antigen receptor T-cell therapy for hematological malignancies. Immunological reviews 263:68-89.
Golshayan, D., S. Jiang, J. Tsang, M.I. Garin, C. Mottet, and R.I. Lechler. 2007. In vitro-expanded donor alloantigen-specific CD4+CD25+ regulatory T cells promote experimental transplantation tolerance. Blood 109:827-835.
Green, E.A., Y.W. Choi, and R.A. Flavell. 2002. Pancreatic lymph node-derived CD4(+)CD25(+) Treg cells: Highly potent regulators of diabetes that require TRANCE-RANK signals. Immunity 16:183-191.
Guillonneau, C., Picarda, E., and I. Anegon. CD8+ regulatory T cells in solid organ transplantation. Curr Opin Organ Transplant. 2010; 15(6):751-6.
Hill GR, Crawford JM, Cooke KR, Brinson YS, Pan L, Ferrara JL. Total body irradiation and acute graft-versus-host disease: the role of gastrointestinal damage and inflammatory cytokines. Blood. 1997;90(8):3204–3213.
Hilton, H. G. & Parham, P. Direct binding to antigen-coated beads refines the specificity and cross-reactivity of four monoclonal antibodies that recognize polymorphic epitopes of HLA class I molecules. Tissue Antigens 81, 212–220 (2013).
Himmel, M.E., K.G. MacDonald, R.V. Garcia, T.S. Steiner, and M.K. Levings. 2013. Helios+ and Helios- cells coexist within the natural FOXP3+ T regulatory cell subset in humans. Journal of immunology 190:2001-2008.
Hombach, A.A., D. Kofler, G. Rappl, and H. Abken. 2009. Redirecting human CD4+CD25+ regulatory T cells from the peripheral blood with pre-defined target specificity. Gene therapy 16:1088-1096.
Hutchinson, J.A., Ahrens, N., and E.K. Geissler. MITAP-compliant characterization of human regulatory macrophages. Transpl Int. 2017 Aug; 30(8):765-775.
Jamnani, F., R., Fatemeh Rahbarizadeh, Mohammad Ali Shokrgozar, Fereidoun Mahboudi, Davoud Ahmadvand, Zahra Sharifzadeh, Ladan Parhamifar, S. Moein Moghimi. 2014. T cells expressing VHH-directed oligoclonal chimeric HER2 antigen receptors: Towards tumor-directed oligoclonal T cell therapy. Biochimica et Biophysica Acta, 1840(1): 378-386.
Joffre, O., T. Santolaria, D. Calise, T. Al Saati, D. Hudrisier, P. Romagnoli, and J.P. van Meerwijk. 2008. Prevention of acute and chronic allograft rejection with CD4+CD25+Foxp3+ regulatory T lymphocytes. Nature medicine 14:88-92.
Jones, B. S., Lamb, L.S., Goldman, F., and A. Di Stasi. 2014. Improving the safety of cell therapy products by suicide gene transfer. Front Pharmacol. Volume 5, 254.
June, C.H., S.R. Riddell, and T.N. Schumacher. 2015. Adoptive cellular therapy: A race to the finish line. Science translational medicine 7:280-287.
Juvet, S.C. and L. Zhang. Double negative regulatory T cells in transplantation and autoimmunity: recent progress and future directions. J Mol Cell Biol. 2012 Feb;4(1):48-58.
Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991).
Kanitakis, J. 2008. The challenge of dermatopathological diagnosis of composite tissue allograft rejection: a review. Journal of cutaneous pathology 35: 738-744.
Khaleghi S. 2012. A caspase 8-based suicide switch induces apoptosis in nanobody-directed chimeric receptor expressing T cells. International journal of hematology, 95(4):434.
Konvalinka, A. and K. Tinckam. Utility of HLA Antibody Testing in Kidney Transplantation. J Am Soc Nephrol. 26(7):1489-502 (2015).
Levings, M.K., R. Sangregorio, and M.G. Roncarolo. 2001. Human cd25(+)cd4(+) t regulatory cells suppress naive and memory T cell proliferation and can be expanded in vitro without loss of function. The Journal of experimental medicine 193:1295-1302.
Li, N., Haiying Fu, Stephen M. Hewitt, Dimiter S. Dimitrov, and Mitchell Ho. 2017. Therapeutically targeting glypican-2 via single-domain antibody-based chimeric antigen receptors and immunotoxins in neuroblastoma. PNAS, 114 (32); E6623-E6631.
MacDonald, K.G., R.E. Hoeppli, Q. Huang, J. Gillies, D.S. Luciani, P.C. Orban, R. Broady, and M.K. Levings. 2016. Alloantigen-specific regulatory T cells generated with a chimeric antigen receptor. The Journal of clinical investigation 126:1413-1424.
Marek-Trzonkowska, N., M. Mysliwiec, A. Dobyszuk, M. Grabowska, I. Techmanska, J. Juscinska, M.A. Wujtewicz, P. Witkowski, W. Mlynarski, A. Balcerska, J. Mysliwska, and P. Trzonkowski. 2012. Administration of CD4+CD25highCD127- regulatory T cells preserves beta-cell function in type 1 diabetes in children. Diabetes care 35:1817-1820.
Massi, D., Franchi, A., Pimpinelli, N., Laszlo, D., Bosi, A., Santucci, M. 1999. A reappraisal of the histopathologic criteria for the diagnosis of cutaneous allogeneic acute graft-vs-host disease. American Journal of Clinical Pathology 112: 791-800.
Masteller, E.L., M.R. Warner, Q. Tang, K.V. Tarbell, H. McDevitt, and J.A. Bluestone. 2005. Expansion of functional endogenous antigen-specific CD4+CD25+ regulatory T cells from nonobese diabetic mice. Journal of immunology 175:3053-3059.
McMurchy, A.N. & Levings, M.K. Suppression assays with human T regulatory cells: a technical guide. Eur. J. Immunol. 42, 27–34 (2012).
Nervi, B., Rettig, M.P., Ritchey, J.K., Wang, H.L., Bauer, G., Walker, J., Bonyhadi, M.L., Berenson, R.J., Prior, J.L., Piwnica-Worms, D., Nolta, J.A., DiPersio, J.F. 2007. Factors affecting human T cell engraftment, trafficking, and associated xenogeneic graft-vs-host disease in NOD/SCID beta2mnull mice. Exp Hematol 35: 1823-1838.
Nishimura, E., T. Sakihama, R. Setoguchi, K. Tanaka, and S. Sakaguchi. 2004. Induction of antigen-specific immunologic tolerance by in vivo and in vitro antigen-specific expansion of naturally arising Foxp3+CD25+CD4+ regulatory T cells. International immunology 16:1189-1201.
Noyan, F., K. Zimmermann, M. Hardtke-Wolenski, A. Knoefel, E. Schulde, R. Geffers, M. Hust, J. Huehn, M. Galla, M. Morgan, A. Jokuszies, M.P. Manns, and E. Jaeckel. 2016. Prevention of Allograft Rejection by Use of Regulatory T Cells With an MHC-Specific Chimeric Antigen Receptor. American journal of transplantation: official journal of the American Society of Transplantation and the American Society of Transplant Surgeons
Papp, G., Boros, P., Nakken, B., Szodoray, P. and M. Zeher. Regulatory immune cells and functions in autoimmunity and transplantation immunology. Autoimmunity Reviews. Volume 16, Issue 5, 2017, Pages 435-444.
Parham, P. and Brodsky, FM (1981) Hum Immunol 3(4): 277-99.
Brian Philip, Evangelia Kokalaki, Leila Mekkaoui, Simon Thomas, Karin Straathof, Barry Flutter, Virna Marin, Teresa Marafioti, Ronjon Chakraverty, David Linch, Sergio A. Quezada, Karl S. Peggs, and Martin Pule. 2014. A highly compact epitope-based marker/suicide gene for easier and safer T-cell therapy. Blood, 124:1277-1287.
Putnam, A.L., N. Safinia, A. Medvec, M. Laszkowska, M. Wray, M.A. Mintz, E. Trotta, G.L. Szot, W. Liu, A. Lares, K. Lee, A. Laing, R.I. Lechler, J.L. Riley, J.A. Bluestone, G. Lombardi, and Q. Tang. 2013. Clinical grade manufacturing of human alloantigen-reactive regulatory T cells for use in transplantation. American journal of transplantation: official journal of the American Society of Transplantation and the American Society of Transplant Surgeons 13:3010-3020.
Robinson, J. et al. The IPD and IMGT/HLA database: allele variant databases. Nucleic Acids Res 43, D423-431, doi:10.1093/nar/gku1161 (2015).
Rosenberg et al, New Eng. J. of Med. 319: 1676, 1988.
Sadelain, M., R. Brentjens, and I. Riviere. 2013. The basic principles of chimeric antigen receptor design. Cancer discovery 3:388-398.
Sagoo, P., N. Ali, G. Garg, F.O. Nestle, R.I. Lechler, and G. Lombardi. 2011. Human regulatory T cells with alloantigen specificity are more potent inhibitors of alloimmune skin graft damage than polyclonal regulatory T cells. Science translational medicine 3:83ra42.
Sambrook et al., 2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York.
Sanchez-Fueyo, A., S. Sandner, A. Habicht, C. Mariat, J. Kenny, N. Degauque, X.X. Zheng, T.B. Strom, L.A. Turka, and M.H. Sayegh. 2006. Specificity of CD4+CD25+ regulatory T cell function in alloimmunity. Journal of immunology 176:329-334.
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. 2012. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9: 671-675.
Schmittgen, T. D. & Livak, K. J. 2008. Analyzing real-time PCR data by the comparative C(T) method. Nature Protocols 3: 1101-1108.
Stephens, L.A., K.H. Malpass, and S.M. Anderton. 2009. Curing CNS autoimmune disease with myelin-reactive Foxp3+ Treg. European journal of immunology 39:1108-1117.
Tang, Q., K.J. Henriksen, M. Bi, E.B. Finger, G. Szot, J. Ye, E.L. Masteller, H. McDevitt, M. Bonyhadi, and J.A. Bluestone. 2004. In vitro-expanded antigen-specific regulatory T cells suppress autoimmune diabetes. The Journal of experimental medicine 199:1455-1465.
Tarbell, K.V., L. Petit, X. Zuo, P. Toy, X. Luo, A. Mqadmi, H. Yang, M. Suthanthiran, S. Mojsov, and R.M. Steinman. 2007. Dendritic cell-expanded, islet-specific CD4+ CD25+ CD62L+ regulatory T cells restore normoglycemia in diabetic NOD mice. The Journal of experimental medicine 204:191-201.
Tarbell, K.V., S. Yamazaki, K. Olson, P. Toy, and R.M. Steinman. 2004. CD25+ CD4+ T cells, expanded with dendritic cells presenting a single autoantigenic peptide, suppress autoimmune diabetes. The Journal of experimental medicine 199:1467-1477.
Trenado, A., M. Sudres, Q. Tang, S. Maury, F. Charlotte, S. Gregoire, M. Bonyhadi, D. Klatzmann, B.L. Salomon, and J.L. Cohen. 2006. Ex vivo-expanded CD4+CD25+ immunoregulatory T cells prevent graft-versus-host-disease by inhibiting activation/differentiation of pathogenic T cells. Journal of immunology 176:1266-1273.
Trzonkowski, P., M. Bieniaszewska, J. Juscinska, A. Dobyszuk, A. Krzystyniak, N. Marek, J. Mysliwska, and A. Hellmann. 2009. First-in-man clinical results of the treatment of patients with graft versus host disease with human ex vivo expanded CD4+CD25+CD127- T regulatory cells. Clinical immunology 133:22-26.
Tsang, J.Y., Y. Tanriver, S. Jiang, S.A. Xue, K. Ratnasothy, D. Chen, H.J. Stauss, R.P. Bucy, G. Lombardi, and R. Lechler. 2008. Conferring indirect allospecificity on CD4+CD25+ Tregs by TCR gene transfer favors transplantation tolerance in mice. The Journal of clinical investigation 118:3619-3628.
Verginis, P., K.A. McLaughlin, K.W. Wucherpfennig, H. von Boehmer, and I. Apostolou. 2008. Induction of antigen-specific regulatory T cells in wild-type mice: visualization and targets of suppression. Proceedings of the National Academy of Sciences of the United States of America 105:3479-3484.
Zhenguang Wang, Zhiqiang Wu, Yang Liu and Weidong Han. 2017. New development in CAR-T cell therapy. Journal of Hematology & Oncology, 10:53.
Watanabe, N., Bajgain, P., Sukumaran, S., Ansari, S., Heslop, H.E., Rooney, C. M., Brenner, M.K., Leen, A.M., Vera, J.F. Fine-tuning the CAR spacer improves T-cell potency. OncoImmunology. 2016, 5(12): e1253656.
Wesch, D., Peters, C., and G.M. Siegers. Human gamma delta T regulatory cells in cancer: fact or fiction? Front Immunol. 2014 Nov 20; 5:598.
Wood, K.J., Bushell, A., and J. Hester. Regulatory immune cells in transplantation. Nat Rev Immunol. 2012, 12(6):417-30.
Zhang, E. and Xu, H. J. 2017. A new insight in chimeric antigen receptor-engineered T cells for cancer immunotherapy. Hematol Oncol, 10:1.
Zhang, N., Schroppel, B., Lal, G., Jakubzick, C., Mao, X., Chen, D., Yin, N., Jessberger, R., Ochando, J.C., Ding, Y., Bromberg, J.S. 2009. Regulatory T cells sequentially migrate from inflamed tissues to draining lymph nodes to suppress the alloimmune response. Immunity. 30: 458-469.
Хотя вышеизложенное изобретение было описано в некоторых деталях для ясности и понимания, специалисту в данной области техники будет понятно из прочтения этого раскрытия, что различные изменения в форме и деталях могут быть сделаны без отклонения от истинного объема изобретения. Например, все методы и устройства, описанные выше, могут использоваться в различных комбинациях. Все публикации, патенты, патентные заявки и/или другие документы, процитированные в этой заявке, полностью включены ссылкой для всех целей в той же степени, как если бы каждая отдельная публикация, патент, заявка на патент и/или другой документ были указаны отдельно для включения ссылкой для всех целей.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> LEVINGS, Megan
ORBAN, Paul
DAWSON, Nicholas
LAMARCHE, Caroline
BERGQVIST, Jan P
<120> ANTI-HLA-A2 ANTIBODIES AND METHODS OF USING THE SAME
<130> CDRD-003WO
<150> US 62/560,574
<151> 2017-09-19
<150> US 62/692,386
<151> 2018-06-29
<160> 224
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 1
Ser Tyr His Ile Gln
1 5
<210> 2
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 2
Gly Tyr Thr Phe Thr Ser Tyr
1 5
<210> 3
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 3
Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 4
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 4
Tyr Pro Gly Asp Gly Ser
1 5
<210> 5
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 5
Trp Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Ser Gln Lys Phe Gln
1 5 10 15
Gly
<210> 6
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 6
Glu Gly Thr Tyr Tyr Ala Met Asp Tyr
1 5
<210> 7
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 7
Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 8
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 8
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 9
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 9
Phe Gln Gly Ser His Val Pro Arg Thr
1 5
<210> 10
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa is Q or K
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa is N or S
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa is E or Q
<220>
<221> misc_feature
<222> (16)..(16)
<223> Xaa is K or Q
<400> 10
Trp Ile Tyr Pro Gly Asp Gly Ser Thr Xaa Tyr Xaa Xaa Lys Phe Xaa
1 5 10 15
Gly
<210> 11
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 11
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser
20 25
<210> 12
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 12
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
20 25 30
<210> 13
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa is R or G
<220>
<221> misc_feature
<222> (15)..(15)
<223> Xaa is I or absent
<400> 13
Trp Val Arg Gln Ala Pro Gly Gln Xaa Leu Glu Trp Met Gly Xaa
1 5 10 15
<210> 14
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 14
Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met Gly
1 5 10
<210> 15
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 15
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ile
1 5 10 15
<210> 16
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 16
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly
1 5 10
<210> 17
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa is R or G
<220>
<221> misc_feature
<222> (15)..(15)
<223> Xaa is I or absent
<400> 17
Trp Val Arg Gln Ala Pro Gly Gln Xaa Leu Glu Trp Met Gly Xaa Trp
1 5 10 15
Ile
<210> 18
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 18
Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met Gly Trp Ile
1 5 10 15
<210> 19
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 19
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ile Trp
1 5 10 15
Ile
<210> 20
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 20
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Trp Ile
1 5 10 15
<210> 21
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa is R or G
<220>
<221> misc_feature
<222> (18)..(18)
<223> Xaa is I or absent
<400> 21
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Xaa Leu Glu Trp Met
1 5 10 15
Gly Xaa Trp Ile
20
<210> 22
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 22
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
1 5 10 15
Gly Trp Ile
<210> 23
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 23
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
1 5 10 15
Gly Ile Trp Ile
20
<210> 24
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 24
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
1 5 10 15
Gly Trp Ile
<210> 25
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa is R or G
<220>
<221> misc_feature
<222> (18)..(18)
<223> Xaa is I or absent
<400> 25
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Xaa Leu Glu Trp Met
1 5 10 15
Gly Xaa
<210> 26
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 26
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
1 5 10 15
Gly
<210> 27
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 27
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
1 5 10 15
Gly Ile
<210> 28
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 28
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
1 5 10 15
Gly
<210> 29
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (1)..(1)
<223> Xaa is R or absent
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa is I or M
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa is R or A
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa is A, T or I
<220>
<221> misc_feature
<222> (16)..(16)
<223> Xaa is E or L
<220>
<221> misc_feature
<222> (19)..(19)
<223> Xaa is S or R
<220>
<221> misc_feature
<222> (23)..(23)
<223> Xaa is E or D
<220>
<221> misc_feature
<222> (25)..(25)
<223> Xaa is T or M
<400> 29
Xaa Val Thr Xaa Thr Xaa Asp Thr Ser Xaa Ser Thr Ala Tyr Met Xaa
1 5 10 15
Leu Ser Xaa Leu Arg Ser Xaa Asp Xaa Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 30
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 30
Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr Met Glu
1 5 10 15
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 31
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 31
Arg Val Thr Ile Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr Met Leu
1 5 10 15
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 32
<211> 31
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 32
Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu
1 5 10 15
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 33
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 33
Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr Met Glu
1 5 10 15
Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 34
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 34
Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr Met Glu
1 5 10 15
Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 35
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa is Q or K
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa is N or S
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa is E or Q
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa is K or Q
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa is R or absent
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa is I or M
<220>
<221> misc_feature
<222> (15)..(15)
<223> Xaa is R or A
<220>
<221> misc_feature
<222> (19)..(19)
<223> Xaa is A, T or I
<220>
<221> misc_feature
<222> (25)..(25)
<223> Xaa is E or L
<220>
<221> misc_feature
<222> (28)..(28)
<223> Xaa is S or R
<220>
<221> misc_feature
<222> (32)..(32)
<223> Xaa is E or D
<220>
<221> misc_feature
<222> (34)..(34)
<223> Xaa is T or M
<400> 35
Thr Xaa Tyr Xaa Xaa Lys Phe Xaa Gly Xaa Val Thr Xaa Thr Xaa Asp
1 5 10 15
Thr Ser Xaa Ser Thr Ala Tyr Met Xaa Leu Ser Xaa Leu Arg Ser Xaa
20 25 30
Asp Xaa Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 36
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa is R or absent
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa is I or M
<220>
<221> misc_feature
<222> (15)..(15)
<223> Xaa is R or A
<220>
<221> misc_feature
<222> (19)..(19)
<223> Xaa is A, T or I
<220>
<221> misc_feature
<222> (25)..(25)
<223> Xaa is E or L
<220>
<221> misc_feature
<222> (28)..(28)
<223> Xaa is S or R
<220>
<221> misc_feature
<222> (32)..(32)
<223> Xaa is E or D
<220>
<221> misc_feature
<222> (34)..(34)
<223> Xaa is T or M
<400> 36
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Xaa Val Thr Xaa Thr Xaa Asp
1 5 10 15
Thr Ser Xaa Ser Thr Ala Tyr Met Xaa Leu Ser Xaa Leu Arg Ser Xaa
20 25 30
Asp Xaa Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 37
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa is R or absent
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa is I or M
<220>
<221> misc_feature
<222> (15)..(15)
<223> Xaa is R or A
<220>
<221> misc_feature
<222> (19)..(19)
<223> Xaa is A, T or I
<220>
<221> misc_feature
<222> (25)..(25)
<223> Xaa is E or L
<220>
<221> misc_feature
<222> (28)..(28)
<223> Xaa is S or R
<220>
<221> misc_feature
<222> (32)..(32)
<223> Xaa is E or D
<220>
<221> misc_feature
<222> (34)..(34)
<223> Xaa is T or M
<400> 37
Thr Lys Tyr Ser Gln Lys Phe Gln Gly Xaa Val Thr Xaa Thr Xaa Asp
1 5 10 15
Thr Ser Xaa Ser Thr Ala Tyr Met Xaa Leu Ser Xaa Leu Arg Ser Xaa
20 25 30
Asp Xaa Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 38
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 38
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
1 5 10 15
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
20 25 30
Asp Thr Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 39
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 39
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Ala Asp
1 5 10 15
Thr Ser Ala Ser Thr Ala Tyr Met Leu Leu Ser Ser Leu Arg Ser Glu
20 25 30
Asp Thr Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 40
<211> 40
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 40
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Val Thr Met Thr Arg Asp Thr
1 5 10 15
Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 41
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 41
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Met Thr Arg Asp
1 5 10 15
Thr Ser Ile Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp
20 25 30
Asp Thr Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 42
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 42
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
1 5 10 15
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
20 25 30
Asp Met Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 43
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 43
Thr Lys Tyr Ser Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Arg Asp
1 5 10 15
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
20 25 30
Asp Thr Ala Val Tyr Tyr Cys Ala Arg
35 40
<210> 44
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 44
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
1 5 10
<210> 45
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (44)..(44)
<223> Xaa is R or G
<220>
<221> misc_feature
<222> (50)..(50)
<223> Xaa is I or absent
<220>
<221> misc_feature
<222> (60)..(60)
<223> Xaa is Q or K
<220>
<221> misc_feature
<222> (62)..(62)
<223> Xaa is N or S
<220>
<221> misc_feature
<222> (63)..(63)
<223> Xaa is E or Q
<220>
<221> misc_feature
<222> (66)..(66)
<223> Xaa is K or Q
<220>
<221> misc_feature
<222> (68)..(68)
<223> Xaa is R or absent
<220>
<221> misc_feature
<222> (71)..(71)
<223> Xaa is I or M
<220>
<221> misc_feature
<222> (73)..(73)
<223> Xaa is R or A
<220>
<221> misc_feature
<222> (77)..(77)
<223> Xaa is A, T or I
<220>
<221> misc_feature
<222> (83)..(83)
<223> Xaa is E or L
<220>
<221> misc_feature
<222> (86)..(86)
<223> Xaa is S or R
<220>
<221> misc_feature
<222> (90)..(90)
<223> Xaa is E or D
<220>
<221> misc_feature
<222> (92)..(92)
<223> Xaa is T or M
<400> 45
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Xaa Leu Glu Trp Met
35 40 45
Gly Xaa Trp Ile Tyr Pro Gly Asp Gly Ser Thr Xaa Tyr Xaa Xaa Lys
50 55 60
Phe Xaa Gly Xaa Val Thr Xaa Thr Xaa Asp Thr Ser Xaa Ser Thr Ala
65 70 75 80
Tyr Met Xaa Leu Ser Xaa Leu Arg Ser Xaa Asp Xaa Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 46
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa is V or I
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa is S or T
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa is L or S
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa is P or S
<220>
<221> misc_feature
<222> (15)..(15)
<223> Xaa is L or P
<220>
<221> misc_feature
<222> (23)..(23)
<223> Xaa is C or F
<400> 46
Asp Xaa Val Met Thr Gln Xaa Pro Leu Ser Xaa Xaa Val Thr Xaa Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Xaa
20
<210> 47
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 47
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys
20
<210> 48
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 48
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys
20
<210> 49
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 49
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys
20
<210> 50
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 50
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Phe
20
<210> 51
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa is F or Y
<220>
<221> misc_feature
<222> (3)..(3)
<223> Xaa is Q or L
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa is R or K
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa is S or P
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa is R or Q
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa is R or L
<400> 51
Trp Xaa Xaa Gln Xaa Pro Gly Gln Xaa Pro Xaa Xaa Leu Ile Tyr
1 5 10 15
<210> 52
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 52
Trp Phe Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile Tyr
1 5 10 15
<210> 53
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 53
Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile Tyr
1 5 10 15
<210> 54
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 54
Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile Tyr
1 5 10 15
<210> 55
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 55
Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile Tyr
1 5 10 15
<210> 56
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa is S or A
<400> 56
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Xaa Gly Thr Asp Phe Thr
1 5 10 15
Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys
20 25 30
<210> 57
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 57
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys
20 25 30
<210> 58
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 58
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr
1 5 10 15
Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys
20 25 30
<210> 59
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 59
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 60
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa is V or I
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa is S or T
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa is L or S
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa is P or S
<220>
<221> misc_feature
<222> (15)..(15)
<223> Xaa is L or P
<220>
<221> misc_feature
<222> (23)..(23)
<223> Xaa is C or F
<220>
<221> misc_feature
<222> (41)..(41)
<223> Xaa is F or Y
<220>
<221> misc_feature
<222> (42)..(42)
<223> Xaa is Q or L
<220>
<221> misc_feature
<222> (44)..(44)
<223> Xaa is R or K
<220>
<221> misc_feature
<222> (48)..(48)
<223> Xaa is S or P
<220>
<221> misc_feature
<222> (50)..(50)
<223> Xaa is R or Q
<220>
<221> misc_feature
<222> (51)..(51)
<223> Xaa is R or L
<220>
<221> misc_feature
<222> (72)..(72)
<223> Xaa is S or A
<400> 60
Asp Xaa Val Met Thr Gln Xaa Pro Leu Ser Xaa Xaa Val Thr Xaa Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Xaa Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Xaa Xaa Gln Xaa Pro Gly Gln Xaa
35 40 45
Pro Xaa Xaa Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Xaa Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 61
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 61
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 62
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 62
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 63
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 63
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 64
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 64
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 65
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 65
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 66
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 66
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Ser Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 67
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 67
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 68
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 68
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 69
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 69
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 70
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 70
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 71
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 71
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 72
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 72
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Phe Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 73
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 73
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 74
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 74
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 75
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 75
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 76
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 76
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 77
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 77
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Phe Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 78
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 78
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 79
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 79
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 80
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 80
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 81
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 81
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 82
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 82
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 83
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 83
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 84
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 84
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 85
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 85
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 86
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 86
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 87
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 87
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 88
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 88
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 89
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 89
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 90
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 90
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 91
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 91
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Ser Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 92
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<220>
<221> misc_feature
<222> (44)..(44)
<223> Xaa is R or G
<220>
<221> misc_feature
<222> (50)..(50)
<223> Xaa is I or absent
<220>
<221> misc_feature
<222> (68)..(68)
<223> Xaa is R or absent
<220>
<221> misc_feature
<222> (71)..(71)
<223> Xaa is I or M
<220>
<221> misc_feature
<222> (73)..(73)
<223> Xaa is R or A
<220>
<221> misc_feature
<222> (77)..(77)
<223> Xaa is A, T or I
<220>
<221> misc_feature
<222> (83)..(83)
<223> Xaa is E or L
<220>
<221> misc_feature
<222> (86)..(86)
<223> Xaa is S or R
<220>
<221> misc_feature
<222> (90)..(90)
<223> Xaa is E or D
<220>
<221> misc_feature
<222> (92)..(92)
<223> Xaa is T or M
<400> 92
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Xaa Leu Glu Trp Met
35 40 45
Gly Xaa Trp Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Ser Gln Lys
50 55 60
Phe Gln Gly Xaa Val Thr Xaa Thr Xaa Asp Thr Ser Xaa Ser Thr Ala
65 70 75 80
Tyr Met Xaa Leu Ser Xaa Leu Arg Ser Xaa Asp Xaa Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 93
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 93
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Phe Gln Gln Arg Pro Gly
195 200 205
Gln Ser Pro Arg Arg Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 94
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 94
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr
165 170 175
Pro Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly
195 200 205
Gln Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 95
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 95
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 96
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 96
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Ser Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 97
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 97
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Phe Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 98
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 98
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Ala Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Leu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Phe Gln Gln Arg Pro Gly
195 200 205
Gln Ser Pro Arg Arg Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 99
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 99
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Ala Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Leu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr
165 170 175
Pro Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly
195 200 205
Gln Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 100
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 100
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Ile Trp Ile Tyr Pro Gly Asp Gly
65 70 75 80
Ser Thr Gln Tyr Asn Glu Lys Phe Lys Gly Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr
165 170 175
Pro Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly
195 200 205
Gln Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 101
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 101
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Ile Trp Ile Tyr Pro Gly Asp Gly
65 70 75 80
Ser Thr Gln Tyr Asn Glu Lys Phe Lys Gly Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 102
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 102
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Ile Trp Ile Tyr Pro Gly Asp Gly
65 70 75 80
Ser Thr Gln Tyr Asn Glu Lys Phe Lys Gly Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Ser Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 103
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 103
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Ile Trp Ile Tyr Pro Gly Asp Gly
65 70 75 80
Ser Thr Gln Tyr Asn Glu Lys Phe Lys Gly Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Phe Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 104
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 104
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Ile Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr
165 170 175
Pro Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly
195 200 205
Gln Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 105
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 105
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Ile Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 106
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 106
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Ile Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Ser Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 107
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 107
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Gly Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Met Thr Arg Asp
85 90 95
Thr Ser Ile Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Phe Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 108
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 108
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Met Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr
165 170 175
Pro Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly
195 200 205
Gln Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 109
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 109
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Met Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 110
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 110
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Met Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Ser Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 111
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 111
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Met Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr
165 170 175
Leu Gly Gln Pro Ala Ser Ile Ser Phe Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Arg Pro Gly
195 200 205
Gln Pro Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 112
<211> 274
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 112
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Val Gln Ser Gly Ala
20 25 30
Glu Val Lys Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Arg Gln Ala Pro
50 55 60
Gly Gln Arg Leu Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Lys Tyr Ser Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Arg Asp
85 90 95
Thr Ser Ala Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr
165 170 175
Pro Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly
195 200 205
Gln Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu
260 265 270
Ile Lys
<210> 113
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 113
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser
20
<210> 114
<211> 21
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 114
Val Asp Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
1 5 10 15
Gly Gly Ser Thr Ser
20
<210> 115
<211> 63
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 115
Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His Phe Val Pro Val
1 5 10 15
Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr
20 25 30
Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala
35 40 45
Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp
50 55 60
<210> 116
<211> 27
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 116
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 117
<211> 41
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 117
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 118
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 118
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 119
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 119
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Phe Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 120
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 120
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 121
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 121
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 122
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 122
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 123
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 123
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 124
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 124
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Phe Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 125
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 125
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 126
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 126
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 127
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 127
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 128
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 128
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 129
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 129
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys
50 55 60
Phe Lys Gly Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 130
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 130
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 131
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 131
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 132
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 132
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 133
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 133
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 134
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 134
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 135
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 135
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 136
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 136
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Val Met Thr
130 135 140
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 137
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 137
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Phe Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Gln Gln Arg Pro Gly Gln Pro Pro Arg Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 138
<211> 510
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 138
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Lys Tyr Ser Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Ile Val Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly Gln Pro Ala Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Asn Arg Ile Arg Gly
245 250 255
Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His
260 265 270
Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro
275 280 285
Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu
290 295 300
Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg
305 310 315 320
Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly Gly Val
325 330 335
Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp
340 345 350
Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val
385 390 395 400
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
405 410 415
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
420 425 430
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
435 440 445
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
450 455 460
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
465 470 475 480
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
485 490 495
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 139
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 139
atggatttcc aagttcaaat cttcagtttc ttgcttatca gtgcttctgt tattatgtca 60
cgagcaagtc aagttcaact cgtacagtct ggagccgagg tgaaaaaacc gggagcgtcc 120
gtgaaagtga gttgcaaggc gagtggatac accttcactt cataccatat acaatgggtt 180
cggcaggcgc ctggtcaacg gctggaatgg atgggctgga tttatcccgg agatggttcc 240
acgcagtaca atgagaaatt caaagggaga gtgacaatca cccgagatac cagtgcctct 300
acggcatata tggaactgag tagtctgcgg tctgaagata cggcggtgta ttattgtgcg 360
agagaaggga cgtactacgc catggattat tggggacaag gaacaacagt cacagtctcc 420
agcgttgatt cctcaggcgg gggcggaagt gggggcggcg gatctggcgg cggcgggtct 480
acgtctgacg tggtcatgac ccaatctcca ttgtctttgc cagttactct gggacagcct 540
gcaagtatca gttgccgatc ctcccaatct atcgtccatt caaacgggaa cacttatttg 600
gaatggtttc aacagagacc tgggcaaagt ccgcgccgac tgatatataa ggtcagtaac 660
cgcttttcag gcgtccccga tcgattcagt ggatctgggt cagggactga cttcactctg 720
aaaatatcaa gagtcgaagc tgaagatgtc ggagtatatt actgtttcca ggggtctcac 780
gtccctcgga cgtttggagg cggaactaag gttgagataa aa 822
<210> 140
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 140
atggattttc aggttcaaat ctttagcttt ctcttgattt ccgcctccgt aataatgagt 60
cgggccagtc aggtacagct cgttcaatct ggggctgaag taaaaaagcc tggagcgtct 120
gtaaaggtat cttgcaaagc gagcggctac acattcacaa gttatcacat ccaatgggtg 180
agacaggccc caggacaacg cttggagtgg atggggtgga tttaccctgg cgacggcagc 240
acacagtaca atgagaaatt taaaggccgg gtgactatca ctcgggacac ctccgccagc 300
acggcttata tggagcttag cagtttgaga tccgaagata cagcggtata ttactgcgcg 360
agagaaggaa cgtactacgc tatggactat tggggtcagg gcacaaccgt tacagtctcc 420
tctgtggaca gctccggagg tgggggttca ggagggggtg gaagcggtgg tggtggcagt 480
acaagcgata tagtaatgac ccaaaccccg ctctctctga gcgtcacgcc aggacaacca 540
gcatcaatct cttgccgcag tagtcaatcc atcgttcact ctaatggaaa cacatacctt 600
gagtggtatc ttcagaaacc aggtcagagc cctcagctcc tcatctataa agtctctaac 660
cggttctcag gtgttccgga ccggttcagt ggttccggct caggaacaga cttcaccttg 720
aagatcagtc gagtagaagc cgaagacgtg ggtgtatatt attgctttca gggttcccac 780
gttccgcgca ccttcggcgg cgggaccaaa gttgagatca aa 822
<210> 141
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 141
atggactttc aagtccaaat cttctcattc ctcctgatct ctgcgtcagt aatcatgtcc 60
agagcgtcac aagtgcaact cgttcaatcc ggagctgagg taaagaagcc cggcgccagc 120
gtgaaagtct cctgcaaagc gagcggctac acgttcacct catatcacat tcaatgggta 180
agacaagcac ctgggcaacg actcgagtgg atggggtgga tctaccctgg ggacgggagc 240
acgcagtata atgagaaatt caaaggcagg gttacaatta cagccgatac cagtgcatct 300
acggcttata tgctcctctc ctcactccgg tctgaggaca cagcggttta ttattgcgca 360
cgggagggaa cgtactacgc gatggactat tgggggcaag gcaccacagt tacagtgagc 420
tcagttgact catcaggagg cggaggatca gggggaggtg gtagtggggg cggtgggagc 480
acatcagatg ttgtcatgac tcagagccca ctttctttgc cggtgacgct ggggcagccc 540
gcttcaatct cttgccgctc atcacagtct atcgttcata gcaatggtaa cacttacttg 600
gaatggttcc aacaaagacc gggtcaaagt ccacggcgct tgatatataa agtatcaaat 660
agattttcag gggtgcctga tcggttcagc ggttctggat ctggcaccga cttcacgctt 720
aaaataagta gggtagaagc cgaagacgtg ggagtgtatt attgtttcca ggggtcacac 780
gtccctcgca cgttcggcgg aggcactaaa gtggagatca aa 822
<210> 142
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 142
atggactttc aagtccaaat cttcagcttt ctccttatat ctgcgtctgt cattatgagt 60
agagcaagtc aagtccagct cgtacaaagt ggagctgagg tgaaaaagcc gggcgcgagt 120
gtgaaagtct catgcaaggc gagtggatac acctttacct cttaccacat tcaatgggtg 180
cggcaggcgc ctgggcagcg cttggaatgg atgggctgga tatatcctgg cgacggaagt 240
acccagtaca acgaaaaatt caaaggtagg gttaccatca ctgctgatac ctccgcgtcc 300
actgcttata tgcttcttag ctccttgcga agcgaggata cagccgtgta ttattgtgcc 360
agagagggga cttattatgc catggactat tggggtcagg gtacaacggt cactgtctca 420
tctgttgaca gtagcggggg aggggggtct ggaggagggg gttccggggg gggaggttcc 480
acgagcgata tagttatgac gcaaacgccc ttgagcctca gtgttacacc cggtcaacct 540
gcctctatta gctgtcgctc ctcccaatca attgtgcata gcaatggaaa tacctacctt 600
gaatggtatc tccaaaagcc cgggcagagt cctcaacttc tcatctacaa agtatccaat 660
cgattcagtg gcgttcctga caggttcagc ggaagtgggt cagggaccga ttttaccctc 720
aaaattagtc gcgtcgaagc tgaggatgtt ggggtgtatt actgcttcca agggtcacac 780
gtaccacgca cattcggggg aggcacgaag gttgaaatta ag 822
<210> 143
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 143
atggatttcc aagtccaaat attcagtttc cttttgataa gtgcttcagt tatcatgtcc 60
cgagcaagcc aggtacagct tgtgcaaagc ggggcggaag ttaagaaacc gggagcctca 120
gttaaagtat cttgcaaagc cagcggttat acattcactt cataccacat acagtgggtg 180
cgccaagcac cggggcagag actggaatgg atgggatgga tttatcccgg tgatggtagt 240
acgcaataca atgaaaaatt caaaggaagg gtgactatca cgcgagacac aagcgcgtcc 300
acggcctata tggaactgag cagtctgaga tccgaggaca ccgctgtgta ttattgcgcc 360
cgagagggga cctactacgc catggattat tggggtcaag ggaccactgt gacagtctct 420
tctgtcgatt ccagcggcgg aggagggagt ggagggggtg ggtccggggg gggagggtct 480
acgagcgaca ttgtcatgac acagacgccg cttagctccc cagttacact cggacagccc 540
gcaagtatta gttgcagaag tagccagtct atcgtacatt caaatggaaa cacctatttg 600
gaatggtatc aacaacggcc tggacagccg cccaggctgc tcatatacaa agtctccaac 660
cgcttcagcg gagtacccga ccgcttttcc ggctccggag caggaactga ctttaccttg 720
aaaattagta gggtcgaagc agaggatgtc ggggtatatt attgtttcca aggtagtcat 780
gtcccacgga cgtttggtgg tgggacgaag gttgagatca aa 822
<210> 144
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 144
atggacttcc aggtgcagat attttccttt cttctcatat ctgcatctgt aataatgtca 60
agggccagcc aggtccagct cgttcaaagc ggagccgagg taaaaaaacc aggcgcttcc 120
gtcaaggtat catgcaaagc gtccggctat accttcacaa gttaccatat ccaatgggtt 180
cgacaggcac ccggacaaag acttgaatgg atgggttgga tataccccgg agacggctcc 240
actcagtata acgaaaagtt taaggggaga gtcacgatca ctagggacac atcagcttct 300
acggcgtata tggaactcag ttctttgcga tccgaggata ctgccgtata ttactgcgcc 360
agagaaggca cgtactacgc aatggattac tgggggcaag ggacaactgt taccgtctca 420
agcgtcgatt catcaggagg cggagggtcc ggaggtgggg gatctggcgg tgggggttct 480
acgtccgatg ttgtgatgac acagtcccca ctctctcttc cagtgacgct gggacagccc 540
gcgagcatct cctgtcgcag ctctcagtcc atagtacaca gtaatggtaa cacctatctt 600
gagtggtatc agcaacgacc cggtcagtct cccaggttgc ttatttataa ggttagtaac 660
cgcttttcag gtgttccaga cagatttagc gggagtggtt ccggtacgga tttcacattg 720
aaaatcagcc gcgtcgaagc cgaggacgtg ggtgtttact actgcttcca gggatctcac 780
gtaccgagga ccttcggcgg aggaacgaaa gtagaaatta ag 822
<210> 145
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 145
atggattttc aagtacaaat cttttccttc ctgcttattt ccgcaagcgt tattatgagt 60
agagccagcc aagtacaact ggtacagtcc ggcgcggagg tgaagaagcc cggagcaagt 120
gtgaaggtat cttgcaaggc gtcaggctat acctttactt cataccatat acagtgggtg 180
cgacaggctc ccggccagcg actcgaatgg atgggctgga tttatcccgg agatggatct 240
acgcagtata atgagaaatt caagggtcgg gtcacgatta cacgagacac gagtgcttcc 300
acagcttata tggaactttc tagcctgagg tctgaggata ctgccgttta ctattgtgca 360
cgagaaggga catactatgc gatggattac tggggacagg gcaccactgt cacagtttcc 420
agcgtggact caagtggagg cggtggatct ggtggtggcg ggtccggggg gggaggcagc 480
accagtgaca ttgtaatgac tcaaacacct ctcagtagcc cagtcactct cggtcagccg 540
gcgagtatct cttttaggtc ctcacaatct atagtgcact ctaacggcaa tacttatctt 600
gaatggtatc aacaaagacc ggggcagcca cctcgccttc tcatctacaa agtaagcaat 660
cgcttctccg gtgtccccga tcgcttctcc ggttcaggag caggaactga cttcacattg 720
aagatttcca gagtggaggc cgaagacgta ggggtatatt attgctttca agggtcccat 780
gtgcccagaa cctttggggg aggaacgaaa gttgagatta aa 822
<210> 146
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 146
atggatttcc aagtgcagat tttctctttt ctcctcataa gcgcctccgt aattatgtct 60
agagctagtc aagtccaatt ggtgcaatcc ggtgccgagg ttaaaaagcc cggcgcaagt 120
gtaaaagtct cctgtaaggc cagtggctac actttcacca gctaccatat acagtgggtg 180
cggcaggcgc ctggtcaggg tctggagtgg atgggtattt ggatttatcc cggagatgga 240
agtactcaat acaatgagaa attcaagggt gtcactatga caagggatac gagcacttct 300
accgtatata tggagttgtc atctttgcga tcagaggata ccgctgtata ttattgcgca 360
cgggaaggta catattatgc catggactac tggggccaag gaaccaccgt gacggtaagc 420
tctgtcgatt ctagcggtgg cgggggctct ggcggtgggg gtagcggggg tggcggatct 480
acatcagata ttgtaatgac acagacccct ctttcacttt ccgtaacgcc aggacagccg 540
gcatcaataa gttgccgatc aagccagtct atcgtacact ccaatggtaa cacatacttg 600
gaatggtatc ttcaaaagcc cggccagagc ccgcagcttt tgatatataa agtgtccaac 660
agattcagtg gggtgccgga ccgctttagt ggatctggtt caggaacgga cttcacattg 720
aaaattagta gagttgaagc ggaagacgtg ggagtctact actgtttcca gggttcacat 780
gtgcctcgga cctttggggg aggcaccaag gttgagataa aa 822
<210> 147
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 147
atggacttcc aagtccaaat cttttctttt ttgttgataa gcgcatcagt tattatgtct 60
cgcgccagtc aagtacaact ggtgcagtcc ggagctgaag tgaaaaaacc aggagcaagc 120
gtgaaagtaa gttgtaaggc aagtggttac actttcacaa gctaccatat tcaatgggtc 180
cgacaggctc ctggacaggg cttggagtgg atgggcatat ggatttaccc tggtgacggg 240
tccacccagt ataatgaaaa gttcaaggga gtcacgatga ccagggacac ctctacatct 300
accgtgtata tggagctctc tagtttgcga tccgaagaca ctgccgttta ttactgtgca 360
agagaaggaa cttattacgc gatggactac tggggtcagg ggacaacagt caccgttagc 420
tccgtcgatt ccagcggggg aggtggctca ggcgggggtg gttctggggg gggcgggagc 480
acttcagata ttgtaatgac ccaaacccca ctgagtagtc cagtcacgct tggtcaaccg 540
gcaagcattt cttgcaggag ctctcagagt attgtccact ctaacgggaa tacatatttg 600
gagtggtatc agcaaagacc gggccaacca ccacgcctct tgatttataa ggtgagcaat 660
aggttttcag gcgtgccaga taggttctca ggctccggag cgggaaccga cttcaccctc 720
aagataagtc gggtggaagc cgaagacgta ggagtttact actgctttca aggatctcat 780
gttccacgaa cgtttggagg aggaaccaag gtggaaataa aa 822
<210> 148
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 148
atggactttc aggtccaaat tttttccttc ttgctcatat ccgcgagtgt catcatgtca 60
agagcaagtc aagttcaact cgttcaatca ggagctgagg tgaaaaaacc aggggcgtct 120
gtcaaagtaa gctgcaaagc atcagggtat acgttcacga gttatcatat ccagtgggtt 180
aggcaggcgc cagggcaggg attggaatgg atgggtatct ggatttaccc gggtgacggc 240
agcactcaat acaatgagaa attcaaaggc gtaacaatga caagggacac gagcacaagc 300
acagtgtaca tggagcttag ctctttgagg tcagaggata ccgctgttta ctattgtgct 360
cgggagggta cttactatgc aatggactac tgggggcaag gcacgaccgt tacagtgagt 420
agcgtagatt cctccggggg tggcggttca ggcggcggag gctcaggcgg aggagggtca 480
acatccgatg tcgtaatgac tcagtcccct ctgtcattgc cggtgacttt gggacagcca 540
gcgtctatat cttgtaggtc ctctcaatca atagtgcatt ccaacggtaa cacctatctg 600
gaatggtatc agcaaaggcc aggacaaagt ccacgcctgc ttatatataa ggtgtctaat 660
cgattcagtg gggttcccga taggttttcc ggctctggta gcgggactga tttcacgttg 720
aaaatatcac gcgtggaagc ggaagatgtt ggggtctatt actgctttca gggtagtcat 780
gtccctcgaa cttttggcgg tggtacaaag gtagaaatca aa 822
<210> 149
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 149
atggattttc aggtacagat attctcattt ctccttatct cagctagtgt cataatgtcc 60
agggcgagtc aagtacaact tgtccagtca ggcgcagagg tcaagaagcc gggcgcaagc 120
gttaaggttt cctgcaaagc atccggctat acattcacgt cctatcacat ccaatgggtc 180
aggcaagcac ccggtcaagg acttgagtgg atgggcatct ggatttaccc tggagatggc 240
agtactcagt acaacgaaaa attcaaaggt gtaaccatga cccgcgacac atctacttcc 300
acagtttata tggaactcag cagtttgcgg agcgaagata ccgctgttta ctactgtgcc 360
cgagagggaa cttactacgc catggactat tggggtcaag gaacgacagt aacagttagt 420
tctgtagatt ccagtggcgg cggtgggagc gggggtgggg gatctggcgg aggcggaagt 480
acaagtgaca tcgttatgac tcagacaccc cttagtagtc ccgttacgtt gggccaaccc 540
gcgagcattt cctttcgatc ctctcagtct atagttcact caaatgggaa tacttatttg 600
gagtggtatc aacagcgccc cggacaacca ccaaggctcc tgatatacaa ggtgtccaat 660
cgattctctg gggtgcctga tagatttagc ggaagtggag ccggtacaga ttttaccctg 720
aaaatatcac gggtagaagc cgaagatgtc ggcgtctact actgtttcca gggttcccat 780
gtaccgcgaa cgttcggggg cggaacaaaa gttgagatca ag 822
<210> 150
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 150
atggatttcc aggttcagat atttagtttc ctcttgattt ctgccagtgt catcatgagc 60
agggcttccc aagttcagtt ggtgcaaagt ggcgctgaag tcaaaaaacc tggggcttcc 120
gttaaagtat cttgcaaggc gtccggctac actttcacat cctaccacat tcaatgggtc 180
cggcaagcgc ccggtcaggg gctcgaatgg atggggtgga tatacccagg agatggatct 240
actcagtaca acgagaaatt taaaggacgg gtgacgatga cgcgcgacac ttcaataagc 300
actgcataca tggaactgtc ccggcttagg tcagatgaca ccgcggtcta ctattgtgcg 360
agagagggta cttactatgc tatggactac tgggggcaag gcacgacggt tacagtttcc 420
tcagtcgata gttcaggcgg aggcggctcc gggggcggtg gtagtggagg gggtggatct 480
acttccgaca ttgtcatgac ccagaccccg ttgagccttt cagtgacgcc cggtcaaccc 540
gccagcataa gttgtcgatc aagccagtct attgtacact ccaatggaaa cacatatttg 600
gagtggtatc tccaaaaacc cggccaaagc cctcaactgc tcatctacaa ggtctcaaac 660
aggtttagcg gggtcccgga tcgcttctca gggtcaggat ctggtacgga ctttacactg 720
aaaatttccc gagtcgaagc ggaagacgtg ggtgtatatt actgcttcca ggggagtcat 780
gttccaagaa cctttggggg aggtacaaag gtcgaaataa aa 822
<210> 151
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 151
atggattttc aggtccaaat tttttccttc ttgcttatca gcgcaagtgt aatcatgtcc 60
cgcgcgtccc aagtacaact tgtgcaatct ggcgcggagg tgaaaaaacc tggagcttcc 120
gtcaaggttt cttgtaaggc ctctggctac accttcacgt cctaccacat tcagtgggtt 180
cgacaggcgc cgggccaagg actggagtgg atgggatgga tatatccagg agatggttct 240
actcagtata atgagaaatt caagggtcgc gtaacaatga cgagggatac atcaatctcc 300
accgcgtaca tggaactttc aagactccgg tcagatgaca cggcggttta ctactgtgct 360
cgggagggca cttactatgc tatggactac tgggggcaag ggacaacggt aacggtatct 420
agtgtggatt ctagtggcgg cggcggttca ggaggaggtg gttcaggcgg ggggggtagt 480
acaagtgata ttgtgatgac ccaaacaccc ctttctagcc ctgttactct gggtcaaccc 540
gcgtccataa gttgtcgaag tagtcaatcc atcgtgcata gcaacggcaa cacttacctt 600
gaatggtatc aacaacgacc cggacagccc ccgcgactgc ttatctataa agtatcaaac 660
aggttcagtg gcgtgccaga tcgattctcc ggctctgggg caggcacaga tttcacgttg 720
aaaatttctc gggtcgaggc cgaggacgtg ggcgtttatt actgtttcca ggggagtcac 780
gtccccagga cgttcggagg aggaactaaa gtcgaaataa ag 822
<210> 152
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 152
atggacttcc aggtccaaat attcagcttc ctcctcattt ccgccagtgt aataatgtcc 60
agagcctcac aagtacagtt ggttcagagc ggggctgagg ttaagaaacc aggcgcgagc 120
gtcaaggtat cctgcaaggc gagtggttat actttcacta gttatcacat tcagtgggtc 180
cgacaggccc ccggtcaagg cctggagtgg atggggtgga tatatccggg agatggttct 240
acccaatata atgagaagtt taaggggaga gtcacaatga caagggacac cagtattagc 300
accgcgtata tggagctttc ccgcctgcga tcagatgaca cggccgtgta ctactgtgct 360
agggagggaa cctattatgc gatggattac tggggacagg gtactacagt cacggtctct 420
agcgtggaca gttccggggg cggtggaagc ggtggtggcg gttcaggtgg aggaggctct 480
acgagtgatg ttgtgatgac tcagtccccg ctttcacttc ccgtcaccct tgggcaaccc 540
gcaagcatct catgccgatc ctcccagtct atagtacata gtaatggcaa cacatatctt 600
gaatggtatc agcagaggcc gggtcagtct ccccgactcc ttatatataa agtgagcaac 660
agattctccg gagtaccgga tagattttcc ggctctggga gcggcaccga ctttacactg 720
aaaatttcac gggttgaagc tgaagatgtt ggggtatact attgtttcca gggttctcac 780
gtcccgagga cattcggggg aggaacgaaa gtcgaaataa ag 822
<210> 153
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 153
atggattttc aggtacaaat cttcagcttc ctgctcatct ccgcgagcgt aatcatgtct 60
agggcgtccc aggtgcagtt ggtgcaatca ggtgcagagg tgaagaagcc tggtgcatcc 120
gttaaagtaa gttgtaaggc aagcggatat acttttacat cctatcatat tcaatgggtc 180
agacaagcac ctggacaggg tcttgagtgg atgggctgga tctatccagg cgatggctca 240
actcaatata acgagaagtt caaggggagg gttactatga ccagggatac gtctatttcc 300
actgcgtaca tggaactctc caggttgaga agtgatgata ccgcggttta ctactgcgct 360
agagaaggaa cgtactacgc tatggattac tgggggcagg gtacaactgt caccgtctca 420
agtgtggatt cttctggggg tgggggatca ggagggggag gctccggtgg gggcgggtca 480
accagcgaca ttgtcatgac tcaaaccccc ctgagcagcc ctgtcaccct gggtcagcct 540
gcctcaatat cctttagaag ctcccaaagc atcgtccatt caaatggtaa tacctatctg 600
gagtggtatc agcaaaggcc tggtcaaccc ccgcgccttc tcatttacaa ggtgtcaaac 660
aggttctccg gcgtaccgga taggttttcc ggaagcggtg ctggaaccga ctttactctc 720
aaaatctcta gggtggaagc tgaggacgtc ggtgtatact attgttttca aggctcccat 780
gttcccagga catttggtgg gggaacgaag gtagaaatca ag 822
<210> 154
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 154
atggactttc aggttcagat tttctctttc ttgttgatct ccgctagtgt cataatgtca 60
cgggcaagtc aggtacaact cgttcagagt ggtgccgaag tgaagaaacc gggtgcctcc 120
gtaaaggtgt catgtaaagc tagtggctat acattcacaa gttatcatat ccaatgggta 180
cgacaagcac cgggacagcg actggaatgg atgggatgga tctatcctgg ggacggatct 240
acacagtaca atgagaaatt taagggacgg gtcacgataa ccagggacac atctgcttcc 300
acggcttaca tggagctttc ctccctgcgg agcgaggaca tggctgttta ctattgcgct 360
cgcgaaggga catactacgc aatggattat tggggccaag gcactaccgt gacggtctct 420
tctgtcgata gttccggagg aggtggttca gggggaggcg gttcaggtgg gggtggatct 480
acctcagata ttgtcatgac acagacacct ttgtccttga gtgtgacacc gggtcaaccg 540
gcgagtataa gctgtcgcag ctcacaatct attgtgcata gcaacgggaa tacatatctc 600
gaatggtatc tccaaaagcc gggccaatcc ccccaacttc tcatttacaa agtttctaat 660
cgattttcag gtgtaccaga tcggttttcc gggtctggct caggtactga cttcaccttg 720
aaaatatcaa gggttgaagc tgaggatgta ggtgtgtact attgcttcca ggggtctcac 780
gttcctcgga cttttggggg gggcacaaaa gtagagatta aa 822
<210> 155
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 155
atggatttcc aggtgcaaat cttctcattt cttttgataa gtgcgtcagt gataatgtct 60
cgggccagtc aagtacagct tgtccaaagt ggcgctgaag tcaagaagcc gggagcctca 120
gttaaggtta gctgcaaggc ctcagggtat acttttacct cctatcatat acagtgggta 180
cgacaagcac cgggacagcg actggagtgg atgggttgga tatatccggg agatggttca 240
acccagtata atgagaagtt caaggggcga gttacgataa cccgcgatac gagtgcatca 300
acagcgtaca tggagttgag ttccctccgc agcgaggaca tggcggtata ctattgtgcc 360
agggagggga cttattatgc catggactac tgggggcagg gcacaaccgt aacagtctct 420
tctgtagaca gttcaggagg gggcggaagt ggaggtggcg gatctggtgg aggtggatct 480
acttccgaca tcgttatgac ccaaacacca ctttcatctc ccgttactct cgggcaacct 540
gctagtattt cctgtagatc ctcacaatct atagtacata gcaatggcaa tacctacctg 600
gagtggtatc aacaacgccc aggccaacca cctcgcctgc ttatctataa agtaagcaat 660
agattcagtg gtgtaccgga taggttctct ggttccggag caggaactga ctttacactc 720
aaaatcagta gggtggaggc ggaagacgtg ggagtatatt attgctttca aggttcacat 780
gtacctcgaa catttggcgg aggaactaag gttgagatta aa 822
<210> 156
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 156
atggatttcc aagtccagat attcagtttt cttttgataa gcgcttctgt aatcatgtct 60
cgggcgtccc aagtacaact ggtgcaatca ggggcagaag tgaaaaaacc aggtgcatcc 120
gttaaggtga gttgcaaggc ttccggctat acctttacat catatcatat tcaatgggtc 180
aggcaagcac ctggtcagcg attggaatgg atgggttgga tatatcctgg tgatgggtct 240
acacaatata acgaaaaatt caaggggcga gtgaccatca caagagatac atcagcgtca 300
acagcgtata tggaactgtc atcccttaga tcagaggaca tggcggtcta ttactgtgcc 360
agagaaggca cttattatgc aatggattat tggggacaag gaaccactgt cactgtttcc 420
agcgtagact cctccggtgg tggtggaagt ggcggcggtg ggtcaggagg gggtgggtca 480
acttctgatg tagtgatgac acagagccct ctgagcttgc ctgtgacctt gggtcagccg 540
gcctcaataa gttgtcgatc tagtcagtca atcgtccata gtaatgggaa cacatacctt 600
gaatggtatc agcaaagacc tggacaatct ccacgactcc ttatatacaa agttagcaac 660
cgatttagcg gagtgccaga ccgcttttct ggttccgggt ctggcacaga ttttaccctt 720
aagatctccc gcgtggaggc ggaagacgtt ggtgtttact attgcttcca ggggtcacac 780
gttccacgca cctttggagg aggtacgaag gtcgagatta ag 822
<210> 157
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 157
atggattttc aagtacagat cttctctttc ttgcttattt cagcgagcgt aatcatgagt 60
agggcatctc aagttcaact cgttcagtca ggtgctgagg taaaaaaacc aggggcttcc 120
gttaaagtta gctgtaaggc atctgggtac acatttacta gctaccatat ccagtgggtg 180
cgacaagccc cggggcagcg cttggaatgg atgggctgga tttacccagg tgacggctcc 240
acgcaatata atgagaaatt taagggtaga gttactatta ccagggacac aagtgcttca 300
actgcctata tggaactgag cagccttcgg agtgaagata tggccgtata ttactgcgca 360
agggagggga cttactatgc aatggactac tggggtcagg gaacgactgt gaccgtgtcc 420
tcagttgact ccagcggtgg tggcggctct ggaggtgggg gttccggcgg aggcggaagc 480
acatctgata tagtgatgac gcaaacgcct ctttcttccc cggtaactct gggacagcca 540
gcgtcaattt catttaggtc ctcccagtca atcgtacata gtaatggaaa tacttacctg 600
gaatggtatc aacaacgacc agggcaaccg ccccgattgt tgatctataa agtgagcaat 660
cgcttttctg gcgtgcccga tcggttctca gggtctggag ctgggactga cttcacattg 720
aaaatttcca gggttgaggc cgaggatgtg ggggtttatt actgcttcca aggctcccac 780
gtcccccgca ccttcggagg gggaaccaaa gtcgaaataa ag 822
<210> 158
<211> 822
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 158
atggattttc aagttcagat attctcattt ttgcttatat cagcctccgt aattatgtca 60
cgggcaagtc aagttcagtt ggtgcagtcc ggagcagaag ttaagaagcc cggtgcttct 120
gtgaaagtct cctgcaaagc gtctgggtac accttcacga gctaccatat acagtgggtc 180
cggcaagcgc ctgggcagag gctggagtgg atgggctgga tttacccagg agatgggagt 240
acaaagtata gtcagaagtt tcaagggcga gtgacgataa ccagagatac gagtgcaagt 300
actgcataca tggaactgag ctccttgagg tccgaggata cagcggtgta ctattgcgct 360
cgggaaggga catattatgc tatggactat tggggacaag ggacaacggt aacggtgagt 420
tccgtcgatt cctcaggtgg cggaggcagt gggggcgggg gttccggcgg tggcgggtcc 480
acgagtgata tagttatgac acagaccccc ctcagccttt ctgtgacccc aggacaaccc 540
gctagtatct cttgccgcag ttctcagtcc atagtacaca gtaacggaaa tacctatctt 600
gagtggtatc ttcaaaagcc cggccagagc cctcaactct tgatatataa agtgtcaaat 660
cgattttcag gtgtgcctga tcgattctca gggtctggtt cagggacaga tttcacgctt 720
aagataagca gagtagaggc tgaagacgtg ggagtctact attgttttca ggggtcacac 780
gttccccgca cttttggtgg gggaaccaag gtggaaatca aa 822
<210> 159
<211> 192
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 159
tcttcagcgc tgagcaactc catcatgtac ttcagccact tcgtgccggt cttcctgcca 60
gcgaagccca ccacgacgcc agcgccgcga ccaccaacac cggcgcccac catcgcgtcg 120
cagcccctgt ccctgcgccc agaggcgtgc cggccagcgg cggggggcgc agtgcacacg 180
agggggctgg ac 192
<210> 160
<211> 81
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 160
ttttgggtgc tggtggtggt tggtggagtc ctggcttgct atagcttgct agtaacagtg 60
gcctttatta ttttctgggt g 81
<210> 161
<211> 123
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 161
aggagtaaga ggagcaggct cctgcacagt gactacatga acatgactcc ccgccgcccc 60
gggcccaccc gcaagcatta ccagccctat gccccaccac gcgacttcgc agcctatcgc 120
tcc 123
<210> 162
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 162
agagtgaagt tcagcaggag cgcagacgcc cccgcgtacc agcagggcca gaaccagctc 60
tataacgagc tcaatctagg acgaagagag gagtacgatg ttttggacaa gagacgtggc 120
cgggaccctg agatgggggg aaagccgaga aggaagaacc ctcaggaagg cctgtacaat 180
gaactgcaga aagataagat ggcggaggcc tacagtgaga ttgggatgaa aggcgagcgc 240
cggaggggca aggggcacga tggcctttac cagggtctca gtacagccac caaggacacc 300
tacgacgccc ttcacatgca ggccctgccc cctcgc 336
<210> 163
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 163
atggatttcc aagttcaaat cttcagtttc ttgcttatca gtgcttctgt tattatgtca 60
cgagcaagtc aagttcaact cgtacagtct ggagccgagg tgaaaaaacc gggagcgtcc 120
gtgaaagtga gttgcaaggc gagtggatac accttcactt cataccatat acaatgggtt 180
cggcaggcgc ctggtcaacg gctggaatgg atgggctgga tttatcccgg agatggttcc 240
acgcagtaca atgagaaatt caaagggaga gtgacaatca cccgagatac cagtgcctct 300
acggcatata tggaactgag tagtctgcgg tctgaagata cggcggtgta ttattgtgcg 360
agagaaggga cgtactacgc catggattat tggggacaag gaacaacagt cacagtctcc 420
agcgttgatt cctcaggcgg gggcggaagt gggggcggcg gatctggcgg cggcgggtct 480
acgtctgacg tggtcatgac ccaatctcca ttgtctttgc cagttactct gggacagcct 540
gcaagtatca gttgccgatc ctcccaatct atcgtccatt caaacgggaa cacttatttg 600
gaatggtttc aacagagacc tgggcaaagt ccgcgccgac tgatatataa ggtcagtaac 660
cgcttttcag gcgtccccga tcgattcagt ggatctgggt cagggactga cttcactctg 720
aaaatatcaa gagtcgaagc tgaagatgtc ggagtatatt actgtttcca ggggtctcac 780
gtccctcgga cgtttggagg cggaactaag gttgagataa aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 164
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 164
atggattttc aggttcaaat ctttagcttt ctcttgattt ccgcctccgt aataatgagt 60
cgggccagtc aggtacagct cgttcaatct ggggctgaag taaaaaagcc tggagcgtct 120
gtaaaggtat cttgcaaagc gagcggctac acattcacaa gttatcacat ccaatgggtg 180
agacaggccc caggacaacg cttggagtgg atggggtgga tttaccctgg cgacggcagc 240
acacagtaca atgagaaatt taaaggccgg gtgactatca ctcgggacac ctccgccagc 300
acggcttata tggagcttag cagtttgaga tccgaagata cagcggtata ttactgcgcg 360
agagaaggaa cgtactacgc tatggactat tggggtcagg gcacaaccgt tacagtctcc 420
tctgtggaca gctccggagg tgggggttca ggagggggtg gaagcggtgg tggtggcagt 480
acaagcgata tagtaatgac ccaaaccccg ctctctctga gcgtcacgcc aggacaacca 540
gcatcaatct cttgccgcag tagtcaatcc atcgttcact ctaatggaaa cacatacctt 600
gagtggtatc ttcagaaacc aggtcagagc cctcagctcc tcatctataa agtctctaac 660
cggttctcag gtgttccgga ccggttcagt ggttccggct caggaacaga cttcaccttg 720
aagatcagtc gagtagaagc cgaagacgtg ggtgtatatt attgctttca gggttcccac 780
gttccgcgca ccttcggcgg cgggaccaaa gttgagatca aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 165
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 165
atggactttc aagtccaaat cttctcattc ctcctgatct ctgcgtcagt aatcatgtcc 60
agagcgtcac aagtgcaact cgttcaatcc ggagctgagg taaagaagcc cggcgccagc 120
gtgaaagtct cctgcaaagc gagcggctac acgttcacct catatcacat tcaatgggta 180
agacaagcac ctgggcaacg actcgagtgg atggggtgga tctaccctgg ggacgggagc 240
acgcagtata atgagaaatt caaaggcagg gttacaatta cagccgatac cagtgcatct 300
acggcttata tgctcctctc ctcactccgg tctgaggaca cagcggttta ttattgcgca 360
cgggagggaa cgtactacgc gatggactat tgggggcaag gcaccacagt tacagtgagc 420
tcagttgact catcaggagg cggaggatca gggggaggtg gtagtggggg cggtgggagc 480
acatcagatg ttgtcatgac tcagagccca ctttctttgc cggtgacgct ggggcagccc 540
gcttcaatct cttgccgctc atcacagtct atcgttcata gcaatggtaa cacttacttg 600
gaatggttcc aacaaagacc gggtcaaagt ccacggcgct tgatatataa agtatcaaat 660
agattttcag gggtgcctga tcggttcagc ggttctggat ctggcaccga cttcacgctt 720
aaaataagta gggtagaagc cgaagacgtg ggagtgtatt attgtttcca ggggtcacac 780
gtccctcgca cgttcggcgg aggcactaaa gtggagatca aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 166
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 166
atggactttc aagtccaaat cttcagcttt ctccttatat ctgcgtctgt cattatgagt 60
agagcaagtc aagtccagct cgtacaaagt ggagctgagg tgaaaaagcc gggcgcgagt 120
gtgaaagtct catgcaaggc gagtggatac acctttacct cttaccacat tcaatgggtg 180
cggcaggcgc ctgggcagcg cttggaatgg atgggctgga tatatcctgg cgacggaagt 240
acccagtaca acgaaaaatt caaaggtagg gttaccatca ctgctgatac ctccgcgtcc 300
actgcttata tgcttcttag ctccttgcga agcgaggata cagccgtgta ttattgtgcc 360
agagagggga cttattatgc catggactat tggggtcagg gtacaacggt cactgtctca 420
tctgttgaca gtagcggggg aggggggtct ggaggagggg gttccggggg gggaggttcc 480
acgagcgata tagttatgac gcaaacgccc ttgagcctca gtgttacacc cggtcaacct 540
gcctctatta gctgtcgctc ctcccaatca attgtgcata gcaatggaaa tacctacctt 600
gaatggtatc tccaaaagcc cgggcagagt cctcaacttc tcatctacaa agtatccaat 660
cgattcagtg gcgttcctga caggttcagc ggaagtgggt cagggaccga ttttaccctc 720
aaaattagtc gcgtcgaagc tgaggatgtt ggggtgtatt actgcttcca agggtcacac 780
gtaccacgca cattcggggg aggcacgaag gttgaaatta agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 167
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 167
atggatttcc aagtccaaat attcagtttc cttttgataa gtgcttcagt tatcatgtcc 60
cgagcaagcc aggtacagct tgtgcaaagc ggggcggaag ttaagaaacc gggagcctca 120
gttaaagtat cttgcaaagc cagcggttat acattcactt cataccacat acagtgggtg 180
cgccaagcac cggggcagag actggaatgg atgggatgga tttatcccgg tgatggtagt 240
acgcaataca atgaaaaatt caaaggaagg gtgactatca cgcgagacac aagcgcgtcc 300
acggcctata tggaactgag cagtctgaga tccgaggaca ccgctgtgta ttattgcgcc 360
cgagagggga cctactacgc catggattat tggggtcaag ggaccactgt gacagtctct 420
tctgtcgatt ccagcggcgg aggagggagt ggagggggtg ggtccggggg gggagggtct 480
acgagcgaca ttgtcatgac acagacgccg cttagctccc cagttacact cggacagccc 540
gcaagtatta gttgcagaag tagccagtct atcgtacatt caaatggaaa cacctatttg 600
gaatggtatc aacaacggcc tggacagccg cccaggctgc tcatatacaa agtctccaac 660
cgcttcagcg gagtacccga ccgcttttcc ggctccggag caggaactga ctttaccttg 720
aaaattagta gggtcgaagc agaggatgtc ggggtatatt attgtttcca aggtagtcat 780
gtcccacgga cgtttggtgg tgggacgaag gttgagatca aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 168
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 168
atggacttcc aggtgcagat attttccttt cttctcatat ctgcatctgt aataatgtca 60
agggccagcc aggtccagct cgttcaaagc ggagccgagg taaaaaaacc aggcgcttcc 120
gtcaaggtat catgcaaagc gtccggctat accttcacaa gttaccatat ccaatgggtt 180
cgacaggcac ccggacaaag acttgaatgg atgggttgga tataccccgg agacggctcc 240
actcagtata acgaaaagtt taaggggaga gtcacgatca ctagggacac atcagcttct 300
acggcgtata tggaactcag ttctttgcga tccgaggata ctgccgtata ttactgcgcc 360
agagaaggca cgtactacgc aatggattac tgggggcaag ggacaactgt taccgtctca 420
agcgtcgatt catcaggagg cggagggtcc ggaggtgggg gatctggcgg tgggggttct 480
acgtccgatg ttgtgatgac acagtcccca ctctctcttc cagtgacgct gggacagccc 540
gcgagcatct cctgtcgcag ctctcagtcc atagtacaca gtaatggtaa cacctatctt 600
gagtggtatc agcaacgacc cggtcagtct cccaggttgc ttatttataa ggttagtaac 660
cgcttttcag gtgttccaga cagatttagc gggagtggtt ccggtacgga tttcacattg 720
aaaatcagcc gcgtcgaagc cgaggacgtg ggtgtttact actgcttcca gggatctcac 780
gtaccgagga ccttcggcgg aggaacgaaa gtagaaatta agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 169
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 169
atggattttc aagtacaaat cttttccttc ctgcttattt ccgcaagcgt tattatgagt 60
agagccagcc aagtacaact ggtacagtcc ggcgcggagg tgaagaagcc cggagcaagt 120
gtgaaggtat cttgcaaggc gtcaggctat acctttactt cataccatat acagtgggtg 180
cgacaggctc ccggccagcg actcgaatgg atgggctgga tttatcccgg agatggatct 240
acgcagtata atgagaaatt caagggtcgg gtcacgatta cacgagacac gagtgcttcc 300
acagcttata tggaactttc tagcctgagg tctgaggata ctgccgttta ctattgtgca 360
cgagaaggga catactatgc gatggattac tggggacagg gcaccactgt cacagtttcc 420
agcgtggact caagtggagg cggtggatct ggtggtggcg ggtccggggg gggaggcagc 480
accagtgaca ttgtaatgac tcaaacacct ctcagtagcc cagtcactct cggtcagccg 540
gcgagtatct cttttaggtc ctcacaatct atagtgcact ctaacggcaa tacttatctt 600
gaatggtatc aacaaagacc ggggcagcca cctcgccttc tcatctacaa agtaagcaat 660
cgcttctccg gtgtccccga tcgcttctcc ggttcaggag caggaactga cttcacattg 720
aagatttcca gagtggaggc cgaagacgta ggggtatatt attgctttca agggtcccat 780
gtgcccagaa cctttggggg aggaacgaaa gttgagatta aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 170
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 170
atggatttcc aagtgcagat tttctctttt ctcctcataa gcgcctccgt aattatgtct 60
agagctagtc aagtccaatt ggtgcaatcc ggtgccgagg ttaaaaagcc cggcgcaagt 120
gtaaaagtct cctgtaaggc cagtggctac actttcacca gctaccatat acagtgggtg 180
cggcaggcgc ctggtcaggg tctggagtgg atgggtattt ggatttatcc cggagatgga 240
agtactcaat acaatgagaa attcaagggt gtcactatga caagggatac gagcacttct 300
accgtatata tggagttgtc atctttgcga tcagaggata ccgctgtata ttattgcgca 360
cgggaaggta catattatgc catggactac tggggccaag gaaccaccgt gacggtaagc 420
tctgtcgatt ctagcggtgg cgggggctct ggcggtgggg gtagcggggg tggcggatct 480
acatcagata ttgtaatgac acagacccct ctttcacttt ccgtaacgcc aggacagccg 540
gcatcaataa gttgccgatc aagccagtct atcgtacact ccaatggtaa cacatacttg 600
gaatggtatc ttcaaaagcc cggccagagc ccgcagcttt tgatatataa agtgtccaac 660
agattcagtg gggtgccgga ccgctttagt ggatctggtt caggaacgga cttcacattg 720
aaaattagta gagttgaagc ggaagacgtg ggagtctact actgtttcca gggttcacat 780
gtgcctcgga cctttggggg aggcaccaag gttgagataa aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 171
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 171
atggacttcc aagtccaaat cttttctttt ttgttgataa gcgcatcagt tattatgtct 60
cgcgccagtc aagtacaact ggtgcagtcc ggagctgaag tgaaaaaacc aggagcaagc 120
gtgaaagtaa gttgtaaggc aagtggttac actttcacaa gctaccatat tcaatgggtc 180
cgacaggctc ctggacaggg cttggagtgg atgggcatat ggatttaccc tggtgacggg 240
tccacccagt ataatgaaaa gttcaaggga gtcacgatga ccagggacac ctctacatct 300
accgtgtata tggagctctc tagtttgcga tccgaagaca ctgccgttta ttactgtgca 360
agagaaggaa cttattacgc gatggactac tggggtcagg ggacaacagt caccgttagc 420
tccgtcgatt ccagcggggg aggtggctca ggcgggggtg gttctggggg gggcgggagc 480
acttcagata ttgtaatgac ccaaacccca ctgagtagtc cagtcacgct tggtcaaccg 540
gcaagcattt cttgcaggag ctctcagagt attgtccact ctaacgggaa tacatatttg 600
gagtggtatc agcaaagacc gggccaacca ccacgcctct tgatttataa ggtgagcaat 660
aggttttcag gcgtgccaga taggttctca ggctccggag cgggaaccga cttcaccctc 720
aagataagtc gggtggaagc cgaagacgta ggagtttact actgctttca aggatctcat 780
gttccacgaa cgtttggagg aggaaccaag gtggaaataa aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 172
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 172
atggactttc aggtccaaat tttttccttc ttgctcatat ccgcgagtgt catcatgtca 60
agagcaagtc aagttcaact cgttcaatca ggagctgagg tgaaaaaacc aggggcgtct 120
gtcaaagtaa gctgcaaagc atcagggtat acgttcacga gttatcatat ccagtgggtt 180
aggcaggcgc cagggcaggg attggaatgg atgggtatct ggatttaccc gggtgacggc 240
agcactcaat acaatgagaa attcaaaggc gtaacaatga caagggacac gagcacaagc 300
acagtgtaca tggagcttag ctctttgagg tcagaggata ccgctgttta ctattgtgct 360
cgggagggta cttactatgc aatggactac tgggggcaag gcacgaccgt tacagtgagt 420
agcgtagatt cctccggggg tggcggttca ggcggcggag gctcaggcgg aggagggtca 480
acatccgatg tcgtaatgac tcagtcccct ctgtcattgc cggtgacttt gggacagcca 540
gcgtctatat cttgtaggtc ctctcaatca atagtgcatt ccaacggtaa cacctatctg 600
gaatggtatc agcaaaggcc aggacaaagt ccacgcctgc ttatatataa ggtgtctaat 660
cgattcagtg gggttcccga taggttttcc ggctctggta gcgggactga tttcacgttg 720
aaaatatcac gcgtggaagc ggaagatgtt ggggtctatt actgctttca gggtagtcat 780
gtccctcgaa cttttggcgg tggtacaaag gtagaaatca aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 173
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 173
atggattttc aggtacagat attctcattt ctccttatct cagctagtgt cataatgtcc 60
agggcgagtc aagtacaact tgtccagtca ggcgcagagg tcaagaagcc gggcgcaagc 120
gttaaggttt cctgcaaagc atccggctat acattcacgt cctatcacat ccaatgggtc 180
aggcaagcac ccggtcaagg acttgagtgg atgggcatct ggatttaccc tggagatggc 240
agtactcagt acaacgaaaa attcaaaggt gtaaccatga cccgcgacac atctacttcc 300
acagtttata tggaactcag cagtttgcgg agcgaagata ccgctgttta ctactgtgcc 360
cgagagggaa cttactacgc catggactat tggggtcaag gaacgacagt aacagttagt 420
tctgtagatt ccagtggcgg cggtgggagc gggggtgggg gatctggcgg aggcggaagt 480
acaagtgaca tcgttatgac tcagacaccc cttagtagtc ccgttacgtt gggccaaccc 540
gcgagcattt cctttcgatc ctctcagtct atagttcact caaatgggaa tacttatttg 600
gagtggtatc aacagcgccc cggacaacca ccaaggctcc tgatatacaa ggtgtccaat 660
cgattctctg gggtgcctga tagatttagc ggaagtggag ccggtacaga ttttaccctg 720
aaaatatcac gggtagaagc cgaagatgtc ggcgtctact actgtttcca gggttcccat 780
gtaccgcgaa cgttcggggg cggaacaaaa gttgagatca agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 174
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 174
atggatttcc aggttcagat atttagtttc ctcttgattt ctgccagtgt catcatgagc 60
agggcttccc aagttcagtt ggtgcaaagt ggcgctgaag tcaaaaaacc tggggcttcc 120
gttaaagtat cttgcaaggc gtccggctac actttcacat cctaccacat tcaatgggtc 180
cggcaagcgc ccggtcaggg gctcgaatgg atggggtgga tatacccagg agatggatct 240
actcagtaca acgagaaatt taaaggacgg gtgacgatga cgcgcgacac ttcaataagc 300
actgcataca tggaactgtc ccggcttagg tcagatgaca ccgcggtcta ctattgtgcg 360
agagagggta cttactatgc tatggactac tgggggcaag gcacgacggt tacagtttcc 420
tcagtcgata gttcaggcgg aggcggctcc gggggcggtg gtagtggagg gggtggatct 480
acttccgaca ttgtcatgac ccagaccccg ttgagccttt cagtgacgcc cggtcaaccc 540
gccagcataa gttgtcgatc aagccagtct attgtacact ccaatggaaa cacatatttg 600
gagtggtatc tccaaaaacc cggccaaagc cctcaactgc tcatctacaa ggtctcaaac 660
aggtttagcg gggtcccgga tcgcttctca gggtcaggat ctggtacgga ctttacactg 720
aaaatttccc gagtcgaagc ggaagacgtg ggtgtatatt actgcttcca ggggagtcat 780
gttccaagaa cctttggggg aggtacaaag gtcgaaataa aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 175
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 175
atggattttc aggtccaaat tttttccttc ttgcttatca gcgcaagtgt aatcatgtcc 60
cgcgcgtccc aagtacaact tgtgcaatct ggcgcggagg tgaaaaaacc tggagcttcc 120
gtcaaggttt cttgtaaggc ctctggctac accttcacgt cctaccacat tcagtgggtt 180
cgacaggcgc cgggccaagg actggagtgg atgggatgga tatatccagg agatggttct 240
actcagtata atgagaaatt caagggtcgc gtaacaatga cgagggatac atcaatctcc 300
accgcgtaca tggaactttc aagactccgg tcagatgaca cggcggttta ctactgtgct 360
cgggagggca cttactatgc tatggactac tgggggcaag ggacaacggt aacggtatct 420
agtgtggatt ctagtggcgg cggcggttca ggaggaggtg gttcaggcgg ggggggtagt 480
acaagtgata ttgtgatgac ccaaacaccc ctttctagcc ctgttactct gggtcaaccc 540
gcgtccataa gttgtcgaag tagtcaatcc atcgtgcata gcaacggcaa cacttacctt 600
gaatggtatc aacaacgacc cggacagccc ccgcgactgc ttatctataa agtatcaaac 660
aggttcagtg gcgtgccaga tcgattctcc ggctctgggg caggcacaga tttcacgttg 720
aaaatttctc gggtcgaggc cgaggacgtg ggcgtttatt actgtttcca ggggagtcac 780
gtccccagga cgttcggagg aggaactaaa gtcgaaataa agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 176
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 176
atggacttcc aggtccaaat attcagcttc ctcctcattt ccgccagtgt aataatgtcc 60
agagcctcac aagtacagtt ggttcagagc ggggctgagg ttaagaaacc aggcgcgagc 120
gtcaaggtat cctgcaaggc gagtggttat actttcacta gttatcacat tcagtgggtc 180
cgacaggccc ccggtcaagg cctggagtgg atggggtgga tatatccggg agatggttct 240
acccaatata atgagaagtt taaggggaga gtcacaatga caagggacac cagtattagc 300
accgcgtata tggagctttc ccgcctgcga tcagatgaca cggccgtgta ctactgtgct 360
agggagggaa cctattatgc gatggattac tggggacagg gtactacagt cacggtctct 420
agcgtggaca gttccggggg cggtggaagc ggtggtggcg gttcaggtgg aggaggctct 480
acgagtgatg ttgtgatgac tcagtccccg ctttcacttc ccgtcaccct tgggcaaccc 540
gcaagcatct catgccgatc ctcccagtct atagtacata gtaatggcaa cacatatctt 600
gaatggtatc agcagaggcc gggtcagtct ccccgactcc ttatatataa agtgagcaac 660
agattctccg gagtaccgga tagattttcc ggctctggga gcggcaccga ctttacactg 720
aaaatttcac gggttgaagc tgaagatgtt ggggtatact attgtttcca gggttctcac 780
gtcccgagga cattcggggg aggaacgaaa gtcgaaataa agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 177
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 177
atggattttc aggtacaaat cttcagcttc ctgctcatct ccgcgagcgt aatcatgtct 60
agggcgtccc aggtgcagtt ggtgcaatca ggtgcagagg tgaagaagcc tggtgcatcc 120
gttaaagtaa gttgtaaggc aagcggatat acttttacat cctatcatat tcaatgggtc 180
agacaagcac ctggacaggg tcttgagtgg atgggctgga tctatccagg cgatggctca 240
actcaatata acgagaagtt caaggggagg gttactatga ccagggatac gtctatttcc 300
actgcgtaca tggaactctc caggttgaga agtgatgata ccgcggttta ctactgcgct 360
agagaaggaa cgtactacgc tatggattac tgggggcagg gtacaactgt caccgtctca 420
agtgtggatt cttctggggg tgggggatca ggagggggag gctccggtgg gggcgggtca 480
accagcgaca ttgtcatgac tcaaaccccc ctgagcagcc ctgtcaccct gggtcagcct 540
gcctcaatat cctttagaag ctcccaaagc atcgtccatt caaatggtaa tacctatctg 600
gagtggtatc agcaaaggcc tggtcaaccc ccgcgccttc tcatttacaa ggtgtcaaac 660
aggttctccg gcgtaccgga taggttttcc ggaagcggtg ctggaaccga ctttactctc 720
aaaatctcta gggtggaagc tgaggacgtc ggtgtatact attgttttca aggctcccat 780
gttcccagga catttggtgg gggaacgaag gtagaaatca agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 178
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 178
atggactttc aggttcagat tttctctttc ttgttgatct ccgctagtgt cataatgtca 60
cgggcaagtc aggtacaact cgttcagagt ggtgccgaag tgaagaaacc gggtgcctcc 120
gtaaaggtgt catgtaaagc tagtggctat acattcacaa gttatcatat ccaatgggta 180
cgacaagcac cgggacagcg actggaatgg atgggatgga tctatcctgg ggacggatct 240
acacagtaca atgagaaatt taagggacgg gtcacgataa ccagggacac atctgcttcc 300
acggcttaca tggagctttc ctccctgcgg agcgaggaca tggctgttta ctattgcgct 360
cgcgaaggga catactacgc aatggattat tggggccaag gcactaccgt gacggtctct 420
tctgtcgata gttccggagg aggtggttca gggggaggcg gttcaggtgg gggtggatct 480
acctcagata ttgtcatgac acagacacct ttgtccttga gtgtgacacc gggtcaaccg 540
gcgagtataa gctgtcgcag ctcacaatct attgtgcata gcaacgggaa tacatatctc 600
gaatggtatc tccaaaagcc gggccaatcc ccccaacttc tcatttacaa agtttctaat 660
cgattttcag gtgtaccaga tcggttttcc gggtctggct caggtactga cttcaccttg 720
aaaatatcaa gggttgaagc tgaggatgta ggtgtgtact attgcttcca ggggtctcac 780
gttcctcgga cttttggggg gggcacaaaa gtagagatta aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 179
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 179
atggatttcc aggtgcaaat cttctcattt cttttgataa gtgcgtcagt gataatgtct 60
cgggccagtc aagtacagct tgtccaaagt ggcgctgaag tcaagaagcc gggagcctca 120
gttaaggtta gctgcaaggc ctcagggtat acttttacct cctatcatat acagtgggta 180
cgacaagcac cgggacagcg actggagtgg atgggttgga tatatccggg agatggttca 240
acccagtata atgagaagtt caaggggcga gttacgataa cccgcgatac gagtgcatca 300
acagcgtaca tggagttgag ttccctccgc agcgaggaca tggcggtata ctattgtgcc 360
agggagggga cttattatgc catggactac tgggggcagg gcacaaccgt aacagtctct 420
tctgtagaca gttcaggagg gggcggaagt ggaggtggcg gatctggtgg aggtggatct 480
acttccgaca tcgttatgac ccaaacacca ctttcatctc ccgttactct cgggcaacct 540
gctagtattt cctgtagatc ctcacaatct atagtacata gcaatggcaa tacctacctg 600
gagtggtatc aacaacgccc aggccaacca cctcgcctgc ttatctataa agtaagcaat 660
agattcagtg gtgtaccgga taggttctct ggttccggag caggaactga ctttacactc 720
aaaatcagta gggtggaggc ggaagacgtg ggagtatatt attgctttca aggttcacat 780
gtacctcgaa catttggcgg aggaactaag gttgagatta aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 180
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 180
atggatttcc aagtccagat attcagtttt cttttgataa gcgcttctgt aatcatgtct 60
cgggcgtccc aagtacaact ggtgcaatca ggggcagaag tgaaaaaacc aggtgcatcc 120
gttaaggtga gttgcaaggc ttccggctat acctttacat catatcatat tcaatgggtc 180
aggcaagcac ctggtcagcg attggaatgg atgggttgga tatatcctgg tgatgggtct 240
acacaatata acgaaaaatt caaggggcga gtgaccatca caagagatac atcagcgtca 300
acagcgtata tggaactgtc atcccttaga tcagaggaca tggcggtcta ttactgtgcc 360
agagaaggca cttattatgc aatggattat tggggacaag gaaccactgt cactgtttcc 420
agcgtagact cctccggtgg tggtggaagt ggcggcggtg ggtcaggagg gggtgggtca 480
acttctgatg tagtgatgac acagagccct ctgagcttgc ctgtgacctt gggtcagccg 540
gcctcaataa gttgtcgatc tagtcagtca atcgtccata gtaatgggaa cacatacctt 600
gaatggtatc agcaaagacc tggacaatct ccacgactcc ttatatacaa agttagcaac 660
cgatttagcg gagtgccaga ccgcttttct ggttccgggt ctggcacaga ttttaccctt 720
aagatctccc gcgtggaggc ggaagacgtt ggtgtttact attgcttcca ggggtcacac 780
gttccacgca cctttggagg aggtacgaag gtcgagatta agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 181
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 181
atggattttc aagtacagat cttctctttc ttgcttattt cagcgagcgt aatcatgagt 60
agggcatctc aagttcaact cgttcagtca ggtgctgagg taaaaaaacc aggggcttcc 120
gttaaagtta gctgtaaggc atctgggtac acatttacta gctaccatat ccagtgggtg 180
cgacaagccc cggggcagcg cttggaatgg atgggctgga tttacccagg tgacggctcc 240
acgcaatata atgagaaatt taagggtaga gttactatta ccagggacac aagtgcttca 300
actgcctata tggaactgag cagccttcgg agtgaagata tggccgtata ttactgcgca 360
agggagggga cttactatgc aatggactac tggggtcagg gaacgactgt gaccgtgtcc 420
tcagttgact ccagcggtgg tggcggctct ggaggtgggg gttccggcgg aggcggaagc 480
acatctgata tagtgatgac gcaaacgcct ctttcttccc cggtaactct gggacagcca 540
gcgtcaattt catttaggtc ctcccagtca atcgtacata gtaatggaaa tacttacctg 600
gaatggtatc aacaacgacc agggcaaccg ccccgattgt tgatctataa agtgagcaat 660
cgcttttctg gcgtgcccga tcggttctca gggtctggag ctgggactga cttcacattg 720
aaaatttcca gggttgaggc cgaggatgtg ggggtttatt actgcttcca aggctcccac 780
gtcccccgca ccttcggagg gggaaccaaa gtcgaaataa agaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 182
<211> 1599
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 182
atggattttc aagttcagat attctcattt ttgcttatat cagcctccgt aattatgtca 60
cgggcaagtc aagttcagtt ggtgcagtcc ggagcagaag ttaagaagcc cggtgcttct 120
gtgaaagtct cctgcaaagc gtctgggtac accttcacga gctaccatat acagtgggtc 180
cggcaagcgc ctgggcagag gctggagtgg atgggctgga tttacccagg agatgggagt 240
acaaagtata gtcagaagtt tcaagggcga gtgacgataa ccagagatac gagtgcaagt 300
actgcataca tggaactgag ctccttgagg tccgaggata cagcggtgta ctattgcgct 360
cgggaaggga catattatgc tatggactat tggggacaag ggacaacggt aacggtgagt 420
tccgtcgatt cctcaggtgg cggaggcagt gggggcgggg gttccggcgg tggcgggtcc 480
acgagtgata tagttatgac acagaccccc ctcagccttt ctgtgacccc aggacaaccc 540
gctagtatct cttgccgcag ttctcagtcc atagtacaca gtaacggaaa tacctatctt 600
gagtggtatc ttcaaaagcc cggccagagc cctcaactct tgatatataa agtgtcaaat 660
cgattttcag gtgtgcctga tcgattctca gggtctggtt cagggacaga tttcacgctt 720
aagataagca gagtagaggc tgaagacgtg ggagtctact attgttttca ggggtcacac 780
gttccccgca cttttggtgg gggaaccaag gtggaaatca aaaaccggat ccgtggggtc 840
accgtctctt cagcgctgag caactccatc atgtacttca gccacttcgt gccggtcttc 900
ctgccagcga agcccaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 960
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 1020
cacacgaggg ggctggaccc ctttgggttt tgggtgctgg tggtggttgg tggagtcctg 1080
gcttgctata gcttgctagt aacagtggcc tttattattt tctgggtgag gagtaagagg 1140
agcaggctcc tgcacagtga ctacatgaac atgactcccc gccgccccgg gcccacccgc 1200
aagcattacc agccctatgc cccaccacgc gacttcgcag cctatcgctc cctcgagaga 1260
gtgagagtga agttcagcag gagcgcagac gcccccgcgt accagcaggg ccagaaccag 1320
ctctataacg agctcaatct aggacgaaga gaggagtacg atgttttgga caagagacgt 1380
ggccgggacc ctgagatggg gggaaagccg agaaggaaga accctcagga aggcctgtac 1440
aatgaactgc agaaagataa gatggcggag gcctacagtg agattgggat gaaaggcgag 1500
cgccggaggg gcaaggggca cgatggcctt taccagggtc tcagtacagc caccaaggac 1560
acctacgacg cccttcacat gcaggccctg ccccctcgc 1599
<210> 183
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 183
Ser Tyr His Ile Gln
1 5
<210> 184
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 184
Gly Tyr Thr Phe Thr Ser Tyr
1 5
<210> 185
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 185
Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 186
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 186
Tyr Pro Gly Asp Gly Ser
1 5
<210> 187
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 187
Glu Gly Thr Tyr Tyr Ala Met Asp Tyr
1 5
<210> 188
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 188
Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 189
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 189
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 190
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 190
Phe Gln Gly Ser His Val Pro Arg Thr
1 5
<210> 191
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 191
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Ile Tyr Phe Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 192
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 192
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Val Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 193
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 193
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Ile Tyr Phe Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Leu Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly Asp Gln Val Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
245 250
<210> 194
<211> 521
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 194
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Leu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Ile Tyr Phe Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Val Asp Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Thr Ser Asp Val Leu Met Thr
130 135 140
Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly Asp Gln Val Ser Ile
145 150 155 160
Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr
165 170 175
Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
180 185 190
Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
210 215 220
Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Arg
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Leu Glu Gln Lys Leu
245 250 255
Ile Ser Glu Glu Asp Leu Asn Arg Ile Arg Gly Val Thr Val Ser Ser
260 265 270
Ala Leu Ser Asn Ser Ile Met Tyr Phe Ser His Phe Val Pro Val Phe
275 280 285
Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro
290 295 300
Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys
305 310 315 320
Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Pro Phe
325 330 335
Gly Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
340 345 350
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg
355 360 365
Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro
370 375 380
Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe
385 390 395 400
Ala Ala Tyr Arg Ser Leu Glu Arg Val Arg Val Lys Phe Ser Arg Ser
405 410 415
Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu
420 425 430
Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg
435 440 445
Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln
450 455 460
Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr
465 470 475 480
Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp
485 490 495
Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala
500 505 510
Leu His Met Gln Ala Leu Pro Pro Arg
515 520
<210> 195
<211> 544
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 195
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Ala Ser Gln Val Gln Leu Gln Gln Ser Gly Pro
20 25 30
Glu Leu Val Lys Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser
35 40 45
Gly Tyr Thr Phe Thr Ser Tyr His Ile Gln Trp Val Lys Gln Arg Pro
50 55 60
Gly Gln Gly Leu Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Gly Ser
65 70 75 80
Thr Gln Tyr Asn Glu Lys Phe Lys Gly Lys Thr Thr Leu Thr Ala Asp
85 90 95
Lys Ser Ser Ser Thr Ala Tyr Met Leu Leu Ser Ser Leu Thr Ser Glu
100 105 110
Asp Ser Ala Ile Tyr Phe Cys Ala Arg Glu Gly Thr Tyr Tyr Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Val Asp Ser
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Thr Ser Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser
165 170 175
Leu Gly Asp Gln Val Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val
180 185 190
His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly
195 200 205
Gln Ser Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly
210 215 220
Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
225 230 235 240
Lys Ile Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe
245 250 255
Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr Lys Leu Glu
260 265 270
Ile Lys Leu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Arg Ile
275 280 285
Arg Gly Val Thr Val Ser Ser Ala Leu Ser Asn Ser Ile Met Tyr Phe
290 295 300
Ser His Phe Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro
305 310 315 320
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
325 330 335
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
340 345 350
Thr Arg Gly Leu Asp Pro Phe Gly Phe Trp Val Leu Val Val Val Gly
355 360 365
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
370 375 380
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
385 390 395 400
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
405 410 415
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Leu Glu Arg Val
420 425 430
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
435 440 445
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
450 455 460
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
465 470 475 480
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
485 490 495
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
500 505 510
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
515 520 525
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
530 535 540
<210> 196
<211> 1635
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 196
atggacttcc aggtgcagat cttcagcttc ctgctgatca gcgccagcgt gatcatgagc 60
cgcgctagcc aggtgcagct gcagcagagc ggccccgagc tggtgaagcc cggcgccagc 120
gtgaagatga gctgcaaggc cagcggctac accttcacca gctaccacat ccagtgggtg 180
aagcagcgcc ccggccaggg cctggagtgg atcggctgga tctaccccgg cgacggcagc 240
acccagtaca acgagaagtt caagggcaag accaccctga ccgccgacaa gagcagcagc 300
accgcctaca tgctgctgag cagcctgacc agcgaggaca gcgccatcta cttctgcgcc 360
cgcgagggca cctactacgc catggactac tggggccagg gcaccagcgt gaccgtgagc 420
agcgtcgaca gcagcggcgg cggcggcagc ggcggcggcg gcagcggcgg cggcggcagc 480
actagtgacg tgctgatgac ccagaccccc ctgagcctgc ccgtgagcct gggcgaccag 540
gtgagcatca gctgccgcag cagccagagc atcgtgcaca gcaacggcaa cacctacctg 600
gagtggtacc tgcagaagcc cggccagagc cccaagctgc tgatctacaa ggtgagcaac 660
cgcttcagcg gcgtgcccga ccgcttcagc ggcagcggca gcggcaccga cttcaccctg 720
aagatcagcc gcgtggaggc cgaggacctg ggcgtgtact actgcttcca gggcagccac 780
gtgccccgca ccttcggcgg cggcaccaag ctggagatca agctcgagca gaagctgatc 840
agcgaggagg acctgaaccg gatccgtggg gtcaccgtct cttcagcgct gagcaactcc 900
atcatgtact tcagccactt cgtgccggtc ttcctgccag cgaagcccac cacgacgcca 960
gcgccgcgac caccaacacc ggcgcccacc atcgcgtcgc agcccctgtc cctgcgccca 1020
gaggcgtgcc ggccagcggc ggggggcgca gtgcacacga gggggctgga cccctttggg 1080
ttttgggtgc tggtggtggt tggtggagtc ctggcttgct atagcttgct agtaacagtg 1140
gcctttatta ttttctgggt gaggagtaag aggagcaggc tcctgcacag tgactacatg 1200
aacatgactc cccgccgccc cgggcccacc cgcaagcatt accagcccta tgccccacca 1260
cgcgacttcg cagcctatcg ctccctcgag agagtgagag tgaagttcag caggagcgca 1320
gacgcccccg cgtaccagca gggccagaac cagctctata acgagctcaa tctaggacga 1380
agagaggagt acgatgtttt ggacaagaga cgtggccggg accctgagat ggggggaaag 1440
ccgagaagga agaaccctca ggaaggcctg tacaatgaac tgcagaaaga taagatggcg 1500
gaggcctaca gtgagattgg gatgaaaggc gagcgccgga ggggcaaggg gcacgatggc 1560
ctttaccagg gtctcagtac agccaccaag gacacctacg acgcccttca catgcaggcc 1620
ctgccccctc gctaa 1635
<210> 197
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 197
ttttctagac gcgtgccacc atggccgtca tggcgcc 37
<210> 198
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 198
aagtcgacgc tagctcacac tttacaagct gtgagagaca 40
<210> 199
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 199
tcaacccgat tgtccaccat 20
<210> 200
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 200
gagtttagtc cgaaatgagg ctg 23
<210> 201
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 201
tccaaagatg tagccgcccc a 21
<210> 202
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 202
ccagtgcctc tttgctgctt tca 23
<210> 203
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 203
ctgagctcgc cagtgaaatg atg 23
<210> 204
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 204
tgctgtagtg gtggtcggag a 21
<210> 205
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 205
acctgcctta agagtggagc ca 22
<210> 206
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 206
acatgtcgca cgtctctgat ga 22
<210> 207
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 207
tgcccagagc atccaaaaga 20
<210> 208
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 208
tgtattgctt tgcgttggac 20
<210> 209
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 209
aggcgctccc caagaagaca 20
<210> 210
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 210
gggctgatta gagagaggtc cct 23
<210> 211
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 211
caagacggac cagagcgaaa 20
<210> 212
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 212
ggcgggtcat gggaataac 19
<210> 213
<211> 468
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 213
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
His Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Thr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu
130 135 140
Ser Val Thr Pro Gly Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln
145 150 155 160
Ser Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln
165 170 175
Lys Pro Gly Gln Ser Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg
180 185 190
Phe Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
195 200 205
Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr
210 215 220
Tyr Cys Phe Gln Gly Ser His Val Pro Arg Thr Phe Gly Gly Gly Thr
225 230 235 240
Lys Val Glu Ile Lys Arg Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr
245 250 255
Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala
260 265 270
Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe
275 280 285
Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val
290 295 300
Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Arg Ser Lys Arg Ser
305 310 315 320
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
325 330 335
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
340 345 350
Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
355 360 365
Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
370 375 380
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
385 390 395 400
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
405 410 415
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
420 425 430
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
435 440 445
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
450 455 460
Leu Pro Pro Arg
465
<210> 214
<211> 1470
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 214
atggctctgc ctgtgacagc tctgctgctg cctctggctc tgcttcttca tgccgccaga 60
ccatctcagg tccagctagt acaaagcggc gccgaagtaa agaaacctgg tgcctctgtg 120
aaggtgagct gcaaggccag cggctacacc ttcaccagct accacatcca gtgggttcga 180
caggcccctg gacagagact agagtggatg ggctggatct atcctggcga cggcagcacc 240
cagtacaacg agaagttcaa gggcagagtt accatcaccc gagacaccag cgccagcaca 300
gcctatatgg agctgagcag cctgcgaagc gaggacacag ctgtttacta ttgtgccaga 360
gagggcacct actacgcaat ggattattgg ggccagggga ccaccgtgac cgtttcttct 420
ggaggcggag gttctggcgg cggaggaagt ggtggcggag gctcagatat tgtaatgacc 480
cagacacctc tgtccctgtc tgtgacacct ggacagcctg caagcatcag ctgtcggagc 540
agccagagca tcgttcacag caacggcaac acctacctgg aatggtatct gcagaagccc 600
ggacagtccc cccagctgct gatctacaag gtgtccaacc gcttcagtgg agtacccgat 660
agattttctg gcagcggctc tggcaccgac ttcaccctga agatctccag agtagaagca 720
gaggacgttg gagtgtacta ctgcttccaa ggcagccatg tgccaagaac ctttggtgga 780
ggcacaaagg tggaaatcaa gcggacaaca acacctgctc ctcggcctcc tacaccagct 840
cctacaattg ccagccagcc actgtctctg aggcccgaag cttgcaggcc tgctgctggc 900
ggagccgtgc atacaagagg actggatttc gcctgcgaca tctacatctg ggcacctctg 960
gctggaacct gtggcgtgct gctgctgagc ctggtcatca ccctgtattg ccggagcaag 1020
agaagcagac tgctgcacag cgactacatg aacatgaccc ctagacggcc cggacctacc 1080
agaaagcact accagcctta cgctcctcct agagacttcg ccgcctacag atccagagtg 1140
aagttcagca gatccgccga cgctcctgcc tatcagcagg gccaaaacca gctctacaac 1200
gagctgaacc tggggagaag agaagagtac gacgtgctgg acaagcggag aggcagagat 1260
cctgaaatgg gcggcaagcc cagacggaag aatcctcaag agggcctgta taatgagcta 1320
cagaaagaca agatggcaga ggcctacagc gagatcggaa tgaagggcga gcgcagaaga 1380
ggcaagggac acgatggact gtaccagggc ctgagcaccg ccaccaagga tacctatgat 1440
gccctgcaca tgcaggccct gcctccaaga 1470
<210> 215
<211> 126
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 215
aaacggggca gaaagaaact cctgtatata ttcaaacaac catttatgag accagtacaa 60
actactcaag aggaagatgg ctgtagctgc cgatttccag aagaagaaga aggaggatgt 120
gaactg 126
<210> 216
<211> 42
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 216
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 217
<211> 135
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 217
accacgacgc cagcgccgcg accaccaaca ccggcgccca ccatcgcgtc gcagcccctg 60
tccctgcgcc cagaggcgtg ccggccagcg gcggggggcg cagtgcacac gagggggctg 120
gacttcgcct gtgat 135
<210> 218
<211> 47
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 218
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr
35 40 45
<210> 219
<211> 45
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 219
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<210> 220
<211> 135
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 220
acaacaacac ctgctcctcg gcctcctaca ccagctccta caattgccag ccagccactg 60
tctctgaggc ccgaagcttg caggcctgct gctggcggag ccgtgcatac aagaggactg 120
gatttcgcct gcgac 135
<210> 221
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 221
Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu
1 5 10 15
Val Ile Thr Leu Tyr Cys
20
<210> 222
<211> 72
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 222
atctacatct gggcgccctt ggccgggact tgtggggtcc ttctcctgtc actggttatc 60
accctttact gc 72
<210> 223
<211> 24
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 223
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr Leu Tyr Cys
20
<210> 224
<211> 72
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic sequence
<400> 224
atctacatct gggcacctct ggctggaacc tgtggcgtgc tgctgctgag cctggtcatc 60
accctgtatt gc 72
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ АНТИ-HLA-А2 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2805674C2 |
| НОВЫЕ КОНСТРУКЦИИ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА, СОДЕРЖАЩИЕ ДОМЕНЫ TNFR2 | 2019 |
|
RU2808254C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА CLL-1 | 2015 |
|
RU2741120C2 |
| ВВЕДЕНИЕ СКОНСТРУИРОВАННЫХ Т-КЛЕТОК ДЛЯ ЛЕЧЕНИЯ РАКА ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ | 2017 |
|
RU2757308C2 |
| Т-КЛЕТКИ С КОСТИМУЛИРУЮЩИМ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ, НАЦЕЛЕННЫЕ НА IL13Rα2 | 2015 |
|
RU2749922C2 |
| ВЫСОКОАФФИННЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP | 2019 |
|
RU2836365C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ M971 | 2018 |
|
RU2770411C1 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
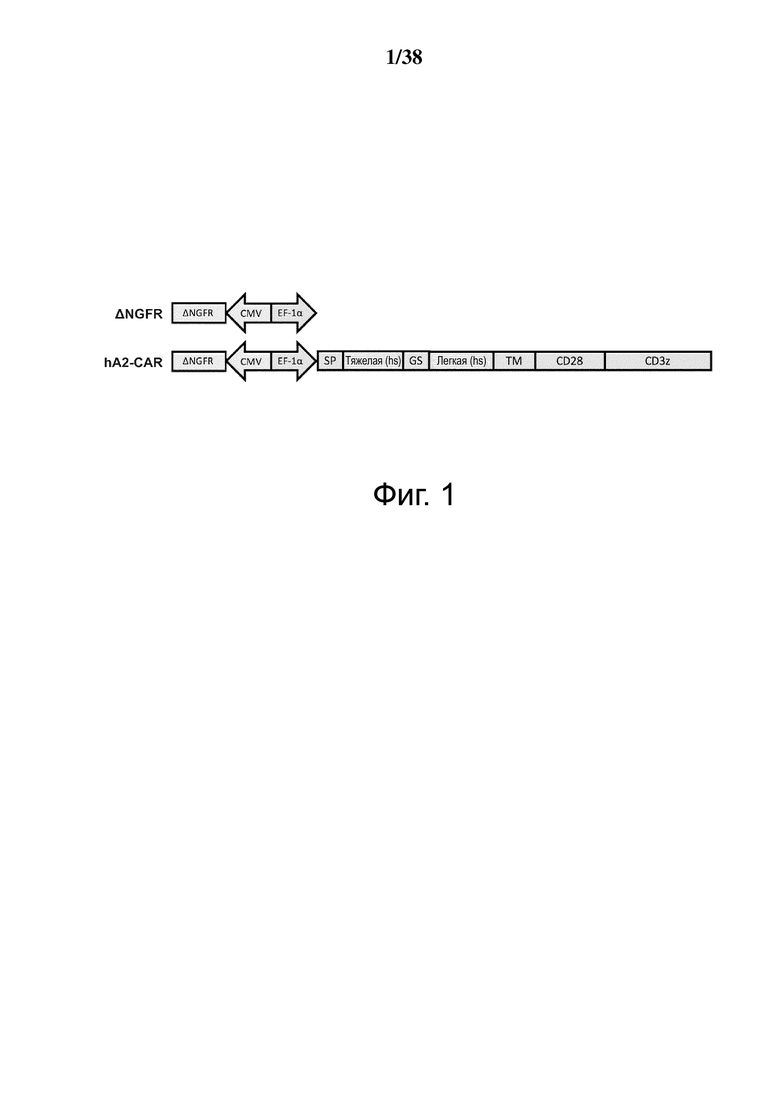
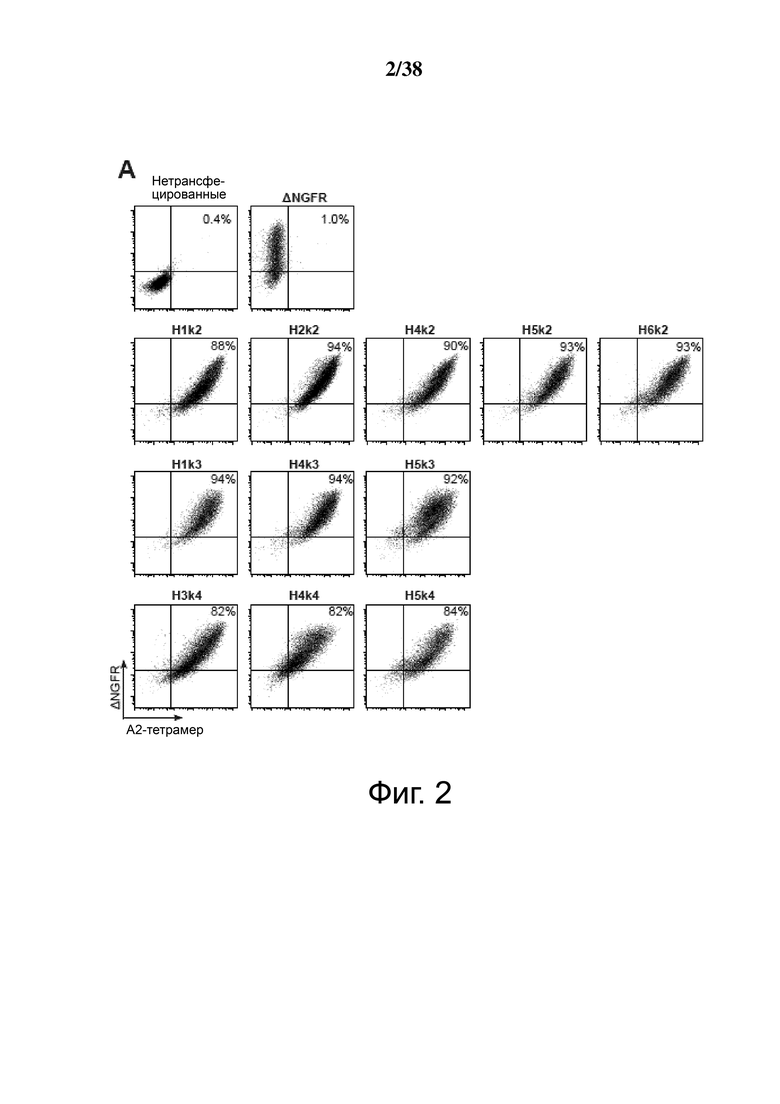
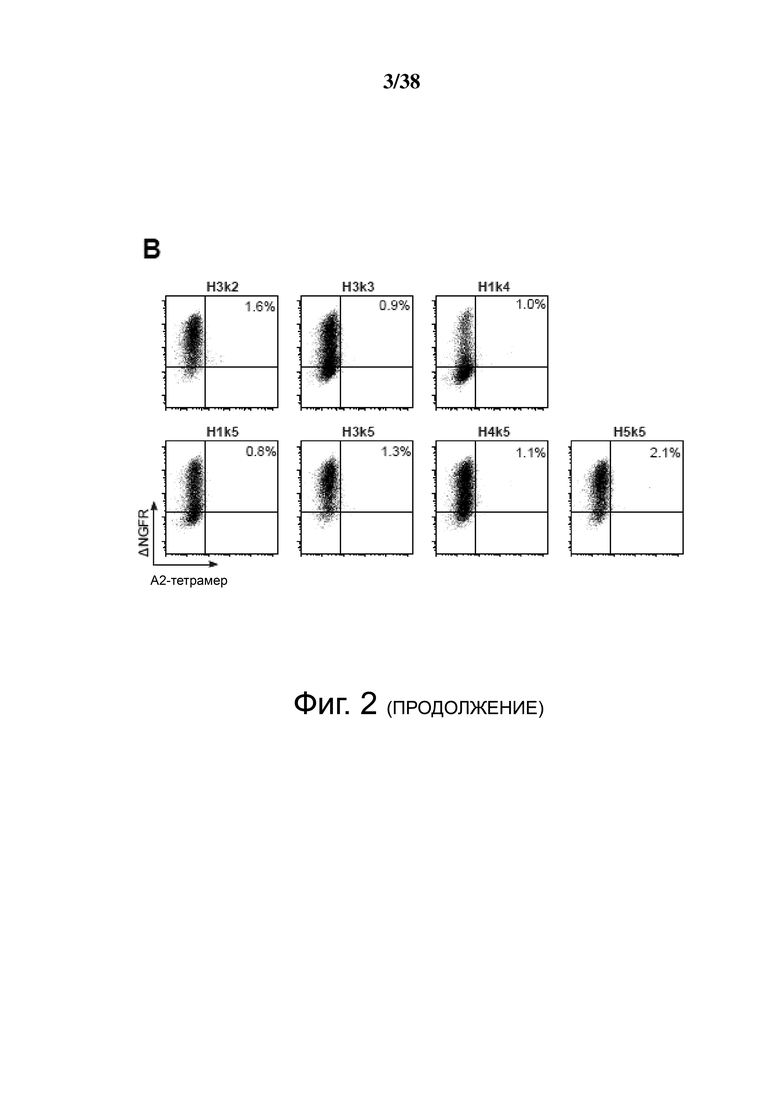
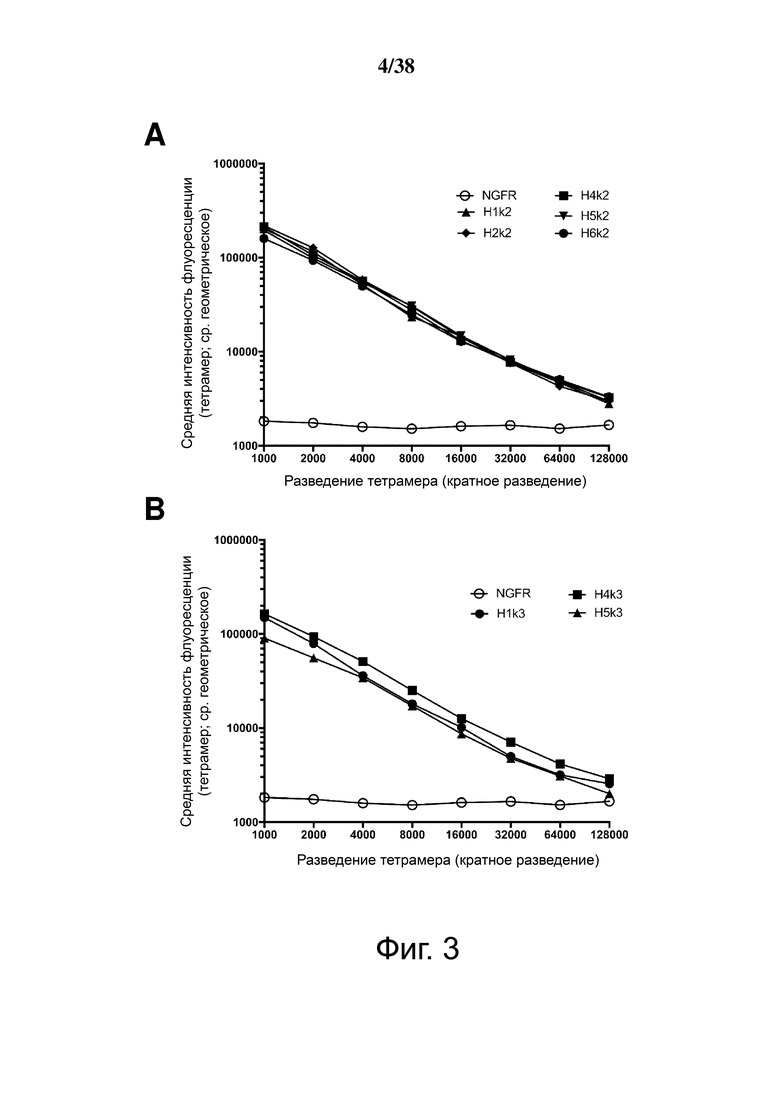
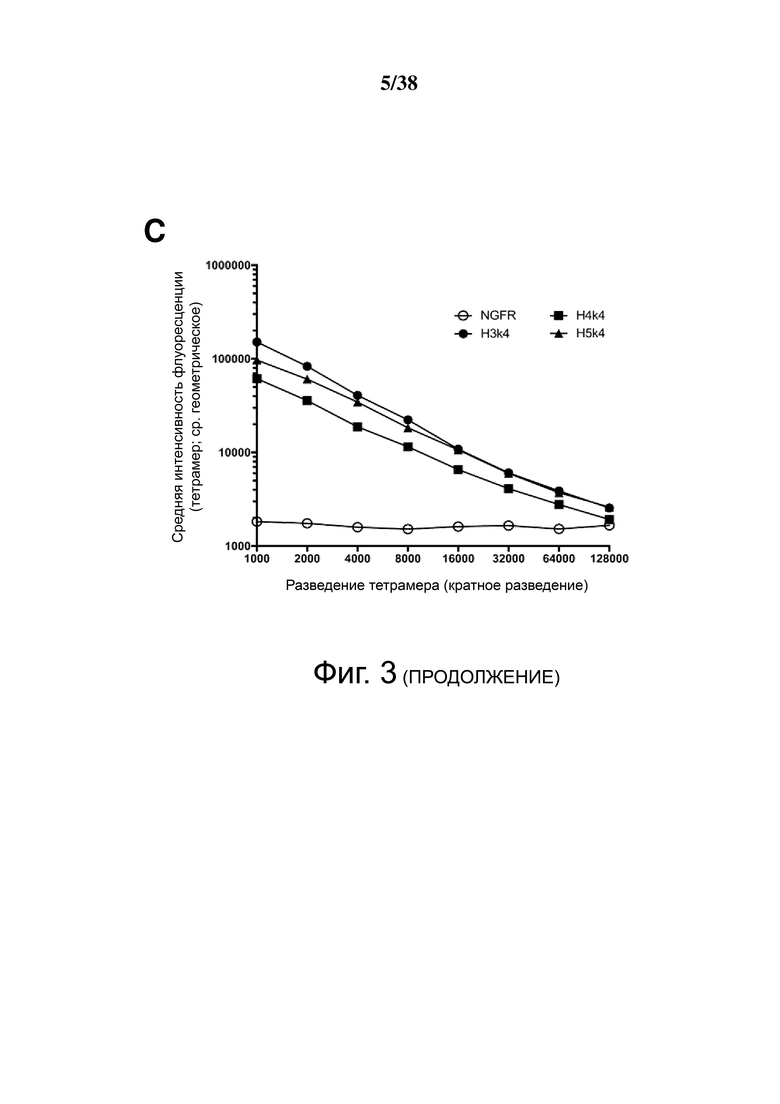
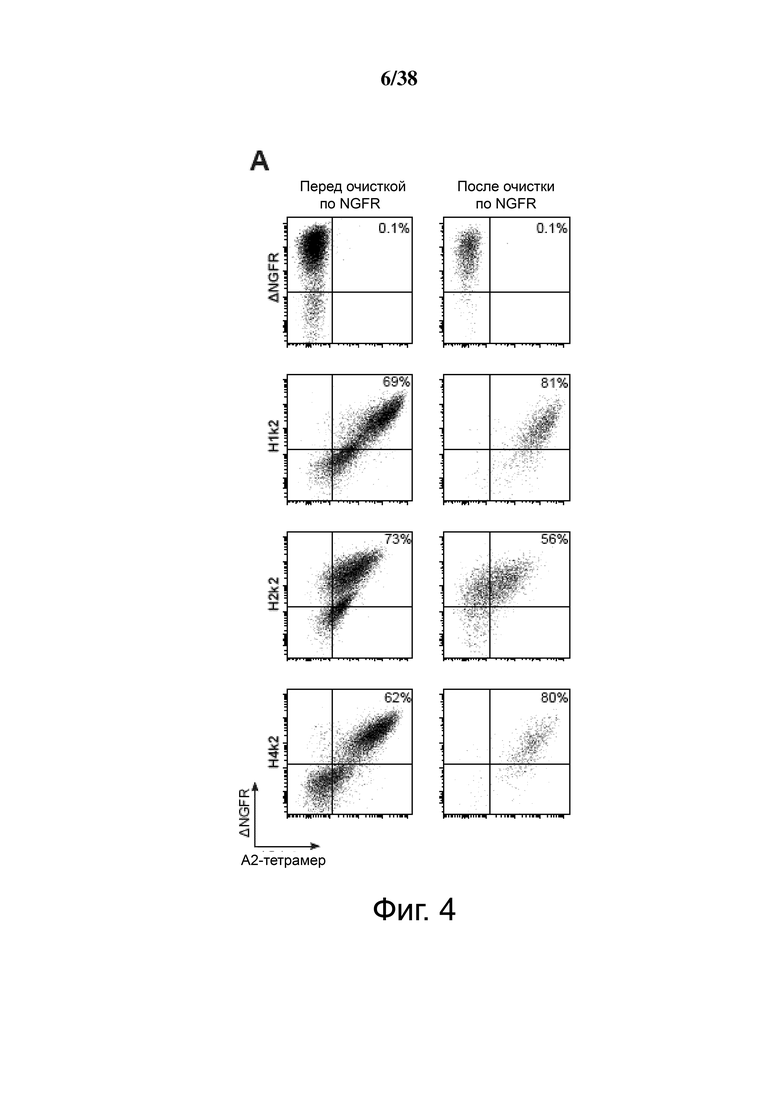
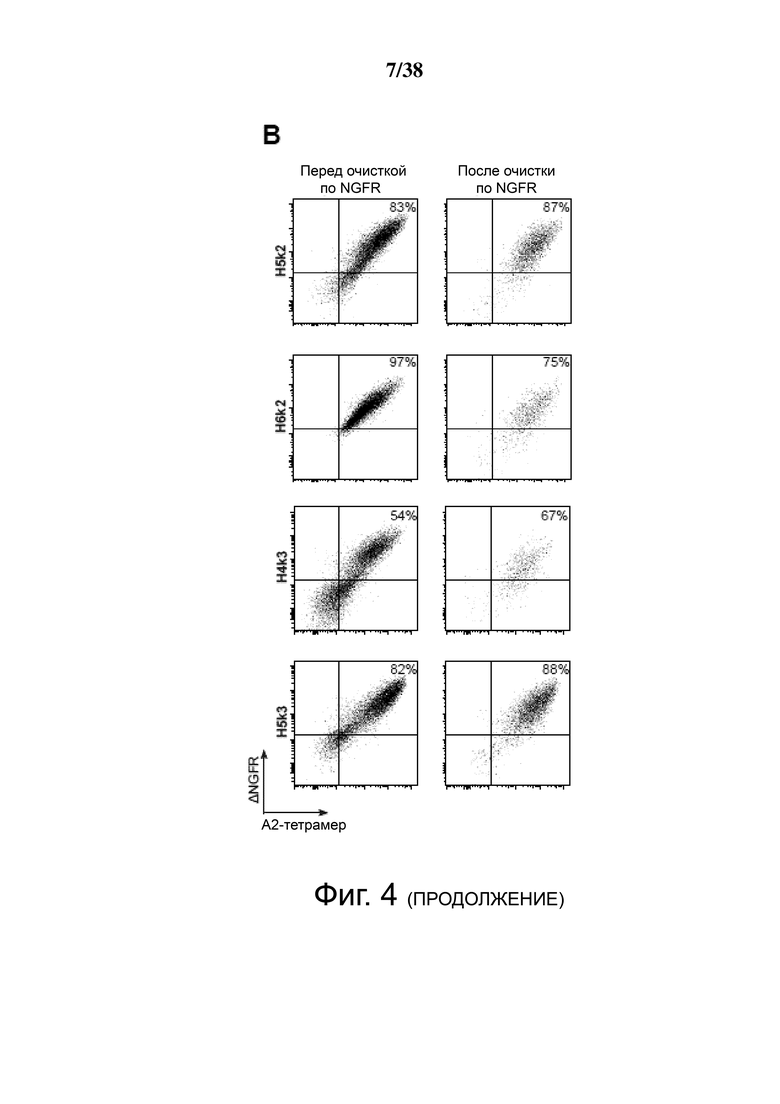
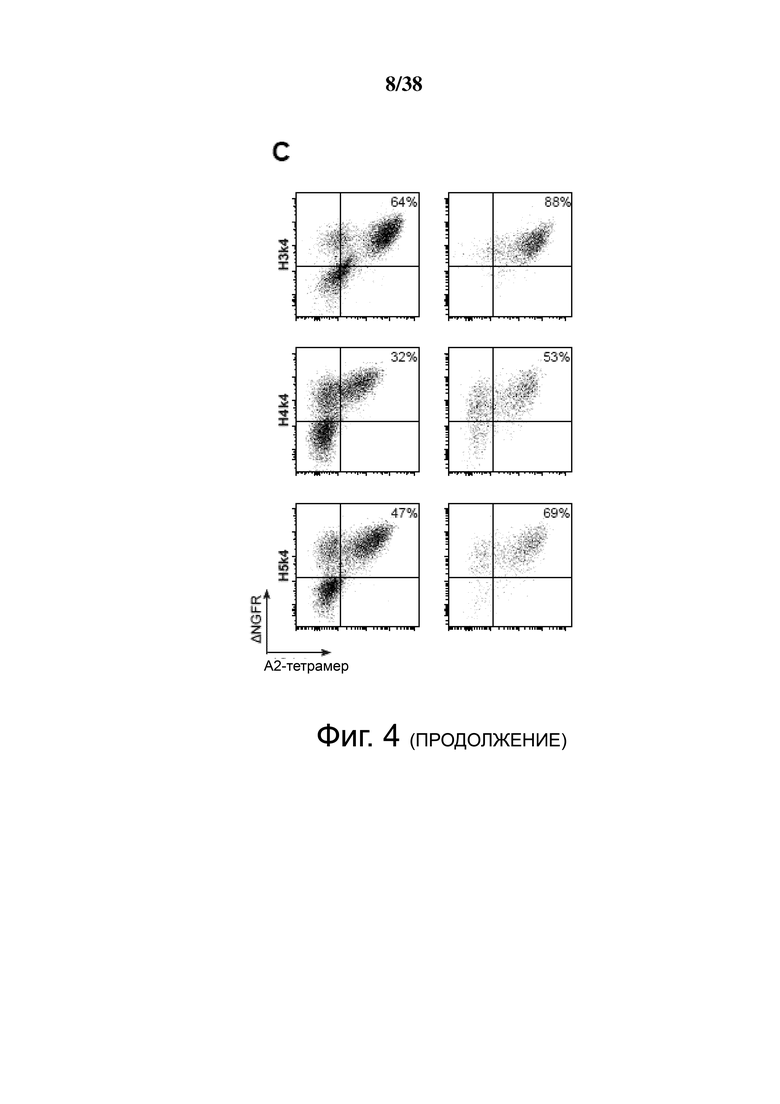
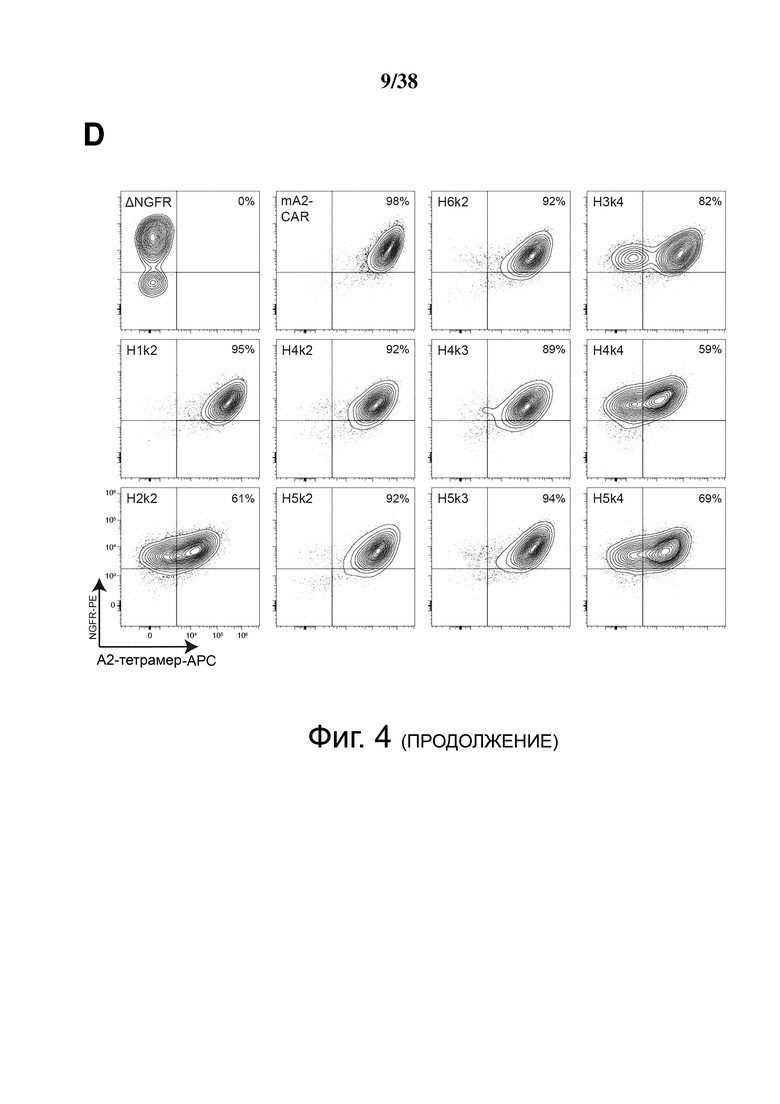
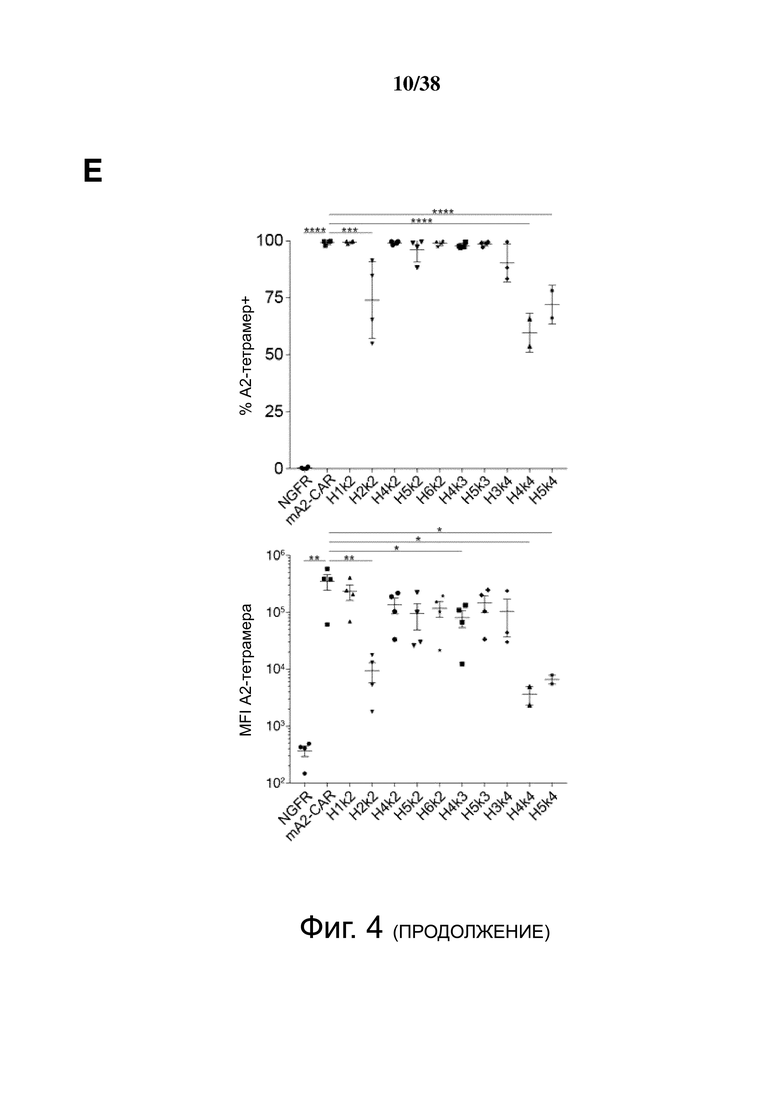
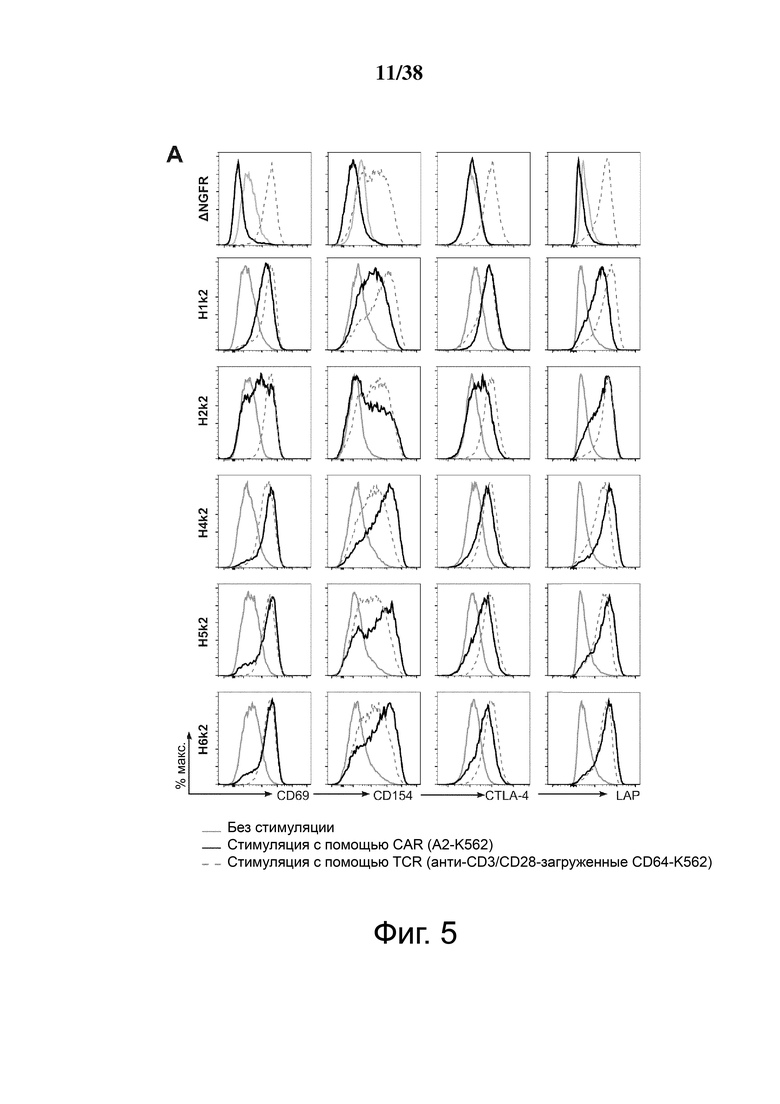
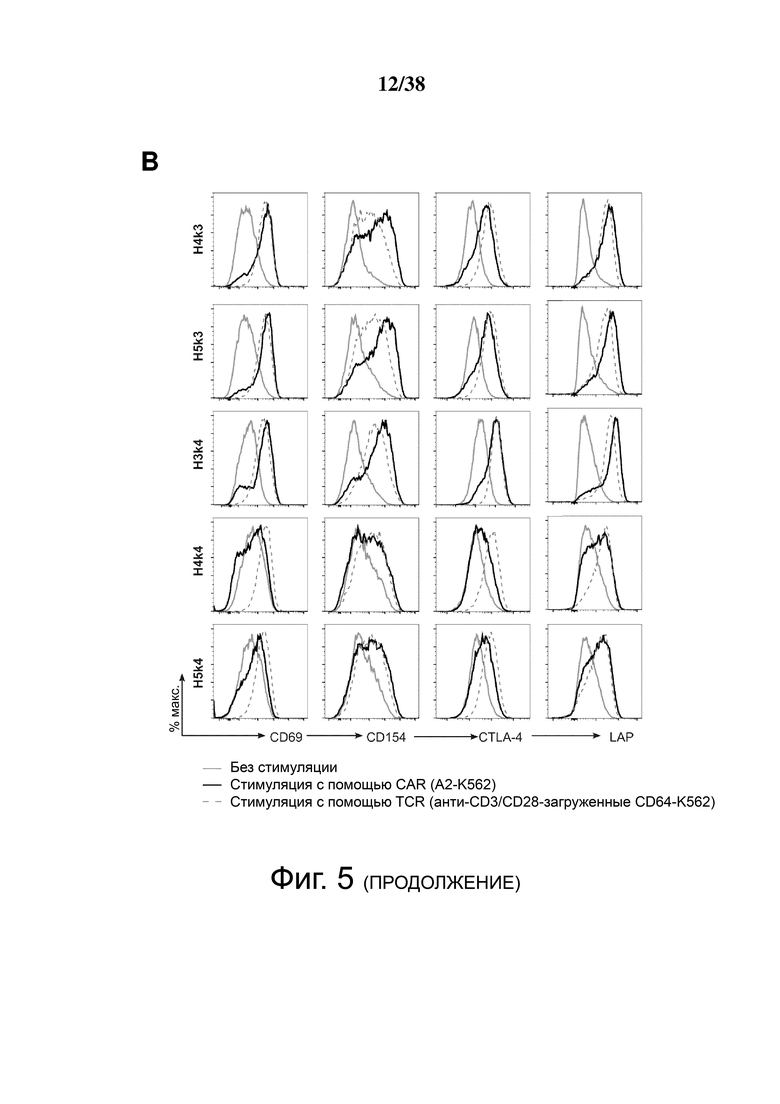
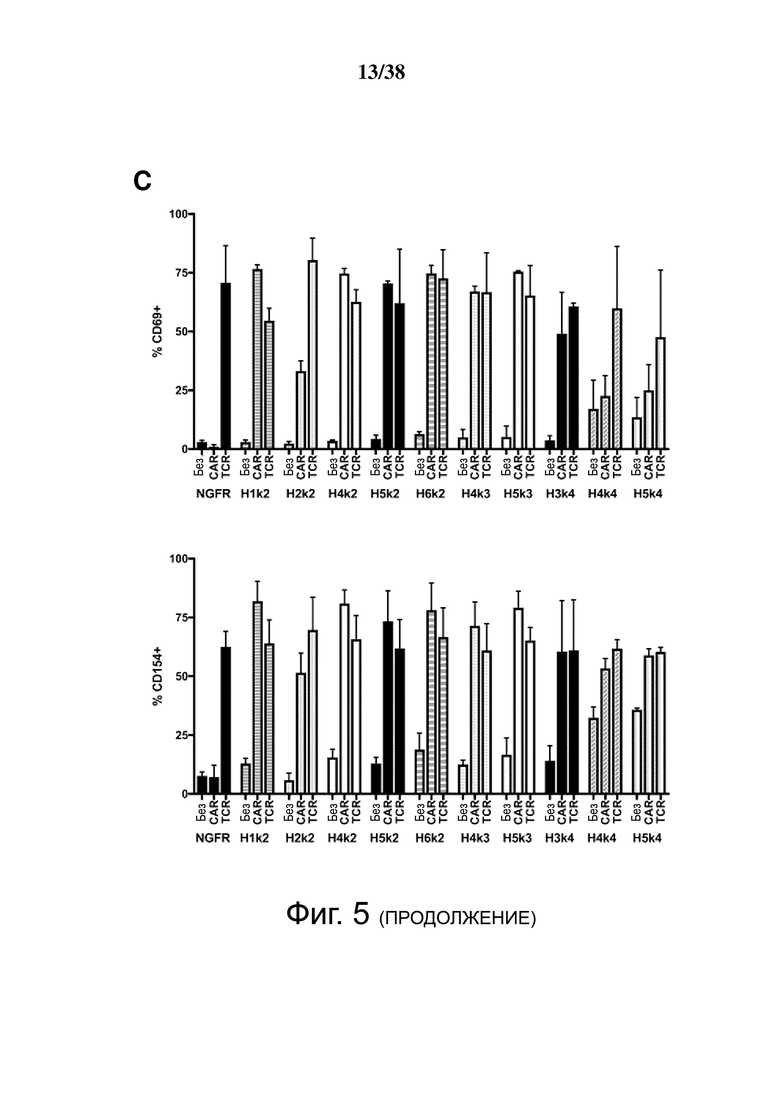
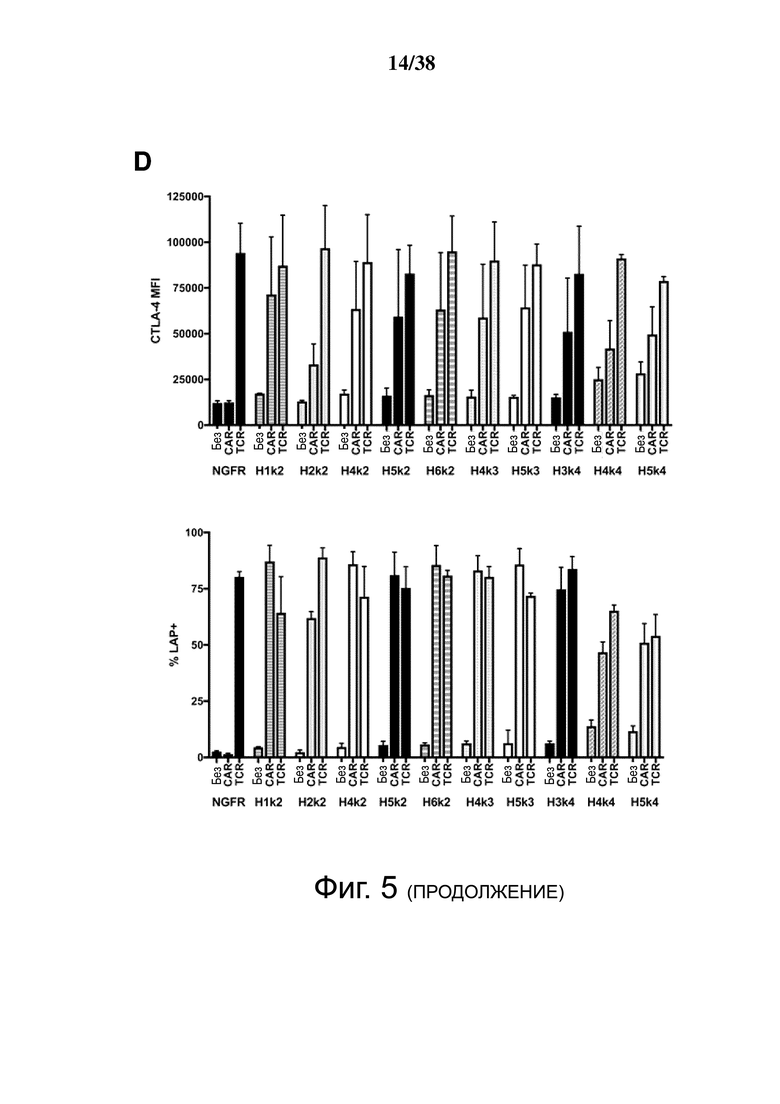
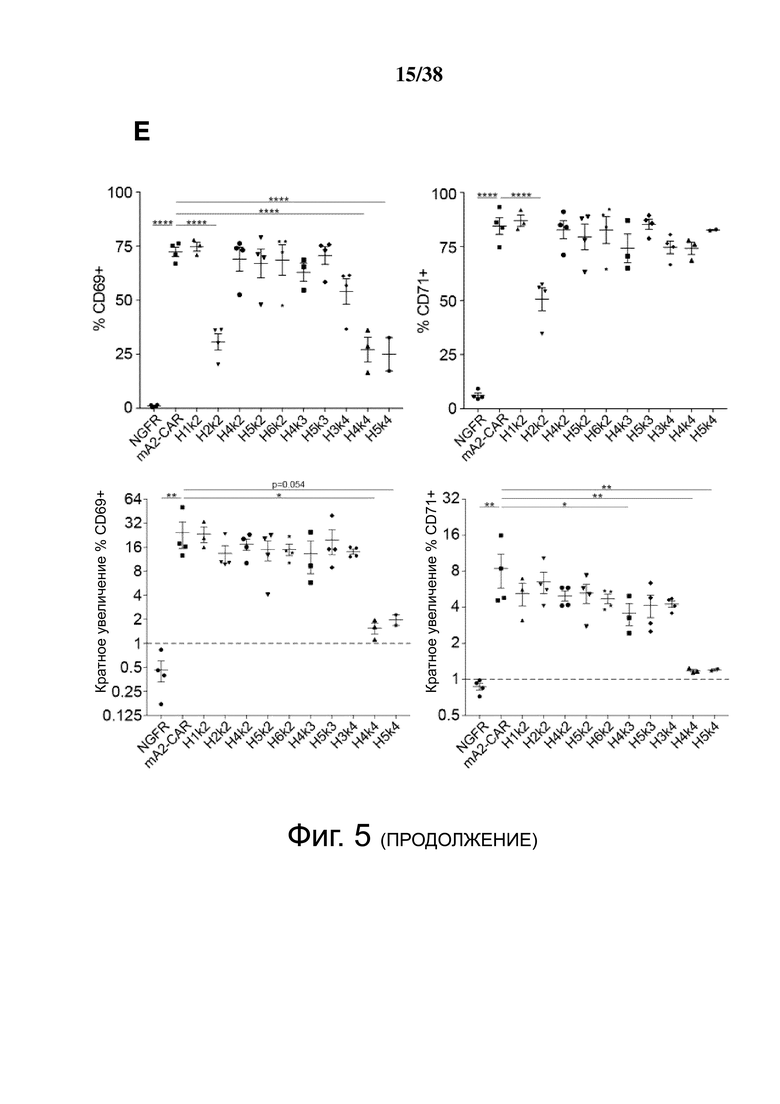
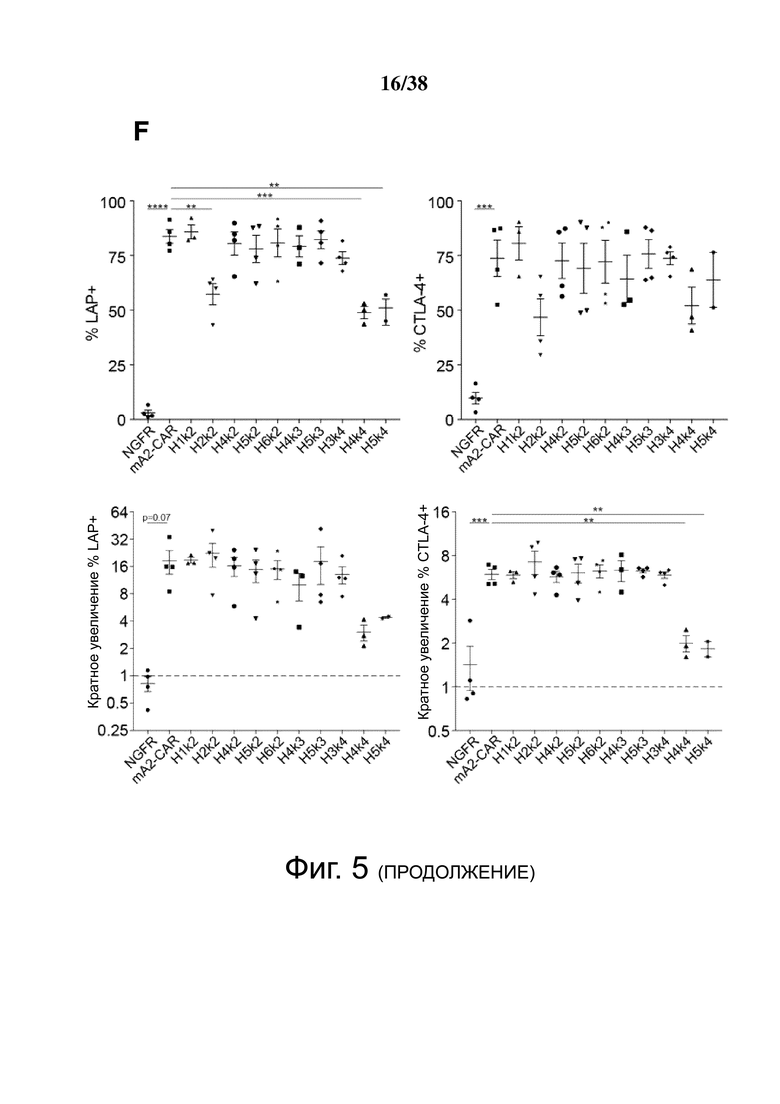
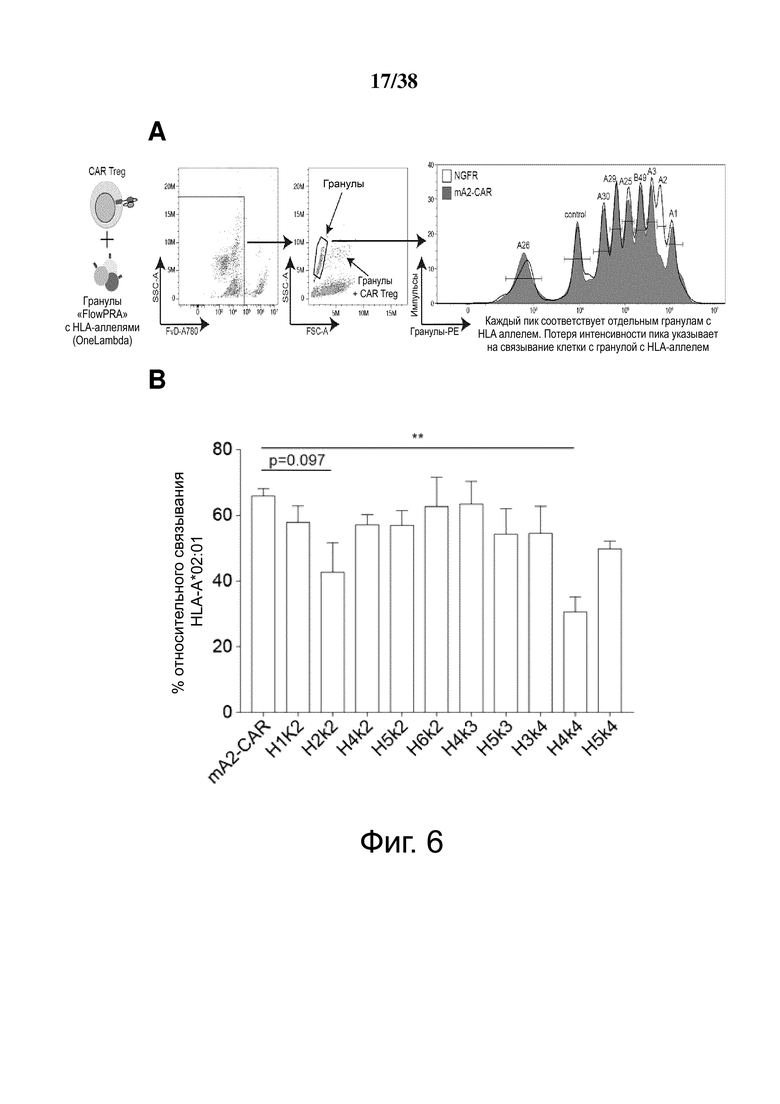
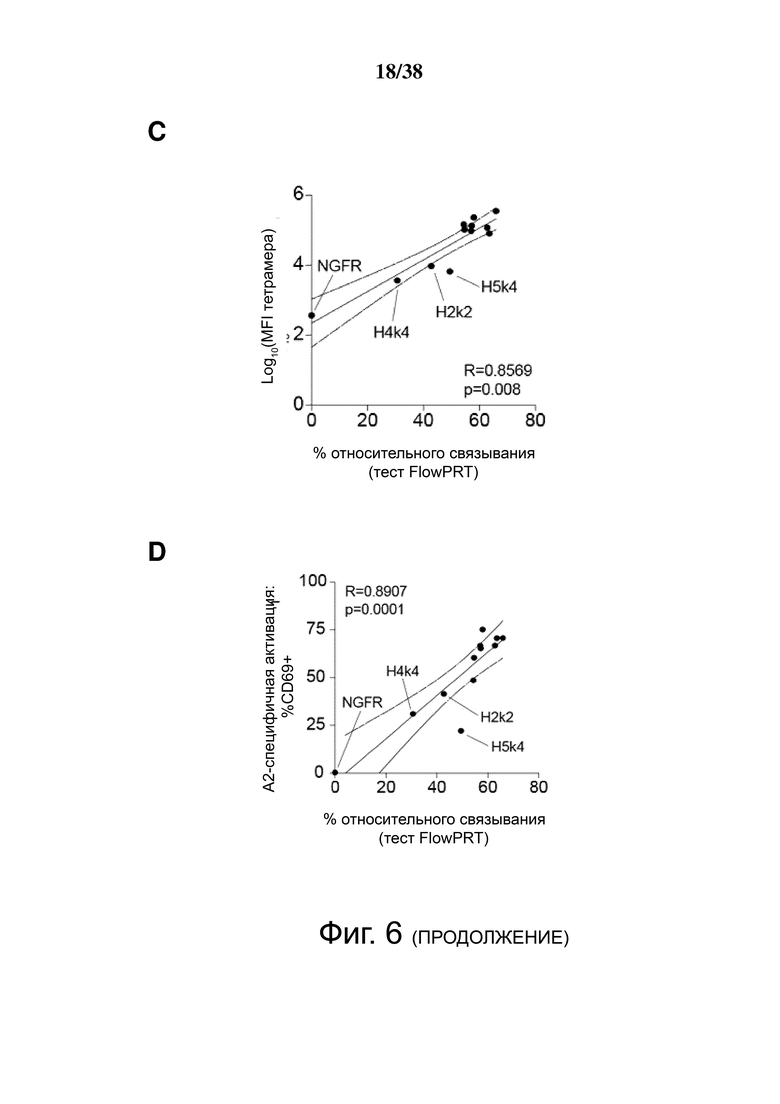
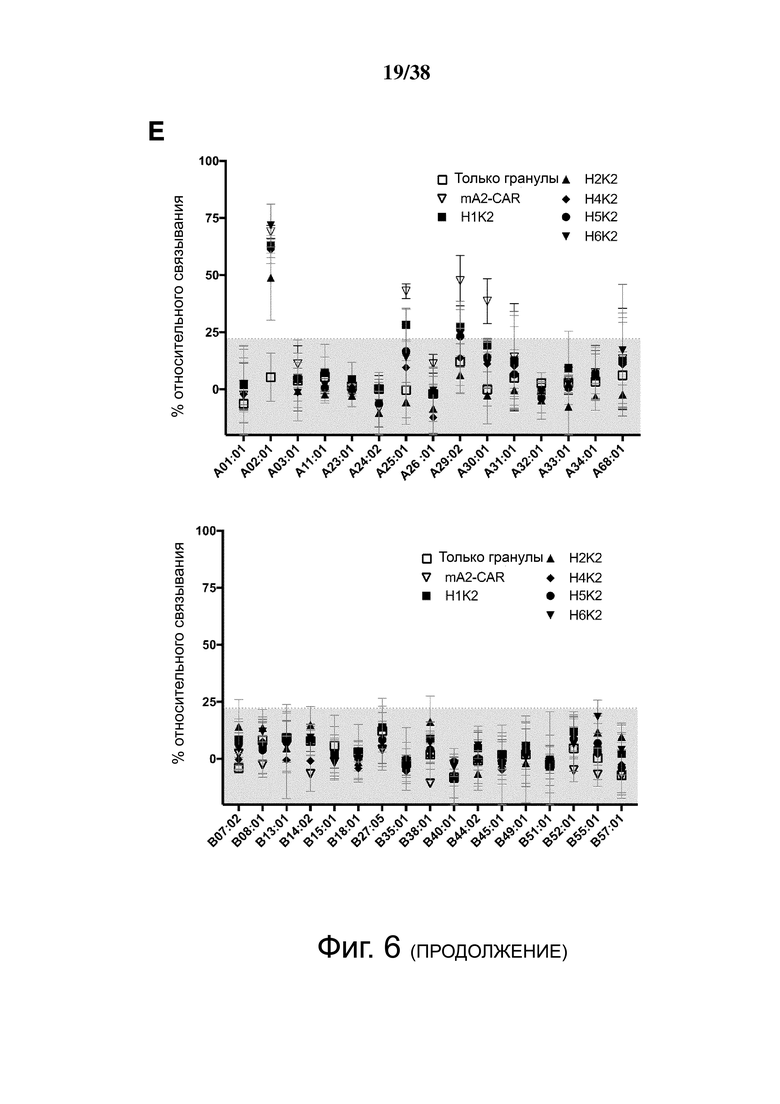
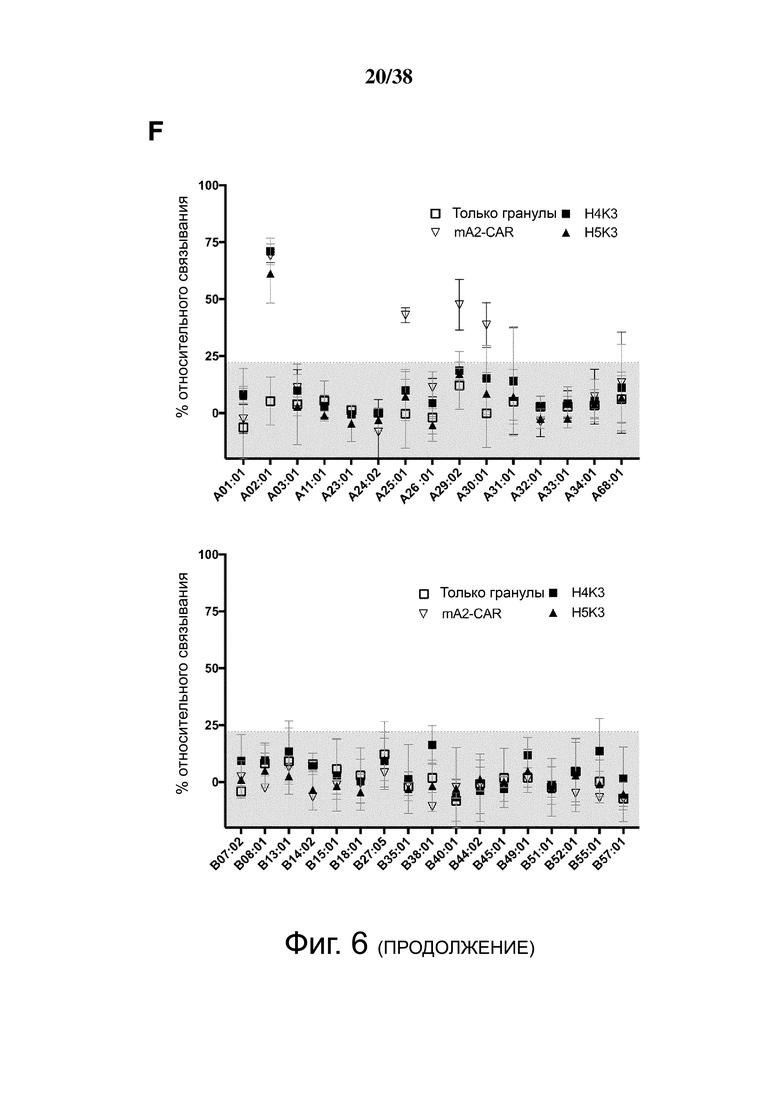
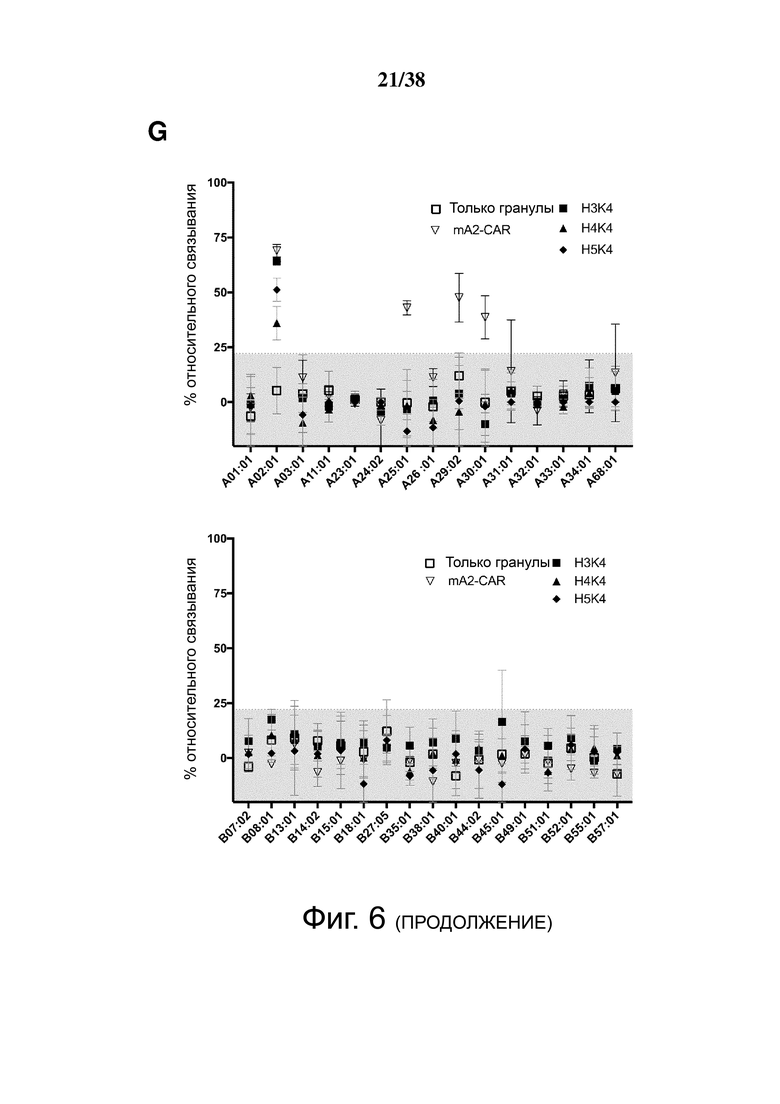
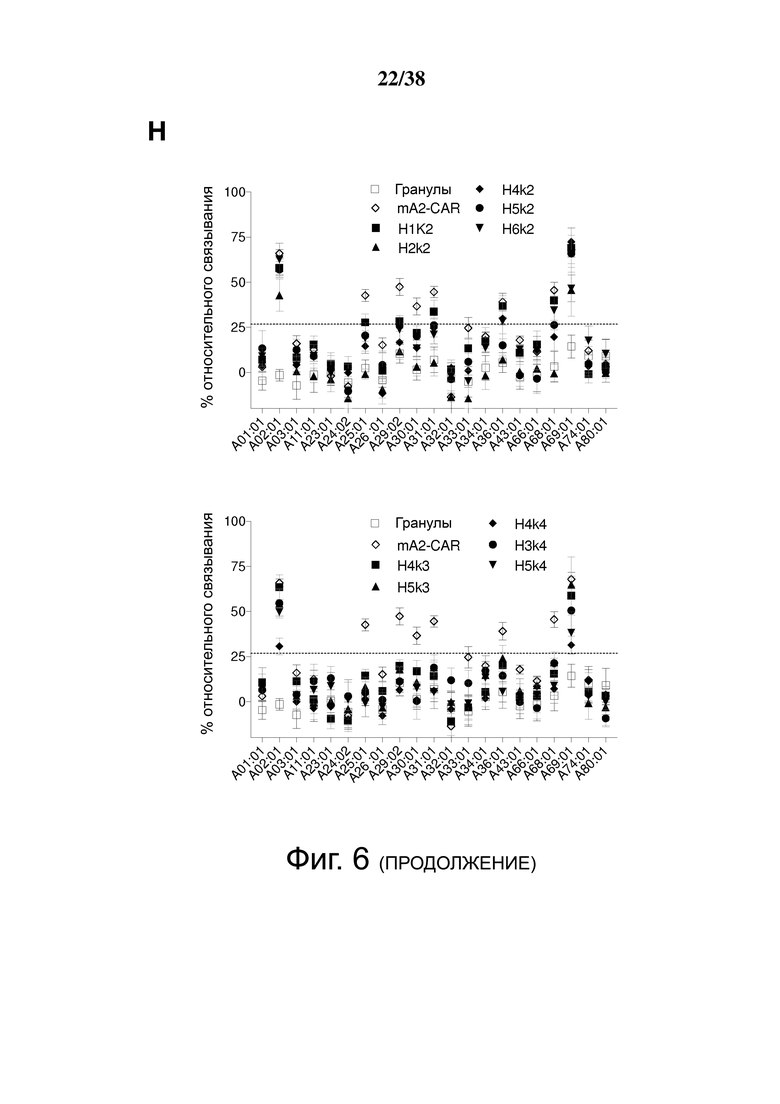

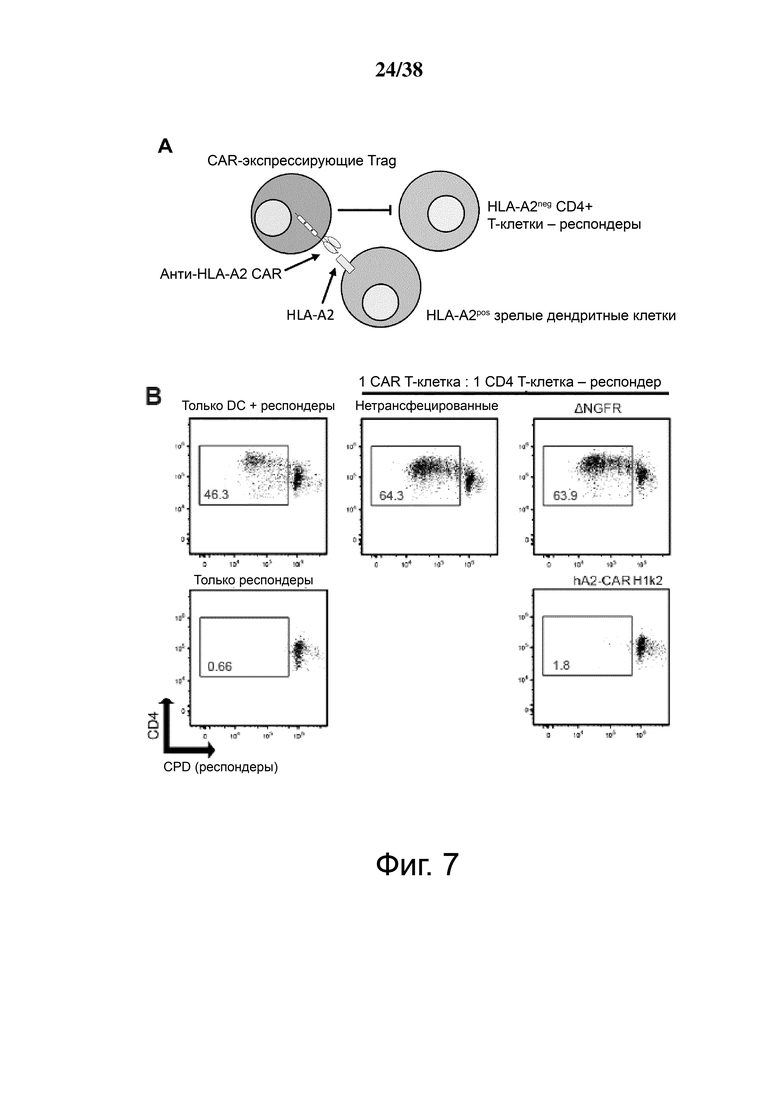
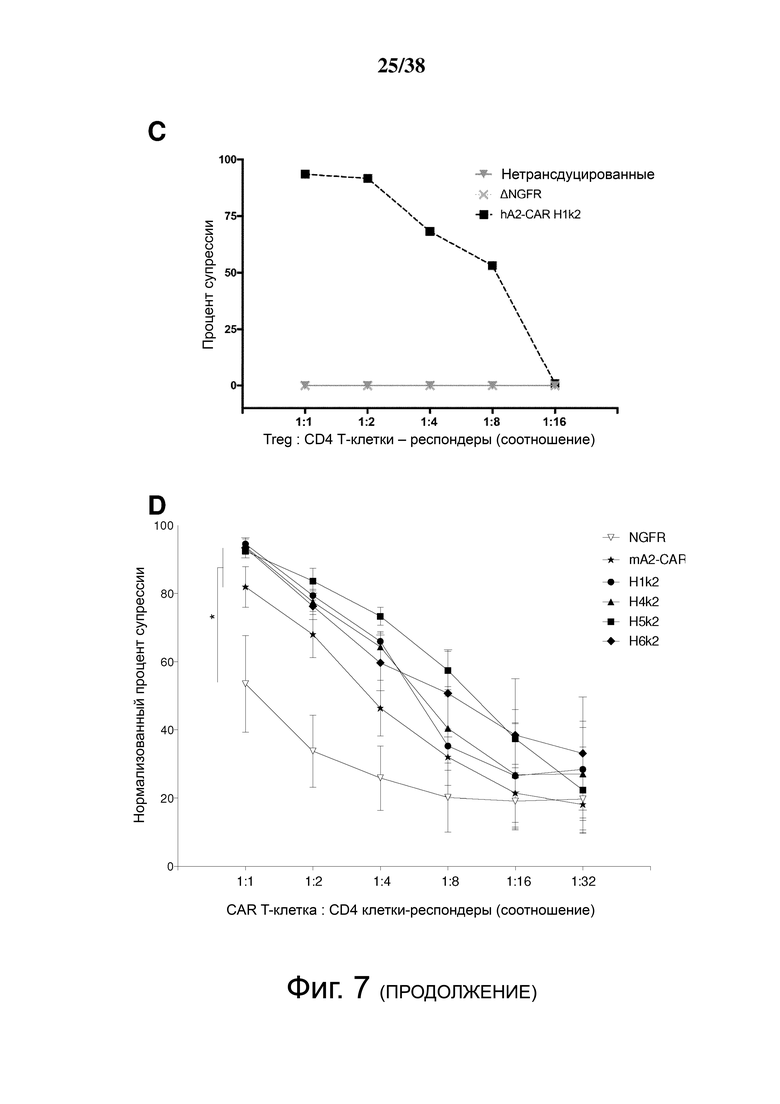
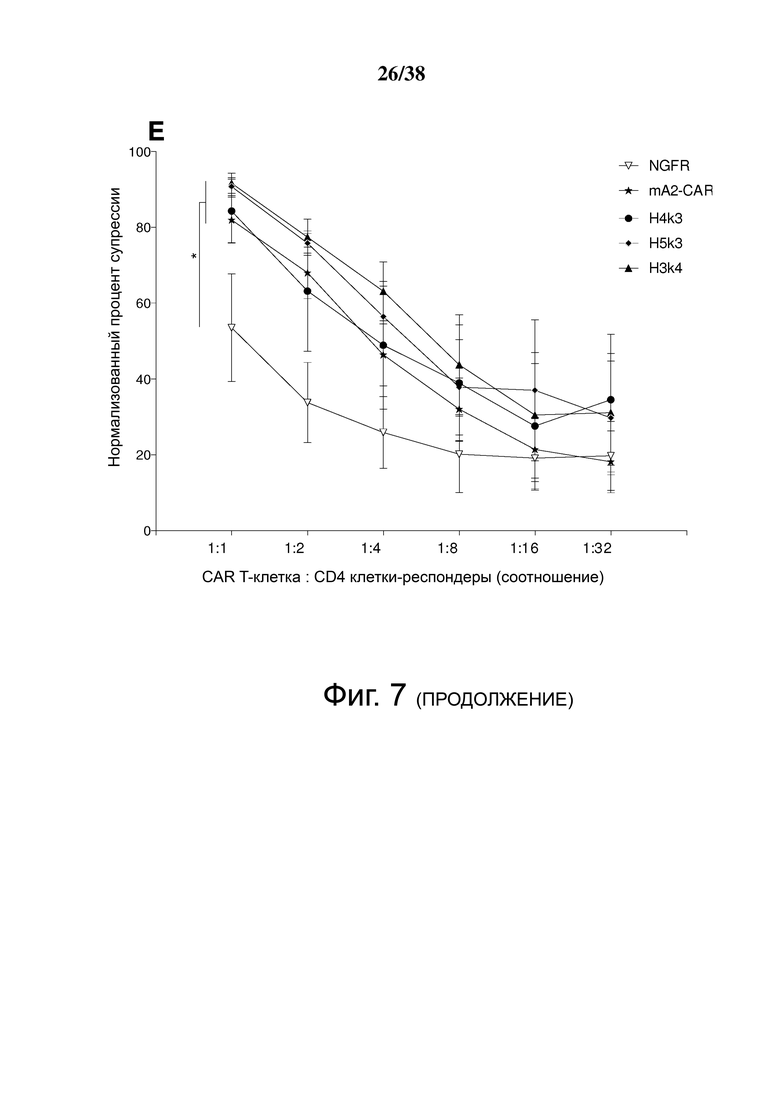
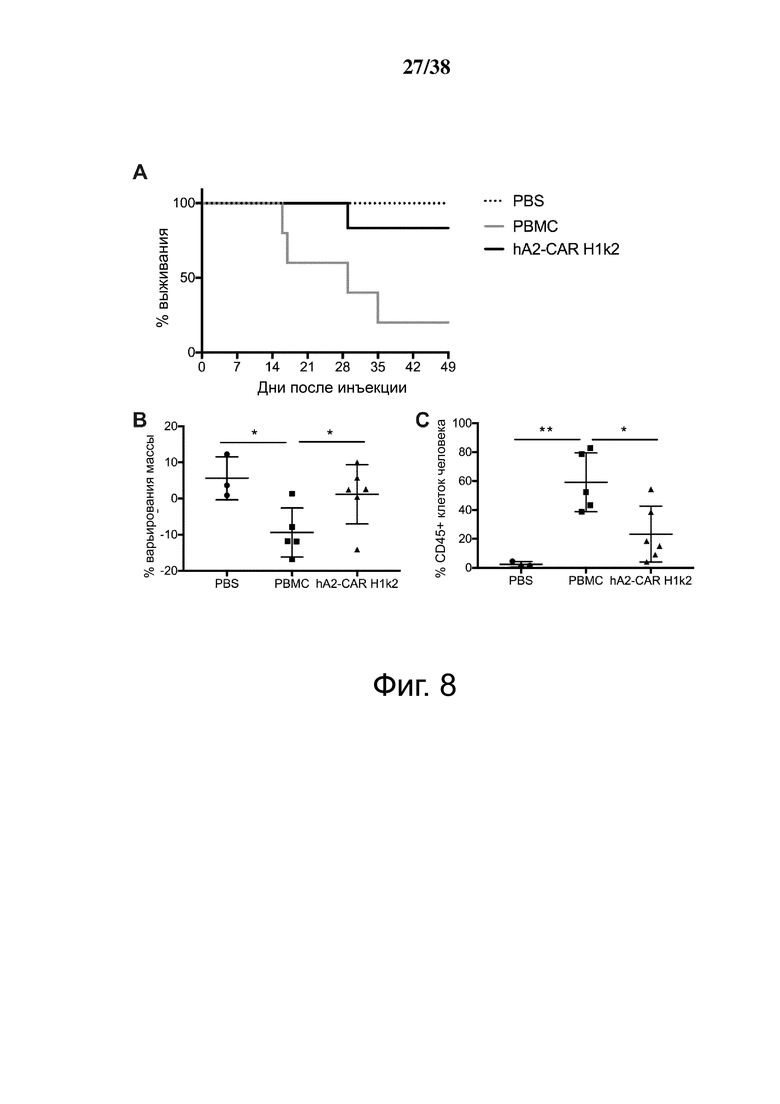
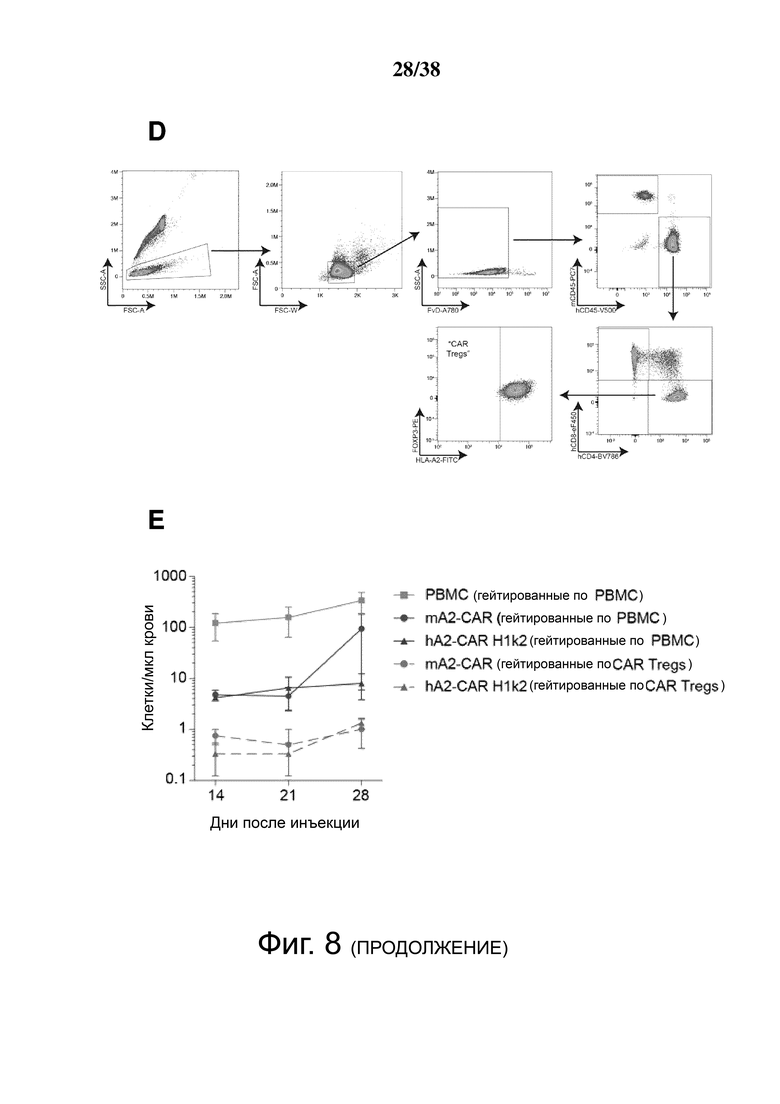
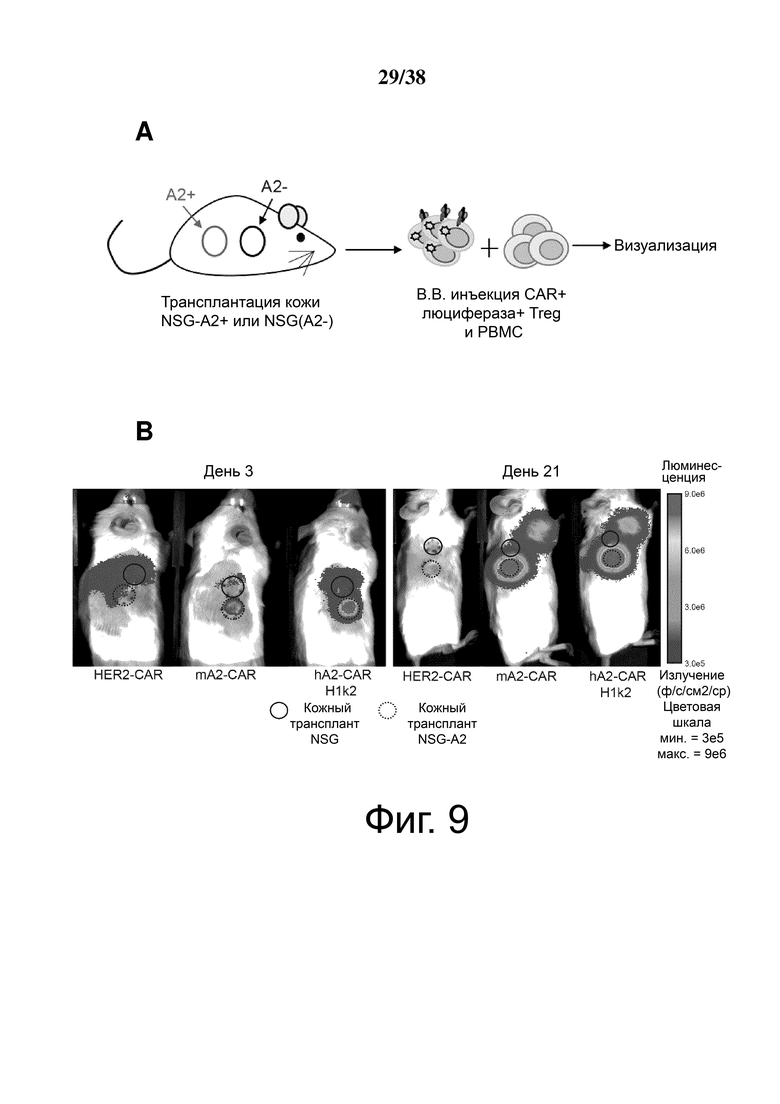
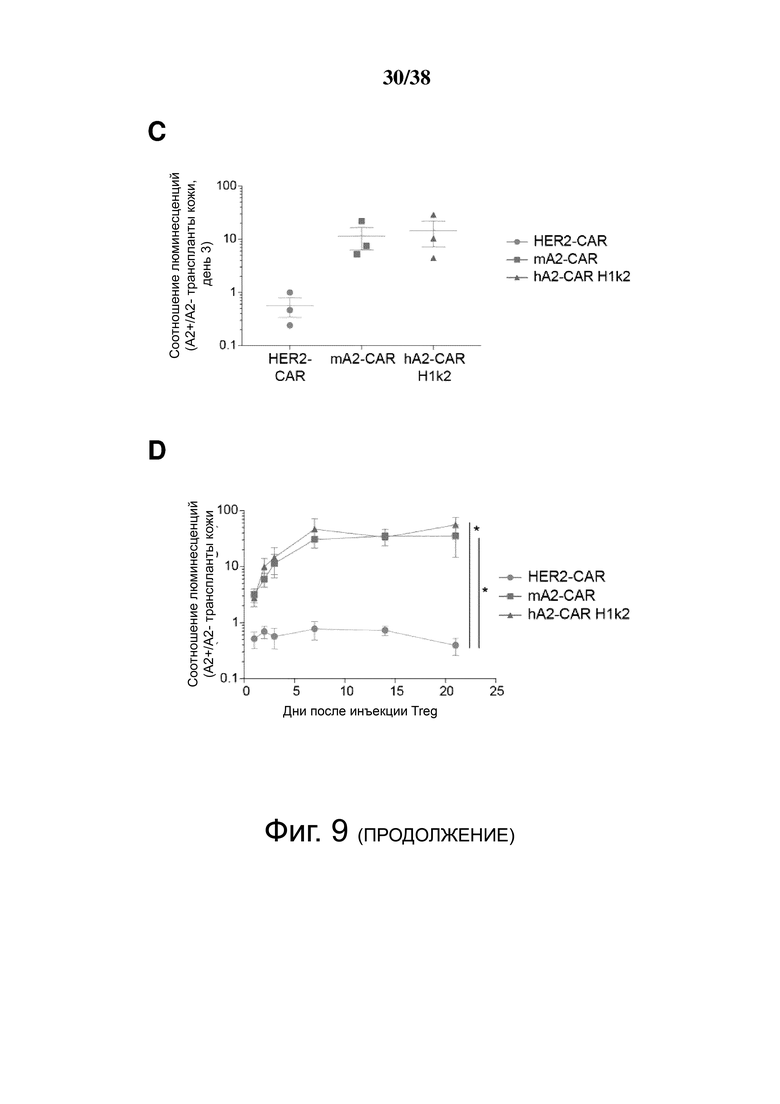
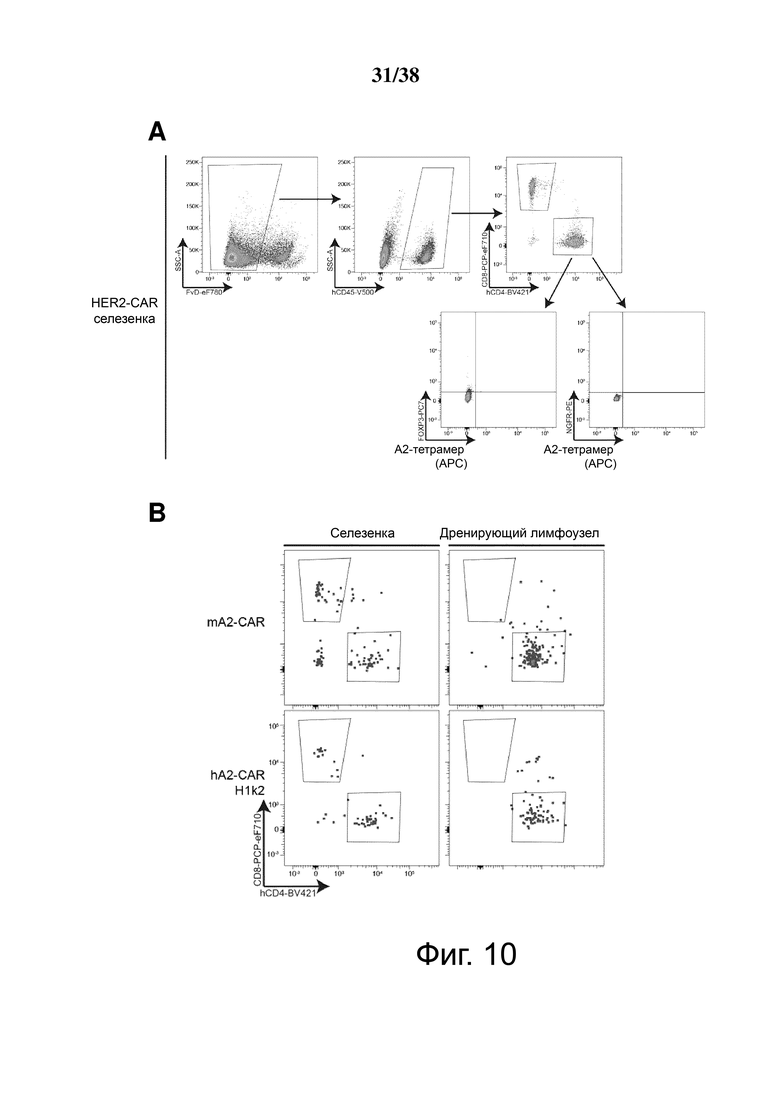

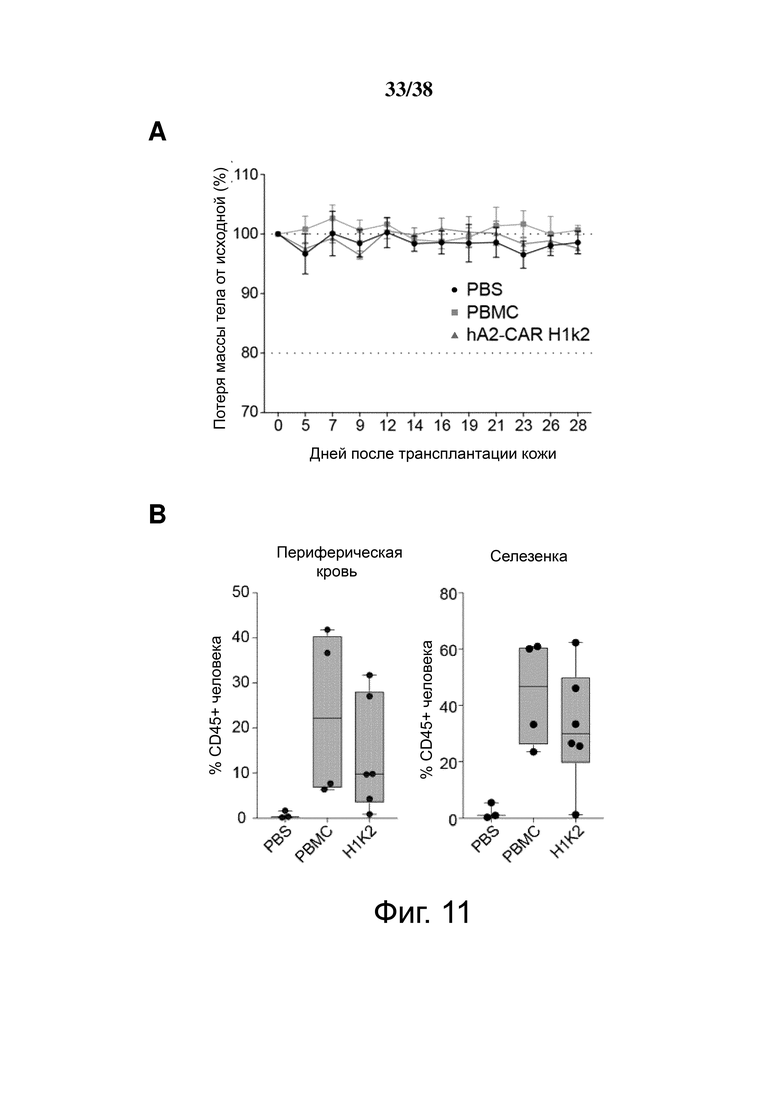
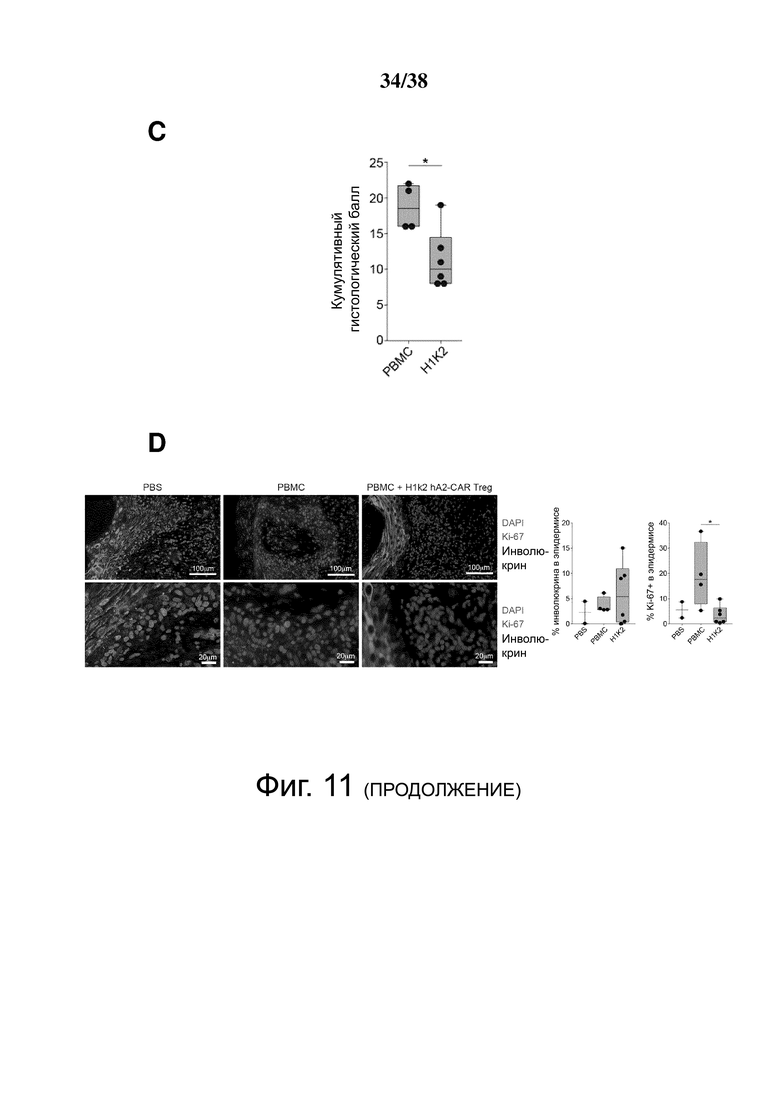
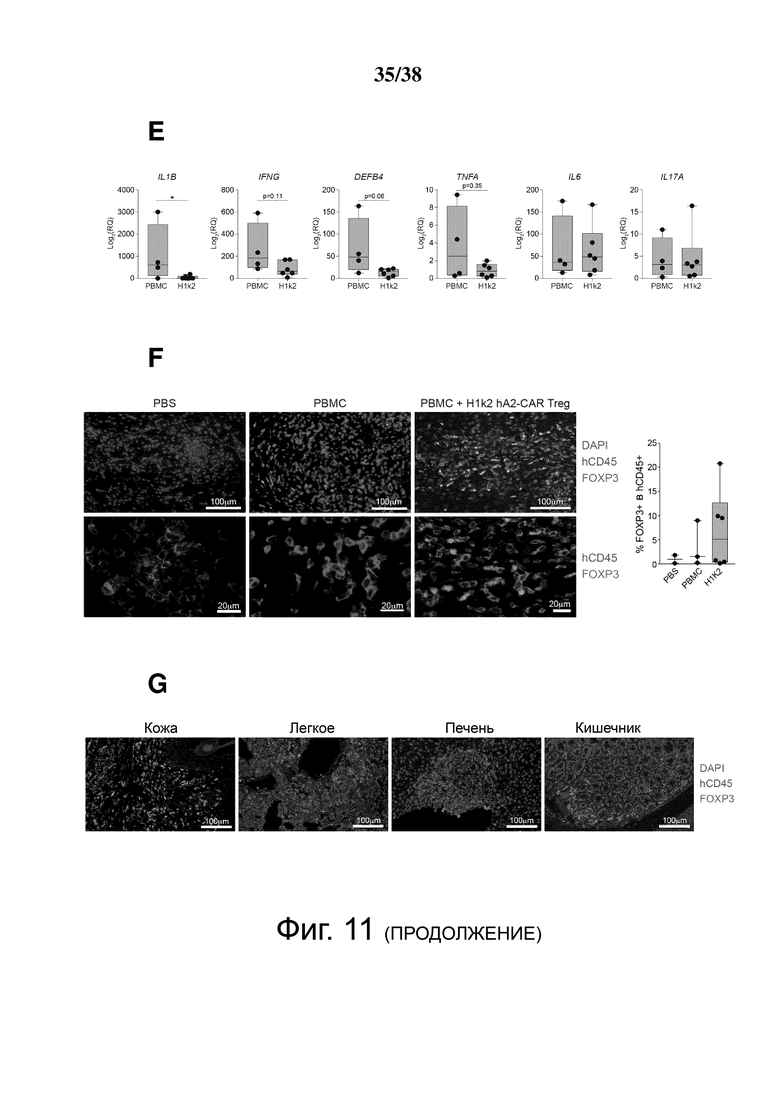
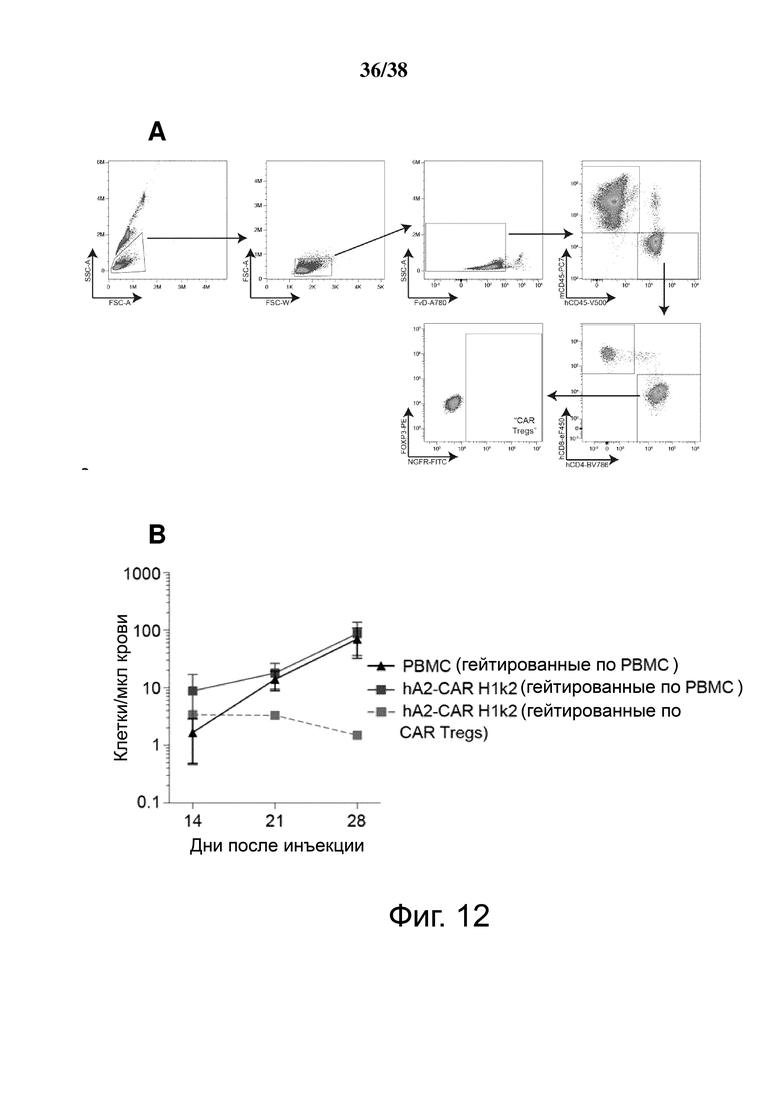
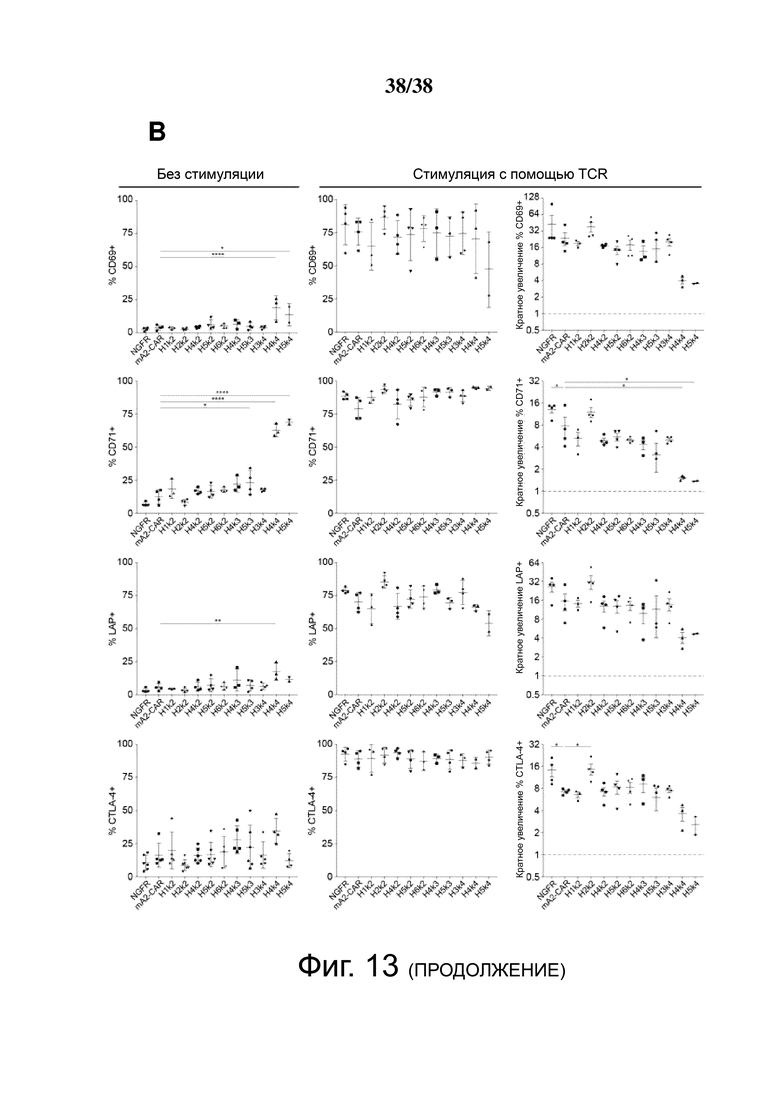
Изобретение относится к области биотехнологии. Предложены гуманизированные анти-HLA-A2 антитела. Также предлагаются CAR, которые включают гуманизированные анти-HLA-A2 антитела. Также предлагаются модифицированные клетки, включающие CAR, и способы применения таких модифицированных клеток для стимулирования иммунной толерантности у субъекта. Изобретение обеспечивает получение CAR со сниженной перекрестной реактивностью к нескольким аллельным вариантам HLA-A. 9 н. и 9 з.п. ф-лы, 13 ил., 9 табл.
1. Гуманизированное анти-HLA-A2 антитело или его антигенсвязывающий фрагмент, где указанное гуманизированное анти-HLA-A2 антитело или его антигенсвязывающий фрагмент содержат:
а) вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность SEQ ID NO: 61, и вариабельный домен легкой цепи (VL), содержащий аминокислотную последовательность SEQ ID NO: 68;
b) VH, содержащий аминокислотную последовательность SEQ ID NO: 64, и VL, содержащий аминокислотную последовательность SEQ ID NO: 68;
c) VH, содержащий аминокислотную последовательность SEQ ID NO: 65, и VL, содержащий аминокислотную последовательность SEQ ID NO: 68;
d) VH, содержащий аминокислотную последовательность SEQ ID NO: 66, и VL, содержащий аминокислотную последовательность SEQ ID NO: 68;
e) VH, содержащий аминокислотную последовательность SEQ ID NO: 64, и VL, содержащий аминокислотную последовательность SEQ ID NO: 69;
f) VH, содержащий аминокислотную последовательность SEQ ID NO: 65, и VL, содержащий аминокислотную последовательность SEQ ID NO: 69;
g) VH, содержащий аминокислотную последовательность SEQ ID NO: 63, и VL, содержащий аминокислотную последовательность SEQ ID NO: 70.
2. Гуманизированное анти-HLA-A2 антитело по п. 1, где указанный вариабельный домен тяжелой цепи содержит аминокислотную последовательность, SEQ ID NO: 61, и/или где указанный вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 68.
3. Гуманизированное анти-HLA-A2 антитело по п. 1, где указанное антитело представляет собой scFv, предпочтительно где указанный scFv, включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 73, 81, 83, 84, 87, 88, и 91.
4. Гуманизированное анти-HLA-A2 антитело или его антигенсвязывающий фрагмент по п. 3, где указанный scFv содержит аминокислотную последовательность SEQ ID NO: 73.
5. Химерный антигенный рецептор (CAR), содержащий:
(i) внеклеточный домен, содержащий гуманизированное анти-HLA-A2 антитело или его антигенсвязывающий фрагмент по п. 1;
(ii) трансмембранный домен; и
(iii) цитоплазматический домен, включающий внутриклеточный сигнальный домен;
где указанный CAR способен экспрессироваться в иммунной клетке, так что указанный CAR специфически связывается с HLA-A2.
6. CAR по п. 5, дополнительно включающий шарнирную область, предпочтительно где указанная шарнирная область включает область стебля CD8α.
7. CAR по п. 5, где указанный трансмембранный домен включает трансмембранный домен белка, выбранного из группы, состоящей из: CD3 гамма, CD3 дельта, CD3 эпсилон, CD3 дзета, альфа-цепи Т-клеточного рецептора, бета-цепи Т-клеточного рецептора, гамма-цепи Т-клеточного рецептора, дельта-цепи Т-клеточного рецептора, CD28, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154 и любой их комбинации, предпочтительно где указанный трансмембранный домен содержит трансмембранный домен CD8.
8. CAR по п. 5, где указанный внутриклеточный сигнальный домен содержит функциональный сигнальный домен белка, выбранного из группы, состоящей из: CD3 гамма, CD3 дельта, CD3 эпсилон, CD3 дзета, FcR гамма, FcR альфа, FcR эпсилон, CD5, CD22, CD79a, CD79b и CD66d и любой их комбинации, предпочтительно где указанный внутриклеточный сигнальный домен содержит функциональный сигнальный домен из CD3 дзета.
9. CAR по п. 5, где указанный внутриклеточный сигнальный домен дополнительно содержит костимулирующий домен, предпочтительно где указанный костимулирующий домен включает функциональный сигнальный домен белка, выбранного из группы, состоящей из OX40, CD27, CD28, антигена-1, связанного с функцией лимфоцитов ((LFA-1) (CD11a/CD18), TNFR1 (CD120a/TNFRSF1A), TNFR2 (CD120b/TNFRSF1B), CTLA-4 (CD152), CD95, ICOS (CD278), 4-1BB (CD137), CD2, CD30, CD40, PD-1, CD7, LIGHT, NKG2C, B7-H3, ICAM-1, лиганд, который специфически связывается с CD83, IL2ra (CD25), IL6Ra (CD126), IL-7Ra (CD127), IL-13RA1, IL-13RA2, IL-33R(IL1RL1), IL-10RA, IL-10RB, IL-4R, IL-5R (CSF2RB), ARHR, рецептор BAFF, IL-21R, TGFbR1, TGFbR2, TGFbR3, общей гамма-цепи и любой их комбинации, более предпочтительно где указанный костимулирующий домен включает функциональный сигнальный домен белка, выбранного из CD28 и 4-1BB, и еще более предпочтительно где указанный костимулирующий домен содержит функциональный сигнальный домен СD28.
10. CAR по п. 5, где указанный внутриклеточный сигнальный домен содержит костимулирующий домен CD28 и первичный сигнальный домен CD3 дзета.
11. Модифицированная иммунная клетка для стимулирования иммунной толерантности у субъекта, включающая CAR по п. 5, предпочтительно где указанная модифицированная иммунная клетка представляет собой регуляторную Т-клетку (Treg).
12. Нуклеиновая кислота, кодирующая CAR по п. 5.
13. Экспрессирующий вектор, включающий нуклеиновую кислоту по п. 12.
14. Иммунная клетка для стимулирования иммунной толерантности у субъекта, включающая экспрессирующий вектор по п. 13, предпочтительно где указанная иммунная клетка представляет собой регуляторную T-клетку (Treg).
15. Фармацевтическая композиция, включающая множество модифицированных иммунных клеток по п. 11.
16. Применение модифицированной иммунной клетки по п. 11 или фармацевтической композиции по п. 15 для стимулирования иммунной толерантности у объекта, предпочтительно где указанная иммунная толерантность представляет собой толерантность к трансплантированному органу или ткани, и/или для предотвращения или лечения болезни «трансплантат против хозяина» (GVHD) у объекта, где предпочтительно указанный объект подвергается или перенес трансплантацию гемопоэтических стволовых клеток и/или для предотвращения или лечения отторжения трансплантата органа или ткани у объекта.
17. Применение по п. 16, где объект дополнительно получает иммуносупрессивный агент.
18. Набор для стимулирования иммунной толерантности у объекта, в первой части которого содержится модифицированная иммунная клетка по п. 11 и во второй части которого содержится другой терапевтический агент, предпочтительно иммуносупрессивный агент.
| US 2013078243 A1, 28.03.2013 | |||
| NAKAUCHI Y | |||
| et al., Effective treatment against severe graft-versus-host disease with allele-specific anti-HLA monoclonal antibody in a humanized mouse model" EXPERIMENTAL HEMATALOGY, 2014, vol | |||
| Зубчатое колесо со сменным зубчатым ободом | 1922 |
|
SU43A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| MACDONALD K.G | |||
| et al., Alloantigen-specific regulatory T cells generated with a chimeric antigen | |||

