[0001] Перечень последовательностей, записанный в файле 10362WO01_ST25.txt, имеет размер 50 килобайт, создан 26 июня 2018 года и включен в данный документ посредством ссылки.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0002] Лечение многих заболеваний является недостаточно эффективным из-за неудовлетворительного нацеливания терапевтических молекул на соответствующие ткань или орган. Доставка биологически активных средств зачастую ограничена из-за трудностей с компонентами, достигающими целевых клетки или ткани.
[0003] Специфические для типа клеток эффекторы интернализации, такие как специфические для типа клеток рецепторы, например, ASGR1 человека, которые опосредованно или прямо интернализуются клеткой-мишенью, могут быть использованы для облегчения и улучшения доставки терапевтических молекул к специфическим клеткам-мишеням in vivo. Аналогично такие эффекторы интернализации, специфические для типа клеток, могут быть использованы с целью терапии для облегчения интернализации рецептора-мишени клеточной поверхности или растворимых белков-мишеней in vivo. Однако остается потребность в подходящих моделях для оценки эффективности таких механизмов доставки и терапевтических механизмов in vivo.
КРАТКОЕ ОПИСАНИЕ
[0004] Представлены животные, отличные от человека, содержащие гуманизированный локус Asgr1 и экспрессирующие гуманизированный или химерный белок ASGR1 в гуманизированном локусе Asgr1, а также способы использования таких животных, отличных от человека (например, грызуна, например, крысы или мыши), клетки и/или ткани, полученные от таких животных, отличных от человека, и нуклеотиды (например, целенаправленно воздействующие векторы, геномы и т.д.), применимые для создания таких животных.
[0005] В одном аспекте представлены животные, отличные от человека, содержащие генетически модифицированный эндогенный локус Asgr1, кодирующий модифицированный белок Asgr1, где модифицированный белок Asgr1 содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, и при этом весь внеклеточный домен или его часть кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. У некоторых таких животных, отличных от человека, внеклеточный домен содержит суперспиральный домен и лектиновый домен С-типа, и весь лектиновый домен С-типа или его часть кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. Необязательно лектиновый домен С-типа представляет собой лектиновый домен С-типа ASGR1 человека. Необязательно лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28.
[0006] У некоторых таких животных внеклеточный домен содержит суперспиральный домен и лектиновый домен С-типа, и весь суперспиральный домен или его часть кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. Необязательно суперспиральный домен представляет собой суперспиральный домен ASGR1 человека. Необязательно суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27.
[0007] У некоторых таких животных как весь суперспиральный домен, так и весь лектиновый домен С-типа или часть и того и другого кодируются сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. Необязательно лектиновый домен С-типа представляет собой лектиновый домен С-типа ASGR1 человека, и суперспиральный домен представляет собой суперспиральный домен ASGR1 человека. Необязательно лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28, и суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27.
[0008] У некоторых таких животных ортологичная последовательность ASGR1 человека содержит экзоны 3 8 гена ASGR1 человека. Необязательно ортологичная последовательность ASGR1 человека кодирует сегмент белка ASGR1, содержащий последовательность, изложенную под SEQ ID NO: 31.
[0009] У некоторых таких животных весь цитоплазматический домен или его часть кодируется эндогенной последовательностью Asgr1 животного, отличного от человека. У некоторых таких животных весь трансмембранный домена или его часть кодируется эндогенной последовательностью Asgr1 животного, отличного от человека. Необязательно как весь цитоплазматический домен, так и весь трансмембранный домен или часть и того и другого кодируются эндогенной последовательностью Asgr1 животного, отличного от человека.
[0010] В другом аспекте представлены клетки или ткани животных, отличных от человека (например, гепатоциты, эмбриональные стволовые клетки (ES) или другие плюрипотентные клетки, отличные от человеческих, такие как клетки зародышевой линии), содержащие гуманизированный локус Asgr1, и композиции in vitro, содержащие их. В одном аспекте представлены клетки животных, отличных от человека, содержащие генетически модифицированный эндогенный локус Asgr1, кодирующий модифицированный белок Asgr1, где модифицированный белок Asgr1 содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, и при этом весь внеклеточный домен или его часть кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. У некоторых таких клеток животных, отличных от человека, внеклеточный домен содержит суперспиральный домен и лектиновый домен С-типа, и весь лектиновый домен С-типа или его часть кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. Необязательно лектиновый домен С-типа представляет собой лектиновый домен С-типа ASGR1 человека. Необязательно лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28.
[0011] У некоторых таких клеток животных, отличных от человека, внеклеточный домен содержит суперспиральный домен и лектиновый домен С-типа, и весь суперспиральный домен или его часть кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. Необязательно суперспиральный домен представляет собой суперспиральный домен ASGR1 человека. Необязательно суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27.
[0012] У некоторых таких клеток животных, отличных от человека, как весь суперспиральный домен, так и весь лектиновый домен С-типа или часть и того и другого кодируются сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека. Необязательно лектиновый домен С-типа представляет собой лектиновый домен С-типа ASGR1 человека, и суперспиральный домен представляет собой суперспиральный домен ASGR1 человека Необязательно лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28, и суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27.
[0013] У некоторых таких клеток животных, отличных от человека, ортологичная последовательность ASGR1 человека содержит экзоны 3-8 гена ASGR1 человека. Необязательно ортологичная последовательность ASGR1 человека кодирует сегмент белка ASGR1, содержащий последовательность, изложенную под SEQ ID NO: 31.
[0014] У некоторых таких клеток животных, отличных от человека, весь цитоплазматический домен или его часть кодируется эндогенной последовательностью Asgr1 животного, отличного от человека. У некоторых таких клеток животных весь трансмембранный домен или его часть кодируется эндогенной последовательностью Asgr1 животного, отличного от человека. Необязательно как весь цитоплазматический домен, так и весь трансмембранный домен или часть и того и другого кодируются эндогенной последовательностью Asgr1 животного, отличного от человека.
[0015] Некоторые такие животные и/или клетки животных являются гетерозиготными по генетически модифицированному эндогенному локусу Asgr1. Некоторые такие животные и/или клетки животных являются гомозиготными по генетически модифицированному эндогенному локусу Asgr1.
[0016] Некоторые такие животные, отличные от человека, и/или клетки животных, отличных от человека, являются млекопитающими и/или клетками млекопитающих. Необязательно млекопитающее является грызуном. Необязательно грызун представляет собой крысу или мышь. Необязательно грызун представляет собой мышь. Необязательно весь цитоплазматический домен или его часть кодируется эндогенной последовательностью Asgr1 мыши. Необязательно цитоплазматический домен содержит последовательность, изложенную под SEQ ID NO: 29. Необязательно весь трансмембранный домен или его часть кодируется эндогенной последовательностью Asgr1 мыши. Необязательно трансмембранный домен содержит последовательность, изложенную под SEQ ID NO: 30. Необязательно как весь цитоплазматический домен, так и весь трансмембранный домен или часть и того и другого кодируются эндогенной последовательностью Asgr1 мыши. Необязательно цитоплазматический домен содержит последовательность, изложенную под SEQ ID NO: 29, и трансмембранный домен содержит последовательность, изложенную под SEQ ID NO: 30. Необязательно внеклеточный домен содержит суперспиральный домен и лектиновый домен С-типа, и где как весь суперспиральный домен, так и весь лектиновый домен С-типа или часть и того и другого кодируются сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека, и при этом как весь цитоплазматический домен, так и весь трансмембранный домен или часть и того и другого кодируются эндогенной последовательностью Asgr1 мыши. Необязательно лектиновый домен С-типа представляет собой лектиновый домен С-типа ASGR1 человека, и суперспиральный домен представляет собой суперспиральный домен ASGR1 человека, цитоплазматический домен представляет собой цитоплазматический домен Asgr1 мыши, а трансмембранный домен представляет собой трансмембранный домен Asgr1 мыши. Необязательно лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28, суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27, цитоплазматический домен содержит последовательность, изложенную под SEQ ID NO: 29, и трансмембранный домен содержит последовательность, изложенную под SEQ ID NO: 30. Необязательно модифицированный белок Asgr1 содержит последовательность, изложенную под SEQ ID NO: 3.
[0017] В другом аспекте представлены способы оценки доставки терапевтического комплекса в печень посредством интернализации, опосредованной ASGR1 человека, in vivo. Некоторые такие способы предусматривают: (а) введение терапевтического комплекса любому из указанных выше животных, отличных от человека, где терапевтический комплекс содержит терапевтическую молекулу и антиген связывающий белок или лиганд, который специфически связывает ASGR1 человека; и (b) оценку доставки терапевтической молекулы в печень животного, отличного от человека. Необязательно терапевтическая молекула представляет собой лизосомальный заместительный белок или фермент или нуклеиновую кислоту, кодирующую лизосомальный заместительный белок или фермент, и стадия (b) предусматривает оценку присутствия или активности лизосомального заместительного белка или фермента в печени животного, отличного от человека. Необязательно терапевтическая молекула представляет собой нуклеиновую кислоту, кодирующую терапевтический секретируемый белок, и стадия (b) предусматривает оценку уровней в сыворотке крови или активности терапевтического секретируемого белка у животного, отличного от человека.
[0018] В другом аспекте представлены способы оценки эффективности терапевтической молекулы, целенаправленно воздействующей на поверхностный белок клеток печени или на растворимый белок в печени, в отношении интернализации посредством ASGR1 человека in vivo. Некоторые такие способы предусматривают: (а) введение терапевтической молекулы любому из указанных выше животных, отличных от человека, где терапевтическая молекула содержит биспецифический антигенсвязывающий белок, который специфически связывает поверхностный белок клеток печени или растворимый белок и специфически связывает ASGR1 человека; и (b) оценку уровней на клеточной поверхности или активности поверхностного белка клеток печени в печени животного, отличного от человека, или оценку экспрессии или активности растворимого белка в печени животного, отличного от человека.
[0019] В другом аспекте представлены нуклеиновые кислоты, например, целенаправленно воздействующие векторы, предназначенные для создания гуманизированного локуса Asgr1, как это описано в данном документе, эндогенный локус Asgr1 животного, отличного от человека, экспрессирующий гуманизированный или химерный белок ASGR1, и/или геном животного, отличного от человека, содержащий локус Asgr1 человека, описанный в данном документе. Некоторые такие нуклеиновые кислоты содержат последовательность, выбранную из группы, включающей SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 23 и их комбинацию. Некоторые такие нуклеиновые кислоты содержат последовательность, которая кодирует аминокислотную последовательность, изложенную под SEQ ID NO: 3, например, содержат последовательность, изложенную под SEQ ID NO: 24.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0020] Файл с патентом или заявкой содержит по меньшей мере один графический материал, выполненный в цвете. Копии данного патента или публикации заявки на патент с цветным (цветными) графическим (графическими) материалом (материалами) будут предоставлены Ведомству после запроса и уплаты необходимой пошлины.
[0021] На фигуре 1А показано схематическое изображение локуса ASGR1 человека. Восемь экзонов обозначены черными прямоугольниками, нетранслируемые участки (UTR) обозначены белыми прямоугольниками, а трансмембранный, суперспиральный домены и лектиновый домен С-типа указаны вверху фигуры. Звездочками обозначены местоположения 5'-праймера (7302hTU) и 3'-праймера (7302hTD), предназначенных для анализа приобретения аллеля. Фрагмент, подлежащий включению в локус Asgr1 мыши для обеспечения гуманизации, показан внизу.
[0022] На фигуре 1В показано схематическое изображение локуса Asgr1 мыши. Восемь экзонов обозначены черными прямоугольниками, UTR обозначены белыми прямоугольниками, а трансмембранный, суперспиральный домены и лектиновый домен С-типа указаны вверху фигуры. Звездочками обозначены местоположения 3'-праймера (7302mTU) и 3'-праймера (7302mTD), предназначенных для анализа потери аллеля. Фрагмент, подлежащий удалению и замене соответствующим фрагментом из локуса ASGR1 человека, показан внизу.
[0023] На фигуре 2А показано схематическое изображение крупного целенаправленно воздействующего вектора, предназначенного для получения гуманизированного аллеля Asgr1 (аллель 7302), содержащего самоудаляющуюся кассету, содержащую ген устойчивости к гигромицину. Экзоны мыши обозначены серыми прямоугольниками, экзоны человека обозначены черными прямоугольниками, a UTR обозначены белыми прямоугольниками. Границы между различными участками (мышь/человек, человек/кассета и кассета/мышь) указаны линиями, соответственно обозначенными А, В и С, в нижней части рисунка.
[0024] На фигуре 2В показано схематическое изображение варианта гуманизированного аллеля Asgr1 с удаленной кассетой на фигуре 2А. Экзоны мыши обозначены серыми прямоугольниками, экзоны человека обозначены черными прямоугольниками, a UTR обозначены белыми прямоугольниками. Границы между различными участками (5'-граница мышь/человек, 3'-граница человек/мышь) указаны линиями, соответственно обозначенными A, D, в нижней части рисунка.
[0025] На фигуре 3 показано выравнивание белка ASGR1 человека (hASGR1; SEQ ID NO: 1), белка Asgr1 мыши (mAsgr1; SEQ ID NO: 2) и гуманизированного белка Asgr1 мыши (7302 humln; SEQ ID NO: 3). Подчеркнутые остатки представляют собой такие остатки, которые кодируются введенными экзонами человека. Остатки, заключенные в рамку, составляют трансмембранный домен. Пунктирная линия обозначает лектиновый домен С-типа. Сплошная жирная линия обозначает суперспиральный участок.
[0026] На фигуре 4 показано, что у гуманизированных по Asgr1 мышей (Asgr1hu/hu) липидные профили в плазме крови (включая общий холестерин, триглицериды, LDL-C и HDL-C) аналогичны таким профилям у их однопометников дикого типа (Asgr1+/+).
[0027] На фигуре 5 показано, что у гуманизированных по Asgr1 мышей (Asgr1hu/hu) вес тела и уровень глюкозы в крови аналогичны таким показателям у их однопометников дикого типа.
[0028] На фигуре 6 показано, что гуманизированный белок Asgr1 гуманизированных по Asgr1 мышей (Asgr1hu/hu) совместно локализуется в мембранах клеток печени, аналогично Asgr1 мыши. Трансферриновый рецептор (TRFR) и глицеральдегид-3-фосфатдегидрогеназу (GAPDH) использовали в качестве загрузочного контроля для мембранных и цитозольных фракций печени соответственно. Показано N=4 на группу.
[0029] На фигуре 7 показано выравнивание белка ASGR1 человека (SEQ ID NO: 1), Macaca fascicularis (макак-крабоед) (SEQ ID NO: 39), белка Asgr1 мыши (SEQ ID NO: 2) и белка Asgr1 крысы (SEQ ID NO: 41). Подчеркнутые остатки представляют собой такие остатки, которые кодируются введенными экзонами человека. Наблюдается 97,6% идентичность последовательности белков ASGR1 человека и макака-крабоеда (98,3% идентичность последовательности во внеклеточном домене), 77% идентичность последовательности белков ASGR1 человека и мыши и 78,4% идентичность последовательности белков ASGR1 человека и крысы.
[0030] На фигурах 8А - 8С представлены изображения, полученные с помощью иммунофлуоресцентной микроскопии, образцов печени (А), селезенки (В) или почки (С), отобранных у мышей C57BL/6, трансгенно модифицированных с целью экспрессии ASGR1 человека клетками печени (i-iv), или мышей C57BL/6 дикого типа (v-viii) через десять дней после внутривенного введения 1×1011 scAAV2-CMV-eGFP дикого типа (i, v), физиологического раствора (ii, vi), 1×1011 вирусных векторов scAAV2-N587myc-CMV-eGFP отдельно (iii, vii) или вирусных векторов scAAV2-N587myc-CMV-eGFP с биспецифическим антителом к myc-ASGR1 (iv, viii).
[0031] На фигурах 9A - 9R представлены изображения, полученные с помощью иммунофлуоресцентной микроскопии, образцов печени, отобранных у мышей C57BL/6, трансгенно модифицированных с целью экспрессии ASGR1 человека клетками печени (фигура 9D - 9F, 9J - 9L, 9Р - 9R), или мышей C57BL/6 дикого типа (фигуры 9А - 9С, 9G - 9I, 9М - 90) через четыре недели после внутривенного введения 2,18×1011 ssAAV2-CAGG-eGFP дикого типа (фигуры 9В, 9С, 9Е, 9F), физиологического раствора (фигуры 9А, 9D), 2,18×1011 вирусных векторов ssAAV2-N587myc-CAGG-eGFP отдельно (фигуры 9G - 9I, 9J - 9L) или вирусных векторов ssAAV2-N587myc-CAGG-eGFP с биспецифическим антителом к myc-ASGR1 (фигуры 9М - 90, 9Р - 9R). Каждое изображение представляет одну мышь.
ОПРЕДЕЛЕНИЯ
[0032] Термины «белок», «полипептид» и «пептид», используемые в данном документе взаимозаменяемо, включают полимерные формы аминокислот любой длины, включая кодируемые и некодируемые аминокислоты и химически или биохимически модифицированные или дериватизированные аминокислоты. Термины также включают полимеры, которые были модифицированы, такие как полипептиды, содержащие модифицированные пептидные остовы. Термин «домен» относится к любой части белка или полипептида, обладающего конкретной функцией или структурой.
[0033] Считается, что белки имеют «N-конец» и «С-конец». Термин «N-конец» относится к началу белка или полипептида, оканчивающемуся аминокислотой со свободной аминогруппой (-NH2). Термин «С-конец» относится к концу аминокислотной цепи (белка или полипептида), оканчивающейся свободной карбоксильной группой (-СООН).
[0034] Термины «нуклеиновая кислота» и «полинуклеотид», используемые в данном документе взаимозаменяемо, включают полимерные формы нуклеотидов любой длины, включая рибонуклеотиды, дезоксирибонуклеотиды или их аналоги или модифицированные варианты. Они включают одно-, двух- и многонитевые ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие природные, химически модифицированные, биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания.
[0035] Считается, что нуклеиновые кислоты имеют «5'-концы» и «3'-концы», потому что мононуклеотиды вступают в реакцию с образованием олигонуклеотидов таким образом, что 5'-фосфат пентозного кольца одного мононуклеотида присоединяется к 3'-кислороду своего соседа в одном направлении через фосфодиэфирную связь. Конец олигонуклеотида называют «5'-концом», если его 5'-фосфат не связан с 3'-кислородом пентозного кольца мононуклеотида. Конец олигонуклеотида называют «3'-концом», если его 3'-кислород не связан с 5'-фосфатом пентозного кольца другого мононуклеотида. Можно также сказать, что последовательность нуклеиновой кислоты, даже если она находится внутри более крупного олигонуклеотида, имеет 5'- и 3'-концы. В молекуле линейной или кольцевой ДНК дискретные элементы называются как «расположенные выше» или в 5'-направлении по отношению к «расположенным ниже» элементам или элементам в 3'-направлении.
[0036] Термин «геномно интегрированный» относится к нуклеиновой кислоте, которая была введена в клетку таким образом, что нуклеотидная последовательность интегрируется в геном клетки и способна наследоваться ее потомством. Любой протокол может быть использован для стабильного включения нуклеиновой кислоты в геном клетки.
[0037] Термин «целенаправленно воздействующий вектор» относится к рекомбинантной нуклеиновой кислоте, которая может быть введена путем гомологичной рекомбинации, лигирования, опосредованного негомологичным соединением концов, или любым другим способом рекомбинации в целевое местоположение в геноме клетки.
[0038] Термин «вирусный вектор» относится к рекомбинантной нуклеиновой кислоте, которая включает по меньшей мере один элемент вирусного происхождения и включает элементы, достаточные для упаковки или допускающие упаковку в частицу вирусного вектора. Вектор и/или частица могут быть использованы с целью переноса ДНК, РНК или других нуклеиновых кислот в клетки ex vivo или in vivo. Известны многочисленные формы вирусных векторов.
[0039] Термин «дикий тип» включает объекты, обладающие структурой и/или активностью, обнаруживаемой в нормальном состоянии (в отличие от мутантного, поврежденного, измененного и т.д.) или окружении. Гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллелях).
[0040] Термин «эндогенный» относится к последовательности нуклеиновой кислоты, которая встречается в природе внутри клетки или животного, отличного от человека. Например, эндогенная последовательность Asgr1 животного, отличного от человека, относится к нативной последовательности Asgr1, которая встречается в природе в локусе Asgr1 животного, отличного от человека.
[0041] «Экзогенные» молекулы или последовательности включают молекулы или последовательности, которые обычно не присутствуют в клетке в такой форме. Присутствие в нормальном состоянии включает присутствие относительно определенной стадии развития и условий окружающей среды клетки. Например, экзогенная молекула или последовательность могут включать мутантный вариант соответствующей эндогенной последовательности в клетке, например, гуманизированный вариант эндогенной последовательности, или могут включать последовательность, соответствующую эндогенной последовательности в клетке, но в другой форме (т.е. не в составе хромосомы). И наоборот, эндогенные молекулы или последовательности включают молекулы или последовательности, которые обычно присутствуют в такой форме в определенной клетке на определенной стадии развития в определенных условиях окружающей среды.
[0042] Термин «гетерологичный» при использовании в контексте нуклеиновой кислоты или белка указывает на то, что нуклеиновая кислота или белок содержат по меньшей мере две части, которые не встречаются в природе вместе в одной и той же молекуле. Например, термин «гетерологичный» при использовании его в отношении частей нуклеиновой кислоты или частей белка указывает, что нуклеиновая кислота или белок содержат две или более подпоследовательности, которые не встречаются в природе в таком же соотношении друг с другом (например, объединенными вместе). В качестве одного примера «гетерологичный» участок вектора, представляющего собой нуклеиновую кислоту, представляет собой сегмент нуклеиновой кислоты в пределах или присоединенный к другой молекуле нуклеиновой кислоты, который не встречается в природе в ассоциации с другой молекулой. Например, гетерологичный участок вектора, представляющего собой нуклеиновую кислоту, может включать кодирующую последовательность, фланкированную последовательностями, которые не встречаются в природе в ассоциации с кодирующей последовательностью. Аналогичным образом «гетерологичный» участок белка представляет собой сегмент аминокислот в пределах или присоединенный к другой пептидной молекуле, который не встречается в природе в ассоциации с другой пептидной молекулой (например, слитый белок или белок с меткой). Аналогичным образом нуклеиновая кислота или белок могут содержать гетерологичную метку или гетерологичную последовательность секреции или последовательность локализации.
[0043] «Оптимизация кодонов» использует преимущества вырожденности кодонов, что проявляется в множественности комбинаций кодонов из трех пар оснований, которые определяют аминокислоту, и, как правило, включает процесс модификации последовательности нуклеиновой кислоты с целью улучшения экспрессии в определенных клетках-хозяевах путем замены по меньшей мере одного кодона нативной последовательности кодоном, который более часто или чаще всего задействован в генах данной клетки-хозяина, сохраняя при этом нативную аминокислотную последовательность. Например, нуклеиновая кислота, кодирующая белок Cas9, может быть модифицирована с целью замены кодонов на другие, имеющие более высокую частоту используемости в данной прокариотической или эукариотической клетке, включая бактериальную клетку, дрожжевую клетку, клетку человека, клетку, отличную от человеческой, клетку млекопитающего, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина, по сравнению со встречающейся в природе последовательностью нуклеиновой кислоты. Списки используемости кодонов являются общедоступными, например, в Базе данных используемости кодонов «Codon Usage Database)). Эти таблицы можно адаптировать в ряде способов. См. Nakamura et al. (2000) Nucleic Acids Research 28:292, включенную в данный документ посредством ссылки во всей своей полноте для всех целей. Также доступны компьютерные алгоритмы оптимизации кодонов определенной последовательности, предназначенную для экспрессии в конкретном хозяине, см., например, Gene Forge.
[0044] Термин «локус» относится к конкретному местоположению гена (или значимой последовательности), последовательности ДНК, последовательности, кодирующей полипептид, или положению на хромосоме генома организма. Например, «локус Asgr1» может относиться к конкретному местоположению гена Asgr1, последовательности ДНК Asgr1, последовательности, кодирующей Asgr1, или к положению Asgr1 на хромосоме генома организма, которое было идентифицировано как такое положение, в котором расположена такая последовательность. «Локус Asgr1» может содержать регуляторный элемент гена Asgr1, включая, например, энхансер, промотор, 5'-концевой и/или 3'-концевой нетранслируемый участок (UTR) или их комбинацию.
[0045] Термин «ген» относится к последовательности ДНК в хромосоме, которая кодирует продукт (например, продукт РНК и/или полипептидный продукт) и включает кодирующий участок, прерываемый некодирующими нитронами, и последовательность, расположенную смежно с кодирующим участком как на 5'-конце, так и на 3'-конце, таким образом, что ген соответствует полноразмерной мРНК (включая 5'- и 3'-нетранслируемые последовательности). Термин «ген» также включает другие некодирующие последовательности, включая регуляторные последовательности (например, промоторы, энхансеры и сайты связывания факторов транскрипции), сигналы полиаденилирования, участки внутренней посадки рибосом, сайленсеры, инсуляторные последовательности и участки прикрепления к матриксу. Эти последовательности могут быть расположены близко к кодирующему участку гена (например, в пределах 10 т.о.) или в отдаленных сайтах, и они влияют на уровень или скорость транскрипции и трансляции гена.
[0046] Термин «аллель» относится к вариантной форме гена. Некоторые гены имеют множество различных форм, которые расположены в одном и том же положении или генетическом локусе на хромосоме. Диплоидный организм имеет два аллеля в каждом генетическом локусе. Каждая пара аллелей представляет генотип определенного генетического локуса. Генотипы описаны как гомозиготные, если в конкретном локусе содержатся два идентичных аллеля, и как гетерозиготные, если эти два аллеля различаются.
[0047] «Промотор» представляет собой регуляторный участок ДНК, обычно содержащий ТАТА-бокс, способный управлять РНК-полимеразой II с целью инициации синтеза РНК в соответствующем сайте инициации транскрипции для конкретной полинуклеотидной последовательности. Промотор может дополнительно содержать другие участки, которые влияют на скорость инициации транскрипции. Описанные в данном документе промоторные последовательности модулируют транскрипцию функционально связанного полинуклеотида. Промотор может быть активным в одном или более типах клеток, раскрытых в данном документе (например, в эукариотической клетке, клетке млекопитающего, отличного от человека, клетке человека, клетке грызуна, плюрипотентной клетке, эмбрионе стадии одной клетки, дифференцированной клетке или их комбинации). Промотор может быть, например, конститутивно активным промотором, условным промотором, индуцируемым промотором, ограниченным во времени промотором (например, промотором, регулируемым развитием) или пространственно ограниченным промотором (например, клеточноспецифическим или тканеспецифическим промотором). Примеры промоторов можно найти, например, в WO 2013/176772, включенной в данный документ посредством ссылки во всей своей полноте для всех целей.
[0048] Термины «функциональная связь» или «функционально связанный» включают сопоставление двух или более компонентов (например, промотора и другого элемента последовательности) таким образом, что оба компонента функционируют нормально и допускают возможность того, что по меньшей мере один из компонентов может опосредовать функцию, которая воздействует по меньшей мере на один из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более транскрипционных регуляторных факторов. Функциональная связь может включать такие последовательности, которые являются смежными друг с другом или проявляют активность в транс-положении (например, регуляторная последовательность может действовать на расстоянии для контроля транскрипции кодирующей последовательности).
[0049] Термин «вариант» относится к нуклеотидной последовательности, отличающейся от последовательности, наиболее распространенной в популяции (например, по одному нуклеотиду), или последовательности белка, отличающейся от последовательности, наиболее распространенной в популяции (например, по одной аминокислоте).
[0050] Термин «фрагмент» применительно к белку означает белок, который короче или содержит меньше аминокислот, чем полноразмерный белок. Термин «фрагмент» применительно к нуклеиновой кислоте означает нуклеиновую кислоту, которая короче или содержит меньше нуклеотидов, чем полноразмерная нуклеиновая кислота. Фрагмент может представлять собой, например, N-концевой фрагмент (т.е. удаление части С-конца белка), С-концевой фрагмент (т.е. удаление части N-конца белка) или внутренний фрагмент.
[0051] «Идентичность последовательности» или «идентичность» в контексте двух полинуклеотидных или полипептидных последовательностей относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия в указанном окне сравнения. Когда процент идентичности последовательности используется в отношении белков, положения остатков, которые не являются идентичными, часто отличаются консервативными аминокислотными заменами, где аминокислотные остатки заменены другими аминокислотными остатками со сходными химическими свойствами (например, зарядом или гидрофобностью) и, следовательно, не изменяют функциональные свойства молекулы. Когда две или более аминокислотных последовательностей отличаются консервативными заменами, процент идентичности можно повышать для коррекции характера консервативности замены. Считается, что последовательности, которые отличаются такими консервативными заменами, имеют «сходство последовательностей» или «сходство». Средства для такой корректировки хорошо известны. Как правило, они включают оценку консервативного замещения как частичного, а не полного несоответствия, за счет чего увеличивается процент идентичности последовательности. Таким образом, например, когда идентичной аминокислоте присваивают оценку 1, а неконсервативной замене присваивают оценку ноль, консервативная замена получает оценку от нуля до 1. Оценка в баллах консервативных замен рассчитывается таким образом, как это, например, реализовано в программе PC/GENE (Intelligenetics, Маунтин-Вью, Калифорния, США)
[0052] «Процент идентичности последовательности» включает значение, определенное путем сравнения двух оптимально выровненных в окне сравнения последовательностей (наибольшее количество идеально совпадающих остатков), где часть полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) по сравнению с эталонной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания двух последовательностей. Процентное значение рассчитывают путем определения количества положений, в которых в обеих последовательностях встречается идентичное основание нуклеиновой кислоты или идентичный аминокислотный остаток, с получением количества положений с совпадениями, деления количества положений с совпадениями на общее количество положений в окне сравнения и умножения результата на 100 с получением процента идентичности последовательности. Если не указано иное (например, более короткая последовательность включает связанную гетерологичную последовательность), окно сравнения представляет собой полную длину более короткой из двух последовательностей сравнения.
[0053] Если не указано иное, значения идентичности/сходства последовательности включают значение, полученное с использованием программы GAP версии 10 с использованием следующих параметров: % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа за открытие гэпа 50 и штрафа за продление гэпа 3 и матрицы замен nwsgapdna.cmp; % идентичности и % сходства для аминокислотной последовательности с использованием штрафа за открытие гэпа 8 и штрафа за продление гэпа 2 и матрицы замен BLOSUM62 или любой эквивалентной программы. «Эквивалентная программа» включает любую программу сравнения последовательностей, которая для любых двух рассматриваемых последовательностей генерирует выравнивание, имеющее совпадения идентичных нуклеотидных или аминокислотных остатков и идентичный процент идентичности последовательностей по сравнению с соответствующим выравниванием, полученным с помощью GAP версии 10.
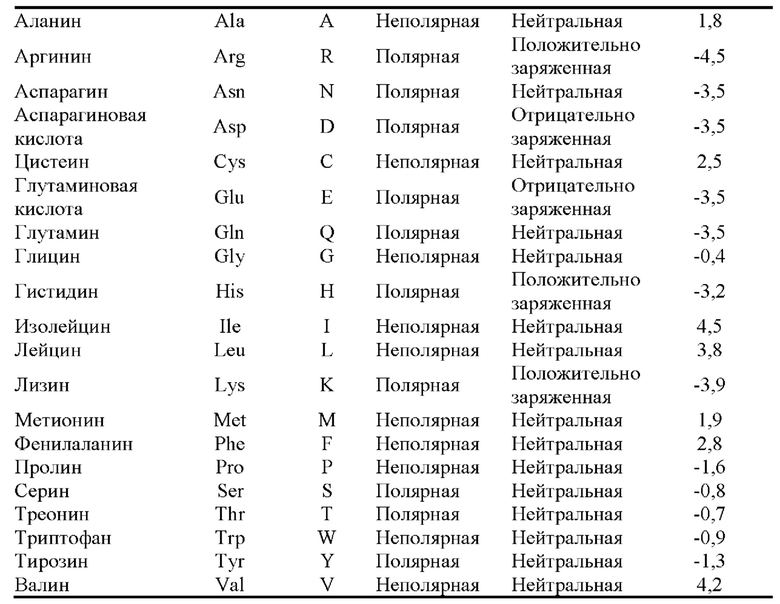
[0054] Термин «консервативная аминокислотная замена» относится к замене аминокислоты, которая обычно присутствует в последовательности, другой аминокислотой с аналогичными размером, зарядом или полярностью. Примеры консервативных замен включают замену неполярного (гидрофобного) остатка, такого как изолейцин, валин или лейцин, другим неполярным остатком. Аналогичным образом примеры консервативных замен включают замену одного полярного (гидрофильного) остатка другим, например, аргинина лизином, глутамина аспарагином или глицина серином. Кроме того, замена основного остатка, такого как лизин, аргинин или гистидин, другим, или замена одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота, другим кислотным остатком являются дополнительными примерами консервативных замен. Примеры неконсервативных замен включают замену неполярного (гидрофобного) аминокислотного остатка, такого как изолейцин, валин, лейцин, аланин или метионин, полярным (гидрофильным) остатком, таким как цистеин, глутамин, глутаминовая кислота или лизин, и/или полярного остатка неполярным остатком. Типичное распределение аминокислот по категориям обобщено ниже.
[0055] «Гомологичная» последовательность (например, последовательность нуклеиновой кислоты) включает последовательность, которая является либо идентичной, либо по сути аналогичной известной эталонной последовательности таким образом, что она, например, на по меньшей мере 50%, на по меньшей мере 55%), на по меньшей мере 60%, на по меньшей мере 65%), на по меньшей мере 70%, на по меньшей мере 75%, на по меньшей мере 80%, на по меньшей мере 85%, на по меньшей мере 90%, на по меньшей мере 95%), на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%), на по меньшей мере 99% или на 100% идентична известной эталонной последовательности. Гомологичные последовательности могут включать, например, ортологичные последовательности и паралогичные последовательности. Например, гомологичные гены обычно передаются от общей предковой последовательности ДНК, либо через событие видообразования (ортологичные гены), либо через событие генетической дупликации (паралогичные гены). «Ортологичные» гены включают гены разных видов, которые произошли от общего предкового гена путем видообразования. Ортологи обычно сохраняют ту же функцию в ходе эволюции. «Паралогичные» гены включают гены, связанные путем дупликации в геноме. У паралогов могут появляться новые функции в ходе эволюции.
[0056] Термин «in vitro» включает искусственную среду и процессы или реакции, которые происходят в искусственной среде (например, в тестовой пробирке). Термин «т vivo» включает природную среду (например, клетку, организм или тело) и процессы или реакции, которые происходят в природной среде. Термин «ех vivo» включает клетки, которые были удалены из организма человека, а также процессы или реакции, которые происходят внутри таких клеток.
[0057] Термин «репортерный ген» относится к нуклеиновой кислоте, имеющей последовательность, кодирующую продукт гена (как правило, фермент), который с легкостью поддается количественному анализу, когда конструкцию, содержащую последовательность репортерного гена, функционально связанную с гетерологичным промоторным и/или энхансерным элементом вводят в клетки, содержащие (или которые могут содержать) факторы, необходимые для активации промоторных и/или энхансерных элементов. Примеры репортерных генов включают без ограничения гены, кодирующие бета-галактозидазу (lacZ), гены бактериальной хлорамфеникол-ацетилтрансферазы (cat), гены люциферазы светлячка, гены, кодирующие бета-глюкуронидазу (GUS), и гены, кодирующие флуоресцентные белки. «Репортерный белою) относится к белку, кодируемому репортерным геном.
[0058] Используемый в данном документе термин «флуоресцентный репортерный белок» означает репортерный белок, который поддается обнаружению за счет флуоресценции, причем флуоресценция может возникать либо непосредственно от репортерного белка, либо в результате активности репортерного белка на флуорогенном субстрате, либо от белка с аффинностью связывания с флуоресцентно меченным соединением. Примеры флуоресцентных белков включают зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, Monomeric Azami Green, CopGFP, AceGFP и ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP и ZsYellowl), синие флуоресцентные белки (например, BFP, eBFP, eBFP2, азурит, mKalamal, GFPuv, Sapphire и T-sapphire), голубые флуоресцентные белки (например, CFP, eCFP, лазурь, CyPet, AmCyanl и Midoriishi-Cyan), красные флуоресцентные белки (например, RFP, mKate, mKate2, mPlum, DsRed monomer, mCherry, mRFPl, DsRed-Express, DsRed2, DsRed-Monomer, HcRed-Tandem, AsRed2, eqFP611, mRaspberry, m Strawberry и Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, Monomeric Kusabira-Orange, mTangerine и tdTomato) и любой другой подходящий флуоресцентный белок, присутствие которого в клетках можно обнаружить с помощью методик проточной цитометрии.
[0059] Термин «рекомбинация» включает любой процесс обмена генетической информацией между двумя полинуклеотидами и может происходить осуществляться посредством любого механизма. Рекомбинация в ответ на двухнитевые разрывы (DSB) происходит главным образом за счет двух консервативных путей репарации ДНК: негомологичное соединение концов (NHEJ) и гомологичная рекомбинация (HR). См. Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22:886-897, включенную в данный документ посредством ссылки во всей своей полноте для всех целей. Аналогичным образом репарация целевой нуклеиновой кислоты, опосредованное экзогенной донорной нуклеиновой кислотой, может включать любой процесс обмена генетической информацией между двумя полинуклеотидами.
[0060] NHEJ включает репарацию двухнитевых разрывов в нуклеиновой кислоте путем прямого лигирования концов разрыва друг с другом или с экзогенной последовательностью без потребности в гомологичной матрице. Лигирование не являющихся смежными последовательностей с помощью NHEJ часто может приводить к делециям, вставкам или транслокациям вблизи сайта двухнитевого разрыва. Например, NHEJ также может приводить к направленной интеграции экзогенной донорной нуклеиновой кислоты посредством прямого лигирования концов разрыва с концами экзогенной донорной нуклеиновой кислоты (т.е. захват на основе NHEJ). Такая NHEJ-опосредованная направленная интеграция может быть предпочтительной для вставки экзогенной донорной нуклеиновой кислоты, если пути гомологичной репарации (HDR) не всегда могут использоваться (например, в неделящихся клетках, первичных клетках и клетках, в которых гомологичная репарация ДНК осуществляется недостаточно эффективно). Кроме того, в отличие от гомологичной репарации, не требуется информация об относительно больших участках идентичности последовательности, фланкирующих сайт расщепления, что может быть полезным при попытке целенаправленной вставки в организмы с геномами, знание о геномной последовательности которых ограничено. Интеграция может происходить посредством лигирования «тупых концов» экзогенной донорной нуклеиновой кислоты и расщепленной геномной последовательности или посредством лигирования «липких концов» (т.е. имеющих 5'- или 3'-свисающие концы) с использованием экзогенной донорной нуклеиновой кислоты, которая фланкирована свисающими концами, которые совместимы с такими концами, образующимся за счет нуклеазного средства в расщепленной геномной последовательности. См., например, US 2011/020722, WO 2014/033644, WO 2014/089290 и Maresca et al. (2013) Genome Res. 23(3): 539-546, каждая из которых включена в данный документ посредством ссылки во всей своей полноте для всех целей. Если «тупые концы» лигированы, может потребоваться эксцизия мишени и/или донора для получения участков микрогомологии, необходимых для соединения фрагментов, что может привести к нежелательным изменениям в целевой последовательности.
[0061] Рекомбинация также может происходить посредством гомологичной репарации (HDR) или гомологичной рекомбинации (HR). HDR или HR включает форму репарации нуклеиновой кислоты, для которая может потребоваться гомология нуклеотидной последовательности, использование «донорной» молекулы в качестве матрицы для репарации «целевой» молекулы (то есть той, в которой произошел двухнитевой разрыв), и которая приводит к передаче генетической информации от донора к мишени. Без ограничения какой-либо конкретной теорией, такой перенос может включать коррекцию несоответствия гетеродуплексной ДНК, которое образуется между поврежденной мишенью и донором, и/или отжиг нитей, зависимый от синтеза, при котором донор используется для повторного синтеза генетической информации, которая станет частью мишени и/или связанных процессов. В ряде случаев донорный полинуклеотид, часть донорного полинуклеотида, копия донорного полинуклеотида или часть копии донорного полинуклеотида интегрируют в ДНК-мишень. См. Wang et al. (2013) Cell 153:910-918; Mandalos et al. (2012) PLOS ONE 7:e45768:1-9; и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждая из которых включена в данный документ посредством ссылки во всей своей полноте для всех целей.
[0062] Термин «антигенсвязывающий белок» включает любой белок, который связывается с антигеном. Примеры антигенсвязывающих белков включают антитело, антигенсвязывающий фрагмент антитела, полиспецифическое антитело (например, биспецифическое антитело), scFV, бис-scFV, диатело, триатело, тетратело, V-NAR, VHH, VL, F(ab), F(ab)2, DVD (антигенсвязывающий белок с двумя вариабельными доменами), SVD (антигенсвязывающий белок с одним вариабельным доменом), биспецифический рекрутер Т-клеток (BiTE) или Davisbody (патент США №8586713, включенный в данный документ посредством ссылки во всей своей полноте для всех целей).
[0063] Термин «полиспецифический» или «биспецифический» в отношении антиген связывающего белка означает, что белок распознает разные эпитопы, либо на одном и том же антигене, либо на разных антигенах. Полиспецифическая антиген связывающая молекула может представлять собой отдельный многофункциональный полипептид или может представлять собой мультимерный комплекс из двух или более полипептидов, ковалентно или нековалентно связанных друг с другом. Например, антитело или его фрагмент могут быть функционально связаны (например, путем химического сочетания, генетического слияния, нековалентной ассоциации или прочих) с одним или более другими молекулярными объектами, такими как белок или его фрагмент, с получением биспецифической или полиспецифической антигенсвязывающей молекулы со второй специфичностью связывания.
[0064] Термин «антиген» относится к веществу, целой молекуле или домену в составе молекулы, которое способно инициировать продуцирование антител со специфичностью связывания с данным веществом. Термин антиген также включает вещества, которые в организмах-хозяевах дикого типа не вызывают продуцирование антител за счет распознавания «своего», но могут вызывать такой ответ у животного-хозяина с соответствующими генноинженерными модификациями, предназначенными для нарушения иммунологической толерантности.
[0065] Термин «эпитоп» относится к сайту на антигене, с которым связывается антигенсвязывающий белок (например, антитело). Эпитоп может образовываться из смежных аминокислот или несмежных аминокислот, сближенных за счет фолдинга одного или более белков в третичную структуру. Эпитопы, образованные смежными аминокислотами (также известные как линейные эпитопы), как правило, сохраняются при воздействии денатурирующими растворителями, тогда как эпитопы, образованные за счет фолдинга в третичную структуру (также известные как конформационные эпитопы), как правило, утрачиваются при обработке денатурирующими растворителями. Как правило, эпитоп включает по меньшей мере 3, а обычно по меньшей мере 5 или 8 10 аминокислот в уникальной пространственной конформации. Способы определения пространственной конформации эпитопов включают, например, рентгеновскую кристаллографию и 2-мерный ядерный магнитный резонанс. См., например, Epitope Mapping Protocols, in Methods in Molecular Biology, Vol.66, Glenn E. Morris, Ed. (1996), включенный в данный документ посредством ссылки во всей своей полноте для всех целей.
[0066] Паратоп антитела, как описано в данном документе, обычно содержит по меньшей мере определяющий комплементарность участок (CDR), который специфически распознает гетерологичный эпитоп (например, участок CDR3 вариабельного домена тяжелой и/или легкой цепи).
[0067] Термин «антитело» предусматривает молекулы иммуноглобулинов, содержащие четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь содержит вариабельный домен тяжелой цепи и константный участок тяжелой цепи (CH). Константный участок тяжелой цепи содержит три домена: CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельный домен легкой цепи и константный участок легкой цепи (CL). Вариабельные домены тяжелой цепи и легкой цепи могут быть дополнительно подразделены на участки гипервариабельности, называемые определяющими комплементарность участками (CDR), чередующиеся с более консервативными участками, называемыми каркасными участками (FR). Каждый вариабельный домен тяжелой и легкой цепей содержит три CDR и четыре FR, расположенных в направлении от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи могут сокращенно называться как HCDR1, HCDR2 и HCDR3; CDR легкой цепи могут сокращенно называться как LCDR1, LCDR2 и LCDR3). Термин «высокоаффинное» антитело относится к антителу, которое характеризуется Kd по отношению к своему целевому эпитопу, составляющей приблизительно 10-9 М или менее (например, приблизительно 1×10-9 М, 1×10-10 М, 1×10-11 М или приблизительно 1×10-12 М). В одном варианте осуществления Kd измеряют с помощью поверхностного плазмонного резонанса, например, BIACORE™; в другом варианте осуществления, Kd измеряют с помощью ELISA.
[0068] Термин «биспецифическое антитело» включает антитело, способное селективно связывать два или более эпитопа. Биспецифические антитела обычно содержат две разные тяжелые цепи, при этом каждая тяжелая цепь специфически связывается с отдельным эпитопом либо на двух разных молекулах (например, на двух разных антигенах), либо на одной и той же молекуле (например, на одном и том же антигене). Если биспецифическое антитело способно селективно связываться с двумя разными эпитопами (первым эпитопом и вторым эпитопом), то аффинность первой тяжелой цепи по отношению к первому эпитопу обычно будет по меньшей мере на один, два, или три, или четыре порядка ниже аффинности первой тяжелой цепи по отношению ко второму эпитопу, и наоборот. Эпитопы, распознаваемые биспецифическим антителом, могут располагаться на одной и той же или на другой мишени (например, на одном и том же или на другом белке). Биспецифические антитела могут быть получены, например, путем объединения тяжелых цепей, которые распознают разные эпитопы одного и того же антигена. Например, последовательности нуклеиновой кислоты, кодирующие те вариабельные последовательности тяжелой цепи, которые распознают разные эпитопы одного и того же антигена, могут быть слиты с последовательностями нуклеиновой кислоты, кодирующими разные константные участки тяжелой цепи, и при этом такие последовательности могут быть экспрессированы в клетке, которая экспрессирует легкую цепь иммуноглобулина. Типичное биспецифическое антитело имеет две тяжелые цепи, каждая из которых содержит три CDR тяжелой цепи, за которыми следует (от N-конца к С-концу) СН1-домен, шарнирный участок, СН2-домен и СН3-домен, и легкую цепь иммуноглобулина, которая либо не наделяет антиген-связывающей специфичностью, но которая при этом может ассоциироваться с каждой тяжелой цепью, либо которая может ассоциироваться с каждой тяжелой цепью и которая может связываться с одним или более эпитопами, связывающимися антиген-связывающими участками тяжелой цепи, либо которая может ассоциироваться с каждой тяжелой цепью и обеспечивать связывание либо одной, либо обеих тяжелых цепей с одним или обоими эпитопами.
[0069] Термин «тяжелая цепь» или «тяжелая цепь иммуноглобулина» включает последовательность тяжелой цепи иммуноглобулина, в том числе последовательность константного участка тяжелой цепи иммуноглобулина, из любого организма. Вариабельные домены тяжелой цепи содержат три CDR тяжелой цепи и четыре FR-участка, если не указано иное. Фрагменты тяжелых цепей включают CDR, CDR и FR и их комбинации. Типичная тяжелая цепь после вариабельного домена (от N-конца к С-концу) содержит CH1-домен, шарнирный участок, CH2-домен и СН3-домен. Функциональный фрагмент тяжелой цепи включает фрагмент, который способен специфически распознавать эпитоп (например, распознавать эпитоп с Kd в микромолярном, наномолярном или пикомолярном диапазоне), который способен к экспрессии и секреции из клетки и который содержит по меньшей мере одну CDR. Вариабельные домены тяжелой цепи кодируются нуклеотидной последовательностью вариабельного участка, который обычно содержит сегменты VH, DH и JH, полученные из репертуара сегментов VH, DH и JH, присутствующих в антителах зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов V, D и J тяжелой цепи для различных организмов можно найти в базе данных IMGT, которая доступна через Интернет во всемирной паутине (www) по адресу ресурса в сети «imgt.org».
[0070] Термин «легкая цепь» включает последовательность легкой цепи иммуноглобулина из любого организма, и, если не указано иное, включает каппа (κ) и лямбда (λ) легкие цепи человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены легкой цепи обычно включают три CDR легкой цепи и четыре каркасных (FR) участка, если не указано иное. Как правило, полноразмерная легкая цепь включает в направлении от аминоконца к карбоксильному концу вариабельный домен, который включает FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и аминокислотную последовательность константного участка легкой цепи. Вариабельные домены легкой цепи кодируются нуклеотидной последовательностью вариабельного участка легкой цепи, который обычно включает сегменты гена VL легкой цепи и JL легкой цепи, полученные из репертуара сегментов гена V и J легкой цепи, присутствующих в антителах зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов гена V и J легкой цепи для различных организмов можно найти в базе данных IMGT, которая доступна через Интернет во всемирной паутине (www) по адресу ресурса в сети Ошибка! Недопустимый объект гиперссылки.. Легкие цепи включают, например, цепи, которые селективно не связывают ни первый, ни второй эпитопы, селективно связанные эпитоп-связывающим белком, в котором они встречаются. Легкие цепи также включают те цепи, которые связывают и распознают или помогают тяжелой цепи в связывании и распознавании одного или более эпитопов, селективно связанных эпитоп-связывающим белком, в котором они находятся.
[0071] Используемый в данном документе термин «определяющий комплементарность участок» или «CDR» предусматривает аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулина организма, которая обычно (т.е. у животного дикого типа) располагается между двумя каркасными участками в вариабельном участке легкой или тяжелой цепей молекулы иммуноглобулина (например, антитела или рецептора Т-клетки). CDR может кодироваться, например, последовательностью зародышевой линии или перегруппированной последовательностью, и, например, наивной или зрелой В-клеткой или Т-клеткой. CDR может быть подвергнут соматической мутации (например, отличаться от последовательности, кодируемой в зародышевой линии у животного), гуманизации и/или модификации с помощью аминокислотных замен, добавлений или делеций. В некоторых случаях (например, в случае CDR3) CDR могут кодироваться двумя или более последовательностями (например, последовательностями зародышевой линии), которые не являются смежными (например, в случае неперегруппированной последовательности нуклеиновой кислоты), но они являются смежными в последовательности нуклеиновой кислоты в В-клетке, например, как результат сплайсинга или соединения последовательностей (например, V-D-J-рекомбинация с образованием CDR3 тяжелой цепи).
[0072] Специфическое связывание антиген связывающего белка с его целевым антигеном включает связывание с аффинностью, составляющей по меньшей мере 106, 107, 108, 109 или 1010 М-1. Специфическое связывание явно выше по значению и отличается от неспецифического связывания, происходящего по меньшей мере с одной неродственной мишенью. Специфическое связывание может быть результатом образования связей между определенными функциональными группами или определенного пространственного соответствия (например, взаимодействия по типу «замок и ключ»), тогда как неспецифическое связывание обычно является результатом действия ван-дер-ваальсовых сил. Однако специфическое связывание не обязательно подразумевает, что антигенсвязывающий белок связывает одну и только одну мишень.
[0073] Композиции или способы, «содержащие» или «включающие» один или более перечисленных элементов, могут включать другие элементы, которые конкретно не указаны. Например, композиция, которая «содержит» или «включает» белок, может содержать белок отдельно или в комбинации с другими ингредиентами. Переходная фраза «состоящий по сути из» означает, что объем формулы изобретения следует интерпретировать как охватывающий указанные элементы, перечисленные в формуле изобретения, и те элементы, которые не оказывают существенного влияния на основную и новую характеристику (характеристики) заявляемого изобретения. Таким образом, выражение «состоящий по сути из» при использовании в формуле настоящего изобретения не предназначен для того, чтобы истолковываться как эквивалент выражению «содержащий».
[0074] «Необязательный» или «необязательно» означает, что описываемое последующее событие или обстоятельство может произойти или может не произойти, и что данное описание включает случаи, когда событие или обстоятельство возникают, и случаи, когда они не возникают.
[0075] Обозначение диапазона значений включает все целые числа определяющие диапазон и все поддиапазоны или в их пределах, определенные целыми числами в пределах диапазона.
[0076] Если иное не очевидно из контекста, выражение «приблизительно» охватывает значения в пределах стандартного предела погрешности измерения (например, SEM) указанного значения.
[0077] Союзы «и/или» относятся к любым возможным комбинациям одного или более связанных перечисленных элементов, а также к отсутствию комбинаций при интерпретации в виде альтернативы («или») и охватывают их.
[0078] Союз «или» относится к любому одному представителю определенного списка, а также включает любую комбинацию представителей этого списка.
[0079] Формы единственного числа включают ссылки на формы множественного числа, если контекст явно не указывает иное. Например, термины «белок» или «по меньшей мере один белок» могут включать множество белков, в том числе их смеси.
[0080] Статистически значимое означает р≤0,05.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Обзор
[0081] В данном документе раскрыты клетки животных, отличных от человека, и животные, отличные от человека, содержащие гуманизированный локус Asgr1, и способы применения таких клеток животных, отличных от человека, и животных, отличных от человека. Клетки животных, отличных от человека, или животные, отличные от человека, содержащие гуманизированный локус Asgr1, экспрессируют белок ASGR1 человека или химерный белок Asgr1, содержащий один или более фрагментов белка ASGR1 человека (например, весь внеклеточный домен ASGR1 человека или его часть). В качестве высокоэффективного специфического для печени рецептора эндоцитоза ASGR1 человека может быть использован для специфической для печени доставки терапевтических средств, таких как антитела, малые молекулы (в качестве части конъюгатов антитело-лекарственное средство) и ДНК. Однако антигенсвязывающие белки или биспецифические антигенсвязывающие белки, которые специфически связывают ASGR1 человека, зачастую не будут связываться с ортологичными белками Asgr1 животного, отличного от человека, такими как Asgr1 мыши, из-за различий между последовательностями ASGR1 человека и Asgr1 животного, отличного от человека. Например, антитела, полученные к внеклеточному домену ASGR1 человека, не связываются с ортологом Asgr1 мыши (данные не показаны). По этой причине эффективность in vivo механизмов доставки, опосредованной ASGR1 человека, или терапевтических механизмов невозможно эффективно оценить у животных, отличных от человека, с немодифицированными эндогенными (т.е. нативными) локусами Asgr1. Гуманизированные по Asgr1 животные, отличные от человека, (например, мыши с гуманизированным Asgr1) можно использовать для валидации специфической для печени доставки различных терапевтических средств посредством интернализации, опосредованной ASGR1 человека, с использованием ряда различных подходов. Например, животные, отличные от человека, содержащие гуманизированный локус Asgr1, могут быть использованы для оценки in vivo эффективности опосредованной ASGR1 человека доставки терапевтических молекул или терапевтических комплексов в печень. Аналогично животные, отличные от человека, содержащие гуманизированный локус Asgr1, могут быть использованы для оценки эффективности терапевтических молекул или терапевтических комплексов, действующих посредством механизмов, опосредованных ASGR1 человека.
II. Животные, отличные от человека, содержащие гуманизированный локус Asgr1
[0082] Клетки и животные, отличные от человека, раскрытые в данном документе, содержат гуманизированный локус Asgr1. Клетки или животные, отличные от человека, содержащие гуманизированный локус Asgr1, экспрессируют белок ASGR1 человека или частично гуманизированный химерный белок Asgr1, в котором один или более фрагментов нативного белка Asgr1 были заменены соответствующими фрагментами из ASGR1 человека (например, весь внеклеточный домен или его часть).
А. Асиалогликопротеиновый рецептор 1 (ASGR1)
[0083] Клетки и животные, отличные от человека, описанные в данном документе, содержат гуманизированный локус Asgr1. Асиалогликопротеиновый рецептор 1 (представитель H1 семейства 4 белков с лектиновым доменом С-типа, печеночный лектин H1, HL-1, ASGP-R 1, ASGPR 1, ASGR1) кодируется геном ASGR1 (CLEC4H1) и является основной субъединицей асиалогликопротеинового рецептора (ASGPR или ASGR). ASGPR представляет собой гетероолигомерный белок, экспрессируемый в основном на клеточной поверхности гепатоцитов, с примерно 1-5×105 сайтов связывания/клетка, который играет важную роль в интернализации и расщеплении десиалилированных гликопротеинов с удалением их из кровотока. ASGPR представляет собой хорошо изученный печеночный лектин С-типа, экспрессирующийся главным образом на синусоидальной поверхности гепатоцитов. ASGPR является ответственным за селективное связывание и интернализацию гликопротеинов с концевыми остатками галактозы или N-ацетилгалактозамина клетками паренхимы печени посредством рецептор-опосредованного эндоцитоза. Он содержит два белка, асиалогликопротеиновые рецепторы 1 и 2 (ASGR1 и ASGR2), кодируемые генами ASGR1 и ASGR2. Обе субъединицы представляют собой однопроходные белки типа II, которые обычно содержат N-концевой цитоплазматический домен, один трансмембранный домен и С-концевой внеклеточный домен распознавания углеводов (CRD). ASGR1 содержит N-концевой цитоплазматический домен (~40 аминокислот), однопроходный трансмембранный домен (~20 аминокислот), внеклеточный суперспиральный стволовой участок (олигомеризация) (~80 аминокислот) и функциональный домен распознавания углеводов С-типа (кальций-зависимый) (лектиновые домены С-типа) (*-140 аминокислот). CRD связывается с гликопротеинами с концевыми галактозными или N-ацетилгалактозаминовыми (GalNac) мотивами. CRD характеризуется низкой аффинностью к десиалилированным гликопротеинам в мономерном состоянии.
[0084] Гены, кодирующие ASGR1 и ASGR2 (ASGR1 и ASGR2 соответственно), расположены на коротком плече хромосомы 17 на расстоянии примерно 58,6 тысяч оснований (т.о.) друг от друга. Гены характеризуются эволюционным родством, но существенно различаются по своей структурной организации: ASGR1 содержит 8 экзонов и имеет длину приблизительно 6 т.о., a ASGR2 содержит 9 экзонов и имеет длину приблизительно 13,5 т.о.
[0085] Иллюстративной кодирующей последовательности для ASGR1 человека присвоен номер доступа NM_001671 (SEQ ID NO: 5) в NCBI. Иллюстративной кодирующей последовательности для Asgr1 мыши присвоен номер доступа NM_009714 (SEQ ID NO: 4) в NCBI. Иллюстративному белку ASGR1 человека присвоен номер доступа Р07306 (SEQ ID NO: 1) в UniProt. Иллюстративному белку Asgr1 мыши присвоен номер доступа Р34927 (SEQ ID NO: 2) в UniProt. Иллюстративный белок Asgr1 мыши с гуманизированными суперспиральным доменом и лектиновым доменом С-типа изложен под SEQ ID NO: 3. Иллюстративному белку Asgr1 крысы присвоен номер доступа: Р02706 в UniProt. Иллюстративному белку Asgr1 орангутана присвоен номер доступа Q5RBQ8 в UniProt.
В. Гуманизированные локусы Asgr1
[0086] Гуманизированный локус Asgr1 может представлять собой локус Asgr1, в котором весь ген Asgr1 заменен соответствующей ортологичной последовательностью ASGR1 человека, или он может представлять собой локус Asgr1, в котором только часть гена Asgr1 заменена соответствующей ортологичной последовательностью ASGR1 человека, (т.е. гуманизирован). Необязательно соответствующую ортологичную последовательность ASGR1 человека подвергают модификации с целью оптимизации кодонов на основе частоты использования кодонов у животных, отличных от человека. Замененные (т.е. гуманизированные) участки могут включать кодирующие участки, такие как экзон, некодирующие участки, такие как интрон, нетранслируемый участок или регуляторный участок (например, промотор, энхансер или элемент, связывающий транскрипционный репрессор) или любую их комбинацию. В качестве одного примера экзоны, соответствующие 1, 2, 3, 4, 5, 6, 7 или всем 8 экзонам гена ASGR1 человека, могут быть гуманизированы. Например, экзоны, соответствующие экзонам 3-8 гена ASGR1 человека, могут быть гуманизированы. Альтернативно участок Asgr1, кодирующий эпитоп, распознаваемый антигенсвязывающим белком, представляющим собой антитело к ASGR1 человека, может быть гуманизирован. В качестве другого примера один или более или все домены, включающие N-концевой цитоплазматический домен, трансмембранный домен, суперспиральный домен или лектиновый домен С-типа, могут быть гуманизированы. Например, весь участок локуса Asgr1 или его часть, кодирующего суперспиральный домен, могут быть гуманизированы, весь участок локуса Asgr1 или его часть, кодирующего лектиновый домен С-типа, могут быть гуманизированы, весь участок локуса Asgr1 или его часть, кодирующего трансмембранный домен, могут быть гуманизированы, и/или весь участок локуса Asgr1 или его часть, кодирующего цитоплазматический домен, могут быть гуманизированы. В одном примере только весь участок локуса Asgr1 или его часть, кодирующего суперспиральный домен, гуманизированы, только весь участок локуса Asgr1 или его часть, кодирующего лектиновый домен С-типа, гуманизированы, или только весь участок локуса Asgr1 или его часть, кодирующего внеклеточный участок (т.е. суперспиральный домен и лектиновый домен С-типа). Например, участки локуса Asgr1, кодирующего суперспиральный домен и лектиновый домен С-типа, могут быть гуманизированы таким образом, что химерный белок Asgr1 будет продуцироваться с эндогенным N-концевым цитоплазматическим доменом, эндогенным трансмембранным доменом, гуманизированным суперспиральным доменом и гуманизированным лектиновым доменом С-типа. Аналогичным образом интроны, соответствующие 1, 2, 3, 4, 5, 6 или всем 7 интронам гена ASGR1 человека, могут быть гуманизированы. Фланкирующие нетранслируемые участки, в том числе регуляторные последовательности, также могут быть гуманизированы. Например, 5'-нетранслируемый участок (UTR), 3'-UTR или и 5'-UTR, и 3'-UTR могут быть гуманизированы, или 5'-UTR, 3'-UTR или и 5'-UTR, и 3-'UTR могут оставаться эндогенными. В одном конкретном примере 3-'UTR гуманизирован, но 5'-UTR остается эндогенным. В зависимости от степени замены ортологичными последовательностями регуляторные последовательности, такие как промотор, могут быть эндогенными или обеспечены замещающей ортологичной последовательностью человека. Например, гуманизированный локус Asgr1 может включать эндогенный промотор Asgr1 животного, отличного от человека.
[0087] Белок Asgr1, кодируемый гуманизированным локусом Asgr1, может содержать один или более доменов, происходящих из белка ASGR1 человека. Например, белок Asgr1 может содержать один или более доменов, включающих суперспиральный домен ASGR1 человека, лектиновый домен С-типа ASGR1 человека, трансмембранный домен ASGR1 человека и цитоплазматический домен ASGR1 человека, или все их. В качестве одного примера белок Asgr1 может содержать только суперспиральный домен ASGR1 человека, только лектиновый домен С-типа ASGR1 человека или только внеклеточный домен ASGR1 человека (т.е. суперспиральный домен и лектиновый домен С-типа). Необязательно белок Asgr1, кодируемый гуманизированным локусом Asgr1, также может содержать один или более доменов, происходящих из эндогенного (т.е. нативного) белка Asgr1 животного, отличного от человека. В качестве одного примера белок Asgr1, кодируемый гуманизированным локусом Asgr1, может содержать суперспиральный домен из белка ASGR1 человека, лектиновый домен С-типа из белка ASGR1 человека, N-концевой цитоплазматический домен из эндогенного (т.е. нативного) белка Asgr1 животного, отличного от человека, и трансмембранный домен из эндогенного (т.е. нативного) белка Asgr1 животного, отличного от человека. Домены из белка ASGR1 человека могут кодироваться полностью гуманизированной последовательностью (т.е. вся последовательность, кодирующая этот домен, заменена ортологичной последовательностью ASGR1 человека), или могут кодироваться частично гуманизированной последовательностью (т.е. некоторые последовательности, кодирующие домен, заменены ортологичной последовательностью ASGR1 человека, а оставшаяся эндогенная (т.е. нативная) последовательность, кодирующая этот домен, кодирует те же аминокислоты, что и ортологичная последовательность ASGR1 человека, таким образом, что кодируемый домен идентичен этому домену в белке ASGR1 человека).
[0088] В качестве одного примера белок Asgr1, кодируемый гуманизированным локусом Asgr1, может содержать суперспиральный домен ASGR1 человека. Необязательно суперспиральный домен ASGR1 человека содержит, состоит по сути из или состоит из последовательности, которая на по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности под SEQ ID NO: 27, и при этом белок Asgr1 сохраняет активность нативного Asgr1 (т.е. сохраняет способность к избирательному связыванию и интернализации гликопротеинов с концевыми остатками галактозы или N-ацетилгалактозамина посредством рецептор-опосредованного эндоцитоза). В качестве другого примера белок Asgr1, кодируемый гуманизированным локусом Asgr1, может содержать лектиновый домен С-типа из ASGR1 человека. Необязательно лектиновый домен С-типа из ASGR1 человека содержит, состоит по сути из или состоит из последовательности, которая на по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности под SEQ ID NO: 28, и при этом белок Asgr1 сохраняет активность нативного Asgr1. Например, участок белка Asgr1, который получен из ASGR1 человека, может содержать, состоять по сути из или состоять из последовательности, которая на по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности под SEQ ID NO: 31, и при этом белок Asgr1 сохраняет активность нативного Asgr1. В качестве другого примера белок Asgr1, кодируемый гуманизированным локусом Asgr1, может содержать эндогенный цитоплазматический домен Asgr1 животного, отличного от человека (например, цитоплазматический домен Asgr1 мыши). Необязательно цитоплазматический домен Asgr1 животного, отличного от человека, содержит, состоит по сути из или состоит из последовательности, которая на по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности под SEQ ID NO: 29, и при этом белок Asgr1 сохраняет активность нативного Asgr1. В качестве другого примера белок Asgr1, кодируемый гуманизированным локусом Asgr1, может содержать эндогенный трансмембранный домен Asgr1 животного, отличного от человека (например, трансмембранный домен Asgr1 мыши). Необязательно трансмембранный домен Asgr1 животного, отличного от человека, состоит по сути из или состоит из последовательности, которая на по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности под SEQ ID NO: 30, и при этом белок Asgr1 сохраняет активность нативного Asgr1. Например, белок Asgr1, кодируемый гуманизированным локусом Asgr1, может содержать, состоять по сути из или состоять из последовательности, которая на по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности под SEQ ID NO: 3, и при белок Asgr1 сохраняет активность нативного Asgr1.
[0089] Необязательно гуманизированный локус Asgr1 может содержать другие элементы. Примеры таких элементов могут включать кассеты с селектируемым маркером, репортерные гены, сайты распознавания рекомбиназ или другие элементы. Альтернативно в гуманизированном локусе Asgr1 могут отсутствовать другие элементы (например, может отсутствовать селектируемый маркер или кассета с селектируемым маркером). Примеры подходящих репортерных генов и репортерных белков раскрыты в другом месте данного документа. Примеры подходящих селектируемых маркеров включают неомицин-фосфотрансферазу (neor), гигромицин-В-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror), бластицидин-S-дезаминазу (bsrr), ксантин/гуанин-фосфорибозилтрансферазу (gpt) и тимидинкиназу вируса простого герпеса (HSV-k). Примеры рекомбиназ включают рекомбиназы Cre, Flp и Dre. Одним примером гена рекомбиназы Cre является Crei, в котором два экзона, кодирующие рекомбиназу Cre, разделены интроном, чтобы предотвратить его экспрессию в прокариотической клетке. Такие рекомбиназы могут дополнительно содержать сигнал внутриядерной локализации для обеспечения локализации в ядре (например, NLS-Crei). Сайты распознавания рекомбиназы включают нуклеотидные последовательности, которые распознаются сайт-специфической рекомбиназой и могут служить в качестве субстрата для события рекомбинации. Примеры сайтов распознавания рекомбиназ включают FRT, FRT11, FRT71, сайты attp, art, rox и lox, такие как loxP, lox511, lox2272, lox66, lox71, loxM2 и lox5171.
[0090] Другие элементы, такие как репортерные гены или кассеты с селектируемым маркером, могут представлять собой самоудаляющиеся кассеты, фланкированные сайтами распознавания рекомбиназ. См., например, US 8697851 и US 2013/0312129, каждый из которых включен в данный документ посредством ссылки во всей своей полноте для всех целей. В качестве примера самоудаляющаяся кассета может содержать ген Crei (содержит два экзона, кодирующих рекомбиназу Cre, которые разделены интроном), функционально связанный с промотором Prm1 мыши, и ген устойчивости к неомицину, функционально связанный с промотором убиквитина человека. При использовании промотора Prm1, самоудаляющаяся кассета может быть специфически удалена в мужских половых клетках зародышевой линии животных F0. Полинуклеотид, кодирующий селектируемый маркер, может быть функционально связан с активным промотором в клетке-мишени. Примеры промоторов описаны в другом месте данного документа. В качестве другого конкретного примера самоудаляющаяся кассета с селектируемым маркером может содержать последовательность, кодирующую ген устойчивости к гигромицину, функционально связанную с одним или более промоторами (например, и с промотором убиквитина человека, и с промоторами ЕМ7), после которых следует сигнал полиаденилирования, после которого следует кодирующая Crei последовательность, функционально связанная с одним или более промоторами (например, с промотором mPrm1), после которого следует другой сигнал полиаденилирования, где вся кассета фланкирована сайтами loxP.
[0091] Гуманизированный локус Asgr1 также может представлять собой условный аллель. Например, условный аллель может представлять собой многофункциональный аллель, описанный в US 2011/0104799, включенном в данный документ посредством ссылки во всей своей полноте для всех целей. Например, условный аллель может содержать: (а) инициирующую последовательность в смысловой ориентации относительно транскрипции гена-мишени; (b) кассету с селектируемым геном устойчивости к лекарственному средству (DSC) в смысловой или антисмысловой ориентации; (с) представляющую интерес нуклеотидную последовательность (NSI) в антисмысловой ориентации; и (d) модуль обусловленный инверсией (COIN, который использует экзон-разделяющий интрон и инвертируемый подобный «генной ловушке» модуль) в обратной ориентации. См., например, US 2011/0104799. Условный аллель может дополнительно содержать рекомбинируемые единицы, которые рекомбинируют при воздействии первой рекомбиназы с образованием условного аллеля, который (i) не имеет инициирующей последовательности и DSC; и (ii) содержит NSI в смысловой ориентации и COIN в антисмысловой ориентации. См., например, US 2011/0104799.
[0092] Один иллюстративный гуманизированный локус Asgr1 (например, гуманизированный локус Asgr1 мыши) представляет собой локус, в котором кодирующие экзоны 3 8 заменены соответствующей последовательностью человека. Данные экзоны кодируют суперспиральный домен и лектиновый домен С-типа Asgr1. Необязательно гуманизированная последовательность может проходить через стоп-кодон и 3-'UTR и, необязательно, в последовательность, расположенную непосредственно ниже 3'-UTR. Необязательно часть интрона, расположенного выше кодирующего экзона 3, также гуманизирована. См. фигуры 2А и 2В и SEQ ID NO: 21 и 24.
С. Клетки, отличные от человеческих, и животные, отличные от человека, содержащие гуманизированный локус Asgr1
[0093] Представлены клетки животных, отличных от человека, и животные, отличные от человека, содержащие гуманизированный локус Asgr1, описанный в другом месте данного документа. Клетки или животные, отличные от человека, могут быть гетерозиготными или гомозиготными по гуманизированному локусу Asgr1. Диплоидный организм имеет два аллеля в каждом генетическом локусе. Каждая пара аллелей представляет генотип определенного генетического локуса. Генотипы описаны как гомозиготные, если в конкретном локусе содержатся два идентичных аллеля, и как гетерозиготные, если эти два аллеля различаются.
[0094] Клетки животных, отличных от человека, представленные в данном документе, могут представлять собой, например, любую клетку, отличную от человеческой, содержащую локус Asgr1 или геномный локус, гомологичный или ортологичный локусу ASGR1 человека. Клетки могут представлять собой эукариотические клетки, которые включают, например, клетки грибов (например, дрожжи), клетки растений, клетки животных, клетки млекопитающих, клетки млекопитающих, отличных от человека, и клетки человека. Термин «животное» включает млекопитающих, рыб и птиц. Клеткой млекопитающего представляет собой, например, клетку млекопитающего, отличного от человека, клетку грызуна, клетку крысы, клетку мыши или клетку хомяка. Другие млекопитающие, отличные от человека, включают, например, приматов, отличных от человека, нечеловекообразных обезьян, человекообразных обезьян, орангутанов, кошек, собак, кроликов, лошадей, быков, оленей, бизонов, домашний скот (например, виды крупного рогатого скота, такие как коровы, быки и т.д.; виды полорогих, такие как овцы, козы и т.д.; и виды свиней, такие как свиньи и кабаны). Птицы включают, например, кур, индеек, страусов, гусей, уток и т.д. Также включены домашние животные и сельскохозяйственные животные. Термин «отличный от человека» исключает людей.
[0095] Клетки также могут быть любого типа в недифференцированном или дифференцированном состоянии. Например, клетка может представлять собой тотипотентную клетку, плюрипотентную клетку (например, плюрипотентную клетку человека или плюрипотентную клетку, отличную от человеческой, такую как эмбрионального стволовую (ES) клетку мыши или ES-клетку крысы), или не являющуюся плюрипотентной клетку. Тотипотентные клетки включают недифференцированные клетки, которые могут давать начало клеткам любого типа, а плюрипотентные клетки включают недифференцированные клетки, которые обладают способностью развиваться в более чем один тип дифференцированных клеток. Такими плюрипотентными и/или тотипотентными клетками могут быть, например, ES-клетки или ES-подобные клетки, такие как индуцированные плюрипотентные стволовые (iPS) клетки. ES-клетки включают тотипотентные или плюрипотентные клетки эмбрионального происхождения, которые способны вносить вклад в любую ткань развивающегося эмбриона при введении в эмбрион. ES-клетки могут быть получены из внутренней клеточной массы бластоцисты и способны дифференцироваться в клетки любого из трех зародышевых листков позвоночных (энтодермы, эктодермы и мезодермы).
[0096] Клетки, представленные в данном документе, также могут представлять собой клетки зародышевой линии (например, сперматозоиды или ооциты). Клетки могут представлять собой митотически компетентные клетки или митотически неактивные клетки, мейотически компетентные клетки или мейотически неактивные клетки. Аналогично клетки также могут представлять собой первичные соматические клетки или клетки, которые не являются первичными соматическими клетками. Соматические клетки включают любую клетку, которая не является гаметой, клеткой зародышевой линии, гаметоцитом или недифференцированной стволовой клеткой. Например, клетки могут представлять собой клетки печени, такие как гепатобласты или гепатоциты.
[0097] Подходящие клетки, представленные в данном документе, также включают первичные клетки. Первичные клетки включают клетки или культуры клеток, которые были выделены непосредственно из организма, органа или ткани. Первичные клетки включают клетки, которые не являются ни трансформированными, ни иммортализованными. Они включают любую клетку, полученную из организма, органа или ткани, которую ранее не подвергали пассированию в культуре ткани или ранее подвергали пассированию в культуре ткани, но не способна бесконечно пассироваться в культуре ткани. Такие клетки могут быть выделены с помощью общепринятых методик и включают, например, гепатоциты.
[0098] Другие подходящие клетки, представленные в данном документе, включают иммортализованные клетки. К иммортализованным клеткам относятся клетки многоклеточного организма, которые в обычных условиях не размножались бы бесконечно, но из-за мутации или изменения избежали нормального клеточного старения и вместо этого могут продолжать дальнейшее деление. Такие мутации или изменения могут происходить естественным путем или могут быть инициированы преднамеренно. Конкретным примером иммортализованной клеточной линии является клеточная линия рака печени человека HepG2. Хорошо известно множество типов иммортализованных клеток. Иммортализованные или первичные клетки включают клетки, которые обычно используются для культивирования или для экспрессии рекомбинантных генов или белков.
[0099] Клетки, представленные в данном документе, также включают эмбрионы на стадии одной клетки (т.е. оплодотворенные ооциты или зиготы). Такие эмбрионы на стадии одной клетки могут быть получены из любого генетического фона (например, BALB/c, C57BL/6, 129 или их комбинации для мышей), могут быть свежими или замороженными, и могут быть получены в результате естественного разведения или оплодотворения in vitro.
[00100] Клетки, представленные в данном документе, могут представлять собой нормальные, здоровые клетки или могут представлять собой пораженные заболеванием или содержащие мутации клетки.
[00101] Животные, отличные от человека, содержащие гуманизированный локус Asgr1, описанный в данном документе, могут быть получены с помощью способов, описанных в другом месте данного документа. Термин «животное» включает млекопитающих, рыб и птиц. Млекопитающие, отличные от человека, включают, например, приматов, отличных от человека, нечеловекообразных обезьян, человекообразных обезьян, орангутанов, кошек, собак, лошадей, быков, оленей, бизонов, овец, кроликов, грызунов (например, мыши, крысы, хомяки и морские свинки) и домашний скот (например, виды крупного рогатого скота, такие как коровы и быки; виды полорогих, такие как овцы и козы; и виды свиней, такие как свиньи и кабаны). Птицы включают, например, кур, индеек, страусов, гусей и уток. Также включены домашние животные и сельскохозяйственные животные. Термин «животное, отличное от человека» исключает людей. Предпочтительные животные, отличные от человека, включают, например, грызунов, таких как мыши и крысы.
[00102] Животные, отличные от человека, могут быть получены из любого генетического фона. Например, подходящие мыши могут быть получены от линии 129, линии C57BL/6, помеси 129 и C57BL/6, линии BALB/c или линии Swiss Webster. Примеры линий 129 включают 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129Sl/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1 и 129Т2. См., например, Festing et al. (1999) Mammalian Genome 10:836, включенную в данный документ посредством ссылки во всей своей полноте для всех целей. Примеры линий C57BL включают C57BL/A, C57BL/An, C57BL/GrFa, C57BL/Kal_wN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. Подходящие мыши также могут быть получены от помеси вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6 (например, 50% 129 и 50% C57BL/6). Аналогичным образом подходящие мыши могут быть получены от помеси вышеупомянутых линий 129 или помеси вышеупомянутых линий BL/6 (например, линия 129S6 (129/SvEvTac)).
[00103] Аналогичным образом крысы могут быть получены от любой линии крыс, в том числе от, например, крыс линии ACI, крыс линии Dark Agouti (DA), крыс линии Wistar, крыс линии LEA, крыс линии Sprague Dawley (SD) или крыс линии Fischer, как, например, Fisher F344 или Fisher F6. Крысы также могут получены из линии, происходящей из помеси двух или более линий, перечисленных выше. Например, подходящая крыса может быть получена из линии DA или линии ACI. Крысы линии ACI характеризуется наличием окраски «черный агути» с белыми брюхом и конечностями и гаплотипом RT1av1. Такие линии доступны из множества источников, в том числе в Harlan Laboratories. Линия крыс Dark Agouti (DA) характеризуется наличием окраски «агути» и гаплотипом RT1av1. Такие крысы доступны из множества источников, в том числе в Charles River и Harlan Laboratories. Некоторые подходящие крысы могут быть получены от инбредных линий крыс. См., например, US 2014/0235933, включенный в данный документ посредством ссылки во всей своей полноте для всех целей.
III. Способы использования животных, отличных от человека, содержащих гуманизированный локус Asgr1, для оценки in vivo эффективности опосредованной ASGR1 человека доставки терапевтических комплексов в печень и терапевтических молекул, действующих через механизмы, опосредованные ASGR1 человека
[00104] Представлены различные способы использования животных, отличных от человека, содержащих гуманизированный локус Asgr1, описанный в другом месте данного документа, для оценки in vivo эффективности опосредованной ASGR1 человека доставки терапевтических молекул или комплексов в печень или опосредованных ASGR1 человека терапевтических механизмов. Поскольку животные, отличные от человека, содержат гуманизированный локус Asgr1, то животные, отличные от человека, будут более точно воспроизводить доставку, опосредованную ASGR1 человека, или опосредованные ASGR1 человека терапевтические механизмы, чем животные, отличные от человека, с негуманизированным локусом Asgr1. В качестве одного примера с помощью данных способов можно оценивать доставку терапевтического комплекса в печень посредством опосредованной ASGR1 человека интернализации in vivo, предусматривающую введение терапевтического комплекса животному, отличному от человека, содержащему гуманизированный локус Asgr1, где терапевтический комплекс содержит терапевтическую молекулу и антигенсвязывающий белок или лиганд, который специфически связывает ASGR1 человека, и затем оценку доставки терапевтической молекулы в печень животного, отличного от человека. В качестве другого примера с помощью данных способов можно оценивать in vivo эффективность терапевтических молекул или комплексов, предназначенных для воздействия посредством опосредованной ASGR1 человека интернализации, таких как терапевтические комплексы, сконструированные для интернализации поверхностного белка-мишени клеток печени посредством ASGR1 человека.
А. Способы оценки in vivo эффективности доставки терапевтических молекул в печень посредством интернализации, опосредованной ASGR1 человека
[00105] Представлены различные способы использования животных, отличных от человека, содержащих гуманизированный локус Asgr1, описанный в другом месте данного документа, для оценки in vivo эффективности доставки терапевтических молекул в печень посредством интернализации, опосредованной ASGR1 человека. Например, такие способы могут предусматривать: (а) введение терапевтического комплекса животному, отличному от человека, содержащему гуманизированный локус Asgr1, где терапевтический комплекс содержит терапевтическую молекулу и антигенсвязывающий белок или лиганд, который специфически связывает ASGR1 человека; и (b) оценку доставки терапевтической молекулы в печень животного, отличного от человека.
[00106] Терапевтическая молекула может представлять собой любое биологическое или химическое средство, используемое в лечении или профилактике любого заболевания или нарушения. Например, терапевтическая молекула может представлять собой терапевтические нуклеиновые кислоты (например, направляющие РНК системы CRISPR/Cas, короткие шпилечные РНК (shRNA) или малые интерферирующие РНК (siRNA)) или нуклеиновые кислоты, кодирующие терапевтические белки (например, белки Cas, как, например, белки Cas9, заместительные ферменты, секретируемые терапевтические белки и т.д.). Альтернативно терапевтическая молекула может представлять собой терапевтический белок, терапевтическое антитело, или антигенсвязывающий белок, или любую другую терапевтическую большую молекулу или малую молекулу.
[00107] Терапевтическая молекула и антигенсвязывающий белок или лиганд ASGR1 человека могут образовывать комплексы друг с другом посредством любых способов. Например, терапевтическая молекула и антигенсвязывающий белок или лиганд ASGR1 человека могут быть соединены посредством прямой ковалентной конъюгации или могут быть соединены посредством линкера, такого как пептидный линкер или химический линкер. Терапевтическая молекула и антигенсвязывающий белок или лиганд ASGR1 человека также могут образовывать комплексы друг с другом посредством образования связей между определенными функциональными группами или за счет определенного пространственного соответствия (например, взаимодействия по типу «замок и ключ»). В качестве конкретного примера антигенсвязывающий белок ASGR1 человека может представлять собой биспецифический антигенсвязывающий белок, который также специфически связывает терапевтическую молекулу.
[00108] Введение терапевтического комплекса может осуществляться посредством любого способа, как это описано более подробно в другом месте данного документа, и с помощью любого пути введения. Средства для доставки терапевтических комплексов и молекул и пути введения более подробно раскрыты в другом месте данного документа.
[00109] Можно использовать антигенсвязывающие белки ASGR1 человека или лиганды ASGR1 человека, которые специфически связывают ASGR1 человека. Поскольку белок ASGR1 человека является трансмембранным белком, который опосредует эндоцитоз определенных гликопротеинов в печени, молекулы, которые образуют комплекс с ASGR1 человека, могут быть интернализованы вместе с ASGR1 человека. Примеры подходящих антигенсвязывающих белков включают слитую с рецептором молекулу, молекулу-ловушку, слитую с рецептором Fc молекулу, антитело, Fab-фрагмент, F(ab')2-фрагмент, Fd-фрагмент, Fv-фрагмент, молекулу одноцепочечного Fv (scFv), dAb-фрагмент, выделенный определяющий комплементарность участок (CDR), пептид CDR3, конформационно ограниченный пептид FR3-CDR3-FR4, домен-специфическое антитело, однодоменное антитело, антитело с удаленным доменом, химерное антитело, антитело с привитым CDR, диатело, триатело, тетратело, минитело, нанотело, одновалентное нанотело, двухвалентное нанотело, иммунофармацевтическое средство на основе модульного белка малого размера (SMIP), верблюжье антитело (гомодимерное антитело на основе VHH тяжелой цепи), вариабельный домен IgNAR акулы и другие. В одном конкретном примере антигенсвязывающий белок представляет собой биспецифическое антитело, которое связывается ASGR1 человека и с терапевтической молекулой (например, с заместительным белком или ферментом или средством для доставки, таким как AAV). Альтернативно можно использовать лиганд или часть лиганда, который специфически взаимодействует с ASGR1 человека (например, асиалооросомукоид (ASOR) или бета-Ga1NAc или его рецептор-связывающая часть для ASGR1).
[00110] Оценку доставки терапевтической молекулы в печень животного, отличного от человека, можно провести при помощи любого известного способа. В качестве одного примера можно провести оценку присутствия терапевтической молекулы в печени. Например, если терапевтическая молекула представляет собой терапевтический белок, то оценку присутствия терапевтического белка в печени животного, отличного от человека, можно провести при помощи известных анализов для выявления белков. Аналогично, если терапевтическая молекула представляет собой нуклеиновую кислоту, кодирующую терапевтический белок, оценку экспрессии нуклеиновой кислоты (например, экспрессии мРНК или экспрессии белка) в печени животного, отличного от человека, можно провести при помощи известных анализов. Если кодируемый терапевтический белок представляет собой секретируемый белок, то с помощью известных анализов могут быть измерены уровни терапевтического белка в сыворотке крови или может быть оценена активность секретируемого терапевтического белка в отношении предусматриваемого типа клетки-мишени, типа ткани-мишени или органа-мишени. Оценку активности терапевтической молекулы в печени животного, отличного от человека, может провести с помощью известных анализов в зависимости от предполагаемой функции терапевтической молекулы. Например, если вводят средство для редактирования генома, такое как CRISPR/Cas, то можно использовать известные анализы для проведения оценки редактирования генома в конкретном целевом геномном локусе.
[00111] В конкретном примере терапевтический комплекс может содержать композиции на основе вирусных векторов (например, композиции на специфических для печени вирусных векторов). Такие композиции на основе вирусных векторов могут характеризоваться пониженным или устраненным природным тропизмом, и они сконструированы так, чтобы быть направленными на ASGR1 человека для целенаправленного воздействия на печень. Например, такой модифицированный вирусный векторный комплекс может содержать: (i) модифицированный вирусный вектор, содержащий нуклеиновую кислоту, кодирующую терапевтическую нуклеиновую кислоту или терапевтический белок, где модифицированный вирусный вектор характеризуется устраненным или пониженным природным тропизмом и содержит гетерологичный эпитоп; и (ii) перенацеливающий фрагмент, содержащий: (1) паратоп антиген-связывающего белка (например, антитела), который специфически связывает гетерологичный эпитоп; и (2) нацеливающий лиганд, который специфически связывает ASGR1 человека.
[00112] В качестве одного примера того, как этого добиться, можно использовать систему для мечения белков, такую как SpyCatcher-SpyTag, для ковалентного связывания антител с поверхностью вируса. Альтернативно могут быть использованы биспецифические антиген связывающие белки (например, биспецифические антитела, в которых одно плечо антитела связывается с вирусом, а другое плечо опосредует связывание с ASGR1 человека). В конкретном примере, перенацеливающий фрагмент представляет собой биспецифический антигенсвязывающий белок (например, биспецифическое антитело), содержащий первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен содержит паратоп, который специфически связывает гетерологичный эпитоп, встроенный в рекомбинантный человеческий вирусный капсидный белок/поверхностно экспрессируемый с его помощью, и второй антигенсвязывающий домен, который специфически связывает ASGR1 человека.
[00113] Примером подходящего гетерологичного эпитопа является Мус-метка. Например, Мус-метка может быть вставлена после N587 в AAV2. Такая вставка устраняет природную лиганд-связывающую активность AAV2, а также позволяет антителу к Мус распознавать модифицированный AAV. Использование биспецифического антитела, которое специфически связывается как с Мус, так и с ASGR1 человека, может затем перенацеливать AAV (например, AAV2 N587 Мус) на клетки печени, экспрессирующие ASGR1 человека (или гуманизированный Asgr1).
[00114] В другом примере терапевтический комплекс содержит лизосомальный заместительный фермент или белок или нуклеиновую кислоту, кодирующую лизосомальный заместительный фермент или белок. Лизосомальные болезни накопления представляют собой класс редких заболеваний, которые влияют на расщепление различных субстратов в лизосоме, включая сфинголипиды, мукополисахариды, гликопротеины, гликоген и олигосахариды, которые могут накапливаться в пораженных клетках, что приводит к гибели клеток. Органы, поражаемые лизосомальными болезнями накопления, включают печень. Патогенез этих заболеваний объясняется накоплением продуктов неполного расщепления в лизосоме, как правило, из-за потери функции белка. Лизосомальные болезни накопления обычно вызываются вариантами с потерей функции или аттенуированными вариантами в белках, нормальная функция которых заключается в расщеплении или координации расщепления содержимого лизосом. Примеры лизосомальных болезней накопления представлены в WO 2017/100467, включенной в данный документ посредством ссылки во всей своей полноте для всех целей. Например, одной из наиболее распространенных лизосомальных болезней накопления является болезнь Помпе. Болезнь Помпе вызывается дефектным лизосомальным ферментом альфа-глюкозидазой (GAA), что приводит к недостаточному процессированию гликогена в лизосомах. Накопление лизосомального гликогена происходит преимущественно в скелетной, сердечной и печеночной тканях.
[00115] Одним из вариантов лечения лизосомальных болезней накопления является заместительная ферментная или белковая терапия. Заместительные ферменты или белки могут быть эффективно доставлены в лизосому специфической клетки-мишени, когда они связаны в терапевтическом комплексе с антиген связывающим белком или лигандом, который специфически связывает ASGR1 человека. См. WO 2017/100467, включенную в данный документ посредством ссылки во всей своей полноте для всех целей. Такой терапевтический комплекс может быть введен субъекту, при этом данный терапевтический комплекс может проникать в лизосому клетки-мишени у субъекта и обеспечивать ферментативную активность, которая замещает ферментативную активность, связанную с лизосомной болезнью накопления. Белок ASGR1 человека представляет собой трансмембранный белок, который опосредует эндоцитоз и лизосомальное расщепление гликопротеинов с доступными концевыми остатками галактозы или N-ацетилгалактозамина в печени. Следовательно, белки, которые связываются и интернализуются вместе с ASGR1 человека, будут нацелены на лизосому. Собственно способы доставки, которые направляют терапевтические молекулы или комплексы на ASGR1 человека, можно использовать для нацеливания терапевтических средств на лизосомы (например, лизосомы в печени).
[00116] Соответственно, некоторые из представленных в данном документе способов представляют собой способы оценки доставки терапевтического комплекса, содержащего лизосомальный заместительный белок или фермент, в печень посредством ASGR1 человека in vivo. Например, такие способы могут предусматривать (а) введение терапевтической молекулы или терапевтического комплекса животным, отличным от человека, описанным в другом месте данного документа, где терапевтическая молекула или терапевтический комплекс содержит лизосомальный заместительный белок или фермент (или нуклеиновую кислоту, кодирующую лизосомальный заместительный белок или фермент) и антигенсвязывающий белок, который специфически связывает ASGR1 человека, где ASGR1 опосредует связывание с клеткой и поглощение лизосомным компартментом; и (b) оценку присутствия или активности лизосомального заместительного белка или фермента в печени животного, отличного от человека. Активность заместительного белка или фермента в печени животного, отличного от человека, можно оценить с помощью известных анализов для конкретного заместительного белка или фермента или путем измерения расщепления различных вовлеченных в заболевание субстратов в лизосоме с помощью известных анализов.
[00117] В качестве другого примера терапевтическая молекула может представлять собой нуклеиновую кислоту, кодирующую терапевтический секретируемый белок. Печень играет важную роль в продуцировании белков, которые секретируются в кровь, включая основные белки плазмы крови, факторы гемостаза и фибринолиза, белки-транспортеры, гормоны, прогормоны и аполипопротеины. Белок ASGR1 человека представляет собой трансмембранный белок, который опосредует эндоцитоз определенных гликопротеинов в печени. Следовательно, молекулы, которые образуют комплекс с ASGR1 человека, могут быть интернализованы вместе с ASGR1 человека. Таким образом, нуклеиновая кислота, кодирующая секретируемый терапевтический белок, может быть нацелена на печень путем ее доставки в комплексе, который нацеливается на ASGR1 человека. Нуклеиновая кислота может быть интернализована посредством ASGR1, и печень может синтезировать и секретировать терапевтический белок.
[00118] Соответственно, некоторые из представленных в данном документе способов представляют собой способы оценки доставки нуклеиновой кислоты, кодирующей терапевтический секретируемый белок, в печень посредством ASGR1 человека in vivo. Например, такие способы могут предусматривать (а) введение нуклеиновой кислоты (например, ДНК), кодирующей терапевтический секретируемый белок, животному, отличному от человека, описанному в другом месте данного документа, где нуклеиновая кислота доставляется в терапевтическом комплексе, который специфически связывается с ASGR1 человека, и при этом ASGR1 опосредует интернализацию комплекса; и (b) оценку секретируемых уровней (например, уровней в сыворотке крови) или активности терапевтического секретируемого белка у животного, отличного от человека.
[00119] Продуцирование и секрецию терапевтического белка можно оценивать при помощи любых известных способов. Например, экспрессия введенной нуклеиновой кислоты может быть оценена путем измерения уровней кодируемой мРНК в печени животного, отличного от человека, или уровней кодируемого терапевтического белка в печени животного, отличного от человека, с использованием известных анализов. Секрецию терапевтического белка можно оценивать путем измерения уровней кодируемого терапевтического белка в сыворотке крови у животного, отличного от человека, с использованием известных анализов. Кроме того, если секретируемый терапевтический белок действует на конкретные тип клеток, ткань или орган, то активность секретируемого терапевтического белка можно оценивать в целевых типе клеток, ткани или органе.
В. Способы оценки in vivo эффективности терапевтического комплекса в отношении интернализации поверхностного белка-мишени клеток печени или растворимого белка-мишени в печени посредством интернализации, опосредованной ASGR1 человека
[00120] Представлены разные способы использования животных, отличных от человека, содержащих гуманизированный локус Asgr1, описанный в другом месте данного документа, для оценки in vivo эффективности терапевтического комплекса, сконструированного для интернализации поверхностного белка-мишени клеток печени или растворимого белка-мишени в печени посредством ASGR1 человека. Способы терапии часто требуют инактивации или блокирования одной или более молекул-мишеней, которые действуют рядом с клеткой или на ее поверхности. Например, терапевтические средства на основе антител зачастую осуществляют свою функцию путем связывания с определенным антигеном, экспрессируемым на поверхности клетки, или с растворимым лигандом, препятствуя тем самым проявлению нормальной биологической активности антигена. Терапевтические средства данного типа обычно осуществляют свою функцию путем блокирования взаимодействия между цитокином и его рецептором для ослабления или подавления клеточной передачи сигнала. Однако в определенных случаях терапевтически преимущественным было бы инактивировать или подавлять активность молекулы-мишени таким образом, который необязательно включает блокирование ее физического взаимодействия с другим компонентом. Одним из способов достижения такого ослабления, не связанного с блокированием молекулы-мишени, может быть уменьшение внеклеточной концентрации молекулы-мишени или на клеточной поверхности. Например, молекула-мишень может быть ослаблена или инактивирована путем содействия или обеспечения физической связи между молекулой-мишенью и интернализующим эффекторным белком, таким как ASGR1. Это может быть достигнуто, например, путем использования полиспецифической (например, биспецифической) антигенсвязывающей молекулы, содержащей первый антигенсвязывающий домен и второй антигенсвязывающий домен. Каждый антигенсвязывающий домен связывает отдельную молекулу: первый специфически связывает молекулу-мишень, а второй специфически связывает ASGR1. Посредством этого типа физического межмолекулярного связывания молекула-мишень может быть принудительно интернализована в клетку вместе с ASGR1 и процессирована посредством внутриклеточного механизма расщепления или иным образом ослаблена, изолируя или инактивирована. См. WO 2013/138400 и US 2013/0243775, которые включены в данный документ посредством ссылки во всей своей полноте для всех целей.
[00121] Поскольку белок ASGR1 человека представляет собой трансмембранный белок, который опосредует эндоцитоз и лизосомальное расщепление гликопротеинов с доступными концевыми остатками галактозы или N-ацетилгалактозамина в печени, белки клеточной поверхности, которые образуют комплекс с ASGR1 человека, могут быть интернализованы вместе с ASGR1 человека, перенаправляя белок-мишень клеточной поверхности или растворимый белок-мишень в расщепляющий компартмент или изолируя белок-мишень клеточной поверхности или растворимый белок-мишень во внутренних компартментах или экзосомах.
[00122] Соответственно, в данном документе представлены способы использования животных, отличных от человека, содержащих гуманизированный локус Asgr1, который описан в другом месте данного документа, для оценки in vivo эффективности терапевтического комплекса, сконструированного для интернализации поверхностного белка-мишени клеток печени или растворимого белка-мишени в печени посредством ASGR1 человека. Например, такие способы могут предусматривать (а) введение терапевтического комплекса животным, отличным от человека, описанным в другом месте данного документа, где терапевтический комплекс содержит биспецифический антигенсвязывающий белок, который специфически связывается с поверхностным белком-мишенью клеток или растворимым белком-мишенью и специфически связывается с ASGR1 человека, где ASGR1 опосредует интернализацию поверхностного белка-мишени клеток или растворимого белка-мишени; и (b) оценку уровней поверхностного белка клеток печени, или активности поверхностного белка-мишени клеток печени, или оценку уровней экспрессии или активности растворимого белка-мишени в печени животного, отличного от человека. Введение терапевтического комплекса может осуществляться с помощью любого подходящего способа, как это описано в другом месте данного документа.
[00123] Белки-мишени клеточной поверхности могут включать любой белок клеточной поверхности, экспрессируемый в печени. Растворимые белки-мишени могут включать любой растворимый белок, экспрессируемый в печени. Уровни поверхностного белка клеток или активность поверхностного белка-мишени клеток печени в печени животного, отличного от человека, могут быть оценены с помощью известных анализов для измерения уровней рецепторов на поверхности клеток и других белков. Аналогичным образом уровни растворимых белков могут быть оценены с помощью известных анализов.
С. Введение молекул животным, отличным от человека
[00124] Способы, раскрытые в данном документе, включают введение животному, отличному от человека, различных молекул (терапевтических молекул или комплексов), в том числе нуклеиновых кислот, белков или белковых комплексов. Введение может быть выполнено с помощью любого способа. Например, такие молекулы могут быть введены животному, отличному от человека, например, посредством доставки с помощью вектора, доставки, опосредованной частицами, доставки, опосредованной экзосомами, доставки, опосредованной липидными наночастицами, доставки, опосредованной проникающим в клетку пептидом, или доставки, опосредованной имплантируемым устройством. В качестве конкретных примеров, молекула может быть введена в клетку или животное, отличное от человека, в носителе, таком как микросфера на основе поли(молочной кислоты) (PLA), микросфера на основе сополимера D,L-молочной кислоты и гликолевой кислоты (PLGA), липосома, мицелла, обратная мицелла, липидный кохлеат или липидная микротрубочка. Некоторые конкретные примеры доставки животному, отличному от человека, включают гидродинамическую доставку, доставку, опосредованную вирусом (например, доставку, опосредованную аденоассоциированным вирусом (AAV)), и доставку, опосредованную липидными наночастицами.
[00125] Введение нуклеиновых кислот, белков или других компонентов животным, отличным от человека, может быть выполнено путем гидродинамической доставки (HDD). Гидродинамическая доставка оказалась идеальным способом внутриклеточной доставки ДНК in vivo. Для доставки гена в паренхиматозные клетки через выбранный кровеносный сосуд следует вводить только требуемые последовательности ДНК, что устраняет проблемы безопасности, связанные с современными вирусными и синтетическими векторами. При введении в кровоток ДНК способна достигать клеток в различных тканях, доступных крови. В гидродинамической доставке используется сила, создаваемая за счет быстрой инъекции большого объема раствора в несжимающуюся кровь в кровотоке, чтобы преодолеть физические барьеры эндотелия и клеточных мембран, которые препятствуют проникновению крупных и не проникающих через мембраны соединений в паренхиматозные клетки. В дополнение к доставке ДНК этот способ применим для эффективной внутриклеточной доставки РНК, белков и других небольших соединений in vivo. См., например, Bonamassa et al. (2011) Pharm. Res. 28(4):694-701, включенную в данный документ посредством ссылки во всей своей полноте для всех целей.
[00126] Введение нуклеиновых кислот также может быть выполнено путем опосредованной вирусом доставки, например, AAV-опосредованной доставки или опосредованной лентивирусом доставки. Другие иллюстративные вирусы/вирусные векторы включают ретровирусы, аденовирусы, вирусы осповакцины, поксвирусы и вирусы простого герпеса. Вирусы могут инфицировать делящиеся клетки, неделящиеся клетки или и делящиеся, и неделящиеся клетки. Вирусы могут интегрироваться в геном хозяина или альтернативно не интегрироваться в геном хозяина. Такие вирусы также могут быть сконструированы с тем, чтобы ослаблять иммунитета. Вирусы могут быть репликационно-компетентными или могут быть репликационно-дефектными (например, дефектными по одному или более генам, необходимым для дополнительных циклов репликации и/или упаковки вириона). Вирусы могут инициировать временную экспрессию, длительную экспрессию (например, по меньшей мере 1 неделю, 2 недели, 1 месяц, 2 месяца или 3 месяца) или постоянную экспрессию (например, Cas9 и/или gRNA). Иллюстративные вирусные титры (например, титры AAV) включают 1012, 1013, 1014, 1015 и 1016 векторных геномов/мл.
[00127] ssDNA генома AAV состоит из двух открытых рамок считывания, Rep и Сар, и фланкирован двумя инвертированными концевыми повторами, которые обеспечивают возможность синтеза комплементарной нити ДНК. При конструировании плазмиды для переноса с помощью AAV трансген помещают между двумя ITR, a Rep и Сар могут обеспечиваться в транс-форме. В дополнение к Rep и Сар для AAV может потребоваться хелперная плазмида, содержащая гены аденовируса. Эти гены (Е4, Е2а и VA) опосредуют репликацию AAV. Например, плазмида для переноса, Rep/Cap и хелперная плазмида могут быть трансфицированы в клетки HEK293, содержащие ген Е1 + аденовируса, для получения инфекционных частиц AAV. Альтернативно Rep, Сар и хелперные гены аденовируса могут быть объединены в одну плазмиду. Аналогичные упаковывающие клетки и способы можно использовать для других вирусов, как, например, для ретровирусов.
[00128] Было идентифицировано множество серотипов AAV. Эти серотипы отличаются по типам клеток, которые они инфицируют (т.е. тропизмом), что обеспечивает возможность преимущественной трансдукции определенных типов клеток. Серотипы для ткани печени включают AAV7, AAV8 и AAV9 и, в частности, AAV8.
[00129] Тропизм можно дополнительно уточнить с помощью псевдотипирования, представляющего собой смешивание капсида и генома различных вирусных серотипов. Использование псевдотипированных вирусов может улучшить эффективность трансдукции, а также изменить тропизм. Гибридные капсиды, полученные от разных серотипов, также могут быть использованы для изменения вирусного тропизма. Например, AAV-DJ содержит гибридный капсид из восьми серотипов и демонстрирует высокую инфицирующую способность в широком спектре типов клеток in vivo. Серотипы AAV также могут быть изменены с помощью мутаций. Другие псевдотипированные/модифицированные варианты AAV включают AAV2/1, AAV2/6, AAV2/7, AAV2/8, AAV2/9, AAV2.5, AAV8.2 и AAV/SASTG.
[00130] Для ускорения экспрессии трансгена могут использоваться самокомплементарные варианты AAV (scAAV). Так как AAV зависит от механизма репликации ДНК клетки, то для синтеза комплементарной нити одноценитевой ДНК генома AAV, экспрессия трансгена может быть замедлена. Для устранения этой задержки можно использовать scAAV, содержащие комплементарные последовательности, которые способны к самопроизвольному отжигу при заражении, что устраняет необходимость синтеза ДНК клетки-хозяина.
[00131] Чтобы увеличить пакующую емкость, более длинные трансгены могут быть разделены между двумя плазмидами для переноса AAV, первая с 3'-донором сплайсинга, а вторая с 5'-акцептором сплайсинга. При совместной инфекции клетки данные вирусы образуют конкатемеры, подвергаются сплайсингу, и может экспрессироваться полноразмерный трансген. Хотя это обеспечивает возможность экспрессии трансгена большей длины, такая экспрессия является менее эффективной. В аналогичных способах увеличения емкости используется гомологичная рекомбинация. Например, трансген может быть разделен между двумя плазмидами для переноса, но с существенным перекрыванием последовательностей с тем, чтобы совместная экспрессия индуцировала гомологичную рекомбинацию и экспрессию полноразмерного трансгена.
[00132] Введение нуклеиновых кислот и белков также может быть выполнено путем доставки, опосредованной липидными наночастицами (LNP). Например, LNP-опосредованная доставка может использоваться для доставки направляющей РНК в форме РНК. Результатом доставки с помощью таких способов является временное присутствие направляющей РНК, а биоразлагаемые липиды улучшают клиренс, улучшают переносимость и снижают иммуногенность. Липидные составы могут защищать биологические молекулы от расщепления при одновременном улучшении их поглощения клетками. Липидные наночастицы представляют собой частицы, содержащие множество молекул липидов, физически связанных друг с другом межмолекулярными силами. Они включают микросферы (в том числе однослойные и многослойные везикулы, например, липосомы), дисперсную фазу в эмульсии, мицеллы или внутреннюю фазу в суспензии. Такие липидные наночастицы могут быть использованы для инкапсуляции одной или более нуклеиновых кислот или белков, предназначенных для доставки. Составы, которые содержат катионные липиды, являются применимыми для доставки полианионов, таких как нуклеиновые кислоты. Другие липиды, которые могут быть включены, представляют собой нейтральные липиды (т.е. незаряженные или цвиттерионные липиды), анионные липиды, хелперные липиды, которые улучшают трансфекцию, и липиды-невидимки, которые увеличивают время существования наночастиц in vivo. Примеры подходящих катионных липидов, нейтральных липидов, анионных липидов, хелперных липидов и липидов-невидимок можно найти в WO 2016/010840 А1, включенной в данный документ посредством ссылки во всей своей полноте для всех целей. Иллюстративная липидная наночастица может содержать катионный липид и один или более других компонентов. В одном примере другой компонент может предусматривать хелперный липид, такой как холестерин. В другом примере другие компоненты могут предусматривать хелперный липид, такой как холестерин, и нейтральный липид, такой как DSPC. В другом примере другие компоненты могут предусматривать хелперный липид, такой как холестерин, необязательный нейтральный липид, такой как DSPC, и липид-невидимку, такой как S010, S024, S027, S031 или S033.
[00133] Способ доставки может быть выбран для снижения иммуногенности. Разные способы могут придавать разные фармакодинамические или фармакокинетические свойства молекуле, доставляемой субъекту. Например, разные способы могут обусловливать разное распределение в тканях, разный период полураспада или разное распределение по времени. Некоторые режимы доставки (например, доставка вектора, представляющего собой нуклеиновую кислоту, который персистирует в клетке за счет автономной репликации или геномной интеграции) приводят к более устойчивой экспрессии и присутствию молекулы, тогда как другие способы режимы являются временными и менее устойчивыми (например, доставка РНК или белка). Доставка компонентов посредством более неустойчивого механизма, например, в виде РНК или белка, может гарантировать, что компоненты присутствуют и будут активными лишь в течение короткого периода времени, и могут снижать иммуногенность.
[00134] Введение in vivo может осуществляться любым подходящим путем, включая, например, парентеральный, внутривенный, подкожный, внутриартериальный или внутрибрюшинный. Системные способы введения включают, например, парентеральные пути, такие как внутривенный, внутриартериальный, подкожный и внутрибрюшинный пути. Конкретным примером является внутривенная инфузия. Также могут быть использованы местные способы введения.
[00135] Композиции, содержащие нуклеиновые кислоты или белки, могут быть составлены с использованием одного или более физиологически и фармацевтически приемлемых носителей, разбавителей, наполнителей или вспомогательных веществ. Состав может зависеть от выбранного пути введения. Термин «фармацевтически приемлемый» означает, что носитель, разбавитель, наполнитель или вспомогательное вещество являются совместимыми с другими ингредиентами состава и, по сути, не вредны для принимающего их пациента.
[00136] Частота введения и количество доз, помимо прочих факторов, могут зависеть от периода полувыведения введенных компонентов и пути введения. Введение нуклеиновых кислот или белков животному, отличному от человека, можно осуществлять один или более раз за определенный период времени. Например, введение можно выполнять по меньшей мере два раза в течение определенного периода времени, по меньшей мере три раза в течение определенного периода времени, по меньшей мере четыре раза в течение определенного периода времени, по меньшей мере пять раз в течение определенного периода времени, по меньшей мере шесть раз в течение определенного периода времени, по меньшей мере семь раз в течение определенного периода времени, по меньшей мере восемь раз в течение определенного периода времени, по меньшей мере девять раз в течение определенного периода времени, по меньшей мере десять раз в течение определенного периода времени, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз в течение определенного периода времени, по меньшей мере тринадцать раз в течение определенного периода времени, по меньшей мере четырнадцать раз в течение определенного периода времени, по меньшей мере пятнадцать раз в течение определенного периода времени, по меньшей мере шестнадцать раз в течение определенного периода времени, по меньшей мере семнадцать раз в течение определенного периода времени, по меньшей мере восемнадцать раз в течение определенного периода времени, по меньшей мере девятнадцать раз в течение определенного периода времени или по меньшей мере двадцать раз в течение определенного периода времени.
IV. Способы получения животных, отличных от человека, содержащих гуманизированный локус Asgr1
[00137] Представлены различные способы получения животного, отличного от человека, содержащего гуманизированный локус Asgr1, раскрытый в другом месте данного документа. Любой общепринятый способ или протокол, предназначенный для получения генетически модифицированного организма, являются подходящими для получения такого генетически модифицированного животного, отличного от человека. См., например, Cho et al. (2009) Current Protocols in Cell Biology 42:19.11:19.11.1-19.11.22 и Gama Sosa et al. (2010) Brain Struct. Funct. 214(2-3):91-109, каждая из которых включена в данный документ посредством ссылки во всей своей полноте для всех целей. Такие генетически модифицированные животные, отличные от человека, могут быть получены, например, посредством нокина гена в целевом локусе Asgr1.
[00138] Например, способ получения животного, отличного от человека, содержащего гуманизированный локус Asgr1, может предусматривать: (1) модификацию генома плюрипотентной клетки со встраиванием гуманизированного локуса Asgr1; (2) идентификацию или отбор генетически модифицированной плюрипотентной клетки, содержащей гуманизированный локус Asgr1; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион животного-хозяина, отличного от человека; и (4) имплантацию и вынашивание эмбриона-хозяина суррогатной матерью. Необязательно эмбрион-хозяин, содержащий модифицированную плюрипотентную клетку (например, ES-клетку, отличную от человеческой), можно инкубировать до стадии бластоцисты перед имплантацией и вынашиванием суррогатной матерью для получения животного F0, отличного от человека. Затем суррогатная мать может произвести животное, отличное от человека, поколения F0, содержащее гуманизированный локус Asgr1.
[00139] Способы могут дополнительно предусматривать идентификацию клетки или животного, имеющих модифицированный целевой геномный локус. Различные способы могут быть использованы для идентификации клеток и животных, имеющих целевую генетическую модификацию.
[00140] Стадия скрининга может предусматривать, например, количественный анализ для оценки модификации аллеля (МОА) в родительской хромосоме. Например, количественный анализ может быть осуществлен с помощью количественной ПЦР, такой как ПЦР в режиме реального времени (qPCR). В ПЦР в режиме реального времени может использоваться первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров может содержать флуоресцентный зонд, который распознает амплифицированную последовательность.
[00141] Другие примеры подходящих количественных анализов включают флуоресцентно-опосредованную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным зондом (иммобилизованными зондами), зонды INVADER®, зонды TAQMAN® от Molecular Beacon или технологию зондов ECLIPSE™ (см., например, US 2005/0144655, включенный в данный документ посредством ссылки во всей своей полноте для всех целей).
[00142] Примером подходящей плюрипотентной клетки является эмбриональная стволовая (ES) клетка (например, ES-клетка мыши или ES-клетка крысы). Модифицированная плюрипотентная клетка может быть получена, например, путем рекомбинации с помощью (а) введения в клетку одного или более нацеливающихся векторов, содержащих вставочную нуклеиновую кислоту, фланкированную 5'- и 3'-плечами гомологии, соответствующими 5'- и 3'-целевым сайтам, где вставочная нуклеиновая кислота содержит гуманизированный локус Asgr1; и (b) идентификации по меньшей мере одной клетки, содержащей в своем геноме вставочную нуклеиновую кислоту, интегрированную в целевой геномный локус. Альтернативно модифицированная плюрипотентная клетка может быть получена путем (а) введения в клетку: (i) средства на основе нуклеаз, где средство на основе нуклеаз индуцирует однонитевый или двухнитевый разрыв в сайте распознавания в пределах целевого геномного локуса; и (ii) одного или более нацеливающихся векторов, содержащих вставочную нуклеиновую кислоту, фланкированную 5'- и 3'-плечами гомологии, соответствующими 5'- и 3'-целевым сайтам, расположенным достаточно близко к сайту распознавания, где вставочная нуклеиновая кислота содержит гуманизированный локус Asgr1; и (с) идентификации по меньшей мере одной клетки, содержащей модификацию (например, интегрированную вставочную нуклеиновую кислоту) в целевом геномном локусе. Можно использовать любое средство на основе нуклеаз, которое индуцирует однонитевый или двухнитевый разрыв в требуемом сайте распознавания. Примеры подходящих нуклеаз включают эффекторную нуклеазу, подобную активаторам транскрипции (TALEN), нуклеазу с «цинковыми пальцами» (ZFN), мегануклеазу и короткие палиндромные повторы, регулярно расположенные группами (CRISPR)/CRISPR-ассоциированные (Cas) системы или компоненты таких системы (например, CRISPR/Cas9). См., например, US 2013/0309670 и US 2015/0159175, каждый из которых включен в данный документ посредством ссылки во всей своей полноте для всех целей.
[00143] Донорную клетку можно вводить в эмбрион-хозяин на любой стадии, как, например, на стадии бластоцисты или стадии ранней морулы (то есть стадии 4 клеток или стадии 8 клеток). Получают потомство, способное передавать генетическую модификацию через зародышевую линию. См., например, патент США №7294754, который включен в данный документ посредством ссылки во всей своей полноте для всех целей.
[00144] Альтернативно способ получения животных, отличных от человека, описанный в другом месте данного документа, может предусматривать: (1) модификацию генома эмбриона на стадии одной клетки со встраиванием гуманизированного локуса Asgr1 с использованием описанных выше способов для модификации плюрипотентных клеток; (2) отбор генетически модифицированных эмбрионов; и (3) имплантацию и вынашивание генетически модифицированного эмбриона суррогатной матерью. Получают потомство, способное передавать генетическую модификацию через зародышевую линию.
[00145] Методики переноса ядер можно также использовать для получения животных-млекопитающих, отличных от человека. Вкратце, способы переноса ядер могут предусматривает стадии: (1) энуклеирования ооцита или обеспечения энуклеированного ооцита; (2) выделения или обеспечения донорных клетки или ядра, подлежащих объединению с энуклеированным ооцитом; (3) вставки клетки или ядра в энуклеированный ооцит с получением реконструированной клетки; (4) имплантирования реконструированной клетки в матку животного, отличного от человека, с получением эмбриона и (5) обеспечения возможности развития эмбриона. В таких способах ооциты обычно получают из умерщвленных животных, несмотря на то, что их также можно выделять из маточных труб и/или яичников живых животных. Перед энуклеированием ооциты можно инкубировать в различных хорошо известных средах с целью их созревания. Энуклеирование ооцита можно осуществлять с помощью ряда хорошо известных способов. Вставка донорной клетки или ядра в энуклеированный ооцит с получением реконструированной клетки может быть достигнута с помощью микроинъекции донорной клетки под блестящую оболочку перед слиянием. Слияние можно индуцировать путем применения импульса электрического постоянного тока по отношению к поверхности контакта/слияния (электрослияние), путем воздействия на клетки химических веществ, способствующих слиянию, таких как полиэтиленгликоль, или путем использования инактивированного вируса, такого как вирус Сендай. Реконструированную клетку можно активировать с помощью способов с использованием электричества и/или без использования электричества до, в ходе и/или после слияния ядерного донора и реципиентного ооцита. Способы активации предусматривают электрические импульсы, химически индуцированный шок, внедрение сперматозоида, повышение уровней двухвалентных катионов в ооците и снижение фосфорилирования клеточных белков (например, с помощью ингибиторов киназ) в ооците. Активированные реконструированные клетки или эмбрионы можно культивировать в хорошо известных средах, а затем переносить в матку животного. См., например, US 2008/0092249, WO 1999/005266, US 2004/0177390 и WO 2008/017234 и патент США №7612250, каждый из которых включен посредством ссылки во всей своей полноте для всех целей.
[00146] Различные способы, представленные в данном документе, позволяют получать генетически модифицированное животное F0, отличное от человека, где клетки генетически модифицированного животного F0 содержат гуманизированный локус Asgr1. Необходимо отметить, что в зависимости от способа, используемого для получения животного F0, количество клеток в животном F0, которые имеют гуманизированный локус Asgr1, будет варьироваться. Введение донорных ES-клеток в эмбрион на стадии ранней морулы из соответствующего организма (например, мышиного эмбриона на стадии 8 клеток), например, с помощью способа VELOCIMOUSE® позволяет получать больший процент клеточной популяции животных F0, содержащих клетки, имеющие представляющую интерес нуклеотидную последовательность, содержащую целевую генетическую модификацию. Например, по меньшей мере 50%, 60%), 65%), 70%, 75%), 85%, 86%, 87%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% клеточного вклада животного F0, отличного от человека, могут содержать клеточную популяцию, имеющую целевую модификацию.
[00147] Клетки генетически модифицированного животного F0 могут быть гетерозиготными по гуманизированному локусу Asgr1 или могут быть гомозиготными по гуманизированному локусу Asgr1.
[00148] Все патентные заявки, веб-сайты, другие публикации, регистрационные номера и им подобные, упомянутые ранее или ниже, включены посредством ссылки во всей их полноте для всех целей в той же степени, как если бы каждый отдельный элемент был специально и индивидуально указан для включения посредством ссылки. Если разные варианты последовательности связаны с номером доступа в разное время, то подразумевается вариант, связанный с номером доступа на действительную дату подачи настоящей заявки. Действительная дата подачи означает наиболее раннюю действительную дату подачи или дату подачи приоритетной заявки со ссылкой на номер доступа, если это необходимо. Аналогичным образом, если разные варианты публикации, веб-сайта и им подобных опубликованы в разное время, то подразумевается вариант, опубликованный раньше всех на действительную дату подачи, если не указано иное. Любые признак, стадия, элемент, вариант осуществления или аспект настоящего изобретения можно использовать в сочетании с любым другим, если специально не указано иное. Несмотря на то, что с целью ясности понимания настоящее изобретение было описано довольно подробно посредством иллюстраций и примеров, будет очевидно, что определенные изменения и модификации можно осуществлять на практике в пределах объема прилагаемой формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[00149] Нуклеотидная и аминокислотная последовательности, перечисленные в прилагаемом перечне последовательностей, показаны с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот. Нуклеотидные последовательности приведены согласно стандартным правилам, начиная с 5'-конца последовательности и продвигаясь вперед (т.е. слева направо в каждой строке) до 3'-конца. Показана только одна нить каждой нуклеотидной последовательности, но считается, что комплементарная цепь включена посредством любой ссылки в представленную последовательность. Аминокислотные последовательности приведены согласно стандартным правилам, начиная с амино-конца последовательности и продвигаясь вперед (то есть слева направо в каждой строке) к карбокси-концу.
[00150] Таблица 1. Описание последовательностей
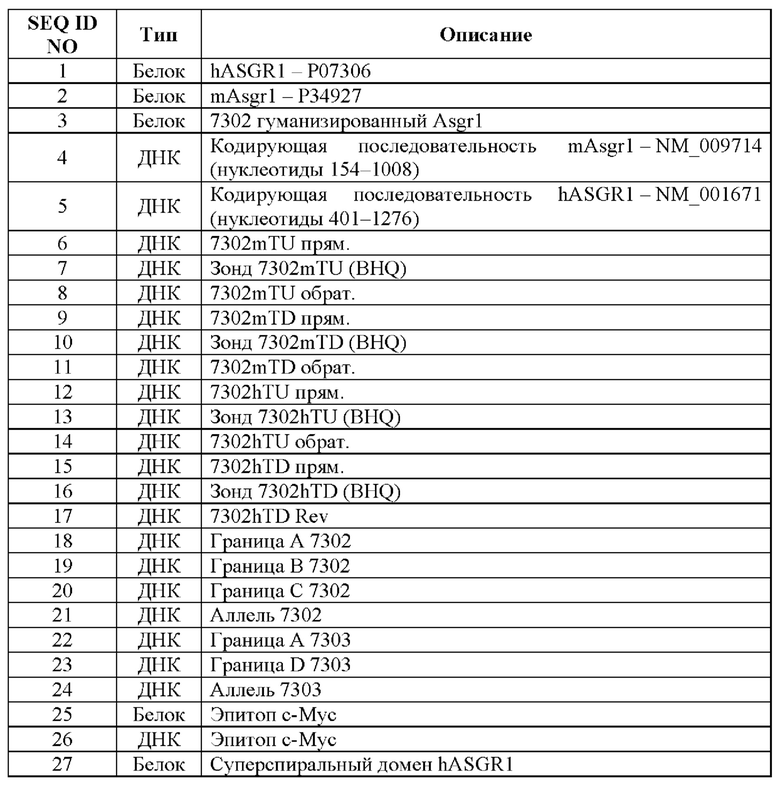

ПРИМЕРЫ
Пример 1. Получение мышей, содержащих гуманизированный локус Asgr1
[00151] Асиалогликопротеиновый рецептор (рецептор Эшвелла, ASGR) представляет собой преимущественно связанный с печеночной мембраной связывающий углеводы белковый рецептор (~33 кДа), относящийся к классу лектиновых рецепторов С-типа. ASGR связывается с гликопротеинами с концевыми галактозными или N-ацетилгалактозаминовыми (GalNac) мотивами. Он может удалять десиалилированные гликопротеины из кровотока с помощью рецептор-опосредованного эндоцитоза. Лиганд подвергается лизосомальному расщеплению, в то время как ASGR возвращается на поверхность клетки. Рецептор представляет собой гетероолигомер из двух субъединиц, ASGR1 и ASGR2 (H1 и Н2). В качестве высокоэффективного специфического для печени рецептора эндоцитоза ASGR1 может быть использован для специфической для печени доставки терапевтических средств, таких как антитела, малые молекулы (в качестве части конъюгатов антитело-лекарственное средство) и ДНК. Однако полученные авторами антитела и биспецифические антитела к внеклеточному домену ASGR1 человека не связываются с ортологом Asgr1 мыши (данные Biacore не показаны). Таким образом, получили гуманизированных по Asgr1 мышей для использования с целью валидации специфической для печени доставки различных терапевтических средств с использованием ряда различных подходов.
[00152] Получили крупный нацеливающийся вектор, содержащий 5'-плечо гомологии, содержащее 24 т.о. из bMQ-69B11, и 3'-плечо гомологии, содержащее 67 т.о. из bMQ-69B11, для замены кодирующих экзонов 3-8 (через стоп-кодон и 3'-UTR в последовательность, расположенную непосредственно после 3'-UTR) Asgr1 мыши соответствующей последовательностью ASGR1 человека. Кодируемый белок Asgr1 будет содержать трансмембранный домен Asgr1 мыши, после которого расположен суперспиральный домен и лектиновый домен С-типа человека. См. фигуры 2А и 3. Для получения мутантного аллеля компоненты системы CRISPR/Cas9 вводили в эмбриональные стволовые клетки мыши F1H4 вместе с крупным нацеливающимся вектором. Для выявления потери эндогенного аллеля мыши проводили анализы потери аллеля с использованием праймеров и зондов, приведенных в таблице 2, а для выявления приобретения гуманизированного аллеля проводили анализы приобретения аллеля с использованием праймеров и зондов, приведенных в таблице 3 Анализы потери аллеля и анализы приобретения аллеля описаны, например, в US 2014/0178879; US 2016/0145646; WO 2016/081923 и Frendewey et al. (2010) Methods Enzymol. 476:295-307, которые включены в данный документ посредством ссылки во всей своей полноте для всех целей.
[00153] Таблица 2. Анализы потери мышиного аллеля TAQMAN®

[00154] Таблица 3. Анализы приобретения аллеля человека TAQMAN®

[00155] Затем получали мышей F0 с использованием способа VELOCIMOUSE®. См., например, US 7576259; US 7659442; US 7294754; US 2008/007800 и Poueymirou et al. (2007) Nature Biotech. 25(1):91-99, которые включены в данный документ посредством ссылки во всей своей полноте для всех целей. Часть интрона, расположенного выше кодирующего экзона 3, была также гуманизирована. Всего 1863 п.о. последовательности Asgr1 мыши было заменено на 3907 п.о. последовательности ASGR1 человека. Кассету loxP-hUb1-em7-ген резистентности к гигромицину-сигнал полиаденилирования-mPrm 1-Crei-сигнал полиаденилирования-1охР (5218 п.о.) также вставили ниже 3'-UTR человека с промежуточной последовательностью размером в примерно 190 п.о. Из 3'-последовательности человека 3'-UTR непосредственно перед кассетой. Полученный частично гуманизированный аллель Asgr1 мыши с самоудаляющейся кассетой резистентности к гигромицину изложен под SEQ ID NO: 21 (обозначен как аллель 7302). Граничные участки последовательности А, В и С на фигуре 2А изложены под SEQ ID NO: 18, 19 и 20 соответственно. Сравнение белка ASGR1 человека (SEQ ID NO: 1), белка Asgr1 мыши (SEQ ID NO: 2) и частично гуманизированного белка Asgr1 мыши (SEQ ID NO: 3) показано на фигуре 3.
[00156] После удаления самоудаляющейся кассеты с рекомбиназой Cre, loxP и сайты клонирования (77 п.о.) остаются ниже 3'-UTR человека с промежуточной последовательностью размером в приблизительно 190 п.о. из 3'-последовательности человека после 3'-UTR непосредственно перед оставшимся loxP. См. фигуру 2В. Полученный частично гуманизированный аллель Asgr1 мыши с удаленной кассетой изложен под SEQ ID NO: 24 (обозначен как аллель 7303). Граничные участки последовательности А и С на фигуре 2В изложены под SEQ ID NO: 22 и 23 соответственно.
Пример 2. Валидация мышей, содержащих гуманизированный локус Asgr1
[00157] Для валидации гуманизированных по Asgr1 мышей (Asgr1hu/hu) в качестве достоверной модели мышей Asgr1hu/hu подвергали фенотипированию, и их фенотип сравнивали с фенотипом однопометников дикого типа. У мышей Asgr1hu/hu не выявляли отличий в уровнях липидов в плазме крови (общий холестерин, триглицериды, HDL-C, LDL-C) по сравнению с однопометниками дикого типа. См. фигуру 4. Аналогичным образом мыши Asgr1hu/hu не демонстрировали отличий в массе тела или уровне глюкозы в крови по сравнению с однопометниками дикого типа. См. фигуру 5. Белок ASGR1 человека был совместно локализован на мембранах клеток печени, аналогично Asgr1 мыши. См. фигуру 6. И в заключение, мыши Asgr1hu/hu экспрессируют белок ASGR1 человека на мембранах клеток печени и характеризуются нормальным липидным профилем в плазме крови.
Способы
[00158] Оценка уровня циркулирующих в крови липидов у гуманизированных по Asgr1 мышей. Плазму крови от самцов гуманизированных по Asgr1 мышей (Asgr1hu/hu) и их однопометников дикого типа (Asgr1+/+) отбирали натощак и анализировали на содержание липидов в сыворотке крови (триглицериды (TG)), общий холестерин, холестерин липопротеинов низкой плотности (LDL)-C)), холестерин липопротеинов высокой плотности (HDL-C)) с использованием биохимического анализатора сыворотки крови ADVIA® Chemistry ХРТ System (Siemens). N=8/группа, возраст 11 недель. Данные, выраженные в виде среднего значения ± SEM для каждого.
[00159] Оценка уровня глюкозы в крови. Глюкозу крови измеряли в крови, отобранной из кончика хвоста, используя глюкометр Accu-Chek (Roche) натощак (16-часовое голодание) и после еды (не натощак).
[00160] Оценка Asgr1 с помощью вестерн-блоттинга. Цельную печень отбирали у гуманизированных по Asgr1 мышей и мышей WT (n=8/генотип) и хранили замороженной при 80°С до обработки. Для каждого образца из цельной печени вырезали кусочек замороженной печени весом ~40 мг, и каждый кусочек помещали в гомогенизатор Даунса и гомогенизировали до получения однородной суспензии. Цитозольные фракции и мембранные фракции каждого образца печени разделяли с использованием коммерческого набора на основе детергента (Thermo #89842) в соответствии с протоколом для «мягких тканей» производителя. После того, как выделили цитозольную фракцию и мембранную фракцию каждого образца, проводили анализ количественного определения белка ВСА (Thermo #23225) для двух фракций из каждого образца в соответствии с протоколом набора для процедуры с микропланшетом. Образцы для вестерн-блоттинга готовили для каждой цитозольной фракции и мембранной фракции каждого образца, используя 5Х восстанавливающий краситель. Все образцы готовили при 0,8 мкг/мкл, и 20 мкг общего белка из каждого образца загружали в гели для вестерн-блоттинга. hASGR1 выявляли с использованием кроличьего поликлонального антитела к hASGR1 (Abgent #АР16133а, в разведении 1:1000 в 2,5% блокирующем молоке в TBS-T). Поскольку антитело проявляло перекрестную реактивность с белком мыши, Asgr1 мыши у однопометных мышей был выявлен с использованием того же самого антитела. Вторичное антитело для выявления hASGR1 представляло собой ослиное антитело к кроличьему IgG-HRP (Jackson #711-035-152, в концентрации 0,1 мкг/мл в 2,5% блокирующем молоке в TBS-T). В качестве загрузочного контроля для цитозольных фракций GAPDH обнаружили во всех образцах с использованием кроличьего моноклонального антитела к GAPDH (Cell Signaling #2118S, в разведении 1:10000 в 2,5% блокирующем молоке в TBS-T). Вторичное антитело для выявления GAPDH представляло собой ослиное антитело к кроличьему IgG-HRP (Jackson #711-035-152, в концентрации 0,1 мкг/мл в 2,5% блокирующем молоке в TBS-T). В качестве загрузочного контроля для мембранных фракций рецептор трансферрина выявляли во всех образцах, используя кроличье поликлональное антитело к рецептору трансферрина (R&D #AF2472, в концентрации 0,25 мкг/мл в 2,5% блокирующем молоке в TBS-T). Вторичное антитело для определения рецептора трансферрина представляло собой ослиное антитело к козьему IgG-HRP (Jackson #705-035-147, в концентрации 0,2 мкг/мл в 2,5%) блокирующем молоке в TBS-T).
Пример 3. Получение векторов на основе аденоассоциированного вируса с гетерологичным эпитопом
[00161] Затем проводили эксперимент с целью определения того, может ли биспецифическое антитело к myc-ASGR1 перенацеливать вирусные частицы scAAV-N587myc на клетки печени, экспрессирующие гуманизированный ASGR1, in vivo у мышей, полученных в примере 1. Чтобы это проверить, сначала получили вирусные частицы.
[00162] Капсидные белки AAV модифицировали таким образом, чтобы они содержали один из нескольких гетерологичных эпитопов: FLAG, c-myc, гексагистидин и т.д., с помощью ПЦР с получением плазмиды, кодирующей рекомбинантный капсидный белок. Вкратце, осуществляли вставку последовательности, кодирующей FLAG, c-myc или гексагистидин, в рамку после кодона, кодирующего N587 AAV2 или Q585 капсидного белка VP1 AAV6.
[00163] Продуцирование аденоассоциированных вирусов осуществляли с использованием способа тройной трансфекции клеток HEK293 (см., например, Erik Arden and Joseph M. Metzger, J Biol Methods. 2016; 3(2)). Клетки высевали за один день до PEFpro-опосредованной (Polyplus transfection, Нью-Йорк, штат Нью-Йорк) трансфекции соответствующими векторами:
- хелперной плазмидой, pHelper (Agilent, кат. №240074);
- плазмидой, кодирующей ген rep/cap AAV дикого типа или модифицированный (pAAV RC2 (Cell biolabs, кат. № VPK-422)), например, pAAV RC2/6 (Cell Biolabs, кат. № VPK-426), pAAV RC2-N587myc, pAAV RC2/6-Q585myc и т.д.; и
- плазмидой, кодирующей представляющую интерес нуклеотидную последовательность и последовательности ITR AAV, например, pscAAV-CMV-eGFP, pAAV-CMVGFP (Agilent, кат. №240074), pAAV-EF 1a-eGFP или pAAV-CAGG-eGFP и т.д.
[00164] Через семьдесят два часа после трансфекции среду собирали и клетки лизировали в буфере [50 мМ Tris-HCl, 150 мМ NaCl и 0,5% дезоксихолата натрия (Sigma, кат. № D6750-100G)]. Затем перед инкубацией при 37°С в течение 60 минут бензоназу (Sigma, Сент-Луис, штат Миссури) добавляли как к среде, так и к клеточному лизату до конечной концентрации 0,5 ед./мкл. Клеточный лизат центрифугировали при 4000 об/мин в течение 30 мин. Клеточный лизат и среду объединяли и осаждали с помощью PEG 8000 (Teknova, кат. № Р4340) в конечной концентрации 8%. Осадок ресуспендировали в 400 мМ NaCl и центрифугировали при 10000 g в течение 10 минут. Вирусы в надосадочной жидкости осаждали ультрацентрифугированием при 149000 g в течение 3 часов и титровали с использованием qPCR.
[00165] При qPCR для титрования геномов AAV образцы AAV обрабатывали ДНКазой I (Thermofisher Scientific, кат. № EN0525) при 37°С в течение одного часа и лизировали с использованием набора DNA extract All Reagents (Thermofisher Scientific, кат. №4403319). Капсидированные вирусные геномы определяли количественно, используя систему для ПЦР в режиме реального времени QuantStudio 3 Real-Time PCR System (Thermofisher Scientific) с применением праймеров для ITR AAV2. Последовательности праймеров для ITR AAV2 представляют собой 5'-GGAACCCCTAGTGATGGAGTT-3' (прямой праймер для ITR; SEQ ID NO: 36) и 5'-CGGCCTCAGTGAGCGA-3' (обратный праймер для ITR; SEQ ID NO: 37) (Aurnhammer et al., 2012), полученные на основе последовательности левого внутреннего инвертированного повтора (ITR) из AAV и последовательности правого внутреннего инвертированного повтора (ITR) из AAV соответственно. Последовательность зонда для ITR AAV2 представляет собой 5'-6-FAM-CACTCCCTCTCTGCGCGCTCG-TAMRA-3' (SEQ ID NO: 38) (Aurnhammer et al. (2012) Hum. Gene Ther. Methods 23:18 28). После стадии активации при 95°С в течение 10 минут проводили двухстадийный цикл ПЦР при 95°С в течение 15 секунд и 60°С в течение 30 секунд в течение 40 циклов. Универсальный мастер-микс для ПЦР TaqMan (Thermofisher Scientific, кат. №4304437) использовали для qPCR. ДНК-плазмиду (Agilent, кат.№240074) использовали в качестве стандарта для определения абсолютных титров.
[00166] Получали векторы на основе аденоассоциированного вируса, содержащие капсид со вставленным эпитопом c-myc. В данном примере эпитоп c-myc (EQKLISEEDL; SEQ ID NO: 25) вставляли между аминокислотами N587 и R588 капсидного белка VP1 AAV2, т.е. нуклеотидную последовательность, кодирующую эпитоп c-myc (GAA CAA AAA CTC АТС TCA GAA GAG GAT CTG; SEQ ID NO: 26), вставляли в плазмиду pAAV RC2 (Cell Biolabs, Inc., Сан-Диего, Калифорния, США), и модифицированную плазмиду pAAV RC2-N587Myc использовали для кодирования модифицированного капсидного белка для вирусных векторов на основе AAV с нейтрализованным тропизмом.
[00167] В частности, первый продукт полимеразной цепной реакции (ПЦР), содержащий (от 5' к 3') сайт рестрикции BsiWl, нуклеотидную последовательность между положениями 3050 и 3773 pAAV RC2 и нуклеотидную последовательность эпитопа c-myc с «липкими» концами, и второй продукт ПЦР, содержащий (от 5' к 3') нуклеотидную последовательность эпитопа c-myc с «липкими» концами, нуклеотидную последовательность между положениями 3774 и 4370 pAAV RC2 и сайт рестрикции Pme1, получали с использованием праймеров, изложенных в таблице 4. Плазмиду pAAV RC2-N587Myc (т.е. плазмиду pAAV RC2, модифицированную для кодирования эпитопа c-myc между аминокислотами N587 и R588 капсидного белка VP1) создавали путем расщепления pAAV RC2 с помощью BsiW1 (New England Biolabs, R0553L) и Pmel (New England Biolabs, R0560L) и вставки двух продуктов ПЦР посредством клонирования, независимого от лигирования, описанного в Li et al. (2012) Methods Mol. Biol. 852:51-59.
[00168] Таблица 4

[00169] В частности, фрагмент ДНК gblock, содержащий положения 3700 и 3940 из pAAV RC2/6 с последовательностью эпитопа c-myc, вставленной между 3757 и 3758, заказали в Integrated DNA Technologies (Коралвилль, Айова). Плазмиду pAAV RC2/6-Q585Myc создавали путем вставки фрагмента gblock в pAAV RC2/6, расщепленную с помощью MscI (New England Biolabs, кат. № R0534L) и AflII (New England Biolabs, кат. № R0520L) посредством клонирования, независимого от лигирования, описанного в Li et al. (2012) Methods Mol. Biol. 852:51-59.
[00170] В частности, pscAAV-CMV-eGFP получали путем введения фрагмента GFP в вектор pscAAV MCS (Cell Biolabs, кат. № VPK-430) с использованием сайта рестрикции для BamHI и NotI. Плазмиду pAAV-EF1a-eGFP и pAAV-CAGG-eGFP синтезировали de novo в Thermofisher Scientific (Уолтем, Массачусетс).
Пример 4. Опосредованная биспецифическими антителами интернализация частиц scAAV-N587Myc у мышей с гуманизированным локусом ASGR1 in vivo
[00171] Чтобы определить, может ли биспецифическое антитело к myc-ASGR1 перенацеливать вирусные векторы scAAV2-N587myc-CMV-eGFP на экспрессирующие hASGR1 клетки печени in vivo, мышам, генетически модифицированным таким образом, что их клетки печени экспрессируют hASGR1 на фоне C57BL/6, и контрольным мышам C57BL/6 дикого типа внутрисосудисто вводили 1×1011 (титрованных с помощью qPCR) только scAAV2-CMV-eGFP дикого типа или вирусных векторов scAAV2-N587myc-CMV-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 при соотношении 1:8 вирусного генома и молекул антитела. Контроль включал мышей с введением им физиологического раствора [250 мМ NaCl] или только вирусного вектора scAAV2-N587myc-CMV-eGFP. Через десять дней после инъекции мышей умерщвляли и проводили транскардиальную перфузию с использованием 4% PFA. Отбирали такие органы, как печень, почки и сердце, и их дегидратировали в 15% сахарозе, а затем в 30% сахарозе. Затем из органов получали криосрезы на предметных стеклах, и их окрашивали с использованием куриного антитела к EGFP (Jackson ImmunoResearch Labs, Inc., Уэст Гроув, Пенсильвания, США) и вторичного антитела к куриному антителу, конъюгированного с Alexa-488 (Jackson ImmunoResearch Labs, Inc., Уэст Гроув, Пенсильвания), (фигуры 8А - 8С). GFP-положительные клетки выявили в печени трансгенных животных, модифицированных для экспрессии ASGR1 в печени и получивших инъекцию scAAV2-CMV-eGFP дикого типа или scAAV2-N587myc-CMV-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигуры 8A(i) и 8A(iv)), и в печени мышей C57BL/6 дикого типа, получивших инъекцию scAAV2-CMV-eGFP дикого типа (фигура 8A(v)). GFP не выявили ни в образцах селезенки или почек (фигуры 8 В и 8С), ни в образцах печени, селезенки или почек, отобранных у любого животного, которым инъецировали физиологический раствор или только вирусные векторы scAAV2-N587myc-CMV-eGFP (фигуры 8A(ii, iii, vi, vii)), ни в образцах печени, отобранных у животных C57BL/6 дикого типа, которым инъецировали scAAV2-N587myc-CMV-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигура 8A(viii)). Таким образом, комбинация вирусных векторов scAAV2-N587myc-CMV-eGFP и биспецифического антитела к myc-ASGR1 инфицировала только те клетки (печени), которые экспрессируют hASGR1, убедительно свидетельствуя о том, что вирусный вектор scAAV2-CMV-eGFP инактивировался модификацией капсидного белка, например, природный тропизм вирусного вектора на основе scAAV может быть нейтрализован, например, с помощью эпитопа c-myc, при этом такие вирусные векторы могут быть специфически повторно активированы, например, специфически перенацелены, например, на клетки печени in vivo, например, с помощью биспецифических антител к myc-ASGR1.
[00172] Аналогично, чтобы определить, могут ли биспецифические антитела к myc-ASGR1 перенацеливать вирусные векторы ssAAV2-N587myc-CAGG-eGFP на экспрессирующие hASGR1 клетки печени in vivo, мышам, генетически модифицированным таким образом, что их клетки печени экспрессируют hASGR1 на фоне C57BL/6, и контрольным мышам C57BL/6 дикого типа внутрисосудисто вводили 2,18×1011 (титрованных с помощью qPCR) только ssAAV2-CAGG-eGFP дикого типа или вирусных векторов ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 при соотношении 1:4 вирусного генома и молекул антитела. Контроли включали мышей с введением им PBS или только вирусного вектора ssAAV2-N587myc-CAGG-eGFP. Через четыре недели после инъекции мышей умерщвляли и проводили транскардиальную перфузию с использованием 4% PFA. Отбирали такие органы, как печень, почки и сердце, и их дегидратировали в 15% сахарозе, а затем в 30% сахарозе. Затем из органов получали криосрезы на предметных стеклах, и их окрашивали с использованием куриного антитела к EGFP (Jackson ImmunoResearch Labs, Inc., Уэст Гроув, Пенсильвания, США) и вторичного антитела к куриному антителу, конъюгированного с Alexa-488 (Jackson ImmunoResearch Labs, Inc., Уэст Гроув, Пенсильвания), (фигуры 9А - 90). GFP-положительные клетки выявили в печени трансгенных животных, модифицированных для экспрессии ASGR1 в печени и получивших инъекцию ssAAV2-CAGG-eGFP дикого типа или ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигуры 9Е - 9F, 9Р - 9R), и в печени мышей C57BL/6 дикого типа, получивших инъекцию ssAAV2-CAGG-eGFP дикого типа (фигура 9В - 9С). Неожиданно, но эффективность инфицирования ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 намного выше, чем у WT ssAAV2-CAGG-GFP (фигуры 9Е - 9F, 9P - 9R). GFP не выявляли или практически не выявляли ни в образцах печени от любого животного, которым инъецировали физиологический раствор или только вирусный вектор ssAAV2-N587myc-CAGG-eGFP (фигуры 9А, 9D, 9G-9L)), ни в образцах печени, отобранных у животных C57BL/6 дикого типа, которым инъецировали ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигура 9М - 90). Таким образом, комбинация вирусных векторов scAAV2-N587myc-CAGG-eGFP и биспецифического антитела к myc-ASGR1 инфицировала только те клетки (печени), которые экспрессируют hASGR1, убедительно свидетельствуя о том, что вирусный вектор ssAAV2-N587myc-CAGG-eGFP инактивировался модификацией капсидного белка, например, природный тропизм вирусного вектора на основе scAAV может быть нейтрализован, например, с помощью эпитопа c-myc, при этом такие вирусные векторы могут быть специфически повторно активированы, например, специфически перенацелены, например, на клетки печени in vivo, например, с помощью биспецифических антител к myc-ASGR1.
[00173] Данный пример демонстрирует, что гуманизированные по ASGR1 мыши могут использоваться для тестирования потенциальных терапевтических средств, которые специфически связываются с ASGR1 человека для целевой доставки в клетки печени.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
110 Регенерон Фармасьютикалс, Инк.
Муджика, Алехо
Гусаров, Виктория
Ванг, Ченг
Киратсус, Христос
Потоки, Терра
Сигнар, Кэтрин
Мартин, Джоел
120 ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ЛОКУС ASGR1
130 009108.362WO1
150 62525,524
151 2017-06-27
160 42
170 Patent In версии 3.5
210 1
211 291
212 БЕЛОК
213 Homosapiens
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (1)..(40)
223 Цитоплазматический домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (41)..(61)
223 Трансмембранный домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (61)..(123)
223 Суперспиральный домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (161)..(278)
223 Лектиновый домен С-типа
400 1
Met Thr Lys Glu Tyr Gln Asp Leu Gln His Leu Asp Asn Glu Glu Ser
1 5 10 15
Asp His His Gln Leu Arg Lys Gly Pro Pro Pro Pro Gln Pro Leu Leu
20 25 30
Gln Arg Leu Cys Ser Gly Pro Arg Leu Leu Leu Leu Ser Leu Gly Leu
35 40 45
Ser Leu Leu Leu Leu Val Val Val Cys Val Ile Gly Ser Gln Asn Ser
50 55 60
Gln Leu Gln Glu Glu Leu Arg Gly Leu Arg Glu Thr Phe Ser Asn Phe
65 70 75 80
Thr Ala Ser Thr Glu Ala Gln Val Lys Gly Leu Ser Thr Gln Gly Gly
85 90 95
Asn Val Gly Arg Lys Met Lys Ser Leu Glu Ser Gln Leu Glu Lys Gln
100 105 110
Gln Lys Asp Leu Ser Glu Asp His Ser Ser Leu Leu Leu His Val Lys
115 120 125
Gln Phe Val Ser Asp Leu Arg Ser Leu Ser Cys Gln Met Ala Ala Leu
130 135 140
Gln Gly Asn Gly Ser Glu Arg Thr Cys Cys Pro Val Asn Trp Val Glu
145 150 155 160
His Glu Arg Ser Cys Tyr Trp Phe Ser Arg Ser Gly Lys Ala Trp Ala
165 170 175
Asp Ala Asp Asn Tyr Cys Arg Leu Glu Asp Ala His Leu Val Val Val
180 185 190
Thr Ser Trp Glu Glu Gln Lys Phe Val Gln His His Ile Gly Pro Val
195 200 205
Asn Thr Trp Met Gly Leu His Asp Gln Asn Gly Pro Trp Lys Trp Val
210 215 220
Asp Gly Thr Asp Tyr Glu Thr Gly Phe Lys Asn Trp Arg Pro Glu Gln
225 230 235 240
Pro Asp Asp Trp Tyr Gly His Gly Leu Gly Gly Gly Glu Asp Cys Ala
245 250 255
His Phe Thr Asp Asp Gly Arg Trp Asn Asp Asp Val Cys Gln Arg Pro
260 265 270
Tyr Arg Trp Val Cys Glu Thr Glu Leu Asp Lys Ala Ser Gln Glu Pro
275 280 285
Pro Leu Leu
290
210 2
211 284
212 PRT
213 Mus musculus
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (1)..(39)
223 Цитоплазматический домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (40)..(60)
223 Трансмембранный домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (60)..(122)
223 Суперспиральный домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (160)..(277)
223 Лектиновый домен С-типа
400 2
Met Thr Lys Asp Tyr Gln Asp Phe Gln His Leu Asp Asn Asp Asn Asp
1 5 10 15
His His Gln Leu Arg Arg Gly Pro Pro Pro Thr Pro Arg Leu Leu Gln
20 25 30
Arg Leu Cys Ser Gly Ser Arg Leu Leu Leu Leu Ser Ser Ser Leu Ser
35 40 45
Ile Leu Leu Leu Val Val Val Cys Val Ile Thr Ser Gln Asn Ser Gln
50 55 60
Leu Arg Glu Asp Leu Leu Ala Leu Arg Gln Asn Phe Ser Asn Leu Thr
65 70 75 80
Val Ser Thr Glu Asp Gln Val Lys Ala Leu Ser Thr Gln Gly Ser Ser
85 90 95
Val Gly Arg Lys Met Lys Leu Val Glu Ser Lys Leu Glu Lys Gln Gln
100 105 110
Lys Asp Leu Thr Glu Asp His Ser Ser Leu Leu Leu His Val Lys Gln
115 120 125
Leu Val Ser Asp Val Arg Ser Leu Ser Cys Gln Met Ala Ala Phe Arg
130 135 140
Gly Asn Gly Ser Glu Arg Thr Cys Cys Pro Ile Asn Trp Val Glu Tyr
145 150 155 160
Glu Gly Ser Cys Tyr Trp Phe Ser Ser Ser Val Arg Pro Trp Thr Glu
165 170 175
Ala Asp Lys Tyr Cys Gln Leu Glu Asn Ala His Leu Val Val Val Thr
180 185 190
Ser Arg Asp Glu Gln Asn Phe Leu Gln Arg His Met Gly Pro Leu Asn
195 200 205
Thr Trp Ile Gly Leu Thr Asp Gln Asn Gly Pro Trp Lys Trp Val Asp
210 215 220
Gly Thr Asp Tyr Glu Thr Gly Phe Gln Asn Trp Arg Pro Glu Gln Pro
225 230 235 240
Asp Asn Trp Tyr Gly His Gly Leu Gly Gly Gly Glu Asp Cys Ala His
245 250 255
Phe Thr Thr Asp Gly Arg Trp Asn Asp Asp Val Cys Arg Arg Pro Tyr
260 265 270
Arg Trp Val Cys Glu Thr Lys Leu Asp Lys Ala Asn
275 280
210 3
211 290
212 БЕЛОК
213 Искусственная последовательность
220
223 Синтетическая
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (1)..(39)
223 Цитоплазматический домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (40)..(60)
223 Трансмембранный домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (60)..(122)
223 Суперспиральный домен
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (62)..(290)
223 Остатки, кодируемые введенным экзоном человека
220
221 ДРУГАЯ_ХАРАКТЕРИСТИКА
222 (160)..(277)
223 Лектиновый домен С-типа
400 3
Met Thr Lys Asp Tyr Gln Asp Phe Gln His Leu Asp Asn Asp Asn Asp
1 5 10 15
His His Gln Leu Arg Arg Gly Pro Pro Pro Thr Pro Arg Leu Leu Gln
20 25 30
Arg Leu Cys Ser Gly Ser Arg Leu Leu Leu Leu Ser Ser Ser Leu Ser
35 40 45
Ile Leu Leu Leu Val Val Val Cys Val Ile Thr Ser Gln Asn Ser Gln
50 55 60
Leu Gln Glu Glu Leu Arg Gly Leu Arg Glu Thr Phe Ser Asn Phe Thr
65 70 75 80
Ala Ser Thr Glu Ala Gln Val Lys Gly Leu Ser Thr Gln Gly Gly Asn
85 90 95
Val Gly Arg Lys Met Lys Ser Leu Glu Ser Gln Leu Glu Lys Gln Gln
100 105 110
Lys Asp Leu Ser Glu Asp His Ser Ser Leu Leu Leu His Val Lys Gln
115 120 125
Phe Val Ser Asp Leu Arg Ser Leu Ser Cys Gln Met Ala Ala Leu Gln
130 135 140
Gly Asn Gly Ser Glu Arg Thr Cys Cys Pro Val Asn Trp Val Glu His
145 150 155 160
Glu Arg Ser Cys Tyr Trp Phe Ser Arg Ser Gly Lys Ala Trp Ala Asp
165 170 175
Ala Asp Asn Tyr Cys Arg Leu Glu Asp Ala His Leu Val Val Val Thr
180 185 190
Ser Trp Glu Glu Gln Lys Phe Val Gln His His Ile Gly Pro Val Asn
195 200 205
Thr Trp Met Gly Leu His Asp Gln Asn Gly Pro Trp Lys Trp Val Asp
210 215 220
Gly Thr Asp Tyr Glu Thr Gly Phe Lys Asn Trp Arg Pro Glu Gln Pro
225 230 235 240
Asp Asp Trp Tyr Gly His Gly Leu Gly Gly Gly Glu Asp Cys Ala His
245 250 255
Phe Thr Asp Asp Gly Arg Trp Asn Asp Asp Val Cys Gln Arg Pro Tyr
260 265 270
Arg Trp Val Cys Glu Thr Glu Leu Asp Lys Ala Ser Gln Glu Pro Pro
275 280 285
Leu Leu
290
210 4
211 855
212 ДНК
213 Mus musculus
400 4
atgacaaagg attatcaaga tttccagcac ctggacaatg ataatgacca tcatcaactc 60
cggagagggc cgcctcccac tccacggctc ttgcagcgac tctgctctgg atcccgcctc 120
ctcctgctct cctcgagcct cagcattctg ttgctggtgg ttgtctgtgt gatcacatcc 180
caaaattccc aactccggga agatctgctg gctctaaggc agaatttcag caacctcact 240
gtgagcactg aggaccaggt caaggccctg agcacccagg gaagtagtgt gggaagaaag 300
atgaagttag tggagtcgaa gctggaaaaa cagcagaagg atctgactga agatcactcc 360
agtttgctac tgcacgtgaa gcagttagtg tctgacgtgc gaagcttgag ctgccagatg 420
gctgcatttc ggggcaatgg ctctgaaagg acctgctgcc ccatcaactg ggtggagtat 480
gaaggcagct gctactggtt ctccagctct gtgaggcctt ggactgaagc tgacaagtac 540
tgccagctgg aaaatgccca tctggtggtg gtgacctcca gggatgagca gaacttcctc 600
cagcgccaca tgggcccctt aaacacttgg attggcctaa ctgaccagaa cgggccctgg 660
aaatgggtgg atggaacaga ctacgagaca ggcttccaga attggagacc agagcagcca 720
gataactggt acggacatgg gcttggagga ggcgaggact gtgcccactt cacgacggat 780
ggccgctgga atgacgacgt ctgcaggagg ccctaccgct gggtctgtga gacaaagttg 840
gataaggcta attag 855
210 5
211 876
212 ДНК
213 Homo sapiens
400 5
atgaccaagg agtatcaaga ccttcagcat ctggacaatg aggagagtga ccaccatcag 60
ctcagaaaag ggccacctcc tccccagccc ctcctgcagc gtctctgctc cggacctcgc 120
ctcctcctgc tctccctggg cctcagcctc ctgctgcttg tggttgtctg tgtgatcgga 180
tcccaaaact cccagctgca ggaggagctg cggggcctga gagagacgtt cagcaacttc 240
acagcgagca cggaggccca ggtcaagggc ttgagcaccc agggaggcaa tgtgggaaga 300
aagatgaagt cgctagagtc ccagctggag aaacagcaga aggacctgag tgaagatcac 360
tccagcctgc tgctccacgt gaagcagttc gtgtctgacc tgcggagcct gagctgtcag 420
atggcggcgc tccagggcaa tggctcagaa aggacctgct gcccggtcaa ctgggtggag 480
cacgagcgca gctgctactg gttctctcgc tccgggaagg cctgggctga cgccgacaac 540
tactgccggc tggaggacgc gcacctggtg gtggtcacgt cctgggagga gcagaaattt 600
gtccagcacc acataggccc tgtgaacacc tggatgggcc tccacgacca aaacgggccc 660
tggaagtggg tggacgggac ggactacgag acgggcttca agaactggag gccggagcag 720
ccggacgact ggtacggcca cgggctcgga ggaggcgagg actgtgccca cttcaccgac 780
gacggccgct ggaacgacga cgtctgccag aggccctacc gctgggtctg cgagacagag 840
ctggacaagg ccagccagga gccacctctc ctttaa 876
210 6
211 19
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 6
tcccaactccgggaagatc 19
210 7
211 24
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 7
tgctggctctaaggcagaatttca 24
210 8
211 20
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 8
tcagtgctcacagtgaggtt 20
210 9
211 22
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 9
gggttggctcatgttaggaagg 22
210 10
211 20
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 10
tcagcagccgagctgtgaaa 20
210 11
211 22
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 11
caggctgtgctacccaaagttc 22
210 12
211 21
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 12
ggaggcaatgtgggaagaaag 21
210 13
211 24
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 13
tgaagtcgctagagtcccagctgg 24
210 14
211 20
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 14
tcaggtccttctgctgtttc 20
210 15
211 20
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 15
gattgggaatccgcccatct 20
210 16
211 29
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 16
cctcttctgctttctcgggaattttcatc 29
210 17
211 20
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 17
aaagcgccacgggtttcaag 20
210 18
211 120
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
220
221 другая_характеристика
222 (1)..(60)
223 Последовательность мыши
220
221 другая_характеристика
222 (61)..(120)
223 Последовательность человека
400 18
catcccaaagtgggtggccagggctgggcagagaaagggggcaacttcgggtgtgtgtga 60
caagggagtggtgggtgcagtggtggcggacacagcgatcccgttttcttctctctgcac 120
210 19
211 160
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
220
221 другая_характеристика
222 (1)..(60)
223 Последовательность человека
220
221 другая_характеристика
222 (61)..(66)
223 XhoI
220
221 другая_характеристика
222 (67)..(100)
223 loxP
220
221 другая_характеристика
222 (101)..(160)
223 Кассета
400 19
tgttatttacagatacgtgagtttgggcaaattattgttctctgtgtcccagctgtaaac 60
ctcgagataacttcgtataatgtatgctatacgaagttatatgcatggcctccgcgccgg 120
gttttggcgcctcccgcgggcgcccccctcctcacggcga 160
210 20
211 191
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
220
221 другая_характеристика
222 (1)..(60)
223 Кассета
220
221 другая_характеристика
222 (61)..(94)
223 loxP
220
221 другая_характеристика
222 (100)..(125)
223 I-CeuI
220
221 другая_характеристика
222 (126)..(131)
223 NheI
220
221 другая_характеристика
222 (132)..(191)
223 Последовательность мыши
400 20
tttcactgcattctagttgtggtttgtccaaactcatcaatgtatcttatcatgtctgga 60
ataacttcgtataatgtatgctatacgaagttatgctagtaactataacggtcctaaggt 120
agcgagctagccgtggacagatacagcaacgtgagctagttattctgtcctaaagtctca 180
gttggaagatg 191
210 21
211 9692
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
220
221 другая_характеристика
222 (1)..(69)
223 Последовательность мыши
220
221 другая_характеристика
222 (70)..(3976)
223 Последовательность человека
220
221 другая_характеристика
222 (3977)..(3982)
223 XhoI
220
221 другая_характеристика
222 (3983)..(4016)
223 loxP
220
221 другая_характеристика
222 (4023)..(5235)
223 Промотор убиквитина человека
220
221 другая_характеристика
222 (5236)..(5302)
223 Промотор EM7
220
221 другая_характеристика
222 (5303)..(6328)
223 Ген устойчивости к гигромицину
220
221 другая_характеристика
222 (6329)..(6813)
223 PGK полиA
220
221 другая_характеристика
222 (6825)..(7506)
223 Промотор Prm
220
221 другая_характеристика
222 (7507)..(8646)
223 Crei
220
221 другая_характеристика
222 (8909)..(9119)
223 SV40 полиA
220
221 другая_характеристика
222 (9124)..(9157)
223 loxP
220
221 другая_характеристика
222 (9163)..(9188)
223 I-CeuI
220
221 другая_характеристика
222 (9189)..(9194)
223 NheI
220
221 другая_характеристика
222 (9195)..(9692)
223 Последовательность мыши
400 21
gtgtgatcacatcccaaagtgggtggccagggctgggcagagaaagggggcaacttcggg 60
tgtgtgtgacaagggagtggtgggtgcagtggtggcggacacagcgatcccgttttcttc 120
tctctgcacgctgtcctggccagactcccagctgcaggaggagctgcggggcctgagaga 180
gacgttcagcaacttcacagcgagcacggaggcccaggtcaagggcttgagcacccaggg 240
tgagggcgct ggggcggggc tggggctggg gctggggctg gggggctgtc gggaacgctg 300
agcgagcctc tcccgcagga ggcaatgtgg gaagaaagat gaagtcgcta gagtcccagc 360
tggagaaaca gcagaaggac ctgagtgaag gtcagagagg gagtgtgtgt gtgtgtgtgt 420
gtgtgtgaaa gagagtgaga atgtgtggat gtgtgtgaga aagtgtgagt gtgtgtggat 480
gtgtgtgaga atgagaggga gtgtgtgtgt gtgtgagtct gtgtgtgaga atgaggggga 540
gtgtgttttg ggtgtgtgta tgagagcctt gtgtggatgt gagaatgaga gggagtgtgt 600
atgtctgtga gtgtgagaat gagatggagt gtgtgtgagt ctgtgtgtga gaatgaggtg 660
tgtgtgtgtg agaatgagat ggtgtgtgtg tgggaatgag agggggtgtg tgtctgagtg 720
tgagaatgag atagagtgtg tgtgagacag tctgtgggaa tgagagggag tgtgtgtgag 780
agtgtgagaa tgacggagtg tgtctgtgag tgtgataatg aggtgtgtgt gagtctgagt 840
gtaagaatga gatggggtgt gtgtgtctgt gagtgtgaga gtgtgagaat gaggggtgtt 900
tgtgtctgag tgtgagtctg tgagtgtgag aatgagatgg ggtgtgtgag tgagtgtgag 960
aatgagatgg ggtgtgtgtg tctgtgagtg tgtgtgtgtt tgtgagtgtg agaatgagat 1020
ggggtgtgtg tgagtgtgag aatgagatgg ggtgtgtgtc tgtgtgtgag aatgagatgg 1080
gtgtgtgtgt gacagagtct gagtgtgaga atgagaggga gtgtgtgtga gtgtgagaat 1140
gagaaggagt ggatgggtgt gtgagtctgt gtgaatgagg gagtgggtgt gtgtacgagt 1200
gtgagtctgt gtttatgtgt gagaatgtgt cagtgtatgt gtgtgagaac gtgtgtatgt 1260
gtgttagtgt gtgttgcgtg tgtgggggaa tgagagggat tgtgtctgtg agtgtgagaa 1320
tgagatggag tgtctgtgag actgtgtgtg aggagtggga gtgtgtgtga gaatgagatg 1380
ggtgtgtgtg tctgagtgtg tgtctgtgag aatgagaggg agtgtgtgtg tgtgtgagag 1440
cctgtgtgaa aatgagaagg agtgtggatg ggtgtttgtg agtgggagag tctgtgtgtt 1500
tatgtgtgtg agaatgaggg agtgtgggtg tgtgtgcgaa tgtgagtctg tgtttatgtg 1560
tgtgagaatg tgtcagtgta tgtgagaacg tgtgtgttag tgtgttgcgt gtgtgagaat 1620
gtaagtatat gtgtaagtgc atgtgagtgt gtgtatgtgc gtgttgtgtg aatgtgcatt 1680
gtgtgtgcat gtgtgaaaga gtatatgtgt gttgtgggtg agtgtgtgtg gtgtgtgtag 1740
tgggtgaggg tgtgttgtat gtgtgggtgt gcgttgtgtg aatgtgtgta tgtgggtgag 1800
ggtgtgtgtg cctgtgtgag ggtgtgttgt ggtttttgtg tgtgtttggg tgagggtgtg 1860
ttgtgtgtgt gtgtgggtga aggtgtgttg tgtgtgtcgt gggtgaaggt gtgttgtgtg 1920
tgtagtgact gtagattagg gtgtgttccg tgtgtgtgtg tgagggtgta tgttgtgggt 1980
gttttgtgtg tgagtgggtg tgtaagggtg tgttgtgtgt atgtgggtta aggtgtgtta 2040
tgcgtgaggg tgtattgtgt gtgtgttttg tgtgtgttgt gtgtatgtgg gttagggtgt 2100
gttgtgtgtt tgtgtgtttt gtgtgttgtc tgtgtatgtg ggttacggtg tgttgtgcgt 2160
gtgagggtgt gttgtgtatg gtgtgttgtg tgtgttgtgt gagtgtgtat gtgagttagg 2220
gtgtgttgtg tctatgtatg tgtgtgtaag ggtgtgttgt gtgtctgtgg gtgtgttttg 2280
tgtatgtggg ttagggtgtg ttgtgtgttc tgtattgtgt gttttatgtg ttgtctgtat 2340
gtgggttatg tgtgttgtgt gtgttgtgga tgtatgtggg ttagggtgtg ttgtgtgtct 2400
ctgtgtgttg tctgcgtttg tgtctgtggg ttagggtgtg ttgtatgtgt tgtgttttgt 2460
gtgttgtccg tgtgtgtgta tgtgggttag gttgtgtgtg tgtgtgttgt atattgtctg 2520
tgtgtgtgtg ttaggatgtg ttgtgtgtct gtgtgagtgt gtgtgtaagg gtgtgttgtg 2580
tgtgtaggag tgtgtgtgtg tgtgtgtatg ggggtctctc aggccaactc cgctgctgtt 2640
tgtggcaatg cgacgggtgt tcgggtccca gcaggaggat gtagggctga cctcgtttcc 2700
cgtttccctc cccgtggttt ccgcatctcc tcccgctccc ctccgcccgg tctccccaga 2760
tcactccagc ctgctgctcc acgtgaagca gttcgtgtct gacctgcgga gcctgagctg 2820
tcagatggcg gcgctccagg gcaatggtaa ggaggccagc ccggcccgct ctctgcctcc 2880
ccccttctct gggcagcgct tagcccctgc gccccgtttc tcccgctcag gctcagaaag 2940
gacctgctgc ccggtcaact gggtggagca cgagcgcagc tgctactggt tctctcgctc 3000
cgggaaggcc tgggctgacg ccgacaacta ctgccggctg gaggacgcgc acctggtggt 3060
ggtcacgtcc tgggaggagc aggtgaggac ccggagggtc tgggaggctg gctggcctcg 3120
gagagatcac cacccgcctt ctctctcctc agaaatttgt ccagcaccac ataggccctg 3180
tgaacacctg gatgggcctc cacgaccaaa acgggccctg gaagtgggtg gacgggacgg 3240
actacgagac gggcttcaag tgagtgcgcg ccctccctcg gcctgggtcc ggccgccttc 3300
gcgccctggg gccctgggct gaggagtctg gagcgacccg cctgcggatc cgacctcctg 3360
gggcccacag ctggctctgt ccccaggaac tggaggccgg agcagccgga cgactggtac 3420
ggccacgggc tcggaggagg cgaggactgt gcccacttca ccgacgacgg ccgctggaac 3480
gacgacgtct gccagaggcc ctaccgctgg gtctgcgaga cagagctgga caaggccagc 3540
caggagccac ctctccttta atttatttct tcaatgcctc gacctgccgc aggggtccgg 3600
gattgggaat ccgcccatct gggggcctct tctgctttct cgggaatttt catctaggat 3660
tttaagggaa ggggaaggat agggtgatgt tccgaaggtg aggagcttga aacccgtggc 3720
gctttctgca gtttgcaggt tatcattgtg aacttttttt ttttaagagt aaaaagaaat 3780
atacctaaac cttctgttag ttgtctggtt attggggatt cggaagcagg agtgggctgg 3840
ttggcattac gaagccttag cgggtgctgt ggcatcatga gaactgtgtg ggctttgggc 3900
cagaatggcc agactttgtt atttacagat acgtgagttt gggcaaatta ttgttctctg 3960
tgtcccagct gtaaacctcg agataacttc gtataatgta tgctatacga agttatatgc 4020
atggcctccg cgccgggttt tggcgcctcc cgcgggcgcc cccctcctca cggcgagcgc 4080
tgccacgtca gacgaagggc gcagcgagcg tcctgatcct tccgcccgga cgctcaggac 4140
agcggcccgc tgctcataag actcggcctt agaaccccag tatcagcaga aggacatttt 4200
aggacgggac ttgggtgact ctagggcact ggttttcttt ccagagagcg gaacaggcga 4260
ggaaaagtag tcccttctcg gcgattctgc ggagggatct ccgtggggcg gtgaacgccg 4320
atgattatat aaggacgcgc cgggtgtggc acagctagtt ccgtcgcagc cgggatttgg 4380
gtcgcggttc ttgtttgtgg atcgctgtga tcgtcacttg gtgagtagcg ggctgctggg 4440
ctggccgggg ctttcgtggc cgccgggccg ctcggtggga cggaagcgtg tggagagacc 4500
gccaagggct gtagtctggg tccgcgagca aggttgccct gaactggggg ttggggggag 4560
cgcagcaaaa tggcggctgt tcccgagtct tgaatggaag acgcttgtga ggcgggctgt 4620
gaggtcgttg aaacaaggtg gggggcatgg tgggcggcaa gaacccaagg tcttgaggcc 4680
ttcgctaatg cgggaaagct cttattcggg tgagatgggc tggggcacca tctggggacc 4740
ctgacgtgaa gtttgtcact gactggagaa ctcggtttgt cgtctgttgc gggggcggca 4800
gttatggcgg tgccgttggg cagtgcaccc gtacctttgg gagcgcgcgc cctcgtcgtg 4860
tcgtgacgtc acccgttctg ttggcttata atgcagggtg gggccacctg ccggtaggtg 4920
tgcggtaggc ttttctccgt cgcaggacgc agggttcggg cctagggtag gctctcctga 4980
atcgacaggc gccggacctc tggtgagggg agggataagt gaggcgtcag tttctttggt 5040
cggttttatg tacctatctt cttaagtagc tgaagctccg gttttgaact atgcgctcgg 5100
ggttggcgag tgtgttttgt gaagtttttt aggcaccttt tgaaatgtaa tcatttgggt 5160
caatatgtaa ttttcagtgt tagactagta aattgtccgc taaattctgg ccgtttttgg 5220
cttttttgtt agacgtgttg acaattaatc atcggcatag tatatcggca tagtataata 5280
cgacaaggtg aggaactaaa ccatgaaaaa gcctgaactc accgcgacgt ctgtcgagaa 5340
gtttctgatc gaaaagttcg acagcgtgtc cgacctgatg cagctctcgg agggcgaaga 5400
atctcgtgct ttcagcttcg atgtaggagg gcgtggatat gtcctgcggg taaatagctg 5460
cgccgatggt ttctacaaag atcgttatgt ttatcggcac tttgcatcgg ccgcgctccc 5520
gattccggaa gtgcttgaca ttggggaatt cagcgagagc ctgacctatt gcatctcccg 5580
ccgtgcacag ggtgtcacgt tgcaagacct gcctgaaacc gaactgcccg ctgttctgca 5640
gccggtcgcg gaggccatgg atgcgattgc tgcggccgat cttagccaga cgagcgggtt 5700
cggcccattc ggaccgcaag gaatcggtca atacactaca tggcgtgatt tcatatgcgc 5760
gattgctgat ccccatgtgt atcactggca aactgtgatg gacgacaccg tcagtgcgtc 5820
cgtcgcgcag gctctcgatg agctgatgct ttgggccgag gactgccccg aagtccggca 5880
cctcgtgcac gcggatttcg gctccaacaa tgtcctgacg gacaatggcc gcataacagc 5940
ggtcattgac tggagcgagg cgatgttcgg ggattcccaa tacgaggtcg ccaacatctt 6000
cttctggagg ccgtggttgg cttgtatgga gcagcagacg cgctacttcg agcggaggca 6060
tccggagctt gcaggatcgc cgcggctccg ggcgtatatg ctccgcattg gtcttgacca 6120
actctatcag agcttggttg acggcaattt cgatgatgca gcttgggcgc agggtcgatg 6180
cgacgcaatc gtccgatccg gagccgggac tgtcgggcgt acacaaatcg cccgcagaag 6240
cgcggccgtc tggaccgatg gctgtgtaga agtactcgcc gatagtggaa accgacgccc 6300
cagcactcgt ccgagggcaa aggaataggg ggatccgctg taagtctgca gaaattgatg 6360
atctattaaa caataaagat gtccactaaa atggaagttt ttcctgtcat actttgttaa 6420
gaagggtgag aacagagtac ctacattttg aatggaagga ttggagctac gggggtgggg 6480
gtggggtggg attagataaa tgcctgctct ttactgaagg ctctttacta ttgctttatg 6540
ataatgtttc atagttggat atcataattt aaacaagcaa aaccaaatta agggccagct 6600
cattcctccc actcatgatc tatagatcta tagatctctc gtgggatcat tgtttttctc 6660
ttgattccca ctttgtggtt ctaagtactg tggtttccaa atgtgtcagt ttcatagcct 6720
gaagaacgag atcagcagcc tctgttccac atacacttca ttctcagtat tgttttgcca 6780
agttctaatt ccatcagacc tcgacctgca gcccctagcc cgggcgccag tagcagcacc 6840
cacgtccacc ttctgtctag taatgtccaa cacctccctc agtccaaaca ctgctctgca 6900
tccatgtggc tcccatttat acctgaagca cttgatgggg cctcaatgtt ttactagagc 6960
ccacccccct gcaactctga gaccctctgg atttgtctgt cagtgcctca ctggggcgtt 7020
ggataatttc ttaaaaggtc aagttccctc agcagcattc tctgagcagt ctgaagatgt 7080
gtgcttttca cagttcaaat ccatgtggct gtttcaccca cctgcctggc cttgggttat 7140
ctatcaggac ctagcctaga agcaggtgtg tggcacttaa cacctaagct gagtgactaa 7200
ctgaacactc aagtggatgc catctttgtc acttcttgac tgtgacacaa gcaactcctg 7260
atgccaaagc cctgcccacc cctctcatgc ccatatttgg acatggtaca ggtcctcact 7320
ggccatggtc tgtgaggtcc tggtcctctt tgacttcata attcctaggg gccactagta 7380
tctataagag gaagagggtg ctggctccca ggccacagcc cacaaaattc cacctgctca 7440
caggttggct ggctcgaccc aggtggtgtc ccctgctctg agccagctcc cggccaagcc 7500
agcaccatgg gtacccccaa gaagaagagg aaggtgcgta ccgatttaaa ttccaattta 7560
ctgaccgtac accaaaattt gcctgcatta ccggtcgatg caacgagtga tgaggttcgc 7620
aagaacctga tggacatgtt cagggatcgc caggcgtttt ctgagcatac ctggaaaatg 7680
cttctgtccg tttgccggtc gtgggcggca tggtgcaagt tgaataaccg gaaatggttt 7740
cccgcagaac ctgaagatgt tcgcgattat cttctatatc ttcaggcgcg cggtctggca 7800
gtaaaaacta tccagcaaca tttgggccag ctaaacatgc ttcatcgtcg gtccgggctg 7860
ccacgaccaa gtgacagcaa tgctgtttca ctggttatgc ggcggatccg aaaagaaaac 7920
gttgatgccg gtgaacgtgc aaaacaggct ctagcgttcg aacgcactga tttcgaccag 7980
gttcgttcac tcatggaaaa tagtgatcgc tgccaggata tacgtaatct ggcatttctg 8040
gggattgctt ataacaccct gttacgtata gccgaaattg ccaggatcag ggttaaagat 8100
atctcacgta ctgacggtgg gagaatgtta atccatattg gcagaacgaa aacgctggtt 8160
agcaccgcag gtgtagagaa ggcacttagc ctgggggtaa ctaaactggt cgagcgatgg 8220
atttccgtct ctggtgtagc tgatgatccg aataactacc tgttttgccg ggtcagaaaa 8280
aatggtgttg ccgcgccatc tgccaccagc cagctatcaa ctcgcgccct ggaagggatt 8340
tttgaagcaa ctcatcgatt gatttacggc gctaaggtaa atataaaatt tttaagtgta 8400
taatgtgtta aactactgat tctaattgtt tgtgtatttt aggatgactc tggtcagaga 8460
tacctggcct ggtctggaca cagtgcccgt gtcggagccg cgcgagatat ggcccgcgct 8520
ggagtttcaa taccggagat catgcaagct ggtggctgga ccaatgtaaa tattgtcatg 8580
aactatatcc gtaacctgga tagtgaaaca ggggcaatgg tgcgcctgct ggaagatggc 8640
gattgatcta gataagtaat gatcataatc agccatatca catctgtaga ggttttactt 8700
gctttaaaaa acctcccaca cctccccctg aacctgaaac ataaaatgaa tgcaattgtt 8760
gttgttaaac ctgccctagt tgcggccaat tccagctgag cgtgcctccg caccattacc 8820
agttggtctg gtgtcaaaaa taataataac cgggcagggg ggatctaagc tctagataag 8880
taatgatcat aatcagccat atcacatctg tagaggtttt acttgcttta aaaaacctcc 8940
cacacctccc cctgaacctg aaacataaaa tgaatgcaat tgttgttgtt aacttgttta 9000
ttgcagctta taatggttac aaataaagca atagcatcac aaatttcaca aataaagcat 9060
ttttttcact gcattctagt tgtggtttgt ccaaactcat caatgtatct tatcatgtct 9120
ggaataactt cgtataatgt atgctatacg aagttatgct agtaactata acggtcctaa 9180
ggtagcgagc tagccgtgga cagatacagc aacgtgagct agttattctg tcctaaagtc 9240
tcagttggaa gatgggagga tttttgacct ctgtctgctg ggggcaggac caaccaccag 9300
ggaactgcag cccccctgtg ctgagtgcat cagagacttg gaatggaaca cactggcctg 9360
cgacactcat cacaacgaac agaaactgct ttgtacactg aataaacgca gtgaataccc 9420
agctcaggat cacagacaca tgaatgcaaa gttatattag tataaccaag ggtgggaatg 9480
agggcaatta cagataactt atagacatga attactaaca aaacagggca aaatgtttgc 9540
tcataaataa catgaaaata caatatatag tcatatgtat atatacatgt atatatataa 9600
atgacataat atgtatatat ttttacaaat acactgcagg aaaataatat ttttcctcta 9660
cagaagagat tggcaaatct gacatctaaa at 9692
210 22
211 160
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
220
221 другая_характеристика
222 (1)..(90)
223 Последовательность мыши
220
221 другая_характеристика
222 (91)..(160)
223 Последовательность человека
400 22
catgatgttt ctttcttagg aaagccaggg catttctcta ttctccaatc tcttggctca 60
atgcccttgg cctctctttt gttccactag tgaagcctct ccagccaggg gctgaggtcc 120
cggtggtgtg ggcccaggag ggggctcctg cccagctccc 160
210 23
211 197
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
220
221 другая_характеристика
222 (1)..(60)
223 Последовательность человека
220
221 другая_характеристика
222 (61)..(66)
223 XhoI
220
221 другая_характеристика
222 (67)..(100)
223 loxP
220
221 другая_характеристика
222 (106)..(131)
223 I-CeuI
220
221 другая_характеристика
222 (132)..(137)
223 NheI
220
221 другая_характеристика
222 (138)..(197)
223 Последовательность мыши
400 23
tgttatttac agatacgtga gtttgggcaa attattgttc tctgtgtccc agctgtaaac 60
ctcgagataa cttcgtataa tgtatgctat acgaagttat gctagtaact ataacggtcc 120
taaggtagcg agctagccgt ggacagatac agcaacgtga gctagttatt ctgtcctaaa 180
gtctcagttg gaagatg 197
210 24
211 4551
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
220
221 другая_характеристика
222 (1)..(69)
223 Последовательность мыши
220
221 другая_характеристика
222 (70)..(3976)
223 Последовательность человека
220
221 другая_характеристика
222 (3977)..(3982)
223 XhoI
220
221 другая_характеристика
222 (3983)..(4016)
223 LoxP
220
221 другая_характеристика
222 (4022)..(4047)
223 I-CeuI
220
221 другая_характеристика
222 (4048)..(4053)
223 NheI
220
221 другая_характеристика
222 (4054)..(4551)
223 Последовательность мыши
400 24
gtgtgatcac atcccaaagt gggtggccag ggctgggcag agaaaggggg caacttcggg 60
tgtgtgtgac aagggagtgg tgggtgcagt ggtggcggac acagcgatcc cgttttcttc 120
tctctgcacg ctgtcctggc cagactccca gctgcaggag gagctgcggg gcctgagaga 180
gacgttcagc aacttcacag cgagcacgga ggcccaggtc aagggcttga gcacccaggg 240
tgagggcgct ggggcggggc tggggctggg gctggggctg gggggctgtc gggaacgctg 300
agcgagcctc tcccgcagga ggcaatgtgg gaagaaagat gaagtcgcta gagtcccagc 360
tggagaaaca gcagaaggac ctgagtgaag gtcagagagg gagtgtgtgt gtgtgtgtgt 420
gtgtgtgaaa gagagtgaga atgtgtggat gtgtgtgaga aagtgtgagt gtgtgtggat 480
gtgtgtgaga atgagaggga gtgtgtgtgt gtgtgagtct gtgtgtgaga atgaggggga 540
gtgtgttttg ggtgtgtgta tgagagcctt gtgtggatgt gagaatgaga gggagtgtgt 600
atgtctgtga gtgtgagaat gagatggagt gtgtgtgagt ctgtgtgtga gaatgaggtg 660
tgtgtgtgtg agaatgagat ggtgtgtgtg tgggaatgag agggggtgtg tgtctgagtg 720
tgagaatgag atagagtgtg tgtgagacag tctgtgggaa tgagagggag tgtgtgtgag 780
agtgtgagaa tgacggagtg tgtctgtgag tgtgataatg aggtgtgtgt gagtctgagt 840
gtaagaatga gatggggtgt gtgtgtctgt gagtgtgaga gtgtgagaat gaggggtgtt 900
tgtgtctgag tgtgagtctg tgagtgtgag aatgagatgg ggtgtgtgag tgagtgtgag 960
aatgagatgg ggtgtgtgtg tctgtgagtg tgtgtgtgtt tgtgagtgtg agaatgagat 1020
ggggtgtgtg tgagtgtgag aatgagatgg ggtgtgtgtc tgtgtgtgag aatgagatgg 1080
gtgtgtgtgt gacagagtct gagtgtgaga atgagaggga gtgtgtgtga gtgtgagaat 1140
gagaaggagt ggatgggtgt gtgagtctgt gtgaatgagg gagtgggtgt gtgtacgagt 1200
gtgagtctgt gtttatgtgt gagaatgtgt cagtgtatgt gtgtgagaac gtgtgtatgt 1260
gtgttagtgt gtgttgcgtg tgtgggggaa tgagagggat tgtgtctgtg agtgtgagaa 1320
tgagatggag tgtctgtgag actgtgtgtg aggagtggga gtgtgtgtga gaatgagatg 1380
ggtgtgtgtg tctgagtgtg tgtctgtgag aatgagaggg agtgtgtgtg tgtgtgagag 1440
cctgtgtgaa aatgagaagg agtgtggatg ggtgtttgtg agtgggagag tctgtgtgtt 1500
tatgtgtgtg agaatgaggg agtgtgggtg tgtgtgcgaa tgtgagtctg tgtttatgtg 1560
tgtgagaatg tgtcagtgta tgtgagaacg tgtgtgttag tgtgttgcgt gtgtgagaat 1620
gtaagtatat gtgtaagtgc atgtgagtgt gtgtatgtgc gtgttgtgtg aatgtgcatt 1680
gtgtgtgcat gtgtgaaaga gtatatgtgt gttgtgggtg agtgtgtgtg gtgtgtgtag 1740
tgggtgaggg tgtgttgtat gtgtgggtgt gcgttgtgtg aatgtgtgta tgtgggtgag 1800
ggtgtgtgtg cctgtgtgag ggtgtgttgt ggtttttgtg tgtgtttggg tgagggtgtg 1860
ttgtgtgtgt gtgtgggtga aggtgtgttg tgtgtgtcgt gggtgaaggt gtgttgtgtg 1920
tgtagtgact gtagattagg gtgtgttccg tgtgtgtgtg tgagggtgta tgttgtgggt 1980
gttttgtgtg tgagtgggtg tgtaagggtg tgttgtgtgt atgtgggtta aggtgtgtta 2040
tgcgtgaggg tgtattgtgt gtgtgttttg tgtgtgttgt gtgtatgtgg gttagggtgt 2100
gttgtgtgtt tgtgtgtttt gtgtgttgtc tgtgtatgtg ggttacggtg tgttgtgcgt 2160
gtgagggtgt gttgtgtatg gtgtgttgtg tgtgttgtgt gagtgtgtat gtgagttagg 2220
gtgtgttgtg tctatgtatg tgtgtgtaag ggtgtgttgt gtgtctgtgg gtgtgttttg 2280
tgtatgtggg ttagggtgtg ttgtgtgttc tgtattgtgt gttttatgtg ttgtctgtat 2340
gtgggttatg tgtgttgtgt gtgttgtgga tgtatgtggg ttagggtgtg ttgtgtgtct 2400
ctgtgtgttg tctgcgtttg tgtctgtggg ttagggtgtg ttgtatgtgt tgtgttttgt 2460
gtgttgtccg tgtgtgtgta tgtgggttag gttgtgtgtg tgtgtgttgt atattgtctg 2520
tgtgtgtgtg ttaggatgtg ttgtgtgtct gtgtgagtgt gtgtgtaagg gtgtgttgtg 2580
tgtgtaggag tgtgtgtgtg tgtgtgtatg ggggtctctc aggccaactc cgctgctgtt 2640
tgtggcaatg cgacgggtgt tcgggtccca gcaggaggat gtagggctga cctcgtttcc 2700
cgtttccctc cccgtggttt ccgcatctcc tcccgctccc ctccgcccgg tctccccaga 2760
tcactccagc ctgctgctcc acgtgaagca gttcgtgtct gacctgcgga gcctgagctg 2820
tcagatggcg gcgctccagg gcaatggtaa ggaggccagc ccggcccgct ctctgcctcc 2880
ccccttctct gggcagcgct tagcccctgc gccccgtttc tcccgctcag gctcagaaag 2940
gacctgctgc ccggtcaact gggtggagca cgagcgcagc tgctactggt tctctcgctc 3000
cgggaaggcc tgggctgacg ccgacaacta ctgccggctg gaggacgcgc acctggtggt 3060
ggtcacgtcc tgggaggagc aggtgaggac ccggagggtc tgggaggctg gctggcctcg 3120
gagagatcac cacccgcctt ctctctcctc agaaatttgt ccagcaccac ataggccctg 3180
tgaacacctg gatgggcctc cacgaccaaa acgggccctg gaagtgggtg gacgggacgg 3240
actacgagac gggcttcaag tgagtgcgcg ccctccctcg gcctgggtcc ggccgccttc 3300
gcgccctggg gccctgggct gaggagtctg gagcgacccg cctgcggatc cgacctcctg 3360
gggcccacag ctggctctgt ccccaggaac tggaggccgg agcagccgga cgactggtac 3420
ggccacgggc tcggaggagg cgaggactgt gcccacttca ccgacgacgg ccgctggaac 3480
gacgacgtct gccagaggcc ctaccgctgg gtctgcgaga cagagctgga caaggccagc 3540
caggagccac ctctccttta atttatttct tcaatgcctc gacctgccgc aggggtccgg 3600
gattgggaat ccgcccatct gggggcctct tctgctttct cgggaatttt catctaggat 3660
tttaagggaa ggggaaggat agggtgatgt tccgaaggtg aggagcttga aacccgtggc 3720
gctttctgca gtttgcaggt tatcattgtg aacttttttt ttttaagagt aaaaagaaat 3780
atacctaaac cttctgttag ttgtctggtt attggggatt cggaagcagg agtgggctgg 3840
ttggcattac gaagccttag cgggtgctgt ggcatcatga gaactgtgtg ggctttgggc 3900
cagaatggcc agactttgtt atttacagat acgtgagttt gggcaaatta ttgttctctg 3960
tgtcccagct gtaaacctcg agataacttc gtataatgta tgctatacga agttatgcta 4020
gtaactataa cggtcctaag gtagcgagct agccgtggac agatacagca acgtgagcta 4080
gttattctgt cctaaagtct cagttggaag atgggaggat ttttgacctc tgtctgctgg 4140
gggcaggacc aaccaccagg gaactgcagc ccccctgtgc tgagtgcatc agagacttgg 4200
aatggaacac actggcctgc gacactcatc acaacgaaca gaaactgctt tgtacactga 4260
ataaacgcag tgaataccca gctcaggatc acagacacat gaatgcaaag ttatattagt 4320
ataaccaagg gtgggaatga gggcaattac agataactta tagacatgaa ttactaacaa 4380
aacagggcaa aatgtttgct cataaataac atgaaaatac aatatatagt catatgtata 4440
tatacatgta tatatataaa tgacataata tgtatatatt tttacaaata cactgcagga 4500
aaataatatt tttcctctac agaagagatt ggcaaatctg acatctaaaa t 4551
210 25
211 10
212 БЕЛОК
213 Искусственная последовательность
220
223 Синтетическая
400 25
Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
1 5 10
210 26
211 30
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 26
gaacaaaaac tcatctcaga agaggatctg 30
210 27
211 63
212 БЕЛОК
213 Homo sapiens
400 27
Ser Gln Asn Ser Gln Leu Gln Glu Glu Leu Arg Gly Leu Arg Glu Thr
1 5 10 15
Phe Ser Asn Phe Thr Ala Ser Thr Glu Ala Gln Val Lys Gly Leu Ser
20 25 30
Thr Gln Gly Gly Asn Val Gly Arg Lys Met Lys Ser Leu Glu Ser Gln
35 40 45
Leu Glu Lys Gln Gln Lys Asp Leu Ser Glu Asp His Ser Ser Leu
50 55 60
210 28
211 118
212 БЕЛОК
213 Homo sapiens
400 28
His Glu Arg Ser Cys Tyr Trp Phe Ser Arg Ser Gly Lys Ala Trp Ala
1 5 10 15
Asp Ala Asp Asn Tyr Cys Arg Leu Glu Asp Ala His Leu Val Val Val
20 25 30
Thr Ser Trp Glu Glu Gln Lys Phe Val Gln His His Ile Gly Pro Val
35 40 45
Asn Thr Trp Met Gly Leu His Asp Gln Asn Gly Pro Trp Lys Trp Val
50 55 60
Asp Gly Thr Asp Tyr Glu Thr Gly Phe Lys Asn Trp Arg Pro Glu Gln
65 70 75 80
Pro Asp Asp Trp Tyr Gly His Gly Leu Gly Gly Gly Glu Asp Cys Ala
85 90 95
His Phe Thr Asp Asp Gly Arg Trp Asn Asp Asp Val Cys Gln Arg Pro
100 105 110
Tyr Arg Trp Val Cys Glu
115
210 29
211 39
212 БЕЛОК
213 Mus musculus
400 29
Met Thr Lys Asp Tyr Gln Asp Phe Gln His Leu Asp Asn Asp Asn Asp
1 5 10 15
His His Gln Leu Arg Arg Gly Pro Pro Pro Thr Pro Arg Leu Leu Gln
20 25 30
Arg Leu Cys Ser Gly Ser Arg
35
210 30
211 21
212 БЕЛОК
213 Mus musculus
400 30
Leu Leu Leu Leu Ser Ser Ser Leu Ser Ile Leu Leu Leu Val Val Val
1 5 10 15
Cys Val Ile Thr Ser
20
210 31
211 229
212 БЕЛОК
213 Homo sapiens
400 31
Asn Ser Gln Leu Gln Glu Glu Leu Arg Gly Leu Arg Glu Thr Phe Ser
1 5 10 15
Asn Phe Thr Ala Ser Thr Glu Ala Gln Val Lys Gly Leu Ser Thr Gln
20 25 30
Gly Gly Asn Val Gly Arg Lys Met Lys Ser Leu Glu Ser Gln Leu Glu
35 40 45
Lys Gln Gln Lys Asp Leu Ser Glu Asp His Ser Ser Leu Leu Leu His
50 55 60
Val Lys Gln Phe Val Ser Asp Leu Arg Ser Leu Ser Cys Gln Met Ala
65 70 75 80
Ala Leu Gln Gly Asn Gly Ser Glu Arg Thr Cys Cys Pro Val Asn Trp
85 90 95
Val Glu His Glu Arg Ser Cys Tyr Trp Phe Ser Arg Ser Gly Lys Ala
100 105 110
Trp Ala Asp Ala Asp Asn Tyr Cys Arg Leu Glu Asp Ala His Leu Val
115 120 125
Val Val Thr Ser Trp Glu Glu Gln Lys Phe Val Gln His His Ile Gly
130 135 140
Pro Val Asn Thr Trp Met Gly Leu His Asp Gln Asn Gly Pro Trp Lys
145 150 155 160
Trp Val Asp Gly Thr Asp Tyr Glu Thr Gly Phe Lys Asn Trp Arg Pro
165 170 175
Glu Gln Pro Asp Asp Trp Tyr Gly His Gly Leu Gly Gly Gly Glu Asp
180 185 190
Cys Ala His Phe Thr Asp Asp Gly Arg Trp Asn Asp Asp Val Cys Gln
195 200 205
Arg Pro Tyr Arg Trp Val Cys Glu Thr Glu Leu Asp Lys Ala Ser Gln
210 215 220
Glu Pro Pro Leu Leu
225
210 32
211 20
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 32
ggagtaccag ctcccgtacg 20
210 33
211 43
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 33
ctcttctgag atgagttttt gttcgttgcc tctctggagg ttg 43
210 34
211 43
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 34
aaactcatct cagaagagga tctgagacaa gcagctaccg cag 43
210 35
211 18
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 35
tccgcccgct gtttaaac 18
210 36
211 21
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 36
ggaaccccta gtgatggagt t 21
210 37
211 16
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 37
cggcctcagt gagcga 16
210 38
211 21
212 ДНК
213 Искусственная последовательность
220
223 Синтетическая
400 38
cactccctct ctgcgcgctc g 21
210 39
211 291
212 БЕЛОК
213 Macaca fascicularis
400 39
Met Thr Lys Glu Tyr Gln Asp Leu Gln His Leu Asp Asn Glu Glu Ser
1 5 10 15
Asp His His Gln Leu Gly Lys Gly Pro Pro Pro Pro Gln Ser Leu Leu
20 25 30
Arg Arg Leu Cys Ser Gly Pro Arg Leu Leu Leu Leu Ser Leu Gly Leu
35 40 45
Ser Leu Leu Leu Leu Val Val Val Cys Val Ile Gly Ser Gln Asn Ala
50 55 60
Gln Leu Gln Arg Glu Leu Arg Gly Leu Arg Glu Thr Leu Ser Asn Phe
65 70 75 80
Thr Ala Ser Thr Glu Ala Gln Val Lys Gly Leu Ser Thr Gln Gly Gly
85 90 95
Asn Val Gly Arg Lys Met Lys Ser Leu Glu Ser Gln Leu Glu Lys Gln
100 105 110
Gln Lys Asp Leu Ser Glu Asp His Ser Ser Leu Leu Leu His Val Lys
115 120 125
Gln Phe Val Ser Asp Leu Arg Ser Leu Ser Cys Gln Met Ala Ala Leu
130 135 140
Gln Gly Asn Gly Ser Glu Arg Ala Cys Cys Pro Val Asn Trp Val Glu
145 150 155 160
His Glu Arg Ser Cys Tyr Trp Phe Ser Arg Ser Gly Lys Ala Trp Ala
165 170 175
Asp Ala Asp Asn Tyr Cys Arg Leu Glu Asp Ala His Leu Val Val Val
180 185 190
Thr Ser Trp Glu Glu Gln Lys Phe Val Gln His His Ile Gly Pro Val
195 200 205
Asn Thr Trp Met Gly Leu His Asp Gln Asn Gly Pro Trp Lys Trp Val
210 215 220
Asp Gly Thr Asp Tyr Glu Thr Gly Phe Lys Asn Trp Arg Pro Glu Gln
225 230 235 240
Pro Asp Asp Trp Tyr Gly His Gly Leu Gly Gly Gly Glu Asp Cys Ala
245 250 255
His Phe Thr Asp Asp Gly Arg Trp Asn Asp Asp Val Cys Gln Arg Pro
260 265 270
Tyr Arg Trp Val Cys Glu Thr Glu Leu Asp Lys Ala Ser Gln Glu Pro
275 280 285
Pro Leu Leu
290
210 40
211 876
212 ДНК
213 Macaca fascicularis
400 40
atgaccaagg agtatcagga cctgcagcat ctggacaatg aggagagtga ccaccatcag 60
ctcggaaaag ggccacctcc tccgcagtcc ctcctgcggc gtctctgctc cggccctcgc 120
ctcctcctgc tctccctggg cctcagcctc ctgctgctgg tggttgtctg tgtgatcgga 180
tcccaaaacg cccagctgca gcgggagctg cggggcctga gagagacgct cagcaacttc 240
acagcgagca ccgaggccca ggtcaagggc ttgagcaccc agggaggcaa tgtgggaaga 300
aagatgaagt cgctggagtc ccagctggag aaacagcaga aggacttgag tgaagatcac 360
tccagcctgc tgctccacgt gaagcagttc gtgtctgacc tgcggagcct gagctgtcag 420
atggcggcgc tccagggcaa tggctcggaa agggcctgct gcccagtcaa ctgggtggag 480
cacgagcgca gctgctactg gttctctcgc tccgggaagg cctgggccga cgccgacaac 540
tactgccggc tggaggacgc gcacctggtg gtggtcacgt cctgggagga gcagaaattt 600
gtccagcacc acataggtcc tgtgaacacc tggatgggcc tccacgacca aaacgggccc 660
tggaagtggg tggacgggac ggactacgag acgggcttca agaactggag accggagcag 720
ccggacgact ggtacggcca cgggctcggg ggaggggagg actgtgccca cttcaccgac 780
gacggccgct ggaacgacga cgtctgccag aggccctacc gctgggtctg cgagacagag 840
ctggacaagg ccagtcagga gccacctctc ctttaa 876
210 41
211 284
212 БЕЛОК
213 Rattus norvegicus
400 41
Met Thr Lys Asp Tyr Gln Asp Phe Gln His Leu Asp Asn Glu Asn Asp
1 5 10 15
His His Gln Leu Gln Arg Gly Pro Pro Pro Ala Pro Arg Leu Leu Gln
20 25 30
Arg Leu Cys Ser Gly Phe Arg Leu Phe Leu Leu Ser Leu Gly Leu Ser
35 40 45
Ile Leu Leu Leu Val Val Val Cys Val Ile Thr Ser Gln Asn Ser Gln
50 55 60
Leu Arg Glu Asp Leu Arg Val Leu Arg Gln Asn Phe Ser Asn Phe Thr
65 70 75 80
Val Ser Thr Glu Asp Gln Val Lys Ala Leu Thr Thr Gln Gly Glu Arg
85 90 95
Val Gly Arg Lys Met Lys Leu Val Glu Ser Gln Leu Glu Lys His Gln
100 105 110
Glu Asp Leu Arg Glu Asp His Ser Arg Leu Leu Leu His Val Lys Gln
115 120 125
Leu Val Ser Asp Val Arg Ser Leu Ser Cys Gln Met Ala Ala Leu Arg
130 135 140
Gly Asn Gly Ser Glu Arg Ile Cys Cys Pro Ile Asn Trp Val Glu Tyr
145 150 155 160
Glu Gly Ser Cys Tyr Trp Phe Ser Ser Ser Val Lys Pro Trp Thr Glu
165 170 175
Ala Asp Lys Tyr Cys Gln Leu Glu Asn Ala His Leu Val Val Val Thr
180 185 190
Ser Trp Glu Glu Gln Arg Phe Val Gln Gln His Met Gly Pro Leu Asn
195 200 205
Thr Trp Ile Gly Leu Thr Asp Gln Asn Gly Pro Trp Lys Trp Val Asp
210 215 220
Gly Thr Asp Tyr Glu Thr Gly Phe Lys Asn Trp Arg Pro Gly Gln Pro
225 230 235 240
Asp Asp Trp Tyr Gly His Gly Leu Gly Gly Gly Glu Asp Cys Ala His
245 250 255
Phe Thr Thr Asp Gly His Trp Asn Asp Asp Val Cys Arg Arg Pro Tyr
260 265 270
Arg Trp Val Cys Glu Thr Glu Leu Gly Lys Ala Asn
275 280
210 42
211 855
212 ДНК
213 Rattus norvegicus
400 42
atgacaaagg attatcaaga tttccagcac ttggacaatg agaacgacca ccatcaactc 60
cagagagggc cacctcccgc tccaaggctc ttgcagcgac tctgctctgg attccgtctc 120
ttcctgcttt ccctgggcct cagcatcctg ctgctggtgg ttgtctgtgt gatcacatcc 180
caaaattccc aactccggga agatctgcgg gttctaaggc agaatttcag caactttacc 240
gtgagcactg aggaccaggt caaggccctg accacccagg gagagagagt gggaagaaag 300
atgaagttag tcgagtcaca gctggaaaaa catcaggagg atctgaggga agaccactct 360
agattgctac tgcatgtaaa gcagttagtg tctgacgtgc gaagcttgag ctgccagatg 420
gccgcacttc ggggcaatgg ctctgaaagg atctgctgcc ccatcaactg ggtggagtat 480
gaaggcagct gctactggtt ctccagctct gtgaagcctt ggacggaagc tgacaagtac 540
tgccagctgg agaacgccca cctggtggtg gtgacttcct gggaggagca gagattcgtc 600
cagcaacaca tgggcccctt aaatacttgg attggcctaa ctgaccagaa cggaccctgg 660
aaatgggtgg atgggacaga ctatgagaca ggcttcaaga actggagacc agggcagcca 720
gatgactggt acggacatgg gcttggaggg ggtgaagact gtgcccactt caccaccgat 780
ggccactgga atgatgacgt ctgcaggagg ccctaccgct gggtctgtga gacagagttg 840
ggcaaggcca attag 855
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ ГУМАНИЗИРОВАННЫЙ КОМПЛЕКС CD3 | 2015 |
|
RU2726446C2 |
| НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ЛОКУС TrkB | 2018 |
|
RU2800428C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ИМЕЮЩИЕ ГУМАНИЗИРОВАННЫЙ ГЕН КЛАСТЕРА ДИФФЕРЕНЦИРОВКИ 47 | 2015 |
|
RU2728412C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ДЛЯ ВЫРАБОТКИ ТЕРАПЕВТИЧЕСКИХ АНТИТЕЛ ПРОТИВ КОМПЛЕКСОВ ПЕПТИД-MHC, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ВАРИАНТЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2819525C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА | 2018 |
|
RU2809246C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ИМЕЮЩИЕ ГУМАНИЗИРОВАННЫЙ ГЕН КЛАСТЕРА ДИФФЕРЕНЦИРОВКИ 47 | 2015 |
|
RU2827833C1 |
| Животные, отличные от человека, содержащие гуманизированный локус альбумина | 2020 |
|
RU2815514C2 |
| ГУМАНИЗИРОВАННЫЕ МЫШИ С НОКИНОМ SIRPA-IL15 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2822370C2 |
| МОДЕЛИ ЗАБОЛЕВАНИЯ DITRA НА ЖИВОТНЫХ, ОТЛИЧНЫХ ОТ ЧЕЛОВЕКА, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2804448C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ЧАСТИЦЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА | 2018 |
|
RU2811426C2 |
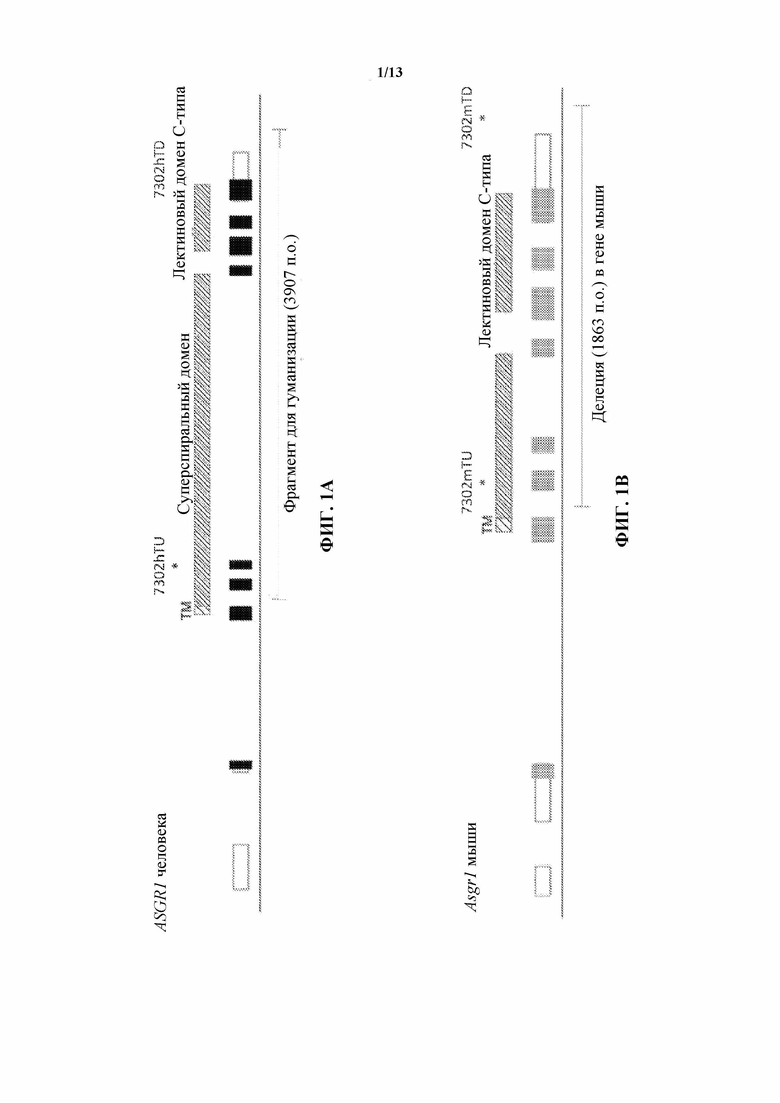

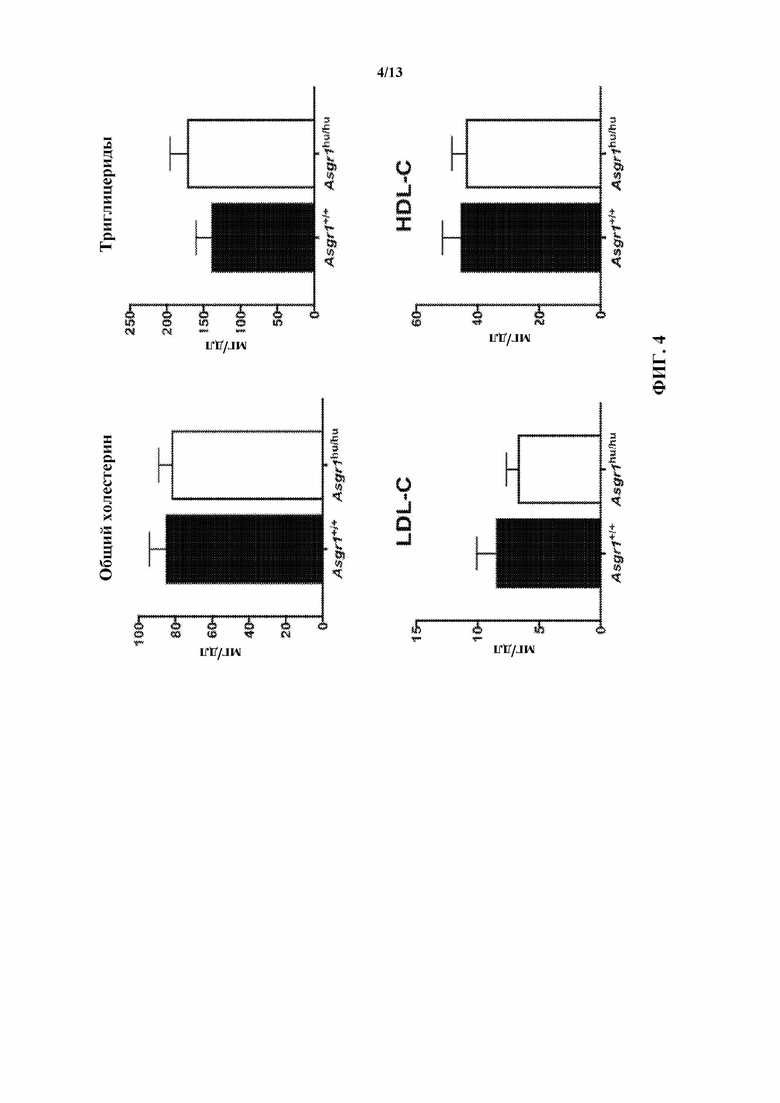
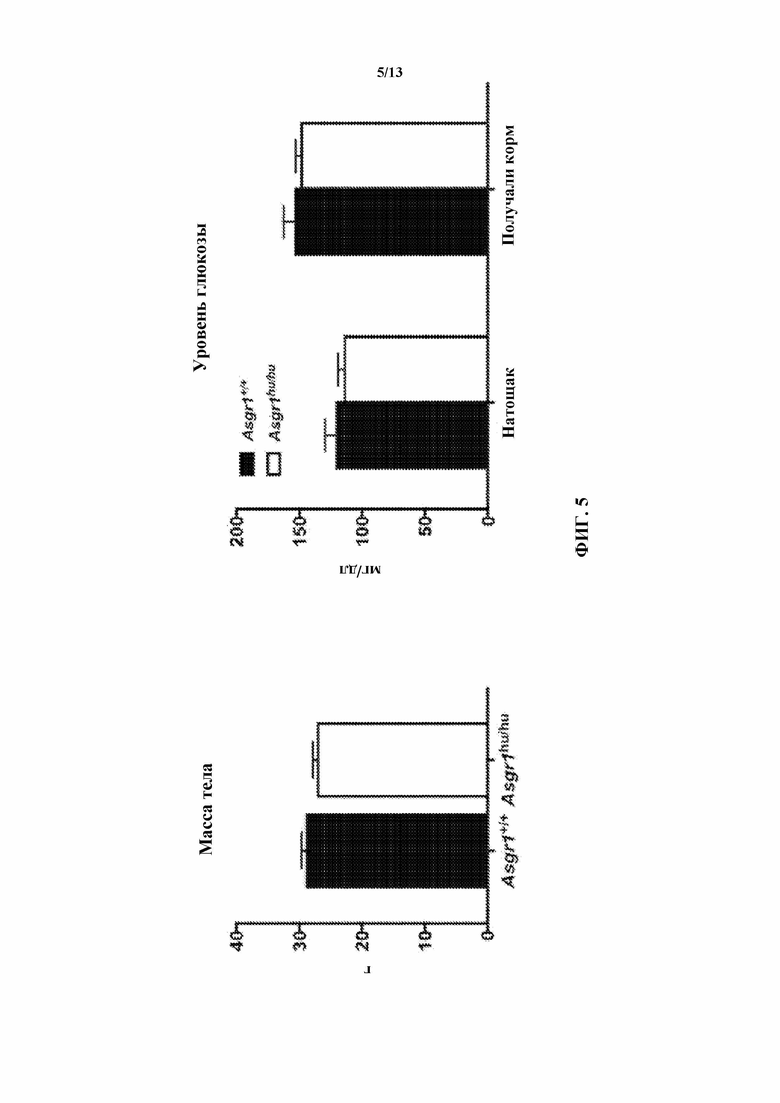
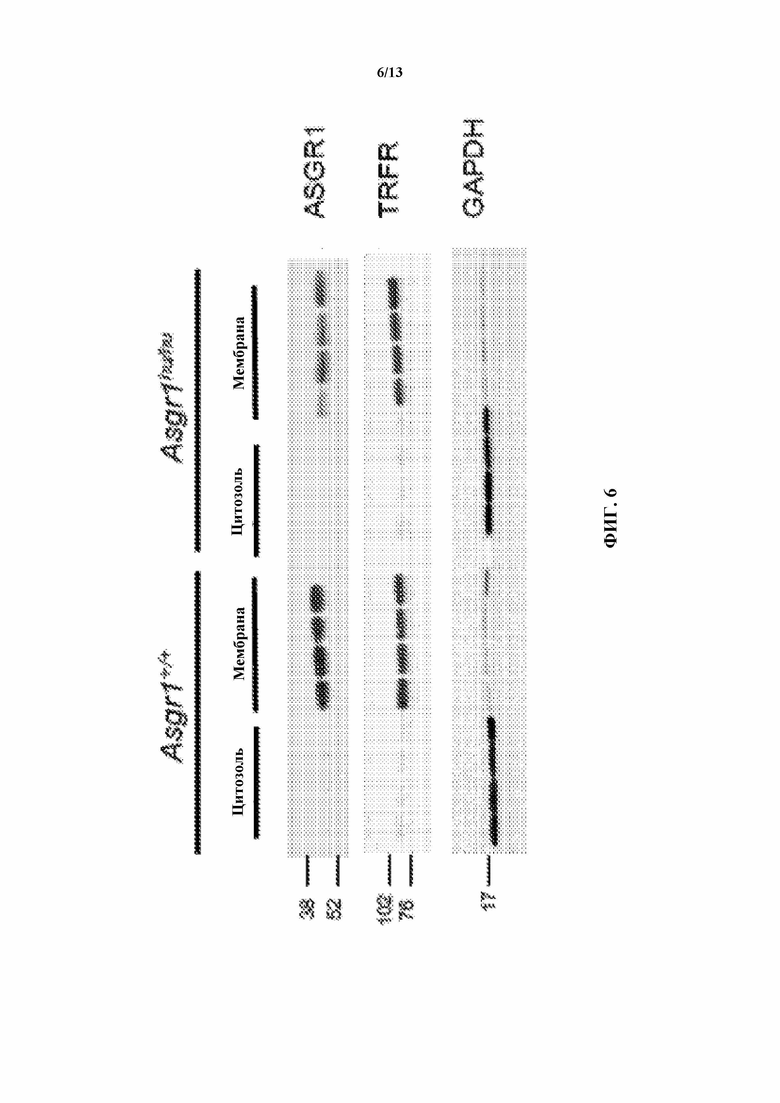







Изобретение относится к области биотехнологии, в частности к животному, отличному от человека, содержащему генетически модифицированный эндогенный локус Asgr1, который содержит эндогенный промотор Asgr1, ри этом животное, отличное от человека, представляет собой грызуна и эндогенный промотор Asgr1 функционально связан с нуклеиновой кислотой, кодирующий модифицированный белок Asgr1, модифицированный белок Asgr1 содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, внеклеточный домен содержит суперспиральный домен. Изобретение эффективно для оценки доставки терапевтического комплекса в печень посредством интернализации, опосредованной ASGR1 человека in vivo, а также для оценки эффективности терапевтической молекулы для целенаправленного воздействия на белок в отношении интернализации в печени посредством ASGR1 человека in vivo. 3 н. и 28 з.п. ф-лы, 9 ил., 4 табл., 4 пр.
1. Животное, отличное от человека, содержащее генетически модифицированный эндогенный локус Asgr1, который содержит эндогенный промотор Asgr1, где животное, отличное от человека представляет собой грызуна, где эндогенный промотор Asgr1 функционально связан с нуклеиновой кислотой, кодирующий модифицированный белок Asgr1, где модифицированный белок Asgr1 содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, где внеклеточный домен содержит суперспиральный домен,
где:
(i) весь внеклеточный домен кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека, или
(ii) весь суперспиральный домен кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека, и
где животное, отличное от человека, экспрессирует модифицированный белок Asgr1.
2. Животное, отличное от человека, по п. 1, где внеклеточный домен содержит суперспиральный домен и лектиновый домен С-типа и весь лектиновый домен С-типа или его часть кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека.
3. Животное, отличное от человека, по п. 2, где лектиновый домен С-типа представляет собой лектиновый домен С-типа ASGR1 человека.
4. Животное, отличное от человека, по п. 2 или 3, где лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28.
5. Животное, отличное от человека, по любому из пп. 1-4, где суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27.
6. Животное, отличное от человека, по любому из пп. 2-5, где лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28, и суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27.
7. Животное, отличное от человека, по любому из предыдущих пунктов, где ортологичная последовательность ASGR1 человека содержит экзоны 3-8 гена ASGR1 человека.
8. Животное, отличное от человека, по п. 7, где ортологичная последовательность ASGR1 человека кодирует сегмент белка ASGR1, содержащий последовательность, изложенную под SEQ ID NO: 31.
9. Животное, отличное от человека, по любому из предыдущих пунктов, где весь цитоплазматический домен или его часть кодируется эндогенной последовательностью Asgr1 животного, отличного от человека.
10. Животное, отличное от человека, по любому из предыдущих пунктов, где весь трансмембранный домен или его часть кодируется эндогенной последовательностью Asgr1 животного, отличного от человека.
11. Животное, отличное от человека, по п. 9 или 10, где как весь цитоплазматический домен, так и весь трансмембранный домен или часть и того и другого кодируются эндогенной последовательностью Asgr1 животного, отличного от человека.
12. Животное, отличное от человека, по любому из предыдущих пунктов, где животное, отличное от человека, является гетерозиготным по генетически модифицированному эндогенному локусу Asgr1.
13. Животное, отличное от человека, по любому из пп. 1-11, где животное, отличное от человека, является гомозиготным по генетически модифицированному эндогенному локусу Asgr1.
14. Животное, отличное от человека, по любому из пп. 1-13, где животное, отличное от человека, представляет собой крысу или мышь.
15. Животное, отличное от человека, по любому из пп. 1-4, где животное, отличное от человека, представляет собой мышь.
16. Животное, отличное от человека, по п. 15, где весь цитоплазматический домен или его часть кодируется эндогенной последовательностью Asgr1 мыши.
17. Животное, отличное от человека, по п. 16, где цитоплазматический домен содержит последовательность, изложенную под SEQ ID NO: 29.
18. Животное, отличное от человека, по п. 15, где весь трансмембранный домен или его часть кодируется эндогенной последовательностью Asgr1 мыши.
19. Животное, отличное от человека, по п. 18, где трансмембранный домен содержит последовательность, изложенную под SEQ ID NO: 30.
20. Животное, отличное от человека, по любому из пп. 16-19, где как весь цитоплазматический домен, так и весь трансмембранный домен или часть и того и другого кодируется эндогенной последовательностью Asgr1 мыши.
21. Животное, отличное от человека, по п. 20, где цитоплазматический домен содержит последовательность, изложенную под SEQ ID NO: 29, и трансмембранный домен содержит последовательность, изложенную под SEQ ID NO: 30.
22. Животное, отличное от человека, по любому из пп. 14-21, где внеклеточный домен содержит суперспиральный домен и лектиновый домен С-типа и где как весь суперспиральный домен, так и весь лектиновый домен С-типа или часть и того и другого кодируется сегментом эндогенного локуса Asgr1, который был удален и заменен ортологичной последовательностью ASGR1 человека, и при этом как весь цитоплазматический домен, так и весь трансмембранный домен или часть и того и другого кодируется эндогенной последовательностью Asgr1 мыши.
23. Животное, отличное от человека, по любому из пп. 14-22, где лектиновый домен С-типа представляет собой лектиновый домен С-типа ASGR1 человека и суперспиральный домен представляет собой суперспиральный домен ASGR1 человека, цитоплазматический домен представляет собой цитоплазматический домен Asgr1 мыши, а трансмембранный домен представляет собой трансмембранный домен Asgr1 мыши.
24. Животное, отличное от человека, по п. 22 или 23, где лектиновый домен С-типа содержит последовательность, изложенную под SEQ ID NO: 28, суперспиральный домен содержит последовательность, изложенную под SEQ ID NO: 27, цитоплазматический домен содержит последовательность, изложенную под SEQ ID NO: 29, и трансмембранный домен содержит последовательность, изложенную под SEQ ID NO: 30.
25. Животное, отличное от человека, по п. 24, где модифицированный белок Asgr1 содержит последовательность, изложенную под SEQ ID NO: 3.
26. Способ оценки доставки терапевтического комплекса в печень посредством интернализации, опосредованной ASGR1 человека, in vivo, предусматривающий:
(a) введение терапевтического комплекса животному, отличному от человека, по любому из пп. 1-25, где терапевтический комплекс содержит терапевтическую молекулу и антигенсвязывающий белок или лиганд, которые специфически связывают ASGR1 человека; и
(b) оценку доставки терапевтической молекулы в печень животного, отличного от человека.
27. Способ по п. 26, где терапевтическая молекула представляет собой лизосомальный заместительный белок, или фермент, или нуклеиновую кислоту, кодирующую лизосомальный заместительный белок или фермент, и стадия (b) предусматривает оценку присутствия или активности лизосомального заместительного белка или фермента в печени животного, отличного от человека.
28. Способ по п. 26, где терапевтическая молекула представляет собой нуклеиновую кислоту, кодирующую терапевтический секретируемый белок, и стадия (b) предусматривает оценку уровней в сыворотке крови или активности терапевтического секретируемого белка у животного, отличного от человека.
29. Способ оценки эффективности терапевтической молекулы для целенаправленного воздействия на белок в отношении интернализации в печени посредством ASGR1 человека in vivo, предусматривающий:
(a) введение терапевтической молекулы животному, отличному от человека, по любому из пп. 1-25, где терапевтическая молекула содержит биспецифический антигенсвязывающий белок, который специфически связывает белок и специфически связывает ASGR1 человека; и
(b) оценку уровней на клеточной поверхности, активности или экспрессии белка в печени животного, отличного от человека.
30. Способ по п. 29, где белок представляет собой поверхностный белок клеток печени и стадия (b) предусматривает оценку уровней на клеточной поверхности или активности поверхностного белка клеток в печени животного, отличного от человека.
31. Способ по п. 29, где белок представляет собой растворимый белок и стадия (b) предусматривает оценку экспрессии или активности растворимого белка в печени животного, отличного от человека.
| WO 2017087780 A1, 26.05.2017 | |||
| WO 2014023709 A1, 13.02.2014 | |||
| WO 2009019312 A2, 12.02.2009 | |||
| SIBYLLE SABRAUTZKI et al | |||
| New mouse models for metabolic bone diseases generated by genome-wide ENU mutagenesis, Mamm Genome, 2012, 23, pp.416-430 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЫШЕЙ | 2009 |
|
RU2425880C2 |

