Область техники
Настоящее изобретение относится к области биотехнологии и, в частности, к Т-клеточному рецептору (TCR), способному распознавать полипептид, происходящий из белка AFP. Изобретение также относится к получению и применениям таких рецепторов.
Предшествующий уровень техники
Существует только два типа молекул, которые могут распознавать антигены специфичным образом. Одним является иммуноглобулин или антитело; и другим является T-клеточный рецептор (TCR), который представляет собой α/β или γ/δ гетеродимерный гликопротеин, находящийся на клеточной мембране. Физический репертуар TCR иммунной системы образуется в тимусе в результате рекомбинации V(D)J с последующими позитивной и негативной селекциями. В периферической окружающей среде TCR опосредуют распознавание T-клетками конкретных комплексов главный комплекс гистосовместимости-пептид (pMHC) и, в связи с этим, являются крайне важными для иммунологического функционирования клеток в иммунной системе.
TCR представляет собой единственный рецептор для презентации конкретных пептидных антигенов в главном комплексе гистосовместимости (MHC). Экзогенный или эндогенный пептиды могут представлять собой единственный признак патологии в клетке. В иммунной системе прямой физический контакт T-клетки и антигенпрезентирующей клетки (APC) будет инициирован связыванием антиген-специфичных TCR с комплексами pMHC. В таком случае, происходит взаимодействие других мембранных молекул в T-клетке и APC, и последующая клеточная сигнализация и другие физиологические реакции инициируются таким образом, что спектр разных антиген-специфичных T-клеток оказывает иммунное действие на свои клетки-мишени.
Молекулярные лиганды MHC класса I и класса II, соответствующие TCR, также представляют собой белки суперсемейства иммуноглобулинов, но являются специфичными в отношении презентации антигена, и разные индивиды имеют разные MHC, презентирующие, таким образом, разные короткие пептиды в одном белковом антигене на поверхности соответствующих клеток APC. Человеческий MHC обычно называют геном HLA или комплексом HLA.
AFP, также известный как α-фетопротеин, представляет собой белок, экспрессируемый во время эмбрионального развития, и главный компонент эмбриональной сыворотки. Во время развития AFP относительно на высоком уровне экспрессируется в желточном мешочке и печени, и впоследствии ингибируется. При гепатоцеллюлярной карциноме экспрессия AFP активируется (Butterfield et al. J Immunol., 2001, Apr 15; 166(8): 5300-8). После продукции AFP в клетке, он деградирует на низкомолекулярные полипептиды и объединяется с молекулами MHC (главный комплекс гистосовместимости) с образованием комплекса, который презентируется на поверхности клетки. FMNKFIYEI представляет собой короткий пептид, происходящий из антигена AFP, и является мишенью в случае лечения заболеваний, связанных с AFP.
Таким образом, маркер, по которому TCR может нацеливаться на опухолевые клетки, предоставляется из комплекса FMNKFIYEI-HLA A0201. TCR, способный связываться с комплексом FMNKFIYEI-HLA A0201, имеет большое прикладное значение для лечения опухолей. Например, TCR, который может нацеливаться на маркер опухолевых клеток, можно применять для доставки цитотоксических агентов или иммуностимуляторов к клеткам-мишеням, или им могут быть трансформированы T-клетки, таким образом, что T-клетки, экспрессирующие этот TCR, могут уничтожать опухолевые клетки и могут быть даны пациентам во время курса лечения, называемого адоптивной иммунотерапией. Для первой цели, идеальный TCR будет обладать высокой аффинностью, таким образом, чтобы TCR мог располагаться на клетках-мишенях длительное время. Для второй цели, предпочтительно применять TCR со средней аффинностью. Таким образом, специалисты в данной области полностью посвящают себя разработке TCR, нацеливающихся на маркеры опухолевых клеток, которые можно было бы применять в разных целях.
Краткое описание изобретения
Одной целью настоящего изобретения является обеспечение TCR, обладающего более высокой аффинностью в отношении комплекса FMNKFIYEI-HLA A0201.
Другой целью настоящего изобретения является обеспечение способа получения TCR указанного выше типа и применений TCR указанного выше типа.
В первом аспекте изобретения предложен T-клеточный рецептор (TCR), который обладает активностью связывания с комплексом FMNKFIYEI-HLA A0201.
В другом предпочтительном воплощении T-клеточный рецептор (TCR) обладает активностью связывания с комплексом FMNKFIYEI-HLA A0201, и данный T-клеточный рецептор содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, причем вариабельный домен цепи α TCR содержит три области CDR (область, определяющая комплементарность), и референсные последовательности данных трех областей CDR вариабельного домена цепи α TCR перечислены ниже,
CDR1α: TSINN
CDR2α: IRSNERE
CDR3α: ATDPRTGANSKLT, и содержит по меньшей мере одну из следующих мутаций:
и/или, вариабельный домен цепи β TCR содержит три области CDR, и референсные последовательности трех областей CDR вариабельного домена цепи β TCR перечислены, как следует ниже:
CDR1β: SEHNR
CDR2β: FQNEAQ
CDR3β: ASEGLAYEQY, и содержит по меньшей мере одну из следующих мутаций:
В еще одном предпочтительном воплощении существуют 1-8 мутаций в 3 областях CDR вариабельного домена цепи α TCR и/или 1-8 мутаций в 3 областях CDR вариабельного домена цепи β TCR.
В еще одном предпочтительном воплощении число мутаций в областях CDR цепи α TCR может составлять 1, 2, 3, 4, 5, 6, 7 или 8.
В еще одном предпочтительном воплощении число мутаций в CDR-областях цепи β TCR может составлять 4, 5, 6, 7 или 8.
В еще одном предпочтительном воплощении аффинность TCR в отношении комплекса FMNKFIYEI-HLA A0201 по меньшей мере в 2 раза выше аффинности TCR дикого типа.
В еще одном предпочтительном воплощении TCR может также связываться с комплексом FMNKFIYEI-HLA A0207.
В еще одном предпочтительном воплощении вариабельный домен цепи α TCR содержит аминокислотную последовательность, имеющую по меньшей мере 90% гомологию последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 1; и/или вариабельный домен цепи β TCR содержит аминокислотную последовательность, имеющую по меньшей мере 90% гомологию последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 2.
В еще одном предпочтительном воплощении мутантное положение в вариабельном домене цепи α TCR включает положение 7 и/или положение 8 CDR3α.
В еще одном предпочтительном воплощении мутация в вариабельном домене цепи α TCR включает следующую группу:
и/или мутация в вариабельном домене цепи β TCR включает следующую группу:
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и указанный вариабельный домен цепи α TCR содержит CDR1α, CDR2a и CDR3α.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и указанный вариабельный домен цепи α TCR содержит CDR1α, CDR2α и CDR3α, где аминокислотная последовательность CDR1α выбрана из группы, состоящей из: TSINN, TPISS, TDIYK, TAVDS, TEISS и TDVSS.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и указанный вариабельный домен цепи α TCR содержит CDR1α, CDR2α и CDR3α, где аминокислотная последовательность CDR2α представляет собой IRSNERE.
В еще одном предпочтительном воплощении CDR1α содержит последовательность:
T[1αX1][1αX2][1αX3][1αX4], где [1αX1], [1αX2], [1αX3], [1αX4] независимо выбраны из любого природного аминокислотного остатка, соответственно.
В еще одном предпочтительном воплощении [1αX1] представляет собой S, или D, или P, или A, или E.
В еще одном предпочтительном воплощении [1αX2] представляет собой I или V.
В еще одном предпочтительном воплощении [1αX3] представляет собой N, или S, или Y, или D.
В еще одном предпочтительном воплощении [1αX4] представляет собой N, или S, или K.
В еще одном предпочтительном воплощении [1αX1] представляет собой S, или D, или P, или A, или E, [1αX2] представляет собой I или V, [1αX3] представляет собой N, или S, или Y, или D, и [1αX4] представляет собой N, или S, или K.
В еще одном предпочтительном воплощении CDR1α содержит последовательность, выбранную из группы, состоящей из: TSINN, TPISS, TDIYK, TAVDS, TEISS и TDVSS.
В еще одном предпочтительном воплощении CDR3α содержит последовательность: ATDP[3αX1][3αX2][3αX3][3αX4][3αX5][3αX6]KLT, где [3αX1], [3αX2], [3αX3], [3αX4], [3αX5], [3αX6] независимо выбраны из любого природного аминокислотного остатка, соответственно.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и указанный вариабельный домен цепи α TCR содержит CDR1a, CDR2a и CDR3a, где CDR3α содержит последовательность: ATDP[3αX1][3αX2][3αX3][3αX4][3αX5][3αX6]KLT, где [3αX1], [3αX2], [3αX3], [3αX4], [3αX5] и [3αX6] независимо выбраны из любого природного аминокислотного остатка, соответственно.
В еще одном предпочтительном воплощении [3αX1] представляет собой R, или A, или S, или D, или E, или Q, или V, или T.
В еще одном предпочтительном воплощении [3αX2] представляет собой T, или M, или R, или L, или A, или N, или S.
В еще одном предпочтительном воплощении [3αX3] представляет собой G, или A, или Q, или S, или E, или N.
В еще одном предпочтительном воплощении [3αX4] представляет собой A, или S, или G o, или P, или W.
В еще одном предпочтительном воплощении [3αX5] представляет собой N, или S, или T.
В еще одном предпочтительном воплощении [3αX6] представляет собой S, или A, или N, или T, или P.
В еще одном предпочтительном воплощении [3αX1] представляет собой R, или A, или S, или D, или E, или Q, или V, или T, [3αX2] представляет собой T, или M, или R, или L, или A, или N, или S, [3αX3] представляет собой G, или A, или Q, или S, или E, или N, [3αX4] представляет собой A, или S, или G, или P, или W, [3αX5] представляет собой N, или S, или T, и [3αX6] представляет собой S, или A, или N, или T, или P.
В еще одном предпочтительном воплощении CDR3α содержит последовательность, выбранную из группы, состоящей из: ATDPSMSPNSKLT, ATDPSLSPNSKLT и ATDPSLQSNSKLT.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и указанный вариабельный домен цепи β TCR содержит CDR1β, CDR2β и CDR3β, где CDR3β содержит последовательность: ASEGLAYEQY.
В еще одном предпочтительном воплощении CDR1β содержит последовательность: [1βX1][1βX2][1βX3][1βX4]R,
где [1βX1], [1βX2], [1βX3], [1βX4] независимо выбраны из любого природного аминокислотного остатка, соответственно.
В еще одном предпочтительном воплощении [1βX1] представляет собой S, или A, или Q, или G.
В еще одном предпочтительном воплощении [1βX2] представляет собой E, или G, или T, или R, или L.
В еще одном предпочтительном воплощении [1βX3] представляет собой H, или W, или R.
В еще одном предпочтительном воплощении [1βX4] представляет собой N, или A, или P.
В еще одном предпочтительном воплощении CDR2β содержит последовательность:
F[2βX1][2βX2][2βX3][2βX4][2βX5], где [2βX1], [2βX2], [2βX3], [2βX4], [2βX5] независимо выбраны из любого природного аминокислотного остатка, соответственно.
В еще одном предпочтительном воплощении [2βX1] представляет собой Q или H.
В еще одном предпочтительном воплощении [2βX2] представляет собой N или G.
В еще одном предпочтительном воплощении [2βX3] представляет собой E или D.
В еще одном предпочтительном воплощении [2βX4] представляет собой A, или M, или L.
В еще одном предпочтительном воплощении [2βX5] представляет собой Q или E.
В еще одном предпочтительном воплощении CDR2β содержит последовательность, выбранную из группы, состоящей из: FQGDME, FQGDLE и FHGDLE.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и указанный вариабельный домен цепи β TCR содержит CDR1β, CDR2β и CDR3β, где аминокислотная последовательность CDR2β выбрана из группы, состоящей из: FQGDME, FQGDLE и FHGDLE.
В еще одном предпочтительном воплощении вариабельный домен цепи α TCR одновременно не содержит следующие CDR: CDR1α: TSINN; CDR2α: IRSNERE; и CDR3α: ATDPRTGANSKLT.
В еще одном предпочтительном воплощении вариабельный домен цепи β TCR одновременно не содержит следующие CDR: CDR1β: SEHNR; CDR2β: FQNEAQ; и CDR3β: ASEGLAYEQY.
В еще одном предпочтительном воплощении мутация встречается в одной или более областях CDR вариабельного домена цепи α и/или цепи β.
В еще одном предпочтительном воплощении мутация встречается в CDR1 и/или CDR3 цепи α, и/или мутация встречается в CDR1 и/или CDR2 цепи β.
В одном предпочтительном воплощении настоящего изобретения аффинность TCR в отношении комплекса FMNKFIYEI-HLA A0201 по меньшей мере в 2 раза выше аффинности TCR дикого типа; предпочтительно по меньшей мере в 5 раз; и более предпочтительно по меньшей мере в 10 раз.
В еще одном предпочтительном воплощении аффинность TCR в отношении комплекса FMNKFIYEI-HLA A0201 по меньшей мере в 50 раз выше аффинности TCR дикого типа; предпочтительно, по меньшей мере в 100 раз; более предпочтительно по меньшей мере в 500 раз; и наиболее предпочтительно, по меньшей мере в 1000 раз.
В еще одном предпочтительном воплощении аффинность TCR в отношении комплекса FMNKFIYEI-HLA A0201 по меньшей мере в 104 раз выше, чем аффинность TCR дикого типа; предпочтительно, по меньшей мере в 105 раз; и более предпочтительно по меньшей мере в 106 раз.
В частности, константа равновесия диссоциации KD TCR в отношении комплекса FMNKFIYEI-HLA A0201 меньше или равна 20 мкМ;
В еще одном предпочтительном воплощении константа равновесия диссоциации TCR в отношении комплекса FMNKFIYEI-HLA A0201 составляет 5 мкМ ≤ KD ≤ 10 мкМ; предпочтительно, 0,1 мкМ ≤ KD ≤ 1 мкМ; и более предпочтительно, 1 нМ ≤ KD ≤ 100 нМ;
В еще одном предпочтительном воплощении константа равновесия диссоциации TCR в отношении комплекса FMNKFIYEI-HLA A0201 составляет 100 пМ ≤ KD ≤ 1000 пМ; и более предпочтительно, 10 пМ ≤ KD ≤ 100 пМ.
В еще одном предпочтительном воплощении TCR имеет CDR, выбранные из группы, состоящей из:
В еще одном предпочтительном воплощении TCR является растворимым.
В еще одном предпочтительном воплощении TCR представляет собой αβ гетеродимерный TCR или одноцепочечный TCR.
В еще одном предпочтительном воплощении TCR по настоящему изобретению представляет собой αβ гетеродимерный TCR, и предпочтительно TCR содержит последовательность константной области цепи α TRAC*01 и последовательность константной области цепи β TRBC1*01 или TRBC2*01. Вариабельный домен цепи α TCR содержит аминокислотную последовательность, обладающую по меньшей мере 90%, предпочтительно по меньшей мере 92%; более предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 90%, 91%, 92% 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 1; и/или вариабельный домен цепи β TCR содержит аминокислотную последовательность, обладающую по меньшей мере 90%, предпочтительно по меньшей мере 92%; более предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 2.
В еще одном предпочтительном воплощении TCR содержит (i) всю или часть цепи α TCR, за исключением ее трансмембранного домена, и (ii) всю или часть цепи β TCR, за исключением ее трансмембранного домена, где обе из (i) и (ii) содержат вариабельный домен и по меньшей мере часть константного домена цепи TCR.
В еще одном предпочтительном воплощении TCR представляет собой αβ гетеродимерный TCR, и искусственная межцепочечная дисульфидная связь содержится между вариабельной областью цепи α и константной областью цепи β TCR.
В еще одном предпочтительном воплощении остатки цистеина, образующие искусственную межцепочечную дисульфидную связь между вариабельной областью цепи α и константной областью цепи β TCR, заменяют одну или более групп аминокислот, выбранных из следующего:
аминокислота в положении 46 TRAV и аминокислота в положении 60 TRBC1*01 или TRBC2*01 экзона 1;
аминокислота в положении 47 TRAV и аминокислота в положении 61 TRBC1*01 или TRBC2*01 экзона 1;
аминокислота в положении 46 TRAV и аминокислота в положении 61 TRBC1*01 или TRBC2*01 экзона 1; или
аминокислота в положении 47 TRAV и аминокислота в положении 60 TRBC1*01 или TRBC2*01 экзона 1.
В еще одном предпочтительном воплощении TCR, содержащий искусственную межцепочечную дисульфидную связь меду вариабельной областью цепи α и константной областью цепи β, содержит вариабельный домен цепи α и вариабельный домен цепи β, а также все или часть константных доменов цепи β, за исключением его трансмембранного домена, однако, он не содержит константный домен цепи α, и вариабельный домен цепи α и цепь β TCR образуют гетеродимер.
В еще одном предпочтительном воплощении TCR, содержащий искусственную межцепочечную дисульфидную связь между вариабельной областью цепи α и константной областью цепи β, содержит (i) всю или часть цепи α TCR, за исключением ее трансмембранного домена, и (ii) всю или часть цепи β TCR, за исключением ее трансмембранного домена, где обе из (i) и (ii) содержат вариабельный домен и по меньшей мере часть константных доменов цепи TCR.
В еще одном предпочтительном воплощении TCR представляет собой αβ гетеродимерный TCR, содержащий (i) всю или часть цепи α TCR, за исключением ее трансмембранного домена, и (ii) всю или часть цепи β TCR, за исключением ее трансмембранного домена, где обе из (i) и (ii) содержат вариабельный домен и по меньшей мере часть константного домена цепи TCR, и искусственная межцепочечная дисульфидная связь содержится между константной областью цепи α и константной областью цепи β.
В еще одном предпочтительном воплощении искусственная межцепочечная дисульфидная связь содержится между константной областью цепи α и константной областью цепи β TCR.
В еще одном предпочтительном воплощении остатки цистеина, образующие искусственную межцепочечную дисульфидную связь между константной областью цепи α TCR и константной областью цепи β, заменены одной или более группами аминокислот, выбранных из нижеследующего:
Thr48 TRAC*01 экзона 1 и Ser57 TRBC1*01 или TRBC2*01 экзона 1;
Thr45 TRAC*01 экзона 1 и Ser77 TRBC1*01 или TRBC2*01 экзона 1;
Tyr10 TRAC*01 экзона 1 и Ser17 TRBC1*01 или TRBC2*01 экзона 1;
Thr45 TRAC*01 экзона 1 и Asp59 TRBC1*01 или TRBC2*01 экзона 1;
Ser15 TRAC*01 экзона 1 и Glu15 TRBC1*01 или TRBC2*01 экзона 1;
Arg53 TRAC*01 экзона 1 и Ser54 TRBC1*01 или TRBC2*01 экзона 1;
Pro89 TRAC*01 экзона 1 и Ala19 TRBC1*01 или TRBC2*01 экзона 1; и
Tyr10 TRAC*01 экзона 1 и Glu20 TRBC1*01 или TRBC2*01 экзона 1.
В еще одном предпочтительном воплощении TCR представляет собой одноцепочечный TCR.
В еще одном предпочтительном воплощении TCR представляет собой одноцепочечный TCR, состоящий из вариабельного домена цепи α и вариабельного домена цепи β, и вариабельный домен цепи α и вариабельный домен цепи β соединены гибкой короткой пептидной последовательностью (линкером).
В еще одном предпочтительном воплощении гидрофобное ядро вариабельного домена цепи α TCR и/или вариабельного домена цепи β является мутантным.
В еще одном предпочтительном воплощении TCR, в котором гидрофобное ядро является мутантным, представляет собой одноцепочечный TCR, состоящий из вариабельного домена α и вариабельного домена β, и вариабельный домен α и вариабельный домен β соединены гибкой короткой пептидной последовательностью (линкером).
В еще одном предпочтительном воплощении TCR по настоящему изобретению представляет собой одноцепочечный TCR, и вариабельный домен цепи α TCR содержит аминокислотную последовательность, обладающую по меньшей мере 85%, предпочтительно по меньшей мере 90%; более предпочтительно по меньшей мере 92%; наиболее предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 3; и/или вариабельный домен цепи β TCR содержит аминокислотную последовательность, обладающую по меньшей мере 85%, предпочтительно по меньшей мере 90%; более предпочтительно по меньшей мере 92%; наиболее предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 4.
В еще одном предпочтительном воплощении аминокислотная последовательность вариабельного домена цепи α TCR выбрана из группы, состоящей из: SEQ ID NO: 9-35, 51-77 и 91-92; и/или аминокислотная последовательность вариабельного домена цепи β TCR выбрана из группы, состоящей из: SEQ ID NO: 36-46, 78-88 и 93-101.
В еще одном предпочтительном воплощении TCR выбран из группы, состоящей из:
В еще одном предпочтительном воплощении TCR выбран из группы, состоящей из:
SEQ ID NO:
SEQ ID NO:
В еще одном предпочтительном воплощении конъюгат связывается с цепью α и/или цепью β TCR на C- или N-конце.
В еще одном предпочтительном воплощении конъюгат, который связывается с TCR, представляет собой детектируемую метку, терапевтическое средство, PK-модифицированную группировку или их комбинацию.
В еще одном предпочтительном воплощении терапевтическое средство, которое связывается с TCR, представляет собой антитело против CD3, связанное с цепью α или β TCR на C- или N-конце.
В одном предпочтительном воплощении T клеточный рецептор (TCR) обладает активностью связывания с комплексом FMNKFIYEI-HLA A2, и содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, вариабельный домен цепи α TCR включает мутацию, как показано в SEQ ID NO: 1, и сайт мутантного аминокислотного остатка включает один или более из 28S, 29I, 30N, 31N, 94R, 95T, 96G, 97A, 98N и 99S, где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 1; и/или вариабельный домен цепи β TCR включает мутацию, как показано в SEQ ID NO: 2, и сайт мутантного аминокислотного остатка включает один или более из 27S, 28E, 29H, 30N, 50Q, 51N, 52E, 53A и 54Q, где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 2;
предпочтительно, при мутации, вариабельный домен цепи α TCR содержит один или более аминокислотных остатков, выбранных из группы, состоящей из: 28D, или 28P, или 28A, или 28E; 29V; 30S, или 30Y, или 30D; 31S, или 31K; 94A, или 94S, или 94V, или 94D, или 94E, или 94Q, или 94T; 95M, или 95R, или 95L, или 95A, или 95N, или 95S; 96A, или 96Q, или 96S, или 96E, или 96N; 97S, или 97G, или 97P, или 97W; 98S, или 98T; 99A, или 99N, или 99T, или 99P; где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 1; и/или при мутации, вариабельный домен цепи β TCR содержит один или более аминокислотных остатков, выбранных из группы, состоящей из: 27A, или 27Q, или 27G; 28G, или 28T, или 28R, или 28L; 29W, или 29R; 30A, или 30P; 50H; 51G; 52D; 53M или 53L; и 54E, где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 2.
Во втором аспекте изобретения предложен поливалентный комплекс TCR, содержащий по меньшей мере две молекулы TCR, и по меньшей мере одна молекула TCR представляет сбой TCR по первому аспекту изобретения.
В третьем аспекте изобретения предложена молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, кодирующую молекулу TCR по первому аспекту изобретения или поливалентный комплекс TCR по второму аспекту изобретения, или ее комплементарную последовательность.
В четвертом аспекте изобретения предложен вектор, содержащий молекулу нуклеиновой кислоты по третьему аспекту изобретения.
В пятом аспекте настоящего изобретения предложена клетка-хозяин, содержащая вектор по четвертому аспекту настоящего изобретения или имеющая экзогенную молекулу нуклеиновой кислоты по третьему аспекту настоящего изобретения, интегрированную в ее геном.
В шестом аспекте изобретения предложена выделенная клетка, экспрессирующая TCR по первому аспекту изобретения.
В седьмом аспекте изобретения предложена фармацевтическая композиция, содержащая фармацевтически приемлемый носитель и TCR по первому аспекту изобретения, или комплекс TCR по второму аспекту изобретения, или клетку по шестому аспекту изобретения.
В восьмом аспекте настоящего изобретения предложен способ лечения заболевания, включающий соответствующее количество TCR по первому аспекту настоящего изобретения, или комплекса TCR по второму аспекту настоящего изобретения, или клетки по шестому аспекту изобретения, или фармацевтической композиции по седьмому аспекту изобретения субъекту, нуждающемуся в этом.
В девятом аспекте изобретения предложено применение TCR по первому аспекту изобретения, или комплекса TCR по второму аспекту изобретения, или клетки по шестому аспекту изобретения, или клетки по шестому аспекту изобретения для получения лекарственного средства для лечения опухоли; и предпочтительно опухоль представляет собой рак печени.
TCR по первому аспекту изобретения или комплекс TCR по второму аспекту изобретения, или клетка по шестому аспекту изобретения для применения в качестве лекарственного средства для лечения опухоли; и предпочтительно опухоль представляет собой рак печени.
В одиннадцатом аспекте изобретения предложен способ получения T-клеточного рецептора по первому аспекту изобретения, включающий следующие стадии:
(i) культивирование клетки-хозяина по пятому аспекту изобретения для экспрессии T-клеточного рецептора по первому аспекту изобретения;
(ii) выделение или очистка T-клеточного рецептора.
Следует понимать, что в объеме настоящего изобретения разные технические характеристики настоящего изобретения и технические характеристики, конкретно описанные далее в данном документе (как в воплощениях), можно объединять друг с другом с образованием нового или предпочтительного технического решения, которые не будут повторяться в данном документе одно за другим.
Описание графических материалов
На Фиг. 1a и Фиг. 1b показаны аминокислотные последовательности вариабельного домена цепи α и β TCR дикого типа, которые способны специфично связываться с комплексом FMNKFIYEI-HLA A0201, соответственно.
На Фиг. 2a и Фиг. 2b показаны аминокислотные последовательности вариабельного домена цепи α и вариабельного домена цепи β одноцепочечной матрицы TCR, сконструированной в настоящем изобретении, соответственно.
На Фиг. 3a и Фиг. 3b показаны ДНК-последовательности вариабельного домена цепи α и вариабельного домена цепи β одноцепочечной матрицы TCR, сконструированной в настоящем изобретении, соответственно.
Фиг. 4a и Фиг. 4b представляют собой аминокислотную последовательность и нуклеотидную последовательность связывающего короткого пептида (линкера) одноцепочечной матрицы TCR, сконструированной в настоящем изобретении, соответственно.
На Фиг. 5(1)-(27) показаны аминокислотные последовательности вариабельного домена цепи α одноцепочечных TCR с высокой аффинностью в отношении комплекса FMNKFIYEI-HLA A0201, соответственно, и подчеркнуты мутантные остатки.
На Фиг. 6(1)-(11) показаны аминокислотные последовательности вариабельного домена цепи β одноцепочечных TCR с высокой аффинностью в отношении комплекса FMNKFIYEI-HLA A0201, соответственно, и подчеркнуты мутантные остатки.
На Фиг. 7a и Фиг. 7b показана аминокислотная последовательность и ДНК-последовательность одноцепочечной матрицы TCR, сконструированной в настоящем изобретении, соответственно.
На Фиг. 8a и 8b показаны аминокислотные последовательности цепей α и β растворимого референсного TCR в настоящем изобретении, соответственно, и подчеркнуты мутантные цистеины.
На Фиг. 9(1)-(29) показаны аминокислотные последовательности вариабельного домена цепи α гетеродимерного TCR с высокой аффинностью в отношении комплекса FMNKFIYEI-HLA A0201, соответственно, и подчеркнуты мутантные остатки.
На Фиг. 10(1)-(20) показаны аминокислотные последовательности вариабельного домена цепи β гетеродимерного TCR с высокой аффинностью в отношении комплекса FMNKFIYEI-HLA A0201, соответственно, и подчеркнуты мутантные остатки.
На Фиг. 11a и Фиг. 11b показаны аминокислотные последовательности цепи α и β TCR дикого типа, которые способны специфично связываться с комплексом FMNKFIYEI-HLA A0201, соответственно.
Фиг. 12 представляет собой кривую связывания растворимого референсного TCR, а именно, TCR дикого типа с комплексом FMNKFIYEI-HLA A0201.
На Фиг. 13a и 13b показаны результаты эксперимента по функции активации эффекторных клеток, трансфицированных TCR с высокой аффинностью, по настоящему изобретению.
На Фиг. 14a, 14b показаны результаты эксперимента по функции «киллинга» эффекторных клеток, трансфицированных TCR с высокой аффинностью, по настоящему изобретению.
На Фиг. 15a, 15b, 15c, 15d, 15e, 15f и 15g показаны результаты эксперимента по верификации функции активации слитого белка, образованного из высокоаффинного TCR по настоящему изобретению и антитела против CD3.
На Фиг. 16 показан результат эксперимента по верификации функции «киллинга» слитого белка, образованного из высокоаффинного TCR по настоящему изобретению и антитела против CD3.
На Фиг. 17a и Фиг. 17b показаны внеклеточные аминокислотные последовательности цепей α и β TCR дикого типа, которые могут специфично связываться с комплексом FMNKFIYEI-HLA A0201, соответственно.
Способы осуществления изобретения
Посредством обширного и интенсивного исследования получали высокоаффинный T-клеточный рецептор (TCR), распознающий короткий пептид FMNKFIYEI (происходящий из белка AFP), и короткий пептид FMNKFIYEI презентирован в форме комплекса пептид-HLA A2. Высокоаффинный TCR имеет мутацию в трех областях CDR его вариабельного домена цепи α:
CDR1α: TSINN
CDR2α: IRSNERE
CDR3α: ATDPRTGANSKLT; и/или имеет мутацию в трех областях CDR его вариабельного домена цепи β:
CDR1β: SEHNR
CDR2β: FQNEAQ
CDR3β: ASEGLAYEQY; и после мутации аффинность и/или время полужизни связывания TCR по настоящему изобретению для упомянутого выше комплекса FMNKFIYEI-HLA A0201 по меньшей мере в 2 раза больше, чем аффинность и/или время полужизни связывания TCR дикого типа.
Перед описанием настоящего изобретения следует понимать, что изобретение не ограничивается конкретными способами и описанными экспериментальными условиями, в связи с этим способы и условия могут варьировать. Также следует понимать, что терминология, используемая в данном документе, предназначена для описания конкретных воплощений и не предназначена для ограничения, и объем настоящего изобретения будет ограничиваться только прилагаемой формулой изобретения.
Все технические и научные термины, используемые в данном документе, имеют то же значение, обычно подразумеваемое специалистом в области, к которой принадлежит данное изобретение, если не определено иное.
Несмотря на то, что любые способы и материалы, аналогичные или эквивалентные способам и материалам, описанным в настоящем изобретении, могут быть использованы на практике или тестировании настоящего изобретения, предпочтительные способы и материалы приведены в качестве примера в данном документе.
Термины
T-клеточный рецептор (TCR)
Международная информационная система по иммуногенетике (IMGT) может использоваться для описания TCR. Нативный αβ гетеродимерный TCR имеет цепь α и цепь β. В общем, каждая цепь содержит вариабельную область, область соединения и константную область, и цепь β обычно также содержит короткую гипервариабельную область между вариабельной областью и областью соединения, которая, однако, часто рассматривается как часть области соединения. Область соединения TCR определяется уникальными TRAJ и TRBJ IMGT, и константная область TCR определяется TACT и TRBC IMGT.
Каждая вариабельная область содержит три CDR (области, определяющие комплементарность), CDR1, CDR2 и CDR3, которые являются химерными в каркасной последовательности. В номенклатуре IMGT разные номера TRAV и TRBV относятся к разным типам Vα и типам Vβ, соответственно. В системе IMGT существуют следующие символы для константного домена цепи α: TRAC*01, где «TR» означает ген T-клеточного рецептора; «A» означает ген цепи α; C означает константную область; «*01» означает аллель гена 1. Имеют место следующие символы для константного домена цепи β: TRBC1*01 или TRBC2*01, где «TR» означает ген T-клеточного рецептора; «B» означает ген цепи β; C означает константную область; «*01» означает аллель гена 1. Константная область цепи α уникально определена, и в виде цепи β, существует два возможных гена константной области «C1» и «C2». Специалист в данной области может получать последовательности генов константной области цепей α и β TCR посредством раскрытой базы данных IMGT.
Цепи α и β TCR обычно рассматриваются как имеющие два «домена», соответственно, а именно, вариабельный домен и константный домен. Вариабельный домен состоит из соединенной вариабельной области и константной области. Таким образом, в описании изобретения и формуле изобретения настоящей заявки, «вариабельный домен цепи α TCR» относится к соединенной области TRAV и TRAJ и т.п., «вариабельный домен цепи β TCR» относится к соединенной области TRBV и TRBD/TRBJ. Три CDR вариабельного домена цепи α представляют собой CDR1α, CDR2α и CDR3α, соответственно; и три CDR вариабельного домена цепи β TCR представляют собой CDR1β, CDR2β и CDR3β, соответственно. Каркасные последовательности вариабельных доменов TCR по изобретению могут происходить из мыши или человека, предпочтительно из человека. Константный домен TCR содержит внутриклеточный участок, трансмембранную область и внеклеточный участок. Для получения растворимого TCR для определения аффинности между TCR и комплексом FMNKFIYEI-HLA A2, TCR по изобретению предпочтительно не содержит трансмембранную область. Более предпочтительно, аминокислотная последовательность TCR по настоящему изобретению относится к внеклеточной аминокислотной последовательности TCR.
Аминокислотная последовательность цепи α и аминокислотная последовательность цепи β «TCR дикого типа», описанные в настоящем изобретении, представляют собой SEQ ID NO: 89 и SEQ ID NO: 90, соответственно, как показано на Фиг. 11a и 11b. Внеклеточные аминокислотные последовательности цепи α и цепи β «TCR дикого типа», описанные в настоящем изобретении, представляют собой SEQ ID NO: 102 и SEQ ID NO: 103, соответственно, как показано на Фиг. 17a и 17b. В настоящем изобретении аминокислотная последовательность цепи α и аминокислотная последовательность цепи β «референсного TCR» представляют собой SEQ ID NO: 49 и SEQ ID NO: 50, соответственно, как показано на Фиг. 8a и 8b. В настоящем изобретении аминокислотные последовательности вариабельного домена цепи α и β TCR дикого типа, способные связываться с комплексом FMNKFIYEI-HLA A0201, представляют SEQ ID NO: 1 и SEQ ID NO: 2, соответственно, как показано на Фиг. 1a и 1b. В настоящем изобретении термины «полипептид по настоящему изобретению», «TCR по настоящему изобретению» и «T-клеточный рецептор по настоящему изобретению» используются взаимозаменяемо.
Природная межцепочечная дисульфидная связь и искусственная межцепочечная дисульфидная связь
Группа дисульфидных связей находится между цепями Cα и Cβ в проксимальной области мембраны нативного TCR, которая называется в данном документе «природной межцепочечной дисульфидной связью». В настоящем изобретении межцепочечная ковалентная дисульфидная связь, которая искусственно введена, и положение которой отличается от положения природной межцепочечной дисульфидной связи, называется «искусственной межцепочечной дисульфидной связью».
Для удобства описания в настоящем изобретении положения аминокислотных последовательностей TRAC*01 и TRBC1*01 или TRBC2*01 последовательно пронумерованы в порядке от N-конца к C-концу. Например, 60ая аминокислота в порядке от N-конца к C-концу в TRBC1*01 или TRBC2*01 представляет собой P (валин), который может быть описан как Pro60 TRBC1*01 или TRBC2*01 экзона 1 в настоящем изобретении, и может также экспрессироваться в виде аминокислоты в положении 60 TRBC1* 01 или TRBC2*01 экзона 1. В качестве еще одного примера, 61ая аминокислота в порядке от N-конца к C-концу в TRBC1*01 или TRBC2*01 представляет собой Q (глутамин), который может быть описан как Gln61 TRBC1*01 или TRBC2*01 экзона 1 в изобретении и может также выражаться как аминокислота в положении 61 TRBC1*01 или TRBC2*01 экзона 1 и т.д. В настоящем изобретении положения аминокислотных последовательностей вариабельных областей TRAV и TRBV пронумерованы в соответствии с положениями, перечисленными в IMGT. В том, что касается аминокислоты в TRAV, положение пронумеровано как 46 в IMGT, которая описана в настоящем изобретении как аминокислота в положении 46 TRAV, и т.д. В настоящем изобретении, если положения в последовательности других аминокислот конкретно описаны, особое описание будет иметь преимущество.
Опухоль
Термин «опухоль» подразумевает включение всех типов роста раковых клеток или процессов канцерогенеза, метастатических тканей или злокачественных трансформированных клеток, тканей или органов, независимо от патологического типа или стадии инфекции. Примеры опухолей включают, без ограничения, солидные опухоли, опухоли мягких тканей и метастатические поражения. Примеры солидных опухолей включают: злокачественные опухоли разных систем органов, такие как саркома, плоскоклеточная карцинома легкого и рак. Например: инфицированные предстательная железа, легкое, молочная железа, лимфа, желудочно-кишечный (например, толстая кишка) и урогенитальный тракт (например, почка, эпителиальные клетки), глотка. Плоскоклеточная карцинома легкого включает злокачественные опухоли, например, большую часть из рака толстой кишки, рака прямой кишки, почечно-клеточной карциномы, рака печени, немелкоклеточного рака легкого, рака тонкой кишки и рака пищевода. Метастатические поражения при упомянутых выше раковых заболеваниях могут также подвергаться лечению и предупреждению с использованием способов и композиций по изобретению.
Подробное описание изобретения
Хорошо известно, что вариабельный домен цепи α и вариабельный домен цепи β TCR содержат три CDR (аналогично областям, определяющим комплементарность, антител), соответственно. CDR3 взаимодействует с коротким пептидом антигена, и CDR1 и CDR2 взаимодействуют с HLA. Таким образом, CDR молекулы TCR определяют ее взаимодействие с комплексом короткий пептид антигена-HLA. Аминокислотные последовательности вариабельного домена цепи α и вариабельного домена цепи β TCR дикого типа, способные связываться с комплексом короткого пептида антигена FMNKFIYEI и HLA-A0201 (а именно, комплексом FMNKFIYEI-HLA-A0201), представляют собой SEQ ID NO: 1 и SEQ ID NO: 2, соответственно. Впервые данные последовательности были открыты авторами изобретения. Он имеет следующие области CDR:
CDR вариабельного домена цепи α
CDR1α: TSINN
CDR2α: IRSNERE
CDR3α: ATDPRTGANSKLT
и CDR вариабельного домена цепи β
CDR1β: SEHNR
CDR2β: FQNEAQ
CDR3β: ASEGLAYEQY.
В настоящем изобретении получают высокоаффинный TCR, подвергая мутации и скринингу в указанных выше областях CDR, который обладает аффинностью в отношении комплекса FMNKFIYEI-HLA-A0201, которая по меньшей мере в 2 раза выше аффинности TCR дикого типа в отношении комплекса FMNKFIYEI-HLA-A0201.
В настоящем изобретении предложен T-клеточный рецептор (TCR), который обладает активностью в связывании с комплексом FMNKFIYEI-HLA-A0201.
T-клеточный рецептор содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, вариабельный домен цепи α TCR содержит три области CDR, и референсные последовательности трех областей CDR вариабельного домена цепи α TCR перечислены ниже:
CDR1α: TSINN
CDR2α: IRSNERE
CDR3α: ATDPRTGANSKLT, и содержит по меньшей мере одну из следующих мутаций:
и/или, вариабельный домен цепи β TCR содержит три области CDR, и референсные последовательности трех областей CDR вариабельного домена цепи β TCR перечислены ниже,
CDR1β: SEHNR
CDR2β: FQNEAQ
CDR3β: ASEGLAYEQY, и содержит по меньшей мере одну из следующих мутаций:
В предпочтительном воплощении настоящего изобретения T-клеточный рецептор по настоящему изобретению содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и вариабельный домен цепи α TCR содержит CDR1α, CDR2α и CDR3α.
В еще одном предпочтительном воплощении CDR1α содержит последовательность:
T[1αX1][1αX2][1αX3][1αX4], где [1αX1], [1αX2], [1αX3], [1αX4] независимо выбраны из любого природного аминокислотного остатка.
В еще одном предпочтительном воплощении [1αX1] представляет собой S, или D, или P, или A, или E.
В еще одном предпочтительном воплощении [1αX2] представляет собой I или V.
В еще одном предпочтительном воплощении [1αX3] представляет собой N, или S, или Y, или D.
В еще одном предпочтительном воплощении [1αX4] представляет собой N или S или K.
В еще одном предпочтительном воплощении [1αX1] представляет собой S, или D, или P, или A, или E, [1αX2] представляет собой I или V, [1αX3] представляет собой N, или S, или Y, или D, и [1αX4] представляет собой N, или S, или K.
В еще одном предпочтительном воплощении CDR1α содержит последовательность, выбранную из группы, состоящей из: TSINN, TPISS, TDIYK, TAVDS, TEISS и TDVSS.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, и вариабельный домен цепи α TCR содержит CDR1α, CDR2α и CDR3α, где аминокислотная последовательность CDR2α представляет собой IRSNERE.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, вариабельный домен цепи α TCR содержит CDR1α, CDR2α и CDR3α, и CDR3α содержит последовательность: ATDP[3αX1][3αX2][3αX3][3αX4][3αX5][3αX6]KLT, где [3αX1], [3αX2], [3αX3], [3αX4], [3αX5] и [3αX6] независимо выбраны из любого природного аминокислотного остатка.
В еще одном предпочтительном воплощении [3αX1] представляет собой R, или A, или S, или D, или E, или Q, или V, или T.
В еще одном предпочтительном воплощении [3αX2] представляет собой T, или M, или R, или L, или A, или N, или S.
В еще одном предпочтительном воплощении [3αX3] представляет собой G, или A, или Q, или S, или E, или N.
В еще одном предпочтительном воплощении [3αX4] представляет собой A, или S, или G, или P, или W.
В еще одном предпочтительном воплощении [3αX5] представляет собой N, или S, или T.
В еще одном предпочтительном воплощении [3αX6] представляет собой S, или A, или N или T или P.
В еще одном предпочтительном воплощении [3αX1] представляет собой R, или A, или S, или D, или E, или Q, или V, или T, [3αX2] представляет собой T, или M, или R, или L, или A, или N, или S, [3αX3] представляет собой G, или A, или Q, или S, или E, или N, [3αX4] представляет собой A, или S, или G, или P, или W, [3αX5] представляет собой N, или S, или T, и [3αX6] представляет собой S, или A, или N, или T, или P.
В еще одном предпочтительном воплощении CDR3α содержит последовательность, выбранную из группы, состоящей из: ATDPSMSPNSKLT, ATDPSLSPNSKLT и ATDPSLQSNSKLT.
В одном предпочтительном воплощении изобретения TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, вариабельный домен цепи β TCR содержит CDR1β, CDR2β и CDR3β, и CDR3β содержит последовательность: ASEGLAYEQY.
В еще одном предпочтительном воплощении CDR1β содержит последовательность: [1βX1][1βX2][1βX3][1βX4]R, где [1βX1], [1βX2], [1βX3], [1βX4] независимо выбраны из любого природного аминокислотного остатка.
В еще одном предпочтительном воплощении [1βX1] представляет собой S, или A, или Q, или G.
В еще одном предпочтительном воплощении [1βX2] представляет собой E, или G, или T, или R, или L.
В еще одном предпочтительном воплощении [1βX3] представляет собой H, или W, или R.
В еще одном предпочтительном воплощении [1βX4] представляет собой N, или A, или P.
В еще одном предпочтительном воплощении CDR2β содержит последовательность:
F[2βX1][2βX2][2βX3][2βX4][2βX5], где [2βX1], [2βX2], [2βX3], [2βX4], [2βX5] независимо выбраны из любого природного аминокислотного остатка.
В еще одном предпочтительном воплощении [2βX1] представляет собой Q или H.
В еще одном предпочтительном воплощении [2βX2] представляет собой N или G.
В еще одном предпочтительном воплощении [2βX3] представляет собой E или D.
В еще одном предпочтительном воплощении [2βX4] представляет собой A, или M, или L.
В еще одном предпочтительном воплощении [2βX5] представляет собой Q или E.
В еще одном предпочтительном воплощении CDR2β содержит последовательность, выбранную из группы, состоящей из: FQGDME, FQGDLE и FHGDLE.
В еще одном предпочтительном воплощении TCR содержит вариабельный домен цепи α TCR и вариабельный домен цепи β TCR, вариабельный домен цепи β TCR содержит CDR1β, CDR2β и CDR3β, и аминокислотная последовательность CDR2β выбрана из группы, состоящей из: FQGDME, FQGDLE и FHGDLE.
В еще одном предпочтительном воплощении вариабельный домен цепи α TCR одновременно не содержит следующие CDR:
CDR1α: TSINN; CDR2α: IRSNERE; и CDR3α: ATDPRTGANSKLT.
В еще одном предпочтительном воплощении вариабельный домен цепи β TCR одновременно не содержит следующие CDR:
CDR1β: SEHNR; CDR2β: FQNEAQ; и CDR3β: ASEGLAYEQY.
В частности, число мутаций в области CDR цепи α TCR может составлять 3, 4, 5, 6, 7 или 8; и/или число мутаций в области CDR цепи β TCR может составлять 4, 5, 6, 7 или 8.
Кроме того, TCR по настоящему изобретению представляет собой αβ гетеродимерный TCR, и вариабельный домен цепи α TCR содержит аминокислотную последовательность, обладающую по меньшей мере 90%, предпочтительно по меньшей мере 92%; более предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 1; и/или вариабельный домен цепи β TCR содержит аминокислотную последовательность, обладающую по меньшей мере 90%, предпочтительно по меньшей мере 92%; более предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 2.
Кроме того, TCR по настоящему изобретению представляет собой одноцепочечный TCR, и вариабельный домен цепи α TCR содержит аминокислотную последовательность, обладающую по меньшей мере 85%, предпочтительно по меньшей мере 90%; более предпочтительно по меньшей мере 92%; наиболее предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 3; и/или вариабельный домен цепи β TCR содержит аминокислотную последовательность, обладающую по меньшей мере 85%, предпочтительно по меньшей мере 90%; более предпочтительно по меньшей мере 92%; наиболее предпочтительно по меньшей мере 94% (например, может иметь место по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомология последовательностей) гомологией последовательностей с аминокислотной последовательностью, показанной в SEQ ID NO: 4.
Предпочтительно, TCR содержит (i) всю или часть цепи α TCR, за исключением ее трансмембранного домена, и (ii) всю или часть цепи β TCR, за исключением ее трансмембранного домена, где обе из (i) и (ii) содержат вариабельный домен и по меньшей мере часть константных доменов цепи TCR.
В настоящем изобретении три CDR вариабельного домена цепи α SEQ ID NO: 1 TCR дикого типа, а именно, CDR1, CDR2 и CDR3 расположены в положениях 27-31, 49-55 и 90-102 SEQ ID NO: 1, соответственно. Соответственно, аминокислотный остаток пронумерован, как показано в SEQ ID NO: 1, 28S представляет собой S во 2м положении CDR1α, 29I представляет собой I в 3м положении CDR1α, 30N представляет собой N в 4м положении CDR1α, 31N представляет собой N в 5м положении CDR1α, 94R представляет собой R в 5м положении CDR3α, 95T представляет собой T в 6м положении CDR3α, 96G представляет собой G в 7м положении CDR3α, 97A представляет собой A в 8м положении CDR3α, 98N представляет собой N в 9м положении CDR3α, и 99S представляет собой S в 10м положении CDR3α.
Аналогично, в настоящем изобретении три CDR вариабельного домена цепи β SEQ ID NO: 2 TCR дикого типа, а именно CDR1, CDR2 и CDR3 расположены в положениях 27-31, 49-54 и 93-102 SEQ ID NO: 2, соответственно. Аналогично, аминокислотный остаток пронумерован, как показано в SEQ ID NO: 2, 27S представляет собой S в 1м положении CDR1β, 28E представляет собой E в 2м положении CDR1β, 29H представляет собой H в 3м положении CDR1β, 30N представляет собой N в 4м положении CDR1β, 50Q представляет собой Q во 2м положении CDR2β, 51N представляет собой N в 3м положении CDR2β, 52E представляет собой E в 4м положении CDR2β, 53A представляет собой A в 5м положении CDR2β, 54Q представляет собой Q в 6м положении CDR2β.
Согласно настоящему изобретению предложен TCR, обладающий свойством связывания с комплексом FMNKFIYEI-HLA-A0201, и содержащий вариабельный домен цепи α и вариабельный домен цепи β, где TCR имеет мутацию в вариабельном домене цепи α, показанную в SEQ ID NO: 1, и участок мутантного аминокислотного остатка включает одну или более из 28S, 29I, 30N, 31N, 94R, 95T, 96G, 97A, 98N и 99S, где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 1; и/или TCR имеет мутацию в вариабельном домене цепи β, показанную в SEQ ID NO: 2, и участок мутантного аминокислотного остатка включает одну или более из 27S, 28E, 29H, 30N, 50Q, 51N, 52E, 53A и 54Q, где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 2;
Предпочтительно, мутантный вариабельный домен цепи α TCR содержит один или более аминокислотных остатков, выбранных из группы, состоящей из: 28D, или 28P, или 28A, или 28E; 29V; 30S, или 30Y, или 30D; 31S, или 31K, 94A, или 94S, или 94V, или 94D, или 94E, или 94Q, или 94T, 95M, или 95R, или 95L, или 95A, или 95N, или 95S, 96A, или 96Q, или 96S, или 96E, или 96N, 97S, или 97G, или 97P, или 97W, 98S, или 98T, 99A, или 99N, или 99T, или 99P, где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 1; и/или мутантный вариабельный домен цепи β содержит один или более аминокислотных остатков, выбранных из группы, состоящей из: 27A или 27Q, или 27G, 28G, или 28T, или 28R, или 28L, 29W, или 29R, 30A, или 30P, 50H, 51G, 52D, 53M, или 53L, 54E, где аминокислотный остаток пронумерован, как показано в SEQ ID NO: 2.
Более конкретно, в вариабельном домене цепи α, конкретные формы мутации включают одну или более групп из S28D/P/A/E, I29V, N30S/Y/D, N31S/K, R94A/S/V/D/E/Q/T, T95M/R/L/A/N/S, G96A/Q/S/E/N, A97S/G/P/W, N98S/T, S99A/N/T/P; и в вариабельном домене цепи β, конкретные формы мутации включают одну или более групп из S27A/Q/G, E28G/T/R/L, H29W/R, N30A/P, Q50H, N51G, E52D, A53M/L, Q54E.
Thr48 константной области цепи α TCR дикого типа TRAC*01 экзона 1 подвергали мутации до цистеина, и Ser57 константной области цепи β TRBC1*01 или TRBC2*01 экзона 1 подвергали мутации до цистеина в соответствии со способом сайт-направленного мутагенеза, хорошо известного специалисту в данной области, таким образом, чтобы получить референсный TCR, аминокислотные последовательности которого показаны на Фиг. 8a и 8b, соответственно, и мутантные остатки цистеина показаны жирными буквами. Приведенные выше цистеиновые замены могут образовывать искусственную межцепочечную дисульфидную связь между константными областями цепи α и β референсного TCR с образованием более стабильного растворимого TCR, таким образом, чтобы легче оценивать аффинность связывания и/или время полужизни связывания у TCR с комплексом FMNKFIYEI-HLA-A2. Следует иметь в виду, что области CDR вариабельной области TCR определяют его аффинность в отношении комплекса pMHC, таким образом, приведенные выше цистеиновые замены в константной области TCR не будет влиять на аффинность связывания и/или время полужизни связывания TCR. Таким образом, в настоящем изобретении считается, что измеренная аффинность связывания референсного TCR с комплексом FMNKFIYEI-HLA-A0201, представляет собой аффинность связывания между TCR дикого типа и комплексом FMNKFIYEI-HLA-A0201. Аналогично, если аффинность связывания TCR по изобретению с комплексом FMNKFIYEI-HLA-A0201, как определено, по меньшей мере в 10 раз выше аффинности связывания референсного TCR с комплексом FMNKFIYEI-HLA-A0201, аффинность связывания TCR по настоящему изобретению с комплексом FMNKFIYEI-HLA-A0201 по меньшей мере в 10 раз выше аффинности связывания TCR дикого типа с комплексом FMNKFIYEI-HLA-A0201.
Аффинность связывания (обратно пропорционально константе равновесия диссоциации KD) и время полужизни связывания (выражено как T1/2) можно определять любым подходящим способом, таким как поверхностный плазмонный резонанс. Следует понимать, что удвоение аффинности TCR будет уменьшать вдвое KD. T1/2 рассчитывают как In2, деленное на скорость диссоциации (Koff). Таким образом, удвоение T1/2 будет уменьшать вдвое Koff. Предпочтительно, аффинность связывания или время полужизни связывания данного TCR выявляют несколько раз посредством использования одного и того же протокола испытаний, например, 3 или более раз, и берут среднее данных результатов. В предпочтительном воплощении выявляют аффинность растворимого TCR методом поверхностного плазмонного резонанса (BIAcore) в Примерах в данном документе со следующими условиями: температура 25 °C и значение pH 7,1-7,5. Константу равновесия диссоциации KD референсного TCR с комплексом FMNKFIYEI-HLA-A2 определили как 4,3E-05M, а именно, 43 мкМ, и в настоящем изобретении, также считается, что константа равновесия диссоциации KD TCR дикого типа с комплексом FMNKFIYEI-HLA-A2 равна 43 мкМ. Поскольку удвоение аффинности TCR будет уменьшать вдвое KD, если константу равновесия диссоциации KD высокоаффинного TCR с комплексом FMNKFIYEI-HLA-A2 определили как 4,3E-06M, а именно, 4,3 мкМ, аффинность высокоаффинного TCR в отношении комплекса FMNKFIYEI-HLA-A2 в 10 раз выше, чем аффинность TCR дикого типа в отношении комплекса FMNKFIYEI-HLA-A2. Специалист в данной области знаком со связью перевода единиц измерения KD, а именно, 1 M равен 106 мкМ, 1 мкМ равен 1000 нМ, и 1 нМ равен 1000 пМ.
В одном предпочтительном воплощении изобретения аффинность TCR в отношении комплекса FMNKFIYEI-HLA-A0201 по меньшей мере в 2 раза выше, чем аффинность TCR дикого типа, предпочтительно по меньшей мере в 5 раз, более предпочтительно по меньшей мере в 10 раз.
В еще одном предпочтительном воплощении аффинность TCR в отношении комплекса FMNKFIYEI-HLA-A0201 по меньшей мере в 50 раз выше аффинности TCR дикого типа, предпочтительно, по меньшей мере в 100 раз, более предпочтительно, по меньшей мере в 500 раз, наиболее предпочтительно по меньшей мере в 1000 раз.
В еще одном предпочтительном воплощении аффинность TCR в отношении комплекса FMNKFIYEI-HLA-A0201 по меньшей мере в 104 раз, предпочтительно по меньшей мере в 105 раз, более предпочтительно по меньшей мере в 106 раз выше аффинности TCR дикого типа.
В частности, константа равновесия диссоциации KD TCR в отношении комплекса FMNKFIYEI-HLA-A0201 составляет 20 мкМ или меньше.
В еще одном предпочтительном воплощении константа равновесия диссоциации TCR в отношении комплекса FMNKFIYEI-HLA-A0201 составляет 5 мкм ≤ KD ≤ 10 мкМ; предпочтительно, 0,1 мкМ ≤ KD ≤ 1 мкМ; и более предпочтительно, 1 нМ ≤ KD ≤ 100 нМ;
В еще одном предпочтительном воплощении константа равновесия диссоциации TCR в отношении комплекса FMNKFIYEI-HLA-A0201 составляет 100 пМ ≤ KD ≤ 1000 пМ; и более предпочтительно 10 пМ ≤ KD ≤ 100 пМ.
Мутации можно осуществлять любым подходящим способом, включая, но не ограничиваясь способами на основе полимеразной цепной реакции (ПЦР), клонирования на основе эндонуклеаз рестрикции или независимого от лигирования клонирования (LIC - ligation-independent cloning). Во многих стандартных учебниках по молекулярной биологии подробно описаны данные способы. Больше подробностей о мутагенезе на основе полимеразной цепной реакции (ПЦР) и клонировании на основе эндонуклеаз рестрикции можно найти в Sambrook and Russell, (2001) Molecular Cloning-A Laboratory Manual (Third Edition) CSHL Publishing house. Больше информации о способе LIC можно найти в Rashtchian, (1995) Curr Opin Biotechnol 6(1): 30-6.
Способ получения TCR по настоящему изобретению может представлять собой скрининг TCR, обладающего высокой аффинностью в отношении комплекса FMNKFIYEI-HLA-A2, из разнообразных библиотек фаговых частиц, экспонирующих такие TCR, как описано в литературе (Li, et al). (2005) Nature Biotech 23(3): 349-354), но не ограничивается им.
Следует понимать, что гены, экспрессирующие аминокислоту вариабельного домена цепи α и β TCR дикого типа, или гены, экспрессирующие аминокислоту вариабельного домена цепи α и β незначительно модифицированного TCR дикого типа, могут быть использованы для получения матрицы TCR. Изменения, необходимые для получения высокоаффинного TCR по изобретению, затем вводят в ДНК, кодирующую вариабельный домен матрицы TCR.
Высокоаффинный TCR по настоящему изобретению содержит одну из аминокислотных последовательностей вариабельного домена цепи α SEQ ID NO: 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 91, 92 и/или одну из аминокислотных последовательностей вариабельного домена цепи β SEQ ID NO: 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 93, 94, 95, 96, 97, 98, 99, 100, 101. Таким образом, цепь α TCR, содержащая аминокислотную последовательность вариабельного домена цепи α TCR дикого типа (SEQ ID NO: 1), может связываться с цепью β TCR, содержащей одну из SEQ ID NO: 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 93, 94, 95, 96, 97, 98, 99, 100, 101, с образованием молекулы гетеродимерного TCR или одноцепочечного TCR. В качестве альтернативы, цепь β TCR, содержащая аминокислотную последовательность вариабельного домена β TCR дикого типа (SEQ ID NO: 2), может связываться с цепью α TCR, содержащей одну из SEQ ID NO: 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 91, 92, с образованием молекулы гетеродимерного TCR или одноцепочечного TCR. В качестве альтернативы, цепь α TCR, содержащая одну из аминокислотных последовательностей вариабельного домена цепи α TCR SEQ ID NO: 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 91, 92, может связываться с цепью β TCR, содержащей одну из аминокислотных последовательностей вариабельного домена цепи β TCR SEQ ID NO: 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 93, 94, 95, 96, 97, 98, 99, 100, 101, с образованием молекулы гетеродимерного TCR или одноцепочечного TCR. В настоящем изобретении аминокислотные последовательности вариабельного домена цепи α и вариабельного домена цепи β, которые образуют молекулу гетеродимерного TCR, предпочтительно выбраны из следующей Таблицы 1:
Таблица 1
В целях настоящего изобретения TCR по изобретению представляет собой группировку, имеющую по меньшей мере один вариабельный домен цепи α TCR и/или β TCR. Они обычно содержат и вариабельный домен цепи α TCR и вариабельный домен цепи β TCR. Они могут представлять собой αβ гетеродимеры или одноцепочечные формы или любые другие стабильные формы. В адоптивной иммунотерапии полноразмерная цепь αβ гетеродимерного TCR (включая цитоплазматические и трансмембранные домены) может быть трансфицирована. TCR по настоящему изобретению можно использовать в качестве средства нацеливания для доставки терапевтического средства к антигенпрезентирующей клетке или в комбинации с другими молекулами для получения бифункционального полипептида для направления эффекторных клеток, когда TCR предпочтительно находится в растворимой форме.
В отношении стабильности в предшествующем уровне техники раскрыто, что растворимая и стабильная молекула TCR может быть получена введением искусственной межцепочечной дисульфидной связи между константными доменами цепи α и β TCR, как описано в PCT/CN2015/093806. Таким образом, TCR по изобретению может представлять собой TCR, в котором искусственную межцепочечную дисульфидную связь вводят между остатками его константных доменов цепи α и β. Цистеиновые остатки образуют искусственную межцепочечную дисульфидную связь между константными доменами цепи α и β TCR. Остаток цистеина может заменять другой аминокислотный остаток в подходящем положении в нативном TCR с образованием искусственной межцепочечной дисульфидной связи. Например, Thr48 TRAC*01 экзона 1 и Ser57 TRBC1*01 или TRBC2*01 экзона 1 могут быть заменены с образованием дисульфидной связи. Другие сайты для введения остатка цистеина с образованием дисульфидной связи могут представлять собой следующие: Thr45 TRAC*01 экзона 1 и Ser77 TRBC1*01 или TRBC2*01 экзона 1; Tyr10 TRAC*01 экзона 1 и Ser17 TRBC1*01 или TRBC2*01 экзона 1; Thr45 TRAC*01 экзона 1 и Asp59 TRBC1*01 или TRBC2*01 экзона 1; Ser15 TRAC*01 экзона 1 и Glu15 TRBC1*01 или TRBC2*01 экзона 1; Arg53 TRAC*01 экзона 1 и Ser54 TRBC1*01 или TRBC2*01 экзона 1; Pro89 TRAC*01 экзона 1 и Ala19 TRBC1*01 или TRBC2*01 экзона 1; или Tyr10 TRAC*01 экзона 1 и Glu20 TRBC1*01 или TRBC2*01 экзона 1. То есть, остатки цистеина заменяют любую группу упомянутых выше сайтов в константных доменах цепи α и β. Максимум 15, или максимум 10, или максимум 8, или меньше аминокислот могут быть усечены на одном или более C-концах константного домена TCR по изобретению таким образом, чтобы он не включал остатки цистеина для достижения цели удаления нативных межцепочечных дисульфидных связей, или остатки цистеина, образующие природную межцепочечную дисульфидную связь, могут быть также подвергнуты мутации до другой аминокислоты для достижения указанной выше цели.
Как описано выше, TCR по настоящему изобретению может содержать искусственную межцепочечную дисульфидную связь, введенную между остатками его константных доменов цепи α и β. Следует отметить, что введенная искусственная дисульфидная связь, как описано выше, может содержаться или не содержаться между константными доменами, и TCR по настоящему изобретению может содержать последовательность константного домена TRAC и последовательность константного домена TRBC1 или TRBC2. Последовательность константного домена TRAC и последовательность константного домена TRBC1 или TRBC2 TCR могут быть соединены природной межцепочечной дисульфидной связью, находящейся в TCR.
Кроме того, относительно стабильности, также в патентной литературе PCT/CN2016/077680 было раскрыто, что введение искусственной межцепочечной дисульфидной связи между вариабельной областью цепи α и константной областью цепи β TCR может значимо улучшать стабильность TCR. Таким образом, искусственная межцепочечная дисульфидная связь может содержаться между вариабельной областью цепи α и константной областью цепи β высокоаффинного TCR по настоящему изобретению. Конкретно, остатки цистеина, образующие искусственную межцепочечную дисульфидную связь между вариабельной областью цепи α и константной областью цепи β TCR, заменены на: аминокислоту в положении 46 TRAV и аминокислоту в положении 60 TRBC1*01 или TRBC2*01 экзона 1; аминокислоту в положении 47 TRAV и аминокислоту в положении 61 TRBC1*01 или TRBC2*01 экзона 1; аминокислоту в положении 46 TRAV и аминокислоту в положении 61 TRBC1*01 или TRBC2*01 экзона 1; или аминокислоту в положении 47 TRAV и аминокислоту в положении 60 TRBC1*01 или TRBC2*01 экзона 1. Предпочтительно, такой TCR может содержать (i) всю или часть цепи α TCR, отличную от ее трансмембранного домена, и (ii) всю или часть цепи β TCR, отличной от ее трансмембранного домена, где обе из (i) и (ii) содержат вариабельный домен и по меньшей мере часть константных доменов цепи TCR, и цепь α и цепь β образуют гетеродимер. Более предпочтительно, такой TCR может содержать вариабельный домен цепи α и вариабельный домен цепи β и всю или часть константного домена цепи β, отличную от трансмембранного домена, которая, однако, не содержит константный домен цепи α, и вариабельный домен цепи α TCR и цепь β образуют гетеродимер.
Для стабильности, в другом аспекте, TCR по настоящему изобретению также включает TCR, имеющий мутацию в своей гидрофобной коровой области, и эти мутации в гидрофобной коровой области предпочтительно представляют собой мутации, способные повышать стабильность TCR по настоящему изобретению, как описано в WO 2014/206304. Такой TCR может иметь мутации в следующих положениях в гидрофобном ядре вариабельного домена: (цепь α и/или β) аминокислоты вариабельной области в положении 11, 13, 19, 21, 53, 76, 89, 91, 94, и/или аминокислота короткого пептида гена J цепи α (TRAJ) в обратных положениях 3, 5, 7 и/или аминокислота короткого пептида гена J цепи β (TRBJ) в обратных положениях 2, 4, 6, где данные положения в аминокислотной последовательности пронумерованы в соответствии с номерами положений, перечисленными в международной информационной системе по иммуногенетике (IMGT). Специалист в данной области будет знаком с описанной выше международной информационной системой по иммуногенетике и может получать номера положений аминокислотных остатков разных TCR в IMGT на основе базы данных.
Более конкретно, в настоящем изобретении TCR, в котором есть мутация в гидрофобной коровой области, может представлять собой высокостабильный одноцепочечный TCR, состоящий из вариабельных доменов цепи α и β TCR, которые связаны гибкой пептидной цепью. Области CDR вариабельной области TCR определяют его аффинность в отношении комплекса короткий пептид-HLA, и мутации в гидрофобном ядре могут повышать стабильность TCR, но не будут влиять на его аффинность в отношении комплекса короткий пептид-HLA. Следует отметить, что гибкая пептидная цепь в настоящем изобретении может представлять собой любую пептидную цепь, подходящую для связывания вариабельных доменов цепи α и β TCR. Цепь-матрица, сконструированная в Примере 1 настоящего изобретения для скрининга высокоаффинных TCR, представляет собой высокостабильный одноцепочечный TCR, содержащий мутации в гидрофобном ядре, как описано выше. Аффинность TCR к комплексу FMNKFIYEI-HLA-A0201 можно легко оценить посредством использования TCR с более высокой стабильностью.
Области CDR вариабельного домена цепи α и вариабельного домена цепи β одноцепочечной матрицы TCR идентичны областям CDR TCR дикого типа. А именно, три CDR вариабельного домена цепи α представляют собой CDR1α: TSINN, CDR2α: IRSNERE и CDR3α: ATDPRTGANSKLT, и три CDR вариабельных доменов цепи β представляют собой CDR1β: SEHNR, CDR2β: FQNEAQ и CDR3β: ASEGLAYEQY, соответственно. Аминокислотная последовательность (SEQ ID NO: 47) и нуклеотидная последовательность (SEQ ID NO: 48) одноцепочечной матрицы TCR показаны на Фиг. 7a и 7b, соответственно, таким образом, осуществляя скрининг одноцепочечного TCR, состоящего из вариабельного домена цепи α и вариабельного домена цепи β, и обладающего высокой аффинностью в отношении комплекса FMNKFIYEI-HLA-A0201.
В настоящем изобретении три CDR вариабельного домена цепи α одноцепочечной матрицы TCR (SEQ ID NO: 3), а именно CDR1, CDR2 и CDR3, расположены в положениях 27-31, 49-55 и 90-102 SEQ ID NO: 3, соответственно. Соответственно, аминокислотные остатки пронумерованы в соответствии с числом, как показано в SEQ ID NO: 3. 28S представляет собой S во 2м положении CDR1α, 29I представляет собой I в 3м положении CDR1α, 30N представляет собой N в 4м положении CDR1α, 31N представляет собой N в 5м положении CDR1α, 94R представляет собой R в 5м положении CDR3α, 95T представляет собой T в 6м положении CDR3α, 96G представляет собой G в 7м положении CDR3α, 97A представляет собой A в 8м положении CDR3α, 98N представляет собой N в 9м положении CDR3α, и 99S представляет собой S в 10м положении CDR3α.
Аналогично, в настоящем изобретении три CDR вариабельного домена цепи β одноцепочечной матрицы TCR (SEQ ID NO: 4), а именно, CDR1, CDR2 и CDR3 расположены в положениях 27-31, 49-54 и 93-102 SEQ ID NO: 2, соответственно. Таким образом, аминокислотные остатки пронумерованы в соответствии с номером, как показано в SEQ ID NO: 4. 27S представляет собой S в 1м положении CDR1β, 28E представляет собой E во 2м положении CDR1β, 29H представляет собой H в 3м положении CDR1β, 30N представляет собой N в 4м положении CDR1β, 50Q представляет собой Q во 2м положении CDR2β, 51N представляет собой N в 3м положении CDR2β, 52E представляет собой E в 4м положении CDR2β, 53A представляет собой A в 5м положении CDR2β, и 54Q представляет собой Q в 6м положении CDR2β.
αβ Гетеродимер по настоящему изобретению, обладающий высокой аффинностью в отношении комплекса FMNKFIYEI-HLA-A0201, получали посредством переноса областей CDR вариабельных доменов цепи α и β отобранного высокоаффинного одноцепочечного TCR в соответствующие положения вариабельного домена цепи α (SEQ ID NO: 1) и вариабельного домена цепи β (SEQ ID NO: 2) TCR дикого типа. Другую часть получали посредством искусственной комбинации на основе сайтов мутации областей CDR, полученных в результате скрининга.
Высокоаффинный TCR по настоящему изобретению дополнительно содержит одну из аминокислотных последовательностей вариабельного домена цепи α SEQ ID NO: 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 и/или одну из аминокислотных последовательностей вариабельного домена цепи β SEQ ID NO: 36, 37, 38, 39, 40, 41, 42, 43, 44, 45 и 46. Таким образом, вариабельный домен цепи α (SEQ ID NO: 3) описанного выше высокостабильного одноцепочечного TCR в качестве цепи матрицы можно объединять с вариабельным доменом цепи β TCR, аминокислотная последовательность которого представляет собой SEQ ID NO: 36, 37, 38, 39, 40, 41, 42, 43, 44, 45 или 46, с образованием молекулы одноцепочечного TCR. В качестве альтернативы, вариабельный домен цепи β (SEQ ID NO: 4) описанного выше высокостабильного одноцепочечного TCR в качестве цепи матрицы можно объединять с вариабельным доменом цепи α TCR, аминокислотная последовательность которого представляет собой SEQ ID NO: 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 или 35, с образованием молекулы одноцепочечного TCR. В качестве альтернативы, один из вариабельных доменов цепи α TCR SEQ ID NO: 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 можно объединять с одним из вариабельных доменов цепи β TCR SEQ ID NO: 36, 37, 38, 39, 40, 41, 42, 43, 44, 45 и 46 с образованием молекулы одноцепочечного TCR. В настоящем изобретении аминокислотные последовательности вариабельного домена цепи α и вариабельного домена цепи β молекулы высокоаффинного одноцепочечного TCR предпочтительно выбраны из следующей Таблицы 2:
Таблица 2
TCR по настоящему изобретению может быть предложен в виде поливалентного комплекса. Поливалентный комплекс TCR по настоящему изобретению содержит полимер, образованный посредством объединения двух, трех, четырех или более TCR по настоящему изобретению, например, тетрамерный домен p53 может быть использован с получением тетрамера. В качестве альтернативы, больше TCR по изобретению можно объединять с другой молекулой с образованием комплекса. Комплексы TCR по изобретению можно использовать для отслеживания или нацеливания на клетки-мишени, которые презентируют конкретный антиген in vitro или in vivo, или продуцируют промежуточные соединения других поливалентных комплексов TCR с такими применениями.
TCR по настоящему изобретению можно использовать отдельно или в комбинации с конъюгатом ковалентно или другим образом, предпочтительно ковалентно. Конъюгат включает детектируемую метку (в целях диагностики, где TCR используют для выявления наличия клетки, презентирующей комплекс FMNKFIYEI-HLA-A2), терапевтическое средство, PK (протеинкиназа) модифицирующую группировку или комбинацию любого из описанных выше веществ.
Детектируемые метки для диагностических целей включают флуоресцентные или люминесцентные метки, радиоактивные метки, MRI (магнитно-резонансная визуализация) или CT (электронная компьютерная томография) с контрастным веществом или ферменты, способные продуцировать выявляемые продукты, но не ограничиваются ими.
Терапевтические средства, которые можно объединять с или связывать с TCR по изобретению, включают, но не ограничиваются следующими средствами: 1. Радионуклиды (Koppe et al., 2005, Cancer metastasis reviews 24, 539); 2. Биотоксин (Chaudhary et al., 1989, Nature 339, 394; Epel et al., 2002, Cancer Immunology and Immunotherapy 51, 565); 3. Цитокины, такие как IL-2, и т.д. (Gillies et al., 1992, National Academy of Sciences (PNAS) 89, 1428; Card et al., 2004, Cancer Immunology and Immunotherapy 53, 345; Halin et al., 2003, Cancer Research 63, 3202); 4. Фрагмент антитела Fc (Mosquera et al., 2005, The Journal Of Immunology 174, 4381); 5. Фрагменты антитела scFv (Zhu et al., 1995, International Journal of Cancer 62, 319); 6. Наночастицы золота/наностержни (Lapotko et al., 2005, Cancer letters 239, 36; Huang et al., 2006, Journal of the American Chemical Society 128, 2115); 7. Вирусные частицы (Peng et al., 2004, Gene therapy 11, 1234); 8. Липосомы (Mamot et al., 2005, Cancer research 65, 11631); 9. Наномагнитные частицы; 10. Пролекарство-активирующие ферменты (например, NADPH-диафораза (DTD - DT-diaphorase) или бифенилгидролаза-подобный белок (BPHL - biphenyl hydrolase-like protein)); 11. Химиотерапевтическое средство (например, цисплатин) или любая форма наночастиц, и т.д.
Антитело, с которым TCR по настоящему изобретению связывается, или его фрагмент включает антитело против T-клеток или антитело, определяющее NK-клетку (естественный киллер), как например, антитело против CD3, или против CD28, или против CD16, и упомянутое выше антитело или его фрагмент связывается с TCR, таким образом, лучше направляя эффекторные клетки к клеткам-мишеням. В предпочтительном воплощении TCR по изобретению связывается с антителом против CD3 или его функциональным фрагментом или вариантом. Конкретно, слитая молекула TCR по настоящему изобретению и одноцепочечного антитела против CD3 содержит вариабельный домен цепи α TCR, аминокислотная последовательность которого выбрана из группы, состоящей из SEQ ID NO: 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 91, 92, и вариабельный домен цепи β TCR, аминокислотная последовательность которого выбрана из группы, состоящей из SEQ ID NO: 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 93, 94, 95, 96, 97, 98, 99, 100, 101.
Изобретение также относится к молекуле нуклеиновой кислоты, кодирующей TCR по изобретению. Молекула нуклеиновой кислоты по изобретению может находиться в виде ДНК или РНК. ДНК может представлять собой кодирующую цепь или некодирующую цепь. Например, последовательность нуклеиновой кислоты, кодирующая TCR по изобретению, может быть такой же, как последовательность нуклеиновой кислоты, изложенная в разделе Графические материалы изобретения, или ее вырожденный вариант. В качестве примера, термин «вырожденный вариант», в том виде, в котором он используется в данном документе, относится к последовательности нуклеиновой кислоты, которая кодирует белок с последовательностью SEQ ID NO: 47, отличается от последовательности SEQ ID NO: 48.
Полноразмерную последовательность молекулы нуклеиновой кислоты по настоящему изобретению или ее фрагмент можно, в общем, получить посредством ПЦР-амплификации, методами генной инженерии или способами синтеза, но не ограничиваясь ими. На данный момент возможно получить ДНК-последовательность, кодирующую TCR (или ее фрагмент или ее производную) по настоящему изобретению, полностью посредством химического синтеза. И затем последовательность ДНК можно вводить в разные существующие молекулы ДНК (или векторы) и клетки, известные в данной области.
Изобретение также относится к векторам, содержащим молекулы нуклеиновой кислоты по изобретению, а также клеткам-хозяевам, генетически сконструированным с использованием векторов или кодирующих последовательностей по изобретению.
Изобретение также охватывает выделенные клетки, в частности, T-клетки, которые экспрессируют TCR по изобретению. Существует целый ряд способов, подходящих для трансфекции T-клеток ДНК или РНК, кодирующих высокоаффинный TCR по изобретению (например, Robbins et al., (2008) J. Immunol. 180: 6116-6131). T-клетки, экспрессирующие высокоаффинный TCR по изобретению, можно использовать в адоптивной иммунотерапии. Специалисту в данной области может быть известно много подходящих способов проведения адоптивной терапии (например, Rosenberg et al., (2008) Nat Rev Cancer 8(4): 299-308).
Согласно изобретению также предложена фармацевтическая композиция, содержащая фармацевтически приемлемый носитель и TCR по изобретению или комплекс TCR по изобретению или клетки, презентирующие TCR по изобретению.
Согласно изобретению также предложен способ лечения заболевания, включающий введение субъекту, нуждающемуся в этом, соответствующего количества TCR по изобретению, или комплекса TCR по изобретению, или клеток, презентирующих TCR по изобретению, или фармацевтической композиции по изобретению.
Следует понимать, что названия аминокислот в данном документе идентифицированы признанными на международном уровне одиночными английскими буквами, и соответствующие названия аминокислот в виде трехбуквенных сокращений выглядят следующим образом: Ala (A), Arg (R), Asn (N), Asp (D), Cys (C), Gln (Q), Glu (E), Gly (G), His (H), Ile (I), Leu (L), Lys (K), Met (M), Phe (F), Pro (P), Ser (S), Thr (T), Trp (W), Tyr (Y), Val (V).
В настоящем изобретении как Pro60, так и 60P означают пролин в положении 60. Кроме того, в отношении экспрессии конкретной формы мутации в настоящем изобретении, как например, «T27G» означает, что T в 27м положении заменен G. Аналогично, «I29A/V» означает, что I в 29м положении заменен A или заменен V, и т п.
В данной области техники, когда аминокислота с похожими свойствами используется для замены, функция белка обычно не меняется. Добавление одной или нескольких аминокислот на C-конце и/или N-конце обычно не меняет структуры и функции белка. Таким образом, TCR по изобретению дополнительно включает TCR, где вплоть до 5, предпочтительно вплоть до 3, более предпочтительно вплоть до 2, наиболее предпочтительно 1 аминокислота (главным образом аминокислота, расположенная за пределами областей CDR) TCR по изобретению заменена аминокислотой с похожими свойствами и все еще способна сохранять свою функцию.
Настоящее изобретение также включает TCR, полученный из TCR по настоящему изобретению в результате незначительной модификации. Форма модификации (обычно без изменения первичной структуры) включает: химически полученные формы TCR по изобретению, как например, ацетилирование или карбоксилирование. Модификации также включают гликозилирование, как например такие TCR, которые получены посредством модификаций на основе гликозилирования на стадии синтеза и обработки или на дополнительных стадиях обработки TCR по изобретению. Такая модификация может быть выполнена посредством подвергания TCR действию фермента, выполняющего гликозилирование (такого как фермент гликозилирования млекопитающего или фермент дегликозилирования). Формы модификации также включают последовательности, имеющие фосфорилированные аминокислотные остатки (такие как фосфотирозин, фосфосерин, фосфотреонин). Также включены TCR, которые были модифицированы для усиления своих антипротеолитических свойств или оптимизации свойств растворимости.
TCR, комплексы TCR по изобретению или T-клетки, трансфицированные TCR по изобретению, могут быть предложены в фармацевтической композиции вместе с фармацевтически приемлемым носителем. TCR, поливалентный комплекс TCR или клетка по изобретению обычно предложены как часть стерильной фармацевтической композиции, которая обычно содержит фармацевтически приемлемый носитель. Фармацевтическая композиция может иметь любую подходящую форму (в зависимости от желательного способа введения пациенту). Она может быть предложена в стандартной лекарственной форме, обычно в герметичном контейнере, и может быть предложена, как часть набора. Такой набор (но не обязательно) включает инструкции. Он может включать множество указанных стандартных лекарственных форм.
Кроме того, TCR по изобретению можно использовать отдельно или в комбинации с другими терапевтическими средствами (например, приготовленными в такой же фармацевтической композиции).
Фармацевтическая композиция может также содержать фармацевтически приемлемый носитель. Термин «фармацевтически приемлемый носитель» относится к носителю для введения терапевтического средства. Термин относится к таким фармацевтическим носителям, которые сами не вызывают продукции антител, вредной для индивидуума, принимающего композицию, и которые не являются избыточно токсичными после введения. Данные носители хорошо известны специалисту в данной области. Полное обсуждение фармацевтически приемлемых эксципиентов можно найти в Remington's Pharmaceutical Sciences (Mack Pub. Co., N. J. 1991). Такие носители включают солевой раствор, буфер, декстрозу, воду, глицерин, этанол, адъюванты и их комбинации, но не ограничиваются ими.
Фармацевтически приемлемый носитель в терапевтической композиции может содержать жидкость, такую как вода, солевой раствор, глицерин и этанол. Кроме того, вспомогательные вещества, такие как увлажнители или эмульгаторы, pH-буферные вещества и тому подобное, могут также находиться в этих носителях.
В общем, терапевтические композиции могут быть приготовлены в виде инъекционных лекарственных средств, таких как жидкие растворы или суспензии; и твердых форм, таких как жидкие носители, которые могут подходить для приготовления в растворе или суспензии перед инъекцией.
Сразу после приготовления композиции по изобретению, ее можно вводить традиционными путями, включая внутриглазное, внутримышечное, внутривенное, подкожное, внутрикожное или местное введение, предпочтительно, парентеральное, включая подкожное, внутримышечное или внутривенное введение, но, не ограничиваясь ими. Субъект, подлежащий предупреждению или лечению, может представлять собой животное; главным образом, человека.
Когда фармацевтическую композицию по настоящему изобретению используют для фактического лечения, можно использовать фармацевтические композиции разных лекарственных форм в зависимости от применений, предпочтительно, инъекцию, пероральный препарат или т.п.
Данные фармацевтические композиции могут быть приготовлены посредством смешивания, разведения или растворения в соответствии с общепринятыми способами, в отдельных случаях, можно добавлять подходящие фармацевтические добавки, такие как эксципиенты, разрыхлители, связующие вещества, смазывающие вещества, разбавители, буферы, агенты, изменяющие изотоничность, консерванты, увлажнители, эмульгаторы, диспергирующие средства, стабилизаторы или сорастворители, и способ приготовления можно осуществлять привычным образом, в зависимости от лекарственной формы.
Фармацевтическую композицию по настоящему изобретению можно также вводить в виде препарата с замедленным высвобождением. Например, TCR по настоящему изобретению может быть включен в пилюлю или микрокапсулу, в которой полимер с замедленным высвобождением используется в качестве носителя, и затем данную пилюлю или микрокапсулу хирургически имплантируют в ткань, подлежащую лечению. Примеры полимера для замедленного высвобождения включают сополимер этилена и винилацетата, полигидрометаакрилат, полиакриламид, поливинилпирролидон, метилцеллюлозу, полимер молочной кислоты, сополимер молочной кислоты и гликолевой кислоты и т.п., предпочтительно биодеградируемый полимер, такой как полимер молочной кислоты и сополимер молочной кислоты и гликолевой кислоты.
Когда фармацевтическую композицию по настоящему изобретению применяют для фактического лечения, количество TCR или комплекса TCR по настоящему изобретению или клетки, презентирующей TCR по настоящему изобретению, в качестве активного ингредиента может быть целесообразно определено в зависимости от массы тела, возраста, пола и степени симптомов каждого пациента, подлежащего лечению, и, в конечном итоге, врачом.
Главные преимущества изобретения:
(1) Аффинность и/или время полужизни связывания TCR по настоящему изобретению в отношении комплекса FMNKFIYEI-HLA-A2 по меньшей мере в 2 раза, предпочтительно по меньшей мере в 10 раз больше аффинности и/или времени полужизни связывания TCR дикого типа.
(2) Аффинность и/или время полужизни связывания TCR по настоящему изобретению в отношении комплекса FMNKFIYEI-HLA-A2 по меньшей мере в 100 раз, предпочтительно по меньшей мере в 1000 раз и более предпочтительно в вплоть до 104-107 раз больше аффинности и/или времени полужизни связывания TCR дикого типа.
(3) Эффекторные клетки, трансдуцированные высокоаффинным TCR по настоящему изобретению, демонстрируют сильное поражающее действие на клетки-мишени.
Изобретение дополнительно проиллюстрировано следующими конкретными примерами. Следует понимать, что данные примеры приведены только в иллюстративных целях и не предназначены для ограничения объема изобретения. Экспериментальные способы в следующих примерах, в которых конкретно не указаны конкретные условия, обычно исполняют в общепринятых условиях, например, условиях, описанных в Sambrook and Russell et al., Molecular Cloning-A Laboratory Manual (Third Edition) (2001) CSHL Publishing company, или в соответствии с условиями, рекомендованными производителем. Проценты и доли являются массовыми, если не установлено иное.
Материалы и методы
Экспериментальные материалы, используемые в примерах настоящего изобретения, могут иметься в продаже, если конкретно не указано иное, среди которых E. coli DH5α приобретали у Tiangen, E. coli BL21 (DE3) приобретали у Tiangen, E. coli Tuner (DE3) приобретали у Novagen, и плазмиду pET28a приобретали у Novagen.
Пример 1. Получение стабильных цепей матрицы одноцепочечного TCR с мутациями в гидрофобном ядре
В настоящем изобретении способ сайт-направленного мутагенеза использовали в соответствии с патентной литературой WO2014/206304 для конструирования стабильной молекулы одноцепочечного TCR, состоящей из вариабельных доменов цепи α и β TCR, соединенных гибким коротким пептидом, и аминокислотные и ДНК-последовательности которых представляют собой SEQ ID NO: 47 и SEQ ID NO: 48, соответственно, как показано на Фиг. 7a и 7b. Молекулу одноцепочечного TCR использовали в качестве матрицы для осуществления скрининга молекул высокоаффинного TCR. Аминокислотные последовательности вариабельного домена α (SEQ ID NO: 3) и вариабельного домена β (SEQ ID NO: 4) цепи матрицы показаны на Фиг. 2a и 2b; соответствующие ДНК-последовательности представляют собой SEQ ID NO: 5 и 6, соответственно, как показано на Фиг. 3a и 3b; и аминокислотная последовательность и ДНК-последовательность гибкого короткого линкера представляют собой SEQ ID NO: 7 и 8, соответственно, как показано на Фиг. 4a и 4b.
Ген-мишень, несущий цепь матрицы, расщепляли NcoI и NotI, и лигировали с вектором pET28a, расщепленным NcoI и NotI. Продуктом лигирования трансформировали E. coli DH5α, помещали на чашку с LB, содержащей канамицин, переворачивали и культивировали при 37 °C в течение ночи, и позитивные клоны отбирали для ПЦР-скрининга. Позитивные рекомбинанты секвенировали для определения правильной последовательности, и рекомбинантную плазмиду выделяли и переносили в E. coli BL21 (DE3) для экспрессии.
Пример 2. Экспрессия, ренатурация и очистка стабильного одноцепочечного TCR, сконструированного в Примере 1
Все колонии BL21(DE3), содержащие рекомбинантную плазмиду pET28a-матричную цепь, полученную в Примере 1, инокулировали в среду LB, содержащую канамицин, и культивировали при 37 °C до тех пор, пока OD600 (оптическая плотность) не составит 0,6-0,8. IPTG добавляли до конечной концентрации 0,5 мМ и культивировали при 37 °C в течение еще 4 часов. Осадки клеток собирали посредством центрифугирования при 5000 об./мин в течение 15 мин, и осадки клеток лизировали с помощью Bugbuster Master Mix (Merck). Тельца включения выделяли посредством центрифугирования при 6000 об./мин в течение 15 мин с последующей промывкой Bugbuster (Merck) для удаления обломков клеток и мембранной фракции. Тельца включения собирали посредством центрифугирования при 6000 об./мин в течение 15 мин и растворяли в буфере (20 мМ Tris-HCl pH 8,0, 8 M мочевина), и нерастворимые вещества удаляли посредством высокоскоростного центрифугирования. Супернатант количественно определяли способом BCA (метод с бицинхониновой кислотой) и затем распределяли и хранили при -80 °C до применения.
К 5 мг растворенного белка телец включения с одноцепочечным TCR добавляли 2,5 мл буфера (6 M Gua-HCl, 50 мМ Tris-HCl, pH 8,1, 100 мМ NaCl, 10 мМ ЭДТА (этилендиаминтетрауксусная кислота)), затем добавляли ДТТ (дитиотреитол) до конечной концентрации 10 мМ и инкубировали при 37 °C в течение 30 мин. Одноцепочечные TCR, как рассмотрено выше, добавляли по каплям к 125 мл ренатурирующего буфера (100 мМ Tris-HCl pH 8,1, 0,4 M L-аргинин, 5 M мочевина, 2 мМ ЭДТА, 6,5 мМ β-меркаптоэтиламин, 1,87 мМ цистамин) посредством шприца и перемешивали при 4 °C в течение 10 мин. Затем, ренатурированный раствор загружали в диализный мешок на основе целлюлозной мембраны с границей отсечения 4 кДа, и диализный мешок помещали в 1 л предварительно охлажденной воды и перемешивали медленно при 4 °C в течение ночи. После 17 часов, диализную жидкость заменяли 1 л предварительно охлажденного буфера (20 мМ Tris-HCl, pH 8,0), и диализ продолжали в течение 8 ч при 4 °C. Диализную жидкость затем заменяли таким же свежим буфером, и диализ продолжали в течение ночи. После 17 часов, образец фильтровали через 0,45 мкм фильтр, подвергали вакуумной дегазации и очищали посредством анионообменной колонки (HiTrap Q HP, GE Healthcare) с элюированием в линейном градиенте 0-1 M NaCl, полученном с помощью 20 мМ Tris-HCl, pH 8,0. Собранные фракции подвергали анализу SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), и данные фракции, содержащие одноцепочечные TCR, концентрировали и дополнительно очищали посредством колонки для гель-фильтрации (Superdex 75 10/300, GE Healthcare), и целевые компоненты также подвергали анализу SDS-PAGE.
Элюированные фракции для анализа BIAcore дополнительно тестировали в отношении чистоты с использованием гель-фильтрации. Условия выглядели следующим образом: хроматографическая колонка Agilent Bio SEC-3 (300 A, ϕ 7,8×300 мм), подвижная фаза 150 мМ фосфатный буфер, скорость потока 0,5 мл/мин, температура колонки 25 °C и длина волны УФ (ультрафиолетовое излучение) при детектирования 214 нм.
Пример 3. Характеристика связывания
Анализ BIAcore
Активность связывания молекулы TCR с комплексом FMNKFIYEI-HLA-A0201 выявляли, используя систему анализа в реальном времени BIAcore T200. Антитело против стрептавидина (GenScript) добавляли к связывающему буферу (10 мМ натрий-ацетатный буфер, pH 4,77), и затем антитело пропускали через чип CM5, предварительно активированный EDC и NHS, для иммобилизации антитела на поверхности чипа. Непрореагировавшую активированную поверхность в конечном итоге блокировали раствором этаноламина в соляной кислоте для завершения процесса связывания на уровне связывания примерно 15000 RU в условиях 25 °C, pH 7,1-7,5.
Низкую концентрацию стрептавидина пропускали через поверхность чипа, покрытого антителом, затем комплекс FMNKFIYEI-HLA-A0201 пропускали через канал выявления, причем другой канал использовали в качестве референсного канала. 0,05 мМ биотина пропускали через чип на протяжении 2 мин со скоростью потока 10 мкл/мин, блокируя, таким образом, оставшиеся сайты связывания для стрептавидина. Аффинность определяли посредством кинетического анализа за один цикл. TCR разводили до нескольких разных концентраций посредством буфера HEPES-EP (10 мМ HEPES, 150 мМ NaCl, 3 мМ ЭДТА, 0,005% P20, pH 7,4) и пропускали через поверхность чипа в свою очередь со скоростью потока 30 мкл/мин, со временем связывания 120 с на инъекцию. После последней инъекции чип помещали для диссоциации на 600 с. В конце каждого раунда анализа чип регенерировали с помощью 10 мМ Gly-HCl, pH 1,75. Кинетические параметры рассчитывали, используя программное обеспечение для оценки BIAcore.
Способ получения указанного выше комплекса FMNKFIYEI-HLA-A0201 описан ниже:
a. Очистка
Собирали 100 мл жидкости E. coli, индуцированной для экспрессии тяжелой или легкой цепи, и центрифугировали при 8000 g в течение 10 мин при 4 °C, и клетки промывали один раз 10 мл PBS (фосфатно-солевой буферный раствор), и затем интенсивно встряхивали в 5 мл экстракционных реагентов BugBuster Master Mix (Merck) для ресуспендирования клеток. Суспензию инкубировали в течение 20 мин при комнатной температуре и затем центрифугировали при 6000 g в течение 15 мин при 4 °C. Супернатант отбрасывали, собирая тельца включения.
Указанные выше тельца включения ресуспендировали в 5 мл BugBuster Master Mix и инкубировали вертикально при комнатной температуре в течение 5 мин. Добавляли 30 мл разведенного в 10 раз BugBuster, смешивали и центрифугировали при 6000 g на протяжении 15 мин при 4°C. Супернатант отбрасывали, 30 мл разведенного в 10 раз BugBuster добавляли для ресуспендирования тельца включения, перемешивали и центрифугировали два раза при 6000 g при 4°C в течение 15 мин. 30 мл 20 мМ Tris-HCl, pH 8,0, добавляли для ресуспендирования телец включения, смешивали и центрифугировали при 6000 g при 4 °C в течение 15 мин. Наконец, тельца включения растворяли в 20 мМ Tris-HCl, 8 M мочевине, и чистоту телец включения определяли посредством SDS-PAGE, и концентрацию измеряли посредством набора BCA.
b. Повторное сворачивание
Синтезированный короткий пептид FMNKFIYEI (Beijing Saibaisheng Gene Technology Co., Ltd.) растворяли в ДМСО (диметилсульфоксид) до концентрации 20 мг/мл. Тельца включения легкой и тяжелой цепей растворяли в 8 M мочевине, 20 мМ Tris pH 8,0, 10 мМ ДТТ, и дополнительно денатурировали посредством добавления 3 M гуанидин гидрохлорида, 10 мМ ацетата натрия, 10 мМ ЭДТА перед повторным сворачиванием. Пептид FMNKFIYEI добавляли к буферу для повторного сворачивания (0,4 M L-аргинин, 100 мМ Tris pH 8,3, 2 мМ ЭДТА, 0,5 мМ окисленный глутатион, 5 мМ восстановленный глутатион, 0,2 мМ PMSF (фенилметилсульфонил фторид), охлажденный до 4°C) в количестве 25 мг/л (конечная концентрация). Затем, 20 мг/л легкой цепи и 90 мг/л тяжелой цепи (конечная концентрация, тяжелую цепь добавляли тремя порциями, 8 ч/порция) последовательно добавляли и подвергали повторному сворачиванию при 4°C на протяжении по меньшей мере 3 суток для завершения повторного сворачивания, и SDS-PAGE использовали для подтверждения повторного сворачивания.
c. Очистка при повторном сворачивании
Буфер для повторного сворачивания заменяли 10 объемами 20 мМ Tris pH 8,0 для диализа, и буфер заменяли по меньшей мере два раза по существу для уменьшения ионной силы раствора. После диализа раствор белка фильтровали через 0,45 мкм фильтр из ацетата целлюлозы и загружали на анионообменную колонку HiTrap Q HP (GE, General Electric Company) (объем слоя 5 мл). Белок элюировали с помощью линейного градиента 0-400 мМ NaCl, полученного в 20 мМ Tris pH 8,0, используя Akta Purifier (GE), и pMHC элюировали приблизительно 250 мМ NaCl. Пиковые фракции собирали, и их чистоту выявляли посредством SDS-PAGE.
d. Биотинилирование
Очищенные молекулы pMHC концентрировали в пробирке для ультрацентрифугирования Millipore, в то время как буфер заменяли 20 мМ Tris, pH 8,0, и затем добавляли реагент для биотинилирования 0,05 M Бицин, pH 8,3, 10 мМ АТФ, 10 мМ MgOAc, 50 мкМ D-Биотин, 100 мкг/мл фермента BirA (GST-BirA). Полученную смесь инкубировали при комнатной температуре в течение ночи, и SDS-PAGE использовали для выявления завершения биотинилирования.
e. Очистка биотинилированного комплекса
Биотинилированные и меченые молекулы pMHC концентрировали до 1 мл в пробирке для ультрацентрифугирования Millipore. Биотинилированный pMHC очищали посредством эксклюзионной хроматографии. 1 мл концентрированных биотинилированных молекул pMHC загружали на колонку HiPrepTM 16/60 S200 HR (GE), предварительно уравновешенную фильтрованным PBS с использованием Akta Purifier (GE), и элюировали PBS при скорости потока 1 мл/мин. Биотинилированные молекулы pMHC элюировали в виде одиночного пика на уровне примерно 55 мл. Фракции, содержащие белок, объединяли и концентрировали в пробирке для ультрацентрифугирования Millipore. Концентрацию белка определяли способом BCA (Thermo), добавляли смесь ингибиторов протеаз (Roche), и биотинилированные молекулы pMHC распределяли и хранили при -80°C.
Пример 4. Получение высокоаффинного одноцепочечного TCR
Технология фагового дисплея представляет собой средство для получения библиотек вариантов высокоаффинного TCR для осуществления скрининга высокоаффинных вариантов. Фаговый дисплей TCR и способ скрининга, описанный Li et al. ((2005) Nature Biotech 23(3): 349-354), применяли в отношении матрицы одноцепочечного TCR Примера 1. Библиотеку высокоаффинных TCR формировали посредством осуществления мутирования областей CDR цепи матрицы и подвергали пэннингу. После нескольких раундов пэннинга фаговая библиотека может специфично связываться с соответствующим антигеном, моноклон отбирали, и проводили анализ последовательностей.
Способ BIAcore Примера 3 использовали для анализа взаимодействия молекулы TCR с комплексом FMNKFIYEI-HLA-A0201, и отбирали высокоаффинный TCR с аффинностью и/или временем полужизни связывания по меньшей мере в 2 раза больше аффинности и/или времени полужизни связывания TCR дикого типа, а именно, константа равновесия диссоциации KD прошедшего скрининг высокоаффинного TCR в отношении связывания комплекса FMNKFIYEI-HLA-A0201 меньше чем или равна половине константны диссоциации KD TCR дикого типа в отношении связывания комплекса FMNKFIYEI-HLA-A0201, и результаты показаны ниже в Таблице 3. Выявляли, что значение KD взаимодействия растворимого референсного TCR с комплексом FMNKFIYEI-HLA-A0201 составляло 43 мкМ посредством использования указанного выше способа, и кривая взаимодействия показана на Фиг. 12, а именно, значение KD взаимодействия TCR дикого типа с комплексом FMNKFIYEI-HLA-A0201 также составляет 43 мкМ (4,3E-05M).
Конкретно, при применении нумерации, показанной в SEQ ID NO: 1, аминокислота в одном или более следующих сайтах в вариабельном домене цепи α данных высокоаффинных TCR-мутантов является мутантной: 28S, 29I, 30N, 31N, 94R, 95T, 96G, 97A, 98N и 99S, и/или при использовании нумерации, показанной в SEQ ID NO: 2, аминокислота в одном или более следующих сайтах в вариабельном домене цепи β данных высокоаффинных TCR-мутантов является мутантной: 27S, 28E, 29H, 30N, 50Q, 51N, 52E, 53A и 54Q.
Более конкретно, при использовании нумерации, показанной в SEQ ID NO: 1, вариабельные домены цепи α данных высокоаффинных TCR содержат один или более аминокислотных остатков, выбранных из группы, состоящей из 28D, или 28P, или 28A, или 28E, 29V, 30S, или 30Y, или 30D, 31S, или 31K, 94A, или 94S, или 94V, или 94D, или 94E, или 94Q, или 94T, 95M, или 95R, или 95L, или 95A, или 95N, или 95S, 96A, или 96Q, или 96S, или 96E, или 96N, 97S, или 97G, или 97P, или 97W, 98S, или 98T, 99A, или 99N, или 99T, или 99P; и/или при использовании нумерации, показанной в SEQ ID NO: 2, вариабельные домены цепи β данных высокоаффинных TCR содержат один или более аминокислотных остатков, выбранных из группы, состоящей из 27A, или 27Q, или 27G, 28G, или 28T, или 28R, или 28L, 29W, или 29R, 30A, или 30P, 50H, 51G, 52D, 53M, или 53L, 54E.
Конкретные аминокислотные последовательности вариабельных доменов цепи α (SEQ ID NO: 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35) и вариабельных доменов цепи β (SEQ ID NO: 36, 37, 38, 39, 40, 41, 42, 43, 44, 45 и 46) высокоаффинных одноцепочечных TCR показаны на Фиг. 5(1)-(27) и Фиг. 6(1)-(11), соответственно.
Таблица 3
Пример 5. Получение высокоаффинного αβ гетеродимерного TCR
Мутации в областях CDR высокоаффинных одноцепочечных TCR, подвергнутых скринингу в Примере 4, вводили в соответствующие сайты вариабельного домена αβ гетеродимерного TCR, и его аффинность в отношении комплекса FMNKFIYEI-HLA-A0201, выявляли посредством BIAcore. Мутантные сайты высокой аффинности могут быть введены в указанных выше областях CDR способом сайт-направленного мутагенеза, хорошо известным специалисту в данной области. Аминокислотные последовательности вариабельного домена цепи α и цепи β указанного выше TCR дикого типа показаны на Фиг. 1a (SEQ ID NO: 1) и 1b (SEQ ID NO: 2), соответственно.
Следует отметить, что для получения более стабильного растворимого TCR для более простой оценки аффинности связывания и/или времени полужизни связывания между TCR и комплексом FMNKFIYEI-HLA A0201, αβ гетеродимерный TCR может представлять собой такой TCR, в котором остаток цистеина соответственно вводят в константный домен цепи α и β с образованием искусственной межцепочечной дисульфидной связи. В данном примере аминокислотные последовательности цепей α и β TCR после введения остатка цистеина показаны на Фиг. 8a (SEQ ID NO: 49) и 8b (SEQ ID NO: 50), и вводимые остатки цистеина показаны буквами, выделенными жирным шрифтом.
В соответствии со стандартными способами, описанными в лабораторном руководстве по молекулярному клонированию (3rd edition, Sambrook and Russell), гены внеклеточных последовательностей цепей α и β TCR, подлежащие экспрессии, синтезируют и вставляют в экспрессионный вектор pET28a+ (Novagene), в котором расположенные выше и расположенные ниже сайты клонирования представляют собой NcoI и NotI, соответственно. Мутации в областях CDR вводят посредством ПЦР с перекрывающимися праймерами, хорошо известной специалисту в данной области. Вставляемый фрагмент секвенировали для подтверждения того, что он правильный.
Пример 6. Экспрессия, повторное сворачивание и очистка αβ гетеродимерного TCR
Экспрессионными векторами для цепи α и β TCR трансформировали экспрессионные бактерии BL21 (DE3) посредством химической трансформации, соответственно. Бактерии выращивали в среде LB и индуцировали конечной концентрацией 0,5 мМ IPTG при OD600, равной 0,6. Тельца включения, образованные после экспрессии цепей α и β TCR, экстрагировали BugBuster Mix (Novagene) и повторно промывали раствором BugBuster. Тельца включения в конечном итоге растворяли в 6 M гуанидин гидрохлориде, 10 мМ дитиотреитоле (ДТТ), 10 мМ этилендиаминтетрауксусной кислоте (ЭДТА) и 20 мМ Tris (pH 8,1).
Растворенные цепи α и β TCR быстро смешивали в 5 M мочевине, 0,4 M аргинине, 20 мМ Tris (pH 8,1), 3,7 мМ цистамине и 6,6 мМ β-меркаптоэтиламине (4°C) в массовом соотношении 1:1. Конечная концентрация составляет 60 мг/мл. После перемешивания раствор подвергали диализу против 10 объемов деионизированной воды (4°C), и через 12 часов деионизированную воду заменяли буфером (20 мМ Tris, pH 8,0), и диализ продолжали при 4°C на протяжении 12 часов. После завершения диализа раствор фильтровали через 0,45 мкМ фильтр и очищали посредством анионообменной колонки (HiTrap Q HP, 5 мл, GE Healthcare). Пик элюирования TCR, содержащего успешно повторно свернутые димеры α и β, подтверждали посредством геля SDS-PAGE. TCR затем дополнительно очищали посредством эксклюзионной хроматографии (HiPrep 16/60, Sephacryl S-100 HR, GE Healthcare). Чистоту очищенного TCR, которую определяли посредством SDS-PAGE, составляла более чем 90%, и его концентрацию определяли способом BCA.
Пример 7. Результаты анализа BIAcore
Аффинность αβ гетеродимерного TCR, в который вводили высокоаффинную область CDR, в отношении комплекса FMNKFIYEI-HLA-A0201 выявляли посредством способа, описанного в Примере 3.
Области CDR, выбранные из цепи α и β высокоаффинного одноцепочечного TCR, переносили в соответствующие положения вариабельного домена цепи α TCR дикого типа SEQ ID NO: 1 и вариабельного домена цепи β SEQ ID NO: 2, соответственно, с образованием αβ гетеродимерного TCR. Дополнительно, αβ гетеродимерные TCR были образованы посредством искусственной комбинации на основе сайтов мутации, подвергавшихся скринингу и полученных в областях CDR. Аминокислотные последовательности полученных новых вариабельных доменов цепи α и β TCR показаны на Фиг. 9(1)-(29) и Фиг. 10(1)-(20), соответственно. Поскольку области CDR молекулы TCR определяют их аффинность в отношении соответствующего комплекса pMHC, специалист в данной области может предвидеть, что αβ гетеродимерный TCR, в который введен сайт мутации высокой аффинности, также обладает высокой аффинностью в отношении комплекса FMNKFIYEI-HLA-A0201. Экспрессионный вектор конструировали способом, описанным в Примере 5, упомянутый выше αβ гетеродимерный TCR с вводимой мутацией высокой аффинности экспрессировали, повторно сворачивали и очищали способом, описанным в Примере 6, и затем аффинность TCR в отношении комплекса FMNKFIYEI-HLA-A0201 определяли посредством BIAcore T200, как показано ниже в Таблице 4.
Таблица 4
SEQ ID NO:
SEQ ID NO:
Как можно видеть из приведенной выше Таблицы 4, αβ гетеродимерный TCR с сайтами мутации, введенными в области CDR, сохраняет высокую аффинность в отношении комплекса FMNKFIYEI-HLA-A0201. Аффинность гетеродимерного TCR в отношении комплекса FMNKFIYEI-HLA-A0201 по меньшей мере в 2 раза выше аффинности TCR дикого типа.
Пример 8. Экспрессия, повторное сворачивание и очистка слитых молекул антител против CD3 с высокоаффинным одноцепочечным TCR
Молекулу высокоаффинного одноцепочечного TCR по настоящему изобретению сливают с одноцепочечной молекулой (scFv) антитела против CD3 с конструированием слитой молекулы. Праймеры конструировали посредством ПЦР с перекрывающимися праймерами, и лигировали гены антител против CD3 и молекулу высокоаффинного одноцепочечного TCR. Промежуточный линкер конструировали в виде GGGGS, и фрагмент гена слитой молекулы имел сайты рестрикции NcoI и NotI. Продукт ПЦР-амплификации расщепляли с помощью NcoI и NotI и лигировали с вектором pET28a, расщепленным NcoI и NotI. Продуктом лигирования трансформировали компетентные клетки E. coli DH5α, и помещали на чашку с LB, содержащей канамицин, и переворачивали и культивировали при 37°C. Позитивные клоны отбирали для ПЦР-скрининга, и позитивные рекомбинантны секвенировали для определения правильной последовательности. Рекомбинантные плазмиды выделяли и ими трансформировали компетентные клетки E. coli BL21 (DE3) для экспрессии.
Экспрессия слитого белка
Экспрессионной плазмидой, содержащей исследуемый ген, трансформировали штамм E. coli BL21 (DE3), помешали на чашку с LB (канамицин, 50 мкг/мл) и культивировали при 37 °C в течение ночи. На следующий день клоны отбирали и инокулировали в 10 мл жидкой среды LB (канамицин, 50 мкг/мл), культивировали в течение 2-3 ч, затем инокулировали в 1 л среды LB (канамицин, 50 мкг/мл) при объемном соотношении 1:100, культивировали до тех, пор пока OD600 не становилась равной 0,5-0,8, и затем индуцировали экспрессию исследуемого белка, используя IPTG в конечной концентрации 0,5 мМ. Спустя 4 часа после индукции, клетки собирали посредством центрифугирования при 6000 об./мин в течение 10 мин. Клетки промывали один раз в буфере PBS и распределяли. Отбирали клетки, соответствующие 200 мл бактериальной культуры, и лизировали посредством 5 мл BugBuster Master Mix (Novagen), и тельца включения собирали посредством центрифугирования при 6000 g в течение 15 минут. Тельца включения 4 раза промывали детергентом для удаления обломков клеток и компонентов мембраны. Тельца включения затем промывают буфером, таким как PBS, для удаления детергента и соли. Наконец, тельца включения растворяли в буферном растворе Tris, содержащем 8 M мочевину, концентрацию телец включения измеряли, и тельца включения распределяли и криоконсервировали при -80°C.
Повторное сворачивание слитых белков
Примерно 10 мг телец включения брали из низкотемпературной морозильной камеры -80°C и размораживали, и дитиотреитол (ДТТ) добавляли до конечной концентрации 10 мМ и инкубировали при 37°C в течение 30 мин - 1 часа для обеспечения разрыва дисульфидной связи. Затем раствор образца телец включения добавляли по каплям в 200 мл 4°C предварительно охлажденного буфера для повторного сворачивания (100 мМ Tris pH 8,1, 400 мМ L-аргинин, 2 мМ ЭДТА, 5 M мочевина, 6,5 мМ β-меркаптоэтиламин, 1,87 мМ цистамин), соответственно, и медленно перемешивали при 4°C в течение примерно 30 минут. Раствор для повторного сворачивания подвергали диализу против 8 объемов предварительно охлажденной H2O на протяжении 16-20 часов. Его дважды дополнительно подвергали диализу с 8 объемами 10 мМ Tris pH 8,0, и диализ продолжали при 4°C в течение 8 часов. После диализа образец фильтровали и подвергали следующему способу очистки.
Первая стадия очистки слитого белка
Подвергнутый диализу и повторному сворачиванию материал (в 10 мМ Tris pH 8,0) элюировали на предварительно наполненной колонке для анионообменной хроматографии POROS HQ/20 (Applied Biosystems) с градиентом 0-600 мМ NaCl, используя очиститель AKTA (GE Healthcare). Каждый компонент анализировали посредством SDS-PAGE, окрашенного Кумасси бриллиантовым синим, и затем объединяли.
Вторая стадия очистки слитого белка
Очищенный и объединенный раствор образца с первой стадии концентрировали для очистки на данной стадии, и слитый белок очищали посредством предварительно наполненной колонки для эксклюзионной хроматографии Superdex 75 10/300 GL (GE Healthcare), предварительно уравновешенной в буфере PBS. Компоненты пиков анализировали посредством SDS-PAGE, окрашенного Кумасси бриллиантовым синим, и затем объединяли.
Пример 9. Экспрессия, повторное сворачивание и очистка слитых белков антитела против CD3 с высокоаффинным αβ гетеродимерным TCR
Слитую молекулу получали посредством слияния одноцепочечного антитела против CD3 (scFv) с αβ гетеродимерным TCR. Антитело против CD3 scFv сливали с цепью β TCR, и цепь β TCR может содержать вариабельный домен цепи β любого из упомянутых выше высокоаффинных αβ гетеродимерных TCR, и цепь α TCR слитой молекулы может содержать вариабельный домен цепи α любого из упомянутых выше высокоаффинных αβ гетеродимерных TCR.
Конструирование экспрессионного вектора для слитой молекулы
1. Конструирование экспрессионного вектора для цепи α
Целевой ген, несущий цепь α αβ гетеродимерного TCR, расщепляли NcoI и NotI, и лигировали с вектором pET28a, расщепленным NcoI и NotI. Продуктом лигирования трансформировали E. coli DH5α, помещали на чашку с LB, содержащую канамицин, и переворачивали и культивировали в течение ночи при 37°C. Позитивные клоны отбирали для ПЦР-скрининга, и позитивные рекомбинанты секвенировали для определения правильной последовательности. Рекомбинантные плазмиды выделяли и ими трансформировали E. coli Tuner (DE3) для экспрессии.
2. Конструирование экспрессионного вектора для антитела против CD3 (scFv)-β цепи
Праймеры конструировали посредством ПЦР с перекрывающимися праймерами с соединением генов антитела против CD3 scFv и цепи β высокоаффинного гетеродимерного TCR.
Антитело против CD3 scFv можно соединять с N-концом или C-концом цепи β TCR. Промежуточный линкер представлял собой GGGGS, и фрагмент гена слитого белка антитела против CD3 scFv и цепи β высокоаффинного гетеродимерного TCR имел сайты эндонуклеаз рестрикции NcoI (CCATGG) и NotI (GCGGCCGC). Продукт ПЦР-амплификации расщепляли NcoI и NotI и лигировали с вектором pET28a, расщепленным NcoI и NotI. Продуктом лигирования трансформировали компетентные клетки E. coli DH5α, помещали на чашку с LB, содержащей канамицин, и переворачивали и культивировали в течение ночи при 37°C. Позитивные клоны отбирали для ПЦР-скрининга, и позитивные рекомбинанты секвенировали для определения правильной последовательности. Рекомбинантные плазмиды выделяли и ими трансформировали компетентные клетки E. coli Tuner (DE3) для экспрессии.
Экспрессия, повторное сворачивание и очистка слитого белка
Экспрессионными плазмидами по отдельности трансформировали компетентные клетки E. coli Tuner (DE3), помещали на чашки с LB (канамицин 50 мкг/мл) и культивировали в течение ночи при 37°C. На следующий день клоны отбирали и инокулировали в 10 мл жидкой среды LB (канамицин 50 мкг/мл) на 2-3 ч, и инокулировали в 1 л среды LB в объемном соотношении 1:100, культивирование продолжали до тех пор, пока OD600 не становилась равной 0,5-0,8, и конечную концентрацию 1 мМ IPTG добавляли для индукции экспрессии исследуемого белка. Спустя 4 часа, клетки собирали посредством центрифугирования при 6000 об./мин в течение 10 мин. Клетки промывали один раз в буфере PBS и распределяли, и клетки, соответствующие 200 мл бактериальной культуры, брали и лизировали 5 мл BugBuster Master Mix (Merck), тельца включения собирали посредством центрифугирования при 6000 g в течение 15 мин и затем 4 раза промывали детергентом для удаления обломков клеток и компонентов мембраны. Тельца включения затем промывали буфером, таким как PBS, для удаления детергента и соли. Наконец, тельца включения растворяли в 6M гуанидин гидрохлориде, 10 мМ дитиотреитоле (ДТТ), 10 мМ этилендиаминтетрауксусной кислоте (ЭДТА), 20 мМ Tris, pH 8,1 буферном растворе, и определяли концентрацию телец включения. Тельца включения распределяли и криоконсервировали при -80°C.
Растворенную цепь α TCR и антитело против CD3 (scFv)-цепь β быстро смешивали в массовом соотношении 2:5 в 5 M мочевине (мочевина), 0,4 M L-аргинине (L-аргинин), 20 мМ Tris pH 8,1, 3,7 мМ цистамине и 6,6 мМ β-меркаптоэтиламине (4°C), и конечные концентрации цепи α и димера антитело против CD3 (scFv)-цепь β составляли 0,1 мг/мл, 0,25 мг/мл, соответственно.
После смешивания раствор подвергали диализу против 10 объемов деионизированной воды (4°C), и после 12 часов деионизированную воду заменяли буфером (10 мМ Tris, pH 8,0) в течение еще 12 часов при 4°C. После завершения диализа раствор фильтровали через фильтр 0,45 мкМ и очищали посредством анионообменной колонки (HiTrap Q HP 5 мл, GE healthcare). Элюированные пики, содержащие восстановленную цепь α TCR и димер антитело против CD3 (scFv)-цепь β TCR, подтверждали посредством геля SDS-PAGE. Слитую молекулу TCR затем очищали посредством эксклюзионной хроматографии (S-100 16/60, GE healthcare) и дополнительно очищали посредством анионообменной колонки (HiTrap Q HP 5 ml, GE healthcare). Чистота очищенной слитой молекулы TCR, как определяли посредством SDS-PAGE, была выше 90%, и концентрацию определяли способом BCA.
Пример 10. Анализ функции активации эффекторных клеток, трансфицированных высокоаффинным TCR по настоящему изобретению
Этот пример демонстрирует, что эффекторные клетки, трансфицированные высокоаффинным TCR по настоящему изобретению, оказывают хорошее специфичное активирующее действие на клетки-мишени.
Функцию и специфичность высокоаффинного TCR по настоящему изобретению в клетках исследовали анализом ELISPOT. Способы выявления клеточной функции с использованием анализов ELISPOT хорошо известны специалисту в данной области. В анализе ELISPOT IFN-γ (interferon-gamma - интерферон гамма) данного примера CD8+ T-клетки, выделенные из крови здоровых добровольцев и трансфицированные высокоаффинным TCR по настоящему изобретению с использованием лентивируса, использовали в качестве эффекторных клеток.
TCR по настоящему изобретению случайным образом отбирали и делили на две группы для эксперимента. TCR и их номера перечислены в Таблице 4.
TCR3 (вариабельный домен цепи α SEQ ID NO: 51, вариабельный домен цепи β SEQ ID NO: 2), TCR2 (вариабельный домен цепи α SEQ ID NO: 52, вариабельный домен цепи β SEQ ID NO: 2), TCR4 (вариабельный домен цепи α SEQ ID NO: 53, вариабельный домен цепи β SEQ ID NO: 2) и TCR1 (вариабельный домен цепи α SEQ ID NO: 54, вариабельный домен цепи β SEQ ID NO: 2), представляли собой первую группу; и эффекторные клетки в контрольной группе метили как WT-TCR (трансфицированные TCR дикого типа) и A6 (трансфицированные другим TCR, который не является TCR по настоящему изобретению). Линии клеток-мишеней представляли собой клетки HepG2, Huh-7, Huh-1, HCCC9810 и SK-HEP-1, среди которых, линия клеток-мишеней HepG2 экспрессирует релевантные антигены, и их генотип также соответствует позитивным клеточным линиям. Huh-7, Huh-1, HCCC9810 и SK-HEP-1 представляют собой негативные клеточные линии и используются в качестве контролей.
TCR7 (вариабельный домен цепи α SEQ ID NO: 57, вариабельный домен цепи β SEQ ID NO: 2), TCR11 (вариабельный домен цепи α SEQ ID NO: 61, вариабельный домен цепи β SEQ ID NO: 2) и TCR13 (вариабельный домен цепи α SEQ ID NO: 63, вариабельный домен цепи β SEQ ID NO: 2) представляли собой вторую группу. Эффекторные клетки в контрольной группе метили как WT-TCR (трансфицированные TCR дикого типа) и A6 (трансфицированные другим TCR, который не является TCR по настоящему изобретению). Линии клеток-мишеней представляли собой HCCC9810-AFP, PLCPRF5-A2, SNU398 и HCCC9810, среди которых линии клеток-мишеней HCCC9810-AFP и PLCPRF5-A2 экспрессируют релевантные антигены, и их генотип также соответствует линиям позитивных клеток. SNU398 и HCCC9810 представляют собой линии негативных клеток и используются в качестве контролей.
Сначала, получали планшет ELISPOT. Планшет ELISPOT активировали и покрывали этанолом в течение ночи при 4°C. В первый день эксперимента покрывающий раствор удаляли, и планшет промывали, блокировали и инкубировали при комнатной температуре в течение 2 часов, и блокирующий раствор удаляли. Компоненты анализа добавляли в планшет ELISPOT в следующем порядке: среду для регуляции числа эффекторных клеток до 1×104 клеток/мл, и среду для регуляции числа каждой линии клеток-мишеней до 2×105 клеток/мл. После однородного смешивания 100 мкл линии клеток-мишеней (т.е., 20000 клеток/лунку) и 100 мкл эффекторных клеток (т.е., 1000 клеток/лунку) добавляли в соответствующие лунки в двухкратной повторности и инкубировали в течение ночи (37°C, 5% CO2). На второй день эксперимента планшет промывали, подвергали вторичному выявлению и проявлению и сушили, и пятна, образованные на пленке, считали, используя ридер immunospot (система ELISPOT READER; компания AID20).
Результаты экспериментов из первой группы и второй группы показаны на Фиг. 13a и Фиг. 13b, в которых эффекторные клетки, трансфицированные высокоаффинным TCR по настоящему изобретению, демонстрируют отсутствие эффектов активации, оказываемых на негативные клетки-мишени, и в то же время демонстрируют превосходные специфичные эффекты активации, оказываемые на позитивные клетки-мишени, которые значимо превосходят эффекты активации эффекторных клеток, трансфицированных WT (дикий тип) TCR.
Пример 11. Анализ функции «киллинга» эффекторных клеток, трансфицированных высокоаффинным TCR по настоящему изобретению
В данном примере высвобождение LDH определяли нерадиоактивным анализом цитотоксичности для проверки функции «киллинга» у клеток, трансдуцирующих TCR по настоящему изобретению. Анализ представляет собой калориметрический анализ, являющийся заменой анализа цитотоксичности по высвобождению 51Cr для количественной оценки лактатдегидрогеназы (LDH), высвобождающейся после лизиса клеток. LDH, высвобождающуюся в среду, выявляли, используя 30 минутную сопряженную ферментативную реакцию, в которой LDH превращает соль тетразолия (INT) в красный формазан. Количество полученного красного продукта прямо пропорционально числу лизированных клеток. Данные по поглощению видимого цвета при 490 нм могут быть собраны, используя стандартный 96-луночный планшет-ридер.
Способы выявления клеточной функции с использованием анализа высвобождения LDH хорошо известны специалисту в данной области. В эксперименте данного примера по LDH клетки PBL, выделенные из крови здоровых добровольцев, трансфицированные TCR посредством лентивируса, использовали в качестве эффекторных клеток. Линии клеток-мишеней представляли собой HepG2, HCCC9810 и SNU-398A375 (3525), среди которых HepG2 представляла собой линию позитивных клеток-мишеней; и HCCC9810 и SNU-398 представляли собой линии негативных клеток-мишеней в качестве контроля.
TCR по настоящему изобретению случайным образом отбирали и делили на две группы для данного эксперимента. TCR и их номера перечислены в Таблице 4: TCR3 (вариабельный домен цепи α SEQ ID NO: 51, вариабельный домен цепи β SEQ ID NO: 2), TCR2 (вариабельный домен цепи α SEQ ID NO: 52, вариабельный домен цепи β SEQ ID NO: 2) и TCR1 (вариабельный домен цепи α SEQ ID NO: 54, вариабельный домен цепи β SEQ ID NO: 2) представляли собой первую группу; и TCR7 (вариабельный домен цепи α SEQ ID NO: 57, вариабельный домен цепи β SEQ ID NO: 2) и TCR13 (вариабельный домен цепи α SEQ ID NO: 63, вариабельный домен цепи β SEQ ID NO: 2) представляли собой вторую группу. Эффекторные клетки в контрольной группе были мечены как WT-TCR (трансфицированы TCR дикого типа) и A6 (трансфицированы другим TCR, который не является TCR по настоящему изобретению).
Сначала, получали планшет LDH. В первый день эксперимента компоненты данного анализа добавляли в планшет в следующем порядке: среду для регулирования числа эффекторных клеток до 3×105 клеток/мл, и среду для регулирования каждой линии клеток-мишеней до 3×105 клеток/мл. После однородного смешивания 100 мкл линии клеток-мишеней (а именно, 30000 клеток/лунка) и 100 мкл эффекторных клеток (а именно, 30000 клеток/лунка) добавляли в соответствующие лунки в трехкратной повторности. Лунки для спонтанных эффекторных клеток, для спонтанных клеток-мишеней, для максимальных клеток-мишеней, для контроля с поправкой на объем и для фонового контроля - среды были одновременно установлены, причем каждая содержала 200 мкл жидкости. Планшет инкубировали в течение ночи (37°C, 5% CO2). На второй день эксперимента выявляли развитие окраски, и после окончания реакции записывали поглощение при 490 нм с использованием микропланшета-ридера (Bioteck).
Результаты экспериментов по первой группе и второй группе показаны на Фиг. 14a и Фиг. 14b, в которых эффекторные клетки, трансфицированные высокоаффинным TCR по настоящему изобретению, демонстрируют сильное поражающее действие на клетки-мишени, экспрессирующие релевантные антигены, в то время как по существу не демонстрируют поражающего действия на клетки-мишени, не экспрессирующие релевантные антигены; и эффекторные клетки, трансфицированные другим TCR, не демонстрируют поражающего действия на клетки-мишени, экспрессирующие релевантные антигены.
Пример 12. Функциональная проверка-анализ активации слитого белка высокоаффинного TCR по настоящему изобретению и антитела против CD3
Данный пример демонстрирует, что слитый белок высокоаффинного TCR по настоящему изобретению и антитела против CD3 способен перенаправлять эффекторные клетки и демонстрирует хорошее активирующее действие.
Способы выявления клеточной функции с использованием анализов ELISPOT хорошо известны специалисту в данной области. Эффекторные клетки, используемые в анализе ELISPOT IFN-γ данного примера, представляли собой CD8+T-клетки, выделенные из крови здоровых добровольцев. Линии клеток-мишеней представляли собой HEPG2, HCCC9810 и SNU-398, где HEPG2 представляла собой линию позитивных клеток-мишеней, и HCCC9810 и SNU-398 представляли собой линии негативных клеток-мишеней. Высокоаффинный TCR по настоящему изобретению случайным образом отбирали, и слитый белок получали, как описано в Примере 9. Конкретно, отобранные TCR и их номера перечислены в Таблице 4, которые представляют собой TCR21 (вариабельный домен цепи α SEQ ID NO: 69, вариабельный домен цепи β SEQ ID NO: 79), TCR24 (вариабельный домен цепи α SEQ ID NO: 71, вариабельный домен цепи β SEQ ID NO: 83), TCR25 (вариабельный домен цепи α SEQ ID NO: 72, вариабельный домен цепи β SEQ ID NO: 84), TCR17 (вариабельный домен цепи α SEQ ID NO: 66, вариабельный домен цепи β SEQ ID NO: 81), TCR23 (вариабельный домен цепи α SEQ ID NO: 71, вариабельный домен цепи β SEQ ID NO: 79), TCR33 (вариабельный домен цепи α SEQ ID NO: 76, вариабельный домен цепи β SEQ ID NO: 79) и TCR34 (вариабельный домен цепи α SEQ ID NO: 72, вариабельный домен цепи β SEQ ID NO: 81), соответственно.
Сначала готовили планшет ELISPOT. Планшет ELISPOT активировали и покрывали этанолом и инкубировали в течение ночи при 4°C. В первый день эксперимента раствор для нанесения покрытия удаляли, и планшет промывали, блокировали и инкубировали при комнатной температуре в течение двух часов, и блокирующий раствор удаляли. Компоненты анализа добавляли в планшет ELISPOT. Клетки-мишени добавляли в количестве 20000 клеток/лунку, эффекторные клетки добавляли в количестве 4000 клеток/лунку, и концентрация слитого белка составляла 0,04 мкМ (серийно разведенные в 10 раз, в общей сложности для 6 градиентов). Устанавливали лунки в двухкратной повторности. Планшет инкубировали в течение ночи (37°C, 5% CO2). На второй день эксперимента, планшет промывали и подвергали вторичному выявлению и развитию окраски. Планшет сушили, и пятна, образованные на пленке, считали, используя планшет-ридер immunospot (система ELISPOT READER; компания AID20).
Результаты экспериментов показаны на Фиг. 15a, 15b и 15c, 15d, 15e, 15f и 15g. Результаты показывают, что слитый белок высокоаффинного TCR по настоящему изобретению и антитела против CD3 может перенаправлять эффекторные клетки, эффекторные клетки демонстрируют сильное активирующее действие на позитивные клетки-мишени, одновременно по существу демонстрируют отсутствие активирующего действия на негативные клетки-мишени, таким образом, что эффекторные клетки демонстрируют хорошую специфичность.
Пример 13. Функциональная проверка-анализ функции «киллинга» слитого белка высокоаффинного TCR по настоящему изобретению и антитела против CD3
В данном примере функцию «киллинга» проверяли нерадиоактивным анализом цитотоксичности. Анализ представляет собой колориметрический анализ, являющийся заменой анализа цитотоксичности по высвобождению 51Cr для количественной оценки лактатдегидрогеназы (LDH), высвобождающейся после лизиса клеток. LDH, высвобождающуюся в среду, выявляли, используя 30-минутную сопряженную ферментативную реакцию, в которой LDH превращает соль тетразолия (INT) в красный формазан. Количество полученного красного продукта прямо пропорционально числу лизированных клеток. Данные по поглощению видимого цвета при 490 нм могут быть собраны, используя стандартный 96-луночный планшет-ридер.
Линии клеток-мишеней представляли собой HepG2, HCCC9810 и SNU-398, среди которых HepG2 представляла собой линию позитивных клеток-мишеней; и HCCC9810 и SNU-398 представляли собой линии негативных клеток-мишеней. Эффекторные клетки представляли собой CD8+ T-клетки. Высокоаффинные TCR по настоящему изобретению отбирали случайным образом. TCR и их номера могут быть найдены в Таблице 4, которые представляют собой TCR23 (вариабельный домен цепи α SEQ ID NO: 71, вариабельный домен цепи β SEQ ID NO: 79). Слитый белок получали в соответствии с Примером 9 и разводили в среде для эксперимента до разных концентраций (10-13-10-8 M). Компоненты анализа добавляли в планшет ELISPOT в следующем порядке: 50 мкл клеток-мишеней, 5×104 клеток/лунка; 100 мкл эффекторных клеток, 5×104 клеток/лунка; и 50 мкл слитого белка в каждую лунку.
Несколько контролей получали следующим образом: спонтанное высвобождение клеток-мишеней: только 200 мкл клеток-мишеней; максимальное высвобождение клеток-мишеней: только 200 мкл клеток-мишеней. Контрольная группа среды: только 200 мкл среды; контрольная группа с поправкой на объем: 200 мкл среды и 20 мкл клеточного лизата. Все лунки получали в трехкратной повторности с конечным объемом 200 мкл. Планшет центрифугировали при 250 g в течение 4 минут и затем инкубировали при 37 °C в течение 20 часов. 20 мкл жидкости с лизатом добавляли к контрольным лункам максимального высвобождения клеток-мишеней и контрольным лункам с поправкой на объем, и супернатант собирали, спустя 45 минут. Планшет центрифугировали при 250 g в течение 4 минут. 50 мкл супернатанта переносили из каждой лунки тестируемого планшета в соответствующую лунку плоскодонного 96-луночного планшета Nunc Vlnxisorb. Смесь субстратов восстанавливали посредством буфера для анализа (12 мл). Затем, 50 мкл восстановленной смеси субстратов добавляли в каждую лунку планшета. Планшет покрывали алюминиевой фольгой и инкубировали при комнатной температуре в течение 30 минут. 50 мкл гасящего раствора добавляли к каждой лунке планшета для гашения реакции. Поглощение записывали при 490 нм посредством ридера Elisa (твердофазный иммуноферментный анализ) за 1 час после добавления гасящего раствора.
Результаты экспериментов показаны на Фиг. 16. Слитый белок высокоаффинного TCR по настоящему изобретению и антитела против CD3 может перенаправлять эффекторные клетки и уничтожать позитивные клетки-мишени, но не оказывает поражающего действия на негативные клетки-мишени, демонстрируя, таким образом, хорошую специфичность.
Все документы, упомянутые в настоящей заявке, включены в данный документ посредством ссылки во всей своей полноте, как если бы каждый был включен посредством ссылки. Кроме того, следует понимать, что после прочтения идей настоящего изобретения, описанного выше, специалист в данной области может вносить разные изменения или модификации изобретения, и данные эквивалентные формы также попадают в объем, как определено прилагаемой формулой изобретения настоящей заявки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ИксЛАЙФЭсСи, ЛТД.
<120> ВЫСОКОАФФИННЫЙ T-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP
<130> P2020-1952
<150> CN2018111096773
<151> 2018-09-21
<160> 103
<170> PatentIn версия 3.5
<210> 1
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 1
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Gly
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 2
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 2
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 3
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 3
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Gly
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 4
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 4
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 5
<211> 339
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный полинуклеотид
<400> 5
tctcaacaag gtgaagaaga tccgcaggca ctgagcattc aggaaggcga aaatgtgacc 60
attaattgta gctataagac cagtattaac aacctgcagt ggtatcgcca gaatagcggt 120
cgcggtctgg ttcatctgat tctgattcgt agcaatgaac gtgaaaaaca tagcggccgt 180
ctgcgtgtga ccctggatac cagtaaaaaa tctagcagcc tggaaattac cgatgtgcgc 240
ccgagcgata ccgcaagtta tttttgtgca accgatccgc gcaccggcgc aaatagcaaa 300
ctgacctttg gtaaaggcat taccctgagt gttcgtccg 339
<210> 6
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный полинуклеотид
<400> 6
gataccggtg ttagtcagga tccgcgccat ctgagcgtta aacgtggcca gaatgttacc 60
ctgcgctgtg atccgattag tgaacataat cgtctgtatt ggtatcgtca gaccccgggt 120
cagggtccgg aatttctgac ctattttcag aatgaagcac agctggaaaa aagccgtctg 180
ctgagtgatc gttttagcgc cgaacgtccg aaaggcagtt ttagtaccct ggaaattcag 240
cgcgtggaac cgggtgacag cgcaatgtat ctgtgcgcca gcgaaggtct ggcctatgaa 300
cagtattttg gcccgggtac ccgcctgacc gttacc 336
<210> 7
<211> 20
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 7
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 8
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный полинуклеотид
<400> 8
ggcggtggtg gcagcggcgg tggtggttca ggcggtggtg gaagcggtgg tggcggtagt 60
<210> 9
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 9
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ala
85 90 95
Ser Ser Ala Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 10
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 10
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Gly
85 90 95
Ser Ser Asn Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 11
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 11
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Met Gln
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 12
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 12
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Arg Gln
85 90 95
Ser Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 13
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 13
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Val Leu Ser
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 14
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 14
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ser
85 90 95
Gly Ser Thr Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 15
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 15
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Asp Arg Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 16
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 16
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ala
85 90 95
Ser Thr Pro Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 17
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 17
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Glu Ala Gln
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 18
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 18
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Glu
85 90 95
Ser Thr Thr Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 19
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 19
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Leu Gln
85 90 95
Ser Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 20
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 20
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Gln Asn Ser
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 21
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 21
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Ser Asn
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 22
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 22
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Asp Val Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Ala Gln
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 23
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 23
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Pro Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Ala Gln
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 24
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 24
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Pro Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Thr Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 25
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 25
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Asp Ile Tyr Lys Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ala
85 90 95
Ser Ser Pro Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 26
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 26
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Leu Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 27
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 27
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Glu Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Leu Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 28
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 28
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 29
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 29
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Glu Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 30
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 30
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Asp Val Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 31
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 31
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Asp Val Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Thr Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 32
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 32
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Thr Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 33
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 33
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Glu Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Thr Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 34
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 34
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Pro Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 35
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 35
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Thr Ser Ser
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 36
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 36
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 37
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 37
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Ala Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 38
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 38
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Gln Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 39
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 39
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Gly Thr Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 40
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 40
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Ala Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 41
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 41
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Gly Thr Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 42
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 42
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Gly Arg Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 43
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 43
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Gln Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 44
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 44
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Ala Arg His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 45
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 45
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Gly Leu Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 46
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 46
Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu Ser Val Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Gln Gly Arg Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 47
<211> 245
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 47
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Val Thr Ile Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Glu Ile Thr Asp Val Arg
65 70 75 80
Pro Ser Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Gly
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Asp Thr Gly Val Ser Gln Asp Pro Arg His Leu
130 135 140
Ser Val Lys Arg Gly Gln Asn Val Thr Leu Arg Cys Asp Pro Ile Ser
145 150 155 160
Glu His Asn Arg Leu Tyr Trp Tyr Arg Gln Thr Pro Gly Gln Gly Pro
165 170 175
Glu Phe Leu Thr Tyr Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg
180 185 190
Leu Leu Ser Asp Arg Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser
195 200 205
Thr Leu Glu Ile Gln Arg Val Glu Pro Gly Asp Ser Ala Met Tyr Leu
210 215 220
Cys Ala Ser Glu Gly Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr
225 230 235 240
Arg Leu Thr Val Thr
245
<210> 48
<211> 735
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный полинуклеотид
<400> 48
tctcaacaag gtgaagaaga tccgcaggca ctgagcattc aggaaggcga aaatgtgacc 60
attaattgta gctataagac cagtattaac aacctgcagt ggtatcgcca gaatagcggt 120
cgcggtctgg ttcatctgat tctgattcgt agcaatgaac gtgaaaaaca tagcggccgt 180
ctgcgtgtga ccctggatac cagtaaaaaa tctagcagcc tggaaattac cgatgtgcgc 240
ccgagcgata ccgcaagtta tttttgtgca accgatccgc gcaccggcgc aaatagcaaa 300
ctgacctttg gtaaaggcat taccctgagt gttcgtccgg gcggtggtgg cagcggcggt 360
ggtggttcag gcggtggtgg aagcggtggt ggcggtagtg ataccggtgt tagtcaggat 420
ccgcgccatc tgagcgttaa acgtggccag aatgttaccc tgcgctgtga tccgattagt 480
gaacataatc gtctgtattg gtatcgtcag accccgggtc agggtccgga atttctgacc 540
tattttcaga atgaagcaca gctggaaaaa agccgtctgc tgagtgatcg ttttagcgcc 600
gaacgtccga aaggcagttt tagtaccctg gaaattcagc gcgtggaacc gggtgacagc 660
gcaatgtatc tgtgcgccag cgaaggtctg gcctatgaac agtattttgg cccgggtacc 720
cgcctgaccg ttacc 735
<210> 49
<211> 208
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 49
Met Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu
1 5 10 15
Gly Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn
20 25 30
Leu Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile
35 40 45
Leu Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val
50 55 60
Thr Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser
65 70 75 80
Arg Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr
85 90 95
Gly Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val
100 105 110
Arg Pro Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp
115 120 125
Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser
130 135 140
Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp
145 150 155 160
Lys Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala
165 170 175
Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn
180 185 190
Asn Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
195 200 205
<210> 50
<211> 243
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 50
Met Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg
1 5 10 15
Gly Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg
20 25 30
Leu Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr
35 40 45
Tyr Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp
50 55 60
Arg Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile
65 70 75 80
Gln Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu
85 90 95
Gly Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val
100 105 110
Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu
115 120 125
Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys
130 135 140
Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val
145 150 155 160
Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu
165 170 175
Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu Ser Ser Arg
180 185 190
Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn His Phe Arg
195 200 205
Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln
210 215 220
Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly
225 230 235 240
Arg Ala Asp
<210> 51
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 51
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ala
85 90 95
Ser Ser Ala Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 52
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 52
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Gly
85 90 95
Ser Ser Asn Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 53
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 53
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Met Gln
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 54
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 54
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Arg Gln
85 90 95
Ser Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 55
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 55
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Val Leu Ser
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 56
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 56
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ser
85 90 95
Gly Ser Thr Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 57
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 57
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Asp Arg Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 58
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 58
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ala
85 90 95
Ser Thr Pro Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 59
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 59
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Glu Ala Gln
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 60
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 60
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Glu
85 90 95
Ser Thr Thr Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 61
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 61
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Leu Gln
85 90 95
Ser Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 62
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 62
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Gln Asn Ser
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 63
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 63
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Ser Asn
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 64
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 64
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Asp Val Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Ala Gln
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 65
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 65
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Pro Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Ala Gln
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 66
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 66
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Pro Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Thr Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 67
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 67
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Asp Ile Tyr Lys Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Ala
85 90 95
Ser Ser Pro Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 68
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 68
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Leu Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 69
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 69
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Glu Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Leu Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 70
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 70
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 71
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 71
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Glu Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 72
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 72
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Asp Val Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 73
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 73
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Asp Val Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Thr Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 74
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 74
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Thr Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 75
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 75
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Glu Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ala Thr Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 76
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 76
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Pro Ile Ser Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Ser Met Ser
85 90 95
Pro Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 77
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 77
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ala Val Asp Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Thr Ser Ser
85 90 95
Trp Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 78
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 78
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 79
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 79
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ala Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 80
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 80
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gln Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 81
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 81
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gly Thr Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 82
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 82
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ala Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 83
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 83
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gly Thr Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 84
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 84
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gly Arg Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 85
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 85
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gln Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 86
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 86
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ala Arg His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 87
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 87
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gly Leu Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 88
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 88
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gln Gly Arg Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Glu Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 89
<211> 254
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 89
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Gly
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys
195 200 205
Asp Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn
210 215 220
Phe Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val
225 230 235 240
Ala Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
245 250
<210> 90
<211> 291
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 90
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
115 120 125
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
130 135 140
Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
145 150 155 160
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys
165 170 175
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
180 185 190
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
195 200 205
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
210 215 220
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
225 230 235 240
Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu Ser
245 250 255
Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala
260 265 270
Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
275 280 285
Ser Arg Gly
290
<210> 91
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 91
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro His Trp Ser
85 90 95
Thr Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 92
<211> 113
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 92
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Asp Ile Ala Ser Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro His Trp Ser
85 90 95
Thr Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro
<210> 93
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 93
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Val Asp Asn Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 94
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 94
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Leu Gly His Ala Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 95
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 95
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gly Trp Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 96
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 96
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Glu Leu Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 97
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 97
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Met Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 98
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 98
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Asp Leu Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 99
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 99
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe His Tyr Thr Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 100
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 100
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Ala Trp Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 101
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 101
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Gly Trp Trp Pro Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Gly Glu Leu Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
<210> 102
<211> 207
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 102
Ser Gln Gln Gly Glu Glu Asp Pro Gln Ala Leu Ser Ile Gln Glu Gly
1 5 10 15
Glu Asn Ala Thr Met Asn Cys Ser Tyr Lys Thr Ser Ile Asn Asn Leu
20 25 30
Gln Trp Tyr Arg Gln Asn Ser Gly Arg Gly Leu Val His Leu Ile Leu
35 40 45
Ile Arg Ser Asn Glu Arg Glu Lys His Ser Gly Arg Leu Arg Val Thr
50 55 60
Leu Asp Thr Ser Lys Lys Ser Ser Ser Leu Leu Ile Thr Ala Ser Arg
65 70 75 80
Ala Ala Asp Thr Ala Ser Tyr Phe Cys Ala Thr Asp Pro Arg Thr Gly
85 90 95
Ala Asn Ser Lys Leu Thr Phe Gly Lys Gly Ile Thr Leu Ser Val Arg
100 105 110
Pro Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
115 120 125
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
130 135 140
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
145 150 155 160
Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
165 170 175
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
180 185 190
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser
195 200 205
<210> 103
<211> 242
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтезированный полипептид
<400> 103
Asp Thr Gly Val Ser Gln Asp Pro Arg His Lys Ile Thr Lys Arg Gly
1 5 10 15
Gln Asn Val Thr Phe Arg Cys Asp Pro Ile Ser Glu His Asn Arg Leu
20 25 30
Tyr Trp Tyr Arg Gln Thr Leu Gly Gln Gly Pro Glu Phe Leu Thr Tyr
35 40 45
Phe Gln Asn Glu Ala Gln Leu Glu Lys Ser Arg Leu Leu Ser Asp Arg
50 55 60
Phe Ser Ala Glu Arg Pro Lys Gly Ser Phe Ser Thr Leu Glu Ile Gln
65 70 75 80
Arg Thr Glu Gln Gly Asp Ser Ala Met Tyr Leu Cys Ala Ser Glu Gly
85 90 95
Leu Ala Tyr Glu Gln Tyr Phe Gly Pro Gly Thr Arg Leu Thr Val Thr
100 105 110
Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
115 120 125
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
130 135 140
Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
145 150 155 160
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys
165 170 175
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
180 185 190
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
195 200 205
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
210 215 220
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
225 230 235 240
Ala Asp
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2017 |
|
RU2775623C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2018 |
|
RU2762255C2 |
| НОВЫЕ БИСПЕЦИФИЧНЫЕ ПОЛИПЕПТИДНЫЕ КОМПЛЕКСЫ ПРОТИВ CD3/CD19 | 2018 |
|
RU2788127C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| КОНСТРУКТЫ Т-КЛЕТОЧНОГО РЕЦЕПТОРА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2785954C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| АНТИ-CEACAM1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2754876C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |







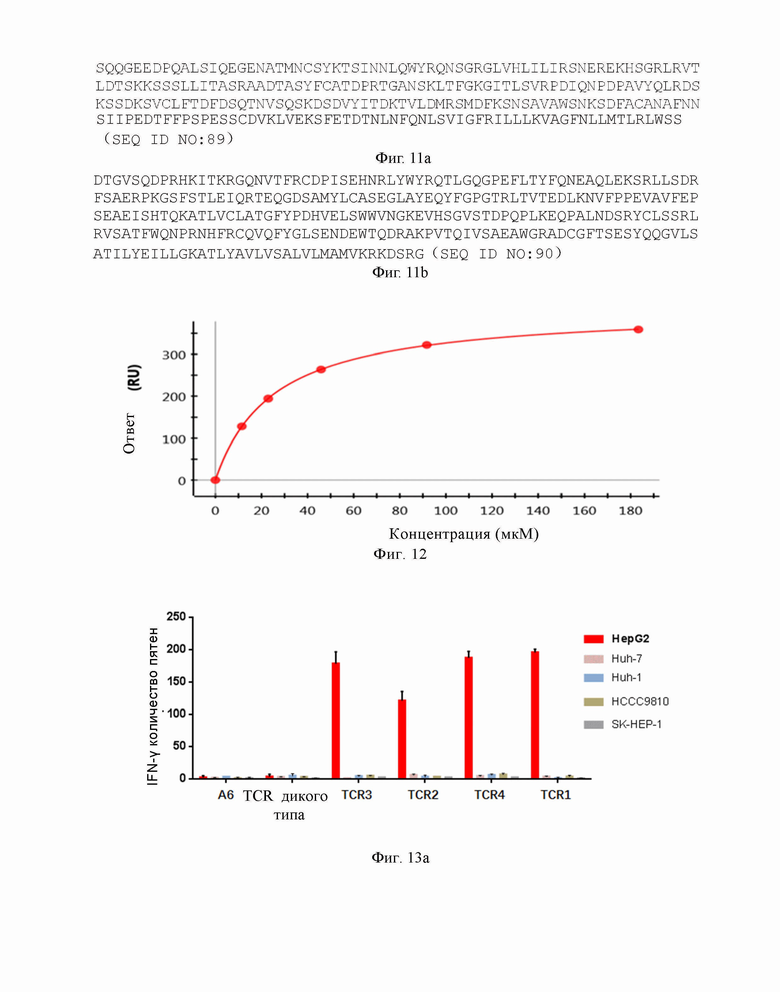
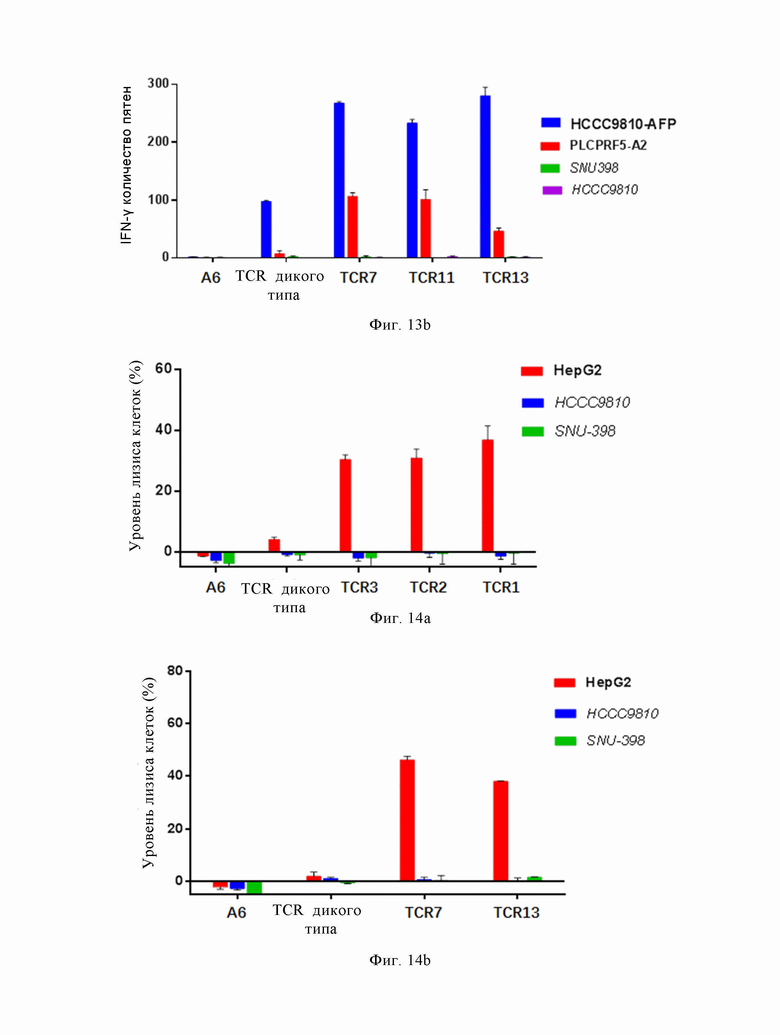
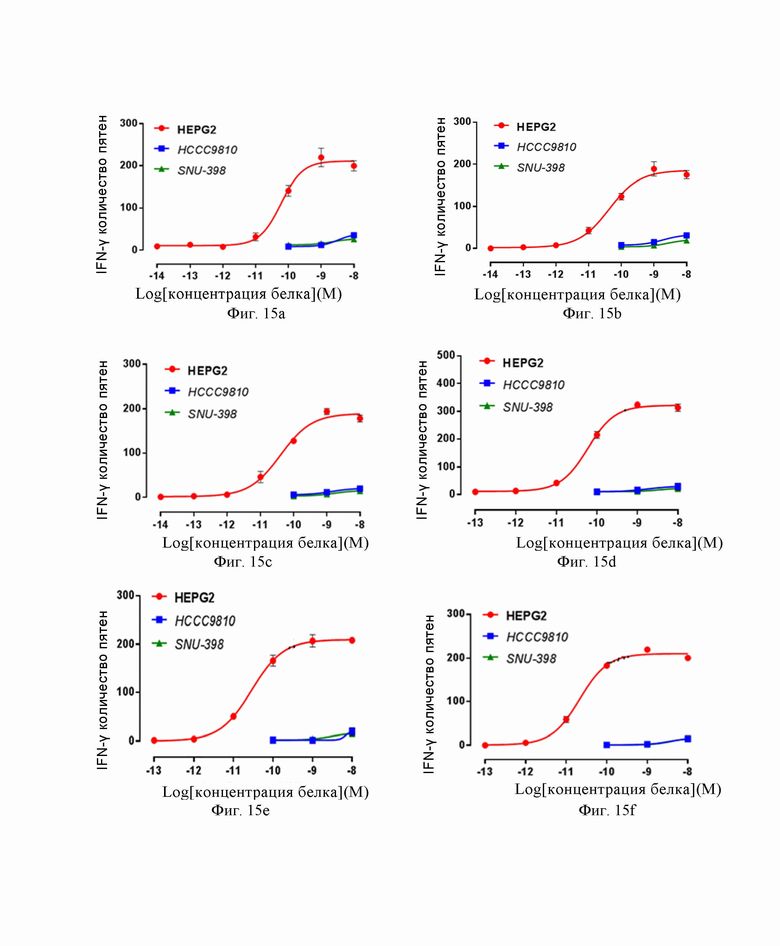
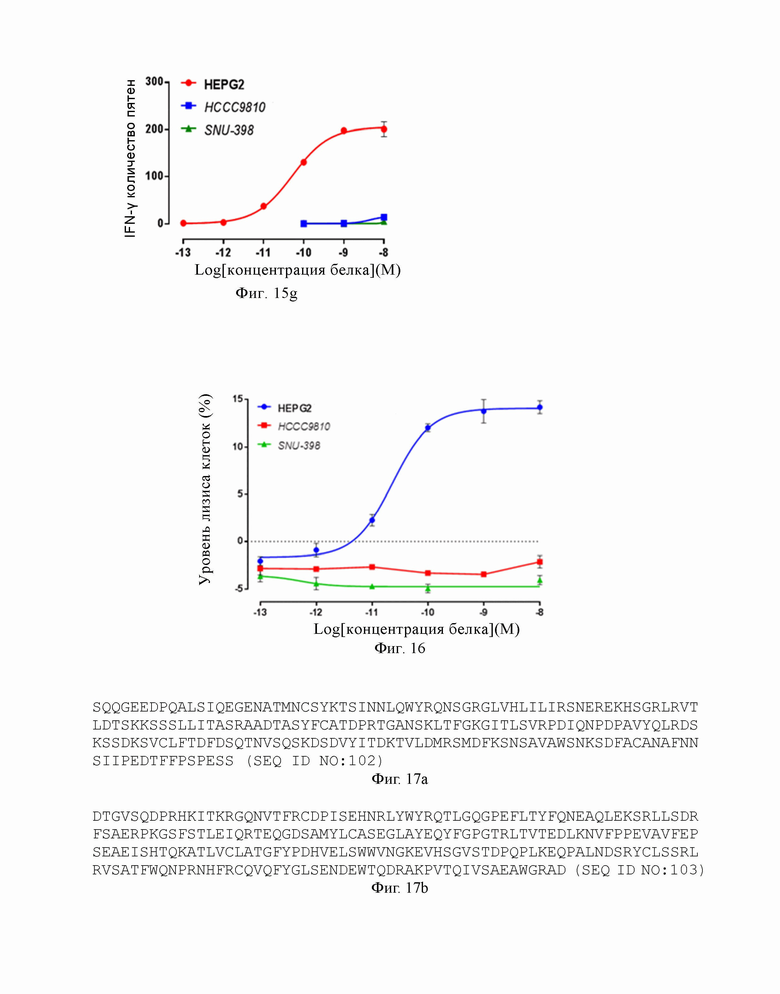
Изобретение относится к области биотехнологии, а именно к T-клеточному рецептору (TCR), обладающему активностью связывания с комплексом FMNKFIYEI-HLA A0201, а также к конъюгату, поливалентному комплексу и композиции, его содержащим. Также раскрыты молекула нуклеиновой кислоты, кодирующая вышеуказанный TCR, а также вектор и клетка, ее содержащие. Изобретение также относится к способу получения TCR, обладающего активностью связывания с комплексом FMNKFIYEI-HLA A0201, который предусматривает культивирование вышеуказанной клетки. Изобретение эффективно для лечения рака печени. 11 н. и 8 з.п. ф-лы, 17 ил., 4 табл., 13 пр.
1. T-клеточный рецептор (TCR), обладающий активностью связывания с комплексом FMNKFIYEI-HLA A0201, содержащий вариабельный домен цепи α TCR и вариабельный домен цепи β TCR; вариабельный домен цепи α TCR содержит три области CDR, и вариабельный домен цепи β TCR содержит три области CDR,
причем TCR имеет CDR, выбранные из группы, состоящей из:
2. TCR по п. 1, который является растворимым.
3. TCR по п. 1, который представляет собой αβ гетеродимерный TCR или одноцепочечный TCR; и предпочтительно, TCR содержит последовательность константной области цепи α TRAC*01 и последовательность константной области цепи β TRBC1*01 или TRBC2*01.
4. TCR по п. 3, содержащий (i) всю или часть цепи α TCR, за исключением ее трансмембранного домена, и (ii) всю или часть цепи β TCR, за исключением ее трансмембранного домена, где обе из (i) и (ii) содержат вариабельный домен и по меньшей мере часть константного домена цепи TCR.
5. TCR по п. 4, где искусственная межцепочечная дисульфидная связь содержится между вариабельной областью цепи α и константной областью цепи β TCR;
предпочтительно остатки цистеина, образующие искусственную межцепочечную дисульфидную связь между вариабельной областью цепи α и константной областью цепи β TCR, заменены на одну или более групп аминокислот, выбранных из следующих:
Thr48 TRAC*01 экзона 1 и Ser57 TRBC1*01 или TRBC2*01 экзона 1;
Thr45 TRAC*01 экзона 1 и Ser77 TRBC1*01 или TRBC2*01 экзона 1;
Tyr10 TRAC*01 экзона 1 и Ser17 TRBC1*01 или TRBC2*01 экзона 1;
Thr45 TRAC*01 экзона 1 и Asp59 TRBC1*01 или TRBC2*01 экзона 1;
Ser15 TRAC*01 экзона 1 и Glu15 TRBC1*01 или TRBC2*01 экзона 1;
Arg53 TRAC*01 экзона 1 и Ser54 TRBC1*01 или TRBC2*01 экзона 1;
Pro89 TRAC*01 экзона 1 и Ala19 TRBC1*01 или TRBC2*01 экзона 1; и
Tyr10 TRAC*01 экзона 1 и Glu20 TRBC1*01 или TRBC2*01 экзона 1.
6. TCR по п. 1, где аминокислотная последовательность вариабельного домена цепи α TCR выбрана из группы, состоящей из: SEQ ID NO: 51-77 и 91-92; и/или аминокислотная последовательность вариабельного домена цепи β TCR выбрана из группы, состоящей из: SEQ ID NO: 78-88 и 93-101.
7. TCR по п. 1, выбранный из группы, состоящей из:
8. TCR по п. 1, представляющий собой одноцепочечный TCR;
предпочтительно представляющий собой одноцепочечный TCR, состоящий из вариабельного домена α и вариабельного домена β, и вариабельный домен α и вариабельный домен β соединены последовательностью гибкого короткого пептида.
9. TCR по п. 1, выбранный из группы, состоящей из:
10. Конъюгат TCR, обладающий активностью связывания с комплексом FMNKFIYEI-HLA A0201, содержащий TCR по любому из пп. 1-9 и детектируемую метку или терапевтическое средство, конъюгированные с цепью α и/или цепью β TCR на C- или N-конце;
предпочтительно терапевтическое средство представляет собой антитело против CD3.
11. Поливалентный комплекс TCR, используемый для отслеживания клеток-мишеней, которые презентируют конкретный антиген, in vitro или in vivo, содержащий по меньшей мере две молекулы TCR, и по меньшей мере одна молекула TCR представляет собой TCR по любому из пп. 1-9.
12. Поливалентный комплекс TCR, используемый для нацеливания на клетки-мишени, которые презентируют конкретный антиген, in vitro или in vivo, содержащий по меньшей мере две молекулы TCR, и по меньшей мере одна молекула TCR представляет собой TCR по любому из пп. 1-9.
13. Молекула нуклеиновой кислоты, используемая для кодирования TCR по любому из пп. 1-9.
14. Вектор для экспрессии TCR по любому из пп. 1-9, содержащий молекулу нуклеиновой кислоты по п. 13.
15. Клетка-хозяин для получения TCR по любому из пп. 1-9, содержащая вектор по п. 14 или имеющая экзогенную молекулу нуклеиновой кислоты по п. 13, интегрированную в ее геном.
16. Выделенная Т-клетка, трансдуцированная TCR по любому из пп. 1-9, для лечения рака печени.
17. Фармацевтическая композиция для лечения рака печени, содержащая фармацевтически приемлемый носитель и TCR по любому из пп. 1-9, или конъюгат TCR по п. 10, или комплекс TCR по п. 11 или 12, или Т-клетку по п. 16.
18. Применение TCR по любому из пп. 1-9, или конъюгата TCR по п. 10, или комплекса TCR по п. 11 или 12, или Т-клетки по п. 16 для получения лекарственного средства для лечения рака печени.
19. Способ получения T-клеточного рецептора по любому из пп. 1-9, включающий стадии:
(i) культивирования клетки-хозяина по п. 15 для экспрессии T-клеточного рецептора по любому из пп. 1-9;
(ii) выделения или очистки T-клеточного рецептора.
| LONGHAO SUN et al | |||
| Engineered cytotoxic T lymphocytes with AFP-specific TCR gene for adoptive immunotherapy in hepatocellular carcinoma, Tumour Biol, 2016 Jan; 37(1):799-806, doi: 10.1007/s13277-015-3845-9 | |||
| WEI ZHU et al | |||
| Identification of α-fetoprotein-specific T-cell receptors for hepatocellular carcinoma immunotherapy, Hepatology, 2018 Aug; |

