ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет заявке США с серийным номером № 62/220196, поданной 17 сентября 2015 года, содержание которой включено в настоящее описание путем ссылки в полном объеме.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка содержит список последовательностей, который предоставлен в электронной форме в формате ASCII и тем самым включен путем ссылки в полном объеме. Указанная копия ASCII, созданная 14 сентября 2016 года, названа N2067-7098WO_SL.txt и имеет размер 507996 байт.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится, главным образом, к применению иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии химерного рецептора антигена (CAR), для лечения заболевания, ассоциированного с экспрессией опухолевого антигена.
УРОВЕНЬ ТЕХНИКИ, К КОТОРОМУ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Терапия посредством адоптивного переноса клеток (ACT) с использованием аутологичных T-клеток, особенно T-клеток, трансдуцированных химерными рецепторами антигенов (CAR), оказалась перспективной в испытаниях на гематологических злокачественных опухолях. Существует медицинская потребность в способах терапии на основе T-клеток, особенно способах терапии на основе CAR T-клеток с повышенной эффективностью.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится композициям и способам, которые нарушают гены метилцитозиндиоксигеназы (например, Tet1, Tet2, Tet3), и к применениям таких композиций и способов для повышения функциональной активности клеток, модифицированных способами инженерии (например, генно-модифицированных антигенспецифических T-клеток, таких как CAR T-клетки). В частности, настоящее изобретение относится к способам и композициям для повышения терапевтической эффективности T-клеток с химерным рецептором антигена (CAR). Без связи с теорией, нарушение одного аллеля в гене Tet (например, Tet1, Tet2 или Tet3) приводит к снижению общих уровней 5-гидроксиметилцитозина совместно с усиленной пролиферацией, регуляцией продукции эффекторных цитокинов и дегрануляций, и тем самым усиливает пролиферацию и/или функцию CAR T-клеток.
Таким образом, настоящее изобретение относится клетке (например, популяции клеток, такой как популяция иммунных эффекторных клеток), модифицированной способами инженерии для экспрессии химерного рецептора антигена (CAR), где CAR содержит антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, и где экспрессия и/или функция Tet1, Tet2 и/или Tet3 в указанной клетке снижена или устранена. В одном варианте осуществления экспрессия и/или функция Tet2 в указанной клетке снижена или устранена. В некоторых вариантах осуществления антигенсвязывающий домен, который связывается с опухолевым антигеном, выбран из группы, состоящей из: TSHR, CD19, CD123, CD22, CD30, CD171, CS-1, CLL-1, CD33, EGFRvIII, GD2, GD3, BCMA, Tn Ag, PSMA, ROR1, FLT3, FAP, TAG72, CD38, CD44v6, CEA, EPCAM, B7H3, KIT, IL-13Ra2, мезотелина, IL-11Ra, PSCA, PRSS21, VEGFR2, LewisY, CD24, PDGFR-бета, SSEA-4, CD20, рецептора фолатов альфа, ERBB2 (Her2/neu), MUC1, EGFR, NCAM, простазы, PAP, ELF2M, эфрина B2, рецептора IGF-I, CAIX, LMP2, gp100, bcr-abl, тирозиназы, EphA2, фукозил-GM1, sLe, GM3, TGS5, HMWMAA, o-ацетил-GD2, рецептора фолатов бета, TEM1/CD248, TEM7R, CLDN6, GPRC5D, CXORF61, CD97, CD179a, ALK, полисиаловой кислоты, PLAC1, GloboH, NY-BR-1, UPK2, HAVCR1, ADRB3, PANX3, GPR20, LY6K, OR51E2, TARP, WT1, NY-ESO-1, LAGE-1a, MAGE-A1, легумаина, HPV E6,E7, MAGE A1, ETV6-AML, белка спермы 17, XAGE1, Tie 2, MAD-CT-1, MAD-CT-2, Fos-родственного антигена 1, p53, мутанта p53, простеина, сурвивина и теломеразы, PCTA-1/галектина 8, MelanA/MART1, мутанта Ras, hTERT, точек разрыва при транслокации саркомы, ML-IAP, ERG (слитый ген TMPRSS2 ETS), NA17, PAX3, рецептора андрогенов, циклина B1, MYCN, RhoC, TRP-2, CYP1B1, BORIS, SART3, PAX5, OY-TES1, LCK, AKAP-4, SSX2, RAGE-1, обратной транскриптазы теломеразы человека, RU1, RU2, кишечной карбоксилэстеразы, mut hsp70-2, CD79a, CD79b, CD72, LAIR1, FCAR, LILRA2, CD300LF, CLEC12A, BST2, EMR2, LY75, GPC3, FCRL5 и IGLL1. В одном варианте осуществления опухолевый антиген представляет собой CD19. В некоторых вариантах осуществления антигенсвязывающий домен представляет собой антитело или фрагмент антитела, как описано, например, в WO2012/079000 или WO2014/153270.
В одном аспекте настоящее изобретение относится к клетке (например, популяции клеток, такой как популяция иммунных эффекторных клеток), модифицированной способами инженерии для экспрессии CAR, и где экспрессия и/или функция Tet1, Tet2 и/или Tet3 в указанной клетке снижена или устранена. В одном варианте осуществления экспрессия и/или функция Tet2 в указанной клетке снижена или устранена. В некоторых вариантах осуществления трансмембранный домен указанного CAR содержит: (i) аминокислотную последовательность, имеющую по меньшей мере одну, две или три модификации, но не более 20, 10 или 5 модификаций аминокислотной последовательности SEQ ID NO: 12, или последовательность, обладающую 95-99% идентичностью с аминокислотной последовательностью SEQ ID NO: 12; или (ii) последовательность SEQ ID NO: 12.
В одном аспекте настоящее изобретение относится к клетке (например, популяции клеток, такой как популяция иммунных эффекторных клеток), модифицированной способами инженерии для экспрессии CAR, и где экспрессия и/или функция Tet1, Tet2 и/или Tet3 в указанной клетке снижена или устранена. В одном варианте осуществления антигенсвязывающий домен указанного CAR связан с трансмембранным доменом шарнирной областью, где указанная шарнирная область содержит SEQ ID NO: 2 или SEQ ID NO: 6, или последовательность, обладающую 95-99% идентичностью с ней. В некоторых вариантах осуществления внутриклеточный сигнальный домен указанного CAR содержит первичный сигнальный домен и/или костимулирующий сигнальный домен, где первичный сигнальный домен содержит функциональный сигнальный домен белка, выбранного из CD3-зета, CD3-гамма, CD3-дельта, CD3, общего FcR-гамма (FCER1G), FcR-бета (Fc-эпсилон R1b), CD79a, CD79b, Fc-гамма RIIa, DAP10 или DAP12.
В некоторых вариантах осуществления первичный сигнальный домен указанного CAR содержит: (i) аминокислотную последовательность, обладающую по меньшей мере одной, двумя или тремя, но не более чем 20, 10 или 5 модификациями аминокислотной последовательности SEQ ID NO: 18 или SEQ ID NO: 20, или последовательность, обладающую 95-99% идентичностью с аминокислотной последовательностью SEQ ID NO: 18 или SEQ ID NO: 20; или (ii) аминокислотную последовательность SEQ ID NO: 18 или SEQ ID NO: 20. В некоторых вариантах осуществления внутриклеточный сигнальный домен указанного CAR содержит костимулирующий сигнальный домен или первичный сигнальный домен и костимулирующий сигнальный домен, где костимулирующий сигнальный домен содержит функциональный сигнальный домен белка, выбранного из группы, состоящей из CD27, CD28, 4-1BB (CD137), OX40, CD30, CD40, PD-1, ICOS, ассоциированного с лимфоцитарной функцией антигена-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, B7-H3, лиганда, который специфически связывается с CD83, CDS, ICAM-1, GITR, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), CD160, CD19, CD4, CD8-альфа, CD8-бета, IL2R-бета, IL2R-гамма, IL7R-альфа, ITGA4, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, TNFR2, TRANCE/RANKL, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Tactile), CEACAM1, CRTAM, Ly9 (CD229), CD160 (BY55), PSGL1, CD100 (SEMA4D), CD69, SLAMF6 (NTB-A, Ly108), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, LAT, GADS, SLP-76, PAG/Cbp, NKp44, NKp30, NKp46 и NKG2D.
В некоторых вариантах осуществления костимулирующий сигнальный домен указанного CAR содержит аминокислотную последовательность, обладающую по меньшей мере одной, двумя или тремя модификациями, но не более чем 20, 10 или 5 модификациями аминокислотной последовательности SEQ ID NO: 14 или SEQ ID NO: 16, или последовательность, обладающую 95-99% идентичностью с аминокислотной последовательностью SEQ ID NO: 14 или SEQ ID NO: 16. В некоторых вариантах осуществления внутриклеточный домен указанного CAR содержит последовательность SEQ ID NO: 14 или SEQ ID NO: 16, и последовательность SEQ ID NO: 18 или SEQ ID NO: 20, где последовательности, содержащие внутриклеточный сигнальный домен, экспрессируются в той же рамке считывания и в качестве одной полипептидной цепи. В некоторых вариантах осуществления CAR по настоящему изобретению, кроме того, содержит лидерную последовательность SEQ ID NO: 2.
В некоторых вариантах осуществления иммунная эффекторная клетка по настоящему изобретению представляет собой T-клетку или NK-клетку. В некоторых вариантах осуществления T-клетка представляет собой CD4+ T-клетку, CD8+ T-клетку или их комбинацию. В одном аспекте клетки по настоящему изобретению представляют собой клетки человека. В одном аспекте клетки (например, модифицированные способами инженерии иммунные эффекторные клетки, например, CAR T-клетки) по настоящему изобретению содержат ингибитор Tet1, Tet2 и/или Tet3. В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой (1) систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 и/или Tet3, или в их регуляторных элементах, например, в Tet2 или его регуляторных элементах; (2) нуклеиновую кислоту, кодирующую один или несколько компонентов указанной системы редактирования генов; или (3) их комбинации.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 и/или Tet3, или в их регуляторных элементах, например, в Tet2 или его регуляторных элементах, и где система редактирования генов выбрана из группы, состоящей из: системы CRISPR/Cas9, системы нуклеазы с цинковыми пальцами, системы TALEN и системы мегануклеазы.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 и/или Tet3, или в их регуляторных элементах, например, в Tet2 или его регуляторных элементах, и где система редактирования генов связывается с последовательностью-мишенью в раннем экзоне или интроне гена, кодирующего Tet1, Tet2 и/или Tet3, например Tet2.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 и/или Tet3, или в их регуляторных элементах, например, в Tet2 или его регуляторных элементах, и где система редактирования генов связывает последовательность-мишень гена, кодирующего tet2, и последовательность-мишень расположена выше по направлению транскрипции относительно экзона 4, например, в экзоне 1, экзоне 2 или экзоне 3, например, в экзоне 3.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 и/или Tet3, или в их регуляторных элементах, например, в Tet2 или его регуляторных элементах, и где система редактирования генов связывается с последовательностью-мишенью в позднем экзоне или интроне гена, кодирующего Tet1, Tet2 и/или Tet3, например Tet2.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 и/или Tet3, или в их регуляторных элементах, например, в Tet2 или его регуляторных элементах, и где система редактирования генов связывает последовательность-мишень гена, кодирующего tet2, и последовательность-мишень расположена ниже по направлению транскрипции относительно экзона 8, например, находится в экзоне 9, экзоне 10 или экзоне 11, например, находится в экзоне 9.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 и/или Tet3, или в их регуляторных элементах, например, в Tet2 или его регуляторных элементах, и где система редактирования генов представляет собой систему CRISPR/Cas, содержащую молекулу гРНК, содержащую нацеливающую последовательность, которая гибридизуется с последовательностью-мишенью гена Tet2. В некоторых вариантах осуществления нацеливающая последовательность представляет собой нацеливающую последовательность, приведенную в таблице 3. В некоторых вариантах осуществления нацеливающая последовательность представляет собой нацеливающую последовательность, приведенную в таблице 5.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой миРНК или кшРНК (siRNA или shRNA), специфичную к Tet1, Tet2, Tet3, или нуклеиновую кислоту, кодирующую указанную миРНК или кшРНК. В некоторых вариантах осуществления миРНК или кшРНК содержит последовательность, комплементарную последовательности мРНК Tet2, например, содержит последовательность-мишень кшРНК, приведенную в таблице 4.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где указанный ингибитор представляет собой низкомолекулярное соединение.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где ингибитор представляет собой белок, например, представляет собой доминантно-негативный партнер по связыванию для Tet1, Tet2 и/или Tet3 (например, деацетилаза гистонов (HDAC), которая взаимодействует с Tet1, Tet2 и/или Tet3), или нуклеиновую кислоту, кодирующую указанный доминантно-негативный партнер по связыванию для Tet1, Tet2 и Tet3.
В некоторых вариантах осуществления клетки по настоящему изобретению содержат CAR и ингибитор Tet1, Tet2 и/или Tet3, где ингибитор представляет собой белок, например, представляет собой доминантно-негативный (например, каталитически неактивный) Tet1, Tet2 или Tet3, или нуклеиновую кислоту, кодирующую указанный доминантно-негативный Tet1, Tet2 или Tet3.
В одном аспекте настоящее изобретение относится к способу повышения терапевтической эффективности экспрессирующей CAR клетки, например, клетки согласно любому из предшествующих положений, например, экспрессирующей CAR19 клетки (например, CTL019), включающему стадию снижения уровня 5-гидроксиметилцитозина в указанной клетке. В некоторых вариантах осуществления указанная стадия включает приведение указанной клетки в контакт с ингибитором Tet (например, Tet1, Tet2 и/или Tet3). В некоторых вариантах осуществления указанный ингибитор Tet представляет собой ингибитор Tet2. В некоторых вариантах осуществления ингибитор Tet (например, Tet1, Tet2 и/или Tet3) по настоящему изобретению выбран из группы, состоящей из: (1) системы редактирования генов, нацеленной на один или несколько участков в гене, кодирующем Tet1, Tet2 или Tet3, или в их соответствующих регуляторных элементах; (2) нуклеиновой кислоты (например, миРНК или кшРНК), которая ингибирует экспрессию Tet1, Tet2 или Tet3; (3) белка (например, доминантно-негативного, например, каталитически неактивного) Tet1, Tet2 или Tet3, или партнера по связыванию для Tet1, Tet2 или Tet3; (4) низкомолекулярного соединения, которое ингибирует экспрессию и/или функцию Tet1, Tet2 или Tet3; (5) нуклеиновой кислоты, кодирующей любой из (1)-(3); и (6) любой комбинации (1)-(5). В некоторых вариантах осуществления ингибитор Tet по настоящему изобретению представляет собой ингибитор Tet2.
В одном аспекте настоящее изобретение относится к способу повышения терапевтической эффективности экспрессирующей CAR клетки, например, клетки согласно любому из предшествующих положений, например, экспрессирующей CAR19 клетки (например, CTL019), включающему стадию снижения уровня 5-гидроксиметилцитозина в указанной клетке. В некоторых вариантах осуществления указанная стадия включает приведение указанной клетки в контакт с ингибитором Tet (например, Tet1, Tet2 и/или Tet3). В некоторых вариантах осуществления указанное приведение в контакт происходит ex vivo. В некоторых вариантах осуществления указанное приведение в контакт происходит in vivo. В некоторых вариантах осуществления указанное приведение в контакт происходит in vivo перед доставкой нуклеиновой кислоты, кодирующей CAR, в клетку. В некоторых вариантах осуществления указанное приведение в контакт происходит in vivo после введения клеток индивидууму, нуждающемуся в этом.
В одном аспекте настоящее изобретение относится к способу повышения терапевтической эффективности экспрессирующей CAR клетки, например, клетки согласно любому из предшествующих положений, например, экспрессирующей CAR19 клетки (например, CTL019), включающему стадию приведения указанной клетки в контакт с ингибитором Tet, например, ингибитором Tet1, Tet2 и/или Tet3. В некоторых вариантах осуществления указанный ингибитор Tet представляет собой ингибитор Tet2. В некоторых вариантах осуществления ингибитор Tet (например, Tet1, Tet2 и/или Tet3) по настоящему изобретению выбран из группы, состоящей из: (1) системы редактирования генов, нацеленной на один или несколько участков в гене, кодирующем Tet1, Tet2 или Tet3, или в их соответствующих регуляторных элементах; (2) нуклеиновой кислоты (например, миРНК или кшРНК), которая ингибирует экспрессию Tet1, Tet2 или Tet3; (3) белка (например, доминантно-негативного, например, каталитически неактивного) Tet1, Tet2 или Tet3, или партнера по связыванию для Tet1, Tet2 или Tet3; (4) низкомолекулярного соединения, которое ингибирует экспрессию и/или функцию Tet1, Tet2 или Tet3; (5) нуклеиновой кислоты, кодирующей любой из (1)-(3); и (6) любой комбинации (1)- (5). В некоторых вариантах осуществления ингибитор Tet по настоящему изобретению представляет собой ингибитор Tet2.
В одном аспекте настоящее изобретение относится к способу повышения терапевтической эффективности экспрессирующей CAR клетки, например, клетки согласно любому из предшествующих положений, например, экспрессирующей CAR19 клетки (например, CTL019), включающему стадию приведения указанной клетки в контакт с ингибитором Tet, например, ингибитором Tet1, Tet2 и/или Tet3. В некоторых вариантах осуществления указанная стадия включает приведение указанных клеток в контакт с ингибитором Tet (например, Tet1, Tet2 и/или Tet3). В некоторых вариантах осуществления указанное приведение в контакт происходит ex vivo. В некоторых вариантах осуществления указанное приведение в контакт происходит in vivo. В некоторых вариантах осуществления указанное приведение в контакт происходит in vivo перед доставкой нуклеиновой кислоты, кодирующей CAR, в клетку. В некоторых вариантах осуществления указанное приведение в контакт происходит in vivo после введения клеток индивидууму, нуждающемуся в этом.
В одном аспекте настоящее изобретение относится к способу лечения индивидуума, нуждающегося в этом, включающему введение указанному индивидууму эффективного количества клеток, как описано в настоящем описании, например, иммунных эффекторных клеток (например, T-клеток или NK-клеток), содержащих CAR, и необязательно введение указанному индивидууму ингибитора Tet1, Tet2 и/или Tet3. В некоторых вариантах осуществления индивидууму проводят предварительное лечение ингибитором Tet1, Tet2 и/или Tet3 перед началом терапии экспрессирующими CAR клетками. В некоторых вариантах осуществления индивидууму проводят сопутствующее лечение ингибитором ингибитором Tet1, Tet2 и/или Tet3 и терапией экспрессирующими CAR клетками. В некоторых вариантах осуществления индивидууму проводят лечение ингибитором Tet1, Tet2 и/или Tet3 после терапии экспрессрующими CAR клетками. В некоторых вариантах осуществления индивидуум имеет заболевание, ассоциированное с экспрессией опухолевого антигена, например, пролиферативное заболевание, предзлокачественное состояние, злокачественную опухоль и не связанное со злокачественной опухолью заболевание, ассоциированное с экспрессией опухолевого антигена. В некоторых вариантах осуществления индивидуум имеет гематологическую злокачественную опухоль, выбранную из одного или нескольких из хронического лимфоцитарного лейкоза (CLL), острых лейкозов, острого лимфоидного лейкоза (ALL), B-клеточного острого лимфоидного лейкоза (B-ALL), T-клеточного острого лимфоидного лейкоза (T-ALL), хронического миелогенного лейкоза (CML), B-клеточного пролимфоцитарного лейкоза, новообразования из бластных плазмацитоидных дендритных клеток, лимфомы Беркитта, диффузной крупноклеточной B-клеточной лимфомы, фолликулярной лимфомы, волосатоклеточного лейкоза, мелкоклеточной или крупноклеточной фолликулярной лимфомы, злокачественных лимфопролиферативных состояний, MALT-лимфомы, лимфомы из клеток мантийной зоны, лимфомы из клеток маргинальной зоны, множественной миеломы, миелодисплазии и миелодиспластического синдрома, неходжскинской лимфомы, лимфомы Ходжкина, плазмабластной лимфомы, новообразования из плазмацитоидных дендритных клеток, макроглобулинемии Валденстрема или предлейкоза.
Настоящее изобретение относится к применению композиций и/или способов, описанных в настоящем описании, для лечения злокачественной опухоли, где злокачественная опухоль выбрана из группы, состоящей из рака толстого кишечника, рака прямой кишки, почечно-клеточного рака, рака печени, немелкоклеточной карциномы легкого, рака тонкого кишечника, рака пищевода, меланомы, рака кости, рака поджелудочной железы, рака кожи, рака головы и шеи, кожной или внутриглазной злокачественной меланомы, рака тела матки, рака яичника, рака прямой кишки, рака анальной области, рака желудка, рака яичка, рака тела матки, карциномы фаллопиевых труб, карциномы эндометрия, карциномы шейки матки, карциномы влагалища, карциномы вульвы, болезни Ходжкина, неходжскинской лимфомы, злокачественной опухоли эндокринной системы, рака щитовидной железы, рака паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака мочеиспускательного канала, рака полового члена, солидных опухолей детского возраста, рака мочевого пузыря, рака почки или мочеточника, рака почечной лоханки, новообразования центральной нервной системы (ЦНС), первичной лимфомы ЦНС, ангиогенеза опухоли, опухоли позвоночника, глиомы ствола головного мозга, аденомы гипофиза, саркомы Капоши, эпидермоидного рака, плоскоклеточного рака, T-клеточной лимфомы, злокачественных опухолей, индуцируемых факторами внешней среды, комбинаций указанных злокачественных опухолей и метастатических очагов указанных опухолей.
Настоящее изобретение относится к ингибиторам Tet1, Tet2 и/или Tet3 для применения для лечения индивидуума, где указанному индивидууму проводили, проводят или намереваются проводить терапию, включающую экспрессирующую CAR клетку.
Кроме того, настоящее изобретение относится к способу получения экспрессирующей CAR клетки, включающему введение нуклеиновой кислоты, кодирующей CAR, в клетку, так чтобы указанная нуклеиновая кислота (или ее кодирующая CAR часть) встраивалась в геном клетки в гене Tet1, Tet2 и/или Tet3 (например, в интроне или экзоне гена Tet1, Tet2 и/или Tet3), так чтобы экспрессия и/или функция Tet1, Tet2 и/или Tet3 снижалась или устранялась.
Кроме того, настоящее изобретение относится к способу получения экспрессирующей CAR клетки, включающему приведение указанной экспрессирующей CAR клетки в контакт с ингибитором Tet1, Tet2 и/или Tet3 ex vivo. В некоторых вариантах осуществления ингибитор представляет собой ингибитор Tet2.
Кроме того, настоящее изобретение относится к вектору, содержащему последовательность, кодирующую CAR, и последовательность, кодирующую ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3. В некоторых вариантах осуществления ингибитор Tet представляет собой (1) систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet1, Tet2 или Tet3, или в их соответствующих регуляторных элементах; (2) нуклеиновую кислоту (например, миРНК или кшРНК), которая ингибирует экспрессию Tet1, Tet2 или Tet3; (3) белок (например, доминантно-негативный, например, каталитически неактивный) Tet1, Tet2 или Tet3, или партнер по связыванию для Tet1, Tet2 или Tet3; и (4) нуклеиновую кислоту, кодирующую любой из (1)-(3), или их комбинации. В некоторых вариантах осуществления последовательность, кодирующая CAR, и последовательность, кодирующая ингибитор Tet, разделены участком 2A.
Кроме того, настоящее изобретение относится к системе редактирования генов, которая является специфичной к последовательности гена Tet или его регуляторных элементов, например, генов Tet1, Tet2 или Tet3 или их регуляторных элементов. В некоторых вариантах осуществления система редактирования генов является специфичной к последовательности гена Tet2. В некоторых вариантах осуществления система редактирования генов представляет собой (1) систему редактирования генов CRISPR/Cas, (2) систему нуклеазы с цинковыми пальцами, систему TALEN и систему мегануклеазы. В некоторых вариантах осуществления система редактирования генов представляет собой систему редактирования генов CRISPR/Cas. В некоторых вариантах осуществления система редактирования генов содержит: молекулу гРНК, содержащую нацеливающую последовательность, специфичную к последовательности гена Tet2 или его регуляторных элементов, и белок Cas9; молекулу гРНК, содержащую нацеливающую последовательность, специфичную к последовательности гена Tet2 или его регуляторных элементов, и нуклеиновую кислоту, кодирующую белок Cas9; нуклеиновую кислоту, кодирующую молекулу гРНК, содержащую нацеливающую последовательность, специфичную к последовательности гена Tet2 или его регуляторных элементов, и белок Cas9; или нуклеиновую кислоту, кодирующую молекулу гРНК, содержащую нацеливающую последовательность, специфичную к последовательности гена Tet2 или его регуляторных элементов, и нуклеиновую кислоту, кодирующую белок Cas9. В некоторых вариантах осуществления система редактирования генов, кроме того, содержит матричную ДНК. В некоторых вариантах осуществления матричная ДНК содержит последовательность нуклеиновой кислоты, кодирующую CAR, например, CAR, как описано в настоящем описании.
Кроме того, настоящее изобретение относится к композиции для получения экспрессирующей CAR клетки ex vivo, содержащей ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3, например, ингибитор Tet2. В некоторых вариантах осуществления ингибитор Tet выбран из N-[3-[7-(2,5-диметил-2H-пиразол-3-иламино)-1-метил-2-оксо-1,4-дигидро-2H-пиримидо[4,5-d]пиримидин-3-ил]-4-метилфенил]-3-трифторметил-бензамида, 2-[(2,6-дихлор-3-метилфенил)амино]бензойной кислоты и 2-гидроксиглутарата.
Кроме того, настоящее изобретение относится к популяции клеток, содержащей одну или несколько клеток, описанных в настоящем описании, где популяция клеток содержит более высокий процент клеток Tscm (например, CD45RA+CD62L+CCR7+CD27+CD95+ T-клеток), чем популяция клеток, которая не содержит одну или несколько клеток, в которых экспрессия и/или функция Tet1, Tet2 и/или Tet3 снижена или устранена.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
ФИГ.1: Экспрессирущие CD19 CART-клетки вводили пациенту (UPCC04409-10) для лечения CLL. Мониторинг CART-клеток у пациента UPCC04409-10 с течением времени проводили путем взятия образцов крови. Определяли уровень экспрессии BBZ в клетках (красный). Определяли количество копий последовательности семейства TCR Vbeta5.1 (синий). Оба измерения проводили для образцов, полученных на указанные сутки после второй инфузии CART-клеток.
ФИГ.2A и 2B: Репертуар T-клеточных рецепторов пациента UPCC04409-10 определяли в образце, полученном на 28 сутки (фиг.2A) или 51 сутки (фиг.2B) после инфузии CART. Это демонстрирует большое количество T-клеточных рецепторов семейства TCRBV05-01 на 51 сутки, что указывает на клональную экспансию с течением времени.
ФИГ.3: Выделенные T-клетки пациента UPCC04409-10 анализировали в отношении одновременной экспрессии CAR19 и 2 различных генов семейства TCR с течением времени (50 сутки и 51 сутки) и сравнивали с исходным дозированным материалом (продукт): на верхней панели представлено семейство TCR Vb13.1; на нижней панели представлено семейство TCR Vb5.1. Данные демонстрируют, что положительные по CAR19 клетки содержат один ген семейства TCR (Vb5.1), уровень которого быстро возрастает между 50 и 51 сутками.
ФИГ.4: Репертуар T-клеточных рецепторов для CD8-положительных клеток от пациента UPCC04409-10 определяли в образце, полученном на 51 сутки после инфузии CART. Это демонстрирует большое количество T-клеточных рецепторов семейства TCRBV05-01 на 51 сутки, что указывает на клональную экспансию CD8-положительных клеток с течением времени.
ФИГ.5: T-клеточный рецептор пациента UPCC04409-10 секвенировали, и представлена последовательность альфа- и бета-цепей (аминокислотные последовательности, представленные в качестве SEQ ID NO: 1297-1298, и нуклеотидные последовательности, представленные в качестве SEQ ID NO: 1299-1301, все, соответственно, в порядке появления).
ФИГ.6: Из T-клеток пациента UPCC04409-10 получали фрагментированную посредством обработки ультразвуковым излучением ДНК. Этот материал используют для амплификации геномных последовательностей, соседних с областью встраивания CAR19. Указанные гены рядом с CAR19 в геноме были идентифицированы как имеющие увеличенную экспрессию относительно инфузированного продукта (D0). В различные моменты времени после указанной инфузии CART (d=сутки; m=месяц), наблюдали различное относительное увеличение в количестве соседних генов, причем увеличение экспрессии Tet2 достигало пика в образцах как периферической крови (PBMC), так и CAR+CD8+ T-клеток на 51 сутки.
ФИГ.7: Участок встраивания гена CAR19 был картирован на гене Tet2. Более конкретно, встраивание происходило между 9 и 10 экзонами гена Tet2. Каталитический домен для Tet2 находится в экзоне 11. Встраивание в этом положении может приводить к экспрессии аберрантных мРНК-транскриптов или снижению экспрессии функционального Tet2 (дикого типа).
ФИГ.8: Транскрипты гена Tet2 из выделенной мРНК пациента UPCC04409-10 оценивали посредством ОТ-ПЦР с использованием праймеров, охватывающих указанные области Tet2 или CAR19, или обоих из них, как указано на фигуре справа. Rxn 3 содержит праймеры, сконструированные для амплификации области транскрипта Tet2, охватывающей экзоны 9 и 10. Rxn, 6, 7, 8, 9 и 10 представляют собой праймеры, сконструированные для амплификации указанных частей лентивируса CAR19. Rxn 12-16 представляют собой пары праймеров, которые содержат последовательность 9 экзона транскрипта Tet2, а также последовательность из лентивирусной конструкции CAR19. Эти данные показывают, что транскрипты получены из локуса Tet2, который содержит как последовательность Tet2, так и последовательность CAR19.
ФИГ.9: Показано схематическое представление транскриптов, происходящих из локуса Tet2, обнаруженных согласно фиг.10A и 10B. На этой фигуре указаны варианты по сплайсингу этой слитой конструкции Tet2/CAR19, которые были обнаружены в образце пациента. Этот анализ показал, что встраивание CAR19 в Tet2 привело к транскриптам, содержащим стоп-кодоны выше экзона 11. Было продемонстрировано, что экзон 11 является важным для функции Tet2. Это указывает на то, что функция Tet2 нарушается посредством встраивания CAR19. Это также указывает на то, что нарушение функции Tet2 приводило к благоприятной экспансии этих индивидуальных клонов CART.
ФИГ.10A и 10B: Схематично представлена ферментативная активность Tet2 (фиг.10A). Белок семейства Tet конвертирует 5-метилцитозин (5-mc) в 5-гидроксиметилцитозин (5-hmc), а затем в 5-формилцитозин (5-fmc), что приводит к деметилированному цитозину. Метилированная ДНК представляет собой эпигенетическое состояние, которое, как известно, влияет на профиль транскрипции. Оценивали состояние метилирования T-клеток от пациента UPCC04409-10 (фиг.10B). T-клетки пациента окрашивали в отношении TCRVb5.1 (который содержит инсерцию CAR19 в Tet2) и 5-hmc и 5-fmc оценивали в положительных по TCRVb5.1 (красный) и отрицательных по TCRVb5.1 (синий) популяциях посредством проточной цитометрии. Эти данные указывают на то, что клетки, содержащие инсерцию CAR19 в гене Tet2, имеют дефект деметилирования.
ФИГ.11: кшРНК против TET2 снижают уровни 5-hmc в нормальных T-клетках человека. TET2 и контрольные конструкции кшРНК с перестановкой, экспрессирующие mCherry, вводили в нормальные T-клетки человека. Уровни 5-hmc определяли по внутриклеточному окрашиванию с помощью FACS на 6 сутки после экспансии с использованием гранул с антителом против CD3/CD28. Нокдаун TET2 снижал общие уровни 5-hmc.
ФИГ.12: кшРНК против TET2 увеличивают в количестве T-клетки Tscm. TET2 и контрольные конструкции кшРНК с перестановкой, экспрессирующие mCherry, вводили в нормальные T-клетки человека. CD45RA+CD62L+CCR7+CD27+CD95+ T-клетки Tscm определяли по окрашиванию с помощью FACS на 11 сутки после экспансии с использованием гранул с антителом против CD3/CD28. Нокдаун TET2 стимулировал экспансию T-клеток с фенотипом Tscm.
ФИГ.13A: Стратегия гейтирования для количественного определения CAR+ клеток.
ФИГ.13B: Уровни экспрессии CAR в клетках, в которые посредством электропорации введены системы CRISPR/Cas, нацеленные на Tet2, по сравнению с нетрансфицированными клетками.
ФИГ.14: Количественное определение CD4+ и CD8+ клеток после трансдукции CAR и редактирования Tet2.
ФИГ.15: Эффект ингибирования Tet2 на экспансию CAR T-клеток с использованием гранул с антителом против CD3/CD28.
ФИГ.16: Эффект ингибирования Tet2 на антигензависимую продукцию интерлейкина-2 (IL-2) CAR T-клетками.
ФИГ.17: Эффект ингибирования Tet2 на антигензависимую продукцию интерферона-гамма CAR T-клетками.
ФИГ.18: Эффект ингибирования Tet2 на запускаемую антигеном пролиферацию CAR+ T-клеток.
ФИГ.19: Эффект ингибирования Tet2 на запускаемую антигеном пролиферацию T-клеток.
ФИГ.20: Эффект ингибирования Tet2 на запускаемую антигеном пролиферацию CD4+ T-клеток.
ФИГ.21: Эффект ингибирования Tet2 на запускаемую антигеном пролиферацию CAR+ CD4+ T-клеток.
ФИГ.22: Эффект ингибирования Tet2 на запускаемую антигеном пролиферацию CD8+ T-клеток.
ФИГ.23: Эффект ингибирования Tet2 на запускаемую антигеном пролиферацию CAR+ CD8+ T-клеток.
ФИГ.24: % редактирование и % редактирование со сдвигом рамки считывания путем систем CRISPR/Cas, нацеленных на Tet2 при определении с использованием NGS.
ФИГ.25: 5 наиболее часто встречающихся инсерций-делеций, наблюдаемых в T-клетках после добавления РНП, который включал указанные гРНК, нацеленные на 3 экзон TET2 (SEQ ID NO: 1302-1326, соответственно, в порядке появления). Представлены изменения относительно немодифицированной последовательности wt, причем инсерции указаны строчными буквами ("a". "t", "g" и "c") и делеции показаны прочерком ("-"). Частота инсерций-делеций показана в крайнем правом столбце.
ФИГ.26: 5 наиболее часто встречающихся инсерций-делеций, наблюдаемых в T-клетках после добавления RNP, который включал указанные гРНК, нацеленные на 9 экзон TET2 (SEQ ID NO: 1327-1356, соответственно, в порядке появления). Представлены изменения относительно немодифицированной последовательности wt, причем инсерции указаны строчными буквами ("a". "t", "g" и "c") и делеции показаны прочерком ("-"). Частота инсерций-делеций показана в крайнем правом столбце.
ФИГ.27: Схематический протокол эксперимента для определения нокдауна TET2 в клетках Jurkat в ответ на лентивирус, кодирующий кшРНК-ингибиторы TET2.
ФИГ.28: Экспрессия RFP в инфицированных кшРНК клетках Jurkat. Определение экспрессии RFP проводили с использованием FACS на 6 сутки после обработки пуромицином. Исходя из экспрессии RFP посредством обработки пуромицином было отобрано более 99% клеток jurkat с введенной кшРНК.
ФИГ.29: Эффективность нокдауна tet2 в инфицированных кшРНК против TET2 клетках Jurkat. Проводили эксперимент с использованием кОТ-ПЦР. Также измеряли уровни экспрессии tet1 и tet3. β-актин служил в качестве внутреннего контроля для количественного определения относительной экспрессии генов среди протестированных образцов. Для повышения надежности кОТ-ПЦР в этом эксперименте использовали два праймера для β-актина и один праймер для RPLP1.
ФИГ.30: Нокдаун белка TET2 в ответ на кшРНК в клетках Jurkat. Проводили эксперимент с использованием вестерн-блоттинга.
ФИГ.31A: Диаграммы Венна для пиков ATAC в CAR+CD8+ T-клетках от пациента с нарушением Tet2 по сравнению с CAR-CD8+ T-клетками от того же пациента в соответствующий момент времени без нарушения Tet2. На коробчатых диаграммах представлены отличия в увеличении уровня ATAC между двумя клеточными популяциями.
ФИГ.31B: Термины GO, ассоциированные с пиками ATAC, наиболее закрытыми в клеточной популяции с нарушением Tet2 по сравнению с его аналогом.
ФИГ.32A: Сайленсинг Tet2 посредством кшРНК в первичных CD8+ T-клетках от здоровых доноров при определении посредством количественной ПЦР. Экспрессия (среднее значение, SEM), нормализованная к GAPDH, представлена в качестве кратности изменения относительно ненацеленной контрольной кшРНК.
ФИГ.32B и 32C: Относительные частоты центральных T-клеток памяти (ФИГ.32B) и эффекторных CD8+ T-клеток (ФИГ.32C) на 14 сутки после увеличения в количестве посредством стимуляции через CD3/CD28 у тех же здоровых доноров, которые представлены на A.
ПОДРОБНОЕ ОПИСАНИЕ
Определения
Если не определено иначе, все технические и научные термины, используемые в настоящем описании, обладают тем же значением, которое обычно подразумевает специалист в области, к которой относится изобретение.
Форма единственного числа относится к одному или более чем к одному (т.е. по меньшей мере к одному) грамматическому объекту в форме единственного числа. В качестве примера "элемент" означает один элемент или более одного элемента.
Термин "приблизительно", когда он относится к поддающейся измерению величине, такой как количество, период времени и т.п., охватывает отклонения±20%, или в некоторых случаях±10%, или в некоторых случаях±5%, или в некоторых случаях±1%, или в некоторых случаях±0,1% от указанной величины, поскольку такие отклонения являются пригодными для выполнения описанных способов.
Термин "химерный рецептор антигена" или альтернативно "CAR" относится к набору полипептидов, как правило, двум в наиболее простых вариантах осуществления, который, когда находится в иммунной эффекторной клетке, обеспечивает специфичность клетки к клетке-мишени, как правило, злокачественной клетке, и индукцию внутриклеточного сигнала. В некоторых вариантах осуществления CAR содержит по меньшей мере внутриклеточный антигенсвязывающий домен, трансмембранный домен и цитоплазматический сигнальный домен (также обозначаемый в настоящем описании как "внутриклеточный сигнальный домен"), содержащий функциональный сигнальный домен, происходящий из стимулирующей молекулы и/или костимулирующей молекулы, как определено ниже. В некоторых аспектах полипептиды в наборе являются соседними друг с другом. В некоторых вариантах осуществления набор полипептидов включает переключатель димеризации, который в присутствии молекулы димеризации может связывать полипептиды друг с другом, например, может связывать антигенсвязывающий домен с внутриклеточным сигнальным доменом. В одном аспекте стимулирующая молекула представляет собой зета-цепь, связанную с T-клеточный рецепторным комплексом. В одном аспекте цитоплазматический сигнальный домен дополнительно содержит один или несколько функциональных сигнальных доменов, происходящих из по меньшей мере одной костимулирующей молекулы, как определено ниже. В одном аспекте костимулирующая молекула выбрана из костимулирующих молекул, описанных в настоящем описании, например, 4-1BB (т.е. CD137), CD27 и/или CD28. В одном аспекте CAR содержит химерный слитый белок, содержащий внеклеточный антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, содержащий функциональный сигнальный домен, происходящий из стимулирующей молекулы. В одном аспекте CAR содержит химерный слитый белок, содержащий внеклеточный антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, содержащий функциональный сигнальный домен, происходящий из костимулирующей молекулы, и функциональный сигнальный домен, происходящий из стимулирующей молекулы. В одном аспекте CAR содержит химерный слитый белок, содержащий внеклеточный антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, содержащий два функциональных сигнальных домена, происходящих из одной или нескольких костсимулирующей молекулы(молекул), и функциональный сигнальный домен, происходящий из стимулирующей молекулы. В одном аспекте CAR содержит химерный слитый белок, содержащий внеклеточный антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, содержащий по меньшей мере два функциональных сигнальных домена, происходящих из одной или нескольких костимулирующей молекулы(молекул), и функциональный сигнальный домен, происходящий из стимулирующей молекулы. В одном аспекте CAR содержит необязательную лидерную последовательность на N-конце (N-ter) слитого белка CAR. В одном аспекте CAR дополнительно содержит лидерную последовательность на N-конце внеклеточного антигенсвязывающего домена, где лидерная последовательность необязательно отщепляется от антигенсвязывающего домена (например, scFv) в ходе клеточного процессинга и локализации CAR на клеточной мембране.
CAR, который содержит антигенсвязывающий домен (например, scFv или TCR), который нацелен на конкретный опухолевый маркер X, такой как опухолевые маркеры, описанные в настоящем описании, также обозначают как XCAR. Например, CAR, который содержит антигенсвязывающий домен, который нацелен на CD19, обозначают как CD19CAR.
Термин "сигнальный домен" относится к функциональной части белка, которая действует, передавая информацию в клетке для регуляции клеточной активности через определенные пути передачи сигнала путем образования вторичных посредников или функционирования в качестве эффекторов посредством ответа на таких посредников.
Термин "антитело", как используют в рамках изобретения, относится к белку, или полипептидной последовательности, происходящим из молекулы иммуноглобулина, которая специфически связывается с антигеном. Антитела могут быть поликлональными или моноклональными, имеющими множество цепей или одноцепочечными, или интактными иммуноглобулинами, и они могут происходить из природных источников или из рекомбинантных источников. Антитела могут представлять собой тетрамеры молекул иммуноглобулинов.
Термин "фрагмент антитела" относится по меньшей мере к одной части антитела, которая сохраняет способность специфически взаимодействовать (например, путем связывания, пространственного препятствования, стабилизации/дестабилизации, пространственного распределения) с эпитопом антигена. Примеры фрагментов антител включают, но не ограничиваются ими, Fab, Fab', F(ab')2, Fv-фрагменты, фрагменты антител scFv, связанные дисульфидной связью Fv (sdFv), Fd-фрагмент, состоящий из доменов VH и CH1, линейные антитела, однодоменные антитела, такие как sdAb (либо VL, либо VH), домены VHH животных семейства верблюжьих, мультиспецифические антитела, образованные фрагментами антител, такими как двухвалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области, и выделенные CDR или другие связывающие эпитоп фрагменты антитела. Антигенсвязывающий фрагмент также может быть включен в однодоменные антитела, максиантитела, миниантитела, наноантитела, интраантитела, диантитела, триантитела, тетраантитела, v-NAR и бис-scFv (см., например, Hollinger and Hudson, Nature Biotechnology 23:1126-1136, 2005). Антигенсвязывающие фрагменты также могут быть пересажены в каркасы на основе полипептидов, таких как фибронектин типа III (Fn3)(см. патент США №: 6703199, в котором описаны миниантитела на основе полипептида фибронектина).
Термин "scFv" относится к слитому белку, содержащему по меньшей мере один фрагмент антитела, содержащий вариабельную область легкой цепи, и по меньшей мере один фрагмент антитела, содержащий вариабельную область тяжелой цепи, где вариабельные области легкой и тяжелой цепей связаны друг с другом, например, через синтетический линкер, например, короткий гибкий полипептидный линкер, и способному экспрессироваться в качестве одноцепочечного полипептида, и где scFv сохраняет специфичность интактного антитела, из которого он происходит. Если не указано, как используют в рамках изобретения, scFv может иметь вариабельные области VL и VH в любом порядке, например, относительно N-конца и C-конца полипептида scFv может содержать VL-линкер-VH или может содержать VH-линкер-VL.
Часть CAR по изобретению, содержащая антитело или фрагмент антитела, может существовать в различных формах, где антигенсвязывающий домен экспрессируется в качестве части непрерывной полипептидной цепи, включая, например, однодоменный фрагмент антитела (sdAb), одноцепочечное антитело (scFv), гуманизированное антитело или биспецифическое антитело (Harlow et al., 1999: Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, NY; Harlow et al., 1989: Antibodies: A Laboratory Manual, Cold Spring Harbor, New York; Houston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-5883; Bird et al., 1988, Science 242:423-426). В одном аспекте антигенсвязывающий домен композиции CAR по изобретению содержит фрагмент антитела. В следующем аспекте CAR содержит фрагмент антитела, который содержит scFv. Точные границы аминокислотных последовательностей данной CDR могут быть определены с использованием любой из ряда хорошо известных схем, включая схемы, описанные Kabat et al. (1991), "Sequences of Proteins of Immunological Interest", 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (схема нумерации "Kabat"), Al-Lazikani et al., (1997) JMB 273,927-948 (схема нумерации "Chothia"), или их комбинации.
Как используют в рамках изобретения, термин "связывающий домен" или "молекула антитела" относится к белку, например, цепи иммуноглобулина или ее фрагменту, содержащему по меньшей мере одну последовательность вариабельного домена иммуноглобулина. Термины "связывающий домен" или "молекула антитела" охватывают антитела и фрагменты антитела. В одном варианте осуществления молекула антитела представляет собой мультиспецифическую молекулу антитела, например, она содержит множество последовательностей вариабельных доменов иммуноглобулинов, где первая последовательность вариабельного домена иммуноглобулина из множества обладает специфичностью связывания в отношении первого эпитопа, а вторая последовательность вариабельного домена иммуноглобулина из множества обладает специфичностью связывания в отношении второго эпитопа. В одном варианте осуществления мультиспецифическая молекула антитела представляет собой биспецифическую молекулу антитела. Биспецифическое антитело обладает специфичностью в отношении не более чем двух антигенов. Биспецифическая молекула антитела характеризуется первой последовательностью вариабельного домена иммуноглобулина, которая обладает специфичностью связывания в отношении первого эпитопа, и второй последовательностью вариабельного домена иммуноглобулина, которая обладает специфичностью связывания в отношении второго эпитопа.
Часть CAR по изобретению, содержащая антитело или фрагмент антитела, может принимать множество форм, где антигенсвязывающий домен экспрессируется в качестве части непрерывной полипептидной цепи, включая, например, однодоменный фрагмент антитела (sdAb), одноцепочечное антитело (scFv), гуманизированное антитело или биспецифическое антитело (Harlow et al., 1999, Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, NY; Harlow et al., 1989, Antibodies: A Laboratory Manual, Cold Spring Harbor, New York; Houston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-5883; Bird et al., 1988, Science 242:423-426). В одном аспекте антигенсвязывающий домен композиции CAR по изобретению содержит фрагмент антитела. В следующем аспекте CAR содержит фрагмент антитела, который содержит scFv.
Термин "тяжелая цепь антитела" относится к более крупной из двух типов полипептидных цепей, присутствующих в молекулах антитела в их встречающихся в природе конформациях, и обычно определяющему класс, к которому относится антитело.
Термин "легкая цепь антитела" относится к меньшей из двух типов полипептидных цепей, присутствующих в молекулах антител в их встречающихся в природе конформациях. Легкие цепи каппа (κ) и лямбда (λ)относятся к двум основным изотипам легких цепей антител.
Термин "рекомбинантное антитело" относится к антителу, которое получают с использованием технологии рекомбинантных ДНК, например, такому как антитело, экспрессируемое бактериофагом или дрожжевой экспрессирующей системой. Термин также следует истолковывать как антитело, полученное посредством синтеза молекулы ДНК, кодирующей антитело, причем эта молекула ДНК экспрессирует белок антитела или аминокислотную последовательность, характеризующую антитело, где ДНК или аминокислотная последовательность получены с использованием технологии рекомбинантных ДНК или аминокислотных последовательностей, которая доступна и хорошо известна в данной области.
Термин "антиген" или "Ag" относится к молекуле, которая индуцирует иммунный ответ. Иммунный ответ может вовлекать либо продукцию антител, либо активацию специфических иммунокомпетентных клеток, или оба из этих вариантов. Квалифицированному специалисту будет понятно, что любая макромолекула, включая практические все белки или пептиды, может выступать в качестве антигена. Более того, антигены могут происходить из рекомбинантной или геномной ДНК. Квалифицированному специалисту будет понятно, что любая ДНК, которая содержит нуклеотидные последовательности или частичную нуклеотидную последовательность, кодирующую белок, который индуцирует иммунный ответ, таким образом, кодирует "антиген", как этот термин используют в настоящем описании. Более того, квалифицированному специалисту будет понятно, что антиген не должен кодироваться исключительно полноразмерной нуклеотидной последовательностью гена. Хорошо понятно, что настоящее изобретение включает, но не ограничивается ими, применение частичных нуклеотидных последовательностей из более чем одного гена, и что эти нуклеотидные последовательности располагаются в различных комбинациях для кодирования полипептидов, которые индуцируют желаемый иммунный ответ. Более того, квалифицированному специалисту будет понятно, что антиген вовсе не должен кодироваться "геном". Хорошо понятно, что антиген можно получать путем синтеза, или он может происходить из биологического образца, или может представлять собой макромолекулу помимо полипептида. Такой биологический образец может включать, но не ограничиваться ими, образец ткани, образец опухоли, клетку или жидкость с другими биологическими компонентами.
Термин "противораковый эффект" относится к биологическому эффекту, который может проявляться различными путями, включая, но не ограничиваясь ими, например, снижение объема опухоли, уменьшение количества злокачественных клеток, уменьшение количества метастазов, увеличение продолжительности жизни, снижение пролиферации злокачественных клеток, снижение выживаемости злокачественных клеток или смягчение различных физиологических симптомов, ассоциированных со злокачественным состоянием. "Противораковый эффект" также может проявляться способностью пептидов, полинуклеотидов, клеток и антител к предупреждению возникновения злокачественной опухоли изначально. Термин "противоопухолевый эффект" относится к биологическому эффекту, который может проявляться различными путями, включая, но не ограничиваясь ими, например, снижение объема опухоли, уменьшение количества опухолевых клеток, снижение пролиферации опухолевых клеток или снижение выживаемости опухолевых клеток.
Термин "аутологичный" относится к любому материалу, происходящему из того же индивидуума, которому его впоследствии обратно вводят.
Термин "аллогенный" относится к любому материалу, происходящему из другого животного того же вида относительно индивидуума, которому материал вводят. Два или более индивидуумов называют аллогенными друг другу, когда гены в одном или нескольких локусов не являются идентичными. В некоторых аспектах аллогенный материал от индивидуумов того же вида может быть достаточно генетически отличающимся, чтобы происходило антигенное взаимодействие.
Термин "ксеногенный" относится к трансплантату, происходящему из животного другого вида.
Термин "злокачественная опухоль" относится к заболеванию, характеризующемуся неконтролируемым ростом аберрантных клеток. Злокачественные клетки могут распространяться локально или через кровоток и лимфатическую систему в другие части организма. Примеры различных злокачественных опухолей описаны в настоящем описании и включают, но не ограничиваются ими, рак молочной железы, рак предстательной железы, рак яичника, рак шейки матки, рак кожи, рак поджелудочной железы, рак ободочной и прямой кишки, рак почки, рак печени, злокачественную опухоль головного мозга, лимфому, лейкоз, рак легкого и т.п. Термины "опухоль" и "злокачественная опухоль" используют в настоящем описании взаимозаменяемо, например, оба термины охватывают солидные и жидкостные, например, диффузные или циркулирующие, опухоли. Как используют в рамках изобретения, термин "злокачественная опухоль" или "опухоль" включает предзлокачественные, а также злокачественные новообразования и опухоли.
Термин "происходящий из", как используют в настоящем описании, указывает на взаимосвязь между первой и второй молекулой. Как правило, он относится к структурному сходству между первой молекулой и второй молекулой и не означает или не включает ограничение процесса или источника в отношении первой молекулы, которая происходит из второй молекулы. Например, в случае внутриклеточного сигнального домена, который происходит из молекулы CD3-зета, внутриклеточный сигнальный домен сохраняет достаточную структуру CD3-зета, чтобы он имел требуемую функцию, а именно, способность генерировать сигнал в соответствующих условиях. Он не означает и не включает ограничение конкретным процессом получения внутриклеточного сигнального домена, например, он не означает, что для предоставления внутриклеточного сигнального домена необходимо начать с последовательности CD3-зета и удалить нежелательную последовательность или внести мутации для получения внутриклеточного сигнального домена.
Выражение "заболевание, ассоциированное с экспрессией опухолевого антигена, как описано в настоящем описании," включает, но не ограничивается ими, заболевание, ассоциированное с экспрессией опухолевого антигена, как описано в настоящем описании, или состояние, ассоциированное с клетками, которые экспрессируют опухолевый антиген, как описано в настоящем описании, включая, например, пролиферативные заболевания, такие как рак, или злокачественное новообразование, или предзлокачественное состояние, такое как миелодисплазия, миелодиспластический синдром или предлейкоз; или незлокачественное состояние, ассоциированное с клетками, которые экспрессируют опухолевый антиген, как описано в настоящем описании. В одном аспекте злокачественная опухоль, ассоциированная с экспрессией опухолевого антигена, как описано в настоящем описании, представляет собой гематологическую злокачественную опухоль. В одном аспекте злокачественная опухоль, ассоциированная с экспрессией опухолевого антигена, как описано в настоящем описании, представляет собой солидную злокачественную опухоль. Следующие заболевания, ассоциированные с экспрессией опухолевого антигена, описанного в настоящем описании, включают, но не ограничиваются ими, например, атипичный и/или неклассический рак, злокачественные новообразования, предзлокачественные состояния или пролиферативные заболевания, ассоциированные с экспрессией опухолевого антигена, как описано в настоящем описании. Незлокачественные состояния, ассоциированные с экспрессией опухолевого антигена, как описано в настоящем описании, включают, но не ограничиваются ими, например, аутоиммунное заболевание (например, волчанка), воспалительные нарушения (аллергия и астма) и трансплантацию. В некоторых вариантах осуществления клетки, экспрессирующие опухолевый антиген, экспрессируют или в какой-либо момент экспрессировали мРНК, кодирующую опухолевый антиген. В одном варианте осуществления клетки, экспрессирующие опухолевый антиген, продуцируют белок опухолевого антигена (например, дикого типа или мутантный), и белок опухолевого антигена может присутствовать на нормальных уровнях или сниженных уровнях. В одном варианте осуществления экспрессирующие опухолевый антиген клетки продуцируют поддающиеся обнаружению уровни белка опухолевого антигена в один момент времени, а затем по существу не продуцируют поддающийся обнаружению белок опухолевого антигена.
Термин "консервативные модификации последовательности" относится к модификациям аминокислот, которые не влияют или не изменяют в значительной степени характеристики связывания антитела или фрагмента антитела, содержащих аминокислотную последовательность. Такие консервативные модификации включают аминокислотные замены, вставки или делеции. Модификации можно вносить в антитело или фрагмент антитела по изобретению стандартными способами, известными в данной области, такими как сайт-направленный мутагенез и ПЦР-опосредуемый мутагенез. Консервативные аминокислотные замены представляют собой замены, в которых аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, определены в данной области. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или несколько аминокислотных остатков в CAR по изобретению могут быть заменены другими аминокислотными остатками из того же семейства боковых цепей, и измененный CAR можно исследовать с использованием функциональных анализов, описанных в настоящем описании.
Термин "стимуляция" относится к первичному ответу, индуцированному посредством связывания стимулирующей молекулы (например, комплекс TCR/CD3 или CAR) с ее собственным лигандом (или опухолевым антигеном в случае CAR), что, тем самым, опосредует событие передачи сигнала, такое как, но не ограничиваясь ими, передача сигнала через комплекс TCR/CD3 или передача сигнала через соответствующий рецептор NK или сигнальные домены CAR. Стимуляция может опосредовать измененную экспрессию определенных молекул.
Термин "стимулирующая молекула" относится к молекуле, экспрессируемой иммунной клеткой (например, T-клеткой, NK-клеткой, B-клеткой), которая предоставляет сигнальную последовательность(и), которая регулирует активацию иммунной клетки стимулирующим образом для по меньшей мере некоторого аспекта каскада передачи сигнала иммунными клетками. В одном аспекте сигнал представляет собой первичный сигнал, который инициируется, например, связыванием комплекса TCR/CD3 с молекулой MHC, нагруженной пептидом, и который приводит к опосредованию T-клеточного ответа, включая, но не ограничиваясь ими, пролиферацию, активацию, дифференцировку и т.п. Первичная цитоплазматическая сигнальная последовательность (также обозначаемая как "первичный сигнальный домен"), которая действует стимулирующим образом, может содержать сигнальный мотив, который известен как иммунорецепторный тирозиновый активирующий мотив или ITAM. Примеры содержащей ITAM цитоплазматической сигнальной последовательности, которая является особенно пригодной в рамках изобретения, включают, но не ограничиваются ими, последовательности, происходящие из CD3-зета, общего FcR-гамма (FCER1G), Fc-гамма RIIa, FcR-бета (Fc-эпсилон R1b), CD3-гамма, CD3-дельта, CD3-эпсилон, CD79a, CD79b, DAP10 и DAP12. В конкретном CAR по изобретению внутриклеточный сигнальный домен в любом одном или нескольких CAR по изобретению содержит внутриклеточную сигнальную последовательность, например, первичную сигнальную последовательность CD3-зета. В конкретном CAR по изобретению первичная сигнальная последовательность CD3-зета представляет собой последовательность, представленную в SEQ ID NO: 17, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, мартышки, человекообразной обезьяны и т.п. В конкретном CAR по изобретению первичная сигнальная последовательность CD3-зета представляет собой последовательность, представленную в SEQ ID NO: 43, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, мартышки, человекообразной обезьяны и т.п.
Термин "антигенпредставляющая клетка" или "APC" относится к клетке иммунной системы, такой как вспомогательная клетка (например, B-клетка, дендритная клетка и т.п.), которая экспонирует чужеродный антиген в комплексе с основным комплексом гистосовместимости (MHC) на ее поверхности. T-клетки могут распознавать эти комплексы с использованием их T-клеточных рецепторов (TCR). APC процессируют антигены и презентируют их T-клеткам.
"Внутриклеточный сигнальный домен", как используют в настоящем описании, относится к внутриклеточной части молекулы. Внутриклеточный сигнальный домен генерирует сигнал, который стимулирует иммунную эффекторную функцию содержащей CAR клетки, например, CART-клетки. Примеры иммунной эффекторной функции, например, в CART-клетке, включают цитолитическую активность и хелперную активность, включая секрецию цитокинов.
В одном варианте осуществления внутриклеточный сигнальный домен может содержать первичный внутриклеточный сигнальный домен. Иллюстративные первичные внутриклеточные сигнальные домены включают домены, происходящие из молекул, ответственных за первичную стимуляцию или антигензависимую стимуляцию. В одном варианте осуществления внутриклеточный сигнальный домен может содержать костисмулирующий внутриклеточный домен. Иллюстративные костимулирующие внутриклеточные сигнальные домены включают домены, происходящие из молекул, ответственных за костисмулирующие сигналы или антиген-независимую стимуляцию. Например, в случае CART первичный внутриклеточный сигнальный домен может содержать цитоплазматическую последовательность T-клеточного рецептора, и костимулирующий внутриклеточный сигнальный домен может содержать цитоплазматическую последовательность из корецептора или костимулирующей молекулы.
Первичный внутриклеточный сигнальный домен может содержать сигнальный мотив, который известен как иммунорецепторный тирозиновый активирующий мотив или ITAM. Примеры содержащих ITAM первичных цитоплазматических сигнальных последовательностей включают, но не ограничиваются ими, последовательности, происходящие из CD3-зета, общего FcR-гамма (FCER1G), Fc-гамма RIIa, FcR-бета (Fc-эпсилон R1b), CD3-гамма, CD3-дельта, CD3-эпсилон, CD79a, CD79b, DAP10 и DAP12.
Термин "зета" или альтернативно "зета-цепь", "CD3-зета" или "TCR-зета" определяют как белок, представленный под номером доступа GenBan № BAG36664.1, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, и т.п., и "зета-стимулирующий домен" или альтернативно "стимулирующий домен CD3-зета" или "стимулирующий домен TCR-зета" определяют как аминокислотные остатки из цитоплазматического домена зета-цепи или их функциональные производные, которые являются достаточными для функциональной передачи первоначального сигнала, необходимого для активации T-клеток. В одном аспекте цитоплазматический домен зета-цепи содержит остатки с 52 по 164 последовательности с номером доступа GenBank № BAG36664.1 или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, мартышки, человекообразной обезьяны и т.п., которые являются их функциональными ортологами. В одном аспекте "зета-стимулирующий домен" или "стимулирующий домен CD3-зета" представляет собой последовательность, предоставленную в качестве SEQ ID NO: 18. В одном аспекте "зета-стимулирующий домен" или "стимулирующий домен CD3-зета" представляет собой последовательность, предоставленную в качестве SEQ ID NO: 20.
Термин "костимулирующая молекула" относится к распознающему связывающему партнеру на T-клетке, который специфически связывается с костимулирующим лигандом, тем самым, опосредуя костимулирующий ответ T-клеткой, такой как, но не ограничиваясь ими, пролиферация. Костимулирующие молекулы представляют собой молекулы клеточной поверхности, отличные от рецепторов антигенов или их лигандов, которые вносят вклад в эффективный иммунный ответ. Костимулирующие молекулы включают, но не ограничиваются ими, молекулу MHC класса I, BTLA и рецептор Toll-лиганда, а также OX40, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278) и 4-1BB (CD137). Следующие примеры таких костимулирующих молекул включают CDS, ICAM-1, GITR, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), NKp44, NKp30, NKp46, CD160, CD19, CD4, CD8-альфа, CD8-бета, IL2R-бета, IL2R-гамма, IL7R альфа, ITGA4, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, NKG2D, NKG2C, TNFR2, TRANCE/RANKL, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Tactile), CEACAM1, CRTAM, Ly9 (CD229), CD160 (BY55), PSGL1, CD100 (SEMA4D), CD69, SLAMF6 (NTB-A, Ly108), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, LAT, GADS, SLP-76, PAG/Cbp, CD19a и лиганд, который специфически связывается с CD83.
Костимулирующий внутриклеточный сигнальный домен может представлять собой внутриклеточную часть костимулирующей молекулы. Костимулирующая молекула может принадлежать одному из следующих семейств белков: белки-рецепторы TNF, иммуноглобулин-подобные белки, рецепторы цитокинов, интегрины, сигнальные молекулы активации лимфоцитов (белки SLAM) и активирующие рецепторы NK-клеток. Примеры таких молекул включают CD27, CD28, 4-1BB (CD137), OX40, GITR, CD30, CD40, ICOS, BAFFR, HVEM, ICAM-1, ассоциированный с функцией лимфоцитов антиген 1 (LFA-1), CD2, CDS, CD7, CD287, LIGHT, NKG2C, NKG2D, SLAMF7, NKp80, NKp30, NKp44, NKp46, CD160, B7-H3 и лиганд, который специфически связывается с CD83, и т.п.
Внутриклеточный сигнальный домен может содержать всю внутриклеточную часть или весь нативный внутриклеточный сигнальный домен молекулы, из которой он происходит, или его функциональный фрагмент или производное.
Термин "4-1BB" относится к представителю суперсемейства TNFR с аминокислотной последовательностью, представленной в качестве последовательности с номером доступа GenBank № AAA62478.2, или эквивалентными остатками из не являющегося человеком вида, например, мыши, грызуна, мартышки, человекообразной обезьяны и т.п.; и "костимулирующий домен 4-1BB" определяют как аминокислотные остатки 214-255 последовательности с номером доступа GenBank № AAA62478.2, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, мартышки, человекообразной обезьяны и т.п. В одном аспекте "костимулирующий домен 4-1BB" представляет собой последовательность, представленную в качестве SEQ ID NO: 14, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, мартышки, человекообразной обезьяны и т.п.
"Иммунная эффекторная клетка", как этот термин используют в настоящем описании, относится к клетке, которая вовлечена в иммунный ответ, например, в стимуляцию ответа иммунных эффекторных клеток. Примеры иммунных эффекторных клеток включают T-клетки, например, альфа/бета T-клетки и гамма/дельта T-клетки, B-клетки, натуральные киллеры (NK), натуральные киллерные T-клетки (NKT), тучные клетки и фагоциты миелоидного происхождения.
"Иммунная эффекторная функция или иммунный эффекторный ответ", как этот термин используют в настоящем описании, относится к функции или ответу, например, иммунной эффекторной клетки, которые усиливают или стимулируют иммунную атаку на клетку-мишень. Например, иммунная эффекторная функция или ответ относятся к свойству T-клеток или NK-клеток, которое стимулирует уничтожение или ингибирование роста или пролиферации клетки-мишени. В случае T-клетки, первичная стимуляция и костимуляция являются примерами иммунной эффекторной функции или ответа.
Термин "кодирующий" относится к свойству, присущему конкретным последовательностям нуклеотидов в полинуклеотиде, таком как ген, кДНК или мРНК, выступать в качестве матрицы для синтеза других полимеров и макромолекул в биологических процессах, имеющих либо определенную последовательность нуклеотидов (например, рРНК, тРНК и мРНК), либо определенную последовательность аминокислот, и к биологическим свойствам, являющимся следствием этого свойства. Таким образом, ген, кДНК или РНК, кодирует белок, если транскрипция и трансляция мРНК, соответствующей этому гену, продуцирует белок в клетке или другой биологической системе. Как кодирующая цепь, нуклеотидная последовательность которой идентична последовательности мРНК и обычно предоставляется в списках последовательностей, так и некодирующая цепь, используемая в качестве матрицы для транскрипции гена или кДНК, могут упоминаться как кодирующие белок или другой продукт этого гена или кДНК.
Если нет иных указаний, "нуклеотидная последовательность, кодирующая аминокислотную последовательность," включает все нуклеотидные последовательности, которые являются вырожденными версиями друг друга и которые кодируют одну и ту же аминокислотную последовательность. Выражение нуклеотидная последовательность, которая кодирует белок или РНК, также может включать интроны, поскольку нуклеотидная последовательность, кодирующая белок, может в некоторой версии содержать интрон(ы).
Термины "эффективное количество" или "терапевтически эффективное количество" используют в настоящем описании взаимозаменяемо, и они относятся к количеству соединения, состава, материала или композиции, как описано в настоящем описании, эффективному для достижения конкретного биологического результата.
Термин "эндогенный" относится к любому материалу из или продуцируемому внутри организма, клетки, ткани или системы.
Термин "экзогенный" относится к любому материалу, внесенному извне или продуцированному вне организма, клетки, ткани или системы.
Термин "экспрессия" относится к транскрипции и/или трансляции конкретной нуклеотидной последовательности, запускаемой промотором.
Термин "вектор для переноса" относится к композиции, которая содержит выделенную нуклеиновую кислоту и которую можно использовать для доставки выделенной нуклеиновой кислоты внутрь клетки. В данной области известны многочисленные векторы, включая, но не ограничиваясь ими, линейные полинуклеотиды, полинуклеотиды, ассоциированные с ионными или амфифильными соединениями, плазмиды и вирусы. Таким образом, термин "вектор для переноса" включает автономно реплицирующуюся плазмиду или вирус. Термин также следует истолковывать как дополнительно включающий неплазмидные и невирусные соединения, которые облегчают перенос нуклеиновой кислоты в клетки, такие как, например, соединение полилизина, липосома и т.п. Примеры вирусных векторов для переноса включают, но не ограничиваются ими, аденовирусные векторы, векторы на основе аденоассоциированного вируса, ретровирусные векторы, лентивирусные векторы и т.п.
Термин "экспрессирующий вектор" относится к вектору, содержащему рекомбинантный полинуклеотид, содержащий последовательности контроля экспрессии, функционально связанные с нуклеотидной последовательностью, подлежащей экспрессии. Экспрессирующий вектор содержит достаточное количество цис-регуляторных элементов для экспрессии; другие элементы для экспрессии могут быть предоставлены клеткой-хозяином или экспрессирующей системой in vitro. Экспрессирующие векторы включают векторы, известные в данной области, включая космиды, плазмиды (например, голые или содержащиеся в липосомах) и вирусы (например, лентивирусы, ретровирусы, аденовирусы и аденоассоциированные вирусы), которые включают рекомбинантный полинуклеотид.
Термин "лентивирус" относится к роду семейства Retroviridae. Лентивирусы являются уникальными среди ретровирусов, поскольку они способны инфицировать неделящиеся клетки; они могут доставлять значительное количество генетической информации в ДНК клетки-хозяина, так что они являются одним из наиболее эффективных способов доставки вектора с геном. Примерами лентивирусов являются ВИЧ, SIV и FIV.
Термин "лентивирусный вектор" относится к вектору, происходящему по меньшей мере из части генома лентивируса, включая, а частности, самоинактивирующийся лентивирусный вектор, представленный в Milone et al., Mol. Ther. 17(8): 1453-1464 (2009). Другие примеры лентивирусных векторов, которые можно использовать в клинике, включают, но не ограничиваются ими, например, технологию доставки генов LENTIVECTOR® от Oxford BioMedica, векторную систему LENTIMAXTM от Lentigen и т.п. Также доступны неклинические типы лентивирусных векторов, и они известны специалисту в данной области.
Термин "гомологичный" или "идентичность" относится к идентичности последовательности субъединиц между двумя полимерными молекулами, например, между двумя молекулами нуклеиновых кислот, таких как две молекулы ДНК или две молекулы РНК, или между двумя полипептидными молекулами. Если положение субъединицы в обеих из двух молекул занимает одна та же мономерная субъединица, например, если положение в каждой из двух молекул ДНК занимает аденин, тогда они являются гомологичными или идентичными в этом положении. Гомология между двумя последовательностями является прямой функцией количества совпадающих или гомологичных положений; например, если половина (например, пять положений в полимере длиной десять субъединиц) из положений в двух последовательностях являются гомологичными, тогда две последовательности являются на 50% гомологичными; если 90% положений (например, 9 из 10) совпадают или гомологичны, тогда две последовательности являются на 90% гомологичными.
"Гуманизированные" формы не являющихся человеческими антител (например, мыши) представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие связывающие антиген подпоследовательности антител), которые содержат минимальную последовательность, происходящую из не являющегося человеческим иммуноглобулина. В основном гуманизированные антитела и их фрагменты представляют собой иммуноглобулины человека (реципиентное антитело или фрагмент антитела), в которых остатки из определяющей комплементарность области (CDR) реципиента заменены остатками из CDR не являющегося человеком вида (донорное антитело), такого как мышь, крыса или кролик, имеющими желаемую специфичность, аффинность и емкость. В некоторых случаях остатки каркасной области Fv (FR) иммуноглобулина человека заменены соответствующими не являющимися человеческими остатками. Более того, гуманизированное антитело/фрагмент антитела может содержать остатки, которые не встречаются ни в реципиентном антителе, ни в импортированных последовательностях CDR или каркасной области. Эти модификации могут далее усовершенствовать и оптимизировать эффективность антитела или фрагмента антитела. Как правило, гуманизированное антитело или его фрагмент содержат по существу весь по меньшей мере один, и, как правило, два, вариабельных домена, в которых все или по существу все из областей CDR соответствуют областям CDR не являющегося человеческим иммуноглобулина и все или значительная часть областей FR представляют собой области FR из последовательности иммуноглобулина человека. Гуманизированное антитело или фрагмент антитела также могут содержать по меньшей мере часть константной области (Fc) иммуноглобулина, как правило, иммуноглобулина человека. Для более подробного описания, см. Jones et al., Nature, 321: 522-525, 1986; Reichmann et al., Nature, 332: 323-329, 1988; Presta, Curr. Op. Struct. Biol., 2: 593-596, 1992.
"Полностью человеческий" относится к иммуноглобулину, такому как антитело или фрагмент антитела, где вся молекула происходит из человека и состоит из аминокислотной последовательности, идентичной человеческой форме антитела или иммуноглобулина.
Термин "выделенный" означает измененный или извлеченный относительно природного состояния. Например, нуклеиновая кислота или пептид, естественным образом присутствующие в живом животном, не являются "выделенными", но те же нуклеиновая кислота или пептид, частично или полностью отделенные от сосуществующих с ними материалов природного состояния, являются "выделенными". Выделенная нуклеиновая кислота или белок могут существовать в по существу очищенной форме или могут существовать в ненативной среде, например, такой как клетка-хозяин.
В контексте настоящего изобретения используют следующие сокращения для обычно встречающихся оснований нуклеиновой кислоты. "A" относится к аденозину, "C" относится к цитозину, "G" относится к гуанозину, "T" относится к тимидину и "U" относится к уридину.
Термин "функционально связанный" или "контроль транскрипции" относится к функциональной связи между регуляторной последовательностью и гетерологичной последовательностью нуклеиновой кислоты, приводящей к экспрессии последней. Например, первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты находится в функциональной взаимосвязи со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с кодирующей последовательностью, если промотор влияет на транскрипцию или экспрессию кодирующей последовательности. Функционально связанные последовательности ДНК могут быть смежными друг с другом и, например, когда необходимо связать две кодирующих белок области, находится в одной рамке считывания.
Термин "парентеральное" введение иммуногенной композиции включает, например, подкожную (п/к), внутривенную (в/в), внутримышечную (в/м) или проводимую внутрь очага повреждения инъекцию, внутриопухолевое введение или способы инфузии.
Термины "нуклеиновая кислота" или "полинуклеотид" относятся к дезоксирибонуклеиновым кислотам (ДНК) или рибонуклеиновым кислотам (РНК) и их полимерам либо в одноцепочечной, либо в двухцепочечной форме. Если конкретно не ограничено, термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые имеют сходные свойства связывания с эталонной нуклеиновой кислотой и метаболизируются аналогично встречающимся в природе нуклеотидам. Если нет иных указаний, конкретная последовательность нуклеиновой кислоты также косвенно охватывает ее консервативно модифицированные варианты (например, вырожденные замены кодонов), аллели, ортологи, SNP и комплементарные последовательности, а также последовательность, прямо указанную. В частности, вырожденные замены кодонов можно осуществлять путем получения последовательностей, в которых третье положение одного или нескольких выбранных (или всех) кодонов замещено смешанными остатками и/или остатками дезоксиинозина (Batzer et al., Нуклеиновая кислота Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); и Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)).
Термины "пептид," "полипептид" и "белок" используют взаимозаменяемо, и они относятся к соединению, состоящему из аминокислотных остатков, ковалентно связанных пептидными связями. Белок или пептид должны содержать по меньшей мере две аминокислоты, и отсутствует ограничение на максимальное количество аминокислот, которые могут содержаться в последовательности белка или пептида. Полипептиды включают любой пептид или белок, содержащие две или более аминокислот, связанных друг с другом пептидными связями. Как используют в рамках изобретения, термин относится к как коротким цепям, которые также обычно называют в данной области, например, пептидами, олигопептидами и олигомерами, так и к более длинным цепям, которые обычно называют в данной области белками, для которых существует множество типов. "Полипептиды" включают, например, биологически активные фрагменты, по существу гомологичные полипептиды, олигопептиды, гомодимеры, гетеродимеры, варианты полипептидов, модифицированные полипептиды, производные, аналоги, слитые белки, среди прочих. Полипептид включает природный пептид, рекомбинантный пептид или их комбинацию.
Термин "промотор" относится к последовательности ДНК, распознаваемой синтетическим аппаратом клетки или введенным синтетическим аппаратом, которая требуется для инициации транскрипции полинуклеотидной последовательности.
Термин "промоторная/регуляторная последовательность" относится к последовательности нуклеиновой кислоты, которая требуется для экспрессии продукта гена, функционально связанного с промоторной/регуляторной последовательностью. В некоторых случаях эта последовательность может представлять собой основную промоторную последовательность и в других случаях эта последовательность также может включать энхансерную последовательность и другие регуляторные элементы, которые требуются для экспрессии продукта гена. Промоторная/регуляторная последовательность, например, может представлять собой последовательность, которая экспрессирует продукт гена тканеспецифическим образом.
Термин "конститутивный" промотор относится к нуклеотидной последовательности, которая, когда она функционально связана с полинуклеотидом, который кодирует или определяет продукт гена, вызывает продукцию продукта гена в клетке при большинстве или всех физиологических условиях в клетке.
Термин "индуцибельный" промотор относится к нуклеотидной последовательности, которая, когда она функционально связана с полинуклеотидом, который кодирует или определяет продукт гена, обеспечивает продукцию продукта гена в клетке, по существу только когда индуктор, который соответствует промотору, присутствует в клетке.
Термин "тканеспецифический" промотор относится к нуклеотидной последовательности, которая, когда она функционально связана с полинуклеотидом, который кодирует или определяет продукт гена, обеспечивает продукцию продукта гена в клетке, по существу только если клетка представляет собой клетку типа ткани, соответствующего промотору.
Термины "ассоциированный со злокачественной опухолью антиген" или "опухолевый антиген" взаимозаменяемо относятся к молекуле (как правило, белок, углевод или липид), которая экспрессируется на поверхности злокачественной клетки, либо полностью, либо в качестве фрагмента (например, MHC/пептид), и которая пригодная для предпочтительного нацеливания фармакологического средства на злокачественную клетку. В некоторых вариантах осуществления опухолевый антиген представляет собой маркер, экспрессируемый как нормальными клетками, так и злокачественными клеткам, например, маркер линии дифференцировки, например, CD19 на B-клетках. В некоторых вариантах осуществления опухолевый антиген представляет собой молекулу клеточной поверхности, которая сверхэкспрессируется в злокачественной клетки по сравнению с нормальной клеткой, например, с 1-кратной сверхэкспрессией, 2-кратной сверхэкспрессией, 3-кратной сверхэкспрессией или более по сравнению с нормальной клеткой. В некоторых вариантах осуществления опухолевый антиген представляет собой молекулу клеточной поверхности, которая ненадлежащим образом синтезируется в злокачественной клетке, например, молекулу, которая содержит делеции, вставки или мутации по сравнению с молекулой, экспрессируемой нормальной клеткой. В некоторых вариантах осуществления опухолевый антиген экспрессируется исключительно на клеточной поверхности злокачественной клетки, полностью или в качестве фрагмента (например, MHC/пептид), и не синтезируется или не экспрессируется на поверхности нормальной клетки. В некоторых вариантах осуществления CAR по настоящему изобретению включают CAR, содержащие антигенсвязывающий домен (например, антитело или фрагмент антитела), который связывается с пептидом, презентируемым MHC. Обычно пептиды, происходящие из эндогенных белков, заполняют карманы молекул основного комплекса гистосовместимости (MHC) класса I и распознаются T-клеточными рецепторами (TCR) на CD8+T-лимфоцитах. Комплексы MHC класса I конститутивно экспрессируются всеми ядросодержащими клетками. В злокачественной опухоли специфические для вируса и/или специфические для опухоли комплексы пептид/MHC соответствуют уникальному классу мишеней, находящихся на клеточной поверхности, для иммунотерапии. Были описаны TCR-подобные антитела, нацеленные на пептиды, происходящие из вирусных или опухолевых антигенов в контексте лейкоцитарного антигена человека (HLA)-A1 или HLA-A2 (см., например, Sastry et al., J Virol. 2011 85(5):1935-1942; Sergeeva et al., Blood, 2011 117(16):4262-4272; Verma et al., J Immunol 2010 184(4):2156-2165; Willemsen et al., Gene Ther 2001 8(21):1601-1608; Dao et al., Sci Transl Med 2013 5(176):176ra33; Tassev et al., Cancer Gene Ther 2012 19(2):84-100). Например, TCR-подобное антитело может быть идентифицировано посредством скрининга библиотеки, такой как библиотека фагового дисплея scFv человека.
Термины "поддерживающий опухоль антиген" или "поддерживающий злокачественную опухоль антиген" взаимозаменяемо относятся к молекуле (как правило, белок, углевод или липид), которая экспрессируется на поверхности клетки, которая сама по себе не является злокачественной, но поддерживает рост злокачественных клеток, например, путем стимуляции их роста или выживания, например, резистентности к иммунным клеткам. Иллюстративные клетки этого типа включают стромальные клетки и супрессорные клетки миелоидного происхождения (MDSC). Поддерживающий опухоль антиген сам по себе не должен играть роль в поддержании опухолевых клеток, если антиген присутствует на клетке, которая поддерживает злокачественные клетки.
Термины "гибкий полипептидный линкер" или "линкер", как используют в контексте scFv, относится к пептидному линкеру, который состоит из аминокислот, таких как остатки глицина и/или серина, используемых отдельно или в комбинации, для связывания вариабельной области тяжелой цепи и вариабельной области легкой цепи вместе. В одном варианте осуществления гибкий полипептидный линкер представляет собой линкер Gly/Ser, и он содержит аминокислотную последовательность (Gly-Gly-Gly-Ser)n, где n представляет собой положительное целое число, равное или превышающее 1. Например, n=1, n=2, n=3, n=4, n=5 и n=6, n=7, n=8, n=9 и n=10 (SEQ ID NO: 28). В одном варианте осуществления гибкие полипептидные линкеры включают, но не ограничиваются ими, (Gly4 Ser)4 (SEQ ID NO: 29) или (Gly4 Ser)3 (SEQ ID NO: 30). В другом варианте осуществления линкеры включают множество повторов (Gly2Ser), (GlySer) или (Gly3Ser) (SEQ ID NO: 31). Также в объем изобретения входят линкеры, описанные в WO2012/138475, включенной в настоящее описание путем ссылки).
Как используют в рамках изобретения, 5'-кэп (также называемый РНК-кэпом, 7-метилгуанозиновым кэпом РНК или m7G-кэпом РНК) представляет собой модифицированный гуаниновый нуклеотид, который добавляется "спереди" или на 5'-конце эукариотической матричной РНК вскоре после начала транскрипции. 5'-кэп состоит из концевой группы, которая связана с первым транскрибируемым нуклеотидом. Его присутствие является важным для распознавания рибосомой и защиты от РНКаз. Присоединение кэпа сопряжено с транскрипцией и происходит котранскрипционно, так что каждое из этих событий влияет на другое из них. Вскоре после начала транскрипции 5'-конец синтезируемой мРНК связывается синтезирующим кэп комплексом, ассоциированным с РНК-полимеразой. Этот ферментативный комплекс катализирует химические реакции, которые требуются для кэпирования мРНК. Синтез протекает в качестве многостадийной биохимической реакции. Кэпирующую часть можно модифицировать для модулирования функциональности мРНК, такой как ее стабильность или эффективность трансляции.
Как используют в рамках изобретения, "транскрибированная in vitro РНК" относится к РНК, предпочтительно мРНК, синтезированной in vitro. Как правило, транскрибированную in vitro РНК получают с помощью вектора для транскрипции in vitro. Вектор для транскрипции in vitro содержит матрицу, которую используют для получения транскрибированной РНК in vitro.
Как используют в рамках изобретения, "поли(A)" представляет собой последовательность остатков аденозина, присоединенную путем полиаденилирования мРНК. В предпочтительном варианте осуществления конструкции для временной экспрессии поли-A имеют от 50 до 5000 (SEQ ID NO: 34), предпочтительно более 64, более предпочтительно более 100, наиболее предпочтительно более 300 или 400 оснований. Последовательности поли(A) можно модифицировать химически или ферментативно для модулирования функциональности мРНК, такой как локализация, стабильность или эффективность трансляции.
Как используют в рамках изобретения, "полиаденилирование" относится к ковалентному присоединению полиаденилильной части или ее модифицированного варианта к молекуле матричной РНК. В эукариотических организмах большинство молекул матричной РНК (мРНК) являются полиаденилированными на 3'-конце. 3'-поли(A)-хвостовая часть представляет собой длинную последовательность адениновых нуклеотидов (часто несколько сотен), присоединяемую к пре-мРНК под действием фермента полиаденилатполимеразы. У высших эукариот поли(A)-хвостовая часть присоединяется к транскриптам, которые содержат конкретную последовательность - сигнал полиаденилирования. Поли(A)-хвостовая часть и белок, связывающийся с ней, способствуют защите мРНК от деградации экзонуклеазами. Полиаденилирование также является важным для терминации транскрипции, экспорта мРНК из ядра и трансляции. Полиаденилирование происходит в ядре непосредственно после транскрипции ДНК в РНК, но, кроме того, также оно происходит позднее в цитоплазме. После завершения транскриции цепь мРНК отщепляется под действием эндонуклеазного комплекса, ассоциированного с РНК-полимеразой. Участок расщепления обычно характеризуется последовательностью оснований AAUAAA вблизи участка расщепления. После отщепления мРНК остатки аденозина являются связанными со свободным 3'-концом участка расщепления.
Как используют в рамках изобретения, "временный" относится к экспрессии невстроенного трансгена в течение периода, составляющего несколько часов, суток или недель, где период экспрессии является меньшим, чем период экспрессии гена, если он встроен в геном или содержится в стабильном плазмидном репликоне в клетке-хозяине.
Как используют в рамках изобретения, термины "лечить", "лечение" и "проведение лечения" относятся к снижению или смягчению прогрессирования, тяжести и/или длительности пролиферативного нарушения, или к смягчению одного или нескольких симптомов (предпочтительно, одного или нескольких заметных симптомов) пролиферативного нарушения в результате введения одного или нескольких терапевтических средств (например, одного или нескольких лекарственных средств, таких как CAR по изобретению). В конкретных вариантах осуществления термины "лечить", "лечение" и "проведение лечения" относятся к смягчению по меньшей мере одного поддающегося количественному определению физического параметра пролиферативного нарушения, такого как рост опухоли, не обязательно заметному для пациента. В других вариантах осуществления термины "лечить", "лечение" и "проведение лечения" относятся к ингибированию прогрессирования пролиферативного нарушения, либо физически, например, путем стабилизации заметного симптома, либо физиологически, например, путем стабилизации физического параметра, либо посредством обоих из них. В других вариантах осуществления термины "treat", "лечение" и "проведение лечения" относятся к снижению или стабилизации размера опухоли или количества злокачественных клеток.
Термин "каскад передачи сигнала" относится к биохимической взаимосвязи между различными молекулами передачи сигнала, которые играют роль в передачи сигнала из одной части клетки в другую часть клетки. Выражение "рецептор клеточной поверхности" включает молекулы и комплексы молекул, способные получать сигнал и передавать сигнал через мембрану клетки.
Термин "индивидуум" включает живые организмы, у которых может быть индуцирован иммунный ответ (например, млекопитающие, человек).
Термин "по существу очищенная" клетка относится к клетке, которая по существу свободна от других типов клеток. По существу очищенная клетка также относится к клетке, которая отделена от других типов клеток, с которыми она обычно ассоциирована в ее природном состоянии. В некоторых случаях популяция по существу очищенных клеток относится к гомогенной популяции клеток. В других случаях этот термин относится просто к клетке, которая отделена от клеток, с которыми она ассоциирована в ее природном состоянии. В некоторых аспектах клетки культивируют in vitro. В других аспектах клетки не культивируют in vitro.
Термин "терапевтический", как используют в рамках изобретения, означает лечение. Терапевтический эффект достигают путем снижения, подавления, обеспечения ремиссии или устранения болезненного состояния.
Термин "профилактика", как используют в рамках изобретения, означает предупреждение или защитное лечение от заболевания или болезненного состояния.
В контексте настоящего изобретения "опухолевый антиген", или "антиген гиперпролиферативного нарушения", или "антиген, ассоциированный с гиперпролиферативным нарушением", относится к антигенам, которые являются общими для конкретных гиперпролиферативных нарушений. В некоторых аспектах антигены гиперпролиферативного нарушения по настоящему изобретению происходят из злокачественных опухолей, включая, но не ограничиваясь ими первичную или метастазирующую меланому, тимому, лимфому, саркому, рак легкого, рак печени, неходжкинскую лимфому, лимфому Ходжкина, лейкозы, рак тела матки, рак шейки матки, рак мочевого пузыря, рак почки и аденокарциномы, такие как рак молочной железы, рак предстательной железы, рак яичника, рак поджелудочной железы и т.п.
Термины "трансфицированный", или "трансформированный", или "трансдуцированный" относятся к процессу, посредством которого экзогенная нуклеиновая кислота переносится или вводится в клетку-хозяина. "Трансфицированная", или "трансформированная", или "трансдуцированная" клетка представляет собой клетку, которая трансфицирована, трансформирована или трансдуцирована экзогенной нуклеиновой кислотой. Клетка включает первичную клетку индивидуума и ее потомство.
Термин "специфически связывается" относится к антителу или лиганду, которые распознают и связываются с белком-партнером по связыванию (например, опухолевым антигеном), присутствующем в образце, но при этом эти антитело или лиганд по существу не распознают или не связывают другие молекулы в образце.
"Регулируемый химерный рецептор антигена (RCAR)", как этот термин используют в настоящем описании, относится к набору полипептидов, как правило, двум в наиболее простых вариантах осуществления, которые, когда он находится в клетке RCARX, обеспечивает специфичность клетке RCARX в отношении клетки-мишени, как правило, злокачественной клетки, и регулируемое генерирование внутриклеточного сигнала или пролиферацию, которые могут оптимизировать иммунное эффекторное свойство клетки RCARX. Клетка RCARX по меньшей мере частично основана на антигенсвязывающем домене для обеспечения специфичности в отношении клетки-мишени, которая содержит антиген, связываемый антигенсвязывающим доменом. В одном варианте осуществления RCAR включает переключатель димеризации, который в присутствии молекулы димеризации может связывать внутриклеточный сигнальный домен с антигенсвязывающим доменом.
"Мембранный якорь" или "связывающий с мембраной домен", как этот термин используют в настоящем описании, относится к полипептиду или части, например, миристоильной группе, достаточным для заякоривания внеклеточного или внутриклеточного домена на плазматической мембране.
"Домен переключения", как этот термин используют в настоящем описании, например, при указании на RCAR, относится к структуре, как правило, структуре на основе полипептида, которая в присутствии молекулы димеризации ассоциирует с другим доменом переключения. Эта ассоциация приводит к функциональному связыванию первой структуры, связанной, например слитой, с первым доменом переключения, и второй структуры, связанной, например слитой, со вторым доменом переключения. Первый и второй домены переключения в совокупности называют в настоящем описании переключателями димеризации. В вариантах осуществления первый и второй домены переключения являются одинаковыми, например, они представляют собой полипептиды, имеющие одинаковую первичную аминокислотную последовательность, и их в совокупности называют переключателями гомодимеризации. В вариантах осуществления первый и второй домены переключения отличаются друг от друга, например, они представляют собой полипептиды, имеющие отличающиеся первичные аминокислотные последовательности, и их в совокупности называют переключателями гетеродимеризации. В вариантах осуществления переключение является внутриклеточным. В вариантах осуществления переключение является внеклеточным. В вариантах осуществления домен переключения представляет собой структуру на основе полипептида, например, на основе FKBP или FRB, и молекула димеризации представляет собой низкомолекулярное соединение, например, рапалог. В вариантах осуществления домен переключения представляет собой структуру на основе полипептида, например scFv, который связывает пептид myc, и молекула димеризации представляет собой полипептид, его фрагмент или мультимер полипептида, например, myc-лиганд или мультимеры myc-лиганда, которые связываются с одним или несколькими scFv к myc. В вариантах осуществления домен переключения представляет собой структуру на основе полипептида, например рецептор myc, и молекула димеризации представляет собой антитело или его фрагменты, например, антитело против myc.
"Молекула димеризации", как этот термин используют в настоящем описании, например, при указании на RCAR, относится к молекуле, которая обеспечивает ассоциацию первого домена переключения со вторым доменом переключения. В вариантах осуществления молекула димеризации не является встречающейся в природе у индивидуума или не встречается в концентрациях, которые привели бы к значительной димеризации. В вариантах осуществления молекула димеризации представляет собой низкомолекулярное соединение, например, рапамицин или рапалог, например RAD001.
Термин "биоэквивалент" относится к количеству средства, отличного от эталонного соединения (например, RAD001), требуемому для обеспечения эффекта, эквивалентного эффекту, обеспечиваемому эталонной дозой или эталонным количеством эталонного соединения (например, RAD001). В одном варианте осуществления эффект представляет собой уровень ингибирования mTOR, например, при измерении по ингибированию киназы P70 S6, например, при оценке в анализе in vivo или in vitro, например, при измерении с использованием анализа, описанного в настоящем описании, например, анализа Boulay. В одном варианте осуществления эффект представляет собой изменение соотношения положительные по PD-1/отрицательные по PD-1 T-клетки при измерении посредством сортировки клеток. В одном варианте осуществления биоэквивалентное количество или доза ингибитора mTOR представляют собой количество или дозу, которые достигают того же уровня ингибирования киназы P70 S6, что и эталонная доза или эталонное количество эталонного соединения. В одном варианте осуществления биоэквивалентное количество или доза ингибитора mTOR представляют собой количество или дозу, которые достигают того же уровня изменения соотношения положительные по PD-1/отрицательные по PD-1 T-клетки, как и эталонная доза или эталонное количество эталонного соединения.
Термин "низкая усиливающая иммунитет доза", когда его используют применительно к ингибитору mTOR, например, аллостерическому ингибитору mTOR, например RAD001 или рапамицину, или каталитическому ингибитору mTOR, относится к дозе ингибитора mTOR, которая частично, но не полностью, ингибирует активность mTOR, например, при измерении по ингибированию активности киназы P70 S6. Способы оценки активности mTOR, например, по ингибированию киназы P70 S6, описаны в настоящем описании. Доза является недостаточной для обеспечения полного подавления иммунного ответа, но является достаточной для усиления иммунного ответа. В одном варианте осуществления низкая усиливающая иммунитет доза ингибитора mTOR приводит к уменьшению количества положительных по PD-1 T-клеток и/или увеличению количества отрицательных по PD-1 T-клеток, или к увеличению соотношения отрицательные по PD-1 T-клетки/положительные по PD-1 T-клетки. В одном варианте осуществления низкая усиливающая иммунитет доза ингибитора mTOR приводит к увеличению количества наивных T-клеток. В одном варианте осуществления низкая усиливающая иммунитет доза ингибитора mTOR приводит к одному или нескольким из следующих:
увеличение экспрессии одного или нескольких из следующих маркеров: CD62Lhigh, CD127high, CD27+ и BCL2, например, на T-клетках памяти, например, предшественниках T-клеток памяти;
снижение экспрессии KLRG1, например, на T-клетках памяти, например, предшественниках T-клеток памяти; и
увеличение количества предшественников T-клеток памяти, например, клеток с любой или комбинацией из следующих характеристик: увеличенный уровень CD62Lhigh, увеличенный уровень CD127high, увеличенный уровень CD27+, сниженный уровень KLRG1 и увеличенный уровень BCL2;
где любое из описанных выше изменений происходит, например, по меньшей мере временно, например, по сравнению с индивидуумом без лечения.
"Рефрактерный", как используют в рамках изобретения, относится к заболеванию, например, злокачественной опухоли, которое не отвечает на лечение. В вариантах осуществления рефрактерная злокачественная опухоль может быть устойчивой к лечению до или в начале лечения. В других вариантах осуществления рефрактерная злокачественная опухоль может стать резистентной в процессе лечения. Рефрактерную злокачественную опухоль также называют резистентной злокачественной опухолью.
"Рецидивирующий", как используют в рамках изобретения, относится к возвращению заболевания (например, злокачественной опухоли) или признаков и симптомов заболевания, такого как злокачественная опухоль, после периода улучшения, например, после предшествующего лечения, например, терапии злокачественной опухоли.
Диапазоны: на протяжении настоящего описания различные аспекты изобретения могут быть представлены в формате диапазонов. Следует понимать, что описание в формате диапазонов предоставлено только для удобства и краткости, и его не следует истолковывать как строгое ограничение объема изобретения. Таким образом, описание диапазона следует считать имеющим конкретно описанные все возможные числовые поддиапазоны, а также индивидуальные числовые величины в этом диапазоне. Например, описание диапазона, такого как от 1 до 6, следует истолковывать как диапазон, имеющий конкретно описанные поддиапазоны, такие как от 1 до 3, от 1 до 4, от 1 до 5, от 2 до 4, от 2 до 6, от 3 до 6 и т.д., а также индивидуальные числа в этом диапазоне, например, 1, 2, 2,7, 3, 4, 5, 5,3 и 6. В качестве другого примера диапазон, такой как идентичность 95-99%, включает что-либо с 95%, 96%, 97%, 98% или 99% идентичностью и включает поддиапазоны, такие как идентичность 96-99%, 96-98%, 96-97%, 97-99%, 97-98% и 98-99%. Это применимо независимо от ширины диапазона.
"Tet", как используют в настоящем описании, относится к семейству генов и к белкам семейства метилцитозиндиоксигеназы, кодируемым указанными генами, с транслокацией десять-одиннадцать. Tet включает, например, Tet1, Tet2 и Tet3.
"Tet2", как используют в настоящем описании, относится к гену tet метилцитозиндиоксигеназы 2 и к белку метилцитозиндиоксигеназе tet2, кодируемому указанным геном, который катализирует конвертирование метилцитозина в 5-гидроксиметилцитозин. Иногда его также называют "KIAA1546", "FLJ20032" и "представителем 2 семейства онкогенов tet". Кодируемый белок вовлечен в миелопоэз и дефекты в этом гене ассоциируют с несколькими миелопролиферативными нарушениями. В геноме человека TET2 расположен на хромосоме 4q24. В настоящее время описано шесть изоформ TET2 и их номера в Genebank представляют собой: NM_001127208.2; XM_005263082.1; XM_006714242.2; NM_017628.4; XM_011532044.1 и XM_011532043.1.
Пример белковой последовательности Tet2 человека приведен под номером доступа UniProt Q6N021:


Ген tet2 расположен на 4 хромосоме, область GRCh38.p2 (GCF_000001405.28) (NC_000004.12 (от 105145875 до 105279803); ID гена 54790.
Примеры последовательностей нуклеиновых кислот, кодирующих Tet2, представлены ниже. Было идентифицировано 6 изоформ Tet2 человека. Последовательности мРНК представлены ниже (В вариантах осуществления в каждой последовательности T может быть заменен на U). В вариантах осуществления Tet2 включает белки, кодируемые каждой из приведенных ниже последовательностей:
[SEQ ID NO: 1358]
[SEQ ID NO: 1359]
[SEQ ID NO: 1360]
[SEQ ID NO: 1361]
[SEQ ID NO: 1362]
[SEQ ID NO: 1363]
Термины "ингибитор Tet" или "ингибитор Tet[x]" (например, "ингибитор Tet1", "ингибитор Tet2" или "ингибитор Tet3"), как используют в настоящем описании, относятся к молекуле или группе молекул (например, система), которые снижают или устраняют функцию и/или экспрессию соответствующего Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В некоторых вариантах осуществления ингибитор Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, представляет собой молекулу, которая ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, например, снижает или устраняет экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В некоторых вариантах осуществления ингибитор Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, представляет собой молекулу, которая ингибирует функцию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Примером ингибитора Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, который ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, является система редактирования генов, например, как описано в настоящем описании, которая нацелена на нуклеиновую кислоту в гене Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, или в его регуляторных элементах, так что модификация нуклеиновой кислоты в или вблизи участка(ов) связывания системы редактирования генов изменяется, снижая или устраняя экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Другим примером ингибитора Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, который ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, является молекула нуклеиновой кислоты, например, молекула РНК, например, короткая шпилечная РНК (кшРНК) или малая интерферирующая РНК (миРНК), способная гибридизоваться с мРНК Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, и вызывать снижение или устранение трансляции Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, также включают нуклеиновые кислоты, кодирующие молекулы, которая ингибируют экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2 (например, нуклеиновая кислота, кодирующая кшРНК или миРНК против Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, или нуклеиновая кислота, кодирующая один или несколько, например, компонентов системы редактирования генов, направленной против Tet, например, Tet1, Tet2 и/или Tet3, например Tet2). Примером молекулы, которая ингибирует функцию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, является молекула, например, белок или низкомолекулярное соединение, которая ингибирует один или несколько видов активности Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Примером является низкомолекулярный ингибитор Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Другим примером является доминантно-негативный белок Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Другим примером является доминантно-негативная версия партнера по связыванию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, например, ассоциированной с ними деацетилазы гистонов (HDAC). Другим примером является молекула, например, низкомолекулярное соединение, которая ингибирует партнер по связыванию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, например, ингибитор ассоциированной с Tet, например, Tet1, Tet2 и/или Tet3, например Tet2 HDAC. Ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, также включают нуклеиновые кислоты, кодирующие ингибиторы функцию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
"Система", как используют в настоящем описании применительно к редактированию генов или ингибированию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, относится к группе молекул, например, к одной или нескольким молекулам, которые действуют вместе, обеспечивая желаемую функцию.
"Система редактирования генов", как используют в настоящем описании, относится к системе, например, к одной или нескольким молекулам, которые обеспечивают и осуществляют изменение, например, делецию, одной или нескольких нуклеиновых кислот в или вблизи участка геномной ДНК, на который нацелена указанная система. Системы редактирования генов известны в данной области и более подробно описаны ниже.
"Партнер по связыванию", как используют в настоящем описании в контексте партнера по связыванию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, относится к молекуле, например, белку, которая взаимодействует, например, связывается, с белком Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Без связи с теорией, полагают, что Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, связывается с одним или несколькими белками HDAC. Такие белки HDAC считаются примерами партнеров по связыванию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
Продукт или белок "доминантно-негативного" гена представляет собой продукт или белок, который препятствует функции другого продукта или белка гена. Другой продукт гена, на который осуществляется воздействие, может быть тем же или может отличаться от доминантно-негативного белка. Продукты доминантно-негативных генов могут иметь множество форм, включая укорочения, полноразмерные белки с точковыми мутациям или их фрагменты, или слитые конструкции полноразмерных белков дикого типа, или мутантных белков, или их фрагментов с другими белками. Наблюдаемый уровень ингибирования моет быть очень низким. Например, чтобы наблюдать эффект может потребоваться большой избыток доминантно-негативного белка по сравнению с функциональным белком или белками, вовлеченными в процесс. Может быть трудно наблюдать эффекты в нормальных условиях биологического анализа. В одном варианте осуществления доминантно-негативный Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, представляет собой каталитически неактивный Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В другом варианте осуществления доминантно-негативный партнер по связыванию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, представляет собой каталитически неактивный ингибитор связывания HDAC с Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
Описание
Настоящее изобретение относится к ингибиторам Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, и к способам их применения. В частности, изобретение относится к экспрессирующим CAR T-клеткам, содержащим ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, и к применению ингибиторов Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, совместно с CART-клетками. Ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, по настоящему изобретению, вместе со способами их применения более подробно описаны ниже. CAR, CART-клетки и способы их применения дополнительно описаны ниже.
Ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2
Настоящее изобретение относится к композициям, например, ингибиторов Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, и к способам усиления функций иммунных эффекторных клеток, например, функций экспрессирующих CAR клеток, с использованием таких композиций и/или других средств, как описано в настоящем описании. Любые ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, известные в данной области, можно использовать в качестве ингибитора Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, в соответствии с настоящим изобретением. Примеры ингибиторов Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, описаны ниже.
Системы редактирования генов
В соответствии с настоящим изобретением, системы редактирования генов можно использовать в качестве ингибиторов Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Также настоящее изобретение предусматривает применение нуклеиновой кислоты, кодирующей один или несколько компонентов системы редактирования генов Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
Системы редактирования генов CRISPR/Cas9
Встречающиеся в природе системы CRISPR/Cas найдены приблизительно в 40% отсеквенированных геномов эубактерий и 90% отсеквенированных геномов архей. Grissa et al. (2007) BMC Bioinformatics 8: 172. Эта система представляет собой тип прокариотической иммунной системы, который сообщает устойчивость к чужеродным генетическим элементам, таким как плазмиды и фаги, и обеспечивает форму приобретенного иммунитета. Barrangou et al. (2007) Science 315: 1709-1712; Marragini et al. ( 2008) Science 322: 1843-1845.
Система CRISPR/Cas была модифицирована для применения в редактировании генов (сайленсинг, усиление или изменение конкретных генов) у эукариот, таких как мыши или приматы. Wiedenheft et al. (2012) Nature 482: 331-8. Это осуществили путем введения в эукариотическую клетку плазмиды, содержащей специально сконструированный CRISPR и один или несколько соответствующих Cas.
Последовательность CRISPR, иногда называемая локусом CRISPR, содержит чередующиеся повторы и спейсеры. Во встречающемся в природе CRISPR спейсеры обычно содержат последовательности, чужеродные для бактерии, такие как последовательность плазмиды или фага; в иллюстративной системе CRISPR/Cas, направленной против Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, спейсеры происходят из последовательности гена Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, или последовательностей их регуляторных элементов.
РНК из локуса CRISPR конститутивно экспрессируется и процессируется в малые РНК. Они содержат спейсер, фланкированный последовательностями повтора. РНК направляют другие белки Cas на подавление экзогенных генетических элементов на уровне РНК или ДНК. Horvath et al. (2010) Science 327: 167-170; Makarova et al. (2006) Biology Direct 1: 7. Таким образом, спейсеры выступают в качестве матриц для молекул РНК, аналогично миРНК. Pennisi (2013) Science 341: 833-836.
Поскольку они встречаются в природе во многих различных типах бактерий, точная организация и структура CRISPR, функция и количество генов Cas и их продуктов отличаются в некоторой степени от вида к виду. Haft et al. (2005) PLoS Comput. Biol. 1: e60; Kunin et al. (2007) Genome Biol. 8: R61; Mojica et al. (2005) J. Mol. Evol. 60: 174-182; Bolotin et al. (2005) Microbiol. 151: 2551-2561; Pourcel et al. (2005) Microbiol. 151: 653-663; и Stern et al. (2010) Trends. Genet. 28: 335-340. Например, белки Cse (подтип Cas, E. coli) (например, CasA) формируют функциональный комплекс, Cascade, который процессирует РНК-транскрипты CRISPR в элементы спейсер-повтор, которые сохраняет Cascade. Brouns et al. (2008) Science 321: 960-964. В других прокариотах Cas6 процессирует транскрипт CRISPR. Инактивация фага на основе CRISPR в E. coli требует Cascade и Cas3, но не Cas1 или Cas2. Белки Cmr (модуль RAMP Cas) в Pyrococcus furiosus и других прокариотах формируют функциональный комплекс с малыми РНК CRISPR, который распознает и расщепляет РНК-мишени. Более простая система CRISPR основана на белке Cas9, который представляет собой нуклеазу с двумя активными участками расщепления, по одному для каждой цепи двойной спирали. Комбинирование Cas9 и модифицированного локуса РНК CRISPR можно использовать в системе редактирования генов. Pennisi (2013) Science 341: 833-836.
Таким образом, систему CRISPR/Cas можно использовать для модификации, например, делеции, одной или нескольких нуклеиновых кислот гена Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, или регуляторного элемента гена Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, или внесения преждевременного стоп-кодона, тем самым уменьшая экспрессию функционального Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Альтернативно систему CRISPR/Cas можно использовать подобно РНК-интерференции, выключая ген Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, обратимым образом. В клетке млекопитающего, например, РНК может направлять белок Cas на промотор Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, пространственно блокируя РНК-полимеразы.
Системы CRISPR/Cas для редактирования генов в эукариотических клетках, как правило, вовлекают (1) молекулу РНК-гида (гРНК), содержащую нацеливающую последовательность (которая способна гибридизоваться с последовательностью-мишенью геномной ДНК), и последовательность, которая способна связываться с ферментом Cas, например, Cas9, и (2) белок Cas, например, Cas9. Нацеливающая последовательность и последовательность, которая способна связываться с ферментом Cas, например, Cas9, могут находиться на одной и той же или на различных молекулах. Если они расположены на различных молекулах, каждая из них включает домен гибридизации, который позволяет молекулам ассоциировать, например, путем гибридизации.
Иллюстративная молекула гРНК по настоящему изобретению содержит, например, состоит из первой нуклеиновой кислоты, имеющей последовательность (где "n" относится к остаткам нацеливающей последовательности (например, как описано в настоящем описании, например, в таблице 3), и может состоять из 15-25 нуклеотидов, например, состоит из 20 нуклеотидов):
nnnnnnnnnnnnnnnnnnnnGUUUUAGAGCUAUGCUGUUUUG (SEQ ID NO: 40);
и второй последовательности нуклеиновой кислоты, имеющей последовательность:
AACUUACCAAGGAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC, необязательно с 1, 2, 3, 4, 5, 6 или 7 (например, 4 или 7, например, 7) дополнительными нуклеотидами U на 3'-конце (SEQ ID NO: 41).
Вторая молекула нуклеиновой кислоты альтернативно может состоять из фрагмента последовательность, приведенной выше, где такой фрагмент способен гибридизоваться с первой нуклеиновой кислотой. Примером такой второй молекулы нуклеиновой кислоты является:
AACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC, необязательно с 1, 2, 3, 4, 5, 6, или 7 (например, 4 или 7, например, 7) дополнительными нуклеотидами U на 3'-конце (SEQ ID NO: 42).
Другая иллюстративная молекула гРНК по настоящему изобретению содержит, например, состоит из первой нуклеиновой кислоты, имеющей последовательность (где "n" относится к остаткам нацеливающей последовательности (например, как описано в настоящем описании, например, в таблице 3), и может состоять из 15-25 нуклеотидов, например, состоит из 20 нуклеотидов):
nnnnnnnnnnnnnnnnnnnGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO: 43), необязательно с 1, 2, 3, 4, 5, 6 или 7 (например, 4 или 7, например, 4) дополнительными нуклеотидами U на 3'-конце. Можно получать искусственные системы CRISPR/Cas, которые ингибируют Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, с использованием технологии, известной в данной области, например, которые описаны в публикации США № 20140068797, WO2015/048577 и Cong (2013) Science 339: 819-823. Также можно получать другие искусственные системы CRISPR/Cas, как известно в данной области, которые ингибируют Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, например, системы, описанные в Tsai (2014) Nature Biotechnol., 32:6 569-576, патенте США № 8871445; 8865406; 8795965; 8771945 и 8697359, содержание которых включено в настоящее описание путем ссылки в полном объеме. Такие системы, которые ингибируют Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, можно получать, например, посредством модификации способами инженерии системы CRISPR/Cas включением молекулы гРНК, содержащей нацеливающую последовательность, которая гибридизуется с последовательностью гена tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В некоторых вариантах осуществления гРНК содержит нацеливающую последовательность, которая полностью комплементарна 15-25 нуклеотидам, например, 20 нуклеотидам, гена tet, например, гена Tet1, Tet2 и/или Tet3, например Tet2. В некоторых вариантах осуществления 15-25 нуклеотидов, например, 20 нуклеотидов, гена tet, например, гена Tet1, Tet2 и/или Tet3, например Tet2, расположены непосредственно с 5'-стороны от последовательностси примыкающего к протоспейсеру мотива (PAM), распознаваемой белком Cas системы CRISPR/Cas (например, когда система содержит белок Cas9 S. pyogenes, последовательность PAM содержит NGG, где N может представлять собой любой из A, T, G или C). В некоторых вариантах осуществления нацеливающая последовательность гРНК содержит, например, состоит из последовательности РНК, комплементарной последовательности, приведенной в таблице 2. В некоторых вариантах осуществления гРНК содержит нацеливающую последовательность, приведенную в таблице 3.
В одном варианте осуществления вместе с системой CRISPR/Cas в клетку можно вводить чужеродную ДНК, например, ДНК, кодирующую CAR, например, как описано в настоящем описании; в зависимости от последовательностей чужеродной ДНК и хромосомной последовательности, этот процесс можно использовать для встраивания ДНК, кодирующей CAR, например, как описано в настоящем описании, в или вблизи участка, на который нацелена система CRISPR/Cas. Как показано в настоящем описании в примерах, но без связи с теорией, такое встраивание может приводить к экспрессии CAR, а также к нарушению гена Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Такую чужеродную молекулу ДНК называют в настоящем описании "матричной ДНК." В некоторых вариантах осуществления матричная ДНК дополнительно содержит плечи гомологии с 5'-стороны, с 3'-стороны или как с 5'-, так и с 3'-стороны от нуклеиновой кислоты матричной ДНК, которая кодирует представляющую интерес молекулу или молекулы (например, которая кодирует CAR, описанный в настоящем описании), где указанные плечи гомологии комплементарны последовательности геномной ДНК, фланкирующей последовательность-мишень.
В одном варианте осуществления система CRISPR/Cas по настоящему изобретению содержит Cas9, например, Cas9 S. pyogenes, и гРНК, содержащую нацеливающую последовательность, которая гибридизуется с последовательностью гена Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В одном варианте осуществления система CRISPR/Cas содержит нуклеиновую кислоту, кодирующую гРНК Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, и нуклеиновую кислоту, кодирующую белок Cas, например, Cas9, например, Cas9 S. pyogenes. В одном варианте осуществления система CRISPR/Cas содержит гРНК Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, и нуклеиновую кислоту, кодирующую белок Cas, например, Cas9, например, Cas9 S. pyogenes.
Примеры геномных последовательностей-мишеней Tet2, для которых можно получать гРНК, содержащие комплементарные нацеливающие последовательности, для применения в рамках настоящего изобретения, приведены в таблице 2 ниже. В некоторых вариантах осуществления гРНК содержит РНК, комплементарную последовательности-мишени, приведенной в таблице ниже (например, для sgTET2_1, гРНК будет содержать CCUUGGACACCUUCUCCUCC (SEQ ID NO: 44)). В некоторых вариантах осуществления гРНК содержит РНК-аналог последовательности-мишени, приведенной в таблице 2 ниже (например, для sgTET2_1, гРНК будет содержать GGAACCUGUGGAAGAGGAGG (SEQ ID NO: 45). В некоторых вариантах осуществления ингибитор Tet2 представляет собой нуклеиновую кислоту, кодирующую молекулу гРНК, специфичную к Tet2, где нуклеиновая кислота содержит последовательность-мишень из таблицы 2 ниже, например, под контролем промотора U6 или H1:
Таблица 2
Примеры нацеливающих последовательностей гРНК, которые пригодны в различных вариантах осуществления настоящего изобретения для ингибирования Tet, например Tet2, приведены ниже в таблице 3. В некоторых вариантах осуществления система CRISPR/Cas по настоящему изобретению содержит молекулу гРНК, содержащую нацеливающую последовательность, содержащую последовательность, приведенную в таблице 3. В некоторых вариантах осуществления система CRISPR/Cas по настоящему изобретению содержит молекулу гРНК, содержащую нацеливающую последовательность, которая представляет собой последовательность, приведенную в таблице 3.
Таблица 3
Системы редактирования генов TALEN
TALEN получают искусственным образом путем слияния эффекторного ДНК-связывающего домена TAL с доменом расщепления ДНК. Эффекты активатора транскрипции (TALE) можно модифицировать способами инженерии для связывания с любой желаемой последовательностью ДНК, включая часть гена HLA или TCR. Путем комбинирования модифицированных способами инженерии TALE с доменом расщепления ДНК можно получать фермент рестрикции, который является специфичным к любой желаемой последовательности ДНК, включая последовательность HLA или TCR. Затем их можно вводить в клетку, где их можно использовать для редактирования генома. Boch (2011) Nature Biotech. 29: 135-6; и Boch et al. (2009) Science 326: 1509-12; Moscou et al. (2009) Science 326: 3501.
TALE представляют собой белки, секретируемые бактериями Xanthomonas. ДНК-связывающий домен содержит повторяющуюся высококонсервативную последовательность из 33-34 аминокислот, за исключением 12-ой и 13-ой аминокислот. Эти два положения являются в высокой степени вариабельными, демонстрируя строгую корреляцию со специфическим распознаванием нуклеотидов. Таким образом, их можно модифицировать способами инженерии для связывания с желаемой последовательностью ДНК.
Для получения TALEN белок TALE подвергают слиянию с нуклеазой (N), которая представляет собой эндонуклеазу FokI дикого типа или мутантную эндонуклеазу FokI. В FokI было внесено несколько мутаций для ее применения в TALEN; они, например, повышают специфичность расщепления или активность. Cermak et al. (2011) Nucl. Acids Res. 39: e82; Miller et al. (2011) Nature Biotech. 29: 143-8; Hockemeyer et al. (2011) Nature Biotech. 29: 731-734; Wood et al. (2011) Science 333: 307; Doyon et al. (2010) Nature Methods 8: 74-79; Szczepek et al. (2007) Nature Biotech. 25: 786-793; и Guo et al. (2010) J. Mol. Biol. 200: 96.
Домен FokI функционирует в качестве димера, требующего две конструкции с уникальными ДНК-связывающими доменами для участков в геноме-мишени с надлежащей ориентаций и расстояниями. Как число аминокислотных остатков между ДНК-связывающим доменом TALE и доменом расщепления FokI, так и число оснований между двумя индивидуальными участками связывания TALEN, по-видимому, являются важными параметрами для достижения высоких уровней активности. Miller et al. (2011) Nature Biotech. 29: 143-8.
TALEN против Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, можно использовать внутри клетки для обеспечения двухцепочечного разрыва (DSB). Мутацию можно вносить в участке разрыва, если механизмы репарации ненадлежащим образом репарируют разрыв посредством негомологичного соединения концов. Например, ненадлежащая репарация может вносить мутацию со сдвигом рамки считывания. Альтернативно вместе с TALEN в клетку можно вводить чужеродную ДНК, например, ДНК, кодирующую CAR, например, как описано в настоящем описании; в зависимости от последовательности чужеродной ДНК и хромосомной последовательности, этот процесс можно использовать для встраивания ДНК, кодирующей CAR, например, как описано в настоящем описании, в или вблизи участка, на который нацелен TALEN. Как показано в настоящем описании в примерах, но без связи с теорией, такое встраивание может приводить экспрессии CAR, а также нарушению гена Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Такую чужеродную молекулу ДНК называют в настоящем описании "матричной ДНК". В некоторых вариантах осуществления матричная ДНК дополнительно содержит плечи гомологии с 5'-стороны, с 3'-стороны или как с 5'-, так и с 3'-стороны от нуклеиновой кислоты матричной ДНК, которая кодирует представляющую интерес молекулу или молекулы (например, которая кодирует CAR, описанный в настоящем описании), где указанные плечи гомологии комплементарны последовательности геномной ДНК, фланкирующей последовательность-мишень.
TALEN, специфичные к последовательностям в Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, можно конструировать с использованием любого способа, известного в данной области, включая различные схемы с использованием модульных компонентов. Zhang et al. (2011) Nature Biotech. 29: 149-53; Geibler et al. (2011) PLoS ONE 6: e19509; US 8420782; US 8470973, содержание которых включено в настоящее описание путем ссылок в полном объеме.
Нуклеаза с цинковыми пальцами для ингибирования Tet, например, Tet1, Tet2 и/или Tet3, например Tet2
"ZFN" или "нуклеаза с цинковыми пальцами" относятся к нуклеазе с цинковыми пальцами, которая представляет собой искусственную нуклеазу, которую можно использовать для модификации, например, делеции, одной или нескольких нуклеиновых кислот с желаемой последовательностью нуклеиновой кислоты, например, Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
Подобно TALEN, ZFN содержит нуклеазный домен FokI (или его производное), слитый с ДНК-связывающим доменом. В случае ZFN ДНК-связывающий домен содержит один или несколько цинковых пальцев. Carroll et al. (2011) Genetics Society of America 188: 773-782; и Kim et al. (1996) Proc. Natl. Acad. Sci. USA 93: 1156-1160.
Цинковый палец представляет собой небольшой белковый структурный мотив, стабилизированный одним или несколькими ионами цинка. Цинковый палец может содержать, например, Cys2His2, и может распознавать последовательность размером приблизительно 3 п.о. Различные цинковые пальцы известной специфичности можно комбинировать для получения полипептидов с несколькими пальцами, которые распознают последовательности размером приблизительно 6, 9, 12, 15 или 18 п.о. Для получения цинковых пальцев (и их комбинаций), распознающих конкретные последовательности, доступны различные способы селекции и модульной сборки, включая фаговый дисплей, дрожжевые моногибридные системы, бактериальные моногибридные и двухгибридные системы и клетки млекопитающих.
Подобно TALEN, ZFN должна димеризоваться для расщепления ДНК. Таким образом, для нацеливания на непалиндромные участки ДНК требуется пара ZFN. Две индивидуальных ZFN должны связывать противоположные цепи ДНК, причем между их нуклеазами должно быть надлежащее пространство. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10570-5.
Также подобно TALEN, ZFN может вносить двухцепочечный разрыв в ДНК, что может приводить к мутации со сдвигом рамки считывания при ненадлежащей репарации, вызывая снижение экспрессии и количества Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, в клетке. ZFN также можно использовать с гомологичной рекомбинацией для внесения мутации в ген Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, или для внесения нуклеиновой кислоты, кодирующей CAR, в участок в или вблизи последоватлеьности-мишени. Как описано выше, нуклеиновая кислота, кодирующая CAR, может быть внесена в качестве части матричной ДНК. В некоторых вариантах осуществления матричная ДНК дополнительно содержит плечи гомологии с 5'-стороны, с 3'-стороны или как с 5'-, так и с 3'-стороны от нуклеиновой кислоты матричной ДНК, которая кодирует представляющую интерес молекулу или молекулы (например, которая кодирует CAR, описанный в настоящем описании), где указанные плечи гомологии комплементарны последовательности геномной ДНК, фланкирующей последовательность-мишень.
ZFN, специфичные к последовательностям в гене Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, можно конструировать с использованием любого способа, известного в данной области. См., например, Provasi (2011) Nature Med. 18: 807-815; Torikai (2013) Blood 122: 1341-1349; Cathomen et al. (2008) Mol. Ther. 16: 1200-7; и Guo et al. (2010) J. Mol. Biol. 400: 96; публикацию патента США 2011/0158957; и публикацию патента США 2012/0060230, содержание которых включено в настоящее описание путем ссылки в полном объеме. В некоторых вариантах осуществления система редактирования генов ZFN также может содержать нуклеиновую кислоту, кодирующую один или несколько компонентов системы редактирования генов ZFN, например, системы редактирования генов ZFN, нацеленной на Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
Без связи с теорией полагают, что использование систем редактирования генов (например, систем редактирования генов CRISPR/Cas), которые нацелены на Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, может позволить ингибирование одной или нескольких функций Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, например, обеспечивая событие редактирования, которое приводит к экспрессии укороченного Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. Вновь, без связи с теорией, такие укороченные белки Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, могут сохранять одну или несколько функций Tet, например, Tet1, Tet2 и/или Tet3, например Tet2 (например, каркасная функция), одновременно ингибируя одну или несколько других функций, например, Tet1, Tet2 и/или Tet3, например Tet2 (например, каталитическая функция), и в силу этого могут быть предпочтительными. Системы редактирования генов, которые нацелены на поздний экзон или интрон гена Tet, например, гена Tet1, Tet2 и/или Tet3, например, гена Tet2, могут быть особенно предпочтительными в этом отношении. В одном аспекте ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3, например, ингибитор Tet2, системы редактирования генов по изобретению нацелен на поздний экзон или интрон гена tet. В одном аспекте ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3, например, ингибитор Tet2 система редактирования генов по изобретению нацелен на экзон или интрон ниже экзона 8. В одном аспекте ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3, например, ингибитор Tet2, системы редактирования генов нацелен на экзон 8 или экзон 9, например, экзон 9, гена tet2.
Без связи с теорией полагают, что в других вариантах осуществления также может быть предпочтительным на ранний экзон или интрон гена Tet, например, гена Tet1, Tet2 и/или Tet3, например, гена Tet2, например, для внесения преждевременного стоп-кодона в ген-мишень, что приводит к отсутствию продукта гена или экспрессии полностью нефункционального продукта гена. Системы редактирования генов, которые нацелены на ранний экзон или интрон гена Tet, например, гена Tet1, Tet2 и/или Tet3, например, гена Tet2, может быть особенно предпочтительной в этом отношении. В одном аспекте ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3, например, ингибитор Tet2, системы редактирования генов по изобретению нацелен на ранний экзон или интрон гена tet. В одном аспекте ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3, например, ингибитор Tet2 система редактирования генов по изобретению нацелен на экзон или интрон выше экзона 4. В некоторых вариантах осуществления ингибитор Tet, например, ингибитор Tet1, Tet2 и/или Tet3, например, ингибитор Tet2 системы редактирования генов, нацелен на экзон 1, экзон 2, или экзон 3, например, экзон 3, гена tet2.
Без связи с теорией, в других вариантах осуществления также может быть предпочтительным нацеливание на последовательность гена Tet, например, гена Tet1, Tet2 и/или Tet3, например, гена Tet2, которое является специфичным к одной или нескольким изоформам tet (например, ген tet2), но не влияет на одну или несколько других изоформ tet (например Tet2). В некоторых вариантах осуществления может быть предпочтительным специфическое нацеливание на изоформы tet (например Tet2), которые содержат каталитический домен.
Ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, на основе дцРНК, например, миРНК или кшРНК
В качестве ингибиторов Tet, например, Tet1, Tet2 и/или Tet3, например Tet2 в соответствии с настоящим изобретением можно использовать двухцепочечную РНК ("дцРНК"), например, миРНК или кшРНК. Также в рамках настоящего изобретения предусматривается применение нуклеиновой кислоты, кодирующей указанные ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, на основе дцРНК.
В одном варианте осуществления ингибитор Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, представляет собой нуклеиновую кислоту, например, дцРНК, например, миРНК или кшРНК, специфичную к нуклеиновой кислоте, кодирующей Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, например, геномной ДНК или мРНК, кодирующей Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
Один аспект изобретения предусматривает композицию, содержащую дцРНК, например, миРНК или кшРНК, содержащую по меньшей мере 15 последовательно расположенных нуклеотидов, например, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 последовательно расположенных нуклеотидов, например, 21 последовательно расположенный нуклеотид, которые комплементарны (например, на 100% комплементарны) последовательности нуклеиновой кислоты Tet, например, Tet1, Tet2 и/или Tet3, например Tet2 (например, геномной ДНК или мРНК, кодирующей Tet, например, Tet1, Tet2 и/или Tet3, например Tet2). В некоторых вариантах осуществления по меньшей мере 15 последовательно расположенных нуклеотидов, например, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 последовательно расположенных нуклеотидов, например, 21 последовательно расположенный нуклеотид, включают последовательно расположенные нуклеотиды последовательности-мишени кшРНК или нуклеиновой кислоты, кодирующей кшРНК Tet2, приведенной в таблице 4. Понятно, что некоторые из последовательностей-мишеней и/или молекул кшРНК присутствуют в качестве ДНК, однако средства на основе дцРНК, нацеленные на эти последовательности или содержащие эти последовательности, могут представлять собой РНК, или любой нуклеотид, модифицированный нуклеотид или их заместитель, описанный в настоящем описании и/или известный в данной области, при условии, что молекула все еще может опосредовать РНК-интерференцию.
В одном варианте осуществления молекула нуклеиновой кислоты, которая кодирует молекулу дцРНК, которая ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, функционально связана с промотором, например, промотором, происходящим из H1 или U6, так что молекула дцРНК, которая ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, экспрессируется в экспрессирующей CAR клетке. См. например, Tiscornia G., "Development of Lentiviral Vectors Expressing siRNA", Chapter 3, Gene Transfer: Delivery and Expression of DNA and RNA (eds. Friedmann и Rossi). Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, USA, 2007; Brummelkamp TR, et al. (2002) Science 296: 550-553; Miyagishi M, et al. (2002) Nat. Biotechnol. 19: 497-500. В одном варианте осуществления молекула нуклеиновой кислоты, которая кодирует молекулу дцРНК, которая ингибирует Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, находится на том же векторе, например, лентивирусном векторе, который содержит молекулу нуклеиновой кислоты, которая кодирует компонент, например, все компоненты, CAR. В таком варианте осуществления молекула нуклеиновой кислоты, которая кодирует молекулу дцРНК, которая ингибирует Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, расположена на векторе, например, лентивирусном векторе с 5'-стороны или с 3'-стороны от нуклеиновой кислоты, которая кодирует компонент, например, все компоненты CAR. Молекула нуклеиновой кислоты, которая кодирует молекулу дцРНК, которая ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, может транскрибироваться в том же или отличающемся положении относительно нуклеиновой кислоты, которая кодирует компонент, например, все компоненты, CAR. В одном варианте осуществления молекула нуклеиновой кислоты, которая кодирует молекулу дцРНК, которая ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, находится на векторе, отличном от вектора, который содержит молекулу нуклеиновой кислоты, которая кодирует компонент, например, все компоненты, CAR. В одном варианте осуществления молекула нуклеиновой кислоты, которая кодирует молекулу дцРНК, которая ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, временно экспрессируется в экспрессирующей CAR клетке. В одном варианте осуществления молекула нуклеиновой кислоты, которая кодирует молекулу дцРНК, которая ингибирует экспрессию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, стабильно встраивается в геном экспрессирующей CAR клетки.
Примеры последовательностей нуклеиновых кислот, которые кодируют последовательности кшРНК, представлены ниже. Последовательность-мишень относится к последовательности в геномной ДНК Tet2 (или окружающей ДНК). Нуклеиновая кислота, кодирующая кшРНК против Tet2, кодирует молекулы кшРНК, пригодные в рамках настоящего изобретения. В одном варианте осуществления ингибитор Tet2 представляет собой миРНК или кшРНК, специфичную к последовательности-мишени, приведенной ниже, или специфичную к комплементарной ей мРНК. В некоторых вариантах осуществления ингибитор Tet2 представляет собой кшРНК, кодируемую нуклеиновой кислотой, кодирующей кшРНК против Tet2, согласно таблице 4 ниже. В некоторых вариантах осуществления ингибитор Tet2 представляет собой нуклеиновую кислоту, содержащую нуклеиновую кислоту, кодирующую кшРНК против Tet2 согласно таблице 4 ниже, например, которая находится под контролем промотора U6 или H1, так чтобы продуцировалась кшРНК против Tet2. В некоторых вариантах осуществления изобретение относится к миРНК или кшРНК, содержащей последовательность, которая является аналогом РНК (т.е. все остатки нуклеиновой кислоты T заменены остатками нуклеиновой кислоты U) последовательности-мишени кшРНК, например, последовательности-мишени кшРНК любой из кшРНК, приведенных в таблице 4.
Таблица 4
(кшРНК против TET2 #1)
(кшРНК против TET2 #2)
(кшРНК против TET2 #3)
(кшРНК против TET2 #4)
(кшРНК против TET2 #5)
(кшРНК против TET2 #9)
Дополнительный ингибитор Tet2 на основе молекул дцРНК, например, кшРНК и миРНК, можно конструировать и тестировать с использованием способов, известных в данной области и описанных в настоящем описании. В некоторых вариантах осуществления ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, нацелен на последовательность SEQ ID NO: 1358. В некоторых вариантах осуществления ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, нацелен на последовательность SEQ ID NO: 1359. В некоторых вариантах осуществления ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, нацелен на последовательность SEQ ID NO: 1360. В некоторых вариантах осуществления ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, нацелен на последовательность SEQ ID NO: 1361. В некоторых вариантах осуществления ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, нацелен на последовательность SEQ ID NO: 1362. В некоторых вариантах осуществления ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, нацелен на последовательность SEQ ID NO: 1363. В некоторых вариантах осуществления ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, нацелен на последовательность мРНК, кодирующей Tet2.
В некоторых вариантах осуществления ингибитор представляет собой нуклеиновую кислоту, например, ДНК, кодирующую ингибитор Tet2 на основе дцРНК, например, кшРНК или миРНК, согласно любому из описанных выше вариантов осуществления. В некоторых вариантах осуществления нуклеиновая кислота, например, ДНК, находится в векторе, например, любой общепринятой экспрессирующей системе, например, как описано в настоящем описании, например, в лентивирусном векторе.
Без связи с теорией, ингибитор TET на основе дцРНК (например, миРНК или кшРНК), нацеленный на последовательность мРНК гена Tet, например, Tet1, Tet2 и/или Tet3, например, мРНК Tet2, который специфичен к одной или нескольким изоформам tet (например Tet2), но не влияет на одну или несколько других изоформ tet (например Tet2) (например, вследствие нацеливания на уникальную точку сплайсинга или нацеливания на домен, который присутствует в одной или нескольких изоформах tet, например Tet2, но не присутствует в одной или нескольких других изоформах tet, например Tet2). В некоторых вариантах осуществления может быть предпочтительным специфическое нацеливание на изоформы tet (например Tet2), которые содержат каталитический домен.
Низкомолекулярные соединения
Ингибиторы Tet
В некоторых вариантах осуществления ингибитор Tet представляет собой низкомолекулярное соединение, которое ингибирует экспрессию и/или функцию Tet, например, Tet1, Tet2 и/или Tet3, например Tet2.
Ингибиторы Tet2
В некоторых вариантах осуществления ингибитор Tet2 представляет собой низкомолекулярное соединение, которое ингибирует экспрессию и/или функцию Tet2. Например, ингибитором Tet2 в соответствии с настоящим изобретением является 2-гидроксиглутарат (CAS #2889-31-8).
В другом примере ингибитор Tet2 в соответствии с настоящим изобретением имеет следующую структуру:
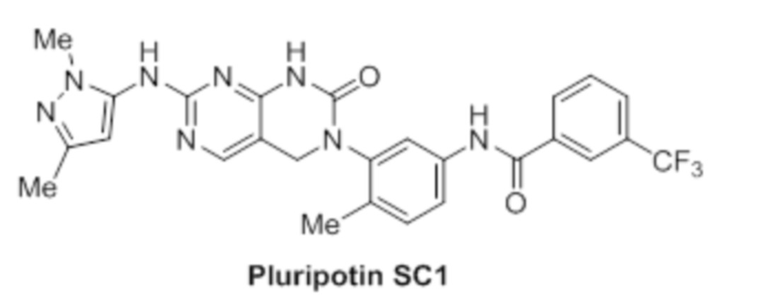
Плурипотин SC1
В другом примере ингибитор Tet2 в соответствии с настоящим изобретением представляет собой N-[3-[7-(2,5-диметил-2H-пиразол-3-иламино)-1-метил-2-оксо-1,4-дигидро-2H-пиримидо[4,5-d]пиримидин-3-ил]-4-метилфенил]-3-трифторметилбензамид (CAS #839707-37-8) и имеет следующую структуру:
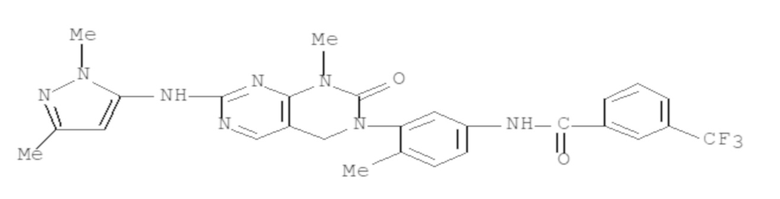
В другом примере ингибитор Tet2 в соответствии с настоящим изобретением представляет собой 2-[(2,6-дихлор-3-метилфенил)амино]бензойную кислоту (CAS # 644-62-2) и имеет следующую структуру:

меклофенаминовая кислота
В некоторых вариантах осуществления ингибитор Tet2 по настоящему изобретению представляет собой фармацевтически приемлемую соль любого из вышеуказанных.
Ингибиторы HDAC
В соответствии с настоящим изобретением можно использовать любые известные ингибиторы HDAC. Неограничивающие примеры ингибиторов HDAC включают Вониностат (Zolinza®); Ромидепсин (Istodax®); Трихостатин A (TSA); Оксамфлатин; Вориностат (Zolinza®, субероиланилидгидроксамовая кислота); Пироксамид (субероил-3-аминопиридинамидгидроксамовая кислота); Трапоксин A (RF-1023A); Трапоксин B (RF-10238); цикло[(αS,2S)-α-амино-η-оксо-2-оксираноктаноил-O-метил-D-тирозил-L-изолейцил-L-пролил] (Cyl-1); цикло[(αS,2S)-α-амино-η-оксо-2-оксираноктаноил-O-метил-D-тирозил-L-изолейцил-(2S)-2-пиперидинкарбонил] (Cyl-2); цикло[L-аланил-D-аланил-(2S)-η-оксо-L-α-аминооксираноктаноил-D-пролил] (HC-токсин); цикло[(αS,2S)-α-амино-η-оксо-2-оксираноктаноил-D-фенилаланил-L-лейцил-(2S)-2-пиперидинкарбонил] (WF-3161); Хламидоцин ((S)-цикло(2-метилаланил-L-фенилаланил-D-пролил-η-оксо-L-α-аминооксираноктаноил); Апидицин (цикло(8-оксо-L-2-аминодеканоил-1-метокси-L-триптофил-L-изолейцил-D-2-пиперидинкарбонил); Ромидепсин (Istodax®, FR-901228); 4-фенилбутират; Спирухостатин A; Милпроин (вальпроевая кислота); Энтиностат (MS-275, N-(2-аминофенил)-4-[N-(пиридин-3-илметоксикарбонил)-амино-метил]-бензамид); Депудецин (4,5:8,9-диангидро-1,2,6,7,11-пентадезокси-D-трео-D-идо-ундека-1,6-диенитол); 4-(ацетиламино)-N-(2-аминофенил)-бензамид (также известный как CI-994); N1-(2-аминофенил)-N8-фенилоктандиамид (также известный как BML-210); 4-(диметиламино)-N-(7-(гидроксиамино)-7-оксогептил)бензамид (также известный как M344); (E)-3-(4-(((2-(1H-индол-3-ил)этил)(2-гидроксиэтил)амино)-метил)фенил)-N-гидроксиакриламид (NVP-LAQ824); Панобиностат (Farydak®); Моцетиностат и Белиностат.
Белки
Доминантно-негативный Tet2
В соответствии с настоящим изобретением, в качестве ингибиторов Tet2 можно использовать доминантно-негативные изоформы Tet2 и нуклеиновую кислоту, кодирующую доминантно-негативный Tet2. В некоторых вариантах осуществления доминантно-негативный Tet2 лишен каталитической функции Tet2. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией R1261G, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией R1262A, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией S1290A, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией WSMYYN (аминокислоты 1291-1296 SEQ ID NO: 1357) на GGSGGS (SEQ ID NNO: 67), в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией M1293A и Y1294A, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией Y1295A, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией S1303N, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией H1382Y, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией D1384A, в соответствии с нумерацией SEQ ID NO: 1357. Примером доминантно-негативного Tet2 является белок, содержащий или состоящий из SEQ ID NO: 1357 с мутацией D1384V, в соответствии с нумерацией SEQ ID NO: 1357. В некоторых вариантах осуществления доминантно-негативный Tet2 может включать комбинации любых из вышеперечисленных мутаций. Такие мутации, кроме того, описаны, например, в Chen et al., Nature, 493:561-564 (2013); Hu et al, Cell, 155:1545-1555 (2013), содержание которой включено в настоящее описание путем ссылки в полном объеме.
Доминантно-негативные партнеры по связыванию Tet2
Без связи с теорией полагают, что Tet2 взаимодействует, например, связывается с одной или несколькими HDAC, например, одной или несколькими HDAC, экспрессируемыми в иммунных эффекторных клетках, например, в T-клетках, и что такие комплексы Tet2:HDAC могут вносить вклад в активность Tet2 в клетке. В некоторых вариантах осуществления ингибитор Tet2 по изобретению представляет собой доминантно-негативный партнер по связыванию Tet2, например, доминантно-негативную Tet2-связывающую HDAC. В других вариантах осуществления ингибитор Tet2 по изобретению содержит нуклеиновую кислоту, кодирующую доминантно-негативный партнер по связыванию Tet2, например, доминантно-негативную Tet2-связывающую HDAC.
Векторы, кодирующие ингибиторы Tet2
Как описано в настоящем описании, изобретение относится к векторам, например, как описано в настоящем описании, которые кодируют ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, такие как системы редактирования генов, ингибиторы на основе кшРНК или миРНК или доминантно-негативные ингибиторы Tet, например, Tet1, Tet2 и/или Tet3, например Tet2 (например, как описано в настоящем описании).
В некоторых вариантах осуществления дополнительно включающих, например, CAR, нуклеиновая кислота может дополнительно содержать последовательность, кодирующую CAR, например, как описано в настоящем описании. В некоторых вариантах осуществления изобретение относится к вектору, содержащему последовательность нуклеиновой кислоты, кодирующую ингибитор Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, описанный в настоящем описании, и содержащему последовательность нуклеиновой кислоты, кодирующую молекулу CAR, описанную в настоящем описании. В некоторых вариантах осуществления последовательности нуклеиновых кислот находятся на отдельных векторах. В других вариантах осуществления две или более последовательности нуклеиновой кислоты кодируются одной молекулой нуклеиновой кислоты в одной рамке считывания и в качестве одной полипептидной цепи. В этом аспекте два или более CAR, например, могут быть одним или несколькими участками расщепления пептидов (например, участок ауторасщепления или субстрат для внутриклеточной протеазы). Примеры участков расщепления пептидов пептид включают следующие, где остатки GSG являются необязательными:
T2A: (GSG) E G R G S L L T C G D V E E N P G P (SEQ ID NO: 68)
P2A: (GSG) A T N F S L L K Q A G D V E E N P G P (SEQ ID NO: 69)
E2A: (GSG) Q C T N Y A L L K L A G D V E S N P G P (SEQ ID NO: 70)
F2A: (GSG) V K Q T L N F D L L K L A G D V E S N P G P (SEQ ID NO: 71).
Эти участки расщепления пептидов в совокупности обозначаются в настоящем описании как "участки 2A". В некоторых вариантах осуществления вектор содержит последовательность нуклеиновой кислоты, кодирующую CAR, описанный в настоящем описании, и последовательность нуклеиновой кислоты, кодирующую ингибитор Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, на основе кшРНК или миРНК, описанный в настоящем описании. В некоторых вариантах осуществления вектор содержит последовательность нуклеиновой кислоты, кодирующую CAR, описанный в настоящем описании, и последовательность нуклеиновой кислоты, кодирующую ингибитор Tet например, Tet1, Tet2 и/или Tet3, например Tet2, на основе системы редактирования генома (например, систему CRISPR/Cas), описанный в настоящем описании.
Способы применения ингибиторов Tet, например, Tet1, Tet2 и/или Tet3, например Tet2
Изобретение относится к способам повышения терапевтической эффективности экспрессирующей CAR клетки, например, клетки, экспрессирующей CAR, как описано в настоящем описании, например, экспрессирующей CAR19 клетки (например, CTL019), включающим стадию снижения уровня 5-гидроксиметилцитозина в указанной клетке. В некоторых вариантах осуществления способ включает снижение или устранение функции или экспрессии Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В некоторых вариантах осуществления способ включает приведение указанных клеток в контакт с ингибитором Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, как описано в настоящем описании.
Кроме того, изобретение относится к способам получения экспрессирующей CAR клетки, например, экспрессирующей CAR клетки, имеющей улучшенную функцию (например, имеющую увеличенную эффективность, например, нацеливание на опухоль или пролиферацию), включающим стадию снижения или устранения экспрессии или функции Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, в указанной клетке. В некоторых вариантах осуществления способ включает приведение указанных клеток в контакт с ингибитором Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, как описано в настоящем описании. В некоторых вариантах осуществления приведение в контакт проводят ex vivo. В некоторых вариантах осуществления приведение в контакт проводят in vivo. В некоторых вариантах осуществления приведение в контакт проводят до, одновременно или после модификации указанных клеток для экспрессии CAR, например, CAR, как описано в настоящем описании.
В некоторых вариантах осуществления изобретение относится к способу ингибирования функции или экспрессии Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, в экспрессирующей CAR клетке, например, клетке, экспрессирующей CAR, как описано в настоящем описании, например, экспрессирующей CAR19 клетке (например, экспрессирующей CTL019 клетке), причем способ включает стадию снижения уровня 5-гидроксиметилцитозина в указанной клетке. В некоторых вариантах осуществления способ включает снижение или устранение функции или экспрессии Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В некоторых вариантах осуществления способ включает приведение указанных клеток в контакт с ингибитором Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, как описано в настоящем описании.
В одном варианте осуществления изобретение относится к способу, например, к способу, описанному выше, который включает введение нуклеиновой кислоты, кодирующей CAR, в клетку, например, иммунную эффекторную клетку, например, T-клетку, в участке в гене Tet или его регуляторных элементах, чтобы экспрессия Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, нарушалась. Встраивание в участке в гене Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, можно проводить, например, с использованием нацеленной на Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, системы редактирования генов как описано выше.
В одном варианте осуществления изобретение относится к способу, например, к способу, описанному выше, включающему стадию введения в клетку системы редактирования генов, например, системы редактирования генов CRISPR/Cas, которая нацелена на Tet, например, Tet1, Tet2 и/или Tet3, например Tet2, например, системы CRISPR/Cas, содержащей гРНК, которая имеет нацеливающую последовательность, комплементарную последовательности-мишени гена Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В некоторых вариантах осуществления систему CRISPR/Cas вводят в указанную клетку в качестве рибонуклеопротеинового комплекса гРНК и фермента, например, вводят посредством электропорации. В одном варианте осуществления способ включает введение нуклеиновой кислоты, кодирующей один или несколько компонентов системы CRISPR/Cas, в указанную клетку. В одном варианте осуществления указанная нуклеиновая кислота находится на векторе, кодирующем CAR, например, CAR, как описано в настоящем описании.
В одном варианте осуществления изобретение относится к способу, например, к способу, описанному выше, включающему стадию введения в клетку ингибиторной дцРНК, например, кшРНК или миРНК, которая нацелена на Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В одном варианте осуществления способ включает введение в указанную клетку нуклеиновой кислоты, кодирующей ингибиторную дцРНК, например, кшРНК или миРНК, которая нацелена на Tet, например, Tet1, Tet2 и/или Tet3, например Tet2. В одном варианте осуществления указанная нуклеиновая кислота находится на векторе, кодирующем CAR, например, CAR, как описано в настоящем описании.
Дополнительные компоненты CAR и CART-клеток и способы, касающиеся изобретения, описаны ниже.
В рамках настоящего изобретения предусматриваются композиции и способы применения для лечения заболевания, такого как злокачественная опухоль, с использованием иммунных эффекторных клеток (например, T-клеток, NK-клеток), сконструированных с CAR по изобретению.
В одном аспекте изобретение относится к ряду химерных рецепторов антигенов (CAR), содержащих антигенсвязывающий домен (например, антитело или фрагмент антитела, TCR или фрагмент TCR), сконструированный для специфического связывания с опухолевым антигеном, например, опухолевым антигеном, описанным в настоящем описании. В одном аспекте изобретение относится к иммунной эффекторной клетке (например, T-клетке, NK-клетке), модифицированной способами инженерии для экспрессии CAR, где модифицированная иммунная эффекторная клетка проявляет противораковое свойство. В одном аспекте клетка трансформирована CAR и CAR экспрессируется на поверхности клетки. В некоторых вариантах осуществления клетка (например, T-клетка, NK-клетка) трансдуцирована вирусным вектором, кодирующим CAR. В некоторых вариантах осуществления вирусный вектор представляет собой ретровирусный вектор. В некоторых вариантах осуществления вирусный вектор представляет собой лентивирусный вектор. В некоторых таких вариантах осуществления клетка может стабильно экспрессировать CAR. В другом варианте осуществления клетка (например, T-клетка, NK-клетка) трансфицирована нуклеиновой кислотой, например, мРНК, кДНК, ДНК, кодирующей CAR. В некоторых таких вариантах осуществления клетка может временно экспрессировать CAR.
В одном аспекте антигенсвязывающий домен CAR, описанного в настоящем описании, представляет собой scFv-фрагмент антитела. В одном аспекте такие фрагменты антител являются функциональными, поскольку они сохраняют эквивалентную аффинность связывания, например, они связывают тот же антиген с аффинностью, сравнимой с IgG-антителом, из которого они происходят. В других вариантах осуществления фрагмент антитела имеет более низкую аффинность связывани, например, он связывает тот же антиген с более низкой аффинностью связывания, чем антитело, из которого он происходит, однако является функциональным в том, что он обеспечивает биологический ответ, описанный в настоящем описании. В одном варианте осуществления молекула CAR содержит фрагмент антитела, который обладает аффинностью связывания KD от 10-4 M до 10-8 M, например, от 10-5 M до 10-7 M, например, 10-6 M или 10-7 M, в отношении антигена-мишени. В одном варианте осуществления фрагмент антитела обладает аффинностью связывания, которая по меньшей мере в пять раз, в 10 раз, в 20 раз, в 30 раз, в 50 раз, в 100 раз или в 1000 раз меньше, чем у эталонного антитела, например, антитела, описанного в настоящем описании.
В одном аспекте такие фрагменты антител являются функциональными в том, что они обеспечивают биологический ответ, который может включать, но не ограничивается ими, активацию иммунного ответа, ингибирование передачи сигнала от антигена-мишени, ингибирование киназной активности и т.п., как будет понятно специалисту в данной области.
В одном аспекте антигенсвязывающий домен CAR представляет собой scFv-фрагмент антитела, который является гуманизированным по сравнению с последовательностью scFv мыши, из которой он происходит.
В одном аспекте антигенсвязывающий домен CAR по изобретению (например, scFv) кодируется молекулой нуклеиновой кислоты, последовательность которой подвергнута оптимизации кодонов для экспрессии в клетке млекопитающего. В одном аспекте вся конструкция CAR по изобретению кодируется молекулой нуклеиновой кислоты, полная последовательность которой подвергнута оптимизации кодонов для экспрессии в клетке млекопитающего. Оптимизация кодонов относится к открытию, что частота встречаемости синонимичных кодонов (т.е. кодонов, которые кодируют одну и ту же аминокислоту) в кодирующей ДНК является смещенной в различных видах. Такая вырожденность кодонов позволяет кодирование идентичных полипептидов различными нуклеотидными последовательностями. Различные способы оптимизации кодонов известны в данной области и включают, например, способы, описанные по меньшей мере в патентах США под номерами 5786464 и 6114148.
В одном аспекте в CAR по изобретению антигенсвязывающий домен специфического антитела комбинирован с внутриклеточной сигнальной молекулой. Например, в некоторых аспектах внутриклеточная сигнальная молекула включает, но не ограничивается ими, сигнальные модули цепи CD3-зета, 4-1BB и CD28 и их комбинации.
Более того, настоящее изобретение относится к CAR и экспрессирующим CAR клеткам и к их применению в лекарственных средствах и способах лечения, среди прочих заболеваний, рака или любого злокачественного новообразования, или аутоиммунных заболеваний, вовлекающих клетки или ткани, которые экспрессируют опухолевый антиген, как описано в настоящем описании.
В одном аспекте CAR по изобретению можно использовать для устранения нормальных клеток, которые экспрессируют опухолевый антиген, как описано в настоящем описании, тем самым, его можно использовать в качестве клеточного кондиционирования перед трансплантацией клеток. В одном аспекте нормальная клетка, которая экспрессирует опухолевый антиген, как описано в настоящем описании, представляет собой нормальную стволовую клетку, и трансплантация клеток представляет собой трансплантацию столовых клеток.
В одном аспекте изобретение относится к иммунной эффекторной клетке (например, T-клетке, NK-клетке), модифицированной способами инженерии для экспрессии химерного рецептора антигена (CAR), где модифицированная способами инженерии иммунная эффекторная клетка демонстрирует противоопухолевое свойство. Предпочтительным антигеном является ассоциированный со злокачественной опухолью антиген (т.е. опухолевый антиген), описанный в настоящем описании. В одном аспекте антигенсвязывающий домен CAR содержит частично гуманизированный фрагмент антитела. В одном аспекте антигенсвязывающий домен CAR содержит частично гуманизированный scFv. Таким образом, изобретение относится к CAR, который содержит гуманизированный антигенсвязывающий домен и введен способами инженерии в клетку, например, T-клетку или NK-клетку, и к способам его применения для адоптивной терапии.
В одном аспекте CAR по изобретению содержат по меньшей мере один внутриклеточный домен, выбранный из группы из сигнального домена CD137 (4-1BB), сигнального домена CD28, сигнального домена CD27, сигнального домена CD3-зета и любой их комбинации. В одном аспекте CAR по изобретению, содержащий по меньшей мере один внутриклеточный сигнальный домен, происходит из одной или нескольких костимулирущей молекулы(молекул), отличной от CD137 (4-1BB) или CD28.
Последовательности некоторых примеров различных компонентов CAR по настоящему изобретению приведены в таблице 1, где а.к. означает аминокислоты, и н.к. означает нуклеиновые кислоты, которые кодируют соответствующий пептид.
Таблица 1. Последовательности различных компонентов CAR (а.к. - аминокислоты, н.к. - нуклеиновые кислоты, которые кодируют соответствующий белок)
AACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTT
TGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGA
AGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGG
AGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGC
ACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGC
CCTTCACATGCAGGCCCTGCCCCCTCGC
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa
Ассоциированные со злокачественной опухолью антигены
Настоящее изобретение относится к иммунным эффекторным клеткам (например, T-клеткам, NK-клеткам), которые модифицированы способами инженерии так, чтобы они содержали один или несколько CAR, которые направляют иммунные эффекторные клетки к злокачественной опухоли. Этого достигают через антигенсвязывающий домен на CAR, который является специфичным к антигену, ассоциированному со злокачественной опухолью. Существует два класса ассоциированных со злокачественной опухолью антигенов (опухолевые антигены), на которые можно нацеливать CAR по настоящему изобретению: (1) ассоциированные со злокачественной опухолью антигены, которые экспрессируются на поверхности злокачественных клеток; и (2) ассоциированные со злокачественной опухолью антигены, которые сами по себе являются внутриклеточными, однако фрагмент такого антигена (пептид) презентируется на поверхности злокачественных клеток посредством MHC (основной комплекс гистосовместимости).
Таким образом, настоящее изобретение относится к CAR, которые нацелены на следующие ассоциированные со злокачественной опухолью антигены (опухолевые антигены): CD19, CD123, CD22, CD30, CD171, CS-1, CLL-1 (CLECL1), CD33, EGFRvIII, GD2, GD3, BCMA, Tn Ag, PSMA, ROR1, FLT3, FAP, TAG72, CD38, CD44v6, CEA, EPCAM, B7H3, KIT, IL-13Ra2, мезотелин, IL-11Ra, PSCA, VEGFR2, LewisY, CD24, PDGFR-beta, PRSS21, SSEA-4, CD20, рецептор фолатов альфа, ERBB2 (Her2/neu), MUC1, EGFR, NCAM, простаза, PAP, ELF2M, эфрин B2, рецептор IGF-I, CAIX, LMP2, gp100, bcr-abl, тирозиназа, EphA2, фукозил-GM1, sLe, GM3, TGS5, HMWMAA, o-ацетил-GD2, рецептор фолатов бета, TEM1/CD248, TEM7R, CLDN6, TSHR, GPRC5D, CXORF61, CD97, CD179a, ALK, полисиаловая кислота, PLAC1, GloboH, NY-BR-1, UPK2, HAVCR1, ADRB3, PANX3, GPR20, LY6K, OR51E2, TARP, WT1, NY-ESO-1, LAGE-1a, легумаин, HPV E6,E7, MAGE-A1, MAGE A1, ETV6-AML, белок спермы 17, XAGE1, Tie 2, MAD-CT-1, MAD-CT-2, Fos-родственный антиген 1, p53, мутантный p53, простеин, сурвивин и теломераза, PCTA-1/галектин 8, MelanA/MART1, мутантный Ras, hTERT, точек разрыва при транслокации саркомы, ML-IAP, ERG (слитый ген TMPRSS2 ETS), NA17, PAX3, рецептор андрогенов, циклин B1, MYCN, RhoC, TRP-2, CYP1B1, BORIS, SART3, PAX5, OY-TES1, LCK, AKAP-4, SSX2, RAGE-1, обратная транскриптаза теломеразы человека, RU1, RU2, кишечная карбоксилэстераза, mut hsp70-2, CD79a, CD79b, CD72, LAIR1, FCAR, LILRA2, CD300LF, CLEC12A, BST2, EMR2, LY75, GPC3, FCRL5 и IGLL1.
Поддерживающие опухоль антигены
CAR, описанный в настоящем описании, может содержать антигенсвязывающий домен (например, антитело или фрагмент антитела, TCR или фрагмент TCR), который связывается с поддерживающим опухоль антигеном (например, поддерживающий опухоль антиген, как описано в настоящем описании). В некоторых вариантах осуществления поддерживающий опухоль антиген представляет собой антиген, присутствующий на стромальной клетке или супрессорной клетке миелоидного происхождения (MDSC). Стромальные клетки могут секретировать факторы роста для стимуляции деления клеток в микроокружении. Клетки MDSC могут ингибировать пролиферацию и активацию T-клеток. Без связи с теорией, в некоторых вариантах осуществления экспрессирующие CAR клетки разрушают поддерживающие опухоль клетки, тем самым непрямо ингибируя рост или выживание опухоли.
В некоторых вариантах осуществления стромальный клеточный антиген выбран из одного или нескольких из: стромального клеточного антигена 2 костного мозга (BST2), белка активации фибробластов (FAP) и тенасцинв. В одном варианте осуществления FAP-специфическое антитело представляет собой, конкурирует за связывание с или имеет те же CDR, что и сибротузумаб. В некоторых вариантах осуществления антиген MDSC выбран из одного или нескольких из: CD33, CD11b, C14, CD15 и CD66b. Таким образом, в некоторых вариантах осуществления поддерживающий опухоль антиген выбран из одного или нескольких из: стромального клеточного антигена 2 костного мозга (BST2), белка активации фибробластов (FAP) или тенасцина, CD33, CD11b, C14, CD15 и CD66b.
Химерный рецептор антигена (CAR)
Настоящее изобретение относится к рекомбинантной конструкции ДНК, содержащей последовательности, кодирующие CAR, где CAR содержит антигенсвязывающий домен (например, антитело или фрагмент антитела, TCR или фрагмент TCR), которые специфически связываются с ассоциированным со злокачественной опухолью антигеном, описанным в настоящем описании, где последовательность антигенсвязывающего фрагмента является соседней с последовательностью нуклеиновой кислоты, кодирующая внутриклеточный сигнальный домен, и находящейся в одной рамке считывания с ней. Внутриклеточный сигнальный домен может содержать костимулирующий сигнальный домен и/или первичный сигнальный домен, например, зета-цепь. Костимулирующий сигнальный домен относится к части CAR, содержащей по меньшей мере часть внутриклеточного домена костимулирующей молекулы.
В конкретных аспектах конструкция CAR по изобретению содержит scFv-домен, где scFv может предшествовать необязательная лидерная последовательность, такая как последовательность, представленная в SEQ ID NO: 2, и за ним может следовать необязательная шарнирная последовательность, такая как последовательность, представленная в SEQ ID NO: 4, или SEQ ID NO: 6, или SEQ ID NO: 8, или SEQ ID NO: 10, трансмембранная область, такая как последовательность, представленная в SEQ ID NO: 12, внутриклеточный сигнальный домен, который включает SEQ ID NO: 14 или SEQ ID NO: 16, и последовательность CD3-зета, которая включает SEQ ID NO: 18 или SEQ ID NO: 20, где домены являются соседними и находятся в одной рамке считывания для формирования слитого белка.
В одном аспекте иллюстративные конструкции CAR содержат необязательную лидерную последовательность (например, лидерную последовательность, описанную в настоящем описании), внеклеточный антигенсвязывающий домен (например, антигенсвязывающий домен, описанный в настоящем описании), шарнирную область (например, шарнирную область, описанную в настоящем описании), трансмембранный домен (например, трансмембранный домен, описанный в настоящем описании) и внутриклеточный стимулирующий домен (например, внутриклеточный стимулирующий домен, описанный в настоящем описании). В одном аспекте иллюстративная конструкция CAR содержит необязательную лидерную последовательность (например, лидерную последовательность, описанную в настоящем описании), внеклеточный антигенсвязывающий домен (например, антигенсвязывающий домен, описанный в настоящем описании), шарнирную область (например, шарнирную область, описанную в настоящем описании), трансмембранный домен (например, трансмембранный домен, описанный в настоящем описании), внутриклеточный костимулирующий сигнальный домен (например, костимулирующий сигнальный домен, описанный в настоящем описании) и/или внутриклеточный первичный сигнальный домен (например, первичный сигнальный домен, описанный в настоящем описании).
Иллюстративная лидерная последовательность предоставлена в качестве SEQ ID NO: 2. Иллюстративная шарнирная/спейсерная последовательность предоставлена в качестве SEQ ID NO: 4 или SEQ ID NO: 6, или SEQ ID NO: 8, или SEQ ID NO: 10. Иллюстративная последовательность трансмембранного домена представлена в качестве SEQ ID NO: 12. Иллюстративная последовательность внутриклеточного сигнального домена белка 4-1BB представлена в качестве SEQ ID NO: 14. Иллюстративная последовательность внутриклеточного сигнального домена CD27 представлена в качестве SEQ ID NO: 16. Иллюстративная последовательность домена CD3-зета представлена в качестве SEQ ID NO: 18 или SEQ ID NO: 20.
В одном аспекте настоящее изобретение относится к рекомбинантной конструкции нуклеиновой кислоты, содержащей молекулу нуклеиновой кислоты, кодирующую CAR, где молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен, например, описанный в настоящем описании, которая является соседней и находится в одной рамке считывания с последовательностью нуклеиновой кислоты, кодирующей внутриклеточный сигнальный домен.
В одном аспекте настоящее изобретение относится к рекомбинантной конструкции нуклеиновой кислоты, содержащей молекулу нуклеиновой кислоты, кодирующую CAR, где молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, кодирующую антигенсвязывающий домен, где последовательность является соседней с последовательностью нуклеиновой кислоты, кодирующей внутриклеточный сигнальный домен, и находится в одной рамке считывания с ней. Иллюстративный внутриклеточный сигнальный домен, который можно использовать в CAR, включает, но не ограничивается ими, один или несколько внутриклеточных сигнальных доменов из, например, CD3-зета, CD28, CD27, 4-1BB и т.п. В некоторых случаях CAR может содержать любую комбинацию CD3-зета, CD28, 4-1BB и т.п.
Последовательности нуклеиновых кислот, кодирующие желаемые молекулы, можно получать с использованием рекомбинантных способов, известных в данной области, например, таких как скрининг библиотек клеток, экспрессирующих молекулу нуклеиновой кислоты, получение молекулы нуклеиновой кислоты из вектора, о котором известно, что он включает его, или выделение непосредственно из клеток и тканей, содержащих его, с использованием стандартных способов. Альтернативно представляющую интерес нуклеиновую кислоту можно получать синтетически, а не клонировать.
Настоящее изобретение включает конструкции ретровирусных и лентивирусных векторов, экспресирующие CAR, которые можно прямо трансдуцировать в клетку.
Настоящее изобретение также включает конструкцию РНК, которая может быть прямо трансфицирована в клетку. Способ получения мРНК для применения в трансфекции вовлекает транскрипцию in vitro (IVT) матрицы с использованием специально сконструированных праймеров, с последующим присоединением поли-A, с получением конструкции, содержащей 3'- и 5'-нетранслируемую последовательность ("UTR") (например, 3'- и/или 5'-UTR, описанную в настоящем описании), 5'-кэп (например, 5'-кэп, описанный в настоящем описании) и/или участок внутренней посадки рибосомы (IRES) (например, IRES, описанный в настоящем описании), нуклеиновую кислоту, подлежащую экспрессии, и поли-A хвостовую часть, как правило, длиной 50-2000 оснований (SEQ ID NO: 32). РНК, полученную таким образом, можно эффективно трансфицировать в различные типы клеток. В одном варианте осуществления матрица включает последовательности CAR. В одном варианте осуществления РНК-вектор CAR трансдуцируют в клетку, например, T-клетку или NK-клетку, способом электропорации.
Антигенсвязывающий домен
В одном аспекте CAR по изобретению содержит специфичный к мишени связывающий элемент, иначе называемый антигенсвязывающим доменом. Выбор части зависит от типа и количества лигандов, которые определяют поверхность клетки-мишени. Например, антигенсвязывающий домен может быть выбран так, чтобы он распознавал лиганд, который выступает в качестве маркера клеточной поверхности на клетках-мишенях, ассоциированных с конкретным болезненным состоянием. Таким образом примеры маркеров клеточной поверхности, которые могут выступать в качестве лигандов для антигенсвязывающего домена в CAR по изобретению, включают маркеры, ассоциированные с вирусными, бактериальными и паразитарными инфекциями, аутоиммунным заболеванием и злокачественными клетками.
В одном аспекте опосредуемый CAR T-клеточный ответ может быть направлен на представляющий интерес антиген посредством встраивания способами инженерии антигенсвязывающего домена, который специфически связывает желаемый антиген, в CAR.
В одном аспекте часть CAR, содержащая антигенсвязывающий домен, содержит антигенсвязывающий домен, который нацелен на опухолевый антиген, например, на опухолевый антиген, описанный в настоящем описании.
Антигенсвязывающий домен может представлять собой любой домен, который связывается с антигеном, включая, но не ограничиваясь ими, моноклональное антитело, поликлональное антитело, рекомбинантное антитело, антитело человека, гуманизированное антитело и его функциональный фрагмент, включая, но не ограничиваясь ими, однодоменное антитело, такое как вариабельный домен тяжелой цепи (VH), вариабельный домен легкой цепи (VL) и вариабельный домен (VHH) наноантитела, происходящего из животного семейства верблюжьих, и альтернативный каркас, о котором в данной области известно, что он функционирует в качестве антигенсвязывающего домена, такой как рекомбинантный домен фибронектина, T-клеточный рецептор (TCR) или их фрагмент, например, одноцепочечный TCR и т.п. В некоторых случаях является полезным, чтобы антигенсвязывающий домен доставлялся тому же виду, в котором в конечном итоге будет использоваться CAR. Например, для применения у человека может быть полезным, чтобы антигенсвязывающий домен CAR содержал человеческие или гуманизированные остатки антигенсвязывающего домена антитела или фрагмента антитела.
В одном варианте осуществления CAR против CD19 представляет собой CAR против CD19, описанный в патенте США № 8399645; патенте США № 7446190; Xu et al., Leuk Lymphoma. 2013 54(2):255-260(2012); Cruz et al., Blood 122(17):2965-2973 (2013); Brentjens et al., Blood, 118(18):4817-4828 (2011); Kochenderfer et al., Blood 116(20):4099-102 (2010); Kochenderfer et al., Blood 122 (25):4129-39(2013); или 16th Annu Meet Am Soc Gen Cell Ther (ASGCT) (May 15-18, Salt Lake City) 2013, Abst 10 (каждая из которых включена в настоящее описание путем ссылки в полном объеме). В одном варианте осуществления антигенсвязывающий домен против CD19 представляет собой антигенсвязывающую часть, например, CDR, из CAR, антитела или их антигенсвязывающего фрагмента, описанных, например, в публикации PCT WO2012/079000 (включенной в настоящее описание путем ссылки в полном объеме). В одном варианте осуществления антигенсвязывающий домен против CD19 представляет собой антигенсвязывающую часть, например, CDR, из CAR, антитела или их антигенсвязывающего фрагмента, описанных, например, в публикации PCT WO2014/153270; Kochenderfer, J.N. et al., J. Immunother. 32 (7), 689-702 (2009); Kochenderfer, J.N., et al., Blood, 116 (20), 4099-4102 (2010); публикации PCT WO2014/031687; Bejcek, Cancer Research, 55, 2346-2351, 1995; или патенте США № 7446190 (каждый из которых включен в настоящее описание путем ссылки в полном объеме).
В одном варианте осуществления антигенсвязывающий домен против мезотелина представляет собой или может происходить из антигенсвязывающего домена, например, CDR, scFv или VH и VL, антитела, антигенсвязывающего фрагмента или CAR, описанного, например, в публикации PCT WO2015/090230 (в одном варианте осуществления CAR представляет собой CAR, описанный в WO2015/090230, содержание которой включено в настоящее описание путем ссылки в полном объеме). В некоторых вариантах осуществления антигенсвязывающий домен против мезотелина представляет собой или происходит из антигенсвязывающей части, например, CDR, scFv или VH и VL, антитела, антигенсвязывающего фрагмента или CAR, описанного, например, в публикациях PCT WO1997/025068, WO1999/028471, WO2005/014652, WO2006/099141, WO2009/045957, WO2009/068204, WO2013/142034, WO2013/040557 или WO2013/063419 (каждая из которых включена в настоящее описание путем ссылки в полном объеме).
В одном варианте осуществления антигенсвязывающий домен против CD123 представляет собой или происходит из антигенсвязывающей части, например, CDR, scFv или VH и VL, антитела, антигенсвязывающего фрагмента или CAR, описанного, например, в публикации PCT WO2014/130635 (включенной в настоящее описание путем ссылки в полном объеме). В одном варианте осуществления антигенсвязывающий домен против CD123 представляет собой или происходит из антигенсвязывающей части, например, CDR, scFv или VH и VL, антитела, антигенсвязывающего фрагмента или CAR, описанного, например, в публикации PCT WO2016/028896 (включенной путем ссылки в полном объеме); в некоторых вариантах осуществления CAR представляет собой CAR, описанный в WO2016/028896. В одном варианте осуществления антигенсвязывающий домен против CD123 представляет собой или происходит из антигенсвязывающей части, например, CDR, scFv или VL и VH, антитела, антигенсвязывающего фрагмента или CAR, описанного, например, в публикации PCT WO1997/024373, WO2008/127735 (например, связывающий CD123 домен из 26292, 32701, 37716 или 32703), WO2014/138805 (например, связывающий CD123 домен из CSL362), WO2014/138819, WO2013/173820, WO2014/144622, WO2001/66139, WO2010/126066 (например, связывающий CD123 домен из любого из Old4, Old5, Old17, Old19, New102 или Old6), WO2014/144622 или US2009/0252742 (каждая из которых включена путем ссылки в полном объеме).
В одном варианте осуществления антигенсвязывающий домен против CD22 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Haso et al., Blood, 121(7): 1165-1174 (2013); Wayne et al., Clin Cancer Res 16(6): 1894-1903 (2010); Kato et al., Leuk Res 37(1):83-88 (2013); Creative BioMart (creativebiomart.net): MOM-18047-S(P).
В одном варианте осуществления антигенсвязывающий домен против CS-1 представляет собой антигенсвязывающую часть, например, CDR, из элотузумаба (BMS), см., например, Tai et al., 2008, Blood 112(4):1329-37; Tai et al., 2007, Blood. 110(5):1656-63.
В одном варианте осуществления антигенсвязывающий домен против CLL-1 представляет собой антигенсвязывающую часть, например, CDR или VH и VL, антитела, антигенсвязывающего фрагмента или CAR, описанного, например, в публикации PCT WO2016/014535, содержание которой включено в настоящее описание путем ссылки в полном объеме. В одном варианте осуществления антигенсвязывающий домен против CLL-1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, доступного от R&D, ebiosciences, Abcam, например, PE-CLL1-hu, каталожный номер № 353604 (BioLegend); и PE-CLL1 (CLEC12A), каталожный номер № 562566 (BD).
В одном варианте осуществления антигенсвязывающий домен против CD33 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Bross et al., Clin Cancer Res 7(6):1490-1496 (2001) (Гемтузумаб озагомицин, hP67.6), Caron et al., Cancer Res 52(24):6761-6767 (1992) (Lintuzumab, HuM195), Lapusan et al., Invest New Drugs 30(3):1121-1131 (2012) (AVE9633), Aigner et al., Leukemia 27(5): 1107-1115 (2013) (AMG330, CD33 BiTE), Dutour et al., Adv hematol 2012:683065 (2012) и Pizzitola et al., Leukemia doi:10,1038/Lue.2014,62 (2014). Иллюстративные молекулы CAR, которые нацелены на CD33, описаны в настоящем описании и представлены в WO2016/014576, например, в таблице 2 WO2016/014576 (включенной в настоящее описание в полном объеме).
В одном варианте осуществления антигенсвязывающий домен против GD2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Mujoo et al., Cancer Res. 47(4):1098-1104 (1987); Cheung et al., Cancer Res 45(6):2642-2649 (1985), Cheung et al., J Clin Oncol 5(9):1430-1440 (1987), Cheung et al., J Clin Oncol 16(9):3053-3060 (1998), Handgretinger et al., Cancer Immunol Immunother 35(3):199-204 (1992). В некоторых вариантах осуществления антигенсвязывающий домен против GD2 представляет собой антигенсвязывающую часть антитела, выбранного из mAb 14.18, 14G2a, ch14.18, hu14.18, 3F8, hu3F8, 3G6, 8B6, 60C3, 10B8, ME36.1 и 8H9, см., например, WO2012033885, WO2013040371, WO2013192294, WO2013061273, WO2013123061, WO2013074916 и WO201385552. В некоторых вариантах осуществления антигенсвязывающий домен против GD2 представляет собой антигенсвязывающую часть антитела, описанного в публикации США № 20100150910 или публикации PCT № WO 2011160119.
В одном варианте осуществления антигенсвязывающий домен против BCMA представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в WO2012163805, WO200112812 и WO2003062401. В некоторых вариантах осуществления дополнительные иллюстративные конструкции CAR против BCMA получают с использованием антигенсвязывающего домена, например, последовательностей CDR, scFv или VH и VL их публикации PCT WO2012/0163805 (содержание которой включено в настоящее описание путем ссылки в полном объеме). В некоторых вариантах осуществления дополнительные иллюстративные конструкции CAR против BCMA получают с использованием антигенсвязывающего домена, например, последовательностей CDRs, scFv или VH и VL из публикации PCT WO2016/014565 (содержание которых включено в настоящее описание путем ссылки в полном объеме). В некоторых вариантах осуществления дополнительные иллюстративные конструкции CAR против BCMA получают с использованием антигенсвязывающего домена, например, последовательностей CDR, scFv или VH и VL из публикации PCT WO2014/122144 (содержание которой включено в настоящее описание путем ссылки в полном объеме). В некоторых вариантах осуществления дополнительные иллюстративные конструкции CAR против BCMA получают с использованием молекул CAR и/или связывающих BCMA доменов (например, последовательностей CDR, scFv или VH и VL) из публикации PCT WO2016/014789 (содержание которой включено в настоящее описание путем ссылки в полном объеме). В некоторых вариантах осуществления дополнительные иллюстративные конструкции CAR против BCMA получают с использованием молекул CAR и/или связывающих BCMA доменов (например, последовательностей CDR, scFv или VH и VL) из публикации PCT WO2014/089335 (содержание которой включено в настоящее описание путем ссылки в полном объеме). В некоторых вариантах осуществления дополнительные иллюстративные конструкции CAR против BCMA получают с использованием молекул CAR и/или связывающих BCMA доменов (например, последовательностей CDR, scFv или VH и VL) из публикации PCT WO2014/140248 (содержание которой включено в настоящее описание путем ссылки в полном объеме).
В одном варианте осуществления антигенсвязывающий домен против антигена Tn представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US 2014/0178365, US8440798, Brooks et al., PNAS 107(22):10056-10061 (2010), и Stone et al., OncoImmunology 1(6):863-873(2012).
В одном варианте осуществления антигенсвязывающий домен против PSMA представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Parker et al., Protein Expr Purif 89(2):136-145 (2013), US 20110268656 (J591 ScFv); Frigerio et al, European J Cancer 49(9):2223-2232 (2013) (scFvD2B); WO 2006125481 (mAb 3/A12, 3/E7 и 3/F11) и одноцепочечных фрагментов антител (scFv A5 и D7).
В одном варианте осуществления антигенсвязывающий домен против ROR1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Hudecek et al., Clin Cancer Res 19(12):3153-3164 (2013); WO 2011159847 и US20130101607.
В одном варианте осуществления антигенсвязывающий домен против FLT3 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в WO2011076922, US5777084, EP0754230, US20090297529, и нескольких коммерческих антител из каталогов (R&D, ebiosciences, Abcam).
В одном варианте осуществления антигенсвязывающий домен против TAG72 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Hombach et al., Gastroenterology 113(4):1163-1170 (1997); и Abcam ab691.
В одном варианте осуществления антигенсвязывающий домен против FAP представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Ostermann et al., Clinical Cancer Research 14:4584-4592 (2008) (FAP5), публикации патента США № 2009/0304718; сибротузумаба (см., например, Hofheinz et al., Oncology Research and Treatment 26(1), 2003); и Tran et al., J Exp Med 210(6):1125-1135 (2013).
В одном варианте осуществления антигенсвязывающий домен против CD38 представляет собой антигенсвязывающую часть, например, CDR, из даратумумаба (см., например, Groen et al., Blood 116(21):1261-1262 (2010); MOR202 (см., например, US8263746); или антител, описанных в US8362211.
В одном варианте осуществления антигенсвязывающий домен против CD44v6 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Casucci et al., Blood 122(20):3461-3472 (2013).
В одном варианте осуществления антигенсвязывающий домен против CEA представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Chmielewski et al., Gastoenterology 143(4):1095-1107 (2012).
В одном варианте осуществления антигенсвязывающий домен против EPCAM представляет собой антигенсвязывающую часть, например, CDR, из антитела, выбранного из MT110, биспецифического Ab против EpCAM-CD3 (см., например, clinicaltrials.gov/ct2/show/NCT00635596); эдреколомаба; 3622W94; ING-1 и адекатумумаба (MT201).
В одном варианте осуществления антигенсвязывающий домен против PRSS21 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного в патенте США №:8080650.
В одном варианте осуществления антигенсвязывающий домен против B7H3 представляет собой антигенсвязывающую часть, например, CDR, из антитела MGA271 (Macrogenics).
В одном варианте осуществления антигенсвязывающий домен против KIT представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US7915391, US20120288506, и нескольких коммерческих антител из каталогов.
В одном варианте осуществления антигенсвязывающий домен против IL-13Ra2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в WO2008/146911, WO2004087758, из нескольких коммерческих антител из каталогов, и из WO2004087758.
В одном варианте осуществления антигенсвязывающий домен против CD30 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US7090843 B1 и EP0805871.
В одном варианте осуществления антигенсвязывающий домен против GD3 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US7253263; US 8207308; US 20120276046; EP1013761; WO2005035577 и US6437098.
В одном варианте осуществления антигенсвязывающий домен против CD171 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Hong et al., J Immunother 37(2):93-104 (2014).
В одном варианте осуществления антигенсвязывающий домен против IL-11Ra представляет собой антигенсвязывающую часть, например, CDR, из антитела, доступного от Abcam (каталожный номер # ab55262) или Novus Biologicals (каталожный номер # EPR5446). В другом варианте осуществления антигенсвязывающий домен против IL-11Ra представляет собой пептид, см., например, Huang et al., Cancer Res 72(1):271-281 (2012).
В одном варианте осуществления антигенсвязывающий домен против PSCA представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Morgenroth et al., Prostate 67(10):1121-1131 (2007) (scFv 7F5); Nejatollahi et al., J of Oncology 2013(2013), article ID 839831 (scFv C5-II); и в публикации патента США № 20090311181.
В одном варианте осуществления антигенсвязывающий домен против VEGFR2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Chinnasamy et al., J Clin Invest 120(11):3953-3968 (2010).
В одном варианте осуществления антигенсвязывающий домен против LewisY представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Kelly et al., Cancer Biother Radiopharm 23(4):411-423 (2008) (hu3S193 Ab (scFvs)); Dolezal et al., Protein Engineering 16(1):47-56 (2003) (NC10 scFv).
В одном варианте осуществления антигенсвязывающий домен против CD24 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Maliar et al., Gastroenterology 143(5):1375-1384 (2012).
В одном варианте осуществления антигенсвязывающий домен против PDGFR-бета представляет собой антигенсвязывающую часть, например, CDR, из антитела Abcam ab32570.
В одном варианте осуществления антигенсвязывающий домен против SSEA-4 представляет собой антигенсвязывающую часть, например, CDR, из антитела MC813 (Cell Signaling), или из других коммерчески доступных антител.
В одном варианте осуществления антигенсвязывающий домен против CD20 представляет собой антигенсвязывающую часть, например, CDR, из антитела ритуксимаба, офатумумаба, окрелизумаба, велтузумаба или GA101.
В одном варианте осуществления антигенсвязывающий домен против рецептора-альфа фолатов представляет собой антигенсвязывающую часть, например, CDR, из антитела IMGN853 или антитела, описанного в US20120009181; US4851332, LK26: US5952484.
В одном варианте осуществления антигенсвязывающий домен против ERBB2 (Her2/neu) представляет собой антигенсвязывающую часть, например, CDR, из антитела трастузумаба или пертузумаба.
В одном варианте осуществления антигенсвязывающий домен против MUC1 представляет собой антигенсвязывающую часть, например, CDR, из антитела SAR566658.
В одном варианте осуществления антигенсвязывающий домен против EGFR представляет собой антигенсвязывающую часть, например, CDR, из антитела цетуксимаба, панитумумаба, залутумумаба, нимотузумаба или матузумаба. В одном варианте осуществления антигенсвязывающий домен против EGFRvIII представляет собой или может происходить из антигенсвязывающего домена, например, CDR, scFv или VH и VL антитела, антигенсвязывающего фрагмента или CAR, описанного, например, в публикации PCT WO2014/130657 (в одном варианте осуществления CAR представляет собой CAR, описанный в WO2014/130657, содержание которой включено в настоящее описание путем ссылки в полном объеме).
В одном варианте осуществления антигенсвязывающий домен против NCAM представляет собой антигенсвязывающую часть, например, CDR, из антитела клона 2-2B: MAB5324 (EMD Millipore)
В одном варианте осуществления антигенсвязывающий домен против эфрина B2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Abengozar et al., Blood 119(19):4565-4576 (2012).
В одном варианте осуществления антигенсвязывающий домен против рецептора IGF-I представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US8344112 B2; EP2322550 A1; WO 2006/138315 или PCT/US2006/022995.
В одном варианте осуществления антигенсвязывающий домен против CAIX представляет собой антигенсвязывающую часть, например, CDR, из антитела клона 303123 (R&D Systems).
В одном варианте осуществления антигенсвязывающий домен против LMP2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US7410640 или US20050129701.
В одном варианте осуществления антигенсвязывающий домен против gp100 представляет собой антигенсвязывающую часть, например, CDR, из антитела HMB45, NKIbetaB, или антитела, описанного в WO2013165940 или US20130295007
В одном варианте осуществления антигенсвязывающий домен против тирозиназы представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US5843674 или US19950504048.
В одном варианте осуществления антигенсвязывающий домен против EphA2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Yu et al., Mol Ther 22(1):102-111 (2014).
В одном варианте осуществления антигенсвязывающий домен против GD3 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US7253263; US 8207308; US 20120276046; EP1013761 A3; 20120276046; WO2005035577 или US6437098.
В одном варианте осуществления антигенсвязывающий домен против фукозил-GM1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US20100297138 или WO2007/067992.
В одном варианте осуществления антигенсвязывающий домен против sLe представляет собой антигенсвязывающую часть, например, CDR, из антитела G193 (for lewis Y), см. Scott AM et al, Cancer Res 60: 3254-61 (2000), как также описано в Neeson et al, J Immunol May 2013 190 (Meeting Abstract Supplement) 177.10.
В одном варианте осуществления антигенсвязывающий домен против GM3 представляет собой антигенсвязывающую часть, например, CDR, из антитела CA 2523449 (mAb 14F7).
В одном варианте осуществления антигенсвязывающий домен против HMWMAA представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Kmiecik et al., Oncoimmunology 3(1):e27185 (2014) (PMID: 24575382) (mAb9.2.27); US6528481; WO2010033866 или US 20140004124.
В одном варианте осуществления антигенсвязывающий домен против o-ацетил-GD2 представляет собой антигенсвязывающую часть, например, CDR, из антитела 8B6.
В одном варианте осуществления антигенсвязывающий домен против TEM1/CD248 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Marty et al., Cancer Lett 235(2):298-308 (2006); Zhao et al., J Immunol Methods 363(2):221-232 (2011).
В одном варианте осуществления антигенсвязывающий домен против CLDN6 представляет собой антигенсвязывающую часть, например, CDR, из антитела IMAB027 (Ganymed Pharmaceuticals), см., например, clinicaltrial.gov/show/NCT02054351.
В одном варианте осуществления антигенсвязывающий домен против TSHR представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US8603466; US8501415 или US8309693.
В одном варианте осуществления антигенсвязывающий домен против GPRC5D представляет собой антигенсвязывающую часть, например, CDR, из антитела FAB6300A (R&D Systems) или LS-A4180 (Lifespan Biosciences).
В одном варианте осуществления антигенсвязывающий домен против CD97 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в US6846911;de Groot et al., J Immunol 183(6):4127-4134 (2009); или антитела из R&D:MAB3734.
В одном варианте осуществления антигенсвязывающий домен против ALK представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Mino-Kenudson et al., Clin Cancer Res 16(5):1561-1571 (2010).
В одном варианте осуществления антигенсвязывающий домен против полисиаловая кислота представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Nagae et al., J Biol Chem 288(47):33784-33796 (2013).
В одном варианте осуществления антигенсвязывающий домен против PLAC1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Ghods et al., Biotechnol Appl Biochem 2013 doi:10.1002/bab.1177.
В одном варианте осуществления антигенсвязывающий домен против GloboH представляет собой антигенсвязывающую часть антитела VK9 или антитела, описанного, например, в Kudryashov V et al, Glycoconj J.15(3):243-9 ( 1998), Lou et al., Proc Natl Acad Sci USA 111(7):2482-2487 (2014); MBr1: Bremer E-G et al. J Biol Chem 259:14773-14777 (1984).
В одном варианте осуществления антигенсвязывающий домен против NY-BR-1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Jager et al., Appl Immunohistochem Mol Morphol 15(1):77-83 (2007).
В одном варианте осуществления антигенсвязывающий домен против WT-1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Dao et al., Sci Transl Med 5(176):176ra33 (2013) или WO2012/135854.
В одном варианте осуществления антигенсвязывающий домен против MAGE-A1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Willemsen et al., J Immunol 174(12):7853-7858 (2005) (TCR-подобный scFv).
В одном варианте осуществления антигенсвязывающий домен против sperm белок 17 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Song et al., Target Oncol 2013 Aug 14 (PMID: 23943313); Song et al., Med Oncol 29(4):2923-2931 (2012).
В одном варианте осуществления антигенсвязывающий домен против Tie 2 представляет собой антигенсвязывающую часть, например, CDR, из антитела AB33 (Cell Signaling Technology).
В одном варианте осуществления антигенсвязывающий домен против MAD-CT-2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в PMID: 2450952; US7635753.
В одном варианте осуществления антигенсвязывающий домен против Fos-родственного антигена 1 представляет собой антигенсвязывающую часть, например, CDR, из антитела 12F9 (Novus Biologicals).
В одном варианте осуществления антигенсвязывающий домен против MelanA/MART1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного в EP2514766 A2 или US 7749719.
В одном варианте осуществления антигенсвязывающий домен против точек разрыва при транслокации саркомы представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Luo et al, EMBO Mol. Med. 4(6):453-461 (2012).
В одном варианте осуществления антигенсвязывающий домен против TRP-2 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Wang et al, J Exp Med. 184(6):2207-16 (1996).
В одном варианте осуществления антигенсвязывающий домен против CYP1B1 представляет собой антигенсвязывающую часть, например, CDR, из антитела, описанного, например, в Maecker et al, Blood 102 (9): 3287-3294 (2003).
В одном варианте осуществления антигенсвязывающий домен против RAGE-1 представляет собой антигенсвязывающую часть, например, CDR, из антитела MAB5328 (EMD Millipore).
В одном варианте осуществления антигенсвязывающий домен против обратной транскриптазы теломеразы человека представляет собой антигенсвязывающую часть, например, CDR, из антитела, имеющего каталожный номер LS-B95-100 (Lifespan Biosciences)
В одном варианте осуществления антигенсвязывающий домен против кишечной карбоксилэстеразы представляет собой антигенсвязывающую часть, например, CDR, из антитело 4F12, каталожный номер: LS-B6190-50 (Lifespan Biosciences).
В одном варианте осуществления антигенсвязывающий домен против mut hsp70-2 представляет собой антигенсвязывающую часть, например, CDR, из антитела Lifespan Biosciences, моноклональное, каталожный номер: LS-C133261-100 (Lifespan Biosciences).
В одном варианте осуществления антигенсвязывающий домен против CD79a представляет собой антигенсвязывающую часть, например, CDR, из антитела против CD79a [HM47/A9] (ab3121), доступного от Abcam; антитела CD79A Antibody #3351, доступного от Cell Signalling Technology; или антитела HPA017748 - антитела против CD79A, продуцируемого в кролике, доступного от Sigma Aldrich.
В одном варианте осуществления антигенсвязывающий домен против CD79b представляет собой антигенсвязывающую часть, например, CDR, из антитела полатузумаба ведотина, антитела против CD79b, описанного в Dornan et al., "Therapeutic potential of an anti-CD79b antibody-drug conjugate, anti-CD79b-vc-MMAE, for the treatment of non-Hodgkin lymphoma" Blood. 2009 Sep 24;114(13):2721-9. doi: 10,1182/blood-2009-02-205500. Epub 2009 Jul 24, или биспецифического антитела против CD79b/CD3, описанного в "4507 Pre-Clinical Characterization of T Cell-Dependent Bispecific Antibody Anti-CD79b/CD3 As a Potential Therapy for B Cell Malignancies" Abstracts of 56th ASH Annual Meeting и Exposition, San Francisco, CA December 6-9 2014.
В одном варианте осуществления антигенсвязывающий домен против CD72 представляет собой антигенсвязывающую часть, например, CDR, из антитела J3-109, описанного в Myers and Uckun, "An anti-CD72 immunotoxin against therapy-refractory B-lineage acute lymphoblastic leukemia". Leuk Lymphoma. 1995 Jun;18(1-2):119-22, или антитела против CD72 (10D6.8.1, mIgG1), описанного в Polson et al., "Antibody-Drug Conjugates for the Treatment of Non-Hodgkin's lymphoma: Target and Linker-Drug Selection" Cancer Res March 15, 2009 69; 2358.
В одном варианте осуществления антигенсвязывающий домен против LAIR1 представляет собой антигенсвязывающую часть, например, CDR, из антитела ANT-301 LAIR1, доступного от ProSpec; или антитела против CD305 человека (LAIR1), доступного от BioLegend.
В одном варианте осуществления антигенсвязывающий домен против FCAR представляет собой антигенсвязывающую часть, например, CDR, из антитела CD89/FCARAntibody (каталожный номер #10414-H08H), доступного Sino Biological Inc.
В одном варианте осуществления антигенсвязывающий домен против LILRA2 представляет собой антигенсвязывающую часть, например, CDR, из моноклонального антитела против LILRA2 (M17), клон 3C7, доступного от Abnova, или антитела мыши против LILRA2, моноклонального (2D7), доступного от Lifespan Biosciences.
В одном варианте осуществления антигенсвязывающий домен против CD300LF представляет собой антигенсвязывающую часть, например, CDR, из антитела мыши против CMRF35-подобной молекулы 1, моноклонального [UP-D2], доступного от BioLegend, или антитела крысы против CMRF35-молекулы 1, моноклонального [234903], доступного от R&D Systems.
В одном варианте осуществления антигенсвязывающий домен против CLEC12A представляет собой антигенсвязывающую часть, например, CDR, из биспецифического scFv-антитела антитело против рекрутера T-клеток (BiTE) и ADC, описанного в Noordhuis et al., "Targeting of CLEC12A In Acute Myeloid Leukemia by Antibody-Drug-Conjugates and Bispecific CLL-1xCD3 BiTE Antibody" 53rd ASH Annual Meeting and Exposition, December 10-13, 2011, и MCLA-117 (Merus).
В одном варианте осуществления антигенсвязывающий домен против BST2 (также называемого CD317) представляет собой антигенсвязывающую часть, например, CDR, из антитела мыши против CD317, моноклонального [3H4], доступного от Antibody-Online, или антитела мыши против CD317, моноклонального [696739], доступного от R&D Systems.
В одном варианте осуществления антигенсвязывающий домен против EMR2 (также называемого CD312) представляет собой антигенсвязывающую часть, например, CDR, из антитела мыши против CD312, моноклонального [LS-B8033], доступного от Lifespan Biosciences, или антитела мыши против CD312, моноклонального [494025], доступного от R&D Systems.
В одном варианте осуществления антигенсвязывающий домен против LY75 представляет собой антигенсвязывающую часть, например, CDR, из антитела мыши против лимфоцитарного антигена 75, моноклонального [HD30], доступного от EMD Millipore, или антитела мыши против лимфоцитарного антигена 75, моноклонального [A15797], доступного от Life Technologies.
В одном варианте осуществления антигенсвязывающий домен против GPC3 представляет собой антигенсвязывающую часть, например, CDR, из антитела hGC33, описанного в Nakano K, Ishiguro T, Konishi H, et al. Generation of a humanized anti-glypican 3 antibody by CDR grafting and stability optimization. Anticancer Drugs. 2010 Nov; 21(10):907-916, или MDX-1414, HN3 или YP7, все три из которых описаны в Feng et al., "Glypican-3 antibodies: a new therapeutic target for liver cancer". FEBS Lett. 2014 Jan 21;588(2):377-82.
В одном варианте осуществления антигенсвязывающий домен против FCRL5 представляет собой антигенсвязывающую часть, например, CDR, из антитела против FcRL5, описанного в Elkins et al., "FcRL5 as a target of antibody-drug conjugates for the treatment of multiple myeloma" Mol Cancer Ther. 2012 Oct;11(10):2222-32.
В одном варианте осуществления антигенсвязывающий домен против IGLL1 представляет собой антигенсвязывающую часть, например, CDR, из антитела мыши против иммуноглобулин лямбда-подобного полипептида 1, моноклонального [AT1G4], доступного от Lifespan Biosciences, антитела против иммуноглобулин лямбда подобного полипептида 1, моноклонального [HSL11], доступного от BioLegend.
В одном варианте осуществления антигенсвязывающий домен содержит одну, две, три (например, все три) CDR тяжелой цепи, CDR1 HC, CDR2 HC и CDR3 HC, из антитела, приведенного выше, и/или одну, две, три (например, все три) CDR легкой цепи, CDR1 LC, CDR2 LC и CDR3 LC, из антитела, приведенного выше. В одном варианте осуществления антигенсвязывающий домен содержит вариабельную область тяжелой цепи и/или вариабельную область легкой цепи антитела, приведенного выше.
В другом аспекте антигенсвязывающий домен содержит гуманизированное антитело или фрагмент антитела. В некоторых аспектах, не являющееся человеческим антитело является гуманизирвоанным, где определенные последовательности или области антитела являются модифицированными для повышения сходства с антителом, естественным образом продуцируемым у человека, или его фрагментом. В одном аспекте антигенсвязывающий домен является гуманизированным.
Гуманизированное антитело можно получать с использованием различных способов, известных в данной области, включая, но не ограничиваясь ими, пересадку CDR (см., например, патент Европы № EP 239400; международную публикацию № WO 91/09967 и патенты США № 5225539, 5530101 и 5585089, каждый из которых включен в настоящее описание путем ссылки в полном объеме), венирование или изменение поверхности см., например, патенты Европы № EP 592106 и EP 519596; Padlan, 1991, Molecular Immunology, 28(4/5):489-498; Studnicka et al., 1994, Protein Engineering, 7(6):805-814; и Roguska et al., 1994, PNAS, 91:969-973, каждый из которых включен в настоящее описание путем ссылки в полном объеме), шаффлинг цепей (см., например, патент США № 5565332, который включен в настоящее описание путем ссылки в полном объеме) и способы, описанные, например, в публикации патентной заявки США № US2005/0042664, публикации патентной заявки США № US2005/0048617, патенте США № 6407213, патенте США № 5766886, международной публикации № WO 9317105, Tan et al., J. Immunol., 169:1119-25 (2002), Caldas et al., Protein Eng., 13(5):353-60 (2000), Morea et al., Methods, 20(3):267-79 (2000), Baca et al., J. Biol. Chem., 272(16):10678-84 (1997), Roguska et al., Protein Eng., 9(10):895-904 (1996), Couto et al., Cancer Res., 55 (23 Supp):5973s-5977s (1995), Couto et al., Cancer Res., 55(8):1717-22 (1995), Sandhu J S, Gene, 150(2):409-10 (1994), и Pedersen et al., J. Mol. Biol., 235(3):959-73 (1994), каждый из которых включен в настоящее описание путем ссылки в полном объеме. Часто, каркасные остатки в каркасных областях заменяют соответствующим остатком из антитела, являющегося донором CDR, для изменения, например, повышения, связывания антигена. Эти замены каркасной области идентифицируют способами, хорошо известными в данной области, например, посредством моделирования взаимодействий CDR и каркасных остатков для идентификации каркасных остатков, важных для связывания антигена, сравнения последовательностей для идентификации необычных каркасных остатков в конкретных положениях (См., например, Queen et al., патент США № 5585089; и Riechmann et al., 1988, Nature, 332:323, которые включены в настоящее описание путем ссылок в полном объеме).
Гуманизированное антитело или фрагмент антитела имеют один или несколько аминокислотных остатков, оставшихся в них из источника, не являющегося человеческим. Эти не являющиеся человеческими аминокислотные остатки часто называют "импортными" остатками, которые, как правило, взяты из "импортного" вариабельного домена. Как описано в настоящем описании, гуманизированные антитела или фрагменты антитела содержат одну или несколько CDR из не являющихся человеческими молекул иммуноглобулинов и каркасных областей, где аминокислотные остатки, содержащие каркасную область, происходят полностью или по большей части из последовательности человека эмбирионального типа. Множество способов гуманизации антител или фрагментов антител хорошо известны в данной области, и ее можно проводить по существу согласно способу Winter и коллег (Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science, 239:1534-1536 (1988)), путем замены CDR или последовательностями CDR грызунов соответствующих последовательностей антитела человека, т.е. пересадки CDR (EP 239400; публикация PCT № WO 91/09967; и патенты США № 4816567; 6331415; 5225539; 5530101; 5585089; 6548640, содержание которых включено в настоящее описание путем ссылок в полном объеме). В таких гуманизированных антителах и фрагментах по существу менее чем интактный вариабельный домен человека заменен соответствующей последовательностью из не являющегося человеком вида. Гуманизированные антитела часто представляют собой антитела человека, в которых некоторые остатки CDR и, возможно, некоторые каркасные (FR) остатки заменены остатками из аналогичных участков антител грызунов. Гуманизацию антител и фрагментов антител также можно осуществлять посредством венирования или изменения поверхности (EP 592,106; EP 519,596; Padlan, 1991, Molecular Immunology, 28(4/5):489-498; Studnicka et al., Protein Engineering, 7(6):805-814 (1994); и Roguska et al., PNAS, 91:969-973 (1994)) или шаффлинга цепей (патент США № 5565332), содержание которых включено в настоящее описание путем ссылки в полном объеме.
Выбор вариабельных доменов человека, как легких, так и тяжелых, для применения для получения гуманизированных антител, проводят так, чтобы снизить антигенность. Согласно так называемому способу "наилучшего соответствия", последовательность вариабельного домена антитела грызуна подвергают скринингу против целой библиотеки известных последовательностей вариабельного домена человека. Последовательность человека, которая является наиболее сходной с последовательностью грызуна, затем принимают в качестве каркасной области человека (FR) для гуманизированного антитела (Sims et al., J. Immunol., 151:2296 (1993); Chothia et al., J. Mol. Biol., 196:901 (1987), содержание которой включено в настоящее описание путем ссылки в полном объеме). В другом способе используют конкретную каркасную область, происходящую из консенсусной последовательности всех антител человека конкретной подгруппы легких или тяжелых цепей. Для нескольких различных гуманизированных антител можно использовано одну и ту же каркасную область (см., например, Nicholson et al. Mol. Immun. 34 (16-17): 1157-1165 (1997); Carter et al., Proc. Natl. Acad. Sci. USA, 89:4285 (1992); Presta et al., J. Immunol., 151:2623 (1993), содержание которых включены в настоящее описание путем ссылок в полном объеме). В некоторых вариантах осуществления каркасная область, например, все четыре каркасных области, вариабельной области тяжелой цепи происходит из последовательности эмбрионального типа VH4_4-59. В одном варианте осуществления каркасная область может содержать одну, две, три, четыре или пять модификаций, например, замен, например, на аминокислоту соответствующей последовательности мыши. В одном варианте осуществления каркасная область, например, все четыре каркасных области вариабельной области легкой цепи, происходит из последовательности эмбрионального типа VK3_1.25. В одном варианте осуществления каркасная область может содержать одну, две, три, четыре или пять модификаций, например, замен, на аминокислоту соответствующей последовательности мыши.
В некоторых аспектах часть композиции CAR по изобретению, которая содержит фрагмент антитела, является гуманизированной с сохранением высокой аффинности в отношении антигена-мишени и других благоприятных биологических свойств. Согласно одному аспекту изобретения гуманизированные антитела и фрагменты антител получают посредством процесса анализа родительских последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются общедоступными и известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и проявляют возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулинов, являющихся кандидатами. Исследование этих изображений позволяют анализ вероятной роли остатков в функционировании последовательности иммуноглобулина, являющейся кандидатом, например, анализ остатков, которые влияют на способность иммуноглобулина, являющегося кандидатом, связывать антиген-мишень. Таким образом, остатки FR можно выбирать и комбинировать из реципиентных и импортных последовательностей, так чтобы достигать желаемых характеристик антитело или фрагмент антитела, таких как увеличенная аффинность в отношении антигена-мишени. Как правило, остатки CDR прямо и наиболее существенно вовлечены во влияние на связывание антигена.
Гуманизированное антитело или фрагмент антитела могут сохранять антигенную специфичность, сходную с исходным антителом, например, в рамках настоящего изобретения, способность связывать ассоциированный со злокачественной опухолью антиген человека, как описано в настоящем описании. В некоторых вариантах осуществления гуманизированное антитело или фрагмент антитела могут иметь увеличенную аффинность и/или специфичность связывания с ассоциированным со злокачественной опухолью антигеном человека, как описано в настоящем описании.
В одном аспекте антигенсвязывающий домен по изобретению характеризуется конкретными функциональными признаками или свойствами антитела или фрагмента антитела. Например, в одном аспекте часть композиции CAR по изобретению, которая содержит антигенсвязывающий домен, специфически связывает опухолеый антиген, как описано в настоящем описании.
В одном аспекте домен, связывающий ассоциированый со злокачетвенной опухолью антиген, как описано в настоящем описании, представляет собой фрагмент, например, одноцепочечный вариабельный фрагмент (scFv). В одном аспекте домен, связывающий ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, представляет собой Fv, Fab, (Fab')2 или бифункциональное (например, биспецифическое) гибридное антитело (например, Lanzavecchia et al., Eur. J. Immunol. 17, 105 (1987)). В одном аспекте антитела и их фрагменты по изобретению связывают ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, с аффинностью дикого типа или усиленной аффинностью.
В некоторых случаях scFv можно получать в соответствии со способом, известным в данной области (см., например, Bird et al., (1988) Science 242:423-426 и Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Молекулы ScFv можно получать путем связывания областей VH и VL вместе с использованием гибких полипептидных линкеров. Молекулы scFv содержат линкер (например, линкер Ser-Gly) с оптимизированной длиной и/или аминокислотным составом. Длина линкера может в значительной степени влиять на то, как вариабельные области scFv будут сворачиваться и взаимодействовать. В действительности, если используют короткий полипептидный линкер (например, 5-10 аминокислот), это препятствует внутрицепочечному фолдингу. Внутрицепочечный фолдинг также требуется для сближения двух вариабельных областей для формирования функционального участка связывания эпитопа. Для примера ориентации и размера линкера см., например, Hollinger et al. 1993 Proc Natl Acad. Sci. U.S.A. 90:6444-6448, публикации патентных заявок США № 2005/0100543, 2005/0175606, 2007/0014794, и публикации PCT № WO2006/020258 и WO2007/024715, которые включены в настоящее описание путем ссылок.
scFv может содержать линкер по меньшей мере из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50 или более аминокислотных остатков между его областями VL и VH. Линкерная последовательность может содержать любую встречающуюся в природе аминокислоту. В некоторых вариантах осуществления линкерная последовательность содержит аминокислоты глицин и серин. В другом варианте осуществления линкерная последовательность содержит наборы глициновых и сериновых повторов, такие как (Gly4Ser)n, где n представляет собой положительное целое число, равное или превышающее 1 (SEQ ID NO: 22). В одном варианте осуществления линкер может представлять собой (Gly4Ser)4 (SEQ ID NO: 29) или (Gly4Ser)3 (SEQ ID NO: 30). Варьирование длины линкера может сохранять или повышать активность, обеспечивая улучшенную эффективность в исследованиях активности.
В другом аспекте антигенсвязывающий домен представляет собой T-клеточный рецептор ("TCR") или его фрагмент, например, одноцепочечный TCR (scTCR). Способы получения таких TCR известны в данной области. См., например, Willemsen RA et al, Gene Therapy 7: 1369-1377 (2000); Zhang T et al, Cancer Gene Ther 11: 487-496 (2004); Aggen et al, Gene Ther. 19(4):365-74 (2012) (ссылки включены в настоящее описание в полном объеме). Например, можно конструировать scTCR, который содержит гены Vα и Vβ из клона T-клеток, связанные линкером (например, гибкий пептид). Этот подход является очень полезным для ассоциированной со злокачественной опухолью мишени, которая сама по себе является внутриклеточной, но фрагмент такого антигена (пептид) презентируется на поверхности клеток посредством MHC.
Биспецифические CAR
В одном варианте осуществления мультиспецифическая молекула антитела представляет собой биспецифическую молекулу антитела. Биспецифическое антитело обладает специфичностью в отношении не более чем двух антигенов. Биспецифическая молекула антитела характеризуется первой последовательностью вариабельного домена иммуноглобулина, которая обладает аффинностью связывания в отношении первого эпитопа, и второй последовательностью вариабельного домена иммуноглобулина, которая обладает специфичностью связывания в отношении второго эпитопа. В одном варианте осуществления первый и второй эпитопы находятся на одном и том же антигене, например, на одном и том же белке (или субъединице мультимерного белка). В одном варианте осуществления первый и второй эпитопы перекрываются. В одном варианте осуществления первый и второй эпитопы не перекрываются. В одном варианте осуществления первый и второй эпитопы находятся на различных антигенах, например, различных белках (или различных субъединицах мультимерного белка). В одном варианте осуществления биспецифическая молекула антитела содержит последовательность вариабельного домена тяжелой цепи и последовательность вариабельного домена легкой цепи, которые обладают специфичностью связывания в отношении первого эпитопа, и последовательность вариабельного домена тяжелой цепи и последовательность вариабельного домена легкой цепи, которые обладают специфичностью связывания в отношении второго эпитопа. В одном варианте осуществления биспецифическая молекула антитела содержит половинное антитело, обладающее специфичностью связывания в отношении первого эпитопа, и половинное антитело, обладающее специфичностью связывания в отношении второго эпитопа. В одном варианте осуществления биспецифическая молекула антитела содержит половинное антитело или его фрагмент, обладающие специфичностью связывания в отношении первого эпитопа, и половинное антитело или его фрагмент, обладающие специфичностью связывания в отношении второго эпитопа. В одном варианте осуществления биспецифическая молекула антитела содержит scFv или его фрагмент, которые обладают специфичностью связывания в отношении первого эпитопа, и scFv или фрагмент, которые обладают специфичностью связывания в отношении второго эпитопа.
В определенных вариантах осуществления молекула антитела является полиспецифической (например, биспецифической или триспецифической) молекулой антитела. Протоколы получения биспецифических или гетеродимерных молекул антител известны в данной области; включая, но не ограничиваясь ими, например, подход "выступа в полости", описанный, например, в US 5731168; образование Fc-пар посредством электростатического наведения, как описано, нарпример, в WO 09/089004, WO 06/106905 и WO 2010/129304; образование гетеодимеров посредством сконструированных путем обмена цепей доменов (SEED), как описано. например, в WO 07/110205; обмен Fab-плечами, как описано, например, в WO 08/119353, WO 2011/131746 и WO 2013/060867; двойной конъюгат антитела, например, путем сшивания антител с образованием биспецифической структуры с использованием гетеробифункционального реагента, имеющего амин-реактивную группу и реакционноспособную сульфгидрильную группу, как описано, например, в US 4433059; детерминанты биспецифических антител, получаемые путем рекомбинации половинных антител (пары тяжелая-легкая цепи или Fab) из различных антител посредством цикла восстановления и окисления дисульфидных связей между двумя тяжелыми цепями, как описано, например, в US 4444878; трифункциональные антитела, например, три Fab'-фрагментыа, сшитых через сульфгидрильные реакционноспособные группы, как описано, например, в US5273743; биосинтетические связывающие белки, например, пару scFv, сшитых через C-концы, предпочтительно посредством дисульфидного или амин-реактивного химического сшивания, как описано, например, в US5534254; бифункциональные антитела, например, Fab-фрагменты, с различающейся специфичностью связывания, димеризованные через лейциновые молнии (например, c-fos и c-jun), которые заменили константный домен, как описано, например, в US5582996; биспецифические и олигоспецифические моно-и олиговалентные рецепторы, например, области VH-CH1 двух антител (два Fab-фрагмента), связанных через полипептидный спейсер между CH1-областью одного антитела и VH-областью другого антитела, как правило, с ассоциированными с ними легкими цепями, как описано, например, в US5591828; биспецифические конъюгаты ДНК, например, сшивание антител или Fab-фрагменты через двухцепочечный фрагмент ДНК, как описано, например, в US5635602; биспецифические слитые белки, например, экспрессирующую конструкцию, содержащую два scFv с гидрофильным спиралевидным пептидным линкером между ними и полную константную область, как описано, например, в US5637481; поливалентные и полиспецифические связывающие белки, например, димер полипептидов, имеющих первый домен со связывающей областью вариабельной области тяжелой цепи Ig и второй домен со связывающей областью вариабельной области легкой цепи Ig, обычно называемые диантителами (также предусматриваются структуры более высокого порядка, образующие биспецифические, триспецифические или тетраспецифические молекулы, как описано, например, в US5837242; конструкции миниантител со связанными VL- и VH-цепями, дополнительно соединенными пептидными спейсерами с шарнирной областью антитела и областью CH3, которые могут димеризоваться с образованием биспецифических/поливалентных молекул, как описано, например, в US5837821; VH- и VL-домены, связанны коротким пептидным линкером (например, 5 или 10 аминокислот) или совсем без линкера в любой ориентации, которые могут образовывать димеры с образованием биспецифических диантител; тримеров и тетрамеров, как описано, например, в US5844094; цепь VH-доменов (или VL-доменов в представителях семейства), соединенных пептидными связями с поддающимися сшиванию группами на C-конце, кроме того, связанные с VL-доменами с образованием серии FV (или scFv), как описано, например, в US5864019; и одноцепочечные связывающие полипептиды как с VH-, так и с VL-доменом, связанными через пептидный линкер, объединенные в поливалентные структуры посредством нековалентного или химического сшивания с образованием, например, гомобивалентных, гетеробивалентных, трехвалентных и четырехвалентных структур с использованием как формата scFV, так и формата диантител, как описано, например, в US5869620. Дополнительные иллюстративные полиспецифические и биспецифические молекулы и способы их получения могут быть найдены, например, в US5910573, US5932448, US5959083, US5989830, US6005079, US6239259, US6294353, US6333396, US6476198, US6511663, US6670453, US6743896, US6809185, US6833441, US7129330, US7183076, US7521056, US7527787, US7534866, US7612181, US2002004587A1, US2002076406A1, US2002103345A1, US2003207346A1, US2003211078A1, US2004219643A1, US2004220388A1, US2004242847A1, US2005003403A1, US2005004352A1, US2005069552A1, US2005079170A1, US2005100543A1, US2005136049A1, US2005136051A1, US2005163782A1, US2005266425A1, US2006083747A1, US2006120960A1, US2006204493A1, US2006263367A1, US2007004909A1, US2007087381A1, US2007128150A1, US2007141049A1, US2007154901A1, US2007274985A1, US2008050370A1, US2008069820A1, US2008152645A1, US2008171855A1, US2008241884A1, US2008254512A1, US2008260738A1, US2009130106A1, US2009148905A1, US2009155275A1, US2009162359A1, US2009162360A1, US2009175851A1, US2009175867A1, US2009232811A1, US2009234105A1, US2009263392A1, US2009274649A1, EP346087A2, WO0006605A2, WO02072635A2, WO04081051A1, WO06020258A2, WO2007044887A2, WO2007095338A2, WO2007137760A2, WO2008119353A1, WO2009021754A2, WO2009068630A1, WO9103493A1, WO9323537A1, WO9409131A1, WO9412625A2, WO9509917A1, WO9637621A2, WO9964460A1. Содержание вышеупомянутых заявок включено в настоящее описание путем ссылки в полном объеме.
В каждом антителе или фрагменте антитела (например, scFv) биспецифической молекулы антитела VH может находиться выше или ниже VL. В некоторых вариантах осуществления вышележащее антитело или фрагмент антитела (например, scFv) организованы так, что их VH (VH1) находится выше их VL (VL1), и нижележащее антитело или фрагмент антитела (например, scFv) организованы так, что их VL (VL2) находится выше их VH (VH2), так что целая биспецифическая молекула антитела имеет порядок VH1-VL1-VL2-VH2. В других вариантах осуществления вышележащее антитело или фрагмент антитела (например, scFv) организованы так, что их VL (VL1) находится выше их VH (VH1) и нижележащее антитело или фрагмент антитела (например, scFv) организованы так, что их VH (VH2) находится выше их VL (VL2), так что целая биспецифическая молекула антитела имеет порядок VL1-VH1-VH2-VL2. Необязательно линкер расположен между двумя антителами или фрагментами антител (например, scFv), например, между VL1 и VL2, если конструкция имеет порядок VH1-VL1-VL2-VH2, или между VH1 и VH2, если конструкция имеет порядок VL1-VH1-VH2-VL2. Линкер может представлять собой линкер, как описано в настоящем описании, например, линкер (Gly4-Ser)n, где n равен 1, 2, 3, 4, 5 или 6, предпочтительно 4 (SEQ ID NO: 72). Как правило, линкер между двумя scFv должен быть достаточно длинным, чтобы избегать неправильного образования пар между доменами двух scFv. Необязательно, линкер расположен между VL и VH первого scFv. Необязательно, линкер расположен между VL и VH второго scFv. В конструкциях, которые имеют множество линкеров, любые два или более из линкеров могут быть одинаковыми или могут различаться. Таким образом, в некоторых вариантах осуществления биспецифический CAR содержит VL, VH и необязательно один или несколько линкеров в таком порядке, который описан в настоящем описании.
Стабильность и мутации
Стабильность антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, например, молекулы scFv (например, растворимые scFv), можно оценивать по сравнению с биофизическими свойствами (например, термической стабильностью) общепринятой контрольной молекулы scFv или полноразмерного антитела. В одном варианте осуществления гуманизированный scFv имеет термическую стабильность, которая более чем приблизительно на 0,1, приблизительно на 0,25, приблизительно на 0,5, приблизительно на 0,75, приблизительно на 1, приблизительно на 1,25, приблизительно на 1,5, приблизительно на 1,75, приблизительно на 2, приблизительно на 2,5, приблизительно на 3, приблизительно на 3,5, приблизительно на 4, приблизительно на 4,5, приблизительно на 5, приблизительно на 5,5, приблизительно на 6, приблизительно на 6,5, приблизительно на 7, приблизительно на 7,5, приблизительно на 8, приблизительно на 8,5, приблизительно на 9, приблизительно на 9,5, приблизительно на 10 градусов, приблизительно на 11 градусов, приблизительно на 12 градусов, приблизительно на 13 градусов, приблизительно на 14 градусов или приблизительно на 15 градусов Цельсия улучшена относительно контрольной связывающей молекулы (например, общепринятой молекулы scFv) в описанных анализах.
Таким образом, улучшенная термическая стабильность антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, сообщается всей конструкции CAR, что приводит к улучшенным терапевтическим свойствам конструкции CAR. Термическая стабильность антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, может быть улучшена по меньшей мере приблизительно на 2°C или 3°C по сравнению с общепринятым антителом. В одном варианте осуществления антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, имеет на 1°C улучшенную термическую стабильность по сравнению с общепринятым антителом. В другом варианте осуществления антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, имеет на 2°C улучшенную термическую стабильность по сравнению с общепринятым антителом. В другом варианте осуществления scFv имеет на 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15°C улучшенную термическую стабильность по сравнению с общепринятым антителом. Сравнения можно проводить, например, между молекулами scFv, описанными в настоящем описании, и молекулами scFv или Fab-фрагментами антитела, из которого происходят VH и VL scFv. Термическую стабильность можно количественно определять с использованием способов, известных в данной области. Например, в одном варианте осуществления можно количественно определять Tm. Способы измерения Tm и другие способы определения стабильности белка более подробно описаны ниже.
Мутации в scFv (вследствие гуманизации или прямого мутагенеза растворимого scFv) могут измерять стабильность scFv и улучшать общую стабильность scFv и конструкции CAR. Стабильность гуманизированного scFv сравнивают с scFv мыши с использованием таких показателей, как Tm, температура денатурации и температура агрегации.
Связывающую способность мутантных scFv можно определять с использованием способов анализа, известных в данной области и описанных в настоящем описании.
В одном варианте осуществления антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, содержит по меньшей мере одну мутацию вследствие процесса гуманизации, так чтобы мутантный scFv сообщал улучшенную стабильность конструкции CAR. В другом варианте осуществления антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, например, scFv, содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 мутаций вследствие процесса гуманизации, так чтобы мутантный scFv сообщал улучшенную стабильность конструкции CAR.
Способы оценки стабильности белка
Стабильность антигенсвязывающего домена можно оценивать с использованием, например, способов, описанных ниже. Такие способы позволяют определение множественных термических переходов разворачивания, где наименее стабильный домен либо разворачивается первым, либо ограничивает общий порог стабильности мультидоменного элемента, который разворачивается кооперативно (например, мультидоменный белок, который демонстрирует единичный переход разворачивания). Наименее стабильный домен можно идентифицировать рядом дополнительных способов. Для изучения того, какой домен ограничивает общую стабильность, можно проводить мутагенез. Кроме того, изучение резистентности мультидоменного белка к протеазам можно проводить в условиях, в которых, как известно, наименее стабильный домен от природы разворачивается, посредством DSC или других спектроскопических способов (Fontana, et al., (1997) Fold. Des., 2: R17-26; Dimasi et al. (2009) J. Mol. Biol. 393: 672-692). После идентификации наименее стабильного домена последовательность, кодирующую этот домен (или ее часть) можно использовать в качестве исследуемой последовательности в способах.
a) Термическая стабильность
Термическую стабильность композиций можно анализировать с использованием ряда неограничивающих биофизических или биохимических способов, известных в данной области. В определенных вариантах осуществления термическую стабильность оценивают посредством аналитической спектроскопии.
Иллюстративным аналитическим способом спектроскопии является дифференциальная сканирующая калориметрия (DSC). В DSC используется калориметр, который является чувствительным к поглощению тепла, сопровождающему разворачивания большинства белков или доменов белков (см., например, Sanchez-Ruiz, et al., Biochemistry, 27: 1648-52, 1988). Для определения термической стабильности белка образец белка помещают в калориметр и температуру повышают до тех пор, пока Fab или scFv не развернется. Температура, при которой белок разворачивается, указывает на общую стабильность белка.
Другим иллюстративным способом аналитической спектроскопии является спектроскопия кругового дихроизма (CD). Спектроскопия CD измеряет оптическую активность композиции в зависимости от повышения температуры. Спектроскопия кругового дихроизма (CD) измеряет различия в поглощении левополяризованного слева провополяризованного света, которая является результатом структурной асимметрии. Нарушенная или развернутая структура приводит к спектру CD, значительно отличающемуся от спектра упорядоченной или свернутой структуры. Спектр CD отражает чувствительность белков к денатурирующим эффектам повышения температуры и, таким образом, указывает на термическую стабильность белка (см. van Mierlo and Steemsma, J. Biotechnol., 79(3):281-98, 2000).
Другим иллюстративным способом аналитической спектроскопии для измерения термической стабильности является флуоресцентная эмиссионная спектроскопия (см. van Mierlo and Steemsma, выше). Другим иллюстративным способом аналитической спектроскопии для измерения термической стабильности является спектроскопия ядерного магнитного резонанса (ЯМР) (см., например, van Mierlo and Steemsma, выше).
Термическую стабильность композиции можно измерять биохимическим путем. Иллюстративным биохимическим способом оценки термической стабильности является анализ с термической нагрузкой. В "анализе с термической нагрузкой" композицию подвергают воздействию диапазона повышенных температур в течение заданного периода времени. Например, в одном варианте осуществления исследуемые молекулы scFv или молекулы, содержащие молекулы scFv, подвергают диапазону возрастающих температур, например, в течение 1-1,5 часа. Затем анализируют активность белка подходящим биохимическим анализом. Например, если белок представляет собой связывающий белок (например, scFv или scFv-содержащий полипептид), активность связывания связывающего белка можно определять функциональным или количественным ELISA.
Такой анализ можно проводить в высокопроизводительном формате и в форматах, описанных в разделе "Примеры" с использованием E. coli и высокопроизводительного скрининга. Можно создавать библиотеку антигенсвязывающих доменов, например, которая включает антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, вариантов scFv, с использованием способов, известных в данной области. Можно индуцировать экспрессию антигенсвязывающего домена, например, против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, и антигенсвязывающий домен, например, против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, можно подвергать термической нагрузке. Исследуемые образцы после нагрузки можно анализировать в отношении связывания, и те антигенсвязывающие домены против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFvs, которые являются стабильными, можно получать в большем количестве и далее охарактеризовывать.
Термическую стабильность оценивают путем измерения температуры плавления (Tm) композиции с использованием любого из описанных выше способов (например, способов аналитической спектроскопии). Температура плавления представляет собой температуру плавления в средней точке кривой температурного перехода, где 50% молекул композиции находятся в свернутом состоянии (см., например, Dimasi et al. (2009) J. Mol Biol. 393: 672-692). В одном варианте осуществления величины Tm для антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, составляют приблизительно 40°C, 41°C, 42°C, 43°C, 44°C, 45°C, 46°C, 47°C, 48°C, 49°C, 50°C, 51°C, 52°C, 53°C, 54°C, 55°C, 56°C, 57°C, 58°C, 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, 65°C, 66°C, 67°C, 68°C, 69°C, 70°C, 71°C, 72°C, 73°C, 74°C, 75°C, 76°C, 77°C, 78°C, 79°C, 80°C, 81°C, 82°C, 83°C, 84°C, 85°C, 86°C, 87°C, 88°C, 89°C, 90°C, 91°C, 92°C, 93°C, 94°C, 95°C, 96°C, 97°C, 98°C, 99°C, 100°C. В одном варианте осуществления величины Tm для IgG составляют приблизительно 40°C, 41°C, 42°C, 43°C, 44°C, 45°C, 46°C, 47°C, 48°C, 49°C, 50°C, 51°C, 52°C, 53°C, 54°C, 55°C, 56°C, 57°C, 58°C, 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, 65°C, 66°C, 67°C, 68°C, 69°C, 70°C, 71°C, 72°C, 73°C, 74°C, 75°C, 76°C, 77°C, 78°C, 79°C, 80°C, 81°C, 82°C, 83°C, 84°C, 85°C, 86°C, 87°C, 88°C, 89°C, 90°C, 91°C, 92°C, 93°C, 94°C, 95°C, 96°C, 97°C, 98°C, 99°C, 100°C. В одном варианте осуществления величины Tm для поливалентного антитела составляют приблизительно 40°C, 41°C, 42°C, 43°C, 44°C, 45°C, 46°C, 47°C, 48°C, 49°C, 50°C, 51°C, 52°C, 53°C, 54°C, 55°C, 56°C, 57°C, 58°C, 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, 65°C, 66°C, 67°C, 68°C, 69°C, 70°C, 71°C, 72°C, 73°C, 74°C, 75°C, 76°C, 77°C, 78°C, 79°C, 80°C, 81°C, 82°C, 83°C, 84°C, 85°C, 86°C, 87°C, 88°C, 89°C, 90°C, 91°C, 92°C, 93°C, 94°C, 95°C, 96°C, 97°C, 98°C, 99°C, 100°C.
Термическую стабильность также оценивают посредством измерения удельного тепла или теплоемкости (Cp) композиции с использованием аналитического калориметрического способа (например, DSC). Удельная теплота композиции представляет собой энергию (например, в ккал/моль), которая требуется для увеличения на 1°C температуры 1 моль воды. Большая Cp является признаком денатурированной или неактивной белковой композиции. Изменение теплоемкости (ΔCp) композиции измеряют посредством определения удельной теплоты композиции до и после ее термического перехода. Термическую стабильность также можно оценивать посредством измерения или определения других параметров термодинамической стабильности, включая свободную энергию Гиббса для разворачивания (ΔG), энтальпию разворачивания (ΔH) или энтропию разворачивания (ΔS). Один или несколько из описанных выше биохимических анализов (например, анализ с термической нагрузкой) используют для определения температуры (т.е. величины TC), при которой 50% композиции сохраняет ее активность (например, активность связывания).
Кроме того, можно вносить мутации антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, для изменения термической стабильности антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv. Когда гуманизированный антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, включен в конструкцию CAR, антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, гуманизированный scFv, сообщает термическую стабильность всему CAR по настоящему изобретению. В одном варианте осуществления антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, содержит одну мутацию, которая сообщает термическую стабильность антигенсвязывающему домену против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv. В другом варианте осуществления антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, содержит множество мутаций, которые сообщают термическую стабильность антигенсвязывающему домену против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv. В одном варианте осуществления множество мутаций в антигенсвязывающем домене против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv, имеет аддитивный эффект на термическую стабильность антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, scFv.
b) % Агрегация
Стабильность композиции можно определять посредством определения ее склонности к агрегации. Агрегацию можно количественно определять с использованием ряда неограничивающих биохимических или биофизических способов. Например, агрегацию композиции можно оценивать с использованием хроматографии, например, эксклюзионной хроматографии (SEC). SEC разделяет молекулы по размеру. Колонку заполняют полутвердыми гранулами из полимерного геля, который пропускает ионы и низкомолекулярные соединения внутрь, но не пропускает крупные. Когда белковую композицию наносят на колонку компактные свернутые белки (т.е. неагрегированные белки) распределяются в большем объеме растворителя, чем доступно для крупных белковых агрегатов. Следовательно, крупные агрегаты двигаются быстрее через колонку и, таким образом, смесь может быть разделена или фракционирована на компоненты. Каждую фракцию можно количественно определять по отдельности (например, по рассеянию света) по мере ее элюирования из геля. Таким образом, % агрегацию композиции можно определять посредством сравнения концентрации фракции с общей концентрацией белка, нанесенного на гель. Стабильные композиции элюируются с колонки по существу в качестве единой фракции и имеют по существу один пик в профиле элюирования или на хроматограмме.
c) Аффинность связывания
Стабильность композиции можно оценивать путем определения ее аффинности связывания с мишенью. Для определения аффинности связывания в данной области известно широкое множество способов. В иллюстративном способе определения аффинности связывания используется поверхностный плазмонный резонанс. Поверхностный плазмонный резонанс представляет собой оптическое явление, которое позволяет анализ биоспецифичексих реакций в реальном времени путем определения изменений концентраций белков на биосенсорной матрице, например, с использованием системы BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, N.J.). Для более подробного описания см. Jonsson, U., et al. (1993) Ann. Biol. Clin. 51:19-26; Jonsson, U., i (1991) Biotechniques 11:620-627; Johnsson, B., et al. (1995) J. Mol. Recognit. 8:125-131; и Johnnson, B., et al. (1991) Anal. Biochem. 198:268-277.
В одном аспекте антигенсвязывающий домен CAR содержит аминокислотную последовательность, которая является гомологичной аминокислотной последовательности антигенсвязывающего домена, описанного в настоящем описании, и антигенсвязывающий домен сохраняет желаемые функциональные свойства антигенсвязывающего домена, описанного в настоящем описании.
В одном конкретном аспекте композиция CAR по изобретению содержит фрагмент антитела. В следующем аспекте фрагмент антитела содержит scFv.
В различных аспектах антигенсвязывающий домен CAR конструируют путем модификации одной или нескольких аминокислот в одной или обеих вариабельных областях (например, VH и/или VL), например, в одной или нескольких областях CDR и/или в одной или нескольких каркасных областях. В одном конкретном аспекте композиция CAR по изобретению содержит фрагмент антитела. В следующем аспекте фрагмент антитела содержит scFv.
Специалисту в данной области будет понятно, что антитело или фрагмент антитела по изобретению можно далее модифицировать так, чтобы их аминокислотная последовательность отличалась (например, относительно дикого типа), но не в отношении желаемой активности. Например, в белок можно вносить дополнительные нуклеотидные замены, ведущие к аминокислотным заменам в "неосновных" аминокислотных остатках. Например, неосновной аминокислотный остаток в молекуле может быть заменен другим аминокислотным остатком из того же семейства боковых цепей. В другом варианте осуществления цепь аминокислот может быть заменена структурно сходной цепью, которая отличается порядком и/или композицией представителей семейства боковых цепей, например, можно вносить консервативную замену, в которой аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь.
Семейства аминокислотных остатков, имеющих сходные боковые цепи, определены в данной области, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислотные боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин).
Процентная идентичность в контексте двух или более последовательностей нуклеиновых кислот или полипептидных последовательностей относится к двум или более последовательностям, которые являются одинаковыми. Две последовательности являются "по существу идентичными", если две последовательности имеют конкретный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (например, 60% идентичность, необязательно 70%, 71%. 72%. 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%,81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичность на протяжении указанной области или, когда не уточняется, на протяжении всей последовательности), при сравнении и выравнивании на максимальное соответствие на протяжении окна сравнения или указанной области, как измеряют с использованием одного из следующих алгоритмов сравнения последовательностей или путем выравнивания вручную или визуального исследования. Необязательно, идентичность существует на протяжении области, которая имеет длину по меньшей мере приблизительно 50 нуклеотидов (или 10 аминокислот), или более предпочтительно на протяжении области, которая имеет длину от 100 до 500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот).
Для сравнения последовательностей, как правило, одна последовательность выступает в качестве эталонной последовательности, с которой сравнивают исследуемые последовательности. При использовании алгоритма сравнения последовательностей, исследуемую и эталонную последовательности вводят в компьютер, координаты подпоследовательностей указывают, если необходимо, и назначают параметры программы с алгоритмом сравнения последовательностей. Можно использовать параметры программы по умолчанию или альтернативно параметры можно назначать. Затем алгоритм сравнения последовательностей вычисляет процентную идентичность последовательностей для исследуемых последовательностей относительно эталонной последовательности, исходя из параметров программы. Способы выравнивания последовательностей для сравнения хорошо известны в данной области. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с использованием алгоритма локальной гомологии Smith and Waterman, (1970) Adv. Appl. Math. 2:482c, с использованием алгоритма выравнивания по гомологии Needleman and Wunsch, (1970) J. Mol. Biol. 48:443, с использованием способа поиска сходства Pearson and Lipman, (1988) Proc. Nat'l. Acad. Sci. USA 85:2444, с использованием компьютерных воплощений этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI), или путем выравнивания вручную и визуального исследования (см., например, Brent et al., (2003) Current Protocols in Molecular Biology).
Двумя примерами алгоритмов, которые пригодны для определения процентной идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., (1977) Nuc. Acids Res. 25:3389-3402; и Altschul et al., (1990) J. Mol. Biol. 215:403-410, соответственно. Программное обеспечение для осуществления анализов BLAST является общедоступным через National Center for Biotechnology Information.
Процентную идентичность между двумя аминокислотными последовательностями также можно определять с использованием алгоритма E. Meyers и W. Miller (Comput. Appl. Biosci., 4:11-17, 1988), который включен в программу ALIGN (версии 2.0), с использованием таблицы веса остатков PAM120, штрафа за продолжение пропуска 12 и штрафа за пропуск 4. Кроме того, процентную идентичность между двумя аминокислотными последовательностями можно определять с использованием алгоритма Needleman и Wunsch (J. Mol, Biol. 48:444-453, 1970), который включен с программу GAP в пакете программ GCG (доступном на http://www.gcg.com), с использованием либо матрицы Blossum 62, либо матрицы PAM250, и штрафа за пропуск 16, 14, 12, 10, 8, 6 или 4 и штрафа за продолжение пропуска 1, 2, 3, 4, 5 или 6.
В одном аспекте настоящее изобретение предусматривает модификации исходной аминокислотной последовательности антитела или фрагмента (например, scFv), которые приводят к функционально эквивалентным молекулам. Например, VH или VL антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, например, scFv, содержащегося в CAR, можно модифицировать для сохранения по меньшей мере приблизительно 70%, 71%, 72%. 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности с исходной каркасной области VH или VL антигенсвязывающего домена против ассоциированного со злокачественной опухолью антигена, например scFv. Настоящее изобретение предусматривает модификации всей конструкции CAR, например, модификации одной или нескольких аминокислотных последовательностей различных доменов конструкции CAR для получения функционально эквивалентных молекул. Конструкцию CAR можно модифицировать, чтобы она сохраняла по меньшей мере приблизительно 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичность с исходной конструкцией CAR.
Трансмембранный домен
Что касается трансмембранного домена, в различных вариантах осуществления CAR можно конструировать так, чтобы он содержал трансмембранный домен, который связан с внеклеточным доменом CAR. Трансмембранный домен может включать одну или несколько дополнительных аминокислот, соседних с трансмембранной областью, например, одну или несколько аминокислот, ассоциированных с внеклеточной областью белка, из которого происходит трансмембранный домен (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 вплоть до 15 аминокислот внеклеточной области) и/или одну или несколько дополнительных аминокислот, ассоциированных с внутриклеточной областью белка, из которой происходит трансмембранный белок (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 вплоть до 15 аминокислот внутриклеточной области). В одном аспекте трансмембранный домен представляет собой домен, который ассоциирован с одним из других доменов, используемых в CAR, например, в одном варианте осуществления трансмембранный домен может происходить из того же белка, что и сигнальный домен, костимулирующий домен или шарнирный домен. В другом аспекте трансмембранный домен не происходит из того же белка, что и любой другой домен CAR. В некоторых случаях трансмембранный домен можно выбирать или модифицировать посредством аминокислотной замены, чтобы избежать связывания таких доменов с трансмембранными доменами тех же или отличающихся поверхностных мембранных белков, например, для минимизации взаимодействий с другими представителями рецепторного комплекса. В одном аспекте трансмембранный домен способен к гомодимеризации с другим CAR на клеточной поверхности экспрессирующей CAR клетки. В другом аспекте аминокислотная последовательность трансмембранного домена может быть модифицирована или замещена, чтобы минимизировать взаимодействия со связывающими доменами нативного связывающего партнера, присутствующего в той же экспрессирующей CAR клетке.
Трансмембранный домен может происходить либо из природного, либо из рекомбинантного источника. Когда источник является природным, домен может происходить из любого мембраносвязанного или трансмембранного белка. В одном аспекте трансмембранный домен способен передавать сигнал на внутриклеточный домен(ы), когда CAR связывается с мишенью. Особенно пригодный в рамках настоящего изобретения трансмембранный домен может включать по меньшей мере трансмембранную область(и), например, альфа-, бета- или зета-цепи T-клеточного рецептора, CD28, CD27, CD3-эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137, CD154. В некоторых вариантах осуществления трансмембранный домен может включать по меньшей мере трансмембранную область(и), например, из KIRDS2, OX40, CD2, CD27, LFA-1 (CD11a, CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD40, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), NKp44, NKp30, NKp46, CD160, CD19, IL2R-бета, IL2R-гамма, IL7Rα, ITGA1, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, TNFR2, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Tactile), CEACAM1, CRTAM, Ly9 (CD229), CD160 (BY55), PSGL1, CD100 (SEMA4D), SLAMF6 (NTB-A, Ly108), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, PAG/Cbp, NKG2D, NKG2C.
В некоторых случаях трансмембранный домен может быть связан с внеклеточной областью CAR, например, антигенсвязывающим доменом CAR, через шарнирную область, например, шарнирную область из белка человека. Например, в одном варианте осуществления шарнирная область может представлять собой шарнирную область Ig (иммуноглобулина) человека (например, шарнирная область IgG4, шарнирная область IgD), линкер GS (например, линкер GS, описанный в настоящем описании), шарнирную область KIR2DS2 или шарнирную область CD8a. В одном варианте осуществления шарнирная область или спейсер содержат (например, состоят из) аминокислотную последовательности SEQ ID NO: 4. В одном аспекте трансмембранный домен содержит (например, состоит из) трансмембранный домен SEQ ID NO: 12.
В одном аспекте шарнирная область или спейсер содержат шарнирную область IgG4. Например, в одном варианте осуществления шарнирная область или спейсер содержат шарнирную область с аминокислотной последовательностью: ESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGKM (SEQ ID NO: 6). В некоторых вариантах осуществления шарнирная область или спейсер содержат шарнирную область, кодируемую нуклеотидной последовательностью: GAGAGCAAGTACGGCCCTCCCTGCCCCCCTTGCCCTGCCCCCGAGTTCCTGGGCGGACCCAGCGTGTTCCTGTTCCCCCCCAAGCCCAAGGACACCCTGATGATCAGCCGGACCCCCGAGGTGACCTGTGTGGTGGTGGACGTGTCCCAGGAGGACCCCGAGGTCCAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCACAACGCCAAGACCAAGCCCCGGGAGGAGCAGTTCAATAGCACCTACCGGGTGGTGTCCGTGCTGACCGTGCTGCACCAGGACTGGCTGAACGGCAAGGAATACAAGTGTAAGGTGTCCAACAAGGGCCTGCCCAGCAGCATCGAGAAAACCATCAGCAAGGCCAAGGGCCAGCCTCGGGAGCCCCAGGTGTACACCCTGCCCCCTAGCCAAGAGGAGATGACCAAGAACCAGGTGTCCCTGACCTGCCTGGTGAAGGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAACGGCCAGCCCGAGAACAACTACAAGACCACCCCCCCTGTGCTGGACAGCGACGGCAGCTTCTTCCTGTACAGCCGGCTGACCGTGGACAAGAGCCGGTGGCAGGAGGGCAACGTCTTTAGCTGCTCCGTGATGCACGAGGCCCTGCACAACCACTACACCCAGAAGAGCCTGAGCCTGTCCCTGGGCAAGATG (SEQ ID NO: 7).
В одном аспекте шарнирная область или спейсер содержат шарнирную область IgD. Например, в одном варианте осуществления шарнирная область или спейсер содержат шарнирную область с аминокислотной последовательностью RWPESPKAQASSVPTAQPQAEGSLAKATTAPATTRNTGRGGEEKKKEKEKEEQEERETKTPECPSHTQPLGVYLLTPAVQDLWLRDKATFTCFVVGSDLKDAHLTWEVAGKVPTGGVEEGLLERHSNGSQSQHSRLTLPRSLWNAGTSVTCTLNHPSLPPQRLMALREPAAQAPVKLSLNLLASSDPPEAASWLLCEVSGFSPPNILLMWLEDQREVNTSGFAPARPPPQPGSTTFWAWSVLRVPAPPSPQPATYTCVVSHEDSRTLLNASRSLEVSYVTDH (SEQ ID NO: 8). В некоторых вариантах осуществления шарнирная область или спейсер содержат шарнирную область, кодируемую нуклеотидной последовательностью AGGTGGCCCGAAAGTCCCAAGGCCCAGGCATCTAGTGTTCCTACTGCACAGCCCCAGGCAGAAGGCAGCCTAGCCAAAGCTACTACTGCACCTGCCACTACGCGCAATACTGGCCGTGGCGGGGAGGAGAAGAAAAAGGAGAAAGAGAAAGAAGAACAGGAAGAGAGGGAGACCAAGACCCCTGAATGTCCATCCCATACCCAGCCGCTGGGCGTCTATCTCTTGACTCCCGCAGTACAGGACTTGTGGCTTAGAGATAAGGCCACCTTTACATGTTTCGTCGTGGGCTCTGACCTGAAGGATGCCCATTTGACTTGGGAGGTTGCCGGAAAGGTACCCACAGGGGGGGTTGAGGAAGGGTTGCTGGAGCGCCATTCCAATGGCTCTCAGAGCCAGCACTCAAGACTCACCCTTCCGAGATCCCTGTGGAACGCCGGGACCTCTGTCACATGTACTCTAAATCATCCTAGCCTGCCCCCACAGCGTCTGATGGCCCTTAGAGAGCCAGCCGCCCAGGCACCAGTTAAGCTTAGCCTGAATCTGCTCGCCAGTAGTGATCCCCCAGAGGCCGCCAGCTGGCTCTTATGCGAAGTGTCCGGCTTTAGCCCGCCCAACATCTTGCTCATGTGGCTGGAGGACCAGCGAGAAGTGAACACCAGCGGCTTCGCTCCAGCCCGGCCCCCACCCCAGCCGGGTTCTACCACATTCTGGGCCTGGAGTGTCTTAAGGGTCCCAGCACCACCTAGCCCCCAGCCAGCCACATACACCTGTGTTGTGTCCCATGAAGATAGCAGGACCCTGCTAAATGCTTCTAGGAGTCTGGAGGTTTCCTACGTGACTGACCATT (SEQ ID NO: 9).
В одном аспекте трансмембранный домен может быть рекомбинантным, и в этом случае он содержит в основном гидрофобные остатки, такие как лейцин и валин. В одном аспекте на каждом конце рекомбинантного трансмембранного домена может находиться триплет из фенилаланина, триптофана и валина.
Необязательно, короткий олиго- или полипептидный линкер длиной от 2 до 10 аминокислот может образовывать связь между трансмембранным доменом и цитоплазматической областью CAR. Глицин-сериновый дублет обеспечивает особенно подходящий линкер. Например, в одном аспекте линкер содержит аминокислотную последовательность GGGGSGGGGS (SEQ ID NO: 10). В некоторых вариантах осуществления линкер кодируется нуклеотидной последовательностью GGTGGCGGAGGTTCTGGAGGTGGAGGTTCC (SEQ ID NO: 11).
В одном аспекте шарнирная область или спейсер содержат шарнирную область KIR2DS2.
Цитоплазматический домен
Цитоплазматический домен или область CAR включает внутриклеточный сигнальный домен. Внутриклеточный сигнальный домен обычно ответственен за активацию по меньшей мере одной из нормальных эффекторных функций иммунной клетки, в которую введен CAR. Термин "эффекторная функция" относится к специализированной функции клетки. Эффекторная функция T-клетки, например, может представлять собой цитолитическую активность или хелперную активность, включая секрецию цитокинов. Таким образом, термин "внутриклеточный сигнальный домен" относится к части белка, которая передает сигнал эффекторной функции и направляет клетку на выполнение специализированной функции. Хотя обычно можно использовать весь внутриклеточный сигнальный домен, во многих случаях не является обязательным использование всей цепи. Когда используют укороченную часть внутриклеточного сигнального домена, такую укороченную часть можно использовать вместо интактной цепи при условии, что она передает сигнал эффекторной функции. Таким образом, подразумевают, что термин "внутриклеточный сигнальный домен" включает любую укороченную часть внутриклеточного сигнального домена, достаточную для передачи сигнала эффекторной функции.
Примеры внутриклеточных сигнальных доменов для применения в CAR по изобретению включают цитоплазматические последовательности T-клеточного рецептора (TCR) и корецепторов, которые действуют совместно для инициации передачи сигнала после связывания рецептором антигена, а также любое производное или вариант этих последовательностей и любую рекомбинантную последовательность, которая имеет ту же функциональную способность.
Известно, что сигналы, индуцируемые TCR отдельно, являются недостаточными для полной активации T-клеток, и что также требуется вторичный и/или костимулирующий сигнал. Таким образом, можно считать, что активация T-клеток опосредуется двумя различными классами цитоплазматических сигнальных последовательностей: последовательности, которые инициируют антигензависимую первичную активацию через TCR (первичные внутриклеточные сигнальные домены), и последовательности, которые действуют независимым от антигена образом, обеспечивая вторичный или костимулирующий (вторичный цитоплазматический домен, например, костимулирующий домен).
Первичный сигнальный домен регулирует первичную активацию комплекса TCR либо стимулирующим путем, либо ингибиторным путем. Первичные внутриклеточные сигнальные домены, которые действуют стимулирующим образом, могут содержать сигнальные мотивы, которые известны как иммунорецепторные тирозиновые активирующие мотивы, или ITAM.
Примеры содержащих ITAM первичных внутриклеточных сигнальных доменов, которые особенно применимы в рамках изобретения, включают сигнальные домены CD3-зета, общего FcR-гамма (FCER1G), Fc-гамма RIIa, FcR-бета (Fc-эпсилон R1b), CD3-гамма, CD3-дельта, CD3-эпсилон, CD79a, CD79b, DAP10 и DAP12. В одном варианте осуществления CAR по изобретению содержит внутриклеточный сигнальный домен, например, первичный сигнальный домен CD3-зета.
В одном варианте осуществления первичный сигнальный домен содержит модифицированный домен ITAM, например, мутантный домен ITAM, который имеет измененную (например, увеличенную или сниженную) активность по сравнению с нативным доменом ITAM. В одном варианте осуществления первичный сигнальный домен содержит модифицированный ITAM-содержащий первичный внутриклеточный сигнальный домен, например, оптимизированный и/или укороченный ITAM-содержащий первичный внутриклеточный сигнальный домен. В одном варианте осуществления первичный сигнальный домен содержит один, два, три, четыре или более мотивов ITAM.
Внутриклеточный сигнальный домен CAR может содержать сигнальный домен CD3-зета сам по себе или в комбинации с любым другим желаемым внутриклеточным сигнальным доменом(ами), пригодным в контексте CAR по изобретению. Например, внутриклеточный сигнальный домен CAR может содержать часть в виде цепи CD3-зета и костимулирующий сигнальный домен. Костсимулирующий сигнальный домен относится к части CAR, содержащей внутриклеточный домен костимулирующей молекулы. Костимулирующая молекула представляет собой молекулу клеточной поверхности, отличную от рецептора антигена или его лигандов, которая требуется для эффективного ответа лимфоцитов на антиген. Примеры таких молекул включают CD27, CD28, 4-1BB (CD137), OX40, CD30, CD40, PD-1, ICOS, ассоциированный с функцией лимфоцитов антиген 1 (LFA-1), CD2, CD7, LIGHT, NKG2C, B7-H3, и лиганд, который специфически связывается с CD83, и т.п. Например, было продемонстрировано, что костимуляция CD27 повышает экспансию, эффекторную функцию и выживаемость клеток CART человека in vitro и усиливает живучесть и противоопухолевую активность T-клеток человека in vivo (Song et al. Blood. 2012; 119(3):696-706). Следующие примеры таких костимулирующих молекул включают CDS, ICAM-1, GITR, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), NKp44, NKp30, NKp46, CD160, CD19, CD4, CD8-альфа, CD8-бета, IL2R-бета, IL2R-гамма, IL7R-альфа, ITGA4, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, TNFR2, TRANCE/RANKL, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Tactile), NKG2D, CEACAM1, CRTAM, Ly9 (CD229), CD160 (BY55), PSGL1, CD100 (SEMA4D), CD69, SLAMF6 (NTB-A, Ly108), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, LAT, GADS, SLP-76, PAG/Cbp и CD19.
Внутриклеточные сигнальные последовательности в цитоплазматической части CAR по изобретению могут быть связаны друг с другом в случайном или определенном порядке. Необязательно короткий олиго- или полипептидный линкер, например, длиной от 2 до 10 аминокислот (например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот), может образовывать связь между внутриклеточными сигнальными последовательностями. В одном варианте осуществления в качестве подходящего линкера можно использовать глицин-сериновый дублет. В одном варианте осуществления в качестве подходящего линкера можно использовать одну аминокислоту, например, аланин, глицин.
В одном аспекте внутриклеточный сигнальный домен сконструирован, чтобы он содержал два или более, например, 2, 3, 4, 5 или более, костимулирующих сигнальных доменов. В одном варианте осуществления два или более, например, 2, 3, 4, 5 или более, костимулирующих сигнальных доменов, разделены линкерной молекулой, например линкерной молекулой, описанной в настоящем описании. В одном варианте осуществления внутриклеточный сигнальный домен содержит два костимулирующих сигнальных домена. В некоторых вариантах осуществления линкерная молекула представляет собой остаток глицина. В некоторых вариантах осуществления линкер представляет собой остаток аланина.
В одном аспекте внутриклеточный сигнальный домен сконструирован так, чтобы он содержал сигнальный домен CD3-зета и сигнальный домен CD28. В одном аспекте внутриклеточный сигнальный домен сконструирован, чтобы он содержал сигнальный домен CD3-зета и сигнальный домен 4-1BB. В одном аспекте сигнальный домен 4-1BB представляет собой сигнальный домен SEQ ID NO: 14. В одном аспекте сигнальный домен CD3-зета представляет собой сигнальный домен SEQ ID NO: 18.
В одном аспекте внутриклеточный сигнальный домен сконструирован так, чтобы он содержал сигнальный домен CD3-зета и сигнальный домен CD27. В одном аспекте сигнальный домен CD27 содержит аминокислотную последовательность QRRKYRSNKGESPVEPAEPCRYSCPREEEGSTIPIQEDYRKPEPACSP (SEQ ID NO: 16). В одном аспекте сигнальный домен CD27 кодируется последовательностью нуклеиновой кислоты AGGAGTAAGAGGAGCAGGCTCCTGCACAGTGACTACATGAACATGACTCCCCGCCGCCCCGGGCCCACCCGCAAGCATTACCAGCCCTATGCCCCACCACGCGACTTCGCAGCCTATCGCTCC (SEQ ID NO: 17).
В одном аспекте экспрессирующая CAR клетка, описанная в настоящем описании, кроме того, может содержать второй CAR, например, второй CAR, который включает отличающийся антигенсвязывающий домен, например, к той же мишени или отличающейся мишени (например, к мишени, отличной от ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, или к отличающемуся ассоциированному со злокачественной опухолью антигену, описанному в настоящем описании). В одном варианте осуществления второй CAR включает антигенсвязывающий домен к мишени, экспрессируемой тем же типом злокачественных клеток, что и ассоциированный со злокачественной опухолью антиген. В одном варианте осуществления экспрессирующая CAR клетка содержит первый CAR, который нацелен на первый антиген и включает внутриклеточный сигнальный домен, имеющий костимулирующий сигнальный домен, но не первичный сигнальный домен, и второй CAR, который нацелен на второй, отличающийся антиген и включает внутриклеточный сигнальный домен, имеющий первичный сигнальный домен, но не костимулирующий сигнальный домен. Без связи с теорией, размещение костимулирующего сигнального домена, например, 4-1BB, CD28, CD27 или OX-40, в первом CAR, и первичного сигнального домен, например, CD3-зета, в первом CAR может ограничить активность CAR клетками, в которых экспрессируются обе мишени. В одном варианте осуществления экспрессирующая CAR клетка содержит первый CAR против ассоциированного со злокачественной опухолью антигена, который включает антигенсвязывающий домен, который связывает антиген-мишень, описанный в настоящем описании, трансмембранный домен и костимулирующий домен, и второй CAR, который нацелен на отличающийся антиген-мишень (например, антиген, экспрессируемый на том же типе злокачественных клеток, что и первый антиген-мишень) и включает антигенсвязывающий домен, трансмембранный домен и первичный сигнальный домен. В другом варианте осуществления экспрессирующая CAR клетка содержит первый CAR, который включает антигенсвязывающий домен, который связывает антиген-мишень, описанный в настоящем описании, трансмембранный домен и первичный сигнальный домен, и второй CAR, который нацелен на антиген, отличный от первого антигена-мишени (например, антиген, экспрессируемый тем же типом злокачественных клеток, что и первый антиген-мишень) и включает антигенсвязывающий домен к антигену, трансмембранный домен и костимулирующий сигнальный домен.
В одном варианте осуществления экспрессирующая CAR клетка содержит XCAR, описанный в настоящем описании, и ингибиторный CAR. В одном варианте осуществления ингибиторный CAR содержит антигенсвязывающий домен, который связывает антиген, встречающийся на нормальных клетках, но не на злокачественных клетках, например, на нормальных клетках, которые также экспрессируют антиген CLL. В одном варианте осуществления ингибиторный CAR содержит антигенсвязывающий домен, трансмембранный домен и внутриклеточный домен ингибиторной молекулы. Например, внутриклеточный домен ингибиторного CAR может представлять собой внутриклеточный домен PD1, PD-L1, CTLA4, TIM3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 или TGF-бета.
В одном варианте осуществления, когда экспрессирующая CAR клетка содержит два или более различных CAR, антигенсвязывающие домены различных CAR могут быть такими, что антигенсвязывающие домены не взаимодействуют друг с другом. Например, клетка, экспрессирующая первый и второй CAR, может иметь антигенсвязывающий домен первого CAR, например, в качестве фрагмента, например scFv, который не образует связи с антигенсвязывающим доменом второго CAR, например, антигенсвязывающий домен второго CAR представляет собой VHH.
В некоторых вариантах осуществления антигенсвязывающий домен включает однодоменные антигенсвязывающие (SDAB) молекулы, которые включают молекулы, чьи определяющие комплементарность области являются частью однодоменного полипептида. Примеры включают, но не ограничиваются ими, вариабельные домены тяжелой цепи, связывающие молекулы, естественным образом лишенные легких цепей, единичные домены, происходящие из обычных антител из 4-цепей, сконструированные домены и однодоменные каркасы, отличные от каркасов, происходящих из антител. Молекулы SDAB могут представлять собой любые молекулы уровня техники или какие-либо будущие однодоменные молекулы. Молекулы SDAB могут происходить из любого вида, включая, но не ограничиваясь ими, мышь, человека, верблюда, ламу, миногу, рыбу, акулу, козу, кролика и быка. Этот термин также включает встречающиеся в природе однодоменные молекулы антител из вида, отличного от Camelidae и акул.
В одном аспекте молекула SDAB может происходить из вариабельной области иммуноглобулина, встречающейся у рыб, например, такой как вариабельная область, которая происходит из изотипа иммуноглобулина, известного как новый рецептор антигенов (NAR), встречающийся в сыворотке акулы. Способы получения однодоменных молекул, происходящих из вариабельной области NAR ("IgNAR"), описаны в WO 03/014161 и Streltsov (2005) Protein Sci. 14:2901-2909.
Согласно другому аспекту молекула SDAB представляет собой встречающуюся в природе однодоменную антигенсвязывающую молекулу, известную как тяжелая цепь, свободная от легких цепей. Такие однодоменные молекулы описаны, например, в WO 9404678 и Hamers-Casterman, C. et al. (1993) Nature 363:446-448. Для ясности, этот вариабельный домен, происходящий из молекулы тяжелой цепи, естественным образом свободной от легкой цепи, известен в настоящем описании как VHH или наноантитела, чтобы отличать его от общепринятого VH иммуноглобулинов из четырех цепей. Такая молекула VHH может происходить из вида Camelidae, например, верблюда, ламы, дромедара, альпаки и гуанако. Другие виды, кроме Camelidae, могут продуцировать молекулы тяжелых цепей, естественным образом лишенные легкой цепи; такие VHH входят в объем изобретения.
Молекулы SDAB могут быть рекомбинантными, имеющими транслантированные CDR, гуманизированными, камелизованными, деимунизированными и/или полученными in vitro (например, отобранными способом фагового дисплея).
Также было обнаружено, что в клетках, имеющих множество химерных погруженных в мембрану рецепторов, содержащих антигенсвязывающий домен, взаимодействия между антигенсвязывающими доменами рецепторов может быть нежелательным, например, поскольку оно ингибирует способность одного или нескольких антигенсвязывающих доменов связывать их антиген. Таким образом, в настоящем описании описаны клетки, имеющие первый и второй не встречающиеся в природе химерные погруженные в мембрану рецепторы, содержащие антигенсвязывающие домены, которые минимизируют такие взаимодействия. Также в настоящем описании описаны нуклеиновые кислоты, кодирующие первый и второй не встречающиеся в природе химерные погруженные в мембрану рецепторы, содержащие антигенсвязывающие домены, которые минимизируют такие взаимодействия, а также способы получения и применения таких клеток и нуклеиновых кислот. В одном варианте осуществления антигенсвязывающий домен одного из указанного первого и указанного второго не встречающихся в природе химерных погруженных в мембрану рецепторов содержит scFv, а другой содержит один домен VH, например, один домен VH животных семейства верблюжьих, акулы или миноги, или один домен VH, происходящий из последовательности человека или мыши.
В некоторых вариантах осуществления заявленное изобретение содержит первый и второй CAR, где антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR не содержит вариабельный домен легкой цепи и вариабельный домен тяжелой цепи. В некоторых вариантах осуществления антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR представляет собой scFv, а другой не является scFv. В некоторых вариантах осуществления антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR содержит один домен VH, например, животного семейства верблюжьих, акулы или миноги, или один домен VH, происходящий из последовательности человека или мыши. В некоторых вариантах осуществления антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR содержит наноантитело. В некоторых вариантах осуществления антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR содержит домен VHH животного семейства верблюжьих.
В некоторых вариантах осуществления антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR содержит scFv, а другой содержит один домен VH, например, один домен VH животного семейства верблюжьих, акулы или миноги, или один домен VH, происходящий из последовательности человека или мыши. В некоторых вариантах осуществления антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR содержит scFv, а другой содержит наноантитело. В некоторых вариантах осуществления антигенсвязывающий домен одного из указанного первого CAR и указанного второго CAR содержит scFv, а другой содержит VHH животного семейства верблюжьих.
В некоторых вариантах осуществления, когда он присутствует на поверхности клетки, связывание антигенсвязывающего домен указанного первого CAR с его антигеном по существу не снижается в результате присутствия указанного второго CAR. В некоторых вариантах осуществления связывание антигенсвязывающего домена указанного первого CAR с его антигеном в присутствии указанного второго CAR составляет 85%, 90%, 95%, 96%, 97%, 98% или 99% от связывания антигенсвязывающего домена указанного первого CAR с его антигеном в отсутствие указанного второго CAR.
В некоторых вариантах осуществления, когда они присутствуют на поверхности клетки, антигенсвязывающие домены указанного первого CAR и указанного второго CAR связываются друг с другом в меньшей степени, чем если бы они оба представляли собой антигенсвязывающие домены scFv. В некоторых вариантах осуществления антигенсвязывающие домены указанного первого CAR и указанного второго CAR связываются друг с другом на 85%, 90%, 95%, 96%, 97%, 98% или 99% меньше, чем если бы они оба представляли собой антигенсвязывающие домены scFv.
В другом аспекте экспрессирующая CAR клетка, описанная в настоящем описании, может дополнительно экспрессировать другое средство, например, средство, которое повышает активность экспрессирующей CAR клетки. Например, в одном варианте осуществления средство может представлять собой средство, которое ингибирует ингибиторную молекулу. Ингибиторные молекулы, например PD1, в некоторых вариантах осуществления могут снижать способность экспрессирующей CAR клетки индуцировать иммунный эффекторный ответ. Примеры ингибиторных молекул включают PD1, PD-L1, CTLA4, TIM3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и TGF-бета. В одном варианте осуществления средство, которое ингибирует ингибирующую молекулу, например, представляет собой молекулу, описанную в настоящем описании, например, средство, которое содержит первый полипептид, например, ингибиторную молекулу, связанный со вторым полипептидом, который дает положительный сигнал клетке, например, внутриклеточным сигнальным доменом, описанным в настоящем описании. В одном варианте осуществления средство содержит первый полипептид, например, из ингибиторной молекулы, такой как PD1, PD-L1, CTLA4, TIM3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 или TGF-бета, или фрагмент любой из них (например, меньшей мере часть внеклеточного домена любой из них), и второй полипептид, который представляет собой внутриклеточный сигнальный домен, описанный в настоящем описании (например, содержащий костимулирующий домен (например, 41BB, CD27 или CD28, например, как описано в настоящем описании) и/или первичный сигнальный домен (например, сигнальный домен CD3-зета, описанный в настоящем описании)). В одном варианте осуществления средство содержит первый полипептид из PD1 или его фрагмента (например, по меньшей мере часть внеклеточного домена PD1), и второй полипептид внутриклеточного сигнального домена, описанный в настоящем описании (например, сигнальный домен CD28, описанный в настоящем описании, и/или сигнальный домен CD3-зета, описанный в настоящем описании). PD1 представляет собой ингибиторный представитель семейства рецепторов CD28, которое также включает CD28, CTLA-4, ICOS и BTLA. PD-1 экспрессируется на активированных B-клетках, T-клетках и миелоидных клетках (Agata et al. 1996 Int. Immunol 8:765-75). Было показано, что два лиганда для PD1, PD-L1 и PD-L2, подавляют активацию T-клеток при связывании с PD1 (Freeman et a. 2000 J Exp Med 192:1027-34; Latchman et al. 2001 Nat Immunol 2:261-8; Carter et al. 2002 Eur J Immunol 32:634-43). PD-L1 распространен в злокачественных опухолях человека (Dong et al. 2003 J Mol Med 81:281-7; Blank et al. 2005 Cancer Immunol. Immunother 54:307-314; Konishi et al. 2004 Clin Cancer Res 10:5094). Иммунную супрессию можно обращать вспять путем ингибирования локального взаимодействия PD1 с PD-L1.
В одном варианте осуществления средство содержит внеклеточный домен (ECD) ингибиторной молекулы, например, Programmed Death 1 (PD1), слитый с трансмембранным доменом и внутриклеточными сигнальными доменами, такими как 41BB и CD3-зета (также обозначаемый в настоящем описании как PD1 CAR). В одном варианте осуществления PD1 CAR, когда его используют в комбинациях с XCAR, описанным в настоящем описании, повышает живучесть T-клетки. В одном варианте осуществления CAR представляет собой PD1 CAR, содержащий внеклеточный домен PD1, указанный подчеркиванием в SEQ ID NO: 26. В одном варианте осуществления PD1 CAR содержит аминокислотную последовательность SEQ ID NO: 26.
Malpvtalllplalllhaarppgwfldspdrpwnpptfspallvvtegdnatftcsfsntsesfvlnwyrmspsnqtdklaafpedrsqpgqdcrfrvtqlpngrdfhmsvvrarrndsgtylcgaislapkaqikeslraelrvterraevptahpspsprpagqfqtlvtttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfacdiyiwaplagtcgvlllslvitlyckrgrkkllyifkqpfmrpvqttqeedgcscrfpeeeeggcelrvkfsrsadapaykqgqnqlynelnlgrreeydvldkrrgrdpemggkprrknpqeglynelqkdkmaeayseigmkgerrrgkghdglyqglstatkdtydalhmqalppr (SEQ ID NO: 26).
В одном варианте осуществления PD1 CAR содержит аминокислотную последовательность, представленную ниже (SEQ ID NO: 39).
pgwfldspdrpwnpptfspallvvtegdnatftcsfsntsesfvlnwyrmspsnqtdklaafpedrsqpgqdcrfrvtqlpngrdfhmsvvrarrndsgtylcgaislapkaqikeslraelrvterraevptahpspsprpagqfqtlvtttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfacdiyiwaplagtcgvlllslvitlyckrgrkkllyifkqpfmrpvqttqeedgcscrfpeeeeggcelrvkfsrsadapaykqgqnqlynelnlgrreeydvldkrrgrdpemggkprrknpqeglynelqkdkmaeayseigmkgerrrgkghdglyqglstatkdtydalhmqalppr (SEQ ID NO: 39).
В одном варианте осуществления средство содержит последовательность нуклеиновой кислоты, кодирующую PD1 CAR, например, PD1 CAR, описанный в настоящем описании. В одном варианте осуществления последовательность нуклеиновой кислоты для PD1 CAR представлена ниже с PD1 ECD, подчеркнутым в SEQ ID NO: 27.
atggccctccctgtcactgccctgcttctccccctcgcactcctgctccacgccgctagaccacccggatggtttctggactctccggatcgcccgtggaatcccccaaccttctcaccggcactcttggttgtgactgagggcgataatgcgaccttcacgtgctcgttctccaacacctccgaatcattcgtgctgaactggtaccgcatgagcccgtcaaaccagaccgacaagctcgccgcgtttccggaagatcggtcgcaaccgggacaggattgtcggttccgcgtgactcaactgccgaatggcagagacttccacatgagcgtggtccgcgctaggcgaaacgactccgggacctacctgtgcggagccatctcgctggcgcctaaggcccaaatcaaagagagcttgagggccgaactgagagtgaccgagcgcagagctgaggtgccaactgcacatccatccccatcgcctcggcctgcggggcagtttcagaccctggtcacgaccactccggcgccgcgcccaccgactccggccccaactatcgcgagccagcccctgtcgctgaggccggaagcatgccgccctgccgccggaggtgctgtgcatacccggggattggacttcgcatgcgacatctacatttgggctcctctcgccggaacttgtggcgtgctccttctgtccctggtcatcaccctgtactgcaagcggggtcggaaaaagcttctgtacattttcaagcagcccttcatgaggcccgtgcaaaccacccaggaggaggacggttgctcctgccggttccccgaagaggaagaaggaggttgcgagctgcgcgtgaagttctcccggagcgccgacgcccccgcctataagcagggccagaaccagctgtacaacgaactgaacctgggacggcgggaagagtacgatgtgctggacaagcggcgcggccgggaccccgaaatgggcgggaagcctagaagaaagaaccctcaggaaggcctgtataacgagctgcagaaggacaagatggccgaggcctactccgaaattgggatgaagggagagcggcggaggggaaaggggcacgacggcctgtaccaaggactgtccaccgccaccaaggacacatacgatgccctgcacatgcaggcccttccccctcgc (SEQ ID NO: 27).
В другом аспекте настоящее изобретение относится популяции экспрессирующих CAR клеток, например, CART-клеток. В некоторых вариантах осуществления популяция экспрессирующих CAR клеток содержит смесь клеток, экспрессирующих различающиеся CAR. Например, в одном варианте осуществления популяция CART-клеток может включать первую клетку, экспрессирующую CAR, имеющий антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, и вторую клетку, экспрессирующую CAR, имеющий отличающийся антигенсвязывающий домен, например, антигенсвязывающий домен против отличающегося ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, например, антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, который отличается от ассоциированного со злокачественной опухолью антигена, связываемого антигенсвязывающим доменом CAR, экспрессируемого первой клеткой. В качестве другого примера популяция экспрессирующих CAR клеток может включать первую клетку, экспрессирующую CAR, которая включает антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, и вторую клетку, экспрессирующую CAR, который включает антигенсвязывающий домен против мишени, отличной от ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании. В одном варианте осуществления популяция экспрессирующих CAR клеток включает, например, первую клетку, экспрессирующую CAR, который включает первичный внутриклеточный сигнальный домен, и вторую клетку, экспрессирующую CAR, который включает вторичный сигнальный домен.
В другом аспекте настоящее изобретение относится популяции клеток, где по меньшей мере одна клетка в популяции экспрессирует CAR, имеющий антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, а вторая клетка экспрессирует другое средство, например, средство, которое повышает активность или функцию экспрессирующей CAR клетки. Например, в одном варианте осуществления средство может представлять собой средство, которое ингибирует ингибиторную молекулу. В некоторых вариантах осуществления ингибиторные молекулы, например PD-1, могут снижать способность экспрессирующей CAR клетки индуцировать иммунный эффекторный ответ. Примеры ингибиторных молекул включают PD-1, PD-L1, CTLA4, TIM3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 или TGF-бета. В одном варианте осуществления средство, которое ингибирует ингибиторную молекулу, например, представляет собой молекулу, описанную в настоящем описании, например, средство, которое содержит первый полипептид, например, ингибиторную молекулу, ассоциированную со вторым полипептидом, которая дает сигнал клетке, например, внутриклеточный сигнальный домен, описанный в настоящем описании. В одном варианте осуществления средство содержит первый полипептид, например, из ингибиторной молекулы, такой как PD-1, PD-L1, CTLA4, TIM3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 или TGF-бета, или фрагмент любого из них, и второй полипептид, который представляет собой внутриклеточный сигнальный домен, описанный в настоящем описании (например, содержащий костимулирующий домен (например, 41BB, CD27, OX40 или CD28, например, как описано в настоящем описании) и/или первичный сигнальный домен (например, сигнальный домен CD3-зета, описанный в настоящем описании). В одном варианте осуществления средство содержит первый полипептид PD-1 или его фрагмент, и второй полипептид внутриклеточного сигнального домена, описанного в настоящем описании (например, сигнальный домен CD28, описанный в настоящем описании, и/или сигнальный домен CD3-зета, описанный в настоящем описании).
В одном аспекте настоящее изобретение относится к способам, включающим введение популяции экспрессирующих CAR клеток, например, CART-клеток, например, к смеси клеток, экспрессирующих различные CAR, в комбинации с другим средством, например, ингибитором киназы, таким как ингибитор киназы, описанный в настоящем описании. В другом аспекте настоящее изобретение относится к способам, включающим введение популяции клеток, где по меньшей мере одна клетка в популяции экспрессирует CAR, имеющий антигенсвязывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, и вторая клетка экспрессирует другое средство, например, средство, которое усиливает активность экспрессирующей CAR клетки, в комбинации с другим средством, например, ингибитором киназы, таким как ингибитор киназы, описанный в настоящем описании.
Регулируемые химерные рецепторы антигенов
В некоторых вариантах осуществления является желательным регулируемый CAR (RCAR), где активность CAR можно контролировать, для оптимизации безопасности и эффективности терапии CAR. Существует много путей регуляции активности CAR. Например, индуцибельный апоптоз с использованием, например, каспазы, слитой с доменом димеризации (см., например, Di et al., N Engl. J. Med. 2011 Nov. 3; 365(18):1673-1683), можно использовать в качестве переключателя безопасности при терапии CAR по настоящему изобретению.
В одном аспекте RCAR содержит набор полипептидов, как правило, два в наиболее простых вариантах осуществления, в которых компоненты стандартного CAR, описанного в настоящем описании, например, антигенсвязывающий домен и внутриклеточный сигнальный домен, разделены на отдельные полипептиды или представители. В некоторых вариантах осуществления набор полипептидов включает переключатель димеризации, который в присутствии молекулы димеризации может связывать полипептиды друг с другом, например, может связывать антигенсвязывающий домен с внутриклеточным сигнальным доменом.
В одном аспекте RCAR содержит два полипептида или представителя: 1) внутриклеточный сигнальный представитель, содержащий внутриклеточный сигнальный домен, например, первичный внутриклеточный сигнальный домен, описанный в настоящем описании, и первый домен переключения; 2) антигенсвязывавющий представитель, содержащий антигенсвязывающий домен, например, который нацелен на опухолевый антиген, описанный в настоящем описании, как описано в настоящем описании, и второй домен переключения. Необязательно, RCAR содержит трансмембранный домен, описанный в настоящем описании. В одном варианте осуществления трансмембранный домен может быть расположен на внутриклеточном сигнальном представителе, на антигенсвязывающем представителе, или на обоих из них. (Если нет иных указаний, когда представители или элементы RCAR описаны в настоящем описании, порядок может быть таким, как представлено, однако также включены другие порядки. Иными словами, в одном варианте осуществления порядок является таким, как указано в тексте, а в других вариантах осуществления порядок может отличаться. Например, порядок элементов на одной стороне трансмембранной области может отличаться от приведенного в качестве примера, например, расположение домена переключения относительно внутриклеточного сигнального домена может отличаться (например, может быть обратным).
В одном варианте осуществления первый и второй домены переключения могут образовывать внутриклеточный или внеклеточный переключатель димеризации. В одном варианте осуществления переключатель димеризации может представлять собой переключатель гомодимеризации, например, где первый и второй домены переключения являются одинаковыми, или переключатель гетеродимеризации, например, где первый и второй домены переключения отличаются друг от друга.
В вариантах осуществления RCAR может содержать "множественный переключатель". Множественный переключатель может содержать домены-переключатели гетеродимеризации или домены-переключатели гомодимеризации. Множественный переключатель содержит множество, например 2, 3, 4, 5, 6, 7, 8, 9 или 10, доменов переключения, независимо, на первом представителе, например, на антигенсвязывающем представителе, и на втором представителе, например, на внутриклеточном сигнальном представителе. В одном варианте осуществления первый представитель может содержать множество первых доменов переключения, например, доменов переключения на основе FKBP, и второй представитель может содержать множество вторых доменов переключения, например, доменов переключения на основе FRB. В одном варианте осуществления первый представитель может содержать первый и второй домены переключения, например, домен переключения на основе FKBP и домен переключения на основе FRB, и второй представитель может содержать первый и второй домены переключения, например, домен переключения на основе FKBP и домен переключения на основе FRB.
В одном варианте осуществления внутриклеточный сигнальный представитель содержит один или несколько внутриклеточных сигнальных доменов, например, первичный внутриклеточный сигнальный домен и один или несколько костимулирующих сигнальных доменов.
В одном варианте осуществления антигенсвязывающий представитель может содержать один или несколько внутриклеточных сигнальных доменов, например, один или несколько костимулирующих сигнальных доменов. В одном варианте осуществления антигенсвязывающий представитель содержит множество, например 2 или 3, костимулирующих сигнальных доменов, описанных в настоящем описании, например, выбранных из 41BB, CD28, CD27, ICOS и OX40, и в вариантах осуществления не содержит первичного внутриклеточного сигнального домена. В одном варианте осуществления антигенсвязывающий представитель содержит следующие костимулирующие сигнальные домены от внеклеточного к внутриклеточному: 41BB-CD27; 41BB-CD27; CD27-41BB; 41BB-CD28; CD28-41BB; OX40-CD28; CD28-OX40; CD28-41BB или 41BB-CD28. В таких вариантах осуществления внутриклеточный связывающий представитель содержит домен CD3-зета. В одном таком варианте осуществления RCAR содержит (1) антигенсвязывающий представитель, содержащий антигенсвязывающий домен, трансмембранный домен, и два костимулирующих домена и первый домен переключения; и (2) внутриклеточный сигнальный домен, содержащий трансмембранный домен или связывающий с мембраной домен, и по меньшей мере один первичный внутриклеточный сигнальный домен, и второй домен переключения.
Вариант осуществления относится к RCAR, где антигенсвязывающий представитель не связан с поверхностью клетки CAR. Это позволяет клетке, имеющей внутриклеточный сигнальный представитель, удобным образом образовывать пару с одним или несколькими антигенсвязывающими доменами без трансформации клетки последовательностью, которая кодирует антигенсвязывающий представитель. В таких вариантах осуществления RCAR содержит: 1) внутриклеточный сигнальный представитель, содержащий: первый домен переключения, трансмембранный домен, внутриклеточный сигнальный домен, например, первичный внутриклеточный сигнальный домен, и первый домен переключения; и 2) антигенсвязывающий представитель, содержащий: антигенсвязывающий домен и второй домен переключения, где антигенсвязывающий представитель не содержит трансмембранный домен или связывающий с мембраной домен, и необязательно не содержит внутриклеточный сигнальный домен. В некоторых вариантах осуществления RCAR может дополнительно содержать 3) второй антигенсвязывающий представитель, содержащий: второй антигенсвязывающий домен, например, второй антигенсвязывающий домен, который связывает отличающийся антиген, чем у антигенсвязывающего домена; и второй домен переключения.
Также в рамках настоящего изобретения предусматриваются RCAR, где антигенсвязывающий представитель обладает способностью биспецифической активации и нацеливания. В этом варианте осуществления антигенсвязывающий представитель может содержать множество, например, 2, 3, 4 или 5 антигенсвязывающих доменов, например, scFv, где каждый антигенсвязывающий домен связывается с антигеном-мишенью, например, различными антигенами или одним антигеном, например, одним и тем же или различными эпитопами на одном антигене. В одном варианте осуществления множество антигенсвязывающих доменов располагаются тандемно, и необязательно между антигенсвязывающими доменами находится линкер или шарнирная область. Пригодные линкеры и шарнирные области описаны в настоящем описании.
Вариант осуществления относится к RCAR, имеющим конфигурацию, которая позволяет переключение пролиферации. В этом варианте осуществления RCAR содержит: 1) внутриклеточный сигнальный представитель, содержащий: необязательно трансмембранный домен или связывающий с мембраной домен; один или несколько костимулирующих сигнальных доменов, например, выбранных из 41BB, CD28, CD27, ICOS и OX40, и домен переключения; и 2) антигенсвязывающий представитель, содержащий: антигенсвязывающий домен, трансмембранный домен, и первичный внутриклеточный сигнальный домен, например, домен CD3-зета, где антигенсвязывающий представитель не содержит домен переключения или не содержит домен переключения, который димеризуется с доменом переключения на внутриклеточном сигнальном представителе. В одном варианте осуществления антигенсвязывающий представитель не содержит костимулирующий сигнальный домен. В одном варианте осуществления внутриклеточный сигнальный представитель содержит домен переключения из переключателя гомодимеризации. В одном варианте осуществления внутриклеточный сигнальный представитель содержит первый домен переключения переключателя гетеродимеризации и RCAR содержит второй внутриклеточный сигнальный представитель, который содержит второй домен переключения переключателя гетеродимеризации. В таких вариантах осуществления второй внутриклеточный сигнальный представитель содержит те же внутриклеточные сигнальные домены, что и внутриклеточный сигнальный представитель. В одном варианте осуществления переключатель димеризации является внутриклеточным. В одном варианте осуществления переключатель димеризации является внеклеточным.
В любой из конфигураций RCAR, описанных в настоящем описании, первый и второй домены переключения содержат переключатель на основе FKBP/FRB, как описано в настоящем описании.
Также в настоящем описании описаны клетки, содержащие RCAR, описанный в настоящем описании. Любую клетку, которая модифицирована способами инженерии для экспрессии RCAR, можно использовать в качестве клетки RCARX. В одном варианте осуществления клетка RCARX представляет собой T-клетку и ее обозначают как RCART-клетка. В одном варианте осуществления RCARX-клетка представляет собой NK-клетку и ее обозначают как RCARN-клетка.
Также в рамках настоящего изобретения предусматриваются нуклеиновые кислоты и векторы, содержащие кодирующие RCAR последовательности. Последовательность, кодирующая различные элементы RCAR, может располагаться на одной молекуле нуклеиновой кислоты, например, на одной плазмиде или векторе, например, вирусном векторе, например, лентивирусном векторе. В одном варианте осуществления (i) последовательность, кодирующая антигенсвязывающий представитель, и (ii) последовательность, кодирующая внутриклеточный сигнальный представитель, может присутствовать на одной нуклеиновой кислоте, например векторе. Продукции соответствующих белков можно достигать, например, с использованием отдельных промоторов или с использованием бицистронного продукта транскрипции (который может обеспечить продукцию двух белков посредством расщепления единого продукта трансляции или посредством трансляции двух отдельных белковых продуктов). В одном варианте осуществления между (i) и (ii) расположена последовательность, кодирующая расщепляемый пептид, например, последовательность P2A или F2A. В одном варианте осуществления между (i) и (ii) расположена последовательность, кодирующая IRES, например, IRES EMCV или EV71. В этих вариантах осуществления (i) и (ii) транскрибируются в качестве единой РНК. В одном варианте осуществления с (i) функционально связан первый промотор и с (ii) функционально связан второй промотор, так что (i) и (ii) транскрибируются в качестве отдельных мРНК. Примеры участков расщепления пептидов включают следующие, где остатки GSG являются необязательными:
T2A: (GSG) E G R G S L L T C G D V E E N P G P (SEQ ID NO: 68)
P2A: (GSG) A T N F S L L K Q A G D V E E N P G P (SEQ ID NO: 69)
E2A: (GSG) Q C T N Y A L L K L A G D V E S N P G P (SEQ ID NO: 70)
F2A: (GSG) V K Q T L N F D L L K L A G D V E S N P G P (SEQ ID NO: 71)
В одном варианте осуществления последовательность, кодирующая IRES, например, IRES EMCV или EV71, находится между (i) и (ii). В этих вариантах осуществления (i) и (ii) транскрибируются в качестве единой РНК. В одном варианте осуществления первый промотор функционально связан с (i) и второй промотор функционально связан с (ii), так что (i) и (ii) транскрибируются в качестве отдельных мРНК.
Альтернативно последовательность, кодирующая различные элементы RCAR, может быть расположена на различных молекулах нуклеиновых кислот, например, различных плазмидах или векторах, например, вирусных векторах, например, лентивирусных векторах. Например, последовательность (i), кодирующая антигенсвязывающий представитель, может присутствовать на первой нуклеиновой кислоте, например, первом векторе, и последовательность (ii), кодирующая внутриклеточный сигнальный представитель, может присутствовать на второй нуклеиновой кислоте, например, втором векторе.
Переключатели димеризации
Переключатели димеризациии могут быть нековалентными или ковалентными. В нековалентном переключателе димеризации молекула димеризации обеспечивает нековалентное взаимодействие между доменами переключения. В ковалентном переключателе димеризации молекула димеризации обеспечивает ковалентное взаимодействие между доменами переключения.
В одном варианте осуществления RCAR содержит переключатель димеризации наоснове FKBP/FRAP или FKBP/FRB. FKBP12 (FKBP, или связывающий FK506 белок) представляет собой распространенный цитоплазматический белок, который служит в качестве первоначальной внутриклеточной мишени для природного иммунодепрессивного лекарственного средства рапамицина. Рапамицин связывается с FKBP и с большим гомологом PI3K FRAP (RAFT, mTOR). FRB представляет собой часть FRAP из 93 аминокислот, которая является достаточной для связывания комплекса FKBP-рапамицин (Chen, J., Zheng, X. F., Brown, E. J. & Schreiber, S. L. (1995) Identification of an 11-kDa FKBP12- rapamycin-binding domain within the 289-kDa FKBP12- rapamycin-associated protein and characterization of a critical serine residue. Proc Natl Acad Sci U S A 92: 4947-51.)
В вариантах осуществления в переключателе на основе FKBP/FRAP, например, FKBP/FRB может использоваться молекула димеризации, например, рапамицин или аналог рапамицина.
Аминокислотная последовательность FKBP является следующей:
D V P D Y A S L G G P S S P K K K R K V S R G V Q V E T I S P G D G R T F P K R G Q T C V V H Y T G M L E D G K K F D S S R D R N K P F K F M L G K Q E V I R G W E E G V A Q M S V G Q R A K L T I S P D Y A Y G A T G H P G I I P P H A T L V F D V E L L K L E T S Y (SEQ ID NO: 54)
В вариантах осуществления домен переключения FKBP может содержать фрагмент FKBP, обладающий способностью связывать FRB, или его фрагмент или аналог в присутствии рапамицина или аналога, например, подчеркнутую часть SEQ ID NO: 54, которая представляет собой:
V Q V E T I S P G D G R T F P K R G Q T C V V H Y T G M L E D G K K F D S S R D R N K P F K F M L G K Q E V I R G W E E G V A Q M S V G Q R A K L T I S P D Y A Y G A T G H P G I I P P H A T L V F D V E L L K L E T S (SEQ ID NO: 55)
Аминокислотная последовательность FRB является следующей:
ILWHEMWHEG LEEASRLYFG ERNVKGMFEV LEPLHAMMER GPQTLKETSF NQAYGRDLME AQEWCRKYMK SGNVKDLTQA WDLYYHVFRR ISK (SEQ ID NO: 56)
"Переключатель на основе FKBP/FRAP, например, FKBP/FRB", как этот термин используют в настоящем описании, относится к переключателю димеризации, содержащему: первый домен переключения, который содержит фрагмент FKBP или его аналог, обладающие способностью связываться с FRB, или его фрагментом или аналогом, в присутствии рапамицина или рапалога, например RAD001, и обладает по меньшей мере 70, 75, 80, 85, 90, 95, 96, 97, 98 или 99% идентичностью с или отличается не более чем на 30, 25, 20, 15, 10, 5, 4, 3, 2 или 1 аминокислотный остаток от последовательности FKBP SEQ ID NO: 54 или 55; и второй домен переключения, который содержит фрагмент FRB или его аналоги, обладающие способностью связываться с FRB, или его фрагментом или аналогом, в присутствии рапамицина или рапалога, и обладает по меньшей мере 70, 75, 80, 85, 90, 95, 96, 97, 98 или 99% идентичностью с или отличается не более чем на 30, 25, 20, 15, 10, 5, 4, 3, 2 или 1 аминокислотный остаток от последовательности FRB SEQ ID NO: 56. В одном варианте осуществления RCAR, описанный в настоящем описании, содержит один домен переключения, который содержит аминокислотные остатки, описанные в SEQ ID NO: 54 (или SEQ ID NO: 55), и один домен переключения, который содержит аминокислотные остатки, описанные в SEQ ID NO: 56.
В вариантах осуществления переключатель димеризации FKBP/FRB содержит модифицированный домен переключения FRB, который проявляет измененное, например усиленное, образование комплекса между доменом переключения на основе FRB, например, модифицированным доменом переключения FRB, доменом переключения на основе FKBP и молекулой димеризации, например, рапамицином или рапалогом, например RAD001. В одном варианте осуществления модифицированный домен переключения FRB содержит одну или несколько мутаций, например 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, выбранных из мутаций в положении(ях) аминокислот L2031, E2032, S2035, R2036, F2039, G2040, T2098, W2101, D2102, Y2105 и F2108, где аминокислота дикого типа заменена любой другой встречающейся в природе аминокислотой. В одном варианте осуществления мутантный FRB содержит мутацию в положении E2032, где E2032 заменен на фенилаланин (E2032F), метионин (E2032M), аргинин (E2032R), валин (E2032V), тирозин (E2032Y), изолейцин (E2032I), например SEQ ID NO: 57, или лейцин (E2032L), например SEQ ID NO: 58. В одном варианте осуществления мутантный FRB содержит мутацию T2098, где T2098 заменен на фенилаланин (T2098F) или лейцин (T2098L), например, SEQ ID NO: 59. В одном варианте осуществления мутантный FRB содержит мутацию в E2032 и в T2098, где E2032 заменен на любую аминокислоту и где T2098 заменен на любую аминокислоту, например, SEQ ID NO: 60. В одном варианте осуществления мутантный FRB содержит мутацию E2032I и T2098L, например, SEQ ID NO: 61. В одном варианте осуществления мутантный FRB содержит мутацию E2032L и T2098L, например, SEQ ID NO: 62.
Таблица 13. Иллюстративный мутантный FRB, обладающий увеличенной аффинностью в отношении молекулы димеризации.
Другие пригодные переключатели димеризации включают переключатель димеризации на основе GyrB-GyrB, переключатель димеризации на основе гибберелина, переключатель димеризации метка/связывающий элемент, переключатель димеризации halo-метка/snap-метка. Согласно руководству, представленному в настоящем описании, такие переключатели и соответствующие молекулы димеризации будут очевидны специалисту в данной области.
Молекула димеризации
Ассоциацию между доменами переключения обеспечивает молекула димеризации. В присутствии молекулы димеризации взаимодействие или ассоциация между такими доменами обеспечивает передачу сигнала между полипептидом, связанным, например слитым, с первым доменом переключения, и полипептидом, связанным, например слитым, со вторым доменом переключения. В присутствии неограничивающих уровней молекулы димеризации передача сигнала возрастает в 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2, 5, 10, 50, 100 раз, например, при измерении в системе, описанной в настоящем описании.
Рапамицин и аналоги рапамицина (иногда называемые рапалогами), например RAD001, можно использовать в качестве молекул димеризации в переключателе димеризации на основе FKBP/FRB, описанном в настоящем описании. В одном варианте осуществления молекула димеризации может быть выбрана из рапамицина (сиролимус), RAD001 (эверолимус), зотапролимуса, темсиролимуса, AP-23573 (ридафоролимус), биолимуса и AP21967. Дополнительные аналоги рапамицина, пригодные для применения с переключателями димеризации на основе FKBP/FRB, описаны ниже в разделе "Комбинированная терапия", или в подразделе под названием "Иллюстративные ингибиторы mTOR".
Расщепленные CAR
В некоторых вариантах осуществления в экспрессирующих CAR клетках используется расщепленный CAR. Подход расщепленных CAR более подробно описан в публикациях WO2014/055442 и WO2014/055657. В кратком изложении, система расщепленного CAR включает клетку, экспрессирующую первый CAR, обладающий первым антигенсвязывающим доменом и костимулирующим доменом (например, 41BB), и клетка также экспрессирует второй CAR, обладающий вторым антигенсвязывающим доменом и внутриклеточным сигнальным доменом (например, CD3-зета). Когда клетка встречает первый антиген, костимулирующий домен активируется, и клетка пролиферирует. Когда клетка встречает второй антиген, внутриклеточный сигнальный домен активируется и начинается активность уничтожения клеток. Таким образом, экспрессирующая CAR клетка полностью активируется в присутствии обоих антигенов.
Трансфекция РНК
В настоящем описании описаны способы получения транскрибируемой in vitro РНК CAR. Настоящее изобретение также включает кодирующую CAR конструкцию РНК, которая может быть прямо трансфицирована в клетку. Способ получения мРНК для применения в трансфекции может вовлекать транскрипцию in vitro (IVT) матрицы с использованием специально сконструированных праймеров с последующим присоединением поли-A, с получением конструкции, содержащей 3'- и 5'-нетранслируемую последовательность ("UTR"), 5'-кэп и/или участок внутренней посадки рибосомы (IRES), нуклеиновую кислоту, подлежащую экспрессии, и поли-A хвостовую часть, как правило, длиной 50-2000 оснований (SEQ ID NO: 32). РНК, полученной таким образом, можно эффективно трансфицировать различные типы клеток. В одном аспекте матрица включает последовательности для CAR.
В одном аспекте CAR по настоящему изобретению кодируется матричной РНК (мРНК). В одном аспекте мРНК, кодирующую CAR, описанный в настоящем описании, вводят в иммунную эффекторную клетку, например, T-клетку или NK-клетку, для получения экспрессирующей CAR клетки, например, CART-клетки или CAR NK-клетки.
В одном варианте осуществления транскрибируемую in vitro РНК CAR можно вводить в клетку посредством временной трансфекции. РНК продуцируют путем транскрипции in vitro с использованием матрицы, полученной посредством полимеразной цепной реакции (ПЦР). Представляющую интерес ДНК из любого источника можно прямо конвертировать способом ПЦР в матрицу для синтеза мРНК in vitro с использованием подходящих праймеров и РНК-полимеразы. Источником ДНК может быть, например, геномная ДНК, плазмидная ДНК, фаговая ДНК, кДНК, синтетическая последовательность ДНК или любой другой подходящий источник ДНК. Желаемой матрицей для транскрипции in vitro является CAR, описанный в настоящем описании. Например, матрица для РНК CAR содержит внеклеточную область, содержащую одноцепочечный вариабельный домен антитела к ассоциированному с опухолью антигену, описанному в настоящем описании; шарнирную область (например, шарнирную область, описанную в настоящем описании), трансмембранный домен (например, трансмембранный домен, описанный в настоящем описании, такой как трансмембранный домен CD8a); и цитоплазматическую область, которая включает внутриклеточный сигнальный домен, например, внутриклеточный ссигнальный домен, описанный в настоящем описании, например, содержащий сигнальный домен CD3-зета и сигнальный домен 4-1BB.
В одном варианте осуществления ДНК для применения в ПЦР содержит открытую рамку считывания. ДНК может происходить из встречающейся в природе последовательности ДНК генома организма. В одном варианте осуществления нуклеиновая кислота может включать некоторые или все из 5'- и/или 3'-нетранслируемых областей (UTR). Нуклеиновая кислота может включать экзоны и интроны. В одном варианте осуществления ДНК для применения в ПЦР представляет собой последовательность нуклеиновой кислоты человека. В другом варианте осуществления ДНК для применения в ПЦР представляет собой последовательность нуклеиновой кислоты человека, включающую 5'- и 3'-UTR. Альтернативно ДНК может представлять собой искусственную последовательность ДНК, которая в норме не экспрессируется во встречающемся в природе организме. Иллюстративная искусственная последовательность ДНК представляет собой последовательность ДНК, которая содержит части генов, которые лигированы с образованием открытой рамки считывания, которая кодирует слитый белок. Части ДНК, которые лигированы вместе, могут быть из одного организма или из более чем одного организма.
ПЦР используют для получения матрицы для транскрипции in vitro мРНК, которую используют для трансфекции. Способы проведения ПЦР хорошо известны в данной области. Праймеры для применения в ПЦР конструируют так, чтобы они имели области, которые по существу комплементарны областям ДНК, используемым в качестве матрицы для ПЦР. "По существу комплементарный", как используют в настоящем описании, относится к последовательностям нуклеотидов, где большинство или все из оснований в последовательности праймеров являются комплементарными, или одно или несколько оснований являются некомплементарными или несоответствующими друг другу. По существу комплементарные последовательности способны к отжигу или гибридизации с предполагаемой ДНК-мишенью в условиях отжига, используемых для ПЦР. Праймеры можно конструировать, чтобы они были по существу комплементарными любой части ДНК-матрицы. Например, праймеры можно конструировать, чтобы они амплифицировали часть нуклеиновой кислоты, которая обычно транскрибируется в клетках (открытая рамка считывания), включая 5'- и 3'-UTR. Праймеры также можно конструировать для амплификации части нуклеиновой кислоты, которая кодирует конкретный представляющий интерес домен. В одном варианте осуществления праймеры конструируют для амплификации кодирующей области кДНК человека, включая все или части 5'- и 3'-UTR. Праймеры, пригодные для ПЦР, можно получать способами синтеза, которые хорошо известны в данной области. "Прямые праймеры" представляют собой праймеры, которые содержат область нуклеотидов, которая по существу комплементарна нуклеотидам на ДНК-матрице, являющейся вышележащей относительно последовательности ДНК, подлежащей амплификации. "Вышележащий" используют в настоящем описании для обозначения положения с 5'-стороны от последовательности ДНК, подлежащей амплификации, относительно кодирующей цепи. "Обратные праймеры" представляют собой праймеры, которые содержат область нуклеотидов, по существу комплементарную двухцепочечной ДНК-матрице, которая являятся нижелещей относительно последовательности ДНК, подлежащей амплификации. "Нижележащий" используют в настоящем описании для обозначения положения с 3'-стороны от последовательности ДНК, подлежащей амплификации, относительно кодирующей цепи.
В способах, описанных в настоящем описании, можно использовать любую ДНК-полимеразу, пригодную для ПЦР. Реагенты и полимераза являются коммерчески доступными из ряда источников.
Также можно использовать химические структуры, обладающие способностью повышать стабильность и/или эффективность трансляции. Предпочтительно РНК имеет 5'- и 3'-UTR. В одном варианте осуществления 5'-UTR имеет длину от одного до 3000 нуклеотидов. Длину последовательностей 5'- и 3'-UTR для присоединения к кодирующей области можно изменять различными способами, включая, но не ограничиваясь ими, конструирование праймеров для ПЦР которые гибридизуются с различными областями UTR. С использованием этого подхода специалист в данной области может модифицировать длину 5'- и 3'-UTR, требуемую для достижения оптимальной эффективности трансляции после трансфекции транскрибированной РНК.
5' и 3'-UTR могут быть представлять собой встречающиеся в природе эндогенные 5'- и 3'-UTR для представляющей интерес нуклеиновой кислоты. Альтернативно последовательности UTR, которые являются эндогенными для представляющей интерес нуклеиновой кислоты, можно добавлять путем включения последовательностей UTR в прямые и обратные праймеры или посредством любых других модификаций матрицы. Использование последовательностей UTR, которые не являются эндогенными для представляющей интерес нуклеиновой кислоты, может быть полезным для модификации стабильности и/или эффективности трансляции РНК. Например, известно, что AU-богатые элементы в последовательностях 3'-UTR могут снижать стабильность мРНК. Таким образом, 3'-UTR можно выбирать или конструировать для повышения стабильности транскрибируемой РНК, исходя из свойств UTR, которые хорошо известны в данной области.
В одном варианте осуществления 5'-UTR может содержать последовательность Козака из эндогенной нуклеиновой кислоты. Альтернативно, когда 5'-UTR, которая не является эндогенной для представляющей интерес нуклеиновой кислоты, добавляют способом ПЦР, как описано выше, консенсусную последовательность Козака можно переконструировать путем добавления последовательности 5'-UTR. Последовательности Козака могут повышать эффективность трансляции некоторых РНК-транскриптов, но, по-видимому, не требуются для обеспечения эффективной трансляции всех РНК. Требование наличия последовательностей Козака для многих мРНК известно в данной области. В других вариантах осуществления 5'-UTR может представлять собой 5'-UTR РНК-вируса, РНК-геном которого стабилен в клетках. В других вариантах осуществления в 3'- или 5'-UTR можно использовать различные нуклеотидные аналоги для препятствования деградации мРНК экзонуклеазами.
Для обеспечения синтеза РНК с ДНК-матрицы без необходимости в клонировании генов промотор транскрипции должен быть связан с ДНК-матрицей выше последовательности, подлежащей транскрипции. Когда последовательность, которая функционирует в качестве промотора РНК-полимеразы, добавляют к 5'-концу прямого праймера, промотор РНК-полимеразы включается в продукт ПЦР выше открытой рамки считывания, подлежащей транскрипции. В одном предпочтительном варианте осуществления промотор представляет собой промотор полимеразы T7, как описано в настоящем описании. Другие пригодные промоторы включают, но не ограничиваются ими, промоторы РНК-полимеразы T3 и SP6. Консенсусные нуклеотидные последовательности для промоторов T7, T3 и SP6, известны в данной области.
В предпочтительном варианте осуществления мРНК имеет как кэп на 5'-конце, так и 3'-поли(A)-хвостовую часть, которая определяет связывание рибосом, инициацию трансляции и стабильность мРНК в клетке. На кольцевой ДНК-матрице, например, плазмидной ДНК, РНК-полимераза продуцирует длинный конкатемерный продукт, который не пригоден для экспрессии в эукариотических клетках. Транскрипция плазмидной ДНК, линеаризованной на конце 3'-UTR, приводит к мРНК нормального размера, которая не является эффективной при эукариотической трансфекции, даже если она полиаденилируется после транскрипции.
На линейной ДНК-матрице РНК-полимераза фага T7 может удлинять 3'-конец транскрипта за пределы последнего основания матрицы (Schenborn and Mierendorf, Nuc Acids Res., 13:6223-36 (1985); Nacheva and Berzal-Herranz, Eur. J. Biochem., 270:1485-65 (2003).
Общепринятым способом встраивания участков поли-A/T в ДНК-матрицу является молекулярное клонирование. Однако последовательность поли-A/T, встроенная в плазмидную ДНК, может приводить к нестабильности плазмиды, поэтому плазмидные ДНК-матрицы, полученные из бактериальных клеток, часто в высокой степени контаминированы делециями и другими аберрациями. Это делает процедуру клонирования не только трудоемкой и времязатратной, но часто и ненадежной. Поэтому является в высокой степени желаемым способ, который позволит конструирование ДНК-матриц с 3'-участками поли-A/T без клонирования.
Сегмент поли-A/T в транскрипционной ДНК-матрице можно продуцировать в процессе ПЦР с использованием обратного праймера, содержащего поли-T-хвостовую часть, такую как 100T-хвостовая часть (SEQ ID NO: 35) (размер может составлять 50-5000 T (SEQ ID NO: 36)), или после ПЦР любым другим способом, включая, но не ограничиваясь ими, лигирование ДНК или рекомбинацию in vitro. Поли-(A)-хвостовые части также обеспечивают стабильность РНК и снижают их деградацию. Как правило, длина поли-(A)-хвостовой части положительно коррелирует со стабильностью транскрибированной РНК. В одном варианте осуществления поли(A)-хвостовая часть имеет от 100 до 5000 остатков аденозина (SEQ ID NO: 37).
Поли(A)-хвостовые части РНК, кроме того, можно удлинять после транскрипции in vitro с использованием поли(A)-полимеразы, такой как поли-A-полимераза E. coli (E-PAP). В одном варианте осуществления увеличение длины поли(A)-хвостовой части от 100 нуклеотидов до 300-400 нуклеотидов (SEQ ID NO: 38) приводит приблизительно к двукратному повышению эффективности трансляции РНК. Кроме того, присоединение различных химических групп к 3'-концу может повышать стабильность мРНК. Такое присоединение может включать модифицированные/искусственные нуклеотиды, аптамеры и другие соединения. Например, ATP аналоги можно встраивать в поли(A)-хвостовую часть с использованием поли(A)-полимеразы. Кроме того, стабильность РНК могут повышать аналоги ATP.
5'-кэпы также обеспечивают стабильность молекул РНК. В предпочтительном варианте осуществления РНК, продуцируемые способами, описанными в настоящем описании, включают 5'-кэп. 5'-кэп предоставляют с использованием способов, известных в данной области и описанных в настоящем описании (Cougot, et al., Trends in Biochem. Sci., 29:436-444 (2001); Stepinski, et al., RNA, 7:1468-95 (2001); Elango, et al., Biochim. Biophys. Res. Commun., 330:958-966 (2005)).
РНК, продуцируемые способами, описанными в настоящем описании, также могут содержать последовательность участка внутренней посадки рибосомы (IRES). Последовательность IRES может представлять собой любую вирусную, хромосомную или искусственно сконструированную последовательность, которая инициирует кэп-независимое связывание рибосомы с мРНК и способствует инициации трансляции. Могут быть включены любые растворенные вещества, пригодные для электропорации клеток, которые могут содержать факторы, способствующие клеточной проницаемости и жизнеспособности, такие как сахара, пептиды, липиды, белки, антиоксиданты и поверхностно-активные вещества.
РНК можно включать в клетки-мишени с использованием любого из ряда различных способов, например коммерчески доступными способами, которые включают, но не ограничиваются ими, электропорацию (Amaxa Nucleofector-II (Amaxa Biosystems, Cologne, Германия)), (ECM 830 (BTX) (Harvard Instruments, Boston, Mass.) или Gene Pulser II (BioRad, Denver, Colo.), Multiporator (Eppendort, Hamburg, Германия), катионную опосредуемую липосомами трансфекцию с использованием липофекции, инкапсулирование в полимер, опосредуемую пептидом трансфекцию, или системы биолистической доставки частиц, такие как "генные пушки" (см., например, Nishikawa, et al. Hum Gene Ther., 12(8):861-70 (2001).
Невирусные способы доставки
В некоторых аспектах невирусные способы можно использовать для доставки нуклеиновой кислоты, кодирующей CAR, описанный в настоящем описании, в клетку, или ткань, или индивидууму.
В некоторых вариантах осуществления невирусный способ включает применение транспозона (также называемого мобильным генетическим элементом). В некоторых вариантах осуществления транспозон представляет собой фрагмент ДНК, который может встраиваться в некотором положении в геноме, например, фрагмент ДНК, который способен самореплицироваться и встраивать его копию в геном, или фрагмент ДНК, который может вырезаться посредством сплайсинга из более длинной нуклеиновой кислоты и встраиваться в другое положение в геноме. Например, транспозон содержит последовательность ДНК, состоящую из инвертированных повторов, фланкирующих гены, для транспозинции.
Иллюстративные способы доставки нуклеиновых кислот с использованием транспозона включают транспозонную систему Sleeping Beauty (SBTS) и транспозонную систему piggyBac (PB). См., например, Aronovich et al. Hum. Mol. Genet. 20.R1(2011): R14-20; Singh et al. Cancer Res. 15(2008):2961-2971; Huang et al. Mol. Ther. 16(2008):580-589; Grabundzija et al. Mol. Ther. 18(2010):1200-1209; Kebriaei et al. Blood. 122,21(2013):166; Williams. Molecular Therapy 16,9(2008):1515-16; Bell et al. Nat. Protoc. 2,12(2007):3153-65; и Ding et al. Cell. 122,3(2005):473-83, все из которых включены в настоящее описание в качестве ссылок.
SBTS включает два компонента: 1) транспозон, содержащий трансген, и 2) источник фермента транспозазы. Транспозаза может переносить транспозон из плазмиды-носителя (или другой донорной ДНК) в ДНК-мишень, такую как хромосома/геном клетки-хозяина. Например, транспозаза связывается с ДНК плазмиды-носителя/донора, вырезает транспозон (включая трансген(ы)) из плазмиды, и встраивает его в геном клетки-хозяина. См., например, Aronovich et al., выше.
Иллюстративные транспозоны включают транспозон на основе pT2. См., например, Grabundzija et al. Nucleic Acids Res. 41,3(2013):1829-47; и Singh et al. Cancer Res. 68,8(2008): 2961-2971, все из которых включены в настоящее описание путем ссылок. Иллюстративные транспозазы включают транспозазу Tc1/mariner-типа, например, транспозазу SB10 или транспозазу SB11 (сверхактивная транспозаза, которая может экспрессироваться, например, из промотора цитомегаловируса). См., например, Aronovich et al.; Kebriaei et al.; и Grabundzija et al., все из которых включены в настоящее описание путем ссылок.
Применение SBTS позволяет эффективное встраивание и экспрессию трансгена, например, нуклеиновой кислоты, кодирующей CAR, описанный в настоящем описании. В рамках настоящего изобретения предусматривается получение клетки, например, T-клетки или NK-клетки, которая стабильной экспрессирует CAR, описанный в настоящем описании, например, с использованием системы транспозона, такой как SBTS.
В соответствии со способами, описанными в настоящем описании, в некоторых вариантах осуществления одну или несколько нуклеиновых кислот, например, плазмид, содержащих компоненты SBTS, доставляют в клетку (например, T- или NK-клетку). Например, нуклеиновую кислоту(ы) доставляют стандартными способами доставки нуклеиновых кислот (например, плазмидной ДНК), например, способами, описанными в настоящем описании, например, электропорацией, трансфекцией или липофекцией. В некоторых вариантах осуществления нуклеиновая кислота содержит транспозон, содержащий трансген, например, нуклеиновую кислоту, кодирующую CAR, описанный в настоящем описании. В некоторых вариантах осуществления нуклеиновая кислота содержит транспозон, содержащий трансген (например, нуклеиновую кислоту, кодирующую CAR, описанный в настоящем описании), а также последовательность нуклеиновой кислоты, кодирующую фермента транспозазу. В других вариантах осуществления предусматривается система с двумя нуклеиновыми кислотами, например, система с двумя плазмидами, например, где первая плазмида содержит транспозон, содержащий трансген, а вторая плазмида содержит последовательность нуклеиновой кислоты, кодирующую фермент транспозазу. Например, первую и вторую нуклеиновые кислоты доставляют в клетку совместно.
В некоторых вариантах осуществления получают клетки, например, T- или NK-клетки, которые экспрессируют CAR, описанный в настоящем описании, с использованием комбинации встраивания гена с использованием SBTS и генетического редактирования с использованием нуклеазы (например, нуклеазы с цинковыми пальцами (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), система CRISPR/Cas или сконструированные мегануклеазы - переконструированные хоминг-эндонуклеазы).
В некоторых вариантах осуществления применение невирусного способа доставки позволяет перепрограммирование клеток, например, T- или NK-клеток, и прямую инфузию клеток индивидууму. Преимущества невирусных векторов включают, но не ограничиваются ими, простоту и относительно низкую стоимость получения достаточных количеств, требуемых для удовлетворения потребностей популяции пациентов, стабильность при хранении и отсутствие иммуногенности.
Конструкции нуклеиновых кислот, кодирующие CAR
Настоящее изобретение также относится к молекулам нуклеиновых кислот, кодирующим одну или несколько конструкций CAR, описанных в настоящем описании. В одном аспекте молекулу нуклеиновой кислоты предоставляют в качестве матричной РНК-транскрипта. В одном аспекте молекулу нуклеиновой кислоты предоставляют в качестве конструкции ДНК.
Таким образом, в одном аспекте изобретение относится к молекуле нуклеиновой кислоты, кодирующей химерный рецептор антигена (CAR), где CAR содержит антигенсвязывающий домен, который связывается с опухолевым антигеном, описанным в настоящем описании, трансмембранный домен (например, трансмембранный домен, описанный в настоящем описании) и внутриклеточный сигнальный домен (например, внутриклеточный сигнальный домен, описанный в настоящем описании), содержащий стимулирующей домен, например, костимулирующий сигнальный домен (например, костимулирующий сигнальный домен, описанный в настоящем описании) и/или первичный сигнальный домен (например, первичный сигнальный домен, описанный в настоящем описании, например, зета-цепь, описанную в настоящем описании). В одном варианте осуществления трансмембранный домен представляет собой трансмембранный домен белка, выбранный из группы, состоящей из альфа-, бета- или зета-цепи T-клеточного рецептора, CD28, CD3-эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154. В некоторых вариантах осуществления трансмембранный домен может включать по меньшей мере трансмембранную область(и), например, из KIRDS2, OX40, CD2, CD27, LFA-1 (CD11a, CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD40, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), NKp44, NKp30, NKp46, CD160, CD19, IL2R-бета, IL2R-гамма, IL7R α, ITGA1, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, NKG2D, NKG2C, TNFR2, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Tactile), CEACAM1, CRTAM, Ly9 (CD229), CD160 (BY55), PSGL1, CD100 (SEMA4D), SLAMF6 (NTB-A, Ly108), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, PAG/Cbp.
В одном варианте осуществления трансмембранный домен содержит последовательность SEQ ID NO: 12 или последовательность, обладающую 95-99% идентичностью с ней. В одном варианте осуществления антигенсвязывающий домен связан с трансмембранным доменом посредством шарнирной области, например, шарнирной области, описанной в настоящем описании. В одном варианте осуществления шарнирная область содержит SEQ ID NO: 4 или SEQ ID NO: 6 или SEQ ID NO: 8 или SEQ ID NO: 10, или последовательность, обладающую 95-99% идентичностью с ними. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кроме того, содержит последовательность, кодирующую костимулирующий домен. В одном варианте осуществления костимулирующий домен представляет собой функциональный сигнальный домен белка, выбранного из группы, состоящей из OX40, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278) и 4-1BB (CD137). Следующие примеры таких костимулирующих молекул включают CDS, ICAM-1, GITR, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), NKp44, NKp30, NKp46, CD160, CD19, CD4, CD8-альфа, CD8-бета, IL2R-бета, IL2R-гамма, IL7R-альфа, ITGA4, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, NKG2D, NKG2C, TNFR2, TRANCE/RANKL, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Tactile), CEACAM1, CRTAM, Ly9 (CD229), CD160 (BY55), PSGL1, CD100 (SEMA4D), CD69, SLAMF6 (NTB-A, Ly108), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, LAT, GADS, SLP-76 и PAG/Cbp. В одном варианте осуществления костимулирующий домен содержит последовательность SEQ ID NO: 16, или последовательность, обладающую 95-99% идентичностью с ней. В одном варианте осуществления внутриклеточный сигнальный домен содержит функциональный сигнальный домен 4-1BB и функциональный сигнальный домен CD3-зета. В одном варианте осуществления внутриклеточный сигнальный домен содержит последовательность SEQ ID NO: 14 или SEQ ID NO: 16, или последовательность, обладающую 95-99% идентичностью с ней, и последовательность SEQ ID NO: 18 или SEQ ID NO: 20, или последовательность, обладающую 95-99% идентичностью с ней, где последовательности, содержащие внутриклеточный сигнальный домен, экспрессируются в одной рамке считывания и в качестве одной полипептидной цепи.
В другом аспекте изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей конструкцию CAR, содержащую лидерную последовательность SEQ ID NO: 2, scFv-домен, как описано в настоящем описании, шарнирную область SEQ ID NO: 4, или SEQ ID NO: 6, или SEQ ID NO: 8, или SEQ ID NO: 10 (или последовательность, обладающую 95-99% идентичностью с ней), трансмембранный домен, обладающий последовательностью SEQ ID NO: 12 (или последовательностью, обладающей 95-99% идентичностью с ней), костимулирующий домен 4-1BB, имеющий последовательность SEQ ID NO: 14, или костимулирующий домен CD27, имеющий последовательность SEQ ID NO: 16 (или последовательность, обладающую 95-99% идентичностью с ней) и стимулирующий домен CD3-зета, обладающий последовательностью SEQ ID NO: 18 или SEQ ID NO: 20 (или последовательностью, обладающей 95-99% идентичностью с ней).
В другом аспекте изобретение относится к молекуле нуклеиновой кислоты, кодирующей молекулу химерного рецептора антигена (CAR), которая содержит антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, содержащий стимулирующий, и где указанный антигенсвязывающий домен связывается с опухолевым антигеном, выбранным из группы, состоящей из: TSHR, CD19, CD123, CD22, CD30, CD171, CS-1, CLL-1, CD33, EGFRvIII, GD2, GD3, BCMA, Tn Ag, PSMA, ROR1, FLT3, FAP, TAG72, CD38, CD44v6, CEA, EPCAM, B7H3, KIT, IL-13Ra2, мезотелина, IL-11Ra, PSCA, PRSS21, VEGFR2, LewisY, CD24, PDGFR-бета, SSEA-4, CD20, рецептора фолатов альфа, ERBB2 (Her2/neu), MUC1, EGFR, NCAM, простазы, PAP, ELF2M, эфрина B2, рецептора IGF-I, CAIX, LMP2, gp100, bcr-abl, тирозиназы, EphA2, фукозил-GM1, sLe, GM3, TGS5, HMWMAA, o-ацетил-GD2, рецептора фолатов бета, TEM1/CD248, TEM7R, CLDN6, GPRC5D, CXORF61, CD97, CD179a, ALK, полисиаловой кислоты, PLAC1, GloboH, NY-BR-1, UPK2, HAVCR1, ADRB3, PANX3, GPR20, LY6K, OR51E2, TARP, WT1, NY-ESO-1, LAGE-1a, MAGE-A1, легумаина, HPV E6,E7, MAGE A1, ETV6-AML, белка спермы 17, XAGE1, Tie 2, MAD-CT-1, MAD-CT-2, Fos-родственного антигена 1, p53, мутанта p53, простеина, сурвивина и теломеразы, PCTA-1/галектина 8, MelanA/MART1, мутанта Ras, hTERT, точек разрыва при транслокации саркомы, ML-IAP, ERG (слитый ген TMPRSS2 ETS), NA17, PAX3, рецептора андрогенов, циклина B1, MYCN, RhoC, TRP-2, CYP1B1, BORIS, SART3, PAX5, OY-TES1, LCK, AKAP-4, SSX2, RAGE-1, обратной транскриптазы теломеразы человека, RU1, RU2, кишечной карбоксилэстераы, mut hsp70-2, CD79a, CD79b, CD72, LAIR1, FCAR, LILRA2, CD300LF, CLEC12A, BST2, EMR2, LY75, GPC3, FCRL5 и IGLL1
В одном варианте осуществления кодируемая молекула CAR, кроме того, содержит последовательность, кодирующую костимулирующий домен. В одном варианте осуществления костимулирующий домен представляет собой функциональный сигнальный домен белка, выбранного из группы, состоящей из OX40, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18) и 4-1BB (CD137). В одном варианте осуществления костимулирующий домен содержит последовательность SEQ ID NO: 14. В одном варианте осуществления трансмембранный домен представляет собой трансмембранный домен белка, выбранного из группы, состоящей из альфа-, бета- или зета-цепи T-клеточного рецептора, CD28, CD3-эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154. В одном варианте осуществления трансмембранный домен содержит последовательность SEQ ID NO: 12. В одном варианте осуществления внутриклеточный сигнальный домен содержит функциональный сигнальный домен 4-1BB и функциональный сигнальный домен зета. В одном варианте осуществления внутриклеточный сигнальный домен содержит последовательность SEQ ID NO: 14 и последовательность SEQ ID NO: 18, где последовательности, содержащие внутриклеточный сигнальный домен, экспрессируются в одной рамке считывания и в качестве единой полипептидной цепи. В одном варианте осуществления связывающий домен против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, связан с трансмембранным доменом шарнирной областью. В одном варианте осуществления шарнирная область содержит SEQ ID NO: 4. В одном варианте осуществления шарнирная область содержит SEQ ID NO: 6, SEQ ID NO: 8 или SEQ ID NO: 10.
Последовательности нуклеиновых кислот, кодирующие желаемые молекулы, можно получать с использованием рекомбинантных способов, известных в данной области, например, таких как скрининг библиотек клеток, экспрессирующих ген, получение гена из вектора, о котором известно, что он его включает, или выделение непосредственно из клеток или тканей, содержащих его, с использованием стандартных способов. Альтернативно представляющий интерес ген может получать способами синтеза, а не клонировать.
Настоящее изобретение также относится к векторам, в которые встроена ДНК по настоящему изобретению. Векторы, происходящие из ретровирусов, таких как лентивирус, являются подходящими инструментами для достижения долговременного переноса генов, поскольку они обеспечивают долговременное стабильное встраивание трансгена и его увеличение в количестве в дочерних клетках. Лентивирусные векторы имеют дополнительное преимущество над векторами, происходящими из онкоретровирусов, таких как вирусы лейкоза мыши, поскольку они могут трансдуцировать непролиферирующие клетки, такие как гепатоциты. Также они имеют дополнительное преимущество, состоящее в низкой иммуногенности. Ретровирусный вектор также может представлять собой, например, гамма-ретровирусный вектор. Гамма-ретровирусный вектор может включать, например, промотор, сигнал упаковывания (ψ), участок связывания праймера (PBS), один или несколько (например, два) длинных концевых повторов (LTR) и представляющий интерес трансген, например, ген, кодирующий CAR. Гамма-ретровирусный вектор может быть лишен структурных генов вируса, таких как gag, pol и env. Иллюстративные гамма-ретровирусные векторы включают вирус лейкоза мышей (MLV), вирус некроза селезенки (SFFV) и вирус миелопролиферативной саркомы (MPSV), и векторы, происходящие из них. Другие гамма-ретровирусные векторы описаны, например, в Tobias Maetzig et al., "Gammaretroviral Vectors: Biology, Technology and Application" Viruses. 2011 Jun; 3(6): 677-713.
В другом варианте осуществления вектор, содержащий нуклеиновую кислоту, кодирующую желаемый CAR по изобретению, представляет собой аденовирусный вектор (A5/35). В другом варианте осуществления экспрессию нуклеиновых кислот, кодирующих CAR, можно проводить с использованием транспозонов, таких как sleeping beauty, crisper, CAS9 и нуклеазы с цинковыми пальцами цинк. См. ниже June et al. 2009 Nature Reviews Immunology 9,10: 704-716, включенную в настоящее описание путем ссылки.
В кратком изложении, экспрессии природных или синтетических нуклеиновых кислот, кодирующих CAR, обычно достигают путем функционального связывания нуклеиновой кислоты, кодирующей полипептид CAR или его части, с промотором, и встраивания конструкции в экспрессирующий вектор. Векторы могут быть пригодными для репликации и встраивания у эукариот. Типичные клонирующие векторы содержат терминаторы транскрипции и трансляции, последовательности инициации и промотор, пригодные для регуляции экспрессии желаемой последовательности нуклеиновой кислоты.
Экспрессирующие конструкции по настоящему изобретению также можно использовать для иммунизации нуклеиновыми кислотами и генной терапии с использованием стандартных протоколов доставки генов. Способы доставки генов известны в данной области. См., например, патенты США № 5399346, 5580859, 5589466, включенные в настоящее описание путем ссылок в полном объеме. В другом варианте осуществления изобретение относится к вектору для генной терапии.
Нуклеиновую кислоту можно клонировать в множество типов векторов. Например, нуклеиновую кислоту можно клонировать в вектор, включающий, но не ограничивающийся ими, плазмиду, фагмиду, производное фага, вирус животных и космиду. Векторы, представляющие особый интерес, включают экспрессирующие векторы, реплицирующиеся векторы, векторы для получения зондов и векторы для секвенирования.
Кроме того, экспрессирующий вектор может быть предоставлен клетке в форме вирусного вектора. Технология вирусных векторов хорошо известна в данной области и описана, например, в Sambrook et al., 2012, MOLECULAR CLONING: A LABORATORY MANUAL, volumes 1-4, Cold Spring Harbor Press, NY), и в других справочниках по вирусологии и молекулярной биологии. Вирусы, которые пригодны в качестве векторов, включают, но не ограничиваются ими, ретровирусы, аденовирусы, аденоассоциированные вирусы, вирусы герпеса и лентивирусы. Как правило, подходящий вектор содержит ориджин репликации, функциональный по меньшей мере в одном организме, промоторную последовательность, удобные участки для эндонуклеаз рестрикции, и один или несколько селективных маркеров (например, WO 01/96584; WO 01/29058 и патент США № 6326193).
Был разработан ряд систем на основе вирусов для переноса генов в клетки млекопитающих. Например, ретровирусы обеспечивают удобную платформу для систем доставки генов. Выбранный ген можно встраивать в вектор и упаковывать ретровирусные частицы с использованием способов, известных в данной области. Затем рекомбинантный вирус можно выделять и доставлять в клетки индивидуума либо in vivo, либо ex vivo. В данной области известен ряд ретровирусных систем. В некоторых вариантах осуществления используют аденовирусные векторы. В данной области известен ряд аденовирусных векторов. В одном варианте осуществления используют лентивирусные векторы.
Дополнительные промоторные элементы, например энхансеры, регулируют частоту инициаций транскрипции. Как правило, они расположены в области на 30-110 п.о. выше участка инициации, хотя было показано, что ряд промоторов также содержат функциональные элементы ниже участка инициации. Спейсер между промоторными элементами часто является гибким, так что промоторная функция сохраняется, когда элементы инвертируются или смещаются друг относительно друга. В промоторе тимидинкиназы (tk) спейсер между промоторными элементами может быть увеличен до 50 п.о. до того, когда активность начнет снижаться. Оказалось, что в зависимости от промотора индивидуальные элементы могут функционировать либо совместно, либо независимо, активируя транскрипцию. Иллюстративные промоторы включают промоторы гена IE CMV, EF-1α, убиквитина C или фосфоглицерокиназы (PGK).
Примером промотора, который способен экпрессировать кодирующую CAR молекулу нуклеиновой кислоты в T-клетке млекопитающего, является промотор EF1a. Нативный промотор EF1a контролирует экспрессию альфа-субъединицы комплекса фактора элонгации 1, который ответственен за ферментативную доставку аминоацил-тРНК в рибосому. Промотор EF1a широко используют в экспрессирующих плазмидах млекопитающих, и было показано, что он является эффективным для запуска экспрессии CAR с молекул нуклеиновых кислот, клонированных в лентивирусный вектор. См., например, Milone et al., Mol. Ther. 17(8): 1453-1464 (2009). В одном аспекте промотор EF1a содержит последовательность, представленную в качестве SEQ ID NO: 1.
Другим примером промотора является последовательность предраннего промотора цитомегаловируса (CMV). Эта промоторная последовательность представляет собой последовательность мощного конститутивного промотора, способного обеспечивать высокие уровни экспрессии любой полинуклеотидной последовательности, функционально связанной с ним. Однако также можно использовать другие конститутивные промоторные последовательности, включая, но не ограничиваясь ими, ранний промотор вируса 40 обезьян (SV40), промотор вируса опухоли молочной железы мыши (MMTV), промотор длинного концевого повтора (LTR) вируса иммунодефицита человека (ВИЧ), промотор MoMuLV, промотор вируса лейкоза птиц, предранний промотор вируса Эпштейна-Барр, промотор вируса саркомы Рауса, а также промоторы генов человека, такие как, но не ограничиваясь ими, промотор актина, промотор миозина, промотор фактора элонгации 1α, промотор гемоглобина и промотор креатинкиназы. Кроме того, изобретение не ограничивается применением конститутивных промоторов. Также в качестве части изобретения предусматриваются индуцибельные промоторы. Применение индуцибельных промоторов обеспечивает молекулярный переключатель, способный запускать экспрессию полинуклеотидной последовательности, с которой он функционально связан, когда такая экспрессия желательна, или выключать экспрессию, когда экспрессия является нежелательной. Примеры индуцибельных промоторов включают, но не ограничиваются ими металлотиониновый промотор, гликокортикоидный промотор, прогестероновый промотор и тетрациклиновый промотор.
Также вектор может включать, например, сигнальную последовательность для облегчения секреции, сигнал полиаденилирования и терминатор транскрипции (например, из гена бычьего гормона роста (BGH)), элемент, позволяющий эписомальную репликацию и репликацию у прокариот (например, ориджин SV40 и ColE1 или другие, известные в данной области) и/или элемент, позволяющие селекцию (например, ген устойчивости к ампициллину и/или зеоциновый).
Для оценки экспрессии полипептида CAR или его частей экспрессирующий вектор, подлежащий введению в клетку, также может содержать либо ген селективного маркера, либо репортерный ген, или оба из них, для облегчения идентификации и селекции экспрессирующих клеток из популяции клеток, которые намереваются трансфицировать или инфицировать вирусными векторами. В других аспектах селективный маркер может содержаться на отдельном фрагменте ДНК и использоваться в методике котрансфекции. Как селективные маркеры, так и репортерные гены, могут фланкироваться соответствующими регуляторными последовательностями для обеспечения экспрессии в клетках-хозяевах. Пригодные селективные маркеры включают, например, гены устойчивости к антибиотикам, такие как neo и т.п.
Репортерные гены используют для идентификации потенциально трансфицированных клеток и для оценки функциональности регуляторных последовательностей. Как правило, репортерный ген представляет собой ген, который не присутствует или не экспрессируется в организме или ткани реципиента и который кодирует полипептид, экспрессия которого проявляется некоторым легко поддающимся обнаружению свойством, например, ферментативной активностью. Экспрессию репортерного гена анализируют в подходящий период времени после встраивания ДНК в реципиентные клетки. Подходящие репортерные гены могут включать гены, кодирующие люциферазу, бета-галактозидазу, хлорамфениколацетилтрансферазу, секретируемую щелочную фосфатазу, или ген зеленого флуоресцентного белка (например, Ui-Tei et al., 2000 FEBS Letters 479: 79-82). Подходящие экспрессирующие системы хорошо известны, и их можно получать с использованием известных способов или приобретать от коммерческих поставщиков. Как правило, в качестве промотора идентифицируют конструкцию с минимальной 5'-фланкирующей областью, демонстрирующей наиболее высокий уровень экспрессии репортерного гена. Такие промоторные области можно связать с репортерным геном и использовать для оценки средств в отношении способности модулировать запускаемую промотором транскрипцию.
Способы введения и экспрессии генов в клетках известны в данной области. В контексте экспрессирующего вектора вектор может быть легко введен в клетку-хозяина, например, клетку млекопитающих, бактерий, дрожжей или насекомых, любым способом, известным в данной области. Например, экспрессирующий вектор можно переносить в клетку-хозяина физическим, химическим или биологическим способами.
Физические способы введения полинуклеотида в клетку-хозяина включают преципитацию с фосфатом кальция, липофекцию, бомбардировку частицами, микроинъекцию, электропорацию и т.п. Способы получения клеток, содержащих векторы и/или экзогенные нуклеиновые кислоты, хорошо известны в данной области. См., например, Sambrook et al., 2012, MOLECULAR CLONING: A LABORATORY MANUAL, volumes 1-4, Cold Spring Harbor Press, NY). Предпочтительным способом введения полинуклеотида в клетку-хозяина является трансфекция с фосфатом кальция.
Биологические способы введения представляющего интерес полинуклеотида в клетку-хозяина включают применение ДНК- и РНК-векторов. Вирусные векторы, и, особенно ретровирусные векторы, стали наиболее широко используемым способом введения генов в клетки млекопитающих, например человека. Другие вирусные векторы могут происходить из лентивирусов, поксвирусов, вируса простого герпеса I, аденовирусов и аденоассоциированных вирусов и т.п. См., например, патенты США № 5350674 и 5585362.
Химические средства для введения полинуклеотида в клетку-хозяина включают коллоидные дисперсные системы, такие как макромолекулярные комплексы, нанокапсулы, микросферы, гранулы и системы на основе липидов, включая эмульсии типа "масло в воде", мицеллы, смешанные мицеллы и липосомы. Иллюстративной колллоидной системой для применения в качестве носителя для доставки in vitro и in vivo является липосома (например, искусственная мембранная везикула). Доступны другие способы уровня техники для направленной доставки нуклеиновых кислот, такие как доставка полинуклеотидов с использованием нацеленных наночастиц или другой подходящей субмикронной системы доставки.
В случае, когда используют невирусную систему доставки, иллюстративным носителем для доставки является липосома. Для введения нуклеиновых кислот в клетку (in vitro, ex vivo или in vivo) предусматривается применение липидных составов. В другом аспекте нуклеиновую кислоту можно связывать с липидом. Нуклеиновая кислота, связанная с липидом, может быть инкапсулирована в водное содержимое липосомы, распределена в липидном бислое липосомы, связана с липосомой через линкерную молекулу, которая связана как с липосомой, так и с олигонуклеотидом, заключена в липосому, быть в комплексе с липосомой, диспергирована в растворе, содержащем липид, смешана с липидом, объединена с липидом, содержаться в качестве суспензии в липиде, содержаться или быть в комплексе с мицеллой, или может быть иным образом ассоциирована с липидом. Композиции, ассоциированные с липидами, липидами/ДНК или липидами/экспрессирующими векторами не ограничиваются какой-либо конкретной структурой в растворе. Например, они могут находиться в бислойной структуре в качестве мицелл или могут быть со "спавшейся" структурой. Они также могут быть просто распределены в растворе, возможно формируя агрегаты, которые не являются однородными по размеру или форме. Липиды представляют собой жирные вещества, которые могут представлять собой встречающиеся в природе или синтетические липиды. Например, липиды включают жировые капли, которые встречаются в природе в цитоплазме, а также класс соединений, который содержит алифатические углеводороды длинной цепи и их производные, такие как жирные кислоты, спирты, амины, аминоспирты и альдегиды.
] Липиды, пригодные для применения, можно получать из коммерческих источников. Например, димиристилфосфатидилхолин ("DMPC") можно получать от Sigma, St. Louis, MO; дицетилфосфат ("DCP") можно получать от K & K Laboratories (Plainview, NY); холестерин ("Choi") можно получать от Calbiochem-Behring; димиристилфосфатидилглицерин ("DMPG") и другие липиды можно получать от Avanti Polar Lipids, Inc. (Birmingham, AL.). Исходные растворы липидов в хлороформе или смеси хлороформ/метанол можно хранить при приблизительно -20°C. Хлороформ используют в качестве единственного растворителя, поскольку он легче испаряется, чем метанол. "Липосома" является общим термином, охватывающим множество однослойных и многослойных липидных везикул, образованных путем формирования заключенных друг в друга липидных бислоев или агрегатов. Липосомы могут быть охарактеризованы как имеющие везикулярные структуры с фосфолипидной бислойной мембраной и внутренней водной средой. Многослойные липосомы имеют множество липидных слоев, разделенных водной средой. Они образуются самопроизвольно, когда фосфолипиды суспендируют в избытке водного раствора. Липидные компоненты претерпевают самоперестройку перед формированием замкнутых структур и заключением воды и растворенных веществ между липидными слоями (Ghosh et al., 1991 Glycobiology 5: 505-10). Однако также предусматриваются композиции, которые имеют в растворе структуры, отличающиеся от нормальной везикулярной структуры. Например, липиды могут принимать мицеллярную структуру или существовать только в качестве неоднородных агрегатов липидных молекул. Также предусматриваются комплексы липофектамин-нуклеиновая кислота.
Независимо от способа, использованного для введения экзогенных нуклеиновых кислот в клетку-хозяина или иного воздействия на клетку ингибитора по настоящему изобретению, для подтверждения присутствия последовательности рекомбинантной ДНК в клетке-хозяине можно проводить различные анализы. Такие анализы включают, например, "молекулярно-биологические" анализы, хорошо известные специалистам в данной области, такие как саузерн- и нозерн-блоттинг, ОТ-ПЦР и ПЦР; "биохимические" анализы, такие как обнаружение присутствия или отсутствия конкретного пептида, например, иммунологическими способами (ELISA и вестерн-блоттинг) или анализами, описанными в настоящем описании, для идентификации средств, входящих в объем изобретения.
Кроме того, настоящее изобретение относится к вектору, содержащему молекулу нуклеиновой кислоты, кодирующую CAR. В одном аспекте вектор CAR можно прямо трансдуцировать в клетку, например T-клетку или NK-клетку. В одном аспекте вектор представляет собой клонирующий или экспрессирующий вектор, например, вектор, включающий, но не ограничивающийся ими, одну или несколько плазмид (например, экспрессирующих плазмид, клонирующих векторов, миниколец, минивекторов, двойных микрохромосом), ретровирусных и лентивирусных векторных конструкций. В одном аспекте вектор способен экспрессировать конструкцию CAR в иммунных эффекторных клетках (например, T-клетках, NK-клетках) млекопитающих. В одном аспекте T-клетка млекопитающего представляет собой T-клетку человека. В одном аспекте NK-клетка млекопитающего представляет собой NK-клетку человека.
Источники клеток
Перед увеличением в количестве и генетической модификации или другой модификации источник клеток, например, T-клеток или натуральных киллерных (NK) клеток) можно получать от индивидуума. Подразумевают, что термин "индивидуум" включает живые организмы, в которых можно индуцировать иммунный ответ (например, млекопитающие). Примеры индивидуумов включают человека, мартышек, шимпанзе, собак, кошек, мышей, крыс и их трансгенные виды. T-клетки можно получать из ряда источников, включая мононуклеарные клетки периферической крови, костный мозг, ткань лимфатических узлов, пуповинную кровь, ткань тимуса, ткань из области инфекции, асцитную жидкость, плевральный выпот, ткань селезенки и опухоли.
В некоторых аспектах по настоящему изобретению иммунные эффекторные клетки, например, T-клетки, можно получать из единицы крови, взятой от индивидуума, с использованием любого из множества способов, известных квалифицированному специалисту, таких как разделение в FicollTM. В одном предпочтительном аспекте клетки из циркулирующей крови индивидуума получают аферезом. Продукт афереза, как правило, содержит лимфоциты, в том числе T-клетки, моноциты, гранулоциты, B-клетки, другие ядросодержащие лейкоциты, эритроциты и тромбоциты. В одном аспекте клетки, полученные аферезом, можно промывать для удаления фракции плазмы и, необязательно, для помещения клеток в соответствующий буфер или среды для последующих стадий обработки. В одном варианте осуществления клетки промывают фосфатно-солевым буфером (PBS). В альтернативном варианте осуществления промывочный раствор лишен кальция и может быть лишен магния или может быть лишен многих, или даже всех, двухвалентных катионов.
Первоначальные стадии активации в отсутствие кальция могут приводить к усиленной активации. Как будет хорошо понятно специалистам в данной области, стадию промывания можно проводить способами, известными в данной области, как например, с использованием полуавтоматической "проточной" центрифуги (например, устройство для обработки клеток Cobe 2991, Baxter CytoMate, или Haemonetics Cell Saver 5) в соответствии с инструкциями изготовителя. После промывания клетки можно ресуспендировать в различных биосовместимых буферах, например, таких как не содержащий Ca, не содержащий Mg PBS, PlasmaLyte A, или другой солевой раствор с буфером или без него. Альтернативно нежелательные компоненты образца для афереза можно удалять и клетки прямо ресуспендировать в культуральной среде.
Является общеизвестным, что в способах применения может быть использованы условиях культуральных сред, включающие 5% или менее, например, 2%, сыворотки AB человека, и могут быть использованы известны условия и композиции культуральных сред, например, описанные в Smith et al., "Ex vivo expansion of human T cells for adoptive immunotherapy using the novel Xeno-free CTS Immune Cell Serum Replacement" Clinical & Translational Immunology (2015) 4, e31; doi:10.1038/cti.2014.31.
В одном аспекте T-клетки выделяют из лимфоцитов периферической крови человека посредством лизиса эритроцитов и устранения моноцитов, например, посредством центрифугирования в градиенте PERCOLLTM или посредством центрифужной элютриации в противотоке.
Способы, описанные в настоящем описании, могут включать, например, селекцию конкретной субпопуляции иммунных эффекторных клеток, например, T-клеток, которые представляют собой популяцию с истощением регуляторных T-клеток, популяцию с истощением CD25+ клеток, с использованием, например, способа отрицательной селекции, например, описанного в настоящем описании. Предпочтительно, популяция клеток с истощением регуляторных T-клеток содержит менее 30%, 25%, 20%, 15%, 10%, 5%, 4%, 3%, 2%, 1% CD25+ клеток.
В одном варианте осуществления регуляторные T-клетки, например, CD25+ T-клетки, устраняют из популяции с использованием антитела против CD25 или его фрагмента, или CD25-связывающего лиганда, IL-2. В одном варианте осуществления антитело против CD25 или его фрагмент, или CD25-связывающий лиганд конъюгированы с подложкой, например, гранулами, или в другом случае нанесены на субстрат, например, гранулы. В одном варианте осуществления антитело против CD25 или его фрагмент конъюгированы с подложкой, как описано в настоящем описании.
В одном варианте осуществления регуляторные T-клетки, например, CD25+ T-клетки, удаляют из популяции с использованием реагента для истощения CD25 от MiltenyiTM. В одном варианте осуществления соотношение клеток и реагента для истощения CD25 составляет 1e7 клеток на 20 мкл, или 1e7 клеток на 15 мкл, или 1e7 клеток на 10 мкл, или 1e7 клеток на 5 мкл, или 1e7 клеток на 2,5 мкл, или 1e7 клеток на 1,25 мкл. В одном варианте осуществления, например, для регуляторных T-клеток, например, используют истощение CD25+ клеток, превышающих 500 миллионов клеток/мл. В следующем аспекте, используют концентрацию клеток, составляющую 600, 700, 800 или 900 миллионов клеток/мл.
В одном варианте осуществления популяция иммунных эффекторных клеток, подлежащих истощению, включает приблизительно 6×109 CD25+ T-клеток. В других аспектах популяция иммунных эффекторных клеток, подлежащая истощению, включает приблизительно от 1×109 до 1×1010 CD25+ T-клеток и любое целое число между ними. В одном варианте осуществления полученная популяция с истощением регуляторных T-клеток имеет 2×109 регуляторных T-клеток, например, CD25+ клеток, или менее (например, 1×109, 5×108, 1×108, 5×107, 1×107 или менее CD25+ клеток).
В одном варианте осуществления регуляторные T-клетки, например, CD25+ клетки, удаляют из популяции с использованием системы CliniMAC с набором трубок для истощения, например, таких как трубки 162-01. В одном варианте осуществления систему CliniMAC используют в настройках для истощения, например, таких как DEPLETION2.1.
Без связи с конкретной теорией, снижение уровня отрицательных регуляторов иммунных клеток (например, снижение количества нежелательных иммунных клеток, например, клеток TREG) у индивидуума перед аферезом или в процессе изготовления продукта с экспрессирующими CAR клетками, может снизить риск рецидива у индивидуума. Например, способы истощения клеток TREG известны в данной области. Способы уменьшения количества клеток TREG включают, но не ограничиваются ими, циклофосфамид, антитело против GITR (антитело против GITR, описанное в настоящем описании), истощение CD25 и их комбинации.
В некоторых вариантах осуществления способы получения включают уменьшение количества (например, истощение) клеток TREG перед получением экспрессирующих CAR клеток. Например, способы получения включают приведение в контакт образца, например, образца для афереза, с антителом против GITR и/или антителом против CD25 (или его фрагментом или CD25-связывающим лигандом), например, для истощения клеток TREG перед изготовлением продукта с экспрессирующими CAR клетками (например, T-клетки, NK-клетки).
В одном варианте осуществления индивидуума предварительно лечат одним или несколькими способами терапии, которые снижают уровень клеток TREG, перед сбором клеток для изготовления продукта с экспрессирующими CAR клетками, тем самым, снижая у индивидуума риск рецидива после лечения экспрессирующими CAR клетками. В одном варианте осуществления способы снижения уровня клеток TREG включают, но не ограничиваются ими, введение индивидууму одного или нескольких из циклофосфамида, антитела против GITR, средства для истощения CD25, или их комбинацию. Введение одного или нескольких из циклофосфамида, антитела против GITR, средства для истощения CD25, или их комбинации может происходить до, в процессе или после инфузии продукта с экспрессирующими CAR клетками.
В одном варианте осуществления индивидуума предварительно лечат циклофосфамидом перед сбором клеток для изготовления продукта с экспрессирующими CAR клетками, тем самым, снижая риск рецидива у индивидуума после лечения экспрессирующими CAR клетками. В одном варианте осуществления индивидуума предварительно лечат антителом против GITR перед сбором клеток для изготовления продукта с экспрессирующими CAR клетками, тем самым, снижая риск рецидива у индивидуума после лечения экспрессирующими CAR клетками.
В одном варианте осуществления популяцией клеток, подлежащей истощению, не являются ни регуляторные T-клетки, не опухолевые клетки, а клетки, которые иным образом отрицательно влияют на экспансию и/или функцию CART-клеток, например, клеток, экспрессирующих CD14, CD11b, CD33, CD15 или другие маркеры, экспрессируемые потенциально иммуносупрессорными клетками. В одном варианте осуществления предусматривается удаление таких клеток одновременно с регуляторными T-клетками и/или опухолевыми клетками, или после указанного истощения, или в другом порядке.
Способы, описанные в настоящем описании, могут включать более одной стадии селекции, на пример, более одной стадии истощения. Увеличение в количестве популяции T-клеток посредством отрицательной селекции можно проводить, например, с использованием комбинации антител, направленных на поверхностные маркеры, уникальные для клеток, подвергнутых отрицательной селекции. Одним способом является сортировка и/или селекция клеток посредством отрицательной магнитной иммунной адгезии или проточной цитометрии, в которых используется смесь моноклональных антител, направленных на маркеры клеточной поверхности, присутствующие на клетках, подвергаемых отрицательной селекции. Например, для увеличения в количестве CD4+ клеток посредством отрицательной селекции, смесь моноклональных антител может включать антитела к CD14, CD20, CD11b, CD16, HLA-DR и CD8.
Способы, описанные в настоящем описании, кроме того, могут включать удаление клеток из популяции, которые экспрессируют опухолевый антиген, например, опухолевый антиген, который не содержит CD25, например, CD19, CD30, CD38, CD123, CD20, CD14 или CD11b, тем самым, получая популяцию с истощенными регуляторными T-клетками, например, истощенными по CD25+ клетками, и истощенными по опухолевому антигену клетками, которая пригодна для экспрессии CAR, например, CAR, описанного в настоящем описании. В одном варианте осуществления клетки, экспрессирующие опухолевый антиген, удаляют одновременно с регуляторными T-клетками, например, CD25+ клетками. Например, антитело против CD25 или его фрагмент и антитело против опухолевого антитела, или его фрагмент, можно связывать с одним и тем же субстратом, например, гранулами, которые можно использовать для удаления клеток, или антитело против CD25 или его фрагмент, или антитело против опухолевого антигена или его фрагмент, можно связывать с отдельными гранулами, смесь которых можно использовать для удаления клеток. В других вариантах осуществления удаление T-регуляторных клеток, например, CD25+ клеток, и удаление клеток, экспрессирующих опухолевый антиген, является последовательным и может происходить, например, в любом порядке.
Также предусматриваются способы, которые включают устранение клеток из популяции, которые экспрессируют ингибитор точки контроля, например, ингибитор точки контроля, описанный в настоящем описании, например, одного или нескольких типов из PD1+ клеток, LAG3+ клеток и TIM3+ клеток, тем самым, обеспечивая популяцию с истощенными регуляторными T-клетками, например, с истощенными CD25+ клетками, и клетками, истощенными по ингибитору точки контроля, например, истощенными PD1+, LAG3+ и/или TIM3+ клетками. Иллюстративные ингибиторы точки контроля включают B7-H1, B7-1, CD160, P1H, 2B4, PD1, TIM3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG3, TIGIT, CTLA-4, BTLA и LAIR1. В одном варианте осуществления клетки, экспрессирующие ингибитор точки контроля, удаляют одновременно с регуляторными T-клетками, например, CD25+ клетками. Например, антитело против CD25 или его фрагмент и антитело против ингибитора точки контроля или его фрагмент можно связывать с одними и теми же гранулами, которые можно использовать для удаления клеток, или антитело против CD25 или его фрагмент, или антитело против ингибитора точки контроля или его фрагмент, можно связывать с отдельными гранулами, смесь которых можно использовать для удаления клеток. В других вариантах осуществления удаление T-регуляторных клеток, например, CD25+ клеток, и удаление клеток, экспрессирующих ингибитор точки контроля, является последовательным и может происходить, например, в любом порядке.
Способы, описанные в настоящем описании, могут включать стадию положительной селекции. Например, T-клетки можно выделять посредством инкубации с гранулами, конъюгированными с антителом против CD3/против CD28 (например, 3×28), такими как DYNABEAD® M-450 CD3/CD28 T, в течение периода времени, достаточного для положительной селекции желаемых T-клеток. В одном варианте осуществления период времени составляет приблизительно 30 минут. В следующем варианте осуществления период времени находится в диапазоне от 30 минут до 36 часов или более, включая все целые числа между ними. В следующем варианте осуществления, период времени составляет по меньшей мере 1, 2, 3, 4, 5 или 6 часов. В другом предпочтительном варианте осуществления период времени составляет от 10 до 24 часов, например, 24 часа. Более длительное время инкубации можно использовать для выделения T-клеток в любой ситуации, где T-клеток мало по сравнению с другими типами клеток, как например, в случае выделения инфильтрирующих опухоль лимфоцитов (TIL) из ткани опухоли или из индивидуумов с иммунодефицитом. Кроме того, использование более длительного времени инкубации может повышать эффективность улавливания CD8+ T-клеток. Таким образом, посредством простого укорочения или удлинения времени T-клеткам позволяют связываться с гранулами CD3/CD28 и/или путем увеличения или снижения соотношения гранул и T-клеток (как дополнительно описано в настоящем описании), можно предпочтительно проводить отбирать в отношении или против субпопуляции T-клеток при инициации культуры или в другие моменты времени процесса. Кроме того, путем увеличения или снижения соотношения антител против CD3 и/или антител против CD28 на гранулах или другой поверхности, можно предпочтительно проводить отбор в отношении или против субпопуляции T-клеток при инициации культуры или в другие желаемые моменты времени.
В одном варианте осуществления можно отбирать популяцию T-клеток, которые экспрессируют один или несколько из IFN-γ, TNFα, IL-17A, IL-2, IL-3, IL-4, GM-CSF, IL-10, IL-13, гранзима B и перфорина, или другие подходящие молекулы, например, другие цитокины. Способы скрининга экспрессии клеток можно определять, например, способами, описанными в публикации PCT № WO 2013/126712.
Для выделения желаемой популяции клеток посредством положительной или отрицательной селекции концентрацию клеток и поверхности (например, частиц, таких как гранулы) можно варьировать. В некоторых аспектах, может быть желательным значительное снижение объема, в котором гранулы и клетки смешивают друг с другом (например, увеличение концентрации клеток), для обеспечения максимального контакта клеток и гранул. Например, в одном аспекте используют концентрацию 10 миллиардов клеток/мл, 9 миллиардов клеток/мл, 8 миллиардов клеток/мл, 7 миллиардов клеток/мл, 6 миллиардов клеток/мл или 5 миллиардов клеток/мл. В одном аспекте используют концентрацию 1 миллиарда клеток/мл. В следующем аспекте используют концентрацию клеток 75, 80, 85, 90, 95 или 100 миллионов клеток/мл. В следующих аспектах можно использовать концентрации, составляющие 125 или 150 миллионов клеток/мл.
Использование высоких концентраций может приводить к увеличению выхода клеток, активации клеток и экспансии клеток. Кроме того, применение высоких концентраций клеток позволяет более эффективное улавливание клеток, которые могут слабо экспрессировать представляющие интерес антигены-мишени, таких как CD28-отрицательные T-клетки, или улавливание из образцов, где присутствует множество опухолевых клеток (например, кровь при лейкозе, опухолевая ткань и т.д.). Такие популяции клеток могут иметь терапевтическую ценность, и их получение может быть желательным. Например, использование высокой концентрации клеток обеспечивает более эффективную селекцию CD8+ T-клеток, которые обычно имеют более слабую экспрессию CD28.
В сходном аспекте может быть желательным снижение концентрации клеток. Путем значительного разбавления смеси T-клеток и поверхности (например, частиц, таких как гранулы), взаимодействия между частицами и клетками минимизируют. Это приводит к селекции клеток, которые экспрессируют высокие количества желаемых антигенов, подлежащих связыванию с частицами. Например, CD4+ T-клетки экспрессируют более высокие уровни CD28 и более эффективно улавливаются, чем CD8+ T-клетки в разбавленных концентрациях. В одном аспекте концентрация используемых клеток составляет 5×10 e6/мл. В других аспектах, используемая концентрация может составлять от приблизительно 1×105/мл до 1×106/мл, включая любое целое число между ними.
В других аспектах клетки можно инкубировать на вращающем устройстве в течение разных периодов времени при различных скоростях либо при 2-10°C, либо при комнатной температуре.
T-клетки, предназначенные для стимуляции, также можно замораживать после стадии промывания. Без связи с теорией полагают, что замораживание и последующее оттаивание обеспечивает более однородный продукт вследствие устранения гранулоцитов и в некоторой степени моноцитов в популяции клеток. После стадии промывания, которая устраняет плазму и тромбоциты, клетки можно суспендировать в растворе для замораживания. Хотя множество растворов и параметров для замораживания известно в данной области и пригодно в этом контексте, один способ вовлекает использование PBS, содержащего 20% DMSO и 8% сывороточный альбумин человека, или культуральной среды, содержащей 10% декстран 40 и 5% декстроза, 20% сывороточный альбумин человека и 7,5% DMSO, или 31,25% Plasmalyte-A, 31,25% декстрозу 5%, 0,45% NaCl, 10% декстран 40 и 5% декстрозу, 20% сывороточный альбумин человека, и 7,5% DMSO или другие походящие среды для замораживания клеток, содержащие, например, Hespan и PlasmaLyte A, затем клетки замораживают до -80°C со скоростью 1° в минуту и хранят в парообразной фазе емкости для хранения с жидком азоте. Можно использовать другие способы контролируемого замораживания, а также неконтролируемого замораживания сразу до -20°C или в жидком азоте.
В некоторых аспектах криосохраненные клетки размораживают и промывают, как описано в настоящем описании, и им позволяют покоиться в течение одного часа при комнатной температуре перед активацией с использованием способов по настоящему изобретению.
Также в контексте изобретения предусматривается получение образцов крови или продукта афереза от индивидуума в период до того, как клетки, описанные в настоящем описании, могут потребоваться. По существу, источник клеток, подлежащих увеличению в количестве, можно собирать в любой требуемый момент времени и желаемые клетки, такие как T-клетки, можно выделять и замораживать для последующего применения в терапии иммунными эффекторными клетками от любого множества заболеваний и состояний, при которых является полезной терапия иммунными эффекторными клетками, таких как заболевания и состояния, описанные в настоящем описании. В одном аспекте получение образца крови или продукта афереза проводят от здорового в основном индивидуума. В некоторых аспектах получение образца крови или продукта афереза проводят от здорового в основном индивидуума, который имеет риск развития заболевания, но у которого заболевание еще не развилось, и представляющие интерес клетки выделяют и замораживают для последующего применения. В некоторых аспектах T-клетки можно увеличивать в количестве, замораживать и использовать позднее. В некоторых аспектах взятие образцов проводят сразу после постановки диагноза конкретного заболевания, как описано в настоящем описании, но перед каким-либо из способов лечения. В следующем аспекте клетки выделяют из образца крови или продукта афереза от индивидуума перед любым из множества соответствующих способов лечения, включая, но не ограничиваясь ими, лечение средствами, такими как натализумаб, эфализумаб, противовирусные средства, химиотерапия, лучевая терапия, иммунодепрессивные средства, такие как циклоспорин, азатиоприн, метотрексат, микофенолат и FK506, антитела или другие иммунодепрессивные средства, такие как CAMPATH, антитела против CD3, цитоксан, флударабин, циклоспорин, FK506, рапамицин, микофеноловая кислота, стероиды, FR901228 и лучевая терапия.
В следующем аспекте настоящего изобретения T-клетки получают от пациента непосредственно после лечения, которое сохраняет у индивидуума функциональные T-клетки. В этом отношении было обнаружено, что в случае определенных способов лечения злокачественной опухоли, в частности, лечения лекарственными средствами, которые повреждают иммунную систему, вскоре после лечения в течение периода, когда пациенты в норме восстанавливаются после лечения, качество полученных T-клеток может быть оптимальным или улучшенным в отношении их способности увеличиваться в количестве ex vivo. Аналогично, после манипулирования ex vivo с использованием способов, описанных в настоящем описании, эти клетки могут быть в предпочтительном состоянии для повышения приживаемости и экспансии in vivo. Таким образом, в контексте изобретения предусматривается сбор клеток крови, в том числе T-клеток, дендритных клеток или других клеток гемопоэтического роста в процессе этой стадии восстановления. Кроме того, в некоторых аспектах, можно использовать режимы мобилизации (например, мобилизация посредством GM-CSF) и кондиционирования у индивидуума, которые способствуют репопуляции, рециркуляции, регенерации и/или экспансии конкретных типов клеток, особенно в течение определенного периода времени после терапии. Иллюстративные типы клеток включают T-клетки, B-клетки, дендритные клетки и другие клетки иммунной системы.
В одном варианте осуществления иммунные эффекторные клетки, экспрессирующие молекулу CAR, например, молекулу CAR, описанную в настоящем описании, получают от индивидуума, которому вводили низкую усиливающую иммунитет дозу ингибитора mTOR. В одном варианте осуществления популяцию иммунных эффекторных клеток, например, T-клеток, подлежащих модификации способами инженерии для экспрессии CAR, собирают после достаточного периода времени или после достаточного дозирования низкой усиливающей иммунитет дозы ингибитора mTOR, так что уровень отрицательных по PD1 иммунных эффекторных клеток, например, T-клеток, или соотношение отрицательные по PD-1 иммунные эффекторные клетки, например, T-клетки/положительные по PD-1 иммунные эффекторные клетки, например, T-клетки, у индивидуума или после взятия у индивидуума по меньшей мере временно увеличивается.
В других вариантах осуществления популяцию иммунных эффекторных клеток, например, T-клеток, которые модифицированы или будут модифицированы способами инженерии для экспрессии CAR, можно обрабатывать ex vivo путем приведения в контакт с количеством ингибитора mTOR, который увеличивает количество отрицательных по PD1 иммунных эффекторных клеток, например, T клеток, или увеличивает соотношение отрицательные по PD-1 иммунные эффекторные клетки, например, T-клетки/положительные по PD-1 иммунные эффекторные клетки, например, T-клетки.
В одном варианте осуществления популяция T-клеток является дефицитной по диаглицеринкиназе (DGK). Клетки с дефицитом DGK включают клетки, которые не экспрессируют РНК или белок DGK, или имеет сниженную или ингибированную активность DGK. Клетки с дефицитом DGK можно получать посредством генетических подходов, например, введения агентов РНК-интерференции, например, миРНК, кшРНК, микроРНК, для снижения или предотвращения экспрессии DGK. Альтернативно клетки с дефицитом DGK можно получать путем обработки ингибиторами DGK, описанными в настоящем описании.
В одном варианте осуществления популяция T-клеток имеет дефицит Ikaros. Клетки с дефицитом Ikaros включают клетки, которые не экспрессируют РНК или белок Ikaros, или обладают сниженной или ингибированной активностью Ikaros. Клетки с дефицитом Ikaros можно получать с использованием генетических подходов, например, введения агентов РНК-интерференции, например, миРНК, кшРНК, микроРНК, для снижения или предотвращения экспрессии Ikaros. Альтернативно клетки с дефицитом Ikaros можно получать путем обработки ингибиторами Ikaros, например, леналидомидом.
В вариантах осуществления популяция T-клеток является дефицитной по DGK и дефицитной по Ikaros, например, не экспрессирует DGK и Ikaros, или имеет сниженную или ингибированную активность DGK и Ikaros. Такие клетки с дефицитом DGK и Ikaros можно получать любыми способами, описанными в настоящем описании.
В одном варианте осуществления NK-клетки получают от индивидуума. В другом варианте осуществления NK-клетки представляют собой линию NK-клеток, например линию клеток NK-92 (Conkwest).
Аллогенный CAR
В вариантах осуществления, описанных в настоящем описании, иммунная эффекторная клетка может представлять собой аллогенную эффекторную клетку, например T-клетку или NK-клетку. Например, клетка может представлять собой аллогенную T-клетку, например аллогенную T-клетку, лишенную экспрессии функционального T-клеточного рецептора (TCR) и/или лейкоцитарного антигена человека (HLA), например, HLA класса I и/или HLA класса II.
Например, T-клетку, лишенную функционального TCR, можно конструировать так, чтобы она не экспрессировала какой-либо функциональный TCR на ее поверхности, можно конструировать так, что она не экспрессировала одной или нескольких субъединиц, которые содержат функциональный TCR, или конструировать так, чтобы она продуцировала очень мало функционального TCR на ее поверхности. Альтернативно T-клетка может экспрессировать по существу нарушенный TCR, например, посредством экспрессии мутантных или укороченных форм одной или нескольких субъединиц TCR. Термин "по существу нарушенный TCR" означает, что этот TCR не индуцирует неблагоприятную иммунную реакцию у хозяина.
T-клетку, описанную в настоящем описании, можно конструировать, например, чтобы она не экспрессировала функциональный HLA на ее поверхности. Например, T-клетку, описанную в настоящем описании, можно модифицировать способами инженерии, чтобы подавлялась экспрессия на клеточной поверхности HLA, например, HLA класса 1 и/или HLA класса II.
В некоторых вариантах осуществления T-клетка может быть лишена функционального TCR и функционального HLA, например, HLA класса I и/или HLA класса II.
Модифицированные T-клетки, которые лишены экспрессии функционального TCR и/или HLA, можно получать любыми подходящими способами, включая нокаут или нокдаун одной или нескольких субъединиц TCR или HLA. Например, T-клетка может включать нокдаун TCR и/или HLA с использованием миРНК, кшРНК, коротких палиндромных повторов, регулярно расположенных кластерами (CRISPR), нуклеазы, подобной активатору транскрипции (TALEN), или эндонуклеазы с цинковыми пальцами (ZFN).
В некоторых вариантах осуществления аллогенная клетка может представлять собой клетку, которая не экспрессирует или экспрессирует на низких уровнях ингибиторную молекулу, например, посредством любого способа, описанного в настоящем описании. Например, клетка может представлять собой клетку, которая не экспрессирует или экспрессирует на низких уровнях ингибиторную молекулу, например, которая может снижать способность экспрессирующей CAR клетки индуцировать иммунный эффекторный ответ. Примеры ингибиторных молекул включают PD1, PD-L1, CTLA4, TIM3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и TGF-бета. Ингибирование ингибиторной молекулы, например, посредством ингибирования на уровне ДНК, РНК или белка, может оптимизировать эффективность экспрессирующей CAR клетки. В вариантах осуществления можно использовать ингибиторную нуклеиновую кислоту, например дцРНК, например, миРНК или кшРНК, короткие палиндромные повторы, регулярно расположенные кластерами (CRISPR), нуклеазу, подобную активатору транскрипции (TALEN) или эндонуклеазу с цинковыми пальцами (ZFN), например, как описано в настоящем описании.
миРНК и кшРНК для ингибирования TCR или HLA
В некоторых вариантах осуществления экспрессию TCR и/или экспрессию HLA можно ингибировать с использованием миРНК или кшРНК, которые нацелены на нуклеиновую кислоту, кодирующую TCR и/или HLA, в T-клетке.
Экспрессии миРНК или кшРНК в T-клетках можно достигать с использованием любой общепринятой экспрессирующей системы, например, такой как лентивирусная экспрессирующая система.
Иллюстративные кшРНК, которые подавляют экспрессию компонентов TCR, описаны, например, в публикации США № 2012/0321667. Иллюстративные миРНК и кшРНК, которые подавляют экспрессию генов HLA класса I и/или HLA класса II, описаны, например, в публикации США № US 2007/0036773.
CRISPR для ингибирования TCR или HLA
"CRISPR" или "CRISPR против TCR и/или HLA" или "CRISPR для ингибирования TCR и/или HLA", как используют в рамках изобретения, относится к набору коротких палиндромных повторов, регулярно расположенных кластерами, или к системе, содержащей такой набор повторов. "Cas", как используют в рамках изобретения, относится к ассоциированному с CRISPR белку. Система "CRISPR/Cas" относится к системе, происходящей из CRISPR и Cas, которую можно использовать для сайленсинга или мутации генов TCR и/или HLA.
Встречающиеся в природе системы CRISPR/Cas найдены приблизительно в 40% отсеквенированных геномов эубактерий и 90% отсеквенированных геномов архей. Grissa et al. (2007) BMC Bioinformatics 8: 172. Эта система представляет собой тип прокариотической иммунной системы, который сообщает устойчивость к чужеродным генетическим элементам, таким как плазмиды и фаги, и обеспечивает форму приобретенного иммунитета. Barrangou et al. (2007) Science 315: 1709-1712; Marragini et al. (2008) Science 322: 1843-1845.
Система CRISPR/Cas была модифицирована для применения в редактировании генов (сайленсинг, усиление или изменение конкретных генов) у эукариот, таких как мыши или приматы. Wiedenheft et al. (2012) Nature 482: 331-8. Это осуществили путем введения в эукариотическую клетку плазмиды, содержащей специально сконструированный CRISPR и один или несколько соответствующих Cas.
Последовательность CRISPR, иногда называемая локусом CRISPR, содержит чередующиеся повторы и спейсеры. Во встречающемся в природе CRISPR спейсеры обычно содержат последовательности, чужеродные для бактерии, такие как последовательность плазмиды или фага; в системе CRISPR/Cas против TCR и/или HLA спейсеры происходят из последовательностей генов TCR или HLA.
РНК из локуса CRISPR конститутивно экспрессируется и процессируется белками Cas в малые РНК. Они содержат спейсер, фланкированный последовательностями повтора. РНК направляют другие белки Cas на подавление экзогенных генетических элементов на уровне РНК или ДНК. Horvath et al. (2010) Science 327: 167-170; Makarova et al. (2006) Biology Direct 1: 7. Таким образом, спейсеры выступают в качестве матриц для молекул РНК, аналогично миРНК. Pennisi (2013) Science 341: 833-836.
Поскольку они встречаются в природе во многих различных типах бактерий, точная организация и структура CRISPR, функция и количество генов Cas и их продуктов отличаются в некоторой степени от вида к виду. Haft et al. (2005) PLoS Comput. Biol. 1: e60; Kunin et al. (2007) Genome Biol. 8: R61; Mojica et al. (2005) J. Mol. Evol. 60: 174-182; Bolotin et al. (2005) Microbiol. 151: 2551-2561; Pourcel et al. (2005) Microbiol. 151: 653-663; и Stern et al. (2010) Trends. Genet. 28: 335-340. Например, белки Cse (подтип Cas, E. coli) (например, CasA) формируют функциональный комплекс, Cascade, который процессирует РНК-транскрипты CRISPR в элементы спейсер-повтор, которые сохраняет Cascade. Brouns et al. (2008) Science 321: 960-964. В других прокариотах Cas6 процессирует транскрипт CRISPR. Инактивация фага на основе CRISPR в E. coli требует Cascade и Cas3, но не Cas1 или Cas2. Белки Cmr (модуль RAMP Cas) в Pyrococcus furiosus и других прокариотах формируют функциональный комплекс с малыми РНК CRISPR, который распознает и расщепляет РНК-мишени. Более простая система CRISPR основана на белке Cas9, который представляет собой нуклеазу с двумя активными участками расщепления, по одному для каждой цепи двойной спирали. Комбинирование Cas9 и модифицированного локуса РНК CRISPR можно использовать в системе редактирования генов. Pennisi (2013) Science 341: 833-836.
Таким образом, систему CRISPR/Cas можно использовать для редактирования гена TCR и/или HLA (присоединение или делеция пары оснований) или внесения преждевременного стоп-кодона, что, таким образом, снижает экспрессию TCR и/или HLA. Альтернативно систему CRISPR/Cas можно использовать подобно РНК-интерференции, выключая гены TCR и/или HLA обратимым образом. В клетке млекопитающего, например, РНК может направлять белок Cas на промотор TCR и/или HLA, пространственно блокируя РНК-полимеразы.
Искусственные системы CRISPR/Cas, которые ингибируют TCR и/или HLA, можно получать с использованием технологии, известной в данной области, например, которая описана в публикации США № 20140068797, и Cong (2013) Science 339: 819-823. Также можно получать другие искусственные системы CRISPR/Cas, известные в данной области, которые ингибируют TCR и/или HLA, например, системы, описанные в Tsai (2014) Nature Biotechnol., 32:6 569-576, патенты США № 8871445; 8865406; 8795965; 8771945 и 8697359.
TALEN для ингибирования TCR и/или HLA
"TALEN" или "TALEN против HLA и/или TCR" или "TALEN для ингибирования HLA и/или TCR" относится к эффекторной нуклеазе, подобной активатору транскрипции, которая представляет собой искусственную нуклеазу, которую можно использовать для редактирования гена HLA и/или TCR.
TALEN получают искусственным образом путем слияния эффекторного ДНК-связывающего домена TAL с доменом расщепления ДНК. Эффекты активатора транскрипции (TALE) можно модифицировать способами инженерии для связывания с любой желаемой последовательностью ДНК, включая часть гена HLA или TCR. Путем комбинирования модифицированных способами инженерии TALE с доменом расщепления ДНК можно получать фермент рестрикции, который является специфичным к любой желаемой последовательности ДНК, включая последовательность HLA или TCR. Затем их можно вводить в клетку, где их можно использовать для редактирования генома. Boch (2011) Nature Biotech. 29: 135-6; и Boch et al. (2009) Science 326: 1509-12; Moscou et al. (2009) Science 326: 3501.
TALE представляют собой белки, секретируемые бактериями Xanthomonas. ДНК-связывающий домен содержит повторяющуюся высококонсервативную последовательность из 33-34 аминокислот, за исключением 12-ой и 13-ой аминокислот. Эти два положения являются в высокой степени вариабельными, демонстрируя строгую корреляцию со специфическим распознаванием нуклеотидов. Таким образом, их можно модифицировать способами инженерии для связывания с желаемой последовательностью ДНК.
Для получения TALEN белок TALE подвергают слиянию с нуклеазой (N), которая представляет собой эндонуклеазу FokI дикого типа или мутантную эндонуклеазу FokI. В FokI было внесено несколько мутаций для ее применения в TALEN; они, например, повышают специфичность расщепления или активность. Cermak et al. (2011) Nucl. Acids Res. 39: e82; Miller et al. (2011) Nature Biotech. 29: 143-8; Hockemeyer et al. (2011) Nature Biotech. 29: 731-734; Wood et al. (2011) Science 333: 307; Doyon et al. (2010) Nature Methods 8: 74-79; Szczepek et al. (2007) Nature Biotech. 25: 786-793; и Guo et al. (2010) J. Mol. Biol. 200: 96.
Домен FokI функционирует в качестве димера, требующего две конструкции с уникальными ДНК-связывающими доменами для участков в геноме-мишени с надлежащей ориентаций и расстояниями. Как число аминокислотных остатков между ДНК-связывающим доменом TALE и доменом расщепления FokI, так и число оснований между двумя индивидуальными участками связывания TALEN, по-видимому, являются важными параметрами для достижения высоких уровней активности. Miller et al. (2011) Nature Biotech. 29: 143-8.
TALEN против HLA или TCR можно использовать внутри клетки для обеспечения двухцепочечного разрыва (DSB). Мутацию можно вносить в участке разрыва, если механизмы репарации ненадлежащим образом репарируют разрыв посредством негомологичного соединения концов. Например, ненадлежащая репарация может вносить мутацию со сдвигом рамки считывания. Альтернативно вместе с TALEN в клетку можно вводить чужеродную ДНК; в зависимости от последовательностей чужеродной ДНК и хромосомной последовательности, этот процесс можно использовать для коррекции дефекта в гене HLA или TCR или внесения такого дефекта в ген HLA или TCR wt, таким образом, снижая экспрессию HLA или TCR.
TALEN, специфичные к последовательностям в HLA или TCR можно конструировать с использованием любого способа, известного в данной области, включая различные схемы с использованием модульных компонентов. Zhang et al. (2011) Nature Biotech. 29: 149-53; Geibler et al. (2011) PLoS ONE 6: e19509.
Нуклеаза с цинковыми пальцами для ингибирования HLA и/или TCR
"ZFN" или "нуклеаза с цинковыми пальцами" или "ZFN против HLA и/или TCR" или "ZFN для ингибирования HLA и/или TCR" относятся к нуклеазе с цинковыми пальцами, которая представляет собой искусственную нуклеазу, которую можно использовать для редактирования гена HLA и/или TCR.
Подобно TALEN, ZFN содержит нуклеазный домен FokI (или его производное), слитый с ДНК-связывающим доменом. В случае ZFN ДНК-связывающий домен содержит один или несколько цинковых пальцев. Carroll et al. (2011) Genetics Society of America 188: 773-782; и Kim et al. (1996) Proc. Natl. Acad. Sci. USA 93: 1156-1160.
Цинковый палец представляет собой небольшой белковый структурный мотив, стабилизированный одним или несколькими ионами цинка. Цинковый палец может содержать, например, Cys2His2, и может распознавать последовательность размером приблизительно 3 п.о. Различные цинковые пальцы известной специфичности можно комбинировать для получения полипептидов с несколькими пальцами, которые распознают последовательности размером приблизительно 6, 9, 12, 15 или 18 п.о. Для получения цинковых пальцев (и их комбинаций), распознающих конкретные последовательности, доступны различные способы селекции и модульной сборки, включая фаговый дисплей, дрожжевые моногибридные системы, бактериальные моногибридные и двухгибридные системы и клетки млекопитающих.
Подобно TALEN, ZFN должна димеризоваться для расщепления ДНК. Таким образом, для нацеливания на непалиндромные участки ДНК требуется пара ZFN. Две индивидуальных ZFN должны связывать противоположные цепи ДНК, причем между их нуклеазами должно быть надлежащее пространство. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10570-5.
Также подобно TALEN, ZFN может вносить двухцепочечный разрыв в ДНК, который может создавать мутацию со сдвигом рамки считывания в случае ненадлежащей репарации, что приводит к снижению экспрессии и количества HLA и/или TCR в клетке. ZFN также можно использовать с гомологичной рекомбинацией для внесения мутаций в гены HLA или TCR.
ZFN, специфичные к последовательностям в HLA и/или TCR, можно конструировать с использованием любого способа, известного в данной области. См., например, Provasi (2011) Nature Med. 18: 807-815; Torikai (2013) Blood 122: 1341-1349; Cathomen et al. (2008) Mol. Ther. 16: 1200-7; и Guo et al. (2010) J. Mol. Biol. 400: 96; публикацию патента США 2011/0158957; публикацию патента США 2012/0060230.
Экспрессия теломеразы
Без связи с какой-либо конкретной теорией, в некоторых вариантах осуществления терапевтическая T-клетка имеет кратковременное нахождение у пациента вследствие укороченных теломер в T-клетках; таким образом, трансфекция гена теломеразы может удлинить теломеры T-клеток и увеличить время существования T-клеток у пациента. См. Carl June, "Adoptive T cell therapy for cancer in the clinic", Journal of Clinical Investigation, 117:1466-1476 (2007). Таким образом, в одном варианте осуществления иммунная эффекторная клетка, например, T-клетка, эктопически экспрессирует субъединицу теломеразы, например, каталитическую субъединицу теломеразы, например, TERT, например, hTERT. В некоторых аспектах настоящее изобретение относится к способу получения экспрессирующей CAR клетки, включающему приведение клетки в контакт с нуклеиновой кислотой, кодирующей субъединицу теломеразы, например, каталитическую субъединицу теломеразы, например, TERT, например, hTERT. Клетку можно приводить в контакт с нуклеиновой кислотой до, одновременно или после приведения в контакт с конструкцией, кодирующей CAR.
В одном аспекте настоящее изобретение относится к способу получения популяции иммунных эффекторных клеток (например, T-клеток, NK-клеток). В одном варианте осуществления способ включает: предоставление популяции иммунных эффекторных клеток (например, T-клеток или NK-клеток), приведение в контакт популяции иммунных эффекторных клеток с нуклеиновой кислотой, кодирующей CAR; и приведение в контакт популяции иммунных эффекторных клеток с нуклеиновой кислотой, кодирующей субъединицу теломеразы, например, hTERT, в условиях, которые позволяют экспрессию CAR и теломеразы.
В одном варианте осуществления нуклеиновая кислота, кодирующая субъединицу теломеразы, представляет собой ДНК. В одном варианте осуществления нуклеиновая кислота, кодирующая субъединицу теломеразы, содержит промотор, способный запускать экспрессию субъединицы теломеразы.
В одном варианте осуществления hTERT имеет аминокислотную последовательность GenBank Protein ID AAC51724.1 (Meyerson et al., "hEST2, the Putative Human Telomerase Catalytic Subunit Gene, Is Up-Regulated in Tumor Cells and during Immortalization" Cell Volume 90, Issue 4, 22 August 1997, Pages 785-795), являющуюся следующей:
MPRAPRCRAVRSLLRSHYREVLPLATFVRRLGPQGWRLVQRGDPAAFRALVAQCLVCVPWDARPPPAAPSFRQVSCLKELVARVLQRLCERGAKNVLAFGFALLDGARGGPPEAFTTSVRSYLPNTVTDALRGSGAWGLLLRRVGDDVLVHLLARCALFVLVAPSCAYQVCGPPLYQLGAATQARPPPHASGPRRRLGCERAWNHSVREAGVPLGLPAPGARRRGGSASRSLPLPKRPRRGAAPEPERTPVGQGSWAHPGRTRGPSDRGFCVVSPARPAEEATSLEGALSGTRHSHPSVGRQHHAGPPSTSRPPRPWDTPCPPVYAETKHFLYSSGDKEQLRPSFLLSSLRPSLTGARRLVETIFLGSRPWMPGTPRRLPRLPQRYWQMRPLFLELLGNHAQCPYGVLLKTHCPLRAAVTPAAGVCAREKPQGSVAAPEEEDTDPRRLVQLLRQHSSPWQVYGFVRACLRRLVPPGLWGSRHNERRFLRNTKKFISLGKHAKLSLQELTWKMSVRGCAWLRRSPGVGCVPAAEHRLREEILAKFLHWLMSVYVVELLRSFFYVTETTFQKNRLFFYRKSVWSKLQSIGIRQHLKRVQLRELSEAEVRQHREARPALLTSRLRFIPKPDGLRPIVNMDYVVGARTFRREKRAERLTSRVKALFSVLNYERARRPGLLGASVLGLDDIHRAWRTFVLRVRAQDPPPELYFVKVDVTGAYDTIPQDRLTEVIASIIKPQNTYCVRRYAVVQKAAHGHVRKAFKSHVSTLTDLQPYMRQFVAHLQETSPLRDAVVIEQSSSLNEASSGLFDVFLRFMCHHAVRIRGKSYVQCQGIPQGSILSTLLCSLCYGDMENKLFAGIRRDGLLLRLVDDFLLVTPHLTHAKTFLRTLVRGVPEYGCVVNLRKTVVNFPVEDEALGGTAFVQMPAHGLFPWCGLLLDTRTLEVQSDYSSYARTSIRASLTFNRGFKAGRNMRRKLFGVLRLKCHSLFLDLQVNSLQTVCTNIYKILLLQAYRFHACVLQLPFHQQVWKNPTFFLRVISDTASLCYSILKAKNAGMSLGAKGAAGPLPSEAVQWLCHQAFLLKLTRHRVTYVPLLGSLRTAQTQLSRKLPGTTLTALEAAANPALPSDFKTILD (SEQ ID NO: 63)
В одном варианте осуществления hTERT имеет последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичную последовательности SEQ ID NO: 63. В одном варианте осуществления hTERT имеет последовательность SEQ ID NO: 63. В одном варианте осуществления hTERT содержит делецию (например, не более чем 5, 10, 15, 20 или 30 аминокислот) на N-конце, C-конце или обоих концах. В одном варианте осуществления hTERT содержи трансгенную аминокислотную последовательность (например, не более чем из 5, 10, 15, 20 или 30 аминокислот) на N-конце, C-конце или обоих концах.
В одном варианте осуществления hTERT кодируется последовательностью нуклеиновой кислоты GenBank под номером доступа № AF018167 (Meyerson et al., "hEST2, the Putative Human Telomerase Catalytic Subunit Gene, Is Up-Regulated in Tumor Cells and during Immortalization" Cell Volume 90, Issue 4, 22 August 1997, Pages 785-795):
1 caggcagcgt ggtcctgctg cgcacgtggg aagccctggc cccggccacc cccgcgatgc
61 cgcgcgctcc ccgctgccga gccgtgcgct ccctgctgcg cagccactac cgcgaggtgc
121 tgccgctggc cacgttcgtg cggcgcctgg ggccccaggg ctggcggctg gtgcagcgcg
181 gggacccggc ggctttccgc gcgctggtgg cccagtgcct ggtgtgcgtg ccctgggacg
241 cacggccgcc ccccgccgcc ccctccttcc gccaggtgtc ctgcctgaag gagctggtgg
301 cccgagtgct gcagaggctg tgcgagcgcg gcgcgaagaa cgtgctggcc ttcggcttcg
361 cgctgctgga cggggcccgc gggggccccc ccgaggcctt caccaccagc gtgcgcagct
н 421 acctgcccaa cacggtgacc gacgcactgc gggggagcgg ggcgtggggg ctgctgttgc
481 gccgcgtggg cgacgacgtg ctggttcacc tgctggcacg ctgcgcgctc tttgtgctgg
541 tggctcccag ctgcgcctac caggtgtgcg ggccgccgct gtaccagctc ggcgctgcca
601 ctcaggcccg gcccccgcca cacgctagtg gaccccgaag gcgtctggga tgcgaacggg
661 cctggaacca tagcgtcagg gaggccgggg tccccctggg cctgccagcc ccgggtgcga
721 ggaggcgcgg gggcagtgcc agccgaagtc tgccgttgcc caagaggccc aggcgtggcg
781 ctgcccctga gccggagcgg acgcccgttg ggcaggggtc ctgggcccac ccgggcagga
841 cgcgtggacc gagtgaccgt ggtttctgtg tggtgtcacc tgccagaccc gccgaagaag
901 ccacctcttt ggagggtgcg ctctctggca cgcgccactc ccacccatcc gtgggccgcc
961 agcaccacgc gggcccccca tccacatcgc ggccaccacg tccctgggac acgccttgtc
1021 ccccggtgta cgccgagacc aagcacttcc tctactcctc aggcgacaag gagcagctgc
1081 ggccctcctt cctactcagc tctctgaggc ccagcctgac tggcgctcgg aggctcgtgg
1141 agaccatctt tctgggttcc aggccctgga tgccagggac tccccgcagg ttgccccgcc
1201 tgccccagcg ctactggcaa atgcggcccc tgtttctgga gctgcttggg aaccacgcgc
1261 agtgccccta cggggtgctc ctcaagacgc actgcccgct gcgagctgcg gtcaccccag
1321 cagccggtgt ctgtgcccgg gagaagcccc agggctctgt ggcggccccc gaggaggagg
1381 acacagaccc ccgtcgcctg gtgcagctgc tccgccagca cagcagcccc tggcaggtgt
1441 acggcttcgt gcgggcctgc ctgcgccggc tggtgccccc aggcctctgg ggctccaggc
1501 acaacgaacg ccgcttcctc aggaacacca agaagttcat ctccctgggg aagcatgcca
1561 agctctcgct gcaggagctg acgtggaaga tgagcgtgcg gggctgcgct tggctgcgca
1621 ggagcccagg ggttggctgt gttccggccg cagagcaccg tctgcgtgag gagatcctgg
1681 ccaagttcct gcactggctg atgagtgtgt acgtcgtcga gctgctcagg tctttctttt
1741 atgtcacgga gaccacgttt caaaagaaca ggctcttttt ctaccggaag agtgtctgga
1801 gcaagttgca aagcattgga atcagacagc acttgaagag ggtgcagctg cgggagctgt
1861 cggaagcaga ggtcaggcag catcgggaag ccaggcccgc cctgctgacg tccagactcc
1921 gcttcatccc caagcctgac gggctgcggc cgattgtgaa catggactac gtcgtgggag
1981 ccagaacgtt ccgcagagaa aagagggccg agcgtctcac ctcgagggtg aaggcactgt
2041 tcagcgtgct caactacgag cgggcgcggc gccccggcct cctgggcgcc tctgtgctgg
2101 gcctggacga tatccacagg gcctggcgca ccttcgtgct gcgtgtgcgg gcccaggacc
2161 cgccgcctga gctgtacttt gtcaaggtgg atgtgacggg cgcgtacgac accatccccc
2221 aggacaggct cacggaggtc atcgccagca tcatcaaacc ccagaacacg tactgcgtgc
2281 gtcggtatgc cgtggtccag aaggccgccc atgggcacgt ccgcaaggcc ttcaagagcc
2341 acgtctctac cttgacagac ctccagccgt acatgcgaca gttcgtggct cacctgcagg
2401 agaccagccc gctgagggat gccgtcgtca tcgagcagag ctcctccctg aatgaggcca
2461 gcagtggcct cttcgacgtc ttcctacgct tcatgtgcca ccacgccgtg cgcatcaggg
2521 gcaagtccta cgtccagtgc caggggatcc cgcagggctc catcctctcc acgctgctct
2581 gcagcctgtg ctacggcgac atggagaaca agctgtttgc ggggattcgg cgggacgggc
2641 tgctcctgcg tttggtggat gatttcttgt tggtgacacc tcacctcacc cacgcgaaaa
2701 ccttcctcag gaccctggtc cgaggtgtcc ctgagtatgg ctgcgtggtg aacttgcgga
2761 agacagtggt gaacttccct gtagaagacg aggccctggg tggcacggct tttgttcaga
2821 tgccggccca cggcctattc ccctggtgcg gcctgctgct ggatacccgg accctggagg
2881 tgcagagcga ctactccagc tatgcccgga cctccatcag agccagtctc accttcaacc
2941 gcggcttcaa ggctgggagg aacatgcgtc gcaaactctt tggggtcttg cggctgaagt
3001 gtcacagcct gtttctggat ttgcaggtga acagcctcca gacggtgtgc accaacatct
3061 acaagatcct cctgctgcag gcgtacaggt ttcacgcatg tgtgctgcag ctcccatttc
3121 atcagcaagt ttggaagaac cccacatttt tcctgcgcgt catctctgac acggcctccc
3181 tctgctactc catcctgaaa gccaagaacg cagggatgtc gctgggggcc aagggcgccg
3241 ccggccctct gccctccgag gccgtgcagt ggctgtgcca ccaagcattc ctgctcaagc
3301 tgactcgaca ccgtgtcacc tacgtgccac tcctggggtc actcaggaca gcccagacgc
3361 agctgagtcg gaagctcccg gggacgacgc tgactgccct ggaggccgca gccaacccgg
3421 cactgccctc agacttcaag accatcctgg actgatggcc acccgcccac agccaggccg
3481 agagcagaca ccagcagccc tgtcacgccg ggctctacgt cccagggagg gaggggcggc
3541 ccacacccag gcccgcaccg ctgggagtct gaggcctgag tgagtgtttg gccgaggcct
3601 gcatgtccgg ctgaaggctg agtgtccggc tgaggcctga gcgagtgtcc agccaagggc
3661 tgagtgtcca gcacacctgc cgtcttcact tccccacagg ctggcgctcg gctccacccc
3721 agggccagct tttcctcacc aggagcccgg cttccactcc ccacatagga atagtccatc
3781 cccagattcg ccattgttca cccctcgccc tgccctcctt tgccttccac ccccaccatc
3841 caggtggaga ccctgagaag gaccctggga gctctgggaa tttggagtga ccaaaggtgt
3901 gccctgtaca caggcgagga ccctgcacct ggatgggggt ccctgtgggt caaattgggg
3961 ggaggtgctg tgggagtaaa atactgaata tatgagtttt tcagttttga aaaaaaaaaa
4021 aaaaaaa (SEQ ID NO: 64)
В одном варианте осуществления hTERT кодируется нуклеиновой кислотой, имеющей последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96, 97%, 98%, или 99% идентичную последовательности SEQ ID NO: 64. В одном варианте осуществления hTERT кодируется нуклеиновой кислотой SEQ ID NO: 64.
Активация и увеличение в количестве иммунных эффекторных клеток (например, T-клеток)
Иммунные эффекторные клетки, такие как T-клетки, можно активировать и увеличивать в количестве, в основном с использованием способов, описанных, например, в патентах США 6352694; 6534055; 6905680; 6692964; 5858358; 6887466; 6905681; 7144575; 7067318; 7172869; 7232566; 7175843; 5883223; 6905874; 6797514; 6867041 и публикации патентной заявки США № 20060121005.
Как правило, популяцию иммунных эффекторных клеток, например, клеток с истощением регуляторных T-клеток, можно увеличивать в количестве путем контакта с поверхностью, имеющей связанное с ней средство, которое стимулирует сигнал, ассоциированный с комплексом CD3/TCR, и лиганд, который стимулирует костимулирующую молекулу на поверхности T-клеток. В частности, популяции T-клеток можно стимулировать, как описано в настоящем описании, например, путем контакта с антителом против CD3 или его антигенсвязывающим фрагментом, или антителом против CD2, иммобилизованным на поверхности, или путем контакта с активатором протеинкиназы C (например, бриостатин) совместно с ионофором кальция. Для стимуляции вспомогательной молекула на поверхности T-клеток можно использовать лиганд, который связывает вспомогательную молекулу. Например, популяцию T-клеток можно приводить в контакт с антителом против CD3 и антителом против CD28 в условиях, пригодных для стимуляции пролиферации T-клеток. Для стимуляции пролиферации либо CD4+ T-клеток, либо CD8+ T-клеток, можно использовать антитело против CD3 и антитело против CD28. Примеры антител против CD28 включают 9.3, B-T3, XR-CD28 (Diaclone,  , Франция), которые можно использовать, а также можно использовать и другие способы, широко известные в данной области (Berg et al., Transplant Proc. 30(8):3975-3977, 1998; Haanen et al., J. Exp. Med. 190(9):13191328, 1999; Garland et al., J. Immunol Meth. 227(1-2):53-63, 1999).
, Франция), которые можно использовать, а также можно использовать и другие способы, широко известные в данной области (Berg et al., Transplant Proc. 30(8):3975-3977, 1998; Haanen et al., J. Exp. Med. 190(9):13191328, 1999; Garland et al., J. Immunol Meth. 227(1-2):53-63, 1999).
В некоторых аспектах первичный стимулирующий сигнал и костимулирующий сигнал для T-клеток может быть предоставлен посредством различных протоколов. Например, средства, обеспечивающие каждый сигнал, могут быть в растворе или могут быть связанными с поверхностью. Когда они связаны с поверхностью, средства могут быть связаны с одной и той же поверхностью (т.е. в "цис"-форме) или с различными поверхностями (т.е. в "транс"-форме). Альтернативно одно средство может быть связано с поверхностью, а другое средство может находиться в растворе. В одном аспекте средство, обеспечивающее костимулирующий сигнал, связано с клеточной поверхностью, и средство, обеспечивающее первичный активирующий сигнал, находится в растворе или связано с поверхностью. В некоторых аспектах оба средства могут находиться в растворе. В одном аспекте средства могут быть в растворимой форме, а затем могут быть сшиты с поверхностью, такой как клетка, экспрессирующая Fc-рецепторы или антитело или другое связывающее соединение, которое будет связываться со средствами. В этом отношении, см., например, публикации патентных заявок США № 20040101519 и 20060034810 для искусственных антигенпредставляющих клеток (aAPC), которые предусматриваются для применения для активации и увеличения в количестве T-клеток в рамках настоящего изобретения.
В одном аспекте два средства иммобилизованы на гранулах, либо на одних и тех же гранулах, т.е. "цис", либо на отдельных гранулах, т.е. "транс". В качестве примера, средство, обеспечивающее первичный сигнал активации, представляет собой антитело против CD3 или его антигенсвязывающий фрагмент и средство, обеспечивающее костимулирующий сигнал, представляет собой антитело против CD28 или его антигенсвязывающий фрагмент; и оба средства совместно иммобилизованы на одних и тех же гранулах в эквивалентных молекулярных количествах. В одном аспекте используют соотношение 1:1 для каждого антитела, связанного с гранулами, для увеличения в количестве CD4+ T-клеток и роста T-клеток. В некоторых аспектах настоящего изобретения используют соотношение антител против CD3:CD28, связанных с гранулами, так чтобы наблюдалось увеличение экспансии T-клеток по сравнению с экспансией, наблюдаемой с использованием соотношения 1:1. В одном конкретном аспекте наблюдается увеличение в от приблизительно 1 до приблизительно 3 раз по сравнению с экспансией, наблюдаемой с использованием соотношения 1:1. В одном аспекте соотношение антител против CD3:CD28, связанных с гранулами, находится в диапазоне от 100:1 до 1:100, включая все целые числа между ними. В одном аспекте с частицами связано больше антитела против CD28, чем антитела против CD3, т.е. соотношение CD3:CD28 меньше единицы. В определенных аспектах соотношение антитела против CD28 и антитела против CD3, связанного с гранулами, превышает 2:1. В одном конкретном аспекте используют соотношение антител против CD3:CD28, связанных с гранулами, равное 1:100. В одном аспекте используют соотношение антител против CD3:CD28, связанных с гранулами, равное 1:75. В следующем аспекте используют соотношение антител против CD3:CD28, связанных с гранулами, равное 1:50. В одном аспекте используют соотношение антител против CD3:CD28, связанных с гранулами, равное 1:30. В одном предпочтительном аспекте используют соотношение антител против CD3:CD28, связанных с гранулами, равное 1:10. В одном аспекте используют соотношение антител против CD3:CD28, связанных с гранулами, равное 1:3. В следующем аспекте используют соотношение антител против CD3:CD28, связанных с гранулами, равное 3:1.
Для стимуляции T-клеток и других клеток-мишеней можно использовать отношения частиц к клеткам от 1:500 до 500:1, включая любые целые числа между ними. Как будет хорошо понятно специалистам в данной области, отношение частиц к клеткам может зависеть от размера частиц относительно клетки-мишени. Например, гранулы малых размеров могут связать только несколько клеток, в то время как гранулы больших размеров могут связывать множество клеток. В некоторых аспектах для стимуляции T-клеток можно использовать соотношение клеток и частиц, которое находится в диапазоне от 1:100 до 100:1, включая любые целые числа между ними, и в следующих аспектах соотношение включает от 1:9 до 9:1, включая любые целые числа между ними. Соотношение частиц с антителом против CD3 и антителом против CD28 и T-клеток, которое приводит к стимуляции T-клеток, может варьироваться, как отмечалось выше, однако определенные предпочтительные величины включают 1:100, 1:50, 1:40, 1:30, 1:20, 1:10, 1:9, 1:8, 1:7, 1:6, 1:5, 1:4, 1:3, 1:2, 1:1, 2:1, 3:1, 4:1, 5:1, 6:1, 7:1, 8:1, 9:1, 10:1 и 15:1, причем одно предпочтительное соотношение частиц на T-клетку составляет по меньшей мере 1:1. В одном аспекте используют соотношение частиц и клеток 1:1 или менее. В одном конкретном аспекте соотношение частица:клетка равно 1:5. В следующих аспектах соотношение частиц и клеток может варьироваться в зависимости от дня стимуляции. Например, в одном аспекте соотношение частиц и клеток составляет от 1:1 до 10:1 на первые сутки, и дополнительные частицы добавляют к клеткам каждые стуки или раз в двое суток после этого в течение вплоть до 10 суток, в конечных соотношениях от 1:1 до 1:10 (исходя из количества клеток в день добавления). В одном конкретном аспекте соотношение частиц и клеток составляет 1:1 в первый день стимуляции и его доводят до 1:5 не третьи и пятые сутки стимуляции. В одном аспекте частицы добавляют раз в сутки или раз в двое суток до конечного соотношения 1:1 на первые сутки и 1:5 на третьи и пятые сутки стимуляции. В одном аспекте соотношение частиц и клеток составляет 2:1 на первые сутки стимуляции и его доводят до 1:10 на третьи и пятые сутки стимуляции. В одном аспекте частицы добавляют раз в стуки или раз в двое суток до конечного соотношения 1:1 на первые сутки, и 1:10 на третьи и пятые сутки стимуляции. Специалисту в данной области будет понятно, что множество других соотношений может быть пригодным для применения в рамках настоящего изобретения. В частности, соотношения будут варьироваться в зависимости от размера частиц и от размера и типа клеток. В одном аспекте наиболее типичные соотношения для применения составляют около 1:1, 2:1 и 3:1 на первые сутки.
В следующих аспектах клетки, такие как T-клетки, комбинируют с покрытыми средством гранулами, затем гранулы и клетки разделяют, а затем клетки культивируют. В альтернативном аспекте перед культивированием покрытые средством гранулы и клетки не отделяют, а культивируют вместе. В следующем аспекте, гранулы и клетки сначала концентрируют с использованием силы, такой как магнитная сила, что приводит к увеличенному лигированию макркеров клеточной поверхности, тем самым индуцируя стимуляцию клеток.
В качестве примера, белки клеточной поверхности можно лигировать, позволяя парамагнитным гранулам, с которыми связаны антитело против CD3 и антитело против CD28 (гранулы 3×28) контактировать с T-клетками. В одном аспекте клетки (например, от 104 дo 109 T-клеток) и гранул (например, парамагнитные гранулы DYNABEAD® M-450 CD3/CD28 T в соотношении 1:1) комбинируют в буфере, например PBS (без двухвалентных катионов, таких как кальций и магний). Вновь, специалисту в данной области будет хорошо понятно, что можно использовать любую концентрацию клеток. Например, клетки-мишени могут быть очень редкими в образце и могут составлять только 0,01% образца, или весь образец (т.е. 100%) может состоять из представляющих интерес клеток-мишеней. Таким образом, в контексте настоящего изобретения предусматривается любое количество клеток. В некоторых аспектах может быть желательным значительное уменьшение объема, в котором частицы и клетки смешивают друг с другом (т.е. увеличение концентрации клеток) для обеспечения максимального контакта клеток и частиц. Например, в одном аспекте используют концентрацию приблизительно 10 миллиардов клеток/мл, 9 миллиардов клеток/мл, 8 миллиардов клеток/мл, 7 миллиардов клеток/мл, 6 миллиардов клеток/мл, 5 миллиардов клеток/мл или 2 миллиарда клеток/мл. В одном аспекте используют более 100 миллионов клеток/мл. В следующем аспекте используют концентрацию клеток, составляющую 10, 15, 20, 25, 30, 35, 40, 45 или 50 миллионов клеток/мл. В другом аспекте используют концентрацию клеток 75, 80, 85, 90, 95 или 100 миллионов клеток/мл. В следующих аспектах можно использовать концентрации 125 или 150 миллионов клеток/мл. Использование высоких концентраций может приводить к увеличению выхода клеток, активации клеток и экспансии клеток. Кроме того, применение высоких концентраций клеток позволяет более эффективное улавливание клеток, которые могут слабо экспрессировать представляющие интерес антигены-мишени, таких как отрицательные по CD28 T-клетки. Такие популяции клеток могут иметь терапевтическую ценность, и в некоторых аспектах может быть желательным их получение. Например, использование высокой концентрации клеток позволяет более эффективную селекцию CD8+ T-клеток, которые обычно имеют более слабую экспрессию CD28.
В одном варианте осуществления клетки, трансдуцированные нуклеиновой кислотой, кодирующей CAR, например, CAR, описанный в настоящем описании, увеличивают в количестве, например, способом, описанным в настоящем описании. В одном варианте осуществления клетки увеличивают в количестве в культуре в течение периода, составляющего от нескольких часов (например, приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 18, 21 час) до приблизительно 14 суток (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 суток). В одном варианте осуществления клетки увеличивают в количестве в течение периода от 4 до 9 суток. В одном варианте осуществления клетки увеличивают в количестве в течение периода 8 суток или менее, например, 7, 6 или 5 суток. В одном варианте осуществления клетки, например, клетки с CAR против CD19, описанные в настоящем описании, увеличивают в количестве в культуре в течение 5 суток, и полученные клетки являются более эффективными, чем те же клетки, которые увеличивали в количестве в культуре в течение 9 суток в тех же условиях культивирования. Эффективность может определяться, например, различными функциями T-клеток, например, пролиферацией, уничтожением клеток-мишеней, продукцией цитокинов, активацией, миграцией и их комбинациями. В одном варианте осуществления клетки, например, клетки с CAR против CD19, описанные в настоящем описании, которые увеличивали в количестве в течение 5 суток, демонстрируют увеличение удвоений клеток по меньшей мере в один, два, три или четыре раза при стимуляции антигеном по сравнению с теми же клетками, которые увеличивали в количестве в течение 9 суток при тех же условиях культивирования. В одном варианте осуществления клетки, например, клетки, экспрессирующие CAR против CD19, описанные в настоящем описании, увеличивают в количестве в культуре в течение 5 суток, и полученные клетки проявляют более высокую продукцию провоспалительных цитокинов, например, уровни IFN-γ и/или GM-CSF, по сравнению с теми же клетками, которые увеличивали в количестве в культуре в течение 9 суток в тех же условиях культивирования. В одном варианте осуществления клетки, например, клетки с CAR против CD19, описанные в настоящем описании, которые увеличивали в количестве в течение 5 суток, демонстрируют увеличение продукции провоспалительных цитокинов в пг/мл, например, уровни IFN-γ и/или GM-CSF, по меньшей мере в один, два, три, четыре, пять, десять раз или более по сравнению с теми же клетками, которые увеличивали в количестве в культуре в течение 9 суток при тех же условиях культивирования.
Также может быть желательным несколько циклов стимуляции, так чтобы время культивирования T-клеток составляло 60 суток или более. Условия, пригодные для культивирования T-клеток, включают соответствующие среды (например, минимальная поддерживающая среда или среда RPMI 1640 или, X-vivo 15, (Lonza)), которые могут содержать факторы, необходимые для пролиферации и жизнеспособности, включая сыворотку (например, эмбриональная сыворотка человек или сыворотка человека), интерлейкин-2 (IL-2), инсулин, IFN-γ, IL-4, IL-7, GM-CSF, IL-10, IL-12, IL-15, TGFβ и TNF-α или любые другие добавки для роста клеток, известные специалисту в данной области. Другие добавки для роста клеток включают, но не ограничиваются ими, поверхностно-активное вещество, плазманат и восстановители, такие как N-ацетилцистеин и 2-меркаптоэтанол. Среды могут включать RPMI 1640, AIM-V, DMEM, MEM, α-MEM, F-12, X-Vivo 15 и X-Vivo 20, Optimizer, с добавлением аминокислот, пирувата натрия и витаминов, либо бессывороточные, либо дополненные соответствующем количеством сыворотки (или плазмы) или определенным набором гормонов, и/или количеством цитокина(ов), достаточным для роста и экспансии T-клеток. Антибиотики, например, пенициллин и стрептомицин, включают только в экспериментальные культуры, но не в культуры клеток, которые подлежат инфузии индивидууму. Клетки-мишени поддерживают в условиях, необходимых для поддержания роста, например, при соответствующей температуре (например, 37°C) и в соответствующей атмосфере (например, воздух плюс 5% CO2).
В одном варианте осуществления клетки увеличивают в количестве в подходящей среде (например, среды, описанные в настоящем описании), которая включает один или несколько интерлейкинов, которые приводят к увеличению количества клеток по меньшей мере в 200 раз (например, в 200 раз, в 250 раз, в 300 раз, в 350раз) в период экспансии в течение 14 суток, например, при измерении способом, описанным в настоящем описании, таким как проточная цитометрия. В одном варианте осуществления клетки увеличивают в количестве в присутствии IL-15 и/или IL-7 (например, IL-15 и IL-7).
В вариантах осуществления способы, описанные в настоящем описании, например, способы получения экспрессирующих CAR клеток, включают удаление регуляторных T-клеток, например, CD25+ T-клеток, из популяции, например, с использованием антитела против CD25 или его фрагмента, или CD25-связывающего лиганда, IL-2. Способы удаления регуляторных T-клеток, например, CD25+ T-клеток, из популяции клеток описаны в настоящем описании. В вариантах осуществления способы, например, способы получения, кроме того, включают приведение в контакт популяции клеток (например, популяции клеток с истощением регуляторных T-клеток, таких как CD25+ T-клеток; или популяции клеток, которую ранее приводили в контакт с антителом против CD25, его фрагментом, или CD25-связывающим лигандом) с IL-15 и/или IL-7. Например, популяцию клеток (например, которую ранее приводили в контакт с антителом против CD25, его фрагментом или CD25-связывающим лигандом) увеличивают в количестве в присутствии IL-15 и/или IL-7.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, приводят в контакт с композицией, содержащей полипептид интерлейкина-15 (IL-15), полипептид альфа-рецептора интерлейкина-15 (IL-15Ra), или комбинацию как полипептида IL-15, так и полипептида IL-15Ra, например, hetIL-15, в процессе получения экспрессирующей CAR клетки, например, ex vivo. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, приводят в контакт с композицией, содержащей полипептид IL-15, в процессе получения экспрессирующей CAR клетки, например, ex vivo. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, приводят в контакт с композицией, содержащей комбинацию как полипептида IL-15, так и полипептида IL-15 Ra, в процессе получения экспрессирующей CAR клетки, например, ex vivo. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, приводят в контакт с композицией, содержащей hetIL-15, в процессе получения экспрессирующей CAR клетки, например, ex vivo.
В одном варианте осуществления экспрессирующую CAR клетку, описанную в настоящем описании, приводят в контакт с композицией, содержащей hetIL-15, в процессе увеличения в количестве ex vivo. В одном варианте осуществления экспрессирующую CAR клетку, описанную в настоящем описании, приводят в контакт с композицией, содержащей полипептид IL-15, в процессе увеличения в количестве ex vivo. В одном варианте осуществления экспрессирующую CAR клетку, описанную в настоящем описании, приводят в контакт с композицией, содержащей как полипептид IL-15, так и полипептид IL-15Ra, в процессе увеличения в количестве ex vivo. В одном варианте осуществления приведение в контакт приводит к выживанию и пролиферации субпопуляции лимфоцитов, например, CD8+ T-клеток.
T-клетки, подвергнутые воздействию в течение различной длительности периодов стимуляции, могут проявлять различные характеристики. Например, типичная кровь или мононуклеарные клеточные продукты подвергнутой аферезу периферической крови имеют популяцию хелперных T-клеток (TH, CD4+), которая превышает популяцию цитотоксических или супрессорных T-клеток (TC, CD8+). Экспансия T-клеток ex vivo посредством стимуляции рецепторов CD3 и CD28 продуцирует популяцию T-клеток, которая до приблизительно 8-9 суток состоит в основном из TH-клеток, в то время как после 8-9 суток, популяция T-клеток содержит значительно более высокую популяцию TC-клеток. Таким образом, в зависимости от цели лечения, инфузия индивидууму популяции T-клеток, содержащей в основном TH-клетки, может быть преимущественной. Аналогично, если антигенспецифическая подгруппа TC-клеток является выделенной, может быть предпочтительным увеличение в количестве этой подгруппы в большей степени.
Кроме того, в дополнение к маркерам CD4 и CD8, другие фенотипические маркеры значительно варьируются, однако по большей части, являются воспроизводимыми в ходе процесса экспансии клеток. Таким образом, такая воспроизводимость обеспечивает возможность адаптации продукта в виде активированных T-клеток к конкретным целям.
После конструирования CAR, описанного в настоящем описании, различные анализы можно использовать для оценки активности молекулы, такой как, но не ограничиваясь ими, способность к экспансии T-клеток после стимуляции антигеном, поддержание экспансии T-клеток в отсутствие рестимуляции и активность против злокачественной опухоли в различных моделях in vitro и на животных. Анализы для оценки эффектов CAR по настоящему изобретению более подробно описаны ниже.
Анализ с использованием вестер-блоттинга экспрессии CAR в первичных T-клетках можно использовать для обнаружения присутствия мономеров и димеров. См., например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). В кратком изложении, T-клетки (смесь 1:1 CD4+ и CD8+ T-клеток), экспрессирующие CAR, увеличивают в количестве in vitro в течение более чем 10 суток с последующим лизисом и SDS-PAGE в восстанавливающих условиях. CAR, содержащие полноразмерный цитоплазматический домен TCR-ζ и эндогенную цепь TCR-ζ, выявляют с помощью вестер-блоттинга с использованием антитела к цепи TCR-ζ. Те же подгруппы T-клеток используют для анализа SDS-PAGE в невосстанавливающих условиях, чтобы иметь возможность оценки образования ковалентного димера.
Экспансию CAR+ T-клеток in vitro после антигенной стимуляции можно измерять проточной цитометрии. Например, смесь CD4+ и CD8+ T-клеток стимулируют APC αCD3/αCD28 с последующей трансдукцией лентивирусными векторами, экспрессирующими GFP под контролем промоторов, подлежащих анализу. Иллюстративные промоторы включают промоторы гена CMV IE, EF-1α, убиквитина C или фосфоглицерокиназы (PGK). Флуоресценцию GFP оценивают на 6 сутки культивирования в подгруппах CD4+ и/или CD8+ T-клеток с использованием проточной цитометрии. См., например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). Альтернативно смесь CD4+ и CD8+ T-клеток стимулируют покрытыми αCD3/αCD28 магнитными гранулами на 0 сутки и трансдуцируют посредством CAR на 1 сутки с использованием бицистронного лентивирусного вектора, экспрессирующего CAR вместе с eGFP, с использованием рибосомальной последовательности пропуска 2A. После промывания культуры повторно стимулируют либо клетками K562, положительными по ассоциированному со злокачественной опухолью антигену (K562, экспрессирующие ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании), либо клетками K562 дикого типа (K562 дикого типа), либо клетками K562, экспрессирующими hCD32 и 4-1BBL, в присутствии антитела против CD3 и антитела против CD28 (K562-BBL-3/28). К культурам раз в двое суток добавляют экзогенный IL-2 в количестве 100 МЕ/мл. GFP+ T-клетки подсчитывают проточной цитометрией с использованием подсчета на основе гранул. См., например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009).
Также можно измерять длительную экспансию CAR+ T-клеток в отсутствие рестимуляции. См., например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). В кратком изложении измеряют средний объем T-клеток (фл) на 8 сутки культивирования с использованием устройства для подсчета частиц Coulter Multisizer III, Nexcelom Cellometer Vision или Millipore Scepter, после стимуляции покрытыми αCD3/αCD28 магнитными гранулами на 0 сутки и трансдукции указанным CAR на 1 сутки.
Также для измерения активности CART можно использовать модели на животных. Например, можно использовать модель с ксенторансплантатом с использованием CAR+ T-клеток, специфичных к ассоциированному со злокачественной опухолью антигену человека, описанному в настоящем описании, для лечения первичного пре-B ALL человека у иммунодефицитных мышей. См., например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). В кратком изложении, после установления ALL мышей случайным образом распределяют в группы лечения. Различные количества модифицированных способами инженерии CART-клеток, специфичных к ассоциированному со злокачественной опухолью антигену, совместно инъецируют в соотношении 1:1 мышам NOD-SCID-γ-/-, имеющим B-ALL. Количество копий вектора с CAR, специфичным к ассоциированному со злокачественной опухолью антигену, в ДНК селезенки от мышей оценивают в различные моменты времени после инъекции T-клеток. Животных оценивают в отношении лейкоза с интервалами раз в неделю. Количества бластных клеток B-ALL, положительных по ассоциированному со злокачественной опухолью антигену, описанному в настоящем описании, в периферической крови измеряют у мышей, которым инъецировали CAR+ T-клетки против ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании, и трансдуцированные имитирующей конструкцией T-клетки. Кривые выживаемости для групп сравнивают с использованием логарифмического рангового критерия. Кроме того, также можно анализировать абсолютное количество CD4+ и CD8+ T-клеток периферической крови через 4 недели после инъекции T-клеток мышам NOD-SCID-γ-/-. Мышам инъецируют лейкозные клетки и через 3 неделе инъецируют T-клетки, модифицированные способами инженерии для экспрессии CAR посредством бицистронного лентивирусного вектора, который кодирует CAR, связанный с eGFP. T-клетки нормализуют к 45-50% GFP+ T-клеток на входе путем смешения с трансдуцированными имитирующей конструкцией клетками перед инъекцией, и подтверждают проточной цитометрией. Животных оценивают в отношении лейкоза с интервалами, составляющими 1 неделю. Кривые выживаемости для групп CAR+ T-клеток сравнивают с использованием логарифмического рангового критерия.
Можно оценивать дозозависимый ответ на введение CAR. См., например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). Например, периферическую кровь получают через 35-70 суток после установления лейкоза у мышей, которым инъецировали на 21 сутки CAR T-клетки, эквивалентное количество трансдуцированных имитирующей конструкцией T-клеток, или не инъецировали T-клетки. У мышей из каждой группы случайным образом проводили взятие крови для определения количеств бластов ALL периферической крови, положительных по ассоциированному со злокачественной опухолью антигену, описанному в настоящем описании, а затем умерщвляли на 35 и 49 сутки. Остальных животных оценивали на 57 и 70 сутки.
Оценка пролиферации клеток и продукции цитокинов была описана ранее, например, в Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). В кратком изложении, оценку опосредуемой CAR пролиферации проводят на микропланшетах для титрования путем смешения промытых T-клеток с клетками K562, экспрессирующими ассоциированный со злокачественной опухолью антиген, описанный в настоящем описании (K19), или CD32 и CD137 (KT32-BBL) в конечном соотношении T-клетки:K562 2:1. Клетки K562 облучают гамма-излучением перед применением. К культурам с клетками KT32-BBL добавляют моноклональные антитела против CD3 (клон OKT3) и против CD28 (клон 9.3), чтобы они служили в качестве положительного контроля стимуляции пролиферации T-клеток, поскольку эти сигналы поддерживают длительную экспансию CD8+ T-клеток ex vivo. T-клетки подсчитывают в культурах с использованием флуоресцентных гранул CountBrightTM (Invitrogen, Carlsbad, CA) и проточной цитометрии, как описано изготовителем. CAR+ T-клетки идентифицируют по экспрессии GFP с использованием T-клеток, которые модифицированы связанными с eGFP-2A экспрессирующими CAR лентивирусными векторами. Для CAR+ T-клеток, не экспрессирующих GFP, CAR+ T-клетки выявляют с использованием биотинлированного рекомбинантного белка ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, и вторичного конъюгата авидин-PE. Также одновременно выявляют экспрессию CD4+ и CD8+ на T-клетках с использованием специфических моноклональных антител (BD Biosciences). Измерение цитокинов проводят в супернатантах, собранных через 24 часа после рестимуляции, с использованием набора цитометрических гранульных матриц для цитокинов TH1/TH2 (BD Biosciences, San Diego, CA) согласно инструкциям изготовителя. Флуоресценцию оценивают с использованием проточного цитометра FACScalibur, и данные анализируют в соответствии с инструкциями изготовителя.
Цитотоксичность можно оценивать способами, описанными в настоящем описании, например в примерах, или с использованием стандартного высвобождения 51Cr. См., например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). В кратком изложении, клетки-мишени (например, клетки BHK или CHO, экспрессирующие мезотелин) нагружают 51Cr (таким как NaCrO4, New England Nuclear, Boston, MA) при 37°C в течение 2 часов при частом встряхивании, промывают дважды в полной RPMI и высевают на микропланшеты для титрования. Эффекторные T-клетки смешивают с клетками-мишенями в лунках в полной RPMI в различных соотношениях эффекторная клетк:клетка-мишень (E:T). Также подготавливают дополнительные лунки, содержащие только среду (самопроизвольное высвобождение, SR) или 1% раствор детергента triton-X 100 (полное высвобождение, TR). После инкубации в течение 4 часов при 37°C супернатант каждой лунки собирают. Затем измеряют высвобждение 51Cr с использованием счетчика гамма-частиц (Packard Instrument Co., Waltham, MA). Каждые условия выполняют по меньшей мере в трех экземплярах, и процентный лизис вычисляют с использованием формулы: % лизис=(ER-SR)/(TR-SR), где ER представляет собой среднее количество 51Cr, высвободившееся для каждых экспериментальных условий. Также можно использовать альтернативные анализы цитотоксичности, такие как проточные анализы цитотоксичности.
Для оценки специфического транспорта и пролиферации CAR в моделях на имеющих опухоль животных можно использовать технологии визуализации. Такие способы анализа описаны, например, в Barrett et al., Human Gene Therapy 22:1575-1586 (2011). В кратком изложении, мышам NOD/SCID/γc-/- (NSG) проводят в/в инъекцию клеток Nalm-6, а затем через 7 суток T-клетки через 4 часа после введения конструкций CAR электропорацией. T-клетки стабильно трансфицируют лентивирусной конструкцией для экспрессии люциферазы светлячка, и мышей визуализируют в отношении биолюминеценции. Альтернативно терапевтическую эффективность однократной инъекции CAR+ T-клеток в модели с ксенотрансплантатом Nalm-6 можно измерять следующим образом: мышам NSG инъецируют Nalm-6, трансдуцированные для стабильной экспрессии люциферазы светлячка, а затем через 7 суток проводят однократную инъекцию в хвостовую вену T-клеток, в которые посредством электропорации был введен CAR по настоящему изобретению. Животных визуализируют в различные моменты времени после инъекции. Например, можно получать тепловые карты плотности фотонов положительных по люциферазе светлячка лейкозных клеток у репрезентативных мышей на 5 сутки (за 2 суток до лечения) и 8 сутки (через 24 ч после CAR+ PBL).
Также для оценки конструкций CAR, описанных в настоящем описании, можно использовать другие анализы, включая анализы, описанные в разделе "Примеры" настоящего писания, а также анализы, известные в данной области.
Терапевтическое применение
В одном аспекте изобретение относится к способам лечения заболевания, ассоциированного с экспрессией ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии XCAR, где X обозначает опухолевый антиген, как описано в настоящем описании, и где злокачественные клетки экспрессируют указанный опухолевый антиген X.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии XCAR, описанного в настоящем описании, где злокачественные клетки экспрессируют X. В одном варианте осуществления X экспрессируется как на нормальных клетках, так и на злокачественных клетках, но на нормальных клетках он экспрессируется на более низких уровнях. В одном варианте осуществления способ дополнительно включает селекцию CAR, который связывает X с аффинностью, которая позволяет XCAR связывать и уничтожать злокачественные клетки, экспрессирующие X, но уничтожается менее 30%, 25%, 20%, 15%, 10%, 5% или менее нормальных клеток, экспрессирующих X, например, как определяют с использованием анализа, описанного в настоящем описании. Например, можно использовать анализ, описанный на фиг.13A и 13B или анализ уничтожения, такой как проточная цитометрия, на основе Cr51 CTL. В одном варианте осуществления отобранный CAR имеет антигенсвязывающий домен, который обладает аффинностью связывания KD от 10-4 M до 10-8 M, например, от 10-5 M до 10-7 M, например, 10-6 M или 10-7 M, в отношении антигена-мишени. В одном варианте осуществления отобранный антигенсвязывающий домен обладает аффинностью связывания, которая по меньшей мере в пять раз, в 10 раз, в 20 раз, в 30 раз, в 50 раз, в 100 раз или в 1000 раз меньше, чем у эталонного антитела, например, антитела, описанного в настоящем описании.
В одном варианте осуществления настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD19 CAR, где злокачественные клетки экспрессируют CD19. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой ALL (острый лимфобластный лейкоз), CLL (хронический лимфоцитарный лейкоз), DLBCL (диффузная крупноклеточная B-клеточная лимфома), MCL (лимфома из клеток мантийной зоны или MM (множественная миелома).
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии EGFRvIIICAR, где злокачественные клетки экспрессируют EGFRvIII. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой глиобластому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против мезотелина, где злокачественные клетки экспрессируют мезотелин. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой мезотелиому, рак поджелудочной железы или рак яичника.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD123CAR, где злокачественные клетки экспрессируют CD123. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой AML.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD22CAR, где злокачественные клетки экспрессируют CD22. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой B-клеточные злокачественные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CS-1CAR, где злокачественные клетки экспрессируют CS-1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой множественную миелому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CLL-1CAR, где злокачественные клетки экспрессируют CLL-1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой AML.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD33CAR, где злокачественные клетки экспрессируют CD33. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой AML.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GD2CAR, где злокачественные клетки экспрессируют GD2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой нейробластому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии BCMACAR, где злокачественные клетки экспрессируют BCMA. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой множественную миелому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TnCAR, где злокачественные клетки экспрессируют антиген Tn. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак яичника.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PSMACAR, где злокачественные клетки экспрессируют PSMA. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак предстательной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии ROR1CAR, где злокачественные клетки экспрессируют ROR1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой B-клеточные злокачественные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии FLT3 CAR, где злокачественные клетки экспрессируют FLT3. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой AML.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TAG72CAR, где злокачественные клетки экспрессируют TAG72. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак желудочно-кишечного тракта.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD38CAR, где злокачественные клетки экспрессируют CD38. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой множественную миелому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD44v6CAR, где злокачественные клетки экспрессируют CD44v6. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак шейки матки, AML или MM.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CEACAR, где злокачественные клетки экспрессируют CEA. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак желудочно-кишечного тракта или рак поджелудочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии EPCAMCAR, где злокачественные клетки экспрессируют EPCAM. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак желудочно-кишечного тракта.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии B7H3CAR, где злокачественные клетки экспрессируют B7H3.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии KITCAR, где злокачественные клетки экспрессируют KIT. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак желудочно-кишечного тракта.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии IL-13Ra2CAR, где злокачественные клетки экспрессируют IL-13Ra2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой глиобластому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PRSS21CAR, где злокачественные клетки экспрессируют PRSS21. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, выбрана из рака яичника, рака поджелудочной железы, рака легкого и рака молочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD30CAR, где злокачественные клетки экспрессируют CD30. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой лимфомы или лейкозы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GD3CAR, где злокачественные клетки экспрессируют GD3. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD171CAR, где злокачественные клетки экспрессируют CD171. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой нейробластому, рак яичника, меланома, рак молочной железы, рак поджелудочной железы, рак толстого кишечника или NSCLC (немелкоклеточный рак легкого).
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии IL-11RaCAR, где злокачественные клетки экспрессируют IL-11Ra. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой остеосаркому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PSCACAR, где злокачественные клетки экспрессируют PSCA. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак предстательной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии VEGFR2CAR, где злокачественные клетки экспрессируют VEGFR2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой солидную опухоль.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии LewisYCAR, где злокачественные клетки экспрессируют LewisY. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак яичника или AML.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD24CAR, где злокачественные клетки экспрессируют CD24. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак поджелудочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против PDGFR-бета, где злокачественные клетки экспрессируют PDGFR-бета. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак молочной железы, рак предстательной железы, GIST (желудочно-кишечная стромальная опухоль), CML, DFSP (возвышающаяся дерматофибросаркома) или глиому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии SSEA-4CAR, где злокачественные клетки экспрессируют SSEA-4. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой глиобластому, рак молочной железы, рак легкого или злокачественную опухоль из стволовых клеток.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD20CAR, где злокачественные клетки экспрессируют CD20. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой B-клеточные злокачественные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против рецептора альфа фолатов, где злокачественные клетки экспрессируют рецептор альфа фолатов. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак яичника, NSCLC, рак эндометрия, рак почки или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии ERBB2CAR, где злокачественные клетки экспрессируют ERBB2 (Her2/neu). В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак молочной железы, рак желудка, рак ободочной и прямой кишки, рак легкого или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии MUC1CAR, где злокачественные клетки экспрессируют MUC1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак молочной железы, рак легкого или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии EGFRCAR, где злокачественные клетки экспрессируют EGFR. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой глиобластому, SCLC (мелкоклеточный рак легкого), SCCHN (плоскоклеточная карцинома головы и шеи), NSCLC или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии NCAMCAR, где злокачественные клетки экспрессируют NCAM. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой нейробластому или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAIXCAR, где злокачественные клетки экспрессируют CAIX. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак почки, CRC, рак шейки матки или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии EphA2CAR, где злокачественные клетки экспрессируют EphA2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой GBM.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GD3CAR, где злокачественные клетки экспрессируют GD3. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против фукозил-GM1, где злокачественные клетки экспрессируют фукозил-GM.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии sLeCAR, где злокачественные клетки экспрессируют sLe. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой NSCLC или AML.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GM3CAR, где злокачественные клетки экспрессируют GM3.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TGS5CAR, где злокачественные клетки экспрессируют TGS5.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии HMWMAACAR, где злокачественные клетки экспрессируют HMWMAA. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланому, глиобластому или рак молочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии o-ацетил-GD2CAR, где злокачественные клетки экспрессируют o-ацетил-GD2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой нейробластому или меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD19CAR, где злокачественные клетки экспрессируют CD19. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой AML, миелому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TEM1/CD248CAR, где злокачественные клетки экспрессируют TEM1/CD248. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой солидную опухоль.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TEM7RCAR, где злокачественные клетки экспрессируют TEM7R. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой солидную опухоль.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CLDN6CAR, где злокачественные клетки экспрессируют CLDN6. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак яичника, рак легкого или рак молочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TSHRCAR, где злокачественные клетки экспрессируют TSHR. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак щитовидной железы или множественную миелому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GPRC5DCAR, где злокачественные клетки экспрессируют GPRC5D. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой множественную миелому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CXORF61CAR, где злокачественные клетки экспрессируют CXORF61.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD97CAR, где злокачественные клетки экспрессируют CD97. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой B-клеточные злокачественные опухоли, рак желудка, рак поджелудочной железы, рак пищевода, глиобластому, рак молочной железы или рак ободочной и прямой кишки.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD179aCAR, где злокачественные клетки экспрессируют CD179a. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой B-клеточные злокачественные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии ALK CAR, где злокачественные клетки экспрессируют ALK. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой NSCLC, ALCL (анапластическая крупноклеточная лимфома), IMT (воспалительная миофибробластная опухоль) или нейробластому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии Полисиаловая кислота CAR, где злокачественные клетки экспрессируют полисиаловую кислоту. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой мелкоклеточный рак легкого.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PLAC1CAR, где злокачественные клетки экспрессируют PLAC1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой HCC (печеночно-клеточная карцинома).
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GloboHCAR, где злокачественные клетки экспрессируют GloboH. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак яичника, рак желудка, рак предстательной железы, рак легкого, рак молочной железы или рак поджелудочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии NY-BR-1CAR, где злокачественные клетки экспрессируют NY-BR-1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак молочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии UPK2CAR, где злокачественные клетки экспрессируют UPK2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак мочевого пузыря.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии HAVCR1CAR, где злокачественные клетки экспрессируют HAVCR1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак почки.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии ADRB3CAR, где злокачественные клетки экспрессируют ADRB3. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой саркому Юинга.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PANX3CAR, где злокачественные клетки экспрессируют PANX3. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой остеосаркому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GPR20CAR, где злокачественные клетки экспрессируют GPR20. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой GIST.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии LY6KCAR, где злокачественные клетки экспрессируют LY6K. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак молочной железы, рак легкого, рак яичника или рак шейки матки.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии OR51E2CAR, где злокачественные клетки экспрессируют OR51E2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак предстательной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TARPCAR, где злокачественные клетки экспрессируют TARP. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак предстательной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии WT1CAR, где злокачественные клетки экспрессируют WT1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии NY-ESO-1CAR, где злокачественные клетки экспрессируют NY-ESO-1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии LAGE-1a CAR, где злокачественные клетки экспрессируют LAGE-1a.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии MAGE-A1CAR, где злокачественные клетки экспрессируют MAGE-A1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии MAGE A1CAR, где злокачественные клетки экспрессируют MAGE A1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии ETV6-AML CAR, где злокачественные клетки экспрессируют ETV6-AML.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии sperm белок 17 CAR, где злокачественные клетки экспрессируют sperm белок 17. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак яичника, HCC или NSCLC.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии XAGE1CAR, где злокачественные клетки экспрессируют XAGE1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой саркому Юинга или рабдомиосаркому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии Tie 2 CAR, где злокачественные клетки экспрессируют Tie 2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой солидную опухоль.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии MAD-CT-1CAR, где злокачественные клетки экспрессируют MAD-CT-1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак предстательной железы или меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии MAD-CT-2CAR, где злокачественные клетки экспрессируют MAD-CT-2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак предстательной железы, меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против Fos-родственного антигена 1, где злокачественные клетки экспрессируют Fos-родственный антиген 1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой глиому, плоскоклеточный рак или рак поджелудочной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии p53CAR, где злокачественные клетки экспрессируют p53.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии prostein CAR, где злокачественные клетки экспрессируют простеин.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против сурвивина и теломеразы, где злокачественные клетки экспрессируют сурвивин и теломеразу.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против PCTA-1/галектина 8, где злокачественные клетки экспрессируют PCTA-1/галектин 8.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии MelanA/MART1CAR, где злокачественные клетки экспрессируют MelanA/MART1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против мутантного Ras, где злокачественные клетки экспрессируют мутантный Ras.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR с мутантным p53, где злокачественные клетки экспрессируют мутантный p53.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии hTERT CAR, где злокачественные клетки экспрессируют hTERT.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против точек разрыва при транслокации саркомы, где злокачественные клетки экспрессируют точки разрыва при транслокации саркомы. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой саркому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии ML-IAP CAR, где злокачественные клетки экспрессируют ML-IAP. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии ERGCAR, где злокачественные клетки экспрессируют ERG (слитый ген TMPRSS2 ETS).
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии NA17CAR, где злокачественные клетки экспрессируют NA17. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PAX3CAR, где злокачественные клетки экспрессируют PAX3. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой альвеолярную рабдомиосаркому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии андроген рецептор CAR, где злокачественные клетки экспрессируют рецептор андрогенов. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой метастазирующий рак предстательной железы.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против циклина B1, где злокачественные клетки экспрессируют циклин B1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии MYCNCAR, где злокачественные клетки экспрессируют MYCN.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии Rh°C CAR, где злокачественные клетки экспрессируют RhoC.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии TRP-2CAR, где злокачественные клетки экспрессируют TRP-2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланому.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CYP1B1CAR, где злокачественные клетки экспрессируют CYP1B1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак молочной железы, рак толстого кишечника, рак легкого, рак пищевода, рак кожи, рак лимфатических узлов, рак головного мозга или рак яичка.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии BORIS CAR, где злокачественные клетки экспрессируют BORIS. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак легкого.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии SART3CAR, где злокачественные клетки экспрессируют SART3.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PAX5CAR, где злокачественные клетки экспрессируют PAX5.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии OY-TES1CAR, где злокачественные клетки экспрессируют OY-TES1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии LCK CAR, где злокачественные клетки экспрессируют LCK.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии AKAP-4CAR, где злокачественные клетки экспрессируют AKAP-4.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии SSX2CAR, где злокачественные клетки экспрессируют SSX2.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии RAGE-1CAR, где злокачественные клетки экспрессируют RAGE-1. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой RCC (почечно-клеточный рак) или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против обратной транскриптазы теломеразы человека, где злокачественные клетки экспрессируют обратную транскриптазу теломеразы человека. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии RU1CAR, где злокачественные клетки экспрессируют RU1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии RU2CAR, где злокачественные клетки экспрессируют RU2.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против кишечной карбоксилэстеразы, где злокачественные клетки экспрессируют кишечную карбоксилэстеразу. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой рак щитовидной железы, RCC, CRC (рак ободочной и прямой кишки), рак молочной железы или другие солидные опухоли.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против простазы, где злокачественные клетки экспрессируют простазу.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии PAPCAR, где злокачественные клетки экспрессируют PAP.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против рецептора IGF-I, где злокачественные клетки экспрессируют рецептор IGF-I.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии gp100 CAR, где злокачественные клетки экспрессируют gp100.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии bcr-abl CAR, где злокачественные клетки экспрессируют bcr-abl.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против тирозиназы, где злокачественные клетки экспрессируют тирозиназу.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CAR против фукозил-GM1, где злокачественные клетки экспрессируют фукозил-GM1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии mut hsp70-2CAR, где злокачественные клетки экспрессируют mut hsp70-2. В одном варианте осуществления злокачественная опухоль, подвергаемая лечению, представляет собой меланома.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD79a CAR, где злокачественные клетки экспрессируют CD79a.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD79b CAR, где злокачественные клетки экспрессируют CD79b.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD72 CAR, где злокачественные клетки экспрессируют CD72.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии LAIR1 CAR, где злокачественные клетки экспрессируют LAIR1.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии FCAR CAR, где злокачественные клетки экспрессируют FCAR.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии LILRA2 CAR, где злокачественные клетки экспрессируют LILRA2.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CD300LF CAR, где злокачественные клетки экспрессируют CD300LF.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии CLEC12A CAR, где злокачественные клетки экспрессируют CLEC12A.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии BST2 CAR, где злокачественные клетки экспрессируют BST2.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии EMR2 CAR, где злокачественные клетки экспрессируют EMR2.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии LY75 CAR, где злокачественные клетки экспрессируют LY75.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии GPC3 CAR, где злокачественные клетки экспрессируют GPC3.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии FCRL5 CAR, где злокачественные клетки экспрессируют FCRL5.
В одном аспекте настоящее изобретение относится к способам лечения злокачественной опухоли путем предоставления индивидууму, нуждающемуся в этом, иммунных эффекторных клеток (например, T-клеток, NK-клеток), модифицированных способами инженерии для экспрессии IGLL1 CAR, где злокачественные клетки экспрессируют IGLL1.
В одном аспекте настоящее изобретение относится к лечению индивидуума in vivo с использованием CAR против PD1, так чтобы происходило ингибирование роста злокачественных опухолей. CAR против PD1 можно использовать отдельно для ингибирования роста злокачественных опухолей. Альтернативно CAR против PD1 можно использовать совместно с другими CAR, иммуногенными средствами, стандартными способами лечения или другими антителами. В одном варианте осуществления индивидуума лечат CAR против PD1 и XCAR, описанным в настоящем описании. В одном варианте осуществления CAR против PD1 используют совместно с другим CAR, например, CAR, описанным в настоящем описании, и ингибитором киназы, например, ингибитором киназы, описанным в настоящем описании.
В другом аспекте предусматривается способ лечения, например, снижения или смягчения, гиперпролиферативного состояния или нарушения (например, злокачественной опухоли), например, солидной опухоли, опухоли мягких тканей или метастатитческого очага, у индивидуума. Как используют в рамках изобретения, термин "злокачественная опухоль" включает все типы злокачественного роста или онкогенных процессов, метастатических тканей или злокачественной трансформированных клеток, тканей или органов, независимо от гистопатологического типа или стадии инвазивности. Примеры солидных опухолей включают злокачественные опухоли, например, саркомы, аденокарциномы и карциномы, различных систем органов, такие как злокачественные опухоли, поражающие печень, легкое, молочную железу, лимфоидную ткань, желудочно-кишечный тракт (например, толстый кишечник), мочеполовой тракт (например, почки, уротелиальные клетки), предстательную железу и глотку. Аденокарциномы включают злокачественные опухоли, такие как большинство злокачественных опухолей толстого кишечника, рак прямой кишки, почечно-клеточную карциному, рак печени, немелкоклеточную карциному легкого, рак тонкого кишечника и рак пищевода. В одном варианте осуществления злокачественная опухоль представляет собой меланому, например, развернутую меланому. Метастатические очаги вышеупомянутых злокачественных опухолей также можно лечить или предупреждать с использованием способов и композиций по изобретению. Примеры других злокачественных опухолей, которые можно лечить, включают рак кости, рак поджелудочной железы, рак кожи, рак головы и шеи, кожную или внутриглазную злокачественную меланому, рак тела матки, рак яичника, рак прямой кишки, рак анальной области, рак желудка, рак яичка, рак тела матки, карциному фаллопиевых труб, карциному эндометрия, карциному шейки матки, карциному влагалища, карциному вульвы, болезнь Ходжкина, неходжкинскую лимфому, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркому мягких тканей, рак уретры, рак полового члена, хронический или острый лейкоз, включая острый миелоидный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, солидные опухоли детского возраста, лимфоцитарную лимфому, рак мочевого пузыря, рак почки или мочеточника, карциному почечной лоханки, новообразование центральной нервной системы (ЦНС), первичную лимфому ЦНС, ангиогенез опухоли, опухоль позвоночника, глиому ствола головного мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак, плоскоклеточный рак, T-клеточную лимфому, индуцируемые факторами внешней среды злокачественные опухоли, включая опухоли, индуцируемые асбестозом, и комбинации указанных злокачественных опухолей. Лечение метастазирующих злокачественных опухолей, например, метастазирующих злокачественных опухолей, которые экспрессируют PD-L1 (Iwai et al. (2005) Int. Immunol. 17:133-144), можно осуществлять с использованием молекул антител, описанных в настоящем описании.
Иллюстративные злокачественные опухоли, рост которых может ингибироваться, включают злокачественные опухоли, обычно отвечающие на иммунотерапию. Неограничивающие примеры злокачественных опухолей, подлежащих лечению, включают меланому (например, метастазирующая злокачественная меланома), рак почки (например, светлоклеточная карцинома), рак предстательной железы (например, рефрактерная к гормонам аденокарцинома предстательной железы), рак молочной железы, рак толстого кишечника и рак легкого (например, немелкоклеточный рак легкого). Кроме того, рефрактерные или рецидивирующие злокачественные опухоли можно лечить с использованием молекул, описанных в настоящем описании.
В одном аспекте изобретение относится к вектору, содержащему CAR, функционально связанный с промотором для экспрессии в иммунных эффекторных клетках млекопитающих (например, T-клетках, NK-клетках). В одном аспекте изобретение относится к рекомбинантной иммунной эффекторной клетке, экспрессирующей CAR по настоящему изобретению, для применения для лечения злокачественной опухоли, экспрессирующей ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании. В одном аспекте экспрессирующие CAR клетки по изобретению способны контактировать с опухолевой клеткой, на поверхности которой экспрессируется по меньшей мере один ассоциированный со злокачественной опухолью антиген, так что экспресирующая CAR клетка нацеливается на злокачественную клетку и рост злокачественной клетки ингибируется.
В одном аспекте изобретение относится к способу ингибирования роста злокачественной опухоли, включающему приведение злокачественной клетки в контакт с экспрессирующей CAR клеткой по настоящему изобретению, так чтобы CART активировалась в ответ на антиген и нацеливалась на злокачественную опухоль, ингибируя рост опухоли.
В одном аспекте изобретение относится к способу лечения злокачественной опухоли у индивидуума. Способ включает введение индивидууму экспрессирующей CAR клетки по настоящему изобретению, так чтобы происходило лечение злокачественной опухоли у индивидуума. В одном аспекте злокачественная опухоль, ассоциированная с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, представляет собой гематологическую злокачественную опухоль. В одном аспекте гематологическая злокачественная опухоль представляет собой лейкоз или лимфому. В одном аспекте злокачественная опухоль, ассоциированная с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, включает злокачественные опухоли и новообразования, включающие, но не ограничивающиеся ими, например, один или несколько из острых лейкозов, включая, но не ограничиваясь ими, например, B-клеточный острый лимфоидный лейкоз ("BALL"), T-клеточный острый лимфоидный лейкоз ("TALL"), острый лимфоидный лейкоз (ALL); один или несколько из хронических лейкозов, включая, но не ограничиваясь ими, например, хронический миелогенный лейкоз (CML), хронический лифмоидный лейкоз (CLL). Дополнительные злокачественные опухоли и гематологические состояния, ассоциированные с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, включают, но не ограничиваются ими, например, B-клеточный пролимфоцитарный лейкоз, новообразование из бластных плазмацитоидных дендритных клеток, лимфому Беркитта, диффузную крупноклеточную B-клеточную лимфому, фолликулярную лимфому, волосатоклеточный лейкоз, мелкоклеточную или крупноклеточную фолликулярную лимфому, злокачественные лимфопролиферативные состояния, лимфому MALT, лимфому из клеток мантийной зоны, лимфому из клеток маргинальной зоны, множественную миелому, миелодисплазию и миелодиспластический синдром, неходжкинскую лимфому, плазмабластную лимфому, новообразование из плазмацитоидных дендритных клеток, макроглобулинемию Вальденстрема и "предлейкоз", который представляет собой разнообразный набор гематологических состояний, характеризующихся неэффективной продукцией (или дисплазией) миелоидных клеток крови, и т.п. Следующее заболевание, ассоциированные с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, включают, но не ограничиваются ими, например, атипичные и/или неклассические злокачественные опухоли, новообразования, предзлокачественные состояния или пролиферативные заболевания, ассоциированные с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании.
В некоторых вариантах осуществления злокачественная опухоль, которую можно лечить экспрессирующей CAR клеткой по настоящему изобретению, представляет собой множественную миелому. Множественная миелома представляет собой злокачественную опухоль крови, характеризующуюся накоплением клона плазмацитов в костном мозге. Современные способы терапии множественной миеломы включают, но не ограничиваются ими, лечение леналидомидом, который является аналогом талидомида. Леналидомид обладает активностью, которая включает противоопухолевую активность, ингибирование ангиогенеза и иммуномодулирование. Как правило, миелоидные клетки считаются отрицательными по экспрессии ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, при определении с использованием проточной цитометрии. Таким образом, в некоторых вариантах осуществления CAR против CD19, например, как описано в настоящем описании, можно использовать для нацеливания на клетки миеломы. В некоторых вариантах осуществления терапию CAR по настоящему изобретению можно использовать в комбинации с одним или несколькими дополнительными способами терапии, например, лечением леналидомидом.
Изобретение относится к типу клеточной терапии, где иммунные эффекторные клетки (например, T-клетки, NK-клетки) генетически модифицированы для экспрессии химерного рецептора антигена (CAR), и экспрессирующую CAR T-клетку или NK-клетку вводят путем инфузии реципиенту, нуждающемуся в этом. Введенная путем инфузии клетка способна уничтожать опухолевые клетки реципиента. В отличие от антительной терапии, модифицированные посредством CAR иммунные эффекторные клетки (например, T-клетки, NK-клетки) способны реплицироваться in vivo, что приводит к длительной персистенции, которая может приводить к длительному контролю опухоли. В различных аспектах иммунные эффекторные клетки (например, T-клетки, NK-клетки), вводимые пациенту, или их потомство, сохраняются у пациента в течение по меньшей мере четырех месяцев, пяти месяцев, шести месяцев, семи месяцев, восьми месяцев, девяти месяцев, десяти месяцев, одиннадцати месяцев, двенадцати месяцев, тринадцати месяцев, четырнадцати месяцев, пятнадцати месяцев, шестнадцати месяцев, семнадцати месяцев, восемнадцати месяцев, девятнадцати месяцев, двадцати месяцев, двадцати одного месяца, двадцати двух месяцев, двадцати трех месяцев, двух лет, трех лет, четырех лет или пяти лет после введения T-клетки или NK-клетки пациенту.
Также изобретение относится к типу клеточной терапии, где иммунные эффекторные клетки (например, T-клетки, NK-клетки) модифицированы, например, транскрибированной in vitro РНК, для временной экспрессии химерного рецептора антигена (CAR), и экспрессирующую CAR T-клетку или NK-клетку вводят путем инфузии реципиенту, нуждающемуся в этом. Введенная путем инфузии клетка способна уничтожать опухолевые клетки реципиента. Таким образом, в различных аспектах иммунные эффекторные клетки (например, T-клетки, NK-клетки), введенные пациенту, сохраняются в течение менее чем одного месяца, например, трех недель, двух недель, одной недели после введения T-клеток пациенту.
Без связи с какой-либо конкретной теорией, иммунный ответ против опухолевых клеток, индуцируемый модифицированными посредством CAR иммунными эффекторными клетками (например, T-клетками, NK-клетками), может представлять собой активный или пассивный иммунный ответ или альтернативно может быть следствием прямого против непрямого иммунного ответа. В одном аспекте трансдуцированные CAR иммунные эффекторные клетки (например, T-клетки, NK-клетки) проявляют специфическую секрецию провоспалительных цитокинов и мощную цитолитическую активность в ответ на злокачественные клетки человека, экспрессирующие ассоциированный со злокачественной опуолью антиген, описанный в настоящем описании, выдерживают ингибирование растворимым ассоциированным со злокачественной опухолью антигеном, описанным в настоящем описании, и опосредуют неспецифический цитолиз и опосредуют регрессию развернутой опухоли человека. Например, опухолевые клетки без антигена в гетерогенной области опухоли, экспрессирующей ассоциированный со злокачественной опухолью антиген, описанный в настоящем описании, могут быть чувствительными к непрямому разрушению перенацеленных на ассоциированный со злокачественной опухолью антиген, описанный в настоящем описании, иммунных эффекторных клеток (например, T-клеток, NK-клеток), которые ранее реагировали с соседними положительными по антигену злокачественными клетками.
В одном аспекте полностью человеческие модифицированные посредством CAR иммунные эффекторные клетки (например, T-клетки, NK-клетки) по изобретению могут представлять собой тип вакцины для иммунизации ex vivo и/или терапии in vivo у млекопитающего. В одном аспекте млекопитающим является человек.
Что касается иммунизации ex vivo, по меньшей мере одно из следующих событий происходит in vitro перед введением клетку в млекопитающего: i) увеличение клеток в количестве, ii) введение нуклеиновой кислоты, кодирующей CAR, в клетки или iii) криоконсервация клеток.
Методики ex vivo хорошо известны в данной области и более подробно рассмотрены ниже. В кратком изложении, выделяют клетки млекопитающего (например, человека) и генетически модифицируют (т.е. трансдуцируют или трансфицируют in vitro) вектором, экспрессирующим CAR, описанный в настоящем описании. Модифицированную посредством CAR клетку можно вводить реципиенту-млекопитающему для обеспечения терапевтической пользы. Реципиент-млекопитающее может представлять собой человека и модифицированная посредством CAR клетка может быть аутологичной для реципиента. Альтернативно клетки могут быть аллогенными, сингенными или ксеногенными в отношении реципиента. В дополнение к использованию клеточной вакцины для иммунизации ex vivo, также в настоящее изобретение включены композиции и способы для иммунизации in vivo для индукции иммунного ответа, направленного против антигена у пациента.
Для клеток по настоящему изобретению можно использовать методику экспансии гематологических стволовых клеток и клеток-предшественников ex vivo, описанную в патенте США № 5199942, включенном в настоящее описание путем ссылки. В данной области известны другие подходящие способы, таким образом, настоящее изобретение не ограничивается каким-либо конкретным способом экспансии клеток ex vivo. В краткому изложении, культивирование и экспансия иммунных эффекторных клеток (например, T-клеток, NK-клеток) ex vivo включает: (1) сбор CD34+ гематологических стволовых клеток и клеток-предшественников из взятой периферической крови или эксплантатов костного мозга; и (2) экспансию таких клеток ex vivo. В дополнение к клеточным факторам роста, описанным в патенте США № 5199942, для культивирования и экспансии клеток можно использовать другие факторы, такие как flt3-L, IL-1, IL-3 и лиганд c-kit.
В дополнение к использованию клеточной вакцины для иммунизации ex vivo, настоящее изобретение также относится к композициям и способам для иммунизации in vivo для индукции иммунного ответа, направленного против антигена у пациента.
Как правило, клетки активированные и увеличенные в количестве, как описано в настоящем описании, можно использовать для лечения и предупреждения заболеваний, которые возникают у индивидуумов с иммунодефицитом. В частности, модифицированные посредством CAR иммунные эффекторные клетки (например, T-клетки, NK-клетки) по изобретению используют для лечения заболеваний, нарушений и состояний, ассоциированных с экспрессией ассоциированного со злокачественной опухолью антигена, описанного в настоящем описании. В определенных аспектах клетки по изобретению используют для лечения пациентов, имеющих риск развития заболеваний, нарушений и состояний, ассоциированных с экспрессией ассоциированного со злокачественной опухолью антигена, описанного внастоящем описании. Таким образом, изобретение относится к способам лечения или предупреждения заболеваний, нарушений и состояний, ассоциированных с экспрессией ассоциированного со злокачественной опухолью антигена, описанного внастоящем описании, включающим введение индивидууму, нуждающемуся в этом, терапевтически эффективного количества модифицированных посредством CAR иммунных эффекторных клеток (например, T-клеток, NK-клеток) по изобретению.
В одном аспекте экспрессирующие CAR клетки по изобретению можно использовать для лечения пролиферативного заболевания, такого как злокачественная опухоль или новобразование, или предзлокачественного состояния, такого как миелодисплазия, миелодиспластический синдром или предлейкоз. Кроме того, заболевание, ассоциированное с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, включает, но не ограничиваясь ими, например, атипичные и/или неклассические злокачественные опухоли, новообразования, предзлокачественные состояния или пролиферативные заболевания, экспрессирующие ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании. Не связанные со злокачественной опухолью заболевания, ассоциированные с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, включают, но не ограничиваются ими, например, аутоиммунное заболевание (например, волчанку), воспалительные нарушения (аллергия и астма) и трансплантацию.
Модифицированные посредством CAR иммунные эффекторные клетки (например, T-клетки, NK-клетки) по настоящему изобретению можно вводить либо отдельно, либо в качестве фармацевтической композиции в комбинации с разбавителями и/или с другими компонентами, такими как IL-2 или другие цитокины или клеточные популяции.
Гематологическая злокачественная опухоль
Гематологические злокачественные состояния представляют собой типы злокачественной опухоли, такие как лейкоз, лимфома и злокачественные лимфоролиферативные состояния, которые поражают кровь, костный мозг и лимфатическую систему.
Лейкоз можно подразделить на острый лейкоз и хронический лейкоз. Острый лейкоз можно далее подразделить на острый миелогенный лейкоз (AML) и острый лимфоидный лейкоз (ALL). Хронический лейкоз включает хронический миелогенный лейкоз (CML) и хронический лимфоидный лейкоз (CLL). Другие родственные состояния включают миелодиспластические синдромы (MDS, ранее известный как "предлейкоз"), которые представляют собой многообразную совокупность гематологических состояний, объединенных неэффективной продукцией (или дисплазией) миелоидных клеток крови и риском трансформации в AML.
Лимфома представляет собой группу опухолей из клеток крови, которая развивается из лимфоцитов. Иллюстративные лимфомы включают неходжкинскую лимфому и лимфому Ходжкина.
Настоящее изобретение относится к композициям и способам для лечения злокачественной опухоли. В одном аспекте злокачественная опухоль представляет собой гематологическую злокачественную опухоль, включая, но не ограничиваясь ими, лейкоз или лимфому. В одном аспекте экспрессирующие CAR клетки по изобретению можно использовать для лечения злокачественных опухолей и новообразований, таких как, но не ограничиваясь ими, например, острые лейкозы, включая, но не ограничиваясь ими, например, B-клеточный острый лимфоидный лейкоз ("BALL"), T-клеточный острый лимфоидный лейкоз ("TALL"), острый лимфоидный лейкоз (ALL); один или несколько из хронических лейкозов, включая, но не ограничиваясь ими, например, хронический миелогенный лейкоз (CML), хронический лимфоцитарный лейкоз (CLL); дополнительные гематологические злокачественные опухоли или гематологические состояния, включая, но не ограничиваясь ими, например, B-клеточный пролимфоцитарный лейкоз, новообразование из бластных плазмацитоидных дендритных клеток, лимфому Беркитта, диффузную крупноклеточную B-клеточную лимфому, фолликулярную лимфому, волосатоклеточный лейкоз, мелкоклеточную или крупноклеточную фолликулярную лимфому, злокачественные лимфопролиферативные состояния, лимфому MALT, лимфому из клеток мантийной зоны, лимфому из клеток маргинальной зоны, множественную миелому, миелодисплазию и миелодиспластический синдром, неходжкинскую лимфому, плазмабластную лимфому, новообразование из плазмацитоидных дендритных клеток, макроглобулинемию Вальденстрема и "предлейкоз", который представляет собой разнообразные гематологические состояния, характеризующихся неэффективной продукцией (или дисплазией) миелоидных клеток крови, и т.п. Следующее заболевание, ассоциированные с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании, включают, но не ограничиваются ими, например, атипичные и/или неклассические злокачественные опухоли, новообразования, предзлокачественные состояния или пролиферативные заболевания, ассоциированные с экспрессией ассоциированного со злокачественной опухолью антигена, как описано в настоящем описании.
Настоящее изобретение также относится к способам ингибирования пролиферации или уменьшения популяции клеток, экспрессирующих ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, причем способы включают приведение в контакт популяции клеток, содержащей клетку, экспрессирующую ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, с экспрессирующей CAR T-клеткой или NK-клеткой по изобретению, которая связывается с клеткой, экспрессирующий ассоциированный злокачественной опухолью антиген, как описано в настоящем описании. В конкретном аспекте настоящее изобретение относится к способам ингибирования пролиферации или уменьшения популяции злокачественных клеток, экспрессирующих ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, причем способы включают приведение в контакт популяции злокачественных клеток, экспрессирующих ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, с экспрессирующей CAR T-клеткой или NK-клеткой по изобретению, которая связывается с клеткой, экспрессирующей ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании. В одном аспекте настоящее изобретение относится к способам ингибирования пролиферации или уменьшения популяции опухолевых клеток, экспрессирующих ассоциированный со злокачественной опухолью антиген как описано в настоящем описании, причем способы включают приведение в контакт популяции злокачественных клеток, экспрессирующих ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, с экспрессирующей CAR T-клеткой или NK-клеткой по изобретению, которая связывается с клеткой, экспрессирующей ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании. В некоторых аспектах, экспрессирующая CAR T-клетка или NK-клетка по изобретению уменьшает количество, число, величину или процент клеток и/или злокачественных клеток по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 65%, по меньшей мере на 75%, по меньшей мере на 85%, по меньшей мере на 95% или по меньшей мере на 99% у индивидуума с миелоидным лейкозом или другой злокачественной опухолью, ассоциированной с клетками, экспрессирующими ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, или в модели на животных для этих заболеваний, относительно отрицательного контроля. В одном аспекте индивидуумом является человек.
Настоящее изобретение также относится к способам предупреждения, лечения и/или управления течением заболевания, ассоциированного с клетками, экспрессирующими ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании (например, гематологическая злокачественная опухоль или атипичная злокачественная опухоль, экспрессирующая ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании), причем способы включают введение индивидууму, нуждающемуся в этом, экспрессирующей CAR T-клетки или NK-летки по изобретению, которая связывается с клеткой, экспрессирующей ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании. В одном аспекте индивидуумом является человек. Неограничивающие примеры нарушений, ассоциированных с клетками, экспрессирующими ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, включают аутоиммунные нарушения (такие как волчанка), воспалительные нарушения (такие как аллергия и астма) и злокачественные опухоли (такие как гематологические злокачественные опухоли или атипичные злокачественные опухоли, экспрессирующие ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании).
Настоящее изобретение также относится к способам предупреждения, лечения и/или управления течением заболевания, ассоциированного с клетками, экспрессирующими ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, причем способы включают введение индивидууму, нуждающемуся в этом, экспрессирующей CAR T-клетки или NK-клетки по изобретению, которая связывается с клеткой, экспрессирующей ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании. В одном аспекте индивидуумом является человек.
Настоящее изобретение относится к способам предупреждения рецидива злокачественной опухоли, ассоциированной с клетками, экспрессирующими ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, причем способы включают введение индивидууму, нуждающегося в этом, экспрессирующей CAR T-клетки или NK-клетки по изобретению, которая связывается с клеткой, экспрессирующей ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании. В одном аспекте способы включают введение индивидууму, нуждающемуся в этом, эффективного количества экспрессирующей CAR T-клетки или NK-клетки, описанной в настоящем описании, которая связывается с клеткой, экспрессирующей ассоциированный со злокачественной опухолью антиген, как описано в настоящем описании, в комбинации с эффективным количеством другого терапевтического средства.
Комбинированные способы терапии
Экспрессирующую CAR клетку, описанную в настоящем описании, можно использовать в комбинации с другими известными средствами и способами терапии. Введение "в комбинации", как используют в рамках изобретения, означает, что два (или более) различных способа лечения осуществляют у индивидуума в процессе наличия у индивидуума нарушения, например, два или более способа лечения проводят после диагностирования у пациента нарушения и пока не произойдет излечение или устранение нарушения, или пока лечение не прекратят по другим причинам. В некоторых вариантах осуществления введение одного лекарственного средства все еще проводят, когда начинается введение второго, так что существует перекрывание с точки зрения введения. Это иногда называют в настоящем описании "одновременной" или "совместной" доставкой. В других вариантах осуществления доставка одного лекарственного средства заканчивается до начала доставки другого лекарственного средства. В некоторых вариантах осуществления в любом случае лечение является более эффективным вследствие комбинированного введения. Например, второе лекарственное средство является более эффективным, например, эквивалентный эффект наблюдают при использовании меньшего количества второго лекарственного средства, или второе лекарственное средство снижает симптомы в большей степени, чем наблюдалось бы, если бы второе лекарственное средство вводили в отсутствие первого лекарственного средства, или аналогичную ситуацию наблюдают для первого лекарственного средства. В некоторых вариантах осуществления доставка является такой, что снижение симптома или другого параметра, связанного с нарушения, является более высоким, чем наблюдалось бы при доставке одного лекарственного средства в отсутствие другого. Эффект двух способов лечения может быть частично аддитивным, полностью аддитивным или более чем аддитивным. Доставка может быть такой, что эффект первого доставляемого лекарственного средства все еще поддается обнаружению, когда доставляют второе лекарственное средство.
Экспрессирующую CAR клетку, описанную в настоящем описании, и по меньшей мере одно дополнительное лекарственное средство можно вводить одновременно, в одной или в отдельных композициях, или последовательно. Для последовательного введения сначала можно вводить экспрессирующую CAR клетку, описанную в настоящем описании, а затем можно вводить дополнительное средство, или порядок введения может быть обратным.
Терапию CAR и/или проведение введения других лекарственных средств, процедур и способов лечения, можно осуществлять в ходе периодов активного нарушения или в ходе периодов ремиссии или менее активного заболевания. Терапию CAR можно проводить до другого способа лечения, одновременно с лечением, после лечения или в ходе ремиссии нарушения.
При введении в комбинации, терапию CAR и введение дополнительного средства (например, второго или третьего средства), или всех из них, можно проводить в количестве или в дозе, которые превышают, являются более низкими или являются такими же, как и количество или доза каждого средства отдельно, например, в качестве монотерапии. В определенных вариантах осуществления введенное количество или доза терапии CAR, дополнительного средства (например, второго или третьего средства), или всех из них, является более низкой (например, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40% или по меньшей мере на 50%) чем количество или доза каждого средства, используемого отдельно, например, в качестве монотерапии. В других вариантах осуществления количество или доза терапии CAR, дополнительного средства (например, второго или третьего средства), или всех из них, которые приводят к желаемому эффекту (например, лечение злокачественной опухоли) являются более низкими (например, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40% или по меньшей мере на 50% более низкими), чем количество или доза каждого средства, используемого отдельно, например, в качестве монотерапии, требуемые для достижения того же терапевтического эффекта.
В следующих аспектах, экспрессирующую CAR клетку, описанную в настоящем описании можно использовать в терапевтическом режиме в комбинации с хирургической операцией, химиотерапией, лучевой терапией, иммунодепрессивными средствами, такими как циклоспорин, азатиоприн, метотрексат, микофенолат и FK506, антителами или другими иммунодеструктивными средствами, такими как CAMPATH, антитела против CD3, или другими способами терапии на основы антител, цитоксином, флударабином, циклоспорином, FK506, рапамицином, микофеноловой кислотой, стероидами, FR901228, цитокинами и лучевой терапией, пептидной вакциной, такой как пептидная вакцина, описанная в Izumoto et al. 2008 J Neurosurg 108:963-971.
В одном варианте осуществления экспрессирующую CAR клетку, описанную в настоящем описании можно использовать в комбинации с другим химиотерапевтическим средством. Иллюстративные химиотерапевтические средства включают антрациклин (например, доксорубицин (например, липосомальный доксорубицин)), алкалоид барвинка (например, винбластин, винкристин, виндезин, винорелбин), алкилирующее средство (например, циклофосфамид, декарбазин, мелфалан, ифосфамид, темозоломид), антитело против иммунных клеток (например, алемтузумаб, гемтузумаб, ритуксимаб, офатумумаб, тозитумомаб, брентуксимаб), антиметаболит (включая, например, антагонисты фолиевой кислоты, аналоги пиримидинов, аналоги пуринов и ингибиторы аденозиндезаминазы (например, флударабин)), ингибитор mTOR, агонист индуцируемого глюкокортикоидами TNFR-родственного белка (GITR), ингибитор протеасом (например, аклациномицин A, глиотоксин или бортезомиб), иммуномодулятор, такой как талидомид или производное талидомида (например, леналидомид).
Основные химические средства, предусматриваемые для применения в комбированных способах терапии, включают анастрозол (Arimidex®), бикалутамид (Casodex®), блеомицин сульфат (Blenoxane®), бусульфан (Myleran®), бусульфан инъекционный (Busulfex®), капецитабин (Xeloda®), N4-пентоксикарбонил-5-дезокси-5-фторцитидин, карбоплатин (Paraplatin®), кармустин (BiCNU®), хлорамбуцил (Leukeran®), цисплатин (Platinol®), кладрибин (Leustatin®), циклофосфамид (Cytoxan® или Neosar®), цитарабин, цитозин арабинозид (Cytosar-U®), инъекционный липосомальный цитарабин (DepoCyt®), дакарбазин (DTIC-Dome®), дактиномицин (актиномицин D, космеган), даунорубицин гидрохлорид (Cerubidine®), инъекционный липосомальный даунорубицина цитрат (DaunoXome®), дексаметазон, доцетаксел (Taxotere®), доксорубицина гидрохлорид (Adriamycin®, Rubex®), этопозид (Vepesid®), флударабина фосфат (Fludara®), 5-фторурацил (Adrucil®, Efudex®), флутамид (Eulexin®), тезацитибин, гемцитабин (дифтордезоксицитидин), гидроксимочевину (Hydrea®), идарубицин (Idamycin®), ифосфамид (IFEX®), иринотекан (Camptosar®), L-аспарагиназу (ELSPAR®), лейковорин кальций, мелфалан (Alkeran®), 6-меркаптопурин (Purinethol®), метотрексат (Folex®), митоксантрон (Novantrone®), милотарг, паклитаксел (Taxol®), феникс (иттрий-90/MX-DTPA), пентостатин, имплантат полифепросана 20 с кармустином (Gliadel®), тамоксифена цитрат (Nolvadex®), тенипозид (Vumon®), 6-тиогуанин, тиотепу, тирапазамин (Tirazone®), топотекана гидрохлорид для инъекций (Hycamptin®), винбластин (Velban®), винкристин (Oncovin®) и винорелбин (Navelbine®).
Иллюстративные алкилирующие средства включают, но не ограничиваются ими, азотистые иприты, производные этиленимина, алкилсульфонаты, нитрозомочевину и триазены: урацил мустард (Aminouracil Mustard®, Chlorethaminacil®, Demethyldopan®, Desmethyldopan®, Haemanthamine®, Nordopan®, Uracil nitrogen mustard®, Uracillost®, Uracilmostaza®, Uramustin®, Uramustine®), хлорметин (Mustargen®), циклофосфамид (Cytoxan®, Neosar®, Clafen®, Endoxan®, Procytox®, RevimmuneTM), ифосфамид (Mitoxana®), мелфалан (Alkeran®), хлорамбуцил (Leukeran®), пипоброман (Amedel®, Vercyte®), триэтиленмеламин (Hemel®, Hexalen®, Hexastat®), триэтилентиофосфорамин, темозоломид (Temodar®), тиотепу (Thioplex®), бусульфан (Busilvex®, Myleran®), кармустин (BiCNU®), ломустин (CeeNU®), стрептозоцин (Zanosar®) и дакарбазин (DTIC-Dome®). Дополнительные иллюстративные алкилирующие средства включают, но не ограничиваются ими, оксалиплатин (Eloxatin®); темозоломид (Temodar® и Temodal®); дактиномицин (также известный как актиномицин-D, Cosmegen®); мелфалан (также известный как L-PAM, L-сарколизин и фенилаланин мустард, Alkeran®); алтретамин (также известный как гексаметилмеламин (HMM), Hexalen®); кармустин (BiCNU®); бендамустин (Treanda®); бусульфан (Busulfex® и Myleran®); карбоплатин (Paraplatin®); ломустин (также известный как CCNU, CeeNU®); цисплатин (также известный как CDDP, Platinol® и Platinol®-AQ); хлорамбуцил (Leukeran®); циклофосфамид (Cytoxan® и Neosar®); дакарбазин (также известный как DTIC, DIC и имидазол карбоксамид, DTIC-Dome®); алтретамин (также известный как гексаметилмеламин (HMM), Hexalen®); ифосфамид (Ifex®); преднумустин; прокарбазин (Matulane®); мехлорэтамин (также известный как азотистый иприт, мустин и мехлорэтамин гидрохлорид, Mustargen®); стрептозоцин (Zanosar®); тиотепу (также известную как тиофосфоамид, TESPA и TSPA, Thioplex®); циклофосфамид (Endoxan®, Cytoxan®, Neosar®, Procytox®, Revimmune®) и бендамустин HCl (Treanda®).
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с флударабином, циклофосфамидом и/или ритуксимабом. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с флударабином, циклофосфамидом и ритуксимабом (FCR). В вариантах осуществления индивидуум имеет CLL. Например, индивидуум имеет делецию в коротком плече хромосомы 17 (del(17p), например, в лейкозной клетке). В других примерах индивидуум не имеет del(17p). В вариантах осуществления индивидуум имеет лейкозную клетку, содержащую мутацию в гене вариабельной области тяжелой цепи иммуноглобулина (IgVH). В других вариантах осуществления индивидуум не имеет лейкозной клетки, содержащей мутацию в гене вариабельной области тяжелой цепи иммуноглобулина (IgVH). В вариантах осуществления флударабин вводят в дозировке приблизительно 10-50 мг/м2 (например, приблизительно 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45 или 45-50 мг/м2), например, внутривенно. В вариантах осуществления циклофосфамид вводят в дозировке приблизительно 200-300 мг/м2 (например, приблизительно 200-225, 225-250, 250-275 или 275-300 мг/м2), например, внутривенно. В вариантах осуществления ритуксимаб вводят в дозировке приблизительно 400-600 мг/м2 (например, 400-450, 450-500, 500-550 или 550-600 мг/м2), например, внутривенно.
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с бендамустином и ритуксимабом. В вариантах осуществления индивидуум имеет CLL. Например, индивидуум имеет делецию в коротком плече хромосомы 17 (del(17p), например, в лейкозной клетке. В других примерах индивидуум не имеет del(17p). В вариантах осуществления индивидуум имеет лейкозную клетку, содержащую мутацию в гене вариабельной области тяжелой цепи иммуноглобулина (IgVH). В других вариантах осуществления индивидуум не имеет лейкозной клетки, содержащей мутацию в гене вариабельной области тяжелой цепи иммуноглобулина (IgVH). В вариантах осуществления бендамустин вводят в дозировке приблизительно 70-110 мг/м2 (например, 70-80, 80-90, 90-100 или 100-110 мг/м2), например, внутривенно. В вариантах осуществления ритуксимаб вводят в дозировке приблизительно 400-600 мг/м2 (например, 400-450, 450-500, 500-550 или 550-600 мг/м2), например, внутривенно.
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ритуксимабом, циклофосфамидом, доксорубицином, винкристином и/или кортикостероидом (например, преднизон). В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ритуксимабом, циклофосфамидом, доксорубицином, винкристином и преднизоном (R-CHOP). В вариантах осуществления индивидуум имеет диффузную крупноклеточную B-клеточную лимфому (DLBCL). В вариантах осуществления индивидуум имеет необъемную DLBCL ограниченной стадии (например, имеет опухоль, имеющую размер/диаметр менее 7 см). В вариантах осуществления индивидуума лечат лучевой терапией в комбинации с R-CHOP. Например, индивидууму проводят терапию R-CHOP (например, 1-6 курсов, например, 1, 2, 3, 4, 5 или 6 курсов R-CHOP), а затем лучевую терапию. В некоторых случаях, индивидууму проводят терапию R-CHOP (например, 1-6 курсов, например, 1, 2, 3, 4, 5 или 6 курсов R-CHOP) после лучевой терапии.
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с этопозидом, преднизоном, винкристином, циклофосфамидом, доксорубицином и/или ритуксимабом. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с этопозидом, преднизоном, винкристином, циклофосфамидом, доксорубицином и ритуксимабом (EPOCH-R). В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации со скорректированной дозой EPOCH-R (DA-EPOCH-R). В вариантах осуществления индивидуум имеет B-клеточную лимфому, например, агрессивную B-клеточную лимфому с реаранжированным Myc.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ритуксимабом и/или леналидомидом. Леналидомид ((RS)-3-(4-амино-1-оксо-1,3-дигидро-2H-изоиндол- 2-ил)пиперидин-2,6-дион) является иммуномодулятором. В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ритуксимабом и леналидомидом. В некоторых вариантах осуществления индивидуум имеет фолликулярную лимфому (FL) или лимфому из клеток мантийной зоны (MCL). В некоторых вариантах осуществления индивидуум имеет FL, и его ранее не лечили терапией против злокачественной опухоли. В некоторых вариантах осуществления леналидомид вводят в дозе приблизительно 10-20 мг (например, 10-15 или 15-20 мг), например, каждые сутки. В некоторых вариантах осуществления ритуксимаб вводят в дозировке приблизительно 350-550 мг/м2 (например, 350-375, 375-400, 400-425, 425-450, 450-475 или 475-500 мг/м2), например, внутривенно.
Иллюстративные ингибиторы mTOR включают, например, темсиролимус; ридафоролимус (ранее известный как деферолимус, (1R,2R,4S)-4-[(2R)-2 [(1R,9S,12S,15R,16E,18R,19R,21R, 23S,24E,26E,28Z,30S,32S,35R)-1,18-дигидрокси-19,30-диметокси-15,17,21,23,29,35-гексаметил-2,3,10,14,20-пентаоксо-11,36-диокса-4-азатрицикло[30.3.1.04,9]гексатриаконта-16,24,26,28-тетраен-12-ил]пропил]-2-метоксициклогексилдиметилфосфинат, также известный как AP23573 и MK8669 и описанный в публикации PCT № WO 03/064383); эверолимус (Afinitor® или RAD001); рапамицин (AY22989, Sirolimus®); симапимод (CAS 164301-51-3); эмсиролимус, (5-{2,4-бис[(3S)-3-метилморфолин-4-ил]пиридо[2,3-d]пиримидин-7-ил}-2-метоксифенил)метанол (AZD8055); 2-амино-8-[транс-4-(2-гидроксиэтокси)циклогексил]-6-(6-метокси-3-пиридинил)-4-метилпиридо[2,3-d]пиримидин-7(8H)-он (PF04691502, CAS 1013101-36-4) и N2-[1,4-диоксо-4-[[4-(4-оксо-8-фенил-4H-1-бензопиран-2-ил)морфолиний-4-ил]метокси]бутил]-L-аргинилглицил-L-α-аспартил-L-серин-, внутренняя соль (SF1126, CAS 936487-67-1) (SEQ ID NO: 1262) и XL765.
Иллюстративные иммуномодуляторы включают, например, афутузумаб (доступный 5от Roche®); пегфилграстим (Neulasta®); леналидомид (CC-5013, Revlimid®); талидомид (Thalomid®), актимид (CC4047) и IRX-2 (смесь цитокинов человека, включая интерлейкин 1, интерлейкин 2 и интерферон γ, CAS 951209-71-5, доступные от IRX Therapeutics).
Иллюстративные антрациклины включают, например, доксорубицин (Adriamycin® и Rubex®); блеомицин (lenoxane®); даунорубицин (даунорубицин гидрохлорид, дауномицин и рубидомицин гидрохлорид, Cerubidine®); липосомальный даунорубицин (липосомальный даунорубицина цитрат, DaunoXome®); митоксантрон (DHAD, Novantrone®); эпирубицин (EllenceTM); идарубицин (Idamycin®, Idamycin PFS®); митомицин C (Mutamycin®); гелданамицин; гербимицин; равидомицин; и дезацетилгавидомицин.
Иллюстративные алкалоиды барвинка включают, например, винорелбина тартрат (Navelbine®), винкристин (Oncovin®) и виндезин (Eldisine®)); винбластин (также известный как винбластина сульфат, винкалейкобластин и VLB, Alkaban-AQ® и Velban®); и винорелбин (Navelbine®).
Иллюстративные ингибиторы протеасом включают бортезомиб (Velcade®); карфилзомиб (PX-171-007, (S)-4-метил-N-((S)-1-(((S)-4-метил-1-((R)-2-метилoксиран-2-ил)-1-оксопентан-2-ил)амино)-1-оксо-3-фенилпропан-2-ил)-2-((S)-2-(2-морфолиноацетамидо)-4-фенилбутанамидо)пентанамид); маризомиб (NPI-0052); иксазомиба цитрат (MLN-9708); деланзомиб (CEP-18770); и O-метил-N-[(2-метил-5-тиазолил)карбонил]-L-серил-O-метил-N-[(1S)-2-[(2R)-2-метил-2-оксиранил]-2-оксо-1-(фенилметил)этил]-L-серинамид (ONX-0912).
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с брентуксимабом. Брентуксимаб представляет собой конъюгат антитело-лекарственное средство между антителом против CD30 и монометилауристатином E. В вариантах осуществления индивидуум имеет лимфому Ходжкина (HL), например, рецидивирующую или рефрактерную HL. В вариантах осуществления индивидуум имеет CD30+ HL. В вариантах осуществления индивидуума подвергали трансплантации аутологичных стволовых клеток (ASCT). В вариантах осуществления индивидуума не подвергали ASCT. В вариантах осуществления брентуксимаб вводят в дозировке приблизительно 1-3 мг/кг (например, приблизительно 1-1,5, 1,5-2, 2-2,5 или 2,5-3 мг/кг), например, внутривенно, например, каждые 3 недели.
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с брентуксимабом и дакарбазином или в комбинации с брентуксимабом и бендамустином. Дакарбазин представляет собой алкилирующее средство, имеющее химическое название 5-(3,3-диметил-1-триазенил)имидазол-4-карбоксамид. Бендамустин представляет собой алкилирующее средство c химическим названием 4-[5-[бис(2-хлорэтил)амино]-1-метилбензимидазол-2-ил]бутановая кислота. В вариантах осуществления индивидуум имеет лимфому Ходжкина (HL). В вариантах осуществления индивидуума ранее не лечили терапией против злокачественной опухоли. В вариантах осуществления индивидуум имеет возраст по меньшей мере 60 лет, например, 60, 65, 70, 75, 80, 85 или старше. В вариантах осуществления дакарбазин вводят в дозировке приблизительно 300-450 мг/м2 (например, приблизительно 300-325, 325-350, 350-375, 375-400, 400-425 или 425-450 мг/м2), например, внутривенно. В вариантах осуществления бендамустин вводят в дозировке приблизительно 75-125 мг/м2 (например, 75-100 или 100-125 мг/м2, например, приблизительно 90 мг/м2), например, внутривенно. В вариантах осуществления брентуксимаб вводят в дозировке приблизительно 1-3 мг/кг (например, приблизительно 1-1,5, 1,5-2, 2-2,5 или 2,5-3 мг/кг), например, внутривенно, например, каждые 3 недели.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором CD20, например, антителом против CD20 (например, моноспецифическое или биспецифическое антитело против CD20) или его фрагментом. Иллюстративные антитела против CD20 включают, но не ограничиваются ими, ритуксимаб, офатумумаб, окрелизумаб, велтузумаб, обинутузумаб, TRU-015 (Trubion Pharmaceuticals), окаратузумаб и Pro131921 (Genentech). См., например, Lim et al. Haematologica. 95.1(2010):135-43.
В некоторых вариантах осуществления антитело против CD20 содержит ритуксимаб. Ритуксимаб представляет собой химерное моноклональное антитело IgG1 каппа мыши/человека, которое связывается с CD20 и вызывает цитолиз CD20-экспрессирующей клетки, например, как описано в www.accessdata.fda.gov/drugsatfda_docs/label/2010/103705s5311lbl.pdf. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ритуксимабом. В вариантах осуществления индивидуум имеет CLL или SLL.
В некоторых вариантах осуществления ритуксимаб вводят внутривенно, например, в качестве внутривенной инфузии. Например, каждая инфузия обеспечивает приблизительно 500-2000 мг (например, приблизительно 500-550, 550-600, 600-650, 650-700, 700-750, 750-800, 800-850, 850-900, 900-950, 950-1000, 1000-1100, 1100-1200, 1200-1300, 1300-1400, 1400-1500, 1500-1600, 1600-1700, 1700-1800, 1800-1900 или 1900-2000 мг) ритуксимаба. В некоторых вариантах осуществления ритуксимаб вводят в дозе от 150 мг/м2 до 750 мг/м2, например, приблизительно 150-175 мг/м2, 175-200 мг/м2, 200-225 мг/м2, 225-250 мг/м2, 250-300 мг/м2, 300-325 мг/м2, 325-350 мг/м2, 350-375 мг/м2, 375-400 мг/м2, 400-425 мг/м2, 425-450 мг/м2, 450-475 мг/м2, 475-500 мг/м2, 500-525 мг/м2, 525-550 мг/м2, 550-575 мг/м2, 575-600 мг/м2, 600-625 мг/м2, 625-650 мг/м2, 650-675 мг/м2 или 675-700 мг/м2, где м2 указывает на площадь поверхности тела индивидуума. В некоторых вариантах осуществления ритуксимаб вводят с интервалом дозирования по меньшей мере 4 суток, например, 4, 7, 14, 21, 28, 35 суток или более. Например, ритуксимаб вводят с интервалом дозирования по меньшей мере 0,5 недели, например, 0,5, 1, 2, 3, 4, 5, 6, 7, 8 недель, или более. В некоторых вариантах осуществления ритуксимаб вводят в дозе и с интервалом дозирования, описанным в настоящем описании, в течение периода времени, например, по меньшей мере 2 недель, например, по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 недель или более. Например, ритуксимаб вводят в дозе и с интервалом дозирования, описанным в настоящем описании, на протяжении всего по меньшей мере 4 доз на курс лечения (например, по меньшей мере 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или более доз на курс лечения).
В некоторых вариантах осуществления антитело против CD20 включает офатумумаб. Офатумумаб представляет собой моноклональное антитело IgG1κ человека против CD20, имеющее молекулярную массу приблизительно 149 кДа. Например, офатумумаб получают с использованием трансгенных мышей и технологии гибридом, и экспрессируют и очищают из рекомбинантной клеточной линии мыши (NS0). См., например, www.accessdata.fda.gov/drugsatfda_docs/label/2009/125326lbl.pdf; и клинические испытания под идентификационным номером NCT01363128, NCT01515176, NCT01626352 и NCT01397591. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с офатумумабом. В вариантах осуществления индивидуум имеет CLL или SLL.
В некоторых вариантах осуществления офатумумаб вводят в качестве внутривенной инфузии. Например, каждая инфузия обеспечивает приблизительно 150-3000 мг (например, приблизительно 150-200, 200-250, 250-300, 300-350, 350-400, 400-450, 450-500, 500-550, 550-600, 600-650, 650-700, 700-750, 750-800, 800-850, 850-900, 900-950, 950-1000, 1000-1200, 1200-1400, 1400-1600, 1600-1800, 1800-2000, 2000-2200, 2200-2400, 2400-2600, 2600-2800 или 2800-3000 мг) офатумумаба. В вариантах осуществления офатумумаб вводят в начальной дозировке приблизительно 300 мг, а затем 2000 мг, например, в течение приблизительно 11 доз, например, в течение 24 недель. В некоторых вариантах осуществления офатумумаб вводят с интервалом дозирования по меньшей мере 4 суток, например, 4, 7, 14, 21, 28, 35 суток или более. Например, офатумумаб вводят с интервалом дозирования, составляющим по меньшей мере 1 недель, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 24, 26, 28, 20, 22, 24, 26, 28, 30 недель или более. В некоторых вариантах осуществления офатумумаб вводят в дозе и с интервалом дозирования, описанными в настоящем описании, в течение периода времени, например, в течение по меньшей мере 1 недели, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 22, 24, 26, 28, 30, 40, 50, 60 недель или более, или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 месяцев или более, или 1, 2, 3, 4, 5 лет или более. Например, офатумумаб вводят в дозе и с интервалом дозирования, описанным в настоящем описании, на протяжении всего по меньшей мере 2 доз на курс лечения (например, по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 18, 20, или более доз на курс лечения).
В некоторых случаях, антитело против CD20 включает окрелизумаб. Окрелизумаб представляет собой гуманизированное моноклональное антитело против CD20, например, как описано в клинических испытаниях под идентификационными номерами № NCT00077870, NCT01412333, NCT00779220, NCT00673920, NCT01194570, и в Kappos et al. Lancet. 19,378(2011):1779-87.
В некоторых случаях антитело против CD20 включает велтузумаб. Велтузумаб представляет собой гуманизированное моноклональное антитело против CD20. См., например, клинические испытания под идентификационными номерами № NCT00547066, NCT00546793, NCT01101581, и Goldenberg et al. Leuk Lymphoma. 51(5)(2010):747-55.
В некоторых случаях, антитело против CD20 включает GA101. GA101 (также называемый обинутузумабом или RO5072759) представляет собой гуманизированное глико-модифицированное моноклональное антитело против CD20. См., например, Robak. Curr. Opin. Investig. Drugs. 10,6(2009):588-96; клинические испытания под идентификационными номерами: NCT01995669, NCT01889797, NCT02229422 и NCT01414205; и www.accessdata.fda.gov/drugsatfda_docs/label/2013/125486s000lbl.pdf.
В некоторых случаях антитело против CD20 включает AME-133v. AME-133v (также называемый LY2469298 или окаратузумабом) представляет собой гуманизированное моноклональное IgG1-антитело против CD20 с увеличенной аффинностью в отношении рецептора FcγRIIIa и усиленной активностью антитело-зависимой клеточной цитотоксичности (ADCC) по сравнению с ритуксимабом. См., например, Robak et al. BioDrugs 25.1(2011):13-25; и Forero-Torres et al. Clin Cancer Res. 18.5(2012):1395-403.
В некоторых случаях антитело против CD20 включает PRO131921. PRO131921 представляет собой гуманизированное моноклональные антитело против CD20, модифицированное способами инженерии, чтобы оно обладало улучшенным связыванием с FcγRIIIa и усиленной ADCC по сравнению с ритуксимабом. См., например, Robak et al. BioDrugs 25.1(2011):13-25; и Casulo et al. Clin Immunol. 154,1(2014):37-46; и клиническое испытание под идентификационным номером № NCT00452127.
В некоторых случаях, антитело против CD20 включает TRU-015. TRU-015 представляет собой слитый белок CD20, образованный доменами антитела против CD20. TRU-015 имеет меньший размер, чем моноклональные антитела, но сохраняет опосредуемые Fc эффекторные функции. См., например, Robak et al. BioDrugs 25.1(2011):13-25. TRU-015 содержит одноцепочечный вариабельный фрагмент (scFv) против CD20, связанный с шарнирной областью IgG1 человека, CH2- и CH3-доменами, но лишен доменов CH1 и CL.
В некоторых вариантах осуществления антитело против CD20, описанное в настоящем описании, конъюгировано или иным образом связано с лекарственным средством, например, химиотерапевтическим средством (например, цитоксан, флударабин, ингибитор деацетилазы гистонов, деметилирующее средство, пептидная вакцина, противоопухолевый антибиотик, ингибитор тирозинкиназы, алкилирующее средство, антитубулиновое или антимитотическое средство), средство против аллергии, средство против тошноты (или противорвотное средство), средство для смягчения боли или цитопротекторное средство, описанное в настоящем описании.
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором B-клеточной лимфомы 2 (BCL-2) (например, венетоклаксом, также называемым ABT-199 или GDC-0199) и/или ритуксимабом. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с венетоклаксом и ритуксимабом. Венетоклакс представляет собой низкомолекулярное соединение, которое ингибирует антиапоптотический белок BCL-2. Структура вентоклакса (4-(4-{[2-(4-хлорфенил)-4,4-диметилциклогекс-1-ен-1-ил]метил}пиперазин-1-ил)-N-({3-нитро-4-[(тетрагидро-2H-пиран-4-илметил)амино]фенил}сульфонил)-2-(1H-пирроло[2,3-b]пиридин-5-илокси)бензамид) представлена ниже.
В вариантах осуществления индивидуум имеет CLL. В вариантах осуществления индивидуум имеет рецидивирующий CLL, например, индивидууму ранее проводили терапию злокачественной опухоли. В вариантах осуществления венетоклакс вводят в дозировке приблизительно 15-600 мг (например, 15-20, 20-50, 50-75, 75-100, 100-200, 200-300, 300-400, 400-500 или 500-600 мг), например, каждые сутки. В вариантах осуществления ритуксимаб вводят в дозировке приблизительно 350-550 мг/м2 (например, 350-375, 375-400, 400-425, 425-450, 450-475, или 475-500 мг/м2), например, внутривенно, например, каждый месяц.
В одном варианте осуществления клетки, экспрессирующие CAR, описанный в настоящем описании, вводят индивидууму в комбинации с молекулой, которая уменьшает популяцию Treg-клеток. Способы, которые уменьшают количество (например, истощают) Treg-клетки, известны в данной области и включают, например, истощение по CD25, введение циклофосфамида, модулирование функции GITR. Без связи с теорией, полагают, что уменьшение количества Treg-клеток у индивидуума перед аферезом или перед введением экспрессирующей CAR клетки, описанной в настоящем описании, уменьшает количество нежелательных иммунных клеток (например, Treg) в микроокружении опухоли и уменьшает риск рецидива у индивидуума. В одном варианте осуществления клетки, экспрессирующие CAR, описанный в настоящем описании, вводят индивидууму в комбинации с молекулой, нацеленной на GITR и/или модулирующей функции GITR, такой как агонист GITR и/или антитело против GITR, которые истощают регуляторные T-клетки (Treg). В вариантах осуществления клетки, экспрессирующие CAR, описанный в настоящем описании, вводят индивидууму в комбинации с циклофосфамидом. В одном варианте осуществления молекулы, связывающие GITR, и/или молекулы, модулирующие функции GITR (например, агонист GITR и/или истощающие Treg антитела против GITR), вводят перед введением экспрессирующей CAR клетки. Например, в одном варианте осуществления агонист GITR можно вводить до афереза клеток. В вариантах осуществления циклофосфамид вводят индивидууму до введения (например, инфузии или реинфузии) экспрессирующей CAR клетки или до афереза клеток. В вариантах осуществления циклофосфамид и антитело против GITR вводят индивидууму перед введением (например, инфузии или реинфузии) экспрессирующей CAR клетки или до афереза клеток. В одном варианте осуществления индивидуум имеет злокачественную опухоль (например, солидную злокачественную опухоль или гематологическую злокачественную опухоль, такую как ALL или CLL). В одном варианте осуществления индивидуум имеет CLL. В вариантах осуществления индивидуум имеет ALL. В вариантах осуществления индивидуум имеет солидную злокачественную опухоль, например, солидную злокачественную опухоль, описанную в настоящем описании. Иллюстративные агонисты GITR включают, например, слитые белки GITR и антитела против GITR (например, двухвалентные антитела против GITR) например, такие как слитый белок GITR, описанный в патенте США № 6111090, патенте Европы № 090505B1, патенте США № 8586023, публикации PCT № WO 2010/003118 и 2011/090754, или антитело против GITR, описанное, например, в патенте США № 7025962, патенте Европы № 1947183B1, патенте США № 7812135, патенте США № 8388967, патенте США № 8591886, патенте Европы № EP 1866339, публикации PCT № WO 2011/028683, публикации PCT № WO 2013/039954, публикации PCT № WO2005/007190, публикации PCT № WO 2007/133822, публикации PCT № WO2005/055808, публикации PCT № WO 99/40196, публикации PCT № WO 2001/03720, публикации PCT № WO99/20758, публикации PCT № WO2006/083289, публикации PCT № WO 2005/115451, патенте США № 7618632 и публикации PCT № WO 2011/051726.
В одном варианте осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором mTOR ингибитор, например, ингибитором mTOR, описанным в настоящем описании, например, рапалогом, таким как эверолимус. В одном варианте осуществления ингибитор mTOR вводят до введения экспрессирующей CAR клетки. Например, в одном варианте осуществления ингибитор mTOR можно вводить до афереза клеток. В одном варианте осуществления индивидуум имеет CLL.
В одном варианте осуществления комбинацию экспрессирующей CAR клетки, описанной в настоящем описании, и ингибитора киназы, описанного в настоящем описании, вводят индивидууму в комбинации с агонистом GITR, например, агонистом GITR, описанным в настоящем описании. В одном варианте осуществления агонист GITR вводят до введения экспрессирующей CAR клетки. Например, в одном варианте осуществления агонист GITR можно вводить до афереза клеток. В одном варианте осуществления индивидуум имеет CLL.
В одном варианте осуществления экспрессирующую CAR клетку, описанную в настоящем описании, можно использовать в комбинации с ингибитором киназы. В одном варианте осуществления ингибитор киназы представляет собой ингибитор CDK4, например, ингибитор CDK4, описанный в настоящем описании, например, ингибитор CD4/6, например, такой как 6-ацетил-8-циклопентил-5-метил-2-(5-пиперазин-1-илпиридин-2-иламино)-8H-пиридо[2,3-d]пиримидин-7-она гидрохлорид (также называемый палбоциклибом или PD0332991). В одном варианте осуществления ингибитор киназы представляет собой ингибитор BTK, например, ингибитор BTK, описанный в настоящем описании, например, такой как ибрутиниб. В одном варианте осуществления ингибитор киназы представляет собой ингибитор mTOR, например, ингибитор mTOR, описанный в настоящем описании, например, такой как рапамицин, аналоги рапамицина, OSI-027. Ингибитор mTOR может представлять собой, например, ингибитор mTORC1 и/или ингибитор mTORC2, например, ингибитор mTORC1 и/или ингибитор mTORC2, описанный в настоящем описании. В одном варианте осуществления ингибитор киназы представляет собой ингибитор MNK, например, ингибитор MNK, описанный в настоящем описании, например, такой как 4-амино-5-(4-фторанилино)пиразолo[3,4-d]пиримидин. Ингибитор MNK может представлять собой, например, ингибитор MNK1a, MNK1b, MNK2a и/или MNK2b. В одном варианте осуществления ингибитор киназы представляет собой двойной ингибитор PI3K/mTOR, описанный в настоящем описании, например, PF-04695102.
В одном варианте осуществления ингибитор киназы представляет собой ингибитор CDK4, выбранный из алоизина A; флавопиридола или HMR-1275, 2-(2-хлорфенил)-5,7-дигидрокси-8-[(3S,4R)-3-гидрокси-1-метил-4-пиперидинил]-4-хроменона; кризотиниба (PF-02341066); 2-(2-хлорфенил)-5,7-дигидрокси-8-[(2R,3S)-2-(гидроксиметил)-1-метил-3-пирролидинил]-4H-1-бензопиран-4-она гидрохлорида (P276-00); 1-метил-5-[[2-[5-(трифторметил)-1H-имидазол-2-ил]-4-пиридинил]окси]-N-[4-(трифторметил)фенил]-1H-бензимидазол-2-амина (RAF265); индисулама (E7070); росковитина (CYC202); палбоциклиба (PD0332991); динациклиба (SCH727965); N-[5-[[(5-трет-бутилоксазол-2-ил)метил]тио]тиазол-2-ил]пиперидин-4-карбоксамида (BMS 387032); 4-[[9-хлор-7-(2,6-дифторфенил)-5H-пиримидо[5,4-d][2]бензазепин-2-ил]амино]бензойной кислоты (MLN8054); 5-[3-(4,6-дифтор-1H-бензимидазол-2-ил)-1H-индазол-5-ил]-N-этил-4-метил-3-пиридинметанамина (AG-024322); 4-(2,6-дихлорбензоиламино)-1H-пиразол-3-карбоновой кислоты N-(пиперидин-4-ил)амида (AT7519); 4-[2-метил-1-(1-метилэтил)-1H-имидазол-5-ил]-N-[4-(метилсульфонил)фенил]-2-пиримидинамина (AZD5438) и XL281 (BMS908662).
В одном варианте осуществления ингибитор киназы представляет собой ингибитор CDK4, например, палбоциклиб (PD0332991), и палбоциклиб вводят в дозе приблизительно 50 мг, 60 мг, 70 мг, 75 мг, 80 мг, 90 мг, 100 мг, 105 мг, 110 мг, 115 мг, 120 мг, 125 мг, 130 мг, 135 мг (например, 75 мг, 100 мг или 125 мг) каждые сутки в течение периода времени, например, каждые сутки в течение 14-21 суток курса из 28 суток, или каждые сутки в течение 7-12 суток курса из 21 суток. В одном варианте осуществления проводят 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более курсов введения палбоциклиба.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором циклин-зависимой киназы (CDK) 4 или 6, например, ингибитором CDK4 или ингибитором CDK6, описанным в настоящем описании. В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором CDK4/6 (например, ингибиторм, который нацелен как на CDK4, так и на CDK6), например, ингибитор CDK4/6, описанный в настоящем описании. В одном варианте осуществления индивидуум имеет MCL. MCL представляет собой агрессивную злокачественную опухоль, которая плохо отвечает на доступные в настоящее время способы терапии, т.е. по существу неизлечима. Во многих случаях MCL, в клетках MCL экспрессируется циклин D1 (регулятор CDK4/6) (например, вследствие хромосомной транслокации, вовлекающей гены иммуноглобулинов и циклина D1). Таким образом, без связи с теорией полагают, что клетки MCL являются в высокой степени чувствительными к ингибированию CDK4/6 с высокой специфичностью (т.е. минимальный эффект на нормальные иммунные клетки). Ингибиторы CDK4/6 отдельно обладают некоторой эффективностью лечения MCL, однако достигают только частичной ремиссии с высокой частотой рецидива. Иллюстративным ингибитором CDK4/6 является LEE011 (также называемый рибоциклибом), структура которого показана ниже.
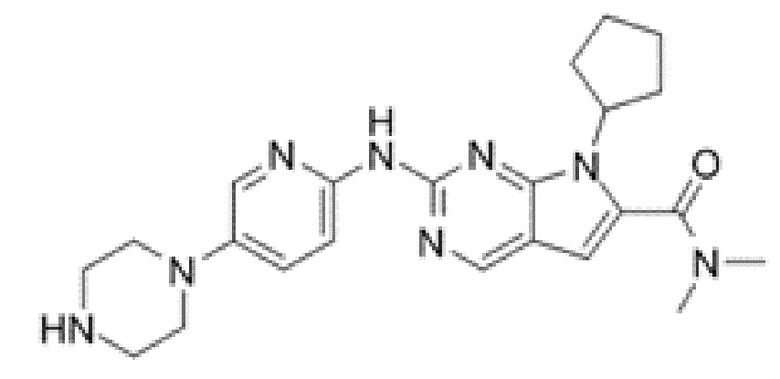
Без связи с теорией, полагают, что введение экспрессирующей CAR клетки, описанной в настоящем описании, с ингибитором CDK4/6 (например, LEE011 или другим ингибитором CDK4/6, описанным в настоящем описании), может достигать более высокой способности к ответу, например, с более высокими показателями ремиссии и/или более низкими показателями рецидива, например, по сравнению с ингибитором CDK4/6 отдельно.
В одном варианте осуществления ингибитор киназы представляет собой ингибитор BTK, выбранный из ибрутиниба (PCI-32765); GDC-0834; RN-486; CGI-560; CGI-1764; HM-71224; CC-292; ONO-4059; CNX-774 и LFM-A13. В одном варианте осуществления ингибитор BTK не снижает или не ингибирует киназную активность индуцируемой интерлейкином-2 киназы (ITK), и он выбран из GDC-0834; RN-486; CGI-560; CGI-1764; HM-71224; CC-292; ONO-4059; CNX-774 и LFM-A13.
В одном варианте осуществления ингибитор киназы представляет собой ингибитор BTK, например, ибрутиниб (PCI-32765). В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором BTK (например, ибрутинибом). В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ибрутинибом (также называемым PCI-32765). Структура ибрутиниба (1-[(3R)-3-[4-амино-3-(4-феноксифенил)-1H-пиразолo[3,4-d]пиримидин-1-ил]пиперидин-1-ил]проп-2-ен-1-он) показана ниже.
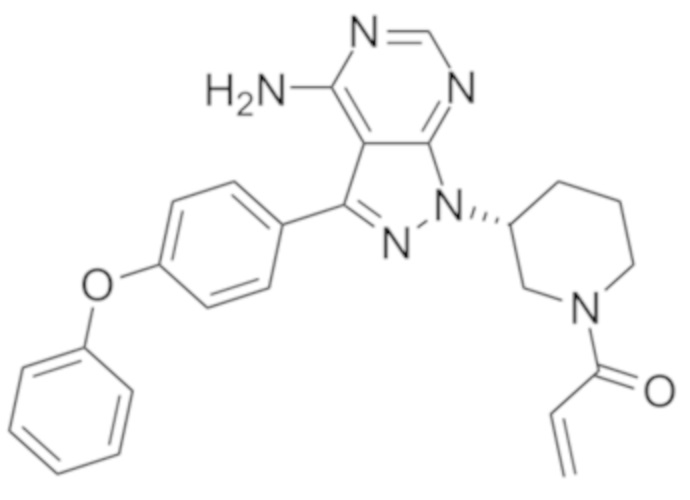
В некоторых вариантах осуществления индивидуум имеет CLL, лимфому из клеток мантийной зоны (MCL) или мелколимфоцитарную лимфому (SLL). Например, индивидуум имеет делецию в коротком плече хромосомы 17 (del(17p), например, в лейкозной клетке). В других примерах индивидуум не имеет del(17p). В некоторых вариантах осуществления индивидуум имеет рецидивирующий CLL или SLL, например, индивидууму ранее проводили терапию злокачественной опухоли (например, ранее проводили одну, две, три или четыре предшествующих терапии злокачественной опухоли). В некоторых вариантах осуществления индивидуум имеет рефрактерный CLL или SLL. В других вариантах осуществления индивидуум имеет фолликулярную лимфому, например, рецидивирующую или рефрактерную фолликулярную лимфому. В некоторых вариантах осуществления ибрутиниб вводят в дозировке приблизительно 300-600 мг/сутки (например, приблизительно 300-350, 350-400, 400-450, 450-500, 500-550 или 550-600 мг/сутки, например, приблизительно 420 мг/сутки или приблизительно 560 мг/сутки), например, перорально. В некоторых вариантах осуществления ибрутиниб вводят в дозе приблизительно 250 мг, 300 мг, 350 мг, 400 мг, 420 мг, 440 мг, 460 мг, 480 мг, 500 мг, 520 мг, 540 мг, 560 мг, 580 мг, 600 мг (например, 250 мг, 420 мг или 560 мг) каждые сутки в течение некоторого периода времени, например, каждые сутки в течение курса из 21 суток или каждые сутки в течение курса из 28 суток. В одном варианте осуществления проводят 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более курсов ибрутиниба. Без связи с теорией полагают, что добавление ибрутиниба усиливают пролиферативный ответ T-клеток и может обеспечить сдвиг T-клеток с фенотипа T-хелперов-2 (Th2) на фенотип T-хелперов-1 (Th1). Th1 и Th2 представляют собой фенотипы хелперных T-клеток, причем Th1 относительно Th2 обеспечивают различные каскады иммунного ответа. Фенотип Th1 ассоциирован с провоспалительными ответами, например, для уничтожения клеток, таких как внутриклеточные патогены/вирусы или злокачественные клетки, или бессрочными аутоиммунными ответами. Фенотип Th2 ассоциирован с накоплением эозинофилов и противовоспалительными ответами.
В одном варианте осуществления ингибитор киназы представляет собой ингибитор mTOR, выбранный из темсиролимуса; ридафоролимуса (1R,2R,4S)-4-[(2R)-2 [(1R,9S,12S,15R,16E,18R,19R,21R, 23S,24E,26E,28Z,30S,32S,35R)-1,18-дигидрокси-19,30-диметокси-15,17,21,23, 29,35-гексаметил-2,3,10,14,20-пентаоксо-11,36-диокса-4-азатрицикло[30,3,1,04,9]гексатриаконта-16,24,26,28-тетраен-12-ил]пропил]-2-метоксициклогексилдиметилфосфината, также известного как AP23573 и MK8669; эверолимуса (RAD001); рапамицина (AY22989); симапимода; (5-{2,4-бис[(3S)-3-метилморфолин-4-ил]пиридо[2,3-d]пиримидин-7-ил}-2-метоксифенил)метанола (AZD8055); 2-амино-8-[транс-4-(2-гидроксиэтокси)циклогексил]-6-(6-метокси-3-пиридинил)-4-метил-пиридо[2,3-d]пиримидин-7(8H)-она (PF04691502); и N2-[1,4-диоксо-4-[[4-(4-оксо-8-фенил-4H-1-бензопиран-2-ил)морфолиний-4-ил]метокси]бутил]-L-аргинилглицил-L-α-аспартил-L-серина-, внутренней соли (SF1126) (SEQ ID NO: 1262) и XL765.
В одном варианте осуществления ингибитор киназы представляет собой ингибитор mTOR, например, рапамицин, и рапамицин вводят в дозе приблизительно 3 мг, 4 мг, 5 мг, 6 мг, 7 мг, 8 мг, 9 мг, 10 мг (например, 6 мг) каждые сутки в течение периода времени, например, каждые сутки в течение курса из 21 суток или каждые сутки в течение курса из 28 суток. В одном варианте осуществления проводят 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более курсов введения рапамицина. В одном варианте осуществления ингибитор киназы представляет собой ингибитор mTOR, например, эверолимус, и эверолимус вводят в дозе приблизительно 2 мг, 2,5 мг, 3 мг, 4 мг, 5 мг, 6 мг, 7 мг, 8 мг, 9 мг, 10 мг, 11 мг, 12 мг, 13 мг, 14 мг, 15 мг (например, 10 мг) каждые сутки в течение периода времени, например, каждые сутки в течение курса из 28 суток. В одном варианте осуществления проводят 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более курсов введения эверолимуса.
В одном варианте осуществления ингибитор киназы представляет собой ингибитор MNK, выбранный из CGP052088; 4-амино-3-(п-фторфениламино)пиразолo[3,4-d]пиримидина (CGP57380); церкоспорамида; ETC-1780445-2 и 4-амино-5-(4-фторанилино)пиразолo[3,4-d]пиримидина.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором фосфоинозитид-3-киназы (PI3K) (например, ингибитор PI3K, описанный в настоящем описании, например, иделалисиб или дувелисиб) и/или ритуксимабом. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с иделалисибом и ритуксимабом. В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с дувелисибом и ритуксимабом. Иделалисиб (также называемый GS-1101 или CAL-101; Gilead) представляет собой низкомолекулярное соединение, которое блокирует дельта-изоформу PI3K. Структура иделалисиба (5-фтор-3-фенил-2-[(1S)-1-(7H-пурин-6-иламино)пропил]-4(3H)-хиназолинона) представлена ниже.
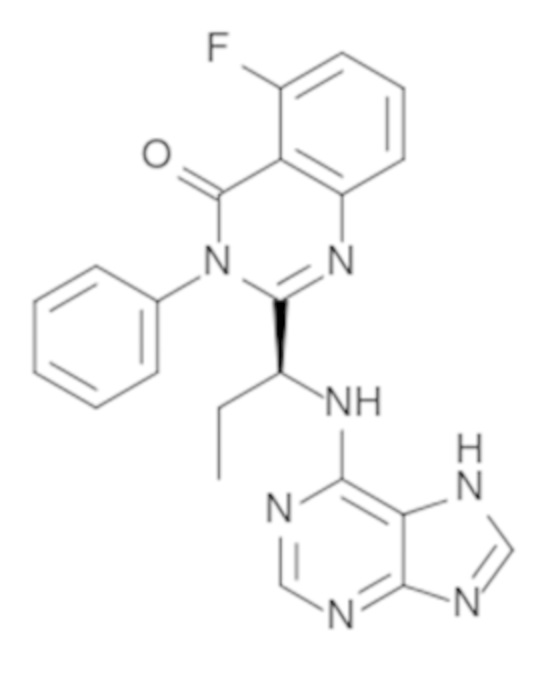
Дувелисиб (также называемый IPI-145; Infinity Pharmaceuticals и Abbvie) представляет собой низкомолекулярное соединение, которое блокирует PI3K-δ,γ. Структура дувелисиба (8-хлор-2-фенил-3-[(1S)-1-(9H-пурин-6-иламино)этил]-1(2H)-изохинолинон) показана ниже.
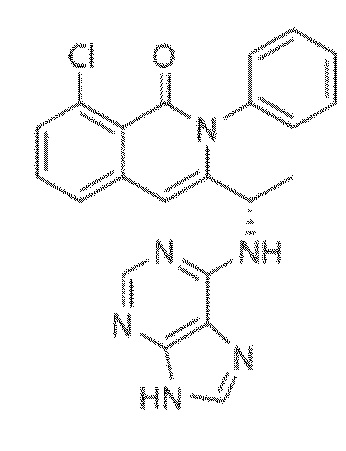
В вариантах осуществления индивидуум имеет CLL. В вариантах осуществления индивидуум имеет рецидивирующий CLL, например, индивидууму ранее проводили терапию злокачественной опухоли (например, ранее вводили антитело против CD20 или ранее вводили ибрутиниб). Например, индивидуум имеет делецию на коротком плече хромосомы 17 (del(17p), например, в лейкозной клетке). В других примерах индивидуум не имеет del(17p). В вариантах осуществления индивидуум имеет лейкозную клетку, содержащую мутацию в гене вариабельной области тяжелой цепи иммуноглобулина (IgVH). В других вариантах осуществления индивидуум не содержит лейкозных клеток. Содержащих мутацию в гене вариабельной области тяжелой цепи иммуноглобулина (IgVH). В вариантах осуществления индивидуум имеет делецию в длинном плече хромосомы 11 (del(11q)). В других вариантах осуществления индивидуум не имеет del(11q). В вариантах осуществления иделалисиб вводят в дозировке приблизительно 100-400 мг (например, 100-125, 125-150, 150-175, 175-200, 200-225, 225-250, 250-275, 275-300, 325-350, 350-375, или 375-400 мг), например, BID. В вариантах осуществления дувелисиб вводят в дозировке приблизительно 15-100 мг (например, приблизительно 15-25, 25-50, 50-75 или 75-100 мг), например, два раза в сутки. В вариантах осуществления ритуксимаб вводят в дозировке приблизительно 350-550 мг/м2 (например, 350-375, 375-400, 400-425, 425-450, 450-475 или 475-500 мг/м2), например, внутривенно.
В одном варианте осуществления ингибитор киназы представляет собой двойной ингибитор фосфатидилинозитол-3-киназы (PI3K) и ингибитор mTOR, выбранный из 2-амино-8-[транс-4-(2-гидроксиэтокси)циклогексил]-6-(6-метокси-3-пиридинил)-4-метилпиридо[2,3-d]пиримидин-7(8H)-она (PF-04691502); N-[4-[[4-(диметиламино)-1-пиперидинил]карбонил]фенил]-N'-[4-(4,6-ди-4-морфолинил-1,3,5-триазин-2-ил)фенил]мочевины (PF-05212384, PKI-587); 2-метил-2-{4-[3-метил-2-оксо-8-(хинолин-3-ил)-2,3-дигидро-1H-имидазо[4,5-c]хинолин-1-ил]фенил}пропаннитрила (BEZ-235); апитолисиба (GDC-0980, RG7422); 2,4-дифтор-N-{2-(метилокси)-5-[4-(4-пиридазинил)-6-хинолинил]-3-пиридинил}бензолсульфонамида (GSK2126458); 8-(6-метоксипиридин-3-ил)-3-метил-1-(4-(пиперазин-1-ил)-3-(трифторметил)фенил)-1H-имидазо[4,5-c]хинолин-2(3H)-она-малеиновой кислоты (NVP-BGT226); 3-[4-(4-морфолинилпиридо[3',2':4,5]фуро[3,2-d]пиримидин-2-ил]фенола (PI-103); 5-(9-изопропил-8-метил-2-морфолино-9H-пурин-6-ил)пиримидин-2-амина (VS-5584, SB2343); и N-[2-[(3,5-диметоксифенил)амино]хиноксалин-3-ил]-4-[(4-метил-3-метоксифенил)карбонил]аминофенилсульфонамида (XL765).
В вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором киназы анапластической лимфомы (ALK). Иллюстративные ингибиторы киназы ALK включают, но не ограничиваются ими, кризотиниб (Pfizer), церитиниб (Novartis), алектиниб (Chugai), бригатиниб (также называемый AP26113; Ariad), энтректиниб (Ignyta), PF-06463922 (Pfizer), TSR-011 (Tesaro) (см., например, клинические испытания под идентификационным номером № NCT02048488), CEP-37440 (Teva), и X-396 (Xcovery). В некоторых вариантах осуществления индивидуум имеет солидную злокачественную опухоль, например, солидную злокачественную опухоль, описанную в настоящем описании, например, рак легкого.
Химическое название кризотиниба представляет собой 3-[(1R)-1-(2,6-дихлор-3-фторфенил)этокси]-5-(1-пиперидин-4-илпиразол-4-ил)пиридин-2-амин. Химическое название церитиниба представляет собой 5-хлор-N2-[2-изопропокси-5-метил-4-(4-пиперидинил)фенил]-N4-[2-(изопропилсульфонил)фенил]-2,4-пиримидиндиамин. Химическое название алектиниба представляет собой 9-этил-6,6-диметил-8-(4-морфолинопиперидин-1-ил)-11-оксо-6,11-дигидро-5H-бензо[b]карбазол-3-карбонитрил. Химическое название бригатиниба представляет собой 5-хлор-N2-{4-[4-(диметиламино)-1-пиперидинил]-2-метоксифенил}-N4-[2-(диметилфосфорил)фенил]-2,4-пиримидиндиамин. Химическое название энтректиниба представляет собой N-(5-(3,5-дифторбензил)-1H-индазол-3-ил)-4-(4-метилпиперазин-1-ил)-2-((тетрагидро-2H-пиран-4-ил)амино)бензамид. Химическое название PF-06463922 представляет собой (10R)-7-амино-12-фтор-2,10,16-триметил-15-оксо-10,15,16,17-тетрагидро-2H-8,4-(метено)пиразолo[4,3-h][2,5,11]-бензоксодиазациклотетрадецин-3-карбонитрил. Химическая структура CEP-37440 представляет собой (S)-2-((5-хлор-2-((6-(4-(2-гидроксиэтил)пиперазин-1-ил)-1-метокси-6,7,8,9-тетрагидро-5H-бензо[7]аннулен-2-ил)амино)пиримидин-4-ил)амино)-N-метилбензамид. Химическое название X-396 представляет собой (R)-6-амино-5-(1-(2,6-дихлор-3-фторфенил)этокси)-N-(4-(4-метилпиперазин-1-карбонил)фенил)пиридазин-3-карбоксамид.
Также можно использовать лекарственные средства, которые либо ингибируют кальцийзависимую фосфатазу кальциневрин (циклоспорин и FK506), либо ингибируют киназу p70S6, которая является важной для индуцируемой факторами роста передачи сигнала (рапамицин). (Liu et al., Cell 66:807-815, 1991; Henderson et al., Immun. 73:316-321, 1991; Bierer et al., Curr. Opin. Immun. 5:763-773, 1993). В следующем аспекте композиции клеток по настоящему изобретению можно вводить пациенту совместно с (например, до, одновременно или после) трансплантации костного мозга, терапией, устраняющей T-клетки, с использованием химиотерапевтических средств, таких как флударабин, лучевая терапия внешним пучком (XRT), циклофосфамид и/или антитела, такие как OKT3 или CAMPATH. В одном аспекте композиции клеток по настоящему изобретению вводят после терапии, устраняющей B-клетки, такой как средства, которые реагируют с CD20, например, ритуксан. Например, в одном варианте осуществления индивидуумы могут подвергаться стандартному лечению химиотерапией в высокой дозе с последующей трансплантацией стволовых клеток периферической крови. В определенных вариантах осуществления после трансплантации индивидуумам проводят инфузию увеличенных в количестве иммунных клеток по настоящему изобретению. В дополнительном варианте осуществления увеличенные в количестве клетки вводят до или после хирургической операции.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с ингибитором индоламин-2,3-диоксигеназы (IDO). IDO представляет собой фермент, который катализирует деградацию аминокислоты L-триптофана до кинуренина. Многие злокачественные опухоли сверхэкспрессируют IDO, например, рак предстательной железы, рак ободочной и прямой кишки, рак поджелудочной железы, рак шейки матки, рак желудка, рак яичника, рак головы и рак легкого. pDC, макрофаги и дендритные клетки (DC) могут экспрессировать IDO. Без связи с теорией, полагают, что снижение уровня L-триптофана (например, катализируемое IDO) приводит к иммунодепрессивной среде путем индукции анергии и апоптоза T-клеток. Таким образом, без связи с теорией, полагают, что ингибитор IDO может повышать эффективность экспрессирующей CAR клетки, описанной в настоящем описании, например, путем уменьшения подавления или гибели экспрессирующих CAR иммунных клеток. В вариантах осуществления индивидуум имеет солидную опухоль, например, солидную опухоль, описанную в настоящем описании, например, рак предстательной железы, рак ободочной и прямой кишки, рак поджелудочной железы, рак шейки матки, рак желудка, рак яичника, рак головы и рак легкого. Иллюстративные ингибиторы IDO включают, но не ограничиваются ими, 1-метилтриптофан, индоксимод (NewLink Genetics) (см., например, клинические испытания под идентификационными номерами № NCT01191216; NCT01792050) и INCB024360 (Incyte Corp.) (см., например, клинические испытания под идентификационными номерами № NCT01604889; NCT01685255).
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с модулятором супрессорных клеток миелоидного происхождения (MDSC). MDSC накапливаются на периферии и в области опухоли многих солидных опухолей. Эти клетки подавляют T-клеточные ответы, тем самым препятствуя эффективности экспрессирующих CAR клеток. Без связи с теорией, полагают, что введение модулятора MDSC повышает эффективность экспрессирующих CAR клеток описанных в настоящем описании. В одном варианте осуществления индивидуум имеет солидную опухоль, например, солидную опухоль, описанную в настоящем описании, например, глиобластому. Иллюстративные модуляторы MDSC включают, но не ограничиваются ими MCS110 и BLZ945. MCS110 представляет собой моноклональное антитело (mAb) против макрофагального колониестимулирующего фактора (M-CSF). См., например, клинические испытания под идентификационным номером № NCT00757757. BLZ945 представляет собой низкомолекулярный ингибитор рецептора колониестимулирующего фактора 1 (CSF1R). См., например, Pyonteck et al. Nat. Med. 19(2013):1264-72. Структура BLZ945 показана ниже.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с CD19 CART-клеткой (например, CTL019, например, как описано в WO2012/079000, включенной в настоящее описание путем ссылки). В некоторых вариантах осуществления индивидуум имеет CD19+ лимфому, например, CD19+ неходжскинскую лимфому (NHL), CD19+ FL или CD19+ DLBCL. В некоторых вариантах осуществления индивидуум имеет рецидивирующую или рефрактерную CD19+ лимфому. В некоторых вариантах осуществления индивидууму проводят истощающую лимфоциты химиотерапию до, одновременно или после введения (например, инфузии) CD19 CART-клеток. В одном примере истощающую лимфоциты химиотерапию проводят у индивидуума до введения CD19 CART-клеток. Например, истощающая лимфоциты химиотерапия оканчивается за 1-4 суток (например, 1, 2, 3 или 4 суток) до инфузии CD19 CART-клеток. В некоторых вариантах осуществления вводят множество доз CD19 CART-клеток, например, как описано в настоящем описании. Например, однократная доза содержит приблизительно 5×108 CD19 CART-клеток. В некоторых вариантах осуществления истощающую лимфоциты химиотерапию проводят у индивидуума до, одновременно или после введения (например, инфузии) экспрессирующей CAR клетки, описанной в настоящем описании, например, экспрессирующей CAR не против CD19 клетки. В некоторых вариантах осуществления CD19 CART вводят индивидууму до, одновременно или после введения (например, инфузии) экспрессирующей CAR клетки не против CD19, например, экспрессирующей CAR клетки не против CD19.
В некоторых вариантах осуществления экспрессирующую CAR клетку, описанную в настоящем описании, вводят индивидууму в комбинации с полипептидом интерлейкина-15 (IL-15), полипептидом альфа-рецептора интерлейкина-15 (IL-15Ra), или с комбинацией как полипептида IL-15, так и полипептида IL-15Ra, например, hetIL-15 (Admune Therapeutics, LLC). hetIL-15 представляет собой гетеродимерный нековалентный комплекс IL-15 и IL-15Ra. hetIL-15 описан, например, в U.S. 8124084, U.S. 2012/0177598, U.S. 2009/0082299, U.S. 2012/0141413 и U.S. 2011/0081311, включенных в настоящее описание путем ссылок. В вариантах осуществления het-IL-15 вводят подкожно. В вариантах осуществления индивидуум имеет злокачественную опухоль, например, солидную злокачественную опухоль, например, меланому или рак толстого кишечника. В вариантах осуществления индивидуум имеет метастазирующую злокачественную опухоль.
В одном варианте осуществления индивидууму можно вводить средство, которое снижает или смягчает побочный эффект, ассоциированный с введением экспрессирующей CAR клетки. Побочные эффекты, ассоциированные с введением экспрессирующей CAR клетки, включают, но не ограничиваются ими CRS и гемофагоцитарный лимфогистиоцитоз (HLH), также называемый синдромом активации макрофагов (MAS). Симптомы CRS включают высокую лихорадку, тошноту, временную гипотензию, гипоксию и т.п. CRS может включать клинические системные признаки и симптомы, такие как лихорадка, усталость, анорексия, миалгии, артралгии, тошнота, рвота и головная боль. CRS может включать клинические кожные признаки и симптомы, такие как сыпь. CRS может включать клинические желудочно-кишечные признаки и симптомы, такие как тошнота, рвота и диарея. CRS может включать клинические респираторные признаки и симптомы, такие как тахипноэ и гипоксемия. CRS может включать клинические сердечно-сосудистые признаки и симптомы, такие как тахикардия, увеличенное пульсовое давление, гипотензия, увеличенный сердечный выброс (ранняя стадия) и потенциально сниженный сердечный выброс (поздняя стадия). CRS может включать клинические признаки и симптомы свертываемости, такие как повышенные уровни d-димера, гипофибриногенемия с кровотечением или без него. CRS может включать клинические почечные признаки и симптомы, такие как азотемия. CRS может включать клинические печеночные признаки и симптомы, такие как трансаминит и гипербилирубинемия. CRS может включать клинические неврологические признаки и симптомы, такие как головная боль, изменения психического состояния, спутанность сознания, делирий, затруднения с подбором слов или афазия Фрэнка, галлюцинации, тремор, дисметрия, изменение походки и судороги.
Таким образом, способы, описанные в настоящем описании, могут включать введение экспрессирующей CAR клетки, описанной в настоящем описании, индивидууму, и, кроме того, введение одного или нескольких средств для контроля повышенных уровней растворимого фактора вследствие лечения экспрессирующей CAR клеткой. В одном варианте осуществления растворимый фактор, повышенный у индивидуума, представляет собой один или несколько из IFN-γ, TNFα, IL-2 и IL-6. В одном варианте осуществления фактор, повышенный у индивидуума, представляет собой один или несколько из IL-1, GM-CSF, IL-10, IL-8, IL-5 и фракталкина. Таким образом, средство, вводимое для лечения этого побочного эффекта, может представлять собой средство, которое нейтрализует один или несколько из этих растворимых факторов. В одном варианте осуществления средство, которое нейтрализует одну или несколько из этих растворимых форм, представляет собой антитело или его антигенсвязывающий фрагмент. Примеры таких средств включают, но не ограничиваются ими, стероид (например, кортикостероид), ингибитор TNFα и ингибитор IL-6. Примером ингибитора TNFα является молекула антитела против TNFα такая как инфликсимаб, адалимумаб, цертолизумаб пегол и голимумаб. Другим примером ингибитора TNFα является слитый белок, такой как этанерцепт. Низкомолекулярные ингибиторы TNFα включают, но не ограничиваются ими, производные ксантина (например, пентоксифиллин) и бупропион. Примером ингибитора IL-6 является молекула антитела против IL-6 или молекула антитела против рецептора IL-6, такая как тоцилизумаб (toc), тоцилизумаб (toc), сарилумаб, элсилимомаб, CNTO 328, ALD518/BMS-945429, CNTO 136, CPSI-2364, CDP6038, VX30, ARGX-109, FE301 и FM101. В одном варианте осуществления молекула антитела против рецептора IL-6 представляет собой тоцилизумаб. Примером ингибитора на основе IL-1R является анакинра.
В одном варианте осуществления индивидууму можно вводить средство, которое повышает активность экспрессирующей CAR клетки. Например, в одном варианте осуществления средство может представлять собой средство, которое ингибирует ингибиторную молекулу. Ингибиторные молекулы, например Programmed Death 1 (PD-1), в некоторых вариантах осуществления могут уменьшать способность экспрессирующей CAR клетки индуцировать иммунный эффекторный ответ. Примеры ингибиторных молекул включают PD-1, PD-L1, CTLA-4, TIM-3, CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5), LAG-3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и TGF-бета. Ингибирование ингибиторной молекулы, например, посредством ингибирования уровня ДНК, РНК или белка, может оптимизировать эффективность экспрессирующей CAR клетки. В вариантах осуществления для ингибирования экспрессии ингибиторной молекулы в экспрессирующей CAR клетке можно использовать ингибиторную нуклеиновую кислоту, например, ингибиторную нуклеиновую кислоту, например дцРНК, например миРНК или кшРНК; короткие палиндромные повторы, регулярно расположенные кластерами (CRISPR), нуклеазу, подобную активатору транскрипции (TALEN) или эндонуклеазу с цинковыми пальцами (ZFN), например, как описано в настоящем описании. В одном варианте осуществления ингибитор представляет собой кшРНК. В одном варианте осуществления ингибиторная молекула ингибируется в экспрессирующей CAR клетке. В этих вариантах осуществления молекула дцРНК, которая ингибирует экспрессию ингибиторнй молекулы, связана с нуклеиновой кислотой, которая кодирует компонент, например, все из компонентов, CAR. В одном варианте осуществления ингибитор ингибиторного сигнала может представлять собой, например, антитело или фрагмент антитела, которые связываются с ингибиторной молекулой. Например, средство может представлять собой антитело или фрагмент антитела, которые связываются с PD1, PD-L1, PD-L2 или CTLA4 (например, ипилимумаб (также называемый MDX-010 и MDX-101, и выпускаемый в продажу как Yervoy®; Bristol-Myers Squibb; тремелимумаб (моноклональное IgG2-антитело, доступное от Pfizer, ранее известное как тицилимумаб, CP-675.206)). В одном варианте осуществления средство представляет собой антитело или фрагмент антитела, которые связываются с TIM3. В одном варианте осуществления средство представляет собой антитело или фрагмент антитела, которые связываются с CEACAM (например, CEACAM-1, CEACAM-3 и/или CEACAM-5). В одном варианте осуществления средство представляет собой антитело или фрагмент антитела, которые связываются с LAG3.
PD-1 представляет собой ингибиторный представитель семейства рецепторов CD28, которое также включает CD28, CTLA-4, ICOS и BTLA. PD-1 экспрессируется на активированных B-клетках, T-клетках и миелоидных клетках (Agata et al. 1996 Int. Immunol 8:765-75). Было показано, что два лиганда для PD-1, PD-L1 и PD-L2, подавляют активацию T-клеток при связывании с PD-1 (Freeman et al. 2000 J Exp Med 192:1027-34; Latchman et al. 2001 Nat Immunol 2:261-8; Carter et al. 2002 Eur J Immunol 32:634-43). PD-L1 распространен в злокачественных опухолях человека (Dong et al. 2003 J Mol Med 81:281-7; Blank et al. 2005 Cancer Immunol. Immunother 54:307-314; Konishi et al. 2004 Clin Cancer Res 10:5094). Иммунную супрессию можно обращать вспять путем ингибирования локального взаимодействия PD-1 с PD-L1. Антитела, фрагменты антител и другие ингибиторы PD-1, PD-L1 и PD-L2 известны в данной области и могут использоваться в комбинации с CAR по настоящему изобретению, описанными в настоящем описании. Например, ниволумаб (также обозначаемый как BMS-936558 или MDX1106; Bristol-Myers Squibb) представляет собой полностью человеческое моноклональное IgG4-антитело, которое специфически блокирует PD-1. Ниволумаб (клон 5C4) и другие моноклональные антитела человека, которые специфически связываются с PD-1, описаны в US 8008449 и WO2006/121168. Пидилизумаб (CT-011; Cure Tech) представляет собой гуманизированное моноклональное IgG1k-антитело, которое связывается с PD-1. Пидилизумаб и другие гуманизированные моноклональные антитела против PD-1 описаны в WO2009/101611. Пембролизумаб (ранее известный как ламбролизумаб и также обозначаемый как MK03475; Merck) представляет собой гуманизированное моноклональное IgG4-антитело, которое связывается с PD-1. Пембролизумаб и другие гуманизированные антитела против PD-1 описаны в US 8354509 и WO2009/114335. MEDI4736 (Medimmune) представляет собой моноклональное антитело человека, которое связывается с PDL1 и ингибирует взаимодействие лиганда с PD1. MDPL3280A (Genentech/Roche) представляет собой моноклональное IgG1-антитело с оптимизированной Fc, которое связывается с PD-L1. MDPL3280A и другие моноклональные антитела человека к PD-L1 описаны в патенте США № 7943743 и публикации США № 20120039906. Другие связывающие PD-L1 средства включают YW243.55.S70 (вариабельные области тяжелой и легкой цепей представлены в SEQ ID NO: 20 и 21 в WO2010/077634) и MDX-1 105 (также обозначаемый как BMS-936559, и, например, связывающие PD-L1 средства, описанные в WO2007/005874). AMP-224 (B7-DCIg; Amplimmune; например, описанный в WO2010/027827 и WO2011/066342), представляет собой слитый растворимый рецептор PD-L2-Fc, который блокирует взаимодействие между PD-1 и B7-H1. Другие антитела против PD-1 включают AMP 514 (Amplimmune), среди прочих, например, антитела против PD-1, описанные в US 8609089, US 2010028330 и/или US 20120114649.
TIM-3 (T-клеточный иммуноглобулин 3) также подавляет T-клеточную функцию, в частности, в секретирующих IFN-γ CD4+ T-хелперных 1 и CD8+ цитотоксических T-клетках 1, и он играет важную роль в устранении T-клеток. Ингибирование взаимодействия между TIM3 и его лигандами, например галектином-9 (Gal9), фосфотидилсерином (PS) и HMGB1, может увеличить иммунный ответ. Антитела, фрагменты антител и другие ингибиторы TIM3 и его лиганды доступны в данной области и могут быть использованы в комбинации с CAR против CD19, описанным в настоящем описании. Например, антитела, фрагменты антитела, низкомолекулярные соединения или пептидные ингибиторы, которые нацелены на TIM3, связываются с IgV-доменом TIM3, ингибируя взаимодействия с его лигандами. Антитела и пептиды, которые ингибируют TIM3, описаны в WO2013/006490 и US20100247521. Другие антитела против TIM3 включают гуманизированные версии RMT3-23 (описанные в Ngiow et al., 2011, Cancer Res, 71:3540-3551) и клон 8B.2C12 (описанный в Monney et al., 2002, Nature, 415:536-541). Биспецифические антитела, которые ингибируют TIM3 и PD-1, описаны в US20130156774.
В других вариантах осуществления средство, которое повышает активность экспрессирующей CAR клетки, представляет собой ингибитор CEACAM (например, ингибитор CEACAM-1, CEACAM-3 и/или CEACAM-5). В одном варианте осуществления ингибитор CEACAM представляет собой молекулу антитела против CEACAM. Иллюстративные антитела против CEACAM-1 описаны в WO 2010/125571, WO 2013/082366 WO 2014/059251 и WO 2014/022332, например моноклональное антитело 34B1, 26H7 и 5F4; или их рекомбинантные формы, как описано, например, в US 2004/0047858, US 7132255 и WO 99/052552. В других вариантах осуществления антитело против CEACAM связывается с CEACAM-5, как описано, например, в Zheng et al. PLoS One. 2010 Sep 2;5(9). pii: e12529 (DOI:10:1371/journal.pone.0021146) или перекрестно реагирует с CEACAM-1 и CEACAM-5, как описано, например, в WO 2013/054331 и US 2014/0271618.
Без связи с теорией, полагают, что канцероэмбриональные антигенные молекулы клеточной адгезии (CEACAM), такие как CEACAM-1 и CEACAM-5, опосредуют, по меньшей мере частично, ингибирование противоопухолевого иммунного ответа (см. например, Markel et al. J Immunol. 2002 Mar 15;168(6):2803-10; Markel et al. J Immunol. 2006 Nov 1;177(9):6062-71; Markel et al. Immunology. 2009 Feb;126(2):186-200; Markel et al. Cancer Immunol Immunother. 2010 Feb;59(2):215-30; Ortenberg et al. Mol Cancer Ther. 2012 Jun;11(6):1300-10; Stern et al. J Immunol. 2005 Jun 1;174(11):6692-701; Zheng et al. PLoS One. 2010 Sep 2;5(9). pii: e12529). Например, CEACAM-1 описан в качестве гетерофильного лиганда для TIM-3, играющего роль в опосредуемой TIM-3 толерантности и устранении T-клеток (см. например, WO 2014/022332; Huang, et al. (2014) Nature doi:10.1038/nature13848). В вариантах осуществления показано, что совместная блокада CEACAM-1 и TIM-3 усиливает противоопухолевый иммунный ответ в моделях с ксенотрансплантатом рака ободочной и прямой кишки (см. например, WO 2014/022332; Huang, et al. (2014), выше). В других вариантах осуществления совместная блокада CEACAM-1 и PD-1 снижает толерантность T-клеток, как описано, например, в WO 2014/059251. Таким образом, ингибиторы CEACAM можно использовать с другими иммуномодуляторами, описанными в настоящем описании (например, ингибиторы PD-1 и/или ингибиторы TIM-3) для усиления иммунного ответа против злокачественной опухоли, например, меланомы, рака легкого (например, NSCLC), рака мочевого пузыря, рака толстого кишечника, рака яичника и других злокачественных опухолей, как описано в настоящем описании.
LAG-3 (ген активации лимфоцитов 3 или CD223) представляет собой молекулу клеточной поверхности, экспрессируемую на активированных T-клетках и B-клетках, для которой было показано, что она играет роль в устранении CD8+ T-клеток. Антитела, фрагменты антитела и другие ингибиторы LAG-3 и его лиганды доступны в данной области, и их можно использовать в комбинации с CAR против CD19, описанным в настоящем описании. Например, BMS-986016 (Bristol-Myers Squib) представляет собой моноклональное антитело, которое нацелено на LAG-3. IMP701 (Immutep) представляет собой антитело-антагонист LAG-3 и IMP731 (Immutep и GlaxoSmithKline) представляет собой истощающее антитело против LAG-3. Другие ингибиторы LAG-3 включают IMP321 (Immutep), который представляет собой рекомбинантный слитый белок растворимой части LAG-3 и Ig, который связывается с молекулами MHC класса II и активирует антигенпредставляющие клетки (APC). Другие антитела описаны, например, в WO2010/019570.
В некоторых вариантах осуществления средство, которое повышает активность экспрессирующей CAR клетки, может представлять собой, например, слитый белок, содержащий первый домен и второй домен, где первый домен представляет собой ингибиторную молекулу или ее фрагмент, и второй домен представляет собой полипептид, который ассоциирован с положительным сигналом, например, полипептид, содержащий внутриклеточный сигнальный домен, как описано в настоящем описании. В некоторых вариантах осуществления полипептид, который ассоциирован с положительным сигналом, может включать костимулирующий домен CD28, CD27, ICOS, например, внутриклеточный сигнальный домен CD28, CD27 и/или ICOS, и/или первичный сигнальный домен, например, из CD3-зета, например, описанный в настоящем описании. В одном варианте осуществления слитый белок экспрессируется той же клеткой, которая экспрессирует CAR. В другом варианте осуществления слитый белок экспрессируется клеткой, например, T-клеткой, которая не экспрессирует CAR по настоящему изобретению.
В одном варианте осуществления средство, которое повышает активность экспрессирующей CAR клетки, описанной в настоящем описании, представляет собой miR-17-92.
В одном варианте осуществления средство, которое усиливает активность CAR, описанного в настоящем описании, представляет собой цитокин. Цитокины имеют важные функции, связанные с экспансией, дифференцировкой, выживаемостью и гомеостазом T-клеток. Цитокины, которые можно вводить индивидууму, которому вводят экспрессирующую CAR клетку, описанную в настоящем описании, включают: IL-2, IL-4, IL-7, IL-9, IL-15, IL-18, и IL-21 или их комбинацию. В предпочтительных вариантах осуществления вводимый цитокин представляет собой IL-7, IL-15 или IL-21, или их комбинацию. Цитокин можно вводить один раз в сутки или более одного раза сутки, например, два раза в сутки, три раза в сутки или четыре раза в сутки. Цитокин можно вводить в течение более чем одних суток, например, цитокин вводят в течение 2 суток, 3 суток, 4 суток, 5 суток, 6 суток, 1 недели, 2 недель, 3 недель или 4 недель. Например, цитокин один раз в сутки в течение 7 суток.
В вариантах осуществления цитокин вводят в комбинации с экспрессирующими CAR T-клетками. Цитокин можно вводить одновременно или совместно с экспрессирующими CAR T-клетками, например, вводить в один и тот же день. Цитокин можно составлять в ту же фармацевтическую композицию, что и экспрессирующие CAR T-клетки, или его можно составлять в отдельную фармацевтическую композицию. Альтернативно цитокин можно вводить вскоре после введения экспрессирующих CAR T-клеток, например, через 1 сутки, 2 суток, 3 суток, 4 суток, 5 суток, 6 суток или 7 суток после введения экспрессирующих CAR T-клеток. В вариантах осуществления, в которых цитокин вводят в режиме дозирования, длящемся более одних суток, первые сутки режима дозирования цитокина могут быть теми же, что и сутки введения экспрессирующих CAR T-клеток, или первые сутки режима дозирования цитокина могут быть через 1 сутки, 2 суток, 3 суток, 4 суток, 5 суток, 6 суток или 7 суток после введения экспрессирующих CAR T-клеток. В одном варианте осуществления на первые сутки экспрессирующие CAR T-клетки вводят индивидууму и на вторые сутки цитокин вводят один раз в сутки в течение последующих 7 суток. В предпочтительном варианте осуществления цитокин, подлежащий введению в комбинации с экспрессирующими CAR T-клетками, представляет собой IL-7, IL-15 или IL-21.
В других вариантах осуществления цитокин вводят после периода времени после введения экспрессирующих CAR клеток, например, по меньшей мере через 2 недели, 3 недели, 4 недели, 6 недель, 8 недель, 10 недель, 12 недель, 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 1 год или более после введения экспрессирующих CAR клеток. В одном варианте осуществления цитокин вводят после оценки ответа индивидуума на экспрессирующие CAR клетки. Например, индивидууму вводят экспрессирующие CAR клетки в соовтетствии с дозировкой и режимами, описанными в настоящем описании. Ответ индивидуума на терапию экспрессирующими CAR клетками оценивают через 2 недели, 3 недели, 4 недели, 6 недель, 8 недель, 10 недель, 12 недель, 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев или 1 год или более после введения экспрессирующих CAR клеток, с использованием любого из способов, описанных в настоящем описании, включая ингибирование роста опухоли, уменьшение количества циркулирующих опухолевых клеток или регрессию опухоли. Индивидуумам, которые не проявляют достаточного ответа на терапию экспрессирущими CAR клетками, можно вводить цитокин. Введение цитокина индивидууму, который имеет субоптимальный ответ на терапию экспрессирующми CAR клетками, повышает эффективность экспрессирующих CAR клеток или противоопухолевую активность. В предпочтительном варианте осуществления цитокин, вводимый после введения экспрессирующих CAR клеток, представляет собой IL-7.
Комбинирование с низкой дозой ингибитора mTOR
В одном варианте осуществления клетки, экспрессирующие молекулу CAR, например, молекулу CAR, описанную в настоящем описании, вводят в комбинации с низкой усиливающей иммунитет дозой ингибитора mTOR.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 5, но не более 90%, по меньшей мере 10, но не более 90%, по меньшей мере 15, но не более 90%, по меньшей мере 20, но не более 90%, по меньшей мере 30, но не более 90%, по меньшей мере 40, но не более 90%, по меньшей мере 50, но не более 90%, по меньшей мере 60, но не более 90%, или по меньшей мере 70, но не более 90%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 5, но не более 80%, по меньшей мере 10, но не более 80%, по меньшей мере 15, но не более 80%, по меньшей мере 20, но не более 80%, по меньшей мере 30, но не более 80%, по меньшей мере 40, но не более 80%, по меньшей мере 50, но не более 80%, или по меньшей мере 60, но не более 80%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 5, но не более 70%, по меньшей мере 10, но не более 70%, по меньшей мере 15, но не более 70%, по меньшей мере 20, но не более 70%, по меньшей мере 30, но не более 70%, по меньшей мере 40, но не более 70%, или по меньшей мере 50, но не более 70%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 5, но не более 60%, по меньшей мере 10, но не более 60%, по меньшей мере 15, но не более 60%, по меньшей мере 20, но не более 60%, по меньшей мере 30, но не более 60%, или по меньшей мере 40, но не более 60%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 5, но не более 50%, по меньшей мере 10, но не более 50%, по меньшей мере 15, но не более 50%, по меньшей мере 20, но не более 50%, по меньшей мере 30, но не более 50%, или по меньшей мере 40, но не более 50%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 5, но не более 40%, по меньшей мере 10, но не более 40%, по меньшей мере 15, но не более 40%, по меньшей мере 20, но не более 40%, по меньшей мере 30, но не более 40%, или по меньшей мере 35, но не более 40%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 5, но не более 30%, по меньшей мере 10, но не более 30%, по меньшей мере 15, но не более 30%, по меньшей мере 20, но не более 30%, или по меньшей мере 25, но не более 30%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 1, 2, 3, 4 или 5, но не более чем 20%, по меньшей мере 1, 2, 3, 4 или 5, но не более чем 30%, по меньшей мере 1, 2, 3, 4 или 5, но не более чем 35, по меньшей мере 1, 2, 3, 4 или 5, но не более чем 40%, или по меньшей мере 1, 2, 3, 4 или 5, но не более чем 45%.
В одном варианте осуществления доза ингибитора mTOR ассоциирована с или обеспечивает ингибирование mTOR, составляющее по меньшей мере 1, 2, 3, 4 или 5, но не более чем 90%.
[0658] Как описано в настоящем описании, степень ингибирования mTOR можно выражать в качестве степени ингибирования P70 S6-киназы, например, степень ингибирования mTOR можно определять по уровню снижения активности P70 S6-киназы, например, по снижению фосфорилирования субстрата P70 S6-киназы. Уровень ингибирования mTOR можно оценивать способом, описанным в настоящем описании, например, с использованием анализа Boulay, или измерения уровней фосфорилированной S6 с помощью вестерн-блоттинга.
ИЛЛЮСТРАТИВНЫЕ ИНГИБИТОРЫ mTOR
Как используют в рамках изобретения, термин "ингибитор mTOR" относится к соединению или лиганду, или их фармацевтически приемлемой соли, которые ингибируют киназу mTOR в клетке. В одном варианте осуществления ингибитор mTOR представляет собой аллостерический ингибитор. В одном варианте осуществления ингибитор mTOR представляет собой каталитический ингибитор.
Аллостерические ингибиторы mTOR включают нейтральное трициклическое соединение рапамицин (сиролимус), родственные рапамицину соединения, которые представляют собой соединения, имеющие структурное и функциональное сходство с рапамицином, включая, например, производные рапамицина, аналоги рапамицина (также называемые рапалогами) и другие макролидные соединения, которые ингибируют активность mTOR.
[0661] Рапамицин представляет собой известный макролидный антибиотик, продуцируемый Streptomyces hygroscopicus, имеющий структуру, показанную в формуле A.
См., например, McAlpine, J.B., et al., J. Antibiotics (1991) 44: 688; Schreiber, S.L., et al., J. Am. Chem. Soc. (1991) 113: 7433; патент США № 3929992. Существуют различные схемы нумерации, предложенные для рапамицина. Во избежание путаницы, когда в настоящем описании упоминаются конкретные аналоги рапамицина, названия приведены относительно рапамицина с использованием схемы нумерации формулы A.
Аналоги рапамицина, пригодные в рамках изобретения, представляют собой, например, O-замещенные аналоги, в которых гидроксильная группа на циклогексильном кольце рапамицина заменена OR1, где R1 представляет собой гидроксиалкил, гидроксиалкоксиалкил, ациламиноалкил или аминоалкил; например RAD001, также известный как эверолимус, как описано в US 5665772 и WO94/09010, содержимое которых включено путем ссылки. Другие пригодные аналоги рапамицина включают аналоги рапамицина, замещенные в положении 26 или 28. Аналог рапамицина может представлять собой эпимер аналога, упомянутого выше, в частности, эпимер аналога, замещенного в положении 40, 28 или 26, и он необязательно может быть далее гидрогенизирован, например, как описано в US 6015815, WO95/14023 и WO99/15530, содержание которых включено в настоящее описание путем ссылки, например ABT578, также известный как зотаролимус, или аналог рапамицина, описанный в US 7091213, WO98/02441 и WO01/14387, содержание которых включено путем ссылки, например AP23573, также известный как ридафоролимус.
Примеры аналогов рапамицина, пригодных для применения в рамках настоящего изобретения, из US 5665772 включают, но не ограничиваются ими, 40-O-бензилрапамицин, 40-O-(4'-гидроксиметил)бензилрапамицин, 40-O-[4'-(1,2-дигидроксиэтил)]бензилрапамицин, 40-O-аллилрапамицин, 40-O-[3'-(2,2-диметил-1,3-диоксолан-4(S)-ил)-проп-2'-ен-1'-ил]-рапамицин, (2'E,4'S)-40-O-(4',5'-дигидроксипент-2'-ен-1'-ил)рапамицин, 40-O-(2-гидрокси)этоксикарбонилметилрапамицин, 40-O-(2-гидрокси)этилрапамицин, 40-O-(3-гидрокси)пропилрапамицин, 40-O-(6-гидрокси)гексилрапамицин, 40-O-[2-(2-гидрокси)этокси]этилрапамицин, 40-O-[(3S)-2,2-диметилдиоксолан-3-ил]метилрапамицин, 40-O-[(2S)-2,3-дигидроксипроп-1-ил]рапамицин, 40-O-(2-ацетокси)этилрапамицин, 40-O-(2-никотиноилокси)этилрапамицин, 40-O-[2-(N-морфолино)ацетокси]этилрапамицин, 40-O-(2-N-имидазолилацетокси)этилрапамицин, 40-O-[2-(N-метил-N'-пиперазинил)ацетокси]этилрапамицин, 39-O-десметил-39,40-O,O-этиленрапамицин, (26R)-26-дигидро-40-O-(2-гидрокси)этилрапамицин, 40-O-(2-аминоэтил)рапамицин, 40-O-(2-ацетаминоэтил)рапамицин, 40-O-(2-никотинамидоэтил)рапамицин, 40-O-(2-(N-метилимидазо-2'-илкарбэтоксамидо)этил)рапамицин, 40-O-(2-этоксикарбониламиноэтил)рапамицин, 40-O-(2-толилсульфонамидоэтил)рапамицин и 40-O-[2-(4',5'-дикарбоэтокси-1',2',3'-триазол-1'-ил)этил]рапамицин.
Другие аналоги рапамицина, пригодные в рамках настоящего изобретения, представляют собой аналоги, в которых гидроксильная группа на циклогексильном кольце рапамицина и/или гидроксигруппа в положении 28 заменены группой сложного гидроксиэфира, например, аналоги рапамицина, описанные в US RE44768, например темсиролимус.
Другие аналоги рапамицина, пригодные в рамках настоящего изобретения, включают аналоги, в которых метоксигруппа в положении 16 заменена другим заместителем, предпочтительно (необязательно гидрокси-замещенным) алкинилокси, бензилом, ортометоксибензил или хлорбензилом и/или где метоксигруппа в положении 39 удалена вместе с углеродом 39, так что циклогексильное кольцо рапамицина становится циклопентильным кольцом, лишенным метоксигруппы в положении 39; например, как описано в WO95/16691 и WO96/41807, содержание которых включено путем ссылки. Аналоги можно далее модифицировать, так что гидрокси в положении 40 рапамицина алкилирован и/или 32-карбонил восстановлен.
Аналоги рапамицина из WO95/16691 включают, но не ограничиваются ими, 16-деметокси-16-(пент-2-инил)оксирапамицин, 16-деметокси-16-(бут-2-инил)оксирапамицин, 16-деметокси-16-(пропаргил)оксирапамицин, 16-деметокси-16-(4-гидроксибут-2-инил)оксирапамицин, 16-деметокси-16-бензилокси-40-O-(2-гидроксиэтил)рапамицин, 16-деметокси-16-бензилоксирапамицин, 16-деметокси-16-ортометоксибензилрапамицин, 16-деметокси-40-O-(2-метоксиэтил)-16-пент-2-инил)оксирапамицин, 39-деметокси-40-дезокси-39-формил-42-нор-рапамицин, 39-деметокси-40-дезокси-39-гидроксиметил-42-норрапамицин, 39-деметокси-40-дезокси-39-карбокси-42-норрапамицин, 39-деметокси-40-дезокси-39-(4-метилпиперазин-1-ил)карбонил-42-норрапамицин, 39-деметокси-40-дезокси-39-(морфолин-4-ил)карбонил-42-норрапамицин, 39-деметокси-40-дезокси-39-[N-метил,N-(2-пиридин-2-илэтил)]карбамоил-42-норрапамицин и 39-деметокси-40-дезокси-39-(п-толуолсульфонилгидразонометил)-42-норрапамицин.
Аналоги рапамицина из WO96/41807 включают, но не ограничиваются ими, 32-деоксорапамицин, 16-O-пент-2-инил-32-деоксорапамицин, 16-O-пент-2-инил-32-деоксо-40-O-(2-гидроксиэтил)рапамицин, 16-O-пент-2-инил-32-(S)-дигидро-40-O-(2-гидроксиэтил)рапамицин, 32(S)-дигидро-40-O-(2-метокси)этилрапамицин и 32(S)-дигидро-40-O-(2-гидроксиэтил)рапамицин.
Другим пригодным аналогом рапамицина является умиролимус, как описано в US2005/0101624, содержание которой включено путем ссылки.
RAD001, иначе известный как эверолимус (Afinitor®), имеет химическое название (1R,9S,12S,15R,16E,18R,19R,21R,23S,24E,26E,28E,30S,32S,35R)-1,18-дигидрокси-12-{(1R)-2-[(1S,3R,4R)-4-(2-гидроксиэтокси)-3-метоксициклогексил]-1-метилэтил}-19,30-диметокси-15,17,21,23,29,35-гексаметил-11,36-диокса-4-азатрицикло[30,3,1,04,9]гексатриаконта-16,24,26,28-тетраен-2,3,10,14,20-пентаон
Следующие примеры аллостерических ингибиторов mTOR включают сиролимус (рапамицин, AY-22989), 40-[3-гидрокси-2-(гидроксиметил)-2-метилпропаноат]рапамицин (также называемый темсиролимусом или CCI-779) и ридафоролимус (AP-23573/MK-8669). Другие примеры аллостерических ингибиторов mTor включают зотаролимус (ABT578) и умиролимус.
Альтернативно или дополнительно, было обнаружено, что каталитические конкурирующие с ATP ингибиторы mTOR нацелены на киназный домен mTOR прямо и нацелены как на mTORC1, так и на mTORC2. Также они являются более эффективными ингибиторами mTORC1, чем такие аллостерические ингибиторы mTOR, как рапамицин, поскольку они модулируют устойчивую к рапамицину mTORC1 активность, такую как фосфорилирование 4EBP1-T37/46 и кэп-зависимая трансляция.
Каталитические ингибиторы включают: BEZ235 или 2-метил-2-[4-(3-метил-2-оксо-8-хинолин-3-ил-2,3-дигидро-имидазо[4,5-c]хинолин-1-ил)фенил]пропионитрил, или его форму монотозилата. Синтез BEZ235 описан в WO2006/122806; CCG168 (иначе известный как AZD-8055, Chresta, C.M., et al., Cancer Res, 2010, 70(1), 288-298), который имеет химическое название {5-[2,4-бис-((S)-3-метилморфолин-4-ил)пиридо[2,3d]пиримидин-7-ил]-2-метоксифенил}метанол; 3-[2,4-бис[(3S)-3-метилморфолин-4-ил]пиридо[2,3-d]пиримидин-7-ил]-N-метилбензамид (WO09104019); 3-(2-аминобензо[d]оксазол-5-ил)-1-изопропил-1H-пиразолo[3,4-d]пиримидин-4-амин (WO10051043 и WO2013023184); N-(3-(N-(3-((3,5-диметоксифенил)амино)хиноксалин-2-ил)сульфамоил)фенил)-3-метокси-4-метилбензамид (WO07044729 и WO12006552); PKI-587 (Venkatesan, A.M., J. Med.Chem., 2010, 53, 2636-2645), который имеет химическое название 1-[4-[4-(диметиламино)пиперидин-1-карбонил]фенил]-3-[4-(4,6-диморфолино-1,3,5-триазин-2-ил)фенил]мочевина; GSK-2126458 (ACS Med. Chem. Lett., 2010, 1, 39-43), который имеет химическое название 2,4-дифтор-N-{2-метокси-5-[4-(4-пиридазинил)-6-хинолинил]-3-пиридинил}бензолсульфонамид; 5-(9-изопропил-8-метил-2-морфолино-9H-пурин-6-ил)пиримидин-2-амин (WO10114484); (E)-N-(8-(6-амино-5-(трифторметил)пиридин-3-ил)-1-(6-(2-цианопропан-2-ил)пиридин-3-ил)-3-метил-1H-имидазо[4,5-c]хинолин-2(3H)-илиден)цианамид (WO12007926).
Следующие примеры каталитических ингибиторов mTOR включают 8-(6-метокси-пиридин-3-ил)-3-метил-1-(4-пиперазин-1-ил-3-трифторметилфенил)-1,3-дигидроимидазо[4,5-c]хинолин-2-он (WO2006/122806) и Ku-0063794 (Garcia-Martinez JM, et al.,Biochem J., 2009, 421(1), 29-42. Ku-0063794 is a specific inhibitor of the mammalian target of rapamycin (mTOR)). WYE-354 является другим примером каталитического ингибитора mTor (Yu K, et al. (2009). Biochemical, Cellular, and In vivo Activity of Novel ATP-Competitive and Selective Inhibitors of the Mammalian Target of Rapamycin. Cancer Res. 69(15): 6232-6240).
Ингибиторы mTOR, пригодные в соответствии с настоящим изобретением, также включают пролекарства, производные, фармацевтически приемлемые соли или аналоги любого из указанных выше средств.
Ингибиторы mTOR, такие как RAD001, можно составлять для доставки на основе хорошо известных в данной области способов на основе конкретных дозировок, описанных в настоящем описании. В частности, в патенте США 6004973 (включенном в настоящее описание путем ссылки) представлены примеры составов, которые могут использоваться с ингибиторами mTOR, описанными в настоящем описании.
ОЦЕНКА ИНГИБИРОВАНИЯ mTOR
mTOR фосфорилирует киназу P70 S6, тем самым активируя киназу P70 S6 и позволяя ей фосфорилировать субстрат. Степень ингибирования mTOR можно выражать как степень ингибирования киназы P70 S6, например, степень ингибирования mTOR можно определять по уровню снижения активности киназы P70 S6, например, по снижению фосфорилирования субстрата киназы P70 S6. Можно определить уровень ингибирования mTOR путем измерения активности киназы P70 S6 (способность киназы P70 S6 фосфорилировать субстрат), в отсутствие ингибитора, например, перед введением ингибитора, и в присутствии ингибитора, или после введения ингибитора. Уровень ингибирования киназы P70 S6 обеспечивает уровень ингибирования mTOR. Таким образом, если киназа P70 S6 ингибируется на 40%, активность mTOR при измерении по активности киназы P70 S6, ингибируется на 40%. Степень или уровень ингибирования, упоминаемые в настоящем описании, представляют собой средний уровень ингибирования на протяжении периода дозирования. В качестве примера, если ингибитор вводят один раз в неделю, уровень ингибирования приводится как средний уровень ингибирования на протяжении этого интервала, а именно, недели.
В Boulay et al., Cancer Res, 2004, 64:252-61, включенной в настоящее описание путем ссылки, описан анализ, который можно использовать для оценки уровня ингибирования mTOR (упоминаемый в настоящем описании как анализ Boulay). В одном варианте осуществления анализ основан на измерении активности киназы P70 S6 из биологических образцов до и после введения ингибитора mTOR, например RAD001. Взятие образцов можно проводить в заданные моменты времени после введения ингибитора mTOR, например, через 24, 48 и 72 часов после введения. Можно использовать биологические образцы, например, из кожи или мононуклеарных клеток периферической крови (PBMC). Из образцов получают экстракты общего белка. Киназу P70 S6 выделяют из белковых экстрактов иммунопреципитацией с использованием антитела, которое специфически распознает киназу P70 S6. Активность выделенной киназы P70 S6 можно измерять в анализе киназы in vitro. Выделенную киназую можно инкубировать с субстратами, представляющими собой рибосомальные субъединицы 40S (которые представляют собой эндогенный субстрат киназы P70 S6) и гамма-32P в условиях, которые обеспечивают фосфорилирование субстрата. Затем реакционную смесь можно разделять на геле SDS-PAGE, и сигнал 32P можно анализировать с использованием FluoroImager. Сигнал 32P, соответствующий размеру рибосомальной субъединицы 40S, указывает на фосфорилированный субстрат и активность киназы P70 S6. Увеличение и снижение активности киназы можно вычислять путем количественного определения площади и интенсивности сигнала 32P у фосфорилированного субстрата (например, с использованием ImageQuant, Molecular Dynamics), присвоения величин произвольных единиц количественно определенному сигналу и сравнения величин после введения с величинами до введения или с эталонной величиной. Например, процентное ингибирование киназной активности можно вычислять по следующей формуле: 1-(величина, полученная после введения/величина, полученная до ведения)×100. Как описано выше, степень или уровень ингибирования, упоминаемые в настоящем описании, представляют собой средний уровень ингибирования на протяжении интервала дозирования.
Способы оценки киназной активности, например, активности киназы P70 S6, также представлены в US 7727950, включенной в настоящее описание путем ссылки.
Уровень ингибирования mTOR также можно оценивать по изменению соотношения отрицательных по PD1 и положительных по PD1 T-клеток. T-клетки из периферической крови можно идентифицировать в качестве положительных или отрицательных по PD1 известными в данной области способами.
Ингибиторы mTOR в низкой дозе
В способах, описанных в настоящем описании, используются низкие усиливающие иммунитет дозы ингибиторов mTOR, например, аллостерических ингибиторов mTOR, включая рапалоги, такие как RAD001. Напротив, уровни ингибитора, которые полностью или практически полностью ингибируют каскад mTOR, являются иммунодепрессивными, и используются, например, для предупреждения отторжения трансплантата органа. Кроме того, высокие дозы рапалогов, которые полностью ингибируют mTOR, также ингибируют рост опухолевых клеток и используются для лечения различных злокачественных опухолей (см., например, Antineoplastic effects of mammalian target of rapamycine inhibitors. Salvadori M. World J Transplant. 2012 Oct 24;2(5):74-83; Current and Future Treatment Strategies for Patients with Advanced Hepatocellular Carcinoma: Role of mTOR Inhibition. Finn RS. Liver Cancer. 2012 Nov;1(3-4):247-256; Emerging Signaling Pathways in Hepatocellular Carcinoma. Moeini A, Cornellà H, Villanueva A. Liver Cancer. 2012 Sep;1(2):83-93; Targeted cancer therapy - Are the days of systemic chemotherapy numbered? Joo WD, Visintin I, Mor G. Maturitas. 2013 Sep 20.; Role of natural and adaptive immunity in renal cell carcinoma response to VEGFR-TKIs and mTOR inhibitor. Santoni M, Berardi R, Amantini C, Burattini L, Santini D, Santoni G, Cascinu S. Int J Cancer. 2013 Oct 2).
Настоящее изобретение основано, по меньшей мере частично, на неожиданном открытии, что дозы ингибиторов mTOR, являющиеся значительно более низкими, чем дозы, используемые в современных клинических условиях, имели улучшенный эффект в отношении повышения иммунного ответа у человека и увеличения соотношения отрицательные по PD-1 T-клетки/положительные по PD-1 T-клетки. Было неожиданным, что низкие дозы ингибиторов mTOR, вызывающие только частичное ингибирование активности mTOR, способны эффективно повышать иммунный ответ у человека и повышать соотношение отрицательные по PD-1 T-клетки/положительные по PD-1 T-клетки.
Альтернативно или дополнительно, без связи с какой-либо теорией полагают, что низкая усиливающая иммунитет доза ингибитора mTOR может увеличивать количества наивных T-клеток, например, по меньшей мере временно, например, по сравнению с индивидуумом без лечения. Альтернативно или дополнительно, вновь без связи с теорией полагают, что такое лечение ингибитором mTOR после достаточного периода времени или достаточного дозирования приводит к одному или нескольким из следующих:
увеличение экспрессии одного или нескольких из следующих маркеров: CD62Lhigh, CD127high, CD27+ и BCL2, например, на T-клетках памяти, например, на предшественниках T-клеток памяти;
снижение экспрессии KLRG1, например, на T-клетках памяти, например, на предшественниках T-клеток памяти; и
увеличение количества предшественников T-клеток памяти, например, клеток с одной или комбинацией следующих характеристик: увеличенный уровень CD62Lhigh, увеличенный уровень CD127high, увеличенный уровень CD27+, сниженный уровень KLRG1, и увеличенный уровень BCL2;
и где любое из описанных выше изменений происходит, например, по меньшей мере временно, например, по сравнению с индивидуумом без лечения (Araki, K et al. (2009) Nature 460:108-112). Предшественники T-клеток памяти представляют собой T-клетки памяти, которые находятся на ранней стадии программы дифференцировки. Например, T-клетки памяти имеют одну или несколько из следующих характеристику: увеличенный уровень CD62Lhigh, увеличенный уровень CD127high, увеличенный уровень CD27+, сниженный уровень KLRG1 и/или увеличенный уровень BCL2.
В одном варианте осуществления изобретение относится к композиции или дозированной форме ингибитора mTOR, например аллостерического ингибитора mTOR, например рапалога, рапамицина или RAD001, или каталитического ингибитора mTOR, которая при введении в выбранном режиме дозирования, например, один раз в сутки или один раз в неделю, ассоциирован с: уровнем ингибирования mTOR, который не ассоциирован с полной или значительной иммунной супрессией, а ассоциирован с усилением иммунного ответа.
Ингибитор mTOR, например, аллостерический ингибитор mTOR, например рапалог, рапамицин или RAD001, или каталитический ингибитор mTOR, может быть предоставлен в составе с замедленным высвобождением. Любая из композиций или единичных дозированных форм, описанных в настоящем описании, может быть предоставлена в составе с замедленным высвобождением. В некоторых вариантах осуществления состав с замедленным высвобождением имеет более низкую биодоступность, чем состав с немедленным высвобождением. Например, в вариантах осуществления для достижения сходного терапевтического эффекта с составом с немедленным высвобождением, состав с замедленным высвобождением будет иметь в от приблизительно 2 до приблизительно 5, в от приблизительно 2,5 до приблизительно 3,5, или приблизительно в 3 раза больше количества ингибитора, чем предоставлено в составе с немедленным высвобождением.
В одном варианте осуществления предусматриваются формы с немедленным высвобождением, например RAD001, как правило, используемые для одного введения в неделю, имеющие от 0,1 до 20, от 0,5 до 10, от 2,5 до 7,5, от 3 до 6 или приблизительно 5 мг на единичную дозированную форму. Для введения один раз в неделю эти составы с немедленным высвобождением соответствуют формам с замедленным высвобождением, имеющим, соответственно, от 0,3 до 60, от 1,5 до 30, от 7,5 до 22,5, от 9 до 18, или приблизительно 15 мг ингибитора mTOR, например аллостерического ингибитора mTOR, например рапамицина или RAD001. В вариантах осуществления обе формы вводят один раз в неделю.
В одном варианте осуществления предусматриваются формы с немедленным высвобождением, например формы RAD001, как правило, используемые для одного введения в сутки, имеющие от 0,005 до 1,5, от 0,01 до 1,5, от 0,1 до 1,5, от 0,2 до 1,5, от 0,3 до 1,5, от 0,4 до 1,5, от 0,5 до 1,5, от 0,6 до 1,5, от 0,7 до 1,5, от 0,8 до 1,5, от 1,0 до 1,5, от 0,3 до 0,6, или приблизительно 0,5 мг на единичную дозированную форму. Для введения один раз в сутки эти формы с немедленным высвобождением соответствуют формам с замедленным высвобождением, имеющим, соответственно, от 0,015 до 4,5, от 0,03 до 4,5, от 0,3 до 4,5, от 0,6 до 4,5, от 0,9 до 4,5, от 1,2 до 4,5, от 1,5 до 4,5, от 1,8 до 4,5, от 2,1 до 4,5, от 2,4 до 4,5, от 3,0 до 4,5, от 0,9 до 1,8, или приблизительно 1,5 мг ингибитора mTOR, например аллостерического ингибитора mTOR, например рапамицина или RAD001. Для введения один раз в неделю эти формы с немедленным высвобождением соответствуют формам с замедленным высвобождением, имеющим, соответственно, от 0,1 до 30, от 0,2 до 30, от 2 до 30, от 4 до 30, от 6 до 30, от 8 до 30, от 10 до 30, от 1,2 до 30, от 14 до 30, от 16 до 30, от 20 до 30, от 6 до 12, или приблизительно 10 мг ингибитора mTOR, например аллостерического ингибитора mTOR, например рапамицина или RAD001.
В одном варианте осуществления предусматриваются формы с немедленным высвобождением, например формы RAD001, как правило, используемые для введения один раз в сутки, имеющие от 0,01 до 1,0 мг на единичную дозированную форму. Для введения один раз в сутки эти формы с немедленным высвобождением соответствуют формам с замедленным высвобождением, имеющим, соответственно, от 0,03 до 3 мг ингибитора mTOR, например аллостерического ингибитора mTOR, например рапамицина или RAD001. Для введения один раз в неделю эти формы с немедленным высвобождением соответствуют формам с замедленным высвобождением, имеющим, соответственно, от 0,2 до 20 мг ингибитора mTOR, например аллостерического ингибитора mTOR, например рапамицина или RAD001.
В одном варианте осуществления предусматриваются формы с немедленным высвобождением, например формы RAD001, как правило, используемые для одного введения в неделю, имеющие от 0,5 до 5,0 мг на единичную дозированную форму. Для введения один раз в неделю эти формы с немедленным высвобождением соответствуют формам с замедленным высвобождением, имеющим, соответственно, от 1,5 до 15 мг ингибитора mTOR, например аллостерического ингибитора mTOR, например, рапамицина или RAD001.
Как описано выше, одной мишенью каскада mTOR является киназа P70 S6. Таким образом, дозы ингибиторов mTOR, которые являются пригодными в способах и композициях, описанных в настоящем описании, представляют собой дозы, которые являются достаточными для достижения не более чем 80% ингибирования активности киназы P70 S6 относительно активности киназы P70 S6 в отсутствие ингибитора mTOR, например, при измерении с использованием анализа, описанного в настоящем описании, например, анализа Boulay. В следующем аспекте изобретение относится к количеству ингибитора mTOR, достаточному для достижения не более чем 38% ингибирования активности киназы P70 S6 относительно активности киназы P70 S6 в отсутствие ингибитора mTOR.
В одном аспекте доза ингибитора mTOR, пригодного в способах и композициях по изобретению, является достаточной для достижения, например, при введении человеку, 90 +/- 5% (т.е., 85-95%), 89 +/- 5%, 88 +/- 5%, 87 +/- 5%, 86 +/- 5%, 85 +/- 5%, 84 +/- 5%, 83 +/- 5%, 82 +/- 5%, 81 +/- 5%, 80 +/- 5%, 79 +/- 5%, 78 +/- 5%, 77 +/- 5%, 76 +/- 5%, 75 +/- 5%, 74 +/- 5%, 73 +/- 5%, 72 +/- 5%, 71 +/- 5%, 70 +/- 5%, 69 +/- 5%, 68 +/- 5%, 67 +/- 5%, 66 +/- 5%, 65 +/- 5%, 64 +/- 5%, 63 +/- 5%, 62 +/- 5%, 61 +/- 5%, 60 +/- 5%, 59 +/- 5%, 58 +/- 5%, 57 +/- 5%, 56 +/- 5%, 55 +/- 5%, 54 +/- 5%, 54 +/- 5%, 53 +/- 5%, 52 +/- 5%, 51 +/- 5%, 50 +/- 5%, 49 +/- 5%, 48 +/- 5%, 47 +/- 5%, 46 +/- 5%, 45 +/- 5%, 44 +/- 5%, 43 +/- 5%, 42 +/- 5%, 41 +/- 5%, 40 +/- 5%, 39 +/- 5%, 38 +/- 5%, 37 +/- 5%, 36 +/- 5%, 35 +/- 5%, 34 +/- 5%, 33 +/- 5%, 32 +/- 5%, 31 +/- 5%, 30 +/- 5%, 29 +/- 5%, 28 +/- 5%, 27 +/- 5%, 26 +/- 5%, 25 +/- 5%, 24 +/- 5%, 23 +/- 5%, 22 +/- 5%, 21 +/- 5%, 20 +/- 5%, 19 +/- 5%, 18 +/- 5%, 17 +/- 5%, 16 +/- 5%, 15 +/- 5%, 14 +/- 5%, 13 +/- 5%, 12 +/- 5%, 11 +/- 5% или 10 +/- 5%, ингибирования активности киназы P70 S6, например, при измерении с использованием анализа, описанного в настоящем описании, например, анализа Boulay.
Активность киназы P70 S6 у индивидуума можно измерять с использованием способов, известных в данной области, например, согласно способам, описанным в патенте США 7727950, посредством анализа с использованием иммуноблоттинга уровней фосфо-P70 S6K и/или уровней фосфо-P70 S6 или посредством анализа киназной активности in vitro.
Как используют в рамках изобретения, термин "приблизительно" применительно к дозе ингибитора mTOR, относится к вплоть до +/-10% вариабельности количества ингибитора mTOR, однако он может включать отсутствие вариабельности указанной дозы.
В некоторых вариантах осуществления изобретение относится к способам, включающим введение индивидууму ингибитора mTOR, например аллостерического ингибитора, например RAD001, в дозировке в пределах заданного остаточного уровня. В некоторых вариантах осуществления остаточный уровень является значительно более низким, чем остаточные уровни, ассоциированные с режимами дозирования, используемыми у пациентов с трансплантатом органа и злокачественной опухолью. В одном варианте осуществления ингибитор mTOR, например RAD001 или рапамицин, вводят для достижения остаточного уровня, который составляет менее 1/2, 1/4, 1/10 или 1/20 от остаточного уровня, который приводит к иммуносупрессии или противораковому эффекту. В одном варианте осуществления ингибитор mTOR, например RAD001 или рапамицин, вводят для достижения остаточного уровня, который составляет менее 1/2, 1/4, 1/10 или 1/20 от остаточного уровня, указанного на одобренном FDA вкладыше в упаковку для применения для иммуносупрессии или противораковых показаний.
В одном варианте осуществления способ, описанный в настоящем описании, включает введение индивидууму ингибитора mTOR, например аллостерического ингибитора, например RAD001, в дозировке, которая обеспечивает заданный остаточный уровень от 0,1 до 10 нг/мл, от 0,1 до 5 нг/мл, от 0,1 до 3 нг/мл, от 0,1 до 2 нг/мл или от 0,1 до 1 нг/мл.
В одном варианте осуществления способ, описанный в настоящем описании, включает введение индивидууму ингибитора mTOR, например аллостерического ингибитора, например RAD001, в дозировке, которая обеспечивает заданный остаточный уровень от 0,2 до 10 нг/мл, от 0,2 до 5 нг/мл, от 0,2 до 3 нг/мл, от 0,2 до 2 нг/мл или от 0,2 до 1 нг/мл.
В одном варианте осуществления способ, описанный в настоящем описании, включает введение индивидууму ингибитора mTOR, например аллостерического ингибитора, например RAD001, в дозировке, которая обеспечивает заданный остаточный уровень от 0,3 до 10 нг/мл, от 0,3 до 5 нг/мл, от 0,3 до 3 нг/мл, от 0,3 до 2 нг/мл или от 0,3 до 1 нг/мл.
В одном варианте осуществления способ, описанный в настоящем описании, включает введение индивидууму ингибитора mTOR, например аллостерического ингибитора, например RAD001, в дозировке, которая обеспечивает заданный остаточный уровень от 0,4 до 10 нг/мл, от 0,4 до 5 нг/мл, от 0,4 до 3 нг/мл, от 0,4 до 2 нг/мл или от 0,4 до 1 нг/мл.
В одном варианте осуществления способ, описанный в настоящем описании, включает введение индивидууму ингибитора mTOR, например аллостерического ингибитора, например RAD001, в дозировке, которая обеспечивает заданный остаточный уровень от 0,5 до 10 нг/мл, от 0,5 до 5 нг/мл, от 0,5 до 3 нг/мл, от 0,5 до 2 нг/мл или от 0,5 до 1 нг/мл.
В одном варианте осуществления способ, описанный в настоящем описании, включает введение индивидууму ингибитора mTOR, например аллостерического ингибитора, например RAD001, в дозировке, которая обеспечивает заданный остаточный уровень от 1 до 10 нг/мл, от 1 до 5 нг/мл, от 1 до 3 нг/мл или от 1 до 2 нг/мл.
Как используют в рамках изобретения, термин "остаточный уровень" относится к концентрации лекарственного средства в плазме непосредственно перед следующей дозой или минимальную концентрацию лекарственного средства между двумя дозами.
В некоторых вариантах осуществления заданный остаточный уровень RAD001 находится в диапазоне приблизительно от 0,1 до 4,9 нг/мл. В одном варианте осуществления заданный остаточный уровень составляет менее 3 нг/мл, например, составляет менее 0,3 или менее 3 нг/мл. В одном варианте осуществления заданный остаточный уровень составляет менее 3 нг/мл, например, составляет приблизительно от 0,3 или менее до 1 нг/мл.
В следующем аспекте в рамках изобретения может использоваться ингибитор mTOR, отличный от RAD001, в количестве, которое ассоциировано с заданным остаточным уровнем, который является биологически эквивалентным указанному заданному остаточному уровню для RAD001. В одном варианте осуществления заданный остаточный уровень для ингибитора mTOR, отличного от RAD001, представляет собой уровень, который обеспечивает тот же уровень ингибирования mTOR (например, при измерении способом, описанным в настоящем описании, например, ингибирование P70 S6-киназы), что и остаточный уровень RAD001, описанный в настоящем описании.
Фармацевтические композиции: ингибиторы mTOR
В одном аспекте настоящее изобретение относится к фармацевтическим композициям, содержащим ингибитор mTOR, например ингибитор mTOR, как описано в настоящем описании, составленный для применения в комбинации с клетками CAR, описанными в настоящем описании.
В некоторых вариантах осуществления ингибитор mTOR составляют для введения в комбинации с дополнительным средством, например, как описано в настоящем описании.
Главным образом, соединения по изобретению можно вводить в терапевтически эффективных количествах, как описано выше, любыми обычными и приемлемыми способами, известными в данной области, либо по отдельности, либо в комбинации с одним или несколькими лекарственными средствами.
Фармацевтические составы можно получать с использованием общепринятых методик растворения и смешения. Например, нерасфасованное лекарственное вещество (например, ингибитор mTOR или стабилизированная форма соединения (например, комплекс с производным циклодекстрина или другим известным комплексообразующим средством)) растворяют в подходящем растворителе в присутствии одного или нескольких эксципиентов, описанных в настоящем описании. Ингибитор mTOR, как правило, составляют в виде фармацевтических дозированных форм для обеспечения легко контролирования дозирования лекарственного средства и для предоставления пациенту удобного и легко используемого продукта.
Соединения по изобретению можно вводить в качестве фармацевтических композиций обычным путем, в частности, энтерально, например перорально, например, в форме таблеток или капсул, или парентерально, например, в форме инъекционных растворов или суспензий, местно, например, в форме лосьонов, гелей, мазей или кремов, или в назальной форме или в форме суппозитория. Когда ингибитор mTOR вводят в комбинации с (либо одновременно, либо отдельно) другим средством, как описано в настоящем описании, в одном аспекте оба компонента можно вводить одним и тем же путем (например, парентерально). Альтернативно другое средство можно вводить отличающимся путем относительно пути введения ингибитора mTOR. Например, ингибитор mTOR можно вводить перорально, а другое средство можно вводить парентерально.
ЗАМЕДЛЕННОЕ ВЫСВОБОЖДЕНИЕ
Ингибиторы mTOR, например, аллостерические ингибиторы mTOR или каталитические ингибиторы mTOR, описанные в настоящем описании, могут быть предоставлены в качестве фармацевтических составов в форме пероральных твердых дозированных форм, содержащих ингибитор mTOR, описанный в настоящем описании, например рапамицин или RAD001, которые удовлетворяют требованиям стабильности продукта и/или имеют благоприятные фармакокинетические свойства относительно таблеток с немедленным высвобождением (IR), такие как сниженные средние пиковые концентрации в плазме, сниженная вариабельность между пациентами и у одного пациента в отношении степени всасывания лекарственного средства и максимальной концентрации в плазме, сниженное соотношение Cmax/Cmin и/или сниженные эффекты питания. Предоставленные фармацевтические составы могут позволить более точный подбор дозы и/или снижение частоты неблагоприятных явлений, таким образом, обеспечивая более безопасное лечение пациентов ингибитором mTOR, описанным в настоящем описании, например рапамицином или RAD001.
В некоторых вариантах осуществления настоящее изобретение относится к стабильным составам с пролонгированным высвобождением ингибитора mTOR, описанного в настоящем описании, например рапамицина или RAD001, которые представляют собой системы из множества частиц и могут иметь функциональные слои и покрытия.
Термин "состав с множеством частиц с пролонгированным высвобождением", как используют в рамках изобретения, относится к составу, который обеспечивает высвобождение ингибитора mTOR, описанного в настоящем описании, например рапамицина или RAD001, в течение пролонгированного периода времени, например, в течение по меньшей мере 1, 2, 3, 4, 5 или 6 часов. Состав с пролонгированным высвобождением может содержать матрицы и покрытия из специальных эксципиентов, например, как описано в настоящем описании, которые составлены так, чтобы активный ингредиент был доступным на протяжении пролонгированного периода времени после приема.
Термин "пролонгированное высвобождение" может быть использован взаимозаменяемо с терминами "замедленное высвобождение" (SR) или "продленное высвобождение". Термин "пролонгированное высвобождение" относится к фармацевтическому составу, который не высвобождает активное вещество сразу после перорального приема, а высвобождает его в течение пролонгированного периода времени в соответствии с определением в статье фармакопеи Ph. Eur. (7-ое издание) о таблетках и капсулах и общей главе USP <1151> о фармацевтических дозированных формах. Термин "немедленное высвобождение" (IR), как используют в рамках изобретения, относится к фармацевтическому составу, который высвобождает 85% активного лекарственного вещества в течение менее чем 60 минут в соответствии с определением "Guidance for Industry: "Dissolution Testing of Immediate Release Solid Oral Dosage Forms" (FDA CDER, 1997). В некоторых вариантах осуществления термин "немедленное высвобождение" означает высвобождение эверолимуса из таблеток в пределах 30 минут, например, при измерении в анализе растворения, описанном в настоящем описании.
Стабильные составы с пролонгированным высвобождением ингибитора mTOR, описанного в настоящем описании, например рапамицина или RAD001, могут быть охарактеризованы посредством профиля высвобождения in vitro с использованием анализов, известных в данной области, таких как анализ растворения, как описано в настоящем описании: емкость для растворения заполняют 900 мл фосфатного буфера, pH 6,8, содержащего додецилсульфат натрия 0,2% при 37°C, и растворение проводят с использованием способ с использованием лопастной мешалки при 75 об/мин в соответствии с USP согласно статье USP о тестировании 711, и статье Ph.Eur. о тестировании 2.9.3, соответственно.
В некоторых вариантах осуществления стабильные составы с пролонгированным высвобождением ингибитора mTOR, описанного в настоящем описании, например рапамицина или RAD001, высвобождают ингибитор mTOR в анализе высвобождения in vitro в соответствии со следующими характеристиками высвобождения:
0,5 ч: <45%, или <40, например <30%
1 ч: 20-80%, например 30-60%
2 ч: >50%, или >70%, например >75%
3 ч: >60%, или >65%, например >85%, например >90%.
В некоторых вариантах осуществления стабильные составы с пролонгированным высвобождением ингибитора mTOR, описанного в настоящем описании, например, рапамицина или RAD001, высвобождают 50% ингибитора mTOR не ранее чем в течение 45, 60, 75, 90, 105 мин или 120 мин в анализе растворения in vitro.
Способы доставки посредством биополимеров
В некоторых вариантах осуществления одну или несколько экспрессирующих CAR клеток, как описано в настоящем описании, можно вводить или доставлять индивидууму через биополимерный каркас, например, биополимерный имплантат. Биополимерные каркасы могут способствовать или усиливать доставку, экспансию и/или распределение экспрессирующих CAR клеток, описанных в настоящем описании. Биополимерный каркас содержит биосовместимый (например, по существу не индуцирует воспалительный или иммунный ответ) и/или биодеградируемый полимер, который может быть встречающимся в природе или синтетическим.
Примеры подходящих биополимеров включают, но не ограничиваются ими, агар, агарозу, альгинат, альгинат/кальций-фосфатный цемент (CPC), бета-галактозидазу (β-GAL), (1,2,3,4,6-пентаацетил a-D-галактоза), целлюлозу, хитин, хитозан, коллаген, эластин, желатин, гиалуроновую кислоту, коллаген, гидроксиапатит, сополимер 3-гидроксибутирата и 3-гидроксигексаноата) (PHBHHx), полилактид, поликапролактон (PCL), сополимер лактида и гликолида (PLG), полиэтиленоксид (PEO), сополимер молочной и гликолевой кислот (PLGA), полипропиленоксид (PPO), поливиниловый спирт (PVA), шелк, соевый белок и изолят соевого белка, отдельно или в комбинации с любой другой полимерной композицией, в любой концентрации и в любом соотношении. Биополимер может быть усилен или модифицирован молекулами, усиливающими адгезию или миграцию, например, пептидами-миметиками коллагена, которые связываются с рецептором коллагена на лимфоцитах, и/или стимулирующие молекулы для повышения доставки, экспансии или функции, например, противоопухолевой активности, клеток, подлежащих доставке. Биополимерный каркас может быть поддающимся инъекции, например, в виде геля или полутвердого вещества, или он может представлять собой твердую композицию.
В некоторых вариантах осуществления экспрессирующие CAR клетки, описанные в настоящем описании, высевают на биополимерный каркас перед доставкой индивидууму. В вариантах осуществления биополимерный каркас, кроме того, содержит одно или несколько дополнительных лекарственных средств, описанных в настоящем описании (например, другая экспрессирующая CAR клетка, антитело или низкомолекулярное соединение), или средств, которые усиливают активность экспрессирующей CAR клетки, например, включенных или конъюгированных с биополимерами каркаса. В вариантах осуществления биополимерный каркас инъецируют, например, внутрь опухоли, или хирургически имплантируют в опухоль или достаточно близко к опухоли, чтобы опосредовать противоопухолевый эффект. Дополнительные примеры биополимерных композиций и способов их доставки описаны в Stephan et al., Nature Biotechnology, 2015, 33:97-101; и WO2014/110591.
Фармацевтические композиции и способы лечения
Фармацевтические композиции по настоящему изобретению могут содержать экспрессирующую CAR клетку, например, множество экспрессирующих CAR клеток, как описано в настоящем описании, в комбинации с одним или несколькими фармацевтически или физиологически приемлемыми носителями, разбавителями или эксципиентами. Такие композиции могут содержать буферы, такие как нейтральный забуференный физиологический раствор, фосфатно-солевой буфер и т.п.; углеводы, такие как глюкоза, манноза, сахароза или декстраны, маннит; белки; полипептиды или аминокислоты, такие как глицин; антиоксиданты; хелатирующие агенты, такие как EDTA или глутатион; адъюванты (например, гидроксид алюминия) и консерванты. В одном аспекте композиции по настоящему изобретению составляют для внутривенного введения.
Фармацевтические композиции по настоящему изобретению можно вводить способом, пригодным для заболевания, подлежащего лечению (или предупреждению). Количество и частота введений будут определяться такими факторами, как состояние пациента и тип и тяжесть заболевания пациента, хотя подходящие дозировки могут быть определены в клинических испытаниях.
В одном варианте осуществления фармацевтическая композиция по существу свободна от примесей, например, отсутствуют поддающиеся обнаружению уровни примесей, например, выбранных из группы, состоящей из эндотоксина, микоплазмы, компетентного по репликации лентивируса (RCL), p24, нуклеиновой кислоты VSV-G, gag ВИЧ, остаточных гранул, покрытых антителом против CD3/антителом против CD28, антител мыши, объединенной сыворотки человека, бычьего сывороточного альбумина, бычьей сыворотки, компонентов культуральной среды, клетки для упаковывания вектора или плазмидных компонентов, бактерий и грибов. В одном варианте осуществления бактерия представляет собой по меньшей мере одну бактерию, выбранную из группы, состоящей из Alcaligenes faecalis, Candida albicans, Escherichia coli, Haemophilus influenza, Neisseria meningitides, Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus pneumoniae и Streptococcus pyogenes группы A.
Когда указано "иммунологически эффективное количество", "эффективное против опухоли количество", "ингибирующее опухоль эффективное количество" или "терапевтическое количество" точное количество композиций по настоящему изобретению, подлежащее введению, может определять врач с учетом индивидуальных отличий возраста, массы тела, размера опухоли, степени инфекции или метастазирования, и состояния пациента (индивидуума). Как правило, можно утверждать, что фармацевтическую композицию, содержащую иммунные эффекторные клетки (например, T-клетки, NK-клетки), описанные в настоящем описании, можно вводить в дозировке от 104 до 109 клеток/кг массы тела, в некоторых случаях от 105 до 106 клеток/кг массы тела, включая все целые числа в этих диапазонах. Композиции T-клеток также можно вводить многократно в этих дозировках. Клетки можно вводить с использованием способов инфузии, которые хорошо известны при иммунотерапии (см., например, Rosenberg et al., New Eng. J. of Med. 319:1676, 1988).
В некоторых аспектах может быть желательным введение активированных иммунных эффекторных клеток (например, T-клеток, NK-клеток) индивидууму, а затем впоследствии повторное взятие крови (или проведение афереза), активация иммунных эффекторных клеток (например, T-клеток, NK-клеток) из нее в соответствии с настоящим изобретением, и повторная инфузия пациенту активированных и увеличенных в количестве иммунных эффекторных клеток (например, T-клеток, NK-клеток). Этот процесс можно проводить многократно каждые несколько недель. В некоторых аспектах иммунные эффекторные клетки (например, T-клетки, NK-клетки) можно активировать из взятой крови в объеме от 10 см3 до 400 см3. В некоторых аспектах иммунные эффекторные клетки (например, T-клетки, NK-клетки) активируют из взятой крови в объеме 20 см3, 30 см3, 40 см3, 50 см3, 60 см3, 70 см3, 80 см3, 90 см3 или 100 см3.
Введение рассматриваемых композиций можно проводить любым удобным способом, включая ингаляцию аэрозоля, инъекцию, прием внутрь, трансфузию, имплантацию или трансплантацию. Композиции, описанные в настоящем описании, можно вводить пациенту трансартериально, подкожно, внутрикожно, внутрь опухоли, внутрь лимфоузла, внутрь костного мозга, внутримышечно, посредством внутривенной (в/в) инъекции или внутрибрюшинно. В одном аспекте композиции T-клеток по настоящему изобретению вводят пациенту посредством внутрикожной или подкожной инъекции. В одном аспекте композиции T-клеток по настоящему изобретению вводят посредством в/в инъекции. Композиции иммунных эффекторных клеток (например, T-клеток, NK-клеток) можно инъецировать прямо в опухоль, лимфатический узел или в область инфекции.
В конкретном иллюстративном аспекте индивидуумов можно подвергать лейкоаферезу, где лейкоциты собирают, увеличивают в количестве или истощают ex vivo для селекции и/или выделения представляющих интерес клеток, например, T-клеток. Эти выделенные T-клетки можно увеличивать в количестве способами, известными в данной области, и обрабатывать так, чтобы можно было вводить одну или несколько конструкций CAR, тем самым, создавая CAR T-клетку по изобретению. Индивидуумов, нуждающихся в этом, можно впоследствии подвергать стандартному лечению химиотерапией в высокой дозе с последующей трансплантацией стволовых клеток периферической крови. В некоторых аспектах после или одновременно с трансплантацией индивидуумам проводят инфузию увеличенных в количестве CAR T-клеток по настоящему изобретению. В дополнительном аспекте увеличенные в количестве клетки вводят до или после хирургической операции.
Дозировка описанных выше лекарственных средств, подлежащая введению пациенту, варьируется в зависимости от точной природы состояния, подвергаемого лечению, и реципиента лечения. Увеличение дозировок в масштабе для введения человеку можно проводить в соответствии с принятой в данной области практикой. Доза CAMPATH, например, как правило, находится в диапазоне от 1 до приблизительно 100 мг для взрослого пациента, обычно вводимых в течение периода от 1 до 30 суток. Предпочтительная суточная доза составляет от 1 до 10 мг в сутки, хотя в некоторых случаях можно использовать более высокие дозы вплоть до 40 мг в сутки (как описано в патенте США № 6120766).
В одном варианте осуществления CAR вводят в иммунные эффекторные клетки (например, T-клетки, NK-клетки), например, с использованием транскрипции in vitro, и индивидууму (например, человеку) проводят первоначальное введение иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) по изобретению, и одно или несколько последующих введений иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) по изобретению, где одно или несколько последующих введений проводят в течение менее чем 15 суток, например 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3 или 2 суток, после предшествующего введения. В одном варианте осуществления индивидууму (например, человеку) проводят более одного введения иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) по изобретению в неделю, например проводят 2, 3 или 4 введений иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) по изобретению в неделю. В одном варианте осуществления индивидууму (например, человек) проводят более одного введения иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) в неделю (например, 2, 3 или 4 введений в неделю) (также называемых в настоящем описании курсом), за которыми следует неделя без введения иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток), а затем индивидууму проводят одно или несколько дополнительных введений иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) (например, более одного введения иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) в неделю). В другом варианте осуществления у индивидуума (например, человеку) проводят более одного курса введения иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток), и время между курсами в каждом случае составляет менее 10, 9, 8, 7, 6, 5, 4 или 3 суток. В одном варианте осуществления иммунные эффекторные клетки с CAR (например, T-клетки, NK-клетки) вводят раз в двое суток на протяжении 3 введений в неделю. В одном варианте осуществления иммунные эффекторные клетки с CAR (например, T-клетки, NK-клетки) по изобретению вводят в течение по меньшей мере двух, трех, четырех, пяти, шести, семи, восьми или более недель.
В одном аспекте экспрессирующие CAR клетки по настоящему изобретению получают с использованием вирусных векторов, таких как лентивирус. Клетки, такие как CART, полученные таким образом, будут иметь стабильную экспрессию CAR.
В одном аспекте экспрессирующие CAR клетки, например, CART, получают с использованием вирусного вектора, такого как гамма-ретровирусный вектор, например, гамма-ретровирусный вектор, описанный в настоящем описании. CART, полученные с использованием этих векторов, могут иметь стабильную экспрессию CAR.
В одном аспекте CART временно экспрессируют векторы CAR в течение 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 суток после трансдукции. Временной экспрессии CAR можно достигать посредством доставки РНК-вектора с CAR. В одном аспекте РНК CAR трансдуцируют в T-клетку посредством электропорации.
Потенциальной проблемой, которая может возникать у пациентов, подвергаемых лечению с использованием иммунных эффекторных клеток с CAR (например, T-клеток, NK-клеток) с временной экспрессией (в частности, CART, содержащих scFv мыши), является анафилаксия после многократных введений.
Без связи с теорией, полагают, что такой анафилактический ответ может быть вызван развитием у пациента гуморального ответа против CAR, т.е. антител против CAR, имеющих анти-IgE изотип. Считается, что антителопродуцирующие клетки пациента претерпевают переключение класса с изотипа IgG (который не вызывает анафилаксию) на изотип IgE, когда существует период перерыва в воздействии антигена, составляющий от десяти до четырнадцати суток.
Если пациент имеет высокий риск индукции антительного ответа против CAR в ходе временной терапии с использованием CAR (такой как терапия, осуществляемая посредством трансдукции РНК), перерывы между инфузиями CART не должны длиться более чем от десяти до четырнадцати суток.
ПРИМЕРЫ
Далее изобретение подробно описано с помощью следующих экспериментальных примеров. Эти примеры предоставлены только для целей иллюстрации и не подразумевается, что они являются ограничивающими, если нет иных указаний. Таким образом, изобретение никоим образом не следует истолковывать как ограниченное следующими примерами, а скорее его следует истолковывать как охватывающее любые и все вариации, которые станут очевидными благодаря указаниям, представленным в настоящем описании.
Лечение пациента с CLL посредством CART19
Пациента UPCC04409-10 лечили аутологичными CART19 T-клетками от CLL. Лечение привело к полной ремиссии CLL.
Анализ популяции CART-клеток
Как показано на фиг.1, мониторинг CART-клеток у пациента UPCC04409-10 с течением времени проводили путем взятия образцов крови. Определяли уровень экспрессии BBZ в клетках (красный). Определяли количество копий последовательности семейства TCR Vbeta5.1 (синий). Оба измерения проводили для образцов, полученных на указанные сутки после второй инфузии CART-клеток. Как показано на фиг.2, репертуар T-клеточных рецепторов пациента UPCC04409-10 определяли в образце, полученном на 28 сутки (фиг.2A) или 51 сутки (фиг.2B) после инфузии CART. Это демонстрирует большое количество T-клеточных рецепторов семейства TCRBV05-01 на 51 сутки, что указывает на клональную экспансию с течением времени. Как показано на фиг.3, выделенные T-клетки пациента UPCC04409-10 анализировали в отношении одновременной экспрессии CAR19 и 2 различных генов семейства TCR с течением времени (50 сутки и 51 сутки) и сравнивали с исходным дозированным материалом (продукт): на верхней панели представлено семейство TCR Vb13.1; на нижней панели представлено семейство TCR Vb5.1. Данные демонстрируют, что положительные по CAR19 клетки содержат один ген семейства TCR (Vb5.1), уровень которого быстро возрастает между 50 и 51 сутками. Как показано на фиг.4, репертуар T-клеточных рецепторов для CD8-положительных клеток от пациента UPCC04409-10 определяли в образце, полученном на 51 сутки после инфузии CART. Это демонстрирует большое количество T-клеточных рецепторов семейства TCRBV05-01 на 51 сутки, что указывает на клональную экспансию CD8-положительных клеток с течением времени.
Анализ персистирования клона CART
Как показано на фиг.6, из T-клеток пациента # получали фрагментированную посредством обработки ультразвуковым излучением ДНК. Этот материал использовали для амплификации геномных последовательностей, соседних с областью встраивания CAR19. Указанные гены рядом с CAR19 в геноме были идентифицированы как имеющие увеличенную экспрессию относительно инфузированного продукта (D0). В различные моменты времени после указанной инфузии CART (d=сутки; m=месяц), наблюдали различное относительное увеличение экспрессии соседних генов, причем увеличение экспрессии Tet2 достигало пика в образцах как периферической крови (PBMC), так и CAR+CD8+ T-клеток на 51 сутки.
Как показано на фиг.7, участок встраивания гена CAR19 был картирован на гене Tet2. Более конкретно, встраивание происходило между 9 и 10 экзонами гена Tet2. Каталитический домен для Tet2 находится в экзоне 11. Встраивание в этом положении может приводить к экспрессии аберрантных мРНК-транскриптов или снижению экспрессии функционального Tet2 (дикого типа).
Как показано на фиг.8, транскрипты гена Tet2 из выделенной мРНК пациента UPCC04409-10 оценивали посредством ОТ-ПЦР с использованием праймеров, охватывающих указанные области Tet2 или CAR19, или обоих из них, как указано на фигуре справа. Rxn 3 содержит праймеры, сконструированные для амплификации области транскрипта Tet2, охватывающей экзоны 9 и 10. Rxn, 6, 7, 8, 9 и 10 представляют собой праймеры, сконструированные для амплификации указанных частей лентивируса CAR19. Rxn 12-16 представляют собой пары праймеров, которые содержат последовательность 9 экзона транскрипта Tet2, а также последовательность из лентивирусной конструкции CAR19. Эти данные показывают, что транскрипты получены из локуса Tet2, который содержит как последовательность Tet2, так и последовательность CAR19.
Анализ функции Tet2
Как показано на фиг.10, на ней схематично представлена ферментативная активность Tet2 (фиг.10A). Белок семейства Tet конвертирует 5-метилцитозин (5-mc) в 5-гидроксиметилцитозин (5-hmc), а затем в 5-формилцитозин (5-fmc), что приводит к деметилированному цитозину. Метилированная ДНК представляет собой эпигенетическое состояние, которое, как известно, влияет на профиль транскрипции. Оценивали состояние метилирования T-клеток от пациента UPCC04409-10 (фиг.10B). T-клетки пациента окрашивали на TCRVb5.1 (который содержит инсерцию CAR19 в Tet2) и 5-hmc и 5-fmc оценивали в положительных по TCRVb5.1 (красный) и отрицательных по TCRVb5.1 (синий) популяциях посредством проточной цитометрии. Эти данные указывают на то, что клетки, содержащие инсерцию CAR19 в гене Tet2, имеют дефект деметилирования.
Обработка T-клеток ингибиторами Tet2 на основе кшРНК
Материалы и способы
Получение лентивируса и инфицирование клеток Jurkat
Лентивирусы получали из клеток 293T на 15-см чашках. В кратком изложении, 10 миллионов клеток 293T высевали на покрытые коллагеном 15-см чашки на сутки -1. На 0 сутки, 15 мкг вектора с кшРНК (т.е. вектор, содержащий последовательность, кодирующую нацеленную на TET2 или контрольную кшРНК), 15 мкг вектора Gag/pol и 5 мкг вектора VSV-G трансфицировали с использованием Lipofectamin 2000 (Invitrogen). Через 24 часа (1 сутки) среду заменяли. После замены среды супернатанты с вирусами собирали на 2 сутки и 3 сутки. Вирусы концентрировали с использованием Lenti-X Concentrator (соотношение по объему 3:1, Clonetech, каталожный номер #: 631231). 100 мкл вирусов добавляли к 0,5 миллиона клеток jurkat в присутствии 6 мкг/мл полибрена. Клетки инфицировали при вращении 2000 об/мин в течение 90 мин при 32°C. После инкубации в течение 1 часа в инкубаторе при 37 градусах добавляли свежую среду RPMI 1640 и переносили в 24-луночный планшет. На 6 сутки клетки переносили в 6-ячеечный планшет в присутствии пуромицина в конечной концентрации 2 мкг/мл на 6 суток.
Антитела
Антитела, использованные для вестерн-блоттинга, были следующими: β-актин (клон #: 8H10D10, Cell Signaling); TET2 (клон #: hT2H21F11, Millipore); IgG мыши (H+L) (конъюгированный с HRP, каталожный #: 115-035-166, Jackson ImmunoResearch); IgG кролика (H+L) (конъюгированный с HRP, каталожный #: 111-035-114, Jackson ImmunoResearch).
Вестерн-блоттинг
Для исследования эффективности нокдауна посредством кшРНК против TET2 на белковом уровне в клетках jurkat получали клеточные лизаты в буфере RIPA, содержавшем коктейль ингибиторов протеаз (Sigma). Концентрацию белка измеряли с использованием наборов для анализа белков BCA (Pierce, Item#: 3603904). 20 мкг белка подвергали SDS-PAGE, а затем белок переносили на нитроцеллюлозную мембрану с использованием системы для переноса iBot (Invitrogen, 20 В, 11 мин 30 с). Мембрану блокировали 5% BSA, содержавшем TBST, в течение 30 мин при комнатной температуре. Антитела инкубировали в течение ночи с мембранами в разведении 1:000 при 4°C. После инкубации с конъюгированными с HRP вторичными антителами проводили детекцию сигнала с использованием устройства для детекции хемилюминесценции (Chemidoc; Bio-Rad).
Количественная ОТ-ПЦР
Для исследования эффективности нокдауна посредством кшРНК против TET2 на уровне ДНК в клетках jurkat проводили количественную полимеразную цепную реакцию с обратной транскрипцией (кОТ-ПЦР). Для экстракции РНК использовали набор RNeasy Micro Kit (Qiagen). мРНК подвергали обратной транскрипции в одноцепочечную комплементарную ДНК (кДНК) с использованием системы SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogen). ПЦР в реальном времени проводили с использованием термоциклера C1000 Touch Thermal Cycler (Biorad). Для обнаружения экспрессии гена использовали протокол на основе SYBR (SsoAdvanced Universal SYBR Green Supermix, Biorad). Реакции ПЦР проводили в 96-луночных планшетах, и использовали рекомендованные изготовителем параметры циклических реакций в трех экземплярах (95°C в течение 3 минут, затем 40 циклов при 95°C в течение 15 секунд и 60°C в течение 30 секунд). Величины порогового цикла (Ct) для представляющих интерес генов нормализовывали к Ct для β-актина. Праймеры, использованные для кОТ-ПЦР, были следующими: β-актин #1 (прямой праймер: CAT GTA CGT TGC TAT CCA GGC (SEQ ID NO: 1263), обратный праймер: CTC CTT AAT GTC ACG CAC GAT (SEQ ID NO: 1264); размер продукта 250 п.н.); β-актин #2 (прямой праймер: CTC ACC ATG GAT GAT GAT ATC GC (SEQ ID NO: 1265), обратный праймер: CCA CAT AGG AAT CCT TCT GAC CC (SEQ ID NO: 1266); размер продукта 169 п.н.); TET1 (прямой праймер: CAG AAC CTA AAC CAC CCG TG (SEQ ID NO: 1267), обратный праймер: TGC TTC GTA GCG CCA TTG TAA (SEQ ID NO: 1268); размер продукта 141 п.н.); TET2 (прямой праймер: ATA CCC TGT ATG AAG GGA AGC C (SEQ ID NO: 1269), обратный праймер: CTT ACC CCG AAG TTA CGT CTT TC (SEQ ID NO: 1270); размер продукта 197 п.н.); TET3 (прямой праймер: CAC CCG GCT CTA TGA AAC CTT (SEQ ID NO: 1271), обратный праймер: CCA GCC ACT CGA GGT AGT CA (SEQ ID NO: 1272); размер продукта 209 п.н.); RPLP1 (каталожный номер #: PPH17813G-200, Qiagen).
Проточная цитометрия
Клетки получали от FACS Fortessa (BD). Обработку данных для представления проводили с использованием программы FlowJo (Treestar Inc.).
Результаты
Валидация эффективности нокдауна Tet2 с использованием кшРНК
Как показано на фиг.27, на ней схематично представлена валидация эффективности нокдауна TET2 посредством кшРНК. кшРНК против TET2 и контрольные конструкции кшРНК с перестановкой, экспрессирующие красный флуоресцентный белок (RPF) и ген резистентности к пуромицину, вводили в клетки jurkat для валидации эффективности нокдауна TET2 посредством кшРНК с использованием кОТ-ПЦР и вестерн-блоттинга.
Как показано на фиг.28, инфицированные кшРНК клетки jurkat экспрессируют RFP. Экспрессию RFP определяли посредством FACS на 6 сутки после обработки пуромицином. Исходя из экспрессии RFP более 99% клеток jurkat с введенной кшРНК было отобрано посредством обработки пуромицином. Следует отметить, что клетки jurkat, инфицированные кшРНК против TET2 #3 и #4, не росли в присутствии пуромицина. Таким образом, клетки jurkat, инфицированные кшРНК против TET2 #3 и #4, далее не использовали. Эти данные указывают на то, что пуромицин является эффективным для селекции клеток jurkat, инфицированных кшРНК.
Как показано на фиг.29, эффективность нокдауна tet2 зависит от кшРНК. Для определения уровня экспрессии мРНК tet1 и tet2 и tet3 в клетках jurkat, инфицированных кшРНК против TET2, проводили эксперимент с использованием кОТ-ПЦР. По сравнению с кшРНК с перестановкой, кшРНК против TET2 #1, #2, #8 и #9 осуществляли нокдаун гена tet2 на уровне 35,6%, 22,7%, 21,6% и 76,7%, соответственно. Неожиданно, хотя кшРНК против TET2 #9 эффективно осуществляет нокдаун tet2, она также подавляет экспрессию tet1 и tet3 на уровне 43,4% и 67,3%, соответственно. β-актин служит в качестве внутреннего контроля для количественного определения относительной экспрессии генов среди протестированных образцов. Для повышения надежности кОТ-ПЦР в этом эксперименте использовали два праймера для β-актина и один праймер для RPLP1. Эти данные указывают на то, что несколько нацеленных на TET2 кшРНК способны снижать уровни мРНК TET2, причем кшРНК #9 демонстрирует наиболее выраженный эффект нокдауна tet2, также влияя на уровни TET1 и TET2.
Как показано на фиг.30, нокдаун белка TET2 в ответ на кшРНК коррелирует с нокдауном уровней мРНК TET2. Для определения уровня экспрессии белка TET2 в клетках jurkat, инфицированных кшРНК против TET2, проводили вестерн-блоттинг. Аналогично данным кОТ-ПЦР, показанным на фиг.29, кшРНК против TET2 #1, #2, #8 и #9 снижали уровень белка TET2 по сравнению с кшРНК с перестановкой, в то время как β-актин конститутивно экспрессировался во всех исследованных образцах. Эти данные указывают на то, что несколько нацеленных на TET2 кшРНК способны снижать уровни белка TET2 в клетках Jurkat, причем кшРНК #9 продемонстрировала наиболее выраженный эффект нокдауна Tet2.
Обработка первичных T-клеток ингибиторами Tet2 на основе кшРНК
Как показано на фиг.11, кшРНК против TET2 снижают уровни 5-hmc в нормальных T-клетках человека. кшРНК против TET2 и контрольные конструкции кшРНК с перестановкой, экспрессирующие mCherry, вводили в нормальные T-клетки человека. Уровни 5-hmc определяли по внутриклеточному окрашиванию с помощью FACS на 6 сутки после экспансии с использованием гранул с антителом против CD3/CD28. Нокдаун TET2 снижал общие уровни 5-hmc.
Как показано на фиг.12, кшРНК против TET2 увеличивают в количестве T-клетки Tscm. кшРНК против TET2 и контрольные конструкции кшРНК с перестановкой, экспрессирующие mCherry, вводили в нормальные T-клетки человека. CD45RA+CD62L+CCR7+CD27+CD95+ T-клетки Tscm определяли по окрашиванию с помощью FACS на 11 сутки после экспансии с использованием гранул с антителом против CD3/CD28. Нокдаун TET2 стимулировал экспансию T-клеток с фенотипом Tscm.
Ингибирование Tet2 в CAR T-клетках с использованием системы редактирования генов CRISPR/Cas
В этом примере исследовали ингибирование TET2 в T-клетках, экспрессирущих химерный рецептор антигена (CAR).
Способы
Молекулы РНК-гида
Для экспериментов, описанных в этом подпримере использовали молекулы гРНК, содержавшие нацеливающие последовательности, приведенные в таблице 5. Если нет иных указаний, все молекулы гРНК исследовали в качестве двойных молекул гРНК, содержащих последовательности tracr и cr-РНК, описанные в этом подпримере.
Таблица 5
Получение CRISPR CAR T-клеток
Выделенные и замороженные тотальные T-клетки размораживали и активировали с использованием гранул с антителом против CD3/CD28 (CD3/CD28 CTS Dynabeads® 43205D) на 0 сутки. Активированные T-клетки трансдуцировали лентивирусом, кодирующим либо BCMA CAR (BCMA-10 (139109), как описано в WO2016/0046724; обозначаемый в настоящем описании как BCMA-10 CAR), либо CD19 CAR (CD19 CAR, имеющий аминокислотную последовательность SEQ ID NO: 12 WO2012/079000; обозначаемый в настоящем описании как CD19 CAR) на 1 сутки. На 3 сутки трансдуцированные CAR T-клетки подвергали электропорации для введения систем CRISPR/Cas в форме предварительного комплексного рибонуклеопротеина гРНК/Cas9 ("РНП"). Для получения РНП все образцы РНК нагревали при 95C. Белок CAS9 S. pyogenes (NLS CAS9 iPROT106154, 37 мкМ) разбавляли в буфере до добавления к нему tracr-РНК (имеющей последовательность: AACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGUUUUUUU (SEQ ID NO: 1296); AXO Labs). После смешения белка CAS9 с tracr-РНК добавляли РНК CRISPR (в каждом случае каждая cr-РНК содержала последовательность nnnnnnnnnnnnnnnnnnnnGUUUUAGAGCUAUGCUGUUUUG (SEQ ID NO: 40), где остатки n соответствуют 20 остаткам рибонуклеиновой кислоты указанной нацеливающей последовательности). Затем предварительные комплексные РНП добавляли всего к 1 миллиону клеток, концентрация РНК составляла 3,2 мкМ. Электропорацию проводили посредством неонового электропоратора с использованием Neon® Transfection System 100 μL Kit (MPK10096) при 1600 В, 10 мс, 3 импульса. Клетки держали в культуре в течение 7 дополнительных суток. Затем клетки разделяли, причем некоторые использовали для проведения проточной цитометрии: окрашивание на CAR (PE), CD3 (PerCP-Cy5.5), CD4 (V450) и CD8 (APC). Остальные T-клетки замораживали и использовали для функциональных анализов и получения образцов для секвенирования нового поколения (NGS).
Получение образца для секвенирования нового поколения (NGS)
Замороженные осадки с отредактированными клетками (описанные выше) размораживали и обрабатывали с использованием DNeasy Blood & Tissue Kit (Qiagen, 69506) для выделения геномной ДНК. Элюированную ДНК использовали для проведения ПЦР с использованием Titan Taq PCR kit (Clontech Laboratories, 639211) и праймеров для TET2 (праймеры, сконструированы для фланкирования ожидаемого участка-мишени гРНК). Продукт ПЦР очищали с использованием QIAquick PCR Purification Kit (Qiagen, 28104). Затем очищенный продукт ПЦР использовали для анализа T7E1 для обнаружения несоответствий оснований и подтверждения редактирования генов. ПЦР-ампликоны подвергали стандартному преобразованию в библиотеку Nextera NGS library prep (Illumina) и секвенировали с использованием считывания спаренных концов на секвенаторе Illumina MiSeq. Отсеквенированные последовательности выравнивали с эталонным геномом и выявляли варианты.
Анализ продукции цитокинов
Эффекторные клетки (CAR T-клетки) размораживали в день анализа и подсчитывали на Cellometer (Nexelcom). Затем эти клетки сокультивировали с различными клетками-мишенями в соотношении клеток эффектор:мишень 2:1. Через 20 ч собирали 100 мкл супернатанта сокультуры. Затем эти супернатанты использовали для количественного определения высвободившихся цитокинов IL-2 и IFN-g с использованием системы Meso Scale Discovery, Proinflammatory Panel 1, каталожный номер # N05049A-1, в соответствии с протоколом изготовителя.
Анализ пролиферации T-клеток
Оценивали пролиферацию CAR T-клеток в ответ на экспрессирующие BCMA или CD19 клетки-мишени. Линии клеток-мишеней представляли собой: положительные по BCMA клеточные линии множественной миеломы, NCI-H929-luc, KMS-11-luc и отрицательные по BCMA Nalm6luc (положительная по CD19 клеточная линия). CAR T-клетки размораживали и инкубировали в течение 2 часов в среде для T-клеток для восстановления. Клетки подсчитывали на Cellometer. Клетки-мишени облучали при 10000 рад. После облучения клетки-мишени промывали два раза в полной среде для T-клеток и подсчитывали. Затем 30000 облученных клеток сокультивировали с CART-клетками в соотношении 1:1. В качестве отрицательного контроля к CAR T-клеткам добавляли только среду.
Сокультуру инкубировали в течение 4 суток при 37°C. На 4 сутки клетки сокультуры окрашивали в течение 20 мин на льду посредством CD3-percp cy5.5(Ebioscience:45-0037), CD4-eflor450(Ebioscience:48-0047) и CD8-APC (Ebioscience 17-0087) и количественно определяли проточной цитометрией относительно CountBright Absolute Counting Beads (Life Technologies) для определения количеств клеток. Экспрессию CAR количественно определяли посредством инкубации в две стадии по 20 мин каждая на льду: биотинилированный белок L+стрептавидин-PE(Jackson immuno research). Данные проточной цитометрии получали с использованием лазера BD 5 Fortessa и анализировали с помощью программного обеспечения FlowJo.
Результаты
На фиг.13A и 13B представлена экспрессия CAR в клетках, подвергнутых электропорации с введением CRISPR против Tet2 или без него. На фиг.13A представлена стратегия гейтирования для определения CAR+ T-клеток. Селекцию лимфоцитов проводили с использованием прямого рассеяния (FSC-A) и бокового рассеяния (SSC-A), как указано обведением в круг. Затем отбирали положительные по CD3 клетки (средняя панель). Затем положительные по CAR (положительные по PE клетки с использованием обнаружения CAR с помощью биотинилированного белка и стрептавидин-PE) клетки, указанные чертой, определяли путем гейтирования относительно пика отрицательного контроля. На фиг.13B представлено количественное определение процента положительных по CAR клеток. Клетки трансдуцировали либо BCMA-10 CAR, либо CD19 CAR, как указано. В клетки посредством электропорации вводили РНП, содержащий белок Cas9, трейсерную РНК и указанную РНК-гид, нацеленную на Tet2 (Ex3-3 нацеленная на экзон 3, или Ex9-6, нацеленная на экзон 9), или электропорацию не проводили. Экспрессию CAR определяли через 10 суток после активации клеток гранулами. Эти данные указывают на то, что редактирование Tet2 не влияет на экспрессию CAR в T-клетках.
На фиг.14 представлено количественное определение положительных по CD4 и CD8 клеток после трансдукции CAR и редактирования Tet2. Клетки окрашивали в отношении экспрессии CD3, CD4, CD8 и CAR в конце экспансии с гранулами в течение 10 суток. На левой панели указан процент положительных по CD4 и CD8 клеток в общей популяции CD3+ клеток. На правой панели указан процент положительных по CD4 и CD8 клеток в популяции CD3+ клеток, которые также являются CAR+. Клетки трансдуцировали либо BCMA-10 CAR, либо CD19 CAR, либо оставляли нетрансдуцированными (UTD). В клетки посредством электропорации вводили РНП, содержащий белок Cas9, трейсерную РНК и указанную РНК-гид, нацеленную на Tet2 (Ex3-3 нацеленная на экзон 3, или Ex9-6, нацеленная на экзон 9), или электропорацию не проводили. Эти данные показали, что редактирование Tet2 вызывает небольшое, но стабильное снижение процента CD8 клеток и увеличение процента CD4 в период экспансии на основе гранул.
На фиг.15 представлен выход и жизнеспособность клеток после экспансии с использованием гранул в течение 10 суток. Количество клеток на мл (левая панель) и жизнеспособность клеток (правая панель) количественно определяли с использованием Cellometer после процесса экспансии с использованием гранул в течение 10 суток. Клетки трансдуцировали либо BCMA-10 CAR, либо CD19 CAR, либо оставляли нетрансдуцированными (UTD). В клетки посредством электропорации вводили РНП, содержащий белок Cas9, трейсерную РНК и указанную РНК-гид, нацеленную на Tet2 (Ex3-3 нацеленная на экзон 3, или Ex9-6, нацеленная на экзон 9), или электропорацию не проводили. Эти данные указывают на то, что редактирование Tet2 приводит к увеличению количества и жизнеспособности клеток относительно клеток без CRISPR/Cas9, причем нацеленный на 9 гид (ex9-6) продемонстрировал наибольшее влияние в нетрансдуцированных, а также в трансдуцированных CAR клетках.
На фиг.16 представлена продукция IL-2 в ответ на клетки, либо положительные, либо отрицательные по антигену, распознаваемому CAR. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA/ отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA /положительными по CD19 клетками (NALM6) и количественно определяли секрецию цитокинов в среду. Представлены уровни IL-2 (пг/мл). Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 вызывает увеличение продукции IL-2 T-клетками в ответ на антиген, причем нацеленный на экзон 9 гид (Ex9-6) продемонстрировал наибольший эффект.
На фиг.17 представлена продукция интерферона-гамма. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA/отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA/положительными по CD19 клетками (NALM6) и определяли секрецию цитокинов в среду. Представлены уровни интерферона-гамма (IFN-g) (пг/мл). Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 приводит к увеличению продукции интерферона-гамма T-клетками в ответ на антиген, причем нацеленный на экзон 9 гид (Ex9-6) продемонстрировал наибольший эффект.
На фиг.18 представлена запускаемая антигеном пролиферация CART-клеток. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA /отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA /положительными по CD19 клетками (NALM6) или культивировали без клеток-мишеней (среда) и количественно определяли пролиферацию CAR-положительных T-клеток. Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 приводит к повышению пролиферации CAR+ T-клеток в ответ на антиген, причем нацеленный на экзон 9 гид (CrEx9-6) продемонстрировал наибольший эффект.
На фиг.19 представлена запускаемая антигеном пролиферация тотальных T-клеток. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA/отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA/положительными по CD19 клетками (NALM6) или культивировали без клеток-мишеней (среда) и количественно определяли пролиферацию всех CD3+ T-клеток. Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 приводит к повышению пролиферации CD3+ T-клеток в ответ на антиген, причем нацеленный на экзон 9 гид (CrEx9-6) продемонстрировал наибольший эффект.
На фиг.20 представлена запускаемая антигеном пролиферация CAR+ T-клеток. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA/отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA/положительными по CD19 клетками (NALM6) или культивировали без клеток-мишеней (среда) и количественно определяли пролиферацию всех CD4+ CD3+ T-клеток. Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 приводит к повышению пролиферации CD4+ T-клеток в ответ на антиген, причем нацеленный на экзон 9 гид (CrEx9-6) продемонстрировал наибольший эффект.
На фиг.21 представлена запускаемая антигеном пролиферация CAR+ CD4+ T-клеток. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA/отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA/положительными по CD19 клетками (NALM6) или культивировали без клеток-мишеней (среда) и количественно определяли пролиферацию всех CAR+CD4+ CD3+ T-клеток. Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 приводит к повышению пролиферации CAR+CD4+ T-клеток в ответ на антиген, причем нацеленный на экзон 9 гид (CrEx9-6) продемонстрировал наибольший эффект.
На фиг.22 представлена запускаемая антигеном пролиферация CD8+ T-клеток. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA/отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA/положительными по CD19 клетками (NALM6) или культивировали без клеток-мишеней (среда) и количественно определяли пролиферацию всех CD8+ CD3+ T-клеток. Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 приводит к повышению пролиферации CD8+ T-клеток в ответ на антиген, причем нацеленный на экзон 9 гид (CrEx9-6) продемонстрировал наибольший эффект.
На фиг.23 представлена запускаемая антигеном пролиферация CAR+ CD8+ T-клеток. CART-клетки или нетрансдуцированные клетки (UTD) сокультивировали с положительными по BCMA/отрицательными по CD19 клетками (KMS11 и NCIH929) или отрицательными по BCMA/положительными по CD19 клетками (NALM6) или культивировали без клеток-мишеней (среда) и количественно определяли пролиферацию всех CAR+CD8+ CD3+ T-клеток. Клетки получали, как описано выше. Эти данные указывают на то, что редактирование Tet2 приводит к повышению пролиферации CAR+CD8+ T-клеток в ответ на антиген, причем нацеленный на экзон 9 гид (CrEx9-6) продемонстрировал наибольший эффект.
На фиг.24 представлено % редактирование и % редактирование со сдвигом рамки считывания путем введения нацеленных на Tet2 систем CRISPR/Cas. Показан уровень редактирования в первичных T-клетках после электропорации РНП, содержавшего белок Cas9, трейсерную РНК и указанные РНК-гиды, нацеленные либо на экзон 3, либо на экзон 9 Tet2. Процент наблюдаемых инсерций или делеций нуклеотидов относительно эталонного генома представлен в среднем столбце (средний % инсерций/делеций). Редактирование, которое приводит к сдвигу открытой рамки считывания, представлено в крайнем правом столбце (средний % со сдвигом рамки считывания). Эти данные представляют собой средние значения для трех повторений эксперимента. Эти данные указывают высокоэффективное редактирование генома в первичных T-клетках с использованием этих последовательностей РНК-гидов.
Профиль инсерций и делеций в или вблизи участка-мишени для каждой гРНК оценивали с использованием секвенирования нового поколения. В кратком изложении, в T-клетки вводили посредством электропорации РНП, содержавший указанную РНГ-гид. Через 48 часов ДНК выделяли и обрабатывали для секвенирования. На фиг.25 представлено 5 наиболее распространенных инсерций-делеций (инсерций и/или делеций), наблюдаемых в первичных T-клетках для РНК-гидов, нацеленных на экзон 3 Tet2. На фиг.26 представлено 5 наиболее распространенных инсерций-делеций (инсерций и/или делеций), наблюдаемых в первичных T-клетках для РНК-гидов, нацеленных на экзон 9 Tet2. Проценты указывают на частоту, с которой наблюдали данный профиль редактирования. Инсерции показаны обозначением нуклеотидов прописными буквами ("a", "g", "c" или "t"), в то время как делеции показаны прочерком ("-").
Эксперименты с ATAC-Seq
CD8+ T-клетки с инсерцией Tet2 или без нее увеличивали в количестве из образца пациента после инфузии. Доступность хроматина оценивали с использованием анализа доступного для транспозазы хроматина с высокопроизводительным секвенированием (ATAC-seq). Он по существу представляет собой способ глобального картирования хроматина на основе транспозиции ДНК-фрагментов со "штрих-кодом". Эти ДНК-фрагменты встраиваются в области открытого хроматина, что позволяет определение того, какие области хроматина открыты, а какие закрыты. Исходя из положения открытых или закрытых областей можно проводить анализ каскадов, предполагая, что "открытый" равноценен "экспрессируемому" и "закрытый" равноценен "репрессируемому". На фиг.30A представлены диаграммы Венна для пиков ATAC в CAR+CD8+ T-клетках от пациента с нарушением Tet2 по сравнению с CAR-CD8+ T-клетками от того же пациента в соответствующий момент времени без нарушения Tet2. На фиг.28B представлены термины GO, ассоциированные с пиками ATAC, наиболее закрытыми в клеточной популяции с нарушением Tet2 по сравнению с его аналогом. Значимость фиг.30B состоит, по меньшей мере частично, в том, что рельеф хроматина CD8+ T-клеток с нарушением Tet2 возможно согласуется с менее дифференцированной клеткой, которая может быть в большей степени "подобной ранней клетке памяти" и в меньшей степени "подобной эффектору". Они представляют собой тип клеток, которые, предположительно, будут длительно сохраняться, обеспечивая устойчивую и длительную противоопухолевую активность.
Исследования с кшРНК
T-клетки от здоровых доноров активировали в течение 24 часов с использованием гранул, покрытых CD3/CD28, а затем проводили трансдукцию лентивирусом либо с ненацеленной (контрольной) кшРНК, либо с кшРНК против Tet2. Как показано на фиг.32A, эффективность нокдауна оценивали посредством кПЦР и показано, что она составляет 50% (может отражать то, что произошло у пациента, у которого Tet2 был разрушен посредством встраивания лентивируса). Фенотип дифференцировки исследовали на 14 посредством исследования CCR7, CD45RO. Центральные клетки памяти определяют как CCR7+CD45RO+, в то время как эффекторные клетки представляют собой CCR7-CD45RO-. Для специфического исследования фенотипа дифференцировки в клетках с нокдауном 50% Tet2, в конструкция кшРНК использовали индикатор GFP. Результаты представлены на фиг.32B и 32C.
Без дальнейшего описания полагают, что специалист в данной области может с использованием предшествующего описания и следующих ниже иллюстративных примеров получать и использовать соединения по настоящему изобретению и применять на практике заявленные способы. Представленные ниже рабочие примеры конкретно указывают на различные аспекты настоящего изобретения, и их не следует считать ограничивающими каким-либо образом остальную часть описания.
ЭКВИВАЛЕНТЫ
Содержание каждого патента, патентной заявки и публикации, цитированных в настоящем описании, включено в настоящее описание путем ссылки в полном объеме. Хотя настоящее изобретение описано в отношении конкретных аспектов, очевидно, что другие аспекты и вариации настоящего изобретения могут быть установлены другими специалистами без отклонения от истинной сущности и объема изобретения. Подразумевают, что прилагаемую формулу изобретения следует истолковывать как включающую все такие аспекты и эквивалентные вариации.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> NOVARTIS AG
THE TRUSTEES OF THE UNIVERSITY OF PENNSYLVANIA
<120> CAR T CELL THERAPIES WITH ENHANCED EFFICACY
<130> N2067-7098WO
<140>
<141>
<150> 62/220,196
<151> 2015-09-17
<160> 1363
<170> PatentIn version 3.5
<210> 1
<211> 1184
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 1
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaag gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacctggctg cagtacgtga ttcttgatcc cgagcttcgg gttggaagtg 360
ggtgggagag ttcgaggcct tgcgcttaag gagccccttc gcctcgtgct tgagttgagg 420
cctggcctgg gcgctggggc cgccgcgtgc gaatctggtg gcaccttcgc gcctgtctcg 480
ctgctttcga taagtctcta gccatttaaa atttttgatg acctgctgcg acgctttttt 540
tctggcaaga tagtcttgta aatgcgggcc aagatctgca cactggtatt tcggtttttg 600
gggccgcggg cggcgacggg gcccgtgcgt cccagcgcac atgttcggcg aggcggggcc 660
tgcgagcgcg gccaccgaga atcggacggg ggtagtctca agctggccgg cctgctctgg 720
tgcctggcct cgcgccgccg tgtatcgccc cgccctgggc ggcaaggctg gcccggtcgg 780
caccagttgc gtgagcggaa agatggccgc ttcccggccc tgctgcaggg agctcaaaat 840
ggaggacgcg gcgctcggga gagcgggcgg gtgagtcacc cacacaaagg aaaagggcct 900
ttccgtcctc agccgtcgct tcatgtgact ccacggagta ccgggcgccg tccaggcacc 960
tcgattagtt ctcgagcttt tggagtacgt cgtctttagg ttggggggag gggttttatg 1020
cgatggagtt tccccacact gagtgggtgg agactgaagt taggccagct tggcacttga 1080
tgtaattctc cttggaattt gccctttttg agtttggatc ttggttcatt ctcaagcctc 1140
agacagtggt tcaaagtttt tttcttccat ttcaggtgtc gtga 1184
<210> 2
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 2
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro
20
<210> 3
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 3
atggccctgc ctgtgacagc cctgctgctg cctctggctc tgctgctgca tgccgctaga 60
ccc 63
<210> 4
<211> 45
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 4
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<210> 5
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 5
accacgacgc cagcgccgcg accaccaaca ccggcgccca ccatcgcgtc gcagcccctg 60
tccctgcgcc cagaggcgtg ccggccagcg gcggggggcg cagtgcacac gagggggctg 120
gacttcgcct gtgat 135
<210> 6
<211> 230
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 6
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys Met
225 230
<210> 7
<211> 690
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 7
gagagcaagt acggccctcc ctgcccccct tgccctgccc ccgagttcct gggcggaccc 60
agcgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagccg gacccccgag 120
gtgacctgtg tggtggtgga cgtgtcccag gaggaccccg aggtccagtt caactggtac 180
gtggacggcg tggaggtgca caacgccaag accaagcccc gggaggagca gttcaatagc 240
acctaccggg tggtgtccgt gctgaccgtg ctgcaccagg actggctgaa cggcaaggaa 300
tacaagtgta aggtgtccaa caagggcctg cccagcagca tcgagaaaac catcagcaag 360
gccaagggcc agcctcggga gccccaggtg tacaccctgc cccctagcca agaggagatg 420
accaagaacc aggtgtccct gacctgcctg gtgaagggct tctaccccag cgacatcgcc 480
gtggagtggg agagcaacgg ccagcccgag aacaactaca agaccacccc ccctgtgctg 540
gacagcgacg gcagcttctt cctgtacagc cggctgaccg tggacaagag ccggtggcag 600
gagggcaacg tctttagctg ctccgtgatg cacgaggccc tgcacaacca ctacacccag 660
aagagcctga gcctgtccct gggcaagatg 690
<210> 8
<211> 282
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 8
Arg Trp Pro Glu Ser Pro Lys Ala Gln Ala Ser Ser Val Pro Thr Ala
1 5 10 15
Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro Ala
20 25 30
Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Lys Glu Lys
35 40 45
Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro
50 55 60
Ser His Thr Gln Pro Leu Gly Val Tyr Leu Leu Thr Pro Ala Val Gln
65 70 75 80
Asp Leu Trp Leu Arg Asp Lys Ala Thr Phe Thr Cys Phe Val Val Gly
85 90 95
Ser Asp Leu Lys Asp Ala His Leu Thr Trp Glu Val Ala Gly Lys Val
100 105 110
Pro Thr Gly Gly Val Glu Glu Gly Leu Leu Glu Arg His Ser Asn Gly
115 120 125
Ser Gln Ser Gln His Ser Arg Leu Thr Leu Pro Arg Ser Leu Trp Asn
130 135 140
Ala Gly Thr Ser Val Thr Cys Thr Leu Asn His Pro Ser Leu Pro Pro
145 150 155 160
Gln Arg Leu Met Ala Leu Arg Glu Pro Ala Ala Gln Ala Pro Val Lys
165 170 175
Leu Ser Leu Asn Leu Leu Ala Ser Ser Asp Pro Pro Glu Ala Ala Ser
180 185 190
Trp Leu Leu Cys Glu Val Ser Gly Phe Ser Pro Pro Asn Ile Leu Leu
195 200 205
Met Trp Leu Glu Asp Gln Arg Glu Val Asn Thr Ser Gly Phe Ala Pro
210 215 220
Ala Arg Pro Pro Pro Gln Pro Gly Ser Thr Thr Phe Trp Ala Trp Ser
225 230 235 240
Val Leu Arg Val Pro Ala Pro Pro Ser Pro Gln Pro Ala Thr Tyr Thr
245 250 255
Cys Val Val Ser His Glu Asp Ser Arg Thr Leu Leu Asn Ala Ser Arg
260 265 270
Ser Leu Glu Val Ser Tyr Val Thr Asp His
275 280
<210> 9
<211> 847
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 9
aggtggcccg aaagtcccaa ggcccaggca tctagtgttc ctactgcaca gccccaggca 60
gaaggcagcc tagccaaagc tactactgca cctgccacta cgcgcaatac tggccgtggc 120
ggggaggaga agaaaaagga gaaagagaaa gaagaacagg aagagaggga gaccaagacc 180
cctgaatgtc catcccatac ccagccgctg ggcgtctatc tcttgactcc cgcagtacag 240
gacttgtggc ttagagataa ggccaccttt acatgtttcg tcgtgggctc tgacctgaag 300
gatgcccatt tgacttggga ggttgccgga aaggtaccca cagggggggt tgaggaaggg 360
ttgctggagc gccattccaa tggctctcag agccagcact caagactcac ccttccgaga 420
tccctgtgga acgccgggac ctctgtcaca tgtactctaa atcatcctag cctgccccca 480
cagcgtctga tggcccttag agagccagcc gcccaggcac cagttaagct tagcctgaat 540
ctgctcgcca gtagtgatcc cccagaggcc gccagctggc tcttatgcga agtgtccggc 600
tttagcccgc ccaacatctt gctcatgtgg ctggaggacc agcgagaagt gaacaccagc 660
ggcttcgctc cagcccggcc cccaccccag ccgggttcta ccacattctg ggcctggagt 720
gtcttaaggg tcccagcacc acctagcccc cagccagcca catacacctg tgttgtgtcc 780
catgaagata gcaggaccct gctaaatgct tctaggagtc tggaggtttc ctacgtgact 840
gaccatt 847
<210> 10
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 10
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 11
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 11
ggtggcggag gttctggagg tggaggttcc 30
<210> 12
<211> 24
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 12
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr Leu Tyr Cys
20
<210> 13
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 13
atctacatct gggcgccctt ggccgggact tgtggggtcc ttctcctgtc actggttatc 60
accctttact gc 72
<210> 14
<211> 42
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 14
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 15
<211> 126
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 15
aaacggggca gaaagaaact cctgtatata ttcaaacaac catttatgag accagtacaa 60
actactcaag aggaagatgg ctgtagctgc cgatttccag aagaagaaga aggaggatgt 120
gaactg 126
<210> 16
<211> 48
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 16
Gln Arg Arg Lys Tyr Arg Ser Asn Lys Gly Glu Ser Pro Val Glu Pro
1 5 10 15
Ala Glu Pro Cys Arg Tyr Ser Cys Pro Arg Glu Glu Glu Gly Ser Thr
20 25 30
Ile Pro Ile Gln Glu Asp Tyr Arg Lys Pro Glu Pro Ala Cys Ser Pro
35 40 45
<210> 17
<211> 123
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 17
aggagtaaga ggagcaggct cctgcacagt gactacatga acatgactcc ccgccgcccc 60
gggcccaccc gcaagcatta ccagccctat gccccaccac gcgacttcgc agcctatcgc 120
tcc 123
<210> 18
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 18
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 19
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 19
agagtgaagt tcagcaggag cgcagacgcc cccgcgtaca agcagggcca gaaccagctc 60
tataacgagc tcaatctagg acgaagagag gagtacgatg ttttggacaa gagacgtggc 120
cgggaccctg agatgggggg aaagccgaga aggaagaacc ctcaggaagg cctgtacaat 180
gaactgcaga aagataagat ggcggaggcc tacagtgaga ttgggatgaa aggcgagcgc 240
cggaggggca aggggcacga tggcctttac cagggtctca gtacagccac caaggacacc 300
tacgacgccc ttcacatgca ggccctgccc cctcgc 336
<210> 20
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 20
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 21
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 21
agagtgaagt tcagcaggag cgcagacgcc cccgcgtacc agcagggcca gaaccagctc 60
tataacgagc tcaatctagg acgaagagag gagtacgatg ttttggacaa gagacgtggc 120
cgggaccctg agatgggggg aaagccgaga aggaagaacc ctcaggaagg cctgtacaat 180
gaactgcaga aagataagat ggcggaggcc tacagtgaga ttgggatgaa aggcgagcgc 240
cggaggggca aggggcacga tggcctttac cagggtctca gtacagccac caaggacacc 300
tacgacgccc ttcacatgca ggccctgccc cctcgc 336
<210> 22
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 22
Gly Gly Gly Gly Ser
1 5
<210> 23
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 23
ggtggcggag gttctggagg tggaggttcc 30
<210> 24
<211> 150
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 24
Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp Asn Pro Pro Thr
1 5 10 15
Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp Asn Ala Thr Phe
20 25 30
Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val Leu Asn Trp Tyr
35 40 45
Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala Ala Phe Pro Glu
50 55 60
Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg Val Thr Gln Leu
65 70 75 80
Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg Ala Arg Arg Asn
85 90 95
Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu Ala Pro Lys Ala
100 105 110
Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val Thr Glu Arg Arg
115 120 125
Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro Arg Pro Ala Gly
130 135 140
Gln Phe Gln Thr Leu Val
145 150
<210> 25
<211> 450
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 25
cccggatggt ttctggactc tccggatcgc ccgtggaatc ccccaacctt ctcaccggca 60
ctcttggttg tgactgaggg cgataatgcg accttcacgt gctcgttctc caacacctcc 120
gaatcattcg tgctgaactg gtaccgcatg agcccgtcaa accagaccga caagctcgcc 180
gcgtttccgg aagatcggtc gcaaccggga caggattgtc ggttccgcgt gactcaactg 240
ccgaatggca gagacttcca catgagcgtg gtccgcgcta ggcgaaacga ctccgggacc 300
tacctgtgcg gagccatctc gctggcgcct aaggcccaaa tcaaagagag cttgagggcc 360
gaactgagag tgaccgagcg cagagctgag gtgccaactg cacatccatc cccatcgcct 420
cggcctgcgg ggcagtttca gaccctggtc 450
<210> 26
<211> 394
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 26
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro
20 25 30
Trp Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly
35 40 45
Asp Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe
50 55 60
Val Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu
65 70 75 80
Ala Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe
85 90 95
Arg Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val
100 105 110
Arg Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser
115 120 125
Leu Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg
130 135 140
Val Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser
145 150 155 160
Pro Arg Pro Ala Gly Gln Phe Gln Thr Leu Val Thr Thr Thr Pro Ala
165 170 175
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
180 185 190
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
195 200 205
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
210 215 220
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
225 230 235 240
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
245 250 255
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
260 265 270
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
275 280 285
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
290 295 300
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
305 310 315 320
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
325 330 335
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
340 345 350
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
355 360 365
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
370 375 380
Ala Leu His Met Gln Ala Leu Pro Pro Arg
385 390
<210> 27
<211> 1182
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 27
atggccctcc ctgtcactgc cctgcttctc cccctcgcac tcctgctcca cgccgctaga 60
ccacccggat ggtttctgga ctctccggat cgcccgtgga atcccccaac cttctcaccg 120
gcactcttgg ttgtgactga gggcgataat gcgaccttca cgtgctcgtt ctccaacacc 180
tccgaatcat tcgtgctgaa ctggtaccgc atgagcccgt caaaccagac cgacaagctc 240
gccgcgtttc cggaagatcg gtcgcaaccg ggacaggatt gtcggttccg cgtgactcaa 300
ctgccgaatg gcagagactt ccacatgagc gtggtccgcg ctaggcgaaa cgactccggg 360
acctacctgt gcggagccat ctcgctggcg cctaaggccc aaatcaaaga gagcttgagg 420
gccgaactga gagtgaccga gcgcagagct gaggtgccaa ctgcacatcc atccccatcg 480
cctcggcctg cggggcagtt tcagaccctg gtcacgacca ctccggcgcc gcgcccaccg 540
actccggccc caactatcgc gagccagccc ctgtcgctga ggccggaagc atgccgccct 600
gccgccggag gtgctgtgca tacccgggga ttggacttcg catgcgacat ctacatttgg 660
gctcctctcg ccggaacttg tggcgtgctc cttctgtccc tggtcatcac cctgtactgc 720
aagcggggtc ggaaaaagct tctgtacatt ttcaagcagc ccttcatgag gcccgtgcaa 780
accacccagg aggaggacgg ttgctcctgc cggttccccg aagaggaaga aggaggttgc 840
gagctgcgcg tgaagttctc ccggagcgcc gacgcccccg cctataagca gggccagaac 900
cagctgtaca acgaactgaa cctgggacgg cgggaagagt acgatgtgct ggacaagcgg 960
cgcggccggg accccgaaat gggcgggaag cctagaagaa agaaccctca ggaaggcctg 1020
tataacgagc tgcagaagga caagatggcc gaggcctact ccgaaattgg gatgaaggga 1080
gagcggcgga ggggaaaggg gcacgacggc ctgtaccaag gactgtccac cgccaccaag 1140
gacacatacg atgccctgca catgcaggcc cttccccctc gc 1182
<210> 28
<211> 40
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (1)..(40)
<223> /примечание="эта последовательность может охватывать 1-10
повторяющихся элементов "Gly-Gly-Gly-Ser""
<400> 28
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
20 25 30
Gly Gly Gly Ser Gly Gly Gly Ser
35 40
<210> 29
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 29
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 30
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 30
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 31
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 31
Gly Gly Gly Ser
1
<210> 32
<211> 2000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<220>
<221> дополнительный признак
<222> (1)..(2000)
<223> /примечание="эта последовательность может охватывать 50-2000
нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 32
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1980
aaaaaaaaaa aaaaaaaaaa 2000
<210> 33
<211> 150
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 33
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 150
<210> 34
<211> 5000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<220>
<221> дополнительный признак
<222> (1)..(5000)
<223> /примечание="эта последовательность может охватывать 50-5000
нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 34
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1980
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2040
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2100
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2160
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2220
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2280
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2340
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2400
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2460
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2520
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2580
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2640
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2700
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2760
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2820
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2880
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2940
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3000
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3060
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4980
aaaaaaaaaa aaaaaaaaaa 5000
<210> 35
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 35
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 60
tttttttttt tttttttttt tttttttttt tttttttttt 100
<210> 36
<211> 5000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<220>
<221> дополнительный признак
<222> (1)..(5000)
<223> /примечание="эта последовательность может охватывать 50-5000
нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 36
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 60
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 120
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 180
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 240
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 300
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 360
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 420
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 480
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 540
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 600
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 660
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 720
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 780
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 840
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 900
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 960
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1020
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1080
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1140
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1200
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1260
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1320
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1380
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1440
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1500
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1560
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1620
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1680
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1740
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1800
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1860
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1920
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1980
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2040
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2100
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2160
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2220
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2280
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2340
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2400
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2460
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2520
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2580
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2640
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2700
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2760
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2820
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2880
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2940
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3000
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3060
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3120
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3180
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3240
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3300
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3360
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3420
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3480
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3540
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3600
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3660
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3720
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3780
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3840
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3900
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3960
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4020
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4080
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4140
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4200
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4260
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4320
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4380
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4440
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4500
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4560
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4620
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4680
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4740
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4800
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4860
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4920
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4980
tttttttttt tttttttttt 5000
<210> 37
<211> 5000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<220>
<221> дополнительный признак
<222> (1)..(5000)
<223> /примечание="эта последовательность может охватывать 100-5000
нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 37
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1980
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2040
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2100
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2160
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2220
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2280
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2340
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2400
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2460
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2520
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2580
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2640
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2700
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2760
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2820
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2880
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2940
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3000
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3060
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4980
aaaaaaaaaa aaaaaaaaaa 5000
<210> 38
<211> 400
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<220>
<221> дополнительный признак
<222> (1)..(400)
<223> /примечание="эта последовательность может охватывать 100-400
нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 38
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 400
<210> 39
<211> 373
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 39
Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp Asn Pro Pro Thr
1 5 10 15
Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp Asn Ala Thr Phe
20 25 30
Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val Leu Asn Trp Tyr
35 40 45
Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala Ala Phe Pro Glu
50 55 60
Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg Val Thr Gln Leu
65 70 75 80
Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg Ala Arg Arg Asn
85 90 95
Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu Ala Pro Lys Ala
100 105 110
Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val Thr Glu Arg Arg
115 120 125
Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro Arg Pro Ala Gly
130 135 140
Gln Phe Gln Thr Leu Val Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr
145 150 155 160
Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala
165 170 175
Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe
180 185 190
Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val
195 200 205
Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly Arg Lys
210 215 220
Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr
225 230 235 240
Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu
245 250 255
Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro
260 265 270
Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly
275 280 285
Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro
290 295 300
Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr
305 310 315 320
Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly
325 330 335
Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln
340 345 350
Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln
355 360 365
Ala Leu Pro Pro Arg
370
<210> 40
<211> 42
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<220>
<221> модифицированное основание
<222> (1)..(20)
<223> a, c, u, g, неизвестные или прочие
<400> 40
nnnnnnnnnn nnnnnnnnnn guuuuagagc uaugcuguuu ug 42
<210> 41
<211> 86
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<220>
<221> дополнительный признак
<222> (80)..(86)
<223> /примечание="Эта область может охватывать 1-7 нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 41
aacuuaccaa ggaacagcau agcaaguuaa aauaaggcua guccguuauc aacuugaaaa 60
aguggcaccg agucggugcu uuuuuu 86
<210> 42
<211> 74
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<220>
<221> дополнительный признак
<222> (68)..(74)
<223> /примечание="Эта область может охватывать 1-7 нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 42
aacagcauag caaguuaaaa uaaggcuagu ccguuaucaa cuugaaaaag uggcaccgag 60
ucggugcuuu uuuu 74
<210> 43
<211> 103
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<220>
<221> модифицированное основание
<222> (1)..(20)
<223> a, c, u, g, неизвестные или прочие
<220>
<221> variation
<222> (97)..(103)
<223> /примечание="Эта область может охватывать 1-7 нуклеотидов"
<220>
<221> источник
<223> /примечание="См. поданное описание для подробного описания замен и
предпочтительных вариантов осуществления"
<400> 43
nnnnnnnnnn nnnnnnnnnn guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc 60
cguuaucaac uugaaaaagu ggcaccgagu cggugcuuuu uuu 103
<210> 44
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 44
ccuuggacac cuucuccucc 20
<210> 45
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 45
ggaaccugug gaagaggagg 20
<210> 46
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 46
ggaacctgtg gaagaggagg 20
<210> 47
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 47
gaaggaagct gaggaacctg 20
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 48
atgacctcca aacaatacac 20
<210> 49
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 49
caagtgctgt ttcaacactg 20
<210> 50
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 50
gggagatgtg aactctggga 20
<210> 51
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 51
ggaggtgatg gtatcaggaa 20
<210> 52
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 52
ggttctgtct ggcaaatggg 20
<210> 53
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 53
ggatgagctc tctcaggcag 20
<210> 54
<211> 132
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 54
Asp Val Pro Asp Tyr Ala Ser Leu Gly Gly Pro Ser Ser Pro Lys Lys
1 5 10 15
Lys Arg Lys Val Ser Arg Gly Val Gln Val Glu Thr Ile Ser Pro Gly
20 25 30
Asp Gly Arg Thr Phe Pro Lys Arg Gly Gln Thr Cys Val Val His Tyr
35 40 45
Thr Gly Met Leu Glu Asp Gly Lys Lys Phe Asp Ser Ser Arg Asp Arg
50 55 60
Asn Lys Pro Phe Lys Phe Met Leu Gly Lys Gln Glu Val Ile Arg Gly
65 70 75 80
Trp Glu Glu Gly Val Ala Gln Met Ser Val Gly Gln Arg Ala Lys Leu
85 90 95
Thr Ile Ser Pro Asp Tyr Ala Tyr Gly Ala Thr Gly His Pro Gly Ile
100 105 110
Ile Pro Pro His Ala Thr Leu Val Phe Asp Val Glu Leu Leu Lys Leu
115 120 125
Glu Thr Ser Tyr
130
<210> 55
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 55
Val Gln Val Glu Thr Ile Ser Pro Gly Asp Gly Arg Thr Phe Pro Lys
1 5 10 15
Arg Gly Gln Thr Cys Val Val His Tyr Thr Gly Met Leu Glu Asp Gly
20 25 30
Lys Lys Phe Asp Ser Ser Arg Asp Arg Asn Lys Pro Phe Lys Phe Met
35 40 45
Leu Gly Lys Gln Glu Val Ile Arg Gly Trp Glu Glu Gly Val Ala Gln
50 55 60
Met Ser Val Gly Gln Arg Ala Lys Leu Thr Ile Ser Pro Asp Tyr Ala
65 70 75 80
Tyr Gly Ala Thr Gly His Pro Gly Ile Ile Pro Pro His Ala Thr Leu
85 90 95
Val Phe Asp Val Glu Leu Leu Lys Leu Glu Thr Ser
100 105
<210> 56
<211> 93
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 56
Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu Glu Ala Ser Arg
1 5 10 15
Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe Glu Val Leu Glu
20 25 30
Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr Leu Lys Glu Thr
35 40 45
Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu Ala Gln Glu Trp
50 55 60
Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp Leu Thr Gln Ala
65 70 75 80
Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser Lys
85 90
<210> 57
<211> 95
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 57
Ile Leu Trp His Glu Met Trp His Glu Gly Leu Ile Glu Ala Ser Arg
1 5 10 15
Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe Glu Val Leu Glu
20 25 30
Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr Leu Lys Glu Thr
35 40 45
Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu Ala Gln Glu Trp
50 55 60
Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp Leu Thr Gln Ala
65 70 75 80
Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser Lys Thr Ser
85 90 95
<210> 58
<211> 95
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 58
Ile Leu Trp His Glu Met Trp His Glu Gly Leu Leu Glu Ala Ser Arg
1 5 10 15
Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe Glu Val Leu Glu
20 25 30
Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr Leu Lys Glu Thr
35 40 45
Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu Ala Gln Glu Trp
50 55 60
Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp Leu Thr Gln Ala
65 70 75 80
Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser Lys Thr Ser
85 90 95
<210> 59
<211> 95
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 59
Ile Leu Trp His Glu Met Trp His Glu Gly Leu Glu Glu Ala Ser Arg
1 5 10 15
Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe Glu Val Leu Glu
20 25 30
Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr Leu Lys Glu Thr
35 40 45
Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu Ala Gln Glu Trp
50 55 60
Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp Leu Leu Gln Ala
65 70 75 80
Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser Lys Thr Ser
85 90 95
<210> 60
<211> 95
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<220>
<221> МОДИФИЦИРОВАННЫЙ ОСТАТОК
<222> (12)..(12)
<223> Любая аминокислота
<220>
<221> МОДИФИЦИРОВАННЫЙ ОСТАТОК
<222> (78)..(78)
<223> Любая аминокислота
<400> 60
Ile Leu Trp His Glu Met Trp His Glu Gly Leu Xaa Glu Ala Ser Arg
1 5 10 15
Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe Glu Val Leu Glu
20 25 30
Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr Leu Lys Glu Thr
35 40 45
Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu Ala Gln Glu Trp
50 55 60
Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp Leu Xaa Gln Ala
65 70 75 80
Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser Lys Thr Ser
85 90 95
<210> 61
<211> 95
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 61
Ile Leu Trp His Glu Met Trp His Glu Gly Leu Ile Glu Ala Ser Arg
1 5 10 15
Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe Glu Val Leu Glu
20 25 30
Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr Leu Lys Glu Thr
35 40 45
Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu Ala Gln Glu Trp
50 55 60
Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp Leu Leu Gln Ala
65 70 75 80
Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser Lys Thr Ser
85 90 95
<210> 62
<211> 95
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 62
Ile Leu Trp His Glu Met Trp His Glu Gly Leu Leu Glu Ala Ser Arg
1 5 10 15
Leu Tyr Phe Gly Glu Arg Asn Val Lys Gly Met Phe Glu Val Leu Glu
20 25 30
Pro Leu His Ala Met Met Glu Arg Gly Pro Gln Thr Leu Lys Glu Thr
35 40 45
Ser Phe Asn Gln Ala Tyr Gly Arg Asp Leu Met Glu Ala Gln Glu Trp
50 55 60
Cys Arg Lys Tyr Met Lys Ser Gly Asn Val Lys Asp Leu Leu Gln Ala
65 70 75 80
Trp Asp Leu Tyr Tyr His Val Phe Arg Arg Ile Ser Lys Thr Ser
85 90 95
<210> 63
<211> 1132
<212> БЕЛОК
<213> Homo sapiens
<400> 63
Met Pro Arg Ala Pro Arg Cys Arg Ala Val Arg Ser Leu Leu Arg Ser
1 5 10 15
His Tyr Arg Glu Val Leu Pro Leu Ala Thr Phe Val Arg Arg Leu Gly
20 25 30
Pro Gln Gly Trp Arg Leu Val Gln Arg Gly Asp Pro Ala Ala Phe Arg
35 40 45
Ala Leu Val Ala Gln Cys Leu Val Cys Val Pro Trp Asp Ala Arg Pro
50 55 60
Pro Pro Ala Ala Pro Ser Phe Arg Gln Val Ser Cys Leu Lys Glu Leu
65 70 75 80
Val Ala Arg Val Leu Gln Arg Leu Cys Glu Arg Gly Ala Lys Asn Val
85 90 95
Leu Ala Phe Gly Phe Ala Leu Leu Asp Gly Ala Arg Gly Gly Pro Pro
100 105 110
Glu Ala Phe Thr Thr Ser Val Arg Ser Tyr Leu Pro Asn Thr Val Thr
115 120 125
Asp Ala Leu Arg Gly Ser Gly Ala Trp Gly Leu Leu Leu Arg Arg Val
130 135 140
Gly Asp Asp Val Leu Val His Leu Leu Ala Arg Cys Ala Leu Phe Val
145 150 155 160
Leu Val Ala Pro Ser Cys Ala Tyr Gln Val Cys Gly Pro Pro Leu Tyr
165 170 175
Gln Leu Gly Ala Ala Thr Gln Ala Arg Pro Pro Pro His Ala Ser Gly
180 185 190
Pro Arg Arg Arg Leu Gly Cys Glu Arg Ala Trp Asn His Ser Val Arg
195 200 205
Glu Ala Gly Val Pro Leu Gly Leu Pro Ala Pro Gly Ala Arg Arg Arg
210 215 220
Gly Gly Ser Ala Ser Arg Ser Leu Pro Leu Pro Lys Arg Pro Arg Arg
225 230 235 240
Gly Ala Ala Pro Glu Pro Glu Arg Thr Pro Val Gly Gln Gly Ser Trp
245 250 255
Ala His Pro Gly Arg Thr Arg Gly Pro Ser Asp Arg Gly Phe Cys Val
260 265 270
Val Ser Pro Ala Arg Pro Ala Glu Glu Ala Thr Ser Leu Glu Gly Ala
275 280 285
Leu Ser Gly Thr Arg His Ser His Pro Ser Val Gly Arg Gln His His
290 295 300
Ala Gly Pro Pro Ser Thr Ser Arg Pro Pro Arg Pro Trp Asp Thr Pro
305 310 315 320
Cys Pro Pro Val Tyr Ala Glu Thr Lys His Phe Leu Tyr Ser Ser Gly
325 330 335
Asp Lys Glu Gln Leu Arg Pro Ser Phe Leu Leu Ser Ser Leu Arg Pro
340 345 350
Ser Leu Thr Gly Ala Arg Arg Leu Val Glu Thr Ile Phe Leu Gly Ser
355 360 365
Arg Pro Trp Met Pro Gly Thr Pro Arg Arg Leu Pro Arg Leu Pro Gln
370 375 380
Arg Tyr Trp Gln Met Arg Pro Leu Phe Leu Glu Leu Leu Gly Asn His
385 390 395 400
Ala Gln Cys Pro Tyr Gly Val Leu Leu Lys Thr His Cys Pro Leu Arg
405 410 415
Ala Ala Val Thr Pro Ala Ala Gly Val Cys Ala Arg Glu Lys Pro Gln
420 425 430
Gly Ser Val Ala Ala Pro Glu Glu Glu Asp Thr Asp Pro Arg Arg Leu
435 440 445
Val Gln Leu Leu Arg Gln His Ser Ser Pro Trp Gln Val Tyr Gly Phe
450 455 460
Val Arg Ala Cys Leu Arg Arg Leu Val Pro Pro Gly Leu Trp Gly Ser
465 470 475 480
Arg His Asn Glu Arg Arg Phe Leu Arg Asn Thr Lys Lys Phe Ile Ser
485 490 495
Leu Gly Lys His Ala Lys Leu Ser Leu Gln Glu Leu Thr Trp Lys Met
500 505 510
Ser Val Arg Gly Cys Ala Trp Leu Arg Arg Ser Pro Gly Val Gly Cys
515 520 525
Val Pro Ala Ala Glu His Arg Leu Arg Glu Glu Ile Leu Ala Lys Phe
530 535 540
Leu His Trp Leu Met Ser Val Tyr Val Val Glu Leu Leu Arg Ser Phe
545 550 555 560
Phe Tyr Val Thr Glu Thr Thr Phe Gln Lys Asn Arg Leu Phe Phe Tyr
565 570 575
Arg Lys Ser Val Trp Ser Lys Leu Gln Ser Ile Gly Ile Arg Gln His
580 585 590
Leu Lys Arg Val Gln Leu Arg Glu Leu Ser Glu Ala Glu Val Arg Gln
595 600 605
His Arg Glu Ala Arg Pro Ala Leu Leu Thr Ser Arg Leu Arg Phe Ile
610 615 620
Pro Lys Pro Asp Gly Leu Arg Pro Ile Val Asn Met Asp Tyr Val Val
625 630 635 640
Gly Ala Arg Thr Phe Arg Arg Glu Lys Arg Ala Glu Arg Leu Thr Ser
645 650 655
Arg Val Lys Ala Leu Phe Ser Val Leu Asn Tyr Glu Arg Ala Arg Arg
660 665 670
Pro Gly Leu Leu Gly Ala Ser Val Leu Gly Leu Asp Asp Ile His Arg
675 680 685
Ala Trp Arg Thr Phe Val Leu Arg Val Arg Ala Gln Asp Pro Pro Pro
690 695 700
Glu Leu Tyr Phe Val Lys Val Asp Val Thr Gly Ala Tyr Asp Thr Ile
705 710 715 720
Pro Gln Asp Arg Leu Thr Glu Val Ile Ala Ser Ile Ile Lys Pro Gln
725 730 735
Asn Thr Tyr Cys Val Arg Arg Tyr Ala Val Val Gln Lys Ala Ala His
740 745 750
Gly His Val Arg Lys Ala Phe Lys Ser His Val Ser Thr Leu Thr Asp
755 760 765
Leu Gln Pro Tyr Met Arg Gln Phe Val Ala His Leu Gln Glu Thr Ser
770 775 780
Pro Leu Arg Asp Ala Val Val Ile Glu Gln Ser Ser Ser Leu Asn Glu
785 790 795 800
Ala Ser Ser Gly Leu Phe Asp Val Phe Leu Arg Phe Met Cys His His
805 810 815
Ala Val Arg Ile Arg Gly Lys Ser Tyr Val Gln Cys Gln Gly Ile Pro
820 825 830
Gln Gly Ser Ile Leu Ser Thr Leu Leu Cys Ser Leu Cys Tyr Gly Asp
835 840 845
Met Glu Asn Lys Leu Phe Ala Gly Ile Arg Arg Asp Gly Leu Leu Leu
850 855 860
Arg Leu Val Asp Asp Phe Leu Leu Val Thr Pro His Leu Thr His Ala
865 870 875 880
Lys Thr Phe Leu Arg Thr Leu Val Arg Gly Val Pro Glu Tyr Gly Cys
885 890 895
Val Val Asn Leu Arg Lys Thr Val Val Asn Phe Pro Val Glu Asp Glu
900 905 910
Ala Leu Gly Gly Thr Ala Phe Val Gln Met Pro Ala His Gly Leu Phe
915 920 925
Pro Trp Cys Gly Leu Leu Leu Asp Thr Arg Thr Leu Glu Val Gln Ser
930 935 940
Asp Tyr Ser Ser Tyr Ala Arg Thr Ser Ile Arg Ala Ser Leu Thr Phe
945 950 955 960
Asn Arg Gly Phe Lys Ala Gly Arg Asn Met Arg Arg Lys Leu Phe Gly
965 970 975
Val Leu Arg Leu Lys Cys His Ser Leu Phe Leu Asp Leu Gln Val Asn
980 985 990
Ser Leu Gln Thr Val Cys Thr Asn Ile Tyr Lys Ile Leu Leu Leu Gln
995 1000 1005
Ala Tyr Arg Phe His Ala Cys Val Leu Gln Leu Pro Phe His Gln
1010 1015 1020
Gln Val Trp Lys Asn Pro Thr Phe Phe Leu Arg Val Ile Ser Asp
1025 1030 1035
Thr Ala Ser Leu Cys Tyr Ser Ile Leu Lys Ala Lys Asn Ala Gly
1040 1045 1050
Met Ser Leu Gly Ala Lys Gly Ala Ala Gly Pro Leu Pro Ser Glu
1055 1060 1065
Ala Val Gln Trp Leu Cys His Gln Ala Phe Leu Leu Lys Leu Thr
1070 1075 1080
Arg His Arg Val Thr Tyr Val Pro Leu Leu Gly Ser Leu Arg Thr
1085 1090 1095
Ala Gln Thr Gln Leu Ser Arg Lys Leu Pro Gly Thr Thr Leu Thr
1100 1105 1110
Ala Leu Glu Ala Ala Ala Asn Pro Ala Leu Pro Ser Asp Phe Lys
1115 1120 1125
Thr Ile Leu Asp
1130
<210> 64
<211> 4027
<212> ДНК
<213> Homo sapiens
<400> 64
caggcagcgt ggtcctgctg cgcacgtggg aagccctggc cccggccacc cccgcgatgc 60
cgcgcgctcc ccgctgccga gccgtgcgct ccctgctgcg cagccactac cgcgaggtgc 120
tgccgctggc cacgttcgtg cggcgcctgg ggccccaggg ctggcggctg gtgcagcgcg 180
gggacccggc ggctttccgc gcgctggtgg cccagtgcct ggtgtgcgtg ccctgggacg 240
cacggccgcc ccccgccgcc ccctccttcc gccaggtgtc ctgcctgaag gagctggtgg 300
cccgagtgct gcagaggctg tgcgagcgcg gcgcgaagaa cgtgctggcc ttcggcttcg 360
cgctgctgga cggggcccgc gggggccccc ccgaggcctt caccaccagc gtgcgcagct 420
acctgcccaa cacggtgacc gacgcactgc gggggagcgg ggcgtggggg ctgctgttgc 480
gccgcgtggg cgacgacgtg ctggttcacc tgctggcacg ctgcgcgctc tttgtgctgg 540
tggctcccag ctgcgcctac caggtgtgcg ggccgccgct gtaccagctc ggcgctgcca 600
ctcaggcccg gcccccgcca cacgctagtg gaccccgaag gcgtctggga tgcgaacggg 660
cctggaacca tagcgtcagg gaggccgggg tccccctggg cctgccagcc ccgggtgcga 720
ggaggcgcgg gggcagtgcc agccgaagtc tgccgttgcc caagaggccc aggcgtggcg 780
ctgcccctga gccggagcgg acgcccgttg ggcaggggtc ctgggcccac ccgggcagga 840
cgcgtggacc gagtgaccgt ggtttctgtg tggtgtcacc tgccagaccc gccgaagaag 900
ccacctcttt ggagggtgcg ctctctggca cgcgccactc ccacccatcc gtgggccgcc 960
agcaccacgc gggcccccca tccacatcgc ggccaccacg tccctgggac acgccttgtc 1020
ccccggtgta cgccgagacc aagcacttcc tctactcctc aggcgacaag gagcagctgc 1080
ggccctcctt cctactcagc tctctgaggc ccagcctgac tggcgctcgg aggctcgtgg 1140
agaccatctt tctgggttcc aggccctgga tgccagggac tccccgcagg ttgccccgcc 1200
tgccccagcg ctactggcaa atgcggcccc tgtttctgga gctgcttggg aaccacgcgc 1260
agtgccccta cggggtgctc ctcaagacgc actgcccgct gcgagctgcg gtcaccccag 1320
cagccggtgt ctgtgcccgg gagaagcccc agggctctgt ggcggccccc gaggaggagg 1380
acacagaccc ccgtcgcctg gtgcagctgc tccgccagca cagcagcccc tggcaggtgt 1440
acggcttcgt gcgggcctgc ctgcgccggc tggtgccccc aggcctctgg ggctccaggc 1500
acaacgaacg ccgcttcctc aggaacacca agaagttcat ctccctgggg aagcatgcca 1560
agctctcgct gcaggagctg acgtggaaga tgagcgtgcg gggctgcgct tggctgcgca 1620
ggagcccagg ggttggctgt gttccggccg cagagcaccg tctgcgtgag gagatcctgg 1680
ccaagttcct gcactggctg atgagtgtgt acgtcgtcga gctgctcagg tctttctttt 1740
atgtcacgga gaccacgttt caaaagaaca ggctcttttt ctaccggaag agtgtctgga 1800
gcaagttgca aagcattgga atcagacagc acttgaagag ggtgcagctg cgggagctgt 1860
cggaagcaga ggtcaggcag catcgggaag ccaggcccgc cctgctgacg tccagactcc 1920
gcttcatccc caagcctgac gggctgcggc cgattgtgaa catggactac gtcgtgggag 1980
ccagaacgtt ccgcagagaa aagagggccg agcgtctcac ctcgagggtg aaggcactgt 2040
tcagcgtgct caactacgag cgggcgcggc gccccggcct cctgggcgcc tctgtgctgg 2100
gcctggacga tatccacagg gcctggcgca ccttcgtgct gcgtgtgcgg gcccaggacc 2160
cgccgcctga gctgtacttt gtcaaggtgg atgtgacggg cgcgtacgac accatccccc 2220
aggacaggct cacggaggtc atcgccagca tcatcaaacc ccagaacacg tactgcgtgc 2280
gtcggtatgc cgtggtccag aaggccgccc atgggcacgt ccgcaaggcc ttcaagagcc 2340
acgtctctac cttgacagac ctccagccgt acatgcgaca gttcgtggct cacctgcagg 2400
agaccagccc gctgagggat gccgtcgtca tcgagcagag ctcctccctg aatgaggcca 2460
gcagtggcct cttcgacgtc ttcctacgct tcatgtgcca ccacgccgtg cgcatcaggg 2520
gcaagtccta cgtccagtgc caggggatcc cgcagggctc catcctctcc acgctgctct 2580
gcagcctgtg ctacggcgac atggagaaca agctgtttgc ggggattcgg cgggacgggc 2640
tgctcctgcg tttggtggat gatttcttgt tggtgacacc tcacctcacc cacgcgaaaa 2700
ccttcctcag gaccctggtc cgaggtgtcc ctgagtatgg ctgcgtggtg aacttgcgga 2760
agacagtggt gaacttccct gtagaagacg aggccctggg tggcacggct tttgttcaga 2820
tgccggccca cggcctattc ccctggtgcg gcctgctgct ggatacccgg accctggagg 2880
tgcagagcga ctactccagc tatgcccgga cctccatcag agccagtctc accttcaacc 2940
gcggcttcaa ggctgggagg aacatgcgtc gcaaactctt tggggtcttg cggctgaagt 3000
gtcacagcct gtttctggat ttgcaggtga acagcctcca gacggtgtgc accaacatct 3060
acaagatcct cctgctgcag gcgtacaggt ttcacgcatg tgtgctgcag ctcccatttc 3120
atcagcaagt ttggaagaac cccacatttt tcctgcgcgt catctctgac acggcctccc 3180
tctgctactc catcctgaaa gccaagaacg cagggatgtc gctgggggcc aagggcgccg 3240
ccggccctct gccctccgag gccgtgcagt ggctgtgcca ccaagcattc ctgctcaagc 3300
tgactcgaca ccgtgtcacc tacgtgccac tcctggggtc actcaggaca gcccagacgc 3360
agctgagtcg gaagctcccg gggacgacgc tgactgccct ggaggccgca gccaacccgg 3420
cactgccctc agacttcaag accatcctgg actgatggcc acccgcccac agccaggccg 3480
agagcagaca ccagcagccc tgtcacgccg ggctctacgt cccagggagg gaggggcggc 3540
ccacacccag gcccgcaccg ctgggagtct gaggcctgag tgagtgtttg gccgaggcct 3600
gcatgtccgg ctgaaggctg agtgtccggc tgaggcctga gcgagtgtcc agccaagggc 3660
tgagtgtcca gcacacctgc cgtcttcact tccccacagg ctggcgctcg gctccacccc 3720
agggccagct tttcctcacc aggagcccgg cttccactcc ccacatagga atagtccatc 3780
cccagattcg ccattgttca cccctcgccc tgccctcctt tgccttccac ccccaccatc 3840
caggtggaga ccctgagaag gaccctggga gctctgggaa tttggagtga ccaaaggtgt 3900
gccctgtaca caggcgagga ccctgcacct ggatgggggt ccctgtgggt caaattgggg 3960
ggaggtgctg tgggagtaaa atactgaata tatgagtttt tcagttttga aaaaaaaaaa 4020
aaaaaaa 4027
<210> 65
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 65
tgaaggagcc cagagagaga 20
<210> 66
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 66
gtaagccaag aaagaaatcc 20
<210> 67
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 67
Gly Gly Ser Gly Gly Ser
1 5
<210> 68
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<220>
<221> VARIANT
<222> (1)..(3)
<223> /замена=" "
<220>
<221> дополнительный признак
<222> (1)..(21)
<223> /примечание="Варианты остатков, приведенные в последовательностях, не
имеют предпочтения относительно остатков в аннотациях для положений
вариантов"
<400> 68
Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu
1 5 10 15
Glu Asn Pro Gly Pro
20
<210> 69
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<220>
<221> VARIANT
<222> (1)..(3)
<223> /замена=" "
<220>
<221> дополнительный признак
<222> (1)..(22)
<223> / примечание="Варианты остатков, приведенные в последовательностях, не
имеют предпочтения относительно остатков в аннотациях для положений
вариантов"
<400> 69
Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val
1 5 10 15
Glu Glu Asn Pro Gly Pro
20
<210> 70
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<220>
<221> VARIANT
<222> (1)..(3)
<223> /замена=" "
<220>
<221> дополнительный признак
<222> (1)..(23)
<223> / примечание="Варианты остатков, приведенные в последовательностях, не
имеют предпочтения относительно остатков в аннотациях для положений
вариантов"
<400> 70
Gly Ser Gly Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp
1 5 10 15
Val Glu Ser Asn Pro Gly Pro
20
<210> 71
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<220>
<221> VARIANT
<222> (1)..(3)
<223> /замена=" "
<220>
<221> дополнительный признак
<222> (1)..(25)
<223> / примечание="Варианты остатков, приведенные в последовательностях, не
имеют предпочтения относительно остатков в аннотациях для положений
вариантов"
<400> 71
Gly Ser Gly Val Lys Gln Thr Leu Asn Phe Asp Leu Leu Lys Leu Ala
1 5 10 15
Gly Asp Val Glu Ser Asn Pro Gly Pro
20 25
<210> 72
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (1)..(30)
<223> /примечание="эта последовательность может охватывать 1-6 повторяющихся
элементов "Gly Gly Gly Gly Ser""
<400> 72
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
<210> 73
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 73
ugucgggucu uuaaaaauac 20
<210> 74
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 74
uacaggcccc uaaagcacua 20
<210> 75
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 75
acaggccccu aaagcacuaa 20
<210> 76
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 76
aagcacuaag ggcaugcccu 20
<210> 77
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 77
gggcaugccc ucggugaaac 20
<210> 78
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 78
ggcaugcccu cggugaaaca 20
<210> 79
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 79
gcaugcccuc ggugaaacag 20
<210> 80
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 80
ugagauuaaa gcgacagaaa 20
<210> 81
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 81
gagauuaaag cgacagaaaa 20
<210> 82
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 82
uaaagcgaca gaaaagggaa 20
<210> 83
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 83
agaaaaggga aaggagagcg 20
<210> 84
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 84
gaaaagggaa aggagagcgc 20
<210> 85
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 85
ggaaaggaga gcgcgggcaa 20
<210> 86
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 86
gaaaggagag cgcgggcaac 20
<210> 87
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 87
gcgcgggcaa cgggaucuaa 20
<210> 88
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 88
cgcgggcaac gggaucuaaa 20
<210> 89
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 89
ucuaaaggga gauagagacg 20
<210> 90
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 90
cuaaagggag auagagacgc 20
<210> 91
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 91
gauagagacg cgggccucug 20
<210> 92
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 92
auagagacgc gggccucuga 20
<210> 93
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 93
gacgcgggcc ucugagggua 20
<210> 94
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 94
gcgggccucu gaggguaagg 20
<210> 95
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 95
cgggccucug aggguaaggu 20
<210> 96
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 96
gaggguaagg ugggcgcaag 20
<210> 97
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 97
gcaugcccuu agugcuuuag 20
<210> 98
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 98
ggcaugcccu uagugcuuua 20
<210> 99
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 99
gggcaugccc uuagugcuuu 20
<210> 100
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 100
gcgcuccccu guuucaccga 20
<210> 101
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 101
agcgcucccc uguuucaccg 20
<210> 102
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 102
ugcgcccacc uuacccucag 20
<210> 103
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 103
agagccggcg guagcggcag 20
<210> 104
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 104
ggcgguagcg gcaguggcag 20
<210> 105
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 105
caguggcagc ggcgagagcu 20
<210> 106
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 106
aguggcagcg gcgagagcuu 20
<210> 107
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 107
ggcagcggcg agagcuuggg 20
<210> 108
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 108
ccucgcgagc gccgcgcgcc 20
<210> 109
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 109
cucgcgagcg ccgcgcgccc 20
<210> 110
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 110
gcaagucacg uccgcccccu 20
<210> 111
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 111
ucacguccgc ccccucggcg 20
<210> 112
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 112
gcgcggccgc cccgagacgc 20
<210> 113
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 113
cugccuuaug aauauugaug 20
<210> 114
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 114
ccuuaugaau auugaugcgg 20
<210> 115
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 115
ugaauauuga ugcggaggcu 20
<210> 116
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 116
ugcuuucgua gagaagcaga 20
<210> 117
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 117
agaagcagaa ggaagcaaga 20
<210> 118
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 118
aagcaagaug gcugcccuuu 20
<210> 119
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 119
cccuuuagga uuuguuagaa 20
<210> 120
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 120
ggagacccga cugcaacugc 20
<210> 121
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 121
gcaacugcug gauugcugca 20
<210> 122
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 122
gcuggauugc ugcaaggcug 20
<210> 123
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 123
cuggauugcu gcaaggcuga 20
<210> 124
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 124
aaggcugagg gacgagaacg 20
<210> 125
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 125
gcugccacug ccgcuaccgc 20
<210> 126
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 126
cgcucgcgag gaggcggcgg 20
<210> 127
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 127
cggcgcucgc gaggaggcgg 20
<210> 128
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 128
gcgcggcgcu cgcgaggagg 20
<210> 129
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 129
ggcgcgcggc gcucgcgagg 20
<210> 130
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 130
ccgggcgcgc ggcgcucgcg 20
<210> 131
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 131
gcgagcggga cccgggcgcg 20
<210> 132
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 132
cuugcaugcg agcgggaccc 20
<210> 133
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 133
acuugcaugc gagcgggacc 20
<210> 134
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 134
gacgugacuu gcaugcgagc 20
<210> 135
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 135
ggacgugacu ugcaugcgag 20
<210> 136
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 136
gggcggccgc gccgaggggg 20
<210> 137
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 137
ucggggcggc cgcgccgagg 20
<210> 138
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 138
cucggggcgg ccgcgccgag 20
<210> 139
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 139
ucucggggcg gccgcgccga 20
<210> 140
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 140
gucucggggc ggccgcgccg 20
<210> 141
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 141
gcggggccgg cgucucgggg 20
<210> 142
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 142
ucagcggggc cggcgucucg 20
<210> 143
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 143
cucagcgggg ccggcgucuc 20
<210> 144
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 144
acucagcggg gccggcgucu 20
<210> 145
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 145
uucucaucac ucagcggggc 20
<210> 146
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 146
ucuguucuca ucacucagcg 20
<210> 147
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 147
gucuguucuc aucacucagc 20
<210> 148
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 148
cgucuguucu caucacucag 20
<210> 149
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 149
ccuccgcauc aauauucaua 20
<210> 150
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 150
ccuuucuaac aaauccuaaa 20
<210> 151
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 151
uccuuucuaa caaauccuaa 20
<210> 152
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 152
gcaauccagc aguugcaguc 20
<210> 153
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 153
agcaauccag caguugcagu 20
<210> 154
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 154
aaacucuguc uucucuaggc 20
<210> 155
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 155
uccuguugag uuacaacgcu 20
<210> 156
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 156
gaguuacaac gcuuggaagc 20
<210> 157
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 157
caacgcuugg aagcaggaga 20
<210> 158
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 158
aacgcuugga agcaggagau 20
<210> 159
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 159
ugggcucagc agcagccaau 20
<210> 160
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 160
cagccaauag gacaugaucc 20
<210> 161
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 161
augauccagg aagagcagua 20
<210> 162
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 162
ugauccagga agagcaguaa 20
<210> 163
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 163
gcaguaaggg acugagcugc 20
<210> 164
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 164
cugagcugcu gguaagacag 20
<210> 165
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 165
gcaaguaaac aaucuugaga 20
<210> 166
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 166
ggcaaguaaa caaucuugag 20
<210> 167
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 167
uuguaacuca acaggagcaa 20
<210> 168
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 168
uccaagcguu guaacucaac 20
<210> 169
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 169
cuuccuggau cauguccuau 20
<210> 170
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 170
cagucccuua cugcucuucc 20
<210> 171
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 171
gcucuuuaga auucaacuag 20
<210> 172
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 172
cucuuuagaa uucaacuaga 20
<210> 173
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 173
ucaacuagag ggcagccuug 20
<210> 174
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 174
cuagagggca gccuugugga 20
<210> 175
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 175
uggccccgaa gcaagccuga 20
<210> 176
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 176
cgaagcaagc cugauggaac 20
<210> 177
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 177
ggauagaacc aaccauguug 20
<210> 178
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 178
gauagaacca accauguuga 20
<210> 179
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 179
cauuugccag acagaaccuc 20
<210> 180
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 180
cuggcuacaa agcuccagaa 20
<210> 181
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 181
agagcucauc cagaaguaaa 20
<210> 182
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 182
aaguaaaugg agacaccaag 20
<210> 183
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 183
cacucuuuca aaaguuauua 20
<210> 184
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 184
uuauggaaua cccuguauga 20
<210> 185
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 185
uauggaauac ccuguaugaa 20
<210> 186
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 186
gacuuuacac aagaaaguag 20
<210> 187
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 187
acuuuacaca agaaaguaga 20
<210> 188
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 188
uauuccaagu guuugcaaaa 20
<210> 189
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 189
uccaaguguu ugcaaaaugg 20
<210> 190
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 190
guuagugaac cuucucucuc 20
<210> 191
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 191
uuagugaacc uucucucucu 20
<210> 192
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 192
gaaauugaaa caagaccaaa 20
<210> 193
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 193
aaacaagacc aaaaggcuaa 20
<210> 194
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 194
aauggagaaa gacguaacuu 20
<210> 195
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 195
auggagaaag acguaacuuc 20
<210> 196
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 196
uggagaaaga cguaacuucg 20
<210> 197
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 197
guaagccaag aaagaaaucc 20
<210> 198
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 198
uuuucaacac auaacugcag 20
<210> 199
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 199
uuucaacaca uaacugcagu 20
<210> 200
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 200
gcuucagauu cugaaugagc 20
<210> 201
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 201
ucagauucug aaugagcagg 20
<210> 202
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 202
cagauucuga augagcagga 20
<210> 203
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 203
agauucugaa ugagcaggag 20
<210> 204
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 204
cauuguauua cuuaaaaaca 20
<210> 205
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 205
aaggcagugc uaaugccuaa 20
<210> 206
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 206
uacaguuucu gccucuuccg 20
<210> 207
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 207
ucuuccgugg aacacacaca 20
<210> 208
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 208
acacacacau ggugaacucc 20
<210> 209
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 209
uccagauugu guuuccauug 20
<210> 210
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 210
cauaaaugcc auuaacaguc 20
<210> 211
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 211
acucacccau cgcauaccuc 20
<210> 212
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 212
cucacccauc gcauaccuca 20
<210> 213
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 213
gccuccaaag ccagcugcag 20
<210> 214
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 214
gccagcugca guggugagug 20
<210> 215
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 215
uccugcagaa aauaacaucc 20
<210> 216
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 216
ccugcagaaa auaacaucca 20
<210> 217
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 217
ggaaccacaa agcuagcguc 20
<210> 218
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 218
ucuggugaag aauucuguuc 20
<210> 219
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 219
agcagcaauu ugcaagcucc 20
<210> 220
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 220
agcaauuugc aagcuccugg 20
<210> 221
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 221
cuccuggugg cagcucugaa 20
<210> 222
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 222
uuaaaacaaa augaaaugaa 20
<210> 223
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 223
gcaaagcuca guguucacua 20
<210> 224
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 224
ucccccuccu ccucuuccac 20
<210> 225
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 225
guuccucagc uuccuucaga 20
<210> 226
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 226
gaaggaaaaa gcacucugaa 20
<210> 227
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 227
ggaaaaagca cucugaaugg 20
<210> 228
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 228
aaaguaacac aacacuuuua 20
<210> 229
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 229
aaguaacaca acacuuuuaa 20
<210> 230
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 230
uuuaagggaa gugaaaauag 20
<210> 231
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 231
uuaagggaag ugaaaauaga 20
<210> 232
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 232
gaaaauagag gguaaaccug 20
<210> 233
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 233
cuucuccgau gcuuucugaa 20
<210> 234
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 234
cucagaauaa uugugugaac 20
<210> 235
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 235
aggaaugaca uacagacugc 20
<210> 236
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 236
ggaaugacau acagacugca 20
<210> 237
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 237
aagcauaacc caccaauuuu 20
<210> 238
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 238
ccaccaauuu uugguagcag 20
<210> 239
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 239
ugguagcagu ggagagcuac 20
<210> 240
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 240
caaagagcaa gagauucuga 20
<210> 241
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 241
aaagagcaag agauucugaa 20
<210> 242
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 242
gauucugaag ggucgagaca 20
<210> 243
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 243
acacagcacu aucugaaacc 20
<210> 244
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 244
agcacuaucu gaaaccagga 20
<210> 245
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 245
accaggaugg auugaauuga 20
<210> 246
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 246
ggccccucgu uuucaccaag 20
<210> 247
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 247
aucccaucua aaacguaaug 20
<210> 248
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 248
augaccucca aacaauacac 20
<210> 249
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 249
acuggaaauu ccaacaugcc 20
<210> 250
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 250
cuggaaauuc caacaugccu 20
<210> 251
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 251
uggaaauucc aacaugccug 20
<210> 252
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 252
ggaaauucca acaugccugg 20
<210> 253
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 253
gaaauuccaa caugccuggg 20
<210> 254
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 254
acaugccugg ggggcuccca 20
<210> 255
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 255
cacccagaaa acaacacagc 20
<210> 256
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 256
uaccaaguug aaaugaauca 20
<210> 257
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 257
accaaguuga aaugaaucaa 20
<210> 258
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 258
augaaucaag ggcaguccca 20
<210> 259
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 259
agggcagucc caagguacag 20
<210> 260
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 260
guuccaaaaa cccucacacc 20
<210> 261
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 261
gcucaugugc agucacugug 20
<210> 262
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 262
gaaacagcac uugaaucaac 20
<210> 263
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 263
gcaacauaag ccucauaaac 20
<210> 264
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 264
auuacaaaua aagaauaaag 20
<210> 265
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 265
aacaaugauc agcaaagaga 20
<210> 266
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 266
caaagagaag gaucauucuu 20
<210> 267
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 267
auucuuuggc cagacuaaag 20
<210> 268
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 268
aaaguggaag aauguuuuca 20
<210> 269
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 269
cgagacucau aauguccaaa 20
<210> 270
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 270
gagacucaua auguccaaau 20
<210> 271
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 271
ucauaauguc caaaugggac 20
<210> 272
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 272
uaauguccaa augggacugg 20
<210> 273
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 273
aucaagugca ugcaaaauac 20
<210> 274
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 274
acacauccug aacuuuuugc 20
<210> 275
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 275
caaagcaaga ucuucuucac 20
<210> 276
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 276
ucacaggugc uuucaagaac 20
<210> 277
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 277
acaacaagcu ucaguucuac 20
<210> 278
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 278
caacaagcuu caguucuaca 20
<210> 279
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 279
aauagaaacc aagauauguc 20
<210> 280
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 280
cugcgcaacu ugcucagcaa 20
<210> 281
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 281
uguuuuuccu gugccugacc 20
<210> 282
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 282
guuuuuccug ugccugacca 20
<210> 283
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 283
uuuccugugc cugaccaggg 20
<210> 284
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 284
cacucagacc ccuccccaga 20
<210> 285
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 285
cucaaaagca ugcugcucua 20
<210> 286
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 286
aaaagcaugc ugcucuaagg 20
<210> 287
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 287
cuugccauag ucagaugcac 20
<210> 288
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 288
ucagaugcac aggccaauua 20
<210> 289
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 289
gaugcacagg ccaauuaagg 20
<210> 290
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 290
aggccaauua agguggaacc 20
<210> 291
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 291
caccaccaga aaacaaaaca 20
<210> 292
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 292
agaaaacaaa acauggaaaa 20
<210> 293
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 293
aaagagcauc auugagacca 20
<210> 294
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 294
caagucguua uuugaccaua 20
<210> 295
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 295
caaguaaaag uugaaauguc 20
<210> 296
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 296
aaguaaaagu ugaaauguca 20
<210> 297
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 297
uacuccuaua aaaaauuuau 20
<210> 298
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 298
uucccaucuu gcagaugugu 20
<210> 299
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 299
cagaaaugua cugagacaca 20
<210> 300
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 300
agcaaauuua ucuucagaua 20
<210> 301
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 301
gcaaauuuau cuucagauau 20
<210> 302
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 302
cuuuuuuuaa aucuugaguc 20
<210> 303
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 303
agucuggcag caauuuguaa 20
<210> 304
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 304
gcucuuugua uauuaucucc 20
<210> 305
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 305
uaucuccugg agagacagcu 20
<210> 306
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 306
aaugagaaaa uaacgaccau 20
<210> 307
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 307
uuuaaauauu uuuuaauuca 20
<210> 308
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 308
uauuaguuuc acaagauuuc 20
<210> 309
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 309
ucacaagauu ucuggcuaau 20
<210> 310
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 310
cacaagauuu cuggcuaaua 20
<210> 311
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 311
uaucuucagu cuucaugagu 20
<210> 312
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 312
aucuucaguc uucaugaguu 20
<210> 313
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 313
ucuucagucu ucaugaguug 20
<210> 314
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 314
cuucagucuu caugaguugg 20
<210> 315
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 315
cuuuucucca uuuauacauu 20
<210> 316
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 316
aaagcuuuuu guuaaaauuc 20
<210> 317
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 317
uaaaauucag gauauguaau 20
<210> 318
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 318
aggauaugua auaggucugu 20
<210> 319
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 319
uagugaaaua uuuuugcuga 20
<210> 320
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 320
gauggaugua gauauauacg 20
<210> 321
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 321
agacaaaugu uaaauuagug 20
<210> 322
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 322
gauaccccac acuguguaga 20
<210> 323
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 323
ccccacacug uguagaagga 20
<210> 324
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 324
cacacugugu agaaggaugg 20
<210> 325
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 325
acacugugua gaaggaugga 20
<210> 326
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 326
cuguguagaa ggauggaggg 20
<210> 327
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 327
cuacuguccc ucuuugcgug 20
<210> 328
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 328
ugguuauuaa guugccucac 20
<210> 329
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 329
gguuauuaag uugccucacu 20
<210> 330
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 330
cacaucucau agauaauauu 20
<210> 331
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 331
ucccacuuuu ccaucuuugu 20
<210> 332
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 332
uucuuuuugc cugacucucc 20
<210> 333
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 333
uucuaaagua cauacuaaua 20
<210> 334
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 334
ucuaaaguac auacuaauau 20
<210> 335
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 335
aguacauacu aauauggguc 20
<210> 336
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 336
aaacagcaau uaaauguuau 20
<210> 337
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 337
aacagcaauu aaauguuaua 20
<210> 338
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 338
auuaaauguu auagggaagu 20
<210> 339
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 339
auagggaagu aggaagaaaa 20
<210> 340
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 340
uagggaagua ggaagaaaaa 20
<210> 341
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 341
agggaaguag gaagaaaaag 20
<210> 342
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 342
caauaaacca agcaauauuc 20
<210> 343
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 343
aauaaaccaa gcaauauucu 20
<210> 344
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 344
auaaaccaag caauauucug 20
<210> 345
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 345
uaaaccaagc aauauucugg 20
<210> 346
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 346
accaagcaau auucuggggg 20
<210> 347
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 347
ccaagcaaua uucugggggu 20
<210> 348
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 348
uucugggggu gggauagagc 20
<210> 349
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 349
ucuuuuaaaa uccaaguaau 20
<210> 350
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 350
uuaaaaucca aguaauaggu 20
<210> 351
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 351
uuuuuuccag cucaaaaaau 20
<210> 352
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 352
uuuguuuagu uucauuuauu 20
<210> 353
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 353
uguacauaua cuuaauuaug 20
<210> 354
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 354
uagagcccuu aauguguagu 20
<210> 355
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 355
agagcccuua auguguaguu 20
<210> 356
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 356
gagcccuuaa uguguaguug 20
<210> 357
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 357
agcccuuaau guguaguugg 20
<210> 358
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 358
uaguuggggg uuaagcuuug 20
<210> 359
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 359
cuuuauauuu aguauaauug 20
<210> 360
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 360
caaauuauug aaaaagauga 20
<210> 361
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 361
gguccuuuuu auacccaucu 20
<210> 362
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 362
uuuuauaccc aucuaggagc 20
<210> 363
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 363
ucuaggagca gguccuaaug 20
<210> 364
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 364
ggcagcuauu agagaaauca 20
<210> 365
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 365
uuagagaaau cauggaagaa 20
<210> 366
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 366
aagaaaggua auuaacgcaa 20
<210> 367
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 367
gguaauuaac gcaaaggcac 20
<210> 368
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 368
guaauuaacg caaaggcaca 20
<210> 369
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 369
uaaauugagu aauuauuagu 20
<210> 370
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 370
uuaguaggcu uagcuauucu 20
<210> 371
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 371
uaguaggcuu agcuauucua 20
<210> 372
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 372
agagagucac aauauuugac 20
<210> 373
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 373
aggacuaaua gucugcuagc 20
<210> 374
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 374
aauagucugc uagcuggcac 20
<210> 375
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 375
acaggcugcc cacuuugcga 20
<210> 376
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 376
cgauggaugc cagaaaaccc 20
<210> 377
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 377
cagaaaaccc aggcaugaac 20
<210> 378
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 378
acccaggcau gaacaggaau 20
<210> 379
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 379
ugaacaggaa ucggccagcc 20
<210> 380
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 380
cagccaggcu gccagccaca 20
<210> 381
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 381
ggcugccagc cacaagguac 20
<210> 382
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 382
cagccacaag guacuggcac 20
<210> 383
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 383
cuggcacagg cuccaacgag 20
<210> 384
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 384
uccaacgaga ggucccacuc 20
<210> 385
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 385
aagugucaaa gcagaaagac 20
<210> 386
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 386
gcagaaagac ugguaaagug 20
<210> 387
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 387
uuuuuuucgc uaucaaucac 20
<210> 388
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 388
ugagcgagau aaugcagaga 20
<210> 389
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 389
cucugagcug uucuucuucu 20
<210> 390
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 390
ucugagcugu ucuucuucua 20
<210> 391
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 391
uagggugccu uuucauuaag 20
<210> 392
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 392
gugccuuuuc auuaagaggu 20
<210> 393
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 393
guauuauuau uaaaguacuu 20
<210> 394
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 394
uuaaaguacu uaggauacau 20
<210> 395
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 395
uaaaguacuu aggauacauu 20
<210> 396
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 396
aaaguacuua ggauacauug 20
<210> 397
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 397
uaggauacau uggggcagcu 20
<210> 398
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 398
uucacuaaau aaucaucuag 20
<210> 399
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 399
uaaauaauca ucuaguggcc 20
<210> 400
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 400
uuguuuuuua aacaagcagu 20
<210> 401
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 401
uuuuuuaaac aagcaguagg 20
<210> 402
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 402
acaagcagua gguggugcuu 20
<210> 403
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 403
uagguggugc uuuggucaua 20
<210> 404
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 404
agguggugcu uuggucauaa 20
<210> 405
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 405
ggaagauaua gucuauuucu 20
<210> 406
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 406
acuauuccau auuuuccaug 20
<210> 407
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 407
uuccauauuu uccauguggc 20
<210> 408
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 408
ucuaaauugu gagacauucu 20
<210> 409
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 409
aaauugugag acauucuugg 20
<210> 410
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 410
uaaaauagcu aaauuuagua 20
<210> 411
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 411
aaaauagcua aauuuaguaa 20
<210> 412
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 412
aucuguacau uuugauauug 20
<210> 413
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 413
uuugauauug aggaaaaaca 20
<210> 414
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 414
aaaccauuau ccaguuugcu 20
<210> 415
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 415
uaauaaaccg uucauuucuc 20
<210> 416
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 416
accguucauu ucucaggaug 20
<210> 417
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 417
uaguugaauu cuaaagagca 20
<210> 418
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 418
uugcuucggg gccauccaca 20
<210> 419
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 419
guuccaucag gcuugcuucg 20
<210> 420
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 420
uguuccauca ggcuugcuuc 20
<210> 421
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 421
cuguuccauc aggcuugcuu 20
<210> 422
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 422
ugguucuauc cuguuccauc 20
<210> 423
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 423
ucuguugccc ucaacauggu 20
<210> 424
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 424
uuagucuguu gcccucaaca 20
<210> 425
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 425
ggaggugaug guaucaggaa 20
<210> 426
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 426
aaaugggagg ugaugguauc 20
<210> 427
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 427
gucuggcaaa ugggagguga 20
<210> 428
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 428
gguucugucu ggcaaauggg 20
<210> 429
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 429
agagguucug ucuggcaaau 20
<210> 430
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 430
cagagguucu gucuggcaaa 20
<210> 431
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 431
uuguagccag agguucuguc 20
<210> 432
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 432
uucuggagcu uuguagccag 20
<210> 433
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 433
caggcagugg gcuuccauuc 20
<210> 434
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 434
gaugagcucu cucaggcagu 20
<210> 435
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 435
ggaugagcuc ucucaggcag 20
<210> 436
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 436
acuucuggau gagcucucuc 20
<210> 437
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 437
uuggugucuc cauuuacuuc 20
<210> 438
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 438
acuuuugaaa gagugccacu 20
<210> 439
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 439
uucuggcuuc ccuucauaca 20
<210> 440
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 440
auucuggcuu cccuucauac 20
<210> 441
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 441
caggacucac acgacuauuc 20
<210> 442
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 442
cuacuuucuu guguaaaguc 20
<210> 443
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 443
uccuccauuu ugcaaacacu 20
<210> 444
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 444
ugaaggagcc cagagagaga 20
<210> 445
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 445
guuucaauuu cuugaucuga 20
<210> 446
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 446
gucuuucucc auuagccuuu 20
<210> 447
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 447
uuucaccugg auuucuuucu 20
<210> 448
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 448
uuugguugac ugcuuucacc 20
<210> 449
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 449
ucacucaaau cggagacauu 20
<210> 450
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 450
uucuuucuua ucacucaaau 20
<210> 451
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 451
aucuuuaacu gcauuuucuu 20
<210> 452
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 452
aaucuuuaac ugcauuuucu 20
<210> 453
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 453
gcaguuaugu guugaaaaac 20
<210> 454
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 454
aucugaagcu cuggauuuuc 20
<210> 455
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 455
ucauucagaa ucugaagcuc 20
<210> 456
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 456
guaauacaau guucuuguca 20
<210> 457
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 457
gcagaaacug uagcaccauu 20
<210> 458
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 458
augugugugu uccacggaag 20
<210> 459
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 459
uucaccaugu guguguucca 20
<210> 460
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 460
auugagacag uguuuuuucc 20
<210> 461
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 461
accgcaaugg aaacacaauc 20
<210> 462
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 462
ugugguuuuc ugcaccgcaa 20
<210> 463
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 463
aauggcauuu augugagaug 20
<210> 464
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 464
auuaguagcc ugacuguuaa 20
<210> 465
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 465
cgauggguga gugaucucac 20
<210> 466
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 466
ucugcccuga gguaugcgau 20
<210> 467
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 467
aucugcccug agguaugcga 20
<210> 468
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 468
ugcggaauug aucugcccug 20
<210> 469
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 469
cucagaguua gaggucugug 20
<210> 470
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 470
uggaggcagc ucagaguuag 20
<210> 471
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 471
accacugcag cuggcuuugg 20
<210> 472
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 472
cucaccacug cagcuggcuu 20
<210> 473
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 473
gccucacuca ccacugcagc 20
<210> 474
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 474
aucagcauca ucagcaucac 20
<210> 475
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 475
uagcauugca gcuaguuuac 20
<210> 476
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 476
uucugguuuc ugaaaggaac 20
<210> 477
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 477
uaguuguucu gguuucugaa 20
<210> 478
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 478
uuuuguuguu guaguuguuc 20
<210> 479
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 479
uguuauuuuc ugcaggagau 20
<210> 480
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 480
auguuauuuu cugcaggaga 20
<210> 481
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 481
cccuggaugu uauuuucugc 20
<210> 482
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 482
acgcuagcuu ugugguuccc 20
<210> 483
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 483
uucaccagac gcuagcuuug 20
<210> 484
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 484
aggagcuugc aaauugcugc 20
<210> 485
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 485
uaccguucag agcugccacc 20
<210> 486
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 486
accacaccau cacccagaaa 20
<210> 487
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 487
accuguggaa gaggaggagg 20
<210> 488
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 488
aaccugugga agaggaggag 20
<210> 489
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 489
gaaccugugg aagaggagga 20
<210> 490
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 490
ggaaccugug gaagaggagg 20
<210> 491
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 491
ugaggaaccu guggaagagg 20
<210> 492
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 492
agcugaggaa ccuguggaag 20
<210> 493
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 493
gaaggaagcu gaggaaccug 20
<210> 494
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 494
uuuccuucug aaggaagcug 20
<210> 495
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 495
agagugcuuu uuccuucuga 20
<210> 496
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 496
uacuuugguu gggguagugg 20
<210> 497
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 497
uguuacuuug guugggguag 20
<210> 498
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 498
guguuguguu acuuugguug 20
<210> 499
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 499
aguguugugu uacuuugguu 20
<210> 500
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 500
aaguguugug uuacuuuggu 20
<210> 501
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 501
uuaaaagugu uguguuacuu 20
<210> 502
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 502
cucugggaag guggugccuc 20
<210> 503
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 503
ggauuaggac ucugggaagg 20
<210> 504
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 504
gauggauuag gacucuggga 20
<210> 505
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 505
uguagaugga uuaggacucu 20
<210> 506
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 506
guguagaugg auuaggacuc 20
<210> 507
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 507
cauacaugug uagauggauu 20
<210> 508
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 508
gggcugcaua cauguguaga 20
<210> 509
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 509
uuucagaaag caucggagaa 20
<210> 510
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 510
cuuucagaaa gcaucggaga 20
<210> 511
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 511
ugaggccuuu cagaaagcau 20
<210> 512
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 512
cuguucacac aauuauucug 20
<210> 513
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 513
cuuguuuucu cagaacacaa 20
<210> 514
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 514
ugcuugaggu guucugacau 20
<210> 515
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 515
aaauuggugg guuaugcuug 20
<210> 516
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 516
cacugcuacc aaaaauuggu 20
<210> 517
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 517
ccacugcuac caaaaauugg 20
<210> 518
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 518
ucuccacugc uaccaaaaau 20
<210> 519
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 519
cuuuguuucu caucaacugc 20
<210> 520
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 520
uucagauagu gcuguguugg 20
<210> 521
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 521
uuucagauag ugcuguguug 20
<210> 522
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 522
guuucagaua gugcuguguu 20
<210> 523
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 523
gguuucagau agugcugugu 20
<210> 524
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 524
gccuucaauu caauccaucc 20
<210> 525
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 525
uuccgcuugg ugaaaacgag 20
<210> 526
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 526
auuccgcuug gugaaaacga 20
<210> 527
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 527
gauuccgcuu ggugaaaacg 20
<210> 528
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 528
guuuuagaug ggauuccgcu 20
<210> 529
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 529
ugccucauua cguuuuagau 20
<210> 530
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 530
augccucauu acguuuuaga 20
<210> 531
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 531
gguugauacu gaagaauuga 20
<210> 532
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 532
gucauuugau uggagagauu 20
<210> 533
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 533
ggucauuuga uuggagagau 20
<210> 534
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 534
uuguuuggag gucauuugau 20
<210> 535
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 535
auuuccagug uauuguuugg 20
<210> 536
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 536
ggaauuucca guguauuguu 20
<210> 537
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 537
ugggagcccc ccaggcaugu 20
<210> 538
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 538
gcuugccuug ggagcccccc 20
<210> 539
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 539
ucugggugua agcuugccuu 20
<210> 540
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 540
uucugggugu aagcuugccu 20
<210> 541
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 541
cuccagcugu guuguuuucu 20
<210> 542
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 542
gcuccagcug uguuguuuuc 20
<210> 543
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 543
gcccuugauu cauuucaacu 20
<210> 544
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 544
auguuggucc acuguaccuu 20
<210> 545
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 545
gauguugguc cacuguaccu 20
<210> 546
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 546
guuuuuggaa cuggagaugu 20
<210> 547
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 547
ggugugaggg uuuuuggaac 20
<210> 548
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 548
gcaccuggug ugaggguuuu 20
<210> 549
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 549
gagaagugca ccugguguga 20
<210> 550
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 550
ggagaagugc accuggugug 20
<210> 551
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 551
cuguuuugga gaagugcacc 20
<210> 552
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 552
uuuugguaaa uggucuguuu 20
<210> 553
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 553
gcacaugagc uuuugguaaa 20
<210> 554
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 554
agugacugca caugagcuuu 20
<210> 555
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 555
ggacauaagu uuuucaguuu 20
<210> 556
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 556
gggacauaag uuuuucaguu 20
<210> 557
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 557
caagugcugu uucaacacug 20
<210> 558
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 558
ucaagugcug uuucaacacu 20
<210> 559
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 559
uucaagugcu guuucaacac 20
<210> 560
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 560
aaaaggugug aguuugaaaa 20
<210> 561
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 561
uaugaggcuu auguugcaaa 20
<210> 562
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 562
guuugugcug ccuguuuaug 20
<210> 563
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 563
gggagaugug aacucuggga 20
<210> 564
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 564
uugagggaga ugugaacucu 20
<210> 565
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 565
uuugagggag augugaacuc 20
<210> 566
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 566
gcugcuguug cugguuuuga 20
<210> 567
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 567
ugcugcuguu gcugguuuug 20
<210> 568
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 568
guaauuuuug cugcuguugc 20
<210> 569
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 569
uuugggggug aggaaaaguc 20
<210> 570
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 570
ucauuguugc uuugggggug 20
<210> 571
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 571
gcugaucauu guugcuuugg 20
<210> 572
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 572
ugcugaucau uguugcuuug 20
<210> 573
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 573
uugcugauca uuguugcuuu 20
<210> 574
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 574
uuugcugauc auuguugcuu 20
<210> 575
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 575
aacauucuuc cacuuuaguc 20
<210> 576
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 576
guacuuccuc cagucccauu 20
<210> 577
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 577
uuucaugguc ugacuauaag 20
<210> 578
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 578
auuucauggu cugacuauaa 20
<210> 579
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 579
gauuucaugg ucugacuaua 20
<210> 580
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 580
uuugcaugca cuugauuuca 20
<210> 581
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 581
guucuuuauu cucugaaacu 20
<210> 582
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 582
uuguuuccug caaaaaguuc 20
<210> 583
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 583
uugcauguga ugcaaguuuu 20
<210> 584
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 584
auugcaugug augcaaguuu 20
<210> 585
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 585
ugcuuuggga ucacauuauu 20
<210> 586
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 586
ugugaagaag aucuugcuuu 20
<210> 587
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 587
cugugaagaa gaucuugcuu 20
<210> 588
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 588
cuuguugacc agacauaucu 20
<210> 589
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 589
gcacaggaaa aacauuugca 20
<210> 590
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 590
cuuccucccu ggucaggcac 20
<210> 591
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 591
gugugacuuc cucccugguc 20
<210> 592
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 592
ucugagugug acuuccuccc 20
<210> 593
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 593
uugagugucc uucuggggag 20
<210> 594
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 594
uuugaguguc cuucugggga 20
<210> 595
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 595
uuuugagugu ccuucugggg 20
<210> 596
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 596
ugcuuuugag uguccuucug 20
<210> 597
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 597
augcuuuuga guguccuucu 20
<210> 598
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 598
caugcuuuug aguguccuuc 20
<210> 599
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 599
cuauggcaag acucaguuug 20
<210> 600
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 600
acuauggcaa gacucaguuu 20
<210> 601
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 601
gacuauggca agacucaguu 20
<210> 602
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 602
uuggccugug caucugacua 20
<210> 603
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 603
cauccagguu ccaccuuaau 20
<210> 604
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 604
caggcaugug gcuugcaucc 20
<210> 605
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 605
gcugugugca uacaggcaug 20
<210> 606
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 606
ugguggugcu gugugcauac 20
<210> 607
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 607
uuccauguuu uguuuucugg 20
<210> 608
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 608
uuuuuccaug uuuuguuuuc 20
<210> 609
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 609
acauuaucac agcuugcagg 20
<210> 610
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 610
ugcacauuau cacagcuugc 20
<210> 611
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 611
cugcuucaga ugcugcucca 20
<210> 612
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 612
cuuaugguca aauaacgacu 20
<210> 613
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 613
auuugagagu aagagccuua 20
<210> 614
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 614
ugucuaguca aaacugugac 20
<210> 615
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 615
gcuaucaagu ucugcagcag 20
<210> 616
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 616
gcugcucuaa agcuggggug 20
<210> 617
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 617
uguuugcugc ucuaaagcug 20
<210> 618
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 618
uuguuugcug cucuaaagcu 20
<210> 619
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 619
guuguuugcu gcucuaaagc 20
<210> 620
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 620
gaagcagcug uucuuuuggu 20
<210> 621
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 621
aacagaagca gcuguucuuu 20
<210> 622
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 622
ggaguaucua guaauuugga 20
<210> 623
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 623
uauaggagua ucuaguaauu 20
<210> 624
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 624
guauccaaua aauuuuuuau 20
<210> 625
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 625
aaaucauauu gagucuugac 20
<210> 626
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 626
uaccuacaca ucugcaagau 20
<210> 627
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 627
uuaccuacac aucugcaaga 20
<210> 628
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 628
caugugucuc aguacauuuc 20
<210> 629
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 629
gaagauaaau uugcuaauuc 20
<210> 630
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 630
acucaagauu uaaaaaaaga 20
<210> 631
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 631
cuuucacaag acacaagcau 20
<210> 632
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 632
gcacgauuau uuaauucuuu 20
<210> 633
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 633
uuuuacagga ucugaagaga 20
<210> 634
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 634
auuuuacagg aucugaagag 20
<210> 635
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 635
cagauacauu caaauuuuac 20
<210> 636
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 636
uaauauacaa agagcuaaau 20
<210> 637
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 637
ugcugccuag cugucucucc 20
<210> 638
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 638
uucguacauu agacugccua 20
<210> 639
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 639
aauggagaaa aggaaacuuu 20
<210> 640
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 640
caaauguaua aauggagaaa 20
<210> 641
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 641
caacauucca aauguauaaa 20
<210> 642
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 642
agaugaaauu uuagagaaaa 20
<210> 643
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 643
aagaugaaau uuuagagaaa 20
<210> 644
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 644
agggaaaaca uggcacgggu 20
<210> 645
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 645
caagagggaa aacauggcac 20
<210> 646
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 646
gcaagaggga aaacauggca 20
<210> 647
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 647
ucauugcaag agggaaaaca 20
<210> 648
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 648
ugggguaucu cauugcaaga 20
<210> 649
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 649
gugggguauc ucauugcaag 20
<210> 650
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 650
ccauccuucu acacagugug 20
<210> 651
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 651
uccauccuuc uacacagugu 20
<210> 652
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 652
cuccauccuu cuacacagug 20
<210> 653
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 653
cacacgcaaa gagggacagu 20
<210> 654
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 654
uuaauaacca cacgcaaaga 20
<210> 655
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 655
cuuaauaacc acacgcaaag 20
<210> 656
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 656
ugugguguuu uagcccagug 20
<210> 657
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 657
aauauuaucu augagaugug 20
<210> 658
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 658
aaaaguggga agauaggggu 20
<210> 659
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 659
gaaaaguggg aagauagggg 20
<210> 660
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 660
auggaaaagu gggaagauag 20
<210> 661
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 661
gauggaaaag ugggaagaua 20
<210> 662
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 662
agauggaaaa gugggaagau 20
<210> 663
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 663
accaacaaag auggaaaagu 20
<210> 664
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 664
aaccaacaaa gauggaaaag 20
<210> 665
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 665
cuguugcaaa ccaacaaaga 20
<210> 666
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 666
gagagucagg caaaaagaag 20
<210> 667
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 667
ggagagucag gcaaaaagaa 20
<210> 668
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 668
uggagaguca ggcaaaaaga 20
<210> 669
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 669
agagaaaauc cuggagaguc 20
<210> 670
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 670
uuuaugauga gagaaaaucc 20
<210> 671
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 671
uauugcuugg uuuauuguca 20
<210> 672
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 672
cccaccccca gaauauugcu 20
<210> 673
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 673
cuggaagccu accuauuacu 20
<210> 674
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 674
aaaaaacauu uaaagcuaac 20
<210> 675
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 675
uacaauccaa uuuuuugagc 20
<210> 676
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 676
cuaaacaaag aauacaguga 20
<210> 677
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 677
acuaaacaaa gaauacagug 20
<210> 678
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 678
auauauuaca uuucagauau 20
<210> 679
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 679
aauauauuac auuucagaua 20
<210> 680
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 680
uauauguaca ugcugguugu 20
<210> 681
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 681
aauuaaguau auguacaugc 20
<210> 682
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 682
cuuuaaaaug aguagauuga 20
<210> 683
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 683
aacccccaac uacacauuaa 20
<210> 684
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 684
uaacccccaa cuacacauua 20
<210> 685
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 685
cucaauuaua cuaaauauaa 20
<210> 686
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 686
gcuccuagau ggguauaaaa 20
<210> 687
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 687
auuaggaccu gcuccuagau 20
<210> 688
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 688
cauuaggacc ugcuccuaga 20
<210> 689
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 689
ucucuaauag cugccacauu 20
<210> 690
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 690
aaaauucuga cauauacaaa 20
<210> 691
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 691
acugcuuugu gugugaaggc 20
<210> 692
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 692
guuuacugcu uuguguguga 20
<210> 693
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 693
aauagcacag uguguagugu 20
<210> 694
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 694
ugucaaauau ugugacucuc 20
<210> 695
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 695
ucuggcaucc aucgcaaagu 20
<210> 696
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 696
uucuggcauc caucgcaaag 20
<210> 697
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 697
cuguucaugc cuggguuuuc 20
<210> 698
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 698
gccgauuccu guucaugccu 20
<210> 699
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 699
ggccgauucc uguucaugcc 20
<210> 700
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 700
cuuguggcug gcagccuggc 20
<210> 701
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 701
guaccuugug gcuggcagcc 20
<210> 702
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 702
cugugccagu accuuguggc 20
<210> 703
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 703
gagccugugc caguaccuug 20
<210> 704
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 704
gccagagugg gaccucucgu 20
<210> 705
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 705
ucagguggga aagccagagu 20
<210> 706
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 706
aucagguggg aaagccagag 20
<210> 707
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 707
uugacacuuu auuaucaggu 20
<210> 708
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 708
uuugacacuu uauuaucagg 20
<210> 709
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 709
ugcuuugaca cuuuauuauc 20
<210> 710
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 710
acuaggugaa uuuaauucag 20
<210> 711
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 711
aaguacucau uugcaacacu 20
<210> 712
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 712
ucacacuugc ucucuuuuua 20
<210> 713
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 713
auagcgaaaa aaaaaaaaaa 20
<210> 714
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 714
ucuucuacau gcaggaguaa 20
<210> 715
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 715
cauaagaguc uucuacaugc 20
<210> 716
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 716
gcuguauaaa uuuauaugaa 20
<210> 717
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 717
cugccuaccu cuuaaugaaa 20
<210> 718
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 718
agaaaugaau aauuuggaaa 20
<210> 719
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 719
uaauuuagaa augaauaauu 20
<210> 720
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 720
ggaaauucac uauuucugcc 20
<210> 721
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 721
guuguuuuuu uuggcacuua 20
<210> 722
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 722
uguuguuuuu uuuggcacuu 20
<210> 723
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 723
uguuuuuuug uuguuuuuuu 20
<210> 724
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 724
auccagccac auggaaaaua 20
<210> 725
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 725
auaguuagua uccagccaca 20
<210> 726
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 726
cacaauuuag aaaaggaggc 20
<210> 727
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 727
gucucacaau uuagaaaagg 20
<210> 728
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 728
aaugucucac aauuuagaaa 20
<210> 729
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 729
uuuagcuauu uuaaaacuug 20
<210> 730
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 730
auuuagcuau uuuaaaacuu 20
<210> 731
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 731
aauuuagcua uuuuaaaacu 20
<210> 732
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 732
uuucacaaag cacaaaauuc 20
<210> 733
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 733
aauuacaugu gggugaaaau 20
<210> 734
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 734
aaauuacaug ugggugaaaa 20
<210> 735
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 735
cuauuuugua aauuacaugu 20
<210> 736
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 736
acuauuuugu aaauuacaug 20
<210> 737
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 737
acgccaagca aacuggauaa 20
<210> 738
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 738
caggucuacg ccaagcaaac 20
<210> 739
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 739
cgguuuauua uuuuuuaaac 20
<210> 740
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 740
accacauccu gagaaaugaa 20
<210> 741
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 741
cuguggguuu cuuuaagguu 20
<210> 742
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 742
uucuuuaagg uuuggacaga 20
<210> 743
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 743
ucuuuaaggu uuggacagaa 20
<210> 744
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 744
gacagaaggg uaaagcuauu 20
<210> 745
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 745
auugaaagag ucaucuauac 20
<210> 746
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 746
gucaucuaua cugguaaaga 20
<210> 747
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 747
uaaagaaggc aaaaguucuc 20
<210> 748
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 748
aaagaaggca aaaguucuca 20
<210> 749
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 749
agggaugucc uauugcuaag 20
<210> 750
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 750
gggauguccu auugcuaagu 20
<210> 751
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 751
acacuuaccc acuuagcaau 20
<210> 752
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 752
ggaaugguga uccacgcagg 20
<210> 753
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 753
ugaagagaag cuacuguguu 20
<210> 754
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 754
agaagcuacu guguuuggug 20
<210> 755
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 755
gaagcuacug uguuuggugc 20
<210> 756
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 756
uguuuggugc gggagcgagc 20
<210> 757
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 757
gcgagcuggc cacaccugug 20
<210> 758
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 758
agugauugug auucucaucc 20
<210> 759
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 759
uugugauucu cauccuggug 20
<210> 760
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 760
ugugauucuc auccuggugu 20
<210> 761
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 761
auucucaucc ugguguggga 20
<210> 762
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 762
ggaaggaauc ccgcugucuc 20
<210> 763
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 763
ucuggcugac aaacucuacu 20
<210> 764
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 764
cggagcuuac cgagacgcug 20
<210> 765
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 765
accgagacgc ugaggaaaua 20
<210> 766
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 766
acggcacgcu caccaaucgc 20
<210> 767
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 767
augaagagua agugaagccc 20
<210> 768
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 768
ugaagaguaa gugaagccca 20
<210> 769
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 769
gcuucugcga accaccugcg 20
<210> 770
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 770
ucacugcagc cucacaggug 20
<210> 771
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 771
cacaaucacu gcagccucac 20
<210> 772
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 772
gcgggauucc uucccacacc 20
<210> 773
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 773
guuugucagc cagagacagc 20
<210> 774
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 774
aguuugucag ccagagacag 20
<210> 775
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 775
gccguauuuc cucagcgucu 20
<210> 776
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 776
auucaaggca caccggcgau 20
<210> 777
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 777
acucuucauu caaggcacac 20
<210> 778
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 778
cuucacuuac ucuucauuca 20
<210> 779
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 779
caggagaacu ugcgccuguc 20
<210> 780
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 780
aggagaacuu gcgccuguca 20
<210> 781
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 781
ggagaacuug cgccugucag 20
<210> 782
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 782
aacuugcgcc ugucaggggc 20
<210> 783
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 783
gggcuggauc cagaaaccug 20
<210> 784
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 784
uguggugccu ccuucucuuu 20
<210> 785
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 785
ccuucucuuu ugguuguuca 20
<210> 786
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 786
ucauggagca uguacuacaa 20
<210> 787
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 787
uugccagaag caagauccca 20
<210> 788
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 788
ccaaggaagu uuaagcugcu 20
<210> 789
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 789
caaggaaguu uaagcugcuu 20
<210> 790
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 790
aaggaaguuu aagcugcuug 20
<210> 791
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 791
gcuuggggau gacccaaaag 20
<210> 792
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 792
uucuggaucc agccccugac 20
<210> 793
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 793
aaggaggcac cacagguuuc 20
<210> 794
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 794
aaaagagaag gaggcaccac 20
<210> 795
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 795
ugaacaacca aaagagaagg 20
<210> 796
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 796
ccaugaacaa ccaaaagaga 20
<210> 797
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 797
cuuccuuggg aucuugcuuc 20
<210> 798
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 798
caagcagcuu aaacuuccuu 20
<210> 799
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 799
ccaagcagcu uaaacuuccu 20
<210> 800
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 800
gaaguaaaca aaccucuuuu 20
<210> 801
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 801
ggaaguaaac aaaccucuuu 20
<210> 802
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 802
cuuuauacag gaagagaaac 20
<210> 803
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 803
gcaaaaccug uccacucuua 20
<210> 804
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 804
accugaugca uauaauaauc 20
<210> 805
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 805
uuggugccau aagaguggac 20
<210> 806
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 806
auauguuggu gccauaagag 20
<210> 807
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 807
ggugcaaguu ucuuauaugu 20
<210> 808
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 808
accugauuau uauaugcauc 20
<210> 809
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 809
cagagcacca gagugccguc 20
<210> 810
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 810
agagcaccag agugccgucu 20
<210> 811
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 811
agagugccgu cugggucuga 20
<210> 812
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 812
ugccgucugg gucugaagga 20
<210> 813
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 813
aaggaaggcc guccauucuc 20
<210> 814
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 814
aggaaggccg uccauucuca 20
<210> 815
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 815
ggaaggccgu ccauucucag 20
<210> 816
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 816
cucagggguc acugcauguu 20
<210> 817
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 817
gacuugcaca acaugcagaa 20
<210> 818
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 818
caugcagaau ggcagcacau 20
<210> 819
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 819
auggcagcac auugguaagu 20
<210> 820
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 820
uggcagcaca uugguaaguu 20
<210> 821
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 821
cacauuggua aguugggcug 20
<210> 822
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 822
uucagaccca gacggcacuc 20
<210> 823
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 823
ggccuuccuu cagacccaga 20
<210> 824
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 824
cagugacccc ugagaaugga 20
<210> 825
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 825
caugcaguga ccccugagaa 20
<210> 826
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 826
cauguugugc aagucucugu 20
<210> 827
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 827
gcauguugug caagucucug 20
<210> 828
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 828
agagaagaca aucgagaauu 20
<210> 829
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 829
gaagacaauc gagaauuugg 20
<210> 830
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 830
agaauuugga ggaaaaccug 20
<210> 831
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 831
uuuauacaaa gucucugacg 20
<210> 832
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 832
gucucugacg uggaugaguu 20
<210> 833
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 833
ucucugacgu ggaugaguuu 20
<210> 834
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 834
cguggaugag uuugggagug 20
<210> 835
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 835
guuugggagu guggaagcuc 20
<210> 836
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 836
ugggagugug gaagcucagg 20
<210> 837
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 837
aagcucagga ggagaaaaaa 20
<210> 838
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 838
caggaggaga aaaaacggag 20
<210> 839
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
<400> 839
aaaacggagu ggugccauuc 20
<210> 840
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 840
uucagguacu gaguucuuuu 20
<210> 841
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 841
guucuuuucg gcgaaaaguc 20
<210> 842
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 842
cagucaagac uugccgacaa 20
<210> 843
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 843
agcugaaaag cuuuccuccc 20
<210> 844
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 844
cagcucaaau aaaaaugaaa 20
<210> 845
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 845
acaaacugaa aacgcaagcc 20
<210> 846
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 846
cgcaagccag gcuaaacagu 20
<210> 847
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 847
agccaggcua aacaguuggc 20
<210> 848
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 848
gugagagugc auaccuggua 20
<210> 849
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 849
agugagagug cauaccuggu 20
<210> 850
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 850
cucuagugag agugcauacc 20
<210> 851
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 851
acgugaagcu gcucauccuc 20
<210> 852
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 852
acgucagaga cuuuguauaa 20
<210> 853
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 853
aaaagaacuc aguaccugaa 20
<210> 854
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 854
cuuugucggc aagucuugac 20
<210> 855
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 855
uggcuucuag uuuccuuugu 20
<210> 856
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 856
cuuuucagcu gcagcuuucu 20
<210> 857
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 857
auuugagcug uucuccaggg 20
<210> 858
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 858
uuuauuugag cuguucucca 20
<210> 859
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 859
uuuuauuuga gcuguucucc 20
<210> 860
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 860
aguuuguuuu guacgugaug 20
<210> 861
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 861
caguuuguuu uguacgugau 20
<210> 862
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 862
ucaguuuguu uuguacguga 20
<210> 863
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 863
uaccugccaa cuguuuagcc 20
<210> 864
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 864
gucaacucuu auucugcuuc 20
<210> 865
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 865
ccaccaaucc auacaugaga 20
<210> 866
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 866
ucacacacuu cagauaucua 20
<210> 867
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 867
uccaccucau cucaagcugc 20
<210> 868
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 868
aaucccauga acccuuaccc 20
<210> 869
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 869
aucccaugaa cccuuacccu 20
<210> 870
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 870
uauccaucau aucaaugcaa 20
<210> 871
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
\олигонуклеотид"
<400> 871
augcaaugga aaccuaucag 20
<210> 872
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 872
ggacaacugc uccccauauc 20
<210> 873
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 873
gacaacugcu ccccauaucu 20
<210> 874
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 874
uucuccccag ucucagccga 20
<210> 875
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 875
cucagccgau ggaucuguau 20
<210> 876
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 876
uccauacacu uuaccagcca 20
<210> 877
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 877
acacuuuacc agccaagguu 20
<210> 878
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 878
aguuuuacau cuaaauacuu 20
<210> 879
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 879
acaucuaaau acuuagguua 20
<210> 880
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 880
uuauggaaac caaaauaugc 20
<210> 881
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 881
uauggaaacc aaaauaugca 20
<210> 882
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 882
aaccaaaaua ugcagggaga 20
<210> 883
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 883
agaccaaaug uacaucaugu 20
<210> 884
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 884
gaccaaaugu acaucaugua 20
<210> 885
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 885
uccuuauccc acucaugaga 20
<210> 886
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 886
uaucccacuc augagaugga 20
<210> 887
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 887
ugagauggau ggccacuuca 20
<210> 888
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 888
gagauggaug gccacuucau 20
<210> 889
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 889
caaucugagc aauccaaaca 20
<210> 890
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 890
ccaaacaugg acuauaaaaa 20
<210> 891
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 891
ccauaacuac agugcagcuc 20
<210> 892
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 892
cauaacuaca gugcagcucc 20
<210> 893
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 893
ugcccugcau cuccaaaaca 20
<210> 894
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 894
augcuuuccc acacagcuaa 20
<210> 895
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 895
ugcuuuccca cacagcuaau 20
<210> 896
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 896
gauagaacug cuugugucca 20
<210> 897
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 897
agaacugcuu guguccaagg 20
<210> 898
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 898
cacaaauuaa gugaugcuaa 20
<210> 899
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 899
auuaagugau gcuaaugguc 20
<210> 900
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 900
uggucaggaa aagcagccau 20
<210> 901
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 901
gcagccauug gcacuagucc 20
<210> 902
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 902
cagccauugg cacuagucca 20
<210> 903
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 903
auuggcacua guccagggug 20
<210> 904
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 904
cuaguccagg guguggcuuc 20
<210> 905
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 905
ggguguggcu ucuggugcag 20
<210> 906
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 906
uggugcagag gacaacgaug 20
<210> 907
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 907
cagaggacaa cgaugagguc 20
<210> 908
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 908
agacagcgag cagagcuuuc 20
<210> 909
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 909
agcuuucugg auccugacau 20
<210> 910
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 910
gcuuucugga uccugacauu 20
<210> 911
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 911
cuuucuggau ccugacauug 20
<210> 912
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 912
uuucuggauc cugacauugg 20
<210> 913
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 913
ggauccugac auugggggag 20
<210> 914
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 914
ugacauuggg ggaguggccg 20
<210> 915
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 915
guggccgugg cuccaacuca 20
<210> 916
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 916
uggccguggc uccaacucau 20
<210> 917
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 917
ccccuuuaaa gaaucccaau 20
<210> 918
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 918
ccaauaggaa ucaccccacc 20
<210> 919
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 919
agcaugaaug agccaaaaca 20
<210> 920
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 920
gaaugagcca aaacauggcu 20
<210> 921
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 921
caaaacaugg cuuggcucuu 20
<210> 922
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 922
aaaacauggc uuggcucuuu 20
<210> 923
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 923
ggcucuuugg gaagccaaaa 20
<210> 924
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 924
ugaaaaagcc cgugagaaag 20
<210> 925
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 925
gaggaagagu gugaaaagua 20
<210> 926
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 926
uaugugccuc agaaauccca 20
<210> 927
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 927
cccauggcaa aaaagugaaa 20
<210> 928
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 928
ccauggcaaa aaagugaaac 20
<210> 929
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 929
ucaucaaguc ucuugccgaa 20
<210> 930
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 930
caucuccaua ugccuucacu 20
<210> 931
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 931
aucuccauau gccuucacuc 20
<210> 932
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 932
uaugccuuca cucgggucac 20
<210> 933
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 933
augccuucac ucgggucaca 20
<210> 934
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 934
augauaucac ccccuuuugu 20
<210> 935
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 935
guaguauagu ucucaugacg 20
<210> 936
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 936
uaguauaguu cucaugacgu 20
<210> 937
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 937
aguucucaug acgugggcag 20
<210> 938
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 938
guucucauga cgugggcagu 20
<210> 939
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 939
uucucaugac gugggcagug 20
<210> 940
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 940
augacguggg caguggggaa 20
<210> 941
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 941
cacaguauuc augacaaaug 20
<210> 942
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 942
aguauucaug acaaaugugg 20
<210> 943
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 943
guauucauga caaauguggu 20
<210> 944
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 944
cagcucacca gcaacaaaag 20
<210> 945
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 945
ccauagcacu uaauuuucac 20
<210> 946
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 946
aauuuucacu ggcucccaag 20
<210> 947
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 947
ggcucccaag uggucacaga 20
<210> 948
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 948
aguggucaca gauggcaucu 20
<210> 949
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 949
aagcauucua ugcaaaaaga 20
<210> 950
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 950
cauucuaugc aaaaagaagg 20
<210> 951
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 951
auucuaugca aaaagaaggu 20
<210> 952
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 952
uucuaugcaa aaagaaggug 20
<210> 953
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 953
caauuuacau uuuuaaacac 20
<210> 954
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 954
uuaaacacug guucuauuau 20
<210> 955
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 955
auaucaaguu ugcauaguca 20
<210> 956
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 956
uacuguagua uuacagugac 20
<210> 957
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 957
aggaaucuua aaauaccauc 20
<210> 958
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 958
uauaugaugu acugaaauac 20
<210> 959
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 959
guacugaaau acuggaauua 20
<210> 960
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 960
uuauuuauca aaauagcuac 20
<210> 961
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 961
cuacaggaaa caugaauagc 20
<210> 962
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 962
aggaaaacac ugaauuuguu 20
<210> 963
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 963
uguuuggaug uucuaagaaa 20
<210> 964
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 964
aagaaauggu gcuaagaaaa 20
<210> 965
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 965
cuccagugcc cuugaauaau 20
<210> 966
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 966
uccagugccc uugaauaaua 20
<210> 967
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 967
ccagugcccu ugaauaauag 20
<210> 968
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 968
caagcuuagu uuuuaaaaug 20
<210> 969
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 969
uuaaaaugug gacauuuuaa 20
<210> 970
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 970
guggacauuu uaaaggccuc 20
<210> 971
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 971
ucauccagug aaguccuugu 20
<210> 972
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 972
ugacaacuug aacaaugcua 20
<210> 973
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 973
augcaaaguu gauuuuuuua 20
<210> 974
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 974
acagccaguu aaauccacca 20
<210> 975
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 975
cagccaguua aauccaccau 20
<210> 976
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 976
agccaguuaa auccaccaug 20
<210> 977
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 977
aaauccacca uggggcuuac 20
<210> 978
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 978
cauggggcuu acuggauuca 20
<210> 979
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 979
auggggcuua cuggauucaa 20
<210> 980
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 980
aguccacaaa acauguuuuc 20
<210> 981
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 981
aagaauuuuc uauuaacugc 20
<210> 982
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 982
cugaagccua ugcuauuuua 20
<210> 983
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 983
uaugcuauuu uauggaucau 20
<210> 984
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 984
aggcucuuca gagaacugaa 20
<210> 985
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 985
uaaguguccu cuuuaacaag 20
<210> 986
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 986
ccugcauaag augaauaaac 20
<210> 987
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 987
cugcauaaga ugaauaaaca 20
<210> 988
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 988
aguuaaaaag aaacaaaaac 20
<210> 989
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 989
aagaaacaaa aacaggcagc 20
<210> 990
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 990
aacaggcagc ugguuugcug 20
<210> 991
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 991
aggcagcugg uuugcugugg 20
<210> 992
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 992
aagcagaauu cacaucauga 20
<210> 993
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 993
cauauaccuc aacacuaguu 20
<210> 994
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 994
cucaacacua guuuggcaau 20
<210> 995
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 995
ccuuuuuguu cuaaaaauuc 20
<210> 996
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 996
cuuuuuguuc uaaaaauuca 20
<210> 997
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 997
uguuuaugua aaauuguugu 20
<210> 998
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 998
uaauaaauau auucuuuguc 20
<210> 999
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 999
aauaaauaua uucuuuguca 20
<210> 1000
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1000
aacuaauuuu guaaaucugu 20
<210> 1001
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1001
aaaagcauuu uaaaaguuug 20
<210> 1002
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1002
aucuuuugac uguuucaagc 20
<210> 1003
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1003
agaaugcacu gaguugauaa 20
<210> 1004
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1004
gaaugcacug aguugauaaa 20
<210> 1005
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1005
ugauaaaggg aaaaauugua 20
<210> 1006
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1006
aaagggaaaa auuguaaggc 20
<210> 1007
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1007
aaaauuguaa ggcaggaguu 20
<210> 1008
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1008
uaaggcagga guuuggcaag 20
<210> 1009
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1009
ggaguuuggc aaguggcugu 20
<210> 1010
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1010
uuugauccug uaaucacuga 20
<210> 1011
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1011
cugaagguac auacuccaug 20
<210> 1012
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1012
cauguggacu ucccuuaaac 20
<210> 1013
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1013
uuaaacaggc aaacaccuac 20
<210> 1014
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1014
caggcaaaca ccuacaggua 20
<210> 1015
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1015
cagauuguac aauuacauuu 20
<210> 1016
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1016
uaaaauaaau ucuuaaucag 20
<210> 1017
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1017
aauaaauucu uaaucagagg 20
<210> 1018
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1018
ucuuaaucag aggaggccuu 20
<210> 1019
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1019
cuuaaucaga ggaggccuuu 20
<210> 1020
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1020
aggaggccuu uggguuuuau 20
<210> 1021
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1021
uuggucaaau cuuuguaagc 20
<210> 1022
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1022
uaaaaaauuu cuugaauuug 20
<210> 1023
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1023
uuugauuacu acaugugcau 20
<210> 1024
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1024
acugucauuu guuaaacugc 20
<210> 1025
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1025
aacugcuggc caacaagaac 20
<210> 1026
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1026
caagaacagg aaguauaguu 20
<210> 1027
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1027
aagaacagga aguauaguuu 20
<210> 1028
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1028
agaacaggaa guauaguuug 20
<210> 1029
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1029
gaacaggaag uauaguuugg 20
<210> 1030
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1030
aacaggaagu auaguuuggg 20
<210> 1031
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1031
ggaaguauag uuuggggggu 20
<210> 1032
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1032
gaaguauagu uugggggguu 20
<210> 1033
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1033
aaguauaguu ugggggguug 20
<210> 1034
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1034
gguuggggag aguuuacaua 20
<210> 1035
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1035
aaggaagaga agaaauugag 20
<210> 1036
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1036
ccugccucag uuagaaugaa 20
<210> 1037
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1037
gaucuacaau uugcuaauau 20
<210> 1038
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1038
uuugcuaaua uaggaauauc 20
<210> 1039
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1039
uacuugaaaa ugcuucugag 20
<210> 1040
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1040
caguucacuu cugaagcuag 20
<210> 1041
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1041
agcuaguggu uaacuugugu 20
<210> 1042
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1042
uuucauuuuc augagauguu 20
<210> 1043
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1043
uguuugguuu auaagaucug 20
<210> 1044
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1044
ugguuuauaa gaucugagga 20
<210> 1045
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1045
uauuguaaug uuaugaaugc 20
<210> 1046
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1046
ucgcaaaagu ucuguggaca 20
<210> 1047
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1047
gucgcaaaag uucuguggac 20
<210> 1048
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1048
acaaagucgc aaaaguucug 20
<210> 1049
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1049
aguugacaga cucugucuga 20
<210> 1050
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1050
gaguugacag acucugucug 20
<210> 1051
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1051
ccgucucaug uauggauugg 20
<210> 1052
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1052
gggccgucuc auguauggau 20
<210> 1053
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1053
ggauugggcc gucucaugua 20
<210> 1054
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1054
ggauaaggac uaacuggauu 20
<210> 1055
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1055
uggauaagga cuaacuggau 20
<210> 1056
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1056
gaguuuggau aaggacuaac 20
<210> 1057
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1057
gugugugaag aguuuggaua 20
<210> 1058
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1058
ucugaagugu gugaagaguu 20
<210> 1059
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1059
ggaauagaag uucauagggc 20
<210> 1060
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1060
agguggaaua gaaguucaua 20
<210> 1061
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1061
gagguggaau agaaguucau 20
<210> 1062
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1062
accugcagcu ugagaugagg 20
<210> 1063
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1063
ugaaccugca gcuugagaug 20
<210> 1064
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1064
agcccagggu aaggguucau 20
<210> 1065
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1065
aagcccaggg uaaggguuca 20
<210> 1066
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1066
gauucaaaag cccaggguaa 20
<210> 1067
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1067
ugauucaaaa gcccagggua 20
<210> 1068
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1068
uauucugauu caaaagccca 20
<210> 1069
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1069
guauucugau ucaaaagccc 20
<210> 1070
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1070
gcauugauau gauggauauu 20
<210> 1071
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1071
ugcauugaua ugauggauau 20
<210> 1072
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1072
uuuccauugc auugauauga 20
<210> 1073
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1073
gggagcaguu guccacugau 20
<210> 1074
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1074
agaauaggaa cccagauaug 20
<210> 1075
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1075
gagaauagga acccagauau 20
<210> 1076
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1076
ggagaauagg aacccagaua 20
<210> 1077
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1077
cggcugagac uggggagaau 20
<210> 1078
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1078
agauccaucg gcugagacug 20
<210> 1079
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1079
cagauccauc ggcugagacu 20
<210> 1080
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1080
acagauccau cggcugagac 20
<210> 1081
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1081
ggauaccuau acagauccau 20
<210> 1082
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1082
uuagacagag ggucuuggcu 20
<210> 1083
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1083
ugagcuuaga cagagggucu 20
<210> 1084
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1084
guagacugag cuuagacaga 20
<210> 1085
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1085
gguagacuga gcuuagacag 20
<210> 1086
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1086
ugguaaagug uauggauggg 20
<210> 1087
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1087
ggcugguaaa guguauggau 20
<210> 1088
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1088
uggcugguaa aguguaugga 20
<210> 1089
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1089
accuuggcug guaaagugua 20
<210> 1090
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1090
ggcuauuucc aaaccuuggc 20
<210> 1091
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1091
cucuggcuau uuccaaaccu 20
<210> 1092
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1092
aguauuuaga uguaaaacuc 20
<210> 1093
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1093
aaccaucucc cugcauauuu 20
<210> 1094
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1094
augauguaca uuuggucuaa 20
<210> 1095
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1095
uucccuacau gauguacauu 20
<210> 1096
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1096
aucucaugag ugggauaagg 20
<210> 1097
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1097
uccaucucau gagugggaua 20
<210> 1098
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1098
uggccaucca ucucaugagu 20
<210> 1099
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1099
guggccaucc aucucaugag 20
<210> 1100
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1100
uagagguggc ucccaugaag 20
<210> 1101
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1101
auuggguggu aaucuagagg 20
<210> 1102
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1102
cagauugggu gguaaucuag 20
<210> 1103
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1103
uuuggauugc ucagauuggg 20
<210> 1104
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1104
auguuuggau ugcucagauu 20
<210> 1105
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1105
cauguuugga uugcucagau 20
<210> 1106
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1106
ccauuuuuau aguccauguu 20
<210> 1107
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1107
uaguuaugga uuaugugaga 20
<210> 1108
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1108
ccggagcugc acuguaguua 20
<210> 1109
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1109
agagagcugu ugaacaugcc 20
<210> 1110
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1110
cuccuuguuu uggagaugca 20
<210> 1111
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1111
ucuccuuguu uuggagaugc 20
<210> 1112
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1112
gcaugucauu cuccuuguuu 20
<210> 1113
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1113
ugauaaccca uuagcugugu 20
<210> 1114
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1114
uugauaaccc auuagcugug 20
<210> 1115
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1115
guucuaucau gguuaagagc 20
<210> 1116
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1116
ggacacaagc aguucuauca 20
<210> 1117
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1117
uuaauuugug uaagccuccu 20
<210> 1118
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1118
acacccugga cuagugccaa 20
<210> 1119
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1119
cugcaccaga agccacaccc 20
<210> 1120
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1120
acggccacuc ccccaauguc 20
<210> 1121
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1121
ugacccauga guuggagcca 20
<210> 1122
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1122
augagaauug acccaugagu 20
<210> 1123
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1123
gggauucuuu aaagggguug 20
<210> 1124
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1124
ccuauuggga uucuuuaaag 20
<210> 1125
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1125
uccuauuggg auucuuuaaa 20
<210> 1126
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1126
uuccuauugg gauucuuuaa 20
<210> 1127
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1127
cugguggggu gauuccuauu 20
<210> 1128
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1128
ccuggugggg ugauuccuau 20
<210> 1129
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1129
agacgaggga gauccuggug 20
<210> 1130
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1130
aagacgaggg agauccuggu 20
<210> 1131
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1131
aaagacgagg gagauccugg 20
<210> 1132
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1132
guaaaagacg agggagaucc 20
<210> 1133
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1133
cuuaugcugg uaaaagacga 20
<210> 1134
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1134
ucuuaugcug guaaaagacg 20
<210> 1135
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1135
gcucauucau gcucuuaugc 20
<210> 1136
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1136
caaagagcca agccauguuu 20
<210> 1137
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1137
acgggcuuuu ucagccauuu 20
<210> 1138
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1138
acacucuucc ucuuucucac 20
<210> 1139
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1139
cacacucuuc cucuuucuca 20
<210> 1140
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1140
auuucugagg cacauagucu 20
<210> 1141
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1141
gauuucugag gcacauaguc 20
<210> 1142
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1142
uuuuugccau gggauuucug 20
<210> 1143
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1143
ccguuucacu uuuuugccau 20
<210> 1144
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1144
cccguuucac uuuuuugcca 20
<210> 1145
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1145
gaaguuucau guggcucagc 20
<210> 1146
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1146
gugggcucug aaguuucaug 20
<210> 1147
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1147
uugaugaaac gcagguaagu 20
<210> 1148
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1148
cuugaugaaa cgcagguaag 20
<210> 1149
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1149
caagagacuu gaugaaacgc 20
<210> 1150
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1150
ggucacggac augguccuuu 20
<210> 1151
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1151
ggagucugug gucacggaca 20
<210> 1152
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1152
uacuguggag ucugugguca 20
<210> 1153
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1153
uguaguuacu guggagucug 20
<210> 1154
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1154
auauggagau guaguuacug 20
<210> 1155
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1155
gugacccgag ugaaggcaua 20
<210> 1156
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1156
aggcccugug acccgaguga 20
<210> 1157
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1157
uaucauauau aucuguugua 20
<210> 1158
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1158
gugagguaac caacaaaagg 20
<210> 1159
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1159
agugagguaa ccaacaaaag 20
<210> 1160
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1160
aagugaggua accaacaaaa 20
<210> 1161
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1161
caagugaggu aaccaacaaa 20
<210> 1162
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1162
gguugugguc uuuucaagug 20
<210> 1163
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1163
uacuacugac agguugguug 20
<210> 1164
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1164
gaacuauacu acugacaggu 20
<210> 1165
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1165
augagaacua uacuacugac 20
<210> 1166
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1166
cuuuuguugc uggugagcug 20
<210> 1167
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1167
aagauaaccu cuuuuguugc 20
<210> 1168
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1168
ccagugaaaa uuaagugcua 20
<210> 1169
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1169
gaugccaucu gugaccacuu 20
<210> 1170
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1170
agaugccauc ugugaccacu 20
<210> 1171
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1171
ucuuuuugca uagaaugcuu 20
<210> 1172
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1172
guguuuaaaa auguaaauug 20
<210> 1173
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1173
agaguuguaa gcgggggggg 20
<210> 1174
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1174
uagaguugua agcggggggg 20
<210> 1175
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1175
guagaguugu aagcgggggg 20
<210> 1176
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1176
uguagaguug uaagcggggg 20
<210> 1177
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1177
guguagaguu guaagcgggg 20
<210> 1178
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1178
uguguagagu uguaagcggg 20
<210> 1179
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1179
auguguagag uuguaagcgg 20
<210> 1180
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1180
gauguguaga guuguaagcg 20
<210> 1181
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1181
agauguguag aguuguaagc 20
<210> 1182
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1182
cagaugugua gaguuguaag 20
<210> 1183
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1183
aaacuugaua uuauuaaaag 20
<210> 1184
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1184
aucauauauu cagcaccaga 20
<210> 1185
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1185
auagcaucuu gaugauauaa 20
<210> 1186
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1186
ccccuauuau ucaagggcac 20
<210> 1187
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1187
aagguacccc uauuauucaa 20
<210> 1188
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1188
aaagguaccc cuauuauuca 20
<210> 1189
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1189
ugauaaaaac uugaaugaaa 20
<210> 1190
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1190
aacuaagcuu guguaagaau 20
<210> 1191
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1191
acuggaugag caaaauccag 20
<210> 1192
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1192
uuguccuaca aggacuucac 20
<210> 1193
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1193
auaucguuua uuguccuaca 20
<210> 1194
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1194
uguucaaguu gucaaagcuu 20
<210> 1195
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1195
auugcucauc agcagaugca 20
<210> 1196
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1196
uuaacuggcu guguuaaaaa 20
<210> 1197
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1197
agccccaugg uggauuuaac 20
<210> 1198
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1198
gaauccagua agccccaugg 20
<210> 1199
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1199
cuugaaucca guaagcccca 20
<210> 1200
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1200
gcaccagaaa acauguuuug 20
<210> 1201
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1201
uaugauccau aaaauagcau 20
<210> 1202
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1202
guacauaauu aucaacacaa 20
<210> 1203
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1203
gcucaauccu cuuguuaaag 20
<210> 1204
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1204
ccuguuuauu caucuuaugc 20
<210> 1205
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1205
uuuuaacuga cagauucaca 20
<210> 1206
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1206
cauuaugaua uauuuguagc 20
<210> 1207
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1207
uguugaggua uaugacaagu 20
<210> 1208
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1208
cuauugccaa acuaguguug 20
<210> 1209
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1209
aaggacuugg aaaaaaauga 20
<210> 1210
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1210
uaacaauaaa aaaaaggacu 20
<210> 1211
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1211
uuuuuuuaac aauaaaaaaa 20
<210> 1212
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1212
agaaaucaag uauugaaaaa 20
<210> 1213
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1213
ccugaauuuu uagaacaaaa 20
<210> 1214
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1214
acaggugaca uguuggcaua 20
<210> 1215
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1215
cacaggugac auguuggcau 20
<210> 1216
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1216
cauaaacaca ggugacaugu 20
<210> 1217
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1217
caacaauuuu acauaaacac 20
<210> 1218
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1218
agaagggauu caaaauaaaa 20
<210> 1219
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1219
uagaagggau ucaaaauaaa 20
<210> 1220
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1220
cauguacaag uaaaauagaa 20
<210> 1221
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1221
acauguacaa guaaaauaga 20
<210> 1222
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1222
gagaguuaca aguaagucuc 20
<210> 1223
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1223
uauguaccuu cagugauuac 20
<210> 1224
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1224
cuguuuaagg gaaguccaca 20
<210> 1225
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1225
guagguguuu gccuguuuaa 20
<210> 1226
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1226
uguagguguu ugccuguuua 20
<210> 1227
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1227
cuguugcaca ccauaccugu 20
<210> 1228
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1228
uaguaagcaa aaauguauuu 20
<210> 1229
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1229
auuugaccaa uaaaacccaa 20
<210> 1230
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1230
uuuuggaaau guuugcaaau 20
<210> 1231
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1231
guaagcaaag caaacauuuu 20
<210> 1232
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1232
caaaaaacau uaaaaucaug 20
<210> 1233
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1233
auguuugggg cuagauauua 20
<210> 1234
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1234
auguaguaau caaauguuug 20
<210> 1235
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1235
cauguaguaa ucaaauguuu 20
<210> 1236
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1236
acauguagua aucaaauguu 20
<210> 1237
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1237
cagaaaucaa auauuaagaa 20
<210> 1238
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1238
gcaguuuaac aaaugacagu 20
<210> 1239
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1239
acuauacuuc cuguucuugu 20
<210> 1240
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1240
ccauucauuc uaacugaggc 20
<210> 1241
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1241
cuuuccauuc auucuaacug 20
<210> 1242
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1242
cucagaagca uuuucaagua 20
<210> 1243
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1243
aacacucaca uagcauuauc 20
<210> 1244
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1244
cacatggcgt ttatccagaa t 21
<210> 1245
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1245
cagatgcaca ggccaattaa g 21
<210> 1246
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1246
gagctgctga attcaactag a 21
<210> 1247
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1247
cagatcgcca taacataaat a 21
<210> 1248
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1248
gaccatggag cagcatctga a 21
<210> 1249
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1249
gccaagtcat tatttgacca t 21
<210> 1250
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1250
cctcagagat attgtgggtt t 21
<210> 1251
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1251
gggtaagcca agaaagaaa 19
<210> 1252
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1252
gggtaagcca agaaagaaa 19
<210> 1253
<211> 112
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 1253
cacatggcgt ttatccagaa tctcgagatt ctggataaac gccatgtgtt ttttgaattc 60
gcaccagcac gctacgcaca cacagtacac acactgacgt ttcgccgtct tc 112
<210> 1254
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 1254
gaagacgcac cggcagatgt acaggctaat taaggttaat attcatagcc ttaattggcc 60
tgtgcatctg ttttttgaat tcgcaccagc acgctacgca acacgtcaac cagtgtcagt 120
gtttcgccgt 130
<210> 1255
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 1255
gaagacgcac cgggagctgc tgaattcaat tagagttaat attcatagct ctagttgaat 60
tcagcagctc ttttttgaat tcgcaccagc acgctacgca tgcagtcaac cagtgtcaac 120
cattcgccgt 130
<210> 1256
<211> 112
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 1256
cagatcgcca taacataaat actcgagtat ttatgttatg gcgatctgtt ttttgaattc 60
gcaccagcac gctacgcatg accagtacac acactgcatg ttcgccgtct tc 112
<210> 1257
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 1257
gaagacgcac cgggaccatg gagtagcatt tgaagttaat attcatagct tcagatgctg 60
ctccatggtc ttttttgaat tcgcaccagc acgctacgca tggtgtcaac cagtgtcagt 120
tgttcgccgt 130
<210> 1258
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1258
gccaagtcat tatttgacca tctcgagatg gtcaaataat gacttggctt ttttga 56
<210> 1259
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1259
cctcagagat attgtgggtt tctcgagaaa cccacaatat ctctgaggtt ttttga 56
<210> 1260
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1260
gggtaagcca agaaagaaac tcgagtttct ttcttggctt accctttttt ga 52
<210> 1261
<211> 126
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 1261
gaagacgcac cgggggtaag ccaagaaaga aagttaatat tcatagcttt ctttcttggc 60
ttaccctttt ttgaattcgc accagcacgc tacgcaacac gtcaaccagt gtcagtgttt 120
cgccgt 126
<210> 1262
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 1262
Arg Gly Asp Ser
1
<210> 1263
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1263
catgtacgtt gctatccagg c 21
<210> 1264
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1264
ctccttaatg tcacgcacga t 21
<210> 1265
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1265
ctcaccatgg atgatgatat cgc 23
<210> 1266
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1266
ccacatagga atccttctga ccc 23
<210> 1267
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1267
cagaacctaa accacccgtg 20
<210> 1268
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1268
tgcttcgtag cgccattgta a 21
<210> 1269
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1269
ataccctgta tgaagggaag cc 22
<210> 1270
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1270
cttaccccga agttacgtct ttc 23
<210> 1271
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1271
cacccggctc tatgaaacct t 21
<210> 1272
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
праймер"
<400> 1272
ccagccactc gaggtagtca 20
<210> 1273
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1273
cagagcacca gagugccguc 20
<210> 1274
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1274
agagcaccag agugccgucu 20
<210> 1275
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1275
uucagaccca gacggcacuc 20
<210> 1276
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1276
auggcagcac auugguaagu 20
<210> 1277
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1277
cacauuggua aguugggcug 20
<210> 1278
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1278
gacuugcaca acaugcagaa 20
<210> 1279
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1279
ucauggagca uguacuacaa 20
<210> 1280
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1280
aacuugcgcc ugucaggggc 20
<210> 1281
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1281
ccaaggaagu uuaagcugcu 20
<210> 1282
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1282
ccaagcagcu uaaacuuccu 20
<210> 1283
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1283
uuggugccau aagaguggac 20
<210> 1284
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1284
gcaaaaccug uccacucuua 20
<210> 1285
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1285
auauguuggu gccauaagag 20
<210> 1286
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1286
aaaacggagu ggugccauuc 20
<210> 1287
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1287
gucucugacg uggaugaguu 20
<210> 1288
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1288
uuuauacaaa gucucugacg 20
<210> 1289
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1289
agagaagaca aucgagaauu 20
<210> 1290
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1290
acgucagaga cuuuguauaa 20
<210> 1291
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1291
ggauagaacc aaccauguug 20
<210> 1292
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1292
uuguagccag agguucuguc 20
<210> 1293
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1293
ucuguugccc ucaacauggu 20
<210> 1294
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1294
gauagaacca accauguuga 20
<210> 1295
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1295
uucuggagcu uuguagccag 20
<210> 1296
<211> 73
<212> РНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1296
aacagcauag caaguuaaaa uaaggcuagu ccguuaucaa cuugaaaaag uggcaccgag 60
ucgguguuuu uuu 73
<210> 1297
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 1297
Cys Leu Val Gly Ala Asn Arg Asp Asp Lys Ile Ile Phe
1 5 10
<210> 1298
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 1298
Cys Ala Ser Ser Leu Asp Gly Ser Gly Gln Gly Ser Asp Tyr Gly Tyr
1 5 10 15
Thr Phe
<210> 1299
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1299
tgtgcagcaa gtagggaggc gacaccgaca agctcatctt t 41
<210> 1300
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1300
tgcctcgtgg gtgcgaacag agatgacaag atcatcttt 39
<210> 1301
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1301
gcgccagcag cttggacggt tcgggacagg gatcggacta tggcta 46
<210> 1302
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1302
aagcaagcct gatggaacag actaagtcca ttcctgatac catcacctcc c 51
<210> 1303
<211> 65
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1303
aagcaagcct gatggaacag gatagaacca acagactaag tccattcctg ataccatcac 60
ctccc 65
<210> 1304
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1304
aagcaagcct gatggaacag gatagaacca accattgagg gcaacagact aagtccattc 60
ctgataccat cacctccc 78
<210> 1305
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1305
aagcaagcct gatggaacag gatagaacca accatgaggg caacagacta agtccattcc 60
tgataccatc acctccc 77
<210> 1306
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1306
aagcaagcct gatggaacag gatagaacag actaagtcca ttcctgatac catcacctcc 60
c 61
<210> 1307
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1307
actaagtcca ttcctgatac catcacctcc catttgccag acagaacgtc tggctacaaa 60
gctccagaat ggaagcccac 80
<210> 1308
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1308
actaagtcca ttcctgatac catcacctcc catttgccag acaagaacct ctggctacaa 60
agctccagaa tggaagccc 79
<210> 1309
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1309
actaagtcca ttcctgatac catcacctcc catttgccag acaagaacgt ctggctacaa 60
agctccagaa tggaagccc 79
<210> 1310
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1310
actaagtcca ttcctgatac catcacctcc catttgcctc tggctacaaa gctccagaat 60
ggaagcccac 70
<210> 1311
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1311
actaagtcca ttcctgatac catcacctcc catttgccag acctctggct acaaagctcc 60
agaatggaag cccac 75
<210> 1312
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1312
atggccccga agcaagcctg atggaacagg atagaaccaa ccaatgttga gggcaacaga 60
ctaagtccat tcctgatac 79
<210> 1313
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1313
atggccccga agcaagcctg atggaacagg atagaaccaa cctgttgagg gcaacagact 60
aagtccattc ctgatacca 79
<210> 1314
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1314
atggccccga agcaagcctg atggaacaga ctaagtccat tcctgatacc a 51
<210> 1315
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1315
atggccccga agcaagcctg atggaacagg atagaaccaa ccgttgaggg caacagacta 60
agtccattcc tgatacca 78
<210> 1316
<211> 65
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1316
atggccccga agcaagcctg atggaacagg atagaaccaa cagactaagt ccattcctga 60
tacca 65
<210> 1317
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1317
agcaagcctg atggaacagg atagaaccaa ccatgtttga gggcaacaga ctaagtccat 60
tcctgatacc atcacctcc 79
<210> 1318
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1318
agcaagcctg atggaacagg atagaaccaa ccatgtgagg gcaacagact aagtccattc 60
ctgataccat cacctccca 79
<210> 1319
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1319
agcaagcctg atggaacaga ctaagtccat tcctgatacc atcacctccc a 51
<210> 1320
<211> 65
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1320
agcaagcctg atggaacagg atagaaccaa cagactaagt ccattcctga taccatcacc 60
tccca 65
<210> 1321
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1321
agcaagcctg atggaacagg atagaaccaa ccatgagggc aacagactaa gtccattcct 60
gataccatca cctccca 77
<210> 1322
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1322
ttcctgatac catcacctcc catttgccag acagaacgtc tggctacaaa gctccagaat 60
ggaagcccac tgcctgagag 80
<210> 1323
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1323
ttcctgatac catcacctcc catttgccag acagaacctc tgggctacaa agctccagaa 60
tggaagccca ctgcctgag 79
<210> 1324
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1324
ttcctgatac catcacctcc catttgccag acagaacctc tgctacaaag ctccagaatg 60
gaagcccact gcctgagag 79
<210> 1325
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1325
ttcctgatac catcacctcc catttgccag acagaatgga agcccactgc ctgagag 57
<210> 1326
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1326
ttcctgatac catcacctcc catttgccag acagaacctc cagaatggaa gcccactgcc 60
tgagag 66
<210> 1327
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1327
ttcagattga atatgaacac agagcaccag agtgcgtctg ggtctgaagg aaggccgtcc 60
attctcaggg gtcactgca 79
<210> 1328
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1328
ttcagattga atatgaacac agagcaccag agtgcccgtc tgggtctgaa ggaaggccgt 60
ccattctcag gggtcactg 79
<210> 1329
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1329
ttcagattga atatgaacac agagcaccag agtgccgtct gggtccgaag gaaggccgtc 60
cattctcagg ggtcactgca 80
<210> 1330
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1330
ttcagattga atatgagcac agagcaccag agtgccgtct gggtctgaag gaaggccgtc 60
cattctcagg ggtcactgca 80
<210> 1331
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1331
ttcagattga atatgaacac agagcaccag agtgccgtct gggtctgaag gagggccgtc 60
cattctcagg ggtcactgca 80
<210> 1332
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1332
tcagattgaa tatgaacaca gagcaccaga gtgcctctgg gtctgaagga aggccgtcca 60
ttctcagggg tcactgcat 79
<210> 1333
<211> 73
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1333
tcagattgaa tatgaacaca gagcaccaga gtgggtctga aggaaggccg tccattctca 60
ggggtcactg cat 73
<210> 1334
<211> 69
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1334
tcagattgaa tatgaacaca gagcaccaga gtctgaagga aggccgtcca ttctcagggg 60
tcactgcat 69
<210> 1335
<211> 76
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1335
tcagattgaa tatgaacaca gagcaccaga gtgctgggtc tgaaggaagg ccgtccattc 60
tcaggggtca ctgcat 76
<210> 1336
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1336
tcagattgaa tatgaacaca gagcaccaga gtgctgaagg aaggccgtcc attctcaggg 60
gtcactgcat 70
<210> 1337
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1337
acttttattt ttcagattga atatgaacac agagcaccgt ctgggtctga aggaaggccg 60
tccattctca gg 72
<210> 1338
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1338
acttttattt ttcagattga atatgaacac agagcaccag agttgccgtc tgggtctgaa 60
ggaaggccgt ccattctca 79
<210> 1339
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1339
acttttattt ttcagattga atatgaacac agagccgtct gggtctgaag gaaggccgtc 60
cattctcagg 70
<210> 1340
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1340
acttttattt ttcagattga atatgaacac agagcaccag agccgtctgg gtctgaagga 60
aggccgtcca ttctcagg 78
<210> 1341
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1341
acttttattt ttcagattga atatgaacac agagcaccag tgccgtctgg gtctgaagga 60
aggccgtcca ttctcagg 78
<210> 1342
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1342
gacttgcaca acatgcagaa tggcagcaca ttgggctgag gacagcttag cagctgttga 60
gtctgttctc ac 72
<210> 1343
<211> 76
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1343
gacttgcaca acatgcagaa tggcagcaca ttggttgggc tgaggacagc ttagcagctg 60
ttgagtctgt tctcac 76
<210> 1344
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1344
gacttgcaca acatgcagaa tggcagcaca ttggtagttg ggctgaggac agcttagcag 60
ctgttgagtc tgttctcac 79
<210> 1345
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1345
gacttgcaca acatgcagaa tggcagctta gcagctgttg agtctgttct cac 53
<210> 1346
<211> 71
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1346
gacttgcaca acatgcagaa tggcagcaca ttggctgagg acagcttagc agctgttgag 60
tctgttctca c 71
<210> 1347
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1347
acaacatgca gaatggcagc acattggtaa gttggctgag gacagcttag cagctgttga 60
gtctgttctc acactgcta 79
<210> 1348
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1348
acaacatgca gaatggcagc acattggtaa gttggggagg acagcttagc agctgttgag 60
tctgttctca cactgcta 78
<210> 1349
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1349
acaacatgca gaatggcagc acattggtaa gtctgttctc acactgcta 49
<210> 1350
<211> 74
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1350
acaacatgca gaatggcagc acattggtaa gttgggacag cttagcagct gttgagtctg 60
ttctcacact gcta 74
<210> 1351
<211> 65
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1351
acaacatgca gaatggcagc acattggtaa gcttagcagc tgttgagtct gttctcacac 60
tgcta 65
<210> 1352
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1352
ctgtgctcat gcccacagag acttgcacaa catgcaagaa tggcagcaca ttggtaagtt 60
gggctgagga cagcttagc 79
<210> 1353
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1353
ctgtgctcat gcccacagag acttgcacat tggtaagttg ggctgaggac agcttagcag 60
<210> 1354
<211> 73
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1354
ctgtgctcat gcccacagag acttgcacaa catggcagca cattggtaag ttgggctgag 60
gacagcttag cag 73
<210> 1355
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1355
ctgtgctcat gcccacagag acttgcagca cattggtaag ttgggctgag gacagcttag 60
cag 63
<210> 1356
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
олигонуклеотид"
<400> 1356
ctgtgctcat gcccacagag acttgcacaa catagaatgg cagcacattg gtaagttggg 60
ctgaggacag cttagcag 78
<210> 1357
<211> 2002
<212> БЕЛОК
<213> Homo sapiens
<400> 1357
Met Glu Gln Asp Arg Thr Asn His Val Glu Gly Asn Arg Leu Ser Pro
1 5 10 15
Phe Leu Ile Pro Ser Pro Pro Ile Cys Gln Thr Glu Pro Leu Ala Thr
20 25 30
Lys Leu Gln Asn Gly Ser Pro Leu Pro Glu Arg Ala His Pro Glu Val
35 40 45
Asn Gly Asp Thr Lys Trp His Ser Phe Lys Ser Tyr Tyr Gly Ile Pro
50 55 60
Cys Met Lys Gly Ser Gln Asn Ser Arg Val Ser Pro Asp Phe Thr Gln
65 70 75 80
Glu Ser Arg Gly Tyr Ser Lys Cys Leu Gln Asn Gly Gly Ile Lys Arg
85 90 95
Thr Val Ser Glu Pro Ser Leu Ser Gly Leu Leu Gln Ile Lys Lys Leu
100 105 110
Lys Gln Asp Gln Lys Ala Asn Gly Glu Arg Arg Asn Phe Gly Val Ser
115 120 125
Gln Glu Arg Asn Pro Gly Glu Ser Ser Gln Pro Asn Val Ser Asp Leu
130 135 140
Ser Asp Lys Lys Glu Ser Val Ser Ser Val Ala Gln Glu Asn Ala Val
145 150 155 160
Lys Asp Phe Thr Ser Phe Ser Thr His Asn Cys Ser Gly Pro Glu Asn
165 170 175
Pro Glu Leu Gln Ile Leu Asn Glu Gln Glu Gly Lys Ser Ala Asn Tyr
180 185 190
His Asp Lys Asn Ile Val Leu Leu Lys Asn Lys Ala Val Leu Met Pro
195 200 205
Asn Gly Ala Thr Val Ser Ala Ser Ser Val Glu His Thr His Gly Glu
210 215 220
Leu Leu Glu Lys Thr Leu Ser Gln Tyr Tyr Pro Asp Cys Val Ser Ile
225 230 235 240
Ala Val Gln Lys Thr Thr Ser His Ile Asn Ala Ile Asn Ser Gln Ala
245 250 255
Thr Asn Glu Leu Ser Cys Glu Ile Thr His Pro Ser His Thr Ser Gly
260 265 270
Gln Ile Asn Ser Ala Gln Thr Ser Asn Ser Glu Leu Pro Pro Lys Pro
275 280 285
Ala Ala Val Val Ser Glu Ala Cys Asp Ala Asp Asp Ala Asp Asn Ala
290 295 300
Ser Lys Leu Ala Ala Met Leu Asn Thr Cys Ser Phe Gln Lys Pro Glu
305 310 315 320
Gln Leu Gln Gln Gln Lys Ser Val Phe Glu Ile Cys Pro Ser Pro Ala
325 330 335
Glu Asn Asn Ile Gln Gly Thr Thr Lys Leu Ala Ser Gly Glu Glu Phe
340 345 350
Cys Ser Gly Ser Ser Ser Asn Leu Gln Ala Pro Gly Gly Ser Ser Glu
355 360 365
Arg Tyr Leu Lys Gln Asn Glu Met Asn Gly Ala Tyr Phe Lys Gln Ser
370 375 380
Ser Val Phe Thr Lys Asp Ser Phe Ser Ala Thr Thr Thr Pro Pro Pro
385 390 395 400
Pro Ser Gln Leu Leu Leu Ser Pro Pro Pro Pro Leu Pro Gln Val Pro
405 410 415
Gln Leu Pro Ser Glu Gly Lys Ser Thr Leu Asn Gly Gly Val Leu Glu
420 425 430
Glu His His His Tyr Pro Asn Gln Ser Asn Thr Thr Leu Leu Arg Glu
435 440 445
Val Lys Ile Glu Gly Lys Pro Glu Ala Pro Pro Ser Gln Ser Pro Asn
450 455 460
Pro Ser Thr His Val Cys Ser Pro Ser Pro Met Leu Ser Glu Arg Pro
465 470 475 480
Gln Asn Asn Cys Val Asn Arg Asn Asp Ile Gln Thr Ala Gly Thr Met
485 490 495
Thr Val Pro Leu Cys Ser Glu Lys Thr Arg Pro Met Ser Glu His Leu
500 505 510
Lys His Asn Pro Pro Ile Phe Gly Ser Ser Gly Glu Leu Gln Asp Asn
515 520 525
Cys Gln Gln Leu Met Arg Asn Lys Glu Gln Glu Ile Leu Lys Gly Arg
530 535 540
Asp Lys Glu Gln Thr Arg Asp Leu Val Pro Pro Thr Gln His Tyr Leu
545 550 555 560
Lys Pro Gly Trp Ile Glu Leu Lys Ala Pro Arg Phe His Gln Ala Glu
565 570 575
Ser His Leu Lys Arg Asn Glu Ala Ser Leu Pro Ser Ile Leu Gln Tyr
580 585 590
Gln Pro Asn Leu Ser Asn Gln Met Thr Ser Lys Gln Tyr Thr Gly Asn
595 600 605
Ser Asn Met Pro Gly Gly Leu Pro Arg Gln Ala Tyr Thr Gln Lys Thr
610 615 620
Thr Gln Leu Glu His Lys Ser Gln Met Tyr Gln Val Glu Met Asn Gln
625 630 635 640
Gly Gln Ser Gln Gly Thr Val Asp Gln His Leu Gln Phe Gln Lys Pro
645 650 655
Ser His Gln Val His Phe Ser Lys Thr Asp His Leu Pro Lys Ala His
660 665 670
Val Gln Ser Leu Cys Gly Thr Arg Phe His Phe Gln Gln Arg Ala Asp
675 680 685
Ser Gln Thr Glu Lys Leu Met Ser Pro Val Leu Lys Gln His Leu Asn
690 695 700
Gln Gln Ala Ser Glu Thr Glu Pro Phe Ser Asn Ser His Leu Leu Gln
705 710 715 720
His Lys Pro His Lys Gln Ala Ala Gln Thr Gln Pro Ser Gln Ser Ser
725 730 735
His Leu Pro Gln Asn Gln Gln Gln Gln Gln Lys Leu Gln Ile Lys Asn
740 745 750
Lys Glu Glu Ile Leu Gln Thr Phe Pro His Pro Gln Ser Asn Asn Asp
755 760 765
Gln Gln Arg Glu Gly Ser Phe Phe Gly Gln Thr Lys Val Glu Glu Cys
770 775 780
Phe His Gly Glu Asn Gln Tyr Ser Lys Ser Ser Glu Phe Glu Thr His
785 790 795 800
Asn Val Gln Met Gly Leu Glu Glu Val Gln Asn Ile Asn Arg Arg Asn
805 810 815
Ser Pro Tyr Ser Gln Thr Met Lys Ser Ser Ala Cys Lys Ile Gln Val
820 825 830
Ser Cys Ser Asn Asn Thr His Leu Val Ser Glu Asn Lys Glu Gln Thr
835 840 845
Thr His Pro Glu Leu Phe Ala Gly Asn Lys Thr Gln Asn Leu His His
850 855 860
Met Gln Tyr Phe Pro Asn Asn Val Ile Pro Lys Gln Asp Leu Leu His
865 870 875 880
Arg Cys Phe Gln Glu Gln Glu Gln Lys Ser Gln Gln Ala Ser Val Leu
885 890 895
Gln Gly Tyr Lys Asn Arg Asn Gln Asp Met Ser Gly Gln Gln Ala Ala
900 905 910
Gln Leu Ala Gln Gln Arg Tyr Leu Ile His Asn His Ala Asn Val Phe
915 920 925
Pro Val Pro Asp Gln Gly Gly Ser His Thr Gln Thr Pro Pro Gln Lys
930 935 940
Asp Thr Gln Lys His Ala Ala Leu Arg Trp His Leu Leu Gln Lys Gln
945 950 955 960
Glu Gln Gln Gln Thr Gln Gln Pro Gln Thr Glu Ser Cys His Ser Gln
965 970 975
Met His Arg Pro Ile Lys Val Glu Pro Gly Cys Lys Pro His Ala Cys
980 985 990
Met His Thr Ala Pro Pro Glu Asn Lys Thr Trp Lys Lys Val Thr Lys
995 1000 1005
Gln Glu Asn Pro Pro Ala Ser Cys Asp Asn Val Gln Gln Lys Ser
1010 1015 1020
Ile Ile Glu Thr Met Glu Gln His Leu Lys Gln Phe His Ala Lys
1025 1030 1035
Ser Leu Phe Asp His Lys Ala Leu Thr Leu Lys Ser Gln Lys Gln
1040 1045 1050
Val Lys Val Glu Met Ser Gly Pro Val Thr Val Leu Thr Arg Gln
1055 1060 1065
Thr Thr Ala Ala Glu Leu Asp Ser His Thr Pro Ala Leu Glu Gln
1070 1075 1080
Gln Thr Thr Ser Ser Glu Lys Thr Pro Thr Lys Arg Thr Ala Ala
1085 1090 1095
Ser Val Leu Asn Asn Phe Ile Glu Ser Pro Ser Lys Leu Leu Asp
1100 1105 1110
Thr Pro Ile Lys Asn Leu Leu Asp Thr Pro Val Lys Thr Gln Tyr
1115 1120 1125
Asp Phe Pro Ser Cys Arg Cys Val Glu Gln Ile Ile Glu Lys Asp
1130 1135 1140
Glu Gly Pro Phe Tyr Thr His Leu Gly Ala Gly Pro Asn Val Ala
1145 1150 1155
Ala Ile Arg Glu Ile Met Glu Glu Arg Phe Gly Gln Lys Gly Lys
1160 1165 1170
Ala Ile Arg Ile Glu Arg Val Ile Tyr Thr Gly Lys Glu Gly Lys
1175 1180 1185
Ser Ser Gln Gly Cys Pro Ile Ala Lys Trp Val Val Arg Arg Ser
1190 1195 1200
Ser Ser Glu Glu Lys Leu Leu Cys Leu Val Arg Glu Arg Ala Gly
1205 1210 1215
His Thr Cys Glu Ala Ala Val Ile Val Ile Leu Ile Leu Val Trp
1220 1225 1230
Glu Gly Ile Pro Leu Ser Leu Ala Asp Lys Leu Tyr Ser Glu Leu
1235 1240 1245
Thr Glu Thr Leu Arg Lys Tyr Gly Thr Leu Thr Asn Arg Arg Cys
1250 1255 1260
Ala Leu Asn Glu Glu Arg Thr Cys Ala Cys Gln Gly Leu Asp Pro
1265 1270 1275
Glu Thr Cys Gly Ala Ser Phe Ser Phe Gly Cys Ser Trp Ser Met
1280 1285 1290
Tyr Tyr Asn Gly Cys Lys Phe Ala Arg Ser Lys Ile Pro Arg Lys
1295 1300 1305
Phe Lys Leu Leu Gly Asp Asp Pro Lys Glu Glu Glu Lys Leu Glu
1310 1315 1320
Ser His Leu Gln Asn Leu Ser Thr Leu Met Ala Pro Thr Tyr Lys
1325 1330 1335
Lys Leu Ala Pro Asp Ala Tyr Asn Asn Gln Ile Glu Tyr Glu His
1340 1345 1350
Arg Ala Pro Glu Cys Arg Leu Gly Leu Lys Glu Gly Arg Pro Phe
1355 1360 1365
Ser Gly Val Thr Ala Cys Leu Asp Phe Cys Ala His Ala His Arg
1370 1375 1380
Asp Leu His Asn Met Gln Asn Gly Ser Thr Leu Val Cys Thr Leu
1385 1390 1395
Thr Arg Glu Asp Asn Arg Glu Phe Gly Gly Lys Pro Glu Asp Glu
1400 1405 1410
Gln Leu His Val Leu Pro Leu Tyr Lys Val Ser Asp Val Asp Glu
1415 1420 1425
Phe Gly Ser Val Glu Ala Gln Glu Glu Lys Lys Arg Ser Gly Ala
1430 1435 1440
Ile Gln Val Leu Ser Ser Phe Arg Arg Lys Val Arg Met Leu Ala
1445 1450 1455
Glu Pro Val Lys Thr Cys Arg Gln Arg Lys Leu Glu Ala Lys Lys
1460 1465 1470
Ala Ala Ala Glu Lys Leu Ser Ser Leu Glu Asn Ser Ser Asn Lys
1475 1480 1485
Asn Glu Lys Glu Lys Ser Ala Pro Ser Arg Thr Lys Gln Thr Glu
1490 1495 1500
Asn Ala Ser Gln Ala Lys Gln Leu Ala Glu Leu Leu Arg Leu Ser
1505 1510 1515
Gly Pro Val Met Gln Gln Ser Gln Gln Pro Gln Pro Leu Gln Lys
1520 1525 1530
Gln Pro Pro Gln Pro Gln Gln Gln Gln Arg Pro Gln Gln Gln Gln
1535 1540 1545
Pro His His Pro Gln Thr Glu Ser Val Asn Ser Tyr Ser Ala Ser
1550 1555 1560
Gly Ser Thr Asn Pro Tyr Met Arg Arg Pro Asn Pro Val Ser Pro
1565 1570 1575
Tyr Pro Asn Ser Ser His Thr Ser Asp Ile Tyr Gly Ser Thr Ser
1580 1585 1590
Pro Met Asn Phe Tyr Ser Thr Ser Ser Gln Ala Ala Gly Ser Tyr
1595 1600 1605
Leu Asn Ser Ser Asn Pro Met Asn Pro Tyr Pro Gly Leu Leu Asn
1610 1615 1620
Gln Asn Thr Gln Tyr Pro Ser Tyr Gln Cys Asn Gly Asn Leu Ser
1625 1630 1635
Val Asp Asn Cys Ser Pro Tyr Leu Gly Ser Tyr Ser Pro Gln Ser
1640 1645 1650
Gln Pro Met Asp Leu Tyr Arg Tyr Pro Ser Gln Asp Pro Leu Ser
1655 1660 1665
Lys Leu Ser Leu Pro Pro Ile His Thr Leu Tyr Gln Pro Arg Phe
1670 1675 1680
Gly Asn Ser Gln Ser Phe Thr Ser Lys Tyr Leu Gly Tyr Gly Asn
1685 1690 1695
Gln Asn Met Gln Gly Asp Gly Phe Ser Ser Cys Thr Ile Arg Pro
1700 1705 1710
Asn Val His His Val Gly Lys Leu Pro Pro Tyr Pro Thr His Glu
1715 1720 1725
Met Asp Gly His Phe Met Gly Ala Thr Ser Arg Leu Pro Pro Asn
1730 1735 1740
Leu Ser Asn Pro Asn Met Asp Tyr Lys Asn Gly Glu His His Ser
1745 1750 1755
Pro Ser His Ile Ile His Asn Tyr Ser Ala Ala Pro Gly Met Phe
1760 1765 1770
Asn Ser Ser Leu His Ala Leu His Leu Gln Asn Lys Glu Asn Asp
1775 1780 1785
Met Leu Ser His Thr Ala Asn Gly Leu Ser Lys Met Leu Pro Ala
1790 1795 1800
Leu Asn His Asp Arg Thr Ala Cys Val Gln Gly Gly Leu His Lys
1805 1810 1815
Leu Ser Asp Ala Asn Gly Gln Glu Lys Gln Pro Leu Ala Leu Val
1820 1825 1830
Gln Gly Val Ala Ser Gly Ala Glu Asp Asn Asp Glu Val Trp Ser
1835 1840 1845
Asp Ser Glu Gln Ser Phe Leu Asp Pro Asp Ile Gly Gly Val Ala
1850 1855 1860
Val Ala Pro Thr His Gly Ser Ile Leu Ile Glu Cys Ala Lys Arg
1865 1870 1875
Glu Leu His Ala Thr Thr Pro Leu Lys Asn Pro Asn Arg Asn His
1880 1885 1890
Pro Thr Arg Ile Ser Leu Val Phe Tyr Gln His Lys Ser Met Asn
1895 1900 1905
Glu Pro Lys His Gly Leu Ala Leu Trp Glu Ala Lys Met Ala Glu
1910 1915 1920
Lys Ala Arg Glu Lys Glu Glu Glu Cys Glu Lys Tyr Gly Pro Asp
1925 1930 1935
Tyr Val Pro Gln Lys Ser His Gly Lys Lys Val Lys Arg Glu Pro
1940 1945 1950
Ala Glu Pro His Glu Thr Ser Glu Pro Thr Tyr Leu Arg Phe Ile
1955 1960 1965
Lys Ser Leu Ala Glu Arg Thr Met Ser Val Thr Thr Asp Ser Thr
1970 1975 1980
Val Thr Thr Ser Pro Tyr Ala Phe Thr Arg Val Thr Gly Pro Tyr
1985 1990 1995
Asn Arg Tyr Ile
2000
<210> 1358
<211> 9796
<212> ДНК
<213> Homo sapiens
<400> 1358
ggcagtggca gcggcgagag cttgggcggc cgccgccgcc tcctcgcgag cgccgcgcgc 60
ccgggtcccg ctcgcatgca agtcacgtcc gccccctcgg cgcggccgcc ccgagacgcc 120
ggccccgctg agtgatgaga acagacgtca aactgcctta tgaatattga tgcggaggct 180
aggctgcttt cgtagagaag cagaaggaag caagatggct gccctttagg atttgttaga 240
aaggagaccc gactgcaact gctggattgc tgcaaggctg agggacgaga acgaggctgg 300
caaacattca gcagcacacc ctctcaagat tgtttacttg cctttgctcc tgttgagtta 360
caacgcttgg aagcaggaga tgggctcagc agcagccaat aggacatgat ccaggaagag 420
cagtaaggga ctgagctgct gaattcaact agagggcagc cttgtggatg gccccgaagc 480
aagcctgatg gaacaggata gaaccaacca tgttgagggc aacagactaa gtccattcct 540
gataccatca cctcccattt gccagacaga acctctggct acaaagctcc agaatggaag 600
cccactgcct gagagagctc atccagaagt aaatggagac accaagtggc actctttcaa 660
aagttattat ggaataccct gtatgaaggg aagccagaat agtcgtgtga gtcctgactt 720
tacacaagaa agtagagggt attccaagtg tttgcaaaat ggaggaataa aacgcacagt 780
tagtgaacct tctctctctg ggctccttca gatcaagaaa ttgaaacaag accaaaaggc 840
taatggagaa agacgtaact tcggggtaag ccaagaaaga aatccaggtg aaagcagtca 900
accaaatgtc tccgatttga gtgataagaa agaatctgtg agttctgtag cccaagaaaa 960
tgcagttaaa gatttcacca gtttttcaac acataactgc agtgggcctg aaaatccaga 1020
gcttcagatt ctgaatgagc aggaggggaa aagtgctaat taccatgaca agaacattgt 1080
attacttaaa aacaaggcag tgctaatgcc taatggtgct acagtttctg cctcttccgt 1140
ggaacacaca catggtgaac tcctggaaaa aacactgtct caatattatc cagattgtgt 1200
ttccattgcg gtgcagaaaa ccacatctca cataaatgcc attaacagtc aggctactaa 1260
tgagttgtcc tgtgagatca ctcacccatc gcatacctca gggcagatca attccgcaca 1320
gacctctaac tctgagctgc ctccaaagcc agctgcagtg gtgagtgagg cctgtgatgc 1380
tgatgatgct gataatgcca gtaaactagc tgcaatgcta aatacctgtt cctttcagaa 1440
accagaacaa ctacaacaac aaaaatcagt ttttgagata tgcccatctc ctgcagaaaa 1500
taacatccag ggaaccacaa agctagcgtc tggtgaagaa ttctgttcag gttccagcag 1560
caatttgcaa gctcctggtg gcagctctga acggtattta aaacaaaatg aaatgaatgg 1620
tgcttacttc aagcaaagct cagtgttcac taaggattcc ttttctgcca ctaccacacc 1680
accaccacca tcacaattgc ttctttctcc ccctcctcct cttccacagg ttcctcagct 1740
tccttcagaa ggaaaaagca ctctgaatgg tggagtttta gaagaacacc accactaccc 1800
caaccaaagt aacacaacac ttttaaggga agtgaaaata gagggtaaac ctgaggcacc 1860
accttcccag agtcctaatc catctacaca tgtatgcagc ccttctccga tgctttctga 1920
aaggcctcag aataattgtg tgaacaggaa tgacatacag actgcaggga caatgactgt 1980
tccattgtgt tctgagaaaa caagaccaat gtcagaacac ctcaagcata acccaccaat 2040
ttttggtagc agtggagagc tacaggacaa ctgccagcag ttgatgagaa acaaagagca 2100
agagattctg aagggtcgag acaaggagca aacacgagat cttgtgcccc caacacagca 2160
ctatctgaaa ccaggatgga ttgaattgaa ggcccctcgt tttcaccaag cggaatccca 2220
tctaaaacgt aatgaggcat cactgccatc aattcttcag tatcaaccca atctctccaa 2280
tcaaatgacc tccaaacaat acactggaaa ttccaacatg cctggggggc tcccaaggca 2340
agcttacacc cagaaaacaa cacagctgga gcacaagtca caaatgtacc aagttgaaat 2400
gaatcaaggg cagtcccaag gtacagtgga ccaacatctc cagttccaaa aaccctcaca 2460
ccaggtgcac ttctccaaaa cagaccattt accaaaagct catgtgcagt cactgtgtgg 2520
cactagattt cattttcaac aaagagcaga ttcccaaact gaaaaactta tgtccccagt 2580
gttgaaacag cacttgaatc aacaggcttc agagactgag ccattttcaa actcacacct 2640
tttgcaacat aagcctcata aacaggcagc acaaacacaa ccatcccaga gttcacatct 2700
ccctcaaaac cagcaacagc agcaaaaatt acaaataaag aataaagagg aaatactcca 2760
gacttttcct cacccccaaa gcaacaatga tcagcaaaga gaaggatcat tctttggcca 2820
gactaaagtg gaagaatgtt ttcatggtga aaatcagtat tcaaaatcaa gcgagttcga 2880
gactcataat gtccaaatgg gactggagga agtacagaat ataaatcgta gaaattcccc 2940
ttatagtcag accatgaaat caagtgcatg caaaatacag gtttcttgtt caaacaatac 3000
acacctagtt tcagagaata aagaacagac tacacatcct gaactttttg caggaaacaa 3060
gacccaaaac ttgcatcaca tgcaatattt tccaaataat gtgatcccaa agcaagatct 3120
tcttcacagg tgctttcaag aacaggagca gaagtcacaa caagcttcag ttctacaggg 3180
atataaaaat agaaaccaag atatgtctgg tcaacaagct gcgcaacttg ctcagcaaag 3240
gtacttgata cataaccatg caaatgtttt tcctgtgcct gaccagggag gaagtcacac 3300
tcagacccct ccccagaagg acactcaaaa gcatgctgct ctaaggtggc atctcttaca 3360
gaagcaagaa cagcagcaaa cacagcaacc ccaaactgag tcttgccata gtcagatgca 3420
caggccaatt aaggtggaac ctggatgcaa gccacatgcc tgtatgcaca cagcaccacc 3480
agaaaacaaa acatggaaaa aggtaactaa gcaagagaat ccacctgcaa gctgtgataa 3540
tgtgcagcaa aagagcatca ttgagaccat ggagcagcat ctgaagcagt ttcacgccaa 3600
gtcgttattt gaccataagg ctcttactct caaatcacag aagcaagtaa aagttgaaat 3660
gtcagggcca gtcacagttt tgactagaca aaccactgct gcagaacttg atagccacac 3720
cccagcttta gagcagcaaa caacttcttc agaaaagaca ccaaccaaaa gaacagctgc 3780
ttctgttctc aataatttta tagagtcacc ttccaaatta ctagatactc ctataaaaaa 3840
tttattggat acacctgtca agactcaata tgatttccca tcttgcagat gtgtagagca 3900
aattattgaa aaagatgaag gtccttttta tacccatcta ggagcaggtc ctaatgtggc 3960
agctattaga gaaatcatgg aagaaaggtt tggacagaag ggtaaagcta ttaggattga 4020
aagagtcatc tatactggta aagaaggcaa aagttctcag ggatgtccta ttgctaagtg 4080
ggtggttcgc agaagcagca gtgaagagaa gctactgtgt ttggtgcggg agcgagctgg 4140
ccacacctgt gaggctgcag tgattgtgat tctcatcctg gtgtgggaag gaatcccgct 4200
gtctctggct gacaaactct actcggagct taccgagacg ctgaggaaat acggcacgct 4260
caccaatcgc cggtgtgcct tgaatgaaga gagaacttgc gcctgtcagg ggctggatcc 4320
agaaacctgt ggtgcctcct tctcttttgg ttgttcatgg agcatgtact acaatggatg 4380
taagtttgcc agaagcaaga tcccaaggaa gtttaagctg cttggggatg acccaaaaga 4440
ggaagagaaa ctggagtctc atttgcaaaa cctgtccact cttatggcac caacatataa 4500
gaaacttgca cctgatgcat ataataatca gattgaatat gaacacagag caccagagtg 4560
ccgtctgggt ctgaaggaag gccgtccatt ctcaggggtc actgcatgtt tggacttctg 4620
tgctcatgcc cacagagact tgcacaacat gcagaatggc agcacattgg tatgcactct 4680
cactagagaa gacaatcgag aatttggagg aaaacctgag gatgagcagc ttcacgttct 4740
gcctttatac aaagtctctg acgtggatga gtttgggagt gtggaagctc aggaggagaa 4800
aaaacggagt ggtgccattc aggtactgag ttcttttcgg cgaaaagtca ggatgttagc 4860
agagccagtc aagacttgcc gacaaaggaa actagaagcc aagaaagctg cagctgaaaa 4920
gctttcctcc ctggagaaca gctcaaataa aaatgaaaag gaaaagtcag ccccatcacg 4980
tacaaaacaa actgaaaacg caagccaggc taaacagttg gcagaacttt tgcgactttc 5040
aggaccagtc atgcagcagt cccagcagcc ccagcctcta cagaagcagc caccacagcc 5100
ccagcagcag cagagacccc agcagcagca gccacatcac cctcagacag agtctgtcaa 5160
ctcttattct gcttctggat ccaccaatcc atacatgaga cggcccaatc cagttagtcc 5220
ttatccaaac tcttcacaca cttcagatat ctatggaagc accagcccta tgaacttcta 5280
ttccacctca tctcaagctg caggttcata tttgaattct tctaatccca tgaaccctta 5340
ccctgggctt ttgaatcaga atacccaata tccatcatat caatgcaatg gaaacctatc 5400
agtggacaac tgctccccat atctgggttc ctattctccc cagtctcagc cgatggatct 5460
gtataggtat ccaagccaag accctctgtc taagctcagt ctaccaccca tccatacact 5520
ttaccagcca aggtttggaa atagccagag ttttacatct aaatacttag gttatggaaa 5580
ccaaaatatg cagggagatg gtttcagcag ttgtaccatt agaccaaatg tacatcatgt 5640
agggaaattg cctccttatc ccactcatga gatggatggc cacttcatgg gagccacctc 5700
tagattacca cccaatctga gcaatccaaa catggactat aaaaatggtg aacatcattc 5760
accttctcac ataatccata actacagtgc agctccgggc atgttcaaca gctctcttca 5820
tgccctgcat ctccaaaaca aggagaatga catgctttcc cacacagcta atgggttatc 5880
aaagatgctt ccagctctta accatgatag aactgcttgt gtccaaggag gcttacacaa 5940
attaagtgat gctaatggtc aggaaaagca gccattggca ctagtccagg gtgtggcttc 6000
tggtgcagag gacaacgatg aggtctggtc agacagcgag cagagctttc tggatcctga 6060
cattggggga gtggccgtgg ctccaactca tgggtcaatt ctcattgagt gtgcaaagcg 6120
tgagctgcat gccacaaccc ctttaaagaa tcccaatagg aatcacccca ccaggatctc 6180
cctcgtcttt taccagcata agagcatgaa tgagccaaaa catggcttgg ctctttggga 6240
agccaaaatg gctgaaaaag cccgtgagaa agaggaagag tgtgaaaagt atggcccaga 6300
ctatgtgcct cagaaatccc atggcaaaaa agtgaaacgg gagcctgctg agccacatga 6360
aacttcagag cccacttacc tgcgtttcat caagtctctt gccgaaagga ccatgtccgt 6420
gaccacagac tccacagtaa ctacatctcc atatgccttc actcgggtca cagggcctta 6480
caacagatat atatgatatc accccctttt gttggttacc tcacttgaaa agaccacaac 6540
caacctgtca gtagtatagt tctcatgacg tgggcagtgg ggaaaggtca cagtattcat 6600
gacaaatgtg gtgggaaaaa cctcagctca ccagcaacaa aagaggttat cttaccatag 6660
cacttaattt tcactggctc ccaagtggtc acagatggca tctaggaaaa gaccaaagca 6720
ttctatgcaa aaagaaggtg gggaagaaag tgttccgcaa tttacatttt taaacactgg 6780
ttctattatt ggacgagatg atatgtaaat gtgatccccc ccccccgctt acaactctac 6840
acatctgtga ccacttttaa taatatcaag tttgcatagt catggaacac aaatcaaaca 6900
agtactgtag tattacagtg acaggaatct taaaatacca tctggtgctg aatatatgat 6960
gtactgaaat actggaatta tggctttttg aaatgcagtt tttactgtaa tcttaacttt 7020
tatttatcaa aatagctaca ggaaacatga atagcaggaa aacactgaat ttgtttggat 7080
gttctaagaa atggtgctaa gaaaatggtg tctttaatag ctaaaaattt aatgccttta 7140
tatcatcaag atgctatcag tgtactccag tgcccttgaa taataggggt accttttcat 7200
tcaagttttt atcataatta cctattctta cacaagctta gtttttaaaa tgtggacatt 7260
ttaaaggcct ctggattttg ctcatccagt gaagtccttg taggacaata aacgtatata 7320
tgtacatata tacacaaaca tgtatatgtg cacacacatg tatatgtata aatattttaa 7380
atggtgtttt agaagcactt tgtctaccta agctttgaca acttgaacaa tgctaaggta 7440
ctgagatgtt taaaaaacaa gtttactttc attttagaat gcaaagttga tttttttaag 7500
gaaacaaaga aagcttttaa aatatttttg cttttagcca tgcatctgct gatgagcaat 7560
tgtgtccatt tttaacacag ccagttaaat ccaccatggg gcttactgga ttcaagggaa 7620
tacgttagtc cacaaaacat gttttctggt gctcatctca catgctatac tgtaaaacag 7680
ttttatacaa aattgtatga caagttcatt gctcaaaaat gtacagtttt aagaattttc 7740
tattaactgc aggtaataat tagctgcatg ctgcagactc aacaaagcta gttcactgaa 7800
gcctatgcta ttttatggat cataggctct tcagagaact gaatggcagt ctgcctttgt 7860
gttgataatt atgtacattg tgacgttgtc atttcttagc ttaagtgtcc tctttaacaa 7920
gaggattgag cagactgatg cctgcataag atgaataaac agggttagtt ccatgtgaat 7980
ctgtcagtta aaaagaaaca aaaacaggca gctggtttgc tgtggtggtt ttaaatcatt 8040
aatttgtata aagaagtgaa agagttgtat agtaaattaa attgtaaaca aaactttttt 8100
aatgcaatgc tttagtattt tagtactgta aaaaaattaa atatatacat atatatatat 8160
atatatatat atatatatat gagtttgaag cagaattcac atcatgatgg tgctactcag 8220
cctgctacaa atatatcata atgtgagcta agaattcatt aaatgtttga gtgatgttcc 8280
tacttgtcat atacctcaac actagtttgg caataggata ttgaactgag agtgaaagca 8340
ttgtgtacca tcattttttt ccaagtcctt ttttttattg ttaaaaaaaa aagcatacct 8400
tttttcaata cttgatttct tagcaagtat aacttgaact tcaacctttt tgttctaaaa 8460
attcagggat atttcagctc atgctctccc tatgccaaca tgtcacctgt gtttatgtaa 8520
aattgttgta ggttaataaa tatattcttt gtcagggatt taaccctttt attttgaatc 8580
ccttctattt tacttgtaca tgtgctgatg taactaaaac taattttgta aatctgttgg 8640
ctctttttat tgtaaagaaa agcattttaa aagtttgagg aatcttttga ctgtttcaag 8700
caggaaaaaa aaattacatg aaaatagaat gcactgagtt gataaaggga aaaattgtaa 8760
ggcaggagtt tggcaagtgg ctgttggcca gagacttact tgtaactctc taaatgaagt 8820
ttttttgatc ctgtaatcac tgaaggtaca tactccatgt ggacttccct taaacaggca 8880
aacacctaca ggtatggtgt gcaacagatt gtacaattac attttggcct aaatacattt 8940
ttgcttacta gtatttaaaa taaattctta atcagaggag gcctttgggt tttattggtc 9000
aaatctttgt aagctggctt ttgtcttttt aaaaaatttc ttgaatttgt ggttgtgtcc 9060
aatttgcaaa catttccaaa aatgtttgct ttgcttacaa accacatgat tttaatgttt 9120
tttgtatacc ataatatcta gccccaaaca tttgattact acatgtgcat tggtgatttt 9180
gatcatccat tcttaatatt tgatttctgt gtcacctact gtcatttgtt aaactgctgg 9240
ccaacaagaa caggaagtat agtttggggg gttggggaga gtttacataa ggaagagaag 9300
aaattgagtg gcatattgta aatatcagat ctataattgt aaatataaaa cctgcctcag 9360
ttagaatgaa tggaaagcag atctacaatt tgctaatata ggaatatcag gttgactata 9420
tagccatact tgaaaatgct tctgagtggt gtcaacttta cttgaatgaa tttttcatct 9480
tgattgacgc acagtgatgt acagttcact tctgaagcta gtggttaact tgtgtaggaa 9540
acttttgcag tttgacacta agataacttc tgtgtgcatt tttctatgct tttttaaaaa 9600
ctagtttcat ttcattttca tgagatgttt ggtttataag atctgaggat ggttataaat 9660
actgtaagta ttgtaatgtt atgaatgcag gttatttgaa agctgtttat tattatatca 9720
ttcctgataa tgctatgtga gtgtttttaa taaaatttat atttatttaa tgcactctaa 9780
aaaaaaaaaa aaaaaa 9796
<210> 1359
<211> 9437
<212> ДНК
<213> Homo sapiens
<400> 1359
aagcagaagg aagcaagatg gctgcccttt aggatttgtt agaaaggaga cccgactgca 60
actgctggat tgctgcaagg ctgagggacg agaacgagaa ttcaactaga gggcagcctt 120
gtggatggcc ccgaagcaag cctgatggaa caggatagaa ccaaccatgt tgagggcaac 180
agactaagtc cattcctgat accatcacct cccatttgcc agacagaacc tctggctaca 240
aagctccaga atggaagccc actgcctgag agagctcatc cagaagtaaa tggagacacc 300
aagtggcact ctttcaaaag ttattatgga ataccctgta tgaagggaag ccagaatagt 360
cgtgtgagtc ctgactttac acaagaaagt agagggtatt ccaagtgttt gcaaaatgga 420
ggaataaaac gcacagttag tgaaccttct ctctctgggc tccttcagat caagaaattg 480
aaacaagacc aaaaggctaa tggagaaaga cgtaacttcg gggtaagcca agaaagaaat 540
ccaggtgaaa gcagtcaacc aaatgtctcc gatttgagtg ataagaaaga atctgtgagt 600
tctgtagccc aagaaaatgc agttaaagat ttcaccagtt tttcaacaca taactgcagt 660
gggcctgaaa atccagagct tcagattctg aatgagcagg aggggaaaag tgctaattac 720
catgacaaga acattgtatt acttaaaaac aaggcagtgc taatgcctaa tggtgctaca 780
gtttctgcct cttccgtgga acacacacat ggtgaactcc tggaaaaaac actgtctcaa 840
tattatccag attgtgtttc cattgcggtg cagaaaacca catctcacat aaatgccatt 900
aacagtcagg ctactaatga gttgtcctgt gagatcactc acccatcgca tacctcaggg 960
cagatcaatt ccgcacagac ctctaactct gagctgcctc caaagccagc tgcagtggtg 1020
agtgaggcct gtgatgctga tgatgctgat aatgccagta aactagctgc aatgctaaat 1080
acctgttcct ttcagaaacc agaacaacta caacaacaaa aatcagtttt tgagatatgc 1140
ccatctcctg cagaaaataa catccaggga accacaaagc tagcgtctgg tgaagaattc 1200
tgttcaggtt ccagcagcaa tttgcaagct cctggtggca gctctgaacg gtatttaaaa 1260
caaaatgaaa tgaatggtgc ttacttcaag caaagctcag tgttcactaa ggattccttt 1320
tctgccacta ccacaccacc accaccatca caattgcttc tttctccccc tcctcctctt 1380
ccacaggttc ctcagcttcc ttcagaagga aaaagcactc tgaatggtgg agttttagaa 1440
gaacaccacc actaccccaa ccaaagtaac acaacacttt taagggaagt gaaaatagag 1500
ggtaaacctg aggcaccacc ttcccagagt cctaatccat ctacacatgt atgcagccct 1560
tctccgatgc tttctgaaag gcctcagaat aattgtgtga acaggaatga catacagact 1620
gcagggacaa tgactgttcc attgtgttct gagaaaacaa gaccaatgtc agaacacctc 1680
aagcataacc caccaatttt tggtagcagt ggagagctac aggacaactg ccagcagttg 1740
atgagaaaca aagagcaaga gattctgaag ggtcgagaca aggagcaaac acgagatctt 1800
gtgcccccaa cacagcacta tctgaaacca ggatggattg aattgaaggc ccctcgtttt 1860
caccaagcgg aatcccatct aaaacgtaat gaggcatcac tgccatcaat tcttcagtat 1920
caacccaatc tctccaatca aatgacctcc aaacaataca ctggaaattc caacatgcct 1980
ggggggctcc caaggcaagc ttacacccag aaaacaacac agctggagca caagtcacaa 2040
atgtaccaag ttgaaatgaa tcaagggcag tcccaaggta cagtggacca acatctccag 2100
ttccaaaaac cctcacacca ggtgcacttc tccaaaacag accatttacc aaaagctcat 2160
gtgcagtcac tgtgtggcac tagatttcat tttcaacaaa gagcagattc ccaaactgaa 2220
aaacttatgt ccccagtgtt gaaacagcac ttgaatcaac aggcttcaga gactgagcca 2280
ttttcaaact cacacctttt gcaacataag cctcataaac aggcagcaca aacacaacca 2340
tcccagagtt cacatctccc tcaaaaccag caacagcagc aaaaattaca aataaagaat 2400
aaagaggaaa tactccagac ttttcctcac ccccaaagca acaatgatca gcaaagagaa 2460
ggatcattct ttggccagac taaagtggaa gaatgttttc atggtgaaaa tcagtattca 2520
aaatcaagcg agttcgagac tcataatgtc caaatgggac tggaggaagt acagaatata 2580
aatcgtagaa attcccctta tagtcagacc atgaaatcaa gtgcatgcaa aatacaggtt 2640
tcttgttcaa acaatacaca cctagtttca gagaataaag aacagactac acatcctgaa 2700
ctttttgcag gaaacaagac ccaaaacttg catcacatgc aatattttcc aaataatgtg 2760
atcccaaagc aagatcttct tcacaggtgc tttcaagaac aggagcagaa gtcacaacaa 2820
gcttcagttc tacagggata taaaaataga aaccaagata tgtctggtca acaagctgcg 2880
caacttgctc agcaaaggta cttgatacat aaccatgcaa atgtttttcc tgtgcctgac 2940
cagggaggaa gtcacactca gacccctccc cagaaggaca ctcaaaagca tgctgctcta 3000
aggtggcatc tcttacagaa gcaagaacag cagcaaacac agcaacccca aactgagtct 3060
tgccatagtc agatgcacag gccaattaag gtggaacctg gatgcaagcc acatgcctgt 3120
atgcacacag caccaccaga aaacaaaaca tggaaaaagg taactaagca agagaatcca 3180
cctgcaagct gtgataatgt gcagcaaaag agcatcattg agaccatgga gcagcatctg 3240
aagcagtttc acgccaagtc gttatttgac cataaggctc ttactctcaa atcacagaag 3300
caagtaaaag ttgaaatgtc agggccagtc acagttttga ctagacaaac cactgctgca 3360
gaacttgata gccacacccc agctttagag cagcaaacaa cttcttcaga aaagacacca 3420
accaaaagaa cagctgcttc tgttctcaat aattttatag agtcaccttc caaattacta 3480
gatactccta taaaaaattt attggataca cctgtcaaga ctcaatatga tttcccatct 3540
tgcagatgtg tagagcaaat tattgaaaaa gatgaaggtc ctttttatac ccatctagga 3600
gcaggtccta atgtggcagc tattagagaa atcatggaag aaaggtttgg acagaagggt 3660
aaagctatta ggattgaaag agtcatctat actggtaaag aaggcaaaag ttctcaggga 3720
tgtcctattg ctaagtgggt ggttcgcaga agcagcagtg aagagaagct actgtgtttg 3780
gtgcgggagc gagctggcca cacctgtgag gctgcagtga ttgtgattct catcctggtg 3840
tgggaaggaa tcccgctgtc tctggctgac aaactctact cggagcttac cgagacgctg 3900
aggaaatacg gcacgctcac caatcgccgg tgtgccttga atgaagagag aacttgcgcc 3960
tgtcaggggc tggatccaga aacctgtggt gcctccttct cttttggttg ttcatggagc 4020
atgtactaca atggatgtaa gtttgccaga agcaagatcc caaggaagtt taagctgctt 4080
ggggatgacc caaaagagga agagaaactg gagtctcatt tgcaaaacct gtccactctt 4140
atggcaccaa catataagaa acttgcacct gatgcatata ataatcagat tgaatatgaa 4200
cacagagcac cagagtgccg tctgggtctg aaggaaggcc gtccattctc aggggtcact 4260
gcatgtttgg acttctgtgc tcatgcccac agagacttgc acaacatgca gaatggcagc 4320
acattggtat gcactctcac tagagaagac aatcgagaat ttggaggaaa acctgaggat 4380
gagcagcttc acgttctgcc tttatacaaa gtctctgacg tggatgagtt tgggagtgtg 4440
gaagctcagg aggagaaaaa acggagtggt gccattcagg tactgagttc ttttcggcga 4500
aaagtcagga tgttagcaga gccagtcaag acttgccgac aaaggaaact agaagccaag 4560
aaagctgcag ctgaaaagct ttcctccctg gagaacagct caaataaaaa tgaaaaggaa 4620
aagtcagccc catcacgtac aaaacaaact gaaaacgcaa gccaggctaa acagttggca 4680
gaacttttgc gactttcagg accagtcatg cagcagtccc agcagcccca gcctctacag 4740
aagcagccac cacagcccca gcagcagcag agaccccagc agcagcagcc acatcaccct 4800
cagacagagt ctgtcaactc ttattctgct tctggatcca ccaatccata catgagacgg 4860
cccaatccag ttagtcctta tccaaactct tcacacactt cagatatcta tggaagcacc 4920
agccctatga acttctattc cacctcatct caagctgcag gttcatattt gaattcttct 4980
aatcccatga acccttaccc tgggcttttg aatcagaata cccaatatcc atcatatcaa 5040
tgcaatggaa acctatcagt ggacaactgc tccccatatc tgggttccta ttctccccag 5100
tctcagccga tggatctgta taggtatcca agccaagacc ctctgtctaa gctcagtcta 5160
ccacccatcc atacacttta ccagccaagg tttggaaata gccagagttt tacatctaaa 5220
tacttaggtt atggaaacca aaatatgcag ggagatggtt tcagcagttg taccattaga 5280
ccaaatgtac atcatgtagg gaaattgcct ccttatccca ctcatgagat ggatggccac 5340
ttcatgggag ccacctctag attaccaccc aatctgagca atccaaacat ggactataaa 5400
aatggtgaac atcattcacc ttctcacata atccataact acagtgcagc tccgggcatg 5460
ttcaacagct ctcttcatgc cctgcatctc caaaacaagg agaatgacat gctttcccac 5520
acagctaatg ggttatcaaa gatgcttcca gctcttaacc atgatagaac tgcttgtgtc 5580
caaggaggct tacacaaatt aagtgatgct aatggtcagg aaaagcagcc attggcacta 5640
gtccagggtg tggcttctgg tgcagaggac aacgatgagg tctggtcaga cagcgagcag 5700
agctttctgg atcctgacat tgggggagtg gccgtggctc caactcatgg gtcaattctc 5760
attgagtgtg caaagcgtga gctgcatgcc acaacccctt taaagaatcc caataggaat 5820
caccccacca ggatctccct cgtcttttac cagcataaga gcatgaatga gccaaaacat 5880
ggcttggctc tttgggaagc caaaatggct gaaaaagccc gtgagaaaga ggaagagtgt 5940
gaaaagtatg gcccagacta tgtgcctcag aaatcccatg gcaaaaaagt gaaacgggag 6000
cctgctgagc cacatgaaac ttcagagccc acttacctgc gtttcatcaa gtctcttgcc 6060
gaaaggacca tgtccgtgac cacagactcc acagtaacta catctccata tgccttcact 6120
cgggtcacag ggccttacaa cagatatata tgatatcacc cccttttgtt ggttacctca 6180
cttgaaaaga ccacaaccaa cctgtcagta gtatagttct catgacgtgg gcagtgggga 6240
aaggtcacag tattcatgac aaatgtggtg ggaaaaacct cagctcacca gcaacaaaag 6300
aggttatctt accatagcac ttaattttca ctggctccca agtggtcaca gatggcatct 6360
aggaaaagac caaagcattc tatgcaaaaa gaaggtgggg aagaaagtgt tccgcaattt 6420
acatttttaa acactggttc tattattgga cgagatgata tgtaaatgtg atcccccccc 6480
cccgcttaca actctacaca tctgtgacca cttttaataa tatcaagttt gcatagtcat 6540
ggaacacaaa tcaaacaagt actgtagtat tacagtgaca ggaatcttaa aataccatct 6600
ggtgctgaat atatgatgta ctgaaatact ggaattatgg ctttttgaaa tgcagttttt 6660
actgtaatct taacttttat ttatcaaaat agctacagga aacatgaata gcaggaaaac 6720
actgaatttg tttggatgtt ctaagaaatg gtgctaagaa aatggtgtct ttaatagcta 6780
aaaatttaat gcctttatat catcaagatg ctatcagtgt actccagtgc ccttgaataa 6840
taggggtacc ttttcattca agtttttatc ataattacct attcttacac aagcttagtt 6900
tttaaaatgt ggacatttta aaggcctctg gattttgctc atccagtgaa gtccttgtag 6960
gacaataaac gtatatatgt acatatatac acaaacatgt atatgtgcac acacatgtat 7020
atgtataaat attttaaatg gtgttttaga agcactttgt ctacctaagc tttgacaact 7080
tgaacaatgc taaggtactg agatgtttaa aaaacaagtt tactttcatt ttagaatgca 7140
aagttgattt ttttaaggaa acaaagaaag cttttaaaat atttttgctt ttagccatgc 7200
atctgctgat gagcaattgt gtccattttt aacacagcca gttaaatcca ccatggggct 7260
tactggattc aagggaatac gttagtccac aaaacatgtt ttctggtgct catctcacat 7320
gctatactgt aaaacagttt tatacaaaat tgtatgacaa gttcattgct caaaaatgta 7380
cagttttaag aattttctat taactgcagg taataattag ctgcatgctg cagactcaac 7440
aaagctagtt cactgaagcc tatgctattt tatggatcat aggctcttca gagaactgaa 7500
tggcagtctg cctttgtgtt gataattatg tacattgtga cgttgtcatt tcttagctta 7560
agtgtcctct ttaacaagag gattgagcag actgatgcct gcataagatg aataaacagg 7620
gttagttcca tgtgaatctg tcagttaaaa agaaacaaaa acaggcagct ggtttgctgt 7680
ggtggtttta aatcattaat ttgtataaag aagtgaaaga gttgtatagt aaattaaatt 7740
gtaaacaaaa cttttttaat gcaatgcttt agtattttag tactgtaaaa aaattaaata 7800
tatacatata tatatatata tatatatata tatatatgag tttgaagcag aattcacatc 7860
atgatggtgc tactcagcct gctacaaata tatcataatg tgagctaaga attcattaaa 7920
tgtttgagtg atgttcctac ttgtcatata cctcaacact agtttggcaa taggatattg 7980
aactgagagt gaaagcattg tgtaccatca tttttttcca agtccttttt tttattgtta 8040
aaaaaaaaag catacctttt ttcaatactt gatttcttag caagtataac ttgaacttca 8100
acctttttgt tctaaaaatt cagggatatt tcagctcatg ctctccctat gccaacatgt 8160
cacctgtgtt tatgtaaaat tgttgtaggt taataaatat attctttgtc agggatttaa 8220
cccttttatt ttgaatccct tctattttac ttgtacatgt gctgatgtaa ctaaaactaa 8280
ttttgtaaat ctgttggctc tttttattgt aaagaaaagc attttaaaag tttgaggaat 8340
cttttgactg tttcaagcag gaaaaaaaaa ttacatgaaa atagaatgca ctgagttgat 8400
aaagggaaaa attgtaaggc aggagtttgg caagtggctg ttggccagag acttacttgt 8460
aactctctaa atgaagtttt tttgatcctg taatcactga aggtacatac tccatgtgga 8520
cttcccttaa acaggcaaac acctacaggt atggtgtgca acagattgta caattacatt 8580
ttggcctaaa tacatttttg cttactagta tttaaaataa attcttaatc agaggaggcc 8640
tttgggtttt attggtcaaa tctttgtaag ctggcttttg tctttttaaa aaatttcttg 8700
aatttgtggt tgtgtccaat ttgcaaacat ttccaaaaat gtttgctttg cttacaaacc 8760
acatgatttt aatgtttttt gtataccata atatctagcc ccaaacattt gattactaca 8820
tgtgcattgg tgattttgat catccattct taatatttga tttctgtgtc acctactgtc 8880
atttgttaaa ctgctggcca acaagaacag gaagtatagt ttggggggtt ggggagagtt 8940
tacataagga agagaagaaa ttgagtggca tattgtaaat atcagatcta taattgtaaa 9000
tataaaacct gcctcagtta gaatgaatgg aaagcagatc tacaatttgc taatatagga 9060
atatcaggtt gactatatag ccatacttga aaatgcttct gagtggtgtc aactttactt 9120
gaatgaattt ttcatcttga ttgacgcaca gtgatgtaca gttcacttct gaagctagtg 9180
gttaacttgt gtaggaaact tttgcagttt gacactaaga taacttctgt gtgcattttt 9240
ctatgctttt ttaaaaacta gtttcatttc attttcatga gatgtttggt ttataagatc 9300
tgaggatggt tataaatact gtaagtattg taatgttatg aatgcaggtt atttgaaagc 9360
tgtttattat tatatcattc ctgataatgc tatgtgagtg tttttaataa aatttatatt 9420
tatttaatgc actctaa 9437
<210> 1360
<211> 9289
<212> ДНК
<213> Homo sapiens
<400> 1360
gtagagaagc agaaggaagc aagatggctg ccctttagga tttgttagaa aggagacccg 60
actgcaactg ctggattgct gcaaggctga gggacgagaa cgaggctggc aaacattcag 120
cagcacaccc tctcaagatt gtttacttgc ctttgctcct gttgagttac aacgcttgga 180
agcaggagat gggctcagca gcagccaata ggacatgatc caggaagagc agtaagggac 240
tgagctgctg aattcaacta gagggcagcc ttgtggatgg ccccgaagca agcctgatgg 300
aacaggatag aaccaaccat gttgagggca acagactaag tccattcctg ataccatcac 360
ctcccatttg ccagacagaa cctctggcta caaagctcca gaatggaagc ccactgcctg 420
agagagctca tccagaagta aatggagaca ccaagtggca ctctttcaaa agttattatg 480
gaataccctg tatgaaggga agccagaata gtcgtgtgag tcctgacttt acacaagaaa 540
gtagagggta ttccaagtgt ttgcaaaatg gaggaataaa acgcacagtt agtgaacctt 600
ctctctctgg gctccttcag atcaagaaat tgaaacaaga ccaaaaggct aatggagaaa 660
gacgtaactt cggggtaagc caagaaagaa atccaggtga aagcagtcaa ccaaatgtct 720
ccgatttgag tgataagaaa gaatctgtga gttctgtagc ccaagaaaat gcagttaaag 780
atttcaccag tttttcaaca cataactgca gtgggcctga aaatccagag cttcagattc 840
tgaatgagca ggaggggaaa agtgctaatt accatgacaa gaacattgta ttacttaaaa 900
acaaggcagt gctaatgcct aatggtgcta cagtttctgc ctcttccgtg gaacacacac 960
atggtgaact cctggaaaaa acactgtctc aatattatcc agattgtgtt tccattgcgg 1020
tgcagaaaac cacatctcac ataaatgcca ttaacagtca ggctactaat gagttgtcct 1080
gtgagatcac tcacccatcg catacctcag ggcagatcaa ttccgcacag acctctaact 1140
ctgagctgcc tccaaagcca gctgcagtgg tgagtgaggc ctgtgatgct gatgatgctg 1200
ataatgccag taaactagct gcaatgctaa atacctgttc ctttcagaaa ccagaacaac 1260
tacaacaaca aaaatcagtt tttgagatat gcccatctcc tgcagaaaat aacatccagg 1320
gaaccacaaa gctagcgtct ggtgaagaat tctgttcagg ttccagcagc aatttgcaag 1380
ctcctggtgg cagctctgaa cggtatttaa aacaaaatga aatgaatggt gcttacttca 1440
agcaaagctc agtgttcact aaggattcct tttctgccac taccacacca ccaccaccat 1500
cacaattgct tctttctccc cctcctcctc ttccacaggt tcctcagctt ccttcagaag 1560
gaaaaagcac tctgaatggt ggagttttag aagaacacca ccactacccc aaccaaagta 1620
acacaacact tttaagggaa gtgaaaatag agggtaaacc tgaggcacca ccttcccaga 1680
gtcctaatcc atctacacat gtatgcagcc cttctccgat gctttctgaa aggcctcaga 1740
ataattgtgt gaacaggaat gacatacaga ctgcagggac aatgactgtt ccattgtgtt 1800
ctgagaaaac aagaccaatg tcagaacacc tcaagcataa cccaccaatt tttggtagca 1860
gtggagagct acaggacaac tgccagcagt tgatgagaaa caaagagcaa gagattctga 1920
agggtcgaga caaggagcaa acacgagatc ttgtgccccc aacacagcac tatctgaaac 1980
caggatggat tgaattgaag gcccctcgtt ttcaccaagc ggaatcccat ctaaaacgta 2040
atgaggcatc actgccatca attcttcagt atcaacccaa tctctccaat caaatgacct 2100
ccaaacaata cactggaaat tccaacatgc ctggggggct cccaaggcaa gcttacaccc 2160
agaaaacaac acagctggag cacaagtcac aaatgtacca agttgaaatg aatcaagggc 2220
agtcccaagg tacagtggac caacatctcc agttccaaaa accctcacac caggtgcact 2280
tctccaaaac agaccattta ccaaaagctc atgtgcagtc actgtgtggc actagatttc 2340
attttcaaca aagagcagat tcccaaactg aaaaacttat gtccccagtg ttgaaacagc 2400
acttgaatca acaggcttca gagactgagc cattttcaaa ctcacacctt ttgcaacata 2460
agcctcataa acaggcagca caaacacaac catcccagag ttcacatctc cctcaaaacc 2520
agcaacagca gcaaaaatta caaataaaga ataaagagga aatactccag acttttcctc 2580
acccccaaag caacaatgat cagcaaagag aaggatcatt ctttggccag actaaagtgg 2640
aagaatgttt tcatggtgaa aatcagtatt caaaatcaag cgagttcgag actcataatg 2700
tccaaatggg actggaggaa gtacagaata taaatcgtag aaattcccct tatagtcaga 2760
ccatgaaatc aagtgcatgc aaaatacagg tttcttgttc aaacaataca cacctagttt 2820
cagagaataa agaacagact acacatcctg aactttttgc aggaaacaag acccaaaact 2880
tgcatcacat gcaatatttt ccaaataatg tgatcccaaa gcaagatctt cttcacaggt 2940
gctttcaaga acaggagcag aagtcacaac aagcttcagt tctacaggga tataaaaata 3000
gaaaccaaga tatgtctggt caacaagctg cgcaacttgc tcagcaaagg tacttgatac 3060
ataaccatgc aaatgttttt cctgtgcctg accagggagg aagtcacact cagacccctc 3120
cccagaagga cactcaaaag catgctgctc taaggtggca tctcttacag aagcaagaac 3180
agcagcaaac acagcaaccc caaactgagt cttgccatag tcagatgcac aggccaatta 3240
aggtggaacc tggatgcaag ccacatgcct gtatgcacac agcaccacca gaaaacaaaa 3300
catggaaaaa ggtaactaag caagagaatc cacctgcaag ctgtgataat gtgcagcaaa 3360
agagcatcat tgagaccatg gagcagcatc tgaagcagtt tcacgccaag tcgttatttg 3420
accataaggc tcttactctc aaatcacaga agcaagtaaa agttgaaatg tcagggccag 3480
tcacagtttt gactagacaa accactgctg cagaacttga tagccacacc ccagctttag 3540
agcagcaaac aacttcttca gaaaagacac caaccaaaag aacagctgct tctgttctca 3600
ataattttat agagtcacct tccaaattac tagatactcc tataaaaaat ttattggata 3660
cacctgtcaa gactcaatat gatttcccat cttgcagatg tgtaggtttg gacagaaggg 3720
taaagctatt aggattgaaa gagtcatcta tactggtaaa gaaggcaaaa gttctcaggg 3780
atgtcctatt gctaagtggg agaacttgcg cctgtcaggg gctggatcca gaaacctgtg 3840
gtgcctcctt ctcttttggt tgttcatgga gcatgtacta caatggatgt aagtttgcca 3900
gaagcaagat cccaaggaag tttaagctgc ttggggatga cccaaaagag gaagagaaac 3960
tggagtctca tttgcaaaac ctgtccactc ttatggcacc aacatataag aaacttgcac 4020
ctgatgcata taataatcag attgaatatg aacacagagc accagagtgc cgtctgggtc 4080
tgaaggaagg ccgtccattc tcaggggtca ctgcatgttt ggacttctgt gctcatgccc 4140
acagagactt gcacaacatg cagaatggca gcacattggt atgcactctc actagagaag 4200
acaatcgaga atttggagga aaacctgagg atgagcagct tcacgttctg cctttataca 4260
aagtctctga cgtggatgag tttgggagtg tggaagctca ggaggagaaa aaacggagtg 4320
gtgccattca ggtactgagt tcttttcggc gaaaagtcag gatgttagca gagccagtca 4380
agacttgccg acaaaggaaa ctagaagcca agaaagctgc agctgaaaag ctttcctccc 4440
tggagaacag ctcaaataaa aatgaaaagg aaaagtcagc cccatcacgt acaaaacaaa 4500
ctgaaaacgc aagccaggct aaacagttgg cagaactttt gcgactttca ggaccagtca 4560
tgcagcagtc ccagcagccc cagcctctac agaagcagcc accacagccc cagcagcagc 4620
agagacccca gcagcagcag ccacatcacc ctcagacaga gtctgtcaac tcttattctg 4680
cttctggatc caccaatcca tacatgagac ggcccaatcc agttagtcct tatccaaact 4740
cttcacacac ttcagatatc tatggaagca ccagccctat gaacttctat tccacctcat 4800
ctcaagctgc aggttcatat ttgaattctt ctaatcccat gaacccttac cctgggcttt 4860
tgaatcagaa tacccaatat ccatcatatc aatgcaatgg aaacctatca gtggacaact 4920
gctccccata tctgggttcc tattctcccc agtctcagcc gatggatctg tataggtatc 4980
caagccaaga ccctctgtct aagctcagtc taccacccat ccatacactt taccagccaa 5040
ggtttggaaa tagccagagt tttacatcta aatacttagg ttatggaaac caaaatatgc 5100
agggagatgg tttcagcagt tgtaccatta gaccaaatgt acatcatgta gggaaattgc 5160
ctccttatcc cactcatgag atggatggcc acttcatggg agccacctct agattaccac 5220
ccaatctgag caatccaaac atggactata aaaatggtga acatcattca ccttctcaca 5280
taatccataa ctacagtgca gctccgggca tgttcaacag ctctcttcat gccctgcatc 5340
tccaaaacaa ggagaatgac atgctttccc acacagctaa tgggttatca aagatgcttc 5400
cagctcttaa ccatgataga actgcttgtg tccaaggagg cttacacaaa ttaagtgatg 5460
ctaatggtca ggaaaagcag ccattggcac tagtccaggg tgtggcttct ggtgcagagg 5520
acaacgatga ggtctggtca gacagcgagc agagctttct ggatcctgac attgggggag 5580
tggccgtggc tccaactcat gggtcaattc tcattgagtg tgcaaagcgt gagctgcatg 5640
ccacaacccc tttaaagaat cccaatagga atcaccccac caggatctcc ctcgtctttt 5700
accagcataa gagcatgaat gagccaaaac atggcttggc tctttgggaa gccaaaatgg 5760
ctgaaaaagc ccgtgagaaa gaggaagagt gtgaaaagta tggcccagac tatgtgcctc 5820
agaaatccca tggcaaaaaa gtgaaacggg agcctgctga gccacatgaa acttcagagc 5880
ccacttacct gcgtttcatc aagtctcttg ccgaaaggac catgtccgtg accacagact 5940
ccacagtaac tacatctcca tatgccttca ctcgggtcac agggccttac aacagatata 6000
tatgatatca cccccttttg ttggttacct cacttgaaaa gaccacaacc aacctgtcag 6060
tagtatagtt ctcatgacgt gggcagtggg gaaaggtcac agtattcatg acaaatgtgg 6120
tgggaaaaac ctcagctcac cagcaacaaa agaggttatc ttaccatagc acttaatttt 6180
cactggctcc caagtggtca cagatggcat ctaggaaaag accaaagcat tctatgcaaa 6240
aagaaggtgg ggaagaaagt gttccgcaat ttacattttt aaacactggt tctattattg 6300
gacgagatga tatgtaaatg tgatcccccc cccccgctta caactctaca catctgtgac 6360
cacttttaat aatatcaagt ttgcatagtc atggaacaca aatcaaacaa gtactgtagt 6420
attacagtga caggaatctt aaaataccat ctggtgctga atatatgatg tactgaaata 6480
ctggaattat ggctttttga aatgcagttt ttactgtaat cttaactttt atttatcaaa 6540
atagctacag gaaacatgaa tagcaggaaa acactgaatt tgtttggatg ttctaagaaa 6600
tggtgctaag aaaatggtgt ctttaatagc taaaaattta atgcctttat atcatcaaga 6660
tgctatcagt gtactccagt gcccttgaat aataggggta ccttttcatt caagttttta 6720
tcataattac ctattcttac acaagcttag tttttaaaat gtggacattt taaaggcctc 6780
tggattttgc tcatccagtg aagtccttgt aggacaataa acgtatatat gtacatatat 6840
acacaaacat gtatatgtgc acacacatgt atatgtataa atattttaaa tggtgtttta 6900
gaagcacttt gtctacctaa gctttgacaa cttgaacaat gctaaggtac tgagatgttt 6960
aaaaaacaag tttactttca ttttagaatg caaagttgat ttttttaagg aaacaaagaa 7020
agcttttaaa atatttttgc ttttagccat gcatctgctg atgagcaatt gtgtccattt 7080
ttaacacagc cagttaaatc caccatgggg cttactggat tcaagggaat acgttagtcc 7140
acaaaacatg ttttctggtg ctcatctcac atgctatact gtaaaacagt tttatacaaa 7200
attgtatgac aagttcattg ctcaaaaatg tacagtttta agaattttct attaactgca 7260
ggtaataatt agctgcatgc tgcagactca acaaagctag ttcactgaag cctatgctat 7320
tttatggatc ataggctctt cagagaactg aatggcagtc tgcctttgtg ttgataatta 7380
tgtacattgt gacgttgtca tttcttagct taagtgtcct ctttaacaag aggattgagc 7440
agactgatgc ctgcataaga tgaataaaca gggttagttc catgtgaatc tgtcagttaa 7500
aaagaaacaa aaacaggcag ctggtttgct gtggtggttt taaatcatta atttgtataa 7560
agaagtgaaa gagttgtata gtaaattaaa ttgtaaacaa aactttttta atgcaatgct 7620
ttagtatttt agtactgtaa aaaaattaaa tatatacata tatatatata tatatatata 7680
tatatatatg agtttgaagc agaattcaca tcatgatggt gctactcagc ctgctacaaa 7740
tatatcataa tgtgagctaa gaattcatta aatgtttgag tgatgttcct acttgtcata 7800
tacctcaaca ctagtttggc aataggatat tgaactgaga gtgaaagcat tgtgtaccat 7860
catttttttc caagtccttt tttttattgt taaaaaaaaa agcatacctt ttttcaatac 7920
ttgatttctt agcaagtata acttgaactt caaccttttt gttctaaaaa ttcagggata 7980
tttcagctca tgctctccct atgccaacat gtcacctgtg tttatgtaaa attgttgtag 8040
gttaataaat atattctttg tcagggattt aaccctttta ttttgaatcc cttctatttt 8100
acttgtacat gtgctgatgt aactaaaact aattttgtaa atctgttggc tctttttatt 8160
gtaaagaaaa gcattttaaa agtttgagga atcttttgac tgtttcaagc aggaaaaaaa 8220
aattacatga aaatagaatg cactgagttg ataaagggaa aaattgtaag gcaggagttt 8280
ggcaagtggc tgttggccag agacttactt gtaactctct aaatgaagtt tttttgatcc 8340
tgtaatcact gaaggtacat actccatgtg gacttccctt aaacaggcaa acacctacag 8400
gtatggtgtg caacagattg tacaattaca ttttggccta aatacatttt tgcttactag 8460
tatttaaaat aaattcttaa tcagaggagg cctttgggtt ttattggtca aatctttgta 8520
agctggcttt tgtcttttta aaaaatttct tgaatttgtg gttgtgtcca atttgcaaac 8580
atttccaaaa atgtttgctt tgcttacaaa ccacatgatt ttaatgtttt ttgtatacca 8640
taatatctag ccccaaacat ttgattacta catgtgcatt ggtgattttg atcatccatt 8700
cttaatattt gatttctgtg tcacctactg tcatttgtta aactgctggc caacaagaac 8760
aggaagtata gtttgggggg ttggggagag tttacataag gaagagaaga aattgagtgg 8820
catattgtaa atatcagatc tataattgta aatataaaac ctgcctcagt tagaatgaat 8880
ggaaagcaga tctacaattt gctaatatag gaatatcagg ttgactatat agccatactt 8940
gaaaatgctt ctgagtggtg tcaactttac ttgaatgaat ttttcatctt gattgacgca 9000
cagtgatgta cagttcactt ctgaagctag tggttaactt gtgtaggaaa cttttgcagt 9060
ttgacactaa gataacttct gtgtgcattt ttctatgctt ttttaaaaac tagtttcatt 9120
tcattttcat gagatgtttg gtttataaga tctgaggatg gttataaata ctgtaagtat 9180
tgtaatgtta tgaatgcagg ttatttgaaa gctgtttatt attatatcat tcctgataat 9240
gctatgtgag tgtttttaat aaaatttata tttatttaat gcactctaa 9289
<210> 1361
<211> 9236
<212> ДНК
<213> Homo sapiens
<400> 1361
aaacagaagg tgggccgggg cggggagaaa cagaactcgg tcaatttccc agtttgtcgg 60
gtctttaaaa atacaggccc ctaaagcact aagggcatgc cctcggtgaa acaggggagc 120
gcttctgctg aatgagatta aagcgacaga aaagggaaag gagagcgcgg gcaacgggat 180
ctaaagggag atagagacgc gggcctctga gggctggcaa acattcagca gcacaccctc 240
tcaagattgt ttacttgcct ttgctcctgt tgagttacaa cgcttggaag caggagatgg 300
gctcagcagc agccaatagg acatgatcca ggaagagcag taagggactg agctgctgaa 360
ttcaactaga gggcagcctt gtggatggcc ccgaagcaag cctgatggaa caggatagaa 420
ccaaccatgt tgagggcaac agactaagtc cattcctgat accatcacct cccatttgcc 480
agacagaacc tctggctaca aagctccaga atggaagccc actgcctgag agagctcatc 540
cagaagtaaa tggagacacc aagtggcact ctttcaaaag ttattatgga ataccctgta 600
tgaagggaag ccagaatagt cgtgtgagtc ctgactttac acaagaaagt agagggtatt 660
ccaagtgttt gcaaaatgga ggaataaaac gcacagttag tgaaccttct ctctctgggc 720
tccttcagat caagaaattg aaacaagacc aaaaggctaa tggagaaaga cgtaacttcg 780
gggtaagcca agaaagaaat ccaggtgaaa gcagtcaacc aaatgtctcc gatttgagtg 840
ataagaaaga atctgtgagt tctgtagccc aagaaaatgc agttaaagat ttcaccagtt 900
tttcaacaca taactgcagt gggcctgaaa atccagagct tcagattctg aatgagcagg 960
aggggaaaag tgctaattac catgacaaga acattgtatt acttaaaaac aaggcagtgc 1020
taatgcctaa tggtgctaca gtttctgcct cttccgtgga acacacacat ggtgaactcc 1080
tggaaaaaac actgtctcaa tattatccag attgtgtttc cattgcggtg cagaaaacca 1140
catctcacat aaatgccatt aacagtcagg ctactaatga gttgtcctgt gagatcactc 1200
acccatcgca tacctcaggg cagatcaatt ccgcacagac ctctaactct gagctgcctc 1260
caaagccagc tgcagtggtg agtgaggcct gtgatgctga tgatgctgat aatgccagta 1320
aactagctgc aatgctaaat acctgttcct ttcagaaacc agaacaacta caacaacaaa 1380
aatcagtttt tgagatatgc ccatctcctg cagaaaataa catccaggga accacaaagc 1440
tagcgtctgg tgaagaattc tgttcaggtt ccagcagcaa tttgcaagct cctggtggca 1500
gctctgaacg gtatttaaaa caaaatgaaa tgaatggtgc ttacttcaag caaagctcag 1560
tgttcactaa ggattccttt tctgccacta ccacaccacc accaccatca caattgcttc 1620
tttctccccc tcctcctctt ccacaggttc ctcagcttcc ttcagaagga aaaagcactc 1680
tgaatggtgg agttttagaa gaacaccacc actaccccaa ccaaagtaac acaacacttt 1740
taagggaagt gaaaatagag ggtaaacctg aggcaccacc ttcccagagt cctaatccat 1800
ctacacatgt atgcagccct tctccgatgc tttctgaaag gcctcagaat aattgtgtga 1860
acaggaatga catacagact gcagggacaa tgactgttcc attgtgttct gagaaaacaa 1920
gaccaatgtc agaacacctc aagcataacc caccaatttt tggtagcagt ggagagctac 1980
aggacaactg ccagcagttg atgagaaaca aagagcaaga gattctgaag ggtcgagaca 2040
aggagcaaac acgagatctt gtgcccccaa cacagcacta tctgaaacca ggatggattg 2100
aattgaaggc ccctcgtttt caccaagcgg aatcccatct aaaacgtaat gaggcatcac 2160
tgccatcaat tcttcagtat caacccaatc tctccaatca aatgacctcc aaacaataca 2220
ctggaaattc caacatgcct ggggggctcc caaggcaagc ttacacccag aaaacaacac 2280
agctggagca caagtcacaa atgtaccaag ttgaaatgaa tcaagggcag tcccaaggta 2340
cagtggacca acatctccag ttccaaaaac cctcacacca ggtgcacttc tccaaaacag 2400
accatttacc aaaagctcat gtgcagtcac tgtgtggcac tagatttcat tttcaacaaa 2460
gagcagattc ccaaactgaa aaacttatgt ccccagtgtt gaaacagcac ttgaatcaac 2520
aggcttcaga gactgagcca ttttcaaact cacacctttt gcaacataag cctcataaac 2580
aggcagcaca aacacaacca tcccagagtt cacatctccc tcaaaaccag caacagcagc 2640
aaaaattaca aataaagaat aaagaggaaa tactccagac ttttcctcac ccccaaagca 2700
acaatgatca gcaaagagaa ggatcattct ttggccagac taaagtggaa gaatgttttc 2760
atggtgaaaa tcagtattca aaatcaagcg agttcgagac tcataatgtc caaatgggac 2820
tggaggaagt acagaatata aatcgtagaa attcccctta tagtcagacc atgaaatcaa 2880
gtgcatgcaa aatacaggtt tcttgttcaa acaatacaca cctagtttca gagaataaag 2940
aacagactac acatcctgaa ctttttgcag gaaacaagac ccaaaacttg catcacatgc 3000
aatattttcc aaataatgtg atcccaaagc aagatcttct tcacaggtgc tttcaagaac 3060
aggagcagaa gtcacaacaa gcttcagttc tacagggata taaaaataga aaccaagata 3120
tgtctggtca acaagctgcg caacttgctc agcaaaggta cttgatacat aaccatgcaa 3180
atgtttttcc tgtgcctgac cagggaggaa gtcacactca gacccctccc cagaaggaca 3240
ctcaaaagca tgctgctcta aggtggcatc tcttacagaa gcaagaacag cagcaaacac 3300
agcaacccca aactgagtct tgccatagtc agatgcacag gccaattaag gtggaacctg 3360
gatgcaagcc acatgcctgt atgcacacag caccaccaga aaacaaaaca tggaaaaagg 3420
taactaagca agagaatcca cctgcaagct gtgataatgt gcagcaaaag agcatcattg 3480
agaccatgga gcagcatctg aagcagtttc acgccaagtc gttatttgac cataaggctc 3540
ttactctcaa atcacagaag caagtaaaag ttgaaatgtc agggccagtc acagttttga 3600
ctagacaaac cactgctgca gaacttgata gccacacccc agctttagag cagcaaacaa 3660
cttcttcaga aaagacacca accaaaagaa cagctgcttc tgttctcaat aattttatag 3720
agtcaccttc caaattacta gatactccta taaaaaattt attggataca cctgtcaaga 3780
ctcaatatga tttcccatct tgcagatgtg taggtaagtg ccagaaatgt actgagacac 3840
atggcgttta tccagaatta gcaaatttat cttcagatat gggattttcc ttcttttttt 3900
aaatcttgag tctggcagca atttgtaaag gctcataaaa atctgaagct tacatttttt 3960
gtcaagttac cgatgcttgt gtcttgtgaa agagaacttc acttacatgc agtttttcca 4020
aaagaattaa ataatcgtgc atgtttattt ttccctctct tcagatcctg taaaatttga 4080
atgtatctgt tttagatcaa ttcgcctatt tagctctttg tatattatct cctggagaga 4140
cagctaggca gcaaaaaaac aatctattaa aatgagaaaa taacgaccat aggcagtcta 4200
atgtacgaac tttaaatatt ttttaattca aggtaaaata tattagtttc acaagatttc 4260
tggctaatag ggaaattatt atcttcagtc ttcatgagtt gggggaaatg ataatgctga 4320
cactcttagt gctcctaaag tttccttttc tccatttata catttggaat gttgtgattt 4380
atattcattt tgattccctt ttctctaaaa tttcatcttt ttgattaaaa aatatgatac 4440
aggcatacct cagagatatt gtgggtttgg ctccatacca caataaaatg aatattacaa 4500
taaagcaagt tgtaaggact ttttggtttc tcactgtatg taaaagttat ttatatacta 4560
tactgtaaca tactaagtgt gcaatagcat tgtgtctaaa aaatatatac tttaaaaata 4620
atttattgtt aaaaaaatgc caacaattat ctgggccttt agtgagtgct aatctttttg 4680
ctggtggagg gtcgtgcttc agtattgatc gctgtggact gatcatggtg gtagttgctg 4740
aaggttgctg ggatggctgt gtgtgtggca atttcttaaa ataagacaac agtgaagtgc 4800
tgtatcaatt gatttttcca ttcacaaaag atttctctgt agcatgcaat gctgtttgat 4860
agcatttaac ccacagcaga atttctttga aaattggact cagtcctctc aaactgtgct 4920
gctgctttat caactaagtt tttgtaattt tctgaatcct ttgttgtcat ttcagcagtt 4980
tacagcatct tcattggaag tatattccat ctcaaacatt ctttgttcat ccataagaag 5040
caacttctta tcaagttttt tcatgacatt gcagtaactc agccccatct tcaggctcta 5100
cttctaattc tggttctctt gctacatctc cctcatctgc agtgacctct ccacggaagt 5160
cttgaactcc tcaaagtaat ccatgagggt tggaatcaac ttctaaactc ctgttaatgt 5220
tgatatattg accccctccc atgaattatg aatgttctta ataacttcta aatggtgata 5280
cctttccaga aggctttcaa tgtactttgc ccggatccat cagaagacta tcttggcagc 5340
tgtagactaa caatatattt cttaaatgat aagacttgaa agtcaaaagt actccttaat 5400
ccataggctg cagaatcaat gttgtattaa caggcacgaa aacagcatta atcttgtgca 5460
tctccatcgg agctcttggg tgactaggtg ccttgagcag taatattttg aaaggaggtt 5520
ttggttttgt tttttgtttt ttttttttgt tttttagcag taagtctcaa cactgggctt 5580
aaaatattca gtaaactatg ttgtaaaaag atgtgttatc atccagactt tgttgttcca 5640
ttactctaca caagcagggt acacttagca taattcttaa gggccttgga attttcagaa 5700
tggtaaatga gtatgggctt caacttaaaa tcatcaactg cattagcctg taacaagaga 5760
gtcagcctgt cctttgaagc aaggcattga cttctatcta tgaaagtctt agatggcacc 5820
ttgtttcaat agtaggctgt ttagtacagc caccttcatc agtgatctta gctagatctt 5880
ctgcataact tgctgcagct tctacatcag cacttgctgc ctcaccttgt ccttttatgt 5940
tatagagaca gctgcgcttc ttaaacttta taaaccaact tctgctagct tccaacttct 6000
cttctgcagc ttcctcattc tcttcataga actgaaggga gtcaaggcct tgctctggat 6060
taagctttgg cttaaggaat gttgtggctg acgtgatctt ctatccagac cactaaagcg 6120
ctctccatat cagcaataag gccgttttgc tttcttacct ttcatgtgtt cactggagta 6180
atttccttca agaatttttc ctttacattc acaacttggc taactggcat gcaaggccta 6240
gctttcagcc tgtcttggct tttgacatgc cttcctcact tagctcgtca tatctagctt 6300
ttgatttaaa gtggcaggca tacaactctt cctttcactt gaacacttag aggccactgt 6360
agggttatta attggcctaa tttcaatatt gttgtgtttt agggaataga gaggcccagg 6420
gagagggaga gagcccaaac ggctggttga tagagcaggc agaatgcaca caacatttat 6480
cagattatgt ttgcaccatt taccagatta tgggtacggt ttgtggcacc ccccaaaaat 6540
tagaatagta acatcaaaga tcactgatca cagatcgcca taacataaat aataataaac 6600
tttaaaatac tgtgagaatt accaaaatgt gatacagaga catgaagtga gcacatgctg 6660
ttgaaaaaaa tgacactgat agacatactt aacacgtggg attgccacaa accttcagtt 6720
tgtaaaagtc acagtaactg tgactcacaa aagaacaaag cacaataaaa cgaggtatgc 6780
ctgtattttt aaaaaaagct ttttgttaaa attcaggata tgtaataggt ctgtaggaat 6840
agtgaaatat ttttgctgat ggatgtagat atatacgtgg atagagatga agatcttaat 6900
tatagctatg cagcatagat ttagtcaaag acatttgaaa agacaaatgt taaattagtg 6960
tggctaatga cctacccgtg ccatgttttc cctcttgcaa tgagataccc cacactgtgt 7020
agaaggatgg agggaggact cctactgtcc ctctttgcgt gtggttatta agttgcctca 7080
ctgggctaaa acaccacaca tctcatagat aatatttggt aagttgtaat cgtcttcact 7140
cttctcttat cacccacccc tatcttccca cttttccatc tttgttggtt tgcaacagcc 7200
ccttcttttt gcctgactct ccaggatttt ctctcatcat aaattgttct aaagtacata 7260
ctaatatggg tctggattga ctattcttat ttgcaaaaca gcaattaaat gttataggga 7320
agtaggaaga aaaaggggta tccttgacaa taaaccaagc aatattctgg gggtgggata 7380
gagcaggaaa ttttattttt aatcttttaa aatccaagta ataggtaggc ttccagttag 7440
ctttaaatgt tttttttttc cagctcaaaa aattggattg tagttgatac tacatataat 7500
acattctaat tccctcactg tattctttgt ttagtttcat ttatttggtt taaaataatt 7560
ttttatccca tatctgaaat gtaatatatt tttatccaac aaccagcatg tacatatact 7620
taattatgtg gcacattttc taatagatca gtccatcaat ctactcattt taaagaaaaa 7680
aaaattttaa agtcactttt agagccctta atgtgtagtt gggggttaag ctttgtggat 7740
gtagccttta tatttagtat aattgaggtc taaaataata atcttctatt atctcaacag 7800
agcaaattat tgaaaaagat gaaggtcctt tttataccca tctaggagca ggtcctaatg 7860
tggcagctat tagagaaatc atggaagaaa ggtaattaac gcaaaggcac agggcagatt 7920
aacgtttatc cttttgtata tgtcagaatt tttccagcct tcacacacaa agcagtaaac 7980
aattgtaaat tgagtaatta ttagtaggct tagctattct agggttgcca acactacaca 8040
ctgtgctatt caccagagag tcacaatatt tgacaggact aatagtctgc tagctggcac 8100
aggctgccca ctttgcgatg gatgccagaa aacccaggca tgaacaggaa tcggccagcc 8160
aggctgccag ccacaaggta ctggcacagg ctccaacgag aggtcccact ctggctttcc 8220
cacctgataa taaagtgtca aagcagaaag actggtaaag tgtggtataa gaaaagaacc 8280
actgaattaa attcacctag tgttgcaaat gagtacttat ctctaagttt tcttttacca 8340
taaaaagaga gcaagtgtga tatgttgaat agaaagagaa acatactatt tacagctgcc 8400
tttttttttt tttttcgcta tcaatcacag gtatacaagt acttgccttt actcctgcat 8460
gtagaagact cttatgagcg agataatgca gagaaggcct ttcatataaa tttatacagc 8520
tctgagctgt tcttcttcta gggtgccttt tcattaagag gtaggcagta ttattattaa 8580
agtacttagg atacattggg gcagctagga catattcagt atcattcttg ctccatttcc 8640
aaattattca tttctaaatt agcatgtaga agttcactaa ataatcatct agtggcctgg 8700
cagaaatagt gaatttccct aagtgccttt tttttgttgt ttttttgttt tgttttttaa 8760
acaagcagta ggtggtgctt tggtcataag ggaagatata gtctatttct aggactattc 8820
catattttcc atgtggctgg atactaacta tttgccagcc tccttttcta aattgtgaga 8880
cattcttgga ggaacagttc taactaaaat ctattatgac tccccaagtt ttaaaatagc 8940
taaatttagt aagggaaaaa atagtttatg ttttagaaga ctgaacttag caaactaacc 9000
tgaattttgt gctttgtgaa attttatatc gaaatgagct ttcccatttt cacccacatg 9060
taatttacaa aatagttcat tacaattatc tgtacatttt gatattgagg aaaaacaagg 9120
cttaaaaacc attatccagt ttgcttggcg tagacctgtt taaaaaataa taaaccgttc 9180
atttctcagg atgtggtcat agaataaagt tatgctcaaa tgttcaaata tttaaa 9236
<210> 1362
<211> 5969
<212> ДНК
<213> Homo sapiens
<400> 1362
tcaggctcta cttctaattc tggttctctt gctacatctc cctcatctgc agtgacctct 60
ccacggaagt cttgaactcc tcaaaagcaa attattgaaa aagatgaagg tcctttttat 120
acccatctag gagcaggtcc taatgtggca gctattagag aaatcatgga agaaaggttt 180
ggacagaagg gtaaagctat taggattgaa agagtcatct atactggtaa agaaggcaaa 240
agttctcagg gatgtcctat tgctaagtgg gtggttcgca gaagcagcag tgaagagaag 300
ctactgtgtt tggtgcggga gcgagctggc cacacctgtg aggctgcagt gattgtgatt 360
ctcatcctgg tgtgggaagg aatcccgctg tctctggctg acaaactcta ctcggagctt 420
accgagacgc tgaggaaata cggcacgctc accaatcgcc ggtgtgcctt gaatgaagag 480
agaacttgcg cctgtcaggg gctggatcca gaaacctgtg gtgcctcctt ctcttttggt 540
tgttcatgga gcatgtacta caatggatgt aagtttgcca gaagcaagat cccaaggaag 600
tttaagctgc ttggggatga cccaaaagag gaagagaaac tggagtctca tttgcaaaac 660
ctgtccactc ttatggcacc aacatataag aaacttgcac ctgatgcata taataatcag 720
attgaatatg aacacagagc accagagtgc cgtctgggtc tgaaggaagg ccgtccattc 780
tcaggggtca ctgcatgttt ggacttctgt gctcatgccc acagagactt gcacaacatg 840
cagaatggca gcacattggt atgcactctc actagagaag acaatcgaga atttggagga 900
aaacctgagg atgagcagct tcacgttctg cctttataca aagtctctga cgtggatgag 960
tttgggagtg tggaagctca ggaggagaaa aaacggagtg gtgccattca ggtactgagt 1020
tcttttcggc gaaaagtcag gatgttagca gagccagtca agacttgccg acaaaggaaa 1080
ctagaagcca agaaagctgc agctgaaaag ctttcctccc tggagaacag ctcaaataaa 1140
aatgaaaagg aaaagtcagc cccatcacgt acaaaacaaa ctgaaaacgc aagccaggct 1200
aaacagttgg cagaactttt gcgactttca ggaccagtca tgcagcagtc ccagcagccc 1260
cagcctctac agaagcagcc accacagccc cagcagcagc agagacccca gcagcagcag 1320
ccacatcacc ctcagacaga gtctgtcaac tcttattctg cttctggatc caccaatcca 1380
tacatgagac ggcccaatcc agttagtcct tatccaaact cttcacacac ttcagatatc 1440
tatggaagca ccagccctat gaacttctat tccacctcat ctcaagctgc aggttcatat 1500
ttgaattctt ctaatcccat gaacccttac cctgggcttt tgaatcagaa tacccaatat 1560
ccatcatatc aatgcaatgg aaacctatca gtggacaact gctccccata tctgggttcc 1620
tattctcccc agtctcagcc gatggatctg tataggtatc caagccaaga ccctctgtct 1680
aagctcagtc taccacccat ccatacactt taccagccaa ggtttggaaa tagccagagt 1740
tttacatcta aatacttagg ttatggaaac caaaatatgc agggagatgg tttcagcagt 1800
tgtaccatta gaccaaatgt acatcatgta gggaaattgc ctccttatcc cactcatgag 1860
atggatggcc acttcatggg agccacctct agattaccac ccaatctgag caatccaaac 1920
atggactata aaaatggtga acatcattca ccttctcaca taatccataa ctacagtgca 1980
gctccgggca tgttcaacag ctctcttcat gccctgcatc tccaaaacaa ggagaatgac 2040
atgctttccc acacagctaa tgggttatca aagatgcttc cagctcttaa ccatgataga 2100
actgcttgtg tccaaggagg cttacacaaa ttaagtgatg ctaatggtca ggaaaagcag 2160
ccattggcac tagtccaggg tgtggcttct ggtgcagagg acaacgatga ggtctggtca 2220
gacagcgagc agagctttct ggatcctgac attgggggag tggccgtggc tccaactcat 2280
gggtcaattc tcattgagtg tgcaaagcgt gagctgcatg ccacaacccc tttaaagaat 2340
cccaatagga atcaccccac caggatctcc ctcgtctttt accagcataa gagcatgaat 2400
gagccaaaac atggcttggc tctttgggaa gccaaaatgg ctgaaaaagc ccgtgagaaa 2460
gaggaagagt gtgaaaagta tggcccagac tatgtgcctc agaaatccca tggcaaaaaa 2520
gtgaaacggg agcctgctga gccacatgaa acttcagagc ccacttacct gcgtttcatc 2580
aagtctcttg ccgaaaggac catgtccgtg accacagact ccacagtaac tacatctcca 2640
tatgccttca ctcgggtcac agggccttac aacagatata tatgatatca cccccttttg 2700
ttggttacct cacttgaaaa gaccacaacc aacctgtcag tagtatagtt ctcatgacgt 2760
gggcagtggg gaaaggtcac agtattcatg acaaatgtgg tgggaaaaac ctcagctcac 2820
cagcaacaaa agaggttatc ttaccatagc acttaatttt cactggctcc caagtggtca 2880
cagatggcat ctaggaaaag accaaagcat tctatgcaaa aagaaggtgg ggaagaaagt 2940
gttccgcaat ttacattttt aaacactggt tctattattg gacgagatga tatgtaaatg 3000
tgatcccccc cccccgctta caactctaca catctgtgac cacttttaat aatatcaagt 3060
ttgcatagtc atggaacaca aatcaaacaa gtactgtagt attacagtga caggaatctt 3120
aaaataccat ctggtgctga atatatgatg tactgaaata ctggaattat ggctttttga 3180
aatgcagttt ttactgtaat cttaactttt atttatcaaa atagctacag gaaacatgaa 3240
tagcaggaaa acactgaatt tgtttggatg ttctaagaaa tggtgctaag aaaatggtgt 3300
ctttaatagc taaaaattta atgcctttat atcatcaaga tgctatcagt gtactccagt 3360
gcccttgaat aataggggta ccttttcatt caagttttta tcataattac ctattcttac 3420
acaagcttag tttttaaaat gtggacattt taaaggcctc tggattttgc tcatccagtg 3480
aagtccttgt aggacaataa acgtatatat gtacatatat acacaaacat gtatatgtgc 3540
acacacatgt atatgtataa atattttaaa tggtgtttta gaagcacttt gtctacctaa 3600
gctttgacaa cttgaacaat gctaaggtac tgagatgttt aaaaaacaag tttactttca 3660
ttttagaatg caaagttgat ttttttaagg aaacaaagaa agcttttaaa atatttttgc 3720
ttttagccat gcatctgctg atgagcaatt gtgtccattt ttaacacagc cagttaaatc 3780
caccatgggg cttactggat tcaagggaat acgttagtcc acaaaacatg ttttctggtg 3840
ctcatctcac atgctatact gtaaaacagt tttatacaaa attgtatgac aagttcattg 3900
ctcaaaaatg tacagtttta agaattttct attaactgca ggtaataatt agctgcatgc 3960
tgcagactca acaaagctag ttcactgaag cctatgctat tttatggatc ataggctctt 4020
cagagaactg aatggcagtc tgcctttgtg ttgataatta tgtacattgt gacgttgtca 4080
tttcttagct taagtgtcct ctttaacaag aggattgagc agactgatgc ctgcataaga 4140
tgaataaaca gggttagttc catgtgaatc tgtcagttaa aaagaaacaa aaacaggcag 4200
ctggtttgct gtggtggttt taaatcatta atttgtataa agaagtgaaa gagttgtata 4260
gtaaattaaa ttgtaaacaa aactttttta atgcaatgct ttagtatttt agtactgtaa 4320
aaaaattaaa tatatacata tatatatata tatatatata tatatatatg agtttgaagc 4380
agaattcaca tcatgatggt gctactcagc ctgctacaaa tatatcataa tgtgagctaa 4440
gaattcatta aatgtttgag tgatgttcct acttgtcata tacctcaaca ctagtttggc 4500
aataggatat tgaactgaga gtgaaagcat tgtgtaccat catttttttc caagtccttt 4560
tttttattgt taaaaaaaaa agcatacctt ttttcaatac ttgatttctt agcaagtata 4620
acttgaactt caaccttttt gttctaaaaa ttcagggata tttcagctca tgctctccct 4680
atgccaacat gtcacctgtg tttatgtaaa attgttgtag gttaataaat atattctttg 4740
tcagggattt aaccctttta ttttgaatcc cttctatttt acttgtacat gtgctgatgt 4800
aactaaaact aattttgtaa atctgttggc tctttttatt gtaaagaaaa gcattttaaa 4860
agtttgagga atcttttgac tgtttcaagc aggaaaaaaa aattacatga aaatagaatg 4920
cactgagttg ataaagggaa aaattgtaag gcaggagttt ggcaagtggc tgttggccag 4980
agacttactt gtaactctct aaatgaagtt tttttgatcc tgtaatcact gaaggtacat 5040
actccatgtg gacttccctt aaacaggcaa acacctacag gtatggtgtg caacagattg 5100
tacaattaca ttttggccta aatacatttt tgcttactag tatttaaaat aaattcttaa 5160
tcagaggagg cctttgggtt ttattggtca aatctttgta agctggcttt tgtcttttta 5220
aaaaatttct tgaatttgtg gttgtgtcca atttgcaaac atttccaaaa atgtttgctt 5280
tgcttacaaa ccacatgatt ttaatgtttt ttgtatacca taatatctag ccccaaacat 5340
ttgattacta catgtgcatt ggtgattttg atcatccatt cttaatattt gatttctgtg 5400
tcacctactg tcatttgtta aactgctggc caacaagaac aggaagtata gtttgggggg 5460
ttggggagag tttacataag gaagagaaga aattgagtgg catattgtaa atatcagatc 5520
tataattgta aatataaaac ctgcctcagt tagaatgaat ggaaagcaga tctacaattt 5580
gctaatatag gaatatcagg ttgactatat agccatactt gaaaatgctt ctgagtggtg 5640
tcaactttac ttgaatgaat ttttcatctt gattgacgca cagtgatgta cagttcactt 5700
ctgaagctag tggttaactt gtgtaggaaa cttttgcagt ttgacactaa gataacttct 5760
gtgtgcattt ttctatgctt ttttaaaaac tagtttcatt tcattttcat gagatgtttg 5820
gtttataaga tctgaggatg gttataaata ctgtaagtat tgtaatgtta tgaatgcagg 5880
ttatttgaaa gctgtttatt attatatcat tcctgataat gctatgtgag tgtttttaat 5940
aaaatttata tttatttaat gcactctaa 5969
<210> 1363
<211> 4594
<212> ДНК
<213> Homo sapiens
<400> 1363
gtagagaagc agaaggaagc aagatggctg ccctttagga tttgttagaa aggagacccg 60
actgcaactg ctggattgct gcaaggctga gggacgagaa cgaggctggc aaacattcag 120
cagcacaccc tctcaagatt gtttacttgc ctttgctcct gttgagttac aacgcttgga 180
agcaggagat gggctcagca gcagccaata ggacatgatc caggaagagc agtaagggac 240
tgagctgctg aattcaacta gagggcagcc ttgtggatgg ccccgaagca agcctgatgg 300
aacaggatag aaccaaccat gttgagggca acagactaag tccattcctg ataccatcac 360
ctcccatttg ccagacagaa cctctggcta caaagctcca gaatggaagc ccactgcctg 420
agagagctca tccagaagta aatggagaca ccaagtggca ctctttcaaa agttattatg 480
gaataccctg tatgaaggga agccagaata gtcgtgtgag tcctgacttt acacaagaaa 540
gtagagggta ttccaagtgt ttgcaaaatg gaggaataaa acgcacagtt agtgaacctt 600
ctctctctgg gctccttcag atcaagaaat tgaaacaaga ccaaaaggct aatggagaaa 660
gacgtaactt cggggtaagc caagaaagaa atccaggtga aagcagtcaa ccaaatgtct 720
ccgatttgag tgataagaaa gaatctgtga gttctgtagc ccaagaaaat gcagttaaag 780
atttcaccag tttttcaaca cataactgca gtgggcctga aaatccagag cttcagattc 840
tgaatgagca ggaggggaaa agtgctaatt accatgacaa gaacattgta ttacttaaaa 900
acaaggcagt gctaatgcct aatggtgcta cagtttctgc ctcttccgtg gaacacacac 960
atggtgaact cctggaaaaa acactgtctc aatattatcc agattgtgtt tccattgcgg 1020
tgcagaaaac cacatctcac ataaatgcca ttaacagtca ggctactaat gagttgtcct 1080
gtgagatcac tcacccatcg catacctcag ggcagatcaa ttccgcacag acctctaact 1140
ctgagctgcc tccaaagcca gctgcagtgg tgagtgaggc ctgtgatgct gatgatgctg 1200
ataatgccag taaactagct gcaatgctaa atacctgttc ctttcagaaa ccagaacaac 1260
tacaacaaca aaaatcagtt tttgagatat gcccatctcc tgcagaaaat aacatccagg 1320
gaaccacaaa gctagcgtct ggtgaagaat tctgttcagg ttccagcagc aatttgcaag 1380
ctcctggtgg cagctctgaa cggtatttaa aacaaaatga aatgaatggt gcttacttca 1440
agcaaagctc agtgttcact aaggattcct tttctgccac taccacacca ccaccaccat 1500
cacaattgct tctttctccc cctcctcctc ttccacaggt tcctcagctt ccttcagaag 1560
gaaaaagcac tctgaatggt ggagttttag aagaacacca ccactacccc aaccaaagta 1620
acacaacact tttaagggaa gtgaaaatag agggtaaacc tgaggcacca ccttcccaga 1680
gtcctaatcc atctacacat gtatgcagcc cttctccgat gctttctgaa aggcctcaga 1740
ataattgtgt gaacaggaat gacatacaga ctgcagggac aatgactgtt ccattgtgtt 1800
ctgagaaaac aagaccaatg tcagaacacc tcaagcataa cccaccaatt tttggtagca 1860
gtggagagct acaggacaac tgccagcagt tgatgagaaa caaagagcaa gagattctga 1920
agggtcgaga caaggagcaa acacgagatc ttgtgccccc aacacagcac tatctgaaac 1980
caggatggat tgaattgaag gcccctcgtt ttcaccaagc ggaatcccat ctaaaacgta 2040
atgaggcatc actgccatca attcttcagt atcaacccaa tctctccaat caaatgacct 2100
ccaaacaata cactggaaat tccaacatgc ctggggggct cccaaggcaa gcttacaccc 2160
agaaaacaac acagctggag cacaagtcac aaatgtacca agttgaaatg aatcaagggc 2220
agtcccaagg tacagtggac caacatctcc agttccaaaa accctcacac caggtgcact 2280
tctccaaaac agaccattta ccaaaagctc atgtgcagtc actgtgtggc actagatttc 2340
attttcaaca aagagcagat tcccaaactg aaaaacttat gtccccagtg ttgaaacagc 2400
acttgaatca acaggcttca gagactgagc cattttcaaa ctcacacctt ttgcaacata 2460
agcctcataa acaggcagca caaacacaac catcccagag ttcacatctc cctcaaaacc 2520
agcaacagca gcaaaaatta caaataaaga ataaagagga aatactccag acttttcctc 2580
acccccaaag caacaatgat cagcaaagag aaggatcatt ctttggccag actaaagtgg 2640
aagaatgttt tcatggtgaa aatcagtatt caaaatcaag cgagttcgag actcataatg 2700
tccaaatggg actggaggaa gtacagaata taaatcgtag aaattcccct tatagtcaga 2760
ccatgaaatc aagtgcatgc aaaatacagg tttcttgttc aaacaataca cacctagttt 2820
cagagaataa agaacagact acacatcctg aactttttgc aggaaacaag acccaaaact 2880
tgcatcacat gcaatatttt ccaaataatg tgatcccaaa gcaagatctt cttcacaggt 2940
gctttcaaga acaggagcag aagtcacaac aagcttcagt tctacaggga tataaaaata 3000
gaaaccaaga tatgtctggt caacaagctg cgcaacttgc tcagcaaagg tacttgatac 3060
ataaccatgc aaatgttttt cctgtgcctg accagggagg aagtcacact cagacccctc 3120
cccagaagga cactcaaaag catgctgctc taaggtggca tctcttacag aagcaagaac 3180
agcagcaaac acagcaaccc caaactgagt cttgccatag tcagatgcac aggccaatta 3240
aggtggaacc tggatgcaag ccacatgcct gtatgcacac agcaccacca gaaaacaaaa 3300
catggaaaaa ggtaactaag caagagaatc cacctgcaag ctgtgataat gtgcagcaaa 3360
agagcatcat tgagaccatg gagcagcatc tgaagcagtt tcacgccaag tcgttatttg 3420
accataaggc tcttactctc aaatcacaga agcaagtaaa agttgaaatg tcagggccag 3480
tcacagtttt gactagacaa accactgctg cagaacttga tagccacacc ccagctttag 3540
agcagcaaac aacttcttca gaaaagacac caaccaaaag aacagctgct tctgttctca 3600
ataattttat agagtcacct tccaaattac tagatactcc tataaaaaat ttattggata 3660
cacctgtcaa gactcaatat gatttcccat cttgcagatg tgtagagcaa attattgaaa 3720
aagatgaagg tcctttttat acccatctag gagcaggtcc taatgtggca gctattagag 3780
aaatcatgga agaaaggtat acaagtactt gcctttactc ctgcatgtag aagactctta 3840
tgagcgagat aatgcagaga aggcctttca tataaattta tacagctctg agctgttctt 3900
cttctagggt gccttttcat taagaggtag gcagtattat tattaaagta cttaggatac 3960
attggggcag ctaggacata ttcagtatca ttcttgctcc atttccaaat tattcatttc 4020
taaattagca tgtagaagtt cactaaataa tcatctagtg gcctggcaga aatagtgaat 4080
ttccctaagt gccttttttt tgttgttttt ttgttttgtt ttttaaacaa gcagtaggtg 4140
gtgctttggt cataagggaa gatatagtct atttctagga ctattccata ttttccatgt 4200
ggctggatac taactatttg ccagcctcct tttctaaatt gtgagacatt cttggaggaa 4260
cagttctaac taaaatctat tatgactccc caagttttaa aatagctaaa tttagtaagg 4320
gaaaaaatag tttatgtttt agaagactga acttagcaaa ctaacctgaa ttttgtgctt 4380
tgtgaaattt tatatcgaaa tgagctttcc cattttcacc cacatgtaat ttacaaaata 4440
gttcattaca attatctgta cattttgata ttgaggaaaa acaaggctta aaaaccatta 4500
tccagtttgc ttggcgtaga cctgtttaaa aaataataaa ccgttcattt ctcaggatgt 4560
ggtcatagaa taaagttatg ctcaaatgtt caaa 4594
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА CLL-1 | 2015 |
|
RU2741120C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2831418C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗБИРАТЕЛЬНОЙ ЭКСПРЕССИИ БЕЛКА | 2017 |
|
RU2795467C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ЛЕЧЕНИЯ РАКА | 2017 |
|
RU2826270C2 |
| CD20 ТЕРАПИЯ, CD22 ТЕРАПИЯ И КОМБИНИРОВАННАЯ ТЕРАПИЯ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) K CD19 | 2016 |
|
RU2752918C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ХИМЕРНЫХ АНТИГЕННЫХ РЕЦЕПТОРОВ И ИНГИБИТОРОВ PD-1 | 2017 |
|
RU2809160C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |

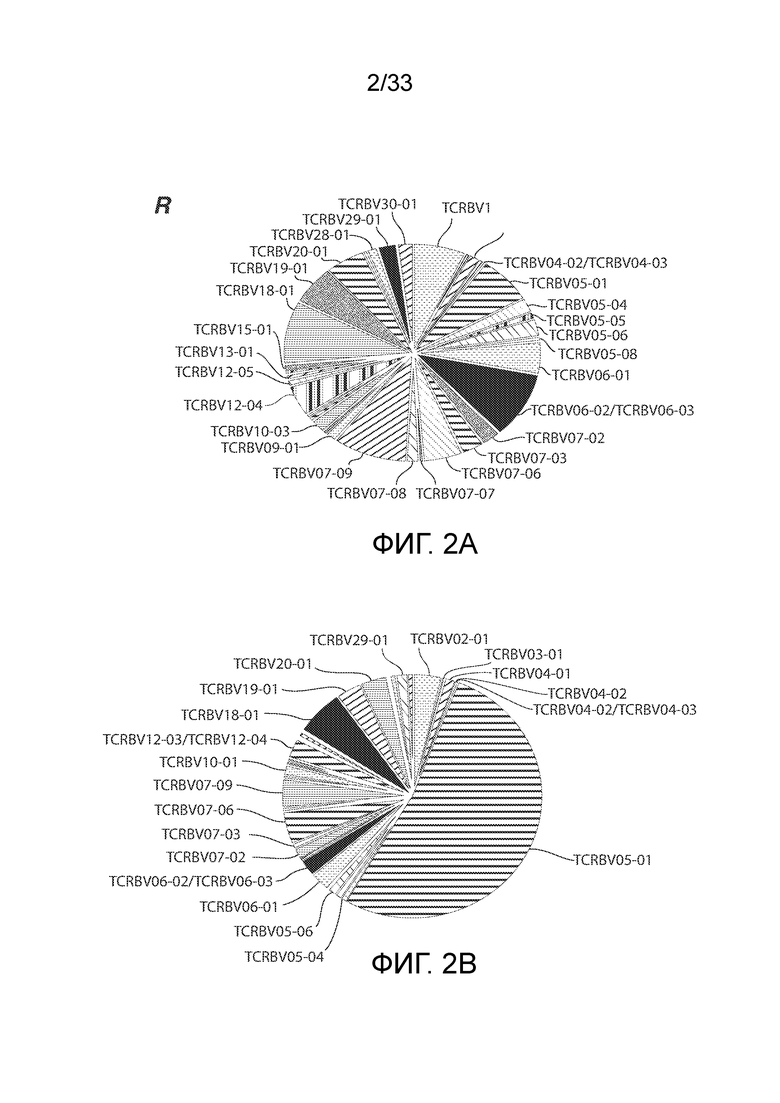

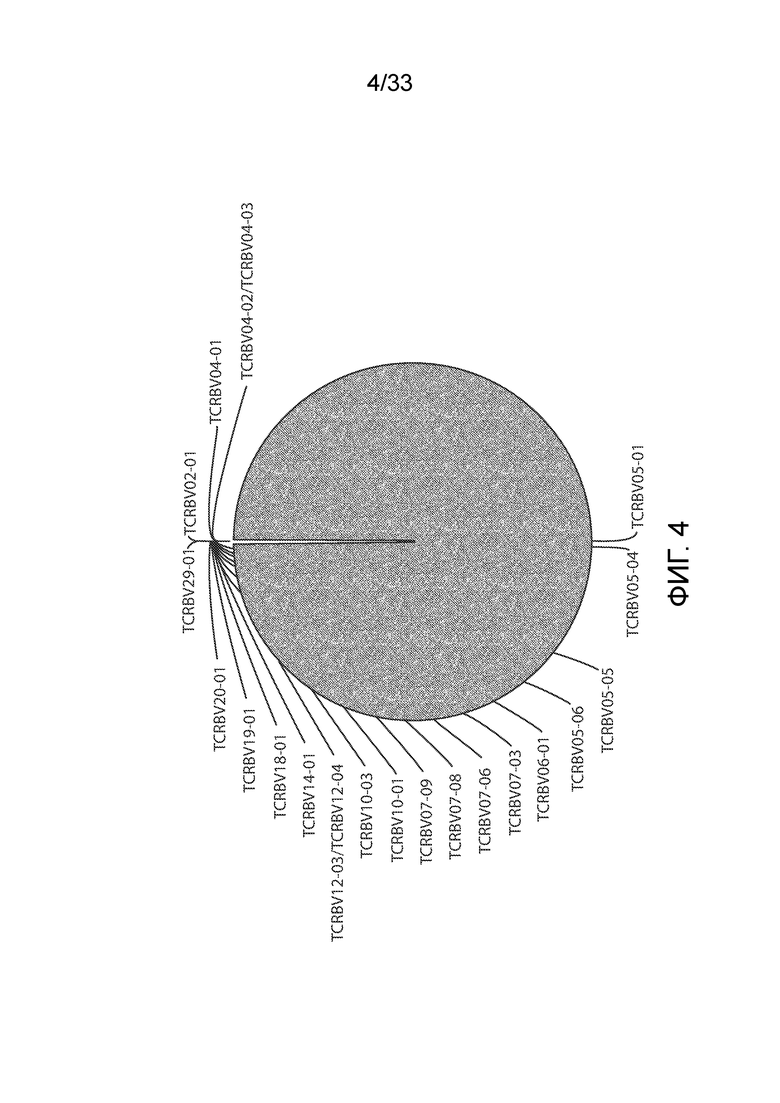
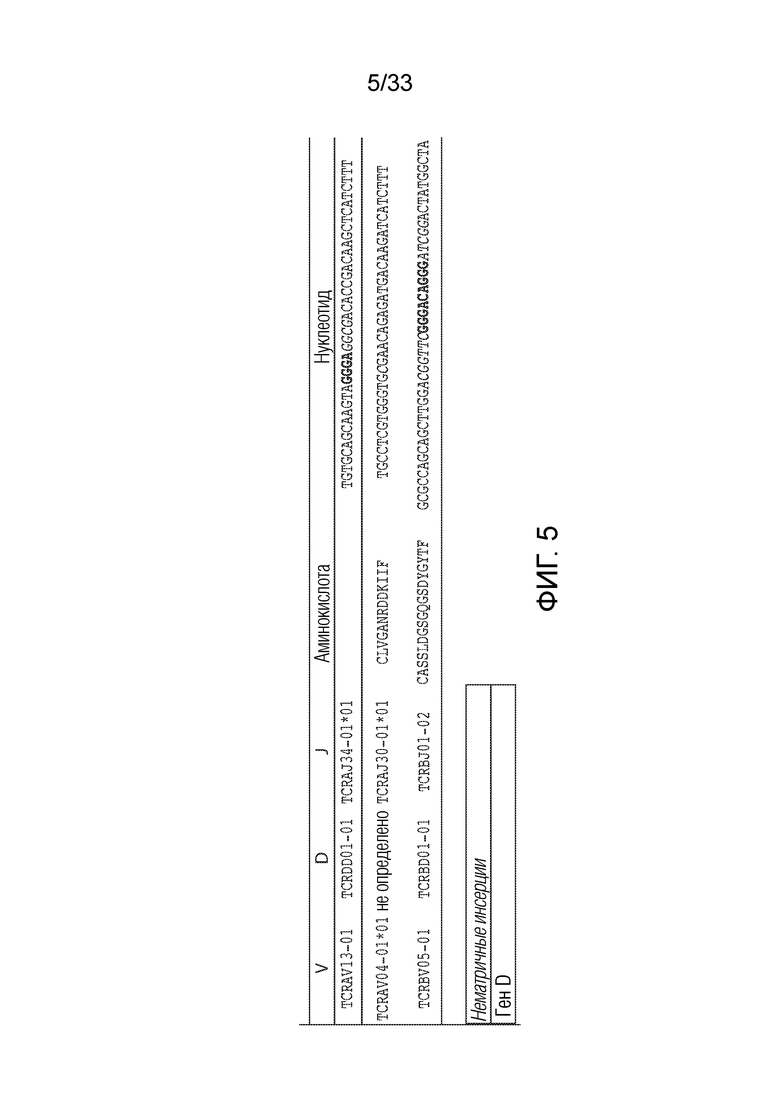
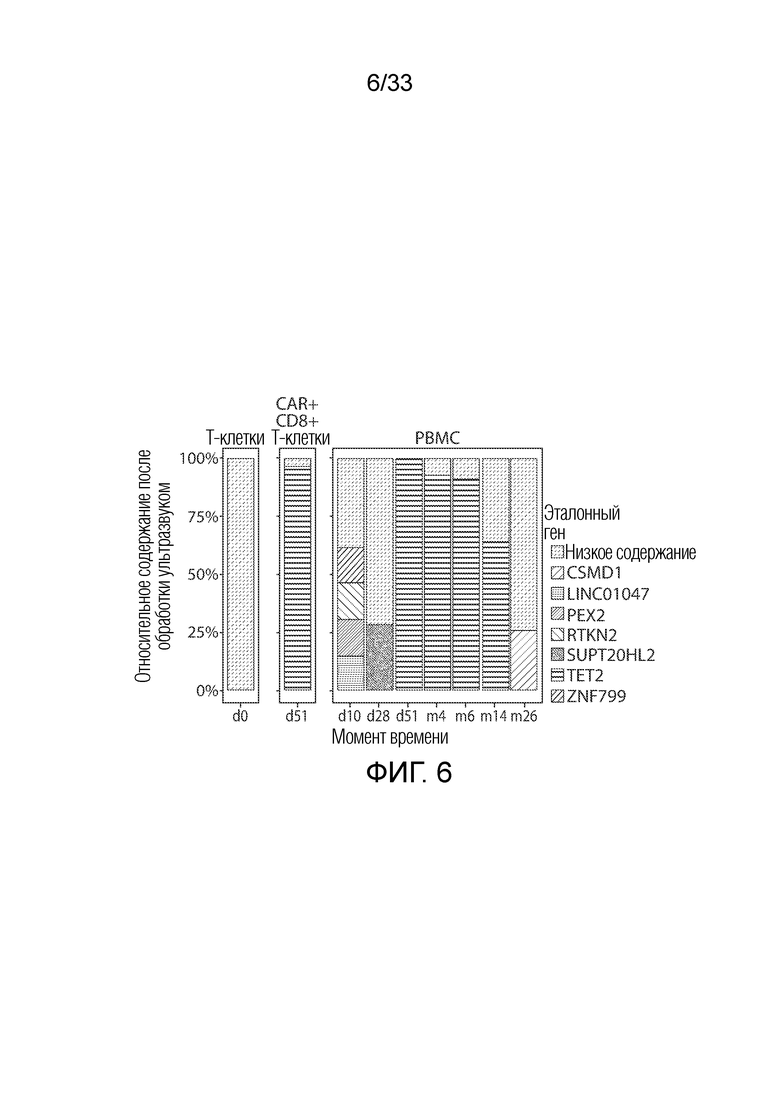
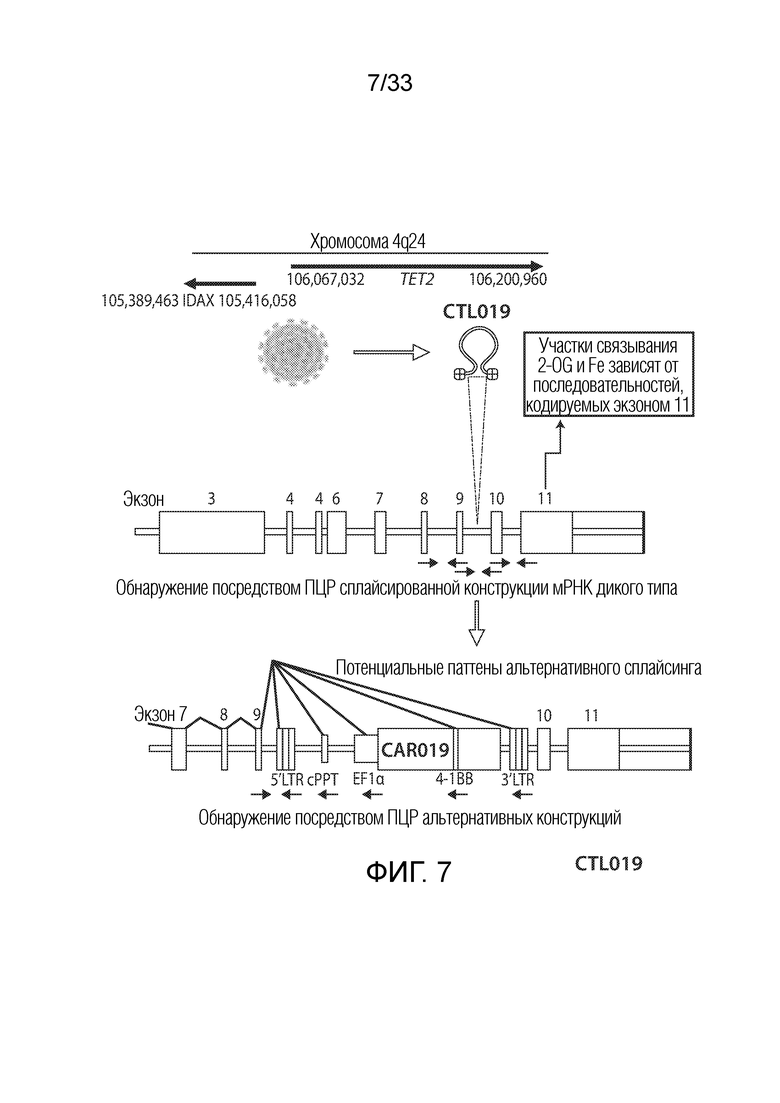
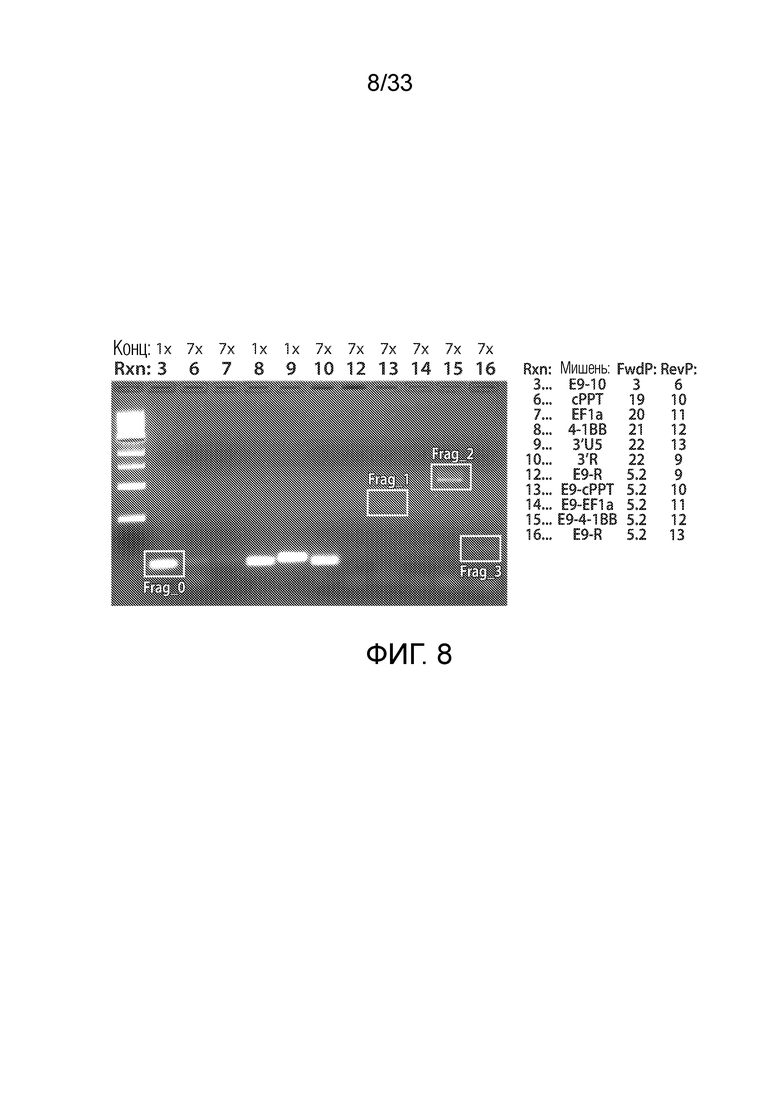
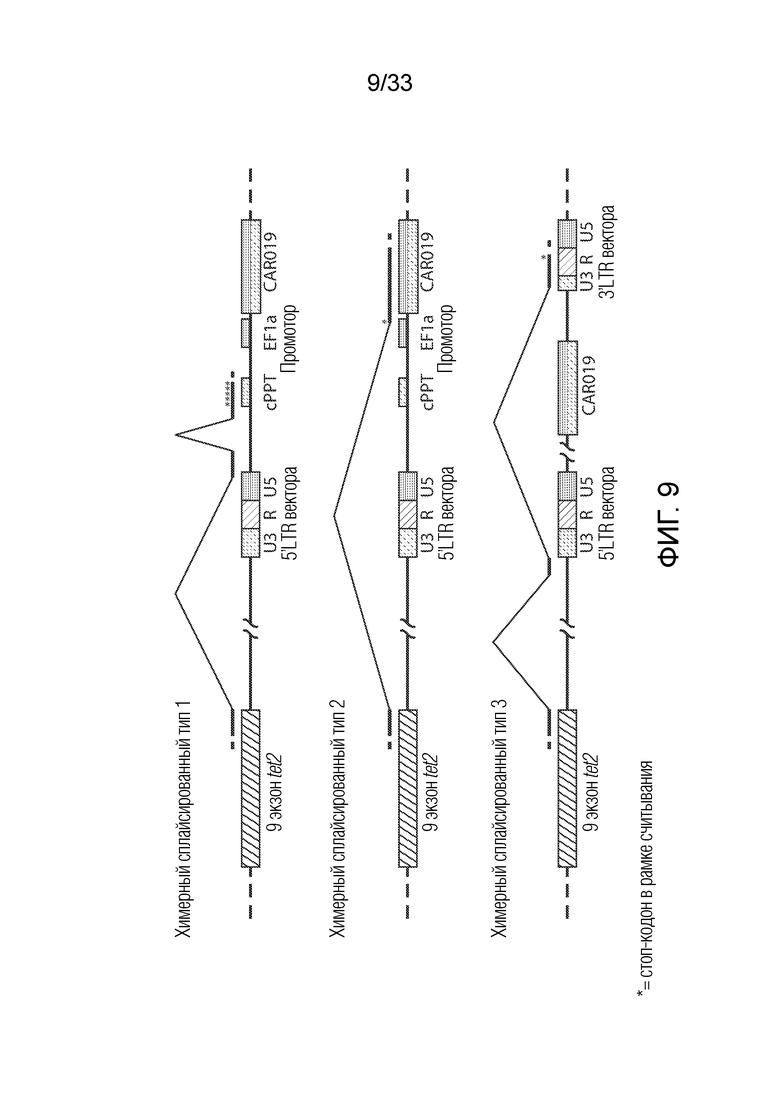
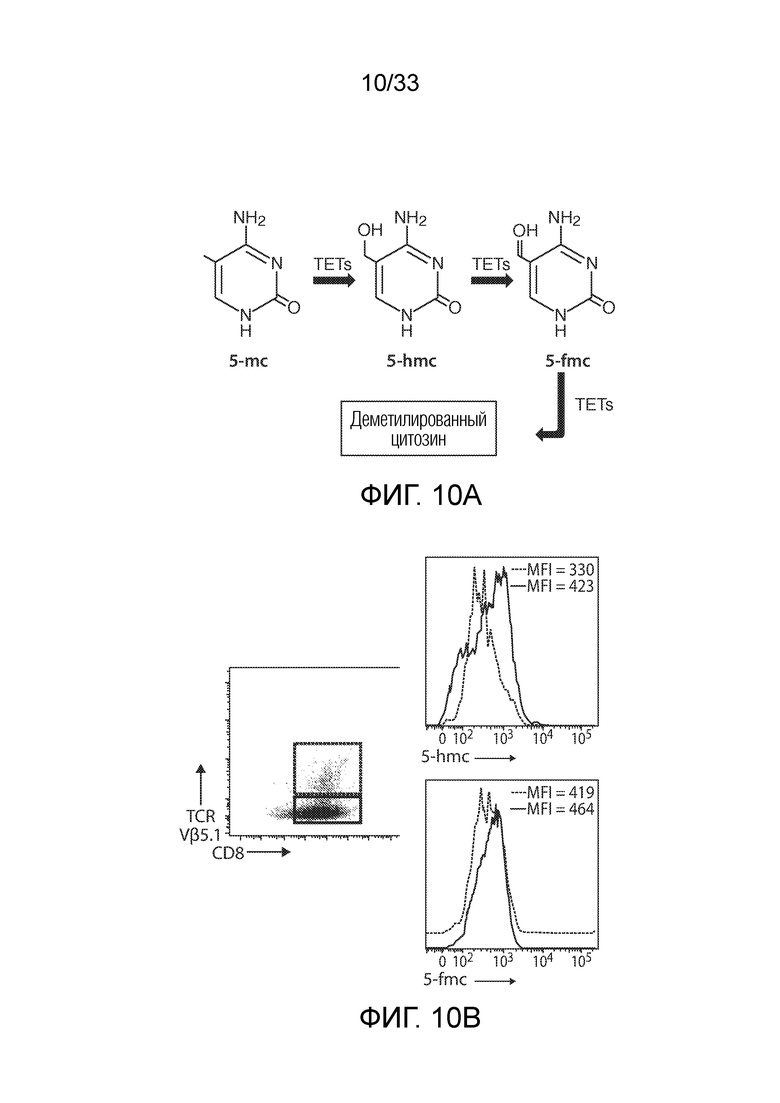
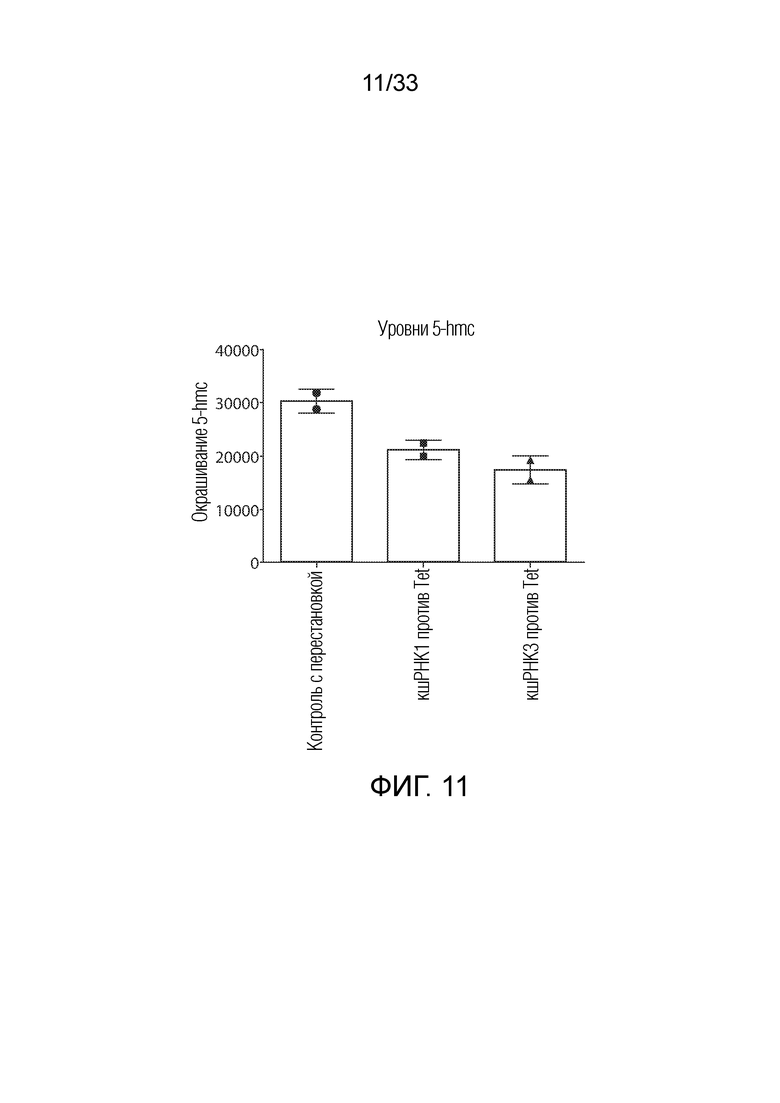
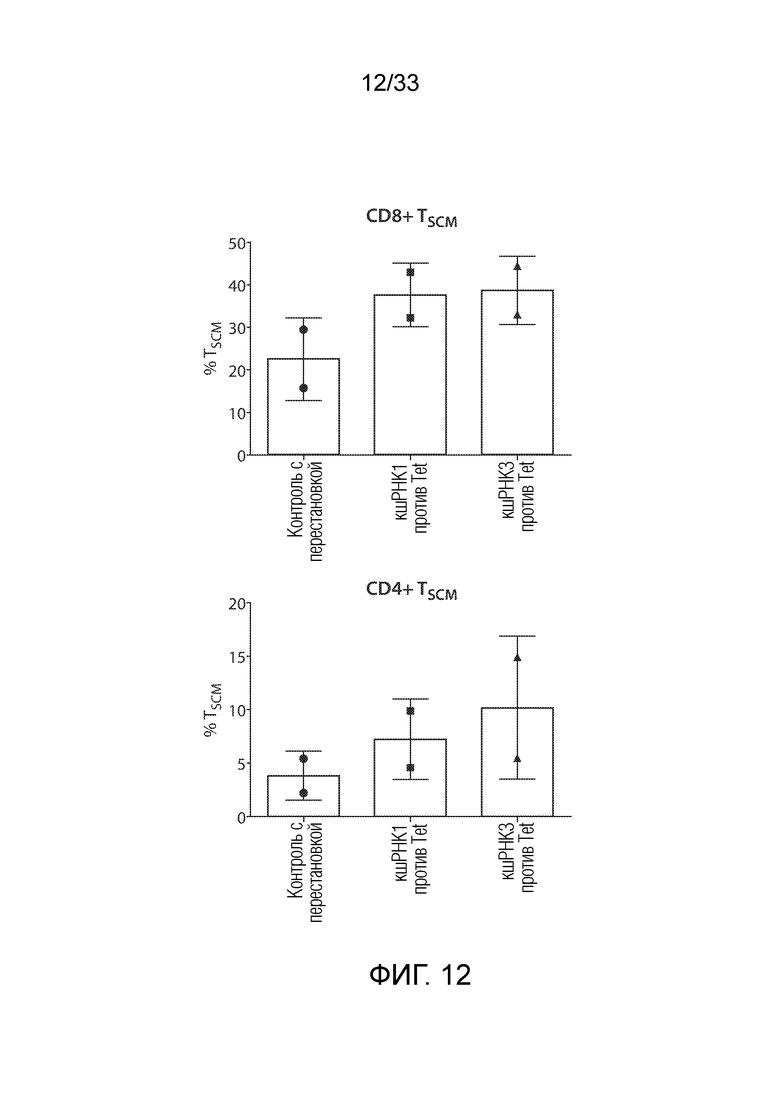

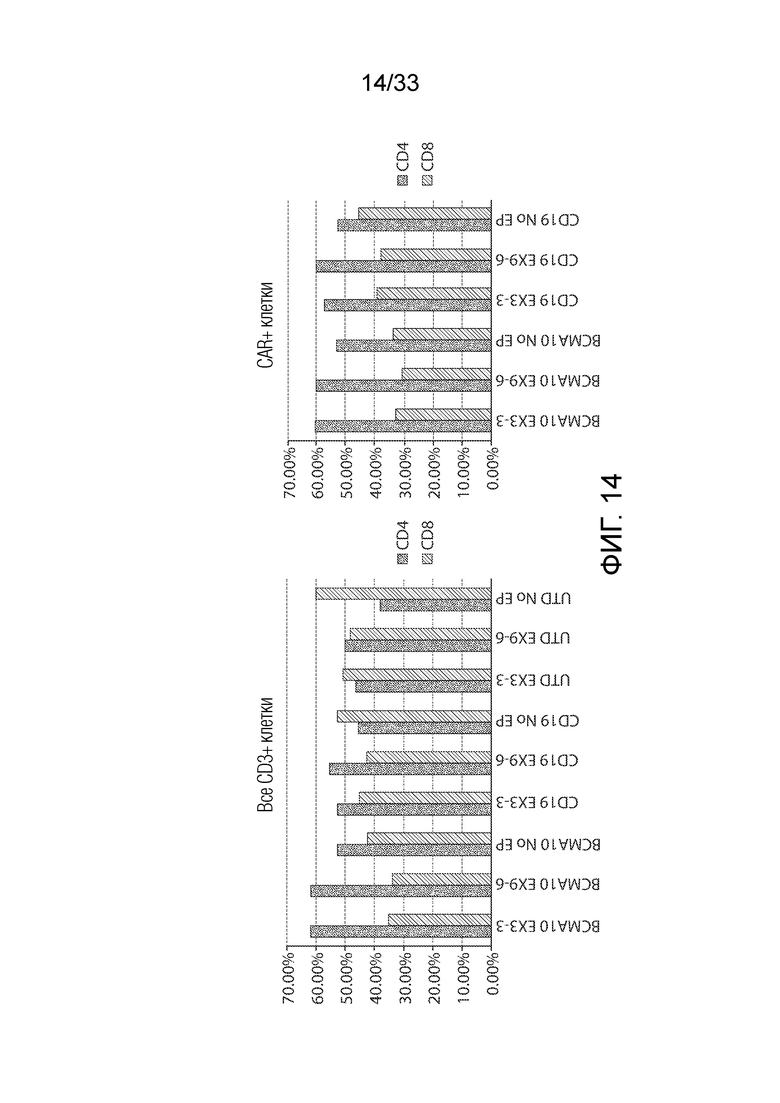

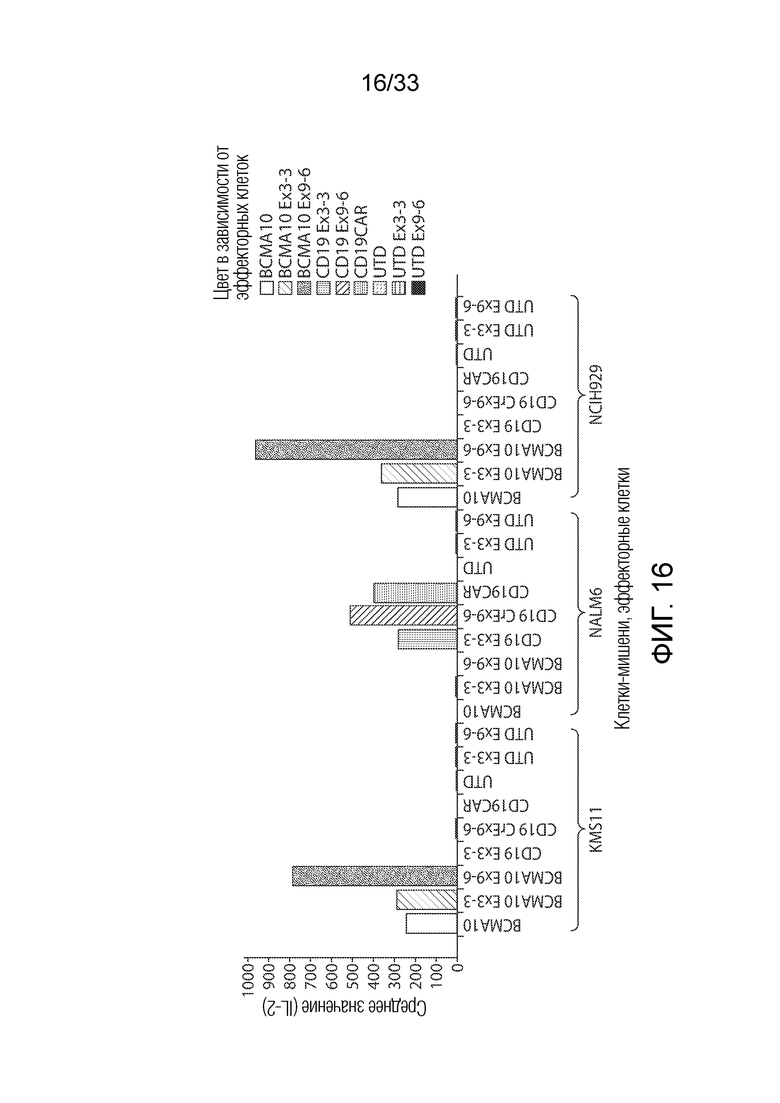
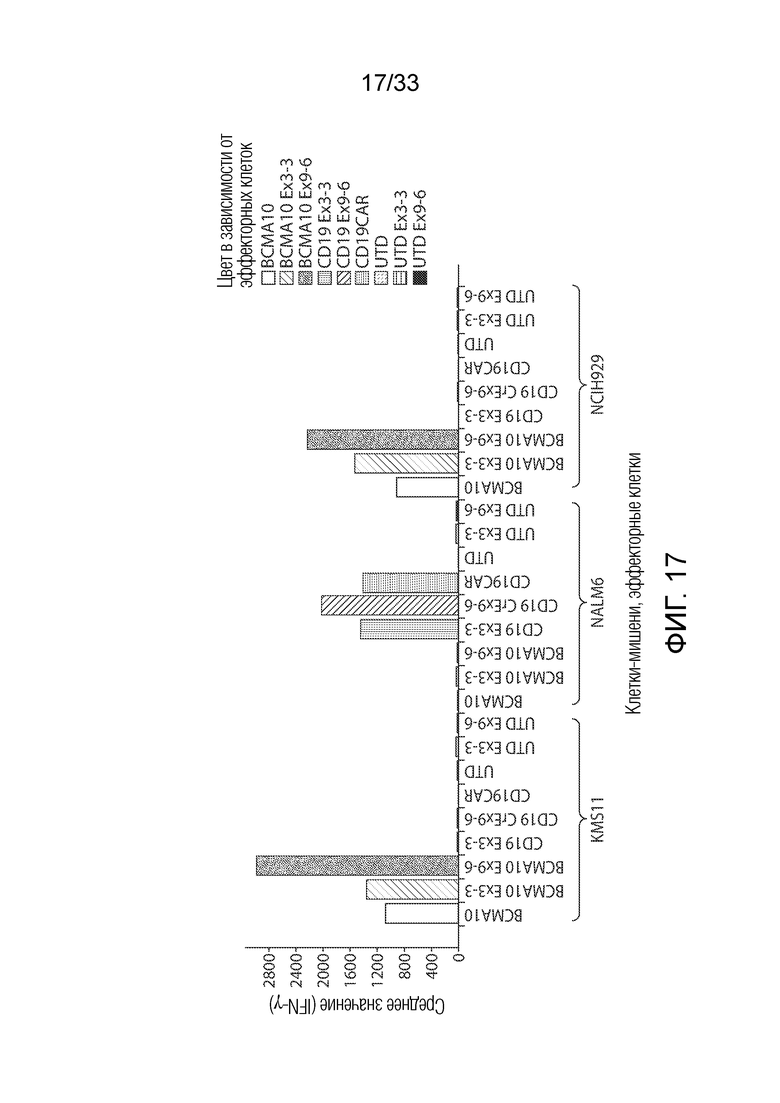
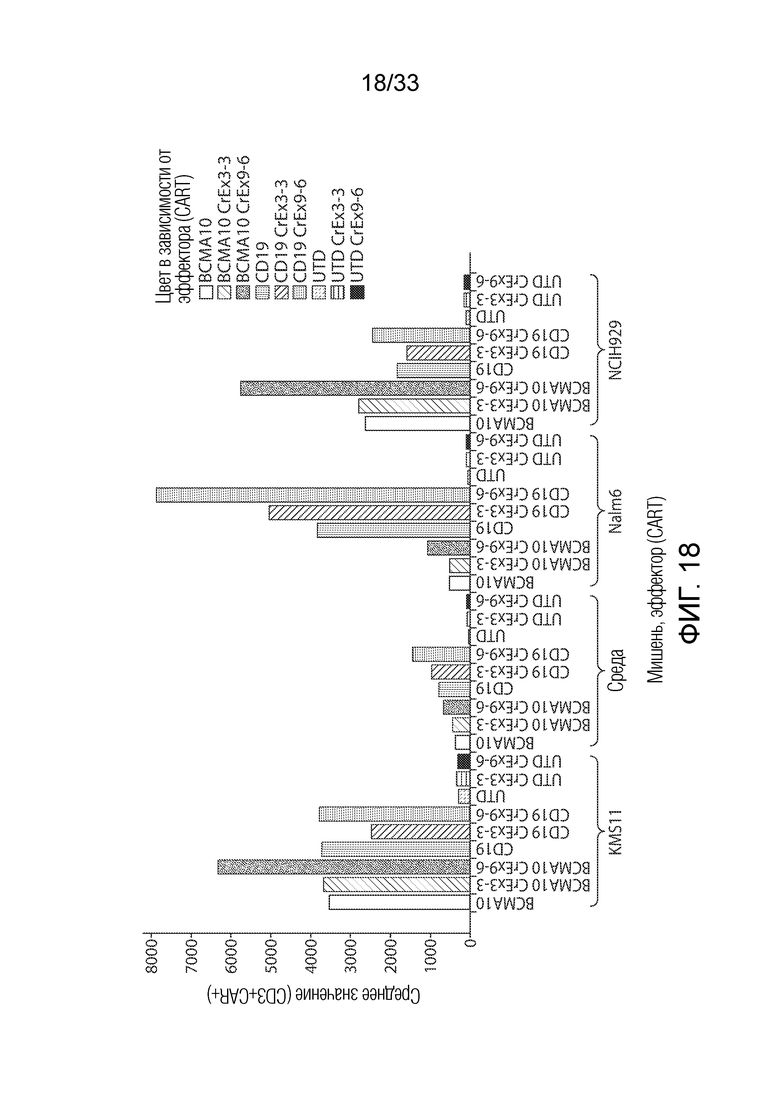
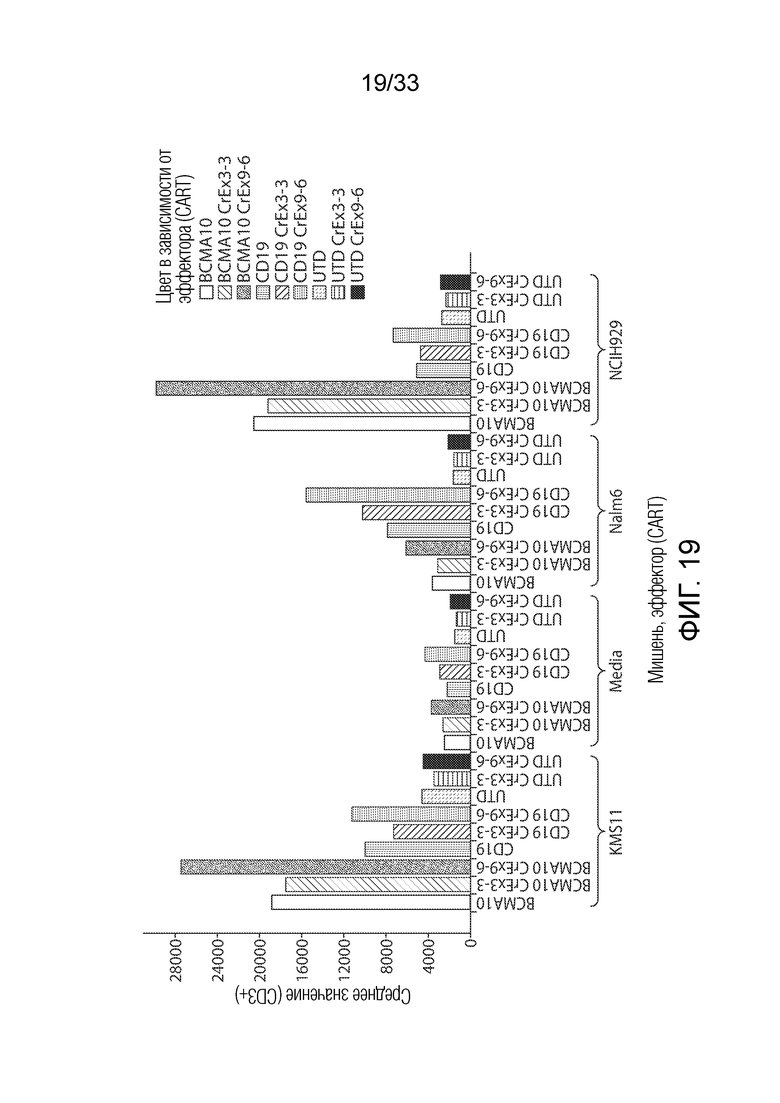
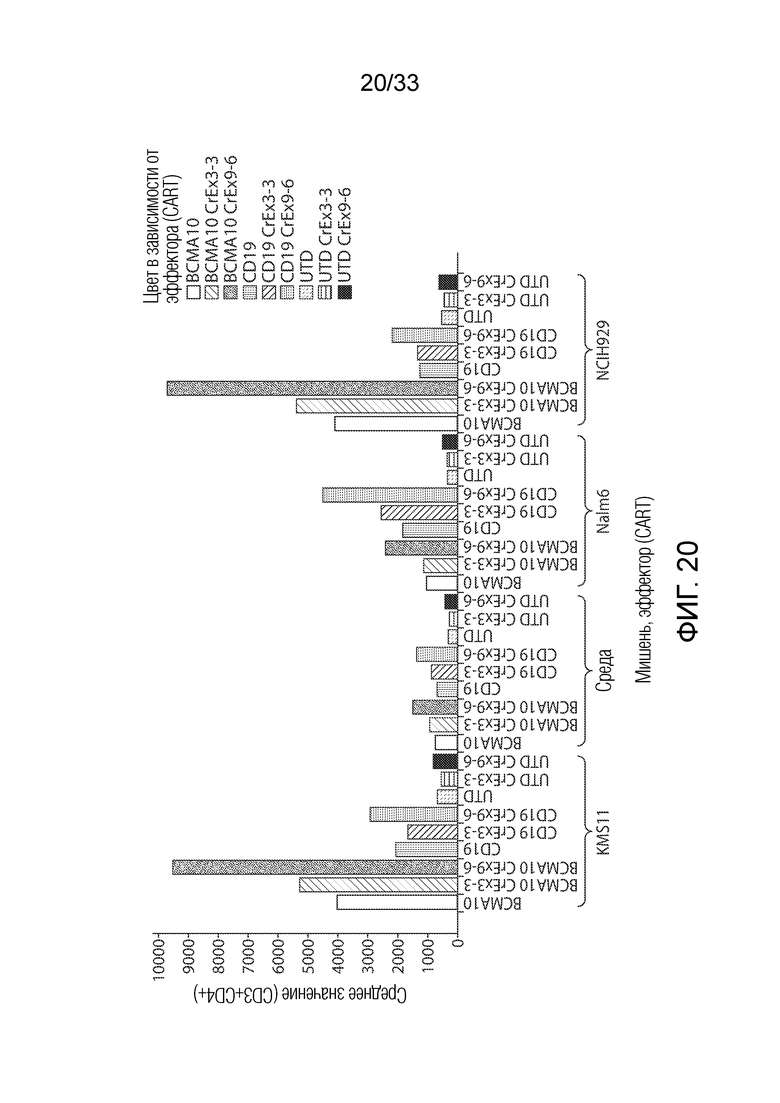
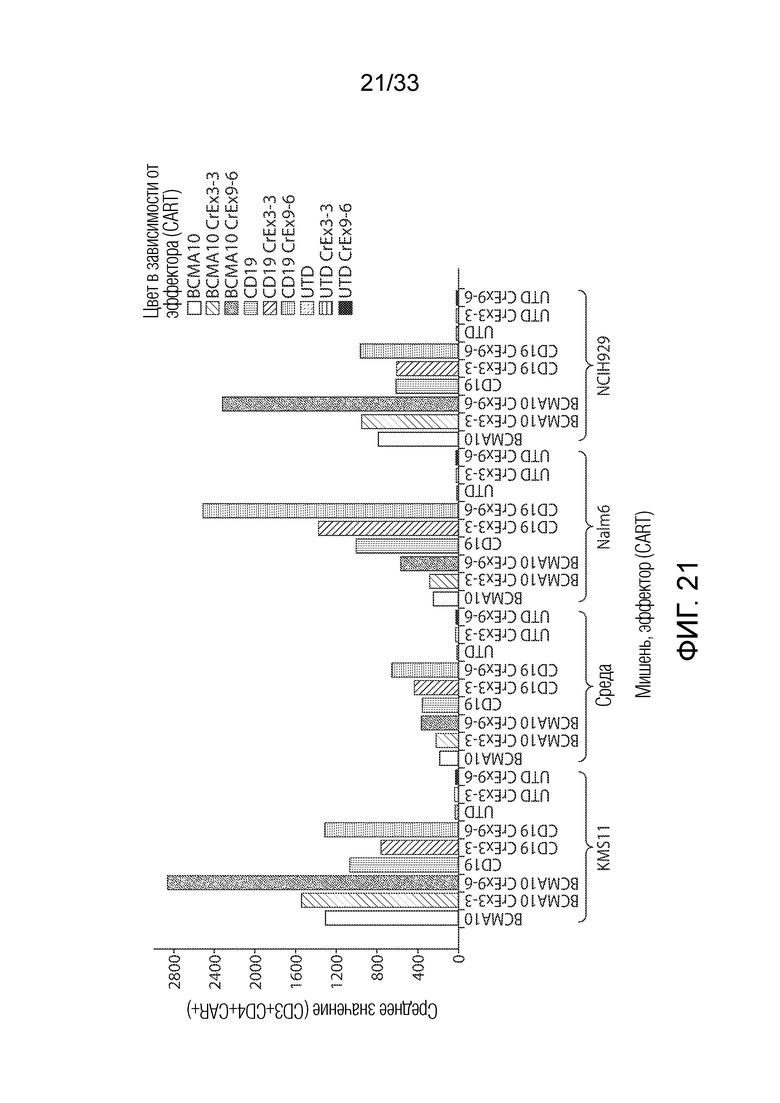
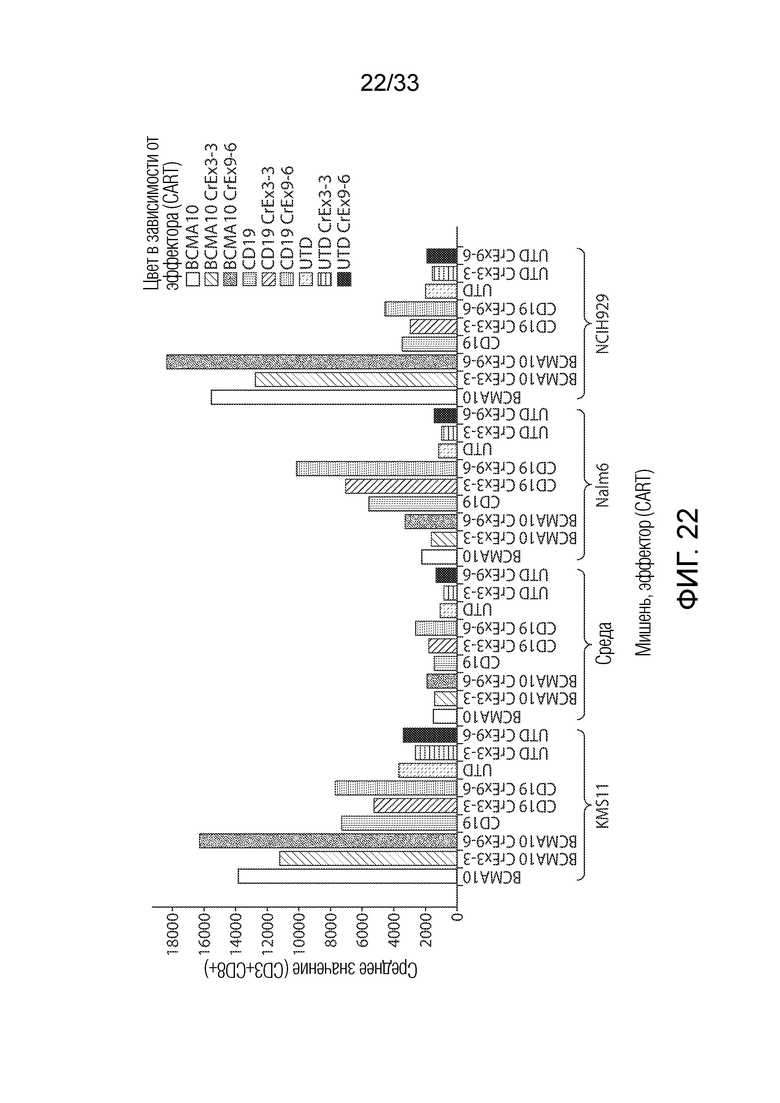
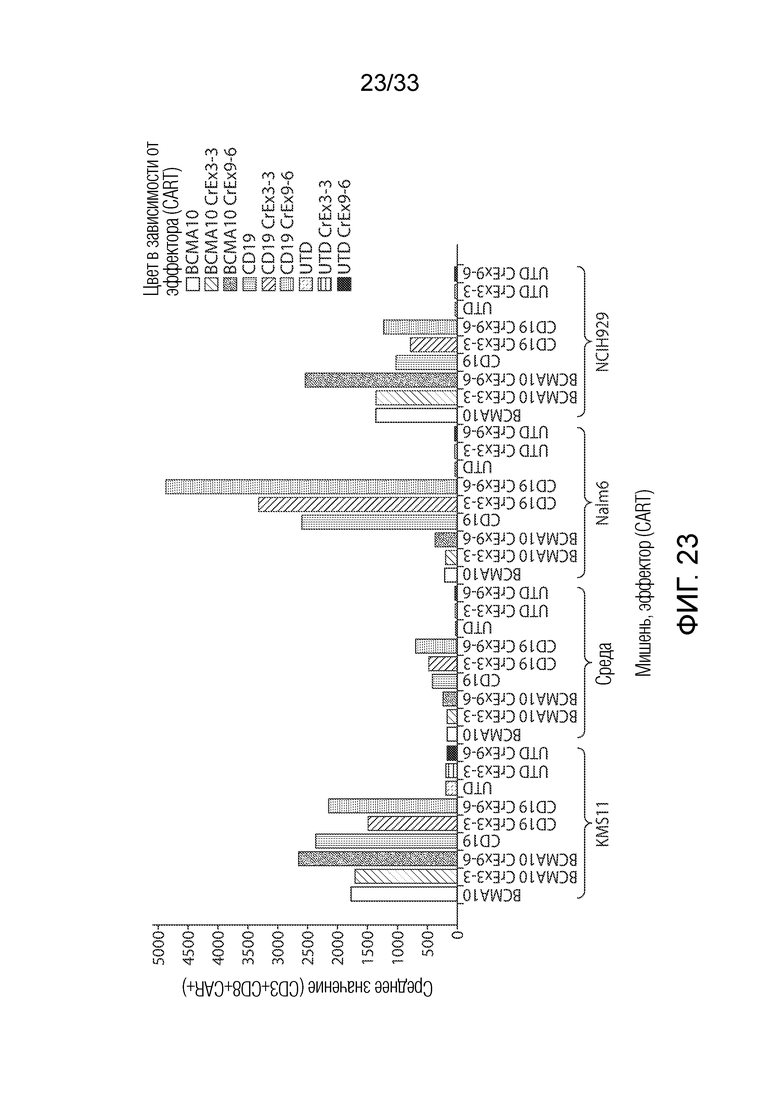
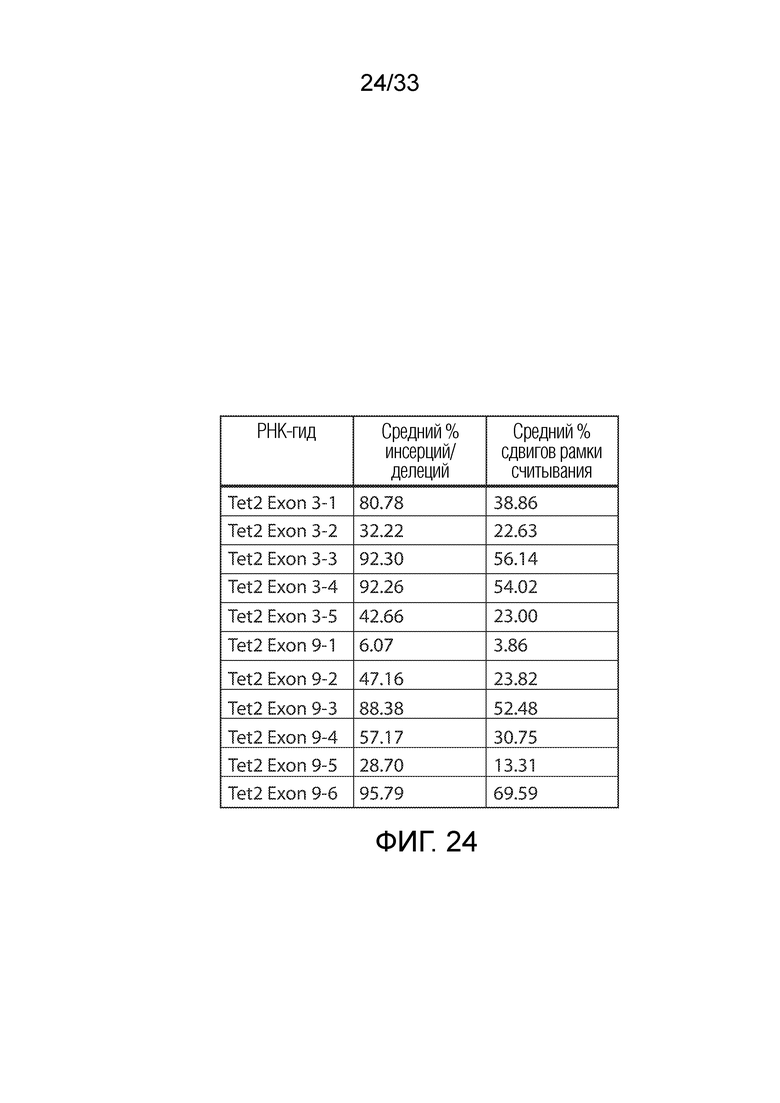
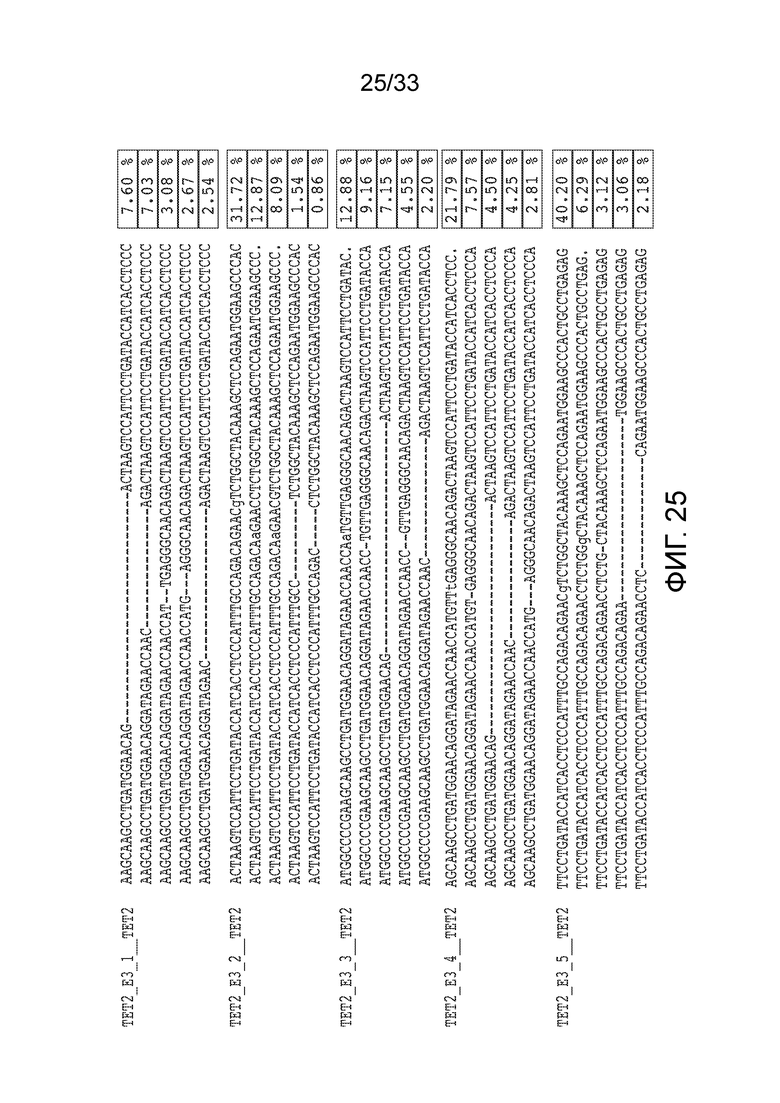
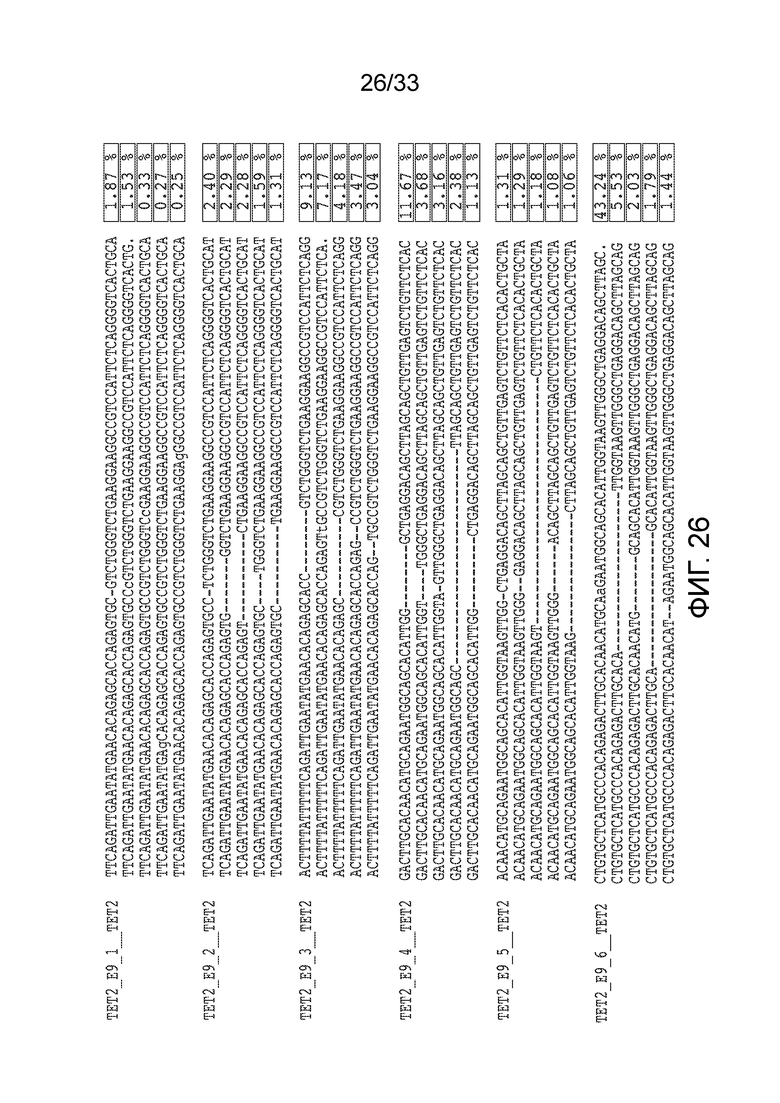
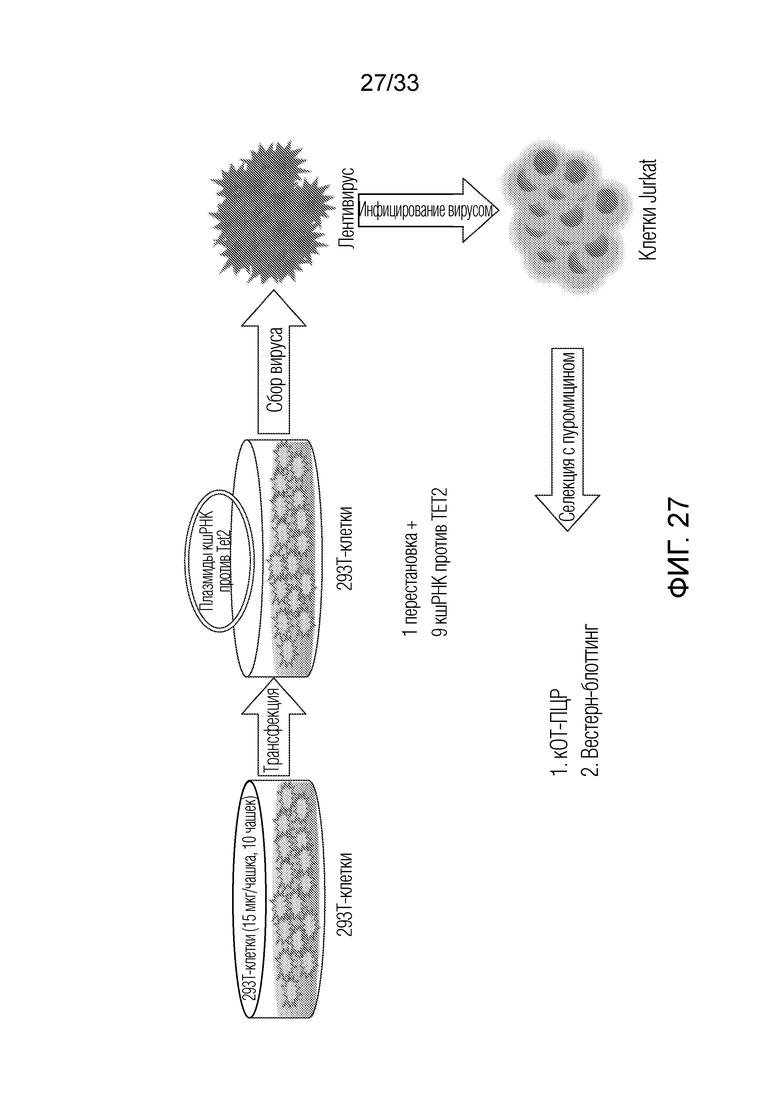
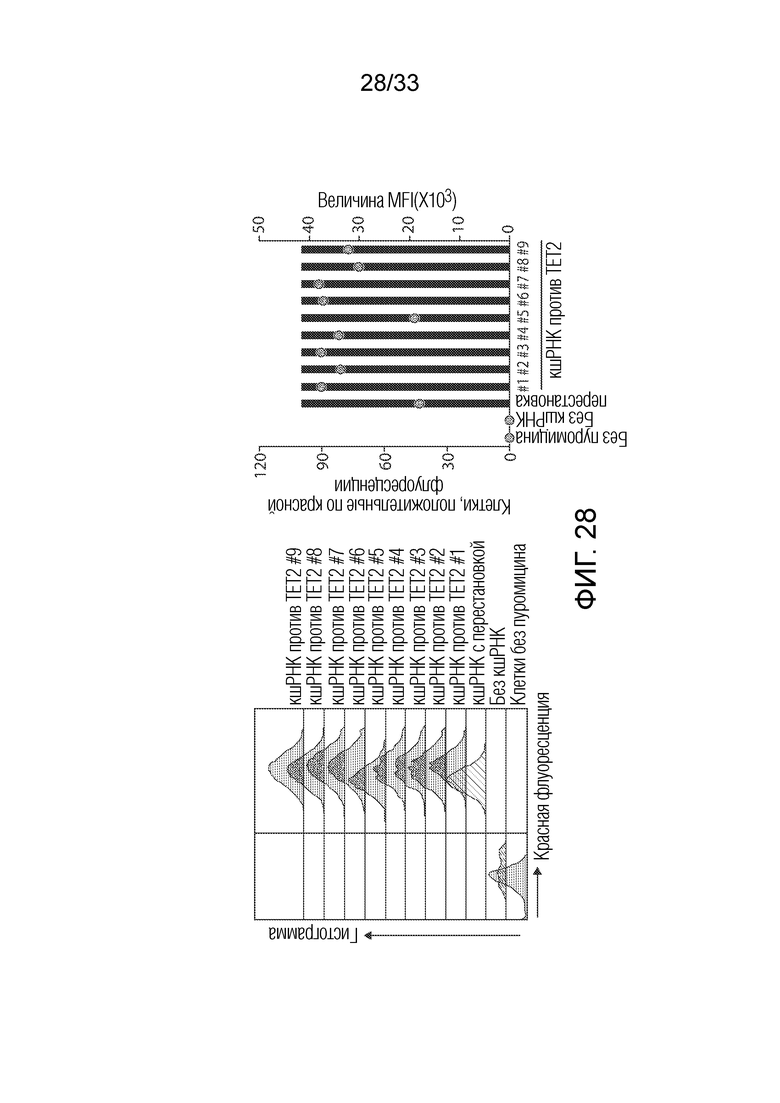

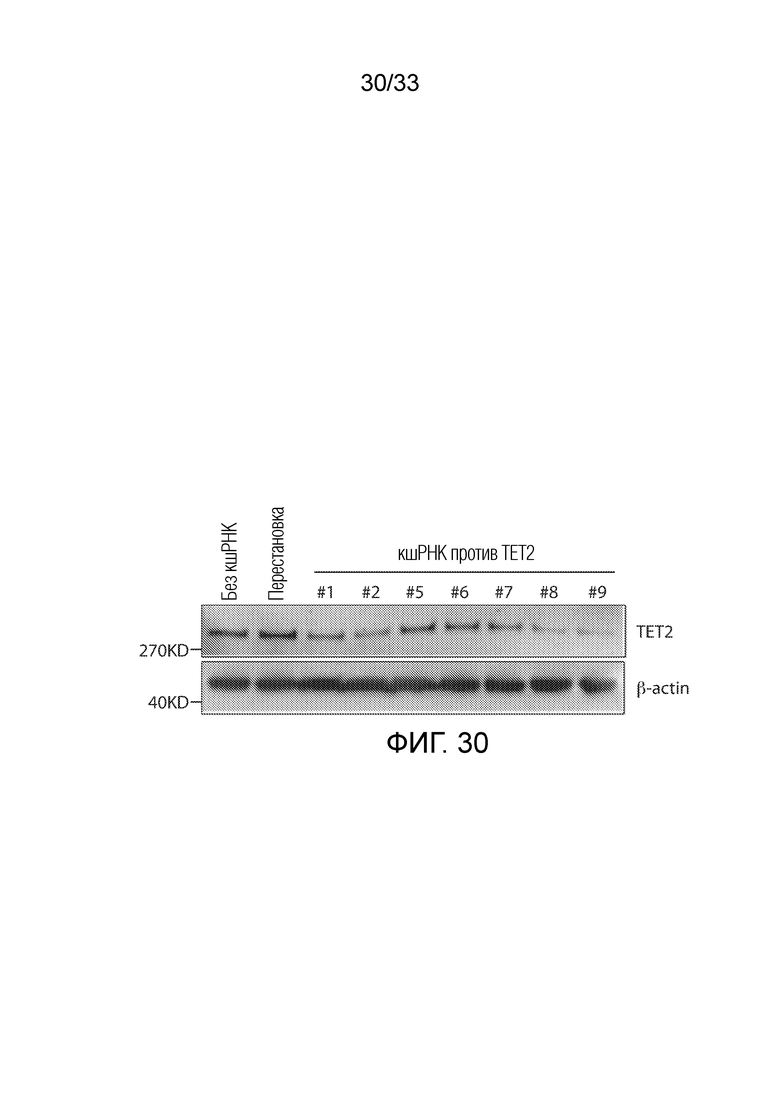

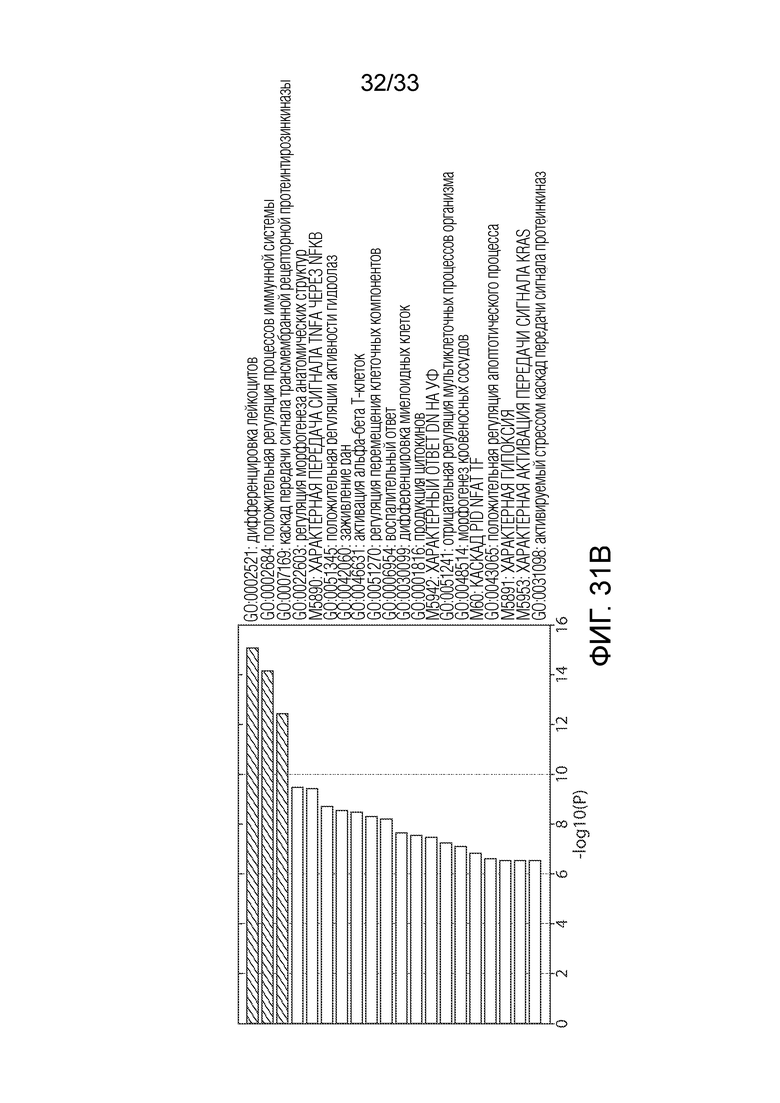
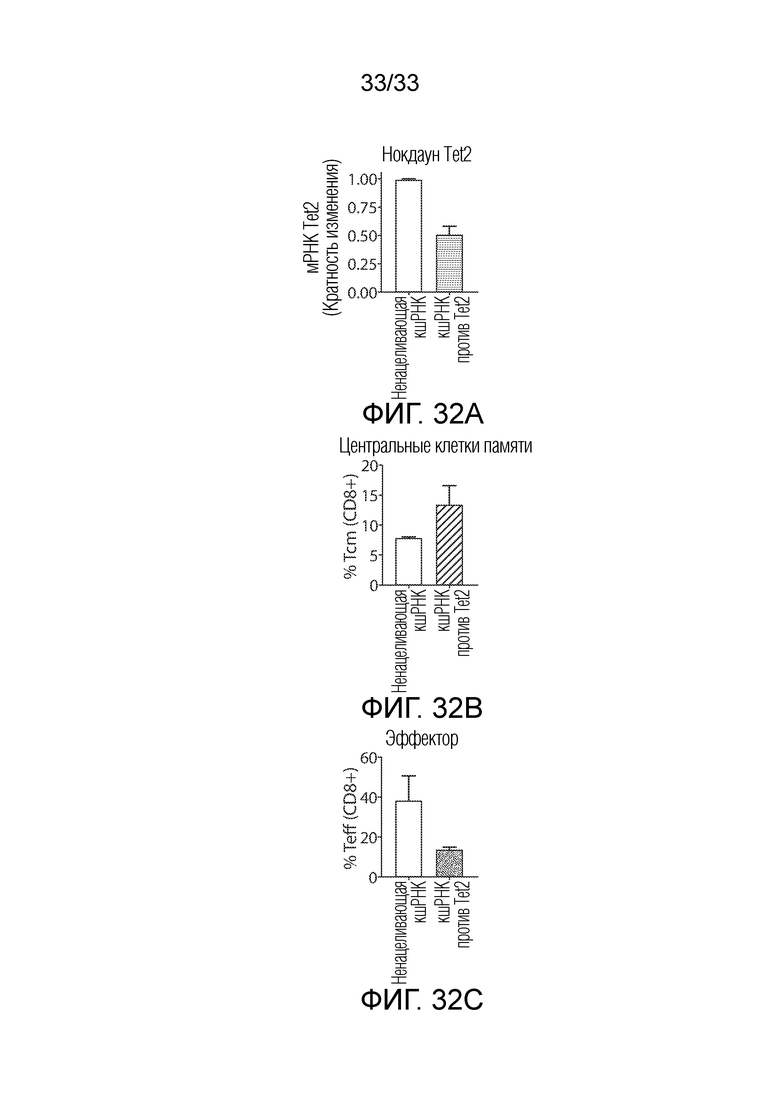
Изобретение относится к области биотехнологии, в частности к иммунной эффекторной клетке, модифицированной способами инженерии для экспрессии химерного рецептора антигена (CAR), содержащей ингибитор Tet2 и нарушение гена Tet2, способу ее производства, а также к содержащей ее композиции и популяции. Также раскрыт способ повышения терапевтической эффективности иммунной эффекторной клетки, включающей стадию приведения указанной иммунной эффекторной клетки в контакт с ингибитором Tet2. Изобретение также относится к вектору, содержащему последовательность, кодирующую CAR, и последовательность, кодирующую ингибитор Tet2. Изобретение эффективно для лечения заболевания, ассоциированного с экспрессией опухолевого антигена, у индивидуума. 9 н. и 35 з.п. ф-лы, 32 ил., 6 табл.
1. Иммунная эффекторная клетка, модифицированная способами инженерии для экспрессии химерного рецептора антигена (CAR), где CAR содержит антигенсвязывающий домен, который связывается с опухолевым антигеном, трансмембранный домен и внутриклеточный сигнальный домен, где иммунная эффекторная клетка представляет собой T-клетку или NK-клетку и где иммунная эффекторная клетка содержит:
(a) ингибитор Tet2, где указанный ингибитор представляет собой:
(i) миРНК или кшРНК, специфичную к Tet2, или нуклеиновую кислоту, кодирующую указанную миРНК или кшРНК; или
(ii) (1) систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet2, или в их регуляторных элементах; (2) нуклеиновую кислоту, кодирующую один или несколько компонентов указанной системы редактирования генов; или (3) их комбинации;
где система редактирования генов представляет собой систему CRISPR/Cas9;
(b) нарушение гена Tet2.
2. Иммунная эффекторная клетка по п. 1, где опухолевый антиген выбран из группы, состоящей из: CD19, TSHR, CD123, CD22, CD30, CD171, CS-1, CLL-1, CD33, EGFRvIII, GD2, GD3, BCMA, Tn Ag, PSMA, ROR1, FLT3, FAP, TAG72, CD38, CD44v6, CEA, EPCAM, B7H3, KIT, IL-13Ra2, мезотелина, IL-11Ra, PSCA, PRSS21, VEGFR2, LewisY, CD24, PDGFR-бета, SSEA-4, CD20, рецептора фолатов альфа, ERBB2 (Her2/neu), MUC1, EGFR, NCAM, простазы, PAP, ELF2M, эфрина B2, рецептора IGF-I, CAIX, LMP2, gp100, bcr-abl, тирозиназы, EphA2, фукозил-GM1, sLe, GM3, TGS5, HMWMAA, o-ацетил-GD2, рецептора фолатов бета, TEM1/CD248, TEM7R, CLDN6, GPRC5D, CXORF61, CD97, CD179a, ALK, полисиаловой кислоты, PLAC1, GloboH, NY-BR-1, UPK2, HAVCR1, ADRB3, PANX3, GPR20, LY6K, OR51E2, TARP, WT1, NY-ESO-1, LAGE-1a, MAGE-A1, легумаина, HPV E6,E7, MAGE A1, ETV6-AML, белка спермы 17, XAGE1, Tie 2, MAD-CT-1, MAD-CT-2, Fos-родственного антигена 1, p53, мутанта p53, простеина, сурвивина и теломеразы, PCTA-1/галектина 8, MelanA/MART1, мутанта Ras, hTERT, точек разрыва при транслокации саркомы, ML-IAP, ERG (слитый ген TMPRSS2 ETS), NA17, PAX3, рецептора андрогенов, циклина B1, MYCN, RhoC, TRP-2, CYP1B1, BORIS, SART3, PAX5, OY-TES1, LCK, AKAP-4, SSX2, RAGE-1, обратной транскриптазы теломеразы человека, RU1, RU2, кишечной карбоксилэстеразы, mut hsp70-2, CD79a, CD79b, CD72, LAIR1, FCAR, LILRA2, CD300LF, CLEC12A, BST2, EMR2, LY75, GPC3, FCRL5 и IGLL1.
3. Иммунная эффекторная клетка по п. 2, где опухолевым антигеном является CD19.
4. Иммунная эффекторная клетка по любому из предшествующих пунктов, где трансмембранный домен содержит:
аминокислотную последовательность, имеющую по меньшей мере одну, две или три модификации, но не более 20, 10 или 5 модификаций аминокислотной последовательности SEQ ID NO: 12 или аминокислотной последовательности, обладающей 95-99% идентичностью с аминокислотной последовательностью SEQ ID NO: 12; или последовательность SEQ ID NO: 12.
5. Иммунная эффекторная клетка по любому из предшествующих пунктов, где антигенсвязывающий домен связан с трансмембранным доменом шарнирной областью, где указанная шарнирная область содержит аминокислотную последовательность SEQ ID NO: 2 или SEQ ID NO: 6 или аминокислотную последовательность, обладающую 95-99% идентичностью с ними.
6. Иммунная эффекторная клетка по любому из предшествующих пунктов, где внутриклеточный сигнальный домен содержит первичный сигнальный домен и/или костимулирующий сигнальный домен, где первичный сигнальный домен содержит функциональный сигнальный домен белка, выбранного из CD3-зета, CD3-гамма, CD3-дельта, CD3, общего FcR-гамма (FCER1G), FcR-бета (Fc-эпсилон R1b), CD79a, CD79b, Fc-гамма RIIa, DAP10 или DAP12.
7. Иммунная эффекторная клетка по п. 6, где первичный сигнальный домен содержит:
аминокислотную последовательность, обладающую по меньшей мере одной, двумя или тремя, но не более чем 20, 10 или 5 модификациями аминокислотной последовательности SEQ ID NO: 18 или SEQ ID NO: 20, или аминокислотную последовательность, обладающую 95-99% идентичностью с аминокислотной последовательностью SEQ ID NO: 18 или SEQ ID NO: 20; или
аминокислотную последовательность SEQ ID NO: 18 или SEQ ID NO: 20.
8. Иммунная эффекторная клетка по любому из предшествующих пунктов, где внутриклеточный сигнальный домен содержит костимулирующий сигнальный домен и/или первичный сигнальный домен, где костимулирующий сигнальный домен содержит функциональный сигнальный домен белка, выбранного из группы, состоящей из CD27, CD28, 4-1BB (CD137), OX40, CD30, CD40, PD-1, ICOS, ассоциированного с лимфоцитарной функцией антигена-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, B7-H3, лиганда, который специфически связывается с CD83, CDS, ICAM-1, GITR, BAFFR, HVEM (LIGHTR), SLAMF7, NKp80 (KLRF1), CD160, CD19, CD4, CD8-альфа, CD8-бета, IL2R-бета, IL2R-гамма, IL7R-альфа, ITGA4, VLA1, CD49a, ITGA4, IA4, CD49D, ITGA6, VLA-6, CD49f, ITGAD, CD11d, ITGAE, CD103, ITGAL, CD11a, LFA-1, ITGAM, CD11b, ITGAX, CD11c, ITGB1, CD29, ITGB2, CD18, LFA-1, ITGB7, TNFR2, TRANCE/RANKL, DNAM1 (CD226), SLAMF4 (CD244, 2B4), CD84, CD96 (Tactile), CEACAM1, CRTAM, Ly9 (CD229), CD160 (BY55), PSGL1, CD100 (SEMA4D), CD69, SLAMF6 (NTB-A, Ly108), SLAM (SLAMF1, CD150, IPO-3), BLAME (SLAMF8), SELPLG (CD162), LTBR, LAT, GADS, SLP-76, PAG/Cbp, NKp44, NKp30, NKp46 и NKG2D
9. Иммунная эффекторная клетка по любому из пп. 6-8, где костимулирующий сигнальный домен указанного CAR содержит аминокислотную последовательность, обладающую по меньшей мере одной, двумя или тремя модификациями, но не более чем 20, 10 или 5 модификациями аминокислотной последовательности SEQ ID NO: 14 или SEQ ID NO: 16, или аминокислотную последовательность, обладающую 95-99% идентичностью с аминокислотной последовательностью SEQ ID NO: 14 или SEQ ID NO: 16.
10. Иммунная эффекторная клетка по любому из пп. 6-8, где костимулирующий сигнальный домен содержит аминокислотную последовательность SEQ ID NO: 14 или SEQ ID NO: 16.
11. Иммунная эффекторная клетка по любому из предшествующих пунктов, где внутриклеточный сигнальный домен содержит аминокислотную последовательность SEQ ID NO: 14 или SEQ ID NO: 16 и аминокислотную последовательность SEQ ID NO: 18 или SEQ ID NO: 20, где последовательности, содержащие внутриклеточный сигнальный домен, экспрессируются в одной рамке считывания и в виде единой полипептидной цепи.
12. Иммунная эффекторная клетка по любому из предшествующих пунктов, дополнительно содержащая лидерную последовательность, которая содержит аминокислотную последовательность SEQ ID NO: 2.
13. Иммунная эффекторная клетка по любому из предшествующих пунктов, где иммунная эффекторная клетка представляет собой T-клетку.
14. Иммунная эффекторная клетка по п. 13, где T-клетка представляет собой CD4+ T-клетку, CD8+ T-клетку или их комбинацию.
15. Иммунная эффекторная клетка по любому из предшествующих пунктов, где иммунная эффекторная клетка представляет собой клетку человека.
16. Иммунная эффекторная клетка по любому из предшествующих пунктов, где система редактирования генов связывается с последовательностью-мишенью в раннем экзоне или интроне гена, кодирующего Tet2.
17. Иммунная эффекторная клетка по п. 16, где последовательность-мишень находится выше по направлению транскрипции относительно экзона 4 или в экзоне 1, экзоне 2 или экзоне 3.
18. Иммунная эффекторная клетка по любому из предшествующих пунктов, где система редактирования генов связывается с последовательностью-мишенью в позднем экзоне или интроне гена, кодирующего Tet2.
19. Иммунная эффекторная клетка по п. 18, где последовательность-мишень расположена ниже по направлению транскрипции относительно экзона 8 или находится в экзоне 9, экзоне 10 или экзоне 11.
20. Иммунная эффекторная клетка по любому из предшествующих пунктов, где система редактирования генов представляет собой систему CRISPR/Cas, содержащую молекулу гРНК, содержащую нацеливающую последовательность, которая гибридизуется с последовательностью-мишенью гена Tet2.
21. Иммунная эффекторная клетка по п. 20, где нацеливающая последовательность представляет собой нацеливающую последовательность, приведенную в любой из SEQ ID NO: 73-1243.
22. Иммунная эффекторная клетка по п. 20, где нацеливающая последовательность представляет собой нацеливающую последовательность, приведенную в любой из SEQ ID NO: 1273-1295.
23. Иммунная эффекторная клетка по любому из предшествующих пунктов, где миРНК или кшРНК содержит последовательность, комплементарную последовательности мРНК Tet2, или содержит последовательность-мишень кшРНК, приведенную в любой из SEQ ID NO: 1244-1252.
24. Способ повышения терапевтической эффективности иммунной эффекторной клетки по любому из предшествующих пунктов, включающий стадию приведения указанной иммунной эффекторной клетки в контакт с ингибитором Tet2, где ингибитор Tet выбран из группы, состоящей из:
(i) миРНК или кшРНК, специфичной к Tet2, или нуклеиновой кислоты, кодирующей указанную миРНК или кшРНК; или
(ii) (1) системы редактирования генов, нацеленной на один или несколько участков в гене, кодирующем Tet2, или в его регуляторных элементах; (2) нуклеиновой кислоты, кодирующей один или несколько компонентов указанной системы редактирования генов; или (3) их комбинации.
25. Способ по п. 24, где указанное приведение в контакт происходит ex vivo.
26. Способ по п. 26, где указанное приведение в контакт происходит in vivo.
27. Способ по п. 26, где приведение в контакт происходит in vivo перед доставкой нуклеиновой кислоты, кодирующей CAR, в клетку.
28. Способ по п. 26, где приведение в контакт происходит in vivo после введения клеток индивидууму, нуждающемуся в этом.
29. Применение иммунной эффекторной клетки по любому из пп. 1-23 для лечения заболевания, ассоциированного с экспрессией опухолевого антигена, у индивидуума.
30. Применение по п. 29, где способ дополнительно включает введение указанному индивидууму ингибитора Tet2.
31. Применение по п. 29 или 30, где заболевание, ассоциированное с экспрессией опухолевого антигена, выбирают из пролиферативного заболевания, предзлокачественного состояния, злокачественной опухоли и не связанного со злокачественной опухолью заболевания, ассоциированного с экспрессией опухолевого антигена.
32. Применение экспрессирующей CAR иммунной эффекторной клетки в комбинации с ингибитором Tet2 для лечения заболевания, ассоциированного с экспрессией опухолевого антигена, у индивидуума, где CAR содержит антигенсвязывающий домен, который связывается с опухолевым антигеном, трансмембранный домен и внутриклеточный сигнальный домен,
где указанный ингибитор Tet2 выбирают из группы, состоящей из
(i) миРНК или кшРНК, специфичной к Tet2, или нуклеиновой кислоты, кодирующей указанную миРНК или кшРНК; или
(ii) (1) системы редактирования генов, нацеленной на один или несколько участков в гене, кодирующем Tet2, или в их регуляторных элементах; (2) нуклеиновой кислоты, кодирующей один или несколько компонентов указанной системы редактирования генов; или (3) их комбинации;
таким образом осуществляя лечение указанного субъекта.
33. Применение по п. 32, где индивидууму проводят предварительное лечение ингибитором Tet2 перед началом введения экспрессирующей CAR иммунной эффекторной клетки.
34. Применение по п. 32, где указанному индивидууму проводят одновременное лечение ингибитором Tet2 и терапию экспрессирующими CAR клетками.
35. Применение по п. 32, где индивидууму проводят лечение ингибитором Tet2 после терапии экспрессирующими CAR клетками.
36. Применение по любому из пп. 31-35, где индивидуум имеет заболевание, ассоциированное с экспрессией опухолевого антигена, или пролиферативное заболевание, предзлокачественное состояние, злокачественную опухоль и не связанное со злокачественной опухолью заболевание, ассоциированное с экспрессией опухолевого антигена.
37. Применение по п. 36, где злокачественная опухоль представляет собой гематологическую злокачественную опухоль, выбранную из одного или нескольких из хронического лимфоцитарного лейкоза (CLL), острых лейкозов, острого лимфобластного лейкоза (ALL), B-клеточного острого лимфобластного лейкоза (B-ALL), T-клеточного острого лимфобластного лейкоза (T-ALL), хронического миелогенного лейкоза (CML), B-клеточного пролимфоцитарного лейкоза, новообразования из бластных плазмацитоидных дендритных клеток, лимфомы Беркитта, диффузной крупноклеточной B-клеточной лимфомы, фолликулярной лимфомы, волосатоклеточного лейкоза, мелкоклеточной или крупноклеточной фолликулярной лимфомы, злокачественных лимфопролиферативных состояний, MALT-лимфомы, лимфомы из клеток мантийной зоны, лимфомы из клеток маргинальной зоны, множественной миеломы, миелодисплазии и миелодиспластического синдрома, неходжскинской лимфомы, лимфомы Ходжкина, плазмабластной лимфомы, новообразования из плазмацитоидных дендритных клеток, макроглобулинемии Валденстрема или предлейкоза.
38. Применение по п. 36, где злокачественная опухоль выбрана из группы, состоящей из рака толстого кишечника, рака прямой кишки, почечно-клеточного рака, рака печени, немелкоклеточной карциномы легкого, рака тонкого кишечника, рака пищевода, меланомы, рака кости, рака поджелудочной железы, рака кожи, рака головы и шеи, кожной или внутриглазной злокачественной меланомы, рака тела матки, рака яичника, рака прямой кишки, рака анальной области, рака желудка, рака яичка, рака тела матки, карциномы фаллопиевых труб, карциномы эндометрия, карциномы шейки матки, карциномы влагалища, карциномы вульвы, болезни Ходжкина, неходжскинской лимфомы, злокачественной опухоли эндокринной системы, рака щитовидной железы, рака паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака мочеиспускательного канала, рака полового члена, солидных опухолей детского возраста, рака мочевого пузыря, рака почки или мочеточника, рака почечной лоханки, новообразования центральной нервной системы (ЦНС), первичной лимфомы ЦНС, ангиогенеза опухоли, опухоли позвоночника, глиомы ствола головного мозга, аденомы гипофиза, саркомы Капоши, эпидермоидного рака, плоскоклеточного рака, T-клеточной лимфомы, злокачественных опухолей, индуцируемых факторами внешней среды, комбинаций указанных злокачественных опухолей и метастатических очагов указанных злокачественных опухолей.
39. Способ производства экспрессирующих CAR клеток, включающий:
(a) введение нуклеиновой кислоты, кодирующей CAR, в клетку, так чтобы указанная нуклеиновая кислота встраивалась в геном клетки в ген Tet2, или указанная интеграция происходит в интрон или экзон гена Tet2, так чтобы экспрессия и/или функция Tet2 снижалась или устранялась; или
(b) обеспечение клетки, полученной у индивидуума, и приведение в контакт клетки ex vivo с ингибитором Tet2, где приведение в контакт проводится до, одновременно или после того, как указанная клетка модифицируется, чтобы экспрессировать CAR,
где указанный CAR содержит антигенсвязывающий домен, который связывается с опухолевым антигеном, трансмембранный домен и внутриклеточный сигнальный домен.
40. Способ производства экспрессирующей CAR клетки, включающий приведение в контакт указанной экспрессирующей CAR клетки ex vivo с ингибитором Tet2, где указанный CAR содержит антигенсвязывающий домен, который связывается с опухолевым антигеном, трансмембранный домен и внутриклеточный сигнальный домен, где указанный ингибитор Tet2 представляет собой:
(i) миРНК или кшРНК, специфичную к Tet2, или нуклеиновую кислоту, кодирующую указанную миРНК или кшРНК; или
(ii) (1) систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet2, или в их регуляторных элементах; (2) нуклеиновую кислоту, кодирующую один или несколько компонентов указанной системы редактирования генов; или (3) их комбинации.
41. Вектор для продуцирования CAR, содержащий последовательность, кодирующую CAR, и последовательность, кодирующую ингибитор Tet2,
где указанный CAR содержит антигенсвязывающий домен, который связывается с опухолевым антигеном, трансмембранный домен и внутриклеточный сигнальный домен, где ингибитор Tet2 представляет собой:
(i) миРНК или кшРНК, специфичную к Tet2; или
(ii) систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet2, или в его соответствующих регуляторных элементах.
42. Вектор по п. 41, где последовательность, кодирующая CAR, и последовательность, кодирующая ингибитор Tet2, разделены участком 2A.
43. Композиция для производства экспрессирующей CAR клетки ex vivo, содержащая ингибитор Tet2,
где указанный ингибитор Tet2 представляет собой:
(i) миРНК или кшРНК, специфичную к Tet2 , или нуклеиновую кислоту, кодирующую указанную миРНК или кшРНК;
(ii) (1) систему редактирования генов, нацеленную на один или несколько участков в гене, кодирующем Tet2, или в его соответствующих регуляторных элементах; (2) нуклеиновую кислоту, кодирующую один или несколько компонентов указанной системы редактирования генов; или (3) их комбинации.
44. Популяция иммунных эффекторных клеток, содержащая одну или несколько клеток по любому из пп. 1-23, для лечения заболевания, ассоциированного с экспрессией опухолевого антигена, где популяция клеток содержит более высокий процент клеток Tscm или CD45RA+CD62L+CCR7+CD27+CD95+ T-клетки, чем популяция клеток, которая не содержит одну или несколько клеток, в которых экспрессия и/или функция Tet2 снижена или устранена.
| WO2012079000 A1, 14.06.2012 | |||
| WO2014190273 A1, 27.11.2014 | |||
| PRONIER, ELODIE, et al., Inhibition of TET2-mediated conversion of 5-methylcytosine to 5-hydroxymethylcytosine disturbs erythroid and granulomonocytic differentiation of human hematopoietic progenitors, Blood, The Journal of the American Society of Hematology, 2011, 118(9): 2551-2555 |

