ВКЛЮЧЕНИЕ ЛЮБЫХ ПРИОРИТЕТНЫХ ЗАЯВОК ПОСРЕДСТВОМ ССЫЛКИ
[0001] Настоящая заявка испрашивает приоритет по предварительной заявке на патент США № 62/791,509, поданной 11 января 2019 г., и предварительной заявке на патент США 62/840,610, поданной 30 апреля 2019 г., которые включены в настоящий документ посредством ссылки во всей своей полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0002] Настоящее изобретение относится к способам, композициям и наборам для создания библиотеки меченых фрагментов нуклеиновых кислот без использования ПЦР-амплификации, включая способы и композиции для фрагментирования и нанесения меток на нуклеиновые кислоты (например, ДНК) с использованием комплексов транспосом, иммобилизованных на твердых подложках.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
[0003] В текущих протоколах секвенирования следующего поколения (NGS) образцов нуклеиновых кислот, как правило, применяют способ получения образца, который преобразует ДНК или РНК в библиотеку фрагментированных матриц, которые можно секвенировать. Для способов получения образцов часто требуется применение множества стадий и переноса материала, а также дорогостоящих приборов для осуществления фрагментации, что может сделать данные способы сложными, трудоемкими, дорогостоящими и неэффективными. Более того, амплификация с использованием праймеров вносит систематическую погрешность в содержимое библиотеки и, таким образом, в полученные данные секвенирования. Например, стадии ПЦР-амплификации могут создавать бреши, которые усугубляются в обогащенных GC областях вследствие неспособности полимераз эффективно копировать обогащенные GC области. Данные бреши создают систематическую погрешность в полученных данных секвенирования из библиотек. Способы получения библиотек со стадиями ПЦР-амплификации могут иметь значительно сниженную эффективность вставки и делеции (индел). Точное секвенирование некоторых областей с множеством расширенных повторов может быть сложным, а ДНК, имеющая обогащенные GC промоторы, может демонстрировать низкое покрытие в пределах генома. Более того, многие способы получения библиотек несовместимы с включением индексов образцов или требуют множества дополнительных стадий для введения таких индексов.
[0004] Для получения библиотек необходимы короткие, эффективные и точные процедуры. В настоящем документе описаны различные способы, композиции и наборы, которые решают данные проблемы и которые поддерживают одинарный и двойной подходы к индексированию.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0005] Настоящее раскрытие изобретения относится к способам, композициям и наборам для создания библиотеки меченых фрагментов нуклеиновых кислот без использования ПЦР-амплификации для включения меток фрагментов, таких как праймерные последовательности и/или индексные последовательности, включая способы и композиции для фрагментации и нанесения меток на нуклеиновые кислоты (например, ДНК) с использованием комплексов транспосом, иммобилизованных на твердых подложках.
[0006] Некоторые варианты осуществления, предложенные в настоящем документе, относятся к комплексу транспосомы, который включает полинуклеотид присоединения. В некоторых аспектах настоящее изобретение относится к комплексу транспосомы, содержащему следующее: (a) транспозаза; (b) первый транспозон, включающий 3’-концевую последовательность транспозона и 5’-адаптерную последовательность; (c) второй транспозон, включающий 5’-концевую последовательность транспозона и 3’-адаптерную последовательность, причем 5’-концевая последовательность комплементарна по меньшей мере участку 3’-концевой последовательности транспозона; и (d) полинуклеотид присоединения, включающий следующее: (i) соединительная адаптерная последовательность, гибридизованная с одной из двух адаптерных последовательностей, и (ii) связывающий элемент. Как правило, концевые транспозонные последовательности ренатурируют вместе с образованием двухцепочечной концевой транспозонной последовательности, которая распознается транспозазой. В некоторых аспектах связывающий элемент иммобилизован на твердой подложке с образованием иммобилизованного комплекса транспосомы.
[0007] В некоторых аспектах настоящее изобретение относится к ренатурированному полинуклеотиду транспозона/присоединения, содержащему первый транспозон, второй транспозон и полинуклеотид присоединения.
[0008] В некоторых аспектах настоящее изобретение относится к способам получения ренатурированного полинуклеотида транспозона/присоединения, содержащего комплекс транспосом, включающий отжиг первого транспозона, второго транспозона и полинуклеотида присоединения. В некоторых аспектах настоящее изобретение относится к способам получения комплекса транспосомы, включающим обработку ренатурированного гибрида полинуклеотида транспозона/присоединения с использованием транспозазы. В некоторых аспектах представлен способ получения иммобилизованного комплекса транспосомы посредством иммобилизации комплекса транспосомы на твердой подложке с использованием связывающего элемента.
[0009] В некоторых аспектах настоящее изобретение относится к комплексу транспосомы, содержащему следующее: (a) транспозаза; (b) первый транспозон, содержащий концевую последовательность 3’-транспозона и 5’-адаптерную последовательность; (c) второй транспозон, содержащий концевую последовательность 5’-транспозона, комплементарную по меньшей мере части 3’-концевой последовательности-транспозона, и 3’-адаптерную последовательность; и (d) связывающий элемент, присоединенный к 5’-адаптерной последовательности посредством расщепляемого линкера. В некоторых аспектах связывающий элемент иммобилизован на твердой подложке с образованием иммобилизованного комплекса транспосомы. В некоторых аспектах настоящее описание относится к ренатурированной олигонуклеотидной конструкции, содержащей первый транспозон, второй транспозон и связывающий элемент.
[0010] В некоторых аспектах настоящее изобретение относится к способам создания библиотеки меченых фрагментов нуклеиновых кислот, включающим приведение целевой нуклеиновой кислоты в контакт с множеством иммобилизованных комплексов транспосом, как описано в настоящем документе, в условиях, достаточных для фрагментации целевой нуклеиновой кислоты во множество целевых фрагментов и для присоединения 3’-концов 3’-концевых последовательностей транспозона к 5’-концам целевых фрагментов для получения множества 5’-меченных целевых фрагментов.
[0011] Некоторые варианты осуществления, предложенные в настоящем документе, относятся к набору для создания библиотеки меченых фрагментов нуклеиновых кислот без применения ПЦР-амплификации. В некоторых вариантах осуществления набор включает в себя иммобилизованный комплекс транспосом, как описано в настоящем документе.
[0012] В некоторых аспектах изобретение относится к способам создания библиотеки меченых фрагментов нуклеиновых кислот, включающим приведение иммобилизованного комплекса транспосомы в контакт с целевой нуклеиновой кислотой в условиях, достаточных для фрагментации целевой нуклеиновой кислоты на множество целевых фрагментов, и присоединение 3’-конца первого транспозона к 5’-концам целевых фрагментов для получения множества 5’-меченных целевых фрагментов; обработку твердой подложки для удаления несвязанных нуклеиновых кислот; или обработку твердой подложки для удаления транспозазы из комплекса, необязательно посредством (a) нагревания твердой подложки и/или (b) мойки твердой подложки ферментативным денатурирующим средством, причем ферментативное денатурирующее средство необязательно содержит додецилсульфат натрия (SDS), гидрохлорид гуанидина, мочевину или протеиназу; обработку множества 5’-меченых целевых фрагментов полимеразой и лигазой для удлинения и лигирования 5’-меченых целевых фрагментов для получения полностью двухцепочечных меченых фрагментов, при этом необязательно обработку полимеразой и лигазой выполняют в присутствии средства разрушения вторичной структуры ДНК, причем средство разрушения необязательно представляет собой ДМСО; удаление полностью двухцепочечных фрагментов с твердой подложки, причем удаление необязательно включает применение нагревания и/или денатурирующего вещества, достаточного для отщепления полностью двухцепочечных меченых фрагментов от твердой подложки, при этом необязательно денатурирующее вещество представляет собой NaOH; и отбор полностью двухцепочечных меченых фрагментов с использованием гранул для захвата, при этом необязательно гранулы для захвата представляют собой магнитные микроносители, и при этом необязательно выполнять две отдельные стадии отбора.
[0013] В некоторых вариантах осуществления иммобилизованный комплекс транспосом содержит твердую подложку и комплекс транспосом, иммобилизованный на твердой подложке, причем комплекс транспосом содержит транспозазу; первый транспозон, содержащий 3’-концевую последовательность транспозона и якорную последовательность (якорь); второй транспозон, содержащий 5’-концевую последовательность транспозона и последовательность B15’; полинуклеотид присоединения, содержащий комплементарную якорную последовательность (якорь’), последовательность A14’, спейсер и последовательность P5’, а также связывающий элемент, содержащий биотин, причем биотин иммобилизован на твердой подложке. В некоторых вариантах осуществления способ дополнительно включает секвенирование одного или более полностью двухцепочечных меченых фрагментов.
[0014] В некоторых вариантах осуществления способ включает приведение в контакт первого иммобилизованного комплекса транспосомы и второго иммобилизованного комплекса транспосомы с целевой нуклеиновой кислотой в условиях, достаточных для фрагментации целевой нуклеиновой кислоты во множество целевых фрагментов, и присоединение 3’-конца каждого первого транспозона к 5’-концам целевых фрагментов для получения множества первых 5’-меченных целевых фрагментов, полученных из первого иммобилизованного комплекса транспосомы, и множества вторых 5’-меченных целевых фрагментов, полученных из второго иммобилизованного комплекса транспосомы. В некоторых вариантах осуществления первый иммобилизованный комплекс транспосом содержит твердую подложку и первый комплекс транспосом, иммобилизованный на твердой подложке, причем комплекс транспосом содержит транспозазу; первый транспозон, содержащий 3’-концевую последовательность транспозона и якорную последовательность; второй транспозон, содержащий 5’-концевую последовательность транспозона и первый полинуклеотид присоединения, содержащий (i) комплементарную якорную последовательность, последовательность A14’, спейсер и последовательность P5’; и (ii) связывающий элемент, содержащий биотин, причем биотин иммобилизован на твердой подложке. В некоторых вариантах осуществления второй иммобилизованный комплекс транспосом содержит твердую подложку и второй комплекс транспосом, иммобилизованный на твердой подложке, причем комплекс транспосом содержит транспозазу; первый транспозон, содержащий 3’-концевую последовательность транспозона и якорную последовательность; второй транспозон, содержащий 5’-концевую последовательность транспозона и второй полинуклеотид присоединения, содержащий (i) комплементарную якорную последовательность, последовательность B15’, спейсер и последовательность P7’; и (ii) связывающий элемент, содержащий биотин, причем биотин иммобилизован на твердой подложке. В некоторых вариантах осуществления способ включает обработку множества 5’-меченных целевых фрагментов лигазой для лигирования каждого 5’-меченого целевого фрагмента либо с первым индексирующим олигонуклеотидом, либо со вторым индексирующим олигонуклеотидом путем приведения 5’-меченных целевых фрагментов в контакт с пулом первого и второго индексирующих олигонуклеотидов, причем каждый первый индексирующий олигонуклеотид содержит последовательность A14, последовательность i5 и последовательность P5 и может связываться с первым меченым 5’-концом; и при этом каждый второй индексирующий олигонуклеотид содержит последовательность B15, последовательность i7 и последовательность P7 и может связываться со вторым меченым 5’-концом для получения множества 5’-меченых целевых фрагментов, лигированных с индексирующими олигонуклеотидами; обработки твердой подложки для удаления транспозаз из комплекса, необязательно посредством (a) нагревания твердой подложки и/или (b) промывки твердой подложки ферментативным денатурирующим средством, причем средство, денатурирующее фермент, необязательно содержит додецилсульфат натрия (SDS), гидрохлорид гуанидина, мочевину или протеиназу; и обработки множества меченных на 5’-конце целевых фрагментов, лигированных с индексированием олигонуклеотидов полимеразой для удлинения с получением полностью двухцепочечных меченых фрагментов. В некоторых вариантах осуществления приведение в контакт первого иммобилизованного комплекса транспосомы со вторым иммобилизованным комплексом транспосомы и обработку множества 5’-меченых целевых фрагментов лигазой проводят в одну реакцию. В некоторых вариантах осуществления двухцепочечные меченые фрагменты получают в растворе.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0015] На Фиг. 1 представлена блок-схема, на которой изображен вариант осуществления способа создания библиотеки меченых фрагментов нуклеиновых кислот без использования ПЦР-амплификации. На первой стадии получают комплекс транспосом, как описано в настоящем документе, иммобилизованный на твердой подложке. Целевую нуклеиновую кислоту наносят на твердую подложку, где протекает реакция тагментации с образованием меченых и фрагментированных нуклеиновых кислот. Применяют смесь индексов, имеющую индексные последовательности, и происходит удлинение и лигирование. Наконец, происходит индексирование меченых фрагментов нуклеиновых кислот. В некоторых вариантах осуществления индексирование происходит одновременно с удлинением и лигированием, а в других вариантах осуществления индексирование происходит после удлинения и лигирования. В других вариантах осуществления схемы расположения стадий различаются, как показано в примере 9.
[0016] На Фиг. 2A-2B представлена принципиальная схема и способ получения библиотеки меченых фрагментов нуклеиновых кислот без применения ПЦР-амплификации. На Фиг. 2A представлен пример конфигурации комплекса транспосомы с биотином (B), присоединенным к одному транспозону посредством расщепляемого линкера. В данном примере осуществления комплекса транспосомы, содержащего транспозазу, первый транспозон, содержащий 3’-концевую последовательность транспозона и 5’-адаптерную последовательность; второй транспозон, содержащий 5’-концевую последовательность транспозона, комплементарную по меньшей мере части 3’-концевой последовательности транспозона и 3’-адаптерной последовательности; и связывающий элемент, присоединенный к 5’-адаптерной последовательности посредством расщепляемого линкера (Фиг. 2A). На Фиг. 2B схематично представлены примеры стадий способа получения библиотеки меченых фрагментов нуклеиновых кислот без применения ПЦР-амплификации с использованием примера комплекса транспосомы, изображенного на Фиг. 2A, иммобилизованного на твердой подложке (твердая подложка не показана), включая стадии нанесения метки и промывки, удлинения и лигирования, а также удаления гранул. На Фиг. 2B показаны стадии тагментации, на которых вставки из целевой нуклеиновой кислоты присоединяются к меткам, удлинению и лигированию, а также отщеплению от твердой подложки.
[0017] На Фиг. 3A-3C представлены примеры стадий способа создания библиотеки меченых фрагментов нуклеиновых кислот без использования ПЦР-амплификации. На Фиг. 3A показан комплекс транспосом, иммобилизованный на твердой подложке посредством полинуклеотида присоединения, несущего биотин (B, твердая подложка не показана). Для упрощения изображение димера показано на Фиг. 3A, но не на Фиг. 3B-3C. На Фиг. 3B показана меченая и фрагментированная нуклеиновая кислота, которая все еще образует комплекс с транспозазой (верхняя панель) и структурой после удаления транспозазы (нижняя панель). На Фиг. 3B представлен комплекс транспосом, имеющий фрагмент нуклеиновой кислоты (вставку), связанный с первым транспозоном, причем 5’-конец вставки присоединен к 3’-концевой последовательности транспозона. Транспозазу удаляют с использованием способов, описанных в настоящем документе, например, с использованием средства для удаления транспозазы, такого как додецилсульфат натрия (SDS). На Фиг. 3C изображена структура после заполнения бреши и удлинения (удлинения и лигирования) (верхняя панель) и после дегибридизации для удаления полученных фрагментов с твердой подложки (нижняя панель). Как показано на Фиг. 3C, индексы добавляют к твердой подложке и гибридизуют с полинуклеотидом присоединения второго транспозона. В частности, как показано в варианте осуществления на Фиг. 3C, индекс i5, имеющий праймерную последовательность (последовательность P5), индексную последовательность (последовательность i5) и якорную последовательность, гибридизуется с полинуклеотидом присоединения в комплементарных последовательностях, так что Р5 гибридизуется с P5’ и якорный участок гибридизуется с якорным участком’. Аналогичным образом в варианте осуществления на Фиг. 3C индекс i7, имеющий праймерную последовательность (P7), индексную последовательность (i7) и адаптерную последовательность (последовательность B15), гибридизуется со вторым транспозоном в комплементарных последовательностях, так что P7 гибридизуется с P7’, i7 гибридизуется с i7’, а B15 гибридизуется с B15’. После приведения твердой подложки в контакт с индексами фрагменты удлиняют и лигируют с использованием смеси для удлинения и лигирования (ELM). Затем твердую подложку обрабатывают средством для денатурации последовательностей цепей, например NaOH, с получением таким образом меченых фрагментов нуклеиновых кислот. Показан вариант осуществления комплекса транспосом, содержащего транспозазу Tn5 с первым и вторым транспозонами. Первый транспозон включает в себя 3’-концевую последовательность транспозона (последовательность МЕ), гибридизованную с 5’-концевой последовательностью транспозона (последовательностью МЕ) второго транспозона. Второй транспозон также включает в себя 3’-адаптерную последовательность (последовательность B15’). Первый транспозон включает в себя 5’-адаптерную последовательность (последовательность A14), которая показана гибридизованной с соединительной адаптерной последовательностью (последовательность A14’) полинуклеотида присоединения. Полинуклеотид присоединения также включает в себя якорную последовательность (якорь’), спейсерную область, праймерную последовательность (последовательность P5’) и линкер, присоединенный к связывающему элементу (B). Комплекс транспосом представляет собой димер с двумя димеризованными мономерами.
[0018] На Фиг. 4A-4E представлена принципиальная схема для сравнения различных конфигураций комплекса транспосомы, иммобилизованного на твердой подложке посредством иллюстративного биотина (B, твердая подложка не показана), включая индексированные гранулы и универсальные гранулы, каждая из которых показана как в режиме единичного индексирования, так и в режиме двойного индексирования, для вариантов осуществления способов создания библиотеки меченых фрагментов нуклеиновых кислот без применения ПЦР-амплификации с использованием вариаций комплексов транспосом, причем компоненты транспозонов и/или полинуклеотидов присоединения изменены, упорядочены или изменены по сравнению друг с другом. На Фиг. 4A представлены различные конфигурации комплекса транспосомы, присоединенного к твердой подложке. На Фиг. 4 A представлены варианты осуществления комплексов транспосом, демонстрирующие транспозазу с первым и вторым транспозоном и присоединенную к твердой подложке посредством полинуклеотида присоединения. На Фиг. 4B показаны различные конфигурации после тагментации. На Фиг. 4B представлена реакция тагментации для индексированных или универсальных гранул с единичным или двойным индексированием, в которой твердую подложку приводят в контакт с фрагментами нуклеиновой кислоты (вставка), которые связываются с 3’-концом первого транспозона. На Фиг. 4C изображены различные конфигурации после удлинения и лигирования. На Фиг. 4C представлено удлинение и лигирование индексированных или универсальных гранул с единичным или двойным индексированием, при котором твердую подложку приводят в контакт со смесью индексов, которая гибридизуется со транспозоном или полинуклеотидом присоединения, и при этом фрагменты нуклеиновых кислот удлинены. На Фиг. 4D показаны различные конфигурации после индексирования (индексированные гранулы для единичного индексирования не показаны, так как индексирование было завершено для данной конфигурации на Фиг. 4C). На Фиг. 4D представлено индексирование гранул с двойным индексированием или универсальных гранул с единичным или двойным индексированием. Как показано на Фиг. 4D, индексирование гранулы с единичным индексированием было завершено на стадии обширного лигирования, показанной на Фиг. 4C. Твердую подложку приводят в контакт со смесью для индексирования, а на фрагменты нуклеиновых кислот наносят метки для индексирования. На Фиг. 4E представлены иллюстративные результаты различных конфигураций, на которых показаны нормализованные частоты считывания в зависимости от размера вставки. На Фиг. 4E представлена нормализованная частота считывания меченых фрагментов нуклеиновых кислот из индексированных или универсальных гранул с единичным или двойным индексированием. На Фиг. 4A-4E, в частности, показаны варианты осуществления комплексов транспосом, расположенных в виде индексированных гранул для единичного индексирования (слева сверху), универсальных гранул для единичного индексирования (справа сверху), индексированных гранул для двойного индексирования (слева снизу) и универсальных гранул для двойного индексирования. Для вариантов осуществления индексированных гранул, показанных на Фиг. 4A-4E, как для одинарного, так и для двойного индексирования (слева), полинуклеотид присоединения гибридизуется со вторым транспозоном в 3’-адаптерной последовательности (последовательность B15’). Для вариантов осуществления универсальных гранул, показанных на Фиг. 4A-4E, как для одинарного, так и для двойного индексирования (справа), полинуклеотид присоединения гибридизуется с первым транспозоном в 5’-адаптерной последовательности (последовательность A14’). В некоторых вариантах осуществления нормализованные библиотеки могут быть получены из исходных образцов, так что нуклеиновую кислоту экстрагируют из исходного образца и непосредственно вводят в систему или способ, описанные в настоящем документе, причем самонормализованный образец обеспечивает прочную CV в диапазоне типов образцов.
[0019] На Фиг. 5A и 5B показана принципиальная схема иллюстративных комплексов транспосом для воздействия, отличного от индексирования, показывающая полинуклеотид присоединения, гибридизованный либо с транспозоном, содержащим последовательность Р7 (Фиг. 5A), либо с транспозоном, содержащим последовательность Р5 (Фиг. 5B). В частности, на Фиг. 5A и 5B показана праймерная последовательность (последовательность P5), присоединенная к 5’-адаптерной последовательности (последовательности A14) первого транспозона, и праймерная последовательность (последовательность P7’), присоединенная к 3’-адаптерной последовательности (последовательности B15’) второго транспозона, с получением таким образом неиндексирующих комплексов транспосом. Полинуклеотид присоединения можно гибридизовать либо на втором транспозоне с использованием 5’-связывающего элемента (B) (Фиг. 5A), либо на первом транспозоне с использованием 3’-связывающего элемента (B) (Фиг. 5B).
[0020] На Фиг. 6A-6D показаны принципиальные схемы примеров комплексов транспосом, включающих в себя последовательность индексирования i5. На Фиг. 6A показан комплекс, в котором полинуклеотид присоединения, содержащий нитропоследовательность и якорную последовательность, может гибридизоваться с индексирующим олигонуклеотидом, который затем может быть лигирован с 5’-концом первого транспозона. На Фиг. 6A комплекс транспосом содержит первый транспозон, содержащий 3’-концевую последовательность транспозона (ME) и 5’-адаптерную последовательность (A14), второй транспозон, содержащий 5’-концевую последовательность транспозона (ME’) и 3’-адаптерую последовательность (B15’), и полинуклеотид присоединения, содержащий соединительную адаптерную последовательность (A14’), гибридизованную с A14 в первом транспозоне, и связывающий элемент (биотин). В данном случае полинуклеотид присоединения дополнительно содержит нитроиндольную последовательность (универсальную последовательность, которая связывается с любой индексной областью i5) и праймерную последовательность (P5’). Область B15’ второго транспозона может быть гибридизована с полинуклеотидом, содержащим комплемент (B15), индексную область и праймерную последовательность P7, которая сама по себе ренатурирована с индексной молекулой P7’-i7’. Лигирование 5’-конца индексной области i7’ к 3’-концу 3’-адаптерной последовательности служит для создания полностью двухцепочечной области.
[0021] На Фиг. 6B показан полинуклеотид присоединения, содержащий якорную последовательность и спейсерную область, который может гибридизоваться с индексирующим олигонуклеотидом, который затем может быть лигирован с 5’-концом первого транспозона. На Фиг. 6B первый транспозон содержит 3’-концевую последовательность транспозона (ME) и 5’-адаптерную последовательность (A14), а второй транспозон содержит 5’-концевую последовательность транспозона (МЕ’) и 3’-адаптерную последовательность (B15’), как показано на Фиг. 6A. Однако в этом случае полинуклеотид присоединения содержит комплементарный адаптер (A14’), фиксированную последовательность, спейсерную область 2 x sp 18, комплемент праймера (P5’) и связывающийся с биотином элемент. Индекс i5 содержит якорную последовательность (комплементарную якорю’), индексную область i5 и праймерную последовательность (P5). Индекс i5 гибридизуется с комплементарными якорными и праймерными последовательностями на полинуклеотиде присоединения по всему спейсеру.
[0022] На Фиг. 6С показан полинуклеотид присоединения, содержащий спейсер и последовательность A14’, который может гибридизоваться с индексирующим олигонуклеотидом, который затем может быть лигирован к последовательности Х у 5’-конца первого транспозона. На Фиг. 6C первый транспозон содержит 3’-концевую последовательность транспозона (ME) и 5’-адаптерную последовательность X, а второй транспозон содержит 5’-концевую последовательность транспозона (МЕ’) и 3’-адаптерную последовательность (B15’). Полинуклеотид присоединения содержит комплемент 5’-адаптерной последовательности (X’), вторую адаптерную последовательность (A14’), спейсерную область (2 x sp18), комплемент праймера (P5’) и связывающий элемент биотин. 5’-адаптерная последовательность (X’) гибридизуется с первой 5’-адаптерной последовательностью транспозона (X). Индекс i5 содержит комплемент второй адаптерной последовательности (A14), спейсерную область 2 x sp18 и праймерную последовательность (P5). Индекс i5 гибридизуется со второй адаптерной последовательностью и комплементом праймера на полинуклеотиде присоединения в спейсерной области, а 3’-конец комплемента второй адаптерной последовательности лигируется с последовательностью X. Кроме того, индекс i7, содержащий 5’-праймерную последовательность (P7), индексную область i7 и комплемент 3’-адаптерной последовательности (B15), отжигают, а 3’-конец 3’-адаптерной последовательности удлиняют с получением двухцепочечной области.
[0023] На Фиг. 6D показан полинуклеотид присоединения, содержащий спейсер и последовательность A14’, который может гибридизоваться с индексирующим олигонуклеотидом, который затем может быть лигирован к последовательности Х у 5’-конца первого транспозона. В данном случае индексирующий олигонуклеотид содержит двухцепочечную праймерную область. На Фиг. 6D первый транспозон, второй транспозон и полинуклеотид присоединения аналогичны представленным на Фиг. 6C. Индекс i7 содержит двухцепочечный праймер (P7/P7’), индексную область i7 и комплемент 3’-адаптерной последовательности (B15). Двухцепочечная область в индексе i7 может быть создана (отожжена) в ходе реакции удлинения и лигирования, например, отсутствует необходимость в отжиге индекса i7 перед самой реакцией. Олигонуклеотид P7 ’может быть включен в реакционную смесь. Индекс i7 отжигают посредством области B15, а удлинение и лигирование от второго транспозона обеспечивают двухцепочечную область. Пример данного способа представлен в примере 5.
[0024] На Фиг. 7A-7B показаны иллюстративные результаты осуществления способа создания библиотеки меченых фрагментов нуклеиновых кислот с использованием ПЦР-амплификации и без нее, включая результаты для улучшенной точности и повторного распознавания инделов (Фиг. 7A) и улучшения покрытия промоторов, обогащенных GC (Фиг. 7B). Для получения этих данных использовали четыре способа: два с помощью ПЦР (TruSeq™ Nano и Nextera™ DNA Flex) и два без ПЦР (настоящий способ и TruSeq™ PCR-Free). Каждый способ предполагал две повторности, причем всего создавали восемь библиотек. После секвенирования данные прореживали 25 раз. На Фиг. 7A показан высокий процент точности индела и повторного распознавания для способа без ПЦР, описанного в настоящем документе. Образцы слева направо включают следующее (в двух повторностях): TruSeq™ Nano (черный); Nextera™ DNA Flex (белый); способ без ПЦР, описанный в настоящем документе (в линию); и TruSeq™ PCR-Free (в клетку). На Фиг. 7B показано улучшение покрытия промоторов, обогащенных GC, для способа без ПЦР, описанного в настоящем документе, по сравнению с другими способами, в том числе: Nextera™ DNA Flex (слева сверху); TruSeq™ Nano (справа сверху); способ без ПЦР, описанный в настоящем документе (слева снизу); и TruSeq™ PCR-Free (справа снизу).
[0025] На Фиг. 8 представлены результаты библиотек секвенирования из образца, содержащего известное 100%-ное повторное размножение GC (FMR1), полученного с использованием ДНК из наборов Nextera™ DNA Flex или TruSeq™ DNA PCR-Free Library Prep, для сравнения со способами, описанными в настоящем документе (в четырех экземплярах образцов 1-4), как описано в примере 3.
[0026] На Фиг. 9A-9C представлены результаты % CV для библиотек, полученных с использованием восьми пар индексов вместе с системами, показанными на Фиг. 6C (данные на Фиг. 9A) и 6D (данные для двух условий реакции на Фиг. 9B и 9C), как описано в примере 5.
[0027] На Фиг. 10 показано графическое представление покрытия секвенированием с пропуском в области гена RNPEPL1 для способов получения библиотек ПЦР (Nextera Flex или Nextera на основе пробирок), но с меньшим количеством пропусков при использовании способов без ПЦР, описанных в настоящем документе (две нижние панели).
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0028] Библиотеки фрагментированных нуклеиновых кислот часто создают из геномных нуклеиновых кислот для применения в областях, связанных с секвенированием следующего поколения (NGS). В настоящем раскрытии изобретения представлены способы, композиции и наборы для создания библиотеки фрагментированных нуклеиновых кислот, которая присоединяет последовательности, необходимые для выполнения операций секвенирования, включая индексы, без использования ПЦР для добавления последовательностей посредством амплификации (также называемой в настоящем документе образованием библиотеки без ПЦР или получением библиотеки без ПЦР). Данный способ получения библиотеки транспосом без ПЦР может уменьшить и/или устранить систематическую ошибку, вызванную ПЦР, в современных подходах тагментации для получения библиотеки.
[0029] «Тагментация» относится к применению транспозазы для фрагментации и нанесения метки на нуклеиновые кислоты. Тагментация включает модификацию ДНК комплексом транспосомы, содержащим фермент транспозазу в комплексе с адаптерами, содержащими концевые последовательности транспозона (называемые в настоящем документе транспозонами). Тагментация обеспечивает одновременную фрагментацию ДНК и лигирование адаптеров к 5’-концам обеих цепей дуплексных фрагментов. Как правило, после стадии очистки для удаления фермента транспозазы дополнительные последовательности добавляют к концам адаптированных фрагментов посредством ПЦР.
[0030] Описанные в настоящем документе способы, композиции, системы и наборы относятся к комплексно-гибридизованным олигонуклеотидам и комплексам, содержащим такие гибриды, включая комплексы, иммобилизованные на поверхности, и к применению комплексов транспосом для образования библиотеки без ПЦР. Как описано в настоящем документе, комплекс транспосом включает в себя транспозазу и связанные с ней транспозоны, которые фрагментируют и наносят метку на целевую молекулу ДНК. В некоторых аспектах комплексно-гибридизованный олигонуклеотид является расщепляемым и включает в себя связывающий элемент, а в других аспектах комплексно-гибридизованный олигонуклеотид содержит полинуклеотид присоединения со связывающим элементом. В некоторых аспектах полинуклеотид присоединения представляет собой нуклеотидную последовательность, которая гибридизуется с транспозоном в комплексе транспосомы и иммобилизована на твердой подложке, такой как слайд, проточная кювета или гранула. Вследствие гибридизации полинуклеотида с транспозоном, комплекс транспосом можно иммобилизовать на твердой подложке опосредованно через полинуклеотид присоединения. Связывание полинуклеотида с твердой подложкой происходит через связывающий элемент на полинуклеотиде присоединения. Целевые нуклеиновые кислоты захватываются комплексами транспосомы, а затем нуклеиновые кислоты фрагментируют и на них наносят метку («тагментация»). Олигонуклеотидная система выполнена с возможностью включения любых меток, необходимых для индексирования и секвенирования, посредством стадий тагментации, удлинения и/или лигирования без ПЦР-амплификации. Таким образом, в некоторых аспектах меченые фрагменты можно удлинить, лигировать и индексировать без амплификации для создания библиотеки фрагментов нуклеиновых кислот без использования ПЦР-амплификации.
[0031] Тагментация на основе раствора имеет недостатки и требует нескольких трудоемких стадий. Кроме того, во время стадий ПЦР-амплификации, применяемых для введения последовательностей тегов, может быть введена систематическая погрешность. Например, снижение индела может быть результатом ПЦР вследствие «проскальзывания» полимеразы. Кроме того, полимеразы ПЦР имеют сложности в некоторых областях, таких как области с высоким уровнем GC или АТ, или области повтора последовательности, что приводит к образованию гэпов, ложных определений структуры в геноме или к потерям экспансии повторов.
[0032] Способы, композиции, системы и наборы, представленные в настоящем документе, позволяют преодолеть данные недостатки и позволяют производить неискаженную подготовку и секвенирование образцов с минимальными требованиями к манипуляциям или переносу образцов. Способы, композиции, системы и наборы, описанные в настоящем документе, относятся к созданию библиотек без применения ПЦР-амплификации. Подход без ПЦР снижает и/или устраняет систематические погрешности, вызванные ПЦР, включая снижение числа и частоты гэпов, в частности, в труднодоступных для ПЦР обогащенных GC областях; повышение эффективности распознавания инделов, в том числе повышение эффективности повторного распознавания инделов и точности инделов; улучшение распознавания размножения повторов; а также улучшение покрытия в обогащенных GC промоторах. В настоящей заявке описаны различные сложные транспосомные конфигурации для выполнения тагментации без ПЦР для улучшения создания библиотек нуклеиновых кислот.
[0033] Кроме того, способы, композиции, системы и наборы, описанные в настоящем документе, могут осуществляться в течение периода меньшего, чем получение и анализ образца нуклеиновой кислоты, с использованием других способов, таких как способы, основанные на ПЦР. Таким образом, в некоторых вариантах осуществления способы образования библиотеки меченых фрагментов нуклеиновых кислот, описанные в данном документе, можно выполнять в течение периода менее приблизительно 5 часов, например, менее 5, менее 4, менее 3 или менее 2 часов. В некоторых вариантах осуществления способы создания библиотеки меченых фрагментов нуклеиновых кислот, описанные в настоящем документе, можно выполнять в течение периода в диапазоне от около 90 минут до около 300 минут, например 90, 105, 120, 135, 150, 165, 180, 195, 210, 225, 240, 255, 270, 285 или 300 минут, или в течение периода в пределах диапазона, определенного любыми двумя из вышеупомянутых значений.
[0034] Более того, в некоторых вариантах осуществления применение способов, композиций, систем и наборов, описанных в настоящем документе, приводит к фрагментации нуклеиновых кислот, которая не зависит от времени, иммобилизация транспосомы приводит к постоянному размеру вставки, а насыщение позволяет интегрировать извлечение и получить библиотеку без количественного определения.
[0035] Дополнительные преимущества способов, композиций, систем и наборов, описанных в настоящем документе, относятся к иммобилизации комплекса транспосомы на твердой поверхности и включают следующее: например, сокращая непосредственное и общее время подготовки библиотеки, стоимость и требования к реагентам, снижая входные требования к образцу и позволяя использовать неочищенные или разложенные образцы в качестве отправной точки для получения библиотеки. Кроме того, описанные в настоящем документе комплексы транспосом также позволяют получать библиотеки с более стабильными размерами вставок по сравнению со способами, в которых применяется фаза растворения, даже при использовании различных входных концентраций образцов.
[0036] В некоторых вариантах осуществления библиотеки нуклеиновых кислот, полученные посредством способов, описанных в настоящем документе, можно секвенировать с использованием любой подходящей платформы секвенирования нуклеиновых кислот для определения последовательности нуклеиновой кислоты целевой последовательности. В некоторых отношениях интересующие последовательности коррелируют или ассоциируют с одним или более врожденными или наследственными расстройствами, патогенностью, резистентностью к антибиотикам или генетическими модификациями. Для определения нуклеотидной последовательности короткого тандемного повтора, однонуклеотидного полиморфизма, гена, экзона, кодирующей области, экзома или их части можно применять секвенирование. Таким образом, способы и композиции, описанные в настоящем документе, относятся к созданию секвенируемых библиотек, применяемых без ограничения в диагностике рака и иных заболеваний, прогнозе и терапевтических средствах, приложениях для ДНК-генотипоскопии (например, для создания баз данных ДНК, в работе по изучению материалов уголовных дел), метагеномных исследованиях и открытиях, сельском хозяйстве, а также для идентификации и мониторинга патогенов.
[0037] В некоторых вариантах осуществления соединительная адаптерная последовательность гибридизована с по меньшей мере частью 5’-адаптерной последовательности, и связывающий элемент находится на 3’-конце полинуклеотида присоединения. В некоторых вариантах осуществления адаптерная последовательность присоединения гибридизуется по меньшей мере с частью 3’-адаптерной последовательности, а связывающий элемент находится на 5’-конце олигонуклеотида присоединения.
[0038] Если не указано иное, все технические и научные термины, применяемые в данном документе, имеют то же значение, которое обычно применяет специалист в данной области. Все патенты, заявки, опубликованные заявки и другие публикации, на которые даны ссылки в настоящем документе, полностью включены в настоящий документ посредством ссылки, если не указано иное. Если не указано иное, при наличии множества определений для термина, представленного в настоящем документе, преимущественными являются определения, приведенные в данном разделе. Применяемые в данном описании и приложенной формуле изобретения формы единственного числа включают упоминания форм множественного числа, если в контексте явно не указано иное. Если не указано иное, применяют традиционные способы масс-спектроскопии, ЯМР, ВЭЖХ, химии белков, биохимии, способы рекомбинантной ДНК и фармакологии. Применение союза «или» или «и» означает «и/или», если не указано иное. Более того, применение термина «включая», а также других форм, таких как «включают», «включает» и «включенный», не имеет ограничительного характера. При использовании в настоящем описании, либо в переходной фразе, либо в структуре формулы изобретения, термины «содержит(ат)» и «содержащий» следует интерпретировать как имеющие неограниченное значение. Другими словами, термины следует интерпретировать как синонимы фраз «имеющий по меньшей мере» или «включающий по меньшей мере». В контексте процесса термин «содержащий» означает, что процесс включает в себя по меньшей мере указанные стадии, но может включать в себя дополнительные стадии. В контексте соединения, композиции или устройства термин «содержащий» означает, что соединение, композиция или устройство включает по меньшей мере указанные особенности или компоненты, но может включать дополнительные особенности или компоненты.
[0039] Заголовки разделов, используемые в настоящем описании, предназначены только для организационных целей и не должны толковаться как ограничивающие описанный предмет.
Комплексы транспосом
[0040] Некоторые варианты осуществления, предложенные в настоящем документе, относятся к композиции для создания библиотеки меченых фрагментов нуклеиновых кислот без ПЦР-амплификации. В некоторых вариантах осуществления композиция включает твердую подложку и комплекс транспосом, иммобилизованный на твердой подложке. В некоторых вариантах осуществления комплекс транспосом включает в себя транспозазу, первый транспозон, полинуклеотид присоединения и второй транспозон. В некоторых вариантах осуществления первый транспозон включает в себя 3’-концевую последовательность транспозона и 5’-адаптерную последовательность. В некоторых вариантах осуществления полинуклеотид присоединения включает в себя адаптерную последовательность присоединения, гибридизованную с 5’-адаптерной последовательностью, и связывающий элемент. В некоторых вариантах осуществления второй транспозон содержит 5’-концевую последовательность транспозона и 3’-адаптерную последовательность. В некоторых вариантах осуществления комплекс транспосом иммобилизуют на твердой подложке посредством полинуклеотида присоединения. В некоторых вариантах осуществления полинуклеотид присоединения дополнительно содержит праймерную последовательность.
[0041] В некоторых вариантах осуществления связывающий элемент содержит или представляет собой необязательно замещенный биотин. В некоторых вариантах осуществления связывающий элемент соединен с полинуклеотидом присоединения посредством линкера. В некоторых вариантах осуществления связывающий элемент содержит или представляет собой линкер биотина. В некоторых вариантах осуществления связывающий элемент содержит или представляет собой 3’, 5’ или внутренний биотин.
[0042] В некоторых вариантах осуществления 3’-концевая последовательность транспозона содержит концевую мозаичную (ME) последовательность, а 5’-концевая последовательность транспозона содержит МЕ’-последовательность. В некоторых вариантах осуществления 5’-адаптерная последовательность содержит последовательность A14, а адаптерная последовательность крепления содержит последовательность A14’. В некоторых вариантах осуществления 3’-адаптерная последовательность содержит последовательность B15’. В некоторых вариантах осуществления 3’-адаптерная последовательность комплементарна по меньшей мере участку индексной адаптерной последовательности. В некоторых вариантах осуществления индексная адаптерная последовательность содержит последовательность В15. В некоторых вариантах осуществления часть полинуклеотида присоединения содержит праймерную последовательность, такую как праймерная последовательность P5’. В некоторых вариантах осуществления праймерная последовательность полинуклеотида присоединения комплементарна по меньшей мере части индексирующей олигонуклеотидной последовательности, такой как праймерная последовательность P5.
[0043] В некоторых вариантах осуществления комплекс транспосом иммобилизуют на твердой подложке посредством связывающего элемента (и необязательного линкера), как описано в настоящем документе. В некоторых вариантах осуществления твердая подложка представляет собой гранулу, парамагнитную гранулу, проточную кювету, поверхность микрожидкостного устройства, пробирку, лунку планшета, предметное стекло, структурированную поверхность или микрочастицу. В некоторых вариантах осуществления твердая подложка содержит или представляет собой гранулу. В одном варианте осуществления гранула представляет собой парамагнитную гранулу. В некоторых вариантах осуществления твердая подложка содержит множество твердых подложек. В некоторых вариантах осуществления комплексы транспосом иммобилизованы на множестве твердых подложек. В некоторых вариантах осуществления множество твердых подложек содержит множество гранул. В некоторых вариантах осуществления множество комплексов транспосом иммобилизуют на твердой подложке с плотностью по меньшей мере 103, 104, 105, 106 комплексов на мМ2. В некоторых вариантах осуществления твердая подложка представляет собой гранулу или парамагнитные гранулы, и с каждой гранулой связано более 10000, 20000, 30000, 40000, 50000 или 60000 комплексов транспосом.
[0044] Для фрагментации ДНК можно применять технологию на основе транспозона, например, как показано в примере рабочего процесса наборов для получения образцов NEXTERA™ FLEX DNA (Illumina, Inc.), причем целевые нуклеиновые кислоты, такие как геномная ДНК, обрабатывают комплексами транспосом, которые одновременно фрагментируют и наносят метки на мишень («тагментация»), создавая таким образом популяцию фрагментированных молекул нуклеиновой кислоты, несущих метку уникальных адаптерных последовательностей на концах фрагментов.
[0045] Реакция транспозиции представляет собой реакцию, в которой один или более транспозонов вводят в целевые нуклеиновые кислоты в случайных сайтах или практически случайных сайтах. Компоненты реакции транспозиции включают транспозазу (или другой фермент, способный фрагментировать и наносить метку на нуклеиновую кислоту, как описано в настоящем документе, такую как интеграза) и транспозонный элемент, который включает двухцепочечную транспозонную концевую последовательность, связывающуюся с транспозазой (или другим ферментом, описанным в настоящем документе), и адаптерную последовательность, присоединенную к одной из двух концевых последовательностей транспозона. Одну цепь двухцепочечной транспозонной концевой последовательности переносят в одну цепь целевой нуклеиновой кислоты, а комплементарную цепь транспозонной концевой последовательности не переносят (транспозонная последовательность без переноса). Адаптерная последовательность может включать в себя одну или более функциональных последовательностей или компонентов (например, праймерные последовательности, якорных последовательностей, универсальных последовательностей, спейсерных участков или последовательностей индексированных меток) по мере необходимости.
[0046] «Комплекс транспосом» состоит по меньшей мере из одной транспозазы (или другого фермента, описанного в настоящем документе) и последовательности распознавания транспозона. В некоторых таких системах транспозаза связывается с последовательностью распознавания транспозона с образованием функционального комплекса, который способен катализировать реакцию транспозиции. В некоторых аспектах последовательность распознавания транспозона представляет собой двухцепочечную концевую последовательность транспозона. Транспозаза связывается с сайтом распознавания транспозазы в целевой нуклеиновой кислоте и вставляет последовательность распознавания транспозона в целевую нуклеиновую кислоту. В некоторых таких явлениях вставки одну цепь последовательности распознавания транспозона (или концевой последовательности) переносят в целевую нуклеиновую кислоту, что приводит к явлению расщепления. Иллюстративные процедуры и системы транспозиции, которые можно легко приспособить для использования с транспозазами.
[0047] Примеры транспозаз, которые можно применять с определенными вариантами осуществления, представленными в настоящем документе, включают следующие (или кодируются следующими): транспозаза Tn5, транспозаза Спящая Красавица (SB), Vibrio harveyi, транспозаза MuA и сайт распознавания транспозазы Mu, содержащий концевые последовательности R1 и R2, транспозаминаза StaphylococcUS aureUS Tn552, Ty1, Tn7, Tn/O и IS10, транспозаза Mariner, Tc1, P-элемент, Tn3, последовательности бактериальной вставки, ретровирусы и ретротранспозон дрожжей. Остальные примеры включают IS5, Tn10, Tn903, IS911 и сконструированные версии ферментов семейства транспозаз. Способы, описанные в настоящем документе, также могут включать комбинации транспозаз, а не только одну транспозазу.
[0048] В некоторых вариантах осуществления транспозаза представляет собой транспозазу Tn5, Tn7, MuA или Vibrio Haryi или ее активный мутант. В других вариантах осуществления транспозаза представляет собой транспозазу Тn5 или ее мутант. В других вариантах осуществления транспозаза представляет собой транспозазу Тn5 или ее мутант. В других вариантах осуществления транспозаза представляет собой транспозазу Тn5 или ее активный мутант. В некоторых вариантах осуществления транспозаза Tn5 представляет собой гиперактивную транспозазу Тn5 или ее активный мутант. В некоторых аспектах транспозаза Tn5 представляет собой транспозазу Tn5, описанную в публикации PCT № WO2015/160895, которая включена в настоящий документ посредством ссылки. В некоторых аспектах транспозаза Tn5 представляет собой гиперактивную транспозазу Tn5 с мутациями в позициях 54, 56, 372, 212, 214, 251 и 338 относительно транспозазы Tn5 дикого типа. В некоторых аспектах транспозаза Tn5 представляет собой гиперактивную транспозазу Tn5 со следующими мутациями относительно транспозазы Tn5 дикого типа: E54K, M56A, L372P, K212R, P214R, G251R и A338V. В некоторых вариантах осуществления транспозаза Tn5 представляет собой белок слияния. В некоторых вариантах осуществления белок слияния транспозазы Tn5 содержит метку слитого фактора удлинения Ts (Tsf). В некоторых вариантах осуществления транспозаза Tn5 представляет собой гиперактивную транспозазу Tn5, содержащую мутации по аминокислотам 54, 56 и 372 относительно последовательности дикого типа. В некоторых вариантах осуществления гиперактивная транспозаза Tn5 представляет собой белок слияния, причем необязательно белок слияния представляет собой фактор удлинения Ts (Tsf). В некоторых вариантах осуществления сайт распознавания представляет собой сайт распознавания транспозазы типа Tn5 (Goryshin and Reznikoff, J. Biol. Chem., 273:7367, 1998). В одном варианте осуществления применяют сайт распознавания транспозазы, образующий комплекс с гиперактивной транспозазой Tn5 (например, транспозаза EZ-Tn5™, Epicentre Biotechnologies, Madison, Wis.). В некоторых вариантах осуществления транспозаза Tn5 представляет собой транспозазу Tn5 дикого типа.
[0049] В некоторых вариантах осуществления комплекс транспосом содержит димер двух молекул транспозазы. В некоторых вариантах осуществления комплекс транспосом представляет собой гомодимер, в котором каждая из двух молекул транспозазы связана с первым и вторым транспозонами одного типа (например последовательности двух транспозонов, связанных с каждым мономером, являются одинаковыми, образуя «гомодимер»). В некоторых вариантах осуществления в композициях и способах, описанных в настоящем документе, применяют две популяции комплексов транспосом. В некоторых вариантах осуществления транспозазы в каждой популяции являются одинаковыми. В некоторых вариантах осуществления комплексы транспомосом в каждой популяции являются гомодимерами, причем первая популяция имеет первую адаптерную последовательность в каждом мономере, а вторая популяция имеет отличную адаптерную последовательность в каждом мономере.
[0050] В некоторых вариантах осуществления комплекс транспозазы содержит димер транспозазы (например, транспозазу Tn5), содержащий первый и второй мономеры. В некоторых аспектах каждый мономер содержит первый транспозон, второй транспозон и полинуклеотид присоединения, причем первый транспозон включает в себя концевую последовательность транспозона на 3’-конце (также называемую 3’ -концевой последовательностью транспозона) и адаптерную последовательность на 5’-конце (также называемую 5’-адаптерной последовательностью); второй транспозон включает в себя концевую последовательность транспозона на 5’-конце (также называемую 5’-концевой последовательностью транспозона) и адаптерную последовательность на 3’-конце (также называемую 3’-адаптерной последовательностью); и полинуклеотид присоединения включает в себя адаптерную последовательность присоединения, гибридизованную с 5’-адаптерной последовательностью первого транспозона, праймерную последовательность и линкер. В некоторых вариантах осуществления 5’-концевая последовательность транспозона второго транспозона по меньшей мере частично комплементарна 3’-концевой последовательности транспозона первого транспозона. В некоторых вариантах осуществления соединительная адаптерная последовательность полинуклеотида присоединения по меньшей мере частично комплементарна 5’-адаптерной последовательности первого транспозона. В некоторых вариантах осуществления линкер полинуклеотида присоединения включает связывающий элемент.
Концевые последовательности
[0051] В любом из вариантов осуществления способа, описанного в настоящем документе, первый транспозон включает в себя 3’-концевую последовательность транспозона, а второй транспозон включает в себя 5’-концевую последовательность транспозона. В некоторых вариантах осуществления 5’-концевая последовательность транспозона по меньшей мере частично комплементарна 3’-концевой последовательности транспозона. В некоторых вариантах осуществления комплементарные концевые последовательности транспозона гибридизуются с образованием двухцепочечной концевой последовательности транспозона, которая связывается с транспозазой (или другим ферментом, как описано в настоящем документе). В некоторых вариантах осуществления концевая последовательность транспозона представляет собой последовательность мозаичного конца (ME). Таким образом, в некоторых вариантах осуществления 3’-концевая последовательность транспозона представляет собой последовательность МЕ, и 5’-концевая последовательность транспозона представляет собой последовательность МЕ.
Адаптерные последовательности
[0052] В любом из вариантов осуществления способа, описанного в настоящем документе, первый транспозон включает в себя 5’-адаптерную последовательность, а второй транспозон включает в себя 3’-адаптерную последовательность. Адаптерные последовательности могут содержать одну или более функциональных последовательностей или компонентов, выбранных из группы, включающей праймерные последовательности, якорные последовательности, универсальные последовательности, спейсерные области, индексные последовательности, последовательности захвата, последовательности банка данных, последовательностей отщепления, последовательности, связанные с секвенированием и их комбинации. В некоторых вариантах осуществления адаптерная последовательность содержит праймерную последовательность. В других вариантах осуществления адаптерная последовательность содержит праймерную последовательность и индексную последовательность или последовательность банка данных. Праймерная последовательность также может представлять собой универсальную последовательность. Данное раскрытие не ограничено типом адаптерных последовательностей, которые можно применять, и квалифицированный специалист распознает дополнительные последовательности, которые можно применять для получения библиотеки и секвенирования следующего поколения. Универсальная последовательность представляет собой область нуклеотидной последовательности, которая является общей для двух или более фрагментов нуклеиновых кислот. Необязательно два или более фрагментов нуклеиновых кислот также имеют области различий последовательности. Универсальная последовательность, которая может присутствовать в разных членах множества фрагментов нуклеиновых кислот, может обеспечивать репликацию или амплификацию множества разных последовательностей с использованием одного универсального праймера, комплементарного универсальной последовательности.
[0053] В некоторых вариантах осуществления полинуклеотид присоединения включает в себя адаптерную последовательность присоединения, гибридизованную с 5’-адаптерной последовательностью. В некоторых вариантах осуществления адаптерная последовательность присоединения по меньшей мере частично комплементарна 5’-адаптерной последовательности. В некоторых вариантах осуществления адаптерная последовательность представляет собой последовательность А14 или последовательность В15. Таким образом, в некоторых вариантах осуществления 5’-адаптерная последовательность представляет собой последовательность A14, а адаптерная последовательность присоединения представляет собой последовательность A14’. В некоторых вариантах осуществления 3’-адаптерная последовательность представляет собой последовательность B15’. В некоторых вариантах осуществления адаптерная последовательность представляет собой любую последовательность для гибридизации (называемую в настоящем документе последовательностью X). В некоторых вариантах осуществления последовательность X содержит 16-20 нуклеотидов. В некоторых вариантах осуществления последовательность X имеет температуру плавления (Tm), аналогичную температуре плавления адаптерной последовательности. В некоторых вариантах осуществления результаты секвенирования улучшаются, если Tm последовательности X имеет температуру плавления, аналогичную температуре плавления адаптерной последовательности. В некоторых вариантах осуществления последовательность X имеет температуру плавления, аналогичную температуре плавления последовательности А14 или последовательности В15. В некоторых вариантах осуществления Tm последовательности X составляет 53°-56°. В некоторых вариантах осуществления адаптерную последовательность переносят к 5’-концам фрагмента нуклеиновой кислоты посредством реакции тагментации.
[0054] В любом из вариантов осуществления адаптерная последовательность или транспозонные концевые последовательности, включая A14-ME, ME, B15-ME, ME’, A14, B15 и ME, представлены ниже:
A14-ME: 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-3′ (SEQ ID NO: 1
B15-ME: 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-3′ (SEQ ID NO: 2).
ME’: 5’-phos-CTGTCTCTTATACACATCT-3’ (SEQ ID NO: 3).
A14: 5′-TCGTCGGCAGCGTC-3′ (SEQ ID NO: 4)
B15: 5′-GTCTCGTGGGCTCGG-3’ (SEQ ID NO: 5)
ME: AGATGTGTATAAGAGACAG (SEQ ID NO: 6)
Полинуклеотид присоединения
[0055] Варианты осуществления описанного в настоящем документе комплекса транспосомы включают в себя полинуклеотид присоединения. В настоящем документе полинуклеотид присоединения представляет собой полинуклеотид, который гибридизуется с транспозоном на одном конце и связывается с поверхностью на втором конце. Таким образом, описанный в настоящем документе комплекс транспосом иммобилизован на твердой подложке посредством полинуклеотида присоединения. В некоторых вариантах осуществления полинуклеотид присоединения включает в себя адаптерную последовательность присоединения, гибридизованную с адаптерной последовательностью первого транспозона или адаптерной последовательностью второго транспозона, последовательностью праймера и линкером. В некоторых вариантах осуществления линкер включает связывающий элемент.
[0056] Как описано в настоящем документе, адаптерная последовательность присоединения может быть по меньшей мере частично комплементарна адаптерной последовательности первого или второго транспозона. В некоторых вариантах осуществления адаптерная последовательность присоединения гибридизуется с 5’-адаптерной последовательностью. В вариантах осуществления, в которых адаптерная последовательность присоединения гибридизуется с 5’-адаптерной последовательностью, где 5’-адаптерная последовательность представляет собой последовательность A14, адаптерная последовательность присоединения представляет собой последовательность A14’. В некоторых вариантах осуществления адаптерная последовательность представляет собой последовательность X. В некоторых вариантах осуществления соединительная адаптерная последовательность гибридизуется с 3’-адаптерной последовательностью. В вариантах осуществления, в которых адаптерная последовательность присоединения гибридизуется с 3’-адаптерной последовательностью, где 3’-адаптерная последовательность представляет собой последовательность B15’, адаптерная последовательность присоединения представляет собой последовательность B15. В любом из данных вариантов осуществления соединительная адаптерная последовательность может быть полностью комплементарна адаптерной последовательности первого или второго транспозона или частично комплементарна адаптерной последовательности первого или второго транспозона.
[0057] В некоторых вариантах осуществления полинуклеотид присоединения содержит праймерную последовательность. В некоторых вариантах осуществления праймерная последовательность представляет собой праймерную последовательность P5, или праймерную последовательность P7, или ее комплемент (например, P5’ или P7’). Праймеры P5 и P7 используют на поверхности коммерческих проточных кювет, поставляемых компанией Illumina, Inc., для секвенирования на различных платформах Illumina. Праймерные последовательности описаны в публикации патента США № 2011/0059865, которая полностью включена в настоящий документ посредством ссылки. Примеры праймеров P5 и P7, которые могут иметь алкиновую концевую группу на 5’-конце, включают в себя следующие:
P5: AATGATACGGCGACCACCGAGAUCTACAC (SEQ ID NO: 7)
P7: CAAGCAGAAGACGGCATACGAG*AT (SEQ ID NO: 8)
и их производные. В некоторых примерах последовательность Р7 включает модифицированный гуанин в положении G*, например, 8-оксогуанин. В других примерах * указывает на то, что связь между G* и смежным 3’ A представляет собой тиофосфатную связь. В некоторых примерах праймеры Р5 и/или Р7 включают линкеры не природного происхождения. Необязательно один или оба праймера P5 и P7 могут включать хвост поли-T. Хвост поли-T по существу расположен на 5’-конце указанной выше последовательности, например, между 5’-основанием и концевым алкиновым звеном, но в некоторых случаях может быть расположен на 3’-конце. Последовательность поли-T может включать любое число Т-нуклеотидов, например от 2 до 20. Несмотря на то, что в качестве примеров приведены праймеры P5 и P7, следует понимать, что в примерах, представленных в настоящем документе, можно использовать любые подходящие праймеры. Индексные последовательности, имеющие праймерные последовательности, включая праймерные последовательности P5 и P7, служат для добавления P5 и P7 для активации библиотеки для секвенирования. Несмотря на то, что в качестве примеров приведены праймеры P5 и P7, следует понимать, что в примерах, представленных в настоящем документе, можно использовать любые подходящие праймеры амплификации.
[0058] В контексте настоящего документа один пример линкера представляет собой функциональную группу, которая ковалентно соединяет связывающий элемент с концом нуклеотидной части полинуклеотида присоединения и может использоваться для иммобилизации полинуклеотида присоединения на твердой подложке. Линкер может представлять собой расщепляемый линкер, например, линкер, способный отщепляться для удаления полинуклеотида присоединения и, таким образом, комплекса транспосомы или продукта мечения с твердой подложки. В контексте настоящего документа отщепляемый линкер представляет собой линкер, который может быть отщеплен химическими или физическими способами, такими как, например, фотолиз, химическое отщепление, термическое отщепление или ферментативное отщепление. В некоторых вариантах осуществления отщепление может осуществляться биохимическими, химическими, ферментативными, нуклеофильными, чувствительными к восстановлению средствами или иными способами. Отщепляемые линкеры могут содержать фрагмент, выбранный из группы, включающей следующее: сайт рестрикционной эндонуклеазы; по меньшей мере один рибонуклеотид, отщепляемый РНКазой; аналоги нуклеотидов, отщепляемые в присутствии определенного(ых) химического(ых) средства(средств); фоторасщепляемое(ые) линкерное(ые) звено (звенья); диольная связь, отщепляемая посредством обработки периодатом (как пример); дисульфидная группа, отщепляемая химическим восстанавливающим средством; отщепляемая функциональная группа, которая может подвергаться фотохимическому отщеплению; и пептид, отщепляемый пептидазой ферментом или другими подходящими способами. Отщепление может быть опосредовано ферментами путем включения отщепляемого нуклеотида или нуклеотидного основания в отщепляемый линкер, такой как урацил или 8-оксогуанин.
[0059] В некоторых вариантах осуществления линкер, описанный в настоящем документе, может быть ковалентно и непосредственно присоединен к полинуклеотиду присоединения, например, с образованием связи -O-, или может быть ковалентно присоединен посредством другой группы, такой как фосфат или сложный эфир. В альтернативном варианте осуществления описанный в настоящем документе линкер может быть ковалентно присоединен к фосфатной группе полинуклеотида присоединения, например, ковалентно присоединен к 3’-гидроксильной группе посредством фосфатной группы, с образованием таким образом связи -O-P(O)3.
[0060] В контексте настоящего документа связывающий элемент представляет собой фрагмент, который можно использовать для связывания, ковалентно или нековалентно, с партнером по связыванию. В некоторых аспектах связывающий элемент находится на комплексе транспосом, а партнер по связыванию находится на твердой подложке. В некоторых вариантах осуществления связывающий элемент может связываться или нековалентно связываться с партнером по связыванию на твердой подложке, нековалентно присоединяя таким образом комплекс транспосом к твердой подложке. В некоторых вариантах осуществления связывающий элемент способен связываться (ковалентно или нековалентно) с партнером по связыванию на твердой подложке. В некоторых аспектах связывающий элемент связан (ковалентно или нековалентно) с партнером по связыванию на твердой подложке, что приводит к образованию иммобилизованного комплекса транспосом.
[0061] В таких вариантах осуществления связывающий элемент содержит или представляет собой, например, биотин, а партнер по связыванию содержит или представляет собой авидин или стрептавидин. В других вариантах осуществления комбинация связывающего элемента/партнера по связыванию содержит или представляет собой FITC/антитело к FITC, дигоксигенин/антитело дигоксигенин или гаптен/антитело. Другие подходящие пары связывания включают без ограничения дитиобиотинавидин, иминобиотинавидин, биотинавидин, дитиобиотинсукцинилированный авидин, иминобиотинсукцинилированный авидин, биотинстрептавидин и биотинсукцинилированный авидин. В некоторых вариантах осуществления связывающий элемент представляет собой биотин, а партнер по связыванию представляет собой стрептавидин.
[0062] В некоторых вариантах осуществления связывающий элемент может связываться с партнером по связыванию посредством химической реакции или ковалентно связываться с партнером по связыванию на твердой подложке, таким образом ковалентно связывая комплекс транспосом с твердой подложкой. В некоторых аспектах комбинация связывающего элемента/партнера по связыванию содержит или представляет собой амин/карбоновую кислоту (например, связывание посредством стандартной реакции сочетания пептидов в условиях, известных специалисту в данной области, таких как EDC или NHS-опосредованное связывание). Реакция двух компонентов соединяет связывающий элемент с партнером по связыванию посредством амидной связи. В альтернативном варианте осуществления связывающий элемент и связывающее вещество могут представлять собой двух партнеров по клик-химии (например, азид/алкин, которые реагируют с образованием триазольной связи).
[0063] В некоторых вариантах осуществления изобретения полинуклеотид присоединения также включает дополнительные последовательности или компоненты, такие как универсальная последовательность, спейсерная область, якорная последовательность или последовательность индексированной метки, или их комбинацию. Универсальная последовательность представляет собой область нуклеотидной последовательности, которая является общей для двух или более фрагментов нуклеиновых кислот. Необязательно два или более фрагментов нуклеиновых кислот также имеют области различий последовательности. Универсальная последовательность, которая может присутствовать в разных членах множества фрагментов нуклеиновых кислот, может обеспечивать репликацию или амплификацию множества разных последовательностей с использованием одного универсального праймера, комплементарного универсальной последовательности.
[0064] В некоторых вариантах осуществления первый транспозон дополнительно содержит 5’-праймерную последовательность 5’-адаптерной последовательности, а полинуклеотид присоединения содержит (i) часть, комплементарную 5’-адаптерной последовательности и гибридизованную с ней, и (ii) комплементарную праймерную последовательность (см. Фиг. 4A, универсальные гранулы с единичным индексированием). Такие конструкции можно применять в качестве универсальных микроносителей в областях применения с единичным индексированием, поскольку отсутствует последовательность индексной метки.
[0065] В некоторых вариантах осуществления первый транспозон дополнительно содержит 5’-праймерную последовательность 5’-адаптерной последовательности, а полинуклеотид присоединения содержит последовательность индексной метки и праймерную последовательность (см., например, Фиг. 4A, индексированные гранулы с единичным индексированием).
[0066] В некоторых вариантах осуществления первый транспозон содержит 5’-адаптерную последовательность, а полинуклеотид присоединения содержит (i) часть, комплементарную 5’-адаптерной последовательности и гибридизованную с ней, (ii) спейсерную область и (iii) праймерную последовательность (см. Фиг. 4A, универсальные гранулы, двойное индексирование).
[0067] В некоторых вариантах осуществления второй транспозон содержит 3’-адаптерную последовательность, а полинуклеотид присоединения содержит (i) часть, комплементарную 3’-адаптерной последовательности и гибридизованную с ней, (ii) последовательность индексной метки и (iii) праймерную последовательность (см. Фиг. 4A, двойное индексирование, индексированные гранулы).
[0068] В некоторых вариантах осуществления изобретения полинуклеотид присоединения содержит спейсерную область (см. например, Фиг. 4A) или спейсерную область и якорную область (см. например, Фиг. 3). В настоящем документе «спейсерная область» представляет собой последовательность нуклеиновой кислоты, не содержащую никакой структурной или кодирующей информации для известных функций гена. Спейсерная область на полинуклеотиде присоединения способна выравниваться с индексирующими олигонуклеотидами с различными последовательностями (например, с диапазоном последовательностей i5). В некоторых вариантах осуществления спейсерная область представляет собой универсальную последовательность. В некоторых вариантах осуществления спейсерная область представляет собой спейсер, отличный от ДНК. В некоторых вариантах осуществления спейсерная область включает универсальные основания, такие как инозины или нитроиндолы. В некоторых вариантах осуществления спейсер включает линкер sp18. В настоящем документе линкер sp18 представляет собой стандартный модифицирующий линкер со спейсером С18 (18-атомный гекса-этиленгликолевый спейсер), который эквивалентен 4 парам оснований по длине. Таким образом, линкер 2 x sp18 эквивалентен по длине 8 парам оснований. В некоторых вариантах осуществления спейсерная область содержит синтетический линкер 2 x sp18. В некоторых вариантах осуществления спейсерная область содержит один или более спейсеров С18, например 1, 2, 3, 4, 5, 6 или более спейсеров С18. В некоторых вариантах осуществления спейсерная область содержит два спейсера С18 (длина которых эквивалентна 8 нуклеотидам). В некоторых вариантах осуществления спейсер представляет собой спейсер C9, эквивалентный по длине 2 парам оснований. В некоторых вариантах осуществления спейсерная область содержит один или более спейсеров C9 (триэтиленгликолевый спейсер), например 1, 2, 3, 4, 5, 6 или более спейсеров C9. В некоторых вариантах осуществления спейсер представляет собой стандартный спейсер, применяемый с существующими индексами, такой как спейсер по 10 паре оснований. В некоторых вариантах осуществления спейсерная область представляет собой комбинацию спейсеров, например комбинацию одного или более спейсеров С18 и одного или более спейсеров С9 или любую комбинацию любых спейсеров, описанных в настоящем документе. В некоторых вариантах осуществления спейсерная область имеет длину, эквивалентную 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 15, 20 или 30 парам оснований. В некоторых вариантах осуществления спейсерная область имеет длину, приблизительно эквивалентную 8 или 10 парам оснований или нуклеотидам. В некоторых вариантах осуществления спейсерная область специфически была такой же длины, что и индексная область. В некоторых вариантах осуществления индексные области имеют длину 8 нуклеотидов, а спейсерная область содержит два спейсера С18. В некоторых вариантах осуществления индексные области имеют длину 10 нуклеотидов, а спейсерная область содержит два спейсера С18 и один спейсер C9.
[0069] В некоторых аспектах полинуклеотид присоединения содержит якорную последовательность. В некоторых вариантах осуществления якорная последовательность представляет собой GGATATGCTCGG (SEQ ID NO: 22). В некоторых вариантах осуществления якорная последовательность представляет собой последовательность A14 (SEQ ID NO: 4). В контексте настоящего документа «якорная область» означает последовательность ДНК, которая комплементарна области комплемента якоря в индексирующем олигонуклеотиде и которая обеспечивает гибридизацию двух компонентов (см. например, Фиг. 3B). В некоторых аспектах якорная область комплементарна участку области комплемента якоря индексирующего олигонуклеотида, причем индексирующий олигонуклеотид содержит комплементарную область якоря и последовательность индексной метки (--якорь’ - последовательность индексной метки -). В некоторых вариантах осуществления якорная последовательность комплементарна области комплемента якоря, общей для множества индексирующих олигонуклеотидов. В некоторых вариантах осуществления каждая последовательность индексной метки во множестве индексирующих олигонуклеотидов является одинаковой (без индексирования) или разной (с индексированием). В некоторых вариантах осуществления последовательность индексной метки представляет собой последовательность i5. Полинуклеотид присоединения может дополнительно включать дополнительные элементы последовательности или компоненты для повышения эффективности и функциональности полинуклеотида присоединения для связывания с индексами, включая, например, праймерные последовательности, якорные последовательности, универсальные последовательности, спейсерные области, индексные последовательности, последовательности захвата, последовательности банка данных, последовательности отщепления, последовательности, связанные с секвенированием, и их комбинации. В некоторых вариантах осуществления полинуклеотид присоединения содержит последовательность A14’.
[0070] Могут быть реализованы варианты комплекса транспосомы, включая транспозазу, транспозоны и полинуклеотид присоединения. Например, могут быть реализованы варианты конфигурации, конструкции, гибридизации, структурных элементов и общего расположения комплекса транспосомы. В описании и графических материалах, представленных в настоящем документе, представлены несколько вариантов, но следует понимать, что могут быть легко реализованы дополнительные варианты в пределах объема настоящего описания изобретения.
7.5. Твердая подложка
[0071] Термины «твердая поверхность»,«твердая подложка» и другие грамматические эквиваленты относятся к любому материалу, подходящему или выполненному с возможностью присоединения комплексов транспосом. Как будет понятно специалистам в данной области, количество возможных подложек является широким. Возможные подложки включают в себя без ограничения стекло и модифицированное или функционализированное стекло, пластмассы (включая акрилы, полистирол и сополимеры стирола и других материалов, полипропилен, полиэтилен, полибутилен, полиуретаны, тефлон и т. п.), полисахариды, материалы на основе полиэдрического органического силсесквиоксана (POSS), нейлон или нитроцеллюлозу, керамику, смолы, кремнезем или материалы на основе кремнезема, в том числе кремний и модифицированный кремний, углерод, металлы, неорганические стекла, пластмассы, волоконно-оптические жгуты, гранулы, парамагнитные гранулы и множество других полимеров.
[0072] Подходящие композиции на основе гранул включают без ограничения пластмассы, керамику, стекло, полистирол, метилстирол, акриловые полимеры, парамагнитные материалы, золь тория, углерод в форме графита, диоксид титана, латекс или сшитый декстран, такой как сефароза, целлюлоза, нейлон, поперечносшитые мицеллы и тефлон, а также любые другие материалы, описанные в настоящем документе применительно к твердым подложкам. В определенных вариантах осуществления микросферы представляют собой магнитные микросферы или гранулы, например парамагнитные частицы, сферы или гранулы. Гранулы не обязательно должны быть сферическими; можно применять частицы неправильной формы. В качестве альтернативы или дополнительно гранулы могут быть пористыми. Размеры гранул варьируют от нанометров, например 100 нМ, до миллиметров, например 1 мМ, предпочтительно от 0,2 микрон до 200 микрон и особенно предпочтительно от 0,5 до 5 микрон, несмотря на то, что в некоторых вариантах осуществления можно применять гранулы меньшего или большего размера. Гранула может быть покрыта партнером по связыванию, например, гранула может быть покрыта стрептавидином. В некоторых вариантах осуществления гранулы представляют собой парамагнитные гранулы, покрытые стрептавидином, например гранулы Dynabeads MyOne streptavidin C1 (Thermo Scientific, № по кат. 65601), парамагнитные частицы стрептавидина (Promega, № по кат. Z5481), магнитные гранулы стрептавидина (NEB, № по кат. S1420S) и MaxBead Streptavidin (Abnova, № по кат. U0087). Твердая подложка также может представлять собой предметное стекло, например проточную кювету или иное предметное стекло, измененное таким образом, чтобы на нем можно было иммобилизовать комплекс транспосом.
[0073] В некоторых вариантах осуществления партнер по связыванию присутствует на твердой подложке или грануле с плотностью от 1000 до около 6000 пМоль/мг, или от около 2000 до около 5000 пМоль/мг, или от около 3000 до около 5000 пМоль/мг, или от около 3500 до около 4500 пМоль/мг.
[0074] В одном варианте осуществления твердая поверхность представляет собой внутреннюю поверхность пробирки для образца. В другом варианте осуществления твердая поверхность представляет собой мембрану для захвата. В одном примере мембрана для захвата представляет собой мембрану, захватывающую биотин (например, поставляемую компанией Promega Corporation). В другом примере мембрана для захвата представляет собой фильтровальную бумагу. В некоторых вариантах осуществления настоящего описания твердые подложки, состоящие из инертного субстрата или матрицы (например, стеклянных предметных стекол, полимерных гранул и т. д.), которые были функционализированы, например, посредством нанесения слоя или покрытия из промежуточного материала, содержащего реакционноспособные группы, которые обеспечивают ковалентное присоединение к молекулам, таким как полинуклеотиды. Примеры таких подложек включают без ограничения полиакриламидные гидрогели, нанесенные на инертный субстрат, такой как стекло, в частности, полиакриламидные гидрогели, описанные в WO2005/065814 и US2008/0280773, содержание которых полностью включено в настоящий документ посредством ссылки. Способы тагментации (фрагментирования и нанесения метки) ДНК на твердой поверхности для конструирования меченой библиотеки ДНК описаны в WO2016/189331 и US2014/0093916A1, которые полностью включены в настоящий документ посредством ссылки. В некоторых вариантах осуществления описанный в данном документе комплекс транспосом иммобилизован на твердой подложке посредством связывающего элемента. В некоторых таких вариантах осуществления твердая подложка содержит стрептавидин в качестве партнера по связыванию, а связывающий элемент представляет собой биотин.
[0075] В некоторых вариантах осуществления комплексы транспосом иммобилизованы на твердой подложке, такой как гранулы, с определенной плотностью или с диапазоном плотностей. В некоторых вариантах осуществления плотность комплексов на твердой подложке относится к концентрации комплексов транспосом в растворе в ходе реакции иммобилизации. Плотность комплекса предполагает, что реакция иммобилизации является количественной. После образования комплексов с конкретной плотностью данная плотность остается постоянной для партии связанных с поверхностью комплексов транспосом. Полученные гранулы могут быть разведены, и полученная концентрация комплексов в разбавленном растворе представляет собой полученную плотность гранул, разделенную на коэффициент разбавления. Маточные растворы разбавленных гранул сохраняют плотность комплекса при их получении, но комплексы присутствуют в более низкой концентрации в разбавленном растворе. На стадии разбавления плотность комплексов на гранулах не изменяется и, следовательно, влияет на выход библиотеки, но не на размер вставки (фрагмента). В некоторых вариантах осуществления плотность составляет от около 5 нМ до около 1000 нМ, или от около 5 до 150 нМ, или от около 10 нМ до 800 нМ. В других вариантах осуществления плотность составляет около 10 нМ, или около 25 нМ, или около 50 нМ, или около 100 нМ, или около 200 нМ, или около 300 нМ, или около 400 нМ, или около 500 нМ, или около 600 нМ, или около 700 нМ, или около 800 нМ, или около 900 нМ, или около 1000 нМ. В некоторых вариантах осуществления плотность составляет приблизительно 100 нМ. В некоторых вариантах осуществления плотность составляет приблизительно 300 нМ. В некоторых вариантах осуществления плотность составляет приблизительно 600 нМ. В некоторых вариантах осуществления плотность составляет приблизительно 800 нМ. В некоторых вариантах осуществления плотность составляет приблизительно 100 нМ. В некоторых вариантах осуществления плотность составляет приблизительно 1000 нМ.
[0076] Некоторые варианты осуществления, предложенные в настоящем документе, относятся к набору, включающему в себя комплекс транспосом, как описано в настоящем документе, и смесь индексов, содержащую индексные последовательности. Некоторые варианты осуществления, предложенные в настоящем документе, относятся к набору, включающему в себя композицию, имеющую твердую подложку с иммобилизованным на ней комплексом транспосом, как описано в настоящем документе, и смесь индексов, содержащую индексные последовательности. В некоторых вариантах осуществления смесь индексов содержит последовательности индексов i5 и последовательности индексов i7, причем последовательности индексов i5 комплементарны полинуклеотиду присоединения и гибридизуются с ним, и причем последовательности индексов i7 комплементарны 3’-адаптерной последовательности и гибридизуются с ней.
7.6. Иммобилизованные комплексы транспосом/способы образования фрагментов нуклеиновых кислот с нанесенной меткой
[0077] В настоящем описании дополнительно предложены способы получения иммобилизованных комплексов транспосом, как описано в настоящем документе. Некоторые варианты осуществления, предложенные в настоящем документе, относятся к способу создания библиотеки меченых фрагментов нуклеиновых кислот. В некоторых вариантах осуществления способ включает получение твердой подложки, имеющей иммобилизованный на ней комплекс транспосом, нанесение целевой нуклеиновой кислоты на твердую подложку в условиях, достаточных для фрагментации целевой нуклеиновой кислоты на множество целевых фрагментов и присоединения 3’-конца первого транспозона к 5’-концам целевых фрагментов для получения множества 5’-меченных целевых фрагментов, и применение смеси индексов, содержащей индексные последовательности, для активации библиотеки для секвенирования. В некоторых вариантах осуществления комплекс транспосом включает транспозазу, связанную с первым и вторым транспозоном, первый транспозон, содержащий 3’-концевую последовательность транспозона и 5’-адаптерную последовательность, полинуклеотид присоединения, содержащий адаптерную последовательность присоединения и связывающий элемент, и второй транспозон, содержащий 5’-концевую последовательность транспозона и 3’-адаптерную последовательность.
[0078] В некоторых вариантах осуществления способ включает создание библиотеки меченых фрагментов нуклеиновых кислот, включающее приведение целевой нуклеиновой кислоты в контакт с множеством иммобилизованных комплексов транспосом, как описано в настоящем документе, в условиях, достаточных для фрагментации целевой нуклеиновой кислоты во множество целевых фрагментов и для присоединения 3’-конца первого транспозона к 5’-концам целевых фрагментов с получением множества 5’-меченных целевых фрагментов.
[0079] В некоторых аспектах способ дополнительно включает следующее: обработка твердой подложки для удаления несвязанных нуклеиновых кислот; или обработка твердой подложки с удалением транспозазы из комплекса, необязательно посредством (a) нагревания твердой подложки и/или (b) промывки твердой подложки ферментным денатурирующим средством, причем ферментное денатурирующее средство необязательно содержит додецилсульфат натрия (SDS), гидрохлорид гуанидина, мочевину или протеиназу.
[0080] В некоторых вариантах осуществления приведение в контакт включает в себя добавление биологического образца к комплексу транспосомы. В некоторых вариантах осуществления биологический образец содержит клеточный лизат или цельные клетки или его выбирают из крови, плазмы, сыворотки, лимфы, слизи, мокроты, мочи, спермы, спинномозговой жидкости, бронхиальной аспирации, фекалий, мацерированной ткани и фиксированной ткани FFPE.
[0081] В некоторых аспектах приведение в контакт дополнительно включает гибридизацию множества индексирующих олигонуклеотидов с полинуклеотидами крепления, причем множество индексирующих олигонуклеотидов содержит одинаковые или разные индексные последовательности.
[0082] В некоторых аспектах способ дополнительно включает обработку множества 5’-меченых целевых фрагментов полимеразой и лигазой для удлинения и лигирования цепей с получением полностью двухцепочечных меченых фрагментов. В некоторых аспектах обработку полимеразой и лигазой выполняют в присутствии средства разрушения вторичной структуры ДНК, причем средство разрушения необязательно представляет собой ДМСО. В некоторых аспектах обработку полимеразой и лигазой проводят в присутствии олигонуклеотида, который комплементарен праймерной последовательности, причем олигонуклеотид представляет собой олигонуклеотид P7’, а праймерная последовательность представляет собой последовательность P7. В некоторых вариантах осуществления полимераза содержит мутант ДНК-полимеразы Т4, не имеющий экзонуклеазной активности.
[0083] В некоторых аспектах способ дополнительно включает удаление из твердой подложки полностью двухцепочечных меченых фрагментов. В некоторых аспектах удаление включает применение нагревания и/или денатурирующего вещества, достаточного для отщепления полностью двухцепочечных меченых фрагментов от твердой подложки.
[0084] В некоторых аспектах способ дополнительно включает секвенирование одного или более 5’-меченных целевых фрагментов полностью двухцепочечных меченых фрагментов. В некоторых аспектах фрагменты не определяют количественно с помощью ПЦР после приведения в контакт и до секвенирования. В некоторых аспектах нуклеиновая кислота представляет собой ДНК или РНК. В некоторых аспектах 3’-адаптерная последовательность комплементарна по меньшей мере участку индексной адаптерной последовательности. В некоторых аспектах праймерная последовательность полинуклеотида присоединения комплементарна по меньшей мере части индексной праймерной последовательности. В некоторых аспектах праймерная последовательность полинуклеотида присоединения комплементарна праймерной последовательности показателя индексирующего полинуклеотида.
[0085] В некоторых вариантах осуществления способ дополнительно включает промывку твердой подложки с удалением несвязанных нуклеиновых кислот. В некоторых вариантах осуществления способ дополнительно включает обработку твердой подложки с удалением транспозазы. В некоторых вариантах осуществления обработка твердой подложки включает промывку твердой подложки ферментативным денатурирующим средством. В некоторых вариантах осуществления ферментативное денатурирующее средство включает в себя уксусную кислоту, диметилсульфоксид (ДМСО), дитиотреитол, этанол, формальдегид, формамид, глутаральдегид, гидрохлорид гуанидина, перхлорат лития, меркаптоэтанол, пропиленгликоль, протеиназу, бикарбонат натрия, додецилсульфат натрия (SDS), салицилат натрия, сульфосалициловую кислоту, трихлоруксусную кислоту, трис(2-карбоксиэтил)фосфин) (TCEP) или мочевину. В некоторых вариантах осуществления обработка твердой подложки включает нагревание твердой подложки с удалением транспозазы. В некоторых вариантах осуществления применение целевой нуклеиновой кислоты включает смешивание биологического образца с комплексом транспосомы. Нуклеиновая кислота не обязательно должна быть полностью очищена или очищена вообще, и может представлять собой часть биологического образца или смеси с белком, другими видами нуклеиновых кислот, другими клеточными компонентами и/или любыми другими загрязнителями. В некоторых вариантах осуществления биологический образец включает в себя клеточный лизат. В некоторых вариантах осуществления биологический образец включает в себя цельные клетки. В некоторых вариантах осуществления биологический образец выбирают из биологической жидкости, крови, плазмы, сыворотки, лимфы, слизи, мокроты, мочи, спермы, спинномозговой жидкости, бронхиальной аспирации, фекалий, мацерированной ткани и фиксированной ткани, зафиксированной в формалине залитой парафином ткани (FFPE).
[0086] В некоторых вариантах осуществления нанесение смеси индексов включает в себя гибридизацию индексной последовательности с полинуклеотидом присоединения и вторым транспозоном. В некоторых вариантах осуществления смесь индексов включает смесь для интенсивного лигирования (ELM). В некоторых вариантах осуществления ELM содержит полимеразу и лигазу. В некоторых вариантах осуществления ELM представляет собой разделенную ELM для лигирования индексов. В некоторых вариантах осуществления полимераза включает ДНК-полимеразу Т4 или ее мутант (например, ДНК-полимеразу Т4 без экзонуклеазной активности, или мутантную ДНК-полимеразу Т4, или мутантную ДНК-полимеразу Т4 без экзонуклеазной активности). В некоторых вариантах осуществления лигаза содержит ДНК-лигазу E. coli. В некоторых вариантах осуществления реакцию удлинения и лигирования проводят в присутствии средства разрушения вторичной структуры ДНК, такого как ДМСО. В некоторых вариантах осуществления способ дополнительно включает денатурацию множества 5’-меченных целевых фрагментов из твердой подложки. В некоторых вариантах осуществления денатурация может быть достигнута посредством применения нагревания и/или денатурирующего вещества, достаточного для отщепления меченных 5’-целевых фрагментов от твердой подложки. В некоторых вариантах осуществления способ дополнительно включает секвенирование одного или более из 5’-меченных целевых фрагментов или продуктов их лигирования. В некоторых вариантах осуществления нуклеиновая кислота представляет собой ДНК или РНК.
[0087] В некоторых аспектах такие способы включают приведение в контакт первого и второго транспозонов, как описано в настоящем документе, с полинуклеотидом присоединения, связанным с поверхностью, в условиях, подходящих для гибридизации полинуклеотида присоединения с первым или вторым транспозоном. В некоторых аспектах способы получения связанного с твердой подложкой комплекса транспосомы включают инкубацию комплекса транспосомы, как описано в настоящем документе, с твердой подложкой, содержащей партнера по связыванию, в условиях, достаточных для связывания (ковалентного или нековалентного) связывающего элемента с партнером по связыванию. Затем иммобилизованный гибридизованный полинуклеотид, который включает полинуклеотид присоединения и первый и второй транспозоны, приводят в контакт с транспозазой в условиях, подходящих для образования комплекса транспосом.
[0088] В некоторых вариантах осуществления первый и второй транспозоны, описанные в настоящем документе, ренатурируют между собой, а первый транспозон ренатурируют с полинуклеотидом присоединения. Затем отожженные полинуклеотиды загружают на транспозазу, такую как транспозаза Tn5, с получением таким образом комплекса транспосом, который затем приводят в контакт и связывают с твердой подложкой, такой как гранула. В некоторых вариантах осуществления отожженные транспозоны связывают с твердой подложкой, такой как гранула, а затем транспозазу связывают в комплекс с транспозонами с получением таким образом транспосомы, которая связывается с твердой подложкой.
[0089] В некоторых аспектах представлены способы получения фрагментов из целевой нуклеиновой кислоты, включающие получение твердой подложки, содержащей иммобилизованный на ней комплекс транспосом, как описано в настоящем документе; применение целевой нуклеиновой кислоты к твердой подложке в условиях, достаточных для фрагментации целевой нуклеиновой кислоты во множество целевых фрагментов и для присоединения 3’-конца первого транспозона к 5’-концам целевых фрагментов с получением множества 5’-меченных целевых фрагментов. В некоторых аспектах способ дополнительно включает применение смеси индексов, включающей в себя индексные последовательности. В некоторых вариантах осуществления состояние фрагмента представляет собой состояние, подходящее для тагментации с использованием комплекса транспосом для фрагмента и нанесения метки на целевую нуклеиновую кислоту.
[0090] В некоторых вариантах осуществления способов, описанных в данном документе, после фрагментации и нанесения метки способы дополнительно включают промывку твердой подложки с удалением несвязанных нуклеиновых кислот. В некоторых вариантах осуществления способов, описанных в настоящем документе, после фрагментации и нанесения метки способы дополнительно включают удаление транспозазы. Удаление транспозазы можно осуществлять в химических условиях, таких как промывка твердой подложки средством для удаления транспозазы. В некоторых вариантах осуществления средство представляет собой додецилсульфат натрия (SDS).
[0091] В некоторых вариантах осуществления способ дополнительно включает приведение твердой подложки в контакт со смесью индексов. Взаимодействие со смесью индексов служит для нанесения меток на фрагменты с определенным индексом и активации библиотеки для секвенирования. В некоторых вариантах осуществления смесь индексов включает в себя олигонуклеотиды, которые гибридизуются с транспозоном или полинуклеотидом присоединения для нанесения метки или индексирования фрагментов нуклеиновых кислот. Таким образом, например, олигонуклеотид включает в себя индекс, а другие области олигонуклеотида комплементарны транспозону или полинуклеотиду присоединения и гибридизуются с ним. В качестве примера, в одном варианте осуществления индекс i5, имеющий праймерную последовательность (последовательность P5), индексную последовательность (последовательность i5) и якорную последовательность, гибридизуется с полинуклеотидом присоединения в комплементарных последовательностях, так что Р5 гибридизуется с P5’ и якорный участок гибридизуется с якорным участком’. В некоторых вариантах осуществления последовательность i5 связывается с последовательностью нитроиндола (нитро) полинуклеотида присоединения. В некоторых вариантах осуществления нитроиндольная последовательность комплементарна любой индексной последовательности i5 и гибридизуется с ней. В одном варианте осуществления индекс i7, имеющий праймерную последовательность (P7), индексную последовательность (i7) и адаптерную последовательность (последовательность B15), гибридизуется со вторым транспозоном в комплементарных последовательностях, так что P7 гибридизуется с P7’, i7 гибридизуется с i7’, а B15 гибридизуется с B15’. В некоторых вариантах осуществления смесь индексов включает в себя двухцепочечный индекс. После приведения твердой подложки в контакт со смесью индексов фрагменты лигируют и удлиняют с использованием смеси для удлинения и лигирования (ELM). ELM может включать, например, ДНК-полимеразу Т4 и ДНК-лигазу E. coli. Примеры полимераз включают без ограничения крупные фрагменты Bst ДНК-полимеразы I Bst, ДНК-полимеразы I E. coli (фрагмент Klenow), фрагмент Klenow (3′-5′ exо-), ДНК-полимеразы T4, ДНК-полимеразы Т7, Deep VentR. (exo-) ДНК-полимеразы, ДНК-полимеразы Deep VentR, ДНК-полимеразы Therminator II, ДНК-полимеразы AmpliTherm, ДНК-полимеразы SP6 или Taq-полимеразы, или мутанты, аналоги или производные любой из вышеупомянутых полимераз. Примеры лигаз включают без ограничения следующее: ДНК-лигаза Т4, РНК-лигаза Т4, ДНК-лигаза Taq, ДНК-лигаза E. Coli, ДНК-лигаза Pfu и Tth ДНК-лигаза. В некоторых вариантах осуществления ELM представляет собой реакцию расщепленного ELM для обеспечения лигирования индекса и удлинения индекса различных индексов, например, для обеспечения отдельного лигирования i5 и удлинения i7. Затем твердую подложку обрабатывают средством для денатурации последовательностей цепей, например NaOH, с получением таким образом меченых фрагментов нуклеиновых кислот.
[0092] В некоторых вариантах осуществления фрагменты депонируют на проточной кювете. В некоторых вариантах осуществления фрагменты гибридизуют с комплементарными праймерами, привитыми на проточную кювету или поверхность. В некоторых вариантах осуществления последовательности фрагментов секвенирования обнаруживают с помощью секвенирования матрицы или способов секвенирования следующего поколения, таких как секвенирование посредством синтеза.
[0093] В таблице 1 представлены примеры последовательностей, применяемых в комплексах транспосом для создания библиотеки меченых фрагментов нуклеиновых кислот.
7.7. Таблица 1: Иллюстративные последовательности
[0094] В некоторых аспектах способы создания библиотеки меченых фрагментов нуклеиновых кислот включают приведение иммобилизованного комплекса транспосомы в контакт с целевой нуклеиновой кислотой в условиях, достаточных для фрагментации целевой нуклеиновой кислоты на множество целевых фрагментов и присоединения 3’-конца первого транспозона к 5’-концам целевых фрагментов для получения множества 5’-меченных целевых фрагментов; обработку твердой подложки для удаления несвязанных нуклеиновых кислот; или обработку твердой подложки для удаления транспозазы из комплекса, необязательно посредством (a) нагревания твердой подложки и/или (b) мойки твердой подложки ферментативным денатурирующим средством, причем ферментативное денатурирующее средство необязательно содержит додецилсульфат натрия (SDS), гидрохлорид гуанидина, мочевину или протеиназу; обработку множества 5’-меченых целевых фрагментов полимеразой и лигазой для удлинения и лигирования 5’-меченых целевых фрагментов для получения полностью двухцепочечных меченых фрагментов, при этом необязательно обработку полимеразой и лигазой выполняют в присутствии средства разрушения вторичной структуры ДНК, причем средство разрушения необязательно представляет собой ДМСО; удаление полностью двухцепочечных фрагментов с твердой подложки, причем удаление необязательно включает применение нагревания и/или денатурирующего вещества, достаточного для отщепления полностью двухцепочечных меченых фрагментов от твердой подложки, при этом необязательно денатурирующее средство представляет собой NaOH; и отбор полностью двухцепочечных меченых фрагментов с использованием гранул для захвата, при этом необязательно гранулы для захвата представляют собой магнитные микроносители, и при этом необязательно выполнять две отдельные стадии отбора.
[0095] В некоторых вариантах осуществления иммобилизованный комплекс транспосом содержит твердую подложку; и комплекс транспосом, иммобилизованный на твердой подложке, причем комплекс транспосом содержит транспозазу; первый транспозон, содержащий 3’-концевую последовательность транспозона и якорную последовательность (якорь); второй транспозон, содержащий 5’-концевую последовательность транспозона и последовательность B15’; и полинуклеотид присоединения, содержащий комплементарную якорную последовательность (якорь’), последовательность A14’, спейсер и последовательность P5’, и связывающий элемент, содержащий биотин, причем биотин иммобилизован на твердой подложке. В некоторых вариантах осуществления способ дополнительно включает секвенирование одного или более полностью двухцепочечных меченых фрагментов.
7.8. Способы создания меченых фрагментов нуклеиновых кислот посредством комбинированной тагментации и индексирования
[0096] В некоторых вариантах осуществления библиотеки с парными концами после двойного индексирования могут быть получены из образца ДНК с использованием комбинированной стадии тагментации и индексирования. Благодаря предотвращению отдельных стадий тагментации и индексирования данный протокол имеет преимущество в виде простого применения и меньшей продолжительности. Более того, данный протокол позволяет избежать стадии денатурации и получить двухцепочечные библиотеки без необходимости отдельной стадии получения двухцепочечных образцов из денатурированных одноцепочечных образцов. Данный протокол также позволяет избежать определенных стадий промывки, что еще больше сокращает время, необходимое для рабочего процесса.
[0097] В данном способе применяют первый иммобилизованный транспозонный комплекс и второй иммобилизованный транспозонный комплекс. В первом иммобилизованном транспозонном комплексе последовательность X в первом транспозоне представляет собой якорную последовательность, а последовательность X’ в полинуклеотиде присоединения представляет собой комплемент якорной последовательности. Во втором иммобилизованном транспозонном комплексе последовательность X в первом транспозоне представляет собой якорную последовательность, а последовательность X’ в полинуклеотиде присоединения представляет собой комплемент якорной последовательности. Якорные последовательности первого и второго иммобилизованных транспозонных комплексов могут быть некомплементарными во избежание перекрестной гибридизации. Первый транспозонный комплекс может содержать иллюстративный первый полинуклеотид присоединения, содержащий (i) комплемент якорной последовательности, последовательность A14’, спейсер и последовательность P5’, и (ii) связывающий элемент, содержащий биотин, а второй транспозонный комплекс может содержать иллюстративный второй полинуклеотид присоединения, содержащий (i) комплемент якорной последовательности, последовательность B15’, спейсер и последовательность P7’, и (ii) связывающий элемент, содержащий биотин.
[0098] При комбинированной тагментации и индексировании в каждую лунку можно добавлять суспензию связанных с твердой подложкой транспозонных комплексов (BLT, т. е. иммобилизованных комплексов транспосом), содержащих первый и второй комплексы транспосом. Затем можно осуществлять стадию индексирования в одном реакционном растворе с адаптерами индекса 1 (i7), адаптерами индекса 2 (i5) и последовательностями, необходимыми для образования кластеров секвенирования, лигированных в том же реакционном растворе, что и для проведения тагментации. Первые индексирующие олигонуклеотиды могут содержать последовательность A14, последовательность i5 и последовательность P5, а вторые индексирующие олигонуклеотиды могут содержать последовательность B15, последовательность i7 и последовательность P7. Условия на стадии тагментации/лигирования включают в себя меченый буфер и ДНК-лигазу E. Coli, добавленные к смеси целевой ДНК, первому и второму комплексам транспосом и первому и второму индексирующим олигонуклеотидам. Данная комбинированная стадия тагментации и индексирования может продолжаться в течение различных периодов, например по меньшей мере от 1 минуты до по меньшей мере 15 минут или по меньшей мере от 5 минут до по меньшей мере 15 минут.
[0099] Реакции тагментации и индексирования можно затем остановить с использованием различных способов, включая нагревание твердой подложки и/или промывку твердой подложки ферментативным денатурирующим средством, например, необязательно с использованием додецилсульфата натрия (SDS), гидрохлорида гуанидина, мочевины или протеиназы. Время остановки реакции как при нагревании, так и/или при использовании стадии промывки может варьироваться от по меньшей мере 1 минуты до по меньшей мере 5 минут.
[0100] Меченные 5’-меткой целевые фрагменты, лигированные для индексирующих олигонуклеотидов, можно обрабатывать полимеразой для удлинения и получения полностью двухцепочечных меченых фрагментов. Реакция удлинения может протекать в течение различных периодов, например по меньшей мере от 1 минуты до по меньшей мере 10 минут или по меньшей мере от 2 минут до по меньшей мере 10 минут.
[0101] Кроме того, при получении библиотеки, включающей комбинированную тагментацию и индексирование, также можно избежать стадии кПЦР, так как конечный продукт представляет собой двухцепочечную библиотеку ДНК в растворе.
[0102] В некоторых вариантах осуществления приведение в контакт первого иммобилизованного комплекса транспосомы со вторым иммобилизованным комплексом транспосомы и обработку множества 5’-меченых целевых фрагментов лигазой проводят в одну реакцию.
[0103] В некоторых вариантах осуществления двухцепочечные меченые фрагменты получают в растворе.
[0104] Некоторые варианты осуществления дополнительно содержат отбор полностью двухцепочечных меченых фрагментов с использованием гранул для захвата, причем необязательно гранулы для захвата представляют собой магнитные микроносители, и необязательно дополнительно выполняют две отдельные стадии отбора.
[0105] В некоторых вариантах осуществления способ дополнительно включает секвенирование одного или более полностью двухцепочечных меченых фрагментов.
Целевая нуклеиновая кислота
[0106] Целевая нуклеиновая кислота может представлять собой любой тип, который содержит ДНК, РНК, кДНК и т. п. Например, целевая нуклеиновая кислота может находиться в различных состояниях очистки, включая очищенную нуклеиновую кислоту. В некоторых вариантах осуществления биологический образец содержит смесь нуклеиновых кислот (таких как ДНК), белка, других видов нуклеиновых кислот, других клеточных компонентов и/или любого другого загрязняющего вещества, приблизительно в той же пропорции, что и in vivo. Например, в некоторых вариантах осуществления компоненты находятся в той же пропорции, что и в интактной клетке. Поскольку предложенные в настоящем документе способы позволяют связывать нуклеиновую кислоту или ДНК с твердой подложкой посредством процесса мечения, другие загрязняющие вещества могут быть удалены посредством промывки твердой подложки после осуществления тагментации. Биологический образец может содержать, например, неочищенный клеточный лизат или цельные клетки. Например, неочищенный клеточный лизат, нанесенный на твердую подложку посредством способа, описанного в настоящем документе, необязательно необходимо подвергнуть одной или более стадиям разделения, которые традиционно применяют для выделения нуклеиновых кислот из других клеточных компонентов.
[0107] Таким образом, в некоторых вариантах осуществления биологический образец может содержать не только очищенные нуклеиновые кислоты из любого источника, но и, например, неочищенные нуклеиновые кислоты, обнаруженные в крови, плазме, сыворотке, лимфе, слизи, мокроте, моче, сперме, спинномозговой жидкости, бронхиальной аспирате, фекалиях и мацерированной ткани или их лизате, или любой другой биологический препарат, содержащий нуклеиновую кислоту или материал ДНК. Целевая нуклеиновая кислота может представлять собой образец ткани, образец опухоли, раковые клетки или образец биопсии. Целевая нуклеиновая кислота может представлять собой бесклеточную ДНК (скДНК).
[0108] Целевая нуклеиновая кислота может быть получена из любого вида или смеси видов. Например, целевая нуклеиновая кислота может быть получена от млекопитающего (такого как человек, собака, кошка, корова, свинья, овца или другое домашнее животное) или от других видов, таких как рыба, бактерии, вирус, грибок или архебактерия. Нуклеиновая кислота может поступать из образцов окружающей среды, таких как почва или вода.
[0109] В некоторых вариантах осуществления целевая нуклеиновая кислота представляет собой ДНК. В одном таком варианте осуществления ДНК является двухцепочечной. В некоторых дополнительных вариантах осуществления двухцепочечная ДНК содержит геномную ДНК. В некоторых других вариантах осуществления целевая нуклеиновая кислота представляет собой РНК или ее производное или кДНК. В некоторых вариантах осуществления целевая нуклеиновая кислота представляет собой продукт реакции выше по потоку, такой как явление амплификации или репликации, например, ампликон. В некоторых вариантах осуществления целевая нуклеиновая кислота представляет собой ДНК, обработанную бисульфитом.
[0110] В некоторых вариантах осуществления биологический образец (необработанный образец или экстракт) обрабатывают для очистки целевых нуклеиновых кислот перед способами тагментации, описанными в настоящем документе. В некоторых вариантах осуществления биологический образец представляет собой необработанный образец или необработанный образец лизата (например, кровь, слюну, клетку или клетки). В некоторых вариантах осуществления способ обработки включает получение исходного образца, лизата исходного образца или предварительно обработанного образца (например, образца крови или слюны), смешивание образца с лизирующим буфером и протеиназой K, инкубирование смеси для лизиса клеток в образце и высвобождение ДНК из клеток, таким образом обеспечивая целевую(ые) нуклеиновую(ые) кислоту(ы) для способов тагментации, описанных в настоящем документе. Количество биологического образца конкретно не указывается при условии, что биологический образец содержит достаточное количество нуклеиновых кислот для анализа. Таким образом, количество биологического образца может включать от приблизительно 1 мкл до приблизительно 500 мкл, например 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80 90, 100, 150, 200, 250, 300, 350, 400, 450 или 500 мкл, или количество в пределах диапазона, определяемого любыми двумя из вышеупомянутых значений.
[0111] Компоненты исходных образцов или лизаты исходных образцов, такие как кровь, или добавки в предварительно обработанных образцах, таких как слюна, собранная в пробирку для сбора генов Oragene (стабилизирующие средства в пробирках для сбора), могут ингибировать реакции тагментации. Таким образом, в настоящем документе предложен способ обработки исходного образца, исходного лизата образца или предварительно обработанного образца для преодоления данной проблемы. В некоторых вариантах осуществления способ включает получение необработанного образца, лизата необработанного образца или предварительно обработанного образца (например, образца крови или слюны), смешивание образа с лизирующим буфером, протеиназой K и гранулами для захвата или очистки ДНК (например, гранулы SPRI, гранулы, содержащие карбоксильные группы, если гранулы необязательно представляют собой магнитные гранулы), инкубацию смеси для лизиса клеток в образце и высвобождения ДНК из клеток, захват таким образом ДНК на гранулы для очистки ДНК или SPRI и отделение гранул, содержащих захваченную ДНК, от смеси. Разделение служит для удаления потенциальных ингибиторов тагментации, присутствующих в супернатанте. Способ дополнительно включает необязательно промывку гранул, содержащих захваченную ДНК, и элюирование ДНК из гранул с образованием целевой(-ых) нуклеиновой(-их) кислоты(кислот).
[0112] В некоторых вариантах осуществления целевая нуклеиновая кислота присутствует в количестве, достаточном для создания библиотеки для секвенирования. В некоторых вариантах осуществления количество целевой нуклеиновой кислоты представляет собой количество гДНК 10-500 нг, например 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400 или 500 нг, или количество в пределах диапазона, определяемого любыми двумя из вышеупомянутых значений. В некоторых вариантах осуществления целевая нуклеиновая кислота представляет собой гДНК, присутствующую в количестве 50 нг.
7.10. Способы секвенирования
[0113] Некоторые из способов, предложенных в настоящем документе, включают способы анализа нуклеиновых кислот. Такие способы включают в себя получение библиотеки матричных нуклеиновых кислот целевой нуклеиновой кислоты, получение данных последовательности из библиотеки матричных нуклеиновых кислот и необязательно сборку представления последовательности целевой нуклеиновой кислоты. Фрагменты ДНК, полученные посредством опосредованной транспосомой тагментации, можно секвенировать в соответствии с любой подходящей методикой секвенирования, такой как прямое секвенирование или секвенирование следующего поколения, включая секвенирование посредством синтеза, секвенирование посредством лигирования, секвенирование посредством гибридизации, секвенирование на основе обнаружения высвобожденных протонов, могут применяться электрический детектор и связанные с ним методики, которые доступны на рынке от компании Ion Torrent (Guilford, CT, дочерняя компания Life Technologies), секвенирование наночастиц и т. п. В некоторых вариантах осуществления фрагменты ДНК секвенируют на твердой подложке, такой как проточная кювета. Примеры проточных кювет и связанных с ними жидкостных систем и платформ обнаружения, которые можно легко применять в рамках данного изобретения, описаны, например, в Bentley et al., Nature 456:53-59 (2008), WO 04/018497; US 7,057,026; WO 91/06678; WO 07/123744; US 7,329,492; US 7,211,414; US 7,315,019; US 7,405,281 и US 2008/0108082, которые включены в настоящий документ посредством ссылки.
[0114] Способы, описанные в настоящем документе, не ограничиваются каким-либо конкретным применяемым типом аппаратуры для секвенирования.
ПРИМЕРЫ
[0115] Следующие примеры служат для иллюстрации, но не ограничивают предложенное в настоящем документе описание.
Пример 1: Олигонуклеотиды, которые не соединяются с отщепляемым пользователем линкером
[0116] Проводили эксперимент для сравнения данных секвенирования без ПЦР с данными секвенирования с использованием ПЦР. Библиотеки получали с применением TruSeq PCR-free, TruSeq Nano (+ПЦР), Nextera Flex (+ПЦР) и способа, составляющего предмет настоящего изобретения, а затем секвенировали до 25-кратного покрытия. Каждый способ предполагал две повторности, причем всего создавали восемь библиотек. На Фиг. 7A показано, что способы без ПЦР (столбцы в линию и столбцы в клетку, которые представляют собой настоящий способ и TruSeq без ПЦР соответственно) имеют улучшенную точность и повторное распознавание индела по сравнению со способами с ПЦР (черные столбцы и белые столбцы, которые представляют собой TruSeq Nano и Nextera Flex соответственно). На Фиг. 7B показано, что покрытие промоторных областей с высоким содержанием GC также улучшено в способах без ПЦР (две нижние панели, причем настоящий способ представлен на нижней левой панели, а TruSeq без ПЦР на нижней правой панели).
Пример 2. Создание библиотеки меченых фрагментов нуклеиновых кислот без использования ПЦР-амплификации
[0117] Стадия А1. Получение иммобилизованных комплексов транспосом - индексированные гранулы. Смесь: (1) первого транспозона, имеющего последовательность 3’-мозаичного конца (ME) (SEQ ID NO: 6), первой последовательности метки (A14) и 5’-праймерной последовательности (P5’) (конечная концентрация 25 мкМ); (2) второго транспозона с 5’-комплементарной концевой мозаичной последовательностью (МЕ’; SEQ ID NO: 3) и второй последовательности метки (B15’) (конечная концентрация 37,5 мкМ); и (3) полинуклеотида присоединения, включающего 3’-последовательность, комплементарную второй последовательности метки (B15), индексную последовательность (в данном случае i701 в таблице 1) и 5’-праймерную последовательность (P7) с биотином на 5’-конце (конечная концентрация 25 мкM), отжигали посредством обработки 10 мМ трис-HCl (pH 8,0), 1 мМ EDTA и 25 мМ NaCl, нагревания при 95°C в течение 10 мин, а затем охлаждения до 10°C в течение приблизительно 15 мин. Затем отожженные транспозоны/полинуклеотид присоединения (конечная концентрация 2 мкМ по отношению к полинуклеотиду присоединения) смешивали с ферментом транспозазой (конечная концентрация 6,1 мкМ) и инкубировали при 37°C в течение ночи с образованием комплексов транспосом в фазе растворения (Фиг. 4A, фигуры с индексированными гранулами). Затем комплексы транспосом в фазе растворения иммобилизовали на твердой подложке посредством смешивания с покрытыми стрептавидином гранулами. В некоторых примерах иммобилизацию можно проводить в присутствии буфера HT1 (Illumina), который представляет собой буфер с высоким содержанием соли, способствующий образованию биотин-стрептавидиновых связей. После перемешивания в HT1 в течение 1 ч гранулы осаждали и однократно промывали в смеси буферного раствора HT1 и 50% глицеринового стандартного буфера для хранения (Illumina) (9:1). Затем гранулы ресуспендировали в 360 мкл буфера, содержащего 50 мМ Tris pH 7,5, 30 мМ NaCl и 0,1% Tween 20. Добавляли 40 мкл EPX2 (Illumina) и гранулы дополнительно вращали в течение 10 мин при комнатной температуре. Гранулы снова осаждали, а затем ресуспендировали в 396 мкл того же буфера, обрабатывали 4 мкл одноцепочечного связывающего белка (5 мг/мл) и снова вращали в течение 10 мин. Гранулы снова осаждали, промывали один раз в смеси 9:1 HT1: стандартный буфер для хранения и ресуспендировали в 15% глицериновом стандартном буфере для хранения (Illumina). Полученные индексированные гранулы применяли в протоколах тагментации с единичным индексированием и двойным индексированием.
[0118] Стадия А2. Формирование иммобилизованных комплексов транспосом - универсальные гранулы. Отожженные транспозоны получали, как описано на стадии А1, за исключением того, что полинуклеотид присоединения включал в себя 5’-последовательность, комплементарную первой последовательности метки (A14’) и 3’-праймерной последовательности (P5’) с биотином на 3’-конце (для одного индексирования), и необязательно промежуточную универсальную последовательность нитроиндола (для двойного индексирования). Транспосомные комплексы получали из отожженных транспозонов и иммобилизовали на твердой подложке, как описано для стадии А1. Полученные универсальные гранулы применяли в протоколах единичного индексирования и двойного индексирования, как указано выше.
[0119] Стадия b. Тагментация. ДНК (например, от приблизительно 50 пг до 5 мкг) смешивали с иммобилизованными комплексами транспосомы и 50 мкл буфера для тагментации (10 мМ трис-ацетата (pH 7,6), 5 мМ ацетата магния и 10% диметилформамида, как описано в патентах США №№ 9,080,211, 9,085,801 и 9,115,396, каждый из которых включен в настоящий документ посредством ссылки, и полученную смесь инкубировали при 55°C в течение 5 мин. Получали библиотеку меченых фрагментов ДНК, иммобилизованных на гранулах. Реакционную смесь для тагментации обрабатывали 10 мкл стоп-буфера, содержащего 5% SDS, 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20, и полученную смесь инкубировали при 37°C в течение 5 мин для денатурации фермента транспозазы из комплексов транспосом. Затем гранулы с иммобилизованными фрагментами ДНК осаждали на магните и три раза промывали промывочным буфером (100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween) для удаления остатков SDS. Гранулы отделяли от супернатанта посредством магнитного захвата и дополнительно промывали с помощью 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20. Полученные меченые фрагменты для всех четырех подходов показаны на Фиг. 4B.
[0120] Стадия С. Удлинение, лигирование и индексирование. Удлинение и лигирование выполняли посредством приведения гранул в контакт со смесью для удлинения и лигирования (ELM), которая включала ДНК-полимеразу Т4 (экзонуклеазу минус) и ДНК-лигазу E. coli, и инкубирования при 30°C в течение 15 мин, а затем при 16°C в течение 15 мин, заполняя таким образом гэп фрагменты ДНК между 3’-концами фрагментов и 5’-концами последовательностей ME’ и удлинения одноцепочечных областей с образованием полностью двухцепочечных продуктов посредством введения оставшихся последовательностей из полинуклеотида присоединения (Фиг. 4C). В определенных случаях на данной стадии также добавляли индексы. Как показано на Фиг. 4D, правая верхняя фигура, к универсальной конструкции гранул/одноиндексных значений добавляли индекс посредством добавления олигонуклеотида с двухцепочечной последовательностью праймера (P7/P7’), двухцепочечной индексной последовательностью (в данном случае i701/i701’) и одноцепочечной выступающей областью с 3’-последовательностью, комплементарной второй последовательности метки (B15). Для подхода с индексированными гранулами/двойным индексированием (Фиг. 4D, левая нижняя фигура) реагент для определения индекса включал в себя двухцепочечную праймерную последовательность (P5/P5’), двухцепочечную индексную последовательность (в данном случае i501/i501’) и одноцепочечную выступающую область с 5’-последовательностью, комплементарной метке первой последовательности (A14’). Для подхода с универсальными гранулами/двойным индексированием (Фиг. 4D, правая нижняя фигура) индексный реагент был таким же, что и в случае универсальной гранулы/единичного индекса, но дополнительный второй индексный реагент, содержащий комплементарную праймерную последовательность (P5) и индексную последовательность (i5), гибридизовали и лигировали с 5’-концом последовательностей A14.
[0121] Реакционную смесь обрабатывали с помощью 10 мкл стоп-буфера, содержащего 5% SDS, 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20, и полученную смесь инкубировали при 37°C в течение 5 мин для денатурации фермента транспозазы из комплексов транспосом. Затем гранулы с иммобилизованными фрагментами ДНК осаждали на магните и три раза промывали промывочным буфером (100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween) для удаления остатков SDS. Гранулы отделяли от супернатанта посредством магнитного захвата и дополнительно промывали с помощью 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20.
[0122] Стадия D. Высвобождение фрагментов с нанесенной меткой. После третьей промывки гранулы ресуспендировали в 100 мкл 0,2 н. NaOH для денатурации фрагментов библиотеки и их высвобождения из гранул. Библиотеку очищали посредством добавления 100 мкл супернатанта непосредственно к 180 мкл гранул SPRI в соответствии со стандартным протоколом очистки SPRI и элюирования в 15 мкл буфера для ресуспендирования Illumina. Библиотеки количественно определяли посредством способа количественной ПЦР, а затем готовили к секвенированию на MiSeq® посредством разведения до 5 мкл около 3200 пМ и денатурации 5 мкл 0,2 н. NaOH при комнатной температуре в течение 5 минут. Добавляли охлажденный буфер Illumina HT1 (990 мкл) и весь 1 мл смеси загружали в картридж MiSeq®.
[0123] Результаты показаны на Фиг. 4E и обобщены в таблице 2 ниже для индексированных гранул или универсальных гранул с единичным или двойным индексированием.
Таблица 2: Результаты получения библиотеки для различных конфигураций комплекса транспосом
[0124] С использованием описанных в данном документе способов удалось преодолеть ошибки в данных секвенирования, которые обычно вносятся посредством ПЦР. Например, при использовании способов без ПЦР, описанных в настоящем документе, полученные библиотеки фрагментов нуклеиновых кислот секвенировали и последовательности не имели значительных гэпов в обогащенных GC областях, что обычно наблюдается при использовании ПЦР.
Пример 3. Сравнение секвенирования испытываемых библиотек
[0125] В данном примере две сравнительные библиотеки из образца, содержащего известное 100%-е повторное размножение GC (FMR1), получали с использованием наборов Nextera™ DNA Flex (который включает ПЦР) и TruSeq™ PCR-Free Library Prep, а четыре библиотеки для испытания получали, как описано в настоящем документе, с использованием способа универсальных гранул/двойного индексирования, как показано на Фиг. 4D. Как показано на Фиг. 8, повторы для данных секвенирования из библиотеки, полученной с использованием набора Nextera™ DNA Flex, не были получены (первая колонка, столбец не показан, так как результат составляет 0). Повторы описывали с использованием TruSeq™ PCR-Free (колонка 2) и четырех библиотек для испытания с использованием способов, описанных в настоящем документе (образцы 1-4, колонки 3-6). Таким образом, настоящие способы демонстрируют улучшение вызова повторных размноженных образцов с 100% GC областей.
Пример 4: Раздвоенные олигонуклеотиды с отщепляемым линкером
[0126] Стадия 1. Образование комплексов транспосом. Биотинилированный олигонуклеотид (50 мкМ), содержащий 5’-биотин, три остатка T, затем три остатка U, P5, A14 и ME, с последовательностью: /5Biosg/TTUUUAATGATACGGCGACCACCGAGATCTACACTCGTCGGCAGCGTCAGATGTGTATAAGAGACA (SEQ ID NO: 20)
и олигонуклеотид, содержащий ME’_B15’_P7’ (75 мкМ), с последовательностью: CTGTCTCTTATACACATCTCCGAGCCCACGAGACATCTCGTATGCCGTCTTCTGCTTG (SEQ ID NO: 21)
обрабатывали 10 мМ трис-HCl (pH 8,0), 1 мМ EDTA и 25 мМ NaCl, нагревали до 95°C в течение 10 мин, а затем охлаждали до 10°C со скоростью -0,1°C в секунду. Затем отожженные транспозоны смешивали с ферментом транспозазой таким образом, чтобы итоговая концентрация биотинилированного олигонуклеотида составляла 2 мкМ, и концентрация транспозазы составляла 4 мкМ. Смесь инкубировали при 37°C в течение ночи с образованием комплексов транспосом в фазе растворения. Затем комплексы транспосом в фазе растворения иммобилизовали на твердой подложке посредством смешивания с покрытыми стрептавидином гранулами и буфером HT1 (Illumina) и вращения при комнатной температуре в течение 1 часа. Затем гранулы трижды промывали в буфере HT1, а затем ресуспендировали в 15%-м глицериновом стандартном буфере для хранения (Illumina) (Фиг. 2A).
[0127] Стадия 2. Тагментация. ДНК (например, от приблизительно 50 пг до 5 мкг) смешивали с иммобилизованными комплексами транспосомы и 50 мкл буфера для тагментации (10 мМ трис-ацетата (pH 7,6), 5 мМ ацетата магния и 10% диметилформамида, как описано в патентах США №№ 9,080,211, 9,085,801 и 9,115,396, каждый из которых включен в настоящий документ посредством ссылки, и полученную смесь инкубировали при 55°C в течение 5 мин. Получали библиотеку меченых фрагментов ДНК, иммобилизованных на гранулах. Реакционную смесь для тагментации обрабатывали 10 мкл стоп-буфера, содержащего 5% SDS, 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20, и полученную смесь инкубировали при 37°C в течение 5 мин для денатурации фермента транспозазы из комплексов транспосом. Затем гранулы с иммобилизованными фрагментами ДНК осаждали на магните и три раза промывали промывочным буфером (100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween) для удаления остатков SDS. Гранулы отделяли от супернатанта посредством магнитного захвата и дополнительно промывали с помощью 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20.
[0128] Стадия 3. Удлинение и лигирование. Удлинение и лигирование выполняли посредством приведения гранул в контакт со смесью для удлинения и лигирования (ELM), которая включала ДНК-полимеразу Т4 (экзонуклеазу минус) и ДНК-лигазу E. coli, и инкубирования при 30°C в течение 15 мин, а затем при 16°C в течение 15 мин, заполняя таким образом гэп фрагменты ДНК между 3’-концами фрагментов и 5’-концами последовательностей ME’. Реакционную смесь обрабатывали с помощью 10 мкл промывочного буфера, содержащего 5% SDS, 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20, и полученную смесь инкубировали при 37°C в течение 5 мин для денатурации фермента транспозазы из комплексов транспосом. Затем гранулы с иммобилизованными фрагментами ДНК осаждали на магните и три раза промывали промывочным буфером (100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween) для удаления остатков SDS. Гранулы отделяли от супернатанта посредством магнитного захвата и дополнительно промывали с помощью 100 мМ трис-HCl (pH 7,5), 100 мМ NaCl и 0,1% Tween 20.
[0129] Стадия 4. Высвобождение фрагментов с нанесенной меткой. После третьей промывки гранулы ресуспендировали в буфере, содержащем 50 мМ ацетата калия, 20 мМ трис-ацетата, 10 мМ ацетата магния и 100 мкг/мл BSA. Добавляли 5 мкл смеси ферментов, содержащей урацил-ДНК-гликозилазу и эндонуклеазу VIII, и библиотеки инкубировали при 37°C в течение 30 мин. Ферменты действовали совместно, расщепляя U’ в олигонуклеотидах присоединения и высвобождая фрагменты библиотеки в раствор. Библиотеку очищали посредством добавления 100 мкл супернатанта непосредственно к гранулам SPRI в соответствии со стандартным протоколом очищения SPRI по размеру (0,5x справа и 0,7-0,8x слева) и элюирования в 15 мкл буфера для ресуспендирования Illumina. Полученные библиотеки количественно определяли посредством кПЦР (Фиг. 2B).
Пример 5: Модифицированный протокол удлинения и лигирования
[0130] Можно применять альтернативный протокол для реакции удлинения и лигирования с индексированными вариациями (пример 2C). В некоторых случаях удлинение индекса i7 неэффективно вследствие вторичной структуры в определенных индексных последовательностях. Таким образом, для конструкции, представленной на Фиг. 6C, исследовали восемь различных пар индексов. Как показано на Фиг. 9A, % CV для пула индексов составлял 76%. Было обнаружено, что добавление олигонуклеотида Р7’ в ходе реакции удлинения и индексирования повышает эффективность определенных пар индексов, возможно, вследствие гибридизации с последовательностью Р7, которая может нарушать или предотвращать образование вторичной структуры. В данном случае реакция удлинения и индексирования также включает лигирование. P7’ (с 5’-фосфатом для лигирования) добавляют в ходе реакции удлинения-лигирования-индексирования при 2,5 мкМ. Для конструкции, представленной на Фиг. 6D, на Фиг. 9B представлен % CV для восьми пар индексов менее 30%.
[0131] Добавление ДМСО к реакции удлинения-лигирования-индексирования олигонуклеотида P7 дополнительно повышает показатель % CV. В данном примере в конструкции, представленной на Фиг. 6D, ДМСО добавляют в реакцию удлинения-лигирования-индексирования на уровне 5%. Реакция удлинения и лигирования содержит все ферменты и компоненты, необходимые для удлинения и лигирования. Реакционную смесь инкубируют при 37°C в течение 30 мин, после чего продолжают работу в нормальном рабочем процессе. Как показано на Фиг. 9C, % CV для восьми разных пар индексов составлял менее 20%. Данный результат позволяет использовать систему без количественной оценки выхода ДНК из библиотеки для каждого образца с целью коррекции вариаций характеристик индексирования.
Пример 6: Иммобилизованная транспосома в пробирке без ПЦР
[0132] Данный пример демонстрирует способ и систему для иммобилизации комплексов транспосом в пробирке, например в пробирке для ПЦР, для секвенирования целого генома без ПЦР. Способы осуществляют аналогично способу, описанному в примере 2, и всю обработку можно выполнять в той же пробирке.
[0133] Получение иммобилизованных комплексов транспосом в пробирке. Смесь: (1) первого транспозона, имеющего последовательность 3’-мозаичного конца (ME) (SEQ ID NO: 6), первой последовательности метки (A14) и 5’-праймерной последовательности (P5) (конечная концентрация 25 мкМ); (2) второго транспозона с 5’-комплементарной концевой мозаичной последовательностью (МЕ’; SEQ ID NO: 3) и второй последовательности метки (B15’) (конечная концентрация 37,5 мкМ); и (3) полинуклеотида присоединения, включающего 3’-последовательность, комплементарную второй последовательности метки (B15), индексную последовательность и 5’-праймерную последовательность (P7) с биотином на 5’-конце (конечная концентрация 25 мкM), отжигали посредством обработки 10 мМ трис-HCl (pH 8,0), 1 мМ EDTA и 25 мМ NaCl, нагревания при 95°C в течение 10 мин, а затем охлаждения до 10°C в течение приблизительно 15 мин. Полинуклеотид присоединения включал в себя 5’-последовательность, комплементарную первой последовательности метки (A14’), и 3’-праймерную последовательность (P5’) с биотином на 3’-конце (для единичного индексирования). В некоторых вариантах осуществления полинуклеотид присоединения включал промежуточную универсальную нитроиндольную последовательность (для двойного индексирования). Затем отожженные транспозоны/полинуклеотиды присоединения (конечная концентрация 2 мкМ на основе полинуклеотида присоединения) смешивали с ферментом транспозазой (конечная концентрация 6,1 мкМ) и инкубировали при 37°C в течение ночи с образованием комплексов транспосом в фазе растворения. Затем комплексы транспосом в фазе растворения иммобилизовали на поверхности пробирки для ПЦР посредством смешивания комплексов (имеющих биотин) в пробирке для ПЦР с поверхностью, покрытой стрептавидином. В некоторых примерах иммобилизацию можно проводить в присутствии буфера HT1 (Illumina, San Diego, CA), который представляет собой буфер с высоким содержанием соли, способствующий образованию биотин-стрептавидиновых связей. После инкубации в течение 1 ч. пробирку для ПЦР промывали в смеси буфера TWB (Illumina, San Diego, CA). Полученные пробирки с иммобилизованными на них транспосомами использовали в протоколах агментации с единичным индексированием и двойным индексированием.
[0134] Рабочий процесс получения библиотеки, включая стадии тагментации, удлинения, лигирования и индексирования, выполняли в соответствии с описанием в примере 2. После завершения стадий, описанных в примере 2, были созданы фрагменты библиотеки, иммобилизованные на поверхности пробирки. Пробирки промывали 50 мкл 0,2 н. NaOH для денатурации фрагментов библиотеки и высвобождения их с поверхности пробирок. Библиотеки количественно определяли посредством способа количественной ПЦР, а затем готовили к секвенированию на MiSeq® посредством разведения до 5 мкл около 3200 пМ и денатурации 5 мкл 0,2 н. NaOH при комнатной температуре в течение 5 минут. Добавляли охлажденный буфер Illumina HT1 (990 мкл) и весь 1 мл смеси загружали в картридж MiSeq®.
[0135] Результаты секвенирования представлены на Фиг. 10, где показана степень покрытия в области генома, которую, как известно, сложно секвенировать (ген RNPEPL1). В двух способах получения библиотек с помощью ПЦР (Nextera Flex и Nextera на основе пробирок) показан гэп, в котором ни одна из последовательностей не покрывала область гена, отмеченную прямоугольной ограничительной рамкой. Было обнаружено, что два способа без ПЦР имеют хорошее покрытие в данной области, как показано в разделах, обозначенных на Фиг. 10 как «PCR-free Flex» и «Nextera без ПЦР на основе пробирок».
Пример 7: Препарат без отщепляемого линкера без использования ПЦР
[0136] Следующий пример демонстрирует способ и систему для иммобилизации комплексов транспосом в пробирке, например, в пробирке для ПЦР, для секвенирования целого генома без ПЦР. Способы осуществляют аналогично способу, описанному в примере 2, но без нуклеотида присоединения, содержащего отщепляемый линкер или стадию отщепления линкера. В данном способе применяют три отдельных олигонуклеотида. Дополнительные компоненты данного способа включают транспозон Tn5, SDS, смесь для интенсивного лигирования (ELM) и покрытую стрептавидином поверхность (такую как гранулы или планшет).
[0137] Смесь (1) первого транспозона, имеющего последовательность ME, первую последовательность метки (A14), последовательность i5 и 5’-праймерную последовательность (P5); (2) второго транспозона с 5’-комплементарной концевой мозаичной последовательностью (МЕ’), второй меткой (B15’), i7’-последовательностью и 3’-праймерной последовательность (P7’); и (3) полинуклеотида присоединения, содержащего последовательность P7 и связывающий элемент (такой как биотин), можно получить и закрепить на поверхности (такой как планшет или гранула, покрытые стрептавидином). Транспосомы можно построить и прикрепить к поверхности в ходе одной стадии в присутствии матричной нуклеиновой кислоты в буфере для тагментации.
[0138] Тагментацию выполняют в течение 5 мин при 55°C. В реакцию тагментации может быть включен ПЭГ. Смесь обрабатывают SDS для удаления транспозазы Tn5. Выполняют ELM и удаляют супернатант. При использовании данного способа не требуются гранулы для захвата SPRI и, следовательно, нет необходимости в настройке размера вставки посредством плотности загрузки гранул. Для подготовки секвенирования фрагменты можно элюировать в NaOH.
Пример 8: Получение библиотек из 96 лунок без ПЦР
[0139] Следующий пример демонстрирует способ и систему для получения до 96 библиотек с парными концами после двойного индексирования из образца ДНК. Способы осуществляют аналогично способу, описанному в примере 2, но используют две стадии инкубации с иммобилизованными гранулами для обеспечения получения без конечной стадии кПЦР. Поскольку кПЦР может занимать много времени (до двух часов), данный протокол имеет преимущество в виде простого применения и меньшую продолжительность применения. Данный способ был совместим с вводимыми количествами ДНК в 25-1000 нг. В образцах человеческой ДНК и других больших сложных геномах вводимое количество ДНК может составлять более 200 нг. Для вводимого количества ДНК от 200 до 1000 нг не требуется количественное определение и нормализация исходного образца ДНК. Перед секвенированием выполняли количественное определение и нормализацию библиотек для вводимого количества от 25 до 200 нг ДНК.
[0140] В данном способе применяют первый транспозон, как описано на Фиг. 6D, причем последовательность X в первом транспозоне представляет собой ATCTGACTATCCCCTGCG (SEQ ID NO: 23), и последовательность X’ в прикрепляемом полинуклеотиде представляет собой CGCAGGGGATAGTCAGAT (SEQ ID NO: 24). Якорная последовательность представляет собой А14, а спейсерная область состоит из 2 углеродных спейсеров C18 и 1 C9 (последовательности, отличные от ДНК).
[0141] Тагментация. В каждую лунку 96-луночного планшета для ПЦР добавляли ДНК в 10 мМ трис-HCl (приблизительно 2-30 мкл) таким образом, чтобы общее вводимое количество (нг) находилось в пределах необходимого диапазона. Если объем ДНК составляет менее 30 мкл, к образцам ДНК добавляют не содержащую нуклеазу воду для доведения общего объема до 30 мкл. В каждую лунку добавляли суспензию связанных с гранулами транспосом (Blt) (10 мкл) с последующим добавлением 10 мкл буфера для тагментации. BLT загружали на гранулы в концентрации от приблизительно 50 до 1000 нМ. Их ресуспендировали в 15% глицериновом стандартном буфере для хранения, который состоит из 15% глицерина, 100 мМ NaCl, 50 мМ Tris HCl pH 7,5, 0,1 мМ EDTA, 1 мМ DTT, 0,1% Triton X-100. Образцы пипетировали для перемешивания до полного ресуспендирования гранул, планшет запечатывали и помещали в термоциклер при 41°C на 5 мин, а затем выдерживали при 10°C. Значения времени тагментации, составляющие 15, 30, 45, 60, 75, 90, 105, 120 и 300 с, обеспечивают сходный выход со сдвигом на большую длину вставки при меньшем времени инкубации (например, 15 с обеспечивали более длинные размеры вставки, чем 60, 120 или 300 с). В некоторых примерах время тагментации сокращено до 1 мин.
[0142] В каждую лунку добавляли 10 мкл стоп-буфера для тагментации, содержащего SDS, полученную смесь пипетировали до полного восстановления гранул и смесь инкубировали в течение 1-5 мин. Планшет помещали на магнитную стойку на 2-5 мин, супернатант извлекали и удаляли. Планшет удаляли с магнита и в каждую лунку непосредственно на гранулы добавляли 150 мкл промывочного буфера для тагментации. Образцы перемешивали пипеткой и планшет помещали на магнитную стойку до тех пор, пока раствор не станет прозрачным (приблизительно 2-5 мин). На данной стадии из образцов удаляли SDS.
[0143] Удлинение/лигирование. Адаптеры индекса 1 (i7), адаптеры индекса 2 (i5) и последовательности, необходимые для генерации кластера секвенирования, добавляют посредством удлинения/лигирования. В то время, как планшет находился на магнитной стойке, супернатант извлекали и удаляли. Планшет извлекали из магнитного штатива и в каждую лунку добавляли по 45 мкл смеси для удлинения и лигирования с последующим повторным суспендированием гранул. Затем к каждому образцу добавляли соответствующие адаптеры индекса (5 мкл), планшет герметизировали и помещали в термоциклер при 37°C на 5 мин, при 50°C на 5 мин, а затем выдерживали при 10°C. Каждый из периодов инкубации при 37°C и 50°C можно выполнять в течение 1, 3 или 5 мин, и все вариации дают аналогичный выход библиотеки. Более короткие периоды инкубации позволяют получать библиотеки с увеличенным размером фрагментов, тогда как более длительные инкубации обеспечивают более высокие концентрации продуктов лигирования. Затем планшет помещали на магнитную стойку на 2-5 мин и удаляли супернатант.
[0144] В лунки добавляли 75 мкл промывочного буфера для тагментации и смесь пипетировали для ресуспендирования гранул для тагментации. Супернатант удаляли, планшет центрифугировали и помещали на магнит на 2-5 мин. В каждую лунку добавляли 45 мкл 0,2 н. NaOH для денатурации фрагментов. Смесь в каждой лунке пипетировали для ресуспендирования гранул для тагментации и планшет инкубировали в течение 1-5 мин при комнатной температуре.
[0145] Очистка библиотеки. В каждую лунку добавляли 36 мкл гранул для захвата (таких как гранулы Ampure XP) и содержимое лунок перемешивали для ресуспендирования гранул, а затем инкубировали в течение 1, 3 или 5 мин. Планшет помещали на магнитную стойку приблизительно на 1-5 мин, до тех пор, пока супернатант не становился прозрачным.
[0146] В каждую лунку второго 96-луночного планшета добавляли 42 мкл гранул для захвата. Затем во второй 96-луночный планшет с гранулами захвата добавляли по 76 мкл супернатанта из каждой лунки первого 96-луночного планшета. Образцы перемешивали пипеткой и инкубировали в течение 1, 3 или 5 мин, а затем помещали на магнитную стойку до тех пор, пока супернатант не становился прозрачным (приблизительно 1-5 мин). В некоторых примерах вторую стадию с гранулами для захвата исключают.
[0147] Подходящими являются протоколы очистки гранул с однократным или двойным захватом и инкубацией в течение 1, 3 или 5 мин для каждой стадии очистки гранул.
[0148] После очистки гранул для захвата супернатант удаляли и гранулы для захвата дважды промывали 180 мкл свежего 80% этанола без перемешивания и инкубировали при комнатной температуре, после чего супернатант удаляли. Остаточный этанол удаляли пипеткой и планшет высушивали на воздухе и удаляли с магнитной стойки.
[0149] В каждую лунку добавляли по 15-22 мкл буфера для ресуспендирования, а содержимое лунки перемешивали пипеткой для ресуспендирования гранул. Планшет инкубировали при комнатной температуре в течение 2 мин, а затем помещали на магнитную стойку до тех пор, пока супернатант не становился прозрачным (приблизительно 2 мин). Всего в третий планшет для ПЦР объемом 96 лунок переносили 14-20 мкл супернатанта из каждой лунки. Данный планшет является конечной библиотекой ДНК, и его подготавливали для секвенирования, как описано выше.
[0150] В данном протоколе применяют две стадии очистки с иммобилизованными гранулами и исключают стадию количественной ПЦР, что позволяет за более короткое время завершить получение библиотеки. Например, образцы получают с применением данных способов в течение менее 30 мин при использовании стадий короткой магнитной инкубации в течение 20 мин.
[0151] Протокол можно осуществлять в других сосудах, таких как пробирки для микроцентрифугирования.
[0152] В некоторых вариантах осуществления для достижения оптимальной плотности кластеров объединяют равные объемы библиотеки и пул перед секвенированием количественно оценивают. При использовании вводимых количеств 200-1000 нг ДНК в некоторых случаях библиотеки объединяют посредством смешивания равных объемов каждой библиотеки в пробирке для микроцентрифугирования объемом 1,7 мл, встряхивания для перемешивания, а затем центрифугирования. Пул библиотеки количественно определяли с использованием набора Qubit ssDNA для определения концентрации пула. При использовании вводимых количеств 25-200 нг ДНК в некоторых примерах каждую библиотеку количественно определяли индивидуально с использованием набора Quibit ssDNA.
[0153] В некоторых примерах секвенирование проводят на картридже NovaSeq6000.
Пример 9: Получение библиотек посредством комбинации тагментации и индексирования
[0154] Следующий пример демонстрирует способ и систему для получения библиотек с парными концами после двойного индексирования из образца ДНК с использованием стадии комбинированной тагментации и индексирования. Способы выполняли аналогично способу, описанному в примере 2, но с комбинированной тагментацией и индексированием за одну стадию. Благодаря предотвращению отдельных стадий тагментации и индексирования данный протокол имеет преимущество в виде простого применения и меньшей продолжительности. Более того, данный протокол позволяет избежать стадии денатурации и получить двухцепочечные библиотеки без необходимости отдельной стадии получения двухцепочечных образцов из денатурированных одноцепочечных образцов. Данный протокол также позволяет избежать определенных стадий промывки, что еще больше сокращает время, необходимое для рабочего процесса.
[0155] Для демонстрации того, что данные библиотеки могут быть получены с использованием реакций, осуществляемых в течение либо долгого, среднего, либо короткого промежутка времени для выполнения, три набора образцов можно обрабатывать с нормальной скоростью рабочего процесса, высокой скоростью рабочего процесса и сверхвысокой скоростью рабочего процесса соответственно.
[0156] В данном способе применяют первый иммобилизованный транспозонный комплекс и второй иммобилизованный транспозонный комплекс. В первом иммобилизованном транспозонном комплексе последовательность X в первом транспозоне представляет собой якорную последовательность, а последовательность X’ в полинуклеотиде присоединения представляет собой комплемент якорной последовательности. Во втором иммобилизованном транспозонном комплексе последовательность X в первом транспозоне представляет собой якорную последовательность, а последовательность X’ в полинуклеотиде присоединения представляет собой комплемент якорной последовательности. Якорная последовательность первого и второго иммобилизованных транспозонных комплексов может быть некомплементарной во избежание перекрестной гибридизации. Первый транспозонный комплекс может содержать иллюстративный первый полинуклеотид присоединения, содержащий (i) комплемент якорной последовательности, последовательность A14’, спейсер и последовательность P5’, и (ii) связывающий элемент, содержащий биотин, а второй транспозонный комплекс может содержать иллюстративный второй полинуклеотид присоединения, содержащий (i) комплемент якорной последовательности, последовательность B15’, спейсер и последовательность P7’, и (ii) связывающий элемент, содержащий биотин.
[0157] Тагментация и индексирование. В каждую лунку 96-луночного планшета для ПЦР добавляли ДНК в 10 мМ трис-HCl (приблизительно 2-30 мкл) таким образом, чтобы общее вводимое количество (нг) находилось в пределах необходимого диапазона. В каждую лунку добавляли суспензию связанных с гранулами транспомосом (BLT, т. е. иммобилизованных транспозоновых комплексов), содержащую первый и второй комплексы транспосом. Стадию индексирования осуществляли в одном реакционном растворе с адаптерами индекса 1 (i7), адаптерами индекса 2 (i5) и последовательностями, необходимыми для образования кластеров секвенирования, лигированных в том же реакционном растворе, что и для проведения тагментации. В данном примере первые индексирующие олигонуклеотиды содержат последовательность A14, последовательность i5 и последовательность P5, а вторые индексирующие олигонуклеотиды содержат последовательность B15, последовательность i7 и последовательность P7. Условия на стадии тагментации/лигирования включают в себя меченый буфер и ДНК-лигазу E. Coli, добавленные к смеси целевой ДНК, первому и второму комплексам транспосом и первому и второму индексирующим олигонуклеотидам при общем объеме 20 мкл. Реакцию тагментации и индексирования проводили при температуре 41°C в течение трех разных временных интервалов (для трех разных образцов): 15 минут (обычный рабочий процесс), 5 минут (быстрый рабочий процесс) и 1 минут (сверхбыстрый рабочий процесс).
[0158] После реакции тагментации и индексирования в каждую лунку добавляли 5 мкл 0,6% SDS с инкубацией при 37°C для остановки реакции агментации посредством промывки SDS для денатурации транспозазы. Стадию остановки проводили в течение трех разных временных интервалов (для трех разных образцов): 5 минут (обычный рабочий процесс), 5 минут (быстрый рабочий процесс) и 1 минута (сверхбыстрый рабочий процесс). Несмотря на то, что стадию остановки выполняют в течение одного и того же периода для обычного и быстрого рабочего процесса, общее время для быстрого рабочего процесса все еще является более коротким вследствие различий по времени на других стадиях.
[0159] Удлинение. В каждую лунку добавляли 75 мкл смеси для удлинения (ДНК-полимераза, dNTP и буфер), и реакцию удлинения проводили при температуре менее 68°C в реакционном объеме 100 мкл. Стадию удлинения выполняли в течение трех разных временных интервалов (для трех разных образцов): 10 минут (обычный рабочий процесс), 2 минут (быстрый рабочий процесс) и 1 минут (сверхбыстрый рабочий процесс). Стадия удлинения позволяет создать библиотеку двухцепочечной ДНК в растворе. Очистку библиотеки можно выполнять с помощью гранул для захвата.
[0160] В целом выходы библиотек обычного и быстрого рабочего процесса сопоставимы со способами, которые включают отдельные стадии тагментации и индексирования, тогда как выходы библиотек сверхбыстрого рабочего процесса, вероятно, достаточны для многих применений, особенно с учетом дополнительного удобства еще более быстрого рабочего процесса. Способы с комбинированной тагментацией и индексированием могут обеспечивать индексирование олигонуклеотидов с транспозазами, которые успешно не фрагментировали образец ДНК (т. е. с побочными продуктами тагментации). Кроме того, комбинированная тагментация и индексирование могут обеспечивать библиотечные продукты, имеющие последовательности P5/P5 или последовательности P7/P7 с обоих концов, и данные библиотечные продукты с гомозиготными концами могут неправильно секвенироваться. Тем не менее, для многих применений количество исходного образца ДНК является достаточным для получения результатов секвенирования, даже если получены некоторые побочные продукты.
[0161] Описан ряд вариантов осуществления. Тем не менее, следует понимать, что возможны различные модификации. Соответственно, нижеследующая формула изобретения охватывает другие варианты осуществления.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 - A14-ME
TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG
SEQ ID NO: 2 - B15-ME
GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG
SEQ ID NO: 3 - ME’
phos-CTGTCTCTTATACACATCT
SEQ ID NO: 4 - A14
TCGTCGGCAGCGTC
SEQ ID NO: 5 - B15
GTCTCGTGGGCTCGG
SEQ ID NO: 6 - ME
AGATGTGTATAAGAGACAG
SEQ ID NO: 7 - P5
AATGATACGGCGACCACCGAGAUCTACAC
SEQ ID NO: 8 - P7
CAAGCAGAAGACGGCATACGAG*A
SEQ ID NO: 9 - P5_A14_ME
AATGATACGGCGACCACCGAGATCTACACTCGTCGGCAGCGTCAGATGTGTATAAGAGACAG
SEQ ID NO: 10 - P_ME’_B15’
5Phos/CTGTCTCTTATACACATCTCCGAGCCCACGAGAC
SEQ ID NO: 11 - Bio_P7_i701_B1_5_ddC
5Biosg/CAAGCAGAAGACGGCATACGAGATTCGCCTTAGTCTCGTGGGCTCGG/3ddC/
SEQ ID NO: 12 - A14’_P5’_bio
GACGCTGCCGACGAGTGTAGATCTCGGTGGTCGCCGTATCATT/3Bio/
SEQ ID NO: 13 - P_A14_ME
5Phos/TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG
SEQ ID NO: 14 - A14’_nitro_P5’_bio
GACGCTGCCGACGACCC/i5NitInd//i5NitInd//i5NitInd//i5NitInd//i5NitInd//i5NitInd//i5NitInd//i5NitInd/GTGTAGATCTCGGTGGTCGCCGTATCATT/3Bio/
SEQ ID NO: 15 - A14’_12anc’_2sp_P5’_bio
GACGCTGCCGACGACCGAGCATATCC/iSp18//iSp18/GTGTAGATCTCGGTGGTCGCCGTATCATT/3BioTEG/
SEQ ID NO: 16 - P5_i501
AATGATACGGCGACCACCGAGATCTACACTAGATCGC
SEQ ID NO: 17 - P_i701’_P7_ddC
5Phos/TAAGGCGAATCTCGTATGCCGTCTTCTGCTTG/3ddC/
SEQ ID NO: 18 - P7_i701_B15_ddC
CAAGCAGAAGACGGCATACGAGATTCGCCTTAGTCTCGTGGGCTCGG/3ddC/
SEQ ID NO: 19 - P5_i501_12anc
AATGATACGGCGACCACCGAGATCTACACTAGATCGCGGATATGCTCGG
SEQ ID NO: 20 - биотинилированный олигонуклеотид
/5Biosg/TTUUUAATGATACGGCGACCACCGAGATCTACACTCGTCGGCAGCGTCAGATGTGTATAAGAGACA
SEQ ID NO: 21 - ME’_B15’_P7’
CTGTCTCTTATACACATCTCCGAGCCCACGAGACATCTCGTATGCCGTCTTCTGCTTG
SEQ ID NO: 22 - иллюстративная якорная последовательность
GGATATGCTCGG
SEQ ID NO: 23: Иллюстративная последовательность X
ATCTGACTATCCCCTGCG
SEQ ID NO: 24: Иллюстративная последовательность X ’
CGCAGGGGATAGTCAGAT
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ILLUMINA CAMBRIDGE LIMITED
<120> СЛОЖНЫЕ КОМПЛЕКСЫ СВЯЗАННОЙ НА ПОВЕРХНОСТИ ТРАНСПОСОМЫ
<130> 01243-0011-00PCT
<150> US 62/840,610
<151> 2019-04-30
<150> US 62/791,509
<151> 2019-01-11
<160> 24
<170> PatentIn версии 3.5
<210> 1
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательности, применяемая в комплексе транспосом
<400> 1
tcgtcggcag cgtcagatgt gtataagaga cag 33
<210> 2
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательности, применяемая в комплексе транспосом
<400> 2
gtctcgtggg ctcggagatg tgtataagag acag 34
<210> 3
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательности, применяемая в комплексе транспосом
<220>
<221> прочие характеристики
<222> (1)..(1)
<223> 5’-фосфатная группа
<400> 3
ctgtctctta tacacatct 19
<210> 4
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная адаптерная последовательность присоединения
<400> 4
tcgtcggcag cgtc 14
<210> 5
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная адаптерная последовательность присоединения
<400> 5
gtctcgtggg ctcgg 15
<210> 6
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательности, применяемая в комплексе транспосом
<400> 6
agatgtgtat aagagacag 19
<210> 7
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный праймер
<400> 7
aatgatacgg cgaccaccga gauctacac 29
<210> 8
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный праймер
<220>
<221> прочие характеристики
<222> (22)..(22)
<223> Необязательно модифицированный гуанин, например 8-оксогуанин
<220>
<221> прочие характеристики
<222> (22)..(23)
<223> Необязательная фосфоротиоатная связь
<400> 8
caagcagaag acggcatacg agat 24
<210> 9
<211> 62
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный 5’-фрагмент транспозона
<400> 9
aatgatacgg cgaccaccga gatctacact cgtcggcagc gtcagatgtg tataagagac 60
ag 62
<210> 10
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный 3’-фрагмент транспозона
<220>
<221> прочие характеристики
<222> (1)..(1)
<223> 5’-фосфатная группа
<400> 10
ctgtctctta tacacatctc cgagcccacg agac 34
<210> 11
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный олигонуклеотид присоединения
<220>
<221> прочие характеристики
<222> (1)..(1)
<223> 5’-биотин
<220>
<221> прочие характеристики
<222> (48)..(48)
<223> 3’-дидезокси-C
<400> 11
caagcagaag acggcatacg agattcgcct tagtctcgtg ggctcggn 48
<210> 12
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный олигонуклеотид присоединения
<220>
<221> прочие характеристики
<222> (43)..(43)
<223> 3’-биотин
<400> 12
gacgctgccg acgagtgtag atctcggtgg tcgccgtatc att 43
<210> 13
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный 5’-фрагмент транспозона
<220>
<221> прочие характеристики
<222> (1)..(1)
<223> 5’-фосфатная группа
<400> 13
tcgtcggcag cgtcagatgt gtataagaga cag 33
<210> 14
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный олигонуклеотид присоединения
<220>
<221> прочие характеристики
<222> (18)..(25)
<223> Int 5-нитроиндол
<220>
<221> прочие характеристики
<222> (54)..(54)
<223> 3’-биотин
<400> 14
gacgctgccg acgacccnnn nnnnngtgta gatctcggtg gtcgccgtat catt 54
<210> 15
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный олигонуклеотид присоединения
<220>
<221> прочие характеристики
<222> (27)..(28)
<223> Int спейсер 18
<220>
<221> прочие характеристики
<222> (57)..(57)
<223> 3’-биотин-ТЭГ
<400> 15
gacgctgccg acgaccgagc atatccnngt gtagatctcg gtggtcgccg tatcatt 57
<210> 16
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный индексирующий олигонуклеотид i501
<400> 16
aatgatacgg cgaccaccga gatctacact agatcgc 37
<210> 17
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный индексирующий олигонуклеотид i701
<220>
<221> прочие характеристики
<222> (1)..(1)
<223> 5’-фосфатная группа
<220>
<221> прочие характеристики
<222> (33)..(33)
<223> 3’-дидезокси-C
<400> 17
taaggcgaat ctcgtatgcc gtcttctgct tgn 33
<210> 18
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный индексирующий олигонуклеотид i701
<220>
<221> прочие характеристики
<222> (48)..(48)
<223> 3’-дидезокси-C
<400> 18
caagcagaag acggcatacg agattcgcct tagtctcgtg ggctcggn 48
<210> 19
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративный индексирующий олигонуклеотид i501
<400> 19
aatgatacgg cgaccaccga gatctacact agatcgcgga tatgctcgg 49
<210> 20
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательности, применяемая в комплексе транспосом
<220>
<221> прочие характеристики
<222> (1)..(1)
<223> 5’-биотин
<400> 20
ttuuuaatga tacggcgacc accgagatct acactcgtcg gcagcgtcag atgtgtataa 60
gagaca 66
<210> 21
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательности, применяемая в комплексе транспосом
<400> 21
ctgtctctta tacacatctc cgagcccacg agacatctcg tatgccgtct tctgcttg 58
<210> 22
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная якорная последовательность
<400> 22
ggatatgctc gg 12
<210> 23
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательность X
<400> 23
atctgactat cccctgcg 18
<210> 24
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Иллюстративная последовательность X’
<400> 24
cgcaggggat agtcagat 18
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТАГМЕНТАЦИЯ С ПРИМЕНЕНИЕМ ИММОБИЛИЗОВАННЫХ ТРАНСПОСОМ С ЛИНКЕРАМИ | 2018 |
|
RU2783536C2 |
| АНАЛИЗ МНОЖЕСТВА АНАЛИТОВ С ИСПОЛЬЗОВАНИЕМ ОДНОГО АНАЛИЗА | 2019 |
|
RU2824049C2 |
| СПОСОБ И КОМПОЗИЦИЯ ДЛЯ АНАЛИЗА КЛЕТОЧНЫХ КОМПОНЕНТОВ | 2016 |
|
RU2717491C2 |
| СПОСОБ И КОМПОЗИЦИЯ ДЛЯ АНАЛИЗА КЛЕТОЧНЫХ КОМПОНЕНТОВ | 2016 |
|
RU2761432C2 |
| ПОЛУЧЕНИЕ БИБЛИОТЕКИ НУКЛЕИНОВЫХ КИСЛОТ С ИСПОЛЬЗОВАНИЕМ ЭЛЕКТРОФОРЕЗА | 2020 |
|
RU2798952C2 |
| СИСТЕМА АНАЛИЗА ДЛЯ ОРТОГОНАЛЬНОГО ДОСТУПА К БИОМОЛЕКУЛАМ И ИХ МЕЧЕНИЯ В КЛЕТОЧНЫХ КОМПАРТМЕНТАХ | 2017 |
|
RU2771892C2 |
| ИДЕНТИФИКАЦИЯ И АНАЛИЗ ОБРАЗЦОВ МИКРООРГАНИЗМОВ ПУТЕМ БЫСТРОГО ИНКУБИРОВАНИЯ И ОБОГАЩЕНИЯ НУКЛЕИНОВЫХ КИСЛОТ | 2020 |
|
RU2809853C2 |
| СПОСОБЫ ИНКАПСУЛИРОВАНИЯ ОДИНОЧНЫХ КЛЕТОК, ИНКАПСУЛИРОВАННЫЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2750567C2 |
| СПОСОБЫ И СРЕДСТВА ПОЛУЧЕНИЯ БИБЛИОТЕКИ ДЛЯ СЕКВЕНИРОВАНИЯ | 2019 |
|
RU2815513C2 |
| СПОСОБЫ ИНКАПСУЛИРОВАНИЯ ОДИНОЧНЫХ КЛЕТОК, ИНКАПСУЛИРОВАННЫЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2793717C2 |
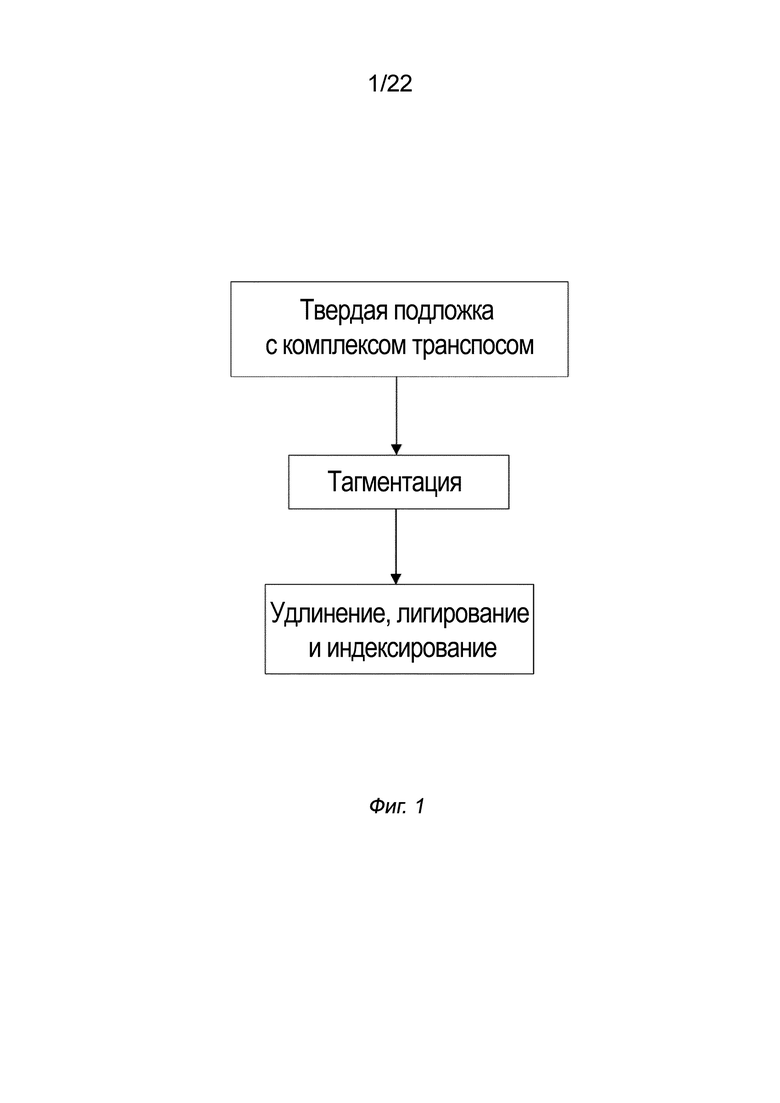
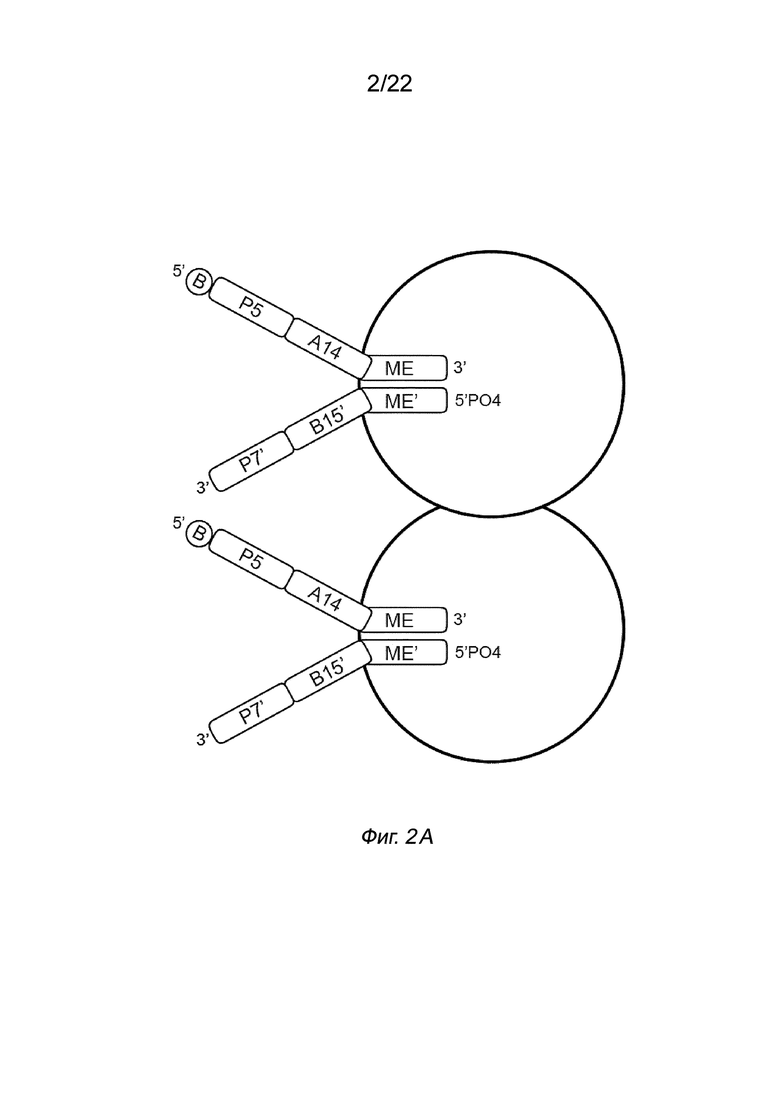
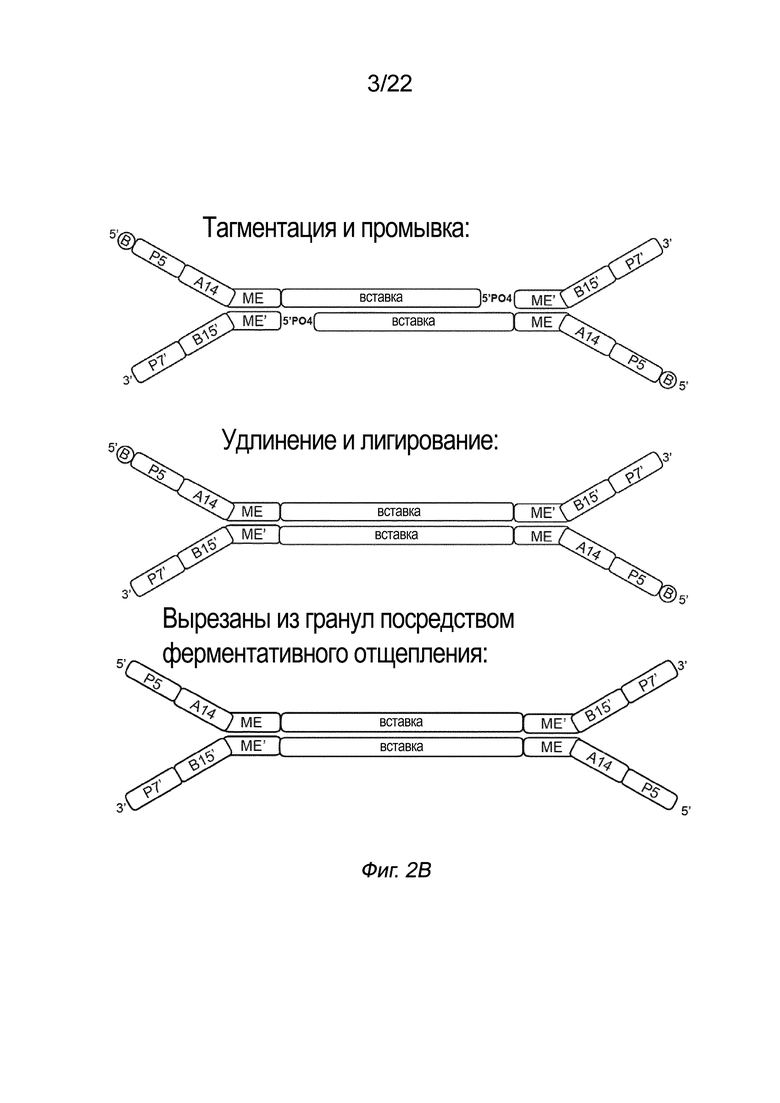
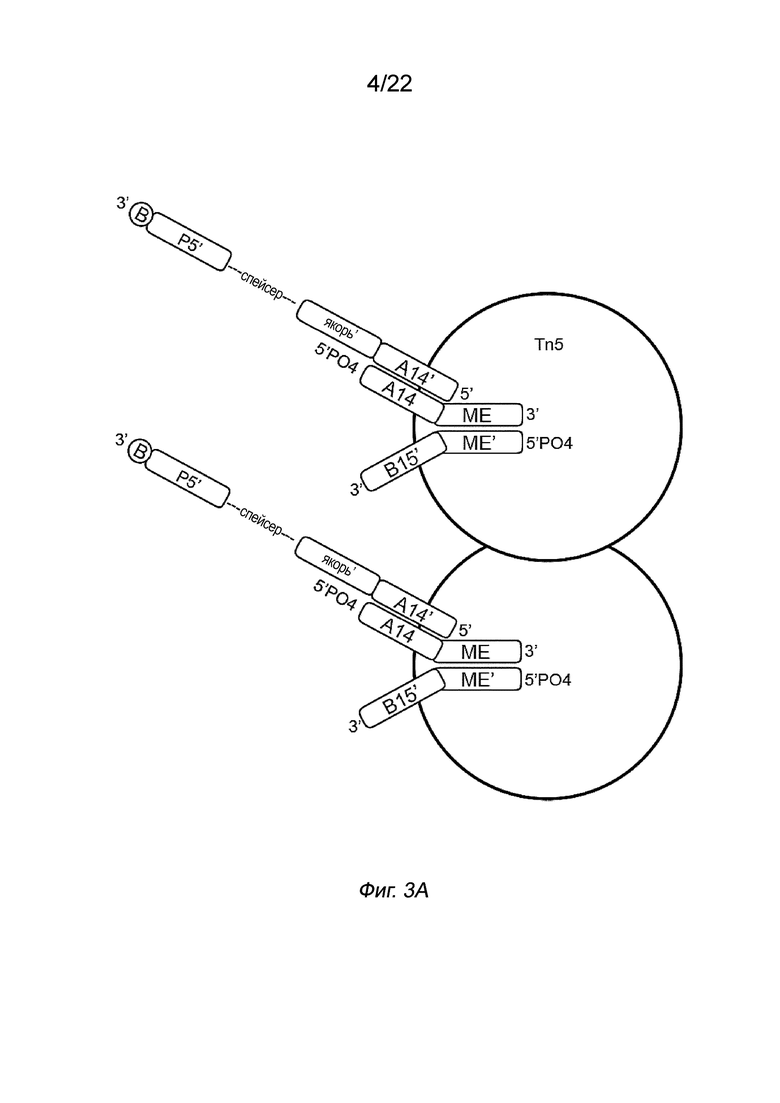
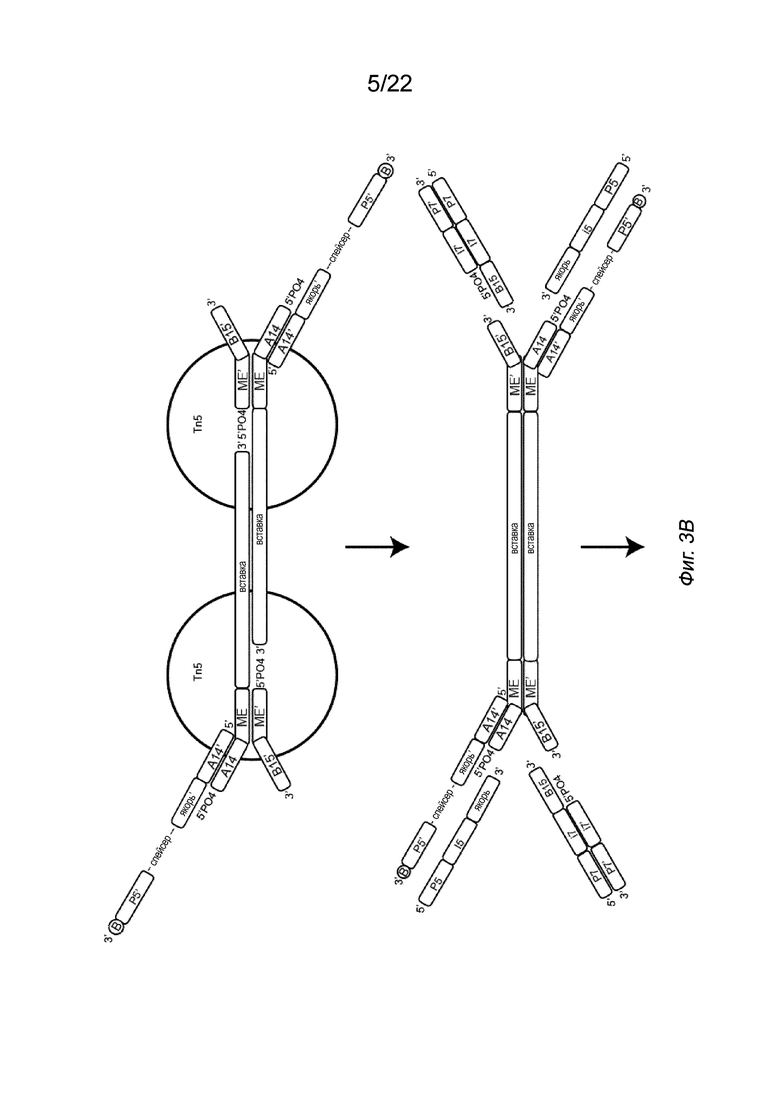
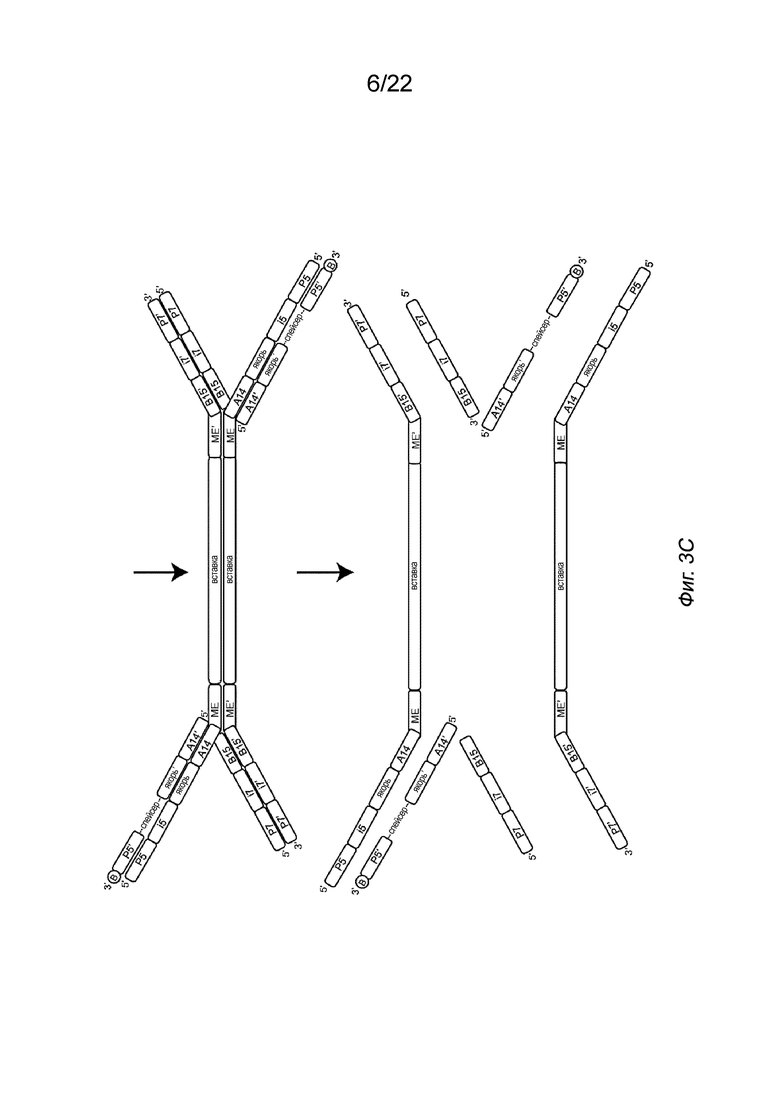
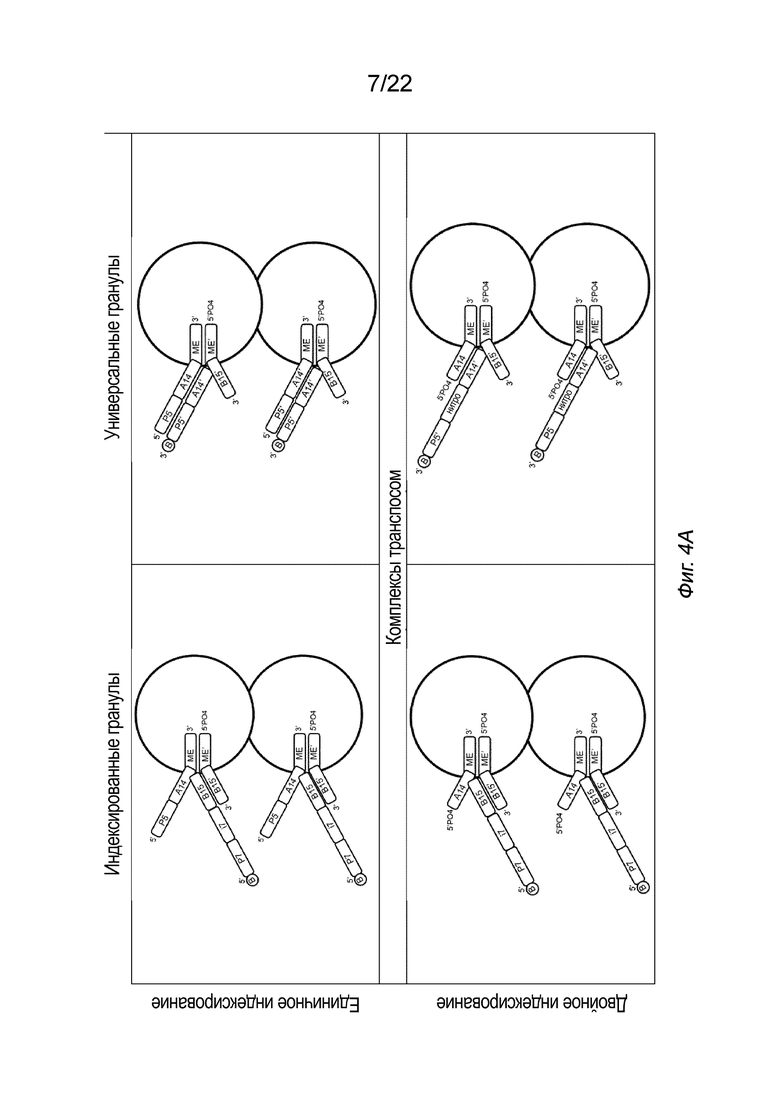
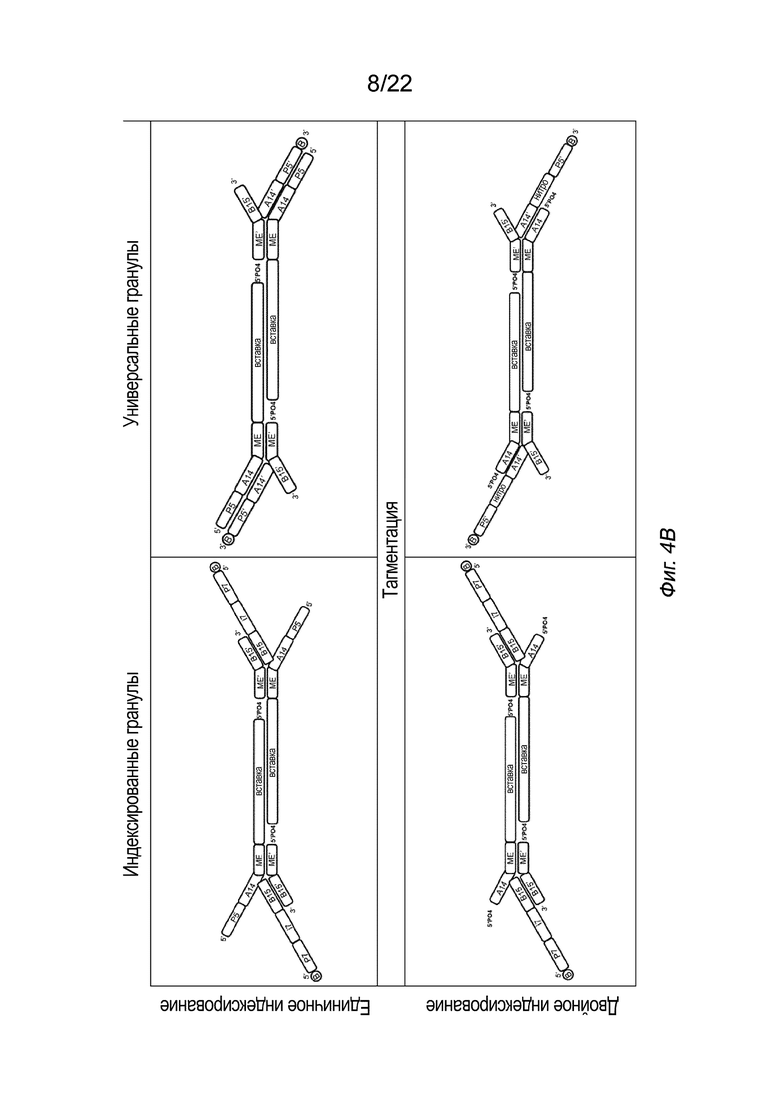
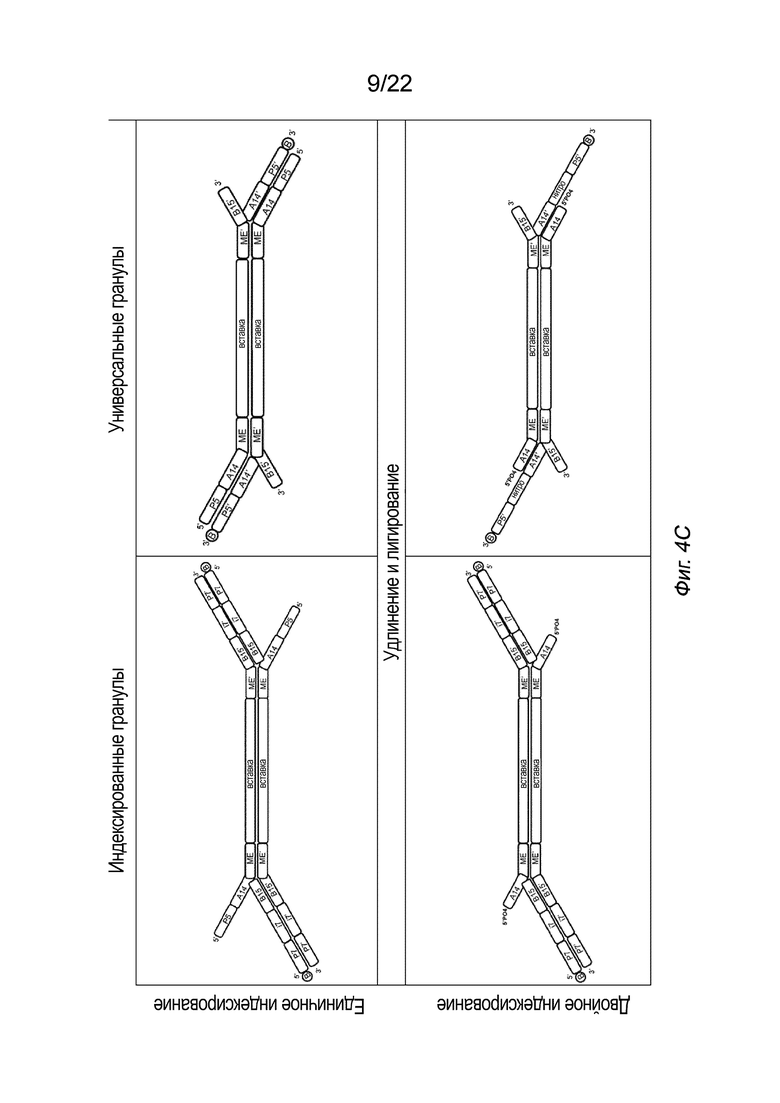
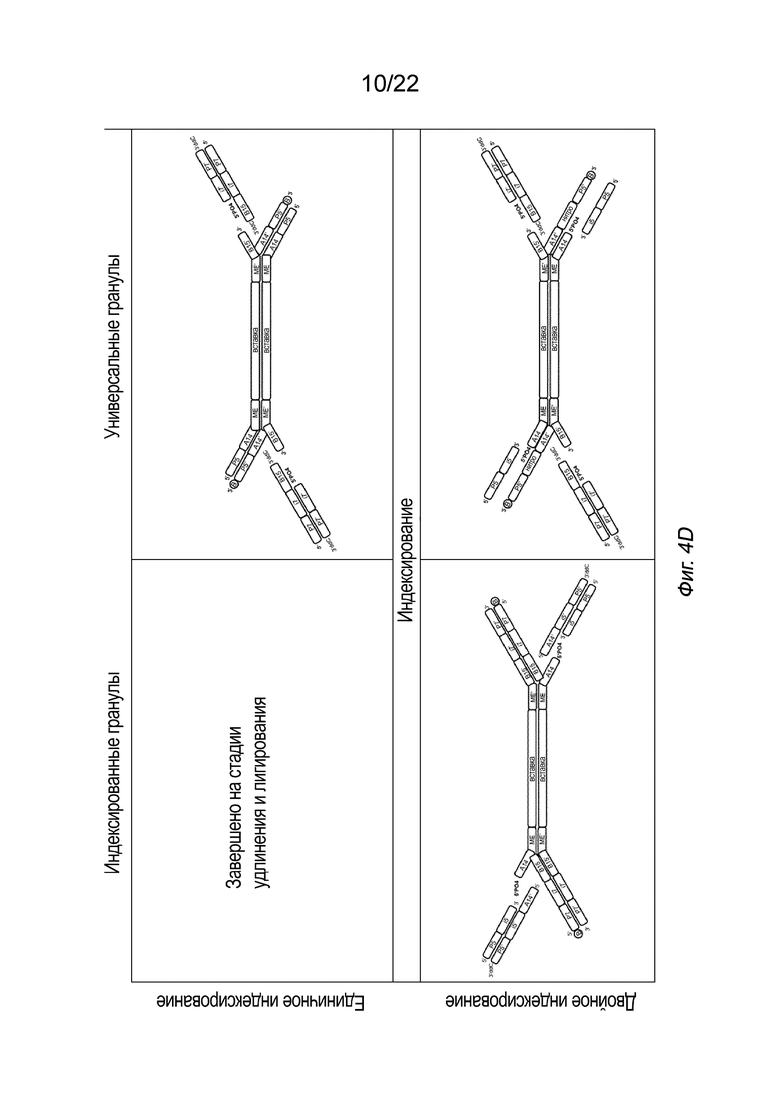
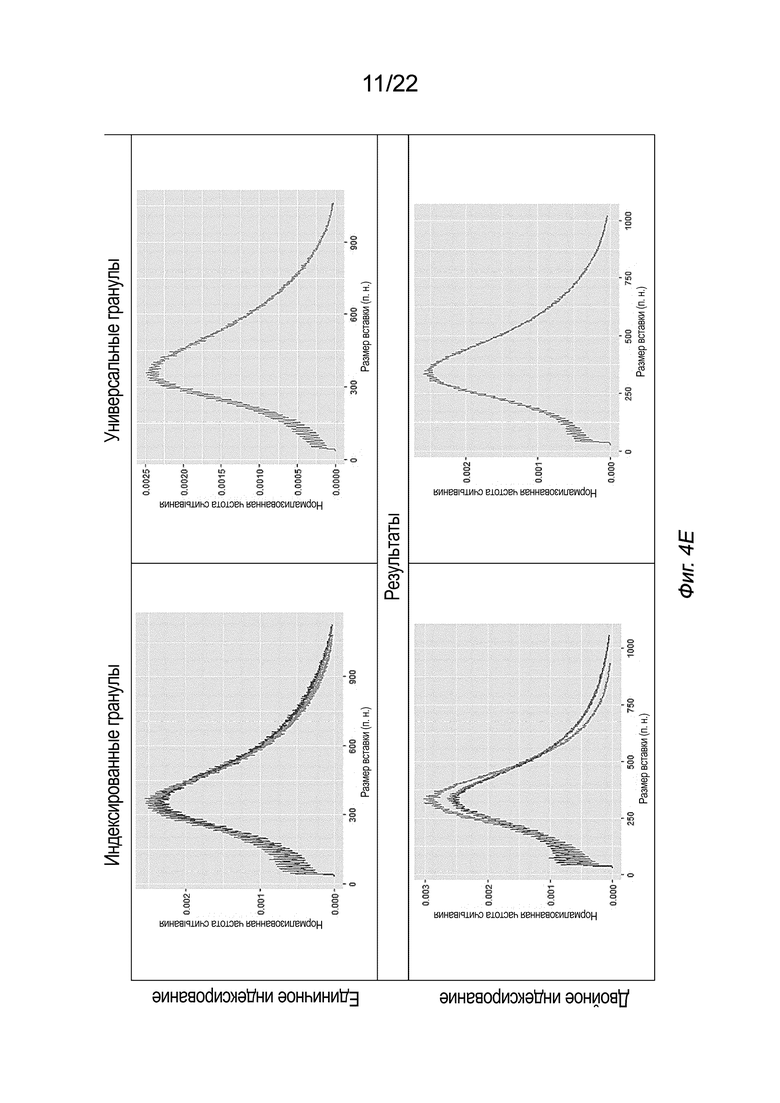
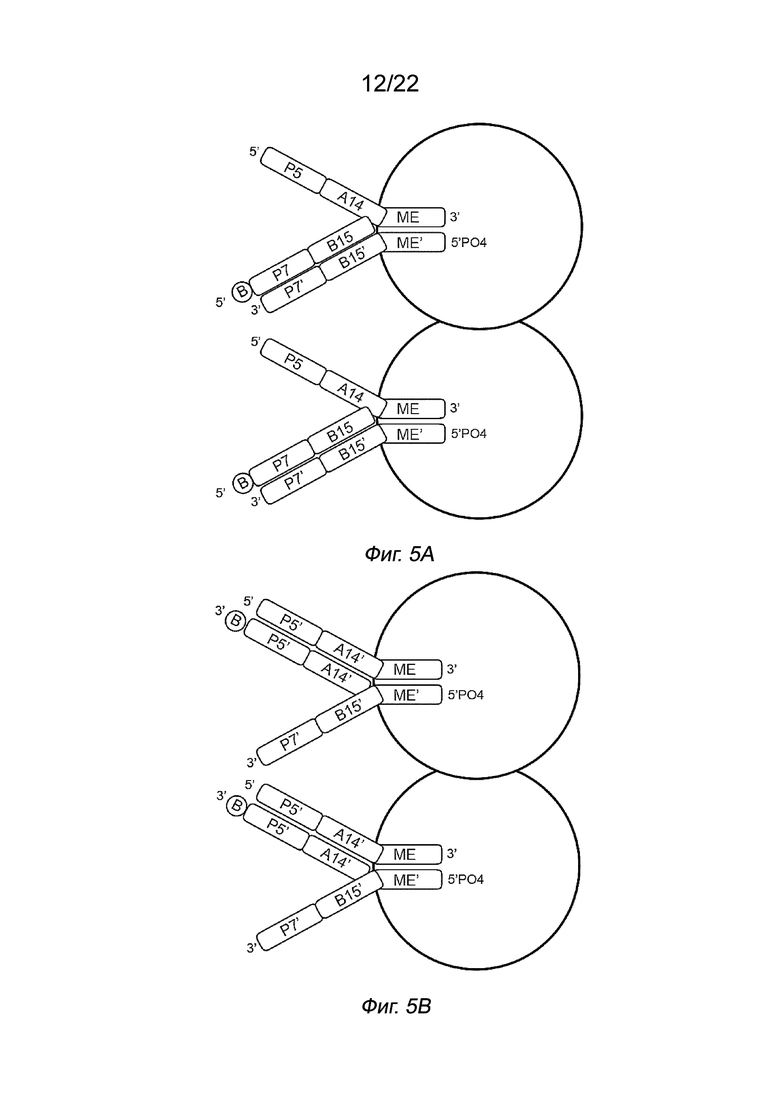
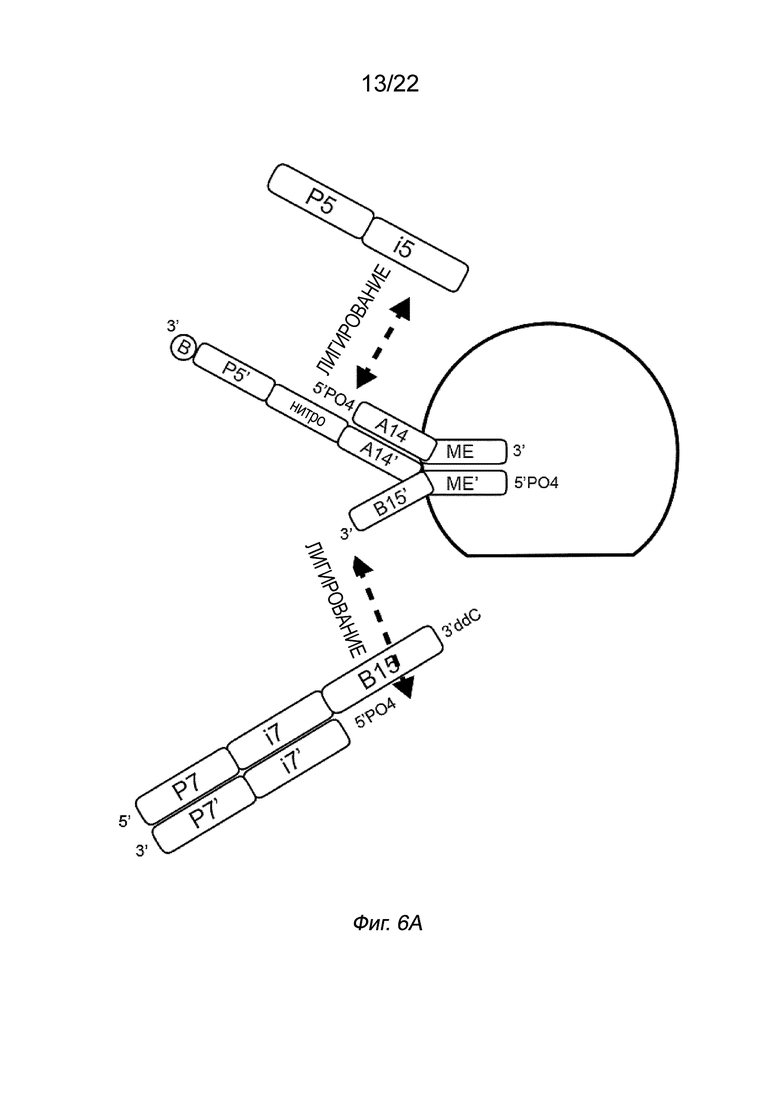
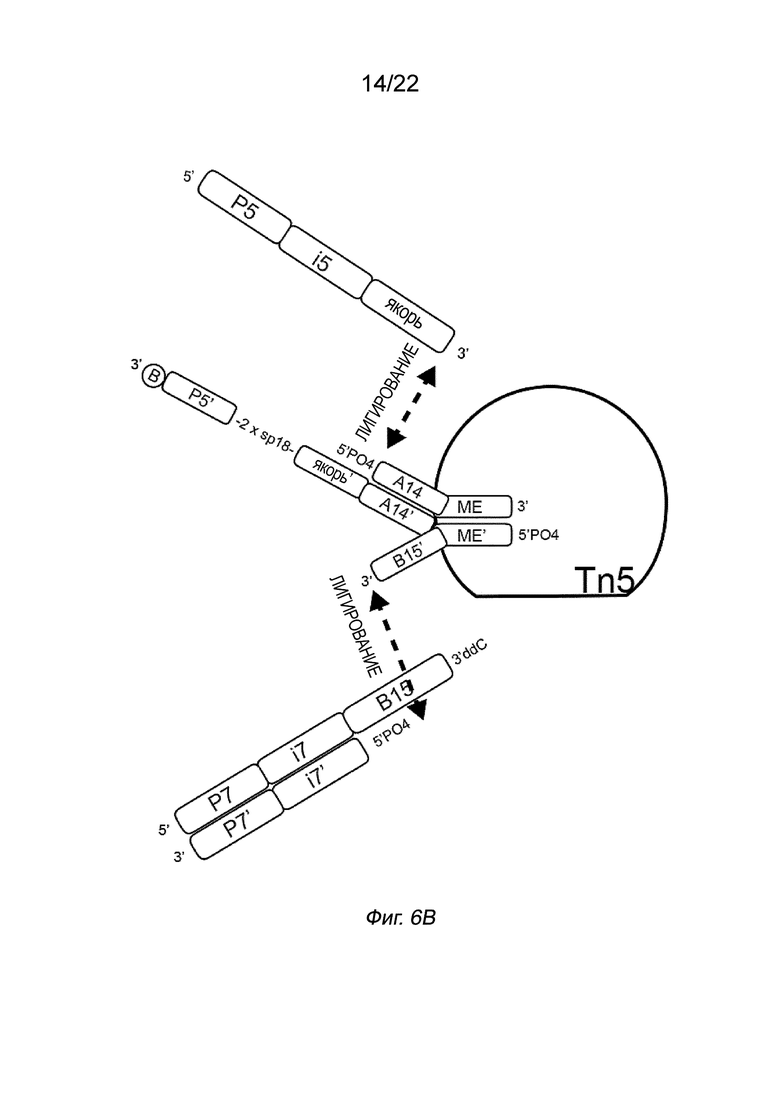
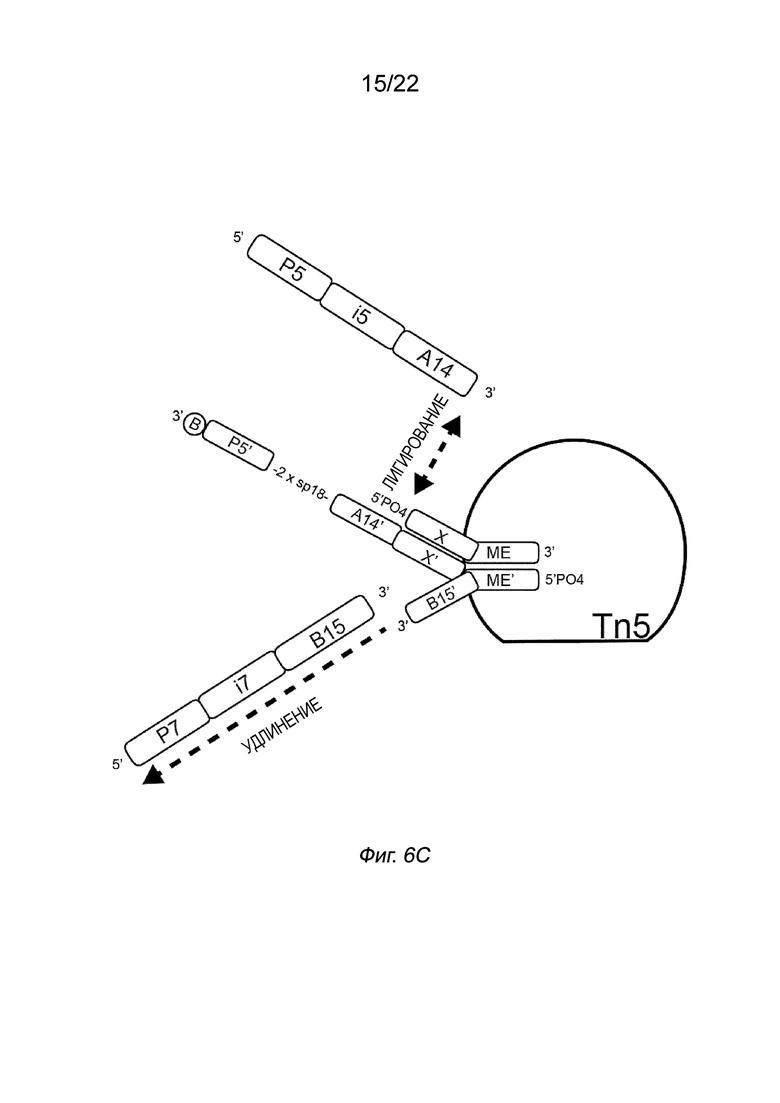
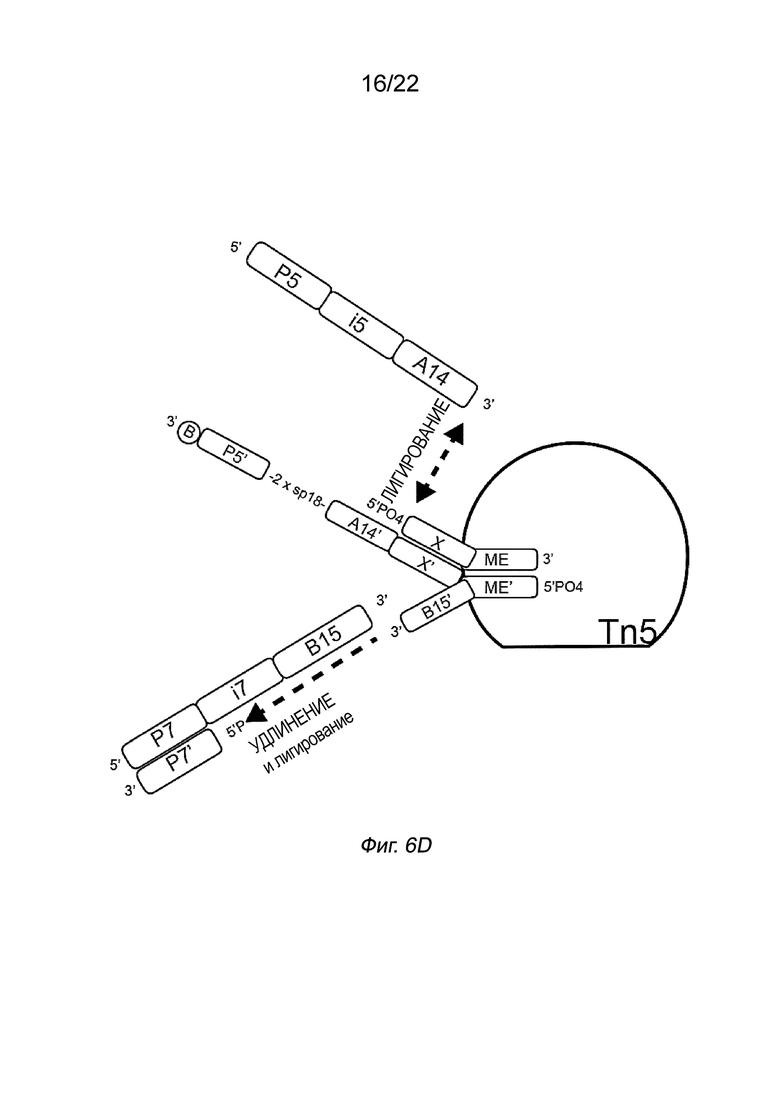
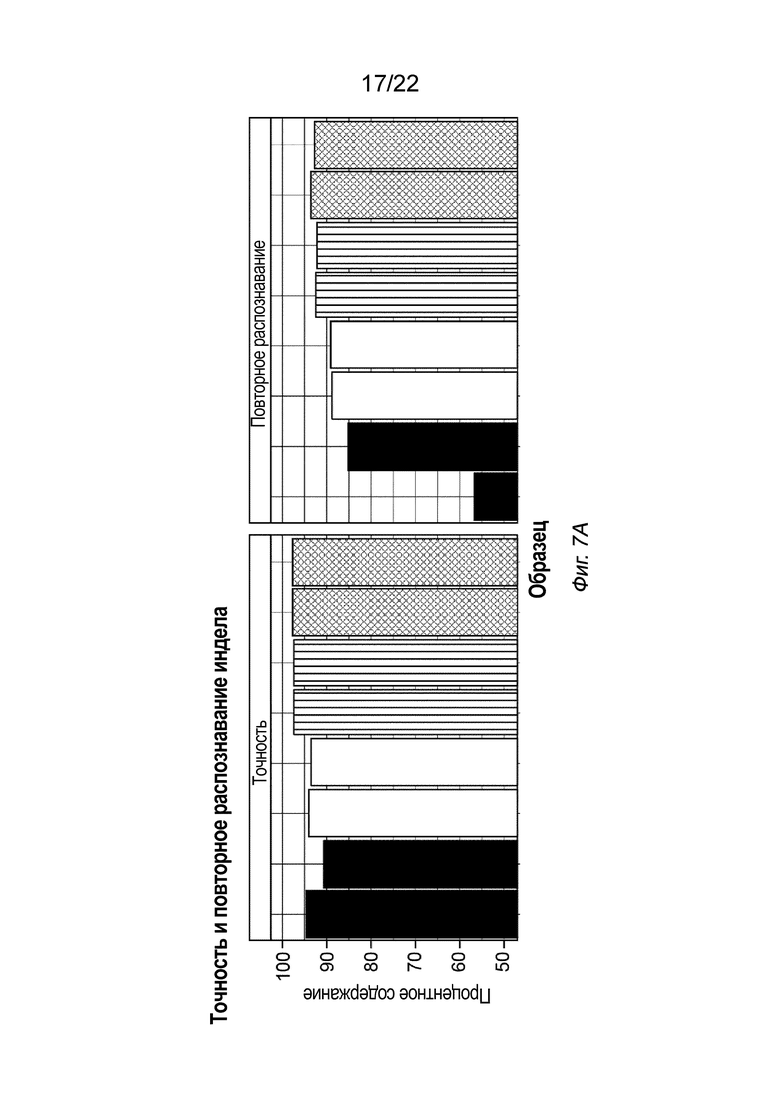
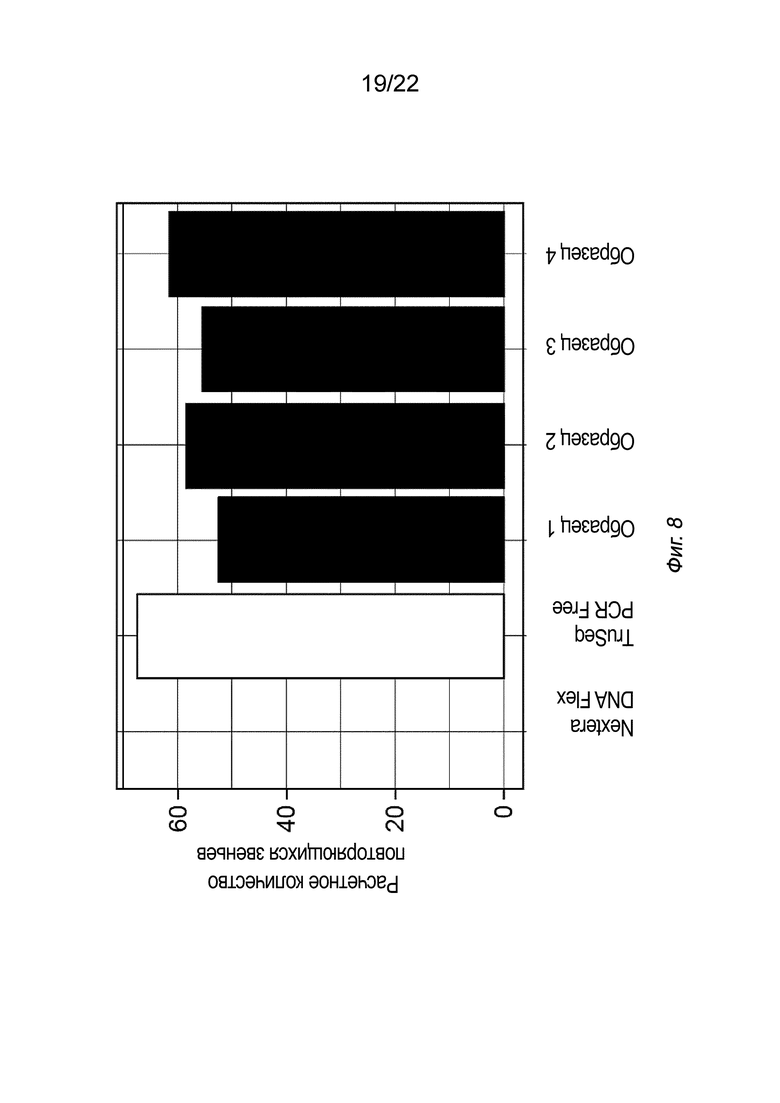
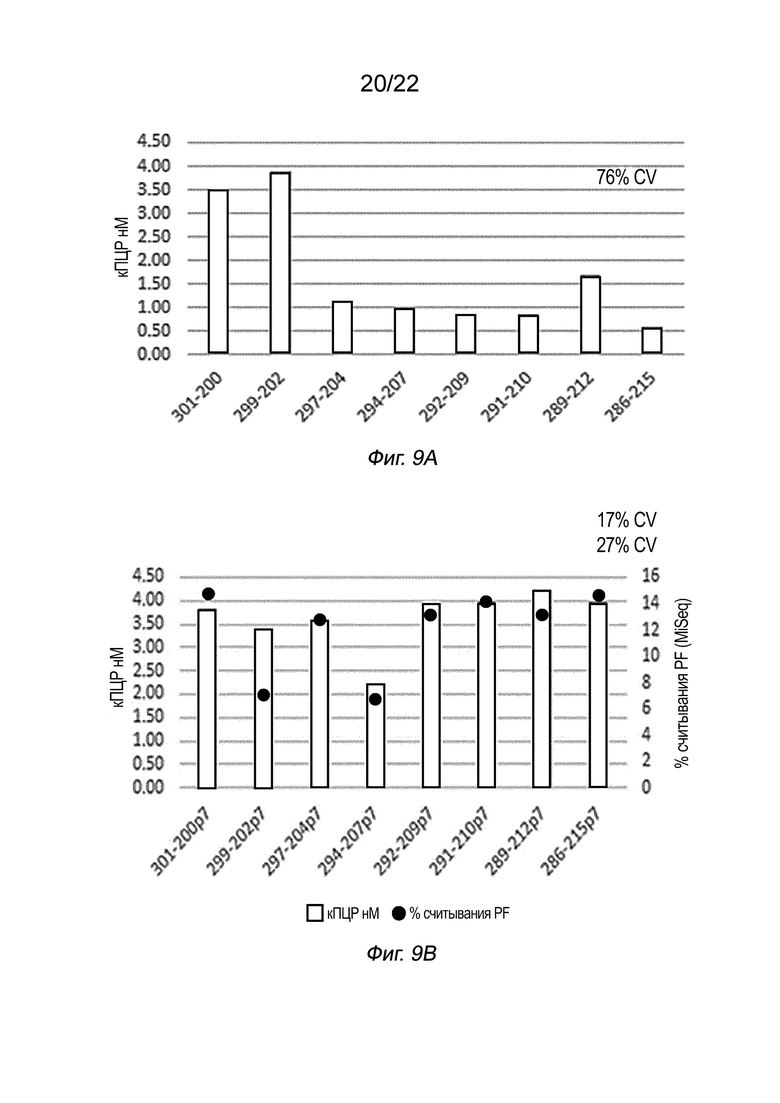
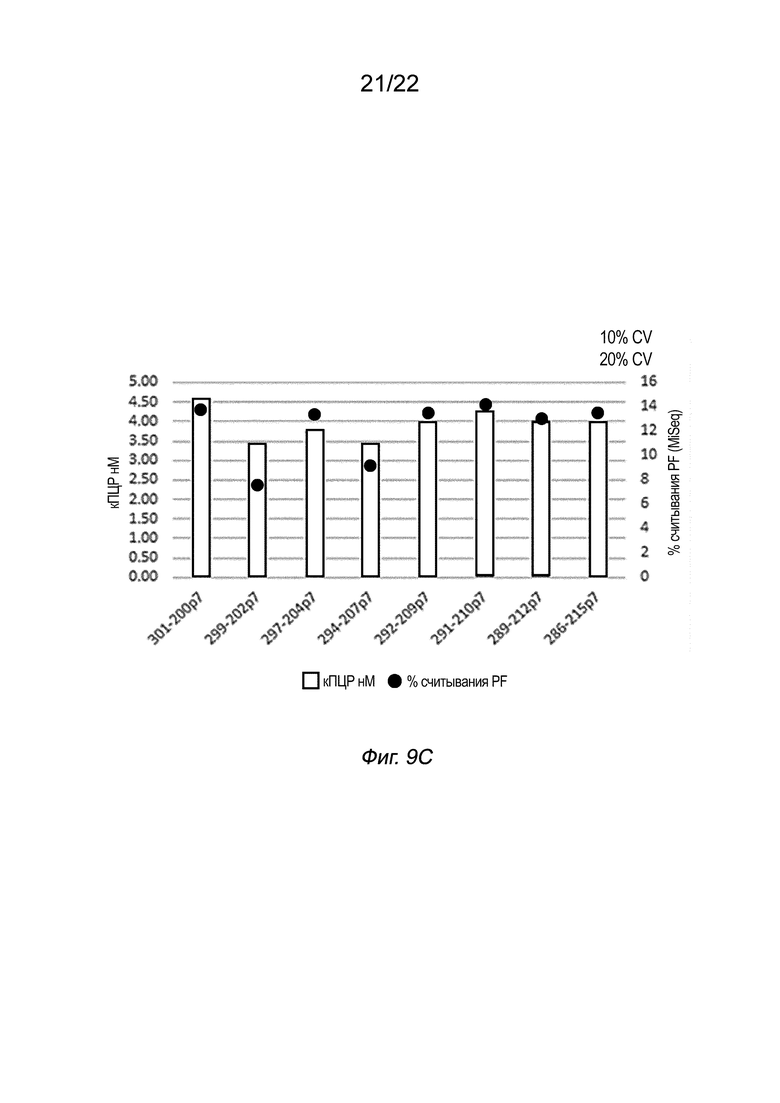

Изобретение относится к области биотехнологии, в частности к отожженному гибриду транспозон/полинуклеотид присоединения, содержащему первый транспозон, второй транспозон и полинуклеотид присоединения. Также раскрыт комплекс транспосом, содержащий транспозазу, первый транспозон, второй транспозон и полинуклеотид присоединения, а также содержащий его набор. Изобретение эффективно для создания библиотеки меченых фрагментов нуклеиновых кислот. 8 н. и 44 з.п. ф-лы, 10 ил., 2 табл., 9 пр.
1. Отожженный гибрид транспозон/полинуклеотид присоединения для создания библиотеки меченых фрагментов нуклеиновых кислот, содержащий следующее:
(а) первый транспозон, содержащий 3’-концевую последовательность транспозона и 5’-адаптерную последовательность;
(b) второй транспозон, содержащий 5’-концевую последовательность транспозона и 3’-адаптерную последовательность, где
(i) 5’-концевая последовательность второго транспозона комплементарна 3’-концевой последовательности первого транспозона,
(ii) 5'-адаптерная последовательность первого транспозона не комплементарна 3'-адаптерной последовательности второго транспозона, и
(iii) первый транспозон и второй транспозон образуют Y-образную форму; и
(c) полинуклеотид присоединения, который гибридизуется с первым транспозоном или вторым транспозоном, содержащий следующее:
(i) соединительная адаптерная последовательность, гибридизованная с 5'-адаптерной последовательностью первого транспозона или 3'-адаптерной последовательностью второго транспозона; и
(ii) связывающий элемент для связывания указанного отожженного гибрида транспозон/полинуклеотид присоединения с твердой подложкой.
2. Комплекс транспосом для создания библиотеки меченых фрагментов нуклеиновых кислот, содержащий следующее:
(a) транспозаза;
(b) первый транспозон, содержащий 3’-концевую последовательность транспозона и 5’-адаптерную последовательность;
(c) второй транспозон, содержащий 5’-концевую последовательность транспозона и 3’-адаптерную последовательность, где
(i) 5’-концевая последовательность второго транспозона комплементарна 3’-концевой последовательности первого транспозона,
(ii) 5'-адаптерная последовательность первого транспозона не комплементарна 3'-адаптерной последовательности второго транспозона, и
(iii) первый транспозон и второй транспозон образуют Y-образную форму; и
(d) полинуклеотид присоединения, который гибридизуется с первым транспозоном или вторым транспозоном, содержащий следующее:
(i) соединительная адаптерная последовательность, гибридизованная с 5'-адаптерной последовательностью первого транспозона или 3'-адаптерной последовательностью второго транспозона, и
(ii) связывающий элемент для связывания указанного комплекса транспосом с твердой подложкой.
3. Иммобилизованный комплекс транспосом для создания библиотеки меченых фрагментов нуклеиновых кислот, содержащий следующее:
твердая подложка; и
комплекс транспосом, иммобилизованный на твердой подложке,
причем комплекс транспосом содержит следующее:
(a) транспозаза;
(b) первый транспозон, содержащий 3’-концевую последовательность транспозона и 5’-адаптерную последовательность;
(c) второй транспозон, содержащий 5’-концевую последовательность транспозона и 3’-адаптерную последовательность, где
(i) 5’-концевая последовательность второго транспозона комплементарна по меньшей мере части 3’-концевой последовательности первого транспозона с образованием двухцепочечной концевой транспозонной последовательности, которая связывается с транспозазой,
(ii) 5'-адаптерная последовательность первого транспозона не комплементарна 3'-адаптерной последовательности второго транспозона, и
(iii) первый транспозон и второй транспозон образуют Y-образную форму; и
(d) полинуклеотид присоединения, который гибридизуется с первым транспозоном или вторым транспозоном, содержащий следующее:
(i) соединительная адаптерная последовательность, гибридизованная с 5'-адаптерной последовательностью первого транспозона или 3'-адаптерной последовательностью второго транспозона, и
(ii) связывающий элемент для связывания указанного комплекса транспосом с твердой подложкой,
причем связывающий элемент иммобилизован на твердой подложке.
4. Иммобилизованный комплекс транспосом для создания библиотеки меченых фрагментов нуклеиновых кислот, содержащий следующее:
(a) транспозаза;
(b) первый транспозон, содержащий 3’-концевую последовательность транспозона и 5’-адаптерную последовательность;
(c) второй транспозон, содержащий 5’-концевую последовательность транспозона и 3’-адаптерную последовательность, где
(i) 5’-концевая последовательность второго транспозона комплементарна по меньшей мере части 3’-концевой последовательности первого транспозона с образованием двухцепочечной концевой транспозонной последовательности, которая связывается с транспозазой,
(ii) 5'-адаптерная последовательность первого транспозона не комплементарна 3'-адаптерной последовательности второго транспозона, и
(iii) первый транспозон и второй транспозон образуют Y-образную форму; и
(d) полинуклеотид присоединения, содержащий:
связывающий элемент для связывания указанного комплекса транспосом с твердой подложкой, где указанный связывающий элемент присоединен к 5’-адаптерной последовательности первого транспозона посредством отщепляемого линкера;
причем связывающий элемент иммобилизован на указанной твердой подложке.
5. Комплекс транспосом по любому из пп. 2-4, в котором полинуклеотид присоединения дополнительно содержит спейсерную область, якорную область, праймерную последовательность, последовательность индексной метки или их комбинацию и необязательно дополнительно содержит последовательность захвата, последовательность банка данных, последовательность отщепления или последовательность, связанную с секвенированием, или их комбинацию.
6. Комплекс транспосом по п. 5, в котором первый транспозон дополнительно содержит 5’-праймерную последовательность 5’-адаптерной последовательности первого транспозона, а полинуклеотид присоединения содержит (i) часть, комплементарную 5’-адаптерной последовательности и гибридизованную с ней, и (ii) комплементарную праймерную последовательность.
7. Комплекс транспосом по п. 5, в котором первый транспозон дополнительно содержит 5’-праймерную последовательность 5’-адаптерной последовательности, а полинуклеотид присоединения содержит последовательность индексной метки и праймерную последовательность.
8. Комплекс транспосом по п. 5, в котором второй транспозон дополнительно содержит 3’-адаптерную последовательность и полинуклеотид присоединения содержит (i) часть, комплементарную 3’-адаптерной последовательности второго транспозона и гибридизованную с ней, и (ii) последовательность индексной метки, и (iii) праймерную последовательность.
9. Комплекс транспосом по п. 5, в котором полинуклеотид присоединения содержит спейсерную область или спейсерную область и якорную область.
10. Комплекс транспосом по п. 9, в котором первый транспозон содержит 5’-адаптерную последовательность и полинуклеотид присоединения содержит (i) часть, комплементарную 5’-адаптерной последовательности первого транспозона и гибридизованную с ней, и (ii) спейсерную область, и (iii) праймерную последовательность.
11. Комплекс транспосом по любому из пп. 2-10, в котором полинуклеотид присоединения содержит последовательность, комплементарную 5’-адаптерной последовательности первого транспозона.
12. Комплекс транспосом по любому из пп. 9-11, в котором спейсерная область не является ДНК-спейсером.
13. Комплекс транспосом по любому из пп. 9-12, в котором спейсерная область имеет длину, приблизительно эквивалентную 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 15, 20 или 30 парам оснований, причем необязательно спейсерная область имеет длину, приблизительно эквивалентную 8 или 10 парам оснований или нуклеотидов.
14. Комплекс транспосом по любому из пп. 9-13, в котором спейсерная область содержит универсальные основания, причем необязательно универсальные основания содержат инозин или нитроиндольную последовательность.
15. Комплекс транспосом по любому из пп. 9-13, в котором спейсерная область содержит один или более 18-атомных гекса-этиленгликолевых спейсеров не из последовательности нуклеиновой кислоты, один или более триэтиленгликолевых спейсеров или их комбинацию.
16. Комплекс транспосом по п. 15, в котором спейсерная область содержит 2 тандемные копии 18-атомного гекса-этиленгликолевого спейсера (синтетический линкер 2 x sp18), два 18-атомных гекса-этиленгликолевых спейсера не из последовательности нуклеиновой кислоты, триэтиленгликолевый спейсер или два 18-атомных гекса-этиленгликолевых спейсера и один триэтиленгликолевый спейсер.
17. Комплекс транспосом по любому из пп. 5-16, в котором якорная область комплементарна якорной области, комплементарной индексирующему олигонуклеотиду, причем индексирующий олигонуклеотид содержит комплемент якорной области и последовательность индексной метки.
18. Комплекс транспосом по п. 17, в котором комплемент якорной области является общим для множества индексирующих олигонуклеотидов.
19. Комплекс транспосом по п. 17, в котором каждая последовательность индексной метки из множества индексирующих олигонуклеотидов является одинаковой или разной.
20. Комплекс транспосом по любому из пп. 2-19, в котором связывающий элемент содержит биотин.
21. Комплекс транспосом по п. 20, в котором связывающий элемент представляет собой линкер 3’-биотин.
22. Комплекс транспосом по любому из пп. 2-10, в котором транспозаза представляет собой транспозазу Tn5, транспозазу Tn5 дикого типа или гиперактивную транспозазу Tn5 или их мутант.
23. Комплекс транспосом по любому из пп. 5-22, в котором праймерная последовательность полинуклеотида присоединения комплементарна индексной праймерной последовательности индексирующего полинуклеотида.
24. Комплекс транспосом по любому из пп. 3-23, в котором твердая подложка представляет собой гранулу, парамагнитную гранулу, проточную кювету, поверхность микрожидкостного устройства, пробирку, лунку планшета, предметное стекло, структурированную поверхность или микрочастицу.
25. Комплекс транспосом по п. 24, в котором твердая подложка включает множество твердых подложек.
26. Комплекс транспосом по п. 25, в котором множество твердых подложек содержит множество гранул.
27. Набор для создания библиотеки меченых фрагментов нуклеиновых кислот, содержащий следующее:
комплекс транспосом по любому из пп. 2-26; и
смесь олигонуклеотидов, содержащая индексные последовательности.
28. Набор по п. 27, в котором смесь олигонуклеотидов содержит последовательности индексов i5 и последовательности индексов i7, причем последовательности индексов i5 гибридизуются с полинуклеотидом присоединения, а последовательности индексов i7 гибридизуются с 3’-адаптерной последовательностью второго транспозона.
29. Способ создания библиотеки меченых фрагментов нуклеиновых кислот, включающий приведение иммобилизованного комплекса транспосом по любому из пп. 3-26 в контакт с целевой нуклеиновой кислотой для фрагментации целевой нуклеиновой кислоты во множество целевых фрагментов и для присоединения 3’-конца первого транспозона к 5’-концам целевых фрагментов с получением множества 5’-меченых целевых фрагментов.
30. Способ по п. 29, дополнительно включающий следующее:
обработка твердой подложки с удалением несвязанных нуклеиновых кислот; или
обработка твердой подложки с удалением транспозазы из комплекса, необязательно посредством следующего:
(a) нагревание твердой подложки и/или
(b) промывка твердой подложки ферментативным денатурирующим средством,
причем ферментативное денатурирующее средство необязательно содержит додецилсульфат натрия (SDS), гидрохлорид гуанидина, мочевину или протеиназу.
31. Способ по п. 29 или 30, в котором приведение в контакт включает смешивание биологического образца с комплексом транспосомы.
32. Способ по п. 31, в котором указанный биологический образец содержит клеточный лизат или цельные клетки или его выбирают из крови, плазмы, сыворотки крови, лимфы, слизи, мокроты, мочи, спермы, спинномозговой жидкости, бронхиальной аспирации, фекалий, мацерированной ткани и фиксированной ткани FFPE.
33. Способ по любому из пп. 29-32, в котором приведение в контакт дополнительно включает гибридизацию множества индексирующих олигонуклеотидов с полинуклеотидами присоединения, причем множество индексирующих олигонуклеотидов содержит одинаковые или разные индексные последовательности.
34. Способ по любому из пп. 29-33, дополнительно включающий обработку множества 5’-меченых целевых фрагментов полимеразой и лигазой для удлинения и лигирования цепей с получением полностью двухцепочечных меченых фрагментов.
35. Способ по п. 34, в котором обработку полимеразой и лигазой выполняют в присутствии средства разрушения вторичной структуры ДНК, причем средство разрушения необязательно представляет собой ДМСО.
36. Способ по п. 34 или 35, в котором обработку полимеразой и лигазой проводят в присутствии олигонуклеотида, который является комплементом праймерной последовательности, причем указанная праймерная последовательность представляет собой последовательность P7, представленную в SEQ ID NO: 8, и указанный олигонуклеотид представляет собой олигонуклеотид, который представляет собой комплмент указанной последовательности P7, представленной в SEQ ID NO: 8.
37. Способ по любому из пп. 34-36, в котором полимераза содержит мутант ДНК-полимеразы Т4, не имеющий экзонуклеазной активности.
38. Способ по любому из пп. 29-37, дополнительно включающий удаление полностью двухцепочечных меченых фрагментов с твердой подложки.
39. Способ по п. 38, в котором удаление включает применение нагревания и/или денатурирующего вещества, достаточного для отщепления полностью двухцепочечных меченых фрагментов от твердой подложки.
40. Способ по любому из пп. 29-39, дополнительно включающий секвенирование одного или более из 5’-меченых целевых фрагментов или полностью двухцепочечных меченых фрагментов.
41. Способ по п. 40, в котором фрагменты не определяют количественно с помощью ПЦР после приведения в контакт и до секвенирования.
42. Способ по любому из пп. 29-41, в котором нуклеиновая кислота представляет собой ДНК или РНК.
43. Способ по п. 29, в котором 3’-адаптерная последовательность второго транспозона комплементарна по меньшей мере участку индексной адаптерной последовательности.
44. Способ по п. 29, в котором праймерная последовательность полинуклеотида присоединения иммобилизованного комплекса транспосом по любому из пп. 5-26 комплементарна по меньшей мере части индексной праймерной последовательности.
45. Способ создания библиотеки меченых фрагментов нуклеиновых кислот, включающий следующее:
a) приведение иммобилизованного комплекса транспосом по любому из пп. 3-26 в контакт с целевой нуклеиновой кислотой для фрагментации целевой нуклеиновой кислоты во множество целевых фрагментов и для присоединения 3’-конца первого транспозона к 5’-концам целевых фрагментов с получением множества 5’-меченых целевых фрагментов;
b) обработка твердой подложки для удаления несвязанных нуклеиновых кислот; или обработка твердой подложки с удалением транспозазы из комплекса, необязательно посредством (a) нагревания твердой подложки и/или (b) промывки твердой подложки ферментным денатурирующим средством, причем ферментное денатурирующее средство необязательно содержит додецилсульфат натрия (SDS), гидрохлорид гуанидина, мочевину или протеиназу;
c) обработка множества 5’-меченых целевых фрагментов полимеразой и лигазой для удлинения и лигирования 5’-меченых целевых фрагментов с получением полностью двухцепочечных меченых фрагментов, причем необязательно обработку полимеразой и лигазой выполняют в присутствии средства разрушения вторичной структуры ДНК, причем средство разрушения необязательно представляет собой ДМСО;
d) удаление полностью двухцепочечных меченых фрагментов с твердой подложки, причем удаление необязательно включает применение нагревания и/или денатурирующего вещества, достаточного для отщепления полностью двухцепочечных меченых фрагментов от твердой подложки, при этом денатурирующее средство необязательно представляет собой NaOH; и
e) отбор полностью двухцепочечных меченых фрагментов с использованием гранул для захвата, необязательно, при этом гранулы для захвата представляют собой магнитные гранулы, причем дополнительно необязательно выполняют две отдельные стадии отбора.
46. Способ по п. 45, в котором иммобилизованный комплекс транспосом включает следующее:
твердая подложка; и
комплекс транспосом, иммобилизованный на твердой подложке,
причем комплекс транспосом содержит следующее:
(a) транспозаза;
(b) первый транспозон, содержащий 3’-концевую последовательность транспозона и якорную последовательность (якорь);
(с) второй транспозон, содержащий 5’-концевую последовательность транспозона и последовательность, комплементарную последовательности B15, представленной в SEQ ID NO: 5, где
(i) 5'-концевая последовательность второго транспозона комплементарна 3'-концевой последовательности первого транспозона,
(ii) 5'-адаптерная последовательность первого транспозона не комплементарна последовательности, комплементарной последовательности B15, представленной в SEQ ID NO: 5, второго транспозона, и
(iii) первый транспозон и второй транспозон образуют Y-образную форму; и
(d) полинуклеотид присоединения, который гибридизуется с первым транспозоном или вторым транспозоном, содержащий следующее:
(i) комплемент якорной последовательности, последовательность, комплементарную последовательности A14, представленной в SEQ ID NO: 4, спейсер и последовательность, комплементарную последовательности P5, представленной в SEQ ID NO: 7; и
(ii) связывающий элемент для связывания указанного транспосомного комплекса с твердой подложкой, содержащий биотин, причем биотин иммобилизован на указанной твердой подложке.
47. Способ по п. 45 или 46, дополнительно включающий секвенирование одного или более полностью двухцепочечных меченых фрагментов.
48. Способ создания библиотеки меченых фрагментов нуклеиновых кислот, включающий следующее:
a) приведение в контакт первого иммобилизованного комплекса транспосомы по любому из пп. 3-26 и второго иммобилизованного комплекса транспосомы по любому из пп. 3-26 с целевой нуклеиновой кислотой для фрагментации целевой нуклеиновой кислоты во множество целевых фрагментов и присоединение 3’-конца каждого первого транспозона к 5’-концам целевых фрагментов для получения множества первых 5’-меченых целевых фрагментов, полученных из первого иммобилизованного комплекса транспосомы, и множества вторых 5’-меченых целевых фрагментов, полученных из второго иммобилизованного комплекса транспосомы,
причем первый иммобилизованный комплекс транспосом содержит твердую подложку и первый комплекс транспосом, иммобилизованный на твердой подложке, и при этом первый комплекс транспосом содержит следующее:
i) транспозаза;
ii) первый транспозон, содержащий 3’-концевую последовательность транспозона и якорную последовательность;
iii) второй транспозон, содержащий 5’-концевую последовательность транспозона, где (1) 5'-концевая последовательность второго транспозона комплементарна 3'-концевой последовательности первого транспозона, (2) якорная последовательность первого транспозона не комплементарна какой-либо части второго транспозона и (3) первый транспозон и второй транспозон образуют Y-образную форму; и
iv) первый полинуклеотид присоединения, который гибридизуется с первым транспозоном или вторым транспозоном, содержащий (i) комплемент якорной последовательности, последовательность, комплементарную последовательности A14, представленной в SEQ ID NO: 4, спейсер и последовательность, комплементарную последовательности P5, представленной в SEQ ID NO: 7, и (ii) связывающий элемент, содержащий биотин, причем биотин иммобилизован на твердой подложке,
и при этом второй иммобилизованный комплекс транспосом содержит твердую подложку и второй комплекс транспосом, иммобилизованный на твердой подложке, причем второй комплекс транспосом содержит следующее:
i) транспозаза;
ii) первый транспозон, содержащий 3’-концевую последовательность транспозона и якорную последовательность;
iii) второй транспозон, содержащий 5’-концевую последовательность транспозона, где (1) 5'-концевая последовательность второго транспозона комплементарна 3'-концевой последовательности первого транспозона, (2) якорная последовательность первого транспозона не комплементарна какой-либо части второго транспозона и (3) первый транспозон и второй транспозон образуют Y-образную форму; и
iv) второй полинуклеотид присоединения, который гибридизуется с первым транспозоном или вторым транспозоном, содержащий (i) комплемент якорной последовательности, последовательность, комплементарную последовательности B15 (SEQ ID NO: 7), спейсер и последовательность, комплементарную последовательности P7 (SEQ ID NO: 8), и (ii) связывающий элемент для связывания второго транспосомного комплекса с твердой подложкой, содержащий биотин, причем биотин иммобилизован на указанной твердой подложке;
b) обработка множества 5’-меченых целевых фрагментов лигазой для лигирования каждого 5’-меченого целевого фрагмента либо с первым индексирующим олигонуклеотидом, либо со вторым индексирующим олигонуклеотидом посредством приведения 5’-меченых целевых фрагментов в контакт с пулом первого и второго индексирующих олигонуклеотидов, причем каждый первый индексирующий олигонуклеотид содержит последовательность A14, последовательность i5 и последовательность Р5 и может связываться с первым 5’-меченым целевым фрагментом; и при этом каждый второй индексирующий олигонуклеотид содержит последовательность В15 (SEQ ID NO: 5), последовательность i7 и последовательность Р7 (SEQ ID NO: 8) и может связываться со вторым 5’-меченым целевым фрагментом для получения множества 5’-меченых целевых фрагментов, лигированных с индексирующими олигонуклеотидами;
c) обработка твердой подложки для удаления транспозаз из комплекса, необязательно посредством (a) нагревания твердой подложки и/или (b) промывки твердой подложки ферментативным денатурирующим средством, причем ферментативное денатурирующее средство необязательно содержит додецилсульфат натрия (SDS), гидрохлорид гуанидина, мочевину или протеиназу; и
d) обработка множества 5’-меченых целевых фрагментов, лигированных с индексирующими олигонуклеотидами, полимеразой для удлинения и получения полностью двухцепочечных меченых фрагментов.
49. Способ по п. 48, в котором приведение в контакт первого иммобилизованного комплекса транспосомы со вторым иммобилизованным комплексом транспосомы и обработку множества 5’-меченых целевых фрагментов лигазой проводят в одну реакцию.
50. Способ по п. 48 или 49, в котором двухцепочечные меченые фрагменты получают в растворе.
51. Способ по любому из пп. 48-50, дополнительно включающий отбор полностью двухцепочечных меченых фрагментов с использованием гранул для захвата, причем гранулы для захвата необязательно представляют собой магнитные гранулы, и дополнительно необязательно выполняют две отдельные стадии отбора.
52. Способ по любому из пп. 48-51, дополнительно включающий секвенирование одного или более из полностью двухцепочечных меченых фрагментов.
| WO 2018156519 A1, 30.08.2018 | |||
| WO 2016061517 A2, 21.04.2016 | |||
| WO 2018208699 A1, 15.11.2018 | |||
| KUAN FENG et al., Next-generation sequencing library construction on a surface, BMC Genomics, 2018, Vol.19, N.416 | |||
| RU 2016107196 A, 26.09.2017. |

