ОБЛАСТЬ ТЕХНИКИ
Настоящее описание относится к новой комбинации терапевтических агентов для лечения опухоли и/или метастаза, предпочтительно метастаза.
УРОВЕНЬ ТЕХНИКИ
Метастатическое распространение основано на способности раковых клеток нарушать межклеточные взаимодействия, мигрировать через внеклеточный матрикс, выживать и размножаться в тканях, отличных от места их происхождения. Физиологический аналог этой сложной программы - известный как «инвазивный рост» - лежит в основе эмбриогенеза и отвечает за заживление ран и регенерацию органов во взрослой жизни. Инвазивный рост жестко регулируется специфическими внеклеточными сигналами, одним из которых является фактор роста гепатоцитов (HGF), лиганд рецептора, кодируемого онкогеном MET. В условиях аберрантной активации передача сигналов HGF/MET управляет возникновением, прогрессированием и метастазированием опухоли в широком спектре злокачественных новообразований человека. В меньшинстве случаев MET ведет себя как «драйвер» онкогена, а рост и выживание опухолевых клеток зависит от конститутивной передачи сигналов MET («зависимость от MET»). Это состояние зависит от наличия генетических повреждений, в основном увеличения числа копий гена или, реже, мутаций2, которые приводят к конститутивной лиганд-независимой активации рецептора. В этом контексте лечение ингибиторами МЕТ является высокоэффективным, вызывая блокировку пролиферации клеток и остановку клеточного цикла in vitro и ингибирование роста опухоли in vivo. Совместная экспрессия лиганда и рецептора в одной и той же клетке является другой стратегией, используемой раком для достижения непрерывной активации MET, и она была описана в основном при неэпителиальных раковых заболеваниях человека, таких как остеосаркомы, глиобластомы и множественные миеломы. Однако в большинстве случаев аберрантная активация МЕТ в опухолях происходит из-за сверхэкспрессии рецептора из-за усиления транскрипции гена дикого типа, запускающего сенсибилизацию раковой клетки к стимуляции лигандом3. В последнем случае передача сигналов MET, которая приводит к проинвазивным и антиапоптотическим ответам, используется раковыми клетками в качестве стратегии обхода стрессовых условий и усиления злокачественного фенотипа («уместность MET»6). В отсутствие специфических генетических повреждений МЕТ не является строго необходимым для роста опухоли, но присутствие лиганда поддерживает активацию рецептора, усиливая злокачественный фенотип. Наконец, МЕТ действует как функциональный маркер раковых «стволовых клеток-предшественников» в глиобластомах8 и поддерживает «стволовой» фенотип при колоректальном раке и раке молочной железы12,15. Более того, было показано, что происходящий из стромы HGF поддерживает путь самообновления WNT стволовых клеток колоректального рака и способствует пролиферации клеток, инициирующих рак толстой кишки, вызывая устойчивость к терапии против EGFR27.
Был разработан и в настоящее время исследуется ряд стратегий, нацеленных на MET или HGF – низкомолекулярные ингибиторы, антитела или рекомбинантные белки. Среди них антитело MvDN30 представляет собой моновалентный химерный Fab-фрагмент, который связывается с внеклеточным доменом MET, вызывая протеолитическое отщепление («шеддинг») рецептора с поверхности клетки21, 28. DecoyMET представляет собой рекомбинантный растворимый рецептор, охватывающий всю внеклеточную область MET; он связывает HGF с высоким сродством и ингибирует лиганд-зависимую биологическую активность in vitro и in vivo при экспрессии с помощью технологии лентивирусных векторов17 или в виде слитого с Fc белка7,26. Как и в большинстве биологических систем, удар по единственному элементу цепи передачи сигнала вряд ли приведет к полному отключению ответа. Таким образом, в случае MET каждая молекула никогда не достигнет 100 % ингибирования, оставляя остаточную активность чувствительной к стимуляции HGF.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является создание новой комбинации противоопухолевых агентов, полезных для лечения онкологических пациентов.
В соответствии с изобретением указанная выше цель достигается благодаря предмету, конкретно упомянутому в приложенной формуле изобретения, которая понимается как составляющая неотъемлемая часть настоящего описания.
Настоящее изобретение относится к фрагменту антитела против рецептора фактора роста гепатоцитов (HGFR) в сочетании с внеклеточной частью человеческого HGFR для использования при лечении пациента, страдающего от опухоли и/или метастаза, предпочтительно метастаза, при этом:
(i) фрагмент антитела против HGFR имеет только один паратоп, способный связываться с эпитопом внеклеточной части HGFR человека, и обладает антагонистической активностью в отношении HGFR,
(ii) внеклеточная часть HGFR человека способна стабильно связываться с фактором роста гепатоцитов (HGF) и содержит по крайней мере одну аминокислотную мутацию в эпитопе, распознаваемом фрагментом антитела против HGFR, для предотвращения связывания с ним фрагмента антитела против HGFR, и
(iii) фрагмент антитела против HGFR и внеклеточная часть HGFR человека подходят для введения пациенту (a) в форме белка или (b) в форме нуклеиновой кислоты.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Далее изобретение будет описано подробно исключительно в качестве иллюстративного и неограничивающего примера со ссылкой на прилагаемые фигуры.
Фиг. 1. Создание мутированного рецептора decoyMET, который не связывает антитело DN30.
(А) Сравнение аминокислотных последовательностей третьего и четвертого доменов IPT MET человека, мыши, крысы и собаки. Остатки, которые изменены исключительно в последовательности мыши, выделены жирным шрифтом, белым на черном фоне, аминокислоты, которые также (или только) изменены в последовательностях крысы или собаки, выделены жирным шрифтом. Показаны только границы IPT-3/IPT-4. (B) Рецепторы decoyMET, несущие одиночные аминокислотные замены, инкубировали с антителом DN30. Комплексы подвергали иммунопреципитации с белком А, который связывается с антителом, и выявляли с помощью конъюгированного с HRP стрептактина, который связывается со стреп-меткой в ловушке (левая панель). 30 мкл нормализованных супернатантов, используемых для иммунопреципитации, обрабатывали на SDS PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) для проверки загрузки рецепторов decoyMET (правая панель). (C) Анализ связывания ELISA (ИФА, иммуноферментный анализ). MAb DN30 находилось в жидкой фазе, decoyMET дикого типа или decoyMETK842E – в твердой фазе. Связывание антител детектировали с использованием конъюгированного с HRP мышиного антитела. OD, оптическая плотность при 450 нм. Каждая точка представляет собой среднее значение трех измерений ± стандартное отклонение SD.
Фиг. 2. DecoyMETK842E связывает HGF с высоким сродством и ингибирует HGF-индуцированное фосфорилирование MET.
(A) Анализ связывания ELISA. DecoyMET дикого типа или decoyMETK842E находились в твердой фазе; увеличивающиеся концентрации HGF добавляли в жидкую фазу. Связывание HGF детектировали с использованием биотинилированного антитела против HGF. Каждая точка представляет собой среднее значение трех измерений ± SD. (B) HGF-индуцированное фосфорилирование МЕТ. Клетки A549 инкубировали с decoyMET дикого типа или decoyMETK842E (2 мкМ) и стимулировали HGF (50 нг/мл). Тотальные клеточные лизаты подвергали иммуноблоттингу с использованием антифосфоМЕТ (верхняя панель), антител против МЕТ (средняя панель) или антител к винкулину (нижняя панель), p145, зрелой формы MET; p190, одноцепочечного предшественника МЕТ; p117, винкулина.
Фиг. 3. MvDN30 и DecoyMETK842E совместно вносят вклад в снижение индуцированного HGF фосфорилирования MET и биологической активности, обусловленной MET.
(A) HGF-индуцированное фосфорилирование МЕТ. Клетки аденокарциномы легких человека A549 инкубировали с 2 мкМ decoyMETK842E, 125 нМ MvDN30 или их комбинацией и стимулировали или не стимулировали 50 нг/мл HGF. Тотальные клеточные лизаты подвергали иммуноблоттингу с использованием антител против фосфоМЕТ (верхняя панель), антител против МЕТ (средняя панель) или антител к винкулину (нижняя панель), p145, зрелой формы MET; p190, одноцепочечного предшественника МЕТ; p117, винкулина. (B) HGF-индуцированное фосфорилирование ERK и AKT. Клетки аденокарциномы легких человека A549 инкубировали с 500 нМ MvDN30, 2 мкМ decoyMETK842E или их комбинацией и стимулировали или не стимулировали 100 нг/мл HGF. Тотальные клеточные лизаты подвергали иммуноблоттингу с анти-фосфоЕРК, анти-ERK, анти-фосфоАКТ, анти-AKT, анти-GAPDH, p42/44, ERK; р60, АКТ; p36, GAPDH. (C) Независимый от закрепления рост, поддерживаемый аутокринной стимуляцией HGF. Клетки глиобластомы человека U87-MG, экспрессирующие белки HGF и MET, обрабатывали 0,5 мкМ MvDN30 или 1 мкМ decoyMETK842E по отдельности или их комбинацией. График представляет процент среднего роста колоний для каждой обработки по сравнению с необработанным контролем. (D) Независимый от закрепления рост, поддерживаемый паракринной стимуляцией HGF. Клетки A549 стимулировали 30 нг/мл HGF и обрабатывали 1 мкМ MvDN30 или 1 мкМ decoyMETK842E, по отдельности или их комбинацией. График показывает процент среднего роста колоний для каждой обработки по сравнению с контролем, стимулированным HGF. (E) Трансвел-анализ инвазии. Клетки аденокарциномы поджелудочной железы человека HPAF-II стимулировали 12,5 нг/мл HGF и обрабатывали 0,5 мкМ MvDN30 или 1 мкМ decoyMETK842E, по отдельности или их комбинацией. График показывает процент инвазии по сравнению с контролем, стимулированным HGF. Правая панель, одно репрезентативное изображение / группа клеток, мигрировавших через слой матригеля; n.t. - необработанные клетки. Увеличение 200×. Каждая точка представляет собой среднее значение в трех экземплярах ± SD. *** означает P ≤ 0,001; ** означает P ≤ 0,01; * означает P ≤ 0,05.
Фиг. 4. MvDN30 и decoyMETK842E в комбинации ингибируют HGF-зависимое фосфорилирование MET.
Клетки U87MG (A), A549 (B) и HPAF-II (C) обрабатывали MvDN30, decoyMETK842E или их комбинацией (COMBO). Клетки HPAF-II и A549 стимулировали или не стримулировали HGF; n.t. - необработанные клетки. Слева: репрезентативные конфокальные срезы, показывающие анти-фосфоМЕТ (верхние ряды) и фаллоидин (нижние ряды). На графиках справа показана средняя интенсивность флуоресценции (MFI) фосфоМЕТ, вычтенная и нормализованная для фаллоидина. Каждая точка представляет собой среднее из 5 значений ± SEM (стандартная ошибка среднего). Масштаб 50 мкм. *** означает P ≤ 0,001; ** означает P ≤ 0,01; * означает P ≤ 0,05.
Фиг. 5. MvDN30 и DecoyMETK842E действуют синергично, подавляя HGF-зависимое рассеяние клеток.
(А) Анализ подвижности клеток. Клетки HPAF-II предварительно инкубировали с различными концентрациями (0,06, 0,25, 1 или 4 мкМ) MvDN30 или decoyMETK842E, отдельно или в комбинации 1:1, а затем стимулировали 6,25 нг/мл HGF. Распределение клеток отслеживали в реальном времени с помощью устройства X-CELLigence RTCA и выражали как нормализованный индекс клеток. Каждый график относится к одной концентрации обработки. (B) Репрезентативные изображения клеток HPAF-II, предварительно инкубированных с 1 мкМ MvDN30 или 1 мкМ decoyMETK842E, отдельно или в комбинации, а затем стимулированных 6,25 нг/мл HGF; n.t. - необработанные клетки. (C) Кривая подвижности клеток. Эффект – это значение индекса клеток, измеренное в конце эксперимента для каждой дозы обработки, нормализованное по значениям, полученным только с HGF, и выраженное как [1-x]. (D) Анализ комбинаций лекарственных средств. Значения кривой подвижности клеток были получены с помощью программного обеспечения Calcusyn для расчета индекса комбинации (CI) для каждой концентрации MvDN30 и decoyMETK842E. CI = 1, взаимодействие; CI < 1, синергизм; CI > 1, антагонизм.
Фиг. 6 Комбинация MvDN30 и decoyMETK842E снижает инвазивный фенотип подкожных опухолей U87-MG.
Клетки U87-MG вводили подкожно мышам NOD-SCID. Когда опухоли достигли объема 80-100 мм3, мыши были разделены на четыре однородные группы: носитель (n = 10); MvDN30 (n = 9); K842E (n = 9); комбинация двух лекарственных средств (n = 10).(A) Объем опухоли при умерщвлении, выраженный как кратное увеличение. Каждая полоса представляет собой среднее значение группы ± SEM. (B) Гистохимический анализ опухолевой нагрузки. Репрезентативные изображения срезов опухоли, окрашенных гематоксилином-эозином. Стрелки указывают на границу между опухолью и окружающей тканью. Увеличение 100×.
Фиг. 7. Влияние MvDN30 или decoyMETK842E, по отдельности или в комбинации, на пролиферацию и апоптоз клеток рака поджелудочной железы, ортотопически инъецированных мышам hHGF-Ki.
Клетки HPAF-II, экспрессирующие люциферазу, вводили в поджелудочную железу мышей hHGF-Ki и разделяли на четыре гомогенные группы: носитель (n = 10), MvDN30 (n = 6), decoyMETK842E (n = 6), комбинация лекарственных средств (n = 6). (A) Биолюминесценция опухоли, обнаруженная IVIS Spectrum. Цифры означают средние значения общего потока биолюминесценции (фотонов/секунду × 108) каждой экспериментальной группы ± SEM. (B) Анализ пролиферации опухолевых клеток, измеренный иммуногистохимическим методом Ki67. Левая панель – репрезентативные изображения каждой экспериментальной группы. Увеличение 200×. График справа показывает средние значения, полученные при анализе пяти изображений на опухоль ± SEM. Индекс пролиферации рассчитывается как отношение Ki67-положительных клеток к общему количеству клеток. (C) Анализ апоптоза опухолевых клеток, измеренный с помощью иммунофлуоресценции расщепленной каспазы-3. Левая панель – репрезентативные изображения каждой экспериментальной группы. Масштаб 50 мкм. График справа показывает средние значения, полученные при анализе 8 изображений на опухоль ± SEM. Индекс апоптоза рассчитывают как отношение количества клеток, положительных на расщепленную каспазу-3, к общему количеству клеток. Масштаб 50 мкм. *** означает значение P < 0,001; ** означает значение P < 0,01; * означает значение P < 0,05.
Фиг. 8. MvDN30 и DecoyMETK842E снижают фосфорилирование МЕТ и метастатическое распространение раковых клеток поджелудочной железы у мышей hHGF-Ki.
Клетки HPAF-II, экспрессирующие люциферазу, вводили в поджелудочную железу мышей hHGF-Ki и разделяли на четыре гомогенные группы: носитель (n = 10), MvDN30 (n = 6), decoyMETK842E (n = 6), комбинация лекарственных средств (n = 6). (A) Статус фосфоMET в опухолях, измеренный с помощью иммунофлуоресцентного анализа. Левая панель, репрезентативные конфокальные срезы каждой экспериментальной группы, показывающие антифосфоМЕТ. График справа показывает среднюю интенсивность флуоресценции (MFI) фосфоMET, фон вычтен и нормализован по DAPI. Каждая точка представляет собой среднее из 12 значений ± SEM. Масштаб 50 мкм. (B) Оценка фенотипа EMT опухолей HPAF-II с помощью иммунофлуоресцентного анализа экспрессии E-кадгерина и виментина. Левая панель, репрезентативные изображения каждой экспериментальной группы. Анти-Е-кадгерин (верхний ряд), антивиментин (нижний ряд). Масштаб 50 мкм. График справа показывает средние значения, полученные при анализе 6 изображений на опухоль ± SEM. Фенотип ЕМТ выражается соотношением виментин/Е-кадгерин. (C) Метастатические узелки в легких, оцениваемые с помощью гистохимического окрашивания HE. График слева: количество метастатических очагов; каждая точка представляет количество поражений, оцененных для каждой мыши. График справа: область метастатического поражения; каждая точка представляет собой среднюю площадь метастазов, измеренную для каждой мыши. Было проанализировано десять слайдов/мышь; метастатические поражения оценивали, а их площадь количественно определяли с помощью ImageJ. *** означает P ≤ 0,001; ** означает P ≤ 0,01; * означает P ≤ 0,05.
Фиг. 9. Схематическое изображение одного из вариантов слитых белков, содержащих фрагмент антитела против HGFR и внеклеточную часть HGFR человека.
(A) Изображение молекулы, образованной слиянием в рамке считывания decoyMETK842E на N-конце и scMvDN30 на C-конце. (B) Изображение молекулы, образованной слиянием в рамке считывания scMvDN30 на N-конце и decoyMETK842E на C-конце. Между двумя группами был вставлен линкер. Последовательность линкера может быть выбрана из последовательностей, перечисленных в (C). SEMA, PSI, IPT 1-4 представляют собой участки decoyMET. Звездочка в IPT 4 показывает мутацию K→Е в позиции 842. Фрагмент антитела против HGFR состоит из двух цепей, соединенных вторым линкером. VL – вариабельная область легкой цепи; CL – константная область легкой цепи; VH – вариабельная область тяжелой цепи; CH – константная область тяжелой цепи. Черный квадрат представляет собой метки Strept-His, включенные в рекомбинантный белок для целей очистки/обнаружения.
Фиг. 10. Анализ связывания слитых белков, содержащих фрагмент антитела против HGFR и внеклеточную часть HGFR человека, с MET или HGF.
(A) Анализ связывания с HGF с помощью ELISA. Левая панель, связывание слитых белков, содержащих decoyMETK842E на N-конце и scMvDN30 на C-конце. Правая панель, связывание слитых белков, содержащих scMvDN30 на N-конце и decoyMETK842E на C-конце. Слитые белки или decoyMETK842E (положительный контроль) находились в твердой фазе. В жидкую фазу добавляли возрастающие концентрации HGF. Связывание HGF детектировали с использованием биотинилированного антитела против HGF. Каждая точка представляет собой среднее значение в трех экземплярах ± SD. (B) Анализ связывания с MET с помощью ELISA. Левая панель, связывание слитых белков, содержащих decoyMETK842E на N-конце и scMvDN30 на C-конце. Правая панель, связывание слитых белков, содержащих scMvDN30 на N-конце и decoyMETK842E на C-конце. DecoyMET дикого типа находился в твердой фазе, а возрастающие концентрации различных слитых белков находились в жидкой фазе; в качестве положительного контроля в анализ был включен scMvDN30. Связывание антител определяли с использованием конъюгированного с HRP антитела против k-цепи человека. OD, оптическая плотность при 450 нм. Каждая точка представляет собой среднее значение в трех измерениях ± SD.
Фиг. 11. Ингибирование роста опухолевых клеток при обработке слитыми белками, содержащими фрагмент антитела против HGFR и внеклеточную часть человеческого HGFR.
(A) Рост клеток карциномы легких EBC-1, обработанных увеличивающимися концентрациями слитых белков, содержащих decoyMETK842E на N-конце и scMvDN30 на C-конце. (B) Рост клеток карциномы легких EBC-1, обработанных увеличивающимися концентрациями слитых белков, содержащих scMvDN30 на N-конце и decoyMETK842E на C-конце. В качестве положительного контроля в анализ были включены эквимолярные комбинации scMvDN30 и decoyMETK842E (Combo). Клетки анализировали через 3 дня после обработки. Образцы представлены в трех экземплярах, столбцы представляют собой стандартное отклонение. (C) Таблица, показывающая значения IC50 для каждого слитого белка, рассчитанные как среднее значение по меньшей мере из трех независимых экспериментов, по сравнению с ингибирующей активностью, проявляемой MvDN30/decoyK842E в комбинации.
Фиг. 12. Ингибирование роста опухолевых клеток при обработке слитыми белками, содержащими фрагмент антитела против HGFR и внеклеточную часть человеческого HGFR.
Клетки HPAF-II предварительно инкубировали с различными слитыми белками (3 мкМ), а затем стимулировали 6,25 нг/мл HGF. Распределение клеток отслеживали в реальном времени с помощью устройства X-CELLigence RTCA и выражали как нормализованный индекс клеток. Испытания образцов проводят в трехкратной повторности. (A) Клетки, обработанные слитыми белками, содержащими decoyMETK842E на N-конце и scMvDN30 на C-конце. (B) Клетки, обработанные слитыми белками, содержащими scMvDN30 на N-конце и decoyMETK842E на C-конце. В качестве положительного контроля в анализ были включены эквимолярные комбинации scMvDN30 и decoyMETK842E (Combo). (C) Таблица, показывающая процент подвижности клеток, рассчитанный как среднее значение из двух независимых экспериментов, по сравнению с клетками, обработанными HGF.
Фиг. 13. Слитые белки, содержащие фрагмент антитела против HGFR и внеклеточную часть человеческого HGFR, уменьшают метастатическое распространение клеток рака поджелудочной железы у мышей hHGF-Ki.
Экспрессирующие люциферазу клетки Capan-1 вводили в поджелудочную железу мышей hHGF-Ki и разделяли на семь гомогенных групп: носитель (n = 14), decoyMETK842E и scMvDN30 в комбинации 1:1 (две группы) и четыре группы для тестирования различных слитых белков, либо содержащие decoyMETK842E на N-конце и scMvDN30 на C-конце, либо содержащие scMvDN30 на N-конце и decoyMETK842E на C-конце. Анализировали два разных линкера (L60 и L134). Combo 1 (n = 4): MvDN30 доставляли 7 раз в неделю, а decoyMETK842E доставляли 2 раза в неделю; combo 2 (n = 6): MvDN30 доставляли 4 раза в неделю, а decoyMETK842E доставляли 2 раза в неделю; K842E_scMvDN30_L60 (n = 4); scMvDN30_K842E_L60 (n = 5); K842E_scMvDN30_L134 (n = 5); K842E_scMvDN30_L134 (n = 6). Все слитые белки доставляли 2 раза в неделю. (A) Таблица, показывающая количество мышей, несущих метастазы в печени, по сравнению с общим количеством проанализированных мышей. Мышей считали положительными, когда биолюминесценция, обнаруженная в печени с помощью IVIS Spectrum, превышала 105 фотонов в секунду. (B) График, показывающий процент биолюминесценции, обнаруженной с помощью IVIS Spectrum в печени каждой экспериментальной группы по сравнению с группой, обработанной носителем.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В нижеследующем описании приведены многочисленные конкретные детали изобретения, чтобы обеспечить полное понимание вариантов осуществления. Варианты осуществления могут быть реализованы без одной или нескольких конкретных деталей или с другими методами, компонентами, материалами и т. д. В других случаях хорошо известные структуры, материалы или операции не показаны или не описаны подробно, чтобы не затруднить понимание аспектов изобретения.
Используемая в настоящем документе отсылка на «один вариант осуществления» или «вариант осуществления» означает, что конкретный элемент, признак, структура или характеристика, описанная в связи с вариантом осуществления, включена в по меньшей мере один вариант осуществления. Таким образом, употребление выражения «в соответствии с одним вариантом осуществления» или «в соответствии с вариантом осуществления» в различных местах настоящего описания не обязательно всегда относится к одному и тому же варианту осуществления. Кроме того, отдельные признаки, структуры или характеристики могут быть объединены любым подходящим образом в одном или более вариантах осуществления.
Названия, представленные здесь, предназначены только для удобства и не интерпретируют объем или значение вариантов осуществления.
Настоящее описание относится к новой комбинации терапевтических агентов для лечения опухоли и/или метастаза, предпочтительно метастаза.
Настоящее изобретение основывается на идее, что более высокая терапевтическая устойчивость при лечении опухолей и/или метастаза, предпочтительно метастаза, может быть достигнута за счет воздействия как на HGF, так и на HGFR. Экспериментальные данные, представленные здесь, на самом деле показывают, что вертикальное ингибирование оси MET/HGF эффективно препятствует росту, подвижности и инвазии опухолевых клеток in vitro и значительно снижает метастатическое распространение in vivo, обеспечивая комбинированную таргетную терапию для широкого спектра раковых заболеваний, экспрессирующих MET дикого типа.
В одном из вариантов осуществления настоящее изобретение относится к комбинации фрагмента антитела против рецептора фактора роста гепатоцитов (HGFR) с внеклеточной частью человеческого HGFR для использования в лечении пациента, страдающего от опухоли и/или метастаза, предпочтительно метастаза, при этом:
(i) фрагмент антитела против HGFR имеет только один паратоп, способный связываться с эпитопом внеклеточной части HGFR человека, и обладает антагонистической активностью в отношении HGFR,
(ii) внеклеточная часть человеческого HGFR способна стабильно связываться с фактором роста гепатоцитов (HGF) и содержит по крайней мере одну аминокислотную мутацию в эпитопе, распознаваемом фрагментом антитела против HGFR, для предотвращения связывания с ним фрагмента антитела против HGFR, и
(iii) фрагмент антитела против HGFR и внеклеточная часть HGFR человека подходят для введения пациенту (а) в форме белка или (b) в форме нуклеиновой кислоты.
В предпочтительном варианте осуществления пациент, подвергаемый терапевтическому лечению с помощью описанной здесь комбинированной терапии, несет онкоген MET дикого типа (т. е. он/она не несет генетических изменений в онкогене MET).
В одном или нескольких вариантах реализации фрагмент антитела против HGFR содержит один вариабельный домен легкой цепи (VL) и один вариабельный домен тяжелой цепи (VH), причем вариабельные домены легкой цепи и тяжелой цепи являются нечеловеческими или гуманизированными, и где вариабельный домен легкой цепи (VL) содержит области, определяющие комплементарность (CDR), имеющие аминокислотные последовательности, указанные в SEQ ID NO: 1-3, и вариабельный домен тяжелой цепи (VH) содержит CDR, имеющие аминокислотные последовательности, указанные в SEQ ID NO: 4-6.
В еще одном варианте осуществления фрагмент антитела против HGFR дополнительно содержит один константный домен легкой цепи человека (CL) и один константный домен CH1 тяжелой цепи человека (CH1), причем вариабельный домен легкой цепи (VL) слит с константным доменом легкой цепи человека (CL) в направлении от N- к C-концу (таким образом образуя легкую цепь VL-CL), причем вариабельный домен тяжелой цепи (VH) слит с константным доменом CH1 тяжелой цепи человека в направлении от N- к C-концу (образуя таким образом тяжелую цепь VH-CH1). В предпочтительном варианте осуществления легкая цепь VL-CL содержит, предпочтительно состоит из аминокислотной последовательности, указанной в SEQ ID NO: 7, а тяжелая цепь VH-CH1 содержит, предпочтительно состоит из аминокислотной последовательности, указанной в SEQ ID NO: 8.
В различных вариантах осуществления, охватываемых настоящим изобретением, фрагмент антитела против HGFR находится в форме нуклеиновой кислоты. В таком случае фрагмент антитела против HGFR кодируется первой и второй молекулами нуклеиновой кислоты, где:
(i) первая молекула нуклеиновой кислоты кодирует один вариабельный домен легкой цепи (VL), содержащий CDR, имеющие последовательности нуклеиновой кислоты, указанные в SEQ ID NO: 9, 10 и 11, и где легкая цепь является нечеловеческой или гуманизированной; и
(ii) вторая молекула нуклеиновой кислоты кодирует один вариабельный домен тяжелой цепи (VH), содержащий CDR, имеющие последовательности нуклеиновой кислоты, указанные в SEQ ID NO: 12, 13 и 14, и где тяжелая цепь является нечеловеческой или гуманизированной.
В еще одном предпочтительном варианте осуществления фрагмент антитела против HGFR кодируется первой и второй молекулами нуклеиновой кислоты, где:
(i) первая молекула нуклеиновой кислоты кодирует (а) один вариабельный домен легкой цепи (VL), содержащий CDR, имеющие последовательности нуклеиновой кислоты, указанные в SEQ ID NO: 9, 10 и 11, где легкая цепь является нечеловеческой или гуманизированной, и (b) один константный домен легкой цепи человека (CL) в направлении от 5'- к 3'-концу (т. е. легкую цепь VL-CL); и
(ii) вторая молекула нуклеиновой кислоты кодирует (а) один вариабельный домен тяжелой цепи (VH), содержащий CDR, имеющие последовательности нуклеиновой кислоты, указанные в SEQ ID NO: 12, 13 и 14, где тяжелая цепь является нечеловеческой или гуманизированной, и (b) один константный домен CH1 тяжелой цепи человека (CH1) в направлении от 5'- к 3'-концу (т. е. тяжелую цепь VH-CH1).
В предпочтительном варианте осуществления первая молекула нуклеиновой кислоты (кодирующая легкую цепь VL-CL) содержит, предпочтительно состоит из последовательности нуклеиновой кислоты, указанной в SEQ ID NO: 15, и вторая молекула нуклеиновой кислоты (кодирующая тяжелую цепь VH-CH1) содержит, предпочтительно состоит из последовательности нуклеиновой кислоты, указанной в SEQ ID NO: 16.
В дополнительном варианте осуществления, охватываемом настоящим изобретением, фрагмент антитела против HGFR представляет собой одноцепочечный Fab-фрагмент против HGFR. Одноцепочечный Fab-фрагмент против HGFR содержит, предпочтительно состоит из аминокислотной последовательности, указанной в SEQ ID NO: 17, или одноцепочечный Fab-фрагмент против HGFR кодируется молекулой нуклеиновой кислоты, содержащей, предпочтительно состоящей из последовательности нуклеиновой кислоты, указанной в SEQ ID NO: 18.
В одном или нескольких вариантах осуществления настоящее изобретение относится к внеклеточной части человеческого HGFR, содержащей домены SEMA, PSI, IPT-1, IPT-2, IPT-3 и IPT-4.
В предпочтительном варианте осуществления внеклеточная часть человеческого HGFR содержит, предпочтительно состоит из аминокислотной последовательности, указанной в SEQ ID NO: 19, где одна или несколько аминокислот между положениями 797 и 875, указанными в SEQ ID NO: 19, мутированы. Согласно настоящему описанию внеклеточная часть человеческого HGFR имеет последовательность, мутированную в соответствии со знаниями, касающимися антител против HGFR и сайтов связывания HGF на HGFR, таким образом, что антитело не взаимодействует с HGFR, в то время как HGF сохраняет свою способность связывания с внеклеточной частью HGFR человека. В другом предпочтительном варианте осуществления внеклеточная часть человеческого HGFR имеет последовательность, указанную в SEQ ID NO: 20.
В дополнительном варианте осуществления, охватываемом настоящим изобретением, внеклеточная часть человеческого HGFR находится в форме молекулы нуклеиновой кислоты, которая содержит, предпочтительно состоит из последовательности нуклеиновой кислоты, как указано в SEQ ID NO: 21, где одна или несколько нуклеиновых кислот между положениями 2391 и 2625, указанными в SEQ ID NO: 21, являются мутированными. Согласно настоящему описанию молекула нуклеиновой кислоты кодирует внеклеточную часть человеческого HGFR, которая имеет последовательность, мутированную в соответствии со знаниями, касающимися антител против HGFR и сайтов связывания HGF на HGFR, таким образом, что антитело не взаимодействует с HGFR, в то время как HGF сохраняет свою способность связывания с внеклеточной частью HGFR человека. В другом предпочтительном варианте осуществления молекула нуклеиновой кислоты, кодирующая мутированную внеклеточную часть человеческого HGFR, имеет последовательность, указанную в SEQ ID NO: 22.
В различных вариантах осуществления, охватываемых настоящим изобретением, фрагмент антитела против HGFR конъюгирован с помощью линкера с внеклеточной частью HGFR человека в направлении от N- к C-концу, таким образом образуя слитый белок фрагмент антитела против HGFR – линкер – внеклеточная часть HGFR человека. Альтернативно, внеклеточная часть человеческого HGFR конъюгирована с помощью линкера с фрагментом антитела против HGFR в направлении от N- к C-концу, таким образом образуя слитый белок внеклеточная часть HGFR человека – линкер – фрагмент антитела против HGFR.
В различных вариантах осуществления, охватываемых настоящим изобретением, молекула нуклеиновой кислоты, кодирующая фрагмент антитела против HGFR, связана с помощью молекулы нуклеиновой кислоты, кодирующей линкер, с молекулой нуклеиновой кислоты, кодирующей внеклеточную часть HGFR человека, в направлении от 5'- к 3'-концу, таким образом кодируя слитый белок фрагмент антитела против HGFR – линкер – внеклеточная часть HGFR человека. Альтернативно, молекула нуклеиновой кислоты, кодирующая внеклеточную часть HGFR человека, связана посредством молекулы нуклеиновой кислоты, кодирующей линкер, с молекулой нуклеиновой кислоты, кодирующей фрагмент антитела против HGFR, в направлении от 5'- к 3'-концу, таким образом кодируя слитый белок внеклеточная часть HGFR человека – линкер – фрагмент антитела против HGFR.
Таким образом, в настоящем изобретении предложен слитый белок, содержащий в направлении от N- к C-концу:
(i) фрагмент антитела против HGFR, линкер и внеклеточную часть HGFR человека; или
(ii) внеклеточную часть HGFR человека, линкер и фрагмент антитела против HGFR.
В предпочтительном варианте осуществления слитый белок, как определено выше, содержит, предпочтительно состоит из аминокислотной последовательности, выбранной из любой из SEQ ID NO: 23-28.
Таким образом, настоящее изобретение охватывает молекулы нуклеиновой кислоты, кодирующие слитый белок, содержащий в направлении от N- к C-концу:
(i) фрагмент антитела против HGFR, линкер и внеклеточную часть HGFR человека; или
(ii) внеклеточную часть HGFR человека, линкер и фрагмент антитела против HGFR.
В предпочтительном варианте осуществления молекула нуклеиновой кислоты, кодирующая слитый белок, как определено выше, содержит, предпочтительно состоит из последовательности нуклеиновой кислоты, выбранной из любой из SEQ ID NO: 29-34.
В дополнительном варианте осуществления настоящее описание касается фармацевтического продукта, содержащего в одном флаконе или в двух флаконах (а) фрагмент антитела против рецептора фактора роста гепатоцитов (HGFR) и фармацевтически приемлемый носитель, и (б) внеклеточную часть человеческого HGFR и фармацевтически приемлемый носитель, где:
(i) фрагмент антитела против HGFR имеет только один паратоп, способный связываться с эпитопом внеклеточной части HGFR человека, и обладает антагонистической активностью в отношении HGFR,
(ii) внеклеточная часть человеческого HGFR способна стабильно связываться с фактором роста гепатоцитов (HGF) и содержит по крайней мере одну аминокислотную мутацию в эпитопе, распознаваемом фрагментом антитела против HGFR, для предотвращения связывания фрагмента антитела против HGFR с эпитопом,
(iii) фрагмент антитела против HGFR и внеклеточная часть человеческого HGFR находятся либо в форме белка, либо в форме нуклеиновой кислоты.
Таким образом, фармацевтический продукт содержит фрагмент антитела против HGFR и внеклеточную часть HGFR человека, имеющую признаки, отдельные или в любой соответствующей комбинации, описанные выше.
В одном аспекте изобретение относится к способу лечения опухоли и/или метастаза у субъекта, который включает введение субъекту эффективного количества комбинации фрагмента антитела против HGFR и внеклеточной части HGFR человека, посредством чего лечат указанное состояние.
В одном аспекте изобретение обеспечивает способ ингибирования роста клетки, экспрессирующей HGFR, который включает контактирование указанной клетки с комбинацией фрагмента антитела против HGFR и внеклеточной части человеческого HGFR по изобретению, вызывая тем самым ингибирование роста указанной клетки.
В одном аспекте изобретение обеспечивает способ терапевтического лечения млекопитающего, имеющего злокачественную опухоль и/или метастаз, где способ включает введение указанному млекопитающему эффективного количества комбинации фрагмента антитела против HGFR и внеклеточной части HGFR человека, обеспечивая тем самым эффективное лечение указанного млекопитающего.
В одном аспекте изобретение обеспечивает способ лечения нарушения пролиферации клеток, который включает введение субъекту, нуждающемуся в таком лечении, эффективного количества комбинации фрагмента анти-HGFR-антитела и внеклеточной части человеческого HGFR, обеспечивая тем самым эффективное лечение или профилактику указанного нарушения пролиферации клеток.
В одном аспекте изобретение обеспечивает способ терапевтического лечения опухоли и/или метастазов у млекопитающего, где рост указанной опухоли по меньшей мере частично зависит от потенцирующего рост эффекта системы HGFR/HGF, обеспечивая таким образом либо увеличение пролиферации клеток, либо защиту от апоптоза, либо и то, и другое. Указанный способ включает контактирование опухолевой клетки с эффективным количеством фрагмента анти-HGFR-антитела в комбинации с эффективным количеством внеклеточной части человеческого HGFR по изобретению, обеспечивая таким образом эффективное лечение указанной опухоли и/или метастаза.
Опухоль, которую можно эффективно лечить с помощью комбинированной терапии по настоящему изобретению, выбирают из опухоли молочной железы, толстой и/или прямой кишки, легких, толстой кишки, поджелудочной железы, предстательной железы, яичников, шейки матки, центральной нервной системы, почек, гепатоцеллюлярной системы, мочевого пузыря, желудка, клетки опухоли головы и шеи, папиллярной карциномы (например, щитовидной железы), меланомы, лимфомы, миеломы, глиомы/глиобластомы (например, анапластической астроцитомы, мультиформной глиобластомы, анапластической олигодендроглиомы, анапластической олигодендроастроцитомы), лейкемической клетки, саркомы, рабдомиосаркомы или опухоли неизвестного первичного происхождения (CUP).
В одном варианте осуществления клетка, которая является терапевтической мишенью в способе по изобретению, представляет собой гиперпролиферативную и/или гиперпластическую клетку. В одном варианте осуществления клетка, на которую нацелен способ по изобретению, представляет собой диспластическую клетку. В еще одном варианте осуществления клетка, на которую нацелен способ по настоящему изобретению, представляет собой метастатическую клетку. В другом варианте осуществления клетка, на которую нацелен способ по настоящему изобретению, представляет собой экспрессирующую HGFR клетку, принадлежащую к микроокружению, поддерживающему опухоль и/или метастаз.
Терапевтические способы по изобретению могут дополнительно включать дополнительные этапы лечения. Например, в одном варианте осуществления терапевтический способ дополнительно включает этап, на котором целевую опухолевую клетку и/или ткань подвергают лучевой терапии или химиотерапевтическому лечению. В дополнительном варианте осуществления в дополнение к фрагменту антитела против HGFR в комбинации с внеклеточной частью человеческого HGFR по изобретению целевую опухолевую клетку и/или ткань обрабатывают молекулами, специфически поражающими другие мишени, имеющие отношение к поддержанию трансформированного фенотипа (т. е. молекулы против EGFR).
Выражение «антагонистическая активность» фрагмента антитела против HGFR в контексте настоящего описания относится к антителу, которое способно гасить внутриклеточную передачу сигналов, вызванную в клетке при активации HGFR. Антагонистическая активность указанного антитела может быть измерена путем оценки уровня экспрессии и/или фосфорилирования HGFR обычными методами, такими как вестерн-блоттинг, иммунофлуоресценция, иммуногистохимия, ELISA, цитофлуориметрический анализ или любой другой метод, который включает использование антитела, которые специфически распознают HGFR, или остатки HGFR Tyr1234-1235, если они фосфорилированы (то есть основной сайт фосфорилирования MET9, или остатки HGFR Tyr1349/1356, если они фосфорилированы (то есть сайт стыковки MET22).
Используемое здесь выражение «внеклеточная часть человеческого HGFR способна стабильно связываться с человеческим HGF» означает, что внеклеточная часть человеческого HGFR связывается с HGF с расчетной Kd не выше 100 нМ.
Выражение «внеклеточная часть человеческого HGFR содержит по меньшей мере одну аминокислотную мутацию в эпитопе, распознаваемом фрагментом антитела против HGFR» в контексте настоящего описания означает наличие одной или нескольких мутаций (т. е. аминокислотных замен, и/или делеций, и/или вставки) во внеклеточной части HGFR, способной индуцировать модификацию во внеклеточной части HGFR, которая предотвращает вовлечение вышеуказанной области вариабельными доменами антитела. Специалист с учетом его общих знаний (представленных, в частности, возможностью создания кДНК, включая изменение одного нуклеотида в заданной последовательности ДНК с использованием специфических праймеров во время дупликации ДНК, см. Maniatis T. Molecular cloning: A laboratory manual Cold Spring Harbor Laboratory (1982)) не нуждается в дополнительных подробностях отностиельно реализации мутантной формы внеклеточной части HGFR человека, сохраняющей способность связываться с HGF человека, но не с фрагментом антитела против HGFR.
Таким образом, настоящее изобретение не следует интерпретировать как охватывающее только мутированную внеклеточную часть человеческого HGFR, как раскрыто в данном документе (т. е. SEQ ID NO: 8), поскольку квалифицированный специалист с точки зрения общих знаний может получить дополнительные мутированные версии внеклеточной части человеческого HGFR, содержащие SEQ ID NO: 19, что предотвращает связывание с ним антитела против HGFR.
Используемый здесь термин «одноцепочечный Fab-фрагмент» относится к одноцепочечному полипептиду, кодирующему домены VL, CL, VH, CH1 антитела, где VL и CL могут быть расположены на N-конце указанного полипептида и присоединены к доменам VH и CH1 с помощью гибкого линкера, или домены VH и CH1 могут быть расположены на N-конце указанного полипептида и связаны с доменами VL и CL гибким линкером.
Термины «SEMA», «PSI», «IPT-1», «IPT-2», «IPT-3» и «IPT-4» относятся к доменам HGFR, составляющим внеклеточную область HGFR. Такие доменные имена лежат в рамках общих знаний специалиста, например, 4, 13. Домен SEMA представляет собой модуль взаимодействия с белками, общий для семафоринов и плексинов, охватывающий область, заключенную между аминокислотами 25-516 MET (SEQ ID NO: 19); PSI является общим доменом с плексинами, семафоринами, интегринами, охватывающим область между аминокислотами 519-561 MET (SEQ ID NO: 19); домен IPT, повторяющийся четыре раза, представляет собой область иммуноглобулина, как и в целом с плексинами и факторами транскрипции, охватывающую область, содержащуюся между аминокислотами 563-934 MET (SEQ ID NO: 19). Более подробно, повтор IPT-1 охватывает аминокислотные положения 563-656 в SEQ ID NO: 19, повтор IPT-2 охватывает аминокислотные положения 657-740 в SEQ ID NO: 19, повтор IPT-3 охватывает аминокислотные положения 741-837 в SEQ ID NO: 19, повтор IPT-4 охватывает аминокислотные положения 838-934 в SEQ ID NO: 19.
Слитые белки, раскрытые в данном документе, могут быть легко изготовлены квалифицированным специалистом либо в форме белков, либо в форме молекул нуклеиновых кислот, кодирующих указанные слитые белки, с учетом общих знаний в области, связанной с технологией рекомбинантной ДНК, как представлено, например, в Maniatis T. Molecular cloning: A laboratory manual Cold Spring Harbor Laboratory (1982). В качестве примера можно использовать следующую стандартную процедуру: (i) синтез соответствующей последовательности кДНК, (ii) вставка кДНК в плазмиду, подходящую для экспрессии у млекопитающих, с помощью обычных методов рекомбинантной ДНК, (iii) временная или стабильная трансфекция линии клеток млекопитающих с помощью упомянутой выше плазмиды, (iv) сбор культурального супернатанта, (v) очистка слитого белка с помощью аффинной хроматографии. Линкеры, используемые для конъюгирования двух белковых последовательностей, то есть фрагмента антитела против HGFR и внеклеточной части человеческого HGFR, могут иметь разную длину и/или аминокислотный состав, быть гибкими, жесткими или представлять собой комбинацию гибких и жестких областей. Линкеры, используемые в получении слитых белков, могут быть выбраны, но не ограничиваются ими, из любой из аминокислотных последовательностей, указанных в SEQ ID NO: 35, 36 и 37. Линкеры, используемые для получения молекул нуклеиновых кислот, кодирующих слитые белки, могут быть выбраны, но не ограничиваются ими, из любой из последовательностей нуклеиновых кислот, указанных в SEQ ID NO: 38, 39 и 40.
Далее настоящее изобретение будет проиллюстрировано ссылкой на комбинацию фрагмента антитела против HGFR, содержащего CDR с SEQ ID NO: 1-6, принадлежащих моноклональному антителу DN30, раскрытому в WO 2007/090807, и внеклеточной части HGFR, содержащей домены SEMA, PSI, IPT-1 - IPT-4, кратко далее обозначенные DecoyMET.
Авторы настоящего изобретения неожиданно обнаружили, что моновалентный фрагмент антитела DN30 (MvDN30) и DecoyMET, используемые в комбинации, позволяют осуществлять двойное нацеливание на лиганд и рецептор, действуя одновременно на MET-экспрессирующие раковые клетки и на секретирующую HGF строму опухоли и оказывая синергетический эффект в лечении опухолей и/или метастаза.
Фармакологическое ингибирование рецептора тирозинкиназы MET в раковых клетках, «зависимых» от онкогенов, подавляет пролиферацию и инвазию клеток. Соответственно, пациенты с усиленным метастазирующим NSCLC (немелкоклеточный рак легких), метастатическим раком желудка или пищевода реагируют на анти-MET терапию5, 14, 19. С другой стороны, раковые клетки без генетических изменений MET используют «физиологическую» программу, запускаемую онкогеном, как «средство» для усиления злокачественного фенотипа6. «Уместность» требует стимуляции МЕТ дикого типа его лигандом HGF. В этом отношении вклад микросреды опухоли в прогрессирование и метастазирование рака становится все более актуальным, поскольку экспериментальные данные показывают, что злокачественный фенотип развивается не строго клеточно-автономным образом, а в довольно сложном взаимодействии между раковыми клетками и стромой хозяина. Микроокружение опухоли является важным источником HGF, секретируемого стромальными клетками мезенхимального происхождения в качестве неактивного предшественника (proHGF). Последний запасается во внеклеточном матриксе благодаря своей авидности к гепарансульфатам и активируется специфическими протеазами, продуцируемыми опухолевыми или стромальными клетками. Следовательно, избыток биологически активного лиганда легко доступен для связывания рецептора MET и запускает инвазивный сигнальный каскад роста в «независимых» клетках. Приведенные здесь данные показывают, что в условиях «уместности» МЕТ сопутствующее вмешательство, затрагивающее обе стороны оси МЕТ/HGF, приводит к улучшенной ингибирующей активности. Одновременное нацеливание было достигнуто путем комбинирования моновалентного антитела против MET, MvDN30, с рекомбинантным растворимым рецептором, decoyMETK842E. Приведенные здесь данные указывают на отсутствие избыточности нацеливания взаимодополняющих средств на один и тот же путь. Два ингибитора были выбраны с учетом их механизмов действия: антитело индуцирует физическое удаление МЕТ с поверхности клетки путем «отщепления» эктодомена. Последний выпускается во внеклеточную среду и действует как «ловушка» для HGF. Экзогенная поставка рекомбинантного decoyMET усиливает HGF-связывающую активность эндогенного decoyMET, генерируемого MvDN30. Чтобы сделать возможным одновременное использование MvDN30 и decoyMET, был создан модифицированный растворимый рецептор (decoyMETK842E), дефицитный по взаимодействию с MvDN30, но наделенный высокими аффинными свойствами связывания с HGF. Эти два агента в комбинации взаимодействуют в различных раковых клетках, что снижает эффективную терапевтическую дозу. Более того, эта «двойная стратегия» демонстрирует сильный синергетический эффект, потенциально проявляя превосходную противоопухолевую эффективность. Экспрессия MET в субпопуляции стволовых/предшественников раковых клеток была определена как «естественная роль» MET, то есть физиологический (естественно присутствующий) HGF-индуцированный внутриклеточный ответ, активированный в раковых стволовых клетках, в отсутствие генетических повреждений ответственен за устойчивость к таргетным способам лечения, таким как использование ингибиторов рецептора эпидермального фактора роста (EGFR) при колоректальном раке. Идея связи раковых стволовых клеток и резистентности к традиционным способам лечения в широко признана, и роль HGF в микросреде в поддержании стволового фенотипа клеток-предшественников, экспрессирующих MET, становится все более и более установленной. Эффективное лечение, нацеленное на MET, в виде комбинации MvDN30 и decoyMETK842E представляет собой терапевтическую поддержку для ослабления раковых стволовых клеток и противодействия возникновению устойчивости к таргетным способам лечения.
Роль микроокружения хозяина трудно исследовать в ксенотрансплантатах мыши из-за ограниченной перекрестной реактивности между факторами, происходящими из стромы мыши, и специфическими мишенями на раковых клетках человека. Это особенно важно в случае системы HGF/MET, поскольку мышиный HGF не активирует человеческий MET. Разработка генетически модифицированных линий мышей, экспрессирующих нокин-ген HGF человека (мыши hHGF-Ki), решила эту проблему. В этой трансгенной модели показано, что сопутствующее нацеливание HGF из окружающей среды и его рецептора на раковые клетки может быть эффективной терапевтической стратегией для предотвращения злокачественного прогрессирования и метастазирования.
Ксенотрансплантаты аденокарциномы поджелудочной железы характеризуются преждевременным метастатическим распространением, возникающим на очень ранней стадии развития опухоли, и поддерживаются обильным стромальным компартментом. Недавно HGF, секретируемый звездчатыми клетками поджелудочной железы, был идентифицирован как фактор, играющий важную роль во взаимодействии опухоль-строма при этом типе злокачественных новообразований. В ортотопической мышиной модели аденокарциномы поджелудочной железы человека, трансплантированной мышам hHGF-Ki, комбинация MvDN30 и decoyMETK842E немного задерживала рост опухоли, как и ожидалось в модели «уместности», где МЕТ не является драйверным онкогеном. С другой стороны, комбинированное лечение оказалось очень эффективным в уменьшении метастатического распространения, что предполагает возможное терапевтическое применение в раковых клетках, независимых от МЕТ, где МЕТ представлен диким типом. Эпидемиологические данные показывают, что только 2-3 % эпителиальных раковых заболеваний основаны на онкогенной зависимости от МЕТ вследствие амплификации, перестройки или мутации генов29. По этой причине ряд клинических испытаний, направленных на неотобранные группы больных раком, провалился. С другой стороны, подавляющее большинство карцином используют лиганд-зависимую активацию МЕТ дикого типа, чтобы запустить инвазивный метастатический фенотип в ответ на гипоксию, ионизирующее излучение или химиотерапию. Таким образом, эти результаты предполагают, что большая группа пациентов, которые в настоящее время не подходят для таргетной терапии MET из-за отсутствия специфического генетического поражения, должны получать пользу от лечения, включающего стратегию двойного антитела-ловушки, которая позволяет оптимально блокировать передачу сигналов MET, управляемую HGF.
Терапевтические композиции, содержащие активные ингредиенты по настоящему изобретению, то есть фрагмент антитела против HGFR и внеклеточную часть человеческого HGFR, могут быть приготовлены либо в виде одного препарата, содержащего два активных ингредиента, смешанных вместе, либо в виде отдельных препаратов, один из которых содержит фрагмент антитела против HGFR и другой - внеклеточный фрагмент человеческого HGFR. Активные ингредиенты готовят для хранения путем смешивания активного ингредиента (-ов), имеющего желаемую степень чистоты, с физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)) в форме водных растворов, лиофилизированных или других высушенных составов. Приемлемые носители, эксципиенты или стабилизаторы нетоксичны для реципиентов в используемых дозировках и концентрациях и включают буферы; антиоксиданты; консерванты; полипептиды с низкой молекулярной массой (менее примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты; моносахариды, дисахариды и другие углеводы; хелатирующие агенты; сахара; солеобразующие противоионы; комплексы металлов и/или неионные поверхностно-активные вещества. Составы, раскрытые в настоящем документе, могут также содержать другое активное соединение (-я), если это необходимо для конкретного показания, подлежащего лечению, предпочтительно соединения с комплементарной активностью, которые не оказывают негативного влияния на терапевтическую активность фрагмента антитела против HGFR и внеклеточной части человеческого HGFR. Такие молекулы соответственно присутствуют в комбинации в количествах, эффективных для предполагаемого назначения.
Активный (-е) ингредиент (-ы) также может быть заключен в микрокапсулы, полученные с помощью методик, раскрытых, например, в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Составы для введения in vivo должны быть стерильными.
Можно приготовить препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащие активные ингредиенты по изобретению, причем эти матрицы имеют форму изделий определенной формы, например пленок или микрокапсул.
Активные ингредиенты по настоящему изобретению можно использовать либо отдельно, либо в комбинации с другим антителом, химиотерапевтическим агентом (-ми) (включая коктейли химиотерапевтических агентов), другим цитотоксическим агентом (-ми), антиангиогенным агентом (-ми), цитокинами и/или агентом (-ы), ингибирующим рост. Такие комбинированные способы лечения, указанные выше, включают комбинированное введение (когда два или более агента включены в один и тот же или отдельные составы) и раздельное введение, и в этом случае введение антитела по изобретению может происходить до и/или после введения дополнительной терапии или способов лечения.
Активные ингредиенты по настоящему изобретению (и дополнительное терапевтическое средство (-а)) вводят любыми подходящими способами, включая парентеральный, подкожный, внутрибрюшинный, внутрилегочный и интраназальный, и внутриочаговое введение, если это желательно для местного лечения. Активные ингредиенты по настоящему изобретению можно подходящим образом вводить пульсирующей инфузией, особенно с уменьшающимися дозами активных ингредиентов. Введение дозы может быть осуществлено любым пригодным способом, например, с помощью инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или длительным. Активные ингредиенты по изобретению также можно доставлять путем переноса генов с помощью вирусных векторов (т. е. лентивирусных векторов), вводить местно или системно, или посредством клеточной терапии, то есть путем внутривенной местной инъекции клеток человека, генетически модифицированных с помощью трансдукции вирусными векторами (т. е. лентивирусными векторами) для экспрессии фрагмента антитела против HGFR и внеклеточной части HGFR человека.
Активные ингредиенты по изобретению будут составляться, дозироваться и вводиться в соответствии с надлежащей медицинской практикой. Факторы, которые следует учитывать в данном контексте, включают конкретное заболевание, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину заболевания, область доставки агента, способ введения, схему введения и другие факторы, известные врачам-практикам. Активные ингредиенты по изобретению необязательно могут быть включены в фармацевтический состав вместе с одним или несколькими агентами, используемых в настоящее время для предотвращения или лечения рассматриваемого заболевания. Эффективное количество таких других агентов зависит от количества активных ингредиентов по изобретению, присутствующих в составе, типа заболевания или лечения и других факторов, обсуждаемых выше. Их обычно используют в тех же дозах и с путями введения, которые были указаны здесь ранее, или примерно от 1 до 99 % от используемых до настоящего момента дозировок.
Для лечения заболевания подходящая дозировка активных ингредиентов по изобретению (при использовании отдельно или в комбинации с другим агентом (-ми), таким как химиотерапевтический агент (-а)), будет зависеть от типа заболевания, подлежащего лечению, степени тяжести и течения заболевания, независимо от того, вводятся ли активные ингредиенты в профилактических или терапевтических целях, при этом должным образом принимается во внимание история болезни пациента и реакция на активные ингредиенты по изобретению, и на усмотрение лечащего врача.
Антитело или иммуноконъюгат соответствующим образом вводят пациенту однократно или в ходе серии сеансов лечения. В зависимости от типа и тяжести заболевания от около 1 до 15 мг/кг антитела является начальной тестовой дозой для введения пациенту, например, путем одного или нескольких отдельных введений или путем непрерывной инфузии. Типичная ежедневная доза может находиться в диапазоне от около 1 до 100 мг/кг или более в зависимости от вышеупомянутых факторов. При повторном введении в течение нескольких дней или дольше, в зависимости от состояния, лечение обычно следует продолжать до желаемого подавления симптомов заболевания. Одна типичная дозировка фрагмента антитела находится в диапазоне от около 0,05 до около 20 мг/кг. Так, пациенту можно вводить одну или более из доз, составляющих около 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любые их комбинации). Такие дозы можно вводить с перерывами, например, каждую неделю или каждые три недели (например, таким образом, что пациент получает от приблизительно двух до приблизительно двадцати, например, приблизительно шесть доз антитела). Можно вводить начальную повышенную нагрузочную дозу, за которой следует одна или более пониженные дозы. Типичный режим дозирования включает введение начальной нагрузочной дозы около 4 мг/кг с последующей еженедельной поддерживающей дозой около 2 мг/кг антитела. Однако, могут быть пригодны другие схемы лечения. Ход такого лечения можно контролировать с помощью обычных методик и анализов. Внеклеточная часть человеческого HGFR должна доставляться одновременно с фрагментом антитела и должна быть дозирована пропорционально фрагменту антитела, предпочтительно, но не ограничиваясь, с молярным соотношением фрагмента антитела к decoyMET 1:1, в зависимости от типа и тяжести заболевания.
РЕЗУЛЬТАТЫ
Сайт-направленный мутагенез ДН30-связывающего эпитопа в decoyMET.
Чтобы использовать активность антитела MvDN30 и decoyMET совместно, необходимо предотвратить взаимодействие между двумя молекулами, которое могло бы привести к взаимной нейтрализации. Ранее было показано, что MvDN30 распознает эпитоп в домене IPT-4 внеклеточной области MET на границе с доменом IPT-328. Предыдущие исследования показали, что исходное антитело DN30, которое связывает человеческий рецептор с пикомолярным сродством, также взаимодействует с МЕТ собак и крыс11,23, хотя оно не реагирует перекрестно с мышью21. После выравнивания аминокислотных последовательностей IPT-3 и IPT-4 вышеупомянутых видов млекопитающих был идентифицирован ряд остатков, которые избирательно изменены у мышей (фиг. 1A). Чтобы проверить, может ли замена отдельных аминокислотных остатков от человека к мыши нарушать связывание антитела, были созданы растворимые рецепторы, несущие точечные мутации в домене IPT 3-4, и протестированы против антитела DN30. Замена лизина 842 на глутаминовую кислоту привела к образованию decoyMETK842E, модифицированного растворимого рецептора, больше не распознаваемого антителами, в то время как все другие мутации не влияли на взаимодействие (фиг. 1B). Неспособность антитела взаимодействовать с decoyMETK842E была подтверждена в анализах ELISA, проведенных с афинно очищенными ловушками в твердой фазе и антителом DN-30 в жидкой фазе (фиг. 1С). Таким образом, замена основной аминокислоты в положении 842 на кислотный остаток приводила к критическому нарушению сайта связывания, препятствуя взаимодействию decoyMETK842E с антителом.
DecoyMETK842E связывает HGF с высоким сродством
Затем авторы настоящего изобретения исследовали, мешает ли аминокислотная замена K842E связыванию HGF. В анализах связывания ELISA константа аффинности decoyMETK842E к HGF (Kd = 1,04 ± 0,05 нМ) совмещалась с Kd = 1,44 ± 0,07 нМ, измеренной для decoyMET дикого типа (фиг. 2A). Наконец, ингибирующая активность decoyMETK842E была протестирована с помощью анализа HGF-индуцированного фосфорилирования МЕТ. Как показано на фиг. 2B, decoyMET дикого типа и decoyMETK842E ингибировали HGF-зависимое фосфорилирование MET в клетках рака легкого A549 с сопоставимой эффективностью. Таким образом, замена K842E не препятствует образованию стабильного комплекса с HGF и не влияет на ингибирующую активность ловушки.
MvDN30 и decoyMETK842E взаимодействуют при ингибировании фосфорилирования MET и последующих биологических ответов
Для оценки ингибирующей активности, вызываемой одновременным нацеливанием лиганда и рецептора на трансдукцию сигнала MET, фосфорилирование MET тестировали в присутствии MvDN30 и decoyMETK842E, по отдельности или в комбинации. С этой целью клетки A549, экспрессирующие МЕТ-рецептор дикого типа, стимулировали наномолярными концентрациями HGF, а активацию МЕТ измеряли с помощью антител к фосфоМЕТ. Обе молекулы проявляли ингибирующую активность, и их комбинация была более эффективной (фиг. 3А). Анализ последующих сигнальных трансдукторов подтвердил, что эта комбинация обеспечивает наиболее эффективное ингибирование активации ERK и AKT (фиг. 3B).
С биологической точки зрения, комбинация MvDN30 и decoyMETK842E сильно ингибировала безъякорный рост колоний в клеточных моделях как аутокринной, так и паракринной стимуляции HGF. В первом случае клетки глиобластомы U87-MG, демонстрирующие очень эффективный рост колоний на мягком агаре за счет аутокринной петли MET/HGF, подавлялись на 75 %, когда MvDN30 и decoyMETK842E использовались в комбинации, тогда как подавление роста колоний никогда не превышало 40 %, когда две молекулы были использованы как отдельные агенты (фиг. 3B). Аналогичным образом, комбинированная обработка полностью блокировала образование колоний A549 на мягком агаре, вызванное наномолярными концентрациями экзогенно введенного HGF, в то время как только MvDN30 и decoyMETK842E обеспечивали частичное, хотя и значительное ингибирование роста колоний (65 % и 74 % соответственно, фиг. 3C). Аналогичные результаты были получены в анализах инвазии, проведенных в камерах, покрытых матригелем: комбинация MvDN30 и decoyMETK842E снижала вызванную HGF инвазию в клетки аденокарциномы поджелудочной железы человека HPAF-II на 85 %, в то время как в качестве отдельных агентов было достигнуто только 59 % и 52 % ингибирования соответственно (фиг. 3D). Во всех этих биологических системах комбинация MvDN30 и decoyMETK842E нарушала фосфорилирование МЕТ более эффективно, чем обработка по отдельности (фиг. 4).
Чтобы оценить, является ли эффект двух ингибиторов аддитивным или синергетическим, был проведен количественный анализ подвижности. Клетки аденокарциномы поджелудочной железы человека HPAF-II, экспрессирующие МЕТ дикого типа, индуцировали для рассеяния действием HGF. Рассеяние клеток определяли количественно путем измерения вариаций электрического импеданса покрытых ячейками электродов (X-CELLigence Real Time Cell Analyzer). Комбинация двух молекул снижала HGF-зависимое рассеяние клеток дозозависимым образом, начиная ингибирование при 250 нМ и достигая полного блокирования в микромолярном диапазоне (фиг. 5 A и B). Значения индекса клеток, измеренные в конце эксперимента (время - 48 ч), были нормализованы по HGF (фиг. 5C) и обработаны с помощью программного обеспечения CalcuSYN для оценки синергизма (фиг. 5D): для всех исследованных концентраций рассчитанный индекс комбинации (CI) был значительно ниже 0,5 (CI = 0,1, 0,09, 0,17 и 0,38 для 0,06, 0,25, 1 и 4 мкМ, соответственно), что указывает на синергетическое отношение MvDN30 и decoyMETK842E10.
MvDN30 и decoyMETK842E ослабляют инвазивный фенотип и уменьшают распространение метастазов
Ингибирующую активность комбинации MvDN30 и decoyMETK842E оценивали in vivo на мышиных моделях лиганд-управляемой стимуляции MET.
Исследовали модель аутокринной стимуляции HGF ксенотрансплантата глиобластомы U87-MG. Клетки вводили подкожно мышам NOD-SCID; когда объем опухолей достигал 80-100 мм3, мышей разделяли на гомогенные группы и случайным образом распределяли по 4 группам обработки: носитель, MvDN30, decoyMETK842E или комбинация последних. После 22 дней обработки мышей умерщвляли и вырезали первичные опухоли для гистологического исследования. В то время как комбинированная обработка вызывала только краевое ингибирование роста (фиг. 6A), наблюдали снижение фенотипических признаков опухолевой инвазии (фиг. 6B).
Комбинированная обработка также была проверена на паракринной модели стимуляции HGF. Как сообщалось ранее, мышиный HGF не активирует человеческий рецептор МЕТ24, 30. Следовательно, чтобы проверить ингибирующую активность decoyMETK842E в ксенотрансплантатах опухолей человека, мы использовали трансгенную мышь SCID, у которой ген HGF мыши был заменен геном человека (hHGF-Ki)20. Клетки аденокарциномы поджелудочной железы человека (HPAF-II) метили путем трансдукции геном люциферазы и вводили ортотопически в поджелудочную железу мышей hHGF-Ki. Приживление проверяли анализом общей люминесценции тела; мышей разделяли на гомогенные группы на основе значений биолюминесценции и случайным образом распределяли на 4 группы обработки: носитель, MvDN30, decoyMETK842E или их комбинация. Рост опухоли контролировали по общей люминесценции тела (фиг. 7A). При умерщвлении через 5 недель после инъекции клетки опухоли вырезали и анализировали на фосфорилирование МЕТ, пролиферацию, апоптоз и экспрессию виментина/Е-кадгерина (маркеры эпителиально-мезенхимального перехода). Одновременно собирали легкие для гистохимической оценки метастатических узелков. Фосфорилированный статус MET по тирозинам 1234-1235 ингибировался обоими агентами, и комбинированная обработка вызывала драматический эффект (фиг. 8A). Как и ожидалось в этой модели «уместности» онкогена MET (т. е. экспрессии MET дикого типа), пролиферация, а также апоптоз были умеренно затронуты, и только при комбинированной обработке (фиг. 7B и C). Анализ соотношения между виментином и Е-кадгерином показал, что комбинированная обработка подтолкнула раковые клетки к более эпителиальному фенотипу (фиг. 8B). Соответственно, одновременное введение MvDN30 и decoyMETK842E значительно ингибировало опосредованное MET метастатическое распространение метастазов в легкие (фиг. 8C).
Создание панели слитых белков, содержащих фрагмент антитела против HGFR и внеклеточную часть HGFR человека
Для создания одной кДНК, кодирующей слитые белки, содержащие фрагмент антитела против HGFR и внеклеточную часть человеческого HGFR, авторы настоящего изобретения сначала разработали одноцепочечный MvDN30 (ScMvDN30), внедрив линкер между легкой цепью (VL-CL) и тяжелая цепь (VH-CH1). Линкер был гибким, богатым остатками глицина/серина и имел длину, которая позволяла предпочтительно генерировать мономерные молекулы, состоящие из ассоциации легкой и тяжелой цепей антитела, принадлежащих одному и тому же полипептиду, а не из отдельных полипепдитов, таким образом генерируя димеры и/или мультимеры. Предпочтительная аминокислотная последовательность линкера представлена в SEQ ID NO: 36; соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 39. Затем кДНК ScMvDN30 сливали в одной рамке считывания с DecoyMETK842E. Были созданы две группы слитых белков: i) decoyMETK842E был расположен на N-конце, а ScMvDN30 - на C-конце, и ii) scMvDN30 был расположен на N-конце, а decoyMetK842E - на C-конце. Чтобы гарантировать высокую степень свободы для всей структуры, авторы настоящего изобретения ввели второй линкер между двумя группами. Использовали три разных линкера, модифицированных по аминокислотному составу и/или длине: i) L45: жесткий линкер из 45 аминокислот, состоящий из двух повторов последовательности, богатой аланином и заряженными остатками, SEQ ID NO: 35, соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 38; ii) L60, гибкий линкер, богатый остатками глицина/серина из 60 аминокислот SEQ ID NO: 36, соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 39; iii) L134, комбинация гибких и жестких областей из 134 аминокислот SEQ ID NO: 37, соответствующая нуклеотидная последовательность представлена в SEQ ID NO: 40. Схематическое изображение слитых белков представлено на фиг. 9.
Слитые белки, содержащие scMvDN30 на N-конце и decoyMETK842E на C-конце, связываются с высокой аффинностью как с MET, так и с HGF.
Авторы настоящего изобретения исследовали с помощью анализов ELISA, сохраняют ли новые созданные гибридные белки способность связывать как HGF, так и MET. Все слитые белки взаимодействовали с HGF с аффинностью, совпадающей с аффинностью decoyMETK842E (фиг. 10A), в то время как слитые белки, содержащие scMvDN30 на С-конце, распознавали MET с более низким сродством по сравнению со слитыми белками с антителом, расположенным на N-конце. Значения Kd, измеренные для этой последней группы молекул, были сопоставимы со значениями, измеренными для одного scMvDN30 (фиг. 10B).
Слитые белки эффективно ингибировали рост опухолевых клеток.
Авторы настоящего изобретения оценивали ингибирующие свойства слитых белков, измеряя нарушение роста клеток после трех дней обработки, по сравнению с scMvDN30 в сочетании с decoyMETK842E в эквимолярном соотношении. Все слитые белки проявляли ингибирующие свойства в зависимости от дозы. В описанном эксперименте слитые с линкером L60 белки были менее эффективны, чем другие молекулы, тогда как белки L45 и L134 проявляли активность, аналогичную комбинации scMvDN30-decoyMETK842E (фиг. 11).
Слитые белки эффективно ингибировали индуцированную HGF подвижность клеток.
Чтобы оценить влияние слитых белков на ингибирование подвижности клеток, управляемой HGF, авторы настоящего изобретения выполнили количественный анализ подвижности с помощью анализатора клеток в реальном времени X-CELLigence, используя в качестве модели клетки аденокарциномы поджелудочной железы человека HPAF-II, обработанные HGF. Все слитые белки были способны снижать HGF-зависимое распространение клеток (фиг. 12).
Слитые белки снижали распространение метастазов сильнее, чем комбинация MvDN30 и decoyMETK842E
Ингибирующую активность слитых белков оценивали in vivo на мышиной модели hHGF-Ki SCID. Клетки аденокарциномы поджелудочной железы человека (Capan-I) метили путем трансдукции геном люциферазы и вводили ортотопически в поджелудочную железу мышей. Приживление проверяли анализом общей люминесценции тела; мыши были разделены на гомогенные группы на основе значений биолюминесценции и обработаны слитыми белками, включающими линкеры L60 или L134. В качестве референсов в экспериментальные группы были включены MvDN30 и decoyMETK842E в комбинации 1:1 и носитель. При умерщвлении через 5 недель после инъекции клеток вырезали печень и анализировали с помощью IVIS Spectrum для подсчета метастазов. Одновременное введение MvDN30 и decoyMETK842E значительно ингибировало метастатическое распространение метастазов в печень; была отмечена высокая эффективность, если введение MvDN30 проводилось ежедневно (группа COMBO 1), тогда как терапевтический ответ был значительно снижен, если применялись менее частые введения (группа COMBO 2). Это ограничение не оценивалось у мышей, получавших слитые белки. Фактически, все они с высокой эффективностью снижали метастатическое распространение в печень, даже если доставку проводили дважды в неделю (фиг. 13).
МАТЕРИАЛЫ И МЕТОДЫ
Культура клеток
Клетки аденокарциномы легких человека A549, клетки аденокарциномы поджелудочной железы человека HPAF-II и Capan-1 и клетки глиобластомы человека U87-MG были получены от ATCC/LGC Standards S.r.l. (Sesto San Giovanni, Италия); клетки карциномы легких человека EBC-1 были из японской исследовательской коллекции биоресурсов. Все клетки культивировали в соответствии с рекомендациями поставщика. Все клеточные культуры были протестированы на заражение микоплазмами.
Создание, экспрессия и очистка фрагмента антитела против HGFR
КДНК, кодирующие легкую цепь (VL-CL) и тяжелую цепь (VH-CH1) фрагмента антитела против HGFR, способного распознавать эпитоп внеклеточной части HGFR, были получены путем синтеза генов, выполненного на аутсорсинге Invitrogen GeneArt. Синтез гена (ThermoFisher) в соответствии с последовательностями, представленными в SEQ ID NO: 7 и SEQ ID NO: 8. Фрагменты антител получали временной трансфекцией клеток HEK-293T плазмидами pcDNA3.1 (кат.# V79020 Invitrogen Corporation, Camarillo, CA), экспрессирующими кДНК, кодирующие легкую цепь (VL-CL) и тяжелую цепь (VH-CH1). Трансфицированные клетки подвергали голоду в течение трех дней и собирали супернатанты клеточных культур, содержащие растворимые рецепторы. Очистку рекомбинантных белков проводили аффинной хроматографией с использованием колонок HisTrap HP (кат.# 17524701 GE Healthcare, Freiburg, Germany) в соответствии с инструкциями производителя. Крупномасштабное производство и очистку белка выполняла компания U-Protein Express BV (Утрехт, Нидерланды).
Создание, экспрессия и очистка мутированных эктодоменов MET
Последовательности кДНК эктодоменов человеческого MET (decoyMET), несущие одиночные аминокислотные замены, были получены с использованием набора для сайт-направленного мутагенеза QuickChange II (кат.# 200524 Agilent Technologies, Санта-Клара, Калифорния) в соответствии с инструкциями производителя. Процедура требует разработки смысловых и антисмысловых олигонуклеотидов, которые включают желаемую точечную мутацию. Были задействованы следующие олигонуклеотиды:
- мутация K842E:
sn. (смысловой)5'-gtacataatcctgtgtttgagccttttgaaaagccagtg-3' (SEQ ID NO: 41)
as. (антисмысловой) 5'-cactggcttttcaaaaggctcaaacacaggattatgtac-3' (SEQ ID NO: 42)
- мутация Q807K:
sn. 5'-ccactccttccctgaaacagctgaatctgcaactcc-3' (SEQ ID NO: 43)
as. 5'-ggagttgcagattcagctgtttcagggaaggagtgg-3' (SEQ ID NO: 44)
- мутация D864N:
sn. 5'-ctggaaattaagggaaataatattgaccctgaagcagttaaagg-3' (SEQ ID NO: 45)
as. 5'-cctttaactgcttcagggtcaatattatttcccttaatttccag-3' (SEQ ID NO: 46).
Сконструированные растворимые рецепторы были получены путем временной трансфекции клеток HEK-293T плазмидами pcDNA3.1 (кат.# V79020 Invitrogen Corporation, Camarillo, CA), экспрессирующие кДНК, кодирующие мутанты decoyMET или decoyMET дикого типа. Трансфицированные клетки подвергали голоду в течение трех дней и собирали супернатанты клеточных культур, содержащие растворимые рецепторы. Очистку рекомбинантных белков проводили аффинной хроматографией с использованием колонок HisTrap HP (кат.# 17524701 GE Healthcare, Freiburg, Germany) в соответствии с инструкциями производителя. Крупномасштабное производство и очистку белка выполняла компания U-Protein Express BV (Утрехт, Нидерланды).
Анализы фосфорилирования МЕТ
A549 после сывороточного голодания инкубировали в течение 24 ч с 125 нМ MvDN30 или 2 мкМ decoyMETK842E, отдельно или в комбинации, а затем стимулировали 50 нг/мл HGF (кат.# 294-HG-025 R&D Systems) в течение 2 часов при 4 °C. Общие клеточные лизаты анализировали с помощью вестерн-блоттинга с использованием следующих первичных антител: фосфо-Tyr1234/1235 против MET (D26, кат.# 3077 Cell Signaling Technology, Беверли, Массачусетс); anti-MET (3D4, кат.№ 08-1366 Invitrogen Corporation); антивинкулин (клон hVIN-1, кат.# V9131 Sigma Life Science, Сент-Луис, Миссури). IgG1 против мыши (кат.# JI115035003) и IgG против кролика (кат.# JI111035003) вторичные антитела, конъюгированные с HRP, были получены от Jackson ImmunoResearch. (West Grove, PA).
Анализы связывания ELISA
Для анализа взаимодействия между decoyMET и mAb DN30 очищенные аффинно-очищенные растворимые рецепторы (decoyMET дикого типа или decoyMETK842E, 100 нг/лунку) иммобилизовали на планшетах для ELISA и добавляли возрастающие концентрации антитела (0-100 нМ) в жидкую фазу. Связывание выявляли с использованием конъюгированных с HRP антител против мыши (кат.# NA931 GE Healthcare). Для анализа взаимодействия между ними и слитыми белками аффинно очищенный decoyMET дикого типа (100 нг/лунку) иммобилизовали на планшетах для ELISA и добавляли возрастающие концентрации слитых белков или scMvDN30 (0-1000 нМ) в жидкой фазе. Связывание выявляли с использованием конъюгированного с HRP антитела против k-цепи человека (кат.# A7164 Sigma-Aldrich). Для анализа связывания decoyMET или слитых белков с HGF растворимые рецепторы (100 нг/лунку) иммобилизовали на планшетах для ELISA и инкубировали с возрастающими концентрациями HGF (0-11 нМ) в растворе. Связывание детектировали с использованием биотинилированного антитела против HGF (кат.# BAF294 R&D Systems) и выявлено с помощью конъюгированного с стрептавидином HRP (кат.№ RPN 1231 GE Healthcare).
Колориметрический анализ был проведен с помощью считывающего устройства для планшетов с несколькими метками VICTOR-X4 (Perkin Elmer Instruments INC., Whaltman, MA). Данные были проанализированы и аппроксимированы с использованием программного обеспечения Prism (GraphPad).
Биологические анализы in vitro
Для анализов роста, не зависящего от закрепления, клетки суспендировали в соответствующей культуральной среде с добавлением 2 % FBS (фетальная бычья сыворотка) и 0,5 % агарозы Seaplaque (кат.# 50100 BMA, Rockland, ME) и высевали в 48-луночные планшеты (500 клеток/лунку) поверх 1 % агарозы. Свежую среду, содержащую препараты, вносили дважды в неделю. Клетки A549 обрабатывали 1 мкМ MvDN30 или 1 мкМ decoyMETK842E, отдельно или в комбинации, в присутствии 30 нг/мл HGF. Клетки U87-MG обрабатывали 0,5 мкМ MvDN30 или 1 мкМ decoyMETK842E, отдельно или в комбинации. Колонии окрашивали солями тетразолия (кат.# T01380 Sigma-Aldrich) через 12 дней культивирования. Рост колоний определяли с использованием программного обеспечения Metamorph (Molecular Devices, Саннивейл, Калифорния). Для анализа клеточной инвазии клетки HPAF-II (1,5 × 105/на лунку) суспендировали в бессывороточной культуральной среде в присутствии 0,5 мкМ MvDN30 или 1 мкМ decoyMETK842E, отдельно или в комбинации, и высевали в верхний отсек камер системы Трансвелл, предварительно покрытых 30 мкг/лунку Matrigel Matrix (кат.# 354234 Corning Incorporated, Нью-Йорк). Культуральная среда с добавлением 2 % FBS и 12,5 нг/мл HGF была добавлена в нижнее отделение камеры. Через 24 ч клетки на верхней стороне фильтров трансвелл механически удаляли, в то время как клетки, мигрировавшие через мембрану, фиксировали 11 % глутаральдегидом (кат.# 340855 Sigma-Aldrich) и окрашивали 0,1 % кристаллическим фиолетовым (кат. # C3886 Sigma-Aldrich). Инвазию клеток количественно оценивали с помощью программного обеспечения Image-J. Для анализа клеточного рассеяния клетки HPAF-II (8000 на лунку) высевали в 96-луночные планшеты в полную культуральную среду. Через 6 ч добавляли увеличивающиеся концентрации (0-4 мкМ) MvDN30 или decoyMETK842E, отдельно или в комбинации 1:1. Еще через 24 часа клетки стимулировали 6,25 нг/мл HGF в течение 20 часов. Клетки фиксировали 11 % глутаровым альдегидом и окрашивали 0,1 % кристаллическим фиолетовым. Для анализа подвижности клеток в реальном времени клетки HPAF-II высевали в E-планшеты (8000 на лунку; Roche Diagnostics, Mannheim, Germany) и обрабатывали, как указано выше; при нанесении слитые белки тестировали в концентрации 3 мкМ. Электрический импеданс контролировали непрерывно в течение 48 часов с записью данных каждые десять минут с использованием устройства X-Celligence RTCA (Roche Diagnostic). Значения выражены в виде индекса клеток, нормализованного в момент добавления HGF.
Для роста клеток, зависимого от закрепления, клетки EBC-1 высевали по 2000 клеток/лунку в 96-луночный планшет в 5 % культуральной среде FBS. Через 24 часа среду заменяли свежей средой с 5 % FBS и тестируемыми молекулами (возрастающие концентрации - от 0,001 до 3 мкМ). Жизнеспособность клеток оценивали через 72 часа с помощью CellTiter-Glo (кат.# G7573 Promega Corp., Мэдисон, Висконсин) в соответствии с инструкциями производителя. Хемолюминесценцию детектировали с помощью VICTOR X4.
Иммунофлуоресцентный анализ
Иммунофлуоресцентный анализ опухолевых клеток и тканей выполняли, как описано16,25. Окрашивание проводили первичным антителом против фосфо-Tyr1234/1235 против MET (D26) и выявляли вторичным антителом, конъюгированным с Alexa Fluor 555 (кат.# A31570 Thermo Fisher Scientific). Клетки контрастировали с 488-конъюгированным фаллоидином (кат.# A12379 Thermo Fisher Scientific). Все изображения были получены с помощью конфокального лазерного сканирующего микроскопа Leica TCS SP5 AOBS (Leica Microsystems). Параметры получения иммунофлуоресценции оставались постоянными для каждой клеточной линии или опухолевой ткани. Средняя интенсивность флуоресценции (MFI) оценивалась с помощью программного обеспечения ImageJ, измеряли среднюю интенсивность пикселей в каждом канале, за вычетом фона. Для клеточных линий MFI фосфоМЕТ нормализовали по фаллоидину, тогда как для опухолей все сигналы нормализовали по DAPI.
Эксперименты in vivo
Все процедуры на животных выполнялись в соответствии с протоколами, утвержденными Комитетом по этике экспериментов на животных Fondazione Piemontese per la Ricerca sul Cancro и Министерством здравоохранения Италии. Мыши NOD-SCID были приобретены у Charles River (Calco, Италия); hHGF-Ki SCID мышей) были получены от AVEO Pharmaceuticals, Кембридж, Массачусетс.
Клетки U87-MG вводили подкожно (2 × 106/мышь) в правый бок самок мышей NOD-SCID. Рост опухоли контролировали измерением штангенциркулем дважды в неделю. Объем опухоли рассчитывали по формуле: V = 4/3 π (x/2)(y/2)(z/2), где x, y и z - высота, ширина и глубина опухолевой массы. Когда опухоли достигли объема 80-100 мм3 (день 0), мышей разделяли на четыре гомогенные группы и дважды в неделю вводили внутриопухолевую инъекцию: носитель (n = 10); MvDN30, 12,5 мкг (n = 9); K842E, 125 мкг (n = 9); комбинация последних (n = 10). После 22 дней обработки мышей умерщвляли, опухоли вырезали и заливали парафином для гистологического анализа. Кратное увеличение объема опухоли рассчитывали как отношение между значением на 22 день и значением на 0 день.
Клетки HPAF-II или Capan-1 трансдуцировали 100 нг/мл p24 лентивирусных векторов, несущих ген люциферазы под контролем промотора CMV, как описано1. Экспрессирующие люциферазу клетки HPAF-II (104/мышь) инъецировали в поджелудочную железу самок мышей hHGF-KI SCID в возрасте от 4 до 6 недель. Через два дня мышам внутрибрюшинно вводили XenoLight D-люциферин (150 мг/кг; кат.# 122799 Perkin Elmer), разделенных на гомогенные группы на основе сигнала биолюминесценции с использованием аппарата IVIS Spectrum CT (Perkin Elmer) и случайным образом распределенных по 4 группам обработки: носитель (n = 10); MvDN30 (10 мг/кг, n = 6); decoyMETK842E (10 мг/кг, n = 6); MvDN30 + decoyMETK842E (10 + 10 мг/кг, n = 6). Обработку проводили ежедневно (MvDN30) или каждые два дня (decoyMETK842E) путем внутрибрюшинной инъекции. При умерщвлении после пяти недель обработки опухоли и легкие были эксплантированы. Опухоли помещали в парафин или реагент ОСТ и обрабатывали для иммуногистохимического или иммунофлуоресцентного анализа соответственно. Пролиферацию опухолевых клеток определяли с использованием моноклонального антитела против Ki67 (MIB-1, кат.# M724001-2 Agilent Technologies), как описано ранее18. Легкие обрабатывали для гистохимического анализа, а микрометастазы оценивали с помощью световой микроскопии на залитых парафином и окрашенных НЕ непоследовательных срезах. Для каждой мыши было проанализировано десять срезов; подсчитывали метастатические очаги и их площадь количественно определяли с помощью программного обеспечения ImageJ.
Экспрессирующий люциферазу Capan-1 (106/мышь) инъецировали в поджелудочную железу самок мышей SCID hHGF-Ki в возрасте от 4 до 6 недель. Стратификацию мышей проводили, как указано выше. Группы обработки: носитель (n= 14), MvDN30 и decoyMETK842E в комбинации (10 + 10 мг/кг; группа 1, n = 4; группа 2, n = 6), scMvDN30_ K842E_L60 (10 мг/кг; n= 5), K842E scMvDN30_L60 (10 мг/кг; n = 4), scMvDN30_K842E_L134 (10 мг/кг; n= 6), K842E_scMvDN30_L134 (10 мг/кг; n = 5). Обработку проводили ежедневно (MvDN30) или каждые два дня (decoyMETK842E) путем внутрибрюшинной инъекции. После пяти недель обработки животных умерщвляли и собирали печень для измерения сигналов биолюминесценции по спектру IVIS. Мышей считали метастазоположительными, когда биолюминесценция, обнаруженная в печени с помощью IVIS Spectrum, превышала 105 фотонов в секунду.
Статистический анализ
Среднее, стандартное отклонение (SD) и стандартная ошибка среднего (SEM) были рассчитаны с использованием программного обеспечения Microsoft Office Excel 2010 (Microsoft Corporation, Редмонд, Вашингтон). Для расчета значений Kd данные анализов ELISA были проанализированы и подогнаны в соответствии с нелинейной регрессией, кривой гиперболы связывания одного сайта, с использованием программного обеспечения GraphPad Prism (GraphPad Software, Сан-Диего, Калифорния). Для расчета значений IC50 данные анализов пролиферации были проанализированы и подогнаны в соответствии с нелинейной регрессией, сигмоидальной кривой зависимости реакции от дозы, с использованием программного обеспечения GraphPad Prism. Статистическую значимость определяли с использованием двустороннего парного критерия Стьюдента. Все эксперименты повторяли не менее трех раз (биологические повторы). Эксперименты in vivo повторяли дважды. На фигурах показан один репрезентативный эксперимент.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> МЕТИС ПРИСИЖН МЕДСИН СБ С.Р.Л.
<120> КОМБИНАЦИЯ АНТИТЕЛА ПРОТИВ HGFR И HEGFR ДЛЯ ЛЕЧЕНИЯ ОПУХОЛИ
И/ИЛИ МЕТАСТАЗА
<130> BIT20544-CF
<150> IT 102018000003875
<151> 2018-03-22
<160> 46
<170> PatentIn версия 3.5
<210> 1
<211> 10
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> VL-CDR1
<400> 1
Gln Ser Val Asp Tyr Asp Gly Gly Ser Tyr
1 5 10
<210> 2
<211> 3
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> VL-CDR2
<400> 2
Ala Ala Ser
1
<210> 3
<211> 9
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> VL-CDR3
<400> 3
Gln Gln Ser Tyr Glu Asp Pro Leu Thr
1 5
<210> 4
<211> 8
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> VH-CDR1
<400> 4
Gly Tyr Thr Phe Thr Ser Tyr Trp
1 5
<210> 5
<211> 8
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> VH-CDR2
<400> 5
Ile Asn Pro Ser Ser Gly Arg Thr
1 5
<210> 6
<211> 5
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> VH-CDR3
<400> 6
Ala Ser Arg Gly Tyr
1 5
<210> 7
<211> 238
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> Аминокислотная последовательность легкой цепи VL-CL
<400> 7
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 8
<211> 281
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи VH-CH1
<400> 8
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Gly His Ser Gln Val Gln Leu Gln Gln Pro Gly Thr Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Glu Ile Asn Pro Ser Ser Gly Arg Thr Asn Tyr Asn
65 70 75 80
Glu Lys Phe Lys Asn Lys Val Thr Val Thr Val Asp Lys Ser Ser Thr
85 90 95
Thr Ala Tyr Met Gln Leu Ser Asn Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Ser Arg Gly Tyr Trp Gly Gln Gly Thr Thr Leu Thr
115 120 125
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
130 135 140
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
145 150 155 160
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
165 170 175
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
180 185 190
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
195 200 205
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
210 215 220
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Gly
225 230 235 240
Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser His
245 250 255
Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser His Pro Gln Phe Glu Lys
260 265 270
Gly Ala Ala His His His His His His
275 280
<210> 9
<211> 30
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность VL-CDR1
<400> 9
caaagtgttg attatgatgg tggtagttat 30
<210> 10
<211> 9
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность VL-CDR2
<400> 10
gctgcatcc 9
<210> 11
<211> 27
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность VL-CDR3
<400> 11
cagcaaagtt atgaagaccc gctcacg 27
<210> 12
<211> 24
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность VH-CDR1
<400> 12
ggctacacct tcaccagtta ctgg 24
<210> 13
<211> 24
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность VH-CDR2
<400> 13
attaatccta gcagcggtcg tact 24
<210> 14
<211> 15
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность VH-CDR3
<400> 14
gcaagtaggg gctac 15
<210> 15
<211> 717
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность легкой цепи VL-CL
<400> 15
atggagacag acacaatcct gctatgggtg ctgctgctct gggttccagg ctccactggt 60
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 120
atctcctgca aggccagcca aagtgttgat tatgatggtg gtagttatat gagttggttc 180
caacagagac caggacagcc acccaaactc ctcatctctg ctgcatccaa ccttgaatct 240
ggcatcccag ccaggtttag tggcagtggc tctgggacag acttcaccct caatatccat 300
cctgtggagg aggaggatgt tgcaacctat tactgtcagc aaagttatga agacccgctc 360
acgttcggtg ctggtaccaa ggtggagatc aaacgaactg tggctgcacc atctgtcttc 420
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 480
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 540
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 600
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 660
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgttaa 717
<210> 16
<211> 846
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность тяжелой цепи VH-CH1
<400> 16
atgggatgga gctatatcat cctctttttg gtagcaacag ctacagatgg ccactcccag 60
gtccaactgc aacagcctgg gactgaactg gtgaagcctg gggcttcagt gaagctgtcc 120
tgcaaggctt ctggctacac cttcaccagt tactggatac actgggtgaa gcagaggcct 180
ggacaaggcc ttgagtggat tggagagatt aatcctagca gcggtcgtac taactacaac 240
gagaaattca agaacaaggt cacagtgact gtagacaaat cttccaccac agcctacatg 300
caactcagca acctgacatc tgaggactct gcggtctatt actgtgcaag taggggctac 360
tggggccaag gcaccactct cacagtctcc tcagctagca cgaagggccc atcggtcttc 420
cccctggcac cctcctccaa gagcacctct gggggcacag cggccctggg ctgcctggtc 480
aaggactact tccccgaacc ggtgacggtg tcgtggaact caggcgccct gaccagcggc 540
gtgcacacct tcccggctgt cctacagtcc tcaggactct actccctcag cagcgtggtg 600
accgtgccct ccagcagctt gggcacccag acctacatct gcaacgtgaa tcacaagccc 660
agcaacacca aggtggacaa gaaagttgag cccaaatctt gtgacaaaac tcacacaggt 720
gccgcatgga gccaccccca gttcgaaaaa ggggccgcat ggagccaccc ccagttcgaa 780
aaaggggccg catggagcca cccccagttc gaaaaagggg ccgcacacca tcaccatcac 840
cattag 846
<210> 17
<211> 562
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> scFab
<400> 17
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly
225 230 235 240
Ser Ser Gly Ser Gly Ser Gly Ser Thr Gly Thr Ser Ser Ser Gly Thr
245 250 255
Gly Thr Ser Ala Gly Thr Thr Gly Thr Ser Ala Ser Thr Ser Gly Ser
260 265 270
Gly Ser Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Gly Gly
275 280 285
Thr Ala Thr Ala Gly Ala Ser Ser Gly Ser Gln Leu Gln Gln Pro Gly
290 295 300
Thr Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala
305 310 315 320
Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His Trp Val Lys Gln Arg
325 330 335
Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Asn Pro Ser Ser Gly
340 345 350
Arg Thr Asn Tyr Asn Glu Lys Phe Lys Asn Lys Val Thr Val Thr Val
355 360 365
Asp Lys Ser Ser Thr Thr Ala Tyr Met Gln Leu Ser Asn Leu Thr Ser
370 375 380
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Ser Arg Gly Tyr Trp Gly Gln
385 390 395 400
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
405 410 415
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
420 425 430
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
435 440 445
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
450 455 460
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
465 470 475 480
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
485 490 495
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
500 505 510
Lys Thr His Thr Thr Gly Thr Arg Gly Ala Ala Trp Ser His Pro Gln
515 520 525
Phe Glu Lys Gly Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala
530 535 540
Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala His His His His
545 550 555 560
His His
<210> 18
<211> 1692
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность scFv
<400> 18
atggagacag acacaatcct gctatgggtg ctgctgctct gggttccagg ctccactggt 60
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 120
atctcctgca aggccagcca aagtgttgat tatgatggtg gtagttatat gagttggttc 180
caacagagac caggacagcc acccaaactc ctcatctctg ctgcatccaa ccttgaatct 240
ggcatcccag ccaggtttag tggcagtggc tctgggacag acttcaccct caatatccat 300
cctgtggagg aggaggatgt tgcaacctat tactgtcagc aaagttatga agacccgctc 360
acgttcggtg ctggtaccaa ggtggagatc aaacgaactg tggctgcacc atctgtcttc 420
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 480
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 540
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 600
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 660
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgtggtggt 720
tcttcgggtt caggctcggg ctccacaggt acatcgtcat cgggtacagg tacttcagct 780
ggcacaacgg gtacctccgc atctacttcc ggttcaggct ccggaggcgg cggcggctcg 840
ggtggtggtg gatctgccgg cggaaccgca accgcaggcg cttcttctgg ttccgtccaa 900
ctgcagcagc ctgggactga actggtgaag cctggggctt cagtgaagct gtcctgcaag 960
gcttctggct acaccttcac cagttactgg atacactggg tgaagcagag gcctggacaa 1020
ggccttgagt ggattggaga gattaatcct agcagcggtc gtactaacta caacgagaaa 1080
ttcaagaaca aggtcacagt gactgtagac aaatcttcca ccacagccta catgcaactc 1140
agcaacctga catctgagga ctctgcggtc tattactgtg caagtagggg ctactggggc 1200
caaggcacca ctctcacagt ctcctcagct agcacgaagg gcccatcggt cttccccctg 1260
gcaccctcct ccaagagcac ctctgggggc acagcggccc tgggctgcct ggtcaaggac 1320
tacttccccg aacccgtgac ggtgtcgtgg aactcaggcg ccctgaccag cggcgtgcac 1380
accttcccgg ctgtcctaca gtcctcagga ctctactccc tcagcagcgt ggtgaccgtg 1440
ccctccagca gcttgggcac ccagacctac atctgcaacg tgaatcacaa gcccagcaac 1500
accaaggtgg acaagaaagt tgagcccaaa tcttgtgaca aaactcacac aaccggtacg 1560
cgtggtgccg catggagcca cccccagttc gaaaaagggg ccgcatggag ccacccccag 1620
ttcgaaaaag gggccgcatg gagccacccc cagttcgaaa aaggggccgc acaccatcac 1680
catcaccatt ag 1692
<210> 19
<211> 976
<212> ПРТ
<213> homo sapiens
<400> 19
Met Lys Ala Pro Ala Val Leu Ala Pro Gly Ile Leu Val Leu Leu Phe
1 5 10 15
Thr Leu Val Gln Arg Ser Asn Gly Glu Cys Lys Glu Ala Leu Ala Lys
20 25 30
Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro Asn Phe Thr Ala
35 40 45
Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu His His Ile Phe Leu
50 55 60
Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn Glu Glu Asp Leu Gln Lys
65 70 75 80
Val Ala Glu Tyr Lys Thr Gly Pro Val Leu Glu His Pro Asp Cys Phe
85 90 95
Pro Cys Gln Asp Cys Ser Ser Lys Ala Asn Leu Ser Gly Gly Val Trp
100 105 110
Lys Asp Asn Ile Asn Met Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp
115 120 125
Gln Leu Ile Ser Cys Gly Ser Val Asn Arg Gly Thr Cys Gln Arg His
130 135 140
Val Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu Val His Cys
145 150 155 160
Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro Asp Cys Val
165 170 175
Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys Asp Arg Phe
180 185 190
Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser Ser Tyr Phe Pro Asp
195 200 205
His Pro Leu His Ser Ile Ser Val Arg Arg Leu Lys Glu Thr Lys Asp
210 215 220
Gly Phe Met Phe Leu Thr Asp Gln Ser Tyr Ile Asp Val Leu Pro Glu
225 230 235 240
Phe Arg Asp Ser Tyr Pro Ile Lys Tyr Val His Ala Phe Glu Ser Asn
245 250 255
Asn Phe Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu Asp Ala Gln
260 265 270
Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu
275 280 285
His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr Glu Lys Arg
290 295 300
Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile Leu Gln Ala Ala
305 310 315 320
Tyr Val Ser Lys Pro Gly Ala Gln Leu Ala Arg Gln Ile Gly Ala Ser
325 330 335
Leu Asn Asp Asp Ile Leu Phe Gly Val Phe Ala Gln Ser Lys Pro Asp
340 345 350
Ser Ala Glu Pro Met Asp Arg Ser Ala Met Cys Ala Phe Pro Ile Lys
355 360 365
Tyr Val Asn Asp Phe Phe Asn Lys Ile Val Asn Lys Asn Asn Val Arg
370 375 380
Cys Leu Gln His Phe Tyr Gly Pro Asn His Glu His Cys Phe Asn Arg
385 390 395 400
Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala Arg Arg Asp Glu Tyr
405 410 415
Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg Val Asp Leu Phe Met Gly
420 425 430
Gln Phe Ser Glu Val Leu Leu Thr Ser Ile Ser Thr Phe Ile Lys Gly
435 440 445
Asp Leu Thr Ile Ala Asn Leu Gly Thr Ser Glu Gly Arg Phe Met Gln
450 455 460
Val Val Val Ser Arg Ser Gly Pro Ser Thr Pro His Val Asn Phe Leu
465 470 475 480
Leu Asp Ser His Pro Val Ser Pro Glu Val Ile Val Glu His Thr Leu
485 490 495
Asn Gln Asn Gly Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys
500 505 510
Ile Pro Leu Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln
515 520 525
Cys Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys
530 535 540
Cys Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
545 550 555 560
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu
565 570 575
Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe Arg Arg
580 585 590
Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu Gly Asn Glu
595 600 605
Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn Thr Leu Lys Cys
610 615 620
Thr Val Gly Pro Ala Met Asn Lys His Phe Asn Met Ser Ile Ile Ile
625 630 635 640
Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr Phe Ser Tyr Val Asp
645 650 655
Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr Gly Pro Met Ala Gly Gly
660 665 670
Thr Leu Leu Thr Leu Thr Gly Asn Tyr Leu Asn Ser Gly Asn Ser Arg
675 680 685
His Ile Ser Ile Gly Gly Lys Thr Cys Thr Leu Lys Ser Val Ser Asn
690 695 700
Ser Ile Leu Glu Cys Tyr Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe
705 710 715 720
Ala Val Lys Leu Lys Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe
725 730 735
Ser Tyr Arg Glu Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser
740 745 750
Phe Ile Ser Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn
755 760 765
Ser Val Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg
770 775 780
Asn Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
785 790 795 800
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu Lys
805 810 815
Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr Phe Asp
820 825 830
Leu Ile Tyr Val His Asn Pro Val Phe Lys Pro Phe Glu Lys Pro Val
835 840 845
Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys Gly Asn Asp
850 855 860
Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys Val Gly Asn Lys
865 870 875 880
Ser Cys Glu Asn Ile His Leu His Ser Glu Ala Val Leu Cys Thr Val
885 890 895
Pro Asn Asp Leu Leu Lys Leu Asn Ser Glu Leu Asn Ile Glu Trp Lys
900 905 910
Gln Ala Ile Ser Ser Thr Val Leu Gly Lys Val Ile Val Gln Pro Asp
915 920 925
Gln Asn Phe Thr Ala Ser Gly Ala Ala Trp Ser His Pro Gln Phe Glu
930 935 940
Lys Gly Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp
945 950 955 960
Ser His Pro Gln Phe Glu Lys Gly Ala Ala His His His His His His
965 970 975
<210> 20
<211> 976
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> Мутированная внеклеточная часть HGFR человека
<400> 20
Met Lys Ala Pro Ala Val Leu Ala Pro Gly Ile Leu Val Leu Leu Phe
1 5 10 15
Thr Leu Val Gln Arg Ser Asn Gly Glu Cys Lys Glu Ala Leu Ala Lys
20 25 30
Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro Asn Phe Thr Ala
35 40 45
Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu His His Ile Phe Leu
50 55 60
Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn Glu Glu Asp Leu Gln Lys
65 70 75 80
Val Ala Glu Tyr Lys Thr Gly Pro Val Leu Glu His Pro Asp Cys Phe
85 90 95
Pro Cys Gln Asp Cys Ser Ser Lys Ala Asn Leu Ser Gly Gly Val Trp
100 105 110
Lys Asp Asn Ile Asn Met Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp
115 120 125
Gln Leu Ile Ser Cys Gly Ser Val Asn Arg Gly Thr Cys Gln Arg His
130 135 140
Val Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu Val His Cys
145 150 155 160
Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro Asp Cys Val
165 170 175
Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys Asp Arg Phe
180 185 190
Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser Ser Tyr Phe Pro Asp
195 200 205
His Pro Leu His Ser Ile Ser Val Arg Arg Leu Lys Glu Thr Lys Asp
210 215 220
Gly Phe Met Phe Leu Thr Asp Gln Ser Tyr Ile Asp Val Leu Pro Glu
225 230 235 240
Phe Arg Asp Ser Tyr Pro Ile Lys Tyr Val His Ala Phe Glu Ser Asn
245 250 255
Asn Phe Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu Asp Ala Gln
260 265 270
Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu
275 280 285
His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr Glu Lys Arg
290 295 300
Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile Leu Gln Ala Ala
305 310 315 320
Tyr Val Ser Lys Pro Gly Ala Gln Leu Ala Arg Gln Ile Gly Ala Ser
325 330 335
Leu Asn Asp Asp Ile Leu Phe Gly Val Phe Ala Gln Ser Lys Pro Asp
340 345 350
Ser Ala Glu Pro Met Asp Arg Ser Ala Met Cys Ala Phe Pro Ile Lys
355 360 365
Tyr Val Asn Asp Phe Phe Asn Lys Ile Val Asn Lys Asn Asn Val Arg
370 375 380
Cys Leu Gln His Phe Tyr Gly Pro Asn His Glu His Cys Phe Asn Arg
385 390 395 400
Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala Arg Arg Asp Glu Tyr
405 410 415
Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg Val Asp Leu Phe Met Gly
420 425 430
Gln Phe Ser Glu Val Leu Leu Thr Ser Ile Ser Thr Phe Ile Lys Gly
435 440 445
Asp Leu Thr Ile Ala Asn Leu Gly Thr Ser Glu Gly Arg Phe Met Gln
450 455 460
Val Val Val Ser Arg Ser Gly Pro Ser Thr Pro His Val Asn Phe Leu
465 470 475 480
Leu Asp Ser His Pro Val Ser Pro Glu Val Ile Val Glu His Thr Leu
485 490 495
Asn Gln Asn Gly Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys
500 505 510
Ile Pro Leu Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln
515 520 525
Cys Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys
530 535 540
Cys Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
545 550 555 560
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu
565 570 575
Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe Arg Arg
580 585 590
Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu Gly Asn Glu
595 600 605
Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn Thr Leu Lys Cys
610 615 620
Thr Val Gly Pro Ala Met Asn Lys His Phe Asn Met Ser Ile Ile Ile
625 630 635 640
Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr Phe Ser Tyr Val Asp
645 650 655
Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr Gly Pro Met Ala Gly Gly
660 665 670
Thr Leu Leu Thr Leu Thr Gly Asn Tyr Leu Asn Ser Gly Asn Ser Arg
675 680 685
His Ile Ser Ile Gly Gly Lys Thr Cys Thr Leu Lys Ser Val Ser Asn
690 695 700
Ser Ile Leu Glu Cys Tyr Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe
705 710 715 720
Ala Val Lys Leu Lys Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe
725 730 735
Ser Tyr Arg Glu Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser
740 745 750
Phe Ile Ser Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn
755 760 765
Ser Val Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg
770 775 780
Asn Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
785 790 795 800
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu Lys
805 810 815
Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr Phe Asp
820 825 830
Leu Ile Tyr Val His Asn Pro Val Phe Glu Pro Phe Glu Lys Pro Val
835 840 845
Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys Gly Asn Asp
850 855 860
Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys Val Gly Asn Lys
865 870 875 880
Ser Cys Glu Asn Ile His Leu His Ser Glu Ala Val Leu Cys Thr Val
885 890 895
Pro Asn Asp Leu Leu Lys Leu Asn Ser Glu Leu Asn Ile Glu Trp Lys
900 905 910
Gln Ala Ile Ser Ser Thr Val Leu Gly Lys Val Ile Val Gln Pro Asp
915 920 925
Gln Asn Phe Thr Ala Ser Gly Ala Ala Trp Ser His Pro Gln Phe Glu
930 935 940
Lys Gly Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp
945 950 955 960
Ser His Pro Gln Phe Glu Lys Gly Ala Ala His His His His His His
965 970 975
<210> 21
<211> 2931
<212> ДНК
<213> homo sapiens
<400> 21
atgaaggccc ccgctgtgct tgcacctggc atcctcgtgc tcctgtttac cttggtgcag 60
aggagcaatg gggagtgtaa agaggcacta gcaaagtccg agatgaatgt gaatatgaag 120
tatcagcttc ccaacttcac cgcggaaaca cccatccaga atgtcattct acatgagcat 180
cacattttcc ttggtgccac taactacatt tatgttttaa atgaggaaga ccttcagaag 240
gttgctgagt acaagactgg gcctgtgctg gaacacccag attgtttccc atgtcaggac 300
tgcagcagca aagccaattt atcaggaggt gtttggaaag ataacatcaa catggctcta 360
gttgtggaca cctactatga tgatcaactc attagctgtg gcagcgtcaa cagagggacc 420
tgccagcgac atgtctttcc ccacaatcat actgctgaca tacagtcgga ggttcactgc 480
atattctccc cacagataga agagcccagc cagtgtcctg actgtgtggt gagcgccctg 540
ggagccaaag tcctttcatc tgtaaaggac cggttcatca acttctttgt aggcaatacc 600
ataaattctt cttatttccc agatcatcca ttgcattcga tatcagtgag aaggctaaag 660
gaaacgaaag atggttttat gtttttgacg gaccagtcct acattgatgt tttacctgag 720
ttcagagatt cttaccccat taagtatgtc catgcctttg aaagcaacaa ttttatttac 780
ttcttgacgg tccaaaggga aactctagat gctcagactt ttcacacaag aataatcagg 840
ttctgttcca taaactctgg attgcattcc tacatggaaa tgcctctgga gtgtattctc 900
acagaaaaga gaaaaaagag atccacaaag aaggaagtgt ttaatatact tcaggctgcg 960
tatgtcagca agcctggggc ccagcttgct agacaaatag gagccagcct gaatgatgac 1020
attcttttcg gggtgttcgc acaaagcaag ccagattctg ccgaaccaat ggatcgatct 1080
gccatgtgtg cattccctat caaatatgtc aacgacttct tcaacaagat cgtcaacaaa 1140
aacaatgtga gatgtctcca gcatttttac ggacccaatc atgagcactg ctttaatagg 1200
acacttctga gaaattcatc aggctgtgaa gcgcgccgtg atgaatatcg aacagagttt 1260
accacagctt tgcagcgcgt tgacttattc atgggtcaat tcagcgaagt cctcttaaca 1320
tctatatcca ccttcattaa aggagacctc accatagcta atcttgggac atcagagggt 1380
cgcttcatgc aggttgtggt ttctcgatca ggaccatcaa cccctcatgt gaattttctc 1440
ctggactccc atccagtgtc tccagaagtg attgtggagc atacattaaa ccaaaatggc 1500
tacacactgg ttatcactgg gaagaagatc acgaagatcc cattgaatgg cttgggttgc 1560
agacatttcc agtcttgcag tcaatgcctc tctgccccac cctttgttca gtgtggctgg 1620
tgccacgaca aatgtgtgcg atcggaggaa tgcctgagcg ggacatggac tcaacagatc 1680
tgtctgcctg caatctacaa ggttttccca aatagtgcac cccttgaagg agggacaagg 1740
ctgaccatat gtggctggga ctttggattt cggaggaata ataaatttga tttaaagaaa 1800
actagagttc tccttggaaa tgagagctgc accttgactt taagtgagag cacgatgaat 1860
acattgaaat gcacagttgg tcctgccatg aataagcatt tcaatatgtc cataattatt 1920
tcaaatggcc acgggacaac acaatacagt acattctcct atgtcgatcc tgtaataaca 1980
agtatttcgc cgaaatacgg tcctatggct ggtggcactt tacttacttt aactggaaat 2040
tacctaaaca gtgggaactc tagacacatt tcaattggtg gaaaaacatg tactttaaaa 2100
agtgtgtcaa acagtattct tgaatgttat accccagccc aaaccatttc aactgagttt 2160
gctgttaaat tgaaaattga cttagccaac cgagagacaa gcatcttcag ttaccgtgaa 2220
gatcccattg tctatgaaat tcatccaacc aaatctttta ttagtggtgg gagcacaata 2280
acaggtgttg ggaaaaacct gaactcagtt agtgtcccga gaatggtcat aaatgtgcat 2340
gaagcaggaa ggaactttac agtggcatgt caacatcgct ctaattcaga gataatctgt 2400
tgtaccactc cttccctgca acagctgaat ctgcaactcc ccctgaaaac caaagccttt 2460
ttcatgttag atggcatcct ttccaaatac tttgatctca tttatgtaca taatcctgtg 2520
tttaagcctt ttgaaaagcc agtgatgatc tcaatgggca atgaaaatgt actggaaatt 2580
aagggaaatg atattgaccc tgaagcagtt aaaggtgaag tgttaaaagt tggaaataag 2640
agctgtgaga atatacactt acattctgaa gccgttttat gcacggtccc caatgacctg 2700
ctgaaattga acagcgagct aaatatagag tggaagcaag caatttcttc aaccgtcctt 2760
ggaaaagtaa tagttcaacc agatcagaat ttcacagcta gcggtgccgc atggagccac 2820
ccccagttcg aaaaaggggc cgcatggagc cacccccagt tcgaaaaagg ggccgcatgg 2880
agccaccccc agttcgaaaa aggggccgca caccatcacc atcaccatta g 2931
<210> 22
<211> 2931
<212> ДНК
<213> Искуственная последовательность
<220>
<223> Нуклеотидная последовательность мутированной внеклеточной части HGFR
человека
<400> 22
atgaaggccc ccgctgtgct tgcacctggc atcctcgtgc tcctgtttac cttggtgcag 60
aggagcaatg gggagtgtaa agaggcacta gcaaagtccg agatgaatgt gaatatgaag 120
tatcagcttc ccaacttcac cgcggaaaca cccatccaga atgtcattct acatgagcat 180
cacattttcc ttggtgccac taactacatt tatgttttaa atgaggaaga ccttcagaag 240
gttgctgagt acaagactgg gcctgtgctg gaacacccag attgtttccc atgtcaggac 300
tgcagcagca aagccaattt atcaggaggt gtttggaaag ataacatcaa catggctcta 360
gttgtggaca cctactatga tgatcaactc attagctgtg gcagcgtcaa cagagggacc 420
tgccagcgac atgtctttcc ccacaatcat actgctgaca tacagtcgga ggttcactgc 480
atattctccc cacagataga agagcccagc cagtgtcctg actgtgtggt gagcgccctg 540
ggagccaaag tcctttcatc tgtaaaggac cggttcatca acttctttgt aggcaatacc 600
ataaattctt cttatttccc agatcatcca ttgcattcga tatcagtgag aaggctaaag 660
gaaacgaaag atggttttat gtttttgacg gaccagtcct acattgatgt tttacctgag 720
ttcagagatt cttaccccat taagtatgtc catgcctttg aaagcaacaa ttttatttac 780
ttcttgacgg tccaaaggga aactctagat gctcagactt ttcacacaag aataatcagg 840
ttctgttcca taaactctgg attgcattcc tacatggaaa tgcctctgga gtgtattctc 900
acagaaaaga gaaaaaagag atccacaaag aaggaagtgt ttaatatact tcaggctgcg 960
tatgtcagca agcctggggc ccagcttgct agacaaatag gagccagcct gaatgatgac 1020
attcttttcg gggtgttcgc acaaagcaag ccagattctg ccgaaccaat ggatcgatct 1080
gccatgtgtg cattccctat caaatatgtc aacgacttct tcaacaagat cgtcaacaaa 1140
aacaatgtga gatgtctcca gcatttttac ggacccaatc atgagcactg ctttaatagg 1200
acacttctga gaaattcatc aggctgtgaa gcgcgccgtg atgaatatcg aacagagttt 1260
accacagctt tgcagcgcgt tgacttattc atgggtcaat tcagcgaagt cctcttaaca 1320
tctatatcca ccttcattaa aggagacctc accatagcta atcttgggac atcagagggt 1380
cgcttcatgc aggttgtggt ttctcgatca ggaccatcaa cccctcatgt gaattttctc 1440
ctggactccc atccagtgtc tccagaagtg attgtggagc atacattaaa ccaaaatggc 1500
tacacactgg ttatcactgg gaagaagatc acgaagatcc cattgaatgg cttgggttgc 1560
agacatttcc agtcttgcag tcaatgcctc tctgccccac cctttgttca gtgtggctgg 1620
tgccacgaca aatgtgtgcg atcggaggaa tgcctgagcg ggacatggac tcaacagatc 1680
tgtctgcctg caatctacaa ggttttccca aatagtgcac cccttgaagg agggacaagg 1740
ctgaccatat gtggctggga ctttggattt cggaggaata ataaatttga tttaaagaaa 1800
actagagttc tccttggaaa tgagagctgc accttgactt taagtgagag cacgatgaat 1860
acattgaaat gcacagttgg tcctgccatg aataagcatt tcaatatgtc cataattatt 1920
tcaaatggcc acgggacaac acaatacagt acattctcct atgtcgatcc tgtaataaca 1980
agtatttcgc cgaaatacgg tcctatggct ggtggcactt tacttacttt aactggaaat 2040
tacctaaaca gtgggaactc tagacacatt tcaattggtg gaaaaacatg tactttaaaa 2100
agtgtgtcaa acagtattct tgaatgttat accccagccc aaaccatttc aactgagttt 2160
gctgttaaat tgaaaattga cttagccaac cgagagacaa gcatcttcag ttaccgtgaa 2220
gatcccattg tctatgaaat tcatccaacc aaatctttta ttagtggtgg gagcacaata 2280
acaggtgttg ggaaaaacct gaactcagtt agtgtcccga gaatggtcat aaatgtgcat 2340
gaagcaggaa ggaactttac agtggcatgt caacatcgct ctaattcaga gataatctgt 2400
tgtaccactc cttccctgca acagctgaat ctgcaactcc ccctgaaaac caaagccttt 2460
ttcatgttag atggcatcct ttccaaatac tttgatctca tttatgtaca taatcctgtg 2520
tttgagcctt ttgaaaagcc agtgatgatc tcaatgggca atgaaaatgt actggaaatt 2580
aagggaaatg atattgaccc tgaagcagtt aaaggtgaag tgttaaaagt tggaaataag 2640
agctgtgaga atatacactt acattctgaa gccgttttat gcacggtccc caatgacctg 2700
ctgaaattga acagcgagct aaatatagag tggaagcaag caatttcttc aaccgtcctt 2760
ggaaaagtaa tagttcaacc agatcagaat ttcacagcta gcggtgccgc atggagccac 2820
ccccagttcg aaaaaggggc cgcatggagc cacccccagt tcgaaaaagg ggccgcatgg 2880
agccaccccc agttcgaaaa aggggccgca caccatcacc atcaccatta g 2931
<210> 23
<211> 1518
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> decoyMET-линкер-антитело
аминокислотная последовательность K842E-линкер L45-scMvDN30
<400> 23
Met Lys Ala Pro Ala Val Leu Ala Pro Gly Ile Leu Val Leu Leu Phe
1 5 10 15
Thr Leu Val Gln Arg Ser Asn Gly Glu Cys Lys Glu Ala Leu Ala Lys
20 25 30
Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro Asn Phe Thr Ala
35 40 45
Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu His His Ile Phe Leu
50 55 60
Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn Glu Glu Asp Leu Gln Lys
65 70 75 80
Val Ala Glu Tyr Lys Thr Gly Pro Val Leu Glu His Pro Asp Cys Phe
85 90 95
Pro Cys Gln Asp Cys Ser Ser Lys Ala Asn Leu Ser Gly Gly Val Trp
100 105 110
Lys Asp Asn Ile Asn Met Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp
115 120 125
Gln Leu Ile Ser Cys Gly Ser Val Asn Arg Gly Thr Cys Gln Arg His
130 135 140
Val Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu Val His Cys
145 150 155 160
Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro Asp Cys Val
165 170 175
Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys Asp Arg Phe
180 185 190
Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser Ser Tyr Phe Pro Asp
195 200 205
His Pro Leu His Ser Ile Ser Val Arg Arg Leu Lys Glu Thr Lys Asp
210 215 220
Gly Phe Met Phe Leu Thr Asp Gln Ser Tyr Ile Asp Val Leu Pro Glu
225 230 235 240
Phe Arg Asp Ser Tyr Pro Ile Lys Tyr Val His Ala Phe Glu Ser Asn
245 250 255
Asn Phe Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu Asp Ala Gln
260 265 270
Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu
275 280 285
His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr Glu Lys Arg
290 295 300
Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile Leu Gln Ala Ala
305 310 315 320
Tyr Val Ser Lys Pro Gly Ala Gln Leu Ala Arg Gln Ile Gly Ala Ser
325 330 335
Leu Asn Asp Asp Ile Leu Phe Gly Val Phe Ala Gln Ser Lys Pro Asp
340 345 350
Ser Ala Glu Pro Met Asp Arg Ser Ala Met Cys Ala Phe Pro Ile Lys
355 360 365
Tyr Val Asn Asp Phe Phe Asn Lys Ile Val Asn Lys Asn Asn Val Arg
370 375 380
Cys Leu Gln His Phe Tyr Gly Pro Asn His Glu His Cys Phe Asn Arg
385 390 395 400
Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala Arg Arg Asp Glu Tyr
405 410 415
Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg Val Asp Leu Phe Met Gly
420 425 430
Gln Phe Ser Glu Val Leu Leu Thr Ser Ile Ser Thr Phe Ile Lys Gly
435 440 445
Asp Leu Thr Ile Ala Asn Leu Gly Thr Ser Glu Gly Arg Phe Met Gln
450 455 460
Val Val Val Ser Arg Ser Gly Pro Ser Thr Pro His Val Asn Phe Leu
465 470 475 480
Leu Asp Ser His Pro Val Ser Pro Glu Val Ile Val Glu His Thr Leu
485 490 495
Asn Gln Asn Gly Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys
500 505 510
Ile Pro Leu Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln
515 520 525
Cys Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys
530 535 540
Cys Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
545 550 555 560
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu
565 570 575
Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe Arg Arg
580 585 590
Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu Gly Asn Glu
595 600 605
Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn Thr Leu Lys Cys
610 615 620
Thr Val Gly Pro Ala Met Asn Lys His Phe Asn Met Ser Ile Ile Ile
625 630 635 640
Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr Phe Ser Tyr Val Asp
645 650 655
Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr Gly Pro Met Ala Gly Gly
660 665 670
Thr Leu Leu Thr Leu Thr Gly Asn Tyr Leu Asn Ser Gly Asn Ser Arg
675 680 685
His Ile Ser Ile Gly Gly Lys Thr Cys Thr Leu Lys Ser Val Ser Asn
690 695 700
Ser Ile Leu Glu Cys Tyr Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe
705 710 715 720
Ala Val Lys Leu Lys Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe
725 730 735
Ser Tyr Arg Glu Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser
740 745 750
Phe Ile Ser Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn
755 760 765
Ser Val Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg
770 775 780
Asn Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
785 790 795 800
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu Lys
805 810 815
Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr Phe Asp
820 825 830
Leu Ile Tyr Val His Asn Pro Val Phe Glu Pro Phe Glu Lys Pro Val
835 840 845
Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys Gly Asn Asp
850 855 860
Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys Val Gly Asn Lys
865 870 875 880
Ser Cys Glu Asn Ile His Leu His Ser Glu Ala Val Leu Cys Thr Val
885 890 895
Pro Asn Asp Leu Leu Lys Leu Asn Ser Glu Leu Asn Ile Glu Trp Lys
900 905 910
Gln Ala Ile Ser Ser Thr Val Leu Gly Lys Val Ile Val Gln Pro Asp
915 920 925
Gln Asn Phe Thr Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu
930 935 940
Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala Ala Ala Glu Ala Ala Ala
945 950 955 960
Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
965 970 975
Ala Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu
980 985 990
Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr
995 1000 1005
Asp Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro Gly Gln
1010 1015 1020
Pro Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser Gly
1025 1030 1035
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1040 1045 1050
Leu Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr
1055 1060 1065
Cys Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr
1070 1075 1080
Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile
1085 1090 1095
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
1100 1105 1110
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln
1115 1120 1125
Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
1130 1135 1140
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
1145 1150 1155
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
1160 1165 1170
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
1175 1180 1185
Ser Phe Asn Arg Gly Glu Cys Gly Gly Ser Ser Gly Ser Gly Ser
1190 1195 1200
Gly Ser Thr Gly Thr Ser Ser Ser Gly Thr Gly Thr Ser Ala Gly
1205 1210 1215
Thr Thr Gly Thr Ser Ala Ser Thr Ser Gly Ser Gly Ser Gly Gly
1220 1225 1230
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Gly Gly Thr Ala Thr
1235 1240 1245
Ala Gly Ala Ser Ser Gly Ser Val Gln Leu Gln Gln Pro Gly Thr
1250 1255 1260
Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala
1265 1270 1275
Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His Trp Val Lys Gln
1280 1285 1290
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Asn Pro Ser
1295 1300 1305
Ser Gly Arg Thr Asn Tyr Asn Glu Lys Phe Lys Asn Lys Val Thr
1310 1315 1320
Val Thr Val Asp Lys Ser Ser Thr Thr Ala Tyr Met Gln Leu Ser
1325 1330 1335
Asn Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Ser Arg
1340 1345 1350
Gly Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser
1355 1360 1365
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
1370 1375 1380
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
1385 1390 1395
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
1400 1405 1410
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
1415 1420 1425
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
1430 1435 1440
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
1445 1450 1455
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
1460 1465 1470
Thr Thr Arg Gly Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly
1475 1480 1485
Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser
1490 1495 1500
His Pro Gln Phe Glu Lys Gly Ala Ala His His His His His His
1505 1510 1515
<210> 24
<211> 1533
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> decoyMET-линкер-антитело
аминокислотная последовательность K842E-линкер L60-scMvDN30
<400> 24
Met Lys Ala Pro Ala Val Leu Ala Pro Gly Ile Leu Val Leu Leu Phe
1 5 10 15
Thr Leu Val Gln Arg Ser Asn Gly Glu Cys Lys Glu Ala Leu Ala Lys
20 25 30
Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro Asn Phe Thr Ala
35 40 45
Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu His His Ile Phe Leu
50 55 60
Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn Glu Glu Asp Leu Gln Lys
65 70 75 80
Val Ala Glu Tyr Lys Thr Gly Pro Val Leu Glu His Pro Asp Cys Phe
85 90 95
Pro Cys Gln Asp Cys Ser Ser Lys Ala Asn Leu Ser Gly Gly Val Trp
100 105 110
Lys Asp Asn Ile Asn Met Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp
115 120 125
Gln Leu Ile Ser Cys Gly Ser Val Asn Arg Gly Thr Cys Gln Arg His
130 135 140
Val Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu Val His Cys
145 150 155 160
Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro Asp Cys Val
165 170 175
Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys Asp Arg Phe
180 185 190
Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser Ser Tyr Phe Pro Asp
195 200 205
His Pro Leu His Ser Ile Ser Val Arg Arg Leu Lys Glu Thr Lys Asp
210 215 220
Gly Phe Met Phe Leu Thr Asp Gln Ser Tyr Ile Asp Val Leu Pro Glu
225 230 235 240
Phe Arg Asp Ser Tyr Pro Ile Lys Tyr Val His Ala Phe Glu Ser Asn
245 250 255
Asn Phe Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu Asp Ala Gln
260 265 270
Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu
275 280 285
His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr Glu Lys Arg
290 295 300
Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile Leu Gln Ala Ala
305 310 315 320
Tyr Val Ser Lys Pro Gly Ala Gln Leu Ala Arg Gln Ile Gly Ala Ser
325 330 335
Leu Asn Asp Asp Ile Leu Phe Gly Val Phe Ala Gln Ser Lys Pro Asp
340 345 350
Ser Ala Glu Pro Met Asp Arg Ser Ala Met Cys Ala Phe Pro Ile Lys
355 360 365
Tyr Val Asn Asp Phe Phe Asn Lys Ile Val Asn Lys Asn Asn Val Arg
370 375 380
Cys Leu Gln His Phe Tyr Gly Pro Asn His Glu His Cys Phe Asn Arg
385 390 395 400
Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala Arg Arg Asp Glu Tyr
405 410 415
Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg Val Asp Leu Phe Met Gly
420 425 430
Gln Phe Ser Glu Val Leu Leu Thr Ser Ile Ser Thr Phe Ile Lys Gly
435 440 445
Asp Leu Thr Ile Ala Asn Leu Gly Thr Ser Glu Gly Arg Phe Met Gln
450 455 460
Val Val Val Ser Arg Ser Gly Pro Ser Thr Pro His Val Asn Phe Leu
465 470 475 480
Leu Asp Ser His Pro Val Ser Pro Glu Val Ile Val Glu His Thr Leu
485 490 495
Asn Gln Asn Gly Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys
500 505 510
Ile Pro Leu Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln
515 520 525
Cys Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys
530 535 540
Cys Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
545 550 555 560
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu
565 570 575
Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe Arg Arg
580 585 590
Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu Gly Asn Glu
595 600 605
Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn Thr Leu Lys Cys
610 615 620
Thr Val Gly Pro Ala Met Asn Lys His Phe Asn Met Ser Ile Ile Ile
625 630 635 640
Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr Phe Ser Tyr Val Asp
645 650 655
Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr Gly Pro Met Ala Gly Gly
660 665 670
Thr Leu Leu Thr Leu Thr Gly Asn Tyr Leu Asn Ser Gly Asn Ser Arg
675 680 685
His Ile Ser Ile Gly Gly Lys Thr Cys Thr Leu Lys Ser Val Ser Asn
690 695 700
Ser Ile Leu Glu Cys Tyr Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe
705 710 715 720
Ala Val Lys Leu Lys Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe
725 730 735
Ser Tyr Arg Glu Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser
740 745 750
Phe Ile Ser Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn
755 760 765
Ser Val Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg
770 775 780
Asn Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
785 790 795 800
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu Lys
805 810 815
Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr Phe Asp
820 825 830
Leu Ile Tyr Val His Asn Pro Val Phe Glu Pro Phe Glu Lys Pro Val
835 840 845
Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys Gly Asn Asp
850 855 860
Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys Val Gly Asn Lys
865 870 875 880
Ser Cys Glu Asn Ile His Leu His Ser Glu Ala Val Leu Cys Thr Val
885 890 895
Pro Asn Asp Leu Leu Lys Leu Asn Ser Glu Leu Asn Ile Glu Trp Lys
900 905 910
Gln Ala Ile Ser Ser Thr Val Leu Gly Lys Val Ile Val Gln Pro Asp
915 920 925
Gln Asn Phe Thr Gly Gly Ser Ser Gly Ser Gly Ser Gly Ser Thr Gly
930 935 940
Thr Ser Ser Ser Gly Thr Gly Thr Ser Ala Gly Thr Thr Gly Thr Ser
945 950 955 960
Ala Ser Thr Ser Gly Ser Gly Ser Gly Gly Gly Gly Gly Ser Gly Gly
965 970 975
Gly Gly Ser Ala Gly Gly Thr Ala Thr Ala Gly Ala Ser Ser Gly Ser
980 985 990
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
995 1000 1005
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr
1010 1015 1020
Asp Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro Gly Gln
1025 1030 1035
Pro Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser Gly
1040 1045 1050
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1055 1060 1065
Leu Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr
1070 1075 1080
Cys Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr
1085 1090 1095
Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile
1100 1105 1110
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
1115 1120 1125
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln
1130 1135 1140
Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
1145 1150 1155
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
1160 1165 1170
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
1175 1180 1185
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
1190 1195 1200
Ser Phe Asn Arg Gly Glu Cys Gly Gly Ser Ser Gly Ser Gly Ser
1205 1210 1215
Gly Ser Thr Gly Thr Ser Ser Ser Gly Thr Gly Thr Ser Ala Gly
1220 1225 1230
Thr Thr Gly Thr Ser Ala Ser Thr Ser Gly Ser Gly Ser Gly Gly
1235 1240 1245
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Gly Gly Thr Ala Thr
1250 1255 1260
Ala Gly Ala Ser Ser Gly Ser Val Gln Leu Gln Gln Pro Gly Thr
1265 1270 1275
Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala
1280 1285 1290
Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His Trp Val Lys Gln
1295 1300 1305
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Asn Pro Ser
1310 1315 1320
Ser Gly Arg Thr Asn Tyr Asn Glu Lys Phe Lys Asn Lys Val Thr
1325 1330 1335
Val Thr Val Asp Lys Ser Ser Thr Thr Ala Tyr Met Gln Leu Ser
1340 1345 1350
Asn Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Ser Arg
1355 1360 1365
Gly Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser
1370 1375 1380
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
1385 1390 1395
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
1400 1405 1410
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
1415 1420 1425
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
1430 1435 1440
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
1445 1450 1455
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
1460 1465 1470
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
1475 1480 1485
Thr Thr Arg Gly Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly
1490 1495 1500
Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser
1505 1510 1515
His Pro Gln Phe Glu Lys Gly Ala Ala His His His His His His
1520 1525 1530
<210> 25
<211> 1607
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> decoyMET-линкер-антитело
аминокислотная последовательность K842E-линкер L134-scMvDN30
<400> 25
Met Lys Ala Pro Ala Val Leu Ala Pro Gly Ile Leu Val Leu Leu Phe
1 5 10 15
Thr Leu Val Gln Arg Ser Asn Gly Glu Cys Lys Glu Ala Leu Ala Lys
20 25 30
Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro Asn Phe Thr Ala
35 40 45
Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu His His Ile Phe Leu
50 55 60
Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn Glu Glu Asp Leu Gln Lys
65 70 75 80
Val Ala Glu Tyr Lys Thr Gly Pro Val Leu Glu His Pro Asp Cys Phe
85 90 95
Pro Cys Gln Asp Cys Ser Ser Lys Ala Asn Leu Ser Gly Gly Val Trp
100 105 110
Lys Asp Asn Ile Asn Met Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp
115 120 125
Gln Leu Ile Ser Cys Gly Ser Val Asn Arg Gly Thr Cys Gln Arg His
130 135 140
Val Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu Val His Cys
145 150 155 160
Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro Asp Cys Val
165 170 175
Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys Asp Arg Phe
180 185 190
Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser Ser Tyr Phe Pro Asp
195 200 205
His Pro Leu His Ser Ile Ser Val Arg Arg Leu Lys Glu Thr Lys Asp
210 215 220
Gly Phe Met Phe Leu Thr Asp Gln Ser Tyr Ile Asp Val Leu Pro Glu
225 230 235 240
Phe Arg Asp Ser Tyr Pro Ile Lys Tyr Val His Ala Phe Glu Ser Asn
245 250 255
Asn Phe Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu Asp Ala Gln
260 265 270
Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu
275 280 285
His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr Glu Lys Arg
290 295 300
Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile Leu Gln Ala Ala
305 310 315 320
Tyr Val Ser Lys Pro Gly Ala Gln Leu Ala Arg Gln Ile Gly Ala Ser
325 330 335
Leu Asn Asp Asp Ile Leu Phe Gly Val Phe Ala Gln Ser Lys Pro Asp
340 345 350
Ser Ala Glu Pro Met Asp Arg Ser Ala Met Cys Ala Phe Pro Ile Lys
355 360 365
Tyr Val Asn Asp Phe Phe Asn Lys Ile Val Asn Lys Asn Asn Val Arg
370 375 380
Cys Leu Gln His Phe Tyr Gly Pro Asn His Glu His Cys Phe Asn Arg
385 390 395 400
Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala Arg Arg Asp Glu Tyr
405 410 415
Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg Val Asp Leu Phe Met Gly
420 425 430
Gln Phe Ser Glu Val Leu Leu Thr Ser Ile Ser Thr Phe Ile Lys Gly
435 440 445
Asp Leu Thr Ile Ala Asn Leu Gly Thr Ser Glu Gly Arg Phe Met Gln
450 455 460
Val Val Val Ser Arg Ser Gly Pro Ser Thr Pro His Val Asn Phe Leu
465 470 475 480
Leu Asp Ser His Pro Val Ser Pro Glu Val Ile Val Glu His Thr Leu
485 490 495
Asn Gln Asn Gly Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys
500 505 510
Ile Pro Leu Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln
515 520 525
Cys Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys
530 535 540
Cys Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
545 550 555 560
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu
565 570 575
Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe Arg Arg
580 585 590
Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu Gly Asn Glu
595 600 605
Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn Thr Leu Lys Cys
610 615 620
Thr Val Gly Pro Ala Met Asn Lys His Phe Asn Met Ser Ile Ile Ile
625 630 635 640
Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr Phe Ser Tyr Val Asp
645 650 655
Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr Gly Pro Met Ala Gly Gly
660 665 670
Thr Leu Leu Thr Leu Thr Gly Asn Tyr Leu Asn Ser Gly Asn Ser Arg
675 680 685
His Ile Ser Ile Gly Gly Lys Thr Cys Thr Leu Lys Ser Val Ser Asn
690 695 700
Ser Ile Leu Glu Cys Tyr Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe
705 710 715 720
Ala Val Lys Leu Lys Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe
725 730 735
Ser Tyr Arg Glu Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser
740 745 750
Phe Ile Ser Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn
755 760 765
Ser Val Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg
770 775 780
Asn Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
785 790 795 800
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu Lys
805 810 815
Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr Phe Asp
820 825 830
Leu Ile Tyr Val His Asn Pro Val Phe Glu Pro Phe Glu Lys Pro Val
835 840 845
Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys Gly Asn Asp
850 855 860
Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys Val Gly Asn Lys
865 870 875 880
Ser Cys Glu Asn Ile His Leu His Ser Glu Ala Val Leu Cys Thr Val
885 890 895
Pro Asn Asp Leu Leu Lys Leu Asn Ser Glu Leu Asn Ile Glu Trp Lys
900 905 910
Gln Ala Ile Ser Ser Thr Val Leu Gly Lys Val Ile Val Gln Pro Asp
915 920 925
Gln Asn Phe Thr Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
930 935 940
Gly Gly Ser Ala Glu Ala Trp Tyr Asn Leu Gly Asn Ala Tyr Tyr Lys
945 950 955 960
Gln Gly Asp Tyr Gln Lys Ala Ile Glu Tyr Tyr Gln Lys Ala Leu Glu
965 970 975
Leu Asp Pro Asn Asn Ala Glu Ala Trp Tyr Asn Leu Gly Asn Ala Tyr
980 985 990
Tyr Lys Gln Gly Asp Tyr Gln Lys Ala Ile Glu Tyr Tyr Gln Lys Ala
995 1000 1005
Leu Glu Leu Asp Pro Asn Asn Ala Glu Ala Trp Tyr Asn Leu Gly
1010 1015 1020
Asn Ala Tyr Tyr Lys Gln Gly Asp Tyr Gln Lys Ala Ile Glu Asp
1025 1030 1035
Tyr Gln Lys Ala Leu Glu Leu Asp Pro Asn Asn Gly Gly Gly Gly
1040 1045 1050
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Asp Ile
1055 1060 1065
Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly Gln
1070 1075 1080
Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp
1085 1090 1095
Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro Gly Gln Pro
1100 1105 1110
Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser Gly Ile
1115 1120 1125
Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
1130 1135 1140
Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr Cys
1145 1150 1155
Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys
1160 1165 1170
Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
1175 1180 1185
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
1190 1195 1200
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
1205 1210 1215
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val
1220 1225 1230
Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
1235 1240 1245
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
1250 1255 1260
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
1265 1270 1275
Phe Asn Arg Gly Glu Cys Gly Gly Ser Ser Gly Ser Gly Ser Gly
1280 1285 1290
Ser Thr Gly Thr Ser Ser Ser Gly Thr Gly Thr Ser Ala Gly Thr
1295 1300 1305
Thr Gly Thr Ser Ala Ser Thr Ser Gly Ser Gly Ser Gly Gly Gly
1310 1315 1320
Gly Gly Ser Gly Gly Gly Gly Ser Ala Gly Gly Thr Ala Thr Ala
1325 1330 1335
Gly Ala Ser Ser Gly Ser Val Gln Leu Gln Gln Pro Gly Thr Glu
1340 1345 1350
Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser
1355 1360 1365
Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His Trp Val Lys Gln Arg
1370 1375 1380
Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Asn Pro Ser Ser
1385 1390 1395
Gly Arg Thr Asn Tyr Asn Glu Lys Phe Lys Asn Lys Val Thr Val
1400 1405 1410
Thr Val Asp Lys Ser Ser Thr Thr Ala Tyr Met Gln Leu Ser Asn
1415 1420 1425
Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Ser Arg Gly
1430 1435 1440
Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr
1445 1450 1455
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
1460 1465 1470
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
1475 1480 1485
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
1490 1495 1500
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
1505 1510 1515
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
1520 1525 1530
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
1535 1540 1545
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
1550 1555 1560
Thr Arg Gly Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala
1565 1570 1575
Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser His
1580 1585 1590
Pro Gln Phe Glu Lys Gly Ala Ala His His His His His His
1595 1600 1605
<210> 26
<211> 1514
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> антитело-линкер-decoyMET
аминокислотная последовательность scMvDN30-линкер L45-K842E
<400> 26
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly
225 230 235 240
Ser Ser Gly Ser Gly Ser Gly Ser Thr Gly Thr Ser Ser Ser Gly Thr
245 250 255
Gly Thr Ser Ala Gly Thr Thr Gly Thr Ser Ala Ser Thr Ser Gly Ser
260 265 270
Gly Ser Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Gly Gly
275 280 285
Thr Ala Thr Ala Gly Ala Ser Ser Gly Ser Val Gln Leu Gln Gln Pro
290 295 300
Gly Thr Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys
305 310 315 320
Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His Trp Val Lys Gln
325 330 335
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Asn Pro Ser Ser
340 345 350
Gly Arg Thr Asn Tyr Asn Glu Lys Phe Lys Asn Lys Val Thr Val Thr
355 360 365
Val Asp Lys Ser Ser Thr Thr Ala Tyr Met Gln Leu Ser Asn Leu Thr
370 375 380
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Ser Arg Gly Tyr Trp Gly
385 390 395 400
Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
405 410 415
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
420 425 430
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
435 440 445
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
450 455 460
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
465 470 475 480
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
485 490 495
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
500 505 510
Asp Lys Thr His Thr Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
515 520 525
Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala Ala Ala Glu Ala Ala
530 535 540
Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala
545 550 555 560
Lys Ala Glu Cys Lys Glu Ala Leu Ala Lys Ser Glu Met Asn Val Asn
565 570 575
Met Lys Tyr Gln Leu Pro Asn Phe Thr Ala Glu Thr Pro Ile Gln Asn
580 585 590
Val Ile Leu His Glu His His Ile Phe Leu Gly Ala Thr Asn Tyr Ile
595 600 605
Tyr Val Leu Asn Glu Glu Asp Leu Gln Lys Val Ala Glu Tyr Lys Thr
610 615 620
Gly Pro Val Leu Glu His Pro Asp Cys Phe Pro Cys Gln Asp Cys Ser
625 630 635 640
Ser Lys Ala Asn Leu Ser Gly Gly Val Trp Lys Asp Asn Ile Asn Met
645 650 655
Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp Gln Leu Ile Ser Cys Gly
660 665 670
Ser Val Asn Arg Gly Thr Cys Gln Arg His Val Phe Pro His Asn His
675 680 685
Thr Ala Asp Ile Gln Ser Glu Val His Cys Ile Phe Ser Pro Gln Ile
690 695 700
Glu Glu Pro Ser Gln Cys Pro Asp Cys Val Val Ser Ala Leu Gly Ala
705 710 715 720
Lys Val Leu Ser Ser Val Lys Asp Arg Phe Ile Asn Phe Phe Val Gly
725 730 735
Asn Thr Ile Asn Ser Ser Tyr Phe Pro Asp His Pro Leu His Ser Ile
740 745 750
Ser Val Arg Arg Leu Lys Glu Thr Lys Asp Gly Phe Met Phe Leu Thr
755 760 765
Asp Gln Ser Tyr Ile Asp Val Leu Pro Glu Phe Arg Asp Ser Tyr Pro
770 775 780
Ile Lys Tyr Val His Ala Phe Glu Ser Asn Asn Phe Ile Tyr Phe Leu
785 790 795 800
Thr Val Gln Arg Glu Thr Leu Asp Ala Gln Thr Phe His Thr Arg Ile
805 810 815
Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu His Ser Tyr Met Glu Met
820 825 830
Pro Leu Glu Cys Ile Leu Thr Glu Lys Arg Lys Lys Arg Ser Thr Lys
835 840 845
Lys Glu Val Phe Asn Ile Leu Gln Ala Ala Tyr Val Ser Lys Pro Gly
850 855 860
Ala Gln Leu Ala Arg Gln Ile Gly Ala Ser Leu Asn Asp Asp Ile Leu
865 870 875 880
Phe Gly Val Phe Ala Gln Ser Lys Pro Asp Ser Ala Glu Pro Met Asp
885 890 895
Arg Ser Ala Met Cys Ala Phe Pro Ile Lys Tyr Val Asn Asp Phe Phe
900 905 910
Asn Lys Ile Val Asn Lys Asn Asn Val Arg Cys Leu Gln His Phe Tyr
915 920 925
Gly Pro Asn His Glu His Cys Phe Asn Arg Thr Leu Leu Arg Asn Ser
930 935 940
Ser Gly Cys Glu Ala Arg Arg Asp Glu Tyr Arg Thr Glu Phe Thr Thr
945 950 955 960
Ala Leu Gln Arg Val Asp Leu Phe Met Gly Gln Phe Ser Glu Val Leu
965 970 975
Leu Thr Ser Ile Ser Thr Phe Ile Lys Gly Asp Leu Thr Ile Ala Asn
980 985 990
Leu Gly Thr Ser Glu Gly Arg Phe Met Gln Val Val Val Ser Arg Ser
995 1000 1005
Gly Pro Ser Thr Pro His Val Asn Phe Leu Leu Asp Ser His Pro
1010 1015 1020
Val Ser Pro Glu Val Ile Val Glu His Thr Leu Asn Gln Asn Gly
1025 1030 1035
Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys Ile Pro Leu
1040 1045 1050
Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln Cys Leu
1055 1060 1065
Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys Cys
1070 1075 1080
Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
1085 1090 1095
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu
1100 1105 1110
Glu Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe
1115 1120 1125
Arg Arg Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu
1130 1135 1140
Gly Asn Glu Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn
1145 1150 1155
Thr Leu Lys Cys Thr Val Gly Pro Ala Met Asn Lys His Phe Asn
1160 1165 1170
Met Ser Ile Ile Ile Ser Asn Gly His Gly Thr Thr Gln Tyr Ser
1175 1180 1185
Thr Phe Ser Tyr Val Asp Pro Val Ile Thr Ser Ile Ser Pro Lys
1190 1195 1200
Tyr Gly Pro Met Ala Gly Gly Thr Leu Leu Thr Leu Thr Gly Asn
1205 1210 1215
Tyr Leu Asn Ser Gly Asn Ser Arg His Ile Ser Ile Gly Gly Lys
1220 1225 1230
Thr Cys Thr Leu Lys Ser Val Ser Asn Ser Ile Leu Glu Cys Tyr
1235 1240 1245
Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe Ala Val Lys Leu Lys
1250 1255 1260
Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe Ser Tyr Arg Glu
1265 1270 1275
Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser Phe Ile Ser
1280 1285 1290
Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn Ser Val
1295 1300 1305
Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg Asn
1310 1315 1320
Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
1325 1330 1335
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu
1340 1345 1350
Lys Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr
1355 1360 1365
Phe Asp Leu Ile Tyr Val His Asn Pro Val Phe Glu Pro Phe Glu
1370 1375 1380
Lys Pro Val Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile
1385 1390 1395
Lys Gly Asn Asp Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu
1400 1405 1410
Lys Val Gly Asn Lys Ser Cys Glu Asn Ile His Leu His Ser Glu
1415 1420 1425
Ala Val Leu Cys Thr Val Pro Asn Asp Leu Leu Lys Leu Asn Ser
1430 1435 1440
Glu Leu Asn Ile Glu Trp Lys Gln Ala Ile Ser Ser Thr Val Leu
1445 1450 1455
Gly Lys Val Ile Val Gln Pro Asp Gln Asn Phe Thr Ala Ser Gly
1460 1465 1470
Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser
1475 1480 1485
His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser His Pro Gln Phe
1490 1495 1500
Glu Lys Gly Ala Ala His His His His His His
1505 1510
<210> 27
<211> 1529
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> антитело-линкер-decoyMET
аминокислотная последовательность scMvDN30-линкер L60-K842E
<400> 27
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly
225 230 235 240
Ser Ser Gly Ser Gly Ser Gly Ser Thr Gly Thr Ser Ser Ser Gly Thr
245 250 255
Gly Thr Ser Ala Gly Thr Thr Gly Thr Ser Ala Ser Thr Ser Gly Ser
260 265 270
Gly Ser Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Gly Gly
275 280 285
Thr Ala Thr Ala Gly Ala Ser Ser Gly Ser Val Gln Leu Gln Gln Pro
290 295 300
Gly Thr Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys
305 310 315 320
Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His Trp Val Lys Gln
325 330 335
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Asn Pro Ser Ser
340 345 350
Gly Arg Thr Asn Tyr Asn Glu Lys Phe Lys Asn Lys Val Thr Val Thr
355 360 365
Val Asp Lys Ser Ser Thr Thr Ala Tyr Met Gln Leu Ser Asn Leu Thr
370 375 380
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Ser Arg Gly Tyr Trp Gly
385 390 395 400
Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
405 410 415
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
420 425 430
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
435 440 445
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
450 455 460
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
465 470 475 480
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
485 490 495
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
500 505 510
Asp Lys Thr His Thr Gly Gly Ser Ser Gly Ser Gly Ser Gly Ser Thr
515 520 525
Gly Thr Ser Ser Ser Gly Thr Gly Thr Ser Ala Gly Thr Thr Gly Thr
530 535 540
Ser Ala Ser Thr Ser Gly Ser Gly Ser Gly Gly Gly Gly Gly Ser Gly
545 550 555 560
Gly Gly Gly Ser Ala Gly Gly Thr Ala Thr Ala Gly Ala Ser Ser Gly
565 570 575
Ser Glu Cys Lys Glu Ala Leu Ala Lys Ser Glu Met Asn Val Asn Met
580 585 590
Lys Tyr Gln Leu Pro Asn Phe Thr Ala Glu Thr Pro Ile Gln Asn Val
595 600 605
Ile Leu His Glu His His Ile Phe Leu Gly Ala Thr Asn Tyr Ile Tyr
610 615 620
Val Leu Asn Glu Glu Asp Leu Gln Lys Val Ala Glu Tyr Lys Thr Gly
625 630 635 640
Pro Val Leu Glu His Pro Asp Cys Phe Pro Cys Gln Asp Cys Ser Ser
645 650 655
Lys Ala Asn Leu Ser Gly Gly Val Trp Lys Asp Asn Ile Asn Met Ala
660 665 670
Leu Val Val Asp Thr Tyr Tyr Asp Asp Gln Leu Ile Ser Cys Gly Ser
675 680 685
Val Asn Arg Gly Thr Cys Gln Arg His Val Phe Pro His Asn His Thr
690 695 700
Ala Asp Ile Gln Ser Glu Val His Cys Ile Phe Ser Pro Gln Ile Glu
705 710 715 720
Glu Pro Ser Gln Cys Pro Asp Cys Val Val Ser Ala Leu Gly Ala Lys
725 730 735
Val Leu Ser Ser Val Lys Asp Arg Phe Ile Asn Phe Phe Val Gly Asn
740 745 750
Thr Ile Asn Ser Ser Tyr Phe Pro Asp His Pro Leu His Ser Ile Ser
755 760 765
Val Arg Arg Leu Lys Glu Thr Lys Asp Gly Phe Met Phe Leu Thr Asp
770 775 780
Gln Ser Tyr Ile Asp Val Leu Pro Glu Phe Arg Asp Ser Tyr Pro Ile
785 790 795 800
Lys Tyr Val His Ala Phe Glu Ser Asn Asn Phe Ile Tyr Phe Leu Thr
805 810 815
Val Gln Arg Glu Thr Leu Asp Ala Gln Thr Phe His Thr Arg Ile Ile
820 825 830
Arg Phe Cys Ser Ile Asn Ser Gly Leu His Ser Tyr Met Glu Met Pro
835 840 845
Leu Glu Cys Ile Leu Thr Glu Lys Arg Lys Lys Arg Ser Thr Lys Lys
850 855 860
Glu Val Phe Asn Ile Leu Gln Ala Ala Tyr Val Ser Lys Pro Gly Ala
865 870 875 880
Gln Leu Ala Arg Gln Ile Gly Ala Ser Leu Asn Asp Asp Ile Leu Phe
885 890 895
Gly Val Phe Ala Gln Ser Lys Pro Asp Ser Ala Glu Pro Met Asp Arg
900 905 910
Ser Ala Met Cys Ala Phe Pro Ile Lys Tyr Val Asn Asp Phe Phe Asn
915 920 925
Lys Ile Val Asn Lys Asn Asn Val Arg Cys Leu Gln His Phe Tyr Gly
930 935 940
Pro Asn His Glu His Cys Phe Asn Arg Thr Leu Leu Arg Asn Ser Ser
945 950 955 960
Gly Cys Glu Ala Arg Arg Asp Glu Tyr Arg Thr Glu Phe Thr Thr Ala
965 970 975
Leu Gln Arg Val Asp Leu Phe Met Gly Gln Phe Ser Glu Val Leu Leu
980 985 990
Thr Ser Ile Ser Thr Phe Ile Lys Gly Asp Leu Thr Ile Ala Asn Leu
995 1000 1005
Gly Thr Ser Glu Gly Arg Phe Met Gln Val Val Val Ser Arg Ser
1010 1015 1020
Gly Pro Ser Thr Pro His Val Asn Phe Leu Leu Asp Ser His Pro
1025 1030 1035
Val Ser Pro Glu Val Ile Val Glu His Thr Leu Asn Gln Asn Gly
1040 1045 1050
Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys Ile Pro Leu
1055 1060 1065
Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln Cys Leu
1070 1075 1080
Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys Cys
1085 1090 1095
Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
1100 1105 1110
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu
1115 1120 1125
Glu Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe
1130 1135 1140
Arg Arg Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu
1145 1150 1155
Gly Asn Glu Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn
1160 1165 1170
Thr Leu Lys Cys Thr Val Gly Pro Ala Met Asn Lys His Phe Asn
1175 1180 1185
Met Ser Ile Ile Ile Ser Asn Gly His Gly Thr Thr Gln Tyr Ser
1190 1195 1200
Thr Phe Ser Tyr Val Asp Pro Val Ile Thr Ser Ile Ser Pro Lys
1205 1210 1215
Tyr Gly Pro Met Ala Gly Gly Thr Leu Leu Thr Leu Thr Gly Asn
1220 1225 1230
Tyr Leu Asn Ser Gly Asn Ser Arg His Ile Ser Ile Gly Gly Lys
1235 1240 1245
Thr Cys Thr Leu Lys Ser Val Ser Asn Ser Ile Leu Glu Cys Tyr
1250 1255 1260
Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe Ala Val Lys Leu Lys
1265 1270 1275
Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe Ser Tyr Arg Glu
1280 1285 1290
Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser Phe Ile Ser
1295 1300 1305
Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn Ser Val
1310 1315 1320
Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg Asn
1325 1330 1335
Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
1340 1345 1350
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu
1355 1360 1365
Lys Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr
1370 1375 1380
Phe Asp Leu Ile Tyr Val His Asn Pro Val Phe Glu Pro Phe Glu
1385 1390 1395
Lys Pro Val Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile
1400 1405 1410
Lys Gly Asn Asp Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu
1415 1420 1425
Lys Val Gly Asn Lys Ser Cys Glu Asn Ile His Leu His Ser Glu
1430 1435 1440
Ala Val Leu Cys Thr Val Pro Asn Asp Leu Leu Lys Leu Asn Ser
1445 1450 1455
Glu Leu Asn Ile Glu Trp Lys Gln Ala Ile Ser Ser Thr Val Leu
1460 1465 1470
Gly Lys Val Ile Val Gln Pro Asp Gln Asn Phe Thr Ala Ser Gly
1475 1480 1485
Ala Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser
1490 1495 1500
His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser His Pro Gln Phe
1505 1510 1515
Glu Lys Gly Ala Ala His His His His His His
1520 1525
<210> 28
<211> 1603
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> антитело-линкер-decoyMET
аминокислотная последовательность scMvDN30-линкер L134-K842E
<400> 28
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Gly Ser Tyr Met Ser Trp Phe Gln Gln Arg Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Ser Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Glu Asp Val Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Tyr Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly
225 230 235 240
Ser Ser Gly Ser Gly Ser Gly Ser Thr Gly Thr Ser Ser Ser Gly Thr
245 250 255
Gly Thr Ser Ala Gly Thr Thr Gly Thr Ser Ala Ser Thr Ser Gly Ser
260 265 270
Gly Ser Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Gly Gly
275 280 285
Thr Ala Thr Ala Gly Ala Ser Ser Gly Ser Val Gln Leu Gln Gln Pro
290 295 300
Gly Thr Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Lys
305 310 315 320
Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His Trp Val Lys Gln
325 330 335
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Glu Ile Asn Pro Ser Ser
340 345 350
Gly Arg Thr Asn Tyr Asn Glu Lys Phe Lys Asn Lys Val Thr Val Thr
355 360 365
Val Asp Lys Ser Ser Thr Thr Ala Tyr Met Gln Leu Ser Asn Leu Thr
370 375 380
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Ser Arg Gly Tyr Trp Gly
385 390 395 400
Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
405 410 415
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
420 425 430
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
435 440 445
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
450 455 460
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
465 470 475 480
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
485 490 495
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
500 505 510
Asp Lys Thr His Thr Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
515 520 525
Gly Gly Gly Ser Ala Glu Ala Trp Tyr Asn Leu Gly Asn Ala Tyr Tyr
530 535 540
Lys Gln Gly Asp Tyr Gln Lys Ala Ile Glu Tyr Tyr Gln Lys Ala Leu
545 550 555 560
Glu Leu Asp Pro Asn Asn Ala Glu Ala Trp Tyr Asn Leu Gly Asn Ala
565 570 575
Tyr Tyr Lys Gln Gly Asp Tyr Gln Lys Ala Ile Glu Tyr Tyr Gln Lys
580 585 590
Ala Leu Glu Leu Asp Pro Asn Asn Ala Glu Ala Trp Tyr Asn Leu Gly
595 600 605
Asn Ala Tyr Tyr Lys Gln Gly Asp Tyr Gln Lys Ala Ile Glu Asp Tyr
610 615 620
Gln Lys Ala Leu Glu Leu Asp Pro Asn Asn Gly Gly Gly Gly Ser Gly
625 630 635 640
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Glu Cys Lys Glu Ala
645 650 655
Leu Ala Lys Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro Asn
660 665 670
Phe Thr Ala Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu His His
675 680 685
Ile Phe Leu Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn Glu Glu Asp
690 695 700
Leu Gln Lys Val Ala Glu Tyr Lys Thr Gly Pro Val Leu Glu His Pro
705 710 715 720
Asp Cys Phe Pro Cys Gln Asp Cys Ser Ser Lys Ala Asn Leu Ser Gly
725 730 735
Gly Val Trp Lys Asp Asn Ile Asn Met Ala Leu Val Val Asp Thr Tyr
740 745 750
Tyr Asp Asp Gln Leu Ile Ser Cys Gly Ser Val Asn Arg Gly Thr Cys
755 760 765
Gln Arg His Val Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu
770 775 780
Val His Cys Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro
785 790 795 800
Asp Cys Val Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys
805 810 815
Asp Arg Phe Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser Ser Tyr
820 825 830
Phe Pro Asp His Pro Leu His Ser Ile Ser Val Arg Arg Leu Lys Glu
835 840 845
Thr Lys Asp Gly Phe Met Phe Leu Thr Asp Gln Ser Tyr Ile Asp Val
850 855 860
Leu Pro Glu Phe Arg Asp Ser Tyr Pro Ile Lys Tyr Val His Ala Phe
865 870 875 880
Glu Ser Asn Asn Phe Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu
885 890 895
Asp Ala Gln Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn
900 905 910
Ser Gly Leu His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr
915 920 925
Glu Lys Arg Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile Leu
930 935 940
Gln Ala Ala Tyr Val Ser Lys Pro Gly Ala Gln Leu Ala Arg Gln Ile
945 950 955 960
Gly Ala Ser Leu Asn Asp Asp Ile Leu Phe Gly Val Phe Ala Gln Ser
965 970 975
Lys Pro Asp Ser Ala Glu Pro Met Asp Arg Ser Ala Met Cys Ala Phe
980 985 990
Pro Ile Lys Tyr Val Asn Asp Phe Phe Asn Lys Ile Val Asn Lys Asn
995 1000 1005
Asn Val Arg Cys Leu Gln His Phe Tyr Gly Pro Asn His Glu His
1010 1015 1020
Cys Phe Asn Arg Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala
1025 1030 1035
Arg Arg Asp Glu Tyr Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg
1040 1045 1050
Val Asp Leu Phe Met Gly Gln Phe Ser Glu Val Leu Leu Thr Ser
1055 1060 1065
Ile Ser Thr Phe Ile Lys Gly Asp Leu Thr Ile Ala Asn Leu Gly
1070 1075 1080
Thr Ser Glu Gly Arg Phe Met Gln Val Val Val Ser Arg Ser Gly
1085 1090 1095
Pro Ser Thr Pro His Val Asn Phe Leu Leu Asp Ser His Pro Val
1100 1105 1110
Ser Pro Glu Val Ile Val Glu His Thr Leu Asn Gln Asn Gly Tyr
1115 1120 1125
Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys Ile Pro Leu Asn
1130 1135 1140
Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln Cys Leu Ser
1145 1150 1155
Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys Cys Val
1160 1165 1170
Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile Cys
1175 1180 1185
Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu
1190 1195 1200
Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe Arg
1205 1210 1215
Arg Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu Gly
1220 1225 1230
Asn Glu Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn Thr
1235 1240 1245
Leu Lys Cys Thr Val Gly Pro Ala Met Asn Lys His Phe Asn Met
1250 1255 1260
Ser Ile Ile Ile Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr
1265 1270 1275
Phe Ser Tyr Val Asp Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr
1280 1285 1290
Gly Pro Met Ala Gly Gly Thr Leu Leu Thr Leu Thr Gly Asn Tyr
1295 1300 1305
Leu Asn Ser Gly Asn Ser Arg His Ile Ser Ile Gly Gly Lys Thr
1310 1315 1320
Cys Thr Leu Lys Ser Val Ser Asn Ser Ile Leu Glu Cys Tyr Thr
1325 1330 1335
Pro Ala Gln Thr Ile Ser Thr Glu Phe Ala Val Lys Leu Lys Ile
1340 1345 1350
Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe Ser Tyr Arg Glu Asp
1355 1360 1365
Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser Phe Ile Ser Gly
1370 1375 1380
Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn Ser Val Ser
1385 1390 1395
Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg Asn Phe
1400 1405 1410
Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys Cys
1415 1420 1425
Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu Lys
1430 1435 1440
Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr Phe
1445 1450 1455
Asp Leu Ile Tyr Val His Asn Pro Val Phe Glu Pro Phe Glu Lys
1460 1465 1470
Pro Val Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys
1475 1480 1485
Gly Asn Asp Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys
1490 1495 1500
Val Gly Asn Lys Ser Cys Glu Asn Ile His Leu His Ser Glu Ala
1505 1510 1515
Val Leu Cys Thr Val Pro Asn Asp Leu Leu Lys Leu Asn Ser Glu
1520 1525 1530
Leu Asn Ile Glu Trp Lys Gln Ala Ile Ser Ser Thr Val Leu Gly
1535 1540 1545
Lys Val Ile Val Gln Pro Asp Gln Asn Phe Thr Ala Ser Gly Ala
1550 1555 1560
Ala Trp Ser His Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser His
1565 1570 1575
Pro Gln Phe Glu Lys Gly Ala Ala Trp Ser His Pro Gln Phe Glu
1580 1585 1590
Lys Gly Ala Ala His His His His His His
1595 1600
<210> 29
<211> 4557
<212> ДНК
<213> Искуственная последовательность
<220>
<223> decoyMET-линкер-антитело
нуклеотидная последовательность K842E-линкер L45-scMvDN30
<400> 29
atgaaggccc ccgctgtgct tgcacctggc atcctcgtgc tcctgtttac cttggtgcag 60
aggagcaatg gggagtgtaa agaggcacta gcaaagtccg agatgaatgt gaatatgaag 120
tatcagcttc ccaacttcac cgcggaaaca cccatccaga atgtcattct acatgagcat 180
cacattttcc ttggtgccac taactacatt tatgttttaa atgaggaaga ccttcagaag 240
gttgctgagt acaagactgg gcctgtgctg gaacacccag attgtttccc atgtcaggac 300
tgcagcagca aagccaattt atcaggaggt gtttggaaag ataacatcaa catggctcta 360
gttgtggaca cctactatga tgatcaactc attagctgtg gcagcgtcaa cagagggacc 420
tgccagcgac atgtctttcc ccacaatcat actgctgaca tacagtcgga ggttcactgc 480
atattctccc cacagataga agagcccagc cagtgtcctg actgtgtggt gagcgccctg 540
ggagccaaag tcctttcatc tgtaaaggac cggttcatca acttctttgt aggcaatacc 600
ataaattctt cttatttccc agatcatcca ttgcattcga tatcagtgag aaggctaaag 660
gaaacgaaag atggttttat gtttttgacg gaccagtcct acattgatgt tttacctgag 720
ttcagagatt cttaccccat taagtatgtc catgcctttg aaagcaacaa ttttatttac 780
ttcttgacgg tccaaaggga aactctagat gctcagactt ttcacacaag aataatcagg 840
ttctgttcca taaactctgg attgcattcc tacatggaaa tgcctctgga gtgtattctc 900
acagaaaaga gaaaaaagag atccacaaag aaggaagtgt ttaatatact tcaggctgcg 960
tatgtcagca agcctggggc ccagcttgct agacaaatag gagccagcct gaatgatgac 1020
attcttttcg gggtgttcgc acaaagcaag ccagattctg ccgaaccaat ggatcgatct 1080
gccatgtgtg cattccctat caaatatgtc aacgacttct tcaacaagat cgtcaacaaa 1140
aacaatgtga gatgtctcca gcatttttac ggacccaatc atgagcactg ctttaatagg 1200
acacttctga gaaattcatc aggctgtgaa gcgcgccgtg atgaatatcg aacagagttt 1260
accacagctt tgcagcgcgt tgacttattc atgggtcaat tcagcgaagt cctcttaaca 1320
tctatatcca ccttcattaa aggagacctc accatagcta atcttgggac atcagagggt 1380
cgcttcatgc aggttgtggt ttctcgatca ggaccatcaa cccctcatgt gaattttctc 1440
ctggactccc atccagtgtc tccagaagtg attgtggagc atacattaaa ccaaaatggc 1500
tacacactgg ttatcactgg gaagaagatc acgaagatcc cattgaatgg cttgggttgc 1560
agacatttcc agtcttgcag tcaatgcctc tctgccccac cctttgttca gtgtggctgg 1620
tgccacgaca aatgtgtgcg atcggaggaa tgcctgagcg ggacatggac tcaacagatc 1680
tgtctgcctg caatctacaa ggttttccca aatagtgcac cccttgaagg agggacaagg 1740
ctgaccatat gtggctggga ctttggattt cggaggaata ataaatttga tttaaagaaa 1800
actagagttc tccttggaaa tgagagctgc accttgactt taagtgagag cacgatgaat 1860
acattgaaat gcacagttgg tcctgccatg aataagcatt tcaatatgtc cataattatt 1920
tcaaatggcc acgggacaac acaatacagt acattctcct atgtcgatcc tgtaataaca 1980
agtatttcgc cgaaatacgg tcctatggct ggtggcactt tacttacttt aactggaaat 2040
tacctaaaca gtgggaactc tagacacatt tcaattggtg gaaaaacatg tactttaaaa 2100
agtgtgtcaa acagtattct tgaatgttat accccagccc aaaccatttc aactgagttt 2160
gctgttaaat tgaaaattga cttagccaac cgagagacaa gcatcttcag ttaccgtgaa 2220
gatcccattg tctatgaaat tcatccaacc aaatctttta ttagtggtgg gagcacaata 2280
acaggtgttg ggaaaaacct gaactcagtt agtgtcccga gaatggtcat aaatgtgcat 2340
gaagcaggaa ggaactttac agtggcatgt caacatcgct ctaattcaga gataatctgt 2400
tgtaccactc cttccctgca acagctgaat ctgcaactcc ccctgaaaac caaagccttt 2460
ttcatgttag atggcatcct ttccaaatac tttgatctca tttatgtaca taatcctgtg 2520
tttgagcctt ttgaaaagcc agtgatgatc tcaatgggca atgaaaatgt actggaaatt 2580
aagggaaatg atattgaccc tgaagcagtt aaaggtgaag tgttaaaagt tggaaataag 2640
agctgtgaga atatacactt acattctgaa gccgttttat gcacggtccc caatgacctg 2700
ctgaaattga acagcgagct aaatatagag tggaagcaag caatttcttc aaccgtcctt 2760
ggaaaagtaa tagttcaacc agatcagaat ttcacagctg aagctgcagc caaagaagct 2820
gcagccaaag aggccgcagc taaggaagcc gcagcaaaag ctgctgctga agctgcagcc 2880
aaagaagctg cagccaaaga ggccgcagct aaggaagccg cagcaaaagc tgacattgtg 2940
ctgacccaat ctccagcttc tttggctgtg tctctagggc agagggccac catctcctgc 3000
aaggccagcc aaagtgttga ttatgatggt ggtagttata tgagttggtt ccaacagaga 3060
ccaggacagc cacccaaact cctcatctct gctgcatcca accttgaatc tggcatccca 3120
gccaggttta gtggcagtgg ctctgggaca gacttcaccc tcaatatcca tcctgtggag 3180
gaggaggatg ttgcaaccta ttactgtcag caaagttatg aagacccgct cacgttcggt 3240
gctggtacca aggtggagat caaacgaact gtggctgcac catctgtctt catcttcccg 3300
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 3360
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 3420
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 3480
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 3540
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgtggtgg ttcttcgggt 3600
tcaggctcgg gctccacagg tacatcgtca tcgggtacag gtacttcagc tggcacaacg 3660
ggtacctccg catctacttc cggttcaggc tccggaggcg gcggcggctc gggtggtggt 3720
ggatctgccg gcggaaccgc aaccgcaggc gcttcttctg gttccgtcca actgcagcag 3780
cctgggactg aactggtgaa gcctggggct tcagtgaagc tgtcctgcaa ggcttctggc 3840
tacaccttca ccagttactg gatacactgg gtgaagcaga ggcctggaca aggccttgag 3900
tggattggag agattaatcc tagcagcggt cgtactaact acaacgagaa attcaagaac 3960
aaggtcacag tgactgtaga caaatcttcc accacagcct acatgcaact cagcaacctg 4020
acatctgagg actctgcggt ctattactgt gcaagtaggg gctactgggg ccaaggcacc 4080
actctcacag tctcctcagc tagcacgaag ggcccatcgg tcttccccct ggcaccctcc 4140
tccaagagca cctctggggg cacagcggcc ctgggctgcc tggtcaagga ctacttcccc 4200
gaacccgtga cggtgtcgtg gaactcaggc gccctgacca gcggcgtgca caccttcccg 4260
gctgtcctac agtcctcagg actctactcc ctcagcagcg tggtgaccgt gccctccagc 4320
agcttgggca cccagaccta catctgcaac gtgaatcaca agcccagcaa caccaaggtg 4380
gacaagaaag ttgagcccaa atcttgtgac aaaactcaca caacgcgtgg tgccgcatgg 4440
agccaccccc agttcgaaaa aggggccgca tggagccacc cccagttcga aaaaggggcc 4500
gcatggagcc acccccagtt cgaaaaaggg gccgcacacc atcaccatca ccattag 4557
<210> 30
<211> 4602
<212> ДНК
<213> Искуственная последовательность
<220>
<223> decoyMET-линкер-антитело
нуклеотидная последовательность K842E-линкер L60-scMvDN30
<400> 30
atgaaggccc ccgctgtgct tgcacctggc atcctcgtgc tcctgtttac cttggtgcag 60
aggagcaatg gggagtgtaa agaggcacta gcaaagtccg agatgaatgt gaatatgaag 120
tatcagcttc ccaacttcac cgcggaaaca cccatccaga atgtcattct acatgagcat 180
cacattttcc ttggtgccac taactacatt tatgttttaa atgaggaaga ccttcagaag 240
gttgctgagt acaagactgg gcctgtgctg gaacacccag attgtttccc atgtcaggac 300
tgcagcagca aagccaattt atcaggaggt gtttggaaag ataacatcaa catggctcta 360
gttgtggaca cctactatga tgatcaactc attagctgtg gcagcgtcaa cagagggacc 420
tgccagcgac atgtctttcc ccacaatcat actgctgaca tacagtcgga ggttcactgc 480
atattctccc cacagataga agagcccagc cagtgtcctg actgtgtggt gagcgccctg 540
ggagccaaag tcctttcatc tgtaaaggac cggttcatca acttctttgt aggcaatacc 600
ataaattctt cttatttccc agatcatcca ttgcattcga tatcagtgag aaggctaaag 660
gaaacgaaag atggttttat gtttttgacg gaccagtcct acattgatgt tttacctgag 720
ttcagagatt cttaccccat taagtatgtc catgcctttg aaagcaacaa ttttatttac 780
ttcttgacgg tccaaaggga aactctagat gctcagactt ttcacacaag aataatcagg 840
ttctgttcca taaactctgg attgcattcc tacatggaaa tgcctctgga gtgtattctc 900
acagaaaaga gaaaaaagag atccacaaag aaggaagtgt ttaatatact tcaggctgcg 960
tatgtcagca agcctggggc ccagcttgct agacaaatag gagccagcct gaatgatgac 1020
attcttttcg gggtgttcgc acaaagcaag ccagattctg ccgaaccaat ggatcgatct 1080
gccatgtgtg cattccctat caaatatgtc aacgacttct tcaacaagat cgtcaacaaa 1140
aacaatgtga gatgtctcca gcatttttac ggacccaatc atgagcactg ctttaatagg 1200
acacttctga gaaattcatc aggctgtgaa gcgcgccgtg atgaatatcg aacagagttt 1260
accacagctt tgcagcgcgt tgacttattc atgggtcaat tcagcgaagt cctcttaaca 1320
tctatatcca ccttcattaa aggagacctc accatagcta atcttgggac atcagagggt 1380
cgcttcatgc aggttgtggt ttctcgatca ggaccatcaa cccctcatgt gaattttctc 1440
ctggactccc atccagtgtc tccagaagtg attgtggagc atacattaaa ccaaaatggc 1500
tacacactgg ttatcactgg gaagaagatc acgaagatcc cattgaatgg cttgggttgc 1560
agacatttcc agtcttgcag tcaatgcctc tctgccccac cctttgttca gtgtggctgg 1620
tgccacgaca aatgtgtgcg atcggaggaa tgcctgagcg ggacatggac tcaacagatc 1680
tgtctgcctg caatctacaa ggttttccca aatagtgcac cccttgaagg agggacaagg 1740
ctgaccatat gtggctggga ctttggattt cggaggaata ataaatttga tttaaagaaa 1800
actagagttc tccttggaaa tgagagctgc accttgactt taagtgagag cacgatgaat 1860
acattgaaat gcacagttgg tcctgccatg aataagcatt tcaatatgtc cataattatt 1920
tcaaatggcc acgggacaac acaatacagt acattctcct atgtcgatcc tgtaataaca 1980
agtatttcgc cgaaatacgg tcctatggct ggtggcactt tacttacttt aactggaaat 2040
tacctaaaca gtgggaactc tagacacatt tcaattggtg gaaaaacatg tactttaaaa 2100
agtgtgtcaa acagtattct tgaatgttat accccagccc aaaccatttc aactgagttt 2160
gctgttaaat tgaaaattga cttagccaac cgagagacaa gcatcttcag ttaccgtgaa 2220
gatcccattg tctatgaaat tcatccaacc aaatctttta ttagtggtgg gagcacaata 2280
acaggtgttg ggaaaaacct gaactcagtt agtgtcccga gaatggtcat aaatgtgcat 2340
gaagcaggaa ggaactttac agtggcatgt caacatcgct ctaattcaga gataatctgt 2400
tgtaccactc cttccctgca acagctgaat ctgcaactcc ccctgaaaac caaagccttt 2460
ttcatgttag atggcatcct ttccaaatac tttgatctca tttatgtaca taatcctgtg 2520
tttgagcctt ttgaaaagcc agtgatgatc tcaatgggca atgaaaatgt actggaaatt 2580
aagggaaatg atattgaccc tgaagcagtt aaaggtgaag tgttaaaagt tggaaataag 2640
agctgtgaga atatacactt acattctgaa gccgttttat gcacggtccc caatgacctg 2700
ctgaaattga acagcgagct aaatatagag tggaagcaag caatttcttc aaccgtcctt 2760
ggaaaagtaa tagttcaacc agatcagaat ttcacaggtg gttcttcggg ttcaggctcg 2820
ggctccacag gtacatcgtc atcgggtaca ggtacttcag ctggcacaac gggtacctcc 2880
gcatctactt ccggttcagg ctccggaggc ggcggcggct cgggtggtgg tggatctgcc 2940
ggcggaaccg caaccgcagg cgcttcttct ggttccgaca ttgtgctgac ccaatctcca 3000
gcttctttgg ctgtgtctct agggcagagg gccaccatct cctgcaaggc cagccaaagt 3060
gttgattatg atggtggtag ttatatgagt tggttccaac agagaccagg acagccaccc 3120
aaactcctca tctctgctgc atccaacctt gaatctggca tcccagccag gtttagtggc 3180
agtggctctg ggacagactt caccctcaat atccatcctg tggaggagga ggatgttgca 3240
acctattact gtcagcaaag ttatgaagac ccgctcacgt tcggtgctgg taccaaggtg 3300
gagatcaaac gaactgtggc tgcaccatct gtcttcatct tcccgccatc tgatgagcag 3360
ttgaaatctg gaactgcctc tgttgtgtgc ctgctgaata acttctatcc cagagaggcc 3420
aaagtacagt ggaaggtgga taacgccctc caatcgggta actcccagga gagtgtcaca 3480
gagcaggaca gcaaggacag cacctacagc ctcagcagca ccctgacgct gagcaaagca 3540
gactacgaga aacacaaagt ctacgcctgc gaagtcaccc atcagggcct gagctcgccc 3600
gtcacaaaga gcttcaacag gggagagtgt ggtggttctt cgggttcagg ctcgggctcc 3660
acaggtacat cgtcatcggg tacaggtact tcagctggca caacgggtac ctccgcatct 3720
acttccggtt caggctccgg aggcggcggc ggctcgggtg gtggtggatc tgccggcgga 3780
accgcaaccg caggcgcttc ttctggttcc gtccaactgc agcagcctgg gactgaactg 3840
gtgaagcctg gggcttcagt gaagctgtcc tgcaaggctt ctggctacac cttcaccagt 3900
tactggatac actgggtgaa gcagaggcct ggacaaggcc ttgagtggat tggagagatt 3960
aatcctagca gcggtcgtac taactacaac gagaaattca agaacaaggt cacagtgact 4020
gtagacaaat cttccaccac agcctacatg caactcagca acctgacatc tgaggactct 4080
gcggtctatt actgtgcaag taggggctac tggggccaag gcaccactct cacagtctcc 4140
tcagctagca cgaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct 4200
gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc cgtgacggtg 4260
tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc 4320
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag 4380
acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gaaagttgag 4440
cccaaatctt gtgacaaaac tcacacaacg cgtggtgccg catggagcca cccccagttc 4500
gaaaaagggg ccgcatggag ccacccccag ttcgaaaaag gggccgcatg gagccacccc 4560
cagttcgaaa aaggggccgc acaccatcac catcaccatt ag 4602
<210> 31
<211> 4824
<212> ДНК
<213> Искуственная последовательность
<220>
<223> decoyMET-линкер-антитело
нуклеотидная последовательность K842E-линкер L134-scMvDN30
<400> 31
atgaaggccc ccgctgtgct tgcacctggc atcctcgtgc tcctgtttac cttggtgcag 60
aggagcaatg gggagtgtaa agaggcacta gcaaagtccg agatgaatgt gaatatgaag 120
tatcagcttc ccaacttcac cgcggaaaca cccatccaga atgtcattct acatgagcat 180
cacattttcc ttggtgccac taactacatt tatgttttaa atgaggaaga ccttcagaag 240
gttgctgagt acaagactgg gcctgtgctg gaacacccag attgtttccc atgtcaggac 300
tgcagcagca aagccaattt atcaggaggt gtttggaaag ataacatcaa catggctcta 360
gttgtggaca cctactatga tgatcaactc attagctgtg gcagcgtcaa cagagggacc 420
tgccagcgac atgtctttcc ccacaatcat actgctgaca tacagtcgga ggttcactgc 480
atattctccc cacagataga agagcccagc cagtgtcctg actgtgtggt gagcgccctg 540
ggagccaaag tcctttcatc tgtaaaggac cggttcatca acttctttgt aggcaatacc 600
ataaattctt cttatttccc agatcatcca ttgcattcga tatcagtgag aaggctaaag 660
gaaacgaaag atggttttat gtttttgacg gaccagtcct acattgatgt tttacctgag 720
ttcagagatt cttaccccat taagtatgtc catgcctttg aaagcaacaa ttttatttac 780
ttcttgacgg tccaaaggga aactctagat gctcagactt ttcacacaag aataatcagg 840
ttctgttcca taaactctgg attgcattcc tacatggaaa tgcctctgga gtgtattctc 900
acagaaaaga gaaaaaagag atccacaaag aaggaagtgt ttaatatact tcaggctgcg 960
tatgtcagca agcctggggc ccagcttgct agacaaatag gagccagcct gaatgatgac 1020
attcttttcg gggtgttcgc acaaagcaag ccagattctg ccgaaccaat ggatcgatct 1080
gccatgtgtg cattccctat caaatatgtc aacgacttct tcaacaagat cgtcaacaaa 1140
aacaatgtga gatgtctcca gcatttttac ggacccaatc atgagcactg ctttaatagg 1200
acacttctga gaaattcatc aggctgtgaa gcgcgccgtg atgaatatcg aacagagttt 1260
accacagctt tgcagcgcgt tgacttattc atgggtcaat tcagcgaagt cctcttaaca 1320
tctatatcca ccttcattaa aggagacctc accatagcta atcttgggac atcagagggt 1380
cgcttcatgc aggttgtggt ttctcgatca ggaccatcaa cccctcatgt gaattttctc 1440
ctggactccc atccagtgtc tccagaagtg attgtggagc atacattaaa ccaaaatggc 1500
tacacactgg ttatcactgg gaagaagatc acgaagatcc cattgaatgg cttgggttgc 1560
agacatttcc agtcttgcag tcaatgcctc tctgccccac cctttgttca gtgtggctgg 1620
tgccacgaca aatgtgtgcg atcggaggaa tgcctgagcg ggacatggac tcaacagatc 1680
tgtctgcctg caatctacaa ggttttccca aatagtgcac cccttgaagg agggacaagg 1740
ctgaccatat gtggctggga ctttggattt cggaggaata ataaatttga tttaaagaaa 1800
actagagttc tccttggaaa tgagagctgc accttgactt taagtgagag cacgatgaat 1860
acattgaaat gcacagttgg tcctgccatg aataagcatt tcaatatgtc cataattatt 1920
tcaaatggcc acgggacaac acaatacagt acattctcct atgtcgatcc tgtaataaca 1980
agtatttcgc cgaaatacgg tcctatggct ggtggcactt tacttacttt aactggaaat 2040
tacctaaaca gtgggaactc tagacacatt tcaattggtg gaaaaacatg tactttaaaa 2100
agtgtgtcaa acagtattct tgaatgttat accccagccc aaaccatttc aactgagttt 2160
gctgttaaat tgaaaattga cttagccaac cgagagacaa gcatcttcag ttaccgtgaa 2220
gatcccattg tctatgaaat tcatccaacc aaatctttta ttagtggtgg gagcacaata 2280
acaggtgttg ggaaaaacct gaactcagtt agtgtcccga gaatggtcat aaatgtgcat 2340
gaagcaggaa ggaactttac agtggcatgt caacatcgct ctaattcaga gataatctgt 2400
tgtaccactc cttccctgca acagctgaat ctgcaactcc ccctgaaaac caaagccttt 2460
ttcatgttag atggcatcct ttccaaatac tttgatctca tttatgtaca taatcctgtg 2520
tttgagcctt ttgaaaagcc agtgatgatc tcaatgggca atgaaaatgt actggaaatt 2580
aagggaaatg atattgaccc tgaagcagtt aaaggtgaag tgttaaaagt tggaaataag 2640
agctgtgaga atatacactt acattctgaa gccgttttat gcacggtccc caatgacctg 2700
ctgaaattga acagcgagct aaatatagag tggaagcaag caatttcttc aaccgtcctt 2760
ggaaaagtaa tagttcaacc agatcagaat ttcacaggtg gtggtggttc tggtggtggt 2820
ggttcaggcg gcggcggctc ggcagaggca tggtataatc tcgggaacgc atactacaaa 2880
cagggcgatt atcagaaggc tatcgaatac taccaaaaag cactcgaact ggaccccaat 2940
aacgcggagg catggtacaa tctcgggaat gcgtactata agcagggtga ctaccagaag 3000
gccattgaat actaccaaaa ggcgcttgaa ctggacccga acaacgccga ggcctggtat 3060
aacctgggga atgcttatta caagcagggg gattatcaaa aggcaataga agactaccag 3120
aaggccctcg aattggaccc taacaatggt ggtggtggtt ctggtggtgg tggttcaggc 3180
ggcggcggct cgggcggaga cattgtgctg acccaatctc cagcttcttt ggctgtgtct 3240
ctagggcaga gggccaccat ctcctgcaag gccagccaaa gtgttgatta tgatggtggt 3300
agttatatga gttggttcca acagagacca ggacagccac ccaaactcct catctctgct 3360
gcatccaacc ttgaatctgg catcccagcc aggtttagtg gcagtggctc tgggacagac 3420
ttcaccctca atatccatcc tgtggaggag gaggatgttg caacctatta ctgtcagcaa 3480
agttatgaag acccgctcac gttcggtgct ggtaccaagg tggagatcaa acgaactgtg 3540
gctgcaccat ctgtcttcat cttcccgcca tctgatgagc agttgaaatc tggaactgcc 3600
tctgttgtgt gcctgctgaa taacttctat cccagagagg ccaaagtaca gtggaaggtg 3660
gataacgccc tccaatcggg taactcccag gagagtgtca cagagcagga cagcaaggac 3720
agcacctaca gcctcagcag caccctgacg ctgagcaaag cagactacga gaaacacaaa 3780
gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc ccgtcacaaa gagcttcaac 3840
aggggagagt gtggtggttc ttcgggttca ggctcgggct ccacaggtac atcgtcatcg 3900
ggtacaggta cttcagctgg cacaacgggt acctccgcat ctacttccgg ttcaggctcc 3960
ggaggcggcg gcggctcggg tggtggtgga tctgccggcg gaaccgcaac cgcaggcgct 4020
tcttctggtt ccgtccaact gcagcagcct gggactgaac tggtgaagcc tggggcttca 4080
gtgaagctgt cctgcaaggc ttctggctac accttcacca gttactggat acactgggtg 4140
aagcagaggc ctggacaagg ccttgagtgg attggagaga ttaatcctag cagcggtcgt 4200
actaactaca acgagaaatt caagaacaag gtcacagtga ctgtagacaa atcttccacc 4260
acagcctaca tgcaactcag caacctgaca tctgaggact ctgcggtcta ttactgtgca 4320
agtaggggct actggggcca aggcaccact ctcacagtct cctcagctag cacgaagggc 4380
ccatcggtct tccccctggc accctcctcc aagagcacct ctgggggcac agcggccctg 4440
ggctgcctgg tcaaggacta cttccccgaa cccgtgacgg tgtcgtggaa ctcaggcgcc 4500
ctgaccagcg gcgtgcacac cttcccggct gtcctacagt cctcaggact ctactccctc 4560
agcagcgtgg tgaccgtgcc ctccagcagc ttgggcaccc agacctacat ctgcaacgtg 4620
aatcacaagc ccagcaacac caaggtggac aagaaagttg agcccaaatc ttgtgacaaa 4680
actcacacaa cgcgtggtgc cgcatggagc cacccccagt tcgaaaaagg ggccgcatgg 4740
agccaccccc agttcgaaaa aggggccgca tggagccacc cccagttcga aaaaggggcc 4800
gcacaccatc accatcacca ttag 4824
<210> 32
<211> 4583
<212> ДНК
<213> Искуственная последовательность
<220>
<223> антитело-линкер-decoyMET
нуклеотидная последовательность scMvDN30-линкер L45-K842E
<400> 32
ggatcccccg gggccaccat ggagacagac acaatcctgc tatgggtgct gctgctctgg 60
gttccaggct ccactggtga cattgtgctg acccaatctc cagcttcttt ggctgtgtct 120
ctagggcaga gggccaccat ctcctgcaag gccagccaaa gtgttgatta tgatggtggt 180
agttatatga gttggttcca acagagacca ggacagccac ccaaactcct catctctgct 240
gcatccaacc ttgaatctgg catcccagcc aggtttagtg gcagtggctc tgggacagac 300
ttcaccctca atatccatcc tgtggaggag gaggatgttg caacctatta ctgtcagcaa 360
agttatgaag acccgctcac gttcggtgct ggtaccaagg tggagatcaa acgaactgtg 420
gctgcaccat ctgtcttcat cttcccgcca tctgatgagc agttgaaatc tggaactgcc 480
tctgttgtgt gcctgctgaa taacttctat cccagagagg ccaaagtaca gtggaaggtg 540
gataacgccc tccaatcggg taactcccag gagagtgtca cagagcagga cagcaaggac 600
agcacctaca gcctcagcag caccctgacg ctgagcaaag cagactacga gaaacacaaa 660
gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc ccgtcacaaa gagcttcaac 720
aggggagagt gtggtggttc ttcgggttca ggctcgggct ccacaggtac atcgtcatcg 780
ggtacaggta cttcagctgg cacaacgggt acctccgcat ctacttccgg ttcaggctcc 840
ggaggcggcg gcggctcggg tggtggtgga tctgccggcg gaaccgcaac cgcaggcgct 900
tcttctggtt ccgtccaact gcagcagcct gggactgaac tggtgaagcc tggggcttca 960
gtgaagctgt cctgcaaggc ttctggctac accttcacca gttactggat acactgggtg 1020
aagcagaggc ctggacaagg ccttgagtgg attggagaga ttaatcctag cagcggtcgt 1080
actaactaca acgagaaatt caagaacaag gtcacagtga ctgtagacaa atcttccacc 1140
acagcctaca tgcaactcag caacctgaca tctgaggact ctgcggtcta ttactgtgca 1200
agtaggggct actggggcca aggcaccact ctcacagtct cctcagcaag cacgaagggc 1260
ccatcggtct tccccctggc accctcctcc aagagcacct ctgggggcac agcggccctg 1320
ggctgcctgg tcaaggacta cttccccgaa cccgtgacgg tgtcgtggaa ctcaggcgcc 1380
ctgaccagcg gcgtgcacac cttcccggct gtcctacagt cctcaggact ctactccctc 1440
agcagcgtgg tgaccgtgcc ctccagcagc ttgggcaccc agacctacat ctgcaacgtg 1500
aatcacaagc ccagcaacac caaggtggac aagaaagttg agcccaaatc ttgtgacaaa 1560
actcacacag ctgaagctgc agccaaagaa gctgcagcca aagaggccgc agctaaggaa 1620
gccgcagcaa aagctgctgc tgaagctgca gccaaagaag ctgcagccaa agaggccgca 1680
gctaaggaag ccgcagcaaa agctgagtgt aaagaggcac tagcaaagtc cgagatgaat 1740
gtgaatatga agtatcagct tcccaacttc accgcggaaa cacccatcca gaatgtcatt 1800
ctacatgagc atcacatttt ccttggtgcc actaactaca tttatgtttt aaatgaggaa 1860
gaccttcaga aggttgctga gtacaagact gggcctgtgc tggaacaccc agattgtttc 1920
ccatgtcagg actgcagcag caaagccaat ttatcaggag gtgtttggaa agataacatc 1980
aacatggctc tagttgtgga cacctactat gatgatcaac tcattagctg tggcagcgtc 2040
aacagaggga cctgccagcg acatgtcttt ccccacaatc atactgctga catacagtcg 2100
gaggttcact gcatattctc cccacagata gaagagccca gccagtgtcc tgactgtgtg 2160
gtgagcgccc tgggagccaa agtcctttca tctgtaaagg accggttcat caacttcttt 2220
gtaggcaata ccataaattc ttcttatttc ccagatcatc cattgcattc gatatcagtg 2280
agaaggctaa aggaaacgaa agatggtttt atgtttttga cggaccagtc ctacattgat 2340
gttttacctg agttcagaga ttcttacccc attaagtatg tccatgcctt tgaaagcaac 2400
aattttattt acttcttgac ggtccaaagg gaaactctag atgctcagac ttttcacaca 2460
agaataatca ggttctgttc cataaactct ggattgcatt cctacatgga aatgcctctg 2520
gagtgtattc tcacagaaaa gagaaaaaag agatccacaa agaaggaagt gtttaatata 2580
cttcaggctg cgtatgtcag caagcctggg gcccagcttg ctagacaaat aggagccagc 2640
ctgaatgatg acattctttt cggggtgttc gcacaaagca agccagattc tgccgaacca 2700
atggatcgat ctgccatgtg tgcattccct atcaaatatg tcaacgactt cttcaacaag 2760
atcgtcaaca aaaacaatgt gagatgtctc cagcattttt acggacccaa tcatgagcac 2820
tgctttaata ggacacttct gagaaattca tcaggctgtg aagcgcgccg tgatgaatat 2880
cgaacagagt ttaccacagc tttgcagcgc gttgacttat tcatgggtca attcagcgaa 2940
gtcctcttaa catctatatc caccttcatt aaaggagacc tcaccatagc taatcttggg 3000
acatcagagg gtcgcttcat gcaggttgtg gtttctcgat caggaccatc aacccctcat 3060
gtgaattttc tcctggactc ccatccagtg tctccagaag tgattgtgga gcatacatta 3120
aaccaaaatg gctacacact ggttatcact gggaagaaga tcacgaagat cccattgaat 3180
ggcttgggtt gcagacattt ccagtcttgc agtcaatgcc tctctgcccc accctttgtt 3240
cagtgtggct ggtgccacga caaatgtgtg cgatcggagg aatgcctgag cgggacatgg 3300
actcaacaga tctgtctgcc tgcaatctac aaggttttcc caaatagtgc accccttgaa 3360
ggagggacaa ggctgaccat atgtggctgg gactttggat ttcggaggaa taataaattt 3420
gatttaaaga aaactagagt tctccttgga aatgagagct gcaccttgac tttaagtgag 3480
agcacgatga atacattgaa atgcacagtt ggtcctgcca tgaataagca tttcaatatg 3540
tccataatta tttcaaatgg ccacgggaca acacaataca gtacattctc ctatgtcgat 3600
cctgtaataa caagtatttc gccgaaatac ggtcctatgg ctggtggcac tttacttact 3660
ttaactggaa attacctaaa cagtgggaac tctagacaca tttcaattgg tggaaaaaca 3720
tgtactttaa aaagtgtgtc aaacagtatt cttgaatgtt ataccccagc ccaaaccatt 3780
tcaactgagt ttgctgttaa attgaaaatt gacttagcca accgagagac aagcatcttc 3840
agttaccgtg aagatcccat tgtctatgaa attcatccaa ccaaatcttt tattagtggt 3900
gggagcacaa taacaggtgt tgggaaaaac ctgaactcag ttagtgtccc gagaatggtc 3960
ataaatgtgc atgaagcagg aaggaacttt acagtggcat gtcaacatcg ctctaattca 4020
gagataatct gttgtaccac tccttccctg caacagctga atctgcaact ccccctgaaa 4080
accaaagcct ttttcatgtt agatggcatc ctttccaaat actttgatct catttatgta 4140
cataatcctg tgtttgagcc ttttgaaaag ccagtgatga tctcaatggg caatgaaaat 4200
gtactggaaa ttaagggaaa tgatattgac cctgaagcag ttaaaggtga agtgttaaaa 4260
gttggaaata agagctgtga gaatatacac ttacattctg aagccgtttt atgcacggtc 4320
cccaatgacc tgctgaaatt gaacagcgag ctaaatatag agtggaagca agcaatttct 4380
tcaaccgtcc ttggaaaagt aatagttcaa ccagatcaga atttcacagc tagcggtgcc 4440
gcatggagcc acccccagtt cgaaaaaggg gccgcatgga gccaccccca gttcgaaaaa 4500
ggggccgcat ggagccaccc ccagttcgaa aaaggggccg cacaccatca ccatcaccat 4560
taggtcgacc tcgaggcggc cgc 4583
<210> 33
<211> 4628
<212> ДНК
<213> Искуственная последовательность
<220>
<223> антитело-линкер-decoyMET
нуклеотидная последовательность scMvDN30-линкер L60-K842E
<400> 33
ggatcccccg gggccaccat ggagacagac acaatcctgc tatgggtgct gctgctctgg 60
gttccaggct ccactggtga cattgtgctg acccaatctc cagcttcttt ggctgtgtct 120
ctagggcaga gggccaccat ctcctgcaag gccagccaaa gtgttgatta tgatggtggt 180
agttatatga gttggttcca acagagacca ggacagccac ccaaactcct catctctgct 240
gcatccaacc ttgaatctgg catcccagcc aggtttagtg gcagtggctc tgggacagac 300
ttcaccctca atatccatcc tgtggaggag gaggatgttg caacctatta ctgtcagcaa 360
agttatgaag acccgctcac gttcggtgct ggtaccaagg tggagatcaa acgaactgtg 420
gctgcaccat ctgtcttcat cttcccgcca tctgatgagc agttgaaatc tggaactgcc 480
tctgttgtgt gcctgctgaa taacttctat cccagagagg ccaaagtaca gtggaaggtg 540
gataacgccc tccaatcggg taactcccag gagagtgtca cagagcagga cagcaaggac 600
agcacctaca gcctcagcag caccctgacg ctgagcaaag cagactacga gaaacacaaa 660
gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc ccgtcacaaa gagcttcaac 720
aggggagagt gtggtggttc ttcgggttca ggctcgggct ccacaggtac atcgtcatcg 780
ggtacaggta cttcagctgg cacaacgggt acctccgcat ctacttccgg ttcaggctcc 840
ggaggcggcg gcggctcggg tggtggtgga tctgccggcg gaaccgcaac cgcaggcgct 900
tcttctggtt ccgtccaact gcagcagcct gggactgaac tggtgaagcc tggggcttca 960
gtgaagctgt cctgcaaggc ttctggctac accttcacca gttactggat acactgggtg 1020
aagcagaggc ctggacaagg ccttgagtgg attggagaga ttaatcctag cagcggtcgt 1080
actaactaca acgagaaatt caagaacaag gtcacagtga ctgtagacaa atcttccacc 1140
acagcctaca tgcaactcag caacctgaca tctgaggact ctgcggtcta ttactgtgca 1200
agtaggggct actggggcca aggcaccact ctcacagtct cctcagcaag cacgaagggc 1260
ccatcggtct tccccctggc accctcctcc aagagcacct ctgggggcac agcggccctg 1320
ggctgcctgg tcaaggacta cttccccgaa cccgtgacgg tgtcgtggaa ctcaggcgcc 1380
ctgaccagcg gcgtgcacac cttcccggct gtcctacagt cctcaggact ctactccctc 1440
agcagcgtgg tgaccgtgcc ctccagcagc ttgggcaccc agacctacat ctgcaacgtg 1500
aatcacaagc ccagcaacac caaggtggac aagaaagttg agcccaaatc ttgtgacaaa 1560
actcacacag gtggttcttc gggttcaggc tcgggctcca caggtacatc gtcatcgggt 1620
acaggtactt cagctggcac aacgggtacc tccgcatcta cttccggttc aggctccgga 1680
ggcggcggcg gctcgggtgg tggtggatct gccggcggaa ccgcaaccgc aggcgcttct 1740
tctggttccg agtgtaaaga ggcactagca aagtccgaga tgaatgtgaa tatgaagtat 1800
cagcttccca acttcaccgc ggaaacaccc atccagaatg tcattctaca tgagcatcac 1860
attttccttg gtgccactaa ctacatttat gttttaaatg aggaagacct tcagaaggtt 1920
gctgagtaca agactgggcc tgtgctggaa cacccagatt gtttcccatg tcaggactgc 1980
agcagcaaag ccaatttatc aggaggtgtt tggaaagata acatcaacat ggctctagtt 2040
gtggacacct actatgatga tcaactcatt agctgtggca gcgtcaacag agggacctgc 2100
cagcgacatg tctttcccca caatcatact gctgacatac agtcggaggt tcactgcata 2160
ttctccccac agatagaaga gcccagccag tgtcctgact gtgtggtgag cgccctggga 2220
gccaaagtcc tttcatctgt aaaggaccgg ttcatcaact tctttgtagg caataccata 2280
aattcttctt atttcccaga tcatccattg cattcgatat cagtgagaag gctaaaggaa 2340
acgaaagatg gttttatgtt tttgacggac cagtcctaca ttgatgtttt acctgagttc 2400
agagattctt accccattaa gtatgtccat gcctttgaaa gcaacaattt tatttacttc 2460
ttgacggtcc aaagggaaac tctagatgct cagacttttc acacaagaat aatcaggttc 2520
tgttccataa actctggatt gcattcctac atggaaatgc ctctggagtg tattctcaca 2580
gaaaagagaa aaaagagatc cacaaagaag gaagtgttta atatacttca ggctgcgtat 2640
gtcagcaagc ctggggccca gcttgctaga caaataggag ccagcctgaa tgatgacatt 2700
cttttcgggg tgttcgcaca aagcaagcca gattctgccg aaccaatgga tcgatctgcc 2760
atgtgtgcat tccctatcaa atatgtcaac gacttcttca acaagatcgt caacaaaaac 2820
aatgtgagat gtctccagca tttttacgga cccaatcatg agcactgctt taataggaca 2880
cttctgagaa attcatcagg ctgtgaagcg cgccgtgatg aatatcgaac agagtttacc 2940
acagctttgc agcgcgttga cttattcatg ggtcaattca gcgaagtcct cttaacatct 3000
atatccacct tcattaaagg agacctcacc atagctaatc ttgggacatc agagggtcgc 3060
ttcatgcagg ttgtggtttc tcgatcagga ccatcaaccc ctcatgtgaa ttttctcctg 3120
gactcccatc cagtgtctcc agaagtgatt gtggagcata cattaaacca aaatggctac 3180
acactggtta tcactgggaa gaagatcacg aagatcccat tgaatggctt gggttgcaga 3240
catttccagt cttgcagtca atgcctctct gccccaccct ttgttcagtg tggctggtgc 3300
cacgacaaat gtgtgcgatc ggaggaatgc ctgagcggga catggactca acagatctgt 3360
ctgcctgcaa tctacaaggt tttcccaaat agtgcacccc ttgaaggagg gacaaggctg 3420
accatatgtg gctgggactt tggatttcgg aggaataata aatttgattt aaagaaaact 3480
agagttctcc ttggaaatga gagctgcacc ttgactttaa gtgagagcac gatgaataca 3540
ttgaaatgca cagttggtcc tgccatgaat aagcatttca atatgtccat aattatttca 3600
aatggccacg ggacaacaca atacagtaca ttctcctatg tcgatcctgt aataacaagt 3660
atttcgccga aatacggtcc tatggctggt ggcactttac ttactttaac tggaaattac 3720
ctaaacagtg ggaactctag acacatttca attggtggaa aaacatgtac tttaaaaagt 3780
gtgtcaaaca gtattcttga atgttatacc ccagcccaaa ccatttcaac tgagtttgct 3840
gttaaattga aaattgactt agccaaccga gagacaagca tcttcagtta ccgtgaagat 3900
cccattgtct atgaaattca tccaaccaaa tcttttatta gtggtgggag cacaataaca 3960
ggtgttggga aaaacctgaa ctcagttagt gtcccgagaa tggtcataaa tgtgcatgaa 4020
gcaggaagga actttacagt ggcatgtcaa catcgctcta attcagagat aatctgttgt 4080
accactcctt ccctgcaaca gctgaatctg caactccccc tgaaaaccaa agcctttttc 4140
atgttagatg gcatcctttc caaatacttt gatctcattt atgtacataa tcctgtgttt 4200
gagccttttg aaaagccagt gatgatctca atgggcaatg aaaatgtact ggaaattaag 4260
ggaaatgata ttgaccctga agcagttaaa ggtgaagtgt taaaagttgg aaataagagc 4320
tgtgagaata tacacttaca ttctgaagcc gttttatgca cggtccccaa tgacctgctg 4380
aaattgaaca gcgagctaaa tatagagtgg aagcaagcaa tttcttcaac cgtccttgga 4440
aaagtaatag ttcaaccaga tcagaatttc acagctagcg gtgccgcatg gagccacccc 4500
cagttcgaaa aaggggccgc atggagccac ccccagttcg aaaaaggggc cgcatggagc 4560
cacccccagt tcgaaaaagg ggccgcacac catcaccatc accattaggt cgacctcgag 4620
gcggccgc 4628
<210> 34
<211> 4850
<212> ДНК
<213> Искуственная последовательность
<220>
<223> антитело-линкер-decoyMET
нуклеотидная последовательность scMvDN30-линкер L134-K842E
<400> 34
ggatcccccg gggccaccat ggagacagac acaatcctgc tatgggtgct gctgctctgg 60
gttccaggct ccactggtga cattgtgctg acccaatctc cagcttcttt ggctgtgtct 120
ctagggcaga gggccaccat ctcctgcaag gccagccaaa gtgttgatta tgatggtggt 180
agttatatga gttggttcca acagagacca ggacagccac ccaaactcct catctctgct 240
gcatccaacc ttgaatctgg catcccagcc aggtttagtg gcagtggctc tgggacagac 300
ttcaccctca atatccatcc tgtggaggag gaggatgttg caacctatta ctgtcagcaa 360
agttatgaag acccgctcac gttcggtgct ggtaccaagg tggagatcaa acgaactgtg 420
gctgcaccat ctgtcttcat cttcccgcca tctgatgagc agttgaaatc tggaactgcc 480
tctgttgtgt gcctgctgaa taacttctat cccagagagg ccaaagtaca gtggaaggtg 540
gataacgccc tccaatcggg taactcccag gagagtgtca cagagcagga cagcaaggac 600
agcacctaca gcctcagcag caccctgacg ctgagcaaag cagactacga gaaacacaaa 660
gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc ccgtcacaaa gagcttcaac 720
aggggagagt gtggtggttc ttcgggttca ggctcgggct ccacaggtac atcgtcatcg 780
ggtacaggta cttcagctgg cacaacgggt acctccgcat ctacttccgg ttcaggctcc 840
ggaggcggcg gcggctcggg tggtggtgga tctgccggcg gaaccgcaac cgcaggcgct 900
tcttctggtt ccgtccaact gcagcagcct gggactgaac tggtgaagcc tggggcttca 960
gtgaagctgt cctgcaaggc ttctggctac accttcacca gttactggat acactgggtg 1020
aagcagaggc ctggacaagg ccttgagtgg attggagaga ttaatcctag cagcggtcgt 1080
actaactaca acgagaaatt caagaacaag gtcacagtga ctgtagacaa atcttccacc 1140
acagcctaca tgcaactcag caacctgaca tctgaggact ctgcggtcta ttactgtgca 1200
agtaggggct actggggcca aggcaccact ctcacagtct cctcagcaag cacgaagggc 1260
ccatcggtct tccccctggc accctcctcc aagagcacct ctgggggcac agcggccctg 1320
ggctgcctgg tcaaggacta cttccccgaa cccgtgacgg tgtcgtggaa ctcaggcgcc 1380
ctgaccagcg gcgtgcacac cttcccggct gtcctacagt cctcaggact ctactccctc 1440
agcagcgtgg tgaccgtgcc ctccagcagc ttgggcaccc agacctacat ctgcaacgtg 1500
aatcacaagc ccagcaacac caaggtggac aagaaagttg agcccaaatc ttgtgacaaa 1560
actcacacag gtggtggtgg ttctggtggt ggtggttcag gcggcggcgg ctcggcagag 1620
gcatggtata atctcgggaa cgcatactac aaacagggcg attatcagaa ggctatcgaa 1680
tactaccaaa aagcactcga actggacccc aataacgcgg aggcatggta caatctcggg 1740
aatgcgtact ataagcaggg tgactaccag aaggccattg aatactacca aaaggcgctt 1800
gaactggacc cgaacaacgc cgaggcctgg tataacctgg ggaatgctta ttacaagcag 1860
ggggattatc aaaaggcaat agaagactac cagaaggccc tcgaattgga ccctaacaat 1920
ggtggtggtg gttctggtgg tggtggttca ggcggcggcg gctcgggcgg agagtgtaaa 1980
gaggcactag caaagtccga gatgaatgtg aatatgaagt atcagcttcc caacttcacc 2040
gcggaaacac ccatccagaa tgtcattcta catgagcatc acattttcct tggtgccact 2100
aactacattt atgttttaaa tgaggaagac cttcagaagg ttgctgagta caagactggg 2160
cctgtgctgg aacacccaga ttgtttccca tgtcaggact gcagcagcaa agccaattta 2220
tcaggaggtg tttggaaaga taacatcaac atggctctag ttgtggacac ctactatgat 2280
gatcaactca ttagctgtgg cagcgtcaac agagggacct gccagcgaca tgtctttccc 2340
cacaatcata ctgctgacat acagtcggag gttcactgca tattctcccc acagatagaa 2400
gagcccagcc agtgtcctga ctgtgtggtg agcgccctgg gagccaaagt cctttcatct 2460
gtaaaggacc ggttcatcaa cttctttgta ggcaatacca taaattcttc ttatttccca 2520
gatcatccat tgcattcgat atcagtgaga aggctaaagg aaacgaaaga tggttttatg 2580
tttttgacgg accagtccta cattgatgtt ttacctgagt tcagagattc ttaccccatt 2640
aagtatgtcc atgcctttga aagcaacaat tttatttact tcttgacggt ccaaagggaa 2700
actctagatg ctcagacttt tcacacaaga ataatcaggt tctgttccat aaactctgga 2760
ttgcattcct acatggaaat gcctctggag tgtattctca cagaaaagag aaaaaagaga 2820
tccacaaaga aggaagtgtt taatatactt caggctgcgt atgtcagcaa gcctggggcc 2880
cagcttgcta gacaaatagg agccagcctg aatgatgaca ttcttttcgg ggtgttcgca 2940
caaagcaagc cagattctgc cgaaccaatg gatcgatctg ccatgtgtgc attccctatc 3000
aaatatgtca acgacttctt caacaagatc gtcaacaaaa acaatgtgag atgtctccag 3060
catttttacg gacccaatca tgagcactgc tttaatagga cacttctgag aaattcatca 3120
ggctgtgaag cgcgccgtga tgaatatcga acagagttta ccacagcttt gcagcgcgtt 3180
gacttattca tgggtcaatt cagcgaagtc ctcttaacat ctatatccac cttcattaaa 3240
ggagacctca ccatagctaa tcttgggaca tcagagggtc gcttcatgca ggttgtggtt 3300
tctcgatcag gaccatcaac ccctcatgtg aattttctcc tggactccca tccagtgtct 3360
ccagaagtga ttgtggagca tacattaaac caaaatggct acacactggt tatcactggg 3420
aagaagatca cgaagatccc attgaatggc ttgggttgca gacatttcca gtcttgcagt 3480
caatgcctct ctgccccacc ctttgttcag tgtggctggt gccacgacaa atgtgtgcga 3540
tcggaggaat gcctgagcgg gacatggact caacagatct gtctgcctgc aatctacaag 3600
gttttcccaa atagtgcacc ccttgaagga gggacaaggc tgaccatatg tggctgggac 3660
tttggatttc ggaggaataa taaatttgat ttaaagaaaa ctagagttct ccttggaaat 3720
gagagctgca ccttgacttt aagtgagagc acgatgaata cattgaaatg cacagttggt 3780
cctgccatga ataagcattt caatatgtcc ataattattt caaatggcca cgggacaaca 3840
caatacagta cattctccta tgtcgatcct gtaataacaa gtatttcgcc gaaatacggt 3900
cctatggctg gtggcacttt acttacttta actggaaatt acctaaacag tgggaactct 3960
agacacattt caattggtgg aaaaacatgt actttaaaaa gtgtgtcaaa cagtattctt 4020
gaatgttata ccccagccca aaccatttca actgagtttg ctgttaaatt gaaaattgac 4080
ttagccaacc gagagacaag catcttcagt taccgtgaag atcccattgt ctatgaaatt 4140
catccaacca aatcttttat tagtggtggg agcacaataa caggtgttgg gaaaaacctg 4200
aactcagtta gtgtcccgag aatggtcata aatgtgcatg aagcaggaag gaactttaca 4260
gtggcatgtc aacatcgctc taattcagag ataatctgtt gtaccactcc ttccctgcaa 4320
cagctgaatc tgcaactccc cctgaaaacc aaagcctttt tcatgttaga tggcatcctt 4380
tccaaatact ttgatctcat ttatgtacat aatcctgtgt ttgagccttt tgaaaagcca 4440
gtgatgatct caatgggcaa tgaaaatgta ctggaaatta agggaaatga tattgaccct 4500
gaagcagtta aaggtgaagt gttaaaagtt ggaaataaga gctgtgagaa tatacactta 4560
cattctgaag ccgttttatg cacggtcccc aatgacctgc tgaaattgaa cagcgagcta 4620
aatatagagt ggaagcaagc aatttcttca accgtccttg gaaaagtaat agttcaacca 4680
gatcagaatt tcacagctag cggtgccgca tggagccacc cccagttcga aaaaggggcc 4740
gcatggagcc acccccagtt cgaaaaaggg gccgcatgga gccaccccca gttcgaaaaa 4800
ggggccgcac accatcacca tcaccattag gtcgacctcg aggcggccgc 4850
<210> 35
<211> 45
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> линкер L45
<400> 35
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
Glu Ala Ala Ala Lys Ala Ala Ala Glu Ala Ala Ala Lys Glu Ala Ala
20 25 30
Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
35 40 45
<210> 36
<211> 60
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> линкер L60
<400> 36
Gly Gly Ser Ser Gly Ser Gly Ser Gly Ser Thr Gly Thr Ser Ser Ser
1 5 10 15
Gly Thr Gly Thr Ser Ala Gly Thr Thr Gly Thr Ser Ala Ser Thr Ser
20 25 30
Gly Ser Gly Ser Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala
35 40 45
Gly Gly Thr Ala Thr Ala Gly Ala Ser Ser Gly Ser
50 55 60
<210> 37
<211> 134
<212> ПРТ
<213> Искуственная последовательность
<220>
<223> линкер L134
<400> 37
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala
1 5 10 15
Glu Ala Trp Tyr Asn Leu Gly Asn Ala Tyr Tyr Lys Gln Gly Asp Tyr
20 25 30
Gln Lys Ala Ile Glu Tyr Tyr Gln Lys Ala Leu Glu Leu Asp Pro Asn
35 40 45
Asn Ala Glu Ala Trp Tyr Asn Leu Gly Asn Ala Tyr Tyr Lys Gln Gly
50 55 60
Asp Tyr Gln Lys Ala Ile Glu Tyr Tyr Gln Lys Ala Leu Glu Leu Asp
65 70 75 80
Pro Asn Asn Ala Glu Ala Trp Tyr Asn Leu Gly Asn Ala Tyr Tyr Lys
85 90 95
Gln Gly Asp Tyr Gln Lys Ala Ile Glu Asp Tyr Gln Lys Ala Leu Glu
100 105 110
Leu Asp Pro Asn Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly
130
<210> 38
<211> 135
<212> ДНК
<213> Искуственная последовательность
<220>
<223> линкер L45
<400> 38
gctgaagctg cagccaaaga agctgcagcc aaagaggccg cagctaagga agccgcagca 60
aaagctgctg ctgaagctgc agccaaagaa gctgcagcca aagaggccgc agctaaggaa 120
gccgcagcaa aagct 135
<210> 39
<211> 180
<212> ДНК
<213> Искуственная последовательность
<220>
<223> линкер L60
<400> 39
ggtggttctt cgggttcagg ctcgggctcc acaggtacat cgtcatcggg tacaggtact 60
tcagctggca caacgggtac ctccgcatct acttccggtt caggctccgg aggcggcggc 120
ggctcgggtg gtggtggatc tgccggcgga accgcaaccg caggcgcttc ttctggttcc 180
<210> 40
<211> 402
<212> ДНК
<213> Искуственная последовательность
<220>
<223> линкер L134
<400> 40
ggtggtggtg gttctggtgg tggtggttca ggcggcggcg gctcggcaga ggcatggtat 60
aatctcggga acgcatacta caaacagggc gattatcaga aggctatcga atactaccaa 120
aaagcactcg aactggaccc caataacgcg gaggcatggt acaatctcgg gaatgcgtac 180
tataagcagg gtgactacca gaaggccatt gaatactacc aaaaggcgct tgaactggac 240
ccgaacaacg ccgaggcctg gtataacctg gggaatgctt attacaagca gggggattat 300
caaaaggcaa tagaagacta ccagaaggcc ctcgaattgg accctaacaa tggtggtggt 360
ggttctggtg gtggtggttc aggcggcggc ggctcgggcg ga 402
<210> 41
<211> 39
<212> ДНК
<213> Искуственная последовательность
<220>
<223> праймер K842E смысловой
<400> 41
gtacataatc ctgtgtttga gccttttgaa aagccagtg 39
<210> 42
<211> 39
<212> ДНК
<213> Искуственная последовательность
<220>
<223> праймер K842E антисмысловой
<400> 42
cactggcttt tcaaaaggct caaacacagg attatgtac 39
<210> 43
<211> 36
<212> ДНК
<213> Искуственная последовательность
<220>
<223> праймер Q807K смысловой
<400> 43
ccactccttc cctgaaacag ctgaatctgc aactcc 36
<210> 44
<211> 36
<212> ДНК
<213> Искуственная последовательность
<220>
<223> праймер Q807K антисмысловой
<400> 44
ggagttgcag attcagctgt ttcagggaag gagtgg 36
<210> 45
<211> 44
<212> ДНК
<213> Искуственная последовательность
<220>
<223> праймер D864N смысловой
<400> 45
ctggaaatta agggaaataa tattgaccct gaagcagtta aagg 44
<210> 46
<211> 44
<212> ДНК
<213> Искуственная последовательность
<220>
<223> праймер D864N антисмысловой
<400> 46
cctttaactg cttcagggtc aatattattt cccttaattt ccag 44
<---
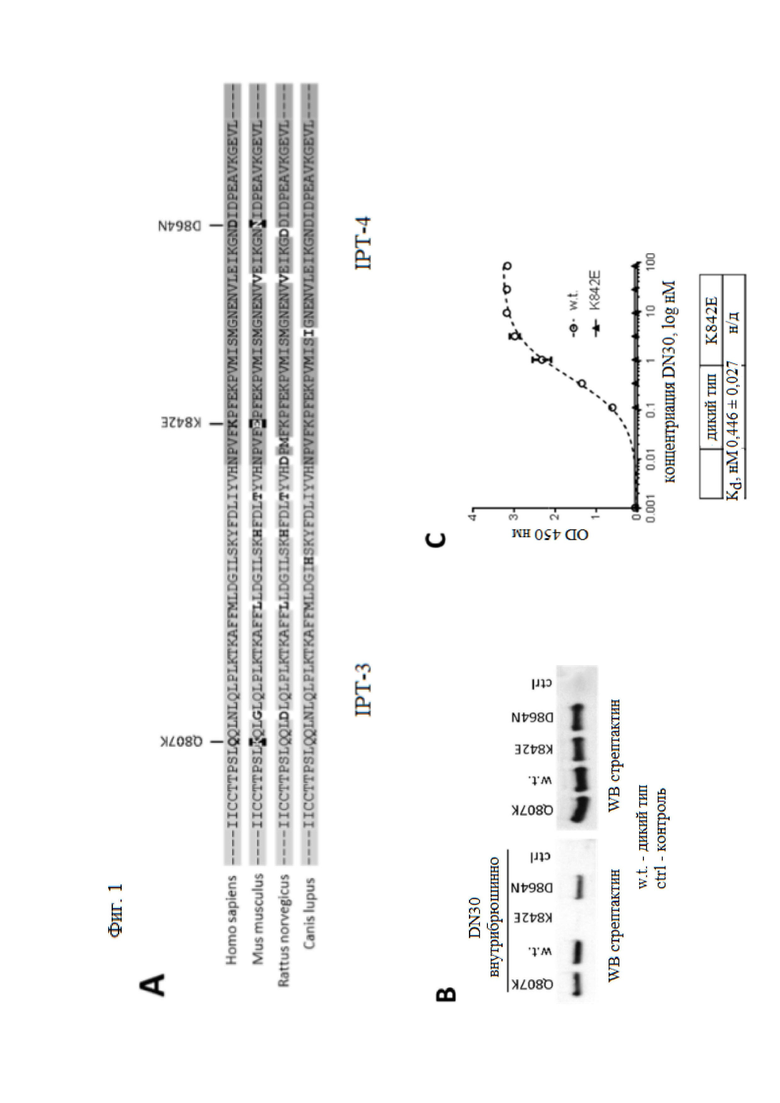
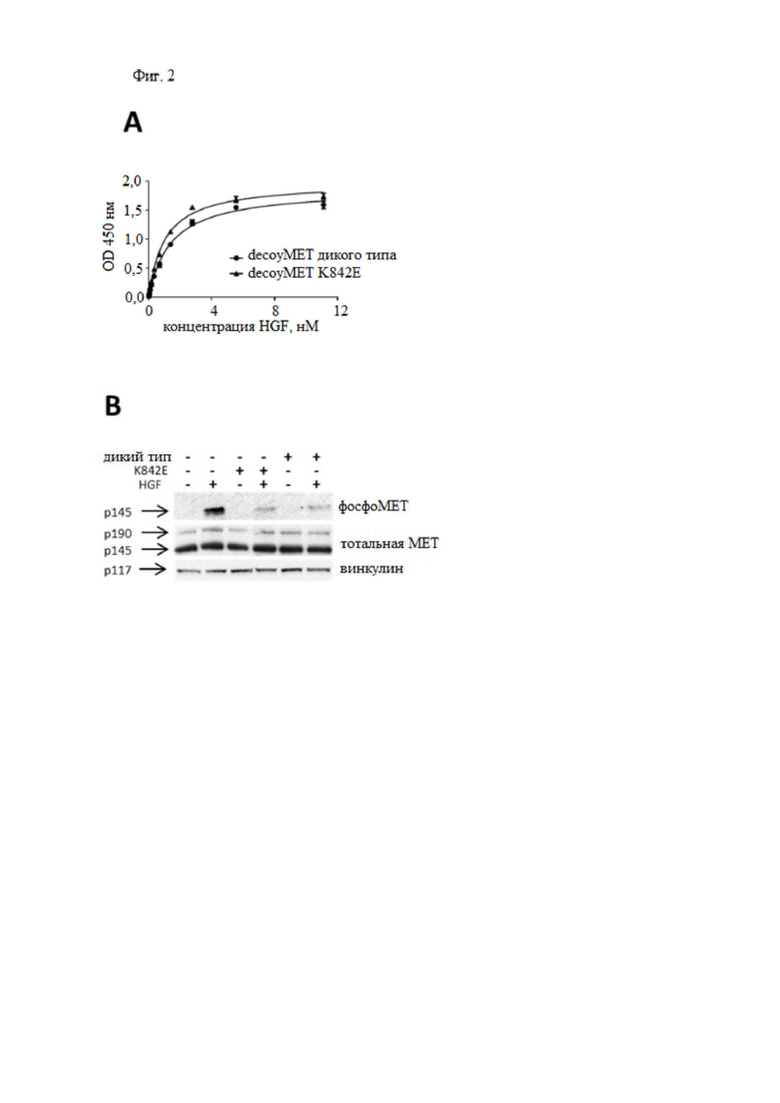
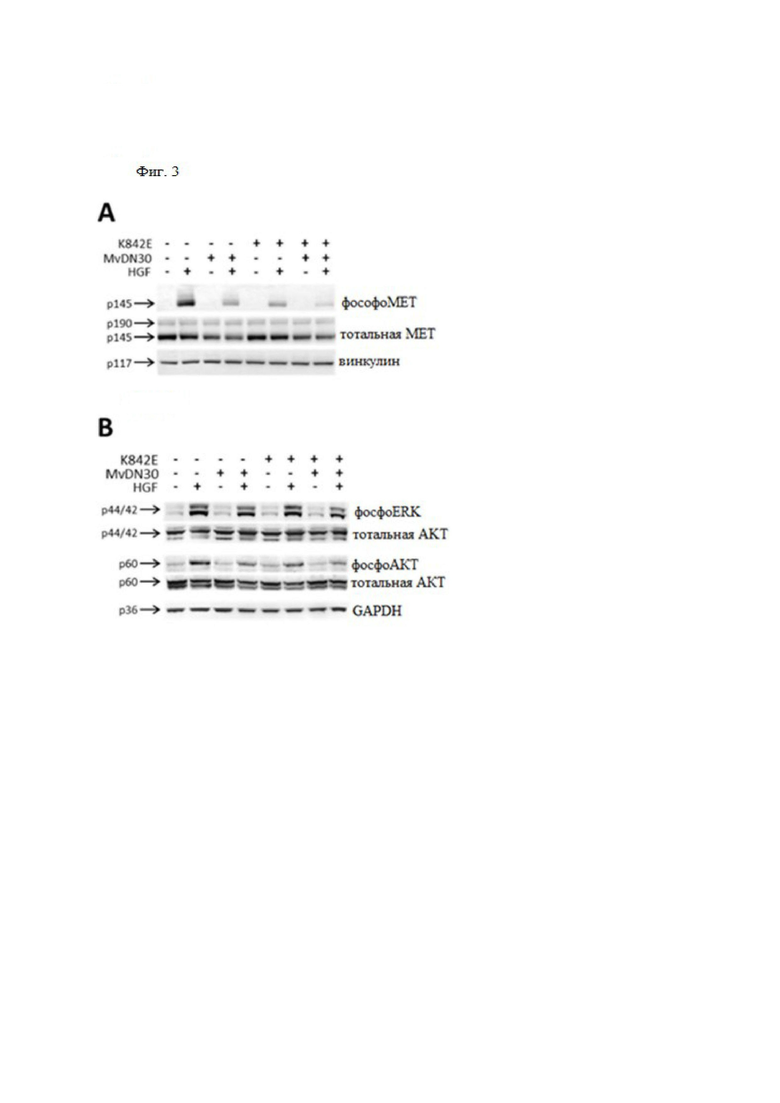
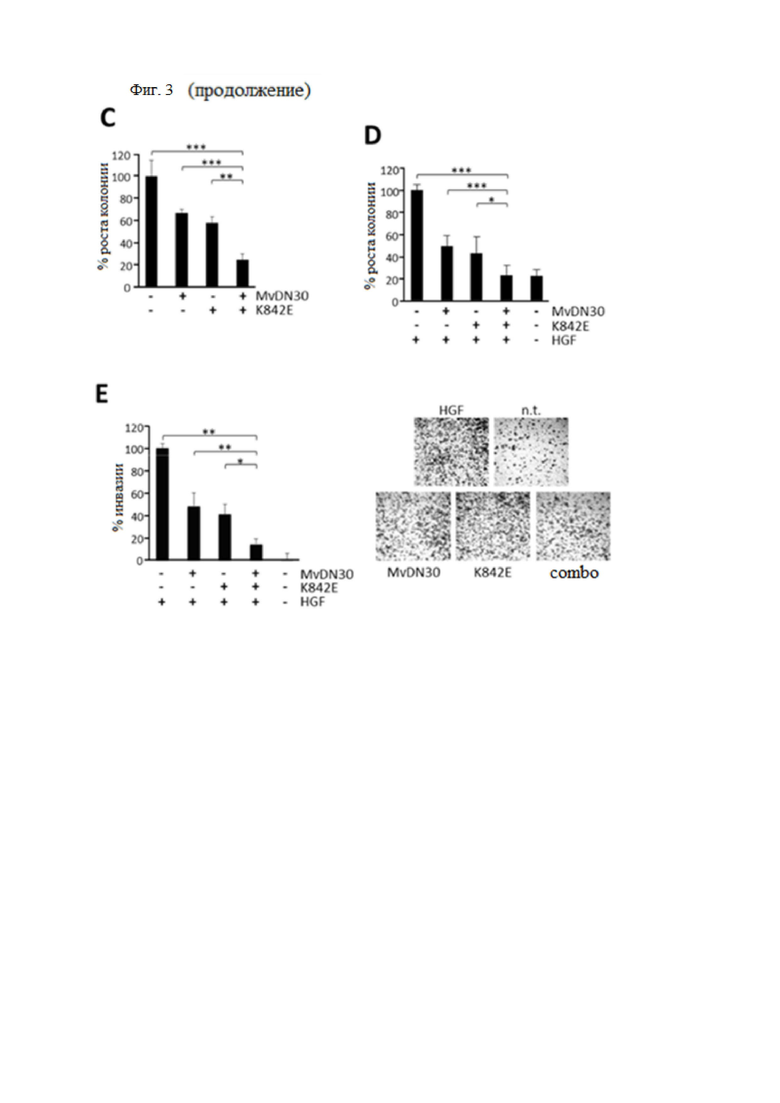
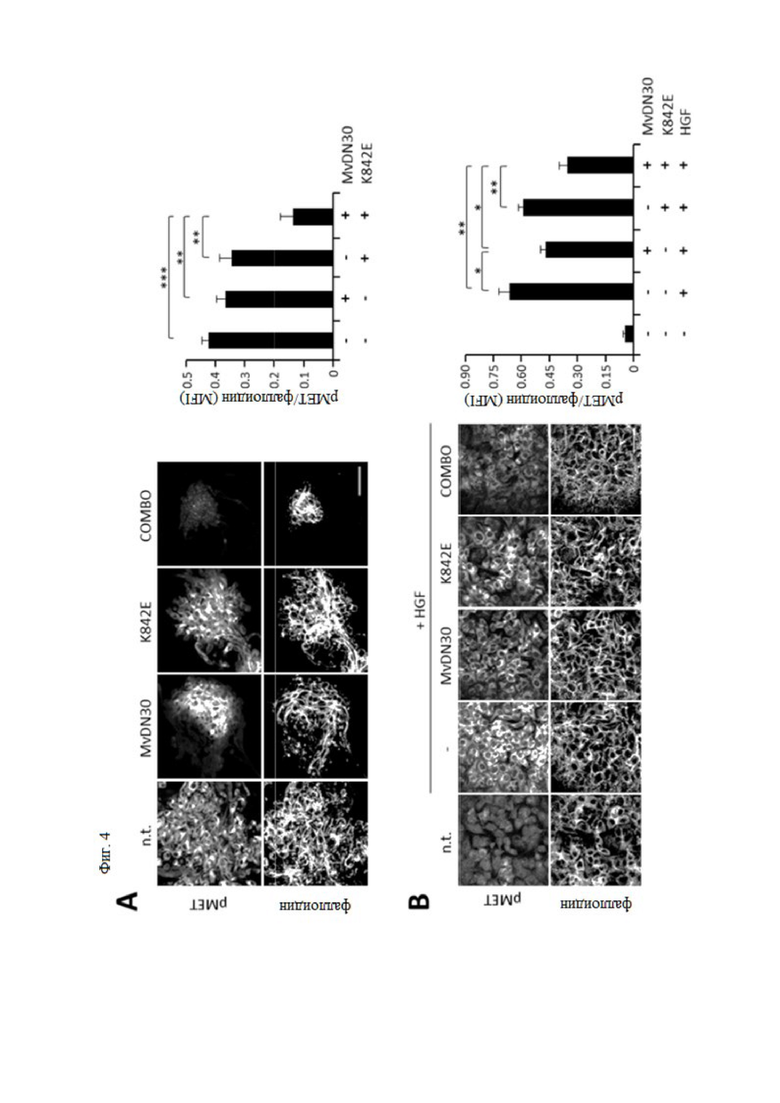
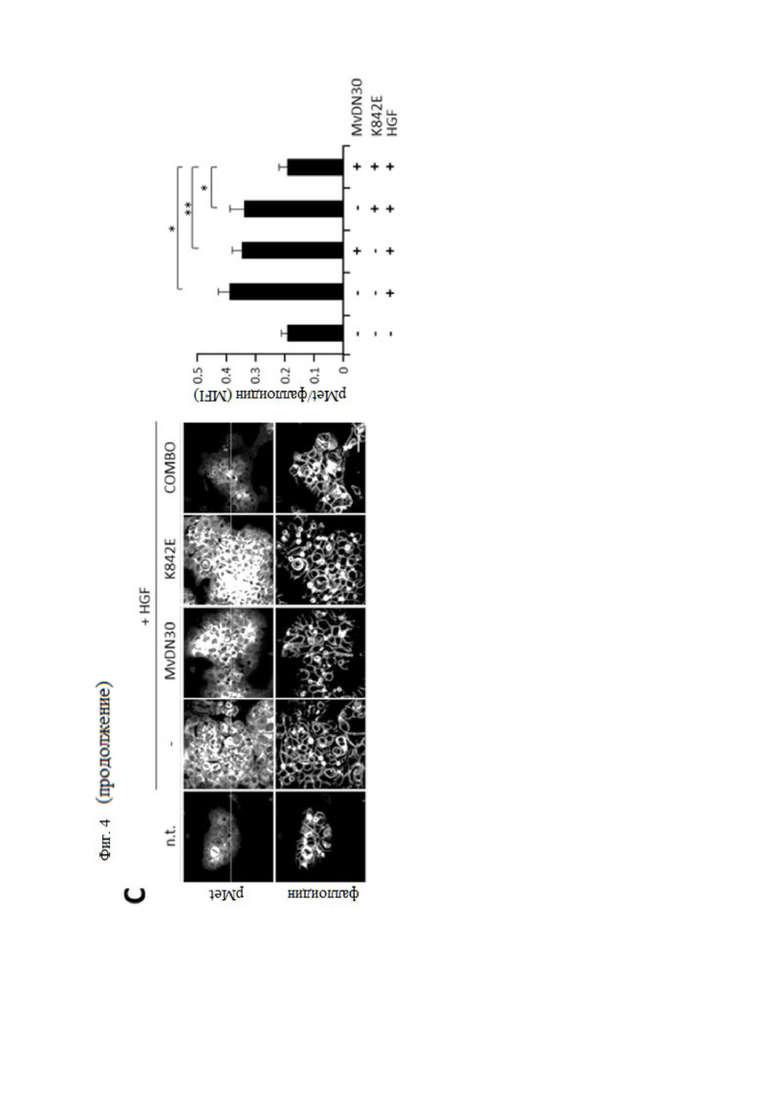
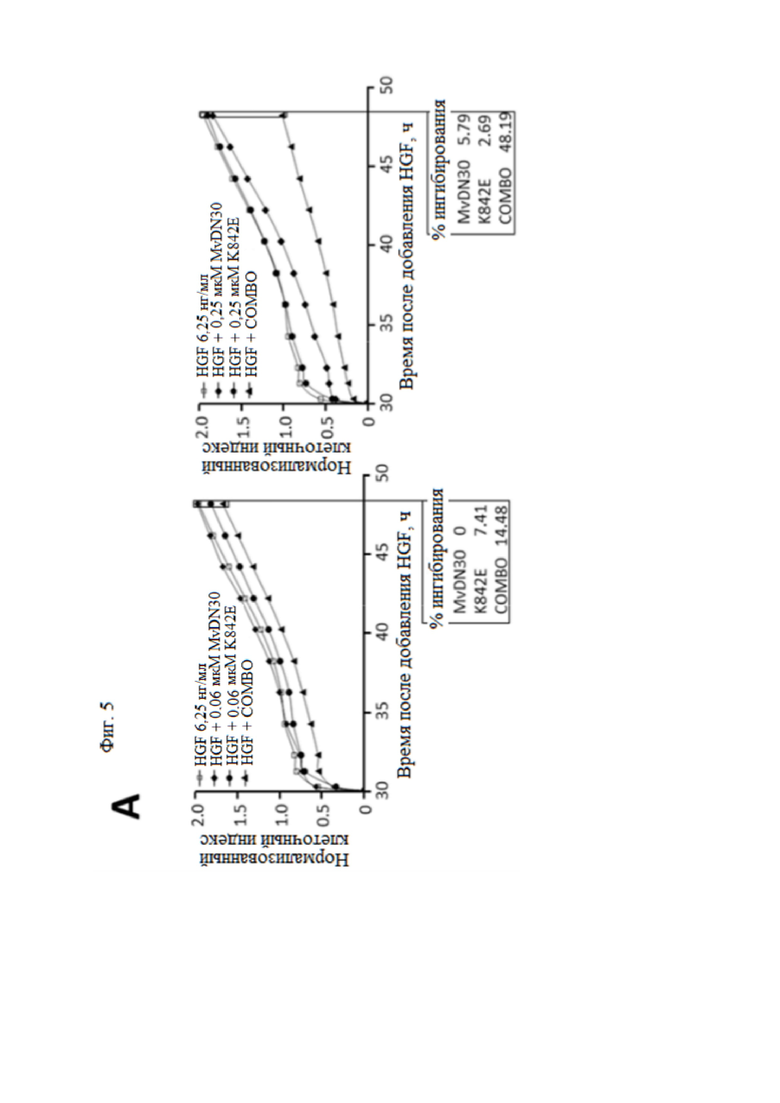
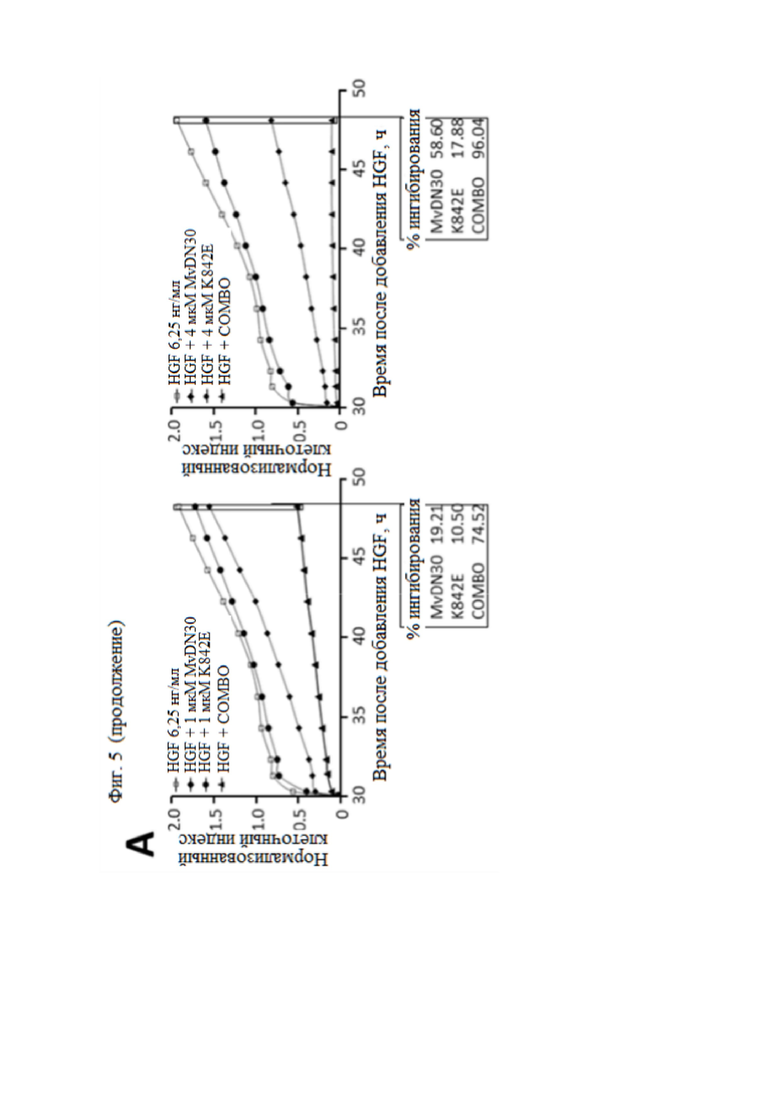
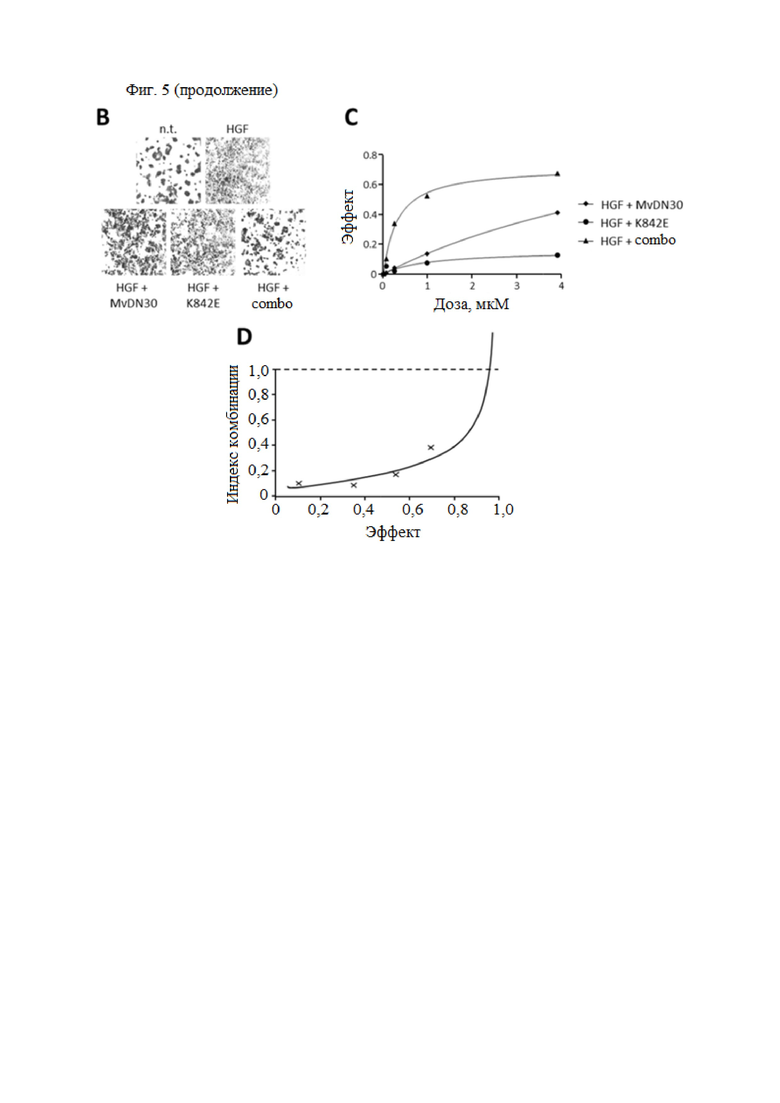
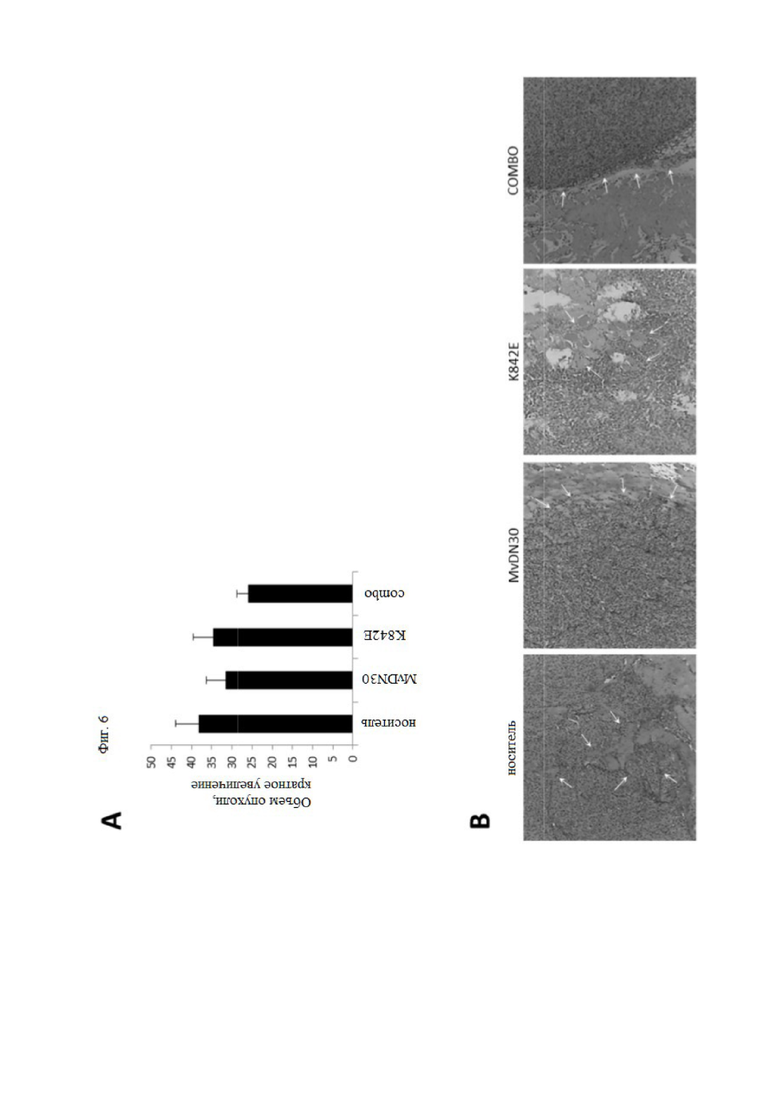
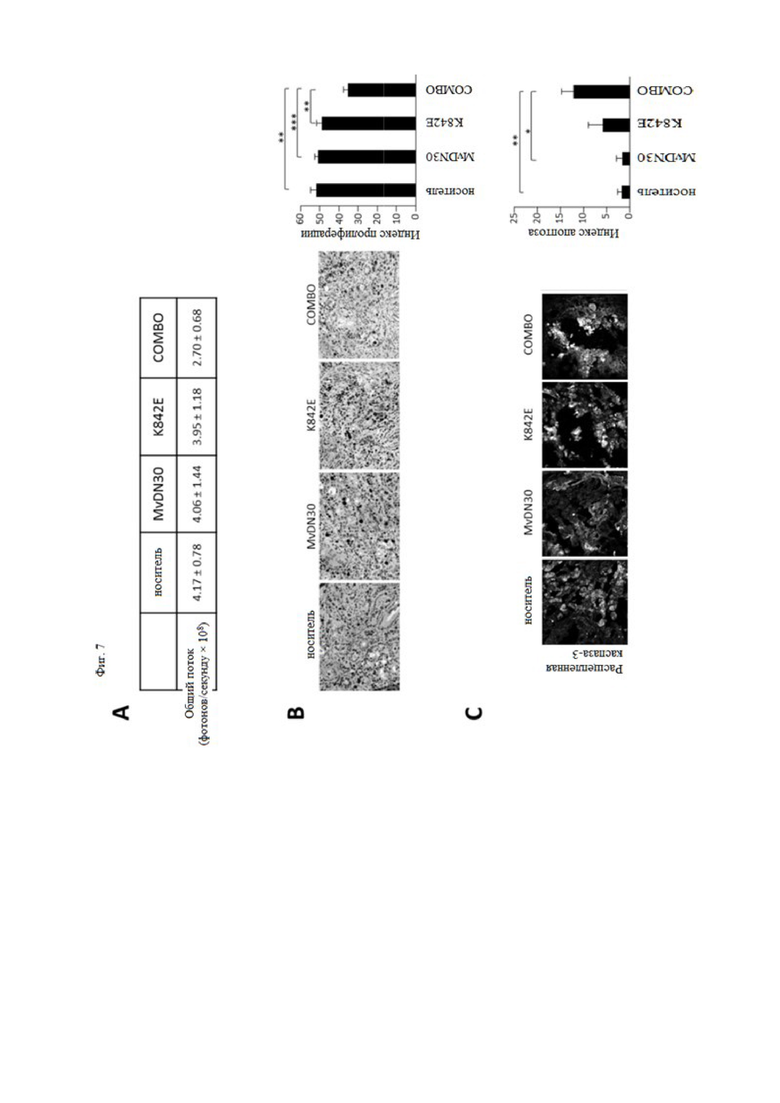
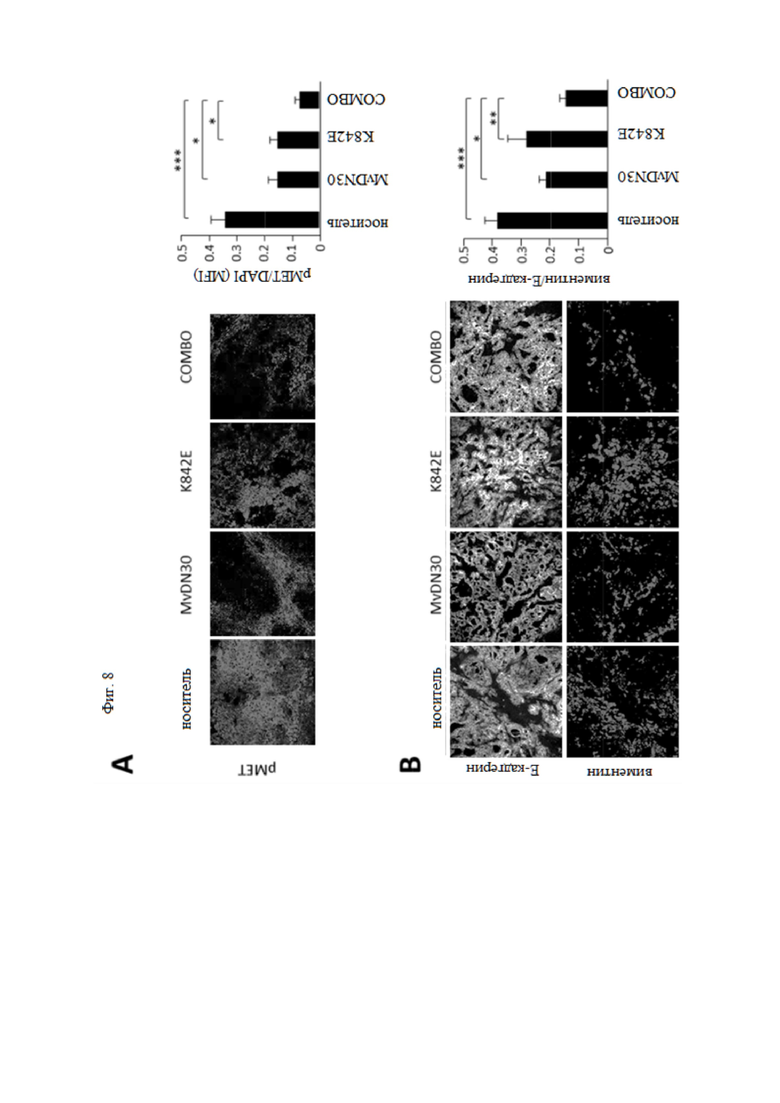
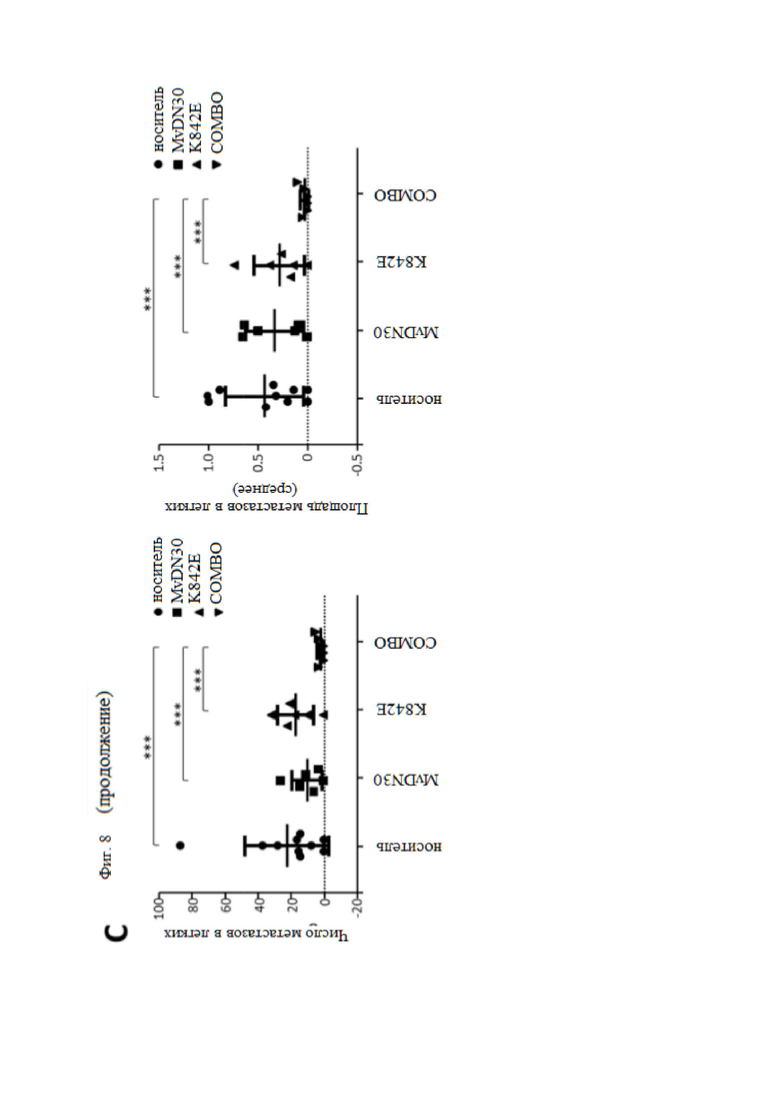
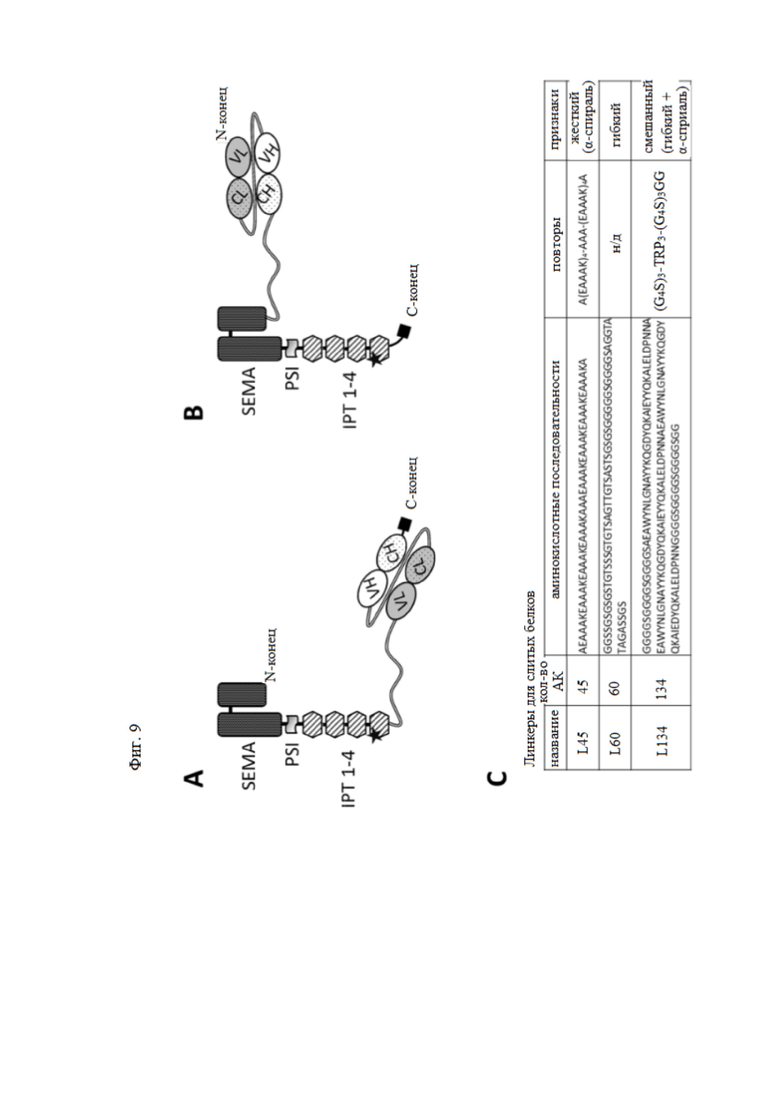
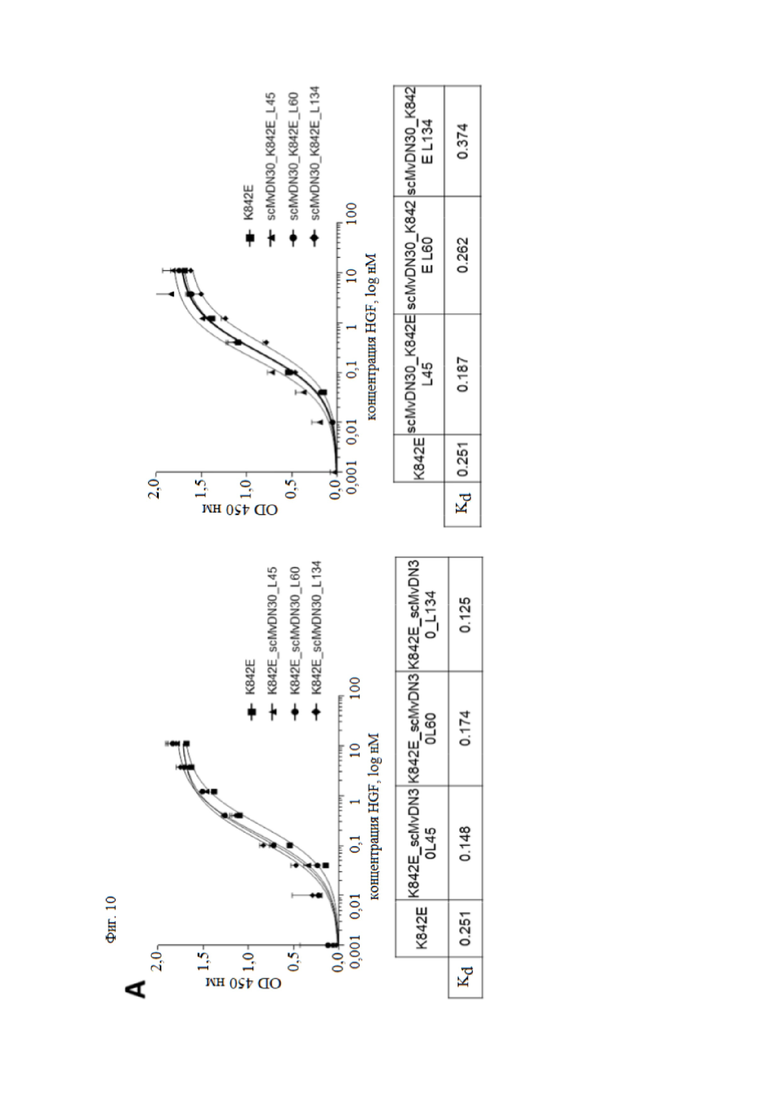
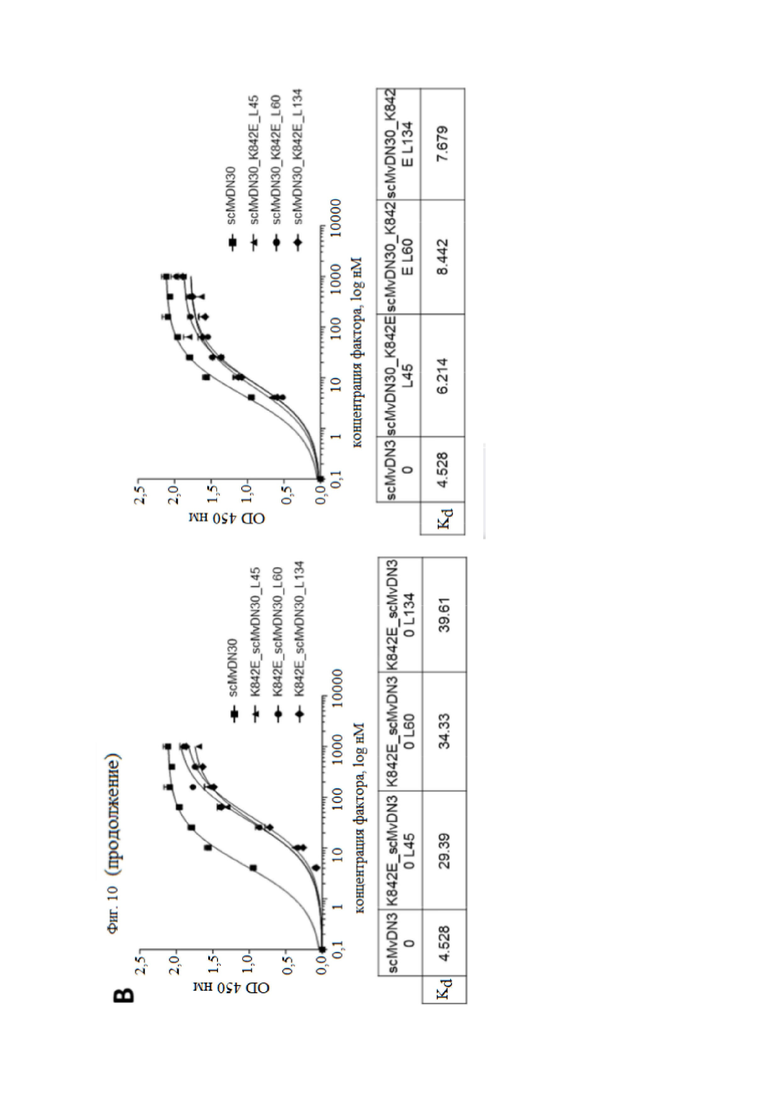
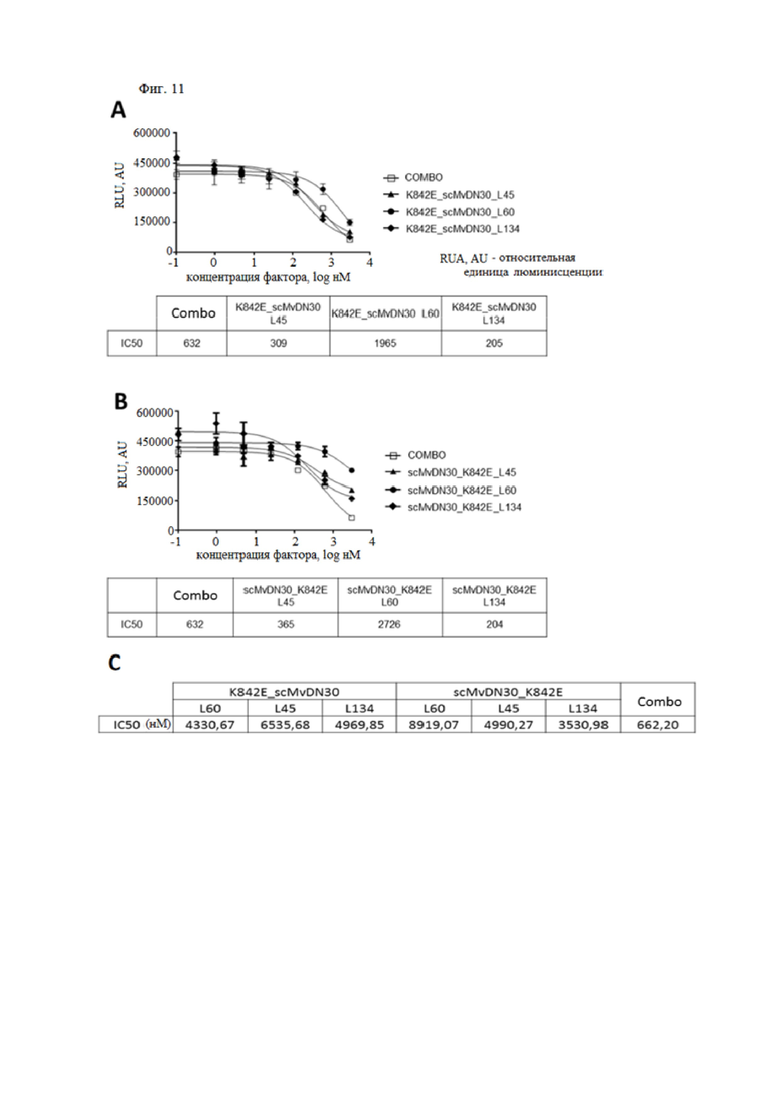
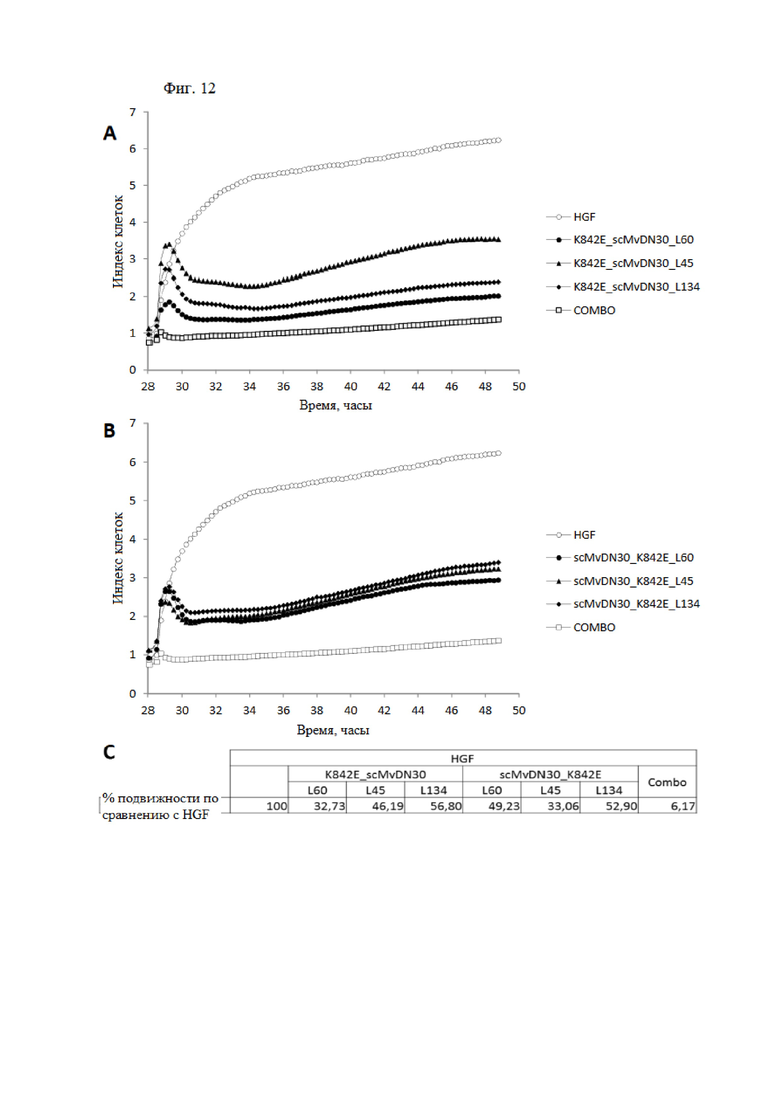
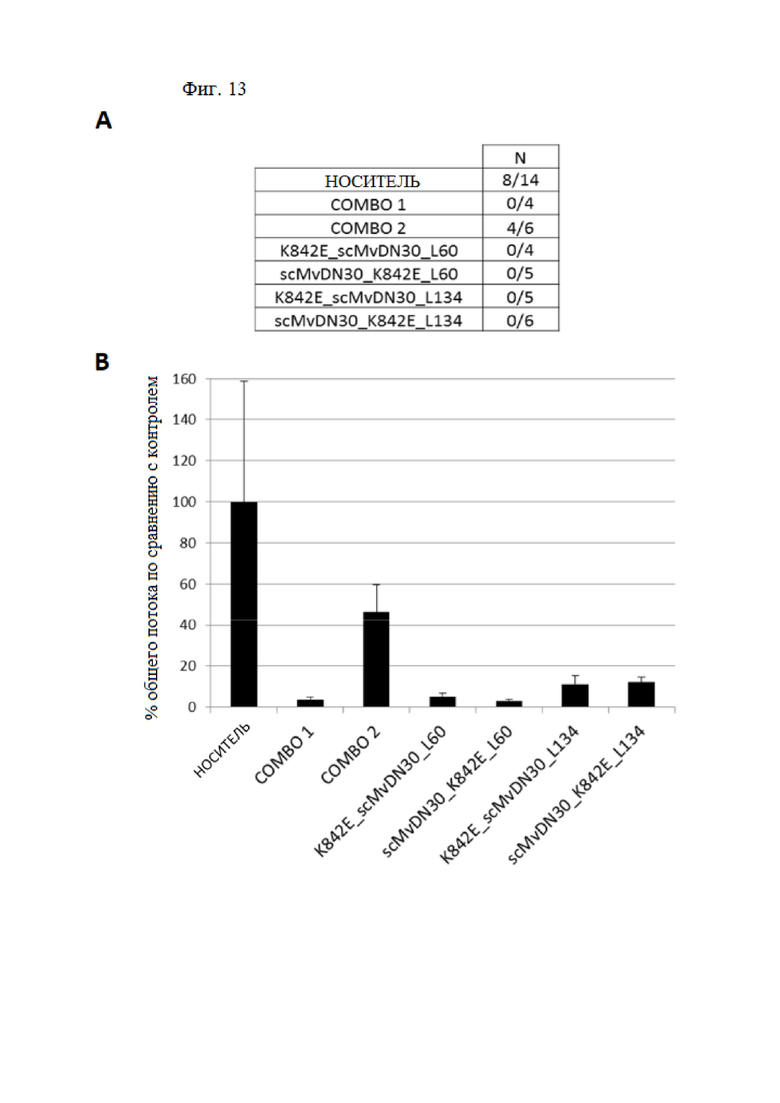
Группа изобретений относится к новой комбинации терапевтических агентов для лечения опухоли и/или метастаза. Предложено применение комбинации фрагмента антитела против рецептора фактора роста гепатоцитов (HGFR) и внеклеточной части HGFR человека в лечении пациента, страдающего от секретирующей HGF опухоли и/или метастаза, предпочтительно секретирующего HGF метастаза. Также предложены слитый белок, обладающий активностью одновременного антагонизма HGFR и связывания HGF, содержащий в направлении от N- к C-концу (i) фрагмент антитела против HGFR, линкер и внеклеточную часть HGFR человека; или (ii) внеклеточную часть HGFR человека, линкер и фрагмент антитела против HGFR, молекула нуклеиновой кислоты, кодирующая слитый белок, и фармацевтическая композиция, содержащая слитый белок и фармацевтически приемлемый носитель. Изобретения позволяют осуществлять двойное нацеливание на фактор роста гепатоцитов (HGF) и рецептор фактора роста гепатоцитов (HGFR), действуя одновременно на MET-экспрессирующие раковые клетки и на секретирующую HGF строму опухоли и оказывая синергетический эффект в лечении опухолей и/или метастаза. 4 н. и 10 з.п. ф-лы, 13 ил.
1. Применение комбинации фрагмента антитела против рецептора фактора роста гепатоцитов (HGFR) и внеклеточной части HGFR человека в лечении пациента, страдающего от секретирующей HGF опухоли и/или метастаза, предпочтительно секретирующего HGF метастаза, где:
(i) фрагмент антитела против HGFR имеет только один паратоп, способный связываться с эпитопом внеклеточной части HGFR человека, и обладает антагонистической активностью в отношении HGFR,
(ii) внеклеточная часть HGFR человека способна стабильно связываться с фактором роста гепатоцитов (HGF) и содержит по меньшей мере одну аминокислотную мутацию в эпитопе, распознаваемом фрагментом антитела против HGFR, для предотвращения связывания с ним фрагмента антитела против HGFR, и
(iii) фрагмент антитела против HGFR и внеклеточная часть HGFR человека подходят для введения пациенту в форме белка,
где фрагмент антитела против HGFR содержит один вариабельный домен легкой цепи (VL) и один вариабельный домен тяжелой цепи (VH), причем вариабельные домены легкой цепи и тяжелой цепи являются нечеловеческими или гуманизированными, и где вариабельный домен легкой цепи (VL) содержит области, определяющие комплементарность (CDR), имеющие аминокислотные последовательности, указанные в SEQ ID NO: 1-3, и вариабельный домен тяжелой цепи (VH) содержит CDR, имеющие аминокислотные последовательности, указанные в SEQ ID NO: 4-6,
где внеклеточная часть HGFR человека содержит последовательность, указанную в SEQ ID NO: 19, где по меньшей мере одна из аминокислот между положениями 797 и 875 последовательности SEQ ID NO: 19 мутирована для предотвращения связывания с ней фрагмента антитела против HGFR, и где:
(a) фрагмент антитела против HGFR конъюгирован с помощью линкера с внеклеточной частью HGFR человека в направлении от N- к C-концу, таким образом образуя слитый белок из фрагмента антитела против HGFR - линкера - внеклеточной части HGFR человека; или
(b) внеклеточная часть HGFR человека конъюгирована с помощью линкера с фрагментом антитела против HGFR в направлении от N- к C-концу, таким образом образуя слитый белок из внеклеточной части HGFR человека - линкера - фрагмента антитела против HGFR,
где пациент несет онкоген МЕТ дикого типа.
2. Применение по п. 1, где фрагмент антитела против HGFR дополнительно содержит один константный домен легкой цепи человека (CL) и один константный домен тяжелой цепи CH1 человека, причем вариабельный домен легкой цепи слит с константным доменом легкой цепи человека в направлении от N- к C-концу и вариабельный домен тяжелой цепи слит с константным доменом тяжелой цепи CH1 человека в направлении от N- к C-концу.
3. Применение по п. 2, где фрагмент антитела против HGFR представляет собой одноцепочечный Fab-фрагмент.
4. Применение по любому из пп. 1-3, где внеклеточная часть HGFR человека содержит домены SEMA, PSI, IPT-1, IPT-2, IPT-3 и IPT-4.
5. Применение по любому из пп. 1-4, где внеклеточная часть HGFR человека имеет последовательность, указанную в SEQ ID NO: 20.
6. Применение по п. 1 или 5, где:
(i) слитый белок из фрагмента антитела против HGFR - линкера - внеклеточной части HGFR человека содержит аминокислотную последовательность, выбранную из SEQ ID NO: 26, 27 и 28; или
(ii) слитый белок из внеклеточной части HGFR человека - линкера - фрагмента антитела против HGFR содержит аминокислотную последовательность, выбранную из SEQ ID NO: 23, 24 и 25.
7. Слитый белок, обладающий активностью одновременного антагонизма HGFR и связывания HGF, содержащий в направлении от N- к C-концу:
(i) фрагмент антитела против HGFR, линкер и внеклеточную часть HGFR человека; или
(ii) внеклеточную часть HGFR человека, линкер и фрагмент антитела против HGFR,
где фрагмент антитела против HGFR имеет только один паратоп, способный связываться с эпитопом внеклеточной части HGFR человека, и обладает антагонистической активностью в отношении HGFR,
где внеклеточная часть HGFR человека способна стабильно связываться с фактором роста гепатоцитов (HGF) и содержит по меньшей мере одну аминокислотную мутацию в эпитопе, распознаваемом фрагментом антитела против HGFR, для предотвращения связывания с ним фрагмента антитела против HGFR,
где фрагмент антитела против HGFR содержит один вариабельный домен легкой цепи (VL) и один вариабельный домен тяжелой цепи (VH), причем вариабельные домены легкой цепи и тяжелой цепи являются нечеловеческими или гуманизированными, и где вариабельный домен легкой цепи (VL) содержит области, определяющие комплементарность (CDR), имеющие аминокислотные последовательности, указанные в SEQ ID NO: 1-3, и вариабельный домен тяжелой цепи (VH) содержит CDR, имеющие аминокислотные последовательности, указанные в SEQ ID NO: 4-6, и
где внеклеточная часть HGFR человека содержит последовательность, указанную в SEQ ID NO: 19, где по меньшей мере одна из аминокислот между положениями 797 и 875 последовательности SEQ ID NO: 19 мутирована для предотвращения связывания с ней фрагмента антитела против HGFR.
8. Слитый белок по п. 7, где фрагмент антитела против HGFR дополнительно содержит один константный домен легкой цепи человека (CL) и один константный домен тяжелой цепи CH1 человека, причем вариабельный домен легкой цепи слит с константным доменом легкой цепи человека в направлении от N- к C-концу и вариабельный домен тяжелой цепи слит с константным доменом тяжелой цепи CH1 человека в направлении от N- к C-концу.
9. Слитый белок по п. 7 или 8, где фрагмент антитела против HGFR представляет собой одноцепочечный Fab-фрагмент.
10. Слитый белок по любому из пп. 7-9, где внеклеточная часть HGFR человека содержит домены SEMA, PSI, IPT-1, IPT-2, IPT-3 и IPT-4.
11. Слитый белок по любому из пп. 7-10, где внеклеточная часть HGFR человека имеет последовательность, указанную в SEQ ID NO: 20.
12. Слитый белок по любому из пп. 7-11, который содержит аминокислотную последовательность, выбранную из любой из SEQ ID NO: 23–28.
13. Молекула нуклеиновой кислоты, кодирующая слитый белок по любому из пп. 7-12.
14. Фармацевтическая композиция, обладающая активностью одновременного антагонизма HGFR и связывания HGF, содержащая слитый белок по любому из пп. 7-12 и фармацевтически приемлемый носитель.
| PETRELLI A | |||
| ET AL | |||
| Ab-induced ectodomain shedding mediates hepatocytes growth factor receptor down-regulation and hampers biological activity, PNAS, 2006, v.103, no.13, p.5090-5095, DOI: 10.1073/PNAS.0508156103 | |||
| MICHIELI P | |||
| ET AL | |||
| Targeting the tumor and its microenvironment by a dual-function decoy Met receptor, CANCER CELL, 2004, v.6, no.1, |

