Ссылка на родственные заявки
[0001] Согласно настоящей заявке испрашивается приоритет в соответствии с заявкой на выдачу патента США №62/539275, поданной 31 июля 2017 г., которая полностью включена в настоящий документ посредством ссылки для всех целей.
Ссылка на перечень последовательностей, представленный в виде текстового файла посредством файловой системы EFS
[0002] Перечень последовательностей, представленный в файле 516569SEQLIST.txt, характеризуется размером, составляющим 178 КБ, был создан 30 июля 2018 г. и включен в настоящий документ посредством ссылки.
Предшествующий уровень техники настоящего изобретения
[0003] Технология CRISPR/Cas является новым многообещающим терапевтическим средством. Однако существует необходимость в более эффективных средствах оценки эффективности создания мутаций или целевой модификации генов введенным агентом CRISPR/Cas in vivo. Одним из ограничений испытания системы in vivo является необходимость одновременного введения всех компонентов в живой организм. Типичный способ внедрения этих компонентов заключается во транзиентной трансфекции ДНК-конструктов в клетки, которые будут генерировать соответствующие РНК и белок. Несмотря на то, что этот подход является эффективным, ему присущим недостаток, поскольку клетки должны полагаться на конструкты плазмидной ДНК, чтобы сначала подвергнуться транскрипции, а затем трансляции, прежде чем белок Cas9 станет доступным для взаимодействия с компонентом sgPHК. Необходимы лучшие способы и инструменты для более эффективной оценки активности введенных агентов CRISPR/Cas и для оценки различных способов и параметров доставки для нацеливания на конкретные ткани или типы клеток in vivo.
[0004] Кроме того, доставке биологически активных агентов, таких как агенты CRISPR/Cas, субъектам часто препятствуют трудности в достижении компонентами целевой клетки или ткани. Эти ограничения могут приводить, например, к необходимости использовать гораздо более высокие концентрации агентов, чем это желательно для достижения результата, что увеличивает риск токсических эффектов и побочных эффектов. Необходимы улучшенные способы доставки и способы оценки таких способов доставки in vivo.
Краткое раскрытие настоящего изобретения
[0005] Предусмотрены Cas9-ready, далее по тексту - экспрессирующие Cas9 (т.е. готовые к использованию системы редактирования генома на основе Cas) не являющиеся человеком животные, а также предусмотрены способы и композиции для оценки способности агентов CRISPR/Cas-нуклеазы модифицировать целевой геномный локус in vivo. Согласно одному аспекту предусмотрены способы испытания способности CRISPR/Cas-нуклеазы модифицировать целевой геномный локус in vivo. Некоторые такие способы предусматривают следующее: (а) введение не являющемуся человеком животному гидовой РНК, предназначенной для нацеливания на целевую последовательность для гидовой РНК в целевом геномном локусе, причем не являющееся человеком животное содержит геномно интегрированную экспрессионную кассету Cas, содержащую кодирующую NLS-Cas последовательность, и причем гидовая РНК введена посредством опосредованной аденоассоциированным вирусом (AAV) доставки; и (b) оценка модификации целевого геномного локуса. Некоторые такие способы предусматривают следующее: (а) введение не являющемуся человеком животному гидовой РНК, предназначенной для нацеливания на целевую последовательность для гидовой РНК в целевом геномном локусе, причем не являющееся человеком животное содержит геномно интегрированную экспрессионную кассету Cas, содержащую кодирующую NLS-Cas последовательность, и причем гидовая РНК введена посредством опосредованной липидными наночастицами (LNP) доставки; и (b) оценка модификации целевого геномного локуса.
[0006] Согласно некоторым таким способам AAV представляет собой AAV7, AAV8 или AAV9, и стадия (b) предусматривает оценку модификации целевого геномного локуса в печени. Необязательно AAV представляет собой AAV8.
[0007] Согласно некоторым таким способам путь введения AAV не являющемуся человеком животному представляет собой внутривенную инъекцию, интрапаренхиматозную инъекцию, интраперитонеальную инъекцию, закапывание в нос или интравитреальную инъекцию.
[0008] Согласно некоторым таким способам экзогенную донорную нуклеиновую кислоту вводят на стадии (а), причем экзогенная донорная нуклеиновая кислота предназначена для рекомбинации с целевым геномным локусом. Необязательно экзогенная донорная нуклеиновая кислота представляет собой одноцепочечный олигодезоксинуклеотид (ssODN).
[0009] Согласно некоторым таким способам не являющееся человеком животное представляет собой крысу или мышь. Необязательно не являющееся человеком животное представляет собой мышь.
[0010] Согласно некоторым таким способам целевой геномный локус содержит целевой ген, и стадия (b) предусматривает измерение экспрессии целевого гена или активности белка, кодируемого целевым геном.
[0011] Согласно некоторым таким способам стадия (b) предусматривает секвенирование целевого геномного локуса в одной или нескольких клетках, выделенных из не являющегося человеком животного.
[0012] Согласно некоторым таким способам стадия (b) предусматривает выделение целевого органа или ткани из не являющегося человеком животного и оценку модификации целевого геномного локуса в целевом органе или ткани. Необязательно стадия (b) предусматривает оценку модификации целевого геномного локуса в двух или больше различных типах клеток в пределах целевого органа или ткани.
[0013] Согласно некоторым таким способам стадия (b) предусматривает выделение нецелевого органа или ткани из не являющегося человеком животного и оценку модификации целевого геномного локуса в нецелевом органе или ткани.
[0014] Согласно некоторым таким способам кодирующая NLS-Cas последовательность представляет собой кодирующую NLS-Cas9 последовательность.
[0015] Согласно некоторым таким способам экспрессионная кассета Cas дополнительно содержит сигнал полиаденилирования, расположенный выше кодирующей NLS-Cas последовательности, причем сигнал полиаденилирования фланкирован сайтами распознавания рекомбиназы, и причем сигнал полиаденилирования в экспрессионной кассете Cas был вырезан тканеспецифическим образом. Необязательно сигнал полиаденилирования, расположенный выше NLS-кодирующей последовательности в экспрессионной кассете Cas, был вырезан в печени. Необязательно рекомбиназа, которая распознает сайты распознавания рекомбиназы в экспрессионной кассете Cas, представляет собой Cre-рекомбиназу. Необязательно не являющееся человеком животное дополнительно содержит геномно интегрированную экспрессионную кассету рекомбиназы Cre, причем экспрессионная кассета рекомбиназы Cre содержит кодирующую Cre-рекомбиназу последовательность, функционально связанную с тканеспецифическим промотором. Необязательно ген Cre-рекомбиназы функционально связан с одним из промоторов, представленных в таблице 2.
[0016] Согласно некоторым таким способам экспрессионная кассета Cas дополнительно содержит сигнал полиаденилирования, расположенный выше кодирующей NLS-Cas последовательности, причем сигнал полиаденилирования фланкирован сайтами распознавания рекомбиназы, и причем способ дополнительно предусматривает введение рекомбиназы не являющемуся человеком животному тканеспецифическим образом. Необязательно рекомбиназу вводят посредством опосредованной аденоассоциированным вирусом (AAV) доставки или опосредованной липидными наночастицами (LNP) доставки. Необязательно рекомбиназу вводят посредством ААУ8-опосредованной доставки. Необязательно рекомбиназу вводят в печень.
[0017] Согласно некоторым таким способам экспрессионная кассета Cas дополнительно содержит кодирующую флуоресцентный белок последовательность. Необязательно экспрессионная кассета Cas содержит мультицистронную нуклеиновую кислоту, содержащую кодирующую NLS-Cas последовательность и кодирующую флуоресцентный белок последовательность, разделенные промежуточным участком внутренней посадки рибосомы (IRES) или промежуточной кодирующей пептид 2А последовательностью. Необязательно мультицистронная нуклеиновая кислота в экспрессионной кассете Cas содержит кодирующую NLS-Cas последовательность и кодирующую зеленый флуоресцентный белок последовательность, разделенные промежуточной кодирующей пептид Р2А последовательностью. Согласно некоторым таким способам экспрессионная кассета Cas дополнительно не содержит кодирующую флуоресцентный белок последовательность. Согласно некоторым таким способам кодирующая NLS-Cas последовательность кодирует белок Cas, содержащий метку белка.
[0018] Согласно некоторым таким способам экспрессионная кассета Cas функционально связана с эндогенным промотором. Согласно некоторым таким способам экспрессионная кассета Cas функционально связана с экзогенным, конститутивным промотором.
[0019] Согласно некоторым таким способам 5' конец экспрессионной кассеты Cas дополнительно содержит 3' последовательность сплайсинга.
[0020] Согласно некоторым таким способам экспрессионная кассета Cas кодирует белок, содержащий последовательность, представленную в SEQ ID NO: 13, 16, 19 или 22. Необязательно экспрессионная кассета Cas содержит последовательность, представленную в SEQ ID NO: 28, 29, 30 или 31. Необязательно экспрессионная кассета Cas содержит последовательность, представленную в SEQ ID NO: 1, 12, 14, 15, 17, 18, 20 или 21.
[0021] Согласно некоторым таким способам экспрессионная кассета Cas интегрирована в локус «безопасная гавань». Необязательно локус «безопасная гавань» представляет собой локус Rosa26. Необязательно экспрессионная кассета Cas интегрирована в первый интрон локуса Rosa26.
[0022] Согласно некоторым таким способам не являющееся человеком животное является гетерозиготным по отношению к экспрессионной кассете Cas. Согласно некоторым таким способам не являющееся человеком животное является гомозиготным по отношению к экспрессионной кассете Cas.
[0023] Согласно некоторым таким способам не являющееся человеком животное представляет собой мышь, AAV представляет собой AAV8, экспрессионная кассета Cas функционально связана с эндогенным промотором Rosa26, вставлена в первый интрон локуса Rosa26 и содержит от 5' к 3' следующее: (i) 3' последовательность сплайсинга; и (ii) кодирующая NLS-Cas9 последовательность, и стадия (b) предусматривает оценку модификации целевого геномного локуса в печени не являющегося человеком животного. Согласно некоторым таким способам не являющееся человеком животное представляет собой мышь, AAV представляет собой AAV8, доставленный не являющемуся человеком животному с помощью внутривенной инъекции, экспрессионная кассета Cas функционально связана с эндогенным промотором Rosa26, вставлена в первый интрон локуса Rosa26 и содержит от 5' к 3' следующее: (i) 3' последовательность сплайсинга; и (ii) кодирующая NLS-Cas9 последовательность, и стадия (b) предусматривает оценку модификации целевого геномного локуса в печени не являющегося человеком животного.
[0024] Согласно одному аспекту предусмотрены способы оптимизации способности CRISPR/Cas-нуклеазы модифицировать целевой геномный локус in vivo. Некоторые такие способы предусматривают следующее: (I) проведение любого из приведенных выше способов испытания способности CRISPR/Cas-нуклеазы модифицировать целевой геномный локус in vivo в первый раз у первого не являющегося человеком животного; (II) изменение переменной и проведение способа согласно стадии (I) во второй раз с измененной переменной у второго не являющегося человеком животного; и (III) сравнение модификации целевого геномного локуса на стадии (I) с модификацией целевого геномного локуса на стадии (II), и выбор способа, приводящего к модификации целевого геномного локуса, характеризующейся одним или несколькими из следующего: повышенная эффективность, повышенная точность, повышенная согласованность, или повышенная специфичность.
[0025] Согласно некоторым таким способам измененная переменная на стадии (II) представляет собой серотип AAV. Согласно некоторым таким способам измененная переменная на стадии (II) представляет собой путь введения для введения гидовой РНК не являющемуся человеком животному. Согласно некоторым таким способам измененная переменная на стадии (II) представляет собой концентрацию или количество гидовой РНК, введенной не являющемуся человеком животному. Согласно некоторым таким способам измененная переменная на стадии (II) представляет собой гидовую РНК (например, форму или последовательность гидовой РНК), введенную не являющемуся человеком животному. Согласно некоторым таким способам способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и причем измененная переменная на стадии (II) представляет собой способ доставки для введения экзогенной донорной нуклеиновой кислоты не являющемуся человеком животному. Согласно некоторым таким способам способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и измененная переменная на стадии (II) представляет собой путь введения для введения экзогенной донорной нуклеиновой кислоты не являющемуся человеком животному. Согласно некоторым таким способам способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и измененная переменная на стадии (II) представляет собой концентрацию или количество экзогенной донорной нуклеиновой кислоты, введенной не являющемуся человеком животному. Согласно некоторым таким способам способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и измененная переменная на стадии (II) представляет собой концентрацию или количество гидовой РНК, введенной не являющемуся человеком животному, по отношению к концентрации или количеству экзогенной донорной нуклеиновой кислоты, введенной не являющемуся человеком животному. Согласно некоторым таким способам измененная переменная на стадии (II) представляет собой экзогенную донорную нуклеиновую кислоту (например, форму экзогенной донорной нуклеиновой кислоты), введенную не являющемуся человеком животному.
Краткое описание фигур
[0026] На фигуре 1 показан аллель Cas9 (MAID2599; без соблюдения масштаба), содержащий от 5' к 3' следующее: 3' последовательность сплайсинга; первый сайт loxP, ген устойчивости к неомицину; сигнал полиаденилирования; второй сайт loxP; кодирующая NLS-Cas9 последовательность; кодирующая пептид Р2А последовательность; и кодирующая GFP последовательность.
[0027] На фигуре 2А показана активность NHEJ в мышиных эмбриональных стволовых клетках F1H4 дикого типа (mESC) и экспрессирующих Cas9 mESC с и без неомициновой кассеты lox-stop-lox (MAID2599 и MAID2600, соответственно) после введения двух sgPHК (в форме плазмиды или в виде РНК), нацеленных на области старт- и стоп-кодона первого целевого гена, необязательно в сочетании с введением плазмиды Cas9. Эффективность 5' разрезания измеряли в левой панели, эффективность 3' разрезания измеряли в средней панели, частоту, с которой промежуточная ДНК подвергалась полной делеции, измеряли в правой панели.
[0028] На фигуре 2В показана эффективность разрезания (левая панель) и эффективность HDR (правая панель) после введения sgPHК (в форме плазмиды или в виде РНК), нацеленной на второй целевой ген вместе с одноцепочечным олигодезоксинуклеотидом (ssODN) в качестве донора точечной мутации, необязательно в комбинации с плазмидой Cas9.
[0029] На фигурах 3A-3F показаны светлопольные изображения тканей печени (фигура 3А), почки (фигура 3В) и головного мозга (фигура 3С) от мышей дикого типа и гетерозиготных экспрессирующих Cas9 мышей (MAID2600), и полученные с помощью GFP флуоресцентные изображения тканей печени (фигура 3D), почки (фигура 3Е) и головного мозга (фигура 3F) у мышей дикого типа и экспрессирующих Cas9 мышей (MAID2600).
[0030] На фигуре 4А показаны уровни экспрессии мРНК Cas9 в различных тканях, выделенных из гетерозиготных экспрессирующих Cas9 мышей (MAID2600), как определено с помощью ОТ-кПЦР. По оси у показано дельта Ct + 1 по сравнению со средним Ct Cas9 из ткани головного мозга.
[0031] На фигуре 4В показана экспрессия белка Cas9 в различных тканях, выделенных из мышей дикого типа и гетерозиготных экспрессирующих Cas9 мышей (MAID2600). Актин использовали в качестве контроля.
[0032] На фигуре 4С показаны средние уровни экспрессии мРНК Cas9 и бета-2-микроглобулина (B2m) в различных тканях, выделенных из гетерозиготных экспрессирующих Cas9 мышей (MAID2600), как определено с помощью ОТ-кПЦР. Число образцов, испытанных из каждого типа ткани, указано выше столбиков.
[0033] На фигурах 5А-5В показана активность NHEJ в процентах (частота инделей) в третьем целевом гене (целевой ген 3) в первичных гепатоцитах, выделенных из мышей дикого типа (фигура 5А) и мышей Cas9 с удаленной кассетой (MAID2600; фигура 5В) после с помощью доставки липидных наночастиц (LNP) любого из следующего: мРНК GFP и контрольная (мертвая) sgPHК, мРНК GFP и sgPHК целевого гена 3 или мРНК Cas9 и sgPHК целевого гена 3. Исследовали концентрации мРНК, составляющие 15,6, 62,5, 250 и 1000 нг/мл.
[0034] На фигурах 6A-6D показаны содержания в сыворотке белка, который секретируется печенью и обнаруживается в сыворотке и кодируется третьим целевым геном (целевой ген 3) после введения sgPHК целевого гена 3 мышам дикого типа (msCas9 -) или экспрессирующим Cas9 мышам с удаленной кассетой (msCas9+; MAID2600) посредством гидродинамической доставки ДНК (HDD), доставки с помощью липидных наночастиц (LNP) или доставки с помощью аденоассоциированного вируса (AAV) с помощью инъекции в хвостовую вену. В некоторых случаях Cas9 также вводят (в форме мРНК для LNP доставки и в форме ДНК для доставки с помощью HDD (плазмида Cas9) и AAV). Не получивших лечение мышей, контрольных мышей LNP, контрольных мышей AAV и контрольных мышей HDD использовали в качестве отрицательных контролей. Для LNP-опосредованной доставки испытывали три группы мышей: (1) экспрессирующие Cas9 мыши (3 самца + 3 самки; 2 мг/кг контрольной гидовой РНК + мРНК GFP); (2) экспрессирующие Cas9 мыши (3 самца + 3 самки; 2 мг/кг гидовой РНК для целевого гена 3 + мРНК GFP); и (3) мыши WT (3 самца + 3 самки; 2 мг/кг гидовой РНК для целевого гена 3 + мРНК Cas9). Для AAV-опосредованной доставки испытывали две группы мышей: (1) экспрессирующие Cas9 мыши (3 самца + 3 самки; AAV8-гидовая РНК для целевого гена 3); и (2) мыши WT (3 самца + 3 самки; AAV8-гидовая РНК для целевого гена 3 + AAV8-Cas9). Для HDD испытывали две группы мышей: (1) экспрессирующие Cas9 мыши (3 самца + 3 самки; гидовая РНК для целевого гена 3); и (2) мыши WT (3 самца + 3 самки; гидовая РНК для целевого гена 3 + Cas9). Содержания в сыворотке белка, кодируемого целевым геном 3, измеряли у самцов мышей (фигуры 6А и 6В) и самок мышей (фигуры 6С и 6D) и измеряли в день 7 (фигуры 6А и 6С) и день 21 (фигуры 6В и 6D).
[0035] На фигуре 7 показана активность NHEJ в процентах (частота инделей) в локусе целевого гена 3 в печени у мышей дикого типа (msCas9 -) и мышей Cas9 с удаленной кассетой (msCas9 +; MAID2600) через один месяц после доставки с помощью липидных наночастиц (LNP) sgPHК отдельно или вместе с мРНК Cas9, гидродинамической доставки (HDD) плазмиды sgPHК отдельно или вместе с плазмидой Cas9 или AAV8-sgPHК отдельно или вместе с AAV8-Cas9.
[0036] На фигуре 8А показана активность NHEJ в процентах (частота инделей) в локусе целевого гена 4 в печени у мышей Cas9 с удаленной кассетой (MAID2600) через 3-4 недели после ААУ8-доставки sgPHК с помощью инъекции в хвостовую вену. UNT = не получивший лечение контроль.
[0037] На фигуре 8В показаны относительные уровни экспрессии целевого гена 4, как определено с помощью анализа TAQMAN, в ткани печени, выделенной из мышей Cas9 с удаленной кассетой (MAID2600) через 3-4 недели после ААУ8-доставки sgPHК с помощью инъекции в хвостовую вену. Мастер-микс WT относится ко всем пяти вирусам sgPHК, смешанным вместе и введенным в виде инъекции мышам дикого типа в качестве отрицательного контроля.
[0038] На фигуре 9 показан вестерн-блоттинг экспрессии Cas9 у мышей LSL-Cas9 (MAID2599) в образцах печени, селезенки и почки, выделенных через одну неделю после того, как LNP-Cre ввели посредством инъекции в хвостовую вену. Мышей без инъекций LNP-Cre использовали в качестве отрицательного контроля. Мышей Cas9 с удаленной кассетой (MAID2600) использовали в качестве положительного контроля.
[0039] На фигуре 10 показаны содержания в сыворотке белка, который секретируется печенью и обнаруживается в сыворотке и кодируется третьим целевым геном (целевой ген 3) через 1 неделю и 3 недели после инъекции sgPHК целевого гена 3 мышам LSL-Cas9 (MAID2599) посредством AAV8, либо отдельно, либо вместе с LNP-Cre. Мышей без введения либо LNP-Cre, либо AAV8-rPHK использовали в качестве отрицательного контроля. Все условия также испытывали на мышах с удаленной кассетой (ROSA Cas9; MAID2600).
[0040] На фигуре 11 показана активность NHEJ в процентах (частота инделей) в локусе целевого гена 3 в образцах печени, выделенных через 3 недели после инъекции sgPHК целевого гена 3 мышам LSL-Cas9 (MAID2599) посредством AAV8, либо отдельно, либо вместе с LNP-Cre. Мышей без введения либо LNP-Cre, либо AAV8-rPHK использовали в качестве отрицательного контроля. Все условия также испытывали на мышах с удаленной кассетой (ROSA Cas9; MAID2600).
[0041] На фигуре 12А показан вестерн-блоттинг для Cas9 в образцах печени, выделенных из мышей LSL-Cas9 (MAID2599) и мышей LSL-Cas9/Alb-Cre. Актин использовали в качестве контроля нагрузки.
[0042] На фигуре 12В показан вестерн-блоттинг для Cas9 в образцах головного мозга, выделенных из мышей LSL-Cas9 (MAID2599) и мышей LSL-Cas9/Alb-Cre. Актин использовали в качестве контроля нагрузки.
[0043] На фигуре 13 показаны содержания в сыворотке белка, который секретируется печенью и обнаруживается в сыворотке и кодируется третьим целевым геном (целевой ген 3) через 1 неделю после инъекции sgPHК целевого гена 3 мышам WT, мышам Cas9 с удаленной кассетой (ROSA Cas9; MAID2600), мышам LSL-Cas9 (MAID2599), мышам Alb-Cre или мышам LSL-Cas9/Alb-Cre посредством AAV8.
[0044] На фигуре 14 показаны четыре различных аллеля Cas9 (без соблюдения масштаба), включая в себя аллель MAID2599 (MAID2600 после удаления кассеты lox-stop-lox (LSL)), аллель MAID2658 (MAID2659 после удаления кассеты LSL), аллель MAID2660 (MAID2661 после удаления кассеты LSL) и аллель MAID2672 (MAID2673 после удаления кассеты LSL).
Определения
[0045] Используемые взаимозаменяемо в настоящем документе термины «белок», «полипептид» и «пептид» включают в себя полимерные формы аминокислот любой длины, включая в себя кодированные и некодированные аминокислоты и химически или биохимически модифицированные или дериватизированные аминокислоты. Термины также включают в себя полимеры, которые были модифицированы, такие как полипептиды, характеризующиеся модифицированными пептидными каркасами.
[0046] Считается, что белки имеют «N-конец» и «С-конец». Термин «N-конец» относится к началу белка или полипептида, оканчивающемуся аминокислотой со свободной аминогруппой (-NH2). Термин «С-конец» относится к концу аминокислотной цепи (белка или полипептида), оканчивающейся свободной карбоксильной группой (-СООН).
[0047] Используемые взаимозаменяемо в настоящем документе термины «нуклеиновая кислота» и «полинуклеотид» включают в себя полимерные формы нуклеотидов любой длины, включая в себя рибонуклеотиды, дезоксирибонуклеотиды или их аналоги или модифицированные версии. Они включают в себя одно-, двух- и многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие природные, химически модифицированные, биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания.
[0048] Считается, что нуклеиновые кислоты характеризуются наличием «5' концов» и «3' концов», потому что мононуклеотиды реагируют с образованием олигонуклеотидов таким образом, что 5'-фосфат одного мононуклеотидпентозного кольца прикреплен к 3' кислороду его соседа в одном направлении через фосфодиэфирную связь. Конец олигонуклеотида называют «5'-концом», если его 5'-фосфат не связан с 3'-кислородом мононуклеотидного пентозного кольца. Конец олигонуклеотида называют «3'-концом», если его 3'-кислород не связан с 5'-фосфатом другого мононуклеотидного пентозного кольца. Можно сказать, что последовательность нуклеиновой кислоты, даже если она находится внутри более крупного олигонуклеотида, характеризуются наличием 5' и 3' концов. В линейной или кольцевой молекуле ДНК дискретные элементы называются «расположенными выше по отношению к ходу транскрипции» или 5' относительно «расположенных ниже» или 3' элементов.
[0049] Термин «геномно интегрированный» относится к нуклеиновой кислоте, которая была введена в клетку так, что нуклеотидная последовательность интегрируется в геном клетки. Любой протокол можно использовать для стабильного включения нуклеиновой кислоты в геном клетки.
[0050] Термин «экспрессионный вектор» или «экспрессионный конструкт» относится к рекомбинантной нуклеиновой кислоте, содержащей требуемую кодирующую последовательность, функционально связанную с соответствующими последовательностями нуклеиновой кислоты, необходимыми для экспрессии функционально связанной кодирующей последовательности в конкретной клетке-хозяине или организме-хозяине. Последовательности нуклеиновых кислот, необходимые для экспрессии в прокариотах, как правило, включают в себя промотор, оператор (необязательно) и сайт связывания рибосомы, а также другие последовательности. Общеизвестно, что эукариотические клетки используют промоторы, энхансеры и сигналы терминации и полиаденилирования, хотя некоторые элементы могут быть удалены, а другие элементы могут быть добавлены без ущерба для необходимой экспрессии.
[0051] Термин «нацеливающий вектор» относится к рекомбинантной нуклеиновой кислоте, которую можно ввести путем гомологичной рекомбинации, лигирования, опосредованного негомологичным соединением концов или любым другим способом рекомбинации в отношении целевого положения в геноме клетки.
[0052] Термин «вирусный вектор» относится к рекомбинантной нуклеиновой кислоте, которая включает в себя по меньшей мере один элемент вирусного происхождения и включает в себя элементы, достаточные для или разрешающие упаковку в частицу вирусного вектора. Вектор и/или частицу можно использовать с целью переноса ДНК, РНК или других нуклеиновых кислот в клетки ex vivo или in vivo. Известны многочисленные формы вирусных векторов.
[0053] Термин «выделенный» в отношении белков, нуклеиновых кислот и клеток включает в себя белки, нуклеиновые кислоты и клетки, которые являются относительно очищенными по отношению к другим клеточным компонентам или компонентам организма, которые обычно могут присутствовать in situ, вплоть до и включая в себя по существу чистый препарат белка, нуклеиновой кислоты или клетки. Термин «выделенный» также включает в себя белки и нуклеиновые кислоты, которые не имеют встречающегося в природе аналога, или белки или нуклеиновые кислоты, которые были химически синтезированы и, таким образом, по существу не загрязнены другими белками или нуклеиновыми кислотами. Термин «выделенный» также включает в себя белки, нуклеиновые кислоты или клетки, которые были отделены или очищены от большинства других клеточных компонентов или компонентов организма, которые им сопутствуют в естественных условиях (например, другие клеточные белки, нуклеиновые кислоты или клеточные или внеклеточные компоненты).
[0054] Термин «дикий тип» включает в себя объекты, характеризующиеся структурой и/или активностью, обнаруживаемой в нормальном (в отличие от мутантного, болезненного, измененного и т.д.) состоянии или контексте. Гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллели).
[0055] Термин «эндогенная последовательность» относится к последовательности нуклеиновой кислоты, которая встречается в природе внутри клетки или животного, не являющегося человеком. Например, эндогенная последовательность Rosa26 не являющегося человеком животного относится к нативной последовательности Rosa26, которая естественным образом встречается в локусе Rosa26 у не являющегося человеком животного.
[0056] «Экзогенные» молекулы или последовательности включают в себя молекулы или последовательности, которые обычно не присутствуют в клетке в этой форме. Нормальное присутствие включает в себя присутствие на конкретной стадии развития и при конкретных условиях окружающей среды клетки. Например, экзогенная молекула или последовательность может включать в себя мутантную версию соответствующей эндогенной последовательности в клетке, такую как гуманизированная версия эндогенной последовательности, или может включать в себя последовательность, соответствующую эндогенной последовательности в клетке, но в другой форма (т.е. не в пределах хромосомы). Напротив, эндогенные молекулы или последовательности включают в себя молекулы или последовательности, которые обычно присутствуют в такой форме в конкретной клетке на определенной стадии развития в определенных условиях окружающей среды.
[0057] Термин «гетерологичный» при использовании в контексте нуклеиновой кислоты или белка указывает на то, что нуклеиновая кислота или белок содержит по меньшей мере два сегмента, которые в природе не встречаются вместе в одной и той же молекуле. Например, термин «гетерологичный» при использовании в отношении сегментов нуклеиновой кислоты или сегментов белка указывает, что нуклеиновая кислота или белок содержит две или более подпоследовательности, которые не находятся в таком же отношении друг к другу (например, объединены) в природе. В качестве одного примера, «гетерологичная» область нуклеиновокислотного вектора представляет собой сегмент нуклеиновой кислоты в пределах или прикрепленный к другой молекуле нуклеиновой кислоты, которая не обнаружена в ассоциации с другой молекулой в природе. Например, гетерологичная область нуклеиновокислотного вектора может включать в себя кодирующую последовательность, фланкированную последовательностями, которые не обнаружены в природе в ассоциации с кодирующей последовательностью. Аналогично, «гетерологичная» область белка представляет собой сегмент аминокислот в пределах или прикрепленный к другой пептидной молекуле, который не обнаружен в природе в ассоциации с другой пептидной молекулой (например, слитый белок или белок с меткой). Аналогично, нуклеиновая кислота или белок могут содержать гетерологичную метку или гетерологичную последовательность секреции или гетерологичную последовательность локализации.
[0058] «Оптимизация ко донов» использует преимущества вырожденности ко донов, что проявляется в множественности комбинаций кодонов с тремя парами оснований, которые определяют аминокислоту, и, как правило, включает в себя процесс модификации последовательности нуклеиновой кислоты для усиленной экспрессии в конкретных клетках-хозяевах путем замены по меньшей мере одного кодона нативной последовательности на кодон, который чаще или чаще всего используется в генах клетки-хозяина при сохранении нативной аминокислотной последовательности. Например, нуклеиновую кислоту, кодирующую белок Cas9, можно модифицировать для замены кодонов, характеризующихся более высокой частотой использования в данной прокариотической или эукариотической клетке, включая в себя бактериальную клетку, дрожжевую клетку, клетку человека, клетку, не относящуюся к человеку, клетку млекопитающего, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина по сравнению с встречающейся в природе последовательностью нуклеиновой кислоты. Таблицы использования кодонов легко доступны, например, в «Базе данных использования кодонов». Эти таблицы можно адаптировать несколькими способами. См. Nakamura et al. (2000)Nucleic Acids Research 28:292, полностью включенный в настоящий документ посредством ссылки для всех целей. Также доступны компьютерные алгоритмы для оптимизации кодонов конкретной последовательности для экспрессии в конкретном хозяине (см., например, Gene Forge).
[0059] «Промотор» представляет собой регуляторную область ДНК, как правило, содержащую ТАТА-бокс, способный направлять РНК-полимеразу II для инициации синтеза РНК в соответствующем сайте инициации транскрипции для конкретной полинуклеотидной последовательности. Промотор может дополнительно содержать другие области, которые влияют на скорость инициации транскрипции. Раскрытые в настоящем документе промоторные последовательности модулируют транскрипцию функционально связанного полинуклеотида. Промотор может являться активным в одном или нескольких типах клеток, раскрытых в настоящем документе (например, эукариотическая клетка, клетка млекопитающего, не являющегося человеком, клетка человека, клетка грызуна, плюрипотентная клетка, эмбрион на стадии одной клетки, дифференцированная клетка или их комбинация). Промотор может являться, например, конститутивно активным промотором, условным промотором, индуцируемым промотором, ограниченным по времени промотором (например, промотором, регулируемым стадией развития) или пространственно ограниченным промотором (например, клеточноспецифическим или тканеспецифическим промотором). Примеры промоторов можно найти, например, в международной патентной публикации WO 2013/176772, полностью включенной в настоящий документ посредством ссылки для всех целей.
[0060] Конститутивный промотор представляет собой промотор, который является активным во всех тканях или конкретных тканях на всех стадиях развития. Примеры конститутивных промоторов включают в себя немедленный ранний промотор цитомегаловируса человека (hCMV), немедленный ранний промотор цитомегаловируса мыши (mCMV), промотор фактора элонгации 1 альфа человека (hEF1α), промотор фактора элонгации 1 альфа мыши (mEF1α), промотор фосфоглицераткиназы мыши (PGK), промотор гибрида бета-актина цыпленка (CAG или CBh), ранний промотор SV40 и промотор бета 2 тубулина.
[0061] Примеры индуцируемых промоторов включают в себя, например, химически регулируемые промоторы и физически регулируемые промоторы. Химически регулируемые промоторы включают в себя, например, регулируемые спиртом промоторы (например, промотор гена алкогольдегидрогеназы (a1cA)), регулируемые тетрациклином промоторы (например, чувствительный к тетрациклину промотор, последовательность оператора тетрациклина (tetO), промотор tet-On или промотор tet-Off), регулируемые стероидами промоторы (например, глюкокортикоидный рецептор крысы, промотор рецептора эстрогена или промотор рецептора экдизона) или регулируемые металлом промоторы (например, промотор металлопротеина). Физически регулируемые промоторы включают в себя, например, регулируемые температурой промоторы (например, промотор теплового шока) и регулируемые светом промоторы (например, светоиндуцируемый промотор или репрессируемый светом промотор).
[0062] Тканеспецифические промоторы могут представлять собой, например, нейрон-специфические промоторы, специфические по отношению к глии промоторы, специфические по отношению к мышечным клеткам промоторы, специфические по отношению к клеткам сердца промоторы, специфические по отношению к клеткам почек промоторы, специфические по отношению к костным клеткам промоторы, специфические по отношению к эндотелиальным клеткам промоторы или специфические по отношению к иммунным клеткам промоторы (например, промотор В-клеток или промотор Т-клеток).
[0063] Регулируемые стадией развития промоторы включают в себя, например, промоторы, активные только во время эмбриональной стадии развития или только во взрослой клетке.
[0064] Термин «функциональная связь» или «функционально связанный» включает в себя смежное положение двух или более компонентов (например, промотора и другого элемента последовательности) так, что оба компонента функционируют нормально и обеспечивают возможность того, что по меньшей мере один из компонентов может опосредовать функцию, на которую действует по крайней мере один из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или нескольких транскрипционных регуляторных факторов. Функциональная связь может включать в себя такие последовательности, которые являются смежными друг с другом или действуют в транс-положении (например, регуляторная последовательность может действовать на расстоянии для управления транскрипцией кодирующей последовательности).
[0065] Термин «комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной цепи нуклеиновой кислоты вследствие ориентации ее групп нуклеиновых оснований образует водородные связи с другой последовательностью на противоположной цепи нуклеиновой кислоты. Комплементарными основаниями в ДНК, как правило, являются А с Т и С с G. В РНК они, как правило, представляют собой С с G и U с А. Комплементарность может являться совершенной или существенной/достаточной. Совершенная комплементарность между двумя нуклеиновыми кислотами означает, что две нуклеиновые кислоты могут образовывать дуплекс, в котором каждое основание в дуплексе связано с комплементарным основанием за счет спаривания Уотсона-Крика. «Существенная» или «достаточная» комплементарность означает, что последовательность в одной цепи не является полностью и/или совершенно комплементарной последовательности в противоположной цепи, но что между основаниями на двух цепях происходит достаточное связывание для образования стабильного гибридного комплекса при наборе условий гибридизации (например, концентрация соли и температура). Такие условия можно предсказать с использованием последовательностей и стандартных математических расчетов для прогнозирования Tm (температуры плавления) гибридизированных нитей или путем эмпирического определения Tm с использованием рутинных способов. Tm включает в себя температуру, при которой популяция гибридизационных комплексов, образованных между двумя цепями нуклеиновой кислоты, денатурируется на 50% (т.е. популяция двухцепочечных молекул нуклеиновой кислоты становится наполовину диссоциированной на отдельные цепи). При температуре ниже Tm предпочтительным является образование гибридизационного комплекса, тогда как при температуре выше Tm предпочтительным является плавление или разделение цепей в гибридизационном комплексе. Tm можно оценить для нуклеиновой кислоты, характеризующейся известным содержанием G+C в водном 1 М растворе NaCl, с использованием, например, Tm=81,5+0,41 (% G+C), хотя другие известные вычисления Tm учитывают структурные характеристики нуклеиновой кислоты.
[0066] «Условие гибридизации» включает в себя совокупную среду, в которой одна цепь нуклеиновой кислоты связывается со второй цепью нуклеиновой кислоты посредством взаимодействия комплементарных цепей и образования водородных связей с образованием гибридизационного комплекса. Такие условия включают в себя химические компоненты и их концентрации (например, соли, хелатирующие агенты, формамид) водного или органического раствора, содержащего нуклеиновые кислоты, и температуру смеси. Другие факторы, такие как продолжительность инкубации или размеры реакционной камеры, могут влиять на окружающую среду. См., например, Sambrook et al., Molecular Cloning, A Laboratory Manual, 2-e изд., С. 1.90-1.91, 9.47-9.51, 1 1.47-11.57 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989), полностью включенный в настоящий документ посредством ссылки для всех целей.
[0067] Гибридизация требует, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несовпадения между основаниями. Условия, подходящие для гибридизации между двумя нуклеиновыми кислотами, зависят от длины нуклеиновых кислот и степени комплементарности, переменные, которые хорошо известны. Чем больше степень комплементарности между двумя нуклеотидными последовательностями, тем больше значение температуры плавления (Tm) для гибридов нуклеиновых кислот, характеризующихся этими последовательностями. Для гибридизации между нуклеиновыми кислотами с короткими участками комплементарности (например, комплементарность более 35 или менее, 30 или менее, 25 или менее, 22 или менее, 20 или менее, или 18 или менее нуклеотидов) положение несовпадений становится важным (см. Sambrook et al., ранее, 11.7-11.8). Как правило, длина гибридизуемой нуклеиновой кислоты составляет по меньшей мере около 10 нуклеотидов. Иллюстративные минимальные длины для гибридизуемой нуклеиновой кислоты включают в себя по меньшей мере приблизительно 15 нуклеотидов, по меньшей мере приблизительно 20 нуклеотидов, по меньшей мере приблизительно 22 нуклеотидов, по меньшей мере приблизительно 25 нуклеотидов и по меньшей мере приблизительно 30 нуклеотидов. Кроме того, температуру и концентрацию соли в отмывающем растворе можно корректировать при необходимости в соответствии с такими факторами, как длина области комплементарности и степень комплементарности.
[0068] Последовательность полинуклеотида не обязательно должна быть на 100% комплементарной последовательности целевой нуклеиновой кислоты, чтобы являться специфически гибридизуемой. Кроме того, полинуклеотид может гибридизоваться с одним или несколькими сегментами, так что промежуточные или смежные сегменты не участвуют в событии гибридизации (например, структура петли или структура шпильки). Полинуклеотид (например, гРНК) может характеризоваться по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 99% или 100% комплементарностью последовательности по отношению к целевой области в пределах целевой последовательности нуклеиновой кислоты, на которую они нацелены. Например, гРНК, в которой 18 из 20 нуклеотидов являются комплементарными целевой области и, следовательно, будут специфически гибридизоваться, будет представлять 90% комплементарность. В этом примере оставшиеся некомплементарные нуклеотиды могут быть кластеризованы или могут перемежаться с комплементарными нуклеотидами и не обязательно должны быть смежными друг с другом или с комплементарными нуклеотидами.
[0069] Комплементарность в процентах между конкретными участками последовательностей нуклеиновых кислот в пределах нуклеиновых кислот можно определить обычным образом с использованием программ BLAST (базовые инструменты поиска локального выравнивания) и программ PowerBLAST (Altschul et al. (1990) J. Mol. Biol. 215:403-410; Zhang and Madden (1997) Genome Res. 7: 649-656) или с помощью программы Gap (пакет анализа последовательности Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, Madison Wis.) с использованием настроек по умолчанию, в которой используют алгоритм Smith и Waterman (Adv. Appl. Math., 1981, 2, 482-489).
[0070] Согласно способам и композициям, представленным в настоящем документе, используют множество различных компонентов. Некоторые компоненты во всем настоящем описании могут характеризоваться наличием активных вариантов и фрагментов. Такие компоненты включают в себя, например, белки Cas, РНК CRISPR, tracrPHK и гидовые РНК. Биологическая активность для каждого из этих компонентов описана в настоящем документе в другом месте. Термин «функциональный» относится к врожденной способности белка или нуклеиновой кислоты (или их фрагмента или варианта) проявлять биологическую активность или функцию. Такие биологические активности или функции могут включать в себя, например, способность белка Cas связываться с гидовой РНК и с целевой последовательностью ДНК. Биологические функции функциональных фрагментов или вариантов могут являться одинаковыми или могут быть фактически изменены (например, в отношении их специфичности, селективности или эффективности) по сравнению с исходными, но с сохранением основной биологической функции.
[0071] Термин «вариант» относится к нуклеотидной последовательности, отличающейся от последовательности, наиболее распространенной в популяции (например, одним нуклеотидом), или последовательности белка, отличающейся от последовательности, наиболее распространенной в популяции (например, одной аминокислотой).
[0072] Термин «фрагмент» применительно к белку означает белок, который является более коротким или содержит меньше аминокислот, чем полноразмерный белок. Термин «фрагмент» в отношении нуклеиновой кислоты означает нуклеиновую кислоту, которая является более короткой или содержит меньше нуклеотидов, чем полноразмерная нуклеиновая кислота. Фрагмент может являться, например, N-концевым фрагментом (т.е. удаление части С-концевого конца белка), С-концевым фрагментом (т.е. удаление части N-концевого конца белка) или внутренним фрагментом.
[0073] Термин «идентичность последовательности» или «идентичность» в контексте двух полинуклеотидных или полипептидных последовательностей относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия в указанном окне сравнения. Когда процент идентичности последовательности используют в отношении белков, положения остатков, которые не являются идентичными, часто отличаются консервативными аминокислотными заменами, где аминокислотные остатки заменены другими аминокислотными остатками со сходными химическими свойствами (например, заряд или гидрофобность) и, следовательно, не меняют функциональные свойства молекулы. Когда последовательности отличаются по консервативным заменам, процентную идентичность последовательности можно повысить для корректировки консервативного характера замены. Говорят, что последовательности, которые отличаются такими консервативными заменами, характеризуются «сходством последовательностей» или «сходством». Средства для такой корректировки хорошо известны. Как правило, это включает в себя оценку консервативного замещения как частичного, а не полного несовпадения, что увеличивает процентную идентичность последовательности. Таким образом, например, когда идентичная аминокислота получает балл 1, а неконсервативная замена получает оценку 0, консервативная замена получает балл от 0 до 1. Рассчитывают балл консервативных замен, например, как это реализовано в программе PC/GENE (Intelligenetics, Mountain View, California).
[0074] «Процентное отношение идентичности последовательности» включает в себя значение, определенное путем сравнения двух оптимально выровненных последовательностей (наибольшее количество идеально совпадающих остатков) в окне сравнения, причем часть полинуклеотидной последовательности в окне сравнения может содержать добавления или удаления (т.е. пропуски) по сравнению с эталонной последовательностью (которая не содержит добавления или удаления) для оптимального выравнивания двух последовательностей. Процентное отношение рассчитывают путем определения количества положений, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток встречается в обеих последовательностях, для получения числа совпадающих положений, деления количества совпадающих положений на общее количество положений в окне сравнения и умножение результата на 100 для получения процентного отношения идентичности последовательности. Если не указано иное (например, более короткая последовательность включает в себя связанную гетерологичную последовательность), окно сравнения представляет собой полную длину более короткой из двух сравниваемых последовательностей.
[0075] Если не указано иное, значения идентичности/сходства последовательности включают в себя значение, полученное с использованием GAP версии 10 с использованием следующих параметров: % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа за внесение пропуска согласно GAP, составляющего 50, и штрафа за удлинение пропуска, составляющего 3, и матрицы замен nwsgapdna. cmp; % идентичности и % сходства для аминокислотной последовательности с использованием штрафа за внесение пропуска согласно GAP, составляющего 8, и штрафа за удлинение пропуска, составляющего 2, и матрицы замен BLOSUM62; или любой эквивалентной им программы. «Эквивалентная программа» включает в себя любую программу сравнения последовательностей, которая для любых двух рассматриваемых последовательностей генерирует выравнивание, характеризующееся идентичными совпадениями нуклеотидных или аминокислотных остатков и идентичным процентным отношением идентичности последовательности по сравнению с соответствующим выравниванием, сгенерированным GAP версии 10.
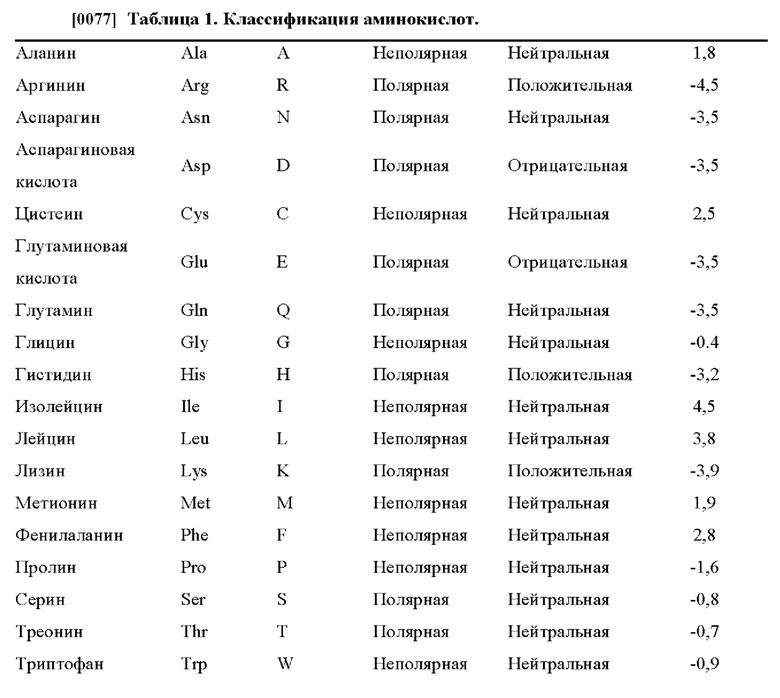
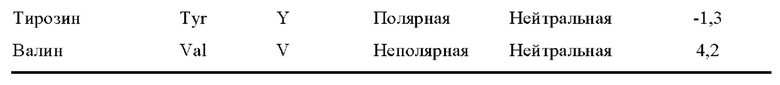
[0076] Термин «консервативная аминокислотная замена» относится к замене аминокислоты, которая обычно присутствует в последовательности, на другую аминокислоту схожего размера, заряда или полярности. Примеры консервативных замен включают в себя замену неполярного (гидрофобного) остатка, такого как изолейцин, валин или лейцин, на другой неполярный остаток. Аналогичным образом, примеры консервативных замен включают в себя замену одного полярного (гидрофильного) остатка на другой, такой как между аргинином и лизином, между глутамином и аспарагином или между глицином и серином. Кроме того, замена основного остатка, такого как лизин, аргинин или гистидин, на другой, или замена одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота, на другой кислотный остаток являются дополнительными примерами консервативных замен. Примеры неконсервативных замен включают в себя замену неполярного (гидрофобного) аминокислотного остатка, такого как изолейцин, валин, лейцин, аланин или метионин, на полярный (гидрофильный) остаток, такой как цистеин, глутамин, глутаминовая кислота или лизин, и/или полярного остатка на неполярный остаток. Типичные классификации аминокислот приведены в таблице 1 ниже.
[0078] Термин «in vitro» включает в себя искусственные среды и процессы или реакции, которые происходят в искусственной среде (например, в пробирке). Термин «in vivo» включает в себя природные среды (например, клетку, организм или организм) и процессы или реакции, которые происходят в естественной среде. Термин «ех vivo» включает в себя клетки, которые были удалены из организма человека, а также процессы или реакции, которые происходят в таких клетках.
[0079] Термин «репортерный ген» относится к нуклеиновой кислоте с последовательностью, кодирующей продукт гена (как правило, фермент), который легко и количественно анализируется, когда конструкт, содержащий последовательность репортерного гена, функционально связан с эндогенным или гетерологичным промоторным и/или энхансерным элементом, введен в клетки, содержащие (или которые могут содержать) факторы, необходимые для активации промоторных и/или энхансерных элементов. Примеры репортерных генов включают в себя без ограничения гены, кодирующие бета-галактозидазу (lacZ), гены бактериальной хлорамфениколацетилтрансферазы (кошка), гены люциферазы светлячка, гены, кодирующие бета-глюкуронидазу (GUS), и гены, кодирующие флуоресцентные белки. «Репортерный белок» относится к белку, кодируемому репортерным геном.
[0080] Используемый в настоящем документе термин «флуоресцентный репортерный белок» означает репортерный белок, который можно обнаружить на основе флуоресценции, где флуоресценция может быть либо непосредственно от репортерного белка, активности репортерного белка на флуорогенном субстрате, либо белка с аффинностью в отношении связывания с флуоресцентным меченым соединением. Примеры флуоресцентных белков включают в себя зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, изумрудно-зеленый, Azami Green, мономерный Azami Green, CopGFP, AceGFP и ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, цитрин, Venus, YPet, PhiYFP и ZsYellowl), синие флуоресцентные белки (например, BFP, eBFP, eBFP2, азурит, mKalamal, GFPuv, сапфир и Т-сапфир), голубые флуоресцентные белки (например, CFP, eCFP, лазурь, CyPet, AmCyanl и Midoriishi-Cyan), красные флуоресцентные белки (например, RFP, mKate, mKate2, mPlum, DsRed-мономер, mCherry, mRFPl, DsRed-Express, DsRed2, DsRed-мономер, HcRed-Tandem, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry и Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine и tdTomato) и любой другой подходящий флуоресцентный белок, присутствие которого в клетках можно обнаружить с помощью способов проточной цитометрии.
[0081] Восстановление в ответ на двухцепочечные разрывы (DSB) происходит главным образом посредством двух консервативных путей восстановления ДНК: гомологичной рекомбинации (HR) и негомологичного соединения концов (NHEJ). См. Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22: 886-897, полностью включенный в настоящий документ посредством ссылки для всех целей. Аналогичным образом, восстановление целевой нуклеиновой кислоты, опосредованной экзогенной донорной нуклеиновой кислотой, может включать в себя любой процесс обмена генетической информацией между двумя полинуклеотидами.
[0082] Термин «рекомбинация» включает в себя любой процесс обмена генетической информацией между двумя полинуклеотидами и может происходить по любому механизму. Рекомбинация может происходить посредством гомологичной репарации (HDR) или гомологичной рекомбинации (HR). HDR или HR включают в себя форму восстановления нуклеиновой кислоты, которая может требовать гомологии нуклеотидной последовательности, использует «донорную» молекулу в качестве матрицы для восстановления «целевой» молекулы (т.е. той, которая испытала двух цепочечный разрыв) и приводит к переносу генетической информации от донора к мишени. Не желая быть связанными какой-либо конкретной теорией, такой перенос может включать в себя коррекцию несовпадения гетеродуплексной ДНК, которая образуется между поврежденной мишенью и донором, и/или отжиг в зависимости от синтеза нитей, при котором донора используют для повторной синтеза генетической информации, которая станет частью мишени, и/или связанные процессы. В некоторых случаях донорный полинуклеотид, часть донорного полинуклеотида, копия донорного полинуклеотида или часть копии донорного полинуклеотида интегрируется в целевую ДНК. См. Wang et al. (2013) Cell 153:910-918; Mandalos et al. (2012) FLOS ONE 7:e45768:1-9; и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей.
[0083] NHEJ включает в себя восстановление двухцепочечных разрывов в нуклеиновой кислоте путем прямого лигирования концов разрывов друг с другом или с экзогенной последовательностью без необходимости гомологичной матрицы. Лигирование несмежных последовательностей с помощью NHEJ часто может приводить к делециям, вставкам или транслокациям вблизи сайта двухцепочечного разрыва. Например, NHEJ также может приводить к целенаправленной интеграции экзогенной донорной нуклеиновой кислоты посредством прямого лигирования концов разрыва с концами экзогенной донорной нуклеиновой кислоты (т.е. захват на основе NHEJ). Такая NHEJ-опосредованная целевая интеграция может быть предпочтительной для вставки экзогенной донорной нуклеиновой кислоты, когда пути гомологичной репарации (HDR) нелегко использовать (например, в неделящихся клетках, первичных клетках и клетках, которые плохо выполняют репарацию ДНК на основе гомологии). Кроме того, в отличие от гомологичной репарации, знание относительно больших областей идентичности последовательностей, фланкирующих сайт расщепления (за пределами свисающих концов, создаваемых Cas-опосредованным расщеплением), не требуется, что может быть благоприятным при попытке целенаправленной вставки в организмы, имеющие геномы, для которых знания о геномной последовательности ограничены. Интеграция может происходить посредством лигирования тупых концов между экзогенной донорной нуклеиновой кислотой и расщепленной геномной последовательностью или посредством лигирования липких концов (т.е. характеризующихся 5' или 3' свисающими концами) с использованием экзогенной донорной нуклеиновой кислоты, которая фланкирована свисающими концами, которые совместимы с таковыми, созданными белком Cas в расщепленной геномной последовательности. См., например, US 2011/020722, WO 2014/033644, WO 2014/089290 и Maresca et al. (2013) Genome Res. 23(3):539-546, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Если тупые концы лигированы, может потребоваться вырезание мишени и/или донора для создания областей микрогомологии, необходимых для соединения фрагментов, что может привести к нежелательным изменениям в целевой последовательности.
[0084] Композиции или способы, «предусматривающие» или «включающие в себя» один или несколько перечисленных элементов, могут включать в себя другие элементы, которые конкретно не указаны. Например, композиция, которая «содержит» или «включает в себя» белок, может содержать белок отдельно или в комбинации с другими ингредиентами. Переходная фраза «состоящий по существу из» означает, что объем формулы изобретения должен интерпретироваться как охватывающий указанные элементы, перечисленные в формуле изобретения, и элементы, которые не оказывают существенного влияния на основную(ые) и новую(ые) характеристику(и) заявленного изобретения. Таким образом, не предусмотрено, что термин «состоящий по существу из» при использовании в формуле изобретения следует интерпретировать как эквивалент «содержащий».
[0085] «Необязательно» или «необязательно» означает, что впоследствии описанное событие или обстоятельство может произойти или может не произойти, и что описание включает в себя случаи, в которых происходит событие или обстоятельство, и случаи, в которых оно не происходит.
[0086] Обозначение диапазона значений включает в себя все целые числа в пределах диапазона или определяющие диапазон и все поддиапазоны, определенные целыми числами в пределах диапазона
[0087] Если иное не очевидно из контекста, термин «приблизительно» охватывает значения в пределах стандартного предела погрешности измерения (например, SEM) установленного значения.
[0088] Термин «и/или» относится и охватывает любые возможные комбинации одного или нескольких связанных перечисленных пунктов, а также отсутствие комбинаций при интерпретации в альтернативе («или»).
[0089] Термин «или» относится к любому одному представителю конкретного перечня, а также включает любую комбинацию представителей этого перечня.
[0090] Формы единственного числа включают в себя формы множественного числа, если контекст явно не предписывает иное. Например, термин «белок Cas» или «по меньшей мере один белок Cas» может включать в себя множество белков Cas, включая в себя их смеси.
[0091] Статистически значимый означает р ≤0,05.
Подробное раскрытие настоящего изобретения
I. Обзор
[0092] Система CRISPR/Cas9 представляет собой мощный инструмент для инженерии генома. Одним из ограничений системы in vivo является необходимость одновременного введения всех компонентов в живой организм. Типичный способ внедрения этих компонентов заключается в транзиентной трансфекции ДНК-конструктов в клетки, которые будут генерировать соответствующие РНК и белок. Хотя этот подход является эффективным, он имеет свойственный недостаток, поскольку клетки должны полагаться на конструкты плазмидной ДНК, чтобы сначала подвергнуться транскрипции, а затем трансляции, прежде чем белок Cas9 станет доступным для взаимодействия с компонентом sgPHК. Необходимы лучшие способы и инструменты для более эффективной оценки активности агентов CRISPR/Cas и для оценки различных способов и параметров доставки для нацеливания на конкретные ткани или типы клеток in vivo.
[0093] В настоящем документе предусмотрены способы и композиции для оценки активности CRISPR/Cas-опосредованного негомологичного соединения концов (NHEJ) и/или CRISPR/Cas-индуцированной рекомбинации целевого геномного локуса с экзогенной донорной нуклеиновой кислотой in vivo и ex vivo. В способах и композициях используют клетки и являющихся человеком животных, содержащих экспрессионную кассету Cas (например, геномно интегрированную Cas экспрессионную кассету), так что белок Cas может быть конститутивно доступным или, например, доступным тканеспецифическим или специфическим для конкретного периода времени образом.
[0094] Не являющиеся человеком животные, содержащие экспрессионные кассеты Cas, упрощают процесс испытания доставки и активности компонентов CRISPR/Cas in vivo, поскольку не являющемуся человеком животному необходимо вводить только гидовые РНК. Кроме того, экспрессионные кассеты Cas могут необязательно являться условными экспрессионными кассетами Cas, которые могут избирательно экспрессироваться в определенных тканях или на определенных стадиях развития, тем самым снижая риск опосредованной Cas токсичности in vivo, или могут быть конститутивно экспрессированы для обеспечения возможности испытания активности в любом и всех типах клеток, тканей и органов.
[0095] Кроме того, предусмотрены способы и композиции для получения и применения этих не являющихся человеком животных для испытания и измерения способности CRISPR/Cas-нуклеазы модифицировать целевой геномный локус in vivo. В некоторых таких способах испытания и измерения способности CRISPR/Cas-нуклеазы модифицировать целевой геномный локус in vivo, гидовую РНК можно доставить в организм эспрессирующего Cas не являющегося человеком животного посредством AAV-опосредованной доставки. Как показано в примере 1, AAV-опосредованная доставка гидовых РНК эспрессирующим Cas9 мышам и, в частности, AAV8-опосредованная доставка в печень, приводит к удивительно более высоким уровням нацеливания CRISPR/Cas, чем доставка гидовых РНК посредством LNP или HDD эспрессирующим Cas9 мышам или доставка как Cas9, так и гидовых РНК мышам дикого типа.
II. Не являющиеся человеком животные, содержащие экспрессионные кассеты Cas
[0096] В раскрытых в настоящем документе способах и композициях используют не являющихся человеком животных или клетки, содержащие экспрессионные кассеты Cas, для оценки способности систем сгруппированных регулярно перемежающихся коротких палиндромных повторов (CRISPR)/ассоциированных с CRISPR белков (Cas) или компонентов таких систем (например, гидовых РНК, введенных организм не являющегося человеком животного или клетки) модифицировать целевой геномный локус in vivo или ex vivo.
[0097] Системы CRISPR/Cas включают в себя транскрипты и другие элементы, участвующие в экспрессии или управлении активностью генов Cas. Система CRISPR/Cas может являться, например, системой типа I, типа II или типа III. Альтернативно, система CRISPR/Cas может представлять собой систему типа V (например, подтип V-A или подтип V-В). Системы CRISPR/Cas, используемые в композициях и способах, раскрытых в настоящем документе, могут являться не встречающимися в природе. «Не встречающаяся в природе» система включает в себя все, что указывает на участие человека, например, один или несколько компонентов системы изменены или мутированы от своего природного состояния, при этом они по меньшей мере по существу не содержат по меньшей мере один другой компонент, с которым они естественным образом ассоциированы в природе, или ассоциированы по меньшей мере с одним другим компонентом, с которым они не ассоциированы естественным образом. Например, не встречающиеся в природе системы CRISPR/Cas могут использовать комплексы CRISPR, содержащие гРНК и белок Cas, которые не встречаются в природе вместе, белок Cas, который не встречается в природе, или гРНК, которую не встречается в природе.
[0098] В раскрытых в настоящем документе способах и композициях используют системы CRISPR/Cas путем испытания способности комплексов CRISPR (содержащих гидовую РНК (гРНК) в комплексе с белком Cas) индуцировать события сайт-направленного расщепления в целевом геномном локусе in vivo, чтобы модифицировать целевой геномный локус посредством не гомологичного соединение концов (NHEJ), посредством гомологичной репарации в присутствии экзогенной донорной нуклеиновой кислоты или с помощью любых других способов репарации или рекомбинации.
А. Экспрессирующие Cas9 не являющиеся человеком животные
[0099] Раскрытые в настоящем документе клетки и не являющиеся человеком животные содержат экспрессионную кассету Cas. Белки Cas, как правило, содержат по меньшей мере один домен распознавания или связывания РНК, который может взаимодействовать с гидовыми РНК (гРНК, более подробно описанными ниже), и нуклеазные домены. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновой кислоты, которое включает в себя разрыв ковалентных связей молекулы нуклеиновой кислоты. Расщепление может привести к образованию тупых концов или расположенных в шахматном порядке концов, и оно может являться одноцепочечным или двухцепочечным. Белок Cas может обладать активностью полного расщепления для создания двухцепочечного разрыва в целевой нуклеиновой кислоте (например, двухцепочечного разрыва с тупыми концами), или может представлять собой никазу, которая создает одноцепочечный разрыв в целевой нуклеиновой кислоте.
[00100] Клетки или не являющиеся человеком животные, содержащие экспрессионную кассету Cas, имеют то преимущество, что им требуется доставка только гидовых РНК для обнаружения CRISPR/Cas-опосредованной модификации целевого геномного локуса.
(1) Экспрессионные кассеты Cas
[00101] Клетки и не являющиеся человеком животные, описанные в настоящем документе, содержат экспрессионную кассету Cas. Экспрессионная кассета Cas может быть стабильно интегрирована в геном (т.е. в хромосому) клетки или не являющегося человеком животного, или она может быть расположена вне хромосомы (например, внехромосомно реплицирующейся ДНК). Необязательно, экспрессионная кассета Cas стабильно интегрирована в геном. Стабильно интегрированная экспрессионная кассета Cas может быть случайным образом интегрирована в геном не являющегося человеком животного (т.е. трансгенного), или она может быть интегрирована в предварительно определенную область генома не являющегося человеком животного (т.е. нокин). Необязательно, экспрессионная кассета Cas стабильно интегрирована в локус «безопасная гавань», как описано в другом месте в настоящем документе. Целевой геномный локус, в котором стабильно интегрирована экспрессионная кассета Cas, может являться гетерозиготным по экспрессионной кассете Cas или гомозиготным по экспрессионной кассете Cas.
[00102] Белок Cas, кодируемый экспрессионной кассетой Cas, может представлять собой любой белок Cas (например, белок Cas9), примеры которого описаны ниже. Кодированный белок Cas может дополнительно содержать один или несколько сигналов ядерной локализации (NLS) (например, N-концевой NLS и С-концевой NLS), и последовательность, кодирующая белок Cas, может являться кодон-оптимизированной для клетки или не являющегося человеком животного, как описано ниже. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка Cas9, представленной в SEQ ID NO: 19. Кодирующая последовательность может содержать, состоять по существу или состоять из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной кодирующей последовательности Cas9, представленной в SEQ ID NO: 30.
[00103] Пример экспрессионной кассеты Cas содержит кодирующую Cas последовательность ниже от сигнала полиаденилирования или терминатора транскрипции, фланкированную сайтами распознавания рекомбиназы, распознаваемыми сайт-специфической рекомбиназой. Сигнал полиаденилирования или терминатор транскрипции предотвращают транскрипцию и экспрессию белка Cas. Однако после воздействия сайт-специфической рекомбиназы сигнал полиаденилирования или терминатор транскрипции будут вырезаны, и белок Cas может быть экспрессирован.
[00104] Такая конфигурация для экспрессионной кассеты Cas может обеспечивать тканеспецифическую экспрессию или специфическую для стадии развития экспрессию у не являющихся человеком животных, содержащих экспрессионную кассету Cas, если сигнал полиаденилирования или терминатор транскрипции вырезан тканеспецифическим или специфическим для стадии развития образом. Это может снизить токсичность из-за пролонгированной экспрессии белка Cas в клетке или организме не являющегося человеком животного, или экспрессии белка Cas на нежелательных стадиях развития или в нежелательных типах клеток или тканей не являющегося человеком животного. Например, токсичность может возникнуть в результате расщепления и разрушения нецелевых участков. См., например, Parikh et at. (2015) PLoS One 10(1):e0116484. Индуцируемая экспрессия также может являться благоприятной, поскольку возможность редактирования некоторых генов в определенных тканях (например, таких как иммунные клетки) может являться вредной, а также потенциально вызывать иммунный ответ. Например, в некоторых случаях, если ген мутирован во всем организме индивидуума, это может быть летальным, но если он мутирует в конкретном типе ткани или клетки, это будет благоприятным. Вырезание сигнала полиаденилирования или терминатора транскрипции может быть достигнуто тканеспецифическим или специфическим для стадии развития способом, если не являющееся человеком животное, содержащее экспрессионную кассету Cas, дополнительно содержит сайт-специфическую рекомбиназу, функционально связанную с тканеспецифическим или специфическим для стадии развития специфический промотором (например, промотор альбумина для специфической в отношении печени экспрессии или промотор инсулина-2 для специфической в отношении поджелудочной железы экспрессии). Аналогичным образом, составы LNP, специфические в отношении печени или других тканей, можно использовать для доставки рекомбиназы, или же для доставки рекомбиназы можно использовать способы доставки AAV или серотипы AAV, специфические для конкретных тканей (например, AAV8 для печени или прямую инъекцию AAV для поджелудочной железы). Сигнал полиаденилирования или терминатор транскрипции затем вырезают только в этих тканях или на этих стадиях развития, что позволяет происходить тканеспецифической или специфической для стадии развития экспрессии белка Cas. В одном примере белок Cas может быть экспрессирован специфическим для печени образом. Примеры таких промоторов, которые были использованы для разработки таких линий не являющихся человеком животных, у которых вырезание участка ДНК осуществляется рекомбиназой ("recombinase deleter"), раскрыты в другом месте в настоящем документе.
[00105] Можно использовать любой терминатор транскрипции или сигнал полиаденилирования. Термин «терминатор транскрипции», используемый в настоящем документе, относится к последовательности ДНК, которая вызывает терминацию транскрипции. У эукариот терминаторы транскрипции распознаются белковыми факторами, а за терминацией следует полиаденилирование, процесс добавления поли(А)-хвоста к транскриптам мРНК в присутствии поли(А)-полимеразы. Сигнал поли(А) млекопитающего, как правило, состоит из сердцевинной последовательности длиной приблизительно 45 нуклеотидов, которая может быть фланкирована различными вспомогательными последовательностями, которые служат для усиления эффективности расщепления и полиаденилирования. Сердцевинная последовательность состоит из высококонсервативного вышележащего элемента (ААТААА или AAUAAA) в мРНК, называемого мотивом распознавания поли-А или последовательностью распознавания поли-А), распознаваемого специфическим в отношении расщепления и полиаденилирования фактором (CPSF), и плохо определенной нижележащей области (богатой Us или Gs и Us), связанной с фактором стимуляции расщепления (CstF). Примеры терминаторов транскрипции, которые можно использовать, включают в себя, например, сигнал полиаденилирования человеческого гормона роста (HGH), поздний сигнал полиаденилирования вируса обезьян 40 (SV40), сигнал полиаденилирования бета-глобина кролика, сигнал полиаденилирования гормона роста крупного рогатого скота (BGH), сигнал полиаденилирования фосфоглицераткиназы (PGK), последовательность терминации транскрипции АОХ1, последовательность терминации транскрипции CYC1 или любая последовательность терминации транскрипции, о которой известно, что она подходит для регуляции экспрессии генов в эукариотических клетках.
[00106] Сайт-специфические рекомбиназы включают в себя ферменты, которые могут облегчать рекомбинацию между сайтами распознавания рекомбиназы, где два сайта рекомбинации физически разделены в одной нуклеиновой кислоте или в отдельных нуклеиновых кислотах. Примеры рекомбиназ включают в себя рекомбиназы Cre, Flp и Dre. Одним примером гена Cre-рекомбиназы является Crei, в котором два экзона, кодирующие Cre-рекомбиназу, разделены интроном для предотвращения его экспрессии в прокариотической клетке. Такие рекомбиназы могут дополнительно содержать сигнал ядерной локализации для облегчения локализации в ядре (например, NLS-Crei). Сайты распознавания рекомбиназы включают в себя нуклеотидные последовательности, которые распознаются сайт-специфической рекомбиназой и могут служить в качестве субстрата для события рекомбинации. Примеры сайтов распознавания рекомбиназы включают в себя FRT, FRT11, FRT71, attp, att, rox и сайты lox, такие как loxP, lox511, lox2272, lox66, lox71, loxM2 и lox5171.
[00107] Экспрессионная кассета Cas может быть функционально связана с любым подходящим промотором для экспрессии in vivo в организме не являющегося человеком животного. Не являющееся человеком животное может представлять собой любое подходящее не являющееся человеком животное, как описано в настоящем документе в другом месте. В качестве одного примера, кассета экспрессии Cas может быть функционально связана с эндогенным промотором в целевом геномном локусе, таким как промотор Rosa26. Альтернативно, экспрессионная кассета Cas может быть функционально связана с экзогенным промотором, таким как конститутивно активный промотор (например, промотор CAG или промотор/энхансер бета-актина курицы в сочетании с немедленно ранним энхансером цитомегаловируса (CMV) (CAGG)), условным промотором, индуцируемым промотором, ограниченным по времени промотором (например, регулируемым стадией развития промотором) или пространственно ограниченным промотором (например, клеточноспецифическим или тканеспецифическим промотором). Такие промоторы хорошо известны и обсуждаются в настоящем документе в другом месте. Иллюстративный промотор CAGG представлен в SEQ ID NO: 38 или содержит, состоит по существу или состоит из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, представленной в SEQ ID NO: 38.
[00108] Экспрессионные кассеты Cas, раскрытые в настоящем документе, могут также содержать другие компоненты. Экспрессионная кассета Cas может дополнительно содержать 3' последовательность сплайсинга на 5' конце экспрессионной кассеты Cas и/или второй сигнал полиаденилирования, следующий за кодирующей последовательностью для белка Cas на 3' конце экспрессионной кассеты Cas. Экспрессионная кассета Cas может дополнительно содержать селекционную кассету, содержащую, например, кодирующую последовательность для белка с устойчивостью лекарственному средству. Примеры подходящих маркеров селекции включают в себя неомицинфосфотрансферазу (neor), гигромицин В-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror), бластицидин S-дезаминазу (bsrr), ксантин/гуанинфосфорибозилтрансферазу (gpt) и тимидинкиназу вируса простого герпеса (HSV-k). Необязательно, селекционная кассета может быть фланкирована сайтами распознавания рекомбиназы для сайт-специфической рекомбиназы. Если экспрессионная кассета Cas также содержит сайты распознавания рекомбиназы, фланкирующие сигнал полиаденилирования выше от кодирующей Cas последовательности, как описано выше, необязательно другой набор сайтов распознавания рекомбиназы, распознаваемых другой рекомбиназой, используют для фланкирования селекционной кассеты. Альтернативно, один и тот же набор сайтов распознавания рекомбиназы может фланкировать как сигнал полиаденилирования перед кодирующей Cas последовательностью, так и селекционную кассету. Иллюстративная последовательность neor-полиаденилирования представлена в SEQ ID NO: 37 или содержит, состоит по существу или состоит из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, представленной в SEQ ID NO: 37.
[00109] Экспрессионная кассета Cas также может содержать нуклеиновую кислоту, кодирующую метку белка, такую как метка 3xFLAG. Пример такой метки представлен в SEQ ID NO: 23, которая необязательно кодируется SEQ ID NO: 34. Например, метка может находиться на N-конце белка Cas, на С-конце белок Cas или внутри белка Cas. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка 3xFLAG-Cas9, представленной в SEQ ID NO: 22, или последовательности белка 3xFLAG-Cas9-P2A-eGFP, представленной в SEQ ID NO: 16. Кодирующая последовательность может содержать, состоять по существу или состоять из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%, идентичной кодирующей 3xFLAG-Cas9 Cas9 последовательности, представленной в SEQ ID NO: 31, или кодирующей 3xFLAG-Cas9-P2A-eGFP последовательности, представленной в SEQ ID NO: 29, соответственно.
[00110] Экспрессионная кассета Cas также может содержать нуклеиновую кислоту, кодирующую один или несколько репортерных белков, таких как флуоресцентный белок (например, зеленый флуоресцентный белок). Можно использовать любой подходящий репортерный белок. Например, можно использовать флуоресцентный репортерный белок, как определено в другом месте в настоящем документе, или не флуоресцентный репортерный белок. Примеры флуоресцентных репортерных белков представлены в другом месте в настоящем документе. Не флуоресцентные репортерные белки включают в себя, например, репортерные белки, которые можно использовать в гистохимических или биолюминесцентных анализах, такие как бета-галактозидаза, люцифераза (например, люцифераза Renilla, люцифераза светлячка и люцифераза NanoLuc) и бета-глюкуронидаза. Экспрессионная кассета Cas может включать в себя репортерный белок, который может быть обнаружен в анализе с помощью проточной цитометрии (например, флуоресцентный репортерный белок, такой как зеленый флуоресцентный белок) и/или репортерный белок, который может быть обнаружен в гистохимическом анализе (например, белок бета-галактозидаза). Одним из примеров такого гистохимического анализа является визуализация гистохимической экспрессии бета-галактозидазы in situ посредством гидролиза X-Gal (5-бром-4-хлор-3-индоил-b-D-галактопиранозида), который дает синий осадок, или с использованием флуорогенных субстратов, таких как бета-метил-умбеллиферилгалактозид (MUG) и флуоресцеиндигалактозид (FDG).
[00111] Экспрессионная кассета Cas в таких случаях может содержать мультицистронную нуклеиновую кислоту. Например, такие нуклеиновые кислоты могут являться кодирующей последовательностью белка Cas и кодирующей последовательностью репортерного белка (в любом порядке), разделенными промежуточным участком внутренней посадки рибосомы (IRES) или промежуточной кодирующей последовательностью пептида 2А. Мультицистронные экспрессионные конструкты одновременно экспрессируют два или более отдельных белков из одной и той же мРНК (т.е. транскрипт, полученный из одного и того же промотора). Подходящие стратегии для мультицистронной экспрессии белков включают в себя, например, использование пептида 2А и использование участка внутренней посадки рибосомы (IRES). Например, такие нуклеиновые кислоты могут содержать кодирующие последовательности для двух или более репортерных белков, разделенных промежуточным участком внутренней посадки рибосомы (IRES) или промежуточной кодирующей пептид 2А последовательностью. В качестве одного примера, такие мультицистронные векторы могут использовать один или несколько участков внутренней посадки рибосомы (IRES), чтобы позволить инициирование трансляции из внутренней области мРНК. В качестве другого примера, такие мультицистронные векторы могут использовать один или несколько пептидов 2А. Эти пептиды представляют собой небольшие «саморасщепляющиеся» пептиды, как правило, характеризующиеся длиной, составляющей 18-22 аминокислоты, и продуцирующие эквимолярные уровни нескольких генов из одной и той же мРНК. Рибосомы пропускают синтез глицил-пролилпептидной связи на С-конце пептида 2А, что приводит к «расщеплению» между пептидом 2А и пептидом, находящимся непосредственно ниже него. См., например, Kim et al. (2011) PLoS One 6(4): e18556, полностью включенный в настоящий документ посредством ссылки для всех целей. «Расщепление» происходит между остатками глицина и пролина, обнаруженными на С-конце, это означает, что находящемся выше цистроне будет добавлено несколько дополнительных остатков, в то время как находящийся ниже по ходу транскрипции цистрон будет начинаться с пролина. В результате «отщепленный» нижележащий пептид содержит пролин на своем N-конце. Опосредованное 2А расщепление является универсальным явлением во всех эукариотических клетках. Пептиды 2А были идентифицированы из пикорнавирусов, вирусов насекомых и ротавирусов типа С. См., например, Szymczak et al. (2005) Expert Opin Biol Ther 5:627-638, полностью включенный в настоящий документ посредством ссылки для всех целей. Примеры пептидов 2А, которые можно использовать, включают в себя 2А вируса Thosea asigna (Т2А); 2А свиного тешовируса-1 (Р2А); 2А вируса лошадиного ринита A (ERAV) (Е2А) и 2А FMDV (F2A). Иллюстративные последовательности Т2А, Р2А, Е2А и F2A включают в себя следующие: Т2А (EGRGSLLTCGDVEENPGP; SEQ ID NO: 2); Р2А (ATNFSLLKQAGDVEENPGP; SEQ ID NO: 3); Е2А (QCTNYALLKLAGDVESNPGP; SEQ ID NO: 4); и F2A (VKQTLNFDLLKLAGDVESNPGP; SEQ ID NO: 5). Остатки GSG могут быть добавлены к 5' концу любого из этих пептидов для улучшения эффективности расщепления. Иллюстративная кодирующая последовательность для Р2А с остатками GSG, добавленными на 5' конце, представлена в SEQ ID NO: 32 или содержит, состоит по существу или состоит из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности, представленной в SEQ ID NO: 32. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 91%, 98%о, 99% или 100% идентичной белковой последовательности Cas9-P2A-eGFP eGFP, представленной в SEQ ID NO: 13, или белковой последовательности 3xFLAG-Cas9-P2A-eGFP, представленной в SEQ ID NO: 16. Кодирующая последовательность может содержать, состоять по существу или состоять из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%) 95%, 96%, 97%, 98%, 99% или 100% идентичной кодирующей Cas9-P2A-eGFP последовательности, представленной в SEQ ID NO: 28, или кодирующей 3xFLAG-Cas9-P2A-eGFP последовательности, представленной в SEQ ID NO: 29, соответственно.
[00112] Экспрессионные кассеты Cas также могут содержать другие элементы, такие как посттранскрипционные регуляторные элементы или сигналы полиаденилирования, расположенные ниже относительно кодирующей Cas последовательности. Иллюстративный посттранскрипционный регуляторный элемент представляет собой посттранскрипционный регуляторный элемент вируса гепатита сурков (WPRE), который представлен в SEQ ID NO: 35 или содержит, состоит по существу или состоит из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 91%, 98%, 99% или 100% идентичной последовательности, представленной в SEQ ID NO: 35. Иллюстративный сигнал полиаденилирования представляет собой сигнал полиаденилирования гормона роста крупного рогатого скота, который представлен в SEQ ID NO: 36 или содержит, состоит по существу или состоит из последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 91%, 98%, 99% или 100% идентичной последовательности, представленной в SEQ ID NO: 36.
[00113]Одна иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) 3' последовательность сплайсинга; (b) сигнал полиаденилирования, фланкированный первым и вторым сайтами распознавания рекомбиназы для рекомбиназы (например, сайты loxP для Cre-рекомбиназы); (с) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце); (d) кодирующая белок 2А последовательность (например, кодирующая Р2А последовательность); и (е) кодирующая последовательность для репортерного белка (например, флуоресцентного репортерного белка, такого как зеленый флуоресцентный белок). См., например, фигуру 1 и MAID2599 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 1. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка Cas9-P2A-eGFP, представленной в SEQ ID NO: 13.
[00114] Другая иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) 3' последовательность сплайсинга; (b) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце); (с) кодирующая белок 2А последовательность (например, кодирующая Р2А последовательность); и (d) кодирующая последовательность для репортерного белка (например, флуоресцентного репортерного белка, такой как зеленый флуоресцентный белок). См., например, MAID2600 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 12. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка Cas9-P2A-eGFP, представленной в SEQ ID NO: 13.
[00115] Другая иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) 3' последовательность сплайсинга; (b) сигнал полиаденилирования, фланкированный первым и вторым сайтами распознавания рекомбиназы для рекомбиназы (например, сайты loxP для Cre-рекомбиназы); и (с) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце). См., например, MAID2660 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 17. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка Cas9, представленной в SEQ ID NO: 19.
[00116] Другая иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) 3' последовательность сплайсинга; и (b) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце). См., например, MAID2661 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 18. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 91%, 98%, 99% или 100% идентичной последовательности белка Cas9, представленной в SEQ ID NO: 19.
[00117] Другая иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) экзогенный промотор (например, конститутивный промотор, такой как промотор CAGG); (b) сигнал полиаденилирования, фланкированный первым и вторым сайтами распознавания рекомбиназы для рекомбиназы (например, сайты loxP для Cre-рекомбиназы); (с) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце, необязательно с меткой на N-концевом или С-концевом конце, такой как метка 3xFLAG на N-концевом конце); (d) кодирующая белок 2А последовательность (например, кодирующая Р2А последовательность); и (е) кодирующая последовательность для репортерного белка (например, флуоресцентного репортерного белка, такого как зеленый флуоресцентный белок). См., например, MAID2658 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 14. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка 3xFLAG-Cas9-P2A-eGFP, представленной в SEQ ГО NO: 16.
[00118] Другая иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) экзогенный промотор (например, конститутивный промотор, такой как промотор CAGG); (b) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце, необязательно с меткой на N-концевом или С-концевом конце, такой как метка 3xFLAG на N-концевом конце); (с) кодирующая белок 2А последовательность (например, кодирующая Р2А последовательность); и (d) кодирующая последовательность для репортерного белка (например, флуоресцентного репортерного белка, такого как зеленый флуоресцентный белок). См., например, MAID2659 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 15. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка 3xFLAG-Cas9-P2A-eGFP, представленной в SEQ ГО NO: 16.
[00119] Другая иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) экзогенный промотор (например, конститутивный промотор, такой как промотор CAGG); (b) сигнал полиаденилирования, фланкированный первым и вторым сайтами распознавания рекомбиназы для рекомбиназы (например, сайты loxP для Cre-рекомбиназы); и (с) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце, необязательно с меткой на N-концевом или С-концевом конце, такой как метка 3xFLAG на N-концевом конце). См., например, MAID2672 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 20. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка 3xFLAG-Cas9, представленной в SEQ ID NO: 22.
[00120] Другая иллюстративная экспрессионная кассета Cas содержит от 5' к 3' следующее: (а) экзогенный промотор (например, конститутивный промотор, такой как промотор CAGG); и (b) кодирующая белок Cas последовательность (например, кодирующая NLS-Cas9 последовательность, например, с NLS на N-концевом конце и NLS на С-концевом конце, необязательно с меткой на N-концевом или С-концевом конце, такой как метка 3xFLAG на N-концевом конце). См., например, MAID2673 на фигуре 14. Такая экспрессионная кассета может содержать, состоять по существу или состоять из последовательности, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, представленной в SEQ ID NO: 21. Например, такая экспрессионная кассета может кодировать белок, содержащий, состоящий по существу или состоящий из аминокислотной последовательности, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности белка 3xFLAG-Cas9, представленной в SEQ ID NO: 22.
[00121] Описанные в настоящем документе экспрессионные кассеты Cas могут находиться в любой форме. Например, экспрессионная кассета Cas может находиться в векторе или плазмиде, такой как вирусный вектор. Экспрессионная кассета Cas может быть функционально связана с промотором в экспрессионной конструкте, способным направлять экспрессию белка Cas после удаления вышележащего сигнала полиаденилирования. Альтернативно, экспрессионная кассета Cas может находиться в нацеливающем векторе, как определено в настоящем документе в другом месте. Например, направляющий вектор может содержать гомологические плечи, фланкирующие экспрессиоиную кассету Cas, причем гомологические плечи подходят для направления рекомбинации с требуемым целевым геномным локусом для облегчения геномной интеграции.
[00122] Описанные в настоящем документе экспрессионные кассеты Cas могут находиться in vitro, они могут находиться внутри клетки (например, эмбриональной стволовой клетки) ex vivo (например, геномно интегрированы или внехромосомно) или они могут находиться в организме (например, не являющегося человеком животного) in vivo (например, геномно интегрированы или внехромосомно). В случае ex vivo экспрессионная кассета Cas может находиться в клетке любого типа из любого организма, такой как тотипотентная клетка, такая как эмбриональная стволовая клетка (например, эмбриональная стволовая клетка мыши или крысы) или индуцированная плюрипотентная стволовая клетка (например, индуцированная плюрипотентная стволовая клетка человека). В случае in vivo экспрессионная кассета Cas может находиться в организме любого типа (например, не являющееся человеком животное, как описано далее в другом месте в настоящем документе).
(2) Белки Cos и полинуклеотиды, кодирующие белки Cas
[00123] Белки Cas, как правило, содержат по меньшей мере один домен распознавания или связывания РНК, который может взаимодействовать с гидовыми РНК (гРНК, более подробно описанные ниже). Белки Cas также могут содержать нуклеазные домены (например, домены ДНКазы или РНКазы), ДНК-связывающие домены, геликазные домены, домены белок-белковых взаимодействий, домены димеризации и другие домены. Некоторые такие домены (например, домены ДНКазы) могут быть из нативного белка Cas. Другие такие домены могут быть добавлены для получения модифицированного белка Cas. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновой кислоты, которое включает в себя разрыв ковалентных связей молекулы нуклеиновой кислоты. Расщепление может привести к образованию тупых концов или расположенных в шахматном порядке концов, и оно может являться одноцепочечным или двухцепочечным. Например, белок Cas9 дикого типа, как правило, создает продукт расщепления с тупым концом. Альтернативно, белок Cpf1 дикого типа (например, FnCpf1) может приводить к продукту расщепления с 5-нуклеотидным 5'-«свисающим» концом, причем расщепление происходит после 18-й пары оснований из последовательности РАМ на нецелевой цепи и после 23-го основания на целевой цепи. Белок Cas может обладать активностью полного расщепления для создания двухцепочечного разрыва в целевом геномном локусе (например, двухцепочечный разрыв с тупыми концами), или это может быть никаза, которая создает одноцепочечный разрыв в целевом геномном локусе.
[00124] Примеры белков Cas включают в себя Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 или Csx12), Cas10, Cas10d, CasF, CasG, CasH, Csy1, Csy2, Csy3, Cse1 (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4 и Cu1966 и их гомологи или модифицированные версии.
[00125] Иллюстративный белок Cas представляет собой белок Cas9 или белок, полученный из Cas9. Белки Cas9 происходят из системы CRISPR/Cas II типа и, как правило, характеризуются общими четырьмя ключевыми мотивами с консервативной архитектурой. Мотивы 1, 2 и 4 представляют собой RuvC-подобные мотивы, и мотив 3 представляет собой мотив HNH. Иллюстративные белки Cas9 происходят из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptomyces viridochromogenes, Streptosporangium roseum, Streptosporangium roseum, Alicyclobacillus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus, Acaryochloris marina, Neisseria meningitidis или Campylobacter jejuni. Дополнительные примеры представителей семейства Cas9 описаны в международной патентной публикации WO 2014/131833, полностью включенной в настоящий документ посредством ссылки для всех целей. Cas9 из S. pyogenes (SpCas9) (присвоенный регистрационный номер SwissProt Q99ZW2) представляет собой иллюстративный белок Cas9. Cas9 из S. aureus (SaCas9) (присвоенный регистрационный номер UniProt J7RUA5) представляет собой другой иллюстративный белок Cas9. Cas9 из Campylobacter jejuni (CjCas9) (присвоенный регистрационный номер UniProt Q0P897) представляет собой еще один иллюстративный белок Cas9. См., например, Kim et al. (2017) Nat. Comm. 8:14500, полностью включенный в настоящий документ посредством ссылки для всех целей. SaCas9 меньше, чем SpCas9, a CjCas9 меньше, чем SaCas9 и SpCas9.
[00126] Другим примером белка Cas является белок Cpf1 (CRISPR из Prevotella и Francisella 1). Cpf1 представляет собой большой белок (приблизительно 1300 аминокислот), который содержит RuvC-подобный нуклеазный домен, гомологичный соответствующему домену Cas9, наряду с аналогом характерного богатого аргинином кластера Cas9. Однако в Cpf1 отсутствует нуклеазный домен HNH, который присутствует в белках Cas9, a RuvC-подобный домен является смежным в последовательности Cpf1, в отличие от Cas9, где он содержит длинные вставки, включая в себя домен HNH. См., например, Zetsche et al. (2015) Cell 163(3):759-771, полностью включенный в настоящий документ посредством ссылки для всех целей. Иллюстративные белки Cpf1 происходят из Francisella tularensis 1, Francisella tularensis subsp.novicida, Prevotella albensis, Lachnospiraceae bacterium MC2017 1, Butyrivibrio proteoclasticus, Peregrinibacteria bacterium GW2011_GWA2_33_10, Parcubacteria bacterium GW2011_GWC2_44_17, Smithella sp. SCADC, Acidaminococcus sp. BV3L6, Lachnospiraceae bacterium MA2020, CandidatusMethanoplasma termitum, Eubacterium eligens, Moraxella bovoculi 237, Leptospira inadai, Lachnospiraceae bacterium ND2006, Porphyromonas crevioricanis 3, Prevotella disiens и Porphyromonas macacae. Cpf1 из Francisella novicida U112 (FnCpf1; присвоенный регистрационный номер UniProt A0Q7Q2) представляет собой иллюстративный белок Cpf1.
[00127] Белки Cas могут представлять собой белки дикого типа (т.е. белки, которые встречаются в природе), модифицированные белки Cas (т.е. варианты белков Cas) или фрагменты белков дикого типа или модифицированных белков Cas. Белки Cas также могут являться активными вариантами или фрагментами в отношении каталитической активности белков Cas дикого типа или модифицированных белков Cas. Активные варианты или фрагменты в отношении каталитической активности могут характеризоваться по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичностью последовательности по отношению к белку Cas дикого типа или модифицированному белку Cas или его части, причем активные варианты сохраняют способность разрезать в требуемом сайте расщепления и, следовательно, сохраняют активность, индуцирующую о дно цепочечный разрыв или двухцепочечный разрыв. Анализы активности в отношении индукции одноцепочечных разрывов или индукции двухцепочечных разрывов являются известными, и, как правило, в них измеряют общую активность и специфичность белка Cas на ДНК-субстратах, содержащих сайт расщепления.
[00128] Белки Cas можно модифицировать для увеличения или уменьшения одного или нескольких из следующего: аффинность связывания нуклеиновой кислоты, специфичность связывания нуклеиновой кислоты и ферментативная активность. Белки Cas также можно модифицировать для изменения любой другой активности или свойства белка, такого как стабильность. Например, один или несколько нуклеазных доменов белка Cas можно модифицировать, удалить или инактивировать, или белок Cas может быть усечен для удаления доменов, которые не являются необходимыми для функции белка или для оптимизации (например, для усиления или снижения активности или свойства белка Cas.
[00129] Одним из примеров модифицированного белка Cas является модифицированный белок SpCas9-HF1, который представляет собой высокоточный вариант Cas9 Streptococcus pyogenes, несущий изменения (N497A/R661A/Q695A/Q926A), предназначенный для уменьшения неспецифических контактов ДНК. См., например, Kleinstiver et al. (2016) Nature 529(7587):490-495, полностью включенный в настоящий документ посредством ссылки для всех целей. Другим примером модифицированного белка Cas является модифицированный вариант eSpCas9 (K848A/K1003A/R1060A), предназначенный для уменьшения нежелательных эффектов. См., например, Slaymaker et al. (2016) Science 351(6268):84-88, полностью включенный в настоящий документ посредством ссылки для всех целей. Другие варианты SpCas9 включают в себя K855A и K810A/K1003A/R1060A.
[00130] Белки Cas могут содержать по меньшей мере один нуклеазный домен, такой как ДНКазный домен. Например, белок Cpf1 дикого типа, как правило, содержит RuvC-подобный домен, который расщепляет обе цепи целевой ДНК, возможно, в димерной конфигурации. Белки Cas также могут содержать по меньшей мере два нуклеазных домена, таких как ДНКазные домены. Например, белок Cas9 дикого типа, как правило, содержит RuvC-подобный нуклеазный домен и HNH-подобный нуклеазный домен. Каждый из доменов RuvC и HNH может разрезать разные нити двухцепочечной ДНК, чтобы сделать двухцепочечный разрыв в ДНК. См., например, Jinek et al. (2012) Science 337:816-821, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00131] Один или несколько нуклеазных доменов могут быть удалены или мутированы, так что они больше не функционируют или характеризуются пониженной нуклеазной активностью. Например, если один из нуклеазных доменов удален или мутирован в белке Cas9, полученный белок Cas9 можно назвать никазой, и он может генерировать однонитевой разрыв в целевой последовательности для гидовой РНК в пределах двухцепочечной ДНК, но не двухцепочечный разрыв (т.е. он может расщеплять комплементарную цепь или не комплементарную цепь, но не обе). Примером мутации, которая превращает Cas9 в никазу, является мутация D10A (аспартат в аланин в положении 10 в Cas9) в RuvC-домене Cas9 из S. pyogenes. Аналогично, Н939А (гистидин в аланин в положении аминокислоты 839), Н840А (гистидин в аланин в положении аминокислоты 840) или N863 А (аспарагин в аланин в положении аминокислоты N863) в домене HNH Cas9 из S. pyogenes могут превращать Cas9 в никазу. Другие примеры мутаций, которые превращают Cas9 в никазу, включают в себя соответствующие мутации в Cas9 из S. thermophilics. См., например, Sapranauskas et al. (2011) Nucleic Acids Research 39:9275-9282 и международную патентную публикацию WO 2013/141680, причем каждый из документов полностью включен в настоящий документ посредством ссылки для всех целей. Такие мутации могут быть получены с использованием таких способов, как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез или общий синтез генов. Примеры других мутаций, создающих никазы, можно найти, например, в международных патентных публикациях WO 2013/176772 и WO 2013/142578, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей.
[00132] Также известны примеры инактивирующих мутаций в каталитических доменах белков Cas9 Staphylococcus aureus. Например, фермент Cas9 Staphyloccocus aureus (SaCas9) может содержать замену в положении N580 (например, замену N580A) и замену в положении D10 (например, замену D10A) для создания белок Cas с неактивной нуклеазой. См., например, международную патентную публикацию WO 2016/106236, полностью включенную в настоящий документ посредством ссылки для всех целей.
[00133] Также известны примеры ин активирующих мутаций в каталитических доменах белков Cpf1. Что касается белков Cpf1 из U112 Francisella novicida (FnCpf1), BV3L6 Acidaminococcus sp. (AsCpf1), ND2006 Lachnospiraceae bacterium (LbCpf1) и 237 Moraxella bovoculi (MbCpf1 Cpf1), такие мутации могут включать в себя мутации в положениях 908, 993 или 1263 в AsCpf1 или в соответствующих положениях в ортологах Cpf1 или 832, 925, 947 или 1180 в LbCpf1 или соответствующих положениях в ортологах Cpf1. Такие мутации могут включать в себя, например, одну или несколько из мутаций D908A, Е993А, и D1263A в AsCpf1 или соответствующие мутации в ортологах Cpf1, или D832A, Е925А, D947A и D1180A в LbCpf1 или соответствующие мутации в ортологах Cpf1. См., например, US 2016/0208243, полностью включенную в настоящий документ посредством ссылки для всех целей.
[00134] Белки Cas также могут быть функционально связаны с гетерологичными полипептидами в качестве слитых белков. Например, белок Cas может быть слит с доменом расщепления или доменом эпигенетической модификации. См. международную патентную публикацию WO 2014/089290, полностью включенную в настоящий документ посредством ссылки для всех целей. Белки Cas также могут быть слиты с гетерологичным полипептидом, обеспечивающим повышенную или пониженную стабильность. Слитый домен или гетерологичный полипептид может быть расположен на N-конце, С-конце или внутри белка Cas.
[00135] В качестве одного примера, белок Cas может быть слит с одним или несколькими гетерологичными полипептидами, которые обеспечивают субклеточную локализацию. Такие гетерологичные полипептиды могут включать в себя, например, один или несколько сигналов ядерной локализации (NLS), таких как одинарный NLS SV40 и/или двойной NLS альфа-импортина для нацеливания на ядро, сигнал митохондриальной локализации для нацеливания на митохондрии, сигнал удерживания в ER и тому подобное. См., например, Lange et al. (2007) J. Biol. Chem. 282:5101-5105, полностью включенный в настоящий документ посредством ссылки для всех целей. Такие сигналы субклеточной локализации могут быть локализованы на N-конце, С-конце или в любом месте белка Cas. NLS может содержать участок основных аминокислот и может представлять собой одинарную последовательность или двойную последовательность. Необязательно белок Cas может содержать два или более NLS, включая в себя NLS (например, NLS альфа-импорта на или одинарный NLS) на N-конце и NLS (например, NLS SV40 или двойной NLS) на С-конце. Белок Cas также может содержать два или более NLS на N-конце и/или два или более NLS на С-конце.
[00136] Белки Cas также могут быть функционально связаны с проникающим в клетку доменом или доменом белковой трансдукции. Например, проникающий в клетку домен может быть получен из белка ТАТ ВИЧ-1, проникающего в клетки TLM мотива вируса гепатита В человека, MPG, Рер-1, VP22, проникающего в клетки пептида вируса простого герпеса или пептидной последовательности полиаргинина. См., например, международные патентные публикации WO 2014/089290 и WO 2013/176772, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей. Проникающий в клетку домен может быть расположен на N-конце, С-конце или в любом месте белка Cas.
[00137] Белки Cas также могут быть функционально связаны с гетерологичным полипептидом для простоты отслеживания или очистки, таким как флуоресцентный белок, метка очистки или метка эпитопа. Примеры флуоресцентных белков включают в себя зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, изумрудно-зеленый, Azami Green, мономерный Azami Green, CopGFP, AceGFP, ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, цитрин, Venus, YPet, PhiYFP, ZsYellowl), голубые флуоресцентные белки (например, eBFP, eBFP2, азурит, mKalamal, GFPuv, сапфир, Т-сапфир), голубые флуоресцентные белки (например, eCFP, лазурь, CyPet, AmCyanl, Midorii), красные флуоресцентные белки (например, mKate, mKate2, mPlum, DsRed-мономер, mCherry, mRFPl, DsRed-Express, DsRed2, DsRed-мономер, HcRed-Tandem, HcRed1, AsRed2, eqFP611, mRespberry, mStrawberry, Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine, tdTomato) и любой другой подходящий флуоресцентный белок. Примеры меток включают в себя глутатион-S-трансферазу (GST), хитин-связывающий белок (СВР), мальтозосвязывающий белок, тиоредоксин (TRX), nonn(NANP), метку тандемной аффинной очистки (ТАР), myc, AcV5, AU1, AU5, Е, ECS, Е2, FLAG, гемагглютинин (НА), nus, Softag 1, Softag 3, Strep, SBP, Glu-Glu, HSV, KT3, S, SI, T7, V5, VSV-G, гистидин (His), белок-носитель биотин-карбоксила (BCCP) и кальмодулин.
[00138] Нуклеиновую кислоту, кодирующую белок Cas, можно оптимизировать по кодонам для эффективной трансляции в белок в конкретной клетке или организме. Например, нуклеиновую кислоту, кодирующую белок Cas, можно модифицировать для замены кодонов, характеризующихся более высокой частотой использования в клетке человека, не относящейся к человеку клетке, клетке млекопитающего, клетке грызуна, клетке мыши, клетке крысы или любой другой представляющей интерес клетке-хозяине по сравнению с встречающейся в природе полинуклеотидной последовательностью.
(3) Гидовые РНК
[00139] «Гидовая РНК» или «гРНК» представляет собой молекулу РНК, которая связывается с белком Cas (например, белком Cas9) и нацеливает белок Cas на специфическое местоположение в пределах целевой ДНК. Гидовые РНК могут содержать два сегмента: «нацеливающий на ДНК сегмент» и «связывающий белок сегмент». «Сегмент» включает в себя участок или область молекулы, такой как непрерывный участок нуклеотидов в РНК. Некоторые гРНК, такие как гРНК для Cas9, могут содержать две отдельные молекулы РНК: «активатор-РНК» (например, tracrPHK) и «targeter-РНК» (например, CRISPR РНК или сrРНК). Другие гРНК представляют собой единую молекулу РНК (полинуклеотид единой РНК), которая также может называться «одномолекулярная гРНК», «единая гидовая РНК» или «sgPHK». См., например, международные патентные публикации WO 2013/176772, WO 2014/065596, WO 2014/089290, WO 2014/093622, WO 2014/099750, WO 2013/142578 и WO 2014/131833, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей. Для Cas9, например, единая гидовая РНК может содержать сrРНК, слитую с tracrPHK (например, посредством линкера). Для Cpf1, например, только сrРНК необходима для достижения связывания и/или расщепления целевой последовательности. Термины «гидовая РНК» и «гРНК» включают в себя как двухмолекулярные (т.е. модульные) гРНК, так и одномолекулярные гРНК.
[00140] Типичная двухмолекулярная гРНК содержит молекулу, подобную сrРНК («CRISPR РНК», или «targeter-РНК», или «сrРНК», или «повтор сrРНК»), и соответствующую tracrPHK-подобную («транс-активная CRISPR РНК» или «активатор-РНК» или «tracrPHK») молекулу. сrРНК содержит как нацеливающий на ДНК сегмент (одноцепочечный) гРНК, так и участок нуклеотидов (т.е. хвост сrРНК), который образует одну половину дцРНК-дуплекса связывающего белок сегмента гРНК. Пример хвоста сrРНК, расположенного ниже (3') от сегмента, нацеливающего на ДНК, содержит, состоит по существу или состоит из GUUUUAGAGCUAUGCU (SEQ ID NO: 25). Любой из сегментов, нацеливающих на ДНК, раскрытых в настоящем документе, можно присоединить к 5' концу SEQ ID NO: 25 с образованием сrРРНК.
[00141] Соответствующая tracrPHK (активатор-РНК) содержит участок нуклеотидов, который образует другую половину дцРНК-дуплекса связывающего белок сегмента гРНК. Участок нуклеотидов сrРНК является комплементарным и гибридизуется с участком нуклеотидов tracrPHK с образованием дцРНК-дуплекса связывающего белок домена гРНК. Таким образом, можно сказать, что каждая сrРНК имеет соответствующую tracrPHK. Пример последовательности tracrPHK содержит, состоит по существу или состоит из AGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGA GUCGGUGCUUU (SEQ ID NO: 26).
[00142] В системах, в которых необходимы как сrРНК, так и tracrPHK, сrРНК и соответствующая tracrPHK гибридизуются с образованием гРНК. В системах, в которых требуется только сrРНК, сrРНК может представлять собой гРНК. Кроме того, сrРНК обеспечивает одноцепочечный нацеливающий на ДНК сегмент, который нацеливается на целевую последовательность для гидовой РНК путем гибридизации с противоположной цепью (т.е. комплементарной цепью). При использовании для модификации внутри клетки точная последовательность данной молекулы сrРНК или tracrPHK может быть разработана так, чтобы она была специфической для видов, в которых будут использоваться молекулы РНК. См., например, Mali et al. (2013) Science 339:823-826; Jinek et al. (2012) Science 337:816-821; Hwang et al. (2013) Nat. Biotechnol. 31:227-229; Jiang et al. (2013) Nat. Biotechnol. 31:233-239; и Cong et al. (2013) Science 339:819-823, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей.
[00143] Нацеливающий на ДНК сегмент (сrРНК) данной гРНК содержит нуклеотидную последовательность, которая комплементарна последовательности (т.е. комплементарная цепь последовательности распознавания гидовой РНК на цепи, противоположной целевой последовательности для гидовой РНК) в целевой ДНК. Нацеливающий на ДНК сегмент гРНК взаимодействует с целевой ДНК специфическим для последовательности образом посредством гибридизации (т.е. спаривания оснований). Как таковая, нуклеотидная последовательность сегмента, нацеливающего на ДНК, может варьироваться и определять местоположение внутри целевой ДНК, с которым будут взаимодействовать гРНК и целевая ДНК. Нацеливающий на ДНК сегмент рассматриваемой гРНК может быть модифицирован для гибридизации с любой требуемой последовательностью в целевой ДНК. Встречающиеся в природе сrРНК различаются в зависимости от системы CRISPR/Cas и организма, но часто содержат нацеливающий сегмент длиной от 21 до 72 нуклеотидов, фланкированный двумя прямыми повторами (DR) длиной от 21 до 46 нуклеотидов (см., например, международную патентную публикацию WO 2014/131833, полностью включенную в настоящий документ посредством ссылки для всех целей). В случае S. pyogenes DR характеризуются длиной, составляющей 36 нуклеотидов, а нацеливающий сегмент составляет 30 нуклеотидов. 3'-расположенный DR является комплементарным и гибридизуется с соответствующей tracrPHK, которая, в свою очередь, связывается с белком Cas.
[00144] Нацеливающий на ДНК сегмент может характеризоваться длиной, составляющей по меньшей мере приблизительно 12 нуклеотидов, по меньшей мере приблизительно 15 нуклеотидов, по меньшей мере приблизительно 17 нуклеотидов, по меньшей мере приблизительно 18 нуклеотидов, по меньшей мере приблизительно 19 нуклеотидов, по меньшей мере приблизительно 20 нуклеотидов, по меньшей мере приблизительно 25 нуклеотидов, по меньшей мере приблизительно 30 нуклеотидов, по меньшей мере приблизительно 35 нуклеотидов или по меньшей мере приблизительно 40 нуклеотидов. Такие нацеливающие на ДНК сегменты могут характеризоваться длиной от приблизительно 12 нуклеотидов до приблизительно 100 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 80 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 50 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 40 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 30 нуклеотидов, от приблизительно 12 нуклеотидов до приблизительно 25 нуклеотидов, или от приблизительно 12 нуклеотидов до приблизительно 20 нуклеотидов. Например, нацеливающий на ДНК сегмент может составлять от приблизительно 15 нуклеотидов до приблизительно 25 нуклеотидов (например, от приблизительно 17 нуклеотидов до приблизительно 20 нуклеотидов или приблизительно 17 нуклеотидов, приблизительно 18 нуклеотидов, приблизительно 19 нуклеотидов или приблизительно 20 нуклеотидов). См., например, US 2016/0024523, полностью включенную в настоящий документ посредством ссылки для всех целей. Для Cas9 из S. pyogenes типичный нацеливающий на ДНК сегмент составляет 16-20 нуклеотидов в длину или 17-20 нуклеотидов в длину. Для Cas9 из S. aureus типичный нацеливающий на ДНК сегмент составляет 21-23 нуклеотида в длину. Для Cpf1 типичный нацеливающий на ДНК сегмент составляет по меньшей мере 16 нуклеотидов в длину или по меньшей мере 18 нуклеотидов в длину.
[00145] tracrPHK могут находиться в любой форме (например, полноразмерные tracrPHK или активные частичные tracrPHK) и могут характеризоваться любой длиной. Они включают в себя первичные транскрипты или процессированные формы. Например, tracrPHK (как часть единой гидовой РНК или как отдельная молекула как часть двухмолекулярной гРНК) могут содержать, состоять по существу или состоять из всей или части последовательности tracrPHK дикого типа (например, приблизительно или больше чем приблизительно 20, 26, 32, 45, 48, 54, 63, 67, 85 или больше нуклеотидов последовательности tracrPHK дикого типа). Примеры последовательностей tracrPHK дикого типа из S. pyogenes включают в себя 171-нуклеотидные, 89-нуклеотидные, 75-нуклеотидные и 65-нуклеотидные версии. См., например, Deltcheva et al. (2011)Nature 471:602-607; международную патентную публикацию WO 2014/093661, причем каждый из этих документов полностью включен в настоящий документ посредством ссылки для всех целей. Примеры tracrPHK в пределах единых гидовых РНК (sgPHK) включают в себя сегменты tracrPHK, обнаруженные в пределах +48, +54, +67 и +85 версий sgPHK, где "+n" указывает на то, что вплоть до +n нуклеотид tracrPHK дикого типа включен в sgPHK. См. патент США №8697359, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00146] Процентное отношение комплементарности между нацеливающей на ДНК последовательностью и комплементарной цепью последовательности распознавания гидовой РНК в пределах целевой ДНК может составлять по меньшей мере 60% (например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%), по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%), по меньшей мере 99% или 100%). Процентное отношение комплементарности между нацеливающей на ДНК последовательностью и комплементарной цепью последовательности распознавания гидовой РНК в пределах целевой ДНК может составлять по меньшей мере 60% по приблизительно 20 смежным нуклеотидам. В качестве примера процентное отношение комплементарности между нацеливающей на ДНК последовательностью и комплементарной цепью последовательности распознавания гидовой РНК в пределах целевой ДНК составляет 100% по 14 смежным нуклеотидам на 5' конце комплементарной цепи последовательности распознавания гидовой РНК в пределах комплементарной цепи целевой ДНК и всего 0% по остальной части. В таком случае может считаться, что нацеливающая на ДНК последовательность составляет 14 нуклеотидов в длину. В качестве другого примера процентное отношение комплементарности между нацеливающей на ДНК последовательностью и комплементарной цепью последовательности распознавания гидовой РНК в пределах целевой ДНК составляет 100% по семи смежным нуклеотидам на 5' конце комплементарной цепи последовательности распознавания гидовой РНК в пределах комплементарной цепи целевой ДНК и только 0% по остальной части. В таком случае может считаться, что нацеливающая на ДНК последовательность составляет 7 нуклеотидов в длину. В некоторых гидовых РНК по меньшей мере 17 нуклеотидов в пределах нацеливающей на ДНК последовательности являются комплементарными целевой ДНК. Например, нацеливающая на ДНК последовательность может составлять 20 нуклеотидов в длину и может содержать 1, 2 или 3 несовпадения с комплементарной цепью последовательности распознавания гидовой РНК. Необязательно несовпадения не являются смежными с последовательностью мотива, прилегающего к протоспейсеру (РАМ) (например, несовпадения находятся на 5' конце нацеливающей на ДНК последовательности или несовпадения находятся по меньшей мере на расстоянии 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 или 19 пар оснований от последовательности РАМ).
[00147] Связывающий белок сегмент гРНК может содержать два участка нуклеотидов, которые являются комплементарными друг Другу. Комплементарные нуклеотиды связывающего белок сегмента гибридизуются с образованием двухцепочечного РНК-дуплекса (дцРНК). Связывающий белок сегмент рассматриваемой гРНК взаимодействует с белком Cas, и гРНК направляет связанный белок Cas на конкретную нуклеотидную последовательность в пределах целевой ДНК посредством нацеливающего на ДНК сегмента.
[00148] Единые гидовые РНК содержат нацеливающий на ДНК сегмент и каркасную последовательность (т.е. связывающую белок или связывающую Cas последовательность гидовой РНК). Например, такие гидовые РНК содержат 5' нацеливающий на ДНК сегмент и 3' каркасную последовательность. Иллюстративные каркасные последовательности содержат, состоят по существу или состоят из следующего: GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAA AAGUGGCACCGAGUCGGUGCU (версия 1; SEQ ID NO: 27); GUUGGAACCAUUCAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAAC UUGAAAAAGUGGCACCGAGUCGGUGC (версия 2; SEQ ID NO: 6); GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAA AAGUGGCACCGAGUCGGUGC (версия 3; SEQ ID NO: 7); и GUUUAAGAGCUAUGCUGGAAACAGCAUAGCAAGUUUAAAUAAGGCUAGUCCGUUAU CAACUUGAAAAAGUGGCACCGAGUCGGUGC (версия 4; SEQ ID NO: 8). Гидовые РНК, нацеливающиеся на любую целевую последовательность для гидовой РНК, могут включать в себя, например, нацеливающий на ДНК сегмент на 5' конце гидовой РНК, слитый с любой из иллюстративных каркасных последовательностей гидовой РНК на 3' конце гидовой РНК. Иными словами, любой из нацеливающих на ДНК сегментов, раскрытых в настоящем документе, может быть соединен с 5' концом любой из SEQ ID NO: 27, 6, 7 или 8 с образованием единой гидовой РНК (химерной гидовой РНК). Версии 1, 2, 3 и 4 гидовой РНК, как раскрыто в другом месте в настоящем документе, носят название нацеливающие на ДНК сегменты, соединенные с каркасными версиями 1, 2, 3 и 4 соответственно.
[00149] Гидовые РНК могут включать в себя модификации или последовательности, которые обеспечивают дополнительные требуемые признаки (например, модифицированная или отрегулированная стабильность; субклеточное нацеливание; отслеживание с помощью флуоресцентной метки; сайт связывания для белка или белкового комплекса; и тому подобное). Примеры таких модификаций включают в себя, например, следующее: 5' кэп (например, 7-метилгуанилатный кэп (m7G)); 3' полиаденилированный хвост (т.е. а 3' поли(А)-хвост); последовательность РНК-переключателя (например, для обеспечения регулируемой стабильности и/или регулируемой доступности белков и/или белковых комплексов); последовательность контроля стабильности; последовательность, которая образует дцРНК дуплекс (т.е. шпилька); модификация или последовательность, которая направляет РНК в субклеточное местоположение (например, ядро, митохондрии, хлоропласты и тому подобное); модификация или последовательность, которая обеспечивает отслеживание (например, прямое конъюгирование с флуоресцентной молекулой, конъюгирование с фрагментом, который облегчает детектирование флуоресценции, последовательность, которая обеспечивает возможность детектирования флуоресценции и т.д.); модификация или последовательность, которая обеспечивает сайт связывания для белков (например, белков, которые действуют на ДНК, включая в себя ДНК-метилтрансферазы, ДНК-деметилазы, гистонацетилтрансферазы, гистондеацетилазы и тому подобное); и их комбинации. Другие примеры модификаций включают в себя сконструированные дуплексные структуры «петля-на-стебле», сконструированные области выпетливания, сконструированные шпильки 3' дуплексной структуры «петля-на-стебле» или любую их комбинацию. См., например, US 2015/0376586, полностью включенную в настоящий документ посредством ссылки для всех целей. Выпетливание может представлять собой неспаренную областью нуклеотидов в дуплексе, состоящей из crPHK-подобной области и минимальной tracrPHK-подобной области. Выпетливание может содержать на одной стороне дуплекса неспаренный 5'-XXXY-3', где X представляет собой любой пурин, a Y может представлять собой нуклеотид, который может образовывать неоднозначную пару с нуклеотидом на противоположной цепи, и неспаренную нуклеотидную область на другой стороне дуплекса
[00150] Немодифицированные нуклеиновые кислоты могут быть склонны к деградации. Экзогенные нуклеиновые кислоты также могут индуцировать врожденный иммунный ответ. Модификации могут помочь ввести стабильность и уменьшить иммуногенность. Гидовые РНК могут содержать модифицированные нуклеозиды и модифицированные нуклеотиды, включая в себя, например, один или несколько из следующего: (1) изменение или замена одного или обоих несвязывающих фосфатных кислородов и/или одного или нескольких связывающих фосфатных кислородов в фосфодиэфирной связи остова; (2) изменение или замена компонента рибозного сахара, например, изменение или замена 2' гидроксила на рибозном сахаре; (3) замена фосфатного фрагмента на дефосфолинкеры; (4) модификация или замена встречающегося в природе нуклеинового основания; (5) замена или модификация рибозофосфатного остова; (6) модификация 3' конца или 5' конца олигонуклеотида (например, удаление, модификация или замена концевой фосфатной группы или конъюгирование фрагмента); и (7) модификация сахара. Другие возможные модификации гидовой РНК включают в себя модификации или замену урацилов или полиурациловых путей. См., например, международную патентную публикацию WO 2015/048577 и US 2016/0237455, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей. Подобные модификации могут быть внесены в кодирующие Cas нуклеиновые кислоты, такие как мРНК Cas.
[00151] В качестве одного примера нуклеотиды на 5' или 3' конце гидовой РНК могут включать в себя фосфоротиоатные связи (например, основания могут содержать модифицированную фосфатную группу, которая является фосфоротиоатной группой). Например, гидовая РНК может включать в себя фосфоротиоатные связи между 2, 3 или 4 концевыми нуклеотидами на 5' или 3' конце гидовой РНК. В качестве другого примера нуклеотиды на 5' и/или 3' конце гидовой РНК могут содержать 2'-O-метильные модификации. Например, гидовая РНК может включать в себя 2'-O-метильные модификации на 2, 3 или 4 концевых нуклеотидах на 5' и/или 3' конце гидовой РНК (например, на 5' конце). См., например, WO 2017/173054 А1 и Finn et al. (2018) Cell Reports 22:1-9, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. В одном конкретном примере гидовая РНК содержит 2'-O-метильные аналоги и 3'-фосфоротиоатные межнуклеотидные связи на первых трех 5' и 3' концевых остатках РНК. В другом конкретном примере гидовая РНК модифицирована таким образом, что все группы 2'ОН, которые не взаимодействуют с белком Cas9, заменены 2'-O-метильными аналогами, а хвостовая область гидовой РНК, которая характеризуется минимальным взаимодействием с Cas9, модифицирована 5' и 3' фосфоротиоатными межнуклеотидными связями. См., например, Yin et al. (2017) Nat. Biotech. 3 5 (12): 1179-1187, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00152] Гидовые РНК могут быть предусмотрены в любой форме. Например, гРНК может быть предусмотрена в форме РНК, либо в виде двух молекул (отдельная сrРНК и tracrPHK), либо в виде одной молекулы (sgPHK) и необязательно в форме комплекса с белком Cas. гРНК также может быть предусмотрена в форме ДНК, кодирующей гРНК. ДНК, кодирующая гРНК, может кодировать одну молекулу РНК (sgPHK) или отдельные молекулы РНК (например, отдельные сrРНК и tracrPHK). В последнем случае ДНК, кодирующая гРНК, может быть предусмотрена в виде одной молекулы ДНК или в виде отдельных молекул ДНК, кодирующих сrРНК и tracrPHK, соответственно.
[00153] Когда гРНК предусмотрена в форме ДНК, гРНК может транзиентно, условно или конститутивно экспрессироваться в клетке. ДНК, кодирующие гРНК, могут быть стабильно интегрированы в геном клетки и функционально связаны с активным в клетке промотором. Альтернативно, ДНК, кодирующие гРНК, могут быть функционально связаны с промотором в экспрессионного конструкта. Например, ДНК, кодирующая гРНК, может находиться в векторе, содержащем гетерологичную нуклеиновую кислоту, такую как нуклеиновая кислота, кодирующая белок Cas. Альтернативно, это может быть вектор или плазмида, которая отделена от вектора, содержащего нуклеиновую кислоту, кодирующую белок Cas. Промоторы, которые можно использовать в таких экспрессионных конструктах, включают в себя промоторы, активные, например, в одной или нескольких из следующего: эукариотическая клетка, клетка человека, не относящаяся к человеку клетка, клетка млекопитающего, клетка млекопитающего, не являющегося человеком, клетка грызуна, клетка крысы, клетка хомяка, клетка кролика, плюрипотентная клетка, эмбриональная клетка стволовая (ES) клетка, стволовая клетка взрослого человека, клетка-предшественник, ограниченная стадией развития, индуцированная плюрипотентная стволовая клетка (iPS) или эмбрион на стадии одной клетки. Такими промоторами могут являться, например, условные промоторы, индуцируемые промоторы, конститутивные промоторы или тканеспецифические промоторы. Такими промоторами также могут являться, например, двунаправленные промоторы. Конкретные примеры подходящих промоторов включают в себя промотор РНК-полимеразы III, такой как промотор U6 человека, промотор U6-полимеразы III крысы или промотор U6-полимеразы III мыши.
[00154] Альтернативно, гРНК можно получить различными другими способами. Например, гРНК можно получить путем транскрипции in vitro с использованием, например, РНК-полимеразы Т7 (см., например, международные патентные публикации WO 2014/089290 и WO 2014/065596, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей). Гидовые РНК также могут представлять собой синтетически полученную молекулу, полученную химическим синтезом.
(4) Последовательности распознавания гидовой РНК и целевые последовательности для гидовой РНК
[00155] Термин «последовательность распознавания гидовой РНК» включает в себя последовательности нуклеиновой кислоты, присутствующие в целевой ДНК, с которой будет связываться с нацеливающим на ДНК сегментом гРНК, при условии наличия достаточных условий для связывания. Используемый в настоящем документе термин «последовательность распознавания гидовой РНК» охватывает обе цепи двухцепочечной целевой ДНК (т.е. последовательность на комплементарной цепи, с которой гибридизуется гидовая РНК, и соответствующую последовательность на некомплементарной цепи, смежную с мотивом, прилегающим к протоспейсеру (РАМ)). Используемый в настоящем документе термин «целевая последовательность для гидовой РНК» относится, в частности, к последовательности на некомплементарной цепи, смежной с РАМ (т.е. выше или 5' от РАМ). То есть целевая последовательность для гидовой РНК относится к последовательности на некомплементарной цепи, соответствующей последовательности, с которой гидовая РНК гибридизуется на комплементарной цепи. Целевая последовательность для гидовой РНК эквивалентна нацеливающему на ДНК сегменту гидовой РНК, но с тиминами вместо урацилов. В качестве одного примера, целевая последовательность для гидовой РНК для фермента Cas9 может относиться к последовательности на некомплементарной цепи, смежной с 5'-NGG-3' РАМ. Последовательности распознавания гидовой РНК включают в себя последовательности, к которым гидовая РНК предназначена обеспечивать комплементарность, где гибридизация между комплементарной цепью последовательности распознавания гидовой РНК и нацеливающей на ДНК последовательностью гидовой РНК способствует образованию комплекса CRISPR. Полная комплементарность не обязательно необходима при условии, что существует достаточная комплементарность, чтобы вызвать гибридизацию и способствовать образованию комплекса CRISPR. Последовательности распознавания гидовой РНК или целевые последовательности для гидовой РНК также включают в себя сайты расщепления для белков Cas, более подробно описанные ниже. Последовательность распознавания гидовой РНК или целевые последовательности для гидовой РНК могут содержать любой полинуклеотид, который может быть расположен, например, в ядре или цитоплазме клетки или в органелле клетки, такой как митохондрия или хлоропласт.
[00156] На последовательность распознавания гидовой РНК в целевой ДНК может быть нацелен (т.е. связан, или может гибридизоваться, или являться комплементарным) белок Cas или гРНК. Подходящие условия связывания ДНК/РНК включают в себя физиологические условия, обычно присутствующие в клетке. Другие подходящие условия связывания ДНК/РНК (например, условия в бесклеточной системе) являются известными (см., например, Molecular Cloning: A Laboratory Manual, 3-е изд. (Sambrook et al., Harbor Laboratory Press 2001), полностью включенный в настоящий документ посредством ссылки для любых целей). Цепь целевой ДНК, которая комплементарна и гибридизуется с белком Cas или гРНК, можно назвать «комплементарной цепью», а цепь целевой ДНК, которая комплементарна «комплементарной цепи» (и, следовательно, не комплементарна белок Cas или гРНК) можно назвать «некомплементарной цепью» или «матричной цепью».
[00157] Белок Cas может расщеплять нуклеиновую кислоту на сайте в пределах или за пределами последовательности нуклеиновой кислоты, присутствующей в целевой ДНК, с которой будет связываться нацеливающий на ДНК сегмент гРНК. «Сайт расщепления» включает в себя положение нуклеиновой кислоты, в котором белок Cas вызывает одноцепочечный разрыв или двухцепочечный разрыв. Например, образование комплекса CRISPR (содержащего гРНК, гибридизованную с комплементарной цепью последовательности распознавания гидовой РНК и образовавшей комплекс с белком Cas), может привести к расщеплению одной или обеих цепей в или вблизи (например, в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар оснований из) последовательности нуклеиновой кислоты, присутствующей в целевой ДНК, с которой будет связываться с нацеливающим на ДНК сегментом гРНК. Если сайт расщепления находится за пределами последовательности нуклеиновой кислоты, с которой будет связываться нацеливающий на ДНК сегмент гРНК, сайт расщепления по-прежнему считается находящимся в «последовательности распознавания гидовой РНК» или целевой последовательности для гидовой РНК. Сайт расщепления может находиться только на одной цепи или на обеих цепях нуклеиновой кислоты. Сайты расщепления могут находиться в одном и том же положении на обеих цепях нуклеиновой кислоты (создавая тупые концы) или могут быть в разных сайтах на каждой цепи (создавая ступенчатые концы (т.е. свисающие концы)). Ступенчатые концы могут быть получены, например, с использованием двух белков Cas, каждый из которых вызывает одноцепочечный разрыв в другом сайте расщепления на другой цепи, тем самым создавая двухцепочечный разрыв. Например, первая никаза может создавать одноцепочечный разрыв на первой цепи двухцепочечной ДНК (дцДНК), а вторая никаза может создавать одноцепочечный разрыв на второй цепи дцДНК, так что создаются свисающие последовательности. В некоторых случаях последовательность распознавания гидовой РНК или целевая последовательность для гидовой РНК никазы на первой цепи отделена от последовательности распознавания гидовой РНК или целевой последовательности для гидовой РНК никазы на второй цепи по меньшей мере 2, 3, 4 г. 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 парами оснований.
[00158] Сайт-специфическое связывание и/или расщепление целевой ДНК белками Cas может происходить в местах, определяемых как (i) комплементарностью спаривания оснований между гРНК и целевой ДНК, так и (ii) коротким мотивом, называемым мотив, прилегающий к протоспейсеру (РАМ), в целевой ДНК. РАМ может фланкировать целевую последовательность для гидовой РНК на некомплементарной цепи, противоположной цепи, с которой гибридизуется гидовая РНК. Необязательно, целевая последовательность для гидовой РНК может быть фланкирована на 3' конце РАМ. Альтернативно, целевая последовательность для гидовой РНК может быть фланкирована на 5' конце РАМ. Например, сайт расщепления белков Cas может составлять от приблизительно 1 до приблизительно 10 или от приблизительно 2 до приблизительно 5 пар оснований (например, 3 пары оснований) выше или ниже последовательности РАМ. В некоторых случаях (например, когда используют Cas9 из S. pyogenes или близкородственный Cas9), последовательность РАМ некомплементарной цепи может представлять собой 5'-NiGG-3', где Ni является любым нуклеотидом ДНК и расположен непосредственно 3' от последовательности распознавания гидовой РНК некомплементарной цепи целевой ДНК (т.е. непосредственно 3' от целевой последовательности для гидовой РНК). Таким образом, последовательность РАМ комплементарной цепи будет представлять собой 5'-CCN2-3', где N2 представляет собой любой нуклеотид ДНК и находится непосредственно 5' от последовательности распознавания гидовой РНК комплементарной цепи целевой ДНК. В некоторых таких случаях N1 и N2 могут являться комплементарными, и пара оснований N1-N2 может быть любой парой оснований (например, N1=C и N2=G; N1=G и N2=C; N1=A и N2=T; или N1=T и N2=А). В случае Cas9 из S. aureus РАМ может представлять собой NNGRRT или NNGRR, где N может представлять собой A, G, С или Т, a R может представлять собой G или А. В случае Cas9 из С. jejuni РАМ может представлять собой, например, NNNNACAC или NNNNRYAC, где N может представлять собой A, G, С или Т, a R может представлять собой G или А. В некоторых случаях (например, для FnCpf1) последовательность РАМ может находиться выше от 5' конца и характеризоваться последовательностью 5'-TTN-3'.
[00159] Примеры целевых последовательностей для гидовой РНК или целевых последовательностей для гидовой РНК в дополнение к последовательности РАМ представлены ниже. Например, целевая последовательность для гидовой РНК может представлять собой 20-нуклеотидную последовательность ДНК, непосредственно предшествующую мотиву NGG, распознаваемому белком Cas9. Примерами таких целевых последовательностей гидовой РНК вместе с последовательностью РАМ являются GN19NGG (SEQ ID NO: 9) или N20NGG (SEQ ID NO: 10). См., например, международную патентную публикацию WO 2014/165825, полностью включенную посредством ссылки для всех целей. Гуанин на 5' конце может облегчать транскрипцию РНК-полимеразой в клетках. Другие примеры целевых последовательностей для гидовой РНК вместе с последовательностью РАМ могут включать в себя два гуаниновых нуклеотида на 5' конце (например, GGN20NGG; SEQ ID NO: 11) для облегчения эффективной транскрипции с помощью Т7-полимеразы in vitro. См., например, международную патентную публикацию WO 2014/065596, полностью включенную посредством ссылки для всех целей. Другие целевые последовательности для гидовой РНК вместе с последовательностью РАМ могут содержать от 4 до 22 нуклеотидов в длину согласно SEQ ID NO: 9-11, включая в себя 5' G или GG и 3' GG или NGG. Еще одна целевая последовательность для гидовой РНК может содержать от 14 до 20 нуклеотидов в длину согласно SEQ ID NO: 9-11.
[00160] Последовательность распознавания гидовой РНК или целевая последовательность для гидовой РНК могут представлять собой любую последовательностью нуклеиновой кислоты, эндогенную или экзогенную для клетки. Последовательность распознавания гидовой РНК или целевая последовательность для гидовой РНК могут представлять собой последовательность, кодирующую продукт гена (например, белок), или некодирующую последовательность (например, регуляторную последовательность) или могут включать в себя обе.
В. Клетки и не являющиеся человеком животные, содержащие экспрессионные кассеты Cas
[00161] Кроме того, предусмотрены клетки и не являющиеся человеком животные, содержащие экспрессионные кассеты Cas, описанные в настоящем документе. Экспрессионная кассета Cas может быть стабильно интегрирована в геном (т.е. в хромосому) клетки или не являющегося человеком животного, или она может быть расположена вне хромосомы (например, внехромосомно реплицирующейся ДНК). Необязательно, экспрессионная кассета Cas стабильно интегрируется в геном. Стабильно интегрированная экспрессионная кассета Cas может быть случайным образом интегрирована в геном животного, не являющегося человеком (т.е. трансгенного), или она может быть интегрирована в предварительно определенную область генома животного, не являющегося человеком (т.е. нокин). Необязательно, экспрессионная кассета Cas стабильно интегрируется в предварительно определенную область генома, такую как локус «безопасная гавань». Целевой геномный локус, в котором стабильно интегрирована экспрессионная кассета Cas, может быть гетерозиготным по экспрессионной кассете Cas или гомозиготным по экспрессионной кассете Cas. Диплоидный организм имеет два аллеля в каждом генетическом локусе. Каждая пара аллелей представляет генотип определенного генетического локуса. Генотипы описаны как гомозиготные, если в конкретном локусе есть присутствуют идентичных аллеля, и как гетерозиготные, если два аллеля различаются.
[00162] Клетки, предусмотренные в настоящем документе, могут представлять собой, например, эукариотические клетки, которые включают в себя, например, клетки грибов (например, дрожжи), клетки растений, клетки животных, клетки млекопитающих, клетки млекопитающих, не являющихся человеком, и клетки человека. Термин «животное» включает в себя млекопитающих, рыб и птиц. Клеткой млекопитающего может являться, например, клетка млекопитающего, не являющегося человеком, клетка человека, клетка грызуна, клетка крысы, клетка мыши или клетка хомяка. Другие млекопитающие, не являющиеся человеком, включают в себя, например, не являющихся человеком приматов, низших обезьян, человекообразных обезьян, кошек, собак, кроликов, лошадей, быков, оленей, бизонов, домашний скот (например, виды крупного рогатого скота, такие как коровы, быки и т.д.; виды овечьих, такие как овцы, козы и т.д.; и виды свинообразных, такие как свиньи и кабаны). Птицы включают в себя, например, кур, индеек, страусов, гусей, уток и так далее. Домашние животные и сельскохозяйственные животные также предусмотрены. Термин «не являющийся человеком» исключает людей.
[00163] Клетки также могут находиться в недифференцированном или дифференцированном состоянии любого типа. Например, клетка может представлять собой тотипотентную клетку, плюрипотентную клетку (например, плюрипотентную клетку человека или плюрипотентную клетку, не относящуюся к человеку, такую как эмбриональная стволовая клетка мыши (ES) или ES клетка крысы), или не плюрипотентную клетку. Тотипотентные клетки включают в себя недифференцированные клетки, которые могут давать клетки любого типа, а плюрипотентные клетки включают в себя недифференцированные клетки, которые обладают способностью развиваться в более чем один тип дифференцированных клеток. Такими плюрипотентными и/или тотипотентными клетками могут являться, например, ES клетки или ES-подобные клетки, такие как индуцированные плюрипотентные стволовые (iPS) клетки. ES клетки включают в себя происходящие из эмбриона тотипотентные или плюрипотентные клетки, которые способны вносить вклад в любую ткань развивающегося эмбриона при введении в эмбрион. ЭС клетки могут быть получены из внутренней клеточной массы бластоцисты и способны дифференцироваться в клетки любого из трех зародышевых слоев позвоночных (энтодерма, эктодерма и мезодерма).
[00164] Примеры плюрипотентных клеток человека включают в себя ES клетки человека, взрослые стволовые клетки человека, ограниченные стадией развития человеческие клетки-предшественники и индуцированные плюрипотентные стволовые (iPS) клетки человека, такие как примированные iPS-клетки человека и наивные iPS-клетки человека. Индуцированные плюрипотентные стволовые клетки включают в себя плюрипотентные стволовые клетки, которые могут быть получены непосредственно из дифференцированной взрослой клетки. Клетки iPS человека могут быть получены путем введения в клетку специфических наборов факторов перепрограммирования, которые могут включать в себя, например, Oct3/4, факторы транскрипции семейства Sox (например, Sox1, Sox2, Sox3, Sox15), факторы транскрипции семейства Мус (например, с-Мус, 1-Мус, п-Мус), факторы транскрипции Krüppel-подобного семейства (KLF) (например, KLF1, KLF2, KLF4, KLF5) и/или родственные факторы транскрипции, такие как NANOG, LIN28 и/или Glis1. iPS клетки человека также могут быть получены, например, путем использования микроРНК, небольших молекул, которые имитируют действие факторов транскрипции или спецификаторов происхождения. iPS клетки человека характеризуются своей способностью дифференцироваться в любую клетку из трех зародышевых слоев позвоночных, например, энтодерму, эктодерму или мезодерму. iPS клетки человека также характеризуются своей способностью размножаться неограниченное время в подходящих условиях культивирования in vitro. См., например, Takahashi and Yamanaka (2006) Cell 126: 663-676, полностью включенный в настоящий документ посредством ссылки для всех целей. Примированные ES клетки человека и примированные iPS клетки человека включают в себя клетки, которые проявляют характеристики, сходные с характеристиками эпибластных клеток после имплантации, и предназначены для спецификации и дифференциации клонов. Наивные ES клетки человека и наивные iPS клетки человека включают в себя клетки, которые проявляют характеристики, сходные с характеристиками ES клеток внутренней клеточной массы предимплантационного эмбриона, и не являются обязательными для характеристики линии. См., например, Nichols and Smith (2009) Cell Stem Cell 4:487-492, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00165] Клетки, предусмотренные в настоящем документе, также могут представлять собой половые клетки (например, сперматозоиды или ооциты). Клетки могут быть митотически компетентными клетками или митотически неактивными клетками, мейотически компетентными клетками или мейотически неактивными клетками. Аналогично, клетки также могут являться первичными соматическими клетками или клетками, которые не являются первичными соматическими клетками. Соматические клетки включают в себя любую клетку, которая не является гаметой, зародышевой клеткой, гаметоцитом или недифференцированной стволовой клеткой. Например, клетки могут представлять собой клетки печени, почек, кроветворные клетки, эндотелиальные клетки, эпителиальные клетки, фибробласты, мезенхимные клетки, кератиноциты, клетки крови, меланоциты, моноциты, мононуклеарные клетки, моноцитарные предшественники, В-клетки, эритроидно-мегакариоцитарные клетки, эозинофилы, макрофаги, Т-клетки, островковые бета-клетки, экзокринные клетки, панкреатические предшественники, эндокринные предшественники, адипоциты, преадипоциты, нейроны, глиальные клетки, нервные стволовые клетки, нейроны, гепатобласты, гепатоциты, кардиомиоциты, скелетные миобласты, клетки гладких мышц, протоковые клетки, ацинарные клетки, альфа-клетки, бета-клетки, дельта-клетки, РР-клетки, холангиоциты, белые или коричневые адипоциты или клетки глаза (например, клетки трабекулярной сети, пигментные эпителиальные клетки сетчатки, микрососудистые эндотелиальные клетки сетчатки, перициты сетчатки, эпителиальные клетки конъюнктивы, фибробласты конъюнктивы, эпителиальные клетки пигмента радужки, кератоциты, эпителиальные клетки хрусталика, непигментные цилиарные эпителиальные клетки, фибробласты глазной сосудистой оболочки, фоторецепторные клетки, ганглиозные клетки, биполярные клетки, горизонтальные клетки или амакринные клетки).
[00166] Подходящие клетки, предусмотренные в настоящем документе, также включают в себя первичные клетки. Первичные клетки включают в себя клетки или культуры клеток, которые были выделены непосредственно из организма, органа или ткани. Первичные клетки включают в себя клетки, которые не являются ни трансформированными, ни бессмертными. Они включают в себя любую клетку, полученную из организма, органа или ткани, которая ранее не была перенесена в культуру ткани или ранее была перенесена в культуру ткани, но не способна бесконечно переноситься в культуру ткани. Такие клетки могут быть выделены с помощью общепринятых техник и включают в себя, например, соматические клетки, кроветворные клетки, эндотелиальные клетки, эпителиальные клетки, фибробласты, мезенхимные клетки, кератиноциты, меланоциты, моноциты, мононуклеарные клетки, адипоциты, преадипоциты, нейроны, глиальные клетки, гепатоциты, скелетные миобласты и гладкомышечные клетки. Например, первичные клетки могут быть получены из соединительных тканей, мышечных тканей, тканей нервной системы или эпителиальных тканей.
[00167] Другие подходящие клетки, предусмотренные в настоящем документе, включают в себя иммортализованные клетки. К иммортализованным клеткам относятся клетки многоклеточного организма, которые в норме не размножаются бесконечно долго, но из-за мутации или изменения избежали нормального клеточного старения и вместо этого могут продолжать подвергаться делению. Такие мутации или изменения могут происходить естественным путем или быть преднамеренно вызванными. Примеры иммортализованных клеток включают в себя клетки яичника китайского хомячка (СНО), клетки эмбриональной почки человека (например, клетки НЕК 293 или клетки 293Т) и мышиные эмбриональные фибробласты (например, клетки 3Т3). Многочисленные типы иммортализованных клеток хорошо известны. Бессмертные или первичные клетки включают в себя клетки, которые, как правило, используют для культивирования или для экспрессии рекомбинантных генов или белков.
[00168] Клетки, предусмотренные в настоящем документе, также включают в себя эмбрионы на стадии одной клетки (т.е. оплодотворенные ооциты или зиготы). Такие одноклеточные эмбрионы могут происходить из любого генетического фона (например, BALB/c, C57BL/6, 129 или их комбинации для мышей), могут являться свежими или замороженными и могут быть получены в результате естественного разведения или оплодотворения in vitro.
[00169] Клетки, предусмотренные в настоящем документе, могут являться нормальными, здоровыми клетками или могут являться патологическими или содержащими мутации клетками.
[00170] Не являющиеся человеком животные, содержащие экспрессионную кассету Cas, как описано в настоящем документе, могут быть получены с помощью способов, описанных в другом месте в настоящем документе. Термин «животное» включает в себя млекопитающих, рыб и птиц. Млекопитающие включают в себя, например, людей, не являющихся человеком приматов, низших обезьян, человекообразных обезьян, кошек, собак, кроликов, лошадей, быков, оленей, бизонов, овец, кроликов, грызунов (например, мышей, крыс, хомяков и морских свинок) и домашний скот (например, виды крупного рогатого скота, такие как коровы и быки; виды овечьих, такие как овцы и козы; и виды свинообразных, такие как свиньи и кабаны). Птицы включают в себя, например, кур, индеек, страусов, гусей и уток. Домашние животные и сельскохозяйственные животные также предусмотрены. Термин «не являющийся человеком» исключает людей. Предпочтительные не являющиеся человеком животные включают в себя, например, грызунов, таких как мыши и крысы.
[00171] Не являющиеся человеком животные могут быть любого генетического происхождения. Например, подходящими мышами могут являться линия 129, линия C57BL/6, смесь 129 и C57BL/6, линия BALB/c или линия Swiss Webster. Примеры линий 129 включают в себя 129Р1, 129Р2, 129РЗ, 129X1, 129S1 (например, 129S1/SV, 129S1/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1 и 129Т2. См., например, Festing et al. (1999)Mammalian Genome 10:836, полностью включенный в настоящий документ посредством ссылки для всех целей. Примеры линий C57BL включают в себя C57BL/A, C57BL/An, C57BL/GrFa, C57BL/Kal wN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. Подходящие мыши также могут быть из смеси вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6 (например, 50% 129 и 50% C57BL/6). Аналогично, подходящие мыши могут быть из смеси вышеупомянутых линий 129 или смеси вышеупомянутых линий BL/6 (например, линия 129S6 (129/SvEvTac)).
[00172] Аналогичным образом, крысы могут быть из любой линии крыс, включая в себя, например, линию крыс ACI, линию крыс Dark Agouti (DA), линию крыс Wistar, линию крыс LEA, линию крыс Sprague Dawley (SD) или линию крыс Fischer, такой как Fisher F344 или Fisher F6. Крысы также могут быть получены из линии, полученного из смеси двух или более линий, указанных выше. Например, подходящая крыса может быть из линии DA или линии ACI. Линия крыс ACI характеризуется тем, что имеет окрас шерсти «черный агути», белый живот и лапы и гаплотип RT1av1. Такие линии доступны из различных источников, включая в себя Harlan Laboratories. Линия крыс Dark Agouti (DA) характеризуется тем, что имеет окрас шерсти «агути» и гаплотип RT1av1. Такие крысы доступны из различных источников, включая в себя Charles River и Harlan Laboratories. В некоторых случаях подходящими крысами могут являться инбредные крысы. См., например, US 2014/0235933, которая полностью включена в настоящий документ посредством ссылки для всех целей.
С. Целевые геномные локусы
[00173] Описанные в настоящем документе экспрессионные кассеты Cas могут являться геномно интегрированными в целевой геномный локус в клетке или организме не являющегося человеком животного. Можно использовать любой целевой геномный локус, способный экспрессировать ген.
[00174] Примером целевого геномного локуса, в который могут стабильно интегрироваться экспрессионные кассеты Cas, описанные в настоящем документе, является локус «безопасная гавань» в геноме не являющегося человеком животного. Взаимодействия между интегрированной экзогенной ДНК и геномом хозяина могут ограничивать надежность и безопасность интеграции и могут приводить к явным фенотипическим эффектам, которые не обусловлены целевой генетической модификацией, а вместо этого обусловлены непреднамеренными эффектами интеграции на окружающие эндогенные гены. Например, случайно вставленные трансгены могут подвергаться эффектам положения и сайленсингу, что делает их экспрессию ненадежной и непредсказуемой. Аналогично, интеграция экзогенной ДНК в хромосомный локус может влиять на окружающие эндогенные гены и хроматин, тем самым изменяя поведение и фенотипы клеток. Локусы «безопасная гавань» включают в себя хромосомные локусы, где трансгены или другие экзогенные вставки нуклеиновой кислоты могут стабильно и надежно экспрессироваться во всех представляющих интерес тканях без явного изменения поведения или фенотипа клетки (т.е. без каких-либо вредных воздействий на клетку-хозяина). См., например, Sadelain et al. (2012) Nat. Rev. Cancer 12:51-58, полностью включенный в настоящий документ посредством ссылки для всех целей. Необязательно, локусом «безопасная гавань» является тот, в котором экспрессия вставленной последовательности гена не нарушается какой-либо сквозной экспрессией от соседних генов. Например, локусы «безопасная гавань» могут включать в себя хромосомные локусы, где экзогенная ДНК может интегрироваться и функционировать предсказуемым образом, не оказывая неблагоприятного влияния на структуру или экспрессию эндогенных генов. Локусы «безопасная гавань» могут включать в себя экстрагенные области или интрагенные области, такие как, например, локусы в генах, которые не являются необходимыми, необязательными или могут быть нарушены без явных фенотипических последствий.
[00175] Например, локус Rosa26 и его эквивалент у человека обеспечивают открытую конфигурацию хроматина во всех тканях и повсеместно экспрессируется во время эмбрионального развития и у взрослых. См., например, Zambrowicz et al. (1997) Proc. Natl. Acad. Sci. USA 94:3789-3794, полностью включенный в настоящий документ посредством ссылки для всех целей. Кроме того, локус Rosa26 может подвергаться нацеленному воздействию с высокой эффективностью, и разрушение гена Rosa26 не дает явного фенотипа. Другие примеры локусов «безопасная гавань» включают в себя CCR5, HPRT, AAVS1 и альбумин. См., например, патенты США №№7888121; 7972854; 7914796; 7951925; 8110379; 8409861; 8586526; и патентные публикации США №№2003/0232410; 2005/0208489; 2005/0026157; 2006/0063231; 2008/0159996; 2010/00218264; 2012/0017290; 2011/0265198; 2013/0137104; 2013/0122591; 2013/0177983; 2013/0177960 и 2013/0122591, причем каждый документ полностью включен в настоящий документ посредством ссылки для всех целей. Двуаллельное нацеливание на локусы «безопасная гавань», такие как локус Rosa26, не имеет отрицательных последствий, поэтому разные гены или репортеры могут быть нацелены на два аллеля Rosa26. В одном примере экспрессионная кассета Cas интегрирована в интрон локуса Rosa26, такой как первый интрон локуса Rosa26.
D. Не являющиеся человеком животные, у которых вырезание участка ДНК осуществляется рекомбиназой
[00176] Клетки или не являющиеся человеком животные, содержащие экспрессионную кассету Cas, содержащую последовательность, кодирующую Cas, ниже от сигнала полиаденилирования или терминатора транскрипции, фланкированную сайтами распознавания рекомбиназы, распознаваемыми сайт-специфической рекомбиназой, как раскрыто в настоящем документе, могут дополнительно содержать экспрессионную кассету рекомбиназы, которая управляет экспрессией сайт-специфической рекомбиназы. Сайт-специфические рекомбиназы включают в себя ферменты, которые могут способствовать рекомбинации между сайтами распознавания рекомбиназы, где два сайта рекомбинации физически разделены в одной нуклеиновой кислоте или в отдельных нуклеиновых кислотах. Примеры рекомбиназ включают в себя рекомбиназы Cre, Flp и Dre. Одним примером гена Cre-рекомбиназы является Crei, в котором два экзона, кодирующие Cre-рекомбиназу, разделены интроном, чтобы предотвратить его экспрессию в прокариотической клетке. Такие рекомбиназы могут дополнительно содержать сигнал ядерной локализации для облегчения локализации в ядре (например, NLS-Crei). Сайты распознавания рекомбиназы включают в себя нуклеотидные последовательности, которые распознаются сайт-специфической рекомбиназой и могут служить в качестве субстрата для события рекомбинации. Примеры сайтов распознавания рекомбиназы включают в себя FRT, FRT11, FRT71, attp, att, rox и сайты lox, такие как loxP, lox511, lox2272, lox66, lox71, loxM2 и lox5171.
[00177] Экспрессионная кассета Cas и экспрессионная кассета рекомбиназы могут быть интегрированы в разные целевые геномные локусы, или они могут быть геномно интегрированы в один и тот же целевой локус (например, локус Rosa26, например, интегрированы в первый интрон локуса Rosa26). Например, клетка или не являющееся человеком животное могут являться гетерозиготными по каждой из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы, причем один аллель целевого геномного локуса содержит экспрессионную кассету Cas, а второй аллель целевого геномного локуса содержит экспрессионную кассету рекомбиназы.
[00178] Ген рекомбиназы в экспрессионной кассете рекомбиназы может быть функционально связан с любым подходящим промотором. Примеры промоторов раскрыты в другом месте в настоящем документе. Например, промотор может являться тканеспецифическим промотором (например, промотор альбумина для специфической для печени экспрессии или промотор инсулина 2 для специфической для поджелудочной железы экспрессии) или специфическим для стадии развития промотором. Преимущество, обеспечиваемое такими промоторами, состоит в том, что экспрессия Cas может активироваться избирательно в требуемой ткани или только на требуемой стадии развития, тем самым уменьшая возможность опосредованной Cas токсичности in vivo. Неограничивающий список иллюстративных промоторов для линий мышей, у которых вырезание участка ДНК осуществляется рекомбиназой, представлен в таблице 2.
[00179]
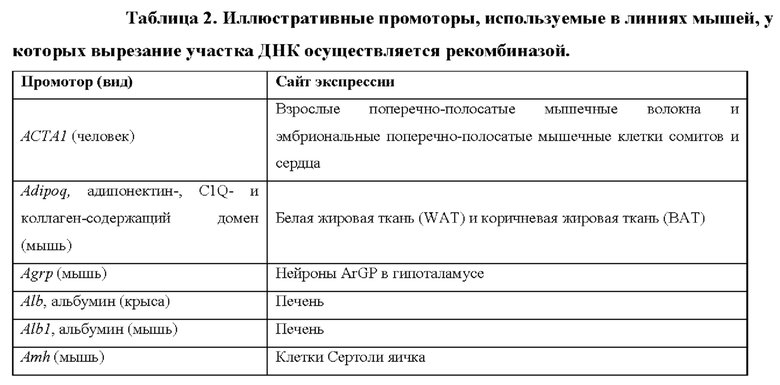
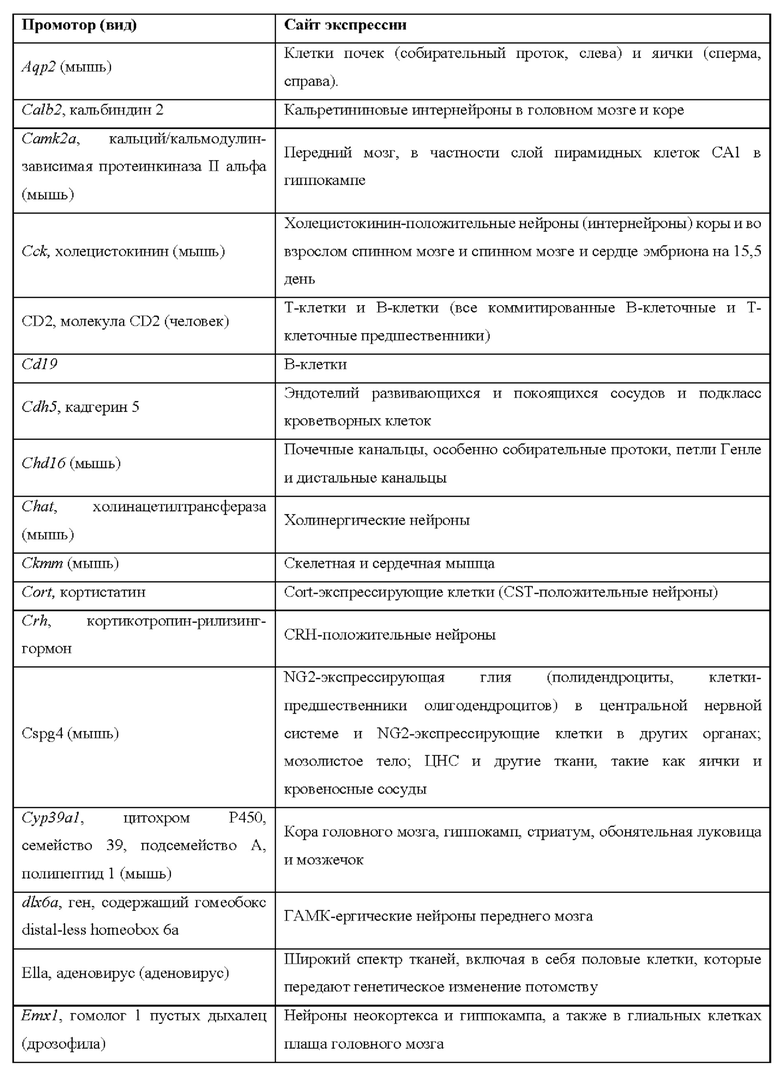
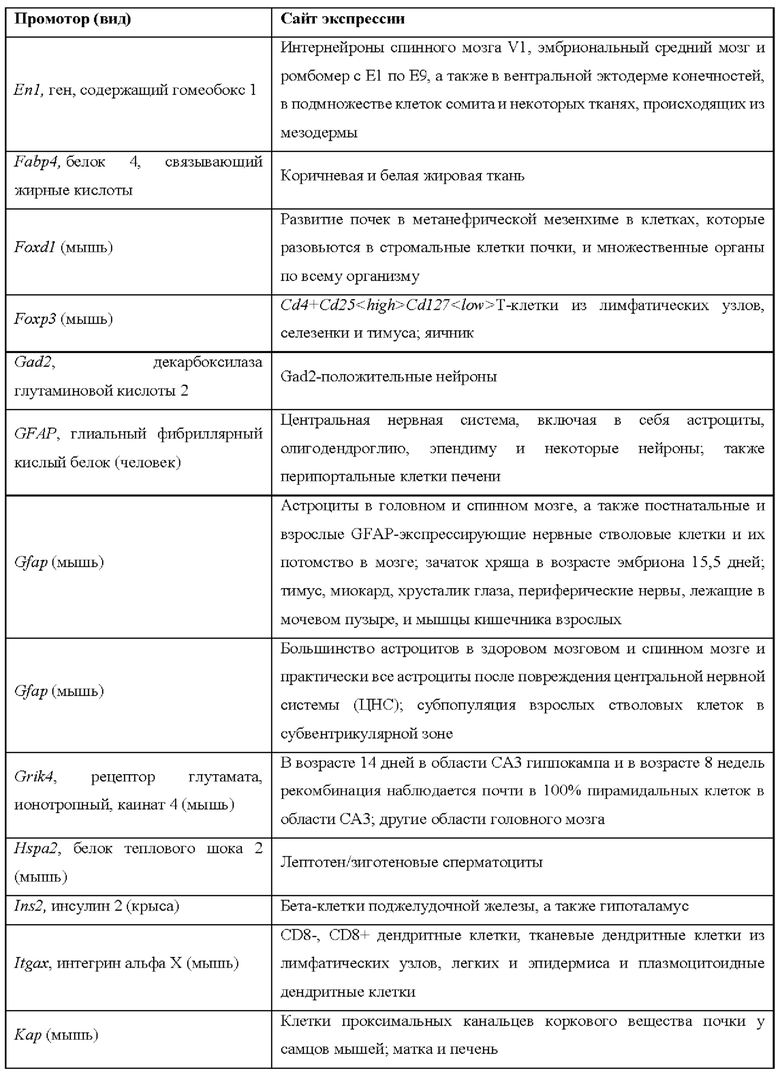
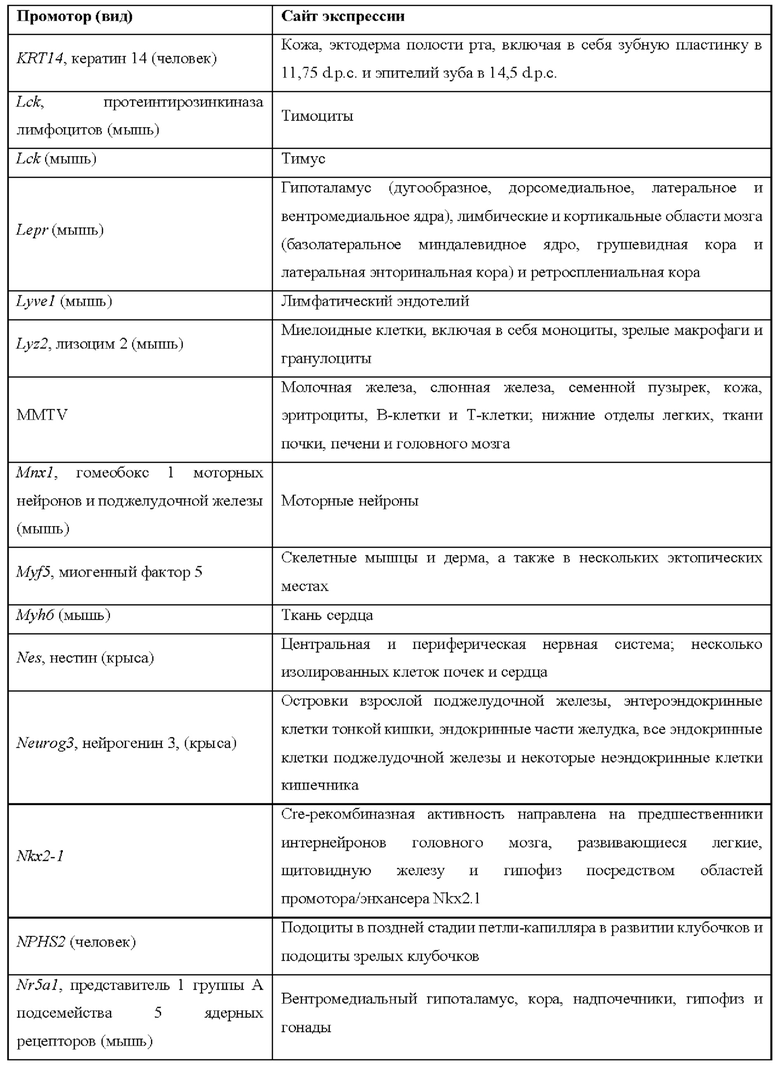
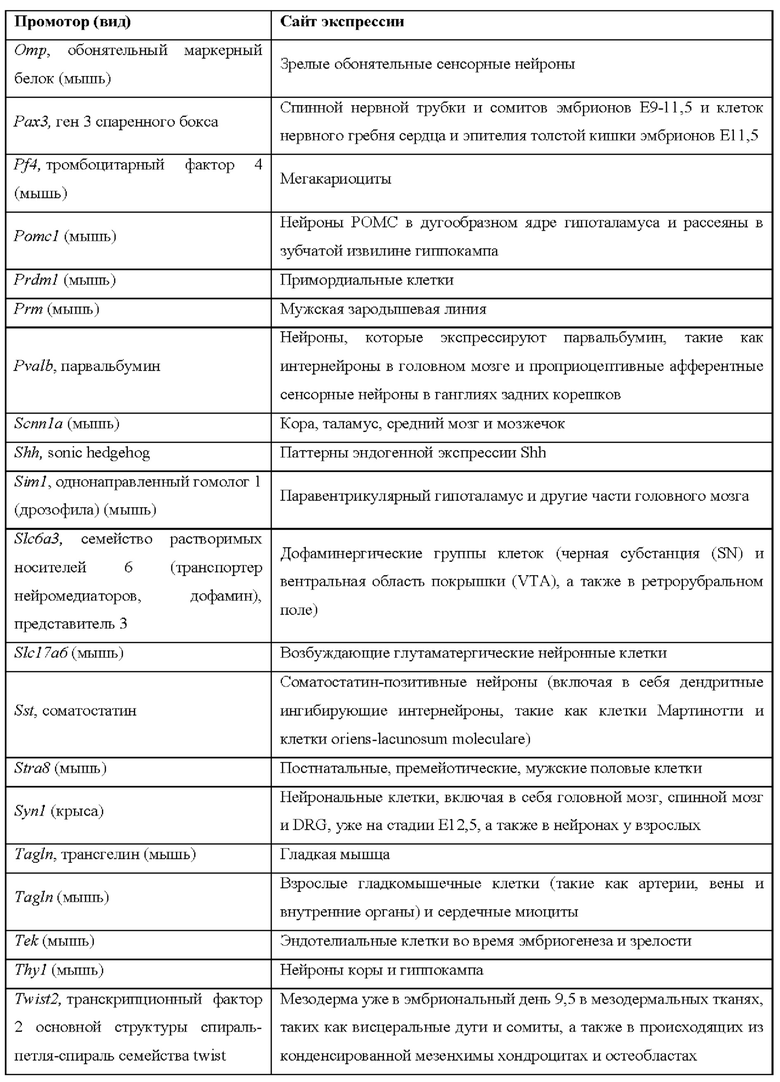
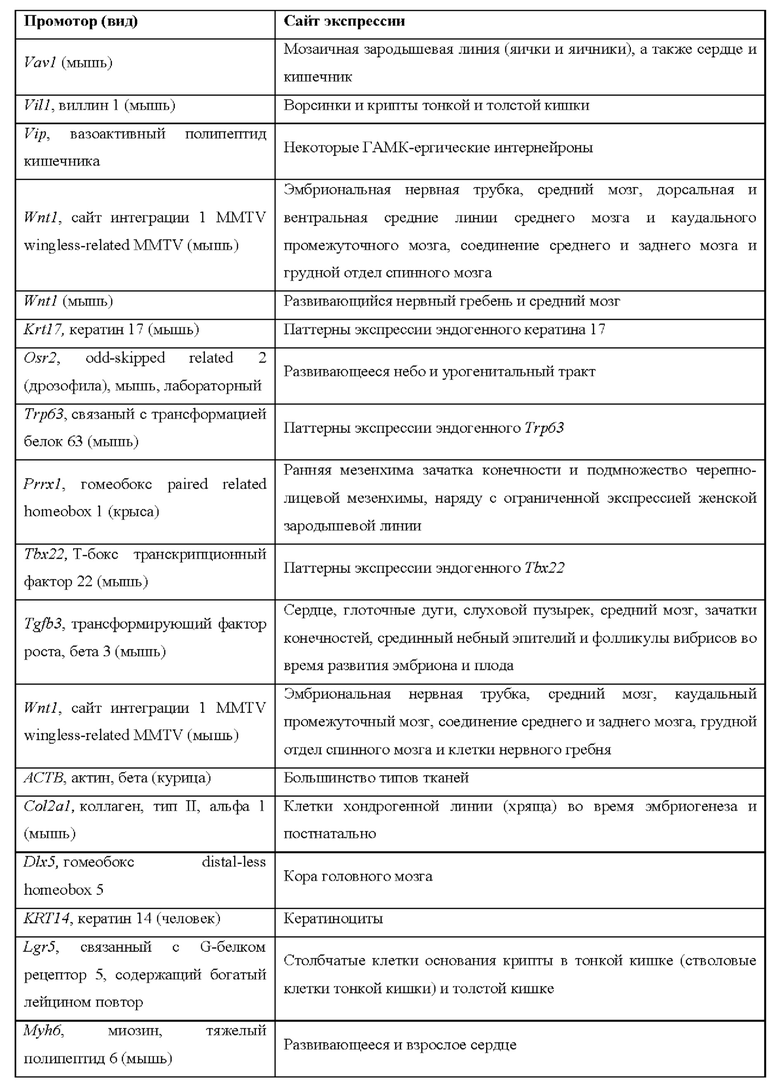
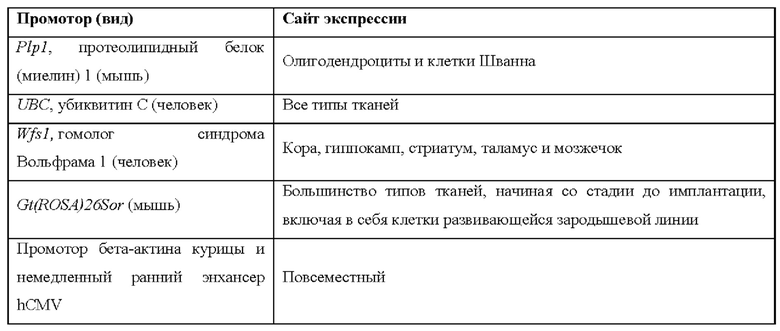
III. Способы оценки активности CRISPR/Cas in vivo
[00180] Предусмотрены различные способы оценки доставки CRISPR/Cas в ткани и органы живого животного и оценки активности CRISPR/Cas в тканях и органах живого животного. Согласно таким способам используют не являющихся человеком животных, содержащих экспрессионную кассету Cas, как описано в другом месте в настоящем документе.
А. Способы испытания способности CRISPR/Cas модифицировать целевой геномный локус in vivo или ex vivo
[00181] Предусмотрены различные способы оценки способности никазы или CRISPR/Cas-нуклеазы модифицировать целевой геномный локус in vivo с использованием не являющихся человеком животных, содержащих экспрессионную кассету Cas, как описано в настоящем документе. Такие способы могут предусматривать следующее: (а) введение не являющемуся человеком животному гидовой РНК, предназначенной для нацеливания на целевую последовательность для гидовой РНК в целевом геномном локусе; и (b) оценка модификации целевого локуса. Модификация целевого геномного локуса будет индуцирована, когда гидовая РНК образует комплекс с белком Cas и направит белок Cas к целевому геномному локусу, и комплекс Cas/гидовая РНК расщепит целевую последовательность для гидовой РНК, запуская репарацию клеткой (например, посредством негомологичного соединения концов (NHEJ), если отсутствует донорная последовательность).
[00182] Необязательно можно вводить две или больше гидовых РНК, причем каждая предназначена для нацеливания на разную целевую последовательность для гидовой РНК в пределах целевого геномного локуса. Например, две гидовые РНК могут быть предназначены для вырезания геномной последовательности между двумя целевыми последовательностями гидовой РНК. Модификация целевого геномного локуса будет индуцирована, когда первая гидовая РНК образует комплекс с белком Cas и направляет белок Cas к целевому геномному локусу, вторая гидовая РНК образует комплекс с белком Cas и направляет белок Cas к целевому геномному локусу, первый комплекс Cas/гидовая РНК расщепляет первую целевую последовательность для гидовой РНК, и второй комплекс Cas/гидовая РНК расщепляет вторую целевую последовательность для гидовой РНК, что приводит к вырезанию промежуточной последовательности. Альтернативно, можно вводить две или больше гидовых РНК, причем каждая предназначенной для нацеливания на разную целевую последовательность для гидовой РНК на разном целевом геномном локусе (т.е. мультиплексирование).
[00183] Необязательно экзогенная донорная нуклеиновая кислота, способная к рекомбинации и модификации целевого геномного локуса, также введена не являющемуся человеком животному. Необязательно белок Cas можно присоединить к экзогенной донорной нуклеиновой кислоте, как описано в другом месте в настоящем документе. Необязательно вводят две или больше экзогенные донорные нуклеиновые кислоты, каждая из которых способна к рекомбинации и модификации различного целевого геномного локуса. Модификация целевого геномного локуса будет индуцирована, например, когда гидовая РНК образует комплекс с белком Cas и направляет белок Cas к целевому геномному локусу, комплекс Cas/гидовая РНК расщепляет целевую последовательность для гидовой РНК, и целевой геномный локус рекомбинируется с экзогенной донорной нуклеиновой кислотой, чтобы модифицировать целевой геномный локус. Экзогенная донорная нуклеиновая кислота может рекомбинироваться с целевым геномным локусом, например, посредством гомологичной репарации (HDR) или посредством NHEJ-опосредованной вставки. Можно использовать любой тип экзогенной донорной нуклеиновой кислоты, примеры которых предусмотрены в другом месте в настоящем документе.
[00184] Аналогично, различные способы, предусмотренные выше для оценки активности CRISPR/Cas in vivo, также можно использовать для оценки активности CRISPR/Cas ex vivo с использованием клеток, содержащих экспрессионную кассету Cas, как описано в другом месте в настоящем документе.
[00185] Гидовые РНК и необязательно экзогенные донорные нуклеиновые кислоты можно вводить в клетку или не являющемуся человеком животному посредством любого способа доставки (например, AAV, LNP, или HDD) и любого пути введения, как раскрыто в другом месте в настоящем документе. Согласно конкретным способам гидовая РНК (или гидовые РНК) доставляют посредством AAV-опосредованной доставки. Например, AAV8 можно использовать, если печень является мишенью.
[00186] Способы оценки модификации целевого геномного локуса предусмотрены в другом месте в настоящем документе и являются хорошо известными. Оценку модификации целевого геномного локуса можно проводить в любом типе клеток, любом типе тканей или любом типе органов, как раскрыто в другом месте в настоящем документе. Согласно некоторым способам модификацию целевого геномного локуса оценивают в клетках печени (например, оценка содержаний в сыворотке секретированного белка, экспрессируемого целевым геномным локусом в клетках печени). Например, целевой геномный локус содержит целевой ген, и оценка может предусматривать измерение экспрессии целевого гена или активности белка, кодируемого целевым геном. Альтернативно или дополнительно, оценка может содержать секвенирование целевого геномного локуса в одной или нескольких клетках, выделенных из не являющегося человеком животного. Оценка может содержать выделение целевого органа или ткани из не являющегося человеком животного, и оценку модификации целевого геномного локуса в целевом органе или ткани. Оценка также может содержать оценку модификации целевого геномного локуса в двух или более различных типах клеток в пределах целевого органа или ткани. Аналогично, оценка может содержать выделение нецелевого органа или ткани (например, двух или больше нецелевых органов или тканей) из не являющегося человеком животного и оценку модификации целевого геномного локуса в нецелевом органе или ткани.
(1) Экзогенные донорные нуклеиновые кислоты
[00187] Согласно раскрытым в настоящем документе способам и композициям используют экзогенные донорные нуклеиновые кислоты, чтобы модифицировать целевой геномный локус после расщепления целевого геномного локуса с помощью белка Cas. В таких способах белок Cas расщепляет целевой геномный локус для создания одноцепочечного разрыва (ник) или двухцепочечного разрыва, и экзогенная донорная нуклеиновая кислота рекомбинируется с целевым геномным локусом посредством опосредованного негомологичным соединением концов (NHEJ) лигирования или посредством явления гомологичной репарации. Необязательно репарация с помощью экзогенной донорной нуклеиновой кислоты удаляет или разрушает целевую последовательность для гидовой РНК или сайт расщепления Cas, так что аллели, на которые осуществлялось нацеливание, не могут вновь подвергнуться нацеливанию с помощью белка Cas.
[00188] Экзогенные донорные нуклеиновые кислоты могут содержать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), они могут являться одноцепочечными или двухцепочечными, и они могут быть в линейной или кольцевой форме. Например, экзогенная донорная нуклеиновая кислота может представлять собой одноцепочечный олигодезоксинуклеотид (ssODN). См., например, Yoshimi et al. (2016) Nat. Commun. 7:10431, полностью включенный в настоящий документ посредством ссылки для всех целей. Иллюстративная экзогенная донорная нуклеиновая кислота составляет от приблизительно 50 нуклеотидов до приблизительно 5 т.п.н. в длину, от приблизительно 50 нуклеотидов до приблизительно 3 т.п.н. в длину или от приблизительно 50 до приблизительно 1000 нуклеотидов в длину. Другие иллюстративные экзогенные донорные нуклеиновые кислоты составляют от приблизительно 40 до приблизительно 200 нуклеотидов в длину. Например, экзогенная донорная нуклеиновая кислота может составлять приблизительно 50-60, 60-70, 70-80,80-90, 90-100, 100-110, 110-120, 120-130, 130-140, 140-150, 150-160, 160-170, 170-180, 180-190 или 190-200 нуклеотидов в длину. Альтернативно, экзогенная донорная нуклеиновая кислота может составлять приблизительно 50-100, 100-200, 200-300, 300-400, 400-500, 500-600, 600-700, 700-800, 800-900 или 900-1000 нуклеотидов в длину. Альтернативно, экзогенная донорная нуклеиновая кислота может составлять приблизительно 1-1,5, 1,5-2, 2-2,5, 2,5-3, 3-3,5, 3,5-4, 4-4,5 или 4,5-5 т.п.н. в длину. Альтернативно, экзогенная донорная нуклеиновая кислота может составлять, например, не более чем 5 т.п.н., 4,5 т.п.н., 4 т.п.н., 3,5 т.п.н., 3 т.п.н., 2,5 т.п.н., 2 т.п.н., 1,5 т.п.н., 1 т.п.н., 900 нуклеотидов, 800 нуклеотидов, 700 нуклеотидов, 600 нуклеотидов, 500 нуклеотидов, 400 нуклеотидов, 300 нуклеотидов, 200 нуклеотидов, 100 нуклеотидов или 50 нуклеотидов в длину. Экзогенные донорные нуклеиновые кислоты (например, нацеливающие векторы) также могут быть длиннее.
[00189] Согласно одному примеру экзогенная донорная нуклеиновая кислота представляет собой ssODN, который составляет от приблизительно 80 нуклеотидов до приблизительно 200 нуклеотидов в длину. Согласно другому примеру экзогенные донорные нуклеиновые кислоты представляют собой ssODN, который составляет от приблизительно 80 нуклеотидов до приблизительно 3 т.п.н. в длину. Такой ssODN может содержать гомологичные плечи, например, каждое из которых составляет от приблизительно 40 нуклеотидов до приблизительно 60 нуклеотидов в длину. Такой ssODN также может содержать гомологичные плечи, например, каждое из которых составляет от приблизительно 30 нуклеотидов до 100 нуклеотидов в длину. Гомологичные плечи могут являться симметричными (например, каждое составляет 40 нуклеотидов или каждое составляет 60 нуклеотидов в длину), или они могут являться асимметричными (например, одно гомологичное плечо, которое составляет 36 нуклеотидов в длину, и одно гомологичное плечо, которое составляет 91 нуклеотид в длину).
[00190] Экзогенные донорные нуклеиновые кислоты могут включать в себя модификации или последовательности, которые обеспечивают дополнительные требуемые признаки (например, измененную или регулируемую стабильность; отслеживание или обнаружение с помощью флуоресцентной метки; сайт связывания для белка или белкового комплекса и т.д.). Экзогенные донорные нуклеиновые кислоты могут содержать одну или несколько флуоресцентных меток, меток очистки, меток эпитопов или их комбинацию. Например, экзогенная донорная нуклеиновая кислота может содержать одну или несколько флуоресцентных меток (например, флуоресцентных белков или других флуорофоров или красителей), таких как по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4 или по меньшей мере 5 флуоресцентных меток. Иллюстративные флуоресцентные метки включают в себя флуорофоры, такие как флуоресцеин (например, 6-карбоксифлуоресцеин (6-FAM)), Texas Red, HEX, Су3, Су5, Су5.5, Pacific Blue, 5-(и-6)-карбокситетраметилродамин (TAMRA) и Су7. Широкий ассортимент флуоресцентных красителей коммерчески доступен для мечения олигонуклеотидов (например, от Integrated DNA Technologies). Такие флуоресцентные метки (например, внутренние флуоресцентные метки) можно использовать, например, для обнаружения экзогенной донорной нуклеиновой кислоты, которая непосредственно интегрирована в расщепленную целевую нуклеиновую кислоту, содержащую выступающие концы, совместимые с концами экзогенной донорной нуклеиновой кислоты. Метка или тег могут находиться на 5' конце, 5' конце или внутри экзогенной донорной нуклеиновой кислоты. Например, экзогенная донорная нуклеиновая кислота может быть конъюгирована на 5' конце с флуорофором IR700 от Integrated DNA Technologies (5'IRDYE®700).
[00191] Экзогенные донорные нуклеиновые кислоты также могут содержать нуклеиновокислотные вставки, включая в себя сегменты ДНК, подлежащие интеграции в целевых геномных локусах. Интеграция нуклеиновокислотной вставки в целевом геномном локусе может привести к добавлению представляющей интерес последовательности нуклеиновой кислоты в целевом геномном локусе, делеции представляющей интерес последовательности нуклеиновой кислоты в целевом геномном локусе или замене представляющей интерес последовательности нуклеиновой кислоты в целевом геномном локусе (т.е. делеция и вставка). Некоторые экзогенные донорные нуклеиновые кислоты предназначены для вставки нуклеиновокислотной вставки в целевом геномном локусе без какой-либо соответствующей делеции в целевом геномном локусе. Другие экзогенные донорные нуклеиновые кислоты предназначенной для делеции представляющей интерес последовательности нуклеиновой кислоты в целевом геномном локусе без какой-либо соответствующей вставки нуклеиновокислотной вставки. Другие же экзогенные донорные нуклеиновые кислоты предназначены для делеции представляющей интерес последовательности нуклеиновой кислоты в целевом геномном локусе и ее замены нуклеиновокислотной вставкой.
[00192] Нуклеиновокислотная вставка или соответствующая нуклеиновая кислота в целевом геномном локусе, подлежащая делеции и/или замене, может характеризоваться различной длиной. Иллюстративная нуклеиновокислотная вставка или соответствующая нуклеиновая кислота в целевом геномном локусе, подлежащая делеции и/или замене, составляет от приблизительно 1 нуклеотида до приблизительно 5 т.п.н. в длину или от приблизительно 1 нуклеотида до приблизительно 1000 нуклеотидов в длину. Например, нуклеиновокислотная вставка или соответствующая нуклеиновая кислота в целевом геномном локусе, подлежащая делеции и/или замене, может составлять приблизительно 1-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140, 140-150, 150-160, 160-170, 170-180, 180-190 или 190-120 нуклеотидов в длину. Аналогично, нуклеиновокислотная вставка или соответствующая нуклеиновая кислота в целевом геномном локусе, подлежащая делеции и/или замене, может составлять 1-100, 100-200, 200-300, 300-400, 400-500, 500-600, 600-700, 700-800, 800-900 или 900-1000 нуклеотидов в длину. Аналогично, нуклеиновокислотная вставка или соответствующая нуклеиновая кислота в целевом геномном локусе, подлежащая делеции и/или замене, может составлять приблизительно 1-1,5, 1,5-2, 2-2,5, 2,5-3, 3-3,5, 3,5-4, 4-4,5 или 4,5-5 т.п.н. в длину или больше.
[00193] Нуклеиновокислотная вставка может содержать последовательность, которая является гомологической или ортологической по отношению ко всей или части последовательности, нацеленной для замены. Например, нуклеиновокислотная вставка может содержать последовательность, которая содержит одну или несколько точечных мутаций (например, 1, 2, 3, 4, 5 или больше) по сравнению с последовательностью, нацеленной для замены в целевом геномном локусе. Необязательно такие точечные мутации могут приводить к консервативной аминокислотной замене (например, замена аспарагиновой кислоты [Asp, D] глутаминовой кислотой [Glu, Е]) в кодируемом полипептиде.
(2) Донорные нуклеиновые кислоты для вставки, опосредованной негомологичным соединением концов
[00194] Некоторые экзогенные донорные нуклеиновые кислоты содержат короткие одноцепочечные области на 5' конце и/или 3' конце, которые являются комплементарными одному или нескольким свисающим концам, созданным с помощью опосредованного белком Cas расщепления в целевом геномном локусе. Эти свисающие концы также могут называться 5' и 3' гомологичные плечи. Например, некоторые экзогенные донорные нуклеиновые кислоты содержат короткие одноцепочечные области на 5' конце и/или 3' конце, которые являются комплементарными одному или нескольким свисающим концам, созданным с помощью опосредованного белком Cas расщепления на 5' и/или 3' целевых последовательностях в целевом геномном локусе. Некоторые такие экзогенные донорные нуклеиновые кислоты содержат комплементарную область только на 5' конце или только на 3' конце. Например, некоторые такие экзогенные донорные нуклеиновые кислоты содержат комплементарную область только на 5' конце, комплементарную свисающему концу, созданному на 5' целевой последовательности в целевом геномном локусе или только на 3' конце, комплементарную свисающему концу, созданному на 3' целевой последовательности в целевом геномном локусе. Другие такие экзогенные донорные нуклеиновые кислоты содержат комплементарные области как на 5', так и на 3' концах. Например, другие такие экзогенные донорные нуклеиновые кислоты содержат комплементарные области как на 5', так и на 3' концах, например, комплементарные первому и второму свисающим концам соответственно, созданным с помощью опосредованного белком Cas расщепления в целевом геномном локусе. Например, если экзогенная донорная нуклеиновая кислота является двухцепочечной, одноцепочечные комплементарные области могут продолжаться от 5' конца верхней цепи донорной нуклеиновой кислоты и 5' конца нижней цепи донорной нуклеиновой кислоты, создавая 5' свисающие концы на каждом конце. Альтернативно, одноцепочечная комплементарная область может продолжаться от 3' конца верхней цепи донорной нуклеиновой кислоты и от 3' конца нижней цепи матрицы, создавая 3' свисающие концы.
[00195] Комплементарные области могут характеризоваться любой длиной, достаточной для стимуляции лигирования между экзогенной донорной нуклеиновой кислотой и целевой нуклеиновой кислотой. Иллюстративные комплементарные области составляют от приблизительно 1 до приблизительно 5 нуклеотидов в длину, между приблизительно 1 до приблизительно 25 нуклеотидов в длину или от приблизительно 5 до приблизительно 150 нуклеотидов в длину. Например, комплементарная область может составлять по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину. Альтернативно, комплементарная область может составлять приблизительно 5-10, 10-20, 20-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-110, 110-120, 120-130, 130-140 или 140-150 нуклеотидов в длину или больше.
[00196] Такие комплементарные области могут являться комплементарными свисающим концам, созданным с помощью двух пар никаз. Два двухцепочечных разрыва с расположенными в шахматном порядке концами можно создать с использованием первой и второй никаз, которые расщепляют противоположные цепи ДНК для создания первого двухцепочечного разрыва, и третей и четвертной никаз, которые расщепляют противоположные цепи ДНК для создания второго двухцепочечного разрыва. Например, белок Cas можно использовать для создания одноцепочечного разрыва в первой, второй, третьей и четвертой целевых последовательностях гидовой РНК, соответствующих первой, второй, третьей и четвертой гидовым РНК. Первая и вторая целевые последовательности для гидовой РНК можно расположить для создания первого сайта расщепления, так что одноцепочечные разрывы, созданные с помощью первой и второй никаз на первой и второй цепях ДНК, создают двухцепочечный разрыв (т.е. первый сайт расщепления содержит одноцепочечные разрывы в пределах первой и второй целевых последовательностей гидовой РНК). Аналогично, третья и четвертая целевые последовательности для гидовой РНК можно расположить для создания второго сайта расщепления, так что одноцепочечные разрывы, созданные с помощью третьей и четвертой никаз на первой и второй цепях ДНК, создают двухцепочечный разрыв (т.е. второй сайт расщепления содержит одноцепочечные разрывы в пределах третьей и четвертой целевых последовательностей гидовой РНК). Необязательно, одноцепочечные разрывы в пределах первой и второй целевых последовательностей гидовой РНК и/или третьей и четвертой целевых последовательностей гидовой РНК могут представлять собой смещенные одноцепочечные разрывы, которые создают свисающие концы. Окно смещения может составлять, например, по меньшей мере приблизительно 5 п.н., 10 п.н., 20 п.н., 30 п.н., 40 п.н., 50 п.н., 60 п.н., 70 п.н., 80 п.н., 90 п.н., 100 п.н. или больше. См. Ran et al. (2013) Cell 154:1380-1389; Mali et al. (2013) Nat. Biotech.31:833-838; и Shen et al. (2014) Nat. Methods 11:399-404, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. В таких случаях двухцепочечная экзогенная донорная нуклеиновая кислота может быть разработана с одноцепочечными комплементарными областями, которые являются комплементарными свисающим концам, созданным с помощью одноцепочечных разрывов в пределах первой и второй целевых последовательностей гидовой РНК и с помощью одноцепочечных разрывов в пределах третьей и четвертой целевых последовательностей гидовой РНК. Такая экзогенная донорная нуклеиновая кислота может быть вставлена с помощью лигирования, опосредованного негомологичным соединением концов.
(3) Донорные нуклеиновые кислоты для вставки с помощью гомологичной репарации [00197] Некоторые экзогенные донорные нуклеиновые кислоты содержат гомологичные плечи. Если экзогенная донорная нуклеиновая кислота также содержит нуклеиновокислотную вставку, гомологичные плечи могут фланкировать нуклеиновокислотную вставку. Для удобства пользования гомологичные плечи в настоящем документе называются 5' и 3' (т.е. вышележащие и нижележащие) гомологичные плечи. Эта терминология относится к относительному положению гомологичных плеч относительно нуклеиновокислотной вставки в пределах экзогенной донорной нуклеиновой кислоты. 5' и 3' гомологичные плечи соответствуют областям в пределах целевого геномного локуса, которые в настоящем документе называются "5' целевая последовательность" и "3' целевая последовательность" соответственно.
[00198] Гомологичное плечо и целевая последовательность "соответствуют" или являются "соответствующими" друг другу, когда две области характеризуются достаточным уровнем идентичности последовательности друг с другом, чтобы действовать в качестве субстратов для реакции гомологичной рекомбинации. Термин "гомология" включает в себя последовательности ДНК, которые либо являются идентичными, либо характеризуются идентичностью последовательности относительно соответствующей последовательности. Идентичность последовательности между данной целевой последовательностью и соответствующим гомологичным плечом, встречающимся в экзогенной донорной нуклеиновой кислоте, может представлять собой любую степень идентичности последовательности, которая позволяет происходить гомологичной рекомбинации. Например, количество идентичности последовательности, которой характеризуются гомологичное плечо экзогенной донорной нуклеиновой кислоты (или его фрагмент) и целевая последовательность (или ее фрагмент), может составлять по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 91%, 98%, 99% или 100% идентичности последовательности, так что последовательности подвергаются гомологичной рекомбинации. Более того, соответствующая область гомологии между гомологичным плечом и соответствующей целевой последовательностью может характеризоваться любой длиной, которая является достаточной для обеспечения гомологичной рекомбинации. Иллюстративные гомологичные плечи составляют от приблизительно 25 нуклеотидов до приблизительно 2,5 т.п.н. в длину, от приблизительно 25 нуклеотидов до приблизительно 1,5 т.п.н. в длину или от приблизительно 25 до приблизительно 500 нуклеотидов в длину. Например, данное гомологичное плечо (или каждое из гомологичных плеч) и/или соответствующая целевая последовательность могут содержать соответствующие области гомологии, которые составляют приблизительно 25-30, 30-40, 40-50, 50-60, 60-70, 70-80, 80-90, 90-100, 100-150, 150-200, 200-250, 250-300, 300-350, 350-400, 400-450 или 450-500 нуклеотидов в длину, так что гомологичные плечи характеризуются достаточной гомологией, чтобы подвергаться гомологичной рекомбинации с соответствующими целевыми последовательностями в пределах целевой нуклеиновой кислоты. Альтернативно, данное гомологичное плечо (или каждое гомологичное плечо) и/или соответствующая целевая последовательность могут содержать соответствующие области гомологии, которые составляют от приблизительно 0,5 т.п.н. до приблизительно 1 т.п.н., от приблизительно 1 т.п.н. до приблизительно 1,5 т.п.н., от приблизительно 1,5 т.п.н. до приблизительно 2 т.п.н. или от приблизительно 2 т.п.н. до приблизительно 2,5 т.п.н. в длину. Например, каждое из гомологичных плеч может составлять приблизительно 750 нуклеотидов в длину. Гомологичные плечи могут являться симметричными (каждое приблизительно одинакового размера в длину) или они могут являться асимметричными (одно длиннее другого).
[00199] Когда систему CRISPR/Cas используют в комбинации с экзогенной донорной нуклеиновой кислотой, 5' и 3' целевые последовательности необязательно расположены в достаточной близости от сайта расщепления Cas (например, в достаточной близости от целевой последовательности для гидовой РНК), так чтобы способствовать возникновению события гомологичной рекомбинации между целевыми последовательностями и гомологичными плечами при одноцепочечном разрыве (ник) или двухцепочечном разрыве на сайте расщепления Cas. Термин "сайт расщепления Cas" включает в себя последовательность ДНК, на которой одноцепочечный разрыв или двухцепочечный разрыв создан с помощью фермента Cas (например, белка Cas9 в комплексе с гидовой РНК). Целевые последовательности в пределах нацеленного локуса, которые соответствуют 5' и 3' гомологичным плечам экзогенной донорной нуклеиновой кислоты, "расположены в достаточной близости" от сайта расщепления Cas, если расстояние является таким, чтобы способствовать возникновению события гомологичной рекомбинации между 5' и 3' целевыми последовательностями и гомологичными плечами при одноцепочечном разрыве или двухцепочечном разрыве на сайте расщепления Cas. Таким образом, целевые последовательности, соответствующие 5' и/или 3' гомологичным плечам экзогенной донорной нуклеиновой кислоты, могут находиться, например, в пределах по меньшей мере 1 нуклеотида от данного сайта расщепления Cas или в пределах по меньшей мере от 10 нуклеотидов до приблизительно 1000 нуклеотидов отданного сайта расщепления Cas. В качестве примера сайт расщепления Cas может являться непосредственно смежным по меньшей мере с одной или обеими целевыми последовательностями.
[00200] Пространственные отношения целевых последовательностей, которые соответствуют гомологичным плечам экзогенной донорной нуклеиновой кислоты, и сайта расщепления Cas могут варьироваться. Например, целевые последовательности могут быть расположены 5' относительно сайта расщепления Cas, целевые последовательности могут быть расположены 3' относительно сайта расщепления Cas, или целевые последовательности могут фланкировать сайт расщепления Cas.
В. Способы оптимизации способности CRISPR/Cas вырезать целевую геномную нуклеиновую кислоту in vivo или ex vivo
[00201] Предусмотрены различные способы оптимизации доставки CRISPR/Cas в клетку или не являющемуся человеком животному или оптимизации активности CRISPR/Cas in vivo. Такие способы могут предусматривать, например, следующее: (а) проведение способа испытания способности CRISPR/Cas модифицировать целевой геномный локус, как описано выше, в первый раз у первого не являющегося человеком животного или в первой клетке; (b) изменение переменной и проведение способа во второй раз у второго не являющегося человеком животного (т.е. того же вида) или во второй клетке с измененной переменной; и (с) сравнение модификации целевого геномного локуса на стадии (а) с модификацией целевого геномного локуса на стадии (b), и выбор способа, приводящего к более эффективной модификации целевого геномного локуса.
[00202] Более эффективная модификация целевого геномного локуса может означать различные вещи в зависимости от требуемого эффекта в пределах не являющегося человеком животного или клетки. Например, более эффективная модификация целевого геномного локуса может означать одно или несколько или все из следующего: повышенная эффективность, повышенная точность, повышенная согласованность или повышенная специфичность. Повышенная эффективность относится к повышенным уровням модификации целевого геномного локуса (например, повышенное процентное отношение клеток нацелено в пределах конкретного целевого типа клеток, в пределах конкретной целевой ткани или в пределах конкретного целевого органа). Повышенная точность относится к более точной модификации целевого геномного локуса (например, повышенное процентное отношение нацеленных клеток с одинаковой модификацией или с требуемой модификацией без избыточных непреднамеренных вставок и делеций (например, инделей NHEJ)). Повышенная согласованность относится к более согласованной модификации целевого геномного локуса среди различных типов нацеленных клеток, тканей или органов, если более одного типа клеток, тканей или органов подвергается нацеливанию (например, модификация большего количества типов клеток в пределах целевого органа). Если конкретный орган подвергается нацеливанию, повышенная согласованность также может относиться к более согласованной модификации во всех местах в пределах органа. Повышенная специфичность может относиться к повышенной специфичности по отношению к нацеленному геномному локусу или локусам, повышенная специфичность по отношению к нацеленному типу клеток, повышенная специфичность по отношению к нацеленному типу ткани или повышенная специфичность по отношению к нацеленному органу. Например, повышенная специфичность в отношении геномного локуса относится к меньшей модификации нецелевых геномных локусов (например, пониженное процентное отношение нацеленных клеток с модификациями на непреднамеренных, нецелевых геномных локусах вместо или в дополнение к модификации целевого геномного локуса). Аналогично, повышенная специфичность в отношении типа клеток, типа ткани или органа относится к меньшей модификации нецелевых типов клеток, типов тканей или типов органов, если конкретный тип клеток, тип тканей или тип органов подлежит нацеливанию (например, когда конкретный орган подлежит нацеливанию (например, печень), существует меньше модификации клеток в органах или тканях, которые не являются предусмотренными мишенями).
[00203] Переменная, которая изменяется, может представлять собой любой параметр. В качестве одного примера измененная переменная может представлять собой способ упаковки или доставки, с помощью которого одну или несколько или все из гидовой РНК (или гидовой РНК, упакованной в AAV) и экзогенной донорной нуклеиновой кислоты вводят в клетку или не являющемуся человеком животному. Примеры способов доставки, таких как LNP, HDD и AAV, раскрыты в настоящем документе в другом месте. Например, измененная переменная может представлять собой серотип AAV. В качестве другого примера измененная переменная может представлять собой путь введения для введения одной или нескольких или всех из гидовой РНК (например, упакованной в AAV) и экзогенной донорной нуклеиновой кислоты в клетку или не являющемуся человеком животному. Примеры способов введения, таких как внутривенный, интравитреальный, интрапаренхимальный способ и закапывание в нос, раскрыты в другом месте в настоящем документе.
[00204] В качестве другого примера измененной переменной может являться концентрация или количество одной или нескольких или всех из введенной гидовой РНК (например, упакованной в AAV) и введенной экзогенной донорной нуклеиновой кислоты. В качестве другого примера измененная переменная может представлять собой концентрацию или количество введенной гидовой РНК (например, упакованной в AAV) относительно концентрации или количества введенной экзогенной донорной нуклеиновой кислоты.
[00205] В качестве другого примера, измененная переменная может представлять собой время введения одной или нескольких или всех из гидовой РНК (например, упакованной в AAV) и экзогенной донорной нуклеиновой кислоты относительно времени измерения экспрессии или активности одного или нескольких репортерных белков. В качестве другого примера, измененной переменной может являться число раз или частота, с которой вводят одну или несколько или все из гидовой РНК (например, упакованной в AAV) и экзогенной донорной нуклеиновой кислоты. В качестве другого примера, измененная переменная может являться временем введения гидовой РНК относительно времени введения экзогенной донорной нуклеиновой кислоты.
[00206] В качестве другого примера измененная переменная может представлять собой форму, в которой вводят одну или несколько или все из гидовой РНК и экзогенной донорной нуклеиновой кислоты. Например, гидовую РНК можно вводить в форме ДНК или в форме РНК. Экзогенная донорная нуклеиновая кислота может являться ДНК, РНК, одноцепочечной, двухцепочечной, линейной, кольцевой и так далее. Аналогично, каждый из компонентов может содержать различные комбинации модификаций для стабильности, для уменьшения нецелевых эффектов, для облегчения доставки и так далее. В качестве другого примера измененная переменная может представлять собой одну или несколько или все из введенной гидовой РНК (например, введение другой гидовой РНК с другой последовательностью) и введенной экзогенной донорной нуклеиновой кислоты (например, введение другой экзогенной донорной нуклеиновой кислоты с другой последовательностью).
С. Введение гидовых РНК и других компонентов в клетки и организм не являющихся человеком животных
[00207] Раскрытые в настоящем документе способы предусматривают введение в клетку или животному, не являющему человеком, одной или нескольких или всех из гидовых РНК и экзогенных донорных нуклеиновых кислот. «Введение» включает в себя представление клетке или не являющемуся человеком животному нуклеиновой кислоты или белка таким образом, что нуклеиновая кислота или белок получает доступ к внутренней части клетки или к внутренней части клеток в организме животного, не являющегося человеком. Введение можно выполнить любыми способами, и два или больше компонентов (например, два из компонентов или все компоненты) можно вводить в клетку или животному, не являющемуся человеком, одновременно или последовательно в любой комбинации. Например, экзогенную донорную нуклеиновую кислоту можно вводить в клетку или животному, не являющемуся человеком, перед введением гидовой РНК, или ее можно вводить после введения гидовой РНК (например, экзогенную донорную нуклеиновую кислоту можно вводить приблизительно за 1, 2, 3, 4, 8, 12, 24, 36, 48 или 72 часа до или после введения гидовой РНК). См., например, US 2015/0240263 и US 2015/0110762, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей. Кроме того, два или более компонентов можно вводить в клетку или животному, не являющемуся человеком, одним и тем же способом доставки или разными способами доставки. Аналогично, два или более компонентов можно вводить животному, не являющемуся человеком, одним и тем же путем введения или различными способами введения.
[00208] Гидовую РНК можно вводить в клетку в форме РНК (например, транскрибированной РНК in vitro) или в форме ДНК, кодирующей гидовую РНК. При введении в форме ДНК кодирующая гидовую РНК ДНК может быть функционально связана с активным в клетке промотором. Например, гидовую РНК можно доставлять посредством AAV, и она может экспрессироваться in vivo под контролем промотора U6. Такие ДНК могут находиться в одной или нескольких экспрессионных конструктах. Например, такие экспрессирующие конструкты могут являться компонентами одной молекулы нуклеиновой кислоты. Альтернативно, они могут быть разделены в любой комбинации среди двух или более молекул нуклеиновой кислоты (т.е. ДНК, кодирующие одну или несколько РНК CRISPR, и ДНК, кодирующие одну или несколько tracrРНК, могут являться компонентами отдельных молекул нуклеиновой кислоты).
[00209] Нуклеиновые кислоты, кодирующие гидовые РНК, могут быть функционально связаны с промотором в экспрессионном конструкте. Экспрессионные конструкты включают в себя любые нуклеиновокислотные конструкты, способные направлять экспрессию гена или другой представляющей интерес последовательности нуклеиновой кислоты и которые могут переносить такую представляющую интерес последовательность нуклеиновой кислоты в целевую клетку. Подходящие промоторы, которые можно использовать в экспрессионном конструкте, включают в себя промоторы, активные, например, в одной или нескольких из следующего: эукариотическая клетка, клетка человека, не относящаяся к человеку клетка, клетка млекопитающего, клетке млекопитающего, не являющегося человеком, клетка грызуна, клетка мыши, клетка крысы, клетка хомяка, клетка кролика, плюрипотентная клетка, эмбриональная стволовая (ES) клетка, взрослая стволовая клетка, клетка-предшественник, ограниченная стадией развития, индуцированная плюрипотентная стволовая клетка (iPS) или эмбрион на стадии одной клетки. Такими промоторами могут являться, например, условные промоторы, индуцируемые промоторы, конститутивные промоторы или тканеспецифические промоторы. Необязательно, промотор может являться двунаправленным промотором, управляющим экспрессией как гидовой РНК в одном направлении, так и другого компонента в другом направлении. Такие двунаправленные промоторы могут состоять из (1) полного общепринятого однонаправленного промотора Pol III, который содержит 3 внешних элемента управления: элемент дистальной последовательности (DSE), элемент проксимальной последовательности (PSE) и ТАТА-бокс; и (2) второй основной промотор Pol III, который включает в себя PSE и ТАТА-бокс, слитый с 5'-концом DSE в обратной ориентации. Например, в промоторе HI DSE соседствует с PSE и ТАТА-боксом, и промотор можно сделать двунаправленным, создав гибридный промотор, в котором транскрипция в обратном направлении контролируется путем добавления PSE и ТАТА-бокса, полученных из промотора U6. См., например, US 2016/0074535, полностью включенную в настоящий документ посредством ссылки для всех целей. Использование двунаправленного промотора для экспрессии генов, кодирующих гидовую РНК и другой компонент одновременно, позволяет создавать компактные экспрессионные кассеты для облегчения доставки.
[00210] Экзогенные донорные нуклеиновые кислоты и гидовые РНК (или нуклеиновые кислоты, кодирующие гидовые РНК) могут быть представлены в композициях, содержащих носитель, повышающий стабильность экзогенной донорной нуклеиновой кислоты или гидовой РНК (например, продлевая период при данных условиях хранения (например, -20°С, 4°С или температура окружающей среды), для которых продукты разложения остаются ниже порогового значения, такого как ниже 0,5% по массе исходной нуклеиновой кислоты или белка, или увеличение стабильности in vivo). Неограничивающие примеры таких носителей включают в себя микросферы поли(молочной кислоты) (PLA), микросферы, микросферы сополимера D,L-молочной и гликолевой кислот (PLGA), липосомы, мицеллы, обратные мицеллы, липидные кохлеаты и липидные микротрубочки.
[00211] В настоящем документе предусмотрены различные способы и композиции, позволяющие вводить нуклеиновую кислоту или белок в клетку или животному, не являющемуся человеком. Способы введения нуклеиновых кислот в клетки различных типов известны в настоящей области техники и включают в себя, например, способы стабильной трансфекции, способы транзиентной трансфекции и способы, опосредованные вирусом.
[00212] Протоколы трансфекции, а также протоколы для введения последовательностей нуклеиновых кислот в клетки могут различаться. Неограничивающие способы трансфекции включают в себя химические способы трансфекции; способы с использованием липосом; наночастиц; фосфата кальция (Graham et al. (1973) Virology 52 (2): 456-67, Bacchetti et al. (1977) Proc. Natl. Acad. Set USA 1A (4): 1590-4, и Kriegler, M (1991). Transfer and Expression: A Laboratory Manual. New York: W. H. Freeman and Company, pp. 96-97); дендримеров или катионных полимеров, таких как DEAE-декстран или полиэтиленимин. Нехимические способы включают в себя электропорацию, сонопорацию и оптическую трансфекцию. Трансфекция на основе частиц включает в себя использование генной пушки или магнитной трансфекции (Bertram (2006) Current Pharmaceutical Biotechnology 7, 277-28). Вирусные техники также можно использовать для трансфекции.
[00213] Введение нуклеиновых кислот или белков в клетку также может быть опосредовано электропорацией, внутрицитоплазматической инъекцией, вирусной инфекцией, аденовирусом, аденоассоциированным вирусом, лентивирусом, ретровирусом, трансфекцией, липид-опосредованной трансфекцией или нуклеофекцией. Нуклеофекция представляет собой усовершенствованную технологию электропорации, которая позволяет доставлять нуклеиновокислотные субстраты не только в цитоплазму, но также через ядерную мембрану и в ядро. Кроме того, использование нуклеофекции в раскрытых в настоящем документе способах, как правило, требует гораздо меньше клеток, чем обычная электропорация (например, только приблизительно 2 миллионов по сравнению с 7 миллионами при обычной электропорации). В одном примере нуклеофекцию выполняют с использованием системы LONZA® NUCLEOFECTOR™.
[00214] Введение нуклеиновых кислот или белков в клетку (например, зиготу) также можно осуществить путем микроинъекции. В зиготах (т.е. эмбрионах на одноклеточной стадии) микроинъекцию можно осуществлять в пронуклеус матери и/или отца или в цитоплазму. Если микроинъекция происходит только в одном пронуклеусе, отцовский пронуклеус предпочтительнее из-за его большего размера. Альтернативно, микроинъекцию можно осуществлять путем инъекции как в ядро/пронуклеус, так и в цитоплазму: сначала можно ввести иглу в ядро/пронуклеус и можно ввести первое количество, при удалении иглы из эмбриона на одноклеточной стадии второе количество может быть введено в цитоплазму. Способы проведения микроинъекции хорошо известны. См., например, Nagy et al. (Nagy A, Gertsenstein M, Vintersten K, Behringer R., 2003, Manipulating the Mouse Embryo. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); см. также Meyer et al. (2010) Proc. Natl. Acad. Sci. USA 107:15022-15026 и Meyer et al. (2012) Proc. Natl. Acad. Sci. USA 109:9354-9359.
[00215] Другие способы введения нуклеиновой кислоты или белков в клетку или животному, не являющемуся человеком, могут включать в себя, например, векторную доставку, опосредованную частицами доставку, опосредованную экзосомами доставку, опосредованную липидными наночастицами доставку, опосредованную проникающим в клетку пептидом доставку или опосредованную имплантируемыми устройствами доставку. В качестве конкретных примеров, нуклеиновую кислоту или белок можно вводить в клетку или животному, не являющемуся человеком, в носителе, таком как микросфера поли(молочной кислоты) (PLA), микросфера сополимера D,L-молочной и гликолевой кислот) (PLGA), липосома, мицелла, обратная мицелла, липидный кохлеат или липидная микротрубочка. Некоторые конкретные примеры доставки животному, не являющемуся человеком, включают в себя гидродинамическую доставку, опосредованную вирусом доставку (например, опосредованную аденоассоциированным вирусом (AAV) доставку) и опосредованную липидными наночастицами доставку.
[00216] Введение нуклеиновых кислот и белков в клетки или организм животных, не являющихся человеком, можно осуществить с помощью гидродинамической доставки (HDD). Гидродинамическая доставка появилась как способ внутриклеточной доставки ДНК in vivo. Для доставки генов в паренхиматозные клетки через выбранный кровеносный сосуд необходимо вводить только необходимые последовательности ДНК, что устраняет проблемы безопасности, связанные с современными вирусными и синтетическими векторами. При введении в кровоток ДНК способна достигать клеток в различных тканях, доступных для крови. Гидродинамическая доставка использует силу, создаваемую быстрой инъекцией большого объема раствора в несжимаемую кровь в кровообращении, чтобы преодолеть физические барьеры эндотелия и клеточных мембран, которые препятствуют проникновению крупных и непроницаемых для мембран соединений в паренхиматозные клетки. В дополнение к доставке ДНК этот способ применим для эффективной внутриклеточной доставки РНК, белков и других небольших соединений in vivo. См., например, Bonamassa et al. (2011) Pharm. Res. 28(4):694-701, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00217] Введение нуклеиновых кислот также можно осуществить путем опосредованной вирусом доставки, такой как AAV-опосредованная доставка или опосредованная лентивирусом доставка. Другие иллюстративные вирусы/вирусные векторы включают в себя ретровирусы, аденовирусы, вирусы коровьей оспы, поксвирусы и вирусы простого герпеса. Вирусы могут инфицировать делящиеся клетки, неделящиеся клетки или как делящиеся, так и неделящиеся клетки. Вирусы могут интегрироваться в геном хозяина или, альтернативно, не интегрироваться в геном хозяина. Такие вирусы также могут быть разработаны для снижения иммунитета. Вирусы могут являться способными к репликации или могут являться дефектными по репликации (например, дефектными в одном или нескольких генах, необходимых для дополнительных циклов репликации и/или упаковки вириона). Вирусы могут вызывать транзиентную экспрессию, длительную экспрессию (например, в течение по меньшей мере 1 недели, 2 недель, 1 месяца, 2 месяцев или 3 месяцев) или постоянную экспрессию (например, Cas9 и/или гРНК). Иллюстративные титры вирусов (например, титры AAV) включают в себя 1012, 1013, 1014, 1015 и 1016 векторных геномов/мл.
[00218] Геном оцДНК AAV состоит из двух открытых рамок считывания, Rep и Сар, фланкированных двумя инвертированными концевыми повторами, которые позволяют синтезировать цепь комплементарной ДНК. При конструировании переносящей плазмиды AAV трансген помещают между двумя ITR, и Rep и Сар могут поставляться в трансположениях. В дополнение к Rep и Cap, AAV может потребовать наличие плазмиды-помощника, содержащей гены аденовируса. Эти гены (Е4, Е2а и VA) опосредуют репликацию AAV. Например, переносящая плазмида, Rep/Cap, и плазмида-помощник могут быть трансфицированы в клетки HEK293, содержащие ген аденовируса Е1+, для получения инфекционных частиц AAV. Альтернативно, Rep, Сар и гены-помощники аденовируса могут быть объединены в одну плазмиду. Подобные упаковывающие клетки и способы можно использовать для других вирусов, таких как ретровирусы.
[00219] Идентифицированы многочисленные серотипы AAV. Эти серотипы различаются по типам клеток, которые они инфицируют (т.е. их тропизм), что позволяет осуществлять преимущественную трансдукцию определенных типов клеток. Серотипы ткани ЦНС включают в себя AAV1, AAV2, AAV4, AAV5, AAV8 и AAV9. Серотипы для ткани сердца включают в себя AAV1, AAV8 и AAV9. Серотипы почечной ткани включают в себя AAV2. Серотипы легочной ткани включают в себя AAV4, AAV5, AAV6 и AAV9. Серотипы ткани поджелудочной железы включают в себя AAV8. Серотипы для фоторецепторных клеток включают AAV2, AAV5 и AAV8. Серотипы ткани пигментного эпителия сетчатки включают в себя AAV1, AAV2, AAV4, AAV5 и AAV8. Серотипы скелетной мышечной ткани включают в себя AAV1, AAV6, AAV7, AAV8 и AAV9. Серотипы ткани печени включают в себя AAV7, AAV8 и AAV9 и, в частности, AAV8.
[00220] Тропизм можно дополнительно уточнить с помощью псевдотипирования, которое представляет собой смешивание капсида и генома из различных вирусных серотипов. Например, AAV2/5 указывает на вирус, содержащий геном серотипа 2, упакованный в капсид из серотипа 5. Использование псевдотипированных вирусов может повысить эффективность трансдукции, а также изменить тропизм. Гибридные капсиды, полученные из разных серотипов, также можно использовать для изменения вирусного тропизма. Например, AAV-DJ содержит гибридный капсид из восьми серотипов и демонстрирует высокую инфекционность в широком диапазоне типов клеток in vivo. AAV-DJ8 является еще одним примером, который проявляет свойства AAV-DJ, но с улучшенным поглощением тканями головного мозга. Серотипы AAV также можно модифицировать с помощью мутаций. Примеры мутационных модификаций AAV2 включают в себя Y444F, Y500F, Y730F и S662V. Примеры мутационных модификаций AAV3 включают в себя Y705F, Y731F и T492V. Примеры мутационных модификаций AAV6 включают в себя S663V и T492V. Другие псевдотипированные/модифицированные варианты AAV включают в себя AAV2/1, AAV2/6, AAV2/7, AAV2/8, AAV2/9, AAV2.5, AAV8.2 и AAV/SASTG.
[00221] Для ускорения экспрессии трансгена можно использовать самокомплементарные варианты AAV (scAAV). Поскольку AAV зависит от механизма репликации ДНК клетки, чтобы синтезировать комплементарную цепь генома одноцепочечной ДНК AAV, экспрессия трансгена может быть отсрочена. Чтобы устранить эту задержку, можно использовать scAAV, содержащие комплементарные последовательности, которые способны к самопроизвольному отжигу при заражении, что устраняет необходимость синтеза ДНК клетки-хозяина. Однако также можно использовать одноцепочечные векторы AAV (ssAAV).
[00222] Чтобы увеличить емкость упаковки, более длинные трансгены можно разделить между двумя переносящими плазмидами AAV, первая с 3'-донорным сайтом сплайсинга, а вторая с 5'-акцепторным сайтом сплайсинга. При коинфекции клетки эти вирусы образуют конкатемеры, сплайсируются вместе, и можно экспрессировать полноразмерный трансген. Хотя это позволяет увеличить экспрессию трансгена, экспрессия является менее эффективной. В подобных способах увеличения емкости используют гомологичную рекомбинацию. Например, трансген может быть разделен между двумя переносящими плазмидами, но с существенным перекрытием последовательности, так что коэкспрессия индуцирует гомологичную рекомбинацию и экспрессию полноразмерного трансгена.
[00223] Введение нуклеиновых кислот и белков также можно осуществить путем доставки, опосредованной липидными наночастицами (LNP). Например, LNP-опосредованную доставку можно использовать для доставки гидовой РНК в форме РНК. Доставка посредством таких способов приводит к временному присутствию гидовой РНК, а биоразлагаемые липиды улучшают клиренс, улучшают переносимость и снижают иммуногенность. Липидные составы могут защищать биологические молекулы от разложения при одновременном улучшении их клеточного поглощения. Липидные наночастицы представляют собой частицы, содержащие множество молекул липидов, физически связанных друг с другом межмолекулярными силами. Они включают в себя микросферы (включая в себя однослойные и многослойные везикулы, например, липосомы), дисперсную фазу в эмульсии, мицеллы или внутреннюю фазу в суспензии. Такие липидные наночастицы можно использовать для инкапсуляции одной или нескольких нуклеиновых кислот или белков для доставки. Составы, которые содержат катионные липиды, являются применимыми для доставки полианионов, таких как нуклеиновые кислоты. Другими липидами, которые могут быть включены, являются нейтральные липиды (т.е. незаряженные или цвиттерионные липиды), анионные липиды, вспомогательные липиды, которые усиливают трансфекцию, и липиды-невидимки, которые увеличивают продолжительность времени, в течение которого наночастицы могут существовать in vivo. Примеры подходящих катионных липидов, нейтральных липидов, анионных липидов, вспомогательных липидов и липидов-невидимок можно найти в международной патентной публикации WO 2016/010840 А1, полностью включенной в настоящий документ посредством ссылки для всех целей. Иллюстративная липидная наночастица может содержать катионный липид и один или несколько других компонентов. В одном примере другой компонент может содержать вспомогательный липид, такой как холестерин. В другом примере другие компоненты могут содержать вспомогательный липид, такой как холестерин, и нейтральный липид, такой как DSPC. В другом примере другие компоненты могут содержать вспомогательный липид, такой как холестерин, необязательный нейтральный липид, такой как DSPC, и липид-невидимку, такой как S010, S024, S027, S031 или S033.
[00224] LNP может содержать одно или несколько или все из следующего: (i) липид для инкапсуляции и для эндосомального высвобождения; (ii) нейтральный липид для стабилизации; (iii) вспомогательный липид для стабилизации; и (iv) липид-невидимка. См., например, Finn et al. (2018) Cell Reports 22:1-9 и WO 2017/173054 Al, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. В некоторых LNP груз может включать в себя гидовую РНК или нуклеиновую кислоту, кодирующую гидовую РНК. В некоторых LNP груз может включать в себя гидовую РНК или нуклеиновую кислоту, кодирующую гидовую РНК и экзогенную донорную нуклеиновую кислоту.
[00225] Липид для инкапсуляции и эндосомального высвобождения может представлять собой катионный липид. Липид также может являться биоразлагаемым липидом, таким как биоразлагаемый ионизируемый липид. Одним примером подходящего липида является липид А или LP01, который представляет собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропилооктадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил(9Z,12Z)-октадека-9,12-диеноат. См., например, Finn et al. (2018) Cell Reports 22:1-9 и международную патентную публикацию WO 2017/173054 A1, причем каждый документ полностью включен в настоящее описание посредством ссылки для всех целей. Другим примером подходящего липида является липид В, который представляет собой ((5-((диметиламино)метил)-1,3-фенилен)бис(окси))бис(октан-8,1-диил)бис(деканоат), который также называется ((5-((диметиламино)метил)-1,3-фенилен)бис(окси))бис(октан-8,1-диил)бис(деканоат). Другим примером подходящего липида является липид С, который представляет собой 2-((4-(((3-(диметиламино)пропокси)карбонил)окси)гексадеканоил)окси)пропан-1,3-диил(9Z,9'Z,12Z,12'Z)-бис(октадека-9,12-диеноат). Другим примером подходящего липида является липид D, который представляет собой 3-(((3-(диметиламино)пропокси)карбонил)окси)-13-(октаноилокси)тридецил-3-октилундеканоат.Другие подходящие липиды включают в себя гептатриаконта-6,9,28,31-тетраен-19-ил-4-(диметиламино)бутаноат (также известный как Dlin-MC3-DMA (МС3))).
[00226] Некоторые такие липиды, подходящие для применения в описанных в настоящем документе LNP, являются биоразлагаемыми in vivo. Например, LNP, содержащие такой липид, включают в себя те, в которых по меньшей мере 75% липида выводится из плазмы в течение 8, 10, 12, 24 или 48 часов или 3, 4, 5, 6, 7 или 10 дней. В качестве другого примера по меньшей мере 50% LNP выводится от плазмы в течение 8, 10, 12, 24 или 48 часов или 3, 4, 5, 6, 7 или 10 дней.
[00227] Такие липиды могут являться ионизируемыми в зависимости от рН среды, в которой они находятся. Например, в слабокислой среде липиды могут протонироваться и, таким образом, нести положительный заряд. И наоборот, в слабоосновной среде, такой как, например, кровь, где рН составляет приблизительно 7,35, липиды могут не протонироваться и, следовательно, нести заряд. Согласно некоторым вариантам осуществления липиды могут протонироваться при рН, по меньшей мере приблизительно 9, 9,5 или 10. Способность такого липида нести заряд связана с его собственной pKa. Например, липид может независимо характеризоваться значением pKa в диапазоне от приблизительно 5,8 до приблизительно 6,2.
[00228] Нейтральные липиды функционируют для стабилизации и улучшения обработки LNP. Примеры подходящих нейтральных липидов включают в себя множество нейтральных, незаряженных или цвиттерионных липидов. Примеры нейтральных фосфолипидов, подходящих для использования в настоящем раскрытии, включают в себя без ограничения 5-гептадецилбензол-1,3-диол (резорцин), дипальмитоилфосфатидилхолин (DPPC), дистеароилфосфатидилхолин (DSPC), фосфохолин (DOPC), димиристоилфосфатидилхолин (DMPC), фосфатидилхолин (PLPC), 1,2-дистеароил-sn-глицеро-3-фосфохолин (DAPC), фосфатидилэтаноламин (РЕ), яичный фосфатидилхолин (РЕС), дилаурилоилфосфатидилхолин (DLPC), димиристоилфосфатидилхолин (DMPC), 1-миристоил-2-пальмитоилфосфатидилхолин (МРРС), 1-пальмитоил-2-миристоилфосфатидилхолин (РМРС), 1-пальмитоил-2-стеароилфосфатидилхолин (PSPC), 1,2-диарахидоил-sn-глицеро-3-фосфохолин (DBPC), 1-стеароил-2-пальмитоилфосфатидилхолин (SPPC), 1,2-диэйкозеноил-sn-глицеро-3-фосфохолин (DEPC), пальмитоилолеоилфосфатидилхолин (РОРС), лизофосфатидилхолин, диолеоилфосфатидилэтаноламин (DOPE), дилинолеоилфосфатидилхолин, дистеароилфосфатидилэтаноламин (DSPE), димиристоилфосфатидилэтаноламин (DMPE), дипальмитоилфосфатидилэтаноламин (DPPE), пальмитоилолеоилфосфатидилэтаноламин (POPE), лизофосфатидилэтаноламин и их комбинации. Например, нейтральный фосфолипид можно выбрать из группы, состоящей из дистеароилфосфатидилхолина (DSPC) и димиристоилфосфатидилэтаноламина (DMPE).
[00229] Вспомогательные липиды включают в себя липиды, которые усиливают трансфекцию. Механизм, с помощью которого вспомогательный липид усиливает трансфекцию, может включать в себя повышение стабильности частиц. В некоторых случаях вспомогательный липид может усиливать фузогенность мембран. Вспомогательные липиды включают в себя стероиды, стеролы и алкилрезорцины. Примеры подходящих вспомогательных липидов, подходящих для использования, включают в себя холестерин, 5-гептадецилрезорцин и холестерингемисукцинат. В одном примере вспомогательный липид может представлять собой холестерин или холестерингемисукцинат.
[00230] Липиды-невидимки включают в себя липиды, которые изменяют продолжительность времени, в течение которого наночастицы могут существовать in vivo. Липиды-невидимки могут содействовать процессу составления, например, путем снижения агрегации частиц и контроля размера частиц. Липиды-невидимки могут модулировать фармакокинетические свойства LNP. Подходящие липиды-невидимки включают в себя липиды с гидрофильной головкой, связанной с липидной частью.
[00231] Гидрофильная головка липида-невидимки может содержать, например, полимерный фрагмент, выбранный из полимеров на основе PEG (которые иногда называются поли(этиленоксид)), поли(оксазолин), поли(виниловый спирт), поли(глицерин), поли(N-винилпирролидон), полиаминокислоты и поли(N-(2-гидроксипропил)метакриламид). Термин PEG означает любой полиэтиленгликоль или другой полиалкиленэфирный полимер. В некоторых составах LNP PEG представляет собой PEG-2K, также называемый PEG 2000, который характеризуется средней молекулярной массой, составляющей приблизительно 2000 дальтон. См., например, международную патентную публикацию WO 2017/173054 А1, полностью включенную в настоящий документ посредством ссылки для всех целей.
[00232] Липидный фрагмент липида-невидимки можно получить, например, из диацилглицерина или диацилгликамида, включая в себя соединения, содержащие диалкилглицериновую или диалкилгликамидную группу с длиной алкильной цепи, независимо содержащей от приблизительно С4 до приблизительно С40 насыщенных или ненасыщенных атомов углерода, где цепь может содержать одну или несколько функциональных групп, таких как, например, амид или сложный эфир. Диалкилглицериновая или диалкилгликамидная группа может дополнительно содержать одну или несколько замещенных алкильных групп.
[00233] В качестве одного примера липид-невидимку можно выбрать из следующего: PEG-дилауроилглицерин, PEG-димиристоилглицерин (PEG-ДМГ), PEG-дипальмитоилглицерин, PEG-дистеароилглицерин (PEG-DSPE), PEG-дилаурилгликамид, PEG-димиристилглимидамид, PEG-дипальмитоилгликамид и PEG-дистеароилгликамид, PEG-холестерин (1-[8'-(холест-5-ен-3-бета-окси)карбоксамидо-3',6'-диоксаоктанил]карбамоил-[омега]-метил-поли(этиленгликоль), PEG-DMB (3,4-дитетрадекоксилбензил-[омега]-метил-поли(этиленгликоль)эфир), 1,2-димиристоил-sn-глицеро-3-фосфоэтаноламин-N-[метокси(полиэтиленгликоль)-2000] (PEG2k-DMG), 1,2-дистеароил-sn-глицеро-3-фосфоэтаноламин-N-[метокси (полиэтиленгликоль)-2000] (PEG2k-DSPE), 1,2-дистеароил-sn-глицерин, метоксиполиэтиленгликоль (PEG2k-DSG), поли(этиленгликоль)-2000-диметакрилат (PEG2k-DMA) и 1,2-дистеарилоксипропил-3-амин-N-[метокси(полиэтиленгликоль)-2000] (PEG2k-DSA). В одном конкретном примере липид-невидимка может представлять собой PEG2k-DMG.
[00234] LNP могут содержать различные соответствующие молярные соотношения компонентов липидов в составе. Соотношение моль% липида CCD может составлять, например, от приблизительно 30 моль% до приблизительно 60 моль%, от приблизительно 35 моль% до приблизительно 55 моль%, от приблизительно 40 моль% до приблизительно 50 моль%, от приблизительно 42 моль% до приблизительно 47 моль% или приблизительно 45%. Соотношение моль% вспомогательного липида может составлять, например, от приблизительно 30 моль% до приблизительно 60 моль%, от приблизительно 35 моль% до приблизительно 55 моль%, от приблизительно 40 моль% до приблизительно 50 моль%, от приблизительно 41 моль% до приблизительно 46 моль%, или приблизительно 44 моль%. Соотношение моль% нейтрального липида может составлять, например, от приблизительно 1 моль% до приблизительно 20 моль%, от приблизительно 5 моль% до приблизительно 15 моль%, от приблизительно 7 моль% до приблизительно 12 моль% или приблизительно 9 моль%. Соотношение моль% липида-невидимки может составлять, например, от приблизительно 1 моль% до приблизительно 10 моль%, от приблизительно 1 моль% до приблизительно 5 моль%, от приблизительно 1 моль% до приблизительно 3 моль%, приблизительно 2 моль% или приблизительно 1 моль%.
[00235] LNP могут характеризоваться различными соотношениями между положительно заряженными аминными группами биоразлагаемого липида (N) и отрицательно заряженными фосфатными группами (Р) нуклеиновой кислоты, которая должна быть инкапсулирована. Это может быть математически представлено соотношением N/P. Например, соотношение N/P может составлять от приблизительно 0,5 до приблизительно 100, от приблизительно 1 до приблизительно 50, от приблизительно 1 до приблизительно 25, от приблизительно 1 до приблизительно 10, от приблизительно 1 до приблизительно 7, от приблизительно 3 до приблизительно 5, от приблизительно 4 до приблизительно 5, приблизительно 4, приблизительно 4,5 или приблизительно 5.
[00236] В некоторых LNP груз может содержать экзогенную донорную нуклеиновую кислоту и гРНК. Экзогенная донорная нуклеиновая кислота и гРНК могут находиться в различных соотношениях. Например, состав LNP может включать в себя соотношение экзогенной донорной нуклеиновой кислоты к нуклеиновой кислоте гРНК, находящееся в диапазоне от приблизительно 25:1 до приблизительно 1:25, находящееся в диапазоне от приблизительно 10:1 до приблизительно 1:10, находящееся в диапазоне от приблизительно 5:1 до приблизительно 1:5, или приблизительно 1:1. Альтернативно, состав LNP может включать в себя соотношение экзогенной донорной нуклеиновой кислоты к нуклеиновой кислоте гРНК от приблизительно 1:1 до приблизительно 1:5, приблизительно 5:1 до приблизительно 1:1, приблизительно 10:1 или приблизительно 1:10. Альтернативно, состав LNP может включать в себя соотношение экзогенной донорной нуклеиновой кислоты к нуклеиновой кислоте гРНК, составляющее приблизительно 1:10, 25:1, 10:1, 5:1, 3:1, 1:1, 1:3, 1:5, 1:10 или 1:25.
[00237] Конкретный пример подходящего LNP характеризуется соотношение азота к фосфату (N/P), составляющим 4,5, и содержит биоразлагаемый катионный липид, холестерин, DSPC и PEG2k-DMG в молярном соотношении 45:44:9:2. Биоразлагаемый катионный липид может представлять собой (9Z,12Z)-3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропилооктадека-9,12-диеноат, также называемый 3-((4,4-бис(октилокси)бутаноил)окси)-2-((((3-(диэтиламино)пропокси)карбонил)окси)метил)пропил-(9Z,12Z)-октадека-9,12-диеноат. См., например, Finn et al. (2018) Cell Reports 22:1-9, полностью включенный в настоящий документ посредством ссылки для всех целей. Другой конкретный пример подходящего LNP содержит Dlin-MC3-DMA (МС3), холестерин, DSPC и PEG-DMG в молярном соотношении 50:38,5:10:1,5.
[00238] Способ доставки можно выбрать для снижения иммуногенности. Например, гРНК и экзогенную донорную нуклеиновую кислоту можно доставлять разными способами (например, бимодальной доставкой). Эти различные способы могут придавать различные фармакодинамические или фармакокинетические свойства изучаемой доставляемой молекуле (например, кодирование гРНК или нуклеиновой кислоты или экзогенная донорная нуклеиновая кислота/матрица для репарации). Например, разные способы могут привести к разному распределению в ткани, разному периоду полужизни или разному распределению по времени. Некоторые способы доставки (например, доставка нуклеиновокислотного вектора, который сохраняется в клетке посредством автономной репликации или геномной интеграции) приводят к более постоянной экспрессии и присутствию молекулы, тогда как другие способы доставки являются временными и менее постоянными (например, доставка РНК или белка). Доставка компонентов более временным способом, например, в виде РНК или белка, может гарантировать, что комплекс Cas/гРНК присутствует и активен только в течение короткого периода времени, и может снизить иммуногенность. Такая временная доставка может также уменьшить вероятность нецелевых модификаций.
[00239] Введение in vivo можно осуществлять любым подходящим путем, включая в себя, например, парентеральный, внутривенный, пероральный, подкожный, внутриартериальный, внутричерепной, интратекальный, интраперитонеальный, местный, интраназальный или внутримышечный. Способы системного введения включают в себя, например, пероральный и парентеральный пути. Примеры парентеральных путей включают в себя внутривенный, внутриартериальный, внутрикостный, внутримышечный, внутрикожный, подкожный, интраназальный и интраперитонеальный пути. Конкретным примером является внутривенная инфузия. Способы местного введения включают в себя, например, интратекальный, интрацеребровентрикулярный, интрапаренхиматозный (например, локализованная интрапаренхиматозная доставку в следующее: стриатум (например, в хвостатое ядро или в путамен), кора головного мозга, прецентральная извилина, гиппокамп (например, зубчатая извилина или область САЗ), височная кора, миндалина, лобная кора, таламус, мозжечок, продолговатый мозг, гипоталамус, крыша среднего мозга, покрышка среднего мозга или черная субстанция), интраокулярный, интраорбитальный, субконъюктивальный, интравитреальный, субретинальный и транссклеральный пути. Значительно меньшие количества компонентов (по сравнению с системными подходами) могут оказывать эффект при местном введении (например, интрапаренхиматозном или интравитреальном) по сравнению с системным введением (например, введением внутривенно). Местные способы введения также могут уменьшать или устранять возникновение потенциально токсических побочных эффектов, которые могут возникнуть, когда терапевтически эффективные количества компонента вводятся системно.
[00240] Введение in vivo можно осуществлять любым подходящим путем, включая в себя, например, парентеральный, внутривенный, пероральный, подкожный, внутриартериальный, внутричерепной, интратекальный, интраперитонеальный, местный, интраназальный или внутримышечный. Конкретным примером является внутривенная инфузия. Закапывание в нос и интравитреальная инъекция являются другими конкретными примерами. Композиции, содержащие гидовые РНК (или нуклеиновые кислоты, кодирующие гидовые РНК), можно составить с использованием одного или нескольких физиологически и фармацевтически приемлемых носителей, разбавителей, вспомогательных веществ или добавок. Состав может зависеть от выбранного пути введения. Термин «фармацевтически приемлемый» означает, что носитель, разбавитель, вспомогательное вещество или добавка совместимы с другими ингредиентами состава и по существу не вредны для их реципиента.
[00241] Частота введения и количество дозировок могут зависеть от периода полужизни экзогенных донорных нуклеиновых кислот или гидовых РНК (или нуклеиновых кислот, кодирующих гидовые РНК) и пути введения среди других факторов. Введение нуклеиновых кислот или белков в клетку или животному, не являющемуся человеком, можно выполнить один раз или несколько раз в течение периода времени. Например, введение можно выполнить по меньшей мере два раза в течение периода времени, по меньшей мере три раза в течение периода времени, по меньшей мере четыре раза в течение периода времени, по меньшей мере пять раз в течение периода времени, по меньшей мере шесть раз в течение периода времени, по меньшей мере семь раз в течение периода времени, по меньшей мере восемь раз в течение периода времени, по меньшей мере девять раз в течение периода времени, по меньшей мере в десять раз в течение периода времени, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз в течение периода времени, по меньшей мере тринадцать раз в течение периода времени, по меньшей мере четырнадцать раз в течение периода времени, по меньшей мере пятнадцать раз в течение периода времени, по меньшей мере шестнадцать раз в течение периода времени, по меньшей мере семнадцать раз в течение периода времени, по меньшей мере восемнадцать раз в течение периода времени, по меньшей мере девятнадцать раз в течение периода времени или по меньшей мере двадцать раз в течение периода времени.
D. Измерение активности CRISPR/Cas in vivo и оценка модификации целевого геномного локуса
[00242] Раскрытые в настоящем документе способы могут дополнительно предусматривать оценку модификации целевого геномного локуса. Способы обнаружения или измерения экспрессии или активности будут зависеть от модифицируемого целевого геномного локуса.
[00243] Например, если целевой геномный локус содержит ген, кодирующий РНК или белок, и предусмотренная модификация заключается в изменении экспрессии кодируемой РНК или белка, способ оценки модификации целевого геномного локуса может предусматривать измерение экспрессии или активности кодируемой РНК или белка. Например, если кодируемый белок представляет собой белок, высвобождающийся в сыворотку, можно измерить содержания в сыворотке кодируемого белка. Анализы для измерения содержаний и активности РНК и белков являются хорошо известными.
[00244] Альтернативно, раскрытые в настоящем документе способы могут дополнительно предусматривать идентификацию клетки, содержащей модифицированный целевой геномный локус, в которой последовательность была модифицирована с помощью негомологичного соединения концов (например, присутствие небольших вставок или делеций (инделей)) после расщепления с помощью CRISPR/Cas, в которой последовательность в целевом геномном локусе между двумя целевыми последовательностями гидовой РНК была вырезана или в которой целевой геномный локус был модифицирован с помощью рекомбинации с экзогенной донорной нуклеиновой кислотой. Различные способы можно использовать для идентификации клеток с целевой генетической модификацией. Скрининг может включать в себя количественный анализ для оценки модификации аллеля (МОА) родительской хромосомы. Например, количественный анализ можно выполнить с помощью количественной ПЦР, такой как ПЦР в режиме реального времени (кПЦР). ПНР в режиме реального времени может использовать первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров может содержать флуоресцентный зонд, который распознает амплифицированную последовательность. Другие примеры подходящих количественных анализов включают в себя флуоресцентно-опосредованную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным(и) зондом(ами), зонды INVADER®, зонды TAQMAN® Molecular Beacon или технологию зондов ECLIPSE™ (см., например, US 2005/0144655, полностью включенную в настоящий документ посредством ссылки для всех целей).
[00245] Секвенирование следующего поколения (NGS) также можно использовать для скрининга. Секвенирование следующего поколения может также носить название «NGS» или «массовое параллельное секвенирование» или «высокопроизводительное секвенирование». NGS можно использовать в качестве инструмента скрининга в дополнение к анализам МОА для определения точного характера целевой генетической модификации и того, является ли это согласованным по типам клеток или типов тканей или типов органов.
[00246] Оценить модификацию целевого геномного локуса у животного, не являющегося человеком, можно в любом типе клеток из любой ткани или органа. Например, модификацию целевого геномного локуса можно оценить в нескольких типах клеток из одной и той же ткани или органа или в клетках из нескольких мест в ткани или органе. Это может предоставить информацию о том, какие типы клеток в целевой ткани или органе модифицируются или какие участки ткани или органа достигаются CRISPR/Cas и модифицируются. В качестве другого примера модификацию целевого геномного локуса можно оценить в нескольких типах тканей или в нескольких органах. В способах, в которых нацеленное воздействие производят на конкретную ткань или орган, это может предоставить информацию о том, насколько эффективно происходит нацеленное воздействие на эту ткань или орган и есть ли нецелевые эффекты в других тканях или органах.
[00247] В некоторых конкретных примерах эспрессирующих Cas9 не являющихся человеком животных можно использовать для оценки частоты редактирования различных гидовых РНК. Гидовые РНК можно вводить либо в виде одной гидовой РНК (модифицированной и немодифицированной), либо дуплексной РНК или экспрессировать под контролем промотора U6 (например, посредством AAV). эспрессирующих Cas9 не являющихся человеком животных также можно скрещивать с не являющимися человеком животными, включая в себя не являющихся человеком животных с гуманизированными аллелями, экспрессирующих гидовые РНК, для оценки при моделировании заболеваний.
IV. Способы получения не являющихся человеком животных, содержащих экспрессионную кассету Cos и/или экспрессионную кассету рекомбиназы
[00248] Предусмотрены различные способы получения не являющегося человеком животного, содержащего одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы, как раскрыто в другом месте в настоящем документе. Любой общепринятый способ или протокол получения генетически модифицированного организма является подходящим для получения такого генетически модифицированного не являющегося человеком животного. См., например, Cho et al. (2009) Current Protocols in Cell Biology 42:19.11:19.11.1-19.11.22 и Gama Sosa et al. (2010) Brain Struct. Funct. 214(2-3):91-109, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Такие генетически модифицированные не являющиеся человеком животные могут быть получены, например, посредством нокина гена в целевом локусе (например, локусе «безопасная гавань», таком как Rosa26) или посредством применения случайно интегрирующегося трансгена. См., например, международные патентные публикации WO 2014/093622 и WO 2013/176772, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей. Способы нацеливания конструкта на локус Rosa26 описаны, например, в US 2012/0017290, US 2011/0265198 и US 2013/0236946, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей.
[00249] Например, способ получения не являющегося человеком животного, содержащего одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы, как раскрыто в другом месте в настоящем документе, может предусматривать следующее: (1) модификация генома плюрипотентной клетки для включения одной или нескольких или всех из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы; (2) идентификация или отбор генетически модифицированной плюрипотентной клетки, содержащей одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион животного-хозяина, не являющегося человеком; и (4) имплантация и вынашивание эмбриона-хозяина в организме суррогатной матери. Необязательно, эмбрион-хозяин, содержащий модифицированную плюрипотентную клетку (например, ES клетку, не относящуюся к человеку), может быть инкубирован до стадии бластоцисты перед имплантацией и гестацией в организме суррогатной матери для получения не являющегося человеком животного F0. Затем суррогатная мать может произвести не являющееся человеком животное поколения F0, содержащее одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы.
[00250] Способы могут дополнительно предусматривать идентификацию клетки или животного с модифицированным целевым геномным локусом. Различные способы можно использовать для идентификации клеток и животных с целевой генетической модификацией.
[00251] Стадия скрининга может предусматривать, например, количественный анализ для оценки модификации аллеля (МОА) родительской хромосомы. Например, количественный анализ можно выполнить с помощью количественной ПЦР, такой как ПЦР в режиме реального времени (кПЦР). В ПЦР в режиме реального времени можно использовать первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров может содержать флуоресцентный зонд, который распознает амплифицированную последовательность.
[00252] Другие примеры подходящих количественных анализов включают в себя флуоресцентно-опосредованную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным(и) зондом(ами), зонды INVADER®, зонды TAQMAN® Molecular Beacon или технологию зондов ECLIPSE™ (см., например, US 2005/0144655, полностью включенную в настоящий документ посредством ссылки для всех целей).
[00253] Примером подходящей плюрипотентной клетки является эмбриональная стволовая (ES) клетка (например, ES клетка мыши или ES клетка крысы). Модифицированную плюрипотентную клетку можно создать, например, путем следующего: (а) введение в клетку одного или нескольких нацеливающих векторов, содержащих нуклеиновую кислоту-вставку, фланкированную 5' и 3' гомологичными плечами, соответствующими 5' и 3' целевым сайтам, причем нуклеиновая кислота-вставка содержит одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы; и (b) идентификация по меньшей мере одной клетки, содержащей в своем геноме нуклеиновую кислоту-вставку, интегрированную в целевом геномном локусе. Альтернативно, модифицированную плюрипотентную клетку можно получить следующим образом: (а) введение в клетку следующего: (i) нуклеазный агент, причем нуклеазный агент индуцирует одноцепочечный разрыв или двухцепочечный разрыв на целевой последовательности в пределах целевого геномного локуса; и (ii) один или несколько нацеливающих векторов, содержащих нуклеиновую кислоту-вставку, фланкированную 5' и 3' гомологичными плечами, соответствующими 5' и 3' целевым сайтам, расположенным в достаточной близости от целевой последовательности, причем нуклеиновая кислота-вставка содержит одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы; и (с) идентификация по меньшей мере одной клетки, содержащей модификацию (например, интеграцию нуклеиновой кислоты-вставки) в целевом геномном локусе. Можно использовать любой нуклеазный агент, который вызывает одноцепочечный разрыв или разрыв двухцепочечной цепи в требуемой целевой последовательности. Примеры подходящих нуклеаз включают в себя эффекторную нуклеазу, подобную активатору транскрипции (TALEN), нуклеазу домена «цинковые пальцы» (ZFN), мегануклеазу и системы сгруппированных регулярно перемежающихся коротких палиндромных повторов (CRISPRyCRISPR-ассоциированных белков (Cas) или компоненты таких системы (например, CRISPR/Cas9). См., например, US 2013/0309670 и US 2015/0159175, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей.
[00254] Донорную клетку можно вводить в эмбрион-хозяин на любой стадии, такой как стадия бластоцисты или стадия предморула (т.е. стадия 4 клеток или стадия 8 клеток). Создается потомство, способное передавать генетическую модификацию через зародышевую линию. См., например, патент США №7294754, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00255] Альтернативно, способ получения не являющихся человеком животных, описанных в другом месте в настоящем документе, может предусматривать следующее: (1) модификация генома эмбриона одноклеточной стадии для включения в него одной или нескольких или всех из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы с использованием способов, описанных выше, для модификации плюрипотентных клеток; (2) отбор генетически модифицированных эмбрионов; и (3) имплантация и вынашивание генетически модифицированного эмбриона в организме суррогатной матери. Создается потомство, способное передавать генетическую модификацию через зародышевую линию.
[00256] Техники ядерного переноса также можно использовать для получения млекопитающих, не являющихся человеком. Вкратце, способы переноса ядра могут предусматривать следующие стадии: (1) энуклеация ооцита или получение энуклеированного ооцита; (2) выделение или получение донорной клетки или ядра для объединения с энуклеированным ооцитом; (3) вставка клетки или ядра в энуклеированный ооцит с образованием реконструированной клетки; (4) имплантация реконструированной клетки в матку животного с образованием эмбриона; и (5) обеспечение развития эмбриона. В таких способах ооциты, как правило, извлекают у умерших животных, хотя они могут быть выделены также из яйцеводов и/или яичников живых животных. Ооциты могут созревать в различных хорошо известных средах до энуклеации. Энуклеацию яйцеклетки можно выполнить с помощью ряда хорошо известных способов. Вставку донорной клетки или ядра в энуклеированный ооцит с образованием реконструированной клетки можно осуществить путем микроинъекции донорной клетки под вителлиновый слой перед слиянием. Слияние может быть индуцировано приложением электрического импульса постоянного тока через плоскость контакта/слияния (электрослияние), воздействием на клетки химических веществ, способствующих слиянию, таких как полиэтиленгликоль, или посредством инактивированного вируса, такого как вирус Сендай. Реконструированная клетка может быть активирована электрическими и/или неэлектрическими способами до, во время и/или после слияния донора-ядра и ооцита-реципиента. Способы активации включают в себя электрические импульсы, химически индуцированный шок, проникновение с помощью сперматозоидов, повышение содержания двухвалентных катионов в ооците и снижение фосфорилирования клеточных белков (как с помощью ингибиторов киназы) в ооците. Активированные реконструированные клетки или эмбрионы можно культивировать в хорошо известных средах и затем переносить в матку животного. См., например, документы US 2008/0092249, WO 1999/005266, US 2004/0177390, WO 2008/017234 и патент США №7612250, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей.
[00257] Различные способы, предусмотренные в настоящем документе, позволяют создавать генетически модифицированное животное F0, не являющееся человеком, причем клетки генетически модифицированного животного F0 содержат одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы. Признано, что в зависимости от способа, используемого для получения животного F0, число клеток в животном F0, которые содержат одну или несколько или все из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы, будет варьироваться. Введение донорных ES клеток в эмбрион на стадии предморула из соответствующего организма (например, эмбриона мыши на стадии 8 клеток), например, с помощью способа VELOCLMOUSE® позволяет получать больший процент клеточной популяции животного F0, которое содержит клетки, содержащие представляющую интерес нуклеотидную последовательность, содержащую целевую генетическую модификацию. Например, по меньшей мере 50%, 60%, 65%, 70%, 75%, 85%, 86%, 87%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% клеточного вклада животного F0, не являющегося человеком, могут содержать клеточную популяцию, содержащую целевую модификацию
[00258] Клетки генетически модифицированного животного F0 могут являться гетерозиготными по одной или нескольким или всем из экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы или могут являться гомозиготными по одной или нескольким или всем экспрессионной кассеты Cas и экспрессионной кассеты рекомбиназы.
[00259] Все патентные заявки, веб-сайты, другие публикации, регистрационные номера и тому подобное, указанные выше или ниже, полностью включены посредством ссылки для всех целей в той же степени, как если бы было специально и индивидуально указано, что каждый отдельный элемент включен в него посредством ссылки. Если разные версии последовательности связаны с регистрационным номером в разное время, подразумевается версия, связанная с регистрационным номером на дату подачи настоящей заявки. Действительная дата подачи означает более раннюю из действительной даты подачи или даты подачи приоритетной заявки со ссылкой на регистрационный номер, если необходимо. Аналогичным образом, если разные версии публикации, веб-сайта или тому подобного публикуются в разное время, подразумевается последняя версия, опубликованная на действительную дату подачи, если не указано иное. Любой признак, стадию, элемент, вариант осуществления или аспект настоящего изобретения можно использовать в сочетании с любым другим, если специально не указано иное. Хотя настоящее изобретение было описано более подробно с помощью иллюстрации и примера для ясности и понимания, будет очевидно, что определенные изменения и модификации можно осуществить в рамках объема прилагаемой формулы изобретения.
Краткое описание последовательностей
[00260] Нуклеотидные и аминокислотные последовательности, перечисленные в прилагаемом перечне последовательностей, показаны с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот. Нуклеотидные последовательности следуют стандартному соглашению, начиная с 5' конца последовательности и продвигаясь вперед (т.е. слева направо в каждой строке) до 3' конца. Показана только одна цепь каждой нуклеотидной последовательности, но считается, что комплементарная цепь включена в любую ссылку на отображаемую цепь. Когда представлена нуклеотидная последовательность, кодирующая аминокислотную последовательность, подразумевается, что также представлены ее кодон-вырожденные варианты, которые кодируют ту же аминокислотную последовательность. Аминокислотные последовательности следуют стандартному соглашению, начиная с амино-конца последовательности и продвигаясь вперед (т.е. слева направо в каждой строке) до карбокси-конца.
[00261] Таблица 3. Описание последовательностей.
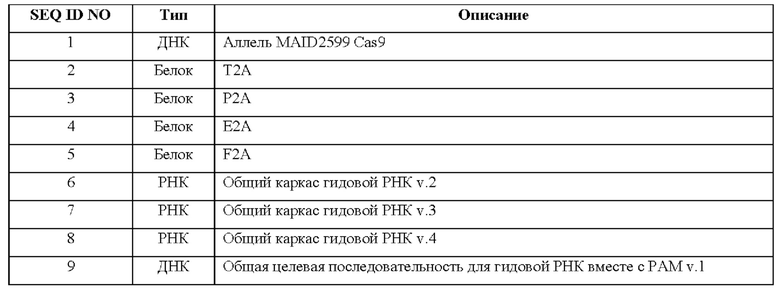

ПРИМЕРЫ
Пример 1. Валидация экспрессирующих Cas9 мышей
[00262] CRISPR/Cas9, ДНК-эндонуклеаза, направляемая РНК, катализирует создание двухцепочечного разрыва (DSB) ДНК в сайте связывания ее РНК-гида. Иллюстративный РНК-гид может состоять из РНК CRISPR из 42 нуклеотидов (сrРНК), которая соединяется с трансактивирующей РНК из 87 нуклеотидов (tracrРНК). tracrРНК комплементарна и спарена по основаниям с сrРНК, образуя функциональный crРНК/tracrРНК-гид. Эта дуплексная РНК становится связанной с белком Cas9 с образованием активного рибонуклеопротеина (RNP), который может детально исследовать геном на предмет комплементарности с 20-нуклеотидной гидовой частью сrРНК. Вторичным требованием для разрыва цепи является то, что белок Cas9 должен распознавать мотив, прилегающий к протоспейсеру (РАМ), непосредственно примыкающий к последовательности, комплементарной гидовой части кРНК (целевая последовательность сrРНК). Альтернативно, активный комплекс RNP также может быть образован путем замены дуплекса crPHК/tracrPHК единой гидовой РНК (sgPHК), образованной путем ковалентного соединения сrРНК и tracrPHК. Такая sgPHК может быть образована, например, путем слияния 20 нуклеотидной гидовой части сrРНК непосредственно с процессированной последовательностью tracrPHК. sgPHК может взаимодействовать как с белком Cas9, так и с ДНК таким же образом и с той же эффективностью, что и дуплекс crPHК/tracrPHК. Было показано, что механизм естественной защиты бактерий CRISPR эффективно функционирует в клетках млекопитающих и активирует пути эндогенной репарации, вызванные разрывом. Когда в геноме происходит разрыв двойной цепи, пути репарации будут пытаться починить ДНК либо каноническими, либо альтернативными путями негомологичного соединения концов (NHEJ) или гомологичной рекомбинацией, также называемой гомологичной репарацией (HDR), если соответствующая матрица доступна. Авторы настоящего изобретения могут использовать эти пути для облегчения сайт-специфической делеции геномных областей или вставки экзогенной ДНК или HDR в клетки млекопитающих.
[00263] Система CRISPR/Cas9 является мощным инструментом для инженерии генома. Тем не менее, одним из ограничений системы для использования in vivo является необходимость одновременного введения всех компонентов в живой организм. Типичный способ введения этих компонентов в клетки заключается во транзиентной трансфекции ДНК-конструктов в клетки, которые будут генерировать соответствующие РНК и белок. Хотя этот подход эффективен, он имеет свойственный недостаток, поскольку клетки должны полагаться на конструкты плазмидной ДНК, чтобы сначала подвергнуться транскрипции, а затем трансляции, прежде чем белок Cas9 станет доступным для взаимодействия с компонентом sgPHК. Авторы настоящего изобретения полагают, что частота мутаций, индуцированных Cas9, и частота рекомбинации могут быть значительно улучшены при наличии конститутивно доступного белка.
[00264] Кодирующая последовательность Cas9 дикого типа (CDS) оптимизировали по кодонам для экспрессии в организме мышей. Затем включали N-концевой одинарный сигнал ядерной локализации (NLS), С-концевой двойной NLS и С-концевой связанный с Р2А флуоресцентный репортер GFP. экспрессионная кассета Cas9 (MAID2599) изображена на фигурах 1 и 14 и SEQ ID NO: 1. P2A-GFP можно использовать для лучшего отслеживания экспрессии Cas9 in vivo. Эти компоненты сконструировали в первый интрон локуса Rosa26 генома мыши вместе с предшествующей фланкированной loxP кассетой устойчивости к неомицину (кассета neo) с соответствующими сигналами сплайсинга и сильным сигналом полиаденилирования (polyA). Компоненты экспрессионной кассеты Cas9 от 5' до 3' показаны в таблице 4, а компоненты аллеля Cas9 после удаления фланкированной 1охР неомициновой кассеты, показаны в таблице 5. Последовательность белка Cas9-P2A-eGFP, кодируемая аллелем, представлена в SEQ ID NO: 13.
[00265] Таблица 4. Компоненты аллеля MAID2599 Cas9.
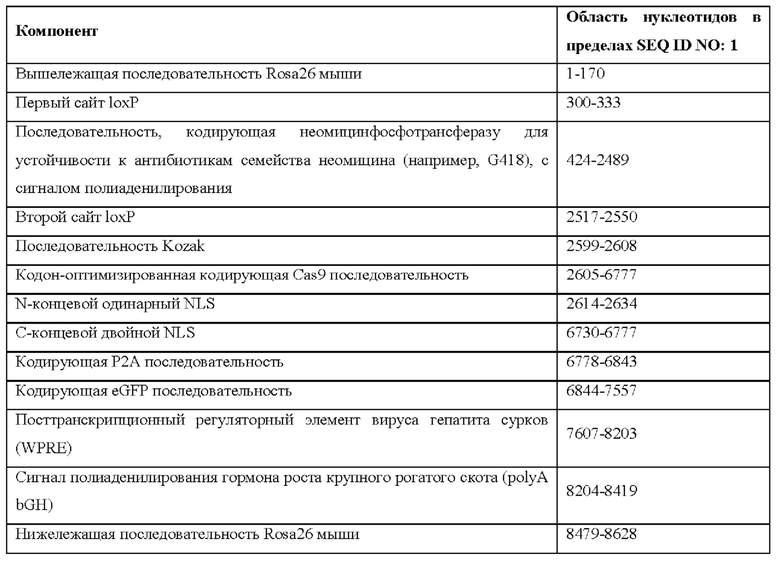
[00266] Таблица 5. Компоненты аллеля MAID2600 Cas9.
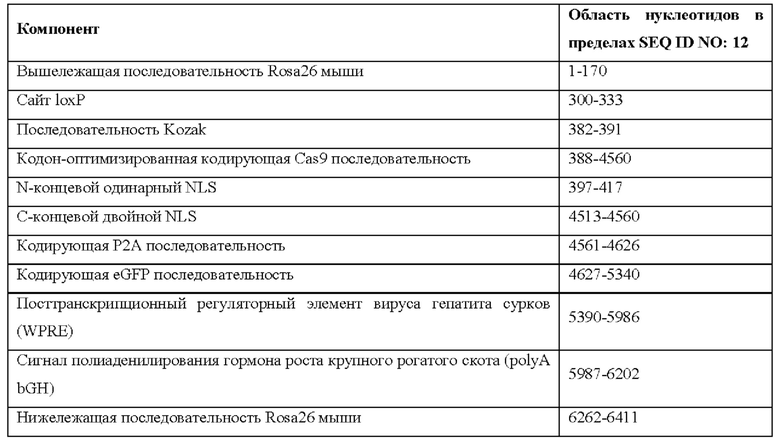
[00267] Перед удалением кассеты под действием Cre-рекомбиназы ген устойчивости к неомицину, как правило, будет эффективно транскрибироваться и транслироваться; однако CDS Cas9 обычно не экспрессируется из-за присутствия сильной области поли(А), которая может эффективно блокировать сквозную транскрипцию. После удаления неомициновой кассеты под действием Cre-рекомбиназы гибридная мРНК для белков Cas9 и GFP, как правило, обычно конститутивно экспрессируется с помощью промотора Rosa26. Нацеленные клетки до и после удаления неомициновой кассеты сначала верифицировали скринингом в отношении потери аллелей для обнаружения единственной сайт-специфической интеграции нацеливающего вектора в локусе Rosa26. Экспрессию Cas9 и GFP валидировали путем выделения общей РНК из нацеленных mESC с последующей обратной транскрипцией для создания кДНК и кПЦР TAQMAN® для обнаружения обратно транскрибированной кДНК (ОТ-кПЦР). В целом, созданная система способна непрерывно или условно экспрессировать постоянные уровни белка Cas9 (требуя удаления кассеты устойчивости к неомицину) в mESC и организме мышей, полученных из них.
[00268] Другую версию разработали без P2A-eGFP (MAID2660). См. фигуру 14. Компоненты экспрессионной кассеты Cas9 от 5' до 3' показаны в таблице 6, а компоненты аллеля Cas9 после удаления фланкированной loxP-сайтами неомициновой кассеты показаны в таблице 7. Последовательность белка Cas9, кодируемая аллелем, представлена в SEQ ID NO: 19.
[00269] Таблица 6. Компоненты аллеля MAID2660 Cas9.
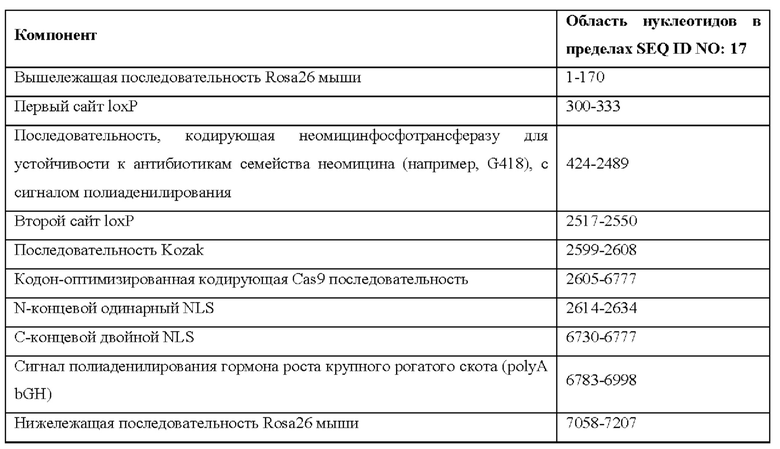 [00270] Таблица 7. Компоненты аллеля MAID2661 Cas9.
[00270] Таблица 7. Компоненты аллеля MAID2661 Cas9.

[00271] Кроме того, разработали две версии с экзогенными промоторами CAGG и последовательностями метки 3xFLAG. Первая версия включала в себя P2A-eGFP (MAID2658), а вторую версию разработали без P2A-eGFP (MAID2672). См. фигуру 14. Компоненты первой версии экспрессионной кассеты CAGG-Cas9 от 5' до 3' показаны в таблице 8, и компоненты первой версии аллеля CAGG-Cas9 после удаления фланкированной loxP-сайтами неомициновой кассеты показаны в таблице 9. Последовательность белка 3xFLAG-Cas9-P2A-eGFP, кодируемая этим аллелем, представлена в SEQ ID NO: 16. Компоненты второй версии экспрессионной кассеты CAGG-Cas9 от 5' до 3' показаны в таблице 10, и компоненты второй версии аллеля CAGG-Cas9 после удаления фланкированной loxP-сайтами кассеты неомицина показаны в таблице 11. Последовательность белка 3xFLAG-Cas9, кодируемая этим аллелем, представлена в SEQ ID NO: 22.
[00272] Таблица 8. Компоненты аллеля MAID2658 Cas9.
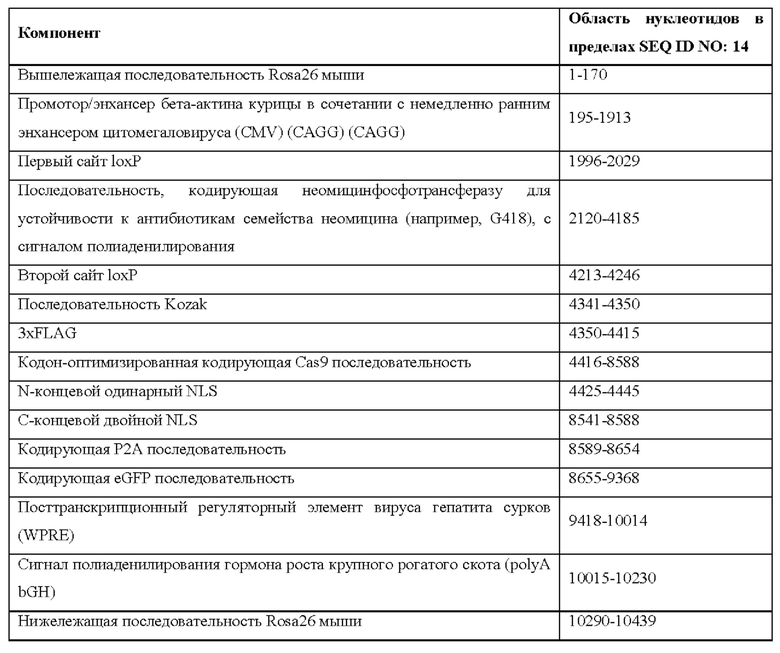
[00273] Таблица 9. Компоненты аллеля MAID2659 Cas9.
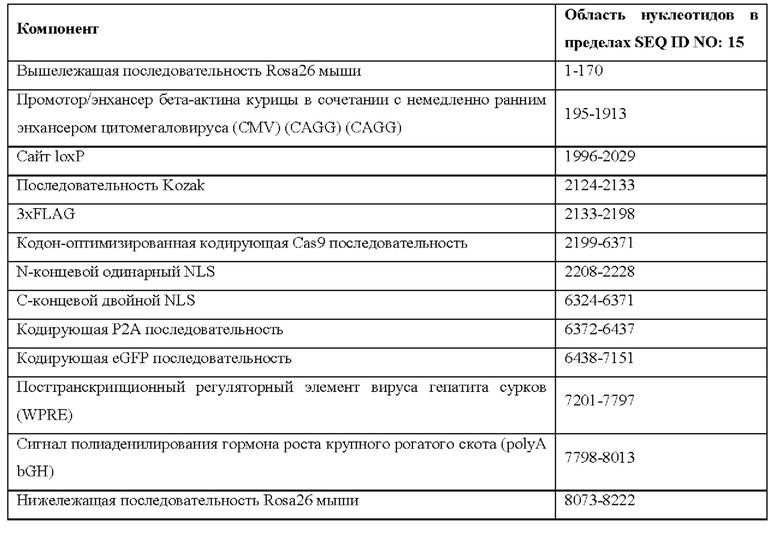
[00274] Таблица 10. Компоненты аллеля MAID2672 Cas9.
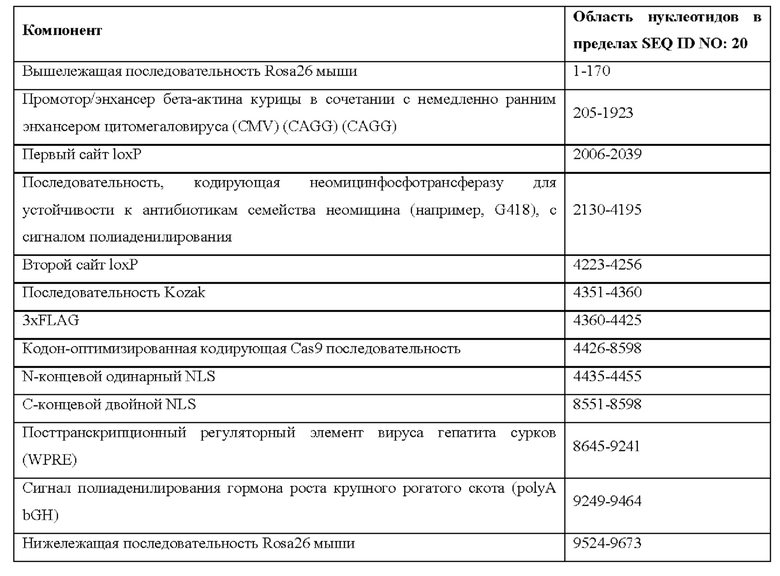
[00275] Таблица 11. Компоненты аллеля MAID2673 Cas9.

[00276] Для валидации системы MAID2599/MAID2600 mESC Cas9 с неомициновой кассетой и без нее (MAID2599 и MAID2600 соответственно) трансфицировали двумя sgPHК, нацеливающимися на области старт- и стоп-кодонов первого целевого гена. См. фигуру 2А. Эффективность расщепления Cas9 затем оценивали с помощью скрининга в отношении потери аллелей для оценки доли клонов mESC с мутациями вставки-делеции в сайтах расщепления Cas9, нацеленных с помощью гРНК. Также определили долю клонов mESC, в которых ДНК между сайтами расщепления Cas9 была удалена на одном или обоих целевых аллелях, вызывая нулевую мутацию. Cas9 был способен вызывать эти геномные изменения только тогда, когда были удалены неомициновая кассета и поли(А) (стоп-последовательность) (MAID2600). Чтобы лучше оценить возможности редактирования генома в отношении гомологичной репарации, вводили sgPHК, нацеленную на второй целевой ген, вместе с одноцепочечный олигодезоксинуклеотидом (ssODN) в качестве донора точечной мутации. См. фигуру 2 В. Описанную в настоящем документе конститутивную систему экспрессии Cas9 сравнивали с традиционными способами введения Cas9 и sgPHК через плазмиды вместе с ssODN. Описанная в настоящем документе система экспрессии Cas9, в сочетании с плазмидой, экспрессирующей sgPHК, была способна активировать индуцированные разрывом пути эндогенной репарации для включения требуемой точечной мутации с частотой, которая была равна той, когда и Cas9 и sgPHК экспрессировались из экзогенной плазмид. Тем не менее, когда описанную в настоящем документе систему экспрессии Cas9 объединяли с напрямую доставленной sgPHК, она индуцировала направляемый гомологией мутагенез методом микроинъекций почти в два раза эффективнее способов доставки с помощью плазмиды.
[00277] Для определения эффективности эндогенно экспрессируемого Cas9 у живых мышей, эти нацеленные mESC вводили с помощью микроинъекции в 8-клеточные эмбрионы мыши с использованием метода VELOCIMOUSE®. См., например, патенты США №№ US 7576259; US 7659442; US 7294754; заявку на выдачу патента США №2008/007800; и Poueymirou et al. (2007) Nature Biotech. 25(l):91-99, причем каждый из этих документов полностью включен в настоящий документ посредством ссылки для всех целей. В частности, в вителлиновом слое создавали небольшое отверстие для облегчения инъекции нацеленной mESC. Эти инъецированные 8-клеточные эмбрионы переносили суррогатным матерям для получения живых мышат, несущих трансген. После беременности у суррогатной матери инъецированные эмбрионы продуцировали мышей F0, которые не несут определяемого вклада эмбрионов хозяина. Мыши, полностью полученные из клеток ES, были нормальными, здоровыми и фертильными (с трансмиссией зародышевой линии). Ткань собирали у мышей F0 с удаленной кассетой (MAID2600) для визуализации GFP, экспрессии мРНК Cas9 и экспрессии белка Cas9. См. фигуры 3A-3F (светлопольная визуализация и визуализация GFP) и фигуры 4А-4С (экспрессия мРНК Cas9 на фигурах 4А и 4С и экспрессия белка на фигуре 4 В). На фигуре 3D показана экспрессия eGFP у гетерозиготных по Rosa26Cas9 мышей (MAID2600), но отсутствует соответствующая экспрессия eGFP у мышей дикого типа в печени. На фигуре 3F показана экспрессия eGFP у гетерозиготных по Rosa26Cas9 мышей, но отсутствует соответствующая экспрессия eGFP у мышей дикого типа в головном мозге. Аналогично, экспрессия мРНК Cas9, анализируемая с помощью ОТ-кПЦР, наблюдалась у гетерозиготных по Rosa26Cas9 мышей в головном мозге, сердце, почках, печени, легких, четырехглавых мышцах, селезенке и тимусе, но в соответствующих тканях из диких животных экспрессия мРНК Cas9 не наблюдалась, типа мышей. См. фигуры 4А и 4С. В экспериментах равные массовые количества РНК из каждой ткани анализировали с помощью ОТ-кПЦР. Данные показывают, что экспрессирующие Cas9 мыши экспрессируют мРНК Cas9 на легко обнаруживаемом уровне во всех тканях. Различные ткани собирали от экспрессирующих Cas9 мышей. Три ткани собирали у четырнадцати мышей и еще пять тканей собирали у четырех мышей для оценки различий от мыши к мыши, а также от ткани к ткани в пределах мыши. В каждой из этих тканей экстрагировали РНК. Геномную ДНК подвергали разложению таким образом, чтобы она не учитывалась в реакции кПЦР. РНК подвергали обратной транскрипции, а затем использовали анализ, специфический для Cas9, для обнаружения транскриптов Cas9. Как и ожидалось, мыши Cas9 показали значительную экспрессию (значения ct ниже 30), тогда как мыши WT показали значения ct 30 и выше, что указывает на отсутствие эндогенной экспрессии белка Cas9.
[00278] Аналогичным образом, экспрессия белка Cas9, определенная с помощью вестерн-блоттинга с использованием антитела ThermFisher Cas9 МА5-23519 в разведении 1:250 и использованием актина в качестве контроля, показала экспрессию белка Cas9 у гетерозиготных по Rosa26Cas9 мышей (MAID2600) в селезенке, печени и головном мозге, тогда как белок Cas9 не наблюдался в тех же тканях у мышей дикого типа. См. фигуру 4 В. Все три теста показали постоянный уровень экспрессии во всех исследуемых тканях.
[00279] Затем был проводили эксперимент по нокауту целевого гена 3, который кодирует белок, секретируемый печенью и обнаруживаемый в сыворотке, путем введения sgРНК в первичные гепатоциты, выделенные из мышей с удаленной кассетой Cas9 (MAID2600), посредством доставки липидными наночастицами (LNP). См. фигуру 5 В. В качестве контроля те же способы введения sgРНК спаривали с экзогенной экспрессией Cas9 в первичных гепатоцитах, выделенных у мышей дикого типа (WT). См. фигуру 5А. Негомологичное соединение концов затем оценивали секвенированием следующего поколения (NGS) для измерения частоты инделей в локусе целевого гена 3. В эксперименте было три условия: (1) опосредованная LNP доставка мРНК GFP и контрольной (т.е. мертвой) sgРНК; (2) опосредованная LNP доставка мРНК GFP и гРНК для целевого гена 3; и (3) опосредованная LNP доставка мРНК Cas9 и гРНК для целевого гена 3). Для каждого условия протестировали четыре концентрации мРНК: 15,6 нг/мл, 62,5 нг/мл, 250 нг/мл и 1000 нг/мл. В первичных гепатоцитах мыши дикого типа дозозависимое увеличение частоты вставок/делеций наблюдали только при введении как мРНК Cas9, так и гРНК для целевого гена 3. В противоположность этому, в первичных гепатоцитах экспрессирующие Cas9 мыши сходный дозозависимый эффект наблюдался при введении гидовой РНК для целевого гена 3 с контрольной мРНК GFP вместо мРНК Cas9, и уровень частоты вставки/делеции являлся по существу идентичным уровням, наблюдаемым, когда также вводят мРНК Cas9.
[00280] Затем проводили эксперимент по нокауту целевого гена 3 in vivo путем введения sgРНК мышам с удаленной кассетой Cas9 (MAID2600) посредством гидродинамической доставки ДНК (HDD), доставки липидными наночастицами (LNP) или введения аденоассоциированного вируса (AAV), несущего последовательность экспрессии sgРНК путем инъекции в хвостовую вену. См. фигуры 6A-6D. В качестве контроля те же способы введения sgРНК спаривали с экзогенной экспрессией Cas9 у мышей дикого типа (WT). Для доставки, опосредованной LNP, протестировали три группы мышей: (1) экспрессирующие Cas9 мыши (3 самца + 3 самки; 2 мг/кг контрольной sgРНК + GFP мРНК); (2) экспрессирующие Cas9 мыши (3 самца + 3 самки; 2 мг/кг sgРНК для целевого гена 3 + GFP мРНК); и (3) WT мыши (3 самца + 3 самки; 2 мг/кг sgРНК для целевого гена 3 + Cas9 мРНК). Для AAV-опосредованной доставки протестировали две группы мышей: (1) экспрессирующие Cas9 мыши (3 самца + 3 самки; AAV8-sgРНК для целевого гена 3); и (2) WT мыши (3 самца + 3 самки; AAV8-sgРНК для целевого гена 3 + AAV8-Cas9). Для HDD протестировали две группы мышей: (1) экспрессирующие Cas9 мыши (3 самца + 3 самки; sgРНК для целевого гена 3); и (2) WT мыши (3 самца + 3 самки; sgРНК для целевого гена 3 + Cas9). экспрессирующие Cas9 мыши характеризовались согласованно и значительно более нацеленной инактивацией гена, чем мыши WT с экзогенной экспрессией Cas9.
[00281] Неожиданно, AAV8-опосредованная доставка sgРНК целевого гена 3 эспрессирующим Cas9 мышам (MAID2600) являлась более эффективной, чем доставка, опосредованная LNP, или HDD при нацеливании на целевой ген 3 печени. См. фигуры 6A-6D. Кроме того, AAV8-опосредованная доставка sgРНК целевого гена 3 эспрессирующим Cas9 мышам, являлась более эффективной, чем AAV8-опосредованная доставка как sgРНК целевого гена, так и Cas9 мышам WT, тогда как между обоими условиями с использованием LNP-опосредованной доставки или HDD не наблюдалось значительного различия доставка. См. фигуры 6A-6D. Эти результаты показывают, что AAV-опосредованная доставка гидовых РНК эспрессирующим Cas9 мышам может быть особенно эффективным средством для испытания активности гРНК in vivo. Содержания в сыворотке белка, кодируемого целевым геном 3 (т.е. целевым белком 3), измеряли у самок и самцов мышей на 7 и 21 день после введения компонентов CRISPR/Cas. Среди испытанных мышей были экспрессирующие Cas9 мыши и мыши дикого типа. Контроли включали в себя экспрессирующих Cas9 мышей и мышей WT, которым не вводили ни sgРНК, ни Cas9, ни целевой ген 3. Гидродинамическая доставка гидовой РНК или комбинации Cas9 и гидовой РНК не снижала содержания в сыворотке целевого белка 3 в значительной степени по сравнению с контрольными мышами WT, которым не вводили ни Cas9, ни гидовую РНК. Для доставки, опосредованной LNP, введение sgРНК эспрессирующим Cas9 мышам приводило к аналогичным содержаниям в сыворотке целевого белка по сравнению с мышами WT, которым вводили как Cas9, так и sgРНК, и каждое из этих условий приводило к пониженным содержаниям в сыворотке целевого белка 3 приблизительно на 50% по сравнению с эспрессирующих Cas9 контрольными мышами, которым не вводили ни Cas9, ни гидовую РНК. Однако при AAV8-опосредованной доставке доставка sgРНК эспрессирующим Cas9 мышам приводила к снижению в несколько раз в содержаниях в сыворотке целевого белка 3 по сравнению с мышами WT, которым вводили и Cas9, и sgРНК, и еще более значительным снижением по сравнению с контрольными мышами WT, которым не вводили ни Cas9, ни гидовую РНК. К 21 дню содержания в сыворотке целевого белка 3 упали почти до предела обнаружения у экспрессирующих Cas9 мышей, которым sgРНК вводили посредством AAV8.
[00282] На фигуре 7 показано процентное отношение активности NHEJ (частота инделей) в целевом локусе гена 3 в печени у мышей дикого типа и у мышей с удаленной кассетой Cas9 (MAID2600) через один месяц после доставки sgРНК с помощью липидных наночастиц (LNP) отдельно или вместе с мРНК Cas9, гидродинамической доставки (HDD) плазмиды sgРНК отдельно или вместе с плазмидой Cas9 или AAV8-sgРНК отдельно или вместе с AAV8-Cas9. Процентное отношение клеток печени со вставками/делециями (инделами) в локусе измеряли с помощью NGS. Гидродинамическая доставка гидовой РНК или комбинации Cas9 и гидовой РНК приводила к низкому процентному отношению инделей. Для доставки, опосредованной LNP, введение sgРНК эспрессирующим Cas9 мышам приводило к аналогичному процентному отношению инделей по сравнению с мышами WT, которым вводили как Cas9, так и sgРНК, и каждое из этих условий приводило к уровню процентного отношения инделей, который составлял приблизительно 40%. Однако для доставки, опосредованной AAV8, доставка sgРНК эспрессирующим Cas9 мышам приводила к гораздо большему проценту инделей (~ 75%) по сравнению с мышами WT, которым вводили как Cas9, так и sgРНК (~ 35%), и еще более значительному увеличению по сравнению с контрольными мышами WT, которым не вводили ни Cas9, ни гидовую РНК.
[00283] Дальнейшее секвенирование следующего поколения (NGS) также выполняли в собранных тканях. Секвенирование ампликона затем использовали для оценки объема редактирования в собранных тканях. Целевые специфические праймеры предназначены для производства продукта ~ 300 п.н., который немного смещен от центра вокруг предполагаемого сайта разреза гида. Затем к праймерам добавляют последовательности «адаптера», которые позволяют штрих-кодировать отдельные образцы во вторичной реакции ПЦР. После добавления штрих-кодов образцы объединяют и загружают в MiSeq. Ожидается от пяти тысяч до десяти тысяч прочтений в представляющей интерес области. Затем запускают программы обработки данных для сопоставления прочтений, чтобы определить точную последовательность каждого редактирования. Затем программа подсчитывает количество WT (неотредактированных) прочтений и предоставляет разбивку типа редактирования, выполненного для всех отредактированных прочтений (оценка количества пар оснований, добавленных и/или удаленных в прогнозируемой области редактирования).
[00284] Затем проводили эксперимент по нокауту целевого гена 4, который кодирует связанный с мембраной гликопротеин II типа, путем введения Cas9 с sgРНК, нацеленной на экзона 2 целевого гена 4 в первичные гепатоциты, выделенные от мышей дикого типа (WT). Протестировали пять различных sgРНК (гиды 1-5). Рибонуклеопротеиновые комплексы Cas9/sgРНК (RNP) вводили в клетки с помощью липофектамина. Негомологичное соединение концов затем оценивали секвенированием следующего поколения (NGS) для измерения частот инделей в локусе целевого гена 4. Процентное редактирование является мерой общего количества событий NHEJ по отношению к общему количеству прочтений. События NHEJ считаются всеми редактированиями (вставка, удаление, изменение основания), которые происходят в 20 п. н. до и после сайта разреза. Процент редактирования для гидов с 1 по 5 являлся следующим: 35,4%, 37,4%, 43,8%, 51,2% и 55,8% соответственно (данные не показаны).
[00285] Затем проводили эксперимент по нокауту целевого гена 4 in vivo путем введения тех же пяти sgРНК отдельным мышам с удаленной кассетой Cas9 (MAID2600) посредством AAV8. В частности, отдельные гиды, экспрессируемые промотором U6, упаковывали в AAV8 и вводили 6-12-недельным мышам с удаленной кассетой Cas9 путем инъекции в хвостовую вену. Введенная вирусная нагрузка составляла 1×1011 - 1×1012, в приблизительном объеме, составляющем 50-100 мкл. Печень собирали через 3-4 недели после инъекции. Процентное редактирование рассчитывали так же, как в первичных гепатоцитах, и оно показано на фигуре 8А. Уровни редактирования соответствовали и фактически превышали уровни редактирования, наблюдаемые в первичных гепатоцитах. Кроме того, протестировали уровни экспрессии мРНК, транскрибированной из целевого гена 4. Как показано на фиг. 8В, каждая гРНК снижала относительные уровни мРНК, транскрибированной из целевого гена 4, в печени, собранной через 3-4 недели после инъекции.
[00286] Кроме того, провели эксперименты по исследованию процентного отношения редактирования в некоторых других целевых генах в печени. В каждом эксперименте возраст мышей составлял приблизительно 6-12 недель. Для каждого целевого гена разработали пять различных гидовых РНК по отношению к критически важным экзонам. Гидовые РНК доставляли посредством AAV8 путем инъекции в хвостовую вену с вирусными нагрузками, составляющими 1×1011 - 1×1012, в приблизительном объеме, составляющем 50-100 мкл. Печень собирали через 3-4 недели после инъекции. Процент редактирования определяли, как объяснено выше. Процент редактирования в печени у мышей с удаленной кассетой Cas9 (MAID2600) посредством доставки AAV8-rРНК показан в таблице 12. Наилучшая гРНК для каждого гена приводила к редактированию, составляющему 48%-70% в печени in vivo.
[00287] Таблица 12. Процентное отношение редактирования в печени.
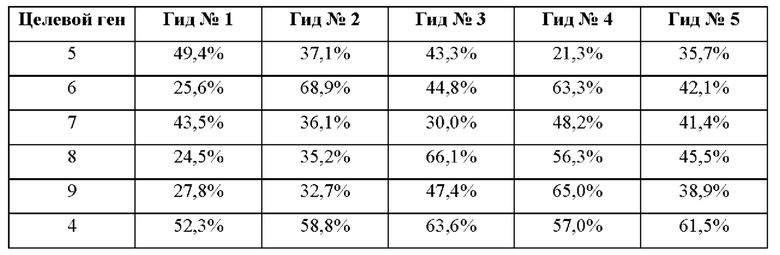
Пример 2. Валидация индуцибельности экспрессирующих Cas9 мышей
[00288] Аллель LSL-Cas9, описанный в примере 1 (MAID2599), включает в себя фланкированную loxP-сайтами область poly(A) (lox-stop-lox, или LSL) выше кодирующей Cas9 последовательности. Перед удалением кассеты под действием Cre-рекомбиназы ген устойчивости к неомицину, как правило, будет эффективно транскрибироваться и транслироваться; однако CDS Cas9, как правило, не экспрессируется из-за присутствия сильной области poly(A), которая может эффективно блокировать сквозную транскрипцию. После удаления неомициновой кассеты под действием Cre-рекомбиназы гибридная мРНК для белков Cas9 и GFP, как правило, конститутивно экспрессируется промотором Rosa26. Это делает индуцируемым аллель Cas9. Это является благоприятным по ряду причин. Возможность редактирования некоторых генов в определенных тканях (например, иммунных клетках) может быть вредной, наряду с тем, что это потенциально вызывает иммунный ответ. Кроме того, в определенных обстоятельствах мутация гена во всем целевом индивидууме может быть летальной, тогда как мутация гена в конкретной ткани или типе клетки будет благоприятной. Индуцируемая природа аллеля MAID2599 обеспечивает большую специфичность в отношении того, какой тип ткани и клетки редактируется путем активации Cas9 только тканеспецифическим или клеточноспецифическим образом.
[00289] Для исследования индуцибельности экспрессии Cas9 в печени in vivo липидные наночастицы (LNP), содержащие мРНК Cre-рекомбиназы, составили на Precision Nanosystems Benchtop NanoAssmblr. мРНК Cre от Trilink (№ по кат. 7211) разбавляли в 10 мМ цитрате натрия и объединяли через кассету NanoAssemblr в соотношении 3:1 с липидной комбинацией катионного липида, DSPC, холестерина и PEG-DMG в молярном соотношении 50:10:38,5:1,5. Этот состав легко усваивается печенью. Полученный LNP-Cre вводили с помощью инъекции через хвостовую вену мышей LSL-Cas9 (MAID2599) в дозе, составляющей 1 мг/кг. Контрольным мышам LNP-Cre не вводили. Через 1 неделю мышей умерщвляли и извлекали органы для анализа вестерн-блоттинга с использованием моноклонального антитела к Cas9 (7А9) (Invitrogen, №по кат. МА5-23519) и моноклонального антитела к актину (С4) (Millipore Sigma, № по кат. МАВ1501). Органы от мышей с удаленной кассетой Cas9 (MAID2600) использовали в качестве положительного контроля. У этих мышей кассета LSL уже была удалена рекомбиназой Сге. Результаты показаны на фигуре 9, на которой показано экспериментальное подтверждение концепции специфической для печени активации Cas9 с доставкой LNP-Cre для редактирования специфичного для печени гена.
[00290] Затем индуцибельность Cas9-опосредованного редактирования генов тестировали in vivo. LNP-Cre составляли, как описано выше. Мышам вводили дозу LNP-Cre и AAV8-rРНК, нацеленную на целевой ген 3 (коинъекция посредством инъекции в хвостовую вену) в следующих группах: (1)3 мыши LSL-Cas9, получавшие лечение с помощью LNP-Cre и AAV8-rРНК; (2) 3 мыши LSL-Cas9, получавшие лечение с помощью LNP-Cre и PBS; (3) 3 мыши LSL-Cas9, получавшие лечение с помощью PBS и AAV8- гРНК; (4) 3 мыши LSL-Cas9, получавшие только PBS; (5) 3 мыши Cas9 с удаленной кассетой, получавшие лечение с помощью LNP-Cre; (6) 3 мыши Cas9 с удаленной кассетой, получавшие лечение с помощью AAV8- гРНК; (7) 3 мыши Cas9 с удаленной кассетой, получавшие лечение с помощью PBS; и (8) 3 мыши WT (не получавшие лечение). В группах, в которые был доставлен LNP-Cre, он был доставлен в концентрации, составляющей 1 мг/кг. В группах, в которые была доставлена AAV8- гРНК, она была доставлена при вирусной нагрузке, составляющей приблизительно 2×1011. Через одну и три недели после инъекции у мышей брали кровь для анализа химического состава сыворотки и для измерения содержаний циркулирующего в сыворотке целевого белка 3. Через три недели ткани также собирали для NGS и для анализа вестерн-блоттинга. Содержания в сыворотке целевого белка 3 измеряли с помощью ELISA. Результаты показаны на фигуре 10. Доставка LNP-Cre мышам LSL-Cas9 вместе с AAV8-rРНК приводила к снижению содержаний в сыворотке целевого белка 3, что согласуется со снижением, наблюдаемым у мышей Cas9 с удаленной кассетой, которым доставляли AAV8-rРНК. Эти результаты ELISA соответствовали результатам NGS в отношении процентного отношения редактирования в целевом гене 3 в печени, выделенной из мышей через 3 недели после инъекции. См. фигуру 11.
[00291] Затем мышей LSL-Cas9 скрещивали с мышами альбумин-Cre от Jax (3601-Tg(Alb-cre)21Mgn; MAID3601), в которых промотор альбумина функционально связан с кодирующей последовательностью Cre-рекомбиназы и управляет его экспрессией в печени. После скрещивания у мышей собирали несколько тканей для вестерн-блоттинга. Соответствующие ткани собирали от мышей LSL-Cas9, которых не скрещивали с мышами альбумина-Cre. Затем проводили вестерн-блоттинг для измерения экспрессии Cas9 в печени и головном мозге. Актин использовали в качестве контроля нагрузки. Предсказанный размер Cas9 составлял 150,48 кДа, а предсказанный размер актина составлял 41,25 кДа. Использовали 17,5 мкг лизатов белка печени и лизатов белка головного мозга. TruCut v2 Cas9 (17,5 мкг) использовали в качестве положительного контроля. Как показано на фигуре 12А, экспрессия Cas9 наблюдалась в печени мышей LSL-Cas9/Alb-Cre, но не в печени мышей LSL-Cas9. Экспрессия Cas9 не наблюдалась в тканях головного мозга ни у одной из мышей (см. фигуру 12 В), подтверждая, что экспрессия Cas9 индуцировалась специфически в печени.
[00292] Проводили эксперимент для испытания Cas9-опосредованного редактирования генов in vivo у этих мышей. Эксперимент включал в себя пять групп мышей в возрасте 8-12-недель, которым вводили инъекцию AAV8-rРНК, нацеленной на целевой ген 3, посредством инъекции в хвостовую вену: (1)3 мыши LSL-Cas9:Alb-Cre (MAID2599 Het (гетерозиготные), MAID3601 Het); (2) 3 мыши WT:Alb-Cre (MAID2599 WT, MAID3601 Het); (3) 3 мыши LSL-Cas9:WT (MAID2599 Het, MAID3601 WT); (4) 3 мыши Cas9 (MAID2600 Het или Horn (гомозиготные)); и (5) 3 мыши 75/25 (50500 WT). Мыши из каждой группы также служили в качестве контролей, которым не вводили инъекцию AAV8-rРНК. В группах, которым доставляли AAV8-rРНК, ее доставляли с вирусной нагрузкой, составляющей приблизительно 2×1011. Через одну неделю после инъекции у мышей проводили забор крови для анализа химического состава сыворотки и анализов ELISA. Через три недели после инъекции у мышей проводили забор крови и собирали ткани. Результаты показаны на фигуре 13. Скрещивание мышей LSL-Cas9 с мышами альбумин-Cre и затем введение с помощью инъекции AAV8-rРНК приводило к снижению содержаний в сыворотке целевого белка 3, что согласуется со снижением, наблюдаемым у мышей Cas9 с удаленной кассетой, которым доставляли AAV8-гРНК.
[00293] Описанная в настоящем документе система экспрессирующих Cas9 мышей способна вызывать более надежное редактирование генов, чем другие способы, основанные на экзогенном введении Cas9. Кроме того, система может условно экспрессировать Cas9 на основе делеции неомициновой кассеты. Комбинируя эту систему с различными линиями мышей, у которых вырезание участка ДНК обеспечивается Cre, можно контролировать время индуцированного Cas9 редактирования генома и обеспечивать тканеспецифическую экспрессию Cas9 in vivo.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>Регенерон Фармасьютикалз, Инк.
<120>ЭКСПРЕССИРУЮЩИЕ CAS МЫШИНЫЕЭМБРИОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ И МЫШИ
И ИХ ПРИМЕНЕНИЯ
<130> 57766-516569
<150>US 62/539275
<151> 2017-07-31
<160> 38
<170>PatentIn версии 3.5
<210> 1
<211> 8628
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221> misc_feature
<222> (300)..(333)
<223> LoxP
<220>
<221> misc_feature
<222> (424)..(2489)
<223> Neo-PolyA
<220>
<221> misc_feature
<222> (2517)..(2550)
<223> LoxP
<220>
<221> misc_feature
<222> (2599)..(2608)
<223> Kozak
<220>
<221> misc_feature
<222> (2605)..(6777)
<223> Cas9
<220>
<221> misc_feature
<222> (2614)..(2634)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (6730)..(6777)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (6778)..(6843)
<223> P2A
<220>
<221> misc_feature
<222> (6844)..(7557)
<223> eGFP
<220>
<221> misc_feature
<222> (7607)..(8203)
<223> WPRE
<220>
<221> misc_feature
<222> (8204)..(8419)
<223>сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (8479)..(8628)
<223>Нижележащая последовательность Rosa26 мыши
<400> 1
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctagttgaccag 180
ctcggcggtgacctgcacgtctagggcgcagtagtccagggtttccttgatgatgtcata 240
cttatcctgtcccttttttttccacagggcgcgccactagtggatccggaacccttaata 300
taacttcgtataatgtatgctatacgaagttattaggtccctcgacctgcaggaattgtt 360
gacaattaatcatcggcatagtatatcggcatagtataatacgacaaggtgaggaactaa 420
accatgggatcggccattgaacaagatggattgcacgcaggttctccggccgcttgggtg 480
gagaggctattcggctatgactgggcacaacagacaatcggctgctctgatgccgccgtg 540
ttccggctgtcagcgcaggggcgcccggttctttttgtcaagaccgacctgtccggtgcc 600
ctgaatgaactgcaggacgaggcagcgcggctatcgtggctggccacgacgggcgttcct 660
tgcgcagctgtgctcgacgttgtcactgaagcgggaagggactggctgctattgggcgaa 720
gtgccggggcaggatctcctgtcatctcaccttgctcctgccgagaaagtatccatcatg 780
gctgatgcaatgcggcggctgcatacgcttgatccggctacctgcccattcgaccaccaa 840
gcgaaacatcgcatcgagcgagcacgtactcggatggaagccggtcttgtcgatcaggat 900
gatctggacgaagagcatcaggggctcgcgccagccgaactgttcgccaggctcaaggcg 960
cgcatgcccgacggcgatgatctcgtcgtgacccatggcgatgcctgcttgccgaatatc 1020
atggtggaaaatggccgcttttctggattcatcgactgtggccggctgggtgtggcggac 1080
cgctatcaggacatagcgttggctacccgtgatattgctgaagagcttggcggcgaatgg 1140
gctgaccgcttcctcgtgctttacggtatcgccgctcccgattcgcagcgcatcgccttc 1200
tatcgccttcttgacgagttcttctgaggggatccgctgtaagtctgcagaaattgatga 1260
tctattaaacaataaagatgtccactaaaatggaagtttttcctgtcatactttgttaag 1320
aagggtgagaacagagtacctacattttgaatggaaggattggagctacgggggtggggg 1380
tggggtgggattagataaatgcctgctctttactgaaggctctttactattgctttatga 1440
taatgtttcatagttggatatcataatttaaacaagcaaaaccaaattaagggccagctc 1500
attcctccca ctcatgatct atagatctat agatctctcg tgggatcatt gtttttctct 1560
tgattcccac tttgtggttc taagtactgt ggtttccaaa tgtgtcagtt tcatagcctg 1620
aagaacgaga tcagcagcct ctgttccaca tacacttcat tctcagtatt gttttgccaa 1680
gttctaattc catcagaagc ttgcagatct gcgactctag aggatctgcg actctagagg 1740
atcataatca gccataccac atttgtagag gttttacttg ctttaaaaaa cctcccacac 1800
ctccccctga acctgaaaca taaaatgaat gcaattgttg ttgttaactt gtttattgca 1860
gcttataatg gttacaaata aagcaatagc atcacaaatt tcacaaataa agcatttttt 1920
tcactgcatt ctagttgtgg tttgtccaaa ctcatcaatg tatcttatca tgtctggatc 1980
tgcgactcta gaggatcata atcagccata ccacatttgt agaggtttta cttgctttaa 2040
aaaacctccc acacctcccc ctgaacctga aacataaaat gaatgcaatt gttgttgtta 2100
acttgtttat tgcagcttat aatggttaca aataaagcaa tagcatcaca aatttcacaa 2160
ataaagcatt tttttcactg cattctagtt gtggtttgtc caaactcatc aatgtatctt 2220
atcatgtctg gatctgcgac tctagaggat cataatcagc cataccacat ttgtagaggt 2280
tttacttgct ttaaaaaacc tcccacacct ccccctgaac ctgaaacata aaatgaatgc 2340
aattgttgtt gttaacttgt ttattgcagc ttataatggt tacaaataaa gcaatagcat 2400
cacaaatttc acaaataaag catttttttc actgcattct agttgtggtt tgtccaaact 2460
catcaatgta tcttatcatg tctggatccc catcaagctg atccggaacc cttaatataa 2520
cttcgtataa tgtatgctat acgaagttat taggtccctc gacctgcagc ccaagctagt 2580
gcccgggaat tcgctagggc caccatggac aagcccaaga aaaagcggaa agtgaagtac 2640
agcatcggcc tggacatcgg caccaactct gtgggctggg ccgtgatcac cgacgagtac 2700
aaggtgccca gcaagaaatt caaggtgctg ggcaacaccg acaggcacag catcaagaag 2760
aacctgatcg gcgccctgct gttcgacagc ggcgaaacag ccgaggccac cagactgaag 2820
agaaccgcca gaagaagata caccaggcgg aagaacagga tctgctatct gcaagagatc 2880
ttcagcaacg agatggccaa ggtggacgac agcttcttcc acagactgga agagtccttc 2940
ctggtggaag aggacaagaa gcacgagaga caccccatct tcggcaacat cgtggacgag 3000
gtggcctacc acgagaagta ccccaccatc taccacctga gaaagaaact ggtggacagc 3060
accgacaagg ccgacctgag actgatctac ctggccctgg cccacatgat caagttcaga 3120
ggccacttcc tgatcgaggg cgacctgaac cccgacaaca gcgacgtgga caagctgttc 3180
atccagctgg tgcagaccta caaccagctg ttcgaggaaa accccatcaa cgccagcggc 3240
gtggacgcca aggctatcct gtctgccaga ctgagcaaga gcagaaggct ggaaaatctg 3300
atcgcccagc tgcccggcga gaagaagaac ggcctgttcg gcaacctgat tgccctgagc 3360
ctgggcctga cccccaactt caagagcaac ttcgacctgg ccgaggatgc caaactgcag 3420
ctgagcaagg acacctacga cgacgacctg gacaacctgc tggcccagat cggcgaccag 3480
tacgccgacc tgttcctggc cgccaagaac ctgtctgacg ccatcctgct gagcgacatc 3540
ctgagagtga acaccgagat caccaaggcc cccctgagcg cctctatgat caagagatac 3600
gacgagcacc accaggacct gaccctgctg aaagctctcg tgcggcagca gctgcctgag 3660
aagtacaaag aaatcttctt cgaccagagc aagaacggct acgccggcta catcgatggc 3720
ggcgctagcc aggaagagtt ctacaagttc atcaagccca tcctggaaaa gatggacggc 3780
accgaggaac tgctcgtgaa gctgaacaga gaggacctgc tgagaaagca gagaaccttc 3840
gacaacggca gcatccccca ccagatccac ctgggagagc tgcacgctat cctgagaagg 3900
caggaagatt tttacccatt cctgaaggac aaccgggaaa agatcgagaa gatcctgacc 3960
ttcaggatcc cctactacgt gggccccctg gccagaggca acagcagatt cgcctggatg 4020
accagaaaga gcgaggaaac catcaccccc tggaacttcg aggaagtggt ggacaagggc 4080
gccagcgccc agagcttcat cgagagaatg acaaacttcg ataagaacct gcccaacgag 4140
aaggtgctgc ccaagcacag cctgctgtac gagtacttca ccgtgtacaa cgagctgacc 4200
aaagtgaaat acgtgaccga gggaatgaga aagcccgcct tcctgagcgg cgagcagaaa 4260
aaggccatcg tggacctgct gttcaagacc aacagaaaag tgaccgtgaa gcagctgaaa 4320
gaggactact tcaagaaaat cgagtgcttc gactccgtgg aaatctccgg cgtggaagat 4380
agattcaacg cctccctggg cacataccac gatctgctga aaattatcaa ggacaaggac 4440
ttcctggata acgaagagaa cgaggacatt ctggaagata tcgtgctgac cctgacactg 4500
tttgaggacc gcgagatgat cgaggaaagg ctgaaaacct acgctcacct gttcgacgac 4560
aaagtgatga agcagctgaa gagaaggcgg tacaccggct ggggcaggct gagcagaaag 4620
ctgatcaacg gcatcagaga caagcagagc ggcaagacaa tcctggattt cctgaagtcc 4680
gacggcttcg ccaaccggaa cttcatgcag ctgatccacg acgacagcct gacattcaaa 4740
gaggacatcc agaaagccca ggtgtccggc cagggcgact ctctgcacga gcatatcgct 4800
aacctggccg gcagccccgc tatcaagaag ggcatcctgc agacagtgaa ggtggtggac 4860
gagctcgtga aagtgatggg cagacacaag cccgagaaca tcgtgatcga gatggctaga 4920
gagaaccaga ccacccagaa gggacagaag aactcccgcg agaggatgaa gagaatcgaa 4980
gagggcatca aagagctggg cagccagatc ctgaaagaac accccgtgga aaacacccag 5040
ctgcagaacg agaagctgta cctgtactac ctgcagaatg gccgggatat gtacgtggac 5100
caggaactgg acatcaacag actgtccgac tacgatgtgg accatatcgt gcctcagagc 5160
tttctgaagg acgactccat cgataacaaa gtgctgactc ggagcgacaa gaacagaggc 5220
aagagcgaca acgtgccctc cgaagaggtc gtgaagaaga tgaagaacta ctggcgacag 5280
ctgctgaacg ccaagctgat tacccagagg aagttcgata acctgaccaa ggccgagaga 5340
ggcggcctga gcgagctgga taaggccggc ttcatcaaga ggcagctggt ggaaaccaga 5400
cagatcacaa agcacgtggc acagatcctg gactcccgga tgaacactaa gtacgacgaa 5460
aacgataagc tgatccggga agtgaaagtg atcaccctga agtccaagct ggtgtccgat 5520
ttccggaagg atttccagtt ttacaaagtg cgcgagatca acaactacca ccacgcccac 5580
gacgcctacc tgaacgccgt cgtgggaacc gccctgatca aaaagtaccc taagctggaa 5640
agcgagttcg tgtacggcga ctacaaggtg tacgacgtgc ggaagatgat cgccaagagc 5700
gagcaggaaa tcggcaaggc taccgccaag tacttcttct acagcaacat catgaacttt 5760
ttcaagaccg aaatcaccct ggccaacggc gagatcagaa agcgccctct gatcgagaca 5820
aacggcgaaa ccggggagat cgtgtgggat aagggcagag acttcgccac agtgcgaaag 5880
gtgctgagca tgccccaagt gaatatcgtg aaaaagaccg aggtgcagac aggcggcttc 5940
agcaaagagt ctatcctgcc caagaggaac agcgacaagc tgatcgccag aaagaaggac 6000
tgggacccca agaagtacgg cggcttcgac agccctaccg tggcctactc tgtgctggtg 6060
gtggctaagg tggaaaaggg caagtccaag aaactgaaga gtgtgaaaga gctgctgggg 6120
atcaccatca tggaaagaag cagctttgag aagaacccta tcgactttct ggaagccaag 6180
ggctacaaag aagtgaaaaa ggacctgatc atcaagctgc ctaagtactc cctgttcgag 6240
ctggaaaacg gcagaaagag aatgctggcc tctgccggcg aactgcagaa gggaaacgag 6300
ctggccctgc ctagcaaata tgtgaacttc ctgtacctgg cctcccacta tgagaagctg 6360
aagggcagcc ctgaggacaa cgaacagaaa cagctgtttg tggaacagca taagcactac 6420
ctggacgaga tcatcgagca gatcagcgag ttctccaaga gagtgatcct ggccgacgcc 6480
aatctggaca aggtgctgtc tgcctacaac aagcacaggg acaagcctat cagagagcag 6540
gccgagaata tcatccacct gttcaccctg acaaacctgg gcgctcctgc cgccttcaag 6600
tactttgaca ccaccatcga ccggaagagg tacaccagca ccaaagaggt gctggacgcc 6660
accctgatcc accagagcat caccggcctg tacgagacaa gaatcgacct gtctcagctg 6720
ggaggcgaca agagacctgc cgccactaag aaggccggac aggccaaaaa gaagaaggga 6780
agcggagcca ctaacttctc cctgttgaaa caagcagggg atgtcgaaga gaatcccggg 6840
ccagtgagca agggcgagga gctgttcacc ggggtggtgc ccatcctggt cgagctggac 6900
ggcgacgtaa acggccacaa gttcagcgtg tccggcgagg gcgagggcga tgccacctac 6960
ggcaagctga ccctgaagtt catctgcacc accggcaagc tgcccgtgcc ctggcccacc 7020
ctcgtgacca ccctgaccta cggcgtgcag tgcttcagcc gctaccccga ccacatgaag 7080
cagcacgact tcttcaagtc cgccatgccc gaaggctacg tccaggagcg caccatcttc 7140
ttcaaggacg acggcaacta caagacccgc gccgaggtga agttcgaggg cgacaccctg 7200
gtgaaccgca tcgagctgaa gggcatcgac ttcaaggagg acggcaacat cctggggcac 7260
aagctggagt acaactacaa cagccacaac gtctatatca tggccgacaa gcagaagaac 7320
ggcatcaagg tgaacttcaa gatccgccac aacatcgagg acggcagcgt gcagctcgcc 7380
gaccactacc agcagaacac ccccatcggc gacggccccg tgctgctgcc cgacaaccac 7440
tacctgagca cccagtccgc cctgagcaaa gaccccaacg agaagcgcga tcacatggtc 7500
ctgctggagt tcgtgaccgc cgccgggatc actctcggca tggacgagct gtacaagtaa 7560
acgcgtatgc atggccggcc ctgcaggaat tcgatatcaa gcttatcgat aatcaacctc 7620
tggattacaa aatttgtgaa agattgactg gtattcttaa ctatgttgct ccttttacgc 7680
tatgtggata cgctgcttta atgcctttgt atcatgctat tgcttcccgt atggctttca 7740
ttttctcctc cttgtataaa tcctggttgc tgtctcttta tgaggagttg tggcccgttg 7800
tcaggcaacg tggcgtggtg tgcactgtgt ttgctgacgc aacccccact ggttggggca 7860
ttgccaccac ctgtcagctc ctttccggga ctttcgcttt ccccctccct attgccacgg 7920
cggaactcat cgccgcctgc cttgcccgct gctggacagg ggctcggctg ttgggcactg 7980
acaattccgt ggtgttgtcg gggaaatcat cgtcctttcc ttggctgctc gcctgtgttg 8040
ccacctggat tctgcgcggg acgtccttct gctacgtccc ttcggccctc aatccagcgg 8100
accttccttc ccgcggcctg ctgccggctc tgcggcctct tccgcgtctt cgccttcgcc 8160
ctcagacgag tcggatctcc ctttgggccg cctccccgca tcgcgacctc gacctcgact 8220
gtgccttcta gttgccagcc atctgttgtt tgcccctccc ccgtgccttc cttgaccctg 8280
gaaggtgcca ctcccactgt cctttcctaa taaaatgagg aaattgcatc gcattgtctg 8340
agtaggtgtc attctattct ggggggtggg gtggggcagg acagcaaggg ggaggattgg 8400
gaagacaatg gcaggcatgc tggggaacta gtggtgccag ggcgtgccct tgggctcccc 8460
gggcgcggcg gccatcgctc gagtaaaatt ggagggacaa gacttcccac agattttcgg 8520
ttttgtcggg aagtttttta ataggggcaa ataaggaaaa tgggaggata ggtagtcatc 8580
tggggtttta tgcagcaaaa ctacaggtta ttattgcttg tgatccgc 8628
<210> 2
<211> 18
<212> PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 2
Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro
1 5 10 15
GlyPro
<210> 3
<211> 19
<212>PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 3
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
1 5 10 15
ProGlyPro
<210> 4
<211> 20
<212>PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 4
GlnCysThrAsnTyrAlaLeuLeuLysLeuAlaGlyAspValGluSer
1 5 10 15
AsnProGlyPro
20
<210> 5
<211> 22
<212>PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 5
ValLysGlnThrLeuAsnPheAspLeuLeuLysLeuAlaGlyAspVal
1 5 10 15
Glu Ser Asn Pro Gly Pro
20
<210> 6
<211> 82
<212> RNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 6
guuggaacca uucaaaacag cauagcaagu uaaaauaagg cuaguccguu aucaacuuga 60
aaaaguggcaccgagucggugc 82
<210> 7
<211> 76
<212>RNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 7
guuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaagu 60
ggcaccgagucggugc 76
<210> 8
<211> 86
<212>RNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 8
guuuaagagcuaugcuggaaacagcauagcaaguuuaaauaaggcuaguccguuaucaac 60
uugaaaaaguggcaccgagucggugc 86
<210> 9
<211> 23
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> misc_feature
<222> (2)..(21)
<223> n = A, T, Cили G
<400> 9
gnnnnnnnnnnnnnnnnnnnngg 23
<210> 10
<211> 23
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> misc_feature
<222> (1)..(21)
<223> n = A, T, Cили G
<400> 10
nnnnnnnnnnnnnnnnnnnnngg 23
<210> 11
<211> 25
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> misc_feature
<222> (3)..(23)
<223> n = A, T, Cили G
<400> 11
ggnnnnnnnnnnnnnnnnnnnnngg 25
<210> 12
<211> 6411
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221> misc_feature
<222> (300)..(333)
<223> LoxP
<220>
<221> misc_feature
<222> (382)..(391)
<223> Kozak
<220>
<221> misc_feature
<222> (388)..(4560)
<223> Cas9
<220>
<221> misc_feature
<222> (397)..(417)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (4513)..(4560)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (4561)..(4626)
<223> P2A
<220>
<221> misc_feature
<222> (4627)..(5340)
<223> eGFP
<220>
<221> misc_feature
<222> (5390)..(5986)
<223> WPRE
<220>
<221> misc_feature
<222> (5987)..(6202)
<223>сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (6262)..(6411)
<223>Нижележащая последовательность Rosa26 мыши
<400> 12
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctagttgaccag 180
ctcggcggtgacctgcacgtctagggcgcagtagtccagggtttccttgatgatgtcata 240
cttatcctgtcccttttttttccacagggcgcgccactagtggatccggaacccttaata 300
taacttcgtataatgtatgctatacgaagttattaggtccctcgacctgcagcccaagct 360
agtgcccgggaattcgctagggccaccatggacaagcccaagaaaaagcggaaagtgaag 420
tacagcatcggcctggacatcggcaccaactctgtgggctgggccgtgatcaccgacgag 480
tacaaggtgcccagcaagaaattcaaggtgctgggcaacaccgacaggcacagcatcaag 540
aagaacctgatcggcgccctgctgttcgacagcggcgaaacagccgaggccaccagactg 600
aagagaaccgccagaagaagatacaccaggcggaagaacaggatctgctatctgcaagag 660
atcttcagcaacgagatggccaaggtggacgacagcttcttccacagactggaagagtcc 720
ttcctggtggaagaggacaagaagcacgagagacaccccatcttcggcaacatcgtggac 780
gaggtggcctaccacgagaagtaccccaccatctaccacctgagaaagaaactggtggac 840
agcaccgacaaggccgacctgagactgatctacctggccctggcccacatgatcaagttc 900
agaggccacttcctgatcgagggcgacctgaaccccgacaacagcgacgtggacaagctg 960
ttcatccagctggtgcagacctacaaccagctgttcgaggaaaaccccatcaacgccagc 1020
ggcgtggacgccaaggctatcctgtctgccagactgagcaagagcagaaggctggaaaat 1080
ctgatcgcccagctgcccggcgagaagaagaacggcctgttcggcaacctgattgccctg 1140
agcctgggcctgacccccaacttcaagagcaacttcgacctggccgaggatgccaaactg 1200
cagctgagcaaggacacctacgacgacgacctggacaacctgctggcccagatcggcgac 1260
cagtacgccgacctgttcctggccgccaagaacctgtctgacgccatcctgctgagcgac 1320
atcctgagagtgaacaccgagatcaccaaggcccccctgagcgcctctatgatcaagaga 1380
tacgacgagcaccaccaggacctgaccctgctgaaagctctcgtgcggcagcagctgcct 1440
gagaagtacaaagaaatcttcttcgaccagagcaagaacggctacgccggctacatcgat 1500
ggcggcgctagccaggaagagttctacaagttcatcaagcccatcctggaaaagatggac 1560
ggcaccgaggaactgctcgtgaagctgaacagagaggacctgctgagaaagcagagaacc 1620
ttcgacaacggcagcatcccccaccagatccacctgggagagctgcacgctatcctgaga 1680
aggcaggaagatttttacccattcctgaaggacaaccgggaaaagatcgagaagatcctg 1740
accttcaggatcccctactacgtgggccccctggccagaggcaacagcagattcgcctgg 1800
atgaccagaaagagcgaggaaaccatcaccccctggaacttcgaggaagtggtggacaag 1860
ggcgccagcgcccagagcttcatcgagagaatgacaaacttcgataagaacctgcccaac 1920
gagaaggtgctgcccaagcacagcctgctgtacgagtacttcaccgtgtacaacgagctg 1980
accaaagtgaaatacgtgaccgagggaatgagaaagcccgccttcctgagcggcgagcag 2040
aaaaaggccatcgtggacctgctgttcaagaccaacagaaaagtgaccgtgaagcagctg 2100
aaagaggactacttcaagaaaatcgagtgcttcgactccgtggaaatctccggcgtggaa 2160
gatagattcaacgcctccctgggcacataccacgatctgctgaaaattatcaaggacaag 2220
gacttcctggataacgaagagaacgaggacattctggaagatatcgtgctgaccctgaca 2280
ctgtttgaggaccgcgagatgatcgaggaaaggctgaaaacctacgctcacctgttcgac 2340
gacaaagtgatgaagcagctgaagagaaggcggtacaccggctggggcaggctgagcaga 2400
aagctgatcaacggcatcagagacaagcagagcggcaagacaatcctggatttcctgaag 2460
tccgacggcttcgccaaccggaacttcatgcagctgatccacgacgacagcctgacattc 2520
aaagaggacatccagaaagcccaggtgtccggccagggcgactctctgcacgagcatatc 2580
gctaacctggccggcagccccgctatcaagaagggcatcctgcagacagtgaaggtggtg 2640
gacgagctcgtgaaagtgatgggcagacacaagcccgagaacatcgtgatcgagatggct 2700
agagagaaccagaccacccagaagggacagaagaactcccgcgagaggatgaagagaatc 2760
gaagagggcatcaaagagctgggcagccagatcctgaaagaacaccccgtggaaaacacc 2820
cagctgcagaacgagaagctgtacctgtactacctgcagaatggccgggatatgtacgtg 2880
gaccaggaactggacatcaacagactgtccgactacgatgtggaccatatcgtgcctcag 2940
agctttctgaaggacgactccatcgataacaaagtgctgactcggagcgacaagaacaga 3000
ggcaagagcgacaacgtgccctccgaagaggtcgtgaagaagatgaagaactactggcga 3060
cagctgctgaacgccaagctgattacccagaggaagttcgataacctgaccaaggccgag 3120
agaggcggcctgagcgagctggataaggccggcttcatcaagaggcagctggtggaaacc 3180
agacagatcacaaagcacgtggcacagatcctggactcccggatgaacactaagtacgac 3240
gaaaacgataagctgatccgggaagtgaaagtgatcaccctgaagtccaagctggtgtcc 3300
gatttccggaaggatttccagttttacaaagtgcgcgagatcaacaactaccaccacgcc 3360
cacgacgcctacctgaacgccgtcgtgggaaccgccctgatcaaaaagtaccctaagctg 3420
gaaagcgagttcgtgtacggcgactacaaggtgtacgacgtgcggaagatgatcgccaag 3480
agcgagcaggaaatcggcaaggctaccgccaagtacttcttctacagcaacatcatgaac 3540
tttttcaagaccgaaatcaccctggccaacggcgagatcagaaagcgccctctgatcgag 3600
acaaacggcgaaaccggggagatcgtgtgggataagggcagagacttcgccacagtgcga 3660
aaggtgctgagcatgccccaagtgaatatcgtgaaaaagaccgaggtgcagacaggcggc 3720
ttcagcaaagagtctatcctgcccaagaggaacagcgacaagctgatcgccagaaagaag 3780
gactgggaccccaagaagtacggcggcttcgacagccctaccgtggcctactctgtgctg 3840
gtggtggctaaggtggaaaagggcaagtccaagaaactgaagagtgtgaaagagctgctg 3900
gggatcaccatcatggaaagaagcagctttgagaagaaccctatcgactttctggaagcc 3960
aagggctacaaagaagtgaaaaaggacctgatcatcaagctgcctaagtactccctgttc 4020
gagctggaaaacggcagaaagagaatgctggcctctgccggcgaactgcagaagggaaac 4080
gagctggccctgcctagcaaatatgtgaacttcctgtacctggcctcccactatgagaag 4140
ctgaagggcagccctgaggacaacgaacagaaacagctgtttgtggaacagcataagcac 4200
tacctggacgagatcatcgagcagatcagcgagttctccaagagagtgatcctggccgac 4260
gccaatctggacaaggtgctgtctgcctacaacaagcacagggacaagcctatcagagag 4320
caggccgagaatatcatccacctgttcaccctgacaaacctgggcgctcctgccgccttc 4380
aagtactttgacaccaccatcgaccggaagaggtacaccagcaccaaagaggtgctggac 4440
gccaccctgatccaccagagcatcaccggcctgtacgagacaagaatcgacctgtctcag 4500
ctgggaggcgacaagagacctgccgccactaagaaggccggacaggccaaaaagaagaag 4560
ggaagcggagccactaacttctccctgttgaaacaagcaggggatgtcgaagagaatccc 4620
gggccagtgagcaagggcgaggagctgttcaccggggtggtgcccatcctggtcgagctg 4680
gacggcgacgtaaacggccacaagttcagcgtgtccggcgagggcgagggcgatgccacc 4740
tacggcaagctgaccctgaagttcatctgcaccaccggcaagctgcccgtgccctggccc 4800
accctcgtgaccaccctgacctacggcgtgcagtgcttcagccgctaccccgaccacatg 4860
aagcagcacgacttcttcaagtccgccatgcccgaaggctacgtccaggagcgcaccatc 4920
ttcttcaaggacgacggcaactacaagacccgcgccgaggtgaagttcgagggcgacacc 4980
ctggtgaaccgcatcgagctgaagggcatcgacttcaaggaggacggcaacatcctgggg 5040
cacaagctggagtacaactacaacagccacaacgtctatatcatggccgacaagcagaag 5100
aacggcatcaaggtgaacttcaagatccgccacaacatcgaggacggcagcgtgcagctc 5160
gccgaccactaccagcagaacacccccatcggcgacggccccgtgctgctgcccgacaac 5220
cactacctgagcacccagtccgccctgagcaaagaccccaacgagaagcgcgatcacatg 5280
gtcctgctggagttcgtgaccgccgccgggatcactctcggcatggacgagctgtacaag 5340
taaacgcgtatgcatggccggccctgcaggaattcgatatcaagcttatcgataatcaac 5400
ctctggattacaaaatttgtgaaagattgactggtattcttaactatgttgctcctttta 5460
cgctatgtggatacgctgctttaatgcctttgtatcatgctattgcttcccgtatggctt 5520
tcattttctcctccttgtataaatcctggttgctgtctctttatgaggagttgtggcccg 5580
ttgtcaggcaacgtggcgtggtgtgcactgtgtttgctgacgcaacccccactggttggg 5640
gcattgccaccacctgtcagctcctttccgggactttcgctttccccctccctattgcca 5700
cggcggaactcatcgccgcctgccttgcccgctgctggacaggggctcggctgttgggca 5760
ctgacaattccgtggtgttgtcggggaaatcatcgtcctttccttggctgctcgcctgtg 5820
ttgccacctggattctgcgcgggacgtccttctgctacgtcccttcggccctcaatccag 5880
cggaccttccttcccgcggcctgctgccggctctgcggcctcttccgcgtcttcgccttc 5940
gccctcagacgagtcggatctccctttgggccgcctccccgcatcgcgacctcgacctcg 6000
actgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgacc 6060
ctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgt 6120
ctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggat 6180
tgggaagacaatggcaggcatgctggggaactagtggtgccagggcgtgcccttgggctc 6240
cccgggcgcggcggccatcgctcgagtaaaattggagggacaagacttcccacagatttt 6300
cggttttgtcgggaagttttttaataggggcaaataaggaaaatgggaggataggtagtc 6360
atctggggttttatgcagcaaaactacaggttattattgcttgtgatccgc 6411
<210> 13
<211> 1651
<212>PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>MISC_FEATURE
<222> (1)..(1391)
<223> Cas9
<220>
<221> MISC_FEATURE
<222> (1392)..(1413)
<223> P2A
<220>
<221> MISC_FEATURE
<222> (1414)..(1651)
<223> eGFP
<400> 13
Met Asp Lys Pro Lys Lys Lys Arg Lys Val Lys Tyr Ser Ile Gly Leu
1 5 10 15
Asp Ile Gly Thr Asn Ser Val Gly Trp Ala Val Ile Thr Asp Glu Tyr
20 25 30
Lys Val Pro Ser Lys Lys Phe Lys Val Leu Gly Asn Thr Asp Arg His
35 40 45
Ser Ile Lys Lys Asn Leu Ile Gly Ala Leu Leu Phe Asp Ser Gly Glu
50 55 60
Thr Ala Glu Ala Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg Tyr Thr
65 70 75 80
Arg Arg Lys Asn Arg Ile Cys Tyr Leu Gln Glu Ile Phe Ser Asn Glu
85 90 95
Met Ala Lys Val Asp Asp Ser Phe Phe His Arg Leu Glu Glu Ser Phe
100 105 110
Leu Val Glu Glu Asp Lys Lys His Glu Arg His Pro Ile Phe Gly Asn
115 120 125
Ile Val Asp Glu Val Ala Tyr His Glu Lys Tyr Pro Thr Ile Tyr His
130 135 140
Leu Arg Lys Lys Leu Val Asp Ser Thr Asp Lys Ala Asp Leu Arg Leu
145 150 155 160
Ile Tyr Leu Ala Leu Ala His Met Ile Lys Phe Arg Gly His Phe Leu
165 170 175
Ile Glu Gly Asp Leu Asn Pro Asp Asn Ser Asp Val Asp Lys Leu Phe
180 185 190
Ile Gln Leu Val Gln Thr Tyr Asn Gln Leu Phe Glu Glu Asn Pro Ile
195 200 205
Asn Ala Ser Gly Val Asp Ala Lys Ala Ile Leu Ser Ala Arg Leu Ser
210 215 220
Lys Ser Arg Arg Leu Glu Asn Leu Ile Ala Gln Leu Pro Gly Glu Lys
225 230 235 240
Lys Asn Gly Leu Phe Gly Asn Leu Ile Ala Leu Ser Leu Gly Leu Thr
245 250 255
Pro Asn Phe Lys Ser Asn Phe Asp Leu Ala Glu Asp Ala Lys Leu Gln
260 265 270
Leu Ser Lys Asp Thr Tyr Asp Asp Asp Leu Asp Asn Leu Leu Ala Gln
275 280 285
Ile Gly Asp Gln Tyr Ala Asp Leu Phe Leu Ala Ala Lys Asn Leu Ser
290 295 300
Asp Ala Ile Leu Leu Ser Asp Ile Leu Arg Val Asn Thr Glu Ile Thr
305 310 315 320
Lys Ala Pro Leu Ser Ala Ser Met Ile Lys Arg Tyr Asp Glu His His
325 330 335
Gln Asp Leu Thr Leu Leu Lys Ala Leu Val Arg Gln Gln Leu Pro Glu
340 345 350
Lys Tyr Lys Glu Ile Phe Phe Asp Gln Ser Lys Asn Gly Tyr Ala Gly
355 360 365
Tyr Ile Asp Gly Gly Ala Ser Gln Glu Glu Phe Tyr Lys Phe Ile Lys
370 375 380
Pro Ile Leu Glu Lys Met Asp Gly Thr Glu Glu Leu Leu Val Lys Leu
385 390 395 400
Asn Arg Glu Asp Leu Leu Arg Lys Gln Arg Thr Phe Asp Asn Gly Ser
405 410 415
Ile Pro His Gln Ile His Leu Gly Glu Leu His Ala Ile Leu Arg Arg
420 425 430
Gln Glu Asp Phe Tyr Pro Phe Leu Lys Asp Asn Arg Glu Lys Ile Glu
435 440 445
Lys Ile Leu Thr Phe Arg Ile Pro Tyr Tyr Val Gly Pro Leu Ala Arg
450 455 460
Gly Asn Ser Arg Phe Ala Trp Met Thr Arg Lys Ser Glu Glu Thr Ile
465 470 475 480
Thr Pro Trp Asn Phe Glu Glu Val Val Asp Lys Gly Ala Ser Ala Gln
485 490 495
Ser Phe Ile Glu Arg Met Thr Asn Phe Asp Lys Asn Leu Pro Asn Glu
500 505 510
Lys Val Leu Pro Lys His Ser Leu Leu Tyr Glu Tyr Phe Thr Val Tyr
515 520 525
Asn Glu Leu Thr Lys Val Lys Tyr Val Thr Glu Gly Met Arg Lys Pro
530 535 540
Ala Phe Leu Ser Gly Glu Gln Lys Lys Ala Ile Val Asp Leu Leu Phe
545 550 555 560
Lys Thr Asn Arg Lys Val Thr Val Lys Gln Leu Lys Glu Asp Tyr Phe
565 570 575
Lys Lys Ile Glu Cys Phe Asp Ser Val Glu Ile Ser Gly Val Glu Asp
580 585 590
Arg Phe Asn Ala Ser Leu Gly Thr Tyr His Asp Leu Leu Lys Ile Ile
595 600 605
Lys Asp Lys Asp Phe Leu Asp Asn Glu Glu Asn Glu Asp Ile Leu Glu
610 615 620
Asp Ile Val Leu Thr Leu Thr Leu Phe Glu Asp Arg Glu Met Ile Glu
625 630 635 640
Glu Arg Leu Lys Thr Tyr Ala His Leu Phe Asp Asp Lys Val Met Lys
645 650 655
Gln Leu Lys Arg Arg Arg Tyr Thr Gly Trp Gly Arg Leu Ser Arg Lys
660 665 670
Leu Ile Asn Gly Ile Arg Asp Lys Gln Ser Gly Lys Thr Ile Leu Asp
675 680 685
Phe Leu Lys Ser Asp Gly Phe Ala Asn Arg Asn Phe Met Gln Leu Ile
690 695 700
His Asp Asp Ser Leu Thr Phe Lys Glu Asp Ile Gln Lys Ala Gln Val
705 710 715 720
Ser Gly Gln Gly Asp Ser Leu His Glu His Ile Ala Asn Leu Ala Gly
725 730 735
Ser Pro Ala Ile Lys Lys Gly Ile Leu Gln Thr Val Lys Val Val Asp
740 745 750
Glu Leu Val Lys Val Met Gly Arg His Lys Pro Glu Asn Ile Val Ile
755 760 765
Glu Met Ala Arg Glu Asn Gln Thr Thr Gln Lys Gly Gln Lys Asn Ser
770 775 780
Arg Glu Arg Met Lys Arg Ile Glu Glu Gly Ile Lys Glu Leu Gly Ser
785 790 795 800
Gln Ile Leu Lys Glu His Pro Val Glu Asn Thr Gln Leu Gln Asn Glu
805 810 815
Lys Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly Arg Asp Met Tyr Val Asp
820 825 830
Gln Glu Leu Asp Ile Asn Arg Leu Ser Asp Tyr Asp Val Asp His Ile
835 840 845
Val Pro Gln Ser Phe Leu Lys Asp Asp Ser Ile Asp Asn Lys Val Leu
850 855 860
Thr Arg Ser Asp Lys Asn Arg Gly Lys Ser Asp Asn Val Pro Ser Glu
865 870 875 880
Glu Val Val Lys Lys Met Lys Asn Tyr Trp Arg Gln Leu Leu Asn Ala
885 890 895
Lys Leu Ile Thr Gln Arg Lys Phe Asp Asn Leu Thr Lys Ala Glu Arg
900 905 910
Gly Gly Leu Ser Glu Leu Asp Lys Ala Gly Phe Ile Lys Arg Gln Leu
915 920 925
Val Glu Thr Arg Gln Ile Thr Lys His Val Ala Gln Ile Leu Asp Ser
930 935 940
Arg Met Asn Thr Lys Tyr Asp Glu Asn Asp Lys Leu Ile Arg Glu Val
945 950 955 960
Lys Val Ile Thr Leu Lys Ser Lys Leu Val Ser Asp Phe Arg Lys Asp
965 970 975
Phe Gln Phe Tyr Lys Val Arg Glu Ile Asn Asn Tyr His His Ala His
980 985 990
Asp Ala Tyr Leu Asn Ala Val Val Gly Thr Ala Leu Ile Lys Lys Tyr
995 1000 1005
Pro Lys Leu Glu Ser Glu Phe Val Tyr Gly Asp Tyr Lys Val Tyr
1010 1015 1020
Asp Val Arg Lys Met Ile Ala Lys Ser Glu Gln Glu Ile Gly Lys
1025 1030 1035
Ala Thr Ala Lys Tyr Phe Phe Tyr Ser Asn Ile Met Asn Phe Phe
1040 1045 1050
Lys Thr Glu Ile Thr Leu Ala Asn Gly Glu Ile Arg Lys Arg Pro
1055 1060 1065
Leu Ile Glu Thr Asn Gly Glu Thr Gly Glu Ile Val Trp Asp Lys
1070 1075 1080
Gly Arg Asp Phe Ala Thr Val Arg Lys Val Leu Ser Met Pro Gln
1085 1090 1095
Val Asn Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly Phe Ser
1100 1105 1110
Lys Glu Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile Ala
1115 1120 1125
Arg Lys Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser
1130 1135 1140
Pro Thr Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys
1145 1150 1155
Gly Lys Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile
1160 1165 1170
Thr Ile Met Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe
1175 1180 1185
Leu Glu Ala Lys Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile
1190 1195 1200
Lys Leu Pro Lys Tyr Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys
1205 1210 1215
Arg Met Leu Ala Ser Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu
1220 1225 1230
Ala Leu Pro Ser Lys Tyr Val Asn Phe Leu Tyr Leu Ala Ser His
1235 1240 1245
Tyr Glu Lys Leu Lys Gly Ser Pro Glu Asp Asn Glu Gln Lys Gln
1250 1255 1260
Leu Phe Val Glu Gln His Lys His Tyr Leu Asp Glu Ile Ile Glu
1265 1270 1275
Gln Ile Ser Glu Phe Ser Lys Arg Val Ile Leu Ala Asp Ala Asn
1280 1285 1290
Leu Asp Lys Val Leu Ser Ala Tyr Asn Lys His Arg Asp Lys Pro
1295 1300 1305
Ile Arg Glu Gln Ala Glu Asn Ile Ile His Leu Phe Thr Leu Thr
1310 1315 1320
Asn Leu Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp Thr Thr Ile
1325 1330 1335
Asp Arg Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp Ala Thr
1340 1345 1350
Leu Ile His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile Asp
1355 1360 1365
Leu Ser Gln Leu Gly Gly Asp Lys Arg Pro Ala Ala Thr Lys Lys
1370 1375 1380
Ala Gly Gln Ala Lys Lys Lys Lys Gly Ser Gly Ala Thr Asn Phe
1385 1390 1395
Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn Pro Gly Pro
1400 1405 1410
Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu
1415 1420 1425
Val Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser
1430 1435 1440
Gly Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Leu Lys
1445 1450 1455
Phe Ile Cys Thr Thr Gly Lys Leu Pro Val Pro Trp Pro Thr Leu
1460 1465 1470
Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg Tyr Pro
1475 1480 1485
Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala Met Pro Glu
1490 1495 1500
Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn
1505 1510 1515
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val
1520 1525 1530
Asn Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn
1535 1540 1545
Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val
1550 1555 1560
Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe
1565 1570 1575
Lys Ile Arg His Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp
1580 1585 1590
His Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro Val Leu Leu
1595 1600 1605
Pro Asp Asn His Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys Asp
1610 1615 1620
Pro Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe Val Thr
1625 1630 1635
Ala Ala Gly Ile Thr Leu Gly Met Asp Glu Leu Tyr Lys
1640 1645 1650
<210> 14
<211> 10439
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221>misc_feature
<222> (195)..(1913)
<223>Промотор CAGG
<220>
<221> misc_feature
<222> (1996)..(2029)
<223> LoxP
<220>
<221> misc_feature
<222> (2120)..(4185)
<223> Neo-PolyA
<220>
<221> misc_feature
<222> (4213)..(4246)
<223> LoxP
<220>
<221> misc_feature
<222> (4341)..(4350)
<223> Kozak
<220>
<221> misc_feature
<222> (4350)..(4415)
<223> 3xFLAG
<220>
<221> misc_feature
<222> (4416)..(8588)
<223> Cas9
<220>
<221> misc_feature
<222> (4425)..(4445)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (8541)..(8588)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (8589)..(8654)
<223> P2A
<220>
<221> misc_feature
<222> (8655)..(9368)
<223> eGFP
<220>
<221> misc_feature
<222> (9418)..(10014)
<223> WPRE
<220>
<221> misc_feature
<222> (10015)..(10230)
<223>сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (10290)..(10439)
<223>Нижележащая последовательность Rosa26 мыши
<400> 14
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctccttaattaa 180
ggcctccaaggcctactagttattaatagtaatcaattacggggtcattagttcatagcc 240
catatatggagttccgcgttacataacttacggtaaatggcccgcctggctgaccgccca 300
acgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaataggga 360
ctttccattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatc 420
aagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaatggcccgcct 480
ggcattatgcccagtacatgaccttatgggactttcctacttggcagtacatctacgtat 540
tagtcatcgctattaccatggtcgaggtgagccccacgttctgcttcactctccccatct 600
cccccccctccccacccccaattttgtatttatttattttttaattattttgtgcagcga 660
tgggggcggggggggggggggggcgcgcgccaggcggggcggggcggggcgaggggcggg 720
gcggggcgaggcggagaggtgcggcggcagccaatcagagcggcgcgctccgaaagtttc 780
cttttatggcgaggcggcggcggcggcggccctataaaaagcgaagcgcgcggcgggcgg 840
ggagtcgctgcgacgctgccttcgccccgtgccccgctccgccgccgcctcgcgccgccc 900
gccccggctctgactgaccgcgttactcccacaggtgagcgggcgggacggcccttctcc 960
tccgggctgtaattagcgcttggtttaatgacggcttgtttcttttctgtggctgcgtga 1020
aagccttgaggggctccgggagggccctttgtgcggggggagcggctcggggggtgcgtg 1080
cgtgtgtgtgtgcgtggggagcgccgcgtgcggctccgcgctgcccggcggctgtgagcg 1140
ctgcgggcgcggcgcggggctttgtgcgctccgcagtgtgcgcgaggggagcgcggccgg 1200
gggcggtgccccgcggtgcggggggggctgcgaggggaacaaaggctgcgtgcggggtgt 1260
gtgcgtgggggggtgagcagggggtgtgggcgcgtcggtcgggctgcaaccccccctgca 1320
cccccctccccgagttgctgagcacggcccggcttcgggtgcggggctccgtacggggcg 1380
tggcgcggggctcgccgtgccgggcggggggtggcggcaggtgggggtgccgggcggggc 1440
ggggccgcctcgggccggggagggctcgggggaggggcgcggcggcccccggagcgccgg 1500
cggctgtcgaggcgcggcgagccgcagccattgccttttatggtaatcgtgcgagagggc 1560
gcagggacttcctttgtcccaaatctgtgcggagccgaaatctgggaggcgccgccgcac 1620
cccctctagcgggcgcggggcgaagcggtgcggcgccggcaggaaggaaatgggcgggga 1680
gggccttcgtgcgtcgccgcgccgccgtccccttctccctctccagcctcggggctgtcc 1740
gcggggggacggctgccttcgggggggacggggcagggcggggttcggcttctggcgtgt 1800
gaccggcggctctagagcctctgctaaccatgttcatgccttcttctttttcctacagct 1860
cctgggcaacgtgctggttattgtgctgtctcatcattttggcaaagaattcgctaggag 1920
aattgatttgataccgcgggccctaagtcgacatttaaatcatttaaatccactagtgga 1980
tccggaacccttaatataacttcgtataatgtatgctatacgaagttattaggtccctcg 2040
acctgcaggaattgttgacaattaatcatcggcatagtatatcggcatagtataatacga 2100
caaggtgaggaactaaaccatgggatcggccattgaacaagatggattgcacgcaggttc 2160
tccggccgcttgggtggagaggctattcggctatgactgggcacaacagacaatcggctg 2220
ctctgatgccgccgtgttccggctgtcagcgcaggggcgcccggttctttttgtcaagac 2280
cgacctgtccggtgccctgaatgaactgcaggacgaggcagcgcggctatcgtggctggc 2340
cacgacgggcgttccttgcgcagctgtgctcgacgttgtcactgaagcgggaagggactg 2400
gctgctattgggcgaagtgccggggcaggatctcctgtcatctcaccttgctcctgccga 2460
gaaagtatccatcatggctgatgcaatgcggcggctgcatacgcttgatccggctacctg 2520
cccattcgaccaccaagcgaaacatcgcatcgagcgagcacgtactcggatggaagccgg 2580
tcttgtcgatcaggatgatctggacgaagagcatcaggggctcgcgccagccgaactgtt 2640
cgccaggctcaaggcgcgcatgcccgacggcgatgatctcgtcgtgacccatggcgatgc 2700
ctgcttgccgaatatcatggtggaaaatggccgcttttctggattcatcgactgtggccg 2760
gctgggtgtggcggaccgctatcaggacatagcgttggctacccgtgatattgctgaaga 2820
gcttggcggcgaatgggctgaccgcttcctcgtgctttacggtatcgccgctcccgattc 2880
gcagcgcatcgccttctatcgccttcttgacgagttcttctgaggggatccgctgtaagt 2940
ctgcagaaattgatgatctattaaacaataaagatgtccactaaaatggaagtttttcct 3000
gtcatactttgttaagaagggtgagaacagagtacctacattttgaatggaaggattgga 3060
gctacgggggtgggggtggggtgggattagataaatgcctgctctttactgaaggctctt 3120
tactattgctttatgataatgtttcatagttggatatcataatttaaacaagcaaaacca 3180
aattaagggccagctcattcctcccactcatgatctatagatctatagatctctcgtggg 3240
atcattgtttttctcttgattcccactttgtggttctaagtactgtggtttccaaatgtg 3300
tcagtttcatagcctgaagaacgagatcagcagcctctgttccacatacacttcattctc 3360
agtattgttttgccaagttctaattccatcagaagcttgcagatctgcgactctagagga 3420
tctgcgactc tagaggatca taatcagcca taccacattt gtagaggttt tacttgcttt 3480
aaaaaacctc ccacacctcc ccctgaacct gaaacataaa atgaatgcaa ttgttgttgt 3540
taacttgttt attgcagctt ataatggtta caaataaagc aatagcatca caaatttcac 3600
aaataaagca tttttttcac tgcattctag ttgtggtttg tccaaactca tcaatgtatc 3660
ttatcatgtc tggatctgcg actctagagg atcataatca gccataccac atttgtagag 3720
gttttacttg ctttaaaaaa cctcccacac ctccccctga acctgaaaca taaaatgaat 3780
gcaattgttg ttgttaactt gtttattgca gcttataatg gttacaaata aagcaatagc 3840
atcacaaatt tcacaaataa agcatttttt tcactgcatt ctagttgtgg tttgtccaaa 3900
ctcatcaatg tatcttatca tgtctggatc tgcgactcta gaggatcata atcagccata 3960
ccacatttgt agaggtttta cttgctttaa aaaacctccc acacctcccc ctgaacctga 4020
aacataaaat gaatgcaatt gttgttgtta acttgtttat tgcagcttat aatggttaca 4080
aataaagcaa tagcatcaca aatttcacaa ataaagcatt tttttcactg cattctagtt 4140
gtggtttgtc caaactcatc aatgtatctt atcatgtctg gatccccatc aagctgatcc 4200
ggaaccctta atataacttc gtataatgta tgctatacga agttattagg tccctcgacc 4260
tgcagcccaa gctagtgccc gggtaggtcc ctcgacctgc agcccaagct agatcgaatt 4320
cggccggcct tcgaacacgt gccaccatgg actataagga ccacgacgga gactacaagg 4380
atcatgatat tgattacaaa gacgatgacg ataagatgga caagcccaag aaaaagcgga 4440
aagtgaagta cagcatcggc ctggacatcg gcaccaactc tgtgggctgg gccgtgatca 4500
ccgacgagta caaggtgccc agcaagaaat tcaaggtgct gggcaacacc gacaggcaca 4560
gcatcaagaa gaacctgatc ggcgccctgc tgttcgacag cggcgaaaca gccgaggcca 4620
ccagactgaa gagaaccgcc agaagaagat acaccaggcg gaagaacagg atctgctatc 4680
tgcaagagat cttcagcaac gagatggcca aggtggacga cagcttcttc cacagactgg 4740
aagagtcctt cctggtggaa gaggacaaga agcacgagag acaccccatc ttcggcaaca 4800
tcgtggacga ggtggcctac cacgagaagt accccaccat ctaccacctg agaaagaaac 4860
tggtggacag caccgacaag gccgacctga gactgatcta cctggccctg gcccacatga 4920
tcaagttcag aggccacttc ctgatcgagg gcgacctgaa ccccgacaac agcgacgtgg 4980
acaagctgtt catccagctg gtgcagacct acaaccagct gttcgaggaa aaccccatca 5040
acgccagcgg cgtggacgcc aaggctatcc tgtctgccag actgagcaag agcagaaggc 5100
tggaaaatct gatcgcccag ctgcccggcg agaagaagaa cggcctgttc ggcaacctga 5160
ttgccctgag cctgggcctg acccccaact tcaagagcaa cttcgacctg gccgaggatg 5220
ccaaactgca gctgagcaag gacacctacg acgacgacct ggacaacctg ctggcccaga 5280
tcggcgacca gtacgccgac ctgttcctgg ccgccaagaa cctgtctgac gccatcctgc 5340
tgagcgacat cctgagagtg aacaccgaga tcaccaaggc ccccctgagc gcctctatga 5400
tcaagagata cgacgagcac caccaggacc tgaccctgct gaaagctctc gtgcggcagc 5460
agctgcctga gaagtacaaa gaaatcttct tcgaccagag caagaacggc tacgccggct 5520
acatcgatgg cggcgctagc caggaagagt tctacaagtt catcaagccc atcctggaaa 5580
agatggacgg caccgaggaa ctgctcgtga agctgaacag agaggacctg ctgagaaagc 5640
agagaacctt cgacaacggc agcatccccc accagatcca cctgggagag ctgcacgcta 5700
tcctgagaag gcaggaagat ttttacccat tcctgaagga caaccgggaa aagatcgaga 5760
agatcctgac cttcaggatc ccctactacg tgggccccct ggccagaggc aacagcagat 5820
tcgcctggat gaccagaaag agcgaggaaa ccatcacccc ctggaacttc gaggaagtgg 5880
tggacaaggg cgccagcgcc cagagcttca tcgagagaat gacaaacttc gataagaacc 5940
tgcccaacga gaaggtgctg cccaagcaca gcctgctgta cgagtacttc accgtgtaca 6000
acgagctgac caaagtgaaa tacgtgaccg agggaatgag aaagcccgcc ttcctgagcg 6060
gcgagcagaa aaaggccatc gtggacctgc tgttcaagac caacagaaaa gtgaccgtga 6120
agcagctgaa agaggactac ttcaagaaaa tcgagtgctt cgactccgtg gaaatctccg 6180
gcgtggaaga tagattcaac gcctccctgg gcacatacca cgatctgctg aaaattatca 6240
aggacaagga cttcctggat aacgaagaga acgaggacat tctggaagat atcgtgctga 6300
ccctgacact gtttgaggac cgcgagatga tcgaggaaag gctgaaaacc tacgctcacc 6360
tgttcgacga caaagtgatg aagcagctga agagaaggcg gtacaccggc tggggcaggc 6420
tgagcagaaa gctgatcaac ggcatcagag acaagcagag cggcaagaca atcctggatt 6480
tcctgaagtc cgacggcttc gccaaccgga acttcatgca gctgatccac gacgacagcc 6540
tgacattcaa agaggacatc cagaaagccc aggtgtccgg ccagggcgac tctctgcacg 6600
agcatatcgc taacctggcc ggcagccccg ctatcaagaa gggcatcctg cagacagtga 6660
aggtggtgga cgagctcgtg aaagtgatgg gcagacacaa gcccgagaac atcgtgatcg 6720
agatggctag agagaaccag accacccaga agggacagaa gaactcccgc gagaggatga 6780
agagaatcga agagggcatc aaagagctgg gcagccagat cctgaaagaa caccccgtgg 6840
aaaacaccca gctgcagaac gagaagctgt acctgtacta cctgcagaat ggccgggata 6900
tgtacgtgga ccaggaactg gacatcaaca gactgtccga ctacgatgtg gaccatatcg 6960
tgcctcagag ctttctgaag gacgactcca tcgataacaa agtgctgact cggagcgaca 7020
agaacagagg caagagcgac aacgtgccct ccgaagaggt cgtgaagaag atgaagaact 7080
actggcgaca gctgctgaac gccaagctga ttacccagag gaagttcgat aacctgacca 7140
aggccgagag aggcggcctg agcgagctgg ataaggccgg cttcatcaag aggcagctgg 7200
tggaaaccag acagatcaca aagcacgtgg cacagatcct ggactcccgg atgaacacta 7260
agtacgacga aaacgataag ctgatccggg aagtgaaagt gatcaccctg aagtccaagc 7320
tggtgtccga tttccggaag gatttccagt tttacaaagt gcgcgagatc aacaactacc 7380
accacgccca cgacgcctac ctgaacgccg tcgtgggaac cgccctgatc aaaaagtacc 7440
ctaagctgga aagcgagttc gtgtacggcg actacaaggt gtacgacgtg cggaagatga 7500
tcgccaagag cgagcaggaa atcggcaagg ctaccgccaa gtacttcttc tacagcaaca 7560
tcatgaactt tttcaagacc gaaatcaccc tggccaacgg cgagatcaga aagcgccctc 7620
tgatcgagac aaacggcgaa accggggaga tcgtgtggga taagggcaga gacttcgcca 7680
cagtgcgaaa ggtgctgagc atgccccaag tgaatatcgt gaaaaagacc gaggtgcaga 7740
caggcggctt cagcaaagag tctatcctgc ccaagaggaa cagcgacaag ctgatcgcca 7800
gaaagaagga ctgggacccc aagaagtacg gcggcttcga cagccctacc gtggcctact 7860
ctgtgctggt ggtggctaag gtggaaaagg gcaagtccaa gaaactgaag agtgtgaaag 7920
agctgctggg gatcaccatc atggaaagaa gcagctttga gaagaaccct atcgactttc 7980
tggaagccaa gggctacaaa gaagtgaaaa aggacctgat catcaagctg cctaagtact 8040
ccctgttcga gctggaaaac ggcagaaaga gaatgctggc ctctgccggc gaactgcaga 8100
agggaaacga gctggccctg cctagcaaat atgtgaactt cctgtacctg gcctcccact 8160
atgagaagct gaagggcagc cctgaggaca acgaacagaa acagctgttt gtggaacagc 8220
ataagcacta cctggacgag atcatcgagc agatcagcga gttctccaag agagtgatcc 8280
tggccgacgc caatctggac aaggtgctgt ctgcctacaa caagcacagg gacaagccta 8340
tcagagagca ggccgagaat atcatccacc tgttcaccct gacaaacctg ggcgctcctg 8400
ccgccttcaa gtactttgac accaccatcg accggaagag gtacaccagc accaaagagg 8460
tgctggacgc caccctgatc caccagagca tcaccggcct gtacgagaca agaatcgacc 8520
tgtctcagct gggaggcgac aagagacctg ccgccactaa gaaggccgga caggccaaaa 8580
agaagaaggg aagcggagcc actaacttct ccctgttgaa acaagcaggg gatgtcgaag 8640
agaatcccgg gccagtgagc aagggcgagg agctgttcac cggggtggtg cccatcctgg 8700
tcgagctgga cggcgacgta aacggccaca agttcagcgt gtccggcgag ggcgagggcg 8760
atgccaccta cggcaagctg accctgaagt tcatctgcac caccggcaag ctgcccgtgc 8820
cctggcccac cctcgtgacc accctgacct acggcgtgca gtgcttcagc cgctaccccg 8880
accacatgaa gcagcacgac ttcttcaagt ccgccatgcc cgaaggctac gtccaggagc 8940
gcaccatctt cttcaaggac gacggcaact acaagacccg cgccgaggtg aagttcgagg 9000
gcgacaccct ggtgaaccgc atcgagctga agggcatcga cttcaaggag gacggcaaca 9060
tcctggggca caagctggag tacaactaca acagccacaa cgtctatatc atggccgaca 9120
agcagaagaa cggcatcaag gtgaacttca agatccgcca caacatcgag gacggcagcg 9180
tgcagctcgc cgaccactac cagcagaaca cccccatcgg cgacggcccc gtgctgctgc 9240
ccgacaacca ctacctgagc acccagtccg ccctgagcaa agaccccaac gagaagcgcg 9300
atcacatggt cctgctggag ttcgtgaccg ccgccgggat cactctcggc atggacgagc 9360
tgtacaagta aacgcgtatg catggccggc cctgcaggaa ttcgatatca agcttatcga 9420
taatcaacct ctggattaca aaatttgtga aagattgact ggtattctta actatgttgc 9480
tccttttacg ctatgtggat acgctgcttt aatgcctttg tatcatgcta ttgcttcccg 9540
tatggctttc attttctcct ccttgtataa atcctggttg ctgtctcttt atgaggagtt 9600
gtggcccgtt gtcaggcaac gtggcgtggt gtgcactgtg tttgctgacg caacccccac 9660
tggttggggc attgccacca cctgtcagct cctttccggg actttcgctt tccccctccc 9720
tattgccacg gcggaactca tcgccgcctg ccttgcccgc tgctggacag gggctcggct 9780
gttgggcact gacaattccg tggtgttgtc ggggaaatca tcgtcctttc cttggctgct 9840
cgcctgtgtt gccacctgga ttctgcgcgg gacgtccttc tgctacgtcc cttcggccct 9900
caatccagcg gaccttcctt cccgcggcct gctgccggct ctgcggcctc ttccgcgtct 9960
tcgccttcgc cctcagacga gtcggatctc cctttgggcc gcctccccgc atcgcgacct 10020
cgacctcgac tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt 10080
ccttgaccct ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat 10140
cgcattgtct gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg 10200
gggaggattg ggaagacaat ggcaggcatg ctggggaact agtggtgcca gggcgtgccc 10260
ttgggctccc cgggcgcggc ggccatcgct cgagtaaaat tggagggaca agacttccca 10320
cagattttcg gttttgtcgg gaagtttttt aataggggca aataaggaaa atgggaggat 10380
aggtagtcat ctggggtttt atgcagcaaa actacaggtt attattgctt gtgatccgc 10439
<210> 15
<211> 8222
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221>misc_feature
<222> (195)..(1913)
<223>Промотор CAGG
<220>
<221> misc_feature
<222> (1996)..(2029)
<223> LoxP
<220>
<221> misc_feature
<222> (2124)..(2133)
<223> Kozak
<220>
<221> misc_feature
<222> (2133)..(2198)
<223> 3xFLAG
<220>
<221> misc_feature
<222> (2199)..(6371)
<223> Cas9
<220>
<221> misc_feature
<222> (2208)..(2228)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (6324)..(6371)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (6372)..(6437)
<223> P2A
<220>
<221> misc_feature
<222> (6438)..(7151)
<223> eGFP
<220>
<221> misc_feature
<222> (7201)..(7797)
<223> WPRE
<220>
<221> misc_feature
<222> (7798)..(8013)
<223>сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (8073)..(8222)
<223>Нижележащая последовательность Rosa26 мыши
<400> 15
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctccttaattaa 180
ggcctccaaggcctactagttattaatagtaatcaattacggggtcattagttcatagcc 240
catatatggagttccgcgttacataacttacggtaaatggcccgcctggctgaccgccca 300
acgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaataggga 360
ctttccattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatc 420
aagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaatggcccgcct 480
ggcattatgcccagtacatgaccttatgggactttcctacttggcagtacatctacgtat 540
tagtcatcgctattaccatggtcgaggtgagccccacgttctgcttcactctccccatct 600
cccccccctccccacccccaattttgtatttatttattttttaattattttgtgcagcga 660
tgggggcggggggggggggggggcgcgcgccaggcggggcggggcggggcgaggggcggg 720
gcggggcgaggcggagaggtgcggcggcagccaatcagagcggcgcgctccgaaagtttc 780
cttttatggcgaggcggcggcggcggcggccctataaaaagcgaagcgcgcggcgggcgg 840
ggagtcgctgcgacgctgccttcgccccgtgccccgctccgccgccgcctcgcgccgccc 900
gccccggctctgactgaccgcgttactcccacaggtgagcgggcgggacggcccttctcc 960
tccgggctgtaattagcgcttggtttaatgacggcttgtttcttttctgtggctgcgtga 1020
aagccttgaggggctccgggagggccctttgtgcggggggagcggctcggggggtgcgtg 1080
cgtgtgtgtgtgcgtggggagcgccgcgtgcggctccgcgctgcccggcggctgtgagcg 1140
ctgcgggcgcggcgcggggctttgtgcgctccgcagtgtgcgcgaggggagcgcggccgg 1200
gggcggtgccccgcggtgcggggggggctgcgaggggaacaaaggctgcgtgcggggtgt 1260
gtgcgtgggggggtgagcagggggtgtgggcgcgtcggtcgggctgcaaccccccctgca 1320
cccccctccccgagttgctgagcacggcccggcttcgggtgcggggctccgtacggggcg 1380
tggcgcggggctcgccgtgccgggcggggggtggcggcaggtgggggtgccgggcggggc 1440
ggggccgcctcgggccggggagggctcgggggaggggcgcggcggcccccggagcgccgg 1500
cggctgtcgaggcgcggcgagccgcagccattgccttttatggtaatcgtgcgagagggc 1560
gcagggacttcctttgtcccaaatctgtgcggagccgaaatctgggaggcgccgccgcac 1620
cccctctagcgggcgcggggcgaagcggtgcggcgccggcaggaaggaaatgggcgggga 1680
gggccttcgtgcgtcgccgcgccgccgtccccttctccctctccagcctcggggctgtcc 1740
gcggggggacggctgccttcgggggggacggggcagggcggggttcggcttctggcgtgt 1800
gaccggcggctctagagcctctgctaaccatgttcatgccttcttctttttcctacagct 1860
cctgggcaacgtgctggttattgtgctgtctcatcattttggcaaagaattcgctaggag 1920
aattgatttgataccgcgggccctaagtcgacatttaaatcatttaaatccactagtgga 1980
tccggaacccttaatataacttcgtataatgtatgctatacgaagttattaggtccctcg 2040
acctgcagcccaagctagtgcccgggtaggtccctcgacctgcagcccaagctagatcga 2100
attcggccggccttcgaacacgtgccaccatggactataaggaccacgacggagactaca 2160
aggatcatgatattgattacaaagacgatgacgataagatggacaagcccaagaaaaagc 2220
ggaaagtgaagtacagcatcggcctggacatcggcaccaactctgtgggctgggccgtga 2280
tcaccgacgagtacaaggtgcccagcaagaaattcaaggtgctgggcaacaccgacaggc 2340
acagcatcaagaagaacctgatcggcgccctgctgttcgacagcggcgaaacagccgagg 2400
ccaccagactgaagagaaccgccagaagaagatacaccaggcggaagaacaggatctgct 2460
atctgcaagagatcttcagcaacgagatggccaaggtggacgacagcttcttccacagac 2520
tggaagagtccttcctggtggaagaggacaagaagcacgagagacaccccatcttcggca 2580
acatcgtggacgaggtggcctaccacgagaagtaccccaccatctaccacctgagaaaga 2640
aactggtggacagcaccgacaaggccgacctgagactgatctacctggccctggcccaca 2700
tgatcaagttcagaggccacttcctgatcgagggcgacctgaaccccgacaacagcgacg 2760
tggacaagctgttcatccagctggtgcagacctacaaccagctgttcgaggaaaacccca 2820
tcaacgccagcggcgtggacgccaaggctatcctgtctgccagactgagcaagagcagaa 2880
ggctggaaaatctgatcgcccagctgcccggcgagaagaagaacggcctgttcggcaacc 2940
tgattgccctgagcctgggcctgacccccaacttcaagagcaacttcgacctggccgagg 3000
atgccaaactgcagctgagcaaggacacctacgacgacgacctggacaacctgctggccc 3060
agatcggcgaccagtacgccgacctgttcctggccgccaagaacctgtctgacgccatcc 3120
tgctgagcgacatcctgagagtgaacaccgagatcaccaaggcccccctgagcgcctcta 3180
tgatcaagagatacgacgagcaccaccaggacctgaccctgctgaaagctctcgtgcggc 3240
agcagctgcctgagaagtacaaagaaatcttcttcgaccagagcaagaacggctacgccg 3300
gctacatcgatggcggcgctagccaggaagagttctacaagttcatcaagcccatcctgg 3360
aaaagatggacggcaccgaggaactgctcgtgaagctgaacagagaggacctgctgagaa 3420
agcagagaaccttcgacaacggcagcatcccccaccagatccacctgggagagctgcacg 3480
ctatcctgagaaggcaggaagatttttacccattcctgaaggacaaccgggaaaagatcg 3540
agaagatcctgaccttcaggatcccctactacgtgggccccctggccagaggcaacagca 3600
gattcgcctggatgaccagaaagagcgaggaaaccatcaccccctggaacttcgaggaag 3660
tggtggacaagggcgccagcgcccagagcttcatcgagagaatgacaaacttcgataaga 3720
acctgcccaacgagaaggtgctgcccaagcacagcctgctgtacgagtacttcaccgtgt 3780
acaacgagctgaccaaagtgaaatacgtgaccgagggaatgagaaagcccgccttcctga 3840
gcggcgagcagaaaaaggccatcgtggacctgctgttcaagaccaacagaaaagtgaccg 3900
tgaagcagctgaaagaggactacttcaagaaaatcgagtgcttcgactccgtggaaatct 3960
ccggcgtggaagatagattcaacgcctccctgggcacataccacgatctgctgaaaatta 4020
tcaaggacaaggacttcctggataacgaagagaacgaggacattctggaagatatcgtgc 4080
tgaccctgacactgtttgaggaccgcgagatgatcgaggaaaggctgaaaacctacgctc 4140
acctgttcgacgacaaagtgatgaagcagctgaagagaaggcggtacaccggctggggca 4200
ggctgagcagaaagctgatcaacggcatcagagacaagcagagcggcaagacaatcctgg 4260
atttcctgaagtccgacggcttcgccaaccggaacttcatgcagctgatccacgacgaca 4320
gcctgacattcaaagaggacatccagaaagcccaggtgtccggccagggcgactctctgc 4380
acgagcatatcgctaacctggccggcagccccgctatcaagaagggcatcctgcagacag 4440
tgaaggtggtggacgagctcgtgaaagtgatgggcagacacaagcccgagaacatcgtga 4500
tcgagatggctagagagaaccagaccacccagaagggacagaagaactcccgcgagagga 4560
tgaagagaatcgaagagggcatcaaagagctgggcagccagatcctgaaagaacaccccg 4620
tggaaaacacccagctgcagaacgagaagctgtacctgtactacctgcagaatggccggg 4680
atatgtacgtggaccaggaactggacatcaacagactgtccgactacgatgtggaccata 4740
tcgtgcctcagagctttctgaaggacgactccatcgataacaaagtgctgactcggagcg 4800
acaagaacagaggcaagagcgacaacgtgccctccgaagaggtcgtgaagaagatgaaga 4860
actactggcgacagctgctgaacgccaagctgattacccagaggaagttcgataacctga 4920
ccaaggccgagagaggcggcctgagcgagctggataaggccggcttcatcaagaggcagc 4980
tggtggaaaccagacagatcacaaagcacgtggcacagatcctggactcccggatgaaca 5040
ctaagtacgacgaaaacgataagctgatccgggaagtgaaagtgatcaccctgaagtcca 5100
agctggtgtccgatttccggaaggatttccagttttacaaagtgcgcgagatcaacaact 5160
accaccacgcccacgacgcctacctgaacgccgtcgtgggaaccgccctgatcaaaaagt 5220
accctaagctggaaagcgagttcgtgtacggcgactacaaggtgtacgacgtgcggaaga 5280
tgatcgccaagagcgagcaggaaatcggcaaggctaccgccaagtacttcttctacagca 5340
acatcatgaactttttcaagaccgaaatcaccctggccaacggcgagatcagaaagcgcc 5400
ctctgatcgagacaaacggcgaaaccggggagatcgtgtgggataagggcagagacttcg 5460
ccacagtgcgaaaggtgctgagcatgccccaagtgaatatcgtgaaaaagaccgaggtgc 5520
agacaggcggcttcagcaaagagtctatcctgcccaagaggaacagcgacaagctgatcg 5580
ccagaaagaaggactgggaccccaagaagtacggcggcttcgacagccctaccgtggcct 5640
actctgtgctggtggtggctaaggtggaaaagggcaagtccaagaaactgaagagtgtga 5700
aagagctgctggggatcaccatcatggaaagaagcagctttgagaagaaccctatcgact 5760
ttctggaagccaagggctacaaagaagtgaaaaaggacctgatcatcaagctgcctaagt 5820
actccctgttcgagctggaaaacggcagaaagagaatgctggcctctgccggcgaactgc 5880
agaagggaaacgagctggccctgcctagcaaatatgtgaacttcctgtacctggcctccc 5940
actatgagaagctgaagggcagccctgaggacaacgaacagaaacagctgtttgtggaac 6000
agcataagcactacctggacgagatcatcgagcagatcagcgagttctccaagagagtga 6060
tcctggccgacgccaatctggacaaggtgctgtctgcctacaacaagcacagggacaagc 6120
ctatcagagagcaggccgagaatatcatccacctgttcaccctgacaaacctgggcgctc 6180
ctgccgccttcaagtactttgacaccaccatcgaccggaagaggtacaccagcaccaaag 6240
aggtgctggacgccaccctgatccaccagagcatcaccggcctgtacgagacaagaatcg 6300
acctgtctcagctgggaggcgacaagagacctgccgccactaagaaggccggacaggcca 6360
aaaagaagaagggaagcggagccactaacttctccctgttgaaacaagcaggggatgtcg 6420
aagagaatcccgggccagtgagcaagggcgaggagctgttcaccggggtggtgcccatcc 6480
tggtcgagctggacggcgacgtaaacggccacaagttcagcgtgtccggcgagggcgagg 6540
gcgatgccacctacggcaagctgaccctgaagttcatctgcaccaccggcaagctgcccg 6600
tgccctggcccaccctcgtgaccaccctgacctacggcgtgcagtgcttcagccgctacc 6660
ccgaccacatgaagcagcacgacttcttcaagtccgccatgcccgaaggctacgtccagg 6720
agcgcaccatcttcttcaaggacgacggcaactacaagacccgcgccgaggtgaagttcg 6780
agggcgacaccctggtgaaccgcatcgagctgaagggcatcgacttcaaggaggacggca 6840
acatcctggggcacaagctggagtacaactacaacagccacaacgtctatatcatggccg 6900
acaagcagaagaacggcatcaaggtgaacttcaagatccgccacaacatcgaggacggca 6960
gcgtgcagctcgccgaccactaccagcagaacacccccatcggcgacggccccgtgctgc 7020
tgcccgacaaccactacctgagcacccagtccgccctgagcaaagaccccaacgagaagc 7080
gcgatcacatggtcctgctggagttcgtgaccgccgccgggatcactctcggcatggacg 7140
agctgtacaagtaaacgcgtatgcatggccggccctgcaggaattcgatatcaagcttat 7200
cgataatcaacctctggattacaaaatttgtgaaagattgactggtattcttaactatgt 7260
tgctccttttacgctatgtggatacgctgctttaatgcctttgtatcatgctattgcttc 7320
ccgtatggctttcattttctcctccttgtataaatcctggttgctgtctctttatgagga 7380
gttgtggcccgttgtcaggcaacgtggcgtggtgtgcactgtgtttgctgacgcaacccc 7440
cactggttggggcattgccaccacctgtcagctcctttccgggactttcgctttccccct 7500
ccctattgccacggcggaactcatcgccgcctgccttgcccgctgctggacaggggctcg 7560
gctgttgggcactgacaattccgtggtgttgtcggggaaatcatcgtcctttccttggct 7620
gctcgcctgtgttgccacctggattctgcgcgggacgtccttctgctacgtcccttcggc 7680
cctcaatccagcggaccttccttcccgcggcctgctgccggctctgcggcctcttccgcg 7740
tcttcgccttcgccctcagacgagtcggatctccctttgggccgcctccccgcatcgcga 7800
cctcgacctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgc 7860
cttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattg 7920
catcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagca 7980
agggggaggattgggaagacaatggcaggcatgctggggaactagtggtgccagggcgtg 8040
cccttgggctccccgggcgcggcggccatcgctcgagtaaaattggagggacaagacttc 8100
ccacagattttcggttttgtcgggaagttttttaataggggcaaataaggaaaatgggag 8160
gataggtagtcatctggggttttatgcagcaaaactacaggttattattgcttgtgatcc 8220
gc 8222
<210> 16
<211> 1673
<212>PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> MISC_FEATURE
<222> (1)..(22)
<223> 3xFLAG
<220>
<221> MISC_FEATURE
<222> (23)..(1413)
<223> Cas9
<220>
<221> MISC_FEATURE
<222> (1414)..(1435)
<223> P2A
<220>
<221> MISC_FEATURE
<222> (1436)..(1673)
<223> eGFP
<400> 16
Asp Tyr Lys Asp His Asp Gly Asp Tyr Lys Asp His Asp Ile Asp Tyr
1 5 10 15
Lys Asp Asp Asp Asp Lys Met Asp Lys Pro Lys Lys Lys Arg Lys Val
20 25 30
Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly Trp Ala
35 40 45
Val Ile Thr Asp Glu Tyr Lys Val Pro Ser Lys Lys Phe Lys Val Leu
50 55 60
Gly Asn Thr Asp Arg His Ser Ile Lys Lys Asn Leu Ile Gly Ala Leu
65 70 75 80
Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala Thr Arg Leu Lys Arg Thr
85 90 95
Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn Arg Ile Cys Tyr Leu Gln
100 105 110
Glu Ile Phe Ser Asn Glu Met Ala Lys Val Asp Asp Ser Phe Phe His
115 120 125
Arg Leu Glu Glu Ser Phe Leu Val Glu Glu Asp Lys Lys His Glu Arg
130 135 140
His Pro Ile Phe Gly Asn Ile Val Asp Glu Val Ala Tyr His Glu Lys
145 150 155 160
Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys Leu Val Asp Ser Thr Asp
165 170 175
Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala Leu Ala His Met Ile Lys
180 185 190
Phe Arg Gly His Phe Leu Ile Glu Gly Asp Leu Asn Pro Asp Asn Ser
195 200 205
Asp Val Asp Lys Leu Phe Ile Gln Leu Val Gln Thr Tyr Asn Gln Leu
210 215 220
Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly Val Asp Ala Lys Ala Ile
225 230 235 240
Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg Leu Glu Asn Leu Ile Ala
245 250 255
Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu Phe Gly Asn Leu Ile Ala
260 265 270
Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys Ser Asn Phe Asp Leu Ala
275 280 285
Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp Thr Tyr Asp Asp Asp Leu
290 295 300
Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln Tyr Ala Asp Leu Phe Leu
305 310 315 320
Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu Leu Ser Asp Ile Leu Arg
325 330 335
Val Asn Thr Glu Ile Thr Lys Ala Pro Leu Ser Ala Ser Met Ile Lys
340 345 350
Arg Tyr Asp Glu His His Gln Asp Leu Thr Leu Leu Lys Ala Leu Val
355 360 365
Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu Ile Phe Phe Asp Gln Ser
370 375 380
Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Gly Ala Ser Gln Glu Glu
385 390 395 400
Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu Lys Met Asp Gly Thr Glu
405 410 415
Glu Leu Leu Val Lys Leu Asn Arg Glu Asp Leu Leu Arg Lys Gln Arg
420 425 430
Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu Gly Glu Leu
435 440 445
His Ala Ile Leu Arg Arg Gln Glu Asp Phe Tyr Pro Phe Leu Lys Asp
450 455 460
Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr Phe Arg Ile Pro Tyr Tyr
465 470 475 480
Val Gly Pro Leu Ala Arg Gly Asn Ser Arg Phe Ala Trp Met Thr Arg
485 490 495
Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn Phe Glu Glu Val Val Asp
500 505 510
Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu Arg Met Thr Asn Phe Asp
515 520 525
Lys Asn Leu Pro Asn Glu Lys Val Leu Pro Lys His Ser Leu Leu Tyr
530 535 540
Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys Tyr Val Thr
545 550 555 560
Glu Gly Met Arg Lys Pro Ala Phe Leu Ser Gly Glu Gln Lys Lys Ala
565 570 575
Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys Val Thr Val Lys Gln
580 585 590
Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys Phe Asp Ser Val Glu
595 600 605
Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser Leu Gly Thr Tyr His
610 615 620
Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe Leu Asp Asn Glu Glu
625 630 635 640
Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr Leu Thr Leu Phe Glu
645 650 655
Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr Tyr Ala His Leu Phe
660 665 670
Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg Arg Tyr Thr Gly Trp
675 680 685
Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile Arg Asp Lys Gln Ser
690 695 700
Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp Gly Phe Ala Asn Arg
705 710 715 720
Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu Thr Phe Lys Glu Asp
725 730 735
Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp Ser Leu His Glu His
740 745 750
Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys Lys Gly Ile Leu Gln
755 760 765
Thr Val Lys Val Val Asp Glu Leu Val Lys Val Met Gly Arg His Lys
770 775 780
Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln Thr Thr Gln
785 790 795 800
Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys Arg Ile Glu Glu Gly
805 810 815
Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu His Pro Val Glu Asn
820 825 830
Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly
835 840 845
Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile Asn Arg Leu Ser Asp
850 855 860
Tyr Asp Val Asp His Ile Val Pro Gln Ser Phe Leu Lys Asp Asp Ser
865 870 875 880
Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg Gly Lys Ser
885 890 895
Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys Asn Tyr Trp
900 905 910
Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys Phe Asp Asn
915 920 925
Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp Lys Ala Gly
930 935 940
Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr Lys His Val
945 950 955 960
Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr Asp Glu Asn Asp
965 970 975
Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu Lys Ser Lys Leu Val
980 985 990
Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys Val Arg Glu Ile Asn
995 1000 1005
Asn Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val Val Gly
1010 1015 1020
Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser Glu Phe Val
1025 1030 1035
Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met Ile Ala Lys
1040 1045 1050
Ser Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr Phe Phe Tyr
1055 1060 1065
Ser Asn Ile Met Asn Phe Phe Lys Thr Glu Ile Thr Leu Ala Asn
1070 1075 1080
Gly Glu Ile Arg Lys Arg Pro Leu Ile Glu Thr Asn Gly Glu Thr
1085 1090 1095
Gly Glu Ile Val Trp Asp Lys Gly Arg Asp Phe Ala Thr Val Arg
1100 1105 1110
Lys Val Leu Ser Met Pro Gln Val Asn Ile Val Lys Lys Thr Glu
1115 1120 1125
Val Gln Thr Gly Gly Phe Ser Lys Glu Ser Ile Leu Pro Lys Arg
1130 1135 1140
Asn Ser Asp Lys Leu Ile Ala Arg Lys Lys Asp Trp Asp Pro Lys
1145 1150 1155
Lys Tyr Gly Gly Phe Asp Ser Pro Thr Val Ala Tyr Ser Val Leu
1160 1165 1170
Val Val Ala Lys Val Glu Lys Gly Lys Ser Lys Lys Leu Lys Ser
1175 1180 1185
Val Lys Glu Leu Leu Gly Ile Thr Ile Met Glu Arg Ser Ser Phe
1190 1195 1200
Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala Lys Gly Tyr Lys Glu
1205 1210 1215
Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys Tyr Ser Leu Phe
1220 1225 1230
Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser Ala Gly Glu
1235 1240 1245
Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys Tyr Val Asn
1250 1255 1260
Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys Gly Ser Pro
1265 1270 1275
Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln His Lys His
1280 1285 1290
Tyr Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe Ser Lys Arg
1295 1300 1305
Val Ile Leu Ala Asp Ala Asn Leu Asp Lys Val Leu Ser Ala Tyr
1310 1315 1320
Asn Lys His Arg Asp Lys Pro Ile Arg Glu Gln Ala Glu Asn Ile
1325 1330 1335
Ile His Leu Phe Thr Leu Thr Asn Leu Gly Ala Pro Ala Ala Phe
1340 1345 1350
Lys Tyr Phe Asp Thr Thr Ile Asp Arg Lys Arg Tyr Thr Ser Thr
1355 1360 1365
Lys Glu Val Leu Asp Ala Thr Leu Ile His Gln Ser Ile Thr Gly
1370 1375 1380
Leu Tyr Glu Thr Arg Ile Asp Leu Ser Gln Leu Gly Gly Asp Lys
1385 1390 1395
Arg Pro Ala Ala Thr Lys Lys Ala Gly Gln Ala Lys Lys Lys Lys
1400 1405 1410
Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp
1415 1420 1425
Val Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu Glu Leu Phe
1430 1435 1440
Thr Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn
1445 1450 1455
Gly His Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr
1460 1465 1470
Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu
1475 1480 1485
Pro Val Pro Trp Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val
1490 1495 1500
Gln Cys Phe Ser Arg Tyr Pro Asp His Met Lys Gln His Asp Phe
1505 1510 1515
Phe Lys Ser Ala Met Pro Glu Gly Tyr Val Gln Glu Arg Thr Ile
1520 1525 1530
Phe Phe Lys Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys
1535 1540 1545
Phe Glu Gly Asp Thr Leu Val Asn Arg Ile Glu Leu Lys Gly Ile
1550 1555 1560
Asp Phe Lys Glu Asp Gly Asn Ile Leu Gly His Lys Leu Glu Tyr
1565 1570 1575
Asn Tyr Asn Ser His Asn Val Tyr Ile Met Ala Asp Lys Gln Lys
1580 1585 1590
Asn Gly Ile Lys Val Asn Phe Lys Ile Arg His Asn Ile Glu Asp
1595 1600 1605
Gly Ser Val Gln Leu Ala Asp His Tyr Gln Gln Asn Thr Pro Ile
1610 1615 1620
Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu Ser Thr
1625 1630 1635
Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His Met
1640 1645 1650
Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
1655 1660 1665
Asp Glu Leu Tyr Lys
1670
<210> 17
<211> 7207
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221>misc_feature
<222> (300)..(333)
<223> LoxP
<220>
<221> misc_feature
<222> (424)..(2489)
<223> Neo-PolyA
<220>
<221> misc_feature
<222> (2517)..(2550)
<223> LoxP
<220>
<221> misc_feature
<222> (2599)..(2608)
<223> Kozak
<220>
<221> misc_feature
<222> (2605)..(6777)
<223> Cas9
<220>
<221> misc_feature
<222> (2614)..(2634)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (6730)..(6777)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (6783)..(6998)
<223>сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (7058)..(7207)
<223>Нижележащая последовательность Rosa26 мыши
<400> 17
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctagttgaccag 180
ctcggcggtgacctgcacgtctagggcgcagtagtccagggtttccttgatgatgtcata 240
cttatcctgtcccttttttttccacagggcgcgccactagtggatccggaacccttaata 300
taacttcgtataatgtatgctatacgaagttattaggtccctcgacctgcaggaattgtt 360
gacaattaatcatcggcatagtatatcggcatagtataatacgacaaggtgaggaactaa 420
accatgggatcggccattgaacaagatggattgcacgcaggttctccggccgcttgggtg 480
gagaggctattcggctatgactgggcacaacagacaatcggctgctctgatgccgccgtg 540
ttccggctgtcagcgcaggggcgcccggttctttttgtcaagaccgacctgtccggtgcc 600
ctgaatgaactgcaggacgaggcagcgcggctatcgtggctggccacgacgggcgttcct 660
tgcgcagctgtgctcgacgttgtcactgaagcgggaagggactggctgctattgggcgaa 720
gtgccggggcaggatctcctgtcatctcaccttgctcctgccgagaaagtatccatcatg 780
gctgatgcaatgcggcggctgcatacgcttgatccggctacctgcccattcgaccaccaa 840
gcgaaacatcgcatcgagcgagcacgtactcggatggaagccggtcttgtcgatcaggat 900
gatctggacgaagagcatcaggggctcgcgccagccgaactgttcgccaggctcaaggcg 960
cgcatgcccgacggcgatgatctcgtcgtgacccatggcgatgcctgcttgccgaatatc 1020
atggtggaaaatggccgcttttctggattcatcgactgtggccggctgggtgtggcggac 1080
cgctatcaggacatagcgttggctacccgtgatattgctgaagagcttggcggcgaatgg 1140
gctgaccgcttcctcgtgctttacggtatcgccgctcccgattcgcagcgcatcgccttc 1200
tatcgccttcttgacgagttcttctgaggggatccgctgtaagtctgcagaaattgatga 1260
tctattaaacaataaagatgtccactaaaatggaagtttttcctgtcatactttgttaag 1320
aagggtgagaacagagtacctacattttgaatggaaggattggagctacgggggtggggg 1380
tggggtgggattagataaatgcctgctctttactgaaggctctttactattgctttatga 1440
taatgtttcatagttggatatcataatttaaacaagcaaaaccaaattaagggccagctc 1500
attcctccca ctcatgatct atagatctat agatctctcg tgggatcatt gtttttctct 1560
tgattcccac tttgtggttc taagtactgt ggtttccaaa tgtgtcagtt tcatagcctg 1620
aagaacgaga tcagcagcct ctgttccaca tacacttcat tctcagtatt gttttgccaa 1680
gttctaattc catcagaagc ttgcagatct gcgactctag aggatctgcg actctagagg 1740
atcataatca gccataccac atttgtagag gttttacttg ctttaaaaaa cctcccacac 1800
ctccccctga acctgaaaca taaaatgaat gcaattgttg ttgttaactt gtttattgca 1860
gcttataatg gttacaaata aagcaatagc atcacaaatt tcacaaataa agcatttttt 1920
tcactgcatt ctagttgtgg tttgtccaaa ctcatcaatg tatcttatca tgtctggatc 1980
tgcgactcta gaggatcata atcagccata ccacatttgt agaggtttta cttgctttaa 2040
aaaacctccc acacctcccc ctgaacctga aacataaaat gaatgcaatt gttgttgtta 2100
acttgtttat tgcagcttat aatggttaca aataaagcaa tagcatcaca aatttcacaa 2160
ataaagcatt tttttcactg cattctagtt gtggtttgtc caaactcatc aatgtatctt 2220
atcatgtctg gatctgcgac tctagaggat cataatcagc cataccacat ttgtagaggt 2280
tttacttgct ttaaaaaacc tcccacacct ccccctgaac ctgaaacata aaatgaatgc 2340
aattgttgtt gttaacttgt ttattgcagc ttataatggt tacaaataaa gcaatagcat 2400
cacaaatttc acaaataaag catttttttc actgcattct agttgtggtt tgtccaaact 2460
catcaatgta tcttatcatg tctggatccc catcaagctg atccggaacc cttaatataa 2520
cttcgtataa tgtatgctat acgaagttat taggtccctc gacctgcagc ccaagctagt 2580
gcccgggaat tcgctagggc caccatggac aagcccaaga aaaagcggaa agtgaagtac 2640
agcatcggcc tggacatcgg caccaactct gtgggctggg ccgtgatcac cgacgagtac 2700
aaggtgccca gcaagaaatt caaggtgctg ggcaacaccg acaggcacag catcaagaag 2760
aacctgatcg gcgccctgct gttcgacagc ggcgaaacag ccgaggccac cagactgaag 2820
agaaccgcca gaagaagata caccaggcgg aagaacagga tctgctatct gcaagagatc 2880
ttcagcaacg agatggccaa ggtggacgac agcttcttcc acagactgga agagtccttc 2940
ctggtggaag aggacaagaa gcacgagaga caccccatct tcggcaacat cgtggacgag 3000
gtggcctacc acgagaagta ccccaccatc taccacctga gaaagaaact ggtggacagc 3060
accgacaagg ccgacctgag actgatctac ctggccctgg cccacatgat caagttcaga 3120
ggccacttcc tgatcgaggg cgacctgaac cccgacaaca gcgacgtgga caagctgttc 3180
atccagctgg tgcagaccta caaccagctg ttcgaggaaa accccatcaa cgccagcggc 3240
gtggacgcca aggctatcct gtctgccaga ctgagcaaga gcagaaggct ggaaaatctg 3300
atcgcccagc tgcccggcga gaagaagaac ggcctgttcg gcaacctgat tgccctgagc 3360
ctgggcctga cccccaactt caagagcaac ttcgacctgg ccgaggatgc caaactgcag 3420
ctgagcaagg acacctacga cgacgacctg gacaacctgc tggcccagat cggcgaccag 3480
tacgccgacc tgttcctggc cgccaagaac ctgtctgacg ccatcctgct gagcgacatc 3540
ctgagagtga acaccgagat caccaaggcc cccctgagcg cctctatgat caagagatac 3600
gacgagcacc accaggacct gaccctgctg aaagctctcg tgcggcagca gctgcctgag 3660
aagtacaaag aaatcttctt cgaccagagc aagaacggct acgccggcta catcgatggc 3720
ggcgctagcc aggaagagtt ctacaagttc atcaagccca tcctggaaaa gatggacggc 3780
accgaggaac tgctcgtgaa gctgaacaga gaggacctgc tgagaaagca gagaaccttc 3840
gacaacggca gcatccccca ccagatccac ctgggagagc tgcacgctat cctgagaagg 3900
caggaagatt tttacccatt cctgaaggac aaccgggaaa agatcgagaa gatcctgacc 3960
ttcaggatcc cctactacgt gggccccctg gccagaggca acagcagatt cgcctggatg 4020
accagaaaga gcgaggaaac catcaccccc tggaacttcg aggaagtggt ggacaagggc 4080
gccagcgccc agagcttcat cgagagaatg acaaacttcg ataagaacct gcccaacgag 4140
aaggtgctgc ccaagcacag cctgctgtac gagtacttca ccgtgtacaa cgagctgacc 4200
aaagtgaaat acgtgaccga gggaatgaga aagcccgcct tcctgagcgg cgagcagaaa 4260
aaggccatcg tggacctgct gttcaagacc aacagaaaag tgaccgtgaa gcagctgaaa 4320
gaggactact tcaagaaaat cgagtgcttc gactccgtgg aaatctccgg cgtggaagat 4380
agattcaacg cctccctggg cacataccac gatctgctga aaattatcaa ggacaaggac 4440
ttcctggata acgaagagaa cgaggacatt ctggaagata tcgtgctgac cctgacactg 4500
tttgaggacc gcgagatgat cgaggaaagg ctgaaaacct acgctcacct gttcgacgac 4560
aaagtgatga agcagctgaa gagaaggcgg tacaccggct ggggcaggct gagcagaaag 4620
ctgatcaacg gcatcagaga caagcagagc ggcaagacaa tcctggattt cctgaagtcc 4680
gacggcttcg ccaaccggaa cttcatgcag ctgatccacg acgacagcct gacattcaaa 4740
gaggacatcc agaaagccca ggtgtccggc cagggcgact ctctgcacga gcatatcgct 4800
aacctggccg gcagccccgc tatcaagaag ggcatcctgc agacagtgaa ggtggtggac 4860
gagctcgtga aagtgatggg cagacacaag cccgagaaca tcgtgatcga gatggctaga 4920
gagaaccaga ccacccagaa gggacagaag aactcccgcg agaggatgaa gagaatcgaa 4980
gagggcatca aagagctggg cagccagatc ctgaaagaac accccgtgga aaacacccag 5040
ctgcagaacg agaagctgta cctgtactac ctgcagaatg gccgggatat gtacgtggac 5100
caggaactgg acatcaacag actgtccgac tacgatgtgg accatatcgt gcctcagagc 5160
tttctgaagg acgactccat cgataacaaa gtgctgactc ggagcgacaa gaacagaggc 5220
aagagcgaca acgtgccctc cgaagaggtc gtgaagaaga tgaagaacta ctggcgacag 5280
ctgctgaacg ccaagctgat tacccagagg aagttcgata acctgaccaa ggccgagaga 5340
ggcggcctga gcgagctgga taaggccggc ttcatcaaga ggcagctggt ggaaaccaga 5400
cagatcacaa agcacgtggc acagatcctg gactcccgga tgaacactaa gtacgacgaa 5460
aacgataagc tgatccggga agtgaaagtg atcaccctga agtccaagct ggtgtccgat 5520
ttccggaagg atttccagtt ttacaaagtg cgcgagatca acaactacca ccacgcccac 5580
gacgcctacc tgaacgccgt cgtgggaacc gccctgatca aaaagtaccc taagctggaa 5640
agcgagttcg tgtacggcga ctacaaggtg tacgacgtgc ggaagatgat cgccaagagc 5700
gagcaggaaa tcggcaaggc taccgccaag tacttcttct acagcaacat catgaacttt 5760
ttcaagaccg aaatcaccct ggccaacggc gagatcagaa agcgccctct gatcgagaca 5820
aacggcgaaa ccggggagat cgtgtgggat aagggcagag acttcgccac agtgcgaaag 5880
gtgctgagca tgccccaagt gaatatcgtg aaaaagaccg aggtgcagac aggcggcttc 5940
agcaaagagt ctatcctgcc caagaggaac agcgacaagc tgatcgccag aaagaaggac 6000
tgggacccca agaagtacgg cggcttcgac agccctaccg tggcctactc tgtgctggtg 6060
gtggctaagg tggaaaaggg caagtccaag aaactgaaga gtgtgaaaga gctgctgggg 6120
atcaccatca tggaaagaag cagctttgag aagaacccta tcgactttct ggaagccaag 6180
ggctacaaag aagtgaaaaa ggacctgatc atcaagctgc ctaagtactc cctgttcgag 6240
ctggaaaacg gcagaaagag aatgctggcc tctgccggcg aactgcagaa gggaaacgag 6300
ctggccctgc ctagcaaata tgtgaacttc ctgtacctgg cctcccacta tgagaagctg 6360
aagggcagcc ctgaggacaa cgaacagaaa cagctgtttg tggaacagca taagcactac 6420
ctggacgaga tcatcgagca gatcagcgag ttctccaaga gagtgatcct ggccgacgcc 6480
aatctggaca aggtgctgtc tgcctacaac aagcacaggg acaagcctat cagagagcag 6540
gccgagaata tcatccacct gttcaccctg acaaacctgg gcgctcctgc cgccttcaag 6600
tactttgaca ccaccatcga ccggaagagg tacaccagca ccaaagaggt gctggacgcc 6660
accctgatcc accagagcat caccggcctg tacgagacaa gaatcgacct gtctcagctg 6720
ggaggcgaca agagacctgc cgccactaag aaggccggac aggccaaaaa gaagaagtga 6780
gtcgacctcg acctcgactg tgccttctag ttgccagcca tctgttgttt gcccctcccc 6840
cgtgccttcc ttgaccctgg aaggtgccac tcccactgtc ctttcctaat aaaatgagga 6900
aattgcatcg cattgtctga gtaggtgtca ttctattctg gggggtgggg tggggcagga 6960
cagcaagggg gaggattggg aagacaatgg caggcatgct ggggaactag tggtgccagg 7020
gcgtgccctt gggctccccg ggcgcggcgg ccatcgctcg agtaaaattg gagggacaag 7080
acttcccaca gattttcggt tttgtcggga agttttttaa taggggcaaa taaggaaaat 7140
gggaggatag gtagtcatct ggggttttat gcagcaaaac tacaggttat tattgcttgt 7200
gatccgc 7207
<210> 18
<211> 4990
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221> misc_feature
<222> (300)..(333)
<223> LoxP
<220>
<221> misc_feature
<222> (382)..(391)
<223> Kozak
<220>
<221> misc_feature
<222> (388)..(4560)
<223> Cas9
<220>
<221> misc_feature
<222> (397)..(417)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (4513)..(4560)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (4566)..(4781)
<223>Сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (4841)..(4990)
<223>Нижележащая последовательность Rosa26 мыши
<400> 18
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctagttgaccag 180
ctcggcggtgacctgcacgtctagggcgcagtagtccagggtttccttgatgatgtcata 240
cttatcctgtcccttttttttccacagggcgcgccactagtggatccggaacccttaata 300
taacttcgtataatgtatgctatacgaagttattaggtccctcgacctgcagcccaagct 360
agtgcccgggaattcgctagggccaccatggacaagcccaagaaaaagcggaaagtgaag 420
tacagcatcggcctggacatcggcaccaactctgtgggctgggccgtgatcaccgacgag 480
tacaaggtgcccagcaagaaattcaaggtgctgggcaacaccgacaggcacagcatcaag 540
aagaacctgatcggcgccctgctgttcgacagcggcgaaacagccgaggccaccagactg 600
aagagaaccgccagaagaagatacaccaggcggaagaacaggatctgctatctgcaagag 660
atcttcagcaacgagatggccaaggtggacgacagcttcttccacagactggaagagtcc 720
ttcctggtggaagaggacaagaagcacgagagacaccccatcttcggcaacatcgtggac 780
gaggtggcctaccacgagaagtaccccaccatctaccacctgagaaagaaactggtggac 840
agcaccgacaaggccgacctgagactgatctacctggccctggcccacatgatcaagttc 900
agaggccacttcctgatcgagggcgacctgaaccccgacaacagcgacgtggacaagctg 960
ttcatccagctggtgcagacctacaaccagctgttcgaggaaaaccccatcaacgccagc 1020
ggcgtggacgccaaggctatcctgtctgccagactgagcaagagcagaaggctggaaaat 1080
ctgatcgcccagctgcccggcgagaagaagaacggcctgttcggcaacctgattgccctg 1140
agcctgggcctgacccccaacttcaagagcaacttcgacctggccgaggatgccaaactg 1200
cagctgagcaaggacacctacgacgacgacctggacaacctgctggcccagatcggcgac 1260
cagtacgccgacctgttcctggccgccaagaacctgtctgacgccatcctgctgagcgac 1320
atcctgagagtgaacaccgagatcaccaaggcccccctgagcgcctctatgatcaagaga 1380
tacgacgagcaccaccaggacctgaccctgctgaaagctctcgtgcggcagcagctgcct 1440
gagaagtacaaagaaatcttcttcgaccagagcaagaacggctacgccggctacatcgat 1500
ggcggcgctagccaggaagagttctacaagttcatcaagcccatcctggaaaagatggac 1560
ggcaccgaggaactgctcgtgaagctgaacagagaggacctgctgagaaagcagagaacc 1620
ttcgacaacggcagcatcccccaccagatccacctgggagagctgcacgctatcctgaga 1680
aggcaggaagatttttacccattcctgaaggacaaccgggaaaagatcgagaagatcctg 1740
accttcaggatcccctactacgtgggccccctggccagaggcaacagcagattcgcctgg 1800
atgaccagaaagagcgaggaaaccatcaccccctggaacttcgaggaagtggtggacaag 1860
ggcgccagcgcccagagcttcatcgagagaatgacaaacttcgataagaacctgcccaac 1920
gagaaggtgctgcccaagcacagcctgctgtacgagtacttcaccgtgtacaacgagctg 1980
accaaagtgaaatacgtgaccgagggaatgagaaagcccgccttcctgagcggcgagcag 2040
aaaaaggccatcgtggacctgctgttcaagaccaacagaaaagtgaccgtgaagcagctg 2100
aaagaggactacttcaagaaaatcgagtgcttcgactccgtggaaatctccggcgtggaa 2160
gatagattcaacgcctccctgggcacataccacgatctgctgaaaattatcaaggacaag 2220
gacttcctggataacgaagagaacgaggacattctggaagatatcgtgctgaccctgaca 2280
ctgtttgaggaccgcgagatgatcgaggaaaggctgaaaacctacgctcacctgttcgac 2340
gacaaagtgatgaagcagctgaagagaaggcggtacaccggctggggcaggctgagcaga 2400
aagctgatcaacggcatcagagacaagcagagcggcaagacaatcctggatttcctgaag 2460
tccgacggcttcgccaaccggaacttcatgcagctgatccacgacgacagcctgacattc 2520
aaagaggacatccagaaagcccaggtgtccggccagggcgactctctgcacgagcatatc 2580
gctaacctggccggcagccccgctatcaagaagggcatcctgcagacagtgaaggtggtg 2640
gacgagctcgtgaaagtgatgggcagacacaagcccgagaacatcgtgatcgagatggct 2700
agagagaaccagaccacccagaagggacagaagaactcccgcgagaggatgaagagaatc 2760
gaagagggcatcaaagagctgggcagccagatcctgaaagaacaccccgtggaaaacacc 2820
cagctgcagaacgagaagctgtacctgtactacctgcagaatggccgggatatgtacgtg 2880
gaccaggaactggacatcaacagactgtccgactacgatgtggaccatatcgtgcctcag 2940
agctttctgaaggacgactccatcgataacaaagtgctgactcggagcgacaagaacaga 3000
ggcaagagcgacaacgtgccctccgaagaggtcgtgaagaagatgaagaactactggcga 3060
cagctgctgaacgccaagctgattacccagaggaagttcgataacctgaccaaggccgag 3120
agaggcggcctgagcgagctggataaggccggcttcatcaagaggcagctggtggaaacc 3180
agacagatcacaaagcacgtggcacagatcctggactcccggatgaacactaagtacgac 3240
gaaaacgataagctgatccgggaagtgaaagtgatcaccctgaagtccaagctggtgtcc 3300
gatttccggaaggatttccagttttacaaagtgcgcgagatcaacaactaccaccacgcc 3360
cacgacgcctacctgaacgccgtcgtgggaaccgccctgatcaaaaagtaccctaagctg 3420
gaaagcgagttcgtgtacggcgactacaaggtgtacgacgtgcggaagatgatcgccaag 3480
agcgagcaggaaatcggcaaggctaccgccaagtacttcttctacagcaacatcatgaac 3540
tttttcaagaccgaaatcaccctggccaacggcgagatcagaaagcgccctctgatcgag 3600
acaaacggcgaaaccggggagatcgtgtgggataagggcagagacttcgccacagtgcga 3660
aaggtgctgagcatgccccaagtgaatatcgtgaaaaagaccgaggtgcagacaggcggc 3720
ttcagcaaagagtctatcctgcccaagaggaacagcgacaagctgatcgccagaaagaag 3780
gactgggaccccaagaagtacggcggcttcgacagccctaccgtggcctactctgtgctg 3840
gtggtggctaaggtggaaaagggcaagtccaagaaactgaagagtgtgaaagagctgctg 3900
gggatcaccatcatggaaagaagcagctttgagaagaaccctatcgactttctggaagcc 3960
aagggctacaaagaagtgaaaaaggacctgatcatcaagctgcctaagtactccctgttc 4020
gagctggaaaacggcagaaagagaatgctggcctctgccggcgaactgcagaagggaaac 4080
gagctggccctgcctagcaaatatgtgaacttcctgtacctggcctcccactatgagaag 4140
ctgaagggcagccctgaggacaacgaacagaaacagctgtttgtggaacagcataagcac 4200
tacctggacgagatcatcgagcagatcagcgagttctccaagagagtgatcctggccgac 4260
gccaatctggacaaggtgctgtctgcctacaacaagcacagggacaagcctatcagagag 4320
caggccgagaatatcatccacctgttcaccctgacaaacctgggcgctcctgccgccttc 4380
aagtactttgacaccaccatcgaccggaagaggtacaccagcaccaaagaggtgctggac 4440
gccaccctgatccaccagagcatcaccggcctgtacgagacaagaatcgacctgtctcag 4500
ctgggaggcgacaagagacctgccgccactaagaaggccggacaggccaaaaagaagaag 4560
tgagtcgacctcgacctcgactgtgccttctagttgccagccatctgttgtttgcccctc 4620
ccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatga 4680
ggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggca 4740
ggacagcaagggggaggattgggaagacaatggcaggcatgctggggaactagtggtgcc 4800
agggcgtgcccttgggctccccgggcgcggcggccatcgctcgagtaaaattggagggac 4860
aagacttcccacagattttcggttttgtcgggaagttttttaataggggcaaataaggaa 4920
aatgggaggataggtagtcatctggggttttatgcagcaaaactacaggttattattgct 4980
tgtgatccgc 4990
<210> 19
<211> 1391
<212>PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 19
Met Asp Lys Pro Lys Lys Lys Arg Lys Val Lys Tyr Ser Ile Gly Leu
1 5 10 15
Asp Ile Gly Thr Asn Ser Val Gly Trp Ala Val Ile Thr Asp Glu Tyr
20 25 30
Lys Val Pro Ser Lys Lys Phe Lys Val Leu Gly Asn Thr Asp Arg His
35 40 45
Ser Ile Lys Lys Asn Leu Ile Gly Ala Leu Leu Phe Asp Ser Gly Glu
50 55 60
Thr Ala Glu Ala Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg Tyr Thr
65 70 75 80
Arg Arg Lys Asn Arg Ile Cys Tyr Leu Gln Glu Ile Phe Ser Asn Glu
85 90 95
Met Ala Lys Val Asp Asp Ser Phe Phe His Arg Leu Glu Glu Ser Phe
100 105 110
Leu Val Glu Glu Asp Lys Lys His Glu Arg His Pro Ile Phe Gly Asn
115 120 125
Ile Val Asp Glu Val Ala Tyr His Glu Lys Tyr Pro Thr Ile Tyr His
130 135 140
Leu Arg Lys Lys Leu Val Asp Ser Thr Asp Lys Ala Asp Leu Arg Leu
145 150 155 160
Ile Tyr Leu Ala Leu Ala His Met Ile Lys Phe Arg Gly His Phe Leu
165 170 175
Ile Glu Gly Asp Leu Asn Pro Asp Asn Ser Asp Val Asp Lys Leu Phe
180 185 190
Ile Gln Leu Val Gln Thr Tyr Asn Gln Leu Phe Glu Glu Asn Pro Ile
195 200 205
Asn Ala Ser Gly Val Asp Ala Lys Ala Ile Leu Ser Ala Arg Leu Ser
210 215 220
Lys Ser Arg Arg Leu Glu Asn Leu Ile Ala Gln Leu Pro Gly Glu Lys
225 230 235 240
Lys Asn Gly Leu Phe Gly Asn Leu Ile Ala Leu Ser Leu Gly Leu Thr
245 250 255
Pro Asn Phe Lys Ser Asn Phe Asp Leu Ala Glu Asp Ala Lys Leu Gln
260 265 270
Leu Ser Lys Asp Thr Tyr Asp Asp Asp Leu Asp Asn Leu Leu Ala Gln
275 280 285
Ile Gly Asp Gln Tyr Ala Asp Leu Phe Leu Ala Ala Lys Asn Leu Ser
290 295 300
Asp Ala Ile Leu Leu Ser Asp Ile Leu Arg Val Asn Thr Glu Ile Thr
305 310 315 320
Lys Ala Pro Leu Ser Ala Ser Met Ile Lys Arg Tyr Asp Glu His His
325 330 335
Gln Asp Leu Thr Leu Leu Lys Ala Leu Val Arg Gln Gln Leu Pro Glu
340 345 350
Lys Tyr Lys Glu Ile Phe Phe Asp Gln Ser Lys Asn Gly Tyr Ala Gly
355 360 365
Tyr Ile Asp Gly Gly Ala Ser Gln Glu Glu Phe Tyr Lys Phe Ile Lys
370 375 380
Pro Ile Leu Glu Lys Met Asp Gly Thr Glu Glu Leu Leu Val Lys Leu
385 390 395 400
Asn Arg Glu Asp Leu Leu Arg Lys Gln Arg Thr Phe Asp Asn Gly Ser
405 410 415
Ile Pro His Gln Ile His Leu Gly Glu Leu His Ala Ile Leu Arg Arg
420 425 430
Gln Glu Asp Phe Tyr Pro Phe Leu Lys Asp Asn Arg Glu Lys Ile Glu
435 440 445
Lys Ile Leu Thr Phe Arg Ile Pro Tyr Tyr Val Gly Pro Leu Ala Arg
450 455 460
Gly Asn Ser Arg Phe Ala Trp Met Thr Arg Lys Ser Glu Glu Thr Ile
465 470 475 480
Thr Pro Trp Asn Phe Glu Glu Val Val Asp Lys Gly Ala Ser Ala Gln
485 490 495
Ser Phe Ile Glu Arg Met Thr Asn Phe Asp Lys Asn Leu Pro Asn Glu
500 505 510
Lys Val Leu Pro Lys His Ser Leu Leu Tyr Glu Tyr Phe Thr Val Tyr
515 520 525
Asn Glu Leu Thr Lys Val Lys Tyr Val Thr Glu Gly Met Arg Lys Pro
530 535 540
Ala Phe Leu Ser Gly Glu Gln Lys Lys Ala Ile Val Asp Leu Leu Phe
545 550 555 560
Lys Thr Asn Arg Lys Val Thr Val Lys Gln Leu Lys Glu Asp Tyr Phe
565 570 575
Lys Lys Ile Glu Cys Phe Asp Ser Val Glu Ile Ser Gly Val Glu Asp
580 585 590
Arg Phe Asn Ala Ser Leu Gly Thr Tyr His Asp Leu Leu Lys Ile Ile
595 600 605
Lys Asp Lys Asp Phe Leu Asp Asn Glu Glu Asn Glu Asp Ile Leu Glu
610 615 620
Asp Ile Val Leu Thr Leu Thr Leu Phe Glu Asp Arg Glu Met Ile Glu
625 630 635 640
Glu Arg Leu Lys Thr Tyr Ala His Leu Phe Asp Asp Lys Val Met Lys
645 650 655
Gln Leu Lys Arg Arg Arg Tyr Thr Gly Trp Gly Arg Leu Ser Arg Lys
660 665 670
Leu Ile Asn Gly Ile Arg Asp Lys Gln Ser Gly Lys Thr Ile Leu Asp
675 680 685
Phe Leu Lys Ser Asp Gly Phe Ala Asn Arg Asn Phe Met Gln Leu Ile
690 695 700
His Asp Asp Ser Leu Thr Phe Lys Glu Asp Ile Gln Lys Ala Gln Val
705 710 715 720
Ser Gly Gln Gly Asp Ser Leu His Glu His Ile Ala Asn Leu Ala Gly
725 730 735
Ser Pro Ala Ile Lys Lys Gly Ile Leu Gln Thr Val Lys Val Val Asp
740 745 750
Glu Leu Val Lys Val Met Gly Arg His Lys Pro Glu Asn Ile Val Ile
755 760 765
Glu Met Ala Arg Glu Asn Gln Thr Thr Gln Lys Gly Gln Lys Asn Ser
770 775 780
Arg Glu Arg Met Lys Arg Ile Glu Glu Gly Ile Lys Glu Leu Gly Ser
785 790 795 800
Gln Ile Leu Lys Glu His Pro Val Glu Asn Thr Gln Leu Gln Asn Glu
805 810 815
Lys Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly Arg Asp Met Tyr Val Asp
820 825 830
Gln Glu Leu Asp Ile Asn Arg Leu Ser Asp Tyr Asp Val Asp His Ile
835 840 845
Val Pro Gln Ser Phe Leu Lys Asp Asp Ser Ile Asp Asn Lys Val Leu
850 855 860
Thr Arg Ser Asp Lys Asn Arg Gly Lys Ser Asp Asn Val Pro Ser Glu
865 870 875 880
Glu Val Val Lys Lys Met Lys Asn Tyr Trp Arg Gln Leu Leu Asn Ala
885 890 895
Lys Leu Ile Thr Gln Arg Lys Phe Asp Asn Leu Thr Lys Ala Glu Arg
900 905 910
Gly Gly Leu Ser Glu Leu Asp Lys Ala Gly Phe Ile Lys Arg Gln Leu
915 920 925
Val Glu Thr Arg Gln Ile Thr Lys His Val Ala Gln Ile Leu Asp Ser
930 935 940
Arg Met Asn Thr Lys Tyr Asp Glu Asn Asp Lys Leu Ile Arg Glu Val
945 950 955 960
Lys Val Ile Thr Leu Lys Ser Lys Leu Val Ser Asp Phe Arg Lys Asp
965 970 975
Phe Gln Phe Tyr Lys Val Arg Glu Ile Asn Asn Tyr His His Ala His
980 985 990
Asp Ala Tyr Leu Asn Ala Val Val Gly Thr Ala Leu Ile Lys Lys Tyr
995 1000 1005
Pro Lys Leu Glu Ser Glu Phe Val Tyr Gly Asp Tyr Lys Val Tyr
1010 1015 1020
Asp Val Arg Lys Met Ile Ala Lys Ser Glu Gln Glu Ile Gly Lys
1025 1030 1035
Ala Thr Ala Lys Tyr Phe Phe Tyr Ser Asn Ile Met Asn Phe Phe
1040 1045 1050
Lys Thr Glu Ile Thr Leu Ala Asn Gly Glu Ile Arg Lys Arg Pro
1055 1060 1065
Leu Ile Glu Thr Asn Gly Glu Thr Gly Glu Ile Val Trp Asp Lys
1070 1075 1080
Gly Arg Asp Phe Ala Thr Val Arg Lys Val Leu Ser Met Pro Gln
1085 1090 1095
Val Asn Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly Phe Ser
1100 1105 1110
Lys Glu Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile Ala
1115 1120 1125
Arg Lys Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser
1130 1135 1140
Pro Thr Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys
1145 1150 1155
Gly Lys Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile
1160 1165 1170
Thr Ile Met Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe
1175 1180 1185
Leu Glu Ala Lys Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile
1190 1195 1200
Lys Leu Pro Lys Tyr Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys
1205 1210 1215
Arg Met Leu Ala Ser Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu
1220 1225 1230
Ala Leu Pro Ser Lys Tyr Val Asn Phe Leu Tyr Leu Ala Ser His
1235 1240 1245
Tyr Glu Lys Leu Lys Gly Ser Pro Glu Asp Asn Glu Gln Lys Gln
1250 1255 1260
Leu Phe Val Glu Gln His Lys His Tyr Leu Asp Glu Ile Ile Glu
1265 1270 1275
Gln Ile Ser Glu Phe Ser Lys Arg Val Ile Leu Ala Asp Ala Asn
1280 1285 1290
Leu Asp Lys Val Leu Ser Ala Tyr Asn Lys His Arg Asp Lys Pro
1295 1300 1305
Ile Arg Glu Gln Ala Glu Asn Ile Ile His Leu Phe Thr Leu Thr
1310 1315 1320
Asn Leu Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp Thr Thr Ile
1325 1330 1335
Asp Arg Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp Ala Thr
1340 1345 1350
Leu Ile His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile Asp
1355 1360 1365
Leu Ser Gln Leu Gly Gly Asp Lys Arg Pro Ala Ala Thr Lys Lys
1370 1375 1380
Ala Gly Gln Ala Lys Lys Lys Lys
1385 1390
<210> 20
<211> 9673
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221>misc_feature
<222> (205)..(1923)
<223>Промотор CAGG
<220>
<221> misc_feature
<222> (2006)..(2039)
<223> LoxP
<220>
<221> misc_feature
<222> (2130)..(4195)
<223> Neo-PolyA
<220>
<221> misc_feature
<222> (4223)..(4256)
<223> LoxP
<220>
<221> misc_feature
<222> (4351)..(4360)
<223> Kozak
<220>
<221> misc_feature
<222> (4360)..(4425)
<223> 3xFLAG
<220>
<221> misc_feature
<222> (4426)..(8598)
<223> Cas9
<220>
<221> misc_feature
<222> (4435)..(4455)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (8551)..(8598)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (8645)..(9241)
<223> WPRE
<220>
<221> misc_feature
<222> (9249)..(9464)
<223>Сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (9524)..(9673)
<223>Нижележащая последовательность Rosa26 мыши
<400> 20
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctccttaattaa 180
acgcgtctggcctcgcgagtgtgtactagttattaatagtaatcaattacggggtcatta 240
gttcatagcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggc 300
tgaccgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacg 360
ccaatagggactttccattgacgtcaatgggtggagtatttacggtaaactgcccacttg 420
gcagtacatcaagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaa 480
tggcccgcctggcattatgcccagtacatgaccttatgggactttcctacttggcagtac 540
atctacgtattagtcatcgctattaccatggtcgaggtgagccccacgttctgcttcact 600
ctccccatctcccccccctccccacccccaattttgtatttatttattttttaattattt 660
tgtgcagcgatgggggcggggggggggggggggcgcgcgccaggcggggcggggcggggc 720
gaggggcggggcggggcgaggcggagaggtgcggcggcagccaatcagagcggcgcgctc 780
cgaaagtttccttttatggcgaggcggcggcggcggcggccctataaaaagcgaagcgcg 840
cggcgggcggggagtcgctgcgacgctgccttcgccccgtgccccgctccgccgccgcct 900
cgcgccgcccgccccggctctgactgaccgcgttactcccacaggtgagcgggcgggacg 960
gcccttctcctccgggctgtaattagcgcttggtttaatgacggcttgtttcttttctgt 1020
ggctgcgtgaaagccttgaggggctccgggagggccctttgtgcggggggagcggctcgg 1080
ggggtgcgtgcgtgtgtgtgtgcgtggggagcgccgcgtgcggctccgcgctgcccggcg 1140
gctgtgagcgctgcgggcgcggcgcggggctttgtgcgctccgcagtgtgcgcgagggga 1200
gcgcggccgggggcggtgccccgcggtgcggggggggctgcgaggggaacaaaggctgcg 1260
tgcggggtgtgtgcgtgggggggtgagcagggggtgtgggcgcgtcggtcgggctgcaac 1320
cccccctgcacccccctccccgagttgctgagcacggcccggcttcgggtgcggggctcc 1380
gtacggggcgtggcgcggggctcgccgtgccgggcggggggtggcggcaggtgggggtgc 1440
cgggcggggcggggccgcctcgggccggggagggctcgggggaggggcgcggcggccccc 1500
ggagcgccggcggctgtcgaggcgcggcgagccgcagccattgccttttatggtaatcgt 1560
gcgagagggcgcagggacttcctttgtcccaaatctgtgcggagccgaaatctgggaggc 1620
gccgccgcaccccctctagcgggcgcggggcgaagcggtgcggcgccggcaggaaggaaa 1680
tgggcggggagggccttcgtgcgtcgccgcgccgccgtccccttctccctctccagcctc 1740
ggggctgtccgcggggggacggctgccttcgggggggacggggcagggcggggttcggct 1800
tctggcgtgtgaccggcggctctagagcctctgctaaccatgttcatgccttcttctttt 1860
tcctacagctcctgggcaacgtgctggttattgtgctgtctcatcattttggcaaagaat 1920
tcgctaggagaattgatttgataccgcgggccctaagtcgacatttaaatcatttaaatc 1980
cactagtggatccggaacccttaatataacttcgtataatgtatgctatacgaagttatt 2040
aggtccctcgacctgcaggaattgttgacaattaatcatcggcatagtatatcggcatag 2100
tataatacgacaaggtgaggaactaaaccatgggatcggccattgaacaagatggattgc 2160
acgcaggttctccggccgcttgggtggagaggctattcggctatgactgggcacaacaga 2220
caatcggctgctctgatgccgccgtgttccggctgtcagcgcaggggcgcccggttcttt 2280
ttgtcaagaccgacctgtccggtgccctgaatgaactgcaggacgaggcagcgcggctat 2340
cgtggctggccacgacgggcgttccttgcgcagctgtgctcgacgttgtcactgaagcgg 2400
gaagggactggctgctattgggcgaagtgccggggcaggatctcctgtcatctcaccttg 2460
ctcctgccgagaaagtatccatcatggctgatgcaatgcggcggctgcatacgcttgatc 2520
cggctacctgcccattcgaccaccaagcgaaacatcgcatcgagcgagcacgtactcgga 2580
tggaagccggtcttgtcgatcaggatgatctggacgaagagcatcaggggctcgcgccag 2640
ccgaactgttcgccaggctcaaggcgcgcatgcccgacggcgatgatctcgtcgtgaccc 2700
atggcgatgcctgcttgccgaatatcatggtggaaaatggccgcttttctggattcatcg 2760
actgtggccggctgggtgtggcggaccgctatcaggacatagcgttggctacccgtgata 2820
ttgctgaagagcttggcggcgaatgggctgaccgcttcctcgtgctttacggtatcgccg 2880
ctcccgattcgcagcgcatcgccttctatcgccttcttgacgagttcttctgaggggatc 2940
cgctgtaagtctgcagaaattgatgatctattaaacaataaagatgtccactaaaatgga 3000
agtttttcctgtcatactttgttaagaagggtgagaacagagtacctacattttgaatgg 3060
aaggattggagctacgggggtgggggtggggtgggattagataaatgcctgctctttact 3120
gaaggctctttactattgctttatgataatgtttcatagttggatatcataatttaaaca 3180
agcaaaaccaaattaagggccagctcattcctcccactcatgatctatagatctatagat 3240
ctctcgtgggatcattgtttttctcttgattcccactttgtggttctaagtactgtggtt 3300
tccaaatgtgtcagtttcatagcctgaagaacgagatcagcagcctctgttccacataca 3360
cttcattctcagtattgttttgccaagttctaattccatcagaagcttgcagatctgcga 3420
ctctagagga tctgcgactc tagaggatca taatcagcca taccacattt gtagaggttt 3480
tacttgcttt aaaaaacctc ccacacctcc ccctgaacct gaaacataaa atgaatgcaa 3540
ttgttgttgt taacttgttt attgcagctt ataatggtta caaataaagc aatagcatca 3600
caaatttcac aaataaagca tttttttcac tgcattctag ttgtggtttg tccaaactca 3660
tcaatgtatc ttatcatgtc tggatctgcg actctagagg atcataatca gccataccac 3720
atttgtagag gttttacttg ctttaaaaaa cctcccacac ctccccctga acctgaaaca 3780
taaaatgaat gcaattgttg ttgttaactt gtttattgca gcttataatg gttacaaata 3840
aagcaatagc atcacaaatt tcacaaataa agcatttttt tcactgcatt ctagttgtgg 3900
tttgtccaaa ctcatcaatg tatcttatca tgtctggatc tgcgactcta gaggatcata 3960
atcagccata ccacatttgt agaggtttta cttgctttaa aaaacctccc acacctcccc 4020
ctgaacctga aacataaaat gaatgcaatt gttgttgtta acttgtttat tgcagcttat 4080
aatggttaca aataaagcaa tagcatcaca aatttcacaa ataaagcatt tttttcactg 4140
cattctagtt gtggtttgtc caaactcatc aatgtatctt atcatgtctg gatccccatc 4200
aagctgatcc ggaaccctta atataacttc gtataatgta tgctatacga agttattagg 4260
tccctcgacc tgcagcccaa gctagtgccc gggtaggtcc ctcgacctgc agcccaagct 4320
agatcgaatt cggccggcct tcgaacacgt gccaccatgg actataagga ccacgacgga 4380
gactacaagg atcatgatat tgattacaaa gacgatgacg ataagatgga caagcccaag 4440
aaaaagcgga aagtgaagta cagcatcggc ctggacatcg gcaccaactc tgtgggctgg 4500
gccgtgatca ccgacgagta caaggtgccc agcaagaaat tcaaggtgct gggcaacacc 4560
gacaggcaca gcatcaagaa gaacctgatc ggcgccctgc tgttcgacag cggcgaaaca 4620
gccgaggcca ccagactgaa gagaaccgcc agaagaagat acaccaggcg gaagaacagg 4680
atctgctatc tgcaagagat cttcagcaac gagatggcca aggtggacga cagcttcttc 4740
cacagactgg aagagtcctt cctggtggaa gaggacaaga agcacgagag acaccccatc 4800
ttcggcaaca tcgtggacga ggtggcctac cacgagaagt accccaccat ctaccacctg 4860
agaaagaaac tggtggacag caccgacaag gccgacctga gactgatcta cctggccctg 4920
gcccacatga tcaagttcag aggccacttc ctgatcgagg gcgacctgaa ccccgacaac 4980
agcgacgtgg acaagctgtt catccagctg gtgcagacct acaaccagct gttcgaggaa 5040
aaccccatca acgccagcgg cgtggacgcc aaggctatcc tgtctgccag actgagcaag 5100
agcagaaggc tggaaaatct gatcgcccag ctgcccggcg agaagaagaa cggcctgttc 5160
ggcaacctga ttgccctgag cctgggcctg acccccaact tcaagagcaa cttcgacctg 5220
gccgaggatg ccaaactgca gctgagcaag gacacctacg acgacgacct ggacaacctg 5280
ctggcccaga tcggcgacca gtacgccgac ctgttcctgg ccgccaagaa cctgtctgac 5340
gccatcctgc tgagcgacat cctgagagtg aacaccgaga tcaccaaggc ccccctgagc 5400
gcctctatga tcaagagata cgacgagcac caccaggacc tgaccctgct gaaagctctc 5460
gtgcggcagc agctgcctga gaagtacaaa gaaatcttct tcgaccagag caagaacggc 5520
tacgccggct acatcgatgg cggcgctagc caggaagagt tctacaagtt catcaagccc 5580
atcctggaaa agatggacgg caccgaggaa ctgctcgtga agctgaacag agaggacctg 5640
ctgagaaagc agagaacctt cgacaacggc agcatccccc accagatcca cctgggagag 5700
ctgcacgcta tcctgagaag gcaggaagat ttttacccat tcctgaagga caaccgggaa 5760
aagatcgaga agatcctgac cttcaggatc ccctactacg tgggccccct ggccagaggc 5820
aacagcagat tcgcctggat gaccagaaag agcgaggaaa ccatcacccc ctggaacttc 5880
gaggaagtgg tggacaaggg cgccagcgcc cagagcttca tcgagagaat gacaaacttc 5940
gataagaacc tgcccaacga gaaggtgctg cccaagcaca gcctgctgta cgagtacttc 6000
accgtgtaca acgagctgac caaagtgaaa tacgtgaccg agggaatgag aaagcccgcc 6060
ttcctgagcg gcgagcagaa aaaggccatc gtggacctgc tgttcaagac caacagaaaa 6120
gtgaccgtga agcagctgaa agaggactac ttcaagaaaa tcgagtgctt cgactccgtg 6180
gaaatctccg gcgtggaaga tagattcaac gcctccctgg gcacatacca cgatctgctg 6240
aaaattatca aggacaagga cttcctggat aacgaagaga acgaggacat tctggaagat 6300
atcgtgctga ccctgacact gtttgaggac cgcgagatga tcgaggaaag gctgaaaacc 6360
tacgctcacc tgttcgacga caaagtgatg aagcagctga agagaaggcg gtacaccggc 6420
tggggcaggc tgagcagaaa gctgatcaac ggcatcagag acaagcagag cggcaagaca 6480
atcctggatt tcctgaagtc cgacggcttc gccaaccgga acttcatgca gctgatccac 6540
gacgacagcc tgacattcaa agaggacatc cagaaagccc aggtgtccgg ccagggcgac 6600
tctctgcacg agcatatcgc taacctggcc ggcagccccg ctatcaagaa gggcatcctg 6660
cagacagtga aggtggtgga cgagctcgtg aaagtgatgg gcagacacaa gcccgagaac 6720
atcgtgatcg agatggctag agagaaccag accacccaga agggacagaa gaactcccgc 6780
gagaggatga agagaatcga agagggcatc aaagagctgg gcagccagat cctgaaagaa 6840
caccccgtgg aaaacaccca gctgcagaac gagaagctgt acctgtacta cctgcagaat 6900
ggccgggata tgtacgtgga ccaggaactg gacatcaaca gactgtccga ctacgatgtg 6960
gaccatatcg tgcctcagag ctttctgaag gacgactcca tcgataacaa agtgctgact 7020
cggagcgaca agaacagagg caagagcgac aacgtgccct ccgaagaggt cgtgaagaag 7080
atgaagaact actggcgaca gctgctgaac gccaagctga ttacccagag gaagttcgat 7140
aacctgacca aggccgagag aggcggcctg agcgagctgg ataaggccgg cttcatcaag 7200
aggcagctgg tggaaaccag acagatcaca aagcacgtgg cacagatcct ggactcccgg 7260
atgaacacta agtacgacga aaacgataag ctgatccggg aagtgaaagt gatcaccctg 7320
aagtccaagc tggtgtccga tttccggaag gatttccagt tttacaaagt gcgcgagatc 7380
aacaactacc accacgccca cgacgcctac ctgaacgccg tcgtgggaac cgccctgatc 7440
aaaaagtacc ctaagctgga aagcgagttc gtgtacggcg actacaaggt gtacgacgtg 7500
cggaagatga tcgccaagag cgagcaggaa atcggcaagg ctaccgccaa gtacttcttc 7560
tacagcaaca tcatgaactt tttcaagacc gaaatcaccc tggccaacgg cgagatcaga 7620
aagcgccctc tgatcgagac aaacggcgaa accggggaga tcgtgtggga taagggcaga 7680
gacttcgcca cagtgcgaaa ggtgctgagc atgccccaag tgaatatcgt gaaaaagacc 7740
gaggtgcaga caggcggctt cagcaaagag tctatcctgc ccaagaggaa cagcgacaag 7800
ctgatcgcca gaaagaagga ctgggacccc aagaagtacg gcggcttcga cagccctacc 7860
gtggcctact ctgtgctggt ggtggctaag gtggaaaagg gcaagtccaa gaaactgaag 7920
agtgtgaaag agctgctggg gatcaccatc atggaaagaa gcagctttga gaagaaccct 7980
atcgactttc tggaagccaa gggctacaaa gaagtgaaaa aggacctgat catcaagctg 8040
cctaagtact ccctgttcga gctggaaaac ggcagaaaga gaatgctggc ctctgccggc 8100
gaactgcaga agggaaacga gctggccctg cctagcaaat atgtgaactt cctgtacctg 8160
gcctcccact atgagaagct gaagggcagc cctgaggaca acgaacagaa acagctgttt 8220
gtggaacagc ataagcacta cctggacgag atcatcgagc agatcagcga gttctccaag 8280
agagtgatcc tggccgacgc caatctggac aaggtgctgt ctgcctacaa caagcacagg 8340
gacaagccta tcagagagca ggccgagaat atcatccacc tgttcaccct gacaaacctg 8400
ggcgctcctg ccgccttcaa gtactttgac accaccatcg accggaagag gtacaccagc 8460
accaaagagg tgctggacgc caccctgatc caccagagca tcaccggcct gtacgagaca 8520
agaatcgacc tgtctcagct gggaggcgac aagagacctg ccgccactaa gaaggccgga 8580
caggccaaaa agaagaagtg ataaatgcat ggccggccct gcaggaattc gatatcaagc 8640
ttatcgataa tcaacctctg gattacaaaa tttgtgaaag attgactggt attcttaact 8700
atgttgctcc ttttacgcta tgtggatacg ctgctttaat gcctttgtat catgctattg 8760
cttcccgtat ggctttcatt ttctcctcct tgtataaatc ctggttgctg tctctttatg 8820
aggagttgtg gcccgttgtc aggcaacgtg gcgtggtgtg cactgtgttt gctgacgcaa 8880
cccccactgg ttggggcatt gccaccacct gtcagctcct ttccgggact ttcgctttcc 8940
ccctccctat tgccacggcg gaactcatcg ccgcctgcct tgcccgctgc tggacagggg 9000
ctcggctgtt gggcactgac aattccgtgg tgttgtcggg gaaatcatcg tcctttcctt 9060
ggctgctcgc ctgtgttgcc acctggattc tgcgcgggac gtccttctgc tacgtccctt 9120
cggccctcaa tccagcggac cttccttccc gcggcctgct gccggctctg cggcctcttc 9180
cgcgtcttcg ccttcgccct cagacgagtc ggatctccct ttgggccgcc tccccgcatc 9240
gataccgtcg acctcgacct cgactgtgcc ttctagttgc cagccatctg ttgtttgccc 9300
ctcccccgtg ccttccttga ccctggaagg tgccactccc actgtccttt cctaataaaa 9360
tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct attctggggg gtggggtggg 9420
gcaggacagc aagggggagg attgggaaga caatggcagg catgctgggg aactagtggt 9480
gccagggcgt gcccttgggc tccccgggcg cggcggccat cgctcgagta aaattggagg 9540
gacaagactt cccacagatt ttcggttttg tcgggaagtt ttttaatagg ggcaaataag 9600
gaaaatggga ggataggtag tcatctgggg ttttatgcag caaaactaca ggttattatt 9660
gcttgtgatccgc 9673
<210> 21
<211> 7456
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221>misc_feature
<222> (1)..(170)
<223>Вышележащая последовательность Rosa26 мыши
<220>
<221> misc_feature
<222> (205)..(1923)
<223>Промотор CAGG
<220>
<221> misc_feature
<222> (2006)..(2039)
<223> LoxP
<220>
<221> misc_feature
<222> (2134)..(2143)
<223> Kozak
<220>
<221> misc_feature
<222> (2143)..(2208)
<223> 3xFLAG
<220>
<221> misc_feature
<222> (2209)..(6381)
<223> Cas9
<220>
<221> misc_feature
<222> (2218)..(2238)
<223>Одинарный NLS
<220>
<221> misc_feature
<222> (6334)..(6381)
<223>Двойной NLS
<220>
<221> misc_feature
<222> (6428)..(7024)
<223> WPRE
<220>
<221> misc_feature
<222> (7032)..(7247)
<223>Сигнал полиаденилирования bGH
<220>
<221>misc_feature
<222> (7307)..(7456)
<223>Нижележащая последовательность Rosa26 мыши
<400> 21
ctgcagtggagtaggcggggagaaggccgcacccttctccggaggggggaggggagtgtt 60
gcaatacctttctgggagttctctgctgcctcctggcttctgaggaccgccctgggcctg 120
ggagaatcccttccccctcttccctcgtgatctgcaactccagtctttctccttaattaa 180
acgcgtctggcctcgcgagtgtgtactagttattaatagtaatcaattacggggtcatta 240
gttcatagcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggc 300
tgaccgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacg 360
ccaatagggactttccattgacgtcaatgggtggagtatttacggtaaactgcccacttg 420
gcagtacatcaagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaa 480
tggcccgcctggcattatgcccagtacatgaccttatgggactttcctacttggcagtac 540
atctacgtattagtcatcgctattaccatggtcgaggtgagccccacgttctgcttcact 600
ctccccatctcccccccctccccacccccaattttgtatttatttattttttaattattt 660
tgtgcagcgatgggggcggggggggggggggggcgcgcgccaggcggggcggggcggggc 720
gaggggcggggcggggcgaggcggagaggtgcggcggcagccaatcagagcggcgcgctc 780
cgaaagtttccttttatggcgaggcggcggcggcggcggccctataaaaagcgaagcgcg 840
cggcgggcggggagtcgctgcgacgctgccttcgccccgtgccccgctccgccgccgcct 900
cgcgccgcccgccccggctctgactgaccgcgttactcccacaggtgagcgggcgggacg 960
gcccttctcctccgggctgtaattagcgcttggtttaatgacggcttgtttcttttctgt 1020
ggctgcgtgaaagccttgaggggctccgggagggccctttgtgcggggggagcggctcgg 1080
ggggtgcgtgcgtgtgtgtgtgcgtggggagcgccgcgtgcggctccgcgctgcccggcg 1140
gctgtgagcgctgcgggcgcggcgcggggctttgtgcgctccgcagtgtgcgcgagggga 1200
gcgcggccgggggcggtgccccgcggtgcggggggggctgcgaggggaacaaaggctgcg 1260
tgcggggtgtgtgcgtgggggggtgagcagggggtgtgggcgcgtcggtcgggctgcaac 1320
cccccctgcacccccctccccgagttgctgagcacggcccggcttcgggtgcggggctcc 1380
gtacggggcgtggcgcggggctcgccgtgccgggcggggggtggcggcaggtgggggtgc 1440
cgggcggggcggggccgcctcgggccggggagggctcgggggaggggcgcggcggccccc 1500
ggagcgccggcggctgtcgaggcgcggcgagccgcagccattgccttttatggtaatcgt 1560
gcgagagggcgcagggacttcctttgtcccaaatctgtgcggagccgaaatctgggaggc 1620
gccgccgcaccccctctagcgggcgcggggcgaagcggtgcggcgccggcaggaaggaaa 1680
tgggcggggagggccttcgtgcgtcgccgcgccgccgtccccttctccctctccagcctc 1740
ggggctgtccgcggggggacggctgccttcgggggggacggggcagggcggggttcggct 1800
tctggcgtgtgaccggcggctctagagcctctgctaaccatgttcatgccttcttctttt 1860
tcctacagctcctgggcaacgtgctggttattgtgctgtctcatcattttggcaaagaat 1920
tcgctaggagaattgatttgataccgcgggccctaagtcgacatttaaatcatttaaatc 1980
cactagtggatccggaacccttaatataacttcgtataatgtatgctatacgaagttatt 2040
aggtccctcgacctgcagcccaagctagtgcccgggtaggtccctcgacctgcagcccaa 2100
gctagatcgaattcggccggccttcgaacacgtgccaccatggactataaggaccacgac 2160
ggagactacaaggatcatgatattgattacaaagacgatgacgataagatggacaagccc 2220
aagaaaaagcggaaagtgaagtacagcatcggcctggacatcggcaccaactctgtgggc 2280
tgggccgtgatcaccgacgagtacaaggtgcccagcaagaaattcaaggtgctgggcaac 2340
accgacaggcacagcatcaagaagaacctgatcggcgccctgctgttcgacagcggcgaa 2400
acagccgaggccaccagactgaagagaaccgccagaagaagatacaccaggcggaagaac 2460
aggatctgctatctgcaagagatcttcagcaacgagatggccaaggtggacgacagcttc 2520
ttccacagactggaagagtccttcctggtggaagaggacaagaagcacgagagacacccc 2580
atcttcggcaacatcgtggacgaggtggcctaccacgagaagtaccccaccatctaccac 2640
ctgagaaagaaactggtggacagcaccgacaaggccgacctgagactgatctacctggcc 2700
ctggcccacatgatcaagttcagaggccacttcctgatcgagggcgacctgaaccccgac 2760
aacagcgacgtggacaagctgttcatccagctggtgcagacctacaaccagctgttcgag 2820
gaaaaccccatcaacgccagcggcgtggacgccaaggctatcctgtctgccagactgagc 2880
aagagcagaaggctggaaaatctgatcgcccagctgcccggcgagaagaagaacggcctg 2940
ttcggcaacctgattgccctgagcctgggcctgacccccaacttcaagagcaacttcgac 3000
ctggccgaggatgccaaactgcagctgagcaaggacacctacgacgacgacctggacaac 3060
ctgctggcccagatcggcgaccagtacgccgacctgttcctggccgccaagaacctgtct 3120
gacgccatcctgctgagcgacatcctgagagtgaacaccgagatcaccaaggcccccctg 3180
agcgcctctatgatcaagagatacgacgagcaccaccaggacctgaccctgctgaaagct 3240
ctcgtgcggcagcagctgcctgagaagtacaaagaaatcttcttcgaccagagcaagaac 3300
ggctacgccggctacatcgatggcggcgctagccaggaagagttctacaagttcatcaag 3360
cccatcctggaaaagatggacggcaccgaggaactgctcgtgaagctgaacagagaggac 3420
ctgctgagaaagcagagaaccttcgacaacggcagcatcccccaccagatccacctggga 3480
gagctgcacgctatcctgagaaggcaggaagatttttacccattcctgaaggacaaccgg 3540
gaaaagatcgagaagatcctgaccttcaggatcccctactacgtgggccccctggccaga 3600
ggcaacagcagattcgcctggatgaccagaaagagcgaggaaaccatcaccccctggaac 3660
ttcgaggaagtggtggacaagggcgccagcgcccagagcttcatcgagagaatgacaaac 3720
ttcgataagaacctgcccaacgagaaggtgctgcccaagcacagcctgctgtacgagtac 3780
ttcaccgtgtacaacgagctgaccaaagtgaaatacgtgaccgagggaatgagaaagccc 3840
gccttcctgagcggcgagcagaaaaaggccatcgtggacctgctgttcaagaccaacaga 3900
aaagtgaccgtgaagcagctgaaagaggactacttcaagaaaatcgagtgcttcgactcc 3960
gtggaaatctccggcgtggaagatagattcaacgcctccctgggcacataccacgatctg 4020
ctgaaaattatcaaggacaaggacttcctggataacgaagagaacgaggacattctggaa 4080
gatatcgtgctgaccctgacactgtttgaggaccgcgagatgatcgaggaaaggctgaaa 4140
acctacgctcacctgttcgacgacaaagtgatgaagcagctgaagagaaggcggtacacc 4200
ggctggggcaggctgagcagaaagctgatcaacggcatcagagacaagcagagcggcaag 4260
acaatcctggatttcctgaagtccgacggcttcgccaaccggaacttcatgcagctgatc 4320
cacgacgacagcctgacattcaaagaggacatccagaaagcccaggtgtccggccagggc 4380
gactctctgcacgagcatatcgctaacctggccggcagccccgctatcaagaagggcatc 4440
ctgcagacagtgaaggtggtggacgagctcgtgaaagtgatgggcagacacaagcccgag 4500
aacatcgtgatcgagatggctagagagaaccagaccacccagaagggacagaagaactcc 4560
cgcgagaggatgaagagaatcgaagagggcatcaaagagctgggcagccagatcctgaaa 4620
gaacaccccgtggaaaacacccagctgcagaacgagaagctgtacctgtactacctgcag 4680
aatggccgggatatgtacgtggaccaggaactggacatcaacagactgtccgactacgat 4740
gtggaccatatcgtgcctcagagctttctgaaggacgactccatcgataacaaagtgctg 4800
actcggagcgacaagaacagaggcaagagcgacaacgtgccctccgaagaggtcgtgaag 4860
aagatgaagaactactggcgacagctgctgaacgccaagctgattacccagaggaagttc 4920
gataacctgaccaaggccgagagaggcggcctgagcgagctggataaggccggcttcatc 4980
aagaggcagctggtggaaaccagacagatcacaaagcacgtggcacagatcctggactcc 5040
cggatgaacactaagtacgacgaaaacgataagctgatccgggaagtgaaagtgatcacc 5100
ctgaagtccaagctggtgtccgatttccggaaggatttccagttttacaaagtgcgcgag 5160
atcaacaactaccaccacgcccacgacgcctacctgaacgccgtcgtgggaaccgccctg 5220
atcaaaaagtaccctaagctggaaagcgagttcgtgtacggcgactacaaggtgtacgac 5280
gtgcggaagatgatcgccaagagcgagcaggaaatcggcaaggctaccgccaagtacttc 5340
ttctacagcaacatcatgaactttttcaagaccgaaatcaccctggccaacggcgagatc 5400
agaaagcgccctctgatcgagacaaacggcgaaaccggggagatcgtgtgggataagggc 5460
agagacttcgccacagtgcgaaaggtgctgagcatgccccaagtgaatatcgtgaaaaag 5520
accgaggtgcagacaggcggcttcagcaaagagtctatcctgcccaagaggaacagcgac 5580
aagctgatcgccagaaagaaggactgggaccccaagaagtacggcggcttcgacagccct 5640
accgtggcctactctgtgctggtggtggctaaggtggaaaagggcaagtccaagaaactg 5700
aagagtgtgaaagagctgctggggatcaccatcatggaaagaagcagctttgagaagaac 5760
cctatcgactttctggaagccaagggctacaaagaagtgaaaaaggacctgatcatcaag 5820
ctgcctaagtactccctgttcgagctggaaaacggcagaaagagaatgctggcctctgcc 5880
ggcgaactgcagaagggaaacgagctggccctgcctagcaaatatgtgaacttcctgtac 5940
ctggcctcccactatgagaagctgaagggcagccctgaggacaacgaacagaaacagctg 6000
tttgtggaacagcataagcactacctggacgagatcatcgagcagatcagcgagttctcc 6060
aagagagtgatcctggccgacgccaatctggacaaggtgctgtctgcctacaacaagcac 6120
agggacaagcctatcagagagcaggccgagaatatcatccacctgttcaccctgacaaac 6180
ctgggcgctcctgccgccttcaagtactttgacaccaccatcgaccggaagaggtacacc 6240
agcaccaaagaggtgctggacgccaccctgatccaccagagcatcaccggcctgtacgag 6300
acaagaatcgacctgtctcagctgggaggcgacaagagacctgccgccactaagaaggcc 6360
ggacaggccaaaaagaagaagtgataaatgcatggccggccctgcaggaattcgatatca 6420
agcttatcgataatcaacctctggattacaaaatttgtgaaagattgactggtattctta 6480
actatgttgctccttttacgctatgtggatacgctgctttaatgcctttgtatcatgcta 6540
ttgcttcccgtatggctttcattttctcctccttgtataaatcctggttgctgtctcttt 6600
atgaggagttgtggcccgttgtcaggcaacgtggcgtggtgtgcactgtgtttgctgacg 6660
caacccccactggttggggcattgccaccacctgtcagctcctttccgggactttcgctt 6720
tccccctccctattgccacggcggaactcatcgccgcctgccttgcccgctgctggacag 6780
gggctcggctgttgggcactgacaattccgtggtgttgtcggggaaatcatcgtcctttc 6840
cttggctgctcgcctgtgttgccacctggattctgcgcgggacgtccttctgctacgtcc 6900
cttcggccctcaatccagcggaccttccttcccgcggcctgctgccggctctgcggcctc 6960
ttccgcgtcttcgccttcgccctcagacgagtcggatctccctttgggccgcctccccgc 7020
atcgataccgtcgacctcgacctcgactgtgccttctagttgccagccatctgttgtttg 7080
cccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaata 7140
aaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggt 7200
ggggcaggacagcaagggggaggattgggaagacaatggcaggcatgctggggaactagt 7260
ggtgccagggcgtgcccttgggctccccgggcgcggcggccatcgctcgagtaaaattgg 7320
agggacaagacttcccacagattttcggttttgtcgggaagttttttaataggggcaaat 7380
aaggaaaatgggaggataggtagtcatctggggttttatgcagcaaaactacaggttatt 7440
attgcttgtgatccgc 7456
<210> 22
<211> 1413
<212>PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> MISC_FEATURE
<222> (1)..(22)
<223> 3xFLAG
<220>
<221> MISC_FEATURE
<222> (23)..(1413)
<223> Cas9
<400> 22
Asp Tyr Lys Asp His Asp Gly Asp Tyr Lys Asp His Asp Ile Asp Tyr
1 5 10 15
Lys Asp Asp Asp Asp Lys Met Asp Lys Pro Lys Lys Lys Arg Lys Val
20 25 30
Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly Trp Ala
35 40 45
Val Ile Thr Asp Glu Tyr Lys Val Pro Ser Lys Lys Phe Lys Val Leu
50 55 60
Gly Asn Thr Asp Arg His Ser Ile Lys Lys Asn Leu Ile Gly Ala Leu
65 70 75 80
Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala Thr Arg Leu Lys Arg Thr
85 90 95
Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn Arg Ile Cys Tyr Leu Gln
100 105 110
Glu Ile Phe Ser Asn Glu Met Ala Lys Val Asp Asp Ser Phe Phe His
115 120 125
Arg Leu Glu Glu Ser Phe Leu Val Glu Glu Asp Lys Lys His Glu Arg
130 135 140
His Pro Ile Phe Gly Asn Ile Val Asp Glu Val Ala Tyr His Glu Lys
145 150 155 160
Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys Leu Val Asp Ser Thr Asp
165 170 175
Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala Leu Ala His Met Ile Lys
180 185 190
Phe Arg Gly His Phe Leu Ile Glu Gly Asp Leu Asn Pro Asp Asn Ser
195 200 205
Asp Val Asp Lys Leu Phe Ile Gln Leu Val Gln Thr Tyr Asn Gln Leu
210 215 220
Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly Val Asp Ala Lys Ala Ile
225 230 235 240
Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg Leu Glu Asn Leu Ile Ala
245 250 255
Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu Phe Gly Asn Leu Ile Ala
260 265 270
Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys Ser Asn Phe Asp Leu Ala
275 280 285
Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp Thr Tyr Asp Asp Asp Leu
290 295 300
Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln Tyr Ala Asp Leu Phe Leu
305 310 315 320
Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu Leu Ser Asp Ile Leu Arg
325 330 335
Val Asn Thr Glu Ile Thr Lys Ala Pro Leu Ser Ala Ser Met Ile Lys
340 345 350
Arg Tyr Asp Glu His His Gln Asp Leu Thr Leu Leu Lys Ala Leu Val
355 360 365
Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu Ile Phe Phe Asp Gln Ser
370 375 380
Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Gly Ala Ser Gln Glu Glu
385 390 395 400
Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu Lys Met Asp Gly Thr Glu
405 410 415
Glu Leu Leu Val Lys Leu Asn Arg Glu Asp Leu Leu Arg Lys Gln Arg
420 425 430
Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu Gly Glu Leu
435 440 445
His Ala Ile Leu Arg Arg Gln Glu Asp Phe Tyr Pro Phe Leu Lys Asp
450 455 460
Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr Phe Arg Ile Pro Tyr Tyr
465 470 475 480
Val Gly Pro Leu Ala Arg Gly Asn Ser Arg Phe Ala Trp Met Thr Arg
485 490 495
Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn Phe Glu Glu Val Val Asp
500 505 510
Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu Arg Met Thr Asn Phe Asp
515 520 525
Lys Asn Leu Pro Asn Glu Lys Val Leu Pro Lys His Ser Leu Leu Tyr
530 535 540
Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys Tyr Val Thr
545 550 555 560
Glu Gly Met Arg Lys Pro Ala Phe Leu Ser Gly Glu Gln Lys Lys Ala
565 570 575
Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys Val Thr Val Lys Gln
580 585 590
Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys Phe Asp Ser Val Glu
595 600 605
Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser Leu Gly Thr Tyr His
610 615 620
Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe Leu Asp Asn Glu Glu
625 630 635 640
Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr Leu Thr Leu Phe Glu
645 650 655
Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr Tyr Ala His Leu Phe
660 665 670
Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg Arg Tyr Thr Gly Trp
675 680 685
Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile Arg Asp Lys Gln Ser
690 695 700
Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp Gly Phe Ala Asn Arg
705 710 715 720
Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu Thr Phe Lys Glu Asp
725 730 735
Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp Ser Leu His Glu His
740 745 750
Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys Lys Gly Ile Leu Gln
755 760 765
Thr Val Lys Val Val Asp Glu Leu Val Lys Val Met Gly Arg His Lys
770 775 780
Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln Thr Thr Gln
785 790 795 800
Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys Arg Ile Glu Glu Gly
805 810 815
Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu His Pro Val Glu Asn
820 825 830
Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly
835 840 845
Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile Asn Arg Leu Ser Asp
850 855 860
Tyr Asp Val Asp His Ile Val Pro Gln Ser Phe Leu Lys Asp Asp Ser
865 870 875 880
Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg Gly Lys Ser
885 890 895
Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys Asn Tyr Trp
900 905 910
Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys Phe Asp Asn
915 920 925
Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp Lys Ala Gly
930 935 940
Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr Lys His Val
945 950 955 960
Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr Asp Glu Asn Asp
965 970 975
Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu Lys Ser Lys Leu Val
980 985 990
Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys Val Arg Glu Ile Asn
995 1000 1005
Asn Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val Val Gly
1010 1015 1020
Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser Glu Phe Val
1025 1030 1035
Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met Ile Ala Lys
1040 1045 1050
Ser Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr Phe Phe Tyr
1055 1060 1065
Ser Asn Ile Met Asn Phe Phe Lys Thr Glu Ile Thr Leu Ala Asn
1070 1075 1080
Gly Glu Ile Arg Lys Arg Pro Leu Ile Glu Thr Asn Gly Glu Thr
1085 1090 1095
Gly Glu Ile Val Trp Asp Lys Gly Arg Asp Phe Ala Thr Val Arg
1100 1105 1110
Lys Val Leu Ser Met Pro Gln Val Asn Ile Val Lys Lys Thr Glu
1115 1120 1125
Val Gln Thr Gly Gly Phe Ser Lys Glu Ser Ile Leu Pro Lys Arg
1130 1135 1140
Asn Ser Asp Lys Leu Ile Ala Arg Lys Lys Asp Trp Asp Pro Lys
1145 1150 1155
Lys Tyr Gly Gly Phe Asp Ser Pro Thr Val Ala Tyr Ser Val Leu
1160 1165 1170
Val Val Ala Lys Val Glu Lys Gly Lys Ser Lys Lys Leu Lys Ser
1175 1180 1185
Val Lys Glu Leu Leu Gly Ile Thr Ile Met Glu Arg Ser Ser Phe
1190 1195 1200
Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala Lys Gly Tyr Lys Glu
1205 1210 1215
Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys Tyr Ser Leu Phe
1220 1225 1230
Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser Ala Gly Glu
1235 1240 1245
Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys Tyr Val Asn
1250 1255 1260
Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys Gly Ser Pro
1265 1270 1275
Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln His Lys His
1280 1285 1290
Tyr Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe Ser Lys Arg
1295 1300 1305
Val Ile Leu Ala Asp Ala Asn Leu Asp Lys Val Leu Ser Ala Tyr
1310 1315 1320
Asn Lys His Arg Asp Lys Pro Ile Arg Glu Gln Ala Glu Asn Ile
1325 1330 1335
Ile His Leu Phe Thr Leu Thr Asn Leu Gly Ala Pro Ala Ala Phe
1340 1345 1350
Lys Tyr Phe Asp Thr Thr Ile Asp Arg Lys Arg Tyr Thr Ser Thr
1355 1360 1365
Lys Glu Val Leu Asp Ala Thr Leu Ile His Gln Ser Ile Thr Gly
1370 1375 1380
Leu Tyr Glu Thr Arg Ile Asp Leu Ser Gln Leu Gly Gly Asp Lys
1385 1390 1395
Arg Pro Ala Ala Thr Lys Lys Ala Gly Gln Ala Lys Lys Lys Lys
1400 1405 1410
<210> 23
<211> 22
<212> PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 23
Asp Tyr Lys Asp His Asp Gly Asp Tyr Lys Asp His Asp Ile Asp Tyr
1 5 10 15
Lys Asp Asp Asp Asp Lys
20
<210> 24
<211> 238
<212> PRT
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 24
Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu Val
1 5 10 15
Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly Glu
20 25 30
Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys
35 40 45
Thr Thr Gly Lys Leu Pro Val Pro Trp Pro Thr Leu Val Thr Thr Leu
50 55 60
Thr Tyr Gly Val Gln Cys Phe Ser Arg Tyr Pro Asp His Met Lys Gln
65 70 75 80
His Asp Phe Phe Lys Ser Ala Met Pro Glu Gly Tyr Val Gln Glu Arg
85 90 95
Thr Ile Phe Phe Lys Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val
100 105 110
Lys Phe Glu Gly Asp Thr Leu Val Asn Arg Ile Glu Leu Lys Gly Ile
115 120 125
Asp Phe Lys Glu Asp Gly Asn Ile Leu Gly His Lys Leu Glu Tyr Asn
130 135 140
Tyr Asn Ser His Asn Val Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly
145 150 155 160
Ile Lys Val Asn Phe Lys Ile Arg His Asn Ile Glu Asp Gly Ser Val
165 170 175
Gln Leu Ala Asp His Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro
180 185 190
Val Leu Leu Pro Asp Asn His Tyr Leu Ser Thr Gln Ser Ala Leu Ser
195 200 205
Lys Asp Pro Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe Val
210 215 220
Thr Ala Ala Gly Ile Thr Leu Gly Met Asp Glu Leu Tyr Lys
225 230 235
<210> 25
<211> 16
<212>RNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 25
guuuuagagcuaugcu 16
<210> 26
<211> 67
<212>RNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 26
agcauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucg 60
gugcuuu 67
<210> 27
<211> 77
<212>RNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 27
guuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaagu 60
ggcaccgagucggugcu 77
<210> 28
<211> 4953
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> misc_feature
<222> (1)..(4173)
<223> Cas9
<220>
<221> misc_feature
<222> (4174)..(4239)
<223> P2A
<220>
<221> misc_feature
<222> (4240)..(4953)
<223> eGFP
<400> 28
atggacaagc ccaagaaaaa gcggaaagtg aagtacagca tcggcctgga catcggcacc 60
aactctgtgg gctgggccgt gatcaccgac gagtacaagg tgcccagcaa gaaattcaag 120
gtgctgggca acaccgacag gcacagcatc aagaagaacc tgatcggcgc cctgctgttc 180
gacagcggcg aaacagccga ggccaccaga ctgaagagaa ccgccagaag aagatacacc 240
aggcggaaga acaggatctg ctatctgcaa gagatcttca gcaacgagat ggccaaggtg 300
gacgacagct tcttccacag actggaagag tccttcctgg tggaagagga caagaagcac 360
gagagacacc ccatcttcgg caacatcgtg gacgaggtgg cctaccacga gaagtacccc 420
accatctacc acctgagaaa gaaactggtg gacagcaccg acaaggccga cctgagactg 480
atctacctgg ccctggccca catgatcaag ttcagaggcc acttcctgat cgagggcgac 540
ctgaaccccg acaacagcga cgtggacaag ctgttcatcc agctggtgca gacctacaac 600
cagctgttcg aggaaaaccc catcaacgcc agcggcgtgg acgccaaggc tatcctgtct 660
gccagactga gcaagagcag aaggctggaa aatctgatcg cccagctgcc cggcgagaag 720
aagaacggcc tgttcggcaa cctgattgcc ctgagcctgg gcctgacccc caacttcaag 780
agcaacttcg acctggccga ggatgccaaa ctgcagctga gcaaggacac ctacgacgac 840
gacctggaca acctgctggc ccagatcggc gaccagtacg ccgacctgtt cctggccgcc 900
aagaacctgt ctgacgccat cctgctgagc gacatcctga gagtgaacac cgagatcacc 960
aaggcccccc tgagcgcctc tatgatcaag agatacgacg agcaccacca ggacctgacc 1020
ctgctgaaag ctctcgtgcg gcagcagctg cctgagaagt acaaagaaat cttcttcgac 1080
cagagcaaga acggctacgc cggctacatc gatggcggcg ctagccagga agagttctac 1140
aagttcatca agcccatcct ggaaaagatg gacggcaccg aggaactgct cgtgaagctg 1200
aacagagagg acctgctgag aaagcagaga accttcgaca acggcagcat cccccaccag 1260
atccacctgg gagagctgca cgctatcctg agaaggcagg aagattttta cccattcctg 1320
aaggacaacc gggaaaagat cgagaagatc ctgaccttca ggatccccta ctacgtgggc 1380
cccctggcca gaggcaacag cagattcgcc tggatgacca gaaagagcga ggaaaccatc 1440
accccctgga acttcgagga agtggtggac aagggcgcca gcgcccagag cttcatcgag 1500
agaatgacaa acttcgataa gaacctgccc aacgagaagg tgctgcccaa gcacagcctg 1560
ctgtacgagt acttcaccgt gtacaacgag ctgaccaaag tgaaatacgt gaccgaggga 1620
atgagaaagc ccgccttcct gagcggcgag cagaaaaagg ccatcgtgga cctgctgttc 1680
aagaccaaca gaaaagtgac cgtgaagcag ctgaaagagg actacttcaa gaaaatcgag 1740
tgcttcgact ccgtggaaat ctccggcgtg gaagatagat tcaacgcctc cctgggcaca 1800
taccacgatc tgctgaaaat tatcaaggac aaggacttcc tggataacga agagaacgag 1860
gacattctgg aagatatcgt gctgaccctg acactgtttg aggaccgcga gatgatcgag 1920
gaaaggctga aaacctacgc tcacctgttc gacgacaaag tgatgaagca gctgaagaga 1980
aggcggtaca ccggctgggg caggctgagc agaaagctga tcaacggcat cagagacaag 2040
cagagcggca agacaatcct ggatttcctg aagtccgacg gcttcgccaa ccggaacttc 2100
atgcagctga tccacgacga cagcctgaca ttcaaagagg acatccagaa agcccaggtg 2160
tccggccagg gcgactctct gcacgagcat atcgctaacc tggccggcag ccccgctatc 2220
aagaagggca tcctgcagac agtgaaggtg gtggacgagc tcgtgaaagt gatgggcaga 2280
cacaagcccg agaacatcgt gatcgagatg gctagagaga accagaccac ccagaaggga 2340
cagaagaact cccgcgagag gatgaagaga atcgaagagg gcatcaaaga gctgggcagc 2400
cagatcctga aagaacaccc cgtggaaaac acccagctgc agaacgagaa gctgtacctg 2460
tactacctgc agaatggccg ggatatgtac gtggaccagg aactggacat caacagactg 2520
tccgactacg atgtggacca tatcgtgcct cagagctttc tgaaggacga ctccatcgat 2580
aacaaagtgc tgactcggag cgacaagaac agaggcaaga gcgacaacgt gccctccgaa 2640
gaggtcgtga agaagatgaa gaactactgg cgacagctgc tgaacgccaa gctgattacc 2700
cagaggaagt tcgataacct gaccaaggcc gagagaggcg gcctgagcga gctggataag 2760
gccggcttca tcaagaggca gctggtggaa accagacaga tcacaaagca cgtggcacag 2820
atcctggact cccggatgaa cactaagtac gacgaaaacg ataagctgat ccgggaagtg 2880
aaagtgatca ccctgaagtc caagctggtg tccgatttcc ggaaggattt ccagttttac 2940
aaagtgcgcg agatcaacaa ctaccaccac gcccacgacg cctacctgaa cgccgtcgtg 3000
ggaaccgccc tgatcaaaaa gtaccctaag ctggaaagcg agttcgtgta cggcgactac 3060
aaggtgtacg acgtgcggaa gatgatcgcc aagagcgagc aggaaatcgg caaggctacc 3120
gccaagtact tcttctacag caacatcatg aactttttca agaccgaaat caccctggcc 3180
aacggcgaga tcagaaagcg ccctctgatc gagacaaacg gcgaaaccgg ggagatcgtg 3240
tgggataagg gcagagactt cgccacagtg cgaaaggtgc tgagcatgcc ccaagtgaat 3300
atcgtgaaaa agaccgaggt gcagacaggc ggcttcagca aagagtctat cctgcccaag 3360
aggaacagcg acaagctgat cgccagaaag aaggactggg accccaagaa gtacggcggc 3420
ttcgacagcc ctaccgtggc ctactctgtg ctggtggtgg ctaaggtgga aaagggcaag 3480
tccaagaaac tgaagagtgt gaaagagctg ctggggatca ccatcatgga aagaagcagc 3540
tttgagaaga accctatcga ctttctggaa gccaagggct acaaagaagt gaaaaaggac 3600
ctgatcatca agctgcctaa gtactccctg ttcgagctgg aaaacggcag aaagagaatg 3660
ctggcctctg ccggcgaact gcagaaggga aacgagctgg ccctgcctag caaatatgtg 3720
aacttcctgt acctggcctc ccactatgag aagctgaagg gcagccctga ggacaacgaa 3780
cagaaacagc tgtttgtgga acagcataag cactacctgg acgagatcat cgagcagatc 3840
agcgagttct ccaagagagt gatcctggcc gacgccaatc tggacaaggt gctgtctgcc 3900
tacaacaagc acagggacaa gcctatcaga gagcaggccg agaatatcat ccacctgttc 3960
accctgacaa acctgggcgc tcctgccgcc ttcaagtact ttgacaccac catcgaccgg 4020
aagaggtaca ccagcaccaa agaggtgctg gacgccaccc tgatccacca gagcatcacc 4080
ggcctgtacg agacaagaat cgacctgtct cagctgggag gcgacaagag acctgccgcc 4140
actaagaagg ccggacaggc caaaaagaag aagggaagcg gagccactaa cttctccctg 4200
ttgaaacaag caggggatgt cgaagagaat cccgggccag tgagcaaggg cgaggagctg 4260
ttcaccgggg tggtgcccat cctggtcgag ctggacggcg acgtaaacgg ccacaagttc 4320
agcgtgtccg gcgagggcga gggcgatgcc acctacggca agctgaccct gaagttcatc 4380
tgcaccaccg gcaagctgcc cgtgccctgg cccaccctcg tgaccaccct gacctacggc 4440
gtgcagtgct tcagccgcta ccccgaccac atgaagcagc acgacttctt caagtccgcc 4500
atgcccgaag gctacgtcca ggagcgcacc atcttcttca aggacgacgg caactacaag 4560
acccgcgccg aggtgaagtt cgagggcgac accctggtga accgcatcga gctgaagggc 4620
atcgacttca aggaggacgg caacatcctg gggcacaagc tggagtacaa ctacaacagc 4680
cacaacgtct atatcatggc cgacaagcag aagaacggca tcaaggtgaa cttcaagatc 4740
cgccacaaca tcgaggacgg cagcgtgcag ctcgccgacc actaccagca gaacaccccc 4800
atcggcgacg gccccgtgct gctgcccgac aaccactacc tgagcaccca gtccgccctg 4860
agcaaagacc ccaacgagaa gcgcgatcac atggtcctgc tggagttcgt gaccgccgcc 4920
gggatcactc tcggcatgga cgagctgtac aag 4953
<210> 29
<211> 5019
<212> DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> misc_feature
<222> (1)..(66)
<223> 3xFLAG
<220>
<221> misc_feature
<222> (67)..(4239)
<223> Cas9
<220>
<221> misc_feature
<222> (4240)..(4305)
<223> P2A
<220>
<221> misc_feature
<222> (4306)..(5019)
<223> eGFP
<400> 29
gactataagg accacgacgg agactacaag gatcatgata ttgattacaa agacgatgac 60
gataagatgg acaagcccaa gaaaaagcgg aaagtgaagt acagcatcgg cctggacatc 120
ggcaccaact ctgtgggctg ggccgtgatc accgacgagt acaaggtgcc cagcaagaaa 180
ttcaaggtgc tgggcaacac cgacaggcac agcatcaaga agaacctgat cggcgccctg 240
ctgttcgaca gcggcgaaac agccgaggcc accagactga agagaaccgc cagaagaaga 300
tacaccaggc ggaagaacag gatctgctat ctgcaagaga tcttcagcaa cgagatggcc 360
aaggtggacg acagcttctt ccacagactg gaagagtcct tcctggtgga agaggacaag 420
aagcacgaga gacaccccat cttcggcaac atcgtggacg aggtggccta ccacgagaag 480
taccccacca tctaccacct gagaaagaaa ctggtggaca gcaccgacaa ggccgacctg 540
agactgatct acctggccct ggcccacatg atcaagttca gaggccactt cctgatcgag 600
ggcgacctga accccgacaa cagcgacgtg gacaagctgt tcatccagct ggtgcagacc 660
tacaaccagc tgttcgagga aaaccccatc aacgccagcg gcgtggacgc caaggctatc 720
ctgtctgcca gactgagcaa gagcagaagg ctggaaaatc tgatcgccca gctgcccggc 780
gagaagaaga acggcctgtt cggcaacctg attgccctga gcctgggcct gacccccaac 840
ttcaagagca acttcgacct ggccgaggat gccaaactgc agctgagcaa ggacacctac 900
gacgacgacc tggacaacct gctggcccag atcggcgacc agtacgccga cctgttcctg 960
gccgccaaga acctgtctga cgccatcctg ctgagcgaca tcctgagagt gaacaccgag 1020
atcaccaagg cccccctgag cgcctctatg atcaagagat acgacgagca ccaccaggac 1080
ctgaccctgc tgaaagctct cgtgcggcag cagctgcctg agaagtacaa agaaatcttc 1140
ttcgaccaga gcaagaacgg ctacgccggc tacatcgatg gcggcgctag ccaggaagag 1200
ttctacaagt tcatcaagcc catcctggaa aagatggacg gcaccgagga actgctcgtg 1260
aagctgaaca gagaggacct gctgagaaag cagagaacct tcgacaacgg cagcatcccc 1320
caccagatcc acctgggaga gctgcacgct atcctgagaa ggcaggaaga tttttaccca 1380
ttcctgaagg acaaccggga aaagatcgag aagatcctga ccttcaggat cccctactac 1440
gtgggccccc tggccagagg caacagcaga ttcgcctgga tgaccagaaa gagcgaggaa 1500
accatcaccc cctggaactt cgaggaagtg gtggacaagg gcgccagcgc ccagagcttc 1560
atcgagagaa tgacaaactt cgataagaac ctgcccaacg agaaggtgct gcccaagcac 1620
agcctgctgt acgagtactt caccgtgtac aacgagctga ccaaagtgaa atacgtgacc 1680
gagggaatga gaaagcccgc cttcctgagc ggcgagcaga aaaaggccat cgtggacctg 1740
ctgttcaaga ccaacagaaa agtgaccgtg aagcagctga aagaggacta cttcaagaaa 1800
atcgagtgct tcgactccgt ggaaatctcc ggcgtggaag atagattcaa cgcctccctg 1860
ggcacatacc acgatctgct gaaaattatc aaggacaagg acttcctgga taacgaagag 1920
aacgaggaca ttctggaaga tatcgtgctg accctgacac tgtttgagga ccgcgagatg 1980
atcgaggaaa ggctgaaaac ctacgctcac ctgttcgacg acaaagtgat gaagcagctg 2040
aagagaaggc ggtacaccgg ctggggcagg ctgagcagaa agctgatcaa cggcatcaga 2100
gacaagcaga gcggcaagac aatcctggat ttcctgaagt ccgacggctt cgccaaccgg 2160
aacttcatgc agctgatcca cgacgacagc ctgacattca aagaggacat ccagaaagcc 2220
caggtgtccg gccagggcga ctctctgcac gagcatatcg ctaacctggc cggcagcccc 2280
gctatcaaga agggcatcct gcagacagtg aaggtggtgg acgagctcgt gaaagtgatg 2340
ggcagacaca agcccgagaa catcgtgatc gagatggcta gagagaacca gaccacccag 2400
aagggacaga agaactcccg cgagaggatg aagagaatcg aagagggcat caaagagctg 2460
ggcagccaga tcctgaaaga acaccccgtg gaaaacaccc agctgcagaa cgagaagctg 2520
tacctgtact acctgcagaa tggccgggat atgtacgtgg accaggaact ggacatcaac 2580
agactgtccg actacgatgt ggaccatatc gtgcctcaga gctttctgaa ggacgactcc 2640
atcgataaca aagtgctgac tcggagcgac aagaacagag gcaagagcga caacgtgccc 2700
tccgaagagg tcgtgaagaa gatgaagaac tactggcgac agctgctgaa cgccaagctg 2760
attacccaga ggaagttcga taacctgacc aaggccgaga gaggcggcct gagcgagctg 2820
gataaggccg gcttcatcaa gaggcagctg gtggaaacca gacagatcac aaagcacgtg 2880
gcacagatcc tggactcccg gatgaacact aagtacgacg aaaacgataa gctgatccgg 2940
gaagtgaaag tgatcaccct gaagtccaag ctggtgtccg atttccggaa ggatttccag 3000
ttttacaaag tgcgcgagat caacaactac caccacgccc acgacgccta cctgaacgcc 3060
gtcgtgggaa ccgccctgat caaaaagtac cctaagctgg aaagcgagtt cgtgtacggc 3120
gactacaagg tgtacgacgt gcggaagatg atcgccaaga gcgagcagga aatcggcaag 3180
gctaccgcca agtacttctt ctacagcaac atcatgaact ttttcaagac cgaaatcacc 3240
ctggccaacg gcgagatcag aaagcgccct ctgatcgaga caaacggcga aaccggggag 3300
atcgtgtggg ataagggcag agacttcgcc acagtgcgaa aggtgctgag catgccccaa 3360
gtgaatatcg tgaaaaagac cgaggtgcag acaggcggct tcagcaaaga gtctatcctg 3420
cccaagagga acagcgacaa gctgatcgcc agaaagaagg actgggaccc caagaagtac 3480
ggcggcttcg acagccctac cgtggcctac tctgtgctgg tggtggctaa ggtggaaaag 3540
ggcaagtcca agaaactgaa gagtgtgaaa gagctgctgg ggatcaccat catggaaaga 3600
agcagctttg agaagaaccc tatcgacttt ctggaagcca agggctacaa agaagtgaaa 3660
aaggacctga tcatcaagct gcctaagtac tccctgttcg agctggaaaa cggcagaaag 3720
agaatgctgg cctctgccgg cgaactgcag aagggaaacg agctggccct gcctagcaaa 3780
tatgtgaact tcctgtacct ggcctcccac tatgagaagc tgaagggcag ccctgaggac 3840
aacgaacaga aacagctgtt tgtggaacag cataagcact acctggacga gatcatcgag 3900
cagatcagcg agttctccaa gagagtgatc ctggccgacg ccaatctgga caaggtgctg 3960
tctgcctaca acaagcacag ggacaagcct atcagagagc aggccgagaa tatcatccac 4020
ctgttcaccc tgacaaacct gggcgctcct gccgccttca agtactttga caccaccatc 4080
gaccggaaga ggtacaccag caccaaagag gtgctggacg ccaccctgat ccaccagagc 4140
atcaccggcc tgtacgagac aagaatcgac ctgtctcagc tgggaggcga caagagacct 4200
gccgccacta agaaggccgg acaggccaaa aagaagaagg gaagcggagc cactaacttc 4260
tccctgttga aacaagcagg ggatgtcgaa gagaatcccg ggccagtgag caagggcgag 4320
gagctgttca ccggggtggt gcccatcctg gtcgagctgg acggcgacgt aaacggccac 4380
aagttcagcg tgtccggcga gggcgagggc gatgccacct acggcaagct gaccctgaag 4440
ttcatctgca ccaccggcaa gctgcccgtg ccctggccca ccctcgtgac caccctgacc 4500
tacggcgtgc agtgcttcag ccgctacccc gaccacatga agcagcacga cttcttcaag 4560
tccgccatgc ccgaaggcta cgtccaggag cgcaccatct tcttcaagga cgacggcaac 4620
tacaagaccc gcgccgaggt gaagttcgag ggcgacaccc tggtgaaccg catcgagctg 4680
aagggcatcg acttcaagga ggacggcaac atcctggggc acaagctgga gtacaactac 4740
aacagccaca acgtctatat catggccgac aagcagaaga acggcatcaa ggtgaacttc 4800
aagatccgcc acaacatcga ggacggcagc gtgcagctcg ccgaccacta ccagcagaac 4860
acccccatcg gcgacggccc cgtgctgctg cccgacaacc actacctgag cacccagtcc 4920
gccctgagca aagaccccaa cgagaagcgc gatcacatgg tcctgctgga gttcgtgacc 4980
gccgccggga tcactctcgg catggacgag ctgtacaag 5019
<210> 30
<211> 4173
<212> DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 30
atggacaagc ccaagaaaaa gcggaaagtg aagtacagca tcggcctgga catcggcacc 60
aactctgtgg gctgggccgt gatcaccgac gagtacaagg tgcccagcaa gaaattcaag 120
gtgctgggca acaccgacag gcacagcatc aagaagaacc tgatcggcgc cctgctgttc 180
gacagcggcg aaacagccga ggccaccaga ctgaagagaa ccgccagaag aagatacacc 240
aggcggaaga acaggatctg ctatctgcaa gagatcttca gcaacgagat ggccaaggtg 300
gacgacagct tcttccacag actggaagag tccttcctgg tggaagagga caagaagcac 360
gagagacacc ccatcttcgg caacatcgtg gacgaggtgg cctaccacga gaagtacccc 420
accatctacc acctgagaaa gaaactggtg gacagcaccg acaaggccga cctgagactg 480
atctacctgg ccctggccca catgatcaag ttcagaggcc acttcctgat cgagggcgac 540
ctgaaccccg acaacagcga cgtggacaag ctgttcatcc agctggtgca gacctacaac 600
cagctgttcg aggaaaaccc catcaacgcc agcggcgtgg acgccaaggc tatcctgtct 660
gccagactga gcaagagcag aaggctggaa aatctgatcg cccagctgcc cggcgagaag 720
aagaacggcc tgttcggcaa cctgattgcc ctgagcctgg gcctgacccc caacttcaag 780
agcaacttcg acctggccga ggatgccaaa ctgcagctga gcaaggacac ctacgacgac 840
gacctggaca acctgctggc ccagatcggc gaccagtacg ccgacctgtt cctggccgcc 900
aagaacctgt ctgacgccat cctgctgagc gacatcctga gagtgaacac cgagatcacc 960
aaggcccccc tgagcgcctc tatgatcaag agatacgacg agcaccacca ggacctgacc 1020
ctgctgaaag ctctcgtgcg gcagcagctg cctgagaagt acaaagaaat cttcttcgac 1080
cagagcaaga acggctacgc cggctacatc gatggcggcg ctagccagga agagttctac 1140
aagttcatca agcccatcct ggaaaagatg gacggcaccg aggaactgct cgtgaagctg 1200
aacagagagg acctgctgag aaagcagaga accttcgaca acggcagcat cccccaccag 1260
atccacctgg gagagctgca cgctatcctg agaaggcagg aagattttta cccattcctg 1320
aaggacaacc gggaaaagat cgagaagatc ctgaccttca ggatccccta ctacgtgggc 1380
cccctggcca gaggcaacag cagattcgcc tggatgacca gaaagagcga ggaaaccatc 1440
accccctgga acttcgagga agtggtggac aagggcgcca gcgcccagag cttcatcgag 1500
agaatgacaa acttcgataa gaacctgccc aacgagaagg tgctgcccaa gcacagcctg 1560
ctgtacgagt acttcaccgt gtacaacgag ctgaccaaag tgaaatacgt gaccgaggga 1620
atgagaaagc ccgccttcct gagcggcgag cagaaaaagg ccatcgtgga cctgctgttc 1680
aagaccaaca gaaaagtgac cgtgaagcag ctgaaagagg actacttcaa gaaaatcgag 1740
tgcttcgact ccgtggaaat ctccggcgtg gaagatagat tcaacgcctc cctgggcaca 1800
taccacgatc tgctgaaaat tatcaaggac aaggacttcc tggataacga agagaacgag 1860
gacattctgg aagatatcgt gctgaccctg acactgtttg aggaccgcga gatgatcgag 1920
gaaaggctga aaacctacgc tcacctgttc gacgacaaag tgatgaagca gctgaagaga 1980
aggcggtaca ccggctgggg caggctgagc agaaagctga tcaacggcat cagagacaag 2040
cagagcggca agacaatcct ggatttcctg aagtccgacg gcttcgccaa ccggaacttc 2100
atgcagctga tccacgacga cagcctgaca ttcaaagagg acatccagaa agcccaggtg 2160
tccggccagg gcgactctct gcacgagcat atcgctaacc tggccggcag ccccgctatc 2220
aagaagggca tcctgcagac agtgaaggtg gtggacgagc tcgtgaaagt gatgggcaga 2280
cacaagcccg agaacatcgt gatcgagatg gctagagaga accagaccac ccagaaggga 2340
cagaagaact cccgcgagag gatgaagaga atcgaagagg gcatcaaaga gctgggcagc 2400
cagatcctga aagaacaccc cgtggaaaac acccagctgc agaacgagaa gctgtacctg 2460
tactacctgc agaatggccg ggatatgtac gtggaccagg aactggacat caacagactg 2520
tccgactacg atgtggacca tatcgtgcct cagagctttc tgaaggacga ctccatcgat 2580
aacaaagtgc tgactcggag cgacaagaac agaggcaaga gcgacaacgt gccctccgaa 2640
gaggtcgtga agaagatgaa gaactactgg cgacagctgc tgaacgccaa gctgattacc 2700
cagaggaagt tcgataacct gaccaaggcc gagagaggcg gcctgagcga gctggataag 2760
gccggcttca tcaagaggca gctggtggaa accagacaga tcacaaagca cgtggcacag 2820
atcctggact cccggatgaa cactaagtac gacgaaaacg ataagctgat ccgggaagtg 2880
aaagtgatca ccctgaagtc caagctggtg tccgatttcc ggaaggattt ccagttttac 2940
aaagtgcgcg agatcaacaa ctaccaccac gcccacgacg cctacctgaa cgccgtcgtg 3000
ggaaccgccc tgatcaaaaa gtaccctaag ctggaaagcg agttcgtgta cggcgactac 3060
aaggtgtacg acgtgcggaa gatgatcgcc aagagcgagc aggaaatcgg caaggctacc 3120
gccaagtact tcttctacag caacatcatg aactttttca agaccgaaat caccctggcc 3180
aacggcgaga tcagaaagcg ccctctgatc gagacaaacg gcgaaaccgg ggagatcgtg 3240
tgggataagg gcagagactt cgccacagtg cgaaaggtgc tgagcatgcc ccaagtgaat 3300
atcgtgaaaa agaccgaggt gcagacaggc ggcttcagca aagagtctat cctgcccaag 3360
aggaacagcg acaagctgat cgccagaaag aaggactggg accccaagaa gtacggcggc 3420
ttcgacagcc ctaccgtggc ctactctgtg ctggtggtgg ctaaggtgga aaagggcaag 3480
tccaagaaac tgaagagtgt gaaagagctg ctggggatca ccatcatgga aagaagcagc 3540
tttgagaaga accctatcga ctttctggaa gccaagggct acaaagaagt gaaaaaggac 3600
ctgatcatca agctgcctaa gtactccctg ttcgagctgg aaaacggcag aaagagaatg 3660
ctggcctctg ccggcgaact gcagaaggga aacgagctgg ccctgcctag caaatatgtg 3720
aacttcctgt acctggcctc ccactatgag aagctgaagg gcagccctga ggacaacgaa 3780
cagaaacagc tgtttgtgga acagcataag cactacctgg acgagatcat cgagcagatc 3840
agcgagttct ccaagagagt gatcctggcc gacgccaatc tggacaaggt gctgtctgcc 3900
tacaacaagc acagggacaa gcctatcaga gagcaggccg agaatatcat ccacctgttc 3960
accctgacaa acctgggcgc tcctgccgcc ttcaagtact ttgacaccac catcgaccgg 4020
aagaggtaca ccagcaccaa agaggtgctg gacgccaccc tgatccacca gagcatcacc 4080
ggcctgtacg agacaagaat cgacctgtct cagctgggag gcgacaagag acctgccgcc 4140
actaagaagg ccggacaggc caaaaagaag aag 4173
<210> 31
<211> 4239
<212> DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<220>
<221> misc_feature
<222> (1)..(66)
<223> 3xFLAG
<220>
<221> misc_feature
<222> (67)..(4239)
<223> Cas9
<400> 31
gactataagg accacgacgg agactacaag gatcatgata ttgattacaa agacgatgac 60
gataagatgg acaagcccaa gaaaaagcgg aaagtgaagt acagcatcgg cctggacatc 120
ggcaccaact ctgtgggctg ggccgtgatc accgacgagt acaaggtgcc cagcaagaaa 180
ttcaaggtgc tgggcaacac cgacaggcac agcatcaaga agaacctgat cggcgccctg 240
ctgttcgaca gcggcgaaac agccgaggcc accagactga agagaaccgc cagaagaaga 300
tacaccaggc ggaagaacag gatctgctat ctgcaagaga tcttcagcaa cgagatggcc 360
aaggtggacg acagcttctt ccacagactg gaagagtcct tcctggtgga agaggacaag 420
aagcacgaga gacaccccat cttcggcaac atcgtggacg aggtggccta ccacgagaag 480
taccccacca tctaccacct gagaaagaaa ctggtggaca gcaccgacaa ggccgacctg 540
agactgatct acctggccct ggcccacatg atcaagttca gaggccactt cctgatcgag 600
ggcgacctga accccgacaa cagcgacgtg gacaagctgt tcatccagct ggtgcagacc 660
tacaaccagc tgttcgagga aaaccccatc aacgccagcg gcgtggacgc caaggctatc 720
ctgtctgcca gactgagcaa gagcagaagg ctggaaaatc tgatcgccca gctgcccggc 780
gagaagaaga acggcctgtt cggcaacctg attgccctga gcctgggcct gacccccaac 840
ttcaagagca acttcgacct ggccgaggat gccaaactgc agctgagcaa ggacacctac 900
gacgacgacc tggacaacct gctggcccag atcggcgacc agtacgccga cctgttcctg 960
gccgccaaga acctgtctga cgccatcctg ctgagcgaca tcctgagagt gaacaccgag 1020
atcaccaagg cccccctgag cgcctctatg atcaagagat acgacgagca ccaccaggac 1080
ctgaccctgc tgaaagctct cgtgcggcag cagctgcctg agaagtacaa agaaatcttc 1140
ttcgaccaga gcaagaacgg ctacgccggc tacatcgatg gcggcgctag ccaggaagag 1200
ttctacaagt tcatcaagcc catcctggaa aagatggacg gcaccgagga actgctcgtg 1260
aagctgaaca gagaggacct gctgagaaag cagagaacct tcgacaacgg cagcatcccc 1320
caccagatcc acctgggaga gctgcacgct atcctgagaa ggcaggaaga tttttaccca 1380
ttcctgaagg acaaccggga aaagatcgag aagatcctga ccttcaggat cccctactac 1440
gtgggccccc tggccagagg caacagcaga ttcgcctgga tgaccagaaa gagcgaggaa 1500
accatcaccc cctggaactt cgaggaagtg gtggacaagg gcgccagcgc ccagagcttc 1560
atcgagagaa tgacaaactt cgataagaac ctgcccaacg agaaggtgct gcccaagcac 1620
agcctgctgt acgagtactt caccgtgtac aacgagctga ccaaagtgaa atacgtgacc 1680
gagggaatga gaaagcccgc cttcctgagc ggcgagcaga aaaaggccat cgtggacctg 1740
ctgttcaaga ccaacagaaa agtgaccgtg aagcagctga aagaggacta cttcaagaaa 1800
atcgagtgct tcgactccgt ggaaatctcc ggcgtggaag atagattcaa cgcctccctg 1860
ggcacatacc acgatctgct gaaaattatc aaggacaagg acttcctgga taacgaagag 1920
aacgaggaca ttctggaaga tatcgtgctg accctgacac tgtttgagga ccgcgagatg 1980
atcgaggaaa ggctgaaaac ctacgctcac ctgttcgacg acaaagtgat gaagcagctg 2040
aagagaaggc ggtacaccgg ctggggcagg ctgagcagaa agctgatcaa cggcatcaga 2100
gacaagcaga gcggcaagac aatcctggat ttcctgaagt ccgacggctt cgccaaccgg 2160
aacttcatgc agctgatcca cgacgacagc ctgacattca aagaggacat ccagaaagcc 2220
caggtgtccg gccagggcga ctctctgcac gagcatatcg ctaacctggc cggcagcccc 2280
gctatcaaga agggcatcct gcagacagtg aaggtggtgg acgagctcgt gaaagtgatg 2340
ggcagacaca agcccgagaa catcgtgatc gagatggcta gagagaacca gaccacccag 2400
aagggacaga agaactcccg cgagaggatg aagagaatcg aagagggcat caaagagctg 2460
ggcagccaga tcctgaaaga acaccccgtg gaaaacaccc agctgcagaa cgagaagctg 2520
tacctgtact acctgcagaa tggccgggat atgtacgtgg accaggaact ggacatcaac 2580
agactgtccg actacgatgt ggaccatatc gtgcctcaga gctttctgaa ggacgactcc 2640
atcgataaca aagtgctgac tcggagcgac aagaacagag gcaagagcga caacgtgccc 2700
tccgaagagg tcgtgaagaa gatgaagaac tactggcgac agctgctgaa cgccaagctg 2760
attacccaga ggaagttcga taacctgacc aaggccgaga gaggcggcct gagcgagctg 2820
gataaggccg gcttcatcaa gaggcagctg gtggaaacca gacagatcac aaagcacgtg 2880
gcacagatcc tggactcccg gatgaacact aagtacgacg aaaacgataa gctgatccgg 2940
gaagtgaaag tgatcaccct gaagtccaag ctggtgtccg atttccggaa ggatttccag 3000
ttttacaaag tgcgcgagat caacaactac caccacgccc acgacgccta cctgaacgcc 3060
gtcgtgggaa ccgccctgat caaaaagtac cctaagctgg aaagcgagtt cgtgtacggc 3120
gactacaagg tgtacgacgt gcggaagatg atcgccaaga gcgagcagga aatcggcaag 3180
gctaccgcca agtacttctt ctacagcaac atcatgaact ttttcaagac cgaaatcacc 3240
ctggccaacg gcgagatcag aaagcgccct ctgatcgaga caaacggcga aaccggggag 3300
atcgtgtggg ataagggcag agacttcgcc acagtgcgaa aggtgctgag catgccccaa 3360
gtgaatatcg tgaaaaagac cgaggtgcag acaggcggct tcagcaaaga gtctatcctg 3420
cccaagagga acagcgacaa gctgatcgcc agaaagaagg actgggaccc caagaagtac 3480
ggcggcttcg acagccctac cgtggcctac tctgtgctgg tggtggctaa ggtggaaaag 3540
ggcaagtcca agaaactgaa gagtgtgaaa gagctgctgg ggatcaccat catggaaaga 3600
agcagctttg agaagaaccc tatcgacttt ctggaagcca agggctacaa agaagtgaaa 3660
aaggacctga tcatcaagct gcctaagtac tccctgttcg agctggaaaa cggcagaaag 3720
agaatgctgg cctctgccgg cgaactgcag aagggaaacg agctggccct gcctagcaaa 3780
tatgtgaact tcctgtacct ggcctcccac tatgagaagc tgaagggcag ccctgaggac 3840
aacgaacaga aacagctgtt tgtggaacag cataagcact acctggacga gatcatcgag 3900
cagatcagcg agttctccaa gagagtgatc ctggccgacg ccaatctgga caaggtgctg 3960
tctgcctaca acaagcacag ggacaagcct atcagagagc aggccgagaa tatcatccac 4020
ctgttcaccc tgacaaacct gggcgctcct gccgccttca agtactttga caccaccatc 4080
gaccggaaga ggtacaccag caccaaagag gtgctggacg ccaccctgat ccaccagagc 4140
atcaccggcc tgtacgagac aagaatcgac ctgtctcagc tgggaggcga caagagacct 4200
gccgccacta agaaggccgg acaggccaaa aagaagaag 4239
<210> 32
<211> 66
<212> DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 32
ggaagcggag ccactaactt ctccctgttg aaacaagcag gggatgtcga agagaatccc 60
gggcca 66
<210> 33
<211> 714
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 33
gtgagcaagggcgaggagctgttcaccggggtggtgcccatcctggtcgagctggacggc 60
gacgtaaacggccacaagttcagcgtgtccggcgagggcgagggcgatgccacctacggc 120
aagctgaccctgaagttcatctgcaccaccggcaagctgcccgtgccctggcccaccctc 180
gtgaccaccctgacctacggcgtgcagtgcttcagccgctaccccgaccacatgaagcag 240
cacgacttcttcaagtccgccatgcccgaaggctacgtccaggagcgcaccatcttcttc 300
aaggacgacggcaactacaagacccgcgccgaggtgaagttcgagggcgacaccctggtg 360
aaccgcatcgagctgaagggcatcgacttcaaggaggacggcaacatcctggggcacaag 420
ctggagtacaactacaacagccacaacgtctatatcatggccgacaagcagaagaacggc 480
atcaaggtgaacttcaagatccgccacaacatcgaggacggcagcgtgcagctcgccgac 540
cactaccagcagaacacccccatcggcgacggccccgtgctgctgcccgacaaccactac 600
ctgagcacccagtccgccctgagcaaagaccccaacgagaagcgcgatcacatggtcctg 660
ctggagttcgtgaccgccgccgggatcactctcggcatggacgagctgtacaag 714
<210> 34
<211> 66
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 34
gactataaggaccacgacggagactacaaggatcatgatattgattacaaagacgatgac 60
gataag 66
<210> 35
<211> 597
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 35
cgataatcaacctctggattacaaaatttgtgaaagattgactggtattcttaactatgt 60
tgctccttttacgctatgtggatacgctgctttaatgcctttgtatcatgctattgcttc 120
ccgtatggctttcattttctcctccttgtataaatcctggttgctgtctctttatgagga 180
gttgtggcccgttgtcaggcaacgtggcgtggtgtgcactgtgtttgctgacgcaacccc 240
cactggttggggcattgccaccacctgtcagctcctttccgggactttcgctttccccct 300
ccctattgccacggcggaactcatcgccgcctgccttgcccgctgctggacaggggctcg 360
gctgttgggcactgacaattccgtggtgttgtcggggaaatcatcgtcctttccttggct 420
gctcgcctgtgttgccacctggattctgcgcgggacgtccttctgctacgtcccttcggc 480
cctcaatccagcggaccttccttcccgcggcctgctgccggctctgcggcctcttccgcg 540
tcttcgccttcgccctcagacgagtcggatctccctttgggccgcctccccgcatcg 597
<210> 36
<211> 216
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 36
cgacctcgacctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccg 60
tgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaa 120
ttgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggaca 180
gcaagggggaggattgggaagacaatggcaggcatg 216
<210> 37
<211> 2066
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 37
atgggatcggccattgaacaagatggattgcacgcaggttctccggccgcttgggtggag 60
aggctattcggctatgactgggcacaacagacaatcggctgctctgatgccgccgtgttc 120
cggctgtcagcgcaggggcgcccggttctttttgtcaagaccgacctgtccggtgccctg 180
aatgaactgcaggacgaggcagcgcggctatcgtggctggccacgacgggcgttccttgc 240
gcagctgtgctcgacgttgtcactgaagcgggaagggactggctgctattgggcgaagtg 300
ccggggcaggatctcctgtcatctcaccttgctcctgccgagaaagtatccatcatggct 360
gatgcaatgcggcggctgcatacgcttgatccggctacctgcccattcgaccaccaagcg 420
aaacatcgcatcgagcgagcacgtactcggatggaagccggtcttgtcgatcaggatgat 480
ctggacgaagagcatcaggggctcgcgccagccgaactgttcgccaggctcaaggcgcgc 540
atgcccgacggcgatgatctcgtcgtgacccatggcgatgcctgcttgccgaatatcatg 600
gtggaaaatggccgcttttctggattcatcgactgtggccggctgggtgtggcggaccgc 660
tatcaggacatagcgttggctacccgtgatattgctgaagagcttggcggcgaatgggct 720
gaccgcttcctcgtgctttacggtatcgccgctcccgattcgcagcgcatcgccttctat 780
cgccttcttgacgagttcttctgaggggatccgctgtaagtctgcagaaattgatgatct 840
attaaacaataaagatgtccactaaaatggaagtttttcctgtcatactttgttaagaag 900
ggtgagaacagagtacctacattttgaatggaaggattggagctacgggggtgggggtgg 960
ggtgggattagataaatgcctgctctttactgaaggctctttactattgctttatgataa 1020
tgtttcatagttggatatcataatttaaacaagcaaaaccaaattaagggccagctcatt 1080
cctcccactcatgatctatagatctatagatctctcgtgggatcattgtttttctcttga 1140
ttcccactttgtggttctaagtactgtggtttccaaatgtgtcagtttcatagcctgaag 1200
aacgagatcagcagcctctgttccacatacacttcattctcagtattgttttgccaagtt 1260
ctaattccatcagaagcttgcagatctgcgactctagaggatctgcgactctagaggatc 1320
ataatcagccataccacatttgtagaggttttacttgctttaaaaaacctcccacacctc 1380
cccctgaacctgaaacataaaatgaatgcaattgttgttgttaacttgtttattgcagct 1440
tataatggttacaaataaagcaatagcatcacaaatttcacaaataaagcatttttttca 1500
ctgcattctagttgtggtttgtccaaactcatcaatgtatcttatcatgtctggatctgc 1560
gactctagaggatcataatcagccataccacatttgtagaggttttacttgctttaaaaa 1620
acctcccacacctccccctgaacctgaaacataaaatgaatgcaattgttgttgttaact 1680
tgtttattgcagcttataatggttacaaataaagcaatagcatcacaaatttcacaaata 1740
aagcatttttttcactgcattctagttgtggtttgtccaaactcatcaatgtatcttatc 1800
atgtctggat ctgcgactct agaggatcat aatcagccat accacatttg tagaggtttt 1860
acttgcttta aaaaacctcc cacacctccc cctgaacctg aaacataaaa tgaatgcaat 1920
tgttgttgtt aacttgttta ttgcagctta taatggttac aaataaagca atagcatcac 1980
aaatttcaca aataaagcat ttttttcact gcattctagt tgtggtttgt ccaaactcat 2040
caatgtatcttatcatgtctggatcc 2066
<210> 38
<211> 1719
<212>DNA
<213>Искусственная последовательность
<220>
<223>Синтетическая
<400> 38
actagttattaatagtaatcaattacggggtcattagttcatagcccatatatggagttc 60
cgcgttacataacttacggtaaatggcccgcctggctgaccgcccaacgacccccgccca 120
ttgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttccattgacgt 180
caatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatg 240
ccaagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccag 300
tacatgacct tatgggactt tcctacttgg cagtacatct acgtattagt catcgctatt 360
accatggtcg aggtgagccc cacgttctgc ttcactctcc ccatctcccc cccctcccca 420
cccccaattt tgtatttatt tattttttaa ttattttgtg cagcgatggg ggcggggggg 480
gggggggggc gcgcgccagg cggggcgggg cggggcgagg ggcggggcgg ggcgaggcgg 540
agaggtgcgg cggcagccaa tcagagcggc gcgctccgaa agtttccttt tatggcgagg 600
cggcggcggc ggcggcccta taaaaagcga agcgcgcggc gggcggggag tcgctgcgac 660
gctgccttcg ccccgtgccc cgctccgccg ccgcctcgcg ccgcccgccc cggctctgac 720
tgaccgcgtt actcccacag gtgagcgggc gggacggccc ttctcctccg ggctgtaatt 780
agcgcttggt ttaatgacgg cttgtttctt ttctgtggct gcgtgaaagc cttgaggggc 840
tccgggaggg ccctttgtgc ggggggagcg gctcgggggg tgcgtgcgtg tgtgtgtgcg 900
tggggagcgc cgcgtgcggc tccgcgctgc ccggcggctg tgagcgctgc gggcgcggcg 960
cggggctttg tgcgctccgc agtgtgcgcg aggggagcgc ggccgggggc ggtgccccgc 1020
ggtgcggggg gggctgcgag gggaacaaag gctgcgtgcg gggtgtgtgc gtgggggggt 1080
gagcaggggg tgtgggcgcg tcggtcgggc tgcaaccccc cctgcacccc cctccccgag 1140
ttgctgagca cggcccggct tcgggtgcgg ggctccgtac ggggcgtggc gcggggctcg 1200
ccgtgccggg cggggggtgg cggcaggtgg gggtgccggg cggggcgggg ccgcctcggg 1260
ccggggaggg ctcgggggag gggcgcggcg gcccccggag cgccggcggc tgtcgaggcg 1320
cggcgagccg cagccattgc cttttatggt aatcgtgcga gagggcgcag ggacttcctt 1380
tgtcccaaat ctgtgcggag ccgaaatctg ggaggcgccg ccgcaccccc tctagcgggc 1440
gcggggcgaa gcggtgcggc gccggcagga aggaaatggg cggggagggc cttcgtgcgt 1500
cgccgcgccg ccgtcccctt ctccctctcc agcctcgggg ctgtccgcgg ggggacggct 1560
gccttcgggg gggacggggc agggcggggt tcggcttctg gcgtgtgacc ggcggctcta 1620
gagcctctgc taaccatgtt catgccttct tctttttcct acagctcctg ggcaacgtgc 1680
tggttattgt gctgtctcat cattttggca aagaattcg 1719
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПЛАТФОРМА CRISPR/CAS ДЛЯ ИСКЛЮЧАЮЩЕГО СКРИНИНГА ДЛЯ ВЫЯВЛЕНИЯ ГЕНЕТИЧЕСКИХ УЯЗВИМОСТЕЙ, СВЯЗАННЫХ С АГРЕГАЦИЕЙ ТАУ-БЕЛКА | 2020 |
|
RU2808829C2 |
| Животные, отличные от человека, содержащие гуманизированный локус альбумина | 2020 |
|
RU2815514C2 |
| СКРИНИНГОВАЯ ПЛАТФОРМА CRISPR/CAS ДЛЯ ИДЕНТИФИКАЦИИ ГЕНЕТИЧЕСКИХ МОДИФИКАТОРОВ ЗАСЕВА ИЛИ АГРЕГАЦИИ ТАУ-БЕЛКА | 2020 |
|
RU2819350C2 |
| МЫШИ, СОДЕРЖАЩИЕ МУТАЦИИ, ВСЛЕДСТВИЕ КОТОРЫХ ЭКСПРЕССИРУЕТСЯ УКОРОЧЕННЫЙ НА С-КОНЦЕ ФИБРИЛЛИН-1 | 2017 |
|
RU2721125C1 |
| НОВЫЕ ФЕРМЕНТЫ И СИСТЕМЫ CRISPR | 2016 |
|
RU2777988C2 |
| КОНСТРУИРОВАНИЕ СИСТЕМ, СПОСОБЫ И ОПТИМИЗИРОВАННЫЕ НАПРАВЛЯЮЩИЕ КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2796017C2 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ DEMEQUINA SEDIMINICOLA | 2019 |
|
RU2722933C1 |
| НОВЫЕ ФЕРМЕНТЫ CRISPR И СИСТЕМЫ | 2016 |
|
RU2771826C2 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA | 2019 |
|
RU2722934C1 |
| НОВЫЕ ФЕРМЕНТЫ И СИСТЕМЫ CRISPR | 2016 |
|
RU2737537C2 |

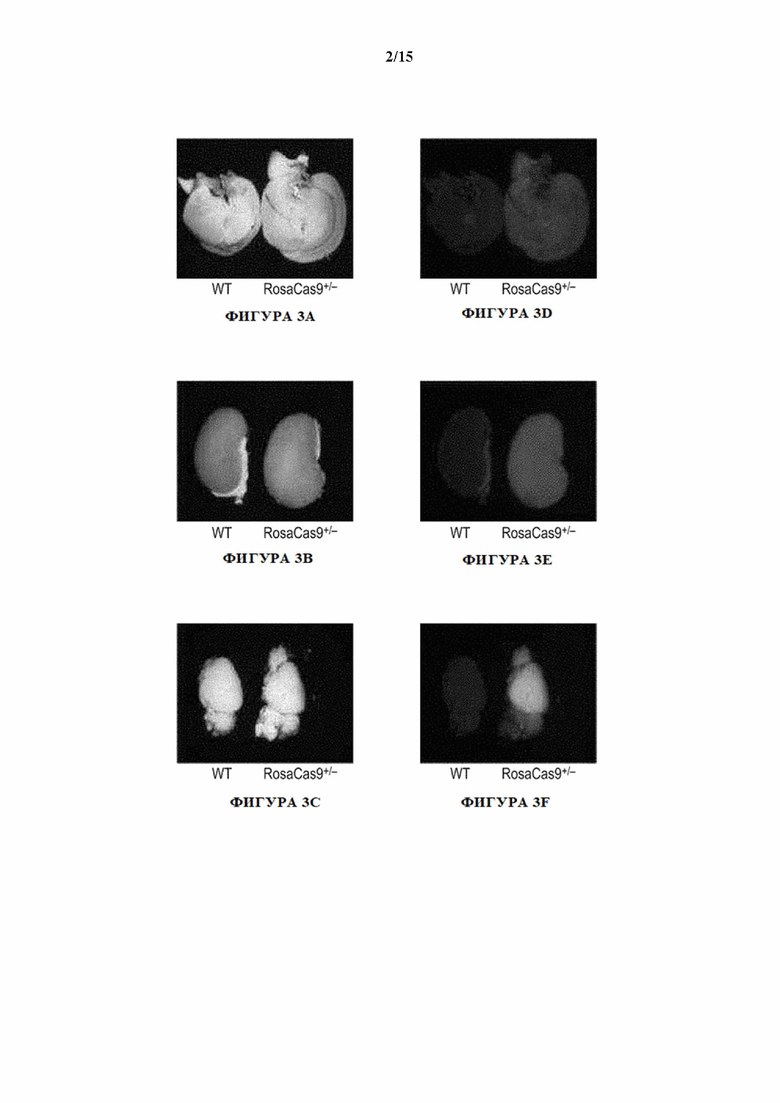

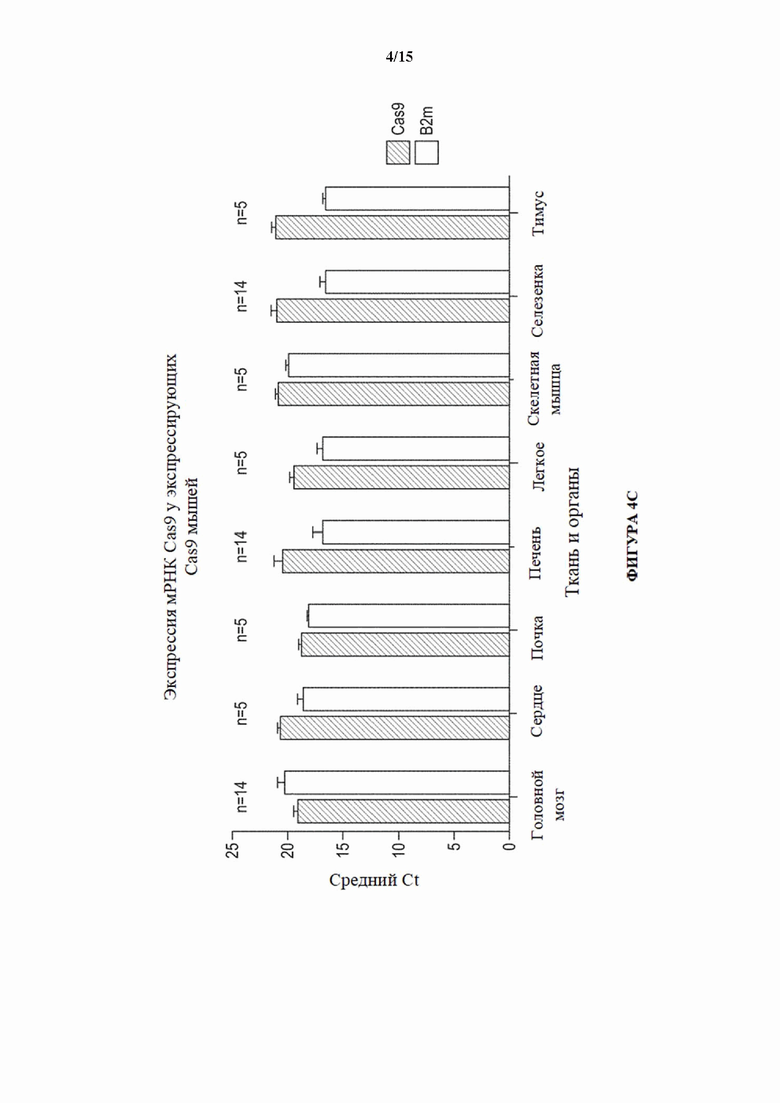
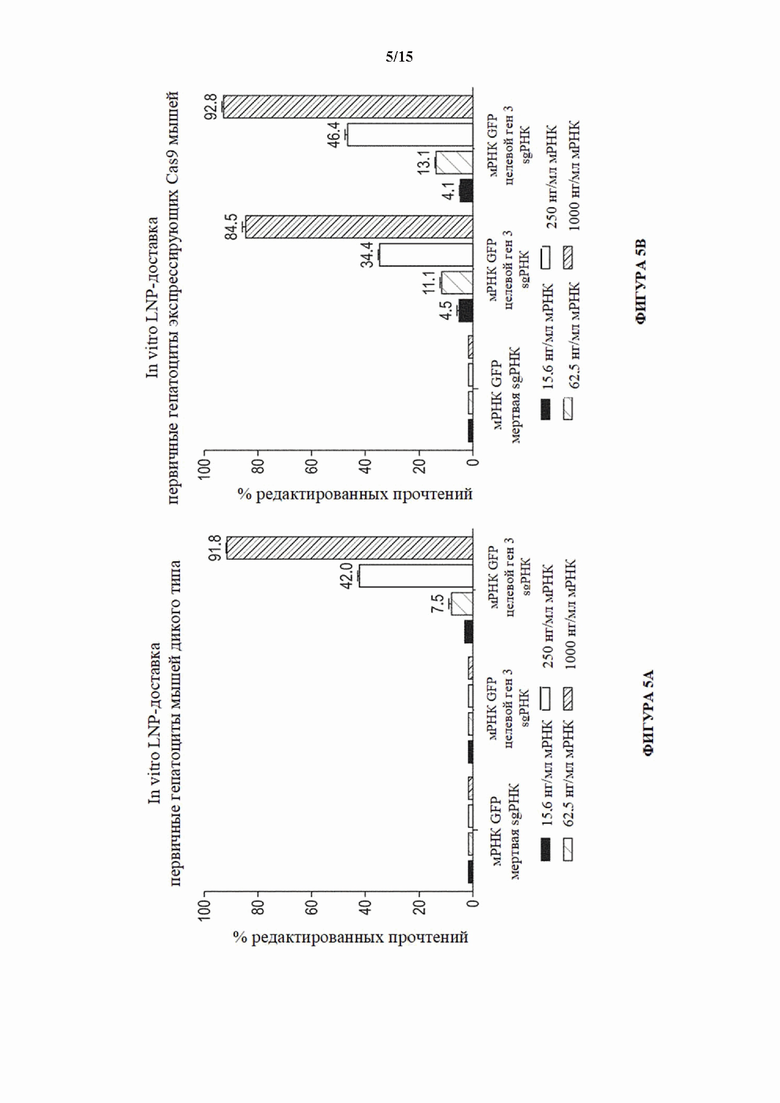

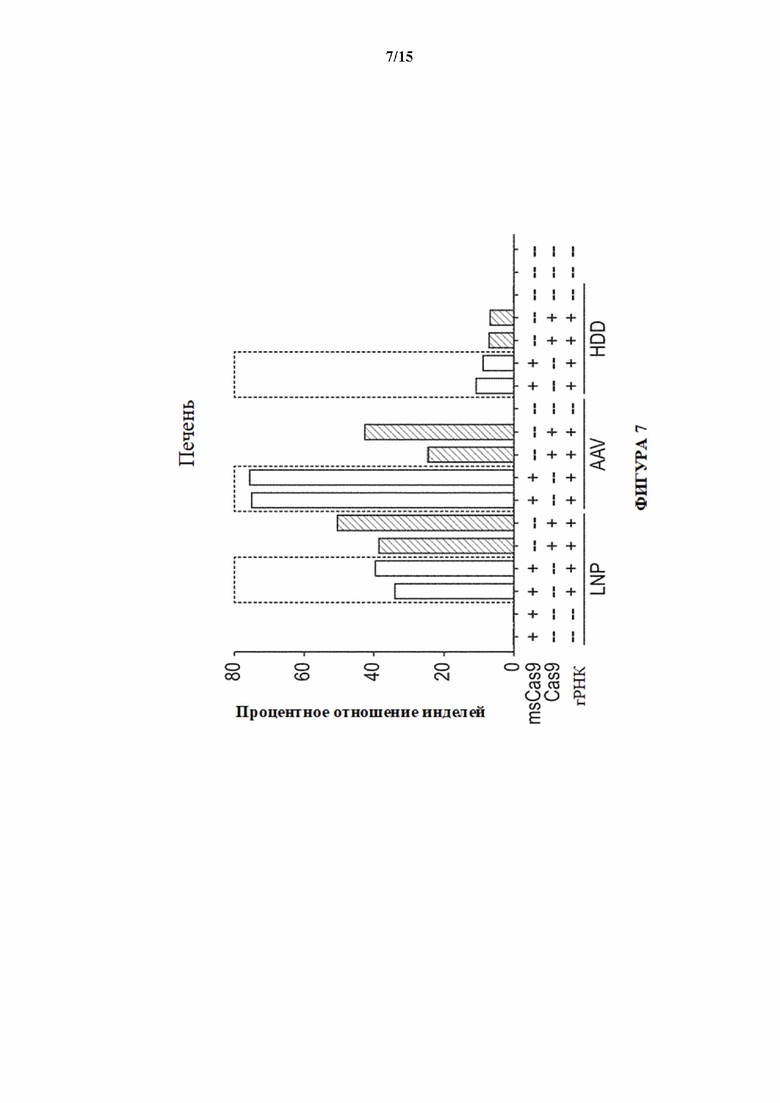
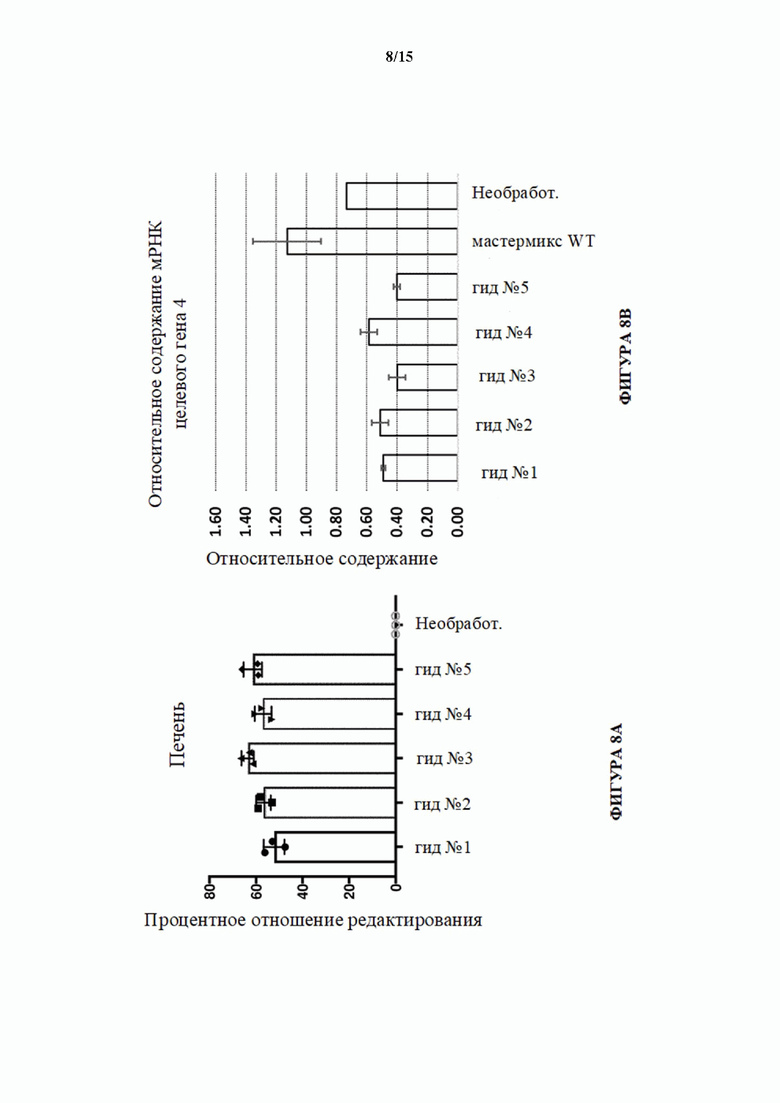
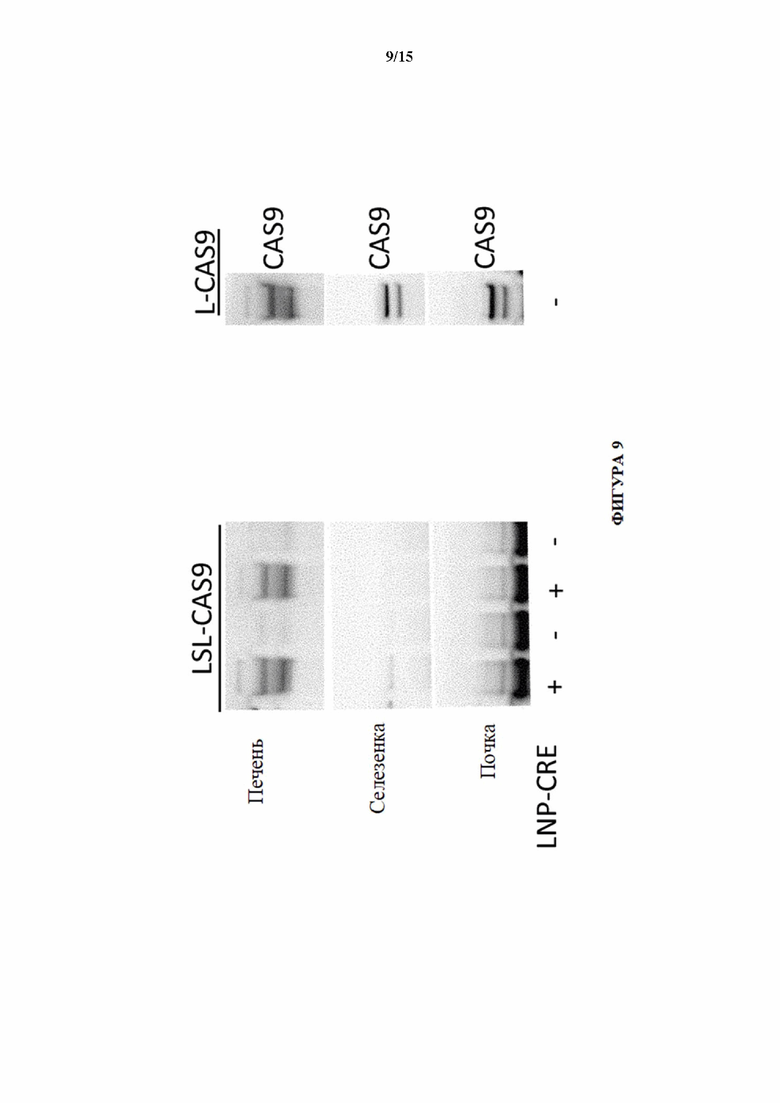
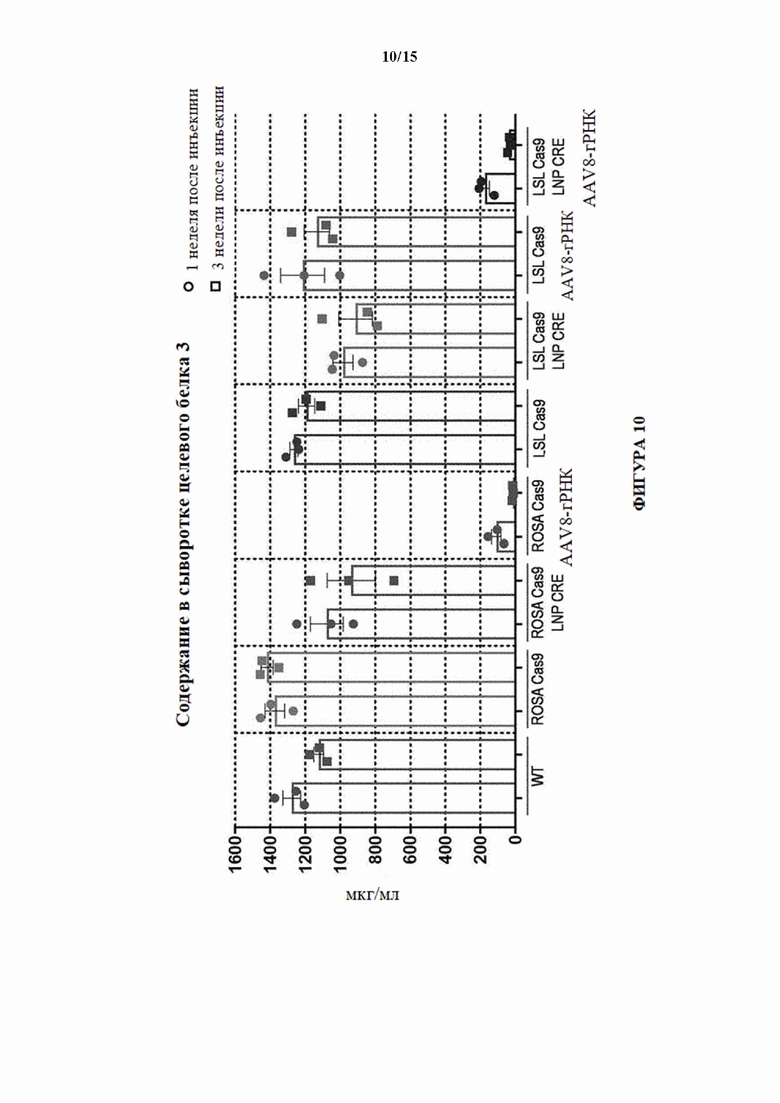
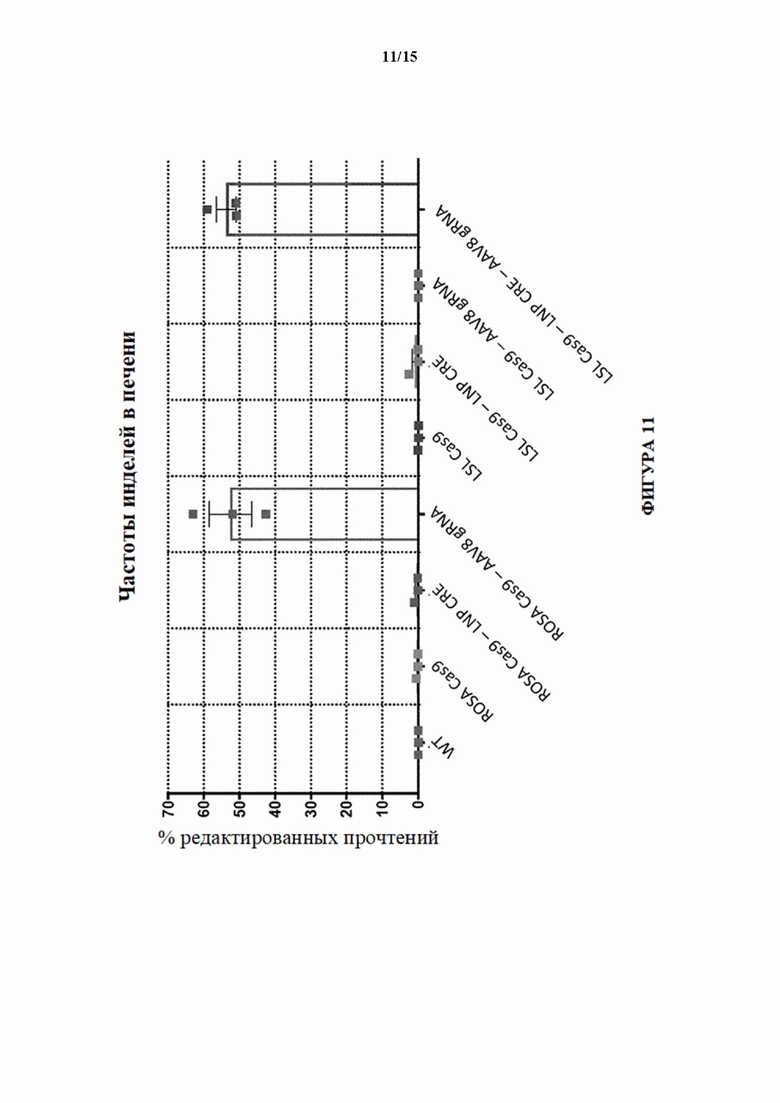
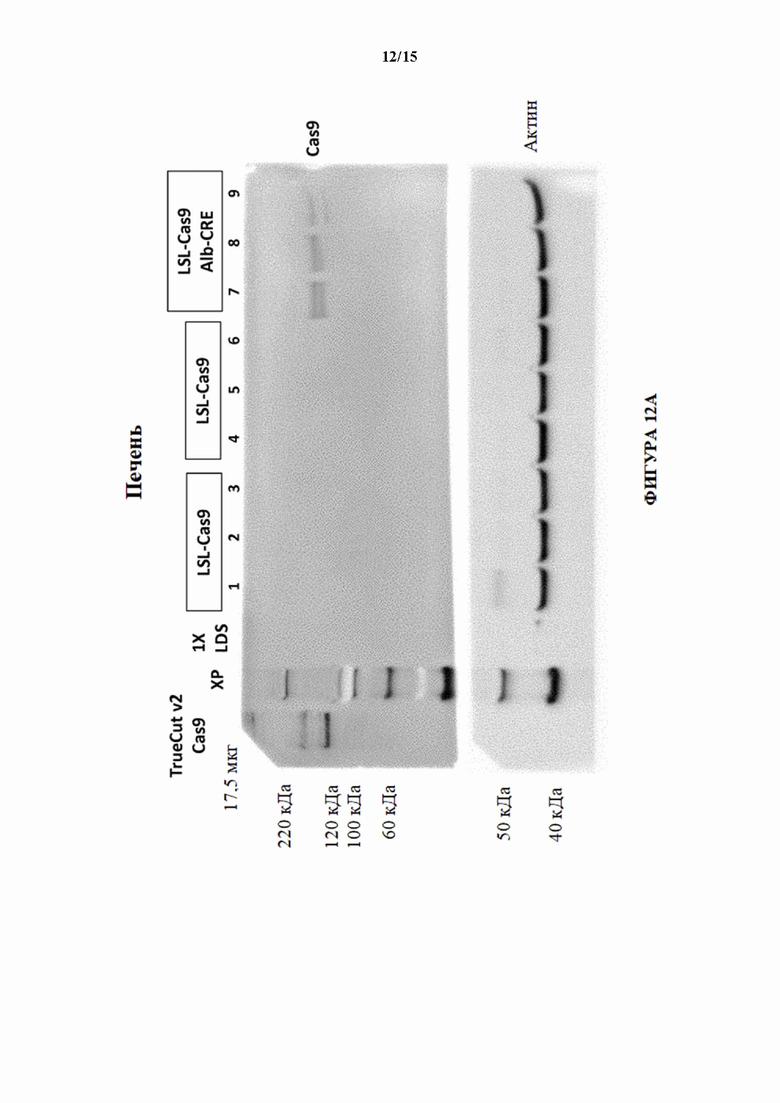
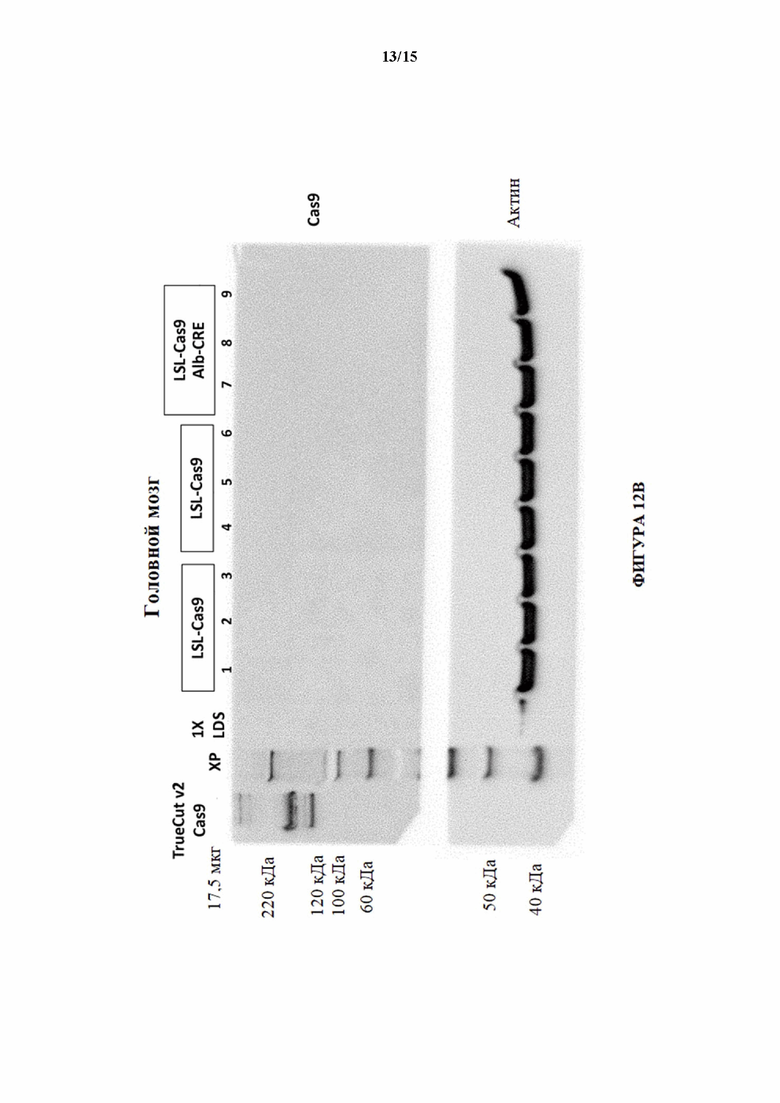
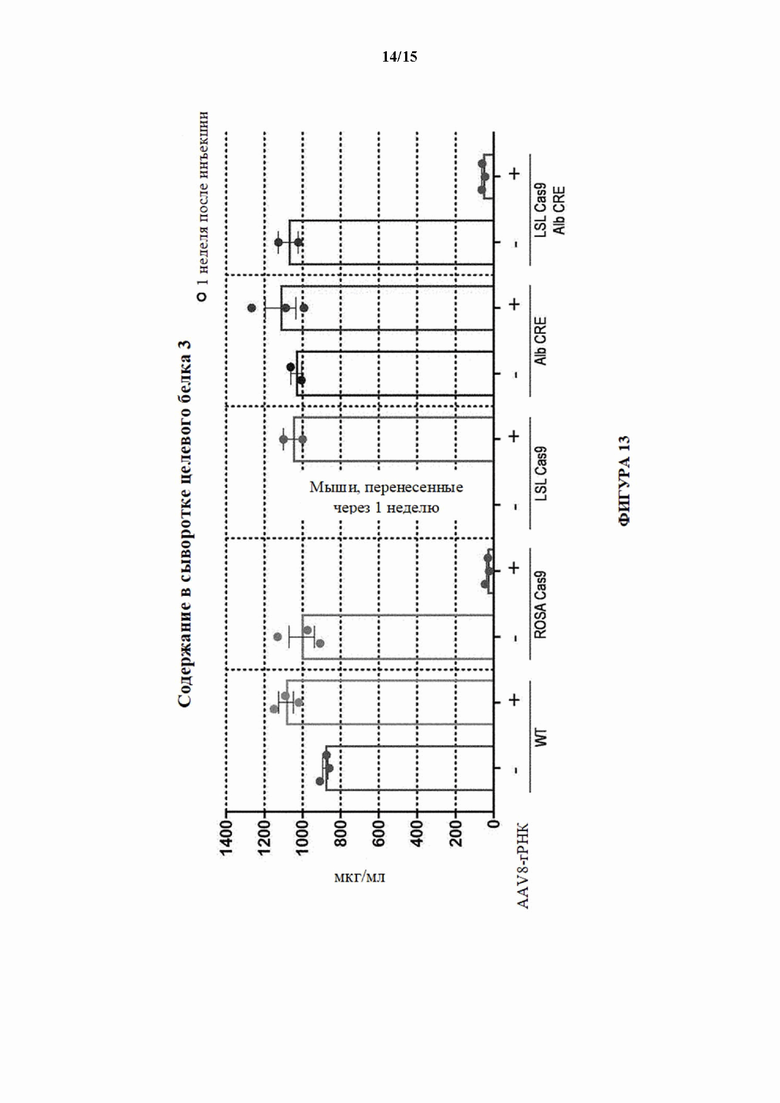
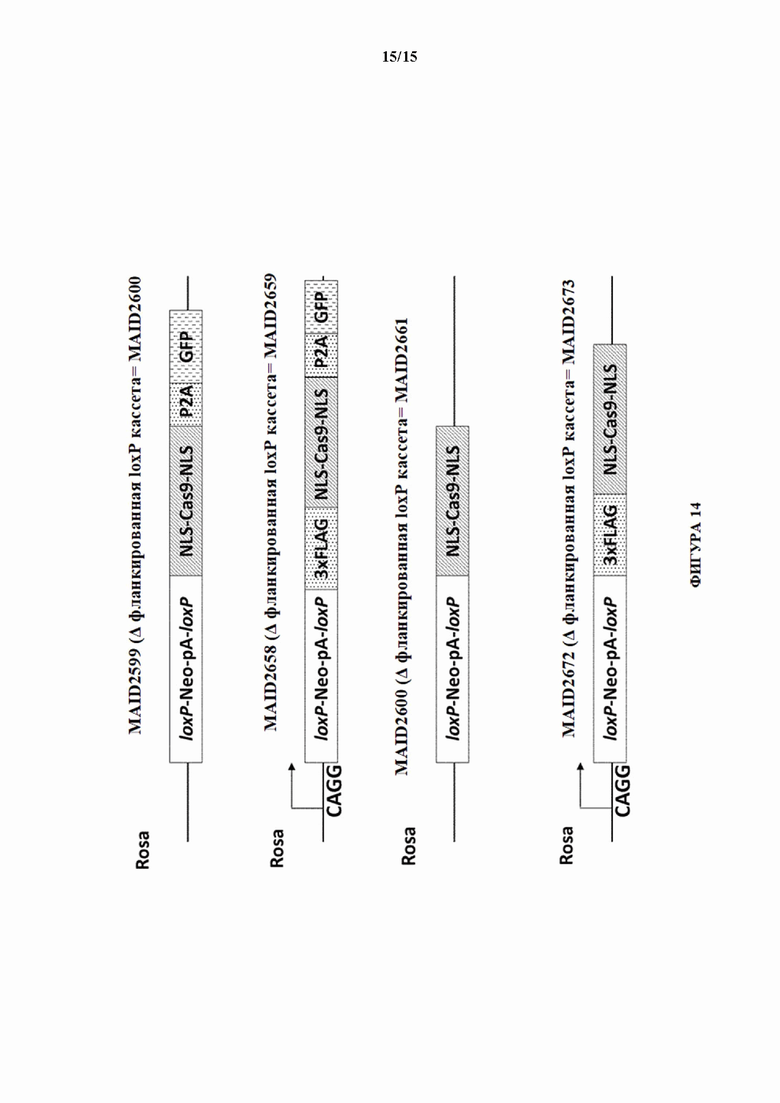
Изобретение относится к области биотехнологии, в частности к мыши, содержащей экспрессионную кассету Cas9, геномно интегрированную в локус Rosa26, причем экспрессионная кассета Cas9 содержит кодирующую последовательность для белка Cas9, к клетке указанной мыши, а также к способу ее получения. Изобретение эффективно для испытания способности CRISPR/Cas9-нуклеазы, содержащей белок Cas9 и гидовую РНК, модифицировать целевой геномный локус in vivo, а также для оптимизации способности CRISPR/Cas9-нуклеазы модифицировать целевой геномный локус in vivo. 6 н. и 61 з.п. ф-лы, 27 ил., 12 табл., 2 пр.
1. Способ испытания способности CRISPR/Cas9-нуклеазы, содержащей белок Cas9 и гидовую РНК, модифицировать целевой геномный локус in vivo, предусматривающий:
(a) введение мыши гидовой РНК, предназначенной для нацеливания на целевую последовательность для гидовой РНК в целевом геномном локусе,
причем мышь в своей зародышевой линии содержит геномно интегрированную экспрессионную кассету Cas9 в локусе Rosa26,
причем экспрессионная кассета Cas9 содержит кодирующую последовательность для белка Cas9, при этом белок Cas9 экспрессируется и содержит полную последовательность, представленную в SEQ ID NO: 19, и
причем гидовую РНК вводят посредством опосредованной аденоассоциированным вирусом (AAV) доставки; и
(b) оценку модификации целевого геномного локуса при помощи CRISPR/Cas9 нуклеазы.
2. Способ по п. 1, в котором AAV представляет собой AAV7, AAV8 или AAV9, и стадия (b) предусматривает оценку модификации целевого геномного локуса в печени.
3. Способ по п. 2, в котором AAV представляет собой AAV8.
4. Способ по любому из предыдущих пунктов, в котором путь введения AAV мыши представляет собой внутривенную инъекцию, интрапаренхиматозную инъекцию, интраперитонеальную инъекцию, закапывание в нос или интравитреальную инъекцию.
5. Способ по любому из предыдущих пунктов, в котором экзогенную донорную нуклеиновую кислоту вводят на стадии (а), причем экзогенная донорная нуклеиновая кислота предназначена для рекомбинации с целевым геномным локусом.
6. Способ по п. 5, в котором экзогенная донорная нуклеиновая кислота представляет собой одноцепочечный олигодезоксинуклеотид (ssODN).
7. Способ по любому из предыдущих пунктов, в котором целевой геномный локус содержит целевой ген, и стадия (b) предусматривает измерение экспрессии целевого гена или активности белка, кодируемого целевым геном.
8. Способ по любому из предыдущих пунктов, в котором стадия (b) предусматривает секвенирование целевого геномного локуса в одной или нескольких клетках, выделенных из мыши.
9. Способ по любому из предыдущих пунктов, в котором стадия (b) предусматривает выделение из мыши целевого органа или ткани и оценку модификации целевого геномного локуса в целевом органе или ткани.
10. Способ по п. 9, в котором стадия (b) предусматривает оценку модификации целевого геномного локуса в двух или более различных типах клеток в пределах целевого органа или ткани.
11. Способ по любому из предыдущих пунктов, в котором стадия (b) предусматривает выделение из мыши нецелевого органа или ткани и оценку модификации целевого геномного локуса в нецелевом органе или ткани.
12. Способ по любому из предыдущих пунктов, в котором экспрессионная кассета Cas9 дополнительно содержит сигнал полиаденилирования выше кодирующей последовательности для белка Cas9, причем сигнал полиаденилирования фланкирован сайтами распознавания рекомбиназы, и причем сигнал полиаденилирования в экспрессионной кассете Cas9 был вырезан тканеспецифическим образом.
13. Способ по п. 12, в котором сигнал полиаденилирования выше кодирующей последовательности для белка Cas9 был вырезан в печени.
14. Способ по п. 12 или 13, в котором рекомбиназа, которая распознает сайты распознавания рекомбиназы в экспрессионной кассете Cas9, представляет собой Cre-рекомбиназу.
15. Способ по п. 14, в котором мышь дополнительно содержит геномно интегрированную экспрессионную кассету Cre-рекомбиназы, содержащую кодирующую Cre-рекомбиназу последовательность, функционально связанную с тканеспецифическим промотором.
16. Способ по п. 14 или 15, в котором ген Cre-рекомбиназы функционально связан с промотором альбумина.
17. Способ по любому из пп. 1-11, в котором экспрессионная кассета Cas9 дополнительно содержит сигнал полиаденилирования выше кодирующей последовательности для белка Cas9, причем сигнал полиаденилирования фланкирован сайтами распознавания рекомбиназы, и причем способ дополнительно предусматривает введение рекомбиназы мыши тканеспецифическим образом.
18. Способ по п. 17, в котором рекомбиназу вводят посредством опосредованной аденоассоциированным вирусом (AAV) доставки или опосредованной липидными наночастицами (LNP) доставки.
19. Способ по любому из предыдущих пунктов, в котором экспрессионная кассета Cas9 дополнительно содержит кодирующую флуоресцентный белок последовательность.
20. Способ по п. 19, в котором экспрессионная кассета Cas9 содержит мультицистронную нуклеиновую кислоту, содержащую кодирующую последовательность для белка Cas9 и кодирующую флуоресцентный белок последовательность, разделенные с помощью промежуточного участка внутренней посадки рибосомы (IRES) или промежуточной кодирующей пептид 2А последовательности.
21. Способ по п. 20, в котором мультицистронная нуклеиновая кислота в экспрессионной кассете Cas9 содержит кодирующую последовательность для белка Cas9 и кодирующую зеленый флуоресцентный белок последовательность, разделенные с помощью промежуточной кодирующей пептид Р2А последовательности.
22. Способ по любому из пп. 1-18, в котором экспрессионная кассета Cas9 дополнительно не содержит кодирующую флуоресцентный белок последовательность.
23. Способ по любому из предыдущих пунктов, в котором белок Cas9 содержит метку белка.
24. Способ по любому из предыдущих пунктов, в котором 5' конец экспрессионной кассеты Cas9 дополнительно содержит 3' последовательность сплайсинга.
25. Способ по любому из предыдущих пунктов, в котором экспрессионная кассета Cas9 функционально связана с эндогенным промотором.
26. Способ по любому из пп. 1-24, в котором экспрессионная кассета Cas9 функционально связана с экзогенным, конститутивным промотором.
27. Способ по любому предыдущему пункту, в котором экспрессионная кассета Cas9 кодирует белок, содержащий последовательность, представленную в SEQ ID NO: 13, 16 или 22.
28. Способ по п. 27, в котором экспрессионная кассета Cas9 содержит последовательность, представленную в SEQ ID NO: 28, 29, 30 или 31.
29. Способ по п. 28, в котором экспрессионная кассета Cas9 содержит последовательность, представленную в SEQ ID NO: 1, 12, 14, 15, 17, 18, 20 или 21.
30. Способ по любому из предыдущих пунктов, в котором экспрессионная кассета Cas9 интегрирована в первый интрон локуса Rosa26.
31. Способ по любому из предыдущих пунктов, в котором мышь является гетерозиготной в отношении экспрессионной кассеты Cas9.
32. Способ по любому из пп. 1-30, в котором мышь является гомозиготной в отношении экспрессионной кассеты Cas9.
33. Способ по любому из предыдущих пунктов,
причем AAV представляет собой AAV8, доставленный в мышь с помощью внутривенной инъекции,
причем экспрессионная кассета Cas9 функционально связана с эндогенным промотором Rosa26, вставлена в первый интрон локуса Rosa26 и содержит от 5' к 3' следующее:
(i) 3' последовательность сплайсинга; и
(ii) кодирующую последовательность для белка Cas9, и
причем стадия (b) предусматривает оценку модификации целевого геномного локуса в печени мыши.
34. Способ оптимизации способности CRISPR/Cas9-нуклеазы модифицировать целевой геномный локус in vivo, предусматривающий:
(I) проведение способа по любому из предыдущих пунктов в первый раз у первой мыши;
(II) изменение переменной и проведение способа согласно стадии (I) во второй раз с измененной переменной у второй мыши; причем:
(А) измененная переменная представляет собой:
(i) серотип AAV;
(ii) путь введения гидовой РНК, вводимой мыши;
(iii) концентрацию или количество гидовой РНК, введенной мыши; или
(iv) последовательность гидовой РНК, введенную мыши, или
(В) стадия (а) дополнительно предусматривает введение экзогенной донорной нуклеиновой кислоты, и измененная переменная представляет собой:
(i) способ доставки или путь введения для введения в мышь экзогенной донорной нуклеиновой кислоты;
(ii) концентрацию или количество экзогенной донорной нуклеиновой кислоты, введенной мыши;
(iii) концентрацию или количество гидовой РНК, введенной мыши, по отношению к концентрации или количеству экзогенной донорной нуклеиновой кислоты, введенной мыши; или
(iv) последовательность экзогенной донорной нуклеиновой кислоты, введенную мыши; и
(III) сравнение модификации целевого геномного локуса на стадии (I) с модификацией целевого геномного локуса на стадии (II), и выбор способа, приводящего к модификации целевого геномного локуса, характеризующейся одним или несколькими из следующего: повышенная эффективность, повышенная точность, повышенная согласованность или повышенная специфичность.
35. Способ по п. 34, в котором измененная переменная на стадии (II) представляет собой серотип AAV.
36. Способ по п. 34, в котором измененная переменная на стадии (II) представляет собой путь введения гидовой РНК, вводимой мыши.
37. Способ по п. 34, в котором измененная переменная на стадии (II) представляет собой концентрацию или количество гидовой РНК, введенной мыши.
38. Способ по п. 34, в котором измененная переменная на стадии (II) представляет собой гидовую РНК, введенную мыши.
39. Способ по п. 34, причем способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и причем измененная переменная на стадии (II) представляет собой способ доставки для введения экзогенной донорной нуклеиновой кислоты мыши.
40. Способ по п. 34, причем способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и измененная переменная на стадии (II) представляет собой путь введения экзогенной донорной нуклеиновой кислоты, вводимой мыши.
41. Способ по п. 34, причем способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и измененная переменная на стадии (II) представляет собой концентрацию или количество экзогенной донорной нуклеиновой кислоты, введенной мыши.
42. Способ по п. 34, причем способ предусматривает введение экзогенной донорной нуклеиновой кислоты, и измененная переменная на стадии (II) представляет собой концентрацию или количество гидовой РНК, введенной мыши, по отношению к концентрации или количеству экзогенной донорной нуклеиновой кислоты, введенной мыши.
43. Способ по п. 34, в котором измененная переменная на стадии (II) представляет собой экзогенную донорную нуклеиновую кислоту, введенную мыши.
44. Мышь для испытания способности CRISPR/Cas9-нуклеазы модифицировать целевой геномный локус in vivo, содержащая экспрессионную кассету Cas9, геномно интегрированную в локус Rosa26, причем экспрессионная кассета Cas9 содержит кодирующую последовательность для белка Cas9, причем белок Cas9 экспрессируется и содержит полную последовательность, представленную в SEQ ID NO: 19.
45. Мышь по п. 44, где экспрессионная кассета Cas9 дополнительно содержит сигнал полиаденилирования выше кодирующей последовательности для белка Cas9, причем сигнал полиаденилирования фланкирован сайтами распознавания рекомбиназы, и причем сигнал полиаденилирования в экспрессионной кассете Cas9 был вырезан тканеспецифическим образом.
46. Мышь по п. 45, где сигнал полиаденилирования выше кодирующей последовательности для белка Cas9 был вырезан в печени.
47. Мышь по п. 45 или 46, где рекомбиназа, которая распознает сайты распознавания рекомбиназы в экспрессионной кассете Cas9, представляет собой Cre-рекомбиназу.
48. Мышь по п. 47, которая дополнительно содержит геномно интегрированную экспрессионную кассету Cre-рекомбиназы, содержащую кодирующую Cre-рекомбиназу последовательность, функционально связанную с тканеспецифическим промотором.
49. Мышь по п. 47 или 48, где ген Cre-рекомбиназы функционально связан с промотором альбумина.
50. Мышь по любому из пп. 44-49, где экспрессионная кассета Cas9 дополнительно содержит кодирующую флуоресцентный белок последовательность.
51. Мышь по п. 50, где экспрессионная кассета Cas9 содержит мультицистронную нуклеиновую кислоту, содержащую кодирующую последовательность для белка Cas9 и кодирующую флуоресцентный белок последовательность, разделенные с помощью промежуточного участка внутренней посадки рибосомы (IRES) или промежуточной кодирующей пептид 2А последовательности.
52. Мышь по п. 51, где мультицистронная нуклеиновая кислота в экспрессионной кассете Cas9 содержит кодирующую последовательность для белка Cas9 и кодирующую зеленый флуоресцентный белок последовательность, разделенные с помощью промежуточной кодирующей пептид Р2А последовательности.
53. Мышь по любому из пп. 44-49, где экспрессионная кассета Cas9 дополнительно не содержит кодирующую флуоресцентный белок последовательность.
54. Мышь по любому из пунктов 44-53, где белок Cas9 содержит метку белка.
55. Мышь по любому из пунктов 44-54, где 5' конец экспрессионной кассеты Cas9 дополнительно содержит 3' последовательность сплайсинга.
56. Мышь по любому из пунктов 44-55, где экспрессионная кассета Cas9 функционально связана с эндогенным промотором.
57. Мышь по любому из пп. 44-55, где экспрессионная кассета Cas9 функционально связана с экзогенным, конститутивным промотором.
58. Мышь по любому из пп. 44-57, где экспрессионная кассета Cas9 кодирует белок, содержащий последовательность, представленную в SEQ ID NO: 13, 16 или 22.
59. Мышь по п. 58, где экспрессионная кассета Cas9 содержит последовательность, представленную в SEQ ID NO: 28, 29, 30 или 31.
60. Мышь по п. 59, где экспрессионная кассета Cas9 содержит последовательность, представленную в SEQ ID NO: 1, 12, 14, 15, 17, 18, 20 или 21.
61. Мышь по любому из пп. 44-60, где экспрессионная кассета Cas9 интегрирована в первый интрон локуса Rosa26.
62. Мышь по любому из пунктов 44-61, которая является гетерозиготной в отношении экспрессионной кассеты Cas9.
63. Мышь по любому из пп. 44-61, которая является гомозиготной в отношении экспрессионной кассеты Cas9.
64. Мышь по любому из пп. 44-63, где экспрессионная кассета Cas9 функционально связана с эндогенным промотором Rosa26, вставлена в первый интрон локуса Rosa26 и содержит от 5' к 3' следующее:
(i) 3' последовательность сплайсинга; и
(ii) кодирующую последовательность для белка Cas9.
65. Клетка мыши для испытания способности CRISPR/Cas9-нуклеазы модифицировать целевой геномный локус, содержащая экспрессионную кассету Cas9, геномно интегрированную в локус Rosa26, причем экспрессионная кассета Cas9 содержит кодирующую последовательность для белка Cas9, и причем белок Cas9 экспрессируется и содержит полную последовательность, представленную в SEQ ID NO: 19.
66. Способ получения мыши по любому из пп. 44-64, предусматривающий:
(a) модификацию генома мышиной эмбриональной стволовой (ES) клетки для содержания экспрессионной кассеты Cas9 геномно интегрированной в локус Rosa26, причем экспрессионная кассета Cas9 содержит кодирующую последовательность для белка Cas9, и причем белок Cas9 экспрессируется и содержит полную последовательность, представленную в SEQ ID NO: 19;
(b) идентификацию или выбор генетически модифицированной мышиной ES клетки, содержащей экспрессионную кассету Cas9;
(c) введение генетически модифицированной мышиной ES клетки в эмбрион-хозяин мыши; и
(d) имплантацию и вынашивание мышиного эмбриона-хозяина в организме суррогатной матери-мыши.
67. Способ получения мыши по любому из пп. 44-64, предусматривающий:
(a) модификацию генома эмбриона мыши на стадии одной клетки для содержания экспрессионной кассеты Cas9 геномно интегрированной в локус Rosa26, причем экспрессионная кассета Cas9 содержит кодирующую последовательность для белка Cas9, и причем белок Cas9 экспрессируется и содержит полную последовательность, представленную в SEQ ID NO: 19;
(b) выбор генетически модифицированного эмбриона мыши на стадии одной клетки, содержащего экспрессионную кассету Cas9; и
(c) имплантацию и вынашивание генетически модифицированного эмбриона мыши на стадии одной клетки в организме суррогатной матери-мыши.
| RANDALL J | |||
| PLATT et al., CRISPR-Cas9 Knockin Mice for Genome Editing and Cancer Modeling, Cell, 2014, 159(2), pp | |||
| Способ обогащения руд | 1915 |
|
SU440A1 |
| US2015020223 A1, 15.01.2015 | |||
| WO2016049024 A2, 31.03.2016 | |||
| CHIANN-MUN CHEN et al., A Comparison of Exogenous Promoter Activity at the ROSA26 Locus Using a PhiC31 Integrase Mediated Cassette Exchange Approach in Mouse ES Cells, | |||

