Изобретение относится к области медицины и биохимии, в частности к молекулярной диагностике, а именно к многопараметрической диагностической тест-системе, и может быть использовано для количественного определения уровней мРНК генов RIG-1, IFIT-1, IFIH-1 человека в биологическом образце.
Первый защитный барьер против вирусной инфекции – это кожные покровы и слизистые оболочки организма хозяина. На первых этапах в ответ на проникновение вирусной инфекции происходит активация экстренного неспецифического иммунного ответа. Существуют различные пути распознавания клеткой вирусных частиц. Наибольшее значение принято отводить толл-подобным рецепторам (TLR). При проникновении вируса в цитоплазму в дело вступают RLR, NLR и цитозольные сенсоры нуклеиновых кислот [1]. Передача сигнала через TLR требует привлечения белковых адаптеров: MyD88 (Myeloid differentiation primary response gene 88), Mal (MyD88 adapter-like), TRIF (TIR-domain-containing adaptor inducing IFNβ) и TRAM (Trif-related adaptor molecule) [2]. Активация молекулярных адаптеров направлена на изменение активности факторов транскрипции (Nuclear factor κВ (NFκВ), IFN-regulatory factor 3 (IRF3), IFN-regulatory factor 7 (IRF7)) и активацию MAPK-зависимых сигнальных путей [1, 3].
Клетка способна к TLR-независимому ответу на проникновение патогена, опосредованному через цитозольные сенсоры. Важнейшим звеном являются РНК-хеликазы, принадлежащие к семейству RLR: Retinoic acid-inducible gene 1 (RIG-I), Melanoma differentiation-associated gene 5 (MDA-5 или IFIH-1) и Laboratory of genetics and physiology-2 (LGP-2). РНК-сенсоры RIG-I и MDA-5 состоят из N-концевого домена CARD (Сaspase activation and recruitment domain), имеющего АТФазную активность DExD/H box домена и С-концевого репрессорного домена. В нормальных условиях RIG-I находится в неактивной форме за счет ингибиторной активности своего регуляторного домена. Наоборот, при проникновении вируса в клетку RIG-I претерпевает конформационные изменения, индуцирующие мультимеризацию данного РНК-сенсора [1]. Активированная мультимерная форма RIG-I или MDA-5 взаимодействует с белковым адаптером Mitochondrial antiviral signaling protein (MAVS), расположенным на оболочке митохондриальной мембраны или в пероксисомах [1]. Активация MAVS-зависимого сигналинга индуцирует активацию транскрипционных факторов IRF-1, IRF-3, IRF-7. Известно, что данные факторы транскрипции необходимы для продукции интерферонов 1-го и 3-го типов, а следовательно, для индукции экспрессии сотен противовирусных ISGs (Interferon-stimulated genes) [3].
РНК-сенсоры RIG-I или MDA-5 играют ключевую роль в антивирусной защите различных типов клеток, включая фибробласты, эпителиальные клетки, дендритные клетки [1]. Вирусная двуцепочечная РНК активирует и RIG-I, и MDA-5, в то время как в качестве активаторов первой РНК-хеликазы могут также выступать молекулы одноцепочечной РНК с 5’-трифосфатным концом без кэп-структуры (РНК такого вида присутствует в геномах многих вирусов). Важно отметить, что RIG-I не способен распознавать одноцепочечную РНК с кэп-структурой, которая продуцируется клеткой. РНК-сенсор MDA-5 отличает вирусную РНК от РНК клетки по длине молекулы. Обычно, длинные двуцепочечные РНК не характерны для здоровой клетки и являются лигандами MDA-5 [1]. Показано, что данные РНК-сенсоры необходимы для распознавания клеткой таких опасных патогенов как вирус бешенства, вирус кори, вирусы гриппа А и В, респираторно-синцитиальный вирус, вирус гепатита С, геморрагическая лихорадка Эбола, менговирус. Все эти вирусы способны продуцировать длинную двуцепочечную РНК, которая является лигандом данных РНК-сенсоров.
Существует взаимосвязь между генами RIG-1, IFIT-1 и IFIH-1. Интерфероны, осуществляя аутокринное или паракринное действие на клетки, активируют JAK/STAT-зависимые сигнальные каскады и индуцируют экспрессию сотен ISGs. Одним из важнейших ISGs является IFIT-1 (ISG-56). IFIT-1 – ген с выраженной противовирусной активностью. Показано, что IFIT-1 ингибирует репликацию РНК- и ДНК-вирусов. Более того, сайленсинг IFIT-1 приводит к повышению репликации вируса гепатита С. С другой стороны, IFIT-1 вовлечен в индукцию клеточного апоптоза. Так, показано, что сайленсинг гена IFIT-1 был ассоциирован с меньшим уровнем апоптоза клеток А549, обработанных 5’-РРР-ssRNA, синтетическим агонистом RIG-1. Следовательно, связанный с действием интерферонов 1-го типа клеточный апоптоз может быть ассоциирован с избыточной экспрессией гена IFIT-1. В силу этого измерение уровня экспрессии гена IFIT-1 можно рассматривать не только как маркер активации врожденного иммунного ответа, но и как маркер ассоциированного с действием интерферонов 1-го типа апоптоза [4].
Совместное определение уровня экспрессии генов RIG-1, IFIT-1 и IFIH-1 позволяет определить уровень активации врожденного иммунного ответа организма и дать оценку активации проапоптотических сигнальных каскадов. Особенную актуальность данная оценка приобретает при анализе патогенеза острых респираторных вирусных инфекций, вызванных вирусами гриппа, пневмовирусами, аденовирусами, коронавирусами. Так, низкий уровень экспрессии РНК-сенсоров MDA-5 и RIG-1 может свидетельствовать о слабой активации врожденного интерферон-опосредованного иммунного ответа на респираторную инфекцию, в то время как избыточная экспрессия IFIT-1 может быть не только сигналом активации интерфероновой защитной системы, но и маркером возможной гибели клеток. Наиболее чувствительным и специфичным методом определения уровня экспрессии генов является ОТ-ПЦР в реальном времени. В данной методике предполагается наличие двух праймеров и флуоресцентно меченного олигонуклеотидного зонда, специфических к интересующей мРНК.
Известен способ идентификации генов, регуляторов экспрессии, рецепторов, рецепторов белковых продуктов и белков, которые могут регулировать риновирусные инфекции [5], включающий: a) контактирование по меньшей мере одного соединения с мишенью, выбранной из определенных генов (группа включает RIG-1, IFIT-1 и IFIH-1), белков, кодируемых этими генами, регуляторов экспрессии генов, рецепторов кодируемых белков, продуктами белков, кодируемых генами, рецепторами продуктов белков генов и их комбинациями; б) определение, связывает ли указанное соединение мишень; и c) идентификация тех соединений, которые связывают мишень, как соединений для регулирования риновирусной инфекции. При этом данное изобретение также относится к способам идентификации терапевтических соединений, которые могут лечить различные расстройства, регулируя экспрессию и активность генов, регуляторов экспрессии, рецепторов, рецепторов белковых продуктов и идентифицированных белков.
Однако в настоящее время не известны тест-системы, которые можно использовать для одновременного определения экспрессии генов RIG-1, IFIT-1 и IFIH-1 методом ОТ-ПЦР в реальном времени. Таким образом, актуальность предлагаемого изобретения обусловлена необходимостью определения уровня экспрессии РНК-сенсоров и гена IFIT-1 для более глубокого понимания патогенеза острых респираторных вирусных инфекций и определения дальнейших стратегий их лечения, определения уровня активации интерферонового ответа и риска клеточной гибели.
Техническая проблема заключается в необходимости разработки тест-системы и способа для количественного определения уровня мРНК генов RIG-1, IFIT-1, IFIH-1 человека, а также в необходимости расширения ассортимента средств диагностики респираторных заболеваний на ранних этапах.
Технический результат состоит в обеспечении возможности быстрого и точного определения уровня мРНК генов RIG-1, IFIT-1, IFIH-1 человека.
Для подбора праймеров и олигонуклеотидных зондов использовали последовательности RIG-1, IFIT-1 и IFIH-1 человека, депонированные в базе данных GenBank (NCBI). Анализируемые гены локализованы на разных хромосомах, длины зрелых транскриптов составляют от 600 до 4700 нуклеотидов. мРНК гена RIG-1 (DDX58) представлена семью, гена IFIT-1 — пятью; а гена IFIH-1 (MDA-5) — одним транскрипционным вариантом.
Оригинальные последовательности праймеров и TaqMan зондов были подобраны к консервативным белок-кодирующим областям и потенциально должны выявлять все транскрипционные варианты заявленных генов. Селекцию необходимых последовательностей проводили исходя из желательных температур плавления праймеров 60°С и выше, а олигонуклеотидного зонда — свыше 63°С, рассчитанных при выбранной концентрации солей 75 мМ. Руководствуясь рациональными критериями дизайна, подбор праймеров проводили так, чтобы они были разделены как минимум одним интроном на последовательности гена и преимущественно были расположены на стыке экзонов. Такой подход позволяет различить геномный продукт ПЦР от продукта ПЦР, полученного с кДНК, и избежать дополнительной процедуры обработки ДНКазами.
Последовательности, сконструированные для выявления мРНК RIG-1, IFIT-1 и IFIH-1 человека были проанализированы в программе BLAST на предмет выявления их возможного спектра неспецифической гибридизации. Последовательностей, высокогомологичных каким-либо другим генам данного вида, выявлено не было.
Технический результат достигается тем, что многопараметрическая диагностическая тест-система для количественного определения уровня мРНК генов RIG-1, IFIT-1, IFIH-1 методом полимеразной цепной реакции с обратной транскрипцией включает набор олигонуклеотидных праймеров и флуоресцентных зондов со следующей структурой:
прямой праймер гена RIG-1 GAGCACTTGTGGACGCTTTA
обратный праймер гена RIG-1 ATACACTTCTGTGCCGGGAG
флуоресцентный зонд гена RIG-1 Rox-CCTGGCATATTGACTGGACGTGGC-BHQ2
прямой праймер гена IFIT-1 AAACTTCGGAGAAAGGCATTAGAT
обратный праймер гена IFIT-1 TGAAATGAAATGTGAAAGTGGCTG
флуоресцентный зонд гена IFIT-1 Hex-CCTGAGACTGGCTGCTGACTTTGAGAAC-BHQ1
прямой праймер гена IFIH-1 AAACCCATGACACAGAATGAACA
обратный праймер гена IFIH-1 TGTGAGCAACCAGGACGTAG
флуоресцентный зонд гена IFIH-1 Cy5.5-CACAGTGGCAGAAGAAGGTCTGGA- BHQ3
Прямой праймер гена RIG-1 представлен в SEQ NO:1; обратный праймер гена RIG-1 представлен в SEQ NO:2; флуоресцентный зонд гена RIG-1 представлен в SEQ NO:3; прямой праймер гена IFIT-1 представлен в SEQ NO:4; обратный праймер гена IFIT-1 представлен в SEQ NO:5; флуоресцентный зонд гена IFIT-1 представлен в SEQ NO:6; прямой праймер гена IFIH-1 представлен в SEQ NO:7; обратный праймер гена IFIH-1 представлен в SEQ NO:8; флуоресцентный зонд гена IFIH-1 представлен в SEQ NO:9.
С помощью указанной тест-системы осуществляют количественное определение уровня мРНК генов RIG-1, IFIT-1, IFIH-1 человека, в ходе которого проводят полимеразную цепную реакцию с обратной транскрипцией, причем первичную денатурацию проводят в течение 5 мин при температуре 95 °С, далее проводят 40 циклов, состоящих из двух этапов: денатурации в течение 10 с при температуре 95°С, затем отжига праймеров и элонгации цепи в течение 30 с при температуре 61,3°С.
Описание фигур
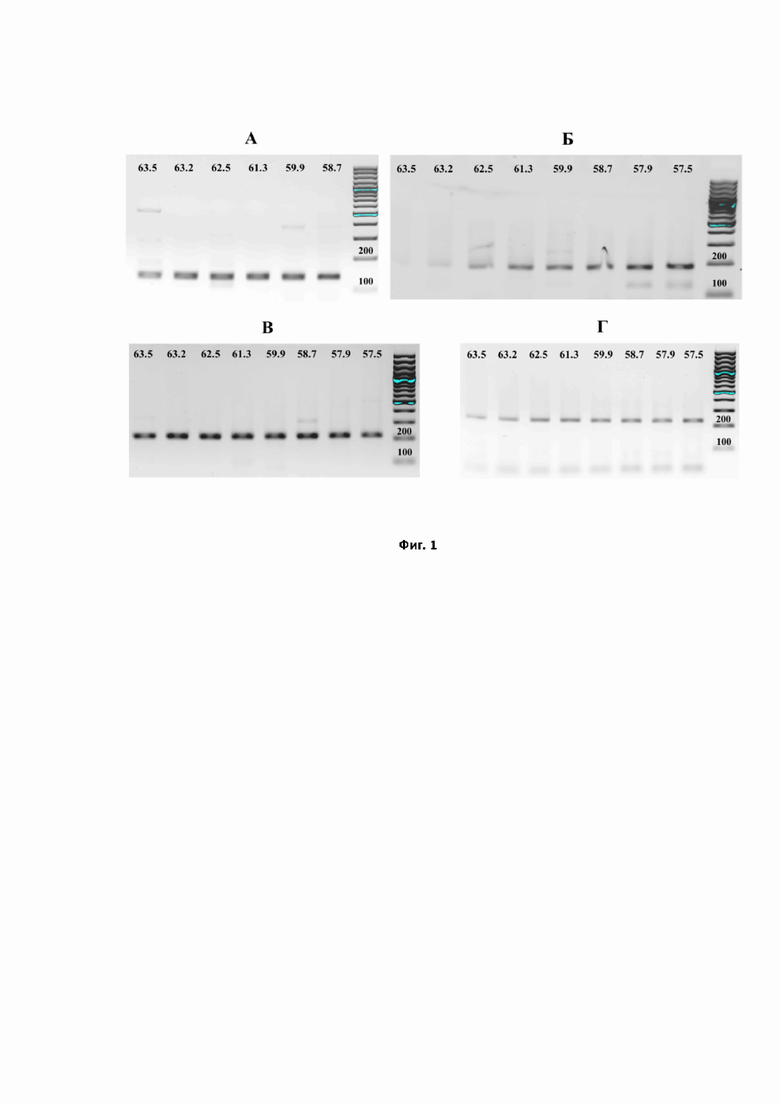
На фиг.1 представлен анализ методом электрофоретического разделения в агарозном геле продуктов ПЦР, полученных при соответствующих температурах отжига (указаны сверху в °С): А – продукты ПЦР для количественной детекции мРНК гена RIG-1; Б – продукты ПЦР для количественной детекции мРНК гена IFIT-1; В – продукты ПЦР для количественной детекции мРНК гена IFIH-1; Г – продукты ПЦР для количественной детекции мРНК гена HPRT-1.
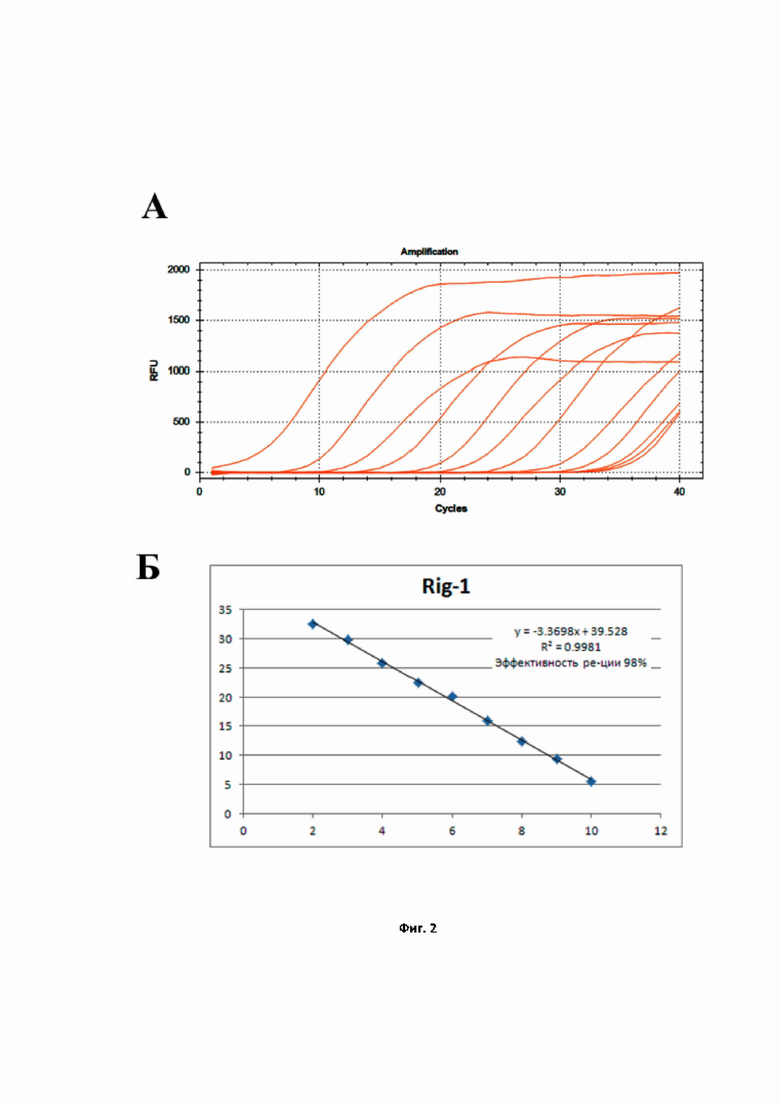
На фиг. 2 представлен расчёт эффективности реакции ПЦР для количественной детекции мРНК гена RIG-1, проведённой в моноплексном режиме: A – кривые роста флуоресценции, полученные для линейки разведений образцов (от –10 до –2) при анализе флуоресценции на канале ROX в режиме реального времени; Б – построенная по полученным значениям Cq кривая, используемая для расчёта эффективности реакции.
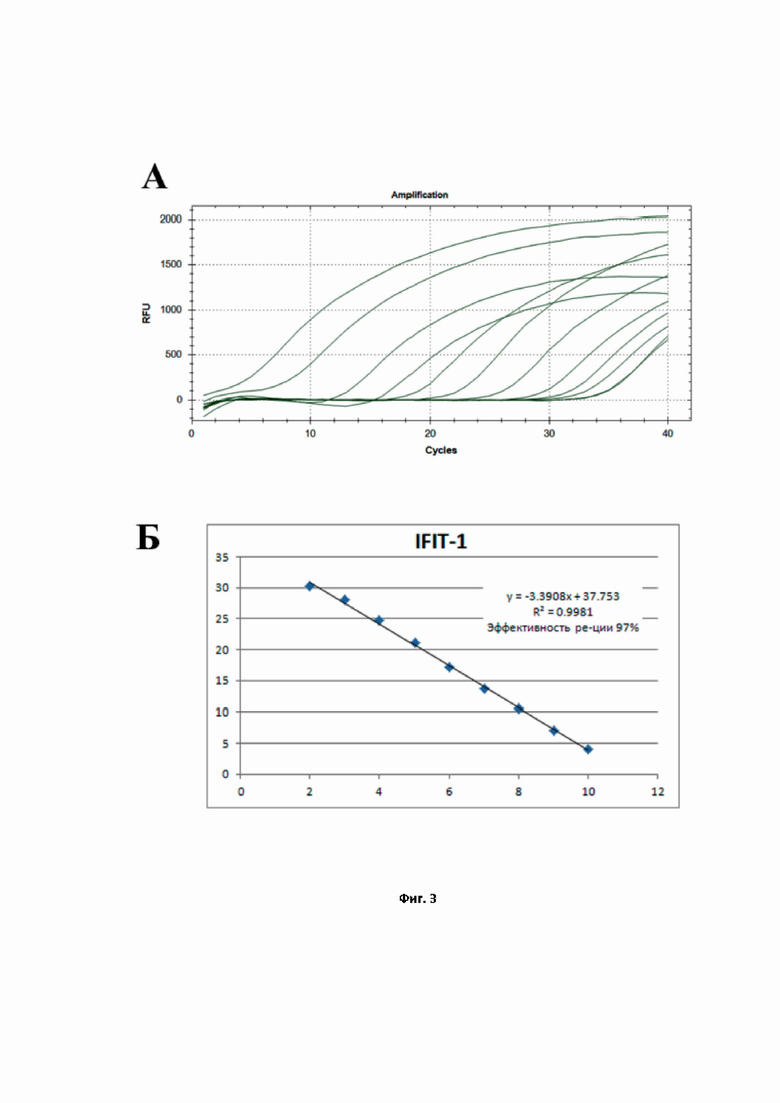
На фиг. 3 представлен Расчёт эффективности реакции ПЦР для количественной детекции мРНК гена IFIT-1, проведённой в моноплексном режиме: A – кривые роста флуоресценции, полученные для линейки разведений образцов (от –10 до –2) при анализе флуоресценции на канале ROX в режиме реального времени; Б – построенная по полученным значениям Cq кривая, используемая для расчёта эффективности реакции.
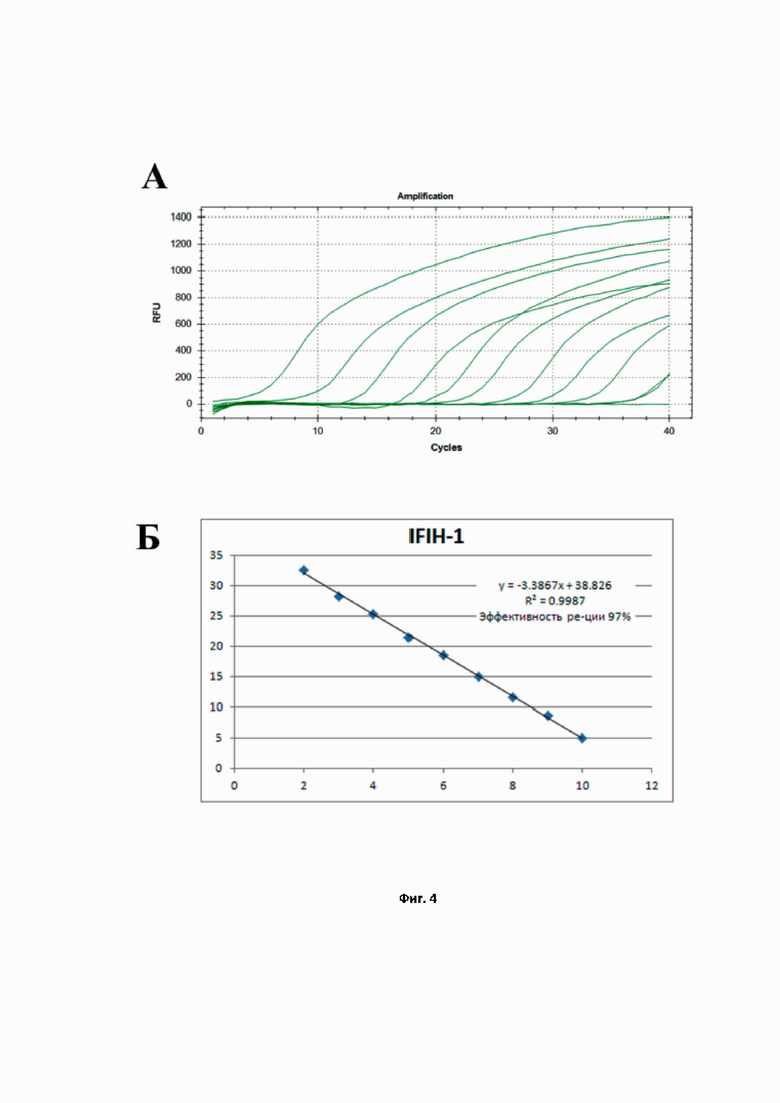
На фиг. 4 представлен расчёт эффективности реакции ПЦР для количественной детекции мРНК гена IFIH-1, проведённой в моноплексном режиме: A – кривые роста флуоресценции, полученные для линейки разведений образцов (от –10 до –2) при анализе флуоресценции на канале Cy5.5 в режиме реального времени; Б – построенная по полученным значениям Cq кривая, используемая для расчёта эффективности реакции.
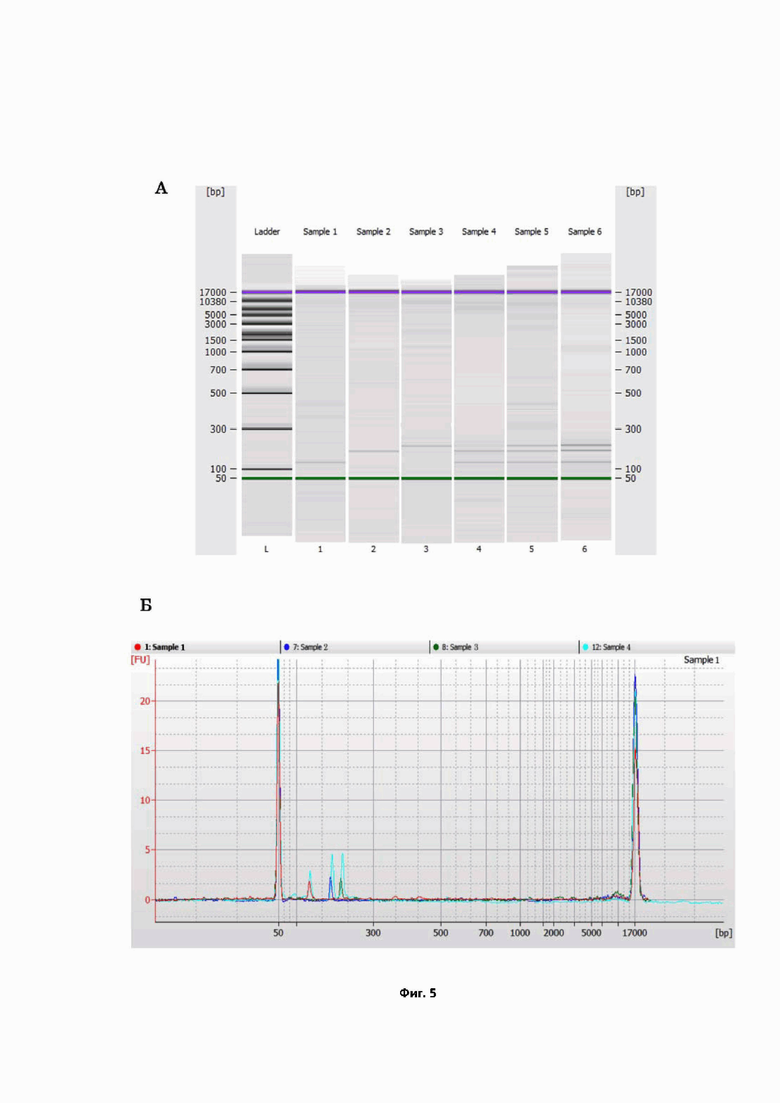
На фиг. 5 представлен анализ методом капиллярного электрофореза с использованием системы Bioanalyzer 2100 (Agilent) продуктов ПЦР, полученных при оптимальных условиях в мультиплексных и моноплексных режимах: A – электрофореграмма продуктов ПЦР: Sample 1–3 содержат продукты моноплексных ПЦР на гены RIG-1, IFIT-1 и IFIH-1, соответственно, Sample 4 – продукты мультиплексной ПЦР на эти гены при одновременной амплификации их в одной пробирке, полученные из образца, выделенного из клеток A549, Sample 5 – смесь продуктов моноплексных ПЦР, полученная путём смешивания образцов Sample 1–3, Sample 6 – продукты мультиплексной ПЦР на гены RIG-1, IFIT-1 и IFIH-1 при одновременной амплификации их в одной пробирке, полученные из образца, выделенного из мононуклеарных клеток периферической крови; Б — спектрофореграмма, отражающая наложение результатов ПЦР в мультиплексном режиме (проба Sample 4, содержащая все три специфических продукта RIG-1, IFIT-1 и IFIH-1) и моноплексных режимах, проба Sample 1 содержит специфический продукт гена RIG-1, Sample 2 и Sample-3 – продукты генов IFIT-1 и IFIH-1, соответственно.
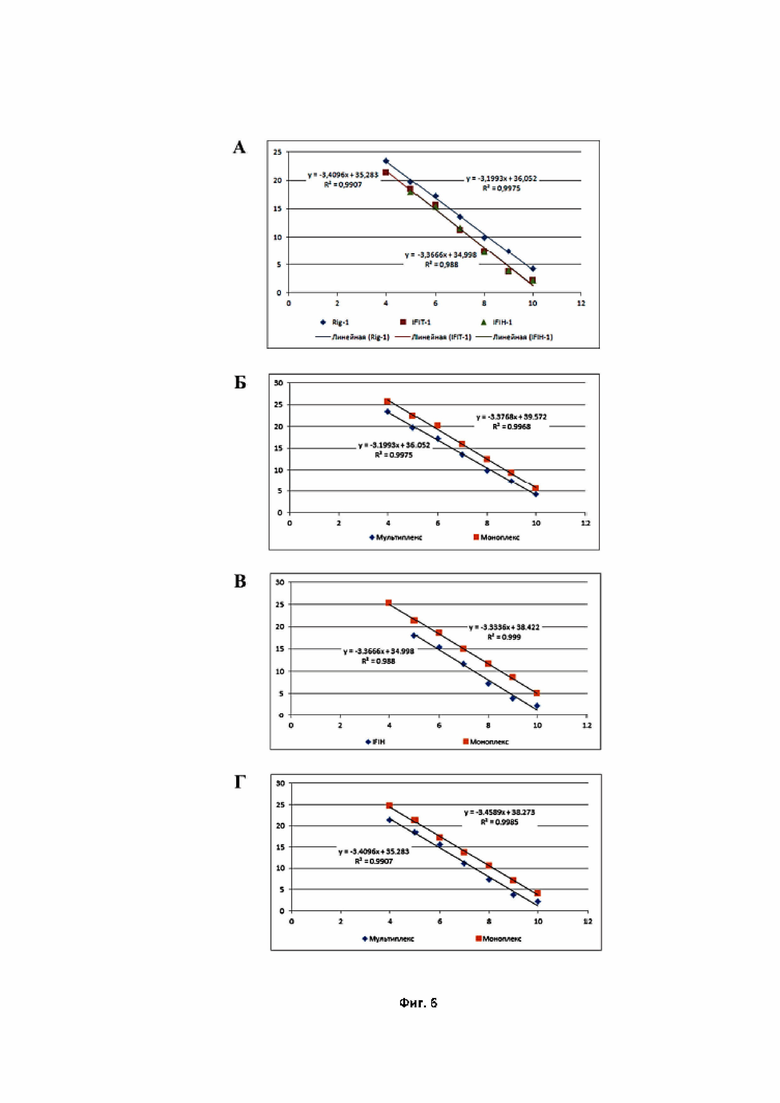
На фиг. 6 представлено сравнение эффективностей реакций ПЦР в моноплексном и мультиплексном форматах: А – представлены стандартные калибровочные кривые, используемые для расчёта эффективностей амплификации RIG-1, IFIT-1 и IFIH-1 в мультиплексном формате (одновременная детекция в одной пробирке); Б – стандартная калибровочная кривая, полученная в мультиплексном формате ПЦР (синие точки) в сравнении калибровочной кривой при моноплексной постановке (оранжевые точки) для гена RIG-1; В – стандартная калибровочная кривая, полученная в мультиплексном формате ПЦР (синие точки), в сравнении с калибровочной кривой при моноплексной постановке (оранжевые точки) для гена IFIH-1; Г – стандартная калибровочная кривая, полученная в мультиплексном формате ПЦР (синие точки), в сравнении калибровочной кривой при моноплексной постановке (оранжевые точки) для гена IFIT-1.
На фиг. 7 представлена специфичность амплификации требуемых продуктов с использованием различных матриц для ПЦР: cDNA – комплементарной ДНК, полученной из препарата мРНК методом обратной транскрипции; RNA – препарата тотальной РНК, выделенной из стимулированных клеток A549, water – используемой в качестве отрицательного контроля ПЦР. Подписи генов RIG-1, IFIT-1 и IFIH-1 означают, что реакция была поставлена в моноплексном формате на соответствующие гены, MIX – мультиплексный формат проведения ПЦР.
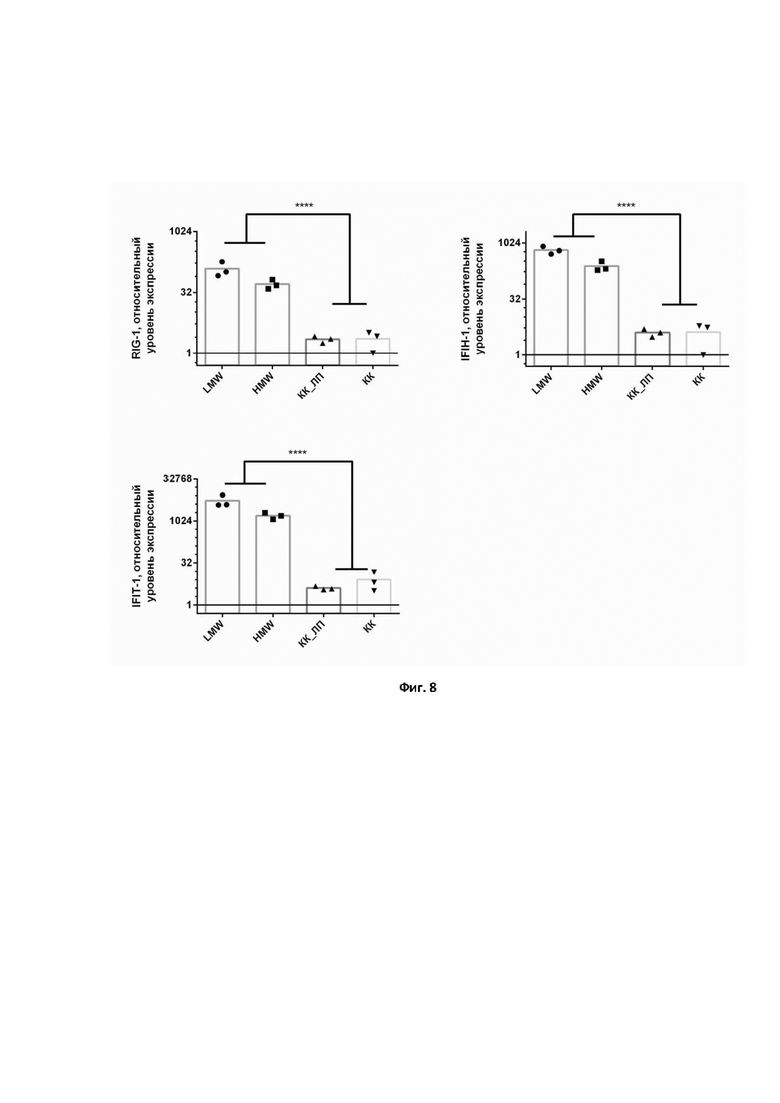
На фиг. 8 представлено определение уровня экспрессии генов RIG-1, IFIT-1, IFIH-1 с использованием разработанной мультиплексной ПЦР в культуре клеток A549, стимулированных низкомолекулярной (LMW) и высокомолекулярной (HMW) poly(I:C) дцРНК. Доставку LMW и HMW в клетки осуществляли с использованием реагента Lipofectamine MessengerMAX, обозначение КК_ЛП – контроль клеток, стимулированных только реагентом Lipofectamine MessengerMAX (без дцРНК), КК – интактный контроль клеток. Достоверность различий определялась с использованием критерия ANOVA (дисперсионный анализ) с множественными сравнениями. **** – P < 0,0001.
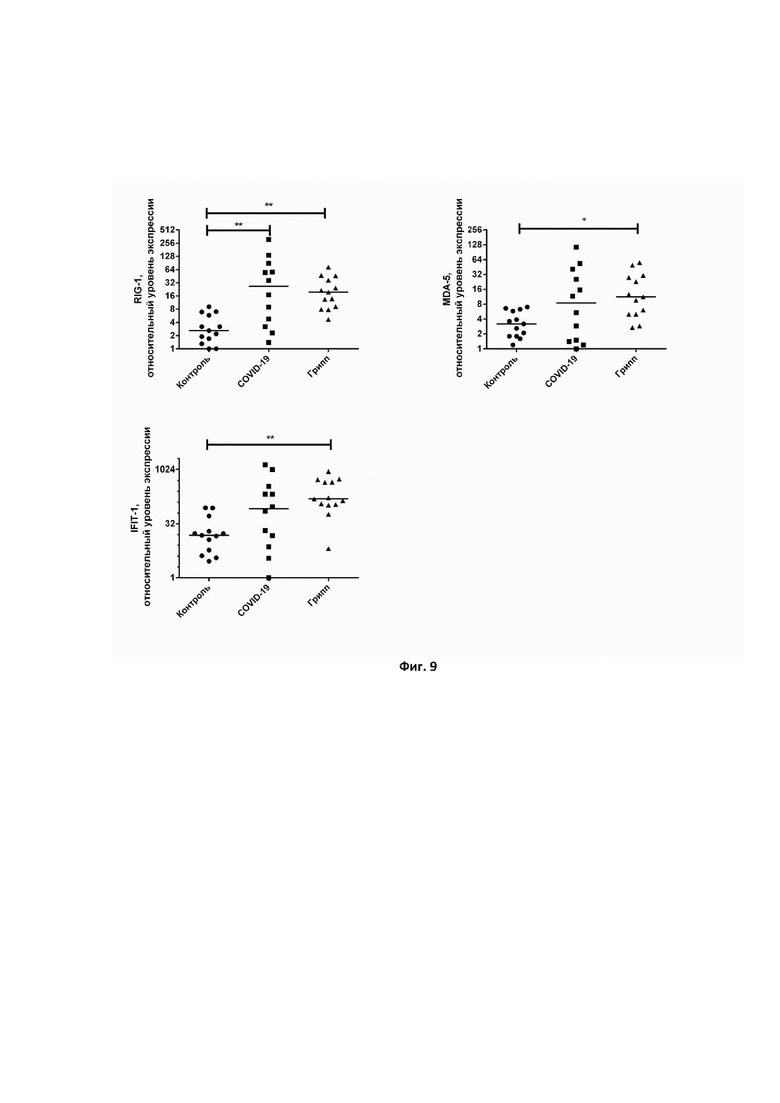
На фиг. 9 представлено определение уровня экспрессии генов RIG-1, IFIT-1, IFIH-1 в периферических мононуклеарных клетках крови здоровых волонтеров и пациентов с гриппозной и новой коронавирусной инфекцией. Достоверность различий определялась с использованием непараметрического критерия Краскела-Уоллиса с множественными сравнениями. * – P < 0,05; ** – P < 0,01.
Для определения уровня экспрессии генов RIG-1, IFIT-1 и IFIH-1 были подобраны праймеры и олигонуклеотидные зонды (таблица 1).
РНК-сенсор RIG-1 человека представлен в базе данных 7 транскрипционными вариантами. Поэтому было проведено множественное выравнивание этих вариантов и к идентичному участку подобрана пара праймеров и олигонуклеотидный зонд.
Ген IFIT-1 представлен 5 транскрипционными вариантами. Поэтому было проведено множественное выравнивание этих вариантов и к идентичному участку подобрана пара праймеров и олигонуклеотидный зонд.
РНК-сенсор MDA-5 (IFIH-1) представлен только одним транскрипционным вариантом.
В качестве референсного гена, подходящего для проведения ОТ-ПЦР в реальном времени, при заражении клеток респираторными инфекциями был выбран ген HPRT-1.
Таблица 1: Подобранные праймеры и TaqMan зонды для определения экспрессии RIG-1, IFIT-1, IFIH-1
Подбор и оптимизация условий ПЦР
На этом этапе работы экспериментально оценили качество подобранных пар праймеров и зондов, а также возможность образования ими гетеро- и гомодимеров и неспецифических продуктов. Материалом для этого исследования служили образцы кДНК, полученные из препаратов тотальной РНК, выделенных из клеток A549, инфицированных вирусом гриппа A. Работа проводилась с использованием набора для проведения ПЦР HS 2× ПЦР-микса («Биолабмикс», Россия). В реакцию объёмом 25 мкл добавляли по 5 пмоль праймеров и зондов (0,5 мкл из стока праймеров и зондов с концентрациями 10 пмоль/мкл).
Для определения оптимального термального профиля были проведены следующие ПЦР с градиентом отжига:
1) первичная денатурация 95°С – 5 мин,
далее следовали 40 трёхступенчатых циклов:
2) денатурация 95°С – 10 с;
3) отжиг праймеров при температурах в диапазоне от 57,5°C до 63,5°С – 30 с (точные температуры составили 63,5°, 63,2°, 62,5°, 61,3°, 59,9°, 58,7°, 57,9°, 57,5°С);
4) элонгация 72°С – 30 с;
Детекцию результатов осуществляли по росту флуоресценции, наличие неспецифических продуктов оценивали электрофоретическим разделением продуктов в агарозном геле. На Фиг. 1 представлены результаты электрофоретического разделения продуктов ПЦР на RIG-1 (Фиг. 1 А), IFIT-1 (Фиг. 1 Б), IFIH-1 (Фиг. 1 В) и на ген HPRT1 (Фиг. 1 Г), предложенный в качестве эндогенного контроля, при проведении реакции при соответствующих температурах отжига. Электрофорез проводили в 1% агарозе на буфере 1× TBE с окрашиванием бромистым этидием.
С учётом полученных результатов в качестве оптимального температурного профиля ПЦР было предложено использовать следующий:
1) первичная денатурация 95°С – 5 мин,
далее 40 двухступенчатых циклов:
2) денатурация 95°С – 10 с;
3) отжиг праймеров и элонгация цепи 61,3°С – 30 c.
Расчет эффективностей ПЦР
Для корректного проведения относительного количественного анализа экспрессии генов RIG-1 подобных рецепторов в мультиплексном формате значения эффективностей амплификации соответствующих генов должны быть идентичными или максимально приближенными друг к другу. Поэтому далее при подобранных оптимальных условиях рассчитали эффективности ПЦР.
Эффективность амплификации продуктов ПЦР рассчитывали по углу наклона кривых, полученных серией последовательных десятикратных разбавлений образцов кДНК.
На Фиг. 2 показаны результаты расчёта эффективности ПЦР при амплификации гена RIG-1 в моноплексном формате. Как видно на Фиг. 2 А, экспрессионные участки кривых накопления, полученных для соответствующих разведений пробы, параллельны, расстояние между последовательными разведениями постоянно, углы наклона кривых одинаковы. Рассчитанная эффективность ПЦР на RIG-1 (Фиг. 2 Б) составила 99,81%, что является практически идеальным параметром амплификации. Аналогичным образом были получены эффективности амплификации ПЦР для IFIT-1 (Фиг. 3) и для IFIH-1 (Фиг. 4). Таким образом, продемонстрировано, что в моноплексном формате эффективности амплификации продуктов трех генов RIG-1, IFIT-1 и IFIH-1 составили: 99,81%, 99,81%, 99,87%, соответственно. Графически это выражается в том, что стандартные калибровочные кривые (представлены на Фиг. 2–4 Б) имеют похожие углы наклона, т.е. параллельны.
Для проведения мультиплексной ПЦР амплификацию всех генов RIG-1, IFIT-1 и IFIH-1 проводили одновременно в одной пробирке. Для этого проба ПЦР, кроме фермента ДНК-зависимой ДНК-полимеразы и прилагаемого к ней буфера, содержала одновременно все подобранные пары праймеров и олигонуклеотидные зонды, специфически выявляющие заявленные гены. Конечные концентрации праймеров и зондов, содержащиеся в пробе при проведении мультиплексной ПЦР, представлены в таблице 2.
Таблица 2: Состав мультиплексной реакции
Ввиду того, что при нахождении в одной пробирке праймеры могу давать неспецифические химерные продукты, анализ продуктов ПЦР в мультиплексном и моноплексном режимах был проведен методом капиллярного электрофореза (Фиг. 5). Как продемонстрировано на Фиг. 5 А, при проведении моноплексной ПЦР в пробирках амплифицировались только продукты, соответствующие заявленным генам RIG-1, IFIT-1, IFIH-1, с длинами 175 п.о., 187 п.о. и 216 п.о. для каждого. При проведении мультиплексной ПЦР в пробирке одновременно выявлялись все три гена, имеющие аналогичные длины. Спектрофореграмма продуктов, полученных в мультиплексном формате, наложенная на спектрофореграммы продуктов ПЦР, образовавшихся в моноплексных форматах, имели идентичные пики (Фиг. 5 Б). Таким образом, мультиплексирование не приводило к образованию каких-либо нежелательных случайных продуктов.
Далее были рассчитаны эффективности амплификации каждого из заявленных генов в мультиплексном формате ПЦР. Как продемонстрировано на Фиг. 6 А, эффективности амплификации всех специфических продуктов при проведении ПЦР в одной пробирке практически идентичны и превышают 98% (для RIG-1, IFIT-1 и IFIH-1 составили: 99,75%, 99,09%, 98,80%, соответственно). Кроме того, мультиплексирование праймеров приводит к снижению амплификации не более чем на 1–2% по сравнению с амплификацией в моноплексном формате для каждого из специфических продуктов (Фиг. 6 В–Г). Такие результаты свидетельствуют, что при проведении мультиплексной ПЦР в каждом её раунде происходит удвоение специфических продуктов.
Для оценки появления возможных неспецифических продуктов, обусловленных геномной контаминацией комплементарных кДНК-проб, разработанные ПЦР в мультиплексном и моноплексном форматах тестировали с использованием различных матриц (РНК или кДНК). Как видно из электрофоретического разделения соответствующих продуктов ПЦР, представленных на Фиг. 7, ПЦР на IFIT-1 даёт положительные специфические продукты, полученные с геномной ДНК (на Фиг. 7 проба RNA). Такие результаты ожидаемы и были предсказаны на этапе подбора последовательностей праймеров и зонда, специфически выявляющего IFIT-1. Тем не менее, такие результаты свидетельствуют о строгой необходимости в предварительной обработке тестируемой в ПЦР пробы РНК ДНКазами для удаления геномной ДНК.
Верификация тест-системы in vitro и на клинических образцах
Разработанная система количественной мультиплексной ПЦР была применена для анализа экспрессии генов RIG-1, IFIT-1 и IFIH-1 в стимулированных культурах клеток A549 и в периферических мононуклеарных клетках крови здоровых волонтеров и пациентов с гриппозной и новой коронавирусной инфекцией.
Известно, что двухцепочечные молекулы РНК (дцРНК) – лиганд, который активирует сенсорные компоненты врожденной иммунной системы. В первую очередь, в ответ на проникновение в клетку дцРНК происходит активация TLR-3 и цитозольных сенсоров IFIH-1 (MDA-5). Длинные дцРНК не характерны для здоровых клеток. Обычно их присутствие связано с проникновением вирусных патогенов [1]. Синтетическим аналогом вирусной дцРНК является poly(I:C) дцРНК. Данная молекула состоит из двух цепей: первая – полимер инозиновой кислоты, вторая — цитидиновой кислоты. Данный агонист используется в исследовательской практике для индукции интерферонов и ISGs. Внесение poly(I:C) дцРНК в клеточную среду моделирует ситуацию вирусной инфекции. Используется два вида синтетических агонистов: высокомолекулярная poly(I:C) дцРНК (High molecular weight poly(I:C) дцРНК, HMW) и низкомолекулярный аналог (Low molecular weight poly(I:C) дцРНК, LMW). Длина HMW составляет от 1500 до 8000 пар оснований, в то время как у LMW – от 200 до 1000 пар азотистых оснований.
Для первого этапа верификации в нашей работе была проведена оценка изменения экспрессии генов RIG-1, IFIT-1, IFIH-1 при стимуляции клеток линии А549 HMW и LMW poly(I:C) дцРНК. Клетки А549 – линия клеток, которая является моделью альвеолярных пневмоцитов II типа. Нами было показано, что HMW и LMW достоверно увеличивают уровень мРНК генов RIG-1, IFIT-1, IFIH-1 (Фиг. 8), что отражено в научной литературе как общеизвестный факт. Следовательно, разрабатываемая нами тест-система пригодна для оценки активации интерферон-опосредованных механизмов врожденной иммунной системы.
На втором этапе верификации разработанная мультиплексная ПЦР применялась для одновременного измерения экспрессии генов RIG-1, IFIT-1, IFIH-1 в периферических мононуклеарных клетках крови здоровых доноров и пациентов с диагнозом COVID-19 или грипп. Диагностика этих инфекций была проведена ПЦР-наборами, сертифицированными для проведения подобных исследований. В результате измерения было показано, что у пациентов, страдающих респираторными инфекциями (грипп, SARS-CoV-2), происходит повышение уровня экспрессии цитозольного РНК-сенсора RIG-1 (Фиг. 9). Таким образом, с использованием мультиплексной ПЦР-системы было показано, что RIG-1 – маркер вирусной инфекции, который неспецифически активируется в ответ на заражение организма гриппозной или новой коронавирусной инфекцией. Наоборот, экспрессия IFIT-1 и IFIH-1 была повышена только у пациентов, зараженных вирусом гриппа. В случае заражения COVID-19 уровень данных маркеров, видимо, зависит от тяжести перенесенного заболевания.
В целом, использование разработанной мультиплексной ПЦР тест-системы позволяет с высокой эффективностью проводить количественный анализ генов RIG-1, IFIT-1, IFIH-1 как в клинических образцах, так и в культуре клеток.
Список литературы
1 Thompson M. R. et al. Pattern recognition receptors and the innate immune response to viral infection//Viruses. – 2011. – Т. 3. – №. 6. – С. 920-940.
2 Kaisho, T., & Akira, S. Toll-like receptor function and signaling//Journal of allergy and clinical immunology. – 2006. – T. 117. – №. 5. – C. 979-987.
3 Randall R. E., Goodbourn S. Interferons and viruses: an interplay between induction, signalling, antiviral responses and virus countermeasures//Journal of General Virology. – 2008. – Т. 89. – №. 1. – С. 1-47.
4 Yap, G. L. et al., Annexin-A1 promotes RIG-I-dependent signaling and apoptosis via regulation of the IRF3–IFNAR–STAT1–IFIT1 pathway in A549 lung epithelial cells//Cell death & disease, – 2020. – Т. 11. – №. 6. – С. 1-12.
5 Methods and targets for identifying compounds for regulating rhinovirus infection: patent appl. CA2679412A1, Canada, appl. CA2679412A, 28.02.2008, publ. 04.09.2009.
| название | год | авторы | номер документа |
|---|---|---|---|
| Тест-система для количественной диагностики мРНК генов MxA, OAS1, EIF2AK2 человека на основе ПЦР | 2022 |
|
RU2811690C1 |
| Тест-система для количественной диагностики мРНК генов OAS1a/g, IL-6, IRF-7, IFNA, IFNB, IFNL мыши методом ПЦР в режиме реального времени | 2022 |
|
RU2796522C1 |
| ТЕСТ-СИСТЕМА ДЛЯ ОПРЕДЕЛЕНИЯ РНК ИНТЕРФЕРОНА λ, ИНТЕРЛЕЙКИНА IL23 И ПРОТИВОВИРУСНОГО БЕЛКА MxA | 2016 |
|
RU2627179C1 |
| Набор олигонуклеотидных праймеров и зондов для идентификации вируса клещевого энцефалита, вируса лихорадки Западного Нила, боррелий и риккетсий методом мультиплексной ПЦР в режиме реального времени | 2016 |
|
RU2629604C1 |
| Тест-система для количественной диагностики мРНК интерферонов I, II и III типов человека на основе ПЦР | 2020 |
|
RU2751791C1 |
| Экспресс-тест на основе ПЦР, позволяющий предсказывать чувствительность опухоли головного мозга конкретного пациента к онколитическим вирусам | 2016 |
|
RU2697412C2 |
| НУКЛЕИНОВАЯ КИСЛОТА, ОБЛАДАЮЩАЯ АКТИВНОСТЬЮ КОРОТКОЙ ИНТЕРФЕРИРУЮЩЕЙ РНК, ВЕКТОР, СОДЕРЖАЩИЙ ДАННУЮ НУКЛЕИНОВУЮ КИСЛОТУ, ФУНКЦИОНАЛЬНО СОЕДИНЕННУЮ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ РЕГУЛЯЦИИ ТРАНСКРИПЦИИ, И СПОСОБ ИХ ПРИМЕНЕНИЯ ДЛЯ ПОДАВЛЕНИЯ ПРОЛИФЕРАЦИИ КЛЕТОК АДНОКАРЦИНОМЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА | 2013 |
|
RU2532842C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ОБНАРУЖЕНИЯ И ИДЕНТИФИКАЦИИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ НУКЛЕИНОВЫХ КИСЛОТ В БИОЛОГИЧЕСКИХ ОБРАЗЦАХ | 2012 |
|
RU2595421C2 |
| Рекомбинантный штамм VV-mNIS-NS1 вируса осповакцины, продуцирующий симпортер йодида натрия мышей mNIS и белок NS1 парвовируса крыс H-1 для тераностики злокачественных опухолей | 2023 |
|
RU2819082C1 |
| Способ диагностики острых лейкозов и наборы для его выполнения | 2021 |
|
RU2806591C2 |

Изобретение относится к области медицины и биохимии, в частности к молекулярной диагностике, а именно к многопараметрической диагностической тест-системе, и может быть использовано для количественного определения уровней мРНК генов RIG-1, IFIT-1, IFIH-1 человека в биологическом образце. Предложена многопараметрическая диагностическая тест-система для количественного определения уровня мРНК генов RIG-1, IFIT-1, IFIH-1 методом полимеразной цепной реакции с обратной транскрипцией. Тест-система включает набор олигонуклеотидных праймеров и флуоресцентных зондов. Тест-система обеспечивает возможность быстрого и точного определения уровня мРНК генов RIG-1, IFIT-1, IFIH-1 человека в биологическом образце. 1 з.п. ф-лы, 2 табл., 9 ил.
1. Многопараметрическая диагностическая тест-система для количественного определения уровня мРНК генов RIG-1, IFIT-1, IFIH-1 методом полимеразной цепной реакции с обратной транскрипцией, включающая набор олигонуклеотидных праймеров и флуоресцентных зондов со следующей структурой:
прямой праймер гена RIG-1 GAGCACTTGTGGACGCTTTA,
обратный праймер гена RIG-1 ATACACTTCTGTGCCGGGAG,
флуоресцентный зонд гена RIG-1
Rox-CCTGGCATATTGACTGGACGTGGC-BHQ2,
прямой праймер гена IFIT-1 AAACTTCGGAGAAAGGCATTAGAT,
обратный праймер гена IFIT-1 TGAAATGAAATGTGAAAGTGGCTG,
флуоресцентный зонд гена IFIT-1
Hex-CCTGAGACTGGCTGCTGACTTTGAGAAC-BHQ1,
прямой праймер гена IFIH-1 AAACCCATGACACAGAATGAACA,
обратный праймер гена IFIH-1 TGTGAGCAACCAGGACGTAG,
флуоресцентный зонд гена IFIH-1
Cy5.5-CACAGTGGCAGAAGAAGGTCTGGA- BHQ3.
2. Тест-система по п. 1, отличающаяся тем, что оптимальная концентрация праймеров и зондов составляет 200 нМ для праймеров и зонда к гену RIG-1, 200 нМ для праймеров и зонда к гену IFIT-1, 750 нМ для праймеров к гену IFIH-1, 250 нМ для зонда к гену IFIH-1.
| US 2009155180 A1, 28.02.2007 | |||
| YAP, G | |||
| L | |||
| et al., Annexin-A1 promotes RIG-I-dependent signaling and apoptosis via regulation of the IRF3-IFNAR-STAT1-IFIT1 pathway in A549 lung epithelial cells, Cell death & disease, 2020, Т | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |

