ОБЛАСТЬ ИЗОБРЕТЕНИЯ
По настоящей заявке испрашивается приоритет в соответствии с патентной заявкой Кореи №10-2018-0111046, поданной 17 сентября 2018 г., содержание которой полностью включено в настоящую заявку посредством ссылки.
Настоящее изобретение относится к антителу, которое специфически связывается с N-концевым доменом лизил-тРНК-синтазы, экспонированным на внеклеточной мембране, более конкретно, оно специфически связывается с N-концевым доменом лизил-тРНК-синтетазы (KRS, лизил-тРНК-синтетаза), экспонированным на внеклеточной мембране, имеющему специфическую последовательность CDR (определяющую комплементарность область), как описано в настоящем документе, и относится к применению для предотвращения, лечения или диагностики рака, метастазов рака или заболеваний, связанных с миграцией иммунных клеток, композиции, содержащей антитело или его фрагмент, обладающее высокой аффинностью и стабильностью, или антитело и его фрагмент в качестве активного ингредиента.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Недавние исследования установили, что лизил-тРНК синтетаза человека (KRS), обычно присутствующая в цитозоле, перемещается к плазматической мембране (клеточной мембране) для взаимодействия с рецептором ламинина массой 67 кДа (67LR), присутствующим на плазматической мембране, тем самым способствуя миграции опухолевых (или раковых) клеток, влияя на метастаз рака (Dae Gyu Kim et al., Chemical inhibition of prometastatic lysyl-tRNA synthetase-laminin receptor interaction, Nat Chem Biol. 2014 Jan; 10(1): 29-34, Dae Gyu Kim et. al. Interaction of two translational components, lysyl-tRNA synthetase and p40/37LRP, in plasma membrane promotes laminin-dependent cell migration, FASEB J. (2012)26, 4142-4159). KRS человека (номер доступа в Genbank NP_005539.1 и т.д.) содержит N-концевое удлинение (1-72), антикодон-связывающий домен (73-209) и каталитический домен (220-597). KRS человека представляет собой фермент, необходимый для синтеза белка, и обычно он находится в составе комплекса мульти-тРНК-синтетазы (MSC) в цитозоле. Однако, после введения сигнала ламинина p38 MAPK (митоген-активируемая протеинкиназа) фосфорилирует KRS по остаткам T52, и KRS перемещается к клеточной мембране, где KRS защищает 67LR от убиквитин-опосредованной деградации. Также сообщалось, что KRS, перемещенная к клеточной мембране, ускоряет метастаз рака за счет стабилизации и взаимодействия с 67LR, ассоциированным с метастазом рака.
В настоящее время тот факт, что Myc-KRS41-597 (ΔN) с делецией 40 концевых остатков в N-концевом удлинении (N-ext) не локализован на плазматической мембране, указывает на то, что область N-ext KRS является важной областью транслокации KRS к клеточной мембране. Что касается метастазов рака, в частности, известно, что область KRS N-ext участвует в связывании KRS и 67LR при их взаимодействии. Чтобы применять этот факт в терапевтических или диагностических целях, необходимо целенаправленно воздействовать на конкретный сайт (особенно, KRS N-ext) в белке KRS в соответствии с характеристиками нескольких доменов, составляющих белок KRS. Соответственно, авторами настоящего изобретения получено антитело, специфически связывающееся с N-концом KRS, которое не проявляет кросс-реакции, а также связывается с ARS (заявка на патент Кореи №10-2018-0035446).
Однако аффинность предыдущих антител, нацеленных на N-конец KRS, ниже, чем у различных антител в полной форме IgG. Следовательно, существует необходимость конструировать антитело, имеющее более высокую аффинность к N-концу KRS.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ТЕХНИЧЕСКАЯ ПРОБЛЕМА
Таким образом, настоящее изобретение было выполнено путем модификации существующей вариабельной области легкой цепи и вариабельной области тяжелой цепи антитела с целью получения антитела, обладающего лучшей аффинностью связывания с N-концевым доменом KRS, по сравнению с существующим антителом, которое специфически связывается с N-концевым доменом KRS, экспонированным на внеклеточной мембране.
Таким образом, цель настоящего изобретения состоит в том, чтобы обеспечить антитело или его фрагмент, специфически связывающееся с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS, лизил-тРНК синтетаза), содержащее:
(а) вариабельную область тяжелой цепи (VH), содержащую
(i) определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность SYDMS,
(ii) определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность X1IX2X3X4X5GX6X7YYADSVKG, где X1 представляет собой A или V, X2 представляет собой S, D или G, X3 представляет собой Y, P, S или A, X4 представляет собой D, Q, L или Y, X5 представляет собой N, M, S или G, X6 представляет собой N, R или P, X7 представляет собой T, V, I или S, и
(iii) определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность X8ALDFDY, где X8 представляет собой M или L, и
(b) вариабельную область легкой цепи (VL), содержащую
(i) определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность TGSSSNIGSNYVT,
(ii) определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность X9NX10X11RPS, где X9 представляет собой D, S или R, X10 представляет собой S или N, и X11 представляет собой N или Q, и
(iii) определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность X12SFSDELGAYV, где X12 представляет собой A или S.
Другой целью настоящего изобретения является обеспечение полинуклеотида, кодирующего антитело или его фрагмент, рекомбинантного вектора экспрессии, содержащего полинуклеотид, и клетки, трансформированной этим вектором.
Другой целью настоящего изобретения является обеспечение способа получения антитела или его фрагмента, специфически связывающегося с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS), включающий:
(а) трансформацию клеток-хозяев рекомбинантным вектором экспрессии,
(b) инкубирование трансформированных клеток-хозяев для продукции антитела или его фрагмента, и
(c) сбор антитела или его фрагмента, продуцируемого в клетках-хозяевах.
Другой целью настоящего изобретения является обеспечение фармацевтической композиции для предотвращения или ингибирования рака и метастазов рака, содержащей антитело или его фрагмент в качестве активного ингредиента.
Кроме того, другой целью настоящего изобретения является обеспечение фармацевтической композиции для предотвращения или ингибирования метастазов рака, состоящей из антитела или его фрагмента в качестве активного ингредиента.
Кроме того, другой целью настоящего изобретения является обеспечение фармацевтической композиции для предотвращения или ингибирования метастазов рака, состоящей по существу из антитела или его фрагмента в качестве активного ингредиента.
Другой целью настоящего изобретения является обеспечение композиции для предотвращения или лечения заболевания, связанного с миграцией иммунных клеток, содержащей антитело или его фрагмент в качестве активного ингредиента.
Кроме того, другой целью настоящего изобретения является обеспечение композиции для предотвращения или лечения заболевания, связанного с миграцией иммунных клеток, состоящей из антитела или его фрагмента в качестве активного ингредиента.
Кроме того, другой целью настоящего изобретения является обеспечение композиции для предотвращения или лечения заболевания, связанного с миграцией иммунных клеток, состоящей по существу из антитела или его фрагмента в качестве активного ингредиента.
Другой целью настоящего изобретения является обеспечение композиции для диагностики заболевания, связанного с миграцией иммунных клеток, содержащей антитело или его фрагмент в качестве активного ингредиента.
Кроме того, другой целью настоящего изобретения является обеспечение композиции для диагностики заболевания, связанного с миграцией иммунных клеток, состоящей из антитела или его фрагмента в качестве активного ингредиента.
Кроме того, другой целью настоящего изобретения является обеспечение композиции для диагностики заболевания, связанного с миграцией иммунных клеток, состоящей по существу из антитела или его фрагмента в качестве активного ингредиента.
Другой целью настоящего изобретения является обеспечение применения антитела или его фрагмента для получения агента для предотвращения или ингибирования рака и метастазов рака.
Другой целью настоящего изобретения является обеспечение способа предотвращения или ингибирования рака и метастазов рака, включающего введение эффективного количества композиции, содержащей антитело или его фрагмент, субъекту, нуждающемуся в этом.
Другой целью настоящего изобретения является обеспечение применения антитела или его фрагмента для получения агента для диагностики рака или метастазов рака.
Другой целью настоящего изобретения является обеспечение способа диагностики рака или метастазов рака, включающего:
а) получение биологического образца от индивидуума (субъекта) с подозрением на метастазы рака;
b) введение композиции, содержащей антитело или его фрагмент, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику рака и метастазов рака, если уровень экспрессии KRS повышен.
Другой целью настоящего изобретения является обеспечение применения антитела или его фрагмента для получения агента для лечения заболевания, связанного с миграцией иммунных клеток.
Другой целью настоящего изобретения является обеспечение способа лечения заболевания, связанного с миграцией иммунных клеток, включающего введение эффективного количества композиции, содержащей антитело или его фрагмент, субъекту, нуждающемуся в этом.
Другой целью настоящего изобретения является обеспечение применения антитела или его фрагмента по п. 1 для получения агента для диагностики заболевания, связанного с миграцией иммунных клеток.
Другой целью настоящего изобретения является обеспечение способа диагностики заболевания, связанного с миграцией иммунных клеток, включающего:
а) получение биологического образца от субъекта с подозрением на заболевание, связанное с миграцией иммунных клеток,
b) введение композиции, содержащей антитело или его фрагмент, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику заболевания, связанного с миграцией иммунных клеток, если уровень экспрессии KRS повышен.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ
Для достижения вышеуказанной цели настоящее изобретение обеспечивает антитело или его фрагмент, специфически связывающееся с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS, лизил-тРНК синтетаза), содержащее:
(а) вариабельную область тяжелой цепи (VH), содержащую
(i) определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность SYDMS,
(ii) определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность X1IX2X3X4X5GX6X7YYADSVKG, где X1 представляет собой A или V, X2 представляет собой S, D или G, X3 представляет собой Y, P, S или A, X4 представляет собой D, Q, L или Y, X5 представляет собой N, M, S или G, X6 представляет собой N, R или P, X7 представляет собой T, V, I или S, и
(iii) определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность X8ALDFDY, где X8 представляет собой M или L, и
(b) вариабельную область легкой цепи (VL), содержащую
(i) определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность TGSSSNIGSNYVT,
(ii) определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность X9NX10X11RPS, где X9 представляет собой D, S или R, X10 представляет собой S или N, и X11 представляет собой N или Q, и
(iii) определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность X12SFSDELGAYV, где X12 представляет собой A или S.
Для достижения вышеуказанной цели настоящее изобретение обеспечивает полинуклеотид, кодирующий антитело или его фрагмент, рекомбинантный вектор экспрессии, содержащий полинуклеотид, и клетку, трансформированную указанным вектором.
В еще одном аспекте настоящего изобретения обеспечен способ получения антитела или его фрагмента, специфически связывающегося с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS), включающий:
(а) трансформацию клеток-хозяев рекомбинантным вектором экспрессии,
(b) инкубирование трансформированных клеток-хозяев для продукции антитела или его фрагмента, и
(c) сбор антитела или его фрагмента, продуцируемого в клетках-хозяевах.
Для достижения другой цели настоящее изобретение обеспечивает фармацевтическую композицию для предотвращения или ингибирования рака и метастазов рака, содержащую антитело или его фрагмент в качестве активного ингредиента.
Кроме того, настоящее изобретение обеспечивает фармацевтическую композицию для предотвращения или ингибирования метастазов рака, состоящую из антитела или его фрагмента в качестве активного ингредиента.
Кроме того, настоящее изобретение обеспечивает фармацевтическую композицию для предотвращения или ингибирования метастазов рака, состоящую по существу из антитела или его фрагмента в качестве активного ингредиента.
Для достижения другой цели настоящее изобретение обеспечивает композицию для предотвращения или лечения заболевания, связанного с миграцией иммунных клеток, содержащую антитело или его фрагмент в качестве активного ингредиента.
Кроме того, настоящее изобретение обеспечивает композицию для предотвращения или лечения заболевания, связанного с миграцией иммунных клеток, состоящую из антитела или его фрагмента в качестве активного ингредиента.
Кроме того, настоящее изобретение обеспечивает композицию для предотвращения или лечения заболевания, связанного с миграцией иммунных клеток, состоящую по существу из антитела или его фрагмента в качестве активного ингредиента.
Для достижения другой цели настоящее изобретение обеспечивает композицию для диагностики заболевания, связанного с миграцией иммунных клеток, содержащую антитело или его фрагмент в качестве активного ингредиента.
Кроме того, настоящее изобретение обеспечивает композицию для диагностики заболевания, связанного с миграцией иммунных клеток, состоящую из антитела или его фрагмента в качестве активного ингредиента.
Кроме того, настоящее изобретение обеспечивает композицию для диагностики заболевания, связанного с миграцией иммунных клеток, состоящую по существу из антитела или его фрагмента в качестве активного ингредиента.
Для достижения другой цели настоящее изобретение обеспечивает применение антитела или его фрагмента для получения агента для предотвращения или ингибирования рака и метастазов рака.
Для достижения другой цели настоящее изобретение обеспечивает способ предотвращения или ингибирования рака и метастазов рака, включающий введение эффективного количества композиции, содержащей антитело или его фрагмент, субъекту, нуждающемуся в этом.
Для достижения другой цели настоящее изобретение обеспечивает применение антитела или его фрагмента для получения агента для диагностики рака или метастазов рака.
Для достижения другой цели настоящее изобретение обеспечивает способ диагностики рака или метастазов рака, включающий:
а) получение биологического образца от индивидуума (субъекта) с подозрением на метастазы рака,
b) введение композиции, содержащей антитело или его фрагмент, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику рака и метастазов рака, если уровень экспрессии KRS повышен.
Для достижения другой цели настоящее изобретение обеспечивает применение антитела или его фрагмента для получения агента для лечения заболевания, связанного с миграцией иммунных клеток.
Для достижения другой цели настоящее изобретение обеспечивает способ лечения заболевания, связанного с миграцией иммунных клеток, включающий введение эффективного количества композиции, содержащей антитело или его фрагмент, субъекту, нуждающемуся в этом.
Для достижения другой цели настоящее изобретение обеспечивает применение антитела или его фрагмента по п. 1 для получения агента для диагностики заболевания, связанного с миграцией иммунных клеток.
Для достижения другой цели настоящее изобретение обеспечивает способ диагностики заболевания, связанного с миграцией иммунных клеток, включающий:
а) получение биологического образца от субъекта с подозрением на заболевание, связанное с миграцией иммунных клеток,
b) введение композиции, содержащей антитело или фрагмент, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику заболевания, связанного с миграцией иммунных клеток, если уровень экспрессии KRS повышен.
Далее настоящее изобретение описано более подробно.
В контексте настоящего изобретения термин "внеклеточно экспонированная N-концевая область лизил-тРНК синтетазы (KRS)" относится к конкретной последовательности, экспонированной во внеклеточную среду или на поверхности клеточной мембраны, когда KRS, продуцируемая в клетках, перемещается к клеточной мембране (или плазматической мембране), и обычно может относиться к частичной или полноразмерной последовательности области из от 1 до 72 аминокислот на N-конце KRS. Кроме того, существует сходство последовательностей у разных видов в N-концевой области KRS, и главным образом, N-концевая область KRS может содержать аминокислотную последовательность, определенную SEQ ID NO: 177.
В контексте настоящего изобретения термин "KRS" относится к полноразмерному полипептиду, известному как лизил-тРНК синтетаза, или к любой последовательности фрагмента KRS, содержащего N-концевую область. Как описано выше, антитела или их фрагменты по настоящему изобретению специфически определяют внеклеточно экспонированную N-концевую область KRS и, таким образом, также могут определять вышеупомянутый полноразмерный полипептид KRS или любую последовательность фрагмента KRS, содержащего N-концевую область. Конкретная последовательность KRS по существу не ограничена при условии, что последовательность содержит полипептид, определенный SEQ ID NO: 117, и известна в данной области техники как лизил-тРНК синтетаза. Например, KRS по настоящему изобретению содержит: последовательность, полученную от человека (Homo sapiens) и известную под номером доступа NP_005539.1 в NCBI (Genbank) или тому подобное; последовательность, полученную от мыши (Mus musculus) и известную под номером доступа NP_444322.1 в NCBI (Genbank) или тому подобное; и последовательность, полученную от крысы (Rattus norvegicus) и известную под номером доступа XP_006255692.1 в NCBI (Genbank) или тому подобное, и, кроме того, последовательность может быть представлена, но не ограничена ими, следующими: XP_005004655.1 (морская свинка: Cavia porcellus), XP_021503253.1 (песчанка, Meriones unguiculatus), XP_002711778.1 (кролик, Oryctolagus cuniculus), XP_536777.2 (собака, Canis lupusiliaris), XP_003126904.2 (свинья, Sus scrofa), XP_011755768.1 (обезьяна, Macaca nemestrina), XP_008984479.1 (мартышка, Callithrix jacchus), XP_019834275.1 (корова, Bos indicus), XP_511115.2 (шимпанзе, Pan troglodytes). Наиболее предпочтительно последовательность может быть известна под номером доступа NP_005539.1 в NCBI (Genbank).
В настоящем изобретении "антитело" также называется иммуноглобулином (Ig) и является общим термином для белков, которые участвуют в биологическом иммунитете путем избирательного воздействия на антигены. Полное антитело, встречающееся в природе, обычно состоит из двух пар легкой цепи (LC) и тяжелой цепи (HC), каждая из которых представляет собой полипептид, состоящий из нескольких доменов, или имеет две пары HC/LC в качестве основной единицы. Существует пять типов тяжелых цепей, составляющих антитела млекопитающих, которые обозначаются греческими буквами: α, δ, ε, γ и μ, и разные типы тяжелых цепей составляют разные типы антител: IgA, IgD, IgE, IgG и IgM, соответственно. Существует два типа легких цепей, составляющих антитела млекопитающих, которые обозначаются λ и κ.
Тяжелые и легкие цепи антител структурно разделены на вариабельную область и константную область в соответствии с вариабельностью аминокислотной последовательности. Константная область тяжелой цепи состоит из трех или четырех константных областей тяжелой цепи, таких как CH1, CH2 и CH3 (антитела IgA, IgD и IgG) и CH4 (антитела IgE и IgM), в зависимости от типа антитела, и легкая цепь имеет одну константную область CL. Каждая из вариабельных областей тяжелой и легкой цепей состоит из одного домена вариабельной области тяжелой цепи (VH) или вариабельной области легкой цепи (VL). Легкая цепь и тяжелая цепь связаны друг с другом одной ковалентной дисульфидной связью, в то время как их вариабельные области и константные области расположены параллельно, и две молекулы тяжелой цепи, которые связаны с легкими цепями, связаны друг с другом двумя ковалентными дисульфидными связями, в результате чего образуется полное антитело. Полное антитело специфически связывается с антигеном через вариабельные области тяжелой и легкой цепей. Полное антитело состоит из двух пар тяжелой и легкой цепей (HC-LC), и, таким образом, одна молекула полного антитела имеет двухвалентную моноспецифичность, где одна молекула полного антитела связывается с двумя одинаковыми антигенами через две вариабельные области.
Каждая из вариабельных областей антитела, которые содержат антигенсвязывающие сайты, подразделяется на каркасные области (FR) с низкой вариабельностью последовательностей и определяющие комплементарность области (CDR), которые представляют собой гипервариабельные области с высокой вариабельностью последовательностей. В VH и VL три CDR и четыре FR расположены в порядке FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4 в направлении от N-конца к C-концу. CDR, которые имеют наивысшую вариабельность последовательностей в вариабельных областях антитела, представляют собой сайты, которые напрямую связываются с антигеном, и очень важны для антигенной специфичности антитела.
Настоящее изобретение относится к антителу или его фрагменту, специфически связывающемуся с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS, лизил-тРНК синтетаза), содержащему:
(а) вариабельную область тяжелой цепи (VH), содержащую
(i) определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность SYDMS,
(ii) определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность X1IX2X3X4X5GX6X7YYADSVKG, где X1 представляет собой A или V, X2 представляет собой S, D или G, X3 представляет собой Y, P, S или A, X4 представляет собой D, Q, L или Y, X5 представляет собой N, M, S или G, X6 представляет собой N, R или P, X7 представляет собой T, V, I или S, и
(iii) определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность X8ALDFDY, где X8 представляет собой M или L, и
(b) вариабельную область легкой цепи (VL), содержащую
(i) определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность TGSSSNIGSNYVT,
(ii) определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность X9NX10X11RPS, где X9 представляет собой D, S или R, X10 представляет собой S или N, и X11 представляет собой N или Q, и
(iii) определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность X12SFSDELGAYV, где X12 представляет собой A или S.
В частности, где (а) вариабельная область тяжелой цепи (VH) содержит определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, и определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23 и SEQ ID NO: 118, и определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5 и SEQ ID NO: 25, и
где (b) вариабельная область легкой цепи (VL) содержит определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, SEQ ID NO: 27 и SEQ ID NO: 29, определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13 и SEQ ID NO: 15.
Антитела, состоящие из последовательностей CDR, обладают превосходной способностью специфически связываться с N-концевой областью KRS, экспонированной на внешней клеточной мембране. Это хорошо раскрыто в примерах описания настоящего изобретения.
В примере настоящего изобретения, для того, чтобы получить антитело, которое специфически связывается с N-концевой областью KRS, экспонированной на внешней клеточной мембране и имеющей высокую аффинность, после улучшения вариабельной области тяжелой цепи и вариабельной области легкой цепи существующего антитела N3 (номер заявки: 10-2018-0035446), улучшенную библиотеку отбирали посредством дрожжевой экспрессии.
Путем первичного, вторичного и третичного скрининга FACS, отбирали N3-1, N3-3 и N3-4 scFv, обладающих высокой аффинностью и специфичностью в N-концевой области KRS, и был выбран новый N3-5 scFv, в котором его VH и VL были объединены друг с другом.
Затем, чтобы выбрать улучшенную библиотеку, которая специфически связывается с GST-связанным N-концом KRS и имеет высокую аффинность, отбирали библиотеку, имеющую улучшенную вариабельную область тяжелой цепи N3-3 scFv. N3-6, N3-7, N3-8 и N3-9 scFv, обладающие высокой аффинностью и специфичностью в N-концевой области KRS, отбирали посредством первичного, вторичного, третичного и четвертого скрининга FACS.
Среди них, для повышения продуктивности и стабильности антитела N3-8 IgG, имеющего наивысшую аффинность, получали 7 видов антител (N3-8-1, N3-8-2, N3-8-3, N3-8- 4, N3-8-5, N3-8-6, N3-8-7) путем введения мутаций в области тяжелой и легкой цепи N3-8.
В результате получили антитела N3-1 IgG, N3-3 IgG, N3-4 IgG, N3-5 IgG, N3-6 IgG, N3-7 IgG, N3-8 IgG, N3-9 IgG, N3-8-1 IgG, N3-8-2 IgG, N3-8-3 IgG, N3-8-4 IgG, N3-8-5 IgG, N3-8-6 IgG и N3-8-7 IgG, и подтвердили, что антитело также проявляет высокую аффинность к N-концу KRS.
"Антитело или его фрагмент, которое специфически связывается с N-концевой областью KRS, экспонированной на внеклеточной мембране" по настоящему изобретению, не ограничивается ими, но предпочтительно представляет собой антитело, содержащее следующие комбинации CDR вариабельной области тяжелой цепи и вариабельной области легкой цепи, где следующие: i, ii, iii, iv, v, vi, vii, viii, ix, x, xi, xii и xiii, соответственно, составляют комбинации CDR антител N3-1, N3-3, N3 -4, N3-5, N3-6, N3-7, N3-8, N3-9, N3-8-1, N3-8-2, N3-8-3, N3-8-4, N3-8-5, N3-8-6 и N3-8-7 в следующих примерах.
Антитело или его фрагмент по п. 1, содержащее
i) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 3, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 13;
ii) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 3, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
iii) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 118, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющая комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 13,
iv) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 118, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность определено SEQ ID NO: 15,
v) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 17, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
vi) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 19, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
vii) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
viii) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 23, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
ix) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 27, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
x) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 29, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
xi) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
xii) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 27, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15, или
xiii) антитело, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 29, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15.
Наиболее предпочтительно, антитело или его фрагмент по настоящему изобретению содержит вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL) следующим образом:
В антителе или его фрагменте, вариабельная область тяжелой цепи содержит одну или более аминокислотных последовательностей, выбранных из группы, состоящей из SEQ ID NO: 31, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45 и SEQ ID NO: 47, и вариабельная область легкой цепи содержит одну или более аминокислотных последовательностей, выбранных из группы, состоящей из SEQ ID NO: 49, SEQ ID NO: 51, SEQ ID NO: 53 и SEQ ID NO: 55.
Предпочтительно, обеспечено антитело, содержащее вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 31, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 49; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 31, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 35, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 49; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 35, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 37, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 39, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 41, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 43, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 45, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 45, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 53; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 45, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 55; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 47, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 51; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 47, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 53; вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 47, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 55.
Конкретнее, антитело типа IgG, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), может представлять собой антитело, состоящее из тяжелой цепи, содержащей по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 89, SEQ ID NO: 93, SEQ ID NO: 95, SEQ ID NO: 97, SEQ ID NO: 99, SEQ ID NO: 101, SEQ ID NO: 103 и SEQ ID NO: 105, и легкой цепи, содержащей по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 107, SEQ ID NO: 109, SEQ ID NO: 111, SEQ ID NO: 113 и SEQ ID NO: 115.
Наиболее предпочтительно, обеспечено антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 89, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 107; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 89, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 93, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 107; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 93, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 95, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 97, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 99, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 101, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 103, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 103, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 113; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 103, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 115; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 105, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 105, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 113; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 105, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 115; обеспечено антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 99, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111.
"Антитело, специфически связывающееся с внеклеточно экспонированной N-концевой областью KRS" по настоящему изобретению, не ограничено его типом, пока антитело имеет вышеуказанные комбинации CDR или комбинации VH и VL. В качестве конкретного примера, антитело может быть выбрано из группы, состоящей из антител IgG, IgA, IgM, IgE и IgD, и предпочтительно может представлять собой антитело IgG.
Антитела по настоящему изобретению могут представлять собой моноклональные антитела или поликлональные антитела при условии, что антитела имеют вышеуказанные комбинации CDR или комбинации VH и VL, которые специфически связываются с N-концевой областью KRS, но предпочтительно представляют собой моноклональные антитела, которые представляют собой группу антител, каждое из которых имеет по существу идентичные аминокислотные последовательности в тяжелой и легкой цепях.
Антитело по настоящему изобретению может быть получено от любых животных, включая млекопитающих, включая людей, и птиц, и предпочтительно может быть получено от человека. Однако антитело по настоящему изобретению может представлять собой химерное антитело, содержащее часть антитела, полученную от человека, и часть антитела, полученную от другого вида животных. То есть настоящее изобретение включает все химерные антитела, гуманизированные антитела и человеческие антитела, и предпочтительно человеческие антитела.
Кроме того, фрагмент антитела по настоящему изобретению относится к фрагменту антитела, который сохраняет антиген-специфическую связывающую способность полного антитела. Предпочтительно фрагмент сохраняет по меньшей мере 20%, 50%, 70%, 80%, 90%, 95% или 100% аффинности связывания с N-концом KRS исходного антитела. В частности, фрагмент может быть в форме Fab, F(ab)2, Fab', F(ab')2, Fv, диатела, scFv или т.п.
Fab (фрагмент, антигенсвязывающий) представляет собой антигенсвязывающий фрагмент антитела и состоит из тяжелой цепи и легкой цепи, причем каждая состоит из одного вариабельного домена и одного константного домена. F(ab')2 представляет собой фрагмент, полученный в результате гидролиза антитела под действием пепсина, и F(ab')2 имеет форму, в которой две молекулы Fab связаны дисульфидными связями в шарнирной области тяжелой цепи. F(ab') представляет собой мономерный фрагмент антитела, в котором шарнир тяжелой цепи добавлен к Fab, отделенному от фрагмента F(ab')2 в результате восстановления его дисульфидных связей. Fv (вариабельный фрагмент) представляет собой фрагмент антитела, состоящий только из соответствующих вариабельных областей тяжелой и легкой цепей. scFv (одноцепочечный вариабельный фрагмент) представляет собой рекомбинантный фрагмент антитела, в котором вариабельная область тяжелой цепи (VH) и вариабельная область легкой цепи (VL) связаны друг с другом через гибкий пептидный линкер. Диатело относится к фрагменту, в котором VH и VL scFv связаны очень коротким линкером и, таким образом, не могут связываться друг с другом, и связываются с VL и VH другого scFv в той же самой форме, соответственно, с образованием димера.
В целях настоящего изобретения фрагмент антитела не ограничен его структурой или конформацией, пока фрагмент антитела сохраняет специфичность связывания с N-концевой областью KRS, но предпочтительно может представлять собой scFv. scFv по настоящему изобретению имеет конформацию CDR или конформацию VH и VL, специфическую для N-концевой области KRS, и ее последовательность по существу не ограничена, при условии, что C-конец VH и N-конец VL связаны через линкер. Линкер конкретно не ограничен его типом, поскольку известен как линкер, применяемый для scFv в данной области техники, но может представлять собой пептид, содержащий аминокислотную последовательность, определенную SEQ ID NO: 57. В частности, scFv по настоящему изобретению может содержать аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 61, SEQ ID NO: 63, SEQ ID NO: 65, SEQ ID NO: 67, SEQ ID NO: 69, SEQ ID NO: 71, SEQ ID NO: 73, SEQ ID NO: 75, SEQ ID NO: 77, SEQ ID NO: 79, SEQ ID NO: 81, SEQ ID NO: 83, SEQ ID NO: 85 и SEQ ID NO: 87.
Антитело или его фрагмент по настоящему изобретению может содержать консервативную аминокислотную замену (также называемую консервативным вариантом антитела), которая по существу не изменяет его биологическую активность.
Кроме того, вышеуказанное антитело или его фрагмент по настоящему изобретению можно конъюгировать с ферментом, флуоресцентным веществом, радиоактивным веществом и белком, но не ограничиваясь ими. Также в данной области техники хорошо известны способы конъюгирования вышеуказанных веществ с антителом.
Кроме того, настоящее изобретение обеспечивает полинуклеотид, кодирующий антитело или его фрагмент.
В описании настоящего изобретения полинуклеотид может быть описан как олигонуклеотид или нуклеиновая кислота и включает: аналоги ДНК или РНК (например, пептидные нуклеиновые кислоты и не встречающиеся в природе нуклеотидные аналоги), полученные с применением молекул ДНК (например, кДНК или геномной ДНК), молекул РНК (например, мРНК) или нуклеотидных аналогов; и их гибриды. Полинуклеотид может быть одноцепочечным или двухцепочечным.
Полинуклеотид относится к нуклеотидной последовательности, кодирующей антитело, состоящее из тяжелой и легкой цепей, причем каждая имеет конформацию CDR или конформацию VH и VL, специфическую к N-концевой области KRS. Полинуклеотид по настоящему изобретению конкретно не ограничен его последовательностью, при условии, что последовательность кодирует антитело или его фрагмент по настоящему изобретению. Полинуклеотиды, кодирующие вышеуказанные последовательности CDR в вышеописанных антителах по настоящему изобретению, конкретно не ограничены их последовательностями, но могут предпочтительно содержать нуклеотидную последовательность, определенную SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 26, SEQ ID NO: 28, SEQ ID NO: 30 или SEQ ID NO: 119.
Кроме того, полинуклеотиды, кодирующие вышеуказанные VH и VL в антителе по настоящему изобретению, конкретно не ограничены их последовательностями, но могут предпочтительно содержать нуклеотидную последовательность, определенную SEQ ID NO: 32 (VH), SEQ ID NO: 34 (VL), SEQ ID NO: 36 (VH), SEQ ID NO: 38 (VH), SEQ ID NO: 40 (VH), SEQ ID NO: 42 (VH), SEQ ID NO: 44 (VH), SEQ ID NO: 46 (VH), SEQ ID NO: 48 (VH), SEQ ID NO: 50 (VL), SEQ ID NO: 52 (VL), SEQ ID NO: 54 (VL) или SEQ ID NO: 56 (VL).
Кроме того, полинуклеотид, кодирующий фрагмент антитела, может предпочтительно содержать нуклеотидную последовательность любой последовательности, выбранной из группы, состоящей из SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66, SEQ ID NO: 68, SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 74, SEQ ID NO: 76, SEQ ID NO: 78, SEQ ID NO: 80, SEQ ID NO: 82, SEQ ID NO: 84, SEQ ID NO: 86 и SEQ ID NO: 88, которые кодируют фрагменты scFv по настоящему изобретению.
Полинуклеотиды, кодирующие антитело или его фрагмент по настоящему изобретению, можно получить способом, известным в данной области техники. Например, на основе последовательностей ДНК, кодирующих часть или полноразмерные тяжелые и легкие цепи антитела или соответствующих аминокислотных последовательностей, полинуклеотиды могут быть получены способами синтеза олигонуклеотидов, которые известны в данной области техники, например, метод полимеразной цепной реакции (ПЦР).
Настоящее изобретение обеспечивает рекомбинантный вектор экспрессии, содержащий полинуклеотид, кодирующий антитело или его фрагмент по настоящему изобретению.
В контексте настоящего описания термин "рекомбинантный" используется как синоним термина "генетическая манипуляция" и относится к конструкции гена в форме, не существующей в природе, с применением экспериментальных способов молекулярного клонирования, таких как генная трансформация, расщепление или связывание.
В контексте настоящего описания термин "экспрессия" относится к продукции белков или нуклеиновых кислот в клетках.
В контексте настоящего описания термин "рекомбинантный вектор экспрессии" представляет собой вектор, который может экспрессировать целевой белок или нуклеиновую кислоту (РНК) в подходящей клетке-хозяине, и относится к генетической конструкции, содержащей основные регуляторные элементы, которые функционально связаны с возможностью экспрессировать вставку полинуклеотида (гена). Термин "функционально связанный" относится к функциональной связи последовательности, регулирующей экспрессию нуклеиновой кислоты, и последовательности нуклеиновой кислоты, кодирующей целевой белок или РНК, так, чтобы выполнять общие функции, что означает связь между ними, с возможностью гена экспрессироваться с помощью данной регуляторной последовательности. Последовательность регуляции экспрессии относится к последовательности ДНК, которая контролирует экспрессию функционально связанной полинуклеотидной последовательности в конкретной клетке-хозяине. Такая последовательность регуляции экспрессии включает промотор транскрипции, любую последовательность оператора для регуляции транскрипции, последовательность для кодирования соответствующего сайта связывания рибосомы мРНК, последовательность регуляции терминации транскрипции и трансляции, кодон инициации, кодон терминации, сигнал полиаденилирования А, энхансер и т.п.
Рекомбинантный вектор экспрессии по настоящему изобретению конкретно не ограничен его типом, в том случае, если вектор обычно применяют в области клонирования, и примеры рекомбинантного вектора экспрессии включают плазмидный вектор, космидный вектор, бактериофаговый вектор, и вирусный вектор, но не ограничиваются ими. Примеры плазмиды могут включать плазмиды, полученные из Escherichia coli (pBR322, pBR325, pUC118, pUC119 и pET-22b(+)), плазмиды, полученные из Bacillus subtilis (pUB110 и pTP5), и плазмиды дрожжевого происхождения (YEp13, YEp24, и YCp50), и примеры вируса могут включать: вирусы животных, такие как ретровирус, аденовирус или вирус коровьей оспы; и вирусы насекомых, такие как бакуловирус.
Рекомбинантный вектор экспрессии по настоящему изобретению означает генетическую конструкцию, которая функционально связана таким образом, чтобы быть способной экспрессировать в подходящей клетке-хозяине полинуклеотид, кодирующий антитело или его фрагмент, состоящий из тяжелой и легкой цепей, имеющих вышеуказанные конформации CDR или VH и VL, способные специфически связывать N-концевую область KRS.
Полинуклеотиды, кодирующие тяжелую и легкую цепи антитела по настоящему изобретению, могут содержаться в отдельных рекомбинантных векторах экспрессии, соответственно, или могут содержаться в одном рекомбинантном векторе экспрессии.
Настоящее изобретение обеспечивает клетки, трансформированные описанным выше рекомбинантным вектором экспрессии.
Клетки по настоящему изобретению конкретно не ограничены их типом при условии, что клетки можно применять для экспрессии полинуклеотида, кодирующего антитело или его фрагмент, содержащийся в рекомбинантном векторе экспрессии по настоящему изобретению. Клетки (клетки-хозяева), трансформированные рекомбинантным вектором экспрессии по настоящему изобретению, могут представлять собой прокариотические клетки (например, E. coli), эукариотические клетки (например, дрожжи или другие грибы), растительные клетки (например, растительные клетки табака или томата), клетки животных (например, клетки человека, клетки обезьяны, клетки хомяка, клетки крысы, клетки мыши или клетки насекомых) или полученные из них гибридомы. Предпочтительно клетки могут происходить от млекопитающих, включая человека.
Типичные прокариоты, подходящие для настоящей цели, включают грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как Escherichia, например, E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescens, и Shigella, а также Bacilli, например, B. subtilis и B. licheniformis, Pseudomonas, например, P. aeruginosa, и Streptomyces. Клетки по настоящему изобретению конкретно не ограничены при условии, что клетки могут экспрессировать вектор по настоящему изобретению, но предпочтительно могут представлять собой E. coli.
Saccharomyces cerevisiae наиболее часто применяют в качестве эукариотических клеток по настоящему изобретению. Однако можно применять ряд других родов, видов и штаммов, но не ограничиваясь ими, например, Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как K. lactis, K. fragilis (ATCC 12,424), K. bulgaricus (ATCC 16,045), K. wickeramii (ATCC 24,178), K. waltii (ATCC 56,500), K. drosophilarum (ATCC 36,906), K. thermotolerans и K. marxianus; Yarrowia (EP 402 226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244 234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитчатые грибы, например, Neurospora, Penicillium, Tolypocladium и Aspergillus хозяева, такие как A. nidulans и A. niger.
Термин "трансформация" относится к модификации генотипа клетки-хозяина посредством введения экзогенных полинуклеотидов, и относится к введению экзогенного полинуклеотида в клетку-хозяина независимо от способа, применяемого для трансформации. Экзогенный полинуклеотид, введенный в клетку-хозяина, встраивается и сохраняется в геноме клетки-хозяина или сохраняется без встраивания в него, и настоящее изобретение включает оба варианта.
Рекомбинантный вектор экспрессии, способный экспрессировать антитело или его фрагмент, специфически связывающийся с N-концевой областью KRS по настоящему изобретению, можно вводить в клетки для получения антитела или его фрагмента способом, известным в данной области техники, например, но не ограничиваясь ими, временная трансфекция, микроинъекция, трансдукция, слияние клеток, осаждение с помощью фосфата кальция, трансфекция, опосредованная липосомами, трансфекция, опосредованная декстраном DEAE (диэтиламиноэтилдекстран), трансфекция, опосредованная полибреном, электропорация, генная пушка и известные способы введения нуклеиновых кислот в клетки, и таким образом можно трансформировать клетки.
В настоящем изобретении обеспечен способ получения антитела или его фрагмента, специфически связывающегося с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS), включающий:
(а) трансформацию клеток-хозяев рекомбинантным вектором экспрессии,
(b) инкубирование трансформированных клеток-хозяев для продукции антитела или его фрагмента, и
(c) сбор антитела или его фрагмента, продуцируемого в клетках-хозяевах.
На стадии (а) для получения антитела или его фрагмента по настоящему изобретению клетки-хозяева трансформируют рекомбинантным вектором экспрессии, в котором полинуклеотид, кодирующий антитело или его фрагмент, функционально связаны.
Специалист в данной области техники может выполнить данную стадию, выбрав подходящий способ трансформации в соответствии с выбранными клетками-хозяевами и рекомбинантным вектором экспрессии, как описано выше. Рекомбинантные векторы экспрессии, содержащие нуклеотидные последовательности тяжелой и легкой цепей, могут быть совместно трансформированы в одной и той же клетке-хозяине, чтобы позволить тяжелой и легкой цепям экспрессироваться в одной клетке, или рекомбинантные векторы экспрессии, содержащие нуклеотидные последовательности тяжелой и легкой цепей, можно трансформировать в отдельные клетки-хозяева, чтобы позволить тяжелой и легкой цепи экспрессироваться по отдельности.
На стадии (b) трансформированные клетки-хозяева инкубируют для получения полипептидов тяжелой и легкой цепей антитела или его фрагмента по настоящему изобретению от рекомбинантного вектора экспрессии, введенного в клетки-хозяева.
Состав среды, условия инкубации и время инкубации для инкубации клеток-хозяев могут быть надлежащим образом выбраны в соответствии со способом, обычно применяемым в данной области техники. Молекулы антитела, продуцируемые в клетке-хозяине, могут накапливаться в клеточной цитоплазме, могут секретироваться наружу из клетки или в культуральную среду с помощью подходящей сигнальной последовательности или могут быть нацелены с помощью периплазмы или тому подобного. Также предпочтительно, чтобы антитело по настоящему изобретению имело функциональную конформацию за счет повторного сворачивания белка с применением способа, известного в данной области техники, таким образом, чтобы сохранялась специфичность связывания с N-концевой областью KRS. В отношении получения антител типа IgG, тяжелые и легкие цепи могут экспрессироваться в отдельных клетках, а затем связываться друг с другом на отдельной стадии с образованием полного антитела, или тяжелые и легкие цепи могут экспрессироваться в одной и той же клетке с образованием полного антитела внутри клетки.
На стадии (c) получают антитело или его фрагмент, продуцируемое в клетках-хозяевах.
Специалист в данной области техники может правильно выбирать и контролировать способ сбора, учитывая характеристики полипептидов антитела или его фрагмента, продуцируемых в клетках-хозяевах, характеристики клеток-хозяев, способ экспрессии или наличие нацеливания или отсутствие нацеливания на полипептид. Например, антитело или его фрагмент, секретируемые в культуральную среду, можно собирать посредством получения культуральной среды, в которой культивируют клетки-хозяева, удаляя примеси центрифугированием и т.п. Для выделения, при необходимости, антитела, находящегося в конкретных органеллах или цитоплазме в клетках, из клеток и сбора антител, клетки можно лизировать в такой степени, которая не влияет на функциональную структуру антитела или его фрагмента. Полученное антитело можно дополнительно подвергнуть процессу дополнительного удаления примесей и осуществления концентрирования с помощью хроматографии, фильтрования с применением фильтра, диализа или тому подобного.
Полипептид в способе получения по настоящему изобретению может представлять собой собственно антитело или его фрагмент по настоящему изобретению и полипептид, с которым дополнительно связана другая аминокислотная последовательность, отличная от антитела или его фрагмента по настоящему изобретению. В данном случае аминокислотная последовательность может быть удалена из антитела или его фрагмента по настоящему изобретению с применением способа, хорошо известного специалисту в данной области техники.
Антитело или его фрагмент по настоящему изобретению специфически связывается с N-концевой областью KRS и, таким образом, может применяться в диагностическом анализе для выявления и количественного определения белков KRS, например, в конкретных клетках, тканях или сыворотке. В частности, внеклеточно экспонированную N-концевую область KRS можно специфически выявлять без лизиса клеток.
Способ выявления по настоящему изобретению может включать стадию получения образца, который подлежит оцениванию на наличие или отсутствие KRS (или внеклеточно экспонированного N-концевого пептида KRS) и измерению его концентрации с применением антитела или его фрагмента по настоящему изобретению (стадия (1)) перед взаимодействием антитела или его фрагмента по настоящему изобретению с образцом.
Специалист в данной области техники может подходящим образом выбрать известный способ выявления белка с применением антитела и получить образец, подходящий для выбранного способа. Кроме того, образец может представлять собой клетки или ткани, полученные с помощью биопсии, кровь, цельную кровь, сыворотку, плазму, слюну, спинномозговую жидкость и т.п., которые взяты у субъекта, подлежащего обследованию на наличие или отсутствие рака (особенно рак молочной железы или рак легкого) или метастазов рака. Примеры способа выявления белка с применением антитела включают, помимо прочего, вестерн-блоттинг, иммуноблоттинг, дот-блоттинг, иммуногистохимию, твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ, анализ конкурентного связывания, иммунопреципитацию и т.п. Например, в случае вестерн-блоттинга получение можно осуществлять путем добавления к образцу или клеточному лизату буфера, подходящего для электрофореза, с последующим кипячением, а в случае иммуногистохимии обработку можно проводить посредством иммобилизации и блокирования клеток или тканевых срезов с последующим блокированием.
Затем осуществляют стадию взаимодействия антитела или его фрагмента по настоящему изобретению с образцом, полученным на вышеописанной стадии (стадия (2)).
Антитело по настоящему изобретению представляет собой антитело или его фрагмент, которые имеют вышеописанные конформации CDR или VH и VL и специфически связываются с N-концевой областью KRS, и их конкретные типы, и организацию последовательности, как описано выше.
Антитело или его фрагмент могут быть помечены обычной детектируемой составляющей для их "выявления". Например, антитело или его фрагмент можно метить радиоизотопной или флуоресцентной меткой, применяя способы, известные в данной области техники. Кроме того, можно применять различные фермент-субстратные метки, и примеры ферментной метки включают: люциферазу, такую как люцифераза дрозофилы и бактериальная люцифераза (патент США №4737456), люциферин, 2,3-дигидрофталазин-дион, малатдегидрогеназу, уреазу, пероксидазу, такую как пероксидаза хрена (HRPO), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазу (например, глюкозооксидаза, галактозоксидаза и глюкозо-6-фосфат дегидрогеназа), гетероциклилоксидазу (например, уриказа и ксантиноксидаза), лактопероксидазу, микропероксидазу и т.д. Способы конъюгирования ферментов с антителами известны в данной области техники. Метки могут быть прямо или непрямо конъюгированы с антителами с применением различных известных способов. Например, антитело можно конъюгировать с биотином, и любые метки, относящиеся к трем классам широко распространенных категорий, перечисленных выше, могут быть конъюгированы с авидином или наоборот. Биотин может избирательно связываться с авидином, и поэтому данная метка может быть конъюгирована с антителом таким непрямым образом. В качестве альтернативы, для достижения непрямой конъюгации метки с антителом, антитело может быть конъюгировано с небольшим гаптеном (например, диоксином), и один из различных типов меток, перечисленных выше, может быть конъюгирован с антителом против гаптена (например, антителом против диоксина). Следовательно, может быть достигнута непрямая конъюгация метки с антителом.
В контексте настоящего документа термин "взаимодействие" применяют в общем смысле и он относится к смешиванию, связыванию или соприкосновению двух или более веществ. Взаимодействие можно осуществлять in vitro или в другом контейнере, или можно осуществлять на месте, in vivo, у субъекта, в ткани или в клетке.
Далее проводят стадию выявления антитела или его фрагмента по настоящему изобретению в образце после выполнения стадии (2) (стадия (3)).
"Выявление" проводят на комплексе антитела или его фрагмента по настоящему изобретению и антигена, где указанный комплекс образуется в образце, и относится к выявлению присутствия или отсутствия N-концевого пептида KRS (или белка, включающего пептид, например, KRS), или измерению (включая качественное измерение, количественное измерение или и то, и другое) уровня пептида. Следовательно, способ выявления по настоящему изобретению может дополнительно включать стадию удаления избыточных антител или их фрагментов, которые не образовали комплекс с N-концевой областью KRS, после осуществления стадии (2) перед стадией (3), описанной позже.
Когда антитело или его фрагмент, применяемое на стадии (2), описанной выше, содержит выявляемый фрагмент, такой как флуоресценция, радиоактивный изотоп или фермент, который прямо метит антитело или его фрагмент, выявление можно осуществлять способом выявления для соответствующего фрагмента, известным в данной области техники. Например, радиоактивность можно измерять, например, посредством сцинтилляционного счета, а флуоресценцию можно количественно оценивать с применением флуорометра.
Когда антитело или его фрагмент, как таковое, применяемое на стадии (2), описанной выше, не содержит вышеупомянутого детектируемого фрагмента, может быть проведено непрямое выявление с применением вторичного антитела, меченного флуоресценцией, радиоактивностью, ферментом или т.п. Вторичное антитело связывается с антителом или его фрагментом (первичным антителом) по настоящему изобретению.
Настоящее изобретение обеспечивает фармацевтическую композицию для предотвращения или ингибирования метастазов рака и композицию для диагностики рака, содержащую антитело или его фрагмент по настоящему изобретению в качестве активного ингредиента.
Рак конкретно не ограничен его типом, в том случае, если рак известен как злокачественная опухоль в данной области техники, и его пример может быть выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака костей, рака поджелудочной железы, рака кожи, рака головы или шеи, кожной или внутриглазной меланомы, рака матки, рака яичника, рака прямой кишки, рака анального канала, рака толстой кишки, рака маточной трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карцинома вульвы, лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака уретры, рака полового члена, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или мочеточника, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли центральной нервной системы, первичной лимфомы центральной нервной системы, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза. Предпочтительно рак может представлять собой рак молочной железы или рак легкого.
Настоящее изобретение может включать только антитело или его фрагмент по настоящему изобретению или может дополнительно включать по меньшей мере один фармацевтически приемлемый носитель. В контексте настоящего документа термин "фармацевтически приемлемый" относится к нетоксичной композиции, которая является физиологически приемлемой, не ингибирует действие активного ингредиента при введении человеку и обычно не вызывает серьезных побочных эффектов.
В фармацевтической композиции по настоящему изобретению антитело или его фрагмент можно вводить в нескольких пероральных и парентеральных лекарственных формах во время клинического введения. Антитело или его фрагмент в составе композиции можно получать с применением обычно используемых разбавителя или эксципиента, такого как наполнитель, разбавитель, связующее вещество, смачивающий агент, разрыхлитель или поверхностно-активное вещество. Твердые композиции для перорального введения включают таблетку, пилюлю, порошок, гранулы, капсулу, пастилку и т.п. Данные твердые композиции могут быть получены путем смешивания арильного производного химической формулы 1 по настоящему изобретению или его фармацевтически приемлемой соли с по меньшей мере одним эксципиентом, например, крахмалом, карбонатом кальция, сахарозой или лактозой или желатином. Кроме того, помимо простых эксципиентов можно применять смазывающие вещества, такие как стеарат магния и тальк. Жидкие композиции для перорального введения включают: суспензию, раствор для внутреннего применения, эмульсию, сироп и т.п. Помимо часто применяемых простых разбавителей, таких как вода и жидкий парафин, в жидких композициях могут содержаться несколько эксципиентов, например, смачивающий агент, подсластитель, ароматизатор, консервант и т.п.
Типичные композиции для парентерального введения включают стерильный водный раствор, неводный растворитель, суспензионный растворитель, эмульсию, лиофилизирующий агент и суппозиторий. Композиция для лечения по настоящему изобретению может быть получена в виде лиофилизата или водного раствора для смешивания и хранения любого физиологически приемлемого носителя, эксципиента или стабилизатора, и антитела с предпочтительной чистотой. Приемлемый носитель, эксципиент или стабилизатор нетоксичен для пользователя в применяемой дозе и концентрации, и их примеры включают: буферы, например фосфорную кислоту, лимонную кислоту и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; низкомолекулярные полипептиды (менее приблизительно 10 остатков); белки, например сывороточный альбумин, желатин или иммуноглобулин; гидрофильные полимеры, например поливинилпирролидон; аминокислоты, например глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрин; хелатирующие агенты, например 1,2-этандитиол; сахароспирты, например маннит или сорбит; солеобразующие противоионы, например натрий; и (или) неионные поверхностно-активные вещества, например твин, плюроники или полиэтиленгликоль (ПЭГ).
Антитело по настоящему изобретению можно вводить в фармацевтически эффективном количестве субъекту, борющемуся с раком или заболеванием, связанным с миграцией иммунных клеток. В контексте настоящего документа термин "фармацевтически эффективное количество" относится к количеству, показывающему более сильный ответ по сравнению с отрицательным контролем, и предпочтительно относится к количеству, достаточному для лечения рака, количеству, достаточному для предотвращения или ингибирования метастазов рака, и количеству, достаточному для лечения заболевания, связанного с миграцией иммунных клеток. Общее эффективное количество антитела или его фрагмента по настоящему изобретению можно вводить пациенту в виде однократной дозы или можно вводить по протоколу фракционированного лечения, при котором многократные дозы вводят в течение длительного периода времени. Доза антитела или его фрагмента по настоящему изобретению для организма человека может обычно составлять от 0,01 до 100 мг/кг/неделю, предпочтительно от 0,1 до 20 мг/кг/неделю, и более предпочтительно от 5 до 10 мг/кг/неделю. Однако, в отношении дозы антитела или его фрагмента по настоящему изобретению, его эффективную дозу для пациента определяют с учетом различных факторов, например, способа введения фармацевтической композиции, количества раз, за которое осуществляется лечение, возраста пациента, массы тела, состояния здоровья и пола, тяжести заболевания, рациона и скорости выведения, и, таким образом, с учетом данных факторов, специалист в данной области техники может определить подходящее эффективное количество антитела или его фрагмента по настоящему изобретению в соответствии с конкретным применением в качестве средства предотвращения или ингибитора метастазов рака. Фармацевтическая композиция по настоящему изобретению конкретно не ограничена лекарственной формой, путем введения и способом ее введения, если композиция демонстрирует действие по настоящему изобретению.
Путь введения композиции по настоящему изобретению может представлять собой известный способ введения антитела, например, инъекцию или инфузию посредством внутривенного, внутрибрюшинного, внутричерепного, подкожного, внутримышечного, внутриглазного, внутриартериального, спинномозгового или внутриочагового пути, или инъекцию или инфузию посредством системы с замедленным высвобождением, как описано ниже. Например, антитело по настоящему изобретению можно вводить в организм системно или местно.
Фармацевтическую композицию по настоящему изобретению можно применять отдельно или в комбинации с хирургическим вмешательством, гормональной терапией, химиотерапией и способами с применением регулятора биологической реакции, для рака или метастазов рака.
Диагностику и прогнозирование рака (или метастазов рака) в соответствии с настоящим изобретением можно осуществлять путем выявления белков KRS (особенно, внеклеточно экспонированной N-концевой области KRS) в биологическом образце.
В контексте настоящего документа термин "диагностика" относится к определению наличия или характеристик патологического состояния. В настоящем изобретении диагностика заключается в определении возникновения или вероятности (риска) возникновения рака или/и метастазов рака или заболевания, связанного с миграцией иммунных клеток.
Термин "выявление" имеет значение, описанное выше, и биологический образец включает кровь и другие жидкие образцы, имеющие биологическое происхождение, образцы биопсии, образцы плотных тканей, такие как культура ткани, или клетки, полученные из нее. В частности, примеры биологического образца могут включать, среди прочего, ткани, экстракты, клеточные лизаты, цельную кровь, плазму, сыворотку, слюну, внутриглазную жидкость, спинномозговую жидкость, пот, мочу, молоко, асцитическую жидкость, синовиальную жидкость, перитонеальную жидкость и т.п. Образец может быть получен от животных, предпочтительно млекопитающих и наиболее предпочтительно человека. Образец можно предварительно обрабатывать перед применением для выявления. Примеры предварительной обработки могут включать фильтрование, дистилляцию, экстракцию, концентрирование, дезактивацию интерференционного вещества, добавление реагента и т.п. Кроме того, для выявления можно применять нуклеиновые кислоты и белки, выделенные из образца.
Антитело или его фрагмент по настоящему изобретению может быть обеспечено в виде набора для диагностики. Набор конкретно не ограничен его типом в том случае, если набор известен в данной области техники, как набор для анализа, который обеспечивает пептид, имеющий антитело или конкретный связывающий домен в качестве компонента, и его примеры включают набор для вестерн-блоттинга, ELISA, радиоиммуноанализа, радиоиммунодиффузии, радиоиммунодиффузии по Оухтерлони, ракетного иммуноэлектрофореза, иммуногистохимия, анализа на основе иммунопреципитации, анализа фиксации комплемента, FACS, белкового чипа и т.п.
Антитело или его фрагмент по настоящему изобретению можно применять в наборе, то есть упакованной комбинации реагентов в заранее определенных количествах с инструкциями по выполнению диагностического анализа. Когда антитело мечено ферментом, набор может включать субстраты и кофакторы, необходимые для фермента (например, предшественник субстрата, который обеспечивает хромофор или флуорофор). Кроме того, могут быть включены другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или буфер для лизиса) и т.п. Относительные количества различных реагентов могут сильно различаться для обеспечения концентраций реагентов в растворе, которые по существу оптимизируют чувствительность анализа. Реагенты могут быть обеспечены в виде сухих порошков, обычно лиофилизированных, включая эксципиенты, которые при растворении будут обеспечивать получение раствора реагента, имеющего подходящую концентрацию.
В настоящем изобретении антитело представляет собой полипептид, содержащий вариант Fc области Fc человеческого IgG дикого типа, и вариант Fc содержит по меньшей мере одну дополнительную аминокислотную замену, такую как L117A, L118A, T182A, P212G области Fc человеческого IgG1 дикого типа, определенную SEQ ID NO: 126 или T179A области Fc человеческого IgG4, определенную SEQ ID NO: 138, где полипептид отличается тем, что он содержит полипептид, имеющий пониженную функцию ADCC/CDC по сравнению с полипептидом, содержащим область Fc IgG дикого типа.
В настоящем изобретении область Fc относится к C-концевой области тяжелой цепи иммуноглобулина, содержащей по меньшей мере часть константной области, и содержит последовательность области Fc дикого типа и вариант области Fc. В настоящем изобретении "вариант Fc" относится к полипептиду, содержащему модификацию в домене Fc. Вариант Fc по настоящему изобретению определяется в соответствии с составляющей его аминокислотной модификацией. В частности, L118A представляет собой вариант Fc, в котором лейцин заменен на аланин в положении 118, T182A представляет собой вариант Fc, в котором треонин заменен на аланин в положении 182, и P212G представляет собой вариант Fc, в котором пролин заменен на глицин в положении 212, по сравнению с исходным полипептидом Fc. Аминокислотные модификации могут представлять собой добавление аминокислот, делеции аминокислот или замены аминокислот. Аминокислотные замены могут включать встречающиеся в природе аминокислоты и не встречающиеся в природе аминокислоты. Варианты могут включать неприродные аминокислоты.
"Аминокислотная замена" относится к замене одного или более существующих аминокислотных остатков другим "замещающим" аминокислотным остатком в данной аминокислотной последовательности.
Замещающие остатки или остатки могут быть "встречающимися в природе аминокислотными остатками" (т.е. кодируемыми генетическим кодом) и могут быть выбраны из группы, состоящей из Аланина (Ala); Аргинина (Arg); Аспарагина (Asn); Аспарагиновой кислоты (Asp); Цистеина (Cys); Глутамина (Gln); Глутаминовой кислоты (Glu); Глицина (Gly); Гистидина (His); Изолейцина (Ile); Лейцина (Leu); Лизина (Lys); Метионина (Met); Фенилаланина (Phe); Пролина (Pro); Серина (Ser); Треонина (Thr); Триптофана (Trp); Тирозина (Tyr); и Валина (Val).
"Функция ADCC/CDC" означает функцию антителозависимой клеточно-опосредованной цитотоксичности (антителозависимая клеточная цитотоксичность, ADCC) и комплемент-зависимой цитотоксичности (комплемент-зависимая цитотоксичность, CDC). "Комплемент-зависимая цитотоксичность" (CDC) относится к лизису антиген-экспрессирующих клеток антителом по настоящему изобретению в присутствии комплемента. "Антителозависимая клеточно-опосредованная цитотоксичность" (ADCC) относится к неспецифическим цитотоксическим клеткам (например, натуральным киллерам (НК), нейтрофилам и макрофагам), экспрессирующим рецепторы Fc (FcR), которые распознают связанные антитела в клетках-мишенях и, следовательно, относится к клеточно-опосредованной реакции, при которой происходит растворение клеток-мишеней. CDC и ADCC можно измерить с помощью анализов, хорошо известных и доступных в данной области техники (Примеры ссылок: US 5,500,362 и 5,821,337, и Clynes et al. (1998) Proc. Natl. Acad. Sci. (USA) 95: 652-656). Константная область антитела важна для способности антитела фиксировать комплемент и опосредовать клеточно-зависимую цитотоксичность. Таким образом, изотип антитела можно выбрать на основании того, желательно ли, чтобы антитело опосредовало цитотоксичность.
В частности, антитело по настоящему изобретению представляет собой антитело, содержащее тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, представленных SEQ ID NO: 140, 142, 144, 146, 148, 150, 152, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 91;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 140, 142, 144, 146, 148, 150, 152, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 107;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 140, 142, 144, 146, 148, 150, 152, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 154, 156, 158, 160, 162, 164, 166, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 107;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 154, 156, 158, 160, 162, 164, 166, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109;
Тяжелую цепь, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из аминокислотных последовательностей, определенных SEQ ID NO: 168, 170, 172, 174, 176, 178, 180, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 182, 184, 186, 188, 190, 192 и 194, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 224, 226, 228, 230, 232, 234, 236, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 210, 212, 214, 216, 218, 220, 222, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 224, 226, 228, 230, 232, 234, 236, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 224, 226, 228, 230, 232, 234, 236, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 113;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 224, 226, 228, 230, 232, 234, 236, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 115;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 238, 240, 242, 244, 246, 248, 250, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111;
Тяжелую цепь, содержащую одну или более аминокислотных последовательностей, выбранных из аминокислотных последовательностей, определенных SEQ ID NO: 238, 240, 242, 244, 246, 248, 250, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 113;
Тяжелую цепь, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из аминокислотных последовательностей, определенных SEQ ID NO: 238, 240, 242, 244, 246, 248, 250, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 115, или
Тяжелую цепь, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из аминокислотных последовательностей, определенных SEQ ID NO: 196, 198, 200, 202, 204, 206, 208, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111.
Настоящее изобретение обеспечивает композицию для предотвращения или лечения заболеваний, связанных с миграцией иммунных клеток, и композицию для диагностики заболеваний, связанных с миграцией иммунных клеток, содержащую антитело или его фрагмент в качестве активного ингредиента.
В настоящем изобретении термин "заболевание, связанное с миграцией иммунных клеток", например, может быть выбрано из группы, состоящей из сердечно-сосудистого заболевания, фиброзного заболевания, воспалительного заболевания и синдрома Альпорта, при этом конкретный тип заболевания не ограничен, если в данной области техники известно, что избыточная миграция (и/или инвазия) иммунных клеток представляет собой основной патогенез заболевания.
Сердечно-сосудистое заболевание может, например, быть выбрано из группы, состоящей из гипертензии (включая воспалительные осложнения, вызванные гипертензией), легочной артериальной гипертензии, атеросклероза, стенокардии, инфаркта миокарда, ишемического цереброваскулярного заболевания, артериосклероза и мезентериального склероза, но конкретно не ограничено ими.
Фиброзное заболевание может быть выбрано из группы, состоящей, например, из склеродермы, ревматоидного артрита, болезни Крона, язвенного колита, миелофиброза, фиброза легкого, фиброза печени, цирроза печени, фиброза почек, гломерулосклероза, миофиброза, фиброза сердца, интерстициального фиброза, фиброза поджелудочной железы, фиброза селезенки, фиброза средостения, фиброза сосудов, фиброза кожи, фиброза глаз, дистрофии желтого пятна, фиброза суставов, фиброза щитовидной железы, эндомиокардиального фиброза, перитонеального фиброза, ретроперитонеального фиброза, массивного прогрессивного фиброза, нефрогенного системного фиброза, системной красной волчанки, наследственного фиброза, инфекционного фиброза, фиброза, вызываемого раздражителем, фиброза, вызываемого хроническим аутоиммунным заболеванием, фиброза, вызываемого антигенной несовместимостью во время трансплантации органов, фиброзные осложнения после операции, фиброза, вызываемого гиперлипидемией, фиброза, вызываемого ожирением, диабетического фиброза, фиброза, вызываемого гипертензией и окклюзии, вызываемой фиброзом при имплантации стента, но конкретно не ограничено ими.
В настоящем изобретении воспалительное заболевание предпочтительно может быть выбрано из группы, состоящей из аутоиммунного заболевания, воспалительного заболевания кишечника, дерматита (например, атопического дерматита, экземы, псориаза и т.д.), диабетического заболевания глаз (диабетической ретинопатии и т.д.), перитонита, остеомиелита, целлюлита, менингита, энцефалита, панкреатита, травматического шока, бронхиальной астмы, ринита, синусита, тимпанита, пневмонии, гастрита, энтерита, муковисцидоза, апоплексии (апоплексия, инсульт и др.), бронхита, бронхиолита, гепатита (цирроз, неалкогольный, стеатогепатит и др.), нефрита (диабетическая почечная недостаточность и др.), протеинурии, артрита (например, псориатического артрита, остеоартрита), неврита (диабетическая нейропатия, рассеянный склероз и др.), подагры, спондилита, синдрома Рейтера, узелкового полиартериита, васкулита, бокового амиотрофического склероза, гранулематоза Вегенера, гиперцитокинемии, ревматической полимиалгии, артикулярно-клеточного артериита, кристаллических артритов с отложением кальция, псевдоподагры, несуставного ревматоида, бурсита, тендосиновита, эпикондилита (теннисный локоть), сустава Шарко, гемартроза, пурпуры Геноха-Шонлейна, гипертрофического остеоартрита, многоцентровой ретикулоцитомы, саркоидоза, гемохроматоза, дрепаноцитоза, гиперлипопроитенемии, гипомагглобулинемии, гиперпаратиреоза, акромегалии, семейной Средиземноморской лихорадки, болезни Бехчета, системной красной волчанки, рецидивирующей лихорадки, псориаза, рассеянного склероза, сепсиса, септического шока, острого респираторного дистресс-синдрома, множественной дисфункции органов, хронической обструктивной болезни легкого, острого повреждения легкого и бронхо-легочной дисплазии, а также включает хронические воспалительные заболевания, но конкретно не ограничено ими.
В настоящем изобретении аутоиммунные заболевания могут быть выбраны из группы, состоящей из ревматоидного артрита, системной склеродермии, системной красной волчанки, псориаза, астмы, язвенного колита, болезни Бехчета, болезни Крона, рассеянного склероза, дерматита, коллагеновой болезни, васкулита, артрита, гранулематоза, органоспецифических аутоиммунных заболеваний, язвенного колита и GvHD (болезнь "трансплантат против хозяина").
Хроническое воспалительное заболевание относится к состоянию, при котором вышеописанные типы воспалительных заболеваний становятся хроническими, и их предпочтительные примеры включают астму, атопический дерматит, экзему, псориаз, остеоартрит, подагру, псориатический артрит, цирроз, неалкогольный стеатогепатит, хроническую обструктивную болезнь легкого, ринит, диабетическую ретинопатию, диабетическую почечную недостаточность, диабетическую невропатию и рассеянный склероз, но не ограничиваются ими.
Кроме того, антитело по настоящему изобретению можно вводить в фармацевтически эффективном количестве субъекту, страдающему заболеванием, связанным с миграцией иммунных клеток. Как описано выше, термин "фармацевтически эффективное количество" относится к количеству, которое демонстрирует более высокий ответ по сравнению с группой отрицательного контроля, и предпочтительно относится к количеству, достаточному для лечения заболеваний, связанных с миграцией иммунных клеток. Общее эффективное количество антитела или его фрагмента по настоящему изобретению, состав композиции, способ введения и путь введения описаны выше.
Фармацевтическую композицию по настоящему изобретению можно применять отдельно или в комбинации с хирургическим вмешательством, гормональной терапией, химиотерапией и способами, в которых применяют модификаторы биологического ответа, для предотвращения или лечения заболеваний, связанных с миграцией иммунных клеток.
Диагностику и прогнозирование заболеваний, связанных с миграцией иммунных клеток в соответствии с настоящим изобретением, можно проводить путем выявления белка KRS (особенно, N-концевой области KRS, экспонированной на внеклеточной мембране) в биологическом образце.
Кроме того, настоящее изобретение обеспечивает применение антитела или его фрагмента для получения агента для предотвращения или ингибирования рака и метастазов рака.
Кроме того, настоящее изобретение обеспечивает способ предотвращения или ингибирования рака и метастазов рака, путем введения эффективного количества композиции, содержащей антитело или его фрагмент, субъекту, нуждающемуся в этом.
Кроме того, настоящее изобретение обеспечивает применение антитела или его фрагмента для получения агента для диагностики рака или метастазов рака.
Настоящее изобретение обеспечивает способ диагностики рака или метастазов рака, включающий:
а) получение биологического образца от индивидуума (субъекта) с подозрением на метастазы рака,
b) введение композиции, содержащей антитело или его фрагмент, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику рака и метастазов рака, если уровень экспрессии KRS повышен.
Настоящее изобретение обеспечивает применение антитела или его фрагмента для получения агента для лечения заболевания, связанного с миграцией иммунных клеток.
Настоящее изобретение обеспечивает применение антитела или его фрагмента для получения агента для диагностики заболевания, связанного с миграцией иммунных клеток.
Настоящее изобретение обеспечивает способ лечения заболевания, связанного с миграцией иммунных клеток, путем введения эффективного количества композиции, содержащей антитело или его фрагмент, субъекту, нуждающемуся в этом.
Настоящее изобретение обеспечивает способ диагностики заболевания, связанного с миграцией иммунных клеток, включающий:
а) получение биологического образца от субъекта с подозрением на заболевание, связанного с миграцией иммунных клеток,
b) введение композиции, содержащей антитело или его фрагмент, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику заболевания, связанного с миграцией иммунных клеток, если уровень экспрессии KRS повышен.
В одном варианте осуществления настоящее изобретение обеспечивает способ диагностики и лечения рака и метастазов рака, включающий следующие стадии:
а) получение биологического образца от индивидуума (субъекта) с подозрением на метастазы рака,
b) введение композиции, содержащей антитело или его фрагмент, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику рака и метастазов рака, если уровень экспрессии KRS повышен, и
e) введение терапевтического лекарственного средства для лечения рака и метастазов рака субъекту с установленным диагнозом или лечение заболевания путем хирургического вмешательства.
В одном варианте осуществления настоящее изобретение обеспечивает способ диагностики и лечения заболеваний, связанных с миграцией иммунных клеток, включающий следующие стадии:
а) получение биологического образца от субъекта с подозрением на заболевание, связанное с миграцией иммунных клеток,
b) введение композиции, содержащей антитело или его фрагмент по п. 1, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b), и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику заболевания, связанного с миграцией иммунных клеток, если уровень экспрессии KRS повышен,
e) введение терапевтического лекарственного средства для лечения заболевания, связанного с миграцией иммунных клеток, субъекту с установленным диагнозом или лечение заболевания путем хирургического вмешательства.
Стадия e) представляет собой стадию лечения заболевания субъекта, с установленным диагнозом на стадии d), с помощью таких средств, как введение терапевтических лекарственных средств или хирургическое вмешательство.
"Лечение" по настоящему изобретению обычно относится к облегчению симптомов рака и метастазов рака или заболевания, связанного с миграцией иммунных клеток, и может включать лечение, по существу предотвращение или облегчение состояния рака и метастазов рака или заболеваний, связанных с миграцией иммунных клеток, и включает, но не ограничивается ими, облегчение, лечение или предотвращение одного или большинства симптомов, возникающих в результате заболевания.
Терапевтическое лекарственное средство конкретно не ограничено, если оно представляет собой вид лекарственного средства, обычно применяемого для лечения рака, метастазов рака или заболеваний, связанных с миграцией иммунных клеток, и в одном варианте осуществления оно может представлять собой одно или более лекарственное средство для лечения, выбранное из группы, состоящей из противоопухолевого агента, противовоспалительного агента (такого как стероидный агент в качестве иллюстративного примера) и терапевтического агента легочной артериальной гипертензии, но не ограничивается ими.
Терапевтическое лекарственное средство вводят субъекту в "терапевтически эффективном количестве", и терапевтически эффективное количество может быть определено специалистами в данной области техники, эффективная доза для пациента может быть определена с учетом различных факторов, таких как тяжесть заболевания пациента, рацион и скорость выведения, а также возраст, масса тела, состояние здоровья, пол и тип заболевания. Путь введения терапевтического лекарственного средства конкретно не ограничен и может представлять собой пероральный или парентеральный путь, и включать как местное, так и системное введение. Парентеральное введение может представлять собой, но не ограничивается ими, интраназальное введение лекарственного средства, подкожную инъекцию и т.п., и, в качестве другого примера, можно применять внутримышечную инъекцию, внутривенную инъекцию или т.п.
В контексте настоящего изобретения термин "содержащий" используется как синоним "включающий" или "характеризуемый" и не исключает дополнительных компонентов или стадий, не упомянутых в композиции или способе. Термин "состоящий из" означает исключение дополнительных элементов, стадий или компонентов, не указанных иным образом. Термин "по существу состоящий из" означает включение упомянутых элементов или стадий, а также любого элемента или стадии, которые существенно не влияют на основные характеристики упомянутых элементов или стадий в объеме композиций или способов.
Преимущественные эффекты
Таким образом, настоящее изобретение обеспечивает антитело, которое имеет специфическую последовательность CDR (определяющей комплементарность области), описанную в настоящем документе, и специфически связывается с N-концевой областью KRS, экспонированной на внеклеточной мембране. Способ по настоящему изобретению можно успешно применять для получения антитела, имеющего более высокую аффинность к N-концу KRS, чем обычное антитело.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 показана схематическая диаграмма стратегии выбора и конструирования двух библиотек, созданных на основе каждой из вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) N3, с целью улучшить аффинность N-конца KRS целевого антитела N3 к N-концу KRS.
На Фиг. 2a показаны результаты анализа пептида KRS (1-72), связанного с 10 нМ, 1 нМ или 0,1 нМ GST, и проанализированная связывающая способность с помощью проточной цитометрии (FACS) для дрожжей, экспрессирующих каждую стадию библиотеки, выбранных с применением FACS (сортировка клеток с активированной флуоресценцией).
На Фиг. 2b показаны результаты анализа пептида KRS (1-72), связанного с 0,1 нМ GST, и связывающей способности с помощью проточного цитометра для дрожжей, экспрессирующих 47 индивидуальных клонов в окончательной выбранной библиотеке.
На Фиг. 3a и 3b показаны результаты ELISA для измерения аффинности к N-концу KRS антитела N3-1, антитела N3-3, антитела N3-4 и антитела N3-5, выбранных как обладающие высокой аффинностью и специфичностью к пептиду N-концу KRS (остатки 1-72).
На Фиг. 4 показаны результаты подтверждения ингибирующего действия на миграцию клеток антитела N3 и антитела N3-1.
На фиг. 5 показан результат сравнения аффинности в отношении KRS антитела N3 и антитела N3-1 способом SPR (поверхностный плазмонный резонанс).
На Фиг. 6 показаны результаты анализа пептида KRS (1-72), связанного с 10 нМ, 1 нМ или 0,1 нМ GST, и способность связывания, проанализированная с помощью проточного цитометра, для дрожжей, экспрессирующих каждую стадию библиотеки, выбранных с применением MACS и FACS (сортировка клеток с активированной флуоресценцией).
На Фиг. 7 показаны результаты ELISA для измерения аффинности антитела N3-1, антитела N3-6, антитела N3-7, антитела N3-8 и антитела N3-9 к N-концу KRS.
На Фиг. 8 показаны результаты сравнения аффинности антитела N3-6, антитела N3-7, антитела N3-8 и антитела N3-9 к KRS способом поверхностного плазмонного резонанса (SPR).
На Фиг. 9 показаны результаты подтверждения ингибирующего действия на миграцию клеток антитела N3, антитела N3-1, антитела N3-6, антитела N3-7, антитела N3-8 и антитела N3-9.
На Фиг. 10 показаны результаты подтверждения эндоцитоза антитела N3 и антитела N3-8 в клетках рака молочной железы способом IHC (иммуногистохимия).
На Фиг. 11 показано изменение конечного систолического давления в правом желудочке (RVESP) в моделях легочной артериальной гипертензии (ЛАГ) при введении антитела N3 по настоящему изобретению (ложный IgG: отрицательный контроль, Ab 1 мг/кг: антитело N3 1 мг/кг, Ab 10 мг/кг: Антитело N3 10 мг/кг, силденафил: положительный контроль).
На Фиг. 12 показаны результаты ИГХ (иммуногистохимического) окрашивания, подтверждающие, что введение антитела N3 по настоящему изобретению уменьшало миграцию и инвазию иммунных клеток в моделях легочной артериальной гипертензии (ЛАГ).
На Фиг. 13 показаны результаты, подтверждающие, что общее количество иммунных клеток, увеличенное в BALF (жидкость бронхоальвеолярного лаважа) на мышиных моделях острого повреждения легкого, было снижено в зависимости от концентрации антитела N3 при обработке (связывание антитела с N-концом KRS).
На Фиг. 14 показаны результаты, подтверждающие, что количество нейтрофилов, которое особенно повышено в жидкости бронхоальвеолярного лаважа (BALF) на мышиных моделях острого повреждения легкого, было снижено в зависимости от концентрации антитела N3 при обработке (связывание антитела с N-концом KRS).
На Фиг. 15 показаны результаты, подтверждающие с помощью FACS, что увеличенная миграция и инвазия макрофагов (IM, CD11b+/F4/80+) в легочную ткань на мышиных моделях острого повреждения легкого снижалась в зависимости от концентрации антитела N3 при обработке (связывание антитела с N-концом KRS).
На Фиг. 16 показан график количественной оценки результатов фиг. 15.
На Фиг. 17 представляет собой изображение ткани, показывающее, что фиброз ткани, прогрессирующий в ткани легкого на мышиных моделях острого повреждения легкого, ингибируется при обработке антителом N3 (антитело, связывающее N-конец KRS). Ткани каждой экспериментальной группы и контрольной группы наблюдали под микроскопом после окрашивания трихромом по Массону.
На Фиг. 18 показаны результаты, подтверждающие, что обработка антителами N3-8, N3-8-1 и производными антителами N3-8-1, из которых были удалены функции ADCC/CDC, ингибировала миграцию клеток.
На Фиг. 19 показаны результаты, подтверждающие, что обработка антителами N3-8, N3-8-1 и производными антителами N3-8-1, из которых были удалены функции ADCC/CDC, ингибировала клеточную миграция раковых клеток.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано подробно.
Однако следующие примеры предназначены только для иллюстрации настоящего изобретения и не предназначены для ограничения объема настоящего изобретения.
Пример 1: Конструирование экспрессионной библиотеки на поверхности дрожжевых клеток для увеличения аффинности
Аффинность к N-концу антитела N3 (номер заявки: 10-2018-0035446), нацеленному на N-конец существующего KRS, составляет приблизительно 150 нМ, что ниже, чем у различных антител в полной форме IgG. Соответственно, чтобы повысить аффинность для получения антитела, имеющего лучший эффект, были улучшены вариабельная область легкой цепи и вариабельная область тяжелой цепи антитела N3.
Модель гомологии применяли для прогнозирования приблизительной структуры N3, посредством которой случайные мутации были введены в области CDR, которые, как предполагается, играют важную роль в связывании антигена. В частности, в библиотеке на основе вариабельной области тяжелой цепи, NNK, применяли вырожденный кодон, который может содержать все 20 аминокислотных последовательностей для остатков CDR3. В библиотеке, основанной на вариабельной области легкой цепи, NNK, применяли вырожденный кодон, который может содержать все 20 аминокислотных последовательностей для остатков CDR3.
В частности, ДНК, кодирующая сконструированную библиотеку, была амплифицирована с применением способа ПЦР, а затем сконцентрирована с применением способа осаждения этанолом.
Вектор поверхностной экспрессии дрожжей (C-aga2), который экспрессирует белок aga2 на С-конце для гомологичной рекомбинации, обрабатывают рестрикционными ферментами NheI и MluI и очищают с применением способа экстракции из агарозного геля и способа осаждения концентрированным этанолом.
Обработанные рестрикционным ферментом 4 мкг векторов для 12 мкг каждой ДНК, кодирующей библиотеку, трансформировали электропорацией в дрожжи EBY100 для экспрессии на поверхности дрожжей, и размер библиотеки подтверждали путем измерения количества колоний, выращенных в селективной среде SD-CAA (20 г/л глюкозы, 6,7 г/л азотистого основания дрожжей без аминокислот, 5,4 г/л Na2HPO4, 8,6 г/л NaH2PO4, 5 г/л казаминокислот) посредством серийных разведений.
Данный способ показан на фиг. 1.
Пример 2: Выбор вариабельной области легкой цепи (VL) и вариабельной области тяжелой цепи (VH) с улучшенной аффинностью к GST-конъюгированному пептиду KRS (1-72)
Два типа библиотек, улучшающих аффинность на основе N3, сконструированных в примере 1, отбирали с применением GST-конъюгированного пептида KRS (1-72) в качестве антигена.
В частности, уровень 10 нМ очищенного GST-конъюгированного пептида KRS (1-72) инкубировали с дрожжами, экспрессирующими библиотеку вариабельной области легкой цепи одноцепочечного Fab (scFab)-типа на поверхности клетки с применением среды SG-CAA (20 мкг/л галактозы, 6,7 г/л основы азотного агара без аминокислот для дрожжей, 5,4 г/л Na2HPO4, 8,6 г/л NaH2PO4, 5 г/л казаминокислот) в течение 1 часа при комнатной температуре для первичного скрининга FACS.
Затем GST-конъюгированный пептид KRS (остатки 1-72) и дрожжи, экспрессирующие библиотеку, подвергали взаимодействию с PE-конъюгированным стрептавидин-R-фикоэритрин конъюгатом (SA-PE) при 4°C в течение 20 минут и суспендировали с помощью FACS (сортировка клеток с активированной флуоресценцией, FACS Caliber; BD biosciences). Впоследствии проводили второй скрининг FACS с 1 нМ пептида KRS (остатки 1-72), конъюгированного с GST, и третий скрининг FACS проводили с 0,5 нМ пептида KRS (остатки 1-72), конъюгированным с GST.
В результате, как показано на фиг. 2a и 2b, посредством отбора с применением FACS, по сравнению с антителом N3, было подтверждено, что клоны, имеющие высокую аффинность к GST-конъюгированному пептиду KRS (1-72) были отобраны, и аффинность зависела от вариабельной области тяжелой цепи (VH) или вариабельной области легкой цепи (VL). Три уникальных клона (N3-1, N3-3 и N3-4), обладающих высокой аффинностью и специфичностью к GST-конъюгированному пептиду KRS (1-72), были отобраны посредством анализа способности связывания отдельных клонов. Кроме того, другой уникальный клон (N3-5) был сконструирован путем комбинирования вариабельной области легкой цепи и вариабельной области тяжелой цепи друг с другом. То есть было отобрано всего четыре уникальных клона (N3-1, N3-3, N3-4, N3-5).
В таблице 1 показаны последовательности CDR вариабельной области легкой цепи и вариабельной области тяжелой цепи четырех отдельных клонов, демонстрирующие высокую способность связывания с GST-конъюгированным пептидом KRS (1-72). В таблице 2 показаны последовательности вариабельной области тяжелой цепи и последовательности вариабельной области легкой цепи.
Таблица 1.
(SEQ ID NO:3)
(SEQ ID NO:5)
(SEQ ID NO:7)
(SEQ ID NO:9)
(SEQ ID NO:11)
(SEQ ID NO:3)
(SEQ ID NO:5)
(SEQ ID NO:7)
(SEQ ID NO:9)
(SEQ ID NO:13)
(SEQ ID NO:3)
(SEQ ID NO:5)
(SEQ ID NO:7)
(SEQ ID NO:9)
(SEQ ID NO:15)
(SEQ ID NO:118)
(SEQ ID NO:5)
(SEQ ID NO:7)
(SEQ ID NO:9)
(SEQ ID NO:13)
(SEQ ID NO:118)
(SEQ ID NO:5)
(SEQ ID NO:7)
(SEQ ID NO:9)
(SEQ ID NO:15)
Таблица 2.
(Название последовательности)
Кроме того, проводили ELISA для измерения аффинности к N-концу KRS, чтобы подтвердить, увеличилась ли аффинность к N-концу KRS.
В частности, N-концевой (остатки 1-72) пептид KRS наносили на 96-луночный планшет EIA/RIA (COSTAR Corning) при 25°C в течение 1 часа, и планшет промывали 3 раза PBS (фосфатно-солевой буфер) (pH 7,4, 137 ммоль/л NaCl, 12 ммоль/л фосфата, 2,7 ммоль/л KCl) (SIGMA) в течение 10 минут. После этого 4% PBS с BSA (4% бычий сывороточный альбумин, pH 7,4, 137 ммоль/л NaCl, 12 ммоль/л фосфат, 2,7 ммоль/л KCl) (SIGMA) обрабатывали в течение 1 часа, а затем промывали 3 раза PBS в течение 10 минут. Затем антитело N3, антитело N3-1, антитело N3-3, антитело N3-4 и антитело N3-5 типа IgG обрабатывали и инкубировали, соответственно, и затем планшет трижды промывали 0,1% PBST (фосфатно-солевой буферный раствор с твином) в течение 10 минут. В качестве меченого антитела применяли mAb (моноклональное антитело) против человека, конъюгированное с пероксидазой хрена (SIGMA). Затем его подвергали взаимодействию с TMB (3,3',5,5'-тетраметилбензидин) (Sigma) и измеряли оптическую плотность при 450 нм для количественного определения связывания антитела.
В результате, как показано на фиг. 3а и 3b, подтверждено, что аффинность мутантных антител N3-1, N3-3, N3-4 и N3-5 была увеличена по сравнению с антителом N3 дикого типа. Было обнаружено, что все клоны не связывались с GST или NRP1-b1b2, применяемыми в качестве отрицательного контроля. Не выявлено значимых различий в способности связывания KRS между мутантными антителами N3-1, N3-3, N3-4 и N3-5.
Пример 3: Сравнение аффинности между антителом N3 и антителом N3-1
3-1. Действие антитела, ингибирующего миграцию клеток
Среди мутантных антител N3, описанных в примере 2, антитело N3-1 было преобразовано в антитело IgG с применением обычного способа. Следующий эксперимент провели с применением преобразованного антитела IgG.
Миграцию клеток измеряли с применением 24-луночной камеры системы Трансвелл, имеющей типичную поликарбонатную мембрану (размер пор 8,0 мкм, Costar). Нижнюю лунку покрывали 10 мкг ламинина в камере системы Трансвелл. Затем клетки A549 суспендировали в бессывороточной среде RPMI и помещали в верхнюю камеру в концентрации 1×105 клеток на лунку. N3, N3-1 IgG и человеческий ложный IgG (контроль) обрабатывали в камере при 10 нМ или 100 нМ, соответственно, и инкубировали в течение 24 часов. Немигрирующие клетки, присутствующие над мембраной, удаляли ватным тампоном. Затем дважды промывали PBS и обрабатывали 70% MeOH (в PBS) в течение 30 минут. После двукратной промывки PBS раствор гематоксилина обрабатывали в течение 30 минут. Затем, после трехкратной промывки камеры дистиллированной водой, мембрану в камере вырезали и помещали на предметное стекло для наблюдения.
В результате, как показано на фиг. 4, было подтверждено, что антитело N3-1 значимо ингибировало миграцию клеток A549 по сравнению с антителом N3.
3-2. KRS-аффинный эффект антитела
Применяя очищенный белок фрагмента KRS (1-207 а.к.) в качестве антигена, анализировали способность связывания с антителами N3 и N3-1 с помощью поверхностного плазменного резонанса (SPR).
Эксперимент SPR проводили с применением Biacore T200 (GE Healthcare), оснащенного сенсорным чипом CM5 серии S (GE Healthcare) при 25°C. После иммобилизации антитела на чипе с применением набора для связывания аминов (GE Healthcare) антиген разводили в 4 раза в растворе PBS в диапазоне от 4,8 до 1250 нМ и давали протечь в течение 60 секунд. После этого в течение 300 секунд пропускали PBS. Полученные данные анализировали с помощью программного обеспечения Biacore T200 Evaluation v2.0 (GE Healthcare).
В результате, как показано на фиг. 5, измеренное значение KD антитела N3-1 составило 31 нМ, что указывает на то, что способность связывания с белком KRS была увеличена по сравнению с антителом N3.
Пример 4: Конструирование библиотеки экспрессии на поверхности дрожжевых клеток для повышения аффинности (антитело N 3-1)
Антитела N3-1, N3-3, N3-4 и N3-5, нацеленные на N-конец KRS, полученные в примере 2, имеют сходную аффинность к KRS, и в результате для антитела N3-1, показано, что оно имеет аффинность приблизительно 31 нМ. Это значение все еще мало по сравнению с аффинностью различных антител в полной форме IgG. Чтобы повысить аффинность и получить более эффективное антитело, была предпринята попытка интенсивного улучшения вариабельной области тяжелой цепи антитела.
Последовательность вариабельной области легкой цепи фиксировали на N3-3, и приблизительную модельную структуру N3-3 предсказывали с применением моделирования гомологии. Благодаря этому в CDR вводили случайную мутацию, которая, как предполагается, играет важную роль в связывании антигена.
В частности, остатки CDR2 и CDR3 вариабельной области тяжелой цепи использованной NNK, вырожденный кодон, который может содержать все 20 аминокислотных последовательностей, и библиотеку сконструировали таким же образом, как в Примере 1.
Пример 5. Выбор вариабельной области легкой цепи (VL) и вариабельной области тяжелой цепи (VH) с улучшенной аффинностью к GST-конъюгированному пептиду KRS (1-72).
Применяя GST-конъюгированный пептид KRS (остатки 1-72) в качестве антигена, были выбраны два типа библиотек, улучшающих аффинность на основе N3-3, сконструированных в Примере 4. Поскольку ожидалось, что сродство N3-3 и N3-1 будет почти одинаковым, и последовательности были почти одинаковыми, сравнительный эксперимент проводили с N3-1.
В частности, дрожжи, экспрессирующие библиотеку, связанную с GTP-конъюгированным пептидом KRS (остатки 1-72), подвергали взаимодействию с микрогранулами Streptavidin Microbead™ (Miltenyi Biotec) при 4°C в течение 20 минут, а дрожжи, экспрессирующие вариабельную область тяжелой цепи с высокой аффинностью к пептиду KRS (1-72 а.к.) суспендировали с применением активируемой магнитным полем сортировки клеток (MACS). Дрожжи, экспрессирующие библиотеку, отобранную с помощью MACS, культивировали в среде SG-CAA (20 г/л галактозы, 6,7 г/л основы азотного агара без аминокислот для дрожжей, 5,4 г/л Na2HPO4, 8,6 г/л NaH2PO4, 5 г/л казаминокислот), чтобы индуцировать экспрессию библиотеки. Впоследствии, таким же образом, как в Примере 2, провели последовательный скрининг с применением FACS.
Первичный скрининг FACS проводили с 10 нМ пептида KRS (1-72), конъюгированного с GST, вторичный скрининг FACS с 1 нМ пептида KRS (1-72), конъюгированного с GST, третий скрининг FACS проводили с 0,5 нМ пептида KRS (1-72), конъюгированного с GST, и четвертый скрининг FACS проводили с 0,1 нМ пептида KRS (1-72), конъюгированного с GST.
В результате, как показано на фиг. 6, посредством отбора с применением MACS и FACS (сортировка клеток с активацией флуоресценции) было подтверждено, что клоны, имеющие высокую аффинность в зависимости от вариабельной области тяжелой цепи (VH) для GST-конъюгированного пептида KRS (1-72), были отобраны для сравнения с антителом N3-1, и четыре уникальных клона (N3-6, N3-7, N3-8, N3-9), обладающих высокой аффинностью и специфичностью к GST-конъюгированному пептиду KRS (1-72), были отобраны путем анализа способностей связывания отдельных клонов.
Последовательности CDR вариабельной области легкой цепи и вариабельной области тяжелой цепи четырех отдельных клонов, которые демонстрируют высокую связывающую способность с пептидом KRS (1-72 а.к.), показаны в таблице 3, а в таблице 4 показаны последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи.
Таблица 3.
(SEQ ID NO: 17)
(SEQ ID NO: 5)
(SEQ ID NO: 7)
(SEQ ID NO: 9)
(SEQ ID NO: 15)
(SEQ ID NO: 19)
(SEQ ID NO: 5)
(SEQ ID NO: 7)
(SEQ ID NO: 9)
(SEQ ID NO: 15)
(SEQ ID NO: 21)
(SEQ ID NO: 5)
(SEQ ID NO: 7)
(SEQ ID NO: 9)
(SEQ ID NO: 15)
(SEQ ID NO: 23)
(SEQ ID NO: 5)
(SEQ ID NO: 7)
(SEQ ID NO: 9)
(SEQ ID NO: 15)
Таблица 4.
(N3 VH мутант 6)
(N3 VL мутант 2)
(N3 VH мутант 5)
(N3 VL мутант 2)
Кроме того, ELISA выполняли таким же образом, как в Примере 2, для измерения аффинности к N-концу KRS, чтобы подтвердить, увеличилась ли аффинность к N-концу KRS.
В частности, N-концевую часть (1-72) KRS наносили в 96-луночный планшет EIA/RIA (COSTAR Corning) при 25°C в течение 1 часа, а затем планшет трижды промывали PBS в течение 10 минут. Затем планшет инкубировали с 4% BSA PBS в течение 1 часа, а затем промывали 3 раза PBS в течение 10 минут. Затем обрабатывали и инкубировали N-концевые целевые антитела-мишени KRS IgG-типа N3-1, N3-6, N3-7, N3-8 и N3-9, и планшет трижды промывали в течение 10 минут 0,1% PBST. Применяли антитело против иммуноглобулина человека, конъюгированное с пероксидазой хрена, и оно реагировало с TMB (3,3',5,5'-тетраметилбензидин), оптическую плотность измеряли при 450 нм и количественно определяли связывание.
В результате, как показано на фиг. 7 было подтверждено, что аффинность мутантных антител N3-6, N3-7, N3-8 и N3-9 была увеличена по сравнению с антителом N3-1. Все антитела не взаимодействовали с NRP1-b1b2, который применяли в качестве отрицательного контроля.
Пример 6: Сравнение аффинности между антителом N3-1 и антителами N3-6, N3-7, N3-8, N3-9
6-1. Сравнение связывания антител с KRS
Применяя эпитоп пептида F4 KRS (EPKLSKNELKRRLKAEKKVAEKEAKQKE: SEQ ID NO: 117) в качестве антигенного эпитопа, анализировали связывающую способность антитела N3, антитела N3-6, антитела N3-7, антитела N3-8 и антитела N3-9 с помощью поверхностного плазменного резонанса (SPR). Эксперимент SPR проводили таким же образом, как в Примере 3-2. Эпитоп разводили в растворе PBS и разбавляли в 2 раза в диапазоне от 15,7 до 4000 нМ и давали протечь в течение 90 секунд. После этого в течение 2400 секунд пропускали PBS. Полученные данные анализировали с помощью программного обеспечения Biacore T200 Evaluation v2.0 (GE Healthcare).
В результате, как показано на фиг. 8, KD антитела N3-8 была отличной, KD антител N3-9 и N3-6 были сходными, а значение KD антитела N3-7 было наибольшим. Время диссоциации антитела N3-6 было больше, чем у антител N3-7 и N3-9, и показало сенсограмму с более длительным связыванием.
Также ELISA выполняли для идентификации остатков, важных для связывания антитела с эпитопом, с применением пептидов, в которых отдельные аминокислоты эпитопного пептида F4 KRS (SEQ ID NO: 117) были заменены аланином (A), соответственно. В результате удалось идентифицировать остатки в эпитопе пептида F4 KRS, которые важны для связывания с каждым антителом.
6-2. Эффект антитела, ингибирующий миграцию клеток
Эксперименты проводили так же, как в Примере 3-1. Антитела N3-6, N3-7, N3-8, N3-9, полученные в приведенном выше примере, были преобразованы в IgG обычным способом. Следующий эксперимент проводили с применением преобразованных антител IgG.
Клетки помещали в верхнюю камеру при концентрации 1×105, а затем N3 IgG обрабатывали при 100 нМ, а N3-1, N3-6, N3-7, N3-8 и N3-9 IgG и человеческий ложный IgG (контроль) каждый обрабатывали в камере при 10 нМ и культивировали в течение 24 часов. Немигрирующие клетки, присутствующие над мембраной, удаляли ватным тампоном.
Затем мембрану дважды промывали PBS и обрабатывали 70% MeOH (в PBS) в течение 30 минут. После двукратной промывки PBS раствор гематоксилина обрабатывали в течение 30 минут. Затем после промывки камеры дистиллированной водой, мембрану в камере устанавливали на предметное стекло и проводили наблюдение.
В результате, как показано на фиг. 9, было подтверждено, что антитела N3-6, N3-7, N3-8 и N3-9 значимо ингибировали миграцию клеток по сравнению с антителом N3-1. Кроме того, не было значимой разницы в эффекте ингибирования миграции клеток среди антител N3-6, N3-7, N3-8 и N3-9.
Пример 7: Уточнение последовательности антитела N3-8
7-1. Мутантное продуцирование последовательности антитела N3-8
В приведенном выше примере было подтверждено, что антитело N3-8 имеет наилучшую аффинность. Таким образом, были проведены следующие эксперименты для подтверждения физических свойств, таких как продуктивность и стабильность антитела N3-8.
Мутацию индуцировали в последовательности, которая, как ожидается, повлияет на стабильность последовательности антитела N3-8. В результате были получены две дополнительные последовательности тяжелой цепи, в которых были введены мутации в последовательность тяжелой цепи (HC) антитела N3-8. Кроме того, можно было получить три дополнительных последовательности легкой цепи, с введенной мутацией. Соответственно, 7 видов последовательностей антител (производных N3-8), в которых последовательность N3-8 была изменена, показаны в таблицах 5 и 6 ниже.
Таблица 5.
(SEQ ID NO: 25)
(SEQ ID NO: 25)
(SEQ ID NO: 27)
(SEQ ID NO: 1)
(SEQ ID NO: 25)
(SEQ ID NO: 29)
(SEQ ID NO: 1)
(SEQ ID NO: 5)
(SEQ ID NO: 7)
(SEQ ID NO: 9)
Таблица 6.
(N3 VH мутант 6)
(N3 VL мутант 2)
(N3 VH мутант 6)
(N3 VH мутант 7)
(N3 VL мутант 2)
(N3 VH мутант 7)
(N3 VL мутант 3)
(N3 VH мутант 7)
(N3 VL мутант 4)
(N3 VH мутант 4)
(N3 VL мутант 2)
7-2. Измерение продуктивности и стабильности производных антитела N3-8 и измерение Tm
Вектор, экспрессирующий производное антитела N3-8, полученный в примере 7-1, экспрессировали и очищали с применением временной трансфекции.
В клетки HEK293-F (Invitrogen), суспендированные в бессывороточной экспрессионной среде FreeStyle 293 (Invitrogen) во встряхиваемой колбе, трансфицировали плазмиду и полиэтиленимин (Polyethylenimine, Polyscience). Во время трансфекции во встряхиваемую колбу на 200 мл клетки HEK293-F засевали в 100 мл среды с плотностью 2×106 клеток/мл и культивировали при 150 об./мин и 37°C при 8% CO2.
Для получения каждого моноклонального антитела, подходящие плазмиды тяжелой и легкой цепей трансфицировали в 10 мл экспрессионной среды FreeStyle 293 (Invitrogen) в соотношении 1:1 или 1:2 ДНК тяжелая цепь: легкая цепь. Когда ДНК тяжелая цепь: легкая цепь используется в соотношении 1:1, 125 мкг тяжелой цепи и 125 мкг легкой цепи, всего 250 мкг (2,5 мкг/мл) ДНК смешивают с 10 мл среды, содержащей 750 мкг PEI (7,5 мкг/мл) при комнатной температуре. Реакцию проводили в течение 10 минут. В случае соотношения 1:2 концентрацию ДНК легкой цепи увеличивали вдвое. После этого смешанную среду обрабатывали клетками, предварительно разделенными на аликвоты по 100 мл, и инкубировали при 150 об./мин и 37°C при 8% CO2 в течение 4 часов, а затем добавляли дополнительные 100 мл экспрессионной среды FreeStyle 293 и культивировали в течение 6 дней.
Затем раствор культуры клеток переносили в пробирки на 50 мл и центрифугировали в течение 5 минут при 3000 об./мин. Затем белок очищали из надосадочной жидкости собранной культуры клеток. Антитело наносили на колонку с протеином А-сефарозой и затем промывали PBS (pH 7,4). После элюирования антитела при pH 3,0 обрабатывали 0,1 моль/л глициновым буфером и образец немедленно нейтрализовали 1 моль/л трис-буфером. Фракцию элюированных антител концентрировали путем замены буфера на PBS (pH 7,4) способом диализа. Очищенный белок количественно определяли на основе измерения оптической плотности и коэффициента поглощения при длине волны 280 нм.
Кроме того, термостабильность антитела измеряли с применением 100 мкл очищенных антител в концентрации 1 мг/мл. Термостабильность измеряли 4 раза с применением набора красителей для анализа плавления белков Protein thermal shift (Thermofisher) и оборудования для ПЦР в реальном времени Quant Studio 3 (Thermofisher).
В результате, как показано в таблице 7 ниже, выход всех протестированных производных антитела N3-8 был улучшен или показал высокий выход при аналогичном уровне антитела N3-8. Кроме того, как показано в таблице 7, было получено значение температуры плавления (Tm). Наблюдали, что тепловой переход составляет 1-2 в зависимости от антитела, но значение Tm увеличивалось во всех производных антитела N3-8.
Таким образом подтверждено, что производные антитела N3-8 имели более высокий выход и их термическая стабильность была улучшена по сравнению с таковой антитела N3-8.
Таблица 7.
В таблице 8 показаны последовательности тяжелой цепи (НС) и легкой цепи (LC) всех антител IgG, применяемых в вышеописанных примерах.
Таблица 8.
7-3. Сравнение аффинности антител N3-8-1 и N3-8-4
Как описано в Примере 6, эпитоп пептида F4 KRS (EPKLSKNELKRRLKAEKKVAEKEAKQKE: SEQ ID NO: 117) применяли в качестве антигенного эпитопа, а силу связывания с антителами N3-8-1 и N3-8-4 анализировали с помощью поверхностного плазмонного резонанса (SPR).
Эксперимент SPR проводили таким же образом, как в Примере 3-2, и эпитоп разводили в растворе PBS, разбавляли дважды в диапазоне от 15,7 до 4000 нМ и давали протечь в течение 90 секунд. После этого в течение 2400 секунд пропускали PBS. Полученные данные анализировали с помощью программного обеспечения Biacore T200 Evaluation v2.0 (GE Healthcare).
В результате, как показано в Таблице 9 ниже, было обнаружено, что KD антитела N3-8-1 оказалась наиболее высокой.
Таблица 9.
Пример 8: Подтверждение механизма действия антител
После конъюгации флуоресцентного зонда с антителом (Ab) и обработки клеток рака молочной железы 4T1 было подтверждено, что антитела против KRS (N3, N3-8) подверглись эндоцитозу.
Антитела против KRS (N3, N3-8), меченные флуоресцентным зондом Alexa fluor 488 (Thermofisher) и 1 мкМ ложного IgG (Thermofisher) в качестве контроля, обрабатывали клетками, и локализацию антител контролировали через 4 часа. В это время Lysotracker (Thermofisher) применяли в качестве маркера лизосом, а DAPI применяли для окрашивания ядра. В отличие от ложного IgG, антитела N3 и N3-8 присутствовали в клетках через 4 часа.
В результате, как показано на фиг. 10, было подтверждено, что антитело против KRS распознает KRS клеточной мембраны и быстро эндоцитируется, тем самым снижая уровень KRS клеточной мембраны.
Пример 9: Последовательность и очистка антитела, из которого удалена функция ADCC/CDC.
9-1. Введение мутации в последовательность антитела для удаления функции ADCC/CDC
Чтобы удалить функцию ADCC/CDC из антитела, эксперимент проводили следующим образом. В каждую из вышеуказанных последовательностей антител вносили мутации в ту часть, которая, как ожидается, несет функцию ADCC/CDC в константной области тяжелой цепи IgG1. Получили пять дополнительных последовательностей тяжелой цепи, в которые внесли мутации. Кроме того, были созданы последовательность тяжелой цепи IgG4 и дополнительная последовательность, в которую ввели мутацию. Соответственно, последовательности мутантных антител, из которых удалили функцию ADCC/CDC каждого антитела, показаны в Таблице 10 соответственно.
Таблица 10.
9-2. Очистка мутантных антител с удаленной функцией ADCC/CDC
Вектор, экспрессирующий мутантное антитело N3-8-1 среди антител, из которых была удалена функция ADCC/CDC, как описано в Примере 9-1, временно трансфицировали в клетки для экспрессии и очистки белка.
Клетки HEK293-F (Invitrogen) трансфицировали во встряхиваемой колбе в соответствии со способом, описанным в Примере 7-2 выше. Затем клетки HEK293-F высевали в среду с плотностью 2×106 клеток/мл и культивировали при 150 об./мин, 8% CO2 и 37°C.
Для получения каждого моноклонального антитела подходящие плазмиды тяжелой и легкой цепей трансфицировали в 10 мл экспрессионной среды FreeStyle 293 (Invitrogen) в соотношении 1:1 или 1:2 ДНК тяжелой цепи:легкой цепи. В случае 1:1 125 мкг тяжелой цепи и 125 мкг легкой цепи, всего 250 мкг (2,5 мкг/мл), смешивали с 10 мл среды, содержащей 750 мкг PEI (7,5 мкг/мл) и оставляли для реакции при комнатной температуре в течение 10 минут. В случае 1:2 концентрацию ДНК легкой цепи увеличивали вдвое. После этого прореагировавшую смешанную среду помещали в 100 мл клеток и инкубировали в течение 4 часов при 150 об./мин, 8% CO2 и 37°C, добавляли дополнительные 100 мл экспрессионной среды FreeStyle 293 и культивировали в течение 6 дней.
Затем раствор культуры клеток переносили в пробирки на 50 мл и центрифугировали при 3000 об./мин в течение 5 минут. Впоследствии белок очищали из надосадочной жидкости собранной культуры клеток. Антитело наносили на колонку с протеином А-сефарозой и промывали PBS (pH 7,4). После элюирования антитела при pH 3,0 с применением 0,1 моль/л глицинового буфера образец немедленно нейтрализовали с применением 1 моль/л трис-буфера. Фракцию элюированных антител концентрировали путем замены буфера на PBS (pH 7,4) способом диализа. Очищенный белок количественно определяли на основе измерения оптической плотности и коэффициента поглощения при длине волны 280 нм.
Измеряли чистоту очищенного антитела и 4 раза исследовали термостабильность с применением оборудования для ПЦР в реальном времени Quant Studio 3 (Thermofisher) и набора красителей для анализа плавления белков Protein thermal shift (Thermofisher).
В результате, как показано в Таблице 11 ниже, выход всех протестированных мутантов антитела N3-8-1 был подобен или выше, чем у дикого типа, из которых мутации LALAPGTA, как было обнаружено, имели самый высокий выход. Кроме того, все мутанты антитела N3-8-1 показали высокую чистоту на уровне, аналогичном таковому у антитела дикого типа.
Таким образом, было подтверждено, что антитела, из которых была удалена функция ADCC/CDC N3-8-1, имеют аналогичный или более высокий выход и аналогичную чистоту по сравнению с антителом N3-8-1.
Таблица 11.
Пример 10: Подтверждение эффективности антител, специфичных к N-концу KRS, на моделях in vivo заболеваний, связанных с миграцией иммунных клеток, модели легочной артериальной гипертензии in vivo
При обработке антителом, которое специфически связывается с N-концом KRS, миграция/инвазия иммунных клеток ингибируется из-за интернализации KRS на участке клеточной мембраны (посредством эндоцитоза и т.д.), и в результате, как видно, он снижает уровень KRS в клеточной мембране. Таким образом, считается, что специфическое к N-концу KRS антитело по настоящему изобретению (обычно антитело N3) будет иметь терапевтический эффект в отношении заболеваний, связанных с миграцией иммунных клеток, что дополнительно продемонстрировано в примерах, описанных ниже.
Способы эксперимента
1) Конструирование моделей легочной артериальной гипертензии (ЛАГ) и введение тестируемого вещества.
Для индукции ЛАГ у крыс SD (Oriental Bio) возраста семь недель подкожно вводили 60 мкл/кг МСТ (монокроталин). После этого крыс разделили на четыре группы (пять животных на группу), и вводили 1 мкг/кг человеческого ложного IgG (Thermo Fisher Scientific, отрицательный контроль), 1 мкг/кг антитела IgG N3, 10 мкг антитела IgG N3 и 25 мг/кг силденафила (положительный контроль) в течение 3 недель. Все антитела внутривенно вводили два раза в неделю, силденафил вводили перорально каждый день.
2) Измерение кровотока и артериального давления
Через три недели крыс анестезировали изофлураном и измеряли кровоток и кровяное давление с помощью системы измерения давления и объема сердечно-сосудистой системы MPVS (название модели: MPVS Ultra, производитель: Millar Instruments). Конечное систолическое давление в правом желудочке (RVESP), конечное диастолическое давление в правом желудочке, конечное систолическое давление в левом желудочке, конечное диастолическое давление в левом желудочке измеряли с помощью эксклюзивного катетера (катетер для измерения давления у крыс Mikro-Tip, производитель: Millar Instruments). Сердечный выброс измеряли с помощью датчика периваскулярного зонда кровотока (зонды Transonic Flow, производитель: Millar Instruments), и данный экспериментальный способ выполняли тем же способом, как описано в следующей публикации: Pacher P, Nagayama T, Mukhopadhyay P, Batkai S, Kass DA. Measurement of cardiac function using pressure-volume conductance catheter technique in mice and rats. Nat Protoc 2008;3(9):1422-34.
3) Иммуногистохимия (ИГХ)
Собранные легкие фиксировали в ПФА (параформальдегиде) в соответствии с обычным способом, а затем заливали парафином путем промывки, дегидратации и очистки. Парафиновые блоки легочной ткани крысы разрезали на толщину 3 мкм и готовили срезы. Образец сначала обрабатывали три раза ксилолом в течение 5 минут, обрабатывали 100% этанолом, 95% этанолом, 90% этанолом и 70% этанолом и дистиллированной водой в указанном порядке в течение 2 минут и промывали PBS в течение 5 минут. После обработки 0,3% H2O2 образец дважды промывали PBS в течение 5 мин. Образец погружали в цитратный буфер 0,01 моль/л, нагревали и промывали PBS-T (0,03% твин 20), а затем проводили блокирование при комнатной температуре в течение 30 минут (2% BSA и 2% козья сыворотка в PBS). Образец окрашивали в течение ночи при 4°C антителом против CD68 (1:200, клон ED1, Abcam). После трехкратной промывки PBS-T в течение 5 минут образец обрабатывали набором антител против мышиных антител, меченных полимером-HRP (DAKO) в течение 1 часа при 4 °C. После трехкратной промывки PBS-T образец проявляли обработкой субстратным буфером DAB и хромогеном DAB 20. Окрашенную ткань обрабатывали гематоксилином Майера (Sigma) в течение 1 минуты, а затем обрабатывали дважды 70% этанолом, 90% этанолом, 95% этанолом и 100% этанолом в указанном порядке в течение 2 мин. Наконец, ткань обрабатывали ксилолом трижды в течение 5 мин, а затем проводили наблюдение под оптическим микроскопом.
Полученные результаты
10-1. Проверка изменений артериального давления и сердечного выброса.
Модели животных с ЛАГ, которая представляет собой заболевание, имеющее тесную связь между инвазией иммунных клеток и патологическими явлениями, обрабатывали антителом N3 IgG (1 мг/кг или 10 мг/кг) в течение 3 недель (внутривенно, два раза в неделю), и затем измеряли конечное систолическое давление в правом желудочке (RVESP), конечное диастолическое давление в правом желудочке (RVEDP), конечное систолическое давление в левом желудочке (LVESP), конечное диастолическое давление в левом желудочке (LVEDP) и сердечный выброс (CO). Результаты представлены в Таблице 12.
Таблица 12.
(отсутствует измерение СО для одного животного группы MCT + ложный IgG и одного животного группы обработки силденафилом, поскольку они умерли от анестезии и во время хирургического вмешательства, соответственно)
Легочная артериальная гипертензия вызывает повышение кровяного давления в правом желудочке вследствие сужения легочной артерии, что приводит к правожелудочковой недостаточности. Кроме того, если компенсаторный механизм нарушается вследствие длительной гипертензии, то это приводит к гипертрофии правого желудочка с последующим увеличением правого желудочка. Это вызывает компрессию левого желудочка из-за движения межжелудочковой перегородки и уменьшение конечного диастолического объема левого желудочка и сердечного выброса (Lee Woo Seok et al., Clinical characteristics and prognostic factors of patients with severe pulmonary hypertension, Korean Circulation J. 2007; 37: 265-270). В результате легочная гипертензия в первую очередь связана с правым желудочком, но также ассоциирована с функцией левого желудочка.
Пациенты с ЛАГ показали увеличение RVESP, которое также наблюдали на животных моделях с ЛАГ в настоящем эксперименте. Напротив, как показано на фиг. 11, антитело N3 (антитело, специфически связывающееся с N-концом KRS) значимо снижало RVESP в обеих концентрациях и, в частности, уменьшало RVESP лучше, чем силденафил, препарат положительного контроля.
Кроме того, не наблюдали снижения конечного систолического давления в левом желудочке (LVESP) после введения антитела N3 (антитела, специфически связывающегося с N-концом KRS). Вместо этого LVESP значительно повышалось в группе, которой вводили антитело по настоящему изобретению, как показано на фиг. 13. Такое действие антитела по настоящему изобретению контрастирует с действием силденафила, который применяют в качестве традиционного средства лечения легочной гипертензии, вызывающим расширение легочной артерии и расширение системной артерии, то есть приводящим к риску снижения системного артериального давления. Таким образом было подтверждено, что антитело по настоящему изобретению показало тенденцию к слабому влиянию на системное артериальное давление по сравнению с силденафилом, и считается, что данное действие является благоприятной характеристикой терапевтического средства, учитывая, что введение силденафила может иметь риск развития гипотензии в клинических учреждениях. Более того, тяжелая легочная артериальная гипертензия вызывает систолическую недостаточность правого желудочка, которая может сопровождаться низким сердечным выбросом и системной гипотензией. При этом ожидается, что лечение для облегчения легочной артериальной гипертензии антителом N3 по настоящему изобретению, увеличит сердечный выброс и системное кровяное давление, тем самым нормализуя кровяное давление.
Таким образом, было подтверждено, что введение антитела, связывающегося с N-концом KRS (в частности, антитела N3) по настоящему изобретению снижает риск побочных эффектов существующих терапевтических лекарственных средств и демонстрирует облегчение симптомов ЛАГ и эффекты лечения.
10-2. Эхокардиография
D-образный левый желудочек, указывающий на перегрузку давлением в правом желудочке, наблюдали у трех животных в группе введения только МСТ (т.е. модели животных без введения антител) и у трех животных в группе введения МСТ и силденафила, но не наблюдали в группах введения терапевтического антитела.
Кроме того, как показано в Таблице 13 ниже, масса тела животных в каждой группе был увеличивалась в одинаковой степени без существенных различий. Не наблюдали никаких признаков отклонения от нормы, включая патологическое снижение массы тела, у животных, получавших терапевтическое антитело.
Таблица 13.
10-3. Проверка миграции и инфильтрации моноцитов/макрофагов
ИГХ окрашивание проводили на тканях легкого каждой экспериментальной группы для выявления CD68, который является маркером моноцитов/макрофагов. Как показано на фиг. 12, было подтверждено, что в группе обработки антителом N3 (антитело, связывающееся с N-концом KRS) по настоящему изобретению явно уменьшалась инфильтрация моноцитов/макрофагов в ткани легкого, и такой эффект был значимо выше, чем у силденафила.
Пример 11: Подтверждение эффективности антител, специфичных к N-концу KRS, на моделях in vivo заболеваний, связанных с миграцией иммунных клеток, модели острого повреждения легкого
Способы
1) Конструирование моделей острого повреждения легкого, вызванного LPS, и введение тестируемого вещества.
Острое повреждение легкого провоцировали мышам путем интратрахеальной инъекции 2,5 мг/кг LPS (Sigma) самцам мышей C57BL/6 в возрасте 7 недель (duothermal bio). Чтобы исследовать влияние ингибиторов KRS на острое повреждение легкого, сначала проводили внутривенную инъекцию антитела N3 IgG мышам C57BL/6 в дозе 1 мг/кг или 10 мг/кг соответственно с последующей эндотрахеальной инъекцией 2,5 мг/кг LPS через 24 часа. Через 24 часа после инъекции LPS каждую мышь умерщвляли для сбора и анализа легочной ткани и BALF (жидкости бронхоальвеолярного лаважа).
2) Подсчет иммунных клеток в жидкости бронхоальвеолярного лаважа (BALF)
BALF, полученную промыванием легкого с помощью PBS, собирали, и осадок клеток собирали центрифугированием при 800×g в течение 10 минут при 4°C. После суспендирования клеток, эритроциты удаляли с применением буфера для лизиса эритроцитов (eBioscience, номер по каталогу 00-4333-57). После остановки реакции с помощью PBS клетки дважды промывали и суспендировали в 400 мкл PBS для измерения количества клеток с помощью гемоцитометра и количества нейтрофилов с помощью окрашивания Hema3.
3) FACS для анализа иммунных клеток в легочной ткани
Ткани легкого собирали и вращали в течение 45 минут при 37°C с применением мягкого MACS Octo Dissociator (MACS Miltenyi Biotec, номер 130-095-937) для раздавливания ткани. После фильтрации с применением сетчатого фильтра для клеток (40 мкм) клетки центрифугировали при комнатной температуре в течение 5 минут при 1500 об./мин. Осадок собирали и эритроциты удаляли с применением буфера для лизиса эритроцитов (eBioscience, номер по каталогу 00-4333-57). Клетки собирали и суспендировали в буфере FACS (PBS, содержащий 1% NaN3 и 3% фетальную бычью сыворотку). Клетки (50 мкл) помещали в пробирку, хорошо перемешивали с таким же количеством смеси антител и окрашивали блокировкой света при 4°C в течение 1 часа. FITC-меченные крысиные антитела к CD11b (BD Pharmingen) и меченные фикоэритрином (РЕ) крысиные антимышиные антитела F4/80 (BD Pharmingen) применяли для анализа интерстициальных макрофагов (IM), инфильтрированных в легкие. После двукратной промывки при 400×g в течение 5 минут с применением буфера FACS клетки анализировали с помощью проточного цитометра Navios (Beckman).
4) Окрашивание легочной ткани трихромом Массона
Ткань легкого заливали парафином оригинальным способом и затем вырезали. После этого срез ткани, с которого парафин удаляли с помощью ксилола, промывали дистиллированной водой, а затем обрабатывали жидкостью Bouin при температуре от 56 до 60°C в течение 1 часа. После окрашивания раствором гематоксилина железа Вейгерта в течение 10 минут срез промывали. После повторного окрашивания раствором фуксина алой кислоты Бибриха в течение от 10 до 15 минут срез промывали. Срез обрабатывали раствором фосфомолибденовой-фосфорновольфрамовой кислоты в течение от 10 до 15 минут, а затем срез переносили в раствор анилинового синего и окрашивали в течение от 5 до 10 минут. После промывания срез обрабатывали 1% раствором уксусной кислоты в течение от 2 до 5 минут. После промывки и дегидратации срез обрабатывали ксилолом и монтировали препарат.
Полученные результаты
11-1. Подтверждение ингибирующего действия на миграцию иммунных клеток в жидкости бронхоальвеолярного лаважа (BALF)
Как показано на фиг. 13, было подтверждено, что общее количество иммунных клеток в BALF увеличивалось у мышей, у которых обработка LPS вызывала острое повреждение легкого. Количество инфильтрированных иммунных клеток снижалось обработкой антителом N3 (антитело, связывающееся с N-концом KRS) в зависимости от концентрации.
В частности, как показано на фиг. 14, было подтверждено, что количество нейтрофилов увеличивалось у мышей с острым повреждением легкого, вызванным обработкой LPS, а обработка антителом N3 (антитело, связывающееся с N-концом KRS) снижало повышенный уровень нейтрофилов. В результате было показано, что инфильтрация иммунных клеток, в частности нейтрофилов, в BALF легких значимо ингибировалась обработкой антителом, специфически связывающегося с N-концом KRS.
11-2. Подтверждение ингибирующего действия антител на миграцию иммунных клеток в легочной ткани
На фиг. 15 и 16 показаны результаты FACS-анализа макрофагов, мигрировавших в легочную ткань с острым повреждением легкого. Интерстициальный макрофаг (IM) - это клетки CD11b+/F4/80+, которые представляют собой мигрирующие макрофаги, которые не находятся в легких, но в определенных ситуациях мигрируют в легкие. Обработка LPS увеличивала инфильтрацию IM в легкие, но обработка антителом N3 уменьшала миграцию IM в легкие в зависимости от концентрации. Благодаря этому было подтверждено, что миграция и инвазия иммунных клеток, таких как макрофаги/моноциты, в ткани легкого ингибировалась обработкой антителами (обычно антителом N3), которые специфически связываются с N-концом KRS.
Чрезмерная миграция и вторжение иммунных клеток, таких как макрофаги/моноциты, являются важным патологическим явлением в тканях при фиброзном заболевании. В результате наблюдения при окрашивании легочной ткани трихромом по Массону модели острого повреждения легкого (фиг. 17) было подтверждено, что фиброз в легочной ткани протекает значительным образом. Напротив, было показано, что обработка антителом N3 (антитело, которое специфически связывается с N-концом KRS) ингибировала такой фиброз.
Пример 12: Анализ миграции иммунных клеток мутантных антител с удаленной функцией ADCC/CDC
Чтобы подтверждения влияния мутантного антитела, из которого была удалена функция ADCC/CDC, на миграцию иммунных клеток, проводили анализ миграции клеток в соответствии с способом, описанным в предшествующей литературе (Park, S. G. et al. Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. U S A 102, 6356-6361 (2005)).
Измерения проводили в камере системы Трансвелл с поликарбонатной мембраной (размер пор 5,0 мкм, Costar). LN421 помещали в нижнюю камеру с концентрацией 2,5 мкг/мл в камеру системы Трансвелл. Затем клетки RAW264.7 помещали в верхнюю камеру в концентрации 5×104 клеток на лунку. Затем каждое антитело помещали в камеру в концентрации 10 нМ, а затем инкубировали в течение 24 часов. Затем его дважды промывали PBS и обрабатывали 70% MeOH (в PBS). После повторной двукратной промывки PBS перенесенные клетки окрашивали кристаллическим фиолетовым (Sigma) и сушили. Затем в верхнюю камеру помещали 33% уксусную кислоту (Merck) и перемешивали в течение 10 минут. Раствор уксусной кислоты, растворенный в кристаллическом фиолетовом, переносили в 96-луночный планшет и измеряли оптическую плотность при 590 нм в считывающем устройстве для микропланшетов (Tecan).
В результате, как показано на фиг. 18, было обнаружено, что все антитела N3-8, N3-8-1, N3-8-1 мутант LALA, N3-8-1 мутант LALATA, N3-8-1 мутант LALAPG, N3-8-1 мутант LALAPGTA ингибировали LN421-зависимую миграцию клеток на уровне, аналогичном таковому в контрольной группе (контроль, C), которую ничем не обрабатывали.
Пример 13: Анализ влияния мутантных антител с удаленной функцией ADCC/CDC на миграцию раковых клеток
Чтобы подтвердить влияние мутантного антитела, из которого была удалена функция ADCC/CDC, на миграцию иммунных клеток, проводили анализ миграции клеток в соответствии со способом, описанным в Примере 12.
Измерения проводили в 24-луночной камере системы Трансвелл с поликарбонатной мембраной (размер пор 8,0 мкм, Costar). Ламинин добавляли в нижнюю камеру в концентрации 1 мг/мл, и анализ миграции проводили с применением стабильных клеток MDA-MB-231, сверхэкспрессирующих T52D KRS, которые имитируют фосфорилированный KRS. Чтобы вызвать экспрессию T52D KRS, стабильные клетки T52D KRS-MDA-MB-231 обрабатывали доксициклином (0,1 мкг/мл) в течение одного дня, а затем высевали в верхнюю камеру в концентрации 4×104 клеток после суспендирования в бессывороточной среде RPMI. Затем добавляли 10 нМ каждого антитела N3-8, N3-8-1, N3-8-1 мутант LALA, N3-8-1 мутант LALATA, N3-8-1 мутант LALAPG, N3-8-1 мутант LALAPGTA в камеру и инкубировали 7 часов. Немигрирующие клетки, существующие над мембраной, удаляли ватным тампоном. Мембрану дважды промывали PBS и обрабатывали 70% MeOH (в PBS) в течение 30 минут. Снова дважды промывали PBS, мембрану окрашивали кристаллическим фиолетовым (SIgma) и сушили. Затем в верхнюю камеру помещали 33% уксусную кислоту (Merck) и перемешивали. Раствор уксусной кислоты, растворенный в кристаллическом фиолетовом, переносили в 96-луночный планшет и измеряли оптическую плотность при 590 нм в считывающем устройстве для микропланшетов (Tecan).
Как показано на фиг. 19, все антитела N3-8, N3-8-1, N3-8-1 мутант LALA, N3-8-1 мутант LALATA, N3-8-1 мутант LALAPG, N3-8-1 мутант LALAPGTA ингибировали ламинин-зависимую миграцию раковых клеток.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Как описано выше, антитела или их фрагменты по настоящему изобретению имеют специфические последовательности CDR (определяющая комплементарность область), описанные в настоящем документе, и обладают абсолютно превосходной способностью к специфическому связыванию и аффинностью к N-концевой области KRS, экспонированной на внеклеточной мембране. Таким образом, их можно применять для диагностики заболеваний, сопровождающихся характерным поведением KRS, таких как рак или заболевания, связанные с миграцией иммунных клеток. И они обладают отличной продуктивностью и стабильностью, а также отличным ингибирующим действием на метастазы рака. Таким образом, их можно с успехом применять в качестве терапевтического средства против рака, а также в качестве агента для предотвращения или ингибирования метастазов рака, а также они могут быть очень полезным для предотвращения, улучшения и лечения заболеваний, связанных с миграцией иммунных клеток.
--->
<110> БайоконТАК Ко., Лтд.
<120> АНТИТЕЛО ДЛЯ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С N-КОНЦЕВЫМ ДОМЕНОМ
ЛИЗИЛ-ТРНК-СИНТЕТАЗЫ, ЭКСПОНИРОВАННЫМ НА ВНЕКЛЕТОЧНОЙ МЕМБРАНЕ
<130> OP21-0031/PCT/RU
<150> KR 10-2018-0111046
<151> 2018-09-17
<150> PCT/KR 2019/011998
<151> 2019-09-17
<160> 251
<170> KoPatentIn 3.0
<210> 1
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H1
<400> 1
Ser Tyr Asp Met Ser
1 5
<210> 2
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H1
<400> 2
agttatgata tgagc 15
<210> 3
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H2
<400> 3
Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 4
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H2
<400> 4
gcgatctctt atgataatgg taatacatat tacgctgatt ctgtaaaagg t 51
<210> 5
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H3
<400> 5
Met Ala Leu Asp Phe Asp Tyr
1 5
<210> 6
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H3
<400> 6
atggcgcttg atttcgacta c 21
<210> 7
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR L1
<400> 7
Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn Tyr Val Thr
1 5 10
<210> 8
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR L1
<400> 8
actggctctt catctaatat tggcagtaat tatgtcacc 39
<210> 9
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR L2
<400> 9
Asp Asn Ser Asn Arg Pro Ser
1 5
<210> 10
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR L2
<400> 10
gataatagta atcggccaag c 21
<210> 11
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR L3
<400> 11
Ala Ser Trp Asp Asp Ser Leu Ser Ala Tyr Val
1 5 10
<210> 12
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR L3
<400> 12
gcttcttggg atgatagcct gagtgcttat gtc 33
<210> 13
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR L3 мутант 1
<400> 13
Ala Ser Phe Ser Asp Glu Leu Gly Ala Tyr Val
1 5 10
<210> 14
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR L3 мутант 1
<400> 14
gcttctttta gtgatgagtt gggggcttat gtc 33
<210> 15
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR L3 мутант 2
<400> 15
Ser Ser Phe Ser Asp Glu Leu Gly Ala Tyr Val
1 5 10
<210> 16
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR L3 мутант 2
<400> 16
tcttctttta gtgatgagtt gggggcttat gtc 33
<210> 17
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H2 мутант 1
<400> 17
Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 18
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H2 мутант 1
<400> 18
gcgatctcgc cgcagatggg tcgggtgtat tacgctgatt ctgtaaaagg t 51
<210> 19
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H2 мутант 2
<400> 19
Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 20
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H2 мутант 2
<400> 20
gcgatcgatc cgttgggggg taatatttat tacgctgatt ctgtaaaagg t 51
<210> 21
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H2 мутант 3
<400> 21
Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 22
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H2 мутант 3
<400> 22
gcgatctctc cgtattcggg taggatttat tacgctgatt ctgtaaaagg t 51
<210> 23
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H2 мутант 4
<400> 23
Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 24
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H2 мутант 4
<400> 24
gcgatcgggg ctgatggggg tccgtcttat tacgctgatt ctgtaaaagg t 51
<210> 25
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H3 мутант 1
<400> 25
Leu Ala Leu Asp Phe Asp Tyr
1 5
<210> 26
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H3 мутант 1
<400> 26
ctggcgcttg atttcgacta c 21
<210> 27
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR L2 мутант 1
<400> 27
Ser Asn Asn Gln Arg Pro Ser
1 5
<210> 28
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR L2 мутант 1
<400> 28
agtaataatc agcggccaag c 21
<210> 29
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR L2 мутант 2
<400> 29
Arg Asn Asn Gln Arg Pro Ser
1 5
<210> 30
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR L2 мутант 2
<400> 30
cggaataatc agcggccaag c 21
<210> 31
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH
<400> 31
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 32
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH
<400> 32
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 33
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VL
<400> 33
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Trp Asp Asp Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 34
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL
<400> 34
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtgct tcttgggatg atagcctgag tgcttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 35
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 1
<400> 35
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 36
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 1
<400> 36
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 37
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 2
<400> 37
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 38
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 2
<400> 38
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 39
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 3
<400> 39
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 40
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 3
<400> 40
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 41
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 4
<400> 41
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 42
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 4
<400> 42
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 43
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 5
<400> 43
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 44
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 5
<400> 44
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 45
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 6
<400> 45
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 46
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 6
<400> 46
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 47
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 7
<400> 47
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 48
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH мутант 7
<400> 48
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 49
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 1
<400> 49
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 50
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 1
<400> 50
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgtgct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 51
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 2
<400> 51
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 52
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 2
<400> 52
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgttct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 53
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 3
<400> 53
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 54
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 3
<400> 54
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat agtaataatc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgttct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 55
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 4
<400> 55
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 56
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL мутант 4
<400> 56
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat cggaataatc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgttct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 57
<211> 15
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Линкер для scFv
<400> 57
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 58
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Линкер для scFv
<400> 58
ggtggaggcg gttcaggcgg aggtggatcc ggcggtggcg gatcg 45
<210> 59
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3 scFv (VH+линкер+VL)
<400> 59
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Trp Asp
210 215 220
Asp Ser Leu Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 60
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 scFv (VH+линкер+VL)
<400> 60
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cggtccgagg atgaggctga ttattactgt 660
gcttcttggg atgatagcct gagtgcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 61
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-1 scFv (VH+линкер+VL)
<400> 61
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 62
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-1 scFv (VH+линкер+VL)
<400> 62
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
gcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 63
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-3 scFv (VH+линкер+VL)
<400> 63
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 64
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-3 scFv (VH+линкер+VL)
<400> 64
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 65
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-4 scFv (VH+линкер+VL)
<400> 65
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 66
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-4 scFv (VH+линкер+VL)
<400> 66
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
gcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 67
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-5 scFv (VH+линкер+VL)
<400> 67
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 68
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-5 scFv (VH+линкер+VL)
<400> 68
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 69
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-6 scFv (VH+линкер+VL)
<400> 69
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 70
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-6 scFv (VH+линкер+VL)
<400> 70
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 71
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-7 scFv (VH+линкер+VL)
<400> 71
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 72
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-7 scFv (VH+линкер+VL)
<400> 72
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 73
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-8-7 scFv (VH+линкер+VL)
<400> 73
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 74
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-8-7 scFv (VH+линкер+VL)
<400> 74
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 75
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-9 scFv (VH+линкер+VL)
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 76
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-9 scFv (VH+линкер+VL)
<400> 76
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 77
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-8 scFv, N3-8-1 scFv (VH+линкер+VL)
<400> 77
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 78
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-8 scFv, N3-8-1 scFv (VH+линкер+VL)
<400> 78
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 79
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-8-2 scFv (VH+линкер+VL)
<400> 79
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 80
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-8-2 scFv (VH+линкер+VL)
<400> 80
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatagtaata atcagcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 81
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-8-3 scFv (VH+линкер+VL)
<400> 81
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 82
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-8-3 scFv (VH+линкер+VL)
<400> 82
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatcggaata atcagcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 83
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-8-4 scFv (VH+линкер+VL)
<400> 83
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 84
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-8-4 scFv (VH+линкер+VL)
<400> 84
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 85
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-8-5 scFv (VH+линкер+VL)
<400> 85
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 86
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-8-5 scFv (VH+линкер+VL)
<400> 86
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatagtaata atcagcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 87
<211> 241
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> N3-8-6 scFv (VH+линкер+VL)
<400> 87
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser
210 215 220
Asp Glu Leu Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 88
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3-8-6 scFv (VH+линкер+VL)
<400> 88
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatcggaata atcagcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cagtccgagg atgaggctga ttattactgt 660
tcttctttta gtgatgagtt gggggcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 89
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG N3
<400> 89
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 90
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG N3
<400> 90
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 91
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG N3
<400> 91
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Trp Asp Asp Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 92
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG N3
<400> 92
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtgct tcttgggatg atagcctgag tgcttatgtc 300
ttcggcggag gcaccaagct gacggtccta cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg t 651
<210> 93
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1
<400> 93
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 94
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1
<400> 94
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 95
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2
<400> 95
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 96
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2
<400> 96
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 97
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3
<400> 97
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 98
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3
<400> 98
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 99
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4
<400> 99
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 100
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4
<400> 100
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 101
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5
<400> 101
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 102
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5
<400> 102
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 103
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6
<400> 103
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 104
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6
<400> 104
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 105
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7
<400> 105
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 106
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7
<400> 106
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 107
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 1
<400> 107
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 108
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 1
<400> 108
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgtgct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg t 651
<210> 109
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 2
<400> 109
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 110
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 2
<400> 110
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgttct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg t 651
<210> 111
<211> 216
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 3
<400> 111
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 112
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 3
<400> 112
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgttct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta ggtcagccca aggctgcccc ctcggtcacg 360
ctcttcccac cctcctctga ggagcttcaa gccaacaagg ccacactggt gtgtctcata 420
agtgacttct acccgggagc cgtgacagtg gcctggaagg cagatagcag ccccgtcaag 480
gcgggagtgg agaccaccac accctccaaa caaagcaaca acaagtacgc ggccagcagc 540
tacctgagcc tgacgcctga gcagtggaag tcccacagaa gctacagctg ccaggtcacg 600
catgaaggga gcaccgtgga gaagacagtg gcccctacag aatgctct 648
<210> 113
<211> 216
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 4
<400> 113
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 114
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 4
<400> 114
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat agtaataatc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgttct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta agtcagccca aggctgcccc ctcggtcacg 360
ctcttcccac cctcctctga ggagcttcaa gccaacaagg ccacactggt gtgtctcata 420
agtgacttct acccgggagc cgtgacagtg gcctggaagg cagatagcag ccccgtcaag 480
gcgggagtgg agaccaccac accctccaaa caaagcaaca acaagtacgc ggccagcagc 540
tacctgagcc tgacgcctga gcagtggaag tcccacagaa gctacagctg ccaggtcacg 600
catgaaggga gcaccgtgga gaagacagtg gcccctacag aatgctct 648
<210> 115
<211> 216
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 5
<400> 115
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Phe Ser Asp Glu Leu
85 90 95
Gly Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 116
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь IgG 5
<400> 116
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat cggaataatc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tccgaggatg aggctgatta ttactgttct tcttttagtg atgagttggg ggcttatgtc 300
ttcggcggag gcaccaagct gacggtccta ggtcagccca aggctgcccc ctcggtcacg 360
ctcttcccac cctcctctga ggagcttcaa gccaacaagg ccacactggt gtgtctcata 420
agtgacttct acccgggagc cgtgacagtg gcctggaagg cagatagcag ccccgtcaag 480
gcgggagtgg agaccaccac accctccaaa caaagcaaca acaagtacgc ggccagcagc 540
tacctgagcc tgacgcctga gcagtggaag tcccacagaa gctacagctg ccaggtcacg 600
catgaaggga gcaccgtgga gaagacagtg gcccctacag aatgctct 648
<210> 117
<211> 28
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> KRS эпитоп F4
<400> 117
Glu Pro Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu
1 5 10 15
Lys Lys Val Ala Glu Lys Glu Ala Lys Gln Lys Glu
20 25
<210> 118
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность CDR H2 мутант 5
<400> 118
Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 119
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК CDR H2 мутант 5
<400> 119
gtgatctctt ctgatggtgg taatacatat tacgctgatt ctgtaaaagg t 51
<210> 120
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область легкой цепи вер.1_аминокислота
<400> 120
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 121
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область легкой цепи вер.1_ДНК
<400> 121
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg t 321
<210> 122
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область легкой цепи вер.2_аминокислота
<400> 122
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 123
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область легкой цепи вер.2_ДНК
<400> 123
ggtcagccca aggctgcccc ctcggtcacg ctcttcccac cctcctctga ggagcttcaa 60
gccaacaagg ccacactggt gtgtctcata agtgacttct acccgggagc cgtgacagtg 120
gcctggaagg cagatagcag ccccgtcaag gcgggagtgg agaccaccac accctccaaa 180
caaagcaaca acaagtacgc ggccagcagc tacctgagcc tgacgcctga gcagtggaag 240
tcccacagaa gctacagctg ccaggtcacg catgaaggga gcaccgtgga gaagacagtg 300
gcccctacag aatgctct 318
<210> 124
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 дикий тип_
аминокислота
<400> 124
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 125
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 дикий тип_ДНК
<400> 125
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtataccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggtaaa 990
<210> 126
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант TA_
аминокислота
<400> 126
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 127
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант TA_ДНК
<400> 127
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcgcctacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtataccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggtaaa 990
<210> 128
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант LALA_
аминокислота
<400> 128
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 129
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант LALA_ДНК
<400> 129
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaagc cgccggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtataccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggtaaa 990
<210> 130
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант
LALATA_аминокислота
<400> 130
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 131
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант
LALATA_ДНК
<400> 131
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaagc cgccggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcgcctacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtataccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggtaaa 990
<210> 132
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант
LALAPG_аминокислота
<400> 132
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 133
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант
LALAPG_ДНК
<400> 133
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaagc cgccggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcggcgccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtataccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggtaaa 990
<210> 134
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант
LALAPGTA_аминокислота
<400> 134
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 135
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG1 мутант
LALAPGTA_ДНК
<400> 135
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaagc cgccggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcgcctacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcggcgccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtataccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggtaaa 990
<210> 136
<211> 327
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG4 дикий
тип_аминокислота
<400> 136
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 137
<211> 981
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG4 дикий тип_ДНК
<400> 137
gccagcacca agggcccttc cgtgtttccc ctggcccctt gctcccggtc cacatctgag 60
agcaccgccg ccctgggctg tctggtgaag gactacttcc cagagcccgt gaccgtgagc 120
tggaacagcg gcgccctgac aagcggcgtg cacacatttc ccgccgtgct gcagagctcc 180
ggcctgtact ccctgtctag cgtggtgaca gtgccttcct ctagcctggg caccaagaca 240
tatacctgta acgtggacca caagccaagc aataccaagg tggataagcg ggtggagtct 300
aagtacggcc ctccttgccc tagctgtcct gctccagagt ttctgggcgg cccttccgtg 360
ttcctgtttc cacccaaacc aaaggacaca ctgatgatct ctagaacacc agaggtgacc 420
tgcgtggtgg tggacgtgag ccaggaggat cccgaggtgc agttcaactg gtacgtggat 480
ggcgtggagg tgcacaatgc caagaccaag ccaagagagg agcagtttaa ctctacatac 540
agggtggtga gcgtgctgac cgtgctgcac caggattggc tcaacggcaa ggagtataag 600
tgcaaggtgt ccaataaggg cctgccctcc tctatcgaga agacaatctc taaggctaag 660
ggccagccaa gagagcctca ggtgtacacc ctgcctccaa gccaggagga gatgacaaag 720
aaccaggtgt ccctgacatg tctggtgaag ggcttctatc cctccgacat cgccgtggag 780
tgggagtcta atggccagcc tgagaacaat tacaagacca caccccctgt gctggactct 840
gatggcagct tctttctgta ttccaggctg accgtggata agtctcggtg gcaggagggc 900
aacgtgttca gctgctctgt gatgcacgaa gccctgcata atcactatac tcagaaaagt 960
ctgtcactgt cactgggaaa g 981
<210> 138
<211> 327
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG4 мутант
TA_аминокислота
<400> 138
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 139
<211> 981
<212> ДНК
<213> Искусственная последовательность
<220>
<223> константная область тяжелой цепи(CH1, CH2, CH3)_IgG4 мутант
TA_ДНК
<400> 139
gccagcacca agggcccttc cgtgtttccc ctggcccctt gctcccggtc cacatctgag 60
agcaccgccg ccctgggctg tctggtgaag gactacttcc cagagcccgt gaccgtgagc 120
tggaacagcg gcgccctgac aagcggcgtg cacacatttc ccgccgtgct gcagagctcc 180
ggcctgtact ccctgtctag cgtggtgaca gtgccttcct ctagcctggg caccaagaca 240
tatacctgta acgtggacca caagccaagc aataccaagg tggataagcg ggtggagtct 300
aagtacggcc ctccttgccc tagctgtcct gctccagagt ttctgggcgg cccttccgtg 360
ttcctgtttc cacccaaacc aaaggacaca ctgatgatct ctagaacacc agaggtgacc 420
tgcgtggtgg tggacgtgag ccaggaggat cccgaggtgc agttcaactg gtacgtggat 480
ggcgtggagg tgcacaatgc caagaccaag ccaagagagg agcagtttaa ctctgcctac 540
agggtggtga gcgtgctgac cgtgctgcac caggattggc tcaacggcaa ggagtataag 600
tgcaaggtgt ccaataaggg cctgccctcc tctatcgaga agacaatctc taaggctaag 660
ggccagccaa gagagcctca ggtgtacacc ctgcctccaa gccaggagga gatgacaaag 720
aaccaggtgt ccctgacatg tctggtgaag ggcttctatc cctccgacat cgccgtggag 780
tgggagtcta atggccagcc tgagaacaat tacaagacca caccccctgt gctggactct 840
gatggcagct tctttctgta ttccaggctg accgtggata agtctcggtg gcaggagggc 900
aacgtgttca gctgctctgt gatgcacgaa gccctgcata atcactatac tcagaaaagt 960
ctgtcactgt cactgggaaa g 981
<210> 140
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант TA_аминокислота
<400> 140
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 141
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант TA_ДНК
<400> 141
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 142
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALA_аминокислота
<400> 142
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 143
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALA_ДНК
<400> 143
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 144
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALATA_аминокислота
<400> 144
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 145
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALATA_ДНК
<400> 145
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 146
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALAPG_аминокислота
<400> 146
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 147
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALAPG_ДНК
<400> 147
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 148
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALAPGTA_аминокислота
<400> 148
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 149
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG1 мутант LALAPGTA_ДНК
<400> 149
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 150
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG4 дикий тип_аминокислота
<400> 150
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 151
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG4 дикий тип_ДНК
<400> 151
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 152
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG4 мутант TA_аминокислота
<400> 152
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 153
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG_IgG4 мутант TA_ДНК
<400> 153
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 154
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант TA_аминокислота
<400> 154
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 155
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант TA_ДНК
<400> 155
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 156
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALA_аминокислота
<400> 156
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 157
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALA_ДНК
<400> 157
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 158
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALATA_аминокислота
<400> 158
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 159
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALATA_ДНК
<400> 159
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 160
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALAPG_аминокислота
<400> 160
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 161
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALAPG_ДНК
<400> 161
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 162
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALAPGTA_аминокислота
<400> 162
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 163
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG1 мутант LALAPGTA_ДНК
<400> 163
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 164
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG4 дикий тип_аминокислота
<400> 164
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 165
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG4 дикий тип_ДНК
<400> 165
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 166
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG4 мутант TA_аминокислота
<400> 166
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Ser Asp Gly Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 167
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 1_IgG4 мутант TA_ДНК
<400> 167
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagtg atctcttctg atggtggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 168
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант TA_аминокислота
<400> 168
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 169
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант TA_ДНК
<400> 169
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 170
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALA_аминокислота
<400> 170
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 171
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALA_ДНК
<400> 171
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 172
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALATA_аминокислота
<400> 172
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 173
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALATA_ДНК
<400> 173
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 174
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALAPG_аминокислота
<400> 174
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 175
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALAPG_ДНК
<400> 175
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 176
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALAPGTA_аминокислота
<400> 176
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 177
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG1 мутант LALAPGTA_ДНК
<400> 177
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 178
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG4 дикий тип_аминокислота
<400> 178
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 179
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG4 дикий тип_ДНК
<400> 179
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 180
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG4 мутант TA_аминокислота
<400> 180
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Gln Met Gly Arg Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 181
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 2_IgG4 мутант TA_ДНК
<400> 181
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcgccgc agatgggtcg ggtgtattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 182
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант TA_аминокислота
<400> 182
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 183
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант TA_ДНК
<400> 183
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 184
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALA_аминокислота
<400> 184
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 185
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALA_ДНК
<400> 185
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 186
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALATA_аминокислота
<400> 186
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 187
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALATA_ДНК
<400> 187
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 188
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALAPG_аминокислота
<400> 188
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 189
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALAPG_ДНК
<400> 189
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 190
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALAPGTA_аминокислота
<400> 190
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 191
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG1 мутант LALAPGTA_ДНК
<400> 191
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 192
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG4 дикий тип_аминокислота
<400> 192
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 193
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG4 дикий тип_ДНК
<400> 193
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 194
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG4 мутант TA_аминокислота
<400> 194
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asp Pro Leu Gly Gly Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 195
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 3_IgG4 мутант TA_ДНК
<400> 195
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcgatccgt tggggggtaa tatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 196
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант TA_аминокислота
<400> 196
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 197
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант TA_ДНК
<400> 197
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 198
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALA_аминокислота
<400> 198
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 199
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALA_ДНК
<400> 199
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 200
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALATA_аминокислота
<400> 200
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 201
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALATA_ДНК
<400> 201
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 202
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALAPG_аминокислота
<400> 202
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 203
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALAPG_ДНК
<400> 203
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 204
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALAPGTA_аминокислота
<400> 204
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 205
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG1 мутант LALAPGTA_ДНК
<400> 205
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 206
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG4 дикий тип_аминокислота
<400> 206
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 207
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG4 дикий тип_ДНК
<400> 207
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 208
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG4 мутант TA_аминокислота
<400> 208
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 209
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 4_IgG4 мутант TA_ДНК
<400> 209
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 210
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант TA_аминокислота
<400> 210
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 211
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант TA_ДНК
<400> 211
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 212
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALA_аминокислота
<400> 212
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 213
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALA_ДНК
<400> 213
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 214
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALATA_аминокислота
<400> 214
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 215
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALATA_ДНК
<400> 215
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 216
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALAPG_аминокислота
<400> 216
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 217
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALAPG_ДНК
<400> 217
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 218
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALAPGTA_аминокислота
<400> 218
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 219
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG1 мутант LALAPGTA_ДНК
<400> 219
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 220
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG4 дикий тип_аминокислота
<400> 220
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 221
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG4 дикий тип_ДНК
<400> 221
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 222
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG4 мутант TA_аминокислота
<400> 222
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Ala Asp Gly Gly Pro Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 223
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 5_IgG4 мутант TA_ДНК
<400> 223
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atcggggctg atgggggtcc gtcttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 224
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант TA_аминокислота
<400> 224
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 225
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант TA_ДНК
<400> 225
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 226
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALA_аминокислота
<400> 226
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 227
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALA_ДНК
<400> 227
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 228
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALATA_аминокислота
<400> 228
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 229
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALATA_ДНК
<400> 229
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 230
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALAPG_аминокислота
<400> 230
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 231
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALAPG_ДНК
<400> 231
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 232
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALAPGTA_аминокислота
<400> 232
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 233
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG1 мутант LALAPGTA_ДНК
<400> 233
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 234
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG4 дикий тип_аминокислота
<400> 234
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 235
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG4 дикий тип_ДНК
<400> 235
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 236
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG4 мутант TA_аминокислота
<400> 236
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 237
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 6_IgG4 мутант TA_ДНК
<400> 237
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 238
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант TA_аминокислота
<400> 238
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 239
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант TA_ДНК
<400> 239
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 240
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALA_аминокислота
<400> 240
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 241
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALA_ДНК
<400> 241
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 242
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALATA_аминокислота
<400> 242
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 243
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALATA_ДНК
<400> 243
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 244
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALAPG_аминокислота
<400> 244
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 245
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALAPG_ДНК
<400> 245
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 246
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALAPGTA_аминокислота
<400> 246
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Ala Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 247
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG1 мутант LALAPGTA_ДНК
<400> 247
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaagccg ccgggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cgcctaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cggcgccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 248
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG4 дикий тип_аминокислота
<400> 248
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 249
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG4 дикий тип_ДНК
<400> 249
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctacatacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<210> 250
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG4 мутант TA_аминокислота
<400> 250
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Pro Tyr Ser Gly Arg Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser
210 215 220
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Ala Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 251
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> тяжелая цепь IgG 7_IgG4 мутант TA_ДНК
<400> 251
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctctccgt attcgggtag gatttattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagactggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc cagcaccaag 360
ggcccttccg tgtttcccct ggccccttgc tcccggtcca catctgagag caccgccgcc 420
ctgggctgtc tggtgaagga ctacttccca gagcccgtga ccgtgagctg gaacagcggc 480
gccctgacaa gcggcgtgca cacatttccc gccgtgctgc agagctccgg cctgtactcc 540
ctgtctagcg tggtgacagt gccttcctct agcctgggca ccaagacata tacctgtaac 600
gtggaccaca agccaagcaa taccaaggtg gataagcggg tggagtctaa gtacggccct 660
ccttgcccta gctgtcctgc tccagagttt ctgggcggcc cttccgtgtt cctgtttcca 720
cccaaaccaa aggacacact gatgatctct agaacaccag aggtgacctg cgtggtggtg 780
gacgtgagcc aggaggatcc cgaggtgcag ttcaactggt acgtggatgg cgtggaggtg 840
cacaatgcca agaccaagcc aagagaggag cagtttaact ctgcctacag ggtggtgagc 900
gtgctgaccg tgctgcacca ggattggctc aacggcaagg agtataagtg caaggtgtcc 960
aataagggcc tgccctcctc tatcgagaag acaatctcta aggctaaggg ccagccaaga 1020
gagcctcagg tgtacaccct gcctccaagc caggaggaga tgacaaagaa ccaggtgtcc 1080
ctgacatgtc tggtgaaggg cttctatccc tccgacatcg ccgtggagtg ggagtctaat 1140
ggccagcctg agaacaatta caagaccaca ccccctgtgc tggactctga tggcagcttc 1200
tttctgtatt ccaggctgac cgtggataag tctcggtggc aggagggcaa cgtgttcagc 1260
tgctctgtga tgcacgaagc cctgcataat cactatactc agaaaagtct gtcactgtca 1320
ctgggaaag 1329
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО, СПЕЦИФИЧНО СВЯЗЫВАЮЩЕЕСЯ С N-КОНЦОМ ЛИЗИЛ-тРНК-СИНТЕТАЗЫ, В КАЧЕСТВЕ ЭФФЕКТИВНОГО ИНГРЕДИЕНТА ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ, ОБУСЛОВЛЕННОГО МИГРАЦИЕЙ КЛЕТОК | 2018 |
|
RU2749591C1 |
| АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ C N-КОНЦЕВОЙ ОБЛАСТЬЮ ЛИЗИЛ-тРНК-СИНТЕТАЗЫ, ЭКСПОНИРОВАННОЙ НА КЛЕТОЧНОЙ МЕМБРАНЕ | 2018 |
|
RU2739393C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИТЕЛО, НАПРАВЛЕННОЕ ПРОТИВ ФАКТОРА СЛИПАНИЯ А (ClfA) S. aureus | 2019 |
|
RU2818805C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
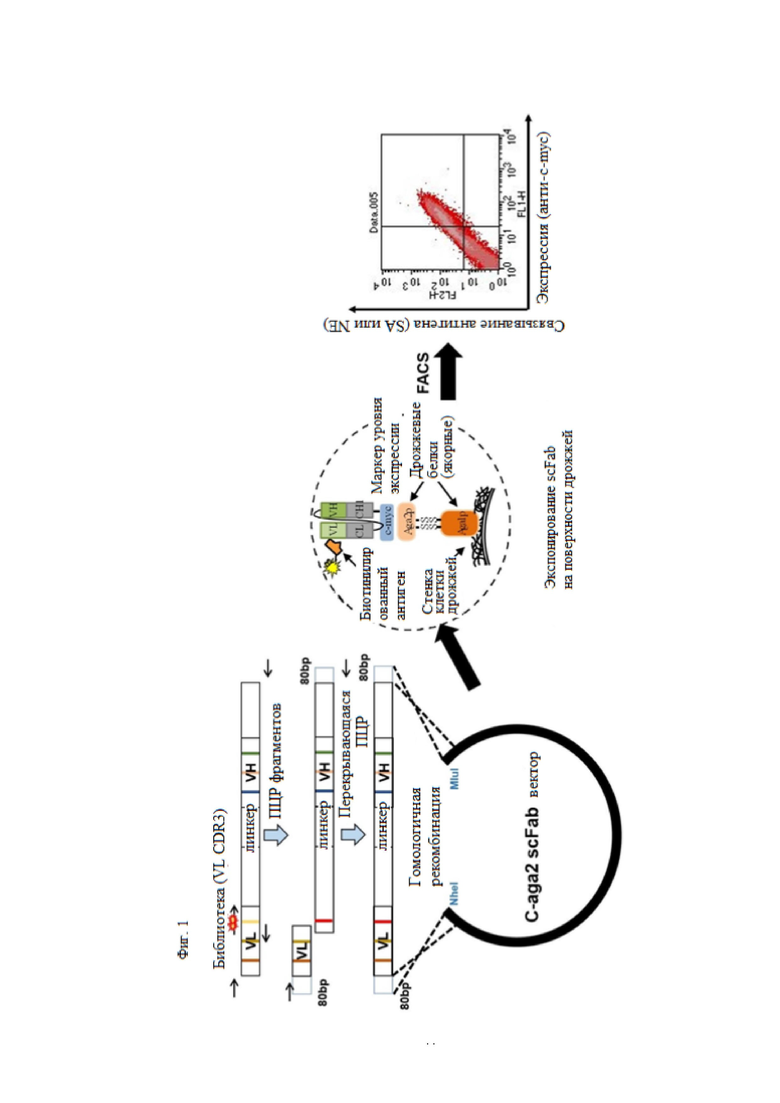
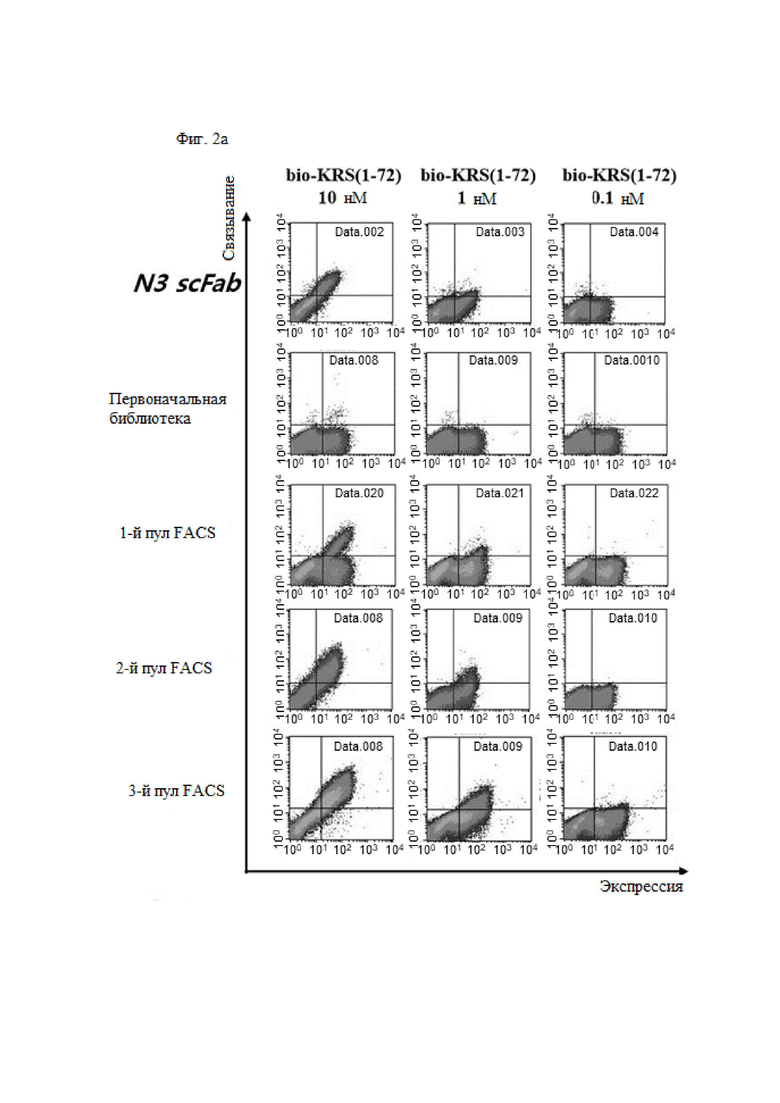

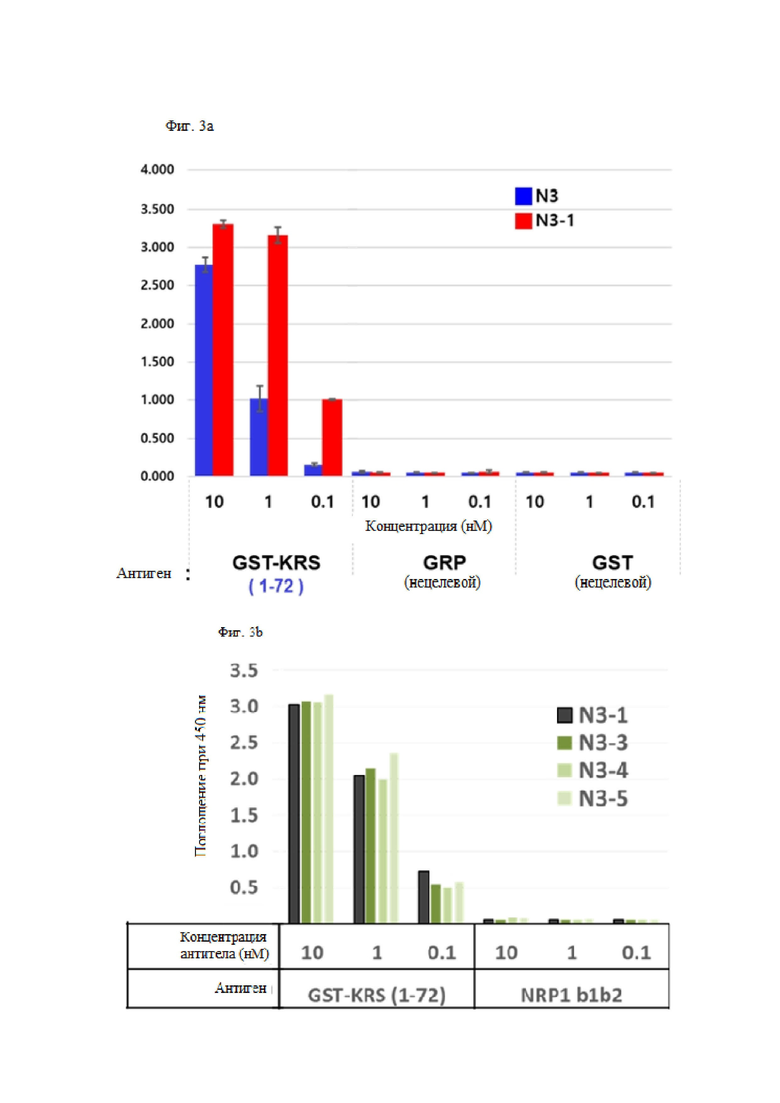
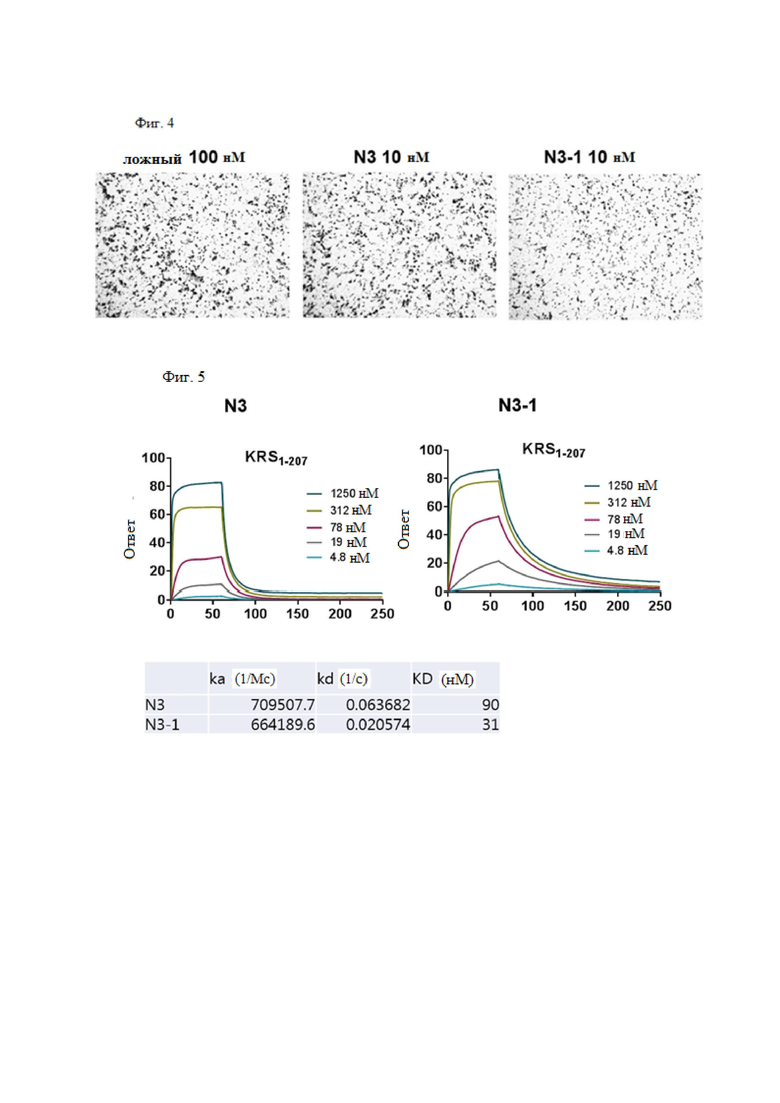
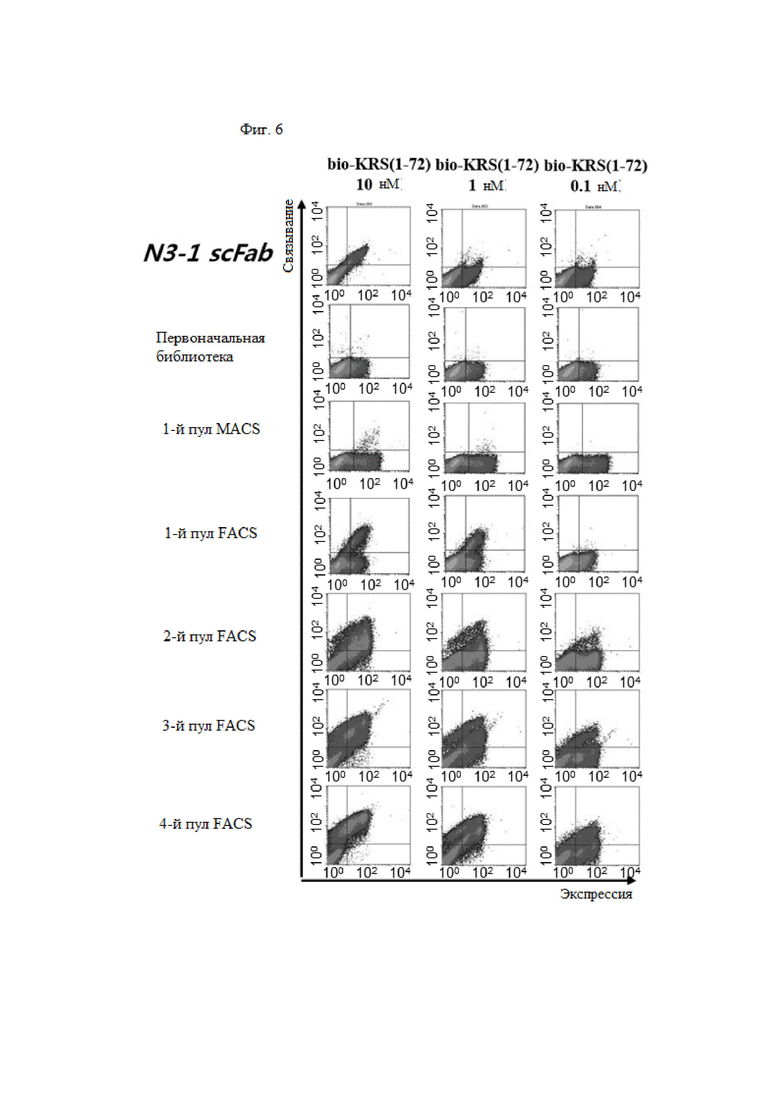
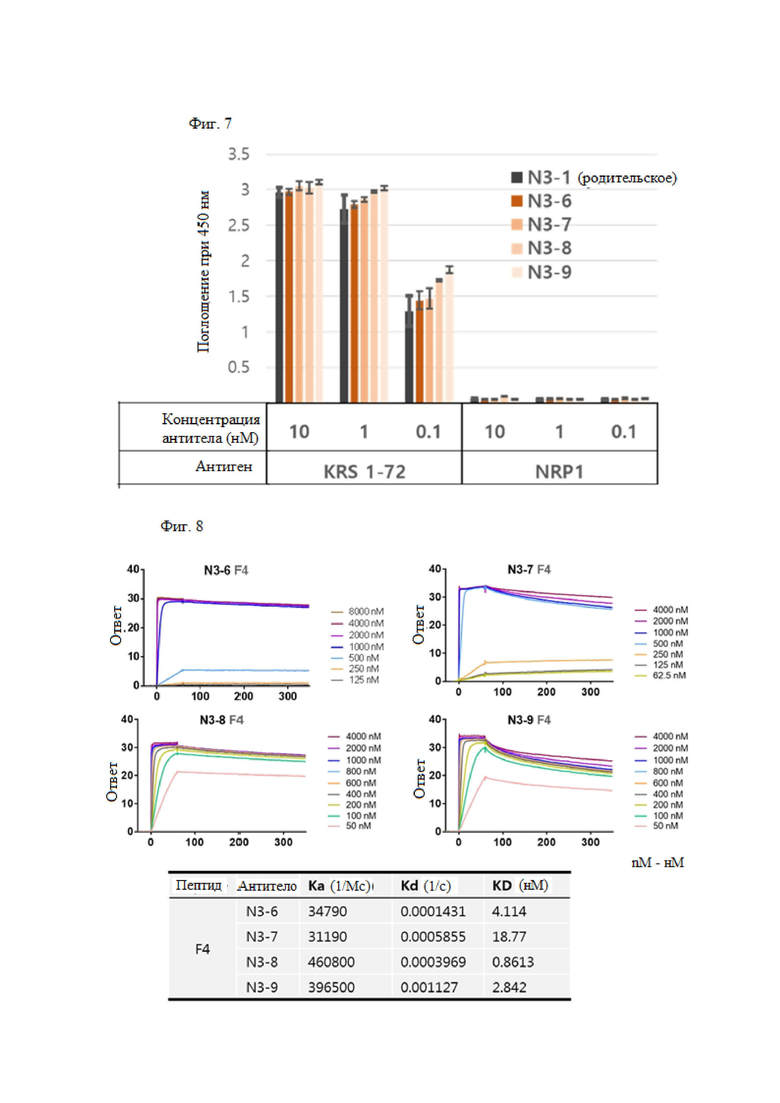
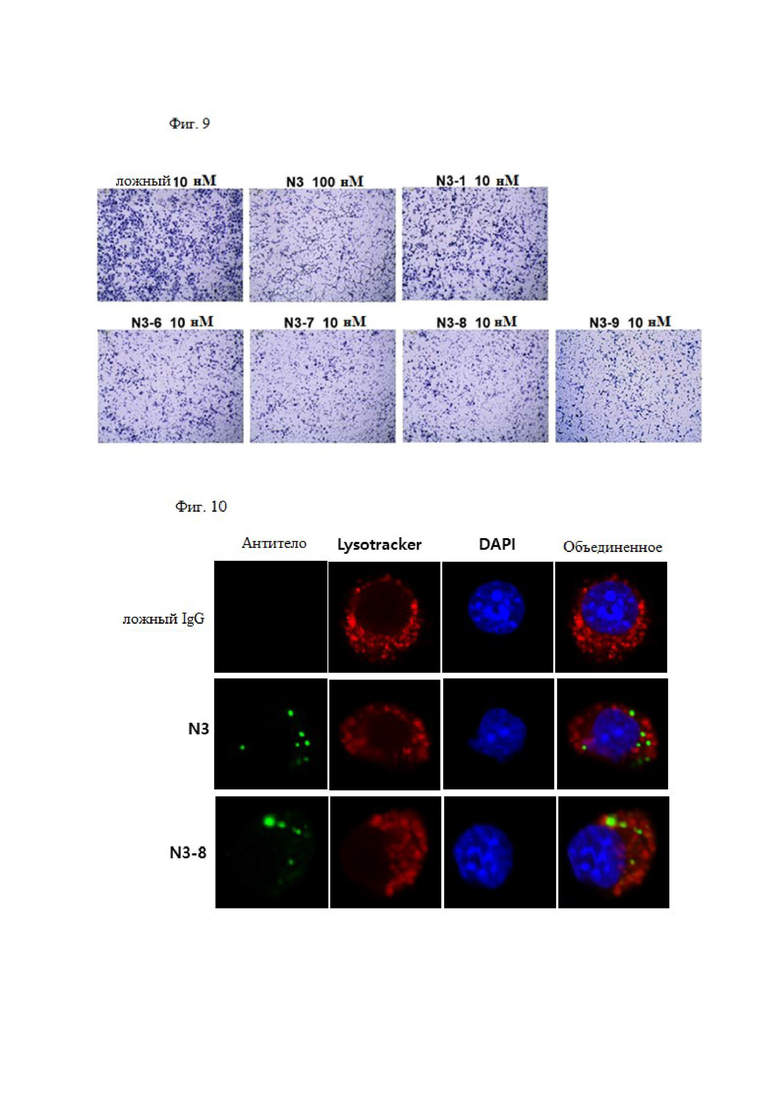
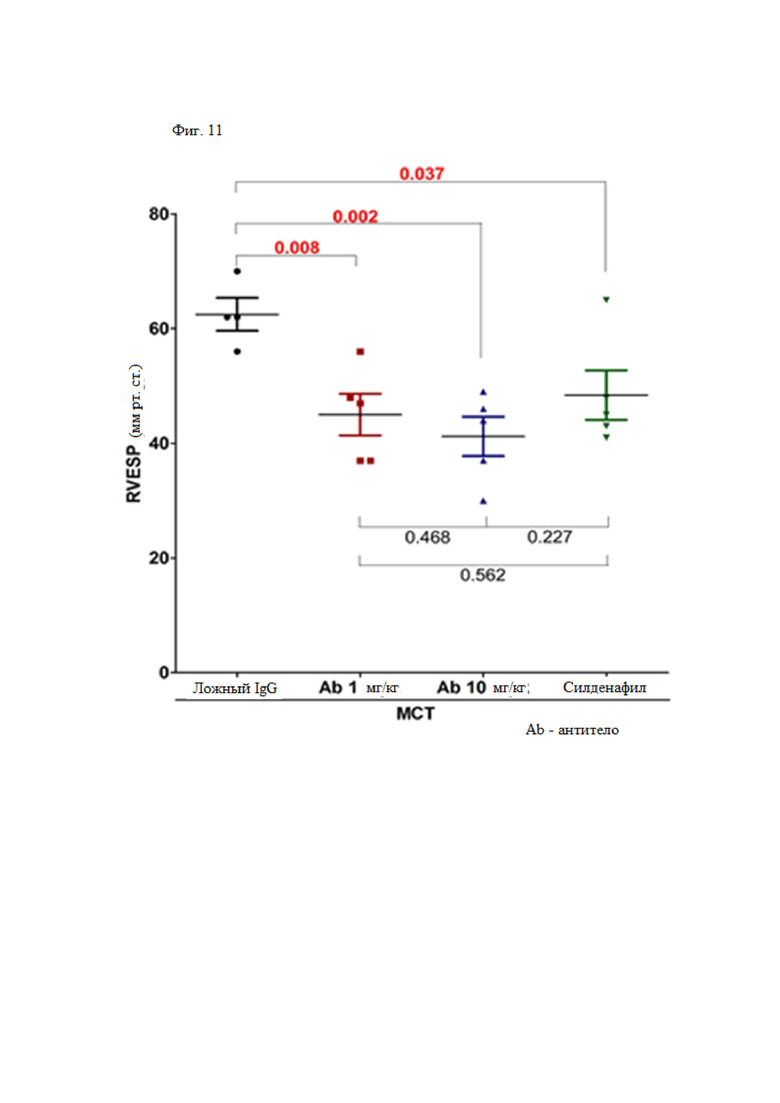
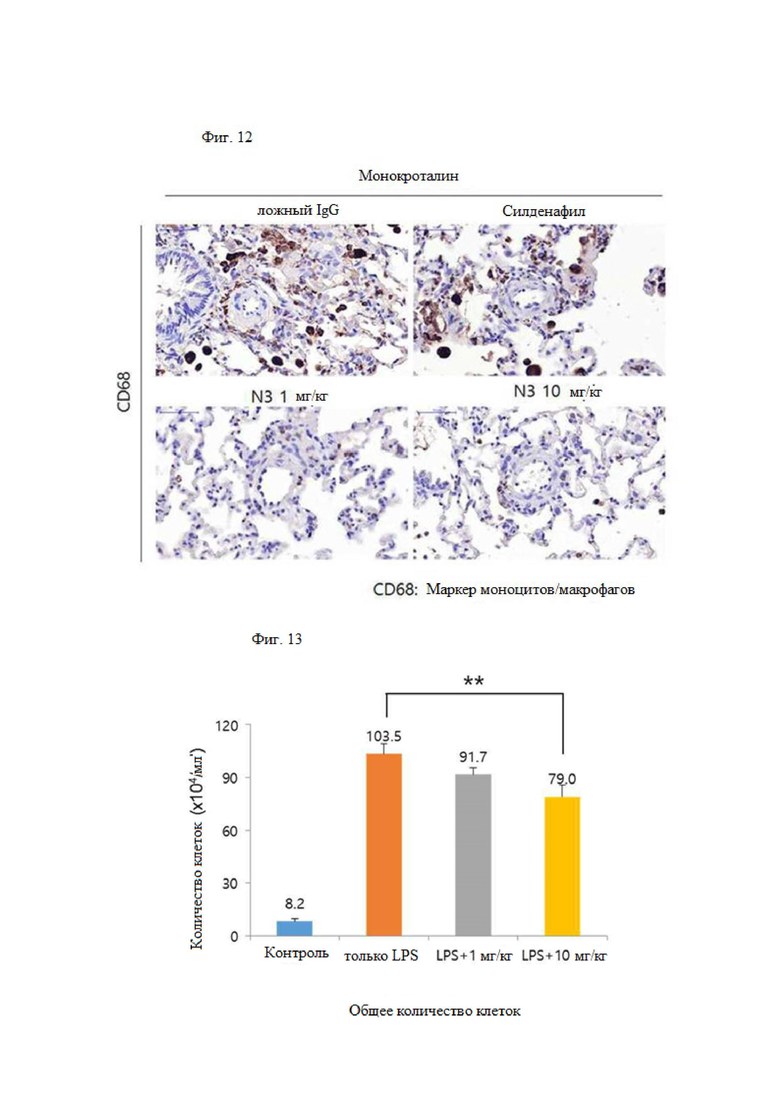
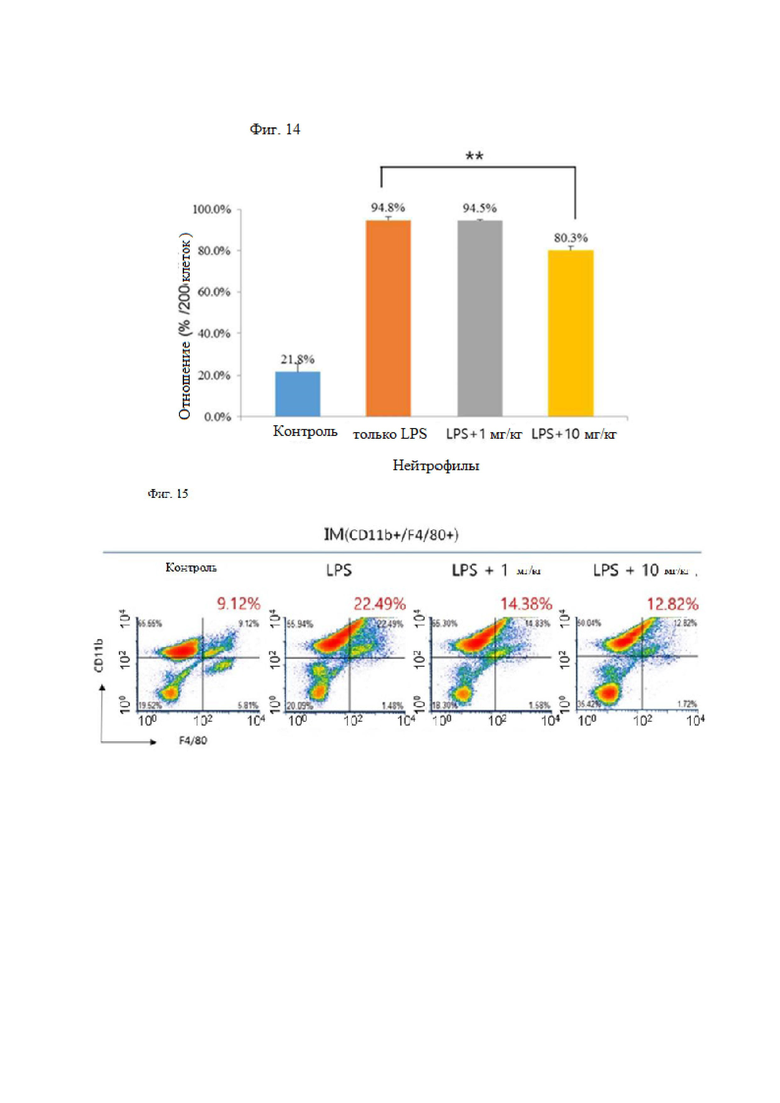
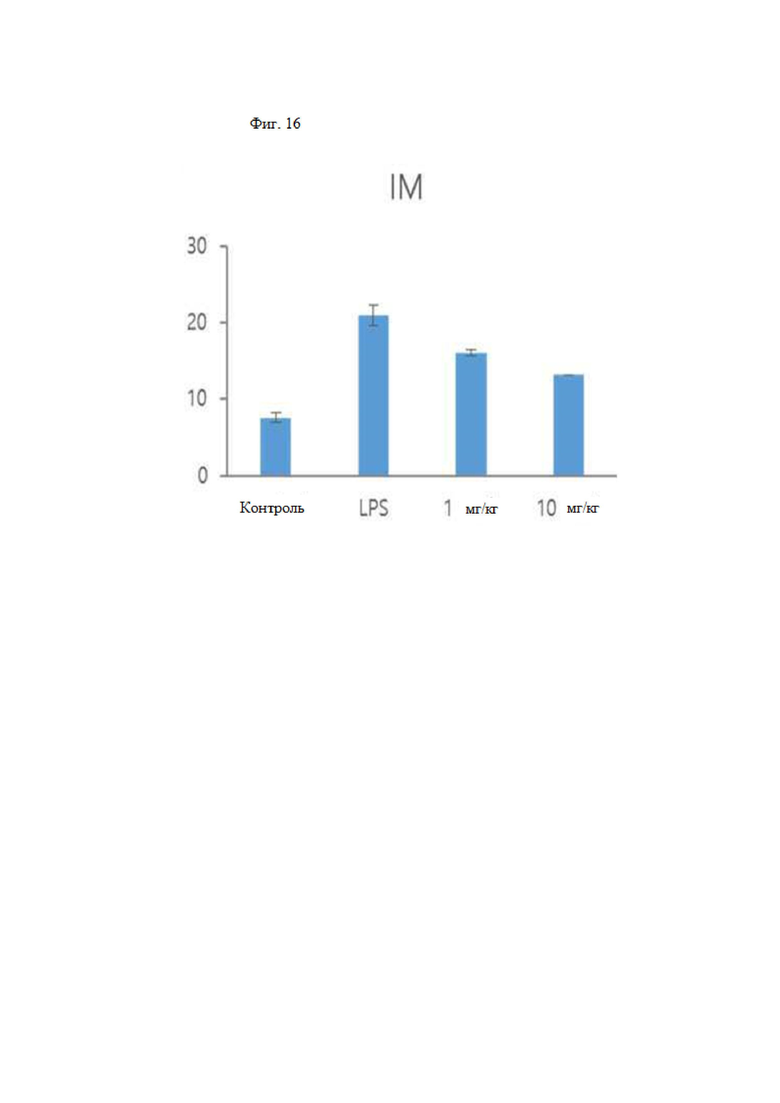
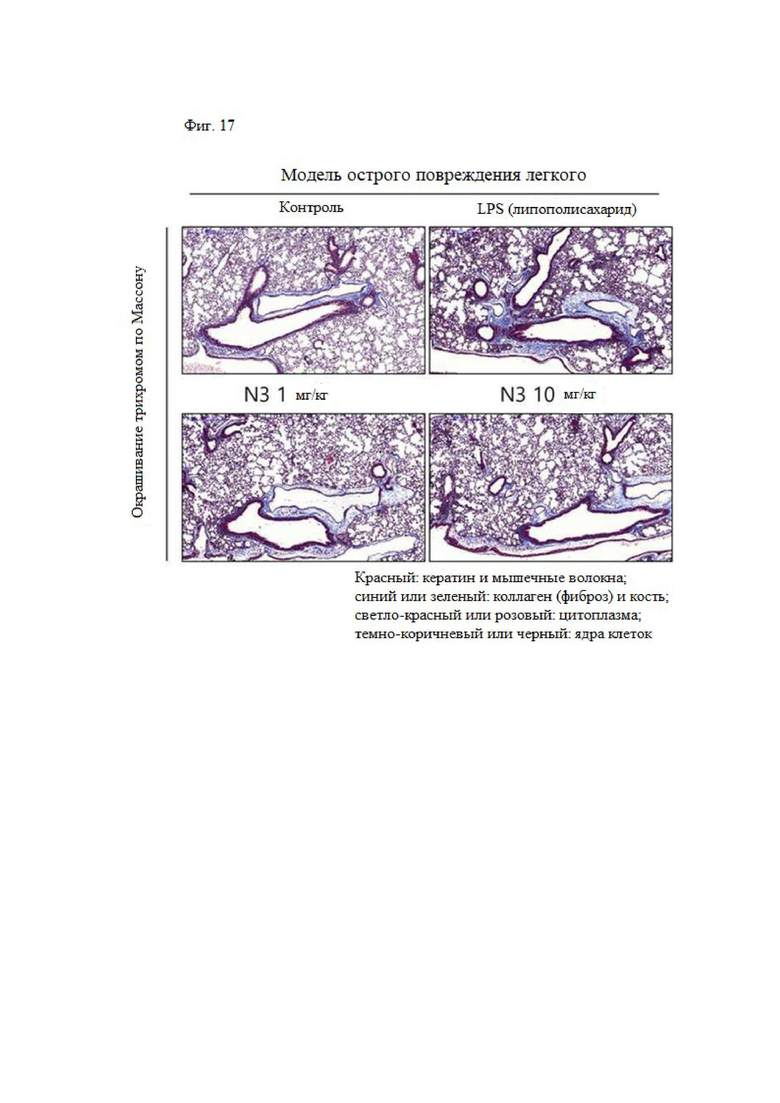

Настоящее изобретение относится к области биотехнологии, в частности к новому антителу, которое специфически связывается с лизил-тРНК-синтетазой, и может быть использовано в медицине. Более конкретно изобретение позволяет получить антитело, которое способно специфически связываться с N-концевым доменом фермента лизил-тРНК-синтетазы (KRS, лизил-тРНК-синтетаза), экспонированным на внеклеточной мембране. Такое антитело может использоваться при лечении или диагностике рака, метастазов при раке или заболеваний, ассоциированных с миграцией иммунных клеток. 17 н. и 9 з.п. ф-лы, 13 пр., 13 табл., 19 ил.
1. Антитело или его фрагмент, специфически связывающееся с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS), содержащее:
(а) вариабельную область тяжелой цепи (VH), содержащую
(i) определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность SYDMS,
(ii) определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность X1IX2X3X4X5GX6X7YYADSVKG, где
X1 представляет собой A или V,
X2 представляет собой S, D или G,
X3 представляет собой Y, P, S или A,
X4 представляет собой D, Q, L или Y,
X5 представляет собой N, M, S или G,
X6 представляет собой N, R или P,
X7 представляет собой T, V, I или S, и
(iii) определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность X8ALDFDY, где
X8 представляет собой M или L, и
(b) вариабельную область легкой цепи (VL), содержащую
(i) определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность TGSSSNIGSNYVT,
(ii) определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность X9NX10X11RPS, где
X9 представляет собой D, S или R,
X10 представляет собой S или N и
X11 представляет собой N или Q, и
(iii) определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность X12SFSDELGAYV, где
X12 представляет собой A или S.
2. Антитело или его фрагмент по п. 1, где (а) вариабельная область тяжелой цепи (VH) содержит определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23 и SEQ ID NO: 118, и определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5 и SEQ ID NO: 25.
3. Антитело или его фрагмент по п. 1, где (b) вариабельная область легкой цепи (VL) содержит определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, SEQ ID NO: 27 и SEQ ID NO: 29, определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13 и SEQ ID NO: 15.
4. Антитело или его фрагмент по п. 1, содержащее:
i) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 3, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 13,
ii) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 3, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
iii) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 118, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 13,
iv) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 118, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность определенную SEQ ID NO: 15,
v) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 17, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
vi) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 19, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
vii) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
viii) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 23, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
ix) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 27, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
x) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 29, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
xi) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15,
xii) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 27, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15, или
xiii) антитело, содержащее вариабельную область тяжелой цепи (VH) антитела, содержащую определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, и вариабельную область легкой цепи (VL) антитела, содержащую определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 29, и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15.
5. Антитело или его фрагмент по п. 1, содержащее
тяжелую цепь (НС), содержащую одну или более аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 89, SEQ ID NO: 93, SEQ ID. NO: 95, SEQ ID NO: 97, SEQ ID NO: 99, SEQ ID NO: 101, SEQ ID NO: 103 и SEQ ID NO: 105, и
легкую цепь (LC), содержащую одну или более аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 107, SEQ ID NO: 109, SEQ ID NO: 111, SEQ ID NO: 113 и SEQ ID NO: 115.
6. Антитело или его фрагмент по п. 1, содержащее:
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 89, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 107,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 89, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 93, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 107,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 93, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 95, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 97, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109,
тяжелую цепь содержащую аминокислотную последовательность, определенную SEQ ID NO: 99, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 101, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 103, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 103, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 113,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 103, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 115,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 105, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 105, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 113,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 105, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 115,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 99, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 111,
тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 103, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 109.
7. Антитело или его фрагмент по п. 1, где антитело выбрано из группы, состоящей из IgG, IgA, IgM, IgE и IgD, и фрагмент выбран из группы, состоящей из диатела, Fab, Fab', F(ab)2, F(ab')2, Fv и scFv.
8. Антитело или его фрагмент по п. 1, где фрагмент содержит одну или более аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 61, SEQ ID NO: 63, SEQ ID NO: 65, SEQ ID NO: 67, SEQ ID NO: 69, SEQ ID NO: 71, SEQ ID NO: 73, SEQ ID NO: 75, SEQ ID NO: 77, SEQ ID NO: 79, SEQ ID NO: 81, SEQ ID NO: 83, SEQ ID NO: 85 и SEQ ID NO: 87.
9. Полинуклеотид, кодирующий антитело или его фрагмент по любому из пп. 1-8.
10. Рекомбинантный вектор экспрессии, содержащий полинуклеотид по п. 9.
11. Клетка-хозяин, трансформированная рекомбинантным вектором по п. 10, для получения антитела по п. 1.
12. Способ получения антитела или его фрагмента, специфически связывающегося с внеклеточно экспонированной N-концевой областью лизил-тРНК синтетазы (KRS), включающий:
(а) трансформацию клеток-хозяев рекомбинантным вектором экспрессии по п. 10,
(b) инкубирование трансформированных клеток-хозяев для продукции антитела или его фрагмента,
(c) сбор антитела или его фрагмента, продуцируемого в клетках-хозяевах;
(d) очистка собранного антитела или его фрагмента; и
(e) выделение очищенного антитела или его фрагмента.
13. Фармацевтическая композиция для предотвращения или ингибирования роста опухоли и метастазов рака, содержащая эффективное количество антитела или его фрагмента по п. 1 в качестве активного ингредиента.
14. Композиция для диагностики рака или метастазов рака, содержащая эффективное количество антитела или его фрагмента по п. 1 в качестве активного ингредиента.
15. Антитело или его фрагмент по п. 1, где указанное антитело представляет собой полипептид, содержащий вариант Fc области Fc человеческого IgG дикого типа, и где вариант Fc содержит L117A, L118A, T182A, P212G области Fc человеческого IgG1 дикого типа, определенной SEQ ID NO: 126, или по меньшей мере одну дополнительную аминокислотную замену, которая представляет собой T179A области Fc человеческого IgG4, определенной SEQ ID NO: 138, и где полипептид имеет пониженную функцию ADCC/CDC (антителозависимая клеточная цитотоксичность/комплементзависимая цитотоксичность) по сравнению с полипептидом, содержащим область Fc IgG дикого типа.
16. Композиция для предотвращения или лечения заболевания, связанного с миграцией иммунных клеток, содержащая эффективное количество антитела или его фрагмента по п. 1 в качестве активного ингредиента.
17. Композиция для диагностики заболевания, связанного с миграцией иммунных клеток, содержащая эффективное количество антитела или его фрагмента по п. 1 в качестве активного ингредиента.
18. Композиция по п. 16 или. 17, где заболевание, связанное с миграцией иммунных клеток, выбрано из группы, состоящей из сердечно-сосудистого заболевания, фиброзного заболевания, воспалительного заболевания и синдрома Альпорта.
19. Применение антитела или его фрагмента по п. 1 для получения агента для предотвращения или ингибирования роста опухоли и метастазов рака.
20. Способ предотвращения или ингибирования роста опухоли и метастазов рака, включающий введение эффективного количества композиции, содержащей антитело или его фрагмент по п. 1, субъекту, нуждающемуся в этом.
21. Применение антитела или его фрагмента по п. 1 для получения агента для диагностики рака или метастазов рака.
22. Способ диагностики рака или метастазов рака, включающий:
а) получение биологического образца от индивидуума (субъекта) с подозрением на метастазы рака,
b) введение композиции, содержащей антитело или его фрагмент по п. 1, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b) и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику рака и метастазов рака, если уровень экспрессии KRS повышен.
23. Применение антитела или его фрагмента по п. 1 для получения агента для лечения заболевания, связанного с миграцией иммунных клеток.
24. Способ лечения заболевания, связанного с миграцией иммунных клеток, включающий введение эффективного количества композиции, содержащей антитело или его фрагмент по п. 1, субъекту, нуждающемуся в этом.
25. Применение антитела или его фрагмента по п. 1 для получения агента для диагностики заболевания, связанного с миграцией иммунных клеток.
26. Способ диагностики заболевания, связанного с миграцией иммунных клеток, включающий:
а) получение биологического образца от субъекта с подозрением на заболевание, связанное с миграцией иммунных клеток,
b) введение композиции, содержащей антитело или его фрагмент по п. 1, в образец или субъекту,
c) определение уровня экспрессии белка KRS в образце или у субъекта со стадии b) и
d) сравнение уровня экспрессии белка KRS с нормальной контрольной группой и диагностику заболевания, связанного с миграцией иммунных клеток, если уровень экспрессии KRS повышен.
| WO 2011153277 A2, 08.12.2011 | |||
| WO 2015102341 A1, 09.07.2015 | |||
| WO 2017146529 A1, 31.08.2017 | |||
| Устройство для прошивки отверстий | 1960 |
|
SU140302A1 |
| СПОСОБ ИНГИБИРОВАНИЯ МЕТАСТАЗИРОВАНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ ИЛИ МИГРАЦИИ ЗЛОКАЧЕСТВЕННЫХ КЛЕТОК ПОСРЕДСТВОМ СНИЖЕНИЯ КЛЕТОЧНОГО УРОВНЯ ЛИЗИЛ-тРНК-СИНТЕТАЗЫ (ВАРИАНТЫ), КОМПОЗИЦИЯ И ПРИМЕНЕНИЕ ВЕКТОРА ЭКСПРЕССИИ ИЛИ АНТИТЕЛА ПРОТИВ KRS ДЛЯ ИНГИБИРОВАНИЯ МЕТАСТАЗИРОВАНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ ИЛИ МИГРАЦИИ ЗЛОКАЧЕСТВЕННЫХ КЛЕТОК | 2008 |
|
RU2465330C1 |

