Область техники
Изобретение относится к области биомедицины, в частности, к улучшенному синтетическому Т-клеточному рецептору и антигенному рецептору (STAR), таргетирующему CD19 и CD20, Т-клетке, содержащей синтетический Т-клеточный рецептор и антигенный рецептор, и их применению.
Уровень техники
Клеточная терапия, особенно терапия, связанная с Т-клетками, быстро развивалась в этом году, и в ее рамках большое внимание привлекла терапия Т-клетками с химерными антигенными рецепторами (CAR-T) и терапия TCR-T.
Терапия CAR-T основана на экспрессии молекул CAR в Т-клетках. Молекула CAR состоит из трех частей: эктодомена, который представляет собой домен распознавания антигена, полученный из антитела и отвечающий за распознавание антигена-мишени; трансмембранный домен; и эндодомен, который представляет собой сигнальную молекулу и костимулирующую сигнальную молекулу, полученную из Т-клеточного рецептора, и отвечает за передачу сигнала активации Т-клетки после получения стимуляции. Когда молекулы CAR связываются с соответствующими их антигенами, они агрегируются, запускают эффекторную функцию Т-клеток и убивают опухолевые клетки-мишени.
Терапия TCR-T основана на Т-клеточном рецепторе (TCR). TCR представляет собой идентичность Т-клеток, которые можно разделить на αβ Т-клетки и γδ Т-клетки в зависимости от типа TCR. В процессе развития, Т-клетка-предшественница претерпевает перегруппировку VDJ в цепях TCRγ и TCRδ, которая, в случае успешной перестройки, развивается в γδ Т-клетку, или, если перестройка заканчивается неудачей, подвергается рекомбинации VDJ в TCRα и TCRβ цепи, и затем развиваются в αβ Т-клетку. αβ Т-клетки составляют 90-95% Т-клеток периферической крови, тогда как γδ Т-клетки составляют 5-10% Т-клеток периферической крови. Два типа Т-клеток распознают антигены MHC-ограниченным и MHC-неограниченным способами, соответственно, которые играют важную роль в иммунитете к патогенам и опухолям.
Молекула комплекса Т-клеточного рецептора (TCR) содержит несколько цепей, в которых TCRα и TCRβ цепи (или TCRγ и TCRδ цепи) отвечают за распознавание молекул MHC-полипептида, и остальные шесть субъединиц CD3 связываются с α/β цепями TCR (или γ/δ цепями TCR), чтобы играть роль трансдукции сигнала. Природный комплекс TCR содержит десять сигнальных последовательностей ITAM, которые теоретически могут передавать более сильные сигналы, чем CAR. Используя функцию трансдукции сигнала природного TCR, можно сконструировать новый рецептор для облегчения нарушения функций Т-клеток, который может играть лучшую роль в борьбе с солидными опухолями. Эктодомен TCR очень похож на Fab домен антитела, поэтому последовательность вариабельной области TCR может быть заменена последовательностью вариабельной области антитела, чтобы получить синтетический TCR и антигенный рецептор (STAR), который обладает не только специфичностью к антителам, но также обладает превосходной функцией передачи сигнала природного TCR при опосредовании активации Т-клеток.
Однако STAR-T, полученный из природного TCR, не обладает костимулирующим сигналом при активации Т-клеток, и его способность к пролиферации и активации часто нарушается. Следовательно, в этой области по-прежнему необходимы улучшенные TCR и соответствующая терапия TCR-T.
Краткое описание чертежей
Фиг. 1 Схематическая диаграмма оптимизации STAR путем модификации цистеина константной области, модификации трансмембранного домена и эндодомена.
Фиг. 2 Схематическая диаграмма оптимизации STAR путем добавления костимулирующих молекул рецепторных эндодоменов к α и/или β цепям.
Фиг. 3. Схематическая диаграмма оптимизации STAR путем добавления костимулирующей молекулы к эндодомену рецептора непосредственно или через линкер после делеции эндодоменов α и/или β цепи.
Фиг. 4 Схематическая диаграмма оптимизации STAR путем добавления эндодоменов костимулирующих молекул рецепторов к субъединицам CD3.
Фиг. 5 Схематическая диаграмма оптимизации STAR путем добавления доменов трансдукции сигнала цитокинового рецептора к α и/или β цепям.
Фиг. 6. Сравнение способности уничтожать опухолевые клетки-мишени Т-клеток WT-STAR и Т-клеток mut-STAR.
Фиг. 7 Влияние различных эндодоменов костимулирующих рецепторов на способность STAR-T-клеток уничтожать мишени при различных соотношениях эффектор/мишень (Е:Т).
Фиг. 8 Влияние различных эндодоменов костимулирующих рецепторов на способность STAR-T-клеток уничтожать мишени при различном времени совместного культивирования.
Фиг. 9 Влияние эндодомена OX40, добавленного к α цепи, β цепи, α цепи и β цепи, на способность STAR-T-клеток уничтожать мишени при различных соотношениях E:T.
Фиг. 10. Влияние эндодомена OX40, добавленного к α цепи, β цепи, α цепи и β цепи, на способность STAR-T-клеток уничтожать мишени при различном времени совместного культивирования.
Фиг. 11 Влияние различных эндодоменов костимулирующих рецепторов на секрецию цитокинов STAR-T-клетками.
Фиг. 12 Влияние различных эндодоменов костимулирующих рецепторов на пролиферацию STAR-T-клеток.
Фиг. 13 Влияние эндодомена OX40, добавленного к α цепи, β цепи, α цепи и β цепи, на способность STAR-T-клеток уничтожать мишени.
Фиг. 14 Способность mut-STAR уничтожать опухоль с различными эндодоменами костимулирующих рецепторов, связанными с α цепью TCR.
Фиг. 15. Влияние различных эндодоменов костимулирующих рецепторов, добавленных к различным субъединицам CD3, на способность STAR-T-клеток уничтожать мишени.
Фиг. 16 Влияние различных эндодоменов костимулирующих рецепторов, добавленных к различным субъединицам CD3, на пролиферацию STAR-T-клеток.
Фиг. 17 Влияние эндодомена OX40, связанного с α цепью с делецией эндодомена через различные линкеры G4S, на способность STAR-T-клеток уничтожать мишени.
Фиг. 18 Влияние эндодомена OX40, связанного с β цепью с делецией эндодомена через различные линкеры G4S, на способность STAR-T-клеток уничтожать мишени.
Фиг. 19. Влияние эндодомена OX40, связанного с α и/или β цепями с делецией эндодомена через различные линкеры G4S, на секрецию IL-2 STAR-T-клетками.
Фиг. 20. Влияние эндодомена OX40, связанного с α и/или β цепями с делецией эндодомена через различные линкеры G4S, на секрецию IFN-γ STAR-T-клетками.
Фиг. 21 Влияние эндодомена OX40, связанного с α цепями с делецией эндодомена через различные линкеры G4S, на дифференциацию Т-клеток центральной памяти.
Фиг. 22. Влияние эндодомена OX40, связанного с α цепями с делецией эндодомена через различные линкеры G4S, на дифференциацию Т-клеток.
Фиг. 23. Влияние модификации лизина в трансмембранном домене или эндодомене на способность STAR-T-клеток уничтожать мишени.
Фиг. 24. Влияние модификации лизина в трансмембранном домене или эндодомене на секрецию IFN-γ STAR-T-клетками.
Фиг. 25 Влияние модификации лизина в трансмембранном домене или эндодомене на секрецию IL-2 STAR-T-клетками.
Фиг. 26 Влияние модификации лизина в трансмембранном или эндодомене на дифференциацию Т-клеток центральной памяти.
Фиг. 27. Влияние модификации лизина в трансмембранном или эндодомене на дифференциацию Т-клеток.
Фиг. 28. Влияние различных доменов передачи сигнала цитокинового рецептора, связанных с α и/или β цепями, на способность STAR-T-клеток уничтожать мишени.
Фиг. 29. Сравнение эффектов уничтожения между мутантным STAR с различными связанными доменами, стимулирующими цитокиновый рецептор, и α-del-(G4S) 3-OX40-STAR.
Фиг. 30. Влияние различных доменов передачи сигнала цитокинового рецептора, связанных с α и/или β цепями, на секрецию IL-2 STAR-T-клетками.
Фиг. 31. Влияние различных доменов передачи сигнала цитокинового рецептора, связанных с α и/или β цепями, на секрецию IFN-γ STAR-T-клетками.
Фиг. 32. Влияние различных доменов передачи сигнала цитокинового рецептора, связанных с α и/или β цепями, на дифференциацию Т-клеток центральной памяти.
Фиг. 33. Влияние различных доменов передачи сигнала цитокинового рецептора, связанных с α и/или β цепями, на дифференциацию Т-клеток.
Фиг. 34. Противоопухолевые влияние in vivo αβ OX40-STAR T, mut-STAR T и CAR-T в мышиной модели опухоли in vivo.
Фиг. 35. Кривые выживания мышей, которым вводят αβ OX40-STAR T, mut-STAR T и CAR-T.
Фиг. 36 Пролиферация in vivo αβ OX40-STAR T, mut-STAR T и CAR-T у мышей.
Фиг. 37. Противоопухолевый эффект in vivo различных структур STAR и CAR-T на мышиной модели опухоли.
Фиг. 38 Схематическая диаграмма оптимизации STAR путем модификации цистеина константной области, трансмембранного домена и N-концевой перегруппировки.
Фиг. 39 Пример положения соединения между доменом костимулирующей молекулы и структурой STAR. Показана только связь с α цепью.
Фиг. 40 Пример структуры STAR, содержащей домен костимулирующей молекулы.
Фиг. 41. Способность уничтожения STAR и STAR, содержащей костимулирующий фактор.
Фиг. 42 Уровень ядерного RELB, связанный с сигналом пролиферации STAR и STAR, содержащих костимулирующие факторы.
Фиг. 43. Результаты анти-CD19 различных STAR.
Фиг. 44. Результаты анти-CD19 и CD20 различных STAR с костимулирующими факторами, добавленными к цепям α и β.
Фиг. 45. Результаты анти-CD19 и CD20 STAR с костимулирующими факторами, добавленными к α цепи.
Сущность изобретения
Если не указано или не определено иное, все используемые термины имеют общепринятое значение в данной области техники, которое будет понятно специалистам в данной области техники. См., например, стандартное руководство, такое как Sambrook et al., “Molecular cloning: a laboratory manual”; Lewin, “Genes VIII” и Roitt et al., “Immunology” (8th edition), и общий предшествующий уровень техники, цитируемый в настоящем документе; кроме того, если не указано иное, все способы, стадии, методы и операции, которые конкретно не описаны, могут и были выполнены известным способом, который будет понятен специалистам в данной области техники. См. также, например, стандартное руководство, приведенный выше общий уровень техники и другие цитируемые в нем ссылки.
Используемый в настоящем документе термин «и/или» охватывает все комбинации элементов, связанных этим термином, и считается, что они перечислены в настоящем документе отдельно. Например, «А и/или В» охватывает «А», «А и В» и «В». Например, «A, B и/или C» охватывает «A», «B», «C», «A и B», «A и C», «B и C» и «A и B и C».
Термин «содержащий» используется в настоящем документе для описания последовательности белка или нуклеиновой кислоты, которая может состоять из указанной последовательности или может иметь дополнительные аминокислоты или нуклеотиды на одном или обоих концах указанного белка или нуклеиновой кислоты, но при этом обладать активностью описан в настоящем документе. Кроме того, специалистам в данной области техники будет понятно, что метионин, кодируемый стартовым кодоном на N-конце полипептида, сохраняется в определенных практических ситуациях (например, при экспрессии в конкретной системе экспрессии), но существенно не влияет на функцию полипептида. Таким образом, при описании аминокислотной последовательности конкретного полипептида, хотя он может и не содержать метионин, кодируемый инициирующим кодоном на его N-конце, но все же охватывает последовательность, содержащую метионин к тому времени, и, соответственно, кодирующие его нуклеотидные последовательности могут также содержать инициирующий кодон; наоборот.
Используемый в настоящем документе термин «номер аминокислоты со ссылкой на SEQ ID NO: x» (SEQ ID NO: x представляет собой конкретную последовательность, указанную в настоящем документе), означает, что номер положения конкретной описанной аминокислоты является номером положения аминокислоты, соответствующей этой аминокислоте в SEQ ID NO:x. Аминокислотное соответствие между различными аминокислотными последовательностями можно определить способами выравнивания последовательностей, известными в данной области техники. Например, соответствие аминокислот можно определить с помощью онлайн-инструмента выравнивания EMBL-EBI (https://www.ebi.ac.uk/Tools/psa/), в котором две последовательности могут быть выровнены с использованием алгоритма Нидлмана-Вунша с параметрами по умолчанию. Например, если аланин в положении 46, начиная с N-конца полипептида, выровнен в выравнивании последовательности с аминокислотой в положении 48 SEQ ID NO: x, тогда аминокислота в полипептиде также может быть описана в настоящем документе как «аланин в положении 48 полипептида, где положение аминокислоты указано со ссылкой на SEQ ID NO: x». В настоящем изобретении делается ссылка на SEQ ID NO: 3 для положения аминокислоты, относящейся к константной области α цепи. В настоящем изобретении делается ссылка на SEQ ID NO: 4 для положения аминокислоты, относящейся к константной области β цепи.
В одном аспекте, в настоящем документе предложен комплекс модифицированного Т-клеточного рецептора (TCR), где,
i) TCR может представлять собой αβ TCR, комплекс αβ TCR, содержащий α цепь TCR, β цепь TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ; где, по меньшей мере, один функциональный домен соединен с С-концом, по меньшей мере, одной из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ;
где α цепь TCR содержит первую константную область и первую антигенсвязывающую область, и β цепь TCR содержит вторую константную область и вторую антигенсвязывающую область;
где первая антигенсвязывающая область специфически связывается с первым антигеном, и вторая антигенсвязывающая область специфически связывается со вторым антигеном; или первая антигенсвязывающая область и вторая антигенсвязывающая область объединены друг с другом для специфического связывания с первым антигеном и вторым антигеном; предпочтительно, первый антиген представляет собой CD19, и второй антиген представляет собой CD20, или первый антиген представляет собой CD20, и второй антиген представляет собой CD19;
или,
ii) TCR может представлять собой γδ TCR, комплекс γδ TCR, содержащий γ цепь TCR, δ цепь TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ; где, по меньшей мере, один функциональный домен соединен с С-концом, по меньшей мере, одной из γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ;
где γ цепь TCR содержит первую константную область и первую антигенсвязывающую область, и δ цепь TCR содержит вторую константную область и вторую антигенсвязывающую область;
где первая антигенсвязывающая область специфически связывается с первым антигеном, и вторая антигенсвязывающая область специфически связывается со вторым антигеном; или первая антигенсвязывающая область и вторая антигенсвязывающая область объединены друг с другом для специфического связывания с первым антигеном и вторым антигеном; предпочтительно, первый антиген представляет собой CD19, и второй антиген представляет собой CD20, или первый антиген представляет собой CD20, и второй антиген представляет собой CD19.
Как правило, в TCR, антигенсвязывающая область расположена на N-конце константной области, обе из которых могут быть связаны напрямую или через линкер.
В одном аспекте, предложен модифицированный Т-клеточный рецептор (TCR), где TCR может быть
i) αβ TCR, содержащим α цепь TCR и β цепь TCR, где, по меньшей мере, один функциональный домен соединен с С-концом α цепи TCR и/или TCR β цепи αβ TCR;
где α цепь TCR содержит первую константную область и первую антигенсвязывающую область, и β цепь TCR содержит вторую константную область и вторую антигенсвязывающую область;
где первая антигенсвязывающая область специфически связывается с первым антигеном, и вторая антигенсвязывающая область специфически связывается со вторым антигеном; или первая антигенсвязывающая область и вторая антигенсвязывающая область объединены друг с другом для специфического связывания с первым антигеном и вторым антигеном; предпочтительно, первый антиген представляет собой CD19, и второй антиген представляет собой CD20, или первый антиген представляет собой CD20, и второй антиген представляет собой CD19;
или,
ii) γδ TCR, содержащим γ цепь TCR и δ цепь TCR, где, по меньшей мере, один функциональный домен соединен с С-концом γ цепи TCR и/или δ цепи TCR γδ TCR;
где γ цепь TCR содержит первую константную область и первую антигенсвязывающую область, и δ цепь TCR содержит вторую константную область и вторую антигенсвязывающую область;
где первая антигенсвязывающая область специфически связывается с первым антигеном, и вторая антигенсвязывающая область специфически связывается со вторым антигеном; или первая антигенсвязывающая область и вторая антигенсвязывающая область объединены друг с другом для специфического связывания с первым антигеном и вторым антигеном; предпочтительно, первый антиген представляет собой CD19, и второй антиген представляет собой CD20, или первый антиген представляет собой CD20, и второй антиген представляет собой CD19.
В некоторых вариантах осуществления, природный эндодомен, по меньшей мере, одной из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе αβ TCR удален, или природный эндодомен, по меньшей мере, одной из γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе γδ TCR удален.
В некоторых вариантах осуществления, природный эндодомен α цепи TCR и/или β цепи TCR в αβ TCR удален, или природный эндодомен γ цепи TCR и/или δ цепи TCR в γδ TCR удален.
В некоторых вариантах осуществления, в комплексе αβ TCR, функциональный домен соединен непосредственно или через линкер с С-концом, по меньшей мере, одной из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ, в которой природный эндодомен удален.
В некоторых вариантах осуществления, в комплексе αβ TCR, функциональный домен соединен непосредственно или через линкер с С-концом α цепи TCR и β цепи TCR, в которой природный эндодомен удален.
В некоторых вариантах осуществления, в комплексе γδ TCR, функциональный домен соединен непосредственно или через линкер с С-концом, по меньшей мере, одной из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ, в которой природный эндодомен удален.
В некоторых вариантах осуществления, в комплексе γδ TCR функциональный домен соединен непосредственно или через линкер с С-концом по меньшей мере, одной из γ цепи TCR, δ цепи TCR, в которой природный эндодомен удален.
В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10. Предпочтительно, n представляет собой целое число от 1 до 6, более предпочтительно, n представляет собой целое число от 2 до 5 и наиболее предпочтительно, n равен 3.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом одной из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе αβ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом α цепи TCR и/или β цепи TCR в αβ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом одной из γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе γδ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом γ цепи TCR и/или δ цепи TCR в γδ TCR.
В некоторых вариантах осуществления, CD3 δ, CD3 γ, CD3 ε и CD3 ζ в комплексе TCR не содержат, по меньшей мере, один функциональный домен, дополнительно связанный с его С-концом.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом α цепи TCR в комплексе αβ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом α цепи TCR в αβ TCR.
В некоторых вариантах осуществления, природный эндодомен α цепи TCR удален.
В некоторых вариантах осуществления, функциональный домен соединен непосредственно или через линкер с С-концом α цепи TCR, в которой природный эндодомен удален. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, от 1 до 6, более предпочтительно, от 2 до 5 и наиболее предпочтительно, n равен 3.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом β цепи TCR в комплексе αβ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом β цепи TCR в αβ TCR.
В некоторых вариантах осуществления, природный эндодомен β цепи TCR удален.
В некоторых вариантах осуществления, функциональный домен соединен непосредственно или через линкер с С-концом β цепи TCR, в которой природный эндодомен удален. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, от 1 до 6, более предпочтительно, от 2 до 5 и наиболее предпочтительно, n равен 3.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом γ цепи TCR в комплексе γδ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом γ цепи TCR в γδ TCR.
В некоторых вариантах осуществления, природный эндодомен γ цепи TCR удален.
В некоторых вариантах осуществления, функциональный домен соединен непосредственно или через линкер с С-концом γ цепи TCR, в которой природный эндодомен удален. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, от 1 до 6, более предпочтительно, от 2 до 5 и наиболее предпочтительно, n равен 3.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом δ цепи TCR в комплексе γδ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концом δ цепи TCR в γδ TCR.
В некоторых вариантах осуществления, природный эндодомен δ цепи TCR удален.
В некоторых вариантах осуществления, функциональный домен соединен непосредственно или через линкер с С-концом дельта цепи TCR, в которой природный эндодомен удален. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, от 1 до 6, более предпочтительно, от 2 до 5 и наиболее предпочтительно, n равен 3.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концами двух из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе αβ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с С-концами двух из γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе γδ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с соответствующими С-концами α цепи TCR и β цепи TCR в комплексе αβ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с соответствующими С-концами α цепи TCR и β цепи TCR в αβ TCR.
В некоторых вариантах осуществления, природный эндодомен каждой из цепей TCRα и β цепи TCR удален.
В некоторых вариантах осуществления, функциональный домен соединен непосредственно или через линкер с С-концом каждой из α цепи TCR и β цепи TCR, в которых природный эндодомен удален. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, от 1 до 6, более предпочтительно, от 2 до 5 и наиболее предпочтительно, n равен 3.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с соответствующими С-концами γ цепи TCR и δ цепи TCR в комплексе γδ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен соединен с соответствующими С-концами γ цепи TCR и δ цепи TCR в γδ TCR.
В некоторых вариантах осуществления, природный эндодомен каждой из γ цепи TCR и δ цепи TCR удален.
В некоторых вариантах осуществления, функциональный домен соединен непосредственно или через линкер с С-концом каждой из γ цепей TCR и δ цепи TCR, в которых природный эндодомен удален. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, от 1 до 6, более предпочтительно, от 2 до 5 и наиболее предпочтительно, n равен 3.
В некоторых вариантах осуществления, две или несколько из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе αβ TCR связаны с одним и тем же или разными функциональными доменами.
В некоторых вариантах осуществления, α цепь TCR и/или β цепь TCR в αβ TCR соединены с одним и тем же или разными функциональными доменами.
В некоторых вариантах осуществления, две или несколько из γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе γδ TCR связаны с одним и тем же или разными функциональными доменами.
В некоторых вариантах осуществления, γ цепь TCR и/или δ цепь TCR в γδ TCR связаны с одним и тем же или разными функциональными доменами.
В некоторых вариантах осуществления, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более функциональных доменов соединены с С-концом, по меньшей мере, одной из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе αβ TCR.
В некоторых вариантах осуществления, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более функциональных доменов соединены с С-концом α цепи TCR и/или β цепи TCR в αβ TCR.
В некоторых вариантах осуществления, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более функциональных доменов соединены с С-концом, по меньшей мере, одной из γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе γδ TCR.
В некоторых вариантах осуществления, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более функциональных доменов соединены с С-концом γ цепи TCR и/или δ цепи TCR в γδ TCR.
В некоторых вариантах осуществления, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединен с С-концом 1, 2, 3, 4, 5 или 6 из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе.
В некоторых предпочтительных вариантах осуществления, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединен с С-концом одной из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе.
Например, в некоторых вариантах осуществления, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединен с С-концом α цепи TCR в комплексе. В некоторых предпочтительных вариантах осуществления, эндодомен костимулирующей молекулы представляет собой OX40 или ICOS. В некоторых вариантах осуществления, цепь TCR β, CD3 δ, CD3 γ, CD3 ε и CD3 ζ может не содержать, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, дополнительно связанный с ее С-концом.
Альтернативно, в некоторых вариантах осуществления, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединен с С-концом CD3δ в комплексе. В некоторых вариантах осуществления, TCR α, TCR β, CD3 γ, CD3 ε и CD3 ζ могут не содержать, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединенный с ее С-концом.
В некоторых предпочтительных вариантах осуществления, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединен с С-концами двух из α цепи TCR, β цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ в комплексе.
Например, в некоторых вариантах осуществления, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединен с С-концами α цепи TCR и β цепи TCR в комплексе. В некоторых предпочтительных вариантах осуществления, эндодомен костимулирующей молекулы представляет собой OX40 или ICOS. В некоторых вариантах осуществления, CD3 δ, CD3 γ, CD3 ε и CD3 ζ не содержат, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, дополнительно связанный с ее С-концом.
В некоторых вариантах осуществления, функциональный домен представляет собой экзогенный функциональный домен. В некоторых вариантах осуществления, функциональный домен представляет собой экзогенный эндодомен, такой как домен, отвечающий за функцию внутриклеточной трансдукции.
Используемый в настоящем документе термин «экзогенный» означает последовательность белка или нуклеиновой кислоты, происходящую от чужеродного вида, или, если она происходит от того же вида, означает последовательность белка или нуклеиновой кислоты, которая претерпела значительные изменения в составе и/или положении по сравнению с ее природной формой в результате преднамеренного вмешательства человека.
Используемый в настоящем документе термин «функциональный домен» выбран из эндодомена костимулирующей молекулы, такой как CD40, OX40, ICOS, CD28, 4-1BB, CD27 и CD137; или эндодомен коингибирующей молекулы, такой как TIM3, PD1, CTLA4 и LAG3; или эндодомен цитокинового рецептора, такого как рецептор интерлейкина (например, рецептор IL-2, рецептор IL-7α или рецептор IL-21), рецептор интерферона, рецептор суперсемейства фактора некроза опухоли, рецептор колониестимулирующего фактора, хемокиновый рецептор, рецептор фактора роста или другие мембранные белки, или домен внутриклеточного белка, такого как NIK. Функциональный домен также может представлять собой слияние эндодомена цитокинового рецептора с группой активации STAT5 человека (аминокислотная последовательность показана в SEQ ID NO: 35) либо непосредственно, либо через линкер (например, (G4S)n, где n представляет собой целое число от 1 до 10).
В некоторых предпочтительных вариантах осуществления, функциональный домен представляет собой эндодомен костимулирующей молекулы, предпочтительно, эндодомен OX40 или ICOS, и более предпочтительно, эндодомен OX40.
Типовой эндодомен CD40 содержит аминокислотную последовательность, показанную в SEQ ID NO: 10. Типовой эндодомен OX40 содержит аминокислотную последовательность, показанную в SEQ ID NO: 11. Типовой эндодомен ICOS содержит аминокислотную последовательность, показанную в SEQ ID NO: 12. Типовой эндодомен CD28 содержит аминокислотную последовательность, показанную в SEQ ID NO: 13. Типовой эндодомен 4-1BB содержит аминокислотную последовательность, показанную в SEQ ID NO: 14. Типовой эндодомен CD27 содержит аминокислотную последовательность, показанную в SEQ ID NO: NO: 15. Типовой эндодомен рецептора IL-2β содержит аминокислотную последовательность, показанную в SEQ ID NO: 32. Типовой эндодомен IL-17α содержит аминокислотную последовательность, показанную в SEQ ID NO: 33. Типовой эндодомен рецептора IL-21 содержит аминокислотную последовательность, показанную в SEQ ID NO: 34. Типовая слитая аминокислотная последовательность эндодомена рецептора IL-2β с группой активации STAT5 человека показана в SEQ ID NO: 36. Типовая слитая аминокислотная последовательность эндодомена рецептора IL-17α с группой активации STAT5 человека показана в SEQ ID NO: 37.
В некоторых вариантах осуществления, первая константная область представляет собой константную область нативной α цепи TCR, например нативную константную область α цепи TCR человека (пример аминокислотной последовательности константной области α цепи TCR человека показан в SEQ ID NO: 1) или нативную константную область α цепи TCR мыши (пример аминокислотной последовательности константной области α цепи TCR мыши показан в SEQ ID NO: 3); или первая константная область представляет собой нативную константную область γ цепи TCR, например, нативную константную область γ цепи TCR человека (пример аминокислотной последовательности константной области γ цепи TCR человека показан в SEQ ID NO: 50) или нативную константную область γ цепи TCR мыши (пример аминокислотной последовательности константной области γ цепи TCR мыши показан в SEQ ID NO: 51).
В некоторых вариантах осуществления, первая константная область представляет собой модифицированную константную область α цепи TCR или модифицированную константную область γ цепи TCR.
В некоторых вариантах осуществления, модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота в положении 48, такая как треонин (Т), заменена на цистеин (С) по сравнению с константной областью α цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота в положении 112, такая как серин (S), заменена на лейцин (L), аминокислота в положении 114, такая как метионин (M), заменена на изолейцин (I), и аминокислота в положении 115, такая как глицин (G), заменена на валин (V), по сравнению с константной областью α цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота (например, E) в положении 6 заменена на D, K в положении 13 заменена на R, и аминокислоты в положениях 15-18 удалены по сравнению с константной областью α цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота в положении 48, такая как треонин (Т), заменена на цистеин (С), аминокислота в положении 112, такая как серин (S), заменена на лейцин (L), аминокислота в положении 114, такая как метионин (M), заменена на изолейцин (I), и аминокислота в положении 115, такая как глицин (G) мутирована в валин (V) по сравнению с константной областью α цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота (например, E) в положении 6 заменена на D, K в положении 13 заменена на R, аминокислоты в положениях 15-18 удалены, аминокислота в положении 48, такая как треонин (Т), заменена на цистеин (С), аминокислота в положении 112, такая как серин (S), заменена на лейцин (L), аминокислота в положении 114, такая как метионин (M), заменена на изолейцин (I), и аминокислота в положении 115, такая как глицин (G), заменена на валин (V), по сравнению с константной областью α цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область α цепи TCR происходит от константной области α цепи TCR мыши, в которой эндодомен константной области удален, например, удалены аминокислоты в положениях 136-137, по сравнению с константной областью α цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, первая константная область содержит аминокислотную последовательность, показанную в одной из SEQ ID No: 1, 3, 5, 7, 8, 26, 41, 42 и 56.
В некоторых вариантах осуществления, вторая константная область представляет собой нативную константную область β цепи TCR, например нативную константную область β цепи TCR человека (пример аминокислотной последовательности константной области β цепи TCR человека показан в SEQ ID NO: 2) или нативную константную область β цепи TCR мыши (пример аминокислотной последовательности константной области β цепи TCR мыши показан в SEQ ID NO: 4); или вторая константная область представляет собой нативную константную область δ цепи TCR, например, нативную константную область δ цепи TCR человека (пример аминокислотной последовательности константной области δ цепи TCR человека показан в SEQ ID NO: 52) или нативную константную область δ цепи TCR мыши (пример аминокислотной последовательности константной области δ цепи TCR мыши показан в SEQ ID NO: 53).
В некоторых вариантах осуществления, вторая константная область представляет собой модифицированную константную область β цепи TCR; или модифицированную константную область δ цепи TCR.
В некоторых вариантах осуществления, модифицированная константная область β цепи TCR получена из константной области β цепи TCR мыши, в которой аминокислота в положении 56, такая как треонин (S), заменена на цистеин (C) по сравнению с константной областью β цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, константная область β цепи модифицированного TCR получена из константной области β цепи TCR мыши, в которой аминокислота (например, R) в положении 3 заменена на K, аминокислота (например, T) в положении 6 заменен на F, K в положении 9 заменена на E, S в положении 11 заменена на A, L в положении 12 заменена на V, и аминокислоты в положениях 17 и 21-25 удалены по сравнению с константной областью β цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область β цепи TCR получена из константной области β цепи TCR мыши, в которой аминокислота в положении 56, такая как серин (S), заменена на цистеин (C), аминокислота ( например, R) в положении 3 заменена на K, аминокислота (например, T) в положении 6 заменена на F, K в положении 9 заменена на E, S в положении 11 заменена на A, L в положении 12 заменена на V, и аминокислоты в положениях 17 и 21-25 удалены по сравнению с константной областью β цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область β цепи TCR происходит от константной области β цепи TCR мыши, в которой эндодомен константной области удален, например, удалены аминокислоты в положениях 167-172, по сравнению с константной областью β цепи TCR мыши дикого типа.
В некоторых вариантах осуществления, модифицированная константная область β цепи TCR содержит аминокислотную последовательность, показанную в одной из SEQ ID No: 2, 4, 6, 9, 27, 43 и 49.
«Антигенсвязывающая область» относится к домену, который сам по себе или в комбинации с другой антигенсвязывающей областью может специфически связываться с антигеном-мишенью.
В некоторых вариантах осуществления, антигенсвязывающая область получена из антитела, которое специфически связывается с антигеном-мишенью.
В некоторых вариантах осуществления, каждая из первой или второй антигенсвязывающей области способна независимо специфически связываться с одним и тем же или разными антигенами-мишенями, например, первая антигенсвязывающая область специфически связывается с первым антигеном, и вторая антигенсвязывающая область специфически связывается с вторым антигеном. В некоторых вариантах осуществления, антигенсвязывающая область представляет собой одноцепочечное антитело (например, scFv) или однодоменное антитело (например, антитело верблюда), предпочтительно, антигенсвязывающая область представляет собой одноцепочечное антитело, такое как scFv. В некоторых вариантах осуществления, одноцепочечное антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, соединенные линкером. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 1 или 3.
В некоторых вариантах осуществления, антигенсвязывающая область, специфически связывающаяся с CD19, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 44, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 45. В некоторых вариантах осуществления, аминокислотная последовательность вариабельной области тяжелой цепи, показанная в SEQ ID NO: 44, и аминокислотная последовательность вариабельной области легкой цепи, показанная в SEQ ID NO: 45, соединены через линкер. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 1 или 3.
В некоторых вариантах осуществления, антигенсвязывающая область, специфически связывающаяся с CD19, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 46, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 47. В некоторых вариантах осуществления, аминокислотная последовательность вариабельной области тяжелой цепи, показанная в SEQ ID NO: 46, и аминокислотная последовательность вариабельной области легкой цепи, показанная в SEQ ID NO: 47, соединены через линкер. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 1 или 3.
В некоторых вариантах осуществления, антигенсвязывающая область, специфически связывающаяся с CD19, содержит аминокислотную последовательность scFv, показанную в SEQ ID NO: 39.
В некоторых вариантах осуществления, антигенсвязывающая область, специфически связывающаяся с CD20, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 54, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 55. В некоторых вариантах осуществления, аминокислотная последовательность вариабельной области тяжелой цепи, показанная в SEQ ID NO: 54, и аминокислотная последовательность вариабельной области легкой цепи, показанная в SEQ ID NO: 55, соединены через линкер. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 1 или 3.
В некоторых вариантах осуществления, антигенсвязывающая область, специфически связывающаяся с CD20, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 56, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 57. В некоторых вариантах осуществления, аминокислотная последовательность вариабельной области тяжелой цепи, показанная в SEQ ID NO: 56, и аминокислотная последовательность вариабельной области легкой цепи, показанная в SEQ ID NO: 57, соединены через линкер. В некоторых вариантах осуществления, линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 1 или 3.
В некоторых вариантах осуществления, антигенсвязывающая область, специфически связывающаяся с CD20, содержит аминокислотную последовательность scFv, показанную в SEQ ID NO: 38.
В некоторых вариантах осуществления, первая антигенсвязывающая область и вторая антигенсвязывающая область комбинируются друг с другом для специфического связывания с антигеном-мишенью. Например, первая антигенсвязывающая область содержит тяжелую цепь антитела, тогда как вторая антигенсвязывающая область содержит легкую цепь антитела, и наоборот.
В некоторых вариантах осуществления, первая антигенсвязывающая область содержит вариабельную область тяжелой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область тяжелой цепи антитела, которая специфически связывается со вторым антигеном, тогда как вторая антигенсвязывающая область содержит вариабельную область легкой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область легкой цепи антитела, которая специфически связывается со вторым антигеном, так что первая антигенсвязывающая область и вторая антигенсвязывающая область объединяются друг с другом для специфического связывания и с первым антигеном и со вторым антигеном.
В некоторых вариантах осуществления, первая антигенсвязывающая область содержит вариабельную область тяжелой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область легкой цепи антитела, которая специфически связывается со вторым антигеном, тогда как вторая антигенсвязывающая область содержит вариабельная область легкой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область тяжелой цепи антитела, которая специфически связывается со вторым антигеном, так что первая антигенсвязывающая область и вторая антигенсвязывающая область объединяются друг с другом для специфического связывания и с первым антигеном и со вторым антигеном.
В некоторых вариантах осуществления, первая антигенсвязывающая область содержит вариабельную область легкой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область легкой цепи антитела, которая специфически связывается со вторым антигеном, тогда как вторая антигенсвязывающая область содержит вариабельную область тяжелой цепи антитела. которая специфически связывается с первым антигеном, и вариабельную область тяжелой цепи антитела, которая специфически связывается со вторым антигеном, так что первая антигенсвязывающая область и вторая антигенсвязывающая область объединяются друг с другом для специфического связывания и с первым антигеном и со вторым антигеном.
В некоторых предпочтительных вариантах осуществления, первый антиген представляет собой CD19, и второй антиген представляет собой CD20. Альтернативно, в некоторых предпочтительных вариантах осуществления, первый антиген представляет собой CD20, и второй антиген представляет собой CD19.
В некоторых вариантах осуществления, вариабельная область тяжелой цепи антитела, которая специфически связывается с CD19, содержит аминокислотную последовательность, показанную в SEQ ID NO: 44, и вариабельная область легкой цепи содержит аминокислотную последовательность, показанную в SEQ ID NO: 45.
В некоторых вариантах осуществления, вариабельная область тяжелой цепи антитела, которая специфически связывается с CD19, содержит аминокислотную последовательность, показанную в SEQ ID NO: 46, и вариабельная область легкой цепи содержит аминокислотную последовательность, показанную в SEQ ID NO: 47.
В некоторых вариантах осуществления, вариабельная область тяжелой цепи антитела, которая специфически связывается с CD20, содержит аминокислотную последовательность, показанную в SEQ ID NO: 54, и вариабельная область легкой цепи содержит аминокислотную последовательность, показанную в SEQ ID NO: 55.
В некоторых вариантах осуществления, вариабельная область тяжелой цепи антитела, которая специфически связывается с CD20, содержит аминокислотную последовательность, показанную в SEQ ID NO: 56, и вариабельная область легкой цепи содержит аминокислотную последовательность, показанную в SEQ ID NO: 57.
В некоторых вариантах осуществления, CD3γ, CD3δ, CD3ε и/или CD3ζ представляют собой CD3γ, CD3δ, CD3ε и/или CD3ζ человека. В некоторых вариантах осуществления, CD3γ человека содержит аминокислотную последовательность, показанную в SEQ ID No: 28. В некоторых вариантах осуществления, CD3γ человека содержит аминокислотную последовательность, показанную в SEQ ID No: 29. В некоторых вариантах осуществления, CD3γ человека содержит аминокислотную последовательность кислотная последовательность, показанную в SEQ ID No: 30. В некоторых вариантах осуществления, CD3γ человека содержит аминокислотную последовательность, показанную в SEQ ID No: 31.
В некоторых вариантах осуществления, модифицированный Т-клеточный рецептор (TCR) или комплекс TCR по настоящему изобретению содержит α цепь TCR, как показано в SEQ ID NO: 59, и β цепь TCR, как показано в SEQ ID NO: 60.
В другом аспекте, в настоящем документе предложена выделенная терапевтическая иммунная клетка, которая содержит модифицированный Т-клеточный рецептор (TCR) или комплекс TCR по настоящему изобретению.
В некоторых вариантах осуществления, иммунная клетка представляет собой Т-клетку. В других вариантах осуществления, иммунная клетка представляет собой NK-клетку.
В другом аспекте, настоящее изобретение относится к выделенному полинуклеотиду, содержащему нуклеотидную последовательность, кодирующую, по меньшей мере, одну из α цепи TCR, β цепи TCR, γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ, как определено выше, где, по меньшей мере, один экзогенный функциональный эндодомен соединен с С-концом, по меньшей мере, одной из α цепи TCR, β цепи TCR, γ цепи TCR, δ цепи TCR, CD3 ε, CD3 γ, CD3 δ и CD3 ζ.
В другом аспекте, настоящее изобретение относится к выделенному полинуклеотиду, содержащему нуклеотидную последовательность, кодирующую TCR, как определено выше.
В некоторых вариантах осуществления, выделенный полинуклеотид содержит нуклеотидную последовательность, кодирующую α цепь TCR и/или β цепь TCR, которая связана, по меньшей мере, с одним эндодоменом костимулирующей молекулы на ее С-конце.
В некоторых вариантах осуществления, полинуклеотид содержит i) нуклеотидную последовательность, кодирующую α цепь, ii) нуклеотидную последовательность, кодирующую β цепь, и iii) нуклеотидную последовательность, кодирующую саморасщепляющийся пептид, расположенный между i) и ii) в той же рамке считывания. Нуклеотидная последовательность, кодирующая α цепь, может быть расположена на 5' конце или на 3' конце нуклеотидной последовательности, кодирующей β цепь.
В некоторых вариантах осуществления, выделенный полинуклеотид содержит нуклеотидную последовательность, кодирующую γ цепь TCR и/или δ цепь TCR, которая связана, по меньшей мере, с одним эндодоменом костимулирующей молекулы на ее С-конце.
В некоторых вариантах осуществления, полинуклеотид содержит i) нуклеотидную последовательность, кодирующую γ цепь, ii) нуклеотидную последовательность, кодирующую δ цепь, и iii) нуклеотидную последовательность, кодирующую саморасщепляющийся пептид, расположенный между i) и ii) в той же рамке считывания. Нуклеотидная последовательность, кодирующая γ цепь, может быть расположена на 5'-конце или на 3'-конце нуклеотидной последовательности, кодирующей δ цепь.
Используемый в настоящем документе термин «саморасщепляющийся пептид» означает пептид, который может осуществлять саморасщепление в клетках. Например, саморасщепляющийся пептид может содержать сайт узнавания протеазой, чтобы его можно было распознать и специфически расщепить протеазами в клетках.
Альтернативно, саморасщепляющийся пептид может представлять собой полипептид 2А. Полипептид 2А представляет собой тип короткого пептида вируса, и его саморасщепление происходит во время трансляции. Когда два разных белка-мишени связаны полипептидом 2А и экспрессируются в одной и той же рамке считывания, два белка-мишени создаются почти в соотношении 1:1. Обычным полипептидом 2А может быть Р2А из тешовируса-1 свиней, Т2А из вируса Thosea asigna, Е2А из вируса ринита А лошадей и F2А из вируса ящура. Среди них, P2A имеет самую высокую эффективность разрезания и поэтому является предпочтительным. В данной области техники также известны различные функциональные варианты этих полипептидов 2А, которые также можно использовать в настоящем изобретении.
В некоторых вариантах осуществления, полинуклеотид содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность, показанную в SEQ ID No: 58.
В другом аспекте, настоящее изобретение относится к вектору экспрессии, содержащему полинуклеотид по настоящему изобретению, функционально связанный с регуляторной последовательностью.
«Вектор экспрессии» по настоящему изобретению может представлять собой линейный фрагмент нуклеиновой кислоты, циклическую плазмиду, вирусный вектор или способную к трансляции РНК (например, мРНК). В некоторых предпочтительных вариантах осуществления, вектор экспрессии представляет собой вирусный вектор, такой как лентивирусный вектор.
Термины «регуляторная последовательность» и «регуляторный элемент» используются взаимозаменяемо для обозначения нуклеотидной последовательности, расположенной выше (5'-некодирующая последовательность), посередине или ниже (3'-некодирующая последовательность) кодирующей последовательности и влияет на транскрипцию, процессинг или стабильность РНК или трансляцию соответствующей кодирующей последовательности. Регуляторный элемент экспрессии относится к нуклеотидной последовательности, которая может контролировать транскрипцию, процессинг или стабильность РНК или трансляцию представляющей интерес нуклеотидной последовательности. Регуляторная последовательность может включать, но не ограничена ими, промотор, лидерную последовательность трансляции, интрон, энхансер и последовательность распознавания полиаденилирования.
Используемый в настоящем документе термин «функционально связанный» означает, что регуляторный элемент (например, но не ограничиваясь ими, промоторная последовательность, последовательность терминации транскрипции и т. д.) связан с последовательностью нуклеиновой кислоты (например, кодирующей последовательностью или открытой рамкой считывания), так что транскрипция нуклеотидной последовательности контролируется и регулируется регуляторным элементом транскрипции. Методы функционального связывания области регуляторного элемента с молекулой нуклеиновой кислоты известны в данной области техники.
В другом аспекте, настоящее изобретение предлагает способ получения терапевтической иммунной клетки по настоящему изобретению, включающий введение полинуклеотида или вектора экспрессии по настоящему изобретению в иммунную клетку.
Иммунная клетка по настоящему изобретению, такая как Т-клетка или NK-клетка, может быть получена различными неограничивающими способами из ряда неограничивающих источников, включая мононуклеарные клетки периферической крови, костный мозг, ткань лимфатических узлов, пуповинную кровь, ткань тимуса, асцит, плевральный выпот, ткань селезенки и опухоль. В некоторых вариантах осуществления, клетка может быть получена от здорового донора или от пациента, у которого диагностирован рак. В некоторых вариантах осуществления, клетка может быть частью смешанной популяции клеток, демонстрирующих разные фенотипические профили. Например, Т-клетки можно получить путем выделения мононуклеарных клеток периферической крови (РВМС), затем активации и амплификации с помощью специфического антитела.
В некоторых вариантах осуществления аспектов изобретения, иммунные клетки, такие как Т-клетки, происходят из аутологичных клеток субъекта. Используемый в настоящем документе термин «аутологичные» означает, что клетки, клеточная линия или популяция клеток, используемые для лечения субъекта, происходят от субъекта. В некоторых вариантах осуществления, иммунные клетки, такие как Т-клетки, происходят из аллогенных клеток, таких как донор, совместимый с лейкоцитарным антигеном человека (HLA). Клетки от доноров могут быть превращены в не аллореактивные клетки с использованием стандартных протоколов, и реплицированы по мере необходимости для продуцирования клеток, которые можно вводить одному или нескольким пациентам.
В другом аспекте, настоящее изобретение предлагает фармацевтическую композицию, которая включает терапевтическую иммунную клетку по настоящему изобретению и фармацевтически приемлемый носитель.
Используемый в настоящем документе термин «фармацевтически приемлемый носитель» включает любые возможные физиологически совместимые растворители, дисперсионную среду, покрытия, антибактериальные и противогрибковые агенты, изотонические агенты и замедлители всасывания и т. д. Предпочтительно, носитель подходит для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии).
В другом аспекте, настоящее изобретение предлагает применение терапевтических иммунных клеток по настоящему изобретению в приготовлении лекарственного средства для лечения заболеваний у субъекта.
Используемый в настоящем документе термин «субъект» относится к организму, который страдает или склонен страдать от заболевания (например, рака), которое можно лечить с помощью клетки, способа или фармацевтической композиции по настоящему изобретению. Неограничивающий пример включает человека, крупный рогатый скот, крысу, мышь, собаку, обезьяну, козу, овцу, корову, оленя и других не млекопитающих. В некоторых предпочтительных вариантах осуществления, субъектом является человек.
В другом аспекте, настоящее изобретение предлагает способ лечения такого заболевания, как рак, у субъекта, где способ включает введение субъекту эффективного количества терапевтических иммунных клеток или фармацевтической композиции по настоящему изобретению.
Используемый в настоящем документе термин «терапевтически эффективное количество» или «терапевтически эффективная доза» или «эффективное количество» относится к количеству вещества, соединения, материала или клетки, которое, по меньшей мере, достаточно для оказания терапевтического эффекта после введения субъекту. Следовательно, это количество необходимо для профилактики, лечения, улучшения, блокирования или частичного блокирования симптомов заболевания или нарушения. Например, «эффективное количество» клетки или фармацевтической композиции по настоящему изобретению, предпочтительно, может приводить к уменьшению тяжести симптомов нарушения, увеличению частоты и продолжительности бессимптомного периода нарушения или профилактике травмы или инвалидности в результате страдания от нарушения. Например, для лечения опухоли, «эффективное количество» клетки или фармацевтической композиции по настоящему изобретению может предпочтительно, ингибировать рост опухолевых клеток или рост опухоли, по меньшей мере, примерно на 10%, предпочтительно, по меньшей мере, примерно на 20%, более предпочтительно, по меньшей мере, примерно на 30%, более предпочтительно, по меньшей мере, примерно на 40%, более предпочтительно, по меньшей мере, примерно на 50%, более предпочтительно, по меньшей мере, примерно на 60%, более предпочтительно, по меньшей мере, примерно на 70% и более предпочтительно, по меньшей мере, примерно на 80% по сравнению с нелеченным субъектом. Способность ингибировать рост опухоли можно оценить в модельной системе животных, которая может спрогнозировать эффективность в отношении опухоли человека. Альтернативно, можно провести оценку путем изучения способности ингибировать рост опухолевых клеток, которую можно определить in vitro с помощью тестов, известных специалистам в данной области техники.
На практике, уровень дозы клеток в фармацевтической композиции по настоящему изобретению может варьироваться для получения количества активного ингредиента, которое позволяет эффективно достичь желаемого терапевтического ответа для конкретного пациента, композиции и пути введения без токсичности для пациента. Выбранный уровень дозы зависит от множества фармакокинетических факторов, включая активность применяемой конкретной композиции изобретения, способ введения, время введения, скорость выведения применяемого конкретного соединения, продолжительность лечения, применяемые другие лекарственные средства, соединения и/или материалы в комбинации с применяемым конкретным составом, возрастом, полом, весом, состоянием, общим состоянием здоровья и историей болезни пациента, подлежащего лечению, и аналогичными факторами, известными в области медицины.
Введение терапевтических иммунных клеток или фармацевтической композиции или лекарственного средства по настоящему изобретению можно осуществлять любым удобным способом, таким как инъекция, инфузия, имплантация или трансплантация. Введение клетки или композиции, описанной в настоящем документе, может быть внутривенным, внутрилимфатическим, интрадермальным, внутриопухолевым, интрамедуллярным, внутримышечным или внутрибрюшинным введением. В одном варианте осуществления, клетку или композицию по настоящему изобретению, предпочтительно, вводят внутривенной инъекцией.
В вариантах осуществления различных аспектов изобретения, заболевание представляет собой заболевание, связанное с CD19 и/или CD20, такое как заболевание, связанное с аномальной экспрессией CD19 и/или CD20, такое как рак, связанный с CD19 и/или CD20. Рак может представлять собой В-клеточную злокачественную опухоль, такую как хронический или острый лейкоз (включая острый миелоидный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, хронический лимфобластный лейкоз), лимфоцитарную лимфому, неходжкинскую лимфому и их комбинации. В некоторых вариантах осуществления, рак представляет собой CD19- и/или CD20-положительный рак.
Примеры
Пример 1: Улучшение STAR
1. Т-клеточный рецептор дикого типа и дизайн его молекулы STAR с мутацией в константной области
1.1 Дизайн прототипа STAR
Секретируемое антитело (антитело, Ab) или В-клеточный рецептор (BCR), продуцируемый В-клетками, имеет большое сходство с Т-клеточным рецептором (TCR) с точки зрения генетической структуры, структуры белка и пространственной конформации. И антитело, и TCR состоят из вариабельной области и константной области, где вариабельная область играет роль распознавания и связывания антигена, а домен константной области играет роль структурного взаимодействия и передачи сигнала. Путем замены вариабельных областей α и β цепей TCR (или γ и δ цепей TCR) вариабельной областью тяжелой цепи (VH) и вариабельной областью легкой цепи (VL) антитела, может быть сконструирована искусственно синтетическая химерная молекула, называемая синтетический Т-клеточный рецептор и рецептор антитела (STAR/WT-STAR), их структура показана на фигуре 1 (слева).
Молекула STAR имеет две цепи, где первая цепь получается путем слияния последовательности распознавания антигена (такой как вариабельная область тяжелой цепи антитела) с константной областью (Cα) α цепи Т-клеточного рецептора (TCRα), и вторую цепь получают путем слияния последовательности распознавания антигена (такой как вариабельная область легкой цепи антитела) с константной областью (Cβ) β цепи Т-клеточного рецептора (TCRβ). Домен распознавания антигена (такой как VH, VL или scFv) и константный домен (константный домен TCR α, β, γ и δ) в конструкции можно расположить и комбинировать с образованием множества конструкций с различными конфигурациями, но схожими функциями.
Первая и вторая цепи молекулы STAR после экспрессии в Т-клетках, будут объединяться с эндогенными цепями CD3 ε δ, CD3 γ ε и CD3 ζ в эндоплазматическом ретикулуме с образованием комплекса из восьми субъединиц, который присутствует на поверхности клеточной мембраны в виде комплекса. Иммунорецепторный тирозиновый активирующий мотив (ITAM) представляет собой мотив трансдукции сигнала в молекуле TCR с его консервативной последовательностью YxxL/V. Эндомены цепей CD3 ε, δ, γ и ε содержат одну последовательность ITAM, и эндодомены цепи CD3 ζ содержат три последовательности ITAM, поэтому полный комплекс STAR имеет всего десять последовательностей ITAM. Когда последовательность распознавания антигена рецептора STAR связывается с его специфическим антигеном, внутриклеточная последовательность ITAM будет последовательно фосфорилироваться, что затем, в свою очередь, активирует нижестоящий сигнальный путь, активируя факторы транскрипции, такие как NF-κ β, NFAT и AP-1, чтобы инициировать активацию Т-клеток и осуществлять эффекторные функции. Предыдущие исследования, проведенные изобретателем, показали, что STAR может активировать Т-клетки лучше, чем обычный химерный антигенный рецептор CAR, и фоновая активация в отсутствие антигенной стимуляции значительно снижается, что имеет значительные преимущества (см. заявку на патент Китая на изобретение № 201810898720.2). Тем не менее, дальнейшее улучшение STAR все еще желательно.
1.2. Дизайн мутантных STAR (mut-STAR) и STAR с модифицированным трансмембранным доменом и эндодоменом (ub-STAR)
В дизайне прототипа STAR используют последовательность константной области α/β цепи гуманизированного TCR (или γ и δ цепей TCR) (константная область TCRα человека дикого типа, SEQ ID NO: 1; константная область TCRβ человека дикого типа, SEQ ID NO: 2). Благодаря константной области последовательностей α/β цепей TCR человека, приматов и мыши (TCRaC-WT мыши, SEQ ID NO: 3; TCRbC-WT мыши, SEQ ID NO: 4) являются высоко консервативными и также имеют одинаковую ключевую аминокислотную последовательность, они могут быть заменены друг другом.
После переноса в Т-клетки, молекулы STAR будут ошибочно спариваться с эндогенным TCR Т-клеток через константную область. С одной стороны, эта проблема ошибочного спаривания может снизить эффективность правильного спаривания молекул STAR и ослабить их функции, что, с другой стороны, может увеличить вероятность неизвестной специфичности из-за ошибочного спаривания и увеличить риск безопасности. Для решения этой проблемы, константную область молекулы STAR изобретатели меняют на последовательность мыши для усиления функций молекулы STAR после переноса в Т-клетки человека. Для дальнейшей оптимизации дизайна молекулы STAR, изобретатели осуществляют мутацию цистеина в молекуле STAR для введения межмолекулярной дисульфидной связи, тем самым усиливая взаимное спаривание между двумя цепями молекулы STAR и уменьшая ошибочно спаривание с эндогенным TCR. В частности, треонин (T) в положении 48 меняют на цистеин (C) (TCRaC-Cys мыши, SEQ ID NO: 5) в константной области α цепи TCR, и серин (S) в положении 56 меняют на цистеин (C) (TCRbC-Cys мыши, SEQ ID NO: 6) в константной области β цепи TCR. Два новых добавленных цистеина будут образовывать дисульфидную связь между двумя цепями STAR, тем самым уменьшая ошибочно спаривание между двумя цепями STAR с эндогенной цепью TCR и помогая молекулам STAR образовывать более стабильные комплексы, тем самым получая лучшие функции. Кроме того, для дальнейшей оптимизации дизайна молекулы STAR, изобретатели осуществляют замену гидрофобной аминокислоты в трансмембранном домене молекулы STAR, чтобы повысить стабильность молекулы STAR и помочь ей выполнять более длительную функцию. В частности, осуществляют три аминокислотные мутации в положениях аминокислот 111-119 в трансмембранном домене константной области α цепи TCR: серин (S) в положении 112 мутируют на лейцин (L), метионин (M) в положении 114 мутируют на изолейцин (I), и глицин (G) в положении 115 мутируют на серин (V). Всю аминокислотную последовательность в этой области изменяют с LSVMGLRIL на LLVIVLRIL, и эту модификацию называют TCRaC-TM9 мыши, которая продуцирует последовательность константной области SEQ ID NO: 7. Этот дизайн увеличивает гидрофобность трансмембранного домена, противодействует нестабильности, вызванной положительными зарядами, которые несет трансмембранный домен TCR, и делает молекулу STAR более стабильной на клеточной мембране, тем самым получая лучшие функции, его структура показана на фигуре 1 (в центре).
После связывания TCR с антигеном и завершения активации, лизин в эндодомене и трансмембранном домене молекулы TCR подвергают модификациям убиквитина посредством серии реакций убиквитинирования с помощью убиквитин-активирующего фермента, убиквитин-связывающего фермента и убиквитинлигазы убиквитината, тем самым продуцируя T клеточный эндоцитоз, приводящий к эндоцитозу молекул TCR в клетки для дальнейшего разложения лизосомами, что снижает концентрацию молекул TCR на поверхности мембраны Т-клеток, что приводит к постоянному снижению эффекта активации Т-клеток. Изобретатели модифицируют аминокислоты в трансмембранном домене или эндодомене α и β цепей в молекуле mut-STAR, что включает: мутацию лизина в эндодомене константной области α цепи и трансмембранном домене константной области β цепи молекулы STAR на аргинин, с получением последовательности константной области TCR α C-Arg mut мыши (SEQ ID NO: 8) и последовательности константной области TCR β C-Arg mut мыши (SEQ ID NO: 9), соответственно, чтобы уменьшить эндоцитоз молекулы STAR, вызванный убиквитинированием лизина. Эта конструкция уменьшает возможность убиквитинирования трансмембранного домена и эндодомена молекулы STAR, тем самым уменьшая эндоцитоз молекул STAR, позволяя молекулам STAR быть более стабильными на клеточной мембране и получить лучшие функции, их структура показана на фигуре 1 (справа).
2. Дизайн дикого типа и мутантных молекул STAR, содержащих эндодомен костимулирующего рецептора.
Чтобы улучшить способность клеток mut-STAR к пролиферации in vivo, время эффекта выживания и способность проникать в микроокружение опухоли для эффективного уничтожения клеток-мишеней, изобретатели разработали новую структуру, в которой mut-STAR комплекс модифицирован, и улучшенная клетка mut-STAR может быть адаптирована, по мере необходимости, чтобы улучшить клинический ответ TCR-T и реализовать длительный лечебный эффект.
2.1. Дизайн молекул mut-STAR (co-STAR), содержащих эндодомен рецептора костимулирующей молекулы
TCR представляет собой особый маркер на поверхности всех Т-клеток, которые можно разделить на αβ TCR и γδ TCR, и соответствующие им Т-клетки представляют собой αβ Т-клетки и γδ Т-клетки, соответственно. Авторы изобретения модифицируют αβ-STAR и tγδ-STAR костимулирующими сигналами, соответственно, для улучшения работы αβ Т-клеток и γδ Т-клеток, соответственно.
TCR αβ Т-клеток состоит из цепей TCRα и TCRβ, которые составляют 90-95% от общего числа Т-клеток. αβ TCR состоит из вариабельной области и константной области, в которой вариабельная область имеет большое разнообразие и играет роль распознавания и связывания антигена, а домен константной области играет роль структурного взаимодействия и трансдукции сигнала. Для повышения токсичности и жизнестойкости пролиферации Т-клеток по настоящему изобретению, в С-конец константной области αβ-STAR (фиг. 2) вводят эндодоменную последовательность гуманизированного костимулирующего рецептора, чтобы изучить влияние на функции Т-клеток STAR. Константная область STAR по настоящему изобретению включает не модифицированную константную область WT-STAR, константную область cys-STAR, содержащую дополнительную межмолекулярную дисульфидную связь, муринизированную константную область hm-STAR и mut-STAR с комбинацией трех модификаций, описанных в подразделе 1. Костимулирующая структура трансдукции сигнала включает внутриклеточные домены передачи сигнала CD40, OX40, ICOS, CD28, 4-1BB или CD27 с последовательностями SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15, соответственно. Костимулирующий эндодомен может быть связан с С-концом α цепи TCR или β цепи TCR или и α, и β цепи TCR (co-STAR). Кроме того, костимулирующий эндодомен может быть соединен напрямую или через линкер G4S/(G4S)n (последовательности G4S линкера представляют собой SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, соответственно) с С-концом константной области TCR или С-концом константной области TCR с удаленной последовательностью эндодомена молекулы TCR (константная область α цепи TCR с делецией эндодомена, содержащая и замену цистеина, и модификацию гидрофобной области (TCRaC-del mut мыши, SEQ ID NO: 26), константная область β цепи TCR делецией эндодомена, содержащая замену цистеина (TCRβC-del mut мыши, SEQ ID NO: 27)) (ко-линкер-STAR, фиг. 3).
TCR γδ Т-клеток состоит из цепей TCR γ и TCR δ, γδ Т-клетки можно разделить на три подгруппы: γδ1, γδ2 и γδ3 в зависимости от типа цепи TCR δ, где разные подгруппы имеют различное распределение в организме человека. γδ Т-клетки распознают антиген MHC-ограниченным способом, что играет важную роль в наблюдении за патогенами и опухолями. Эксперименты показали, что CD28 или 4-1BB и аналогичные костимулирующие сигналы играют важную роль в активации и пролиферации γδ Т-клеток. Эндоменная последовательность рецептора костимулирующей молекулы человека введена изобретателями в С-конец TCR γ и TCR δ, соответственно (фиг. 2, справа) для улучшения работы γδ Т-клеток.
2.2. Дизайн молекулы CD3 (co-CD3-STAR), включающей эндодомен рецептора костимулирующей молекулы
Субъединица CD3 включает γ цепь, δ цепь, ε цепь и ζ цепь и образует комплекс Т-клеточного рецептора с молекулой TCR, которая передает сигналы от эктодомена к эндодомену, чтобы регулировать состояние клеток и ответ на стимулы. Чтобы сконструировать Т-клетки с усиленным TCR и улучшить способность уничтожать опухоли, способность к пролиферации и время выживания Т-клеток in vivo, изобретатели модифицируют молекулу CD3 путем введения эндодомена рецептора костимулирующей молекулы человека в С-конец γ цепи (SEQ ID NO: 28), δ цепи (SEQ ID NO: 29), ε цепи (SEQ ID NO: 30) и ζ цепи (SEQ ID NO: 31) CD3 (фиг. 4). Модифицированную молекулу CD3 экспрессируют в Т-клетках mut-STAR для улучшения ее функции.
2.3. Дизайн молекулы CD3, включающей стимулирующую область цитокинового рецептора (цитокин-STAR, CK-STAR)
Цитокины играют важную роль в пролиферации, противоопухолевом действии и дифференциации Т-клеток. Различные цитокины объединяются с соответствующими рецепторами для передачи сигналов от эктодомена к эндодомену, чтобы регулировать состояние клеток и ответ на стимулы. Кроме того, исследования показали, что нижестоящая молекула STAT5 (SEQ ID NO: 35) активируется каскадной реакцией на эндодомене рецептора IL-2, тем самым усиливая транскрипцию молекул, связанных с пролиферацией Т-клеток, и повышая способность к пролиферации CAR-T-клеток. Чтобы сконструировать улучшенные STAR-T-клетки и улучшить способность к уничтожению опухолей, способность к пролиферации и время выживания T-клеток in vivo, изобретатели модифицируют молекулу STAR путем связывания внутриклеточного домена трансдукции сигнала цитокинового рецептора человека (например, эндодомена рецептора IL-2β IL2Rb, SEQ ID NO: 32; эндодомена рецептора IL-7α, SEQ ID NO: 33; эндодомена рецептора IL-21, SEQ ID NO: 34 и т. д.)) с С-концом α цепи или β цепи TCR или обеих, α и β цепей, или путем дальнейшего связывания активирующей части STAT5 с эндодоменом рецептора IL-2β или IL-7Rα через G4S (IL-2RbQ, SEQ ID NO: 36; IL-7RbQ, SEQ ID NO: 37) (фиг. 5).
3. Конструирование вектора дикого типа и мутантных молекул STAR, содержащих эндодомен рецептора костимулирующей молекулы.
3.1 Источники вектора
Векторы, используемые в настоящем изобретении, включая вирусные векторы, плазмидные векторы и т.д., приобретают у или синтезируют в коммерческих компаниях, и получают полноразмерные последовательности этих векторов, где известны специфические сайты расщепления.
3.2. Источники фрагмента
TCR, упомянутый в изобретении, может быть любым функциональным TCR, включая WT-STAR, mut-STAR, ub-STAR, co-STAR, co-линкер-STAR, CK-STAR, co-CD3-STAR и т.п., используемые в изобретении. Все фрагменты гена, такие как вариабельная область TCR, константная область TCR, эндодомен рецептора костимулирующей молекулы, область трансдукции внутриклеточного сигнала рецептора цитокина, последовательность метки, линкер и подобные, используемые в настоящем изобретении, синтезируют в коммерческих компаниях. Эти фрагменты генов связаны ПЦР.
В этом примере, оптимизацию комплекса TCR подтверждают с использованием STAR, содержащего ScFv, показанного в SEQ ID NO: 38 (получен из антитела, таргетирующего CD20, OFA), и/или ScFv, показанного в SEQ ID NO: 39 (получен из антитела, таргетирующего CD19, FMC63), и сравнивают с пустой контрольной группой «Имитация», экспрессирующей только белок RFP, показанный в SEQ ID NO: 40 (красный флуоресцентный белок, RFP).
3.3. Конструирование вектора
Используемый в настоящем документе лентивирусный вектором представляет собой pHAGE-EF1 α-IRES-RFP, где линейный вектор получают с помощью рестрикционного фермента Not I/Nhe I, фрагмент гена получают синтезом и ПЦР, и полный вектор получают гомологичной рекомбинацией.
4 Конструирование клеточных линий:
4.1 Конструирование плазмиды pHAGE-люцифераза-GFP
Лентивирусный вектор pHAGE может стабильно вставлять гены-мишени в геном клетки-мишени, что является важным способом создания стабильной клеточной линии. Люцифераза представляет собой вид фермента с каталитической активностью, который может катализировать химическую аутолюминесценцию субстрата, чтобы клетка-мишень стабильно экспрессировала люциферазу, количество клеток-мишеней может быть указано после добавления субстрата, что отражает влияние функциональных клеток на клетки-мишени. Вектор pHAGE-EF1A, несущий сайты расщепления эндонуклеазой рестрикции NotI/ClaI, расщепляют двумя ферментами, и последовательности люциферазы и GFP получают посредством NCBI, фрагменты синтезируют в коммерческой компании Ruiboxingke путем объединения гена люциферазы с геном GFP с использованием перекрывающей ПЦР, и затем фрагмент люцифераза-GFP соединяют с вектором pHAGE гомологичной рекомбинацией.
4.2 Конструирование линии клеток-мишеней, несущих люциферазу
После успешного конструирования лентивирусного вектора, несущего люциферазу и GFP, лентивирус упаковывают с помощью Lenti-X-293T, и раствор лентивируса концентрируют с помощью PEG8000, измеряют титр вируса способом градиентного разведения, и затем клеточную линию лимфомы Raji заражают, через 72 часа заражения, наблюдают наличие GFP-положительных клеток с помощью флуоресцентного микроскопа, и затем GFP-положительные клетки сортируют с помощью проточного сортировщика, и моноклональные клетки отбирают для создания и сохранения библиотеки. В то же время, субстрат люциферазы используют для инкубации с клетками-мишенями, и определяют уровни экспрессии и обнаружения люциферазы для определения уровня экспрессии.
4.3 Конструирование клеточной линии Jurkat с нокаутом TCR α-β
Основываясь на характеристиках структуры и последовательности TCR, конструируют направляющую последовательность в константных областях α и β цепей для конструирования клеточной линии TCR α-β-Jurkat. Последовательности экзонов константных областей α и β цепей TCR получают в NCBI, и последовательности экзона 1 константных областей α и β цепей TCR отправляют на сайт tools.genome-engineering.org для разработки направляющих последовательностей, на основании результата, синтезируют олиго последовательность, и затем конструируют лентивирусный вектор sgRNA-LentiCRISPR (купленный у Aidi gene). Направляющую последовательность α цепи связывают с LentiCRISPR-puro, и направляющую последовательность β цепи связывают с LentiCRISPR-BSD.
Упаковка лентивируса sgRNA-LentiCRISPR: HEK-293T заранее выкладывают на 10 см чашку, и когда клетки вырастают до 80%-90%, к HEK-293T добавляют систему трансфекции, и клетки помещали обратно в инкубатор при 37° для культивирования. Это время считают за 0 часов; через 12 часов после трансфекции добавляют свежую 10% FBS-DMEM. Вирус собирают через 48 часов и 72 часа после трансфекции. Культуральную среду, содержащую вирус, центрифугируют и фильтруют, смешивают с PEG8000, выдерживают при 4° более 12 часов, затем центрифугируют при 3500 об/мин в течение 30 минут, ресуспендируют и осаждают с соответствующим объемом среды после удаления супернатанта. Полученный продукт замораживают при -80° или используют напрямую.
Заражение и скрининг Jurkat Т-клеток и идентификация моноклональной клеточной линии: Jurkat Т-клетки инокулируют в 12- или 24-луночный планшет с последующим добавлением sgRNA-LentiCRISPR вируса α цепи и β цепи в соответствующих объемах одновременно, а также полибрена (добавляют в соотношении 1:1000 к общему объему) и хорошо перемешивают. Заражение центрифугированием проводят при 1000 об/мин при 32° в течение 90 мин. Полученную смесь помещают в инкубатор при 37°, считая за 0 часов; жидкость меняют через 10-12 часов; через 48 часов добавляют пуромицин и BSD до соответствующей конечной концентрации, и после обработки в течение дополнительных 48 часов, как показано, все клетки в не зараженной контрольной группе погибают. Выжившие клетки отсасывают, центрифугируют и культивируют в полной среде для получения банка TCR α-β-Jurkat клеток. Отдельные клетки из банка TCR α-β-Jurkat клеток сортируют в 96-луночный планшет с помощью проточного сортировщика Aria, через две недели культивирования, выращенные моноклоны отсасывают для культуры амплификации. Моноклональные клеточные линии идентифицируют с антителами α цепи и β цепи TCR, соответственно, и клеточные линии с дефицитом обеих цепей амплифицируют для получения клеточной линии Jurkat-T с нокаутом эндогенного TCR.
5. Конструкция системы упаковки вируса для трансдукции Т-клеток
5.1 Лентивирусная система и способ упаковки (разные поколения)
Клетки Lentix-293T инокулируют в 10 см чашку для культивирования в количестве 5х105/мл и культивируют в инкубаторе при 37°C с 5% CO2, трансфекцию проводят, когда плотность клеток достигает примерно 80% (наблюдают под микроскопом). Три плазмиды смешивают с 500 мкл бессывороточной DMEM в соотношении 1:2:3 PMD2.G:PSPAX:трансферная плазмида. 54 мкл PEI-Max и 500 мкл бессывороточной DMEM однородно смешивают и оставляют при комнатной температуре на 5 мин (в отношении PEI-Max к плазмиде 3:1 объем/массу). Смесь PEI-max медленно добавляют к смеси плазмид, осторожно продувают, равномерно перемешивают и затем оставляют при комнатной температуре на 15 мин. Конечную смесь медленно добавляют в культуральную среду и равномерно перемешивают, затем снова помещают в инкубатор для другого культивирования в течение 12-16 часов, затем меняют на среду 6% FBS DMEM для другого культивирования, и раствор вируса собирают через 48 ч и 72 ч.
5.2 Измерение титра вируса
Клетки Jurkat-C5 инокулируют в плоскодонный 96-луночный планшет в концентрации 1,5х105 клеток/мл, и в каждую лунку добавляют 100 мкл среды 1640, содержащей 10% FBS и 0,2 мкл 1000х полибрена. Разведение вируса проводят с полной средой 1640 при 10-кратном разведении, доза вируса в первой лунке составляет 100 мкл при определении в виде маточного раствора вируса, или 1 мкл при определении в виде концентрированного раствора. Разведенные клетки добавляют в лунки с вирусом по 100 мкл/лунку, перемешивают при 32°С, центрифугируют при 1500 об/мин в течение 90 мин, культивируют в инкубаторе при 37°С с 5% СО2 в течение 72 часов. Клетки из плоскодонного 96-луночного планшета отсасывают в круглодонный 96-луночный планшет, центрифугируют при 4°С и 1800 об/мин в течение 5 мин, и супернатант отбрасывают. После добавления 200 мкл 1хPBS, центрифугирование проводят при 4°С и 1800 об/мин в течение 5 мин, супернатант отбрасывают. Добавляют 200 мкл 4% раствора для фиксации тканей, полученный раствор хранят вдали от света и измеряют с помощью проточной цитометрии. Эффективность заражения измеряют способом проточной цитометрии, лунку со скоростью воздействия 2-30% выбирают при расчете титра по формуле: титр (TU/мл)=1,5 х 104 х положительная скорость/объем вируса (мкл) х 1000. Вышеуказанным способом упаковывают вирусы следующих плазмид: pHAGE-EF1A-IRES-RFP, WT-STAR, mut-STAR, co-WT-STAR (αβ-4-1BB-WT, αβ-CD27-WT, αβ-CD28-WT, αβ-ICOS-WT, αβ-OX40-WT, αβ-OX40-WT), co-STAR (αβ-4-1BB, αβ-CD27, αβ-CD28, αβ-ICOS, αβ-OX40, αβ-OX40), co-CD3-STAR (CD3 δ-4-1BB, CD3 δ-CD28, CD3 δ-ICOS, CD3 δ-OX40, CD3 ε-4-1BB, CD3 ε-CD28, CD3 ε-ICOS, CD3 ε-OX40, CD3 γ-OX40, CD3 γ-ICOS, CD3 γ-OX40, CD3 ζ-41BB, CD3 ζ-CD28, CD3 ζ-ICOS, CD3 ζ-OX40), co-линкер-STAR (TCR β-del-OX40, TCR α-del-G4S-OX40, TCR α-del-G4S-OX40, TCR β-del-(G4S) 3-OX40, TCR β-del-(G4S) 7-OX40), C K-STAR (β-IL-2Rb STAR, β-IL-2RbQ STAR, α-IL-2RbQ STAR, α-IL-7RA STAR, α-IL-7RAQ STAR, α-IL-21R STAR) и так далее.
6. Создание культуры Т-клеток и способ заражения
6.1 Культивирование Т-клеточной линии Jurkat
Т-клеточную линию Jurkat культивируют в среде RPMI1640, содержащей 10% FBS, с плотностью культуры от 3х105/мл до 3х106/мл и субкультивируют каждые 1-2 дня. После подсчета клеток, берут необходимое количество клеток и добавляют культуральную среду для доведения до вышеуказанной плотности и помещают в инкубатор с СО2 для культивирования.
6.2 Заражение Т-клеточной линии Jurkat
Клетки подсчитывают, берут 1х106 клеток/мл, центрифугируют и заменяют жидкостью, ресуспендируют в 1 мл среды RPMI 1640, содержащей 10% FBS, и добавляют в 24-луночный планшет с добавлением необходимого количества вирусного раствора, центрифугируют при 1500 об/мин в течение 90 мин и помещают в инкубатор с СО2 для культивирования. Жидкость полностью заменяли свежей средой RPMI 1640, содержащей 10% FBS, через 12 часов после заражения, и положительный результат определяют через 72 часа.
6.3 Первичная культура Т-клеток человека
Первичные Т-клетки выделяют способом Ficoil и культивируют в среде X-VIVO, содержащей 10% FBS и 100 МЕ/мл IL-2, при исходной плотности культуры 1х106/мл, и затем добавляют в планшет с лунками, предварительно покрытый CD3 и RetroNectin r-фибронектином (с конечной концентрацией 5 мкг/мл каждого). Плотность анафазной культуры составляет от 5х105/мл до 3х106/мл, субкультивирование проводят каждые 1-2 дня.
6.4 Заражение первичных Т-клеток человека
После культивирования в течение 48 ч, первичные Т-клетки добавляют к раствору вируса с MOI=20, центрифугируют при 1500 об/мин в течение 90 мин, и затем помещают в СО2 инкубатор для культивирования. Через 24 часа после заражения добавляют среду X-VIVO, содержащую 10% FBS и 100 МЕ/мл IL-2, и лунки подвергают ротации, через 72 часа эффективность заражения определяют с помощью меченого белка или антитела.
6.5. Способ определения эффективности заражения
Через 72 часа после заражения клетки равномерно продувают и подсчитывают, собирают при 5х105/мл, центрифугируют, затем супернатант отбрасывают, используют раствор для окрашивания PBS+2% FBS+2 мМ ЭДТА, соответствующее антитело добавляют для инкубации в течение 30 мин, затем дважды добавляют PBS для промывания и проводят определение на компьютере.
7. Функциональный анализ in vitro для рецептора WT-STAR и его мутантных Т-клеток mut-STAR.
7.1. Способ совместного культивирования in vitro Т-клеток и клеток-мишеней
Клетки-мишени Raji-люцифераза, RajiCD19KO-люцифераза, RajiCD20KO-люцифераза и первичные Т-клетки представляют собой суспендированные клетки, для совместной инкубации берут соответствующее количество клеток и смешивают со средой клеток-мишеней и центрифугируют для культивирования. Конкретные стадии следующие: первичные Т-клетки заражают упакованными и очищенными вирусами WT-STAR и mut-STAR T, и за день до совместного культивирования эффективность заражения определяют с помощью проточной цитометрии, и соотношение эффекторной клетки к клетке-мишени определяют и обычно используют в соотношении 1:1, и общее количество Т-клеток рассчитывают в соответствии с эффективностью заражения, обычное использование клеток-мишеней составляет 1х105/лунку (96-луночный планшет).
7.2. Стимуляция Т-клеток антигеном-мишенью
Антиген-мишень по настоящему изобретению, как правило, представляет собой белок клеточной поверхности, который можно непосредственно использовать для активации Т-клеток для определения функции Т-клеток. Положительные Т-клетки обычно добавляют в количестве 1х105 на лунку, центрифугируют и активируют в течение 24 часов, затем собирают Т-клетки или клетки-мишени для определения функции Т-клеток.
7.3. Подтверждение функции уничтожения Т-клеток: анализ люциферазы
Т-клетки совместно культивируют с клетками-мишенями в течение 24 ч, затем клеточную суспензию осторожно равномерно продувают, берут по 150 мкл клеточной суспензии на лунку и добавляют в белый 96-луночный планшет, центрифугируют при 1500 об/мин в течение 5 мин, и супернатант берут и добавляют клеточный лизат для лизиса при комнатной температуре в течение 15 мин, затем центрифугируют при 4° при 4000 об/мин в течение 15 мин, затем супернатант берут в 2 параллельные лунки на каждую лунку, и добавляют субстрат люциферазы (реагент для анализа люциферазы светлячка), затем определяют с помощью многофункционального микропланшетного ридера с фиксированным значением усиления, равным 100, для получения значения хемилюминесценции. Расчет уничтожения клеток: эффективность уничтожения=100% - (эффекторные клетки и клетки-мишени на лунку/контрольные клетки и клетки-мишени на лунку).
7.4. Результаты обнаружения функции уничтожения Т-клеток с помощью анализа люциферазы показывают, что мутантные Т-клетки mut-STAR проявляют более сильную способность Т-клеток уничтожать опухоли, как показано на фигуре 6 и в таблице 1, после того, как Т-клетки mut-STAR, таргетирующие CD19 и CD20, уничтожают Raji-люциферазу, RajiCD19KO-люциферазу и RajiCD20KO-люциферазу, показатели выживаемости остаточных опухолей составляют 2,41%, 13,94% и 24,40%, соответственно, что значительно лучше, чем у WT-STAR на 45,73%, 77,00% и 76,16%, что указывает на то, что мутант mut-STAR обладает более значительным эффектом уничтожения опухоли.
8. Влияние костимулирующего эндодомена, связанного с С-концами как α цепи TCR, так и β цепи TCR, или С-концом α цепи TCR или β цепи TCR, на функцию STAR-T-клеток.
8.1. Способ совместного культивирования in vitro Т-клеток и клеток-мишеней
Клетки-мишени Raji-люцифераза и первичные Т-клетки представляют собой суспензионные клетки, для совместной инкубации берут соответствующее количество клеток, смешивают со средой клеток-мишеней и центрифугируют для культивирования. Конкретные стадии включают следующие: первичные Т-клетки заражают упакованными и очищенными вирусами WT-STAR и mut-STAR T, и за один день до совместного культивирования, эффективность заражения определяют с помощью проточной цитометрии, и определяют соотношение эффекторной клетки к клетке-мишени, и совместную инкубацию проводят обычно в соотношении 8:1, 4:1, 2:1, 1:1, 1:2, 1:4 и 1:8, а разницу совместной инкубации во времени также определяют , обычно через 6 ч, 12 ч, 24 ч, 36 ч и 48 ч. Для определения пролиферации co-STAR Т-клеток, клетки-мишени Raji-люцифераза инкубируют с первичными Т-клетками в течение 7 дней для наблюдения за изменениями количества пролиферации клеток и секреции IL-2, затем положительные Т-клетки сортируют с помощью проточной цитометрии и обрабатывают до покоящейся культуры без антигенной стимуляции в течение двух дней, затем снова совместно культивируют с клетками-мишенями в течение 24 часов для обнаружения уничтожения Т-клеток, где обычное использование клеток-мишеней составляет 1х105/лунку (96-луночный планшет).
8.2. Способ стимуляции Т-клеток антигеном-мишенью
Антиген-мишень по настоящему изобретению, как правило, представляет собой белок клеточной поверхности, который можно непосредственно использовать для активации Т-клеток для определения функции Т-клеток, в частности, антиген-мишень обычно добавляют с 1х105/лунку положительными Т-клетками, центрифугируют и активируют в течение 24 часов для сбора клеточной суспензии или культурального супернатанта для определения функции Т-клеток, или активируют в течение 6 часов, 12 часов, 24 часов, 36 часов, 48 часов или 7 дней для определения функции уничтожения Т-клеток.
8.3. Подтверждение функции уничтожения Т-клеток: анализ люциферазы
Т-клетки совместно культивируют с клетками-мишенями в течение разного времени, затем клеточную суспензию осторожно равномерно продувают, берут 150 мкл клеточной суспензии на лунку и добавляют в белый 96-луночный планшет, центрифугируют при 1500 об/мин в течение 5 мин, и супернатант берут и добавляют клеточный лизат для лизиса при комнатной температуре в течение 15 мин, затем центрифугируют при 4° при 4000 об/мин в течение 15 мин, затем берут супернатант, отбирая по 2 параллельные лунки на каждую лунку, и добавляют субстрат люциферазы (реагент для анализа люциферазы светлячка), затем определяют с помощью многофункционального микропланшетного ридера с фиксированным значением усиления, равным 100, с получением значения хемилюминесценции. Расчет уничтожения клеток: эффективность уничтожения=100% - (эффекторные клетки и клетки-мишени на лунку/контрольные клетки и клетки-мишени на лунку).
Результаты обнаружения функции уничтожения Т-клеток с помощью люциферазного анализа показали, что mut-STAR с костимулирующим эндодоменом, связанным с С-концами и α, и β цепи TCR, демонстрируют сходный эффект уничтожения опухоли при различных соотношениях E:T T-клеток и клеток-мишеней, и нет существенной разницы в остаточной выживаемости опухоли при различных соотношениях E:T, как показано на фиг. 7 и в таблице 2. Однако, с точки зрения результатов обнаружения уничтожения опухоли в разное время, mut-STAR с эндодоменами OX40, связанными с С-концами и α, и β цепей TCR (αβ-OX40), и mut-STAR с инкубацией с Raji-люцифераза в течение 48 ч демонстрируют аналогичное влияние на уничтожение опухоли, примерно 24% и 19%, соответственно, которые значительно превосходят mut-STAR с другими костимулирующими эндодоменами, связанными с С-концами и α, и β цепей TCR (αβ-41BB, αβ-CD27, αβ-CD28, αβ-ICOS), как показано на фиг. 8 и таблице 3. На основании приведенных выше результатов установлено, что связывание эндодомена костимулирующей молекулы OX40 с С-концами и α, и β цепей TCR не влияет на эффект уничтожения mut-STAR. Кроме того, основываясь на приведенных выше результатах, mut-STAR с эндодоменом OX40, связанным с C-концом α цепи (α-OX40) или β цепи (β-OX40) TCR, отдельно обнаружен на функции уничтожения Т-клеток с помощью люциферазного анализа, и результаты обнаружения показывают, что при различных соотношениях E:T Т-клеток к совместно инкубируемым опухолевым клеткам или при совместном инкубировании в течение 24 часов, α-OX40 и β-OX40 не демонстрируют значительных отличий от α-β-OX40, как показано на фиг. 9 и 10, и в таблицах 4 и 5. На основании приведенных выше результатов уничтожения, не обнаружено существенной разницы между mut-STAR с эндодоменом костимулирующей молекулы OX40, связанным с С-концами и α-, и β цепи TCR (αβ-OX40) и mut-STAR с эндодоменом костимулирующей молекулы OX40, связанным с С-концом α цепи (α-OX40) или β цепи (β-OX40) TCR, по отдельности, но их способность уничтожать опухоль значительно лучше, чем у mut-STAR.
8.4. Анализ секреции цитокинов Т-клетками: ELISA
Во время активации Т-клеток высвобождается большое количество цитокинов, таких как TNF-α, IFN-γ и IL-2, которые помогают Т-клеткам убивать клетки-мишени или способствуют размножению самих Т-клеток. После стимуляции Т-клеток клетками-мишенями или антигенами, Т-клетки собирают, центрифугируют и сортируют отбирают супернатант. Используемые наборы ELISA для TNF-α, IFN-γ и IL-2 представляют собой наборы ELISA без покрытия IL-2 человека, ELISA без покрытия TNF-α человека и ELISA без покрытия IFN-γ человека (арт. № 88-7025, 88-7346, 88-7316, соответственно). Конкретные стадии включают следующие: разведение 10X буфера для покрытия до 1X с ddH2O, добавление антитела с покрытием (250X), тщательное перемешивание и добавление в 96-луночный планшет по 100 мкл/лунку. После герметичного закрытия пластиковой пленкой и выдерживания в течение ночи при 4°C, 1X PBST (1X PBS с добавлением 0,05% Tween 20) используют для промывки 3 раза по 260 мкл/лунку каждый раз, 5X ELISA/ELISPOT разбавитель разводят до 1X с ddH2O, и затем добавляют в 96-луночный планшет по 200 мкл/лунку с последующим выдерживанием при комнатной температуре в течение 1 часа. PBST используют для однократной промывки, и разведение проводят по стандартной кривой (в диапазоне: от 2 до 250, от 4 до 500, от 4 до 500, соответственно), образцы разводят в 20-50 раз 1x разбавителем. Образцы, разведенные в соответствии со стандартной кривой, добавляют по 100 мкл на лунку, берут две параллельные лунки и инкубируют при комнатной температуре в течение 2 часов, PBST используют для промывки три раза, затем разводят антитело для обнаружения, разведенное 1x разбавителем, после инкубации в течение 1 ч, PBST используют для промывки три раза, затем добавляют HRP, разведенный 1x разбавителем, после инкубации в течение 30 минут раствор промывают шесть раз, добавляют TMB для проявления цвета со временем проявления цвета менее 15 мин, и 2N H2SO4 добавляют для терминации, обнаруживают абсорбцию света при 450 нм.
Результаты ELISA показали, что после совместной инкубации Т-клеток с клетками-мишенями в течение 24 ч, секреция IL-2 mut-STAR (αβ-OX40) с эндодоменом OX40, связанным с С-концами и α цепи, и β цепи TCR, составляет примерно 10000 пг/мл, что значительно выше, чем у STAR с другими структурами, в которых значения для mut-STAR, αβ-41BB, αβ-CD27, αβ-CD28 и αβ-ICOS составляют примерно 7700 пг/мл, 6450 пг/мл, 6690 пг/мл, 6000 пг/мл и 6050 пг/мл, соответственно; с точки зрения секреции TNF α и IFN-γ, αβ-OX40 также показывает аналогичные результаты с mut-STAR, в то время как другие структуры показали другое снижение, как показано на фигуре 11 и в таблице 6. На основании результатов ELISA, показано, что секреция IL-2 с помощью mut-STAR с эндодоменом костимулирующей молекулы OX40, связанным с С-концами и α, и β цепи TCR (αβ-OX40), значительно выше, чем у mut-STAR.
8.5. Обнаружение изменений пролиферации Т-клеток: подсчет проточной цитометрией
Во время активации Т-клеток высвобождается большое количество цитокинов, которые помогают Т-клеткам убивать клетки-мишени или способствовать размножению самих Т-клеток, и наиболее очевидным явлением в пролиферации Т-клеток является значительное изменение числа Т-клеток. Т-клетки инкубируют с клетками-мишенями в течение 7 дней, затем центрифугируют, ресуспендируют до 200 мкл с PBS, и количество положительных Т-клеток подсчитывают проточной цитометрией. Изменения пролиферации Т-клеток: кратность пролиферации=количество положительных Т-клеток через 7 дней/начальное количество добавленных положительных Т-клеток.
После сортировки, клетки mut-STAR-T с различной структурой совместно культивируют с клетками Raji-люцифераза в соотношении 1:3 E:T, считая за 0 день, и затем клетки собирают в 1 и 7 день для проточного анализа, соответственно. Среди них, используемая среда представляет собой 1640 полных сред без IL-2, и начальное количество Т-клеток TCR составляет 1х105 клеток, образцы в каждый момент времени инкубируют независимо, и оставшиеся совместно инкубированные образцы были частично заменяют жидкостью на следующий день и дополняют клетками-мишенями. Клетки, используемые для проточного анализа, заранее окрашивают антителом против CD3 человека, и их определенный объем собирают и регистрируют при проведении анализа на машине, количество и доля Т-клеток в системе известны путем преобразования. Как показано на фиг. 12 и в таблице 7, и можно увидеть из кривой кратности пролиферации абсолютного числа клеток mut-STAR, mut-STAR с эндодоменом OX40, соединенным с С-концами и α, и β цепи TCR (αβ-OX40) показали лучшую активацию и пролиферацию после распознавания эпитопа клетки-мишени, которые значительно выше, чем у mut-STAR. Пролиферация mut-STAR с эндодоменом OX40, соединенным с С-концом α цепи TCR (α-OX40) или β цепи (β-ОХ40), по отдельности, показана на фиг. 13 и в таблице 8, где пролиферация Т-клеток mut-STAR с эндодоменом OX40, соединенным с С-концом α цепи TCR (α-OX40), отдельно, через 7 дней является 11,75-кратной, тогда как у других структур, таких как mut-STAR, β-OX40 и αβ-OX40, является 2,755-кратной, 4,128-кратной и 6,744-кратной, соответственно, таким образом, эффект пролиферации α-OX40 значительно выше, чем у других структур. Объединяя приведенные выше результаты уничтожения опухоли, результаты ELISA и результаты пролиферации Т-клеток, mut-STAR с эндодоменом OX40, соединенным с C-концом α цепи TCR (α-OX40), показывает более повышенную способность к пролиферации и уничтожению опухоли, чем другие структуры.
В соответствии с приведенными выше результатами, обнаружено, что наилучший эффект пролиферации получают путем связывания костимулирующей молекулы OX40 с цепью TCR-α, не влияя также на эффект уничтожения Т-клеток. Таким образом, влияние mu-STAR с эндодоменом различных костимулирующих молекул, связанных с α цепью TCR, на эффект уничтожения при различных соотношениях E:T Т-клетки к клетке-мишени показывают, что эффект уничтожения mut-STAR, связанного с эндодоменом костимулирующей молекулы OX40, лучше, чем у STAR с другим связанным костимулирующим доменом. При соотношениях Е:Т 1:2 и 1:4, эффект Т-клеток mut-STAR с присоединенным эндодоменом костимулирующей молекулы OX40 (α-OX40) является таким же, как эффект αβ-OX40, и лучше, чем эффект Т-клеток mut-STAR с другими связанными костимулирующими молекулами. Результаты показаны на фиг. 14 и в таблице 9. На основании приведенных выше результатов уничтожения опухоли, способности к уничтожению опухоли и пролиферации, полученные с помощью mut-STAR с эндодоменом OX40, связанным с α цепью TCR (α-OX40), значительно выше, чем способности mut-STAR.
Таблица 2. Выживаемость клеток-мишеней после совместного культивирования с ко-STAR при различных соотношениях E:T (%)
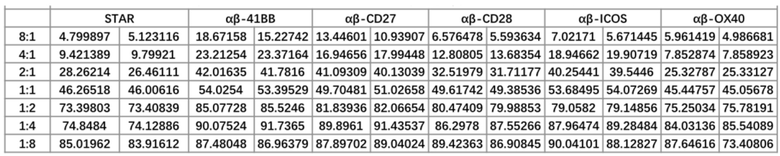
Таблица 3. Выживаемость клеток-мишеней после совместного культивирования с ко-STAR в разное время (%)
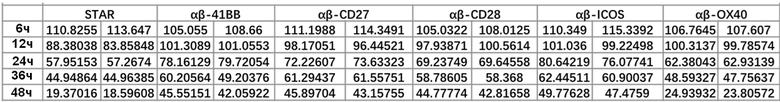
Таблица 4. Степень выживаемости клеток-мишеней после совместного культивирования со STAR с добавлением OX40 к α цепи и β цепи, соответственно, при различных соотношениях E:T (%)
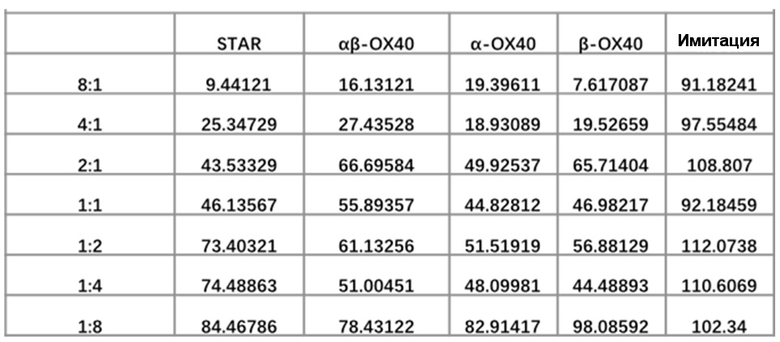
Таблица 5. Выживаемость клеток-мишеней после совместного культивирования со STAR с добавлением OX40 к α цепи и β цепи, соответственно, в разное время (%)
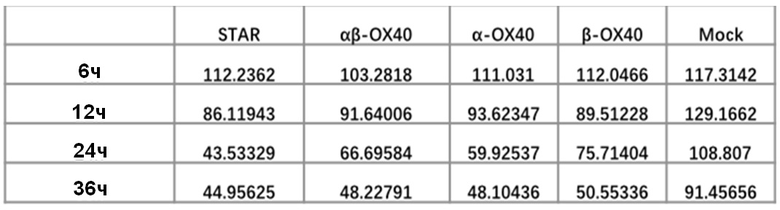
Таблица 6.
Секреция IL-2 после совместного культивирования co-STAR с клетками-мишенями
 Секреция IFN-α после совместного культивирования co-STAR с клетками-мишенями
Секреция IFN-α после совместного культивирования co-STAR с клетками-мишенями
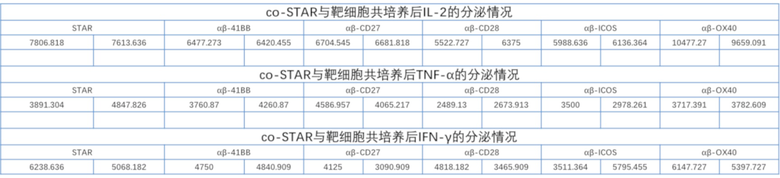
Секреция IFN-γ после совместного культивирования co-STAR с клетками-мишенями

Таблица 7. Пролиферация после совместного культивирования co-STAR с клетками-мишенями
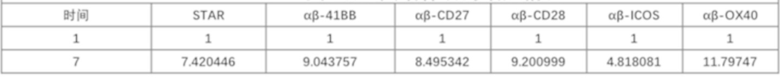
Таблица 8. Сравнение пролиферации после совместного культивирования клеток-мишеней со STAR, содержащего OX40, связанный с его α или β цепями
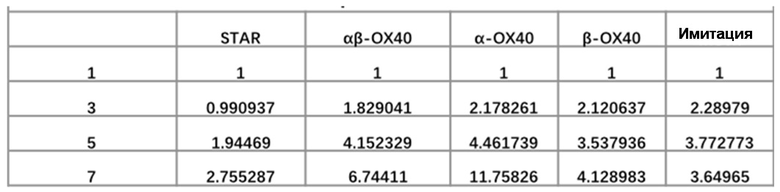
9. Влияние костимуляторной структуры, связанной с различными цепями CD3 (co-CD3-STAR), на функцию STAR-T-клеток.
9.1. Способ совместного культивирования in vitro Т-клеток и клеток-мишеней
Клетки-мишени Raji-люцифераза и первичные Т-клетки представляют собой суспензионные клетки, для совместной инкубации берут соответствующее количество клеток, смешивают со средой клеток-мишеней и центрифугируют для культивирования. Конкретные стадии включают следующие: первичные Т-клетки заражают упакованными и очищенными вирусами WT-STAR и mut-STAR T, и за день до совместного культивирования, эффективность заражения определяют с помощью проточной цитометрии, и определяют соотношение эффекторной клетке к клетке-мишени, совместную инкубацию проводят обычно в соотношении 1:1 или 2:1, кроме того, клетки-мишени Raji-люцифераза и первичные Т-клетки совместно инкубируют в течение 7 дней, чтобы наблюдать изменения в количестве пролифераций клеток. Общее количество Т-клеток рассчитывают на основе эффективности заражения, и обычное использование клеток-мишеней составляет 1х105/лунку (96-луночный планшет).
9.2. Стимуляция Т-клеток антигеном-мишенью
Антиген-мишень по настоящему изобретению, как правило, представляет собой белок клеточной поверхности, который можно непосредственно использовать для активации Т-клеток для определения функции Т-клеток. Положительные Т-клетки обычно добавляют в количестве 1х105/лунку, центрифугируют и активируют в течение 24 часов, клеточную суспензию или супернатант культуры собирают для определения функции Т-клеток.
9.3. Подтверждение функции уничтожения Т-клеток: анализ люциферазы
Т-клетки совместно культивируют с клетками-мишенями в течение разного времени, затем клеточную суспензию осторожно равномерно продувают, 150 мкл клеточной суспензии на лунку переносят в белый 96-луночный планшет, центрифугируют при 1500 об/мин в течение 5 мин, отбирают супернатант и добавляют клеточный лизат для лизиса при комнатной температуре в течение 15 мин, затем центрифугируют при 4° при 4000 об/мин в течение 15 мин, затем отбирают супернатант, отбирая 2 параллельные лунки на каждую лунку, и добавляют субстрат люциферазы (реагент для анализа люциферазы светлячка), затем обнаруживают с помощью многофункционального микропланшетного ридера с фиксированным значением усиления, равным 100, для получения значения хемилюминесценции. Расчет уничтожения клеток: эффективность уничтожения=100% - (эффекторные клетки и клетки-мишени на лунку/контрольные клетки и клетки-мишени на лунку).
Результаты обнаружения функции уничтожения Т-клеток с помощью анализа люциферазы показали, что, когда костимулирующий эндодомен соединен с С-концом цепи CD3 δ, CD3 γ, CD3 ε или CD3 ζ и коэкспрессирован на mut-STAR T, результаты при соотношении 1:1 E:T различных Т-клеток к клеткам-мишеням показывают, что все co-CD3-STAR не демонстрируют лучший эффект уничтожения клеток-мишеней, чем αβ-OX40, но по сравнению с mut-STAR, CD3δ-OX40, CD3ζ-41BB, CD3ζ-CD28, CD3ζ-ICOS, CD3ζ-OX40 демонстрируют аналогичный эффект уничтожения опухоли, их остаточная выживаемость опухоли существенно не различается при соотношении E:T 1:1, как показано на фиг.15 и в таблице 10. Основываясь на приведенных выше результатах, путем связывания костимулирующего эндодомена с С-концом различных цепей CD3, эффект уничтожения опухоли mut-STAR значительно не улучшается, но эффект mut-STAR с ко-стимулирующим эндодоменом, связанным с С-концом ζ цепи CD3, существенно не уменьшается.
9.4. Обнаружение изменений пролиферации Т-клеток: подсчет методом проточной цитометрии
Во время активации Т-клеток высвобождается большое количество цитокинов, которые помогают Т-клеткам убивать клетки-мишени или способствовать размножению самих Т-клеток, и наиболее очевидным явлением в пролиферации Т-клеток является значительное изменение числа Т-клеток. Т-клетки инкубируют с клетками-мишенями в течение 7 дней, затем центрифугируют, ресуспендируют до 200 мкл с PBS, и количество положительных Т-клеток подсчитывают с помощью проточной цитометрии. Изменения пролиферации Т-клеток: кратность пролиферации=количество положительных Т-клеток через 7 дней/начальное количество добавленных положительных Т-клеток.
После сортировки, клетки mut-STAR-T с различной структурой совместно культивируют с клетками Raji-люцифераза в соотношении 1:3 E:T, считая за 0 день, и затем клетки собирают в 1 и 7 день для проточного анализа, соответственно. Среди них, используемая среда представляет собой 1640 полную среду без IL-2, и начальное количество Т-клеток TCR составляет 1х105 клеток, образцы в каждый момент времени инкубируют независимо, и оставшиеся совместно инкубированные образцы частично заменяют жидкостью на следующий день и дополняют клетками-мишенями. Клетки, используемые для проточного анализа, заранее окрашивают антителом против CD3 человека, и их определенный объем собирают и регистрируют при проведении анализа на машине, количество и доля Т-клеток в системе известны путем преобразования. Как показано на фиг. 16 и в таблице 11, из кривой кратности абсолютного числа клеток mut-STAR видно, что все ко-CD3-STAR не демонстрируют лучший эффект уничтожения клеток-мишеней по сравнению с αβ-OX40-mut-STAR, но CD3 ε-CD28, CD3 δ-OX40, CD3 ζ-CD28, CD3 ζ-ICOS, CD3 ζ-OX40 и подобные демонстрируют аналогичные эффекты пролиферации. Объединяя результаты обнаружения уничтожения с помощью анализа люциферазы и результаты пролиферации Т-клеток, структура с костимулирующим эндодоменом, соединенным с С-концом цепи CD3δ или CD3γ, или CD3ε, или CD3ζ, и коэкспрессируемая на mut-STAR T не демонстрирует лучших эффектов уничтожения и пролиферации опухоли по сравнению с T-клетками αβ-OX40-mut-STAR.
Таблица 10. Выживаемость клеток-мишеней после совместного культивирования со STAR с костимулирующим доменом, связанным с другими эндодоменами CD3
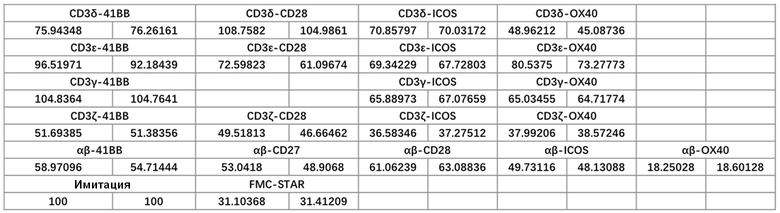
Таблица 11. Пролиферация после совместного культивирования STAR с костимулирующим доменом, связанным с другими эндодоменами CD3, с клетками-мишенями в течение 7 дней
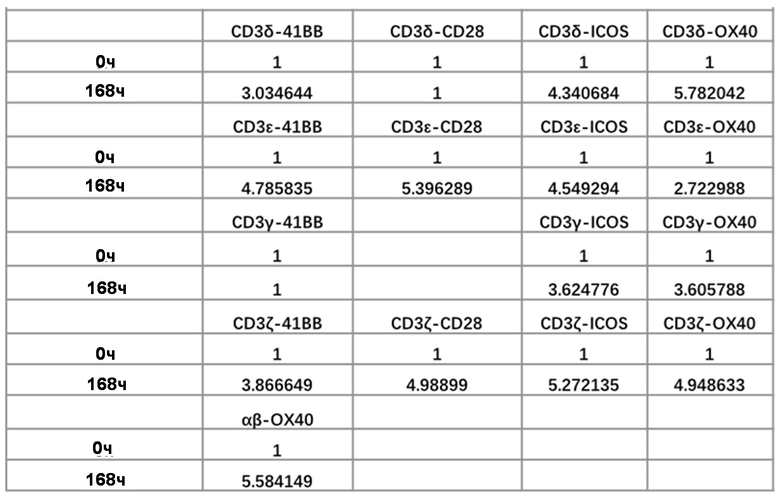
10. Влияние костимулирующего эндодомена, содержащего линкер G4S, соединенный с константной областью α или β с делецией эндодомена, на функцию STAR-T-клеток.
10.1. Способ совместного культивирования in vitro Т-клеток и клеток-мишеней
Клетки-мишени Raji-люцифераза и первичные Т-клетки представляют собой суспензионные клетки, для совместной инкубации берут соответствующее количество клеток, смешивают со средой клеток-мишеней и центрифугируют для культивирования. Конкретные стадии включают следующие: первичные Т-клетки заражают упакованными и очищенными вирусами WT-STAR и mut-STAR T, и за день до совместного культивирования, эффективность заражения определяют с помощью проточной цитометрии, и соотношение определяют эффекторной клетки к клетке-мишени, и проводят совместную инкубацию, как правило, в соотношении 1:1 или 2:1, кроме того, клетки-мишени Raji-люцифераза и первичные Т-клетки совместно инкубируют в течение 7 дней, чтобы наблюдать изменения в количестве пролифераций клеток. Общее количество Т-клеток рассчитывают на основе эффективности заражения, и обычное использование клеток-мишеней составляет 1х105/лунку (96-луночный планшет).
10.2. Стимуляция Т-клеток антигеном-мишенью
Антиген-мишень по настоящему изобретению, как правило, представляет собой белок клеточной поверхности, который можно непосредственно использовать для активации Т-клеток для определения функции Т-клеток, в частности, антиген-мишень обычно добавляют с 1х105/лунку положительных Т-клеток, центрифугируют и активируют в течение 24 часов для сбора клеточной суспензии или супернатанта культуры для определения функции Т-клеток.
10.3. Подтверждение функции уничтожения Т-клеток: анализ люциферазы
Т-клетки совместно культивируют с клетками-мишенями в течение разного времени, затем клеточную суспензию осторожно равномерно продувают, 150 мкл клеточной суспензии на лунку переносят в белый 96-луночный планшет, центрифугируют при 1500 об/мин в течение 5 мин, отбирают супернатант и добавляют клеточный лизат для лизиса при комнатной температуре в течение 15 мин, затем центрифугируют при 4° при 4000 об/мин в течение 15 мин, затем отбирают супернатант, отбирая 2 параллельные лунки на каждую лунку, и добавляют с субстратом люциферазы (реагент для анализа люциферазы светлячка), затем обнаруживают с помощью многофункционального микропланшетного ридера с фиксированным значением усиления, равным 100, для получения значения хемилюминесценции. Расчет уничтожения клеток: эффективность уничтожения=100% - (эффекторные клетки и клетки-мишени на лунку/контрольные клетки и клетки-мишени на лунку).
Результаты обнаружения функции уничтожения Т-клеток с помощью люциферазного анализа показали, что костимулирующий эндодомен с линкером G4S различной длины был связан с α или β константной областью с удаленным эндодоменом, результаты при 1:1 E:T соотношении Т-клетки к клетке-мишени показывают, что, когда линкер соединен с эндодоменом константной области α, α-del-OX40, α-OX40, α-del-G4S-OX40, α-del-(G4S) 3-OX40 и α-β-ox40 демонстрируют аналогичный эффект уничтожения, который выше, чем у α-del-(G4S) 7-OX40 и α-del-(G4S) 10-OX40, и чем длиннее линкер, тем слабее эффект уничтожения Т-клеток, когда линкер соединен с эндодоменом β-константной области, β-del-OX40, β-ox40, β-del-(G4S) 3-OX40 и α-β-ox40 демонстрируют аналогичный эффект уничтожения, и еще, чем длиннее линкер, тем слабее эффект уничтожения Т-клеток. Соединение с OX40 (α-del-OX40 или β-del-OX40) после удаления эндодомена константной области демонстрирует небольшое различие в эффекте по сравнению с таковым без такого удаления, но после добавления линкера (количество линкеров не более 3), эффект α-del-(G4S) 1-3-OX40 или β-del-(G4S) 1-3-OX40 лучше, чем эффект α-del-OX40 или β-del-OX40. Сравнение эффекта, когда линкер соединен с эндодоменом константной области α, и эффекта, когда линкер соединен с эндодоменом константной области β, показывает, что эффект, когда линкер соединен с эндодоменом константной области α, лучше, чем эффект, когда линкер соединен с эндодоменом константной области β. Однако существенной разницы в выживаемости остаточной опухоли при соотношении E:T 2:1, как показано на фигурах 17 и 18 и в таблицах 12 и 13, нет. На основании приведенных выше результатов, mut-STAR с костимулирующим эндодоменом, содержащим линкер G4S, связанный с константной областью α или β с удаленным эндодоменом, не влияет на эффект уничтожения опухоли mut-STAR, и эффект при связывании с эндодоменом с константной областью α лучше, чем при связывании с константной областью β эндодомена, но чем больше длина линкера, тем слабее эффект уничтожения опухоли.
10.4. Анализ секреции цитокинов Т-клетками: ELISA
Во время активации Т-клеток высвобождается большое количество цитокинов, таких как TNF-α, IFN-γ и IL-2, которые помогают Т-клеткам убивать клетки-мишени или способствуют размножению самих Т-клеток. После стимуляции Т-клеток клетками-мишенями или антигенами, Т-клетки собирают, центрифугируют и отбирают супернатант. Используемые наборы ELISA для TNF-α, IFN-γ и IL-2 представляют собой наборы ELISA без покрытия IL-2 человека, ELISA без покрытия TNF-α человека и ELISA без покрытия IFN-γ человека (арт. № 88-7025, 88-7346, 88-7316, соответственно). Конкретные стадии включают следующие: разведение 10Х буфера для покрытия ddH2O до 1Х, добавление антитела с покрытием (250Х), тщательное перемешивание и добавление в 96-луночный планшет по 100 мкл/лунку. После герметичного закрытия полиэтиленовой пленкой и выдерживания в течение ночи при 4°C, 1X PBST (также называемый промывочным буфером, 1X PBS с добавлением 0,05% Tween 20) используют для промывки 3 раза по 260 мкл/лунку, 5X ELISA/ELISPOT разбавитель разводят до 1X с ddH2O и затем добавляют в 96-луночный планшет по 200 мкл/лунку с последующим выдерживанием при комнатной температуре в течение 1 часа. PBST используют для однократной промывки, и разведение проводят по стандартной кривой (в диапазоне: от 2 до 250, от 4 до 500, от 4 до 500, соответственно), образцы разводят в 20-50 раз 1x разбавителем. Образцы в соответствии со стандартной кривой добавляют по 100 мкл на лунку, берут две параллельные лунки и инкубируют при комнатной температуре в течение 2 часов, PBST используют для промывки 3 раза, затем добавляют антитело для обнаружения, разведенное 1x разбавителем, после инкубации в течение 1 ч, PBST используют для промывки 3 раза, затем добавляют HRP, разведенный 1x разбавителем, после инкубации в течение 30 минут раствор промывают шесть раз, добавляют TMB для проявления цвета со временем проявления цвета менее 15 мин и 2N H2SO4 добавляют для терминации, определяют поглощение света при 450 нм.
Результаты ELISA показали, что после совместной инкубации Т-клеток с клетками-мишенями в течение 24 ч, секреция IL-2 и IFN-γ mut-STAR со структурой α-del-(G4S)7-OX40 OX40 значительно ниже, чем у mut-STAR, в то время как секреция IL-2 других структур β-del-OX40, α-del-OX40, α-del-G4S-OX40 и α-del-(G4S) 3-OX40, каждая, является аналогичной с mut-STAR; только β-del-OX40 демонстрирует секрецию IFN-γ, аналогичную секреции mut-STAR, в то время как секреция IFN-γ других структур демонстрирует другое снижение, как показано на фигурах 19 и 20 и таблицах 14 и 15. Согласно по результатам ELISA, в структуре с делецией эндодомена константной области α или β, делеция эндодомена α цепи влияет на секрецию IFN-γ, но не влияет на секрецию IL-2, в то время как делеция эндодомена β цепи не влияет на секрецию ни IL-2, ни IFN-γ; в то же время, после делеции эндодомена α цепи, линкер используют для связывания с эндодоменом OX40, что, в случае линкера длиной менее 7, не влияет на секрецию IL-2, но снижает секрецию IFN-γ.
10.5. Обнаружение изменений дифференциации Т-клеток: анализ проточной цитометрией
Во время активации Т-клеток высвобождается большое количество цитокинов и других хемокинов, и сигналы трансдуцируются в ядро через цитокины или хемокиновые рецепторы для регуляции дифференциации Т-клеток. Т-клетки дифференцируют из примитивных Т-клеток (наивных) в Т-клетки центральной памяти (Tcm), в эффекторные Т-клетки памяти (Tem) и, наконец, в эффекторные Т-клетки (Teff). Однако на пролиферацию и жизнестойкость Т-клеток in vivo влияет количество Т-клеток, дифференцировавших в центральные Т-клетки памяти (Tcm) в эффекторные Т-клетки памяти (Tem). Т-клетки памяти можно классифицировать на Т-клетки стволовых клеток, Т-клетки центральной памяти и Т-клетки эффекторной памяти. Коэффициент дифференциации Т-клеток центральной памяти влияет на стойкий эффект уничтожения Т-клеток in vivo. Соотношение примитивных Т-клеток и эффекторных Т-клеток влияет на эффект уничтожения опухоли и жизнестойкость Т-клеток in vivo. Экспрессия CD45RA и CCR7 на поверхности Т-клеток обнаружена с помощью проточной цитометрии, поэтому можно определить дифференциацию Т-клеток. Т-клетки инкубируют с клетками-мишенями в течение 7 дней, центрифугируют, окрашивают потоком антител против CD45RA-Percp-cy5.5 человека и против CCR7-APC человека в течение 30 минут, снова центрифугируют, промывают PBS и фиксируют 4% раствора параформальдегида, и дифференциацию Т-клеток обнаруживают с помощью проточной цитометрии.
Результаты проточной цитометрии показали, что при присоединении костимулирующего эндодомена, содержащего линкер G4S, к структуре с удаленным эндодоменом константной области α или β, полученный α-del-(G4S)3-OX40 демонстрирует значительную дифференциацию Т-клеток центральной памяти, в то время как структура с OX40, непосредственно связанная с эндодоменом константной области α (α-del-OX40) или β (β-del-OX40), также способствует дифференциации Т-клеток центральной памяти, как показано на фиг. 21 и в таблице 16. В то же время, хотя количество эффекторных Т-клеток mut-STAR Т-клеток с различными модифицированными структурами значительно ниже, чем у mut-STAR, доля наивных Т-клеток значительно выше, чем у mut-STAR (28,3%), как показано на фигуре 22 и в таблице 17. Комбинируя результаты уничтожения опухоли, результаты ELISA и обнаружение дифференциации клеток проточной цитометрией, mut-STAR с удаленным эндодоменом константной области α или β и связанным с эндодоменом OX40 через линкер (G4S)3, может значительно улучшать дифференциацию популяции клеток памяти Т-клеток, не влияя на эффект уничтожения опухоли и секрецию IL-2.
Таблица 12. Эффекты уничтожения клеток-мишеней после совместного культивирования клеток-мишеней со STAR с OX40, связанным с его α цепью через разные линкеры
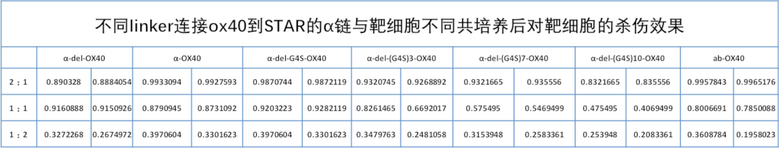
Таблица 13. Эффект уничтожения клеток-мишеней после совместного культивирования клеток-мишеней со STAR с OX40, присоединенным к его β цепи через различные линкеры
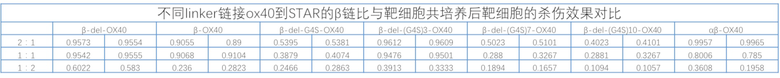
Таблица 14. Секреция IL-2 после совместного культивирования клеток-мишеней со STAR с OX40, связанным с его α цепью через разные линкеры
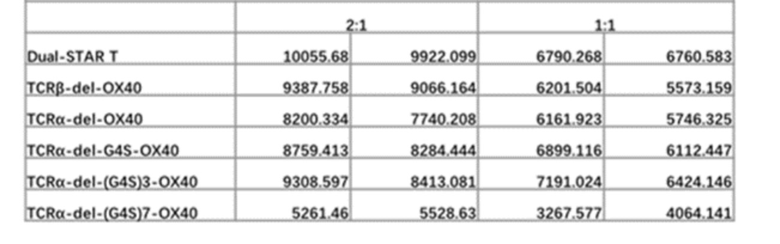
Таблица 15. Секреция IFN-α после совместного культивирования клеток-мишеней со STAR с OX40, присоединенным к его α цепи через разные линкеры
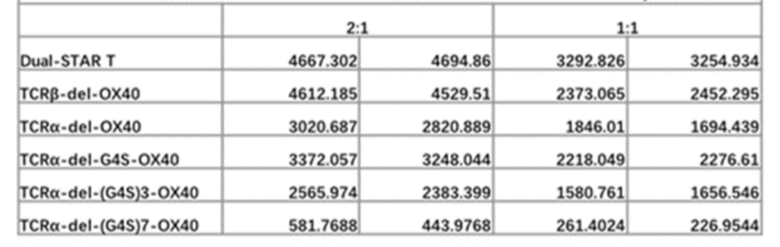
Таблица 16. Влияние STAR с OX40, связанным с его α цепью через различные линкеры, на дифференциацию Т-клеток центральной памяти
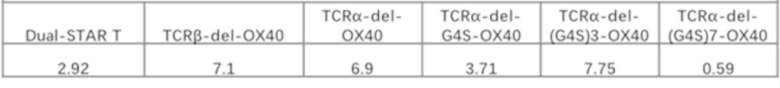
Таблица 17. Влияние STAR с OX40, соединенным с его α цепью через различные линкеры, на дифференциацию Т-клеток
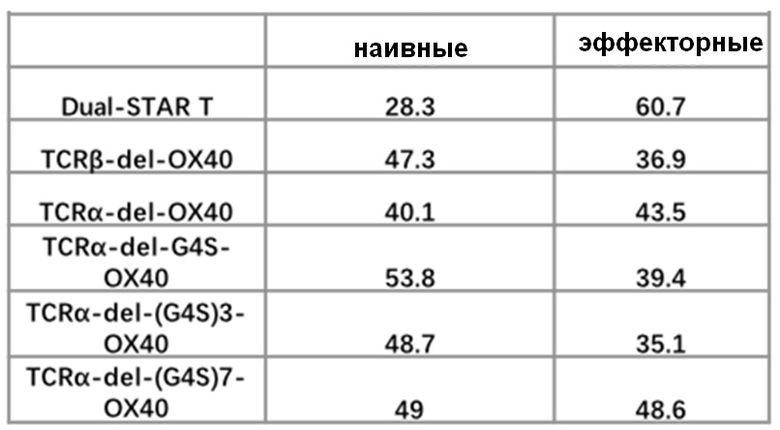
11. Влияние связанной с эндоцитозом TCR модификации лизина в аргинин в трансмембранном домене или эндодомене на функцию мутантных STAR-T-клеток.
11.1. Способ совместного культивирования in vitro Т-клеток и клеток-мишеней
Клетки-мишени Raji-люцифераза и первичные Т-клетки представляют собой суспензионные клетки, для совместной инкубации берут соответствующее количество клеток, смешивают со средой клеток-мишеней и центрифугируют для культивирования. Конкретные стадии включают следующие: первичные Т-клетки заражают упакованными и очищенными вирусами WT-STAR и mut-STAR T, и за день до совместного культивирования, эффективность заражения определяют с помощью проточной цитометрии, и определяют соотношение эффекторной клетки к клетке-мишени, совместную инкубацию проводят обычно в соотношении 1:1 или 2:1, кроме того, клетки-мишени Raji-люцифераза и первичные Т-клетки совместно инкубируют в течение 7 дней, чтобы наблюдать изменения в количестве пролифераций клеток. Общее количество Т-клеток рассчитывают на основе эффективности заражения, и обычное использование клеток-мишеней составляет 1х105/лунку (96-луночный планшет).
11.2. Стимуляция Т-клеток антигеном-мишенью
Мишенью настоящего изобретения, как правило, является белок клеточной поверхности, который можно непосредственно использовать для активации Т-клеток для определения функции Т-клеток, в частности, антиген-мишень обычно добавляют с 1х105/лунку положительных Т-клеток, центрифугируют и активируют в течение 24 часов для сбора клеточной суспензии или супернатанта культуры для определения функции Т-клеток.
11.3. Подтверждение функции уничтожения Т-клеток: анализ люциферазы
Т-клетки совместно культивируют с клетками-мишенями в течение разного времени, затем клеточную суспензию осторожно равномерно продувают, 150 мкл клеточной суспензии на лунку переносят в белый 96-луночный планшет, центрифугируют при 1500 об/мин в течение 5 мин, отбирают супернатант и добавляют клеточный лизат для лизиса при комнатной температуре в течение 15 мин, затем центрифугируют при 4° при 4000 об/мин в течение 15 мин, затем отбирают супернатант, отбирая 2 параллельные лунки на каждую лунку, и добавляют с субстратом люциферазы (реагент для анализа люциферазы светлячка), затем обнаруживают с помощью многофункционального микропланшетного ридера с фиксированным значением усиления, равным 100, для получения значения хемилюминесценции. Расчет уничтожения клеток: эффективность уничтожения=100% - (эффекторные клетки и клетки-мишени на лунку/контрольные клетки и клетки-мишени на лунку).
Результаты обнаружения функции уничтожения Т-клеток с помощью анализа люциферазы показали, что эффект уничтожения опухолевых клеток ub-STAR, mut-STAR со связанным с эндоцитозом TCR лизином, модифицированным в аргинин в трансмембранном домене или эндодомене, в соотношении E:T 2:1 или 1:1 Т-клетки к клетке-мишени, значительно ниже, чем у Т-клеток mut-STAR, как показано на фигуре 23 и в таблице 18.
11.4. Анализ цитокинов, секретируемых Т-клетками: ELISA
Во время активации Т-клеток высвобождается большое количество цитокинов, таких как TNF-α, IFN-γ и IL-2, которые помогают Т-клеткам убивать клетки-мишени или способствуют размножению самих Т-клеток. После стимуляции Т-клеток клетками-мишенями или антигенами, Т-клетки собирают, центрифугируют и отбирают супернатант. Используемые наборы ELISA для TNF-α, IFN-γ и IL-2 представляют собой наборы ELISA без покрытия IL-2 человека, ELISA без покрытия TNF-α человека и ELISA без покрытия IFN-γ человека (арт. № 88-7025, 88-7346, 88-7316, соответственно). Конкретные стадии включают следующие: разведение 10Х буфера для покрытия ddH2O до 1Х, добавление антитела с покрытием (250Х), тщательное перемешивание и добавление в 96-луночный планшет по 100 мкл/лунку. После герметичного закрытия полиэтиленовой пленкой и выдерживания в течение ночи при 4°C, 1X PBST (также называемый промывочным буфером, 1X PBS с добавлением 0,05% Tween 20) используют для промывки 3 раза по 260 мкл/лунку, 5X ELISA/ELISPOT разбавитель разводят до 1X с ddH2O и затем добавляют в 96-луночный планшет по 200 мкл/лунку с последующим выдерживанием при комнатной температуре в течение 1 часа. PBST используют для однократной промывки, и разведение проводят по стандартной кривой (в диапазоне: от 2 до 250, от 4 до 500, от 4 до 500, соответственно), образцы разводят в 20-50 раз 1x разбавителем. Образцы в соответствии со стандартной кривой добавляют по 100 мкл на лунку, берут две параллельные лунки и инкубируют при комнатной температуре в течение 2 часов, PBST используют для промывки 3 раза, затем добавляют антитело для обнаружения, разведенное 1x разбавителем, после инкубации в течение 1 ч, PBST используют для промывки 3 раза, затем добавляют HRP, разведенный 1x разбавителем, после инкубации в течение 30 минут раствор промывают шесть раз, добавляют TMB для проявления цвета со временем проявления цвета менее 15 мин и 2N H2SO4 добавляют для терминации, определяют поглощение света при 450 нм.
Результаты ELISA показали, что секреция IL-2 и IFN-γ с помощью ub-STAR, mut-STAR со связанным с эндоцитозом TCR лизином, модифицированным в аргинин в трансмембранном домене или эндодомене, который совместно инкубируют с клетками-мишенями в соотношение E:T 2:1 или 1:1 или 1:2 Т-клетки к клетке-мишени в течение 24 часов, значительно ниже, чем у Т-клеток mut-STAR, как показано на фигурах 24, 25 и в таблицах 19, 20.
11.5. Обнаружение изменений дифференциации Т-клеток: анализ проточной цитометрией
Во время активации Т-клеток высвобождается большое количество цитокинов и других хемокинов, и сигналы трансдуцируются в ядро через цитокины или хемокиновые рецепторы для регуляции дифференциации Т-клеток. На пролиферацию и жизнестойкость Т-клеток in vivo влияет количество Т-клеток, дифференцировавших в Т-клетки памяти. Т-клетки памяти можно разделить на Т-клетки стволовых клеток, Т-клетки центральной памяти и Т-клетки эффекторной памяти. Экспрессия CD45RA и CCR7 на поверхности Т-клеток обнаружена с помощью проточной цитометрии, поэтому можно определить дифференциацию Т-клеток. Т-клетки инкубируют с клетками-мишенями в течение 7 дней, центрифугируют, окрашивали потоком антител против CD45RA-Percp-cy5.5 человека и против CCR7-APC человека в течение 30 минут, снова центрифугируют, промывают PBS и фиксируют 4% раствором параформальдегида, и дифференциацию Т-клеток обнаруживают с помощью проточной цитометрии.
Результаты проточной цитометрии показали, что не обнаружено существенной разницы ни в дифференциации Т-клеток центральной памяти, ни в отношении наивных Т-клеток к эффекторным Т-клеткам между mut-STAR и ub-STAR, mut-STAR со связанным с эндоцитозом TCR лизином, модифицированным в аргинин в трансмембранном домене или эндодомене, что указывает на то, что связанная с эндоцитозом TCR модификация лизина в аргинин в трансмембранном домене или эндодомене не оказывает существенного влияния на дифференциацию Т-клеток mut-STAR, как показано на фиг. 26 и 27 и в таблицах 21 и 22. Комбинируя результаты уничтожения опухоли, результаты ELISA и тест дифференциации клеток проточной цитометрией, ub-STAR, mut-STAR со связанным с эндоцитозом TCR лизином, модифицированным в аргинин в трансмембранном домене или эндодомене, не оказывает эффективный стимулирующий эффект на модификацию mut-STAR.
Таблица 18. Эффект уничтожения при совместном культивировании клеток-мишеней со STAR с модификацией трансмембранного домена
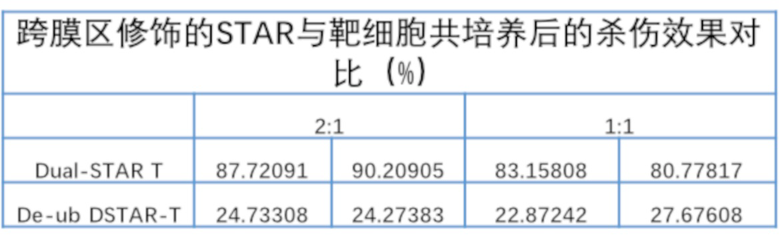
Таблица 19. Секреция IL-2 после совместного культивирования клеток-мишеней со STAR с модификацией трансмембранного домена
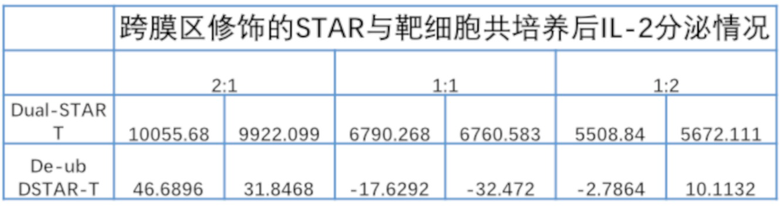
Таблица 20. Секреция IFN-γ после совместного культивирования клеток-мишеней со STAR с модификацией трансмембранного домена
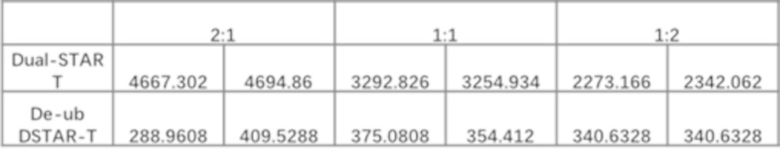
Таблица 21. Влияние модификации трансмембранного домена на дифференциацию STAR-T-клеток центральной памяти
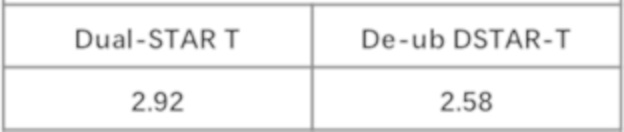
Таблица 22. Влияние модификации трансмембранного домена на дифференциацию STAR-T-клеток
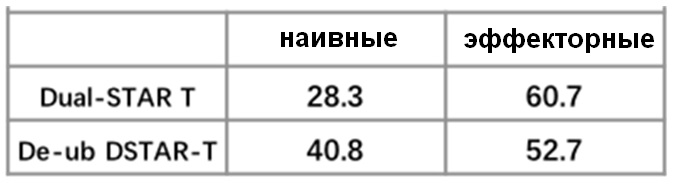
12. Влияние мутантного STAR с различными стимулирующими областями цитокинового рецептора, связанными с эндодоменом константной области α или β, на функцию STAR-T-клеток.
12.1. Способ совместного культивирования in vitro Т-клеток и клеток-мишеней
Клетки-мишени Raji-люцифераза и первичные Т-клетки представляют собой суспензионные клетки, для совместной инкубации берут соответствующее количество клеток, смешивают со средой клеток-мишеней и центрифугируют для культивирования. Конкретные стадии включают следующие: первичные Т-клетки заражают упакованными и очищенными вирусами WT-STAR и mut-STAR T, и за день до совместного культивирования, эффективность заражения определяют с помощью проточной цитометрии, и определяют соотношение эффекторной клетки к клетке-мишени, совместную инкубацию проводят обычно в соотношении 1:1 или 2:1, кроме того, клетки-мишени Raji-люцифераза и первичные Т-клетки совместно инкубируют в течение 7 дней, чтобы наблюдать изменения в количестве пролифераций клеток. Общее количество Т-клеток рассчитывают на основе эффективности заражения, и обычное использование клеток-мишеней составляет 1х105/лунку (96-луночный планшет).
12.2. Стимуляция Т-клеток антигеном-мишенью
Мишенью настоящего изобретения, как правило, является белок клеточной поверхности, который можно непосредственно использовать для активации Т-клеток для определения функции Т-клеток, в частности, антиген-мишень обычно добавляют с 1х105/лунку положительных Т-клеток, центрифугируют и активируют в течение 24 часов для сбора клеточной суспензии или супернатанта культуры для определения функции Т-клеток.
12.3. Подтверждение функции уничтожения Т-клеток: анализ люциферазы
Т-клетки совместно культивируют с клетками-мишенями в течение разного времени, затем клеточную суспензию осторожно равномерно продувают, 150 мкл клеточной суспензии на лунку переносят в белый 96-луночный планшет, центрифугируют при 1500 об/мин в течение 5 мин, отбирают супернатант и добавляют клеточный лизат для лизиса при комнатной температуре в течение 15 мин, затем центрифугируют при 4° при 4000 об/мин в течение 15 мин, затем отбирают супернатант, отбирая 2 параллельные лунки на каждую лунку, и добавляют с субстратом люциферазы (реагент для анализа люциферазы светлячка), затем обнаруживают с помощью многофункционального микропланшетного ридера с фиксированным значением усиления, равным 100, для получения значения хемилюминесценции. Расчет уничтожения клеток: эффективность уничтожения=100% - (эффекторные клетки и клетки-мишени на лунку/контрольные клетки и клетки-мишени на лунку).
Результаты обнаружения функции уничтожения Т-клеток с помощью анализа люциферазы показали, что, когда различные области, стимулирующие цитокиновые рецепторы, связаны с эндодоменом α или β области мутантного STAR, при соотношении E:T 2:1 Т-клетки к клетке-мишени, β-IL-2Rb, α-IL-2Rb, α-IL-7RA, α-IL21R демонстрируют эффект уничтожения, аналогичный эффекту mut-STAR, в то время как β-IL2RbQ, α-IL2RbQ и α-IL7RAQ демонстрируют эффект уничтожения опухоли значительно ниже, чем у mut-STAR, как показано на фигуре 28 и в таблице 23. Кроме того, эффект уничтожения мутантных STAR с различными связанными стимулирующими областями цитокинового рецептора обнаруживают и сравнивают с эффектом α-del-(G4S) 3-OX40-STAR, как показано на фигуре 29 и в таблице 24, при соотношениях E:T 2:1 и 1:1 эффект уничтожения α-IL7RA несколько ниже, чем у α-del-(G4S) 3-OX40-STAR, в то время как эффект уничтожения STAR с другими связанными структурами цитокиновых рецепторов значительно ниже, чем у α-del-(G4S) 3-OX40-STAR.
12.4. Анализ цитокинов, секретируемых Т-клетками: ELISA
Во время активации Т-клеток высвобождается большое количество цитокинов, таких как TNF-α, IFN-γ и IL-2, которые помогают Т-клеткам убивать клетки-мишени или способствуют размножению самих Т-клеток. После стимуляции Т-клеток клетками-мишенями или антигенами, Т-клетки собирают, центрифугируют и отбирают супернатант. Используемые наборы ELISA для TNF-α, IFN-γ и IL-2 представляют собой наборы ELISA без покрытия IL-2 человека, ELISA без покрытия TNF-α человека и ELISA без покрытия IFN-γ человека (арт. № 88-7025, 88-7346, 88-7316, соответственно). Конкретные стадии включают следующие: разведение 10Х буфера для покрытия ddH2O до 1Х, добавление антитела с покрытием (250Х), тщательное перемешивание и добавление в 96-луночный планшет по 100 мкл/лунку. После герметичного закрытия полиэтиленовой пленкой и выдерживания в течение ночи при 4°C, 1X PBST (также называемый промывочным буфером, 1X PBS с добавлением 0,05% Tween 20) используют для промывки 3 раза по 260 мкл/лунку, 5X ELISA/ELISPOT разбавитель разводят до 1X с ddH2O и затем добавляют в 96-луночный планшет по 200 мкл/лунку с последующим выдерживанием при комнатной температуре в течение 1 часа. PBST используют для однократной промывки, и разведение проводят по стандартной кривой (в диапазоне: от 2 до 250, от 4 до 500, от 4 до 500, соответственно), образцы разводят в 20-50 раз 1x разбавителем. Образцы в соответствии со стандартной кривой добавляют по 100 мкл на лунку, берут две параллельные лунки и инкубируют при комнатной температуре в течение 2 часов, PBST используют для промывки 3 раза, затем добавляют антитело для обнаружения, разведенное 1x разбавителем, после инкубации в течение 1 ч, PBST используют для промывки 3 раза, затем добавляют HRP, разведенный 1x разбавителем, после инкубации в течение 30 минут раствор промывают шесть раз, добавляют TMB для проявления цвета со временем проявления цвета менее 15 мин и 2N H2SO4 добавляют для терминации, определяют поглощение света при 450 нм.
Результаты ELISA показали, что при инкубации Т-клеток с клетками-мишенями в течение 24 часов при соотношении Е:Т 1:1 или 1:2, секреция IL-2 в β-IL-2Rb значительно выше, чем у мутантных. STAR, в то время как α-IL-2Rb, β-IL-2RbQ и α-IL-7RAQ демонстрируют аналогичную секрецию IL-2, однако секреция IL-2 в α-IL-2RbQ значительно ниже, чем у mut-STAR, как показано на фигуре 30 и в таблице 25. Секреция IFN-γ в β-IL-2Rb значительно выше, чем в mut-STAR, в то время как секреция IFN-γ в других структурах значительно ниже, чем в mut-STAR, как показано на фигуре 31 и в таблице 26.
12.5. Обнаружение изменений дифференциации Т-клеток: анализ проточной цитометрией
Во время активации Т-клеток высвобождается большое количество цитокинов и других хемокинов, и сигналы трансдуцируются в ядро через цитокины или хемокиновые рецепторы для регуляции дифференциации Т-клеток. На пролиферацию и жизнестойкость Т-клеток in vivo влияет количество Т-клеток, дифференцировавших в Т-клетки памяти. Т-клетки памяти можно разделить на Т-клетки стволовых клеток, Т-клетки центральной памяти и Т-клетки эффекторной памяти. Экспрессия CD45RA и CCR7 на поверхности Т-клеток обнаружена с помощью проточной цитометрии, поэтому можно определить дифференциацию Т-клеток. Т-клетки инкубируют с клетками-мишенями в течение 7 дней, центрифугируют, окрашивали потоком антител против CD45RA-Percp-cy5.5 человека и против CCR7-APC человека в течение 30 минут, снова центрифугируют, промывают PBS и фиксируют 4% раствором параформальдегида, и дифференциацию Т-клеток обнаруживают с помощью проточной цитометрии.
Мутантные STAR с различными стимулирующими областями цитокиновых рецепторов, связанными с эндодоменом константной области α или β, демонстрируют различные различия в дифференциации центральных Т-клеток памяти и соотношении наивных Т-клеток к эффекторным Т-клеткам, где α-IL-2Rb, β- IL-2Rb, α-IL-2RbQ, β-IL-2RbQ не демонстрируют существенной разницы по сравнению с mut-STAR, в то время как α-IL-7RA, α-IL-7RAQ и α-IL-21R не демонстрируют существенной разницы по сравнению с mut-STAR, как показано на фиг. 32 и 33 и в таблицах 27 и 28. На основании приведенных выше результатов можно сказать, что на дифференциацию мутантных Т-клеток STAR подвергается значительному влиянию, когда различные области, стимулирующие цитокиновый рецептор, соединяют с эндодоменом константной области α или β мутантного STAR.
Таблица 23. Эффект уничтожения после совместного культивирования клеток-мишеней со STAR с различными стимулирующими доменами цитокиновых рецепторов, связанными с их α цепью
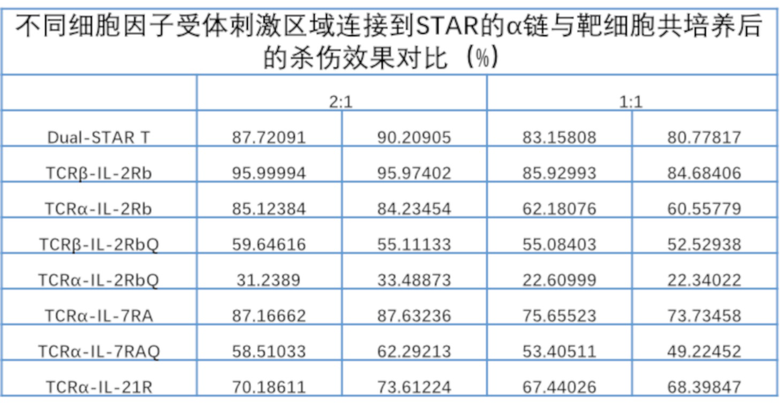
Таблица 24. Эффект уничтожения после совместного культивирования клеток-мишеней со STAR с различными стимулирующими доменами цитокиновых рецепторов, связанными с их α цепью
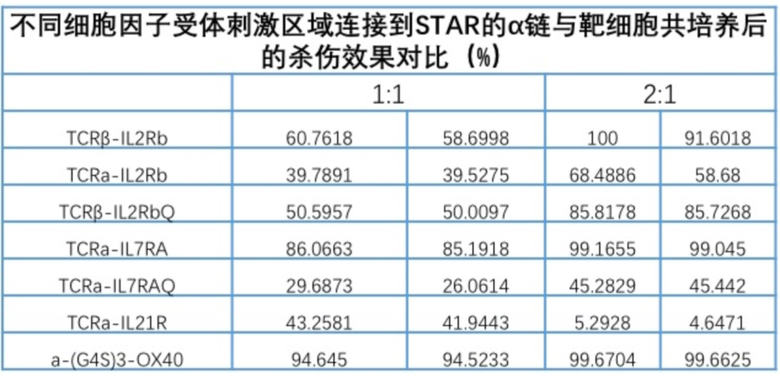
Таблица 25. Секреция IL-2 после совместного культивирования клеток-мишеней со STAR с различными стимулирующими доменами цитокиновых рецепторов, связанными с их α цепью
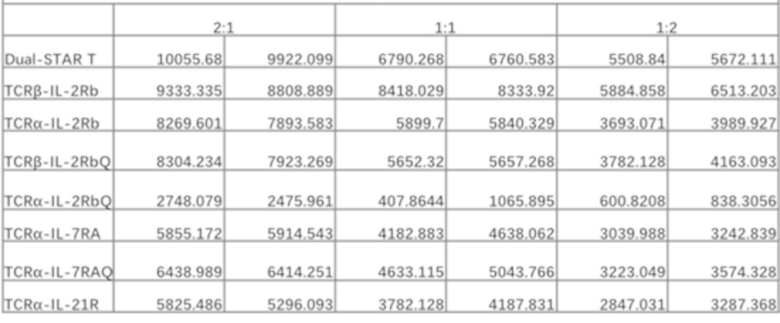
Таблица 26. Секреция IFN-γ после совместного культивирования клеток-мишеней со STAR с различными стимулирующими доменами цитокиновых рецепторов, связанными с их α цепью
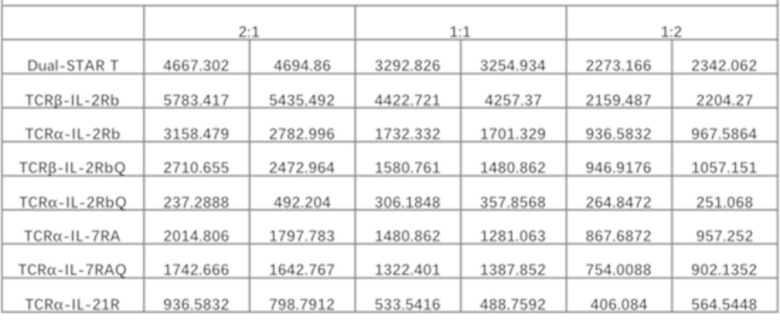
Таблица 27. Влияние STAR с различными стимулирующими доменами цитокинового рецептора, связанными с его α цепью, на дифференциацию Т-клеток центральной памяти
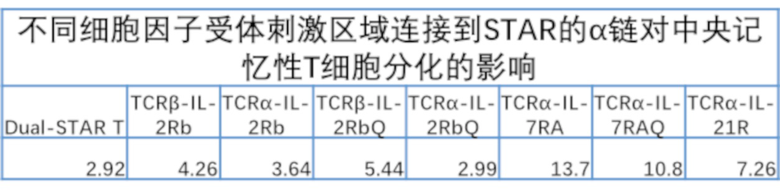
Таблица 28. Влияние STAR с различными стимулирующими доменами цитокиновых рецепторов, связанными с их α цепью, на дифференциацию Т-клеток
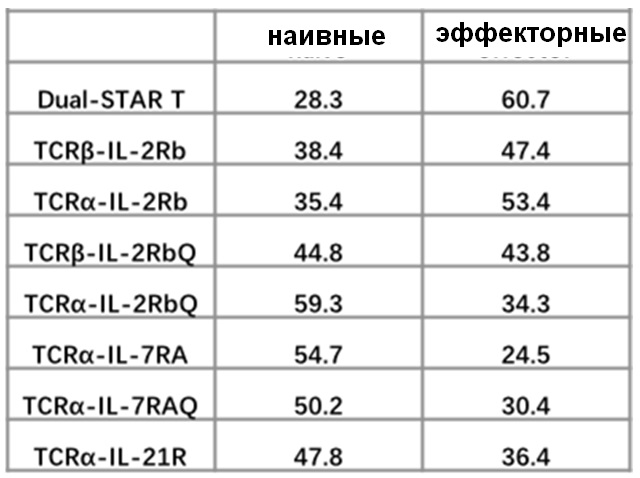
13. Оценка in vivo пролиферации и противоопухолевой способности различных STAR-подобных Т-клеток.
1) Экспериментальная животная модель
В этом эксперименте, в качестве модели используют иммунодефицитную мышь NSG. Генотип мыши NOD-Prkdcem26Il2rgem26/Nju с отсутствием Т-клеток, В-клеток и NK-клеток, и их макрофаги и дендритные клетки также отсутствуют. В настоящее время, наиболее иммунодефицитной линией мышей является NSG мышь, которую можно широко использовать в доклинических исследованиях Т-клеточной терапии, поскольку она не будет отторгать трансплантированные опухоли и Т-клетки. В этом эксперименте используют самок мышей NSG в возрасте 6~8 недель, и разница в весе мышей в каждой партии контролируют в пределах 2 г. Мышей содержат в отдельных вентилируемых клетках в пределах беспатогенного (SPF) барьера, и обеспечивают нормальным кормом и питьевой водой со слегка кислым pH для профилактики заражения патогенами.
2) Конструирование модели опухоли
При построении гематологической модели опухоли, для ксенотрансплантации используют клетки Raji линии клеток лимфомы Беркитта человека. Клетки Raji представляют собой клеточные штаммы, экспрессирующие ген люциферазы с помощью лентивирусного вектора, и за развитием и изменениями опухоли Raji следят в режиме реального времени с помощью флуоресцентной хемилюминесценции и визуализации in vivo у мышей. В этой модели, различные дозы (обычно от 1 до 3х106 клеток) клеток Raji-люцифераза смешивают с матричным гелем и инокулируют самкам мышей NSG в возрасте от 6 до 8 недель путем внутрибрюшинной инъекции. Через три дня, мышам внутрибрюшинно вводят раствор калиевой соли флуоресцеина, и с помощью визуализации in vivo обнаруживают сигналы флуоресценции опухолевых клеток in vivo. Клетки Raji быстро растут у мышей, которые продуцируют солидные опухоли в брюшной полости, вызывая такие симптомы, как потеря веса у мышей; в отсутствие терапевтического лечения, опухолевое бремя Raji приводит к смерти мыши примерно через 40 дней.
3) Эксперимент на животных
Все операции на животных проводят после утверждения Протокола для животных.
4) Средства мониторинга роста опухоли
В этом эксперименте, флуоресцентную визуализацию in vivo в основном используют путем: инъекции опухолевых клеток с геном люциферазы животным для колонизации. Мышам внутрибрюшинно вводят раствор калиевой соли флуоресцеина, которая, в качестве субстрата, испускает свет с определенной длиной волны в присутствии фермента, и с помощью визуализации in vivo регистрируют сигналы флуоресценции опухолевых клеток in vivo. Количественный анализ сигналов флуоресценции проводят и строят тепловую карту для количественного отражения роста опухоли.
5) Способ определения активности и амплификации Т-клеток у животных.
Выживание и размножение Т-клеток in vivo напрямую связаны с их конечным противоопухолевым эффектом. Для обнаружения активности и пролиферации Т-клеток у животных, регулярно собирают образцы крови у мышей и анализируют долю, состояние клеток и группировку STAR-T-клеток в периферической крови. Конкретная операция заключается в следующем: каждые 3-4 дня мышей анестезируют изофлураном и собирают примерно 100 мкл крови из глазницы мышей. Образец крови подвергают антикоагуляции, сбору плазмы и расщеплению эритроцитов, затем оставшиеся клетки подвергают проточному окрашиванию для определения соотношения CD4 к CD8 и молекул, таких как CCR7, CD45RA, PD-1, LAG-3 и TIM-3, которые используют для анализа подмножества Т-клеток и анализа состояния клеток. В то же время, абсолютное количество STAR-T-клеток в периферической крови мышей получают с помощью проточной цитометрии или цифровой ПЦР. Кроме того, в конце эксперимента, мыши могут быть вскрыты для определения доли Т-клеток в других иммунных органах мышей.
6) Способ оценки безопасности Т-клеток у животных.
Чтобы оценить токсичность и безопасность STAR-T-клеток, исследуют, вызывают ли STAR-T-клетки побочные эффекты у экспериментальных животных. Наблюдая за поведением мышей, анализируя патологию мышей и анализируя срезы, взятые из важных органов мышей, можно оценить, обладают ли повторно инфузированные Т-клетки значительной токсичностью. В то же время, анализируя инфильтрацию Т-клеток в неопухолевых тканях мышей, можно определить, оказывают ли Т-клетки действие уничтожения, не связанное с мишенью, на неопухолевые ткани мышей. Кроме того, определяя уровень цитокинов, таких как IL-2, IFN-γ, TNF α или IL-6 в крови мышей, можно определить, могут ли Т-клетки вызывать систематический цитокиновый шторм.
7) Способ оценки способности Т-клеток к инфильтрации опухоли
Способность Т-клеток проникать в опухоли является их основной способностью противостоять солидным опухолям. Чтобы определить способность Т-клеток к инфильтрации, опухолевые ткани можно сначала отделить, а затем переварить и измельчить, чтобы получить отдельные клетки, которые подвергают проточному окрашиванию для определения доли Т-клеток в опухолевых тканях. В то же время, опухолевые клетки, опухолевые стромальные клетки и иммунные клетки в опухолевой суспензии могут быть дополнительно разделены центрифугированием в градиенте плотности (например, в градиенте Перколла, градиенте Фиколла и т. д.), чтобы получить очищенные инфильтрирующие опухоль Т-клетки, и их характеристики, такие как экспрессия хемокинового рецептора, истощение Т-клеток и так далее, могут быть подробно проанализированы секвенированием и другими способами.
8) Результаты
В соответствии с результатами описанной выше функции in vitro, 5х105 опухолевых клеток Raji-люцифераза инокулируют самкам мышей NCG в возрасте 6-8 недель через хвостовую вену, чтобы сконструировать модель опухоли мыши (фигура 34), и in vivo дополнительно оценивают функцию экспрессии αβ OX40-STAR или mut-STAR Т-клеток. На 6 день, мышей с опухолями делят на четыре группы: A: группа инъекции PBS (инъецируют равный объем PBS); B: группа инъекции mut-STAR Т-клеток; C: группа инъекции αβ OX40-STAR Т-клеток; D: группа инъекции BBz-CAR-T-клеток. Мышам в группе B/C/D вводят 5х105 Т-клеток TCR через хвостовую вену, и мышам в группе A вводят равный объем 200 мкл PBS. В течение следующих нескольких недель наблюдают за ростом опухолевых клеток, пролиферацией Т-клеток TCR in vivo и выживаемостью мышей. Как показано на фигурах 34 и 35, по сравнению с контрольной группой, αβ OX40-STAR T и mut-STAR T клетки, сконструированные в этом примере, могут значительно продлить время выживания мышей с опухолью, хотя после исчезновения опухоли и повторной инфузии опухолевых клеток мышам в разные моменты времени, αβ OX40-STAR T и mut-STAR T могут должным образом уничтожать опухолевые клетки, и эффект αβ OX40-STAR T лучше, чем у mut-STAR T, и время выживания мышей в группе αβ OX40-STAR T выше, чем в группе STAR-T и группе CAR-T. Кроме того, как показано на фиг. 36, по сравнению с группой STAR-T, эффект пролиферации in vivo у αβ OX40-STAR T-клеток лучше, чем у STAR-T-клеток, и в случае повторной инфузии опухоли возникает незначительная пролиферация в αβ OX40-STAR T-клетках, чего, однако, не происходит в STAR-T-клетках. На основании результатов вышеуказанных экспериментов на животных можно обнаружить, что in vitro и in vivo эффект структуры αβ OX40-STAR лучше, чем у mut-STAR.
В соответствии с приведенными выше результатами функции in vitro, 2х106 опухолевых клеток Raji-люцифераза инокулируют внутрибрюшинно самкам мышей NCG в возрасте 6-8 недель для создания мышиной модели опухоли (фигура 37), и далее оценивают in vivo функцию mut-STAR Т-клеток, экспрессирующих различные эффекты. На 8 день, мышей с опухолями делят на семь групп: A: группа инъекции PBS (инъецируют равный объем PBS); B: dual-car; C: группа инъекции dual-STAR-T-клеток; D: группа инъекции αβOX40-STAR Т-клеток; E: группа инъекции α-del-OX40-STAR-T-клеток; F: группа инъекции α-del-(G4S)3-OX40-STAR-T-клеток; и G: группа инъекции α-IL7R-STAR-T-клеток. Мышам группы B/C/D/E/F/G вводят 2х106 Т-клеток TCR через хвостовую вену, мышам группы A вводят равный объем 200 мкл PBS. В течение следующих нескольких недель наблюдают за ростом опухолевых клеток, пролиферацией STAR-T-клеток in vivo и выживаемостью мышей. Как показано на фиг. 37, по сравнению с контрольной группой, α-del-(G4S) 3-OX40-STAR-T-клетки, сконструированные в этом примере, могут значительно уничтожать опухолевые клетки мышей с опухолями, и их эффект выше, чем в других группах. На основании результатов экспериментов на животных можно обнаружить, что in vitro и in vivo эффект структуры α-del-(G4S)3-OX40-STAR лучше, чем у mut-STAR.
Пример 2. Влияние модификации N-конца константной области на STAR.
1. Дизайн домена константной области рецептора STAR
В этом примере, hSTAR относится к STAR, содержащему константную область TCR человека. hmct STAR относится к STAR, содержащему константную область с цистеиновой заменой и модификацией трансмембранного домена, как показано в примере 1. Константная область α цепи TCR мыши представляет собой hmct STAR TCR α (SEQ ID NO: 41), и константная область β цепи TCR мыши представляет собой hmct STAR TCRb (SEQ ID NO: 6) с конкретными структурами, показанными на фиг. 38.
Для дальнейшей оптимизации конструкции молекулы STAR на основе муринизации константной области, мутации цистеина и мутации гидрофобной аминокислоты в константной области α цепи, N-конец константной области молекулы STAR специально перегруппировывают для получения лучших результатов. Перегруппировка означает, что некоторые последовательности удалены, при этом проведены гуманизированные мутации некоторых последовательностей. Значение гуманизированной мутации заключается в уменьшении нечеловеческой последовательности в молекуле STAR, насколько это возможно, при сохранении функции молекулы STAR, чтобы избежать возможности отторжения STAR-T-клеток рецепторами при клиническом применении до наибольшей степени.
Осуществляют схему из 18 аминокислотных перегруппировок (последовательность мыши представляет собой DIQNPEPAVYQLKDPRSQ) на N-конце константной области α цепи TCR, на основе анализа аминокислот обнаруживают, что E6D, K13R, R16K и Q18S на мышиной и гуманизированной последовательностях принадлежат гомологичной аминокислотной замене, в то время как замена P15S принадлежит неполярной аминокислоте замене полярной аминокислоты, поэтому можно считать, что белок вблизи этого сайта не является консервативным по своей природе и может быть модифицирован без влияния на его функцию. Таким образом, аминокислотные последовательности в положениях 1-14 сохраняют и гуманизируют, и аминокислоты в положениях 15-18 удаляют.
Осуществляют схему из 25 аминокислотных перегруппировок (последовательность мыши представляет собой DLRNVTPPKVSLFEPSKAEIANKQK) на N-конце константной области β цепи TCR, на основе аминокислотного анализа обнаруживают, что только R3K и L12V в мышиной и гуманизированной последовательностях являются гомологичными аминокислотными заменами, T6F, K9E, S11A, K17E, A21S, N22H и K23T являются гетерогенными аминокислотными заменами. Поэтому можно считать, что свойства белков вблизи этих сайтов не являются консервативными по своей природе и могут быть модифицированы без влияния на их функции. Таким образом, аминокислотные последовательности в положениях 1-16 сохраняют и гуманизируют, и аминокислоты в положениях 17 и 21-25 удаляют.
После N-концевой перегруппировки, константная область α цепи TCR представляет собой Nrec STAR TCRa (SEQ ID NO: 42), и константная область β цепи TCR представляет собой Nrec STAR TCRb (SEQ ID NO: 43) со специфическими структурами, показанными на фиг. 38.
2. Дизайн костимулирующего фактора STAR
1) Комбинация различных способов оптимизации STAR
Для проверки влияния костимулирующих факторов на различные функции STAR используют оригинальную неоптимизированную структуру STAR (TCR α/β STAR человека, hSTAR), hmct STAR на основе hSTAR с муринизацией C-области, модификацией цистина и трансмембранной модификацией, и выбирают Nrec STAR на основе hmct STAR с N-концевой модификацией.
2 Дизайн структуры STAR, включающей костимулирующие факторы
Для усиления токсичности STAR-T-клеток и сохранения пролиферации Т-клеток по настоящему изобретению, в С-конец константной области STAR вводят эндодоменную последовательность гуманизированного костимулирующего рецептора (фиг. 39) для изучения влияния на функции STAR-Т-клеток. Оригинальную не оптимизированную структуру STAR (TCR α/β STAR человека, hSTAR), hmct STAR на основе hSTAR и Nrec STAR на основе hmct STAR с N-концевой модификацией выбирают в качестве структуры константной области STAR настоящего изобретения. Структура трансдукции костимулирующего сигнала включает внутриклеточные домены передачи сигнала CD40, OX40, ICOS, CD28, 4-1BB и CD27. В изобретении, модификация может происходить в α, β цепи и αβ цепи TCR. В изобретении, костимулирующая молекулярная модификация происходит в αβ цепи TCR, как показано на фиг. 40.
3. Функция FMC63-STAR-T, таргетирующая CD19.
1) Определение последовательности CD19-таргетирующего антитела
Опубликованную последовательность scFv FMC63 выбирают для вариабельной области тяжелой цепи антитела, таргетирующего CD19 (анти-CD19 FMC63-VH, SEQ ID NO: 44), и вариабельной области легкой цепи антитела (анти-CD19 FMC63-VL, SEQ ID NO: 45).
2) Конструирование STAR, таргетирующего CD19, и вектора, содержащего костимулирующие факторы
STAR содержит две полипептидных цепи: первую полипептидную цепь, которая образована путем слияния анти-CD19 FMC63-VL с цепями TCR bC hSTAR/hmct STAR/Nrec STAR, соответственно, и вторую полипептидную цепь, которая образована путем слияния анти-CD19 FMC63 VH с аС цепями TCR hSTAR/hmct STAR/Nrec STAR, соответственно. В обеих цепях используют сигнальный пептид GM-CSFR. Двухцепочечные последовательности hSTAR/hmct STAR/Nrec STAR соединяют полипептидным фрагментом сайта расщепления протеазой Furin-SGSG-p2A, транскрибируют и транслируют в белок вместе, затем расщепляют соответствующей протеазой фурина и p2A, в результате чего образуются две независимые белковые цепи. Целый ген вставляют в лентивирусный вектор экспрессии pHAGE через сайты эндонуклеаз рестрикции NheI и NotI. Вектор переносят с геном резистентности к ампицину, промотором EF 1α и флуоресцентным репортерным геном IRES-RFP. Путем клонирования, сборки, трансформации, секвенирования и плазмидного выделения фрагментов генов получают следующие плазмиды: FMC63-hSTAR, FMC63-hmct STAR, FMC63-Nrec STAR.
Конструирование вектора STAR, содержащего костимулирующие факторы: на основе трех векторов STAR, таргетирующих CD19, FMC63-hSTAR, FMC63-hmct STAR и FMC63-Nrec STAR, на них конструируют CD40, OX40, ICOS, CD28, 41BB и CD27, и вышеуказанные последовательности получают путем генного синтеза. К векторам FMC63-hSTAR, FMC63-hmct STAR и FMC63-Nrec STAR добавляют костимулирующие факторы с помощью ПЦР и гомологичной рекомбинации путем одновременного добавления одного и того же костимулирующего фактора к α и β цепям TCR. В конце создают FMC63-hSTAR-CD40, FMC63-hSTAR-OX40, FMC63-hSTAR-ICOS, FMC63-hSTAR-CD28, FMC63-hSTAR-41BB, FMC63-hSTAR-CD27, FMC63-hmct STAR-CD40, FMC63-hmct STAR- OX40, FMC63-hmct STAR-ICOS, FMC63-hmct STAR-CD28, FMC63-hmct STAR-41BB, FMC63-hmct STAR-CD27, FMC63-Nrec STAR-CD40, FMC63-Nrec STAR-OX40, FMC63-Nrec STAR-ICOS, FMC63-Nrec STAR-CD28, FMC63-Nrec STAR-41BB и FMC63-Nrec STAR-CD27.
3) Обнаружение способности уничтожения STAR, таргетирующих CD19, и STAR, содержащих костимулирующий фактор.
Не зараженные Т-клетки (группа NC), FMC63-hSTAR, FMC63-hmct STAR, FMC63-Nrec STAR, FMC63-hSTAR-CD40, FMC63-hSTAR-OX40, FMC63-hSTAR-ICOS, FMC63-hSTAR-CD28, FMC63-hSTAR-41BB, FMC63-hSTAR-CD27, FMC63-hmct STAR-CD40, FMC63-hmct STAR- OX40, FMC63-hmct STAR-ICOS, FMC63-hmct STAR-CD28, FMC63-hmct STAR-41BB, FMC63-hmct STAR-CD27, FMC63-Nrec STAR-CD40, FMC63-Nrec STAR-OX40, FMC63-Nrec STAR-ICOS, FMC63-Nrec STAR-CD28, FMC63-Nrec STAR-41BB и FMC63-Nrec STAR-CD27 совместно культивируют с клетками RAJI-luc. в течение 24 ч в 24-луночном планшете. Количество положительных клеток Т-клеток корректируют в соответствии с количеством положительных результатов RFP до 4E5 для каждой, и количество клеток RAJI-luc составляет 4E5 с общим объемом системы совместного культивирования 1 мл. Через 24 часа совместного культивирования, совместно культивируемые клетки равномерно перемешивают, отсасывают 150 мкл суспензии и добавляют 70 мкл субстрата люциферазы, после встряхивания в течение 10 минут на низкой скорости в темноте значение флуоресценции определяют с помощью многофункционального микропланшетного ридера, и рассчитывают эффект уничтожения клеток-мишеней для каждой группы. Результаты показали, что эффективность уничтожения hSTAR значительно ниже, чем у hmct STAR и Nrec STAR, среди них Nrec STAR был самым высоким. Результаты того же STAR, включающего костимулирующие факторы, показали, что OX40 и ICOS могут значительно повышать эффект уничтожения STAR, в то время как другие костимулирующие факторы не оказывают существенного влияния на способность уничтожения STAR, 41BB снижет функцию уничтожения STAR, однако, без существенной разницы (фигура 41).
Фиг. 4. Обнаружение уровня ядерного RelB в CD19-таргетирующем STAR и STAR, содержащем костимулирующий фактор.
Следующие вирусы FMC63-hSTAR, FMC63-hmct STAR, FMC63-Nrec STAR, FMC63-hSTAR-CD40, FMC63-hSTAR-OX40, FMC63-hSTAR-ICOS, FMC63-hSTAR-CD28, FMC63-hSTAR-41BB, FMC63-hSTAR-CD27, FMC63-hmct STAR-CD40, FMC63-hmct STAR- OX40, FMC63-hmct STAR-ICOS, FMC63-hmct STAR-CD28, FMC63-hmct STAR-41BB, FMC63-hmct STAR-CD27, FMC63-Nrec STAR-CD40, FMC63-Nrec STAR-OX40, FMC63-Nrec STAR-ICOS, FMC63-Nrec STAR-CD28, FMC63-Nrec STAR-41BB и FMC63-Nrec STAR-CD27 заражают клетками Jurkat в титре MOI=1, через 4 дня после заражения, Т-клеточные линии и белок CD19 совместно культивируют в 12-луночном планшете (2 мкг/мл, 500 мкл наносят на 12-луночный планшет и оставляют на ночь в холодильнике при 4°C), в котором клетки STAR-T составляют 4E6, и клетки-мишени составляют 2E5 (клетки-мишени инокулируют в 12-луночный планшет за один день), культивируют в течение 6 часов, затем клетки собирают для экстракции ядерных белков, которые обнаруживают с помощью Вестерн-блоттинга для уровня ядерного RelB. Результаты показали, что уровень ядерного RelB является очень низким в группах FMC63-hSTAR, FMC63-hmct STAR и FMC63-Nrec STAR без костимулирующих факторов, что согласуется с не зараженной группой STAR-T, после добавления костимулирующих факторов, другие костимулирующие факторы, кроме CD28, значительно повышают уровень ядерного RelB, среди которых 41BB больше всего улучшает уровень ядерного RelB, как показано на фигуре 42.
4. Функция 334-STAR-T, таргетирующего CD19.
1) Определение последовательности CD19-таргетирующего антитела
Последовательность антитела 334, разработанную авторами настоящего изобретения, выбирают для вариабельной области тяжелой цепи антитела, таргетирующего CD19 (анти-CD19 334-VH, SEQ ID NO: 46), и вариабельной области легкой цепи антитела (анти-CD19 334-VL, SEQ ID №: 47).
2) Конструирование STAR, таргетирующего CD19, и вектора, содержащего костимулирующие факторы.
STAR содержит две полипептидных цепи: первую полипептидную цепь, которая образована путем слияния анти-CD19 334-VL с bC цепями TCR hSTAR/hmct STAR/Nrec STAR, соответственно, и вторую полипептидную цепь, которая была образована путем слияния анти-CD19 334 VH с аС цепями TCR hSTAR/hmct STAR/Nrec STAR, соответственно. В обеих цепях используют сигнальный пептид GM-CSFR. Двухцепочечные последовательности hmct STAR/Nrec STAR соединяют полипептидным фрагментом сайта расщепления протеазой Furin-SGSG-p2A, транскрибируют и транслируют в белок вместе, затем расщепляют соответствующей протеазой фурина и p2A, в результате чего образуются две независимые цепи белка. Целый ген был вставляют в лентивирусный вектор экспрессии pHAGE через сайты эндонуклеаз рестрикции NheI и NotI. Вектор переносят с геном резистентности к ампицину, промотором EF 1α и флуоресцентным репортерным геном IRES-RFP. Путем клонирования, сборки, трансформации, секвенирования и выделения плазмид фрагментов генов получают следующие плазмиды: 334-hmct STAR, 334-Nrec STAR.
Конструирование вектора STAR, включающего костимулирующие факторы: на основе векторов STAR, таргетирующих CD19, 334-hmct STAR и 334-Nrec STAR конструируют костимулирующий фактор OX40, и вышеуказанные последовательности получают путем генного синтеза. К векторам 334-hmct STAR и 334-Nrec STAR добавляют костимулирующий фактор с помощью ПЦР и гомологичной рекомбинации путем одновременного добавления того же костимулирующего фактора к α и β цепям TCR. Наконец, создают 334-hmct STAR-OX40 и 334-Nrec STAR-OX40.
3) Обнаружение способности уничтожения STAR, таргетирующих CD19, и STAR, содержащих костимулирующий фактор.
Не зараженные Т-клетки (группа NC), 334-hmct STAR, 334-Nrec STAR, 334-hmct STAR-OX40 и 334-Nrec STAR-OX40 совместно культивируют с клетками RAJI-luc. в течение 24 ч в 24-луночном планшете. Количество положительных клеток Т-клеток корректируют в соответствии с количеством положительных результатов RFP до 4E5 для каждой, и количество клеток RAJI-luc составляет 4E5 с общим объемом системы совместного культивирования 1 мл. Через 24 часа совместного культивирования, совместно культивируемые клетки равномерно перемешивают, отсасывают 150 мкл суспензии и добавляют 70 мкл субстрата люциферазы, после встряхивания в течение 10 минут на низкой скорости в темноте значение флуоресценции определяют с помощью многофункционального микропланшетного ридера, и рассчитывают эффект уничтожения клеток-мишеней для каждой группы. Результаты показали, что OX40 может значительно повысить эффективность уничтожения по сравнению с hmct STAR и Nrec STAR. Кроме того, OX40 может значительно увеличить способность к пролиферации и уровень ядерного RelB STAR-T-клеток. См. фиг. 43.
5 Функцию STAR-T по таргетированию CD19 и CD 20
1) Определение последовательности антител, таргетирующих CD19 и CD20.
Вариабельная область тяжелой цепи антитела, таргетирующего CD19 (анти-CD19 FMC63-VH, SEQ ID NO: 44) и вариабельная область легкой цепи антитела (анти-CD19 FMC63-VL, SEQ ID NO: 45); вариабельная область тяжелой цепи антитела, таргетирующего CD20 (анти-CD20 2C6-VH, SEQ ID NO: 54), и вариабельная область легкой цепи антитела (анти-CD20 2C6-VL, SEQ ID NO: 55).
2) Конструирование STAR, таргетирующего CD19 и CD20, и вектора, содержащего костимулирующие факторы.
STAR содержит две полипептидные цепи: первую полипептидную цепь, которая образована путем слияния анти-CD20 2C6 VL-(G4S)3-VH с bC цепями TCR hmct STAR/Nrec STAR, соответственно, и вторую полипептидную цепь, которая образована путем слияния анти-CD19 FMC63 VL-(G4S)3-VH с aC цепями TCR hmct STAR/Nrec STAR, соответственно. В обеих цепях используют сигнальный пептид GM-CSFR. Две последовательности цепей hmct STAR/Nrec STAR соединяют полипептидным фрагментом сайта расщепления протеазой Furin-SGSG-p2A, транскрибируют и транслируют в белок вместе, затем расщепляют соответствующей протеазой фурина и p2A, в результате чего образуются две независимые цепи белка. Целый ген вставляют в лентивирусный вектор экспрессии pHAGE через сайты эндонуклеаз рестрикции NheI и NotI. Вектор переносят с геном резистентности к ампицину, промотором EF 1α и флуоресцентным репортерным геном IRES-RFP. Путем клонирования, сборки, трансформации, секвенирования и плазмидного выделения фрагментов генов получают следующие плазмиды: FMC63-2C6-hmct STAR и FMC63-2C6-Nrec STAR.
Конструирование вектора STAR, содержащего костимулирующие факторы: на основе векторов STAR, таргетирующих CD19 и CD20, FMC63-2C6-hmct STAR и FMC63-2C6-Nrec STAR, на них конструируют костимулирующий фактор OX40, и указанные выше последовательности получают путем генного синтеза. К векторам FMC63-2C6-hmct STAR и FMC63-2C6-Nrec STAR добавляют костимулирующие факторы с помощью ПЦР и гомологичной рекомбинации путем одновременного добавления одного и того же костимулирующего фактора к α и β цепям TCR. Наконец, конструируют FMC63-2C6-hmct STAR-OX40 и FMC63-2C6-Nrec STAR-OX40.
3) Обнаружение способности уничтожения STAR, таргетирующих CD19 и CD20, и STAR, включающих костимулирующий фактор.
Не зараженные Т-клетки (группа NC), FMC63-2C6-hmct STAR, FMC63-2C6-Nrec STAR, FMC63-2C6-hmct STAR-OX40 и FMC63-2C6-Nrec STAR-OX40 совместно культивируют с клетками RAJI-luc. в течение 24 ч в 24-луночном планшете. Количество положительных клеток Т-клеток корректируют в соответствии с количеством положительных результатов RFP до 4E5 для каждой, и количество клеток RAJI-luc составляет 4E5 с общим объемом системы совместного культивирования 1 мл. Через 24 часа совместного культивирования, совместно культивируемые клетки равномерно перемешивают, отсасывают 150 мкл суспензии и добавляют 70 мкл субстрата люциферазы, после встряхивания в течение 10 минут на низкой скорости в темноте значение флуоресценции определяют с помощью многофункционального микропланшетного ридера, и рассчитывают эффект уничтожения клеток-мишеней для каждой группы. Результаты показали, что OX40 может значительно повысить эффективность уничтожения по сравнению с hmct STAR и Nrec STAR. Кроме того, OX40 может значительно увеличить способность к пролиферации и уровень ядерного RelB STAR-T-клеток. См. фиг. 44.
4) Улучшение пролиферационных способностей STAR, таргетирующих CD19 и CD20, при добавлении костимулирующего фактора только к α цепи TCR.
Первую полипептидную цепь получают путем слияния анти-CD20 2C6 VL-(G4S)3-VH с TCR βC (SEQ ID NO: 6) структуры hmct STAR, вторую полипептидную цепь получают путем слияния анти-CD19 FMC63 VL-(G4S)3-V с константной областью TCR αC (SEQ ID NO: 26) hmct STAR, в которой природный эндодомен удален, и костимулирующий домен OX40 добавляют к С-концу константной области. В обеих цепях используют сигнальный пептид GM-CSFR. Двухцепочечные последовательности STAR соединяют полипептидным фрагментом сайта расщепления протеазой фурин-SGSG-p2A, транскрибируют и транслируют в белок (SEQ ID NO: 58) вместе, затем расщепляют соответствующей протеазой фурина и p2A, в результате чего получают две независимые белковые цепи. Таким образом, получают CD19 и CD20-таргетирующий STAR с костимулирующими факторами, связанными только с α цепью TCR: a(G4S)3 OX40. Сравнение способности к уничтожению и пролиферации проводят между такими STAR с CD19 и CD20-таргетирующими hmct STAR abOX40, в которых костимулирующий фактор связан как с α, так и с β цепями TCR. Результаты представлены на фигуре 45: a(G4S)3 OX40 превосходит abOX40.
Список последовательностей
SEQ ID NO: 1 Аминокислотная последовательность константной области TCR α человека дикого типа
DIQNPDPAVYQLRDSKSSDKSVCLFTDFDSQTNVSQSKDSDVYITDKTVLDMRSMDF KSNSAVAWSNKSDFACANAFNNSIIPEDTFFPSPESSCDVKLVEKSFETDTNLNFQNLS VIGFRILLLKVAGFNLLMTLRLWSS*
SEQ ID NO: 2 Аминокислотная последовательность константной области TCR β человека дикого типа
DLKNVFPPEVAVFEPSEAEISHTQKATLVCLATGFYPDHVELSWWVNGKEVHSGVST DPQPLKEQPALNDSRYCLSSRLRVSATFWQNPRNHFRCQVQFYGLSENDEWTQDRAK PVTQIVSAEAWGRADCGFTSESYQQGVLSATILYEILLGKATLYAVLVSALVLMAMVK RKDF*
SEQ ID NO: 3 Аминокислотная последовательность константной области TCR α мыши дикого типа
DIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKTVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLSVMGLRILLLKVAGFNLLMTLRLWSS
SEQ ID NO: 4 Аминокислотная последовательность константной области TCR β мыши дикого типа
DLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGKEVHSGVSTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGKATLYAVLVSTLVVMAMVKRKNS
SEQ ID NO: 5 α цепь константной области Т-клеточного рецептора мыши с цистеиновой заменой (TCRaC-Cys мыши)
DIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLSVMGLRILLLKVAGFNLLMTLR LWSS
SEQ ID NO: 6 β цепь константной области Т-клеточного рецептора мыши с цистеиновой заменой (TCRβC-Cys мыши)
DLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGKEVHSGVCTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGKATLYAVLVSTLVVMAMVKRKNS
SEQ ID NO: 7 α цепь константной области Т-клеточного рецептора мыши с заменой гидрофобной аминокислотой (TCRaC-TM9 мыши)
DIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKTVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLKVAGFNLLMTLRLWSS
SEQ ID NO: 8 α цепь константной области Т-клеточного рецептора мыши с цистеиновой заменой в трансмембранном домене (TCRaC-Arg mut мыши)
DIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLRVAGFNLLMTLRLWSS
SEQ ID NO: 9 β цепь константной области Т-клеточного рецептора мыши с цистеиновой заменой в трансмембранном домене и эндодомене (TCRβC-Arg mut мыши)
DLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGKEVHSGVCTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGRATLYAVLVSTLVVMAMVRRRNS
SEQ ID NO: 10 Аминокислотная последовательность CD40 эндодомена
KKVAKKPTNKAPHPKQEPQEINFPDDLPGSNTAAPVQETLHGCQPVTQEDGKESRISVQERO
SEQ ID NO: 11 Аминокислотная последовательность OX40 эндодомена
RRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKI
SEQ ID NO:12 Аминокислотная последовательность ICOS эндодомена
KKKYSSSVHDPNGEYMFMRAVNTAKKSRLTDVTL
SEQ ID NO: 13 Аминокислотная последовательность CD28 эндодомена
RSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS
SEQ ID NO:14 Аминокислотная последовательность 4-1BB эндодомена
KRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL
SEQ ID NO: 15 Аминокислотная последовательность CD27 эндодомена
QRRKYRSNKGESPVEPAEPCRYSCPREEEGSTIPIQEDYRKPEPACSP
SEQ ID NO: 16 Аминокислотная последовательность G4S линкера
GGGGS
SEQ ID NO: 17 Аминокислотная последовательность (G4S)2 линкера
GGGGSGGGGS
SEQ ID NO: 18 Аминокислотная последовательность (G4S)3 линкера
GGGGSGGGGSGGGGS
SEQ ID NO: 19 Аминокислотная последовательность (G4S)4 линкера
GGGGSGGGGSGGGGSGGGGS
SEQ ID NO: 20 Аминокислотная последовательность (G4S)5 линкера
GGGGSGGGGSGGGGSGGGGSGGGGS
SEQ ID NO: 21 Аминокислотная последовательность (G4S)6 линкера
GGGGSGGGGSGGGGSGGGGSGGGGSGGGGS
SEQ ID NO: 22 Аминокислотная последовательность (G4S)7 линкера
GGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGS
SEQ ID NO: 23 Аминокислотная последовательность (G4S)8 линкера
GGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGS
SEQ ID NO: 24 Аминокислотная последовательность (G4S)9 линкера
GGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGS
SEQ ID NO: 25 Аминокислотная последовательность (G4S)10 линкера
GGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGSGGGGS
SEQ ID NO: 26 α цепь константной области Т-клеточного рецептора мыши с удаленным эндодоменом с цистеиновой заменой и гидрофобной модификацией (TCRαC-del mut мыши)
DIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLKVAGFNLLMTLRLW
SEQ ID NO: 27 β цепь константной области Т-клеточного рецептора мыши с удаленным эндодоменом с цистеиновой заменой (TCRβC-del mut мыши)
DLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGKEVHSGVCTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGKATLYAVLVSTLVVMAM
SEQ ID NO: 28 Аминокислотная последовательность CD3γ человека
MEQGKGLAVLILAIILLQGTLAQSIKGNHLVKVYDYQEDGSVLLTCDAEAKNITWFKDGKMIGFLTEDKKKWNLGSNAKDPRGMYQCKGSQNKSKPLQVYYRMCQNCIELNAATISGFLFAEIVSIFVLAVGVYFIAGQDGVRQSRASDKQTLLPNDQLYQPLKDREDDQYSHLQGNQLRRN
SEQ ID NO: 29 Аминокислотная последовательность CD3δ человека
MEHSTFLSGLVLATLLSQVSPFKIPIEELEDRVFVNCNTSITWVEGTVGTLLSDITRLDLGKRILDPRGIYRCNGTDIYKDKESTVQVHYRMCQSCVELDPATVAGIIVTDVIATLLLALGVFCFAGHETGRLSGAADTQALLRNDQVYQPLRDRDDAQYSHLGGNWARNK
SEQ ID NO: 30 Аминокислотная последовательность CD3ε человека
MQSGTHWRVLGLCLLSVGVWGQDGNEEMGGITQTPYKVSISGTTVILTCPQYPGSEILWQHNDKNIGGDEDDKNIGSDEDHLSLKEFSELEQSGYYVCYPRGSKPEDANFYLYLRARVCENCMEMDVMSVATIVIVDICITGGLLLLVYYWSKNRKAKAKPVTRGAGAGGRQRGQNKERPPPVPNPDYEPIRKGQRDLYSGLNQRRI
SEQ ID NO: 31 Аминокислотная последовательность CD3ζ человека
MKWKALFTAAILQAQLPITEAQSFGLLDPKLCYLLDGILFIYGVILTALFLRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
SEQ ID NO: 32 Аминокислотная последовательность эндодомена IL-2β рецептора человека
NCRNTGPWLKKVLKCNTPDPSKFFSQLSSEHGGDVQKWLSSPFPSSSFSPGGLAPEISPLEVLERDKVTQLLPLNTDAYLSLQELQGQDPTHLV
SEQ ID NO: 33 Аминокислотная последовательность эндодомена IL-7α рецептора человека
KKRIKPIVWPSLPDHKKTLEHLCKKPRKNLNVSFNPESFLDCQIHRVDDIQARDEVEGFLQDTFPQQLEESEKQRLGGDVQSPNCPSEDVVITPESFGRDSSLTCLAGNVSACDAPILSSSRSLDCRESGKNGPHVYQDLLLSLGTTNSTLPPPFSLQSGILTLNPVAQGQPILTSLGSNQEEAYVTMSSFYQNQ
SEQ ID NO: 34 Аминокислотная последовательность IL-21 рецептора человека эндодомена
SLKTHPLWRLWKKIWAVPSPERFFMPLYKGCSGDFKKWVGAPFTGSSLELGPWSPEVPSTLEVYSCHPPRSPAKRLQLTELQEPAELVESDGVPKPSFWPTAQNSGGSAYSEERDRPYGLVSIDTVTVLDAEGPCTWPCSCEDDGYPALDLDAGLEPSPGLEDPLLDAGTTVLSCGCVSAGSPGLGGPLGSLLDRLKPPLADGEDWAGGLPWGGRSPGGVSESEAGSPLAGLDMDTFDSGFVGSDCSSPVECDFTSPGDEGPPRSYLRQWVVIPPPLSSPGPQAS
SEQ ID NO: 35 Аминокислотная последовательность активирующей группы STAT5 человека
YRHQ
SEQ ID NO: 36 Аминокислотная последовательность эндодомена IL-2β рецептора человека и активирующей группы STAT5 человека
NCRNTGPWLKKVLKCNTPDPSKFFSQLSSEHGGDVQKWLSSPFPSSSFSPGGLAPEISPLEVLERDKVTQLLPLNTDAYLSLQELQGQDPTHLVGGGGSYRHQ
SEQ ID NO: 37 Аминокислотная последовательность эндодомена IL-7α рецептора человека и активирующей группы STAT5 человека
KKRIKPIVWPSLPDHKKTLEHLCKKPRKNLNVSFNPESFLDCQIHRVDDIQARDEVEGFLQDTFPQQLEESEKQRLGGDVQSPNCPSEDVVITPESFGRDSSLTCLAGNVSACDAPILSSSRSLDCRESGKNGPHVYQDLLLSLGTTNSTLPPPFSLQSGILTLNPVAQGQPILTSLGSNQEEAYVTMSSFYQNQGGGGSYRHQ
SEQ ID NO: 38 Аминокислотная последовательность анти-CD20 OFA ScFv
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPITFGQGTRLEIKGGGGSGGGGSGGGGSEVQLVESGGGLVQPGRSLRLSCAASGFTFNDYAMHWVRQAPGKGLEWVSTISWNSGSIGYADSVKGRFTISRDNAKKSLYLQMNSLRAEDTALYYCAKDIQYGNYYYGMDVWGQGTTVTVSS
SEQ ID NO: 39 Аминокислотная последовательность анти-CD19 FMC63 ScFv
DIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSS
SEQ ID NO: 40 Аминокислотная последовательность красного флуоресцентного белка
MASSEDVIKEFMRFKVRMEGSVNGHEFEIEGEGEGRPYEGTQTAKLKVTKGGPLPFAWDILSPQFQYGSKAYVKHPADIPDYLKLSFPEGFKWERVMNFEDGGVVTVTQDSSLQDGEFIYKVKLRGTNFPSDGPVMQKKTMGWEASTERMYPEDGALKGEIKMRLKLKDGGHYDAEVKTTYMAKKPVQLPGAYKTDIKLDITSHNEDYTIVEQYERAEGRHSTGA*
SEQ ID NO: 41 TCRaC-Cys-TM9 мыши (hmct STAR TCRaC)
DIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLKVAGFNLLMTLRLWSS
SEQ ID NO: 42 TCRaC-Cys-TM9-N.Rec мыши (Nrec STAR TCRaC)
DIQNPDPAVYQLRDDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLKVAGFNLLMTLRLWSS
SEQ ID NO: 43 β цепь константной области Т-клеточного рецептора мыши с N-концевой модификацией и цистеиновой заменой (TCRbC-Cys-N.Rec мыши, Nrec STAR TCRbC)
DLKNVFPPEVAVFEPSAEIATLVCLARGFFPDHVELSWWVNGKEVHSGVCTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGKATLYAVLVSTLVVMAMVKRKNS
SEQ ID NO:44 анти-CD19 FMC63 VH
EVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSS
SEQ ID NO:45 анти-CD19 FMC63 VL
DIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEIT
SEQ ID NO:46 анти-CD19 334 VH
QVQLQQSGAELVRPGASVKLSCKALGFIFTDYEIHWVKQTPVHGLEWIGAFHPGSGGSAYNQKFKGKATLTADKSSSTAYMELSSLTFEDSAVYHCTRQLGPDWGQGTLVTVS
SEQ ID NO:47 анти-CD19 334 VL
DVVMTQTPLTLSVTIGQPASISCKSSQSLLESDGKTYLNWLLQRPGQSPKRLIYLVSKLDSGVPDRFTGSGSGTDFTLRISRVEAEDLGVYYCWQGTQFPWTFGGGTKLEIK
SEQ ID NO: 48 α цепь константной области Т-клеточного рецептора мыши с удаленным эндодоменом с N-концевой модификацией, цистеиновой заменой и гидрофобной модификацией в трансмембранном домене
DIQNPDPAVYQLRDDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLKVAGFNLLMTLRLW
SEQ ID NO: 49 β цепь константной области Т-клеточного рецептора мыши с удаленным эндодоменом с N-концевой модификацией и цистеиновой заменой
DLKNVFPPEVAVFEPSAEIATLVCLARGFFPDHVELSWWVNGKEVHSGVCTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGKATLYAVLVSTLVVMAM
SEQ ID NO: 50 Аминокислотная последовательность γ цепи константной области TCR человека дикого типа:
DKQLDADVSPKPTIFLPSIAETKLQKAGTYLCLLEKFFPDVIKIHWQEKKSNTILGSQEGNTMKTNDTYMKFSWLTVPEKSLDKEHRCIVRHENNKNGVDQEIIFPPIKTDVITMDPKDNCSKDANDTLLLQLTNTSAYYMYLLLLLKSVVYFAIITCCLLRRTAFCCNGEKS
SEQ ID NO: 51 Аминокислотная последовательность γ цепи константной области TCR мыши дикого типа:
XKRLDADISPKPTIFLPSVAETNLHKTGTYLCLLEKFFPDVIRVYWKEKDGNTILDSQEGDTLKTNDTYMKFSWLTVPERAMGKEHRCIVKHENNKGGADQEIFFPSIKKVAVSTKPTTCWQDKNDVLQLQFTITSAYYTYLLLLLKSVIYLAIISFSLLRRTSVCGNEKKS
SEQ ID NO: 52 Аминокислотная последовательность δ цепи константной области TCR человека дикого типа:
XSQPHTKPSVFVMKNGTNVACLVKEFYPKDIRINLVSSKKITEFDPAIVISPSGKYNAVKLGKYEDSNSVTCSVQHDNKTVHSTDFEVKTDSTDHVKPKETENTKQPSKSCHKPKAIVHTEKVNMMSLTVLGLRMLFAKTVAVNFLLTAKLFFL*
SEQ ID NO: 53 Аминокислотная последовательность δ цепи константной области TCR мыши дикого типа
XSQPPAKPSVFIMKNGTNVACLVKDFYPKEVTISLRSSKKIVEFDPAIVISPSGKYSAVKLGQYGDSNSVTCSVQHNSETVHSTDFEPYANSFNNEKLPEPENDTQISEPCYGPRVTVHTEKVNMMSLTVLGLRLLFAKTIAINFLLTVKLFF*
SEQ ID NO: 54 анти-CD20 2C6 VH
AVQLVESGGGLVQPGRSLRLSCAASGFTFGDYTMHWVRQAPGKGLEWVSGISWNSGSIGYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCTKDNQYGSGSTYGLGVWGQGTLVTVSS
SEQ ID NO: 55 анти-CD20 2C6 VL
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSG TDFTLTISSLEPEDFAVYYCQQRSNWPLTFGGGTKVEIK
SEQ ID NO: 56 анти-CD20 OFA VH
EVQLVESGGGLVQPGRSLRLSCAASGFTFNDYAMHWVRQAPGKGLEWVSTISWNSGSIGYADSVKGRFTISRDNAKKSLYLQMNSLRAEDTALYYCAKDIQYGNYYYGMDVWGQGTTVTVSS
SEQ ID NO: 57 анти-CD20 OFA VL
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPITFGQGTRLEIK
SEQ ID NO: 58 CD19 и CD20-таргетирующий a(G4S)3OX40
MLLLVTSLLLCELPHPAFLLIPEIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPLTFGGGTKVEIKGGGGSGGGGSGGGGSAVQLVESGGGLVQPGRSLRLSCAASGFTFGDYTMHWVRQAPGKGLEWVSGISWNSGSIGYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCTKDNQYGSGSTYGLGVWGQGTLVTVSSEDLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGKEVHSGVCTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGKATLYAVLVSTLVVMAMVKRKNSRRKRSGSGATNFSLLKQAGDVEENPGPMLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLKVAGFNLLMTLRLWGGGGSGGGGSGGGGSRRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKI*
SEQ ID NO: 59 α цепь TCR CD19 и CD20-таргетирующего a(G4S)3OX40
DIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKCVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVPCDATLTEKSFETDMNLNFQNLLVIVLRILLLKVAGFNLLMTLRLWGGGGSGGGGSGGGGSRRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKI
SEQ ID NO: 60 β цепь TCR CD19 и CD20-таргетирующего a(G4S)3OX40
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPLTFGGGTKVEIKGGGGSGGGGSGGGGSAVQLVESGGGLVQPGRSLRLSCAASGFTFGDYTMHWVRQAPGKGLEWVSGISWNSGSIGYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCTKDNQYGSGSTYGLGVWGQGTLVTVSSEDLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGKEVHSGVCTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRADCGITSASYQQGVLSATILYEILLGKATLYAVLVSTLVVMAMVKRKNS
--->
Перечень последовательностей
<110> China Immunotech (Beijing) Biotechnology Co., Ltd.
Tsinghua University
<120> Улучшенный синтетический Т-клеточный рецептор и антигенный рецептор
<130> P2021TC1626
<150> 202011549176.4
<151> 2020-12-24
<150> 202010449454.2
<151> 2020-05-25
<160> 60
<170> PatentIn version 3.5
<210> 1
<211> 141
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 2
<211> 176
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro Ser
1 5 10 15
Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys Glu
50 55 60
Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu Arg
65 70 75 80
Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys Gln
85 90 95
Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp Arg
100 105 110
Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg Ala
115 120 125
Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu Ser Ala
130 135 140
Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val
145 150 155 160
Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp Phe
165 170 175
<210> 3
<211> 137
<212> БЕЛОК
<213> Mus musculus
<400> 3
Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Ser
100 105 110
Val Met Gly Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 4
<211> 172
<212> БЕЛОК
<213> Mus musculus
<400> 4
Asp Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro Ser
1 5 10 15
Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Ala Tyr Lys Glu
50 55 60
Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala Thr
65 70 75 80
Phe Trp His Asn Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe His
85 90 95
Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro Val
100 105 110
Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile
115 120 125
Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr
130 135 140
Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val Leu Val Ser Thr
145 150 155 160
Leu Val Val Met Ala Met Val Lys Arg Lys Asn Ser
165 170
<210> 5
<211> 137
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 5
Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Ser
100 105 110
Val Met Gly Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 6
<211> 172
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 6
Asp Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro Ser
1 5 10 15
Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Ala Tyr Lys Glu
50 55 60
Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala Thr
65 70 75 80
Phe Trp His Asn Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe His
85 90 95
Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro Val
100 105 110
Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile
115 120 125
Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr
130 135 140
Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val Leu Val Ser Thr
145 150 155 160
Leu Val Val Met Ala Met Val Lys Arg Lys Asn Ser
165 170
<210> 7
<211> 137
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 7
Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu
100 105 110
Val Ile Val Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 8
<211> 137
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 8
Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu
100 105 110
Val Ile Val Leu Arg Ile Leu Leu Leu Arg Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 9
<211> 172
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 9
Asp Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro Ser
1 5 10 15
Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Ala Tyr Lys Glu
50 55 60
Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala Thr
65 70 75 80
Phe Trp His Asn Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe His
85 90 95
Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro Val
100 105 110
Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile
115 120 125
Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr
130 135 140
Glu Ile Leu Leu Gly Arg Ala Thr Leu Tyr Ala Val Leu Val Ser Thr
145 150 155 160
Leu Val Val Met Ala Met Val Arg Arg Arg Asn Ser
165 170
<210> 10
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 10
Lys Lys Val Ala Lys Lys Pro Thr Asn Lys Ala Pro His Pro Lys Gln
1 5 10 15
Glu Pro Gln Glu Ile Asn Phe Pro Asp Asp Leu Pro Gly Ser Asn Thr
20 25 30
Ala Ala Pro Val Gln Glu Thr Leu His Gly Cys Gln Pro Val Thr Gln
35 40 45
Glu Asp Gly Lys Glu Ser Arg Ile Ser Val Gln Glu Arg
50 55 60
<210> 11
<211> 37
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 11
Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly Gly
1 5 10 15
Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His Ser
20 25 30
Thr Leu Ala Lys Ile
35
<210> 12
<211> 34
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 12
Lys Lys Lys Tyr Ser Ser Ser Val His Asp Pro Asn Gly Glu Tyr Met
1 5 10 15
Phe Met Arg Ala Val Asn Thr Ala Lys Lys Ser Arg Leu Thr Asp Val
20 25 30
Thr Leu
<210> 13
<211> 41
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 13
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 14
<211> 42
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 14
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 15
<211> 48
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 15
Gln Arg Arg Lys Tyr Arg Ser Asn Lys Gly Glu Ser Pro Val Glu Pro
1 5 10 15
Ala Glu Pro Cys Arg Tyr Ser Cys Pro Arg Glu Glu Glu Gly Ser Thr
20 25 30
Ile Pro Ile Gln Glu Asp Tyr Arg Lys Pro Glu Pro Ala Cys Ser Pro
35 40 45
<210> 16
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 16
Gly Gly Gly Gly Ser
1 5
<210> 17
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 17
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 18
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 18
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 19
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 19
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 20
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 20
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 21
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 21
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
<210> 22
<211> 35
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 22
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser
35
<210> 23
<211> 40
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 23
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser
35 40
<210> 24
<211> 45
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 24
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
35 40 45
<210> 25
<211> 50
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 25
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
35 40 45
Gly Ser
50
<210> 26
<211> 135
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 26
Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu
100 105 110
Val Ile Val Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp
130 135
<210> 27
<211> 166
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 27
Asp Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro Ser
1 5 10 15
Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Ala Tyr Lys Glu
50 55 60
Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala Thr
65 70 75 80
Phe Trp His Asn Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe His
85 90 95
Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro Val
100 105 110
Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile
115 120 125
Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr
130 135 140
Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val Leu Val Ser Thr
145 150 155 160
Leu Val Val Met Ala Met
165
<210> 28
<211> 182
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Met Glu Gln Gly Lys Gly Leu Ala Val Leu Ile Leu Ala Ile Ile Leu
1 5 10 15
Leu Gln Gly Thr Leu Ala Gln Ser Ile Lys Gly Asn His Leu Val Lys
20 25 30
Val Tyr Asp Tyr Gln Glu Asp Gly Ser Val Leu Leu Thr Cys Asp Ala
35 40 45
Glu Ala Lys Asn Ile Thr Trp Phe Lys Asp Gly Lys Met Ile Gly Phe
50 55 60
Leu Thr Glu Asp Lys Lys Lys Trp Asn Leu Gly Ser Asn Ala Lys Asp
65 70 75 80
Pro Arg Gly Met Tyr Gln Cys Lys Gly Ser Gln Asn Lys Ser Lys Pro
85 90 95
Leu Gln Val Tyr Tyr Arg Met Cys Gln Asn Cys Ile Glu Leu Asn Ala
100 105 110
Ala Thr Ile Ser Gly Phe Leu Phe Ala Glu Ile Val Ser Ile Phe Val
115 120 125
Leu Ala Val Gly Val Tyr Phe Ile Ala Gly Gln Asp Gly Val Arg Gln
130 135 140
Ser Arg Ala Ser Asp Lys Gln Thr Leu Leu Pro Asn Asp Gln Leu Tyr
145 150 155 160
Gln Pro Leu Lys Asp Arg Glu Asp Asp Gln Tyr Ser His Leu Gln Gly
165 170 175
Asn Gln Leu Arg Arg Asn
180
<210> 29
<211> 171
<212> БЕЛОК
<213> Homo sapiens
<400> 29
Met Glu His Ser Thr Phe Leu Ser Gly Leu Val Leu Ala Thr Leu Leu
1 5 10 15
Ser Gln Val Ser Pro Phe Lys Ile Pro Ile Glu Glu Leu Glu Asp Arg
20 25 30
Val Phe Val Asn Cys Asn Thr Ser Ile Thr Trp Val Glu Gly Thr Val
35 40 45
Gly Thr Leu Leu Ser Asp Ile Thr Arg Leu Asp Leu Gly Lys Arg Ile
50 55 60
Leu Asp Pro Arg Gly Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys
65 70 75 80
Asp Lys Glu Ser Thr Val Gln Val His Tyr Arg Met Cys Gln Ser Cys
85 90 95
Val Glu Leu Asp Pro Ala Thr Val Ala Gly Ile Ile Val Thr Asp Val
100 105 110
Ile Ala Thr Leu Leu Leu Ala Leu Gly Val Phe Cys Phe Ala Gly His
115 120 125
Glu Thr Gly Arg Leu Ser Gly Ala Ala Asp Thr Gln Ala Leu Leu Arg
130 135 140
Asn Asp Gln Val Tyr Gln Pro Leu Arg Asp Arg Asp Asp Ala Gln Tyr
145 150 155 160
Ser His Leu Gly Gly Asn Trp Ala Arg Asn Lys
165 170
<210> 30
<211> 207
<212> БЕЛОК
<213> Homo sapiens
<400> 30
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr
20 25 30
Gln Thr Pro Tyr Lys Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Pro Gln Tyr Pro Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys
50 55 60
Asn Ile Gly Gly Asp Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp
65 70 75 80
His Leu Ser Leu Lys Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr
85 90 95
Val Cys Tyr Pro Arg Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu
100 105 110
Tyr Leu Arg Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp Val Met
115 120 125
Ser Val Ala Thr Ile Val Ile Val Asp Ile Cys Ile Thr Gly Gly Leu
130 135 140
Leu Leu Leu Val Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys
145 150 155 160
Pro Val Thr Arg Gly Ala Gly Ala Gly Gly Arg Gln Arg Gly Gln Asn
165 170 175
Lys Glu Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg
180 185 190
Lys Gly Gln Arg Asp Leu Tyr Ser Gly Leu Asn Gln Arg Arg Ile
195 200 205
<210> 31
<211> 163
<212> БЕЛОК
<213> Homo sapiens
<400> 31
Met Lys Trp Lys Ala Leu Phe Thr Ala Ala Ile Leu Gln Ala Gln Leu
1 5 10 15
Pro Ile Thr Glu Ala Gln Ser Phe Gly Leu Leu Asp Pro Lys Leu Cys
20 25 30
Tyr Leu Leu Asp Gly Ile Leu Phe Ile Tyr Gly Val Ile Leu Thr Ala
35 40 45
Leu Phe Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr
50 55 60
Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg
65 70 75 80
Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met
85 90 95
Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu
100 105 110
Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys
115 120 125
Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu
130 135 140
Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu
145 150 155 160
Pro Pro Arg
<210> 32
<211> 94
<212> БЕЛОК
<213> Homo sapiens
<400> 32
Asn Cys Arg Asn Thr Gly Pro Trp Leu Lys Lys Val Leu Lys Cys Asn
1 5 10 15
Thr Pro Asp Pro Ser Lys Phe Phe Ser Gln Leu Ser Ser Glu His Gly
20 25 30
Gly Asp Val Gln Lys Trp Leu Ser Ser Pro Phe Pro Ser Ser Ser Phe
35 40 45
Ser Pro Gly Gly Leu Ala Pro Glu Ile Ser Pro Leu Glu Val Leu Glu
50 55 60
Arg Asp Lys Val Thr Gln Leu Leu Pro Leu Asn Thr Asp Ala Tyr Leu
65 70 75 80
Ser Leu Gln Glu Leu Gln Gly Gln Asp Pro Thr His Leu Val
85 90
<210> 33
<211> 195
<212> БЕЛОК
<213> Homo sapiens
<400> 33
Lys Lys Arg Ile Lys Pro Ile Val Trp Pro Ser Leu Pro Asp His Lys
1 5 10 15
Lys Thr Leu Glu His Leu Cys Lys Lys Pro Arg Lys Asn Leu Asn Val
20 25 30
Ser Phe Asn Pro Glu Ser Phe Leu Asp Cys Gln Ile His Arg Val Asp
35 40 45
Asp Ile Gln Ala Arg Asp Glu Val Glu Gly Phe Leu Gln Asp Thr Phe
50 55 60
Pro Gln Gln Leu Glu Glu Ser Glu Lys Gln Arg Leu Gly Gly Asp Val
65 70 75 80
Gln Ser Pro Asn Cys Pro Ser Glu Asp Val Val Ile Thr Pro Glu Ser
85 90 95
Phe Gly Arg Asp Ser Ser Leu Thr Cys Leu Ala Gly Asn Val Ser Ala
100 105 110
Cys Asp Ala Pro Ile Leu Ser Ser Ser Arg Ser Leu Asp Cys Arg Glu
115 120 125
Ser Gly Lys Asn Gly Pro His Val Tyr Gln Asp Leu Leu Leu Ser Leu
130 135 140
Gly Thr Thr Asn Ser Thr Leu Pro Pro Pro Phe Ser Leu Gln Ser Gly
145 150 155 160
Ile Leu Thr Leu Asn Pro Val Ala Gln Gly Gln Pro Ile Leu Thr Ser
165 170 175
Leu Gly Ser Asn Gln Glu Glu Ala Tyr Val Thr Met Ser Ser Phe Tyr
180 185 190
Gln Asn Gln
195
<210> 34
<211> 285
<212> БЕЛОК
<213> Homo sapiens
<400> 34
Ser Leu Lys Thr His Pro Leu Trp Arg Leu Trp Lys Lys Ile Trp Ala
1 5 10 15
Val Pro Ser Pro Glu Arg Phe Phe Met Pro Leu Tyr Lys Gly Cys Ser
20 25 30
Gly Asp Phe Lys Lys Trp Val Gly Ala Pro Phe Thr Gly Ser Ser Leu
35 40 45
Glu Leu Gly Pro Trp Ser Pro Glu Val Pro Ser Thr Leu Glu Val Tyr
50 55 60
Ser Cys His Pro Pro Arg Ser Pro Ala Lys Arg Leu Gln Leu Thr Glu
65 70 75 80
Leu Gln Glu Pro Ala Glu Leu Val Glu Ser Asp Gly Val Pro Lys Pro
85 90 95
Ser Phe Trp Pro Thr Ala Gln Asn Ser Gly Gly Ser Ala Tyr Ser Glu
100 105 110
Glu Arg Asp Arg Pro Tyr Gly Leu Val Ser Ile Asp Thr Val Thr Val
115 120 125
Leu Asp Ala Glu Gly Pro Cys Thr Trp Pro Cys Ser Cys Glu Asp Asp
130 135 140
Gly Tyr Pro Ala Leu Asp Leu Asp Ala Gly Leu Glu Pro Ser Pro Gly
145 150 155 160
Leu Glu Asp Pro Leu Leu Asp Ala Gly Thr Thr Val Leu Ser Cys Gly
165 170 175
Cys Val Ser Ala Gly Ser Pro Gly Leu Gly Gly Pro Leu Gly Ser Leu
180 185 190
Leu Asp Arg Leu Lys Pro Pro Leu Ala Asp Gly Glu Asp Trp Ala Gly
195 200 205
Gly Leu Pro Trp Gly Gly Arg Ser Pro Gly Gly Val Ser Glu Ser Glu
210 215 220
Ala Gly Ser Pro Leu Ala Gly Leu Asp Met Asp Thr Phe Asp Ser Gly
225 230 235 240
Phe Val Gly Ser Asp Cys Ser Ser Pro Val Glu Cys Asp Phe Thr Ser
245 250 255
Pro Gly Asp Glu Gly Pro Pro Arg Ser Tyr Leu Arg Gln Trp Val Val
260 265 270
Ile Pro Pro Pro Leu Ser Ser Pro Gly Pro Gln Ala Ser
275 280 285
<210> 35
<211> 4
<212> БЕЛОК
<213> Homo sapiens
<400> 35
Tyr Arg His Gln
1
<210> 36
<211> 103
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 36
Asn Cys Arg Asn Thr Gly Pro Trp Leu Lys Lys Val Leu Lys Cys Asn
1 5 10 15
Thr Pro Asp Pro Ser Lys Phe Phe Ser Gln Leu Ser Ser Glu His Gly
20 25 30
Gly Asp Val Gln Lys Trp Leu Ser Ser Pro Phe Pro Ser Ser Ser Phe
35 40 45
Ser Pro Gly Gly Leu Ala Pro Glu Ile Ser Pro Leu Glu Val Leu Glu
50 55 60
Arg Asp Lys Val Thr Gln Leu Leu Pro Leu Asn Thr Asp Ala Tyr Leu
65 70 75 80
Ser Leu Gln Glu Leu Gln Gly Gln Asp Pro Thr His Leu Val Gly Gly
85 90 95
Gly Gly Ser Tyr Arg His Gln
100
<210> 37
<211> 204
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 37
Lys Lys Arg Ile Lys Pro Ile Val Trp Pro Ser Leu Pro Asp His Lys
1 5 10 15
Lys Thr Leu Glu His Leu Cys Lys Lys Pro Arg Lys Asn Leu Asn Val
20 25 30
Ser Phe Asn Pro Glu Ser Phe Leu Asp Cys Gln Ile His Arg Val Asp
35 40 45
Asp Ile Gln Ala Arg Asp Glu Val Glu Gly Phe Leu Gln Asp Thr Phe
50 55 60
Pro Gln Gln Leu Glu Glu Ser Glu Lys Gln Arg Leu Gly Gly Asp Val
65 70 75 80
Gln Ser Pro Asn Cys Pro Ser Glu Asp Val Val Ile Thr Pro Glu Ser
85 90 95
Phe Gly Arg Asp Ser Ser Leu Thr Cys Leu Ala Gly Asn Val Ser Ala
100 105 110
Cys Asp Ala Pro Ile Leu Ser Ser Ser Arg Ser Leu Asp Cys Arg Glu
115 120 125
Ser Gly Lys Asn Gly Pro His Val Tyr Gln Asp Leu Leu Leu Ser Leu
130 135 140
Gly Thr Thr Asn Ser Thr Leu Pro Pro Pro Phe Ser Leu Gln Ser Gly
145 150 155 160
Ile Leu Thr Leu Asn Pro Val Ala Gln Gly Gln Pro Ile Leu Thr Ser
165 170 175
Leu Gly Ser Asn Gln Glu Glu Ala Tyr Val Thr Met Ser Ser Phe Tyr
180 185 190
Gln Asn Gln Gly Gly Gly Gly Ser Tyr Arg His Gln
195 200
<210> 38
<211> 244
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 38
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
115 120 125
Ser Gly Gly Gly Leu Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys
130 135 140
Ala Ala Ser Gly Phe Thr Phe Asn Asp Tyr Ala Met His Trp Val Arg
145 150 155 160
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Thr Ile Ser Trp Asn
165 170 175
Ser Gly Ser Ile Gly Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asn Ala Lys Lys Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Lys Asp Ile Gln Tyr
210 215 220
Gly Asn Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
225 230 235 240
Thr Val Ser Ser
<210> 39
<211> 242
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 39
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr His Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu
115 120 125
Ser Gly Pro Gly Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys
130 135 140
Thr Val Ser Gly Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg
145 150 155 160
Gln Pro Pro Arg Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser
165 170 175
Glu Thr Thr Tyr Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile
180 185 190
Lys Asp Asn Ser Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln
195 200 205
Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly
210 215 220
Gly Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val
225 230 235 240
Ser Ser
<210> 40
<211> 225
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 40
Met Ala Ser Ser Glu Asp Val Ile Lys Glu Phe Met Arg Phe Lys Val
1 5 10 15
Arg Met Glu Gly Ser Val Asn Gly His Glu Phe Glu Ile Glu Gly Glu
20 25 30
Gly Glu Gly Arg Pro Tyr Glu Gly Thr Gln Thr Ala Lys Leu Lys Val
35 40 45
Thr Lys Gly Gly Pro Leu Pro Phe Ala Trp Asp Ile Leu Ser Pro Gln
50 55 60
Phe Gln Tyr Gly Ser Lys Ala Tyr Val Lys His Pro Ala Asp Ile Pro
65 70 75 80
Asp Tyr Leu Lys Leu Ser Phe Pro Glu Gly Phe Lys Trp Glu Arg Val
85 90 95
Met Asn Phe Glu Asp Gly Gly Val Val Thr Val Thr Gln Asp Ser Ser
100 105 110
Leu Gln Asp Gly Glu Phe Ile Tyr Lys Val Lys Leu Arg Gly Thr Asn
115 120 125
Phe Pro Ser Asp Gly Pro Val Met Gln Lys Lys Thr Met Gly Trp Glu
130 135 140
Ala Ser Thr Glu Arg Met Tyr Pro Glu Asp Gly Ala Leu Lys Gly Glu
145 150 155 160
Ile Lys Met Arg Leu Lys Leu Lys Asp Gly Gly His Tyr Asp Ala Glu
165 170 175
Val Lys Thr Thr Tyr Met Ala Lys Lys Pro Val Gln Leu Pro Gly Ala
180 185 190
Tyr Lys Thr Asp Ile Lys Leu Asp Ile Thr Ser His Asn Glu Asp Tyr
195 200 205
Thr Ile Val Glu Gln Tyr Glu Arg Ala Glu Gly Arg His Ser Thr Gly
210 215 220
Ala
225
<210> 41
<211> 137
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 41
Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu
100 105 110
Val Ile Val Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 42
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 42
Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Asp Ser
1 5 10 15
Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile Asn Val Pro Lys
20 25 30
Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys Val Leu Asp Met
35 40 45
Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala Trp Ser Asn Gln
50 55 60
Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr Asn Ala Thr Tyr
65 70 75 80
Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr Glu Lys Ser Phe
85 90 95
Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu Val Ile Val Leu
100 105 110
Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu Leu Met Thr Leu
115 120 125
Arg Leu Trp Ser Ser
130
<210> 43
<211> 166
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 43
Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro Ser
1 5 10 15
Ala Glu Ile Ala Thr Leu Val Cys Leu Ala Arg Gly Phe Phe Pro Asp
20 25 30
His Val Glu Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly
35 40 45
Val Cys Thr Asp Pro Gln Ala Tyr Lys Glu Ser Asn Tyr Ser Tyr Cys
50 55 60
Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp His Asn Pro Arg
65 70 75 80
Asn His Phe Arg Cys Gln Val Gln Phe His Gly Leu Ser Glu Glu Asp
85 90 95
Lys Trp Pro Glu Gly Ser Pro Lys Pro Val Thr Gln Asn Ile Ser Ala
100 105 110
Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile Thr Ser Ala Ser Tyr Gln
115 120 125
Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys
130 135 140
Ala Thr Leu Tyr Ala Val Leu Val Ser Thr Leu Val Val Met Ala Met
145 150 155 160
Val Lys Arg Lys Asn Ser
165
<210> 44
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 44
Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln
1 5 10 15
Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu Pro Asp Tyr
20 25 30
Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 45
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 45
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr His Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr
100 105
<210> 46
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 46
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Leu Gly Phe Ile Phe Thr Asp Tyr
20 25 30
Glu Ile His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Phe His Pro Gly Ser Gly Gly Ser Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Phe Glu Asp Ser Ala Val Tyr His Cys
85 90 95
Thr Arg Gln Leu Gly Pro Asp Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser
<210> 47
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 47
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr Gln Phe Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 48
<211> 131
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 48
Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Asp Ser
1 5 10 15
Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile Asn Val Pro Lys
20 25 30
Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys Val Leu Asp Met
35 40 45
Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala Trp Ser Asn Gln
50 55 60
Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr Asn Ala Thr Tyr
65 70 75 80
Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr Glu Lys Ser Phe
85 90 95
Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu Val Ile Val Leu
100 105 110
Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu Leu Met Thr Leu
115 120 125
Arg Leu Trp
130
<210> 49
<211> 160
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 49
Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro Ser
1 5 10 15
Ala Glu Ile Ala Thr Leu Val Cys Leu Ala Arg Gly Phe Phe Pro Asp
20 25 30
His Val Glu Leu Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly
35 40 45
Val Cys Thr Asp Pro Gln Ala Tyr Lys Glu Ser Asn Tyr Ser Tyr Cys
50 55 60
Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp His Asn Pro Arg
65 70 75 80
Asn His Phe Arg Cys Gln Val Gln Phe His Gly Leu Ser Glu Glu Asp
85 90 95
Lys Trp Pro Glu Gly Ser Pro Lys Pro Val Thr Gln Asn Ile Ser Ala
100 105 110
Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile Thr Ser Ala Ser Tyr Gln
115 120 125
Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys
130 135 140
Ala Thr Leu Tyr Ala Val Leu Val Ser Thr Leu Val Val Met Ala Met
145 150 155 160
<210> 50
<211> 173
<212> БЕЛОК
<213> Homo sapiens
<400> 50
Asp Lys Gln Leu Asp Ala Asp Val Ser Pro Lys Pro Thr Ile Phe Leu
1 5 10 15
Pro Ser Ile Ala Glu Thr Lys Leu Gln Lys Ala Gly Thr Tyr Leu Cys
20 25 30
Leu Leu Glu Lys Phe Phe Pro Asp Val Ile Lys Ile His Trp Gln Glu
35 40 45
Lys Lys Ser Asn Thr Ile Leu Gly Ser Gln Glu Gly Asn Thr Met Lys
50 55 60
Thr Asn Asp Thr Tyr Met Lys Phe Ser Trp Leu Thr Val Pro Glu Lys
65 70 75 80
Ser Leu Asp Lys Glu His Arg Cys Ile Val Arg His Glu Asn Asn Lys
85 90 95
Asn Gly Val Asp Gln Glu Ile Ile Phe Pro Pro Ile Lys Thr Asp Val
100 105 110
Ile Thr Met Asp Pro Lys Asp Asn Cys Ser Lys Asp Ala Asn Asp Thr
115 120 125
Leu Leu Leu Gln Leu Thr Asn Thr Ser Ala Tyr Tyr Met Tyr Leu Leu
130 135 140
Leu Leu Leu Lys Ser Val Val Tyr Phe Ala Ile Ile Thr Cys Cys Leu
145 150 155 160
Leu Arg Arg Thr Ala Phe Cys Cys Asn Gly Glu Lys Ser
165 170
<210> 51
<211> 172
<212> БЕЛОК
<213> Mus musculus
<220>
<221> прочий_признак
<222> (1)..(1)
<223> Xaa может быть любой существующей в природе аминокислотой
<400> 51
Xaa Lys Arg Leu Asp Ala Asp Ile Ser Pro Lys Pro Thr Ile Phe Leu
1 5 10 15
Pro Ser Val Ala Glu Thr Asn Leu His Lys Thr Gly Thr Tyr Leu Cys
20 25 30
Leu Leu Glu Lys Phe Phe Pro Asp Val Ile Arg Val Tyr Trp Lys Glu
35 40 45
Lys Asp Gly Asn Thr Ile Leu Asp Ser Gln Glu Gly Asp Thr Leu Lys
50 55 60
Thr Asn Asp Thr Tyr Met Lys Phe Ser Trp Leu Thr Val Pro Glu Arg
65 70 75 80
Ala Met Gly Lys Glu His Arg Cys Ile Val Lys His Glu Asn Asn Lys
85 90 95
Gly Gly Ala Asp Gln Glu Ile Phe Phe Pro Ser Ile Lys Lys Val Ala
100 105 110
Val Ser Thr Lys Pro Thr Thr Cys Trp Gln Asp Lys Asn Asp Val Leu
115 120 125
Gln Leu Gln Phe Thr Ile Thr Ser Ala Tyr Tyr Thr Tyr Leu Leu Leu
130 135 140
Leu Leu Lys Ser Val Ile Tyr Leu Ala Ile Ile Ser Phe Ser Leu Leu
145 150 155 160
Arg Arg Thr Ser Val Cys Gly Asn Glu Lys Lys Ser
165 170
<210> 52
<211> 154
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> прочий_признак
<222> (1)..(1)
<223> Xaa может быть любой существующей в природе аминокислотой
<400> 52
Xaa Ser Gln Pro His Thr Lys Pro Ser Val Phe Val Met Lys Asn Gly
1 5 10 15
Thr Asn Val Ala Cys Leu Val Lys Glu Phe Tyr Pro Lys Asp Ile Arg
20 25 30
Ile Asn Leu Val Ser Ser Lys Lys Ile Thr Glu Phe Asp Pro Ala Ile
35 40 45
Val Ile Ser Pro Ser Gly Lys Tyr Asn Ala Val Lys Leu Gly Lys Tyr
50 55 60
Glu Asp Ser Asn Ser Val Thr Cys Ser Val Gln His Asp Asn Lys Thr
65 70 75 80
Val His Ser Thr Asp Phe Glu Val Lys Thr Asp Ser Thr Asp His Val
85 90 95
Lys Pro Lys Glu Thr Glu Asn Thr Lys Gln Pro Ser Lys Ser Cys His
100 105 110
Lys Pro Lys Ala Ile Val His Thr Glu Lys Val Asn Met Met Ser Leu
115 120 125
Thr Val Leu Gly Leu Arg Met Leu Phe Ala Lys Thr Val Ala Val Asn
130 135 140
Phe Leu Leu Thr Ala Lys Leu Phe Phe Leu
145 150
<210> 53
<211> 153
<212> БЕЛОК
<213> Mus musculus
<220>
<221> прочий_признак
<222> (1)..(1)
<223> Xaa может быть любой существующей в природе аминокислотой
<400> 53
Xaa Ser Gln Pro Pro Ala Lys Pro Ser Val Phe Ile Met Lys Asn Gly
1 5 10 15
Thr Asn Val Ala Cys Leu Val Lys Asp Phe Tyr Pro Lys Glu Val Thr
20 25 30
Ile Ser Leu Arg Ser Ser Lys Lys Ile Val Glu Phe Asp Pro Ala Ile
35 40 45
Val Ile Ser Pro Ser Gly Lys Tyr Ser Ala Val Lys Leu Gly Gln Tyr
50 55 60
Gly Asp Ser Asn Ser Val Thr Cys Ser Val Gln His Asn Ser Glu Thr
65 70 75 80
Val His Ser Thr Asp Phe Glu Pro Tyr Ala Asn Ser Phe Asn Asn Glu
85 90 95
Lys Leu Pro Glu Pro Glu Asn Asp Thr Gln Ile Ser Glu Pro Cys Tyr
100 105 110
Gly Pro Arg Val Thr Val His Thr Glu Lys Val Asn Met Met Ser Leu
115 120 125
Thr Val Leu Gly Leu Arg Leu Leu Phe Ala Lys Thr Ile Ala Ile Asn
130 135 140
Phe Leu Leu Thr Val Lys Leu Phe Phe
145 150
<210> 54
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 54
Ala Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Asp Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Asn Ser Gly Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Lys Asp Asn Gln Tyr Gly Ser Gly Ser Thr Tyr Gly Leu Gly Val
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 55
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 55
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 56
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 56
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Ser Trp Asn Ser Gly Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys Asp Ile Gln Tyr Gly Asn Tyr Tyr Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 57
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 57
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 58
<211> 918
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 58
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Glu Ile Val Leu Thr Gln Ser Pro Ala Thr
20 25 30
Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
35 40 45
Gln Ser Val Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
50 55 60
Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile
65 70 75 80
Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
100 105 110
Arg Ser Asn Trp Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
115 120 125
Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Ala Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
145 150 155 160
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Asp Tyr
165 170 175
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
180 185 190
Ser Gly Ile Ser Trp Asn Ser Gly Ser Ile Gly Tyr Ala Asp Ser Val
195 200 205
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
210 215 220
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
225 230 235 240
Thr Lys Asp Asn Gln Tyr Gly Ser Gly Ser Thr Tyr Gly Leu Gly Val
245 250 255
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Asp Leu Arg Asn
260 265 270
Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro Ser Lys Ala Glu Ile
275 280 285
Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu Ala Arg Gly Phe Phe
290 295 300
Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly Lys Glu Val His
305 310 315 320
Ser Gly Val Cys Thr Asp Pro Gln Ala Tyr Lys Glu Ser Asn Tyr Ser
325 330 335
Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp His Asn
340 345 350
Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe His Gly Leu Ser Glu
355 360 365
Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro Val Thr Gln Asn Ile
370 375 380
Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile Thr Ser Ala Ser
385 390 395 400
Tyr Gln Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu
405 410 415
Gly Lys Ala Thr Leu Tyr Ala Val Leu Val Ser Thr Leu Val Val Met
420 425 430
Ala Met Val Lys Arg Lys Asn Ser Arg Arg Lys Arg Ser Gly Ser Gly
435 440 445
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
450 455 460
Pro Gly Pro Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu
465 470 475 480
Pro His Pro Ala Phe Leu Leu Ile Pro Asp Ile Gln Met Thr Gln Thr
485 490 495
Thr Ser Ser Leu Ser Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys
500 505 510
Arg Ala Ser Gln Asp Ile Ser Lys Tyr Leu Asn Trp Tyr Gln Gln Lys
515 520 525
Pro Asp Gly Thr Val Lys Leu Leu Ile Tyr His Thr Ser Arg Leu His
530 535 540
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr
545 550 555 560
Ser Leu Thr Ile Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe
565 570 575
Cys Gln Gln Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys
580 585 590
Leu Glu Ile Thr Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
595 600 605
Gly Gly Ser Glu Val Lys Leu Gln Glu Ser Gly Pro Gly Leu Val Ala
610 615 620
Pro Ser Gln Ser Leu Ser Val Thr Cys Thr Val Ser Gly Val Ser Leu
625 630 635 640
Pro Asp Tyr Gly Val Ser Trp Ile Arg Gln Pro Pro Arg Lys Gly Leu
645 650 655
Glu Trp Leu Gly Val Ile Trp Gly Ser Glu Thr Thr Tyr Tyr Asn Ser
660 665 670
Ala Leu Lys Ser Arg Leu Thr Ile Ile Lys Asp Asn Ser Lys Ser Gln
675 680 685
Val Phe Leu Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr
690 695 700
Tyr Cys Ala Lys His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr
705 710 715 720
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Asp Ile Gln Asn Pro
725 730 735
Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg Ser Gln Asp Ser Thr
740 745 750
Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile Asn Val Pro Lys Thr
755 760 765
Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys Val Leu Asp Met Lys
770 775 780
Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala Trp Ser Asn Gln Thr
785 790 795 800
Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr Asn Ala Thr Tyr Pro
805 810 815
Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr Glu Lys Ser Phe Glu
820 825 830
Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu Val Ile Val Leu Arg
835 840 845
Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu Leu Met Thr Leu Arg
850 855 860
Leu Trp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
865 870 875 880
Ser Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly
885 890 895
Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His
900 905 910
Ser Thr Leu Ala Lys Ile
915
<210> 59
<211> 429
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 59
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr His Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Thr Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Lys Leu Gln Glu
115 120 125
Ser Gly Pro Gly Leu Val Ala Pro Ser Gln Ser Leu Ser Val Thr Cys
130 135 140
Thr Val Ser Gly Val Ser Leu Pro Asp Tyr Gly Val Ser Trp Ile Arg
145 150 155 160
Gln Pro Pro Arg Lys Gly Leu Glu Trp Leu Gly Val Ile Trp Gly Ser
165 170 175
Glu Thr Thr Tyr Tyr Asn Ser Ala Leu Lys Ser Arg Leu Thr Ile Ile
180 185 190
Lys Asp Asn Ser Lys Ser Gln Val Phe Leu Lys Met Asn Ser Leu Gln
195 200 205
Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Ala Lys His Tyr Tyr Tyr Gly
210 215 220
Gly Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val
225 230 235 240
Ser Ser Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp
245 250 255
Pro Arg Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser
260 265 270
Gln Ile Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp
275 280 285
Lys Cys Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala
290 295 300
Ile Ala Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys
305 310 315 320
Glu Thr Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr
325 330 335
Leu Thr Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn
340 345 350
Leu Leu Val Ile Val Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe
355 360 365
Asn Leu Leu Met Thr Leu Arg Leu Trp Gly Gly Gly Gly Ser Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Arg Arg Asp Gln Arg Leu Pro Pro
385 390 395 400
Asp Ala His Lys Pro Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln
405 410 415
Glu Glu Gln Ala Asp Ala His Ser Thr Leu Ala Lys Ile
420 425
<210> 60
<211> 418
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Искусственная последовательность
<400> 60
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Val Gln Leu Val Glu
115 120 125
Ser Gly Gly Gly Leu Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys
130 135 140
Ala Ala Ser Gly Phe Thr Phe Gly Asp Tyr Thr Met His Trp Val Arg
145 150 155 160
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Gly Ile Ser Trp Asn
165 170 175
Ser Gly Ser Ile Gly Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys Thr Lys Asp Asn Gln Tyr
210 215 220
Gly Ser Gly Ser Thr Tyr Gly Leu Gly Val Trp Gly Gln Gly Thr Leu
225 230 235 240
Val Thr Val Ser Ser Glu Asp Leu Arg Asn Val Thr Pro Pro Lys Val
245 250 255
Ser Leu Phe Glu Pro Ser Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala
260 265 270
Thr Leu Val Cys Leu Ala Arg Gly Phe Phe Pro Asp His Val Glu Leu
275 280 285
Ser Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp
290 295 300
Pro Gln Ala Tyr Lys Glu Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg
305 310 315 320
Leu Arg Val Ser Ala Thr Phe Trp His Asn Pro Arg Asn His Phe Arg
325 330 335
Cys Gln Val Gln Phe His Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu
340 345 350
Gly Ser Pro Lys Pro Val Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly
355 360 365
Arg Ala Asp Cys Gly Ile Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu
370 375 380
Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr
385 390 395 400
Ala Val Leu Val Ser Thr Leu Val Val Met Ala Met Val Lys Arg Lys
405 410 415
Asn Ser
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР | 2016 |
|
RU2826122C1 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2016 |
|
RU2752880C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2810092C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2017 |
|
RU2775623C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) С АНТИГЕНСВЯЗЫВАЮЩИМИ ДОМЕНАМИ К КОНСТАНТНОЙ ОБЛАСТИ β Т-КЛЕТОЧНОГО РЕЦЕПТОРА | 2015 |
|
RU2830064C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) С АНТИГЕНСВЯЗЫВАЮЩИМИ ДОМЕНАМИ К КОНСТАНТНОЙ ОБЛАСТИ β Т-КЛЕТОЧНОГО РЕЦЕПТОРА | 2015 |
|
RU2744046C2 |
| СВЯЗЫВАЮЩИЙ ДОМЕН | 2019 |
|
RU2829934C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2018 |
|
RU2762255C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| ВЫСОКОАФФИННЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP | 2019 |
|
RU2836365C2 |
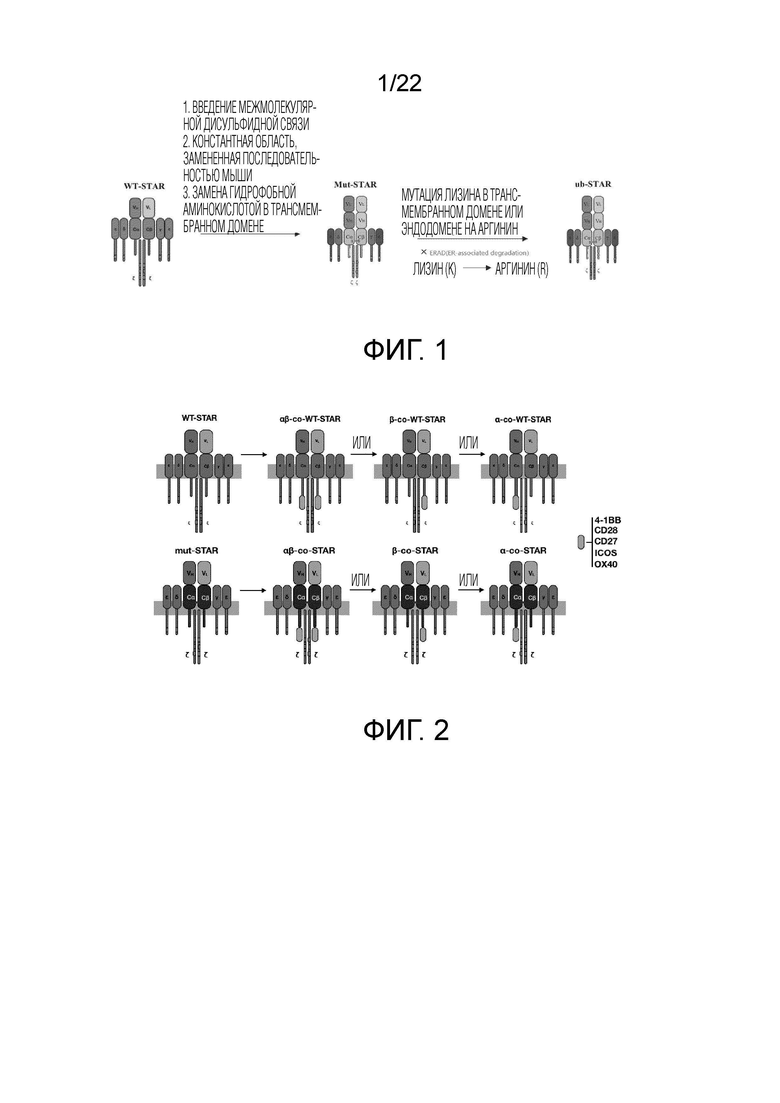
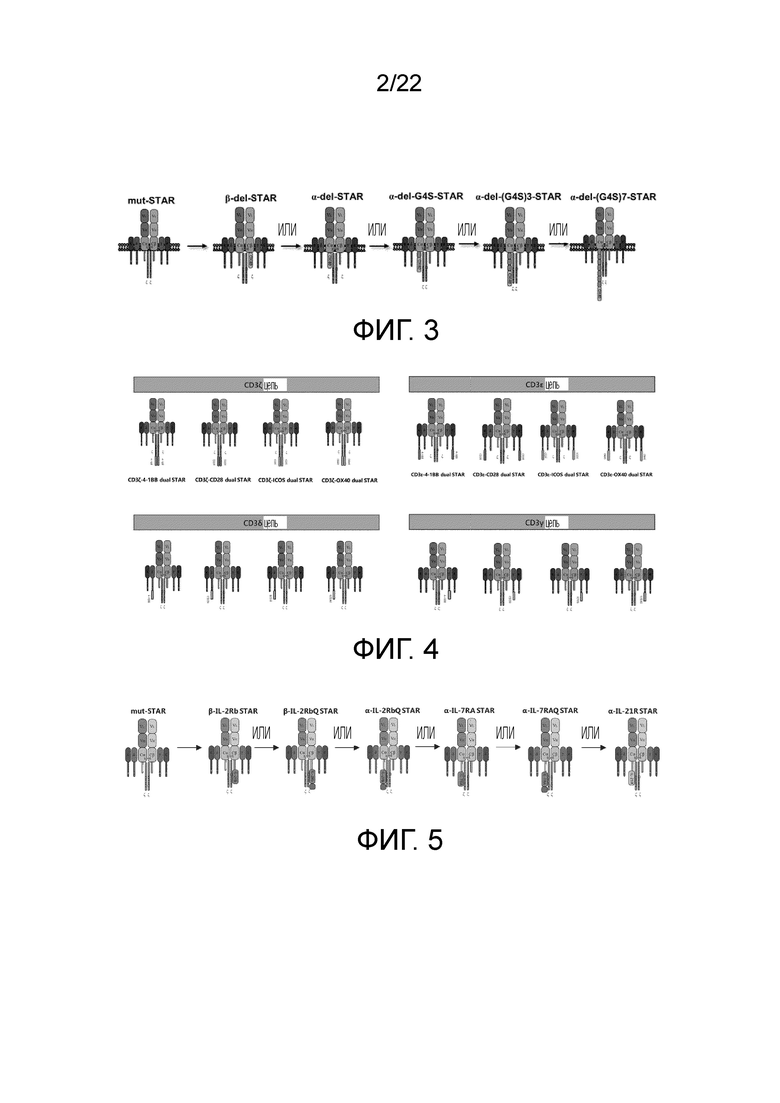
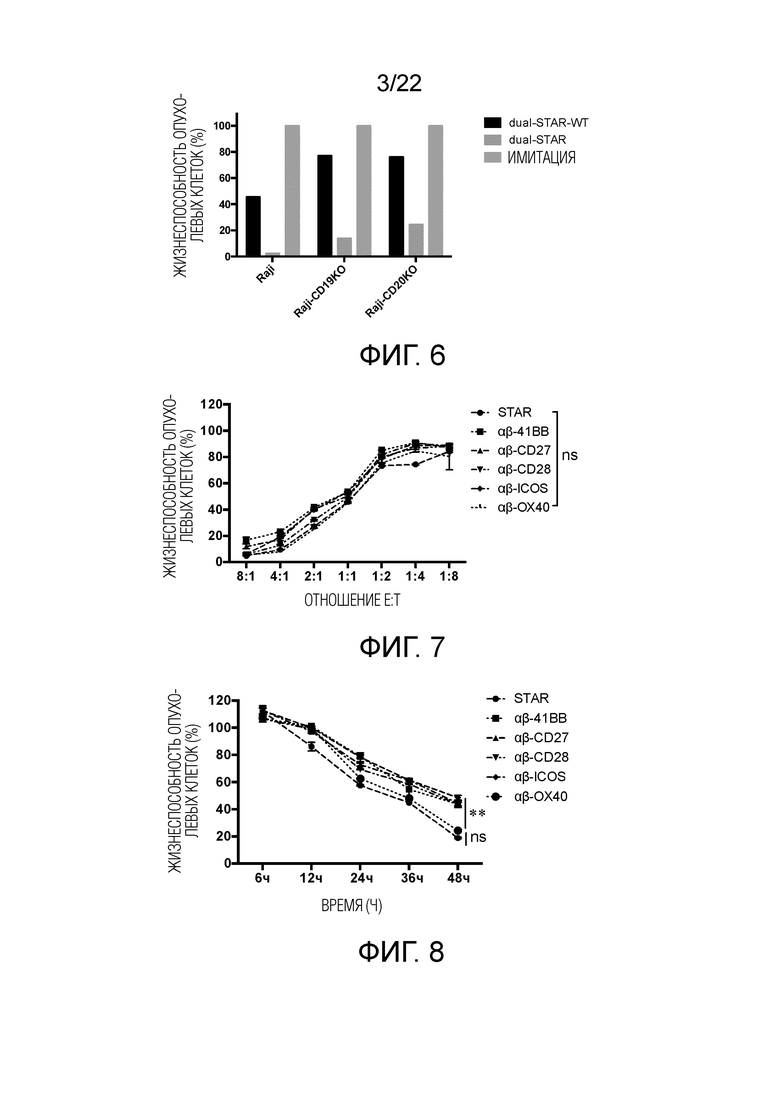
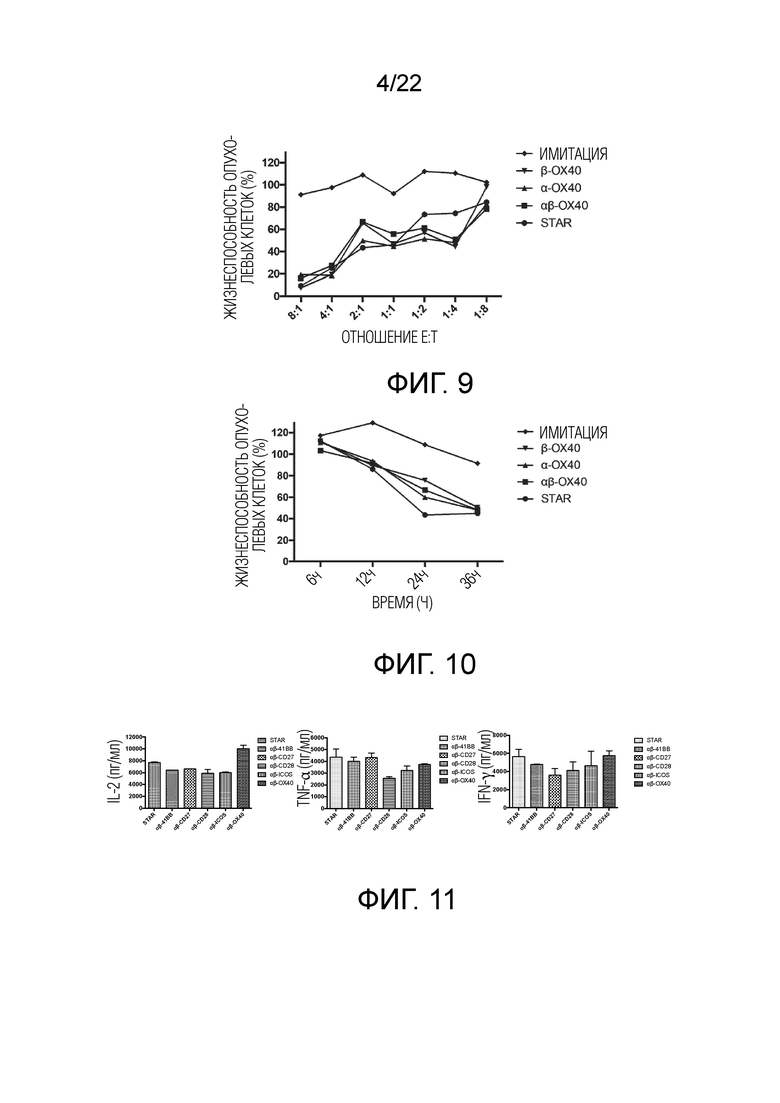
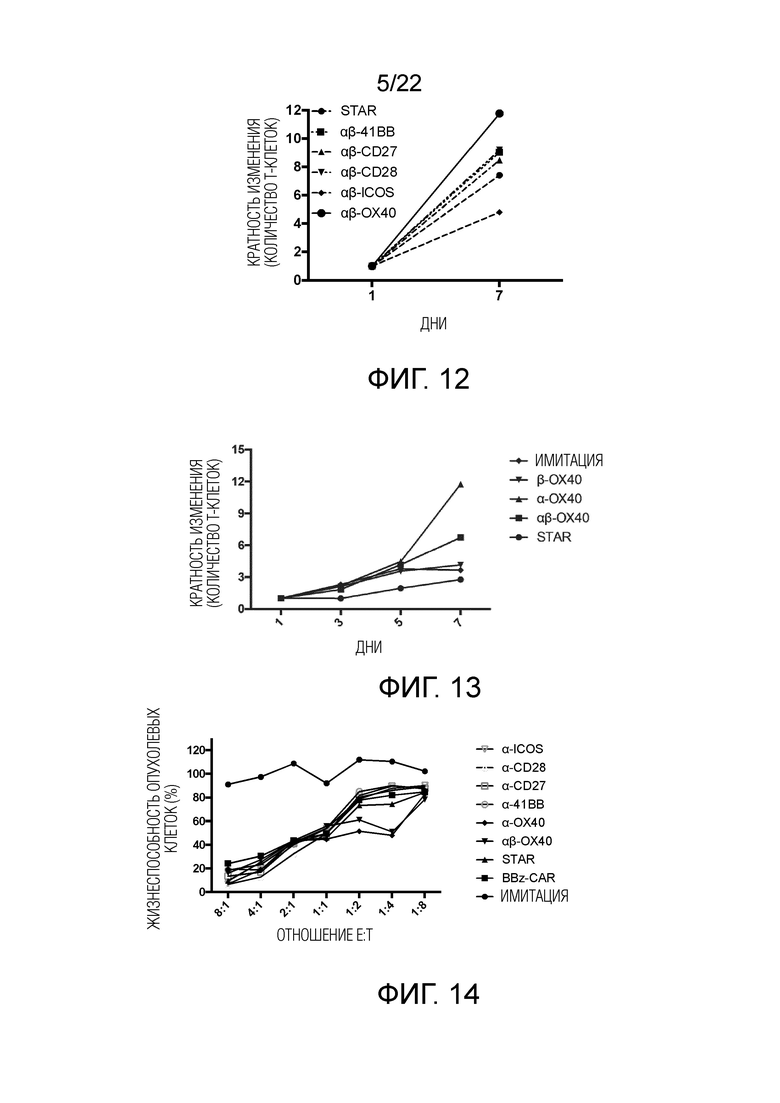
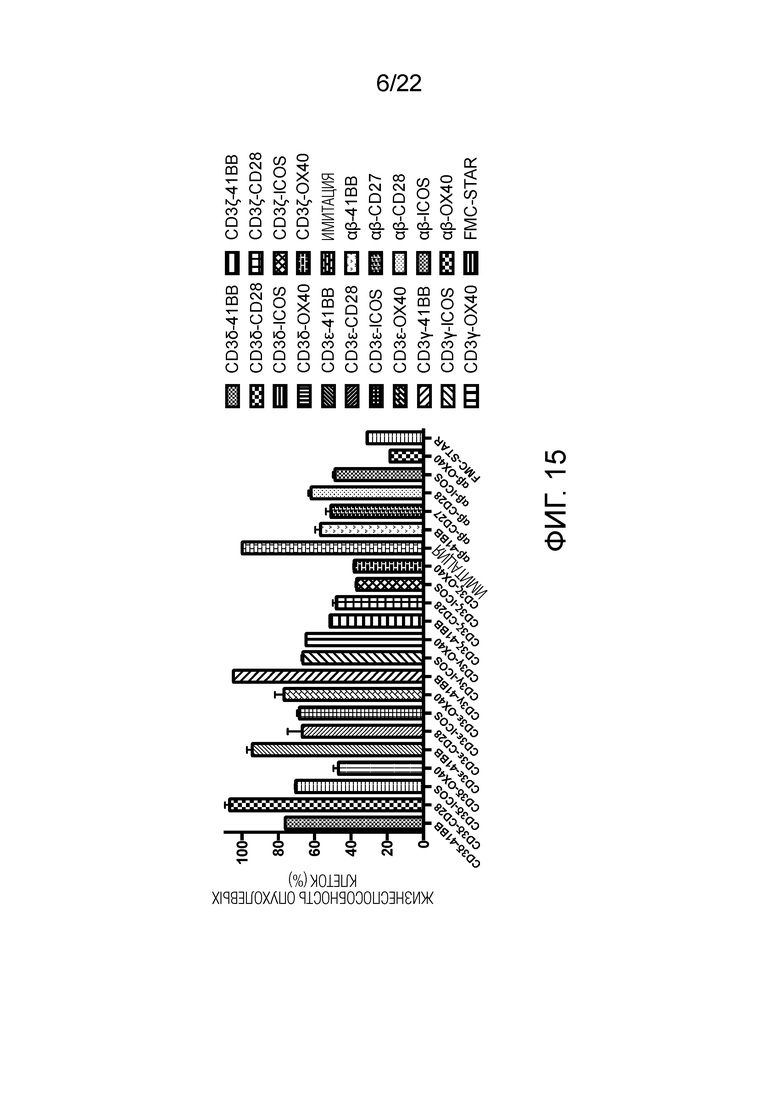
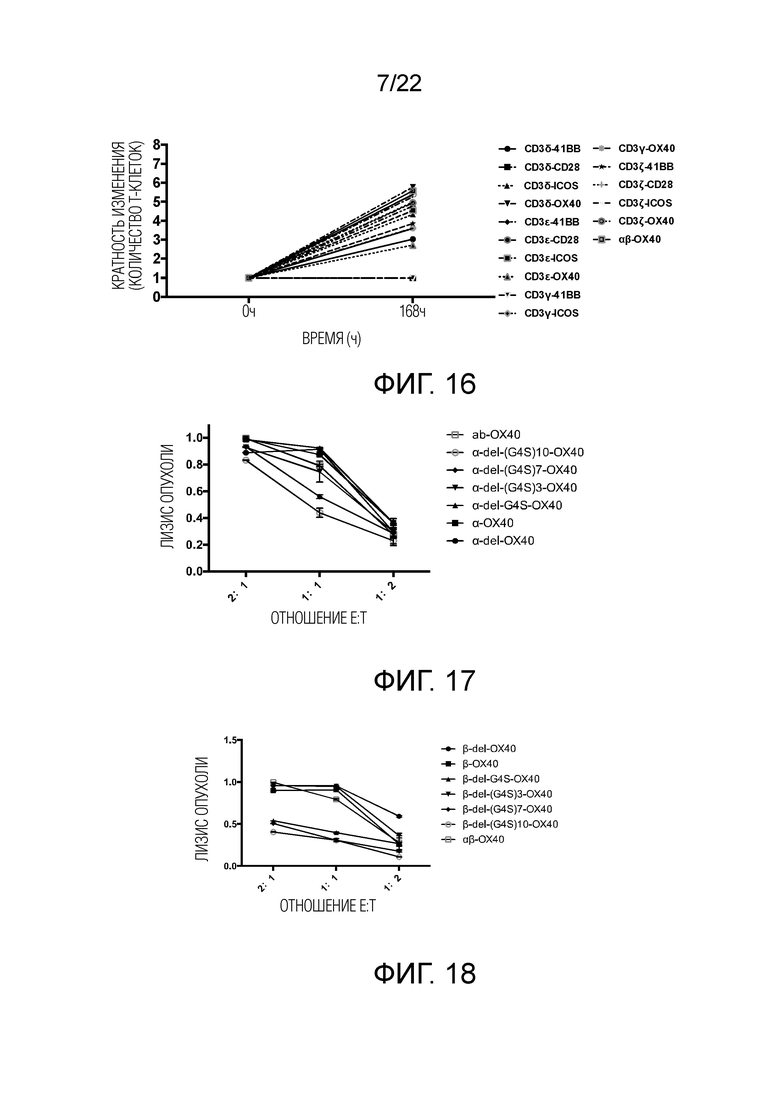
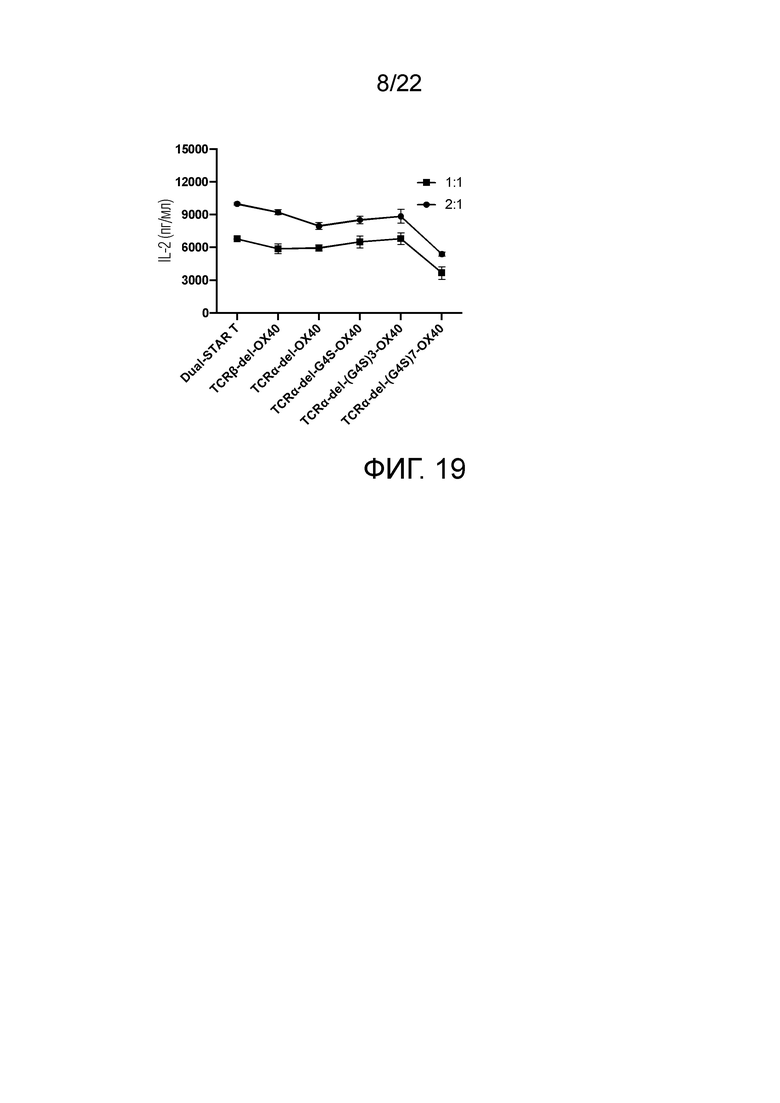
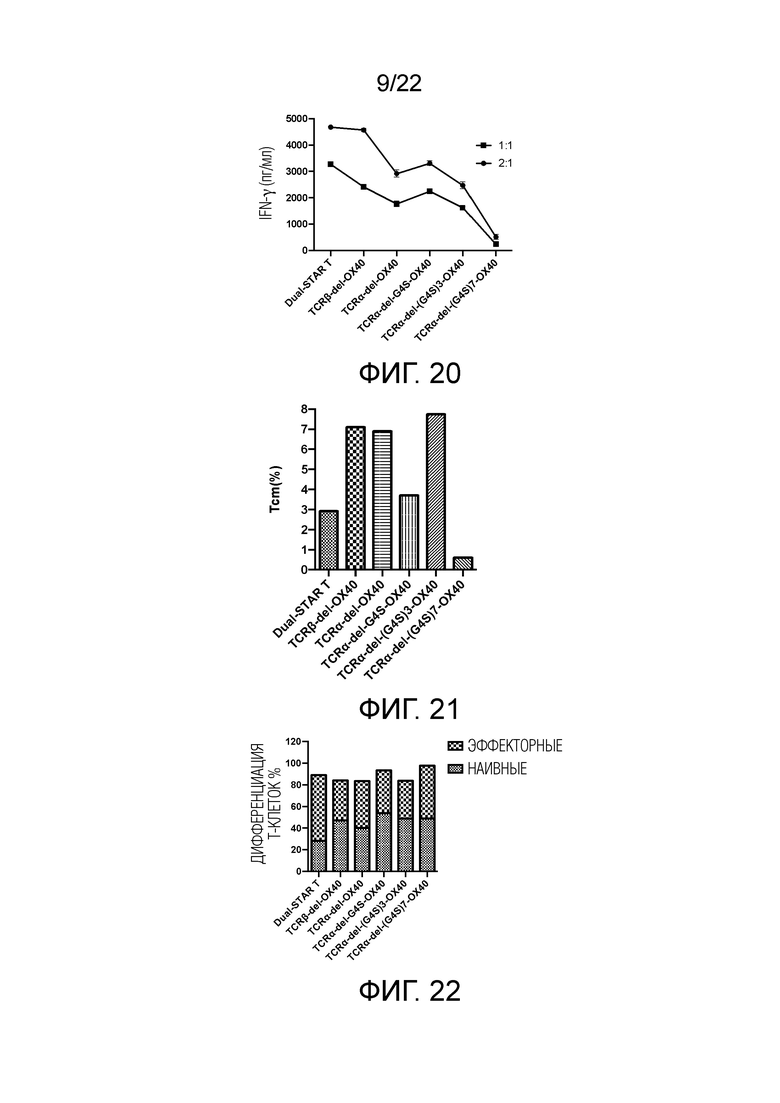
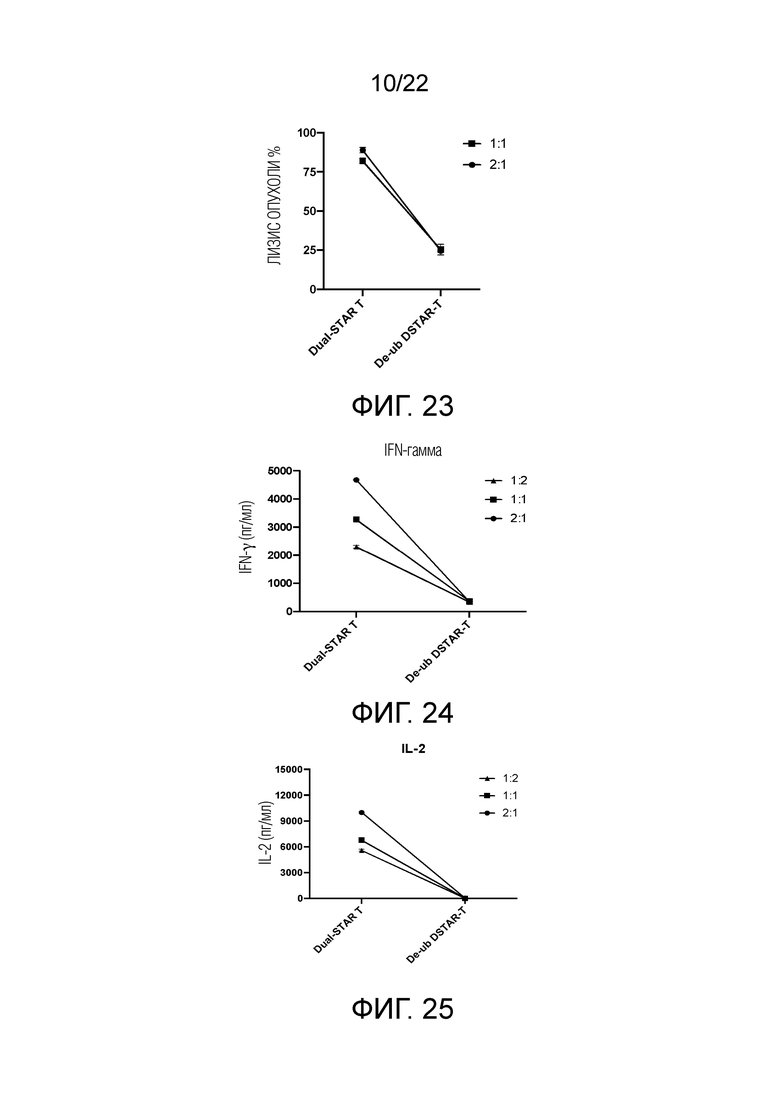
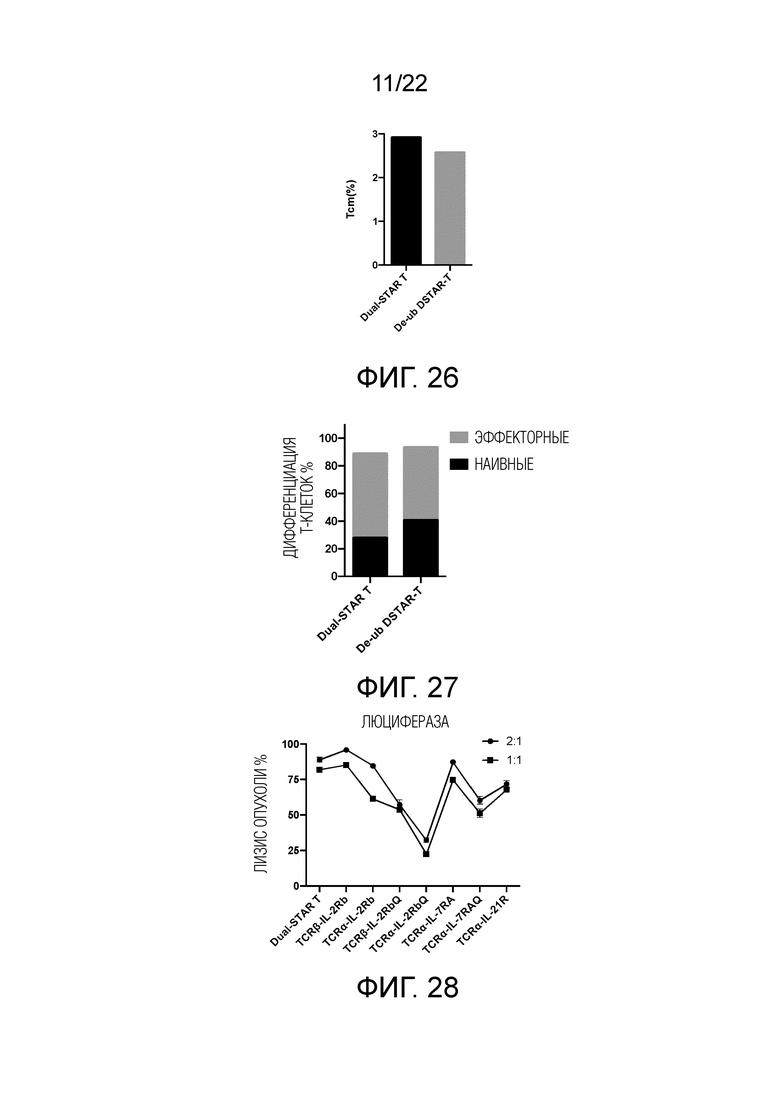
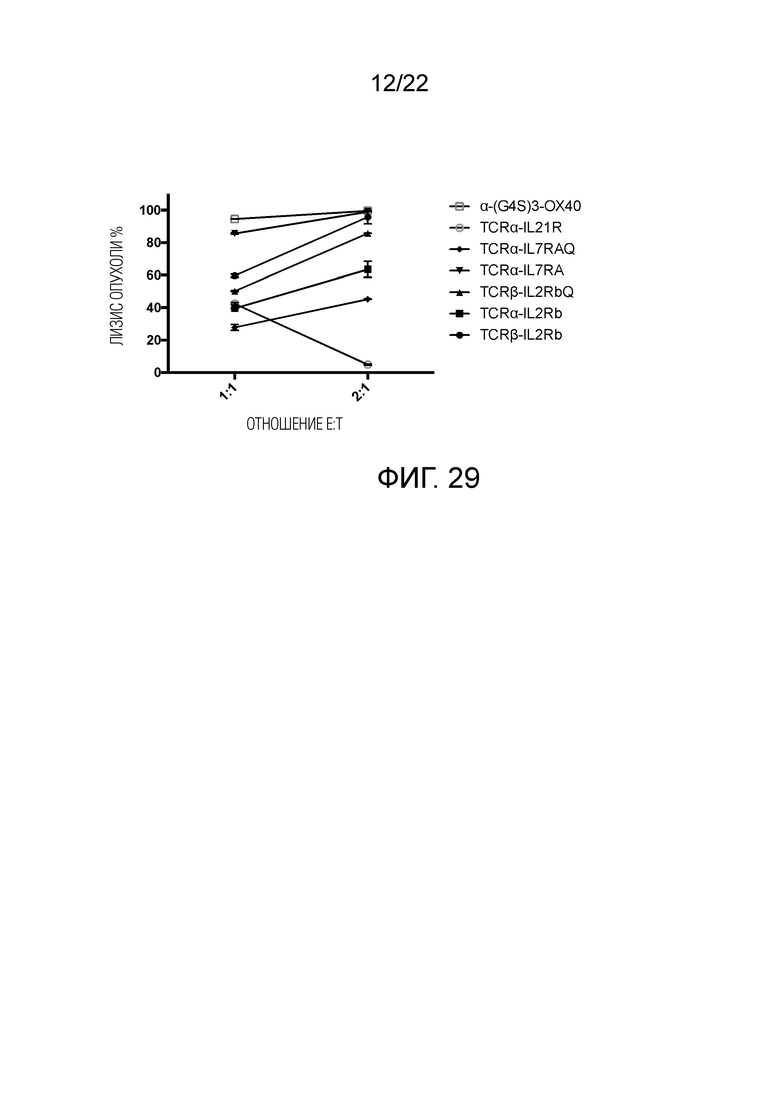
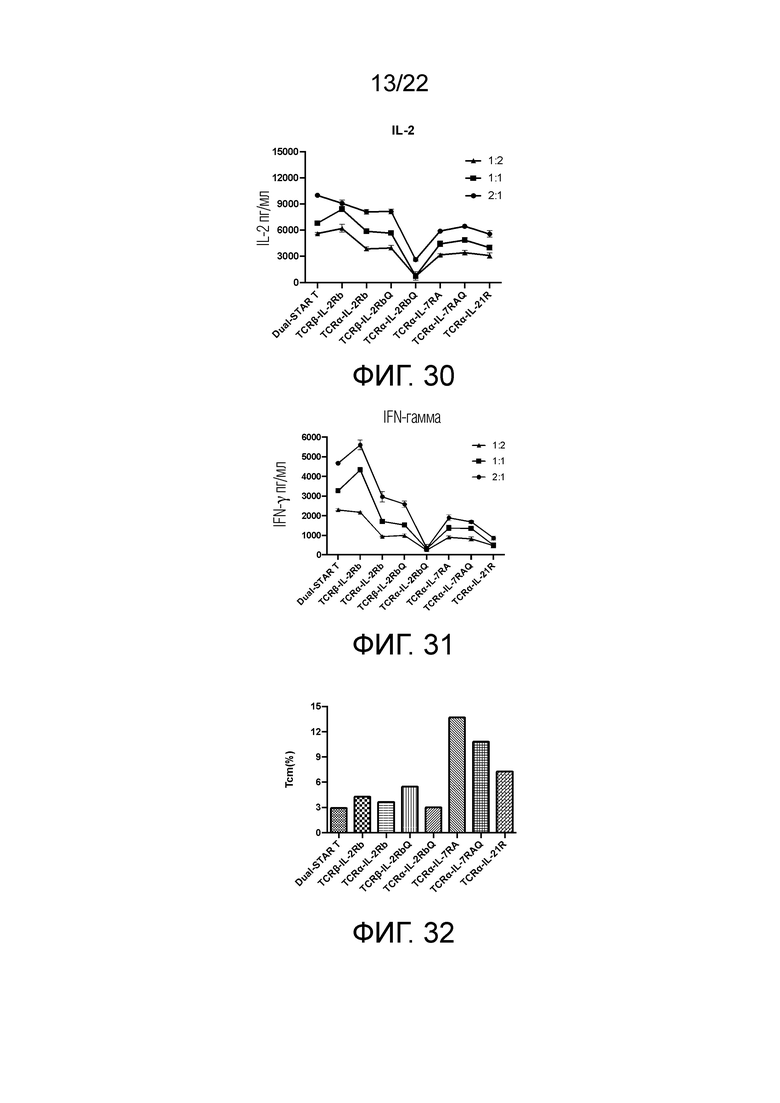
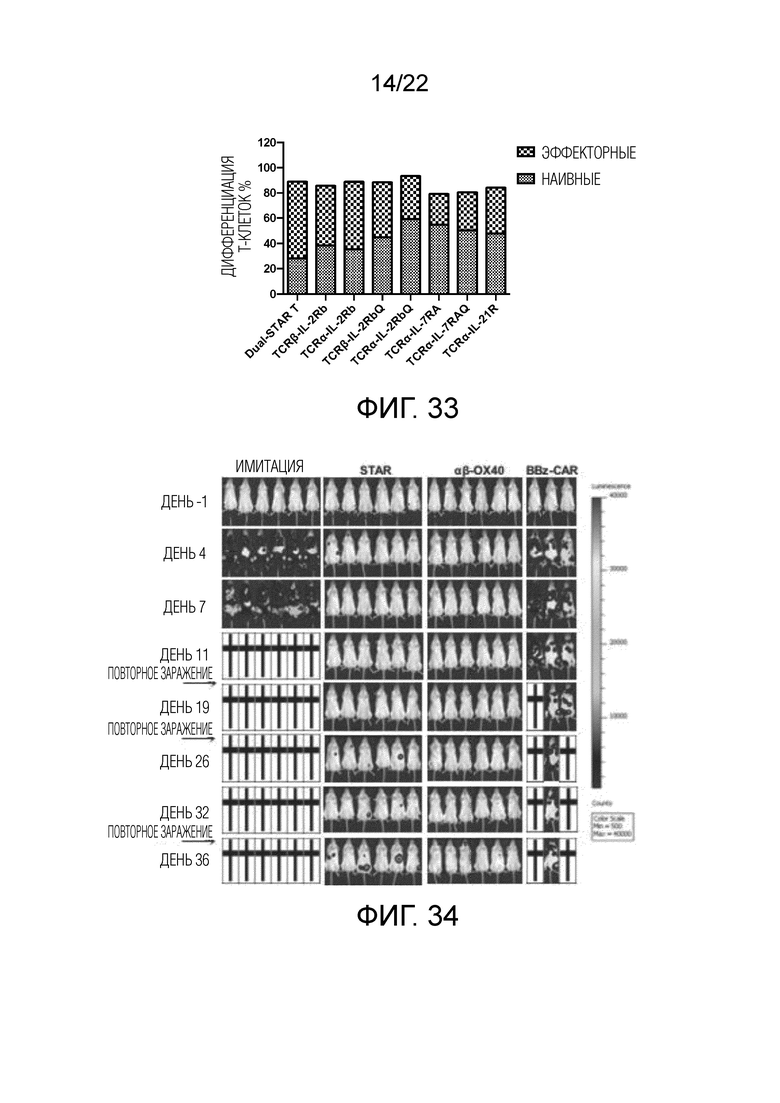
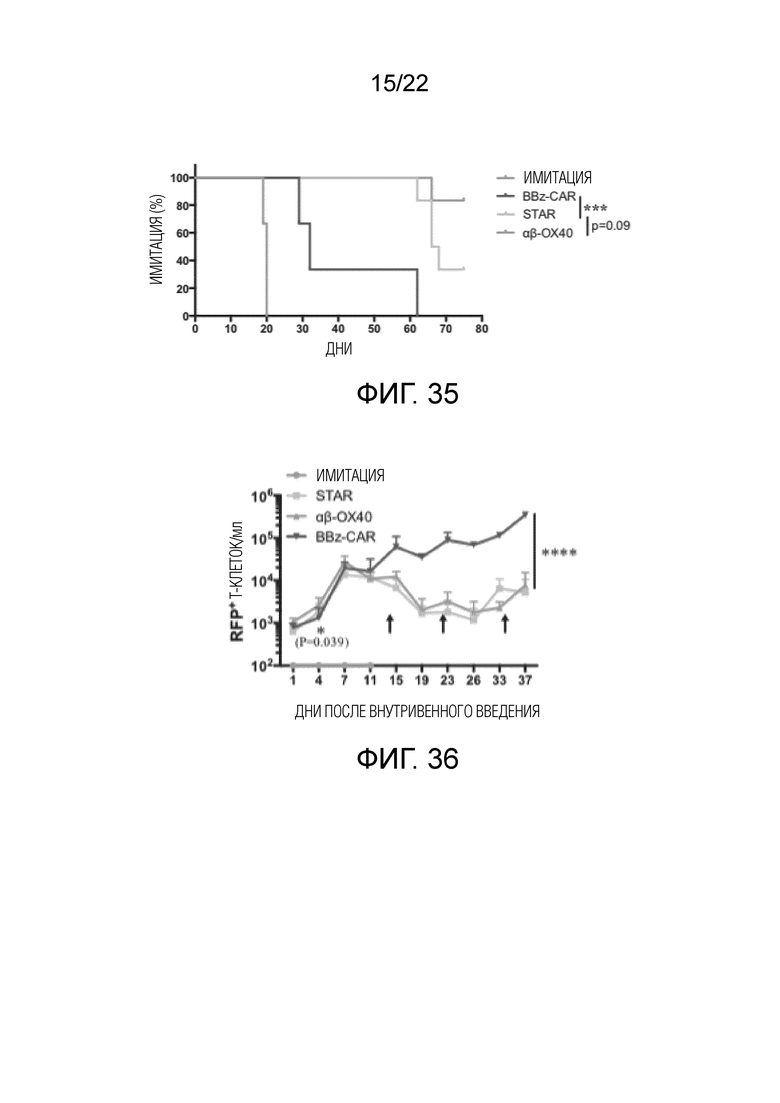
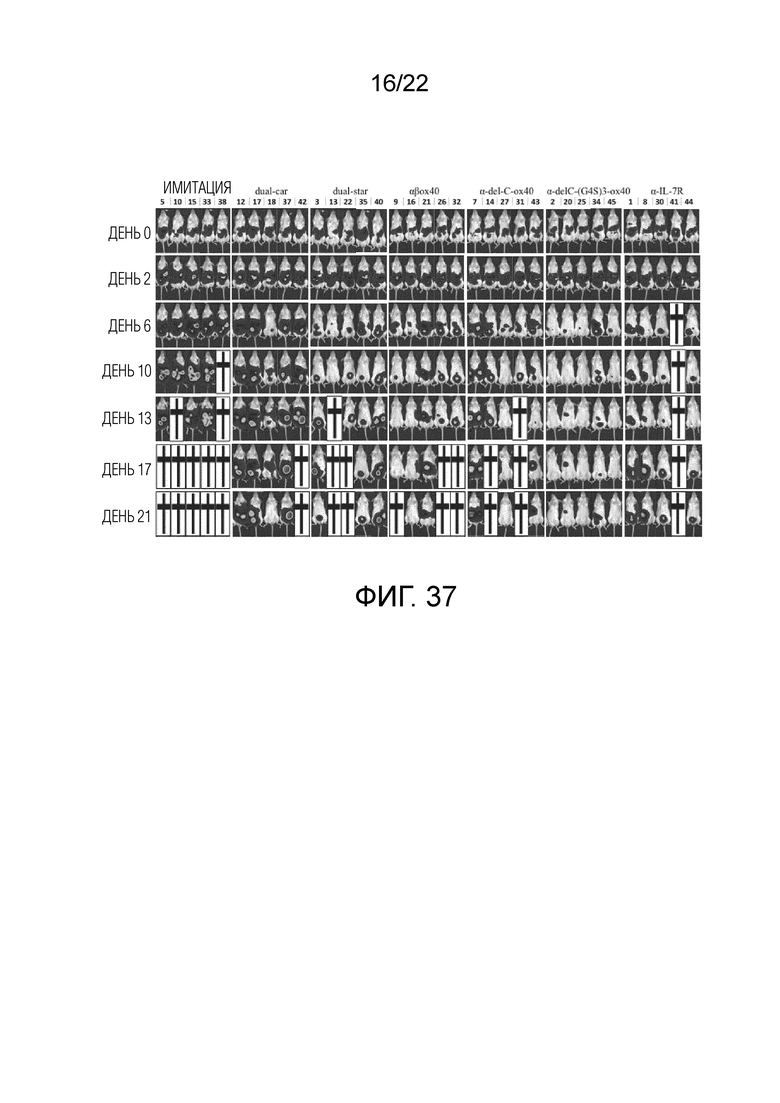
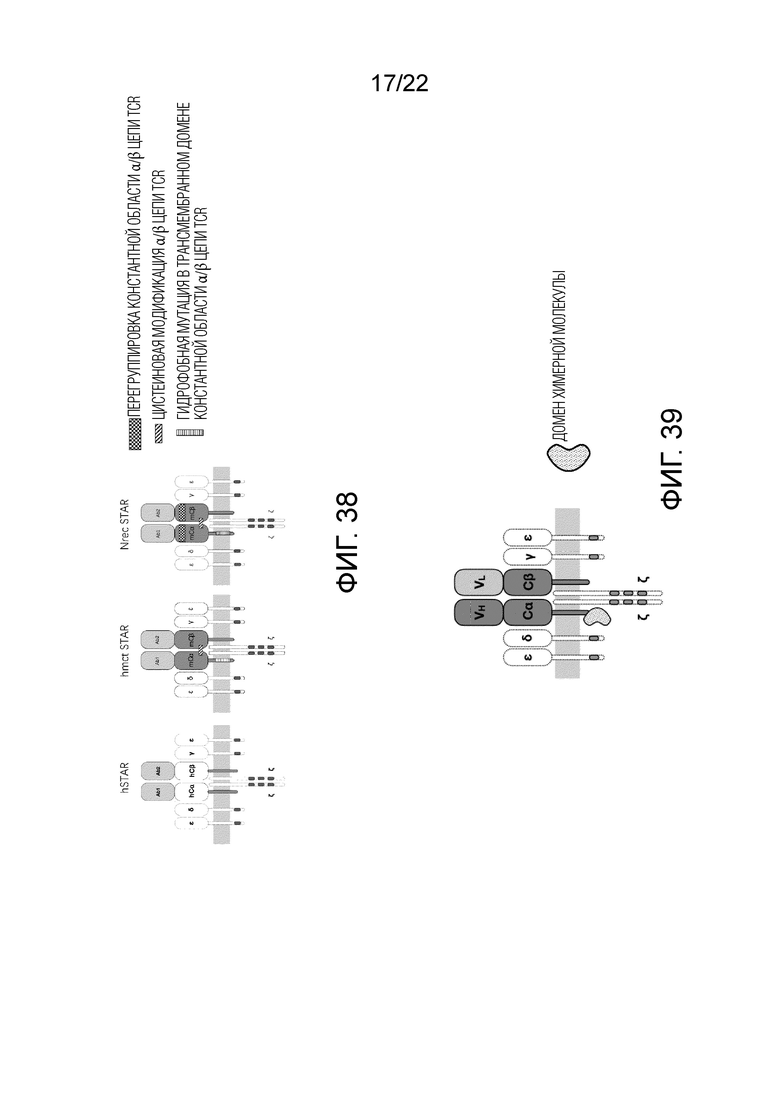
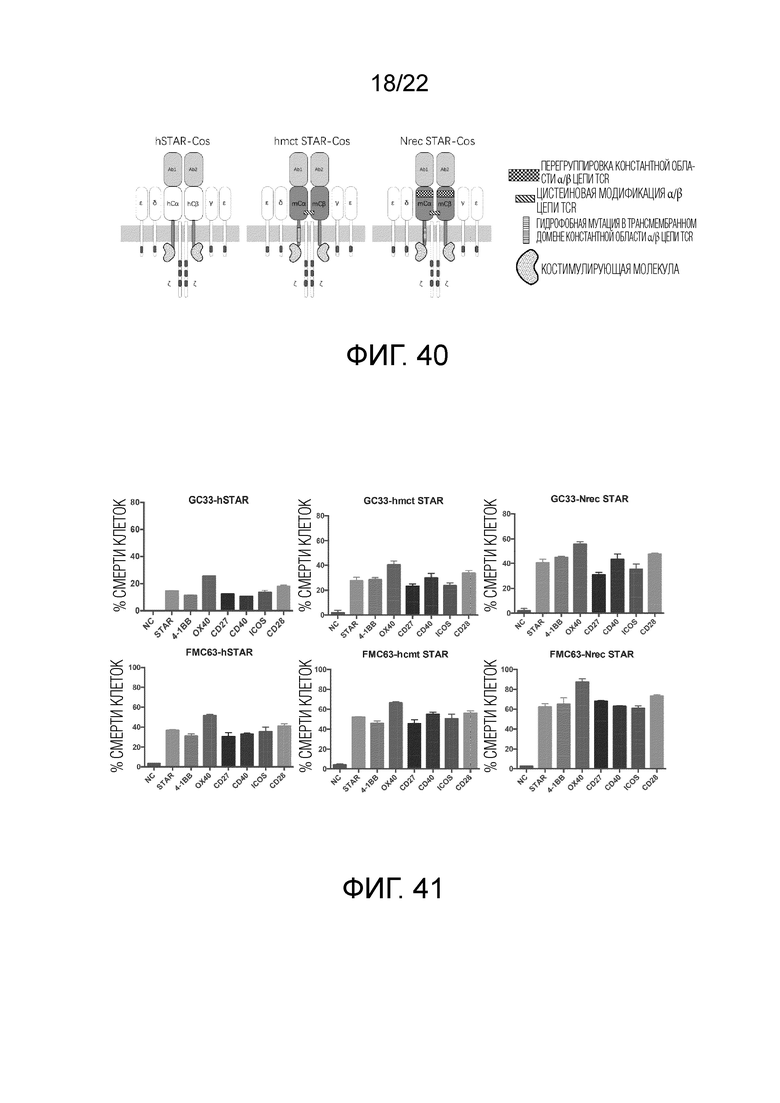
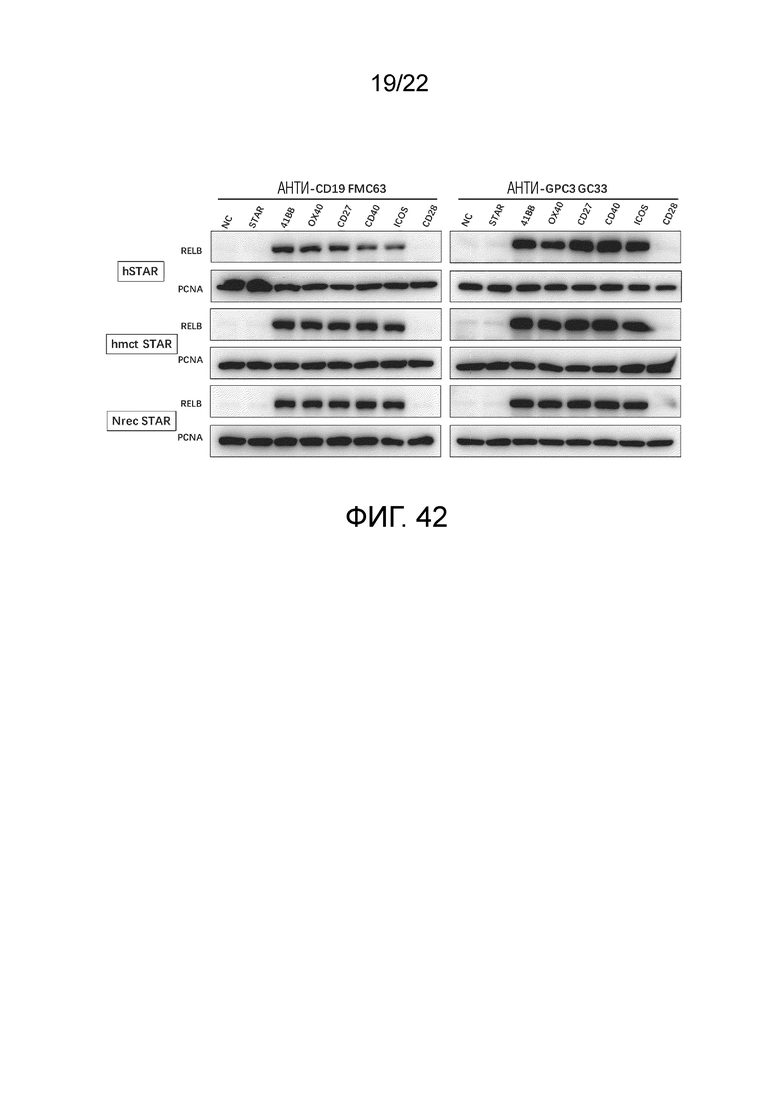
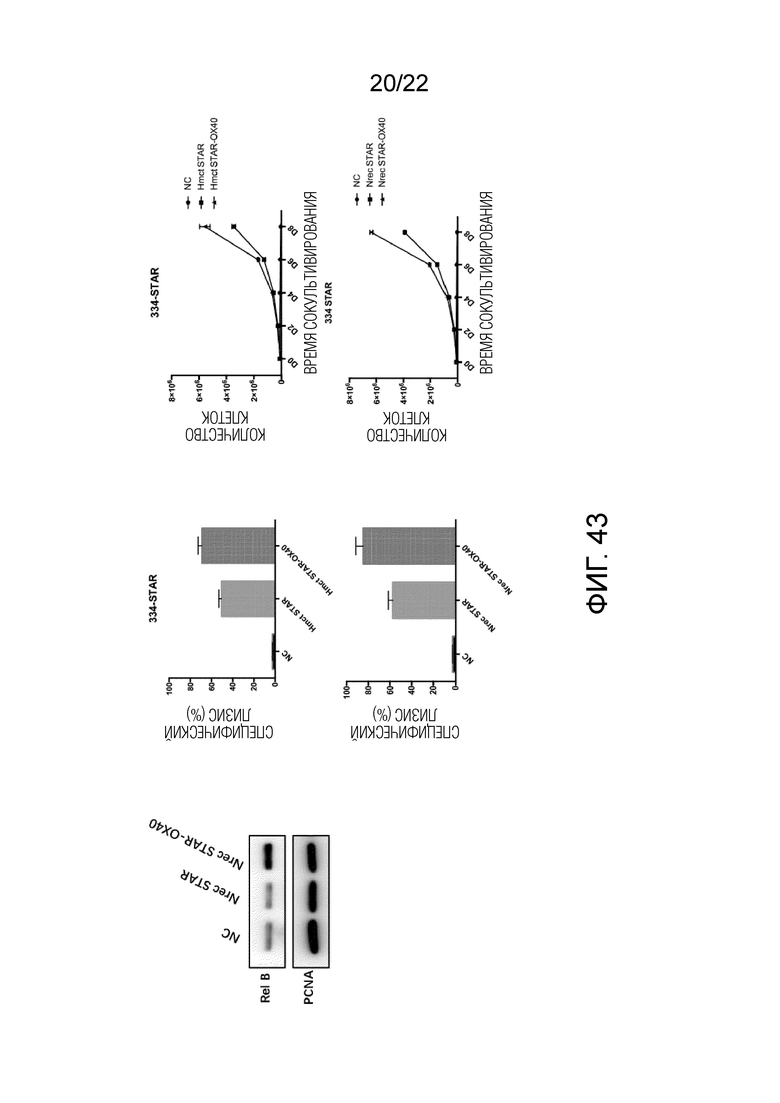
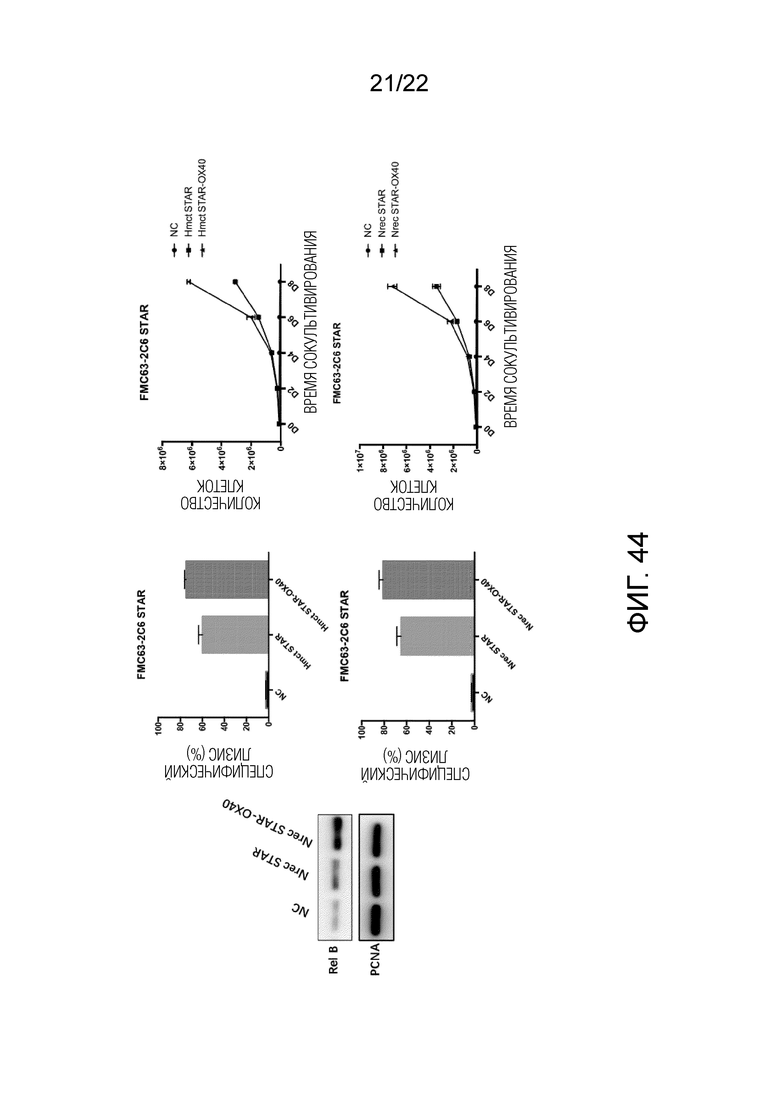
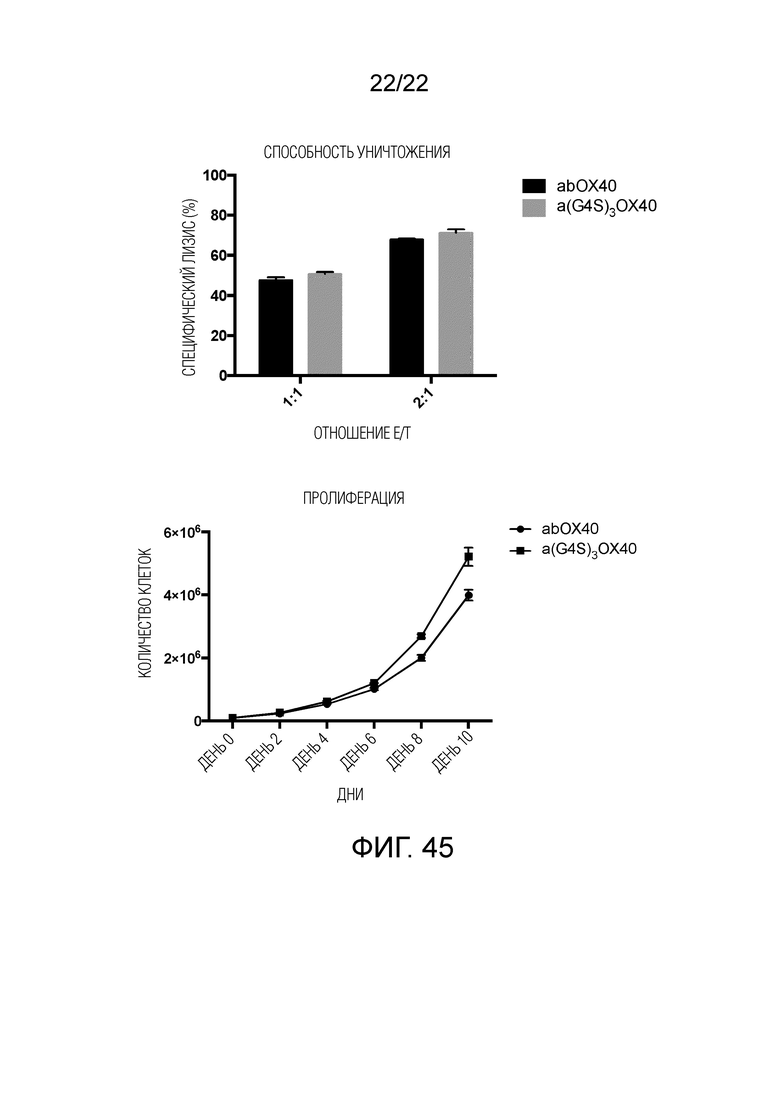
Изобретение относится к области биотехнологии, иммунологии и биомедицины, в частности к улучшенному синтетическому Т-клеточному рецептору и антигенному рецептору (STAR), таргетирующему CD19 и CD20, Т-клетке, содержащей синтетический Т-клеточный рецептор и антигенный рецептор, и их применению. Изобретение предлагает получение улучшенных TCR и соответствующей терапии TCR-T. 4 н. и 39 з.п. ф-лы, 45 ил., 28 табл., 2 пр.
1. Модифицированный Т-клеточный рецептор (TCR) для лечения CD19 и/или CD20-положительного рака у субъекта, где TCR представляет собой
αβ TCR, содержащий α цепь TCR и β цепь TCR, где, по меньшей мере, один функциональный домен соединен с С-концом α цепи TCR и/или β цепи TCR αβ TCR;
где α цепь TCR содержит первую константную область и первую антигенсвязывающую область, и β цепь TCR содержит вторую константную область и вторую антигенсвязывающую область;
где первая константная область представляет собой константную область α цепи природного TCR человека, представленную последовательностью SEQ ID NO:1 или константную область α цепи природного TCR мыши, представленную последовательностью SEQ ID NO:3, или где первая константная область представляет собой модифицированную константную область α цепи TCR, выделенную из константной области α цепи TCR мыши, представленную последовательностью SEQ ID NO:3, в которой аминокислота в положении 48, такая как треонин (T), мутирована на цистеин (C) по сравнению с константной областью α цепи TCR мыши дикого типа;
где вторая константная область представляет собой константную область β цепи природного TCR человека, представленную последовательностью SEQ ID NO:2, или константную область β цепи природного TCR мыши, представленную последовательностью SEQ ID NO:4, или где вторая константная область представляет собой модифицированную константную область β цепи TCR, выделенную из константной области β цепи TCR мыши, представленную последовательностью SEQ ID NO:4, в которой аминокислота в положении 56, такая как серин (S), мутирована на цистеин (C) по сравнению с константной областью β цепи TCR мыши дикого типа;
где первая антигенсвязывающая область специфически связывается с первым антигеном, и вторая антигенсвязывающая область специфически связывается со вторым антигеном; или первая антигенсвязывающая область и вторая антигенсвязывающая область объединены друг с другом для специфического связывания с первым антигеном и вторым антигеном; и
первый антиген представляет собой CD19, и второй антиген представляет собой CD20, или первый антиген представляет собой CD20, и второй антиген представляет собой CD19.
2. Модифицированный Т-клеточный рецептор по п. 1, отличающийся тем, что антигенсвязывающая область получена из антитела.
3. Модифицированный Т-клеточный рецептор по п. 1 или 2, отличающийся тем, что природный эндодомен α цепи TCR и/или β цепи TCR αβ-TCR удален.
4. Модифицированный Т-клеточный рецептор по п. 3, отличающийся тем, что в αβ TCR функциональный домен соединен непосредственно или через линкер с С-концом α цепи TCR и/или β цепи TCR, в которой природный эндодомен удален.
5. Модифицированный Т-клеточный рецептор по п. 4, отличающийся тем, что линкер представляет собой (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 3.
6. Модифицированный Т-клеточный рецептор по любому из пп. 1-5, отличающийся тем, что, по меньшей мере, один функциональный домен, такой как эндодомен костимулирующей молекулы, соединен с С-концом только одной из α цепи TCR и β цепи TCR αβ TCR.
7. Модифицированный Т-клеточный рецептор по п. 6, отличающийся тем, что, по меньшей мере, один функциональный домен соединен с С-концом α цепи TCR αβ TCR.
8. Модифицированный Т-клеточный рецептор по п. 7, отличающийся тем, что природный эндодомен α цепи TCR удален.
9. Модифицированный Т-клеточный рецептор по п. 8, отличающийся тем, что функциональный домен соединен непосредственно или через линкер с С-концом α цепи TCR, в которой природный эндодомен удален.
10. Модифицированный Т-клеточный рецептор по п. 9, отличающийся тем, что линкер представляет собой, например, (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 3.
11. Модифицированный Т-клеточный рецептор по п. 6, отличающийся тем, что, по меньшей мере, один функциональный домен соединен с С-концом β цепи TCR αβ TCR.
12. Модифицированный Т-клеточный рецептор по п. 11, отличающийся тем, что природный эндодомен β цепи TCR удален.
13. Модифицированный Т-клеточный рецептор по п. 12, отличающийся тем, что функциональный домен соединен непосредственно или через линкер с С-концом β цепи TCR, в которой природный эндодомен удален.
14. Модифицированный Т-клеточный рецептор по п. 13, отличающийся тем, что линкер представляет собой, например, (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 3.
15. Модифицированный Т-клеточный рецептор по любому из пп. 1-5, отличающийся тем, что, по меньшей мере, один функциональный домен соединен с соответствующими С-концами α цепи TCR и β цепи TCR αβ TCR.
16. Модифицированный Т-клеточный рецептор по п. 15, отличающийся тем, что природные эндодомены как α цепи TCR, так и β цепи TCR удалены.
17. Модифицированный Т-клеточный рецептор по п. 16, отличающийся тем, что функциональный домен соединен непосредственно или через линкер с С-концами цепи TCR α и цепи TCR β, в каждой из которых природный эндодомен удален.
18. Модифицированный Т-клеточный рецептор по п. 17, отличающийся тем, что линкер представляет собой, например, (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 3.
19. Модифицированный Т-клеточный рецептор по любому из пп. 1-5, отличающийся тем, что и α цепь TCR, и β цепь TCR αβ TCR связаны с одними и теми же или разными функциональными доменами.
20. Модифицированный Т-клеточный рецептор по любому из пп. 1-19, отличающийся тем, что 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более функциональных доменов соединены с С-концами α цепи TCR и/или β цепи TCR αβ TCR.
21. Модифицированный Т-клеточный рецептор по любому из пп. 1-20, отличающийся тем, что, по меньшей мере, один функциональный домен выбран из эндодомена костимулирующей молекулы, такой как CD40, OX40, ICOS, CD28, 4-1BB, CD27, и CD137, или эндодомена коингибирующей молекулы, такой как TIM3, PD1, CTLA4 и LAG3, или эндодомена рецептора цитокинов, такого как рецептор интерлейкина, рецептора интерферона, рецептора суперсемейства фактора некроза опухоли, рецептора колониестимулирующего фактора, хемокинового рецептора, рецептора фактора роста или других мембранных белков, или домена внутриклеточного белка, такого как NIK.
22. Модифицированный Т-клеточный рецептор по любому из пп. 1-21, отличающийся тем, что эндодомен костимулирующей молекулы представляет собой OX40 или ICOS, предпочтительно, OX40.
23. Модифицированный Т-клеточный рецептор по любому из пп. 1-22, отличающийся тем, что модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота в положении 112, такая как серин (S), заменена на лейцин (L), аминокислота в положении 114, такая как метионин (M), заменена на изолейцин I, и аминокислота в положении 115, такая как глицин (G), заменена на валин (V), по сравнению с константной областью α цепи TCR мыши дикого типа.
24. Модифицированный Т-клеточный рецептор по любому из пп. 1-22, отличающийся тем, что модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота, такая как E, в положении 6, заменена на D, K в положении 13 заменена на R, и аминокислоты в положениях 15-18 удалены по сравнению с константной областью α цепи TCR мыши дикого типа.
25. Модифицированный Т-клеточный рецептор по любому из пп. 1-22, отличающийся тем, что модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота в положении 48, такая как треонин (Т), заменена на цистеин (C), аминокислота в положении 112, такая как серин (S), заменена на лейцин (L), аминокислота в положении 114, такая как метионин (M), заменена на изолейцин (I), и аминокислота в положении 115, такая как глицин (G), заменена на валин (V) по сравнению с константной областью α цепи TCR мыши дикого типа.
26. Модифицированный Т-клеточный рецептор по любому из пп. 1-22, отличающийся тем, что модифицированная константная область α цепи TCR получена из константной области α цепи TCR мыши, в которой аминокислота, такая как E, в положении 6, заменена на D, K в положении 13 заменена на R, аминокислоты в положениях 15-18 удалены, аминокислота в положении 48, такая как треонин (T), заменена на цистеин (C), аминокислота в положении 112, такая как серин (S), заменена на лейцин (L), аминокислота в положении 114, такая как метионин (M), заменена на изолейцин (I), и аминокислота в положении 115, такая как глицин (G), заменена на валин (V) по сравнению с константной областью α цепи TCR мыши дикого типа.
27. Модифицированный Т-клеточный рецептор по любому из пп. 1-22, отличающийся тем, что первая константная область содержит нуклеотидную последовательность, показанную в одной из SEQ ID NO: 1, 3, 5, 7, 8, 26, 41, 42 и 48.
28. Модифицированный Т-клеточный рецептор по любому из пп. 1-27, отличающийся тем, что модифицированная константная область β цепи TCR получена из константной области β цепи TCR мыши, в которой аминокислота в положении 3, такая как R, заменена на K, аминокислота кислота в положении 6, такая как T, заменена на F, K в положении 9 заменена на E, S в положении 11 заменена на A, L в положении 12 заменена на V, и аминокислоты в положениях 17 и 21-25 удалены по сравнению с константной областью β цепи TCR мыши дикого типа.
29. Модифицированный Т-клеточный рецептор по любому из пп. 1-27, отличающийся тем, что модифицированная константная область β цепи TCR получена из константной области β цепи TCR мыши, в которой аминокислота в положении 56, такая как серин (S), заменена на цистеин (C), аминокислота в положении 3, такая как R, заменена на K, аминокислота в положении 6, такая как T, заменена на F, K в положении 9 заменена на E, S в положении 11 заменена на A, L в положении 12 заменена на V, и аминокислоты в положениях 17 и 21-25 удалены по сравнению с константной областью β цепи TCR мыши дикого типа.
30. Модифицированный Т-клеточный рецептор по любому из пп. 1-27, отличающийся тем, что модифицированный Т-клеточный рецептор содержит нуклеотидную последовательность, показанную в одной из SEQ ID NO: 2, 4, 6, 9, 27, 43 и 49.
31. Модифицированный Т-клеточный рецептор по любому из пп. 1-30, в котором антигенсвязывающая область получена из антитела.
32. Модифицированный Т-клеточный рецептор по любому из пп. 1-31, отличающийся тем, что антигенсвязывающая область содержит одноцепочечное антитело или однодоменное антитело,
например, одноцепочечное антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, связанные линкером, таким как (G4S)n, где n представляет собой целое число от 1 до 10, предпочтительно, n равно 1 или 3.
33. Модифицированный Т-клеточный рецептор по любому из пп. 1-32, отличающийся тем, что антигенсвязывающая область, специфически связывающаяся с CD19, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 44, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 45.
34. Модифицированный Т-клеточный рецептор по любому из пп. 1-32, отличающийся тем, что антигенсвязывающая область, специфически связывающаяся с CD19, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 46, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 47.
35. Модифицированный Т-клеточный рецептор по любому из пп. 1-32, отличающийся тем, что антигенсвязывающая область, специфически связывающаяся с CD19, содержит аминокислотную последовательность scFv, показанную в SEQ ID NO: 39.
36. Модифицированный Т-клеточный рецептор по любому из пп. 1-35, отличающийся тем, что антигенсвязывающая область, специфически связывающаяся с CD20, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 54, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 55.
37. Модифицированный Т-клеточный рецептор по любому из пп. 1-35, отличающийся тем, что антигенсвязывающая область, специфически связывающаяся с CD20, содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в SEQ ID NO: 56, и аминокислотную последовательность вариабельной области легкой цепи, показанную в SEQ ID NO: 57.
38. Модифицированный Т-клеточный рецептор по любому из пп. 1-35, отличающийся тем, что антигенсвязывающая область, специфически связывающаяся с CD20, содержит аминокислотную последовательность scFv, показанную в SEQ ID NO: 38.
39. Модифицированный Т-клеточный рецептор по п.1, отличающийся тем, что α цепь TCR содержит аминокислотную последовательность, показанную в SEQ ID NO: 59, β цепь TCR содержит аминокислотную последовательность, показанную в SEQ ID NO: 60.
40. Модифицированный Т-клеточный рецептор по любому из пп. 1-31, отличающийся тем, что
i) первая антигенсвязывающая область содержит вариабельную область тяжелой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область тяжелой цепи антитела, которая специфически связывается со вторым антигеном, тогда как вторая антигенсвязывающая область содержит вариабельную область легкой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область легкой цепи антитела, которая специфически связывается со вторым антигеном, так что первая антигенсвязывающая область и вторая антигенсвязывающая область объединяются друг с другом для специфического связывания и с первым антигеном и со вторым антигеном; или
ii) первая антигенсвязывающая область содержит вариабельную область тяжелой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область легкой цепи антитела, которая специфически связывается со вторым антигеном, тогда как вторая антигенсвязывающая область содержит вариабельную область легкой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область тяжелой цепи антитела, которая специфически связывается со вторым антигеном, так что первая антигенсвязывающая область и вторая антигенсвязывающая область объединяются друг с другом для специфического связывания и с первым антигеном и со вторым антигеном; или
iii) первая антигенсвязывающая область содержит вариабельную область легкой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область легкой цепи антитела, которая специфически связывается со вторым антигеном, тогда как вторая антигенсвязывающая область содержит вариабельную область тяжелой цепи антитела, которая специфически связывается с первым антигеном, и вариабельную область тяжелой цепи антитела, которая специфически связывается со вторым антигеном, так что первая антигенсвязывающая область и вторая антигенсвязывающая область объединяются друг с другом для специфического связывания и с первым антигеном и со вторым антигеном;
первый антиген представляет собой CD19, и второй антиген представляет собой CD20, или первый антиген представляет собой CD20, и второй антиген представляет собой CD19.
41. Выделенная терапевтическая иммунная клетка для лечения CD19 и/или CD20-положительного рака у субъекта, содержащая модифицированный Т-клеточный рецептор по любому из пп. 1-40, где терапевтическая иммунная клетка представляет собой Т-клетку или NK-клетку.
42. Фармацевтическая композиция для лечения CD19 и/или CD20-положительного рака у субъекта, содержащая терапевтическую иммунную клетку по п. 41 и фармацевтически приемлемый носитель.
43. Применение терапевтических иммунных клеток по п. 41 или фармацевтической композиции по п.42 в приготовлении лекарственного средства для лечения CD19 и/или CD20-положительного рака у субъекта.
| CN 110741016 A, 31.01.2018 | |||
| MARTYNISZYN A | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| ШАХТНАЯ ТОПКА ДЛЯ ДРОВ С НАКЛОННЫМИ И ГОРИЗОНТАЛЬНЫМИ КОЛОСНИКАМИ | 1919 |
|
SU1147A1 |
| MA Q | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

