Настоящее изобретение относится к способу размножения негемопоэтических тканерезидентных γδ T-клеток ex vivo. Термин "негемопоэтические тканерезидентные γδ T-клетки" относится к субпопуляциям Т-лимфоцитов, которые постоянно находятся в негемопоэтических тканях, но не в лимфоидных органах и крови. Такие клетки включают в себя клетки, отличные от Vδ2, например. Vδ1, Vδ3 и Vδ5. Изобретение также относится к применению таких клеток в адоптивной терапии с использованием Т-клеток и в терапии, опосредованной химерными антигенными рецепторами, а также к их применению в способе скрининга модуляторов, контролирующих ключевые точки. Изобретение также относится к клеткам, полученным в результате размножения ex vivo негемопоэтических тканерезидентных γδ Т-клеток.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Растущий интерес к Т-клеточной иммунотерапии рака сосредоточен на очевидной способности субпопуляций Т-клеток CD8+ (1-4) и CD4+ αβ (5,6) распознавать раковые клетки и опосредовать защитный функциональный потенциал хозяина, особенно после дерепрессии под действием клинически опосредованного антагонизма ингибиторных путей, опосредуемых PD1 (7,8), CTLA4 (9,10) и другими рецепторами (11). Тем не менее, остается много вопросов. Например, существует множество основных клинических сценариев, при которых эффективность таких методов лечения оказывается низкой (11); часто возникают тяжелые побочные эффекты (AE) (12); возможность прогнозировать либо эффективность, либо AE чрезвычайно ограничена (13); и существует очень мало объяснений взаимодействий, позволяющих хозяину чувствовать опухолевые клетки (так называемая "иммуногенность"), что обязательно должно предшествовать активации ответов, опосредуемых обычными антигенспецифическими αβ T-клетками CD8+ и CD4+.
В этом отношении многие ученые и клиницисты также переоценивают потенциал γδ T-клеток, третьей линии лимфоцитов с соматически генерируемыми рецепторами, которые эволюционно являются также высоко консервативными, как αβ T-клетки и B-клетки. Существуют две основные субпопуляции человеческих γδ T-клеток: одна из них преобладает в периферической крови человека, экспрессируя, главным образом, рецептор T-клеток Vδ2 (TCR); а другая преобладает в негемопоэтических тканях, большинство из клеток этой субпопуляции экспрессирует TCR Vδ1, также существуют более маленькие популяции, экспрессирующие TCR, которые содержат цепь Vδ3, или Vδ5, или какую-либо другую цепь, отличную от Vδ2 (14).
У большинства взрослых клетки Vδ2 составляют в стационарном состоянии лишь небольшую и высоко вариабельную часть Т-клеток крови (0,01-5%), однако эти клетки быстро размножаются, временно достигая ~25% от клеток CD3+, после заражения широким спектром агентов, включающих в себя многочисленные бактерии и паразиты (14). Основой такого ответа является способность Vδ2 распознавать при посредстве TCR низкомолекулярные "фосфо-фрагменты", в том числе гидроксилметилбут-2-енилпирофосфат (HMBPP) (15), промежуточное соединение в ключевом микробном пути синтеза холестерина и других липидов, которые используются для модификации белков, например, путем геранилирования или фарнезилирования. У приматов этот синтез протекает через мевалонатный путь, одно из промежуточных соединений которого - изопентенилпирофосфат (IPP) - экспрессируется на очень высоких уровнях в инфицированных вирусом и трансформированных клетках и также является мишенью распознавания Vδ2, опосредованного TCR (16).
Кроме того, большинство T-клеток Vδ2 экспрессируют высокие уровни рецептора NKG2D, который может активировать или совместно стимулировать (вместе с TCR) цитолитические потенциалы клеток в присутствии лигандов NKG2D, например, MICA, MICB и ULBP. Эти лиганды представляют собой белки хозяина, которые подвергаются повышающей регуляции при воздействии на клетки таких факторов, как окислительный или осмотический стресс или ультрафиолетовый свет. Данные факторы стимулируют гиперактивный сигнальный путь рецептора эпидермального фактора роста (EGFR), регуляция которого также обычно нарушена в солидных опухолях человека (17).
Способность T-клеток Vδ2 обнаруживать трансформированные клетки с использованием их TCR и/или NKG2D (18-20), наряду с их мощной цитолитической способностью и выраженной способностью презентировать антигены T-клеткам CD8+ (21), обусловливает мнение, что T-клетки Vδ2 можно использовать в клинике для иммунотерапии рака. Этого можно достигнуть путем адоптивного переноса клеток, в связи с чем неспособность МНС подавлять γδ Т-клетки в значительной и благоприятной мере ограничивает возможность возникновения заболевания трансплантата против хозяина (GvHD) (22). Для этого постоянно присутствующие в крови клетки Vγ9Vδ2 γδ Т-клетки можно размножить ex vivo путем добавления цитокинов, таких как интерлейкин (IL)-2, наряду с экзогенными TCR-активирующими средствами, такими как фосфо-фрагменты (например, BrHPP), или наряду с клинически одобренными бисфосфонатами (такими как золедроновая кислота), которые ингибируют фарнезилпирофосфатсинтазу в мевалонатном пути, тем самым индуцируя накопление TCR-активирующего фрагмента, IPP. Однако длительная активация клеток Vγ9Vδ2 под действием таких средств, как BrHPP, может привести к постепенному истощению клеток и уменьшению потенциала цитотоксичности.
Альтернативно собственные γδ Т-клетки пациентов можно активировать in situ с использованием либо фармакологически модифицированных форм HMBPP, либо клинически одобренных аминобензофосфонатов. Указанные подходы использовали для лечения более 250 больных раком, по-видимому, они являются безопасными, но полной ремиссии позволяют достичь только в редких случаях. Одной из основных проблем, обуславливающих ограниченную клиническую эффективность клеток, является их склонность к необратимому истощению при хроническом воздействии антигена. Вторая основная проблема заключается в их кажущейся неэффективности при возвращении в солидные опухоли и ткани, несущие такие опухоли (23).
Терапия с использованием Т-клеток, несущих химерные антигенные рецепторы (CAR-T), является перспективным клиническим методом лечения В-клеточных злокачественных опухолей. Однако при лечении солидных опухолей эффективность клеток CAR-T до настоящего времени была ниже ожиданий, наблюдалась более низкая эффективность при индуцировании полных реакций опухолей и высокая встречаемость внеопухолевой цитотоксичности (24). Что касается γδ T-клеток периферической крови, основным препятствием для успешного применения подходов с использованием CAR-T к солидным опухолям является вероятная неспособность системных клеток CAR-T мигрировать в участки злокачественной опухоли и оставаться там в функционально-эффективном состоянии (25). Кроме того, будучи зависимыми от обычных αβ Т-клеток, клетки CAR-T должны преодолевать иммуносупрессорные сигналы в микроокружении опухоли, например, сигналы, которые передаются через рецептор PD1.
Способы CAR-T с использованием γδ T-клеток могут иметь преимущества, поскольку эти клетки можно трансдуцировать активными в отношении опухоли антигенспецифичными химерными TCR и сохранить при этом их способность распознавать трансформированные клетки при посредстве таких рецепторов, как NKG2D. Таким образом, данные клетки могут одновременно обладать опухоль-адаптивными (TCR) и собственными (NKG2D)-опосредованными свойствами. Однако остается проблема кажущейся неспособности γδ Т-клеток человеческой крови возвращаться в опухоли твердых тканей и оставаться там в активной форме. Данная проблема инициировала более подробные исследования γδ T-клеток, которые обычно находятся в негемопоэтических тканях.
Такие Т-клетки мигрируют в негемопоэтические ткани, что является частью их развития, и, как таковые, отличаются от Т-клеток, например, тканерезидентных Т-клеток памяти TCRαβ+ (так называемых клеток TRM), которые инфильтруют ткань после системного примирования. Тканерезидентные γδ Т-клетки наиболее хорошо изучены у мышей, у которых они, как показано, распространены в коже, кишечнике и репродуктивных тканях, а также в других участках организма. Было показано, что многие такие клетки обладают функциональными механизмами, подобными врожденным механизмам, которые обуславливают их способность отвечать на заражение посредством активации рецептора NKG2D. Авторы настоящего изобретения недавно получили результаты, свидетельствующие о том, что кожа и кишечник человека также содержат большие популяции не гемопоэтических тканерезидентных γδ Т-клеток с активностью, подобной врожденной. И еще следует отметить, что в исследованиях злокачественных новообразований, воспаления, атопии, аллергии и других патологий, формирующихся в негемопоэтических тканях, не учитывалось потенциальное воздействие указанных врожденных человеческих Т-клеток, постоянно находящихся в тканях, в которых присутствуют патологические поражения.
Человеческие γδ Т-клетки, постоянно присутствующие в негемопоэтических тканях, гораздо менее изучены, поскольку отбор образцов клеток осложнен вследствие их локализации и, кроме того, отсутствуют разработанные способы их культивирования. Из относительно скудной существующей информации можно заключить, что данный подтип включает в себя разнообразные клетки с MHC-неограниченным цитолитическим потенциалом, совсем не способные взаимодействовать с низкомолекулярными фосфо-фрагментами, поскольку они не экспрессируют TCR, содержащие Vγ2. Хотя для таких клеток известно очень мало TCR с конкретной специфичностью, существующие данные свидетельствуют о том, что данные клетки способны взаимодействовать с собственными антигенами, такими как эндотелиальный рецептор белка C (EPCR), который экспрессируется на повышенном уровне клетками, инфицированными цитомегаловирусом (CMV), а также клетками многих солидных опухолей (32). Негемопоэтические ассоциированные с тканями γδ Т-клетки также обычно экспрессируют NKG2D (14). Учитывая эти свойства и физиологическое местонахождение клеток в негемопоэтических тканях, таких как кожа и кишечник, адоптивный перенос таких клеток раковым пациентам может быть значительно более эффективным в отношении солидных опухолей, а, возможно, и других иммунопатологий.
Чтобы использовать клетки, отличные от Vδ2, для иммунотерапии, нужно либо разработать способ размножения клеток in situ, либо собрать клетки и размножить их ex vivo с последующей повторной инфузией. Обычно используют последний подход, поскольку отсутствуют известные TCR-активирующие средства, обладающие доказанной способностью стимулировать размножение большого числа отличных от Vδ2 клеток in situ. Чтобы преодолеть проблему ограниченного доступа к негемопоэтическим тканям, некоторые исследователи предприняли попытки размножить очень небольшое число отличных от Vδ2 клеток, взятых из крови, где Vδ2-экспрессирующие клетки являются доминирующей субпопуляцией, сделав предположение, что эти клетки эквивалентны тканерезидентным клеткам, отличным от Vδ2. Небольшое число отличных от Vδ2 γδ Т-клеток, взятых из крови, размножают по существу в процессе активной инфекции CMV, причем полученные клетки демонстрируют превосходную реакционную способность по отношению к CMV по сравнению с Т-клетками Vδ2 и, по-видимому, обладают способностью защищать человеческий плод в случаях инфекции CMV в утробе. Кроме того, реакционноспособные в отношении CMV отличные от Vδ2 γδ Т-клетки, по-видимому, защищают пациентов после трансплантации от повторной активации CMV в процессе иммуносупрессии, и, посредством перекрестных взаимодействий с трансформированными клетками, уменьшают риск вторичных злокачественных новообразований (26). Подобным образом, существуют данные, свидетельствующие о том, что γδ T-клетки играют полезную роль в борьбе с инфекцией HIV, в данном случае отличные от Vδ2 γδ Т-клетки размножают в крови по сравнению с Т-клетками Vδ2 (24).
Присутствующие в крови отличные от Vδ2 клетки размножают ex vivo либо путем добавления экзогенных средств, которые непосредственно активируют сигнальный путь TCR, например, таких средств, как антитело против CD3, pan γδ-TCR-специфичное антитело или фитогемагглютинин (PHA), либо путем совместного культивирования стимулированных отличных от Vδ2 Т-клеток с искусственными антигенпрезентирующими клетками (aAPC), где непосредственный контакт между γδ Т-клетками и aAPC необходим для размножения отличных от Vδ2 T-клеток ex vivo (41-44). Альтернативно размножение клеток осуществляют путем стимуляции сигнального пути рецептора NKG2D с помощью иммобилизованного рекомбинантного MICA (лиганд NKG2D), например, как при поддержании пролиферации ex vivo культур γδ T-клеток, полученных из эпителиальных лимфоцитов, инфильтрующих опухоли (TIL) (28). Таким образом, существующие в настоящее время способы размножения ex vivo Vδ2-экспрессирующих γδ Т-клеток крови, или отличных от Vδ2 γδ Т-клеток крови, требуют добавления средств, заведомо способствующих активации рецепторов TCR и/или NKG2D, наряду с дополнительными цитокинами, такими как интерлейкин-2 (IL-2) (41-44). Указанное сочетание рецептор-активирующих сигналов и цитокинов отражает стандартный подход к культивированию и выращиванию Т-клеток, широко используемый научным сообществом. На сегодняшний день не описаны способы эффективного размножения γδ Т-клеток, находящихся в негемопоэтической ткани. Такой способ описан в настоящем документе.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В рамках фенотипической и функциональной характеристики Т-клеток человеческой кожи авторы настоящего изобретения выделили особую и большую популяцию γδ Т-клеток, обычно присутствующих в негемопоэтических тканях и обладающих уникальными свойствами по сравнению с αβ Т-клетками и присутствующими в крови γδ T клетками. Авторы обнаружили, что клетки способны давать сильные, TCR-независимые ответы, подобные врожденным, на NKG2D-лиганды и цитокины. Авторы настоящего изобретения неожиданно обнаружили, что, хотя для размножения первичных αβ T-клеток обычно требуется совместное культивирование с другими поддерживающими клетками в качестве источника полезных факторов роста (29), размножение γδ Т-клеток, присутствующих в коже и других негемопоэтических тканях, кардинально и специфично подавляется совместным культивированием этих клеток в контакте с аутологичными дермальными фибробластами и, возможно, с другими стромальными компонентами, такими как кератиноциты и эндотелиальные клетки. Устранение таких взаимодействий позволяет быстро размножать клетки в больших количествах для потенциальных клинических применений.
Кроме того, в отличие от предпринятых на сегодняшний день усилий по размножению γδ Т-клеток, полученных из крови и опухоли, авторы настоящего изобретения показали, что такие негемопоэтические тканерезидентные γδ Т-клетки можно размножать без преднамеренного добавления каких-либо экзогенных средств, активирующих сигнальные пути, опосредуемые TCR или NKG2D.
В данном документе раскрываются новые способы, позволяющие эффективно и воспроизводимо выделять и размножать γδ Т-клетки, полученные из негемопоэтической ткани человека или отличного от человека животного, такой как кожа и кишечник. Размножение инициируют путем нарушения контакта γδ Т-клеток, полученных из негемопоэтической ткани, с аутологичными фибробластами и, возможно, с другими стромальными компонентами и поддерживают путем культивирования в присутствии интерлейкина-2 (IL-2) и/или интерлейкина-15 (IL-15).
Размножение является высоко селективным, поскольку размножение αβ Т-клеток, или Vδ2-экспрессирующих Т-клеток, или NK-клеток не индуцируется путем нарушения их контактов с аутологичными фибробластами (фиг.3А, 3С и 3D). Указанное размножение негемопоэтических тканерезидентных γδ Т-клеток в результате освобождения от фибробласт-опосредованной модуляции контрольных точек также приводит к "спонтанной" активации эффекторных потенциалов клеток (фиг.5А и 5В), которые весьма желательны в контексте противоопухолевой активности. Эти разработки позволяют размножать в культуре и активировать негемопоэтические тканерезидентные γδ Т-клетки для потенциального применения в виде "готовых" клеточных инфузий пациентам. В то же время разработка антитела или другого ингибитора модуляции контрольной точки тканерезидентных γδ T-клеток фибробластами (или другими стромальными или эпителиальными клетками) должна обеспечить активацию негемопоэтических тканерезидентных γδ Т-клеток in situ, например, у ракового пациента, посредством блокады контрольной точки.
Возможность получать и выращивать γδ Т-клетки, полученные из негемопоэтической ткани (например, кожи), позволила выявить четкие отличия от Т-клеток Vδ1, полученных из крови. Например, в Т-клетках Vδ1, полученных из кожи, наблюдаются маркеры предшествующей активации Т-клеток, такие как экспрессия CD69, положительная реакция на ICOS и TIM3, а также незначительная или не выраженная экспрессия классической костимуляторной молекулы CD28 (фиг.10А). Кроме того, в них наблюдается высокая экспрессия NKG2D. И наоборот, Т-клетки Vδ1, полученные из человеческой крови, не экспрессируют CD69 или TIM3, экспрессируют лишь незначительные уровни ICOS и также являются в некоторой степени положительными по экспрессии CD28. Более того, экспрессия NKG2D под действием Т-клеток Vδ1, полученных из крови, намного ниже, чем экспрессия под действием Т-клеток Vδ1, полученных из кожи, и, в отличие от Т-клеток Vδ1, полученных из крови, T-клетки Vδ1, полученные из кожи, обладают врожденной способностью взаимодействовать с лигандами NKG2D, такими как рекомбинантный MICA, в отсутствии стимуляции Т-клеточного рецептора (фиг.10В). Описанные здесь γδ Т-клетки из негемопоэтических тканей также могут характеризоваться более высокой экспрессией CCR3, CD39, CD11b, IL-13 и/или CD9, чем полученные из крови T-клетки Vδ1 и другие популяции лимфоцитов.
В первом аспекте изобретение предлагает способ размножения негемопоэтических тканерезидентных γδ Т-клеток in vitro, который включает в себя культивирование лимфоцитов, полученных из негемопоэтической ткани человека или отличного от человека животного, в присутствии IL-2 и/или интерлейкина-15 (IL-15), где лимфоциты не находятся в непосредственном контакте со стромальными или эпителиальными клетками в процессе культивирования.
Предпочтительно лимфоциты не находятся в непосредственном контакте с фибробластами в процессе культивирования.
γδ Т-клетки изначально присутствуют в негемопоэтической ткани in vivo.
Предпочтительно способ включает в себя культивирование лимфоцитов, полученных из негемопоэтической ткани человека или отличного от человека животного, в присутствии IL-2 и IL-15.
В некоторых вариантах осуществления лимфоциты, полученные из негемопоэтической ткани человека или отличного от человека животного, можно выращивать в отсутствии активаторов TCR или костимуляторов, которые индуцируют активацию Т-клеток. Например, лимфоциты можно выращивать в отсутствии агонистов пути TCR, например, активаторов CD3, таких как антитела против CD3, в присутствии или отсутствии активаторов CD28.
Добавление таких экзогенных активаторов TCR-сигнализации не требуется для размножения негемопоэтических тканерезидентных γδ Т-клеток при использовании способов настоящего изобретения. Как таковая, среда для размножения γδ Т-клеток, подходящая для применения в способах настоящего изобретения, может быть лишена способности активировать Т-клетки, например, способности активировать αβ Т-клетки или γδ Т-клетки крови, и может не обладать способностью активировать или костимулировать TCR.
Например, среда для размножения γδ Т-клеток может не содержать, или практически не содержать средства или факторы, активирующие Т-клеточный сигнальный путь, такие как активаторы или костимуляторы TCR, включающие в себя экзогенные агонисты пути TCR. Среда для размножения γδ Т-клеток может содержать IL-2 и/или IL-15. В некоторых вариантах осуществления среда для размножения γδ Т-клеток помимо IL-2 и/или IL-15 может содержать один или несколько дополнительных факторов роста, таких как цитокины. Подходящие факторы роста не обладают способностью активировать Т-клетки. В других вариантах осуществления среда для размножения γδ Т-клеток может быть лишена факторов роста, отличных от IL-2 и/или IL-15; например, среда для размножения γδ Т-клеток может состоять из основной среды, дополненной IL-2 и/или IL-15.
В одном варианте осуществления лимфоциты, полученные из негемопоэтической ткани человека или отличного от человека животного, можно выращивать в отсутствии стромальных или эпителиальных клеток. Например, стромальные или эпителиальные клетки можно удалить до культивирования. Предпочтительно, чтобы лимфоциты, полученные из негемопоэтической ткани человека или отличного от человека животного, можно культивировать в отсутствии фибробластов. Например, фибробласты можно удалить до культивирования.
Лимфоциты можно получить из любой подходящей негемопоэтической ткани человека или отличного от человека животного, такой как ткань кожи, желудочно-кишечного тракта (например, толстой кишки или подвздошной кишки), молочной железы, легких, печени, поджелудочной железы, жировой ткани или предстательной железы.
Негемопоэтические тканерезидентные γδ Т-клетки предпочтительно представляют собой клетки, отличные от Vδ2, наиболее предпочтительно клетки, экспрессирующие TCR, содержащие цепи Vδ1, т.е. клетки Vδ1. Негемопоэтические тканерезидентные γδ Т-клетки также могут включать в себя так называемые двойные отрицательные (DN) γδ Т-клетки, определенные как клетки, экспрессирующие γδ TCR, не содержащие ни цепи Vδ1, ни цепи Vδ2.
Способ необязательно включает в себя стадию получения лимфоцитов из негемопоэтической ткани человека или отличного от человека животного. Например, лимфоциты можно получить из образца негемопоэтической ткани человека или отличного от человека животного. Способ может включать в себя получение образца негемопоэтической ткани человека или отличного от человека животного и отделение лимфоцитов от негемопоэтических клеток указанного образца с получением популяции лимфоцитов, которая практически не содержит стромальных клеток.
Во втором аспекте изобретение предлагает способ размножения γδ Т-клеток, включающий в себя (i) получение популяции γδ Т-клеток из негемопоэтической ткани; и (ii) выращивание γδ Т-клеток, по существу не контактирующих со стромальными клетками, с получением увеличенной популяции γδ Т-клеток.
Популяция γδ Т-клеток, полученных из негемопоэтической ткани, может представлять собой практически чистую популяцию γδ Т-клеток.
Популяция γδ Т-клеток, полученных из негемопоэтической ткани, предпочтительно представляет собой клетки, отличные от Vδ2, наиболее предпочтительно клетки, экспрессирующие TCR, содержащие цепи Vδ1, т.е. клетки Vδ1. Популяция γδ Т-клеток может также содержать γδ T-клетки DN.
Популяция γδ Т-клеток, полученных из негемопоэтической ткани, может экспрессировать один или несколько дополнительных маркеров тканерезидентных γδ Т-клеток, таких как CLA, IL13, CCL1, CD103 и CCR8. В некоторых вариантах осуществления популяция γδ Т-клеток, полученных из негемопоэтической ткани, может содержать γδ Т-клетки Vδ1+ CCR8+.
γδ Т-клетки можно выращивать в отсутствии контакта со стромальными клетками с получением увеличенной популяции γδ Т-клеток (то есть при отсутствии контакта γδ клеток и стромальных клеток в культуре).
γδ Т-клетки можно выращивать в отсутствии сигналов активации TCR или костимуляторных сигналов. В некоторых вариантах осуществления стадию выращивания можно проводить в среде, кондиционированной по стромальным клеткам, или в присутствии IL-2, IL-15 или их сочетания. Например, γδ T-клетки можно выращивать в среде для культивирования γδ, содержащей IL-2 и/или IL-15. Подходящая среда для выращивания γδ не способна активировать или костимулировать TCR. Например, среда для выращивания γδ может не содержать или практически не содержать средств или факторов, которые активируют Т-клеточный сигнальный путь, таких как активаторы или костимуляторы TCR, в том числе агонисты пути TCR. Среда для выращивания γδ может содержать IL-2 и/или IL-15. В некоторых вариантах осуществления среда для выращивания γδ помимо IL-2 и/или IL-15 может содержать один или несколько дополнительных факторов роста, таких как цитокины. Подходящие факторы роста не способны активировать Т-клетки. В других вариантах осуществления среда для выращивания γδ может не содержать факторы роста, отличные от IL-2 и/или IL-15; например, среда для выращивания γδ может состоять из основной среды, дополненной IL-2 и/или IL-15.
В третьем аспекте изобретение предлагает способ размножения γδ Т-клеток, включающий в себя: (i) получение негемопоэтической ткани, содержащей негемопоэтические клетки и γδ Т-клетки; (ii) отделение γδ Т-клеток от негемопоэтических клеток с получением популяции γδ Т-клеток, которая по существу не содержит стромальных клеток; и (iii) выращивание популяции, полученной на стадии (ii), в отсутствии сигналов активации TCR или костимуляторных сигналов с получением увеличенной популяции γδ Т-клеток.
Например, γδ Т-клетки можно отделить от αβ Т-клеток.
Популяция, полученная на стадии (ii), может представлять собой практически чистую популяцию γδ Т-клеток.
γδ Т-клетки в популяции, полученной на стадии (ii), могут содержать γδ Т-клетки, отличные от Vδ2, наиболее предпочтительно экспрессирующие TCR, содержащие цепи Vδ1, т.е. γδ Т-клетки Vδ1. γδ Т-клетки в популяции, полученной на стадии (ii), также могут содержать γδ Т-клетки DN.
γδ Т-клетки в популяции, полученной на стадии (ii), также могут содержать γδ Т-клетки, которые экспрессируют один или несколько дополнительных маркеров тканерезидентных γδ Т-клеток, таких как CLA, CD103 и CCR8. В некоторых вариантах осуществления γδ Т-клетки в популяции, полученной на стадии (ii), могут содержать γδ Т-клетки Vδ1+CCR8+.
Выращивание на стадии (iii) можно проводить по существу в отсутствии контакта стромальных клеток с популяцией, полученной на стадии (ii), и/или в отсутствии сигналов активации TCR или костимулирующих сигналов. Например, выращивание можно проводить в отсутствии контакта γδ Т-клеток со стромальными клетками. В некоторых вариантах осуществления выращивание на стадии (iii) проводят в среде, кондиционированной по стромальным клеткам, или в присутствии IL-2, IL-15, или их сочетания.
В некоторых вариантах осуществления γδ Т-клетки можно выращивать в среде для культивирования γδ, содержащей IL-2 и/или IL-15. Подходящая среда для культивирования γδ не может активировать или костимулировать TCR. Например, среда для выращивания γδ может не содержать, или по существу не содержать средства или факторы, которые активируют Т-клеточный сигнальный путь, например, активаторы или костимуляторы TCR, такие как агонисты пути TCR. В некоторых вариантах осуществления среда для выращивания γδ помимо IL-2 и/или IL-15 может содержать один или несколько дополнительных факторов роста, таких как цитокины. Подходящие факторы роста не способны активировать Т-клетки. В других вариантах осуществления среда для выращивания γδ может состоять из основной среды, дополненной IL-2 и/или IL-15.
Число γδ Т-клеток в увеличенной популяции по второму или третьему аспекту может по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 15 раз, по меньшей мере в 20 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз, по меньшей мере в 50 раз, по меньшей мере в 100 раз, по меньшей мере в 500 раз, по меньшей мере в 1000 раз или, по меньшей мере, в 10000 раз превышает число γδ Т-клеток в негемопоэтической ткани, или отделенных от негемопоэтических клеток. Увеличенную популяцию можно получить путем выращивания в течение 3 дней, 5 дней, 7 дней, 10 дней, 14 дней, 21 дня или 28 дней.
γδ Т-клетки в увеличенной популяции предпочтительно представляют собой Т-клетки Vδ2-. Увеличенная популяция γδ Т-клеток может содержать по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% клеток Vδ1+. Увеличенная популяция γδ Т-клеток может быть по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, или по меньшей мере на 90% положительной по одному, двум или всем трем из CCR4, CCR8 и CD103. Например, увеличенная популяция может быть по меньшей мере на 10% положительной по CD103, по меньшей мере на 30% положительной по CCR4 и по меньшей мере на 60% положительной по CCR8.
В четвертом аспекте изобретение относится к негемопоэтическим тканерезидентным γδ Т-клеткам или их популяциям, полученным по способу, описанному в первом, втором или третьем аспекте настоящего изобретения.
В пятом аспекте изобретение относится к способу скрининга модуляторов контрольных точек с использованием негемопоэтических тканерезидентных γδ Т-клеток, где способ включает в себя:
(i) выращивание негемопоэтических тканерезидентных γδ Т-клеток in vitro в непосредственном контакте со стромальными или эпителиальными клетками (например, фибробластами) в присутствии и отсутствии тестируемого соединения, или выращивание негемопоэтических тканерезидентных γδ Т-клеток in vitro при непосредственном контакте со стромальными или эпителиальными клетками (например, фибробластами) в условиях изменения экспрессии тестируемого гена в γδ Т-клетках и/или в стромальных или эпителиальных клетках (например, фибробластах); и
(ii) определение скорости пролиферации или активации негемопоэтических тканерезидентных γδ Т-клеток в присутствии и отсутствии тестируемого соединения, или в присутствии и отсутствии изменения экспрессии тестируемого гена в стромальных или эпителиальных клетках (таких как фибробласты) и/или γδ Т-клетках, или определение скорости уничтожения стромальных или эпителиальных клеток (например, фибробластов) в присутствии и отсутствии тестируемого соединения, или в присутствии и отсутствии изменения экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, в фибробластах) и/или γδ Т-клетках,
причем, если скорость пролиферации или активации Т-клеток в присутствии тестируемого соединения выше, чем в отсутствии тестируемого соединения, или в присутствии изменения экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах) и/или γδ Т-клетках, выше, чем в отсутствии изменения экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах) и/или γδ Т-клетках, и/или если скорость уничтожения стромальных или эпителиальных клеток (например, фибробластов) в присутствии тестируемого соединения выше, чем в отсутствии тестируемого соединения, или в присутствии изменения экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах) и/или γδ Т-клетках, выше, чем в отсутствии изменения экспрессии тестируемого гена в фибробластах и/или γδ Т-клетках, то тестируемое соединение, скорее всего, является модулятором контрольной точки, или тестируемый ген, по всей вероятности, является геном-кандидатом контрольной точки или его регулятором.
Экспрессию тестируемого гена в γδ Т-клетках и/или в стромальных или эпителиальных клетках (таких как фибробласты) можно изменить, например, с помощью РНК-направленного средства, такого как малая интерферирующая РНК (миРНК) или малая шпилечная РНК (мшРНК), или путем редактирования гена, например, с использованием системы CRISPR/Cas.
В шестом аспекте изобретение относится к способу лечения индивидуума с помощью адоптивной терапии Т-клетками, где способ включает в себя введение негемопоэтических тканерезидентных γδ Т-клеток, полученных по способу, описанному в первом, втором или третьем аспекте настоящего изобретения, индивидууму, нуждающемуся в этом. Индивидуум предпочтительно представляет собой человека.
Индивидуум предпочтительно представляет собой человека-пациента, страдающего от рака или вирусной инфекции, такого как CMV-инфицированный пациент или ВИЧ-инфицированный пациент.
В седьмом аспекте изобретение относится к применению негемопоэтических тканерезидентных γδ Т-клеток, полученных по способу, описанному в первом, втором или третьем аспекте настоящего изобретения, в способе лечения человека или отличного от человека животного с помощью адоптивной Т-клеточной терапии. Негемопоэтическая тканерезидентная γδ Т-клетка может обладать одним или несколькими из следующих свойств:
(i) имеет фенотип CD69высокий, ICOSвысокий, TIM3высокий и CD28низкий/отсутствует
(ii) осуществляет повышающую регуляцию одного или нескольких из CCR3, CD11b, CD9 и CD39,
(iii) продуцирует IFN-γ в ответ на лиганд NKG2D в отсутствии агонистов TCR,
(iv) продуцирует IL-13 в отсутствии агонистов TCR,
(v) продуцирует один или несколько из IFN-γ, TNF-α и GM-CSF в ответ на активацию TCR,
(vi) не продуцирует или практически не продуцирует IL-17 в ответ на активацию TCR,
(vii) растет в культуральной среде, содержащей IL-2 в отсутствии дополнительных факторов роста,
(viii) обеспечивает цитотоксический Т-клеточный ответ в отсутствии агонистов TCR и/или
(ix) обладает селективной цитотоксичностью в отношении опухолевых клеток по сравнению с нормальными клетками.
В предпочтительном варианте осуществления человек представляет собой человека-пациента, страдающего от рака или вирусной инфекции, например, CMV-инфицированного или ВИЧ-инфицированного пациента, где инфекция CMV или ВИЧ связана с MICA.
В восьмом аспекте изобретение предлагает способ лечения индивидуума с помощью терапии, опосредованной химерным антигенным рецептором, где способ включает в себя введение негемопоэтических тканерезидентных γδ Т-клеток, полученных по способу, описанному в первом, втором или третьем аспекте изобретения, индивидууму, нуждающемуся в этом. Индивидуум предпочтительно представляет собой человека.
В предпочтительном варианте осуществления индивидуум представляет собой человека-пациента, страдающего от рака.
В девятом аспекте изобретение относится к применению негемопоэтических тканерезидентных γδ Т-клеток, полученных по способу, описанному в первом, втором или третьем аспекте настоящего изобретения, в способе лечения человека или отличного от человека животного с использованием химерного антигенного рецептора.
В предпочтительном варианте осуществления человек представляет собой человека-пациента, страдающего от рака.
Каждый из описанных выше вариантов осуществления можно объединить с одним или несколькими из любых других вариантов осуществления.
Эти и другие аспекты изобретения более подробно описаны ниже.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1A-1D показано, что кожа человека содержит заметную популяцию резидентных γδ Т-клеток. Фиг. 1A: Резидентные лимфоциты кожи выделяют с использованием органотипической клеточной культуры, описанной Clark et al. (29), "метод Кларка". В применении к клеткам CD45+ антитела против CD3 используют для окрашивания Т-клеток, а антитела против CD56 используют для идентификации NK-клеток CD3-CD56+ соответственно. В применении к клеткам CD3+ антитела против рецептора γδ Т-клеток pan используют для идентификации резидентных γδ T-клеток кожи, а антитела против CD8альфа используют для идентификации соотношений обычных CD4- и CD8-положительных αβ Т-клеток в воротах CD3+, pan γδ TCR. На фиг. 1B показаны результаты указанных экспериментов для 7-10 доноров с использованием метода Кларка. В данном методе лимфоциты кожи человека все еще находятся в контакте с дермальными фибробластами, либо в отсутствии каких-либо цитокинов, либо в присутствии интерлейкина-2 (IL-2), интерлейкина-15 (IL-15), или IL-2 и IL-15, и полученные результаты свидетельствуют о том, что добавление цитокинов не влияет на состав резидентных лимфоцитов кожи, за исключением небольшого увеличения популяции γδ T-клеток при добавлении к культуре IL-15, или IL-2 и IL-15, в соответствии с методом Clark et al. Состав лимфоцитов после 3-недельного органотипического культивирования кожи показан с указанием цитокинов в сумме для 4 доноров. Фиг. 1C: Резидентные γδ клетки кожи состоят преимущественно из Vδ1-экспрессирующих γδ Т-клеток (76,24%±17,3), небольшой популяции Vδ2-экспрессирующих Т-клеток (3,06%±6,1) и популяции pan-γδ TCR-положительных клеток, окрашиваемых отрицательно на Vδ1 или Vδ2, также называемых здесь двойные отрицательные (DN) γδ Т-клетки (20,7%±13,97). Контрольное окрашивание крови здоровых добровольцев демонстрирует сильную компартментализацию человеческих γδ T-клеток, поскольку внутри крови доминирует популяция γδ T-клеток, экспрессирующих цепь Vδ2 TCR. Фиг. 1D: Резидентные γδ Т-клетки кожи содержат маркеры, как показано ранее, ассоциированные с хронически активированными Т-клетками, хотя эти маркеры скорее являются характерными показателями постоянного нахождения в ткани, чем хронической активации. Гистограммы демонстрируют окрашивание указанных маркеров на γδ Т-клетках (закрашенная гистограмма) по сравнению с соответствующим изотипическим контролем для каждого антитела (незакрашенная гистограмма).
На фиг. 2А-2D показано, что резидентные γδ T-клетки кожи, полученные непосредственно из кожи человека по методу Кларка, дают так называемый TH1-смещенный ответ при активации под действием средств, обычно активирующих Т-клетки, и подобным образом дают TH1-смещенный ответ при активации только под действием лигандов NKG2D. Фиг. 2A: В резидентных γδ T-клетках кожи наблюдается высокий уровень экспрессии активаторного и ассоциированного с NK-клетками рецептора NKG2D (закрашенная гистограмма по сравнению с изотипом, представленным незакрашенной гистограммой). При активации под действием связанного с планшетом рекомбинантного MICA, одного из известных лигандов рецепторов NKG2D, γδ Т-клетки кожи отвечают в отсутствии какой-либо другой стимуляции, причем ответ не зависит от TCR, поскольку он отменяется в присутствии антител, блокирующих NKG2D. Клетки стимулируют в течение 6 часов в присутствии брефелдина А и 100 единиц IL-2/мл и затем анализируют дегрануляцию путем окрашивания на CD107a. Продукцию TNFα и INF-γ анализируют по пермеабилизации после поверхностного окрашивания и последующего окрашивания внутриклеточных цитокинов. Форбол 12-миристат 13-ацетат (P) в сочетании с иономицином (I) используют в качестве положительного контроля для активации Т-клеток. Фиг. 2B: Резидентные γδ T-клетки кожи дают TH1-смещенный ответ. γδ Т-клетки извлекают с помощью метода Кларка и стимулируют PMA и иономицином в течение 6 ч в присутствии брефелдина А, после чего окрашивают на внутриклеточные цитокины. γδ Т-клетки, свежевыделенные из человеческой кожи, продуцируют TNFα и IFN-γ после стимуляции, но лишь небольшие или недетектируемые количества цитокинов, например, IL-4, IL-17A, IL-13, IL-22, которые связаны с клетками TH2 или TH-17, тогда как обычные αβ Т-клетки CD4+ продуцируют гораздо более широкий спектр цитокинов. Фиг. 2C: Среди лимфоцитов, полученных непосредственно из кожи человека, γδ Т-клетки, обычные αβ Т-клетки CD8a+ и NK-клетки экспрессируют разные уровни рецептора NKG2D. Среди указанных клеток NK-клетки отвечают на воздействие только лигандов NKG2D, однако из Т-клеток лишь популяция γδ Т-клеток дает цитокиновый ответ при стимуляции лигандами NKG2D в отсутствии какой-либо стимуляции TCR (см. верхний ряд точечной диаграммы проточной цитометрии). Растворимые блокирующие антитела против NKG2D блокируют ответ, свидетельствуя о том, что ответ опосредуется исключительно рецептором NKG2D. Фиг. 2D: Среди резидентных γδ T-клеток кожи только γδ T-клетки Vδ1 и DN обладают врожденной способностью к активации под действием только рекомбинантного MICA (обозначены символом *). Vδ2-экспрессирующие Т-клетки, обнаруженные в небольших количествах в коже, не дают такого ответа.
На фиг.3А-3D показано, что резидентные γδ Т-клетки кожи исключительно отвечают на выделение из дермальной стромы сильной активацией и пролиферацией. Фиг. 3A: Резидентные лимфоциты кожи выделяют по методу Кларка. После органотипического культивирования в течение 3 недель лимфоциты кожи собирают, отделяют от остальных клеток кожи, включающих в себя фибробласты, и помещают в лунки для культивирования тканей при плотности 1 млн лимфоцитов/мл и добавляют 100 ед/мл IL-2. Еще через 3 недели резидентные γδ Т-клетки сильно увеличиваются в числе и обогащают культуру лимфоцитов кожи. Данное явление интенсивной пролиферации свойственно только резидентным γδ Т-клеткам, представленным в основном Т-клетками Vδ1+, число которых через 3 недели увеличивается в среднем в 127,18 раза, тогда как число обычных αβ T-клеток в среднем увеличивается только в 5,21 раза; что более чем в 20 раз меньше. Фиг. 3B: Резидентные Т-клетки кожи Vδ1+ реагируют на потерю ткани путем интенсивной повышающей регуляции маркера Ki-67 (указывающего на клеточный цикл) в течение 14 дней (изотипический контроль представлен незакрашенной гистограммой, ограниченной пунктирной линией, экспрессия Ki-67 на 0 день представлена незакрашенной гистограммой, экспрессия Ki-67 на 7 день представлена светло-серой гистограммой, экспрессия Ki-67 при окрашивании на 14 день представлена темно-серой гистограммой). Кроме того, резидентные T-клетки кожи Vδ1, которые в большинстве своем являются отрицательными по рецептору IL-2 альфа (CD25) при контакте с дермальной стромой, осуществляют повышающую регуляцию CD25 после выделения из ткани (изотипический контроль: пунктирная гистограмма, окрашивание на 0 день: светло-серая гистограмма, окрашивание на 7 день: темно-серая гистограмма).
Фиг. 3C: Высокие скорости клеточного цикла, о которых свидетельствуют значения средней интенсивности флуоресценции (MFI) Ki-67, наблюдаются только в резидентных T-клетках кожи, представленных Т-клетками Vδ1+, и не наблюдаются ни в обычных αβ Т-клетках, ни в NK-клетках, где MFI фактически уменьшается в течение 14 дней. Фиг. 3D: После выращивания в течение 3 недель лимфоциты кожи, отделенные от стромальных клеток, сильно обогащены популяцией γδ T-клеток. Эта популяция γδ T-клеток состоит в основном из Vδ1-положительных клеток (77,49%±17,04) и pan γδ TCR-положительных T-клеток DN (21,46%±16,92). Небольшая популяция Vδ2 T-клеток, присутствующая изначально в свежесобранных лимфоцитах кожи, полученных по методу Кларка, уменьшается и почти исчезает (0,6%±1,204) после выращивания тканевых γδ T-клеток в течение 3 недель.
На фиг.4А и 4В показано, что резидентные γδ Т-клетки кожи отвечают на потерю ткани и удерживаются под контролем клеток дермальной стромы, особенно фибробластов, посредством зависимого от контакта механизма. Фиг. 4А: Смешанные лимфоциты кожи собирают после органотипического культивирования по методу Кларка в течение 3 недель. Затем смешанные лимфоциты высевают на поверхность достигшего слияния слоя аутологичных фибробластов кожи и в трансвел, чтобы контролировать наличие растворимых ингибиторов, продуцируемых фибробластами. Через 14 дней измеряют интенсивность размножения путем определения абсолютного числа γδ T-клеток и обычных αβ T-клеток. После отделения от ткани и в присутствии фибробластов резидентные γδ T-клетки кожи дают сильный пролиферативный ответ, но только в том случае, если они не находятся в непосредственном контакте с аутологичными фибробластами. Обычные αβ Т-клетки не дают такого ответа ни в каком тестируемом состоянии. Фиг. 4B: Смешанные лимфоциты, полученные из органотипической культуры, высевают на монослой аутологичных фибробластов (светло-серые гистограммы) или в пустые лунки (темно-серые гистограммы), добавляют IL-2 и выращивают в течение 7 дней. Резидентные Т-клетки кожи Vδ1+(левые панели), а также pan γδ TCR+ Т-клетки DN (правые панели) остаются спокойными в непосредственном присутствии фибробластов, но сильно активируются при отделении от дермальной органотипической культуры и в отсутствии фибробластов, на что указывает повышенная экспрессия (MFI) CD25, TH-ассоциированного фактора транскрипции T-bet и маркера клеточного цикла Ki-67 (пунктирные, незакрашенные гистограммы обозначают соответствующий изотипический контроль).
На фиг. 5А и 5В показано, что при размножении γδ Т-клеток кожи проявляются признаки дерепрессии и увеличения сильного цитотоксического потенциала. Фиг. 5A: Резидентные γδ Т-клетки кожи оставляют размножаться в течение 14 дней после отделения от органотипической клеточной культуры. Затем γδ Т-клетки подвергают отрицательному сортингу методом проточной цитометрии, исключая все обычные Т-клетки, окрашенные моноклональным антителом pan αβ TCR. Затем 150000 отсортированных γδ Т-клеток высевают в плоский 96-луночный культуральный планшет с двойными повторами и оставляют культивироваться в течение 24 часов без добавления цитокинов, или без добавления какого-либо активирующего лиганда. Супернатанты собирают и анализируют на наличие цитокинов с использованием цитокиновой матрицы Affymetrix LUMINEX®. Фиг. 5B: Полученные путем отрицательного сортинга γδ Т-клетки также высевают на линии раковых клеток, высеянных за 1 день до этого в концентрации 10000 клеток на лунку. В качестве контроля используют полученные путем отрицательного сортинга обычные αβ Т-клетки кожи. Т-клетки высевают в следующих эффекторных условиях: целевые соотношения указаны в присутствии и отсутствии блокирующего антитела NKG2D в присутствии 100 ед/мл IL-2. Резидентные γδ Т-клетки кожи обладают превосходной способностью к уничтожению, что демонстрируют по высвобождению злокачественными клеточными линиями, измеряемому методом ELISA, расщепленного каспазой эпителиального специфического цитокератина 18 (CK18), по сравнению с обычными αβ Т-клетками. Цитотоксичность по меньшей мере частично опосредуется рецептором NKG2D, что демонстрируется уменьшением его уровня в культурах, содержащих антитело, блокирующее рецептор NKG2D.
На фиг. 6A-6D показан анализ тканерезидентных γδ T-клеток в кишечнике человека. Фиг. 6A: Резидентные лимфоциты кишечника выделяют с помощью адаптированного метода Кларка. Смешанные лимфоциты кишечника содержат большую популяцию тканерезидентных γδ Т-клеток, которые в основном состоят из клеток Vδ1, но также содержат клетки Vδ2 и двойные отрицательные γδ Т-клетки. Фиг. 6B: γδ Т-клетки, выделенные из органотипической культуры клеток кишечника, дают ответы, подобные ответам γδ Т-клеток кожи, поскольку они после отделения от стромы кишечника активируют Ki-67 с течением времени. Фиг. 6C: γδ Т-клетки, полученные из кишечника, отвечают на стимулы, подобные врожденным, такие как рекомбинантный MICA, путем продукции IFN-γ и дегрануляции, что измеряют по регуляции CD107a. Фиг. 6D: γδ Т-клетки, выделенные из органотипической культуры клеток кишечника, дают ответы, подобные ответам γδ Т-клеток кожи, и растут со временем в культуре клеток, что можно видеть по общему обогащению культур лимфоцитов, лишенных контакта со стромой кишечника.
На фиг. 7А и 7В показан тканевой фенотип полученных после культивирования γδ Т-клеток кожи. Фиг. 7A: γδ Т-клетки кожи характеризуются положительным окрашиванием на кожный лимфоцитарный антиген (CLA), кожные хоминговые рецепторы хемокинов CCR4 и CCR8. Фиг. 7B: Уровни экспрессии в культивированных γδ Т-клетках, полученных из кожи или крови, различаются.
На фиг. 8 показано, что дерепрессия γδ Т-клеток кожи, без какой-либо стимуляции TCR, приводит к спонтанной продукции цитокина TH-1, и, что интересно, в отличие от свежих TCR-активированных γδ Т-клеток, к продукции атопического цитокина IL-13. Как и свеже полученные γδ Т-клетки, дерепрессированные и культивированные γδ Т-клетки продуцируют незначительное количество TH-2-ассоциированных цитокинов, таких как IL-4 и IL-5. γδ Т-клетки кожи оставляют расти в течение 14 дней и затем подвергают отрицательному сортингу путем исключения обычных αβ Т-клеток. 150000 смешанных γδ Т-клеток культивируют при плотности 1 млн клеток/мл в 96-луночном плоскодонном планшете с двойными повторами для 4 доноров без какой-либо стимуляции или добавления цитокинов. Супернатанты собирают через 24 часа и анализируют с использованием матрицы цитокинов на основе LUMINEX®, Affymetrix.
На фиг. 9A-9C показано, что полученные после культивирования и отрицательного сортинга γδ Т-клетки кожи обладают сильной цитотоксичностью в отношении разных линий опухолевых клеток человека, с которыми их совместно культивируют (фиг.9A: HCT1954, фиг.9B: HCT1 16, фиг.9C: MD231), где цитотоксичность измеряют методом ELISA по высвобождению клетками-мишенями расщепленного каспазой цитокератина 18.
На фиг. 10А и 10В показано, что в свежих, некультивированных Т-клетках кожи Vδ1 маркеры присутствуют до активации Т-клеток. Фиг. 10A: Т-клетки кожи Vδ1 экспрессируют CD69, ICOS и TIM3 на высоких уровнях и CD28 на низком уровне. Кроме того, они экспрессируют на высоком уровне маркер активации NKG2D. Данный фенотип поддерживается Т-клетками кожи Vδ1 в процессе культивирования in vitro. И наоборот, T-клетки Vδ1, полученные из человеческой крови, не имеют этих признаков активации, не экспрессируют CD69 или TIM3 и экспрессируют только незначительные уровни ICOS. Экспрессия NKG2D в Т-клетках крови Vδ1 значительно ниже, чем в Т-клетках кожи Vδ1, однако Т-клетки крови Vδ1 экспрессируют костимулирующую молекулу CD28. Фиг. 10B: Только Т-клетки кожи Vδ1 взаимодействуют с лигандами NKG2D, такими как рекомбинантный MICA, в отсутствие каких-либо других стимулов, таких как лиганд Т-клеточного рецептора. Т-клетки крови Vδ1 или Vδ2 не проявляют такой реакции на естественные стимулы. Клетки высевают в 96-луночные планшеты, содержащие рекомбинантный MICA или антитела против CD3, или то и другое, как указано. Клетки выращивают в течение 6 часов в присутствии 100 ед/мл IL-2 и BFA в течение последних 4 часов с последующим окрашиванием поверхностного антигена, пермеабилизацией и внутриклеточным окрашиванием IFN-γ.
На фиг. 11 показано, что Т-клетки кожи Vδ1 экспрессируют минорные уровни CD16, но экспрессируют высокоаффинный рецептор IgG CD64. Следовательно, помимо непосредственной цитотоксической активности тканевые T-клетки Vδ1 также можно использовать для повышения эффективности терапии моноклональными антителами, такой как терапия, направленная на CD20 или Her2, поскольку они определяются антителом со стороны злокачественных новообразований и метастазов, распознают опсонизированные опухолевые клетки и уничтожают их посредством антителозависимой клеточно-опосредованной цитотоксичности (ADCC). Результаты показаны для одного типичного донора (из четырех).
На фиг. 12 показано культивирование Т-клеток Vδ1 в присутствии IL-2 (левая панель), IL-15 (центральная панель) и IL-2+IL-5 (правая панель). Свежевыделенные лимфоциты кожи культивируют в течение 7 дней в 96-луночных плоскодонных планшетах в среде RPMI, содержащей 10% FCS и 1% Pen/Strep, в которую добавляют соответственно IL-2, IL-15 или IL-2+IL-15. И IL-2, и IL-15, а также сочетание обоих цитокинов индуцирует пролиферацию T-клеток Vδ1, что показывает сдвиг окраски KI-67 по сравнению с изотипическим (истинно отрицательным) окрашиванием в отсутствии каких-либо стромальных клеток. Ki-67 окрашивает только клетки, которые вышли из GO клеточного цикла и обычно ассоциируется с пролиферацией.
На фиг. 13 показаны результаты проточной цитометрии, которые свидетельствуют об экспрессии CD9, CCR3 и CD39 на поверхности культивированных γδ Т-клеток Vδ1 на 21-й день. В культивированных T-клетках кожи Vδ1 поддерживаются высокие уровни клеточных поверхностных маркеров, CCR3, CD39 и CD9, как указано (темная гистограмма) по сравнению с эквивалентным изотипическим окрашиванием (истинная отрицательная, незакрашенная гистограмма).
На фиг.14 показана экспрессия мРНК CCR3 и CD9 в T-клетках кожи Vδ1 (темные столбики) и T-клетках крови Vδ1 (светлые столбики). T-клетки кожи Vδ1 выращивают по описанному здесь способу, а Т-клетки крови Vδ1 выращивают с использованием связанных с планшетом антител против рецептора Т-клеток Vδ (20 мкг/мл). После выращивания Т-клетки Vδ1 получают методом сортировки клеток с активацией флуоресценции (FACS) от трех доноров из обеих групп и выделяют РНК (кровь=серый, кожа=черный). Целую мРНК секвенируют, уровни экспрессии указанных мРНК нормализуют и трансформируют в log2. Все уровни экспрессии показаны в непосредственном сравнении и в отношении к GAPDH, распространенному гену домашнего хозяйства, экспрессируемому на высоких уровнях в большинстве человеческих клеток.
На фиг. 15 показана экспрессия мРНК IL-13 в T-клетках кожи Vδ1 (темные столбики) и тромбоцитах Vδ1, полученных из крови (светлые столбики). T-клетки кожи Vδ1 выращивают по описанному здесь способу, а Т-клетки крови Vδ1 выращивают с использованием высоких доз связанных с планшетом антител против рецептора Т-клеток Vδ (20 мкг/мл). После выращивания Т-клетки Vδ1 получают методом FACS от трех доноров из обеих групп и выделяют РНК (кровь=серый, кожа=черный). Целую мРНК секвенируют, уровни экспрессии мРНК IL-13 нормализуют и трансформируют в log2. Все уровни экспрессии показаны в непосредственном сравнении и в отношении к GAPDH.
На фиг. 16А и 16В показана продукция цитокинов в T-клетках кожи Vδ1 после стимуляции TCR PMA/иономицином (фиг.16А) или антителом против CD3 (фиг.16В). После выделения и выращивания T-клетки кожи Vδ1 очищают методом сортировки клеток с активацией флуоресценции (FACS). 150000 Т-клеток Vδ1 высевают в 96-луночный плоскодонный планшет с двойными повторами для трех доноров и стимулируют либо связанным с планшетом антителом против CD3 (5 мкг/мл), либо PMA/иономицином в течение 24 часов. Супернатанты анализируют на абсолютные количества указанных цитокинов с использованием платформы LUMINEX®.
ПОДРОБНОЕ ОПИСАНИЕ
Гамма дельта Т-клетки (γδ Т-клетки) представляют собой небольшую субпопуляцию Т-клеток, которые экспрессируют на поверхности особый, определяющий Т-клетки рецептор (TCR). Этот TCR состоит из одной гамма (γ) и одной дельта (δ) цепи.
Существует два основных подтипа человеческих γδ Т-клеток: один из них преобладает в периферической крови, а второй преобладает в негемопоэтических тканях.
Локализация второго подтипа человеческих γδ T-клеток в негемопоэтической ткани затрудняет получение образцов, кроме того, отсутствуют стандартные способы культивирования таких клеток. Для удовлетворения данной потребности настоящее изобретение предлагает способ выращивания резидентных γδ Т-клеток негемопоэтической ткани, альтернативно называемых здесь нативные γδ Т-клетки негемопоэтической ткани. Указанные γδ Т-клетки обычно находятся в негемопоэтических тканях. Резидентные γδ Т-клетки негемопоэтической ткани, подходящие для применения в описанном здесь способе, могут происходить или быть получены из негемопоэтической ткани. Негемопоэтические ткани могут содержать негемопоэтические клетки и γδ Т-клетки.
С помощью описанного здесь способа можно выращивать γδ Т-клетки из любой негемопоэтической ткани человека или отличного от человека животного, которую можно удалить из организма пациента, такой как ткань кожи, желудочно-кишечного тракта (например, ободочной кишки), молочной железы, легкого, простаты, печени, селезенки и поджелудочной железы. γδ Т-клетки также могут постоянно находиться в человеческих раковых тканях, например, в ткани опухоли молочной железы или предстательной железы. В некоторых вариантах осуществления γδ Т-клетки могут быть получены из человеческих раковых тканей. В других вариантах осуществления γδ Т-клетки могут быть получены из негемопоэтической ткани, отличной от раковой ткани человека.
γδ Т-клетки, которые преобладают в крови, представляют собой, в основном, Т-клетки Vδ2, тогда как γδ Т-клетки, которые преобладают в негемопоэтических тканях, представляют собой, в основном, Т-клетки Vδ1, так что Т-клетки Vδ1 составляют примерно 70-80% от резидентной популяции γδ Т-клеток негемопоэтической ткани. Однако некоторые Т-клетки Vδ2 также присутствуют в негемопоэтических тканях, например, в кишечнике, где они могут составлять примерно 10-20% от γδ Т-клеток (фиг.6). Некоторые γδ Т-клетки, постоянно присутствующие в негемопоэтических тканях, не экспрессируют ни Vδ1, ни Vδ2 TCR, и авторы называют их двойные отрицательные (DN) γδ Т-клетки. Указанные γδ Т-клетки DN, скорее всего, преимущественно экспрессируют Vδ3, однако небольшое количество Т-клеток экспрессируют Vδ5.
Следовательно, γδ Т-клетки, которые обычно находятся в негемопоэтических тканях, и которые выращивают по способу настоящего изобретения, предпочтительно представляют собой Т-клетки, отличные от Vδ2, например, Т-клетки Vδ1, содержащие небольшое число γδ T-клеток DN.
В используемом здесь значении "двойные отрицательные" γδ Т-клетки (γδ T-клетки DN) представляют собой γδ Т-клетки, которые экспрессируют рецепторы γδ (то есть положительные окрашиваемые на pan-TCR), но являются отрицательными по рецепторам Vδ1 и Vδ2. γδ Т-клетки DN включают в себя клетки, которые экспрессируют рецепторы Vδ, отличные от Vδ1 и Vδ2 (например, Vδ3, Vδ4, Vδ5 или Vδ8). Клетку можно охарактеризовать как положительную по маркеру (например, как Vδ1+), если она экспрессирует маркер на уровне, превышающем уровень экспрессии в клетке, используемой в качестве отрицательного контроля, как определено стандартными методами пропускания FACS.
Описанный здесь способ может включать в себя культивирование in vitro лимфоцитов, полученных из негемопоэтической ткани человека или отличного от человека животного.
Лимфоциты могут быть получены из любой подходящей негемопоэтической ткани человека или отличного от человека животного. Негемопоэтическая ткань представляет собой ткань, отличную от ткани крови, костного мозга или тимуса. В некоторых вариантах осуществления γδ Т-клетки не получены из конкретных типов образцов биологических жидкостей, таких как кровь или синовиальная жидкость. Примерами таких подходящих негемопоэтических тканей человека или отличного от человека животного являются ткани кожи или ее части (например, дермы, эпидермиса), желудочно-кишечного тракта (например, желудочно-кишечного эпителия, толстой кишки, тонкой кишки, желудка, аппендикса, слепой кишки или прямой кишки), молочной железы, легкого (предпочтительно, если ткань не получена путем бронхоальвеолярного лаважа), простаты, печени, селезенки и поджелудочной железы. γδ Т-клетки также могут постоянно присутствовать в человеческих раковых тканях, например, в тканях рака молочной железы и предстательной железы. В некоторых вариантах осуществления γδ Т-клетки не получают из раковой ткани человека. Образцы негемопоэтических тканей можно получить с помощью стандартных методов, таких как эксплантация (например, биопсийного образца).
Лимфоциты можно получить любым подходящим способом, который позволяет выделить лимфоциты из негемопоэтической ткани человека или отличного от человека животного. Один из таких способов предложен в Clark et al. (29), где описана методика трехмерного культивирования эксплантата кожи для выделения лимфоцитов из кожи человека. Эксплантат можно прикрепить к синтетическому каркасу для облегчения выхода лимфоцитов из эксплантатов на каркас. Синтетический каркас представляет собой неприродную трехмерную структуру, подходящую для поддержания роста клеток. Синтетические каркасы можно изготовить из таких материалов, как полимеры (например, натуральные или синтетические полимеры, такие как поливинилпирролидоны, полиметилметакрилат, метилцеллюлоза, полистирол, полипропилен, полиуретан), керамические материалы (например, трикальция фосфат, алюминат кальция, гидроксиапатит кальция), или металлы (тантал, титан, платина и металлы из той же группы элементов, что и платина, ниобий, гафний, вольфрам, а также сочетания их сплавов). Биологические факторы (такие как коллагены (например, коллаген I или коллаген II), фибронектины, ламинины, интегрины, ангиогенные факторы, противовоспалительные факторы, гликозаминогликаны, витрогены, антитела и их фрагменты, цитокины (например, интерлейкин-2 (IL-2) или интерлейкин-15 (IL-15) и их сочетания) можно нанести на поверхность каркаса или инкапсулировать в каркасном материале для улучшения адгезии, миграции, выживания или пролиферации клеток в соответствии со способами, известными в данной области. Данный способ и другие способы можно использовать для выделения лимфоцитов из ряда других негемопоэтических тканей, таких как ткани кишечника, предстательной железы и молочной железы. Другие примеры подходящих способов включают в себя ферментативное расщепление ткани и метод "выползания", описанный Carrasco et al. (30), в котором ткань измельчают и добавляют IL-2, чтобы лимфоциты "выползали".
Как указано выше, можно использовать любые подходящие негемопоэтические ткани, такие как ткани кожи, желудочно-кишечного тракта (например, ободочной кишки), молочной железы, легкого, простаты, печени, селезенки и поджелудочной железы.
Резидентные γδ Т-клетки негемопоэтической ткани предпочтительно получают из ткани человека. Однако они могут быть получены из негемопоэтической ткани любого подходящего отличного от человека животного, такого как мышь, крыса, собака, лошадь и свинья.
Критической стадией является целенаправленное разделение, например, после культивирования в течение нескольких дней или недель, резидентных Т-клеток негемопоэтической ткани (например, в смешанной популяции лимфоцитов, которая может содержать, например, αβ, γδ2 и отличные от γδ2 Т-клетки) и негемопоэтических клеток (таких как стромальные клетки, особенно фибробласты) ткани, из которой были получены Т-клетки, с последующим культивированием лимфоцитов в присутствии цитокинов, как описано ниже. Это обеспечивает селективный и значительный рост в течение следующих дней и недель тканевых Т-клеток γδ 1 и γδ DN.
В настоящем описании термины "разделение", "разделенные" или "разделять" относятся к акту нарушения или прекращения физического контакта между разными популяциями клеток. Разделение можно осуществить, например, путем интенсивного пипетирования смешанной популяции клеток с целью разрушения межмембранных ассоциаций или путем индуцирования "выползания" популяции клеток, например, из тканевой матрицы, при культивировании, например, в присутствии хемокинов или цитокинов, как описано Carrasco et al. (30). Разделение можно поддерживать в процессе культивирования с использованием системы Трансвел или аналогичных методов культивирования, которые позволяют предотвратить физический контакт между разными популяциями клеток.
Используемый здесь термин "практически чистый" относится к чистоте более 90% по количеству, массе или объему. Термин "практически не содержит" означает наличие менее 5% по количеству, массе или объему указанного компонента. Лимфоциты, полученные из негемопоэтической ткани человека или отличного от человека животного, можно культивировать в течение по меньшей мере 3 дней, по меньшей мере 4 дней, по меньшей мере 5 дней, по меньшей мере 6 дней, по меньшей мере 7 дней, по меньшей мере 8 дней, по меньшей мере 9 дней, по меньшей мере 10 дней, по меньшей мере 2 недель, по меньшей мере 3 недель, или по меньшей мере 4 недель.
Способ включает в себя культивирование лимфоцитов, полученных из негемопоэтической ткани человека или отличного от человека животного, в присутствии IL-2. Концентрация IL-2 предпочтительно составляет по меньшей мере 10 международных единиц/мл (МЕ/мл или ед/мл), по меньшей мере, 20 ед/мл, по меньшей мере 30 ед/мл, по меньшей мере 40 ед/мл, по меньшей мере 50 ед/мл, по меньшей мере, 60 ед/мл, по меньшей мере, 70 ед/мл, по меньшей мере, 80 ед/мл, по меньшей мере 90 ед/мл или по меньшей мере 100 ед/мл.
Использование IL-2 для стимулирования экспрессии γδ Т-клеток кожи не является очевидным, поскольку клетки экспрессируют очень низкие уровни высокоаффинного рецептора IL-2, известного как CD25 (фиг. 1D). Однако данный рецептор подвергается повышающей регуляции в отдельных субпопуляциях γδ T-клеток в результате отделения от клеток других типов, таких как стромальные или эпителиальные клетки (например, фибробласты) (см. фиг.3B и 4B), что делает клетки высокочувствительными к IL-2.
Используемый здесь термин "IL-2" относится к IL-2 дикого типа (например, нативному или рекомбинантному), или к средству, которое действует как агонист на одну или нескольких субъединиц рецептора IL-2 (IL-2R) (такому как мутеины IL-2, аналоги IL-2 длительного действия, их субъединицы, комплексы с рецепторами). Такие средства могут поддерживать пролиферацию IL-2-зависимой клеточной линии, CTLL-2 (33, Американская коллекция типовых культур (ATCC®), TIB 214). Зрелый IL-2 человека встречается в виде последовательности из 133 аминокислот (меньше сигнального пептида, который дополнительно содержит 20 N-концевых аминокислот), как описано в Fujita, et al. (34). Мутеин IL-2 представляет собой полипептид, полученный в результате специфических замен в последовательности белка интерлейкина-2 и сохраняющий способность связывать IL-2Rβ, как описано в US 2014/0046026. Мутеины IL-2 могут содержать аминокислотные вставки, делеции, замены и модификации в одном или нескольких положениях нативной полипептидной цепи IL-2, или по другим остаткам нативной полипептидной цепи IL-2. В соответствии с настоящим описанием любые такие вставки, делеции, замены и модификации позволяют получить мутеин IL-2, который сохраняет способность к связыванию IL-2Rβ. Типичные мутеины можно получить путем замены 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот.
Нуклеиновую кислоту, кодирующую человеческий IL-2, можно получить с помощью традиционных способов, таких как полимеразная цепная реакция (ПЦР). Аминокислотную последовательность человеческого IL-2 (Gene ID 3558) можно найти в Genbank под номером доступа NP_000577.2 GI: 28178861. Аминокислотную последовательность мышиного (Mus musculus) IL-2 (Gene ID 16183) можно найти в Genbank под номером доступа NP_032392.1 GI: 7110653.
Лимфоциты предпочтительно культивируют в присутствии IL-2 и IL-15, так как добавление IL-15 в сочетании с IL-2 приводит к усилению роста пролиферирующих негемопоэтических тканерезидентных γδ Т-клеток по сравнению с одним IL-2. Концентрация IL-15 предпочтительно составляет по меньшей мере 10 нг/мл.
IL-15, как и IL-2, является известным фактором роста Т-клеток, который может поддерживать пролиферацию IL-2-зависимой клеточной линии CTLL-2. IL-15 впервые описан Grabstein et al. (35) как зрелый белок, содержащий 114 аминокислот. Используемый здесь термин "IL-15" относится к нативному или рекомбинантному IL-15, а также к его мутеинам, аналогам, субъединицам или комплексам (например, рецепторным комплексам, таким как пептиды sushi, описанные в WO2007/046006), способным стимулировать пролиферацию клеток CTLL-2. Анализы пролиферации CTLL-2 демонстрируют, что супернатанты клеток, трансфицированных экспрессированным рекомбинантным методом предшественником и гибридами зрелых форм IL-15, полученными с сохранением рамки считывания, могут индуцировать пролиферацию клеток CTLL-2.
Используемый здесь термин IL-15 также относится к IL-15, полученному из разных видов млекопитающих, включающих в себя, например, человека, обезьян, быков, свиней, лошадей и мышей. "Мутеин" или "вариант" IL-15, в соответствии с данным документом, представляет собой полипептид, практически гомологичный последовательности нативного IL-15 млекопитающих, но имеющий аминокислотную последовательность, отличающуюся от последовательности нативного полипептида IL-15 млекопитающих вследствие аминокислотных удалений, вставок или замен. Варианты могут содержать консервативно замещенные последовательности, то есть последовательности, в которых конкретный аминокислотный остаток заменен остатком, имеющим сходные физико-химические характеристики. Примеры консервативных замен включают замены одного алифатического остатка на другой, например, замены Ile, Val, Leu или Ala друг на друга, или замены одного полярного остатка на другой, например, замены между Lys и Arg; Glu и Asp; или Gln и Asn. Хорошо известны другие консервативные замены, например, замены целых участков, обладающих сходными характеристиками гидрофобности. Природные варианты IL-15 также входят в объем изобретения. Примерами таких вариантов являются белки, образовавшиеся в результате событий альтернативного сплайсинга мРНК или протеолитического расщепления белка IL-15, которые сохраняют связывающие свойства IL-15. Альтернативный сплайсинг мРНК может приводить к образованию укороченного, но биологически активного белка IL-15. Вариации, обусловленные протеолизом, включают в себя, например, различия в N- или С-концах при экспрессии в клетках-хозяевах разных типов вследствие протеолитического удаления одной или нескольких концевых аминокислот из белка IL-15 (обычно от 1 до 10 аминокислот).
Человеческий IL-15 можно получить с помощью способов, описанных Grabstein et al (35), или с помощью традиционных способов, таких как полимеразная цепная реакция (ПЦР). кДНК человеческого IL-15 помещена в ATCC® 19 февраля 1993 года с присвоением номера доступа 69245.
Аминокислотная последовательность человеческого IL-15 (Gene ID 3600) находится в Genbank под номером доступа NP000576.1 GI: 10835153 (изоформа 1) и NP_751915.1 GI: 26787986 (изоформа 2). Аминокислотная последовательность мышиного IL-15 (Mus musculus) (Gene ID 16168) находится в Genbank под номером доступа NP_001241676.1 GI: 363000984.
Лимфоциты можно культивировать в отсутствии IL-6, IL-23 и IL-1B, или в присутствии низких концентраций указанных цитокинов (например, менее 20 нг/мл), поскольку добавление этого сочетания цитокинов, по-видимому, снижает пролиферацию негемопоэтических тканерезидентных γδ Т-клеток. Это является неожиданным, так как считается, что указанные цитокины способствуют пролиферации.
Лимфоциты, полученные из негемопоэтической ткани, можно культивировать в отсутствии средств, активирующих Т-клеточный сигнальный путь (например, агонистов пути Т-клеточного рецептора (TCR)). Например, лимфоциты, полученные из негемопоэтической ткани, можно культивировать в среде, которая не поддерживает или не индуцирует пролиферацию или активацию αβ Т-клеток и резидентных γδ Т-клеток крови. Подходящая среда может не содержать или практически не содержать агонисты TCR или другие средства, активирующие Т-клеточный сигнальный путь. И наоборот, для культивирования γδ T-клеток из гемопоэтических тканей требуется присутствие средства, активирующего Т-клеточный сигнальный путь, такого как золедронат (41, 42) или антитело против CD3, такое как OKT3 (43).
Средства, активирующие Т-клеточный сигнальный путь, относятся к соединениям, которые индуцируют пролиферацию или активацию Т-клеток, таких как αβ Т-клетки и/или резидентные γδ Т-клетки крови, через сигнальный путь или костимуляцию TCR. Модуляторы Т-клеточного сигнального пути действуют путем последовательной активации Src-родственных тирозин-специфичных протеинкиназ (PTK), LcK и Fyn и связанной с зета-цепью (TCR) протеинкиназы размером 70 кДа (ZAP70). Указанные PTK фосфорилируют полипептиды, включая линкерный активатор Т-клеток (LAT), что приводит к нижестоящей стимуляции через киназу, регулируемую внеклеточными сигналами (ERK), c-Jun N-концевой киназы (JNK) и ядерного фактора активированных Т-клеток (NFAT). Совместная стимуляция, например, через CD28 и CD45, может усиливать фосфорилирование и сигнальные пути TCR. Таким образом, любое средство, направленное на часть TCR или костимуляторный путь, может активировать Т-клеточный сигнальный путь. Средства, активирующие Т-клеточный сигнальный путь, могут быть растворимыми или связанными с мембраной, и могут, например, презентироваться на клетках, таких как искусственные антигенпрезентирующие клетки (aAPC). aAPC, подходящие для активации Т-клеточного сигнального пути, известны в данной области (44).
В некоторых вариантах осуществления лимфоциты можно культивировать в отсутствии экзогенно добавленных агонистов пути Т-клеточного рецептора, таких как активаторы CD3 и/или CD28 (например, моноклональные антитела против CD3 и/или против CD28); фитогемагглютинин (PHA); конканавалин А, синтетические фосфоантигены, такие как BrHPP (бромгидринпирофосфат), 2M3B1PP (2-метил-3-бутенил-1-пирофосфат), HMBPP ((E)-4-гидрокси-3-метилбут-2-енила пирофосфат) или IPP (изопентенилпирофосфат); N-бисфосфонаты, такие как золедронат; рекомбинантный CD70; моноклональные антитела против CD2; моноклональные антитела против CD27; моноклональные антитела против pan-TCRγδ; моноклональные антитела против CD277; или искусственные антигенпрезентирующие клетки (aAPC). Средства, активирующие Т-клеточный сигнальный путь, также включают в себя молекулы, связанные с клеточной поверхностью, такие как комплексы MHC или HLA, связанные с антигенпрезентирующими клетками (APC) или искусственными APC. Подходящие способы активации Т-клеток путем экзогенного добавления агонистов пути TCR хорошо известны в данной области и приведены на фиг.1 в Deniger et al. (44).
Например, лимфоциты можно культивировать в среде, которая не содержит или практически не содержит экзогенно добавленных агонистов пути Т-клеточного рецептора. При использовании способа настоящего изобретения добавление таких средств, активирующих сигнальный путь Т-клеточного рецептора, не требуется для роста негемопоэтических резидентных γδ Т-клеток. И наоборот, для роста гемопоэтических тканевых γδ T-клеток требуется присутствие как IL-2, так и средства, активирующего сигнальный путь Т-клеточного рецептора, такого как золедронат (41, 42).
В некоторых вариантах осуществления лимфоциты можно культивировать в кондиционированных средах из культур стромальных клеток, чтобы обеспечить добавки, необходимые для роста γδ T-клеток.
В некоторых вариантах осуществления γδ Т-клетки можно культивировать в среде для выращивания γδ, содержащей IL-2 и/или IL-15. Подходящая среда для выращивания γδ лишена средств, активирующих Т-клетки, например, средств, активирующих αβ Т-клетки или γδ Т-клетки крови, и, например, может не содержать или практически не содержать агонисты TCR или костимулирующие средства. В некоторых вариантах осуществления среда для выращивания γδ может содержать один или несколько дополнительных факторов роста, таких как цитокины, в дополнение к IL-2 и/или IL-15. Подходящие факторы роста не обладают способностью активировать Т-клетки. В других вариантах осуществления среда для выращивания γδ может не содержать факторы роста, отличных от IL-2 и/или IL-15; например, среда для выращивания γδ может состоять из основной среды, дополненной IL-2 и/или IL-15.
Существуют многочисленные основные культуральные среды, подходящие для пролиферации γδ T-клеток, в частности полные среды, такие как AIM-V, среда Искова и RPMI-1640 (Life Technologies). Среда может быть дополнена другими компонентами среды, такими как сыворотка, сывороточные белки и средства селекции, такие как антибиотики. Например, в некоторых вариантах осуществления можно использовать среду RPMI-1640, содержащую 2 мМ глутамин, 10% FBS, 10 мМ HEPES, pH 7,2, 1% пенициллина-стрептомицина, пируват натрия (1 мМ, Life Technologies), незаменимые аминокислоты (например, 100 мкМ Gly, Ala, Asn, Asp, Glu, Pro и Ser; 1X MEM незаменимые аминокислоты Life Technologies) и 10 мкл/л β-меркаптоэтанола. Основная среда может быть дополнена IL-2 и/или IL-15 в стандартных концентрациях, которые может легко определить квалифицированный специалист путем рутинного эксперимента.
Обычно клетки культивируют при 37°С во влажной атмосфере, содержащей 5% СО2, в подходящей культуральной среде.
Т-клетки γδ можно культивировать с помощью описанного здесь способа в любой подходящей системе, включая ферментеры с механическим перемешиванием, эрлифтные ферментеры, роллерные флаконы, культуральные мешки или чашки и другие биореакторы, в частности половолоконные биореакторы. Применение таких систем хорошо известно в данной области.
Способы и методы культивирования лимфоцитов хорошо известны в данной области (36-39).
В процессе культивирования лимфоциты не находятся в непосредственном контакте со стромальными или эпителиальными клетками. Это связано с тем, что непосредственный контакт лимфоцитов со стромальными или эпителиальными клетками, по-видимому, препятствует росту тканерезидентных γδ T-клеток.
Стромальные клетки представляют собой негемопоэтические клетки соединительной ткани любого органа и поддерживают функцию паренхимных клеток этого органа. Примеры стромальных клеток включают в себя фибробласты, перициты, мезенхимальные клетки, кератиноциты, эндотелиальные клетки и клетки негематологических опухолей. Предпочтительно лимфоциты не находятся в непосредственном контакте с фибробластами во время культивирования.
Эпителиальные клетки представляют собой негемопоэтические клетки, которые выстилают полости и поверхности кровеносных сосудов и органов по всему организму. Обычно они плоские, столбчатые или кубовидные по форме и могут находиться в виде одного слоя клеток или в виде двух или более слоев клеток.
Фибробласты и/или другие стромальные или эпителиальные клетки предпочтительно присутствуют во время культивирования лимфоцитов, поскольку факторы, секретируемые этими клетками, могут способствовать росту негемопоэтических тканерезидентных γδ Т-клеток, но не находятся в непосредственном контакте с лимфоцитами, поскольку непосредственный контакт препятствует росту негемопоэтических тканерезидентных γδ Т-клеток. Например, лимфоциты можно культивировать в трансвелах, которые обеспечивают физическое разделение лимфоцитов и фибробластов. Примеры фибробластных клеточных линий, подходящих для применения, включают в себя фибробласты крайней плоти человека (например, BJ (ATCC® CRL-2522™)), нормальные фибробласты кожи (например, CCD-1059Sk (ATCC® CRL-2072™)) и фибробласты легких (например, HEL 299 (ATCC® CRL-137)).
С помощью метода Кларка негемопоэтические тканерезидентные лимфоциты можно собрать и отделить от стромальных клеток, таких как фибробласты кожи, например, путем жесткого пипетирования. Собранные лимфоциты можно затем промыть через нейлоновую сетку 40 мкл, чтобы сохранить агрегированные фибробласты, которые могут стать свободными во время процесса. Лимфоциты также можно выделить путем флуоресцентной или магнитной сортировки клеток с использованием, например, антител против CD45. Чтобы свести к минимуму активацию Т-клеток, их также можно отсортировать исключительно по параметрам прямого и бокового рассеяния. Затем лимфоциты можно выращивать отдельно от стромальных клеток (например, фибробластов), или в их присутствии, но не в непосредственном контакте. Например, лимфоциты можно выращивать в корзинке системы трансвел так, чтобы слившийся монослой фибробластов в лунке для культивирования клеток находился ниже, чтобы обеспечить обмен растворимыми факторами роста, продуцируемыми фибробластами, при отсутствии непосредственного контакта. Альтернативно фибробласты можно культивировать в корзинке системы трансвел, так, чтобы лимфоциты, растущие в лунке для культивирования клеток, находились значительно ниже. Можно также использовать среды, кондиционированные негемопоэтическими клетками (например, фибробластами), чтобы дополнить рост лимфоцитов.
Кондиционированная среда содержит растворимые факторы, секретируемые негемопоэтическими клетками (например, стромальными клетками, такими как фибробласты). Кондиционированные среды могут содержать или не содержать клетки, которые секретируют факторы. Например, γδ Т-клетки можно культивировать в присутствии клеток, которые секретируют кондиционирующие факторы в процессе культивирования γδ T-клеток. Альтернативно негемопоэтические клетки можно удалить из среды до культивирования γδ T-клеток, оставив в среде секретируемые ими факторы. Кондиционированные среды также включают в себя среды, дополненные предварительно полученными факторами негемопоэтических клеток (например, в виде концентрата или лиофилизированного порошка).
Хотя стромальные или эпителиальные клетки предпочтительно присутствуют во время культивирования лимфоцитов (но не находятся в непосредственном контакте с лимфоцитами), их можно удалить, так чтобы культивировать лимфоциты в отсутствии стромальных или эпителиальных клеток, например, в отсутствии фибробластов.
В некоторых вариантах осуществления после роста негемопоэтических тканерезидентных γδ Т-клеток в отсутствии средств, активирующих Т-клеточный сигнальный путь, таких как экзогенно добавленные агонисты пути Т-клеточного рецептора, описанные выше, размноженные γδ Т-клетки можно дополнительно культивировать в присутствии одного или нескольких средств, активирующих Т-клеточный сигнальный путь, таких как экзогенные агонисты пути Т-клеточного рецептора, и/или одного или нескольких факторов роста, таких как цитокины. После выращивания по описанному здесь способу и необязательно дополнительного культивирования негемопоэтические тканерезидентные γδ Т-клетки можно выделять или дополнительно очищать, хранить, смешивать с другими реагентами, такими как фармацевтически приемлемые вспомогательные вещества и/или использовать по мере необходимости.
Негемопоэтические тканерезидентные γδ Т-клетки, полученные по способу настоящего изобретения, можно отличить от других γδ Т-клеток, полученных из крови, поскольку они отвечают на лиганд NKG2D (MICA), который в значительной степени ассоциируется со злокачественными новообразованиями, в отсутствии какого-либо лиганда, стимулирующего T-клеточный рецептор, например, по увеличению продукции TNFα, IFNγ и CD107a (фиг.2A-2D, 10A и 10B). Они также дают цитотоксический Т-клеточный ответ, не подвергаясь какой-либо экзогенной фармакологической или лиганд-опосредованной активации Т-клеточного рецептора и, следовательно, являются цитотоксичными в отсутствии стимуляции (фиг.3 и фиг.5). Это означает, что в отличие от других γδ Т-клеток, αβ Т-клеток или NK-клеток, негемопоэтические тканерезидентные γδ Т-клетки, полученные по способу настоящего изобретения, обладают уникальной способностью отвечать и размножаться в отсутствии добавления каких-либо экзогенных средств, активирующих сигнальный путь Т-клеточного рецептора (фиг. 3). Негемопоэтические тканерезидентные γδ Т-клетки, полученные по способу настоящего изобретения, также положительно окрашиваются по CD69 и PD-1, не экспрессируют CD28 и содержат только низкие уровни CD25 (см. фиг.1D). Данное сочетание маркеров не экспрессируется γδ Т-клетками, полученными из крови. Кроме того, они экспрессируют тканевые хоминг-рецепторы, такие как CCR4 и CCR8, на более высоком уровне, чем полученные из крови культивированные Vd2 γδ Т-клетки (фиг. 7B). Негемопоэтические тканерезидентные γδ Т-клетки, полученные по способу настоящего изобретения, можно культивировать в присутствии IL2 и/или IL15 и в отсутствии агонистов TCR или других факторов роста. Например, негемопоэтическая тканерезидентная γδ Т-клетка может расти в среде RPMI 1640, дополненной IL-2.
Таким образом, негемопоэтическая тканерезидентная γδ Т-клетка, полученная по способу настоящего изобретения, может обладать одним или несколькими из следующих свойств:
(i) имеет фенотип CD69высокий, ICOSвысокий, TIM3высокий и CD28низкий/отсутствует
(ii) осуществляет повышающую регуляцию одного или нескольких из CCR3, CD39, CD11b и CD9
(iii) продуцирует IFN-γ в ответ на лиганд NKG2D в отсутствии агонистов TCR,
(iv) продуцирует IL-13 в отсутствии агонистов TCR,
(v) продуцирует один или несколько из IFN-γ, TNF-α и GM-CSF в ответ на активацию TCR,
(vi) не продуцирует, или практически не продуцирует IL-17 в ответ на активацию TCR,
(vii) растет в культуральной среде, содержащей IL-2 в отсутствии дополнительных факторов роста,
(viii) дает цитотоксический Т-клеточный ответ в отсутствии агонистов TCR и/или
(ix) обладает избирательной цитотоксичностью в отношении опухолевых клеток в отличие от нормальных клеток.
Предпочтительно негемопоэтическая тканерезидентная γδ Т-клетка, полученная по способу настоящего изобретения, продуцирует IL-13 в отсутствии агонистов TCR, и/или продуцирует IFN-γ в ответ на лиганд NKG2D в отсутствии агонистов TCR.
Т-клетки γδ, полученные по способу настоящего изобретения, можно использовать в способе скрининга на модулятор контрольной точки негемопоэтических тканерезидентных γδ T-клеток. Идентификацию таких модуляторов контрольных точек можно использовать для разработки противораковых иммунотерапевтических средств, поскольку модулятор контрольной точки является потенциальной мишенью для противораковой терапии.
Чтобы определить, является ли тестируемое соединение модулятором контрольной точки, негемопоэтические тканерезидентные γδ Т-клетки можно культивировать in vitro при непосредственном контакте со стромальными или эпителиальными клетками (например, фибробластами) в присутствии или отсутствии тестируемого соединения. Скорость пролиферации или степень активации негемопоэтических тканерезидентных γδ Т-клеток определяют в присутствии и в отсутствии тестируемого соединения. Если скорость пролиферации или степень активации в присутствии тестируемого соединения выше, чем в отсутствии тестируемого соединения, то тестируемое соединение, скорее всего, является возможным модулятором контрольной точки. Это связано с тем, что тестируемое соединение способно ослаблять контактное ингибирование стромальными или эпителиальными клетками (например, фибробластами).
Тестируемое соединение выбирают на основе его способности модулировать контрольные точки тканерезидентных Т-клеток.
Кажущееся увеличение пролиферации негемопоэтических тканерезидентных γδ Т-клеток, которое также может быть обусловлено ингибированием гибели клеток, можно измерить по увеличению количества Т-клеток или их маркеров и по уменьшению экспрессии маркеров программируемой гибели клеток. Повышенную активацию можно оценить путем измерения высвобождения цитокинов, таких как IFN-γ, которые секретируются активными тканерезидентными γδ Т-клетками.
Альтернативно негемопоэтические тканерезидентные γδ Т-клетки можно культивировать in vitro в непосредственном контакте со стромальными или эпителиальными клетками (например, фибробластами) в условиях изменения экспрессии тестируемого гена в γδ Т-клетках, и/или стромальных или эпителиальных клетках (например, фибробластах), или в клетках обоих типов. Экспрессию тестируемого гена в γδ Т-клетках, и/или в стромальных или эпителиальных клетках (например, фибробластах), можно изменить, например, с использованием средства, специфичного к РНК, такого как малая интерферирующая РНК (миРНК) или короткая шпилечная РНК (кшРНК), или путем редактирования генов, например, с использованием системы CRISPR/Cas. Скорость пролиферации или степень активации негемопоэтических тканерезидентных γδ Т-клеток определяют в присутствии и в отсутствии изменения экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах), и/или в γδ Т-клетках. Если при изменениях экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах) и/или в γδ Т-клетках скорость пролиферации или активации выше, чем при отсутствии изменения тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах) и/или в γδ Т-клетках, то тестируемый ген, скорее всего, является возможным геном контрольной точки.
Альтернативно, если скорость уничтожения стромальных или эпителиальных клеток (например, фибробластов) в присутствии тестируемого соединения выше, чем в отсутствии тестируемого соединения, или при изменении экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах), и/или в γδ Т-клетках выше, чем при отсутствии изменения тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах), и/или в γδ Т-клетках, то тестируемое соединение, скорее всего, является модулятором контрольной точки, или тестируемый ген, скорее всего, является возможным геном контрольной точки. Скорость уничтожения можно измерить, например, путем количественного определения молекул, высвобождаемых умирающими клетками. Более высокая скорость гибели клеток в присутствии тестируемого соединения или при изменении экспрессии тестируемого гена в стромальных или эпителиальных клетках (например, фибробластах) и/или в γδ Т-клетках указывает на то, что контрольная точка, которая подавляла уничтожение клеток, была выключена, и поэтому можно сделать вывод, что тестируемое соединение, скорее всего, является модулятором контрольной точки, или тестируемый ген, скорее всего, является возможным геном контрольной точки.
Примеры линий фибробластов, которые можно использовать в каждом варианте осуществления, включают фибробласты крайней плоти человека (например, BJ (ATCC® CRL-2522™)), нормальные фибробласты кожи (например, CCD-1059Sk (ATCC® CRL-2072™)) и фибробласты легких (например, HEL 299 (ATCC® CRL-137™)).
Чтобы идентифицировать тестируемое соединение как возможный модулятор контрольной точки, или чтобы идентифицировать тестируемый ген как возможный ген контрольной точки, скорость пролиферации и/или активация γδ Т-клеток в присутствии тестируемого соединения или в присутствии изменения тестируемого гена должна быть по меньшей мере в 1,5 раза, по меньшей мере в 2 раза, по меньшей мере в 3 раза, по меньшей мере в 4 раза, или по меньшей мере в 5 раз выше, чем в отсутствии тестируемого соединения, или в отсутствии изменения тестируемого гена. Клеточный цикл можно определить с помощью ряда способов, например, путем определения абсолютного числа клеток в 0 день и 7 день, уровней экспрессии Ki-67 и CD25 (которые являются маркерами клеточного цикла), и путем использования клеточных культуральных красителей, таких как CFSE или CELLTRACE™ фиолетовый. Активацию клеток можно измерить по продукции эффекторных белков, таких как IFN-γ. Кажущееся увеличение пролиферации негемопоэтических тканерезидентных γδ T-клеток, которое также может быть обусловлено ингибированием гибели клеток, можно измерить по увеличению числа Т-клеток или их маркеров, и по уменьшению экспрессии маркеров программируемой гибели клеток.
γδ Т-клетки, полученные по способу настоящего изобретения, можно использовать в качестве лекарственного средства, например, для адоптивной терапии Т-клетками. Такая терапия включает в себя перенос пациенту γδ Т-клеток, полученных по способу настоящего изобретения. Терапия может быть аутологичной, когда γδ Т-клетки переносят обратно тому же пациенту, из организма которого они были получены, или терапия может быть аллогенной, когда γδ Т-клетки, полученные от одного индивидуума, переносят другому пациенту. Способ лечения может включать в себя:
получение образца негемопоэтической ткани от индивидуума-донора,
культивирование γδ Т-клеток из описанного выше образца с получением увеличенной популяции и
введение увеличенной популяции γδ T-клеток индивидууму-реципиенту.
Индивидуум-донор и индивидуум-реципиент могут быть одинаковыми или разными.
γδ Т-клетки можно вводить пациенту или индивидууму, нуждающемуся в лечении, любым подходящим способом. Например, γδ Т-клетки можно вводить пациенту или индивидууму, нуждающемуся в лечении, внутривенно или внутритуморально.
Пациент или индивидуум, подлежащий лечению, предпочтительно представляет собой пациента-человека, страдающего от рака или вирусной инфекции, такой как инфекция CMV или HIV.
Поскольку γδ Т-клетки не связаны с МНС, они не распознают хозяина, которому их переносят как чужеродные, следовательно, они менее склонны вызывать болезнь трансплантат против хозяина. Это означает, что их можно использовать "в готовом виде" и переносить любому реципиенту, например, с целью аллогенной адоптивной терапии Т-клетками.
Поскольку они обычно находятся в негемопоэтических тканях, тканерезидентные Vδ1 T и DN γδ Т-клетки также с большей вероятностью будут возвращаться в опухолевые массы и оставаться там, чем их аналоги, обычно присутствующие в системной крови, и при адоптивном переносе указанные клетки с большей вероятностью будут эффективно направляться к солидным опухолям и, возможно, к другим негемопоэтическим тканеспецифическим иммунопатологиям.
В некоторых вариантах осуществления способ лечения индивидуума, страдающего от опухоли в негемопоэтической ткани, может включать в себя:
получение образца указанной негемопоэтической ткани от индивидуума-донора,
культивирование γδ Т-клеток из описанного выше образца с получением увеличенной популяции и
введение увеличенной популяции γδ T-клеток индивидууму, несущему опухоль.
Негемопоэтические тканерезидентные γδ Т-клетки, полученные по способу настоящего изобретения, экспрессируют NKG2D и отвечают на лиганд NKG2D (например, MICA), тесно связананый со злокачественными новообразованиями. Они также экспрессируют цитотоксический профиль в отсутствии какой-либо активации и, следовательно, могут эффективно уничтожать опухолевые клетки. Например, негемопоэтические тканерезидентные γδ Т-клетки, полученные по описанному здесь способу, могут экспрессировать один или несколько, предпочтительно все из IFN-γ, TNF-α, GM-CSF, CCL4, IL-13, гранулизина, гранзимов A и B и перфорина в отсутствии какой-либо активации. IL-17A может не экспрессироваться.
Таким образом, описанные здесь результаты являются убедительным доказательством практической пригодности негемопоэтических тканерезидентных γδ Т-клеток, полученных по способу настоящего изобретения, для клинического применения в виде "готового" иммунотерапевтического реагента. Указанные клетки обладают врожденной способностью к уничтожению других клеток, не связаны с МНС и обладают улучшенной способностью достигать опухоли и/или оставаться в опухолях, по сравнению с другими Т-клетками.
Негемопоэтические тканерезидентные γδ Т-клетки, полученные по способу настоящего изобретения, также можно использовать для терапии CAR-T. Данная терапия включает в себя получение рекомбинантных Т-клеточных рецепторов (TCR), чтобы перепрограммировать Т-клетку на новую специфичность, например, специфичность моноклонального антитела. Рекомбинантный TCR может придавать Т-клеткам специфичность к злокачественным клеткам и, следовательно, может использоваться для иммунотерапии рака. Например, Т-клетки могут распознавать раковые клетки, экспрессирующие опухолевый антиген, такой как ассоциированный с опухолью антиген (ТАА), который не экспрессируется нормальными соматическими клетками ткани индивидуума. Таким образом, CAR-модифицированные Т-клетки можно использовать для адоптивной терапии Т-клетками, например, раковых пациентов.
Описано применение резидентных γδ Т-клеток крови в терапии CAR. Однако негемопоэтические тканерезидентные γδ Т-клетки, полученные по способу настоящего изобретения, вероятно, будут особенно хорошими средствами для подходов CAR-T, поскольку их можно трансдуцировать химерными антиген-специфическими TCR, и при этом они сохраняют свою врожденную способность распознавать трансформированные клетки и, по всей вероятности, обладают улучшенной способностью проникать в опухоль и оставаться там по сравнению с резидентными γδ Т-клетками крови или обычными системными αβ Т-клетками. Кроме того, отсутствие у них MHC-зависимой презентации антигена снижает вероятность заболевания трансплантат против хозяина и обеспечивает специфичность к опухолям, экспрессирующим низкие уровни МНС. Подобным образом, их нечувствительность к обычной костимуляции, например, посредством CD28, усиливает направленность на опухоли, экспрессирующие низкие уровни лигандов костимуляторных рецепторов.
Рак может характеризоваться аномальной пролиферацией злокачественных раковых клеток и может включать в себя лейкозы, такие как острый миелоидный лейкоз (AML), хронический миелолейкоз (CML), острый лимфобластный лейкоз (ALL) и хронический лимфолейкоз (CLL), лимфомы, такие как лимфома Ходжкина, неходжкинская лимфома и множественная миелома, а также солидные раковые опухоли, такие как саркомы, рак кожи, меланома, рак мочевого пузыря, рак головного мозга, рак молочной железы, рак матки, рак яичников, рак предстательной железы, рак легкого, колоректальный рак, рак шейки матки, рак печени, рак головы и шеи, рак пищевода, рак поджелудочной железы, рак почки, рак надпочечников, рак желудка, рак яичек, рак желчного пузыря и желчных путей, рак щитовидной железы, рак тимуса, рак кости и церебральный рак.
Раковые клетки у больного раком могут иммунологически отличаться от нормальных соматических клеток индивидуума (то есть раковая опухоль может быть иммуногенной). Например, раковые клетки могут вызывать системный иммунный ответ у ракового пациента в отношении одного или нескольких антигенов, экспрессируемых раковыми клетками. Антигены, которые вызывают иммунный ответ, могут представлять собой опухолевые антигены, или они могут также являться антигенами нормальных клеток. Раковый пациент может иметь по меньшей мере один идентифицируемый признак, симптом или результат лабораторного анализа, который является достаточным для постановки диагноза рака в соответствии с клиническими стандартами, известными в данной области. Примеры таких клинических стандартов можно найти в учебниках медицины, таких как "Принципы лечения внутренних болезней" Харрисона (40). В некоторых случаях диагностика рака у индивидуума может включать в себя идентификацию клетки конкретного типа (например, раковой клетки) в образце жидкости или ткани организма, полученном от индивидуума.
Пациент, субъект или индивидуум, подходящий для лечения по описанному выше способу может представлять собой млекопитающее, например, относящееся к семейству грызунов (например, морская свинка, хомяк, крыса, мышь), мышиных (например, мышь), собачьих (например, собака), кошачьих (например, кошка), лошадиных (например, лошадь), приматов, обезьян (например, мартышка или человекообразная обезьяна), мартышек (например, мартышка или бабуин), человекообразных обезьян (например, горилла, шимпанзе, орангутан или гиббон), или человека.
В некоторых предпочтительных вариантах осуществления пациент, субъект или индивидуум представляет собой человека. В других предпочтительных вариантах осуществления пациент может представлять собой отличное от человека млекопитающее, особенно млекопитающее, которое обычно используют в качестве модели для демонстрации терапевтической эффективности у людей (такое как мышь, примат, свинья, собака или кролик).
В некоторых вариантах осуществления у пациента, субъекта или индивидуума может присутствовать минимальное остаточное заболевание (MRD) после первичного лечения рака.
Лечение может представлять собой любое лечение и терапию, человека или животного (например, в ветеринарных применениях), при котором достигается некоторый желаемый терапевтический эффект, например, ингибирование или задержка развития состояния, включая уменьшение скорости развития, прекращение развития, улучшение состояния, излечение или ремиссию (частичную или полную) состояния, предотвращение, отсрочку, устранение или прекращение одного или нескольких симптомов и/или признаков состояния, или увеличение продолжительности жизни индивидуума или пациента сверх ожидаемой в отсутствии лечения.
Изобретение также охватывает лечение как профилактическую меру (например, профилактику). Например, пациента, субъекта или индивидуума, предрасположенного или подверженного риску возникновения или повторного появления рака, можно лечить с помощью описанного здесь способа. Такое лечение может предотвратить или задержать возникновение или повторное появление рака у пациента, субъекта или индивидуума.
В частности, лечение может включать в себя ингибирование роста раковой опухоли, включая полную ремиссию рака, и/или ингибирование метастазирования рака. Рост раковой опухоли обычно характеризуется любым из нескольких показателей, которые свидетельствуют об изменении рака на более развитую форму. Так, показатели, используемые для измерения ингибирования роста раковой опухоли, включают в себя снижение выживаемости раковых клеток, уменьшение объема или морфологии опухоли (например, определяемое с помощью компьютерной томографии (КТ), сонографии или другого метода визуализации), замедление роста опухоли, разрушение сосудистой сети опухоли, улучшение показателей при кожном тесте на гиперчувствительность замедленного типа, увеличение активности цитолитических Т-лимфоцитов и уменьшение уровней опухолевых специфических антигенов. Снижение подавления иммунного ответа в раковых опухолях индивидуума может улучшить способность индивидуума противостоять росту раковой опухоли, в частности, росту уже присутствующей у индивидуума раковой опухоли, и/или уменьшить предрасположенность к росту рака у индивидуума.
Другие аспекты и варианты осуществления изобретения включают в себя описанные выше аспекты и варианты осуществления, где термин "содержащий" заменяется термином "состоящий из", а также описанные выше аспекты и варианты осуществления, где термин "содержащий" заменяется термином "состоящий по существу из".
Следует понимать, что, если контекст не указывает иначе, настоящая заявка раскрывает любые сочетания вышеописанных аспектов и вариантов осуществления. Подобным образом, если контекст не указывает иначе, настоящая заявка раскрывает любые сочетания предпочтительных и/или необязательных признаков либо отдельно, либо вместе с любым другим аспектом.
После прочтения настоящего описания для квалифицированного специалиста станут очевидны модификации вышеупомянутых вариантов осуществления, другие варианты осуществления и их модификации, которые также входят в объем настоящего изобретения.
Все документы и записи в базе данных последовательностей, упомянутые в настоящем описании, включены в данный документ посредством ссылки во всей их полноте для всех целей.
Используемый здесь термин "и/или" следует рассматривать как конкретное раскрытие каждого из двух указанных признаков или компонентов в присутствии или в отсутствии другого. Например, выражение "A и/или B" следует понимать как конкретное раскрытие каждого из (i) A, (ii) B и (iii) A и B, как если бы каждый из них был описан здесь отдельно.
Некоторые аспекты и варианты осуществления изобретения далее иллюстрируются посредством примера и со ссылкой на фигуры, описанные выше.
МЕТОДЫ
Выделение лимфоцитов из кожи человека путем трехмерного культивирования эксплантата
Разработан метод трехмерного культивирования эксплантата кожи, как описано в данном документе (29). Матрицы Cellfoam 9 мм×9 мм×1 мм (Cytomatrix Pty Ltd, Victoria, Australia) подвергают автоклавированию, затем инкубируют в растворе коллагена из сухожилий крысиного хвоста I (BD Biosciences) в PBS, 100 мг/мл, в течение 30 минут при комнатной температуре, затем один раз промывают PBS. Образцы кожи взрослого человека получают в течение 3-6 часов после кожной хирургии. Подкожный жир удаляют, а оставшуюся ткань кожи измельчают с получением фрагментов размером примерно 1 мм×1 мм. Примерно пять фрагментов/эксплантатов кожи помещают на поверхность каждой матрицы и прижимают. Каждую матрицу помещают в отдельную лунку 24-луночного планшета (Corning), содержащую 2 мл среды "Skin-T" (среда Дульбекко в модификации Искова (IMDM, Life Technologies), содержащая 10% инактивированной нагреванием фетальной бычьей сыворотки (Life Technologies ), L-глутамин (292 мг/мл, Life Technologies), пенициллин (100 единиц/мл, Life Technologies), стрептомицин (100 мг/мл, Life Technologies), N-2-гидроксиэтилпиперазин-N-2-этансульфокислоту (HEPES, 0,01 М, Life Technologies), пируват натрия (1 мМ, Life Technologies), минимальную эссенциальную среду (MEM), неэсенциальные аминокислоты (1×, Life Technologies) и 3,5 мкл/л 2-меркаптоэтанола (Life Technologies). В течение первых 7 дней культивирования в среду добавляют амфотерицин (2,5 мг/мл, Life Technologies). Среду заменяют три раза в неделю. Подпитку проводят путем отсасывания сверху 1 мл среды из каждой лунки и замены свежей средой. IL-2 и IL-15 добавляют вначале культивирования и при каждой замене среды до выделения лимфоцитов на 21 день. Человеческий рекомбинантный IL-2 (Proleukin, Novartis Pharmaceutical UK Ltd) добавляют в концентрации 100 ме/мл. Человеческий рекомбинантный IL-15 (Biolegend) добавляют в концентрации 20 нг/мл. До 96 лунок (четыре 24-луночных планшета) используют для культивирования для каждого донора.
Чтобы выделить лимфоциты, матрицы переносят в центрифужную пробирку (Corning) объемом 50 мл, содержащую 10 мл сбалансированного солевого раствора Хэнкса (HBSS, Life Technologies) и 0,01 мМ HEPES (до 12 матриц/пробирку). Матрицы промывают клеточной суспензией с помощью пипетки на 10 мл, клеточную суспензию пропускают через фильтр 70 мкм (BD Biosciences) в свежую центрифужную пробирку (Corning) объемом 50 мл. "Промывание" матриц повторяют еще два раза. Среду из культуральной лунки также отсасывают и пропускают через фильтр 70 мкм (BD Biosciences) в свежую центрифужную пробирку (Corning) объемом 50 мл. Лунки промывают еще два раза 1 мл 0,01 мМ HEPES/HBSS и пропускают через фильтр 70 мкм (BD Biosciences). Затем клетки выделяют центрифугированием (1600 об/мин в течение 15 минут). Осадок ресуспендируют в среде "Skin-T". Конечный осадок клеток ресуспендируют в среде "Skin-T" для последующего анализа методом проточной цитометрии или функциональных исследований. Если нужно определить число клеток, лейкоциты считают на данной стадии либо (1) путем окрашивания трипановым синим (0,4%) (Life Technologies) с последующим использованием гемоцитометра, либо (2) с помощью цитометра CASY®, модель TT, и анализатора (Roche).
Чтобы выделить лимфоциты, матрицы переносят в центрифужную пробирку (Corning) объемом 50 мл, содержащую 10 мл сбалансированного солевого раствора Хэнкса (HBSS, Life Technologies) и 0,01 мМ HEPES (до 12 матриц/пробирки). Матрицы промывают клеточной суспензией с помощью пипетки на 10 мл, клеточную суспензию пропускают через фильтр 70 мкм (BD Biosciences) в свежую центрифужную пробирку (Corning) объемом 50 мл. "Промывание" матриц повторяют еще два раза. Среду из культуральной лунки также отсасывают и пропускают через фильтр 70 мкм (BD Biosciences) в свежую центрифужную пробирку (Corning) объемом 50 мл. Лунки промывают еще два раза 1 мл 0,01 мМ HEPES/HBSS и пропускают через фильтр 70 мкм (BD Biosciences). Затем клетки выделяют центрифугированием (1600 об/мин в течение 15 минут). Осадок ресуспендируют в среде "Skin-T". Конечный осадок клеток ресуспендируют в среде "Skin-T" для последующего анализа методом проточной цитометрии или функциональных исследований. Если нужно определить число клеток, лейкоциты считают на данной стадии либо (1) путем окрашивания трипановым синим (0,4%) (Life Technologies) с последующим использованием гемоцитометра, либо (2) с помощью цитометра CASY®, модель TT, и анализатора (Roche).
Поскольку первичные образцы кишечника более подвержены загрязнению, полученные биопсийные образцы сначала дважды промывают IMDM, содержащей 10% FCS, пенициллин 500 ед/мл, стрептомицин 500 мг/мл, гентамицин 100 мг/мл, амфотерицин B 12,5 мкг/мл и метронидазол 5 мкг/мл, затем измельчают и помещают на сетки. Культуры клеток кишечника на сетках выращивали в среде Gut-T (IMDM, 10% FCS, пенициллина 100 ед/мл, стрептомицина 100 мг/мл, гентамбина 20 мг/мл, метронидазола 1 мг/мл. В течение первой недели выращивания также используют амфотерицин В, 2,5 мг/мл, как и при выращивании клеток кожи. Среда содержит IL-2 (100IU/мл) и IL-15 (20 нг/мл), замену среды проводят 3 раза в неделю. Поскольку структура кишечника является более рыхлой, чем структура кожи, лимфоциты можно собирать через неделю.
Выращивание тканевых γδ T-клеток
Для выращивания γδ T-клеток человеческой кожи смешанные лимфоциты, собранные после культивирования на сетке в течение 3-4 недель, промывают PBS, центрифугируют и повторно суспендируют в среде 1640 Мемориального института Розуэлла Парка (RPMI-1640, Life Technologies), содержащей 10% инактивированной нагреванием фетальной бычьей сыворотки (Life Technologies), L-глутамин (292 мг/мл, Life Technologies), пенициллин (100 единиц/мл, Life Technologies), стрептомицин (100 мг/мл, Life Technologies), N-2-гидроксиэтилпиперазин-N-2-этансульфокислоту (HEPES, 0,01 М, Life Technologies), пируват натрия (1 мМ, Life Technologies), неэссенциальные аминокислоты минимальной эссенциальной среды (MEM) (1×, Life Technologies) и 10 мл/л 2-меркаптоэтанола Life Technologies), при плотности 1×106/мл и добавляют либо один IL-2 (100 ме/мл), либо IL-2 плюс IL-15 (20 нг/мл). Клетки высевают в количестве 2×105/лунку в 96-луночные плоскодонные планшеты (Corning) или в количестве 2×106/лунку в 12-луночные планшеты (Corning) для выращивания. Клетки ежедневно контролируют с помощью микроскопии, свежую среду и цитокины добавляют 3 раза в неделю. По достижению полного слияния и после того, как клеточные агрегаты станут видимыми, клетки разделяли 1:1 в дополнительные лунки и планшеты соответственно. Клетки собирают и анализируют методом проточной цитометрии, или используют для функциональных анализов через 7, 14 или 21 день, в зависимости от анализа. Если нужны чистые γδ Т-клетки, клетки ресуспендируют в 1 мл буфера FACS (PBS, содержащем 2% инактивированной нагреванием фетальной телячьей сыворотки и 0,01 М EDTA) и окрашивают на рецептор αβ Т-клеток (Biolegend, клон IP26, 1:50) в течение 30 минут в темноте на льду, после чего все отрицательные клетки сортируют с помощью сортера Aria, работающего с использованием DIVA (BD Biosciences).
Совместное культивирование с фибробластами
Для каждой культуры на сетке готовят две чашки Петри (100×25 мм, Corning), которые процарапывают в нескольких местах с помощью скальпеля. Измельченные кусочки кожи помещают на царапины. После высыхания на воздухе в течение 5-10 минут кусочки кожи обычно прилипают к чашке, после чего к ним добавляют 10 мл среды Skin-T. Среду меняют один раз в неделю, а первичные фибробласты собирают после обработки ACCUTASE® (Life Technologies) через 3 недели после начала выращивания. Фибробласты высевают либо в 48-луночные планшеты в количестве 1×104, либо в нижние камеры 24-луночных планшетов в количестве 2×104 в случае экспериментов с использованием системы трансвел. Через 2-3 дня, когда фибробласты достигнут слияния, начинают эксперименты по совместному культивированию с использованием RPMI и указанных цитокинов, путем добавления 2×105 смешанных лимфоцитов кожи в случае 48-луночных планшетов или 3×105 лимфоцитов в случае 24-луночных планшетов, донных лунок, а также трансвелов.
Проточная цитометрия
Проточную цитометрию проводят с использованием следующих антител, связанных с указанными флуорохромами: Ki-67-BV421, CD3-BV510, Vδ1-PeVio770, TIM-3-PE, CD9-PE, CCR3-BV421, CD39-BV421. Все образцы также всегда окрашивают на жизнеспособность помощью eFluor770NIR. Коммерческие антитела приобретают у Biolegend или Miltenyi. Краситель для подтверждения жизнеспособности (вблизи ИК) получают от eBioscience. Окрашивание Ki-67 проводят на клетках, фиксированных и пермеабилизированных с помощью набора для окрашивания Foxp3 (eBioscience). После завершения каждого эксперимента популяцию клеток промывают PBS и разделяют пополам. Клетки окрашивали eFluor770 NIR на жизнеспособность и промывают, а затем TrueStain (Biolegend), чтобы избежать неспецифического связывания окрашивающих антител. Половину образца окрашивают на указанные поверхностные маркеры, тогда как другую половину окрашивают только на маркеры линии (CD3, Vδ1) с использованием эквивалентного изотипического контроля для используемых поверхностных маркеров. Это означает, что соответствующее изотипическое мышиное антитело, конъюгированное с тем же флуорохромом, используют в такой же концентрации. Изотипические контроли не связываются с какими-либо известными человеческими антигенами и поэтому свидетельствуют о неспецифическом связывании или ложно-положительном ответе. Его также называют реально отрицательным. Каждая гистограмма (закрашенная) показана в сравнении с соответствующим изотипическим контролем (незакрашенная, пунктирные линии). Приведенные данные отражают процент клеток, положительно окрашенных по указанному маркеру, и, следовательно, характеризующихся уровнем, превышающем уровень в изотипическом контроле. Анализ результатов проточной цитометрии проводят на FlowJo (версия 10.1).
Секвенирование РНК
Т-клетки Vδ1 человеческой кожи и Т-клетки Vδ1 человеческой крови (после роста, инициированного Т-клеточным рецептором) сортируют (FACS), центрифугируют и клеточный осадок ресуспендируют в буфере RLT. РНК получают с помощью набора RNA-Micro-plus (QIAGEN). Библиотеки РНК получают с использованием набора KAPA Stranded RNA-seq с RiboErase (HMR) (KAPA BIOSYSTEMS). Секвенирование спаренных концов проводят на HiSeq 2500 (подсветка) с использованием химии быстрого запуска (длина прочтения: 100 п.о.). Прочтенные спаренные концы длиной 101 пара оснований выравнивают и подвергают количественному анализу с использованием RSEM (v1.2.1 1) и Bowtie2. Прочтенные последовательности выравнивают с транскриптомом человека, рассчитанные значения преобразовывают в log2 и подвергают квантильной нормализации.
Количественный анализ цитокинов
Клетки Vδ1 кожи человека стимулируют PMA и иономицином или связанным с планшетом mAb против CD3 (OKT3, 5 мкг/мл) в течение 24 часов. Затем супернатанты анализируют с использованием панели 1A ProcartaPlex Human Cytokine & Chemokine (34 компонента), eBioscience. Анализы проводят с помощью Luminex FlexMap3D (Luminex Corp). Результаты анализируют в Microsoft Excel, показаны средние значения от трех доноров (анализы проводят с двойными повторами). Планки погрешностей обозначают стандартное отклонение.
Выращивание γδ T-клеток крови
γδ T-клетки крови в популяции РВМС могут расти только после стимуляции лигандами TCR (в случае Vδ2 такими лигандами могут являться, например, IPP, HMBPP, бисфосфонаты) (41, 42), или после добавления антител, перекрестно связывающихся либо с рецептором TCR (mAb), либо с TCR-ассоциированной киназой CD3 (43). Такой же эффект перекрестного связывания TCR также можно достичь с использованием лектинов, таких как PHA. В отсутствии добавления таких TCR-стимулирующих средств γδ T-клетки в РВМС выживают в течение нескольких дней, но не могут размножаться и остаются в исходном составе субпопуляций Т-клеток с небольшими вариациями.
Кровь здоровых добровольцев используют для выделения РВМС путем наслоения целой крови на Фиколл с последующим центрифугированием при 400 g в течение 20 минут для разделения красных кровяных телец, плазмы крови и белых лимфоцитов/моноцитов. Белые кровяные клетки тщательно собирают пипеткой и промывают четыре раза холодным PBS. Клетки ресуспендируют в среде RPMI-1640 (Life Technologies), содержащей 10% инактивированной нагреванием фетальной телячьей сыворотки (Life Technologies), L-глутамин (292 мг/мл, Life Technologies), пенициллин (100 единиц/мл, Life Technologies), стрептомицин (100 Мг/мл, Life Technologies), N-2-гидроксиэтилпиперазин-N-2-этансульфокислоту (HEPES, 0,01 М, Life Technologies), пируват натрия (1 мМ, Life Technologies), неэссенциальные аминокислоты минимальной эсенциальной среды (MEM) (1×; Life Technologies), при плотности 1×106/мл и добавляют IL-2 (100 ме/мл). Клетки переносят в 24-луночный планшет, покрытый моноклональным антителом pan γδ TCR (20 мкг/мл, клон B1, Biolegend) за 90 минут до переноса клеток. Клетки выращивают в течение 14 дней, среду заменяют каждые 2-3 дня и добавляют свежие цитокины. По достижении слияния клетки разделяют 1:1. В указанных условиях через 14 дней первоначально минорная популяция γδ T-клеток обычно находится в высоко активированном через TCR (на что указывает повышающая регуляция CD69 и CD25) и в значительной степени обогащенном состоянии и состоит в основном из T-клеток Vδ2, но также содержит T-клетки Vδ1 (которые составляют до 30% от всех γδ Т-клеток). Т-клетки Vδ1 можно затем выделить методом FACS для функционального, фенотипического или генетического анализа.
РЕЗУЛЬТАТЫ
Человеческие γδ Т-клетки присутствуют в коже в большом количестве, экспрессируют TCR, отличный от Vδ2, и участвуют в контролирующем ответе лимфоидной ткани человека на стресс
По методу Кларка (29) культивируют образцы остаточной кожи человека, к которым добавляют IL-2 и IL-15, чтобы обеспечить разрастание тканерезидентных лимфоцитов в течение 3 недель, с получением популяции лимфоцитов, содержащей в среднем 240000 клеток на сетку. В соответствии с предыдущими сообщениями (29) авторы идентифицируют отдельные субпопуляции резидентных лимфоцитов кожи, причем большинство клеток экспрессирует обычный αβ TCR, в основном из тканерезидентного типа "TRM". В целом 59,9% (±8,6) клеток CD45+ являются CD4+, а 18,3% (±2,8) представляют собой αβ Т-клетки CD8+ с фракцией NK-клеток 8,7% (±3,6). Кроме того, авторы обнаружили у доноров значительную популяцию γδ T-клеток (в среднем 8,513% клеток CD45+, ±6,564%) (фиг. 1А и 3D). Данное представление субпопуляции лимфоцитов после органотипического культивирования является высоко воспроизводимым примерно у 100 доноров и сопоставимым со свеже расщепленными образцами кожи, отличающимися только по слегка увеличенной популяции γδ, но с практической точки зрения более полезное, поскольку оно предлагает гораздо более крупные и более чистые популяции лимфоцитов, чем стандартные методы расщепления ткани. В соответствии с описанными в литературе данными, касающимися тканевой компартментализации человеческих γδ T-клеток на основе дельта-цепи их TCR, большинство человеческих γδ Т-клеток кожи экспрессируют цепь TCR Vδ1 в паре с разными γ-цепями, идентифицированными методом проточной цитометрии. Этим они отличаются от большинства γδ Т-клеток периферической крови, которые содержат один специфический гетеродимер TCR с цепью Vδ2, соединенной с Vγ9, и практически отсутствуют в образцах кожи человека. Тем не менее, важно отметить, что значительная субпопуляция не экспрессирует ни Vδ1 TCR, ни Vδ2 TCR, что позволило ее назвать "двойные отрицательные" γδ T-клетки (фиг. 1С).
Резидентные γδ T-клетки кожи, выращенные по указанному способу, имеют фенотип с неполной дифференцировкой памяти, который характеризуется отсутствием экспрессии CD45RA и экспрессией разных уровней костимуляторной молекулы CCR7. Аналогично обычным системным Т-клеткам, интенсивная экспрессия поверхностного белка CD69 наряду с экспрессией рецептора программируемой гибели 1 (PD-1); низкий уровень рецептора IL-2 α (CD25) или его отсутствие; и отсутствие костимуляторной молекулы CD28 дают картину ранее активированных или хронически активированных Т-клеток (фиг. 1D). В соответствии с их локализацией в ткани клетки Vδ1 и DN экспрессируют кожные и тканевые хоминг-маркеры, такие как CLA, CCR4, CCR8 и интегрин αE (CD103) (см. Фиг. 7). Указанный набор тканевых хоминг-маркеров, по-видимому, может оказаться полезным при применении в иммунотерапии. Кроме того, резидентные γδ Т-клетки кожи экспрессируют на высоком уровне активаторный рецептор NKG2D (фиг. 2А), что позволяет предположить возможность участия этих клеток в контролирующем ответе лимфоидной ткани на стресс. Лиганды NKG2D, такие как MICA, MICB и ULBP, соответственно, подвергаются повышающей регуляции клетками в ответ на повреждение ДНК, активацию EGF-рецептора и окислительный стресс и, следовательно, могут обеспечивать экспрессию Т-клетками NKG2D для идентификации и устранения стрессированных или трансформированных клеток, тем самым поддерживая тканевый гомеостаз. В соответствии с этим принципом авторы обнаружили, что резидентные γδ T-клетки кожи, выращенные по способу настоящего изобретения, активируются при воздействии рекомбинантных лигандов рецептора NKG2D (MICA, ULBP2), демонстрируя дегрануляцию, измеряемую по повышающей регуляция ассоциированного с лизосомами мембранного белка CD107a (фиг. 2А). Данная врожденная функция присуща исключительно γδ Т-клеткам Vδ1+ и DN, так как другие тканерезидентные Т-клетки (фиг. 2C) и системные γδ T-клетки не способны давать такой ответ (фиг. 10B).
Таким образом, активированные резидентные γδ Т-клетки кожи Vδ1+ и DN выполняют провоспалительную TH1-смещенную цитокиновую программу (положительное окрашивание на IFN-γ, TNF-α и GM-CSF) после активации PMA/иономцином или лигандами NKG2D, например. рекомбинантным белком MICA (фиг.2А и 2В), тем самым подтверждая врожденные ответы клеток. И действительно, ответы на MICA почти полностью прекращаются в результате блокирования рецептора NKG2D под действием антитела (фиг. 2В и 2С).
Известно, что γδ Т-клетки секретируют IL-17 при определенных заболеваниях, таких как псориаз, и в некоторых типах опухолей. γδ Т-клетки, выращенные по способу настоящего изобретения, продуцируют низкий уровень или не продуцируют IL-17, даже при экстенсивной активации (фиг. 2В и 8). И наоборот, тканерезидентные CD4-экспрессирующие αβ T-клетки продуцируют IL-17 при активации TCR (фиг. 2B). Как правило, αβ T-клетки в ответ на PMA/иономицин продуцируют репертуар цитокинов, сильно отличающийся от продуцируемого γδ Т-клетками Vδ1+ и DN, которые ограничены TH1-смещенной программой, связанной с защитой хозяина.
Отделение от ткани вызывает активацию и массивную пролиферацию человеческих тканевых γδ Т-клеток
Чтобы дополнительно исследовать человеческую ткань, γδ Т-клетки и их биологию, смешанные лимфоциты кожи переносят в лунки для культивирования клеток и добавляют IL-2, чтобы поддерживать жизнеспособность с течением времени. Интересно, что авторы быстро обнаружили, что после отделения от стромальных и эпителиальных клеток, присутствующих в органотипической культуре, T-клетки Vδ1 демонстрируют уникальные признаки активации и пролиферации в отсутствии каких-либо других добавленных стимулов. В течение менее 7 дней в γδ Т-клетках Vδ1+ и DN наблюдается уникальная и массовая повышающая регуляция ядерного фактора Ki-67 и увеличение поверхностной экспрессии рецептора IL-2 α (CD25) (фиг. 3B и 4B). Удивительно, что после выращивания в течение 3 недель только в присутствии IL-2 тканевые γδ Т-клетки Vδ1+ и DN превосходят по росту все другие субпопуляции T-клеток, составляя до 65% от всех лимфоцитов кожи, причем их число увеличивается в среднем в 127,18 раз, тогда как число αβ T-клеток увеличивается только в 5,21 раза (р=0,0124), что определяют по абсолютному числу клеток (фиг.3А). MFI ассоциированного с клеточным циклом ядерного фактора KI-67 в γδ Т-клетках Vδ1+ и DN увеличивается с 2664,5 (±1876,1) до 8457,7 (±4574,2) за 14 дней, тогда как в αβ Т-клетках MFI уменьшается с 592,8 (±390,5) до 284,7 (±140,1) за тот же период времени (фиг.3C). Такой селективный рост резидентных γδ T-клеток кожи можно дополнительно поддерживать путем добавления рекомбинантного IL-15, который повышает жизнеспособность и общее число лимфоцитов.
γδ Т-клетки кожи активно подавляются фибробластами в контакт-зависимой манере
Удивительный рост γδ T-клеток Vδ1+ и DN, описанный в предыдущем абзаце, никогда не встречается в системах органотипического культивирования, в которых наблюдается экстенсивное разрастание фибробластов. Поэтому авторы выращивают аутологичные фибробласты, чтобы непосредственно проверить, действительно ли они при совместном культивировании с γδ T-клетками Vδ1+ и DN ингибируют рост Т-клеток. После культивирования на сетке в течение 3 недель смешанные лимфоциты кожи высевают в лунки, которые либо являются пустыми, либо содержат ранее полученный слившийся монослой фибробластов, причем в каждом случае в среду добавляют экзогенный IL-2, чтобы поддерживать рост Т-клеток. Кроме того, используют транслунки, которые препятствуют непосредственному контакту Т-лимфоцитов с фибробластами внутри одних и тех же лунок, но позволяют любым растворимым факторам, продуцируемым фибробластами, оказывать влияние на лимфоциты. После совместного культивирования в течение 14 дней γδ T-клетки Vδ1+ и DN начинают пролиферировать в лунках, в которых фибробласты отсутствуют, а также в лунках, в которых Т-клетки не могут непосредственно контактировать с фибробластами. В соответствии с полученными ранее результатами, пролиферация αβ T-клеток является низкой при любых условиях. Следует отметить, что если Т-клетки вступают в непосредственный контакт с фибробластами, скорость роста γδ T-клеток Vδ1+ и DN в течение двух недель значительно снижается с 22,6 (SEM 8,07) раз в лунках с отсутствием контакта с фибробластами до 3,3 (SEM 0,17) раз (фиг. 4А). Такое опосредуемое контактом ингибирование дополнительно подтверждается отсутствием повышающей регуляции CD25, Ki-67 и фактора транскрипции T-bet в клетках Vδ1 в течение 7 дней, в отличие от лимфоцитов, выращенных отдельно (фиг. 4В). Некоторая форма опосредованного тканью контроля иммунной системы, по-видимому, имеет фундаментальное значение для поддержания гомеостаза тканей, поскольку без такого контроля может существовать риск стойкого воспаления. Примером такого контроля может служить супрессорная регуляция γδ T-клеток Vδ1+ и DN стромальными фибробластами.
Таким образом, фенотип резидентных γδ T-клеток Vδ1+ и DN кожи и их удивительные функциональные потенциалы свидетельствуют о предварительно активированных Т-клетках, которые обычно контролируются соседними фибробластами кожи посредством зависимого от контакта механизма, который еще подлежит изучению. Инактивация этого механизма путем освобождения Т-клеток от контакта с фибробластами селективно обеспечивает выраженный рост γδ T-клеток Vδ1+ и DN, в отличие от других Т-клеток кожи.
Освобождение от опосредуемого контактом ингибирования вызывает цитотоксический TH1-смещенный цитокиновый ответ γδ T-клеток Vδ1+ и DN
Смешанные лимфоциты кожи выращивают в течение 14 дней и с помощью сортировки клеток, связанной с флуоресценцией, αβ Т-клетки отделяют от γδ Т-клеток, которые таким образом можно получить с чистотой до 90%. Полученные высокообогащенные клетки помещают в лунки для культивирования клеток в концентрации 150000 клеток/лунку в среде RPMI, содержащей 10% FCS, через 24 часа супернатанты собирают и анализируют на широкий спектр эффекторных цитокинов с использованием матрицы на основе LUMINEX®. Совершенно неожиданно обнаружено, что растущие γδ Т-клетки (рост которых инициируется только их отделением от фибробластов) спонтанно продуцируют большие количества TH1-ассоциированных цитокинов, таких как IFN-γ (12383,46±16,618,90 пг/мл), GM-CSF (4316,73±4,534,96 пг/мл), и провоспалительных хемокинов CCL4 (14877,34±10935,64 пг/мл) и CCL3 (1303,07±757,23 пг/мл) (фиг.5A).
Кроме того, в процессе роста клетки продуцируют большое количество спонтанного IL-13, связанного с атопическими ответами, в отличие от свежевыделенных TCR-активированных γδ Т-клеток кожи; другие цитокины продуцируются на гораздо более низких уровнях или совсем не продуцируются, например, IL-17A (фиг. 8). Высокие эффекторные потенциалы клеток можно дополнительно увеличить путем стимуляции рекомбинантным MICA (лигандом NKG2D), перекрестно сшитым антителом против CD3, или PMA/иономицином. Чтобы оценить цитотоксический потенциал выращенных γδ Т-клеток в отношении злокачественных клеток-мишеней, в 24-часовых экспериментах по совместному культивированию используют стандартные трансформированные клеточные линии. γδ T-клетки Vδ1+ и DN обладают очень высокой дозозависимой цитотоксической активностью по отношению к клеткам Hela (шейки матки) и Caco2 (толстой кишки), намного превосходя традиционные тканевые αβ T-клетки (фиг. 5B). Кроме того, цитотоксичность, опосредованную γδ-клетками, можно эффективно ингибировать путем блокирования рецептора NKG2D растворимыми моноклональными антителами, а это свидетельствует о том, что указанный рецептор по меньшей мере отчасти участвует в контролировании опухоли посредством дерепрессии человеческих γδ Т-клеток кожи. Цитотоксический потенциал этих клеток также подтверждают с использованием других мишеней: HCT1954, MDAMB231 (карцинома молочной железы), и HCT116 (толстая кишка) (фиг. 9).
Тканерезидентные γδ Т-клетки в кишечнике человека
Авторы также идентифицировали популяцию негемопоэтических тканерезидентных γδ Т-клеток, экспрессирующих Т-клеточный рецептор Vδ1 в культурах на сетке, полученных из толстой кишки человека (фиг. 6). В случае 3 доноров авторам удалось размножить эти клетки с помощью методов, используемых 15 для клеток кожи, в течение 4-5 недель. Во время экспрессии γδ T-клетки Vδ1+ и DN, полученные из толстой кишки, характеризуются подобной картиной повышающей регуляции Ki-67 после их отделения от обогащенной фибробластами органотипической клеточной культуры. Аналогично γδ T-клетки Vδ1+ и DN толстой кишки подвергаются эффективной стимуляции под действием лигандов рецептора NKG2D. Ранее было описано, что полученные из крови γδ Т-клетки хорошо приспособлены для выполнения антителозависимой клеточно-опосредованной цитотоксичности посредством экспрессии CD16, что подтверждает повышенную цитотоксичность, направленную против CD20-положительной лимфомы B-линии в сочетании с ритуксимабом. Подобным образом, клетки хронического лимфолейкоза (CLL) и HER2-положительного рака молочной железы уничтожаются более эффективно при использовании моноклональных антител (31). Чтобы оценить потенциал Т-клеток кожи Vδ1 для нацеливания на опсонизированные антителами клетки-мишени, проверяют уровни экспрессии трех IgG1-ассоциированных рецепторов Fc CD16, CD32 и CD64. Полученные из кожи T-клетки Vδ1 экспрессируют небольшие уровни рецептора FC CD16, но высокие уровни высокоаффинного рецептора IgG CD64 (фиг. 11). Следовательно, полученные из ткани T-клетки Vδ1 могут быть пригодными для применения в качестве адъюванта в терапии с использованием моноклональных антител, такой как CD20- или Her2-терапия, поскольку они могут направляться антителом к злокачественным опухолям и метастазам, распознавать опсонизированные опухолевые клетки и уничтожать мишени посредством ADCC.
СПИСОК ЛИТЕРАТУРЫ
1. Sensi M et al. Cancer Research. 2005; 65(2):632-40.
2. Gaudin C et al. Journal of Immunology. 1999; 162(3):1730-8.
3. Echchakir H et al. Cancer Research. 2001; 61(10):4078-83.
4. Karanikas V et al. Cancer Research. 2001; 61(9):3718-24.
5. Pieper R et al. Journal of Experimental Medicine 1999; 189(5):757-66.
6. Wang RF et al. Science. 1999; 284(5418):1351-4.
7. Topalian SL et al. The New England Journal of Medicine. 2012; 366(26):2443-54.
8. Brahmer JR et al. The New England Journal of Medicine. 2012; 366(26):2455-65.
9. Hodi FS et al. The New England Journal of Medicine. 2010; 363(8):711-23.
10. Robert C et al. The New England Journal of Medicine. 2011; 364(26):2517-26.
11. Pardoll DM. Nature Reviews Cancer. 2012; 12(4):252-64.
12. Fecher LA et al. The Oncologist. 2013; 18(6):733-43.
13. Ribas A. The New England Journal of Medicine. 2012; 366(26):2517-9.
14. Vantourout P, Hayday A. Nature Reviews Immunology. 2013; 13(2):88-100.
15. Hintz M et al. FEBS Letters. 2001; 509(2):317-22.
16. Freed-Pastor WA et al. Cell. 2012; 148(1-2):244-58.
17. Vantourout P et al. Science Translational Medicine. 2014; 6(231):231ra49.
18. Wrobel P et al. Scandinavian Journal of Immunology. 2007; 66(2-3):320-8.
19. Todaro M et al. Journal of Immunology. 2009; 182(11):7287-96.
20. Gertner-Dardenne J et al. Journal of Immunology. 2012; 188(9):4701-8.
21. Mao C et al. Journal of Immunol Research. 2014; 2014:593562.
22. Godder KT et al. Bone Marrow Transplantation. 2007; 39(12):751-7.
23. Fournie JJ et al. Cellular & Molecular Immunology. 2013; 10(1):35-41.
24. Curran KJ et al. Journal of Clinical Oncology. 2015; 33(15):1703-6.
25. Hillerdal V et al. BioDrugs. 2015; 29(2):75-89.
26. Couzi L et al. Journal of the American Society of Nephrology. 2010; 21(1):181-8.
27. Zheng J et al. Cellular & Molecular Immunology. 2013; 10(1):50-57.
28. Deniger DC et al. Molecular Therapy. 2013; 21(3):638-47.
29. Clark RA et al. 2006. Journal of Investigational Dermatology. 126(5):1059-70.
30. Carrasco A. et al. 2013. Journal of Immunological Methods. 389(1-2):29-37.
31. Tokuyama H. et al. International Journal of Cancer. 2008; 122(11):2526-34.
32. Willcox CR et al. Nature Immunology. 2012; 13(9):872-9.
33. Gillis and Smith. Nature. 1977; 268(5616):154-6.
34. Fujita et al. Proceedings of the National Academy of Sciences USA. 1984; 80(24):7437-41.
35. Grabstein et al. Science. 1994; 264(5161):965-8.
36. Helgason C et al. Methods in Molecular Biology. Humana Press Inc. U.S. (15 Oct 2004) ISBN: 1588295451.
37. Picot J. Methods in Molecular Medicine. Humana Press Inc., U.S. (9 Dec 2004) ISBN: 1588292223.
38. Freshney R. Culture of Animal Cells: A Manual of Basic Technique. John Wiley & Sons Inc., (2 Aug 2005) ISBN: 0471453293.
39. Ho WY et al. Journal of Immunological Methods. 2006;310:40-52.
40. Fauci AS et al. eds. Harrison's Principles of Internal Medicine. 2001; 15th Ed., McGraw-Hill, New York.
41. Petrini I. et al. European Journal of Clinical Investigation. 2009; 39(9):813-8.
42. Gruenbacher et al. Blood. 2009; 114(20):4422-31.
43. Almeida et al. Clin Cancer Res (2016) 10.1158/1078-0432.CCR-16-0597.
44. Deniger D. et al. Frontiers in Immunology. 2014:5(636):1-10.
| название | год | авторы | номер документа |
|---|---|---|---|
| Биомедицинский клеточный продукт со специфической противоопухолевой активностью, представленный популяциями лимфокин-активированных киллеров и анти-HER2 CAR-γδΤ-ОИЛ и анти-HER2 CAR-T-NK | 2022 |
|
RU2786210C1 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ГАММА ДЕЛЬТА Т-КЛЕТКИ | 2015 |
|
RU2756247C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР | 2013 |
|
RU2665548C2 |
| АНТИГЕННЫЙ РЕЦЕПТОР | 2020 |
|
RU2832189C2 |
| КОМПОЗИЦИИ Т-КЛЕТОК С НЕДОСТАТОЧНОСТЬЮ РЕЦЕПТОРОВ Т-КЛЕТОК | 2013 |
|
RU2653761C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| СПОСОБЫ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ИСТОЩЕНИЯ TCRαβ КЛЕТОК | 2020 |
|
RU2829624C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ T-КЛЕТОК ДЛЯ T-КЛЕТОЧНОЙ ТЕРАПИИ | 2020 |
|
RU2834232C1 |
| ИММУНОКОМПЕТЕНТНАЯ КЛЕТКА И ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, ЭКСПРЕССИРУЮЩИЕ РЕГУЛЯТОРНЫЕ ФАКТОРЫ ИММУННОЙ ФУНКЦИИ | 2017 |
|
RU2770812C2 |
| КОМПОЗИЦИИ Т-КЛЕТОК С НЕДОСТАТОЧНОСТЬЮ РЕЦЕПТОРОВ Т-КЛЕТОК | 2013 |
|
RU2781979C2 |
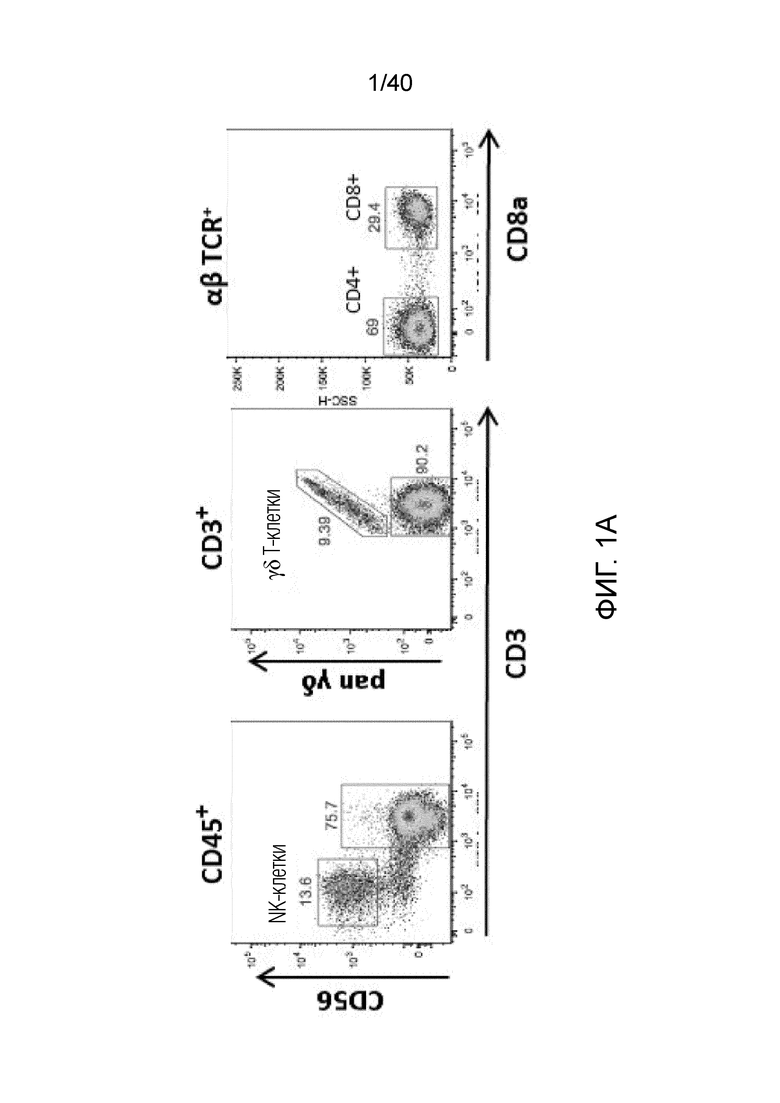
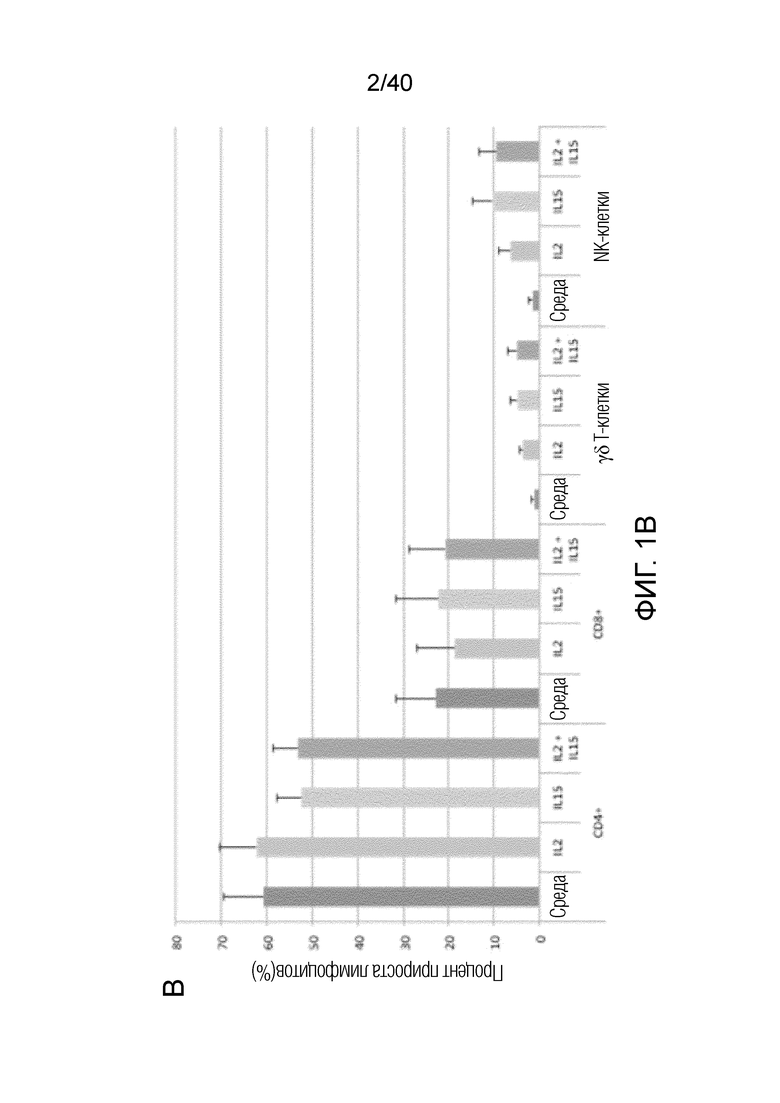
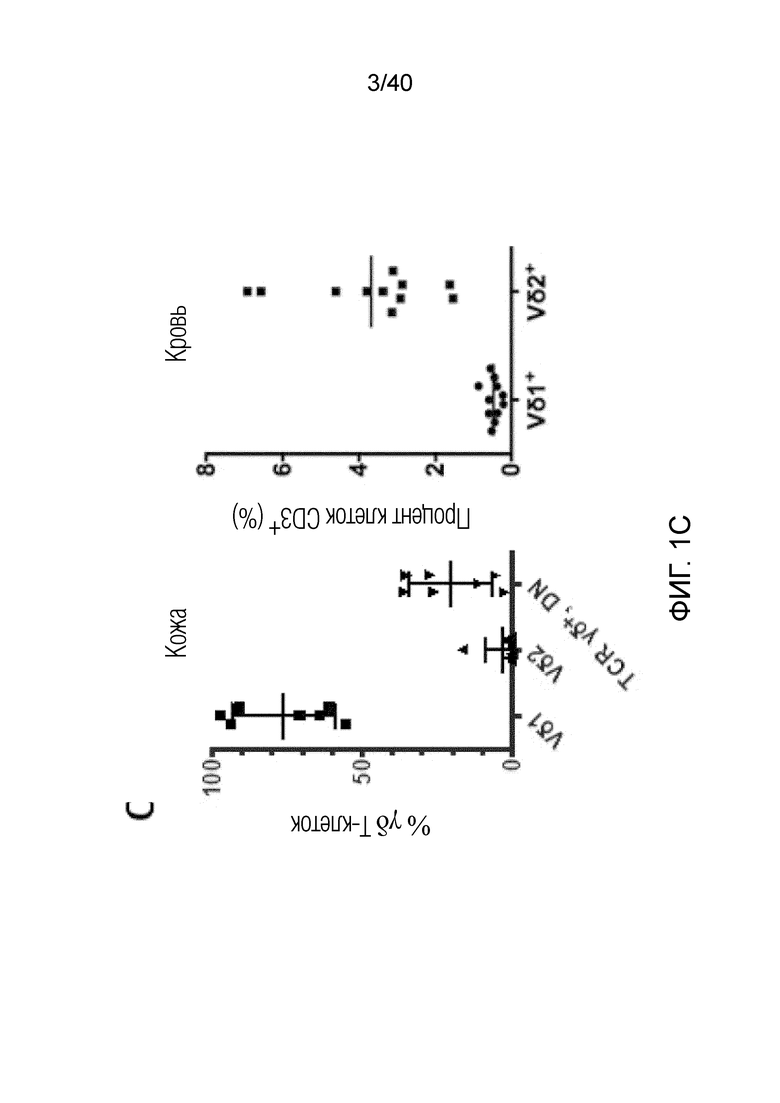
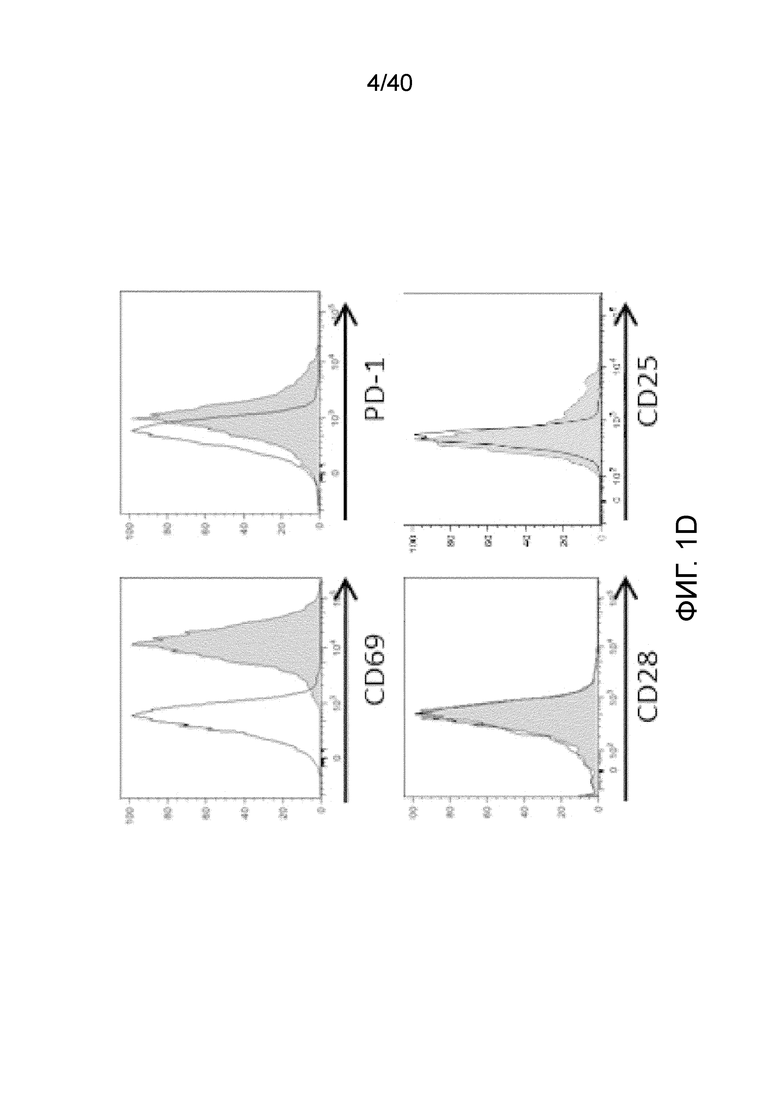
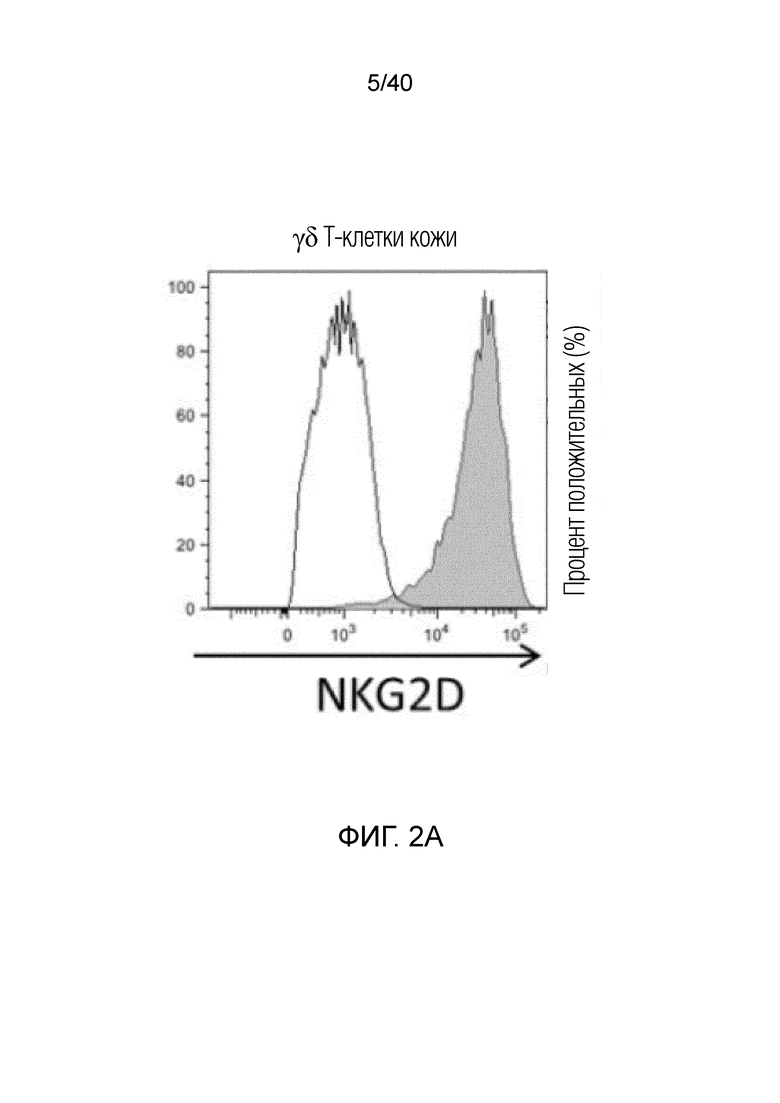
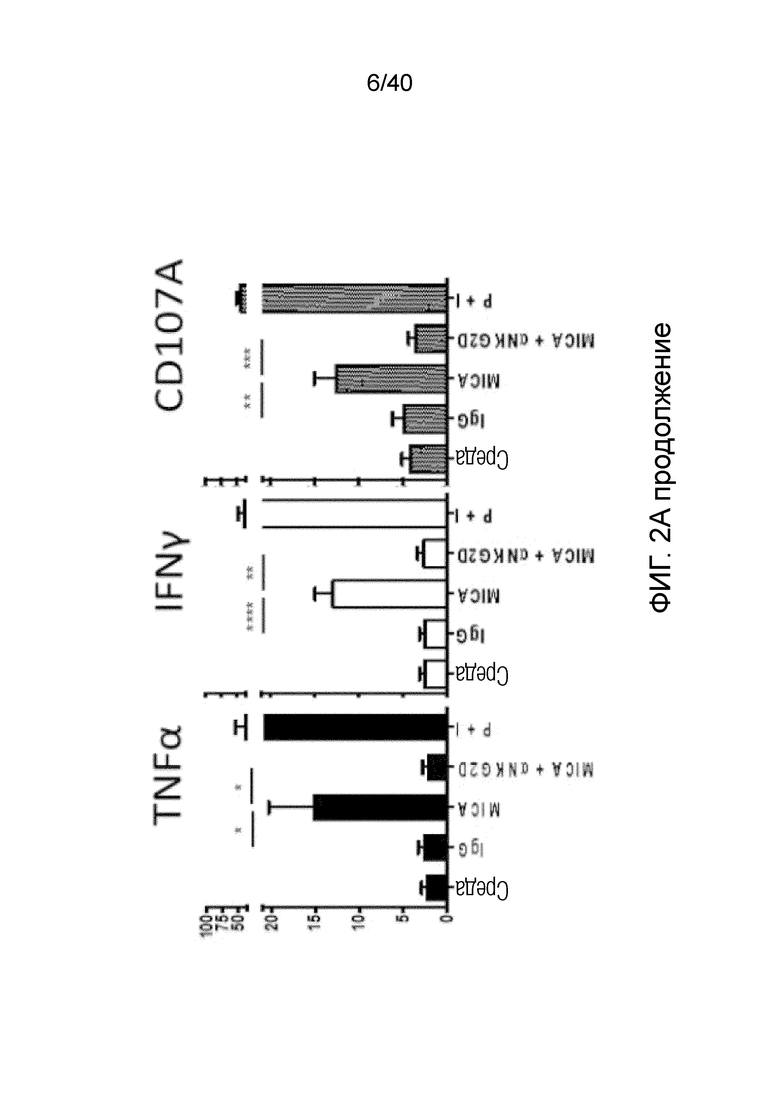
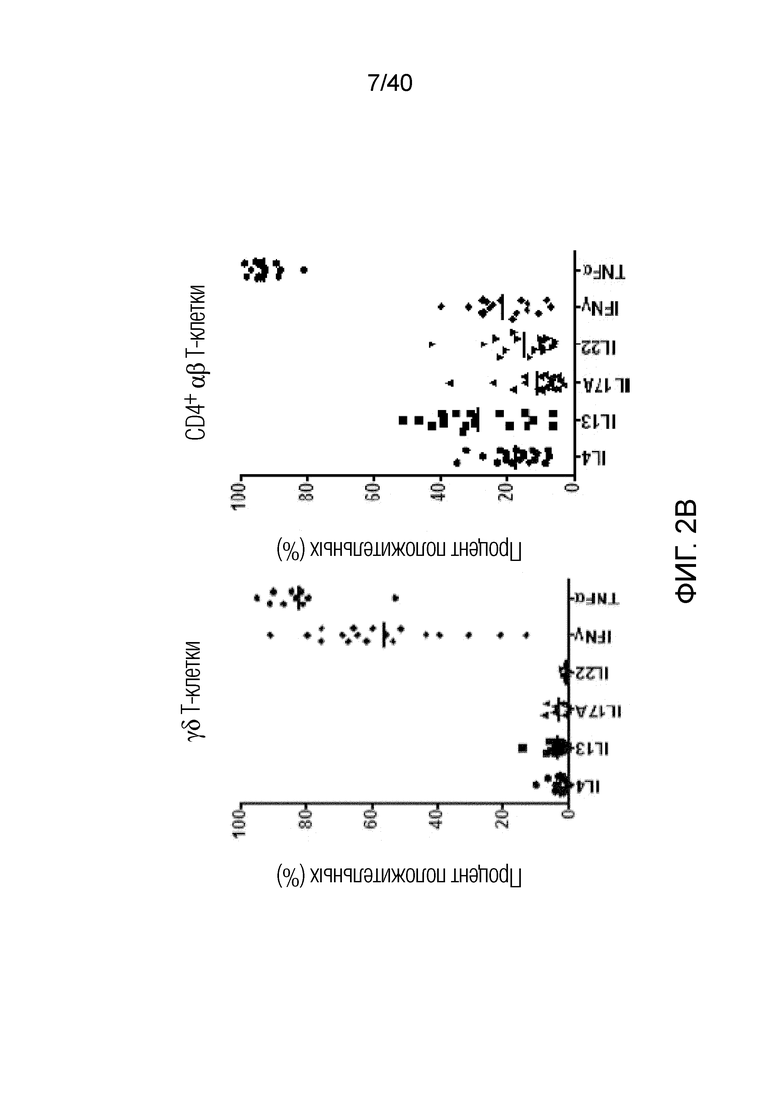

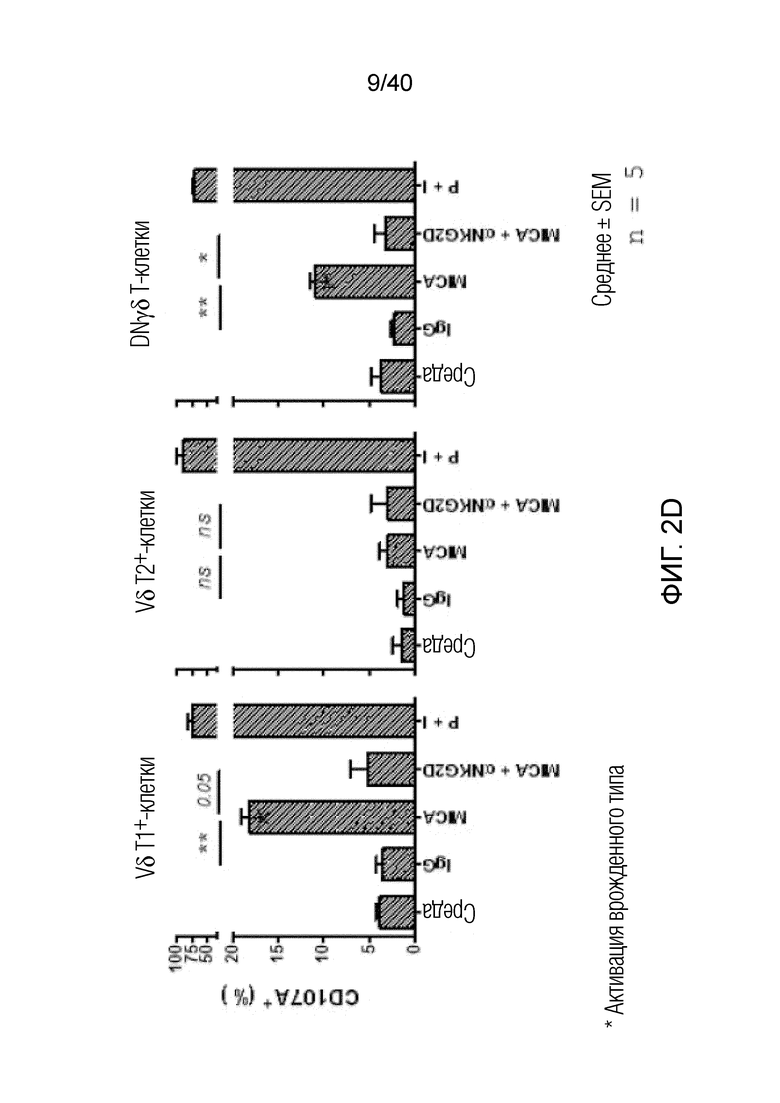
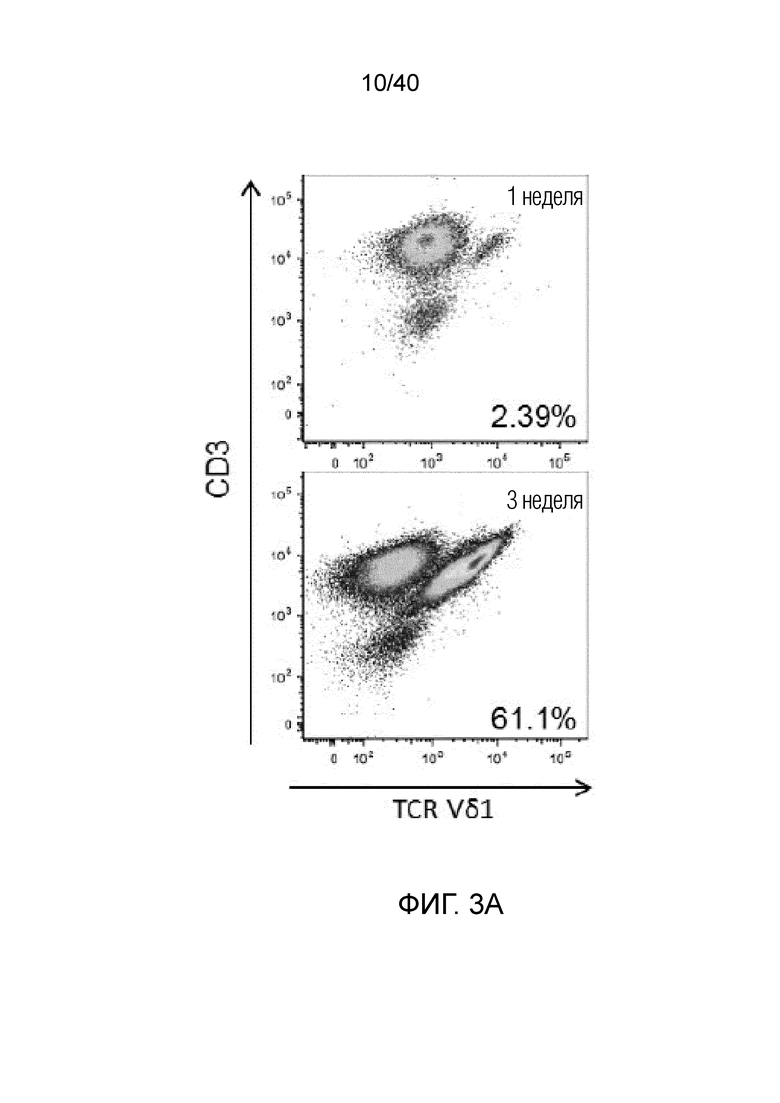
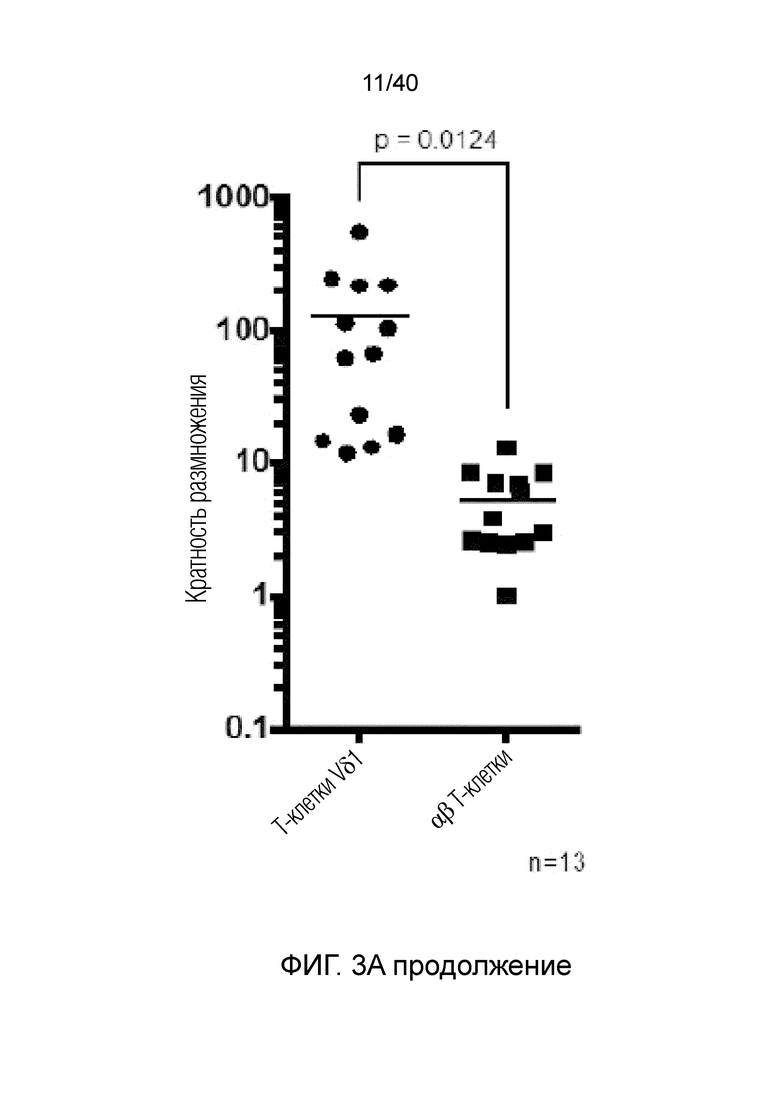
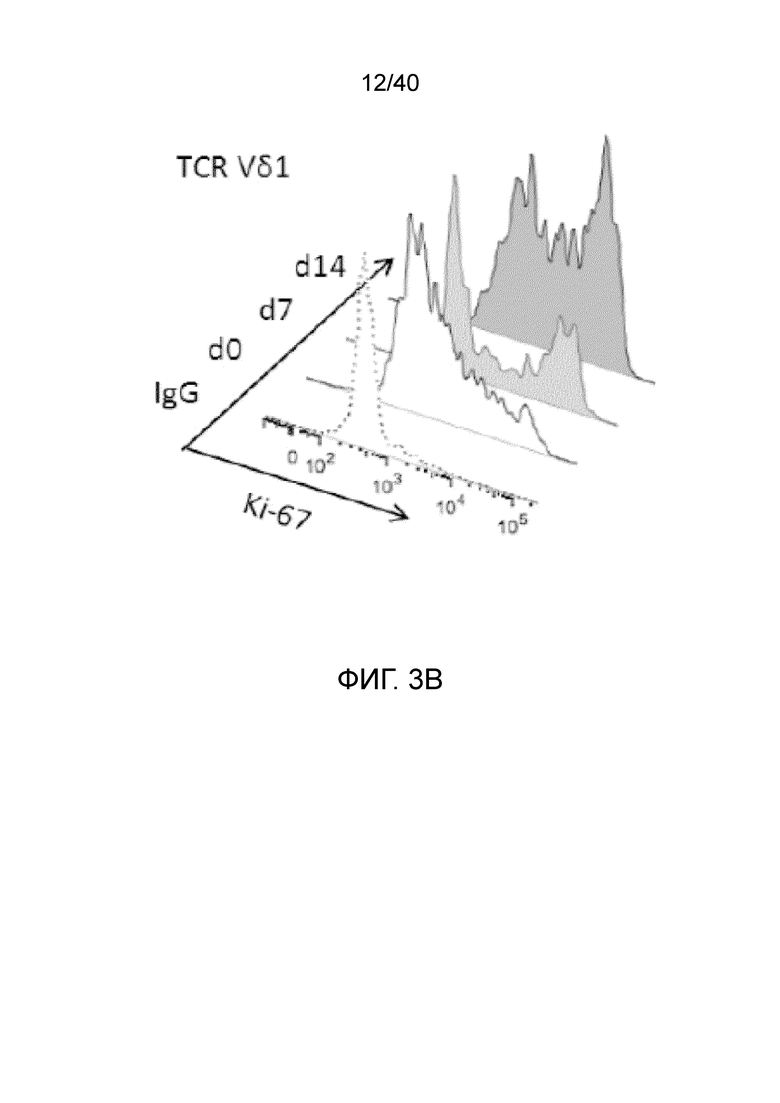
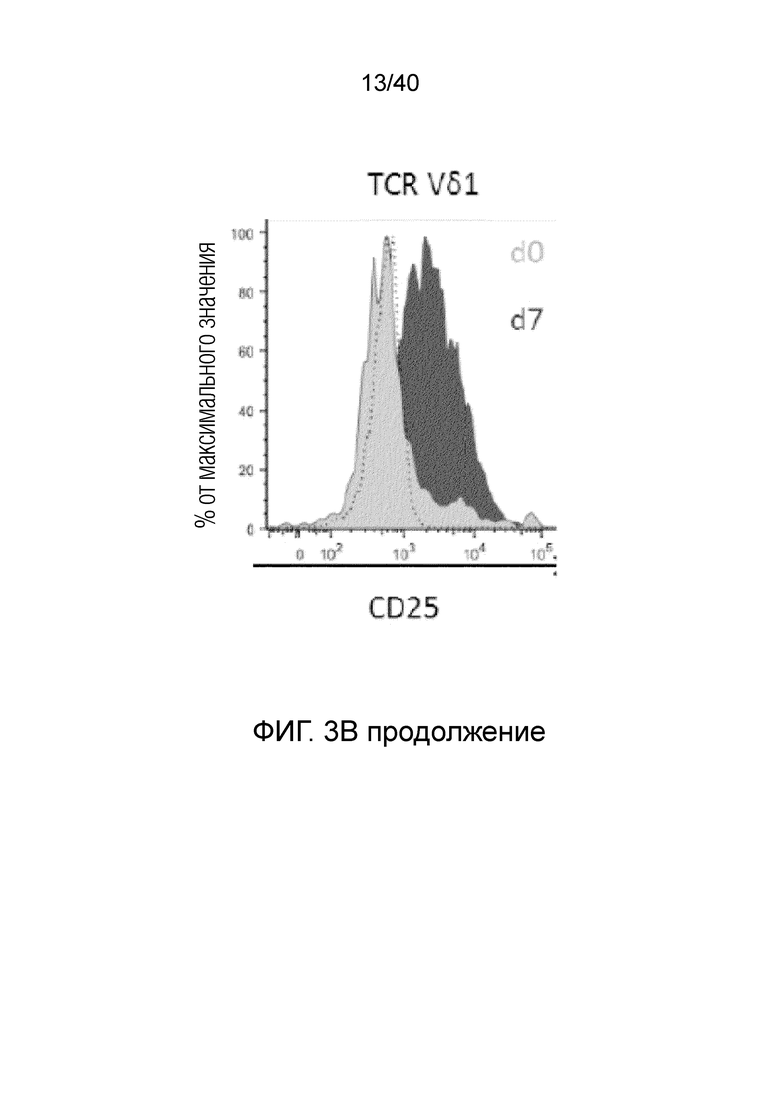
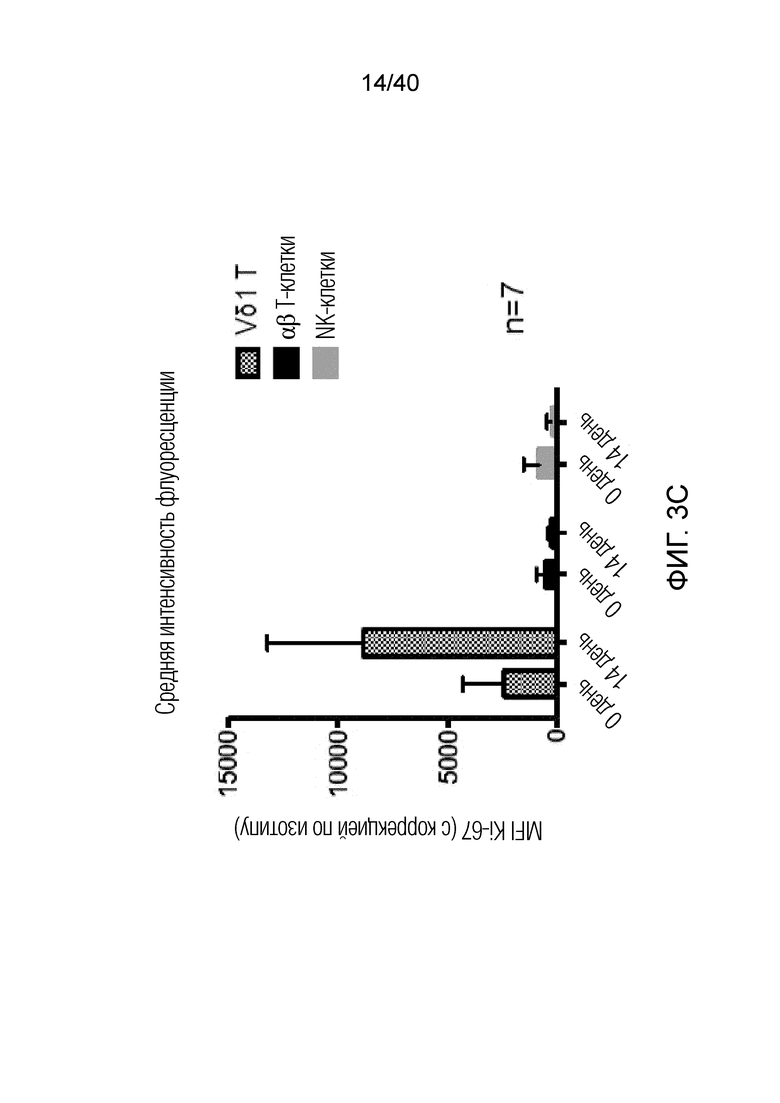
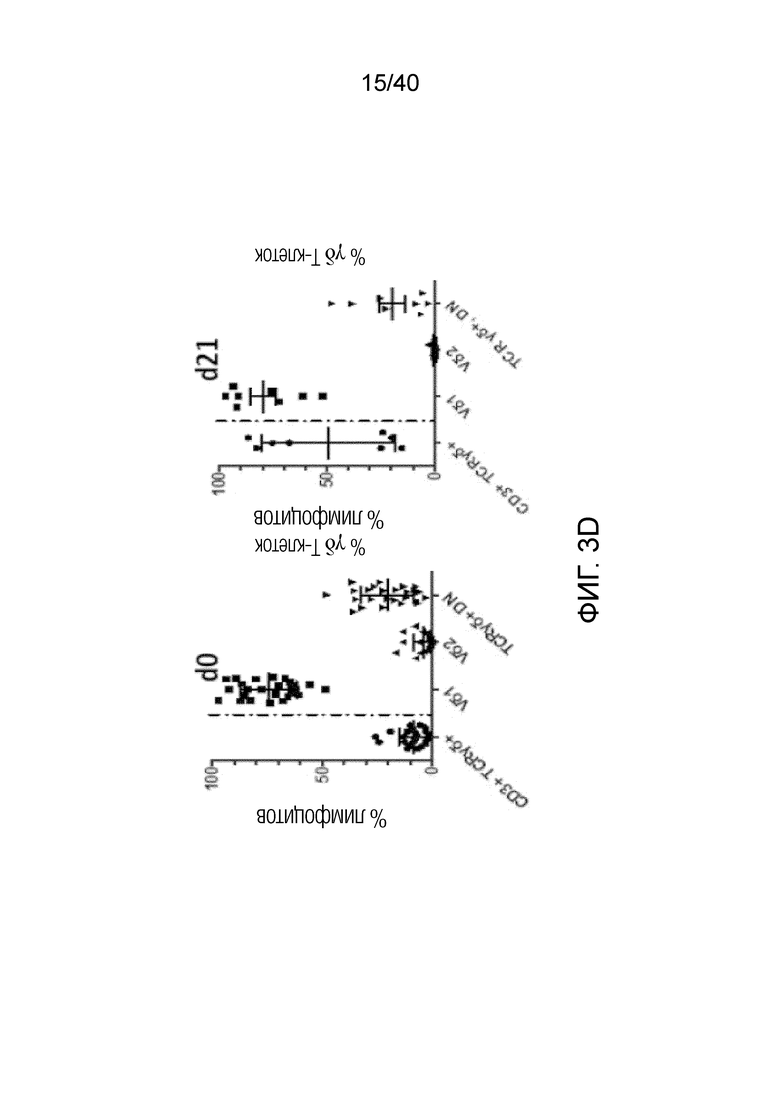
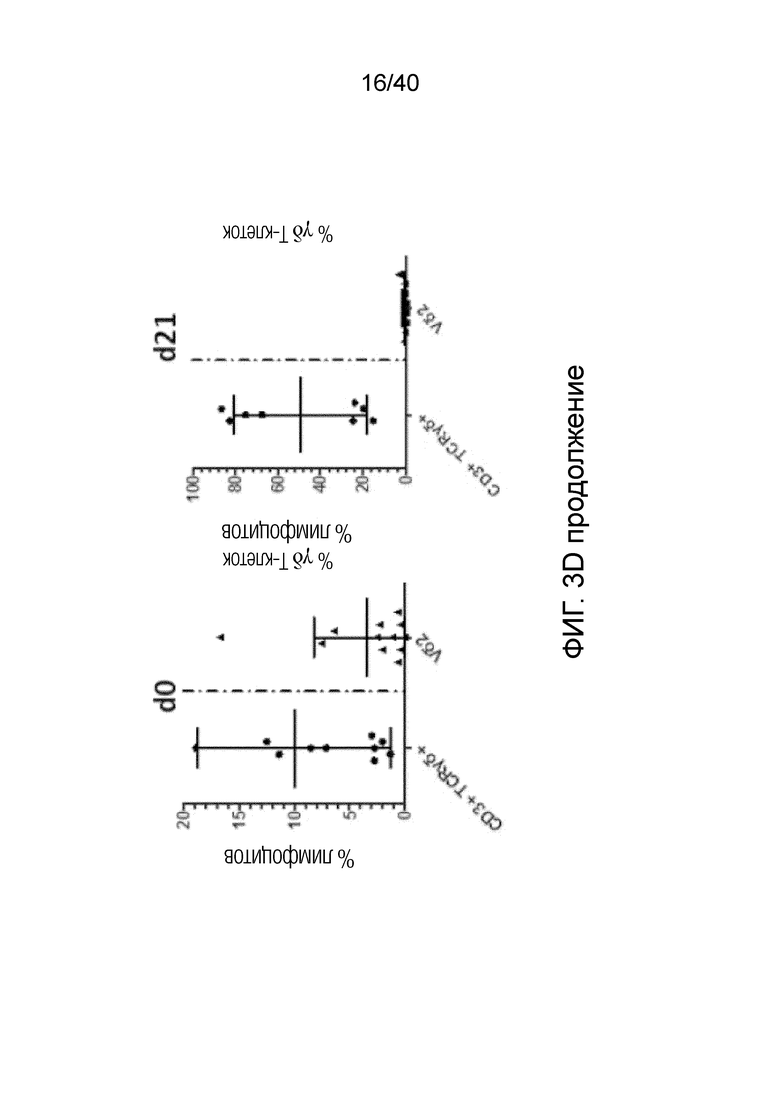
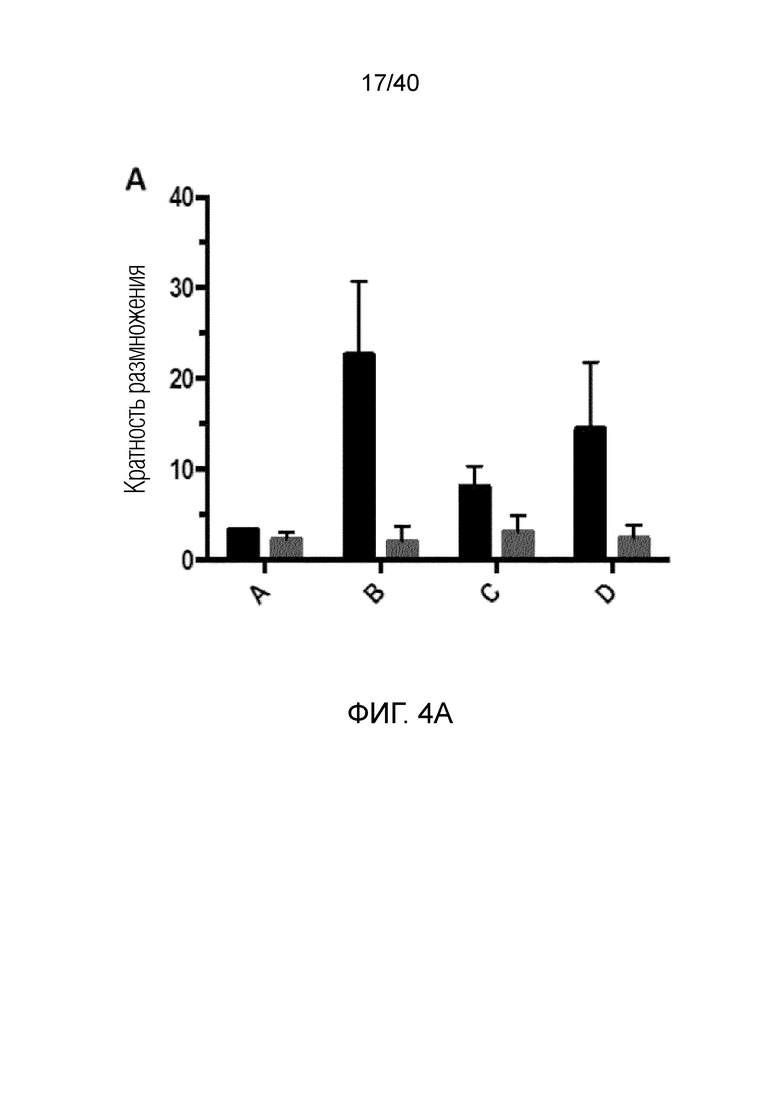
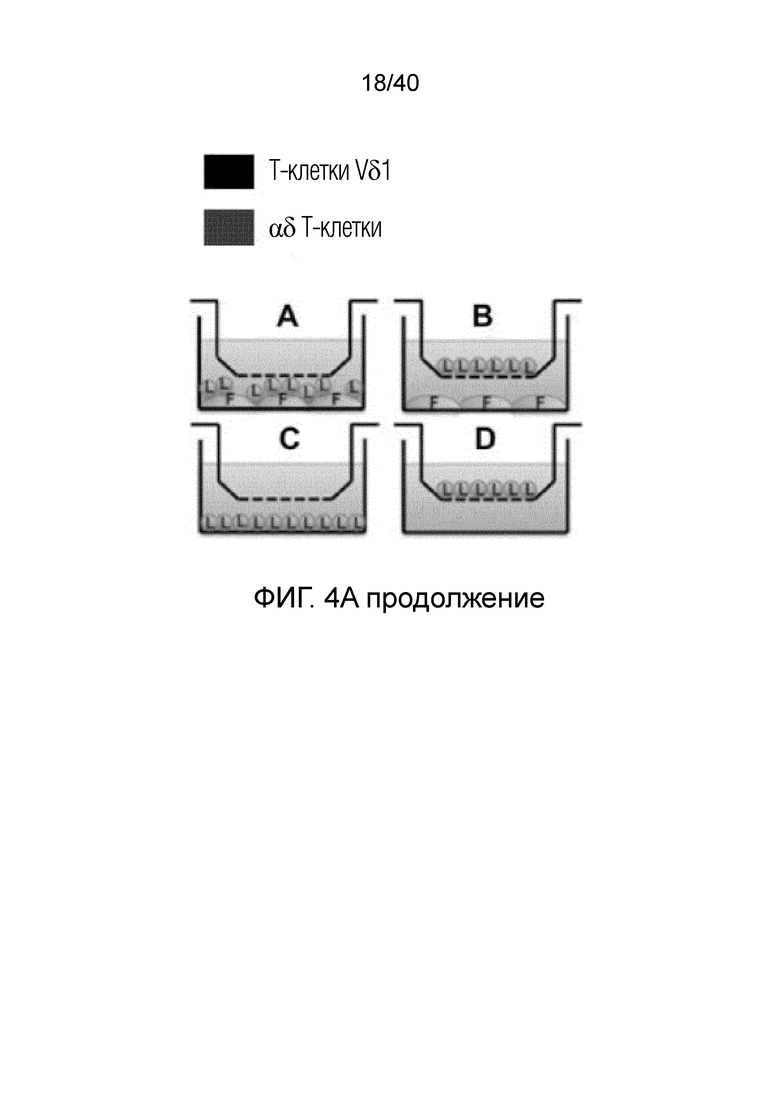
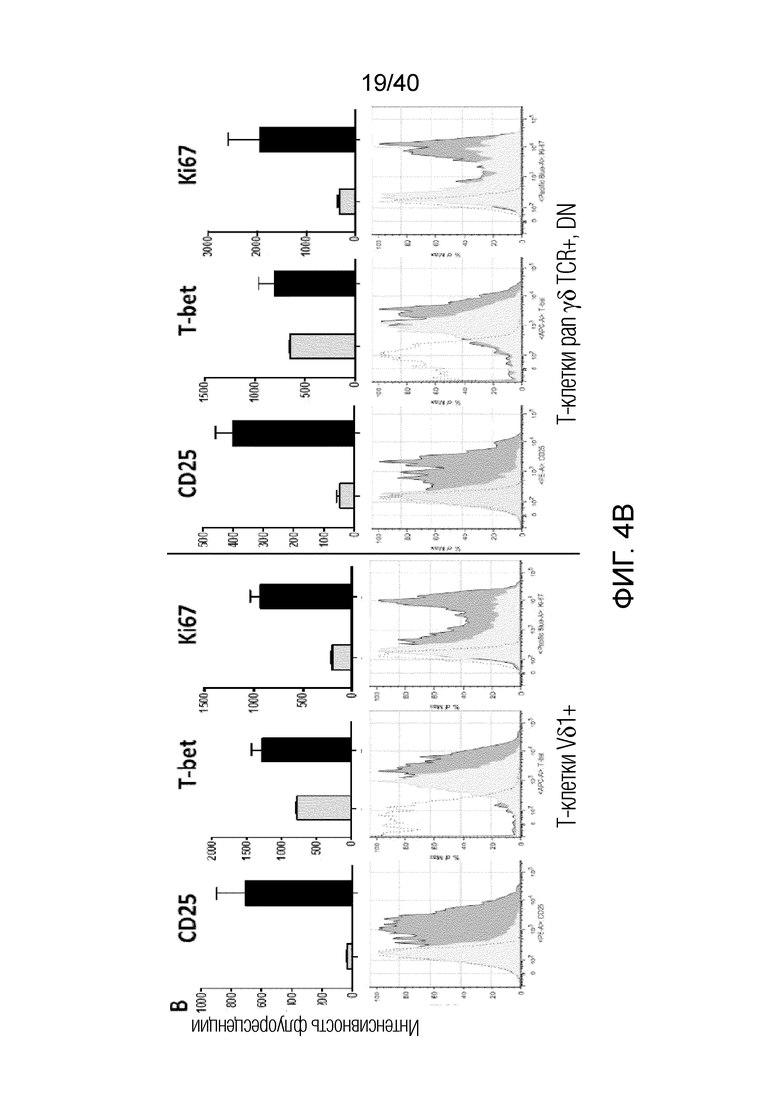
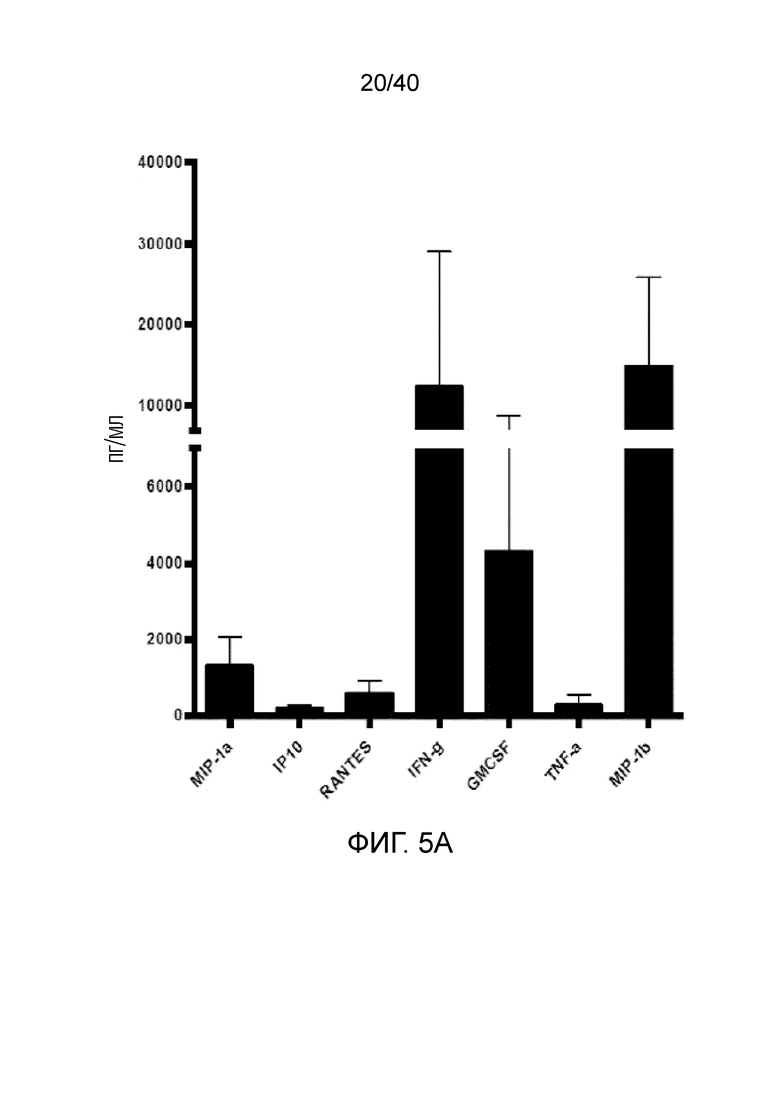

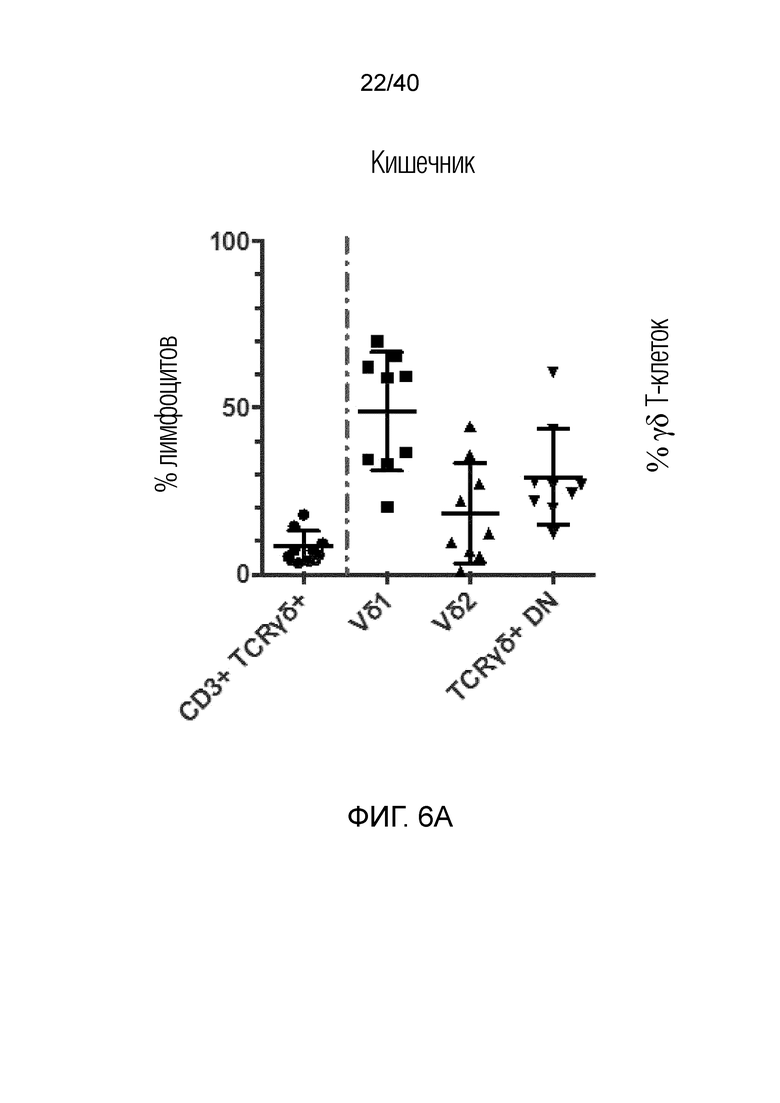
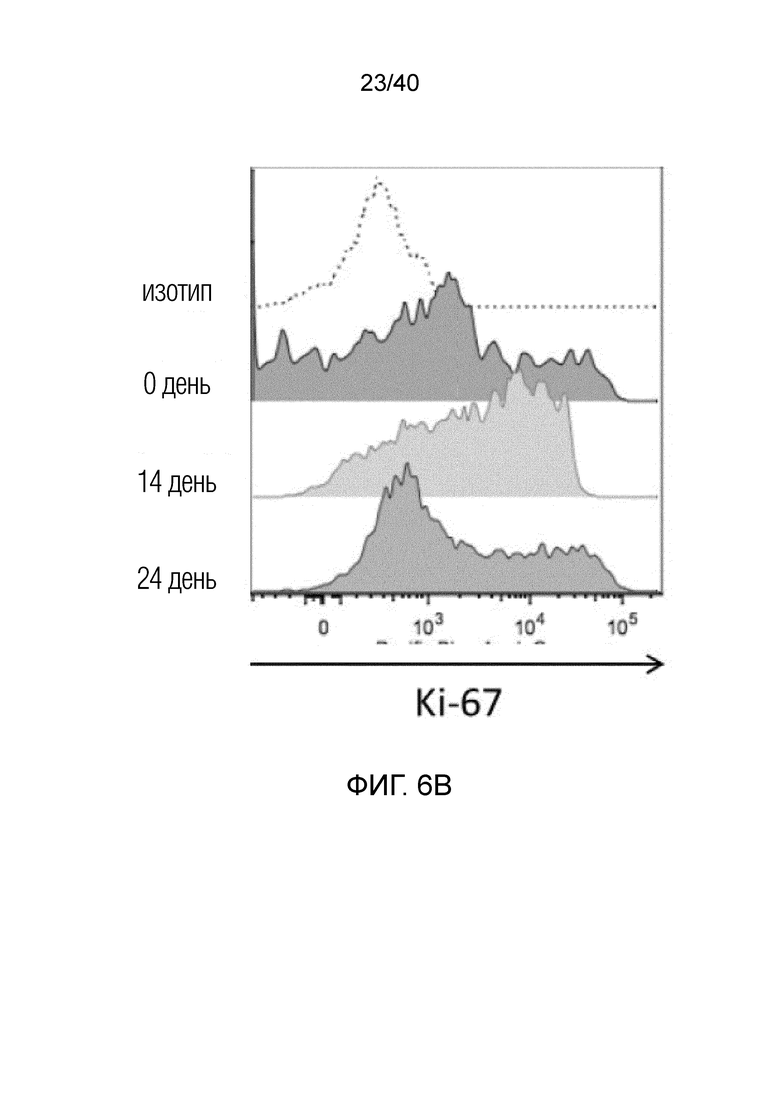
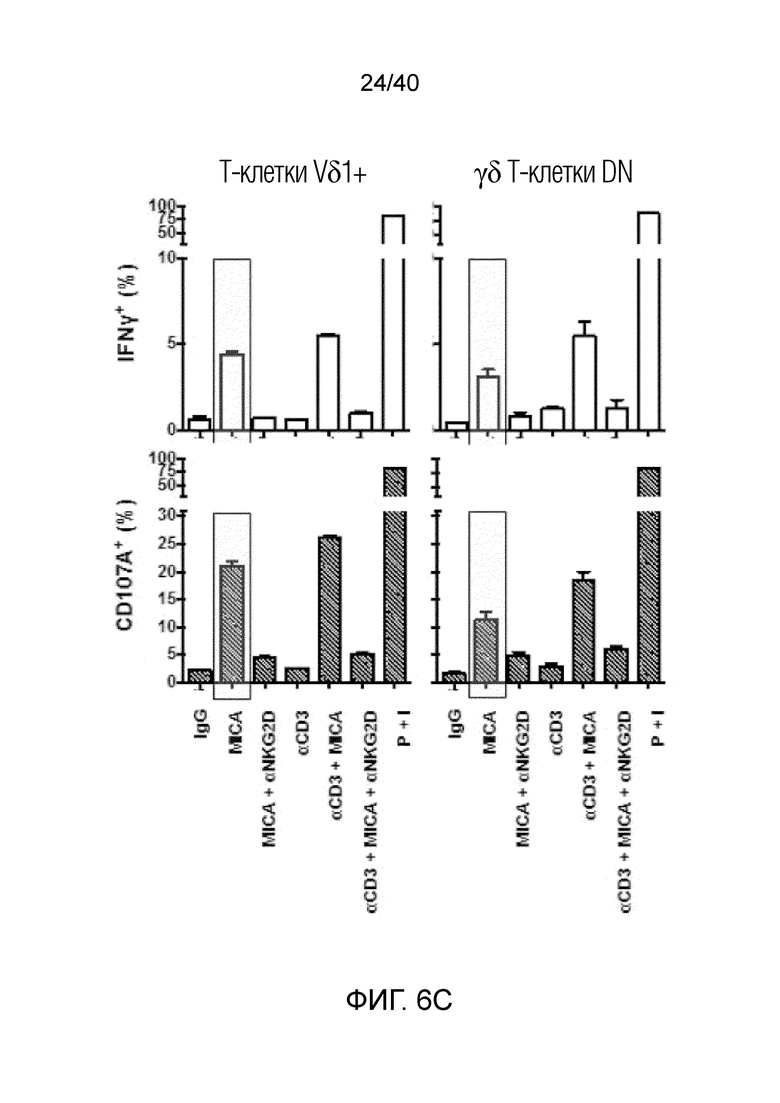
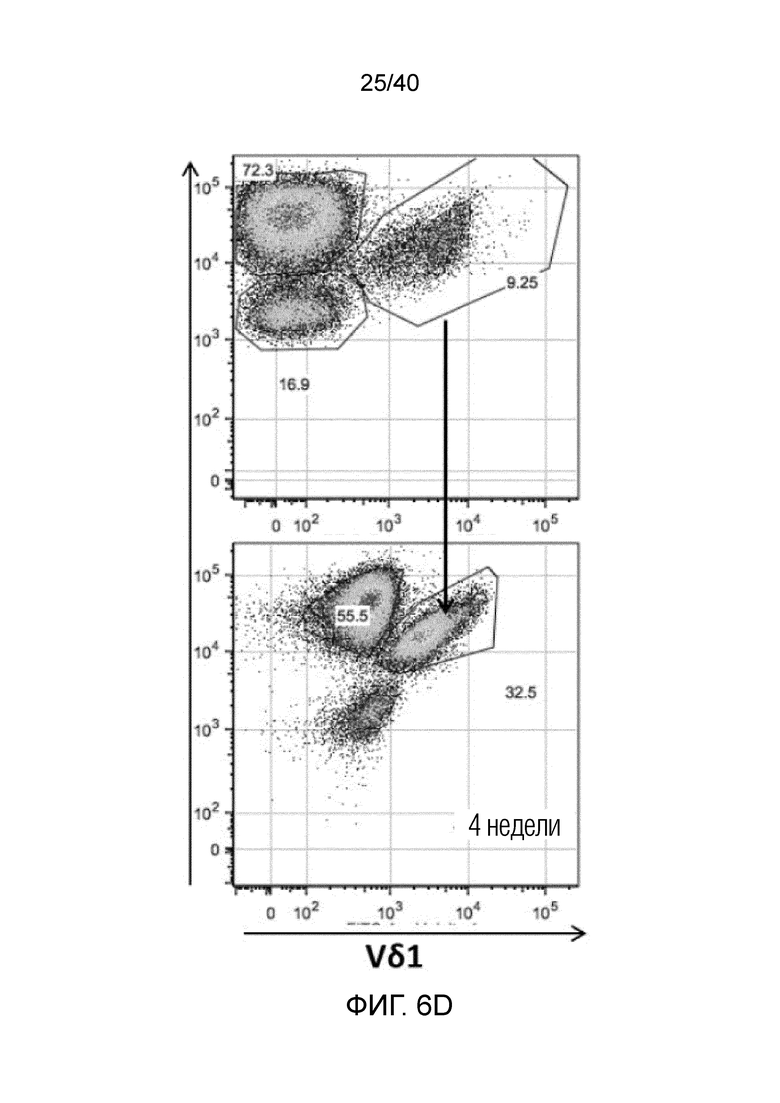
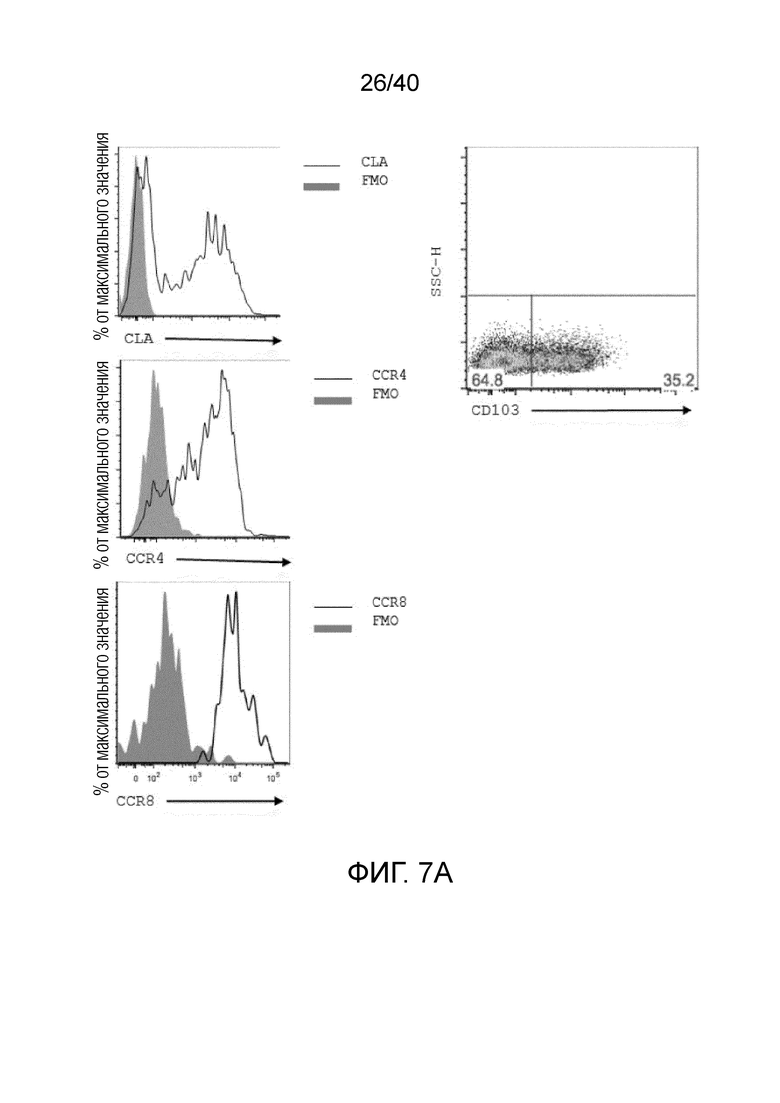
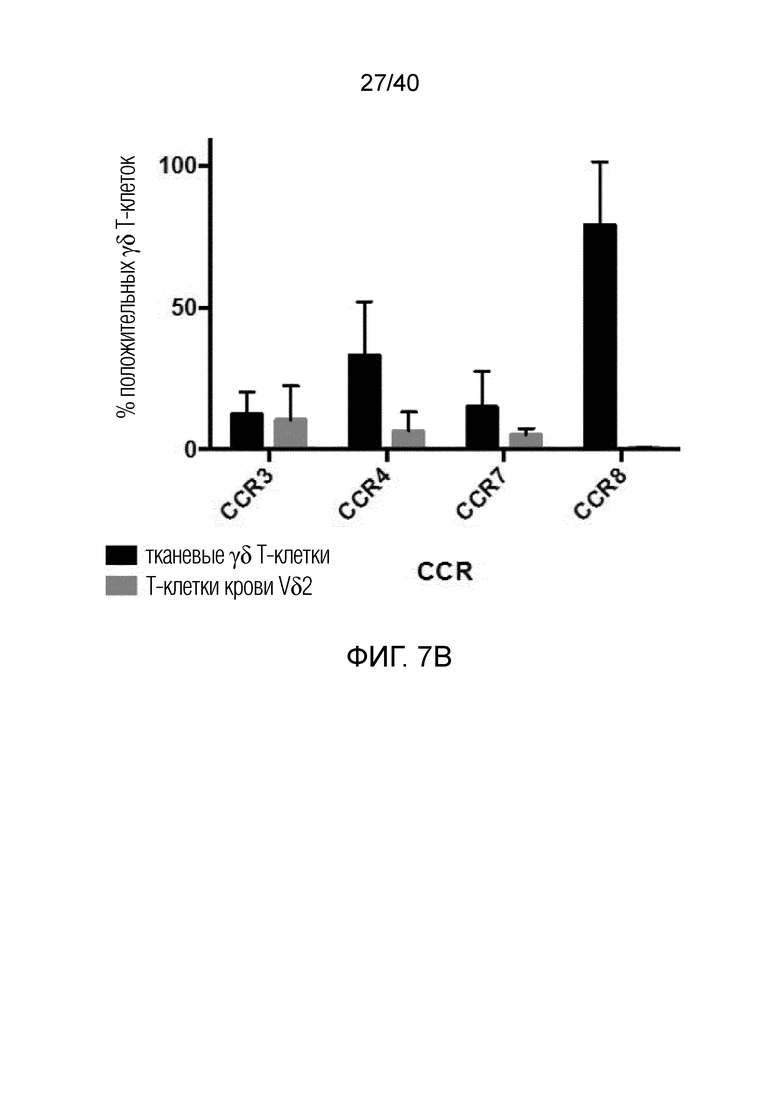
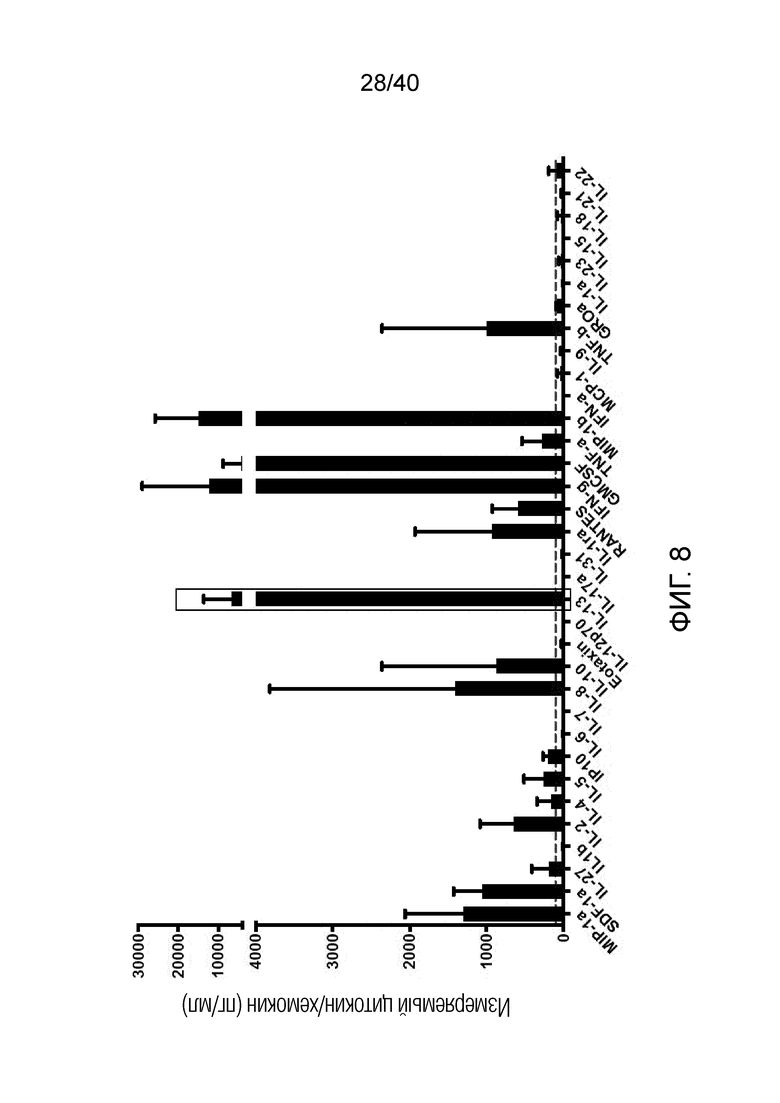
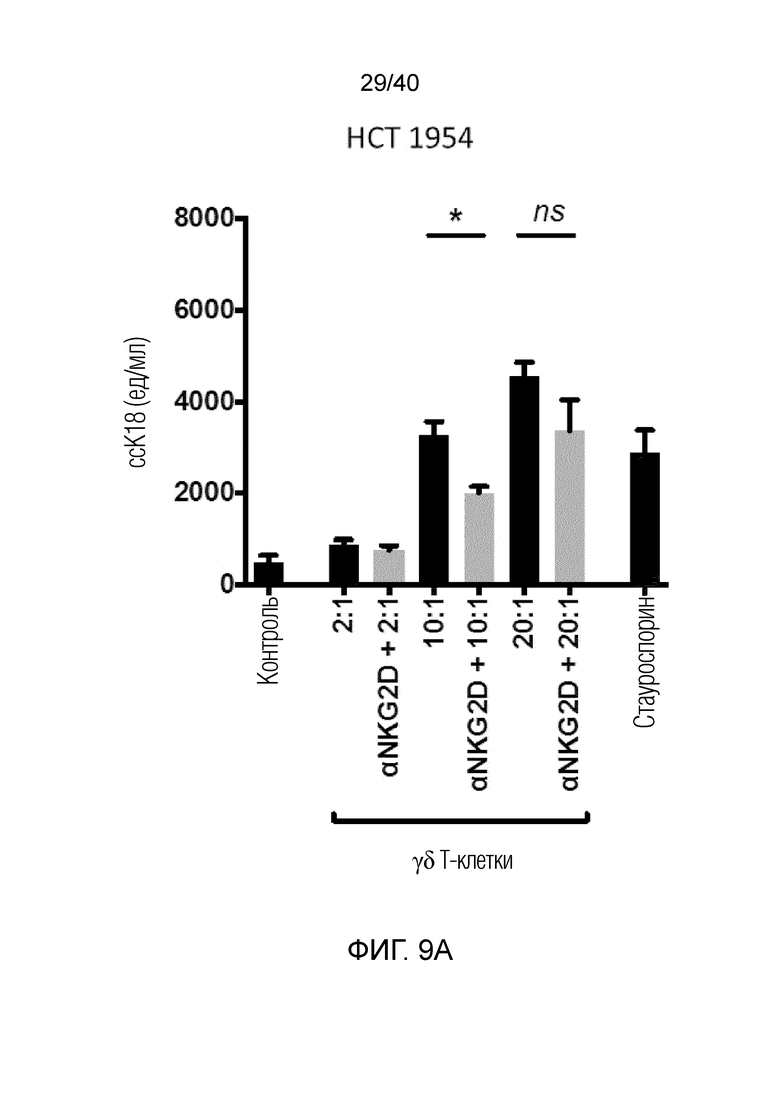
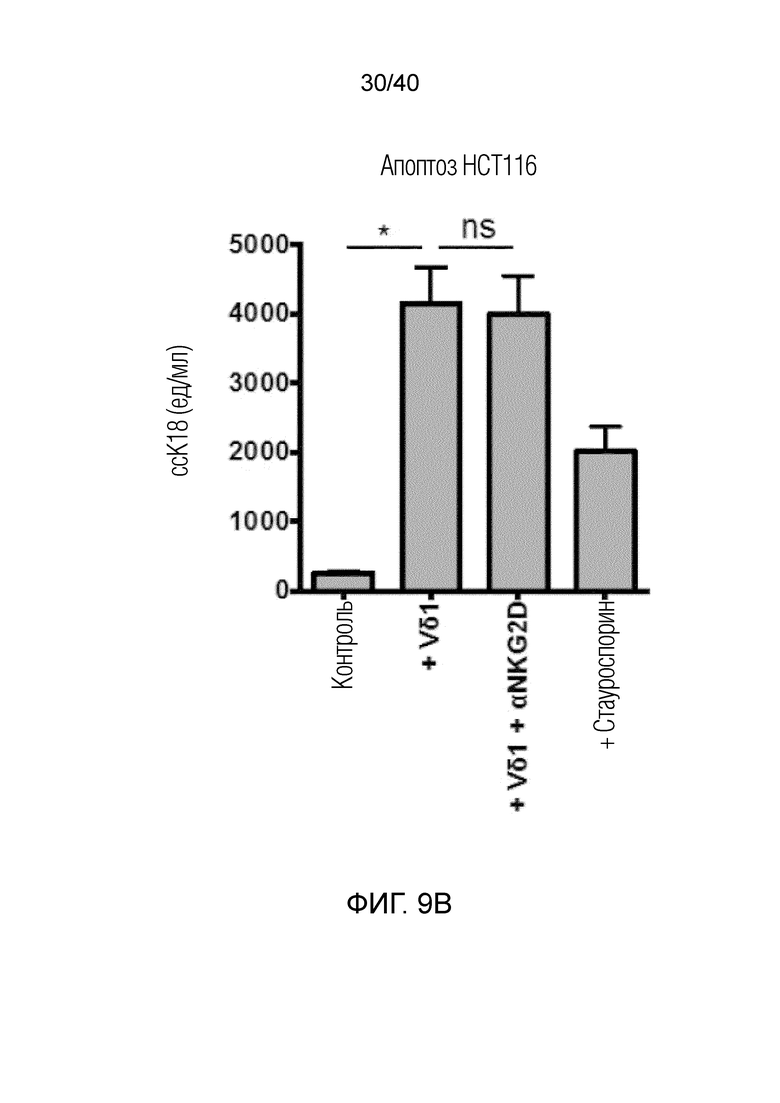
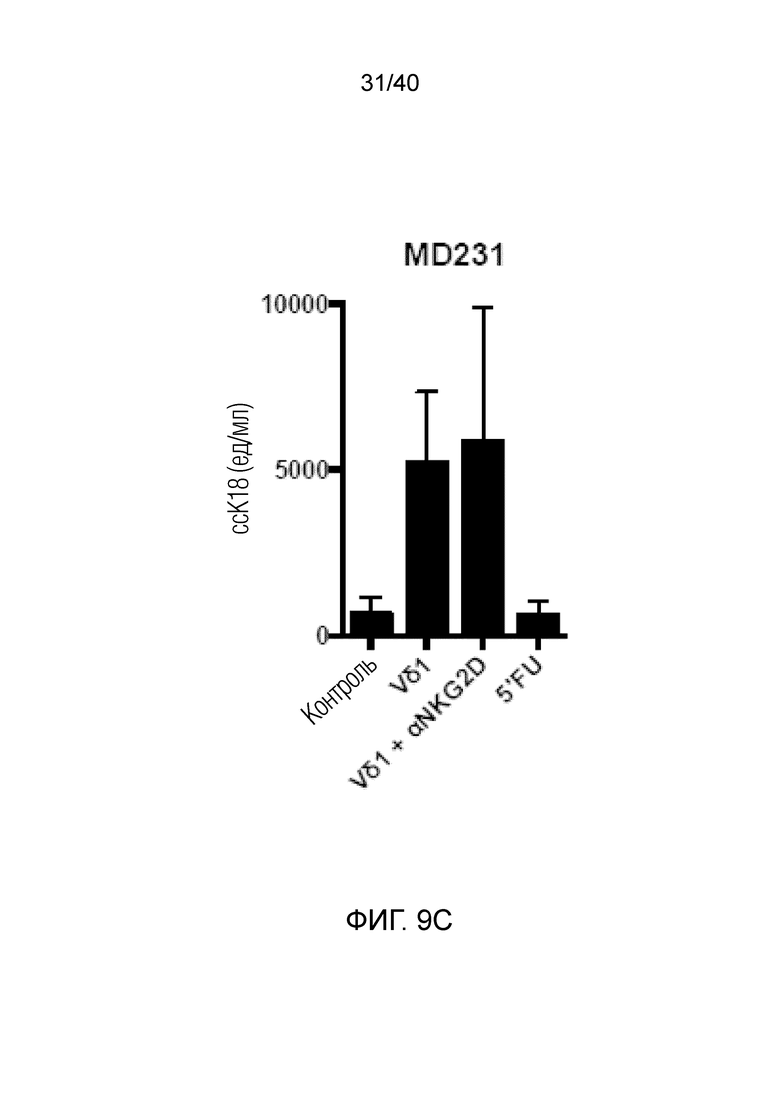
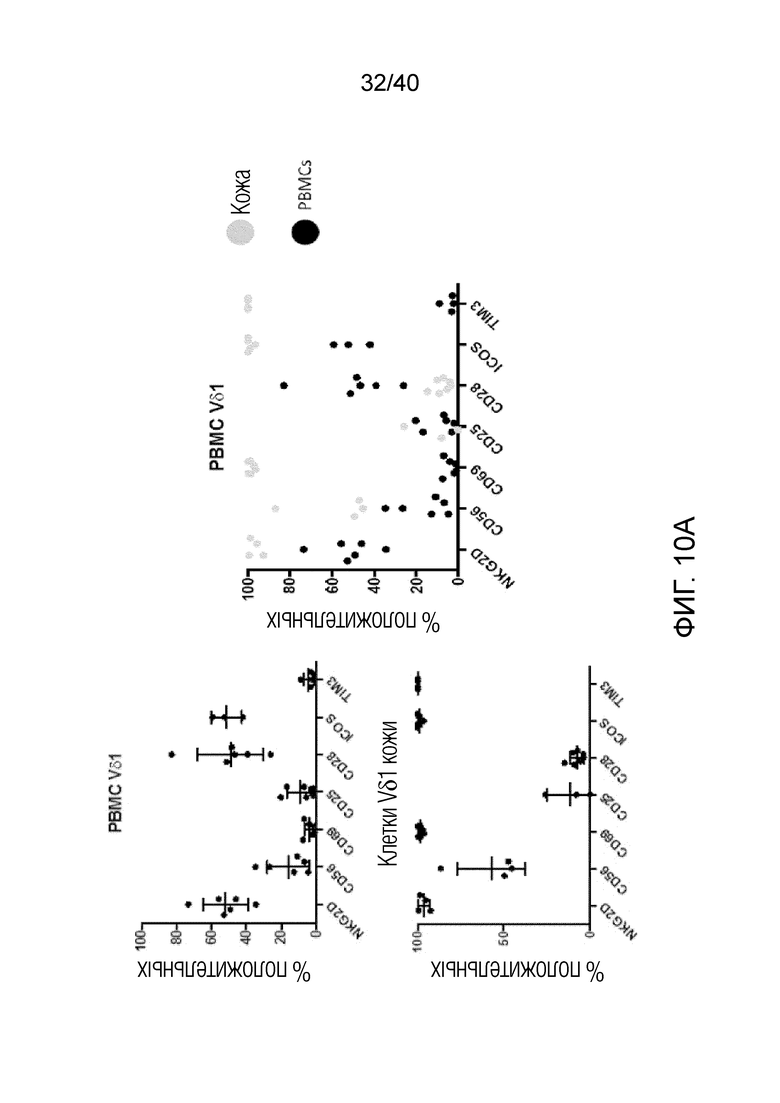
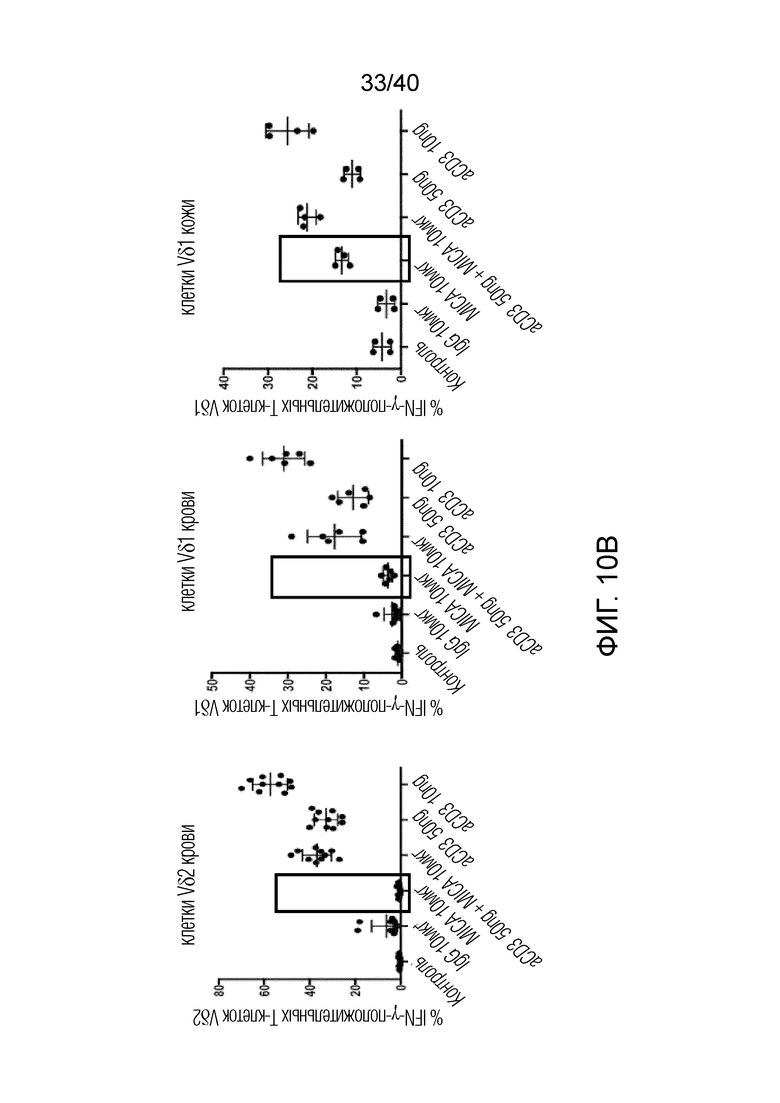
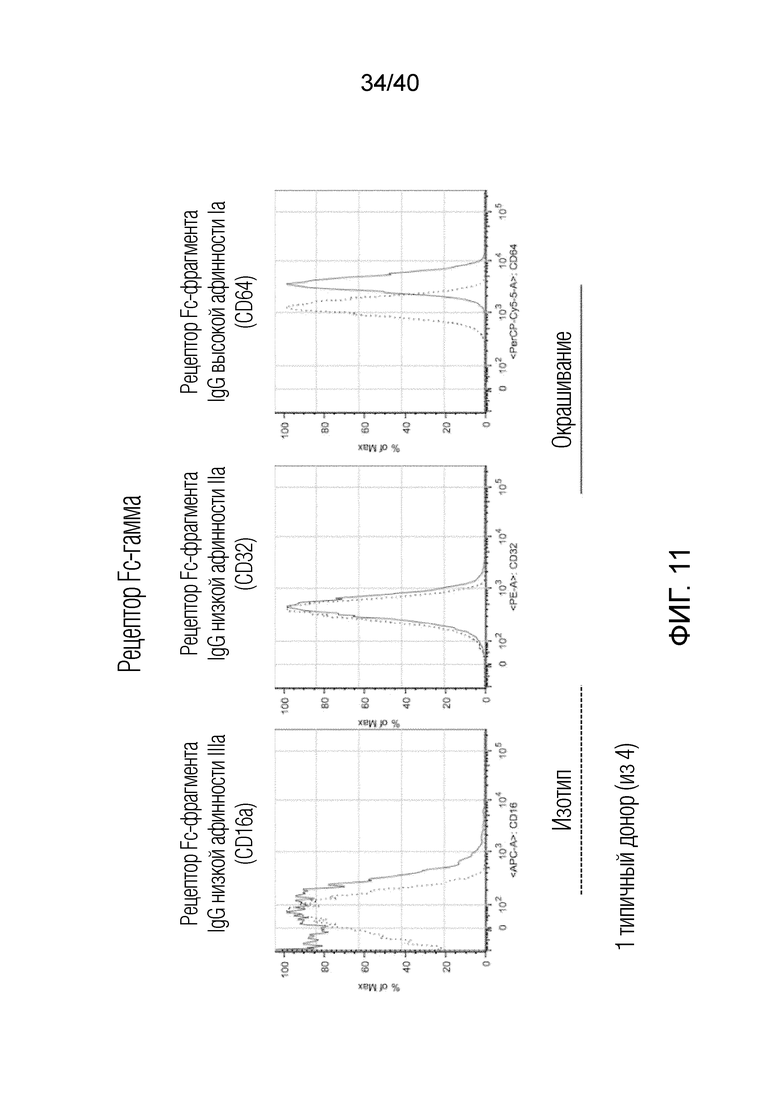
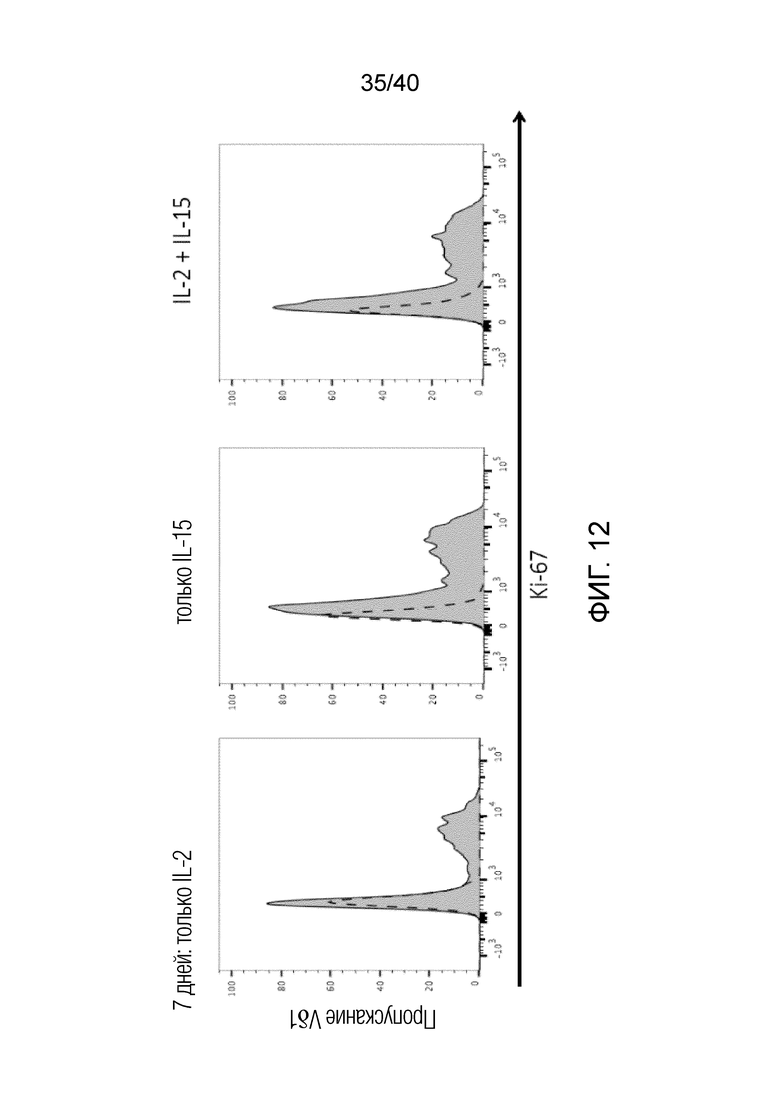
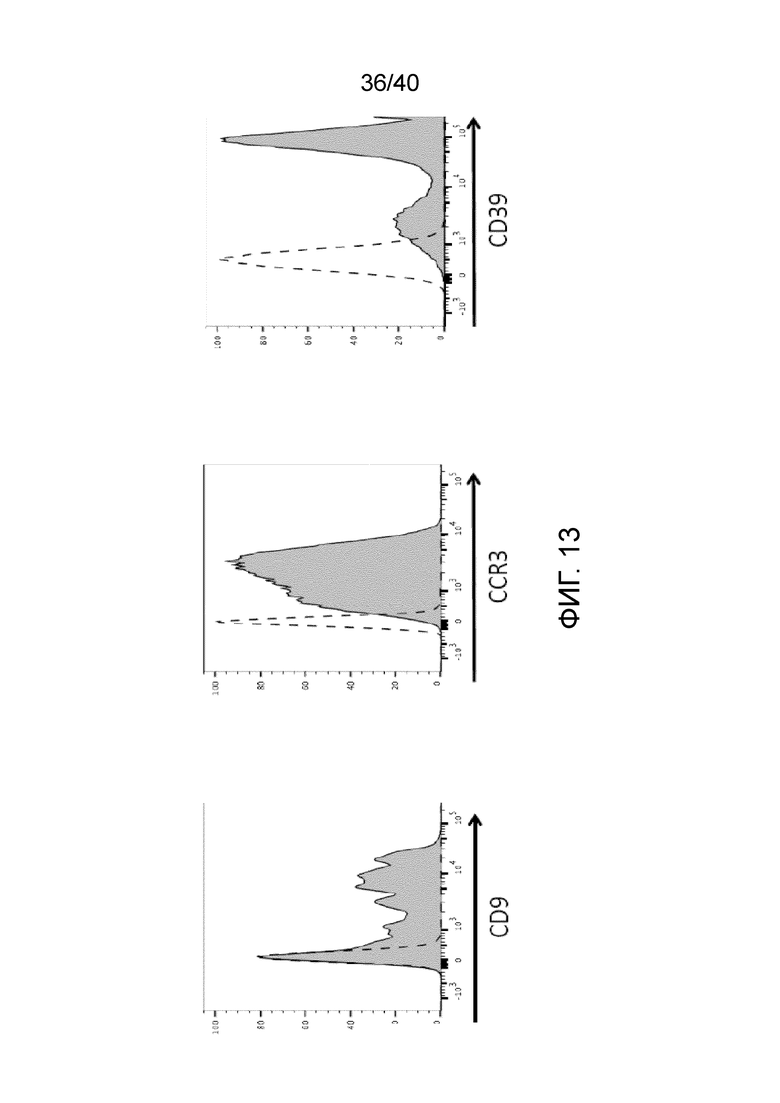
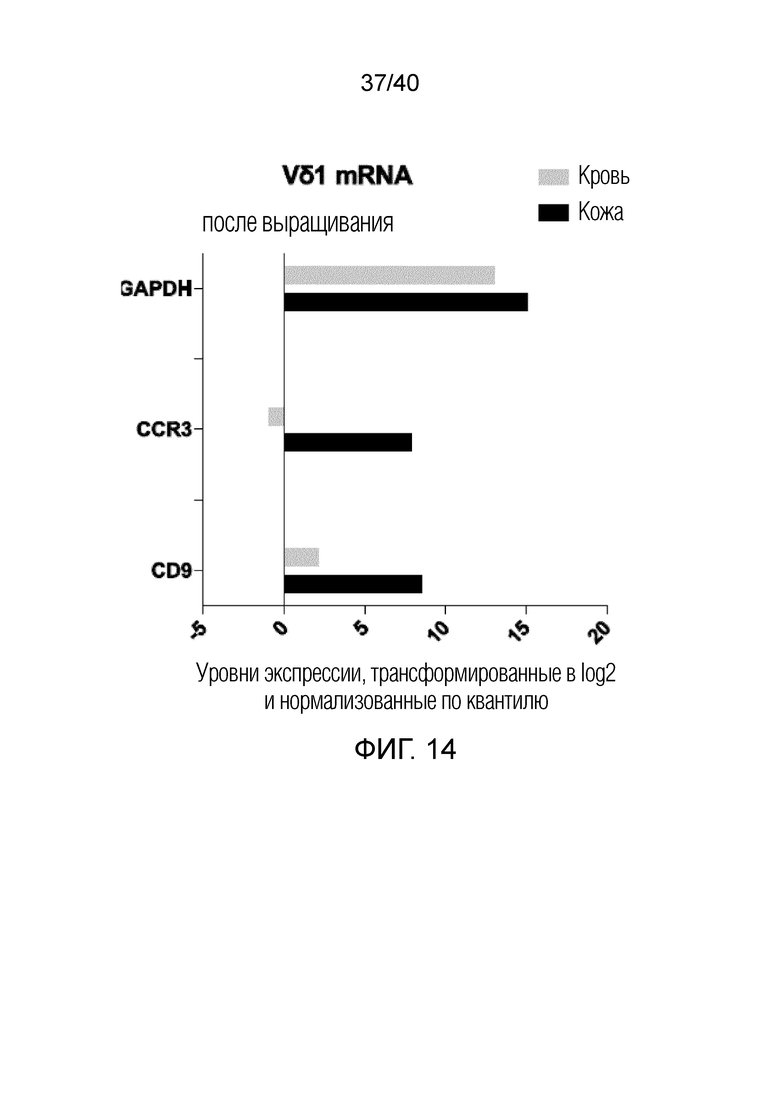
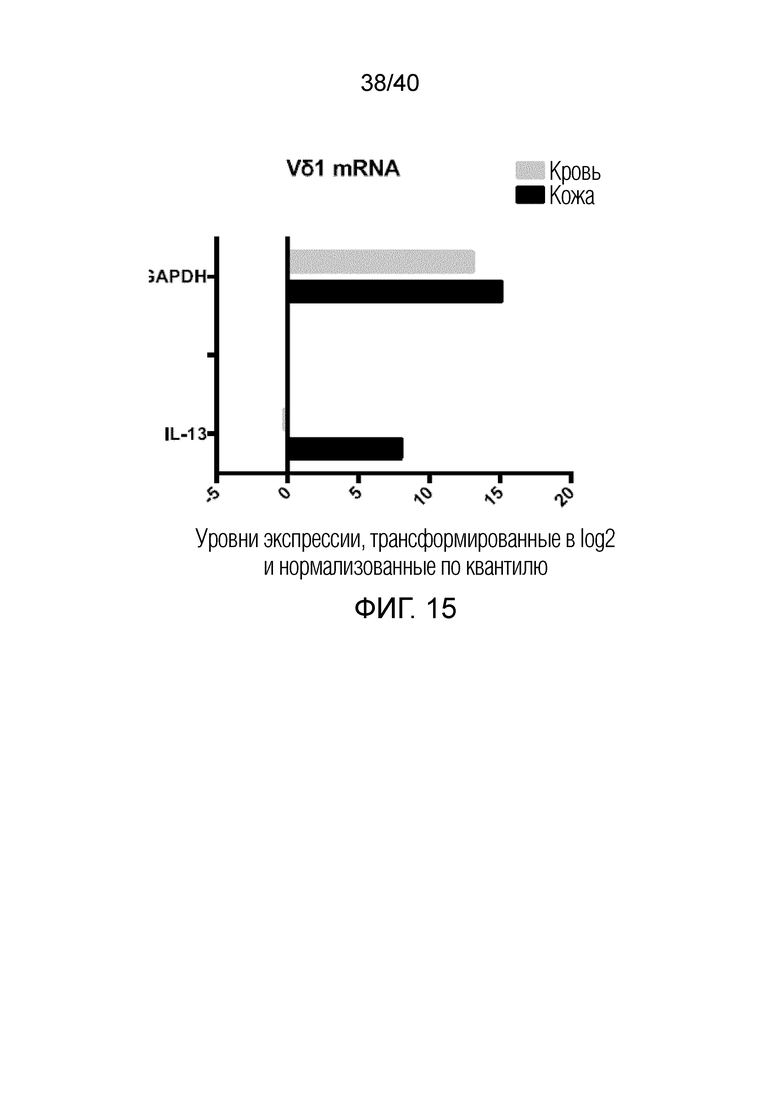
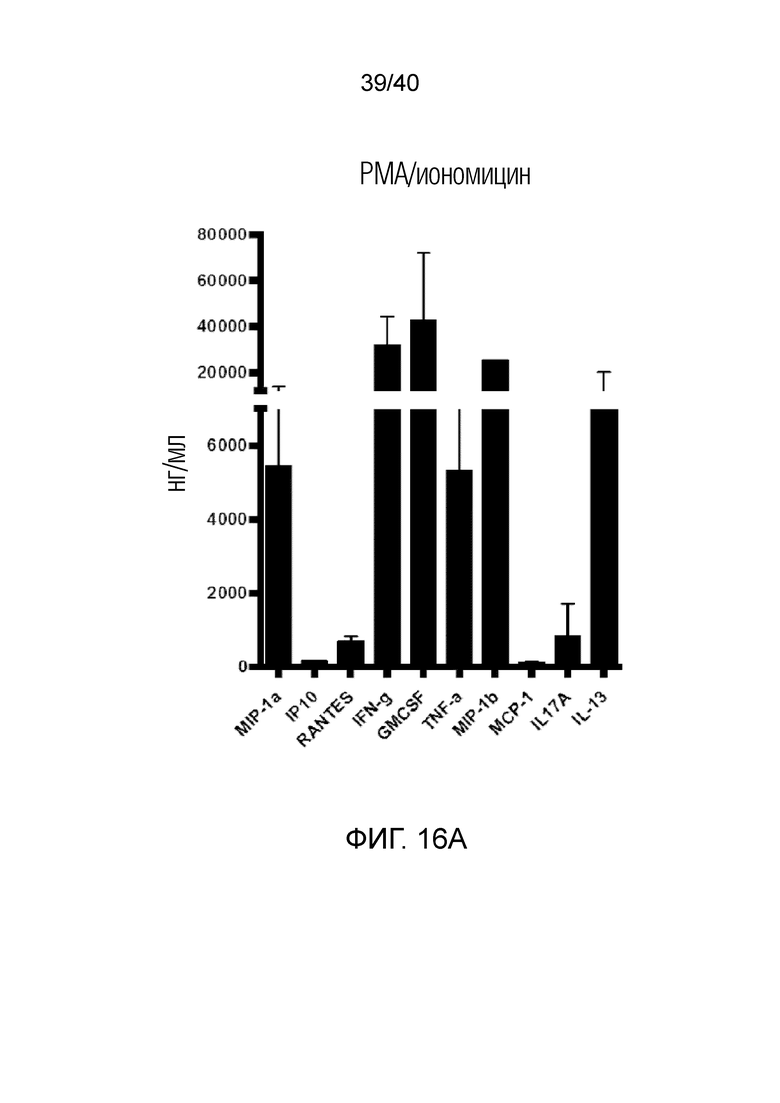
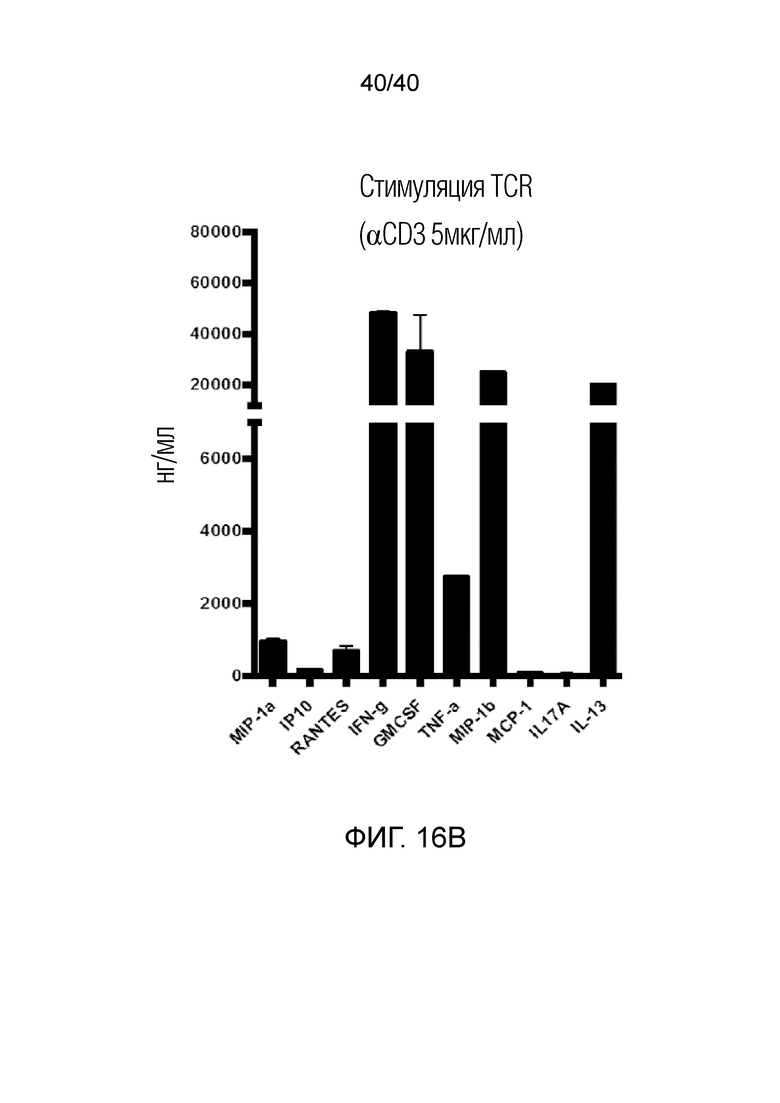
Настоящее изобретение относится к клеточной биологии и иммунологии, в частности к способу размножения негемопоэтических тканерезидентных γδ T-клеток in vitro. Для осуществления способа культивируют лимфоциты, полученные из негемопоэтической ткани человека или отличного от человека животного, в присутствии IL-15 или IL-15 и IL-2. При этом лимфоциты не находятся в непосредственном контакте со стромальными или эпителиальными клетками в процессе культивирования. При это лимфоциты культивируются в отсутствии экзогенно добавленного агониста пути Т-клеточного рецептора. Настоящее изобретение позволяет повысить эффективность способа размножения γδ Т-клеток, находящихся в негемопоэтической ткани. 14 з.п. ф-лы, 1 пр., 35 ил.
1. Способ размножения негемопоэтических тканерезидентных γδ Т-клеток in vitro, включающий в себя культивирование лимфоцитов, полученных из негемопоэтической ткани человека или отличного от человека животного, в присутствии интерлейкина-15 (IL-15), где лимфоциты не находятся в непосредственном контакте со стромальными или эпителиальными клетками в процессе культивирования, и где лимфоциты культивируются в отсутствии экзогенно добавленного агониста пути Т-клеточного рецептора.
2. Способ по п.1, где лимфоциты не находятся в непосредственном контакте с фибробластами в процессе культивирования.
3. Способ по п.1 или 2, где способ включает в себя культивирование лимфоцитов, полученных из негемопоэтической ткани человека или отличного от человека животного, в присутствии IL-2 и IL-15.
4. Способ по любому из предшествующих пунктов, где способ включает в себя культивирование лимфоцитов в отсутствии сигналов активации TCR.
5. Способ по любому из предшествующих пунктов, где лимфоциты культивируют в отсутствии стромальных или эпителиальных клеток.
6. Способ по п.5, где стромальные или эпителиальные клетки удаляют перед культивированием.
7. Способ по п.5, где лимфоциты культивируют в отсутствии фибробластов.
8. Способ по п.7, где фибробласты удаляют перед культивированием.
9. Способ по любому из предшествующих пунктов, где лимфоциты культивируют в среде для выращивания γδ, содержащей IL-2 и IL-15.
10. Способ по п.9, где среде для выращивания γδ не активирует или не стимулирует Т-клеточные рецепторы.
11. Способ по любому из предшествующих пунктов, где лимфоциты получают из ткани кожи, желудочно-кишечного тракта, молочной железы, легкого, печени, поджелудочной железы или предстательной железы.
12. Способ по п.11, где лимфоциты получают из толстой кишки.
13. Способ по любому из предшествующих пунктов, где γδ Т-клетки представляют собой клетки, отличные от Vδ2.
14. Способ по п.13, где γδ Т-клетки представляют собой клетки Vδ1.
15. Способ по п.13, где γδ Т-клетки представляют собой двойные отрицательные (DN) γδ Т-клетки.
| KIM K | |||
| et al., Comparative analysis of human epidermal and peripheral blood γδ T cell cytokine profiles, Ann Dermatol., 2014, 26(3), pp | |||
| Распределительный механизм для паровых машин | 1921 |
|
SU308A1 |
| GARCIA V.E | |||
| et al., IL-15 enhances the response of human γδ T cells to nonpetide microbial antigens, J Immunol., 1998, 160(9), pp | |||
| МНОГОКАМЕРНАЯ ПЕЧЬ КОЛЬЦЕОБРАЗНОЙ ФОРМЫ ДЛЯ ОБЖИГА ДИНАСА И ДР. КЕРАМИЧЕСКИХ ИЗДЕЛИЙ | 1924 |
|
SU4322A1 |
| VAN ACKER H.H | |||
| et al., | |||

