Область техники, к которой относится изобретение
Настоящее изобретение касается антител против рецептора IGF-I, в частности, антитела против рецептора IGF-I, которое специфически связывается с рецептором IGF-I позвоночных.
Уровень техники
1. IGF-I
IGF-I – инсулиноподобный фактор роста, который секретируется главным образом из печени и воздействует на рецептор IGF-I, вызывая проявление разнообразных физиологических функций в различных органах. Поэтому ожидается, что IGF-I будет применяться для лечения различных заболеваний. Поскольку аминокислотная последовательность IGF-I имеет высокое сходство примерно на 40% с таковой у проинсулина, IGF-I может связываться с рецептором инсулина и при этом проявлять инсулиноподобные эффекты (непатентная ссылка 1). Кроме того, поскольку аминокислотная последовательность рецептора IGF-I имеет высокое сходство примерно на 60% с таковой у рецептора инсулина, эти рецепторы могут образовывать гетеродимеры (непатентная ссылка 1). Инсулин может воздействовать на рецептор инсулина, проявляя при этом сильный эффект снижения уровня глюкозы в крови, поэтому он применяется в качестве гипогликемического средства.
2. Рецептор IGF-I
Рецептор IGF-I – трансмембранный белок, состоящий из α-цепи и β-цепи, который содержит 6 внеклеточных доменов (L1, CR, L2, Fn1, Fn2 и Fn3), трансмембранный домен и внутриклеточный домен (непатентная ссылка 2). Внутриклеточный домен рецептора IGF-I включает тирозинкиназу. Внеклеточный домен представляет собой домен CR (богатый цистеином) и участвует в активации внутриклеточной тирозинкиназы, связанной с конформационным изменением рецептора IGF-I, которое происходит при связывании IGF-I с рецептором IGF-I. Рецептор IGF-I образует гомодимерный комплекс (гомоформу). Связывание IGF-I с рецептором IGF-I (гомоформой) запускает сигнализацию посредством активации рецепторной киназы. Рецептор IGF-I также образует гетеродимерный комплекс (гетероформу) с рецептором инсулина. Связывание инсулина или IGF-I с рецептором IGF-I (гетероформой) запускает сигнализацию посредством активации рецепторной киназы (непатентные ссылки 3 и 4).
3. Физиологические эффекты IGF-I
Было показано, что IGF-I проявляет эффекты усиления роста типа увеличения длины и массы тела и инсулиноподобные метаболические эффекты типа ускорения метаболизма глюкозы и гипогликемические эффекты. Оказалось, что мекасермин, рекомбинантный IGF-I человека, улучшает симптомы, связанные с аномалией рецептора инсулина, типа гипергликемии, гиперинсулинемии, черного акантоза и гирсутизма. IGF-I также улучшает состояние при расстройствах роста типа карликовости, резистентных к гормону роста (непатентная ссылка 5).
Наряду с эффектами усиления роста, IGF-I также повышает способность к синтезу ДНК у хрящевых клеток человека. Также известно, что введение IGF-I крысам с гипофизэктомией повышает у них массу тела и длину бедренных костей (непатентная ссылка 5).
4. Влияние IGF-I на возрастание мышечной массы
Усиление активности пролиферации клеток у IGF-I требует непрерывной активации рецепторов IGF-I (непатентная ссылка 6). У генно-инженерных животных, гиперэкспрессирующих рецептор IGF-I, проявляется возрастание мышечной массы (непатентная ссылка 7). Продолжительное введение IGF-I/IGFBP3 пациентам с переломом проксимального отдела бедренной кости повышает у них силу захвата и улучшает способность вставать из сидячего положения без посторонней помощи (непатентная ссылка 8). Известно, что уровни IGF-I в мышцах у пожилых людей и мышей ниже, чем у молодых (непатентные ссылки 9 и 10). Гиперэкспрессия IGF-I конкретно в мышечных тканях пожилых мышей улучшала их мышечную массу по сравнению с мышами дикого типа (непатентная ссылка 11).
5. Предшествующие препараты для увеличения мышечной массы
Анаморелин, агонист рецептора грелина, повышал безжировую массу тела при клинических испытаниях по поводу кахексии, которая представляет собой мышечную атрофию от бездействия. Однако он имеет отрицательные эффекты типа тошноты и гипергликемии (непатентная ссылка 12).
Миостатин, фактор отрицательного контроля миогенеза скелетных мышц, влияет на рецептор II активина (ActRII) и тем самым ингибирует Akt/mTOR (непатентные ссылки 13-15).
LY2495655, антитело против миостатина, повышает мышечную массу пациентов, перенесших тотальное эндопротезирование тазобедренного сустава, а также у пожилых пациентов (непатентные ссылки 16 и 17).
Бимагрумаб, антитело против ActRII, повышает мышечную массу у пациентов с нервно-мышечными заболеваниями (непатентная ссылка 18).
Однако до сих пор нет лекарственных средств, способствующих формированию скелетных мышц, которые могли бы применяться для лечения нуждающихся в этом субъектов.
6. Предшествующие препараты для усиления роста
Состав с рекомбинантным гормоном роста (GH) человека активирует рецепторы GH и индуцирует секрецию IGF-I, проявляя при этом эффекты усиления роста. Однако, поскольку состав требует ежедневного введения посредством подкожной инъекции, он часто вызывает плохое соблюдение приема препарата (напр., непреднамеренный пропуск препарата) и приводит к снижению эффектов роста (непатентная ссылка 19). В настоящее время предпринимаются попытки разработать препарат GH пролонгированного действия с улучшенной кинетикой, который можно будет вводить раз в одну или две недели.
Однако до сих пор нет лекарственных средств, проявляющих стимулирующее рост действие, которые могли бы применяться для лечения нуждающихся в этом субъектов, с лучшим соблюдение приема препарата. Кроме того, оказалось, что состав GH проявляет снижение эффектов роста у пациентов с аномалией рецепторов GH с пониженной чувствительностью к активации рецепторов GH или у пациентов, невосприимчивых к лечению GH (непатентная ссылка 20).
IGF-I является единственным терапевтическим средством, обладающим стимулирующим рост действием на пациентов с пониженной чувствительностью к активации рецепторов GH, так как он действует в любой точке по нисходящей от рецепторов GH. Однако препарат IGF-I представляет собой парентеральный раствор для введения два раза в день, поэтому он может приводить к плохому соблюдению приема препарата. Кроме того, было показано, что он вызывает гипогликемию в качестве побочного действия (непатентная ссылка 21). До сих пор нет лекарственных средств с лучшим соблюдением приема препарата и меньшей частотой возникновения гипогликемии, чем IGF-I, которые могли бы применяться в качестве альтернативного терапевтического средства.
7. Гипогликемическое действие IGF-I
Известно, что IGF-I обладает гипогликемическим действием в качестве инсулиноподобного действия. IGF-I усиливает эффект поглощения глюкозы у мышечных клеток крыс (непатентная ссылка 5). Введение IGF-I также снижает уровень глюкозы в крови у крыс (непатентная ссылка 5).
Сообщалось, что снижение IGF-I уровня глюкозы вызывает гипогликемию в качестве клинического отрицательного эффекта (непатентная ссылка 21). Точно так же введение IGF-I больным людям вызывает гипогликемию. Поэтому с самого начала терапии IGF-I необходимо продолжать контролировать дозировку, начиная с низкой дозы, и отслеживать различные клинические показатели, включая уровень глюкозы в крови после введения (непатентная ссылка 5).
IGF-I проявляет гипогликемическое действие посредством стимуляции фосфорилирования Akt, что является нисходящим сигналом рецептора IGF-I. Активный вариант Akt усиливает поглощение глюкозы клетками 3T3-L1 (непатентная ссылка 22). С другой стороны, у мышей с дефицитом Akt2 проявляется повышение уровня глюкозы в крови (непатентная ссылка 23). Ингибитор Akt ингибирует индуцированное инсулином поглощение глюкозы мышечными клетками крыс (непатентная ссылка 24). Также известно, что IGF-I активирует рецептор инсулина, который играет роль в гипогликемическом действии. Эти данные свидетельствуют о том, что гипогликемическое действие IGF-I включает гиперактивацию Akt и активацию инсулиновых рецепторов.
8. Короткий период полужизни IGF-I в крови
IGF-I имеет короткий период полужизни в крови, поэтому он требует частого введения при применении в терапии. Так, мекасермин, рекомбинантный IGF-I человека, имеет период полужизни в крови от 11 часов до 16 часов, поэтому при лечении карликовости его нужно вводить один-два раза в день (непатентная ссылка 5).
Примерно 70-80% IGF-I связывается с IGFBP3 в крови, тогда как физиологическое действие проявляет свободная форма IGF-I. Связывание IGF-I с IGFBP3 поддерживает период его полужизни в крови от 10 часов до 16 часов (непатентная ссылка 1).
IPLEX, комбинированный препарат IGF-I с IGFBP3, проявляет повышение периода полужизни IGF-I в крови до 21-26 часов и тем самым позволяет уменьшить частоту введения до одного раза в день (непатентная ссылка 23). Однако IPLEX уже снят с продажи.
Также предпринималась попытка разработать ПЭГилированный IGF-I с улучшенной кинетикой IGF-I, но до сих пор еще ни один препарат не был успешно разработан и доступен в настоящее время (патентная ссылка 1).
9. Ожидаемые терапевтические эффекты под действием IGF-I
Известно, что IGF-I влияет на различные органы и выполняет широкий спектр физиологических функций (непатентная ссылка 21).
Сообщалось, что IGF-I оказывает нейропротекторное действие на центральную нервную систему, защищая митохондрии, и антиоксидантное действие путем активации рецептора IGF-I (непатентные ссылки 26 и 27). IGF-I способствует регенерации поврежденных нейритов (непатентная ссылка 28).
IGF-I является главным фактором стимулирования роста (непатентные ссылки 29 и 30). Так, мекасермин, рекомбинантный IGF-I человека, имеет клиническое применение в качестве препарата для лечения карликовости.
IGF-I считается эффективным при лечении цирроза печени, который возникает при повреждениях печени или хронических заболеваниях печени и включает фиброз печени. Введение IGF-I улучшает состояние при фиброзе печени на модели у животных с циррозом печени (непатентная ссылка 31).
IGF-I также играет роль в развитии и функциях почек. IGF-I оказывает защитное действие против окислительного стресса и апоптоза при глюкотоксичности у мезангиальных клеток почек (непатентная ссылка 32). IGF-I предполагается в качестве средства для лечения нефропатии.
Примеры состояний, при которых ожидаются улучшения при введении IGF-I, включают: карликовость, синдром Ларона, цирроз печени, фиброз печени, старение, ограничение внутриутробного развития (IUGR), неврологические заболевания, инсульт, травмы спинного мозга, защита сердечно-сосудистой системы, инсулинорезистентный диабет, метаболический синдром, нефропатия, остеопороз, муковисцидоз, заживление ран, миотоническая дистрофия, связанная со СПИД саркопения, связанный с ВИЧ синдром перераспределения жира, ожоги, болезнь Крона, синдром Вернера, сцепленный с Х-хромосомой комбинированный иммунодефицит, потеря слуха, нервная анорексия и ретинопатия недоношенных (непатентная ссылка 21).
Таким образом, IGF-I предполагается в качестве средства для лечения различных заболеваний вследствие широкого спектра его физиологических эффектов. Однако такие проблемы, как неблагоприятный гипогликемический эффект и короткий период полужизни, требующий многократного введения, препятствуют его клиническому применению.
10. Антитела – агонисты рецептора IGF-I
Как правило, препараты антител имеют длительный период полужизни и оказываются эффективными при введении один-два раза в месяц. Хотя сообщалось, что некоторые антитела-агонисты рецептора IGF-I эффективны в активации рецептора in vitro, не было сообщений об антителах, проявляющих агонистическую активность против рецептора IGF-I in vivo (непатентные ссылки 33-37).
В частности, антитела 3B7 и 2D1 усиливают синтез клеточной ДНК in vitro (непатентная ссылка 34).
Антитела 11A1, 11A4, 11A11 и 24-57 усиливают фосфорилирование тирозина у рецептора IGF-I in vitro (непатентная ссылка 35).
Антитела 16-13, 17-69, 24-57, 24-60, 24-31 и 26-3 оказались эффективными в стимулировании синтеза клеточной ДНК и поглощения глюкозы in vitro и способны проявлять гипогликемический эффект (непатентные ссылки 36 и 37).
Однако ни одно из антител-агонистов рецептора IGF-I не было отмечено как проявляющее эффекты пролиферации клеток в эксперименте in vitro на первичных культурах клеток, в частности, миобластов человека, уж не говоря об эффектах увеличения мышечной массы in vivo.
11. Антитела – антагонисты рецептора IGF-I
Предпринимались попытки применения антител, связывающихся с рецептором IGF-I, для лечения злокачественных новообразований, исходя из их антагонистического действия – ингибирования связывания IGF-I с рецептором IGF-I. Однако существующие антагонистические антитела к рецептору IGF-I имеют различные побочные эффекты типа гипергликемии при монотерапии (непатентная ссылка 38) и проявляют повышенную частоту гипергликемии при использовании в сочетании с другими противораковыми средствами (непатентная ссылка 39). Соответственно ожидается, что их терапевтическое применение будет ограниченным.
Список цитированной литературы
Патентная литература
[1] Use of PEGylated Igf-I Variants for the Treatment of Neuromuscular Disorders. JP 2011-518778A (WO 2009/121759A) (2011).
Непатентная литература
[1] Ohlsson C. et al. The role of liver-derived insulin-like growth factor-I. Endocr Rev., 2009, 30(5): p. 494-535.
[2] Kavran J.M. et al. How IGF-I activates its receptor. Elife, 2014, 3.
[3] Bailyes E.M. et al. Insulin receptor/IGF-I receptor hybrids are widely distributed in mammalian tissues: quantification of individual receptor species by selective immunoprecipitation and immunoblotting. Biochem J, 1997, 327 (Pt 1): p. 209-15.
[4] Pandini G. et al. Insulin/insulin-like growth factor I hybrid receptors have different biological characteristics depending on the insulin receptor isoform involved. J Biol Chem., 2002, 277(42): p. 39684-95.
[5] OrphanPacific, IF, 2015.
[6] Fukushima T. et al. Phosphatidylinositol 3-kinase (PI3K) activity bound to insulin-like growth factor-I (IGF-I) receptor, which is continuously sustained by IGF-I stimulation, is required for IGF-I-induced cell proliferation. J Biol Chem., 2012, 287(35): p. 29713-21.
[7] Schiaffino S. and C. Mammucari. Regulation of skeletal muscle growth by the IGF-I-Akt/PKB pathway: insights from genetic models. Skelet Muscle, 2011, 1(1): p. 4.
[8] Boonen S. et al. Musculoskeletal effects of the recombinant human IGF-I/IGF binding protein-3 complex in osteoporotic patients with proximal femoral fracture: a double-blind, placebo-controlled pilot study. J Clin Endocrinol Metab., 2002, 87(4): p. 1593-9.
[9] Barton-Davis E.R. et al. Viral mediated expression of insulin-like growth factor I blocks the aging-related loss of skeletal muscle function. Proc Natl Acad Sci. USA, 1998, 95 (26): p. 15603-7.
[10] Lamberts S.W., A.W. van den Beld, and A.J. van der Lely. The endocrinology of aging. Science, 1997, 278(5337): p. 419-24.
[11] Musaro A. et al. Localized IGF-I transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat Genet., 2001, 27(2): p. 195-200.
[12] Temel J.S. et al. Anamorelin in patients with non-small-cell lung cancer and cachexia (ROMANA 1 and ROMANA 2): results from two randomized, double-blind, phase 3 trials. Lancet Oncol., 2016, 17(4): p. 519-31.
[13] Glass D.J. Signaling pathways perturbing muscle mass. Curr Opin Clin Nutr Metab Care, 2010, 13(3): p. 225-9.
[14] Lee S.J. and A.C. McPherron. Regulation of myostatin activity and muscle growth. Proc Natl Acad Sci USA, 2001, 98(16): p. 9306-11.
[15] Amirouche A. et al. Down-regulation of Akt/mammalian target of rapamycin signaling pathway in response to myostatin overexpression in skeletal muscle. Endocrinology, 2009, 150(1): p. 286-94.
[16] Woodhouse L. et al. A Phase 2 Randomized Study Investigating the Efficacy and Safety of Myostatin Antibody LY2495655 versus Placebo in Patients Undergoing Elective Total Hip Arthroplasty. J Frailty Aging, 2016, 5(1): p. 62-70.
[17] Becker C. et al. Myostatin antibody (LY2495655) in older weak fallers: a proof-of-concept, randomized, phase 2 trial. Lancet Diabetes Endocrinol., 2015, 3(12): p. 948-57.
[18] Amato A.A. et al. Treatment of sporadic inclusion body myositis with bimagrumab. Neurology, 2014, 83(24): p. 2239-46.
[19] Cutfield W.S. et al. Non-compliance with growth hormone treatment in children is common and impairs linear growth. PLos One, 2011, 6(1):e16223
[20] Bang P. et al. Identification and management of poor response to growth-promoting therapy in children with short stature. Clin Endocrinol. (Oxf.), 2012, 77(2):p.169-181.
[21] Puche J.E. and I. Castilla-Cortazar. Human conditions of insulin-like growth factor-I (IGF-I) deficiency. J Transl Med., 2012, 10: p. 224.
[22] Kohn A.D. et al. Expression of a constitutively active Akt Ser/Thr kinase in 3T3-L1 adipocytes stimulates glucose uptake and glucose transporter 4 translocation. J Biol Chem., 1996, 271(49): p. 31372-8.
[23] Cho H. et al. Insulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt2 (PKB beta). Science, 2001, 292(5522): p. 1728-31.
[24] Green C.J. et al. Use of Akt inhibitor and a drug-resistant mutant validates a critical role for protein kinase B/Akt in the insulin-dependent regulation of glucose and system A amino acid uptake. J Biol Chem., 2008, 283(41): p. 27653-67.
[25] Submission for marketing application to FDA, APPLICATION NUMBER, 21-884.
[26] Garcia-Fernandez M. et al. Low doses of insulin-like growth factor I improve insulin resistance, lipid metabolism, and oxidative damage in aging rats. Endocrinology, 2008, 149(5): p. 2433-42.
[27] Puche J.E. et al. Low doses of insulin-like growth factor-I induce mitochondrial protection in aging rats. Endocrinology, 2008, 149(5): p. 2620-7.
[28] Joseph D’Ercole A. and P. Ye. Expanding the mind: insulin-like growth factor I and brain development. Endocrinology, 2008, 149(12): p. 5958-62.
[29] Abuzzahab M.J. et al. IGF-I receptor mutations resulting in intrauterine and postnatal growth retardation. N Engl J Med., 2003, 349(23): p. 2211-22.
[30] Woods K.A. et al. Intrauterine growth retardation and postnatal growth failure associated with deletion of the insulin-like growth factor I gene. N Engl J Med., 1996, 335(18): p. 1363-7.
[31] Perez R. et al. Mitochondrial protection by low doses of insulin-like growth factor-I in experimental cirrhosis. World J Gastroenterol., 2008, 14(17): p. 2731-9.
[32] Kang B.P. et al. IGF-I inhibits the mitochondrial apoptosis program in mesangial cells exposed to high glucose. Am J Physiol Renal Physiol., 2003, 285(5): p. F1013-24.
[33] Bhaskar V. et al. A fully human, allosteric monoclonal antibody that activates the insulin receptor and improves glycemic control. Diabetes, 2012, 61(5): p. 1263-71.
[34] Xiong L. et al. Growth-stimulatory monoclonal antibodies against human insulin-like growth factor I receptor. Proc Natl Acad Sci USA, 1992, 89(12): p. 5356-60.
[35] Runnels H.A. et al. Human monoclonal antibodies to the insulin-like growth factor 1 receptor inhibit receptor activation and tumor growth in preclinical studies. Adv Ther., 2010, 27(7): p. 458-75.
[36] Soos M.A. et al. A panel of monoclonal antibodies for the type I insulin-like growth factor receptor. Epitope mapping, effects on ligand binding, and biological activity. J Biol Chem, 1992, 267(18): p. 12955-63.
[37] Kato H. et al. Role of tyrosine kinase activity in signal transduction by the insulin-like growth factor-I (IGF-I) receptor. Characterization of kinase-deficient IGF-I receptors and the action of an IGF-I-mimetic antibody (alpha IR-3). J Biol Chem., 1993, 268(4): p. 2655-61.
[38] Atzori F. et al. A Phase I Pharmacokinetic and Pharmacodynamic Study of Dalotuzumab (MK-0646), an Anti-Insulin-like Growth Factor-1 Receptor Monoclonal Antibody, in Patients with Advanced Solid Tumors. Clin Cancer Res., 2011, 17(19): p.6304-12.
[39] de Bono J.S. et al. Phase II randomized study of figitumumab plus docetaxel and docetaxel alone with crossover for metastatic castration-resistant prostate cancer. Clin Cancer Res., 2014, 20(7): p. 1925-34.
Cущность изобретения
Задача, решаемая изобретением
Целью настоящего изобретения является получение такого антитела против рецептора IGF-I либо его фрагмента или производного, которое специфически связываются с рецептором IGF-I позвоночных. Другой целью настоящего изобретения является получение такого антитела, которое повышает мышечную массу или толщину хряща ростовой пластинки через рецептор IGF-I, не снижая при этом уровень глюкозы в крови.
Средства для решения задачи
Настоящее изобретение касается следующих аспектов.
Аспект [1] Антитело против рецептора IGF-I либо его фрагмент или производное, которое специфически связывается с рецептором IGF-I позвоночных и проявляет активность усиления роста клеток от позвоночных.
Аспект [2] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 1, при этом активность антитела, фрагмента или производного в усилении роста клеток от позвоночных равна или выше, чем соответствующая активность IGF-I дикого типа.
Аспект [3] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 1 или 2, при этом значение EC50 антитела, фрагмента или производного для усиления роста клеток от позвоночных in vitro составляет 1/20 или меньше от соответствующего значения IGF-I дикого типа.
Аспект [4] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-3, при этом при контактировании антитела, фрагмента или производного с культивируемыми клетками от позвоночных продолжительность действия антитела, фрагмента или производного по усилению роста культивируемых клеток относительно продолжительности контакта лучше, чем у IGF-I дикого типа.
Аспект [5] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 2-4, при этом IGF-I дикого типа представляет собой IGF-I человека, имеющий аминокислотную последовательность, приведенную в SEQ ID NO:1.
Аспект [6] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-5, при этом значение EC50 антитела, фрагмента или производного для усиления роста клеток от позвоночных in vitro составляет 0,1 нМ или меньше.
Аспект [7] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-6, которое проявляет активность, индуцирующую увеличение мышечной массы и/или длины тела позвоночных при парентеральном введении позвоночным.
Аспект [8] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-7, которое вводится позвоночным с частотой раз в неделю или реже.
Аспект [9] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-8, при этом позвоночные представлены человеком; другими животными, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак или птиц; или же другими животными, подвергнутыми инженерии для экспрессии рецептора IGF-I человека.
Аспект [10] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-9, которое не индуцирует поглощения глюкозы дифференцированными мышечными клетками при введении в дозе, достаточной для индуцирования роста клеток от позвоночных.
Аспект [11] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 10, которое не индуцирует поглощения глюкозы дифференцированными мышечными клетками при введении в дозе, в 100 или больше раз превышающей значение EC50 для индуцирования роста клеток от позвоночных in vitro.
Аспект [12] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 10 или 11, при этом клетки от позвоночных представляют собой миобласты, полученные от человека или другого млекопитающего.
Аспект [13] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 7-12, которое не снижает уровень глюкозы в крови позвоночных при парентеральном введении позвоночным в дозе, достаточной для индуцирования увеличения мышечной массы и/или длины тела у позвоночных.
Аспект [14] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 13, которое не изменяет уровень глюкозы в крови позвоночных при парентеральном введении позвоночным в дозе, которая в 10 или больше раз превышает эффективную дозу, достаточную для индуцирования увеличения мышечной массы и/или длины тела у позвоночных.
Аспект [15] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-14, которое связывается с доменом CR рецептора IGF-I.
Аспект [16] Антитело против рецептора IGF-I либо его фрагмент или производное, которое связывается с доменом CR рецептора IGF-I и ингибирует связывание IGF-I или IGF-II с рецептором IGF-I.
Аспект [17] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 16, которое связывается с эпитопом, содержащим ProSerGlyPheIleArgAsnX1X2GlnSerMet (где X1 означает Gly или Ser, а X2 означает Ser или Thr) или его часть по соседству в последовательности домена CR рецептора IGF-I.
Аспект [18] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 17, которое связывается с эпитопом, содержащим ProSerGlyPheIleArgAsnGlySerGlnSerMet или его часть по соседству в последовательности домена CR рецептора IGF-I.
Аспект [19] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-18, которое обладает перекрестной реактивностью с рецептором IGF-I человека или других животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак или птиц.
Аспект [20] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-19, которое вызывает реакцию антиген-антитело со сродством по равновесной константе диссоциации (KD) 1×10−8 M или меньше.
Аспект [21] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 16-20, которое обладает по меньшей мере одним из следующих признаков:
1) проявляет активность, вызывающую рост клеток от позвоночных;
2) проявляет активность, вызывающую увеличение мышечной массы и/или длины тела позвоночных при парентеральном введении позвоночным;
3) не вызывает поглощения глюкозы дифференцированными мышечными клетками при введении в дозе, достаточной для того, чтобы вызвать рост клеток от позвоночных; и
4) не изменяет уровень глюкозы в крови позвоночных при парентеральном введении позвоночным в дозе, достаточной для того, чтобы вызвать увеличение мышечной массы и/или длины тела у позвоночных.
Аспект [22] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 16-21, которое обладает по меньшей мере одним из следующих признаков:
1) ингибирует рост клеток от позвоночных, вызванный IGF-I;
2) ингибирует индуцированную IGF-I пролиферацию клеток у позвоночных, страдающих связанным с пролиферацией клеток заболеванием, при парентеральном введении позвоночным;
3) не влияет на поглощение глюкозы дифференцированными мышечными клетками в дозе, достаточной для ингибирования роста клеток от позвоночных, индуцированного IGF-I; и
4) не изменяет уровень глюкозы в крови позвоночных, страдающих связанным с пролиферацией клеток заболеванием, при парентеральном введении позвоночным в дозе, достаточной для ингибирования индуцированной IGF-I пролиферации клеток у позвоночных.
Аспект [23] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-22, которое представляет собой Fab, scFv, диатело или биспецифичное антитело либо его производное.
Аспект [24] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-23, состоящее из аминокислотной последовательности, включающей:
в качестве последовательности CDR-1 вариабельной области тяжелой цепи (CDR-H1) – аминокислотную последовательность, приведенную в SEQ ID NO: 3, или аминокислотную последовательность, полученную из SEQ ID NO: 3 путем замены, делеции или вставки любого аминокислотного остатка;
в качестве последовательности CDR-2 вариабельной области тяжелой цепи (CDR-H2) – аминокислотную последовательность, приведенную в SEQ ID NO: 4, или аминокислотную последовательность, полученную из SEQ ID NO: 4 путем замены, делеции или вставки одного или двух аминокислотных остатков;
в качестве последовательности CDR-3 вариабельной области тяжелой цепи (CDR-H3) – аминокислотную последовательность, приведенную в SEQ ID NO: 5, или аминокислотную последовательность, полученную из SEQ ID NO: 5 путем замены, делеции или вставки одного или двух аминокислотных остатков;
в качестве последовательности CDR-1 вариабельной области легкой цепи (CDR-L1) – аминокислотную последовательность, приведенную в SEQ ID NO: 6, или аминокислотную последовательность, полученную из SEQ ID NO: 6 путем замены, делеции или вставки одного или двух аминокислотных остатков;
в качестве последовательности CDR-2 вариабельной области легкой цепи (CDR-L2) – аминокислотную последовательность, приведенную в SEQ ID NO: 7, или аминокислотную последовательность, полученную из SEQ ID NO: 7 путем замены, делеции или вставки любого аминокислотного остатка;
в качестве последовательности CDR-3 вариабельной области легкой цепи (CDR-L3) – аминокислотную последовательность, приведенную в SEQ ID NO: 8, или аминокислотную последовательность, полученную из SEQ ID NO: 8 путем замены, делеции или вставки одного или двух аминокислотных остатков;
Аспект [25] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 24, дополнительно содержащее каркасную последовательность иммуноглобулина.
Аспект [26] Антитело против рецептора IGF-I либо его фрагмент или производное по аспекту 25, при этом каркасная последовательность иммуноглобулина представляет собой каркасную последовательность каждого класса иммуноглобулина человека или других животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак, птиц, мышей или крыс.
Аспект [27] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-26, состоящее из аминокислотной последовательности, включающей:
в качестве вариабельной области тяжелой цепи – аминокислотную последовательность, приведенную в SEQ ID NO: 9, или аминокислотную последовательность, имеющую сходство на 90% или больше с SEQ ID NO: 9; и
в качестве вариабельной области легкой цепи – аминокислотную последовательность, приведенную в SEQ ID NO: 10, или аминокислотную последовательность, имеющую сходство на 90% или больше с SEQ ID NO: 10.
Аспект [28] Антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-27, дополнительно содержащее константную область каждого класса иммуноглобулина человека или других животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак, птиц, мышей или крыс.
Аспект [29] Молекула нуклеиновой кислоты, состоящая из последовательности полинуклеотида, кодирующего антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-28.
Аспект [30] Клонирующий вектор или экспрессирующий вектор, содержащий по меньшей мере одну молекулу нуклеиновой кислоты по аспекту 29.
Аспект [31] Рекомбинантная клетка, полученная из клетки хозяина путем трансфекции вектором по аспекту 30.
Аспект [32] Способ получения антитела против рецептора IGF-I либо его фрагмента или производного по любому из аспектов 1-28, включающий:
культивирование рекомбинантной клетки по аспекту 31; и
очистку антитела против рецептора IGF-I, его фрагмента или производного, полученного из рекомбинантной клетки.
Аспект [33] Фармацевтическая композиция, содержащая антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-28, молекулы нуклеиновой кислоты по аспекту 29, вектор по аспекту 30 или рекомбинантные клетки по аспекту 31.
Аспект [34] Фармацевтическая композиция по аспекту 33, также содержащая дополнительный активный ингредиент, отличный от антитела, его фрагмента или производного по любому из аспектов 1-28, молекул нуклеиновой кислоты по аспекту 29, вектора по аспекту 30 или рекомбинантных клеток по аспекту 31.
Аспект [35] Фармацевтическая композиция по аспекту 34, при этом активным ингредиентом является одно или несколько из числа гормона роста или его аналогов, инсулина или его аналогов, IGF-II или его аналогов, антител против миостатина, антагонистов миостатина, антител против рецептора активина типа IIB, антагонистов рецептора активина типа IIB, растворимого рецептора активина типа IIB или его аналогов, грелина или его аналогов, фоллистатина или его аналогов, β2-агонистов и селективных модуляторов андрогеновых рецепторов.
Аспект [36] Фармацевтическая композиция по аспекту 34 или 35, при этом активный ингредиент включает ингредиент, выбранный из группы, состоящей из: кортикостероидов, противорвотных средств, ондансетрона гидрохлорида, гранисетрона гидрохлорида, метоклопрамида, домперидона, галоперидола, циклизина, лоразепама, прохлорперазина, дексаметазона, левомепромазина, трописетрона, противораковой вакцины, ингибиторов GM-CSF, ДНК-вакцины против GM-CSF, клеточной вакцины, вакцины против дендритных клеток, рекомбинантной вирусной вакцины, вакцины против белка теплового шока (HSP), гомологичной противоопухолевой вакцины, аутологичной противоопухолевой вакцины, анальгетиков, ибупрофена, напроксена, трисалицилат холина магния, оксикодона гидрохлорида, антиангиогенных, антитромботических средств, антител против PD-1, ниволумаба, пембролизумаба, антител против PD-L1, атезолизумаба, антител против CTLA4, ипилимумаба, антител против CD20, ритуксимаба, антител против HER2, трастузумаба, антител против CCR4, могамулизумаба, антител против VEGF, бевацизумаба, антител против рецептора VEGF, растворимого фрагмента рецептора VEGF, антител против TWEAK, антител против рецептора TWEAK, растворимого фрагмента рецептора TWEAK, AMG 706, AMG 386, антипролиферативных средств, ингибиторов фарнезил-протеинтрансферазы, ингибиторов αVβ3, ингибиторов αVβ5, ингибиторов р53, ингибиторов Kit-рецептора, ингибиторов RET-рецептора, ингибиторов PDGFR, ингибиторов секреции гормона рос-
та, ингибиторов ангиопоэтина, ингибиторов инфильтрирующих опухоли макрофагов, ингибиторов c-FMS, антител против c-FMS, ингибиторов CSF-1, антител против CSF-1, растворимого фрагмента c-FMS, пегвисоманта, гемцитабина, панитумумаба, иринотекана и SN-38.
Аспект [37] Лекарственный препарат для применения при лечении или профилактике состояния, связанного с IGF-I или IGF-II, содержащий антитело против рецептора IGF-I либо его фрагмент или производное по любому из аспектов 1-28, молекулы нуклеиновой кислоты по аспекту 29, вектор по аспекту 30 или рекомбинантные клетки по аспекту 31.
Аспект [38] Лекарственный препарат по аспекту 37, при этом состояние, связанное с IGF-I, выбрано из числа мышечной атрофии от бездействия, карликовости, диабетической нефропатии, хронической почечной недостаточности, синдрома Ларона, цирроза печени, фиброза печени, старения, ограничения внутриутробного развития (IUGR), неврологических заболеваний, инсульта, травмы спинного мозга, защиты сердечно-сосудистой системы, инсулинорезистентного диабета, метаболического синдрома, остеопороза, муковисцидоза, заживления ран, миотонической дистрофии, связанной со СПИД саркопении, связанного с ВИЧ синдрома перераспределения жира, ожогов, болезни Крона, синдрома Вернера, сцепленного с Х-хромосомой комбинированного иммунодефицита, потери слуха, нервной анорексии и ретинопатии недоношенных, синдрома Тернера, синдрома Прадера-Вилли, синдрома Сильвера-Рассела, идиопатической карликовости, ожирения, рассеянного склероза, фибромиалгии, язвенного колита, низкой мышечной массы, ишемии миокарда и снижения плотности костей.
Аспект [39] Лекарственный препарат по аспекту 37 или 38, который вводится парентерально.
Аспект [40] Лекарственный препарат по любому из аспектов 37-39, который представляет собой ветеринарный препарат для введения животным, а не людям.
Аспект [41] Лекарственный препарат по аспекту 40, при этом ветеринарный препарат вводится с целью повышения мышечной массы и/или длины тела, усиления роста, повышения продукции молока, стимулирования размножения или предотвращения старения.
Аспект [42] Лекарственный препарат по аспекту 40 или 41, при этом животные представляют собой морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак или птиц;
Аспект [43] Лекарственный препарат по любому из аспектов 37-42 для лечения или профилактики заболевания, вызванного действием IGF-I или IGF-II на рецептор IGF-I.
Аспект [44] Лекарственный препарат по аспекту 43, при этом заболевание, вызванное действием IGF-I или IGF-II на рецептор IGF-I, выбрано из группы, состоящей из: рака печени, нейробластомы, саркомы поперечно-полосатых мышц, рака костей, детского рака, акромегалии, рака яичников, рака поджелудочной железы, доброкачественной гипертрофии предстательной железы, рака молочной железы, рака простаты, рака костей, рака легких, колоректального рака, рака шейки матки, синовиальной саркомы, рака мочевого пузыря, рака желудка, опухоли Вильмса, диареи, связанной с метастатическими карциноидными и секретирующими вазоактивный кишечный пептид опухолями, випомы, синдрома Вернера-Моррисона, синдрома Беквита-Видеманна, рака почек, почечно-клеточного рака, переходно-клеточного рака, саркомы Юинга, лейкемии, острой лимфобластной лейкемии, опухоли головного мозга, глиобластомы, неглиобластомной опухоли головного мозга, менингиомы, аденомы гипофиза, вестибулярной шванномы, примитивной нейроэктодермальной опухоли, медуллобластомы, астроцитомы, олигодендроглиомы, эпендимомы, папилломы хороидного сплетения, гигантизма, псориаза, атеросклероза, рестеноза гладких мышц сосудов, неадекватного роста микрососудов, диабетической ретинопатии, болезниГрейвса, рассеянного склероза, системной красной волчанки, хронического тиреоидита, миастении, аутоиммунного тиреоидита и болезни Бехчета.
Аспект [45] Способ культивирования клеток от позвоночных in vitro, включающий контактирование клеток от позвоночных с антителом против рецептора IGF-I либо его фрагментом или производным по любому из аспектов 1-28, молекулой нуклеиновой кислоты по аспекту 29, вектором по аспекту 30 или рекомбинантными клетками по аспекту 31 при культивировании клеток.
Аспект [46] Способ по аспекту 45, при этом контактирование проводится с целью усиления роста или индуцирования дифференцировки клеток от позвоночных.
Аспект [47] Способ по аспекту 45 или 46, при этом антитело против рецептора IGF-I, его фрагмент или производное адсорбировано или иммобилизовано на твердой фазе.
Аспект [48] Трансгенное животное, содержащее ген рецептора IGF-I, который подвергался мутации в своем домене CR посредством генной инженерии с тем, чтобы домен CR включал в себя аминокислотную последовательность, представленную ProSerGlyPheIleArgAsnGlySerGlnSerMet.
Аспект [49] Трансгенное животное, в которое был трансфецирован гетерологичный ген рецептора IGF-I, причем аминокислотная последовательность, кодируемая гетерологичным геном рецептора IGF-I, отличается от аминокислотной последовательности, кодируемой собственным геном рецептора IGF-I животного, по аминокислотным остаткам X1 и/или X2 в последовательности, представленной ProSerGlyPheIleArgAsnX1X2GlnSerMet в домене CR.
Действие изобретения
Антитело против рецептора IGF-I либо его фрагменты или производные по настоящему изобретению обладают эффектом специфического связывания с рецептором IGF-I позвоночных.
Краткое описание фигур
На фиг. 1 представлено выравнивание аминокислотных последовательностей домена CR у рецепторов IGF-I мыши, крысы, человека, морской свинки и кролика, причем аминокислотные последовательности приведены с использованием однобуквенного кода.
На фиг. 2 представлен график, показывающий результаты ELISA с использованием вариантов предположительного эпитопа IGF11-16.
На фиг. 3 представлен график, показывающий активность роста миобластов человека после удаления IGF11-16 и IGF-I.
На фиг. 4 представлен график, показывающий эффект поглощения глюкозы дифференцированными мышечными клетками человека после добавления IGF-I и IGF11-16.
На фиг. 5 представлен график, показывающий вес мышц длинных разгибателей пальцев (extensor digitorum longus) у морских свинок, получавших продолжительное введение IGF-I с помощью осмотического насоса в течение двух недель либо однократное подкожное или внутривенное введение дозы IGF11-16 за две недели до этого.
На фиг. 6 представлен график, показывающий временную зависимость уровня глюкозы в крови морских свинок натощак после однократного подкожного введения дозы IGF-I.
На фиг. 7 представлен график, показывающий временную зависимость уровня глюкозы в крови морских свинок натощак после однократного подкожного введения дозы IGF11-16.
На фиг. 8 представлен график, показывающий временную зависимость уровня глюкозы в крови морских свинок натощак после однократного внутривенного введения дозы IGF11-16.
На фиг. 9 представлен график, показывающий влияние IGF11-16 на увеличение толщины хряща ростовой пластинки у гипофизэктомированных морских свинок (HPX).
На фиг. 10 представлен график, показывающий влияние IGF11-16 на увеличение длины большеберцовой кости у гипофизэктомированных морских свинок (HPX).
На фиг. 11 представлен график, показывающий кинетику IGF-I в крови морских свинок натощак после однократного подкожного введения дозы.
На фиг. 12 представлен график, показывающий кинетику IGF11-16 в крови морских свинок натощак после однократного подкожного введения дозы.
Осуществление изобретения
В последующем описании настоящее изобретение будет раскрываться со ссылкой на конкретные воплощения, хотя настоящее изобретение никоим образом не должно ограничиваться этими воплощениями. Все документы, цитируемые в настоящем описании, включая патентные публикации, публикации нерассмотренных заявок и непатентные ссылки, тем самым включены сюда путем ссылки во всей полноте для всех целей.
IGF
IGF означает инсулиноподобный фактор роста, который может быть представлен IGF-I или IGF-II. IGF-I и IGF-II – биологические лиганды с активностью агонистов, которые связываются с рецептором IGF-I (рецептором инсулиноподобного фактора роста-I) и передают сигналы типа деления клеток и метаболизма внутрь клетки. IGF-I и IGF-II также обладают активностью перекрестного связывания с рецептором инсулина (INSR), который структурно близок рецептору IGF-I. В настоящем описании будет обсуждаться в основном IGF-I, так как его свойства типа физиологических функций известны лучше, чем таковые IGF-II. Однако при обсуждении различных эффектов и заболеваний, опосредованных связыванием лигандов с рецептором IGF-I, могут собирательно упоминаться и IGF-I, и IGF-II.
IGF-I, который также именуется соматомедином C, представляет собой гормон из одного полипептида, состоящего из 70 аминокислот. Последовательность IGF-I человека доступна, напр., в EMBL-EBI с номером доступа P50919 в UniProtKB. Аминокислотная последовательность зрелого IGF-I приведена в SEQ ID NO: 1 из прилагаемого перечня последовательностей. Эта последовательность из 70 аминокислот консервативна у многих видов. В настоящем изобретении термин “IGF-I” без каких-либо ограничений означает белок IGF-I, обладающий активностью такого гормона, если не указано иначе.
IGF-I вырабатывается различными клетками в живом организме, включая клетки печени, и существует в крови и других жидкостях организма. Следовательно, IGF-I дикого типа может быть получен путем очистки из жидкой среды организма животных или из первичной культуры клеток или культуры линии клеток, происходящих из животных. Поскольку гормон роста индуцирует выработку IGF-I клетками, IGF-I также можно выделить из жидкой среды организма животных, которым вводили гормон роста, или же из первичной культуры клеток животных или линии клеток животных, проинкубированных в присутствии гормона роста. В качестве другого метода IGF-I также может быть получен из рекомбинантных клеток, полученных при трансфекции экспрессирующим вектором, несущим молекулы нуклеиновой кислоты, кодирующей аминокислотную последовательность IGF-I, в организме хозяина типа прокариотического организма (напр., E. coli) или эукариотических клеток, включая дрожжевые, клетки насекомых или культуры клеток млекопитающих, или же из трансгенных животных или трансгенных растений, в которые был трансфецирован ген IGF-I. IGF-I человека также доступен в виде реагента для исследований (Enzo Life Sciences, кат. № ADI-908-059-0100, Abnova, кат. № P3452 и др.) или же в виде фармацевтического препарата (мекасермин Somazon®, Increlex® и др.). Активность IGF-I для применения in vivo и in vitro можно оценить в виде удельной активности относительно стандартной субстанции IGF-I с кодом NIBSC 91/554, активность которой соответствует 1 международной единице на мкг. Стандартную субстанцию можно получить из Национального института биологических стандартов и контроля (NIBSC) Всемирной организации здравоохранения. В контексте настоящего изобретения считается, что IGF-I обладает удельной активностью, эквивалентной IGF-I с кодом NIBSC 91/554.
Рецептор IGF-I
Термин “рецептор IGF-I” относится к рецептору инсулиноподобного фактора роста -I. При этом термин “рецептор IGF-I” означает белок рецептора IGF-I, если не указано иначе. Рецептор IGF-I представляет собой белок, образованный двумя субъединицами, каждая из которых состоит из α-цепи и β-цепи. Аминокислотная последовательность рецептора IGF-I человека приведена в SEQ ID NO: 2, в которой подпоследовательность, состоящая из аминокислотных остатков 31-735, представляет α-цепь, а подпоследовательность, начинающаяся с аминокислотного остатка 740, представляет β-цепь. α-Цепь рецептора IGF-I содержит участок, с которым связывается IGF-I, тогда как β-цепь имеет трансмембранную структуру и обладает функцией передачи сигналов внутрь клетки. α-Цепь рецептора IGF-I подразделяется на домены L1, CR, L2, FnIII-1 и FnIII-2a/ID/FnIII-2b. В соответствии с аминокислотной последовательностью рецептора IGF-I человека, приведенной в SEQ ID NO: 2, домену L1 соответствуют остатки 31-179, домену CR соответствуют остатки 180-32, домену L2 соответствуют остатки 329-491, домену FnIII-1 соответствуют остатки 492-607, а домену FnIII-2a/ID/FnIII-2b соответствуют остатки 608-735. Из них домен CR (богатый цистеином) участвует в активации внутриклеточной тирозинкиназы в β-цепи, которая связана с конформационным изменением рецептора IGF-I, происходящим при связывании IGF-I с рецептором. Аминокислотная последовательность рецептора IGF-I человека доступна, напр., в EMBL-EBI с номером доступа UniProtKB P08069, а также приведена в перечне последовательностей как SEQ ID NO: 2.
Известно, что рецептор IGF-I экспрессируется в широком спектре тканей и клеток в живом организме и получает различные стимулы через IGF-I типа индукции пролиферации клеток и активации внутриклеточных сигналов. В частности, эффекты IGF-I на миобласты через рецептор IGF-I можно оценить по активности пролиферации клеток в качестве индикатора. По этой причине миобласты полезны при анализе эффектов связывания антител с рецептором IGF-I. Клетки, экспрессирующие рецептор IGF-I, полученный из человека или любых других позвоночных, могут быть получены искусственно путем трансфекции экспрессирующего вектора, несущего молекулу нуклеиновой кислоты, кодирующей аминокислотную последовательность рецептора IGF-I человека или какого-либо другого позвоночного, в эукариотические клетки хозяина типа культуры клеток насекомых или клеток млекопитающих, получая рекомбинантные клетки, экспрессирующие кодируемый трансфецированной нуклеиновой кислотой рецептор IGF-I на своей клеточной мембране. Полученные клетки, экспрессирующие рецептор IGF-I, можно использовать для анализа способности к связыванию и передачи внутриклеточных сигналов антител.
Антитела против рецептора IGF-I
Антитело представляет собой гликопротеин, содержащий по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, соединенные вместе дисульфидными связями. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно VH) и константную область тяжелой цепи. Константная область тяжелой цепи содержит три домена, а именно CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельную область легкой цепи (сокращенно VL) и константную область легкой цепи. Константная область легкой цепи содержит один домен, т.е. CL. Существует два типа константных областей легкой цепи, а именно цепь λ (лямбда) и цепь κ (каппа). Константные области тяжелой цепи подразделяются на цепь γ (гамма), цепь μ (мю), цепь α (альфа), цепь δ (дельта) и цепь ε (эпсилон), а различные типы константной области тяжелой цепи дают различные изотипы антител, т.е. IgG, IgM, IgA, IgD и IgE, соответственно. Каждая из областей VH и VL также подразделяется на четыре относительно консервативных участка (FR-1, FR-2, FR-3 и FR-4), совокупно именуемые каркасными участками (FR), и три очень вариабельных участка (CDR-1, CDR-2 и CDR-3), совокупно именуемые участками, определяющими комплементарность (CDR). Область VH включает в себя три CDR и четыре FR, расположенные в следующем порядке: FR-1, CDR-1 (CDR-H1), FR-2, CDR-2 (CDR-H2), FR-3, CDR-3 (CDR-H3) и FR-4 от N-конца к C-концу. VL включает в себя три CDR и четыре FR, расположенные в порядке FR-1, CDR-1 (CDR-L1), FR-2, CDR-2 (CDR-L2), FR-3, CDR-3 (CDR -L3) и FR-4 от N-конца к C-концу. Вариабельная область каждой тяжелой цепи и легкой цепи включает в себя домен связывания, который взаимодействует с антигеном.
Антитела по настоящему изобретению могут представлять собой фрагменты и/или производные антител. Примеры фрагментов антител включают F(ab′)2, Fab и Fv. Примеры производных антител включают антитела, в которые была введена аминокислотная мутация в константной области; антитела, в которых было модифицировано расположение доменов константных областей; антитела, содержащие два или больше Fc на 1 молекулу; антитела, состоящие только из тяжелой цепи либо только из легкой цепи; антитела с модифицированным гликозилированием; биспецифичные антитела; конъюгаты антител или фрагментов антител с соединениями или белками, отличными от антител; антитела-ферменты; нанотела; тандемные scFv; биспецифичные тандемные scFv; диатела; и VHHs. В настоящем изобретении термин “антитело” охватывает такие фрагменты и/или производные антител, если не указано иначе.
Термин “моноклональное антитело” обычно означает молекулы антитела, полученные из клона, происходящего из одной вырабатывающей антитело клетки, то есть одну разновидность молекул антитела, содержащих комбинацию VH и VL с определенными аминокислотными последовательностями. Моноклональное антитело также можно получить методами генной инженерии, получая молекулу нуклеиновой кислоты, имеющую последовательность гена, кодирующего аминокислотную последовательность белка моноклонального антитела. Специалистам в данной области также должны быть известны методики модификации моноклональных антител с использованием генетической информации, напр., об H-цепях, L-цепях, их вариабельных областях и последовательностях их CDR с тем, чтобы улучшить способность к связыванию и специфичность антител, а также способы получения антител, подходящих для терапевтических средств, путем преобразования антител животных типа антител мыши в человеческие антитела. Моноклональные антитела человеческого типа также можно получить путем сенсибилизации трансгенного животного (не человека), несущего ген антитела человека, к антигену. Другой способ, не требующий сенсибилизации животных, который включает: приготовление фаговой библиотеки, экспрессирующей антигенсвязывающий участок человеческого антитела или его часть (фаговый дисплей человеческого антитела); получение фагового клона, экспрессирующего пептид, который специфически связывается с соответствующим антигеном или антителом, имеющим нужную аминокислотную последовательность; и получение требуемого человеческого антитела на основе информации о выбранном фаговом клоне. Специалисты в данной области могут использовать такую методику по обстоятельствам (напр., см. обзор Taketo Tanaka et al., Keio J. Med., Vol. 60, pp. 37-46). Специалисты в данной области также могут разработать антитела для введения животным таким же способом, как и гуманизованные антитела, используя информацию об аминокислотных последовательностях участков CDR и вариабельных областей, как потребуется.
Термин “реакция антиген-антитело” в настоящем изобретении означает, что антитело связывается с рецептором IGF-I со сродством, представленным равновесной константой диссоциации (KD) в 1×10−8 M или меньше. Антитело по настоящему изобретению должно предпочтительно связываться с рецептором IGF-I со значением KD в 1×10−8 M или меньше, предпочтительно 1×10−9 M или меньше, более предпочтительно 1×10−10 M или меньше.
Термин “специфичность” антитела в настоящем изобретении означает, что антитело вызывает прочное связывание на основе реакции антиген-антитело с конкретным антигеном. В контексте настоящего изобретения специфичное к рецептору IGF-I антитело означает такое антитело, которое при использовании в концентрации, достаточной для запуска значительной реакции антиген-антитело с клетками, экспрессирующими рецептор IGF-I, вызывает реакцию антиген-антитело с INSR с иммунореактивностью, которая в 1,5 раза или меньше реактивности с посторонними клетками. INSR имеет большое сходство
с рецептором IGF-I по первичной структуре (аминокислотной последовательности) и по структуре более высокого порядка.
Специалисты в данной области смогут провести измерение реакции антиген-антитело, выбрав подходящий метод связывания в твердофазной системе или в жидкой фазе. Примеры таких методов включают, без ограничения: твердофазный иммуноферментный анализ (ELISA), ферментный иммуноанализ (EIA), поверхностный плазмонный резонанс (SPR), флуоресцентно-резонансный перенос энергии (FRET) и люминесцентно-резонансный перенос энергии (LRET). Измерение сродства связывания антитела с антигеном может проводиться, напр., путем мечения антитела и/или антигена, напр., с помощью фермента, флуоресцентного вещества, люминесцентного вещества или радиоизотопа, и детектирования реакции антиген-антитело методом, подходящим для измерения физических и/или химических свойств, характерных для используемой метки.
Антитела против рецептора IGF-I по настоящему изобретению охватывают как антитела-агонисты, так и антитела-антагонисты. При использовании по отдельности антитела-агонисты рецептора IGF-I по настоящему изобретению обладают эффектом усиления активности роста миобластов. При использовании в комбинации с IGF-I антитела-антагонисты рецептора IGF-I по настоящему изобретению обладают эффектом ингибирования индуцированной IGF-I активности роста миобластов.
Антитела-агонисты рецептора IGF-I по настоящему изобретению, которые прочно связываются со специфическим доменом рецептора IGF-I, обладают эффектом усиления активности роста миобластов in vitro.
Антитела-агонисты рецептора IGF-I по настоящему изобретению не обладают эффектом усиления поглощения глюкозы дифференцированными мышечными клетками in vitro в эффективной концентрации, достаточной для усиления активности роста миобластов, предпочтительно в концентрации, в 10 раз превышающей эффективную концентрацию, более предпочтительно в концентрации, в 100 раз превышающей эффективную концентрацию.
Хотя IGF-I обладает заметным гипогликемическим действием в дозах, достаточных для проявления эффекта увеличения мышечной массы, антитела-агонисты рецептора IGF-I по настоящему изобретению не обладают гипогликемическим действием в эффективных дозах, достаточных для проявления эффекта увеличения мышечной массы, предпочтительно в дозах, в 10 раз превышающих эффективные дозы.
Кроме того, антитела-агонисты рецептора IGF-I при введении морским свинкам в однократной дозе проявляют активность увеличения мышечной массы in vivo, которая соответствует активности, достигаемой при продолжительном введении IGF-I. Антитела-агонисты рецептора IGF-I по настоящему изобретению также имеют длительный период полужизни в крови и проявляют эффект увеличения мышечной массы при однократном введении дозы животным.
Таким образом, антитела-агонисты рецептора IGF-I по настоящему изобретению обладают потенциалом в качестве терапевтических или профилактических средств для многих заболеваний, связанных с рецептором IGF-I, типа мышечной атрофии от бездействия и карликовости, при которых IGF-I также должен быть эффективным. Кроме того, антитела-агонисты рецептора IGF-I по настоящему изобретению могут решить проблемы, связанные с IGF-I, напр., путем преодоления гипогликемического эффекта и увеличения периода полужизни в крови.
Антитела-антагонисты рецептора IGF-I по настоящему изобретению ингибируют связывание IGF-I с рецептром IGF-I. Согласно одному воплощению антител-антагонистов рецептора IGF-I по настоящему изобретению, эти антитела активируют рецептор IGF-I и в то же время ингибируют действие IGF-I на рецептор IGF-I. В этом воплощении антитела обладают эффектом устранения аддитивной агонистической активности с IGF-I, напр., эффектом устранения активности IGF-I по индукции роста миобластов. В другом воплощении антител-антагонистов рецептора IGF-I по настоящему изобретению они связываются, но не активируют рецептор IGF-I. Примеры таких антагонистических антител, которые не вызывают активации рецептора IGF-I посредством поперечной сшивки, включают, без ограничения: антитела с моновалентной антигенсвязывающей способностью типа Fab и scFv; и антитела с бивалентными сайтами связывания типа биспецифичных антител, у которых только один из сайтов связывания связывается со специфичным доменом рецептора IGF-I или же сайты связывания разнесены с контролируемым интервалом с помощью линкера. При получении антител-антагонистов рецептора IGF-I по настоящему изобретению можно проверить, связывается ли антитело с рецептором IGF-I, но не обладает агонистической активностью, путем: определения того, что антитело обладает способностью связываться с рецептором IGF-I методом измерения реактивности антиген-антитело между антителом и рецептором IGF-I; или определения того, что антитело не обладает активностью вызывать пролиферацию клеток с помощью теста на пролиферацию клеток, напр., миобластов. С другой стороны, антитела-антагонисты рецептора IGF-I не влияют на поглощение глюкозы дифференцированными мышечными клетками in vitro или на уровень глюкозы в крови in vivo. Следовательно, антитела-антагонисты рецептора IGF-I по настоящему изобретению обладают потенциалом в качестве терапевтических или профилактических средств без побочных эффектов типа гипергликемии и могут применяться для лечения злокачественных опухолей типа рака молочной железы, рака кишечника, саркомы, рака легких, рака простаты, рака щитовидной железы и миеломы.
Связывающая способность антител против рецептора IGF-I
Антитела против рецептора IGF-I по настоящему изобретению связываются с доменом CR рецептора IGF-I в качестве эпитопа. С другой стороны, антитела-агонисты рецептора IGF-I не обладают способностью связываться с INSR, который имеет большое сходство с рецептором IGF-I по первичной структуре (аминокислотной последовательности) и структуре более высокого порядка.
При связывании с доменом CR рецептора IGF-I считается, что антитела против рецептора IGF-I по настоящему изобретению активируют рецептор гомотипа, который является димером из двух копий рецептора IGF-I, или же рецептор гетеротипа, который является димером между рецептором IGF-I и INSR.
Последовательность антител против рецептора IGF-I
Последовательность антител против рецептора IGF-I по настоящему изобретению не имеет особых ограничений при условии, что они специфически связываются с рецептором IGF-I позвоночных и обладают активностью вызывать пролиферацию клеток.
Однако антитела предпочтительно должны иметь специфические аминокислотные последовательности в качестве последовательностей CDR, как будет подробно объяснено ниже. В контексте настоящего изобретения термин “идентичность” аминокислотных последовательностей означает долю идентичных аминокислотных остатков между последовательностями, тогда как термин “сходство” аминокислотных последовательностей означает долю идентичных или сходных аминокислотных остатков между последовательностями. Сходство и идентичность аминокислотных последовательностей можно определить, напр., методом BLAST (с параметрами PBLAST по умолчанию, предусмотренными NCBI).
Термин “сходные аминокислотные остатки” в настоящем изобретении означает группу аминокислотных остатков с боковыми цепями, имеющими близкие химические свойства (напр., электрический заряд или гидрофобность). Группы сходных аминокислотных остатков включают:
1) аминокислотные остатки с алифатическими боковыми цепями: остатки глицина, аланина, валина, лейцина и изолейцина;
2) аминокислотные остатки с алифатическими гидроксильными боковыми цепями: остатки серина и треонина;
3) аминокислотные остатки с амидсодержащими боковыми цепями: остатки аспарагина и глутамина;
4) аминокислотные остатки с ароматическими боковыми цепями: остатки фенилаланина, тирозина и триптофана;
5) аминокислотные остатки с основными боковыми цепями: остатки лизина, аргинина и гистидина;
6) аминокислотные остатки с кислотными боковыми цепями: остатки аспарагиновой кислоты и глутаминовой кислоты; и
7) аминокислотные остатки с серосодержащими боковыми цепями: остатки цистеина и метионина.
Согласно настоящему изобретению, последовательность CDR-1 вариабельной области тяжелой цепи (CDR-H1) предпочтительно должна составлять аминокислотная последовательность, приведенная в SEQ ID NO: 3 (SerTyrTrpMetHis), либо аминокислотная последовательность, полученная из SEQ ID NO: 3 путем замены, делеции или вставки какого-либо одного аминокислотного остатка. Последовательность CDR-H1 также предпочтительно должна иметь сходство на 80% или больше с SEQ ID NO: 3. В контексте настоящего изобретения, если какой-то аминокислотный остаток (в дальнейшем “первый аминокислотный остаток”) аминокислотной последовательности заменяется на другой аминокислотный остаток (в дальнейшем “второй аминокислотный остаток”), то первый аминокислотный остаток до замены и второй аминокислотный остаток после замены весьма предпочтительно должны быть близкими друг другу по структуре и/или характеристикам.
Последовательность CDR-2 вариабельной области тяжелой цепи (CDR-H2) предпочтительно должна составлять аминокислотная последовательность, приведенная в SEQ ID NO: 4 (GluThrAsnProSerAsnSerValThrAsnTyrAsnGluLysPheLysSer), либо аминокислотная последовательность, полученная из SEQ ID NO: 4 путем замены, делеции или вставки какого-либо одного аминокислотного остатка. Последовательность CDR-H2 также предпочтительно должна иметь сходство на 82% или больше, предпочтительно на 88% или больше, более предпочтительно на 94% или больше с SEQ ID NO: 4.
Последовательность CDR-3 вариабельной области тяжелой цепи (CDR-H3) предпочтительно должна составлять аминокислотная последовательность, приведенная в SEQ ID NO: 5 (GlyArgGlyArgGlyPheAlaTyr), либо аминокислотная последовательность, полученная из SEQ ID NO: 5 путем замены, делеции или вставки какого-либо одного аминокислотного остатка. Последовательность CDR-H3 также предпочтительно должна иметь сходство на 75% или больше, предпочтительно на 87% или больше с SEQ ID NO: 5.
Последовательность CDR-1 вариабельной области легкой цепи (CDR-L1) предпочтительно должна составлять аминокислотная последовательность, приведенная в SEQ ID NO: 6 (ArgAlaSerGlnAsnIleAsnPheTrpLeuSer), либо аминокислотная последовательность, полученная из SEQ ID NO: 6 путем замены, делеции или вставки какого-либо одного аминокислотного остатка. Последовательность CDR-L1 также предпочтительно должна иметь сходство на 81% или больше, предпочтительно на 90% или больше с SEQ ID NO: 6.
Последовательность CDR-2 вариабельной области легкой цепи (CDR-L2) предпочтительно должна составлять аминокислотная последовательность, приведенная в SEQ ID NO: 7 (LysAlaSerAsnLeuHisThr), либо аминокислотная последовательность, полученная из SEQ ID NO: 7 путем замены, делеции или вставки какого-либо одного аминокислотного остатка. Последовательность CDR-L2 также предпочтительно должна иметь сходство на 85% или больше с SEQ ID NO: 7.
Последовательность CDR-3 вариабельной области легкой цепи (CDR-L3) предпочтительно должна составлять аминокислотная последовательность, приведенная в SEQ ID NO: 8 (LeuGlnGlyGlnSerTyrProTyrThr), либо аминокислотная последовательность, полученная из SEQ ID NO: 8 путем замены, делеции или вставки какого-либо одного аминокислотного остатка. Последовательность CDR-L3 также предпочтительно должна иметь сходство на 77% или больше, предпочтительно на 88% или больше с SEQ ID NO: 8.
Еще более предпочтительно антитела против рецептора IGF-I по настоящему изобретению должны иметь комбинацию последовательностей CDR:
в качестве последовательности CDR-H1 – аминокислотная последовательность, приведенная в SEQ ID NO: 3;
в качестве последовательности CDR-H2 – аминокислотная последовательность, приведенная в SEQ ID NO: 4;
в качестве последовательности CDR-H3 – аминокислотная последовательность, приведенная в SEQ ID NO: 5;
в качестве последовательности CDR-L1 – аминокислотная последовательность, приведенная в SEQ ID NO: 6;
в качестве последовательности CDR-L2 – аминокислотная последовательность, приведенная в SEQ ID NO: 7; и
в качестве последовательности CDR-L3 – аминокислотная последовательность, приведенная в SEQ ID NO: 8.
Методы идентификации последовательности каждого из CDR-H1, CDR-H2, CDR-H3, CDR-L1, CDR-L2 и CDR-L3 антител включают: метод Kabat (Kabat et al., Journal of Immunology, 1991, Vol. 147, No. 5, pp. 1709-1719) и метод Chothia (Al-Lazikani et al., Journal of Molecular Biology, 1997, Vol. 273, No. 4, pp. 927-948). Эти методы общеизвестны специалистам в данной области, их краткое изложение доступно, напр., на веб-сайте группы Dr. Andrew C.R. Martin’s Group (http://www.bioinf.org.uk/abs/).
Каркасные последовательности иммуноглобулина для антител по настоящему изобретению предпочтительно должны составлять каркасные последовательности каждого класса иммуноглобулинов позвоночных, более предпочтительно каркасные последовательности каждого класса иммуноглобулина человека или животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак, птиц, мышей или крыс.
Антитела против рецептора IGF-I по настоящему изобретению предпочтительно должны иметь специфические аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи, как описано ниже.
Вариабельная область тяжелой цепи предпочтительно должна иметь аминокислотную последовательность, приведенную в SEQ ID NO: 9, аминокислотную последовательность, полученную из SEQ ID NO: 9 путем замены, делеции или вставки какого-либо одного или двух аминокислотных остатков, или же аминокислотную последовательность, имеющую сходство на 90% или больше с SEQ ID NO: 9. Вариабельная область легкой цепи предпочтительно должна иметь аминокислотную последовательность, приведенную в SEQ ID NO: 10, аминокислотную последовательность, полученную из SEQ ID NO: 10 путем замены, делеции или вставки какого-либо одного или двух аминокислотных остатков, или же аминокислотную последовательность, имеющую сходство на 90% или больше с SEQ ID NO: 10. Антителом против рецептора IGF-I по настоящему изобретению более предпочтительно должно быть IGF11-16, то есть оно должно включать комбинацию SEQ ID NO: 9 в качестве вариабельной области тяжелой цепи и SEQ ID NO: 10 в качестве вариабельной области легкой цепи.
Специалисты в данной области смогут разработать гуманизованные антитела против рецептора IGF-I по настоящему изобретению путем выбора аминокислотных последовательностей CDR и/или вариабельных областей тяжелой цепи и легкой цепи из числа приведенных выше и комбинирования их с аминокислотными последовательностями каркасных участков и/или константных областей тяжелой цепи и легкой цепи человеческих антител надлежащим образом. Аминокислотные последовательности каркасных участков и/или константных областей тяжелой цепи и легкой цепи гуманизованных антител могут быть выбраны, напр., из классов IgG, IgA, IgM, IgE и IgD человека либо их вариантов.
Когда антитело против рецептора IGF-I по настоящему изобретению представляет собой антитело-агонист рецептора IGF-I, то антитело по настоящему изобретению или его антигенсвязывающий фрагмент предпочтительно должны быть из класса IgG человека либо его варианта, более предпочтительно из подкласса IgG4 человека или подкласса IgG1 человека либо его варианта. Согласно одному примеру, стабилизованная константная область IgG4 содержит пролин в положении 241 в шарнирной области по системе нумерации Кабата. Это положение соответствует положению 228 в шарнирной области по системе нумерации EU (Kabat et al., Sequences of Proteins of Immunological Interest, DIANE Publishing, 1992; Edelman et al., Proc. Natl. Acad. Sci. USA, 63, 78-85, 1969). Остаток в этом положении в IgG4 человека обычно представлен серином, тогда как замена серина на пролин может вызвать стабилизацию полученного антитела. Согласно другому примеру, введение мутации N297A в константную область IgG1 служит для минимизации способности к связыванию с Fc-рецептором и/или способности к фиксации комплемента.
Конкурентное связывание
Гуманизованные антитела, которые вызывают конкуренцию за связывание с рецептором IGF-I с гуманизованными антителами против рецептора IGF-I по настоящему изобретению, также входят в рамки настоящего изобретения. Термин “конкурентное связывание” в настоящем изобретении означает такое явление, когда при наличии двух или более моноклональных антител вместе с антигеном связывание одного из антител с антигеном ингибируется при связывании другого антитела с антигеном. Конкурентное связывание обычно измеряют, напр., путем добавления к одному моноклональному антителу при его постоянном количестве (концентрации) другого моноклонального антитела в различных количествах (концентрациях) и определения такого количества (концентрации) последнего моноклонального антитела, при котором снижается уровень связывания первого моноклонального антитела, находящегося при постоянном количестве. Степень ингибирования выражается в единицах IC50 или Ki. Моноклональное антитело, которое вызывает конкуренцию за связывание с гуманизованным антителом против рецептора IGF-I по настоящему изобретению, означает такое антитело, у которого IC50 составляет 1000 нМ или меньше, предпочтительно 100 нМ или меньше, более предпочтительно 10 нМ или меньше при измерении связывания антиген-антитело с использованием гуманизованного антитела против рецептора IGF-I по настоящему изобретению, напр., антитела IGF11-16, при 10 нМ. Измерение конкурентного связывания также может проводиться путем мечения используемого антитела, напр., с помощью фермента, флуоресцентного вещества, люминесцентного вещества, радиоизотопа и т.д., и детектирования метки методом измерения, подходящим для выявления физических и/или химических свойств метки.
Перекрестные реакции
Антитела против рецептора IGF-I по настоящему изобретению предпочтительно должны давать перекрестные реакции с рецептором IGF-I других позвоночных. Термин “перекрестная реакция” означает, что, если антитело вызывает реакцию антиген-антитело с рецептором IGF-I из целевого животного (как-то человека), то антитело также обладает способностью связываться с антигеном, полученным от других животных, отличных от целевого животного. Антитело предпочтительно должно обладать перекрестной реактивностью с рецептором IGF-I животных, отличных от целевого животного, чей рецептор IGF-I является мишенью реакции антиген-антитело у данного антитела, как-то человека или других животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак или птиц. В примере 7 показано, что антитело IGF11-16 против рецептора IGF-I связывается с последовательностью ProSerGlyPheIleArgAsnGlySerGlnSerMet в домене CR рецептора IGF-I человека. Поскольку эта последовательность ProSerGlyPheIleArgAsnGlySerGlnSerMet консервативна в гомологичных частях рецепторов IGF-I обезьян (макаки-крабоеда), кроликов, морских свинок, коров, овец, лошадей и собак, то это антитело обладает способностью к перекрестному связыванию с рецепторами IGF-I из этих видов. Кроме того, поскольку аминокислотные последовательности гомологичных частей у мышей и крыс представлены ProSerGlyPheIleArgAsnSerThrGlnSerMet, то скрининг на антитела против рецептора IGF-I, которые связываются с данной частью, дает возможность получить антитела, которые связываются с рецепторами IGF-I, напр., мышей и крыс, а также имеют характеристики и функции, аналогичные IGF11-16.
С другой стороны, клетки или животные того вида, который не дает перекрестной реакции с антителами против рецептора IGF-I по настоящему изобретению, методами генной инженерии могут быть преобразованы в клетки или животные, экспрессирующие рецептор IGF-I, с которым антитела против рецептора IGF-I по настоящему изобретению дают перекрестную реакцию.
Активность, индуцирующая рост клеток от позвоночных, и активность, индуцирующая увеличение мышечной массы и/или длины тела
Антитела против рецептора IGF-I согласно одному воплощению настоящего изобретения обладают активностью индуцирования роста клеток от позвоночных. Хотя антитела-агонисты рецептора IGF-I уже были известны, однако не было отмечено ни одно антитело, проявляющее активность, индуцирующую рост первичных культур клеток, в частности миобластов. Также пока еще не известно ни одно антитело, проявляющее индуцирующее рост клеток действие при меньших дозах, чем значение ЕС50 IGF-I in vitro. Клетки от позвоночных в контексте настоящего изобретения предпочтительно должны представлять собой клетки, происходящие из млекопитающих, птиц, рептилий, амфибий или рыб, более предпочтительно клетки, происходящие из млекопитающих или птиц, еще более предпочтительно клетки, происходящие из человека, обезьян, кроликов, морских свинок, коров, свиней, овец, лошадей или собак. Клетки, происходящие из тех видов, которые экспрессируют рецептор IGF-I, с которым дают перекрестные реакции антитела против рецептора IGF-I по настоящему изобретению, можно индуцировать для пролиферации антителами против рецептора IGF-I по настоящему изобретению. Клетки от позвоночных по настоящему изобретению также включают клетки и животных, подвергнутых генной инженерии для экспрессии рецептора IGF-I того вида, с которым дают перекрестные реакции антитела против рецептора IGF-I по настоящему изобретению; и модифицированные клетки животных, полученные из таких генно-инженерных клеток и животных.
Активность антител по индуцированию роста клеток от позвоночных можно анализировать in vitro, используя первичные культуры клеток, установленные линии клеток или трансформантов, полученных из таких клеток. Термин “первичная культура клеток” означает клетки, которые были выделены из органа или ткани живого организма и которые обычно можно субкультивировать в течение нескольких пассажей. Первичные культуры клеток от позвоночных могут быть получены из органа или ткани позвоночных путем обработки ферментами, диспергирования физическими средствами или методом эксплантации. Для анализа приведенной выше активности антител также можно использовать органы или ткани либо их фрагменты, полученные от позвоночных. Предпочтительные примеры органов и тканей, из которых получают первичные клетки, включают: эндокринные ткани, как-то щитовидная железа, околощитовидная железа и надпочечники; иммунные ткани, как-то аппендикс, миндалины, лимфатические узлы и селезенка; органы дыхания, как-то трахеи и легкие; пищеварительные органы, как-то желудок, двенадцатиперстная кишка, тонкая кишка и толстая кишка; мочевыделительные органы, как-то почки и мочевой пузырь; мужские половые органы, как-то семявыносящие протоки, яички и предстательная железа; женские половые органы, как-то молочные железы и фаллопиевы трубы; и мышечные ткани, как-то сердечные мышцы и скелетные мышцы. Более предпочтительные примеры включают печень, почки или органы пищеварения или мышечные ткани, из которых наиболее предпочтительны мышечные ткани. Первичные культуры клеток, которые можно использовать для анализа индуцирующей рост активности антител против рецептора IGF-I в контексте настоящего изобретения, – это клетки, которые экспрессируют рецептор IGF-I и могут индуцироваться для пролиферации при связывании IGF-I с рецептором IGF-I. Типичными примерами таковых являются миобласты скелетных мышц, которые представляют собой первичные культуры клеток, выделенных из мышечной ткани. Также можно получить и использовать первичные культуры клеток от человека или животных, доступные по поручению или коммерческим путем на рынке. Первичные культуры клеток человека доступны из различных учреждений и компаний, напр., ATCC®, ECACC, Lonza, Gibco®, Cell Applications, ScienCell Research Laboratories и PromoCell.
Термин “клеточная линия” означает линию культивируемых клеток, которые были получены из живого организма, а затем иммортализованы с тем, чтобы они могли почти перманентно размножаться с сохранением своих специфических свойств. Клеточные линии подразделяются на нераковые линии клеток и раковые линии клеток. Линии клеток от позвоночных, которые можно использовать для анализа индуцирующей рост активности антител против рецептора IGF-I по настоящему изобретению, представляют собой клетки, которые экспрессируют рецептор IGF-I и могут индуцироваться для пролиферации при связывании IGF-I с рецептором IGF-I. Примеры клеточных линий, которые экспрессируют рецептор IGF-I и могут индуцироваться к пролиферации под действием IGF-I, включают, без ограничения: нейробластому SH-SY5Y человека, линию эпидермальных кератиноцитов HaCaT человека, линию клеток альвеолярной базально-эпителиальной аденокарциномы A549 человека, линию клеток Caco-2 аденокарциномы толстой кишки, линию клеток HepG2 гепатоцеллюлярного рака человека, линию раковых клеток Hela шейки матки человека, линию раковых клеток SiHa шейки матки человека, линию раковых клеток MCF7 молочной железы человека, плюрипотентную линию NTERA-2 эмбриональной карциномы человека и линию раковых клеток U-2-OS костей человека.
Другие клетки, которые можно использовать для анализа индуцирующей рост активности антител против рецептора IGF-I по настоящему изобретению, – это трансформанты, происходящие из первичных культур клеток и линий клеток. Примеры таких трансформантов включают: клетки iPS, полученные из первичной культуры клеток; и клетки и ткани, прошедшие дифференцировку от таких клеток iPS. Другие трансформанты включают первичные культуры клеток и линии клеток, подвергнутые генной инженерии для включения генов с тем, чтобы кратковременно или перманентно экспрессировать эти гены. Примеры генов для введения и экспрессирования такими клетками включают гены рецептора IGF-I человека и других видов.
Способы определения способности антител против IGF-I-рецептора по настоящему изобретению индуцировать пролиферацию клеток от позвоночных включают: подсчет клеток, измерение синтеза ДНК и измерение изменений активности метаболических ферментов. Методы подсчета клеток включают методы с использованием пластинок для подсчета клеток крови или устройств для подсчета клеток типа счетчика Коултера. Методы измерения синтеза ДНК включают методы на основе поглощения [3H]-тимидина или 5-бром-2′-дезоксиуридина (BrdU). Методы измерения изменений активности метаболических ферментов включают метод МТТ, метод XTT и метод WST. Специалисты в данной области также могут использовать и другие методы, если потребуется. Активность по индуцированию пролиферации клеток обнаруживаются, если рост культуры клеток после реакции с антителом против рецептора IGF-I по настоящему изобретению будет выше, чем рост клеток в отсутствие антитела. При этом удобно также измерять индуцирующую активность в тех же условиях с использованием IGF-I, который является исходным лигандом рецептора IGF-I, в качестве контроля.
Культуры клеток для тестирования подвергают реакции либо с антителом против рецептора IGF-I по настоящему изобретению, либо с IGF-I при различных концентрациях, а концентрацию, при которой проявляется 50% от максимальной активности роста, принимают за значение EC50. При использовании миобластов скелетных мышц человека для оценки активности роста значение EC50 у антител против рецептора IGF-I по настоящему изобретению для запуска пролиферации клеток предпочтительно должно быть сравнимо с таковым у IGF-I, более предпочтительно оно составляет 1/10 или меньше от такового у IGF-I, еще более предпочтительно 1/20 или меньше и еще более предпочтительно 1/50 или меньше от такового у IGF-I. При использовании миобластов скелетных мышц человека для оценки активности роста значение EC50 у антител против рецептора IGF-I по настоящему изобретению предпочтительно должно составлять 0,5 нМ или меньше, более предпочтительно 0,3 нМ или меньше и еще более предпочтительно 0,1 нМ или меньше.
Способы измерения активности индуцирования роста клеток от позвоночных in vivo включают: способ, включающий парентеральное введение антитела против рецептора IGF-I по настоящему изобретению позвоночным и измерение изменений массы, размера, количества клеток и т.п. для всего тела индивида, которому его вводили, или для органа или ткани, выделенной из индивида; и способ, включающий использование животного с пересадкой клеток позвоночных и измерение изменений веса, размера, количества клеток и т.п. у трансплантата, содержащего клетки позвоночных. Измерения для всего тела индивида включают: измерения массы тела, длины тела и окружностей четырех конечностей; измерение состава организма методом импеданса; и измерение креатинина и коэффициента роста. Измерения для органа, ткани или трансплантата от индивида включают: в случае животных, но не человека – способ, включающий непосредственное извлечение целевого органа, ткани или трансплантата и измерение его веса, размера или количества содержащихся в нем клеток. Неинвазивные измерения для органа, ткани или трансплантата от индивида включают: анализ изображений по рентгеновским снимкам, КТ и МРТ; и контрастные методы с использованием индикаторов с изотопами или флуоресцентными веществами. Если целевой тканью являются скелетные мышцы, то в качестве показателя также можно использовать изменение силы мышц. Специалисты в данной области также могут использовать и другие подходящие методы для анализа активности антител против рецептора IGF-I по настоящему изобретению по индуцированию роста клеток от позвоночных in vivo. Способы измерения активности антител против рецептора IGF-I по настоящему изобретению по индуцированию роста клеток от позвоночных in vivo включают: проведение измерений с использованием, напр., приведенных выше методов для индивидов, которым вводились антитела против рецептора IGF-I по настоящему изобретению, и индивидов, которым вводили другие антитела, отличные от антител против рецептора IGF-I по настоящему изобретению, или какие-либо другие контрольные вещества, и сравнение полученных измерений между этими индивидами.
Антитела против рецептора IGF-I по настоящему изобретению характеризуются большей продолжительностью индуцирующего рост клеток эффекта относительно времени контакта с клетками по сравнению с продолжительностью у IGF-I дикого типа, тем самым проявляя лучшую устойчивость. Согласно примеру 12, в котором представлена индуцирующая пролиферацию клеток активность in vitro, при контактировании клеток с IGF-I дикого типа, а затем промывании культуральной средой без IGF-I индуцирующая пролиферацию клеток активность IGF-I дикого типа исчезает после промывки. С другой стороны, при контактировании клеток с IGF11-16 (антителом против рецептора IGF-I по настоящему изобретению), а затем промывании культуральной средой без IGF11-16 индуцирующая рост клеток активность сохраняется даже после промывки. Согласно примеру 16, в котором сравнивается кинетика IGF-I и IGF11-16 (антитела против рецептора IGF-I по настоящему изобретению) в крови, 50% или больше IGF-I дикого типа, введенного животным, исчезало из крови в пределах 24 часов после введения, тогда как 60% или больше антител IGF11-16, введенных животным, оставалось в крови даже через 48 часов после введения. Таким образом, было показано, что антитело IGF11-16 остается в крови в течение длительного времени. Эти результаты показывают, что антитела против рецептора IGF-I по настоящему изобретению проявляют долгосрочный эффект индуцирования пролиферации клеток как in vitro, так и in vivo.
Также ожидается, что антитела против рецептора IGF-I по настоящему изобретению проявляют эффект увеличения мышечной массы и/или длины тела in vivo. В частности, IGF-I обладает эффектом индуцирования роста и дифференцировки миобластов в скелетных мышцах, как указано выше, а также эффектом расширения мышечных волокон. Ожидается, что эти эффекты в совокупности приводят к эффекту повышения мышечной массы. Подобно IGF-I, при введении животным антител против рецептора IGF-I по настоящему изобретению они также проявляют эффект увеличения мышечной массы у животных. Антитела против рецептора IGF-I по настоящему изобретению являются первыми антителами-агонистами рецептора IGF-I, которые, как было показано, проявляют эффект увеличения мышечной массы in vivo.
Способы измерения активности антител против рецептора IGF-I по настоящему изобретению по повышению мышечной массы включают: для всего тела индивида, которому проводилось введение – измерение массы тела, длины тела и окружностей четырех конечностей; измерение состава организма методом импеданса; и измерение креатинина и коэффициента роста. Другие методы включают: анализ изображений по рентгеновским снимкам, КТ и МРТ; контрастные методы с использованием индикаторов с изотопами или флуоресцентными веществами; и измерение изменения силы мышц. В случае животных, но не человека, также можно использовать способ, включающий непосредственное извлечение целевого органа, ткани или трансплантата и измерение его веса и/или размера. Эффект увеличения мышечной массы можно оценить путем: сравнения увеличения мышечной массы между индивидом, которому вводилось антитело против рецептора IGF-I по настоящему изобретению, и индивидом, которому антитело не вводилось; или же сравнения мышечной массы индивида до и после введения антитела против рецептора IGF-I по настоящему изобретению. Эффект увеличения мышечной массы выявляется, если имеется какое-либо увеличение мышечной массы индивида до и после введения антитела против рецептора IGF-I по настоящему изобретению. Предпочтительно эффект, достигаемый при введении антител против рецептора IGF-I по настоящему изобретению, выявляется, если существует разница, предпочтительно в 103% или больше, более предпочтительно в 104% или больше мышечной массы между индивидом, которому вводили антитело против рецептора IGF-I по настоящему изобретению, и индивидом, которому не вводили антитело, либо у одного и того же индивида до и после введения антитела против рецептора IGF-I по настоящему изобретению. IGF-I также играет роль в росте костей и обладает эффектом увеличения длины тела (или роста в случае человека). Таким образом, антитела против рецептора IGF-I по настоящему изобретению также проявляют эффект увеличения длины тела при введении животным. Эффект антител против рецептора IGF-I по настоящему изобретению в увеличении длины тела индивида можно определить путем измерения массы тела, длины тела и окружностей четырех конечностей индивида.
Влияние на поглощение глюкозы клетками от позвоночных и/или на уровень глюкозы в крови животных
Антитела против рецептора IGF-I согласно одному воплощению настоящего изобретения характеризуются тем, что они не влияют на поглощение глюкозы внутрь клетки дифференцированными мышечными клетками от позвоночных и/или на уровень глюкозы в крови позвоночных. В частности, известно, что IGF-I обладает эффектом повышения поглощения глюкозы внутрь клетки и эффектом снижения уровня глюкозы в крови как составной части его агонистического воздействия на рецепторы IGF-I. С другой стороны, хотя антитела против рецептора IGF-I по настоящему изобретению функционируют в качестве антител-агонистов рецептора IGF-I, вне ожидания они не индуцируют поглощения глюкозы дифференцированными мышечными клетками даже в дозах, в 100 и более раз превышающих значение ЕС50 in vitro для индуцирующей рост клеток активности. В частности, при парентеральном введении животным в дозах, в 10 и более раз превышающих эффективную дозу, достаточную для индуцирования повышения мышечной массы, антитела вне ожидания не изменяют уровень глюкозы в крови. Кроме того, обладая еще и свойствами антител-антагонистов рецептора IGF-I, их характеристики, не влияющие на внутриклеточное поглощение глюкозы дифференцированными мышечными клетками от позвоночных и/или на уровень глюкозы в крови позвоночных, являются полезными и служат для устранения проблемы возникновения гипергликемии, которую нужно было решить для того, чтобы эти антитела применялись для лечения людей, но которая не была решена обычными антителами-антагонистами рецептора IGF-I. Клетки от позвоночных по настоящему изобретению предпочтительно должны представлять собой клетки, происходящие из млекопитающих, птиц, рептилий, амфибий или рыб, более предпочтительно это клетки, происходящие из млекопитающих или птиц, еще более предпочтительно это клетки, происходящие из человека, обезьян, кроликов, морских свинок, коров, свиней, овец, лошадей или собак. Клетки от позвоночных по настоящему изобретению также охватывают клетки и животных, подвергнутых генной инженерии для экспрессии рецептора IGF-I того вида позвоночных, с которым дают перекрестные реакции антитела против рецептора IGF-I по настоящему изобретению; клетки и животных, подвергнутых генной инженерии для экспрессии мутантного рецептора IGF-I, обладающего способностью к связыванию; и клетки, полученные из таких генно-инженерных животных.
При анализе отсутствия влияния антител против рецептора IGF-I по настоящему изобретению на внутриклеточное поглощение глюкозы клетками от позвоночных in vitro можно использовать первичные культуры клеток, линии клеток и трансформантов, полученных из таких клеток. Термин “первичная культура клеток” означает клетки, которые были выделены из органа или ткани живого организма и которые обычно можно субкультивировать в течение нескольких пассажей. Первичные культуры клеток от позвоночных могут быть получены из органа или ткани позвоночных путем обработки ферментами, диспергирования физическими средствами или методом эксплантации. Предпочтительные примеры органов и тканей, из которых получают первичные клетки, включают: эндокринные ткани, как-то щитовидная железа, околощитовидная железа и надпочечники; иммунные ткани, как-то аппендикс, миндалины, лимфатические узлы и селезенка; органы дыхания, как-то трахеи и легкие; пищеварительные органы, как-то желудок, двенадцатиперстная кишка, тонкая кишка и толстая кишка; мочевыделительные органы, как-то почки и мочевой пузырь; мужские половые органы, как-то семявыносящие протоки, яички и предстательная железа; женские половые органы, как-то молочные железы и фаллопиевы трубы; и мышечные ткани, как-то сердечные мышцы и скелетные мышцы. Более предпочтительные примеры включают печень, почки или органы пищеварения или мышечные ткани, из которых наиболее предпочтительны мышечные ткани.
Первичные культуры клеток, которые можно использовать для анализа отсутствия влияния антител против рецептора IGF-I на внутриклеточное поглощение глюкозы в контексте настоящего изобретения, – это клетки, которые экспрессируют рецептор IGF-I и могут индуцироваться для внутриклеточного поглощения глюкозы при связывании IGF-I с рецептором IGF-I. Типичными примерами таковых являются мышечные клетки, прошедшие дифференцировку из миобластов скелетных мышц, которые представляют собой первичные культуры клеток, выделенных из мышечной ткани.
“Дифференцированные мышечные клетки” в настоящем изобретении представляют собой мышечные клетки, прошедшие дифференцировку, и включают клетки, которые не полностью дифференцированы. Для удобства термин “дифференцированные мышечные клетки” в контексте настоящего изобретения относится к клеткам, которые претерпели дифференцировку в течение по меньшей мере шести дней от начала дифференцировки. Также можно получить и использовать первичные культуры клеток от человека или животных, доступные по поручению или коммерческим путем на рынке. Первичные культуры клеток человека доступны из различных учреждений и компаний, напр., ATCC®, ECACC, Lonza, Gibco®, Cell Applications, ScienCell Research Laboratories и PromoCell.
Термин “клеточная линия” означает линию культивируемых клеток, которые были получены из живого организма, а затем иммортализованы с тем, чтобы они могли почти перманентно размножаться с сохранением своих специфических свойств. Клеточные линии подразделяются на нераковые линии клеток и раковые линии клеток. Линии клеток от позвоночных, которые можно использовать для анализа действия антител против рецептора IGF-I по настоящему изобретению на внутриклеточное поглощение глюкозы, представляют собой клетки, которые экспрессируют рецептор IGF-I и могут индуцироваться для внутриклеточного поглощения глюкозы при связывании IGF-I с рецептором IGF-I. Примеры клеточных линий, которые экспрессируют рецептор IGF-I и могут индуцироваться для внутриклеточного поглощения глюкозы под действием IGF-I, включают, без ограничения: клетки скелетных мышц, жировые клетки и эпидермальные кератиноциты.
Другие клетки, которые можно использовать для анализа эффекта антител против рецептора IGF-I по настоящему изобретению на внутриклеточное поглощение глюкозы, – это трансформанты, происходящие из первичных культур клеток и линий клеток. Примеры таких трансформантов включают: клетки iPS, полученные из первичной культуры клеток; и клетки или ткани, прошедшие дифференцировку из таких клеток iPS. Другие трансформанты включают первичные культуры клеток и линии клеток, подвергнутые генной инженерии для включения генов с тем, чтобы кратковременно или перманентно экспрессировать эти гены. Примеры генов для введения и экспрессирования такими клетками включают гены рецептора IGF-I человека и других видов.
Способы определения эффекта антител против рецептора IGF-I по настоящему изобретению на поглощение глюкозы клетками от позвоночных включают: измерение внутриклеточной концентрации глюкозы; измерение внутриклеточного поглощения вещества-индикатора – аналога глюкозы; и измерение изменений уровня переносчика глюкозы. Методы измерения концентрации глюкозы включают методы измерения оптического поглощения типа ферментных методов. Методы измерения степени внутриклеточного поглощения вещества-индикатора – аналога глюкозы включают измерение степени поглощения [3H]-2′-дезоксиглюкозы. Методы измерения изменений уровня переносчика глюкозы включают иммуноцитохимическое окрашивание и вестерн-блоттинг. Специалисты в данной области также могут использовать и другие методы, если потребуется. Факт отсутствия влияния на внутриклеточное поглощение глюкозы подтверждается, если внутриклеточное поглощение глюкозы культурой клеток после реакции с антителом против рецептора IGF-I по настоящему изобретению будет почти таким же, как и внутриклеточное поглощение глюкозы культурой клеток в отсутствие антител. При этом удобно также проводить измерения в тех же условиях с использованием IGF-I, который является исходным лигандом рецептора IGF-I, в качестве контроля.
Культуры клеток для тестирования обрабатывают либо антителом против рецептора IGF-I по настоящему изобретению, либо IGF-I при различных концентрациях, а поглощение глюкозы в опытной группе выражают в процентах, принимая за 100% внутриклеточное поглощение глюкозы в группе без обработки. При использовании для оценки поглощения глюкозы дифференцированных мышечных клеток человека поглощение глюкозы под действием антитела против рецептора IGF-I по настоящему изобретению предпочтительно должно быть равно или меньше, чем поглощение глюкозы под действием IGF-I при такой же концентрации. Более предпочтительно поглощение глюкозы под действием антитела против рецептора IGF-I по настоящему изобретению должно составлять 110% или меньше, еще более предпочтительно 100% от степени поглощения глюкозы в группе без обработки. При использовании для оценки поглощения глюкозы дифференцированных мышечных клеток человека поглощение глюкозы под действием антитела против рецептора IGF-I по настоящему изобретению, добавленного в количестве 100 нМ, предпочтительно должно составлять 110% или меньше, более предпочтительно 105% или меньше, еще более предпочтительно от 95% до 100%.
Способы определения поглощения глюкозы клетками от позвоночных in vivo включают: способы, включающие парентеральное введение антитела против рецептора IGF-I по настоящему изобретению позвоночным и определение изменения содержания глюкозы в органе или ткани индивида. Способы измерения для всего тела индивида, которому проводилось введение, включают: измерение уровня глюкозы в крови; и гемоглобина A1C с использованием гликозилированных белков в качестве индикаторов. Способы измерения поглощения глюкозы для органа или ткани индивида включают: в случае животных, но не человека – непосредственное извлечение целевого органа или ткани и вычисление концентрации глюкозы или индикатора. Неинвазивные методы измерения поглощения глюкозы для органа или ткани индивида включают: анализ изображений по рентгеновским снимкам, КТ и МРТ; и контрастные методы с использованием индикаторов с изотопами или флуоресцентными веществами. Если целевой тканью являются скелетные мышцы, то в качестве показателя также можно использовать глюкозный клэмп (glucose clamp). Специалисты в данной области также могут использовать и другие подходящие методы для анализа эффекта антител против рецептора IGF-I по настоящему изобретению на поглощение глюкозы клетками от позвоночных in vivo.
Антитела против рецептора IGF-I по настоящему изобретению также отличаются тем, что при парентеральном введении позвоночным даже в эффективных дозах, достаточных для увеличения мышечной массы у позвоночных, предпочтительно в дозах, превышающих эффективные дозы в 10 и более раз, они не изменяют уровень глюкозы в крови позвоночных. При оценке влияния антител против рецептора IGF-I по настоящему изобретению на изменение уровня глюкозы в крови позвоночных предпочтительно используются животные, принадлежащие к млекопитающим, птицам, рептилиям, амфибиям или рыбам, более предпочтительно животные, принадлежащие к млекопитающим или птицам, еще более предпочтительно люди, обезьяны, кролики, морские свинки, коровы, свиньи, овцы, лошади или собаки. Для оценки влияния антител против рецептора IGF-I по настоящему изобретению на изменение уровня глюкозы в крови также можно использовать животных, подвергнутых генной инженерии для экспрессии рецептора IGF-I того вида, с которым дают перекрестные реакции антитела против рецептора IGF-I по настоящему изобретению. Инвазивные методы измерения уровня глюкозы в крови включают колориметрические методы и электродные методы. Примеры ферментных методов, применяемых для обнаружения, включают глюкозооксидазный метод (метод GOD) и глюкозо-дегидрогеназный метод (метод GDH). Неинвазивные методы включают оптические методы измерения. Специалисты в данной области также могут выбрать какой-либо другой метод, если потребуется. В случае человека нормальный уровень глюкозы в крови натощак составляет от 100 мг/дл до 109 мг/дл. В отношении неблагоприятных событий по уровню глюкозы в крови, возникающих при приеме лекарственных средств (Common Terminology Criteria for Adverse Events v4.0), уровень глюкозы в крови ниже, чем в пределах от 77 мг/дл до 55 мг/дл, определяется как показатель низкого уровня глюкозы в крови, тогда как уровень глюкозы в крови выше, чем в пределах от 109 мг/дл до 160 мг/дл, определяется как показатель высокого уровня глюкозы в крови. Считается, что введение лекарственных средств не влияет на уровень глюкозы в крови, если уровень глюкозы в крови после введения препарата будет выше 55 мг/дл или ниже 160 мг/дл, более предпочтительно выше 77 мг/дл или ниже 109 мг/дл. Однако нормальные значения уровня глюкозы в крови и их диапазон колебаний варьируются в зависимости от животных, которым вводится препарат, и даже у людей уровень глюкозы в крови не всегда может быть в пределах нормального диапазона во время введения препарата. Соответственно, в контексте настоящего изобретения предпочтительно следует считать, что антитело против рецептора IGF-I по настоящему изобретению не изменяет уровень глюкозы в крови того позвоночного, которому вводится антитело, если изменение уровня глюкозы в крови позвоночного предпочтительно составляет 30% или меньше, более предпочтительно 20% или меньше, еще более предпочтительно 10% или меньше.
Способ получения антител против рецептора IGF-I
Антитела по настоящему изобретению могут быть получены различными методами, хорошо известными специалистам. В частности, антитела по настоящему изобретению могут представлять собой поликлональные антитела или моноклональные антитела (Milstein et al., Nature (England), October 6, 1983, Vol.305, No.5934, pp.537-540). Поликлональные антитела по настоящему изобретению могут быть получены, к примеру, путем сенсибилизации млекопитающих пептидом рецептора IGF-I, приведенным в SEQ ID NO: 2 в качестве антигена, и извлечения полученных антител, напр., из сыворотки крови животных. При использовании пептида в качестве антигена пептид может быть связан с белком-носителем типа BSA или KLH либо соединен с полилизином. Конкретные примеры пептидов, которые можно использовать в качестве антигена, включают, без ограничения, ProSerGlyPheIleArgAsnGlySerGlnSerMet, часть последовательности SEQ ID NO: 2. Моноклональные антитела по настоящему изобретению могут быть получены, к примеру, путем сенсибилизации млекопитающих таким антигеном, извлечения из них иммунных клеток и слияния их с клетками миеломы для получения гибридомы, клонирования и культивирования гибридомы и выделения полученных антител из культуры гибридомы. Пример такого способа получения моноклональных антител описан в Примере 1, а примеры полученных таким образом моноклональных антител включают, без ограничения, моноклональное антитело с аминокислотной последовательностью VH, приведенной в SEQ ID NO: 9, и аминокислотной последовательностью VL, приведенной в SEQ ID NO: 10 (IGF11-16).
После получения такого моноклонального антитела, а затем и молекул нуклеиновой кислоты, содержащей генную последовательность, кодирующую аминокислотную последовательность белка антитела, такие молекулы нуклеиновой кислоты тоже можно использовать для получения антител методами генной инженерии. Специалистам в данной области должны быть известны различные методы использования информации о генах антител типа информации об H-цепях и L-цепях, их вариабельных областях и последовательностях CDR для модификации антител с целью улучшения их способности к связыванию или специфичности либо преобразования антител животных типа антител мыши в человеческие антитела, получая при этом антитела со структурой, подходящей в качестве терапевтических средств для человека. Человеческие моноклональные антитела также можно получить, используя в качестве животных для сенсибилизации антигеном трансгенных животных, в которые был введен ген человеческого антитела. Другой метод, который не требует сенсибилизации животных, включает использование фаговой библиотеки, экспрессирующей антигенсвязывающую область антитела человека или её часть (фаговый дисплей антител человека), и получение фагового клона, экспрессирующего пептид, который специфически связывается с соответствующим антигеном, или антитело с требуемой аминокислотной последовательностью, и получение требуемого человеческого антитела на основании информации о выбранном фаговом клоне. Специалисты в данной области техники могут использовать такой метод, который подойдет (напр., см. обзор Taketo Tanaka et al., Keio J. Med., Vol.60, pp.37-46).
Способ получения моноклональных антител, который приведен выше, включает культивирование гибридомы, вырабатывающей требуемое антитело, и выделение полученных антител из супернатанта культуры стандартными способами. Другой способ получения моноклональных антител, приведенный выше, включает получение гибридомы, вырабатывающей требуемое антитело, или получение фагового клона из фагового дисплея человеческих антител, получение гена, кодирующего такое антитело, более конкретно гена, кодирующего тяжелую цепь и/или легкую цепь иммуноглобулина, получение вектора, экспрессирующего ген, и введение вектора в клетки хозяина (клетки млекопитающих, клетки насекомых, микроорганизмов и т.д.) для получения антитела. Специалисты в данной области также могут модифицировать этот способ путем генетической инженерии гена, кодирующего тяжелую цепь и/или легкую цепь иммуноглобулина, для введения требуемого признака, и получения гуманизованного антитела, химерного белка антитела, низкомолекулярного антитела или каркаса антитела, используя информацию о структуре вариабельных областей или участков CDR тяжелой цепи и/или легкой цепи иммуноглобулина с помощью известных методов. Для улучшения характеристик антител или устранения побочных эффектов специалисты в данной области также могут вносить изменения в структуру константных областей или сахаридных цепей антител с помощью методов, хорошо известных специалистам в данной области.
Антитела против рецептора IGF-I по настоящему изобретению могут быть получены методами, хорошо известными специалистам в данной области. В частности, поскольку гуманизованные антитела против рецептора IGF-I по настоящему изобретению обычно представляют собой моноклональные антитела (Milstein et al., Nature, 1983, Vol. 305, No.5934, pp.537-540), то такие моноклональные антитела могут быть получены, напр., следующим способом.
Этот способ начинается, к примеру, с получения молекул нуклеиновой кислоты, кодирующей аминокислотную последовательность тяжелой цепи и/или легкой цепи иммуноглобулина в антителах против рецептора IGF-I по настоящему изобретению. Затем молекулы нуклеиновой кислоты можно клонировать в различные векторы или плазмиды для получения вектора или плазмиды, содержащей молекулу нуклеиновой кислоты. Затем молекулы нуклеиновой кислоты, векторы или плазмиды используют для трансформации клеток хозяина, которые можно выбрать, напр., из эукариотических клеток типа клеток млекопитающих, клеток насекомых, дрожжевых клеток, растительных клеток и клеток бактерий. Трансформированные клетки хозяина затем культивируют в соответствующих условиях, способствующих вырабатыванию антител против рецептора IGF-I по настоящему изобретению. Если нужно, полученные антитела против рецептора IGF-I по настоящему изобретению можно выделить из клеток хозяина. Различные способы, которые можно использовать для этой процедуры, хорошо известны специалистам в данной области.
Способ на основе иммунизации животных включает получение трансгенного животного, в которое был введен ген антитела человека, в качестве подлежащего иммунизации, иммунизацию животного рецептором IGF-I и/или его частичным пептидом в качестве антигена, выделение иммунных клеток из животного и слияние их с клетками миеломы с образованием гибридомы, которую затем клонируют для получения антитела, которое затем выделяют из супернатанта культуры по стандартной процедуре очистки. Пример такого способа получения моноклональных антител описан, напр, в WO 2013/180238A.
Другой доступный способ включает использование фаговой библиотеки, экспрессирующей вариабельную область требуемого гуманизованного антитела или её часть (фаговый дисплей антител человека), получая при этом антитело, которое специфически связывается с соответствующим антигеном, или фаговый клон со специфической аминокислотной последовательностью, информация о которой затем используется для получения требуемого гуманизованного антитела (напр., см. обзор Taketo Tanaka et al., The Keio Journal of Medicine, Vol.60, pp.37-46).
В этой связи специалисты в данной области могут получать различные антитела, как-то химерные белки антител, низкомолекулярные антитела и каркасы антител, используя известные методы, напр., путем генетической модификации гена, кодирующего тяжелую цепь и/или легкую цепь иммуноглобулина для введения нужного признака, или с помощью информации о структуре вариабельных областей или участков CDR тяжелой цепи и/или легкой цепи иммуноглобулина. Кроме того, для улучшения характеристик антитела или устранения побочных эффектов можно вносить модификации в структуру константной области антител или вводить сайты гликозилирования антител с помощью методов, хорошо известных специалистам в данной области, как потребуется.
Препараты, содержащие антитела против рецептора IGF-I
Антитела против рецептора IGF-I по настоящему изобретению могут применяться в качестве терапевтических или профилактических средств при состояниях, связанных с IGF-I, или состояниях, вызванных каким-либо воздействием на рецептор IGF-I. В частности, состояния, связанные с IGF-I, либо состояния, которые могут быть мишенью для терапии или профилактики с помощью антител-агонистов рецептора IGF-I, включают: мышечную атрофию от бездействия, карликовость, цирроз печени, фиброз печени, диабетическую нефропатию, хроническую почечную недостаточность, синдром Ларона, старение, ограничения внутриутробного развития (IUGR), защиты сердечно-сосудистой системы, инсулинорезистентный диабет, метаболический синдром, остеопороз, муковисцидоз, миотоническую дистрофию, связанную со СПИД саркопению, связанный с ВИЧ синдром перераспределения жира, болезнь Крона, синдром Вернера, сцепленный с Х-хромосомой комбинированный иммунодефицит, потерю слуха, нервную анорексию и ретинопатию недоношенных, синдром Тернера, синдром Прадера-Вилли, синдром Сильвера-Рассела, идиопатическую карликовость, ожирение, рассеянный склероз, язвенный колит, низкую мышечную массу, ишемию миокарда и снижение плотности костей. Заболевания, которые могут быть мишенью для терапии или профилактики с помощью антител-антагонистов рецептора IGF-I, включают: нейробластому, саркому поперечно-полосатых мышц, рак костей, детский рак, акромегалию, рак яичников, рак поджелудочной железы, доброкачественную гипертрофию предстательной железы, рак молочной железы, рак простаты, рак костей, рак легких, колоректальный рак, рак шейки матки, синовиальную саркому, рак мочевого пузыря, рак желудка, опухоль Вильмса, диарею, связанную с метастатическими карциноидными и секретирующими вазоактивный кишечный пептид опухолями, випому, синдром Вернера-Моррисона, синдром Беквита-Видеманна, рак почек, почечно-клеточный рак, переходно-клеточный рак, саркому Юинга, лейкемию, острую лимфобластную лейкемию, опухоли головного мозга, глиобластому, неглиобластомные опухоли головного мозга, менингиому, аденому гипофиза, вестибулярную шванному, примитивные нейроэктодермальные опухоли, медуллобластому, астроцитому, олигодендроглиому, эпендимому, папиллому хороидного сплетения, гигантизм, псориаз, атеросклероз, рестеноз гладких мышц сосудов, неадекватный рост микрососудов, диабетическую ретинопатию, болезнь Грейвса, системную красную волчанку, хронический тиреоидит, миастению, аутоиммунный тиреоидит и болезнь Бехчета. Особенно предпочтительным применением антител против рецептора IGF-I по настоящему изобретению является применение в качестве терапевтических или профилактических средств против мышечной атрофии от бездействия и/или карликовости. Антитела против рецептора IGF-I по настоящему изобретению имеют то преимущество, что они не изменяют уровень глюкозы в крови после введения.
Препараты, содержащие антитела против рецептора IGF-I по настоящему изобретению, могут быть составлены в виде фармацевтической композиции, содержащей, наряду с антителом против рецептора IGF-I по настоящему изобретению, фармацевтически приемлемый носитель и/или любой другой наполнитель. Получение лекарственных форм с использованием фармацевтически приемлемых носителей и/или других наполнителей может проводиться в соответствии, напр., с методами, описанными в The University of the Sciences in Philadelphia, “Remington: The Science and Practice of Pharmacy, 20th edition”, Lippincott, Williams & Wilkins, 2000. Такие терапевтические или профилактические средства могут быть представлены в виде жидкой формы, приготовленной путем растворения, суспендирования или эмульгирования ингредиентов в стерильной водной среде или в масляной среде, либо в виде лиофилизованной формы. Средой или растворителем в качестве разбавителя для приготовления таких форм может быть водная среда, примеры которой включают дистиллированную воду для инъекций и физиологический солевой раствор, который необязательно может использоваться с добавлением осморегулирующего вещества (напр., D-глюкозы, D-сорбита, D-маннита или хлорида натрия) и/или в сочетании с подходящим растворяющим средством типа спирта (напр., этанола), полиспирта (напр., пропиленгликоля или полиэтиленгликоля) или неионного поверхностно-активного вещества (напр., полисорбата 80 или полиоксиэтилена с гидрогенизированным касторовым маслом 50). Такие формы также могут быть получены с масляной средой или растворителем, примеры которых включают кунжутное масло и соевое масло, которые необязательно можно использовать в сочетании с растворяющим средством типа бензилбензоата или бензилового спирта. Такие жидкие препараты зачастую готовят с соответствующими добавками типа буферных веществ (напр., фосфатных буферных веществ и ацетатных буферных веществ), смягчающих средств (напр., бензалкония хлорида и прокаина гидрохлорида), стабилизаторов (напр., сывороточного альбумина человека и полиэтиленгликоля), консервантов (напр., аскорбиновой кислоты, эриторбиновой кислоты и их солей), красителей (напр., хлорофилл-β-каротина меди, Red #2 и Blue #1), антисептических средств (напр., сложных эфиров параоксибензойной кислоты, фенола, бензетония хлорида и бензалкония хлорида), загустителей (напр., гидроксипропилцеллюлозы, карбоксиметилцеллюлозы и их солей), стабилизаторов (напр., сывороточного альбумина человека, маннита и сорбита) и средств, устраняющих запахи (напр., ментоловых и цитрусовых ароматизаторов). Другие альтернативные формы включают терапевтические средства или профилактические средства для нанесения на слизистые оболочки, причем такие формы часто содержат такие добавки, как чувствительные к давлению адгезивы, чувствительные к давлению усилители, регуляторы вязкости, загустители и др. (напр., муцин, агар, желатин, пектин, каррагинан, альгинат натрия, камедь рожкового дерева, ксантановую камедь, трагакантовую камедь, гуммиарабик, хитозан, пуллулан, воскообразный крахмал, сукральфат, целлюлозу и её производные типа гидроксипропилметилцеллюлозы, сложные эфиры полиглицерина и жирных кислот, сополимеры акриловой кислоты-алкил(мет)акрилата либо их соли и сложные эфиры полиглицерина и жирных кислот), главным образом для придания им свойств всасывания или удержания на слизистой оболочке. Однако формы, растворители и добавки для терапевтических средств или профилактических средств, предназначенных для введения в организм, этим не ограничиваются, и специалисты в данной области могут сделать соответствующий выбор.
Препараты, содержащие антитела против рецептора IGF-I по настоящему изобретению, также могут содержать, наряду с антителами против рецептора IGF-I по настоящему изобретению, и другие известные средства (активные ингредиенты). Препараты, содержащие антитела против рецептора IGF-I по настоящему изобретению, можно комбинировать с другими известными средствами в виде набора. Примеры активных ингредиентов для комбинирования с антителами-агонистами рецептора IGF-I включают: гормон роста или его аналоги, инсулин или его аналоги, IGF-II или его аналоги, антитела против миостатина, антагонисты миостатина, антитела против рецептора активина типа IIB, антагонисты рецептора активина типа IIB, растворимый рецептор активина типа IIB или его аналоги, грелин или его аналоги, фоллистатин или его аналоги, β2-агонисты и селективные модуляторы андрогеновых рецепторов. Примеры активных ингредиентов для комбинирования с антителами-антагонистами рецептора IGF-I включают: кортикостероиды, противорвотные средства, ондансетрон гидрохлорид, гранисетрон гидрохлорид, метоклопрамид, домперидон, галоперидол, циклизин, лоразепам, прохлорперазин, дексаметазон, левомепромазин, трописетрон, противораковые вакцины, ингибиторы GM-CSF, ДНК-вакцины против GM-CSF, клеточные вакцины, вакцины против дендритных клеток, рекомбинантные вирусные вакцины, вакцины против белка теплового шока (HSP), гомологичные противоопухолевые вакцины, аутологичные противоопухолевые вакцины, анальгетики, ибупрофен, напроксен, трисалицилат холина магния, оксикодон гидрохлорид, антиангиогенные, антитромботические средства, антитела против PD-1, ниволумаб, пембролизумаб, антитела против PD-L1, атезолизумаб, антитела против CTLA4, ипилимумаб, антитела против CD20, ритуксимаб, антитела против HER2, трастузумаб, антитела против CCR4, могамулизумаб, антитела против VEGF, бевацизумаб, антитела против рецептора VEGF, растворимые фрагменты рецептора VEGF, антитела против TWEAK, антитела против рецептора TWEAK, растворимые фрагменты рецептора TWEAK, AMG 706, AMG 386, антипролиферативные средства, ингибиторы фарнезил-протеинтрансферазы, ингибиторы αVβ3, ингибиторы αVβ5, ингибиторы р53, ингибиторы Kit-рецептора, ингибиторы RET-рецептора, ингибиторы PDGFR, ингибиторы секреции гормона роста, ингибиторы ангиопоэтина, ингибиторы инфильтрирующих опухоли макрофагов, ингибиторы c-FMS, антитела против c-FMS, ингибиторы CSF-1, антитела против CSF-1, растворимые фрагменты c-FMS, пегвисомант, гемцитабин, панитумумаб, иринотекан и SN-38. Дозировка других средств, используемых в комбинации с антителами против рецептора IGF-I, может находиться в пределах дозировок, используемых для нормальной терапии, но может повышаться или уменьшаться в зависимости от ситуации.
Терапевтические или профилактические средства по настоящему изобретению можно вводить парентерально с целью улучшения симптомов. Для парентерального введения можно приготовить трансназальные средства и выбрать жидкий препарат, суспензию или твердую форму. В качестве другой формы для парентерального введения можно приготовить инъекции, причем инъекции выбирают в виде подкожных инъекций, внутривенных инъекций, вливаний, внутримышечных инъекций, интрацеребровентрикулярных инъекций или внутрибрюшинных инъекций. Другие формы, используемые для парентерального введения, включают свечи, сублингвальные формы, чрескожные формы и формы для трансмукозального введения, отличные от трансназальных форм. Кроме того, возможно и внутрисосудистое местное введение посредством добавления или нанесения на стент или внутрисосудистый обтуратор.
Дозировка средств для лечения или профилактики по изобретению будет отличаться в зависимости от возраста, пола, массы тела и симптомов пациента, терапевтического эффекта, способа введения, времени лечения или типа активных ингредиентов в лекарственной композиции, но обычно их можно вводить в диапазоне от 0,1 мг до 1 г, предпочтительно в диапазоне от 0,5 мг до 300 мг активного соединения на 1 введение для взрослых, один раз от одной до четырех недель или один раз в один-два месяца. Таким образом, введение предпочтительно должно проводиться менее одного раза в неделю. Однако, поскольку доза и частота введения будут варьироваться в зависимости от целого ряда условий, может быть достаточной более низкая доза и меньшая частота введения, чем приведенные выше, или же может потребоваться доза и частота введения, превышающая эти диапазоны.
Применения для животных, но не человека
Антитела против рецептора IGF-I согласно одному воплощению настоящего изобретения могут применяться в животноводстве или ветеринарии на животных, а не на людях. Животные как мишени антител против рецептора IGF-I по настоящему изобретению для применения в животноводстве или ветеринарии предпочтительно должны быть представлены животными, принадлежащими к млекопитающим, птицам, рептилиям, амфибиям или рыбам, более предпочтительно животными, принадлежащими к млекопитающим или птицам, еще более предпочтительно животными из числа обезьян, кроликов, морских свинок, коров, свиней, овец, лошадей или собак. Хотя гормоны роста коров и гормоны роста свиней в настоящее время применяются для повышения молочной продуктивности коров и для усиления роста поросят, соответственно, однако считается, что эти эффекты возникают под действием IGF-I, экспрессия которого индуцируется гормоном роста (см. H. Jiang and X. Ge, Journal of Animal Science, Vol. 92, pp. 21-29, 2014). Таким образом, агонистические эффекты антител против рецептора IGF-I по настоящему изобретению можно использовать в целях повышения молочной продуктивности животных и усиления роста плодов или новорожденных животных. Примеры других применений, для которых можно использовать антитела против рецептора IGF-I по настоящему изобретению, включают, без ограничения: увеличение мышечной массы животных, увеличение отношения массы мышц к жиру животных, повышение эффективности преобразования корма в ткани организма, повышение репродуктивной эффективности, повышение репродуктивной способности вида для его сохранения и лечение травм и симптомов истощения, связанных с изнурительными заболеваниями. Антагонистический эффект, возникающий при другом воплощении антител против рецептора IGF-I по настоящему изобретению, можно использовать для лечения злокачественных опухолей у животных, контролирования частоты размножения животных, контролирования роста особей и других применений. Специалисты в данной области также могут модифицировать структуру антител против рецептора IGF-I по настоящему изобретению надлежащим образом, чтобы изменить аминокислотные последовательности каркасных участков или константных областей антител и тем самым уменьшить их иммуногенность в зависимости от того вида животных, которым вводятся антитела.
Способ культивирования клеток с использованием антител против рецептора IGF-I
IGF-I и его производные широко применяются в методах культивирования клеток для поддержания, выращивания и/или дифференцировки клеток от позвоночных in vitro и коммерчески доступны в качестве реагентов для культивирования клеток. Однако, поскольку IGF-I может терять свои эффекты при продолжительном культивировании, напр., вследствие его недостаточной стабильности, то необходимо, напр., все время регулировать его концентрацию с тем, чтобы стабильно проводить культивирование клеток. Кроме того, поскольку IGF-I индуцирует поглощение глюкозы клетками, то существует вероятность того, что метаболизм и характеристики клеток могут изменяться из-за повышения внутриклеточной концентрации глюкозы, а также что условия культивирования могут изменяться из-за уменьшения концентрации глюкозы в культуральной среде. По сравнению с IGF-I антитела против рецептора IGF-I по настоящему изобретению отличаются тем, что они более стабильны, сохраняют свой эффект пролиферации клеток даже после контакта с клетками, проявляют активность, индуцирующую пролиферацию клеток даже при низких концентрациях и не вызывают внутриклеточного поглощения глюкозы. Антитела против рецептора IGF-I по настоящему изобретению можно использовать для культивирования клеток путем добавления соответствующего количества антител в культуральную среду или путем адсорбции или иммобилизации соответствующего количества антител на твердой фазе сосуда для культивирования. Таким образом, антитела против рецептора IGF-I по настоящему изобретению дают возможность уменьшить используемое количество и эффективно индуцировать пролиферацию клеток, адгезированных к твердой фазе. Клетки от позвоночных по настоящему изобретению предпочтительно должны представлять собой клетки, происходящие из млекопитающих, птиц, рептилий, амфибий или рыб, более предпочтительно клетки, происходящие из млекопитающих или птиц, еще более предпочтительно клетки, происходящие из человека, обезьян, кроликов, морских свинок, коров, свиней, овец, лошадей или собак. Используемые клетки могут представлять собой первичные культуры клеток, линии клеток, трансформантов, полученных из таких клеток, или клетки, полученные из трансгенных животных. В частности, примеры субъектов, которые можно культивировать с использованием антител против рецептора IGF-I по настоящему изобретению, также включают органы или ткани позвоночных или трансгенных животных, полученных из таких позвоночных. Антитела против рецептора IGF-I по настоящему изобретению можно использовать для культивирования клеток в целях клеточного производства веществ или клеточной терапии и восстановительной медицины с помощью таких клеток.
ПРИМЕРЫ
Пример 1. Получение мышиного моноклонального антитела
Мышиные моноклональные антитела можно получить методом гибридомы, разработанным Kohler et al. (Nature 256: 495-497, 1975). Антитело-агонист рецептора IGF-I получали путем иммунизации мышей клетками, экспрессирующими рецептор IGF-I человека, по стандартной технологии гибридом. Все эксперименты на животных проводились в соответствии с правилами учреждения. Выполняли стандартный метод, включающий слияние клеток, полученных из селезенки мыши, с линией клеток миеломы мыши (P3U1). Гибридомы отбирали на среде, содержащей гипоксантин, аминоптерин и тимидин. Для оценки сродства методом Cell ELISA на клетках, экспрессирующих рецептор IGF-I, использовали бульон от гибридомы, а для отбора положительной содержащей гибридомы лунки использовали определение активации внутриклеточной тирозинкиназы рецептора IGF-I методом PathHunter®. Из гибридом, содержащихся в этой лунке, выделяли одиночные клоны методом предельного разведения. Эти положительные одиночные клоны гибридомы культивировали без сыворотки и выделяли моноклональные антитела из бульона на колонке с белком A (Ab-Capcher, ProteNova). При оценке пролиферационной активности этих моноклональных антител на миобластах человека было обнаружено антитело-агонист рецептора IGF-I, которое назвали IGF11-16.
Пример 2. Определение изотипа антитела
Для определения изотипа антитела-агониста рецептора IGF-I проводили ELISA с использованием антител, специфичных к соответствующим изотипам антител. В 96-луночный планшет (Nunc, MaxiSorp) добавляли антитело против IgG мыши (TAGO, 6150), разведенное PBS в 2000 раз, в количестве 50 мкл на лунку и оставляли на всю ночь при 4°C. Раствор в 96-луночном планшете заменяли на 3% BSA/PBS и использовали планшет при ELISA. В 96-луночный планшет с иммобилизованным антителом против IgG мыши добавляли антитело-агонист рецептора IGF-I в количестве 30 мкл на лунку и проводили реакцию при комнатной температуре в течение 1,5 ч. Каждую лунку отмывали промывочной жидкостью, а затем добавляли антитела, специфически реагирующие с соответствующими изотипами IgG мыши: конъюгат антитела против IgG1 мыши с ALP (SBA, 1070-04), конъюгат антитела против IgG2a мыши с ALP (SBA, 1080-04), конъюгат антитела против IgG2b мыши с ALP (SBA, 1090-04) и конъюгат антитела против IgG3 мыши с ALP (SBA, 1100-04) в количестве 30 мкл на лунку и проводили реакцию при комнатной температуре в течение 1 часа. Добавляли субстрат (PNPP) в количестве 100 мкл на лунку и проводили реакцию при комнатной температуре в течение 45 мин. Вычисляли разность между значениями поглощения при 405 и 550 нм в качестве оценки авидности.
Поскольку IGF11-16 проявляло реактивность с антителом против IgG1 мыши, то изотип антитела был IgG1.
Пример 3. Определение последовательности антитела
Для определения последовательностей генов легкой цепи и тяжелой цепи антитела-агониста рецептора IGF-I применяли метод SMARTer® RACE. Из РНК, полученной из гибридомы, вырабатывающей антитело, методом SMARTer® RACE получали фрагменты генов, кодирующих тяжелую цепь и легкую цепь антитела и содержащих кодоны инициации и терминации, и определяли их нуклеотидные последовательности. Используя в качестве матрицы общую РНК, полученную из гибридомы, синтезировали первую нить кДНК с помощью набора SMARTer® RACE 5′/3′ Kit (634859, Clontech), а затем амплифицировали её с помощью реакции ПЦР. Используя кДНК в качестве матрицы, проводили реакцию ПЦР с помощью универсальных праймеров для секвенирования, прилагаемых к набору и специфичных для тяжелой цепи и легкой цепи антител, соответственно. Праймеры для легкой цепи (каппа) антитела мыши и IgG1 антитела мыши были разработаны по номерам доступа BC080787 и LT160966, соответственно. Нуклеотидная последовательность праймера, составленного для легкой цепи антитела мыши: GGTGAAGTTGATGTCTTGTGAGTGG, а нуклеотидная последовательность праймера, составленного для тяжелой цепи антитела мыши: GCTCTTCTCAGTATGGTGGTTGTGC. Эти праймеры использовались в экспериментах. Полученные продукты использовали в качестве 5′-RACE при TA-клонировании.
При TA-клонировании ПЦР-продукты 5'-RACE подвергали электрофорезу и выделяли кДНК с целевой последовательностью с помощью набора для экстракции гелей QIAEX II (20021, Qiagen). Очищенную кДНК подвергали реакции с помощью TaKaRa-Taq (R001A, Takara) при 72°C в течение 5 мин для присоединения аденина к 5′- и 3′-концам. кДНК клонировали в активированный топоизомеразой I вектор pCR®II-TOPO® (в дальнейшем именуется вектором TOPO) с помощью набора TOPO® TA Cloning® kit (450641, Thermo Fisher) по методике, прилагаемой к набору. Вектором TOPO, клонированным с целевой кДНК, трансформировали E. coli TOP10, после чего культивировали в агаровой среде, содержащей канамицин 50 мкг/мл. Встраивание целевой кДНК в вектор TOPO проверяли методом ПЦР колоний. Идентифицировали нуклеотидную последовательность клонированной кДНК. Таким же образом идентифицировали нуклеотидную последовательность ПЦР-продукта 3′-RACE для определения полноразмерной последовательности гена антитела. Полноразмерная нуклеотидная и аминокислотная последовательность легкой цепи IGF11-16 приведена в SEQ ID NO: 27 и SEQ ID NO: 28, соответственно, а полноразмерная нуклеотидная и аминокислотная последовательность тяжелой цепи IGF11-16 приведена в SEQ ID NO: 29 и SEQ ID NO: 30, соответственно. Аминокислотные последовательности CDR-H1, CDR-H2, CDR-H3, CDR-L1, CDR-L2, CDR-L3, вариабельной области тяжелой цепи и вариабельной области легкой цепи IGF11-16 приведены в SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10, соответственно.
Пример 4. Авидность к рецептору IGF-I (ELISA)
Для исследования авидности антитела-агониста рецептора IGF-I к рецептору IGF-I человека (SEQ ID NO: 2, NP_000866), морской свинки (SEQ ID NO: 11, XP_003475316), макаки-крабоеда (SEQ ID NO: 12, NP_001248281), кролика (SEQ ID NO: 13, XP_017193273), крысы (SEQ ID NO: 14, NP_494694) и мыши (SEQ ID NO: 15, NP_034643) проводили Cell ELISA на клетках, экспрессирующих соответствующие рецепторы IGF-I.
В клетки P3U1 трансфецировали экспрессирующий вектор pEF1 (Thermo Fisher), содержащий ген рецептора IGF-I человека (SEQ ID NO: 16), морской свинки (SEQ ID NO: 17), макаки-крабоеда (SEQ ID NO: 18), кролика (SEQ ID NO: 19), крысы (SEQ ID NO: 20) или мыши (SEQ ID NO: 21) методом липофекции. После липофекции клетки P3U1 культивировали как минимум в течение ночи и вносили в 96-луночный планшет (покрытый поли-D-лизином) по 0,8×105 клеток на лунку и иммобилизировали 10% забуференным формалином (Mildform® 10NM, Wako), после чего блокировали фосфатным буфером, содержащим 3% BSA. Полученный планшет использовали для ELISA.
При ELISA в каждую лунку добавляли 30 мкл раствора антитела IGF11-16, доведенного до 10 нМ с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1,5 ч. После двукратной отмывки промывочной жидкостью в соответствующие лунки добавляли растворы конъюгата антитела против IgG мыши с HRP (30 мкл), доведенные до заданных концентраций с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1 часа. После двукратной отмывки промывочной жидкостью в каждую лунку добавляли 50 мкл субстрата (TMB) для запуска реакции. Примерно через 20 мин в каждую лунку добавляли 50 мкл 0,5 М серной кислоты. Измеряли поглощение при 450 нм и 550 нм и вычисляли разность между значениями поглощения при 450 нм и 550 нм. Рассчитывали авидность, исходя из разности между значениями поглощения при 450 нм и 550 нм для клеток (холостых клеток, SEQ ID NO: 22), трансфецированных вектором, не содержащим гена рецептора IGF-I, которую принимали за 1 (табл. 1).
Таблица 1
IGF11-16 повышал авидность к клеткам, экспрессирующим рецепторы IGF-I человека, морской свинки, макаки-крабоеда и кролика, более чем в 5 раз по сравнению с холостыми клетками. Напротив, авидность IGF11-16 к клеткам, экспрессирующим рецепторы IGF-I крысы и мыши, была почти эквивалентна авидности к холостым клеткам и не повышалась. Эти результаты свидетельствуют, что IGF11-16 связывается с рецепторами IGF-I человека, морской свинки, макаки-крабоеда и кролика, но не связывается с рецепторами IGF-I крысы и мыши.
Пример 5. Авидность к рецептору инсулина (ELISA)
Для исследования авидности антитела-агониста рецептора IGF-I к рецептору инсулина проводили Cell ELISA на клетках, экспрессирующих человеческий рецептор инсулина.
В клетки HEK 293T трансфецировали экспрессирующий вектор pEF1 (Thermo Fisher), содержащий ген человеческого рецептора инсулина, методом липофекции. После липофекции клетки HEK 293T вносили в 96-луночный планшет (покрытый поли-D-лизином) по 0,8×105 клеток на лунку (около 180 мкл на лунку) и иммобилизировали 10% забуференным формалином (Mildform® 10NM, Wako), после чего блокировали фосфатным буфером, содержащим 3% BSA. Планшет использовали для ELISA.
При ELISA в соответствующие лунки добавляли растворы антитела (30 мкл), доведенные до заданных концентраций с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1 часа. После двукратной отмывки промывочной жидкостью (трис-буфер, содержащий Tween) в соответствующие лунки добавляли растворы конъюгата антитела против IgG мыши с ALP (30 мкл), доведенные до заданных концентраций с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1 часа. После двукратной отмывки промывочной жидкостью в каждую лунку добавляли 100 мкл субстрата (PNPP) для запуска реакции. Примерно через 30 мин измеряли поглощение при 450 нм и 550 нм и вычисляли разность между значениями поглощения при 450 и 550 нм. Авидность рассчитывали, исходя из разности между значениями поглощения при 450 и 550 нм для клеток (холостых клеток, SEQ ID NO: 22), трансфецированных вектором, не содержащим гена рецептора IGF-I и гена рецептора инсулина, которую принимали за 1 (табл. 2).
Таблица 2
При ELISA на иммобилизованных клетках, экспрессирующих человеческий рецептор IGF-I, разность между значениями поглощения при 405 и 550 нм при 0,5 нМ и 5 нМ IGF11-16 возрастала примерно в 3 раза или больше по сравнению с холостыми клетками. Напротив, при ELISA на иммобилизованных клетках, экспрессирующих инсулиновый рецептор человека, разность между значениями поглощения при 405 и 550 нм при 0,5 нМ и 5 нМ IGF11-16 возрастала не более чем в 1,5 раза. Эти результаты свидетельствуют, что IGF11-16 сильнее связывается с рецептором IGF-I, чем с рецептором инсулина.
Пример 6. Анализ сайта связывания у рецептора IGF-I (ELISA)
Для того, чтобы идентифицировать эпитоп у антитела против рецептора IGF-I – агониста рецептора IGF-I, у антитела-агониста рецептора IGF-I измеряли авидность к вариантам, полученным путем замены каждого домена рецептора IGF-I на домен рецептора инсулина, имеющий структуру, близкую структуре рецептора IGF-I.
Внеклеточный домен человеческого рецептора IGF-I (NP_000866) заменяли на внеклеточный домен рецептора инсулина или же внеклеточный домен человеческого рецептора инсулина (NP_000199) заменяли на внеклеточный домен рецептора IGF-I. При этом получали следующие четыре замены:
замена 1: замена hIGFIR[L1-L2]/hINSR, при которой домен L1 и домен L2 человеческого рецептора инсулина заменен на домен L1 и домен L2 человеческого рецептора IGF-I;
замена 2: замена hINSR[L1-L2]/hIGFIR, при которой домен L1 и домен L2 человеческого рецептора IGF-I заменен на домен L1 и домен L2 человеческого рецептора инсулина;
замена 3: замена hINSR[L1]/hIGFIR, при которой домен L1 человеческого рецептора IGF-I заменен на домен L1 человеческого рецептора инсулина;
замена 4: замена hINSR[L2]/hIGFIR, при которой домен L2 человеческого рецептора IGF-I заменен на домен L2 человеческого рецептора инсулина.
В клетки P3U1 трансфецировали экспрессирующие векторы pEF1 (Thermo Fisher), содержащие соответствующие гены рецептора IGF-I человека с вышеприведенными четырьмя заменами методом липофекции. Ген hIGFIR[L1-L2]/hINSR в качестве замены 1 приведен в SEQ ID NO: 23; ген hINSR[L1-L2]/hIGFIR в качестве замены 2 приведен в SEQ ID NO: 24; ген hINSR[L1]/hIGFIR в качестве замены 3 приведен в SEQ ID NO: 25; и ген hINSR[L2]/hIGFIR в качестве замены 4 приведен в SEQ ID NO: 26. После липофекции клетки P3U1 культивировали как минимум в течение ночи и вносили в 96-луночный планшет (покрытый поли-D-лизином) по 0,8×105 клеток на лунку и иммобилизировали 10% забуференным формалином (Mildform® 10NM, Wako), после чего блокировали фосфатным буфером, содержащим 3% BSA. Планшет использовали для ELISA.
При ELISA в каждую лунку добавляли 30 мкл раствора антитела, доведенного до 10 нМ с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1,5 ч. После двукратной отмывки промывочной жидкостью в каждую лунку добавляли 30 мкл раствора конъюгата антитела против IgG мыши с HRP, доведенного до 5 нМ с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1 часа. После двукратной отмывки промывочной жидкостью в каждую лунку добавляли 50 мкл субстрата (TMB) для запуска реакции. Примерно через 20 мин в каждую лунку добавляли 50 мкл 0,5 М серной кислоты для остановки реакции. Измеряли поглощение при 450 нм и 550 нм и вычисляли разность между значениями поглощения при 450 нм и 550 нм. Рассчитывали авидность, исходя из разности между значениями поглощения при 450 нм и 550 нм для клеток (холостых клеток, SEQ ID NO: 22), трансфецированных вектором, не содержащим никаких генов с заменами, которую принимали за 1 (табл. 1).
Таблица 3
При ELISA на иммобилизованных клетках, экспрессирующих hIGFIR[L1-L2]/hINSR, hINSR[L1]/hIGFIR или hINSR[L2]/hIGFIR, разность между поглощением при 405 и 550 нм с IGF11-16 возрастала в 5 раз или больше по сравнению с холостыми клетками. Напротив, авидность IGF11-16 к клеткам, экспрессирующим hINSR[L1-L2]/hIGFIR, была слабой. Эти результаты свидетельствуют, что IGF11-16 связывается с доменом CR рецептора IGF-I.
Пример 7. Определение эпитопа IGF11-16
Для того, чтобы более подробно идентифицировать эпитоп из домена CR в качестве эпитопа IGF11-16, оценивали связывающуюся последовательность по отличиям между видами в авидности IGF11-16 к рецептору IGF-I. На фиг. 1 представлены аминокислотные последовательности домена CR рецептора IGF-I соответствующих видов.
IGF11-16 связывается с рецепторами IGF-I человека, морской свинки и кролика, но не связывается с рецепторами IGF-I мыши и крысы. Исходя из этого, в качестве эпитопа IGF11-16 принимали аминокислотную последовательность, общую для человека, морской свинки и кролика, но не для мыши и крысы, исходя из аминокислотных последовательностей домена CR рецептора IGF-I.
Для определения аминокислотного сайта в домене CR рецептора IGF-I, с которым связывается IGF11-16, измеряли авидность при замене каждой аминокислоты домена CR методом ELISA.
Проводили Cell ELISA на клетках, экспрессирующих рецептор IGF-I, у которого в домене CR была модифицирована аминокислотная последовательность, предположительно связывающаяся с IGF11-16.
В качестве аминокислотных замен в домене CR использовали три замены, представленные ниже. Кроме того, в качестве положительного контроля и отрицательного контроля, соответственно, использовали рецептор IGF-I дикого типа человека и рецептор IGF-I дикого типа крысы, каждый из которых был встроен в экспрессирующий вектор pEF1 (Thermo Fisher). Определяли уровень экспрессии каждого рецептора IGF-I по реакции антитела FLAG M2 с меткой FLAG (AspTyrLysAspAspAspAspLys), присоединенной к внутриклеточному домену рецептора IGF-I, в качестве показателя.
Замена 1 в домене CR: в аминокислотной последовательности рецептора IGF-I человека (NP_000866, SEQ ID NO: 2) аспарагиновая кислота в положении 245 и аланин в положении 247 были заменены на аспарагин и треонин, соответственно.
Замена 2 в домене CR: в аминокислотной последовательности рецептора IGF-I человека (NP_000866, SEQ ID NO: 2) глутаминовая кислота в положении 294 была заменена на аспарагиновую кислоту.
Замена 3 в домене CR: в аминокислотной последовательности рецептора IGF-I человека (NP_000866, SEQ ID NO: 2) глицин в положении 315 и серин в положении 316 были заменены на серин и треонин, соответственно.
В чашку на 10 см, покрытую поли-D-лизином, высеивали клетки HEK 293T по 9×106 клеток на лунку. На следующий день каждую плазмидную ДНК трансфецировали в клетки методом липофекции. На следующий день клетки HEK 293T отделяли с помощью 0,25% трипсина/ЭДТА и суспендировали в бульоне. Клетки HEK 293T вносили в 96-луночный планшет (покрытый поли-D-лизином) по 0,8×105 клеток на лунку и инкубировали при 37°C в присутствии 5% CO2 в течение ночи. Из 96-луночного планшета удаляли среду и иммобилизировали клетки 10% забуференным формалином (Mildform® 10NM, Wako). 10% забуференный формалин заменяли блокирующим буфером (3% BSA/PBS/азид натрия) и использовали планшет при ELISA.
При ELISA в каждую лунку добавляли 50 мкл раствора антитела IGF11-16 или антитела FLAG M2, доведенного до 1 нМ с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1 часа. После двукратной отмывки промывочной жидкостью в соответствующие лунки добавляли растворы конъюгата антитела против IgG мыши с HRP (50 мкл), доведенные до заданных концентраций с помощью 0,1% обезжиренного молока/3% BSA/PBS, и проводили реакцию при комнатной температуре в течение 1 часа. После двукратной отмывки промывочной жидкостью в каждую лунку добавляли 100 мкл субстрата (TMB) для запуска реакции. Примерно через 30 мин в каждую лунку добавляли 100 мкл 0,5 М серной кислоты для остановки реакции и измеряли поглощение при 450 нм. В качестве сродства принимали значение поглощения при 450 нм.
Результаты представлены на фиг. 2. Было подтверждено, что реакции антитела FLAG M2 с клетками, экспрессирующими соответствующие замены в домене CR, практически эквивалентны друг другу, и уровни экспрессии отдельных замен в домене CR практически одинаковы. IGF11-16 повышало значение поглощения при 450 нм до 2 и более с рецептором IGF-I дикого типа человека, у которого не было модификаций в домене CR, и проявляло усиление авидности. IGF11-16 повышало значение поглощения при 450 нм до 2 и более при заменах 1 и 2 в домене CR и проявляло усиление авидности. Напротив, значение поглощения при 450 нм при замене 3 в домене CR было около 1 и было на том же уровне, что и поглощение с рецептором IGF-I крысы в качестве отрицательного контроля, и авидность не проявлялась. Эти результаты свидетельствуют, что аминокислоты в положениях 315 и 316 рецептора IGF-I важны для авидности IGF11-16 к домену CR рецептора IGF-I.
Результаты свидетельствуют, что сайт связывания IGF11-16 с рецептором IGF-I человека находится вблизи глицина (Gly) в положении 315 и серина (Ser) в положении 316. В общем, распознающая последовательность антител состоит из восьми аминокислотных остатков (в среднем от 6 до 10 остатков), а IGF11-16 обладает перекрестной реактивностью, но не проявляет авидности к рецептору IGF-I крысы, а проявляет авидность к рецепторам IGF-I кролика и человека; поэтому последовательность сайта связывания IGF11-16 с рецептором IGF-I человека оценивается как ProSerGlyPheIleArgAsnGly*Ser*GlnSerMet (Gly*Ser* означает аминокислотную последовательность в положениях 315 и 316).
Пример 8. Сродство к рецептору IGF-I при определении методом поверхностного плазмонного резонанса
Измеряли сродство (скорость связывания и скорость диссоциации) веществ к рецептору IGF-I методом поверхностного плазмонного резонанса (SPR).
На сенсорном чипе CM3 (GE) иммобилизировали моноклональное антитело против His с помощью набора Amine Coupling kit (BR-1000-50, GE) и набора His Capture kit (28-9950-56, GE). Условия иммобилизации: NHS/EDC: 7 мин; моноклональное антитело против His 50 мкг/мл: 3 мин; этаноламин: 7 мин; и мишень: 3000 относ. ед. (RU). В качестве аналитов использовали вещества в заданных концентрациях. В качестве лиганда использовали рекомбинантный рецептор IGF-I человека с His-меткой (305-GR-050, R&D Systems, в дальнейшем именуется IGF-IR-His). В качестве отрицательного контроля использовали очищенный IgG2a κ мыши контрольного изотипа, клон: MG2a-53 (401502, BioLegend, в дальнейшем именуется Ctrl IgG2a).
Сенсорный чип CM3 с иммобилизованным моноклональным антителом против His устанавливали на прибор Biacore T200, выставляли температуру реакции 36°C и скорость подачи рабочего буфера (HBS-EP+, BR-1006-69, GE) в 30 мкл/мин. Количество связывающегося лиганда составляло около 100 RU, а на сенсорный чип подавали IGF-IR-His (от 0,5 до 2×10−8 М) для захвата моноклональным антителом против His. Ctrl IgG2a (10 нМ) подавали для реакции на 1 мин, а HBS-EP+ подавали со скоростью в 30 мкл/мин не менее 10 мин. Аналит и HBS-EP+ вводили в проточные ячейки 1 и 2 и проточные ячейки 3 и 4, соответственно.
Условия реакции были установлены на время связывания 600 сек и время диссоциации 600 сек. По завершении реакции проводили промывку регенерационным буфером 1 (0,2% SDS), регенерационным буфером 2 (100 мМ трис-HCl (pH 8,5), 1 М NaCl, 15 мМ MgCl2) и регенерационным буфером 3 (10 мМ глицин-HCl (рН 1,5)) по 1 минуте при скорости потока 30 мкл/мин. Рассчитывали константу скорости диссоциации (ka, 1/M·с), константу скорости связывания (kd, 1/с) и константу диссоциации (KD, M) при анализе на модели связывания 1:1 с помощью программы Biacore T200 Evaluation software (ver. 2.0). Результаты представлены в табл. 4.
Таблица 4
* Значение меньше нижнего предела измерения на приборе
Значение ka у IGF11-16 против рецептора IGF-I человека составляло примерно одну пятую от такового у IGF-I, что указывает на низкую скорость связывания. Напротив, значение kd у IGF11-16 против рецептора IGF-I человека было меньше нижнего предела измерения на приборе и составляло менее 1/1000 от такового у IGF-I, что указывает на значительно меньшую скорость диссоциации и несущественную диссоциацию IGF11-16, связанного с рецептором IGF-I. Значение KD у IGF11-16 против рецептора IGF-I человека составляло менее 1/50 от такового у IGF-I, что указывает на высокую прочность связывания. Это значит, что сродство IGF11-16 к рецептору IGF-I более высокое, чем у IGF-I.
Пример 9. Активирующее действие на рецептор IGF-I или рецептор инсулина при определении методом PathHunter®
Для того, чтобы установить активирующее действие антитела-агониста рецептора IGF-I на рецептор IGF-I, измеряли активацию нижележащего сигнала рецептора IGF-I с помощью функционального анализа PathHunter® IGF1R (DiscoverX).
Использовали линию клеток, в которых принудительно экспрессируется внутриклеточно слитый с адаптерным белком SHC1-Enzyme Acceptor (EA) белок, включающий рецептор IGF-I и домен SH2, связывающийся с внутриклеточной тирозинкиназой рецептора IGF-I. Для выявления активирующего действия антитела-агониста рецептора IGF-I на рецептор инсулина измеряли активацию нижележащего сигнала рецептора инсулина с помощью функционального анализа PathHunter® INSR (DiscoverX). Использовали еще одну линию клеток, в которых принудительно экспрессируется внутриклеточно слитый с адаптерным белком PLCG1-EA белок, включающий рецептор инсулина и домен SH2, связывающийся с внутриклеточной тирозинкиназой рецептора инсулина. В каждой линии клеток лиганд связывается с рецептором IGF-I или рецептором инсулина, что вызывает димеризацию рецептора; фосфорилирование рецептора с рекрутированием адаптерного белка, содержащего домен SH2; формирование рецепторного комплекса передачи сигналов; ускорение связывания между пространственно смежной тирозинкиназой и EA; и реконструкцию активированной β-галактозидазы. Влияние веществ на рецепторную тирозинкиназу можно определить путем измерения уровня хемилюминесцентного сигнала субстрата, гидролизованного под действием реконструированной β-галактозидазы.
В 96-луночный планшет (черный/прозрачный или белый/прозрачный), покрытый поли-D-лизином или коллагеном-I, высеивали клетки, экспрессирующие рецептор IGF-I или рецептор инсулина, по 90 мкл на лунку (2×104 клеток на лунку или 5×103 клеток на лунку) и инкубировали при 37°C в присутствии 5% CO2. На следующий день добавляли вещества в заданных концентрациях по 10 мкл на лунку, а затем инкубировали при 37°C в присутствии 5% CO2. На следующий день по 30 мкл супернатантов культур добавляли к 15 мкл раствора субстрата с последующей реакцией в течение 60 мин и измеряли люминесцентный сигнал на люминометре (Tristar, Berthold Japan K.K.). Рассчитывали активацию рецептора IGF-I, принимая за 100% активность в группе, в которой был только растворитель. Результаты представлены в табл. 5.
Таблица 5
Рассчитывали активацию рецептора инсулина, принимая за 100% активность в группе, в которой был только растворитель. Результаты представлены в табл. 6.
Таблица 6
Активацию рецептора IGF-I веществами измеряли на линии клеток, экспрессирующих рецептор IGF-I. На линии клеток, экспрессирующих рецептор IGF-I, IGF-I и IGF11-16 проявляли активирующее действие на рецептор IGF-I по сравнению с контролем.
Активацию рецептора инсулина веществами измеряли на линии клеток, экспрессирующих рецептор инсулина. На линии клеток, экспрессирующих рецептор инсулина, наблюдалось активирующее действие инсулина на рецептор инсулина. IGF-I зависимым от концентрации образом активировал рецептор инсулина, причем значительный эффект активации наблюдался при 50 нМ. Напротив, антитело IGF11-16 не активировало рецептор инсулина.
Известно, что IGF-I способен реагировать с рецептором инсулина. Также известно, что активация рецептора инсулина вызывает гипогликемический эффект. Было показано, что IGF11-16 специфически действует на рецептор IGF-I и не обладает гипогликемическим действием через рецептор инсулина.
Пример 10. Активность по пролиферации клеток на миобластах человека
Для того, чтобы исследовать пролиферационную активность антитела-агониста рецептора IGF-I на миобластах человека, к миобластам человека добавляли вещества и через 4 дня измеряли содержание АТФ в клетках.
В 96-луночный планшет (покрытый коллагеном типа I) высеивали нормальные миобласты скелетных мышц человека (HSMM, Lonza) по 0,1 мл на лунку (2×103 клеток на лунку), используя среду SkBM-2 (Lonza, CC-3246), содержащую 1% BSA, и инкубировали при 37°C в присутствии 5% CO2. На следующий день после высева клеток добавляли вещества по 25 мкл на лунку и инкубировали при 37°C в присутствии 5% CO2 в течение 4 дней. В качестве показателя пролиферации клеток измеряли внутриклеточное содержание АТФ с помощью набора люминесцентного анализа жизнеспособности клеток CellTiter-Glo® (Promega). Из 96-луночного планшета после инкубации в течение 4 дней удаляли супернатанты так, чтобы бульон в каждой лунке составлял 50 мкл, а затем оставляли планшет при комнатной температуре как минимум на 30мин. Добавляли реагент CellTiter-Glo® по 50 мкл на лунку и проводили реакцию в течение по меньшей мере 10 мин. Затем измеряли люминесцентный сигнал на люминометре (Tristar, Berthold Japan K.K.). Рассчитывали активность, принимая за 100% активность в группе, в которой был только растворитель. Результаты представлены в табл. 7.
Таблица 7
* Контроли на растворитель 1 и 2 содержат азид натрия в количестве 0,005 нМ и 0,5 нМ, соответственно, что равно его содержанию в антителах 16-13 и 26-3.
IGF-I и IGF11-16 усиливали активность по пролиферации клеток по сравнению с контрольным антителом (FLAG M2, Sigma-Aldrich).
Активность по пролиферации миобластов человека повышалась зависимым от концентрации образом при 0,00005, 0,0005, 0,005, 0,05, 0,5, 5, 50 и 500 нМ IGF11-16. Значения ЕС50 по пролиферативной активности миобластов для IGF11-16 и IGF-I составляли 0,004 нМ и 0,61 нМ, соответственно. Результаты показывают, что активность IGF11-16 была более чем в 100 раз выше активности IGF-I.
Антитела 16-13 и 26-3, описанные в непатентной ссылке 35, не проявляли заметной активности по пролиферации клеток по сравнению с контролем на растворитель (содержащим азид натрия), причем активность была слабой по сравнению с IGF11-16.
Пример 11. Активность по пролиферации клеток на миобластах морских свинок
В 96-луночный планшет (покрытый коллагеном типа I) высеивали миобласты морской свинки (Cell Applications) по 0,1 мл на лунку (4×103 клеток на лунку), используя среду SkBM-2 (Lonza, CC-3246), содержащую 1% BSA, и инкубировали при 37°C в присутствии 5% CO2. На следующий день после высева клеток добавляли вещества по 25 мкл на лунку и инкубировали при 37°C в присутствии 5% CO2 в течение 4 дней. В качестве показателя пролиферации клеток измеряли внутриклеточное содержание АТФ с помощью набора для люминесцентного анализа жизнеспособности клеток CellTiter-Glo® (Promega). Из 96-луночного планшета после инкубации в течение 4 дней удаляли супернатанты так, чтобы бульон в каждой лунке составлял 50 мкл, а затем оставляли планшет при комнатной температуре как минимум на 30мин. Добавляли реагент CellTiter-Glo® по 50 мкл на лунку и проводили реакцию в течение по меньшей мере 10 мин. Затем измеряли люминесцентный сигнал на люминометре (Tristar, Berthold Japan K.K.).
Активность по пролиферации миобластов морской свинки повышалась зависимым от концентрации образом при 0,00005, 0,0005, 0,005, 0,05, 0,5, 5, 50 и 500 нМ IGF11-16. Значения ЕС50 по пролиферативной активности миобластов для IGF11-16 и IGF-I составляли 0,004 нМ и 0,76 нМ, соответственно. Результаты показывают, что активность IGF11-16 была более чем в 100 раз выше активности IGF-I.
Пример 12. Сравнение по стойкости действия с IGF-I in vitro
Для того, чтобы сравнить стойкость действия IGF11-16 и IGF-I, через 18 часов после добавления IGF11-16 или IGF-I заменяли среду и измеряли активность по пролиферации миобластов человека в условиях после удаления IGF11-16 и IGF-I.
В 96-луночный планшет (покрытый коллагеном типа I) высеивали нормальные миобласты скелетных мышц человека (клетки миобластов скелетных мышц человека, HSMM, Lonza) по 0,1 мл на лунку (2×103 клеток на лунку), используя среду SkBM-2 (Lonza, CC-3246), содержащую 1% BSA, и инкубировали при 37°C в присутствии 5% CO2. На следующий день после высева клеток добавляли IGF11-16 или IGF-I по 25 мкл на лунку. Через 18 часов после добавления среду заменяли средой, не содержащей IGF11-16 или IGF-I, либо средой, содержащей их, и инкубировали при 37°C в присутствии 5% CO2 в течение 4 дней. В качестве показателя пролиферации клеток измеряли внутриклеточное содержание АТФ с помощью набора для люминесцентного анализа жизнеспособности клеток CellTiter-Glo® (Promega). Из 96-луночного планшета после инкубации в течение 4 дней удаляли супернатанты так, чтобы бульон в каждой лунке составлял 50 мкл, а затем оставляли планшет при комнатной температуре как минимум на 30 мин. Добавляли реагент CellTiter-Glo® по 50 мкл на лунку и проводили реакцию в течение по меньшей мере 10 мин. Затем измеряли люминесцентный сигнал на люминометре (Tristar, Berthold Japan K.K.). В качестве активности пролиферации клеток рассчитывали соотношение (контрольная группа: 0%) относительно контрольной группы, содержащей только растворитель. Результаты представлены на фиг. 3.
Активность пролиферации клеток повышалась до 39% и 75%, соответственно, в группах, в которых добавляли 1 нМ и 5 нМ IGF-I, соответственно, на 4 дня. Активность пролиферации клеток повышалась до 8% и 10%, соответственно, в группах, в которых добавляли 1 нМ и 5 нМ IGF-I, соответственно, на 18 часов, а затем отмывали, причем активность составляла менее 1/5 от таковой в группах, в которых IGF-I добавляли на 4 дня, указывая на заметное снижение эффекта.
В группе, в которой добавляли 0,5 нМ IGF11-16 на 4 дня, активность пролиферации клеток повышалась до 49%. Активность пролиферации клеток в группе, в которой добавляли 0,5 нМ IGF11-16 на 18 часов, а затем отмывали, повышалась до 30%, что соответствует более 60% от активности в группе, в которой IGF11-16 добавляли на 4 дня.
Сравнивали активность по пролиферации клеток в группе, обработанной 0,5 нМ IGF11-16, в которой проводилось отмывание после добавления агента, с активностью по пролиферации клеток в группах, обработанных 1 нМ и 5 нМ IGF-I. Активность IGF11-16 была высокой при статистической значимости. Эти результаты свидетельствуют, что IGF11-16 сохраняет активность по пролиферации миобластов человека даже после отмывания агента и обладает сильным действием по сравнению с IGF-I. IGF11-16 сохраняет активность пролиферации клеток даже после вымывания. Результаты показывают, что в отличие от действия IGF-I, IGF11-16 прочно связывается с рецептором IGF-I и оказывает стойкое активирующее действие на рецептор IGF-I.
Пример 13. Поглощение глюкозы в дифференцированных мышечных клетках человека
Для того, чтобы исследовать действие IGF11-16 на поглощение глюкозы, измеряли степень поглощения помеченной радиоактивной меткой 3H-2-дезоксиглюкозы в дифференцированных мышечных клетках человека и сравнивали с эффектом IGF-I.
В 24-луночный планшет (Costar, 3526) высеивали нормальные миобласты скелетных мышц человека (клетки миобластов скелетных мышц человека, HSMM, Lonza) по 0,5 мл на лунку (2×104 клеток на лунку) и инкубировали при 37°C в присутствии 5% CO2. Среду SkBM-2 (Lonza, CC-3246) с добавлением FBS (Lonza, CC-4423W), L-глутамина (Lonza, CC-4422W), дексаметазона (Lonza, CC-4421W), rhEGF (Lonza, CC-4420W) и GA-1000 (Lonza, CC-4419W) заменяли свежей средой вплоть до слияния клеток. Среду по 0,5 мл на лунку заменяли средой для дифференцировки (DMEM/F12 (1:1) (Gibco, 11320), содержащей 2% лошадиной сыворотки (Sigma, H1270), 50 ед./мл пенициллина, 50 мкг/мл стрептомицина (Gibco, 15070-063)) и инкубировали слившиеся клетки HSMM при 37°C в присутствии 5% CO2 для запуска дифференцировки в мышечные клетки. Через 6 дней от начала дифференцировки клетки использовали в качестве дифференцированных мышечных клеток человека в эксперименте по поглощению глюкозы.
Среду по 0,5 мл на лунку заменяли истощающей средой (DMEM, содержащая 1 г/л глюкозы (Gibco, 11885), с добавлением 0,1% BSA без жирных кислот (Seikagaku Corporation, 82-002-5), 50 ед./мл пенициллина и 50 мкг/мл стрептомицина (Gibco, 15070-063)) и инкубировали дифференцированные мышечные клетки человека при 37°C в присутствии 5% CO2 в течение ночи. На следующий день среду по 0,5 мл на лунку заменяли истощающей средой и инкубировали клетки при 37°C в присутствии 5% CO2 в течение 2 часов. Лунки промывали PBS по 1 мл на лунку и добавляли в них по 0,5 мл на лунку среды для обработки, содержащей соответствующие вещества, после чего инкубировали при 37°C в присутствии 5% CO2 в течение 2 часов. Среду для обработки готовили так, чтобы конечные концентрации составляли 0,1 мМ глюкозы, 0,1% BSA, 1 мкКи/мл 3H-2-дезоксиглюкозы и различные концентрации рекомбинантного IGF-I человека или антитела-агониста рецептора IGF-I человека в буфере для поглощения глюкозы (содержащем 20 мМ HEPES (Dojindo, 342-01375), 150 мМ NaCl (Sigma, S5150), 5 мМ KCl (Wako, 163-03545), 5 мМ MgSO4 (Wako, 131-00405), 1,2 мМ KH2PO4 (Wako, 169-04245), 25 мМ CaCl2 (Fluka, 21114) и 2мМ пирувата (Wako, 190-14881) в воде для инъекций, доведенном до рН 7,4 с помощью NaOH). Лунки трижды промывали добавлением охлажденного PBS по 1 мл на лунку для остановки поглощения глюкозы. Клетки лизировали добавлением 1N NaOH по 0,25 мл на лунку. Весь объем клеточного лизата вносили во флакон, содержащий 3 мл жидкого сцинтиллятора Ultima Gold (PerkinElmer Japan), и перемешивали. Измеряли радиоактивность (dpm) 3H в течение 3 мин на жидкостном сцинтилляционном счетчике. Рассчитывали относительное поглощение глюкозы в обработанной группе, принимая за 100% средний уровень поглощения глюкозы (dpm) в группе без обработки (контрольной группе). Результаты представлены на фиг. 4.
При 0,8, 4, 20 и 100 нМ IGF-I поглощение глюкозы зависело от концентрации и значительно повышалось. Напротив, IGF11-16 не проявлял значительного эффекта вплоть до 100 нМ. Эти результаты свидетельствуют, что IGF11-16 проявляет чрезвычайно слабое действие на поглощение глюкозы в дифференцированных мышечных клетках человека.
Пример 14. Эффективность in vivo (эффект увеличения мышечной массы у морских свинок)
Для проверки эффективности антитела-агониста рецептора IGF-I in vivo морским свинкам однократно вводили IGF11-16 и через 2 недели измеряли мышечную массу для сравнения с эффектом при непрерывном введении IGF-I. Эффект увеличения мышечной массы определяется как эффект возрастания массы мышц у морских свинок на 5% или больше по сравнению с контрольной группой.
Нормальным морским свинкам однократно вводили IGF11-16 (0,03, 0,1 или 0,3 мг/кг) подкожно или внутривенно. В качестве положительного контроля вводили рекомбинантный IGF-I человека (Mecasermin) непрерывно по 1 мг/кг/день с помощью вставленного подкожно осмотического насоса (Alzet). Через две недели после введения препарата морских свинок забивали путем обескровливания под наркозом и измеряли массу мышц длинных разгибателей пальцев (extensor digitorum longus). Результаты представлены на фиг. 5.
В группе внутривенного (iv) введения IGF11-16 в дозе 0,03, 0,1 или 0,3 мг/кг мышечная масса дозозависимым образом значительно возрастала по сравнению с контрольной группой, получавшей только растворитель. Даже в группе подкожного (sc) введения IGF11-16 по 0,3 мг/кг мышечная масса значительно возрастала по сравнению с контрольной группой.
Увеличение массы мышц в группах введения от 0,03 до 0,3 мг/кг IGF11-16 при однократном введении было эквивалентно таковому в группе непрерывного введения (инфузии) 1 мг/кг/день рекомбинантного IGF-I человека. Результаты показывают, что IGF11-16 проявляет эффективность даже in vivo при однократном внутривенном или подкожном введении.
Было показано, что IGF11-16 при однократном введении проявляет эффективность, эквивалентную таковой при непрерывном введении IGF-I. При клиническом применении IGF-I (Mecasermin) вводится один или два раза в день. Напротив, IGF11-16 при введении раз в две недели проявляет эффективность in vivo, эквивалентную таковой при непрерывном введении IGF-I, указывая на то, что IGF11-16 обладает отличной стойкостью действия по сравнению с IGF-I.
Пример 15. Гипогликемический эффект in vivo (гипогликемический эффект у морских свинок)
Для проверки того, обладает ли антитело-агонист рецептора IGF-I гипогликемическим действием in vivo или нет, морским свинкам однократно вводили IGF11-16 и измеряли уровень глюкозы в крови по времени и сравнивали с гипогликемическим эффектом IGF при однократном введении. Гипогликемический эффект определяется как эффект снижения уровня глюкозы в крови до 50 мг/дл или меньше либо вызывающий гипогликемию.
Морским свинкам подкожно вводили IGF-I один раз и изучали гипогликемический эффект. Морских свинок не кормили в течение 12 часов, а затем подкожно вводили рекомбинантный IGF-I человека (Mecasermin) в дозе 0,3, 1, 3 или 10 мг/кг один раз. Морских свинок не кормили в течение 24 часов после введения. У бодрствующих морских свинок брали кровь перед введением (0 часов) и через 1, 2, 4, 8, 10 и 24 часа после введения и проводили измерение уровня глюкозы в крови с помощью датчика Glutest (Sanwa Kagaku Kenkyusyo). Результаты представлены на фиг. 6.
IGF-I проявлял значительный эффект снижения глюкозы при 0,3 мг/кг и более. Гипогликемия наблюдалась при 1 мг/кг и более. Смерть наступала при 3 мг/кг и более.
Морским свинкам подкожно вводили IGF11-16 один раз и изучали гипогликемическое действие. Морских свинок не кормили в течение 12 часов, а затем подкожно вводили IGF11-16 в дозе 10, 30 или 100 мг/кг один раз. Морских свинок не кормили в течение 24 часов после введения. У бодрствующих морских свинок брали кровь перед введением (0 часов) и через 2, 4, 8, 10 и 24 часа после введения и проводили измерение уровня глюкозы в крови с помощью датчика Glutest (Sanwa Kagaku Kenkyusyo). Результаты представлены на фиг. 7.
IGF11-16 не вызывал каких-либо существенных отличий в уровне глюкозы в крови даже в группе введения 100 мг/кг по сравнению с контрольной группой, в которой вводили только растворитель. Результаты показывают, что подкожное введение IGF11-16 не оказывает гипогликемического действия и не влияет на уровень глюкозы в крови.
Морским свинкам внутривенно вводили IGF11-16 один раз и изучали гипогликемическое действие. Морских свинок не кормили в течение 12 часов, а затем внутривенно вводили IGF11-16 в дозе 0,1, 1,5, 6 или 20 мг/кг один раз. Морских свинок не кормили в течение 24 часов после введения. У бодрствующих морских свинок брали кровь перед введением (0 часов) и через 0,5, 1, 2, 4, 8 и 24 часа после введения и проводили измерение уровня глюкозы в крови с помощью датчика Glutest (Sanwa Kagaku Kenkyusyo). Результаты представлены на фиг. 8.
IGF11-16 не вызывал каких-либо существенных отличий в уровне глюкозы в крови даже в группе введения 20 мг/кг по сравнению с контрольной группой, в которой вводили только растворитель. Результаты показывают, что внутривенное введение IGF11-16 не оказывает гипогликемического действия и не влияет на уровень глюкозы в крови.
IGF11-16 не обладает заметным гипогликемическим действием при подкожном и внутривенном введении, в отличие от IGF-I, и не влияет на уровень глюкозы в крови, указывая на то, что IGF11-16 обладает потенциалом в качестве средства, преодолевающего побочный эффект IGF-I – гипогликемию.
Пример 16. Эффективность in vivo (эффект усиления роста у морских свинок)
Для проверки эффективности антитела-агониста рецептора IGF-I in vivo на костях сравнивали действие IGF-I при непрерывном введении и гормона роста (GH) при многократном введении по одному разу в день. IGF11-16 вводили один раз морским свинкам с гипофизэктомией. Через две недели измеряли длину большеберцовой кости и толщину хряща ростовой пластинки как показатели эффекта усиления роста. IGF11-16 (0,3 мг/кг и 1 мг/кг) вводили подкожно морским свинкам с гипофизэктомией один раз. В качестве контроля вводили рекомбинантный IGF-I человека (Mecasermin) непрерывно по 1 мг/кг/день с помощью вставленного подкожно осмотического насоса (Alzet). Другой контроль, рекомбинантный GH человека (Genotropin®), вводили подкожно в дозе 1 мг/кг многократно по одному разу в день. Через две недели после введения препарата морских свинок забивали путем обескровливания под наркозом и измеряли толщину хряща ростовой пластинки и длину большеберцовой кости. Результаты представлены на фиг. 9 и 10.
В группах (IGF11-16), в которых IGF11-16 вводили подкожно в дозах 0,3 мг/кг и 1 мг/кг, толщина хряща ростовой пластинки и длина большеберцовой кости дозозависимым образом значительно возрастали, свидетельствуя об эффекте усиления роста по сравнению с контрольной группой (носителя), в которой морские свинки с гипофизэктомией получали только растворитель.
Эффект усиления роста в группе однократного введения IGF11-16 при 0,3 мг/кг был эквивалентным таковому в группе (IGF-I) непрерывного введения рекомбинантного IGF-I человека при 1 мг/кг/день. Эффект усиления роста в группе однократного введения IGF11-16 при 1 мг/кг был эквивалентен таковому в группе (GH) многократного введения рекомбинантного GH человека при 1 мг/кг/день. Результаты показывают, что однократное введение IGF11-16 проявляет эффективность, эквивалентную таковой при непрерывном введения IGF-I и при многократном введения GH по одному разу в день. При клиническом применении рекомбинантный IGF-I человека (Mecasermin) и рекомбинантный GH человека (Genotropin®) вводятся посредством подкожной инъекции один или два раза в день и шесть или семь раз в неделю, соответственно. Напротив, IGF11-16 при введении один раз в две недели проявляет эффективность in vivo, эквивалентную таковой при непрерывном введении IGF-I и при многократном введения GH по одному разу в день, указывая на то, что IGF11-16 обладает отличной стойкостью действия по сравнению с IGF-I и GH.
Пример 17. Кинетика IGF-I и IGF11-16 в крови
Кинетика IGF-I в крови
Морских свинок не кормили в течение 12 часов, а затем подкожно вводили рекомбинантный IGF-I человека (Mecasermin) в дозе 0,3, 1, 3 или 10 мг/кг. Морских свинок не кормили в течение 24 часов после введения. У бодрствующих морских свинок брали кровь перед введением (0 часов) и через 1, 2, 4, 8, 10 и 24 часа после введения. Измеряли концентрацию IGF-I человека в плазме методом ELISA (DG100, R&D). Результаты представлены на фиг. 11.
Концентрация IGF-I в плазме повышалась в зависимости от вводимой дозы, а через 24 часа после введения снижалась примерно до 50% от максимальной концентрации IGF-I в плазме. В группе введения 0,3 мг/кг концентрация IGF-I через 24 часа после введения была меньше нижнего предела измерения. В группе введения 10 мг/кг морские свинки умирали вследствие гипогликемии через 4 часа после введения, поэтому не удалось получить плазму.
Кинетика IGF11-16 в крови
Морских свинок не кормили в течение 12 часов, а затем подкожно вводили антитело-агонист рецептора IGF-I в дозе 0,3, 1, 3, 10, 30 или 100 мг/кг. Морских свинок не кормили в течение 24 часов после введения, а затем покормили. У бодрствующих морских свинок брали кровь перед введением (0 часов) и через 2, 4, 8, 10, 24, 48 и 72 часа после введения. Измеряли концентрацию IGF11-16 в плазме методом ELISA. Результаты представлены на фиг. 12.
Концентрация IGF11-16 в плазме повышалась в зависимости от вводимой дозы, а через 48 часов после введения она оставалась на уровне примерно 50% или больше от таковой через 24 часа после введения, указывая на то, что кинетика IGF11-16 в крови превосходна по стойкости по сравнению с таковой IGF-I.
Промышленная применимость
Настоящим изобретением предусмотрено антитело, которое специфически связывается с рецептором IGF-I позвоночных и при этом увеличивает мышечную массу или толщину хряща ростовой пластинки через рецептор IGF-I, но не снижает уровень глюкозы в крови. Следовательно, настоящее изобретение может применяться для лечения, профилактики или диагностики заболеваний, связанных с антителами против рецептора IGF-I.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО ПРОТИВ РЕЦЕПТОРА IGF-1 | 2019 |
|
RU2784079C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО ПРОТИВ РЕЦЕПТОРА IGF-1 | 2021 |
|
RU2836869C1 |
| УЛУЧШЕННЫЕ АНТИТЕЛА-АНТАГОНИСТЫ ПРОТИВ GDF-8 И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2630634C2 |
| СТАБИЛИЗИРОВАННЫЕ ПОЛИПЕПТИДЫ ИНСУЛИНОПОДОБНОГО ФАКТОРА РОСТА | 2007 |
|
RU2477287C2 |
| АНТАГОНИСТЫ РЕЦЕПТОРОВ ДЛЯ ЛЕЧЕНИЯ МЕТАСТАТИЧЕСКОГО РАКА | 2009 |
|
RU2455026C2 |
| АНТИТЕЛА ПРОТИВ PDGFRα ДЛЯ ЛЕЧЕНИЯ ВТОРИЧНОЙ ОПУХОЛИ КОСТИ | 2006 |
|
RU2502523C2 |
| СПОСОБЫ РАЗРУШЕНИЯ КЛЕТОК С ИСПОЛЬЗОВАНИЕМ ЭФФЕКТОРНЫХ ФУНКЦИЙ АНТИ-EphA4 АНТИТЕЛ | 2007 |
|
RU2429246C2 |
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА ПРОТИВ GDF-8 И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2360925C2 |
| ХИМЕРНЫЕ ФАКТОРЫ РОСТА ФИБРОБЛАСТОВ С ИЗМЕНЕННОЙ РЕЦЕПТОРНОЙ СПЕЦИФИЧНОСТЬЮ | 2010 |
|
RU2573896C2 |
| PSCAXCD3, CD19XCD3, C-METXCD3, ЭНДОСИАЛИНXCD3, EPCAMXCD3, IGF-1RXCD3 ИЛИ FAP-АЛЬФАXCD3 БИСПЕЦИФИЧЕСКОЕ ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2009 |
|
RU2547600C2 |
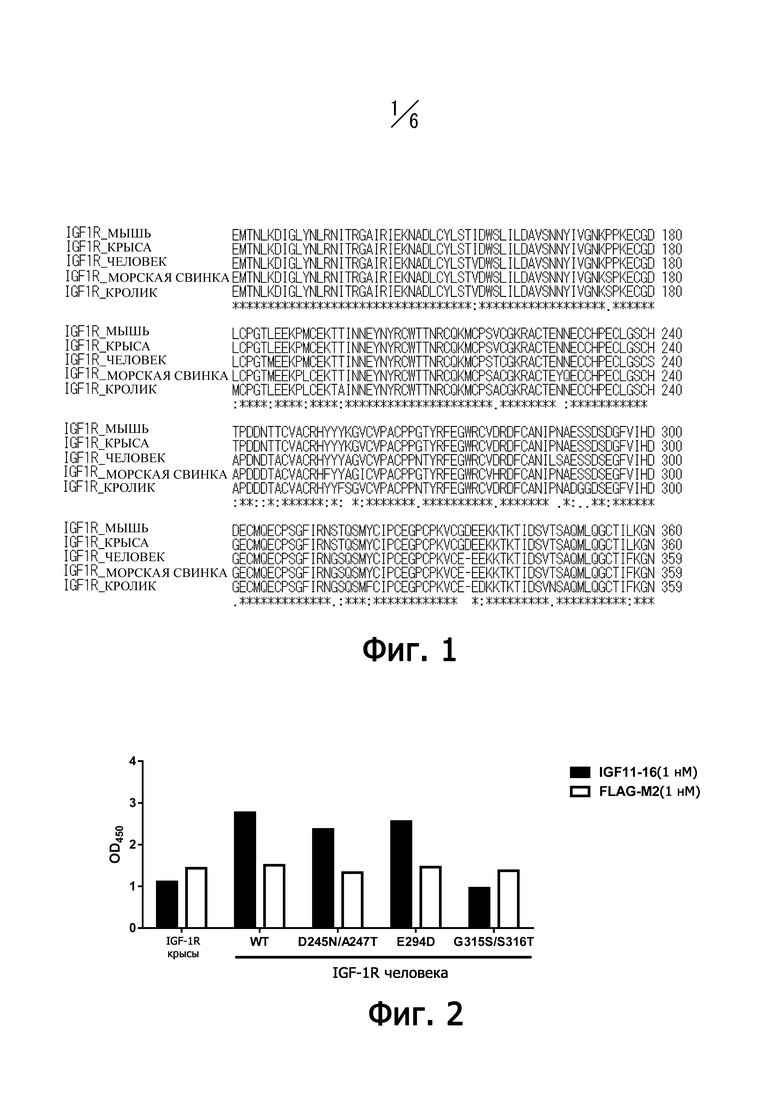
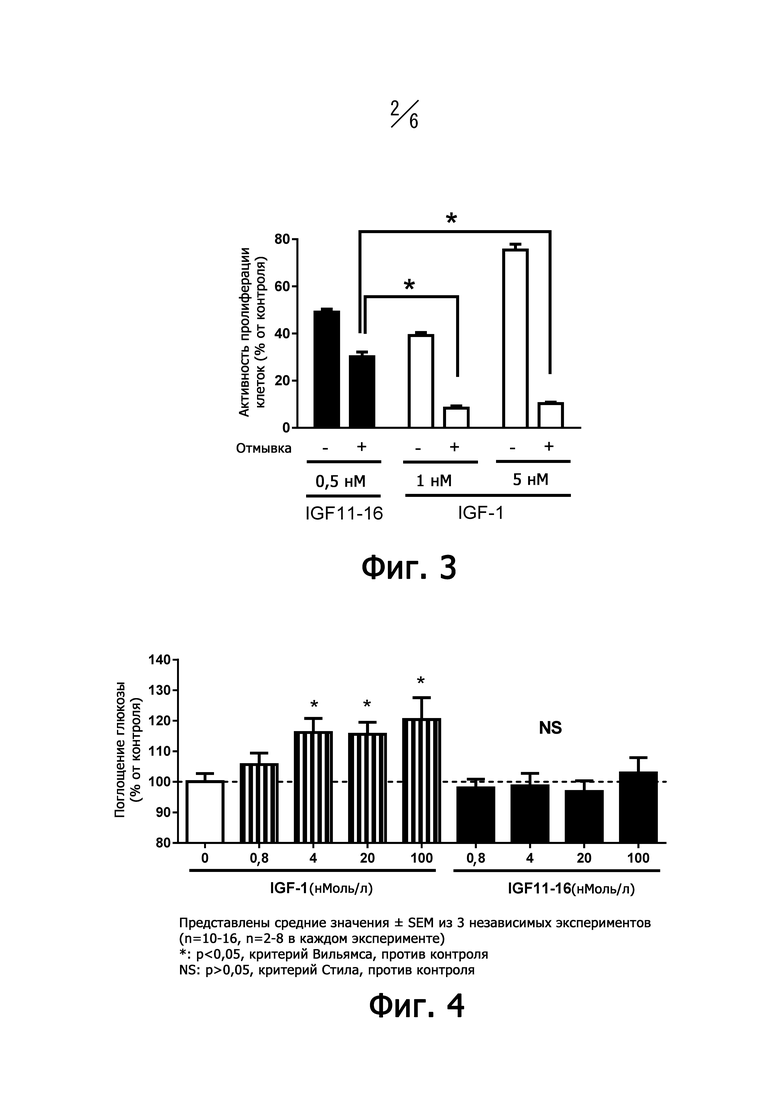
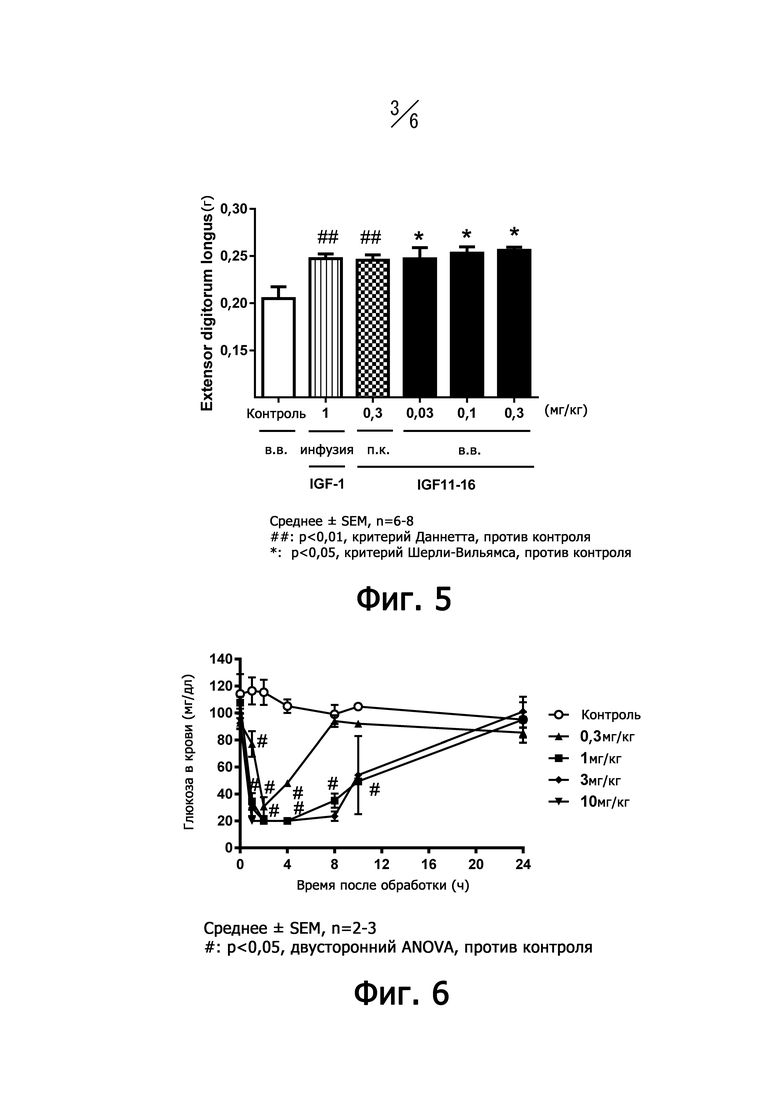
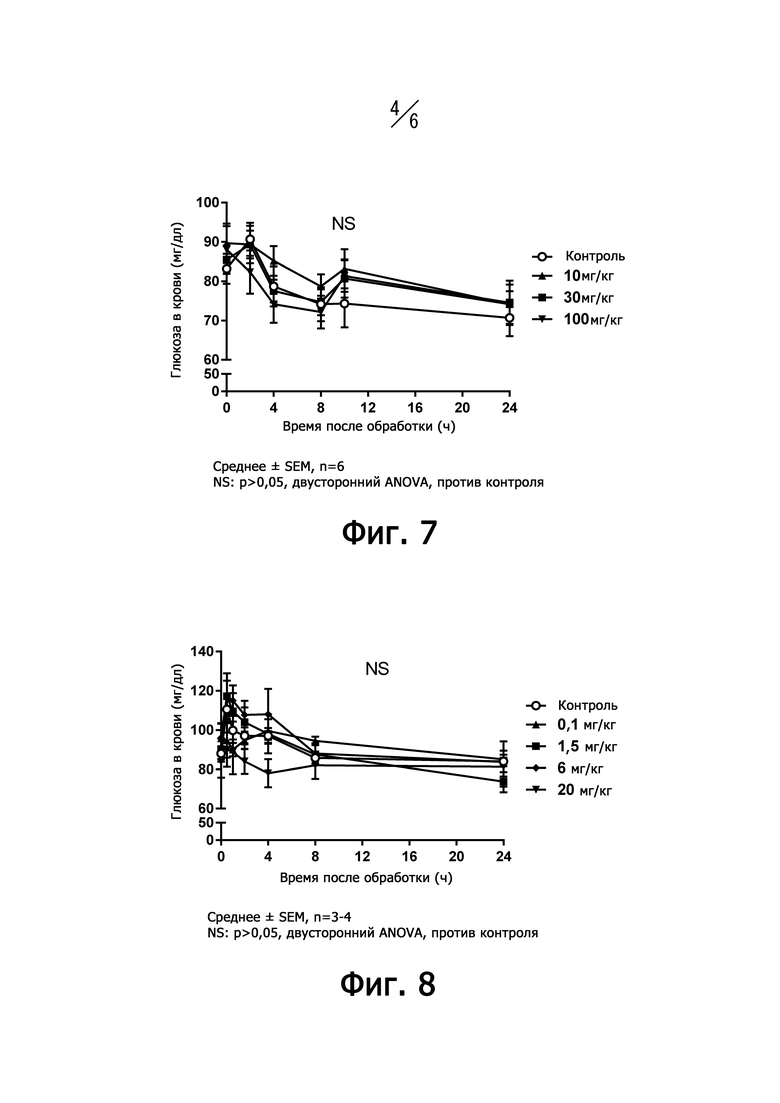
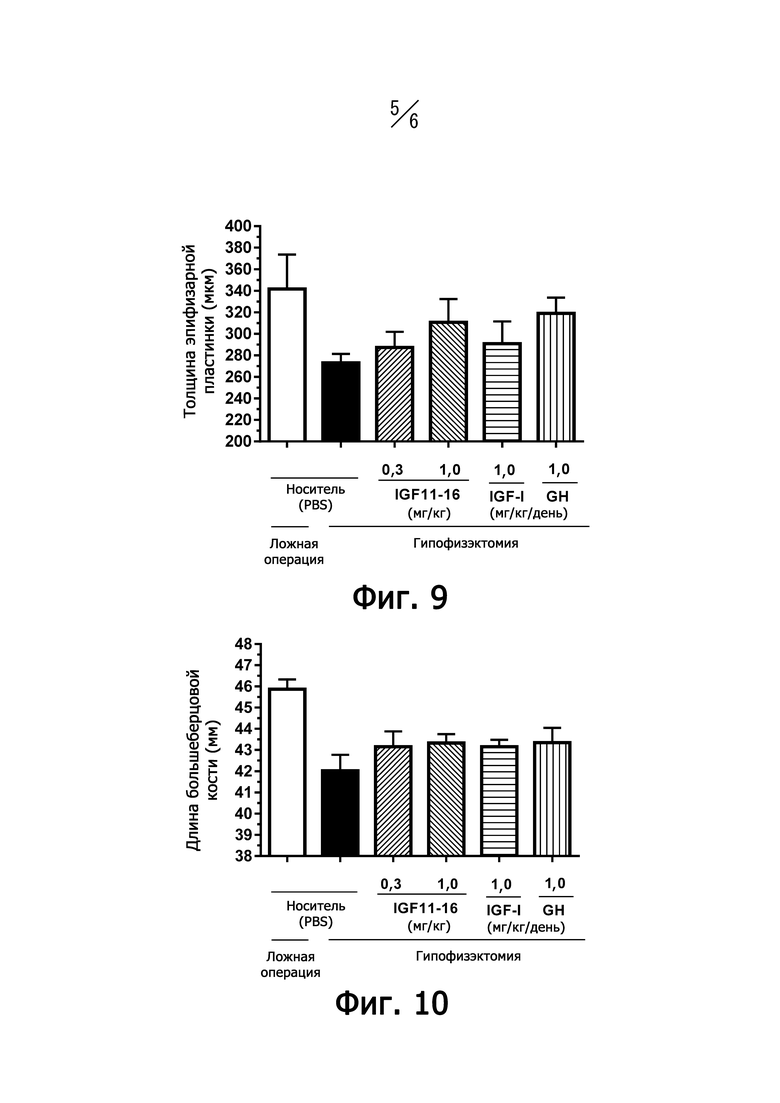

Группа изобретений относится к антителу против рецептора IGF-I. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент, которые специфически связываются с рецептором IGF-I позвоночных и проявляют активность усиления роста клеток от позвоночных, связываются с эпитопом ProSerGlyPheIleArgAsnGlySerGlnSerMet и содержат вариабельную область тяжелой цепи, содержащую три участка, определяющих комплементарность (CDR H1-H3) - SEQ ID NO: 3-5, и вариабельную область легкой цепи, содержащую три участка, определяющих комплементарность (CDR L1-L3) - SEQ ID NO: 6-8. Также раскрыты производное антитела, молекула нуклеиновой кислоты, клонирующий вектор, экспрессирующий вектор, рекомбинантная клетка, способы получения антитела, фармацевтическая композиция и лекарственный препарат для лечения или профилактики состояния, связанного с IGF-I или IGF-II, способ культивирования клеток от позвоночных in vitro, способ анализа связывания антитела, способ исследования внутриклеточной передачи сигнала. Группа изобретений обеспечивает повышение мышечной массы или толщины хряща ростовой пластинки через рецептор IGF-I, не снижая при этом уровень глюкозы в крови. 14 н. и 29 з.п. ф-лы, 12 ил., 7 табл., 17 пр.
1. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент, которые специфически связываются с рецептором IGF-I позвоночных и проявляют активность усиления роста клеток от позвоночных, где указанное антитело или его антигенсвязывающий фрагмент связываются с эпитопом ProSerGlyPheIleArgAsnGlySerGlnSerMet, где указанное антитело или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую три участка, определяющих комплементарность (CDR H1-H3), и далее содержат вариабельную область легкой цепи, содержащую три участка, определяющих комплементарность (CDR L1-L3),
где CDR-H1 содержит аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность SEQ ID NO: 3, в которой заменена, вставлена или удалена одна аминокислота;
где CDR-H2 содержит аминокислотную последовательность SEQ ID NO: 4 или аминокислотную последовательность SEQ ID NO: 4, в которой вплоть до двух аминокислотных остатков заменены, делетированы или вставлены;
где CDR-H3 содержит аминокислотную последовательность SEQ ID NO: 5 или аминокислотную последовательность SEQ ID NO: 5, в которой вплоть до двух аминокислотных остатков заменены, делетированы или вставлены;
где CDR-L1 содержит аминокислотную последовательность SEQ ID NO: 6 или аминокислотную последовательность SEQ ID NO: 6, в которой вплоть до двух аминокислотных остатков заменены, делетированы или вставлены;
где CDR-L2 содержит аминокислотную последовательность SEQ ID NO: 7 или аминокислотную последовательность SEQ ID NO: 7, в которой заменена, вставлена или удалена одна аминокислота;
где CDR-L3 содержит аминокислотную последовательность SEQ ID NO: 8 или аминокислотную последовательность SEQ ID NO: 8, в которой вплоть до двух аминокислотных остатков заменены, делетированы или вставлены.
2. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по п. 1, при этом значение EC50 антитела или фрагмента для усиления роста клеток от позвоночных in vitro составляет 1/20 или меньше от соответствующего значения IGF-I дикого типа.
3. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1 или 2, при этом при контактировании антитела или фрагмента с культивируемыми клетками от позвоночных продолжительность действия антитела или фрагмента по усилению роста культивируемых клеток относительно продолжительности контакта лучше, чем у IGF-I дикого типа.
4. Антитело против рецептора IGF-I либо его антигенсвязывающий по любому из пп. 1-3, при этом значение EC50 антитела или фрагмента для усиления роста клеток от позвоночных in vitro составляет 0,1 нМ/л или меньше.
5. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-4, которое проявляет активность, индуцирующую увеличение мышечной массы и/или длины тела позвоночных при парентеральном введении позвоночным.
6. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-5, которое имеет длительный период полужизни в крови.
7. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-6, при этом позвоночные представлены человеком; другими животными, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак или птиц; или же животными, отличными от человека, подвергнутыми инженерии для экспрессии рецептора IGF-I человека.
8. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-7, которое не индуцирует поглощение глюкозы дифференцированными мышечными клетками при введении в дозе, достаточной для индуцирования роста клеток от позвоночных.
9. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по п. 8, которое не индуцирует поглощения глюкозы дифференцированными мышечными клетками при введении в дозе, в 100 или больше раз превышающей значение EC50 для индуцирования роста клеток от позвоночных in vitro.
10. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по п. 8 или 9, при этом клетки от позвоночных представляют собой миобласты, полученные от человека или другого млекопитающего.
11. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 5-10, которое не снижает уровень глюкозы в крови позвоночных при парентеральном введении позвоночным в дозе, достаточной для индуцирования увеличения мышечной массы и/или длины тела у позвоночных.
12. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по п. 11, которое не изменяет уровень глюкозы в крови позвоночных при парентеральном введении позвоночным в дозе, которая в 10 или больше раз превышает эффективную дозу, достаточную для индуцирования увеличения мышечной массы и/или длины тела у позвоночных.
13. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-12, которое обладает перекрестной реактивностью с рецептором IGF-I человека или других животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак или птиц.
14. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-13, которое вызывает реакцию антиген-антитело со сродством по равновесной константе диссоциации (KD) 1×10−8 M или меньше.
15. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-14, которое обладает по меньшей мере одним из следующих признаков:
1) проявляет активность, вызывающую рост клеток от позвоночных;
2) проявляет активность, вызывающую увеличение мышечной массы и/или длины тела позвоночных при парентеральном введении позвоночным;
3) не вызывает поглощения глюкозы дифференцированными мышечными клетками при введении в дозе, достаточной для того, чтобы вызвать рост клеток от позвоночных; и
4) не изменяет уровень глюкозы в крови позвоночных при парентеральном введении позвоночным в дозе, достаточной для того, чтобы вызвать увеличение мышечной массы и/или длины тела у позвоночных.
16. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-14, которое обладает по меньшей мере одним из следующих признаков:
1) ингибирует рост клеток от позвоночных, вызванный IGF-I;
2) ингибирует индуцированную IGF-I пролиферацию клеток у позвоночных, страдающих связанным с пролиферацией клеток заболеванием, при парентеральном введении позвоночным;
3) не влияет на поглощение глюкозы дифференцированными мышечными клетками в дозе, достаточной для ингибирования роста клеток от позвоночных, индуцированного IGF-I; и
4) не изменяет уровень глюкозы в крови позвоночных, страдающих связанным с пролиферацией клеток заболеванием, при парентеральном введении позвоночным в дозе, достаточной для ингибирования индуцированной IGF-I пролиферации клеток у позвоночных.
17. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-16, которое представляет собой Fab, scFv, диатело или биспецифичное антитело.
18. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по п. 1, при этом каркасная последовательность иммуноглобулина представляет собой каркасную последовательность каждого класса иммуноглобулина человека или других животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак, птиц, мышей или крыс.
19. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-18, состоящее из аминокислотной последовательности, включающей:
в качестве вариабельной области тяжелой цепи – аминокислотную последовательность, приведенную в SEQ ID NO: 9, или аминокислотную последовательность, имеющую сходство на 90% или больше с SEQ ID NO: 9; и
в качестве вариабельной области легкой цепи – аминокислотную последовательность, приведенную в SEQ ID NO: 10, или аминокислотную последовательность, имеющую сходство на 90% или больше с SEQ ID NO: 10.
20. Антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-19, дополнительно содержащее константную область каждого класса иммуноглобулина человека или других животных, включая морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак, птиц, мышей или крыс.
21. Производное антитела против рецептора IGF-I или его антигенсвязывающего фрагмента, которое связывает рецептор IGF-I и выбрано из:
антитела против рецептора IGF-I или его антигенсвязывающего фрагмента по любому из пп. 1-20, у которых была введена мутация в константную область;
антитела против рецептора IGF-I или его антигенсвязывающего фрагмента по любому из пп. 1-20, у которых было модифицировано расположение доменов константных областей;
антитела против рецептора IGF-I или его антигенсвязывающего фрагмента по любому из пп. 1-20, содержащих два или больше Fc на 1 молекулу;
антигенсвязывающего фрагмента антитела против рецептора IGF-I по любому из пп.1-20, состоящего только из тяжелой цепи либо только из легкой цепи;
антитела против рецептора IGF-I или его антигенсвязывающего фрагмента по любому из пп. 1-20 с модифицированным гликозилированием;
биспецифического антитела, содержащего антитело против рецептора IGF-I или его антигенсвязывающий фрагмент по любому из пп. 1-20;
конъюгата антитела против рецептора IGF-I или его антигенсвязывающего фрагмента по любому из пп. 1-20 с соединением или белком, отличным от антител;
антитела-фермента из антитела против рецептора IGF-I или его антигенсвязывающего фрагмента по любому из пп. 1-20;
нанотела из антитела против рецептора IGF-I по любому из пп. 1-20;
тандемного scFv из антитела против рецептора IGF-I по любому из пп. 1-20;
биспецифичного тандемного scFv из антитела против рецептора IGF-I по любому из пп. 1-20;
диатела из антитела против рецептора IGF-I по любому из пп. 1-20; или
VHH из антитела против рецептора IGF-I по любому из пп. 1-20,
где указанное производное антитела или его антигенсвязывающего фрагмента связываются с эпитопом ProSerGlyPheIleArgAsnGlySerGlnSerMet.
22. Молекула нуклеиновой кислоты, состоящая из последовательности полинуклеотида, кодирующего антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-20.
23. Клонирующий вектор, содержащий по меньшей мере одну молекулу нуклеиновой кислоты по п. 22.
24. Экспрессирующий вектор, содержащий по меньшей мере одну молекулу нуклеиновой кислоты по п. 22.
25. Рекомбинантная клетка, полученная из клетки хозяина путем трансфекции вектором по п. 23 или 24.
26. Способ получения антитела против рецептора IGF-I либо его антигенсвязывающего фрагмента по любому из пп. 1-20, включающий:
культивирование рекомбинантной клетки по п. 25; и
очистку антитела против рецептора IGF-I или его антигенсвязывающего фрагмента, полученного из рекомбинантной клетки.
27. Фармацевтическая композиция для лечения или профилактики состояния, связанного с IGF-I или IGF-II, содержащая антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-20, производное антитела по п. 21, молекулу нуклеиновой кислоты по п. 22, вектор по п. 23 или 24 или рекомбинантную клетку по п. 25.
28. Фармацевтическая композиция по п. 27, также содержащая дополнительный активный ингредиент, отличный от антитела или фрагмента по любому из пп. 1-20, производного антитела по п. 21, молекулы нуклеиновой кислоты по п. 22, вектора по п. 23 или 24 или рекомбинантной клетки по п. 25.
29. Фармацевтическая композиция по п. 28, при этом активным ингредиентом является одно или несколько из числа гормона роста или его аналогов, инсулина или его аналогов, IGF-II или его аналогов, антител против миостатина, антагонистов миостатина, антител против рецептора активина типа IIB, антагонистов рецептора активина типа IIB, растворимого рецептора активина типа IIB или его аналогов, грелина или его аналогов, фоллистатина или его аналогов, β2-агонистов и селективных модуляторов андрогеновых рецепторов.
30. Фармацевтическая композиция по п. 28 или 29, при этом активный ингредиент включает ингредиент, выбранный из группы, состоящей из: кортикостероидов, противорвотных средств, ондансетрона гидрохлорида, гранисетрона гидрохлорида, метоклопрамида, домперидона, галоперидола, циклизина, лоразепама, прохлорперазина, дексаметазона, левомепромазина, трописетрона, противораковой вакцины, ингибиторов GM-CSF, ДНК-вакцины против GM-CSF, клеточной вакцины, вакцины из дендритных клеток, рекомбинантной вирусной вакцины, вакцины против белка теплового шока (HSP), гомологичной противоопухолевой вакцины, аутологичной противоопухолевой вакцины, анальгетиков, ибупрофена, напроксена, трисалицилат холина магния, оксикодона гидрохлорида, антиангиогенных, антитромботических средств, антител против PD-1, ниволумаба, пембролизумаба, антител против PD-L1, атезолизумаба, антител против CTLA4, ипилимумаба, антител против CD20, ритуксимаба, антител против HER2, трастузумаба, антител против CCR4, могамулизумаба, антител против VEGF, бевацизумаба, антител против рецептора VEGF, растворимого фрагмента рецептора VEGF, антител против TWEAK, антител против рецептора TWEAK, растворимого фрагмента рецептора TWEAK, AMG 706, AMG 386, антипролиферативных средств, ингибиторов фарнезил-протеинтрансферазы, ингибиторов αVβ3, ингибиторов αVβ5, ингибиторов р53, ингибиторов Kit-рецептора, ингибиторов RET-рецептора, ингибиторов PDGFR, ингибиторов секреции гормона роста, ингибиторов ангиопоэтина, ингибиторов инфильтрирующих опухоли макрофагов, ингибиторов c-FMS, антител против c-FMS, ингибиторов CSF-1, антител против CSF-1, растворимого фрагмента c-FMS, пегвисоманта, гемцитабина, панитумумаба, иринотекана и SN-38.
31. Лекарственный препарат для лечения или профилактики состояния, связанного с IGF-I, содержащий антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-20, производное антитела по п. 21, молекулу нуклеиновой кислоты по п. 22, вектор по п. 23 или 24 или рекомбинантную клетку по п. 25, при этом состояние, связанное с IGF-I, выбрано из числа мышечной атрофии от бездействия, карликовости, диабетической нефропатии, хронической почечной недостаточности, синдрома Ларона, цирроза печени, фиброза печени, старения, ограничения внутриутробного развития (IUGR), неврологических заболеваний, инсульта, травмы спинного мозга, инсулинорезистентного диабета, метаболического синдрома, остеопороза, муковисцидоза, ран, миотонической дистрофии, связанной со СПИД саркопении, связанного с ВИЧ синдрома перераспределения жира, ожогов, болезни Крона, синдрома Вернера, сцепленного с Х-хромосомой комбинированного иммунодефицита, потери слуха, нервной анорексии и ретинопатии недоношенных, синдрома Тернера, синдрома Прадера-Вилли, синдрома Сильвера-Рассела, идиопатической карликовости, ожирения, рассеянного склероза, фибромиалгии, язвенного колита, низкой мышечной массы, ишемии миокарда и снижения плотности костей.
32. Лекарственный препарат по п. 31, который представлен в форме для парентерального введения.
33. Лекарственный препарат по любому из пп.31, 32, который представляет собой ветеринарный препарат для введения животным, а не людям.
34. Лекарственный препарат по п. 33, при этом ветеринарный препарат вводится с целью повышения мышечной массы и/или длины тела, усиления роста, повышения продукции молока, стимулирования размножения или предотвращения старения.
35. Лекарственный препарат по п. 33 или 34, при этом животные представляют собой морских свинок, обезьян, кроликов, коров, свиней, лошадей, овец, собак или птиц.
36. Лекарственный препарат для лечения или профилактики заболевания, вызванного действием IGF-I или IGF-II на рецептор IGF-I, содержащий антитело против рецептора IGF-I либо его антигенсвязывающий фрагмент по любому из пп. 1-20, производное антитела по п. 21, молекулу нуклеиновой кислоты по п. 22, вектор по п. 23 или 24 или рекомбинантную клетку по п. 25.
37. Лекарственный препарат по п. 36, при этом заболевание, вызванное действием IGF-I или IGF-II на рецептор IGF-I, выбрано из группы, состоящей из: рака печени, нейробластомы, саркомы поперечно-полосатых мышц, рака костей, детского рака, акромегалии, рака яичников, рака поджелудочной железы, доброкачественной гипертрофии предстательной железы, рака молочной железы, рака простаты, рака костей, рака легких, колоректального рака, рака шейки матки, синовиальной саркомы, рака мочевого пузыря, рака желудка, опухоли Вильмса, диареи, связанной с метастатическими карциноидными и секретирующими вазоактивный кишечный пептид опухолями, випомы, синдрома Вернера-Моррисона, синдрома Беквита-Видеманна, рака почек, почечно-клеточного рака, переходно-клеточного рака, саркомы Юинга, лейкемии, острой лимфобластной лейкемии, опухоли головного мозга, глиобластомы, неглиобластомной опухоли головного мозга, менингиомы, аденомы гипофиза, вестибулярной шванномы, примитивной нейроэктодермальной опухоли, медуллобластомы, астроцитомы, олигодендроглиомы, эпендимомы, папилломы хороидного сплетения, гигантизма, псориаза, атеросклероза, рестеноза гладких мышц сосудов, неадекватного роста микрососудов, диабетической ретинопатии, болезни Грейвса, рассеянного склероза, системной красной волчанки, хронического тиреоидита, миастении, аутоиммунного тиреоидита и болезни Бехчета.
38. Способ культивирования клеток от позвоночных in vitro, включающий контактирование клеток от позвоночных с антителом против рецептора IGF-I либо его антигенсвязывающим фрагментом по любому из пп. 1-20, производным по п. 21, молекулой нуклеиновой кислоты по п. 22, вектором по п. 23 или 24 или рекомбинантной клеткой по п. 25 при культивировании клеток.
39. Способ по п.38, при этом контактирование проводится с целью усиления роста или индуцирования дифференцировки клеток от позвоночных.
40. Способ по п. 38 или 39, при этом антитело против рецептора IGF-I, его фрагмент или производное адсорбировано или иммобилизовано на твердой фазе.
41. Способ анализа связывания антитела против рецептора IGF-I, его фрагмента или производного, указанный способ включает использование трансгенного животного, чей ген рецептора IGF-I был генетически модифицирован путем введения мутации, так чтобы экспрессировался IGF-I рецептор, имеющий CR домен, содержащий аминокислотную последовательность ProSerGlyPheIleArgAsnGlySerGlnSerMet.
42. Способ исследования внутриклеточной передачи сигнала посредством антитела против рецептора IGF-I, его фрагмента или производного, указанный способ включает использование трансгенного животного, чей ген рецептора IGF-I был генетически модифицирован путем введения мутации, так чтобы экспрессировался IGF-I рецептор, имеющий CR домен, содержащий аминокислотную последовательность ProSerGlyPheIleArgAsnGlySerGlnSerMet.
43. Способ получения антитела против рецептора IGF-I, включающий стадии:
(а) иммунизации трансгенного животного рецептором IGF-1 или частичным пептидом из него и выделения антитело-продуцирующих клеток из иммунизированного животного, где ген рецептора IGF-I трансгенного животного был генетически модифицирован путем введения мутации, так чтобы экспрессировался IGF-I рецептор, имеющий CR домен, содержащий аминокислотную последовательность ProSerGlyPheIleArgAsnGlySerGlnSerMet;
(б) слияния антитело-продуцирующих клеток животного с клетками миеломы и подобными, чтобы получить гибридомы; и
(в) клонирования гибридомы и выделения антитела против рецептора IGF-I из их культуры.
| SOOS M.A | |||
| et al | |||
| A Panel of Monoclonal Antibodies for the Type I Insulin-Like Growth Factor Receptor | |||
| Epitope Mapping, Effects on Ligand Binding, and Biological Activity // J Biol Chem., 1992, Jun 25; 267(18): 12955-63 | |||
| XIONG L | |||
| et al | |||
| Growth-stimulatory monoclonal antibodies against human insulin-like growth factor I receptor // Proc Natl Acad |

