ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка испрашивает приоритет на основании заявки на патент США №62/858,589, поданной 7 июня 2019 г., и заявки на патент США №62/916,666, поданной 17 октября 2019 г., каждая из которых включена в настоящий документ посредством ссылки в полном объеме для всех целей.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ,
ПОДАННЫЙ В ВИДЕ ТЕКСТОВОГО ФАЙЛА ПОСРЕДСТВОМ EFS WEB
[0002] Перечень последовательностей, записанный в файле 548157SEQLIST.txt, имеет объем 158 килобайт, был создан 27 мая 2020 г. и включен в настоящий документ посредством ссылки.
УРОВЕНЬ ТЕХНИКИ
[0003] Генная терапия - многообещающий терапевтический подход для лечения ряда заболеваний человека. Одним из подходов в генной терапии является вставка трансгена в локус «безопасной гавани» в геноме. Локусы «безопасной гавани» включают хромосомные локусы, в которых трансгены или другие экзогенные вставки нуклеиновых кислот могут стабильно и надежно экспрессироваться во всех представляющих интерес тканях без непосредственного изменения клеточного поведения или фенотипа. Часто локус «безопасной гавани» представляет собой локус, в котором экспрессия вставленной последовательности гена не нарушается какой-либо сквозной экспрессией соседних генов. Например, локусы «безопасной гавани» могут включать хромосомные локусы, в которых экзогенная ДНК может интегрироваться и функционировать предсказуемым образом без неблагоприятного воздействия на структуру или экспрессию эндогенного гена. Локусы «безопасной гавани» могут включать внегенные области или внутригенные области, такие как, например, локусы в генах, которые не относятся к незаменимым, являются необязательными или могут быть нарушены без явных фенотипических последствий.
[0004] Одним из примеров локуса «безопасной гавани» является альбумин. Тем не менее, остается потребность в подходящих животных, отличных от человека, обеспечивающих истинную или близко приближенную к истинной человеческую геномную ДНК-мишень для реагентов, нацеленных на человеческий альбумин, в эндогенном локусе альбумина in vivo, чтобы обеспечить возможность тестирования эффективности и принципа действия таких агентов на живых животных, а также проведения фармакокинетических и фармакодинамических исследований в условиях, когда гуманизированный ген является единственной доступной версией альбумина.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0005] Предложены животные, отличные от человека, содержащие гуманизированный локус альбумина (ALB), а также способы создания и применения таких животных, отличных от человека. Также предложены геномы или клетки животных, отличных от человека, содержащие гуманизированный локус альбумина (ALB). Также предложены гуманизированные гены альбумина.
[0006] В одном аспекте предложены геномы животных, отличных от человека, клетки животных, отличных от человека, или животные, отличные от человека, содержащие гуманизированный локус альбумина (ALB). Такие геномы животных, отличных от человека, клетки животных, отличных от человека, или животные, отличные от человека, могут содержать в своих геномах гуманизированный эндогенный локус альбумина, в котором сегмент эндогенного локуса альбумина был удален и заменен соответствующей последовательностью человеческого альбумина.
[0007] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина кодирует белок, содержащий пептид человеческого сывороточного альбумина. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина кодирует белок, содержащий пропептид человеческого альбумина. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина кодирует белок, содержащий сигнальный пептид человеческого альбумина.
[0008] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, область эндогенного локуса альбумина, содержащая как кодирующую последовательность, так и некодирующую последовательность, была удалена и заменена соответствующей последовательностью человеческого альбумина, содержащей как кодирующую последовательность, так и некодирующую последовательность. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина содержит эндогенный промотор альбумина, где последовательность человеческого альбумина функционально связана с эндогенным промотором альбумина. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, по меньшей мере один интрон и по меньшей мере один экзон эндогенного локуса альбумина был удален и заменен соответствующей последовательностью человеческого альбумина.
[0009] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, вся кодирующая последовательность альбумина эндогенного локуса альбумина была удалена и заменена соответствующей последовательностью человеческого альбумина. Необязательно, область эндогенного локуса альбумина от старт-кодона до стоп-кодона была удалена и заменена соответствующей последовательностью человеческого альбумина.
[0010] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина содержит 3’-нетранслируемую область человеческого альбумина. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, эндогенная 5’-нетранслируемая область альбумина не была удалена и заменена соответствующей последовательностью человеческого альбумина.
[0011] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, область эндогенного локуса альбумина от старт-кодона до стоп-кодона была удалена и заменена последовательностью человеческого альбумина, содержащей соответствующую последовательность человеческого альбумина и 3’-нетранслируемую область человеческого альбумина, и эндогенная 5’-нетранслируемая область альбумина не была удалена и заменена соответствующей последовательностью человеческого альбумина, и эндогенный промотор альбумина не был удален и заменен соответствующей последовательностью человеческого альбумина.
[0012] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, последовательность человеческого альбумина в гуманизированном эндогенном локусе альбумина содержит последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 35. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина кодирует белок, содержащий последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 5. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина содержит кодирующую последовательность, содержащую последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 13. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина содержит последовательность, по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную последовательности, приведенной в SEQ ID NO: 17 или 18. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, последовательность человеческого альбумина в гуманизированном эндогенном локусе альбумина содержит последовательность, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичную последовательности, приведенной в SEQ ID NO: 35. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина кодирует белок, содержащий последовательность, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичную последовательности, приведенной в SEQ ID NO: 5. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина содержит кодирующую последовательность, содержащую последовательность, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичную последовательности, приведенной в SEQ ID NO: 13. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина содержит последовательность, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичную последовательности, приведенной в SEQ ID NO: 17 или 18.
[0013] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, гуманизированный эндогенный локус альбумина не содержит кассету селекции или репортерный ген.
[0014] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, указанное животное, отличное от человека, является гомозиготным по гуманизированному эндогенному локусу альбумина. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, указанное животное, отличное от человека, содержит гуманизированный эндогенный локус альбумина в своей зародышевой линии.
[0015] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, указанное животное, отличное от человека, представляет собой млекопитающее. Необязательно, указанное животное, отличное от человека, представляет собой крысу или мышь. Необязательно, указанное животное, отличное от человека, представляет собой мышь.
[0016] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, уровни сывороточного альбумина у указанного животного, отличного от человека, составляют по меньшей мере приблизительно 10 мг/мл. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, уровни сывороточного альбумина у указанного животного, отличного от человека, по меньшей мере столь же высоки, как уровни сывороточного альбумина у контрольного животного, отличного от человека, содержащего локус альбумина дикого типа.
[0017] В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, указанные геном, клетка или животное являются гетерозиготными по гуманизированному эндогенному локусу альбумина. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, указанные геном, клетка или животное являются гомозиготными по гуманизированному эндогенному локусу альбумина. В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, указанные геном, клетка или животное дополнительно содержат кодирующую последовательность экзогенного белка, интегрированную в по меньшей мере один аллель гуманизированного эндогенного локуса альбумина в одной или более клетках указанного животного, отличного от человека. Необязательно, кодирующая последовательность экзогенного белка интегрирована в интрон 1 по меньшей мере одного аллеля гуманизированного эндогенного локуса альбумина (например, в одной или более клетках указанного животного, отличного от человека). В некоторых таких геномах животных, отличных от человека, клетках животных, отличных от человека, или животных, отличных от человека, указанные геном, клетка или животное дополнительно содержат инактивированный эндогенный локус, который не является эндогенным локусом альбумина. Необязательно, геном животного, отличного от человека, клетка животного, отличного от человека, или животное, отличное от человека, дополнительно содержит кодирующую последовательность экзогенного белка, интегрированную в по меньшей мере один аллель гуманизированного эндогенного локуса альбумина (например, в одной или более клетках указанного животного, отличного от человека), где указанный экзогенный белок заменяет функцию указанного инактивированного эндогенного локуса. Необязательно, указанный инактивированный эндогенный локус представляет собой инактивированный локус F9.
[0018] В еще одном аспекте предложены нацеливающие векторы для создания геномов животных, отличных от человека, клеток животных, отличных от человека, или животных, отличных от человека, описанных выше. Такие нацеливающие векторы могут быть предназначены для создания гуманизированного эндогенного локуса альбумина, в котором сегмент эндогенного локуса альбумина был удален и заменен соответствующей последовательностью человеческого альбумина, где нацеливающий вектор содержит вставку нуклеиновой кислоты, содержащую соответствующую последовательность человеческого альбумина, фланкированную 5’-гомологичным плечом, нацеленным на 5’-последовательность-мишень в эндогенном локусе альбумина, и 3’-гомологичным плечом, нацеленным на 3’-последовательность-мишень в эндогенном локусе альбумина.
[0019] В еще одном аспекте предложены способы оценки активности реагента, нацеленного на человеческий альбумин, in vivo. Некоторые такие способы включают: (a) введение указанного реагента, нацеленного на человеческий альбумин, животному, отличному от человека, описанному выше; и (b) оценку активности указанного реагента, нацеленного на человеческий альбумин, у указанного животного, отличного от человека.
[0020] В некоторых таких способах введение включает доставку, опосредованную аденоассоциированным вирусом (AAV), доставку, опосредованную липидными наночастицами (LNP), или гидродинамическую доставку (HDD). Необязательно, введение включает доставку, опосредованную LNP. Необязательно, доза LNP составляет от приблизительно 0,1 мг/кг до приблизительно 2 мг/кг. В некоторых таких способах введение включает опосредованную AAV8 доставку.
[0021] В некоторых таких способах стадия (b) включает выделение печени указанного животного, отличного от человека, и оценку активности указанного реагента, нацеленного на человеческий альбумин, в указанной печени.
[0022] В некоторых таких способах указанный реагент, нацеленный на человеческий альбумин, представляет собой агент для редактирования генома, и указанная оценка включает оценку модификации гуманизированного эндогенного локуса альбумина. Необязательно, указанная оценка включает измерение частоты вставок или делеций в гуманизированном эндогенном локусе альбумина.
[0023] В некоторых таких способах указанная оценка включает измерение экспрессии матричной РНК альбумина, кодируемой гуманизированным эндогенным локусом альбумина. В некоторых таких способах указанная оценка включает измерение экспрессии белка альбумина, кодируемого гуманизированным эндогенным локусом альбумина. Необязательно, оценка экспрессии белка альбумина включает измерение сывороточных уровней белка альбумина у указанного животного, отличного от человека. Необязательно, оценка экспрессии белка альбумина включает измерение экспрессии белка альбумина в печени указанного животного, отличного от человека.
[0024] В некоторых таких способах указанный реагент, нацеленный на человеческий альбумин, содержит нуклеазный агент, разработанный для нацеливания на область гена человеческого альбумина. В некоторых таких способах указанный реагент, нацеленный на человеческий альбумин, содержит нуклеазный агент или нуклеиновую кислоту, кодирующую указанный нуклеазный агент, где указанный нуклеазный агент разработан для нацеливания на область гена человеческого альбумина. Необязательно, указанный нуклеазный агент содержит белок Cas и гидовую РНК, разработанную для нацеливания на последовательность-мишень гидовой РНК в гене человеческого альбумина. Необязательно, указанная последовательность-мишень гидовой РНК находится в интроне 1 гена человеческого альбумина. Необязательно, белок Cas представляет собой белок Cas9.
[0025] В некоторых таких способах указанный реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту, которая разработана для нацеливания на ген человеческого альбумина, и, необязательно, где указанная экзогенная донорная нуклеиновая кислота доставляется посредством AAV. Необязательно, указанная экзогенная донорная нуклеиновая кислота представляет собой одноцепочечный олигодезоксинуклеотид (оцОДН). Необязательно, указанная экзогенная донорная нуклеиновая кислота способна вставляться в гуманизированный локус альбумина путем негомологичного соединения концов.
[0026] В некоторых способах указанная экзогенная донорная нуклеиновая кислота не содержит гомологичных плеч. В некоторых способах указанная экзогенная донорная нуклеиновая кислота содержит вставку нуклеиновой кислоты, фланкированную 5’-гомологичным плечом, нацеленным на 5’-последовательность-мишень в гуманизированном эндогенном локусе альбумина, и 3’-гомологичным плечом, нацеленным на 3’-последовательность-мишень в гуманизированном эндогенном локусе альбумина. Необязательно, каждая из 5’-последовательности-мишени и 3’-последовательности-мишени содержит сегмент интрона 1 гена человеческого альбумина.
[0027] В некоторых таких способах указанная экзогенная донорная нуклеиновая кислота кодирует экзогенный белок. Необязательно, указанный белок, кодируемый гуманизированным эндогенным локусом альбумина, на который осуществляли нацеливание с помощью указанной экзогенной донорной нуклеиновой кислоты, представляет собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с указанным экзогенным белком. Необязательно, указанный экзогенный белок представляет собой белок фактора IX. Необязательно, указанная оценка включает измерение сывороточных уровней белка фактора IX у указанного животного, отличного от человека, и/или включает оценку активированного частичного тромбопластинового времени или проведение теста генерации тромбина. Необязательно, указанное животное, отличное от человека, дополнительно содержит инактивированный локус F9, и указанная оценка включает измерение сывороточных уровней белка фактора IX у указанного животного, отличного от человека, и/или включает оценку активированного частичного тромбопластинового времени или проведение теста генерации тромбина. Необязательно, указанный реагент, нацеленный на человеческий альбумин, содержит (1) нуклеазный агент, разработанный для нацеливания на область гена человеческого альбумина, и (2) экзогенную донорную нуклеиновую кислоту, которая разработана для нацеливания на ген человеческого альбумина, где указанная экзогенная донорная нуклеиновая кислота кодирует экзогенный белок, и указанный белок, кодируемый гуманизированным эндогенным локусом альбумина, на который осуществляли нацеливание с помощью указанной экзогенной донорной нуклеиновой кислоты, представляет собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с указанным экзогенным белком. Необязательно, указанная оценка включает измерение экспрессии матричной РНК, кодируемой указанной экзогенной донорной нуклеиновой кислотой. Необязательно, указанная оценка включает измерение экспрессии указанного экзогенного белка. Необязательно, оценка экспрессии указанного гетерологичного белка включает измерение сывороточных уровней указанного гетерологичного белка у указанного животного, отличного от человека. Необязательно, оценка экспрессии указанного гетерологичного белка включает измерение экспрессии в печени указанного животного, отличного от человека.
[0028] В еще одном аспекте предложены способы оптимизации активности реагента, нацеленного на человеческий альбумин, in vivo. Некоторые такие способы включают: (I) осуществление любого из вышеуказанных способов оценки активности реагента, нацеленного на человеческий альбумин, in vivo первый раз у первого животного, отличного от человека, содержащего в своем геноме гуманизированный эндогенный локус альбумина; (II) изменение переменной и осуществление способа со стадии (I) второй раз с измененной переменной у второго животного, отличного от человека, содержащего в своем геноме гуманизированный эндогенный локус альбумина; и (III) сравнение активности указанного реагента, нацеленного на человеческий альбумин, на стадии (I) с активностью указанного реагента, нацеленного на человеческий альбумин, на стадии (II), и выбор способа, обеспечивающего более высокую активность.
[0029] В некоторых таких способах указанная изменяемая переменная на стадии (II) представляет собой способ доставки для введения указанного реагента, нацеленного на человеческий альбумин, указанному животному, отличному от человека. Необязательно, указанное введение включает опосредованную LNP доставку, и указанная изменяемая переменная на стадии (II) представляет собой препарат LNP. В некоторых таких способах указанная изменяемая переменная на стадии (II) представляет собой путь введения указанного реагента, нацеленного на человеческий альбумин, указанному животному, отличному от человека. В некоторых таких способах указанная изменяемая переменная на стадии (II) представляет собой концентрацию или количество указанного реагента, нацеленного на человеческий альбумин, введенного указанному животному, отличному от человека. В некоторых таких способах указанная изменяемая переменная на стадии (II) представляет собой форму указанного реагента, нацеленного на человеческий альбумин, введенного указанному животному, отличному от человека. В некоторых таких способах указанная изменяемая переменная на стадии (II) представляет собой указанный реагент, нацеленный на человеческий альбумин, введенный указанному животному, отличному от человека.
[0030] В некоторых таких способах указанный реагент, нацеленный на человеческий альбумин, содержит белок Cas и гидовую РНК, разработанную для нацеливания на последовательность-мишень гидовой РНК в гене человеческого альбумина. В некоторых таких способах указанный реагент, нацеленный на человеческий альбумин, содержит белок Cas или нуклеиновую кислоту, кодирующую белок Cas, и гидовую РНК или ДНК, кодирующую указанную гидовую РНК, где указанная гидовая РНК разработана для нацеливания на последовательность-мишень гидовой РНК в гене человеческого альбумина. Необязательно, указанная изменяемая переменная на стадии (II) представляет собой последовательность указанной гидовой РНК или указанную последовательность-мишень гидовой РНК. Необязательно, и указанный белок Cas, и указанную гидовую РНК вводят в форме РНК, и указанная изменяемая переменная на стадии (II) представляет собой отношение мРНК Cas к гидовой РНК. Необязательно, указанная изменяемая переменная на стадии (II) представляет собой модификации гидовой РНК. Необязательно, указанный реагент, нацеленный на человеческий альбумин, содержит матричную РНК (мРНК), кодирующую указанный белок Cas, и указанную гидовую РНК, и указанная изменяемая переменная на стадии (II) представляет собой отношение мРНК Cas к гидовой РНК.
[0031] В некоторых таких способах указанный реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту. Необязательно, указанная изменяемая переменная на стадии (II) представляет собой форму указанной экзогенной донорной нуклеиновой кислоты. Необязательно, указанная экзогенная донорная нуклеиновая кислота содержит вставку нуклеиновой кислоты, фланкированную 5’-гомологичным плечом, нацеленным на 5’-последовательность-мишень в гуманизированном эндогенном локусе альбумина, и 3’-гомологичным плечом, нацеленным на 3’-последовательность-мишень в гуманизированном эндогенном локусе альбумина, и указанная изменяемая переменная на стадии (II) представляет собой последовательность или длину указанного 5’-гомологичного плеча и/или последовательность или длину указанного 3’-гомологичного плеча.
[0032] В еще одном аспекте предложены способы создания любых из вышеуказанных животных, отличных от человека. Некоторые такие способы включают: (a) введение в эмбриональную стволовую (ЭС) клетку животного, отличного от человека: (i) нуклеазного агента, нацеленного на последовательность-мишень в эндогенном локусе альбумина; и (ii) нацеливающего вектора, содержащего вставку нуклеиновой кислоты, содержащую последовательность человеческого альбумина, фланкированную 5’-гомологичным плечом, соответствующим 5’-последовательности-мишени в эндогенном локусе альбумина, и 3’-гомологичным плечом, соответствующим 3’-последовательности-мишени в эндогенном локусе альбумина, где указанный нацеливающий вектор рекомбинирует с эндогенным локусом альбумина с получением генетически модифицированной ЭС клетки, отличной от человеческой, содержащей в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина; (b) введение указанной генетически модифицированной ЭС клетки, отличной от человеческой, в эмбрион животного-хозяина, отличного от человека; и (c) вынашивание указанного эмбриона животного-хозяина, отличного от человека, у суррогатной матери, где указанная суррогатная мать производит потомство F0 генетически модифицированного животного, отличного от человека, содержащего в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина. В еще одном аспекте предложены способы создания любых из вышеуказанных животных, отличных от человека. Некоторые такие способы включают: (a) введение в эмбриональную стволовую (ЭС) клетку животного, отличного от человека: (i) нуклеазного агента или нуклеиновой кислоты, кодирующей указанный нуклеазный агент, где указанный нуклеазный агент нацелен на последовательность-мишень в эндогенном локусе альбумина; и (ii) нацеливающего вектора, содержащего вставку нуклеиновой кислоты, содержащую последовательность человеческого альбумина, фланкированную 5’-гомологичным плечом, соответствующим 5’-последовательности-мишени в эндогенном локусе альбумина, и 3’-гомологичным плечом, соответствующим 3’-последовательности-мишени в эндогенном локусе альбумина, где указанный нацеливающий вектор рекомбинирует с эндогенным локусом альбумина с получением генетически модифицированной ЭС клетки, отличной от человеческой, содержащей в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина; (b) введение указанной генетически модифицированной ЭС клетки, отличной от человеческой, в эмбрион животного-хозяина, отличного от человека; и (c) вынашивание указанного эмбриона животного-хозяина, отличного от человека, у суррогатной матери, где указанная суррогатная мать производит потомство F0 генетически модифицированного животного, отличного от человека, содержащего в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина. Необязательно, указанный нацеливающий вектор представляет собой большой нацеливающий вектор длиной по меньшей мере 10 кб, или в котором указанные 5’- и 3’-гомологичные плечи в совокупности составляют по меньшей мере 10 кб.
[0033] Некоторые такие способы включают: (a) введение в находящийся на одноклеточной стадии эмбрион животного, отличного от человека: (i) нуклеазного агента, нацеленного на последовательность-мишень в эндогенном локусе альбумина; и (ii) нацеливающего вектора, содержащего вставку нуклеиновой кислоты, содержащую последовательность человеческого альбумина, фланкированную 5’-гомологичным плечом, соответствующим 5’-последовательности-мишени в эндогенном локусе альбумина, и 3’-гомологичным плечом, соответствующим 3’-последовательности-мишени в эндогенном локусе альбумина, где указанный нацеливающий вектор рекомбинирует с эндогенным локусом альбумина с получением генетически модифицированного находящегося на одноклеточной стадии эмбриона, отличного от человеческого, содержащего в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина; (b) вынашивание указанного генетически модифицированного находящегося на одноклеточной стадии эмбриона животного, отличного от человека, у суррогатной матери с получением поколения F0 генетически модифицированного животного, отличного от человека, содержащего в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина. Некоторые такие способы включают: (a) введение в находящийся на одноклеточной стадии эмбрион животного, отличного от человека: (i) нуклеазного агента или нуклеиновой кислоты, кодирующей указанный нуклеазный агент, где указанный нуклеазный агент нацелен на последовательность-мишень в эндогенном локусе альбумина; и (ii) нацеливающего вектора, содержащего вставку нуклеиновой кислоты, содержащую последовательность человеческого альбумина, фланкированную 5’-гомологичным плечом, соответствующим 5’-последовательности-мишени в эндогенном локусе альбумина, и 3’-гомологичным плечом, соответствующим 3’-последовательности-мишени в эндогенном локусе альбумина, где указанный нацеливающий вектор рекомбинирует с эндогенным локусом альбумина с получением генетически модифицированного находящегося на одноклеточной стадии эмбриона, отличного от человеческого, содержащего в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина; (b) вынашивание указанного генетически модифицированного находящегося на одноклеточной стадии эмбриона животного, отличного от человека, у суррогатной матери с получением поколения F0 генетически модифицированного животного, отличного от человека, содержащего в своем геноме гуманизированный эндогенный локус альбумина, содержащий последовательность человеческого альбумина.
[0034] В некоторых таких способах указанный нуклеазный агент содержит белок Cas и гидовую РНК. Необязательно, белок Cas представляет собой белок Cas9. Необязательно, стадия (a) дополнительно включает введение второй гидовой РНК, нацеленной на вторую последовательность-мишень в эндогенном локусе альбумина.
[0035] В некоторых таких способах указанное животное, отличное от человека, представляет собой мышь или крысу. Необязательно, указанное животное, отличное от человека, представляет собой мышь.
[0036] В еще одном аспекте предложены способы создания любых из вышеуказанных животных, отличных от человека. Некоторые такие способы включают: (a) модификацию генома плюрипотентной клетки животного, отличного от человека, для включения гуманизированного эндогенного локуса альбумина; (b) идентификацию или отбор генетически модифицированной плюрипотентной клетки животного, отличного от человека, содержащей гуманизированный эндогенный локус альбумина; (c) введение генетически модифицированной плюрипотентной клетки животного, отличного от человека, в эмбрион животного-хозяина, отличного от человека; и (d) вынашивание указанного эмбриона животного-хозяина, отличного от человека, у суррогатной матери. Некоторые такие способы включают: (a) модификацию генома находящегося на одноклеточной стадии эмбриона животного, отличного от человека, для включения гуманизированного эндогенного локуса альбумина; (b) отбор генетически модифицированного находящегося на одноклеточной стадии эмбриона животного, отличного от человека, содержащего гуманизированный эндогенный локус альбумина; и (c) вынашивание указанного генетически модифицированного находящегося на одноклеточной стадии эмбриона животного, отличного от человека, у суррогатной матери.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0037] На фиг. 1A (не в масштабе) показано схематическое изображение гуманизированного мышиного локуса альбумина (Alb) с кассетой селекции неомицином (MAID 7626). Последовательности соединений A, B и C приведены в SEQ ID NO: 19-21 соответственно.
[0038] На фиг. 1B (не в масштабе) показано схематическое изображение гуманизированного мышиного локуса альбумина (Alb) после удаления кассеты селекции неомицином (MAID 7627). Последовательности соединений A и D приведены в SEQ ID NO: 19 и 22 соответственно.
[0039] На фиг. 2 (не в масштабе) показано расположение зондов TAQMAN® для скрининговой гуманизации мышиного локуса альбумина (Alb). Зонды на приобретение аллеля (GOA) включают 7626hU и 7626hD. Зонды на потерю аллеля (LOA) включают 7626mTU и 7626mTD.
[0040] На фиг. 3A и 3B показано выравнивание белков мышиного (мышиный Alb), человеческого (человеческий ALB) и гуманизированного (7626 HumIn Prot) альбумина. Остатки, обведенные рамкой, составляют сигнальный пептид. Пунктирные линии обозначают пептидную последовательность сывороточного альбумина. Жирная сплошная линия обозначает пропептидную последовательность. Все остатки в белке гуманизированного альбумина кодируются введенными человеческими экзонами.
[0041] На фиг. 4 показаны уровни человеческого альбумина в образцах плазмы мышей с гуманизированным альбумином (ALBhu/hu) и мышей дикого типа (WT). В качестве положительного контроля использовали объединенную нормальную человеческую плазму (George King-Biomedical Inc.). В качестве отрицательного контроля использовали мышей VelocImmune (VI).
[0042] На фиг. 5 показаны уровни мышиного альбумина в образцах плазмы мышей с гуманизированным альбумином (ALBhu/hu) и мышей дикого типа (WT). В качестве отрицательного контроля использовали объединенную нормальную человеческую плазму (George King-Biomedical Inc.). Мышей VI использовали в качестве положительного контроля.
[0043] На фиг. 6A и 6B показаны уровни человеческого фактора IX в плазме в результате вставки AAV-hF9 у мышей с гуманизированным альбумином.
[0044] На фиг. 7 показаны уровни человеческого фактора IX в плазме на 7 неделе после инъекции донора AAV-hF9 и LNP-CRISPR/Cas9, нанесенные на график в виде зависимости от процента клеток, положительных на мРНК hALB-hFIX, как определено с помощью BASESCOPE™.
[0045] На фиг. 8 показаны уровни человеческого фактора IX в плазме в результате вставки AAV-hF9 у мышей ALBm/hu x F9-/-.
[0046] На фиг. 9 показаны эффекты АЧТВ в образцах человеческой и мышиной плазмы в результате вставки AAV-hF9 у мышей ALBm/hu x F9-/-.
[0047] На фиг. 10A и 10B показаны профили ТГТ-EA. На фиг. 10A показан профиль ТГТ-EA образцов человеческой нормальной плазмы и образцов человеческой плазмы с дефицитом фактора IX. На фиг. 10B показан профиль ТГТ-EA мышиной плазмы в результате вставки AAV-hF9 у мышей ALBm/hu x F9-/-.
[0048] На фиг. 11 показано образование тромбина в образцах мышиной плазмы в результате вставки AAV-hF9 у мышей ALBm/hu x F9-/-.
ОПРЕДЕЛЕНИЯ
[0049] Термины «белок», «полипептид» и «пептид», используемые в контексте настоящего документа взаимозаменяемо, включают полимерные формы аминокислот любой длины, включая кодируемые и некодируемые аминокислоты, а также химически или биохимически модифицированные или дериватизированные аминокислоты. Данные термины также включают модифицированные полимеры, такие как полипептиды, имеющие модифицированные пептидные остовы. Термин «домен» относится к любой части белка или полипептида, имеющей конкретную функцию или структуру.
[0050] Термины «нуклеиновая кислота» и «полинуклеотид», используемые в контексте настоящего документа взаимозаменяемо, включают полимерные формы нуклеотидов любой длины, включая рибонуклеотиды, дезоксирибонуклеотиды или их аналоги или модифицированные версии. Они включают одно-, двух- и многоцепочечную ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие встречающиеся в природе, химически модифицированные, биохимически модифицированные, не встречающиеся в природе или дериватизированные нуклеотидные основания.
[0051] Термин «интегрированная в геном» относится к нуклеиновой кислоте, которая была введена в клетку таким образом, что нуклеотидная последовательность интегрировалась в геном клетки. Для стабильного включения нуклеиновой кислоты в геном клетки может быть использован любой протокол.
[0052] Термин «нацеливающий вектор» относится к рекомбинантной нуклеиновой кислоте, которая может быть введена в целевое положение в геноме клетки путем гомологичной рекомбинации, лигирования, опосредованного негомологичным соединением концов, или любым другим способом рекомбинации.
[0053] Термин «вирусный вектор» относится к рекомбинантной нуклеиновой кислоте, которая включает по меньшей мере один элемент вирусного происхождения и включает элементы, достаточные для упаковки в частицу вирусного вектора или допускающие указанную упаковку. Вектор и/или частица могут быть использованы для переноса ДНК, РНК или других нуклеиновых кислот в клетки in vitro, ex vivo или in vivo. Известно множество форм вирусных векторов.
[0054] Термин «выделенный» применительно к клетке, ткани (например, образцам печени), белкам и нуклеиновым кислотам включает клетки, ткани (например, образцы печени), белки и нуклеиновые кислоты, которые относительно очищены по сравнению с другими бактериальными, вирусными, клеточными или другими компонентами, которые обычно могут присутствовать in situ, вплоть до по существу чистого препарата клеток, тканей (например, образцов печени), белков и нуклеиновых кислот, и включая такой препарат. Термин «выделенный» также включает: клетки, ткани (например, образцы печени), белки и нуклеиновые кислоты, не имеющие аналогов в природе, были химически синтезированы и, таким образом, по существу не загрязнены другими клетками, тканями (например, образцами печени), белками и нуклеиновыми кислотами, или были отделены или очищены от большинства других компонентов (например, клеточных компонентов), с которыми они естественным образом связаны (например, другими клеточными белками, полинуклеотидами или клеточными компонентами).
[0055] Термин «дикий тип» включает объекты, имеющие структуру и/или активность, которые наблюдаются в нормальном (в отличие от мутантного, болезненного, измененного и т.д.) состоянии или контексте. Гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллелях).
[0056] Термин «эндогенная последовательность» относится к последовательности нуклеиновой кислоты, которая встречается в природе внутри клетки или животного, отличного от человека. Например, эндогенная последовательность альбумина животного, отличного от человека, относится к нативной последовательности альбумина, которая естественным образом встречается в локусе альбумина у указанного животного, отличного от человека.
[0057] «Экзогенные» молекулы или последовательности включают молекулы или последовательности, которые обычно не присутствуют в клетке в этой форме или локализации (например, в геномном локусе). Нормальное присутствие включает наличие в зависимости от конкретной стадии развития и условий окружающей среды клетки. Экзогенная молекула или последовательность, например, может включать мутированную версию соответствующей эндогенной последовательности внутри клетки, такую как гуманизированная версия эндогенной последовательности, или может включать последовательность, соответствующую эндогенной последовательности внутри клетки, но в другой форме (т.е. не в хромосоме). Напротив, эндогенные молекулы или последовательности включают молекулы или последовательности, которые обычно присутствуют в этой форме и локализации в определенной клетке на определенной стадии развития в определенных условиях окружающей среды.
[0058] Термин «гетерологичный» при использовании в контексте нуклеиновой кислоты или белка указывает на то, что нуклеиновая кислота или белок содержит по меньшей мере два сегмента, которые в природе не встречаются вместе в одной и той же молекуле. Например, термин «гетерологичный», когда он используется применительно к сегментам нуклеиновой кислоты или сегментам белка, означает, что данная нуклеиновая кислота или данный белок содержит две или более субпоследовательностей, которые не находятся в такой взаимосвязи друг с другом (например, соединенными вместе) в природе. В качестве одного примера, «гетерологичная» область нуклеиново-кислотного вектора представляет собой сегмент нуклеиновой кислоты внутри другой молекулы нуклеиновой кислоты или присоединенный к другой молекуле нуклеиновой кислоты, который не находится в ассоциации с указанной другой молекулой в природе. Например, гетерологичная область нуклеиново-кислотного вектора может включать кодирующую последовательность, фланкированную последовательностями, не находящимися в ассоциации с указанной кодирующей последовательностью в природе. Аналогичным образом, «гетерологичная» область белка представляет собой сегмент из аминокислот внутри другой молекулы пептида или присоединенный к другой молекуле пептида, который не находится в ассоциации с указанной другой молекулой пептида в природе (например, слитый белок или белок с меткой). Аналогичным образом, нуклеиновая кислота или белок могут содержать гетерологичную метку или гетерологичную последовательность секреции или локализации.
[0059] При «оптимизации кодонов» используется свойство вырожденности кодонов, что демонстрируется множеством комбинаций кодонов из трех пар оснований, которые определяют аминокислоту, и, как правило, она включает процесс модификации последовательности нуклеиновой кислоты для усиления экспрессии в конкретных клетках-хозяевах путем замены по меньшей мере одного кодона нативной последовательности на кодон, который чаще или наиболее часто используется в генах клетки-хозяина, при сохранении нативной аминокислотной последовательности. Например, нуклеиновая кислота, кодирующая белок Cas9, может быть модифицирована для замены кодонов, имеющих более высокую частоту использования в данной прокариотической или эукариотической клетке, включая бактериальную клетку, дрожжевую клетку, человеческую клетку, клетку, отличную от человеческой, клетку млекопитающего, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина, по сравнению с встречающейся в природе последовательностью нуклеиновой кислоты. Таблицы частот использования кодонов легко доступны, например, в базе данных «Codon Usage Database». Эти таблицы можно адаптировать разными способами. См. источник Nakamura et al. (2000) Nucleic Acids Research 28:292, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Также доступны компьютерные алгоритмы для оптимизации кодонов конкретной последовательности для экспрессии в конкретном хозяине (см., например, Gene Forge).
[0060] Термин «локус» относится к конкретному местоположению гена (или значимой последовательности), последовательности ДНК, последовательности, кодирующей полипептид, или положению на хромосоме генома организма. Например, «локус альбумина» или «локус Alb» может относиться к конкретному местоположению гена альбумина (Alb), последовательности ДНК альбумина, кодирующей альбумин последовательности, или положению альбумина на хромосоме генома организма, которое было идентифицировано как место, где находится такая последовательность. «Локус альбумина» может содержать регуляторный элемент гена альбумина, включая, например, энхансер, промотор, 5’- и/или 3’-нетранслируемую область (UTR) или их комбинацию.
[0061] Термин «ген» относится к последовательности ДНК в хромосоме, которая кодирует продукт (например, РНК-продукт и (или) продукт-полипептид) и включает в себя кодирующую область, прерванную некодирующими интронами, и последовательность, расположенную смежно с указанной кодирующей областью как на 5’-конце, так и на 3’-конце, таким образом, что ген соответствует полноразмерной мРНК (включая 5’- и 3’-нетранслируемые последовательности). Термин «ген» также включает в себя другие некодирующие последовательности, включая регуляторные последовательности (например, промоторы, энхансеры и сайты связывания факторов транскрипции), сигналы полиаденилирования, участок внутренней посадки рибосомы, сайленсеры, изолирующие последовательности и области прикрепления к матриксу. Эти последовательности могут быть расположены вблизи кодирующей области гена (например, в пределах 10 кб) или в отдаленных сайтах, и они влияют на уровень или скорость транскрипции и трансляции данного гена.
[0062] Термин «аллель» относится к вариантной форме гена. Некоторые гены имеют множество различных форм, которые расположены в одном и том же месте, или генетическом локусе, на хромосоме. Диплоидный организм имеет два аллеля в каждом генетическом локусе. Каждая пара аллелей отражает генотип конкретного генетического локуса. Генотипы описываются как гомозиготные, если в определенном локусе находятся два идентичных аллеля, и как гетерозиготные, если эти два аллеля различаются.
[0063] «Промотор» представляет собой регуляторную область ДНК, обычно содержащую ТАТА-бокс, способную управлять РНК-полимеразой II, чтобы инициировать синтез РНК в соответствующем сайте инициации транскрипции для конкретной полинуклеотидной последовательности. Промотор может дополнительно содержать другие области, которые влияют на скорость инициации транскрипции. Раскрытые в настоящем документе промоторные последовательности модулируют транскрипцию функционально связанного полинуклеотида. Промотор может быть активным в одном или более типах клеток, раскрытых в настоящем документе (например, эукариотической клетке, клетке млекопитающего, отличного от человека, человеческой клетке, клетке грызуна, плюрипотентной клетке, находящемся на одноклеточной стадии эмбрионе, дифференцированной клетке или их комбинации). Промотор может представлять собой, например, конститутивно активный промотор, условный промотор, индуцибельный промотор, временно ограниченный промотор (например, промотор, регулируемый развитием) или пространственно ограниченный промотор (например, клеточноспецифический или тканеспецифический промотор). Примеры промоторов можно найти, например, в источнике WO 2013/176772, включенном в настоящий документ посредством ссылки в полном объеме для всех целей.
[0064] Примеры индуцибельных промоторов включают, например, химически регулируемые промоторы и физически регулируемые промоторы. Химически регулируемые промоторы включают, например, алкогольрегулируемые промоторы (например, промотор гена алкогольдегидрогеназы (alcA)), тетрациклин-регулируемые промоторы (например, тетрациклин-чувствительный промотор, последовательность оператора тетрациклина (tetO), промотор tet-On или промотор tet-Off), регулируемые стероидами промоторы (например, промотор глюкокортикоидного рецептора крысы, промотор рецептора эстрогена или промотор рецептора экдизона), или регулируемые металлами промоторы (например, промотор металлопротеина). Физически регулируемые промоторы включают, например, терморегулируемые промоторы (например, промотор теплового шока) и светорегулируемые промоторы (например, индуцируемый светом промотор или репрессируемый светом промотор).
[0065] Тканеспецифические промоторы могут представлять собой, например, нейрон-специфические промоторы, специфичные для глиальных клеток промоторы, специфичные для мышечных клеток промоторы, специфичные для клеток сердца промоторы, специфичные для клеток почек промоторы, специфичные для костных клеток промоторы, специфичные для эндотелиальных клеток промоторы или специфичные для иммунных клеток промоторы (например, промотор В-клеток или промотор Т-клеток).
[0066] Промоторы, регулируемые развитием, включают, например, промоторы, активные только на эмбриональной стадии развития или только во взрослой клетке.
[0067] Термин «функциональная связь» или «функционально связанный» включает в себя контактное расположение двух или более компонентов (например, промотора и другого элемента последовательности) таким образом, что оба компонента функционируют нормально, и допускается возможность того, что по меньшей мере один из компонентов может опосредовать функцию, которая воздействует на по меньшей мере один другой из компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более факторов регуляции транскрипции. Функциональная связь может включать в себя случаи, когда такие последовательности расположены смежно друг с другом или оказывают действие в транс-положении (например, регуляторная последовательность может оказывать действие на расстоянии, контролируя транскрипцию кодирующей последовательности).
[0068] «Комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной цепи нуклеиновой кислоты из-за ориентации ее групп азотистых оснований образует водородные связи с другой последовательностью на противоположной цепи нуклеиновой кислоты. Комплементарные основания в ДНК обычно представляют собой A с T и C с G. В РНК они обычно представляют собой C с G и U с A. Комплементарность может быть абсолютной или существенной/достаточной. Абсолютная комплементарность между двумя нуклеиновыми кислотами означает, что эти две нуклеиновые кислоты могут образовывать дуплекс, в котором каждое основание связано с комплементарным основанием путем спаривания по Уотсону-Крику. «Существенная» или «достаточная» комплементарность означает, что последовательность в одной цепи не полностью и/или не абсолютно комплементарна последовательности в противоположной цепи, но что между основаниями на двух цепях имеет место достаточное связывание для образования стабильного гибридного комплекса в наборе условий гибридизации (например, концентрации соли и температура). Такие условия можно спрогнозировать с помощью последовательностей и стандартных математических расчетов для прогнозирования Tm (температуры плавления) гибридизованных цепей, или путем эмпирического определения Tm с использованием обычных способов. Tm включает температуру, при которой популяция гибридизационных комплексов, образованных между двумя цепями нуклеиновой кислоты, денатурируется на 50% (т.е. популяция двухцепочечных молекул нуклеиновой кислоты становится наполовину диссоциированной на одноцепочечные). При температуре ниже Tm преимущественно происходит образование гибридизационного комплекса, тогда как при температуре выше Tm преимущественно происходит плавление или разделение цепей в гибридизационном комплексе. Tm может быть оценена для нуклеиновой кислоты, имеющей известное содержание G+C, в водном 1M растворе NaCl, используя, например, Tm=81,5+0,41(% G+C), хотя другие известные методы вычисления Tm учитывают структурные характеристики нуклеиновой кислоты.
[0069] Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несовпадения между основаниями. Условия, подходящие для гибридизации двух нуклеиновых кислот, зависят от длины нуклеиновых кислот и степени комплементарности - переменных, которые хорошо известны. Чем выше степень комплементарности между двумя нуклеотидными последовательностями, тем больше будет значение температуры плавления (Tm) для гибридов нуклеиновых кислот, имеющих эти последовательности. Для гибридизации нуклеиновых кислот с короткими участками комплементарности (например, комплементарность 35 или менее, 30 или менее, 25 или менее, 22 или менее, 20 или менее или 18 или менее нуклеотидов) положение несовпадений становится важным (см. Sambrook et al., цитируется выше, 11.7-11.8). Обычно длина поддающийся гибридизации нуклеиновой кислоты составляет по меньшей мере приблизительно 10 нуклеотидов. Иллюстративные минимальные длины поддающийся гибридизации нуклеиновой кислоты включают по меньшей мере приблизительно 15 нуклеотидов, по меньшей мере приблизительно 20 нуклеотидов, по меньшей мере приблизительно 22 нуклеотида, по меньшей мере приблизительно 25 нуклеотидов и по меньшей мере приблизительно 30 нуклеотидов. Кроме того, температуру и концентрацию соли в промывочном растворе можно регулировать по мере необходимости в соответствии с такими факторами, как длина области комплементарности и степень комплементарности.
[0070] Последовательность полинуклеотида не обязательно должна быть на 100% комплементарной последовательности его нуклеиновой кислоты-мишени для специфической гибридизации. Более того, полинуклеотид может гибридизоваться с одним или более сегментами, так что промежуточные или прилегающие сегменты не участвуют в событии гибридизации (например, петлевая структура или шпилечная структура). Полинуклеотид (например, гРНК) может обладать по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 99% или 100% комплементарностью последовательности с целевой областью в последовательности нуклеиновой кислоты-мишени, на которую они нацелены. Например, гРНК, в которой 18 из 20 нуклеотидов комплементарны целевой области и поэтому специфически гибридизуются, будет иметь 90% комплементарности. В этом примере оставшиеся некомплементарные нуклеотиды могут быть сгруппированы или могут перемежаться с комплементарными нуклеотидами и не обязательно должны быть смежными друг с другом или с комплементарными нуклеотидами.
[0071] Процент комплементарности между конкретными участками последовательностей нуклеиновых кислот в нуклеиновых кислотах может быть определен обычным способом с использованием программ BLAST (средство поиска основного локального выравнивания) и программ PowerBLAST (см. источники Altschul et al. (1990) J. Mol. Biol. 215:403-410; Zhang and Madden (1997) Genome Res. 7:649-656, каждый из которых включен в настоящий документ в полном объеме посредством ссылки для всех целей), или с использованием программы Gap (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, Мэдисон, Висконсин), используя настройки по умолчанию, в которых используется алгоритм, раскрытый в источнике Smith and Waterman (1981) Adv. Appl. Math. 2:482-489, который включен в настоящий документ в полном объеме посредством ссылки для всех целей.
[0072] В способах и композициях, предложенных в настоящем документе, используется множество различных компонентов. Некоторые компоненты, упомянутые в описании, могут иметь активные варианты и фрагменты. Такие компоненты включают, например, белки Cas, CRISPR-РНК, tracrРНК и гидовые РНК. Биологическая активность каждого из этих компонентов описана в других местах настоящего документа. Термин «функциональный» относится к присущей белку или нуклеиновой кислоте (или их фрагменту или варианту) способности проявлять биологическую активность или функцию. Такие биологические активности или функции могут включать, например, способность белка Cas связываться с гидовой РНК и с последовательностью ДНК-мишени. Биологические функции функциональных фрагментов или вариантов могут быть такими же или могут, напротив, быть изменены (например, в отношении их специфичности, селективности или эффективности) по сравнению с исходной молекулой, но с сохранением основной биологической функции молекулы.
[0073] Термин «вариант» относится к нуклеотидной последовательности, отличающейся от последовательности, наиболее распространенной в популяции (например, на один нуклеотид), или к последовательности белка, отличной от последовательности, наиболее распространенной в популяции (например, на одну аминокислоту).
[0074] Термин «фрагмент» применительно к белку означает белок, который имеет меньшую длину или содержит меньше аминокислот, чем полноразмерный белок. Термин «фрагмент» применительно к нуклеиновой кислоте означает нуклеиновую кислоту, которая имеет меньшую длину или содержит меньше нуклеотидов, чем полноразмерная нуклеиновая кислота. Фрагмент может представлять собой, например, когда речь идет о фрагменте белка, N-концевой фрагмент (т.е. образован путем удаления части C-конца белка), C-концевой фрагмент (т.е. образован путем удаления части N-конца белка) или внутренний фрагмент (т.е. образован путем удаления части каждого из N-конца и C-конца белка). Фрагмент может представлять собой, например, когда речь идет о фрагменте нуклеиновой кислоты, 5’-фрагмент (т.е. образован путем удаления части 3’-конца нуклеиновой кислоты), 3’-фрагмент (т.е. образован путем удаления части 5’-конца нуклеиновой кислоты) или внутренний фрагмент (т.е. образован путем удаления части каждого из 5’-конца и 3’-конца нуклеиновой кислоты).
[0075] «Идентичность последовательностей» или «идентичность» в контексте двух полинуклеотидных или полипептидных последовательностей относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании последовательностей для максимального соответствия в пределах установленного окна сравнения. Когда процент идентичности последовательностей используется в отношении белков, положения остатков, которые не являются идентичными, часто отличаются консервативными аминокислотными заменами, при которых аминокислотные остатки заменены на другие аминокислотные остатки с аналогичными химическими свойствами (например, зарядом или гидрофобностью) и, следовательно, не меняют функциональных свойств данной молекулы. Когда последовательности отличаются консервативными аминокислотными заменами, процент идентичности последовательностей может быть увеличен для поправки на консервативный характер замены. Последовательности, отличающиеся такими консервативными заменами, называют обладающими «сходством последовательностей» или «сходством». Средства для выполнения данной поправки хорошо известны. Обычно оно включает в себя оценку консервативной замены как частичного, а не полного несоответствия, что тем самым увеличивает процент идентичности последовательностей. Таким образом, например, если идентичной аминокислоте присваивается оценка, равная 1, а неконсервативной замене присваивается оценка, равная нулю, то консервативной замене присваивается оценка в интервале от нуля до 1. Оценка консервативных замен рассчитывается, например, так, как реализовано в программе PC/GENE (Intelligenetics, Маунтин-Вью, Калифорния, США).
[0076] «Процент идентичности последовательностей» включает значение, определяемое путем сравнения двух оптимально выровненных последовательностей (наибольшее количество полностью совпадающих остатков) в пределах окна сравнения, где часть полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) по сравнению с референсной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания данных двух последовательностей. Данный процент рассчитывают путем определения количества положений, в которых в обеих последовательностях присутствует одно и то же основание нуклеиновой кислоты или один и тот же аминокислотный остаток, чтобы получить количество совпадающих положений, деления количества совпадающих положений на общее количество положений в окне сравнения и умножения полученного результата на 100, с получением процента идентичности последовательностей. Если не указано иное (например, более короткая последовательность включает связанную гетерологичную последовательность), окно сравнения представляет собой полную протяженность более короткой из двух сравниваемых последовательностей.
[0077] Если не указано иное, значения идентичности/сходства последовательностей включают в себя значение, полученное с использованием программного обеспечения GAP версии 10 с использованием следующих параметров: % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа в 50 за открытие гэпа и штрафа в 3 за удлинение гэпа, а также весовой матрицы nwsgapdna.cmp; % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа в 8 за открытие гэпа и штрафа в 2 за удлинение гэпа, а также весовой матрицы BLOSUM62; или с помощью любой эквивалентной программы. «Эквивалентная программа» включает в себя любую программу для сравнения последовательностей, которая для любых двух рассматриваемых последовательностей генерирует выравнивание, имеющее идентичные совпадения нуклеотидных или аминокислотных остатков и идентичный процент идентичности последовательностей по сравнению с соответствующим выравниванием, созданным в программе GAP версии 10.
[0078] Термин «консервативная аминокислотная замена» относится к замене аминокислоты, которая обычно присутствует в данной последовательности, на другую аминокислоту аналогичного размера, заряда или полярности. Примеры консервативных замен включают в себя замену неполярного (гидрофобного) остатка, такого как изолейцин, валин или лейцин, на другой неполярный остаток. Аналогичным образом, примеры консервативных замен включают в себя замену одного полярного (гидрофильного) остатка на другой, например, замену аргинина на лизин и наоборот, замену глутамина на аспарагин и наоборот или замену глицина на серин и наоборот. Кроме того, дополнительными примерами консервативных замен являются замена основного остатка, такого как лизин, аргинин или гистидин, на другой остаток, или замена одного кислого остатка, такого как аспарагиновая кислота или глутаминовая кислота, на другой кислый остаток. Примеры неконсервативных замен включают в себя замену неполярного (гидрофобного) аминокислотного остатка, такого как изолейцин, валин, лейцин, аланин или метионин, на полярный (гидрофильный) остаток, такой как цистеин, глутамин, глутаминовая кислота или лизин, и/или замену полярного остатка на неполярный остаток. Стандартная классификация аминокислот обобщенно представлена в Таблице 1 ниже.
[0079] Таблица 1. Классификация аминокислот.
[0080] Термин «гомологичная» последовательность (например, последовательность нуклеиновой кислоты) включает в себя последовательность, которая либо идентична, либо по существу сходна с известной референсной последовательностью таким образом, что она, например, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична известной референсной последовательности. Гомологичные последовательности могут включать в себя, например, ортологичную последовательность и паралогичные последовательности. Гомологичные гены, например, обычно происходят от общей предковой последовательности ДНК либо в результате видообразующего события (ортологичные гены), либо в результате события генетической дупликации (паралогичные гены). «Ортологичные» гены включают в себя гены разных видов, которые произошли от общего предкового гена путем видообразования. Ортологи обычно сохраняют одну и ту же функцию в ходе эволюции. «Паралогичные» гены включают в себя гены, связанные с дупликацией в геноме. У паралогов в ходе эволюции могут развиться новые функции.
[0081] Термин «in vitro» включает в себя искусственные среды и процессы либо реакции, которые происходят в искусственной среде (например, в пробирке, в выделенной клетке или клеточной линии). Термин «in vivo» включает в себя естественные среды (например, клетку, или организм, или тело) и процессы либо реакции, которые происходят в естественной среде. Термин «ex vivo» включает в себя клетки, которые были удалены из организма индивидуума, а также процессы или реакции, которые происходят в таких клетках.
[0082] Термин «репортерный ген» относится к нуклеиновой кислоте, имеющей последовательность, кодирующую продукт гена (обычно фермент), которая поддается легкому и количественному анализу при введении конструкции, содержащей последовательность репортерного гена, функционально связанную с гетерологичным промотором и/или энхансерным элементом, в клетки, которые содержат (или могут быть модифицированы так, чтобы они содержали) факторы, необходимые для активации промоторных и/или энхансерных элементов. Примеры репортерных генов включают, не ограничиваясь перечисленным, гены, кодирующие бета-галактозидазу (lacZ), гены бактериальной хлорамфениколацетилтрансферазы (cat), гены люциферазы светлячка, гены, кодирующие бета-глюкуронидазу (GUS), и гены, кодирующие флуоресцентные белки. «Репортерный белок» относится к белку, кодируемому репортерным геном.
[0083] Термин «флуоресцентный репортерный белок», используемый в контексте настоящего документа, означает репортерный белок, который может быть обнаружен посредством флуоресценции, при этом флуоресценция может исходить либо непосредственно от репортерного белка, либо в результате действия репортерного белка на флуорогенный субстрат, либо от белка с аффинностью связывания с флуоресцентно меченым соединением. Примеры флуоресцентных белков включают в себя зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, мономерный Azami Green, CopGFP, AceGFP и ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP и ZsYellowl), синие флуоресцентные белки (например, BFP, eBFP, eBFP2, Azurite, mKalamal, GFPuv, Sapphire и T-sapphire), голубые флуоресцентные белки (например, CFP, eCFP, Cerulean, CyPet, AmCyanl и Midoriishi-Cyan), красные флуоресцентные белки (например, RFP, mKate, mKate2, mPlum, мономер DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, мономер DsRed, тандем HcRed, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry и Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine и tdTomato) и любой другой подходящий флуоресцентный белок, присутствие которого в клетках можно определить методами проточной цитометрии.
[0084] Репарация в ответ на двухцепочечные разрывы (DSB) происходит главным образом посредством двух консервативных путей репарации ДНК: гомологичной рекомбинации (HR) и негомологичного соединения концов (NHEJ). См. источник Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22:886-897, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Точно так же репарация нуклеиновой кислоты-мишени, опосредованная экзогенной донорной нуклеиновой кислотой, может включать в себя любой процесс обмена генетической информацией между двумя указанными полинуклеотидами.
[0085] Термин «рекомбинация» включает в себя любой процесс обмена генетической информацией между двумя полинуклеотидами и может происходить по любому механизму. Рекомбинация может происходить посредством гомологически направленной репарации (HDR) или гомологичной рекомбинации (HR). HDR или HR включает в себя вид репарации нуклеиновой кислоты, для которого может требоваться гомология нуклеотидной последовательности, в котором используется «донорная» молекула в качестве матрицы для репарации молекулы-«мишени» (т.е. той, в которой произошел двухцепочечный разрыв), и приводящий к передаче генетической информации от донора к молекуле-мишени. Не ограничиваясь какой-либо конкретной теорией, такой перенос может включать в себя исправление ошибки спаривания нуклеотидов гетеродуплексной ДНК, которая образуется между поврежденной мишенью и донором, и/или синтез-зависимый отжиг цепей, при котором донор используется для повторного синтеза генетической информации, которая станет частью мишени, и/или связанные процессы. В некоторых случаях донорный полинуклеотид, часть донорного полинуклеотида, копия донорного полинуклеотида или часть копии донорного полинуклеотида интегрируется в ДНК-мишень. См. источники Wang et al. (2013) Cell 153:910-918; Mandalos et al. (2012) PLOS ONE 7:e45768:1-9; и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей.
[0086] Негомологичное соединение концов (NHEJ) включает репарацию двухцепочечных разрывов в нуклеиновой кислоте путем прямого лигирования концов разрывов друг с другом или с экзогенной последовательностью без необходимости в гомологичной матрице. Лигирование несмежных последовательностей с помощью NHEJ часто может приводить к делециям, вставкам или транслокациям вблизи сайта двухцепочечного разрыва. Например, NHEJ может также привести к направленной интеграции экзогенной донорной нуклеиновой кислоты посредством непосредственного лигирования концов разрывов с концами экзогенной донорной нуклеиновой кислоты (т.е. захвату на основе NHEJ). Такая направленная NHEJ-опосредованная интеграция может быть предпочтительной для вставки экзогенной донорной нуклеиновой кислоты, когда использование путей репарации, опосредованной гомологичной рекомбинацией (HDR), затруднительно (например, в неделящихся клетках, первичных клетках и клетках, в которых плохо осуществляется репарация ДНК на основе гомологии). Кроме того, в отличие от репарации, опосредованной гомологичной рекомбинацией, не требуется информация о больших областях идентичности последовательностей, фланкирующих сайт расщепления, что может быть выгодно при попытке направленной вставки в организмы, о геномной последовательности геномов которых имеются лишь ограниченные сведения. Интеграция может происходить посредством лигирования тупых концов между экзогенной донорной нуклеиновой кислотой и расщепленной геномной последовательностью или посредством лигирования липких концов (т.е. концов, имеющих 5’- или 3’-выступы) с использованием экзогенной донорной нуклеиновой кислоты, фланкированной выступами, которые совместимы с теми, которые генерируются нуклеазным агентом в указанной расщепленной геномной последовательности. См., например, источники US 2011/020722, WO 2014/033644, WO 2014/089290 и Maresca et al. (2013) Genome Res. 23(3):539-546, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. При лигировании тупых концов может потребоваться обрезание последовательности-мишени и/или донорной последовательности для создания областей микрогомологии, необходимых для соединения фрагментов, что может создать нежелательные изменения в последовательности-мишени.
[0087] Композиции или способы, «содержащие» или «включающие» один или более перечисленных элементов, могут включать другие элементы, не перечисленные специально. Например, композиция, которая «содержит» или «включает» белок, может содержать белок отдельно или в комбинации с другими ингредиентами. Переходная фраза «по состоящий существу из» означает, что объем пункта формулы изобретения следует интерпретировать как включающий в себя конкретные указанные в данном пункте формулы изобретения элементы, а также элементы, которые не оказывают существенного влияния на основную и новую характеристику(ки) заявленного изобретения. Таким образом, термин «по существу состоящий из» при использовании в пункте формулы настоящего изобретения не предусматривает интерпретации как эквивалента термина «содержащий».
[0088] «Необязательный» или «необязательно» означает, что описанное далее событие или обстоятельство может произойти или не произойти, и что описание включает случаи, в которых это событие или обстоятельство происходит, и случаи, при которых это событие или обстоятельство не происходит.
[0089] Обозначение диапазона значений включает все целые числа в пределах данного диапазона или все целые числа, определяющие данный диапазон, а также все поддиапазоны, определяемые целыми числами в пределах данного диапазона.
[0090] Если иное явным образом не следует из контекста, термин «приблизительно» охватывает значения в пределах стандартной погрешности измерения (например, SEM (стандартной ошибки среднего)) для указанного значения.
[0091] Термин «и/или» относится и охватывает все возможные комбинации одного или более связанных перечисленных элементов, а также отсутствие комбинаций при интерпретации в части альтернативы («или»).
[0092] Термин «или» относится к любому члену конкретного списка, а также включает любую комбинацию элементов этого списка.
[0093] Формы единственного числа включают в себя соответствующие им формы множественного числа, если из контекста явным образом не следует иное. Например, термины «белок» или «по меньшей мере один белок» могут включать в себя множество белков, включая их смеси.
[0094] «Статистически значимое» означает p ≤ 0,05.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Обзор
[0095] В настоящем документе раскрыты геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, содержащие гуманизированный локус альбумина (ALB), и способы применения клеток животных, отличных от человека, и животных, отличных от человека. Клетки животных, отличных от человека, или животные, отличные от человека, содержащие гуманизированный локус альбумина, экспрессируют белок человеческого альбумина или белок химерного альбумина, содержащий один или более фрагментов белка человеческого альбумина. Такие клетки животных, отличных от человека, и животные, отличные от человека, могут применяться для оценки доставки или эффективности агентов, нацеленных на человеческий альбумин (например, агентов для редактирования генома CRISPR/Cas9) in vitro, ex vivo или in vivo, и могут применяться в способах оптимизации доставки или эффективности таких агентов in vitro, ex vivo или in vivo.
[0096] В некоторых клетках животных, отличных от человека, и животных, отличных от человека, раскрытых в настоящем документе, большая часть или вся геномная ДНК человеческого альбумина вставлена в соответствующий ортологичный локус альбумина животного, отличного от человека. В некоторых клетках животных, отличных от человека, и животных, отличных от человека, раскрытых в настоящем документе, большая часть или вся геномная ДНК человеческого альбумина заменена один к одному соответствующей геномной ДНК ортологичного человеческого альбумина. По сравнению с животными, отличными от человека, со вставками кДНК, уровни экспрессии должны быть выше при сохранении структуры интрон-экзон и механизма сплайсинга, поскольку консервативные регуляторные элементы с большей вероятностью останутся нетронутыми, а сплайсированные транскрипты, которые подвергаются процессингу РНК, более стабильны, чем кДНК. Напротив, вставка кДНК человеческого альбумина в локус альбумина животного, отличного от человека, приведет к устранению консервативных регуляторных элементов, таких как содержащиеся в первом экзоне и интроне альбумина животного, отличного от человека. Замена геномной последовательности животного, отличного от человека, соответствующей ортологичной человеческой геномной последовательностью, или вставка геномной последовательности человеческого альбумина в соответствующий ортологичный локус альбумина, отличного от человеческого, с большей вероятностью приведет к верной экспрессии трансгена из эндогенного локуса альбумина. Аналогичным образом, трансгенные животные, отличные от человека, с трансгенной вставкой последовательностей, кодирующих человеческий альбумин, в случайный геномный локус, а не в эндогенный локус альбумина животного, отличного от человека, не будут столь точно отражать эндогенную регуляцию экспрессии альбумина. Гуманизированный аллель альбумина, возникающий в результате замены большей части или всей геномной ДНК животного, отличного от человека, один к одному соответствующей ортологичной человеческой геномной ДНК, или вставки геномной последовательности человеческого альбумина в соответствующий ортологичный локус альбумина, отличный от человеческого, обеспечит истинную человеческую мишень или близкое приближение к истинной человеческой мишени для реагентов, нацеленных на человеческий альбумин (например, реагентов CRISPR/Cas9, разработанных для нацеливания на человеческий альбумин), тем самым позволяя тестировать эффективность и принцип действия таких агентов на живых животных, а также проводить фармакокинетические и фармакодинамические исследования в условиях, когда гуманизированный белок и гуманизированный ген являются единственной доступной версией альбумина.
II. Животные, отличные от человека, содержащие гуманизированный локус альбумина (ALB)
[0097] Раскрытые в настоящем документе геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, содержат гуманизированный локус альбумина (ALB). Клетки или животные, отличные от человека, содержащие гуманизированный локус альбумина, экспрессируют белок человеческого альбумина или белок частично гуманизированного химерного альбумина, в котором один или более фрагментов белка нативного альбумина были заменены соответствующими фрагментами из человеческого альбумина. В настоящем документе также раскрыты гуманизированные гены альбумина животного, отличного от человека, в которых сегмент гена альбумина, отличного от человеческого, был удален и заменен соответствующей последовательностью человеческого альбумина.
[0098] Раскрытые в настоящем документе геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, могут дополнительно содержать инактивированный (нокаутированный) эндогенный ген, который не является локусом альбумина. Такие геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, могут применяться, например, для скрининга реагентов для генной терапии (например, трансгенов) на предмет вставки в гуманизированный локус альбумина для замены инактивированного эндогенного гена. Вставка в гуманизированный локус альбумина для замены инактивированного эндогенного гена может, например, восстановить нокаут. В одном конкретном примере геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, могут дополнительно содержать инактивированный (нокаутированный) эндогенный F9 (кодирует фактор свертывания крови IX). Инактивированный (нокаутированный) эндогенный ген F9 - это ген, который не экспрессирует какой-либо фактор свертывания крови IX (также известный как фактор Кристмаса, плазменный тромбопластиновый компонент или ПТК). Человеческому белку фактора свертывания крови IX дикого типа был присвоен регистрационный номер UniProt P00740, а человеческому гену F9 был присвоен GeneID 2158. Мышиному белку фактора свертывания крови IX дикого типа был присвоен регистрационный номер UniProt P16294, а мышиному гену F9 был присвоен GeneID 14071. Крысиному белку фактора свертывания крови IX дикого типа был присвоен регистрационный номер UniProt P16296, а крысиному гену F9 был присвоен GeneID 24946.
[0099] Раскрытые в настоящем документе геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, могут дополнительно содержать кодирующую последовательность экзогенного белка, интегрированного в по меньшей мере один аллель гуманизированного локуса альбумина (например, в одной или более клетках животного, отличного от человека, например, в одной или более клетках печени животного, отличного от человека). Кодирующая последовательность может быть интегрирована, например, в интрон 1, интрон 12 или интрон 13 гуманизированного локуса альбумина. В некоторых случаях экспрессия человеческого альбумина из гуманизированного локуса альбумина поддерживается на том же уровне после интеграции кодирующей последовательности экзогенного белка в по меньшей мере один аллель гуманизированного локуса альбумина (например, в одной или более клетках животного, отличного от человека, например, в одной или более клетках печени животного, отличного от человека). В одном примере геном животного, отличного от человека, клетка животного, отличного от человека, или животное, отличное от человека, дополнительно содержат инактивированный (нокаутированный) эндогенный ген, который не является локусом альбумина, и экзогенный белок заменяет функцию инактивированного эндогенного гена (например, восстанавливает нокаут). В одном конкретном примере экзогенный белок представляет собой фактор свертывания крови IX (например, человеческий фактор свертывания крови IX).
A. Альбумин
[00100] Описанные в настоящем документе клетки и животные, отличные от человека, содержат гуманизированный локус альбумина (ALB). Альбумин кодируется геном ALB (также известным как альбумин, сывороточный альбумин, PRO0883, PRO0903, HSA, GIG20, GIG42, PRO1708, PRO2044, PRO2619, PRO2675 и UNQ696/PRO1341). Альбумин синтезируется в печени в виде препроальбумина, имеющего N-концевой пептид, который удаляется до того, как образующийся белок высвобождается из гранулярного эндоплазматического ретикулума. Продукт - проальбумин - в свою очередь, расщепляется в пузырьках Гольджи с образованием секретируемого альбумина (сывороточного альбумина). Человеческий сывороточный альбумин представляет собой сывороточный альбумин, присутствующий в крови человека. Он является самым распространенным белком в плазме крови человека; он составляет около половины сывороточного белка. Вырабатывается в печени. Растворим в воде и является мономерным. Альбумин, помимо других функций, переносит гормоны, жирные кислоты и другие соединения, буферизует pH и поддерживает онкотическое давление. Концентрации человеческого альбумина в сыворотке обычно составляет примерно 35-50 г/л (3,5-5,0 г/дл). Его период полувыведения из сыворотки составляет примерно 20 дней. Он имеет молекулярную массу 66,5 кДа.
[00101] Альбумин считается геномным локусом «безопасной гавани» из-за его очень высокого уровня экспрессии и удобства использования печени для доставки генов и редактирования in vivo по сравнению с другими тканями. Локусы «безопасной гавани» включают хромосомные локусы, в которых трансгены или другие экзогенные вставки нуклеиновых кислот могут стабильно и надежно экспрессироваться во всех представляющих интерес тканях без непосредственного изменения клеточного поведения или фенотипа. Часто локус «безопасной гавани» представляет собой локус, в котором экспрессия вставленной последовательности гена не нарушается какой-либо сквозной экспрессией соседних генов. Например, локусы «безопасной гавани» могут включать хромосомные локусы, в которых экзогенная ДНК может интегрироваться и функционировать предсказуемым образом без неблагоприятного воздействия на структуру или экспрессию эндогенного гена. Локусы «безопасной гавани» могут включать внегенные области или внутригенные области, такие как, например, локусы в генах, которые не относятся к незаменимым, являются необязательными или могут быть нарушены без явных фенотипических последствий.
[00102] Структура гена альбумина подходит для нацеливания трансгена на интронные последовательности, поскольку его первый экзон кодирует секреторный пептид (сигнальный пептид), который отщепляется от конечного белкового продукта. Например, интеграция кассеты без промотора, несущей акцептор сплайсинга и терапевтический трансген, будет обеспечивать экспрессию и секрецию многих различных белков.
[00103] Человеческий ALB картирован в область 4q13.3 человека на хромосоме 4 (NCBI RefSeq Gene ID 213; Assembly GRCh38.p12 (GCF_000001405.38); положение NC_000004.12 (73404239..73421484 (+))). Сообщается, что данный ген имеет 15 экзонов. Из них 14 экзонов являются кодирующими экзонами, а экзон 15 является некодирующим экзоном, который является частью 3’-нетранслируемой области (UTR). Белку человеческого альбумина дикого типа был присвоен регистрационный номер UniProt P02768. Известны по меньшей мере три изоформы (с P02768-1 по P02768-3). Последовательность для одной изоформы, P02768-1 (идентична регистрационному номеру NCBI NP_000468.1), приведена в SEQ ID NO: 5. мРНК (кДНК), кодирующей каноническую изоформу, присвоен регистрационный номер NCBI NM_000477.7, и она приведена в SEQ ID NO: 37. Иллюстративной кодирующей последовательности (CDS) присвоен идентификатор CCDS CCDS3555.1, и она приведена в SEQ ID NO: 13. Полноразмерный белок человеческого альбумина, приведенный в SEQ ID NO: 5, содержит 609 аминокислот, включая сигнальный пептид (аминокислоты 1-18), пропептид (аминокислоты 19-24) и сывороточный альбумин (аминокислоты 25-609). Границы между этими доменами соответствуют указанным в UniProt. Ссылка на человеческий альбумин включает канонические формы (дикого типа), а также все аллельные формы и изоформы. В любых других формах человеческого альбумина аминокислоты пронумерованы для максимального выравнивания с формой дикого типа, выровненные аминокислоты обозначены одним и тем же номером.
[00104] Мышиный Alb картирован в область 5 E1; 5 44.7 cM мыши на хромосоме 5 (NCBI RefSeq Gene ID 11657; Assembly GRCm38.p4 (GCF_000001635.24); положение NC_000071.6 (90,460,870..90,476,602 (+))). Сообщается, что данный ген имеет 15 экзонов. Из них 14 экзонов являются кодирующими экзонами, а экзон 15 является некодирующим экзоном, который является частью 3’-нетранслируемой области (UTR). Белку мышиного альбумина дикого типа был присвоен регистрационный номер UniProt P07724. Последовательность мышиного альбумина (идентична регистрационному номеру NCBI NP_033784.2) приведена в SEQ ID NO: 1. Иллюстративной изоформе мРНК (кДНК), кодирующей каноническую изоформу, присвоен регистрационный номер NCBI NM_009654.4, и она приведена в SEQ ID NO: 36. Иллюстративная кодирующая последовательность (CDS) (идентификатор CCDS CCDS19412.1) приведена в SEQ ID NO: 9. Канонический полноразмерный белок мышиного альбумина, приведенный в SEQ ID NO: 1, содержит 608 аминокислот, включая сигнальный пептид (аминокислоты 1-18), пропептид (аминокислоты 19-24) и сывороточный альбумин (аминокислоты 25-608). Границы между этими доменами соответствуют указанным в UniProt. Ссылка на мышиный альбумин включает канонические формы (дикого типа), а также все аллельные формы и изоформы. В любых других формах мышиного альбумина аминокислоты пронумерованы для максимального выравнивания с формой дикого типа, выровненные аминокислоты обозначены одним и тем же номером.
[00105] Также известны последовательности альбумина для многих других животных, отличных от человека. К ним относятся, например, крупный рогатый скот (регистрационный номер UniProt P02769; идентификатор гена NCBI RefSeq 280717), крыса (регистрационный номер UniProt P02770; идентификатор гена NCBI RefSeq 24186), курица (регистрационный номер UniProt P19121), суматранский орангутан (регистрационный номер UniProt Q5NVH5; идентификатор гена NCBI RefSeq 100174145), лошадь (регистрационный номер UniProt P35747; идентификатор гена NCBI RefSeq 100034206), кошка (регистрационный номер UniProt P49064; идентификатор гена NCBI RefSeq 448843), кролик (регистрационный номер UniProt P49065; идентификатор гена NCBI RefSeq 100009195), собака (регистрационный номер UniProt P49822; идентификатор гена NCBI RefSeq 403550), свинья (регистрационный номер UniProt P08835; идентификатор гена NCBI RefSeq 396960), монгольская песчанка (регистрационный номер UniProt O35090), макак-резус (регистрационный номер UniProt Q28522; идентификатор гена NCBI RefSeq 704892), осел (регистрационный номер UniProt Q5XLE4; идентификатор гена NCBI RefSeq 106835108), овца (регистрационный номер UniProt P14639; идентификатор гена NCBI RefSeq 443393), американская лягушка-бык (регистрационный номер UniProt P21847), золотистый хомячок (регистрационный номер UniProt A6YF56; идентификатор гена NCBI RefSeq 101837229) и коза (регистрационный номер UniProt P85295).
B. Гуманизированные локусы альбумина
[00106] Гуманизированный локус альбумина - это локус альбумина, в котором сегмент эндогенного локуса альбумина был удален и заменен ортологичной последовательностью человеческого альбумина. Гуманизированный локус альбумина может представлять собой локус альбумина, в котором весь ген альбумина заменен соответствующей ортологичной последовательностью человеческого альбумина, или он может представлять собой локус альбумина, в котором только часть гена альбумина заменена соответствующей ортологичной последовательностью человеческого альбумина (т.е. гуманизирована). Например, вся кодирующая последовательность альбумина в эндогенном локусе альбумина может быть удалена и заменена соответствующей последовательностью человеческого альбумина. Последовательность человеческого альбумина, соответствующая конкретному сегменту последовательности эндогенного альбумина, относится к области человеческого альбумина, которая совпадает с конкретным сегментом последовательности эндогенного альбумина, когда человеческий альбумин и эндогенный альбумин оптимально выровнены. Оптимально выровненный относится к наибольшему количеству полностью совпадающих остатков. Соответствующая ортологичная человеческая последовательность может содержать, например, комплементарную ДНК (кДНК) или геномную ДНК. Необязательно, соответствующая последовательность ортологичного человеческого альбумина модифицирована для оптимизации кодонов, исходя из использования кодонов у животного, отличного от человека. Замененные или вставленные (т.е. гуманизированные) области могут включать кодирующие области, такие как экзон, некодирующие области, такие как интрон, нетранслируемую область или регуляторную область (например, промотор, энхансер или элемент, связывающий репрессор трансляции), или любую их комбинацию. Например, экзоны, соответствующие 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или всем 15 экзонам гена человеческого альбумина, могут быть гуманизированы. Например, экзоны, соответствующие всем экзонам (т.е. экзонам 1-15) гена человеческого альбумина, могут быть гуманизированы. В качестве еще одного примера экзоны, соответствующие 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или всем 14 экзонам гена человеческого альбумина, могут быть гуманизированы. Например, экзоны, соответствующие всем кодирующим экзонам (т.е. экзонам 1-14) гена человеческого альбумина, могут быть гуманизированы. Аналогичным образом, интроны, соответствующие 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или всем 14 интронам гена человеческого альбумина, могут быть гуманизированы или могут оставаться эндогенными. Например, интроны, соответствующие всем интронам (т.е. интронам 1-14) человеческого альбумина, могут быть гуманизированы. Аналогичным образом, интроны, соответствующие 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или всем 13 интронам между кодирующими экзонами гена человеческого альбумина, могут быть гуманизированы или могут оставаться эндогенными. Например, интроны, соответствующие всем интронам между кодирующими экзонами (т.е. интронам 1-13) гена человеческого альбумина, могут быть гуманизированы. Фланкирующие нетранслируемые области, включая регуляторные последовательности, также могут быть гуманизированы или могут оставаться эндогенными. Например, 5’-нетранслируемая область (UTR), 3’ UTR или обе из 5’ UTR и 3’ UTR могут быть гуманизированы, или 5’ UTR, 3’ UTR или обе из 5’ UTR и 3’ UTR могут оставаться эндогенными. Одна или обе из человеческих 5’ и 3’ UTR могут быть вставлены, и/или одна или обе из эндогенных 5’ и 3’ UTR могут быть удалены. В конкретном примере обе из 5’ UTR и 3’ UTR остаются эндогенными. В другом конкретном примере 5’ UTR остается эндогенной, а 3’ UTR гуманизируют. В зависимости от степени замены ортологичными последовательностями регуляторные последовательности, такие как промотор, могут быть эндогенными или могут обеспечиваться замещающей человеческой ортологичной последовательностью. Например, гуманизированный локус альбумина может включать эндогенный промотор альбумина животного, отличного от человека (т.е. последовательность человеческого альбумина может быть функционально связана с эндогенным промотором животного, отличного от человека).
[00107] Одна или более, или все области, кодирующие сигнальный пептид, пропептид или сывороточный альбумин, могут быть гуманизированы, или одна или более таких областей могут оставаться эндогенными. Примеры кодирующих последовательностей сигнального пептида, пропептида мышиного альбумина и мышиного сывороточного альбумина приведены в SEQ ID NO: 10-12 соответственно. Примеры кодирующих последовательностей сигнального пептида, пропептида и сывороточного альбумина человеческого альбумина приведены в SEQ ID NO: 14-16 соответственно.
[00108] Например, вся или часть области локуса альбумина, кодирующей сигнальный пептид, может быть гуманизирована, и/или вся или часть области локуса альбумина, кодирующей пропептид, может быть гуманизирована, и/или вся или часть области локуса альбумина, кодирующей сывороточный альбумин, может быть гуманизирована. В качестве альтернативы или дополнительно, вся или часть области локуса альбумина, кодирующей сигнальный пептид, может оставаться эндогенной, и/или вся или часть области локуса альбумина, кодирующей пропептид, может оставаться эндогенной, и/или вся или часть области локуса альбумина, кодирующей сывороточный альбумин, может оставаться эндогенной. В одном примере все или часть областей локуса альбумина, кодирующих сигнальный пептид, пропептид и сывороточный альбумин, гуманизированы. Необязательно, CDS гуманизированной области локуса альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 13 (или ее вырожденным вариантам). Необязательно, CDS гуманизированной области локуса альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 13 (или ее вырожденным вариантам). Необязательно, гуманизированная область локуса альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 35. Необязательно, гуманизированная область локуса альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 35. Необязательно, гуманизированный локус альбумина кодирует белок, содержащий, по существу состоящий из или состоящий из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 5. Необязательно, гуманизированный локус альбумина кодирует белок, содержащий, по существу состоящий из или состоящий из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 5. Необязательно, гуманизированный локус альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 17 или 18. Необязательно, гуманизированный локус альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 17 или 18.
[00109] Белок альбумина, кодируемый гуманизированным локусом альбумина, может содержать один или более доменов, происходящих из белка человеческого альбумина, и/или один или более доменов, происходящих из белка эндогенного (т.е. нативного) альбумина. Примеры аминокислотных последовательностей сигнального пептида, пропептида мышиного альбумина и мышиного сывороточного альбумина приведены в SEQ ID NO: 2-4 соответственно. Примеры аминокислотных последовательностей сигнального пептида, пропептида человеческого альбумина и сывороточного человеческого альбумина приведены в SEQ ID NO: 6-8 соответственно.
[00110] Белок альбумина может содержать один или более из, или все из сигнального пептида человеческого альбумина, пропептида человеческого альбумина и сывороточного человеческого альбумина. В качестве альтернативы или дополнительно, белок альбумина может содержать один или более доменов, происходящих из белка эндогенного (т.е. нативного) альбумина животного, отличного от человека. Например, белок альбумина может содержать сигнальный пептид из белка эндогенного (т.е. нативного) альбумина животного, отличного от человека, и/или пропептид из белка эндогенного (т.е. нативного) альбумина животного, отличного от человека, и/или сывороточный альбумин из белка эндогенного (т.е. нативного) альбумина животного, отличного от человека. В качестве одного из примеров белок альбумина может содержать человеческий сигнальный пептид, пропептид и сывороточный альбумин.
[00111] Домены в химерном белке альбумина, происходящие из белка человеческого альбумина, могут кодироваться полностью гуманизированной последовательностью (т.е. вся последовательность, кодирующая этот домен, заменена последовательностью ортологичного человеческого альбумина) или могут кодироваться частично гуманизированной последовательностью (т.е. часть последовательности, кодирующей этот домен, заменена последовательностью ортологичного человеческого альбумина, а оставшаяся эндогенная (т.е. нативная) последовательность, кодирующая этот домен, кодирует те же аминокислоты, что и последовательность ортологичного человеческого альбумина, так что кодируемый домен идентичен этому домену в белке человеческого альбумина). Аналогичным образом, домены в химерном белке, происходящие из белка эндогенного альбумина, может кодироваться полностью эндогенной последовательностью (т.е. вся последовательность, кодирующая этот домен, представляет собой последовательность эндогенного альбумина) или может кодироваться частично гуманизированной последовательностью (т.е. часть последовательности, кодирующей этот домен, заменена последовательностью ортологичного человеческого альбумина, но последовательность ортологичного человеческого альбумина кодирует те же аминокислоты, что и замененная последовательность эндогенного альбумина, так что кодируемый домен идентичен этому домену в белке эндогенного альбумина).
[00112] Например, белок альбумина, кодируемый гуманизированным локусом альбумина, может содержать сигнальный пептид человеческого альбумина. Необязательно, сигнальный пептид человеческого альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 6. Необязательно, сигнальный пептид человеческого альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 6. В качестве еще одного примера, белок альбумина, кодируемый гуманизированным локусом альбумина, может содержать пропептид человеческого альбумина. Необязательно, пропептид человеческого альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 7. Необязательно, пропептид человеческого альбумина содержит, по существу состоит из или состоит из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 7. В качестве еще одного примера, белок альбумина, кодируемый гуманизированным локусом альбумина, может содержать человеческий сывороточный альбумин. Необязательно, человеческий сывороточный альбумин содержит, по существу состоит из или состоит из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 8. Необязательно, человеческий сывороточный альбумин содержит, по существу состоит из или состоит из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 8. Например, белок альбумина, кодируемый гуманизированным локусом альбумина, может содержать, по существу состоять из или состоять из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 5. Например, белок альбумина, кодируемый гуманизированным локусом альбумина, может содержать, по существу состоять из или состоять из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 5. Необязательно, CDS альбумина, кодируемая гуманизированным локусом альбумина, может содержать, по существу состоять из или состоять из последовательности, по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентичной SEQ ID NO: 13 (или ее вырожденным вариантам). Необязательно, CDS альбумина, кодируемая гуманизированным локусом альбумина, может содержать, по существу состоять из или состоять из последовательности, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99% или приблизительно на 100% идентичной SEQ ID NO: 13 (или ее вырожденным вариантам).
[00113] Белок гуманизированного альбумина может сохранять активность белка нативного альбумина и/или белка человеческого альбумина.
[00114] Необязательно, гуманизированный локус альбумина может содержать другие элементы. Примеры таких элементов могут включать кассеты селекции, репортерные гены, сайты распознавания рекомбиназы или другие элементы. В качестве альтернативы, в гуманизированном локусе альбумина могут отсутствовать другие элементы (например, может отсутствовать маркер селекции или кассета селекции). Примеры подходящих репортерных генов и репортерных белков раскрыты в других местах настоящего документа. Примеры подходящих маркеров селекции включают неомицинфосфотрансферазу (neor), гигромицин-B-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror), бластицидин-S-дезаминазу (bsrr), ксантин-гуанинфосфорибозилтрансферазу (gpt) и тимидинкиназу вируса простого герпеса (HSV-k). Примеры рекомбиназ включают рекомбиназы Cre, Flp и Dre. Одним из примеров гена рекомбиназы Cre является Crei, в котором два экзона, кодирующие рекомбиназу Cre, разделены интроном для предотвращения его экспрессии в прокариотической клетке. Такие рекомбиназы могут дополнительно содержать сигнал ядерной локализации для облегчения локализации в ядре (например, NLS-Crei). Сайты распознавания рекомбиназы включают нуклеотидные последовательности, которые распознаются сайт-специфической рекомбиназой и могут служить субстратом для события рекомбинации. Примеры сайтов распознавания рекомбиназы включают сайты FRT, FRT11, FRT71, attp, att, rox и lox, такие как loxP, lox511, lox2272, lox66, lox71, loxM2 и lox5171.
[00115] Другие элементы, такие как репортерные гены или кассеты селекции, могут представлять собой самоудаляющиеся кассеты, фланкированные сайтами распознавания рекомбиназы. См., например, источники US 8697851 и US 2013/0312129, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. В качестве примера, самоудаляющаяся кассета может содержать ген Crei (содержит два экзона, кодирующих рекомбиназу Cre, которые разделены интроном), функционально связанный с промотором мышиного Prm1 и геном устойчивости к неомицину, функционально связанным с промотором убиквитина человека. За счет использования промотора Prm1 самоудаляющаяся кассета может быть специфично удалена в мужских половых клетках животных F0. Полинуклеотид, кодирующий маркер селекции, может быть функционально связан с промотором, активным в клетке, выступающей в качестве мишени. Примеры промоторов описаны в других местах настоящего документа. В качестве еще одного конкретного примера, самоудаляющаяся кассета селекции может содержать кодирующую последовательность гена устойчивости к гигромицину, функционально связанную с одним или более промоторами (например, одновременно с промоторами убиквитина человека и EM7), за которой следует сигнал полиаденилирования, за которым следует кодирующая последовательность Crei, функционально связанная с одним или более промоторами (например, промотором mPrm1), за которой следует еще один сигнал полиаденилирования, где вся кассета фланкирована сайтами loxP.
[00116] Гуманизированный локус альбумина также может представлять собой условный аллель. Например, нсловный аллель может быть многофункциональным аллелем, как описано в заявке US 2011/0104799, включенной в настоящий документ посредством ссылки в полном объеме для всех целей. Например, условный аллель может содержать: (a) активирующую последовательность в смысловой ориентации относительно транскрипции целевого гена; (b) кассету для отбора по чувствительности к лекарственному препарату (DSC) в смысловой или антисмысловой ориентации; (c) представляющую интерес нуклеотидную последовательность (NSI) в антисмысловой ориентации; и (d) условный по инверсии модуль (COIN, в котором используется разделяющий экзон интрон и инвертируемый модуль, подобный генной ловушке) в обратной ориентации. См., например, US 2011/0104799. Условный аллель может дополнительно содержать рекомбинируемые блоки, которые рекомбинируются при воздействии первой рекомбиназы с образованием условного аллеля, который (i) лишен активирующей последовательности и DSC; и (ii) содержит NSI в смысловой ориентации и COIN в антисмысловой ориентации. См., например, US 2011/0104799.
[00117] Одним из примеров гуманизированного локуса альбумина (например, гуманизированный мышиный локус альбумина) является локус, в котором область от старт-кодона до стоп-кодона заменена соответствующей человеческой последовательностью. См. фиг. 1A и 1B и SEQ ID NO: 17 и 18. В конкретном примере область от старт-кодона ATG до стоп-кодона (т.е. кодирующие экзоны 1-14) может быть удалена из локуса альбумина (Alb) животного (например, мыши), отличного от человека, и вместо удаленной эндогенной области может быть вставлена соответствующая область человеческого альбумина (ALB) от старт-кодона ATG до около 100 п.н. ниже стоп-кодона.
C. Геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, содержащие гуманизированный локус альбумина (ALB)
[00118] Предложены геномы животных, отличных от человека, клетки животных, отличных от человека, и животные, отличные от человека, содержащие гуманизированный локус альбумина (ALB), как описано в других местах настоящего документа. Геномы, клетки или животные, отличные от человека, могут быть мужского или женского пола. Геномы, клетки или животные, отличные от человека, могут быть гетерозиготными или гомозиготными по гуманизированному локусу альбумина. Диплоидный организм имеет два аллеля в каждом генетическом локусе. Каждая пара аллелей отражает генотип конкретного генетического локуса. Генотипы описываются как гомозиготные, если в определенном локусе находятся два идентичных аллеля, и как гетерозиготные, если эти два аллеля различаются. Животное, отличное от человека, содержащее гуманизированный локус альбумина, может содержать гуманизированный эндогенный локус альбумина в своей зародышевой линии.
[00119] Предложенные в настоящем документе геномы или клетки животного, отличного от человека, могут представлять собой, например, любой геном или клетку животного, отличного от человека, содержащего локус альбумина или геномный локус, гомологичный или ортологичный человеческому локусу альбумина. Геномы могут происходить из или клетки могут представлять собой эукариотические клетки, которые включают, например, клетки грибов (например, дрожжевые), клетки растений, клетки животных, клетки млекопитающих, клетки млекопитающих, отличных от человека, и человеческие клетки. Термин «животное» включает любого представителя царства животных, включая, например, млекопитающих, рыб, рептилий, земноводных, птиц и червей. Клетка млекопитающего может представлять собой, например, клетку млекопитающего, отличного от человека, клетку грызуна, крысиную клетку, мышиную клетку или клетку хомяка. К другим млекопитающим, отличным от человека, относятся, например, отличные от человека приматы, мартышки, обезьяны, орангутаны, кошки, собаки, кролики, лошади, быки, олени, бизоны, домашний скот (например, такие виды крупного рогатого скота, как коровы, кастрированные бычки и т.д.; виды овечьих, такие как овцы, козы и т.д., и виды свинообразных, такие как свиньи и кабаны). К птицам относятся, например, куры, индюки, страусы, гуси, утки и т.д. Включены также домашние животные и сельскохозяйственные животные. Термин «отличный от человека» исключает людей.
[00120] Клетки также могут находиться в недифференцированном или дифференцированном состоянии любого типа. Например, клетка может быть тотипотентной клеткой, плюрипотентной клеткой (например, человеческой плюрипотентной клеткой или плюрипотентной клеткой, отличной от человеческой, такой как мышиная эмбриональная стволовая (ЭС) клетка или крысиная ЭС клетка) или неплюрипотентной клеткой. Тотипотентные клетки включают недифференцированные клетки, которые могут давать начало любому типу клеток, а плюрипотентные клетки включают недифференцированные клетки, которые обладают способностью развиваться в больше чем один тип дифференцированных клеток. Такие плюрипотентные и/или тотипотентные клетки могут представлять собой, например, ЭС клетки или ЭС-подобные клетки, такие как индуцированные плюрипотентные стволовые (иПС) клетки. ЭС клетки включают тотипотентные или плюрипотентные клетки эмбрионального происхождения, которые способны вносить вклад в клеточную популяцию любой ткани развивающегося эмбриона при введении в эмбрион. ЭС клетки могут происходить из внутренней клеточной массы бластоцисты и способны дифференцироваться в клетки любого из трех зародышевых листков позвоночных (энтодермы, эктодермы и мезодермы).
[00121] Предложенные в настоящем документе клетки также могут быть зародышевыми клетками (например, сперматозоидами или ооцитами). Клетки могут быть митотически компетентными клетками или митотически неактивными клетками, мейотически компетентными клетками или мейотически неактивными клетками. Точно так же клетки также могут быть первичными соматическими клетками или клетками, которые не являются первичными соматическими клетками. Соматические клетки включают любые клетки, не являющиеся гаметами, зародышевыми клетками, гаметоцитами или недифференцированными стволовыми клетками. Например, клетки могут быть клетками печени, такими как гепатобласты или гепатоциты.
[00122] Предложенные в настоящем документе подходящие клетки также включают первичные клетки. Первичные клетки включают клетки или культуры клеток, которые были выделены непосредственно из организма, органа или ткани. Первичные клетки включают клетки, которые не являются ни трансформированными, ни иммортализованными. Они включают в себя любую клетку, полученную из организма, органа или ткани, которая ранее не пассировалась в культуре клеток или ранее пассировалась в культуре клеток, но неспособна к бесконечно долгому пассированию в культуре клеток. Такие клетки могут быть выделены обычными методами и включают, например, гепатоциты.
[00123] Другие подходящие клетки, предложенные в настоящем документе, включают иммортализованные клетки. Иммортализованные клетки включают клетки из многоклеточного организма, которые в обычных условиях неспособны бесконечно долго пролиферировать, но из-за мутации или изменения избежали нормального клеточного старения и вместо этого могут продолжать делиться. Такие мутации или изменения могут происходить естественным образом или быть вызваны намеренно. Конкретным примером иммортализованной клеточной линии является клеточная линия рака печени человека HepG2. Хорошо известно множество типов иммортализованных клеток. Иммортализованные или первичные клетки включают клетки, которые обычно используются для культивирования или для экспрессии рекомбинантных генов или белков.
[00124] Предложенные в настоящем документе клетки также включают находящиеся на одноклеточной стадии эмбрионы (т.е. оплодотворенные ооциты или зиготы). Такие находящиеся на одноклеточной стадии эмбрионы могут иметь любое генетическое происхождение (например, для мышей - BALB/c, C57BL/6, 129 или их комбинация), могут быть свежими или замороженными и могут быть получены в результате естественного размножения или оплодотворения in vitro.
[00125] Предложенные в настоящем документе клетки могут быть нормальными, здоровыми клетками или могут быть больными или несущими мутации клетками.
[00126] Животные, отличные от человека, содержащие гуманизированный локус альбумина, как описано в настоящем документе, могут быть получены способами, описанными в других местах настоящего документа. Термин «животное» включает любого представителя царства животных, включая, например, млекопитающих, рыб, рептилий, земноводных, птиц и червей. В конкретном примере животное, отличное от человека, представляет собой млекопитающее, отличное от человека. Млекопитающие, отличные от человека, включают, например, отличных от человека приматов, мартышек, обезьян, орангутанов, кошек, собак, лошадей, быков, оленей, бизонов, овец, кроликов, грызунов (например, мышей, крыс, хомяков и морских свинок) и домашний скот (например, такие виды крупного рогатого скота, как коровы и кастрированные бычки; виды овечьих, такие как овцы и козы; и виды свинообразных, такие как свиньи и кабаны). К птицам относятся, например, куры, индюки, страусы, гуси и утки. Включены также домашние животные и сельскохозяйственные животные. Термин «животное, отличное от человека» исключает людей. Предпочтительные животные, отличные от человека, включают, например, грызунов, таких как мыши и крысы.
[00127] Животные, отличные от человека, могут иметь любое генетическое происхождение. Например, подходящие мыши могут происходить из линии 129, линии C57BL/6, линии 129 и C57BL/6, линии BALB/c или линии Swiss Webster. Примеры линий 129 включают 129P1, 129P2, 129P3, 129X1, 129S1 (e.g., 129S1/SV, 129S1/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1 и 129T2. См., например, источник Festing et al. (1999) Mammalian Genome 10:836, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Примеры линий C57BL включают C57BL/A, C57BL/An, C57BL/GrFa, C57BL/Kal_wN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. Подходящие мыши также могут происходить из комбинации вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6 (например, 50% 129 и 50% C57BL/6). Аналогичным образом, подходящие мыши могут происходить из комбинации вышеупомянутых линий 129 или из комбинации вышеупомянутых линий BL/6 (например, линия 129S6 (129/SvEvTac)).
[00128] Аналогичным образом крысы могут быть из любой линии крыс, включая, например, линию крыс ACI, линию крыс Dark Agouti (DA), линию крыс Wistar, линию крыс LEA, линию крыс Sprague Dawley (SD) или линию крыс Fischer, например, Fisher F344 или Fisher F6. Крысы также могут быть получены из линии, происходящей из комбинации двух или более линий, перечисленных выше. Например, подходящая крыса может происходить из линии DA или линии ACI. Линия крыс ACI характеризуется черным агути с белыми животом и лапами и гаплотипом RT1av1. Такие линии доступны из различных источников, включая Harlan Laboratories. Линия крыс Dark Agouti (DA) характеризуется наличием окраса агути и гаплотипом RT1av1. Такие крысы доступны из различных источников, включая Charles River и Harlan Laboratories.. В некоторых случаях подходящие крысы могут происходить из инбредной линии крыс. См., например, источник US 2014/0235933, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00129] Животные, отличные от человека (например, мыши или крысы), содержащие гуманизированный локус альбумина (например, гомозиготный гуманизированный локус альбумина), могут экспрессировать альбумин из гуманизированного локуса альбумина, так что уровни сывороточного альбумина (например, уровни человеческого сывороточного альбумина) сопоставимы с уровнями сывороточного альбумина у контрольных животных дикого типа, отличных от человека. В одном примере животные, отличные от человека, содержащие гуманизированный локус альбумина (например, гомозиготный гуманизированный локус альбумина), могут иметь уровни сывороточного альбумина (например, уровни человеческого сывороточного альбумина), которые составляют по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, при по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 100% от уровней сывороточного альбумина у контрольного животного дикого типа, отличного от человека. В еще одном примере животные, отличные от человека, содержащие гуманизированный локус альбумина (например, гомозиготный гуманизированный локус альбумина), могут иметь уровни сывороточного альбумина (например, уровни человеческого сывороточного альбумина), по меньшей мере столь же высокие, как уровни сывороточного альбумина у контрольного животного дикого типа, отличного от человека. В еще одном примере животные, отличные от человека, содержащие гуманизированный локус альбумина (например, гомозиготный гуманизированный локус альбумина), могут иметь уровни сывороточного альбумина (например, уровни человеческого сывороточного альбумина), превышающие уровни сывороточного альбумина у контрольного животного дикого типа, отличного от человека. Например, животное, отличное от человека, содержащее гуманизированный локус альбумина (например, гомозиготный гуманизированный локус альбумина), может иметь уровни сывороточного альбумина (например, уровни человеческого сывороточного альбумина), равные по меньшей мере приблизительно 1 мг/мл, по меньшей мере приблизительно 2 мг/мл, по меньшей мере приблизительно 3 мг/мл, по меньшей мере приблизительно 4 мг/мл, по меньшей мере приблизительно 5 мг/мл, по меньшей мере приблизительно 6 мг/мл, по меньшей мере приблизительно 7 мг/мл, по меньшей мере приблизительно 8 мг/мл, по меньшей мере приблизительно 9 мг/мл, по меньшей мере приблизительно 10 мг/мл, по меньшей мере приблизительно 11 мг/мл, по меньшей мере приблизительно 12 мг/мл, по меньшей мере приблизительно 13 мг/мл, по меньшей мере приблизительно 14 мг/мл или по меньшей мере приблизительно 15 мг/мл. В более конкретном примере животное, отличное от человека, содержащее гуманизированный локус альбумина (например, гомозиготный гуманизированный локус альбумина), может представлять собой мышь и может иметь уровни сывороточного альбумина (например, уровни человеческого сывороточного альбумина), равные по меньшей мере приблизительно 1 мг/мл, по меньшей мере приблизительно 2 мг/мл, по меньшей мере приблизительно 3 мг/мл, по меньшей мере приблизительно 4 мг/мл, по меньшей мере приблизительно 5 мг/мл, по меньшей мере приблизительно 6 мг/мл, по меньшей мере приблизительно 7 мг/мл, по меньшей мере приблизительно 8 мг/мл, по меньшей мере приблизительно 9 мг/мл, по меньшей мере приблизительно 10 мг/мл, по меньшей мере приблизительно 11 мг/мл, по меньшей мере приблизительно 12 мг/мл, по меньшей мере приблизительно 13 мг/мл, по меньшей мере приблизительно 14 мг/мл или по меньшей мере приблизительно 15 мг/мл. В конкретном примере животное, отличное от человека (например, мышь), содержащее гуманизированный локус альбумина (например, гомозиготный гуманизированный локус альбумина), может иметь уровни сывороточного альбумина (например, уровни человеческого сывороточного альбумина) от приблизительно 10 мг/мл до приблизительно 15 мг/мл. В любом из приведенных выше примеров альбумин, кодируемый гуманизированным локусом альбумина, может содержать, например, сигнальный пептид человеческого альбумина. Например, в одном примере вся кодирующая последовательность альбумина эндогенного локуса альбумина была удалена и заменена соответствующей последовательностью человеческого альбумина, или область эндогенного локуса альбумина от старт-кодона до стоп-кодона была удалена и заменена соответствующей последовательностью человеческого альбумина.
III. Способы применения животных, отличных от человека, содержащих гуманизированный локус альбумина, для оценки эффективности реагентов, нацеленных на человеческий альбумин, in vivo или ex vivo
[00130] Предложены различные способы применения животных, отличных от человека, содержащих гуманизированный локус альбумина, как описано в других местах настоящего документа, для оценки или оптимизации доставки или эффективности реагентов, нацеленных на человеческий альбумин (например, терапевтических молекул или комплексов) in vivo или ex vivo. Поскольку животные, отличные от человека, содержат гуманизированный локус альбумина, указанные животные, отличные от человека, будут более точно отражать эффективность реагента, нацеленного на человеческий альбумин. Такие животные, отличные от человека, особенно полезны для тестирования реагентов для редактирования генома, разработанных для нацеливания на ген человеческого альбумина, поскольку животные, отличные от человека, раскрытые в настоящем документе, содержат гуманизированные эндогенные локусы альбумина, а не трансгенные вставки последовательности человеческого альбумина в случайных геномных локусах, и гуманизированные эндогенные локусы альбумина могут содержать ортологичную человеческую геномную последовательность альбумина как из кодирующих, так и из некодирующих областей, а не искусственную последовательность кДНК.
A. Способы тестирования эффективности реагентов, нацеленных на человеческий альбумин, in vivo или ex vivo
[00131] Предложены различные способы для оценки доставки или эффективности реагентов, нацеленных на человеческий альбумин, in vivo с использованием животных, отличных от человека, содержащих гуманизированный локус альбумина, как описано в других местах настоящего документа. Такие способы могут включать: (a) введение животному, отличному от человека, реагента, нацеленного на человеческий альбумин (т.е. введение реагента, нацеленного на человеческий альбумин, животному, отличному от человека); и (b) оценку активности указанного реагента, нацеленного на человеческий альбумин.
[00132] Реагент, нацеленный на человеческий альбумин, может представлять собой любой биологический или химический агент, нацеленный на человеческий локус альбумина (гена человеческого альбумина), мРНК человеческого альбумина или белок человеческого альбумина. Примеры реагентов, нацеленных на человеческий альбумин, раскрыты в других местах настоящего документа и включают, например, агенты для редактирования генома. Например, реагент, нацеленный на человеческий альбумин, может представлять собой нуклеиновую кислоту, нацеленную на альбумин (например, CRISPR/гидовые РНК Cas, короткие шпилечные РНК (кшРНК) или малые интерферирующие РНК (миРНК)), или нуклеиновую кислоту, кодирующую белок, нацеленный на альбумин (например, белок Cas, такой как Cas9, ZFN или TALEN). В качестве альтернативы, реагент, нацеленный на человеческий альбумин, может представлять собой антитело или антигенсвязывающий белок, нацеленные на альбумин, или любую другую большую молекулу или малую молекулу, нацеленную на человеческий альбумин. В одном примере реагент, нацеленный на человеческий альбумин, представляет собой агент для редактирования генома, такой как нуклеазный агент и/или экзогенная донорная нуклеиновая кислота (например, нацеливающий вектор). В конкретном примере агент для редактирования генома может нацеливаться на интрон 1, интрон 12 или интрон 13 гена человеческого альбумина. Например, агент для редактирования генома может нацеливаться на интрон 1 гена человеческого альбумина.
[00133] Такие нацеленные на человеческий альбумин реагенты могут быть введены с помощью любого способа доставки (например, AAV, LNP или HDD), как более подробно описано в других местах настоящего документа, и любого пути введения. Способы доставки терапевтических комплексов и молекул, а также пути введения более подробно раскрыты в других местах настоящего документа. В частных вариантах способов реагенты доставляют посредством опосредованной AAV доставки. Например, для нацеливания на печень может быть использован AAV8. В других частных вариантах способов реагенты доставляют с помощью опосредованной LNP доставки. В других частных вариантах способов реагенты доставляют с помощью гидродинамической доставки (HDD). Доза может представлять собой любую подходящую дозу. Например, в некоторых способах, в которых реагенты (например, мРНК и гРНК Cas9) доставляют с помощью опосредованной LNP доставки, доза может составлять от приблизительно 0,01 до приблизительно 10 мг/кг, от приблизительно 0,01 до приблизительно 5 мг/кг, от приблизительно 0,01 до приблизительно 4 мг/кг, от приблизительно 0,01 до приблизительно 3 мг/кг, от приблизительно 0,01 до приблизительно 2 мг/кг, от приблизительно 0,01 до приблизительно 1 мг/кг, от приблизительно 0,1 до приблизительно 10 мг/кг, от приблизительно 0,1 до приблизительно 6 мг/кг; от приблизительно 0,1 до приблизительно 5 мг/кг, от приблизительно 0,1 до приблизительно 4 мг/кг, от приблизительно 0,1 до приблизительно 3 мг/кг, от приблизительно 0,1 до приблизительно 2 мг/кг, от приблизительно 0,1 до приблизительно 1 мг/кг, от приблизительно 0,3 до приблизительно 10 мг/кг, от приблизительно 0,3 до приблизительно 6 мг/кг; от приблизительно 0,3 до приблизительно 5 мг/кг, от приблизительно 0,3 до приблизительно 4 мг/кг, от приблизительно 0,3 до приблизительно 3 мг/кг, от приблизительно 0,3 до приблизительно 2 мг/кг, от приблизительно 0,3 до приблизительно 1 мг/кг, приблизительно 0,1 мг/кг, приблизительно 0,3 мг/кг, приблизительно 1 мг/кг, приблизительно 2 мг/кг или приблизительно 3 мг/кг. В конкретном примере доза составляет от приблизительно 0,1 до приблизительно 6 мг/кг; от приблизительно 0,1 до приблизительно 3 мг/кг или от приблизительно 0,1 до приблизительно 2 мг/кг.
[00134] Способы оценки активности реагента, нацеленного на человеческий альбумин, хорошо известны и приведены в других местах настоящего документа. Оценка активности может проводиться в любом типе клеток, любом типе ткани или любом типе органа, как раскрыто в других местах настоящего документа. В некоторых способах оценку активности проводят в клетках печени. Если нацеленный на альбумин реагент представляет собой реагент для редактирования генома (например, нуклеазный агент), такие способы могут включать оценку модификации гуманизированного локуса альбумина. В качестве примера, указанная оценка может включать измерение активности негомологичного соединения концов (NHEJ) в гуманизированном локусе альбумина. Оно может включать, например, измерение частоты вставок или делеций в гуманизированном локусе альбумина. Например, указанная оценка может включать секвенирование гуманизированного локуса альбумина в одной или более клетках, выделенных от животного, отличного от человека (например, секвенирование нового поколения). Оценка может включать выделение целевого органа или ткани (например, печени) или ткани у животного, отличного от человека, и оценку модификации гуманизированного локуса альбумина в целевом органе или ткани. Оценка также может включать оценку модификации гуманизированного локуса альбумина в двух или более различных типах клеток в целевом органе или ткани. Аналогичным образом, оценка может включать выделение нецелевого органа или ткани (например, двух или более нецелевых органов или тканей) от животного, отличного от человека, и оценку модификации гуманизированного локуса альбумина в нецелевом органе или ткани.
[00135] Такие методы также могут включать измерение уровней экспрессии мРНК, продуцируемой гуманизированным локусом альбумина, или путем измерения уровней экспрессии белка, кодируемого гуманизированным локусом альбумина. Например, могут быть измерены уровни белка в конкретном типе клетки, ткани или органа (например, печени), или могут быть измерены секретированные уровни в сыворотке. Методы оценки экспрессии мРНК альбумина или белка, экспрессируемого из гуманизированного локуса альбумина, приведены в других местах настоящего документа и хорошо известны. В качестве примера может быть использован анализ in situ гибридизации РНК BASESCOPE™ (ISH), например, для количественной оценки клеточно-специфичных отредактированных транскриптов.
[00136] В некоторых способах реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту (например, нацеливающий вектор). Такие экзогенные донорные нуклеиновые кислоты могут кодировать экзогенный белок, не кодируемый или не экспрессируемый эндогенным локусом альбумина дикого типа (например, могут содержать вставку нуклеиновой кислоты, кодирующую экзогенный белок). В одном примере экзогенный белок может представлять собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с белком, не кодируемым или не экспрессируемым эндогенным локусом альбумина дикого типа. В одном примере экзогенный белок, кодируемый экзогенной донорной нуклеиновой кислотой после интегрирования в гуманизированный локус альбумина может представлять собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с белком, не кодируемым или не экспрессируемым эндогенным локусом альбумина дикого типа. В некоторых способах оценка может включать измерение экспрессии матричной РНК, кодируемой указанной экзогенной донорной нуклеиновой кислотой. Оценка также может включать измерение экспрессии экзогенного белка. Например, экспрессия экзогенного белка может быть измерена в печени животного, отличного от человека, или могут быть измерены сывороточные уровни экзогенного белка.
[00137] В некоторых способах животные, отличные от человека, содержащие гуманизированный локус альбумина, как описано в других местах настоящего документа, дополнительно содержат инактивированный (нокаутированный) эндогенный ген, который не является локусом альбумина, и, необязательно, реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту (например, нацеливающий вектор), кодирующий экзогенный белок для замены функции инактивированного эндогенного гена. В конкретном примере инактивированным эндогенным геном является F9, а экзогенным белком является фактор свертывания крови IX (например, человеческий фактор свертывания крови IX).
[00138] В некоторых способах реагент, нацеленный на человеческий альбумин, содержит (1) нуклеазный агент, разработанный для нацеливания на область гена человеческого альбумина, и (2) экзогенную донорную нуклеиновую кислоту, которая разработана для нацеливания на ген человеческого альбумина. Экзогенная донорная нуклеиновая кислота может, например, кодировать экзогенный белок, необязательно где указанный белок, кодируемый гуманизированным эндогенным локусом альбумина, на который осуществляли нацеливание с помощью указанной экзогенной донорной нуклеиновой кислоты, представляет собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с указанным экзогенным белком.
[00139] В качестве одного конкретного примера, если реагент, нацеленный на человеческий альбумин, представляет собой реагент для редактирования генома (например, нуклеазный агент), может быть оценен процент редактирования (например, общее количество вставок или делеций, получаемое из общего количества последовательностей, считанных в реакции ПЦР из пула лизированных клеток) в гуманизированном локусе альбумина (например, в клетках печени).
[00140] Различные способы, приведенные выше для оценки активности in vivo, также могут быть использованы для оценки активности реагентов, нацеленных на человеческий альбумин, ex vivo, как описано в других местах настоящего документа.
[00141] В некоторых способах реагент, нацеленный на человеческий альбумин, представляет собой нуклеазный агент, такой как нуклеазный агент CRISPR/Cas, нацеленный на гена человеческого альбумина. Такие способы могут включать, например: (a) введение животному, отличному от человека, нуклеазного агента, разработанного для расщепления гена человеческого альбумина (например, белка Cas, такого как Cas9, и гидовой РНК, разработанной для нацеливания на последовательность-мишень гидовой РНК в ген человеческого альбумина); и (b) оценку модификации гуманизированного локуса альбумина.
[00142] Например, в случае нуклеазы CRISPR/Cas модификация гуманизированного локуса альбумина будет вызвана при образовании гидовой РНК комплекса с белком Cas и направлении белка Cas в гуманизированный локус альбумина, и комплекс Cas/гидовая РНК расщепляет последовательность-мишень гидовой РНК, запуская репарацию в клетке (например, посредством негомологичного соединения концов (NHEJ), при отсутствии донорной последовательности).
[00143] Необязательно, могут быть введены две или более гидовых РНК, каждая из которых разработана для нацеливания на отличающуюся последовательность-мишень гидовой РНК в гене человеческого альбумина. Например, могут быть разработаны две гидовые РНК для вырезания геномной последовательности между двумя последовательностями-мишенями гидовой РНК. Модификация гуманизированного локуса альбумина будет вызвана, когда первая гидовая РНК образует комплекс с белком Cas и направляет белок Cas к гуманизированному локусу альбумина, вторая гидовая РНК образует комплекс с белком Cas и направляет белок Cas к гуманизированному локусу альбумина, первый комплекс Cas/гидовая РНК расщепляет первую последовательность-мишень гидовой РНК, а второй комплекс Cas/гидовая РНК расщепляет вторую последовательность-мишень гидовой РНК, что приводит к вырезанию промежуточной последовательности.
[00144] Дополнительно или в качестве альтернативы, животному, отличному от человека, также вводят экзогенную донорную нуклеиновую кислоту (например, нацеливающий вектор), способную вступать в рекомбинацию с геном человеческого альбумина и модифицировать его. Необязательно, нуклеазный агент или белок Cas могут быть присоединены к экзогенной донорной нуклеиновой кислоте, как описано в других местах настоящего документа. Модификация гуманизированного локуса альбумина будет вызвана, например, при образовании гидовой РНК комплекса с белком Cas и направлении белка Cas в гуманизированный локус альбумина, комплекс Cas/гидовая РНК расщепляет последовательность-мишень гидовой РНК, и гуманизированный локус альбумина вступает в рекомбинацию с экзогенной донорной нуклеиновой кислотой с модификацией гуманизированного локуса альбумина. Экзогенная донорная нуклеиновая кислота может вступать в рекомбинацию с гуманизированным локусом альбумина, например, посредством репарации, опосредованной гомологичной рекомбинацией (HDR), или посредством NHEJ-опосредованной вставки. Может быть использован любой тип экзогенной донорной нуклеиновой кислоты, примеры которых приведены в других местах настоящего документа.
[00145] В некоторых способах реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту (например, нацеливающий вектор). Такие экзогенные донорные нуклеиновые кислоты могут кодировать экзогенный белок, не кодируемый или не экспрессируемый эндогенным локусом альбумина дикого типа (например, могут содержать вставку нуклеиновой кислоты, кодирующую экзогенный белок). В одном примере экзогенный белок может представлять собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с белком, не кодируемым или не экспрессируемым эндогенным локусом альбумина дикого типа. Например, экзогенная донорная нуклеиновая кислота может представлять собой кассету без промотора, содержащую акцептор сплайсинга, и экзогенная донорная нуклеиновая кислота может быть нацелена на первый интрон человеческого альбумина.
B. Способы оптимизации доставки или эффективности реагента, нацеленного на человеческий альбумин, in vivo или ex vivo
[00146] Предложены различные способы оптимизации доставки реагентов, нацеленных на человеческий альбумин, в клетку или в организм животного, отличного от человека, или оптимизации активности или эффективности реагентов, нацеленных на человеческий альбумин, in vivo. Такие способы могут включать, например: (a) осуществление способа тестирования эффективности реагента, нацеленного на человеческий альбумин, как описано выше, в первый раз у первого животного, отличного от человека, или в первой клетке, содержащей гуманизированный локус альбумина; (b) изменение переменной и осуществление способа второй раз у второго животного, отличного от человека (т.е. того же вида), или во второй клетке, содержащей гуманизированный локус альбумина с измененной переменной; и (c) сравнение активности указанного реагента, нацеленного на человеческий альбумин, на стадии (a) с активностью указанного реагента, нацеленного на человеческий альбумин, на стадии (b), и выбор способа, обеспечивающего более высокую активность.
[00147] Способы измерения доставки, эффективности или активности реагентов, нацеленных на человеческий альбумин, раскрыты в других местах настоящего документа. Например, такие способы могут включать измерение модификации гуманизированного локуса альбумина. Более эффективная модификация гуманизированного локуса альбумина может означать разные вещи в зависимости от желаемого эффекта у животного, отличного от человека, или в клетке, отличной от человеческой. Например, более эффективная модификация гуманизированного локуса альбумина может означать одно или более, или все из более высоких уровней модификации, более высокой точности, более высокого единообразия или более высокой специфичности. Более высокие уровни модификации (т.е. более высокая эффективность) гуманизированного локуса альбумина относятся к более высокому проценту клеток, на которые осуществляется нацеливание, в определенном целевом типе клеток, в определенной целевой ткани или в определенном целевом органе (например, печени). Более высокая точность относится к более точной модификации гуманизированного локуса альбумина (например, более высокому проценту целевых клеток, имеющих одинаковую модификацию или имеющих желаемую модификацию без дополнительных непреднамеренных вставок и делеций (например, инделей NHEJ)). Более высокое единообразие означает более единообразную модификацию гуманизированного локуса альбумина среди различных типов целевых клеток, тканей или органов, если нацеливание осуществляется более чем на один тип клетки, ткани или органа (например, модификация большего количества типов клеток в печени). Если мишенью является конкретный орган, более высокое единообразие может также относиться к более единообразным модификациям во всех местах органа (например, в печени). Более высокая специфичность может относиться к более высокой специфичности в отношении целевого геномного локуса или локусов, более высокой специфичности в отношении целевого типа клеток, более высокой специфичности в отношении целевого типа ткани или более высокой специфичности в отношении целевого органа. Например, повышенная специфичность к геномному локусу относится к меньшей модификации нецелевых геномных локусов (например, более низкому проценту целевых клеток, имеющих модификации в непреднамеренных, нецелевых геномных локусах вместо или в дополнение к модификации целевого геномного локуса). Аналогичным образом, повышенная специфичность к типу клеток, ткани или органа относится к меньшим нецелевым модификациям типов клеток, типов тканей или типов органов, если мишенью является конкретный тип клеток, тип ткани или тип органа (например, когда мишенью является конкретный орган (например, печень), имеет место меньше изменений в клетках в органах или тканях, которые не являются предполагаемыми мишенями).
[00148] Изменяемая переменная может быть любым параметром. В качестве примера, изменяемой переменной может быть упаковка или способ доставки, с помощью которых реагент или реагенты, нацеленные на человеческий альбумин, вводятся в клетку или животное, отличное от человека. Примеры способов доставки, таких как LNP, HDD и AAV, раскрыты в других местах в настоящей заявке. Например, изменяемой переменной может быть серотип AAV. Аналогичным образом, введение может включать опосредованную LNP доставку, и изменяемая переменная может представлять собой препарат LNP. В качестве еще одного примера, изменяемая переменная может представлять собой путь введения реагента или реагентов, нацеленных на человеческий альбумин, в клетку или животное, отличное от человека. Примеры путей введения, таких как внутривенный, интравитреальный, интрапаренхиматозный и назальная инстилляция, раскрыты в других местах настоящего документа.
[00149] В качестве еще одного примера, изменяемая переменная может представлять собой концентрацию или количество введенного реагента или реагентов, нацеленных на человеческий альбумин. В качестве еще одного примера, изменяемая переменная может представлять собой концентрацию или количество одного введенного реагента, нацеленного на человеческий альбумин (например, гидовой РНК, белка Cas или экзогенной донорной нуклеиновой кислоты), по отношению к концентрации или количеству другого реагента, нацеленного на человеческий альбумин (например, гидовой РНК, белка Cas или экзогенной донорной нуклеиновой кислоты).
[00150] В качестве еще одного примера, изменяемая переменная может представлять собой время введения реагента или реагентов, нацеленных на человеческий альбумин, относительно времени оценки активности или эффективности указанных реагентов. В качестве еще одного примера, изменяемая переменная может представлять собой количество раз или частота введения реагента или реагентов, нацеленных на человеческий альбумин. В качестве еще одного примера, изменяемая переменная может представлять собой время введения одного введенного реагента, нацеленного на человеческий альбумин (например, гидовой РНК, белка Cas или экзогенной донорной нуклеиновой кислоты), по отношению ко времени введения другого реагента, нацеленного на человеческий альбумин (например, гидовой РНК, белка Cas или экзогенной донорной нуклеиновой кислоты).
[00151] В качестве еще одного примера, изменяемая переменная может представлять собой форму, в которой вводят реагент или реагенты, нацеленные на человеческий альбумин. Например, гидовая РНК может быть введена в форме ДНК или в форме РНК. Белок Cas (например, Cas9) может быть введен в форме ДНК, в форме РНК или в форме белка (например, в комплексе с гидовой РНК). Экзогенная донорная нуклеиновая кислота может быть ДНК, РНК, одноцепочечной, двухцепочечной, линейной, кольцевой и так далее. Точно так же каждый из компонентов может содержать различные комбинации модификаций для стабильности, для уменьшения нецелевых эффектов, для облегчения доставки и так далее.
[00152] В качестве еще одного примера, изменяемая переменная может представлять собой вводимые реагент или реагенты, нацеленные на человеческий альбумин. Например, если реагент, нацеленный на человеческий альбумин, содержит гидовую РНК, изменяемая переменная может представлять собой введение другой гидовой РНК с другой последовательностью (например, нацеленной на другую последовательность-мишень гидовой РНК). Аналогичным образом, если реагент, нацеленный на человеческий альбумин, содержит белок Cas, изменяемая переменная может представлять собой введение другого белка Cas (например, введение другого белка Cas с другой последовательностью) или нуклеиновой кислоты с другой последовательностью (например, кодон-оптимизированной), но кодирующей ту же аминокислотную последовательность белка Cas. Аналогичным образом, если реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту, изменяемая переменная может представлять собой введение другой экзогенной донорной нуклеиновой кислоты с другой последовательностью (например, другой вставленной нуклеиновую кислотой или другими гомологичными плечами (например, более длинными или более короткими гомологичными плечами или гомологичными плечами, нацеленными на другую область гена человеческого альбумина)).
[00153] В конкретном примере реагент, нацеленный на человеческий альбумин, содержит белок Cas и гидовую РНК, разработанную для нацеливания на последовательность-мишень гидовой РНК в гене человеческого альбумина. В таких способах изменяемая переменная может представлять собой последовательность гидовой РНК и/или последовательность-мишень гидовой РНК. В некоторых таких способах и белок Cas, и гидовая РНК могут быть введены в форме РНК, и изменяемая переменная может представлять собой отношение мРНК Cas к гидовой РНК (например, в препарате LNP). В некоторых таких способах изменяемая переменная может представлять собой модификации гидовой РНК (например, гидовая РНК с модификацией сравнивается с гидовая РНК без модификации).
C. Реагенты, нацеленные на человеческий альбумин
[00154] Реагентом, нацеленным на человеческий альбумин, может быть любой реагент, нацеленный на гена человеческого альбумина, мРНК человеческого альбумина или белок человеческого альбумина. Например, он может представлять собой реагент для редактирования генома, такой как нуклеазный агент, который расщепляет последовательность-мишень в гене человеческого альбумина, и/или экзогенную донорную последовательность, которая вступает в рекомбинацию с геном человеческого альбумина, он может представлять собой антисмысловой олигонуклеотид, нацеленный на мРНК человеческого альбумина, он может представлять собой антигенсвязывающий белок, нацеленный на эпитоп белка человеческого альбумина, или он может представлять собой малую молекулу, нацеленную на человеческий альбумин. Реагенты, нацеленные на человеческий альбумин, в раскрытых в настоящем документе способах могут представлять собой известные реагенты, нацеленные на человеческий альбумин, могут представлять собой реагенты, предположительно нацеленные на альбумин (например, реагенты-кандидаты, разработанные для нацеливания на человеческий альбумин), или могут представлять собой реагенты, проверяемые на предмет активности нацеливания на человеческий альбумин.
(1) Нуклеазные агенты, нацеленные на гена человеческого альбумина
[00155] Реагент, нацеленный на человеческий альбумин, может представлять собой реагент для редактирования генома, такой как нуклеазный агент, который расщепляет последовательность-мишень в гене человеческого альбумина. Последовательность-мишень нуклеазы содержит последовательность ДНК, в которой нуклеазный агент вызывает ник или двухцепочечный разрыв. Последовательность-мишень для нуклеазного агента может быть эндогенной (или нативной) по отношению к клетке, или последовательность-мишень может быть экзогенной по отношению к клетке. Последовательность-мишень, экзогенная по отношению к клетке, в природе не встречается в геноме клетки. Последовательность-мишень также может быть экзогенной по отношению к представляющим интерес полинуклеотидам, которые требуется поместить в локусе-мишени. В некоторых случаях последовательность-мишень присутствует в виде только одной копии в геноме клетки-хозяина. В конкретном примере последовательность-мишень нуклеазы может находиться в интроне 1, интроне 12 или интроне 13 гена человеческого альбумина. Например, последовательность-мишень нуклеазы может находиться в интроне 1 гена человеческого альбумина.
[00156] Длина последовательности-мишени может варьироваться и включает, например, последовательности-мишени, которые составляют приблизительно 30-36 п.н. для пары нуклеаз с «цинковыми пальцами» (ZFN) (т.е. приблизительно 15-18 п.н. для каждой ZFN), приблизительно 36 п.н. для эффекторной нуклеазы, подобной активатору транскрипции (TALEN), или приблизительно 20 п.н. для гидовой РНК CRISPR/Cas9.
[00157] В способах и композициях, описанных в данном документе, может быть использован любой нуклеазный агент, который вызывает ник или двухцепочечный разрыв в желаемой последовательности-мишени. Может быть использован встречающийся в природе или нативный нуклеазный агент при условии, что нуклеазный агент вызывает ник или двухцепочечный разрыв в желаемой последовательности-мишени. В качестве альтернативы, может быть использован модифицированный или сконструированный нуклеазный агент. «Сконструированный нуклеазный агент» включает нуклеазу, которая сконструирована (модифицирована или получена) из ее нативной формы таким образом, чтобы она могла специфически распознавать и вызывать ник или двухцепочечный разрыв в желаемой последовательности-мишени. Таким образом, сконструированный нуклеазный агент может быть получен из нативного природного нуклеазного агента, или может быть создан или синтезирован искусственно. Сконструированная нуклеаза может вызывать ник или двухцепочечный разрыв в последовательности-мишени, например, когда последовательность-мишень не является последовательностью, которая была бы распознана нативным (несконструированным или немодифицированным) нуклеазным агентом. Модификация нуклеазного агента может представлять собой изменение всего одной аминокислоты в белке расщепляющего агента или одного нуклеотида в нуклеиновой кислоте расщепляющего агента. Создание ника или двухцепочечного разрыва в последовательности-мишени или другой ДНК может называться в настоящем документе «разрезанием» или «расщеплением» последовательности-мишени или другой ДНК.
[00158] Также предложены активные варианты и фрагменты приведенных в качестве примеров последовательностей-мишеней. Такие активные варианты могут обладать по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью последовательности с рассматриваемой последовательностью-мишенью, при этом активные варианты сохраняют биологическую активность и, следовательно, способны распознаваться и расщепляться нуклеазным агентом сиквенс-специфичным образом. Способы измерения двухцепочечного разрыва последовательности-мишени нуклеазным агентом хорошо известны. См., например, источник Frendewey et al. (2010) Methods in Enzymology 476:295-307, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00159] Последовательность-мишень нуклеазного агента может быть расположена в любом месте или рядом с локусом алюбумина. Последовательность-мишень может быть расположена в кодирующей области гена альбумина или в регуляторных областях, которые влияют на экспрессию гена. Последовательность-мишень нуклеазного агента может быть расположена в интроне, экзоне, промоторе, энхансере, регуляторной области или любой небелковой кодирующей области.
[00160] Одним из типов нуклеазных агентов является эффекторная нуклеаза, подобная активатору транскрипции (TALEN). Эффекторные нуклеазы TAL представляют собой класс сиквенс-специфичных нуклеаз, которые могут быть использованы для создания двухцепочечных разрывов в определенных последовательностях-мишенях в геноме прокариотического или эукариотического организма. Эффекторные нуклеазы TAL создают путем слияния природного или сконструированного эффектора, подобного активатору транскрипции (TAL), или его функциональной части, с каталитическим доменом эндонуклеазы, такой как, например, FokI. Уникальный модульный эффекторный ДНК-связывающий домен TAL позволяет разрабатывать белки, которые потенциально могут обладать специфичностью распознавания любой рассматриваемой ДНК. Таким образом, ДНК-связывающие домены эффекторных нуклеаз TAL могут быть разработаны для распознавания специфических сайтов-мишеней ДНК и, таким образом, использованы для создания двухцепочечных разрывов в желаемых последовательностях-мишенях. См. источники WO 2010/079430; Morbitzer et al. (2010) PNAS 10.1073/pnas.1013133107; Scholze & Boch (2010) Virulence 1:428-432; Christian et al. Genetics (2010) 186:757-761; Li et al. (2010) Nuc. Acids Res. (2010) doi:10.1093/nar/gkq704; и Miller et al. (2011) Nature Biotechnology 29:143-148, каждый из которых включен в настоящий документ посредством ссылки в полном объеме.
[00161] Примеры подходящих нуклеаз TAL и способы получения подходящих нуклеаз TAL раскрыты, например, в источниках US 2011/0239315 A1, US 2011/0269234 A1, US 2011/0145940 A1, US 2003/0232410 A1, US 2005/0208489 A1, US 2005/0026157 A1, US 2005/0064474 A1, US 2006/0188987 A1 и US 2006/0063231 A1, каждый из которых включен в настоящий документ посредством ссылки в полном объеме. В различных вариантах осуществления эффекторные нуклеазы TAL сконструированы таким образом, что они создают разрез в последовательности нуклеиновой кислоты-мишени или рядом с ней, например, в представляющем интерес локусе или представляющем интерес геномном локусе, причем последовательность нуклеиновой кислоты-мишени находится в последовательности, подлежащей модификации посредством нацеливающего вектора, или рядом с ней. Нуклеазы TAL, подходящие для применения с различными способами и композициями, предложенными в настоящем документе, включают специально разработанные для связывания с последовательностями нуклеиновых кислот-мишеней, подлежащих модификации нацеливающими векторами, как описано в настоящем документе, или рядом с ними.
[00162] В некоторых TALEN каждый мономер TALEN содержит 33-35 повторов TAL, которые распознают одну пару оснований посредством двух гипервариабельных остатков. В некоторых TALEN нуклеазный агент представляет собой химерный белок, содержащий ДНК-связывающий домен на основе TAL-повтора, функционально связанный с независимой нуклеазой, такой как эндонуклеаза FokI. Например, нуклеазный агент может содержать ДНК-связывающий домен на основе первого TAL-повтора и второй ДНК-связывающий домен на основе TAL-повтора, при этом каждый из первого и второго ДНК-связывающего домена на основе TAL-повторов функционально связан с нуклеазой FokI, при этом первый и второй ДНК-связывающий домен на основе TAL-повтора распознает две смежные последовательности ДНК-мишени в каждой цепи последовательности ДНК-мишени, разделенные спейсерной последовательностью различной длины (12-20 п.н.), и при этом субъединицы нуклеазы FokI димеризуются с образованием активной нуклеазы, которая делает двухцепочечный разрыв в последовательности-мишени.
[00163] Нуклеазный агент, используемый в различных раскрытых в настоящем документе способах и композициях, может дополнительно содержать нуклеазу с «цинковыми пальцами» (ZFN). В некоторых ZFN каждый мономер ZFN содержит 3 или более ДНК-связывающих домена с «цинковыми пальцами», при этом каждый ДНК-связывающий домен с «цинковыми пальцами» связывается с субсайтом размером 3 п.н. В других ZFN, ZFN представляет собой химерный белок, содержащий ДНК-связывающий домен с «цинковыми пальцами», функционально связанный с независимой нуклеазой, такой как эндонуклеаза FokI. Например, нуклеазный агент может содержать первую ZFN и первую ZFN, при этом каждая из первой ZFN и второй ZFN функционально связана с субъединицей нуклеазы FokI, при этом первая и вторая ZFN распознают две смежные последовательности ДНК-мишени в каждой цепи последовательности ДНК-мишени, разделенные спейсером размером приблизительно 5-7 п.н., и при этом субъединицы нуклеазы FokI димеризуются с образованием активной нуклеазы, которая делает двухцепочечный разрыв. См., например, источники US 20060246567; US 20080182332; US 20020081614; US 20030021776; WO/2002/057308A2; US 20130123484; US 20100291048; WO/2011/017293A2; и Gaj et al. (2013) Trends in Biotechnology, 31(7):397-405, каждый из которых включен в настоящий документ посредством ссылки.
[00164] Другой тип подходящего нуклеазного агента представляет собой сконструированную мегануклеазу. На основе консервативных мотивов последовательностей мегануклеазы были классифицированы на четыре семейства, которые представляют собой семейства LAGLIDADG, GIY-YIG, H-N-H и His-Cys бокс. Эти мотивы участвуют в координации ионов металлов и гидролизе фосфодиэфирных связей. Мегануклеазы отличаются своими длинными последовательностями-мишенями и устойчивостью к некоторым полиморфизмам последовательностей в их ДНК-субстратах. Домены, структура и функция мегануклеаз известны, см., например, источники Guhan and Muniyappa (2003) Crit Rev Biochem Mol Biol 38:199-248; Lucas et al., (2001) Nucleic Acids Res 29:960-9; Jurica and Stoddard, (1999) Cell Mol Life Sci 55:1304-26; Stoddard, (2006) Q Rev Biophys 38:49-95; и Moure et al., (2002) Nat Struct Biol 9:764. В некоторых примерах используют встречающийся в природе вариант и/или сконструированную производную мегануклеазу. Известны способы модификации кинетики, взаимодействий с кофакторами, экспрессии, оптимальных условий и/или специфичности к последовательности-мишени, а также скрининга на активность. См., например, источники Epinat et al., (2003) Nucleic Acids Res 31:2952-62; Chevalier et al., (2002) Mol Cell 10:895-905; Gimble et al., (2003) Mol Biol 334:993-1008; Seligman et al., (2002) Nucleic Acids Res 30:3870-9; Sussman et al., (2004) J Mol Biol 342:31-41; Rosen et al., (2006) Nucleic Acids Res 34:4791-800; Chames et al., (2005) Nucleic Acids Res 33:e178; Smith et al., (2006) Nucleic Acids Res 34:e149; Gruen et al., (2002) Nucleic Acids Res 30:e29; Chen and Zhao, (2005) Nucleic Acids Res 33:e154; WO 2005105989; WO 2003078619; WO 2006097854; WO 2006097853; WO 2006097784; и WO 2004031346, каждый из которых включен в настоящий документ посредством ссылки в полном объеме.
[00165] Может быть использована любая мегануклеаза, включая, например, I-SceI, I-SceII, I-SceIII, I-SceIV, I-SceV, I-SceVI, I-SceVII, I-CeuI, I-CeuAIIP, I-CreI, I-CrepsbIP, I-CrepsbIIP, I-CrepsbIIIP, I-CrepsbIVP, I-TliI, I-PpoI, PI-PspI, F-SceI, F-SceII, F-SuvI, F-TevI, F-TevII, I-AmaI, I-AniI, I-ChuI, I-CmoeI, I-CpaI, I-CpaII, I-CsmI, I-CvuI, I-CvuAIP, I-DdiI, I-DdiII, I-DirI, I-DmoI, I-HmuI, I-HmuII, I-HsNIP, I-LlaI, I-MsoI, I-NaaI, I-NanI, I-NcIIP, I-NgrIP, I-NitI, I-NjaI, I-Nsp236IP, I-PakI, I-PboIP, I-PcuIP, I-PcuAI, I-PcuVI, I-PgrIP, I-PobIP, I-PorI, I-PorIIP, I-PbpIP, I-SpBetaIP, I-ScaI, I-SexIP, I-SneIP, I-SpomI, I-SpomCP, I-SpomIP, I-SpomIIP, I-SquIP, I-Ssp6803I, I-SthPhiJP, I-SthPhiST3P, I-SthPhiSTe3bP, I-TdeIP, I-TevI, I-TevII, I-TevIII, I-UarAP, I-UarHGPAIP, I-UarHGPA13P, I-VinIP, I-ZbiIP, PI-MtuI, PI-MtuHIP PI-MtuHIIP, PI-PfuI, PI-PfuII, PI-PkoI, PI-PkoII, PI-Rma43812IP, PI-SpBetaIP, PI-SceI, PI-TfuI, PI-TfuII, PI-ThyI, PI-TliI, PI-TliII, или любые их активные варианты или фрагменты.
[00166] Мегануклеазы могут распознавать, например, двухцепочечные последовательности ДНК длиной от 12 до 40 пар оснований. В некоторых случаях мегануклеаза распознает одну идеально соответствующую последовательность-мишень в геноме.
[00167] Некоторые мегануклеазы являются хоминг-нуклеазами. Одним из типов хоминг-нуклеазы является семейство хоминг-нуклеаз LAGLIDADG, включающее, например, I-SceI, I-CreI и I-Dmol.
[00168] Нуклеазные агенты могут дополнительно содержать системы CRISPR/Cas, как более подробно описано ниже.
[00169] Также предложены активные варианты и фрагменты нуклеазных агентов (т.е. сконструированный нуклеазный агент). Такие активные варианты могут обладать по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью последовательности с нативным нуклеазным агентом, при этом активные варианты сохраняют способность разрезать желаемую последовательность-мишень и, следовательно, сохраняют активность вызывания ников или двухцепочечных разрывов. Например, любой из описанных в данном документе нуклеазных агентов может быть модифицирован относительно последовательности нативной эндонуклеазы и разработан для распознавания и вызывания ников или двухцепочечных разрывов в последовательности-мишени, которая не распознавалась нативным нуклеазным агентом. Таким образом, некоторые сконструированные нуклеазы обладают специфичностью вызывания ников или двухцепочечных разрывов в последовательности-мишени, которая отличается от соответствующей последовательности-мишени нативного нуклеазного агента. Анализы на активность вызывания ников или двухцепочечных разрывов известны, и обычно в них измеряют общую активность и специфичность эндонуклеазы на ДНК-субстратах, содержащих последовательность-мишень.
[00170] Нуклеазный агент может быть введен в клетку или животное, отличное от человека, любым известным способом. Полипептид, кодирующий нуклеазный агент, может быть введен непосредственно в клетку или животное, отличное от человека. В качестве альтернативы, в клетку может быть введен полинуклеотид, кодирующий нуклеазный агент. При введении в клетку полинуклеотида, кодирующего нуклеазный агент, нуклеазный агент может временно, условно или конститутивно экспрессироваться внутри клетки. Таким образом, полинуклеотид, кодирующий нуклеазный агент, может содержаться в кассете экспрессии и быть функционально связан с условным промотором, индуцибельным промотором, конститутивным промотором или тканеспецифическим промотором. Примеры промоторов обсуждаются более подробно в других местах настоящего документа. В качестве альтернативы, нуклеазный агент вводят в клетку в виде мРНК, кодирующей нуклеазный агент.
[00171] Полинуклеотид, кодирующий нуклеазный агент, может быть стабильно интегрирован в геном клетки и функционально связан с активным в клетке промотором. В качестве альтернативы, полинуклеотид, кодирующий нуклеазный агент, может находиться в нацеливающем векторе.
[00172] Когда нуклеазный агент вводят в клетку посредством внесения полинуклеотида, кодирующего нуклеазный агент, такой полинуклеотид, кодирующий нуклеазный агент, может быть модифицирован для замены кодонов, имеющих более высокую частоту использования в представляющей интерес клетке, по сравнению с встречающейся в природе полинуклеотидной последовательностью, кодирующей нуклеазный агент. Например, полинуклеотид, кодирующий нуклеазный агент, может быть модифицирован для замены кодонов, имеющих более высокую частоту использования в данной представляющей интерес эукариотической клетке, включая человеческую клетку, клетку, отличную от человеческой, клетку млекопитающего, клетку грызуна, мышиную клетку, крысиную клетку или любую другую представляющую интерес клетку-хозяина, по сравнению с встречающейся в природе полинуклеотидной последовательностью.
(2) Системы CRISPR/Cas, нацеленные на гена человеческого альбумина
[00173] Особым типом реагента, нацеленного на человеческий альбумин, может быть система кластерные короткие палиндромные повторы, разделенные регулярными промежутками (CRISPR)/CRISPR-ассоциированная (Cas) система, нацеленная на гена человеческого альбумина. Системы CRISPR/Cas включают в себя транскрипты и другие элементы, участвующие в экспрессии генов Cas или управляющие активностью генов Cas. Система CRISPR/Cas может представлять собой, например, систему типа I, типа II, типа III или систему типа V (например, подтип V-A или подтип V-B). Системы CRISPR/Cas, используемые в раскрытых в настоящем документе композициях и способах, могут быть не встречающимися в природе. «Не встречающаяся в природе» система включает в себя то, что было создано деятельностью человека, например, один или более компонентов системы, измененных или мутированных по сравнению с тем состоянием, в котором они встречаются в природе, по меньшей мере по существу не содержащих по меньшей мере одного другого компонента, с которым они естественным образом связаны в природе, или связанных с по меньшей мере одним другим компонентом, с которым они не связаны естественным образом. Например, в некоторых системах CRISPR/Cas используются не встречающиеся в природе комплексы CRISPR, содержащие гРНК и белок Cas, которые вместе не встречаются в природе, используется белок Cas, который не встречается в природе, или используется гРНК, которая не встречается в природе.
[00174] Белки Cas и полинуклеотиды, кодирующие белки Cas. Белки Cas обычно содержат по меньшей мере один РНК-распознающий или связывающий домен, который может взаимодействовать с гидовыми РНК (гРНК). Белки Cas также могут содержать нуклеазные домены (например, ДНКазные домены или РНКазные домены), ДНК-связывающие домены, хеликазные домены, домены белок-белкового взаимодействия, домены димеризации и другие домены. Некоторые такие домены (например, ДНКазные домены) могут происходить из природного белка Cas. Другие такие домены могут быть добавлены для получения модифицированного белка Cas. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновой кислоты, которое включает разрыв ковалентных связей молекулы нуклеиновой кислоты. Расщепление может приводить к образованию тупых или ступенчатых концов, и оно может быть одноцепочечным или двухцепочечным. Например, белок Cas9 дикого типа обычно создает продукт расщепления с тупыми концами. В качестве альтернативы, белок Cpf1 дикого типа (например, FnCpf1) может приводить к образованию продукта расщепления с 5’-выступом из 5 нуклеотидов, при этом расщепление происходит после 18-й пары оснований от последовательности PAM в нецелевой цепи и после 23-го основания в целевой цепи. Белок Cas может обладать полной активностью расщепления для создания двухцепочечного ника в целевом геномном локусе (например, двухцепочечного разрыва с тупыми концами), или может представлять собой никазу, которая создает одноцепочечный разрыв в целевом геномном локусе.
[00175] Примеры белков Cas включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 или Csx12), Cas10, Cas10d, CasF, CasG, CasH, Csy1, Csy2, Csy3, Cse1 (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4 и Cu1966, а также их модифицированные версии и их гомологи.
[00176] Иллюстративный белок Cas представляет собой белок Cas9 или белок, производный из белка Cas9. Белки Cas9 происходят из системы CRISPR/Cas II типа и обычно имеют четыре общих ключевых мотива с консервативной структурой. Мотивы 1, 2 и 4 представляют собой RuvC-подобные мотивы, а мотив 3 представляет собой мотив HNH. Иллюстративные белки Cas9 происходят из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptomyces viridochromogenes, Streptosporangium roseum, Streptosporangium roseum, Alicyclobacillus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus, Acaryochloris marina, Neisseria meningitidis или Campylobacter jejuni. Дополнительные примеры представителей семейства Cas9 описаны в источнике WO 2014/131833, включенном в настоящий документ посредством ссылки в полном объеме для всех целей. Cas9 из S. pyogenes (SpCas9) (которой присвоен регистрационный номер SwissProt Q99ZW2) представляет собой пример белка Cas9. Cas9 из S. aureus (SaCas9) (которой присвоен регистрационный номер UniProt J7RUA5) представляет собой еще один пример белка Cas9. Cas9 из Campylobacter jejuni (CjCas9) (которой присвоен регистрационный номер UniProt Q0P897) представляет собой еще один иллюстративный белок Cas9. См., например, источник Kim et al. (2017) Nat. Comm. 8:14500, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. SaCas9 имеет меньший размер, чем SpCas9, а CjCas9 имеет меньший размер, чем оба белка SaCas9 и SpCas9. Иллюстративная последовательность белка Cas9 может содержать, по существу состоять из или по состоять из SEQ ID NO: 38. Иллюстративная ДНК, кодирующая белок Cas9, может содержать, по существу состоять из или по состоять из SEQ ID NO: 39.
[00177] Еще одиним примером белка Cas является белок Cpf1 (CRISPR из Prevotella и Francisella 1). Cpf1 представляет собой большой белок (приблизительно 1300 аминокислот), содержащий RuvC-подобный нуклеазный домен, гомологичный соответствующему домену Cas9, вместе с аналогом характерного богатого аргинином кластера Cas9. Тем не менее, в Cpf1 отсутствует нуклеазный домен HNH, который присутствует в белках Cas9, а RuvC-подобный домен в последовательности Cpf1 является непрерывным, в отличие от Cas9, где он содержит длинные вставки, включая домен HNH. См., например, источник Zetsche et al. (2015) Cell 163(3):759-771, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Иллюстративные белки Cpf1 происходят из Francisella tularensis 1, Francisella tularensis subsp. novicida, Prevotella albensis, Lachnospiraceae bacterium MC2017 1, Butyrivibrio proteoclasticus, Peregrinibacteria bacterium GW2011_GWA2_33_10, Parcubacteria bacterium GW2011_GWC2_44_17, Smithella sp. SCADC, Acidaminococcus sp. BV3L6, Lachnospiraceae bacterium MA2020, Candidatus Methanoplasma termitum, Eubacterium eligens, Moraxella bovoculi 237, Leptospira inadai, Lachnospiraceae bacterium ND2006, Porphyromonas crevioricanis 3, Prevotella disiens и Porphyromonas macacae. Cpf1 из Francisella novicida U112 (FnCpf1; присвоен регистрационный номер UniProt A0Q7Q2) представляет собой пример белка Cpf1.
[00178] Белки Cas могут представлять собой белки дикого типа (т.е. белки, которые встречаются в природе), модифицированные белки Cas (т.е. вариантные белки Cas) или фрагменты белков Cas дикого типа или модифицированных белков Cas. Белки Cas также могут быть активными вариантами или фрагментами в отношении каталитической активности белков Cas дикого типа или модифицированных белков Cas. Активные варианты или фрагменты в отношении каталитической активности могут обладать по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью последовательности с белком Cas дикого типа или модифицированным белком Cas, или его частью, при этом активные варианты сохраняют способность делать разрез в желаемом сайте расщепления и, следовательно, сохраняют активность вызывания ников или двухцепочечных разрывов. Анализы на активность вызывания ников или двухцепочечных разрывов известны, и обычно в них измеряют общую активность и специфичность белка Cas на ДНК-субстратах, содержащих сайт расщепления.
[00179] Белки Cas могут быть модифицированы для увеличения или уменьшения одного или более из аффинности связывания нуклеиновой кислоты, специфичности связывания нуклеиновой кислоты и ферментативной активности. Белки Cas также могут быть модифицированы для изменения любой другой активности или свойства белка, например, стабильности. Например, один или более нуклеазных доменов белка Cas могут быть модифицированы, удалены или инактивированы, или белок Cas может быть усечен для удаления доменов, которые не являются существенными для функции белка, или для оптимизации (например, увеличения или уменьшения) активности или свойства белка Cas.
[00180] Одним из примеров модифицированного белка Cas является модифицированный белок SpCas9-HF1, который представляет собой высокоточный вариант Cas9 Streptococcus pyogenes, несущий изменения (N497A/R661A/Q695A/Q926A), предназначенные для уменьшения неспецифических контактов с ДНК. См., например, источник Kleinstiver et al. (2016) Nature 529(7587):490-495, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Еще одним примером модифицированного белка Cas является модифицированный вариант eSpCas9 (K848A/K1003A/R1060A), разработанный для уменьшения нецелевых воздействий. См., например, источник Slaymaker et al. (2016) Science 351(6268):84-88, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Другие варианты SpCas9 включают K855A и K810A/K1003A/R1060A.
[00181] Белки Cas могут содержать по меньшей мере один нуклеазный домен, такой как ДНКазный домен. Например, белок Cpf1 дикого типа обычно содержит RuvC-подобный домен, который расщепляет обе цепи ДНК-мишени, возможно, в димерной конфигурации. Белки Cas также могут содержать по меньшей мере два нуклеазных домена, таких как ДНКазные домены. Например, белок Cas9 дикого типа обычно содержит RuvC-подобный нуклеазный домен и HNH-подобный нуклеазный домен. Каждый из доменов RuvC и HNH может разрезать разные цепи двухцепочечной ДНК с образованием двухцепочечного разрыва в ДНК. См., например, источник Jinek et al. (2012) Science 337:816-821, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00182] Один или более, или все нуклеазные домены могут быть удалены или мутированы таким образом, что они больше не функциональны или имеют пониженную нуклеазную активность. Например, если один из нуклеазных доменов в белке Cas9 удален или мутирован, то полученный в результате белок Cas9 может называться никазой и может генерировать одноцепочечный разрыв в двухцепочечной ДНК-мишени, но не двухцепочечный разрыв (т.е. он может расщеплять комплементарную цепь или некомплементарную цепь, но не обе из них). Если оба нуклеазных домена удалены или мутированы, то полученный в результате белок Cas (например, Cas9) будет иметь пониженную способность расщеплять обе цепи двухцепочечной ДНК (например, белок Cas с нулевой нуклеазной активностью или белок Cas с инактивированной нуклеазной активностью, или каталитически мертвый белок Cas (dCas)). Примером мутации, которая превращает Cas9 в никазу, является мутация D10A (аспартат в аланин в положении 10 Cas9) в домене RuvC Cas9 из S. pyogenes. Аналогичным образом, H939A (гистидин в аланин в аминокислотном положении 839), H840A (гистидин в аланин в аминокислотном положении 840) или N863A (аспарагин в аланин в аминокислотном положении N863) в домене HNH белка Cas9 из S. pyogenes могут превращать Cas9 в никазу. Другие примеры мутаций, которые превращают Cas9 в никазу, включают соответствующие мутации белка Cas9 из S. thermophilus. См., например, источники Sapranauskas et al. (2011) Nucleic Acids Research 39:9275-9282 и WO 2013/141680, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Такие мутации могут быть получены с использованием таких способов, как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез или полный генный синтез. Примеры других мутаций, создающих никазы, можно найти, например, в источниках WO 2013/176772 и WO 2013/142578, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Если в белке Cas удалены или мутированы все нуклеазные домены (например, в белке Cas9 удалены или мутированы оба нуклеазных домена), то полученный в результате белок Cas (например, Cas9) будет иметь пониженную способность расщеплять обе цепи двухцепочечной ДНК (например, белок Cas с нулевой нуклеазной активностью или с белок Cas с инактивированной нуклеазной активностью). Одним из конкретных примеров является двойной мутант D10A/H840A Cas9 из S. pyogenes или соответствующий двойной мутант Cas9 из другого вида при оптимальном выравнивании с последовательностью Cas9 из S. pyogenes. Еще одним конкретным примером является двойной мутант D10A/N863A Cas9 из S. pyogenes или соответствующий двойной мутант Cas9 из другого вида при оптимальном выравнивании с Cas9 S. pyogenes.
[00183] Также известны примеры инактивирующих мутаций в каталитических доменах белков Cas9 Staphylococcus aureus. Например, фермент Cas9 Staphylococcus aureus (SaCas9) может содержать замену в положении N580 (например, замену N580A) и замену в положении D10 (например, замену D10A) для создания белка Cas с инактивированной нуклеазной активностью. См., например, источник WO 2016/106236, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00184] Также известны примеры инактивирующих мутаций в каталитических доменах белков Cpf1. Для белков Cpf1 из Francisella novicida U112 (FnCpf1), Acidaminococcus sp. BV3L6 (AsCpf1), Lachnospiraceae bacterium ND2006 (LbCpf1) и Moraxella bovoculi 237 (MbCpf1 Cpf1) такие мутации могут включать мутации в положениях 908, 993 или 1263 в AsCpf1, или в соответствующих положениях в ортологах Cpf1, либо в положениях 832, 925, 947 или 1180 в LbCpf1, или в соответствующих положениях в ортологах Cpf1. Такие мутации могут включать, например, одну или более из мутаций D908A, E993A и D1263A в AsCpf1, или соответствующие мутации в ортологах Cpf1, или D832A, E925A, D947A и D1180A в LbCpf1, или соответствующие мутации в ортологах Cpf1. См., например, источник US 2016/0208243, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00185] Белки Cas (например, белки Cas с нуклеазной активностью или белки Cas с инактивированной нуклеазной активностью) также могут быть функционально связаны с гетерологичными полипептидами в качестве слитых белков. Например, белок Cas может быть слит с доменом расщепления или доменом эпигенетической модификации. См. источник WO 2014/089290, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Белки Cas также могут быть слиты с гетерологичным полипептидом, обеспечивающим повышенную или пониженную стабильность. Слитый домен или гетерологичный полипептид может быть расположен на N-конце, C-конце или внутри белка Cas.
[00186] В качестве примера, белок Cas может быть слит с одним или более гетерологичными полипептидами, которые обеспечивают субклеточную локализацию. Такие гетерологичные полипептиды могут включать, например, один или более сигналов ядерной локализации (NLS), таких как одинарный NLS SV40 и/или двойной NLS импортина-альфа, для нацеливания на ядро, сигнал митохондриальной локализации для нацеливания на митохондрии, сигнал ER-удержания и т.п. См., например, источник Lange et al. (2007) J. Biol. Chem. 282:5101-5105, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Такие сигналы субклеточной локализации могут быть расположены на N-конце, C-конце или в любой области в пределах белка Cas. NLS может содержать отрезок основных аминокислот и может быть монопартитной одинарной или двойной последовательностью. Необязательно, белок Cas может содержать два или более NLS, включая NLS (например, NLS импортина-альфа или одинарный NLS) на N-конце и NLS (например, NLS SV40 или двойной NLS) на C-конце. Белок Cas также может содержать два или более NLS на N-конце и/или два или более NLS на C-конце.
[00187] Белки Cas также могут быть функционально связаны с доменом проникновения в клетку или доменом белковой трансдукции. Например, проникающий в клетку домен может происходить из белка ТАТ ВИЧ-1, проникающего в клетку мотива TLM из вируса гепатита В человека, MPG, Pep-1, VP22, проникающего в клетку белка из вируса простого герпеса, или из полиаргининовой пептидной последовательности. См., например, источники WO 2014/089290 и WO 2013/176772, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Проникающий в клетку домен может быть расположен на N-конце, C-конце или в любой области в пределах белка Cas.
[00188] Белки Cas также могут быть функционально связаны с гетерологичным полипептидом для облегчения отслеживания или очистки, например, с флуоресцентным белком, меткой очистки или эпитопной меткой. Примеры флуоресцентных белков включают зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, мономерный Azami Green, CopGFP, AceGFP, ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP, ZsYellowl), синие флуоресцентные белки (например, eBFP, eBFP2, Azurite, mKalamal, GFPuv, Sapphire, T-sapphire), голубые флуоресцентные белки (например, eCFP, Cerulean, CyPet, AmCyanl, Midoriishi-Cyan), красные флуоресцентные белки (например, mKate, mKate2, mPlum, мономер DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, мономер DsRed, тандем HcRed, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry, Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine, tdTomato) и любой другой подходящий флуоресцентный белок. Примеры меток включают в себя глутатион-S-трансферазу (GST), хитин-связывающий белок (CBP), мальтозо-связывающий белок, тиоредоксин (TRX), поли(NANP), метку для тандемной аффинной очистки (TAP), myc, AcV5, AU1, AU5, E, ECS, E2, FLAG, гемагглютинин (HA), nus, Softag 1, Softag 3, Strep, SBP, Glu-Glu, HSV, KT3, S, S1, T7, V5, VSV-G, гистидин (His), белок-носитель карбоксила биотина (BCCP) и кальмодулин.
[00189] Белки Cas также могут быть присоединены к экзогенным донорным нуклеиновым кислотам или меченым нуклеиновым кислотам. Такое присоединение (т.е. физическое связывание) может быть достигнуто посредством ковалентных взаимодействий или нековалентных взаимодействий, и присоединение может быть прямым (например, посредством прямого слияния или химической конъюгации, что может быть достигнуто путем модификации остатков цистеина или лизина в белке или интеиновой модификации), или может быть достигнуто с помощью одного или более промежуточных линкеров или адаптерных молекул, таких как стрептавидин или аптамеры. См., например, источники Pierce et al. (2005) Mini Rev. Med. Chem. 5(1):41-55; Duckworth et al. (2007) Angew. Chem. Int. Ed. Engl. 46(46):8819-8822; Schaeffer and Dixon (2009) Australian J. Chem. 62(10):1328-1332; Goodman et al. (2009) Chembiochem. 10(9):1551-1557; и Khatwani et al. (2012) Bioorg. Med. Chem. 20(14):4532-4539, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Нековалентные стратегии синтеза конъюгатов белок-нуклеиновая кислота включают биотин-стрептавидиновые и никель-гистидиновые методы. Ковалентные конъюгаты белок-нуклеиновая кислота могут быть синтезированы путем соединения соответствующим образом функционализированных нуклеиновых кислот и белков с использованием широкого разнообразия химических процессов. Некоторые из этих химических процессов включают прямое присоединение олигонуклеотида к аминокислотному остатку на поверхности белка (например, амину лизина или тиолу цистеина), в то время как другие более сложные схемы требуют посттрансляционной модификации белка или участия каталитического или реакционноспособного белкового домена. Способы ковалентного присоединения белков к нуклеиновым кислотам могут включать, например, химическое сшивание олигонуклеотидов с остатками лизина или цистеина белка, лигирование синтезированных белков, химиоферментные методы и использование фотоаптамеров. Экзогенная донорная нуклеиновая кислота или меченая нуклеиновая кислота может быть присоединена к С-концу, N-концу или к внутренней области в белке Cas. В одном примере экзогенная донорная нуклеиновая кислота или меченая нуклеиновая кислота присоединена к С-концу или N-концу белка Cas. Аналогичным образом, белок Cas может быть присоединен к 5’-концу, 3’-концу или к внутренней области в экзогенной донорной нуклеиновой кислоте или меченой нуклеиновой кислоте. То есть экзогенная донорная нуклеиновая кислота или меченая нуклеиновая кислота может быть присоединена в любой ориентации и полярности. Например, белок Cas может быть присоединен к 5’-концу или 3’-концу экзогенной донорной нуклеиновой кислоты или меченой нуклеиновой кислоты.
[00190] Белки Cas могут быть предоставлены в любой форме. Например, белок Cas может быть предоставлен в форме белка, такого как белок Cas в комплексе с гРНК. В качестве альтернативы, белок Cas может быть предоставлен в форме нуклеиновой кислоты, кодирующей белок Cas, такой как РНК (например, матричная РНК (мРНК)) или ДНК. Необязательно, нуклеиновая кислота, кодирующая белок Cas, может быть кодон-оптимизирована для эффективной трансляции в белок в конкретной клетке или организме. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована для замены кодонов, которые чаще используются в бактериальной клетке, дрожжевой клетке, человеческой клетке, клетке, отличной от человеческой, клетке млекопитающего, клетке грызуна, мышиной клетке, крысиной клетке или любой другой представляющей интерес клетке-хозяине по сравнению со встречающейся в природе полинуклеотидной последовательностью. Когда нуклеиновая кислота, кодирующая белок Cas, вводится в клетку, белок Cas может временно, условно или конститутивно экспрессироваться в клетке.
[00191] Белки Cas, предоставленные в виде мРНК, могут быть модифицированы для улучшения свойств стабильности и/или иммуногенности. Модификации могут быть внесены в один или большее число нуклеозидов в мРНК. Примеры химических модификаций нуклеиновых оснований мРНК включают псевдоуридин, 1-метилпсевдоуридин и 5-метилцитидин. Например, можно использовать кэпированную и полиаденилированную мРНК Cas, содержащую N1-метилпсевдоуридин. Аналогичным образом, мРНК Cas могут быть модифицированы за счет обеднения уридином с использованием синонимичных кодонов.
[00192] Нуклеиновые кислоты, кодирующие белки Cas, могут быть стабильно интегрированы в геном клетки и функционально связаны с активным в клетке промотором. В качестве альтернативы, нуклеиновые кислоты, кодирующие белки Cas, могут быть функционально связаны с промотором в экспрессирующей конструкции. Экспрессирующие конструкции включают любые конструкции нуклеиновых кислот, способные регулировать экспрессию гена или другой представляющей интерес последовательности нуклеиновой кислоты (например, гена Cas), и которые могут переносить такую представляющую интерес последовательность нуклеиновой кислоты в целевую клетку. Например, нуклеиновая кислота, кодирующая белок Cas, может находиться в нацеливающем векторе, содержащем вставку нуклеиновой кислоты, и/или в векторе, содержащем ДНК, кодирующую нРНК. В качестве альтернативы, она может находиться в векторе или плазмиде, отдельных от нацеливающего вектора, содержащего вставку нуклеиновой кислоты, и/или отдельных от вектора, содержащего ДНК, кодирующую нРНК. Промоторы, которые могут быть использованы в экспрессирующей конструкции, включают промоторы, активные, например, в одной или более из эукариотической клетки, человеческой клетки, клетки, отличной от человеческой, клетки млекопитающего, клетки млекопитающего, отличного от человека, клетки грызуна, мышиной клетки, крысиной клетки, клетки хомяка, клетки кролика, плюрипотентной клетки, эмбриональной стволовой (ЭС) клетки или зиготы. Такие промоторы могут представлять собой, например, условные промоторы, индуцибельные промоторы, конститутивные промоторы или тканеспецифические промоторы. Необязательно, промотор может быть двунаправленным промотором, управляющим экспрессией как белка Cas в одном направлении, так и гидовой РНК в другом направлении. Такие двунаправленные промоторы могут состоять из (1) полного обычного однонаправленного промотора Pol III, который содержит 3 внешних контрольных элемента: дистальный промоторный элемент (DSE), проксимальный промоторный элемент (PSE) и TATA-бокс; и (2) второго базового промотора Pol III, включающего PSE и TATA-бокс, слитый с 5’-концом DSE в обратной ориентации. Например, в промоторе H1 DSE прилегает к PSE и TATA-боксу, и промотор можно сделать двунаправленным путем создания гибридного промотора, в котором транскрипция в обратном направлении контролируется путем добавления PSE и TATA-бокса, полученного из промотора U6. См., например, источник US 2016/0074535, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Применение двунаправленного промотора для одновременной экспрессии генов, кодирующих белок Cas и гидовую РНК, позволяет создавать компактные кассеты экспрессии для облегчения доставки.
[00193] Гидовые РНК. «Гидовая РНК» или «гРНК» представляет собой молекулу РНК, которая связывается с белком Cas (например, с белком Cas9) и нацеливает белок Cas на определенное место в ДНК-мишени. Гидовые РНК могут содержать два сегмента: «ДНК-нацеливающий сегмент» и «белок-связывающий сегмент». «Сегмент» включает часть или область молекулы, такую как непрерывный участок нуклеотидов в РНК. Некоторые гРНК, например, гРНК для Cas9, могут содержать две отдельные молекулы РНК: «активирующую РНК» (например, tracrРНК) и «нацеливающую РНК» (например, CRISPR РНК или crРНК). Другие гРНК представляют собой одинарную молекулу РНК (полинуклеотид одинарной РНК), которую также можно назвать «одномолекулярной гРНК», «единственной гидовой РНК» или «sgРНК». См., например, источники WO 2013/176772, WO 2014/065596, WO 2014/089290, WO 2014/093622, WO 2014/099750, WO 2013/142578 и WO 2014/131833, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Для Cas9, например, единственная гидовая РНК может содержать crРНК, слитую с tracrРНК (например, с помощью линкера). Для Cpf1, например, для достижения связывания и/или расщепления последовательности-мишени необходима только crРНК. Термины «гидовая РНК» и «гРНК» включают в себя как двухмолекулярные (т.е. модульные) гРНК, так и одномолекулярные гРНК.
[00194] Иллюстративная двухмолекулярная гРНК содержит crРНК-подобную («CRISPR РНК» или «нацеливающую РНК», или «crРНК», или «crРНК-повтор») молекулу и соответствующую tracrРНК-подобную («трансактивирующая CRISPR РНК» или «активирующая РНК», или «tracrРНК») молекулу. crРНК содержит как ДНК-нацеливающий сегмент (одноцепочечный) гРНК, так и участок нуклеотидов (т.е. хвост crРНК), который формирует половину дуплекса дцРНК белок-связывающего сегмента гРНК. Пример хвоста crРНК, расположенного по ходу транскрипции (с 3’-конца) от ДНК-нацеливающего сегмента, содержит, по существу состоит из или состоит из GUUUUAGAGCUAUGCU (SEQ ID NO: 40). Любой из описанных в настоящем документе ДНК-нацеливающих сегментов может быть присоединен к 5’-концу SEQ ID NO: 40 с образованием crРНК.
[00195] Соответствующая tracrРНК (активирующая РНК) содержит участок нуклеотидов, который формирует другую половину дуплекса дцРНК сегмента гРНК для связывания с белком. Участок нуклеотидов crРНК комплементарен участку нуклеотидов tracrРНК и гибридизуется с ним с образованием дуплекса дцРНК белок-связывающего сегмента гРНК. Таким образом, можно сказать, что каждая crРНК имеет соответствующую tracrРНК. Пример последовательности tracrРНК содержит, по существу состоит из или состоит из AGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUU (SEQ ID NO: 41).
[00196] В системах, в которых необходимы как crРНК, так и tracrРНК, crРНК и соответствующая tracrРНК гибридизуются с образованием гРНК. В системах, в которых необходима только crРНК, crРНК может представлять собой гРНК. crРНК дополнительно обеспечивает одноцепочечный ДНК-нацеливающий сегмент, который гибридизуется с комплементарной цепью ДНК-мишени. При использовании для модификации в клетке точная последовательность отдельно взятой молекулы crРНК или tracrРНК может быть разработана таким образом, чтобы быть специфичной для видов, в которых будут использоваться данные молекулы РНК. См., например, источники Mali et al. (2013) Science 339:823-826; Jinek et al. (2012) Science 337:816-821; Hwang et al. (2013) Nat. Biotechnol. 31:227-229; Jiang et al. (2013) Nat. Biotechnol. 31:233-239; и Cong et al. (2013) Science 339:819-823, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей.
[00197] ДНК-нацеливающий сегмент (crРНК) отдельно взятой гРНК содержит нуклеотидную последовательность, комплементарную последовательности на комплементарной цепи ДНК-мишени, как более подробно описано ниже. ДНК-нацеливающий сегмент гРНК взаимодействует с ДНК-мишенью сиквенс-специфическим образом посредством гибридизации (т.е. спаривания оснований). Как таковая, нуклеотидная последовательность ДНК-нацеливающего сегмента может варьировать и определяет местоположение в ДНК-мишени, с которым будут взаимодействовать гРНК и ДНК-мишень. ДНК-нацеливающий сегмент рассматриваемой гРНК может быть модифицирован путем гибридизации с любой желаемой последовательностью в ДНК-мишени. Встречающиеся в природе crРНК различаются в зависимости от системы CRISPR/Cas и организма, но часто содержат нацеливающий сегмент длиной от 21 до 72 нуклеотидов, фланкированный двумя прямыми повторами (DR) длиной от 21 до 46 нуклеотидов (см., например, источник WO 2014/131833, включенный в настоящий документ посредством ссылки в полном объеме для всех целей). В случае S. pyogenes длина DR составляет 36 нуклеотидов, а длина нацеливающего сегмента составляет 30 нуклеотидов. Расположенный с 3’-конца DR комплементарен соответствующей tracrРНК и гибридизуется с ней, а она, в свою очередь, связывается с белком Cas.
[00198] ДНК-нацеливающий сегмент может иметь длину, например, по меньшей мере приблизительно 12, 15, 17, 18, 19, 20, 25, 30, 35 или 40 нуклеотидов. Такие ДНК-нацеливающие сегменты могут иметь длину, например, от приблизительно 12 до приблизительно 100, от приблизительно 12 до приблизительно 80, от приблизительно 12 до приблизительно 50, от приблизительно 12 до приблизительно 40, от приблизительно 12 до приблизительно 30, от приблизительно 12 до приблизительно 25 или от приблизительно 12 до приблизительно 20 нуклеотидов. Например, ДНК-нацеливающий сегмент может состоять из от приблизительно 15 до приблизительно 25 нуклеотидов (например, из от приблизительно 17 до приблизительно 20 нуклеотидов, или приблизительно 17, 18, 19 или 20 нуклеотидов). См., например, источник US 2016/0024523, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Для Cas9 из S. pyogenes типичный ДНК-нацеливающий сегмент имеет от 16 до 20 нуклеотидов в длину или от 17 до 20 нуклеотидов в длину. Для Cas9 из S. aureus типичный ДНК-нацеливающий сегмент имеет от 21 до 23 нуклеотидов в длину. Для Cpf1 типичный ДНК-нацеливающий сегмент имеет по меньшей мере 16 нуклеотидов в длину или по меньшей мере 18 нуклеотидов в длину.
[00199] TracrРНК могут быть в любой форме (например, полноразмерных tracrРНК или активных частичных tracrРНК) и различной длины. Они могут включать первичные транскрипты или процессированные формы. Например, tracrРНК (как часть единственной гидовой РНК или как отдельная молекула, представляющая собой часть двухмолекулярной гРНК) могут содержать, по существу состоять из, или состоять из всей или части последовательности tracrРНК дикого типа (например, из приблизительно или более чем приблизительно 20, 26, 32, 45, 48, 54, 63, 67, 85 или более нуклеотидов из последовательности tracrРНК дикого типа). Примеры последовательностей tracrРНК дикого типа из S. pyogenes включают 171-нуклеотидную, 89-нуклеотидную, 75-нуклеотидную и 65-нуклеотидную версии. См., например, источники Deltcheva et al. (2011) Nature 471:602-607; WO 2014/093661, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Примеры tracrРНК в единственных гидовых РНК (sgРНК) включают сегменты tracrРНК, присутствующие в +48, +54, +67 и +85 версиях sgРНК, где «+n» означает, что в sgРНК включено вплоть до +n нуклеотидов из tracrРНК дикого типа. См. источник US 8697359, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00200] Процент комплементарности между ДНК-нацеливающим сегментом гидовой РНК и комплементарной цепью ДНК-мишени может составлять по меньшей мере 60% (например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%). Процент комплементарности между ДНК-нацеливающим сегментом и комплементарной цепью ДНК-мишени может составлять по меньшей мере 60% по приблизительно 20 смежным нуклеотидам. Например, процент комплементарности между ДНК-нацеливающим сегментом и комплементарной цепью ДНК-мишени может составлять 100% по 14 смежным нуклеотидам на 5’-конце комплементарной цепи ДНК-мишени и до 0% по оставшимся нуклеотидам. В таком случае можно считать, что ДНК-нацеливающий сегмент имеет 14 нуклеотидов в длину. В качестве еще одного примера, процент комплементарности между ДНК-нацеливающим сегментом и комплементарной цепью ДНК-мишени может составлять 100% по семи смежным нуклеотидам на 5’-конце комплементарной цепи ДНК-мишени и до 0% по оставшимся нуклеотидам. В таком случае можно считать, что ДНК-нацеливающий сегмент имеет 7 нуклеотидов в длину. В некоторых гидовых РНК по меньшей мере 17 нуклеотидов в ДНК-нацеливающем сегменте комплементарны комплементарной цепи ДНК-мишени. Например, ДНК-нацеливающий сегмент может иметь 20 нуклеотидов в длину и может содержать 1, 2 или 3 несовпадения с комплементарной цепью ДНК-мишени. В одном примере несовпадения не прилегают к области комплементарной цепи, соответствующей последовательности мотива, прилегающего к протоспейсеру (PAM) (т.е. обратному комплементу последовательности PAM) (например, несовпадения находятся на 5’-конце ДНК-нацеливающего сегмента гидовой РНК, или несовпадения находятся по меньшей мере в 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 или 19 парах оснований от области комплементарной цепи, соответствующей последовательности PAM).
[00201] Белок-связывающий сегмент гРНК может содержать два участка нуклеотидов, которые комплементарны друг другу. Комплементарные нуклеотиды белок-связывающего сегмента гибридизуются с образованием дуплекса двухцепочечной РНК (дцРНК). Белок-связывающий сегмент рассматриваемой гРНК взаимодействует с белком Cas, и гРНК направляет связанный белок Cas к определенной нуклеотидной последовательности в ДНК-мишени посредством ДНК-нацеливающего сегмента.
[00202] Единственные гидовые РНК могут содержать ДНК-нацеливающий сегмент, присоединенный к каркасной последовательности (т.е. белок-связывающую или Cas-связывающую последовательность гидовой РНК). Например, такие единственные гидовые РНК могут иметь 5’-ДНК-нацеливающий сегмент и 3’-каркасную последовательность. Иллюстративные каркасные последовательности содержат, по существу состоят из или состоят из: GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCU (версия 1; SEQ ID NO: 42); GUUGGAACCAUUCAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (версия 2; SEQ ID NO: 43); GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (версия 3; SEQ ID NO: 44); и GUUUAAGAGCUAUGCUGGAAACAGCAUAGCAAGUUUAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (версия 4; SEQ ID NO: 45). Гидовые РНК, нацеленные на любую последовательность-мишень гидовой РНК, могут включать, например, ДНК-нацеливающий сегмент на 5’-конце гидовой РНК, слитый с любой из иллюстративных каркасных последовательностей гидовой РНК на 3’-конце гидовой РНК. Это значит, что любой из раскрытых в настоящем документе ДНК-нацеливающих сегментов может быть присоединен к 5’-концу любой из SEQ ID NO: 42-45 с образованием единственной гидовой РНК (химерной гидовой РНК). Версии 1, 2, 3 и 4 гидовой РНК, как раскрыто в других местах настоящего документа, относятся к ДНК-нацеливающим сегментам (т.е. гидовым последовательностям или направляющим), соединенным с каркасами версий 1, 2, 3 и 4, соответственно.
[00203] Гидовые РНК могут включать модификации или последовательности, которые обеспечивают дополнительные желательные свойства (например, модифицированную или регулируемую стабильность; субклеточное нацеливание; отслеживание с помощью флуоресцентной метки; сайт связывания для белка или белкового комплекса и т.п.). Примеры таких модификаций включают, например, 5’-кэп (например, 7-метилгуанилатный кэп (m7G)); 3’-полиаденилированный хвост (т.е. 3’-поли(А) хвост); последовательность рибопереключателя (например, для обеспечения регулируемой стабильности и/или регулируемой доступности для белков и/или белковых комплексов); последовательность контроля устойчивости; последовательность, которая образует дуплекс дцРНК (т.е. шпильку); модификацию или последовательность, которая нацеливает РНК на субклеточную локализацию (например, ядро, митохондрии, хлоропласты и тому подобное); модификацию или последовательность, которая обеспечивает отслеживание (например, прямая конъюгация с флуоресцентной молекулой, конъюгация с фрагментом, который облегчает флуоресцентное детектирование, последовательность, которая позволяет проводить флуоресцентное детектирование и тому подобное); модификацию или последовательность, которая обеспечивает формирование сайта связывания для белков (например, белков, которые действуют на ДНК, включая ДНК-метилтрансферазы, ДНК-деметилазы, гистоновые ацетилтрансферазы, гистоновые деацетилазы и тому подобное); и их комбинации. Другие примеры модификаций включают сконструированные дуплексные структуры «петля на стебле», сконструированные области петель, сконструированные шпильки на 3’-конце дуплексной структуры «петля на стебле» или любую их комбинацию. См., например, источник US 2015/0376586, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Петля может представлять собой область неспаренных нуклеотидов внутри дуплекса, состоящую из crРНК-подобной области и минимальной tracrРНК-подобной области. Петля может содержать на одной стороне дуплекса неспаренный 5’-XXXY-3’, где X представляет собой любой пурин, а Y может представлять собой нуклеотид, который может формировать неоднозначную пару с нуклеотидом на противоположной цепи, и область неспаренных нуклеотидов на другой стороне дуплекса.
[00204] Немодифицированные нуклеиновые кислоты могут быть подвержены деградации. Экзогенные нуклеиновые кислоты могут также вызывать врожденный иммунный ответ. Модификации могут способствовать повышению стабильности и снижению иммуногенности. Гидовые РНК могут содержать модифицированные нуклеозиды и модифицированные нуклеотиды, включая, например, одно или более из следующего: (1) изменение или замена одного или обоих несвязывающих атомов кислорода из фосфата и/или одного или более связывающих атомов кислорода из фосфата в связи фосфодиэфирного остова; (2) изменение или замена компонента рибозного сахара, такие как изменение или замена 2’-гидроксила в рибозном сахаре; (3) замена фосфатного фрагмента дефосфорилированными линкерами; (4) модификация или замена встречающегося в природе азотистого основания; (5) замена или модификация рибозо-фосфатного остова; (6) модификация 3’-конца или 5’-конца олигонуклеотида (например, удаление, модификация или замена концевой фосфатной группы или конъюгация фрагмента); и (7) модификация сахара. Другие возможные модификации гидовой РНК включают в себя модификации или замену урацилов или полиурациловых трактов. См., например, источники WO 2015/048577 и US 2016/0237455, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Похожие модификации могут быть внесены в нуклеиновые кислоты, кодирующие Cas, такие как мРНК Cas.
[00205] В качестве одного примера, нуклеотиды на 5’- или 3’-конце гидовой РНК могут содержать тиофосфатные связи (например, основания могут содержать модифицированную фосфатную группу, которая представляет собой тиофосфатную группу). Например, гидовая РНК может включать тиофосфатные связи между 2, 3 или 4 концевыми нуклеотидами на 5’- или 3’-конце гидовой РНК. В качестве другого примера, нуклеотиды на 5’- и/или 3’-конце гидовой РНК могут иметь 2’-O-метильные модификации. Например, гидовая РНК может включать 2’-O-метильные модификации на 2, 3 или 4 концевых нуклеотидах на 5’- и/или 3’-конце гидовой РНК (например, на 5’-конце). См., например, источники WO 2017/173054 A1 и Finn et al. (2018) Cell Reports 22:1-9, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. В одном конкретном примере гидовая РНК содержит 2’-O-метильные аналоги и 3’-тиофосфатные межнуклеотидные связи в первых трех 5’- и 3’-концевых остатках РНК. В другом конкретном примере гидовая РНК модифицирована таким образом, что все группы 2’OH, которые не взаимодействуют с белком Cas9, заменены 2’-O-метильными аналогами, а хвостовая часть гидовой РНК, которая характеризуется минимальным взаимодействием с белком Cas9, модифицирована с помощью 5’- и 3’-тиофосфатных межнуклеотидных связей. См., например, источник Yin et al. (2017) Nat. Biotech. 35(12):1179-1187, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Другие примеры модифицированных гидовых РНК приведены, например, в источнике WO 2018/107028 A1, включенном в настоящий документ посредством ссылки в полном объеме для всех целей.
[00206] Гидовые РНК могут быть предоставлены в любой форме. Например, гРНК может быть предоставлена в форме РНК, либо в виде двух молекул (отдельные crРНК и tracrРНК), либо в виде одной молекулы (sgРНК), и, необязательно, в форме комплекса с белком Cas. гРНК также может быть предоставлена в форме ДНК, кодирующей гРНК. ДНК, кодирующая гРНК, может кодировать одну молекулу РНК (sgРНК) или отдельные молекулы РНК (например, отдельные crРНК и tracrРНК). В последнем случае ДНК, кодирующая гРНК, может быть предоставлена в виде одной молекулы ДНК или в виде отдельных молекул ДНК, кодирующих crРНК и tracrРНК, соответственно.
[00207] Когда гРНК предоставляется в форме ДНК, гРНК может кратковременно, условно или конститутивно экспрессироваться в клетке. ДНК, кодирующие гРНК, могут быть стабильно интегрированы в геном клетки и функционально связаны с активным в клетке промотором. В качестве альтернативы, ДНК, кодирующие гРНК, могут быть функционально связаны с промотором в экспрессирующей конструкции. Например, ДНК, кодирующая гРНК, может находиться в векторе, содержащем гетерологичную нуклеиновую кислоту, такую как нуклеиновая кислота, кодирующая белок Cas. В качестве альтернативы, она может находиться в векторе или плазмиде, отдельной от вектора, содержащего нуклеиновую кислоту, кодирующую белок Cas. Подходящие промоторы, которые могут быть использованы в таких экспрессирующих конструкциях, включают промоторы, активные, например, в одной или более из эукариотической клетки, человеческой клетки, клетка, отличной от человеческой, клетки млекопитающего, клетки млекопитающего, отличного от человека, клетки грызуна, мышиной клетки, крысиной клетки, клетки хомяка, клетки кролика, плюрипотентной клетки, эмбриональной стволовой (ЭС) клетки, взрослой стволовой клетки, ограниченной стадией развития клетки-предшественника, индуцированной плюрипотентной стволовой (иПС) клетки или находящегося на одноклеточной стадии эмбриона. Такие промоторы могут представлять собой, например, условные промоторы, индуцибельные промоторы, конститутивные промоторы или тканеспецифические промоторы. Такие промоторы также могут представлять собой, например, двунаправленные промоторы. Конкретные примеры подходящих промоторов включают промотор РНК-полимеразы III, такой как промотор U6 человека, промотор U6 полимеразы III крысы или промотор U6 полимеразы III мыши.
[00208] В качестве альтернативы, гРНК могут быть получены различными другими способами. Например, гРНК могут быть получены путем транскрипции in vitro с использованием, например, РНК-полимеразы T7 (см., например, источники WO 2014/089290 и WO 2014/065596, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей). Гидовые РНК также могут представлять собой синтетически полученные молекулы, полученные с помощью химического синтеза.
[00209] Гидовые РНК (или нуклеиновые кислоты, кодирующие гидовые РНК) могут содержаться в композициях, содержащих одну или более гидовых РНК (например, 1, 2, 3, 4 или более гидовых РНК) и носитель, повышающий стабильность гидовой РНК (например, носитель, пролонгирующий период при заданных условиях хранения (например, -20°C, 4°C или температуре окружающей среды), в течение которого продукты разложения остаются ниже порогового значения, например, ниже 0,5 мас.% относительно исходной нуклеиновой кислоты или белка; или повышающий стабильность in vivo). Неограничивающие примеры таких носителей включают микросферы полимолочной кислоты (PLA), микросферы сополимера D,L-молочной и гликолевой кислот (PLGA), липосомы, мицеллы, обратные мицеллы, липидные кохлеаты и липидные микротрубочки. Такие композиции могут дополнительно содержать белок Cas, такой как белок Cas9, или нуклеиновую кислоту, кодирующую белок Cas.
[00210] Последовательности-мишени гидовой РНК. ДНК-мишени для гидовых РНК включают последовательности нуклеиновых кислот, присутствующие в ДНК, с которыми будет связываться ДНК-нацеливающий сегмент гРНК, при условии, что обеспечены достаточные условия для связывания. Подходящие условия для связывания ДНК/РНК включают в себя физиологические условия, обычно присутствующие в клетке. Другие подходящие условия для связывания ДНК/РНК (например, условия в бесклеточной системе) известны в данной области техники (см., например, источник Molecular Cloning: A Laboratory Manual, 3rd Ed. (Sambrook et al., Harbor Laboratory Press 2001), включенный в настоящий документ посредством ссылки в полном объеме для всех целей). Цепь ДНК-мишени, которая комплементарна гРНК и гибридизуется с ней, может быть названа «комплементарной цепью», а цепь ДНК-мишени, которая комплементарна «комплементарной цепи» (и, следовательно, не комплементарна белку Cas или гРНК), может быть названа «некомплементарной цепью» или «матричной цепью».
[00211] ДНК-мишень содержит как последовательность на комплементарной цепи, с которой гибридизуется гидовая РНК, так и соответствующую последовательность на некомплементарной цепи (например, смежную с мотивом, прилегающим к протоспейсеру (PAM)). В контексте настоящего документа термин «последовательность-мишень гидовой РНК» относится конкретно к последовательности на некомплементарной цепи (т.е. обратному комплементу), соответствующей последовательности, с которой гибридизируется гидовая РНК на комплементарной цепи. То есть последовательность-мишень гидовой РНК относится к последовательности на некомплементарной цепи, прилегающей к PAM (например, расположенной против хода транскрипции или с 5’-конца относительно PAM в случае Cas9). Последовательность-мишень гидовой РНК эквивалентна ДНК-нацеливающему сегменту гидовой РНК, но с тиминами вместо урацилов. В качестве одного примера, последовательность-мишень гидовой РНК для фермента SpCas9 может относиться к последовательности, расположенной против хода транскрипции относительно PAM 5’-NGG-3’ на некомплементарной цепи. Гидовая РНК разработана таким образом, чтобы быть комплементарной к комплементарной цепи ДНК-мишени, при этом гибридизация между ДНК-нацеливающим сегментом гидовой РНК и комплементарной цепью ДНК-мишени способствует образованию комплекса CRISPR. Полная комплементарность не обязательно требуется при условии, что комплементарность достаточна, чтобы вызвать гибридизацию и способствовать образованию комплекса CRISPR. Если гидовая РНК упоминается в настоящем документе как нацеленная на последовательность-мишень гидовой РНК, это означает, что гидовая РНК гибридизируется с последовательностью комплементарной цепи ДНК-мишени, которая представляет собой обратный комплемент последовательности-мишени гидовой РНК на некомплементарной цепи.
[00212] ДНК-мишень или последовательность-мишень гидовой РНК может содержать любой полинуклеотид и может располагаться, например, в ядре или цитоплазме клетки или в органелле клетки, такой как митохондрия или хлоропласт. ДНК-мишень или последовательность-мишень гидовой РНК может представлять собой любую последовательность нуклеиновой кислоты, эндогенную или экзогенную для клетки. Последовательность-мишень гидовой РНК может представлять собой последовательность, кодирующую продукт гена (например, белок), или некодирующую последовательность (например, регуляторную последовательность), или может содержать и то, и другое. В частном примере последовательность-мишень гидовой РНК может находиться в интроне 1, интроне 12 или интроне 13 гена человеческого альбумина. Например, последовательность-мишень гидовой РНК может находиться в интроне 1 гена человеческого альбумина.
[00213] Сайт-специфическое связывание и расщепление ДНК-мишени белком Cas может происходить в местах, определяемых как (i) комплементарностью спаривания оснований между гидовой РНК и комплементарной цепью ДНК-мишени, так и (ii) коротким мотивом, называемым мотивом, прилегающим к протоспейсеру (PAM), в некомплементарной цепи ДНК-мишени. PAM может фланкировать последовательность-мишень гидовой РНК. Необязательно, последовательность-мишень гидовой РНК может быть фланкирована на 3’-конце посредством PAM (например, в случае Cas9). В качестве альтернативы, последовательность-мишень гидовой РНК может быть фланкирована на 5’-конце посредством PAM (например, в случае Cpf1). Например, сайт расщепления белков Cas может располагаться на расстоянии от приблизительно 1 до приблизительно 10 или от приблизительно 2 до приблизительно 5 пар оснований (например, 3 пар оснований) против хода транскрипции или по ходу транскрипции от последовательности PAM (например, в пределах последовательности-мишени гидовой РНК). В случае SpCas9 последовательность PAM (т.е. на некомплементарной цепи) может представлять собой 5’-N1GG-3’, где N1 представляет собой любой нуклеотид ДНК, и где PAM расположен непосредственно с 3’-конца от последовательности-мишени гидовой РНК на некомплементарной цепи ДНК-мишени. Таким образом, последовательность, соответствующая РАМ на комплементарной цепи (т.е., обратный комплемент) будет представлять собой 5’-CCN2-3’, где N2 представляет собой любой нуклеотид ДНК и расположен непосредственно с 5’-конца от последовательности, с которой ДНК-нацеливающий сегмент гидовой РНК гибридизуется на комплементарной цепи ДНК-мишени. В некоторых таких случаях N1 и N2 могут быть комплементарными, и пара оснований N1- N2 может представлять собой любую пару оснований (например, N1=C и N2=G; N1=G и N2=C; N1=A и N2=T; или N1=T и N2=A). В случае Cas9 из S. aureus последовательность PAM может представлять собой NNGRRT или NNGRR, где N может представлять собой A, G, C или T, а R может представлять собой G или A. В случае Cas9 из C. jejuni последовательность PAM может представлять собой, например, NNNNACAC или NNNNRYAC, где N может представлять собой A, G, C или T, а R может представлять собой G или A. В некоторых случаях (например, для FnCpf1) последовательность PAM может быть расположена против хода транскрипции от 5’-конца и имеет последовательность 5’-TTN-3’.
[00214] Примером последовательности-мишени гидовой РНК является последовательность ДНК из 20 нуклеотидов, непосредственно предшествующая мотиву NGG, распознаваемому белком SpCas9. Например, двумя примерами последовательностей-мишеней для гидовой РНК плюс PAM являются GN19NGG (SEQ ID NO: 46) или N20NGG (SEQ ID NO: 47). См., например, источник WO 2014/165825, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Гуанин на 5’-конце может способствовать транскрипции РНК-полимеразой в клетках. Другие примеры последовательностей-мишеней гидовой РНК плюс PAM могут включать два гуаниновых нуклеотида на 5’-конце (например, GGN20NGG; SEQ ID NO: 48) для облегчения эффективной транскрипции полимеразой Т7 in vitro. См., например, источник WO 2014/065596, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Другие последовательности-мишени гидовой РНК плюс PAM могут иметь длину от 4 до 22 нуклеотидов последовательностей SEQ ID NO: 46-48, включая 5’ G или GG и 3’ GG или NGG. Другие последовательности-мишени гидовой РНК плюс PAM могут иметь длину от 14 до 20 нуклеотидов последовательностей SEQ ID NO: 46-48.
[00215] Образование комплекса CRISPR, гибридизованного с ДНК-мишенью, может приводить к расщеплению одной или обеих цепей ДНК-мишени в пределах или вблизи области, соответствующей последовательности-мишени гидовой РНК (т.е. последовательности-мишени гидовой РНК на некомплементарной цепи ДНК-мишени и обратному комплементу на комплементарной цепи, с которой гибридизуется гидовая РНК). Например, сайт расщепления может располагаться в последовательности-мишени гидовой РНК (например, в определенном месте относительно последовательности PAM). «Сайт расщепления» включает положение в ДНК-мишени, в котором белок Cas вызывает одноцепочечный разрыв или двухцепочечный разрыв. Сайт расщепления может находиться только на одной цепи (например, когда используется никаза) или на обеих цепях двухцепочечной ДНК. Сайты расщепления могут находиться в одном и том же положении на обеих цепях (образуя тупые концы; например, Cas9)) или могут находиться в разных сайтах на каждой цепи (образуя ступенчатые концы (т.е. выступы); например, Cpf1). Ступенчатые концы могут быть образованы, например, с помощью использования двух белков Cas, каждый из которых вызывает одноцепочечный разрыв в отличающемся сайте расщепления на другой цепи, тем самым вызывая двухцепочечный разрыв. Например, первая никаза может создавать одноцепочечный разрыв в первой цепи двухцепочечной ДНК (дцДНК), а вторая никаза может создавать одноцепочечный разрыв во второй цепи дцДНК, и, таким образом, создаются последовательности с выступами. В некоторых случаях последовательность-мишень гидовой РНК или сайт расщепления для никазы в первой цепи отделены от последовательности-мишени гидовой РНК или от сайта расщепления для никазы во второй цепи по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 пар оснований.
(3) Экзогенные донорные нуклеиновые кислоты, нацеленные на гена человеческого альбумина
[00216] В раскрытых в настоящем документе способах и композициях могут быть использованы экзогенные донорные нуклеиновые кислоты для модификации гуманизированного локуса альбумина после расщепления гуманизированного локуса альбумина нуклеазным агентом или независимо от расщепления гуманизированного локуса альбумина нуклеазным агентом. В таких способах с использованием нуклеазного агента белок нуклеазного агента расщепляет гуманизированный локус альбумина, создавая одноцепочечный разрыв (ник) или двухцепочечный разрыв, а экзогенная донорная нуклеиновая кислота рекомбинирует гуманизированный локус альбумина посредством лигирования, опосредованного негомологичным соединением концов (NHEJ) или посредством события репарации, опосредованной гомологичной рекомбинацией. Необязательно, репарация экзогенной донорной нуклеиновой кислотой удаляет или разрушает последовательность-мишень нуклеазы, так чтобы на аллели, выступавшие в качестве мишеней, не мог быть повторно нацелен нуклеазный агент.
[00217] Экзогенная донорная нуклеиновая кислота может нацеливаться на любую последовательность гена человеческого альбумина. Некоторые экзогенные донорные нуклеиновые кислоты содержат гомологичные плечи. Другие экзогенные донорные нуклеиновые кислоты не содержат гомологичные плечи. Экзогенные донорные нуклеиновые кислоты могут быть способны вставляться в гуманизированный локус альбумина посредством репарации, опосредованной гомологичной рекомбинацией, и/или они могут быть способны вставляться в гуманизированный локус альбумина посредством негомологичного соединения концов. В одном примере экзогенная донорная нуклеиновая кислота (например, нацеливающий вектор) может быть нацелена на интрон 1, интрон 12 или интрон 13 гена человеческого альбумина. Например, экзогенная донорная нуклеиновая кислота может быть нацелена на интрон 1 гена человеческого альбумина.
[00218] Экзогенные донорные нуклеиновые кислоты могут включать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), они могут быть одноцепочечными или двухцепочечными, и они могут иметь линейную или кольцевую форму. Например, экзогенная донорная нуклеиновая кислота может представлять собой одноцепочечный олигодезоксинуклеотид (оцОДН). См., например, источник Yoshimi et al. (2016) Nat. Commun. 7:10431, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Экзогенные донорные нуклеиновые кислоты могут представлять собой голые нуклеиновые кислоты или могут доставляться вирусами, такими как AAV. В конкретном примере экзогенная донорная нуклеиновая кислота может быть доставлена посредством AAV и может быть способной к вставке в гуманизированный локус альбумина посредством негомологичного соединения концов (например, экзогенная донорная нуклеиновая кислота может представлять собой такую нуклеиновую кислоту, которая не содержит гомологичные плечи).
[00219] Иллюстративная экзогенная донорная нуклеиновая кислота имеет от приблизительно 50 нуклеотидов до приблизительно 5 кб, от приблизительно 50 нуклеотидов до приблизительно 3 кб, или от приблизительно 50 до приблизительно 1000 нуклеотидов в длину. Другие иллюстративные экзогенные донорные нуклеиновые кислоты имеют от приблизительно 40 до приблизительно 200 нуклеотидов в длину. Например, экзогенная донорная нуклеиновая кислота может иметь приблизительно 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140, 140-150, 150-160, 160-170, 170-180, 180-190 или 190-200 нуклеотидов в длину. В качестве альтернативы, экзогенная донорная нуклеиновая кислота может иметь приблизительно 50-100, 100-200, 200-300, 300-400, 400-500, 500-600, 600-700, 700-800, 800-900 или 900-1000 нуклеотидов в длину. В качестве альтернативы, экзогенная донорная нуклеиновая кислота может иметь приблизительно 1-1,5, 1,5-2, 2-2,5, 2,5-3, 3-3,5, 3,5-4, 4-4,5 или 4,5-5 кб в длину. В качестве альтернативы, экзогенная донорная нуклеиновая кислота может иметь, например, не более 5 кб, 4,5 кб, 4 кб, 3,5 кб, 3 кб, 2,5 кб, 2 кб, 1,5 кб, 1 кб, 900 нуклеотидов, 800 нуклеотидов, 700 нуклеотидов, 600 нуклеотидов, 500 нуклеотидов, 400 нуклеотидов, 300 нуклеотидов, 200 нуклеотидов, 100 нуклеотидов или 50 нуклеотидов в длину. Экзогенные донорные нуклеиновые кислоты (например, нацеливающие векторы) также могут иметь большую длину.
[00220] В одном примере экзогенная донорная нуклеиновая кислота представляет собой оцОДН, имеющий от приблизительно 80 нуклеотидов до приблизительно 200 нуклеотидов в длину. В еще одном примере экзогенная донорная нуклеиновая кислота представляет собой оцОДН, имеющий от приблизительно 80 нуклеотидов до приблизительно 3 кб в длину. Такой оцОДН может иметь гомологичные плечи или комплементарные области, например, каждая из которых имеет от приблизительно 40 нуклеотидов до приблизительно 60 нуклеотидов в длину. Такой оцОДН также может иметь гомологичные плечи или комплементарные области, например, каждая из которых имеет от приблизительно 30 нуклеотидов до 100 нуклеотидов в длину. Гомологичные плечи могут быть симметричными (например, каждое длиной 40 нуклеотидов или каждое длиной 60 нуклеотидов), или они могут быть асимметричными (например, одно гомологичное плечо длиной 36 нуклеотидов и одно гомологичное плечо длиной 91 нуклеотид).
[00221] Экзогенные донорные нуклеиновые кислоты могут включать модификации или последовательности, которые обеспечивают дополнительные желательные свойства (например, модифицированную или регулируемую стабильность; отслеживание или обнаружение с помощью флуоресцентной метки; сайт связывания для белка или белкового комплекса и так далее). Экзогенные донорные нуклеиновые кислоты могут содержать одну или более флуоресцентных меток, меток очистки, эпитопных меток или их комбинации. Например, экзогенная донорная нуклеиновая кислота может содержать одну или более флуоресцентных меток (например, флуоресцентные белки или другие флуорофоры или красители), например, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 флуоресцентных меток. Примеры флуоресцентных меток включают флуорофоры, такие как флуоресцеин (например, 6-карбоксифлуоресцеин (6-FAM)), Texas Red, HEX, Cy3, Cy5, Cy5.5, Pacific Blue, 5-(и-6)-карбокситетраметилродамин (TAMRA) и Cy7. Для мечения олигонуклеотидов коммерчески доступен широкий спектр флуоресцентных красителей (например, от Integrated DNA Technologies). Такие флуоресцентные метки (например, внутренние флуоресцентные метки) могут быть использованы, например, для обнаружения экзогенной донорной нуклеиновой кислоты, которая была непосредственно интегрирована в расщепленную нуклеиновую кислоту-мишень, имеющую выступающие концы, совместимые с концами экзогенной донорной нуклеиновой кислоты. Маркер или метка может находиться на 5’-конце, 3’-конце или внутри экзогенной донорной нуклеиновой кислоты. Например, экзогенная донорная нуклеиновая кислота может быть конъюгирована на 5’-конце с флуорофором IR700 от Integrated DNA Technologies (5’IRDYE®700).
[00222] Экзогенные донорные нуклеиновые кислоты могут также содержать вставки нуклеиновых кислот, включая сегменты ДНК, которые должны быть интегрированы в гуманизированный локус альбумина. Интеграция вставки нуклеиновой кислоты в гуманизированный локус альбумина может привести к добавлению представляющей интерес последовательности нуклеиновой кислоты к гуманизированному локусу альбумина, делеции представляющей интерес последовательности нуклеиновой кислоты в гуманизированном локусе альбумина или замене представляющей интерес последовательности нуклеиновой кислоты интерес в гуманизированном локусе альбумина (т.е. делеция и вставка). Некоторые экзогенные донорные нуклеиновые кислоты разработаны для встраивания вставки нуклеиновой кислоты в гуманизированный локус альбумина без какой-либо соответствующей делеции в гуманизированном локусе альбумина. Другие экзогенные донорные нуклеиновые кислоты разработаны для удаления представляющей интерес последовательности нуклеиновой кислоты в гуманизированном локусе альбумина без какой-либо соответствующей вставки нуклеиновой кислоты. Другие экзогенные донорные нуклеиновые кислоты разработаны для удаления представляющей интерес последовательности нуклеиновой кислоты в гуманизированном локусе альбумина и замены ее вставкой нуклеиновой кислоты.
[00223] Вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в удаляемом и/или заменяемом гуманизированном локусе альбумина может иметь различную длину. Иллюстративная вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в удаляемом и/или заменяемом гуманизированном локусе альбумина имеет от приблизительно 1 нуклеотида до приблизительно 5 кб в длину или от приблизительно 1 нуклеотида до приблизительно 1000 нуклеотидов в длину. Например, вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в удаляемом и/или заменяемом гуманизированном локусе альбумина может иметь приблизительно 1-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140, 140-150, 150-160, 160-170, 170-180, 180-190 или 190-120 нуклеотидов в длину. Аналогичным образом, вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в удаляемом и/или заменяемом гуманизированном локусе альбумина может иметь 1-100, 100-200, 200-300, 300-400, 400-500, 500-600, 600-700, 700-800, 800-900 или 900-1000 нуклеотидов в длину. Аналогичным образом, вставка нуклеиновой кислоты или соответствующая нуклеиновая кислота в удаляемом и/или заменяемом гуманизированном локусе альбумина может иметь приблизительно 1-1,5, 1,5-2, 2-2,5, 2,5-3, 3-3,5, 3,5-4, 4-4,5 или 4,5-5 кб, или более, в длину.
[00224] Вставка нуклеиновой кислоты может содержать последовательность, гомологичную или ортологичную всей или части последовательности, выступающей в качестве мишени для замены. Например, вставка нуклеиновой кислоты может содержать последовательность, которая содержит одну или более точечных мутаций (например, 1, 2, 3, 4, 5 или более) по сравнению с последовательностью, выступающей в качестве мишени для замены в гуманизированном локусе альбумина. Необязательно, такие точечные мутации могут приводить к консервативной аминокислотной замене (например, замене аспарагиновой кислоты [Asp, D] на глутаминовую кислоту [Glu, E]) в кодируемом полипептиде.
[00225] Некоторые экзогенные донорные нуклеиновые кислоты могут кодировать экзогенный белок, не кодируемый или не экспрессируемый эндогенным локусом альбумина дикого типа (например, могут содержать вставку нуклеиновой кислоты, кодирующую экзогенный белок). В одном примере гуманизированный локус альбумина, на который нацелена экзогенная донорная нуклеиновая кислота, может кодировать гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с белком, не кодируемым или не экспрессируемым эндогенным локусом альбумина дикого типа. Например, экзогенная донорная нуклеиновая кислота может представлять собой кассету без промотора, содержащую акцептор сплайсинга, и экзогенная донорная нуклеиновая кислота может быть нацелена на первый интрон человеческого альбумина.
[00226] Донорные нуклеиновые кислоты для вставки, опосредованной негомологичным соединением концов. Некоторые экзогенные донорные нуклеиновые кислоты способны вставляться в гуманизированный локус альбумина путем негомологичного соединения концов. В некоторых случаях такие экзогенные донорные нуклеиновые кислоты не содержат гомологичные плечи. Например, такие экзогенные донорные нуклеиновые кислоты могут быть вставлены в двухцепочечный разрыв с тупым концом после расщепления нуклеазным агентом. В конкретном примере экзогенная донорная нуклеиновая кислота может быть доставлена посредством AAV и может быть способной к вставке в гуманизированный локус альбумина посредством негомологичного соединения концов (например, экзогенная донорная нуклеиновая кислота может представлять собой такую нуклеиновую кислоту, которая не содержит гомологичные плечи). В конкретном примере экзогенная донорная нуклеиновая кислота может быть вставлена посредством независимой от гомологии направленной интеграции. Например, последовательность вставки в экзогенной донорной нуклеиновой кислоте, которая должна быть вставлена в гуманизированный локус альбумина, может быть фланкирована с каждой стороны сайтом-мишенью для нуклеазного агента (например, тем же сайтом-мишенью, что и в гуманизированном локусе альбумина, и тем же самым нуклеазным агентом, который используется для расщепления сайта-мишени в гуманизированном локусе альбумина). Затем нуклеазный агент может расщеплять сайты-мишени, фланкирующие вставленную последовательность. В конкретном примере, экзогенную донорную нуклеиновую кислоту доставляют с помощью опосредованной AAV доставки, и расщепление сайтов-мишеней, фланкирующих вставленную последовательность, может удалить инвертированные концевые повторы (ITR) AAV. В некоторых способах сайт-мишень в гуманизированном локусе альбумина (например, последовательность-мишень гРНК, включая фланкирующий мотив, прилегающий к протоспейсеру) удаляется, если последовательность вставки вставлена в гуманизированный локус альбумина в правильной ориентации, но восстанавливается, если последовательность вставки вставлена в гуманизированный локус альбумина в противоположной ориентации. Это может помочь гарантировать, что последовательность вставки вставлена в правильной ориентации для экспрессии.
[00227] Другие экзогенные донорные нуклеиновые кислоты имеют короткие одноцепочечные области на 5’-конце и/или 3’-конце, которые комплементарны одному или более выступам, созданным в результате опосредованного нуклеазным агентом расщепления в гуманизированном локусе альбумина. Эти выступы также могут называться 5’- и 3’-гомологичными плечами. Например, некоторые экзогенные донорные нуклеиновые кислоты имеют короткие одноцепочечные области на 5’-конце и/или 3’-конце, которые комплементарны одному или более выступам, созданным в результате опосредованного нуклеазным агентом расщепления в 5’- и/или 3’-последовательностях-мишенях в гуманизированном локусе альбумина. Некоторые из таких экзогенных донорных нуклеиновых кислот имеют комплементарную область только на 5’-конце или только на 3’-конце. Например, некоторые такие экзогенные донорные нуклеиновые кислоты иметь комплементарную область только на 5’-конце, комплементарном выступу, созданному на 5’-последовательности-мишени в гуманизированном локусе альбумина, или только на 3’-конце, комплементарном выступу, созданному на 3’-последовательности-мишени в гуманизированном локусе альбумина. Другие подобные экзогенные донорные нуклеиновые кислоты имеют комплементарные области как на 5’-, так и на 3’-концах. Например, другие подобные экзогенные донорные нуклеиновые кислоты имеют комплементарные области как на 5’-, так и на 3’-концах, например, комплементарные первому и второму выступам, соответственно, генерируемые в результате опосредованного нуклеазным агентом расщепления в гуманизированном локусе альбумина. Например, если экзогенная донорная нуклеиновая кислота является двухцепочечной, одноцепочечные комплементарные области могут выступать с 5’-конца верхней цепи донорной нуклеиновой кислоты и 5’-конца нижней цепи донорной нуклеиновой кислоты, создавая 5’-выступы на каждом конце. В качестве альтернативы, одноцепочечная комплементарная область может выступать с 3’-конца верхней цепи донорной нуклеиновой кислоты и с 3’-конца нижней цепи матрицы, создавая 3’-выступы.
[00228] Комплементарные области могут иметь любую длину, достаточную для облегчения лигирования между экзогенной донорной нуклеиновой кислотой и нуклеиновой кислотой-мишенью. Иллюстративные комплементарные области имеют от приблизительно 1 до приблизительно 5 нуклеотидов в длину, от приблизительно 1 до приблизительно 25 нуклеотидов в длину или от приблизительно 5 до приблизительно 150 нуклеотидов в длину. Например, комплементарная область может иметь по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину. В качестве альтернативы, комплементарная область может иметь приблизительно 5-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140 или 140-150, или больше, нуклеотидов в длину.
[00229] Такие комплементарные области могут быть комплементарными выступам, созданным двумя парами никаз. Два двухцепочечных разрыва со ступенчатыми концами могут быть созданы с использованием первой и второй никаз, которые расщепляют противоположные цепи ДНК, создавая первый двухцепочечный разрыв, и третьей и четвертой никаз, которые расщепляют противоположные цепи ДНК, создавая второй двухцепочечный разрыв. Например, белок Cas может быть использован для никования первой, второй, третьей и четвертой последовательностей-мишеней гидовой РНК, соответствующих первой, второй, третьей и четвертой гидовой РНК. Первая и вторая последовательности-мишени гидовой РНК могут быть расположены таким образом, чтобы создать первый сайт расщепления, так что ники, созданные первой и второй никазами на первой и второй цепях ДНК, создают двухцепочечный разрыв (т.е. первый сайт расщепления содержит ники в первой и второй последовательностях-мишенях гидовой РНК). Аналогичным образом, третья и четвертая последовательности-мишени гидовой РНК могут быть расположены таким образом, чтобы создать второй сайт расщепления, так что ники, созданные третьей и четвертой никазами на первой и второй цепях ДНК, создают двухцепочечный разрыв (т.е. первый сайт расщепления содержит ники в третьей и четвертой последовательностях-мишенях гидовой РНК). Предпочтительно ники в первой и второй последовательностях-мишенях гидовой РНК и/или третьей и четвертой последовательностях-мишенях гидовой РНК могут представлять собой смещенные ники, которые создают выступы. Окно смещения может составлять, например, по меньшей мере приблизительно 5 п.н., 10 п.н., 20 п.н., 30 п.н., 40 п.н., 50 п.н., 60 п.н., 70 п.н., 80 п.н., 90 п.н., 100 п.н. или более. См. источники Ran et al. (2013) Cell 154:1380-1389; Mali et al. (2013) Nat. Biotech. 31:833-838; и Shen et al. (2014) Nat. Methods 11:399-404, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. В таких случаях двухцепочечная экзогенная донорная нуклеиновая кислота может быть разработана с одноцепочечными комплементарными областями, которые комплементарны выступам, создаваемым никами в первой и второй последовательностях-мишенях гидовой РНК и никами в третьей и четвертой последовательностях-мишенях гидовой РНК. Такая экзогенная донорная нуклеиновая кислота затем может быть вставлена путем лигирования, опосредованного негомологичным соединением концов.
[00230] Донорные нуклеиновые кислоты для введения путем репарации, опосредованной гомологичной рекомбинацией. Некоторые экзогенные донорные нуклеиновые кислоты содержат гомологичные плечи. Если экзогенная донорная нуклеиновая кислота также содержит вставку нуклеиновой кислоты, гомологичные плечи могут фланкировать указанную вставку нуклеиновой кислоты. Для простоты ссылки гомологичные плечи обозначаются в настоящем документе как 5’- и 3’- (т.е. расположенные по ходу и против хода транскрипции) гомологичные плечи. Эта терминология относится к относительному положению гомологичных плеч относительно вставки нуклеиновой кислоты внутри экзогенной донорной нуклеиновой кислоты. 5’-и 3’-гомологичные плечи соответствуют областям в гуманизированном локусе альбумина, которые обозначаются в настоящем документе как «5’-последовательность-мишень» и «3’-последовательность-мишень», соответственно.
[00231] Гомологичное плечо и последовательность-мишень «соответствуют» или являются «соответствующими» друг другу тогда, когда две области обладают достаточным уровнем идентичности последовательностей друг к другу, чтобы действовать в качестве субстратов для реакции гомологичной рекомбинации. Термин «гомология» включает последовательности ДНК, которые либо идентичны, либо обе обладают определенной идентичностью соответствующей последовательности. Идентичность последовательностей между данной последовательностью-мишенью и соответствующим гомологичным плечом, содержащимся в донорной нуклеиновой кислоте, может иметь любую степень идентичности последовательностей, которая допускает гомологичную рекомбинацию. Например, степень идентичности последовательностей, общая для гомологичного плеча экзогенной донорной нуклеиновой кислоты (или ее фрагмента) и последовательности-мишени (или ее фрагмента), может представлять собой по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей, так что последовательности подвергаются гомологичной рекомбинации. Кроме того, соответствующая область гомологии между гомологичным плечом и соответствующей последовательностью-мишенью может иметь любую длину, достаточную для способствования гомологичной рекомбинации. Иллюстративные гомологичные плечи имеют длину от приблизительно 25 нуклеотидов до приблизительно 2,5 кб, от приблизительно 25 нуклеотидов до приблизительно 1,5 кб или от приблизительно 25 до приблизительно 500 нуклеотидов. Например, отдельно взятое гомологичное плечо (или каждое из гомологичных плеч) и/или соответствующая последовательность-мишень могут содержать соответствующие области гомологии, имеющие приблизительно 25-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-150, 150-200, 200-250, 250-300, 300-350, 350-400, 400-450 или 450-500 нуклеотидов в длину, так что гомологичные плечи обладают достаточной гомологией для гомологичной рекомбинации с соответствующими последовательностями-мишенями в нуклеиновой кислоте-мишени. В качестве альтернативы, отдельно взятое гомологичное плечо (или каждое из гомологичных плеч) и/или соответствующая последовательность-мишень могут содержать соответствующие области гомологии, имеющие от приблизительно 0,5 кб до приблизительно 1 кб, от приблизительно 1 кб до приблизительно 1,5 кб, от приблизительно 1,5 кб до приблизительно 2 кб или от приблизительно 2 кб до приблизительно 2,5 кб в длину. Например, каждое гомологичное плечо может иметь приблизительно 750 нуклеотидов в длину. Гомологичные плечи могут быть симметричными (каждое плечо приблизительно одинакового размера по длине) или асимметричными (одно плечо длиннее другого).
[00232] Когда нуклеазный агент используется в комбинации с экзогенной донорной нуклеиновой кислотой, 5’- и 3’-последовательности-мишени предпочтительно расположены в достаточной близости от сайта расщепления нуклеазой (например, в достаточной близости от последовательности-мишени нуклеазы), чтобы способствовать возникновению события гомологичной рекомбинации между последовательностями-мишенями и гомологичными плечами при одноцепочечном разрыве (нике) или двухцепочечном разрыве в сайте расщепления нуклеазой. Термин «сайт расщепления нуклеазой» включает последовательность ДНК, в которой нуклеазный агент создает ник или двухцепочечный разрыв (например, белок Cas9 в комплексе с гидовой РНК). Последовательности-мишени в локусе-мишени, которые соответствуют 5’- и 3’-гомологичным плечам экзогенной донорной нуклеиновой кислоты, «расположены в достаточной близости» к сайту расщепления нуклеазой, если расстояние таково, что способствует возникновению события гомологичной рекомбинации между 5’- и 3’-последовательностями-мишенями и гомологичными плечами при одноцепочечном или двухцепочечном разрыве в сайте расщепления нуклеазой. Таким образом, последовательности-мишени, соответствующие 5’- и/или 3’-гомологичным плечам экзогенной донорной нуклеиновой кислоты, могут находиться, например, в пределах по меньшей мере 1 нуклеотида от данного сайта расщепления нуклеазой или в пределах от по меньшей мере 10 нуклеотидов до приблизительно 1000 нуклеотидов от данного сайта расщепления нуклеазой. Например, сайт расщепления нуклеазой может непосредственно прилегать к по меньшей мере одной или обеим последовательностям-мишеням.
[00233] Пространственное взаимное расположение последовательностей-мишеней, которые соответствуют гомологичным плечам экзогенной донорной нуклеиновой кислоты и сайту расщепления нуклеазой, может варьироваться. Например, последовательности-мишени могут быть расположены с 5’-конца от сайта расщепления нуклеазой, последовательности-мишени могут быть расположены с 3’-конца от сайта расщепления нуклеазой, или последовательности-мишени могут фланкировать сайт расщепления нуклеазой.
(4) Другие реагенты, нацеленные на человеческий альбумин
[00234] Активность любого другого известного или предполагаемого реагента, нацеленного на человеческий альбумин, также может быть оценена с использованием раскрытых в настоящем документе животных, отличных от человека. Аналогичным образом, любая другая молекула может быть проверена на активность нацеливания на человеческий альбумин с использованием раскрытых в настоящем документе животных, отличных от человека.
[00235] Примеры других нацеленных на человеческий альбумин реагентов включают антисмысловые олигонуклеотиды (например, миРНК или кшРНК), которые действуют посредством РНК-интерференции (РНКи). Антисмысловые олигонуклеотиды (АСО) или антисмысловые РНК представляют собой короткие синтетические цепочки нуклеотидов, предназначенные для предотвращения экспрессии целевого белка путем избирательного связывания с РНК, кодирующей целевой белок, и предотвращения тем самым трансляции. Эти соединения связываются с РНК с высокой аффинностью и селективностью за счет хорошо охарактеризованного спаривания оснований по Уотсону-Крику (гибридизации). РНК-интерференция (РНКи) представляет собой эндогенный клеточный механизм для контроля экспрессии генов, в котором малые интерферирующие РНК (миРНК), которые связаны с РНК-индуцированным комплексом сайленсинга (RISC), опосредуют расщепление матричной РНК (мРНК)-мишени.
[00236] Другие нацеленные на человеческий альбумин реагенты включают антитела или антигенсвязывающие белки, разработанные для специфического связывания эпитопа человеческого альбумина. Другие реагенты, нацеленные на человеческий альбумин, включают низкомолекулярные реагенты.
D. Введение реагентов, нацеленных на человеческий альбумин, животным, отличным от человека, или в клетки, отличные от человеческих
[00237] Раскрытые в настоящем документе способы могут включать введение животному, отличному от человека, или в клетку, отличную от человеческой, различных молекул (например, реагентов, нацеленных на человеческий альбумин, таких как терапевтические молекулы или комплексы), включая, например, нуклеиновые кислоты, белки, комплексы нуклеиновая кислота-белок или белковые комплексы. «Введение» включает представление клетке или животному, отличному от человека, молекулы (например, нуклеиновой кислоты или белка) таким образом, что она получает доступ внутрь клетки или внутрь клеток в организме животного, отличного от человека. Введение может быть осуществлено любыми способами, и два или более компонентов (например, два компонента или все компоненты) могут быть введены в клетку или животное, отличное от человека, одновременно или последовательно в любой комбинации. Например, белок Cas может быть введен в клетку или животное, отличное от человека, до введения гидовой РНК, или он может быть введен после введения гидовой РНК. В качестве еще одного примера, экзогенная донорная нуклеиновая кислота может быть введена до введения белка Cas и гидовой РНК, или она может быть введена после введения белка Cas и гидовой РНК (например, экзогенная донорная нуклеиновая кислота может быть введена приблизительно в пределах 1, 2, 3, 4, 8, 12, 24, 36, 48 или 72 часов до или после введения белка Cas и гидовой РНК). См., например, источники US 2015/0240263 и US 2015/0110762, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Кроме того, два или более компонентов могут быть введены в клетку или животному, отличному от человека, одним и тем же способом доставки или разными способами доставки. Точно так же два или более компонентов могут быть введены животному, отличному от человека, одним и тем же путем введения или разными путями введения.
[00238] В некоторых способах компоненты системы CRISPR/Cas вводят животному, отличному от человека, или в клетку, отличную от человеческой. Гидовая РНК может быть введена животному, отличному от человека, или в клетку, отличную от человеческой, в форме РНК (например, РНК, транскрибируемой in vitro) или в форме ДНК, кодирующей гидовую РНК. При введении в форме ДНК, указанная ДНК, кодирующая гидовую РНК, может быть функционально связана с промотором, активным в клетке животного, отличного от человека. Например, гидовая РНК может быть доставлена при помощи AAV и экспрессироваться in vivo под управлением промотора U6. Такие ДНК могут присутствовать в одной или более экспрессирующих конструкциях. Например, такие экспрессирующие конструкции могут быть компонентами одной молекулы нуклеиновой кислоты. В качестве альтернативы, они могут быть разделены в любой комбинации между двумя или более молекулами нуклеиновой кислоты (т.е. ДНК, кодирующие одну или более РНК CRISPR, и ДНК, кодирующие одну или более tracrРНК, могут быть компонентами отдельных молекул нуклеиновой кислоты).
[00239] Аналогичным образом, белки Cas могут быть предоставлены в любой форме. Например, белок Cas может быть предоставлен в форме белка, такого как белок Cas в комплексе с гРНК. В качестве альтернативы, белок Cas может быть предоставлен в форме нуклеиновой кислоты, кодирующей белок Cas, такой как РНК (например, матричная РНК (мРНК)) или ДНК. Необязательно, нуклеиновая кислота, кодирующая белок Cas, может быть кодон-оптимизирована для эффективной трансляции в белок в конкретной клетке или организме. Например, нуклеиновая кислота, кодирующая белок Cas, может быть модифицирована для замены кодонов, которые чаще используются в клетке млекопитающего, клетке грызуна, мышиной клетке, крысиной клетке или любой другой представляющей интерес клетке-хозяине по сравнению со встречающейся в природе полинуклеотидной последовательностью.Когда нуклеиновая кислота, кодирующая белок Cas, вводится животному, отличному от человека, белок Cas может временно, условно или конститутивно экспрессироваться в организме животного, отличного от человека.
[00240] Нуклеиновые кислоты, кодирующие белки Cas или гидовые РНК, могут быть функционально связаны с промотором в экспрессирующей конструкции. Экспрессирующие конструкции включают любые конструкции нуклеиновых кислот, способные регулировать экспрессию гена или другой представляющей интерес последовательности нуклеиновой кислоты (например, гена Cas), и которые могут переносить такую представляющую интерес последовательность нуклеиновой кислоты в целевую клетку. Например, нуклеиновая кислота, кодирующая белок Cas, может находиться в векторе, содержащем ДНК, кодирующую одну или более гРНК. В качестве альтернативы, она может находиться в векторе или плазмиде, отдельных от вектора, содержащего ДНК, кодирующую одну или более гРНК. Подходящие промоторы, которые могут быть использованы в экспрессирующей конструкции, включают промоторы, активные, например, в одной или более из эукариотической клетки, человеческой клетки, клетка, отличной от человеческой, клетки млекопитающего, клетки млекопитающего, отличного от человека, клетки грызуна, мышиной клетки, крысиной клетки, клетки хомяка, клетки кролика, плюрипотентной клетки, эмбриональной стволовой (ЭС) клетки, взрослой стволовой клетки, ограниченной стадией развития клетки-предшественника, индуцированной плюрипотентной стволовой (иПС) клетки или находящегося на одноклеточной стадии эмбриона. Такие промоторы могут представлять собой, например, условные промоторы, индуцибельные промоторы, конститутивные промоторы или тканеспецифические промоторы. Необязательно, промотор может быть двунаправленным промотором, управляющим экспрессией как белка Cas в одном направлении, так и гидовой РНК в другом направлении. Такие двунаправленные промоторы могут состоять из (1) полного обычного однонаправленного промотора Pol III, который содержит 3 внешних контрольных элемента: дистальный промоторный элемент (DSE), проксимальный промоторный элемент (PSE) и TATA-бокс; и (2) второго базового промотора Pol III, включающего PSE и TATA-бокс, слитый с 5’-концом DSE в обратной ориентации. Например, в промоторе H1 DSE прилегает к PSE и TATA-боксу, и промотор можно сделать двунаправленным путем создания гибридного промотора, в котором транскрипция в обратном направлении контролируется путем добавления PSE и TATA-бокса, полученного из промотора U6. См., например, источник US 2016/0074535, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. Применение двунаправленного промотора для одновременной экспрессии генов, кодирующих белок Cas и гидовую РНК, позволяет создавать компактные кассеты экспрессии для облегчения доставки.
[00241] Молекулы (например, белки Cas или гидовые РНК), введенные животному, отличному от человека, или в клетку, отличную от человеческой, могут быть представлены в композициях, содержащих носитель, повышающий стабильность вводимых молекул (например, носитель, пролонгирующий период при заданных условиях хранения (например, -20°C, 4°C или температуре окружающей среды), в течение которого продукты разложения остаются ниже порогового значения, например, ниже 0,5 мас.% относительно исходной нуклеиновой кислоты или белка; или повышающий стабильность in vivo). Неограничивающие примеры таких носителей включают микросферы полимолочной кислоты (PLA), микросферы сополимера D,L-молочной и гликолевой кислот (PLGA), липосомы, мицеллы, обратные мицеллы, липидные кохлеаты и липидные микротрубочки.
[00242] В настоящем документе представлены различные способы и композиции, позволяющие ввести молекулу (например, нуклеиновую кислоту или белок) в клетку или животному, отличному от человека. Способы введения молекул в различные типы клеток известны и включают, например, способы стабильной трансфекции, способы временной трансфекции и опосредованные вирусами способы.
[00243] Протоколы трансфекции, а также протоколы введения молекул в клетки могут быть различными. Неограничивающие способы трансфекции включают химические способы трансфекции с использованием липосом; наночастиц; фосфата кальция (Graham et al. (1973) Virology 52 (2): 456-67, Bacchetti et al. (1977) Proc. Natl. Acad. Sci. USA 74 (4): 1590-4, и Kriegler, M (1991). Transfer and Expression: A Laboratory Manual. New York: W.H. Freeman and Company. pp. 96-97); дендримеров; или катионных полимеров, такие как ДЭАЭ-декстран или полиэтиленимин. Нехимические способы включают электропорацию, сонопорацию и оптическую трансфекцию. Трансфекция на основе частиц включает применение генной пушки или трансфекцию с помощью магнита (Bertram (2006) Current Pharmaceutical Biotechnology 7, 277-28). Для трансфекции также могут применяться вирусные способы.
[00244] Введение молекул (например, нуклеиновых кислот или белков) в клетку также может быть опосредовано электропорацией, интрацитоплазматической инъекцией, вирусной инфекцией, аденовирусом, аденоассоциированным вирусом, лентивирусом, ретровирусом, трансфекцией, липид-опосредованной трансфекцией или нуклеофекцией. Нуклеофекция представляет собой усовершенствованную технологию электропорации, которая позволяет доставлять субстраты нуклеиновых кислот не только в цитоплазму, но также через ядерную мембрану и в ядро. Кроме того, для применения нуклеофекции в описанных в настоящем документе способах обычно требуется гораздо меньше клеток, чем при обычной электропорации (например, всего около 2 миллионов по сравнению с 7 миллионами при обычной электропорации). В одном из примеров нуклеофекцию проводят с использованием системы LONZA® NUCLEOFECTOR™.
[00245] Введение молекул (например, нуклеиновых кислот или белков) в клетку (например, зиготу) также можно осуществить с помощью микроинъекции. В зиготах (т.е. находящихся на одноклеточной стадии эмбрионах) микроинъекция может осуществляться в материнский и/или отцовский пронуклеус или в цитоплазму. При осуществлении микроинъекции только в один пронуклеус отцовский пронуклеус более предпочтителен из-за его большего размера. Микроинъекция мРНК предпочтительно осуществляется в цитоплазму (например, для доставки мРНК непосредственно в аппарат трансляции), тогда как микроинъекция белка Cas или полинуклеотида, кодирующего белок Cas или кодирующего РНК, предпочтительно осуществляется в ядро/пронуклеус. В качестве альтернативы, микроинъекция может быть осуществлена с помощью инъекции как в ядро/пронуклеус, так и в цитоплазму: сначала игла может быть введена в ядро/пронуклеус, и может быть произведена инъекция первого количества, а при извлечении иглы из находящегося на одноклеточной стадии эмбриона может быть введено второе количество - в цитоплазму. При введении белка Cas вводят в цитоплазму белок Cas предпочтительно содержит сигнал ядерной локализации для обеспечения доставки в ядро/пронуклеус. Способы осуществления микроинъекций хорошо известны. См., например, источник Nagy et al. (Nagy A, Gertsenstein M, Vintersten K, Behringer R., 2003, Manipulating the Mouse Embryo. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); см. также источники Meyer et al. (2010) Proc. Natl. Acad. Sci. USA 107:15022-15026 и Meyer et al. (2012) Proc. Natl. Acad. Sci. USA 109:9354-9359.
[00246] Другие способы введения молекул (например, нуклеиновой кислоты или белков) в клетку или животному, отличному от человека, могут включать, например, векторную доставку, опосредованную частицами доставку, опосредованную экзосомами доставку, опосредованную липидными наночастицами доставку, опосредованную проникающим пептидом доставку или опосредованную имплантируемым устройством доставку. В качестве конкретных примеров нуклеиновая кислота или белок могут быть введены в клетку или животному, отличному от человека, в носителе, таком как микросфера из полимолочной кислоты) (PLA), микросфера из сополимера D,L-молочной и гликолевой кислот (PLGA), липосома, мицелла, обратная мицелла, липидный кохелат или липидная микротрубочка. Некоторые конкретные примеры доставки животному, отличному от человека, включают гидродинамическую доставку, доставку, опосредованную вирусом (например, доставку, опосредованную аденоассоциированным вирусом (AAV)) и доставку, опосредованную липидными наночастицами.
[00247] Введение нуклеиновых кислот и белков в клетки или животным, отличным от человека, может осуществляться при помощи гидродинамической доставки (HDD). Для доставки генов в паренхимные клетки необходимо вводить только необходимые последовательности ДНК через выбранный кровеносный сосуд, что устраняет проблемы безопасности, связанные с существующими вирусными и синтетическими векторами. При попадании в кровоток ДНК способна достигать клеток в различных тканях, доступных для крови. В гидродинамической доставке используется сила, создаваемая быстрой инъекцией большого объема раствора в несжимаемую кровь в кровотоке, чтобы преодолеть физические барьеры эндотелия и клеточных мембран, которые предотвращают попадание крупных и непроницаемых для мембран соединений в паренхимные клетки. Помимо доставки ДНК, этот способ подходит для эффективной внутриклеточной доставки РНК, белков и других небольших соединений in vivo. См., например, источник Bonamassa et al. (2011) Pharm. Res. 28(4):694-701, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00248] Введение нуклеиновых кислот также может осуществляться посредством опосредованной вирусом доставки, такой как опосредованная AAV доставка или опосредованная лентивирусом доставка. Другие иллюстративные вирусы/вирусные векторы включают ретровирусы, аденовирусы, вирусы осповакцины, поксвирусы и вирусы простого герпеса. Вирусы могут инфицировать делящиеся клетки, неделящиеся клетки или как делящиеся, так и неделящиеся клетки. Вирусы могут интегрироваться в геном хозяина или, в качестве альтернативы, не интегрироваться в геном хозяина. Такие вирусы также могут быть сконструированы таким образом, чтобы они имели сниженный иммунитет. Вирусы могут быть репликационно-компетентными или могут быть репликационно-дефектными (например, дефектными в одном или нескольких генах, необходимых для дополнительных раундов репликации и/или упаковки вириона). Вирусы могут вызывать временную экспрессию, длительную экспрессию (например, по меньшей мере 1 неделю, 2 недели, 1 месяц, 2 месяца или 3 месяца) или постоянную экспрессию (например, Cas9 и/или гРНК). Иллюстративные вирусные титры (например, титры AAV) включают 1012, 1013, 1014, 1015, и 1016 геномов вектора/мл.
[00249] Геном оцДНК AAV состоит из двух открытых рамок считывания, Rep и Cap, фланкированных двумя инвертированными концевыми повторами, которые обеспечивают синтез комплементарной цепи ДНК. При конструировании плазмиды для переноса AAV трансген помещают между двумя ITR, и Rep и Cap могут быть представлены в транс-положении. Помимо Rep и Cap для AAV может потребоваться хелперная плазмида, содержащая гены аденовируса. Эти гены (E4, E2a и VA) опосредуют репликацию AAV. Например, плазмида для переноса, Rep/Cap и хелперная плазмида могут быть трансфицированы в клетки HEK293, содержащие ген аденовируса E1+, для получения инфекционных частиц AAV. В качестве альтернативы, Rep, Cap и хелперные гены аденовируса могут быть объединены в одной плазмиде. Аналогичные упаковывающие клетки и способы могут быть использованы для других вирусов, например, ретровирусов.
[00250] Идентифицировано несколько серотипов AAV. Эти серотипы различаются типами клеток, которые они инфицируют (т.е. их тропизмом), что позволяет осуществлять избирательную трансдукцию определенных типов клеток. Серотипы для тканей ЦНС включают AAV1, AAV2, AAV4, AAV5, AAV8 и AAV9. Серотипы для сердечной ткани включают AAV1, AAV8 и AAV9. Серотипы для ткани почек включают AAV2. Серотипы для легочной ткани включают AAV4, AAV5, AAV6 и AAV9. Серотипы для ткани поджелудочной железы включают AAV8. Серотипы для фоторецепторных клеток включают AAV2, AAV5 и AAV8. Серотипы для пигментного эпителия сетчатки включают AAV1, AAV2, AAV4, AAV5 и AAV8. Серотипы для скелетной мышечной ткани включают AAV1, AAV6, AAV7, AAV8 и AAV9. Серотипы для ткани печени включают AAV7, AAV8 и AAV9, и особенно AAV8.
[00251] Тропизм может быть дополнительно уточнен с помощью псевдотипирования, которое представляет собой смешение капсида и генома разных вирусных серотипов. Например, AAV2/5 означает вирус, содержащий геном серотипа 2, упакованный в капсид серотипа 5. Применение псевдотипированных вирусов может улучшить эффективность трансдукции, а также изменить тропизм. Гибридные капсиды, полученные из разных серотипов, также могут быть использованы для изменения вирусного тропизма. Например, AAV-DJ содержит гибридный капсид восьми серотипов и демонстрирует высокую инфицирующую способность в широком диапазоне типов клеток in vivo. AAV-DJ8 представляет собой еще один пример, который демонстрирует свойства AAV-DJ, но с улучшенным захватом головным мозгом. Серотипы AAV также могут быть изменены посредством мутаций. Примеры мутационных модификаций AAV2 включают Y444F, Y500F, Y730F и S662V. Примеры мутационных модификаций AAV3 включают Y705F, Y731F и T492V. Примеры мутационных модификаций AAV6 включают S663V и T492V. Другие псевдотипированные/модифицированные варианты AAV включают AAV2/1, AAV2/6, AAV2/7, AAV2/8, AAV2/9, AAV2.5, AAV8.2 и AAV/SASTG.
[00252] Для ускорения экспрессии трансгена можно использовать самокомплементарные варианты AAV (scAAV). Поскольку AAV зависит от механизма репликации ДНК клетки для синтеза комплементарной цепи генома в виде одноцепочечной ДНК AAV, экспрессия трансгена может быть отсрочена. Чтобы устранить эту задержку, можно использовать scAAV, содержащие комплементарные последовательности, которые способны спонтанно отжигаться при инфицировании, что устраняет потребность в синтезе ДНК клетки-хозяина. Однако, также можно использовать одноцепочечные векторы AAV (ssAAV).
[00253] Чтобы увеличить упаковочную способность, более длинные трансгены могут быть разделены между двумя плазмидами для переноса AAV, первая с 3’-донором сплайсинга, а вторая с 5’-акцептором сплайсинга. При совместном инфицировании клетки эти вирусы образуют конкатемеры, стыкуются и обеспеяивают возможность экспрессии полноразмерного трансгена. Хотя это обеспечивает более длительную экспрессию трансгена, экспрессия менее эффективна. В аналогичных способах увеличения способности используется гомологичная рекомбинация. Например, трансген может быть разделен между двумя плазмидами для переноса, но со значительным перекрытием последовательностей, так что совместная экспрессия вызывает гомологичную рекомбинацию и экспрессию полноразмерного трансгена.
[00254] Введение нуклеиновых кислот и белков также может осуществляться путем доставки, опосредованной липидными наночастицами (LNP). Например, опосредованная LNP доставка может быть использована для доставки комбинации мРНК Cas и гидовой РНК или комбинации белка Cas и гидовой РНК. Доставка такими способами приводит к временной экспрессии Cas, а биоразлагаемые липиды улучшают клиренс, улучшают переносимость и снижают иммуногенность. Липидные составы могут защищать биологические молекулы от деградации, улучшая их клеточный захват. Липидные наночастицы представляют собой частицы, содержащие множество липидных молекул, физически связанных друг с другом межмолекулярными силами. К ним относятся микросферы (включая однослойные и многослойные везикулы, например, липосомы), дисперсную фазу в эмульсии, мицеллы или внутреннюю фазу в суспензии. Такие липидные наночастицы могут быть использованы для инкапсуляции одной или более нуклеиновых кислот или белков для доставки. Составы, содержащие катионные липиды, пригодны для доставки полианионов, таких как нуклеиновые кислоты. Другие липиды, которые могут быть включены, представляют собой нейтральные липиды (т.е. незаряженные или цвиттерионные липиды), анионные липиды, вспомогательные липиды, которые усиливают трансфекцию, и липиды-невидимки, которые увеличивают продолжительность существования наночастиц in vivo. Примеры подходящих катионных липидов, нейтральных липидов, анионных липидов, вспомогательных липидов и липидов-невидимок можно найти в источнике WO 2016/010840 A1, включенном в настоящий документ посредством ссылки в полном объеме для всех целей. Иллюстративная липидная наночастица может содержать катионный липид и один или более других компонентов. В одном примере другой компонент может включать вспомогательный липид, такой как холестерин. В еще одном примере другие компоненты могут включать вспомогательный липид, такой как холестерин, и нейтральный липид, такой как DSPC (дипальмитоилфосфатидилхолин). В еще одном примере другие компоненты могут содержать вспомогательный липид, такой как холестерин, необязательный нейтральный липид, такой как DSPC, и липид-невидимку, такой как S010, S024, S027, S031 или S033.
[00255] LNP может содержать одно или более, или все из следующего: (i) липид для инкапсуляции и для эндосомального высвобождения; (ii) нейтральный липид для стабилизации; (iii) вспомогательный липид для стабилизации; и (iv) липид-невидимка. См., например, источники Finn et al. (2018) Cell Reports 22:1-9 и WO 2017/173054 A1, каждый из которых включен в данный документ посредством ссылки в полном объеме для всех целей. В некоторых LNP груз может включать гидовую РНК или нуклеиновую кислоту, кодирующую гидовую РНК. В некоторых LNP груз может включать мРНК, кодирующую нуклеазу Cas, такую как Cas9, и гидовую РНК или нуклеиновую кислоту, кодирующую гидовую РНК.
[00256] Липид для инкапсуляции и эндосомального высвобождения может представлять собой катионный липид. Липид также может представлять собой биоразлагаемый липид, такой как биоразлагаемый ионизируемый липид. Одним из примеров подходящего липида является липид A или LP01, который представляет собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропилоктадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил(9Z,12Z)-октадека-9,12-диеноат. См., например, источники Finn et al. (2018) Cell Reports 22:1-9 и WO 2017/173054 A1, каждый из которых включен в данный документ посредством ссылки в полном объеме для всех целей. Еще одним примером подходящего липида является липид B, который представляет собой ((5-((диметиламино)метил)-1,3-фенилен)бис(окси))бис(октан-8,1-диил)бис(деканоат), также называемый ((5-((диметиламино)метил)-1,3-фенилен)бис(окси))бис(октан-8,1-диил)бис(деканоатом). Еще одним примером подходящего липида является липид C, который представляет собой 2-(4-(((3-(диметиламино)пропокси)карбонил)окси)гексадеканоил)окси)пропан-1,3-диил(9Z,9'Z,12Z,12'Z)-бис(октадека-9,12-диеноат). Еще одним примером подходящего липида является липид D, который представляет собой 3-(((3-(диметиламино)пропокси)карбонил)окси)-13-(октаноилокси)тридецил-3-октилундеканоат. Другие подходящие липиды включают: гептатриаконта-6,9,28,31-тетраен-19-ил 4-(диметиламино)бутаноат (также известный как Dlin-MC3-DMA (MC3))).
[00257] Некоторые такие липиды, подходящие для применения в описанных в настоящем документе LNP, являются биоразлагаемыми in vivo. Например, LNP, содержащие такой липид, включают те, в которых по меньшей мере 75% липида выводится из плазмы в течение 8, 10, 12, 24 или 48 часов, или 3, 4, 5, 6, 7 или 10 дней. В качестве еще одного примера, по меньшей мере 50% LNP выводится из плазмы в течение 8, 10, 12, 24 или 48 часов, или 3, 4, 5, 6, 7 или 10 дней.
[00258] Такие липиды могут быть ионизируемыми в зависимости от pH среды, в которой они находятся. Например, в слабокислой среде липиды могут быть протонированы и, таким образом, нести положительный заряд. И наоборот, в слабощелочной среде, такой как, например, кровь, где pH составляет приблизительно 7,35, липиды не могут протонироваться и, таким образом, не имеют заряда. В некоторых вариантах осуществления липиды могут быть протонированы при pH по меньшей мере приблизительно 9, 9,5 или 10. Способность такого липида нести заряд связана с его собственной pKa. Например, липид может независимо иметь pKa в диапазоне от приблизительно 5,8 до приблизительно 6,2.
[00259] Нейтральные липиды стабилизируют и улучшают процессинг LNP. Примеры подходящих нейтральных липидов включают множество нейтральных, незаряженных или цвиттерионных липидов. Примеры нейтральных фосфолипидов, подходящих для применения в настоящем изобретении, включают, не ограничиваясь перечисленным, 5-гептадецилбензол-1,3-диол (резорцин), дипальмитоилфосфатидилхолин (DPPC), дистеароилфосфатидилхолин (DSPC), фосфохолин (DOPC), димиристоилфосфатидилхолин (DMPC), фосфатидилхолин (PLPC), 1,2-диарахидоноил-sn-глицеро-3-фосфохолин (DAPC), фосфатидилэтаноламин (PE), яичный фосфатидилхолин (EPC), дилаурилоилфосфатидилхолин (DLPC), димиристоилфосфатидилхолин (DMPC), 1-миристоил-2-пальмитоилфосфатилхолин (MPPC), 1-пальмоитил-2-миристоилфосфатидилхолин (PMPC), 1-пальмитоил-2-стеароилфосфатидилхолин (PSPC), 1,2-диарахидоил-sn-глицеро-3-фосфохолин (DBPC), 1-стеароил-2-пальмитоилфосфатидилхолин (SPPC), 1,2-диэйкозеноил-sn глицеро-3-фосфохолин (DEPC), пальмитоилолеоилфосфатидилхолин (POPC), лизофосфатидилхолин, диолеоилфосфатидилэтаноламин (DOPE), дилинолеоилфосфатидилхолин дистеароилфосфатидилэтаноламин (DSPE), димиристоилфосфатидилэтаноламин (DMPE), дипальмитоилфосфатидилэтаноламин (DPPE), пальмитоилолеилфосфатидилэтаноламин (POPE), лизофосфатидилэтаноламин и их комбинации. Например, нейтральный фосфолипид может быть выбран из группы, состоящей из дистеароилфосфатидилхолина (DSPC) и димиристоилфосфатидилэтаноламина (DMPE).
[00260] Вспомогательные липиды включают липиды, которые усиливают трансфекцию. Механизм, с помощью которого вспомогательный липид усиливает трансфекцию, может включать повышение стабильности частиц. В некоторых случаях вспомогательный липид может усиливать фузогенность мембран. Вспомогательные липиды включают стероиды, стерины и алкилрезорцины. Примеры подходящих вспомогательных липидов включают холестерин, 5-гептадецилрезорцин и холестерин гемисукцинат. В одном примере вспомогательный липид может представлять собой холестерин или холестерин гемисукцинат.
[00261] Липиды-невидимки включают липиды, которые изменяют продолжительность существования наночастиц in vivo. Липиды-невидимки могут способствовать процессу приготовления препарата, например, уменьшая агрегацию частиц и контролируя размер частиц. Липиды-невидимки могут модулировать фармакокинетические свойства LNP. Подходящие липиды-невидимки включают липиды, имеющие гидрофильную головную группу, связанную с липидным фрагментом.
[00262] Гидрофильная головная группа липида-невидимки может содержать, например, полимерный фрагмент, выбранный из полимеров на основе PEG (иногда называемый поли(этиленоксидом)), поли(оксазолином), поли(виниловым спиртом), поли(глицерином), поли(N-винилпирролидоном), полиаминокислотами и поли-N-(2-гидроксипропил)метакриламидом). Термин «PEG» означает любой полиэтиленгликоль или другой полимер на основе простого полиалкиленового эфира. В некоторых препаратах LNP PEG представляет собой PEG-2К, также называемый PEG 2000, который имеет среднюю молекулярную массу приблизительно 2000 Дальтон. См., например, источник WO 2017/173054 A1, включенный в настоящий документ посредством ссылки в полном объеме для всех целей.
[00263] Липидный фрагмент липида-невидимки может быть получен, например, из диацилглицерина или диацилгликамида, включая группы, содержащие диалкилглицериновую или диалкилгликамидную группу, имеющую длину алкильной цепи, независимо содержащую от приблизительно C4 до приблизительно C40 насыщенных или ненасыщенных атомов углерода, где цепь может содержать одну или более функциональных групп, таких как, например, амид или сложный эфир. Диалкилглицериновая или диалкилгликамидная группа может дополнительно содержать одну или более замещенных алкильных групп.
[00264] В качестве одного примера, липид-невидимка может быть выбран из PEG-дилауроилглицерина, PEG-димиристоилглицерина (PEG-DMG), PEG-дипальмитоилглицерина, PEG-дистеароилглицерина (PEG-DSPE), PEG-дилаурилгликамида, PEG-димиристилгликамида, PEG-дипальмитоилгликамида и PEG-дистеароилгликамида, PEG-холестерина (1-[8’-(холест-5-ен-3[бета]-окси)карбоксамидо-3’,6’-диоксаоктанил]карбамоил-[омега]-метилполи(этиленгликоль), PEG-DMB (3,4-дитетрадекоксилбензил-[омега]-метилполи(этиленгликоль)эфир), 1,2-димиристоил-sn-глицеро-3-фосфоэтаноламин-N-[метокси(полиэтиленгликоль)-2000] (PEG2k-DMG), 1,2-дистеароил-sn-глицеро-3-фосфоэтаноламин-N-[метокси(полиэтиленгликоль)-2000] (PEG2k-DSPE), 1,2-дистеароил-sn-глицерин метоксиполиэтиленгликоля (PEG2k-DSG), поли(этиленгликоль)-2000-диметакрилата (PEG2k-DMA) и 1,2-дистеарилоксипропил-3-амин-N-[метокси(полиэтиленгликоль)-2000] (PEG2k-DSA). В одном конкретном примере липид-невидимка может представлять собой PEG2k-DMG.
[00265] LNP могут содержать различные соответствующие молярные соотношения входящих в состав препарата липидов. Мол.% липида CCD может составлять, например, от приблизительно 30 мол.% до приблизительно 60 мол.%, от приблизительно 35 мол.% до приблизительно 55 мол.%, от приблизительно 40 мол.% до приблизительно 50 мол.%, от приблизительно 42 мол.% до приблизительно 47 мол.%, или приблизительно 45%. Мол.% вспомогательного липида может составлять, например, от приблизительно 30 мол.% до приблизительно 60 мол.%, от приблизительно 35 мол.% до приблизительно 55 мол.%, от приблизительно 40 мол.% до приблизительно 50 мол.%, от приблизительно 41 мол.% до приблизительно 46 мол.%, или приблизительно 44 мол.%. Мол.% нейтрального липида может составлять, например, от приблизительно 1 мол.% до приблизительно 20 мол.%, от приблизительно 5 мол.% до приблизительно 15 мол.%, от приблизительно 7 мол.% до приблизительно 12 мол.%, или приблизительно 9 мол.%. Мол.% липида-невидимки может составлять, например, от приблизительно 1 мол.% до приблизительно 10 мол.%, от приблизительно 1 мол.% до приблизительно 5 мол.%, от приблизительно 1 мол.% до приблизительно 3 мол.%, приблизительно 2 мол.%, или приблизительно 1 мол.%.
[00266] LNP могут иметь разные соотношения между положительно заряженными аминогруппами биоразлагаемого липида (N) и отрицательно заряженными фосфатными группами (P) нуклеиновой кислоты, которую необходимо инкапсулировать. Это может быть математически представлено уравнением N/P. Например, соотношение N/P может составлять от приблизительно 0,5 до приблизительно 100, от приблизительно 1 до приблизительно 50, от приблизительно 1 до приблизительно 25, от приблизительно 1 до приблизительно 10, от приблизительно 1 до приблизительно 7, от приблизительно 3 до приблизительно 5, от приблизительно 4 до приблизительно 5, приблизительно 4, приблизительно 4,5 или приблизительно 5. Соотношение N/P также может составлять от приблизительно 4 до приблизительно 7 или от приблизительно 4,5 до приблизительно 6. В конкретных примерах соотношение N/P может составлять 4,5 или может составлять 6.
[00267] В некоторых LNP груз может содержать мРНК Cas и гРНК. мРНК Cas и гРНК могут находиться в разных соотношениях. Например, препарат LNP может включать соотношение мРНК Cas и нуклеиновой кислоты гРНК в диапазоне от приблизительно 25:1 до приблизительно 1:25, в диапазоне от приблизительно 10:1 до приблизительно 1:10, в диапазоне от приблизительно 5:1 до приблизительно 1:5, или приблизительно 1:1. В качестве альтернативы, препарат LNP может включать соотношение мРНК Cas и нуклеиновой кислоты гРНК от приблизительно 1:1 до приблизительно 1:5, или приблизительно 10:1. В качестве альтернативы, препарат LNP может включать соотношение мРНК Cas и нуклеиновой кислоты гРНК, равное приблизительно 1:10, 25:1, 10:1, 5:1, 3:1, 1:1, 1:3, 1:5, 1:10 или 1:25. В качестве альтернативы, препарат LNP может включать соотношение мРНК Cas и нуклеиновой кислоты гРНК от приблизительно 1:1 до приблизительно 1:2. В конкретных примерах соотношение мРНК Cas и гРНК может составлять приблизительно 1:1 или приблизительно 1:2.
[00268] В некоторых LNP груз может содержать экзогенную донорную нуклеиновую кислоту и гРНК. Экзогенная донорная нуклеиновая кислота и гРНК могут находиться в разных соотношениях. Например, препарат LNP может включать соотношение экзогенной донорной нуклеиновой кислоты и нуклеиновой кислоты гРНК в диапазоне от приблизительно 25:1 до приблизительно 1:25, в диапазоне от приблизительно 10:1 до приблизительно 1:10, в диапазоне от приблизительно 5:1 до приблизительно 1:5, или приблизительно 1:1. В качестве альтернативы, препарат LNP может включать соотношение экзогенной донорной нуклеиновой кислоты и нуклеиновой кислоты гРНК от приблизительно 1:1 до приблизительно 1:5, от приблизительно 5:1 до приблизительно 1:1, приблизительно 10:1 или приблизительно 1:10. В качестве альтернативы, препарат LNP может включать соотношение экзогенной донорной нуклеиновой кислоты и нуклеиновой кислоты гРНК, равное приблизительно 1:10, 25:1, 10:1, 5:1, 3:1, 1:1, 1:3, 1:5, 1:10 или 1:25.
[00269] Конкретный пример подходящей LNP имеет соотношение азота и фосфата (N/P) 4,5 и содержит биоразлагаемый катионный липид, холестерин, DSPC и PEG2k-DMG в молярном соотношении 45:44:9:2. Биоразлагаемый катионный липид может представлять собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропилоктадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил(9Z,12Z)-октадека-9,12-диеноатом. См., например, источник Finn et al. (2018) Cell Reports 22:1-9, включенный в настоящий документ посредством ссылки в полном объеме для всех целей. мРНК Cas9 может находиться в массовом соотношении 1:1 с гидовой РНК. Еще один конкретный пример подходящей LNP содержит Dlin-MC3-DMA (MC3), холестерин, DSPC и PEG-DMG в молярном соотношении 50:38,5:10:1,5.
[00270] Еще один конкретный пример подходящей LNP имеет соотношение азота и фосфата (N/P), равное 6, и содержит биоразлагаемый катионный липид, холестерин, DSPC и PEG2k-DMG в молярном соотношении 50:38:9:3. Биоразлагаемый катионный липид может представлять собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропилоктадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил(9Z,12Z)-октадека-9,12-диеноатом. мРНК Cas9 может находиться в массовом соотношении 1:2 с гидовой РНК.
[00271] Способ доставки может быть выбран для снижения иммуногенности. Например, белок Cas и гРНК могут быть доставлены разными способами (например, бимодальная доставка). Эти разные способы могут придавать различные фармакодинамические или фармакокинетические свойства отдельно взятой доставляемой молекуле (например, Cas или кодирующей нуклеиновой кислоте, гРНК или кодирующей нуклеиновой кислоте, или экзогенной донорной нуклеиновой кислоте/матрице репарации). Например, разные способы могут приводить к разному распределению в тканях, разному периоду полувыведения или разному распределению по времени. Некоторые способы доставки (например, доставка нуклеиново-кислотного вектора, который сохраняется в клетке путем автономной репликации или геномной интеграции) приводят к более устойчивой экспрессии и присутствию молекулы, тогда как другие способы доставки являются временными и менее устойчивыми (например, доставка РНК или белка). Доставка белков Cas более временным способом, например в виде мРНК или белка, может гарантировать, что комплекс Cas/гРНК присутствует и активен только в течение короткого периода времени, и может снизить иммуногенность, вызываемую пептидами из фермента Cas, полученного из бактерий, экспонируемыми на поверхности клетки молекулами MHC. Такая временная доставка может также снизить вероятность нецелевых модификаций.
[00272] Введение in vivo может осуществляться любым подходящим путем, включая, например, парентеральный, внутривенный, пероральный, подкожный, внутриартериальный, интракраниальный, интратекальный, внутрибрюшинный, местный, интраназальный или внутримышечный. Системные способы введения включают, например, пероральный и парентеральный пути. Примеры парентеральных путей включают внутривенный, внутриартериальный, внутрикостный, внутримышечный, внутрикожный, подкожный, интраназальный и внутрибрюшинный пути. Конкретный пример представляет собой внутривенную инфузию. Другими конкретными примерами являются закапывание в нос и интравитреальная инъекция. Локальные способы введения включают, например, интратекальный, интрацеребровентрикулярный, интрапаренхиматозный (например, локализованную интрапаренхиматозную доставку в полосатое тело (например, в хвостатое ядро или в дорсолатеральную часть), кору головного мозга, прецентральную извилину, гиппокамп (например, в зубчатую извилину или область CA3), височную кору, миндалину, лобную кору, таламус, мозжечок, продолговатый мозг, гипоталамус, перемычку, тегментум или черную субстанцию), внутриглазной, внутриглазничный, субконъюнктивальный, интравитреальный, субретинальный и трансклеральный пути. Значительно меньшие количества компонентов (по сравнению с системными подходами) могут оказывать эффект при локальном введении (например, интрапаренхиматозном или интравитреальном) по сравнению с системным введением (например, внутривенным). Способы местного введения могут также снижать или устранять частоту потенциально токсичных побочных действий, которые могут возникать при системном введении терапевтически эффективных количеств компонента.
[00273] Введение in vivo может осуществляться любым подходящим путем, включая, например, парентеральный, внутривенный, пероральный, подкожный, внутриартериальный, интракраниальный, интратекальный, внутрибрюшинный, местный, интраназальный или внутримышечный. Конкретный пример представляет собой внутривенную инфузию. Композиции, содержащие гидовые РНК и/или белки Cas (или нуклеиновые кислоты, кодирующие гидовые РНК и/или белки Cas), могут быть составлены с использованием одного или более физиологически и фармацевтически приемлемых носителей, разбавителей, эксципиентов или вспомогательных веществ. Состав может зависеть от выбранного пути введения. Термин «фармацевтически приемлемый» означает, что носитель, разбавитель, эксципиент или вспомогательное вещество совместимы с другими ингредиентами препарата и по существу не являются вредными для их реципиента.
[00274] Частота введения и количество доз могут зависеть от периода полувыведения экзогенных донорных нуклеиновых кислот, гидовых РНК или белков Cas (или нуклеиновых кислот, кодирующих гидовые РНК или белки Cas) и пути введения среди других факторов. Введение нуклеиновых кислот или белков в клетку или животному, отличному от человека, может быть проведено один или более раз в течение определенного периода времени. Например, введение может быть проведено по меньшей мере два раза в течение определенного периода времени, по меньшей мере три раза в течение определенного периода времени, по меньшей мере четыре раза в течение определенного периода времени, по меньшей мере пять раз в течение определенного периода времени, по меньшей мере шесть раз в течение определенного периода времени, по меньшей мере семь раз в течение определенного периода времени, по меньшей мере восемь раз в течение определенного периода времени, по меньшей мере девять раз в течение определенного периода времени, по меньшей мере десять раз в течение определенного периода времени, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз в течение определенного периода времени, по меньшей мере тринадцать раз в течение определенного периода времени, по меньшей мере пятнадцать раз в течение определенного периода времени, по меньшей мере шестнадцать раз в течение определенного периода времени, по меньшей мере семнадцать раз в течение определенного периода времени, по меньшей мере восемнадцать раз в течение определенного периода времени, по меньшей мере девятнадцать раз в течение определенного периода времени или по меньшей мере двадцать раз в течение определенного периода времени.
E. Измерение доставки, активности или эффективности реагентов, нацеленных на человеческий альбумин, in vivo или ex vivo
[00275] Раскрытые в настоящем документе способы могут дополнительно включать обнаружение или измерение активности реагентов, нацеленных на человеческий альбумин. Например, если реагент, нацеленный на человеческий альбумин, представляет собой реагент для редактирования генома (например, CRISPR/Cas, разработанный для нацеливания на локус человеческого альбумина), измерение может включать оценку гуманизированного локуса альбумина на предмет модификаций.
[00276] Для идентификации клеток, имеющих целевую генетическую модификацию, могут быть использованы различные способы. Скрининг может включать количественный анализ для оценки модификации аллеля (MOA) исходной хромосомы. См., например, источники US 2004/0018626; US 2014/0178879; US 2016/0145646; WO 2016/081923; и Frendewey et al. (2010) Methods Enzymol. 476:295-307, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Например, количественный анализ может быть проведен с помощью количественной ПЦР, такой как ПЦР в реальном времени (кПЦР). В ПЦР в реальном времени может использоваться первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой референсный локус. Набор праймеров может включать флуоресцентный зонд, распознающий амплифицированную последовательность. Другие примеры подходящих количественных анализов включают флуоресцентную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным(и) зондом(ами), зонды INVADER®, зонды типа «молекулярный маяк» (Molecular Beacon) TAQMAN® или технологию зондов ECLIPSE™ (см., например, источник US 2005/0144655, включенный в настоящий документ посредством ссылки в полном объеме для всех целей).
[00277] Для скрининга также может быть использовано секвенирование нового поколения (NGS). Секвенирование нового поколения также может называться «NGS», «массовое параллельное секвенирование» или «высокопроизводительное секвенирование». NGS может быть использовано в качестве инструмента скрининга в дополнение к анализам MOA для определения точной природы целевой генетической модификации и ее соответствия между типами клеток, типами тканей или типами органов.
[00278] Оценка модификации гуманизированного локуса альбумина у животного, отличного от человека, может быть осуществлена в любом типе клеток из любой ткани или органа. Например, оценка может быть осуществлена в нескольких типах клеток из одной и той же ткани или органа или в клетках из нескольких мест в ткани или органе. Она может предоставить информацию о том, какие типы клеток в целевой ткани или органе выступают в качестве мишеней, или какие участки ткани или органа достигаются реагентом, нацеленным на человеческий альбумин. В качестве еще одного примера, оценка может быть осуществлена в нескольких типах тканей или в нескольких органах. В способах, в которых нацеливание осуществляется на конкретную ткань, орган или тип клеток, это может предоставить информацию о том, насколько эффективно осуществляется нацеливание на эту ткань или орган, и есть ли побочные действия в других тканях или органах.
[00279] Если реагент предназначен для инактивации гуманизированного локуса альбумина, воздействия на экспрессию гуманизированного локуса альбумина, предотвращения трансляции мРНК гуманизированного альбумина или устранения белка гуманизированного альбумина, измерение может включать оценку экспрессии мРНК или белка гуманизированного альбумина. Это измерение может быть осуществлено в печени или определенных типах клеток или областях в печени, или оно может включать измерение сывороточных уровней секретируемого белка гуманизированного альбумина.
[00280] Если реагент представляет собой экзогенную донорную нуклеиновую кислоту, кодирующую экзогенный белок, не кодируемый или не экспрессируемый эндогенным локусом альбумина дикого типа, измерение может включать оценку экспрессии мРНК, кодируемой экзогенной донорной нуклеиновой кислотой, или оценку экспрессии экзогенного белка. Это измерение может быть осуществлено в печени или определенных типах клеток или областях в печени, или оно может включать измерение сывороточных уровней секретируемого экзогенного белка. В конкретном примере экзогенный белок представляет собой белок фактора IX. Необязательно, указанная оценка включает измерение сывороточных уровней белка фактора IX у указанного животного, отличного от человека, и/или включает оценку активированного частичного тромбопластинового времени или проведение теста генерации тромбина. Необязательно, животное, отличное от человека, дополнительно содержит инактивированный локус F9, и указанная оценка включает измерение сывороточных уровней белка фактора IX у указанного животного, отличного от человека, и/или включает оценку активированного частичного тромбопластинового времени (АЧТВ) или проведение теста генерации тромбина (ТГТ). Эти анализы более подробно описаны в примерах.
[00281] Одним из примеров анализа, который может быть использован, является анализ in situ гибридизации РНК BASESCOPE™ (ISH), который представляет собой метод, позволяющий количественно оценить клеточно-специфичные отредактированные транскрипты. ISH анализ РНК BASESCOPE™ может дополнять NGS и кПЦР при определения характеристик редактирования генов. В то время как NGS/кПЦР могут предоставить количественные средние значения последовательностей дикого типа и отредактированных последовательностей, они не предоставляют информации о неоднородности или процентном содержании отредактированных клеток в ткани. ISH анализ BASESCOPE™ может обеспечить обзор всей ткани и количественную оценку транскриптов дикого типа по сравнению с отредактированными транскриптами с разрешением на уровне отдельных клеток, где можно количественно оценить фактическое количество клеток в целевой ткани, содержащих отредактированный транскрипт мРНК. Анализ BASESCOPE™ обеспечивает обнаружение одиночной молекулы РНК с использованием парных олигозондов («ZZ») для усиления сигнала без неспецифического фона. Тем не менее, конструкция зонда BASESCOPE™ и система амплификации сигнала позволяют обнаруживать одиночные молекулы РНК с помощью зонда ZZ и способны дифференцированно обнаруживать единичные нуклеотидные изменения и мутации в интактной фиксированной ткани.
[00282] Получение и секреция белка гуманизированного альбумина или экзогенного белока могут быть оценены любыми известными способами. Например, экспрессия может быть оценена путем измерения уровней кодируемой мРНК в печени животного, отличного от человека, или уровней кодируемого белка в печени животного, отличного от человека, с использованием известных анализов. Секреция белка гуманизированного альбумина или экзогенного белка может быть оценена путем измерения уровней кодируемого белка гуманизированного альбумина или экзогенного белка в плазме или сыворотке животного, отличного от человека, с использованием известных анализов.
IV. Способы создания животных, отличных от человека, содержащих гуманизированный локус альбумина
[00283] Предложены различные способы создания генома животного, отличного от человека, клетки животного, отличного от человека, или животного, отличного от человека, содержащих гуманизированный локус альбумина (ALB), как раскрыто в других местах настоящего документа. Любой удобный способ или протокол получения генетически модифицированного организма подходит для получения такого генетически модифицированного животного, отличного от человека. См., например, источники Cho et al. (2009) Current Protocols in Cell Biology 42:19.11:19.11.1-19.11.22 и Gama Sosa et al. (2010) Brain Struct. Funct. 214(2-3):91-109, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Такие генетически модифицированные животные, отличные от человека, могут быть созданы, например, посредством нокина гена в целевом локусе альбумина.
[00284] Например, способ получения животного, отличного от человека, содержащего гуманизированный локус альбумина, может включать: (1) модификацию генома плюрипотентной клетки для включения гуманизированного локуса альбумина; (2) идентификацию или отбор генетически модифицированной плюрипотентной клетки, содержащей гуманизированный локус альбумина; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион животного-хозяина, отличного от человека; и (4) имплантацию и вынашивание эмбриона-хозяина в организме суррогатной матери. Например, способ получения животного, отличного от человека, содержащего гуманизированный локус альбумина, может включать: (1) модификацию генома плюрипотентной клетки для включения гуманизированного локуса альбумина; (2) идентификацию или отбор генетически модифицированной плюрипотентной клетки, содержащей гуманизированный локус альбумина; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион животного-хозяина, отличного от человека; и (4) вынашивание эмбриона-хозяина у суррогатной матери. Необязательно, эмбрион-хозяин, содержащий модифицированную плюрипотентную клетку (например, ЭС клетку, отличную от человеческой), может быть инкубирован до стадии бластоцисты перед имплантацией и вынашиванием в организме суррогатной матери для получения животного, отличного от человека, поколения F0. Затем суррогатная мать может произвести на свет животное, отличное от человека, поколения F0, содержащее гуманизированный локус альбумина.
[00285] Способы могут дополнительно включать идентификацию клетки или животного, имеющего модифицированный целевой геномный локус. Для идентификации клеток и животных, имеющих целевую генетическую модификацию, могут быть использованы различные способы.
[00286] На стадии модификации генома можно, например, использовать экзогенные донорные нуклеиновые кислоты (например, нацеливающие векторы) для модификации локуса альбумина, чтобы он содержал гуманизированный локус альбумина, раскрытый в настоящем документе. В качестве одного примера, нацеливающий вектор может быть предназначен для создания гуманизированного гена альбумина в эндогенном локусе альбумина (например, эндогенном локусе альбумина животного, отличного от человека), где нацеливающий вектор содержит 5’-гомологичное плечо, нацеленное на 5’-последовательность-мишень в эндогенном локусе альбумина, и 3’-гомологичное плечо, нацеленное на 3’-последовательность-мишень в эндогенном локусе альбумина. Экзогенные донорные нуклеиновые кислоты могут также содержать вставки нуклеиновых кислот, включая сегменты ДНК, которые должны быть интегрированы в локус альбумина. Интеграция вставки нуклеиновой кислоты в локус альбумина может привести к добавлению представляющей интерес последовательности нуклеиновой кислоты в локусе альбумина, делеции представляющей интерес последовательности нуклеиновой кислоты в локусе альбумина или замене представляющей интерес последовательности нуклеиновой кислоты интерес в локусе альбумина (т.е. делеция и вставка). Гомологичные плечи могут фланкировать вставку нуклеиновой кислоты, содержащую последовательность человеческого альбумина, для создания гуманизированного локуса альбумина (например, для удаления сегмента эндогенного локуса альбумина и замены ортологичной последовательностью человеческого альбумина).
[00287] Экзогенные донорные нуклеиновые кислоты могут быть предназначены для вставки, опосредованной негомологичным соединением концов, или для гомологичной рекомбинации. Экзогенные донорные нуклеиновые кислоты могут включать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), они могут быть одноцепочечными или двухцепочечными, и они могут иметь линейную или кольцевую форму. Например, матрица репарации может представлять собой одноцепочечный олигодезоксинуклеотид (оцОДН).
[00288] Экзогенные донорные нуклеиновые кислоты могут также содержать гетерологичную последовательность, которая не присутствует в нецелевом эндогенном локусе альбумина. Например, экзогенные донорные нуклеиновые кислоты могут содержать кассету селекции, такую как кассета селекции, фланкированная сайтами распознавания рекомбиназы.
[00289] Некоторые экзогенные донорные нуклеиновые кислоты содержат гомологичные плечи. Если экзогенная донорная нуклеиновая кислота также содержит вставку нуклеиновой кислоты, гомологичные плечи могут фланкировать указанную вставку нуклеиновой кислоты. Для простоты ссылки гомологичные плечи обозначаются в настоящем документе как 5’- и 3’- (т.е. расположенные по ходу и против хода транскрипции) гомологичные плечи. Эта терминология относится к относительному положению гомологичных плеч относительно вставки нуклеиновой кислоты внутри экзогенной донорной нуклеиновой кислоты. 5’- и 3’-плечи гомологии соответствуют областям в геномном локусе-мишени, которые обозначаются в настоящем документе как «5’-последовательность-мишень» и «3’-последовательность-мишень», соответственно.
[00290] Плечо гомологии и последовательность-мишень «соответствуют» или являются «соответствующими» друг другу тогда, когда две области обладают достаточным уровнем идентичности последовательностей друг к друг, чтобы действовать в качестве субстратов для реакции гомологичной рекомбинации. Термин «гомология» включает последовательности ДНК, которые либо идентичны, либо обе обладают определенной идентичностью соответствующей последовательности. Идентичность последовательностей между данной последовательностью-мишенью и соответствующим гомологичным плечом, содержащимся в донорной нуклеиновой кислоте, может иметь любую степень идентичности последовательностей, которая допускает гомологичную рекомбинацию. Например, степень идентичности последовательностей, общая для гомологичного плеча экзогенной донорной нуклеиновой кислоты (или ее фрагмента) и последовательности-мишени (или ее фрагмента), может представлять собой по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей, так что последовательности подвергаются гомологичной рекомбинации. Кроме того, соответствующая область гомологии между гомологичным плечом и соответствующей последовательностью-мишенью может иметь любую длину, достаточную для способствования гомологичной рекомбинации. В некоторых нацеливающих векторах предполагаемая мутация в эндогенном локусе альбумина включена во вставку нуклеиновой кислоты, фланкированную гомологичными плечами.
[00291] В клетках, отличных от находящхися на одноклеточной стадии эмбрионов, экзогенная донорная нуклеиновая кислота может представлять собой «большой нацеливающий вектор» или «LTVEC», который включает нацеливающие векторы, содержащие гомологичные плечи, которые соответствуют последовательностям нуклеиновых кислот, более крупными, чем обычно используемые в других подходах, предназначенных для осуществления гомологичной рекомбинации в клетках, и получены из них. LTVEC также включают нацеливающие векторы, содержащие вставки нуклеиновых кислот, имеющие последовательности нуклеиновых кислот, более длинные, чем обычно используемые в других подходах, предназначенных для осуществления гомологичной рекомбинации в клетках. Например, LTVEC делают возможной модификацию больших локусов, которые не могут быть охвачены традиционными нацеливающими векторами на основе плазмид из-за их ограничений по размеру. Например, целевой локус может представлять собой (т.е. 5’- и 3’-гомологичные плечи гомологии могут соответствовать) локус клетки, на который не может быть осуществлено нацеливание с помощью обычного способа, или на который может быть осуществлено нацеливание только некорректным образом или только с крайне низкой эффективностью при отсутствии ника или двухцепочечного разрыва, вызванного нуклеазным агентом (например, белком Cas). LTVEC могут иметь любую длину и обычно имеют длину по меньшей мере 10 кб. 5’-гомологичное плечо и 3’-гомологичное плечо в LTVEC в совокупности обычно составляет по меньшей мере 10 кб.
[00292] Этап скрининга может включать, например, количественный анализ для оценки модификации аллеля (MOA) исходной хромосомы. Например, количественный анализ может быть проведен с помощью количественной ПЦР, такой как ПЦР в реальном времени (кПЦР). В ПЦР в реальном времени может использоваться первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой референсный локус. Набор праймеров может включать флуоресцентный зонд, распознающий амплифицированную последовательность.
[00293] Другие примеры подходящих количественных анализов включают флуоресцентную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным(и) зондом(ами), зонды INVADER®, зонды типа «молекулярный маяк» (Molecular Beacon) TAQMAN® или технологию зондов ECLIPSE™ (см., например, источник US 2005/0144655, включенный в настоящий документ посредством ссылки в полном объеме для всех целей).
[00294] Примером подходящей плюрипотентной клетки является эмбриональная стволовая (ЭС) клетка (например, мышиная ЭС клетка или крысиная ЭС клетка). Модифицированная плюрипотентная клетка может быть получена, например, с помощью рекомбинации путем (a) введения в клетку одной или более экзогенных донорных нуклеиновых кислот (например, нацеливающих векторов), содержащих вставку нуклеиновой кислоты, фланкированную, например, 5’ и 3’-гомологичными плечами, соответствующими 5’ и 3’-сайтам-мишеням, где указанная вставка нуклеиновой кислоты содержит последовательность человеческого альбумина для создания гуманизированного локуса альбумина; и (b) идентификации по меньшей мере одной клетки, содержащей в своем геноме указанную вставку нуклеиновой кислоты, интегрированную в эндогенном локусе альбумина (т.е. идентификацию по меньшей мере одной клетки, содержащей гуманизированный локус альбумина). Модифицированная плюрипотентная клетка может быть получена, например, с помощью рекомбинации путем (a) введения в клетку одного или более нацеливающих векторов, содержащих вставку нуклеиновой кислоты, фланкированную 5’ и 3’-гомологичными плечами, соответствующими 5’ и 3’-сайтам-мишеням, где указанная вставка нуклеиновой кислоты содержит гуманизированный локус альбумина; и (b) идентификации по меньшей мере одной клетки, содержащей в своем геноме указанную вставку нуклеиновой кислоты, интегрированную в целевом геномном локусе.
[00295] В качестве альтернативы, модифицированная плюрипотентная клетка может быть получена путем (а) введения в клетку: (i) нуклеазного агента, где нуклеазный агент вызывает ник или двухцепочечный разрыв в сайте-мишени в эндогенном локусе альбумина; и (ii) одной или более экзогенных донорных нуклеиновых кислот (например, нацеливающих векторов), необязательно содержащих вставку нуклеиновой кислоты, фланкированную, например, 5’ и 3’-гомологичными плечами, соответствующими 5’ и 3’-сайтам-мишеням, расположенным в достаточной близости от сайта-мишени нуклеазы, где указанная вставка нуклеиновой кислоты содержит последовательность человеческого альбумина для создания гуманизированного локуса альбумина; и (c) идентификации по меньшей мере одной клетки, содержащей в своем геноме указанную вставку нуклеиновой кислоты, интегрированную в эндогенном локусе альбумина (т.е. идентификацию по меньшей мере одной клетки, содержащей гуманизированный локус альбумина). В качестве альтернативы, модифицированная плюрипотентная клетка может быть получена путем (а) введения в клетку: (i) нуклеазного агента или нуклеиновой кислоты, кодирующей указанный нуклеазный агент, где нуклеазный агент вызывает ник или двухцепочечный разрыв в сайте-мишени в эндогенном локусе альбумина; и (ii) одной или более экзогенных донорных нуклеиновых кислот (например, нацеливающих векторов), необязательно содержащих вставку нуклеиновой кислоты, фланкированную, например, 5’ и 3’-гомологичными плечами, соответствующими 5’ и 3’-сайтам-мишеням, расположенным в достаточной близости от сайта-мишени нуклеазы, где указанная вставка нуклеиновой кислоты содержит последовательность человеческого альбумина для создания гуманизированного локуса альбумина; и (c) идентификации по меньшей мере одной клетки, содержащей в своем геноме указанную вставку нуклеиновой кислоты, интегрированную в эндогенном локусе альбумина (т.е. идентификацию по меньшей мере одной клетки, содержащей гуманизированный локус альбумина). В качестве альтернативы, модифицированная плюрипотентная клетка может быть получена путем (а) введения в клетку: (i) нуклеазного агента, где нуклеазный агент вызывает ник или двухцепочечный разрыв в сайте распознавания в целевом геномном локусе; и (ii) одного или более нацеливающих векторов, содержащих вставку нуклеиновой кислоты, фланкированную 5’ и 3’-гомологичными плечами, соответствующими 5’ и 3’-сайтам-мишеням, расположенным в достаточной близости от указанного сайта распознавания, где указанная вставка нуклеиновой кислоты содержит гуманизированный локус альбумина; и (c) идентификации по меньшей мере одной клетки, содержащей модификацию (например, интеграцию указанной вставки нуклеиновой кислоты) в целевом геномном локусе. Может быть использован любой нуклеазный агент, который вызывает ник или двухцепочечный разрыв в желаемом сайте распознавания. В качестве альтернативы, модифицированная плюрипотентная клетка может быть получена путем (а) введения в клетку: (i) нуклеазного агента или нуклеиновой кислоты, кодирующей указанный нуклеазный агент, где указанный нуклеазный агент вызывает ник или двухцепочечный разрыв в сайте распознавания в целевом геномном локусе; и (ii) одного или более нацеливающих векторов, содержащих вставку нуклеиновой кислоты, фланкированную 5’ и 3’-гомологичными плечами, соответствующими 5’ и 3’-сайтам-мишеням, расположенным в достаточной близости от указанного сайта распознавания, где указанная вставка нуклеиновой кислоты содержит гуманизированный локус альбумина; и (c) идентификации по меньшей мере одной клетки, содержащей модификацию (например, интеграцию указанной вставки нуклеиновой кислоты) в целевом геномном локусе. Может быть использован любой нуклеазный агент, который вызывает ник или двухцепочечный разрыв в желаемом сайте распознавания. Примеры подходящих нуклеаз включают эффекторную нуклеазу, подобную активатору транскрипции (TALEN), нуклеазу с «цинковыми пальцами» (ZFN), мегануклеазу и системы кластерные короткие палиндромные повторы, разделенные регулярными промежутками (CRISPR)/CRISPR-ассоциированная (Cas) (например, системы CRISPR/Cas9) или компоненты таких систем (например, CRISPR/Cas9). См., например, источники US 2013/0309670 и US 2015/0159175, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей.
[00296] Донорская клетка может быть введена в эмбрион-хозяин на любой стадии, такой как стадия бластоцисты или стадия предморулы (т.е. 4-клеточная стадия или 8-клеточная стадия). Получают потомство, способное передавать генетическую модификацию через зародышевую линию. См., например, патент США №7294754, включенный в настоящий документ в полном объеме посредством ссылки для всех целей.
[00297] В качестве альтернативы, способ получения животных, отличных от человека, описанный в других местах настоящего документа, может включать: (1) модификацию генома находящегося на одноклеточной стадии эмбриона для включения гуманизированного локуса альбумина с применением описанных выше способов модификации плюрипотентных клеток; (2) отбор генетически модифицированного эмбриона; и (3) имплантацию и вынашивание генетически модифицированного эмбриона в организме суррогатной матери. В качестве альтернативы, способ получения животных, отличных от человека, описанный в других местах настоящего документа, может включать: (1) модификацию генома находящегося на одноклеточной стадии эмбриона для включения гуманизированного локуса альбумина с применением описанных выше способов модификации плюрипотентных клеток; (2) отбор генетически модифицированного эмбриона; и (3) вынашивание генетически модифицированного эмбриона у суррогатной матери. Получают потомство, способное передавать генетическую модификацию через зародышевую линию.
[00298] Для создания животных-млекопитающих, отличных от человека также могут быть использованы методы ядерного транспорта. Вкратце, способы ядерного транспорта могут включать стадии: (1) удаление ядра из ооцита или обеспечение безъядерного ооцита; (2) выделение или обеспечение донорской клетки или ядра для объединения с безъядерным ооцитом; (3) вставка клетки или ядра в безъядерный ооцит с образованием восстановленной клетки; (4) имплантация восстановленной клетки в матку животного с образованием эмбриона; и (5) обеспечение развития эмбриона. В таких способах ооциты обычно получают от умерших животных, хотя их можно также выделить из маточных труб и/или яичников живых животных. До удаления ядер ооциты могут созревать в различных хорошо известных средах. Удаление ядра из ооцита может быть проведено рядом хорошо известных способов. Вставка донорской клетки или ядра в безъядерный ооцит с образованием восстановленной клетки может быть осуществлено путем микроинъекции донорской клетки под блестящую оболочку перед слиянием. Слияние может быть вызвано приложением электрического импульса постоянного тока через плоскость контакта/слияния (электрослияние), воздействием на клетки химических веществ, способствующих слиянию, таких как полиэтиленгликоль, или посредством инактивированного вируса, такого как вирус Сендай. Восстановленная клетка может быть активирована электрическими и/или неэлектрическими средствами до, во время и/или после слияния донорского ядра и ооцита-реципиента. Способы активации включают электрические импульсы, химически индуцированный шок, проникновение сперматозоидов, повышение уровней двухвалентных катионов в ооците и снижение фосфорилирования клеточных белков (например, с помощью ингибиторов киназ) в ооците. Активированные восстановленные клетки или эмбрионы могут быть культивированы в хорошо известных средах, а затем перенесены в матку животного. См., например, источники US 2008/0092249, WO 1999/005266, US 2004/0177390, WO 2008/017234 и патент США №7612250, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей.
[00299] Различные способы, предложенные в настоящем документе, позволяют получить генетически модифицированное животное, отличное от человека, поколения F0, где клетки генетически модифицированного животного поколения F0 содержат гуманизированный локус альбумина. Признано, что в зависимости от метода, используемого для создания животного поколения F0, количество клеток в животном поколения F0, которые имеют гуманизированный локус альбумина, будет варьироваться. Введение донорских ЭС клеток эмбриону на стадии морулы из соответствующего организма (например, мышиному эмбриону на 8-клеточной стадии) например, методом VELOCIMOUSE®, обеспечивает более высокий процент клеточной популяции животного поколения F0, содержащей клетки, имеющие представляющую интерес нуклеотидную последовательность, содержащую целевую генетическую модификацию. Например, по меньшей мере 50%, 60%, 65%, 70%, 75%, 85%, 86%, 87%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% клеточного вклада животного, отличного от человека, поколения F0 может составлять клеточная популяция, имеющая целевую модификацию.
[00300] Клетки генетически модифицированного животного поколения F0 могут быть гетерозиготными по гуманизированному локусу альбумина или могут быть гомозиготными по гуманизированному локусу альбумина.
[00301] Все патентные документы, веб-сайты, другие публикации, регистрационные номера и т.п., цитируемые выше или ниже, включены посредством ссылки в полном объеме для всех целей в той же степени, как если бы каждый отдельный элемент был специально и отдельно указан как включенный посредством ссылки. Если в разное время разные версии последовательности связаны с определенным регистрационным номером, то подразумевается та версия, которая связана с регистрационным номером на действительную дату подачи данной заявки. Действительная дата подачи означает более раннюю дату из даты фактической подачи или даты подачи приоритетной заявки, содержащей ссылку на регистрационный номер, если это применимо. Аналогичным образом, если в разное время опубликованы разные версии публикации, веб-сайта и т.п., то подразумевается та версия, которая была опубликована последней на действительную дату подачи данной заявки, если не указано иное. Любые признак, стадия, элемент, вариант осуществления или аспект данного изобретения могут быть использованы в комбинации с любым другим, если специально не указано иное. Несмотря на то, что данное изобретение было описано довольно подробно с помощью иллюстраций и примеров для ясности и понимания, очевидно, что могут быть осуществлены определенные изменения и модификации в пределах объема прилагаемой формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[00302] Нуклеотидные и аминокислотные последовательности, перечисленные в прилагаемом перечне последовательностей, представлены с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот. Нуклеотидные последовательности соответствуют стандартному соглашению: они приведены, начиная с 5’-конца последовательности, вперед (т.е. слева направо в каждой строке), до 3’-конца. Приведена только одна цепь каждой нуклеотидной последовательности, но подразумевается, что комплементарная ей цепь включена посредством любой ссылки на представленную цепь. Когда представлена нуклеотидная последовательность, кодирующая аминокислотную последовательность, подразумевается, что также представлены ее варианты с вырожденными кодонами, кодирующие ту же аминокислотную последовательность. Аминокислотные последовательности соответствуют стандартному соглашению: они приведены, начиная с аминоконца последовательности, вперед (т.е. слева направо в каждой строке), до карбоксиконца.
[00303] Таблица 2. Описание последовательностей.
ПРИМЕРЫ
Пример 1. Получение мышей, содержащих гуманизированный локус альбумина (ALB)
[00304] Был создан большой нацеливающий вектор (LTVEC), содержащий 5’-гомологичное плечо, содержащее 20 кб мышиного локуса альбумина (Alb) (из bMQ-127G8), и 3’-гомологичное плечо, содержащее 127 кб мышиного локуса альбумина (Alb) (из bMQ-127G8), для замены области 14,4 кб (14 376 п.н.) из гена мышиного альбумина (Alb) посредством 17,3 кб (17 335 п.н.) соответствующей человеческой последовательности альбумина (ALB) (из RP11-31P12). Сведения о мышином и человеческом альбумине представлены в таблице 3. Получение и применение больших нацеливающих векторов (LTVEC), полученных из ДНК бактериальной искусственной хромосомы (BAC) посредством реакций бактериальной гомологичной рекомбинации (BHR) с использованием технологии генной инженерии VELOCIGENE®, описано, например, в источниках US 6586251 и Valenzuela et al. (2003) Nat. Biotechnol. 21(6):652-659, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Создание LTVEC с помощью способов сборки in vitro описано, например, в источниках US 2015/0376628 и WO 2015/200334, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей.
[00305] Таблица 3. Мышиный и человеческий альбумин (ALB).
[00306] В частности, из мышиного локуса альбумина (Alb) была удалена область от старт-кодона ATG до стоп-кодона (т.е. кодирующие экзоны 1-14). Вместо удаленной мышиной области была вставлена соответствующая область человеческого альбумина (ALB) от старт-кодона ATG до 100 п.н. по ходу транскрипции от стоп-кодона. По ходу транскрипции от человеческой 3’-UTR была вставлена кассета loxP-mPrm1-Crei-pA-hUb1-em7-Neo-pA-loxP (4766 п.н.) с буфером примерно 100 п.н. из человеческой 3’-последовательности после 3’-UTR непосредственно перед кассетой. Эта область представляет собой аллель MAID 7626. См. фиг. 1A. После делеции кассеты loxP и сайты клонирования (38 п.н.) остались по ходу транскрипции от человеческой 3’-UTR, с буфером из 100 п.н. человеческой 3’-последовательности после 3’-UTR непосредственно перед оставшимся сайтом loxP. Эта область представляет собой аллель MAID 7627. См. фиг. 1B.
[00307] Последовательности сигнального пептида, пропептида и сывороточного альбумина мышиного альбумина приведены в SEQ ID NO: 2-4, соответственно, а соответствующие кодирующие последовательности приведены в SEQ ID NO: 10-12, соответственно. Последовательности сигнального пептида, пропептида и сывороточного альбумина человеческого альбумина приведены в SEQ ID NO: 6-8, соответственно, а соответствующие кодирующие последовательности приведены в SEQ ID NO: 14-16, соответственно. Ожидаемый кодируемый белок гуманизированного альбумина идентичен белку человеческого альбумина. См. фиг. 1A и 1B. Выравнивание белков мышиного и человеческого альбумина с белком гуманизированного альбумина представлено на фиг. 3A-3B. Кодирующие последовательности мышиного и человеческого Alb/ALB приведены в SEQ ID NO: 9 и 13 соответственно. Последовательности белка мышиного и человеческого альбумина приведены в SEQ ID NO: 1 и 5 соответственно. Последовательности ожидаемой кодирующей последовательности гуманизированного ALB и ожидаемого белка гуманизированного альбумина приведены в SEQ ID NO: 13 и 5 соответственно.
[00308] Для создания мутантного аллеля в мышиные эмбриональные стволовые клетки F1H4 вводили большой нацеливающий вектор, описанный выше. Мышиные ЭС клетки F1H4 были получены из гибридных эмбрионов, полученных путем скрещивания самки мыши C57BL/6NTac с самцом мыши 12956/SvEvTac. См., например, источники US 2015-0376651 и WO 2015/200805, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. После селекции по чувствительности к антибиотику колонии собирали, размножали и подвергали скринингу с помощью TAQMAN®. См. фиг. 2. Анализы потери аллеля проводили для обнаружения потери эндогенного мышиного аллеля, а анализы приобретения аллеля проводили для обнаружения приобретения гуманизированного аллеля с использованием праймеров и зондов, приведенных в таблице 4.
[00309] Таблица 4. Скрининговые анализы.
[00310] Анализы модификации аллеля (MOA), включая анализы потери аллеля (LOA) и приобретения аллеля (GOA), описаны, например, в источниках US 2014/0178879; US 2016/0145646; WO 2016/081923; и Frendewey et al. (2010) Methods Enzymol. 476:295-307 каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. Анализ потери аллеля (LOA) имеет логику, обратную стандартной логике скрининга, и в нем определяют количество копий в образце геномной ДНК нативного локуса, на который была направлена мутация. В правильно нацеленном гетерозиготном клоне клетки анализ LOA определяет один из двух нативных аллелей (для генов, не принадлежащих X или Y хромосоме), а другой аллель нарушен целевой модификацией. Тот же принцип может быть применен в обратном порядке в качестве анализа приобретения аллеля (GOA) для количественного определения количества копий вставленного нацеливающего вектора в образце геномной ДНК.
[00311] Мыши поколения F0 были получены из модифицированных ЭС клеток с использованием метода VELOCIMOUSE®. В частности, клоны мышиных ЭС клеток, содержащие гуманизированный локус альбумина, описанный выше, которые были выбраны с помощью анализа МОА, описанного выше, инъецировали в эмбрионы на 8-клеточной стадии с использованием метода VELOCIMOUSE®. См., например, источники US 7,576,259; US 7,659,442; US 7,294,754; US 2008/0078000; и Poueymirou et al. (2007) Nat. Biotechnol. 25(1):91-99, каждый из которых включен в настоящий документ посредством ссылки в полном объеме для всех целей. В методе VELOCIMOUSE® осуществляют инъекцию целевых мышиных эмбриональных стволовых (ЭС) клеток посредством осуществляемой под воздействием лазера инъекции в эмбрионы на стадии предморулы, например, эмбрионы на восьмиклеточной стадии, что эффективно дает мыши поколения F0, полностью полученные из ЭС клеток. В методе VELOCIMOUSE® инъецированные эмбрионы на стадии предморулы культивировали до стадии бластоцисты, и эмбрионы на стадии бластоцисты вводили суррогатным матерям и вынашивали в них для получения мышей поколения F0. Используя в качестве исходного материала клоны мышиных ЭС клеток, гомозиготных по целевой модификации, получают мышей поколения F0, гомозиготных по целевой модификации. Используя в качестве исходного материала клоны мышиных ЭС клеток, гетерозиготных по целевой модификации, может быть проведено последующее размножение с получением мышей, гомозиготных по целевой модификации.
Пример 2. Проверка мышей, содержащих гуманизированный локус альбумина (ALB)
[00312] Для проверки гуманизированного альбумина у мышей измеряли уровни мышиного и человеческого альбумина в образцах плазмы с использованием наборов ELISA для человеческого и мышиного сывороточного альбумина (Abcam ab179887 и ab207620, соответственно). Гуманизированные мыши, использованные для проверки, были мышами F1, у которых самоудаляющаяся кассета селекции самоудалилась. Белок человеческого альбумина был обнаружен в образцах нормальной плазмы человека и мышей с гуманизированным альбумином, но не в образцах плазмы мышей дикого типа (WT) или мышей VelocImmune (VI). См. фиг. 4. Белок мышиного альбумина был обнаружен в образцах плазмы мышей дикого типа и образцах плазмы мышей VI, но не в образцах плазмы мышей с гуманизированным альбумином. См. фиг. 5. В частности, объединенная нормальная человеческая плазма (приобретенная у George King-Biomedical Inc.) содержала приблизительно 30-40 мг/мл человеческого альбумина. Плазма мышей с гуманизированным альбумином содержала приблизительно 10-15 мг/мл человеческого альбумина, но мышиный альбумин не определялся. Нормальная плазма мышей VI и WT содержала приблизительно 7-13 мг/мл мышиного альбумина.
Пример 3. Проверка мышей, содержащих гуманизированный локус альбумина (ALB) - гидовые РНК, нацеленные на человеческий альбумин, для вставки F9
[00313] Для дальнейшей проверки мышей с гуманизированным альбумином использовали мышей с гуманизированным альбумином для оценки использования технологии CRISPR/Cas9 для встраивания трансгена F9 в локус альбумина. В частности, авторы тестировали встраивание и экспрессию встроенного человеческого F9 варианта Padua (hF9-R338L) на гомозиготных мышах с гуманизированным альбумином. Были сконструированы различные гидовые РНК против интрона 1 человеческого локуса альбумина. Были проведены два отдельных эксперимента на мышах с использованием мышей ALBhu/hu для скрининга в общей сложности 11 гидовых РНК, каждая из которых была нацелена на первый интрон человеческого локуса альбумина. Всех мышей взвешивали и осуществляли инъекцию через хвостовую вену в день 0 эксперимента. Кровь собирали на 1, 3, 4 и 6 неделях посредством надреза хвоста и отделяли плазму. Мышей умерщвляли на 7 неделе. Собирали кровь из полой вены и отделяли плазму. Также иссекали печень и селезенку. Гидовые последовательности (ДНК-нацеливающие сегменты) этих гидовых РНК приведены в таблице 5.
[00314] Таблица 5. Последовательности гРНК человеческого альбумина и хромосомные координаты.
[00315] В первом эксперименте были протестированы LNP, содержащие мРНК Cas9 и каждую из следующих шести гидовых РНК по отдельности: G009852, G009859, G009860, G009864, G009874 и G012764. LNP разбавляли до 0,3 мг/кг (используя среднюю массу 30 г) и вводили совместно с AAV8, упакованным матрицей двунаправленной вставки hF9 (SEQ ID NO: 63; ITR-акцептор сплайсинга-hF9 (экзоны 2-8)-bGH-полиA SV40-кодон-оптимизированный hF9-pLac-pMB-акцептор сплайсинга-устойчивость к Kan) в дозе 3E11 вирусных геномов на мышь. Инъекцию осуществляли пяти самцам мышей ALBhu/hu на группу в возрасте от 12 до 14 недель. Пяти мышам из той же когорты инъецировали AAV8, упакованный промотором CAGG, функционально связанным с hF9 (SEQ ID NO: 64; CAGG-ITR-hF9-WPRE-bGH-ITR-pLac-pMB-устойчивость к Amp), что приводит к эписомальной экспрессии hF9 (в количестве 3E11 вирусных геномов на мышь). Имелось три группы отрицательного контроля с тремя мышами на группу, которым вводили только буфер, AAV8, упакованный только матрицей двунаправленной вставки hF9, или только LNP-G009874.
[00316] Во втором эксперименте были протестированы LNP, содержащие мРНК Cas9 и каждую из следующих шести гидовых РНК по отдельности: G009860, G012764, G009844, G009857, G012752, G012753 и G012761. LNP разбавляли до 0,3 мг/кг (используя среднюю массу 40 г) и вводили совместно с AAV8, упакованным матрицей двунаправленной вставки hF9 (SEQ ID NO: 63), в дозе 3E11 вирусных геномов на мышь. Инъекцию осуществляли пяти самцам мышей ALBhu/hu на группу в возрасте 30 недель. Пяти мышам из той же когорты инъецировали AAV8, упакованный промотором CAGG, функционально связанным с hF9 (SEQ ID NO: 64), что приводит к эписомальной экспрессии hF9 (в количестве 3E11 вирусных геномов на мышь). Имелось три группы отрицательного контроля с тремя мышами на группу, которым вводили только буфер, AAV8, упакованный только матрицей двунаправленной вставки hF9, или только LNP-G009874.
[00317] Для анализа проводили ELISA для измерения уровней hFIX в кровообращении у мышей в каждой временной точке. Для этой цели использовали наборы для ELISA человеческого фактора IX (ab188393), и все планшеты анализировали с человеческой объединенной нормальной плазмой от George King Bio-Medical в качестве положительного контроля для анализа. Уровни экспрессии человеческого фактора IX в образцах плазмы в каждой группе на 6 неделе после инъекции показаны на фиг. 6A и 6B. В соответствии с данными о вставке in vitro, низкие или нулевые уровни фактора IX в сыворотке не были обнаружены при использовании гидовой РНК G009852. В соответствии с отсутствием прилегающей последовательности PAM в человеческом альбумине уровни фактора IX в сыворотке не были обнаружены при использовании гидовой РНК G009864. Гидовая последовательность (ДНК-нацеливающий сегмент) G009864 представляет собой UACUUUGCACUUUCCUUAGU (SEQ ID NO: 61), и она нацелена на геномные координаты яванского макака (mf5) chr5:61199187-61199207. Экспрессия фактора IX в сыворотке наблюдалась для нескольких других гидовых РНК, включая G009857, G009859, G009860, G009874 и G0012764.
[00318] Селезенки и часть левой боковой доли всех образцов печени были направлены на анализ методом секвенирования нового поколения (NGS). NGS использовали для оценки процента клеток печени со вставками/делециями (инделами) в гуманизированном локусе альбумина на 7 неделе после инъекции донора AAV-hF9 и LNP-CRISPR/Cas9. В соответствии с отсутствием прилегающей последовательности PAM в человеческом альбумине, при использовании гидовой РНК G009864 в печени не было обнаружено никакого редактирования. Редактирование в печени наблюдалось для групп, где использовали гидовые РНК G009859, G009860, G009874 и G012764 (данные не показаны).
[00319] Оставшуюся печень фиксировали в течение 24 часов в 10% нейтральном забуференном формалине, а затем переносили в 70% этанол. От четырех до пяти образцов из отдельных долей были вырезаны и отправлены в HistoWisz, где они были обработаны и заключены в парафиновые блоки. Затем каждый парафиновый блок нарезали на срезы толщиной пять мкм и проводили BASESCOPE™ на Ventana Ultra Discovery (Roche) с использованием универсальной процедуры BASESCOPE™ и реагентов от Advanced Cell Diagnostics, а также специально разработанного зонда, нацеленного на уникальное соединение мРНК, образованное между сигнальной последовательностью человеческого альбумина от первого интрона локуса альбумина ALBhu/hu и трансгеном hF9 при успешной интеграции и транскрипции. Затем использовали программное обеспечение для визуализации HALO (Indica Labs) для количественной оценки процента положительных клеток в каждом образце. Затем средний процент положительных клеток в нескольких долях для каждого животного коррелировали с уровнями hFIX в сыворотке на 7 неделе. Результаты представлены на фиг. 7 и в таблице 6. Сывороточные уровни на 7 неделе и % положительных на мРНК hALB-hFIX клеток сильно коррелировали (r = 0,89; R2 = 0,79).
[00320] Таблица 6. hFIX и данные BASESCOPE™ на 7 неделе.
(7 неделя)
(4-5 срезов)
Пример 4. Проверка мышей, содержащих гуманизированный локус альбумина (ALB) - вставка F9 у мышей F9 KO
[00321] Для дальнейшей проверки мышей с гуманизированным альбумином указанных мышей скрещивали с мышами с нокаутом F9 для получения мышей ALBm/hu x F9-/- (гетерозиготных по гуманизации локуса альбумина и гомозиготных по нокауту F9), чтобы использовать их для оценки использования технологии CRISPR/Cas9 для встраивания трансгена F9 в локус альбумина.
[00322] Затем мышей F9 KO с гуманизированным альбумином использовали для тестирования вставки трансгена человеческого F9 варианта Padua (hF9-R338L) в интрон 1 гуманизированного локуса альбумина. Всех мышей взвешивали и осуществляли инъекцию через хвостовую вену в день 0 эксперимента. Кровь собирали на 1-й и 3-й неделях посредством надреза хвоста и отделяли плазму. Мышей умерщвляли на 4 неделе. Собирали кровь из полой вены и отделяли плазму. Также иссекали печень и селезенку.
[00323] Были протестированы LNP, содержащие мРНК Cas9 и следующие две гидовые РНК по отдельности: G009860 (нацеленная на первый интрон человеческого локуса альбумина) и G000666 (нацеленная на первый интрон мышиного локуса альбумина). Гидовая последовательность (ДНК-нацеливающий сегмент) G009860 приведена в таблице 5. Гидовой последовательностью G000666 является CACUCUUGUCUGUGGAAACA (SEQ ID NO: 62), и она нацелена на мышиные геномные координаты (mm10) chr5:90461709-90461729. G009860 разбавляли до 0,3 мг/кг, а G000666 разбавляли до 1,0 мг/кг (используя среднюю массу 31,2 г) и вводили обе из них совместно с AAV8, упакованным матрицей двунаправленной вставки hF9 (SEQ ID NO: 63), в дозе 3E11 вирусных геномов на мышь. Инъекцию осуществляли пяти самцам мышей ALBms/hu x F9-/- на группу (в возрасте 16 недель). Пяти мышам из той же когорты инъецировали AAV8, упакованный промотором CAGG, функционально связанным с hF9 (SEQ ID NO: 64), что приводит к эписомальной экспрессии hF9 (в количестве 3E11 вирусных геномов на мышь). Имелось шесть животных отрицательного контроля, по одной мыши на группу, которым вводили только буфер или AAV8, упакованный только матрицей двунаправленной вставки hF9, и две мыши на группу, которым вводили только LNP-G009860 или LNP-G000666 в дозе 0,3 мг/кг и 1,0 мг/кг, соответственно.
[00324] Для анализа проводили ELISA для измерения уровней hFIX в кровообращении у мышей в каждой временной точке. Для этой цели использовали наборы для ELISA человеческого фактора IX (ab188393), и все планшеты анализировали с человеческой объединенной нормальной плазмой от George King Bio-Medical в качестве положительного контроля для анализа. Селезенки и часть левой латеральной доли всех образцов печени были направлены на анализ методом NGS.
[00325] Уровни экспрессии человеческого фактора IX в образцах плазмы в каждой группе через 1, 2 и 4 недели после инъекции приведены на фиг. 8 и в таблице 7. Кроме того, результаты NGS, показывающие уровни вставки и делеции (индел) в локусе альбумина в печени и селезенке, приведены в таблице 7. Как показано на фиг. 8 и в таблице 7, hFIX был обнаружен в плазме подвергшихся воздействию мышей Alb+/hu/F9-/- через 1, 3 и 4 недели, причем ELISA показал значения экспрессии 0,5-10 мкг/мл через 1, 3 и 4 недели.
[00326] Таблица 7. Уровни человеческого FIX в плазме и результаты NGS.
(мкг/мл)
(мкг/мл)
(мкг/мл)
Печень
Селезенка
[00327] Оставшуюся печень фиксировали в течение 24 часов в 10% нейтральном забуференном формалине, а затем переносили в 70% этанол. От четырех до пяти образцов из отдельных долей были вырезаны и отправлены в HistoWiz, где они были обработаны и заключены в парафиновые блоки. Затем каждый парафиновый блок нарезали на срезы толщиной пять мкм для анализа посредством BASESCOPE™ на Ventana Ultra Discovery (Roche) с использованием универсальной процедуры BASESCOPE™ и реагентов от Advanced Cell Diagnostics, а также специально разработанного зонда, нацеленного на уникальное соединение мРНК, образованное между сигнальной последовательностью человеческого или мышиного альбумина от первого интрона каждого соответствующего локуса альбумина у мышей ALBms/hu и трансгеном hF9 при успешной интеграции и транскрипции. Использовали программное обеспечение для визуализации HALO (Indica Labs) для количественной оценки процента положительных клеток в каждом образце.
[00328] Затем взятую после умерщвления кровь использовали для оценки функциональной коагуляционной активности с помощью активированного частичного тромбопластинового времени (АЧТВ) и теста генерации тромбина (ТГТ). Активированное частичное тромбопластиновое время (АЧТВ) является клинической мерой внутреннего пути активации свёртывания в плазме. Свертывание плазмы индуцируется добавлением эллаговой кислоты или каолина, оба из которых активируют фактор свертывания XII во внутреннем пути (также известном как контактный путь) свертывания, что впоследствии приводит к образованию фибрина из фибриногена после активации тромбина. Анализ АЧТВ обеспечивает оценку способности индивидуума образовывать сгусток, и эту информацию можно использовать для определения риска кровотечения или тромбоза. Для тестирования АЧТВ использовалась полуавтоматическая настольная система (Diagnostica Stago STart 4) с электромеханическим методом обнаружения сгустков (система обнаружения на основе вязкости) для оценки свертывания в плазме. В каждую кювету со стальным шариком добавляли 50 мкл цитратной плазмы и инкубировали при 37°C в течение 5 мин, а затем инициировали свертывание добавлением 50 мкл эллаговой кислоты (конечная концентрация 30 мкМ) при 37°C на 300 секунд. После окончательной активации свертывания путем добавления 50 мкл 0,025 М хлорида кальция (конечная концентрация 8 мМ) в каждую кювету стальной шарик начал колебаться вперед и назад между двумя катушками возбуждения. Движение шарика фиксировалось измерительной катушкой. Образование фибрина увеличивало вязкость плазмы до тех пор, пока шарик не переставал двигаться, что регистрировалось как время свертывания. Единственным измеряемым параметром было время свертывания. Анализы проводили в двух экземплярах.
[00329] Тест генерации тромбина (ТГТ) является неклинической оценкой кинетики образования тромбина в активированной плазме. Образование тромбина является важным процессом свертывания крови, поскольку тромбин отвечает за активацию других факторов свертывания и распространение дополнительного тромбина (через активацию FXI) для превращения фибриногена в фибрин. Тест генерации тромбина позволяет оценить способность человека вырабатывать тромбин, и эту информацию можно использовать для определения риска кровотечения или тромбоза. Для проведения ТГТ использовали калиброванную автоматическую тромбограмму для оценки уровней образования тромбина на спектрофотометре (Thrombinograph™, Thermo Scientific). Для высокопроизводительных экспериментов использовали 96-луночные планшеты (Immulon II HB). В каждую лунку добавляли 55 мкл цитратной плазмы (4-кратно разбавленной физиологическим раствором для мышиной плазмы) и инкубировали при 37°C в течение 30 мин. Образование тромбина запускалось добавлением 15 мкл 2 мкМ эллаговой кислоты (конечная концентрация 0,33 мкМ) при 37°C в течение 45 мин. Образование тромбина определяли после автоматической инъекции 15 мкл флуорогенного субстрата с 16 мМ CaCl2 (FluCa; Thrombinoscope BV) в каждую лунку. Флуорогенный субстрат реагировал с образовавшимся тромбином, который непрерывно измеряли в плазме каждые 33 секунды в течение 90 минут при 460 нм. Интенсивность флуоресценции была пропорциональна протеолитической активности тромбина. Основными параметрами, измеренными при отслеживании, были время задержки, пик образования тромбина, время до достижения пика образования тромбина и эндогенный тромбиновый потенциал (ЭТП). Время задержки позволяет оценить время, необходимое для первоначального обнаружения тромбина в плазме. Пик - это максимальное количество тромбина, образовавшееся в данный момент времени после активации. Время до достижения пика образования тромбина - это время от начала каскада коагуляции до пика образования тромбина. ЭТП - это общее количество тромбина, образовавшееся в течение 60 минут измерения. Анализы проводили в двух экземплярах.
[00330] Как показано на фиг. 9 и в таблице 8, вставка трансгена hF9 с использованием либо гРНК мышиного альбумина, либо гРНК человеческого альбумина показала восстановление функции свертывания в анализе АЧТВ. Образцы отрицательного контроля с физиологическим раствором, только AAV и только LNP показали увеличенное время АЧТВ, равное 45-60 секундам. Положительный контроль CAGG и тестируемые образцы (AAV8+LNP) были ближе к нормальному человеческому АЧТВ, равному 28-34 секундам.
Таблица 8. АЧТВ и ТГТ-EA.
[00331] Как показано на фиг. 10A, 10B и 11 и в таблице 8, вставка трансгена hF9 с использованием либо гРНК мышиного альбумина, либо гРНК человеческого альбумина показала повышенное образование тромбина в анализе ТГТ-EA. Концентрации тромбина были выше в образцах положительного контроля CAGG и AAV8+LNP по сравнению с образцами отрицательного контроля.
[00332] Таким образом, hFIX был обнаружен в плазме мышей Alb+/hu/F9-/- через 1, 3 и 4 недели, и было обнаружено, что экспрессированный hFIX-R338L является функциональным, поскольку в анализе ТГТ образовывался тромбин, и время свертывания АЧТВ было улучшено.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Regeneron Pharmaceuticals, Inc.
<120> ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ЛОКУС АЛЬБУМИНА
<130> 057766/548157
<150> US 62/858,589
<151> 2019-06-07
<150> US 62/916,666
<151> 2019-10-17
<160> 64
<170> PatentIn версии 3.5
<210> 1
<211> 608
<212> БЕЛОК
<213> Mus musculus
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (1)..(18)
<223> Сигнальный пептид
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (19)..(24)
<223> Пропептид
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (25)..(608)
<223> Сывороточный альбумин
<400> 1
Met Lys Trp Val Thr Phe Leu Leu Leu Leu Phe Val Ser Gly Ser Ala
1 5 10 15
Phe Ser Arg Gly Val Phe Arg Arg Glu Ala His Lys Ser Glu Ile Ala
20 25 30
His Arg Tyr Asn Asp Leu Gly Glu Gln His Phe Lys Gly Leu Val Leu
35 40 45
Ile Ala Phe Ser Gln Tyr Leu Gln Lys Cys Ser Tyr Asp Glu His Ala
50 55 60
Lys Leu Val Gln Glu Val Thr Asp Phe Ala Lys Thr Cys Val Ala Asp
65 70 75 80
Glu Ser Ala Ala Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp
85 90 95
Lys Leu Cys Ala Ile Pro Asn Leu Arg Glu Asn Tyr Gly Glu Leu Ala
100 105 110
Asp Cys Cys Thr Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln
115 120 125
His Lys Asp Asp Asn Pro Ser Leu Pro Pro Phe Glu Arg Pro Glu Ala
130 135 140
Glu Ala Met Cys Thr Ser Phe Lys Glu Asn Pro Thr Thr Phe Met Gly
145 150 155 160
His Tyr Leu His Glu Val Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro
165 170 175
Glu Leu Leu Tyr Tyr Ala Glu Gln Tyr Asn Glu Ile Leu Thr Gln Cys
180 185 190
Cys Ala Glu Ala Asp Lys Glu Ser Cys Leu Thr Pro Lys Leu Asp Gly
195 200 205
Val Lys Glu Lys Ala Leu Val Ser Ser Val Arg Gln Arg Met Lys Cys
210 215 220
Ser Ser Met Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val
225 230 235 240
Ala Arg Leu Ser Gln Thr Phe Pro Asn Ala Asp Phe Ala Glu Ile Thr
245 250 255
Lys Leu Ala Thr Asp Leu Thr Lys Val Asn Lys Glu Cys Cys His Gly
260 265 270
Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Glu Leu Ala Lys Tyr Met
275 280 285
Cys Glu Asn Gln Ala Thr Ile Ser Ser Lys Leu Gln Thr Cys Cys Asp
290 295 300
Lys Pro Leu Leu Lys Lys Ala His Cys Leu Ser Glu Val Glu His Asp
305 310 315 320
Thr Met Pro Ala Asp Leu Pro Ala Ile Ala Ala Asp Phe Val Glu Asp
325 330 335
Gln Glu Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly
340 345 350
Thr Phe Leu Tyr Glu Tyr Ser Arg Arg His Pro Asp Tyr Ser Val Ser
355 360 365
Leu Leu Leu Arg Leu Ala Lys Lys Tyr Glu Ala Thr Leu Glu Lys Cys
370 375 380
Cys Ala Glu Ala Asn Pro Pro Ala Cys Tyr Gly Thr Val Leu Ala Glu
385 390 395 400
Phe Gln Pro Leu Val Glu Glu Pro Lys Asn Leu Val Lys Thr Asn Cys
405 410 415
Asp Leu Tyr Glu Lys Leu Gly Glu Tyr Gly Phe Gln Asn Ala Ile Leu
420 425 430
Val Arg Tyr Thr Gln Lys Ala Pro Gln Val Ser Thr Pro Thr Leu Val
435 440 445
Glu Ala Ala Arg Asn Leu Gly Arg Val Gly Thr Lys Cys Cys Thr Leu
450 455 460
Pro Glu Asp Gln Arg Leu Pro Cys Val Glu Asp Tyr Leu Ser Ala Ile
465 470 475 480
Leu Asn Arg Val Cys Leu Leu His Glu Lys Thr Pro Val Ser Glu His
485 490 495
Val Thr Lys Cys Cys Ser Gly Ser Leu Val Glu Arg Arg Pro Cys Phe
500 505 510
Ser Ala Leu Thr Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Lys Ala
515 520 525
Glu Thr Phe Thr Phe His Ser Asp Ile Cys Thr Leu Pro Glu Lys Glu
530 535 540
Lys Gln Ile Lys Lys Gln Thr Ala Leu Ala Glu Leu Val Lys His Lys
545 550 555 560
Pro Lys Ala Thr Ala Glu Gln Leu Lys Thr Val Met Asp Asp Phe Ala
565 570 575
Gln Phe Leu Asp Thr Cys Cys Lys Ala Ala Asp Lys Asp Thr Cys Phe
580 585 590
Ser Thr Glu Gly Pro Asn Leu Val Thr Arg Cys Lys Asp Ala Leu Ala
595 600 605
<210> 2
<211> 18
<212> БЕЛОК
<213> Mus musculus
<400> 2
Met Lys Trp Val Thr Phe Leu Leu Leu Leu Phe Val Ser Gly Ser Ala
1 5 10 15
Phe Ser
<210> 3
<211> 6
<212> БЕЛОК
<213> Mus musculus
<400> 3
Arg Gly Val Phe Arg Arg
1 5
<210> 4
<211> 584
<212> БЕЛОК
<213> Mus musculus
<400> 4
Glu Ala His Lys Ser Glu Ile Ala His Arg Tyr Asn Asp Leu Gly Glu
1 5 10 15
Gln His Phe Lys Gly Leu Val Leu Ile Ala Phe Ser Gln Tyr Leu Gln
20 25 30
Lys Cys Ser Tyr Asp Glu His Ala Lys Leu Val Gln Glu Val Thr Asp
35 40 45
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Ala Asn Cys Asp Lys
50 55 60
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Ala Ile Pro Asn Leu
65 70 75 80
Arg Glu Asn Tyr Gly Glu Leu Ala Asp Cys Cys Thr Lys Gln Glu Pro
85 90 95
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Ser Leu
100 105 110
Pro Pro Phe Glu Arg Pro Glu Ala Glu Ala Met Cys Thr Ser Phe Lys
115 120 125
Glu Asn Pro Thr Thr Phe Met Gly His Tyr Leu His Glu Val Ala Arg
130 135 140
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Tyr Tyr Ala Glu Gln
145 150 155 160
Tyr Asn Glu Ile Leu Thr Gln Cys Cys Ala Glu Ala Asp Lys Glu Ser
165 170 175
Cys Leu Thr Pro Lys Leu Asp Gly Val Lys Glu Lys Ala Leu Val Ser
180 185 190
Ser Val Arg Gln Arg Met Lys Cys Ser Ser Met Gln Lys Phe Gly Glu
195 200 205
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Thr Phe Pro
210 215 220
Asn Ala Asp Phe Ala Glu Ile Thr Lys Leu Ala Thr Asp Leu Thr Lys
225 230 235 240
Val Asn Lys Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
245 250 255
Arg Ala Glu Leu Ala Lys Tyr Met Cys Glu Asn Gln Ala Thr Ile Ser
260 265 270
Ser Lys Leu Gln Thr Cys Cys Asp Lys Pro Leu Leu Lys Lys Ala His
275 280 285
Cys Leu Ser Glu Val Glu His Asp Thr Met Pro Ala Asp Leu Pro Ala
290 295 300
Ile Ala Ala Asp Phe Val Glu Asp Gln Glu Val Cys Lys Asn Tyr Ala
305 310 315 320
Glu Ala Lys Asp Val Phe Leu Gly Thr Phe Leu Tyr Glu Tyr Ser Arg
325 330 335
Arg His Pro Asp Tyr Ser Val Ser Leu Leu Leu Arg Leu Ala Lys Lys
340 345 350
Tyr Glu Ala Thr Leu Glu Lys Cys Cys Ala Glu Ala Asn Pro Pro Ala
355 360 365
Cys Tyr Gly Thr Val Leu Ala Glu Phe Gln Pro Leu Val Glu Glu Pro
370 375 380
Lys Asn Leu Val Lys Thr Asn Cys Asp Leu Tyr Glu Lys Leu Gly Glu
385 390 395 400
Tyr Gly Phe Gln Asn Ala Ile Leu Val Arg Tyr Thr Gln Lys Ala Pro
405 410 415
Gln Val Ser Thr Pro Thr Leu Val Glu Ala Ala Arg Asn Leu Gly Arg
420 425 430
Val Gly Thr Lys Cys Cys Thr Leu Pro Glu Asp Gln Arg Leu Pro Cys
435 440 445
Val Glu Asp Tyr Leu Ser Ala Ile Leu Asn Arg Val Cys Leu Leu His
450 455 460
Glu Lys Thr Pro Val Ser Glu His Val Thr Lys Cys Cys Ser Gly Ser
465 470 475 480
Leu Val Glu Arg Arg Pro Cys Phe Ser Ala Leu Thr Val Asp Glu Thr
485 490 495
Tyr Val Pro Lys Glu Phe Lys Ala Glu Thr Phe Thr Phe His Ser Asp
500 505 510
Ile Cys Thr Leu Pro Glu Lys Glu Lys Gln Ile Lys Lys Gln Thr Ala
515 520 525
Leu Ala Glu Leu Val Lys His Lys Pro Lys Ala Thr Ala Glu Gln Leu
530 535 540
Lys Thr Val Met Asp Asp Phe Ala Gln Phe Leu Asp Thr Cys Cys Lys
545 550 555 560
Ala Ala Asp Lys Asp Thr Cys Phe Ser Thr Glu Gly Pro Asn Leu Val
565 570 575
Thr Arg Cys Lys Asp Ala Leu Ala
580
<210> 5
<211> 609
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (1)..(18)
<223> Сигнальный пептид
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (19)..(24)
<223> Пропептид
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (25)..(609)
<223> Сывороточный альбумин
<400> 5
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Arg Gly Val Phe Arg Arg Asp Ala His Lys Ser Glu Val Ala
20 25 30
His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu
35 40 45
Ile Ala Phe Ala Gln Tyr Leu Gln Gln Cys Pro Phe Glu Asp His Val
50 55 60
Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp
65 70 75 80
Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp
85 90 95
Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala
100 105 110
Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln
115 120 125
His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val
130 135 140
Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys
145 150 155 160
Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro
165 170 175
Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys
180 185 190
Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu
195 200 205
Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys
210 215 220
Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val
225 230 235 240
Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser
245 250 255
Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly
260 265 270
Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile
275 280 285
Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu
290 295 300
Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp
305 310 315 320
Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser
325 330 335
Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly
340 345 350
Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val
355 360 365
Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys
370 375 380
Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu
385 390 395 400
Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys
405 410 415
Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu
420 425 430
Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val
435 440 445
Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His
450 455 460
Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val
465 470 475 480
Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg
485 490 495
Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe
500 505 510
Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala
515 520 525
Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu
530 535 540
Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys
545 550 555 560
Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala
565 570 575
Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe
580 585 590
Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly
595 600 605
Leu
<210> 6
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser
<210> 7
<211> 6
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Arg Gly Val Phe Arg Arg
1 5
<210> 8
<211> 585
<212> БЕЛОК
<213> Homo sapiens
<400> 8
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
1 5 10 15
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
20 25 30
Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
35 40 45
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
50 55 60
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
65 70 75 80
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
85 90 95
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
100 105 110
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
115 120 125
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
130 135 140
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
145 150 155 160
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
165 170 175
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
180 185 190
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
195 200 205
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
210 215 220
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
225 230 235 240
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
245 250 255
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
260 265 270
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
275 280 285
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
290 295 300
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
305 310 315 320
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
325 330 335
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
340 345 350
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
355 360 365
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
370 375 380
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
385 390 395 400
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
405 410 415
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
420 425 430
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
435 440 445
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
450 455 460
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
465 470 475 480
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
485 490 495
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
500 505 510
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
515 520 525
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
530 535 540
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
545 550 555 560
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
565 570 575
Ala Ala Ser Gln Ala Ala Leu Gly Leu
580 585
<210> 9
<211> 1827
<212> ДНК
<213> Mus musculus
<220>
<221> прочий_признак
<222> (1)..(72)
<223> Сигнальный пептид
<220>
<221> прочий_признак
<222> (73)..(90)
<223> Пропептид
<220>
<221> прочий_признак
<222> (91)..(1824)
<223> Сывороточный альбумин
<400> 9
atgaagtggg taacctttct cctcctcctc ttcgtctccg gctctgcttt ttccaggggt 60
gtgtttcgcc gagaagcaca caagagtgag atcgcccatc ggtataatga tttgggagaa 120
caacatttca aaggcctagt cctgattgcc ttttcccagt atctccagaa atgctcatac 180
gatgagcatg ccaaattagt gcaggaagta acagactttg caaagacgtg tgttgccgat 240
gagtctgccg ccaactgtga caaatccctt cacactcttt ttggagataa gttgtgtgcc 300
attccaaacc tccgtgaaaa ctatggtgaa ctggctgact gctgtacaaa acaagagccc 360
gaaagaaacg aatgtttcct gcaacacaaa gatgacaacc ccagcctgcc accatttgaa 420
aggccagagg ctgaggccat gtgcacctcc tttaaggaaa acccaaccac ctttatggga 480
cactatttgc atgaagttgc cagaagacat ccttatttct atgccccaga acttctttac 540
tatgctgagc agtacaatga gattctgacc cagtgttgtg cagaggctga caaggaaagc 600
tgcctgaccc cgaagcttga tggtgtgaag gagaaagcat tggtctcatc tgtccgtcag 660
agaatgaagt gctccagtat gcagaagttt ggagagagag cttttaaagc atgggcagta 720
gctcgtctga gccagacatt ccccaatgct gactttgcag aaatcaccaa attggcaaca 780
gacctgacca aagtcaacaa ggagtgctgc catggtgacc tgctggaatg cgcagatgac 840
agggcggaac ttgccaagta catgtgtgaa aaccaggcga ctatctccag caaactgcag 900
acttgctgcg ataaaccact gttgaagaaa gcccactgtc ttagtgaggt ggagcatgac 960
accatgcctg ctgatctgcc tgccattgct gctgattttg ttgaggacca ggaagtgtgc 1020
aagaactatg ctgaggccaa ggatgtcttc ctgggcacgt tcttgtatga atattcaaga 1080
agacaccctg attactctgt atccctgttg ctgagacttg ctaagaaata tgaagccact 1140
ctggaaaagt gctgcgctga agccaatcct cccgcatgct acggcacagt gcttgctgaa 1200
tttcagcctc ttgtagaaga gcctaagaac ttggtcaaaa ccaactgtga tctttacgag 1260
aagcttggag aatatggatt ccaaaatgcc attctagttc gctacaccca gaaagcacct 1320
caggtgtcaa ccccaactct cgtggaggct gcaagaaacc taggaagagt gggcaccaag 1380
tgttgtacac ttcctgaaga tcagagactg ccttgtgtgg aagactatct gtctgcaatc 1440
ctgaaccgtg tgtgtctgct gcatgagaag accccagtga gtgagcatgt taccaagtgc 1500
tgtagtggat ccctggtgga aaggcggcca tgcttctctg ctctgacagt tgatgaaaca 1560
tatgtcccca aagagtttaa agctgagacc ttcaccttcc actctgatat ctgcacactt 1620
ccagagaagg agaagcagat taagaaacaa acggctcttg ctgagctggt gaagcacaag 1680
cccaaggcta cagcggagca actgaagact gtcatggatg actttgcaca gttcctggat 1740
acatgttgca aggctgctga caaggacacc tgcttctcga ctgagggtcc aaaccttgtc 1800
actagatgca aagacgcctt agcctaa 1827
<210> 10
<211> 72
<212> ДНК
<213> Mus musculus
<400> 10
atgaagtggg taacctttct cctcctcctc ttcgtctccg gctctgcttt ttccaggggt 60
gtgtttcgcc ga 72
<210> 11
<211> 18
<212> ДНК
<213> Mus musculus
<400> 11
gaagcacaca agagtgag 18
<210> 12
<211> 1734
<212> ДНК
<213> Mus musculus
<400> 12
atcgcccatc ggtataatga tttgggagaa caacatttca aaggcctagt cctgattgcc 60
ttttcccagt atctccagaa atgctcatac gatgagcatg ccaaattagt gcaggaagta 120
acagactttg caaagacgtg tgttgccgat gagtctgccg ccaactgtga caaatccctt 180
cacactcttt ttggagataa gttgtgtgcc attccaaacc tccgtgaaaa ctatggtgaa 240
ctggctgact gctgtacaaa acaagagccc gaaagaaacg aatgtttcct gcaacacaaa 300
gatgacaacc ccagcctgcc accatttgaa aggccagagg ctgaggccat gtgcacctcc 360
tttaaggaaa acccaaccac ctttatggga cactatttgc atgaagttgc cagaagacat 420
ccttatttct atgccccaga acttctttac tatgctgagc agtacaatga gattctgacc 480
cagtgttgtg cagaggctga caaggaaagc tgcctgaccc cgaagcttga tggtgtgaag 540
gagaaagcat tggtctcatc tgtccgtcag agaatgaagt gctccagtat gcagaagttt 600
ggagagagag cttttaaagc atgggcagta gctcgtctga gccagacatt ccccaatgct 660
gactttgcag aaatcaccaa attggcaaca gacctgacca aagtcaacaa ggagtgctgc 720
catggtgacc tgctggaatg cgcagatgac agggcggaac ttgccaagta catgtgtgaa 780
aaccaggcga ctatctccag caaactgcag acttgctgcg ataaaccact gttgaagaaa 840
gcccactgtc ttagtgaggt ggagcatgac accatgcctg ctgatctgcc tgccattgct 900
gctgattttg ttgaggacca ggaagtgtgc aagaactatg ctgaggccaa ggatgtcttc 960
ctgggcacgt tcttgtatga atattcaaga agacaccctg attactctgt atccctgttg 1020
ctgagacttg ctaagaaata tgaagccact ctggaaaagt gctgcgctga agccaatcct 1080
cccgcatgct acggcacagt gcttgctgaa tttcagcctc ttgtagaaga gcctaagaac 1140
ttggtcaaaa ccaactgtga tctttacgag aagcttggag aatatggatt ccaaaatgcc 1200
attctagttc gctacaccca gaaagcacct caggtgtcaa ccccaactct cgtggaggct 1260
gcaagaaacc taggaagagt gggcaccaag tgttgtacac ttcctgaaga tcagagactg 1320
ccttgtgtgg aagactatct gtctgcaatc ctgaaccgtg tgtgtctgct gcatgagaag 1380
accccagtga gtgagcatgt taccaagtgc tgtagtggat ccctggtgga aaggcggcca 1440
tgcttctctg ctctgacagt tgatgaaaca tatgtcccca aagagtttaa agctgagacc 1500
ttcaccttcc actctgatat ctgcacactt ccagagaagg agaagcagat taagaaacaa 1560
acggctcttg ctgagctggt gaagcacaag cccaaggcta cagcggagca actgaagact 1620
gtcatggatg actttgcaca gttcctggat acatgttgca aggctgctga caaggacacc 1680
tgcttctcga ctgagggtcc aaaccttgtc actagatgca aagacgcctt agcc 1734
<210> 13
<211> 1830
<212> ДНК
<213> Homo sapiens
<220>
<221> прочий_признак
<222> (1)..(72)
<223> Сигнальный пептид
<220>
<221> прочий_признак
<222> (73)..(90)
<223> Пропептид
<220>
<221> прочий_признак
<222> (91)..(1827)
<223> Сывороточный альбумин
<400> 13
atgaagtggg taacctttat ttcccttctt tttctcttta gctcggctta ttccaggggt 60
gtgtttcgtc gagatgcaca caagagtgag gttgctcatc ggtttaaaga tttgggagaa 120
gaaaatttca aagccttggt gttgattgcc tttgctcagt atcttcagca gtgtccattt 180
gaagatcatg taaaattagt gaatgaagta actgaatttg caaaaacatg tgttgctgat 240
gagtcagctg aaaattgtga caaatcactt catacccttt ttggagacaa attatgcaca 300
gttgcaactc ttcgtgaaac ctatggtgaa atggctgact gctgtgcaaa acaagaacct 360
gagagaaatg aatgcttctt gcaacacaaa gatgacaacc caaacctccc ccgattggtg 420
agaccagagg ttgatgtgat gtgcactgct tttcatgaca atgaagagac atttttgaaa 480
aaatacttat atgaaattgc cagaagacat ccttactttt atgccccgga actccttttc 540
tttgctaaaa ggtataaagc tgcttttaca gaatgttgcc aagctgctga taaagctgcc 600
tgcctgttgc caaagctcga tgaacttcgg gatgaaggga aggcttcgtc tgccaaacag 660
agactcaagt gtgccagtct ccaaaaattt ggagaaagag ctttcaaagc atgggcagta 720
gctcgcctga gccagagatt tcccaaagct gagtttgcag aagtttccaa gttagtgaca 780
gatcttacca aagtccacac ggaatgctgc catggagatc tgcttgaatg tgctgatgac 840
agggcggacc ttgccaagta tatctgtgaa aatcaagatt cgatctccag taaactgaag 900
gaatgctgtg aaaaacctct gttggaaaaa tcccactgca ttgccgaagt ggaaaatgat 960
gagatgcctg ctgacttgcc ttcattagct gctgattttg ttgaaagtaa ggatgtttgc 1020
aaaaactatg ctgaggcaaa ggatgtcttc ctgggcatgt ttttgtatga atatgcaaga 1080
aggcatcctg attactctgt cgtgctgctg ctgagacttg ccaagacata tgaaaccact 1140
ctagagaagt gctgtgccgc tgcagatcct catgaatgct atgccaaagt gttcgatgaa 1200
tttaaacctc ttgtggaaga gcctcagaat ttaatcaaac aaaattgtga gctttttgag 1260
cagcttggag agtacaaatt ccagaatgcg ctattagttc gttacaccaa gaaagtaccc 1320
caagtgtcaa ctccaactct tgtagaggtc tcaagaaacc taggaaaagt gggcagcaaa 1380
tgttgtaaac atcctgaagc aaaaagaatg ccctgtgcag aagactatct atccgtggtc 1440
ctgaaccagt tatgtgtgtt gcatgagaaa acgccagtaa gtgacagagt caccaaatgc 1500
tgcacagaat ccttggtgaa caggcgacca tgcttttcag ctctggaagt cgatgaaaca 1560
tacgttccca aagagtttaa tgctgaaaca ttcaccttcc atgcagatat atgcacactt 1620
tctgagaagg agagacaaat caagaaacaa actgcacttg ttgagctcgt gaaacacaag 1680
cccaaggcaa caaaagagca actgaaagct gttatggatg atttcgcagc ttttgtagag 1740
aagtgctgca aggctgacga taaggagacc tgctttgccg aggagggtaa aaaacttgtt 1800
gctgcaagtc aagctgcctt aggcttataa 1830
<210> 14
<211> 72
<212> ДНК
<213> Homo sapiens
<400> 14
atgaagtggg taacctttat ttcccttctt tttctcttta gctcggctta ttccaggggt 60
gtgtttcgtc ga 72
<210> 15
<211> 18
<212> ДНК
<213> Homo sapiens
<400> 15
gatgcacaca agagtgag 18
<210> 16
<211> 1737
<212> ДНК
<213> Homo sapiens
<400> 16
gttgctcatc ggtttaaaga tttgggagaa gaaaatttca aagccttggt gttgattgcc 60
tttgctcagt atcttcagca gtgtccattt gaagatcatg taaaattagt gaatgaagta 120
actgaatttg caaaaacatg tgttgctgat gagtcagctg aaaattgtga caaatcactt 180
catacccttt ttggagacaa attatgcaca gttgcaactc ttcgtgaaac ctatggtgaa 240
atggctgact gctgtgcaaa acaagaacct gagagaaatg aatgcttctt gcaacacaaa 300
gatgacaacc caaacctccc ccgattggtg agaccagagg ttgatgtgat gtgcactgct 360
tttcatgaca atgaagagac atttttgaaa aaatacttat atgaaattgc cagaagacat 420
ccttactttt atgccccgga actccttttc tttgctaaaa ggtataaagc tgcttttaca 480
gaatgttgcc aagctgctga taaagctgcc tgcctgttgc caaagctcga tgaacttcgg 540
gatgaaggga aggcttcgtc tgccaaacag agactcaagt gtgccagtct ccaaaaattt 600
ggagaaagag ctttcaaagc atgggcagta gctcgcctga gccagagatt tcccaaagct 660
gagtttgcag aagtttccaa gttagtgaca gatcttacca aagtccacac ggaatgctgc 720
catggagatc tgcttgaatg tgctgatgac agggcggacc ttgccaagta tatctgtgaa 780
aatcaagatt cgatctccag taaactgaag gaatgctgtg aaaaacctct gttggaaaaa 840
tcccactgca ttgccgaagt ggaaaatgat gagatgcctg ctgacttgcc ttcattagct 900
gctgattttg ttgaaagtaa ggatgtttgc aaaaactatg ctgaggcaaa ggatgtcttc 960
ctgggcatgt ttttgtatga atatgcaaga aggcatcctg attactctgt cgtgctgctg 1020
ctgagacttg ccaagacata tgaaaccact ctagagaagt gctgtgccgc tgcagatcct 1080
catgaatgct atgccaaagt gttcgatgaa tttaaacctc ttgtggaaga gcctcagaat 1140
ttaatcaaac aaaattgtga gctttttgag cagcttggag agtacaaatt ccagaatgcg 1200
ctattagttc gttacaccaa gaaagtaccc caagtgtcaa ctccaactct tgtagaggtc 1260
tcaagaaacc taggaaaagt gggcagcaaa tgttgtaaac atcctgaagc aaaaagaatg 1320
ccctgtgcag aagactatct atccgtggtc ctgaaccagt tatgtgtgtt gcatgagaaa 1380
acgccagtaa gtgacagagt caccaaatgc tgcacagaat ccttggtgaa caggcgacca 1440
tgcttttcag ctctggaagt cgatgaaaca tacgttccca aagagtttaa tgctgaaaca 1500
ttcaccttcc atgcagatat atgcacactt tctgagaagg agagacaaat caagaaacaa 1560
actgcacttg ttgagctcgt gaaacacaag cccaaggcaa caaaagagca actgaaagct 1620
gttatggatg atttcgcagc ttttgtagag aagtgctgca aggctgacga taaggagacc 1680
tgctttgccg aggagggtaa aaaacttgtt gctgcaagtc aagctgcctt aggctta 1737
<210> 17
<211> 23484
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(46)
<223> Мышиная последовательность
<220>
<221> прочий_признак
<222> (47)..(17381)
<223> Человеческая последовательность
<220>
<221> прочий_признак
<222> (17382)..(17387)
<223> XhoI
<220>
<221> прочий_признак
<222> (17388)..(17421)
<223> LoxP
<220>
<221> прочий_признак
<222> (17422)..(18108)
<223> Prm1
<220>
<221> прочий_признак
<222> (18109)..(19249)
<223> Crei
<220>
<221> прочий_признак
<222> (19250)..(19545)
<223> Поли(А) SV40
<220>
<221> прочий_признак
<222> (19546)..(20758)
<223> hUbi
<220>
<221> прочий_признак
<222> (20759)..(20825)
<223> em7
<220>
<221> прочий_признак
<222> (20826)..(21629)
<223> Neo
<220>
<221> прочий_признак
<222> (21630)..(22119)
<223> ПолиA PGK
<220>
<221> прочий_признак
<222> (22120)..(22153)
<223> LoxP
<220>
<221> прочий_признак
<222> (22160)..(22185)
<223> I-CeuI
<220>
<221> прочий_признак
<222> (22186)..(22191)
<223> NheI
<220>
<221> прочий_признак
<222> (22192)..(23484)
<223> Мышиная последовательность
<400> 17
tgcacacaga tcacctttcc tatcaacccc actagcctct ggcaaaatga agtgggtaac 60
ctttatttcc cttctttttc tctttagctc ggcttattcc aggggtgtgt ttcgtcgaga 120
tgcacgtaag aaatccattt ttctattgtt caacttttat tctattttcc cagtaaaata 180
aagttttagt aaactctgca tctttaaaga attattttgg catttatttc taaaatggca 240
tagtattttg tatttgtgaa gtcttacaag gttatcttat taataaaatt caaacatcct 300
aggtaaaaaa aaaaaaaggt cagaattgtt tagtgactgt aattttcttt tgcgcactaa 360
ggaaagtgca aagtaactta gagtgactga aacttcacag aatagggttg aagattgaat 420
tcataactat cccaaagacc tatccattgc actatgcttt atttaaaaac cacaaaacct 480
gtgctgttga tctcataaat agaacttgta tttatattta ttttcatttt agtctgtctt 540
cttggttgct gttgatagac actaaaagag tattagatat tatctaagtt tgaatataag 600
gctataaata tttaataatt tttaaaatag tattcttggt aattgaatta ttcttctgtt 660
taaaggcaga agaaataatt gaacatcatc ctgagttttt ctgtaggaat cagagcccaa 720
tattttgaaa caaatgcata atctaagtca aatggaaaga aatataaaaa gtaacattat 780
tacttcttgt tttcttcagt atttaacaat cctttttttt cttcccttgc ccagacaaga 840
gtgaggttgc tcatcggttt aaagatttgg gagaagaaaa tttcaaagcc ttgtaagtta 900
aaatattgat gaatcaaatt taatgtttct aatagtgttg tttattattc taaagtgctt 960
atatttcctt gtcatcaggg ttcagattct aaaacagtgc tgcctcgtag agttttctgc 1020
gttgaggaag atattctgta tctgggctat ccaataaggt agtcactggt cacatggcta 1080
ttgagtactt caaatatgac aagtgcaact gagaaacaaa aacttaaatt gtatttaatt 1140
gtagttaatt tgaatgtata tagtcacatg tggctaatgg ctactgtatt ggacagtaca 1200
gctctggaac ttgcttggtg gaaaggactt taatataggt ttcctttggt ggcttaccca 1260
ctaaatcttc tttacatagc aagcattcct gtgcttagtt gggaatattt aatttttttt 1320
tttttttaag acagggtctc gctctgtcgc ccaggctgga gtgcagtggc gcaatctcgg 1380
ctcactgcaa actccgcctc ccgggttcac gccattctcc tgcctcagcc tcccgagtag 1440
ctgggactac aggcgcccgc catcacgccc ggctaatctt ttgtattttt agtagagatg 1500
gggtttcacc gtgtgccagg atggtctcaa tctcctgaca tcgtgatctg cccacctcgg 1560
cctcccaaag tgctgggatt acaggagtga gccaccgcgc ccggcctatt taaatgtttt 1620
ttaatctagt aaaaaatgag aaaattgttt ttttaaaagt ctacctaatc ctacaggcta 1680
attaaagacg tgtgtgggga tcaggtgcgg tggttcacac ctgtaatccc agcactttgg 1740
aaggctgatg caggaggatt gcttgagccc aggagttcaa gaccagcctg ggcaagtctc 1800
tttaaaaaaa acaaaacaaa caaacaaaaa aattaggcat ggtggcacat gcctgtagtc 1860
ctagctactt aggaggctga cgtaggagga tcgtttggac ctgagaggtc aaggctacag 1920
tgagccatga ttgtgccact gcactccagc ctgggtgaca gagtgagact ctgtctcaaa 1980
aaagaaaaag gaaatctgtg gggtttgttt tagttttaag taattctaag gactttaaaa 2040
atgcctagtc ttgacaatta gatctatttg gcatacaatt tgcttgctta atctatgtgt 2100
gtgcatagat ctactgacac acgcatacat ataaacatta gggaactacc attctctttg 2160
cgtaggaagc cacatatgcc tatctaggcc tcagatcata cctgatatga ataggctttc 2220
tggataatgg tgaagaagat gtataaaaga tagaacctat acccatacat gatttgttct 2280
ctagcgtagc aacctgttac atattaaagt tttattatac tacatttttc tacatccttt 2340
gtttcagggt gttgattgcc tttgctcagt atcttcagca gtgtccattt gaagatcatg 2400
taaaattagt gaatgaagta actgaatttg caaaaacatg tgttgctgat gagtcagctg 2460
aaaattgtga caaatcactt gtaagtacat tctaattgtg gagattcttt cttctgtttg 2520
aagtaatccc aagcatttca aaggaatttt ttttaagttt tctcaattat tattaagtgt 2580
cctgatttgt aagaaacact aaaaagttgc tcatagactg ataagccatt gtttcttttg 2640
tgatagagat gctttagcta tgtccacagt tttaaaatca tttctttatt gagaccaaac 2700
acaacagtca tggtgtattt aaatggcaat ttgtcattta taaacacctc tttttaaaat 2760
ttgaggtttg gtttcttttt gtagaggcta atagggatat gatagcatgt atttatttat 2820
ttatttatct tattttatta tagtaagaac ccttaacatg agatctaccc tgttatattt 2880
ttaagtgtac aatccattat tgttaactac gggtacactg ttgtatagct tactcatctt 2940
gctgtattaa aactttgtgc ccattgatta gtaacccctc gtttcgtcct cccccagcca 3000
ctggcaacca gcattatact ctttgattct atgagtttga ctactttagc taccttatat 3060
aagtggtatt atgtactgtt tatcttttta tgactgactt atttccctta gcatagtgca 3120
ttcaaagtcc aaccatgttg ttgcctattg cagaatttcc ttcttttcaa ggctgaataa 3180
tattccagtg catgtgtgta ccacattttc tttatccatt aatttgttga ttgatagaca 3240
tttaggttgg ttttctacat cttgactatc atgaatagtg ttgcaatgaa cacaggagag 3300
ctactatctc ttagagatga tatcatggtt tttatcatca gaaaacaccc actgatttct 3360
atgctaattt tgttacctgg gtggaataat agtacagcta tatattcctc attttagata 3420
tctttgtatt tctacataca ataaaaaagc agagtactta gtcatgttga agaactttaa 3480
acttttagta tttccagatc aatcttcaaa acaaggacag gtttatcttt ctctcaccac 3540
tcaatctata tatacctctt gtgggcaagg ccagttttta tcactggagc ctttcccctt 3600
tttattatgt acctctccct cacagcagag tcaggacttt aactttacac aatactatgg 3660
ctctacatat gaaatcttaa aaatacataa aaattaataa attctgtcta gagtagtata 3720
ttttccctgg ggttacagtt actttcataa taaaaattag agataaggaa aggactcatt 3780
tattggaaag tgattttagg taacatttct ggaagaaaaa tgtctatatc ttaatagtca 3840
cttaatatat gatggattgt gttactcctc agttttcaat ggcatatact aaaacatggc 3900
cctctaaaaa gggggcaaat gaaatgagaa actctctgaa tgtttttctc ccctaggtga 3960
attcacctgc tgcttagaag cttattttct cttgatttct gttataatga ttgctcttac 4020
cctttagttt taagtttcaa aataggagtc atataacttt ccttaaagct attgactgtc 4080
tttttgtcct gttttattca ccatgagtta tagtgtgaca gttaattctt atgaaaatta 4140
tatagagatg gttaaatcat cagaaactgt aaacctcgat tgggagggga agcggatttt 4200
taaatgattt cctgaccaag cttaaccagt atattaaatc ctttgtactg ttctttggct 4260
ataaagaaaa aaggtactgt ccagcaactg aaacctgctt tcttccattt agcataccct 4320
ttttggagac aaattatgca cagttgcaac tcttcgtgaa acctatggtg aaatggctga 4380
ctgctgtgca aaacaagaac ctgagagaaa tgaatgcttc ttgcaacaca aagatgacaa 4440
cccaaacctc ccccgattgg tgagaccaga ggttgatgtg atgtgcactg cttttcatga 4500
caatgaagag acatttttga aaaagtaagt aatcagatgt ttatagttca aaattaaaaa 4560
gcatggagta actccatagg ccaacactct ataaaaatta ccataacaaa aatattttca 4620
acattaagac ttggaagttt tgttatgatg attttttaaa gaagtagtat ttgataccac 4680
aaaattctac acagcaaaaa atatgatcaa agatattttg aagtttattg aaacaggata 4740
caatctttct gaaaaattta agatagacaa attatttaat gtattacgaa gatatgtata 4800
tatggttgtt ataattgatt tcgttttagt cagcaacatt atattgccaa aatttaacca 4860
tttatgcaca cacacacaca cacacacaca cttaaccctt ttttccacat acttaaagaa 4920
tgacagagac aagaccatca tgtgcaaatt gagcttaatt ggttaattag atatctttgg 4980
aatttggagg ttctggggag aatgtcgatt acaattattt ctgtaatatt gtctgctata 5040
gaaaagtgac tgtttttctt tttcaaaatt tagatactta tatgaaattg ccagaagaca 5100
tccttacttt tatgccccgg aactcctttt ctttgctaaa aggtataaag ctgcttttac 5160
agaatgttgc caagctgctg ataaagctgc ctgcctgttg ccaaaggtat tatgcaaaag 5220
aatagaaaaa aagagttcat tatccaacct gattttgtcc attttgtggc tagatttagg 5280
gaacctgagt gtctgataca aactttccga catggtcaaa aaagccttcc ttttatctgt 5340
cttgaaaatc tttcatcttt gaaggcctac actctcgttt cttcttttaa gatttgccaa 5400
tgatgatctg tcagaggtaa tcactgtgca tgtgtttaaa gatttcacca ctttttatgg 5460
tggtgatcac tatagtgaaa tactgaaact tgtttgtcaa attgcacagc aaggggccac 5520
agttcttgtt tatcttttca tgataatttt tagtagggag ggaattcaaa gtagagaatt 5580
ttactgcatc tagatgcctg agttcatgca ttcattccat aaatatatat tatggaatgc 5640
tttattttct tttctgagga gtttactgat gttggtggag gagagactga aatgaattat 5700
acacaaaatt taaaaattag caaaattgca gcccctggga tattagcgta ctctttctct 5760
gacttttctc ccacttttaa ggctcttttt cctggcaatg tttccagttg gtttctaact 5820
acatagggaa ttccgctgtg accagaatga tcgaatgatc tttccttttc ttagagagca 5880
aaatcattat tcgctaaagg gagtacttgg gaatttaggc ataaattatg ccttcaaaat 5940
ttaatttggc acagtctcat ctgagcttat ggaggggtgt ttcatgtaga atttttcttc 6000
taattttcat caaattattc ctttttgtag ctcgatgaac ttcgggatga agggaaggct 6060
tcgtctgcca aacagagact caagtgtgcc agtctccaaa aatttggaga aagagctttc 6120
aaagcatggt aaatactttt aaacatagtt ggcatcttta taacgatgta aatgataatg 6180
cttcagtgac aaattgtaca tttttatgta ttttgcaaag tgctgtcaaa tacatttctt 6240
tggttgtcta acaggtagaa ctctaataga ggtaaaaatc agaatatcaa tgacaatttg 6300
acattatttt taatcttttc ttttctaaat agttgaataa tttagaggac gctgtccttt 6360
ttgtcctaaa aaaagggaca gatatttaag ttctatttat ttataaaatc ttggactctt 6420
attctaatgg ttcattattt ttatagagct gtaggcatgg ttctttattt aattttttaa 6480
agttattttt aatttttgtg gatacagagt aggtatacat atttacgggg tatatgagat 6540
attttgatat aagtatacaa catatataat ccctttattt aattttatct tccccccaat 6600
gatctaaaac tatttgcttg tccttttatg tcttatagtt aaattcagtc accaactaag 6660
ttgaagttac ttcttatttt tgcatagctc cagctctgat cttcatctca tgtttttgcc 6720
tgagcctctg ttttcatatt acttagttgg ttctgggagc atactttaat agccgagtca 6780
agaaaaatac tagctgcccc gtcacccaca ctcctcacct gctagtcaac agcaaatcaa 6840
cacaacagga aataaaatga aaataataga cattatgcat gctctctaga aactgtcaat 6900
tgaactgtat ttgctcatca ttcctaccat ctacaccacc aaaatcaacc aaatttatga 6960
aaaaaaacag ccccaacata aaattataca cagataaaca ggctatgatt ggttttggga 7020
aagaagtcac ctttacctga tttaggcaac tgtgaaatga ctagagaatg aagaaaatta 7080
gacgtttaca tcttgtcata gagtttgaag atagtgctgg atctttcttt ttataagtaa 7140
gatcaataaa aactccctca ttctgtagaa gttatgattt cttttctaag agacctttag 7200
aagtcagaaa aaatgtgttt caattgagaa aaaagataac tggagtttgt gtagtacttc 7260
ccagattata aaatgctttt gtatgtatta tctaatttaa tcctcaaaac ttcttcaatt 7320
tagcatgttg tcatgacact gcagaggctg aagctcagag aggctgagcc ctctgctaac 7380
aagtcctact gctaacaagt gataaagcca gagctggaag tcacatctgg actccaaacc 7440
tgatgcttct cagcctgttg ccccttttag agttcctttt taatttctgc ttttatgact 7500
tgctagattt ctacctacca cacacactct taaatggata attctgccct aaggataagt 7560
gattaccatt tggttcagaa ctagaactaa tgaattttaa aaattatttc tgtatgtcca 7620
ttttgaattt tcttatgaga aatagtattt gcctagtgtt ttcatataaa atatcgcatg 7680
ataataccat tttgattggc gattttcttt ttagggcagt agctcgcctg agccagagat 7740
ttcccaaagc tgagtttgca gaagtttcca agttagtgac agatcttacc aaagtccaca 7800
cggaatgctg ccatggagat ctgcttgaat gtgctgatga cagggtaaag agtcgtcgat 7860
atgctttttg gtagcttgca tgctcaagtt ggtagaatgg atgcgtttgg tatcattggt 7920
gatagctgac agtgggttga gattgtcttc tgtgctttcg tctgtcctat cttcaatctt 7980
tccctgccta tggtggtggt acctttctgt ttttaacctg gctataaatt accagataaa 8040
cccattcact gatttgtaac tcctttcagt catgctctaa ctgtaaatga aggcttaaac 8100
tgaagtagaa cagttacaag gttttacttg gcagaacatc ttgcaaggta gatgtctaag 8160
aagatttttt tttctttttt taagacagag tttcgctctt gtttcccagg ctggggtgca 8220
atggtgtgat cttggctcag cgcaacctct gcctcctggg ttcaagtgat tctcatgcct 8280
cagcctccca agtagctggg attacaggca tgcgccacca cacctggcta attttgtatt 8340
tttagtagag gcggggtttc accatattgt ccagactggt ctcgaactcc tgacctcagg 8400
tgatccaccc gccttggcct cccaaagtgc tgggattaca ggcatgagcc accttgccca 8460
gcctaagaag attttttgag ggaggtaggt ggacttggag aaggtcacta cttgaagaga 8520
tttttggaaa tgatgtattt ttcttctcta tattccttcc cttaattaac tctgtttgtt 8580
agatgtgcaa atatttggaa tgatatctct tttctcaaaa cttataatat tttctttctc 8640
cctttcttca agattaaact tatgggcaaa tactagaatc ctaatctctc atggcacttt 8700
ctggaaaatt taaggcggtt attttatata tgtaagcagg gcctatgact atgatcttga 8760
ctcatttttc aaaaatcttc tatattttat ttagttattt ggtttcaaaa ggcctgcact 8820
taattttggg ggattatttg gaaaaacagc attgagtttt aatgaaaaaa acttaaatgc 8880
cctaacagta gaaacataaa attaataaat aactgagctg agcacctgct actgattagt 8940
ctattttaat taagtgggaa tgtttttgta gtcctatcta catctccagg tttaggagca 9000
aacagagtat gttcatagaa ggaatatgtg tatggtctta gaatacaatg aatatgttct 9060
gccaacttaa taaaggtctg aggagaaagt gtagcaatgt caattcgtgt tgaacaattt 9120
ccaccaactt acttataggc ggaccttgcc aagtatatct gtgaaaatca agattcgatc 9180
tccagtaaac tgaaggaatg ctgtgaaaaa cctctgttgg aaaaatccca ctgcattgcc 9240
gaagtggaaa atgatgagat gcctgctgac ttgccttcat tagctgctga ttttgttgaa 9300
agtaaggatg tttgcaaaaa ctatgctgag gcaaaggatg tcttcctggg catgtaagta 9360
gataagaaat tattctttta tagctttggc atgacctcac aacttaggag gatagcctag 9420
gcttttctgt ggagttgcta caatttccct gctgcccaga atgtttcttc atccttccct 9480
ttcccaggct ttaacaattt ttgaaatagt taattagttg aatacattgt cataaaataa 9540
tacatgttca tggcaaagct caacattcct tactccttag gggtatttct gaaaatacgt 9600
ctagaaacat tttgtgtata tataaattat gtatacttca gtcattcatt ccaagtgtat 9660
ttcttgaaca tctataatat atgtgtgtga ctatgtattg cctgtctatc taactaatct 9720
aatctaatct agtctatcta tctaatctat gcaatgatag caaagaagta taaaaagaaa 9780
tatagagtct gacaccaggt gctttatatt tggtgaaaag accagaagtt cagtataatg 9840
gcaatatggt aggcaactca attacaaaat aaatgtttac atattgtcag aagttgtggt 9900
gataaactgc atttttgttg ttggattatg ataatgcact aaataatatt tcctaaaatt 9960
atgtacccta caagatttca ctcatacaga gaagaaagag aatattttaa gaacatatct 10020
ctgcccatct atttatcaga atccttttga gatgtagttt aaatcaaaca aaatgttaat 10080
aaaaataaca agtatcattc atcaaagact tcatatgtgc caagcagtgt gtgctttgtg 10140
tagattatgt catatagttc tcataatcca ccttccgaga cagatactat ttattttttg 10200
agacagagtt ttactcttgt tgcccaggct ggagtgcaat ggtgccatct cggctcacca 10260
caacctccgc ctcccaggtt caagcgattc tcctgcctca gcctcctggg attacaggca 10320
tgcaccacca tgcctggcta attttgtatt tttagtagag atggggtttc accatgttgg 10380
tcagactggt ctcaaactcc tgacctctgg tgatatgcct gcctcagcct cctaaagtgc 10440
tgggattaca ggcatgagcc actgtgccca gccgacagat actattatta tttccattct 10500
accgagaagg agactaaggc tctgatcatt taaataagtt gcctaaggtg atgcagtgat 10560
ataagtagca gagctaggaa ttgagccttg gtaactttaa ctctggaccc caagtcctta 10620
gctactaagc tttactgcat ggggtttagt caaattaaga cttttggaat atgagttact 10680
tttgagatta gctttgtgat attttttgtg ctcatttgtc caacaaagtc tattttattt 10740
tcatcttaat taggtttttg tatgaatatg caagaaggca tcctgattac tctgtcgtgc 10800
tgctgctgag acttgccaag acatatgaaa ccactctaga gaagtgctgt gccgctgcag 10860
atcctcatga atgctatgcc aaagtggtag gtttattgtt ggaaaaaaat gtagttcttt 10920
gactgatgat tccaataatg agaaagaaaa ataatgcaag aatgtaaaat gatatacagt 10980
gcaatttaga tcttttcttg agatggtttc aattctggaa tcttaaacat gaaagaaaaa 11040
gtagccttag aatgattaac aaaatttaga ctagttagaa tagaaagatc tgaatagagc 11100
aatctctaaa aaattttgat ctttttttct ctttttcaca atcctgagaa caaaaaaaaa 11160
ttaaatttaa atgttaatta gaagatattt aacttagatg taaagtgagt taacctgatt 11220
ccaggattaa tcaagtacta gaattagtat cttatggcaa attatagaac ctatcccttt 11280
agaatatttt caaatctttt tgaggatgtt taggaatagt tttacaagaa attaagttag 11340
gagaggaaat ctgttctgga ggatttttag ggttcccact agcatatgta atggtttctg 11400
aactattcag aatcagagaa aactcatttt tcctgctttc aagaagctac tgtatgccag 11460
gcaccatgca caaacaatga ccaacgtaaa atctctcatt ttggagagcc tggaatctaa 11520
ctggaaaggt gaactaataa taataatatg tacaatcata gccatcattt attaaacttt 11580
tattatatgc aaggcactgt ttaatttcat tagcttacct ggtttacaga gcagctctat 11640
gagatgagtg ccatctttgc ccctatttta gggataagga ttctgaaatg tggagatggt 11700
aagtaaaatt gcacaactga agaatgagtt acatgacttg gctcaaatac tggtcattga 11760
actccagagc ctgaatattc ttaaccactt acatgatgca agctcaccaa ataaatagtt 11820
cgaatgtatt gtgacagagc ggcattgata ttcatctatt catgtggctt tgagtaggaa 11880
gaagaaagga tatcattctg accagagggg tgaaaaacaa cctgcatctg atcctgaggc 11940
ataatactat taacacaatt cttttatgtt tcagttcgat gaatttaaac ctcttgtgga 12000
agagcctcag aatttaatca aacaaaattg tgagcttttt gagcagcttg gagagtacaa 12060
attccagaat gcgtaagtaa tttttattga ctgatttttt ttatcaattt gtaattattt 12120
aagacttaat atatgagcca cctagcatag aacttttaag aatgaaaata cattgcatat 12180
ttctaatcac tctttgtcaa gaaagatagg agaggagaga taaaatagtt gatggggtgg 12240
agaggtctat atttgaatgt agtctaaaaa ttgttctctt aagattggaa gtatgtaggc 12300
tgggagggta aataccaaat cttggtatat cagaactgag catgtccctt gaaggttaag 12360
aaatagttaa tgggcaaata gagcatggca atattttgta gagcagcaag tagtaggcct 12420
tgaatagatg tcgctcaaaa agtaatatgt aagctgaaca caaaaatgta acaaatgaat 12480
ttagatacat atttgaatat taaattcagg ttgtttggga gatgcaccta gtctttgatg 12540
gttaaacctt tccctccata gaagagacag agacagaatg gcttgctgga ctaatgtccc 12600
aattcaatag agtcttatct atgaaggtta aaaacaagaa gagacatatt atacagtaga 12660
tatttattgt gtggctcata cacatggtgc tcttctgatt atggatttta gagataataa 12720
cagtgaacaa gacatagttt ctttcctcga gtagattaaa gtcatacatt gacttttaat 12780
ggtgactggc attcttaata catgattatt atatattagg taccatgtca gattaattat 12840
aatactttac tacttttaat ttaacccttg aactatccct attgagtcag atatatttcc 12900
ttccattttc tacttgtatc tttcaagttt agcatatgct gatacatatg aagctctctc 12960
caggttttat tgaaagaaga aattaataaa tttattaatg tcactgaatt aggcaactca 13020
ctttcccaag attatgcaag tggtacaggt ggaactcaaa gccaagttta actagttgtt 13080
caggagaatg ttttctaccc tccactaacc cactactctg cagatggaga taatatgatg 13140
aatggaacat agcaacatct tagttgattc cggccaagtg ttctctgttt tatctactat 13200
gttagacagt ttcttgcctt gctgaaaaca catgacttct ttttttcagg ctattagttc 13260
gttacaccaa gaaagtaccc caagtgtcaa ctccaactct tgtagaggtc tcaagaaacc 13320
taggaaaagt gggcagcaaa tgttgtaaac atcctgaagc aaaaagaatg ccctgtgcag 13380
aagactatgt gagtctttaa aaaaatataa taaattaata atgaaaaaat tttaccttta 13440
gatattgata atgctagctt tcataagcag aaggaagtaa tgtgtgtgtg tgcatgtttg 13500
tgtgcatgtg tgtgtgcatg cacgtgtgtg tatgtgtgat attggcagtc aaggccccga 13560
ggatgataat tttttttttt tttttgagac ggagtctcgc tttgttgtcc aggctggagt 13620
gcagtggtgc catctcggct cactgcaacc tccgcctccc aggttcaagc cattctcctg 13680
cctcagcctc ccaagtagct gggactacag gtgcatgcca ccatgcctgg ctaatttttt 13740
gtatttttag tagaaaattt tcagcttcac ctcttttgaa tttctgctct cctgcctgtt 13800
ctttagctat ccgtggtcct gaaccagtta tgtgtgttgc atgagaaaac gccagtaagt 13860
gacagagtca ccaaatgctg cacagaatcc ttggtgaaca ggcgaccatg cttttcagct 13920
ctggaagtcg atgaaacata cgttcccaaa gagtttaatg ctgaaacatt caccttccat 13980
gcagatatat gcacactttc tgagaaggag agacaaatca agaaacaaac gtgaggagta 14040
tttcattact gcatgtgttt gtagtcttga tagcaagaac tgtcaattca agctagcaac 14100
tttttcctga agtagtgatt atatttctta gaggaaagta ttggagtgtt gcccttatta 14160
tgctgataag agtacccaga ataaaatgaa taacttttta aagacaaaat cctctgttat 14220
aatattgcta aaattattca gagtaatatt gtggattaaa gccacaatag aataacatgt 14280
taggccatat tcagtagaaa aagatgaaca attaactgat aaatttgtgc acatggcaaa 14340
ttagttaatg ggaaccatag gagaatttat ttctagatgt aaataattat tttaagtttg 14400
ccctatggtg gccccacaca tgagacaaac ccccaagatg tgacttttga gaatgagact 14460
tggataaaaa acatgtagaa atgcaagccc tgaagctcaa ctccctattg ctatcacagg 14520
ggttataatt gcataaaatt tagctataga aagttgctgt catctcttgt gggctgtaat 14580
catcgtctag gcttaagagt aatattgcaa aacctgtcat gcccacacaa atctctccct 14640
ggcattgttg tctttgcaga tgtcagtgaa agagaaccag cagctcccat gagtttggat 14700
agccttattt tctatagcct ccccactatt agctttgaag ggagcaaagt ttaagaacca 14760
aatataaagt ttctcatctt tatagatgag aaaaatttta aataaagtcc aagataatta 14820
aatttttaag gatcattttt agctctttaa tagcaataaa actcaatatg acataatatg 14880
gcacttccaa aatctgaata atatataatt gcaatgacat acttcttttc agagatttac 14940
tgaaaagaaa tttgttgaca ctacataacg tgatgagtgg tttatactga ttgtttcagt 15000
tggtcttccc accaactcca tgaaagtgga ttttattatc ctcatcatgc agatgagaat 15060
attgagactt atagcggtat gcctgagccc caaagtactc agagttgcct ggctccaaga 15120
tttataatct taaatgatgg gactaccatc cttactctct ccatttttct atacgtgagt 15180
aatgtttttt ctgttttttt tttttctttt tccattcaaa ctcagtgcac ttgttgagct 15240
cgtgaaacac aagcccaagg caacaaaaga gcaactgaaa gctgttatgg atgatttcgc 15300
agcttttgta gagaagtgct gcaaggctga cgataaggag acctgctttg ccgaggaggt 15360
actacagttc tcttcatttt aatatgtcca gtattcattt ttgcatgttt ggttaggcta 15420
gggcttaggg atttatatat caaaggaggc tttgtacatg tgggacaggg atcttatttt 15480
acaaacaatt gtcttacaaa atgaataaaa cagcactttg tttttatctc ctgctctatt 15540
gtgccatact gttaaatgtt tataatgcct gttctgtttc caaatttgtg atgcttatga 15600
atattaatag gaatatttgt aaggcctgaa atattttgat catgaaatca aaacattaat 15660
ttatttaaac atttacttga aatgtggtgg tttgtgattt agttgatttt ataggctagt 15720
gggagaattt acattcaaat gtctaaatca cttaaaattg ccctttatgg cctgacagta 15780
actttttttt attcatttgg ggacaactat gtccgtgagc ttccgtccag agattatagt 15840
agtaaattgt aattaaagga tatgatgcac gtgaaatcac tttgcaatca tcaatagctt 15900
cataaatgtt aattttgtat cctaatagta atgctaatat tttcctaaca tctgtcatgt 15960
ctttgtgttc agggtaaaaa acttgttgct gcaagtcaag ctgccttagg cttataacat 16020
cacatttaaa agcatctcag gtaactatat tttgaatttt ttaaaaaagt aactataata 16080
gttattatta aaatagcaaa gattgaccat ttccaagagc catatagacc agcaccgacc 16140
actattctaa actatttatg tatgtaaata ttagctttta aaattctcaa aatagttgct 16200
gagttgggaa ccactattat ttctattttg tagatgagaa aatgaagata aacatcaaag 16260
catagattaa gtaattttcc aaagggtcaa aattcaaaat tgaaaccaaa gtttcagtgt 16320
tgcccattgt cctgttctga cttatatgat gcggtacaca gagccatcca agtaagtgat 16380
ggctcagcag tggaatactc tgggaattag gctgaaccac atgaaagagt gctttatagg 16440
gcaaaaacag ttgaatatca gtgatttcac atggttcaac ctaatagttc aactcatcct 16500
ttccattgga gaatatgatg gatctacctt ctgtgaactt tatagtgaag aatctgctat 16560
tacatttcca atttgtcaac atgctgagct ttaataggac ttatcttctt atgacaacat 16620
ttattggtgt gtccccttgc ctagcccaac agaagaattc agcagccgta agtctaggac 16680
aggcttaaat tgttttcact ggtgtaaatt gcagaaagat gatctaagta atttggcatt 16740
tattttaata ggtttgaaaa acacatgcca ttttacaaat aagacttata tttgtccttt 16800
tgtttttcag cctaccatga gaataagaga aagaaaatga agatcaaaag cttattcatc 16860
tgtttttctt tttcgttggt gtaaagccaa caccctgtct aaaaaacata aatttcttta 16920
atcattttgc ctcttttctc tgtgcttcaa ttaataaaaa atggaaagaa tctaatagag 16980
tggtacagca ctgttatttt tcaaagatgt gttgctatcc tgaaaattct gtaggttctg 17040
tggaagttcc agtgttctct cttattccac ttcggtagag gatttctagt ttcttgtggg 17100
ctaattaaat aaatcattaa tactcttcta agttatggat tataaacatt caaaataata 17160
ttttgacatt atgataattc tgaataaaag aacaaaaacc atggtatagg taaggaatat 17220
aaaacatggc ttttacctta gaaaaaacaa ttctaaaatt catatggaat caaaaaagag 17280
cctgcagaac caaagtaaga ctaagcaaaa agaacaaatt acctgatttc aaactacact 17340
ataaggccat agtcaccgaa acagcaaggt actggtataa actcgagata acttcgtata 17400
atgtatgcta tacgaagtta tatgcatgcc agtagcagca cccacgtcca ccttctgtct 17460
agtaatgtcc aacacctccc tcagtccaaa cactgctctg catccatgtg gctcccattt 17520
atacctgaag cacttgatgg ggcctcaatg ttttactaga gcccaccccc ctgcaactct 17580
gagaccctct ggatttgtct gtcagtgcct cactggggcg ttggataatt tcttaaaagg 17640
tcaagttccc tcagcagcat tctctgagca gtctgaagat gtgtgctttt cacagttcaa 17700
atccatgtgg ctgtttcacc cacctgcctg gccttgggtt atctatcagg acctagccta 17760
gaagcaggtg tgtggcactt aacacctaag ctgagtgact aactgaacac tcaagtggat 17820
gccatctttg tcacttcttg actgtgacac aagcaactcc tgatgccaaa gccctgccca 17880
cccctctcat gcccatattt ggacatggta caggtcctca ctggccatgg tctgtgaggt 17940
cctggtcctc tttgacttca taattcctag gggccactag tatctataag aggaagaggg 18000
tgctggctcc caggccacag cccacaaaat tccacctgct cacaggttgg ctggctcgac 18060
ccaggtggtg tcccctgctc tgagccagct cccggccaag ccagcaccat gggaaccccc 18120
aagaagaaga ggaaggtgcg taccgattta aattccaatt tactgaccgt acaccaaaat 18180
ttgcctgcat taccggtcga tgcaacgagt gatgaggttc gcaagaacct gatggacatg 18240
ttcagggatc gccaggcgtt ttctgagcat acctggaaaa tgcttctgtc cgtttgccgg 18300
tcgtgggcgg catggtgcaa gttgaataac cggaaatggt ttcccgcaga acctgaagat 18360
gttcgcgatt atcttctata tcttcaggcg cgcggtctgg cagtaaaaac tatccagcaa 18420
catttgggcc agctaaacat gcttcatcgt cggtccgggc tgccacgacc aagtgacagc 18480
aatgctgttt cactggttat gcggcggatc cgaaaagaaa acgttgatgc cggtgaacgt 18540
gcaaaacagg taaatataaa atttttaagt gtataatgat gttaaactac tgattctaat 18600
tgtttgtgta ttttaggctc tagcgttcga acgcactgat ttcgaccagg ttcgttcact 18660
catggaaaat agcgatcgct gccaggatat acgtaatctg gcatttctgg ggattgctta 18720
taacaccctg ttacgtatag ccgaaattgc caggatcagg gttaaagata tctcacgtac 18780
tgacggtggg agaatgttaa tccatattgg cagaacgaaa acgctggtta gcaccgcagg 18840
tgtagagaag gcacttagcc tgggggtaac taaactggtc gagcgatgga tttccgtctc 18900
tggtgtagct gatgatccga ataactacct gttttgccgg gtcagaaaaa atggtgttgc 18960
cgcgccatct gccaccagcc agctatcaac tcgcgccctg gaagggattt ttgaagcaac 19020
tcatcgattg atttacggcg ctaaggatga ctctggtcag agatacctgg cctggtctgg 19080
acacagtgcc cgtgtcggag ccgcgcgaga tatggcccgc gctggagttt caataccgga 19140
gatcatgcaa gctggtggct ggaccaatgt aaatattgtc atgaactata tccgtaacct 19200
ggatagtgaa acaggggcaa tggtgcgcct gctggaagat ggcgattagg cggccggccg 19260
ctaatcagcc ataccacatt tgtagaggtt ttacttgctt taaaaaacct cccacacctc 19320
cccctgaacc tgaaacataa aatgaatgca attgttgttg ttaacttgtt tattgcagct 19380
tataatggtt acaaataaag caatagcatc acaaatttca caaataaagc atttttttca 19440
ctgcattcta gttgtggttt gtccaaactc atcaatgtat cttatcatgt ctggatcccc 19500
cggctagagt ttaaacacta gaactagtgg atcccccggg atcatggcct ccgcgccggg 19560
ttttggcgcc tcccgcgggc gcccccctcc tcacggcgag cgctgccacg tcagacgaag 19620
ggcgcagcga gcgtcctgat ccttccgccc ggacgctcag gacagcggcc cgctgctcat 19680
aagactcggc cttagaaccc cagtatcagc agaaggacat tttaggacgg gacttgggtg 19740
actctagggc actggttttc tttccagaga gcggaacagg cgaggaaaag tagtcccttc 19800
tcggcgattc tgcggaggga tctccgtggg gcggtgaacg ccgatgatta tataaggacg 19860
cgccgggtgt ggcacagcta gttccgtcgc agccgggatt tgggtcgcgg ttcttgtttg 19920
tggatcgctg tgatcgtcac ttggtgagta gcgggctgct gggctggccg gggctttcgt 19980
ggccgccggg ccgctcggtg ggacggaagc gtgtggagag accgccaagg gctgtagtct 20040
gggtccgcga gcaaggttgc cctgaactgg gggttggggg gagcgcagca aaatggcggc 20100
tgttcccgag tcttgaatgg aagacgcttg tgaggcgggc tgtgaggtcg ttgaaacaag 20160
gtggggggca tggtgggcgg caagaaccca aggtcttgag gccttcgcta atgcgggaaa 20220
gctcttattc gggtgagatg ggctggggca ccatctgggg accctgacgt gaagtttgtc 20280
actgactgga gaactcggtt tgtcgtctgt tgcgggggcg gcagttatgg cggtgccgtt 20340
gggcagtgca cccgtacctt tgggagcgcg cgccctcgtc gtgtcgtgac gtcacccgtt 20400
ctgttggctt ataatgcagg gtggggccac ctgccggtag gtgtgcggta ggcttttctc 20460
cgtcgcagga cgcagggttc gggcctaggg taggctctcc tgaatcgaca ggcgccggac 20520
ctctggtgag gggagggata agtgaggcgt cagtttcttt ggtcggtttt atgtacctat 20580
cttcttaagt agctgaagct ccggttttga actatgcgct cggggttggc gagtgtgttt 20640
tgtgaagttt tttaggcacc ttttgaaatg taatcatttg ggtcaatatg taattttcag 20700
tgttagacta gtaaattgtc cgctaaattc tggccgtttt tggctttttt gttagacgtg 20760
ttgacaatta atcatcggca tagtatatcg gcatagtata atacgacaag gtgaggaact 20820
aaaccatggg atcggccatt gaacaagatg gattgcacgc aggttctccg gccgcttggg 20880
tggagaggct attcggctat gactgggcac aacagacaat cggctgctct gatgccgccg 20940
tgttccggct gtcagcgcag gggcgcccgg ttctttttgt caagaccgac ctgtccggtg 21000
ccctgaatga actgcaggac gaggcagcgc ggctatcgtg gctggccacg acgggcgttc 21060
cttgcgcagc tgtgctcgac gttgtcactg aagcgggaag ggactggctg ctattgggcg 21120
aagtgccggg gcaggatctc ctgtcatctc accttgctcc tgccgagaaa gtatccatca 21180
tggctgatgc aatgcggcgg ctgcatacgc ttgatccggc tacctgccca ttcgaccacc 21240
aagcgaaaca tcgcatcgag cgagcacgta ctcggatgga agccggtctt gtcgatcagg 21300
atgatctgga cgaagagcat caggggctcg cgccagccga actgttcgcc aggctcaagg 21360
cgcgcatgcc cgacggcgat gatctcgtcg tgacccatgg cgatgcctgc ttgccgaata 21420
tcatggtgga aaatggccgc ttttctggat tcatcgactg tggccggctg ggtgtggcgg 21480
accgctatca ggacatagcg ttggctaccc gtgatattgc tgaagagctt ggcggcgaat 21540
gggctgaccg cttcctcgtg ctttacggta tcgccgctcc cgattcgcag cgcatcgcct 21600
tctatcgcct tcttgacgag ttcttctgag gggatccgct gtaagtctgc agaaattgat 21660
gatctattaa acaataaaga tgtccactaa aatggaagtt tttcctgtca tactttgtta 21720
agaagggtga gaacagagta cctacatttt gaatggaagg attggagcta cgggggtggg 21780
ggtggggtgg gattagataa atgcctgctc tttactgaag gctctttact attgctttat 21840
gataatgttt catagttgga tatcataatt taaacaagca aaaccaaatt aagggccagc 21900
tcattcctcc cactcatgat ctatagatct atagatctct cgtgggatca ttgtttttct 21960
cttgattccc actttgtggt tctaagtact gtggtttcca aatgtgtcag tttcatagcc 22020
tgaagaacga gatcagcagc ctctgttcca catacacttc attctcagta ttgttttgcc 22080
aagttctaat tccatcagac ctcgacctgc agcccctaga taacttcgta taatgtatgc 22140
tatacgaagt tatgctaggt aactataacg gtcctaaggt agcgagctag cacacatcac 22200
aaccacaacc ttctcaggta actatacttg ggacttaaaa aacataatca taatcatttt 22260
tcctaaaacg atcaagactg ataaccattt gacaagagcc atacagacaa gcaccagctg 22320
gcactcttag gtcttcacgt atggtcatca gtttgggttc catttgtaga taagaaactg 22380
aacatataaa ggtctaggtt aatgcaattt acacaaaagg agaccaaacc agggagagaa 22440
ggaaccaaaa ttaaaaattc aaaccagagc aaaggagtta gccctggttt tgctctgact 22500
tacatgaacc actatgtgga gtcctccatg ttagcctagt caagcttatc ctctggatga 22560
agttgaaacc atatgaagga atatttgggg ggtgggtcaa aacagttgtg tatcaatgat 22620
tccatgtggt ttgacccaat cattctgtga atccatttca acagaagata caacgggttc 22680
tgtttcataa taagtgatcc acttccaaat ttctgatgtg ccccatgcta agctttaaca 22740
gaatttatct tcttatgaca aagcagcctc ctttgaaaat atagccaact gcacacagct 22800
atgttgatca attttgttta taatcttgca gaagagaatt ttttaaaata gggcaataat 22860
ggaaggcttt ggcaaaaaaa ttgtttctcc atatgaaaac aaaaaactta tttttttatt 22920
caagcaaaga acctatagac ataaggctat ttcaaaatta tttcagtttt agaaagaatt 22980
gaaagttttg tagcattctg agaagacagc tttcatttgt aatcataggt aatatgtagg 23040
tcctcagaaa tggtgagacc cctgactttg acacttgggg actctgaggg accagtgatg 23100
aagagggcac aacttatatc acacatgcac gagttggggt gagagggtgt cacaacatct 23160
atcagtgtgt catctgccca ccaagtaaca gatgtcagct aagactaggt catgtgtagg 23220
ctgtctacac cagtgaaaat cgcaaaaaga atctaagaaa ttccacattt ctagaaaata 23280
ggtttggaaa ccgtattcca ttttacaaag gacacttaca tttctctttt tgttttccag 23340
gctaccctga gaaaaaaaga catgaagact caggactcat cttttctgtt ggtgtaaaat 23400
caacacccta aggaacacaa atttctttaa acatttgact tcttgtctct gtgctgcaat 23460
taataaaaaa tggaaagaat ctac 23484
<210> 18
<211> 17768
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(46)
<223> Мышиная последовательность
<220>
<221> прочий_признак
<222> (47)..(17381)
<223> Человеческая последовательность
<220>
<221> прочий_признак
<222> (17382)..(17387)
<223> XhoI
<220>
<221> прочий_признак
<222> (17388)..(17421)
<223> LoxP
<220>
<221> прочий_признак
<222> (17428)..(17453)
<223> I-CeuI
<220>
<221> прочий_признак
<222> (17454)..(17459)
<223> NheI
<220>
<221> прочий_признак
<222> (17460)..(17768)
<223> Мышиная последовательность
<400> 18
tgcacacaga tcacctttcc tatcaacccc actagcctct ggcaaaatga agtgggtaac 60
ctttatttcc cttctttttc tctttagctc ggcttattcc aggggtgtgt ttcgtcgaga 120
tgcacgtaag aaatccattt ttctattgtt caacttttat tctattttcc cagtaaaata 180
aagttttagt aaactctgca tctttaaaga attattttgg catttatttc taaaatggca 240
tagtattttg tatttgtgaa gtcttacaag gttatcttat taataaaatt caaacatcct 300
aggtaaaaaa aaaaaaaggt cagaattgtt tagtgactgt aattttcttt tgcgcactaa 360
ggaaagtgca aagtaactta gagtgactga aacttcacag aatagggttg aagattgaat 420
tcataactat cccaaagacc tatccattgc actatgcttt atttaaaaac cacaaaacct 480
gtgctgttga tctcataaat agaacttgta tttatattta ttttcatttt agtctgtctt 540
cttggttgct gttgatagac actaaaagag tattagatat tatctaagtt tgaatataag 600
gctataaata tttaataatt tttaaaatag tattcttggt aattgaatta ttcttctgtt 660
taaaggcaga agaaataatt gaacatcatc ctgagttttt ctgtaggaat cagagcccaa 720
tattttgaaa caaatgcata atctaagtca aatggaaaga aatataaaaa gtaacattat 780
tacttcttgt tttcttcagt atttaacaat cctttttttt cttcccttgc ccagacaaga 840
gtgaggttgc tcatcggttt aaagatttgg gagaagaaaa tttcaaagcc ttgtaagtta 900
aaatattgat gaatcaaatt taatgtttct aatagtgttg tttattattc taaagtgctt 960
atatttcctt gtcatcaggg ttcagattct aaaacagtgc tgcctcgtag agttttctgc 1020
gttgaggaag atattctgta tctgggctat ccaataaggt agtcactggt cacatggcta 1080
ttgagtactt caaatatgac aagtgcaact gagaaacaaa aacttaaatt gtatttaatt 1140
gtagttaatt tgaatgtata tagtcacatg tggctaatgg ctactgtatt ggacagtaca 1200
gctctggaac ttgcttggtg gaaaggactt taatataggt ttcctttggt ggcttaccca 1260
ctaaatcttc tttacatagc aagcattcct gtgcttagtt gggaatattt aatttttttt 1320
tttttttaag acagggtctc gctctgtcgc ccaggctgga gtgcagtggc gcaatctcgg 1380
ctcactgcaa actccgcctc ccgggttcac gccattctcc tgcctcagcc tcccgagtag 1440
ctgggactac aggcgcccgc catcacgccc ggctaatctt ttgtattttt agtagagatg 1500
gggtttcacc gtgtgccagg atggtctcaa tctcctgaca tcgtgatctg cccacctcgg 1560
cctcccaaag tgctgggatt acaggagtga gccaccgcgc ccggcctatt taaatgtttt 1620
ttaatctagt aaaaaatgag aaaattgttt ttttaaaagt ctacctaatc ctacaggcta 1680
attaaagacg tgtgtgggga tcaggtgcgg tggttcacac ctgtaatccc agcactttgg 1740
aaggctgatg caggaggatt gcttgagccc aggagttcaa gaccagcctg ggcaagtctc 1800
tttaaaaaaa acaaaacaaa caaacaaaaa aattaggcat ggtggcacat gcctgtagtc 1860
ctagctactt aggaggctga cgtaggagga tcgtttggac ctgagaggtc aaggctacag 1920
tgagccatga ttgtgccact gcactccagc ctgggtgaca gagtgagact ctgtctcaaa 1980
aaagaaaaag gaaatctgtg gggtttgttt tagttttaag taattctaag gactttaaaa 2040
atgcctagtc ttgacaatta gatctatttg gcatacaatt tgcttgctta atctatgtgt 2100
gtgcatagat ctactgacac acgcatacat ataaacatta gggaactacc attctctttg 2160
cgtaggaagc cacatatgcc tatctaggcc tcagatcata cctgatatga ataggctttc 2220
tggataatgg tgaagaagat gtataaaaga tagaacctat acccatacat gatttgttct 2280
ctagcgtagc aacctgttac atattaaagt tttattatac tacatttttc tacatccttt 2340
gtttcagggt gttgattgcc tttgctcagt atcttcagca gtgtccattt gaagatcatg 2400
taaaattagt gaatgaagta actgaatttg caaaaacatg tgttgctgat gagtcagctg 2460
aaaattgtga caaatcactt gtaagtacat tctaattgtg gagattcttt cttctgtttg 2520
aagtaatccc aagcatttca aaggaatttt ttttaagttt tctcaattat tattaagtgt 2580
cctgatttgt aagaaacact aaaaagttgc tcatagactg ataagccatt gtttcttttg 2640
tgatagagat gctttagcta tgtccacagt tttaaaatca tttctttatt gagaccaaac 2700
acaacagtca tggtgtattt aaatggcaat ttgtcattta taaacacctc tttttaaaat 2760
ttgaggtttg gtttcttttt gtagaggcta atagggatat gatagcatgt atttatttat 2820
ttatttatct tattttatta tagtaagaac ccttaacatg agatctaccc tgttatattt 2880
ttaagtgtac aatccattat tgttaactac gggtacactg ttgtatagct tactcatctt 2940
gctgtattaa aactttgtgc ccattgatta gtaacccctc gtttcgtcct cccccagcca 3000
ctggcaacca gcattatact ctttgattct atgagtttga ctactttagc taccttatat 3060
aagtggtatt atgtactgtt tatcttttta tgactgactt atttccctta gcatagtgca 3120
ttcaaagtcc aaccatgttg ttgcctattg cagaatttcc ttcttttcaa ggctgaataa 3180
tattccagtg catgtgtgta ccacattttc tttatccatt aatttgttga ttgatagaca 3240
tttaggttgg ttttctacat cttgactatc atgaatagtg ttgcaatgaa cacaggagag 3300
ctactatctc ttagagatga tatcatggtt tttatcatca gaaaacaccc actgatttct 3360
atgctaattt tgttacctgg gtggaataat agtacagcta tatattcctc attttagata 3420
tctttgtatt tctacataca ataaaaaagc agagtactta gtcatgttga agaactttaa 3480
acttttagta tttccagatc aatcttcaaa acaaggacag gtttatcttt ctctcaccac 3540
tcaatctata tatacctctt gtgggcaagg ccagttttta tcactggagc ctttcccctt 3600
tttattatgt acctctccct cacagcagag tcaggacttt aactttacac aatactatgg 3660
ctctacatat gaaatcttaa aaatacataa aaattaataa attctgtcta gagtagtata 3720
ttttccctgg ggttacagtt actttcataa taaaaattag agataaggaa aggactcatt 3780
tattggaaag tgattttagg taacatttct ggaagaaaaa tgtctatatc ttaatagtca 3840
cttaatatat gatggattgt gttactcctc agttttcaat ggcatatact aaaacatggc 3900
cctctaaaaa gggggcaaat gaaatgagaa actctctgaa tgtttttctc ccctaggtga 3960
attcacctgc tgcttagaag cttattttct cttgatttct gttataatga ttgctcttac 4020
cctttagttt taagtttcaa aataggagtc atataacttt ccttaaagct attgactgtc 4080
tttttgtcct gttttattca ccatgagtta tagtgtgaca gttaattctt atgaaaatta 4140
tatagagatg gttaaatcat cagaaactgt aaacctcgat tgggagggga agcggatttt 4200
taaatgattt cctgaccaag cttaaccagt atattaaatc ctttgtactg ttctttggct 4260
ataaagaaaa aaggtactgt ccagcaactg aaacctgctt tcttccattt agcataccct 4320
ttttggagac aaattatgca cagttgcaac tcttcgtgaa acctatggtg aaatggctga 4380
ctgctgtgca aaacaagaac ctgagagaaa tgaatgcttc ttgcaacaca aagatgacaa 4440
cccaaacctc ccccgattgg tgagaccaga ggttgatgtg atgtgcactg cttttcatga 4500
caatgaagag acatttttga aaaagtaagt aatcagatgt ttatagttca aaattaaaaa 4560
gcatggagta actccatagg ccaacactct ataaaaatta ccataacaaa aatattttca 4620
acattaagac ttggaagttt tgttatgatg attttttaaa gaagtagtat ttgataccac 4680
aaaattctac acagcaaaaa atatgatcaa agatattttg aagtttattg aaacaggata 4740
caatctttct gaaaaattta agatagacaa attatttaat gtattacgaa gatatgtata 4800
tatggttgtt ataattgatt tcgttttagt cagcaacatt atattgccaa aatttaacca 4860
tttatgcaca cacacacaca cacacacaca cttaaccctt ttttccacat acttaaagaa 4920
tgacagagac aagaccatca tgtgcaaatt gagcttaatt ggttaattag atatctttgg 4980
aatttggagg ttctggggag aatgtcgatt acaattattt ctgtaatatt gtctgctata 5040
gaaaagtgac tgtttttctt tttcaaaatt tagatactta tatgaaattg ccagaagaca 5100
tccttacttt tatgccccgg aactcctttt ctttgctaaa aggtataaag ctgcttttac 5160
agaatgttgc caagctgctg ataaagctgc ctgcctgttg ccaaaggtat tatgcaaaag 5220
aatagaaaaa aagagttcat tatccaacct gattttgtcc attttgtggc tagatttagg 5280
gaacctgagt gtctgataca aactttccga catggtcaaa aaagccttcc ttttatctgt 5340
cttgaaaatc tttcatcttt gaaggcctac actctcgttt cttcttttaa gatttgccaa 5400
tgatgatctg tcagaggtaa tcactgtgca tgtgtttaaa gatttcacca ctttttatgg 5460
tggtgatcac tatagtgaaa tactgaaact tgtttgtcaa attgcacagc aaggggccac 5520
agttcttgtt tatcttttca tgataatttt tagtagggag ggaattcaaa gtagagaatt 5580
ttactgcatc tagatgcctg agttcatgca ttcattccat aaatatatat tatggaatgc 5640
tttattttct tttctgagga gtttactgat gttggtggag gagagactga aatgaattat 5700
acacaaaatt taaaaattag caaaattgca gcccctggga tattagcgta ctctttctct 5760
gacttttctc ccacttttaa ggctcttttt cctggcaatg tttccagttg gtttctaact 5820
acatagggaa ttccgctgtg accagaatga tcgaatgatc tttccttttc ttagagagca 5880
aaatcattat tcgctaaagg gagtacttgg gaatttaggc ataaattatg ccttcaaaat 5940
ttaatttggc acagtctcat ctgagcttat ggaggggtgt ttcatgtaga atttttcttc 6000
taattttcat caaattattc ctttttgtag ctcgatgaac ttcgggatga agggaaggct 6060
tcgtctgcca aacagagact caagtgtgcc agtctccaaa aatttggaga aagagctttc 6120
aaagcatggt aaatactttt aaacatagtt ggcatcttta taacgatgta aatgataatg 6180
cttcagtgac aaattgtaca tttttatgta ttttgcaaag tgctgtcaaa tacatttctt 6240
tggttgtcta acaggtagaa ctctaataga ggtaaaaatc agaatatcaa tgacaatttg 6300
acattatttt taatcttttc ttttctaaat agttgaataa tttagaggac gctgtccttt 6360
ttgtcctaaa aaaagggaca gatatttaag ttctatttat ttataaaatc ttggactctt 6420
attctaatgg ttcattattt ttatagagct gtaggcatgg ttctttattt aattttttaa 6480
agttattttt aatttttgtg gatacagagt aggtatacat atttacgggg tatatgagat 6540
attttgatat aagtatacaa catatataat ccctttattt aattttatct tccccccaat 6600
gatctaaaac tatttgcttg tccttttatg tcttatagtt aaattcagtc accaactaag 6660
ttgaagttac ttcttatttt tgcatagctc cagctctgat cttcatctca tgtttttgcc 6720
tgagcctctg ttttcatatt acttagttgg ttctgggagc atactttaat agccgagtca 6780
agaaaaatac tagctgcccc gtcacccaca ctcctcacct gctagtcaac agcaaatcaa 6840
cacaacagga aataaaatga aaataataga cattatgcat gctctctaga aactgtcaat 6900
tgaactgtat ttgctcatca ttcctaccat ctacaccacc aaaatcaacc aaatttatga 6960
aaaaaaacag ccccaacata aaattataca cagataaaca ggctatgatt ggttttggga 7020
aagaagtcac ctttacctga tttaggcaac tgtgaaatga ctagagaatg aagaaaatta 7080
gacgtttaca tcttgtcata gagtttgaag atagtgctgg atctttcttt ttataagtaa 7140
gatcaataaa aactccctca ttctgtagaa gttatgattt cttttctaag agacctttag 7200
aagtcagaaa aaatgtgttt caattgagaa aaaagataac tggagtttgt gtagtacttc 7260
ccagattata aaatgctttt gtatgtatta tctaatttaa tcctcaaaac ttcttcaatt 7320
tagcatgttg tcatgacact gcagaggctg aagctcagag aggctgagcc ctctgctaac 7380
aagtcctact gctaacaagt gataaagcca gagctggaag tcacatctgg actccaaacc 7440
tgatgcttct cagcctgttg ccccttttag agttcctttt taatttctgc ttttatgact 7500
tgctagattt ctacctacca cacacactct taaatggata attctgccct aaggataagt 7560
gattaccatt tggttcagaa ctagaactaa tgaattttaa aaattatttc tgtatgtcca 7620
ttttgaattt tcttatgaga aatagtattt gcctagtgtt ttcatataaa atatcgcatg 7680
ataataccat tttgattggc gattttcttt ttagggcagt agctcgcctg agccagagat 7740
ttcccaaagc tgagtttgca gaagtttcca agttagtgac agatcttacc aaagtccaca 7800
cggaatgctg ccatggagat ctgcttgaat gtgctgatga cagggtaaag agtcgtcgat 7860
atgctttttg gtagcttgca tgctcaagtt ggtagaatgg atgcgtttgg tatcattggt 7920
gatagctgac agtgggttga gattgtcttc tgtgctttcg tctgtcctat cttcaatctt 7980
tccctgccta tggtggtggt acctttctgt ttttaacctg gctataaatt accagataaa 8040
cccattcact gatttgtaac tcctttcagt catgctctaa ctgtaaatga aggcttaaac 8100
tgaagtagaa cagttacaag gttttacttg gcagaacatc ttgcaaggta gatgtctaag 8160
aagatttttt tttctttttt taagacagag tttcgctctt gtttcccagg ctggggtgca 8220
atggtgtgat cttggctcag cgcaacctct gcctcctggg ttcaagtgat tctcatgcct 8280
cagcctccca agtagctggg attacaggca tgcgccacca cacctggcta attttgtatt 8340
tttagtagag gcggggtttc accatattgt ccagactggt ctcgaactcc tgacctcagg 8400
tgatccaccc gccttggcct cccaaagtgc tgggattaca ggcatgagcc accttgccca 8460
gcctaagaag attttttgag ggaggtaggt ggacttggag aaggtcacta cttgaagaga 8520
tttttggaaa tgatgtattt ttcttctcta tattccttcc cttaattaac tctgtttgtt 8580
agatgtgcaa atatttggaa tgatatctct tttctcaaaa cttataatat tttctttctc 8640
cctttcttca agattaaact tatgggcaaa tactagaatc ctaatctctc atggcacttt 8700
ctggaaaatt taaggcggtt attttatata tgtaagcagg gcctatgact atgatcttga 8760
ctcatttttc aaaaatcttc tatattttat ttagttattt ggtttcaaaa ggcctgcact 8820
taattttggg ggattatttg gaaaaacagc attgagtttt aatgaaaaaa acttaaatgc 8880
cctaacagta gaaacataaa attaataaat aactgagctg agcacctgct actgattagt 8940
ctattttaat taagtgggaa tgtttttgta gtcctatcta catctccagg tttaggagca 9000
aacagagtat gttcatagaa ggaatatgtg tatggtctta gaatacaatg aatatgttct 9060
gccaacttaa taaaggtctg aggagaaagt gtagcaatgt caattcgtgt tgaacaattt 9120
ccaccaactt acttataggc ggaccttgcc aagtatatct gtgaaaatca agattcgatc 9180
tccagtaaac tgaaggaatg ctgtgaaaaa cctctgttgg aaaaatccca ctgcattgcc 9240
gaagtggaaa atgatgagat gcctgctgac ttgccttcat tagctgctga ttttgttgaa 9300
agtaaggatg tttgcaaaaa ctatgctgag gcaaaggatg tcttcctggg catgtaagta 9360
gataagaaat tattctttta tagctttggc atgacctcac aacttaggag gatagcctag 9420
gcttttctgt ggagttgcta caatttccct gctgcccaga atgtttcttc atccttccct 9480
ttcccaggct ttaacaattt ttgaaatagt taattagttg aatacattgt cataaaataa 9540
tacatgttca tggcaaagct caacattcct tactccttag gggtatttct gaaaatacgt 9600
ctagaaacat tttgtgtata tataaattat gtatacttca gtcattcatt ccaagtgtat 9660
ttcttgaaca tctataatat atgtgtgtga ctatgtattg cctgtctatc taactaatct 9720
aatctaatct agtctatcta tctaatctat gcaatgatag caaagaagta taaaaagaaa 9780
tatagagtct gacaccaggt gctttatatt tggtgaaaag accagaagtt cagtataatg 9840
gcaatatggt aggcaactca attacaaaat aaatgtttac atattgtcag aagttgtggt 9900
gataaactgc atttttgttg ttggattatg ataatgcact aaataatatt tcctaaaatt 9960
atgtacccta caagatttca ctcatacaga gaagaaagag aatattttaa gaacatatct 10020
ctgcccatct atttatcaga atccttttga gatgtagttt aaatcaaaca aaatgttaat 10080
aaaaataaca agtatcattc atcaaagact tcatatgtgc caagcagtgt gtgctttgtg 10140
tagattatgt catatagttc tcataatcca ccttccgaga cagatactat ttattttttg 10200
agacagagtt ttactcttgt tgcccaggct ggagtgcaat ggtgccatct cggctcacca 10260
caacctccgc ctcccaggtt caagcgattc tcctgcctca gcctcctggg attacaggca 10320
tgcaccacca tgcctggcta attttgtatt tttagtagag atggggtttc accatgttgg 10380
tcagactggt ctcaaactcc tgacctctgg tgatatgcct gcctcagcct cctaaagtgc 10440
tgggattaca ggcatgagcc actgtgccca gccgacagat actattatta tttccattct 10500
accgagaagg agactaaggc tctgatcatt taaataagtt gcctaaggtg atgcagtgat 10560
ataagtagca gagctaggaa ttgagccttg gtaactttaa ctctggaccc caagtcctta 10620
gctactaagc tttactgcat ggggtttagt caaattaaga cttttggaat atgagttact 10680
tttgagatta gctttgtgat attttttgtg ctcatttgtc caacaaagtc tattttattt 10740
tcatcttaat taggtttttg tatgaatatg caagaaggca tcctgattac tctgtcgtgc 10800
tgctgctgag acttgccaag acatatgaaa ccactctaga gaagtgctgt gccgctgcag 10860
atcctcatga atgctatgcc aaagtggtag gtttattgtt ggaaaaaaat gtagttcttt 10920
gactgatgat tccaataatg agaaagaaaa ataatgcaag aatgtaaaat gatatacagt 10980
gcaatttaga tcttttcttg agatggtttc aattctggaa tcttaaacat gaaagaaaaa 11040
gtagccttag aatgattaac aaaatttaga ctagttagaa tagaaagatc tgaatagagc 11100
aatctctaaa aaattttgat ctttttttct ctttttcaca atcctgagaa caaaaaaaaa 11160
ttaaatttaa atgttaatta gaagatattt aacttagatg taaagtgagt taacctgatt 11220
ccaggattaa tcaagtacta gaattagtat cttatggcaa attatagaac ctatcccttt 11280
agaatatttt caaatctttt tgaggatgtt taggaatagt tttacaagaa attaagttag 11340
gagaggaaat ctgttctgga ggatttttag ggttcccact agcatatgta atggtttctg 11400
aactattcag aatcagagaa aactcatttt tcctgctttc aagaagctac tgtatgccag 11460
gcaccatgca caaacaatga ccaacgtaaa atctctcatt ttggagagcc tggaatctaa 11520
ctggaaaggt gaactaataa taataatatg tacaatcata gccatcattt attaaacttt 11580
tattatatgc aaggcactgt ttaatttcat tagcttacct ggtttacaga gcagctctat 11640
gagatgagtg ccatctttgc ccctatttta gggataagga ttctgaaatg tggagatggt 11700
aagtaaaatt gcacaactga agaatgagtt acatgacttg gctcaaatac tggtcattga 11760
actccagagc ctgaatattc ttaaccactt acatgatgca agctcaccaa ataaatagtt 11820
cgaatgtatt gtgacagagc ggcattgata ttcatctatt catgtggctt tgagtaggaa 11880
gaagaaagga tatcattctg accagagggg tgaaaaacaa cctgcatctg atcctgaggc 11940
ataatactat taacacaatt cttttatgtt tcagttcgat gaatttaaac ctcttgtgga 12000
agagcctcag aatttaatca aacaaaattg tgagcttttt gagcagcttg gagagtacaa 12060
attccagaat gcgtaagtaa tttttattga ctgatttttt ttatcaattt gtaattattt 12120
aagacttaat atatgagcca cctagcatag aacttttaag aatgaaaata cattgcatat 12180
ttctaatcac tctttgtcaa gaaagatagg agaggagaga taaaatagtt gatggggtgg 12240
agaggtctat atttgaatgt agtctaaaaa ttgttctctt aagattggaa gtatgtaggc 12300
tgggagggta aataccaaat cttggtatat cagaactgag catgtccctt gaaggttaag 12360
aaatagttaa tgggcaaata gagcatggca atattttgta gagcagcaag tagtaggcct 12420
tgaatagatg tcgctcaaaa agtaatatgt aagctgaaca caaaaatgta acaaatgaat 12480
ttagatacat atttgaatat taaattcagg ttgtttggga gatgcaccta gtctttgatg 12540
gttaaacctt tccctccata gaagagacag agacagaatg gcttgctgga ctaatgtccc 12600
aattcaatag agtcttatct atgaaggtta aaaacaagaa gagacatatt atacagtaga 12660
tatttattgt gtggctcata cacatggtgc tcttctgatt atggatttta gagataataa 12720
cagtgaacaa gacatagttt ctttcctcga gtagattaaa gtcatacatt gacttttaat 12780
ggtgactggc attcttaata catgattatt atatattagg taccatgtca gattaattat 12840
aatactttac tacttttaat ttaacccttg aactatccct attgagtcag atatatttcc 12900
ttccattttc tacttgtatc tttcaagttt agcatatgct gatacatatg aagctctctc 12960
caggttttat tgaaagaaga aattaataaa tttattaatg tcactgaatt aggcaactca 13020
ctttcccaag attatgcaag tggtacaggt ggaactcaaa gccaagttta actagttgtt 13080
caggagaatg ttttctaccc tccactaacc cactactctg cagatggaga taatatgatg 13140
aatggaacat agcaacatct tagttgattc cggccaagtg ttctctgttt tatctactat 13200
gttagacagt ttcttgcctt gctgaaaaca catgacttct ttttttcagg ctattagttc 13260
gttacaccaa gaaagtaccc caagtgtcaa ctccaactct tgtagaggtc tcaagaaacc 13320
taggaaaagt gggcagcaaa tgttgtaaac atcctgaagc aaaaagaatg ccctgtgcag 13380
aagactatgt gagtctttaa aaaaatataa taaattaata atgaaaaaat tttaccttta 13440
gatattgata atgctagctt tcataagcag aaggaagtaa tgtgtgtgtg tgcatgtttg 13500
tgtgcatgtg tgtgtgcatg cacgtgtgtg tatgtgtgat attggcagtc aaggccccga 13560
ggatgataat tttttttttt tttttgagac ggagtctcgc tttgttgtcc aggctggagt 13620
gcagtggtgc catctcggct cactgcaacc tccgcctccc aggttcaagc cattctcctg 13680
cctcagcctc ccaagtagct gggactacag gtgcatgcca ccatgcctgg ctaatttttt 13740
gtatttttag tagaaaattt tcagcttcac ctcttttgaa tttctgctct cctgcctgtt 13800
ctttagctat ccgtggtcct gaaccagtta tgtgtgttgc atgagaaaac gccagtaagt 13860
gacagagtca ccaaatgctg cacagaatcc ttggtgaaca ggcgaccatg cttttcagct 13920
ctggaagtcg atgaaacata cgttcccaaa gagtttaatg ctgaaacatt caccttccat 13980
gcagatatat gcacactttc tgagaaggag agacaaatca agaaacaaac gtgaggagta 14040
tttcattact gcatgtgttt gtagtcttga tagcaagaac tgtcaattca agctagcaac 14100
tttttcctga agtagtgatt atatttctta gaggaaagta ttggagtgtt gcccttatta 14160
tgctgataag agtacccaga ataaaatgaa taacttttta aagacaaaat cctctgttat 14220
aatattgcta aaattattca gagtaatatt gtggattaaa gccacaatag aataacatgt 14280
taggccatat tcagtagaaa aagatgaaca attaactgat aaatttgtgc acatggcaaa 14340
ttagttaatg ggaaccatag gagaatttat ttctagatgt aaataattat tttaagtttg 14400
ccctatggtg gccccacaca tgagacaaac ccccaagatg tgacttttga gaatgagact 14460
tggataaaaa acatgtagaa atgcaagccc tgaagctcaa ctccctattg ctatcacagg 14520
ggttataatt gcataaaatt tagctataga aagttgctgt catctcttgt gggctgtaat 14580
catcgtctag gcttaagagt aatattgcaa aacctgtcat gcccacacaa atctctccct 14640
ggcattgttg tctttgcaga tgtcagtgaa agagaaccag cagctcccat gagtttggat 14700
agccttattt tctatagcct ccccactatt agctttgaag ggagcaaagt ttaagaacca 14760
aatataaagt ttctcatctt tatagatgag aaaaatttta aataaagtcc aagataatta 14820
aatttttaag gatcattttt agctctttaa tagcaataaa actcaatatg acataatatg 14880
gcacttccaa aatctgaata atatataatt gcaatgacat acttcttttc agagatttac 14940
tgaaaagaaa tttgttgaca ctacataacg tgatgagtgg tttatactga ttgtttcagt 15000
tggtcttccc accaactcca tgaaagtgga ttttattatc ctcatcatgc agatgagaat 15060
attgagactt atagcggtat gcctgagccc caaagtactc agagttgcct ggctccaaga 15120
tttataatct taaatgatgg gactaccatc cttactctct ccatttttct atacgtgagt 15180
aatgtttttt ctgttttttt tttttctttt tccattcaaa ctcagtgcac ttgttgagct 15240
cgtgaaacac aagcccaagg caacaaaaga gcaactgaaa gctgttatgg atgatttcgc 15300
agcttttgta gagaagtgct gcaaggctga cgataaggag acctgctttg ccgaggaggt 15360
actacagttc tcttcatttt aatatgtcca gtattcattt ttgcatgttt ggttaggcta 15420
gggcttaggg atttatatat caaaggaggc tttgtacatg tgggacaggg atcttatttt 15480
acaaacaatt gtcttacaaa atgaataaaa cagcactttg tttttatctc ctgctctatt 15540
gtgccatact gttaaatgtt tataatgcct gttctgtttc caaatttgtg atgcttatga 15600
atattaatag gaatatttgt aaggcctgaa atattttgat catgaaatca aaacattaat 15660
ttatttaaac atttacttga aatgtggtgg tttgtgattt agttgatttt ataggctagt 15720
gggagaattt acattcaaat gtctaaatca cttaaaattg ccctttatgg cctgacagta 15780
actttttttt attcatttgg ggacaactat gtccgtgagc ttccgtccag agattatagt 15840
agtaaattgt aattaaagga tatgatgcac gtgaaatcac tttgcaatca tcaatagctt 15900
cataaatgtt aattttgtat cctaatagta atgctaatat tttcctaaca tctgtcatgt 15960
ctttgtgttc agggtaaaaa acttgttgct gcaagtcaag ctgccttagg cttataacat 16020
cacatttaaa agcatctcag gtaactatat tttgaatttt ttaaaaaagt aactataata 16080
gttattatta aaatagcaaa gattgaccat ttccaagagc catatagacc agcaccgacc 16140
actattctaa actatttatg tatgtaaata ttagctttta aaattctcaa aatagttgct 16200
gagttgggaa ccactattat ttctattttg tagatgagaa aatgaagata aacatcaaag 16260
catagattaa gtaattttcc aaagggtcaa aattcaaaat tgaaaccaaa gtttcagtgt 16320
tgcccattgt cctgttctga cttatatgat gcggtacaca gagccatcca agtaagtgat 16380
ggctcagcag tggaatactc tgggaattag gctgaaccac atgaaagagt gctttatagg 16440
gcaaaaacag ttgaatatca gtgatttcac atggttcaac ctaatagttc aactcatcct 16500
ttccattgga gaatatgatg gatctacctt ctgtgaactt tatagtgaag aatctgctat 16560
tacatttcca atttgtcaac atgctgagct ttaataggac ttatcttctt atgacaacat 16620
ttattggtgt gtccccttgc ctagcccaac agaagaattc agcagccgta agtctaggac 16680
aggcttaaat tgttttcact ggtgtaaatt gcagaaagat gatctaagta atttggcatt 16740
tattttaata ggtttgaaaa acacatgcca ttttacaaat aagacttata tttgtccttt 16800
tgtttttcag cctaccatga gaataagaga aagaaaatga agatcaaaag cttattcatc 16860
tgtttttctt tttcgttggt gtaaagccaa caccctgtct aaaaaacata aatttcttta 16920
atcattttgc ctcttttctc tgtgcttcaa ttaataaaaa atggaaagaa tctaatagag 16980
tggtacagca ctgttatttt tcaaagatgt gttgctatcc tgaaaattct gtaggttctg 17040
tggaagttcc agtgttctct cttattccac ttcggtagag gatttctagt ttcttgtggg 17100
ctaattaaat aaatcattaa tactcttcta agttatggat tataaacatt caaaataata 17160
ttttgacatt atgataattc tgaataaaag aacaaaaacc atggtatagg taaggaatat 17220
aaaacatggc ttttacctta gaaaaaacaa ttctaaaatt catatggaat caaaaaagag 17280
cctgcagaac caaagtaaga ctaagcaaaa agaacaaatt acctgatttc aaactacact 17340
ataaggccat agtcaccgaa acagcaaggt actggtataa actcgagata acttcgtata 17400
atgtatgcta tacgaagtta tgctaggtaa ctataacggt cctaaggtag cgagctagca 17460
cacatcacaa ccacaacctt ctcaggtaac tatacttggg acttaaaaaa cataatcata 17520
atcatttttc ctaaaacgat caagactgat aaccatttga caagagccat acagacaagc 17580
accagctggc actcttaggt cttcacgtat ggtcatcagt ttgggttcca tttgtagata 17640
agaaactgaa catataaagg tctaggttaa tgcaatttac acaaaaggag accaaaccag 17700
ggagagaagg aaccaaaatt aaaaattcaa accagagcaa aggagttagc cctggttttg 17760
ctctgact 17768
<210> 19
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(60)
<223> Мышиная последовательность
<220>
<221> прочий_признак
<222> (61)..(120)
<223> Человеческая последовательность
<400> 19
agagcgagtc tttctgcaca cagatcacct ttcctatcaa ccccactagc ctctggcaaa 60
atgaagtggg taacctttat ttcccttctt tttctcttta gctcggctta ttccaggggt 120
<210> 20
<211> 160
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(60)
<223> Человеческая последовательность
<220>
<221> прочий_признак
<222> (61)..(66)
<223> XhoI
<220>
<221> прочий_признак
<222> (67)..(100)
<223> LoxP
<220>
<221> прочий_признак
<222> (101)..(160)
<223> Кассета
<400> 20
cctgatttca aactacacta taaggccata gtcaccgaaa cagcaaggta ctggtataaa 60
ctcgagataa cttcgtataa tgtatgctat acgaagttat atgcatgcca gtagcagcac 120
ccacgtccac cttctgtcta gtaatgtcca acacctccct 160
<210> 21
<211> 192
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(60)
<223> Кассета
<220>
<221> прочий_признак
<222> (61)..(94)
<223> LoxP
<220>
<221> прочий_признак
<222> (101)..(126)
<223> I-CeuI
<220>
<221> прочий_признак
<222> (127)..(132)
<223> NheI
<220>
<221> прочий_признак
<222> (133)..(192)
<223> Мышиная последовательность
<400> 21
cattctcagt attgttttgc caagttctaa ttccatcaga cctcgacctg cagcccctag 60
ataacttcgt ataatgtatg ctatacgaag ttatgctagg taactataac ggtcctaagg 120
tagcgagcta gcacacatca caaccacaac cttctcaggt aactatactt gggacttaaa 180
aaacataatc at 192
<210> 22
<211> 198
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(60)
<223> Человеческая последовательность
<220>
<221> прочий_признак
<222> (61)..(66)
<223> XhoI
<220>
<221> прочий_признак
<222> (67)..(100)
<223> LoxP
<220>
<221> прочий_признак
<222> (107)..(132)
<223> I-CeuI
<220>
<221> прочий_признак
<222> (133)..(138)
<223> NheI
<220>
<221> прочий_признак
<222> (139)..(198)
<223> Мышиная последовательность
<400> 22
cctgatttca aactacacta taaggccata gtcaccgaaa cagcaaggta ctggtataaa 60
ctcgagataa cttcgtataa tgtatgctat acgaagttat gctaggtaac tataacggtc 120
ctaaggtagc gagctagcac acatcacaac cacaaccttc tcaggtaact atacttggga 180
cttaaaaaac ataatcat 198
<210> 23
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 23
gtaaccttta tttcccttct ttttctctt 29
<210> 24
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 24
agctcggctt attc 14
<210> 25
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 25
cgtgcatctc gacgaaacac 20
<210> 26
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 26
gcagaaccaa agtaagacta agcaaa 26
<210> 27
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 27
agaacaaatt acctgatttc 20
<210> 28
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 28
tgtttcggtg actatggcct tat 23
<210> 29
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 29
gccgagaagc acgtaagagt tt 22
<210> 30
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 30
atgttttttc atctctgctt gt 22
<210> 31
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 31
aataccaggc ttccattact agaaaaa 27
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 32
ccctcccatg gcctaacaac 20
<210> 33
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 33
ttgggcacaa cagatgtcag agagc 25
<210> 34
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 34
acgtgccttg cattgctta 19
<210> 35
<211> 17335
<212> ДНК
<213> Homo sapiens
<400> 35
atgaagtggg taacctttat ttcccttctt tttctcttta gctcggctta ttccaggggt 60
gtgtttcgtc gagatgcacg taagaaatcc atttttctat tgttcaactt ttattctatt 120
ttcccagtaa aataaagttt tagtaaactc tgcatcttta aagaattatt ttggcattta 180
tttctaaaat ggcatagtat tttgtatttg tgaagtctta caaggttatc ttattaataa 240
aattcaaaca tcctaggtaa aaaaaaaaaa aggtcagaat tgtttagtga ctgtaatttt 300
cttttgcgca ctaaggaaag tgcaaagtaa cttagagtga ctgaaacttc acagaatagg 360
gttgaagatt gaattcataa ctatcccaaa gacctatcca ttgcactatg ctttatttaa 420
aaaccacaaa acctgtgctg ttgatctcat aaatagaact tgtatttata tttattttca 480
ttttagtctg tcttcttggt tgctgttgat agacactaaa agagtattag atattatcta 540
agtttgaata taaggctata aatatttaat aatttttaaa atagtattct tggtaattga 600
attattcttc tgtttaaagg cagaagaaat aattgaacat catcctgagt ttttctgtag 660
gaatcagagc ccaatatttt gaaacaaatg cataatctaa gtcaaatgga aagaaatata 720
aaaagtaaca ttattacttc ttgttttctt cagtatttaa caatcctttt ttttcttccc 780
ttgcccagac aagagtgagg ttgctcatcg gtttaaagat ttgggagaag aaaatttcaa 840
agccttgtaa gttaaaatat tgatgaatca aatttaatgt ttctaatagt gttgtttatt 900
attctaaagt gcttatattt ccttgtcatc agggttcaga ttctaaaaca gtgctgcctc 960
gtagagtttt ctgcgttgag gaagatattc tgtatctggg ctatccaata aggtagtcac 1020
tggtcacatg gctattgagt acttcaaata tgacaagtgc aactgagaaa caaaaactta 1080
aattgtattt aattgtagtt aatttgaatg tatatagtca catgtggcta atggctactg 1140
tattggacag tacagctctg gaacttgctt ggtggaaagg actttaatat aggtttcctt 1200
tggtggctta cccactaaat cttctttaca tagcaagcat tcctgtgctt agttgggaat 1260
atttaatttt tttttttttt taagacaggg tctcgctctg tcgcccaggc tggagtgcag 1320
tggcgcaatc tcggctcact gcaaactccg cctcccgggt tcacgccatt ctcctgcctc 1380
agcctcccga gtagctggga ctacaggcgc ccgccatcac gcccggctaa tcttttgtat 1440
ttttagtaga gatggggttt caccgtgtgc caggatggtc tcaatctcct gacatcgtga 1500
tctgcccacc tcggcctccc aaagtgctgg gattacagga gtgagccacc gcgcccggcc 1560
tatttaaatg ttttttaatc tagtaaaaaa tgagaaaatt gtttttttaa aagtctacct 1620
aatcctacag gctaattaaa gacgtgtgtg gggatcaggt gcggtggttc acacctgtaa 1680
tcccagcact ttggaaggct gatgcaggag gattgcttga gcccaggagt tcaagaccag 1740
cctgggcaag tctctttaaa aaaaacaaaa caaacaaaca aaaaaattag gcatggtggc 1800
acatgcctgt agtcctagct acttaggagg ctgacgtagg aggatcgttt ggacctgaga 1860
ggtcaaggct acagtgagcc atgattgtgc cactgcactc cagcctgggt gacagagtga 1920
gactctgtct caaaaaagaa aaaggaaatc tgtggggttt gttttagttt taagtaattc 1980
taaggacttt aaaaatgcct agtcttgaca attagatcta tttggcatac aatttgcttg 2040
cttaatctat gtgtgtgcat agatctactg acacacgcat acatataaac attagggaac 2100
taccattctc tttgcgtagg aagccacata tgcctatcta ggcctcagat catacctgat 2160
atgaataggc tttctggata atggtgaaga agatgtataa aagatagaac ctatacccat 2220
acatgatttg ttctctagcg tagcaacctg ttacatatta aagttttatt atactacatt 2280
tttctacatc ctttgtttca gggtgttgat tgcctttgct cagtatcttc agcagtgtcc 2340
atttgaagat catgtaaaat tagtgaatga agtaactgaa tttgcaaaaa catgtgttgc 2400
tgatgagtca gctgaaaatt gtgacaaatc acttgtaagt acattctaat tgtggagatt 2460
ctttcttctg tttgaagtaa tcccaagcat ttcaaaggaa ttttttttaa gttttctcaa 2520
ttattattaa gtgtcctgat ttgtaagaaa cactaaaaag ttgctcatag actgataagc 2580
cattgtttct tttgtgatag agatgcttta gctatgtcca cagttttaaa atcatttctt 2640
tattgagacc aaacacaaca gtcatggtgt atttaaatgg caatttgtca tttataaaca 2700
cctcttttta aaatttgagg tttggtttct ttttgtagag gctaataggg atatgatagc 2760
atgtatttat ttatttattt atcttatttt attatagtaa gaacccttaa catgagatct 2820
accctgttat atttttaagt gtacaatcca ttattgttaa ctacgggtac actgttgtat 2880
agcttactca tcttgctgta ttaaaacttt gtgcccattg attagtaacc cctcgtttcg 2940
tcctccccca gccactggca accagcatta tactctttga ttctatgagt ttgactactt 3000
tagctacctt atataagtgg tattatgtac tgtttatctt tttatgactg acttatttcc 3060
cttagcatag tgcattcaaa gtccaaccat gttgttgcct attgcagaat ttccttcttt 3120
tcaaggctga ataatattcc agtgcatgtg tgtaccacat tttctttatc cattaatttg 3180
ttgattgata gacatttagg ttggttttct acatcttgac tatcatgaat agtgttgcaa 3240
tgaacacagg agagctacta tctcttagag atgatatcat ggtttttatc atcagaaaac 3300
acccactgat ttctatgcta attttgttac ctgggtggaa taatagtaca gctatatatt 3360
cctcatttta gatatctttg tatttctaca tacaataaaa aagcagagta cttagtcatg 3420
ttgaagaact ttaaactttt agtatttcca gatcaatctt caaaacaagg acaggtttat 3480
ctttctctca ccactcaatc tatatatacc tcttgtgggc aaggccagtt tttatcactg 3540
gagcctttcc cctttttatt atgtacctct ccctcacagc agagtcagga ctttaacttt 3600
acacaatact atggctctac atatgaaatc ttaaaaatac ataaaaatta ataaattctg 3660
tctagagtag tatattttcc ctggggttac agttactttc ataataaaaa ttagagataa 3720
ggaaaggact catttattgg aaagtgattt taggtaacat ttctggaaga aaaatgtcta 3780
tatcttaata gtcacttaat atatgatgga ttgtgttact cctcagtttt caatggcata 3840
tactaaaaca tggccctcta aaaagggggc aaatgaaatg agaaactctc tgaatgtttt 3900
tctcccctag gtgaattcac ctgctgctta gaagcttatt ttctcttgat ttctgttata 3960
atgattgctc ttacccttta gttttaagtt tcaaaatagg agtcatataa ctttccttaa 4020
agctattgac tgtctttttg tcctgtttta ttcaccatga gttatagtgt gacagttaat 4080
tcttatgaaa attatataga gatggttaaa tcatcagaaa ctgtaaacct cgattgggag 4140
gggaagcgga tttttaaatg atttcctgac caagcttaac cagtatatta aatcctttgt 4200
actgttcttt ggctataaag aaaaaaggta ctgtccagca actgaaacct gctttcttcc 4260
atttagcata ccctttttgg agacaaatta tgcacagttg caactcttcg tgaaacctat 4320
ggtgaaatgg ctgactgctg tgcaaaacaa gaacctgaga gaaatgaatg cttcttgcaa 4380
cacaaagatg acaacccaaa cctcccccga ttggtgagac cagaggttga tgtgatgtgc 4440
actgcttttc atgacaatga agagacattt ttgaaaaagt aagtaatcag atgtttatag 4500
ttcaaaatta aaaagcatgg agtaactcca taggccaaca ctctataaaa attaccataa 4560
caaaaatatt ttcaacatta agacttggaa gttttgttat gatgattttt taaagaagta 4620
gtatttgata ccacaaaatt ctacacagca aaaaatatga tcaaagatat tttgaagttt 4680
attgaaacag gatacaatct ttctgaaaaa tttaagatag acaaattatt taatgtatta 4740
cgaagatatg tatatatggt tgttataatt gatttcgttt tagtcagcaa cattatattg 4800
ccaaaattta accatttatg cacacacaca cacacacaca cacacttaac ccttttttcc 4860
acatacttaa agaatgacag agacaagacc atcatgtgca aattgagctt aattggttaa 4920
ttagatatct ttggaatttg gaggttctgg ggagaatgtc gattacaatt atttctgtaa 4980
tattgtctgc tatagaaaag tgactgtttt tctttttcaa aatttagata cttatatgaa 5040
attgccagaa gacatcctta cttttatgcc ccggaactcc ttttctttgc taaaaggtat 5100
aaagctgctt ttacagaatg ttgccaagct gctgataaag ctgcctgcct gttgccaaag 5160
gtattatgca aaagaataga aaaaaagagt tcattatcca acctgatttt gtccattttg 5220
tggctagatt tagggaacct gagtgtctga tacaaacttt ccgacatggt caaaaaagcc 5280
ttccttttat ctgtcttgaa aatctttcat ctttgaaggc ctacactctc gtttcttctt 5340
ttaagatttg ccaatgatga tctgtcagag gtaatcactg tgcatgtgtt taaagatttc 5400
accacttttt atggtggtga tcactatagt gaaatactga aacttgtttg tcaaattgca 5460
cagcaagggg ccacagttct tgtttatctt ttcatgataa tttttagtag ggagggaatt 5520
caaagtagag aattttactg catctagatg cctgagttca tgcattcatt ccataaatat 5580
atattatgga atgctttatt ttcttttctg aggagtttac tgatgttggt ggaggagaga 5640
ctgaaatgaa ttatacacaa aatttaaaaa ttagcaaaat tgcagcccct gggatattag 5700
cgtactcttt ctctgacttt tctcccactt ttaaggctct ttttcctggc aatgtttcca 5760
gttggtttct aactacatag ggaattccgc tgtgaccaga atgatcgaat gatctttcct 5820
tttcttagag agcaaaatca ttattcgcta aagggagtac ttgggaattt aggcataaat 5880
tatgccttca aaatttaatt tggcacagtc tcatctgagc ttatggaggg gtgtttcatg 5940
tagaattttt cttctaattt tcatcaaatt attccttttt gtagctcgat gaacttcggg 6000
atgaagggaa ggcttcgtct gccaaacaga gactcaagtg tgccagtctc caaaaatttg 6060
gagaaagagc tttcaaagca tggtaaatac ttttaaacat agttggcatc tttataacga 6120
tgtaaatgat aatgcttcag tgacaaattg tacattttta tgtattttgc aaagtgctgt 6180
caaatacatt tctttggttg tctaacaggt agaactctaa tagaggtaaa aatcagaata 6240
tcaatgacaa tttgacatta tttttaatct tttcttttct aaatagttga ataatttaga 6300
ggacgctgtc ctttttgtcc taaaaaaagg gacagatatt taagttctat ttatttataa 6360
aatcttggac tcttattcta atggttcatt atttttatag agctgtaggc atggttcttt 6420
atttaatttt ttaaagttat ttttaatttt tgtggataca gagtaggtat acatatttac 6480
ggggtatatg agatattttg atataagtat acaacatata taatcccttt atttaatttt 6540
atcttccccc caatgatcta aaactatttg cttgtccttt tatgtcttat agttaaattc 6600
agtcaccaac taagttgaag ttacttctta tttttgcata gctccagctc tgatcttcat 6660
ctcatgtttt tgcctgagcc tctgttttca tattacttag ttggttctgg gagcatactt 6720
taatagccga gtcaagaaaa atactagctg ccccgtcacc cacactcctc acctgctagt 6780
caacagcaaa tcaacacaac aggaaataaa atgaaaataa tagacattat gcatgctctc 6840
tagaaactgt caattgaact gtatttgctc atcattccta ccatctacac caccaaaatc 6900
aaccaaattt atgaaaaaaa acagccccaa cataaaatta tacacagata aacaggctat 6960
gattggtttt gggaaagaag tcacctttac ctgatttagg caactgtgaa atgactagag 7020
aatgaagaaa attagacgtt tacatcttgt catagagttt gaagatagtg ctggatcttt 7080
ctttttataa gtaagatcaa taaaaactcc ctcattctgt agaagttatg atttcttttc 7140
taagagacct ttagaagtca gaaaaaatgt gtttcaattg agaaaaaaga taactggagt 7200
ttgtgtagta cttcccagat tataaaatgc ttttgtatgt attatctaat ttaatcctca 7260
aaacttcttc aatttagcat gttgtcatga cactgcagag gctgaagctc agagaggctg 7320
agccctctgc taacaagtcc tactgctaac aagtgataaa gccagagctg gaagtcacat 7380
ctggactcca aacctgatgc ttctcagcct gttgcccctt ttagagttcc tttttaattt 7440
ctgcttttat gacttgctag atttctacct accacacaca ctcttaaatg gataattctg 7500
ccctaaggat aagtgattac catttggttc agaactagaa ctaatgaatt ttaaaaatta 7560
tttctgtatg tccattttga attttcttat gagaaatagt atttgcctag tgttttcata 7620
taaaatatcg catgataata ccattttgat tggcgatttt ctttttaggg cagtagctcg 7680
cctgagccag agatttccca aagctgagtt tgcagaagtt tccaagttag tgacagatct 7740
taccaaagtc cacacggaat gctgccatgg agatctgctt gaatgtgctg atgacagggt 7800
aaagagtcgt cgatatgctt tttggtagct tgcatgctca agttggtaga atggatgcgt 7860
ttggtatcat tggtgatagc tgacagtggg ttgagattgt cttctgtgct ttcgtctgtc 7920
ctatcttcaa tctttccctg cctatggtgg tggtaccttt ctgtttttaa cctggctata 7980
aattaccaga taaacccatt cactgatttg taactccttt cagtcatgct ctaactgtaa 8040
atgaaggctt aaactgaagt agaacagtta caaggtttta cttggcagaa catcttgcaa 8100
ggtagatgtc taagaagatt tttttttctt tttttaagac agagtttcgc tcttgtttcc 8160
caggctgggg tgcaatggtg tgatcttggc tcagcgcaac ctctgcctcc tgggttcaag 8220
tgattctcat gcctcagcct cccaagtagc tgggattaca ggcatgcgcc accacacctg 8280
gctaattttg tatttttagt agaggcgggg tttcaccata ttgtccagac tggtctcgaa 8340
ctcctgacct caggtgatcc acccgccttg gcctcccaaa gtgctgggat tacaggcatg 8400
agccaccttg cccagcctaa gaagattttt tgagggaggt aggtggactt ggagaaggtc 8460
actacttgaa gagatttttg gaaatgatgt atttttcttc tctatattcc ttcccttaat 8520
taactctgtt tgttagatgt gcaaatattt ggaatgatat ctcttttctc aaaacttata 8580
atattttctt tctccctttc ttcaagatta aacttatggg caaatactag aatcctaatc 8640
tctcatggca ctttctggaa aatttaaggc ggttatttta tatatgtaag cagggcctat 8700
gactatgatc ttgactcatt tttcaaaaat cttctatatt ttatttagtt atttggtttc 8760
aaaaggcctg cacttaattt tgggggatta tttggaaaaa cagcattgag ttttaatgaa 8820
aaaaacttaa atgccctaac agtagaaaca taaaattaat aaataactga gctgagcacc 8880
tgctactgat tagtctattt taattaagtg ggaatgtttt tgtagtccta tctacatctc 8940
caggtttagg agcaaacaga gtatgttcat agaaggaata tgtgtatggt cttagaatac 9000
aatgaatatg ttctgccaac ttaataaagg tctgaggaga aagtgtagca atgtcaattc 9060
gtgttgaaca atttccacca acttacttat aggcggacct tgccaagtat atctgtgaaa 9120
atcaagattc gatctccagt aaactgaagg aatgctgtga aaaacctctg ttggaaaaat 9180
cccactgcat tgccgaagtg gaaaatgatg agatgcctgc tgacttgcct tcattagctg 9240
ctgattttgt tgaaagtaag gatgtttgca aaaactatgc tgaggcaaag gatgtcttcc 9300
tgggcatgta agtagataag aaattattct tttatagctt tggcatgacc tcacaactta 9360
ggaggatagc ctaggctttt ctgtggagtt gctacaattt ccctgctgcc cagaatgttt 9420
cttcatcctt ccctttccca ggctttaaca atttttgaaa tagttaatta gttgaataca 9480
ttgtcataaa ataatacatg ttcatggcaa agctcaacat tccttactcc ttaggggtat 9540
ttctgaaaat acgtctagaa acattttgtg tatatataaa ttatgtatac ttcagtcatt 9600
cattccaagt gtatttcttg aacatctata atatatgtgt gtgactatgt attgcctgtc 9660
tatctaacta atctaatcta atctagtcta tctatctaat ctatgcaatg atagcaaaga 9720
agtataaaaa gaaatataga gtctgacacc aggtgcttta tatttggtga aaagaccaga 9780
agttcagtat aatggcaata tggtaggcaa ctcaattaca aaataaatgt ttacatattg 9840
tcagaagttg tggtgataaa ctgcattttt gttgttggat tatgataatg cactaaataa 9900
tatttcctaa aattatgtac cctacaagat ttcactcata cagagaagaa agagaatatt 9960
ttaagaacat atctctgccc atctatttat cagaatcctt ttgagatgta gtttaaatca 10020
aacaaaatgt taataaaaat aacaagtatc attcatcaaa gacttcatat gtgccaagca 10080
gtgtgtgctt tgtgtagatt atgtcatata gttctcataa tccaccttcc gagacagata 10140
ctatttattt tttgagacag agttttactc ttgttgccca ggctggagtg caatggtgcc 10200
atctcggctc accacaacct ccgcctccca ggttcaagcg attctcctgc ctcagcctcc 10260
tgggattaca ggcatgcacc accatgcctg gctaattttg tatttttagt agagatgggg 10320
tttcaccatg ttggtcagac tggtctcaaa ctcctgacct ctggtgatat gcctgcctca 10380
gcctcctaaa gtgctgggat tacaggcatg agccactgtg cccagccgac agatactatt 10440
attatttcca ttctaccgag aaggagacta aggctctgat catttaaata agttgcctaa 10500
ggtgatgcag tgatataagt agcagagcta ggaattgagc cttggtaact ttaactctgg 10560
accccaagtc cttagctact aagctttact gcatggggtt tagtcaaatt aagacttttg 10620
gaatatgagt tacttttgag attagctttg tgatattttt tgtgctcatt tgtccaacaa 10680
agtctatttt attttcatct taattaggtt tttgtatgaa tatgcaagaa ggcatcctga 10740
ttactctgtc gtgctgctgc tgagacttgc caagacatat gaaaccactc tagagaagtg 10800
ctgtgccgct gcagatcctc atgaatgcta tgccaaagtg gtaggtttat tgttggaaaa 10860
aaatgtagtt ctttgactga tgattccaat aatgagaaag aaaaataatg caagaatgta 10920
aaatgatata cagtgcaatt tagatctttt cttgagatgg tttcaattct ggaatcttaa 10980
acatgaaaga aaaagtagcc ttagaatgat taacaaaatt tagactagtt agaatagaaa 11040
gatctgaata gagcaatctc taaaaaattt tgatcttttt ttctcttttt cacaatcctg 11100
agaacaaaaa aaaattaaat ttaaatgtta attagaagat atttaactta gatgtaaagt 11160
gagttaacct gattccagga ttaatcaagt actagaatta gtatcttatg gcaaattata 11220
gaacctatcc ctttagaata ttttcaaatc tttttgagga tgtttaggaa tagttttaca 11280
agaaattaag ttaggagagg aaatctgttc tggaggattt ttagggttcc cactagcata 11340
tgtaatggtt tctgaactat tcagaatcag agaaaactca tttttcctgc tttcaagaag 11400
ctactgtatg ccaggcacca tgcacaaaca atgaccaacg taaaatctct cattttggag 11460
agcctggaat ctaactggaa aggtgaacta ataataataa tatgtacaat catagccatc 11520
atttattaaa cttttattat atgcaaggca ctgtttaatt tcattagctt acctggttta 11580
cagagcagct ctatgagatg agtgccatct ttgcccctat tttagggata aggattctga 11640
aatgtggaga tggtaagtaa aattgcacaa ctgaagaatg agttacatga cttggctcaa 11700
atactggtca ttgaactcca gagcctgaat attcttaacc acttacatga tgcaagctca 11760
ccaaataaat agttcgaatg tattgtgaca gagcggcatt gatattcatc tattcatgtg 11820
gctttgagta ggaagaagaa aggatatcat tctgaccaga ggggtgaaaa acaacctgca 11880
tctgatcctg aggcataata ctattaacac aattctttta tgtttcagtt cgatgaattt 11940
aaacctcttg tggaagagcc tcagaattta atcaaacaaa attgtgagct ttttgagcag 12000
cttggagagt acaaattcca gaatgcgtaa gtaattttta ttgactgatt ttttttatca 12060
atttgtaatt atttaagact taatatatga gccacctagc atagaacttt taagaatgaa 12120
aatacattgc atatttctaa tcactctttg tcaagaaaga taggagagga gagataaaat 12180
agttgatggg gtggagaggt ctatatttga atgtagtcta aaaattgttc tcttaagatt 12240
ggaagtatgt aggctgggag ggtaaatacc aaatcttggt atatcagaac tgagcatgtc 12300
ccttgaaggt taagaaatag ttaatgggca aatagagcat ggcaatattt tgtagagcag 12360
caagtagtag gccttgaata gatgtcgctc aaaaagtaat atgtaagctg aacacaaaaa 12420
tgtaacaaat gaatttagat acatatttga atattaaatt caggttgttt gggagatgca 12480
cctagtcttt gatggttaaa cctttccctc catagaagag acagagacag aatggcttgc 12540
tggactaatg tcccaattca atagagtctt atctatgaag gttaaaaaca agaagagaca 12600
tattatacag tagatattta ttgtgtggct catacacatg gtgctcttct gattatggat 12660
tttagagata ataacagtga acaagacata gtttctttcc tcgagtagat taaagtcata 12720
cattgacttt taatggtgac tggcattctt aatacatgat tattatatat taggtaccat 12780
gtcagattaa ttataatact ttactacttt taatttaacc cttgaactat ccctattgag 12840
tcagatatat ttccttccat tttctacttg tatctttcaa gtttagcata tgctgataca 12900
tatgaagctc tctccaggtt ttattgaaag aagaaattaa taaatttatt aatgtcactg 12960
aattaggcaa ctcactttcc caagattatg caagtggtac aggtggaact caaagccaag 13020
tttaactagt tgttcaggag aatgttttct accctccact aacccactac tctgcagatg 13080
gagataatat gatgaatgga acatagcaac atcttagttg attccggcca agtgttctct 13140
gttttatcta ctatgttaga cagtttcttg ccttgctgaa aacacatgac ttcttttttt 13200
caggctatta gttcgttaca ccaagaaagt accccaagtg tcaactccaa ctcttgtaga 13260
ggtctcaaga aacctaggaa aagtgggcag caaatgttgt aaacatcctg aagcaaaaag 13320
aatgccctgt gcagaagact atgtgagtct ttaaaaaaat ataataaatt aataatgaaa 13380
aaattttacc tttagatatt gataatgcta gctttcataa gcagaaggaa gtaatgtgtg 13440
tgtgtgcatg tttgtgtgca tgtgtgtgtg catgcacgtg tgtgtatgtg tgatattggc 13500
agtcaaggcc ccgaggatga taattttttt tttttttttg agacggagtc tcgctttgtt 13560
gtccaggctg gagtgcagtg gtgccatctc ggctcactgc aacctccgcc tcccaggttc 13620
aagccattct cctgcctcag cctcccaagt agctgggact acaggtgcat gccaccatgc 13680
ctggctaatt ttttgtattt ttagtagaaa attttcagct tcacctcttt tgaatttctg 13740
ctctcctgcc tgttctttag ctatccgtgg tcctgaacca gttatgtgtg ttgcatgaga 13800
aaacgccagt aagtgacaga gtcaccaaat gctgcacaga atccttggtg aacaggcgac 13860
catgcttttc agctctggaa gtcgatgaaa catacgttcc caaagagttt aatgctgaaa 13920
cattcacctt ccatgcagat atatgcacac tttctgagaa ggagagacaa atcaagaaac 13980
aaacgtgagg agtatttcat tactgcatgt gtttgtagtc ttgatagcaa gaactgtcaa 14040
ttcaagctag caactttttc ctgaagtagt gattatattt cttagaggaa agtattggag 14100
tgttgccctt attatgctga taagagtacc cagaataaaa tgaataactt tttaaagaca 14160
aaatcctctg ttataatatt gctaaaatta ttcagagtaa tattgtggat taaagccaca 14220
atagaataac atgttaggcc atattcagta gaaaaagatg aacaattaac tgataaattt 14280
gtgcacatgg caaattagtt aatgggaacc ataggagaat ttatttctag atgtaaataa 14340
ttattttaag tttgccctat ggtggcccca cacatgagac aaacccccaa gatgtgactt 14400
ttgagaatga gacttggata aaaaacatgt agaaatgcaa gccctgaagc tcaactccct 14460
attgctatca caggggttat aattgcataa aatttagcta tagaaagttg ctgtcatctc 14520
ttgtgggctg taatcatcgt ctaggcttaa gagtaatatt gcaaaacctg tcatgcccac 14580
acaaatctct ccctggcatt gttgtctttg cagatgtcag tgaaagagaa ccagcagctc 14640
ccatgagttt ggatagcctt attttctata gcctccccac tattagcttt gaagggagca 14700
aagtttaaga accaaatata aagtttctca tctttataga tgagaaaaat tttaaataaa 14760
gtccaagata attaaatttt taaggatcat ttttagctct ttaatagcaa taaaactcaa 14820
tatgacataa tatggcactt ccaaaatctg aataatatat aattgcaatg acatacttct 14880
tttcagagat ttactgaaaa gaaatttgtt gacactacat aacgtgatga gtggtttata 14940
ctgattgttt cagttggtct tcccaccaac tccatgaaag tggattttat tatcctcatc 15000
atgcagatga gaatattgag acttatagcg gtatgcctga gccccaaagt actcagagtt 15060
gcctggctcc aagatttata atcttaaatg atgggactac catccttact ctctccattt 15120
ttctatacgt gagtaatgtt ttttctgttt tttttttttc tttttccatt caaactcagt 15180
gcacttgttg agctcgtgaa acacaagccc aaggcaacaa aagagcaact gaaagctgtt 15240
atggatgatt tcgcagcttt tgtagagaag tgctgcaagg ctgacgataa ggagacctgc 15300
tttgccgagg aggtactaca gttctcttca ttttaatatg tccagtattc atttttgcat 15360
gtttggttag gctagggctt agggatttat atatcaaagg aggctttgta catgtgggac 15420
agggatctta ttttacaaac aattgtctta caaaatgaat aaaacagcac tttgttttta 15480
tctcctgctc tattgtgcca tactgttaaa tgtttataat gcctgttctg tttccaaatt 15540
tgtgatgctt atgaatatta ataggaatat ttgtaaggcc tgaaatattt tgatcatgaa 15600
atcaaaacat taatttattt aaacatttac ttgaaatgtg gtggtttgtg atttagttga 15660
ttttataggc tagtgggaga atttacattc aaatgtctaa atcacttaaa attgcccttt 15720
atggcctgac agtaactttt ttttattcat ttggggacaa ctatgtccgt gagcttccgt 15780
ccagagatta tagtagtaaa ttgtaattaa aggatatgat gcacgtgaaa tcactttgca 15840
atcatcaata gcttcataaa tgttaatttt gtatcctaat agtaatgcta atattttcct 15900
aacatctgtc atgtctttgt gttcagggta aaaaacttgt tgctgcaagt caagctgcct 15960
taggcttata acatcacatt taaaagcatc tcaggtaact atattttgaa ttttttaaaa 16020
aagtaactat aatagttatt attaaaatag caaagattga ccatttccaa gagccatata 16080
gaccagcacc gaccactatt ctaaactatt tatgtatgta aatattagct tttaaaattc 16140
tcaaaatagt tgctgagttg ggaaccacta ttatttctat tttgtagatg agaaaatgaa 16200
gataaacatc aaagcataga ttaagtaatt ttccaaaggg tcaaaattca aaattgaaac 16260
caaagtttca gtgttgccca ttgtcctgtt ctgacttata tgatgcggta cacagagcca 16320
tccaagtaag tgatggctca gcagtggaat actctgggaa ttaggctgaa ccacatgaaa 16380
gagtgcttta tagggcaaaa acagttgaat atcagtgatt tcacatggtt caacctaata 16440
gttcaactca tcctttccat tggagaatat gatggatcta ccttctgtga actttatagt 16500
gaagaatctg ctattacatt tccaatttgt caacatgctg agctttaata ggacttatct 16560
tcttatgaca acatttattg gtgtgtcccc ttgcctagcc caacagaaga attcagcagc 16620
cgtaagtcta ggacaggctt aaattgtttt cactggtgta aattgcagaa agatgatcta 16680
agtaatttgg catttatttt aataggtttg aaaaacacat gccattttac aaataagact 16740
tatatttgtc cttttgtttt tcagcctacc atgagaataa gagaaagaaa atgaagatca 16800
aaagcttatt catctgtttt tctttttcgt tggtgtaaag ccaacaccct gtctaaaaaa 16860
cataaatttc tttaatcatt ttgcctcttt tctctgtgct tcaattaata aaaaatggaa 16920
agaatctaat agagtggtac agcactgtta tttttcaaag atgtgttgct atcctgaaaa 16980
ttctgtaggt tctgtggaag ttccagtgtt ctctcttatt ccacttcggt agaggatttc 17040
tagtttcttg tgggctaatt aaataaatca ttaatactct tctaagttat ggattataaa 17100
cattcaaaat aatattttga cattatgata attctgaata aaagaacaaa aaccatggta 17160
taggtaagga atataaaaca tggcttttac cttagaaaaa acaattctaa aattcatatg 17220
gaatcaaaaa agagcctgca gaaccaaagt aagactaagc aaaaagaaca aattacctga 17280
tttcaaacta cactataagg ccatagtcac cgaaacagca aggtactggt ataaa 17335
<210> 36
<211> 2076
<212> ДНК
<213> Mus musculus
<400> 36
atattagagc gagtctttct gcacacagat cacctttcct atcaacccca ctagcctctg 60
gcaaaatgaa gtgggtaacc tttctcctcc tcctcttcgt ctccggctct gctttttcca 120
ggggtgtgtt tcgccgagaa gcacacaaga gtgagatcgc ccatcggtat aatgatttgg 180
gagaacaaca tttcaaaggc ctagtcctga ttgccttttc ccagtatctc cagaaatgct 240
catacgatga gcatgccaaa ttagtgcagg aagtaacaga ctttgcaaag acgtgtgttg 300
ccgatgagtc tgccgccaac tgtgacaaat cccttcacac tctttttgga gataagttgt 360
gtgccattcc aaacctccgt gaaaactatg gtgaactggc tgactgctgt acaaaacaag 420
agcccgaaag aaacgaatgt ttcctgcaac acaaagatga caaccccagc ctgccaccat 480
ttgaaaggcc agaggctgag gccatgtgca cctcctttaa ggaaaaccca accaccttta 540
tgggacacta tttgcatgaa gttgccagaa gacatcctta tttctatgcc ccagaacttc 600
tttactatgc tgagcagtac aatgagattc tgacccagtg ttgtgcagag gctgacaagg 660
aaagctgcct gaccccgaag cttgatggtg tgaaggagaa agcattggtc tcatctgtcc 720
gtcagagaat gaagtgctcc agtatgcaga agtttggaga gagagctttt aaagcatggg 780
cagtagctcg tctgagccag acattcccca atgctgactt tgcagaaatc accaaattgg 840
caacagacct gaccaaagtc aacaaggagt gctgccatgg tgacctgctg gaatgcgcag 900
atgacagggc ggaacttgcc aagtacatgt gtgaaaacca ggcgactatc tccagcaaac 960
tgcagacttg ctgcgataaa ccactgttga agaaagccca ctgtcttagt gaggtggagc 1020
atgacaccat gcctgctgat ctgcctgcca ttgctgctga ttttgttgag gaccaggaag 1080
tgtgcaagaa ctatgctgag gccaaggatg tcttcctggg cacgttcttg tatgaatatt 1140
caagaagaca ccctgattac tctgtatccc tgttgctgag acttgctaag aaatatgaag 1200
ccactctgga aaagtgctgc gctgaagcca atcctcccgc atgctacggc acagtgcttg 1260
ctgaatttca gcctcttgta gaagagccta agaacttggt caaaaccaac tgtgatcttt 1320
acgagaagct tggagaatat ggattccaaa atgccattct agttcgctac acccagaaag 1380
cacctcaggt gtcaacccca actctcgtgg aggctgcaag aaacctagga agagtgggca 1440
ccaagtgttg tacacttcct gaagatcaga gactgccttg tgtggaagac tatctgtctg 1500
caatcctgaa ccgtgtgtgt ctgctgcatg agaagacccc agtgagtgag catgttacca 1560
agtgctgtag tggatccctg gtggaaaggc ggccatgctt ctctgctctg acagttgatg 1620
aaacatatgt ccccaaagag tttaaagctg agaccttcac cttccactct gatatctgca 1680
cacttccaga gaaggagaag cagattaaga aacaaacggc tcttgctgag ctggtgaagc 1740
acaagcccaa ggctacagcg gagcaactga agactgtcat ggatgacttt gcacagttcc 1800
tggatacatg ttgcaaggct gctgacaagg acacctgctt ctcgactgag ggtccaaacc 1860
ttgtcactag atgcaaagac gccttagcct aaacacatca caaccacaac cttctcaggc 1920
taccctgaga aaaaaagaca tgaagactca ggactcatct tttctgttgg tgtaaaatca 1980
acaccctaag gaacacaaat ttctttaaac atttgacttc ttgtctctgt gctgcaatta 2040
ataaaaaatg gaaagaatct aaaaaaaaaa aaaaaa 2076
<210> 37
<211> 2285
<212> ДНК
<213> Homo sapiens
<400> 37
ctagcttttc tcttctgtca accccacacg cctttggcac aatgaagtgg gtaaccttta 60
tttcccttct ttttctcttt agctcggctt attccagggg tgtgtttcgt cgagatgcac 120
acaagagtga ggttgctcat cggtttaaag atttgggaga agaaaatttc aaagccttgg 180
tgttgattgc ctttgctcag tatcttcagc agtgtccatt tgaagatcat gtaaaattag 240
tgaatgaagt aactgaattt gcaaaaacat gtgttgctga tgagtcagct gaaaattgtg 300
acaaatcact tcataccctt tttggagaca aattatgcac agttgcaact cttcgtgaaa 360
cctatggtga aatggctgac tgctgtgcaa aacaagaacc tgagagaaat gaatgcttct 420
tgcaacacaa agatgacaac ccaaacctcc cccgattggt gagaccagag gttgatgtga 480
tgtgcactgc ttttcatgac aatgaagaga catttttgaa aaaatactta tatgaaattg 540
ccagaagaca tccttacttt tatgccccgg aactcctttt ctttgctaaa aggtataaag 600
ctgcttttac agaatgttgc caagctgctg ataaagctgc ctgcctgttg ccaaagctcg 660
atgaacttcg ggatgaaggg aaggcttcgt ctgccaaaca gagactcaag tgtgccagtc 720
tccaaaaatt tggagaaaga gctttcaaag catgggcagt agctcgcctg agccagagat 780
ttcccaaagc tgagtttgca gaagtttcca agttagtgac agatcttacc aaagtccaca 840
cggaatgctg ccatggagat ctgcttgaat gtgctgatga cagggcggac cttgccaagt 900
atatctgtga aaatcaagat tcgatctcca gtaaactgaa ggaatgctgt gaaaaacctc 960
tgttggaaaa atcccactgc attgccgaag tggaaaatga tgagatgcct gctgacttgc 1020
cttcattagc tgctgatttt gttgaaagta aggatgtttg caaaaactat gctgaggcaa 1080
aggatgtctt cctgggcatg tttttgtatg aatatgcaag aaggcatcct gattactctg 1140
tcgtgctgct gctgagactt gccaagacat atgaaaccac tctagagaag tgctgtgccg 1200
ctgcagatcc tcatgaatgc tatgccaaag tgttcgatga atttaaacct cttgtggaag 1260
agcctcagaa tttaatcaaa caaaattgtg agctttttga gcagcttgga gagtacaaat 1320
tccagaatgc gctattagtt cgttacacca agaaagtacc ccaagtgtca actccaactc 1380
ttgtagaggt ctcaagaaac ctaggaaaag tgggcagcaa atgttgtaaa catcctgaag 1440
caaaaagaat gccctgtgca gaagactatc tatccgtggt cctgaaccag ttatgtgtgt 1500
tgcatgagaa aacgccagta agtgacagag tcaccaaatg ctgcacagaa tccttggtga 1560
acaggcgacc atgcttttca gctctggaag tcgatgaaac atacgttccc aaagagttta 1620
atgctgaaac attcaccttc catgcagata tatgcacact ttctgagaag gagagacaaa 1680
tcaagaaaca aactgcactt gttgagctcg tgaaacacaa gcccaaggca acaaaagagc 1740
aactgaaagc tgttatggat gatttcgcag cttttgtaga gaagtgctgc aaggctgacg 1800
ataaggagac ctgctttgcc gaggagggta aaaaacttgt tgctgcaagt caagctgcct 1860
taggcttata acatcacatt taaaagcatc tcagcctacc atgagaataa gagaaagaaa 1920
atgaagatca aaagcttatt catctgtttt tctttttcgt tggtgtaaag ccaacaccct 1980
gtctaaaaaa cataaatttc tttaatcatt ttgcctcttt tctctgtgct tcaattaata 2040
aaaaatggaa agaatctaat agagtggtac agcactgtta tttttcaaag atgtgttgct 2100
atcctgaaaa ttctgtaggt tctgtggaag ttccagtgtt ctctcttatt ccacttcggt 2160
agaggatttc tagtttcttg tgggctaatt aaataaatca ttaatactct tctaagttat 2220
ggattataaa cattcaaaat aatattttga cattatgata attctgaata aaagaacaaa 2280
aacca 2285
<210> 38
<211> 1391
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (4)..(10)
<223> 5' NLS
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (1376)..(1391)
<223> 3' NLS
<400> 38
Met Asp Lys Pro Lys Lys Lys Arg Lys Val Lys Tyr Ser Ile Gly Leu
1 5 10 15
Asp Ile Gly Thr Asn Ser Val Gly Trp Ala Val Ile Thr Asp Glu Tyr
20 25 30
Lys Val Pro Ser Lys Lys Phe Lys Val Leu Gly Asn Thr Asp Arg His
35 40 45
Ser Ile Lys Lys Asn Leu Ile Gly Ala Leu Leu Phe Asp Ser Gly Glu
50 55 60
Thr Ala Glu Ala Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg Tyr Thr
65 70 75 80
Arg Arg Lys Asn Arg Ile Cys Tyr Leu Gln Glu Ile Phe Ser Asn Glu
85 90 95
Met Ala Lys Val Asp Asp Ser Phe Phe His Arg Leu Glu Glu Ser Phe
100 105 110
Leu Val Glu Glu Asp Lys Lys His Glu Arg His Pro Ile Phe Gly Asn
115 120 125
Ile Val Asp Glu Val Ala Tyr His Glu Lys Tyr Pro Thr Ile Tyr His
130 135 140
Leu Arg Lys Lys Leu Val Asp Ser Thr Asp Lys Ala Asp Leu Arg Leu
145 150 155 160
Ile Tyr Leu Ala Leu Ala His Met Ile Lys Phe Arg Gly His Phe Leu
165 170 175
Ile Glu Gly Asp Leu Asn Pro Asp Asn Ser Asp Val Asp Lys Leu Phe
180 185 190
Ile Gln Leu Val Gln Thr Tyr Asn Gln Leu Phe Glu Glu Asn Pro Ile
195 200 205
Asn Ala Ser Gly Val Asp Ala Lys Ala Ile Leu Ser Ala Arg Leu Ser
210 215 220
Lys Ser Arg Arg Leu Glu Asn Leu Ile Ala Gln Leu Pro Gly Glu Lys
225 230 235 240
Lys Asn Gly Leu Phe Gly Asn Leu Ile Ala Leu Ser Leu Gly Leu Thr
245 250 255
Pro Asn Phe Lys Ser Asn Phe Asp Leu Ala Glu Asp Ala Lys Leu Gln
260 265 270
Leu Ser Lys Asp Thr Tyr Asp Asp Asp Leu Asp Asn Leu Leu Ala Gln
275 280 285
Ile Gly Asp Gln Tyr Ala Asp Leu Phe Leu Ala Ala Lys Asn Leu Ser
290 295 300
Asp Ala Ile Leu Leu Ser Asp Ile Leu Arg Val Asn Thr Glu Ile Thr
305 310 315 320
Lys Ala Pro Leu Ser Ala Ser Met Ile Lys Arg Tyr Asp Glu His His
325 330 335
Gln Asp Leu Thr Leu Leu Lys Ala Leu Val Arg Gln Gln Leu Pro Glu
340 345 350
Lys Tyr Lys Glu Ile Phe Phe Asp Gln Ser Lys Asn Gly Tyr Ala Gly
355 360 365
Tyr Ile Asp Gly Gly Ala Ser Gln Glu Glu Phe Tyr Lys Phe Ile Lys
370 375 380
Pro Ile Leu Glu Lys Met Asp Gly Thr Glu Glu Leu Leu Val Lys Leu
385 390 395 400
Asn Arg Glu Asp Leu Leu Arg Lys Gln Arg Thr Phe Asp Asn Gly Ser
405 410 415
Ile Pro His Gln Ile His Leu Gly Glu Leu His Ala Ile Leu Arg Arg
420 425 430
Gln Glu Asp Phe Tyr Pro Phe Leu Lys Asp Asn Arg Glu Lys Ile Glu
435 440 445
Lys Ile Leu Thr Phe Arg Ile Pro Tyr Tyr Val Gly Pro Leu Ala Arg
450 455 460
Gly Asn Ser Arg Phe Ala Trp Met Thr Arg Lys Ser Glu Glu Thr Ile
465 470 475 480
Thr Pro Trp Asn Phe Glu Glu Val Val Asp Lys Gly Ala Ser Ala Gln
485 490 495
Ser Phe Ile Glu Arg Met Thr Asn Phe Asp Lys Asn Leu Pro Asn Glu
500 505 510
Lys Val Leu Pro Lys His Ser Leu Leu Tyr Glu Tyr Phe Thr Val Tyr
515 520 525
Asn Glu Leu Thr Lys Val Lys Tyr Val Thr Glu Gly Met Arg Lys Pro
530 535 540
Ala Phe Leu Ser Gly Glu Gln Lys Lys Ala Ile Val Asp Leu Leu Phe
545 550 555 560
Lys Thr Asn Arg Lys Val Thr Val Lys Gln Leu Lys Glu Asp Tyr Phe
565 570 575
Lys Lys Ile Glu Cys Phe Asp Ser Val Glu Ile Ser Gly Val Glu Asp
580 585 590
Arg Phe Asn Ala Ser Leu Gly Thr Tyr His Asp Leu Leu Lys Ile Ile
595 600 605
Lys Asp Lys Asp Phe Leu Asp Asn Glu Glu Asn Glu Asp Ile Leu Glu
610 615 620
Asp Ile Val Leu Thr Leu Thr Leu Phe Glu Asp Arg Glu Met Ile Glu
625 630 635 640
Glu Arg Leu Lys Thr Tyr Ala His Leu Phe Asp Asp Lys Val Met Lys
645 650 655
Gln Leu Lys Arg Arg Arg Tyr Thr Gly Trp Gly Arg Leu Ser Arg Lys
660 665 670
Leu Ile Asn Gly Ile Arg Asp Lys Gln Ser Gly Lys Thr Ile Leu Asp
675 680 685
Phe Leu Lys Ser Asp Gly Phe Ala Asn Arg Asn Phe Met Gln Leu Ile
690 695 700
His Asp Asp Ser Leu Thr Phe Lys Glu Asp Ile Gln Lys Ala Gln Val
705 710 715 720
Ser Gly Gln Gly Asp Ser Leu His Glu His Ile Ala Asn Leu Ala Gly
725 730 735
Ser Pro Ala Ile Lys Lys Gly Ile Leu Gln Thr Val Lys Val Val Asp
740 745 750
Glu Leu Val Lys Val Met Gly Arg His Lys Pro Glu Asn Ile Val Ile
755 760 765
Glu Met Ala Arg Glu Asn Gln Thr Thr Gln Lys Gly Gln Lys Asn Ser
770 775 780
Arg Glu Arg Met Lys Arg Ile Glu Glu Gly Ile Lys Glu Leu Gly Ser
785 790 795 800
Gln Ile Leu Lys Glu His Pro Val Glu Asn Thr Gln Leu Gln Asn Glu
805 810 815
Lys Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly Arg Asp Met Tyr Val Asp
820 825 830
Gln Glu Leu Asp Ile Asn Arg Leu Ser Asp Tyr Asp Val Asp His Ile
835 840 845
Val Pro Gln Ser Phe Leu Lys Asp Asp Ser Ile Asp Asn Lys Val Leu
850 855 860
Thr Arg Ser Asp Lys Asn Arg Gly Lys Ser Asp Asn Val Pro Ser Glu
865 870 875 880
Glu Val Val Lys Lys Met Lys Asn Tyr Trp Arg Gln Leu Leu Asn Ala
885 890 895
Lys Leu Ile Thr Gln Arg Lys Phe Asp Asn Leu Thr Lys Ala Glu Arg
900 905 910
Gly Gly Leu Ser Glu Leu Asp Lys Ala Gly Phe Ile Lys Arg Gln Leu
915 920 925
Val Glu Thr Arg Gln Ile Thr Lys His Val Ala Gln Ile Leu Asp Ser
930 935 940
Arg Met Asn Thr Lys Tyr Asp Glu Asn Asp Lys Leu Ile Arg Glu Val
945 950 955 960
Lys Val Ile Thr Leu Lys Ser Lys Leu Val Ser Asp Phe Arg Lys Asp
965 970 975
Phe Gln Phe Tyr Lys Val Arg Glu Ile Asn Asn Tyr His His Ala His
980 985 990
Asp Ala Tyr Leu Asn Ala Val Val Gly Thr Ala Leu Ile Lys Lys Tyr
995 1000 1005
Pro Lys Leu Glu Ser Glu Phe Val Tyr Gly Asp Tyr Lys Val Tyr
1010 1015 1020
Asp Val Arg Lys Met Ile Ala Lys Ser Glu Gln Glu Ile Gly Lys
1025 1030 1035
Ala Thr Ala Lys Tyr Phe Phe Tyr Ser Asn Ile Met Asn Phe Phe
1040 1045 1050
Lys Thr Glu Ile Thr Leu Ala Asn Gly Glu Ile Arg Lys Arg Pro
1055 1060 1065
Leu Ile Glu Thr Asn Gly Glu Thr Gly Glu Ile Val Trp Asp Lys
1070 1075 1080
Gly Arg Asp Phe Ala Thr Val Arg Lys Val Leu Ser Met Pro Gln
1085 1090 1095
Val Asn Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly Phe Ser
1100 1105 1110
Lys Glu Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile Ala
1115 1120 1125
Arg Lys Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser
1130 1135 1140
Pro Thr Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys
1145 1150 1155
Gly Lys Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile
1160 1165 1170
Thr Ile Met Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe
1175 1180 1185
Leu Glu Ala Lys Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile
1190 1195 1200
Lys Leu Pro Lys Tyr Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys
1205 1210 1215
Arg Met Leu Ala Ser Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu
1220 1225 1230
Ala Leu Pro Ser Lys Tyr Val Asn Phe Leu Tyr Leu Ala Ser His
1235 1240 1245
Tyr Glu Lys Leu Lys Gly Ser Pro Glu Asp Asn Glu Gln Lys Gln
1250 1255 1260
Leu Phe Val Glu Gln His Lys His Tyr Leu Asp Glu Ile Ile Glu
1265 1270 1275
Gln Ile Ser Glu Phe Ser Lys Arg Val Ile Leu Ala Asp Ala Asn
1280 1285 1290
Leu Asp Lys Val Leu Ser Ala Tyr Asn Lys His Arg Asp Lys Pro
1295 1300 1305
Ile Arg Glu Gln Ala Glu Asn Ile Ile His Leu Phe Thr Leu Thr
1310 1315 1320
Asn Leu Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp Thr Thr Ile
1325 1330 1335
Asp Arg Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp Ala Thr
1340 1345 1350
Leu Ile His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile Asp
1355 1360 1365
Leu Ser Gln Leu Gly Gly Asp Lys Arg Pro Ala Ala Thr Lys Lys
1370 1375 1380
Ala Gly Gln Ala Lys Lys Lys Lys
1385 1390
<210> 39
<211> 4176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(3)
<223> Старт-кодон
<220>
<221> прочий_признак
<222> (10)..(30)
<223> 5' NLS
<220>
<221> прочий_признак
<222> (4126)..(4173)
<223> 3' NLS
<220>
<221> прочий_признак
<222> (4174)..(4176)
<223> Стоп-кодон
<400> 39
atggacaagc ccaagaaaaa gcggaaagtg aagtacagca tcggcctgga catcggcacc 60
aactctgtgg gctgggccgt gatcaccgac gagtacaagg tgcccagcaa gaaattcaag 120
gtgctgggca acaccgacag gcacagcatc aagaagaacc tgatcggcgc cctgctgttc 180
gacagcggcg aaacagccga ggccaccaga ctgaagagaa ccgccagaag aagatacacc 240
aggcggaaga acaggatctg ctatctgcaa gagatcttca gcaacgagat ggccaaggtg 300
gacgacagct tcttccacag actggaagag tccttcctgg tggaagagga caagaagcac 360
gagagacacc ccatcttcgg caacatcgtg gacgaggtgg cctaccacga gaagtacccc 420
accatctacc acctgagaaa gaaactggtg gacagcaccg acaaggccga cctgagactg 480
atctacctgg ccctggccca catgatcaag ttcagaggcc acttcctgat cgagggcgac 540
ctgaaccccg acaacagcga cgtggacaag ctgttcatcc agctggtgca gacctacaac 600
cagctgttcg aggaaaaccc catcaacgcc agcggcgtgg acgccaaggc tatcctgtct 660
gccagactga gcaagagcag aaggctggaa aatctgatcg cccagctgcc cggcgagaag 720
aagaacggcc tgttcggcaa cctgattgcc ctgagcctgg gcctgacccc caacttcaag 780
agcaacttcg acctggccga ggatgccaaa ctgcagctga gcaaggacac ctacgacgac 840
gacctggaca acctgctggc ccagatcggc gaccagtacg ccgacctgtt cctggccgcc 900
aagaacctgt ctgacgccat cctgctgagc gacatcctga gagtgaacac cgagatcacc 960
aaggcccccc tgagcgcctc tatgatcaag agatacgacg agcaccacca ggacctgacc 1020
ctgctgaaag ctctcgtgcg gcagcagctg cctgagaagt acaaagaaat cttcttcgac 1080
cagagcaaga acggctacgc cggctacatc gatggcggcg ctagccagga agagttctac 1140
aagttcatca agcccatcct ggaaaagatg gacggcaccg aggaactgct cgtgaagctg 1200
aacagagagg acctgctgag aaagcagaga accttcgaca acggcagcat cccccaccag 1260
atccacctgg gagagctgca cgctatcctg agaaggcagg aagattttta cccattcctg 1320
aaggacaacc gggaaaagat cgagaagatc ctgaccttca ggatccccta ctacgtgggc 1380
cccctggcca gaggcaacag cagattcgcc tggatgacca gaaagagcga ggaaaccatc 1440
accccctgga acttcgagga agtggtggac aagggcgcca gcgcccagag cttcatcgag 1500
agaatgacaa acttcgataa gaacctgccc aacgagaagg tgctgcccaa gcacagcctg 1560
ctgtacgagt acttcaccgt gtacaacgag ctgaccaaag tgaaatacgt gaccgaggga 1620
atgagaaagc ccgccttcct gagcggcgag cagaaaaagg ccatcgtgga cctgctgttc 1680
aagaccaaca gaaaagtgac cgtgaagcag ctgaaagagg actacttcaa gaaaatcgag 1740
tgcttcgact ccgtggaaat ctccggcgtg gaagatagat tcaacgcctc cctgggcaca 1800
taccacgatc tgctgaaaat tatcaaggac aaggacttcc tggataacga agagaacgag 1860
gacattctgg aagatatcgt gctgaccctg acactgtttg aggaccgcga gatgatcgag 1920
gaaaggctga aaacctacgc tcacctgttc gacgacaaag tgatgaagca gctgaagaga 1980
aggcggtaca ccggctgggg caggctgagc agaaagctga tcaacggcat cagagacaag 2040
cagagcggca agacaatcct ggatttcctg aagtccgacg gcttcgccaa ccggaacttc 2100
atgcagctga tccacgacga cagcctgaca ttcaaagagg acatccagaa agcccaggtg 2160
tccggccagg gcgactctct gcacgagcat atcgctaacc tggccggcag ccccgctatc 2220
aagaagggca tcctgcagac agtgaaggtg gtggacgagc tcgtgaaagt gatgggcaga 2280
cacaagcccg agaacatcgt gatcgagatg gctagagaga accagaccac ccagaaggga 2340
cagaagaact cccgcgagag gatgaagaga atcgaagagg gcatcaaaga gctgggcagc 2400
cagatcctga aagaacaccc cgtggaaaac acccagctgc agaacgagaa gctgtacctg 2460
tactacctgc agaatggccg ggatatgtac gtggaccagg aactggacat caacagactg 2520
tccgactacg atgtggacca tatcgtgcct cagagctttc tgaaggacga ctccatcgat 2580
aacaaagtgc tgactcggag cgacaagaac agaggcaaga gcgacaacgt gccctccgaa 2640
gaggtcgtga agaagatgaa gaactactgg cgacagctgc tgaacgccaa gctgattacc 2700
cagaggaagt tcgataacct gaccaaggcc gagagaggcg gcctgagcga gctggataag 2760
gccggcttca tcaagaggca gctggtggaa accagacaga tcacaaagca cgtggcacag 2820
atcctggact cccggatgaa cactaagtac gacgaaaacg ataagctgat ccgggaagtg 2880
aaagtgatca ccctgaagtc caagctggtg tccgatttcc ggaaggattt ccagttttac 2940
aaagtgcgcg agatcaacaa ctaccaccac gcccacgacg cctacctgaa cgccgtcgtg 3000
ggaaccgccc tgatcaaaaa gtaccctaag ctggaaagcg agttcgtgta cggcgactac 3060
aaggtgtacg acgtgcggaa gatgatcgcc aagagcgagc aggaaatcgg caaggctacc 3120
gccaagtact tcttctacag caacatcatg aactttttca agaccgaaat caccctggcc 3180
aacggcgaga tcagaaagcg ccctctgatc gagacaaacg gcgaaaccgg ggagatcgtg 3240
tgggataagg gcagagactt cgccacagtg cgaaaggtgc tgagcatgcc ccaagtgaat 3300
atcgtgaaaa agaccgaggt gcagacaggc ggcttcagca aagagtctat cctgcccaag 3360
aggaacagcg acaagctgat cgccagaaag aaggactggg accccaagaa gtacggcggc 3420
ttcgacagcc ctaccgtggc ctactctgtg ctggtggtgg ctaaggtgga aaagggcaag 3480
tccaagaaac tgaagagtgt gaaagagctg ctggggatca ccatcatgga aagaagcagc 3540
tttgagaaga accctatcga ctttctggaa gccaagggct acaaagaagt gaaaaaggac 3600
ctgatcatca agctgcctaa gtactccctg ttcgagctgg aaaacggcag aaagagaatg 3660
ctggcctctg ccggcgaact gcagaaggga aacgagctgg ccctgcctag caaatatgtg 3720
aacttcctgt acctggcctc ccactatgag aagctgaagg gcagccctga ggacaacgaa 3780
cagaaacagc tgtttgtgga acagcataag cactacctgg acgagatcat cgagcagatc 3840
agcgagttct ccaagagagt gatcctggcc gacgccaatc tggacaaggt gctgtctgcc 3900
tacaacaagc acagggacaa gcctatcaga gagcaggccg agaatatcat ccacctgttc 3960
accctgacaa acctgggcgc tcctgccgcc ttcaagtact ttgacaccac catcgaccgg 4020
aagaggtaca ccagcaccaa agaggtgctg gacgccaccc tgatccacca gagcatcacc 4080
ggcctgtacg agacaagaat cgacctgtct cagctgggag gcgacaagag acctgccgcc 4140
actaagaagg ccggacaggc caaaaagaag aagtga 4176
<210> 40
<211> 16
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 40
guuuuagagc uaugcu 16
<210> 41
<211> 67
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 41
agcauagcaa guuaaaauaa ggcuaguccg uuaucaacuu gaaaaagugg caccgagucg 60
gugcuuu 67
<210> 42
<211> 77
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 42
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcu 77
<210> 43
<211> 82
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 43
guuggaacca uucaaaacag cauagcaagu uaaaauaagg cuaguccguu aucaacuuga 60
aaaaguggca ccgagucggu gc 82
<210> 44
<211> 76
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 44
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugc 76
<210> 45
<211> 86
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 45
guuuaagagc uaugcuggaa acagcauagc aaguuuaaau aaggcuaguc cguuaucaac 60
uugaaaaagu ggcaccgagu cggugc 86
<210> 46
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (2)..(21)
<223> n представляет собой a, c, g или t
<400> 46
gnnnnnnnnn nnnnnnnnnn ngg 23
<210> 47
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (1)..(21)
<223> n представляет собой a, c, g или t
<400> 47
nnnnnnnnnn nnnnnnnnnn ngg 23
<210> 48
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> прочий_признак
<222> (3)..(23)
<223> n представляет собой a, c, g или t
<400> 48
ggnnnnnnnn nnnnnnnnnn nnngg 25
<210> 49
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 49
gagcaaccuc acucuugucu 20
<210> 50
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 50
ugcauuuguu ucaaaauauu 20
<210> 51
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 51
auuuaugaga ucaacagcac 20
<210> 52
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 52
uuaaauaaag cauagugcaa 20
<210> 53
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 53
uaaagcauag ugcaauggau 20
<210> 54
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 54
uaauaaaauu caaacauccu 20
<210> 55
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 55
ugacugaaac uucacagaau 20
<210> 56
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 56
gacugaaacu ucacagaaua 20
<210> 57
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 57
agugcaaugg auaggucuuu 20
<210> 58
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 58
ccucacucuu gucugggcaa 20
<210> 59
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 59
accucacucu ugucugggca 20
<210> 60
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 60
ugagcaaccu cacucuuguc 20
<210> 61
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 61
uacuuugcac uuuccuuagu 20
<210> 62
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 62
cacucuuguc uguggaaaca 20
<210> 63
<211> 6150
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 63
aacacccctt gtattactgt ttatgtaagc agacagtttt attgttcatg atgatatatt 60
tttatcttgt gcaatgtaac atcagagatt ttgagacacg ggccagagct gcatcgcgcg 120
tttcggtgat gacggtgaaa acctctgaca catgcagctc ccggagacgg tcacagcttg 180
tctgtaagcg gatgccggga gcagacaagc ccgtcagggc gcgtcagcgg gtgttggcgg 240
gtgtcggggc tggcttaact atgcggcatc agagcagatt gtactgagag tgcaccatat 300
gcggtgtgaa ataccgcaca gatgcgtaag gagaaaatac cgcatcaggc gccattcgcc 360
attcaggctg cgcaactgtt gggaagggcg atcggtgcgg gcctcttcgc tattacgcca 420
gctggcgaaa gggggatgtg ctgcaaggcg attaagttgg gtaacgccag ggttttccca 480
gtcacgacgt tgtaaaacga cggccagaga attcgagctc ggtacctcgc gaatacatct 540
agataggaac ccctagtgat ggagttggcc actccctctc tgcgcgctcg ctcgctcact 600
gaggccgccc gggcaaagcc cgggcgtcgg gcgacctttg gtcgcccggc ctcagtgagc 660
gagcgagcgc gcagagaggg agtggccaaa gatctcttag gtcagtgaag agaagaacaa 720
aaagcagcat attacagtta gttgtcttca tcaatcttta aatatgttgt gtggtttttc 780
tctccctgtt tccacagttt ttcttgatca tgaaaacgcc aacaaaattc tgaatcggcc 840
aaagaggtat aattcaggta aattggaaga gtttgttcaa gggaaccttg agagagaatg 900
tatggaagaa aagtgtagtt ttgaagaagc acgagaagtt tttgaaaaca ctgaaagaac 960
aactgaattt tggaagcagt atgttgatgg agatcagtgt gagtccaatc catgtttaaa 1020
tggcggcagt tgcaaggatg acattaattc ctatgaatgt tggtgtccct ttggatttga 1080
aggaaagaac tgtgaattag atgtaacatg taacattaag aatggcagat gcgagcagtt 1140
ttgtaaaaat agtgctgata acaaggtggt ttgctcctgt actgagggat atcgacttgc 1200
agaaaaccag aagtcctgtg aaccagcagt gccatttcca tgtggaagag tttctgtttc 1260
acaaacttct aagctcaccc gtgctgagac tgtttttcct gatgtggact atgtaaattc 1320
tactgaagct gaaaccattt tggataacat cactcaaagc acccaatcat ttaatgactt 1380
cactcgggtt gttggtggag aagatgccaa accaggtcaa ttcccttggc aggttgtttt 1440
gaatggtaaa gttgatgcat tctgtggagg ctctatcgtt aatgaaaaat ggattgtaac 1500
tgctgcccac tgtgttgaaa ctggtgttaa aattacagtt gtcgcaggtg aacataatat 1560
tgaggagaca gaacatacag agcaaaagcg aaatgtgatt cgaattattc ctcaccacaa 1620
ctacaatgca gctattaata agtacaacca tgacattgcc cttctggaac tggacgaacc 1680
cttagtgcta aacagctacg ttacacctat ttgcattgct gacaaggaat acacgaacat 1740
cttcctcaaa tttggatctg gctatgtaag tggctgggga agagtcttcc acaaagggag 1800
atcagcttta gttcttcagt accttagagt tccacttgtt gaccgagcca catgtcttct 1860
atctacaaag ttcaccatct ataacaacat gttctgtgct ggcttccatg aaggaggtag 1920
agattcatgt caaggagata gtgggggacc ccatgttact gaagtggaag ggaccagttt 1980
cttaactgga attattagct ggggtgaaga gtgtgcaatg aaaggcaaat atggaatata 2040
taccaaggta tcccggtatg tcaactggat taaggaaaaa acaaagctca cttaacctcg 2100
actgtgcctt ctagttgcca gccatctgtt gtttgcccct cccccgtgcc ttccttgacc 2160
ctggaaggtg ccactcccac tgtcctttcc taataaaatg aggaaattgc atcgcattgt 2220
ctgagtaggt gtcattctat tctggggggt ggggtggggc aggacagcaa gggggaggat 2280
tgggaagaca atagcaggca tgctggggat gcggtgggct ctatggcttc tgaggcggaa 2340
agaaccagct ggggctctag ggggtatccc caaaaaacct cccacacctc cccctgaacc 2400
tgaaacataa aatgaatgca attgttgttg ttaacttgtt tattgcagct tataatggtt 2460
acaaataaag caatagcatc acaaatttca caaataaagc atttttttca ctgcattcta 2520
gttgtggttt gtccaaactc atcaatgtat cttatcatgt ctgttaggtg agcttagtct 2580
tttcttttat ccaattcacg tagcgagaga ccttcgtata gatgccatat ttccccttca 2640
tcgcacattc ctccccccaa cttattatcc cggtcaagaa acttgttcct tcgacttcag 2700
tgacgtgtgg tccacctgaa tcaccttggc atgagtcgcg accgccctcg tgaaacccag 2760
cacaaaacat gttattgtaa atcgtaaatt tcgtggacag aagacaggtc gctctatcga 2820
ccaacgggac gcgcaaatat tgcagaacga gggctgatcg acctttgtgg aagacccgcc 2880
cccacccact cacatatccg ctcccaaatt tcaagaagat atttgtatat tctttatcgg 2940
ctatacaaat cggggtaaca taggagttaa gtacgagtgg ctcgtccagc tccaggaggg 3000
ctatatcatg gttgtacttg tttatagcgg cattataatt gtgatggggt atgatcctga 3060
taacattcct tttctgttca gtatgctcag tttcttcaat gttgtgttcg ccagccacga 3120
ccgtaatctt aacccccgtc tcgacacagt gtgcggccgt tacaatccac ttttcattga 3180
ctatggagcc cccacaaaac gcgtcgactt ttccgttgag caccacctgc catggaaatt 3240
ggccaggttt agcgtcctcg cccccgacaa ccctagtaaa gtcattaaat gactgtgtgg 3300
attgtgttat attatcaaga atcgtttcgg cttcagtaga gttaacgtag tccacatcgg 3360
gaaaaactgt ctcggccctt gtcaactttg atgtctggga cacacttacc cgaccgcacg 3420
ggaagggcac cgccggttca cagctctttt gattctcagc gagccggtag ccctcagtgc 3480
aactacacac aactttgttg tcggcggaat ttttacagaa ttgctcgcat cgtccatttt 3540
taatgttgca ggtgacgtcc aactcgcagt tttttccttc aaaaccaaaa gggcaccaac 3600
actcgtagga atttatatcg tctttacaac tccccccatt cagacatgga ttagattcgc 3660
attggtcccc atcgacatat tgcttccaga actcagtggt ccgttctgta ttctcaaaca 3720
cctcgcgcgc ttcttcaaaa ctgcattttt cctccataca ctctcgctcc aagttccctt 3780
gcacgaattc ttcaagcttt cctgagttat accttttagg ccggttaagt atcttattcg 3840
cgttttcgtg gtccagaaaa actgtggaaa cagggagaga aaaaccacac aacatattta 3900
aagattgatg aagacaacta actgtaatat gctgcttttt gttcttctct tcactgacct 3960
aagagatcta ggaaccccta gtgatggagt tggccactcc ctctctgcgc gctcgctcgc 4020
tcactgaggc cgcccgggca aagcccgggc gtcgggcgac ctttggtcgc ccggcctcag 4080
tgagcgagcg agcgcgcaga gagggagtgg ccaaactcgg atcccgggcc cgtcgactgc 4140
agaggcctgc atgcaagcgt ggtgtaatca tggtcatagc tgtttcctgt gtgaaattgt 4200
tatccgctca caattccaca caacatacga gccggaagca taaagtgtaa agcctggggt 4260
gcctaatgag tgagctaact cacattaatt gcgttgcgct cactgcccgc tttccagtcg 4320
ggaaacctgt cgtgccagct gcattaatga atcggccaac gcgcggggag aggcggtttg 4380
cgtattgggc gctcttccgc ttcctcgctc actgactcgc tgcgctcggt cgttcggctg 4440
cggcgagcgg tatcagctca ctcaaaggcg gtaatacggt tatccacaga atcaggggat 4500
aacgcaggaa agaacatgtg agcaaaaggc cagcaaaagg ccaggaaccg taaaaaggcc 4560
gcgttgctgg cgtttttcca taggctccgc ccccctgacg agcatcacaa aaatcgacgc 4620
tcaagtcaga ggtggcgaaa cccgacagga ctataaagat accaggcgtt tccccctgga 4680
agctccctcg tgcgctctcc tgttccgacc ctgccgctta ccggatacct gtccgccttt 4740
ctcccttcgg gaagcgtggc gctttctcat agctcacgct gtaggtatct cagttcggtg 4800
taggtcgttc gctccaagct gggctgtgtg cacgaacccc ccgttcagcc cgaccgctgc 4860
gccttatccg gtaactatcg tcttgagtcc aacccggtaa gacacgactt atcgccactg 4920
gcagcagcca ctggtaacag gattagcaga gcgaggtatg taggcggtgc tacagagttc 4980
ttgaagtggt ggcctaacta cggctacact agaagaacag tatttggtat ctgcgctctg 5040
ctgaagccag ttaccttcgg aaaaagagtt ggtagctctt gatccggcaa acaaaccacc 5100
gctggtagcg gtggtttttt tgtttgcaag cagcagatta cgcgcagaaa aaaaggatct 5160
caagaagatc ctttgatctt ttctacgggg tctgacgctc agtggaacga aaactcacgt 5220
taagggattt tggtcatgag attatcaaaa aggatcttca cctagatcct tttaaattaa 5280
aaatgaagtt ttaaatcaag cccaatctga ataatgttac aaccaattaa ccaattctga 5340
ttagaaaaac tcatcgagca tcaaatgaaa ctgcaattta ttcatatcag gattatcaat 5400
accatatttt tgaaaaagcc gtttctgtaa tgaaggagaa aactcaccga ggcagttcca 5460
taggatggca agatcctggt atcggtctgc gattccgact cgtccaacat caatacaacc 5520
tattaatttc ccctcgtcaa aaataaggtt atcaagtgag aaatcaccat gagtgacgac 5580
tgaatccggt gagaatggca aaagtttatg catttctttc cagacttgtt caacaggcca 5640
gccattacgc tcgtcatcaa aatcactcgc atcaaccaaa ccgttattca ttcgtgattg 5700
cgcctgagcg agacgaaata cgcgatcgct gttaaaagga caattacaaa caggaatcga 5760
atgcaaccgg cgcaggaaca ctgccagcgc atcaacaata ttttcacctg aatcaggata 5820
ttcttctaat acctggaatg ctgtttttcc ggggatcgca gtggtgagta accatgcatc 5880
atcaggagta cggataaaat gcttgatggt cggaagaggc ataaattccg tcagccagtt 5940
tagtctgacc atctcatctg taacatcatt ggcaacgcta cctttgccat gtttcagaaa 6000
caactctggc gcatcgggct tcccatacaa gcgatagatt gtcgcacctg attgcccgac 6060
attatcgcga gcccatttat acccatataa atcagcatcc atgttggaat ttaatcgcgg 6120
cctcgacgtt tcccgttgaa tatggctcat 6150
<210> 64
<211> 6901
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 64
gaggcccttt cgtctcgcgc gtttcggtga tgacggtgaa aacctctgac acatgcagct 60
cccggagacg gtcacagctt gtctgtaagc ggatgccggg agcagacaag cccgtcaggg 120
cgcgtcagcg ggtgttggcg ggtgtcgggg ctggcttaac tatgcggcat cagagcagat 180
tgtactgaga gtgcaccata tgcggtgtga aataccgcac agatgcgtaa ggagaaaata 240
ccgcatcagg cgccattcgc cattcaggct gcgcaactgt tgggaagggc gatcggtgcg 300
ggcctcttcg ctattacgcc agctggcgaa agggggatgt gctgcaaggc gattaagttg 360
ggtaacgcca gggttttccc agtcacgacg ttgtaaaacg acggccagtg aattcgagct 420
cggtacccct gcaggcagct gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc 480
gggcgtcggg cgacctttgg tcgcccggcc tcagtgagcg agcgagcgcg cagagaggga 540
gtggccaact ccatcactag gggttcctcg ggcaaagcca cgcgtactag ttattaatag 600
taatcaatta cggggtcatt agttcatagc ccatatatgg agttccgcgt tacataactt 660
acggtaaatg gcccgcctgg ctgaccgccc aacgaccccc gcccattgac gtcaataatg 720
acgtatgttc ccatagtaac gccaataggg actttccatt gacgtcaatg ggtggagtat 780
ttacggtaaa ctgcccactt ggcagtacat caagtgtatc atatgccaag tacgccccct 840
attgacgtca atgacggtaa atggcccgcc tggcattatg cccagtacat gaccttatgg 900
gactttccta cttggcagta catctacgta ttagtcatcg ctattaccat ggtcgaggtg 960
agccccacgt tctgcttcac tctccccatc tcccccccct ccccaccccc aattttgtat 1020
ttatttattt tttaattatt ttgtgcagcg atgggggcgg gggggggggg ggggcgcgcg 1080
ccaggcgggg cggggcgggg cgaggggcgg ggcggggcga ggcggagagg tgcggcggca 1140
gccaatcaga gcggcgcgct ccgaaagttt ccttttatgg cgaggcggcg gcggcggcgg 1200
ccctataaaa agcgaagcgc gcggcgggcg gggagtcgct gcgacgctgc cttcgccccg 1260
tgccccgctc cgccgccgcc tcgcgccgcc cgccccggct ctgactgacc gcgttactcc 1320
cacaggtgag cgggcgggac ggcccttctc ctccgggctg taattagcgc ttggtttaat 1380
gacggcttgt ttcttttctg tggctgcgtg aaagccttga ggggctccgg gagggccctt 1440
tgtgcggggg gagcggctcg gggggtgcgt gcgtgtgtgt gtgcgtgggg agcgccgcgt 1500
gcggctccgc gctgcccggc ggctgtgagc gctgcgggcg cggcgcgggg ctttgtgcgc 1560
tccgcagtgt gcgcgagggg agcgcggccg ggggcggtgc cccgcggtgc ggggggggct 1620
gcgaggggaa caaaggctgc gtgcggggtg tgtgcgtggg ggggtgagca gggggtgtgg 1680
gcgcgtcggt cgggctgcaa ccccccctgc acccccctcc ccgagttgct gagcacggcc 1740
cggcttcggg tgcggggctc cgtacggggc gtggcgcggg gctcgccgtg ccgggcgggg 1800
ggtggcggca ggtgggggtg ccgggcgggg cggggccgcc tcgggccggg gagggctcgg 1860
gggaggggcg cggcggcccc cggagcgccg gcggctgtcg aggcgcggcg agccgcagcc 1920
attgcctttt atggtaatcg tgcgagaggg cgcagggact tcctttgtcc caaatctgtg 1980
cggagccgaa atctgggagg cgccgccgca ccccctctag cgggcgcggg gcgaagcggt 2040
gcggcgccgg caggaaggaa atgggcgggg agggccttcg tgcgtcgccg cgccgccgtc 2100
cccttctccc tctccagcct cggggctgtc cgcgggggga cggctgcctt cgggggggac 2160
ggggcagggc ggggttcggc ttctggcgtg tgaccggcgg ctctagagcc tctgctaacc 2220
atgttcatgc cttcttcttt ttcctacagc tcctgggcaa cgtgctggtt attgtgctgt 2280
ctcatcattt tggcaaagaa ttcctcgaga tgcagcgcgt gaacatgatc atggcagaat 2340
caccaggcct catcaccatc tgccttttag gatatctact cagtgctgaa tgtacagttt 2400
ttcttgatca tgaaaacgcc aacaaaattc tgaatcggcc aaagaggtat aattcaggta 2460
aattggaaga gtttgttcaa gggaaccttg agagagaatg tatggaagaa aagtgtagtt 2520
ttgaagaagc acgagaagtt tttgaaaaca ctgaaagaac aactgaattt tggaagcagt 2580
atgttgatgg agatcagtgt gagtccaatc catgtttaaa tggcggcagt tgcaaggatg 2640
acattaattc ctatgaatgt tggtgtccct ttggatttga aggaaagaac tgtgaattag 2700
atgtaacatg taacattaag aatggcagat gcgagcagtt ttgtaaaaat agtgctgata 2760
acaaggtggt ttgctcctgt actgagggat atcgacttgc agaaaaccag aagtcctgtg 2820
aaccagcagt gccatttcca tgtggaagag tttctgtttc acaaacttct aagctcaccc 2880
gtgctgagac tgtttttcct gatgtggact atgtaaattc tactgaagct gaaaccattt 2940
tggataacat cactcaaagc acccaatcat ttaatgactt cactcgggtt gttggtggag 3000
aagatgccaa accaggtcaa ttcccttggc aggttgtttt gaatggtaaa gttgatgcat 3060
tctgtggagg ctctatcgtt aatgaaaaat ggattgtaac tgctgcccac tgtgttgaaa 3120
ctggtgttaa aattacagtt gtcgcaggtg aacataatat tgaggagaca gaacatacag 3180
agcaaaagcg aaatgtgatt cgaattattc ctcaccacaa ctacaatgca gctattaata 3240
agtacaacca tgacattgcc cttctggaac tggacgaacc cttagtgcta aacagctacg 3300
ttacacctat ttgcattgct gacaaggaat acacgaacat cttcctcaaa tttggatctg 3360
gctatgtaag tggctgggga agagtcttcc acaaagggag atcagcttta gttcttcagt 3420
accttagagt tccacttgtt gaccgagcca catgtcttcg atctacaaag ttcaccatct 3480
ataacaacat gttctgtgct ggcttccatg aaggaggtag agattcatgt caaggagata 3540
gtgggggacc ccatgttact gaagtggaag ggaccagttt cttaactgga attattagct 3600
ggggtgaaga gtgtgcaatg aaaggcaaat atggaatata taccaaggta tcccggtatg 3660
tcaactggat taaggaaaaa acaaagctca cttaagcggc cgcgtttaaa ctcaacctct 3720
ggattacaaa atttgtgaaa gattgactgg tattcttaac tatgttgctc cttttacgct 3780
atgtggatac gctgctttaa tgcctttgta tcatgctatt gcttcccgta tggctttcat 3840
tttctcctcc ttgtataaat cctggttgct gtctctttat gaggagttgt ggcccgttgt 3900
caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca acccccactg gttggggcat 3960
tgccaccacc tgtcagctcc tttccgggac tttcgctttc cccctcccta ttgccacggc 4020
ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg gctcggctgt tgggcactga 4080
caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct tggctgctcg cctgtgttgc 4140
cacctggatt ctgcgcggga cgtccttctg ctacgtccct tcggccctca atccagcgga 4200
ccttccttcc cgcggcctgc tgccggctct gcggcctctt ccgcgtcttc gccttcgccc 4260
tcagacgagt cggatctccc tttgggccgc ctccccgcag aattcctgca gctagttgcc 4320
agccatctgt tgtttgcccc tcccccgtgc cttccttgac cctggaaggt gccactccca 4380
ctgtcctttc ctaataaaat gaggaaattg catcgcattg tctgagtagg tgtcattcta 4440
ttctgggggg tggggtgggg caggacagca agggggagga ttgggaagac aatagcaggc 4500
atgctgggga tgcggtgggc tctatggggt aaccaggaac ccctagtgat ggagttggcc 4560
actccctctc tgcgcgctcg ctcgctcact gaggccgggc gaccaaaggt cgcccgacgc 4620
ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc gcagctgcct gcaggaagct 4680
tggcgtaatc atggtcatag ctgtttcctg tgtgaaattg ttatccgctc acaattccac 4740
acaacatacg agccggaagc ataaagtgta aagcctgggg tgcctaatga gtgagctaac 4800
tcacattaat tgcgttgcgc tcactgcccg ctttccagtc gggaaacctg tcgtgccagc 4860
tgcattaatg aatcggccaa cgcgcgggga gaggcggttt gcgtattggg cgctcttccg 4920
cttcctcgct cactgactcg ctgcgctcgg tcgttcggct gcggcgagcg gtatcagctc 4980
actcaaaggc ggtaatacgg ttatccacag aatcagggga taacgcagga aagaacatgt 5040
gagcaaaagg ccagcaaaag gccaggaacc gtaaaaaggc cgcgttgctg gcgtttttcc 5100
ataggctccg cccccctgac gagcatcaca aaaatcgacg ctcaagtcag aggtggcgaa 5160
acccgacagg actataaaga taccaggcgt ttccccctgg aagctccctc gtgcgctctc 5220
ctgttccgac cctgccgctt accggatacc tgtccgcctt tctcccttcg ggaagcgtgg 5280
cgctttctca tagctcacgc tgtaggtatc tcagttcggt gtaggtcgtt cgctccaagc 5340
tgggctgtgt gcacgaaccc cccgttcagc ccgaccgctg cgccttatcc ggtaactatc 5400
gtcttgagtc caacccggta agacacgact tatcgccact ggcagcagcc actggtaaca 5460
ggattagcag agcgaggtat gtaggcggtg ctacagagtt cttgaagtgg tggcctaact 5520
acggctacac tagaaggaca gtatttggta tctgcgctct gctgaagcca gttaccttcg 5580
gaaaaagagt tggtagctct tgatccggca aacaaaccac cgctggtagc ggtggttttt 5640
ttgtttgcaa gcagcagatt acgcgcagaa aaaaaggatc tcaagaagat cctttgatct 5700
tttctacggg gtctgacgct cagtggaacg aaaactcacg ttaagggatt ttggtcatga 5760
gattatcaaa aaggatcttc acctagatcc ttttaaatta aaaatgaagt tttaaatcaa 5820
tctaaagtat atatgagtaa acttggtctg acagttacca atgcttaatc agtgaggcac 5880
ctatctcagc gatctgtcta tttcgttcat ccatagttgc ctgactcccc gtcgtgtaga 5940
taactacgat acgggagggc ttaccatctg gccccagtgc tgcaatgata ccgcgagacc 6000
cacgctcacc ggctccagat ttatcagcaa taaaccagcc agccggaagg gccgagcgca 6060
gaagtggtcc tgcaacttta tccgcctcca tccagtctat taattgttgc cgggaagcta 6120
gagtaagtag ttcgccagtt aatagtttgc gcaacgttgt tgccattgct acaggcatcg 6180
tggtgtcacg ctcgtcgttt ggtatggctt cattcagctc cggttcccaa cgatcaaggc 6240
gagttacatg atcccccatg ttgtgcaaaa aagcggttag ctccttcggt cctccgatcg 6300
ttgtcagaag taagttggcc gcagtgttat cactcatggt tatggcagca ctgcataatt 6360
ctcttactgt catgccatcc gtaagatgct tttctgtgac tggtgagtac tcaaccaagt 6420
cattctgaga atagtgtatg cggcgaccga gttgctcttg cccggcgtca atacgggata 6480
ataccgcgcc acatagcaga actttaaaag tgctcatcat tggaaaacgt tcttcggggc 6540
gaaaactctc aaggatctta ccgctgttga gatccagttc gatgtaaccc actcgtgcac 6600
ccaactgatc ttcagcatct tttactttca ccagcgtttc tgggtgagca aaaacaggaa 6660
ggcaaaatgc cgcaaaaaag ggaataaggg cgacacggaa atgttgaata ctcatactct 6720
tcctttttca atattattga agcatttatc agggttattg tctcatgagc ggatacatat 6780
ttgaatgtat ttagaaaaat aaacaaatag gggttccgcg cacatttccc cgaaaagtgc 6840
cacctgacgt ctaagaaacc attattatca tgacattaac ctataaaaat aggcgtatca 6900
c 6901
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЭКСПРЕССИРУЮЩИЕ CAS МЫШИНЫЕ ЭМБРИОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ И МЫШИ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2782358C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ВКЛЮЧАЮЩИЕ В СЕБЯ ГУМАНИЗИРОВАННЫЙ TTR ЛОКУС, И СПОСОБЫ ПРИМЕНЕНИЯ | 2018 |
|
RU2784927C1 |
| МЫШИ, СОДЕРЖАЩИЕ МУТАЦИИ, ВСЛЕДСТВИЕ КОТОРЫХ ЭКСПРЕССИРУЕТСЯ УКОРОЧЕННЫЙ НА С-КОНЦЕ ФИБРИЛЛИН-1 | 2017 |
|
RU2721125C1 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ЛОКУС ASGR1 | 2018 |
|
RU2796949C2 |
| НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ЛОКУС TrkB | 2018 |
|
RU2800428C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ХАРАКТЕРИЗУЮЩИЕСЯ ЭКСПАНСИЕЙ ГЕКСАНУКЛЕОТИДНЫХ ПОВТОРОВ В ЛОКУСЕ C9ORF72 | 2017 |
|
RU2760877C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОЦЕНКИ ОПОСРЕДОВАННОГО CRISPR/CAS РАЗРУШЕНИЯ ИЛИ ВЫРЕЗАНИЯ И ИНДУЦИРОВАННОЙ CRISPR/CAS РЕКОМБИНАЦИИ С ЭКЗОГЕННОЙ ДОНОРСКОЙ НУКЛЕИНОВОЙ КИСЛОТОЙ IN VIVO | 2018 |
|
RU2782356C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ДЛЯ ВЫРАБОТКИ ТЕРАПЕВТИЧЕСКИХ АНТИТЕЛ ПРОТИВ КОМПЛЕКСОВ ПЕПТИД-MHC, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ВАРИАНТЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2819525C2 |
| НЕДЕСТРУКТИВНАЯ ГЕННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ MMA | 2018 |
|
RU2820602C2 |
| СТРАТЕГИИ CRISPR И AAV ДЛЯ ТЕРАПИИ Х-СЦЕПЛЕННОГО ЮВЕНИЛЬНОГО РЕТИНОШИЗИСА | 2020 |
|
RU2833486C1 |






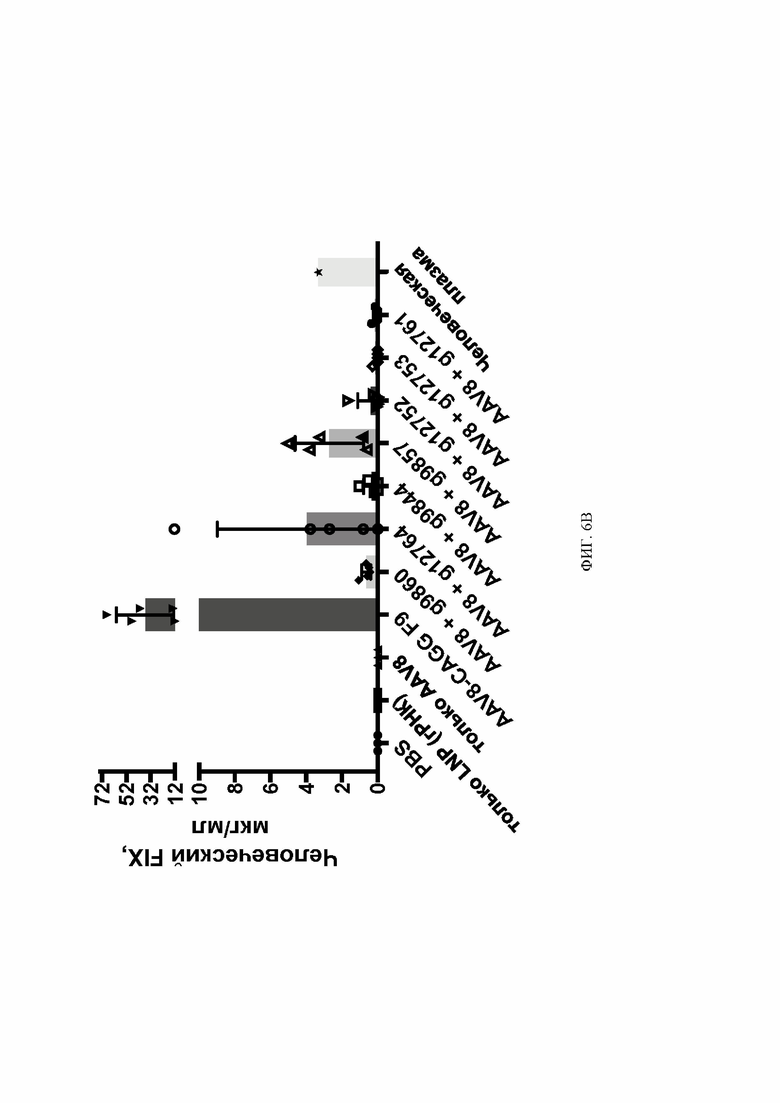





Изобретение относится к области биотехнологии, а именно к животному, отличному от человека, для экспрессии белка человеческого альбумина, содержащему в своем геноме гуманизированный эндогенный локус альбумина, в котором область эндогенного локуса альбумина от старт-кодона до стоп-кодона удалена и заменена соответствующей последовательностью человеческого альбумина, содержащей область от старт-кодона до стоп-кодона гена человеческого альбумина, к его клетке, а также к способу получения указанного животного. Изобретение эффективно для оценки активности реагента, нацеленного на человеческий альбумин, а также для выбора способа, обеспечивающего более высокую активность реагента, нацеленного на человеческий альбумин. 5 н. и 45 з.п. ф-лы, 11 ил., 8 табл., 4 пр.
1. Животное, отличное от человека, для экспрессии белка человеческого альбумина, при этом животное, отличное от человека, содержит в своем геноме гуманизированный эндогенный локус альбумина, в котором область эндогенного локуса альбумина от старт-кодона до стоп-кодона удалена и заменена соответствующей последовательностью человеческого альбумина, содержащей область от старт-кодона до стоп-кодона гена человеческого альбумина,
где гуманизированный эндогенный локус альбумина содержит эндогенный промотор альбумина,
где последовательность человеческого альбумина функционально связана с эндогенным промотором альбумина, и
причем указанное животное, отличное от человека, представляет собой мышь или крысу.
2. Животное, отличное от человека, по п.1, где гуманизированный эндогенный локус альбумина содержит 3’-нетранслируемую область человеческого альбумина.
3. Животное, отличное от человека, по любому из предшествующих пунктов, где эндогенная 5’-нетранслируемая область альбумина не была удалена и заменена соответствующей последовательностью человеческого альбумина.
4. Животное, отличное от человека, по любому из предшествующих пунктов, где область эндогенного локуса альбумина от старт-кодона до стоп-кодона удалена и заменена последовательностью человеческого альбумина, содержащей соответствующую последовательность человеческого альбумина и 3’-нетранслируемую область человеческого альбумина, и
где эндогенная 5’-нетранслируемая область альбумина не была удалена и заменена соответствующей последовательностью человеческого альбумина, и
где эндогенный промотор альбумина не был удален и заменен соответствующей последовательностью человеческого альбумина.
5. Животное, отличное от человека, по любому из предшествующих пунктов, где:
(i) последовательность человеческого альбумина в гуманизированном эндогенном локусе альбумина содержит последовательность, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичную последовательности, приведенной в SEQ ID NO: 35; или
(ii) гуманизированный эндогенный локус альбумина кодирует белок, содержащий последовательность, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или по меньшей мере на 100% идентичную последовательности, приведенной в SEQ ID NO: 5;
(iii) гуманизированный эндогенный локус альбумина содержит кодирующую последовательность, содержащую последовательность, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичную последовательности, приведенной в SEQ ID NO: 13; или
(iv) гуманизированный эндогенный локус альбумина содержит последовательность, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичную последовательности, приведенной в SEQ ID NO: 17 или 18.
6. Животное, отличное от человека, по любому из предшествующих пунктов, где гуманизированный эндогенный локус альбумина не содержит кассету селекции или репортерный ген.
7. Животное, отличное от человека, по любому из предшествующих пунктов, причем указанное животное, отличное от человека, содержит гуманизированный эндогенный локус альбумина в своей зародышевой линии.
8. Животное, отличное от человека, по любому из предшествующих пунктов, причем указанное животное, отличное от человека, представляет собой мышь.
9. Животное, отличное от человека, по любому из предшествующих пунктов, причем указанное животное, отличное от человека, является гомозиготным по гуманизированному эндогенному локусу альбумина.
10. Животное, отличное от человека, по любому из пп. 1-8, причем указанное животное, отличное от человека, является гетерозиготным по гуманизированному эндогенному локусу альбумина.
11. Животное, отличное от человека, по любому из предшествующих пунктов, причем указанное животное, отличное от человека, содержит уровни сывороточного альбумина, составляющие по меньшей мере приблизительно 10 мг/мл.
12. Животное, отличное от человека, по любому из предшествующих пунктов, причем уровни сывороточного альбумина у указанного животного, отличного от человека, по меньшей мере столь же высоки, как уровни сывороточного альбумина у контрольного животного дикого типа, отличного от человека, содержащего локус альбумина дикого типа.
13. Животное, отличное от человека, по любому из предшествующих пунктов, причем указанное животное, отличное от человека, дополнительно содержит кодирующую последовательность экзогенного белка, интегрированную в по меньшей мере один аллель гуманизированного эндогенного локуса альбумина в одной или более клетках указанного животного, отличного от человека.
14. Животное, отличное от человека, по п. 13, где кодирующая последовательность экзогенного белка интегрирована в интрон 1 по меньшей мере одного аллеля гуманизированного эндогенного локуса альбумина в одной или более клетках указанного животного, отличного от человека.
15. Животное, отличное от человека, по любому из предшествующих пунктов, причем указанное животное, отличное от человека, дополнительно содержит инактивированный эндогенный локус, который не является эндогенным локусом альбумина.
16. Животное, отличное от человека, по п. 15, причем указанное животное, отличное от человека, дополнительно содержит кодирующую последовательность экзогенного белка, интегрированную в по меньшей мере один аллель гуманизированного эндогенного локуса альбумина в одной или более клетках указанного животного, отличного от человека, где указанный экзогенный белок заменяет функцию указанного инактивированного эндогенного локуса.
17. Животное, отличное от человека, по п. 15 или 16, где указанный инактивированный эндогенный локус представляет собой инактивированный локус F9.
18. Клетка животного, отличного от человека, для экспрессии белка человеческого альбумина, при этом клетка животного, отличного от человека, содержит в своем геноме гуманизированный эндогенный локус альбумина, в котором область эндогенного локуса альбумина от старт-кодона до стоп-кодона удалена и заменена соответствующей последовательностью человеческого альбумина, содержащей область от старт-кодона до стоп-кодона гена человеческого альбумина,
где гуманизированный эндогенный локус альбумина содержит эндогенный промотор альбумина,
где последовательность человеческого альбумина функционально связана с эндогенным промотором альбумина, и
причем указанное животное, отличное от человека, представляет собой мышь или крысу.
19. Способ оценки активности реагента, нацеленного на человеческий альбумин, in vivo, включающий:
(a) введение указанного реагента, нацеленного на человеческий альбумин, животному, отличному от человека, по любому из пп. 1-17; и
(b) оценку активности указанного реагента, нацеленного на человеческий альбумин, у указанного животного, отличного от человека.
20. Способ по п. 19, где введение включает доставку, опосредованную аденоассоциированным вирусом (AAV), доставку, опосредованную липидными наночастицами (LNP), или гидродинамическую доставку (HDD).
21. Способ по п. 20, где введение включает опосредованную LNP доставку.
22. Способ по п. 21, где доза LNP составляет от приблизительно 0,1 мг/кг до приблизительно 2 мг/кг.
23. Способ по п. 22, где введение включает опосредованную AAV8 доставку.
24. Способ по любому из пп. 19-23, где стадия (b) включает выделение печени указанного животного, отличного от человека, и оценку активности указанного реагента, нацеленного на человеческий альбумин, в указанной печени.
25. Способ по любому из пп. 19-24, где указанный реагент, нацеленный на человеческий альбумин, представляет собой агент для редактирования генома, и указанная оценка включает оценку модификации гуманизированного эндогенного локуса альбумина.
26. Способ по п. 25, где указанная оценка включает измерение частоты вставок или делеций в гуманизированном эндогенном локусе альбумина.
27. Способ по любому из пп. 19-26, где указанная оценка включает измерение экспрессии матричной РНК альбумина, кодируемой гуманизированным эндогенным локусом альбумина или указанная оценка включает измерение экспрессии белка альбумина, кодируемого гуманизированным эндогенным локусом альбумина.
28. Способ по п. 27, где оценка экспрессии белка альбумина включает измерение сывороточных уровней белка альбумина у указанного животного, отличного от человека или оценка экспрессии белка альбумина включает измерение экспрессии белка альбумина в печени указанного животного, отличного от человека.
29. Способ по любому из пп. 19-28, где указанный реагент, нацеленный на человеческий альбумин, содержит нуклеазный агент или нуклеиновую кислоту, кодирующую указанный нуклеазный агент, где указанный нуклеазный агент разработан для нацеливания на область гена человеческого альбумина.
30. Способ по п. 29, где указанный нуклеазный агент содержит белок Cas и гидовую РНК, разработанную для нацеливания на последовательность-мишень гидовой РНК в гене человеческого альбумина.
31. Способ по п. 30, где указанная последовательность-мишень гидовой РНК находится в интроне 1 гена человеческого альбумина.
32. Способ по п. 30 или 31, где указанный белок Cas представляет собой белок Cas9.
33. Способ по любому из пп. 19-32, где указанный реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту, причем указанная экзогенная донорная нуклеиновая кислота разработана для нацеливания на ген человеческого альбумина, и при этом указанная экзогенная донорная нуклеиновая кислота доставляется посредством AAV.
34. Способ по п. 33, где указанная экзогенная донорная нуклеиновая кислота представляет собой одноцепочечный олигодезоксинуклеотид (оцОДН).
35. Способ по п. 33 или 34, где указанная экзогенная донорная нуклеиновая кислота не содержит гомологичных плеч.
36. Способ по п. 33 или 34, где указанная экзогенная донорная нуклеиновая кислота содержит вставку нуклеиновой кислоты, фланкированную 5’-гомологичным плечом, нацеленным на 5’-последовательность-мишень в гуманизированном эндогенном локусе альбумина, и 3’-гомологичным плечом, нацеленным на 3’-последовательность-мишень в гуманизированном эндогенном локусе альбумина.
37. Способ по п. 36, где каждая из 5’-последовательности-мишени и 3’-последовательности-мишени содержит сегмент интрона 1 гена человеческого альбумина.
38. Способ по любому из пп. 33-37, где указанная экзогенная донорная нуклеиновая кислота кодирует экзогенный белок.
39. Способ по п. 38, где указанный белок, кодируемый гуманизированным эндогенным локусом альбумина, на который осуществляли нацеливание с помощью указанной экзогенной донорной нуклеиновой кислоты, представляет собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с указанным экзогенным белком.
40. Способ по п. 38 или 39, где указанный экзогенный белок представляет собой белок фактора IX.
41. Способ по п. 40, где указанная оценка включает измерение сывороточных уровней белка фактора IX у указанного животного, отличного от человека, и/или включает оценку активированного частичного тромбопластинового времени или проведение теста генерации тромбина.
42. Способ по любому из пп. 19-41, где указанный реагент, нацеленный на человеческий альбумин, содержит (1) нуклеазный агент, разработанный для нацеливания на область гена человеческого альбумина, и (2) экзогенную донорную нуклеиновую кислоту,
где указанная экзогенная донорная нуклеиновая кислота разработана для нацеливания на ген человеческого альбумина,
где указанная экзогенная донорная нуклеиновая кислота кодирует экзогенный белок, и
где указанный белок, кодируемый гуманизированным эндогенным локусом альбумина, на который осуществляли нацеливание с помощью указанной экзогенной донорной нуклеиновой кислоты, представляет собой гетерологичный белок, содержащий сигнальный пептид человеческого альбумина, слитый с указанным экзогенным белком.
43. Способ по любому из пп. 38-42, где указанная оценка включает измерение экспрессии матричной РНК, кодируемой указанной экзогенной донорной нуклеиновой кислотой.
44. Способ по п. 43, где указанная оценка включает анализ гибридизации in situ для количественного определения экспрессии матричной РНК, кодируемой указанной экзогенной донорной нуклеиновой кислотой, с разрешением на уровне отдельных клеток.
45. Способ по п. 43 или 44, где указанная оценка включает измерение экспрессии матричной РНК, кодируемой указанной экзогенной донорной нуклеиновой кислотой, в нескольких долях печени животного, отличного от человека.
46. Способ по любому из пп. 38-45, где указанная оценка включает измерение экспрессии указанного экзогенного белка.
47. Способ по п. 46, где оценка экспрессии указанного гетерологичного белка включает измерение сывороточных уровней указанного гетерологичного белка у указанного животного, отличного от человека, или оценка экспрессии указанного гетерологичного белка включает измерение экспрессии в печени указанного животного, отличного от человека.
48. Способ выбора способа, обеспечивающего более высокую активность реагента, нацеленного на человеческий альбумин, in vivo, включающий:
(a) осуществление способа по любому из пп. 19-47 первый раз у первого животного, отличного от человека, содержащего в своем геноме гуманизированный эндогенный локус альбумина;
(b) изменение переменной и осуществление способа со стадии (a) второй раз с измененной переменной у второго животного, отличного от человека, содержащего в своем геноме, гуманизированный эндогенный локус альбумина; и
(c) сравнение активности указанного реагента, нацеленного на человеческий альбумин, на стадии (a) с активностью указанного реагента, нацеленного на человеческий альбумин, на стадии (b), и выбор способа, обеспечивающего более высокую активность.
49. Способ по п. 48, где:
(I) указанная изменяемая переменная на стадии (b) представляет собой способ доставки для введения указанного реагента, нацеленного на человеческий альбумин, указанному животному, отличному от человека;
(II) указанное введение включает опосредованную LNP доставку, и указанная изменяемая переменная на стадии (b) представляет собой препарат LNP;
(III) указанная изменяемая переменная на стадии (b) представляет собой путь введения указанного реагента, нацеленного на человеческий альбумин, указанному животному, отличному от человека;
(IV) указанная изменяемая переменная на стадии (b) представляет собой концентрацию или количество указанного реагента, нацеленного на человеческий альбумин, введенного указанному животному, отличному от человека;
(V) указанная изменяемая переменная на стадии (b) представляет собой форму указанного реагента, нацеленного на человеческий альбумин, введенного указанному животному, отличному от человека;
(VI) указанная изменяемая переменная на стадии (b) представляет собой указанный реагент, нацеленный на человеческий альбумин, введенный указанному животному, отличному от человека;
(VII) указанный реагент, нацеленный на человеческий альбумин, содержит белок Cas или нуклеиновую кислоту, кодирующую белок Cas, и гидовую РНК или ДНК, кодирующую гидовую РНК, где указанная гидовая РНК разработана для нацеливания на последовательность-мишень гидовой РНК в гене человеческого альбумина, и где (1) указанная изменяемая переменная на стадии (b) представляет собой последовательность указанной гидовой РНК или указанную последовательность-мишень гидовой РНК; (2) указанный реагент, нацеленный на человеческий альбумин, содержит матричную РНК (мРНК), кодирующую указанный белок Cas, и указанную гидовую РНК, и указанная изменяемая переменная на стадии (b) представляет собой отношение мРНК Cas к гидовой РНК или (3) изменяемая переменная на стадии (b) представляет собой модификации гидовой РНК; или
(VIII) указанный реагент, нацеленный на человеческий альбумин, содержит экзогенную донорную нуклеиновую кислоту, и где (1) указанная изменяемая переменная на стадии (b) представляет собой форму указанной экзогенной донорной нуклеиновой кислоты, или (2) указанная экзогенная донорная нуклеиновая кислота содержит вставку нуклеиновой кислоты, фланкированную 5’-гомологичным плечом, нацеленным на 5’-последовательность-мишень в гуманизированном эндогенном локусе альбумина, и 3’-гомологичным плечом, нацеленным на 3’-последовательность-мишень в гуманизированном эндогенном локусе альбумина, и указанная изменяемая переменная на стадии (b) представляет собой последовательность или длину указанного 5’-гомологичного плеча и/или последовательность или длину указанного 3’-гомологичного плеча.
50. Способ создания животного, отличного от человека, по любому из пп. 1-17, включающий:
(I) (a) модификацию генома эмбриональной стволовой (ЭС) клетки мыши или крысы, для включения гуманизированного эндогенного локуса альбумина, в котором область эндогенного локуса альбумина от старт-кодона до стоп-кодона удалена и заменена соответствующей последовательностью человеческого альбумина, содержащей область от старт-кодона до стоп-кодона гена человеческого альбумина, где гуманизированный эндогенный локус альбумина содержит эндогенный промотор альбумина, и где последовательность человеческого альбумина функционально связана с эндогенным промотором альбумина;
(b) идентификацию или отбор генетически модифицированной ЭС клетки мыши или крысы, содержащей гуманизированный эндогенный локус альбумина;
(c) введение генетически модифицированной ЭС клетки мыши или крысы в эмбрион хозяина мыши или крысы; и
(d) вынашивание указанного эмбриона хозяина, мыши или крысы у суррогатной матери мыши или крысы; или
(II) (a) модификацию генома находящегося на одноклеточной стадии эмбриона мыши или крысы для включения гуманизированного эндогенного локуса альбумина, в котором область эндогенного локуса альбумина от старт-кодона до стоп-кодона удалена и заменена соответствующей последовательностью человеческого альбумина, содержащей область от старт-кодона до стоп-кодона гена человеческого альбумина, где гуманизированный эндогенный локус альбумина содержит эндогенный промотор альбумина, и где последовательность человеческого альбумина функционально связана с эндогенным промотором альбумина;
(b) отбор генетически модифицированного находящегося на одноклеточной стадии эмбриона мыши или крысы, содержащего гуманизированный эндогенный локус альбумина; и
(c) вынашивание генетически модифицированного находящегося на одноклеточной стадии эмбриона мыши или крысы у суррогатной матери мыши или крысы.
| VIUFF D | |||
| et al | |||
| Generation of a double transgenic humanized neonatal Fc receptor (FcRn)/albumin mouse to study the pharmacokinetics of albumin-linked drugs | |||
| Journal of Controlled Release, 2016, Vol.223, pp.22-30, doi:10.1016/j.jconrel.2015.12.019 | |||
| URANO Y.et al | |||
| The human albumin gene | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |

