РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет на основании предварительной заявки США №62/473595, поданной 20 марта 2017 г. под названием «METHODS AND COMPOISTIONS FOR PREPARING NUCLEIC ACID LIBRARIES)) ("СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ БИБЛИОТЕК НУКЛЕИНОВЫХ КИСЛОТ"), описание которой полностью включено в настоящий документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка включает перечень последовательностей в электронном формате. Указанный перечень последовательностей представлен в виде файла с именем ILLINC362WOSEQLIST, созданного 15 марта 2018 г., размером приблизительно 4 кБ. Информация, содержащаяся перечне последовательностей в электронном формате, полностью включена в настоящий документ посредством ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0003] Системы, способы и композиции, предложенные в настоящем документе, относятся к получению библиотек нуклеиновых кислот. Некоторые варианты реализации включают получение библиотек нуклеиновых кислот посредством лигирования одноцепочечных нуклеиновых кислот.
УРОВЕНЬ ТЕХНИКИ
[0004] Для быстрого и экономичного определения полной последовательности генома существует несколько технологий секвенирования следующего поколения. Обычно до секвенирования получают библиотеку матричных нуклеиновых кислот из образца двуцепочечной целевой геномной ДНК. Получение образца обычно включает этап фрагментирования ДНК, на котором происходит разрыв крупных цепей ДНК на фрагменты меньшего размера, более подходящие для технологий секвенирования следующего поколения. Часто к концам фрагментов ДНК присоединяют адаптеры, что можно выполнять путем репарации концов ДНК с последующим лигированием адаптеров, или, в последнее время, с использованием системы транспосом. Использование транспосом, представляющих собой комплекс транспозазы и последовательностей транспозонов, позволяет одновременно фрагментировать геном и лигировать адаптеры к фрагментам, тем самым упрощая получение библиотеки. Способы получения библиотек обычно трудоемки и требуют выполнения нескольких этапов, выполняемых оператором, на различных стадиях.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0005] Некоторые варианты реализации включают способ получения библиотеки нуклеиновых кислот, включающий: (а) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой одноцепочечные нуклеиновые кислоты; (b) дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот; (с) лигирование первого адаптера с 3'-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, причем 3'-конец первого адаптера содержит блокирующую группу; (d) фосфорилирование 5'-концов лигированных одноцепочечных нуклеиновых кислот; и (е) лигирование второго адаптера с 5'-концами фосфорилированных лигированных одноцепочечных нуклеиновых кислот в присутствии лигазы, за счет чего получают библиотеку нуклеиновых кислот.
[0006] В некоторых вариантах реализации 5'-конец второго адаптера нефосфорилирован.
[0007] В некоторых вариантах реализации второй адаптер прикреплен к подложке. В некоторых вариантах реализации подложка включает гранулу или проточную ячейку.
[0008] Некоторые варианты реализации также включают удаление нелигированных одноцепочечных первых адаптеров до этапа (е).
[0009] Некоторые варианты реализации также включают гибридизацию нелигированных одноцепочечных первых адаптеров с зондом захвата. В некоторых вариантах реализации зонд захвата содержит последовательность, комплементарную по меньшей мере части первого адаптера. В некоторых вариантах реализации 3'-конец зонда захвата содержит блокирующую группу. В некоторых вариантах реализации 5'-конец зонда захвата содержит блокирующую группу.
[0010] Некоторые варианты реализации также включают гидролиз нелигированных одноцепочечных первых адаптеров. В некоторых вариантах реализации гидролиз нелигированных одноцепочечных первых адаптеров включает приведение нелигированных одноцепочечных первых адаптеров в контакт с 5'-фосфат-зависимой экзонуклеазой. В некоторых вариантах реализации гидролиз нелигированных одноцепочечных первых адаптеров включает приведение нелигированных одноцепочечных первых адаптеров в контакт с 5'-фосфат-зависимой экзонуклеазой и 5'-деаденилазой.
[0011] В некоторых вариантах реализации фосфорилирование 5'-конца лигированных одноцепочечных нуклеиновых кислот включает приведение лигированных одноцепочечных нуклеиновых кислот в контакт с киназой.
[0012] Некоторые варианты реализации включают способ получения библиотеки нуклеиновых кислот, включающий: (а) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой двуцепочечные нуклеиновые кислоты; (b) приведение двуцепочечных нуклеиновых кислот в контакт с 5'-экзонуклеазой с получением множества модифицированных двуцепочечных нуклеиновых кислот с одноцепочечными 3'-выступающими концами; (с) лигирование первого адаптера с 3'-концами модифицированных двуцепочечных нуклеиновых кислот в присутствии лигазы, причем 3'-конец первого адаптера содержит блокирующую группу; (d) денатурацию модифицированных двуцепочечных нуклеиновых кислот, лигированных с первыми адаптерами, с получением множества одноцепочечных нуклеиновых кислот; и (е) лигирование второго адаптера с 5'-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, за счет чего получают библиотеку нуклеиновых кислот.
[0013] В некоторых вариантах реализации 5'-конец второго адаптера нефосфорилирован.
[0014] Некоторые варианты реализации включают способ получения библиотеки нуклеиновых кислот, включающий: (а) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой одноцепочечные нуклеиновые кислоты; (b) дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот; (с) лигирование первого адаптера с 3'-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, причем 3'-конец первого адаптера содержит блокирующую группу; (d) гибридизацию лигированного первого адаптера с зондом захвата; и (е) удлинение зонда захвата и лигирование второго адаптера с 3'-концом удлиненного зонда захвата в присутствии лигазы, причем 3'-конец второго адаптера содержит блокирующую группу, за счет чего получают библиотеку нуклеиновых кислот.
[0015] Некоторые варианты реализации также включают удаление гибридизованного лигированного первого адаптера с удлиненного зонда захвата.
[0016] В некоторых вариантах реализации зонд захвата прикреплен к подложке. В некоторых вариантах реализации подложка включает гранулу или проточную ячейку. В некоторых вариантах реализации зонд захвата содержит индекс зонда захвата. В некоторых вариантах реализации зонд захвата содержит расщепляемый линкер. Некоторые варианты реализации также включают расщепление расщепляемого линкера.
[0017] Некоторые варианты реализации включают способ получения библиотеки нуклеиновых кислот, включающий: (а) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой одноцепочечные нуклеиновые кислоты; (b) дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот; (с) лигирование линкера с 3'-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, причем указанный линкер содержит первый адаптер и второй адаптер, и 3'-конец линкера содержит блокирующую группу; (d) фосфорилирование 5'-концов лигированных одноцепочечных нуклеиновых кислот; и (е) снятие защиты с 3'-концов фосфорилированных нуклеиновых кислот и замыкание нуклеиновых кислот со снятой защитой в кольцо, за счет чего получают библиотеку нуклеиновых кислот.
[0018] В некоторых вариантах реализации линкер содержит расщепляемый сайт между первым адаптером и вторым адаптером.
[0019] Некоторые варианты реализации также включают линеаризацию кольцевых нуклеиновых кислот путем расщепления по расщепляемому линкеру. В некоторых вариантах реализации расщепляемый сайт содержит остаток урацила. Некоторые варианты реализации также включают приведение кольцевых нуклеиновых кислот в контакт с урацил-специфическим эксцизионным реагентом. В некоторых вариантах реализации урацил-специфический эксцизионный реагент содержит фермент, выбранный из урацил-ДНК-гликозилазы (UDG) и ДНК-гликозилазы-лиазы эндонуклеазы VTII.
[0020] Некоторые варианты реализации также включают амплификацию кольцевых нуклеиновых кислот способом, включающим гибридизацию по меньшей мере праймера с первым адаптером или вторым адаптером. В некоторых вариантах реализации амплификация выбрана из ПНР и амплификации по типу катящегося кольца (RCA). В некоторых вариантах реализации амплификация включает приведение кольцевых нуклеиновых кислот в контакт с полимеразой, которая образует линейный продукт при контакте с остатком урацила в матрице.
[0021] В некоторых вариантах реализации дефосфорилирование 5'-конца одноцепочечных нуклеиновых кислот включает приведение одноцепочечных нуклеиновых кислот в контакт с фосфатазой.
[0022] В некоторых вариантах реализации этапы (b)-(е) выполняют в одном реакционном объеме.
[0023] В некоторых вариантах реализации этапы (b)-(e) выполняют в одном реакционном сосуде.
[0024] В некоторых вариантах реализации первый адаптер и/или второй адаптер содержит сайт связывания праймера для секвенирования.
[0025] В некоторых вариантах реализации первый адаптер содержит последовательность Р7, последовательность Р5 или их комплемент или обратный комплемент. В некоторых вариантах реализации второй адаптер содержит последовательность Р7, последовательность Р5 или их комплемент или обратный комплемент.
[0026] В некоторых вариантах реализации первый адаптер и/или второй адаптер содержит индекс адаптера (adaptor index). В некоторых вариантах реализации индекс первого адаптера отличается от индекса второго адаптера. В некоторых вариантах реализации индекс адаптера указывает на источник множества нуклеиновых кислот.
[0027] В некоторых вариантах блокирующая группа содержит 3'-спейсер С3 или дидезоксинуклеотид.
[0028] В некоторых вариантах реализации изобретения лигаза содержит лигазу одноцепочечной нуклеиновой кислоты.
В некоторых вариантах реализации лигирование первого адаптера и/или лигирование второго адаптера выполняют в присутствии агента, вызывающего эффект исключения объема (volume excluding agent). В некоторых вариантах реализации агент, вызывающий эффект исключения объема, выбран из группы, состоящей из полиэтиленгликоля (ПЭГ), декстрана, гидроксиэтилкрахмала, фиколла и поливинилпирролидона. В некоторых вариантах реализации первый этап лигирования и/или второй этап лигирования выполняют в реакционном объеме, содержащем по меньшей мере приблизительно 37% (вес/об.) ПЭГ. В некоторых вариантах реализации первый этап лигирования и/или второй этап лигирования выполняют в реакционном объеме, содержащем по меньшей мере приблизительно 60% (вес/об.) ПЭГ.
[0030] В некоторых вариантах реализации множество нуклеиновых кислот содержат РНК. В некоторых вариантах реализации множество нуклеиновых кислот содержат кДНК или геномную ДНК.
[0031] В некоторых вариантах реализации средний размер нуклеиновой кислоты из множества нуклеиновых кислот составляет менее приблизительно 200 нуклеотидов.
[0032] В некоторых вариантах реализации множество нуклеиновых кислот получают из низкокачественного источника нуклеиновой кислоты. В некоторых вариантах реализации множество нуклеиновых кислот получают из фиксированного образца.
[0033] Некоторые варианты реализации также включают амплификацию библиотеки нуклеиновых кислот.
[0034] Некоторые варианты реализации также включают получение данных о последовательностях из библиотеки нуклеиновых кислот.
[0035] Некоторые варианты реализации включают библиотеку нуклеиновых кислот, полученную способом согласно любому из вышеуказанных вариантов реализации.
[0036] Некоторые варианты реализации включают набор, содержащий: первый адаптер, причем 3'-конец указанного первого адаптера содержит блокирующую группу, причем первый адаптер содержит последовательность Р7, последовательность Р5 или их комплемент или обратный комплемент; лигазу; и компонент, выбранный из группы, состоящей из агента, вызывающего эффект исключения объема, киназы, фосфатазы, 5'-фосфат-зависимой экзонуклеазы, 5'-деаденилазы, полимеразы, образующей линейный продукт при контакте с остатком урацила в матрице, урацил-специфического эксцизионного реагента, и второй адаптер, причем указанный второй адаптер содержит последовательность Р7, последовательность Р5 или их комплемент или обратный комплемент. В некоторых вариантах реализации линкер содержит первый и второй адаптеры.
[0037] Некоторые варианты реализации включают реакционный сосуд, содержащий реакционный объем, содержащий: множество нуклеиновых кислот; первый адаптер, причем 3'-конец первого адаптера содержит блокирующую группу; лигазу; и агент, вызывающий эффект исключения объема.
[0038] В некоторых вариантах реализации множество нуклеиновых кислот представляет собой одноцепочечные нуклеиновые кислоты.
[0039] В некоторых вариантах реализации первый адаптер лигирован с 3'-концами множества одноцепочечных нуклеиновых кислот, тем самым образуя множество модифицированных одноцепочечных нуклеиновых кислот.
[0040] В некоторых вариантах реализации нелигированный первый адаптер гибридизован с зондом захвата. В некоторых вариантах реализации зонд захвата содержит последовательность, комплементарную по меньшей мере части первого адаптера.
[0041] В некоторых вариантах реализации множество нуклеиновых кислот представляет собой двуцепочечные нуклеиновые кислоты с 3'-выступающими концами. В некоторых вариантах реализации первый адаптер лигирован с 3'-концами множества двуцепочечных нуклеиновых кислот с 3'-выступающими концами, тем самым образуя множество модифицированных двуцепочечных нуклеиновых кислот.
[0042] Некоторые варианты реализации также включают второй адаптер. В некоторых вариантах реализации 5'-конец второго адаптера нефосфорилирован.
[0043] Некоторые варианты реализации также включают дефосфорилирующий агент. В некоторых вариантах реализации дефосфорилирующий агент содержит фосфатазу. В некоторых вариантах реализации указанная фосфатаза инактивирована.
[0044] В некоторых вариантах реализации агент, вызывающий эффект исключения объема, выбран из группы, состоящей из полиэтиленгликоля (ПЭГ), декстрана, гидроксиэтилкрахмала, фиколла и поливинилпирролидона. В некоторых вариантах реализации реакционный объем содержит по меньшей мере приблизительно 37% (вес/об.) ПЭГ. В некоторых вариантах реализации реакционный объем содержит по меньшей мере приблизительно 60% (вес/об.) ПЭГ.
[0045] В некоторых вариантах реализации первый адаптер и/или второй адаптер содержит сайт связывания праймера для секвенирования.
[0046] В некоторых вариантах реализации первый адаптер и/или второй адаптер содержит индекс адаптера. В некоторых вариантах реализации индекс первого адаптера отличается от индекса второго адаптера. В некоторых вариантах реализации индекс адаптера указывает на источник множества одноцепочечных нуклеиновых кислот.
[0047] В некоторых вариантах блокирующая группа содержит 3'-спейсер С3 или дидезоксинуклеотид. В некоторых вариантах реализации блокирующая группа содержит 3'-спейсер С3.
[0048] В некоторых вариантах реализации лигаза содержит лигазу одноцепочечной нуклеиновой кислоты.
[0049] В некоторых вариантах реализации множество нуклеиновых кислот содержат РНК. В некоторых вариантах реализации множество нуклеиновых кислот содержат кДНК или геномную ДНК.
[0050] В некоторых вариантах реализации множество одноцепочечных нуклеиновых кислот получают из низкокачественного источника нуклеиновой кислоты. В некоторых вариантах реализации множество нуклеиновых кислот получают из фиксированного образца.
[0051] Некоторые варианты реализации включают проточную ячейку, содержащую реакционный сосуд согласно любому из вышеуказанных вариантов реализации.
[0052] Некоторые варианты реализации включают систему, содержащую реакционный сосуд согласно любому из вышеуказанных вариантов реализации, и детектор для получения данных секвенирования.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0053] Фиг. 1 представляет собой схему способа получения библиотеки одноцепочечных нуклеиновых кислот в соответствии с одним вариантом реализации. Двуцепочечную нуклеиновую кислоту денатурируют с образованием одноцепочечной нуклеиновой кислоты, и библиотеку можно получить, дефосфорилируя 5'-конец одноцепочечной нуклеиновой кислоты, лигируя Р7'-адаптер, содержащий 3'-блокирующую группу, с 3'-концом одноцепочечной нуклеиновой кислоты, повторно фосфорилируя 5'-конец лигированной одноцепочечной нуклеиновой кислоты и лигируя адаптер Р5, содержащий нефосфорилированный 5'-конец, с 5'-концом лигированной одноцепочечной нуклеиновой кислоты.
[0054] Фиг. 2 представляет собой схему способа получения библиотеки одноцепочечных нуклеиновых кислот в соответствии с одним вариантом реализации. Двуцепочечную нуклеиновую кислоту частично гидролизуют 5'-экзонуклеазой с образованием двуцепочечной нуклеиновой кислоты с 3'-выступающими концами, денатурируют с образованием одноцепочечной нуклеиновой кислоты, адаптер Р7', содержащий 3'-блокирующую группу, можно лигировать с 3'-концами двуцепочечной нуклеиновой кислоты с 3'-выступающими концами, лигированную двуцепочечную нуклеиновую кислоту с 3'-выступающими концами можно денатурировать с образованием одноцепочечной нуклеиновой кислоты, а адаптер Р5, содержащий нефосфорилированный 5'-конец, можно лигировать с 5'-концом лигированной одноцепочечной нуклеиновой кислоты.
[0055] На фиг.3 изображена схема способа получения библиотеки одноцепочечных нуклеиновых кислот с использованием гранул в соответствии с одним вариантом реализации. Библиотеку можно получить путем дефосфорилирования 5'-конца одноцепочечной нуклеиновой кислоты, лигирования Р5'-адаптера, содержащего 3'-блокирующую группу, с 3'-концом одноцепочечной нуклеиновой кислоты, гибридизации первого продукта лигирования с зондом захвата Р5, присоединенным к грануле, удлинения зонда захвата, удаления гибридизированного первого продукта лигирования с удлиненного зонда захвата и лигирования адаптера Р7', содержащего 3'-блокирующую группу, с 3'-концом удлиненного зонда захвата.
[0056] Фиг. 4А представляет собой схему способа получения библиотеки кольцевых одноцепочечных нуклеиновых кислот в соответствии с одним вариантом реализации. Двуцепочечную нуклеиновую кислоту денатурируют с образованием одноцепочечной нуклеиновой кислоты, и можно получить библиотеку, дефосфорилируя 5'-конец одноцепочечной нуклеиновой кислоты, лигируя линкер, содержащий Р7'-адаптер, расщепляемый сайт ("U"), адаптер Р5, содержащий 3'-блокирующую группу ("R"), с 3'-концом одноцепочечной нуклеиновой кислоты; повторно фосфорилируя 5'-конец лигированной одноцепочечной нуклеиновой кислоты; снимая защиту с 3'-конца лигированной одноцепочечной нуклеиновой кислоты и замыкая нуклеиновые кислоты со снятой защитой в кольцо с образованием члена библиотеки, содержащей кольцевые одноцепочечные нуклеиновые кислоты.
[0057] Фиг. 4В представляет собой схему, на которой кольцевую нуклеиновую кислоту согласно фиг. 4В можно перевести в линейную форму, расщепляя расщепляемый сайт ("U"), или амплифицировать посредством ПНР или амплификации по принципу катящегося кольца (RCA).
[0058] Фиг. 5 представляет собой схему способа получения библиотеки одноцепочечных нуклеиновых кислот, включающего вариант реализации, показанный на фиг. 1. На фиг. 5 зонд захвата, содержащий последовательность Р7, гибридизуют с адаптером Р7', лигированным с нуклеиновой кислотой-мишенью, и с избытком нелигированных адаптеров Р7'. Гибридизация зонда захвата с нелигированными адаптерами Р7' ингибирует лигирование нелигированных адаптеров Р7' с другими нуклеиновыми кислотами.
[0059] Фиг. 6 представляет собой схему способа получения библиотеки одноцепочечных нуклеиновых кислот в соответствии с одним вариантом реализации. Двуцепочечную нуклеиновую кислоту денатурируют с образованием одноцепочечной нуклеиновой кислоты, и можно получить библиотеку, дефосфорилируя 5'-конец одноцепочечной нуклеиновой кислоты, лигируя Р7'-адаптер, содержащий 3'-блокирующую группу, с 3'-концом одноцепочечной нуклеиновой кислоты, удаляя избыток нелигированных одноцепочечных адаптеров Р7' посредством гидролиза 5'-фосфат-зависимой экзонуклеазы и 5'-деаденилазы, повторно фосфорилируя 5'-конец лигированной одноцепочечной нуклеиновой кислоты и лигируя адаптер Р5, содержащий нефосфорилированный 5'-конец, с 5'-концом лигированной одноцепочечной нуклеиновой кислоты.
[0060] На фиг. 7 изображены результаты эксперимента по дефосфорилированию 5'-конца 60-членной одноцепочечной нуклеиновой кислоты (3'ОН-60-Phos 5') и подтверждение того, что продукты (3'ОН-60-OH 5') не могут образовывать конкатемеры. Верхняя панель представляет собой схему, на которой показано дефосфорилирование. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0061] На фиг. 8 изображены результаты эксперимента по лигированию 64-членного адаптера Р7' (3'C3-P7-phos 5'), содержащего 3'-блокирующую группу, с 3'-концом дефосфорилированной 60-членной одноцепочечной нуклеиновой кислоты (3'ОН-60-OH 5'). Верхняя панель представляет собой схему, на которой показано лигирование. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0062] На фиг. 9 изображены результаты эксперимента по повторному фосфорилированию 5'-конца 124-членной дефосфорилированной одноцепочечной нуклеиновой кислоты, содержащей лигированный 64-членный адаптер с 3' блокирующей группой (3'С3-Р7-60-ОН). Верхняя панель представляет собой схему, на которой показана киназная реакция. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0063] На фиг. 10 изображены результаты эксперимента по лигированию 60-членного адаптера Р5, содержащего нефосфорилированный 5'-конец (5'P5-OH), с 5'-концом 124-членной одноцепочечной нуклеиновой кислоты (3'C3-60-B2-Phos). 124-членная одноцепочечная нуклеиновая кислота содержит лигированный 64-членный адаптер с 3'-блокирующей группой (3'C3-P7-phos 5'). Верхняя панель представляет собой схему, на которой показано лигирование. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0064] На фиг. 11 изображены результаты эксперимента по лигированию 64-членного адаптера Р7', содержащего 3'-блокирующую группу (3'C3-P7-phos 5'), с 3'-концом одноцепочечной дефосфорилированной бесклеточной ДНК (бкДНК). В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0065] На фиг. 12 изображены результаты эксперимента, в котором дефосфорилированную бкДНК (бкДНК [Apex]) обрабатывали киназой (PNK) и лигировали 64-членный адаптер Р7', содержащий 3'-блокирующую группу (3'C3-P7-phos 5'), с 3'-концом повторно фосфорилированной бкДНК. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0066] На фиг. 13 изображены результаты эксперимента, в котором 64-членный адаптер Р7', содержащий 3'-блокирующую группу (3'C3-P7-phos 5'), лигировали с 3'-концом дефосфорилированной бкДНК, полученный первый лигированный продукт (3'С3-Р7-бкДНК-phos 5') обрабатывали киназой (PNK) и лигировали 60-членный адаптер Р5 с нефосфорилированным 5'-концом (5P5'-OH) с 3'-концом первого продукта лигирования с образованием второго продукта лигирования (Р7'-бкДНК-Р5). В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0067] На фиг. 14 изображены гели, демонстрирующие продукты лигирования из примера 2, и продукты амплификации продуктов лигирования «Экстракт В1» и «Экстракт В2». На дорожке 'Ref экстракт В1 обозначен двумя черными прямоугольниками, а экстракт В2 - одним белым прямоугольником.
[0068] На фиг. 15 изображена электрофореграмма амплифицированных продуктов лигирования «Экстракт В1» и «Экстракт В2». Пики представляют собой праймер-димер длиной 126 п.о. (Р7'-Р5), разновидности длиной 292 п.о. (Р7'-бкДНК-Р5) и 480 п.о. (Р7'-бкДНК-Р5).
[0069] На фиг. 16 изображены гели, демонстрирующие результаты использования грануд SPRI в различных соотношениях для экстракции продуктов амплификации.
[0070] На фиг.17 изображены результаты эксперимента по лигированию 63-членного адаптера Р7', содержащего 3'-блокирующую группу (3'C3-P7-Phos 5'), с 3'-концом одноцепочечной дефосфорилированной бкДНК. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0071] На фиг. 18 изображены результаты эксперимента по лигированию 60-членного адаптера Р5, содержащего нефосфорилированный 5'-конец (5'P5-OH), с 3'-концом первого продукта лигирования (3'С3-Р7'-бкДНК-phos5') в присутствии 63-членного зонда захвата Р7 (3'С3-Р7-С35'), содержащего 5'- и 3'-блокирующие группы. Верхняя панель представляет собой схему, на которой показано лигирование. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0072] На фиг. 19 изображен анализ электрофореграммы амплифицированных продуктов лигирования «Экстракт В1» и «Экстракт В2». Пики представляют собой праймер-димер длиной 126 п.о. (Р7'-Р5), разновидности длиной 292 п.о. (Р7'-бкДНК-Р5) и 480 п.о. (Р7'-бкДНК-Р5).
[0073] На фиг. 20 изображены гели продуктов лигирования до амплификации, после амплификации и после экстракции с гранулами SPRI.
[0074] На фиг. 21 изображены результаты эксперимента по лигированию 39-членного олигонуклеотида 1 с дефосфорилированным 5'-концом (5'OH-олиго 1-OH3'), с 37-членным олигонуклеотидом 2 с фосфорилированной 3'-блокирующей группой (5'phos-Oligo 2-phos3') в присутствии различных концентраций полиэтиленгликоля (PEG). В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0075] На фиг. 22 изображены результаты эксперимента по лигированию олигонуклеотида 1 длиной 39 п.о. с дефосфорилированным 5'-концом (5'OH-олиго 1-OH3'), с олигонуклеотидом 2 длиной 37 п.о. с фосфорилированной 3'-блокирующей группой (5'phos-Oligo 2-phos3') при различных соотношениях «олигонуклеотид 1 : олигонуклеотид 2». В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0076] На фиг. 23 изображены результаты эксперимента по лигированию олигонуклеотида 1 длиной 39 п.о. с дефосфорилированным 5'-концом (5'OH-олиго 1-OH3'), с олигонуклеотидом 2 длиной 37 п.о. с фосфорилированной 3'-блокирующей группой (5'phos-Oligo 2-phos3') при различном количестве лигазы (CIRCLIGASE). В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0077] На фиг. 24 изображены результаты эксперимента по лигированию 37-членного олигонуклеотида 2 с фосфорилированной 3'-блокирующей группой (5'phos-Oligo 2-phos3') с 39-членным олигонуклеотидом 1 с дефосфорилированным 5'-концом (5'OH-олиго 1-OH3'), конъюгированным с гранулами. Продукты лигирования анализировали путем отщепления от гранул с использованием урацил-ДНК-гликозилазы (UDG), которая расщепляла сайт урацила в олигонуклеотиде 1. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0078] На фиг. 25 изображены результаты эксперимента по лигированию 37-членного олигонуклеотида 2 с фосфорилированной 3'-блокирующей группой (5'phos-Oligo 2-phos3') с 39-членным олигонуклеотидом 1 с дефосфорилированным 5'-концом (5'OH-олиго 1-OH3'), конъюгированным с гранулами, в конечном реакционном объеме, содержащем 45% ПЭГ. В таблице приведены условия проведения реакций и анализа в геле, показанном на нижней панели.
[0079] На фиг. 26 изображена схема способа получения библиотеки одноцепочечных нуклеиновых кислот с использованием гранул в соответствии с одним вариантом реализации. Библиотеку можно получить путем дефосфорилирования 5'-конца одноцепочечной нуклеиновой кислоты, лигирования Р5'-адаптера, содержащего 3'-блокирующую группу, с 3'-концом одноцепочечной нуклеиновой кислоты, гибридизации первого продукта лигирования с зондом захвата Р5, присоединенным к проточной ячейке, удлинения зонда захвата, удаления гибридизированного первого продукта лигирования с удлиненного зонда захвата, гибридизации комплементарных праймеров, лигирования адаптера Р7', содержащего 3'-блокирующую группу, с 3'-концом удлиненного зонда захвата, и денатурирования комплементарных праймеров.
ПОДРОБНОЕ ОПИСАНИЕ
[0080] Варианты реализации систем, способов и композиций, предложенные в настоящем документе, относятся к получению библиотек нуклеиновых кислот. Некоторые варианты реализации включают лигирование одноцепочечных нуклеиновых кислот с адаптерами для получения библиотек нуклеиновых кислот из небольших количеств исходных нуклеиновых кислот. Некоторые варианты реализации включают лигирование двухцепочечных нуклеиновых кислот с 3'-выступающими концами с адаптерами для получения библиотек нуклеиновых кислот из небольших количеств исходных нуклеиновых кислот. Некоторые варианты реализации включают лигирование одноцепочечных нуклеиновых кислот с линкерами, содержащими адаптеры, для получения библиотек кольцевых нуклеиновых кислот из небольших количеств исходных нуклеиновых кислот.
[0081] В одном варианте реализации одноцепочечные нуклеиновые кислоты получают путем уменьшения вероятности самоконкатемеризации реагирующих соединений. Например, для лигирования 3'-адаптера с 3'-концом одноцепочечной нуклеиновой кислоты 3'-адаптер может содержать блокирующую группу, которая ингибирует самоконкатемеризацию. Аналогичным образом, 5'-конец одноцепочечной нуклеиновой кислоты можно дефосфорилировать для ингибирования самоконкатемеризации. Продукты данной реакции лигирования могут включать лигированную одноцепочечную нуклеиновую кислоту, содержащую 3'-блокирующую группу. В некоторых вариантах реализации обнаружено, что лигирование 3'-адаптера с 3'-концом одноцепочечной нуклеиновой кислоты является высокоэффективным и выполняется в присутствии агента, вызывающего эффект исключения объема. 5'-конец лигированной одноцепочечной нуклеиновой кислоты, содержащей 3'-блокирующую группу, можно дефосфорилировать, так что с ее 5'-концом можно лигировать 5'-адаптер. Для лигирования 5'-адаптера с 5'-концом повторно фосфорилированной лигированной одноцепочечной нуклеиновой кислоты, содержащей 3'-блокирующую группу, 5'-адаптер может содержать дефосфорилированный 5'-конец для ингибирования самоконкатемеризации. Продукт данной реакции лигирования может включать одноцепочечную нуклеиновую кислоту, содержащую 5'-адаптер и 3'-адаптер. Адаптеры могут содержать сайты праймеров для секвенирования, сайты праймеров для амплификации и индексы. В настоящем документе термин «индекс» может включать последовательность нуклеотидов, которую можно использовать в качестве молекулярного идентификатора и/или штрих-кода для маркировки нуклеиновой кислоты и/или для идентификации источника нуклеиновой кислоты. В некоторых вариантах реализации индекс можно использовать для идентификации одиночной нуклеиновой кислоты или субпопуляции нуклеиновых кислот. В некоторых вариантах реализации библиотеки одноцепочечных нуклеиновых кислот можно получить в одном реакционном сосуде в одном объеме. Кроме того, варианты реализации, предложенные в настоящем документе, могут снизить вероятность образования праймер-димерных продуктов в последующих реакциях амплификации.
[0082] Пример способа согласно одному варианту реализации приведен на фиг. 1. Как проиллюстрировано, 5'-концы одноцепочечных нуклеиновых кислот-мишеней дефосфорилируют во избежание образования конкатемеров на последующих стадиях лигирования. Первые адаптеры лигируют с 3'-концами дефосфорилированных мишеней с использованием одноцепочечной лигазы. Во избежание образования конкатемеров первых адаптеров можно заблокировать 3'-концы первых адаптеров. 5'-концы лигированных мишеней повторно фосфорилируют с помощью киназы, и лигируют второй адаптер с 5'-концами дефосфорилированных мишеней с использованием одноцепочечной лигазы, тем самым формируя библиотеку нуклеиновых кислот. Во избежание образования конкатемеров вторых адаптеров можно не фосфорилировать 5'-концы вторых адаптеров.
[0083] Еще один пример способа согласно одному варианту реализации приведен на фиг. 2, где адаптеры лигируют с двуцепочечными нуклеиновыми кислотами с 3'-выступающими концами. В некоторых вариантах реализации первый адаптер можно лигировать с 3'-концами двуцепочечных нуклеиновых кислот с 3'-выступающими концами. Во избежание образования конкатемеров первых адаптеров можно заблокировать 3'-концы первых адаптеров. Двуцепочечные нуклеиновые кислоты, лигированные с первыми адаптерами, можно денатурировать с получением множества одноцепочечных нуклеиновых кислот. Второй адаптер можно лигировать с 5'-концами одноцепочечных нуклеиновых кислот.
[0084] Еще один пример способа согласно одному варианту реализации приведен на фиг .3, на котором библиотеку нуклеиновых кислот можно получить с использованием адаптеров, присоединенных к гранулам. В некоторых таких вариантах реализации первый адаптер лигируют с 3'-концами дефосфорилированных мишеней с использованием одноцепочечной лигазы. Цитированные первые адаптеры можно гибридизовать с зондами захвата, присоединенными к гранулам. Зонды захвата можно удлинить, и с удлиненным зондом захвата можно лигировать второй адаптер. Нелигированные одноцепочечные адаптеры можно удалить, отмыв от гранул. Некоторые варианты реализации получения библиотеки нуклеиновых кислот, включающие использование адаптеров, присоединенных к гранулам, можно применять для добавления индексов к библиотекам нуклеиновых кислот. Например, первый адаптер, содержащий первый индекс, можно присоединить к грануле, а второй адаптер, содержащий второй индекс и зонд захвата, можно лигировать с первым адаптером. Нуклеиновую кислоту-мишень, гибридизованную с зондом захвата, можно удлинить путем включения первого и второго индекса.
[0085] Еще один пример способа согласно одному варианту реализации приведен на фиг. 4А, на котором библиотеку одноцепочечных кольцевых нуклеиновых кислот можно получить с использованием линкера, содержащего первый и второй адаптеры. В некоторых таких вариантах реализации линкер может содержать расщепляемый сайт между первым адаптером и вторым адаптером, так что расщепление расщепляемого сайта кольцевой нуклеиновой кислоты может привести к получению линейной нуклеиновой кислоты, содержащей первый адаптер на одном конце, а второй адаптер на другом конце линейной нуклеиновой кислоты.
[0086] В некоторых вариантах реализации библиотеку нуклеиновых кислот можно амплифицировать с использованием сайтов праймеров в последовательностях адаптеров. В некоторых вариантах реализации эффективность последующих этапов амплификации может быть снижена за счет образования праймеров-димеров. Для повышения эффективности последующих этапов амплификации можно удалить нелигированные одноцепочечные адаптеры из продуктов лигирования. Пример, приведенный на фиг. 5 и описанный далее в настоящем документе, включает удаление нелигированных одноцепочечных первых адаптеров путем гибридизации с зондами захвата. Другой пример, показанный на фиг. 6 и описанный далее в настоящем документе, включает удаление нелигированных одноцепочечных первых адаптеров путем гидролиза с использованием 5'-фосфат-зависимой экзонуклеазы и 5'-деаденилазы.
Получение одноцепочечных библиотек
[0087] Некоторые варианты реализации систем, способов и композиций, предложенных в настоящем документе, включают способы, в которых адаптеры лигированы с нуклеиновыми кислотами-мишенями. Некоторые варианты реализации, предложенные в настоящем документе для получения библиотек нуклеиновых кислот, предложенных в настоящем документе, можно с успехом выполнять в одном реакционном объеме.
[0088] Адаптеры содержат нуклеиновые кислоты, например, одноцепочечные нуклеиновые кислоты. Адаптеры могут содержать короткие нуклеиновые кислоты, длина которых менее, более или равна приблизительно 5 нуклеотидам, 10 нуклеотидам, 20 нуклеотидам, 30 нуклеотидам, 40 нуклеотидам, 50 нуклеотидам, 60 нуклеотидам, 70 нуклеотидам, 80 нуклеотидам, 90 нуклеотидам, 100 нуклеотиды или находится в диапазоне между любыми двумя из вышеуказанных размеров. Адаптеры могут содержать сайты связывания праймеров для секвенирования, сайты связывания праймеров для амплификации и/или индексы. Например, адаптер может содержать последовательность Р5, последовательность Р7 или их комплемент. Индексы можно применять для идентификации источника молекулы нуклеиновой кислоты. Примеры использования и получения индексов, которые можно применять в вариантах реализации, предложенных в настоящем документе, можно найти в международных публикациях № WO 2012/061832 и WO 2014/142850; и патентах США №9074251, 8829171, каждый из которых полностью включен в настоящее описание посредством ссылки. В некоторых вариантах реализации адаптер можно модифицировать во избежание образования конкатемеров, например, путем добавления блокирующих групп, которые предотвращают удлинение адаптера на одном или обоих концах. Блокирующая группа представляет собой любую химическую группу, которая блокирует реакцию соединения, к которому она присоединена, на последующих этапах рабочего процесса, например, группу, блокирующую активацию адаптера лигазой, реакцию с фосфатом и реакцию с самим собой. Примеры 3'-блокирующих групп включают 3'-спейсер С3, дидезоксинуклеотид и присоединение к подложке. Дополнительные примеры 3'-блокирующих групп включают алкил, необязательно замещенный одной или несколькими гидроксильными группами, тиол, азид или алкин. В некоторых аспектах блокирующие группы могут обладать функциональными возможностями, которые можно применять при последующих операциях, например, обогащении, очистке, дальнейшей функционализации и т.п. Примеры 5'-блокирующих групп включают дефосфорилированный 5'-нуклеотид и присоединение к подложке (например, где блокирующая группа представляет собой подложку, необязательно присоединенную посредством линкера).
[0089] Нуклеиновые кислоты-мишени включают одноцепочечные нуклеиновые кислоты и двуцепочечные нуклеиновые кислоты. Способы денатурации двуцепочечных нуклеиновых кислот с образованием одноцепочечных нуклеиновых кислот хорошо известны в данной области техники и включают термические или химические способы. Примеры нуклеиновых кислот-мишеней включают ДНК, например, геномную ДНК или кДНК; РНК, например, мРНК, короткую РНК или рРНК, или гибриды ДНК и РНК. Нуклеиновая кислота может содержать фосфодиэфирные связи и может включать другие типы каркаса, включая, например, фосфорамидные, тиофосфатные, тиодифосфатные, О-метилфосфороамидитные и пептидные каркасы и связи нуклеиновых кислот. Нуклеиновая кислота может содержать любую комбинацию дезоксирибо- и рибонуклеотидов и любую комбинацию оснований, включая урацил, аденин, тимин, цитозин, гуанин, инозин, ксантанин, гипоксантанин, изоцитозин, изогуанин и аналоги оснований, например, нитропиррол (включая 3-нитропиррол) и нитроиндол (включая 5-нитроиндол) и т.д. В некоторых вариантах реализации нуклеиновая кислота может включать по меньшей мере одно случайное основание. Случайное основание может являться парой оснований, содержащей основания более чем одного из различных типов и может быть полезным, например, при его включении в олигонуклеотидные праймеры или вставки, применяемые для случайной гибридизации в образцах нуклеиновых кислот сложного состава, например, образцах геномной ДНК. Пример случайного основания включает инозин, который может обрзовывать пару с аденином, тимином или цитозином. Другие примеры включают гипоксантин, 5-нитроиндол, ацильный 5-нитроиндол, 4-нитропиразол, 4-нитроимидазол и 3-нитропиррол. Можно использовать случайные основания, которые могут образовывать пару с по меньшей мере двумя, тремя, четырьмя или более типами оснований
[0090] Нуклеиновые кислоты-мишени могут включать образец, в котором средний размер двуцепочечной нуклеиновой кислоты меньше, больше или равен примерно 2 т.п.о., 1 т.п.о., 500 п.о., 400 п.о., 200 п.о., 100 п.о., 50 п.о. или находится в диапазоне между любыми двумя из вышеупомянутых размеров. В некоторых вариантах реализации средний размер одноцепочечной нуклеиновой кислоты в образце меньше, больше или равен приблизительно 2000 нуклеотидов, 1000 нуклеотидов, 500 нуклеотидов, 400 нуклеотидов, 200 нуклеотидов, 100 нуклеотидов, 50 нуклеотидов или находится в диапазоне между любыми двумя из вышеупомянутых значений размера. Образец может содержать нуклеиновые кислоты-мишени в количестве, меньшем, большем или равном приблизительно 50 мкг, 10 мкг, 5 мкг, 1 мкг, 500 нг, 400 нг, 200 нг, 100 нг, 50 нг, 10 нг, или в диапазоне между любыми двумя из вышеупомянутых количеств. Нуклеиновые кислоты-мишени можно получить из низкокачественных источников нуклеиновых кислот, например, образца, подвергавшегося разложению, например, фиксированного образца и древнего образца. Примеры фиксированных образцов включают образцы, фиксированные с помощью таких соединений, как формалин, глутаральдегид, спирт, осмиевая кислота и параформальдегид. Фиксированный образец может включать образец, фиксированный формалином и включенный в парафин (FFPE). Древние образцы могут включать образцы, возраст которых превышает 5 лет, 10 лет, 20 лет, 50 лет, 100 лет, 500 лет, 1000 лет или находится в диапазоне между любыми двумя из вышеуказанных значений возраста.
[0091] Способы дефосфорилирования нуклеиновых кислот, например, 5'-нуклеотида нуклеиновой кислоты, включают приведение нуклеиновой кислоты в контакт с фосфатазой. Примеры фосфатаз включают кишечную фосфатазу теленка, щелочную фосфатазу креветки, антарктическую фосфатазу и щелочную фосфатазу APEX (Epicenter, Мэдисон, штат Висконсин, США).
[0092] Способы лигирования одноцепочечных нуклеиновых кислот включают приведение одноцепочечных нуклеиновых кислот в контакт с лигазой. Примеры одноцепочечных лигаз включают РНК-лигазу 1 Т4, РНК-лигазу 2 Т4, RtcB-лигазу, РНК-лигазу Methanobacterium и РНК-лигазу TS2126 (CIRCLIGASE; Epicenter, Мэдисон, штат Висконсин, США). Способы лигирования двух нуклеиновых кислот друг с другом можно осуществлять в реакционном объеме в присутствии агента, вызывающего эффект исключения объема. В настоящем документе термин «агент, вызывающий эффект исключения объема» может включать агенты, уменьшающие эффективный объем, в котором может происходить реакция, например, реакция лигирования. Примеры агентов, вызывающих эффект исключения объема, включают полимеры, например, инертные полимеры, например, полиэтиленгликоль (ПЭГ), фиколл, декстран, гидроксиэтилкрахмал и поливинилпирролидон. Примеры ПЭГ, которые можно примерять в вариантах реализации, предложенных в настоящем документе, включают ПЭГ 600, ПЭГ-800, ПЭГ-1000, ПЭГ-6000 и ПЭГ-8000. Реакционный объем может содержать концентрацию агента, вызывающего эффект исключения объема, например, в массо-объемных процентах, которая меньше, больше или равна приблизительно 5%, 10%, 20%, 30%, 40%, 50% 60%, 70%, 80%, 90%, 95% или находится в диапазоне между любыми двумя из вышеупомянутых процентных значений.
[0093] Способы фосфорилирования нуклеиновых кислот, например, 5'-нуклеотида нуклеиновой кислоты, включают приведение нуклеиновой кислоты в контакт с киназой. Примеры киназ включают полинуклеотидкиназу Т4.
[0094] Некоторые варианты реализации, предложенные в настоящем документе, можно осуществить в одном реакционном объеме. В некоторых вариантах реализации способ получения нуклеиновой кислоты можно осуществить в течение сокращенного периода времени, например, менее приблизительно 12 часов, менее приблизительно 6 часов или менее приблизительно 3 часов.
[0095] На фиг. 1 изображен вариант реализации, в котором двуцепочечный фрагмент ДНК-мишени дефосфорилирован и денатурирован с образованием одноцепочечной ДНК-мишени. Первый адаптер размером 64 п.о., содержащий фосфорилированный 5'-конец, последовательность Р7' и заблокированный 3'-конец, лигируют с 3'-концом одноцепочечной ДНК-мишени с образованием лигированной ДНК с использованием лигазы, например, CIRCLIGASE™ (Lucigen Corporation, Миддлтон, штат Висконсин, США). 5'-конец лигированной ДНК фосфорилируют с использованием киназы, например, полинуклеотидкиназы Т4. Второй адаптер длиной 60 п.о., содержащий последовательность Р5, лигируют с 5'-фосфорилированной лигированной ДНК с образованием члена (5'-Р5-оцДНК-инсерт-Р7-3') библиотеки ДНК. Примеры последовательностей праймеров могут включать: последовательность праймера P5 (SEQ ID NO: 01: aatgatacggcgaccaccga); последовательность праймера  (SEQ ID NO: 02: tcggtggtcgccgtatcatt); последовательность праймера Р7 (SEQ ID NO: 03: caagcagaagacggcatacga); и последовательность праймера
(SEQ ID NO: 02: tcggtggtcgccgtatcatt); последовательность праймера Р7 (SEQ ID NO: 03: caagcagaagacggcatacga); и последовательность праймера  (SEQ ID NO: 04: tcgtatgccgtcttctgcttg).
(SEQ ID NO: 04: tcgtatgccgtcttctgcttg).
[0096] В некоторых вариантах реализации способ получения библиотеки нуклеиновых кислот может включать дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот-мишеней во избежение образования конкатемеров на последующих стадиях лигирования; лигирование первых адаптеров с 3'-концами дефосфорилированных мишеней с использованием одноцепочечной лигазы, причем 3'-концы первых адаптеров блокированы; повторное фосфорилирование 5'-концов лигированных мишеней; лигирование второго адаптера с 5'-концами дефосфорилированных мишеней с использованием одноцепочечной лигазы, причем 5'-концы вторых адаптеров нефосфорилированы, в результате чего получают библиотеку нуклеиновых кислот.В некоторых вариантах реализации второй адаптер прикреплен к подложке. В некоторых вариантах реализации подложка включает гранулу.
[0097] Некоторые варианты реализции включают удаление нелигированных одноцепочечных первых адаптеров перед лигированием второго адаптера с 5'-концами дефосфорилированных мишеней. В некоторых вариантах реализации нелигированные одноцепочечные первые адаптеры можно гибридизовать с зондом захвата. В некоторых вариантах реализации зонд захвата содержит последовательность, комплементарную по меньшей мере части первого адаптера. В некоторых таких вариантах реализации гибридизация с зондом захвата ингибирует лигирование нелигированных первых адаптеров с одноцепочечными вторыми адаптерами. В некоторых вариантах реализации 3'-конец зонда захвата содержит блокирующую группу. В некоторых вариантах реализации 5'-конец зонда захвата содержит блокирующую группу. В некоторых вариантах реализации нелигированные одноцепочечные первые адаптеры можно удалить путем гидролиза. В некоторых вариантах реализации гидролиз нелигированных одноцепочечных первых адаптеров включает приведение нелигированных одноцепочечных первых адаптеров в контакт с 5'-фосфат-зависимой экзонуклеазой, а также, необязательно, 5'-деаденилазой.
[0098] На фиг. 2 изображен еще один вариант реализации, в котором двуцепочечную нуклеиновую кислоту-мишень частично гидролизуют 5'-экзонуклеазой с образованием двуцепочечной нуклеиновой кислоты с одноцепочечными 3'-выступающими концами. Первый адаптер размером 64 п.о., содержащий фосфорилированный 5'-конец, последовательность Р7' и заблокированный 3'-конец, лигируют с 3'-концами двуцепочечной ДНК-мишени с образованием лигированной ДНК с использованием лигазы, например, CIRCLIGASE™. Лигированную ДНК денатурируют с образованием одноцепочечных нуклеиновых кислот. Второй адаптер длиной 60 п.о., содержащий последовательность Р5, лигируют с 5'-фосфорилированной лигированной ДНК с образованием члена (5'-Р5-оцДНК-инсерт-Р7-3') библиотеки ДНК.
[0099] В некоторых вариантах реализации способ получения библиотеки нуклеиновых кислот может включать приведение двуцепочечных нуклеиновых кислот-мишеней в контакт с 5'-экзонуклеазой с получением множества модифицированных двуцепочечных нуклеиновых кислот с одноцепочечными 3'-выступающими концами; лигирование первого адаптера с 3'-концами модифицированных двуцепочечных нуклеиновых кислот в присутствии лигазы и агента, вызывающего эффект исключения объема, причем 3'-конец первого адаптера содержит блокирующую группу; денатурацию модифицированных двуцепочечных нуклеиновых кислот, лигированных с первыми адаптерами, с получением множества одноцепочечных нуклеиновых кислот; и лигирование второго адаптера с 5'-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, за счет чего получают библиотеку нуклеиновых кислот.
[0100] На фиг. 3 изображен вариант реализации, в котором одноцепочечную ДНК-мишень дефосфорилируют с использованием щелочной фосфатазы. Первый адаптер, содержащий фосфорилированный 5'-конец, последовательность Р5' и заблокированный 3'-конец, лигируют с 3'-концом одноцепочечной ДНК-мишени с образованием лигированной ДНК с использованием лигазы, например, CIRCLIGASE™. Лигированную ДНК гибридизуют через первый адаптер с зондом захвата, содержащим последовательность Р5 и присоединенным к грануле. Зонд захвата удлиняют с образованием удлиненной ДНК, а игированную ДНК удаляют с удлиненной ДНК. Второй адаптер, содержащий фосфорилированный 5'-конец, последовательность Р7' и заблокированный 3'-конец, лигируют с 3'-концом удлиненной ДНК с использованием лигазы, например, CIRCLIGASE™, и с образованием члена (Р5-инсерт'-Р7') библиотеки ДНК, присоединенного к грануле.
[0101] В некоторых вариантах реализации способ получения библиотеки нуклеиновых кислот может включать дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот-мишеней; лигирование первого адаптера с 3'-концами одноцепочечных нуклеиновых кислот в присутствии лигазы и агента,, вызывающего эффект исключения объема, причем 3'-конец первого адаптера содержит блокирующую группу; гибридизацию лигированного первого адаптера с зондом захвата; удлинение зонда захвата и лигирование второго адаптера с 3'-концом удлиненного зонда захвата в присутствии лигазы, причем 3'-конец второго адаптера содержит блокирующую группу, за счет чего получают библиотеку нуклеиновых кислот. Некоторые варианты реализации также включают удаление гибридизованного лигированного первого адаптера с удлиненного зонда захвата. В некоторых вариантах реализации зонд захвата присоединен к подложке. В некоторых вариантах реализации подложка с зондом захвата представляет собой проточную ячейку, гранулу, стекло, стекло с контролируемым размером пор, пластмассу, кремний, кварцевое стекло, диоксид кремния, нитрид кремния, производные кремния, арсенид галлия, фосфид индия, алюминий, керамику, полиимид, кварц, смолы или полимер или сополимер (например, полистирол). Подложки могут состоять из слоев из нескольких материалов или смеси материалов. Подложки могут быть жесткими или полужесткими. Подложки могут быть плоскими, круглыми или текстурированными. В некоторых вариантах реализации подложки могут представлять собой жесткий материал, например, производные стекла и/или диоксида кремния, с нанесенным одним или несколькими полимерными материалами. В некоторых вариантах реализации подложка включает гранулу. В некоторых вариантах реализации зонд захвата содержит индекс зонда захвата. В некоторых вариантах реализации индекс зонда захвата указывает на источник зонда захвата. В некоторых вариантах реализации зонд захвата содержит расщепляемый линкер. Некоторые варианты реализации также включают расщепление расщепляемого линкера. В некоторых вариантах реализации зонд захвата присоединен к подложке, например, проточной ячейке (см. фиг. 26). и его последующее удлинение происходит на подложке. Если подложка представляет собой проточную ячейку, полученную библиотеку можно непосредственно секвенировать. В некоторых вариантах реализации способ дополнительно включает гибридизацию комплементарных праймеров с удлиненным зондом захвата, лигирование адаптера Р7', содержащего 3'-блокирующую группу, с 3'-концом удлиненного зонда захвата и денатурацию комплементарных праймеров, как показано, например, на фиг. 26.
[0102] На фиг. 4А изображен вариант реализации, в котором двуцепочечный фрагмент ДНК-мишени дефосфорилирован и денатурирован с образованием одноцепочечной ДНК-мишени. Предложен адаптер, содержащий от 5'-конца к 3-концу': фосфорилированный 5'-конец, последовательность Р7', расщепляемый сайт ("U"), последовательность Р5 и заблокированный 3'-конец ("R"). Адаптер лигируют с помощью лигазы с 3'-концом одноцепочечной ДНК-мишени с образованием лигированной ДНК. 5'-конец лигированной ДНК фосфорилируют с помощью киназы. С заблокированного 3'-конца лигированной ДНК снимают защиту, и лигированную ДНК замыкают в кольцо путем лигирования с помощью лигазы с образованием члена библиотеки, содержащей кольцевую одноцепочечную ДНК. В некоторых вариантах реализации кольцевую нуклеиновую кислоту можно использовать для получения одноцепочечных нуклеиновых кислот любым из множества способов. Некоторые типичные варианты реализации изображены на фиг. 4В, на котором кольцевую нуклеиновую кислоту можно перевести в линейную форму путем расщепления расщепляемого сайта, который может содержать остаток урацила. Кольцевую нуклеиновую кислоту также можно применять для получения одноцепочечных нуклеиновых кислот путем амплификации, например, ПЦР, с использованием сайтов праймеров внутри адаптера. Кольцевую нуклеиновую кислоту также можно амплифицировать путем амплификации по принципу катящегося кольца (RCA).
[0103] В некоторых вариантах реализации способ получения библиотеки нуклеиновых кислот может включать: дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот-мишеней; лигирование линкера с 3'-концами одноцепочечных нуклеиновых кислот, причем линкер содержит первый адаптер и второй адаптер, а 3'-конец линкера содержит блокирующую группу; фосфорилирование 5'-концов лигированных одноцепочечных нуклеиновых кислот; снятие защиты с 3'-концов фосфорилированных нуклеиновых кислот и замыкание нуклеиновых кислот со снятой защитой в кольцо путем лигирования, за счет чего получают библиотеку кольцевых нуклеиновых кислот.
[0104] В некоторых вариантах реализации линкер содержит расщепляемый сайт между первым адаптером и вторым адаптером. В некоторых таких вариантах реализации расщепление расщепляемого сайта кольцевой нуклеиновой кислоты может привести к получению линейной нуклеиновой кислоты, содержащей первый адаптер на одном конце, а второй адаптер на другом конце линейной нуклеиновой кислоты. В некоторых вариантах реализации расщепляемые сайты включают сайты рестрикции, сайты, содержащие по меньшей мере один остаток урацила, и сайты, содержащие атипичные основания, например, 8-оксогуанин и дитиольные группы. Некоторые варианты реализации включают перевод кольцевых нуклеиновых кислот в линейную форму путем расщепления по расщепляемому сайту. Примеры расщепления расщепляемого сайта включают приведение указанного сайта в контакт с урацил-специфическим эксцизионным реагентом, например, ферментом, выбранным из урацил-ДНК-гликозилазы (UDG) и ДНК-гликозилазы-лиазы-эндонуклеазы VTII или их смеси, или приведение указанного сайта в контакт с эндонуклеазой рестрикции.
[0105] Некоторые варианты реализации также включают амплификацию кольцевых нуклеиновых кислот путем включения гибридизующихся праймеров в первый и второй адаптеры. Примеры способов амплификации включают ПЦР, амплификацию по принципу катящегося кольца (RCA) и кластерную амплификацию. В некоторых таких вариантах реализации амплификацию по принципу RCA можно выполнять в растворе или можно выполнять захват кольцевых одноцепочечных нуклеиновых кислот на поверхности, например, на поверхности проточной ячейки, и амплифицировать их. В некоторых вариантах реализации амплификация включает приведение кольцевых нуклеиновых кислот в контакт с полимеразой, которая образует линейный продукт при контакте с остатком урацила в матрице. Примеры полимераз включают корректирующую полимеразу, подобную полимеразе Pyrococcus, например, ДНК-полимеразу PHUSION.
[0106] Некоторые варианты реализации включают способ получения библиотеки одноцепочечных нуклеиновых кислот, включающий: (а) получение множества одноцепочечных нуклеиновых кислот; (b) приведение одноцепочечных нуклеиновых кислот в контакт с щелочной фосфатазой APEX, за счет чего происходит дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот; (с) лигирование первого адаптера с 3'-концами одноцепочечных нуклеиновых кислот в реакционном объеме, содержащем РНК-лигазу TS2126 (Lucigen Corporation, Миддлтон, штат Висконсин, США) и по меньшей мере 45% (вес/об.) полиэтиленгликоля (ПЭГ), причем 3'-конец первого адаптера содержит 3'-спейсерную блокирующую группу С3; (d) приведение лигированных одноцепочечных нуклеиновых кислот в контакт с полинуклеотидкиназой Т4, за счет чего происходит фосфорилирование 5'-концов лигированных одноцепочечных нуклеиновых кислот; и (е) лигирование второго адаптера с 5'-концами фосфорилированных лигированных одноцепочечных нуклеиновых кислот в присутствии лигазы и ПЭГ, причем 5'-конец второго адаптера не фосфорилирован, за счет чего получают библиотеку нуклеиновых кислот.
[0107] Некоторые варианты реализации включают способ получения библиотеки одноцепочечных нуклеиновых кислот, включающий: (а) получение множества двуцепочечных нуклеиновых кислот; (b) приведение двуцепочечных нуклеиновых кислот в контакт с 5'-экзонуклеазой с получением множества модифицированных двуцепочечных нуклеиновых кислот с одноцепочечными 3'-выступающими концами; (с) лигирование первого адаптера с 3'-концами модифицированных двуцепочечных нуклеиновых кислот в реакционном объеме, содержащем РНК-лигазу TS2126 и по меньшей мере 45% (вес/об.) полиэтиленгликоля (ПЭГ), причем 3'-конец первого адаптера содержит 3'-спейсерную блокирующую группу С3; (d) денатурацию модифицированных двуцепочечных нуклеиновых кислот, лигированных с первыми адаптерами, с получением множества одноцепочечных нуклеиновых кислот; и (е) лигирование второго адаптера с 5'-концами одноцепочечных нуклеиновых кислот в присутствии лигазы и ПЭГ, причем 5'-конец второго адаптера не фосфорилирован, за счет чего получают библиотеку нуклеиновых кислот.
[0108] Некоторые варианты реализации включают способ получения библиотеки одноцепочечных нуклеиновых кислот, включающий: (а) получение множества одноцепочечных нуклеиновых кислот; (b) приведение одноцепочечных нуклеиновых кислот в контакт с щелочной фосфатазой APEX, за счет чего происходит дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот; (с) лигирование первого адаптера с 3'-концами одноцепочечных нуклеиновых кислот в реакционном объеме, содержащем РНК-лигазу TS2126 и по меньшей мере 45% (вес/об.) полиэтиленгликоля (ПЭГ), причем 3'-конец первого адаптера содержит 3'-спейсерную блокирующую группу С3; (d) приведение лигированных одноцепочечных нуклеиновых кислот в контакт с полинуклеотидкиназой Т4, за счет чего происходит фосфорилирование 5'-концов лигированных одноцепочечных нуклеиновых кислот; (е) удаление нелигированных одноцепочечных первых адаптеров из лигированных одноцепочечных нуклеиновых кислот путем гибридизации одноцепочечных первых адаптеров с зондом захвата, причем зонд захвата содержит: 3' конец, содержащий 3'-спейсерную блокирующую группу С3, нефосфорилированный 5'-конец и последовательность, комплементарную по меньшей мере части первого адаптера; и (f) лигирование второго адаптера с 5'-концами фосфорилированных лигированных одноцепочечных нуклеиновых кислот в присутствии лигазы и ПЭГ, причем 5'-конец второго адаптера не фосфорилирован, за счет чего получают библиотеку нуклеиновых кислот.
[0109] Некоторые варианты реализации включают способ получения библиотеки нуклеиновых кислот, присоединенных к гранулам, включающий: (а) получение множества одноцепочечных нуклеиновых кислот; (b) приведение одноцепочечных нуклеиновых кислот в контакт с щелочной фосфатазой APEX, за счет чего происходит дефосфорилирование 5'-концов одноцепочечных нуклеиновых кислот; (с) лигирование первого адаптера с 3'-концами одноцепочечных нуклеиновых кислот в реакционном объеме, содержащем РНК-лигазу TS2126 и по меньшей мере 45% (вес/об.) полиэтиленгликоля (ПЭГ), причем 3'-конец первого адаптера содержит 3'-спейсерную блокирующую группу С3; (d) гибридизацию лигированного первого адаптера с зондом захвата, причем зонд захвата присоединен к грануле; (е) удлинение зонда захвата; и (f) лигирование второго адаптера с 3'-концом удлиненного зонда захвата в присутствии лигазы и ПЭГ, причем 5'-конец второго адаптера не фосфорилирован, за счет чего получают библиотеку нуклеиновых кислот.
[0110] Некоторые варианты реализации включают способ получения библиотеки индексированных нуклеиновых кислот, включающий: (а) получение множества первых одноцепочечных нуклеиновых кислот, содержащих первый индекс, причем 5'-концы первых одноцепочечных нуклеиновых кислот присоединены к множеству гранул, и множества вторых одноцепочечных нуклеиновых кислот, содержащих второй индекс и зонд захвата, причем 3'-концы вторых одноцепочечных нуклеиновых кислот содержат 3'-спейсерную блокирующую группу С3; (b) лигирование 3'-концов первых одноцепочечных нуклеиновых кислот с 5'-концами вторых одноцепочечных нуклеиновых кислот в реакционном объеме, содержащем РНК-лигазу TS2126 и по меньшей мере 45% (вес/об.) полиэтиленгликоля (ПЭГ); (с) гибридизацию нуклеиновой кислоты-мишени с зондом захвата; и (d) удлинение нуклеиновой кислоты-мишени с получением библиотеки индексированных нуклеиновых кислот.
Удаление нелигированных одноцепочечных адаптеров
[0111] Некоторые варианты реализации систем, способов и композиций, предложенных в настоящем документе, включают удаление одноцепочечных нелигированных адаптеров от лигированных адаптеров, например, адаптеров, лигированных с одноцепочечными нуклеиновыми кислотами. В некоторых вариантах реализации удаление одноцепочечных нелигированных адаптеров от лигированных адаптеров может повысить эффективность амплификации полученной библиотеки нуклеиновых кислот, например, уменьшить вероятность образования или количество праймеров-димеров на последующих этапах амплификации.
[0112] Некоторые варианты реализации включают системы или способы удаления одноцепочечных нелигированных адаптеров и могут включать гибридизацию одноцепочечных нелигированных адаптеров с зондом захвата. На фиг. 5 изображен вариант реализации, в котором двуцепочечный фрагмент ДНК-мишени дефосфорилирован и денатурирован с образованием одноцепочечной ДНК-мишени. Первый адаптер размером 64 п.о., содержащий фосфорилированный 5'-конец, последовательность Р7' и заблокированный 3'-конец, лигируют с 3'-концом одноцепочечной ДНК-мишени с образованием лигированной ДНК. 5'-конец лигированной ДНК фосфорилируют с использованием киназы, например, полинуклеотидкиназы Т4. Нелигированные первые адаптеры удаляют от лигированной ДНК посредством гибридизации с зондом захвата, содержащим последовательность Р7. Второй адаптер длиной 60 п.о., содержащий последовательность Р5, лигируют с 5'-фосфорилированной лигированной ДНК с образованием члена (5'-Р5-оцДНК-инсерт-Р7-3') библиотеки ДНК. В некоторых таких вариантах реализации гибридизация с зондом захвата банка может ингибировать лигирование нелигированных первых адаптеров с одноцепочечными вторыми адаптерами. В некоторых вариантах реализации зонд захвата может содержать комплементарную последовательность по меньшей мере фрагмента одноцепочечного нелигированного адаптера. В некоторых вариантах реализации зонд захвата может включать 3' и/или 5'-блокирующую группу во избежание образования конкатемеров. В некоторых вариантах реализации зонд захвата можно присоединить к подложке, например, грануле.
[0113] Некоторые варианты реализации с удалением одноцепочечных нелигированных адаптеров могут включать гидролиз одноцепочечных нелигированных адаптеров. Некоторые такие варианты реализации могут включать приведение одноцепочечных нелигированных адаптеров в контакт с нуклеазой, например, 5'-фосфат-зависимой экзонуклеазой, например, TERMINATOR (Epicentre, Мэдисон, штат Висконсин, США). На фиг. 6 изображен вариант реализации, в котором двуцепочечный фрагмент ДНК-мишени дефосфорилирован и денатурирован с образованием одноцепочечной ДНК-мишени. Первый адаптер размером 64 п.о., содержащий фосфорилированный 5'-конец, последовательность Р7' и заблокированный 3'-конец, лигируют с 3'-концом одноцепочечной ДНК-мишени с образованием лигированной ДНК. Нелигированные первые адаптеры удаляют гидролизом с помощью 5'-фосфат-зависимой экзонуклеазы и 5'-деаденилазы. 5'-конец лигированной ДНК фосфорилируют с использованием киназы, например, полинуклеотидкиназы Т4. Второй адаптер длиной 60 п.о., содержащий последовательность Р5, лигируют с 5'-фосфорилированной лигированной ДНК с образованием члена (5'-Р5-оцДНК-инсерт-Р7-3') библиотеки ДНК.
[0114] Некоторые варианты реализации с удалением одноцепочечных нелигированных адаптеров могут включать применение гранул. Предпочтительное применение гранул может включать отмывание компонентов реакции от гранул и компонентов, присоединенных к гранулам. В некоторых вариантах реализации первый адаптер можно лигировать с 3'-концами дефосфорилированных одноцепочечных нуклеиновых кислот. Первый адаптер может содержать 3'-блокирующую группу и может содержать последовательности-мишени. Последовательности-мишени можно гибридизовать с зондами захвата, присоединенными к подложке. В некоторых вариантах реализации подложка может включать гранулу. Зонд захвата может содержать сайты праймеров для секвенирования, индексы и сайты праймеров для амплификации. Одноцепочечные нелигированные адаптеры можно отмывать от подложки и гибридизованных лигированных одноцепочечных нуклеиновых кислот. Зонд захвата можно удлинять. В некоторых вариантах реализации гибридизованные нелигированные одноцепочечные нуклеиновые кислоты можно удалить от удлиненного зонда захвата. Второй адаптер можно лигировать с 5'-концами удлиненных зондов захвата. В некоторых вариантах реализации вторые адаптеры содержат блокирующую группу на 3'-концах. Второй адаптер может содержать сайты праймеров для секвенирования, индексы и сайты праймеров для амплификации.
Получение индексированных библиотек
[0115] Некоторые варианты реализации систем, способов и композиций, предложенных в настоящем документе, включают способы получения индексированной библиотеки нуклеиновых кислот. Адаптеры и/или зонды захвата могут содержать индексы. Примеры использования и получения индексов, которые можно применять в вариантах реализации, предложенных в настоящем документе, можно найти в международных публикациях № WO 2012/061832 и WO 2014/142850; и патентах США №9074251, 8829171, каждый из которых полностью включен в настоящее описание посредством ссылки. В некоторых вариантах реализации индекс можно включить в нуклеиновую кислоту-мишень посредством лигирования. В некоторых вариантах реализации индекс можно включить в нуклеиновую кислоту посредством удлинения зонда захвата, адаптера или нуклеиновой кислоты-мишени. Таким образом, индексированные библиотеки легко получить способами, предложенными в настоящем документе.
[0116] В некоторых вариантах реализации можно получить популяцию зондов захвата. В некоторых вариантах реализации зонд захвата можно получить путем лигирования первого адаптера со вторым адаптером. Каждый адаптер может содержать индекс. Первый адаптер можно присоединить к подложке, например, грануле. Второй адаптер может содержать гибридизационный зонд. Гибридизационный зонд можно гибридизовать с одноцепочечными нуклеиновыми кислотами-мишенями. Одноцепочечные нуклеиновые кислоты-мишени можно гибридизовать с гибридизационным зондом и удлинять, тем самым включая последовательности, комплементарные индексу одного или более адаптеров.
[0117] Некоторые варианты реализации могут включать получение множества первых адаптеров, содержащих первый индекс, причем 5'-концы первых адаптеров содержат 5'-блокирующую группу, и множества вторых адаптеров, содержащих второй индекс и зонд захвата, причем 3'-концы вторых адаптеров содержат 3'-блокирующую группу. Некоторые варианты реализации также включают лигирование 3'-концов первых одноцепочечных нуклеиновых кислот с 5'-концами вторых одноцепочечных нуклеиновых кислот в присутствии лигазы. Некоторые варианты реализации также включают гибридизацию нуклеиновой кислоты-мишени с зондом захвата. Некоторые варианты реализации также включают удлинение нуклеиновых кислот-мишеней, за счет чего получают библиотеку индексированных нуклеиновых кислот. Некоторые варианты реализации также включают амплификацию удлиненных нуклеиновых кислот-мишеней. В некоторых вариантах реализации 5'-блокирующая группа содержит дефосфорилированный нуклеотид или присоединена к подложке. В некоторых вариантах реализации подложка включает множество гранул.
Наборы, реакционные сосуды и проточные ячейки
[0118] Варианты реализации систем и способов, предложенных в настоящем документе, включают наборы, реакционные сосуды и проточные ячейки, содержащие любой один или более из компонентов, пригодных для получения библиотек нуклеиновых кислот путем лигирования адаптеров с нуклеиновыми кислотами, например, одноцепочечными нуклеиновыми кислотами и/или двуцепочечными нуклеиновыми кислотами. Типичные компоненты включают первый адаптер, причем 3'-конец первого адаптера содержит блокирующую группу, лигазу, агент, вызывающий эффект исключения объема, дефосфорилирующий агент, второй адаптер, причем 3'-конец второго адаптера содержит блокирующую группу, праймеры для секвенирования, праймеры для амплификации, нуклеиновые кислоты, содержащие индексы, и 5'-экзонуклеазы. В некоторых вариантах реализации набор может включать реагент для любого способа, предложенного в настоящем документе. В некоторых вариантах реализации набор может включать реакционный сосуд. В некоторых вариантах реализации набор может включать проточную ячейку, содержащую реакционный сосуд.
[0119] Некоторые варианты реализации включают реакционный сосуд, содержащий реакционный объем, в котором можно осуществить способ, предложенный в настоящем документе. В некоторых таких вариантах реализации реакционный сосуд может включать компоненты любой стадии или этапа способа, предложенного в настоящем документе. В некоторых вариантах реализации реакционный сосуд может содержать множество нуклеиновых кислот, например, одноцепочечных нуклеиновых кислот и/или двуцепочечных нуклеиновых кислот; первый адаптер, причем 3'-конец первого адаптера содержит блокирующую группу; лигазу; и агент, вызывающий эффект исключения объема. В некоторых вариантах реализации реакционный сосуд также может содержать второй адаптер. В некоторых вариантах реализации реакционный сосуд также может содержать дефосфорилирующий агент.
[0120] В некоторых вариантах реализации реакционный сосуд может содержать реакционный объем. В некоторых вариантах реализации реакционный объем может содержать множество нуклеиновых кислот, например, одноцепочечных нуклеиновых кислот и/или двуцепочечных нуклеиновых кислот; первый адаптер, причем 3'-конец первого адаптера содержит блокирующую группу; второй адаптер; дефосфорилирующий агент; лигазу; и агент, вызывающий эффект исключения объема. В некоторых вариантах реализации реакционный объем может содержать множество первых одноцепочечных нуклеиновых кислот, содержащих первый индекс, причем 5'-концы первых одноцепочечных нуклеиновых кислот содержат 5'-блокирующую группу, множество вторых одноцепочечных нуклеиновых кислот, содержащих второй индекс и зонд захвата, причем 3'-концы вторых одноцепочечных нуклеиновых кислот содержат 3'-блокирующую группу; и лигазу.
[0121] В некоторых вариантах реализации первый адаптер лигирован с 3'-концами множества одноцепочечных нуклеиновых кислот, тем самым образуя множество модифицированных одноцепочечных нуклеиновых кислот. В некоторых вариантах реализации нелигированный первый адаптер гибридизован с зондом захвата. В некоторых вариантах реализации зонд захвата содержит последовательность, комплементарную по меньшей мере части первого адаптера.
[0122] В некоторых вариантах реализации 5'-конец второго адаптера нефосфорилирован. В некоторых вариантах реализации второй адаптер лигируют с 3'-концами множества модифицированных одноцепочечных нуклеиновых кислот. В некоторых вариантах реализации дефосфорилирующий агент содержит фосфатазу. В некоторых вариантах реализации указанная фосфатаза инактивирована. Примеры фосфатаз включают кишечную фосфатазу теленка, щелочную фосфатазу креветки, антарктическую фосфатазу и щелочную фосфатазу APEX.
[0123] В некоторых вариантах реализации первый адаптер и/или второй адаптер может содержать сайт связывания праймера для секвенирования. Примеры сайтов связывания праймеров для секвенирования включают последовательность Р7, ее комплемент или обратный комплемент; и последовательность Р5, ее комплемент или обратный комплемент.
[0124] В некоторых вариантах реализации первый адаптер и/или второй адаптер может содержать индекс адаптера. В некоторых вариантах реализации индекс первого адаптера отличается от индекса второго адаптера. В некоторых вариантах реализации индекс адаптера указывает на источник множества одноцепочечных нуклеиновых кислот.
[0125] Некоторые варианты реализации, предложенные в настоящем документе, включают реакционный сосуд, содержащий реакционный объем, содержащий: множество первых одноцепочечных нуклеиновых кислот, содержащих первый индекс, причем 5'-концы первых одноцепочечных нуклеиновых кислот содержат 5'-блокирующую группу, множество вторых одноцепочечных нуклеиновых кислот, содержащих второй индекс и зонд захвата, причем 3'-концы вторых одноцепочечных нуклеиновых кислот содержат 3'-блокирующую группу; и лигазу.
[0126] В некоторых вариантах блокирующая группа содержит 3'-спейсер С3, дидезоксинуклеотид или фосфатную группу. В некоторых вариантах реализации второй адаптер может включать присоединение к подложке. Примеры подложки включают гранулы.
[0127] В некоторых вариантах реализации агент, вызывающий эффект исключения объема, выбран из группы, состоящей из полиэтиленгликоля (ПЭГ), декстрана, гидроксиэтилкрахмала, фиколла и поливинилпирролидона. Примеры ПЭГ, которые можно примерять в вариантах реализации, предложенных в настоящем документе, включают ПЭГ 600, ПЭГ-800, ПЭГ-1000, ПЭГ-6000 и ПЭГ-8000. Реакционный объем может содержать концентрацию агента, вызывающего эффект исключения объема, например, в массо-объемных процентах, которая меньше, больше или равна приблизительно 5%, 10%, 20%, 30%, 40%, 50% 60%, 70%, 80%, 90%, 95% или находится в диапазоне между любыми двумя из вышеупомянутых процентных значений.
[0128] В некоторых вариантах реализации изобретения лигаза содержит лигазу одноцепочечной нуклеиновой кислоты. Примеры лигаз одноцепочечных нуклеиновых кислот включают РНК-лигазу 1 Т4, РНК-лигазу 2 Т4, RtcB-лигазу, РНК-лигазу Methanobacterium и РНК-лигазу TS2126 (CIRCLIGASE).
[0129] Некоторые варианты реализации, предложенные в настоящем документе, включают проточную ячейку, содержащую реакционный сосуд, предложенный в настоящем документе. Некоторые варианты реализации включают систему, содержащую реакционный сосуд, предложенный в настоящем документе, и детектор для получения данных секвенирования.
[0130] Проточная ячейка может содержать камеру с поверхностью, через которую можно направлять поток одного или более из жидких реагентов. В общем случае в проточной ячейке есть входное отверстие и выходное отверстие для облегчения потока жидкости. Примеры проточных ячеек и связанных с ними жмдкостных систем и платформ обнаружения, которые легко применять в способах согласно настоящему изобретению, описаны, например, в Bentley et al, Nature 456:53-59 (2008), WO 04/018497; US 7057026; WO 91/06678; WO 071123744; US 7329492; US 7211414; US 7315019; US 7405281 и US 2008/0108082, каждый из которых полностью включен в настоящий документ посредством ссылки.
ПРИМЕРЫ
Пример 1 - лигирование адаптеров с одноцепочечными нуклеиновыми кислотами-мишенями
[0131] Четырехэтапный способ получения библиотеки одноцепочечной ДНК (оцДНК) выполняли с использованием одноцепочечной ДНК в качестве нуклеиновой кислоты-мишени. На фиг. 1 представлен обзор четырех этапов, которые включали 5'-дефосфорилирование нуклеиновой кислоты-мишени; лигирование 3'-адаптера с 3'-концом нуклеиновой кислоты-мишени, причем 3'-конец 3'-адаптера блокировали для ингибирования самоконкатемеризации; повторное фосфорилирование 5'-дефосфорилированной лигированной нуклеиновой кислоты-мишени; и лигирование 5'-адаптера с 5'-концом лигированной нуклеиновой кислоты-мишени, причем 5'-конец 5'-адаптера блокировали для ингибирования самоконкатемеризации.
[0132] Для проверки этапа дефосфорилирования 5'-конец 60-членной одноцепочечной нуклеиновой кислоты (3'ОН-60-Phos 5') дефосфорилировали с использованием щелочной фосфатазы, фосфатазы APEX (Epicenter, Мэдисон, штат Висконсин, США) в реакционном объеме с 25% полиэтиленгликоля (ПЭГ). Степень 5'-дефосфорилирования анализировали с помощью одноцепочечной лигазы - лигазы CIRCLIGASE™ (Lucigen Corporation, Миддлтон, штат Висконсин, США) для подтверждения того, что дефосфорилированные продукты не образуют конкатемеры. Как изображено на фиг. 7, эффективность дефосфорилирования с помощью щелочной фосфатазы составляла приблизительно 100%.
[0133] Для проверки этапа начального лигирования 64-членный адаптер Р7' (3'C3-P7-phos 5'), содержащий 3'-блокирующую группу, лигировали с 3'-концом дефосфорилированной 60-членной одноцепочечной нуклеиновой кислоты (3'ОН-60-OH 5') с помощью CIRCLIGASE в реакционном объеме, содержащем 25% ПЭГ. 3'-конец адаптера Р7' блокировали для ингибирования самоконкатемеризации. Как показано на фиг. 8, выход этого этапа лигирования (дорожка 2) составлял 97.5%.
[0134] Для проверки киназного этапа 5'-конец 124-членной дефосфорилированной одноцепочечной нуклеиновой кислоты, содержащей лигированный 64-членный адаптер с 3' блокирующей группой (3'С3-Р7-60-ОН), повторно фосфорилировали с помощью полинуклеотидкиназы Т4 (PNK). Как изображено на фиг. 9, выход повторного фосфорилирования составлял приблизительно 99%.
[0135] Для проверки второго этапа лигирования 60-членный адаптер Р5 с нефосфорилированным 5'-концом (5'Р5-ОН), лигировали с 5'-концом 124-членной одноцепочечной нуклеиновой кислоты (3'C3-60-B2-Phos) с помощью CIRCLIGASE в реакционном объеме, содержащем 1,6% ПЭГ. 124-членная одноцепочечная нуклеиновая кислота содержала лигированный 64-членный адаптер с 3'-блокирующей группой (3'C3-P7-phos 5'). Как показано на фиг.10, выход этого этапа лигирования (дорожка 2) составлял >85%. Таким образом, общий выход 4-этапного подхода к получению библиотеки оцДНК составлял >80%.
Пример 2 - лигирование адаптеров с бесклеточными нуклеиновыми кислотами-мишенями
[0136] Схему, представленную на фиг. 1, осуществляли с использованием бесклеточной ДНК (бкДНК). Двуцепочечные нуклеиновые кислоты-мишени дефосфорилировали с использованием щелочной фосфатазы APEX и денатурировали с образованием одноцепочечных нуклеиновых кислот-мишеней. Для проверки этапа начального лигирования 64-членный адаптер Р7', содержащий 3'-блокирующую группу (3'C3-P7-phos 5'), лигировали с 3'-концом одноцепочечной дефосфорилированной бесклеточной ДНК (бкДНК) с помощью CIRCLIGASE в реакционном объеме, содержащем 22,5% ПЭГ, в течение 3 часов. Как показано на фиг. 11, выход этого этапа лигирования (дорожка 2) составлял более 95%.
[0137] Для проверки киназного этапа дефосфорилированную бкДНК (бкДНК [Apex]) обрабатывали полинуклеотидкиназой Т4 (PNK) и лигировали 64-членный адаптер Р7', содержащий 3'-блокирующую группу (3'C3-P7-phos 5'), с 3'-концом повторно фосфорилированной бкДНК. Реакцию выполняли в объеме, содержащем 11.25% ПЭГ, в течение 1 часа (фиг. 12).
[0138] Для проверки второго этапа лигирования 64-членный адаптер Р7', содержащий 3'-блокирующую группу (3'C3-P7-phos 5'), лигировали с 3'-концом дефосфорилированной бкДНК, полученный первый лигированный продукт (3'С3-Р7-бкДНК-phos 5') обрабатывали киназой (PNK) и лигировали 60-членный адаптер Р5 с нефосфорилированным 5'-концом (5'Р5-ОН) с 5'-концом первого продукта лигирования с образованием второго продукта лигирования (Р7'-бкДНК-Р5). Реакцию выполняли с использованием CIRCLIGASE в реакционном объеме, содержащем 3.8% ПЭГ. На фиг. 13 показаны продукты лигирования (Р7'-бкДНК-Р5) и (Р7'-Р5).
Пример 3 - анализ продуктов лигирования
[0139] Продукты лигирования из примера 2 разделяли в геле, полосы продуктов лигирования экстрагировали из геля, экстрагированные продукты лигирования амплифицировали, а амплифицированные продукты разделяли с помощью капиллярного электрофореза (BioAnalyzer, Agilent).
[0140] Продукты лигирования разделяли в геле, полосы «Экстракт В1» и «Экстракт В2» вырезали из геля. Как показано на фиг. 14, на дорожке 'REF' экстракт В1 обозначен двумя черными прямоугольниками, а экстракт В2 - одним белым прямоугольником. Экстракты амплифицировали посредством ПЦР с использованием праймеров Р7, не предназначенных для ПЦР (не амплифицируемых), в соотношении с ПЦР-праймерами Р7 от 1:40 до 1:20 в течение 24 циклов. Продукты ПЦР экстрагировали с помощью твердофазной обратимой иммобилизации (SPRI) на гранулах в соотношении 2:1. Результаты показаны на фиг. 14.
[0141] Затем амплифицированные продукты экстракта В1 и В2 анализировали посредством капиллярного электрофореза (BioAnalyser, Agilent). Как показано на фиг. 15, экстракт В2 преимущественно содержал продукты длиной 127 п.о. (Р7'-Р5), а экстракт В1 содержал разновидности продуктов длиной 292 п.о. (Р7'-бкДНК-Р5) и 480 п.о. (Р7'-бкДНК-Р5).
[0142] Количество гранул SPRI, использованных для очистки экстрактов из геля, меняли для определения соотношения, позволявшего эффективно удалять праймеры-димеры (фиг. 16). Соотношение гранулы SPRI : образец (об : об), равное 1.2:1, наиболее эффективно позволяло удалять праймеры-димеры без потери существенной части библиотеки бкДНК.
Пример 4 - захват нелигированных праймеров Р7'
[0143] Избыток нелигированных праймеров Р7' удаляли по схеме, приведенной на фиг. 5. Вкратце, праймер Р7, комплементарный праймеру адаптера Р7', добавляли к реакционному объему после лигирования праймера адаптера Р7' с 3'-концом нуклеиновой кислоты-мишени. Праймер Р7 содержал блокирующие группы на 5'- и 3'-концах. После добавления праймера Р7 адаптер Р5 лигировали с 5'-концом повторно фосфорилированной лигированной нуклеиновой кислоты-мишени.
[0144] 63-членный праймер Р7' (3'C3-P7'-phos 5') лигировали с 3'-концами бкДНК в реакционном объеме, содержавшем 25% ПЭГ. Как показано на фиг.17, выход лигированного продукта составлял >95%. Для проверки лигирования в присутствии зонда захвата 60-членный адаптер Р5, содержащий нефосфорилированный 5'-конец (5'Р5-ОН), лигировали с 5'-концом первого продукта лигирования (3'С3-Р7'-бкДНК-phos5') в присутствии 63-членного зонда захвата Р7 (3'С3-Р7-С35'), содержащего 5'- и 3'-блокирующие группы. Реакцию выполняли в реакционном объеме, содержащем 6,5% ПЭГ. На фиг. 18 показаны продукты лигирования (Р7'-бкДНК-Р5) и (Р7'-Р5).
[0145] Затем анализировали продукты лигирования. Как показано на фиг. 19, продукты лигирования разделяли по размеру в геле, некоторые полосы экстрагировали из геля, экстракт В1 и экстракт В2 амплифицировали. Амплифицированные продукты анализировали посредством капиллярного электрофореза; они содержали разновидности продуктов длиной 127 п.о. (Р7'-Р5), 292 п.о. (Р7'-бкДНК-Р5) и 480 п.о. (Р7'-бкДНК-Р5). На фиг. 20 изображены гели продуктов лигирования до амплификации, после амплификации и после экстракции с гранулами SPRI.
[0146] Включение Р7, не предназначенных для ПЦР, для захвата избытка Р7', существенно снижало образование праймеров-димеров. Кроме того, образование праймеров-димеров существенно снижалось при удалении с использованием 1.2Х SPRI без существенной потери библиотеки бкДНК.
Пример 5-лигирование одноцепочечных нуклеиновых кислот, конъюгированных с гранулами
[0147] Выполнили проверку эффективности лигирования двух олигомеров (олигонуклеотида 1 и олигонуклеотида 2) при различных концентрациях ПЭГ. Олигонуклеотид 1 содержал блокированный 3'-конец (ОН), а олигонуклеотид 2 содержал блокированный 5'-конец (phos), таким образом, лигирование приводило к получению единственного лигированного продукта олигонуклеотид 1-олигонуклеотид 2. Условия реакции включали: 10 пмоль каждого олигонуклеотида + 2 мкл 10 × буфера + 1 мкл 1 мМ АТФ, 1 мкл 50 мМ MnCl2, 1 мкл CIRCLIGASE (100 ед/мкл) + 15 мкл 0, 25, 50 или 60% ПЭГ в конечном реакционном объеме 20 мкл при концентрации ПЭГ 0, 19, 38 и 45%, соответственно. Результаты показаны на фиг. 21. Реакционный объем, содержавший 45% ПЭГ, обеспечивал 55.9% выход лигированных олигонуклеотида 1 и олигонуклеотида 2.
[0148] Выполнили проверку эффективности лигирования олигонуклеотида 1 и олигонуклеотида 2 при различных концентрациях олигонуклеотида 2. Условия реакции включали: 10 пмоль олигонуклеотида 1 + 10 × пмоль олигонуклеотида 2 + 2 мкл 10 × буфера + 1 мкл 1 мМ АТФ, 1 мкл 50 мМ MnCl2, 2 мкл CIRCLIGASE (100 ед/мкл) + 15 мкл 60% ПЭГ, что обеспечивало конечный реакционный объем 20 мкл при концентрации ПЭГ 45%. Результаты показаны на фиг. 22. Реакционный объем, содержавший 4Х избыток олигонуклеотида 2 и 2Х CIRCLIGASE, обеспечивал 79,6% выход лигированных олигонуклеотида 1 и олигонуклеотида 2.
[0149] Выполнили проверку эффективности лигирования олигонуклеотида 1 и олигонуклеотида 2 при различных концентрациях CIRCLIGASE. Условия реакции включали: 10 пмоль каждого олигонуклеотида + 2 мкл 10 × буфера + 1 мкл 1 мМ АТФ, 1 мкл 50 мМ MnCl2, X мкл CIRCLIGASE (100 ед/мкл) + 15 мкл 60% ПЭГ, что обеспечивало конечный реакционный объем 20 мкл при концентрации ПЭГ 45%. Результаты показаны на фиг. 23. Реакционный объем, содержавший 8 X концентрацию CIRCLIGASE, обеспечивал 75.3% выход лигированных олигонуклеотида 1 и олигонуклеотида 2.
[0150] Олигонуклеотид 1 конъюгировали с гранулами посредством гидразин-альдегидной реакции присоединения. 39-членный олигонуклеотид 1 содержал два урациловых основания, которые можно было расщеплять с использованием урацил-ДНК-гликозилазы (UDG), получая более короткий 27-членный олигонуклеотид (олигонуклеотид 1*).
[0151] В ходе реакции лигирования проверяли различные концентрации олигонуклеотида 1, конъюгированного с гранулами. Лигирование олигонуклеотида 2 с конъюгированным олигонуклеотидом 1 анализировали посредством последующей обработки UDG. Условия реакции лигирования предполагали, что количество олигонуклеотида 1, использованного для присоединения к гранулам, было равно количеству олигонуклеотида 1 на гранулах. Проверили четыре начальные концентрации общего олигонуклеотида 1: 0.5 нмоль, 1 нмоль, 2 нмоль и 4 нмоль. Использовали 10% общего раствора гранул, таким образом, предполагаемое количество олигонуклеотида 1 в каждой реакции составляло 0.1 нмоль, 0.2 нмоль, 0.3 нмоль и 0.4 нмоль. Реакционная смесь: 10 мкмоль олигонуклеотида раствора гранул (2 × концентрации), олигонуклеотид 2 (0,4 нмоль), 2 мкл 10 × буфера, 1 мкл 1 мМ АТФ, 1 мкл 50 мМ MnCl2, 1 мкл CIRCLIGASE (100 ед/мкл) + 5 мкл 90% ПЭГ, что обеспечивало конечный реакционный объем 20 мкл при концентрации ПЭГ 22,5%. Реакционную смесь инкубировали при 60°C в течение 3 ч и инактивировали при 80°C в теечние 10 мин. Соотношения олигонуклеотид 1: олигонуклеотид 2 в реакциях: 0.05 нмоль (1:8); 0.1 нмоль (1:4), 0.2 нмоль (1:2), 0.4 нмоль (1:1). Расщепление по урацилу выполняли с использованием 10 мкл LMX1 в конечном реакционном объеме 10 мкл и инкубировали при 37°C в течение 1 часа. Результаты показаны на фиг. 24. Реакционный объем, содержавший соотношение олигонуклеотид 1: олигонуклеотид 2, равное 1:1, обеспечивал 85,2% выход лигированных олигонуклеотида 1 и олигонуклеотида 2. Анализ повторяли для конечных реакционных объемов, содержащих 45% ПЭГ. Результаты показаны на фиг. 25. Реакционный объем, содержавший соотношение олигонуклеотид 1 : олигонуклеотид 2, равное 1:1, обеспечивал 84.5% выход лигированных олигонуклеотида 1 и олигонуклеотида 2.
[0152] Термин «содержащий», используемый в настоящем документе как синоним терминов «включающий», «состоящий» или «характеризующийся», является открытым и не исключает наличия дополнительных, не указанных элементов или этапов способа.
[0153] В вышеприведенном описании изложены несколько способов и материалов согласно настоящему изобретению. Настоящее изобретение подвержено модификациям в части способов и материалов, а также изменениям в части способов изготовления и оборудования. Такие модификации очевидны для специалистов в данной области техники после рассмотрения описания или реализации настоящего изобретения, описанного в настоящем документе. Соответственно, настоящее изобретение не должно ограничиваться конкретными вариантами реализации, описанными в настоящем документе, но охватывает все модификации и альтернативы, исходящие из истинной сущности настоящего изобретения.
[0154] Все источники, процитированные в настоящем документ, включая опубликованные и неопубликованные заявки, патенты и литературные источники, но не ограничиваясь ими, полностью включены в настоящий документ посредством ссылок, и являются частью настоящего документа. В той степени, в которой публикации и патенты или заявки на патенты, включенные посредством ссылки, противоречат описанию, содержащемуся в настоящем документе, настоящий документ должен не принимать во внимание любые такие противоречащие материалы и/или иметь преимущество перед ними.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДУЛЯЦИЯ ПОЛИМЕРНЫХ ГРАНУЛ ДЛЯ ОБРАБОТКИ ДНК | 2019 |
|
RU2822154C2 |
| АНАЛИЗ МНОЖЕСТВА АНАЛИТОВ С ИСПОЛЬЗОВАНИЕМ ОДНОГО АНАЛИЗА | 2019 |
|
RU2824049C2 |
| ПРЯМОЙ ЗАХВАТ, АМПЛИФИКАЦИЯ И СЕКВЕНИРОВАНИЕ ДНК-МИШЕНИ С ИСПОЛЬЗОВАНИЕМ ИММОБИЛИЗИРОВАННЫХ ПРАЙМЕРОВ | 2011 |
|
RU2565550C2 |
| НЕИНВАЗИВНАЯ ДИАГНОСТИКА ПУТЕМ СЕКВЕНИРОВАНИЯ 5-ГИДРОКСИМЕТИЛИРОВАННОЙ БЕСКЛЕТОЧНОЙ ДНК | 2017 |
|
RU2742355C2 |
| ЗОНД НУКЛЕИНОВОЙ КИСЛОТЫ И СПОСОБ ДЕТЕКТИРОВАНИЯ ГЕНОМНЫХ ФРАГМЕНТОВ | 2014 |
|
RU2733888C2 |
| СПОСОБЫ ИНКАПСУЛИРОВАНИЯ ОДИНОЧНЫХ КЛЕТОК, ИНКАПСУЛИРОВАННЫЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2750567C2 |
| СПОСОБЫ И СИСТЕМЫ ДЛЯ ПОЛУЧЕНИЯ НАБОРОВ УНИКАЛЬНЫХ МОЛЕКУЛЯРНЫХ ИНДЕКСОВ С ГЕТЕРОГЕННОЙ ДЛИНОЙ МОЛЕКУЛ И КОРРЕКЦИИ В НИХ ОШИБОК | 2018 |
|
RU2766198C2 |
| БИБЛИОТЕКИ ДЛЯ СЕКВЕНИРОВАНИЯ НОВОГО ПОКОЛЕНИЯ | 2014 |
|
RU2698125C2 |
| СЛОЖНЫЕ КОМПЛЕКСЫ СВЯЗАННОЙ НА ПОВЕРХНОСТИ ТРАНСПОСОМЫ | 2020 |
|
RU2790295C2 |
| МУЛЬТИПЛЕКСНОЕ ДЕТЕКТИРОВАНИЕ НУКЛЕИНОВЫХ КИСЛОТ | 2014 |
|
RU2733887C2 |


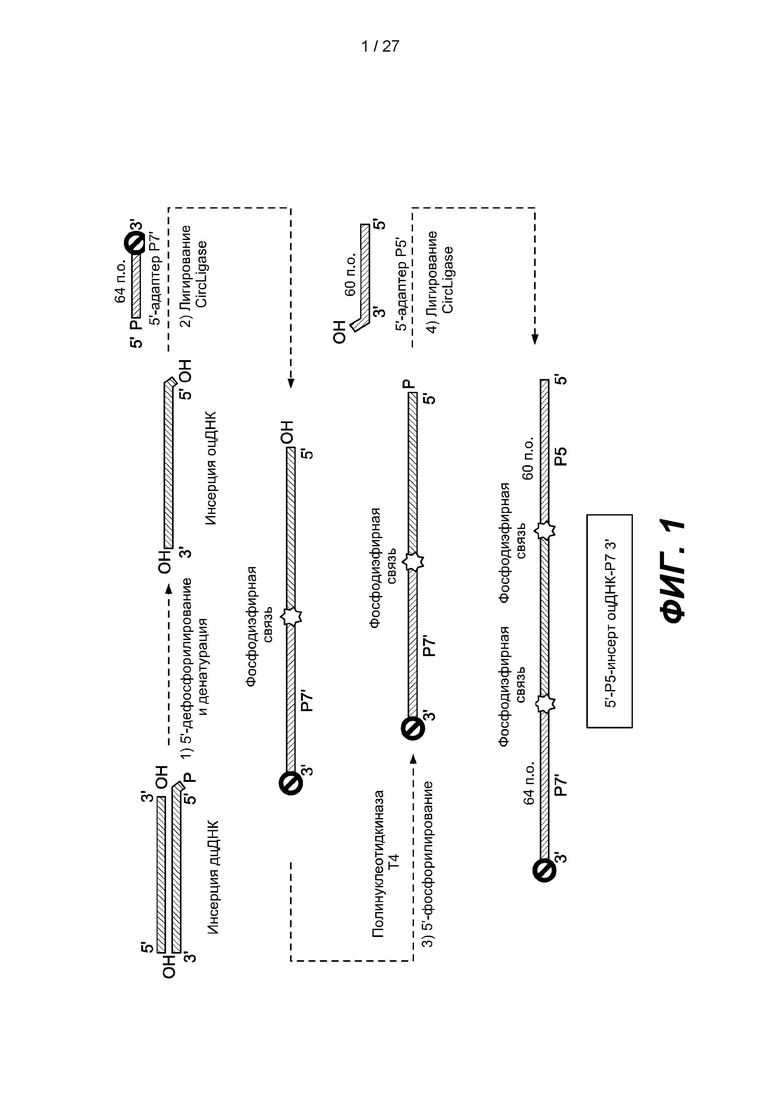
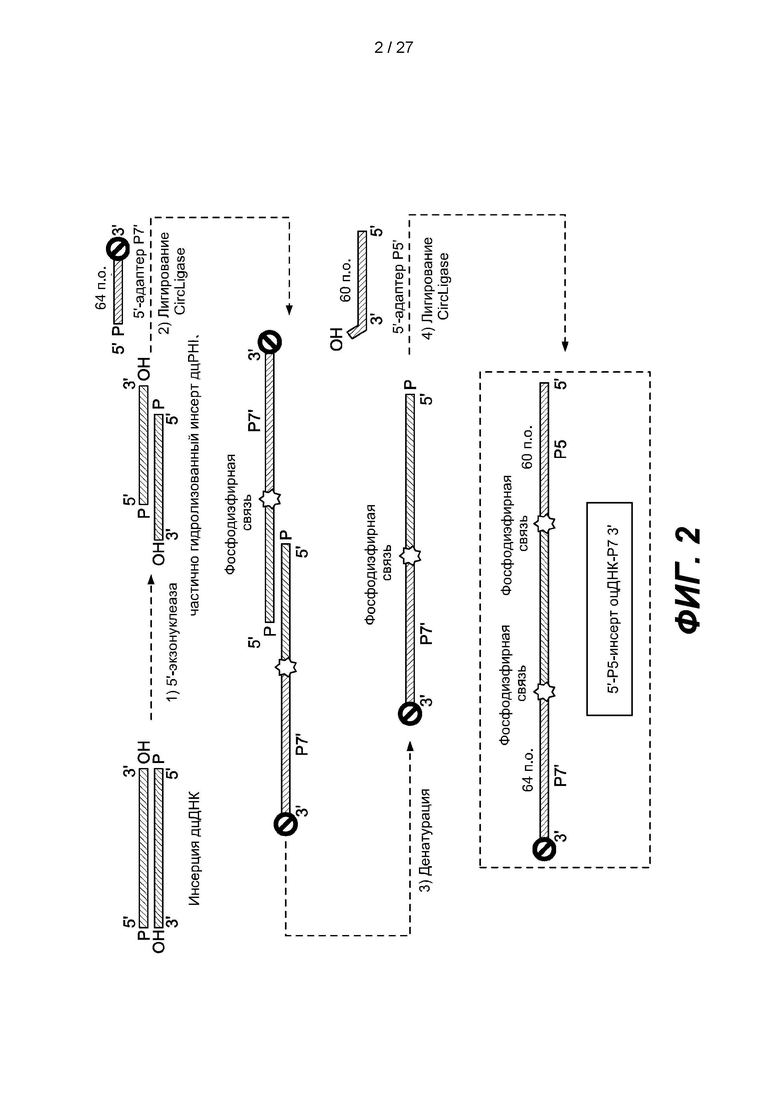
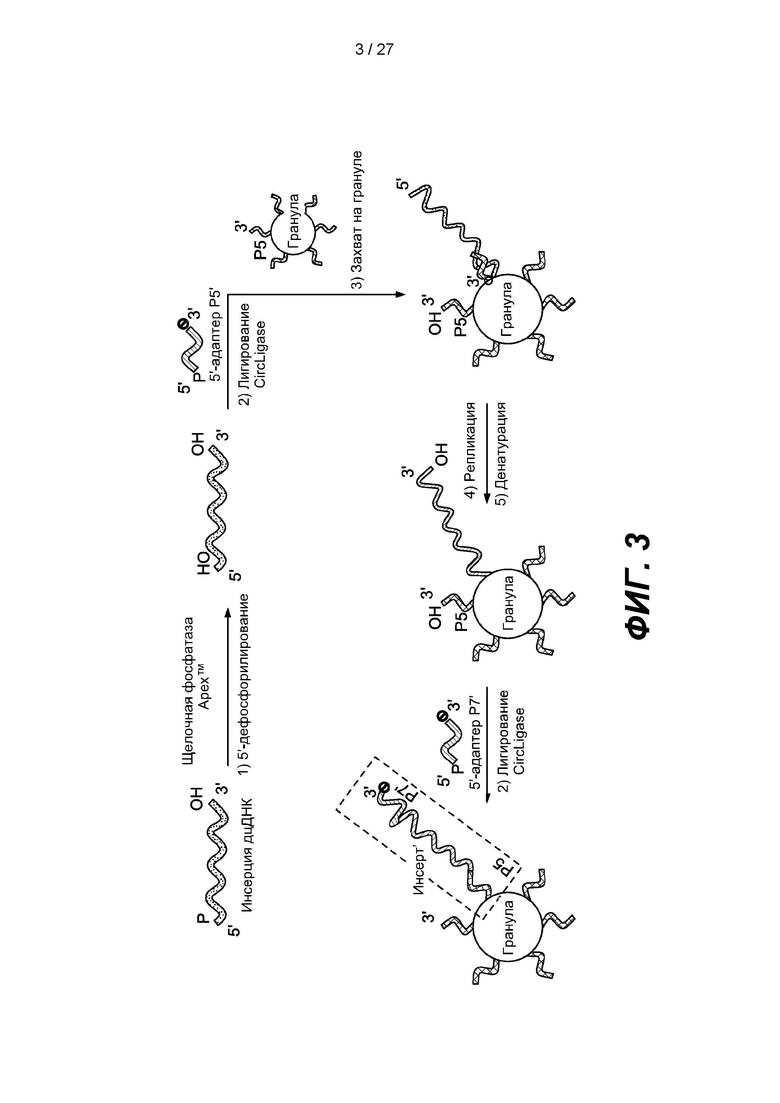
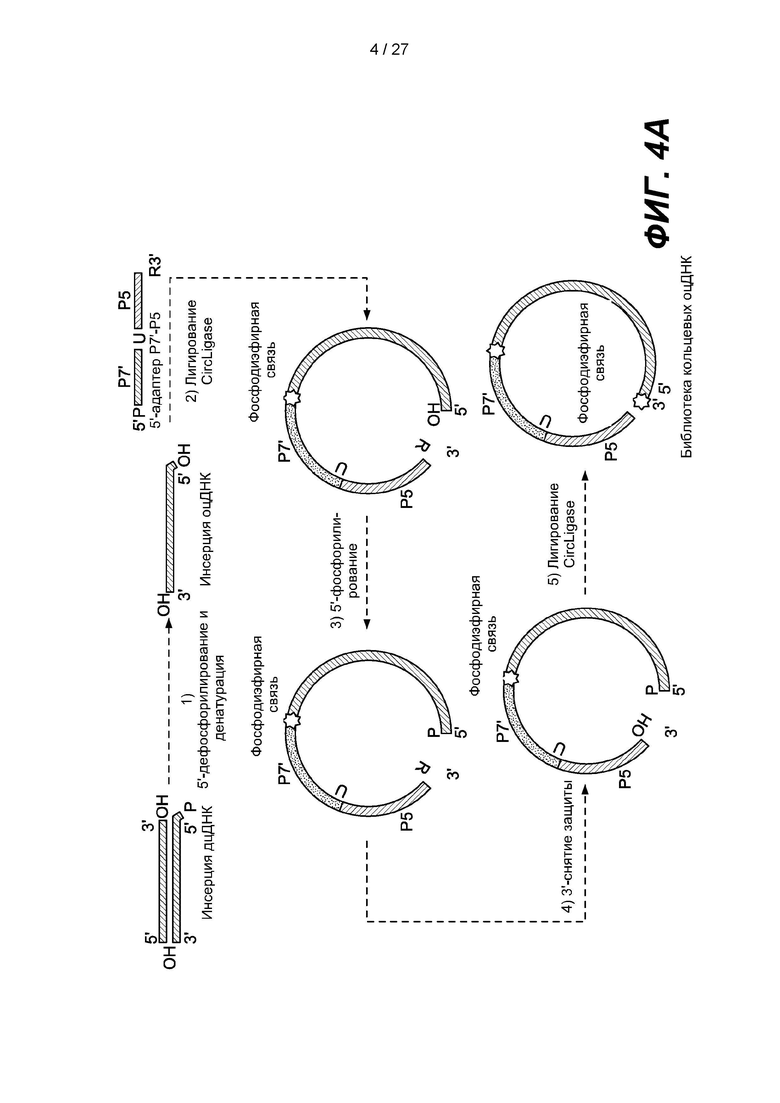
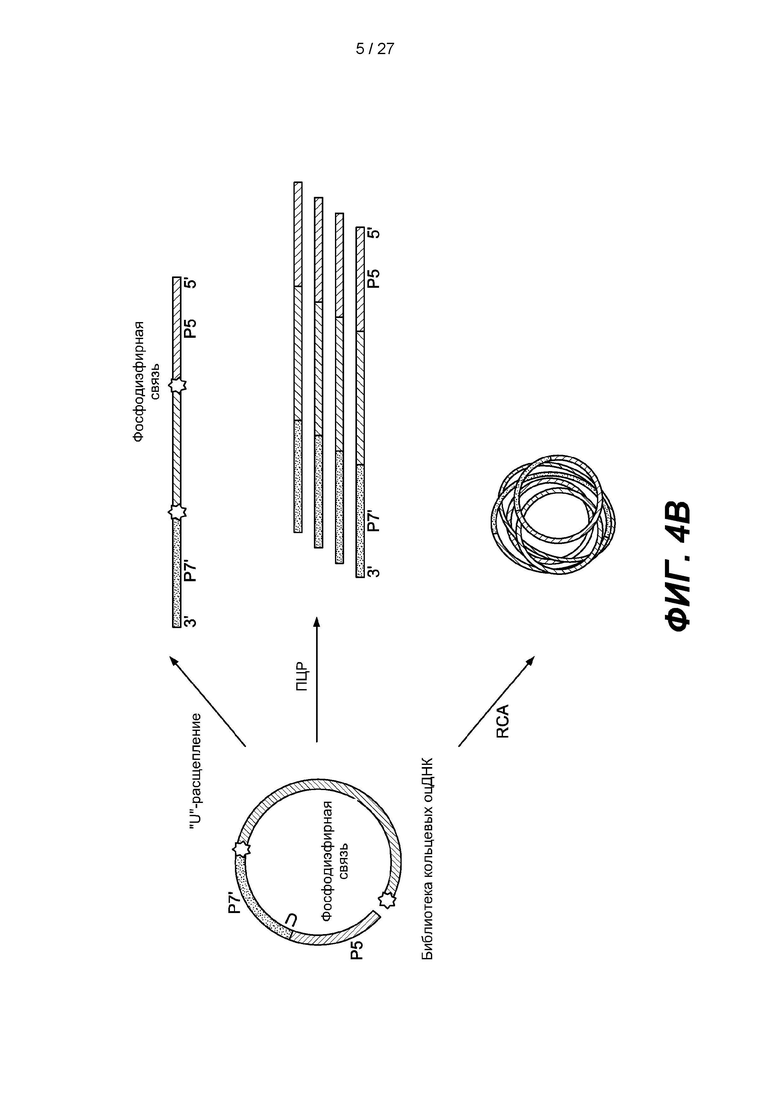
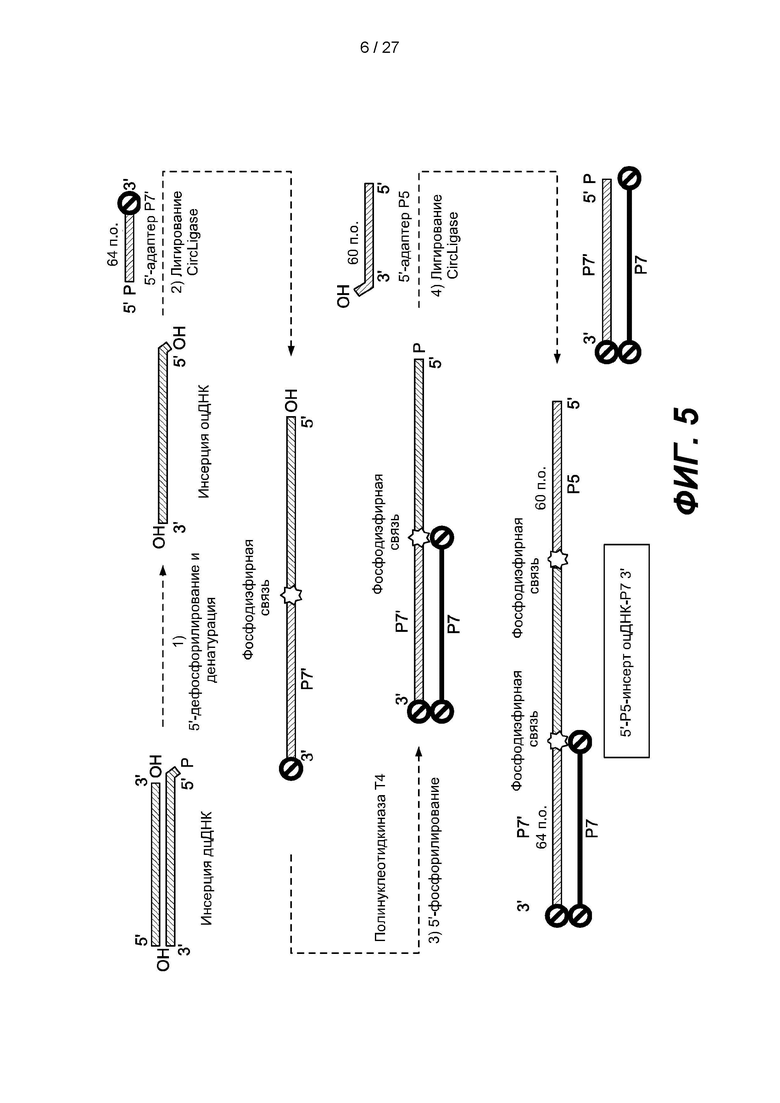
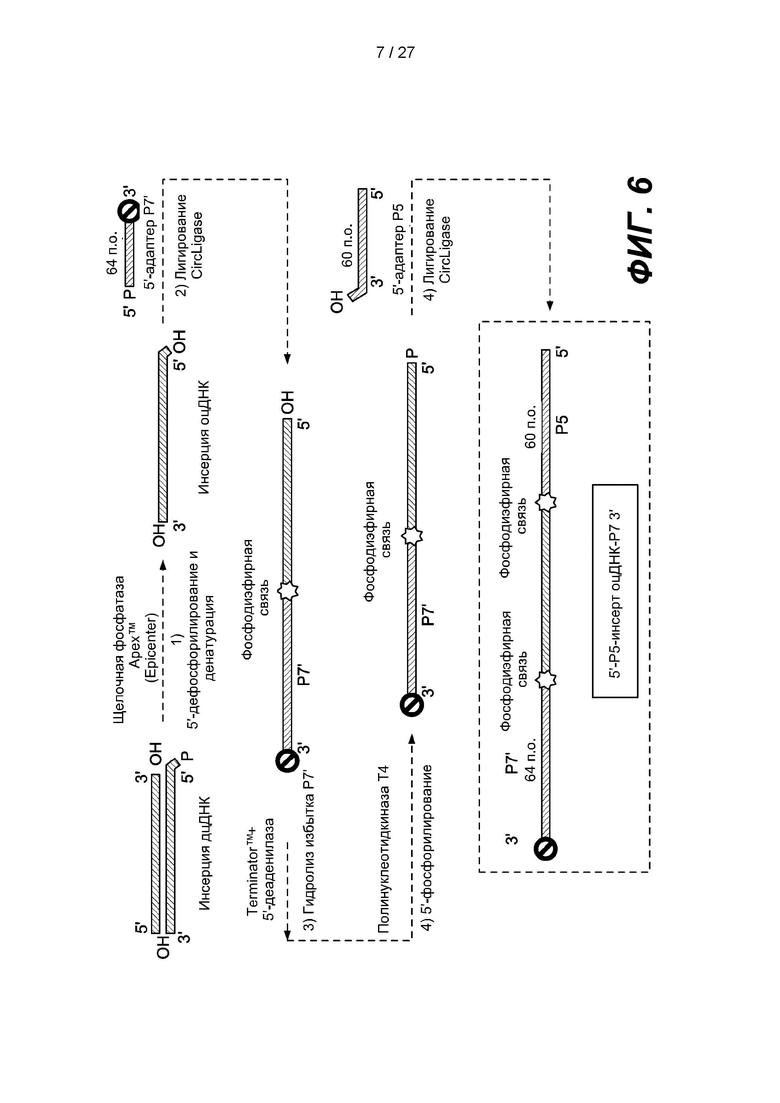
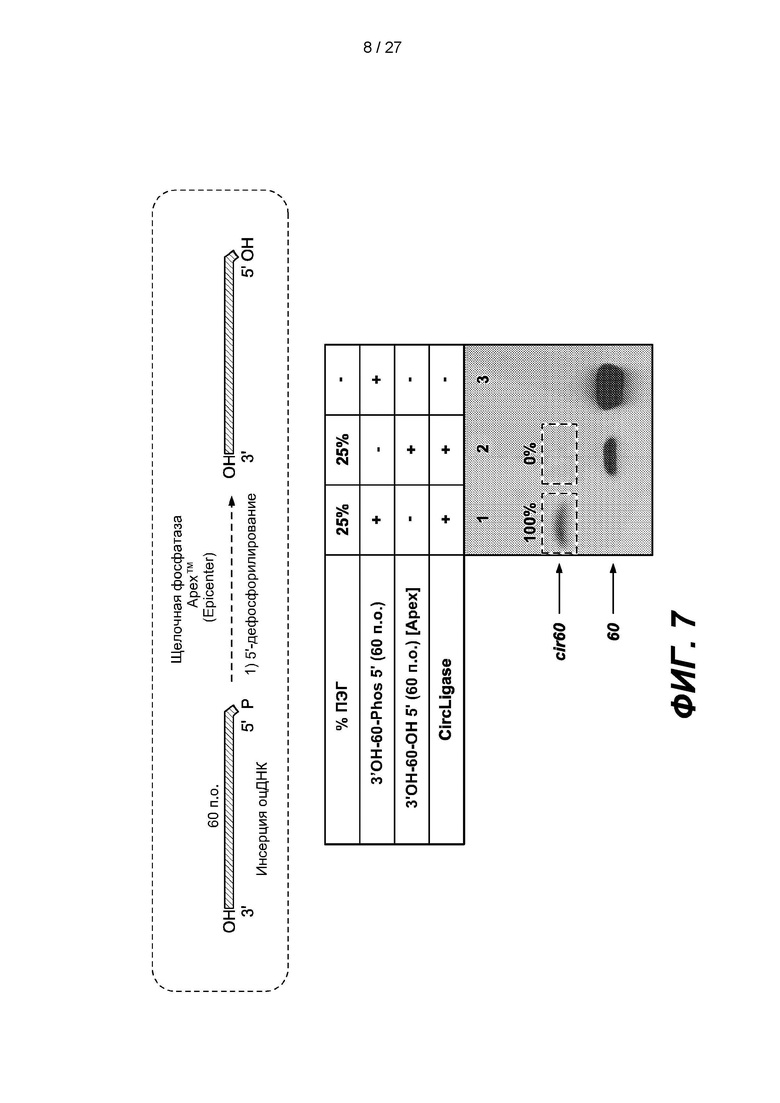
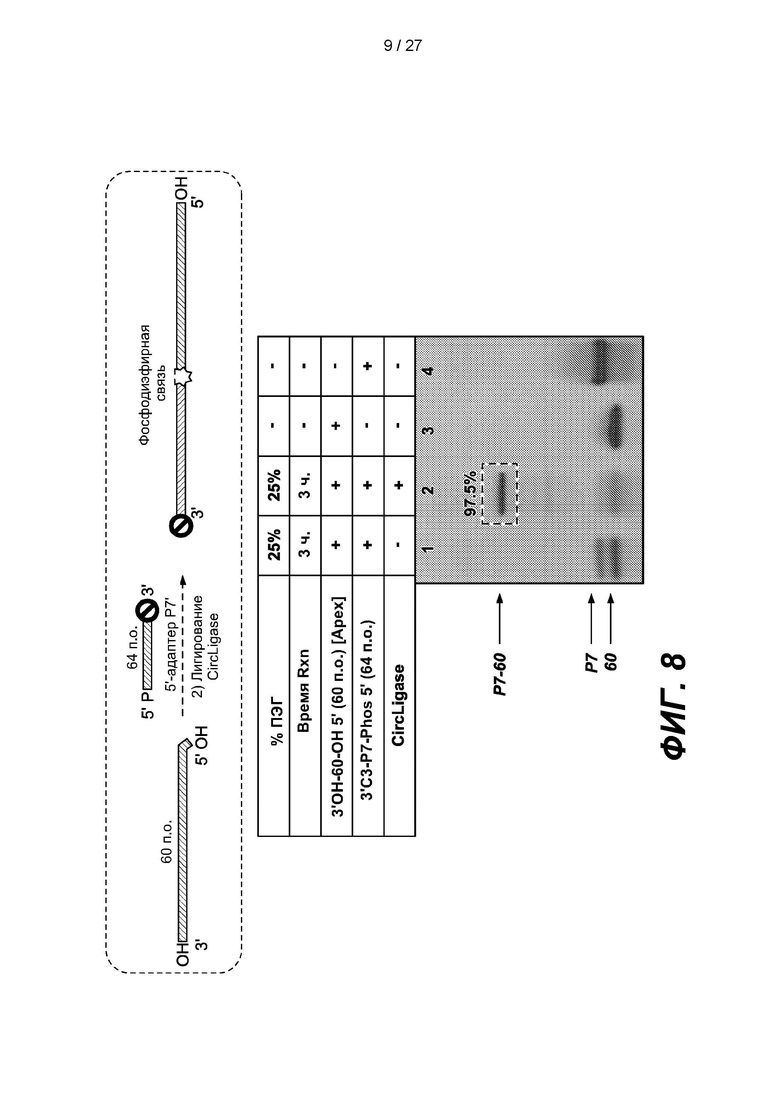
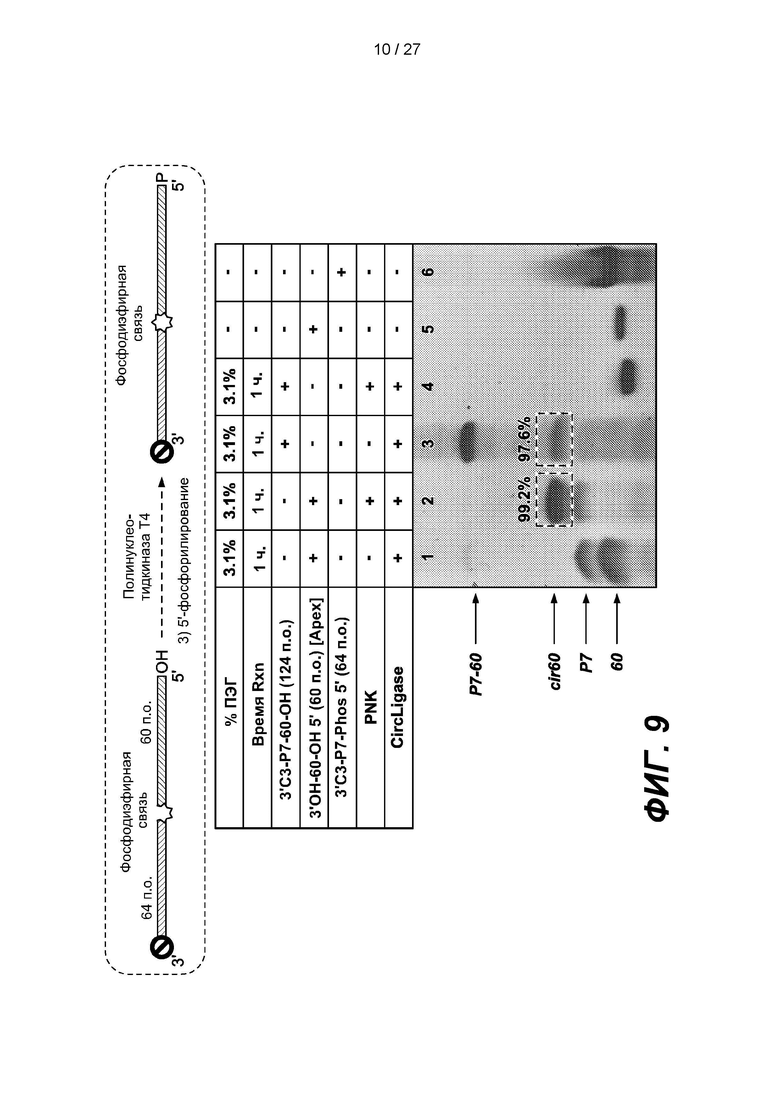
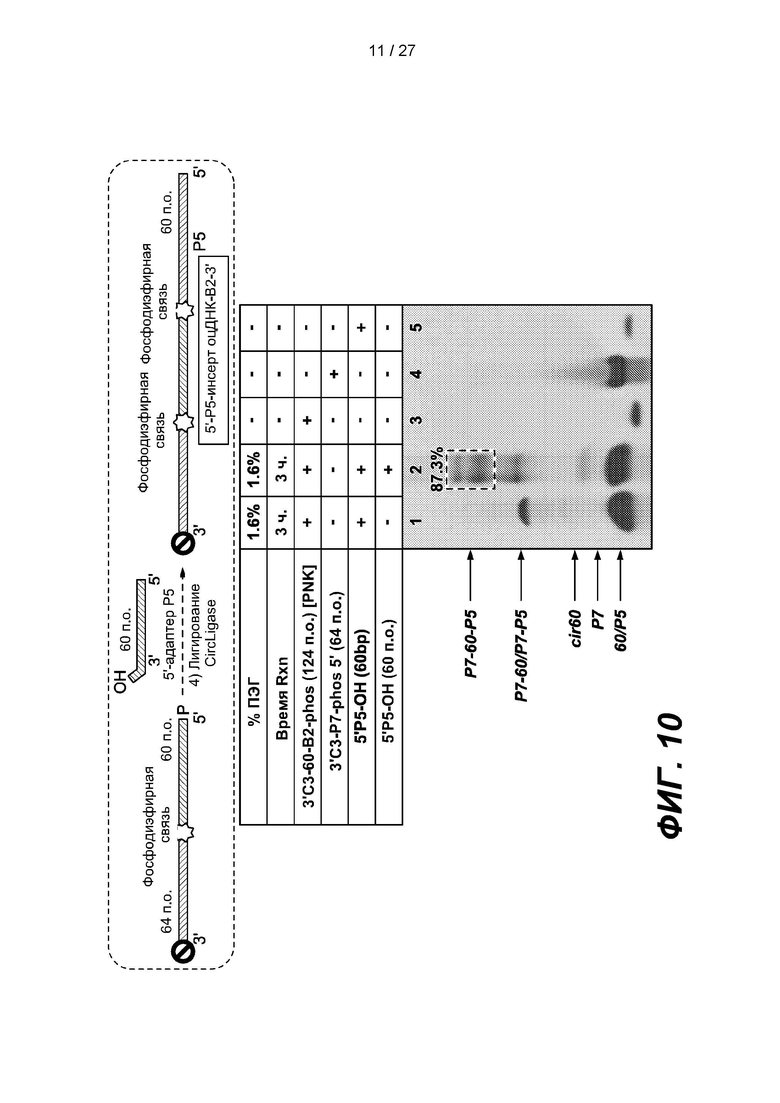
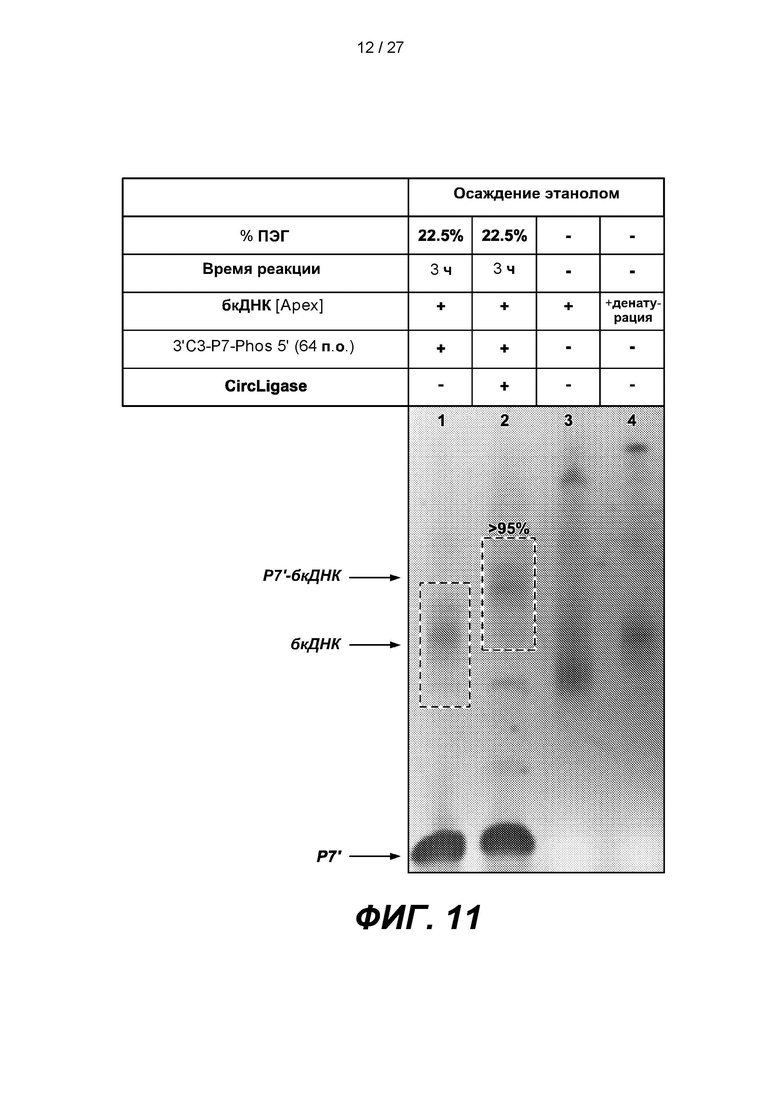
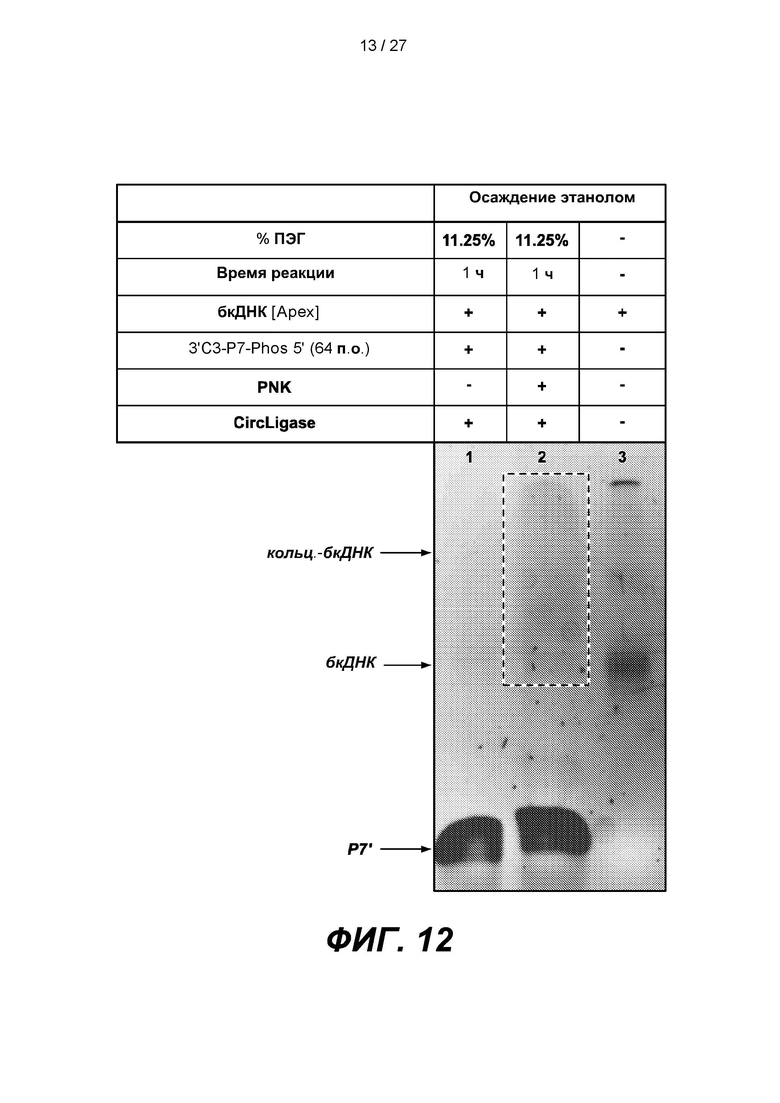
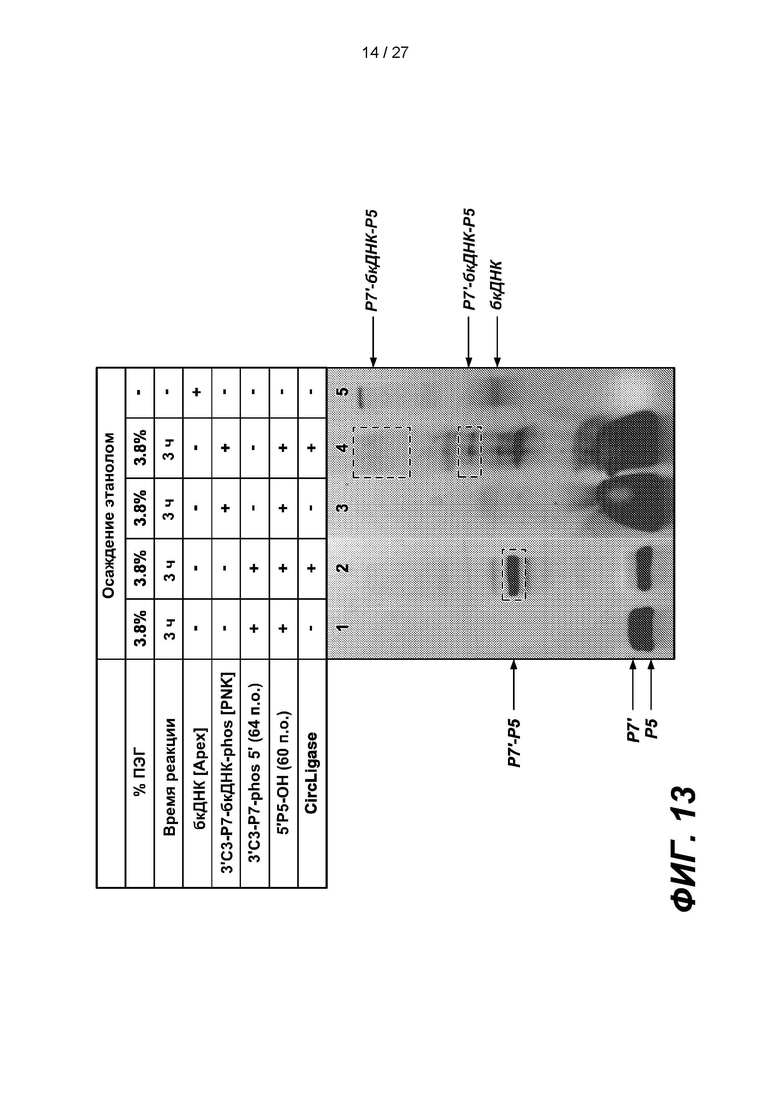
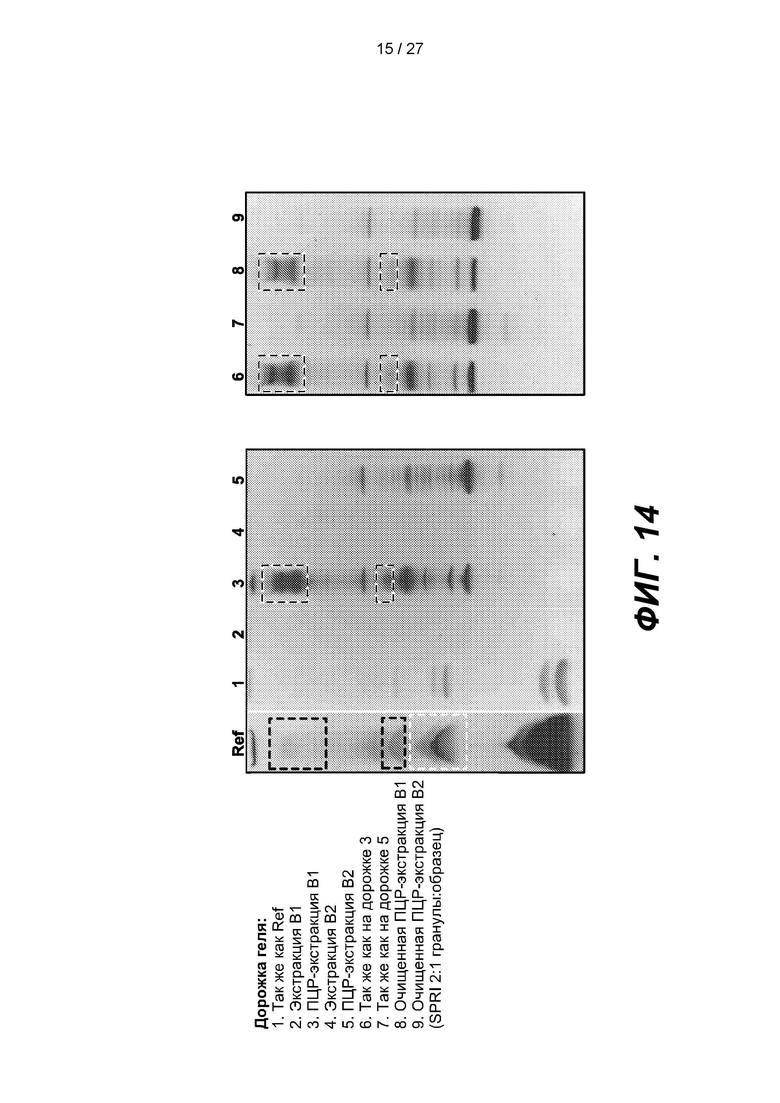
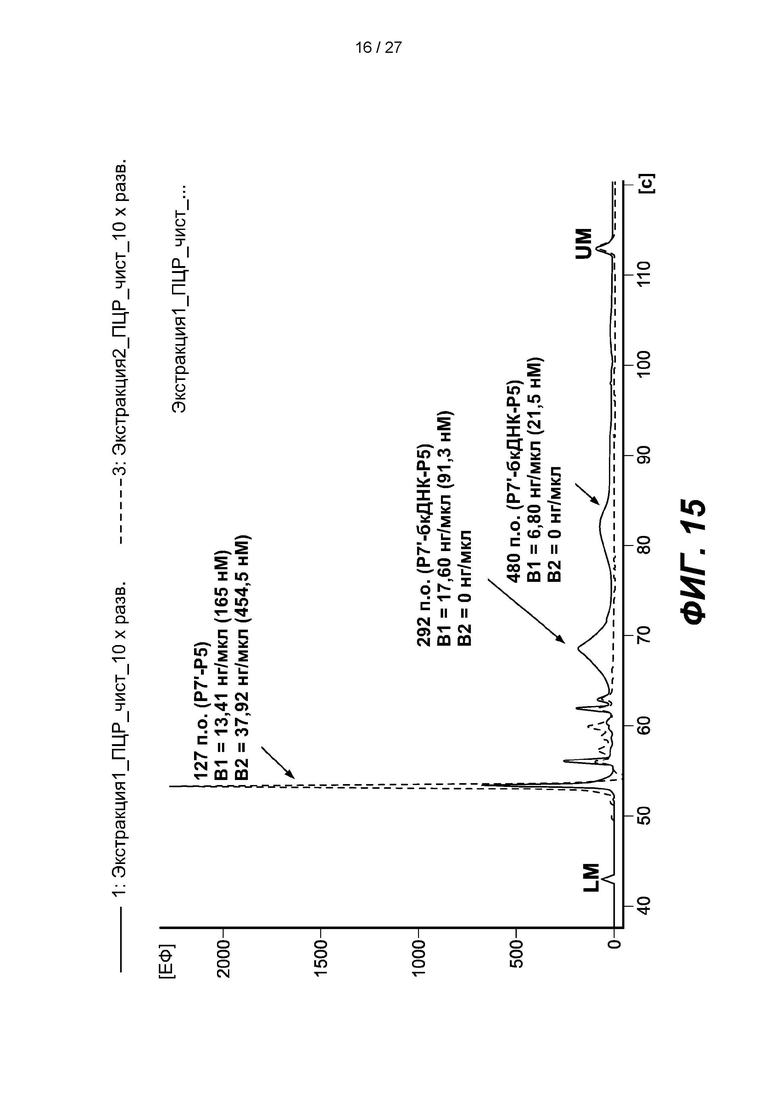
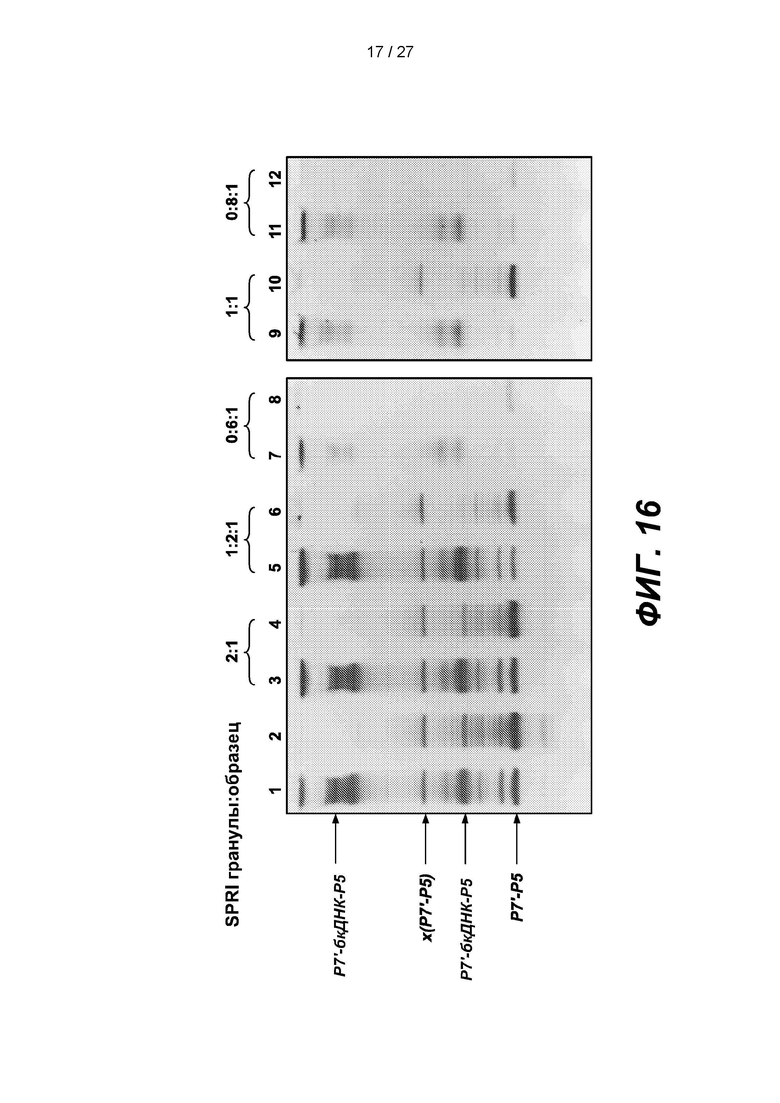
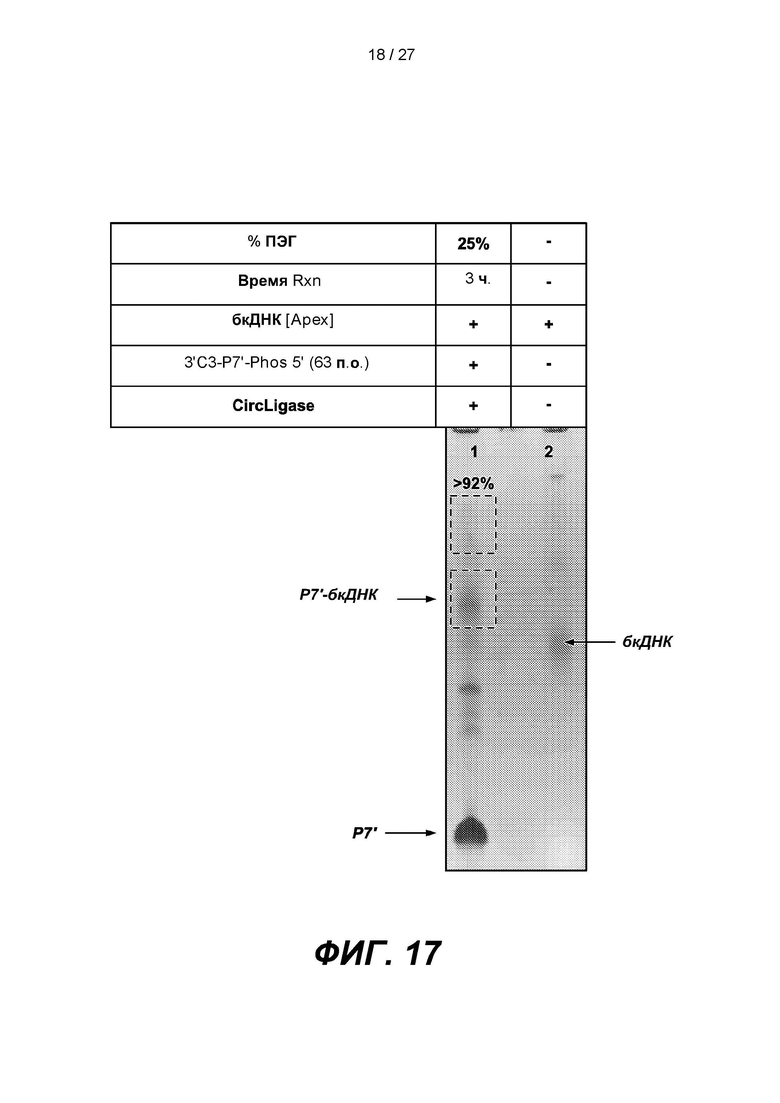
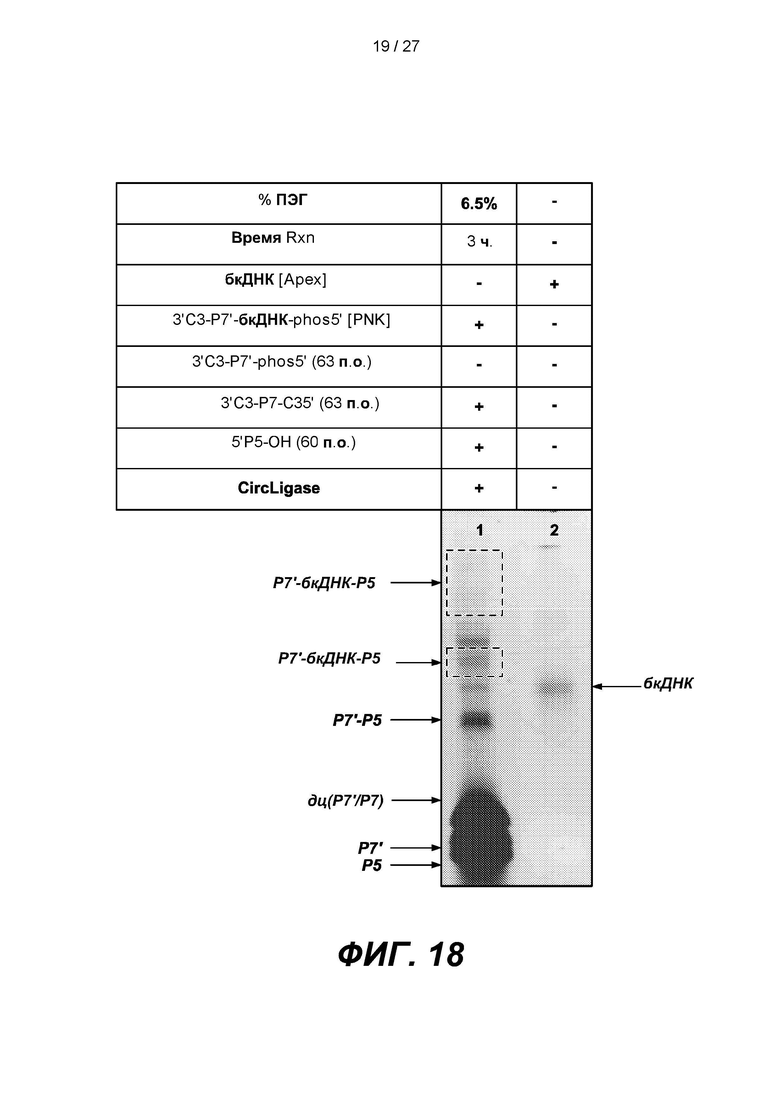
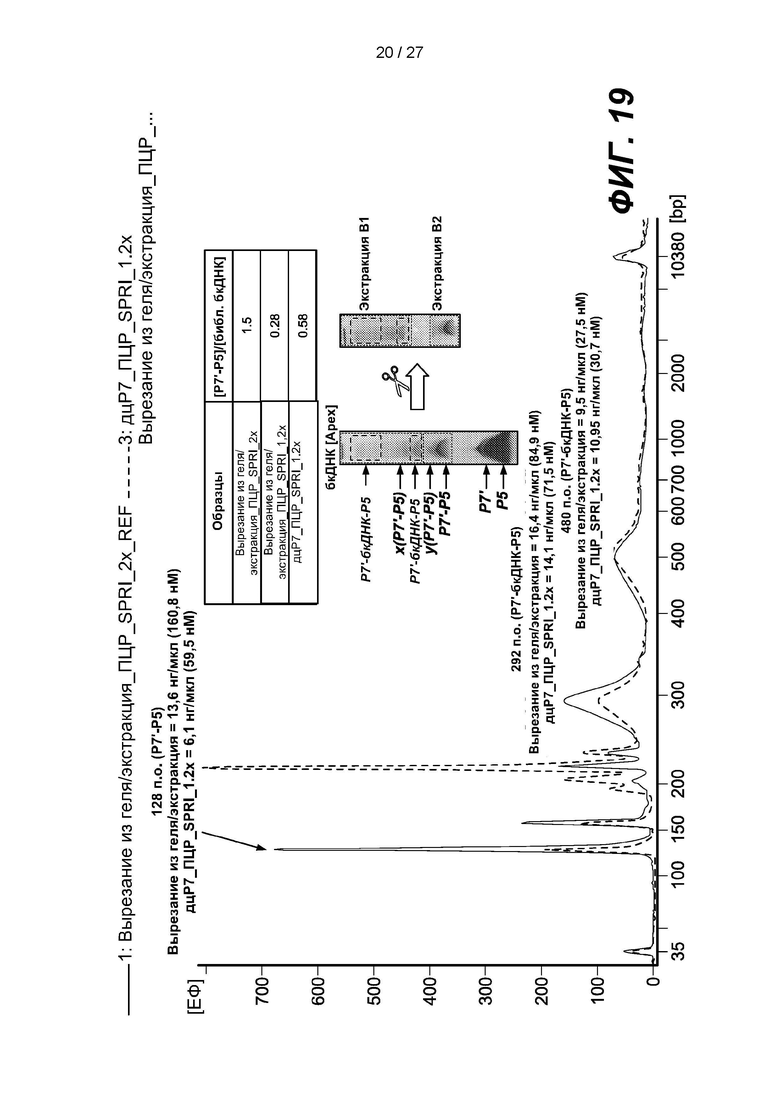
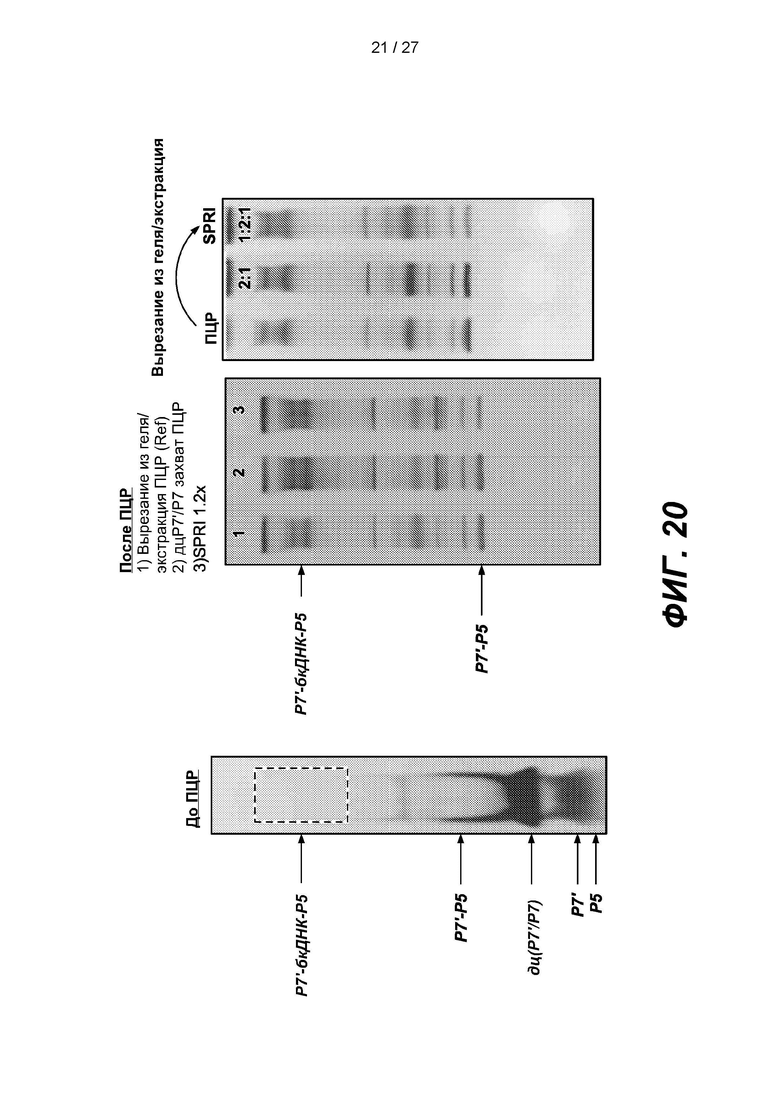

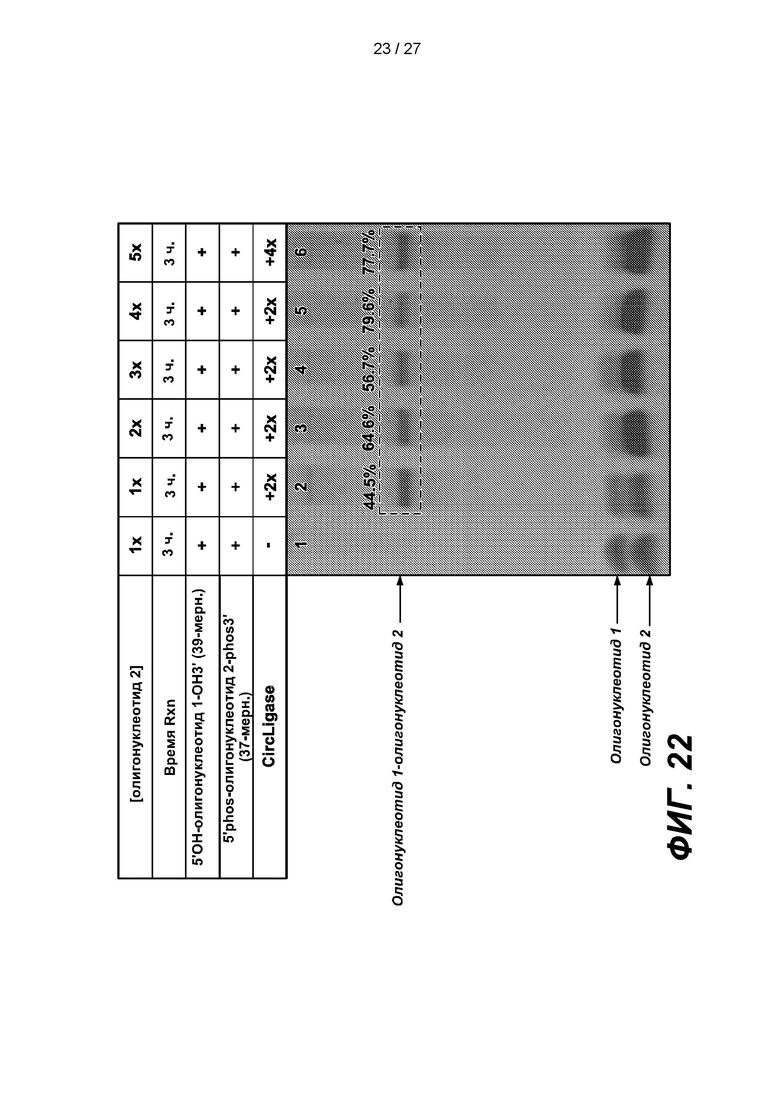
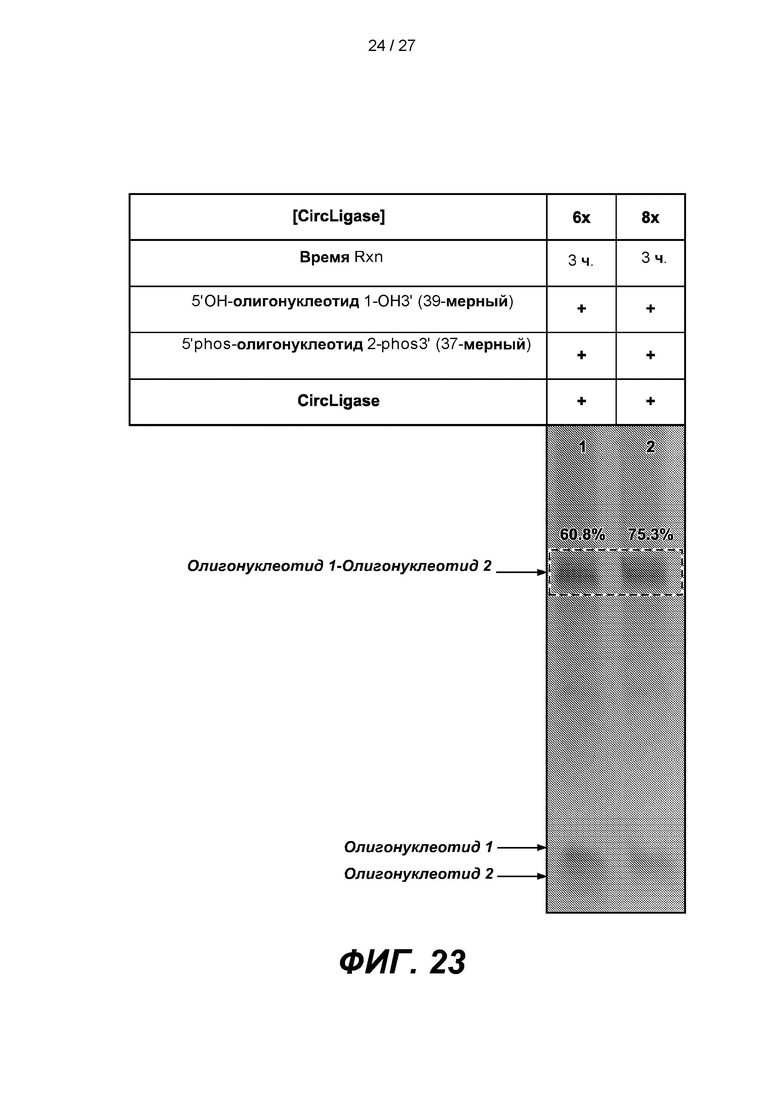


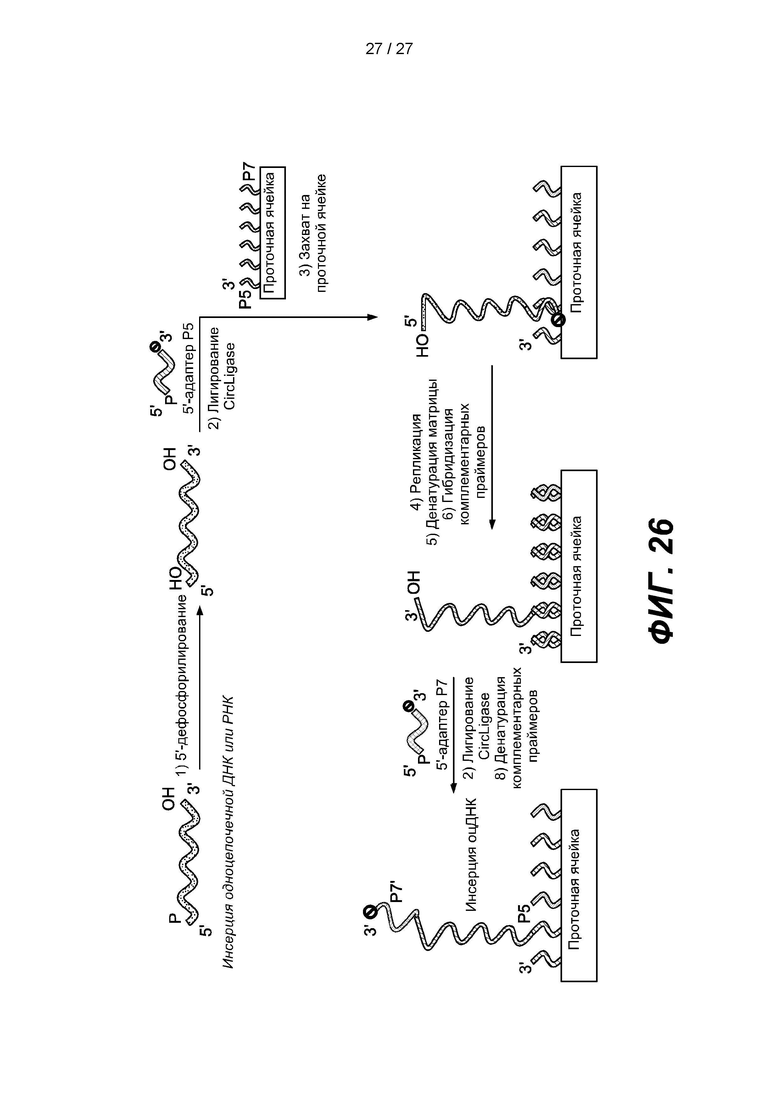
Группа изобретений относится к получению библиотек нуклеиновых кислот. Способ получения библиотеки нуклеиновых кислот включает получение множества одноцепочечных нуклеиновых кислот или множества двуцепочечных нуклеиновых кислот с последующей денатурацией последних с получением множества одноцепочечных нуклеиновых кислот, и осуществление лигирования первого и второго адаптеров. Осуществление группы изобретений позволяет одновременно фрагментировать геном и лигировать адаптеры к фрагментам, тем самым упрощая получение библиотеки нуклеиновых кислот. 4 н. и 46 з.п. ф-лы, 27 ил., 4 пр.
1. Способ получения библиотеки нуклеиновых кислот, включающий:
(a) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой одноцепочечные нуклеиновые кислоты;
(b) дефосфорилирование 5’-концов одноцепочечных нуклеиновых кислот;
(c) лигирование первого адаптера с 3′-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, причем 3′-конец первого адаптера содержит блокирующую группу;
(d) гибридизацию нелигированных одноцепочечных первых адаптеров с зондом захвата;
(e) фосфорилирование 5’-концов лигированных одноцепочечных нуклеиновых кислот; и
(f) лигирование второго адаптера с 5′-концами фосфорилированных лигированных одноцепочечных нуклеиновых кислот в присутствии лигазы, за счет чего получают библиотеку нуклеиновых кислот.
2. Способ по п. 1, характеризующийся тем, что зонд захвата находится в растворе.
3. Способ по п. 1 или 2, характеризующийся тем, что этапы (b)–(f) выполняют в едином реакционном объеме.
4. Способ по любому из пп. 1-3, характеризующийся тем, что 5'-конец второго адаптера нефосфорилирован.
5. Способ по любому из пп. 1-4, характеризующийся тем, что второй адаптер присоединен к подложке.
6. Способ по п. 5, характеризующийся тем, что подложка включает гранулу или проточную ячейку.
7. Способ по любому из пп. 1-6, характеризующийся тем, что зонд захвата содержит последовательность, комплементарную по меньшей мере части первого адаптера.
8. Способ по любому из пп. 1-7, характеризующийся тем, что 3'-конец зонда захвата содержит блокирующую группу.
9. Способ по любому из пп. 1-8, характеризующийся тем, что 5′-конец зонда захвата содержит блокирующую группу.
10. Способ по любому из пп. 1-9, характеризующийся тем, что фосфорилирование 5'-конца лигированных одноцепочечных нуклеиновых кислот включает приведение лигированных одноцепочечных нуклеиновых кислот в контакт с киназой.
11. Способ получения библиотеки нуклеиновых кислот, включающий:
(a) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой двуцепочечные нуклеиновые кислоты;
(b) приведение двуцепочечных нуклеиновых кислот в контакт с 5'-экзонуклеазой с получением множества модифицированных двуцепочечных нуклеиновых кислот с одноцепочечными 3'-выступающими концами;
(c) лигирование первого адаптера с 3′-концами модифицированных двуцепочечных нуклеиновых кислот в присутствии лигазы, причем 3′-конец первого адаптера содержит блокирующую группу;
(d) денатурацию модифицированных двуцепочечных нуклеиновых кислот, лигированных с первыми адаптерами, с получением множества одноцепочечных нуклеиновых кислот; и
(e) лигирование второго адаптера с 5′-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, за счет чего получают библиотеку нуклеиновых кислот.
12. Способ по п. 11, характеризующийся тем, что 5'-конец второго адаптера нефосфорилирован.
13. Способ получения библиотеки нуклеиновых кислот, включающий:
(a) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой одноцепочечные нуклеиновые кислоты;
(b) дефосфорилирование 5’-концов одноцепочечных нуклеиновых кислот;
(c) лигирование первого адаптера с 3′-концами одноцепочечных нуклеиновых кислот в присутствии лигазы, причем 3′-конец первого адаптера содержит блокирующую группу;
(d) гибридизацию лигированного первого адаптера с зондом захвата;
(e) удлинение зонда захвата и лигирование второго адаптера с 3′-концом удлиненного зонда захвата в присутствии лигазы, причем 3′-конец второго адаптера содержит блокирующую группу, за счет чего получают библиотеку нуклеиновых кислот.
14. Способ по п. 13, включающий удаление гибридизованного лигированного первого адаптера с удлиненного зонда захвата.
15. Способ по п. 13 или 14, характеризующийся тем, что зонд захвата присоединен к подложке.
16. Способ по п. 15, характеризующийся тем, что подложка включает гранулу или проточную ячейку.
17. Способ по любому из пп. 13-16, характеризующийся тем, что зонд захвата содержит индекс зонда захвата.
18. Способ по любому из пп. 13-17, характеризующийся тем, что зонд захвата содержит отщепляемый линкер.
19. Способ по п. 18, включающий отщепление отщепляемого линкера.
20. Способ получения библиотеки нуклеиновых кислот, включающий:
(a) получение множества нуклеиновых кислот, причем указанное множество нуклеиновых кислот представляет собой одноцепочечные нуклеиновые кислоты;
(b) дефосфорилирование 5’-концов одноцепочечных нуклеиновых кислот;
(c) лигирование линкера с 3′-концами одноцепочечных нуклеиновых кислот, причем линкер содержит первый адаптер и второй адаптер, а 3′-конец линкера содержит блокирующую группу;
(d) фосфорилирование 5’-концов лигированных одноцепочечных нуклеиновых кислот;
(e) снятие защиты с 3′-концов фосфорилированных нуклеиновых кислот и замыкание нуклеиновых кислот со снятой защитой в кольцо путем лигирования, за счет чего получают библиотеку кольцевых нуклеиновых кислот.
21. Способ по п. 20, характеризующийся тем, что линкер содержит расщепляемый сайт между первым адаптером и вторым адаптером.
22. Способ по п. 21, дополнительно включающий перевод кольцевых нуклеиновых кислот в линейную форму путем расщепления по расщепляемому линкеру.
23. Способ по п. 21 или 22, характеризующийся тем, что расщепляемый сайт содержит остаток урацила.
24. Способ по п. 23, включающий приведение кольцевых нуклеиновых кислот в контакт с урацил-специфическим эксцизионным реагентом.
25. Способ по п. 24, характеризующийся тем, что урацил-специфический эксцизионный реагент содержит фермент, выбранный из урацил-ДНК-гликозилазы (UDG) и ДНК-гликозилазы-лиазы-эндонуклеазы VIII.
26. Способ по любому из пп. 20-25, дополнительно включающий амплификацию кольцевых нуклеиновых кислот способом, включающим гибридизацию по меньшей мере праймера с первым адаптером или вторым адаптером.
27. Способ по п. 26, характеризующийся тем, что амплификацию выбирают из ПЦР и амплификации по принципу катящегося кольца (RCA).
28. Способ по п. 26, характеризующийся тем, что амплификация включает приведение кольцевых нуклеиновых кислот в контакт с полимеразой, которая образует линейный продукт при контакте с остатком урацила в матрице.
29. Способ по любому из пп. 20-28, характеризующийся тем, что дефосфорилирование 5'-конца одноцепочечных нуклеиновых кислот включает приведение одноцепочечных нуклеиновых кислот в контакт с фосфатазой.
30. Способ по любому из пп. 11-29, характеризующийся тем, что этапы (b)–(e) выполняют в едином реакционном объеме.
31. Способ по любому из пп. 11-30, характеризующийся тем, что этапы (b)–(e) выполняют в едином реакционном сосуде.
32. Способ по любому из пп. 1-31, характеризующийся тем, что первый адаптер и/или второй адаптер содержат сайт связывания праймера для секвенирования.
33. Способ по п. 32, характеризующийся тем, что первый адаптер содержит последовательность P7, последовательность P5 или их комплемент или обратный комплемент.
34. Способ по п. 32, характеризующийся тем, что второй адаптер содержит последовательность P7, последовательность P5 или их комплемент или обратный комплемент.
35. Способ по любому из пп. 1-34, характеризующийся тем, что первый адаптер и/или второй адаптер содержат индекс адаптера.
36. Способ по п. 35, характеризующийся тем, что индекс первого адаптера отличается от индекса второго адаптера.
37. Способ по п. 35 или 36, характеризующийся тем, что индекс адаптера указывает на источник множества нуклеиновых кислот.
38. Способ по любому из пп. 1-37, характеризующийся тем, что блокирующая группа содержит 3'-спейсер C3 или дидезоксинуклеотид.
39. Способ по любому из пп. 1-38, характеризующийся тем, что лигаза содержит лигазу одноцепочечных нуклеиновых кислот.
40. Способ по любому из пп. 1-39, характеризующийся тем, что лигирование первого адаптера и/или лигирование второго адаптера выполняют в присутствии агента, вызывающего эффект исключения объема.
41. Способ по п. 40, характеризующийся тем, что агент, вызывающий эффект исключения объема, выбран из группы, состоящей из полиэтиленгликоля (ПЭГ), декстрана, гидроксиэтилкрахмала, фиколла и поливинилпирролидона.
42. Способ по п. 40, характеризующийся тем, что первый этап лигирования и/или второй этап лигирования выполняют в реакционном объеме, содержащем по меньшей мере приблизительно 37 % (вес./об.) ПЭГ.
43. Способ по п. 40, характеризующийся тем, что первый этап лигирования и/или второй этап лигирования выполняют в реакционном объеме, содержащем по меньшей мере приблизительно 60 % (вес./об.) ПЭГ.
44. Способ по любому из пп. 1-43, характеризующийся тем, что множество нуклеиновых кислот содержит РНК.
45. Способ по любому из пп. 1-43, характеризующийся тем, что множество нуклеиновых кислот содержит кДНК или геномную ДНК.
46. Способ по любому из пп. 1-45, характеризующийся тем, что средний размер нуклеиновой кислоты из множества нуклеиновых кислот составляет менее приблизительно 200 нуклеотидов.
47. Способ по любому из пп. 1-46, характеризующийся тем, что множество нуклеиновых кислот получают из низкокачественного источника нуклеиновых кислот.
48. Способ по п. 47, характеризующийся тем, что множество нуклеиновых кислот получают из фиксированного образца.
49. Способ по любому из пп. 1-48, дополнительно включающий амплификацию библиотеки нуклеиновых кислот.
50. Способ по любому из пп. 1-49, дополнительно включающий получение данных о последовательности из библиотеки нуклеиновых кислот.
| ANA P | |||
| VIVANCOS ET AL | |||
| "Strand-specific deep sequencing of the transcriptome", Genome Research, vol | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| J.J | |||
| SCHWARTZ ET AL | |||
| "Capturing native long-range contiguity by in situ library construction and optical sequencing" PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES, vol | |||
| Шкив для канатной передачи | 1920 |
|
SU109A1 |

