ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка истребует преимущества патентной заявки Китая 202010351280.6, поданной 28 апреля 2021, содержание которой целиком включено в настоящий документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области противоопухолевых иммунотерапевтических агентов. В частности, настоящее изобретение относится к молекуле TGFβR2 (фактор иммуномодуляции) с укороченным внеклеточным доменом, слитому белку, содержащему молекулу TGFβR2 с укороченным внеклеточным доменом и нацеливающую часть. Настоящее изобретение включает фармацевтические композиции и их применение в качестве противоопухолевых лекарственных средств.
УРОВЕНЬ ТЕХНИКИ
Трансформирующий фактор роста-β (TGF-β) принадлежит к надсемейству TGF-β, которое регулирует рост и дифференцировку клеток, и представляет собой плейотропный и многофункциональный цитокин, который регулирует пролиферацию, дифференцировку и апоптоз клеток через сигнальные пути рецепторов клеточной поверхности аутокринным или паракринным образом. Он также играет важную регуляторную роль в синтезе внеклеточного матрикса, восстановлении травм, иммунной функции и т.д. Три изоформы, TGF-β1, TGF-β2 и TGF-β3, присутствуют у млекопитающих, и наиболее распространенной и экспрессируемой изоформой является TGF-β1. TGF-β инициирует классические пути Smad и не-Smad для передачи сигналов вниз по цепочке путем связывания с рецепторами серин-треонинкиназы TGFβR1 и ΤGFβR2 на поверхности клеточной мембраны[1]. При нормальном гомеостазе in vivo TGF-β регулирует передачу сигналов ключевых процессов, такие как рост, регенерация, дифференцировка и т.д. различных тканей. В иммунной системе TGF-β вызывает толерантность и подавляет воспаление. Генетические мутации могут изменить передачу сигналов TGF-β в клетках, инициирующих опухоль. На начальных стадиях онкогенеза TGF-β играет ключевую роль в ингибировании рака, ингибируя пролиферацию клеток и запуская программу апоптоза. Однако по мере прогрессирования опухоли селективное давление опухолевых клеток приводит к утрате у TGF-β функции подавления опухоли посредством различных механизмов. Приобретенные инактивирующие мутации в сигнальном пути TGF-β позволяют множеству злокачественных клеток расти в среде, обогащенной TGF-β. Дополнительно опухолевые клетки каким-то образом преобразуют проапоптотическую способность TGF-β в проопухолевые функции развития, такие как способность к инвазии и миграции, а также содействие мезенхимальному переходу[2-4]. TGF-β регулирует различные типы и функции иммунных клеток. TGF-β контролирует адаптивный иммунитет, напрямую стимулируя пролиферацию Treg-клеток, тем самым подавляя продукцию и функцию эффекторных Т-клеток и антигенпрезентирующих дендритных клеток. TGF-β также ингибирует функцию NK-клеток и превращает макрофаги и нейтрофилы в подтипы проопухолевого роста, способствуя формированию опухолевого микроокружения с негативной иммунной регуляцией опухоли[4]. TGF-β1 часто экспрессируется с более высокими уровнями, чем в нормальной паранеопластической ткани, в различных солидных опухолях, включая EGFR-положительный рак толстой и прямой кишки, немелкоклеточный рак легких, плоскоклеточную карциному головы и шеи. Клинические данные свидетельствуют о том, что блокада пути TGF-β сама по себе недостаточна для полного восстановления иммунной системы для подавления онкогенеза, и поэтому антитела к TGF-β еще не получены.
Рецептор эпидермального фактора роста (EGFR) представляет собой продукт экспрессии протоонкогена C-ErbB1, и он является рецептором клеточной пролиферации и передачи сигнала эпителиального фактора роста (EGF), членом семейства рецепторов эпидермального фактора роста (HER), и рецептором тирозинкиназы с молекулярной массой 170 кДа[5]. EGFR состоит из внеклеточного лиганд-связывающего участка, трансмембранного участка и внутриклеточного киназного участка. Внеклеточный домен EGFR трансформируется из мономера в димер после связывания с лигандом, активирует внутриклеточный киназный участок и несколько нижестоящих сигнальных путей и играет важную роль в росте, пролиферации и дифференцировке клеток[6]. Высокая экспрессия EGFR вызывает усиленную передачу сигналов вниз по цепочке повышенная экспрессия мутантных рецепторов или лигандов EGFR приводит к устойчивой активации EGFR, повышенной активности секреторной петли и нарушению механизма подавления рецептора и т.д., что, в свою очередь, активирует гены, связанные с пролиферацией и дифференцировкой опухоли, и играет важную роль в образовании и развитии опухоли[7]. Повышенная экспрессия EGFR связана со снижением выживаемости при некоторых типах рака, включая рак головы и шеи, мочевого пузыря, яичников, шейки матки и рак пищевода. Дополнительно было показано, что препараты против EGFR эффективны при лечении нескольких типов солидных опухолей, таких как колоректальные опухоли, опухоли головы и шеи, немелкоклеточный рак легкого (НМРЛ) и рак поджелудочной железы, включая общую выживаемость, выживаемость без прогрессирования, и общие показатели ответов.[8] Таким образом, в качестве четкой мишени, связанной с пролиферацией опухоли, агенты, нацеленные на EGFR, стали вариантом лечения первой линии для различных злокачественных новообразований.
Хотя сигналы, инициируемые TGF-β, отличаются от сигналов, инициируемых путем EGF/EGFR, пути передачи сигналов между ними могут взаимодействовать. Было обнаружено, что пути передачи сигналов TGF-β и EGFR взаимодействуют между собой и совместно способствуют прогрессированию опухолей при различных типов. TGF-β может индуцировать транс-активацию EGFR в зависимости от типа клеток и контекста. Например, TGF-β способствует миграции и инвазии клеток рака молочной железы (MDA-МВ-231, T47D, 4Т1) путем активизации EGFR через классические сигнальные пути Smad и ERK/Sp1[9,10]. В плоскоклеточной карциноме (А431, SCC13), TGF-β активирует путь EGFR через H2O2-зависимые механизмы для повышения уровня фосфорилирования Erk1/2[11]. EGF и связанные с ним нижестоящие сигнальные пути также могут регулировать передачу сигналов TGF-β в различных типах клеток. Например, в клетках первичного рака яичников человека EGF снижает чувствительность клеток рака яичников к антипролиферативному действию TGF-β за счет уменьшения экспрессии мРНК TCF-β-индуцируемого регулятора клеточного цикла p15INK4B[12].Онкогенный Ras в эпителиальных клетках молочной железы и легких снижает эффект TGF-β по ингибированию роста клеток за счет ингибирования TGF-β-опосредованной передачи сигналов посредством негативной регуляции Smad2 и Smad3[13,14]. EGF также положительно регулирует передачу сигналов Smad2 в клетках COS7, увеличивая фосфорилирование Smad2 через путь ERK[15].
TGF-β и EGF синергически способствуют злокачественному фенотипу опухолей, и исследования на различных тканях показали, что EGF в сочетании с TGF-β усиливает эпителиально-мезенхимальный переход (ЭМП). Например, EGF и TGF-β1 способствуют экспрессии ламинина-332 (Laminin-332), что синергетически способствует ЭМП при раке эпителия полости рта[16]. EGF и TGF-β1 способствуют ЭМП в эпителиальных клетках кишечника путем подавления Е-кадгерина по пути MAPK, а не по пути PI3K, p38MAPK, JNK или AP-1[17]. EGF и TGF-β1 индуцируют экспрессию Slug и Snail через Smad и MEK1/2-зависимые сигнальные пути, подавляют Е-кадгерин и способствуют ЭМП в эпителиальных клетках яичников[18]. EGF и TGF-β1 активируют сигнальный путь ERK1/2, синергически усиливают экспрессию белка Snail и способствуют ЭМП и миграции в эпителиальных клетках проксимальных канальцев коры почек человека (HK-2)[19]. EGF усиливает TGF-β-индуцированный ЭМП в клетках рака легких (Н322, Н358) и рака поджелудочной железы (HPAF-II, CAPAN-2), способствуя связыванию SHP2 с GAB1[20].
Несколько клинических исследований показали, что повышенные уровни TGF-β тесно связаны с развитием лекарственной устойчивости и неблагоприятным прогнозом. ЭМП, индуцированный TGF-β1, в раковых стволовых клетках остеосаркомы снижает экспрессию miR-499a, что приводит к увеличению экспрессии SHKBP1, происходящему одновременно с TGFβ-индуцированным переключением киназы, связанной с ЕМТ, в АКТ-активируемое EGFR-независимое состояние, тем самым снижая активность EGFR и индуцируя резистентность остеосаркомы к ингибиторам киназы EGFR (Фиг. 5)[21]. Клетки Treg являются одними из основных клеток, продуцирующих TGF-β. Повышенное количество Treg в опухолях больных плоскоклеточным раком головы и шеи, получавших цетуксимаб, сопровождается повышением содержания TGF-β, в то время как содержание TGF-β у больных с низкой эффективностью цетуксимаба даже выше[22]. Повышенный уровень TGF-β может индуцировать резистентность к терапии антителами против EGFR путем ингибирования экспрессии соответствующих молекулярных эффекторов эффекторно-клеточной цитотоксичности, активации EGFR-независимого пути АКТ и усиления индукции ЭМП[23]. В мутантном EGFR немелкоклеточного рака легких классический сигнальный путь TGF-βSmad участвует в развитии PD-L1-индуцированной резистентности опухоли к ингибиторам киназы EGFR[24]. В тканях рака молочной железы экспрессия TGF-β положительно коррелировала с экспрессией EGFR, а повышенные уровни TGF-β и EGFR были связаны с плохим прогнозом у пациентов с раком молочной железы[9]. Таким образом, EGFR и TGF-β играют относительно независимые, но близкородственные роли в указанном пути развития опухоли. Дополнительно TGF-β представляет собой ключевую молекулу в развитии приобретенной резистентности опухолей к терапии, направленной на EGFR. Исследования на животных показали, что ингибирование TGF-β улучшает противоопухолевый эффект цетуксимаба in vivo на ксенотрансплантаты плоскоклеточных опухолей головы и шеи[23]. Эти данные обеспечивают теоретическую основу для комбинированного нацеливания TGF-β для повышения терапевтической эффективности антитела против EGFR в отношении EGFR-положительных опухолей.
Настоящее изобретение обеспечивает новые слитые белки, содержащие укороченный TGFβR2, которые могут специфически нацеливаться как на EGFR, так и на TGF-β, два относительно независимых, но тесно связанных сигнальных пути. Они используются для лечения солидных опухолей, включая рак желудка, но не ограничиваясь им.
КРАТКОЕ ОПИСАНИЕ
В одном аспекте настоящее изобретение обеспечивает молекулу TGFβR2 с укороченным внеклеточным доменом, в которой, по сравнению с ее природной формой,
a) по меньшей мере остатки аминокислот в положениях 6-16 удалены, и дополнительно, необязательно, остатки аминокислот в положениях 17-17+n удалены, где n представляет собой целое число от 1 до 10; предпочтительно, n равно 2, 4, 8, 9 или 10; более предпочтительно, n равно 9; или
b) на базе делеции остатков аминокислот в положениях 6-26, в дополнение, остатки аминокислот в положениях 5, 4-5, 3-5, 2-5, 1, 1-2, 1-3, или 1-4 удалены; или
c) остатки аминокислот в положениях 7-26 удалены.
В одном варианте реализации указанная последовательность аминокислот включает любую из последовательностей SEQ ID NO: 48-62.
В другом аспекте настоящее изобретение обеспечивает слитой белок, содержащий молекулы, описанные выше.
В одном варианте реализации указанный слитой белок содержит
a) указанную молекулу TGFβR2 с укороченным внеклеточным доменом и
b) нацеливающую часть.
В одном варианте реализации указанного слитого белка нацеливающая часть представляет собой специфическую к раковым клеткам нацеливающую часть, выбранную из антитела или его антиген-связывающего фрагмента, функционального лиганда или его слитого белка с Fc, и рецепторного белка или его слитого белка с Fc.
В одном варианте реализации указанного слитого белка нацеливающая часть представляет собой антитело против EGFR или его антиген-связывающий фрагмент.
В одном варианте реализации N-конец указанной молекулы TGFβR2 с укороченным внеклеточным доменом в указанном слитом белке соединен с С-концом указанной тяжелой цепи нацеливающей части, необязательно, посредством линкера.
В одном варианте реализации указанный линкер представляет собой предпочтительно G4S гибкий пептидный линкер, предпочтительно (G4S)4 пептидный линкер.
В другом аспекте настоящее изобретение обеспечивает выделенное связывающее антитело или его антиген-связывающий фрагмент против EGFR, содержащее
(а) вариабельный участок тяжелой цепи, содержащий домены CDR1 тяжелой цепи, CDR2 тяжелой цепи и CDR3 тяжелой цепи, содержащие SEQ ID NO: 19, 20, и 21, соответственно, и/или
(b) а вариабельный участок легкой цепи, содержащий а домены CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, содержащему SEQ ID NO: 16, 17, и 18, соответственно.
В одном варианте реализации указанное антитело или его антиген-связывающий фрагмент содержит
а) вариабельный участок тяжелой цепи, содержащий последовательность, содержащую SEQ ID NO: 28 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью; и/или
(b) вариабельный участок легкой цепи, содержащий последовательность, содержащую SEQ ID NO: 29 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью.
В одном варианте реализации указанное антитело дополнительно содержит:
a) константный участок тяжелой цепи, содержащий последовательность, содержащую SEQ ID NO: 30 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью; и/или
b) константный участок легкой цепи, содержащий последовательность, содержащую SEQ ID NO: 31 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью.
В одном варианте реализации нацеливающая часть указанного слитого белка выбрана из антитела против EGFR, трастузумаба, бевацизумаба, рамуцирумаба, ипилимумаба или панитумумаба.
В одном варианте реализации указанный слитой белок содержит
a) последовательность аминокислот тяжелой цепи, содержащую SEQ ID NO: 141 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью; и
b) последовательность аминокислот легкой цепи, содержащую SEQ ID NO: 23 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью,
которые содержат две тяжелых цепи и две легких цепи; дисульфидная связь образована между его первой легкой цепью и первой тяжелой цепью, дисульфидная связь образована между его второй легкой цепью и второй тяжелой цепью, и дисульфидная связь образована между его первой тяжелой цепью и второй тяжелой цепью.
В одном варианте реализации указанный слито белок имеет значение KD в диапазоне 2,92 пМ - 26,3 пМ, предпочтительно 7 пМ - 9 пМ, более предпочтительно 8,77 пМ, по аффинности связывания с белком EGFR человека; и
имеет значение KD в диапазоне 23 пМ - 288,3 пМ, предпочтительно 64 пМ - 144 пМ, более предпочтительно 96,1 пМ по аффинности связывания с белком TGF-β1 человека.
В другом аспекте настоящее изобретение обеспечивает конъюгат, содержащий молекулу TGFβR2 с укороченным внеклеточным доменом, описанную в настоящем документе, слитой белок, описанный в настоящем документе, антитело или его антиген-связывающий фрагмент, описанный в настоящем документе, и дополнительный терапевтический агент, при этом предпочтительно указанное антитело или его антиген-связывающий фрагмент и указанный дополнительный терапевтический агент соединены посредством линкера.
В другом аспекте настоящее изобретение обеспечивает нуклеиновую кислоту, кодирующую указанную молекулу TGFβR2 с укороченным внеклеточным доменом, описанную в настоящем документе, слитой белок, описанный в настоящем документе, антитело или его антиген-связывающий фрагмент, описанный в настоящем документе, которая представляет собой мРНК и/или ДНК.
В одном варианте реализации указанная нуклеиновая кислота содержит
любую из последовательностей SEQ ID NO: 32-39;
любую из последовательностей SEQ ID NO: 67-84; или
любую из последовательностей SEQ ID NO: 148-163, или их функциональные варианты.
В другом аспекте настоящее изобретение обеспечивает вектор экспрессии, содержащий указанную нуклеиновую кислоту, описанную в настоящем документе.
В другом аспекте настоящее изобретение обеспечивает клетку-хозяина, содержащую указанную нуклеиновую кислоту, описанную в настоящем документе или указанный вектор экспрессии, описанный в настоящем документе.
В другом аспекте настоящее изобретение обеспечивает способ получения указанной молекулы TGFβR2 с укороченным внеклеточным доменом, описанной в настоящем документе, слитого белка, описанного в настоящем документе, антитела или его антиген-связывающего фрагмента, описанного в настоящем документе, включающий выращивание указанной клетки-хозяина, описанной в настоящем документе в условиях, подходящих для экспрессии молекулы описанного выше белка, и выделение экспрессированного продукта из культуральной жидкости.
В другом аспекте настоящее изобретение обеспечивает фармацевтическую композицию, содержащую
а) молекулу TGFβR2 с укороченным внеклеточным доменом, описанную в настоящем документе, слитой белок, описанный в настоящем документе, антитело или его антиген-связывающий фрагмент, описанный в настоящем документе, конъюгат, описанный в настоящем документе, нуклеиновую кислоту, описанную в настоящем документе, или вектор экспрессии, описанный в настоящем документе; и
b) фармацевтически приемлемый носитель; необязательно
c) один или более других терапевтических агентов.
В другом аспекте согласно настоящему изобретению предложено применение указанной молекулы TGFβR2 с укороченным внеклеточным доменом, описанной в настоящем документе, слитого белка, описанного в настоящем документе, антитела или его антиген-связывающего фрагмента, описанного в настоящем документе, конъюгата описанного в настоящем документе, нуклеиновой кислоты, описанной в настоящем документе, вектора экспрессии, описанного в настоящем документе, или фармацевтической композиции, описанной в настоящем документе для предотвращения и лечения рака, предпочтительно для лечения рака желудка.
В другом аспекте согласно настоящему изобретению предложено применение указанной молекулы TGFβR2 с укороченным внеклеточным доменом, описанной в настоящем документе, слитого белка, описанного в настоящем документе, антитела или его антиген-связывающего фрагмента, описанного в настоящем документе, конъюгата описанного в настоящем документе, нуклеиновой кислоты, описанной в настоящем документе, вектора экспрессии, описанного в настоящем документе, или фармацевтической композиции, описанной в настоящем документе для получения лекарственного средства для предотвращения и лечения рака, предпочтительно для лечения рака желудка.
В другом аспекте настоящее изобретение обеспечивает фармацевтическую комбинацию, содержащую молекулу TGFβR2 с укороченным внеклеточным доменом, описанный в настоящем документе, слитой белок, описанный в настоящем документе, антитело или его антиген-связывающий фрагмент, описанный в настоящем документе, конъюгат, описанный в настоящем документе, нуклеиновую кислоту, описанную в настоящем документе, вектор экспрессии, описанный в настоящем документе, или фармацевтическую композицию, описанную в настоящем документе, и один или более дополнительных терапевтических агентов.
В другом аспекте настоящее изобретение обеспечивает набор, содержащий молекулу TGFβR2 с укороченным внеклеточным доменом, описанный в настоящем документе, слитой белок, описанный в настоящем документе, антитело или его антиген-связывающий фрагмент, описанный в настоящем документе, конъюгат, описанный в настоящем документе, нуклеиновую кислоту, описанную в настоящем документе, вектор экспрессии, описанный в настоящем документе, или фармацевтическую композицию, описанную в настоящем документе, предпочтительно, дополнительно содержащий устройство для введения лекарственного средства.
В другом аспекте настоящее изобретение обеспечивает способ предотвращения и лечения новообразований, включающий введение субъекту указанной молекулы TGFβR2 с укороченным внеклеточным доменом, описанной в настоящем документе, слитого белка, описанного в настоящем документе, антитела или его антиген-связывающего фрагмента, описанного в настоящем документе, конъюгата, описанного в настоящем документе, нуклеиновой кислоты, описанной в настоящем документе, вектора экспрессии, описанного в настоящем документе, или фармацевтической композиции, описанной в настоящем документе.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На Фиг. 1 показан анализ связывания антитела мыши EGFR-mhPA8 с рекомбинантным белком EGFR-His человека.
На Фиг. 2 показана блокировка связывания EGF с EGFR посредством антитела мыши EGFR-mhPA8.
На Фиг. 3 показано ингибирование пролиферации клеток MDA-MB-468 посредством антитела мыши EGFR-mhPA8.
На Фиг. 4 показан анализ связывания гуманизированного антитела EGFR-HPA8 с рекомбинантным белком EGFR-His человека.
На Фиг. 5 показана блокировка связывания EGF с EGFR посредством гуманизированного антитела EGFR-HPA8.
На Фиг. 6 показано ингибирование пролиферации клеток MDA-MB-468 посредством гуманизированного антитела EGFR-HPA8 в различных условиях лиганда.
На Фиг. 7 показано ингибирование пролиферации клеток Fadu посредством гуманизированного антитела EGFR-HPA8 в различных условиях лиганда.
На Фиг. 8 показан эффект ADCC, опосредованный гуманизированным антителом EGFR-HPA8.
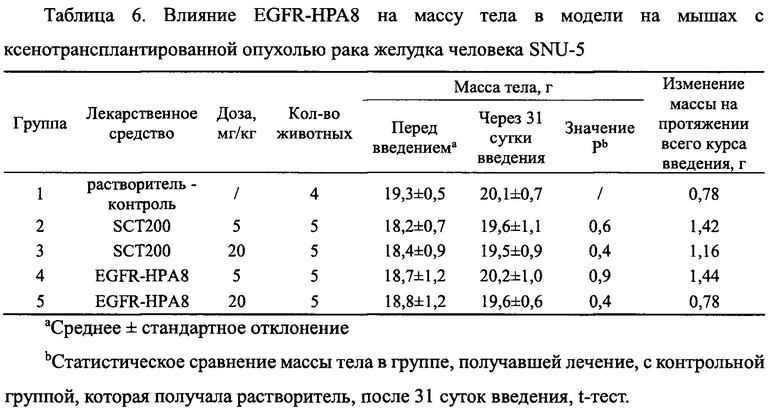
На Фиг. 9 показано влияние EGFR-HPA8 на массу тела мышей с подкожно ксенотрансплантрированными клетками рака желудка SNU-5.
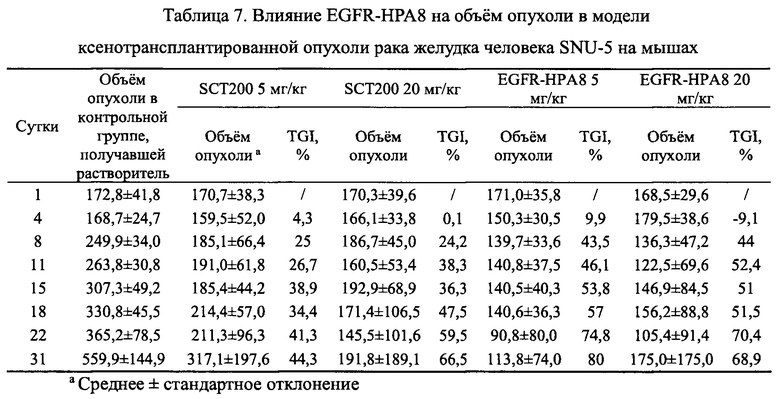
На Фиг. 10 показано влияние EGFR-HPA8 на объем опухоли и результаты TGI у мышей с подкожно ксенотрансплантрированными клетками рака желудка SNU-5.
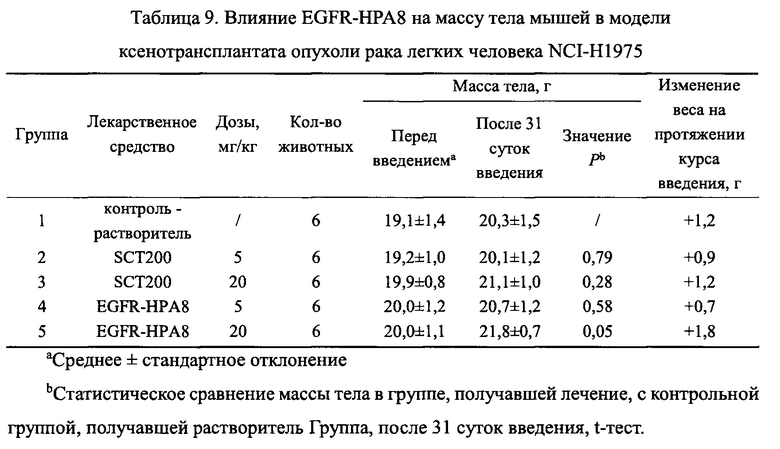
На Фиг. 11 показано влияние EGFR-HPA8 на массу тела у мышей с подкожно ксенотрансплантрированными клетками немелкоклеточного рака легких NCI-H1975.
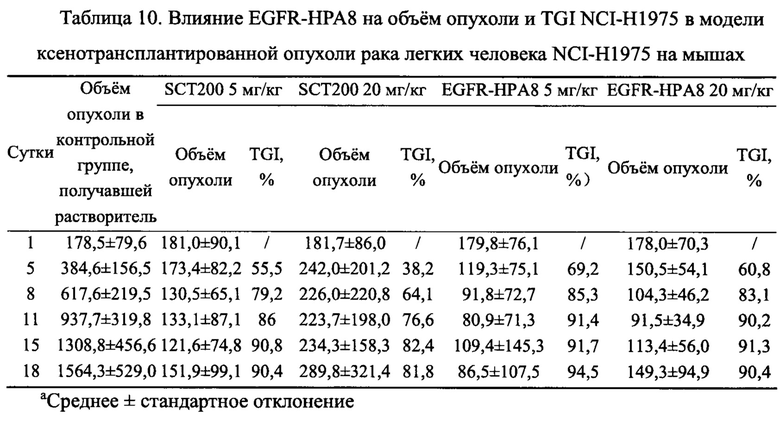
На Фиг. 12 показано влияние EGFR-HPA8 на объем опухоли и результаты TGI у мышей с подкожно ксенотрансплантрированными клетками немелкоклеточного рака легких NCI-H1975.
На Фиг. 13 показана схематическая структура слитого белка EGFR антитело/TGFβR2.
На Фиг. 14 показана детекция деградации слитого белка EGFR антитело/TGFβR2.
На Фиг. 15 показана детекция склонности к деградации слитого белка EGFR антитело/TGFβR2.
На Фиг. 16 показана детекция способности к связыванию различных укороченных форм слитого белка EGFR антиттело/TGFβR2 с TGF-β1 и EGFR.
На Фиг. 17 показана детекция способности укороченного TGFβR2 слитого белка EGFR антитело/TGFβR2 к нейтрализации TGF-β1.
На Фиг. 18 показана детекция способности укороченного TGFβR2 слитого белка EGFR антитело/TGFβR2 к нейтрализации TGF-β3.
На Фиг. 19 показан анализ способности слитого белка 6 связываться с белками TGF-β1 и TGF-β3, соответственно.
На Фиг. 20 показан анализ способности слитого белка 6 блокировать связывание белка TGF-β1 или белка TGF-β3 с TGFβR2-Fc, соответственно.
На Фиг. 21 показан анализ связывания и способности к конкурированию слитого белка 6.
На Фиг. 22 показан анализ аффинности слитого белка 6 к рекомбинантному белку EGFR человека.
На Фиг. 23 показан анализ аффинности слитого белка 6 к рекомбинантному белок TGF-β1 человека.
На Фиг. 24 показан нейтрализующий эффект TGF-β1 посредством слитого белка 6 на клетках Mv-1-lu.
На Фиг. 25 показан нейтрализующий эффект TGF-β1 посредством слитого белка 6, проанализированный с использованием системы с репортерным геном.
На Фиг. 26 показано ингибирование пролиферации клеток MDA-MB-468 посредством слитого белка 6.
На Фиг. 27 показан эффект ADCC, опосредованной слитым белком 6.
На Фиг. 28 показано влияние слитого белка 6 на массу тела у мышей с подкожно ксенотрансплантрированными клетками немелкоклеточного рака легких NCI-H1975.
На Фиг. 29 показано влияние слитого белка 6 на объем опухоли мышей с подкожно ксенотрансплантрированными клетками немелкоклеточного рака легких NCI-H1975.
На Фиг. 30 показан ультрафильтрационный анализ стабильности слитого белка 6.
На Фиг. 31 показан анализ деградации слитого белка X/TGFβR2.
На Фиг. 32 показан анализ склонности к деградации слитого белка X/TGFβR2.
На Фиг. 33 показан анализ способности связывания слитого белка X/TGFβR2 с TGF-β1.
На Фиг. 34 показан анализ способности слитого белка X/TGFβR2 к нейтрализации TGF-β1 и TGF-β3.
На Фиг. 35 показан анализ связывания слитого белка X/TGFβR2 с мишенью части X.
ПОДРОБНОЕ ОПИСАНИЕ
Определения
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют значение, обычно понятное специалистам в данной области техники, к которой относится настоящее изобретение. Для целей настоящего изобретения дополнительно определены следующие термины.
При использовании в настоящем описании и в прилагаемой формуле изобретения формы единственного числа «один», «другой» и «указанный» включают обозначение объекта во множественном числе, если контекст явно не указывает на иное.
Термин «укороченная» форма белковой молекулы означает, что в модифицированном виде отсутствуют один или более остатков аминокислот по сравнению с природной формой белковой молекулы.
Термин «слитой белок» относится к белковой молекуле, объединяющей два или более белков. Обычно ее получают путем экспрессии гибридного гена, объединяющего последовательности двух или более генов, которые встраивают в вектор экспрессии в форме с соответствующей совпадающей рамкой считывания.
Термин «антитело» относится к молекуле иммуноглобулина и относится к любой форме антитела, которая проявляет желаемую биологическую активность. Он включает моноклональное антитело (включая полноразмерное моноклональное антитело), поликлональные антитела и полиспецифическое антитела (например, биспецифические антитела), и даже фрагменты антител, но не ограничивается ими.
Термин «вариабельный участок» относится к участку в тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные участки тяжелой и легкой цепи природного антитела (VH и VL, соответственно) в целом имеют сходную структуру и могут дополнительно подразделяться на сильно вариабельные участки (называемые участками, определяющими комплементарность, (CDR)) с вкраплениями более консервативных областей (называемых каркасными участками (FR)).
Участок, определяющий комплементарность, (CDR) и каркасный участок (FR) конкретного антитела могут быть идентифицированы с использованием системы Кабата (Kabat и др.: Sequences of Proteins of Immunological Interest, 5th edition, U.S. Department of Health и Human Services, PHS, NIH, NIH Publication No. 91-3242, 1991).
Термин «константный участок» относится к таким последовательностям аминокислот в легких и тяжелых цепях антитела, которые непосредственно не участвуют в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как вызываемая антителом цитотоксичность (ADCC).
«Антиген-связывающий фрагмент антитела» включает часть интактной молекулы антитела, которая сохраняет по меньшей мере некоторую специфичность связывания родительского антитела и обычно включает по меньшей мере часть антиген-связывающего участка или вариабельного участка (например, один или более CDR) родительского антитела. Примеры антиген-связывающих фрагментов включают, Fv, Fab, Fab', Fab'-SH, F(ab')2, Fd фрагменты, Fd' фрагменты, молекулы одноцепочечных антител (например, scFv, ди-scFv или три-scFv, двудольные антитела или scFab), и однодоменные антитела, но не ограничиваются ими.
Термин «конъюгат» относится к биологически активным белковым или пептидным молекулам, образующим ковалентную или нековалентную связь с другой молекулой, будь то малая молекула или биомолекула.
Термин «моноклональное антитело» относится к антителу, полученному из по существу однородной популяции антител, т.е. популяции, содержащей гомогенные антитела, идентичные друг другу, за исключением возможных мутаций (например, естественных мутаций), которые могут присутствовать в очень малых количествах. Соответственно, термин «моноклональное» указывает на природу указанного антитела, а не на смесь неродственных антител. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела против разных кластеров (эпитопов), каждое моноклональное антитело препарата моноклонального антитела направлено против конкретного кластера антигена. В дополнение к их специфичности препараты моноклонального антитела имеют то преимущество, что они обычно не контаминированы другими антителами. Термин «моноклональное» не следует толковать как требующее какого-либо конкретного метода получения определенного антитела. Термин «моноклональное антитело» конкретно включает химерные антитела, гуманизированные антитела и антитела человека.
Антитело «специфически связывается» с антигеном-мишенью, таким как белок антигена, ассоциированного с опухолью (в данном документе, EGFR), т.е., связывает указанный антиген с достаточной аффинностью, чтобы позволить использовать указанное антитело в качестве терапевтического агента, нацеленного на ткань или клетку, экспрессирующую указанный антиген, и без значительной перекрестной реактивности с другими белками или с белками, отличными от гомологов и вариантов (например, мутантных форм, сплайс-вариантов или гидролизованных укороченных форм белка) антигенов-мишеней, упомянутых выше.
Термин «аффинность связывания» относится к силе суммы нековалентных взаимодействий между отдельными сайтами связывания молекулы и ее партнерами по связыванию. Если не указано иное, термин «аффинность связывания», используемый в настоящем документе, относится к внутренней аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающейся пары (например, антитело и антиген). Такие параметры как «KD», «константа скорости связывания kon» и «константа скорости диссоциации koff» обычно используют для описания сродства между молекулой (например, антителом) и ее партнером по связыванию (например, антигеном, т.е., насколько прочно лиганд связывается с конкретным белком. На аффинность связывания влияют нековалентные межмолекулярные взаимодействия, такие как водородная связь, электростатические взаимодействия, а также гидрофобные и ван-дер-ваальсовые силы между двумя молекулами. Дополнительно на аффинность связывания между лигандом и его молекулой-мишенью может влиять присутствие других молекул. Аффинность может быть проанализирована обычными методами, известными в данной области техники, включая ИФА, описанный в настоящем документе.
Термин «нацеливающая часть» относится к части слитого белка, которая выполняет функцию специфического связывания с клеткой-мишенью. Термин включает антитела и другие природные (например, рецепторы, лиганды) или синтетические (например, DARPin) молекулы, которые специфически связываются с клеткой-мишенью. В настоящем тексте «специфически связывается с клеткой-мишенью» указывает на, что указанная часть предпочтительно связывает клетки-мишени в сложной смеси.
«Выделенная» биомолекула представляет собой биомолекулу, которая была идентифицирована и выделена из клетки, которая естественным образом экспрессирует такие молекулы. Выделенные биомолекулы включают биомолекулы in situ в рекомбинантных клетках, а также биомолекулы, которые обычно получают по меньшей мере за одну стадию очистки.
Термин «рецептор» представляет собой биохимическое понятие, относящееся к классу молекул, которые специфически распознают и связывают внеклеточные сигналы (т.е., термин «лиганд») и производят специфический эффект внутри клетки. Произведенные эффекты могут длиться только в течение короткого периода времени, например, изменение клеточного метаболизма или подвижности клеток. Это также может иметь длительный эффект, такой как повышение или понижение экспрессии гена или генов.
«Антитело-зависимая клеточно-опосредованная цитотоксичность» или «ADCC» относится к форме цитотоксичности, при которой связывание секретируемого Ig с рецепторами Fcγ, присутствующими на некоторых цитотоксических клетках (например, NK-клетках, нейтрофилах и макрофагах), позволяет этим цитотоксическим эффекторным клеткам специфически связываться с клетками-мишенями, несущими антиген, и впоследствии убивать указанные клетки-мишени с использованием, например, цитотоксического агента. Для оценки ADCC-активности целевого антитела может быть проведен анализ ADCC in vitro, такой как анализ ADCC in vitro, задокументированный в патенте США №5,500,362 или 5,821,337 или патенте США №6,737,056 (Presta) и способы, описанные в вариантах осуществления настоящей заявки. Полезные эффекторные клетки для таких анализов включают клетки РВМС и NK-клетки.
Молекула TGFβR2 с укороченным внеклеточным доменом согласно настоящему изобретению, слитые белки, содержащие молекулу TGFβR2 с укороченным внеклеточным доменом и нацеливающую часть
Между сайтами 7-15 N-конца полноразмерного внеклеточного домена TGFβR2 существует множество сайтов чувствительности. Чтобы улучшить структурную стабильность слитого белка, авторы настоящего изобретения модифицировали N-конец последовательности аминокислот внеклеточного домена TGFβR2 путем делеции различного количества аминокислот.Было получено пятнадцать молекул TGFβR2 с укороченным внеклеточным доменом.
Авторы настоящего изобретения использовали свои недавно выделенные антитела против EGFR EGFR-HPA8, трастузумаб, бевацизумаб, рамуцирумаб, ипилимумаб или панитумумаб в качестве нацеливающей части слитого белка и молекулу белка TGFβR2 с укороченным внеклеточным доменом в качестве иммуномодуляторной части слитого белка. Молекулу белка TGFβR2 с укороченным внеклеточным доменом, соединенную с С-концом тяжелой цепи антитела против EGFR, получали методом гомологичной рекомбинации, то есть получали слитой белок (EGFR/TGFβR2), содержащий две цепи, т.е., легкую цепь и тяжелую цепь антитела против EGFR, и внеклеточный домен TGFβR2, структура которого показана на Фиг. 13. В слитом белке EGFR антитело/TGFβR2, аминокислота тяжелой цепи С-конца антитела против EGFR связана с внеклеточным доменом TGFβR2 с различными формами делеции аминокислот посредством (G4S)4 линкера (SEQ ID NO: 66). Дополнительно остаток лизина С-конца тяжелой цепи антитела против EGFR был удален для снижения риска протеолиза.
В одном из предпочтительных вариантов реализации настоящего изобретения, слитой белок 6 сохраняет высокую аффинность связывания с белком EGFR человека, имея значение KD 8,77 пМ по аффинности связывания с белком EGFR человека, значение константы связывания kon 1,68Е+06 М-1 с-1, и значение константы диссоциации kdis 1,47Е-05 с-1. Дополнительно слитой белок 6, который содержит укороченный TGFβR2, обладает схожей аффинностью с белком TGF-β1 человека, что и слитой белок 1, который содержит полноразмерный TGFβR2, со значением KD 96,1 пМ, значением константы связывания kon 1,53Е+06 M-1 с-1, и значением константы диссоциации kdis 1,47Е-04 с-1.
Нуклеиновые кислоты согласно настоящему изобретению
Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим молекулы TGFβR2 с укороченным внеклеточным доменом согласно настоящему изобретению, молекулы, включая слитые белки, содержащие укороченную молекулу и нацеливающую часть, или или их части. Некоторые примеры последовательностей этих нуклеиновых кислот приведены в Перечне последовательностей.
Молекулы нуклеиновых кислот согласно настоящему изобретению не ограничены последовательностями, описанными в настоящем описании, но также включают их варианты и другие соответствующие формы нуклеиновых кислот, такие как мРНК, к ДНК, и их варианты. Варианты согласно настоящему изобретению могут быть описаны со ссылкой на их физические свойства в гибридизации. Специалист в данной области техники понимает, что использование методик гибридизации нуклеиновых кислот может применяться идентифицировать их комплементы, а также их эквиваленты или конгенеры.
Рекомбинантные векторы и экспрессия
Согласно настоящему изобретению также предложены рекомбинантные конструкции, содержащие одну или более последовательностей нуклеотидов согласно настоящему изобретению. Рекомбинантные конструкции согласно настоящему изобретению могут использоваться в векторах, например, плазмидах, фагмидах, фагах, или вирусных векторах, а молекулы нуклеиновых кислот, кодирующих антитела согласно настоящему изобретению, могут вводиться в указанные векторы.
Указанная молекула TGFβR2 с укороченным внеклеточным доменом, нацеливающие части, слитые белки, содержащие указанные укороченную молекулу и нацеливающая часть, могут быть получены посредством рекомбинантной экспрессии последовательности нуклеотидов, кодирующей указанную молекулу или белок в клетке-хозяине. Указанные молекула или белок могут содержать более одной последовательности аминокислот, и для того, чтобы экспрессировать множество последовательностей аминокислот рекомбинантными методами, один или более рекомбинантных векторов экспрессии, содержащих кодирующие последовательности нуклеотидов, могут быть трансфицированы в указанную клетку-хозяина так, что указанное множество последовательностей аминокислот экспрессируется в указанной клетке-хозяине. Стандартные методологии рекомбинантных ДНК используют для получения нуклеиновых кислот, кодирующих указанные молекулы или белки, включают эти нуклеиновые кислоты в рекомбинантные векторы экспрессии и вводят указанные векторы в клетки-хозяева, смотри, например, Sambrook, Fritsch, и Maniatis (eds.), Molecular Cloning; A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y., (1989), Ausubel, F. M. и др. (eds.) Current Protocols in Molecular Biology, Greene Publishing Associates, (1989) и методологии, задокументированные в патенте США №4,816,397 (Boss и др.).
Примерами прокариотических клеток-хозяев являются бактерии, а примерами эукариотических клеток-хозяев являются клетки дрожжей, насекомых или млекопитающих. Следует понимать, что на дизайн векторной экспрессии, включая выбор регуляторной последовательности, влияют различные факторы, такие как выбор клетки-хозяина, уровень экспрессии желаемого белка и является ли экспрессия конститутивной или индуцируемой.
Молекулы TGFβR2 с укороченным внеклеточным доменом, нацеливающая часть и слитые белки согласно настоящему изобретению могут быть выделены и очищены от культур рекомбинантных клеток хорошо известными способами, указанные хорошо известные способы включают преципитацию сульфатом аммония или этанолом, кислотную экстракцию, аффинную хроматографию на белке А, аффинную хроматографию на белке G, анионо- или катионообменную хроматографию, хроматографию на фосфатах целлюлозы, хроматографию гидрофобного взаимодействия, аффинную хроматографию, хроматографию на гидроксиапатите, хроматографию на лектинах, но не ограничиваются ими. Высокоэффективная жидкостная хроматография ("ВЭЖХ") также может быть использована для очистки.
Применение
Молекулы TGFβR2 с укороченным внеклеточным доменом, антитела и слитые белки согласно настоящему изобретению могут применяться для лечения рака, такого как рак желудка.
Фармацевтические композиции
Один или более видов укороченного белка(ов), слитые белки, антитела и антиген-связывающий фрагменты, конъюгаты укороченного белка и лекарственного средства, конъюгаты слитого белка лекарственного средства, конъюгаты, нуклеиновые кислоты и носители согласно настоящему изобретению могут быть приготовлены с по меньшей мере одним другим химическим агентом с образованием фармацевтической композиции, содержащей предшествующий активный ингредиент(ы), описанный выше, и один или более фармацевтически приемлемых носителей, разбавителей или вспомогательных веществ; они, необязательно, также могут включать один или более других терапевтических агентов.
Наборы реагентов
Настоящее изобретение также относится к фармацевтическим упаковкам и наборам, содержащим один или более контейнеров, где указанные контейнеры содержат фармацевтические композиции согласно настоящему изобретению, упомянутые выше. Ассоциированные с такими контейнерами, они могут быть представлены в форме, установленной государственным органом, регулирующим производство и использование или распространение лекарственного препарата или биологического продукта, что отражает одобрение для введения их человеку указанным органом государства, в котором указанный продукт производится, используется или распространяется.
Приготовление и хранение
Фармацевтические композиции настоящего изобретения могут быть получены способами, известными в данной области техники, например, с помощью обычных методов смешивания, растворения, гранулирования, измельчения, эмульгирования, инкапсулирования, инкапсулирования или лиофилизации.
После приготовления фармацевтической формы композиция, содержащая соединения согласно настоящему изобретению, изготовленные в виде лекарственной формы в приемлемом носителе, может быть помещена в соответствующие дозаторы и помечена для лечения указанного состояния. Такая маркировка должна включать количество, частоту и способ введения лекарственного средства.
Комбинации лекарственных средств
Комбинации, содержащие антитела согласно настоящему изобретению, описанные выше, также комбинируются с одним или более другими терапевтическими агентами, если при этом полученная комбинация не вызывает неприемлемый неблагоприятный эффект.
Последующие варианты осуществления используются для иллюстрации настоящего изобретения в качестве примеров и не предназначены для ограничения настоящего изобретения.
Примеры
Пример 1: Скрининг антител мыши, блокирующих связывание EGF с EGFR, с использованием фаговой библиотеки антител
1.1 Иммунизация мышей и скрининг фаговой библиотеки антител
Мышей иммунизировали плазмидами pCMV3-mFlt3L (G12FE5S01-D, сконструирована Sinocelltech Limited, SEQ ID NO: 142) и pCMV3-mCSF2 (G12FE5S02-D, сконструирована Sinocelltech Limited, SEQ ID NO: 143), содержащими указанные нуклеиновые кислоты. Конкретный метод заключается в следующем: мышей иммунизировали кардиотоксином 10 M внутримышечно за двое суток до первой иммунизации. Вводили внутримышечно 20 г в ногу смеси pCMV3-mFlt3L и pCMV3-mCSF2 в соотношении 1:1 и внутримышечно 30 г в ногу pGS6-EGFR-TT-WPRE (сконструирована Sinocelltech Limited, SEQ ID NO: 144) через трое суток после иммунизации, внутрибрюшинная инъекция 5×106 клеток насекомых, сверхэкспрессирующих EGFR, для четвертой и пятой иммунизации с интервалами иммунизации 2 недели, 2 недели, 2 недели и 2 недели последовательно. Начиная с третьей иммунизации, кровь собирали через медиальное кантальное сплетение глаза через семь дней после каждой иммунизации. Делали покрытие рекомбинантного белка EGFR человека (источник: Sino Biological, Inc., кат. №10001-Н08В, в дальнейшем то же самое) для определения титра анти-EGFR в сыворотке мышей. Сыворотки пятой иммунизации достигли критериев, при которых титры иммунной сыворотки достигают 8000-кратного значения и титр сыворотки OD>1, затем проводили внутрибрюшинную бустерную инъекцию с использованием 5×106 клеток MDA-MB468 с интервалом 20 дней, через 7 дней мышей умерщвляли, а ткани селезенки замораживали в жидком азоте.
Ткань селезенки мыши экстрагировали реагентом TriPure Isolation Reagent (источник: Roche кат. №11 667 165 001) для экстракции РНК, и кДНК получали обратной транскрипцией с помощью набора для обратной транскрипции TriPure Isolation Reagent (источник: Invitrogen кат.№18080-051), осуществляли ПЦР-амплификацию, в результате чего получали последовательности нуклеотидов, кодирующих вариабельные участки легкой и тяжелой цепи антитела мыши, указанная последовательность нуклеотидов, кодирующих легкую и тяжелую цепь вариабельных участков антитела мыши сплайсировали в последовательность нуклеотидов, кодирующих scFv, путем ПЦР сплайсинга удлинения с перекрытием, вариабельные участки легкой и тяжелой цепи связывали линкером [25]:
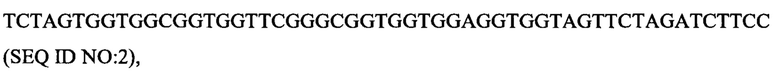
затем лигировали в фаговый вектор pComb3x (источник: Sino Biological, Inc.) с помощью эндонуклеазы рестрикции SfiI (источник: Fermentas), и библиотеку фагового дисплея антител scFv для иммунизации мышей сконструировали электротрансформацией компетентных клеток X-Blue. Планшет для ИФА покрывали рекомбинантным белком EGFR человека, и фаговую библиотеку, обогащенную положительными антителами против EGFR, получали методом пэннинга фаговых антител. Затем, полученную библиотеку смешивали с клетками MDA-MB-468 (источник: Центр клеточных ресурсов, Шанхайский институт биологических наук, академия наук Китая), и фаговые библиотеки, обогащенные положительными антителами против клеток MDA-MB-468, отбирали из вышеуказанных библиотек методом пэннинга фаговых антител (O'Brien, Р. М., & Aitken, R. (Eds.), Springer Science & Business Media. 2002; ISBN: 9780896037113). Колонии фага выбирали из обогащенной библиотеки, экспрессировали, связывание с рекомбинантным белком EGFR человека выявляли методом ИФА, а клоны с высокосвязывающим антителом EGFR-mhPA8 scFv со специфическим связыванием с рекомбинантным EGFR человека подвергали скринингу и отправляли в компанию, занимающуюся секвенированием, в результате чего получили последовательность нуклеотидов антитела EGFR-mhPA8 scFv (SEQ ID NO: 3).
1.2 Функциональный анализ антител мыши, нацеленных на EGFR
1.2.1 Функции связывания антител мыши и конкурирования лиганда Для определения способности антител мыши связываться с рекомбинантным белком EGFR человека использовали ИФА. Рекомбинантный белок EGFR-His человека (источник: Sino Biological, Inc., в дальнейшем тот же самый) в разных концентрациях (1,37 нг/мл, 4,12 нг/мл, 12,35 нг/мл, 37,04 нг/мл, 111,11 нг/мл, 333,33 нг/мл, 1000 нг/мл и 3000 нг/мл) наносили в 96-луночные планшеты, 100 мкл на лунку, и оставляли в течение ночи при 4°С. Планшеты промывали на следующий день и блокировали при комнатной температуре в течение 1 ч. EGFR-mhPA8 и антитело H7N9-R1 в качестве отрицательного контроля (источник: Sinocelltech Limited, далее такое же) добавляли в концентрации 13,89 нМ и инкубировали в течение 1 ч, после чего планшеты промывали для удаления несвязавшихся антител. Вторичное антитело козы против hIgG F(ab)2/HRP (источник: Jackson ImmunoResearch, кат. №109-036-006, далее такое же) добавляли и инкубировали в течение 1 ч, планшеты многократно промывали и добавляли раствор хромогенного субстрата для проявления окраски, затем измеряли OD450 с помощью микропланшетного ридера после завершения реакции. Используя концентрацию рекомбинантного белка EGFR-His человека в качестве горизонтальной координаты и значение OD450 в качестве вертикальной координаты, строили S-образную кривую и ЕС50 связывания антитела с рекомбинантным белком EGFR-His человека анализировали с помощью программного обеспечения GraphPad Prism 6.0. Результаты приведены на Фиг. 1. Химерное антитело EGFR-mhPA8 человека и мыши может эффективно связывать рекомбинантный EGFR-His человека с ЕС50, равной 128,7 нг/мл, и R2=1.000. Отрицательный контроль, H7N9-R1, не связывался с рекомбинантным белком EGFR-His человека.
В этом Примере дополнительно анализировали способность EGFR-mhPA8 блокировать связывание лиганда и рецептора EGFR с помощью FACS. Клетки MDA-MB-468 в количестве 3×105 добавляли к 10 мкл меченого биотином белка EGF-Fc в конечной концентрация 217,1 нМ (источник: Sino Biological, Inc.). Антитела EGFR-mhPA8 в конечных концентрациях 306,74 нМ, 102,25 нМ, 34,08 нМ и 11,36 нМ добавляли после 30 мин инкубации при 2-8°С, и H7N9-R1 использовали в качестве антитела отрицательного контроля. Перемешивали и инкубировали при 2-8°С в течение 20 мин, а затем промывали БСА и центрифугировали для удаления несвязавшихся антител и лигандов. Добавляли вторичное антитело Стрептавидин-488-FITC (источник: Sino Biological, Inc., далее такое же) и инкубировали в течение 1 ч при 2-8°С. Повторяли отмывку и центрифугирование для удаления несвязавшихся вторичных антител. Наконец добавляли 200 мкл БСА к ресуспендированным клеткам и отфильтровывали через фильтр 400 меш в проточную трубку для проточной детекции. Результаты приведены на Фиг. 2, EGFR-mhPA8 может эффективно блокировать связывание белка EGF-Fc с EGFR на клетках MDA-MB-468 в градиенте концентраций.
1.2.2 Ингибирование пролиферации клеток MDA-MB-468 антителами мыши
Клетки рака молочной железы MDA-MB-468 на высоком уровне экспрессируют EGFR, а также различные аутокринные лигандные факторы EGFR. В этом Примере функцию антител мыши по ингибированию роста определяли на клетках MDA-MB-468 с помощью анализа WST-8.
Клетки MDA-MB-468 равномерно инокулировали в 96-луночные планшеты в количестве 5×103 на лунку. Клетки инкубировали в инкубаторе с СО2 в течение 3 ч и добавляли различные концентрации антител мыши EGFR-mhPA8 (66,7 нМ, 22,2 нМ, 7,4 нМ, 2,5 нМ, 0,82 нМ, 0,27 нМ, 0,093 нМ, 0,033 нМ и 0,013 нМ). Также использовали группу M отрицательного контроля (содержащую клетки) и контрольную группу В (только среда без клеток). Клетки инкубировали в инкубаторе с СО2 при 37°С и 5% СО2 в течение 5 суток, а затем добавляли WST-8 в количестве 15 мкл на лунку. Через 240 мин инкубации измеряли OD450-OD630 с помощью микропланшетного ридера и степень ингибирования полученного из мыши антитела рассчитывали путем вычитания детектированного значения показаний лунки контроля группы В. Степень ингибирования=(OD группы M - OD образца)/(OD группы М) × 100%, количественную кривую эффективности анализировали и строили с помощью программного обеспечения GraphPad Prism, где горизонтальной координатой являлась концентрация антитела и вертикальной координатой являлась степень ингибирования. Как показано на Фиг. 3, EGFR-mhPA8 эффективно ингибировал пролиферацию MDA-MB-468 и степень ингибирования повышалась с повышением концентрации лекарственного средства на S-образной кривой. EGFR-mhPA8 ингибировал клетки MDA-MB-468 с EC50, равной 2,78 hM и R2=0,991.
Пример 2: Гуманизация антитела мыши EGFR-mhPA8 и определение характеристик гуманизированного антитела ex vivo
2.1 Гуманизация и получение антитела мыши mhPA8
Последовательность нуклеотидов антитела EGFR-mhPA8 была определена, в результате чего получили последовательность аминокислот тяжелой цепи (SEQ ID NO: 8) и вариабельного участка легкой цепи (SEQ ID NO: 9) антитела EGFR-mhPA8 scFv. Последовательность аминокислот каждого из трех CDR для легкой и тяжелой цепей EGFR-mhPA8 scFv определили со ссылкой на подходы Кабата [26] и нумерацию IMGT, смотрите SEQ ID NO: 10-15. Согласно нумерации Кабата, за исключением мутации N на D в положении 52 в LCDR2, соответствующие три CDR вышеупомянутых легких и тяжелых цепей были трансплантированы на последующем этапе гуманизации и сохранены в конечном гуманизированном антителе EGFR-HPA8 scFv.
Гуманизацию антител мыши осуществляли с использованием классического метода трансплантации CDR[27,28]. Антитела, у которых имеется более чем 50% сходство последовательности аминокислот с вариабельными участками как легкой цепи, так и тяжелой цепи антитела мыши и более чем 50% сходство последовательности аминокислот с каркасными участками вариабельных участков легкой цепи и тяжелой цепи антитела мыши, которые предполагалось модифицировать, соответственно, были выбраны в качестве библиотеки шаблонов для гуманизации. Антитело человека с наивысшим пространственным сходством с указанным вариабельным участком антитела, которые предполагалось модифицировать, было выбрано в качестве матрицы для гуманизации. Тремя последовательностями CDR легкой или тяжелой цепи антитела мыши были заменены соответствующие последовательности аминокислот CDR в матрице гуманизации и аминокислоты NG, NS, NA и NT, которые подвергаются высокому риску дезамидирования в мутированной последовательности, подверглись мутации с целью улучшения химической стабильности и сохранения биологической функции указанного антитела. Аффинность гуманизированного антитела определяют с помощью ИФА и отбирают гуманизированные антитела, чья аффинность сохранена. Матрицей для гуманизации, используемой для трансплантации вариабельного участка легкой цепи из EGFR-mhPA8, в этом примере является IGKV1-NL1*01, который имеет 68,4% гомологии с легкой цепью EGFR-mhPA8, а матрицей для гуманизации, используемой для трансплантации вариабельного участка тяжелой цепи, является IGHV1-69-2*01, который имеет 64,9% гомологии с тяжелой цепью EGFR-mhPA8. N52 вариабельного участка LCDR2 гуманизированного антитела EGFR-HPA8, склонный к дезамидированию, мутирован в D.
Поскольку ключевой сайт каркасного участка антитела мыши необходим для поддержания стабильности пространственной структуры CDR, ключевой сайт должен быть обратно мутирован в соответствующие аминокислоты антитела мыши. По нумерации Кабата легкая цепь была обратно мутирована в Q в положении 45, в I в положении 48, в К в положении 74 и в D в положении 76, а тяжелая цепь была обратно мутирована в К в положении 38, в I в положении 48 и в L в положении 70. Гуманизированное антитело EGFR-HPA8 получали посредством гуманизационной трансплантации CDR и реверсивных мутаций каркасной области, а последовательности аминокислот его тяжелой цепи и легкой цепь показаны в SEQ ID NO: 22/23, соответственно; последовательности аминокислот его тяжелой цепь и легкой цепи, содержащих сигнальный пептид, показаны в SEQ ID NO: 24/25, соответственно, содержащих последовательности аминокислот сигнального пептида, последовательно соединенного с тяжелой цепью/легкой цепью, показаны в (SEQ ID NO: 26/27), вариабельный участок тяжелой цепи/легкой цепи гуманизированного антитела (SEQ ID NO: 28/29) и константный участок гуманизированного антитела, а именно константный участок тяжелой цепи/ константный участок легкой цепи каппа IgG1 человека (SEQ ID NO: 30/31); и последовательности его гуманизированных CDR показаны в Таблице 2.

Последовательность нуклеотидов, содержащую сигнальный пептид легкой цепи антитела EGFR-HPA8 (SEQ ID NO: 33), последовательно соединенную с последовательностью нуклеотидов легкой цепи (SEQ ID NO: 35), последовательность нуклеотидов вариабельного участка легкой цепи гуманизированного антитела (SEQ ID NO: 37) и последовательность нуклеотидов константного участка легкой цепи каппа человека (SEQ ID NO: 39) амплифицировали путем сплайсинга ПЦР. Указанный выше продукт ПЦР вставляли в вектор pSTEP2 (источник: сконструирована Sinocelltech Limited, такая же далее) инфузионным методом (двойное расщепление HindIII+XbaI), и правильность последовательности плазмиды проверяли секвенированием. Последовательность нуклеотидов вариабельного участка тяжелой цепи антитела EGFR-НРА8 (SEQ ID NO: 36) получали путем синтеза полного гена и вставляли в вектор pSTEP2 (после двойного расщепления SeaI+NheI), содержащий последовательность нуклеотидов сигнального пептида тяжелой цепи (SEQ ID NO: 34) и последовательность нуклеотидов константного участка тяжелой цепи IgGl человека (SEQ ID NO: 38) инфузионным методом и правильность последовательности вектора экспрессии легкой и тяжелой цепей EGFR-HPA8 проверяли секвенированием. После выделения плазмиды и трансфекции клеток HEK-293 (нокаут по fut8), указанные клетки культивировали и экспрессировали в течение 7 суток, клеточный супернатант очищали с использованием аффинной хроматографии после центрифугирования, при этом хроматографическая среда представляла собой упаковку белка А, которой взаимодействует с Fc. После уравновешивания хроматографической колонки с белком А буфером состава 50 мМ Трис, 10 мМ NaCl, рН 8,0 5-10 объемами колонки, отфильтрованный культуральный супернатант добавляли в хроматографическую колонку для связывания, и колонку наполняли буфером состава 20 мМ Трис, 0,3 M Arg, рН 6,5 5-10 объемами колонки, а затем колонку промывали элюирующим буфером состава 0,1 M Gly, 10 мМ NaCl, рН 3,5, а собранные образцы нейтрализовали 2 M Трис (рН 8,0), в результате чего получали высокочистое и высококачественное ADCC-усиленное антитело EGFR-HPA8.
Последовательности праймеров для сплайсинга для ПЦР-амплификации легкой цепи антитела EGFR-HPA8, содержащей сигнальный пептид:

Последовательности праймеров для полногеномного синтеза вариабельного участка тяжелой цепи антитела EGFR-HPA8:

2.2. Определение характеристик гуманизированного антитела EGFR-HPA8 in vitro
2.2.1. Анализ специфичности гуманизированного антитела и конкурентности лиганда
Ссылаясь на Пример 1.2.1, связывающая способность антитела человека с рекомбинантным белком EGFR человека была обнаружена методом ИФА с использованием SCT200 (задокументировано в CN200610012002.8, далее так же), Erbitux (MERCK, 201621, далее так же) и отрицательного контроля. Как показано на Фиг. 4, ЕС50 для гуманизированного антитела EGFR-HPA8 специфическое связывание с рекомбинантным EGFR-His человека составляла 116,6 нг/мл, R2=1,000; ЕС50 для SCT200 составляла 166,5 нг/мл, R2=1,000; ЕС50 для Erbitux составляла 253,6 нг/мл, R2=1,000; отрицательный контроль H7N9-R1 не связывался. Эти результаты указывают на то, что EGFR-HPA8 обладает лучшей способностью блокировать рекомбинантный EGFR-his человека, чем SCT200 и Erbitux.
В то же время, способность гуманизированного антитела EGFR-HPA8 блокировать связывание лиганда и рецептора EGFR анализировали с помощью FACS со ссылкой на Пример 1.2.1, используя SCT200, Erbitux и отрицательный контроль. Результаты приведены на Фиг. 5, способность EGFR-HPA8 блокировать связывание белка EGF-Fc с EGFR на клетках MDA-MB-468 является более сильной, чем у SCT200 и Erbitux.
2.2.2. Ингибирование пролиферации различных опухолевых клеток гуманизированными антителами
2.2.2.1. Способность гуманизированных антител ингибировать пролиферацию клеток MDA-MB-468
Клетки MDA-MB-468 равномерно инокулировали в 96-луночные планшеты в количестве 5×103 на лунку. Клетки инкубировали в инкубаторе с СО2 в течение 3 ч, и добавляли различные концентрации гуманизированного антитела EGFR-HPA8 (666,7 нМ, 222,2 нМ, 74,1 нМ, 24,7 нМ, 8,23 нМ, 2,74 нМ, 0,91 нМ, 0,31 нМ и 0,1 нМ), a SCT200 и Erbitux использовали в качестве контроля. Затем добавляли лиганды HB-EGF (источник: Sino Biological, Inc., далее такой же) в конечной концентрации 8 нг/мл, BTC-Fc (источник: Sino Biological, Inc., далее такой же) в конечной концентрации 200 нг/мл или Fc-EREG (источник: Sino Biological, Inc., далее такой же) в конечной концентрации 1 мкг/мл, соответственно. Клетки инкубировали в инкубаторе с СО2 при 37°С и 5% СО2 в течение 5 суток, затем добавляли WST-8 в количестве 15 мкл на лунку. Через 240 минут инкубации измеряли OD450-OD630 с помощью микропланшетного ридера, и рассчитывали рост степени ингибирования клеток антителом. Добавление лиганда без антитела использовали в качестве группы М. Степень ингибирования=(OD группы M - OD образца)/(OD группы M) × 100%, а количественную кривую эффективности анализировали и строили с помощью программного обеспечения GraphPad Prism, горизонтальной координатой являлась концентрация антитела, а вертикальной координатой являлась степень ингибирования. Результаты приведены на Фиг. 6А и в Таблице 3. EGFR-HPA8 показало лучшее ингибирование роста клеток MDA-MB-468, чем контрольные антитела SCT200 и цетуксимаб в условиях без лиганда. EGFR-FJPA8 обладает максимальной степенью ингибирования, схожей с SCT200, но имеет меньшую ЕС50 ингибирования роста. Результаты Примера 2.2.1 демонстрируют, что способность EGFR-HPA8 блокировать связывание белка EGF-Fc с EGFR на клетках MDA-MB-468 была более сильной, чем у SCT200 и Erbitux. В этом Примере добавляли различные лиганды EGFR в анализе ингибирования роста клеток MDA-MB-468, чтобы дополнительно оценить способность EGFR-HPA8 блокировать связывание лиганда и рецептора EGFR на клеточном функциональном уровне. Результаты приведены на Фиг. 6B-6D и в Таблице 3. EGFR-HPA8 ингибировало пролиферацию клеток MDA-MB-468 лучше, чем SCT200 и цетуксимаб в условиях с различными лигандами.
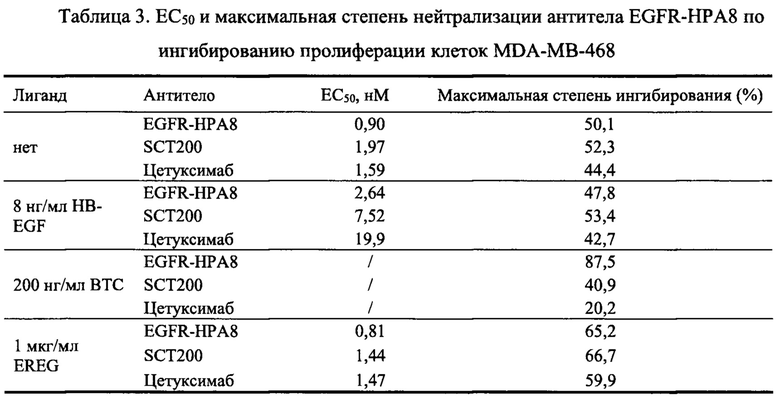
2.2.2.2. Способность гуманизированных антител ингибировать пролиферацию клеток Fadu
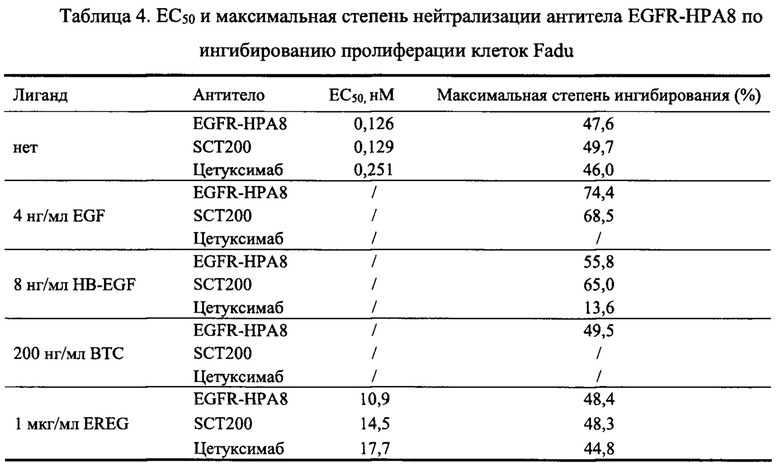
Fadu представляют собой штамм клеток плоскоклеточной карциномы глотки человека, которые сильно экспрессируют EGFR, а также аутокринирует различные лигандные факторы EGFR. Со ссылкой на Пример 2.2.2.1, ингибирование пролиферации клеток Fadu (источник: Центр клеточных ресурсов, Шанхайский институт биологических наук, академия наук Китая) антителом EGFR-HPA8 в условиях с различными лигандами измеряли с помощью метода WST-8. Результаты приведены на Фиг. 7 и в Таблице 4. Способность EGFR-HPA8 ингибировать рост клеток Fadu была сходной с SCT200 и лучшей, чем у цетуксимаба в условиях без лиганда, в то время как способность EGFR-НРА8 ингибировать рост клеток Fadu была значительно лучшей, чем у SCT200 и цетуксимаба в условиях с различными лигандами. Эти результаты продемонстрировали, что антитело EGFR-HPA8 обладало лучшей способностью к ингибированию связывания лиганда с рецептором EGFR по сравнению с SCT200 и цетуксимабом.
2.2.3. Действие гуманизированного антитела ADCC
В этом Примере использована система с рекомбинантным репортерным геном CD16a для определения эффектов ADCC, опосредованных гуманизированным антителом EGFR-HPA8. Система с рекомбинантным репортерным геном CD16a включает эффекторные клетки Jurkat-NFAT-Luc2p-CD16A и целевые клетки, экспрессирующие EGFR. При совместном культивировании двух типов клеток и одновременном добавлении Fab-фрагмента антитела EGFR, антитело EGFR связывается с EGFR, экспрессируемым на поверхности клеток-мишеней, а его Fc-фрагмент может связываться с эффектором клетки, сверхэкспрессирующей FCy рецептор CD16a, тем самым активируя эффекторные клетки Jurkat-NFAT-Luc2p-CD16A и стимулируя NFAT-RE-опосредованную биолюминесценцию.
Целевые клетки А431 (источник: Центр клеточных ресурсов, Шанхайский институт биологических наук, академия наук Китая) равномерно инокулировали в 96-луночные планшеты в количестве 1×104 на лунку. После инкубации в течение ночи добавляли различные концентрации антител (2,67 нМ, 0,53 нМ, 0,11 нМ, 0,021 нМ, 0,0043 нМ, 0,00085 нМ, 0,00017 нМ и 0,000034 нМ) в количестве 40 мкл на лунку, а затем 1×105 эффекторных клеток Jurkat-NFAT-Luc2p-CD16A (Источник: Sinocelltech Limited, далее тот же), 40 мкл на лунку, для каждого анализа использовали 3 повторные лунки. Также использовали лунки с целевыми клетками, эффекторными клетками и антителом отрицательного контроля. Клетки инкубировали в течение 4 ч в инкубаторе с СО2 при 37°С и 5% СО2, и добавляли 5 х буфер пассивного лизирования, 20 мкл на лунку. Клетки один раз замораживали и оттаивали, и 20 мкл супернатанта из каждой лунки переносили в 96-луночный планшет с белым дном, затем встряхивали планшет и перемешивали, детекцию люминесценции осуществляли с помощью детектора LB960-Microplate Luminol Detector. Количественные кривые эффективности были проанализированы и построены с использованием программного обеспечения GraphPad Prism, где горизонтальная координата представляла собой концентрацию образца, а вертикальная координата представляла собой значение RLU. Множитель индукции интенсивности биолюминесценции = RLU образца/отрицательный контроль RLU. Результаты приведены на Фиг. 8, EGFR-HPA8, положительные контроли Erbitux и SCT200 могут опосредовать эффективное действие ADCC на опухолевые клетки А431, экспрессирующие EGFR. Среди них, EGFR-HPA8 имело преимущество в полуэффективной концентрации и индукционной кратности с ЕС50, равной 0,008 нМ и R2=0,999 в отношении индукции ADCC.
2.3 Эффективность EGFR-HPA-8 в отношении подкожно ксенотрансплантированных опухолей линии клеток рака желудка человека SNU-5 и линии немелкоклеточного рака легких человека NCI-H1975 у мышей
Клетки SNU-5 на логарифмической стадии роста (банк клеток АТСС) промывали PBS и расщепляли 0,25% трипсином, продукт расщепления собирали с последующим центрифугированием при 800 об/мин в течение 5 мин. Клетки ресуспендировали в PBS, и концентрацию клеток доводили до 5,0×107 клеток/мл (с 50% матриксным гелем). Мышам Balb/c-nude (Beijing Viton Lever Laboratory Animal Technology Co.) подкожно иннокулировали суспензию клеток SNU-5 в количестве 5,0×106 в правую часть спины, 100 мкл на мышь. После того как объем опухоли достигал примерно 170 мм3, мышей случайным образом делили на семь групп в соответствии с объемом опухоли, по пять мышей в каждой из групп. Лекарственное средство вводили внутрибрюшинно (IP) в день разделения на группы, а затем два раза в неделю в виде 7 последовательных доз. Конкретный режим дозирования показан в Таблице 5 ниже.

Примечание: Объем введения рассчитывали на основе массы тела мышей в количестве 10 мл/кг.
Значение ингибирования роста опухоли (TGI) рассчитывали следующим образом: Т/С(%)=TRTV/CRTV×100% (Trtv представляет собой RTV группы, получавшей лечения; CRTV представляет собой RTV группы отрицательного контроля), относительный объем опухоли (RTV)=VT/V0, V0 - объем опухоли, измеренный в день 0 (D0), а именно день распределения на группы и начала введения дозы, VT - объем опухоли, измеряемый за определенное время. TGI(%)=1-Т/С(%).
Все подопытные животные были в хорошем состоянии и показали некоторую прибавку массы тела на протяжении всего курса введения. Существенной разницы в массе тела мышей в каждой из групп, получавшей дозу лекарственного средства, не было по сравнению с контрольной группой, получавшей растворитель (Р>0,05). Изменения массы тела всех животных показаны на Фиг. 9 и в Таблице 6.
Объем опухоли и результаты TGI для каждой из групп в этом исследовании показаны в Таблице 7 и на Фиг. 10. Через 31 сутки введения доз средний объем опухоли в группе, получавшей носитель, составлял 559,9±144,9 мм3 и в группе положительного контроля с низкой дозой SCT200 5 мг/кг составлял 317,1±197,6 мм3 со значением TGI, равным 44,3%, которое не сильно отличалось группы, получавшей носитель, (Р=11,28%). Группа, получавшая низкую дозу EGFR-HPA8 в 5 мг/кг показала лучшую эффективность в отношении объема опухоли, равной 113,8±74,0 мм3 и TGI 80,0%, это значительная разница по сравнению с группой, получавшей носитель, (Р<0,05), что указывает на то, что EGFR-HPA8 продемонстрировало несколько лучшие свойства ингибирования опухоли, чем положительный контроль SCT200 при этой дозе (Р=0,09). Объем опухоли в группе положительного контроля SCT200 20 мг/кг составлял 191,8±189,1 мм3 со значением TGI, равным 66,5%, что существенно отличалось от группы, получавшей носитель (Р<0,05). Объем опухоли в группе, получавшей большую дозу EGFR-HPA8 20 мг/кг, составлял 175,0±175,0 мм3 со значением TGI, равным 68,9%, что существенно отличалось от группы, получавшей носитель (Р<0,05). Группы, получавшие EGFR-HPA8 и положительный контроль SCT200 20 мг/кг, показали существенный эффект по ингибированию опухоли, и не было существенной разницы в объеме опухолей между двумя группами, получавшими лечение (Р=0,9). В заключение, молекулы EGFR-HPA8 продемонстрировали значительную противоопухолевую эффективность в модели ксенотрансплантированной опухоли рака желудка человека SNU-5 как при уровне дозировки 5 мг/кг, так и 20 мг/кг, и эффект по ингибированию опухоли был немного лучше, чем у SCT200 при низких дозах.
EGFR-HPA8 также показало значительное ингибирующее действие на клеточную линию NCI-H1975 опухоли немелкоклеточного рака легкого человека, в модели подкожной ксенотрансплантации на мышах. Мышам Balb/c-nude подкожно инокулировали клетки NCI-H1975 в правую сторону спины (Центр клеточных ресурсов, Шанхайский институт биологических наук, академия наук Китая), и распределяли по группам и вводили дозы как показано в Таблице 8.

Все подопытные животные были в хорошем состоянии и показали некоторое увеличение массы тела на протяжении всего курса введения, и не было значимой разницы в массе тела мышей в каждой группе введения по сравнению с контрольной группой, получавшей растворитель (Р>0,05). Изменения массы тела всех животных показаны на Фиг. 11 и в Таблице 9.
Объем опухоли и результаты TGI для каждой из групп в этом исследовании показаны в Таблице 10 и на Фиг. 12. Через 18 суток после распределения по группам и введения доз, группы положительного контроля SCT200 и группы EGFR-HPA8 с высокой и низкой дозировкой показали существенные эффекты по ингибированию роста опухоли и статистически значимые различия в объеме опухоли между каждой из групп, получавшей лечение, и группой, получавшей носитель. Средний объем опухоли в группе, получавшей носитель, на 18 сутки составлял 1564,3±529,0 мм3. Объем опухолей после лечения SCT200 5 мг/кг и EGFR-HPA8 5 мг/кг составлял 151,9±99,1 мм3 и 86,5±107,5 мм3, со значениями TGI, равными 90,4% и 94,5%, соответственно. Объем опухолей после лечения SCT200 20 мг/кг и EGFR-HPA8 20 мг/кг составлял 289,8±321,4 мм3 и 149,3±94,9 мм3, со значениями TGI, равными 81,8% и 90,4%, соответственно. EGFR-HPA8 как в дозировке 5 мг/кг, так и 20 мг/кг показало несколько лучший противоопухолевой эффект, чем положительный контроль SCT200, но статистически значимой разницы не было. В совокупности эти результаты позволяют предположить, что молекула EGFR-HPA8 обладает значительной противоопухолевой эффективностью в модели ксенотрансплантированной опухоли немелкоклеточного рака легких человека NCI-H1975.
Пример 3: Конструирование и исследование слитых белков EGFR антитело/TGFβR2, содержащего различные укороченные формы TGFβR2
3.1 Конструирование векторов экспрессии слитого белка EGFR антитело/TGFβR2 с различными укороченными формами TGFβR2, экспрессия белка и очистка
В этом варианте осуществления использовали антитело против EGFR в качестве нацеливающей части слитого белка и внеклеточный домен TGFβR2 в качестве иммуномодуляторной части слитого белка. Внеклеточный домен TGFβR2 соединен с С-концом тяжелой цепи антитела EGFR посредством гомологичной рекомбинации, а слитой белок EGFR антитело/внеклеточный домен TGFβR2 (EGFR/TGFβR2) образован двумя цепями, легкой цепью и тяжелой цепью. Структура слитого белка показана на Фиг. 13. Результаты масс-спектрометрии показали наличие множества сайтов чувствительности между сайтами 7-15 N-конца полноразмерного внеклеточного домена TGFβR2. Чтобы улучшить структурную стабильность белка, последовательность аминокислот N-конца внеклеточного домена TGFβR2 модифицировали делециями аминокислот в различных количествах аминокислот в этом Примере (SEQ ID NO: 47-65). В слитом белке EGFR антитело/TGFβR2 аминокислота С-конца тяжелой цепи антитела EGFR связаны с внеклеточным доменом TGFβR2 с различными формами делегированных аминокислот посредством линкера (G4S)4 (SEQ ID NO: 66). Дополнительно лизин С-конца тяжелой цепи антитела EGFR удаляли для снижения риска протеолиза. Конкретный протокол конструирования слитого белка EGFR антитело/TGFβR2 показан в Таблице 11.
В приведенном выше протоколе целевой ген амплифицировали с помощью ПЦР или ПЦР с перекрытием, лигировали в вектор экспрессии путем инфузии, а плазмиды экстрагировали отдельно после проверки последовательностей и транзиентно переносили в клетки HEK-293 (нокаут fut8) и культивировали до 7-го дня. Центрифугировали и собрали супернатант.Клеточный супернатант после центрифугирования очищали с помощью аффинной хроматографии с белком А, в результате чего получали слитой белок EGFR антитело/ТСРβR2 с усиленной ADCC.
3.2 Расщепление слитых белков EGFR антитело/TGFβR2, содержащих различные укороченные формы TGFβR2
Чистоту, а также расщепление продуктов экспрессии анализировали с помощью SDS-ПААГ в восстанавливающих условиях. Различные укороченные слитые белки EGFR антитело/TGFβR2 1-16, очищенные в Примере 3.1, использовали в SDS- ПААГ в восстанавливающих условиях. Конкретные стадии SDS- ПААГ в восстанавливающих условиях: (1) подготовка SDS- ПААГ: 3,9% концентрирующий гель, 13% разделяющий гель; (2) образцы кипятили при 100°С в течение 2 мин, центрифугировали, а затем наносили образцы в количестве 8 мкг; (3) постоянный ток 40 мА, время электрофореза -1 ч. Результаты приведены на Фиг. 14. Молекулярная масса указанной легкой цепи слитого белка EGFR антитело/TGFβR2 составляла примерно 25 кДа, молекулярная масса указанной тяжелой цепи составляла примерно 66 кДа, и молекулярная масса укороченных вариантов была в диапазоне 45-66 кДа. Результаты показали, что во внеклеточном домене TGFβR2 присутствовали явные расщепленные фрагменты, в то время как у различных укороченных форм слитых белков TGFβR2 внеклеточный домен имел значительно меньшее количество полос его расщепления, чем у полноразмерных форм внеклеточного домена TGFβR2 в слитом белке. Соответственно, различные укороченные формы внеклеточного домена TGFβR2, полученные в настоящем изобретении, существенно повышали стабильность слитого белка, содержащего рецептор TGF-β и антитело.
3.3 тенденция к деградации слитых белков EGFR антитело/TGFβR2, содержащих различные укороченные формы TGFβR2
Супернатант клеток 293Е, который ускоряет процессинг слитых белков, дополнительно использовали для оценки стабильности различных укороченных форм слитых белков EGFR антитело/TGFβR2. Система клеточной экспрессии 293Е, которую часто используют для экспрессии антитела, экспрессирует различные белки клетки-хозяина (НСР) и протеазы, необходимые для роста клеток. Таким образом, устойчивость антител можно оценить, наблюдая за склонностью к деградации слитого белка в супернатанте клеток 293Е.
Очищенный слитой белок смешивали с супернатантом клеток 293Е, культивированных в течение 10 дней при объемном соотношении 1:0,3, и конечная концентрация слитого белка составляла примерно 1 мг/мл. Смешанные образцы встряхивали и хорошо перемешивали и инкубировали при 37°С в течение 48 часов. Также использовали контроль инкубации без супернатанта клеток. Чистоту образцов и содержание укороченных форм определяли с помощью SDS- ПААГ, а чистоту тяжелой цепи слитого белка определяли с помощью программного обеспечения BandScan.
Результаты анализа показаны на Фиг. 15, а процент расщепления в каждом образце показан в Таблице 12. Результаты показали, что процент расщепленных форм TGFβR2 укороченной формы слитого белка (слитые белки 2-16) во всех контрольных группах был менее 4,0%, что значительно ниже, чем у полноразмерной формы TGFβR2 слитого белка (слитой белок 1), у которой этот процент составлял 24,8%. После инкубации с тем же клеточным супернатантом в течение 48 часов при 37°С все полноразмерные формы TGFβR2 слитого белка (слитой белок 1) в экспериментальной группе оказались чувствительными с процентом 100%. Укороченные формы TGFβR2 слитых белков (слитые белки 2-16) содержали различный процент расщепленных форм, но они были значительно лучше, чем полноразмерный контрольный белок, при этом слитой белок 2, слитой белок 6, слитой белок 9, слитой белок 10 и слитой белок 13 продемонстрировали наилучшие качества с процентом содержания расщепленных форм менее 3,0%.

Показанные выше результаты указывают на то что слитые белки EGFR антитело/TGFβR2 с различными укороченными формами TGFβR2 являются более устойчивыми к расщеплению протеазами, чем слитой белок 1 с полноразмерной формой TGFβR2.
3.4 Анализ связывания слитого белка EGFR антитело/TGFβR2 с различными укороченными формами TGFβR2 с TGF-β
100 нг/мл белка TGF-β1 и 40 нг/мл белка EGFR-His наносили в 96-луночные планшеты в количестве 100 мкл на лунку, соответственно, и выдерживали в течение ночи при 4°С. Планшеты промывали на следующий день, блокировали при комнатной температуре в течение 1 ч, и добавляли 2 мкг/мл слитого белка EGFR антитело/TGFβR2 с различными укороченными формами TGFβR2 в количестве 100 мкл на лунку. Планшеты промывали после инкубации в течение 1 часа для удаления несвязавшихся антител, добавляли вторичное антитело козы против hIgG Fc/HRP, инкубировали и повторяли отмывку. Наконец, добавляли раствор хромогенного субстрата для развития окраски, и значение OD450 считывали с помощью микропланшетного ридера после окончания реакции. Результаты приведены на Фиг. 16. Способность к связыванию слитого белка EGFR антитело/TGFβR2 с различными укороченными формами TGFβR2 с TGF-β1 была разной, но способность к связыванию с EGFR была одинаковой.
3.5 Анализ нейтрализации TGF-β слитым белком EGFR антитело/TGFβR2 с различными укороченными формами TGFβR2
TGF-β регулирует клеточные функции, регулируя транскрипцию нескольких генов-мишеней. Ингибитор активатора плазминогена 1 (PAI-1) представляет собой важную мишень нижестоящего сигнального пути TGF-β1/Smad и активирует связывание Smad3 с цис-действующим элементом промоторной области PAI-1, чтобы регулировать экспрессию PAI-1. Элемент, содержащий участок промоторной области PAI-1, встраивали в специфической форме в вектор, содержащий люциферазу, и переносили в клетки HepG2. В этой системе репортерных генов добавление экзогенного белка TGF-β инициирует экспрессию репортерного гена люциферазы и люминесценцию в присутствии субстрата. При добавлении экзогенного антитела TGF-β оно нейтрализует белок TGF-β, блокирует связывание TGF-β с TGFβR2, ингибирует нижестоящий сигнальный путь и, наконец, ингибирует экспрессию репортерного гена люциферазы. Следовательно, эффективность in vitro TGF-β антитела, нейтрализующего TGF-β, может быть определена путем определения интенсивности светового сигнала.
В 96-луночные планшеты равномерно наносили клетки HepG2-3TP-Luc2p-puro (источник: Sinocelltech Limited, далее такой же) в количестве 30,000 клеток на лунку. После выдерживания монослойной культуры в течение ночи питательную среду удаляли из 96-луночного планшета и замещали средой DMEM, содержащей 0,5% ФБС, и инкубировали в течение 6 часов при 37°С с инкубаторе с 5% СО2. Среду в 96-луночном планшете удаляли и добавляли 4 нг/мл белка TGF-β1 вместе со слитым белком EGFR антитело/TGFβR2 в конечной концентрации 0,.02 мкг/мл и инкубировали в течение 18 ч при 37°С в инкубаторе с 5% СО2. Одновременно использовали группу отрицательного контроля M (содержащую клетки и TGF-β1) и группу отрицательного контроля М' (содержащую клетки без TGF-β1). Наконец добавляли 5х лизирующий буфер и отбирали 10 мкл образцов клеток для определения значения интенсивности биолюминесценции (RLU) и рассчитывали степень нейтрализации слитого белка EGFR антитело/TGFβR2. Степень нейтрализации, %=(значение RLU группы Μ - значение RLU образца)/(значение OD группы M - значение OD группы М') × 100%. В качестве горизонтальной координаты использовали концентрацию антител, а в качестве вертикальной координаты использовали уровень нейтрализации антителом, и количественные кривые эффективности были проанализированы и построены с использованием программного обеспечения GraphPad Prism. Как показано на Фиг. 17, слитой белок EGFR антитело/TGFβR2 2, слитой белок 6, слитой белок 8, слитой белок 13, слитой белок 14 и слитой белок 16, содержащие укороченные формы TGFβR2 обладали определенной способностью к нейтрализации TGF-β1, среди которых слитой белок 2 имел сходную нейтрализующую способность со слитым белком 1, содержащим полноразмерную форму TGFβR2, в то время как слитой белок 5, слитой белок 6 и слитой белок 8 показали лучшую нейтрализующую способность, чем слитой белок 1 при такой же концентрации. Слитой белок 6 продемонстрировал наиболее сильную способность к нейтрализации TGF-β1. Оставшиеся слитые белки EGFR антитело/TGFβR2, содержащие укороченные формы TGFβR2, по существу не обладали или обладали слабой нейтрализующей способностью.
Белок TGF-β3 обладает высокой аффинностью к TGFβR2 и может активировать нижестоящую передачу сигналов TGF-β. Способность слитого белка к нейтрализации TGF-β3 (конечная концентрация 20 нг/мл) была проанализирована с применением системы репортерного гена в этом примере. Результаты представлены на Фиг. 18. Слитой белок 2, слитой белок 4 и слитой белок 13 обладали такой же способностью к нейтрализации TGF-β3, как и слитой белок 1, в то время как слитой белок 3, слитой белок 5, слитой белок 6 и слитой белок 8 обладали лучшей склонностью к нейтрализации TGF-β3, чем слитой белок 1. Слитой белок 6 также показал самую высокую способность к нейтрализации TGF-β3.
Основываясь на приведенном выше анализе состава и способности к нейтрализации слитых белков, содержащих укороченные формы TGFβR2, в настоящем изобретении предпочтительными оказались формы слитого белка 2 ~ 6, 8, 13, 14 и 16 с укороченным TGFβR2, более предпочтительными - формы слитого белка 2, 5, 6 и 8 с укороченным TGFβR2, и наиболее предпочтительной - форма слитого белка 6 с укороченным TGFβR2.
Пример 4: In vitro биологические свойства слитого белка 6, слитого белка EGFR антитело/TGFβR2 с укороченной формой TGFβR2
4.1 Анализ связывающей способности слитого белка EGFR антитело/TGFβR2 4.1.1 Свойства слитого белка EGFR антитело/TGFβR2 по связыванию и конкуренции с TGF-β
Белок TGF-β1 и белок TGF-β3 в конечной концентрации 2 мкг/мл вносили в 96-луночные планшеты в количестве 100 мкл на лунку, соответственно, и оставляли образовывать покрытие в течение ночи при 4°С. Планшеты промывали на следующий день, блокировали при комнатной температуре в течение 1 часа и инкубировали с различными концентрациями (1,22 пМ, 4,88 пМ, 19,53 пМ, 78,13 пМ, 312,5 пМ, 1250 пМ, 500 пМ, 2000 пМ) слитого белка EGFR антитело/TGFβR2 6 в течение 1 часа. После этого планшеты промывали для удаления несвязавшихся антител, и инкубировали со вторичным антителом козы против hIgG502F(ab)2/HRP и повторно промывали, добавляли раствор хромогенного субстрата для развития окраски, и считывали значение OD450 с помощью микропланшетного ридера после окончания реакции. Результаты приведены на Фиг. 19. Способность к связыванию слитого белка 6 с белками TGF-β1 и TGF-β3 была сходной со способностью слитого белка 1, с ЕС50, равной 91 пМ и R2=0,998 по связыванию с белком TGF-β1, и с ЕС50, равной 102 пМ и R2=0,998 по связыванию с белком TGFβ3.
В этом Примере дополнительно анализировали способность слитого белка 6 конкурировать с белком TGF-β1 или белком TGF-β3 за связывание с белком TGFβR2-Fc на уровне белка.
Белок TGF-β1 в конечной концентрации 0,2 мкг/мл или белок TGF-β3 в концентрации 0,5 мкг/мл наносили на 96-луночный планшет в количестве 100 мкл на лунку и инкубировали в течение ночи при 4°С. Планшеты промывали на следующий день, блокировали при комнатной температуре в течение 1 часа, и добавляли 100 мкл слитого белка EGFR антитело/TGFβR2 в разных концентрациях (0,05 нМ, 0,14 нМ, 0,42 нМ, 1,25 нМ, 3,75 нМ, 11,24 нМ, 33,71 нМ, 101,12 нМ) со 100 мкл конечной концентрации 0,2 мкг/мл (конкуренция с TGF-β1) или 1 мкг/мл (конкуренция с TGF-β3) меченого биотином белка TGFβR2-Fc (источник белка: Sino Biological, Inc., а меченого биотином белка: Sinocelltech Limited, далее такой же). Также использовали лунки с белком TGFβR2-Fc в качестве положительного контроля. Планшет промывали после 1 часа инкубации, а затем планшет несколько раз промывали после инкубации в течение 1 часа после добавления вторичного детектирующего антитела стрептавидин/пероксидаза хрена. Наконец, добавляли раствор хромогенного субстрата для развития окраски, и после окончания реакции, микропланшетный ридер считывал значение OD450. Степень конкуренции ингибирования PI% слитого белка рассчитывали на основании значения OD450, и степень ингибирования PI(%)=(значение OD450 лунки с положительным контролем - значение OD450 лунки с образцом)/значение OD450 лунки с положительным контролем × 100%. Результаты приведены на Фиг. 20, а слитой белок 6 обладал схожей способностью блокировать связывание белка TGF-β1 или белка TGF-β3 с TGFβR2-Fc, что и слитой белок 1.
4.1.2. Связывающие свойства слитого белка EGFR антитело/TGFβR2 с EGFR
Со ссылкой на Пример 1.2.1. способность к связыванию слитого белка с рекомбинантным белок EGFR человека измеряли с помощью ИФА. Как показано на Фиг. 21, способность к связыванию слитого белка 6 с EGFR и способность конкурировать с EGF в связывании с EGFR была схожей с такой способностью у EGFR-HPA8, когда ЕС50 была равной 133,1 нг/мл и R2=1,000.
4.2 Анализ аффинности связывания слитого белка EGFR антитело/TGFβR2
В этом Примере аффинность слитого белка EGFR антитело/TGFβR2 в связывании биотинилированного рекомбинантного белка EGFR человека и белка TGF-β1 определяли с помощью системы анализа биомолекулярных взаимодействий (модель: OctetRED96e, производитель: Fortebio) с EGFR-HPA8 и H7N9-R1, соответственно, в качестве отрицательных контролей. Параметры аффинности были получены путем подгонки кривых связывания и диссоциации множества точек концентрации, результаты представлены в Таблицах 13 и 14, а специфические кривые параметров кинетических характеристик показаны на Фиг. 22 и 23.
Эти результаты показывают, что слитой белок 6 сохранил высокую аффинность связывания с белком EGFR человека по сравнению с моноклональным антителом EGFR-НРА8, со значением KD, равным 8,77 пМ, и значением константы связывания kon, равным 1,68Е+06 М-1 с-1, и значением константы диссоциации kdis, равным 1,47Е-05 с-1. В дополнение, слитой белок 6 с укороченной формой TGFβR2 обладает схожей аффинностью с белком EGFR человека, что и слитой белок 1 с полноразмерной формой TGFβR2 со значение KD, равным 96,1 пМ, значением константы связывания kon, равным 1,53Е+06 M с-1, и значением константы диссоциации kdis, равным 1,47Е-04 с-1.
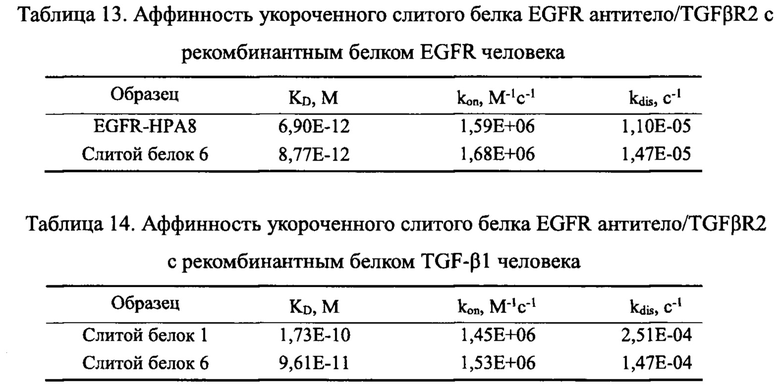
Приведенные выше результаты показывают, что слитой белок 6 имеет хорошее сродство как с EGFR человека, так и с TGF-β1.
4.3. Анализ слитого белка EGFR антитело/TGFβR2 по нейтрализации TGF-β
TGF-β1 может ингибировать пролиферацию клеток Mv-1-lu, поэтому способность слитого белка EGFR антитело/TGFβR2 по нейтрализации TGF-β1 может быть обнаружена с помощью анализа WST-8.
Клетки Mv-1-lu (источник: Центр клеточных ресурсов, Шанхайский институт биологических наук, академия наук Китая) равномерно инокулировали в количестве 50 мкл на лунку в 96-луночный планшет при плотности клеток 1×103 на лунку. Клетки инкубировали в инкубаторе с СО2 в течение примерно 3 ч для их прилипания к стенкам лунки, а затем образцы слитого белка EGFR антитело/TGFβR2 в разных концентрациях (0,0078 нМ, 0,0156 нМ, 0,0313 нМ, 0,0625 нМ, 0,125 нМ, 0,25 нМ, 0,5 нМ, 1 нМ, 2 нМ), разбавленных в среде 1640, содержащей 10% ФБС, добавляли в количестве 50 мкл на лунку. Наконец, добавляли фактор TGF-β1 в конечной концентрации 1 нг/мл в количестве 10 мкл на лунку. Также использовали группу M в качестве положительного контроля (содержащую клетки и TGF-β1), группу М' в качестве отрицательного контроля (содержащую клетки без TGF-β1), и группу В в качестве пустого контроля (содержащую только питательную среду без клеток). Клетки инкубировали в инкубаторе с СО2 при 37°С и 5% СО2 в течение 5 суток, а затем добавляли WST-8 в количестве 10 мкл на лунку. Образцы оставляли в течение 180 мин и значение OD450-OD630 измеряли с помощью микрогшаншетаого ридера, а степень нейтрализации слитого белка EGFR антитело/TGFβR2 рассчитывали путем вычитания значения пустого контроля В. Степень нейтрализации, %=(OD группы М' - OD образца)/(OD группы М' - OD группы M) × 100%, и количественную кривую эффективности анализировали и построили с помощью программного обеспечения GraphPad Prism, горизонтальной координатой являлась концентрация антитела и вертикальной координатой являлась степень ингибирования. Как показано на Фиг. 24, как слитой белок 6, так и контрольный продукт H7N9-R1-43-IgG1 (L9) для TGFβR2 (источник: Sinocelltech Limited для мечения биотином, далее такой же) могли эффективно нейтрализовать ингибирование пролиферации Mv-1-lu посредством TGF-β1 в дозозависимом виде, а полуэффективная концентрация слитого белка 6 была меньше, чем у контрольного продукта H7N9- R1-43-IgG1 (L9), что указывает на лучшую нейтрализующую активность указанной молекулы. EGFR-HPA8 не нейтрализовал TGF-β1, что позволяет предположить, что именно часть TGFβR2 слитого белка 6 проявляет нейтрализующее действие в отношении TGF-β1.
Способность слитого белка 6 к нейтрализации TGF-β была дополнительно проанализирована с использованием системы репортерного гена в этом Примере, со ссылкой на Пример 3.5. Результаты приведены на Фиг. 25, и как слитой белок 6, так и контрольный продукт H7N9-R1-43-IgG1 (L9) для TGFβR2 может эффективно нейтрализовать TGF-β1 в дозозависимом виде, а максимальная скорость нейтрализации слитого белка 6 (74,8%) значительно выше, чем у контрольного продукта H7N9-R1-43-IgG1 (L9) (55.3%), что дополнительно указывает на превосходную нейтрализующую активность слитого белка 6.
4.4. Анализ активности слитого белка EGFR антитело/TGFβR2 по ингибированию пролиферации клеток
Ингибирование слитым белком EGFR антитело/TGFβR2 роста клеток MDA-MB-468 анализировали с помощью метода WST-8, в соответствии с Примером 1.2.2 для определения свойств части EGFR. Как показано на Фиг. 26, способность слитого белка 6 ингибировать пролиферацию клеток MDA-MB-468 была сходной с EGFR-HPA8, а степень ингибирования повышалась с повышением концентрации лекарственного средства по S-образной кривой. Контрольный продукт H7N9-R1-43-IgG1 (L9) с функцией в отношении TGFβR2 не ингибировал пролиферацию клеток MDA-MB-468, указывая на то, что слитой белок 6 ингибировал пролиферацию опухолевых клеток посредством его части EGFR.
4.5. Влияние слитого белка EGFR антитело/TGFβR2 на ADCC
Влияние на ADCC, опосредованное слитым белком EGFR антитело/TGFβR2, клеток, экспрессирующих EGFR,- анализировали со ссылкой на Пример 2.2.3. Результаты приведены на Фиг. 27. В диапазоне концентраций 0,00004-3 нМ, слитой белок 6 и антитело против EGFR EGFR-HPA8 могут вызывать сходные эффекты ADCC на опухолевые клетки А431, экспрессирующие EGFR. Контрольный продукт H7N9-R1-43-IgG1 (L9) с функцией в отношении TGFβR2 на оказывал влияния на ADCC клеток А431, указывая на то, что именно часть EGFR слитого белка 6 вызывала ADCC в этой экспериментальной системе.
Пример 5: Фармакодинамическое исследование слитого белка EGFR антитело/TGFβR2 с укороченной формой TGFβR2 (слитой белок 6) в модели опухоли подкожного трансплантата NCI-H1975
Мышах Balb/c-nu подкожно вводили 1×106 клеток NCI-H1975 в правую сторону грудной клетки. Когда объем опухоль достиг примерно 300 мм3, животных случайным образом группировали по объему опухоли по 6 животных в каждой группе и вводили дозу. Вводить дозу начинали в день распределения по группам, препарат вводили посредством внутрибрюшинной инъекцией (LP) дважды в неделю в течение 10 последовательных доз и прекращали введение после последней дозы для наблюдения за рецидивом опухоли. Конкретный режим дозирования введения показан в Таблице 15 ниже.

Животные в каждой группе находились в хорошем общем состоянии, таком как активность и питание в течение курса введения, масса тела у них несколько увеличилась. Не было существенной разницы в массе тела между группой, которой вводили дозы, и контрольной группой, получавшей растворитель, после введенных доз (Р>0,05). Изменение массы тела всех животных показаны в Таблице 16 и на Фиг. 28.


Результаты измерения объема опухоли для каждой из групп в этом исследовании показаны в Таблице 17 и на Фиг. 29. Через 35 суток лечения в группах, средний объем опухоли контрольной группе, получавшей растворитель, был 7150,78±780,4 мм3. У пяти из шести мышей, которым вводили слитой белок 6, опухоль полностью исчезла (CR) с средним объемом опухоли 4,55±4,55 мм3 и TGI, равным 99,9%, что значимо отличалось от такового у контрольной группы, получавшей растворитель (Р<0,001). В противоположность этому, только у одной мыши из группы, получавшей EGFR-HPA8, обнаружилось полное исчезновение опухоли, со средним объемом опухоли 79,44±28,65 мм3 и TGI, равным 98,9%. Результаты показали, что слитой белок 6 оказывает значительное ингибирующее действие на опухоли немелкоклеточного рака легких NCI-Н1975, трансплантированные подкожно, и эффект ингибирования опухоли превосходит эффект EGFR-HPA8 при той же молярной дозе (Р=0,237).


Пример 6: Анализ стабильности слитого белка EGFR антитело/TGFβR2 с укороченной формой TGFβR2 (слитой белок 6)
6.1. Ультрафильтрационный анализ стабильности слитого белка EGFR антитело/TGFβR2
Образцы слитого белка EGFR антитело/TGFβR2 концентрировали до концентрации примерно 10 мг/мл посредством ультрафильтрации в буфере 100 мМ глицина, 10 мМ NaCl, 50 мМ Трис, рН 7.5. Концентрированные образцы проверяли на чистоту и стабильность ультрафильтрованных образцов методами SDS-ПААГ в восстанавливающих условиях и молекулярно-ситовой хроматографии (SEC-ВЭЖХ, система жидкостной хроматографии Agilent 1260, колонка TSK-G3000SWXL). Стадии SEC-ВЭЖХ: (1) подвижная фаза: 200 мМ NaH2PO4, 100 мМ аргинин, рН 6.5; (2) объем загрузки 80 мкг; (3) время анализа - 30 мин, скорость потока - 0,5 мл/мин, и температура колонки 25°С; (4) чистоту рассчитывали по методу нормализации площади пика.
Результаты теста на чистоту после концентрирования образцов показаны на Фиг. 30, а чистота образцов и соотношение фрагментов показаны в Таблице 18. Результаты показали, что слитой белок 6 с укороченным TGFβR2 с меньшей вероятностью разрушался после концентрирования и имел более высокую степень ультрафильтрации по сравнению со слитым белком с полноразмерным TGFβR2.
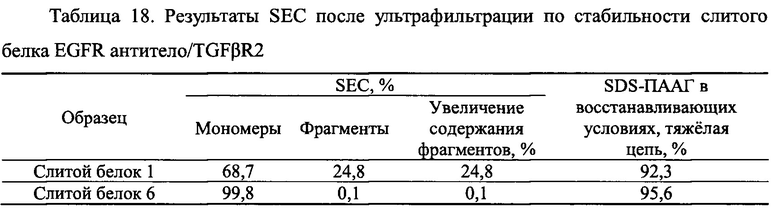
6.2. Анализ термической стабильности слитого белка EGFR антитело/TGFβR2
Термическую стабильность образцов измеряли методом дифференциальной сканирующей флуориметрии (ДСФ) с использованием системы UNcle (Unchained Labs, модель: UNCLE-0330). Стадии процесса: (1) объем образца составлял 9 мкл; (2) экспериментальные параметры установлены следующими: диапазон температур с 25°С по 95°С, а скорость нагревания составляла 0,3°С/мин; (3) для анализа данных использовали программное обеспечение UNcle Analysis, среднее значение кривой изменения внутренней флуоресценции под УФ266 принимали за Tm, а температуру начала агрегации кривой изменения агрегации, формируемой сигналом статического светорассеяния под действием УФ266/Синий473, принимали за Tagg266 и Tagg473.
В Таблице 19 показаны результаты анализа термической стабильность слитого белка 6, который показал хорошую термическую стабильность.

6.3. Анализ термоускоренной стабильности слитого белка EGFR антитело/TGFβR2
После хранения образцов при 45°С в течение 1 недели, термоускоренную стабильность образцов анализировали с помощью SEC-ВЭЖХ и SDS-ПААГ, и процедура была такой же, как и в 6.1.
Результаты анализа термоускоренной стабильности слитого белка 6 показаны в Таблице 20. После 1 недели хранения при 45°С, SEC чистота слитого белка 6 снизилась на 0,7%, но чистота осталась высокой, уровень агрегатов увеличился в меньшей степени, а уровень фрагментов не изменился, что показало хорошую термоускоренную стабильность.

6.4. Анализ стабильности при замораживании/оттаивании слитого белка EGFR антитело/TGFβR2
Образцы хранили при -80°С в течение 3 часов, а затем помещали при 45°С на 1 час для оттаивания, и так осуществляли пять повторных замораживаний и оттаиваний. Стабильность образцов при замораживании/оттаивании анализировали с помощью SEC-ВЭЖХ, и процедура была такой же как и в 6.1.
Результаты стабильности слитого белка 6 при замораживании/оттаивании показаны в Таблице 21. После пяти последовательных замораживаний/оттаиваний, SEC чистота слитого белка 6 существенно не изменилась, а уровни агрегатов и фрагментов существенно не повысился, что продемонстрировало хорошую стабильность при замораживании/оттаивании.

6.5. Стабильность слитого белка EGFR антитело/TGFβR2 при встряхивании Образцы помещали в глубоколуночные планшеты и встряхивали на вортексном шейкере при 800 об./мин в течение 24 часов. Образцы анализировали с помощью SEC-ВЭЖХ по той же методике, что и в 6.1. Результаты приведены в Таблице 22, которые показывают, что чистота SEC образцов в отношении мономеров существенно не изменилась после 24 часов встряхивания, и уровни агрегатов и фрагментов существенно не изменились, указывая на то, что слитой белок 6 обладает хорошей стабильностью при встряхивании.

Пример 7: Конструирование и анализ свойств слитых белков антител X, направленных на антигены солидных опухолей, и TGFβR2, где часть TGFβR2 представляет собой укороченную форму TGFβR2 (слитой белок 6)
7.1 Конструирование вектора экспрессии слитого белка антитело Х/укороченный TGFβR2, экспрессия и очистка
Чтобы дополнительно оценить структурную стабильность и способность к нейтрализации TGF-β1 выбранных укороченных форм TGFβR2 в Примере 3.2, в этом Примере использовали различные антигены твердых опухолей в качестве нацеливающей части слитого белка и внеклеточный домен TGFβR2 (полноразмерный и делегированный ECD (6-26), слитой белок 6) в качестве иммуномодуляторной части слитого белка для образования слитого белка антитело Х/внеклеточный домен TGFβR2 (слитой белок X/TGβPR2). Аналогично описанному выше, в слитом белке X/TGFβR2 С-концевая аминокислота тяжелой цепи антитела X связана с внеклеточным доменом TGFβR2 посредством линкера (G4S)4. Дополнительно С-концевой лизин тяжелой цепи антитела X удаляли для снижения риска протеолитического расщепления. Протокол конструирования слитого белка X/TGFβR2 показан в Таблице 23.
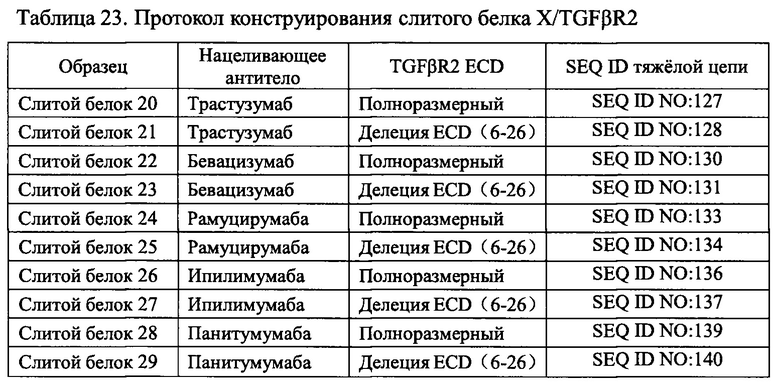
С помощью ПЦР или ПЦР с перекрыванием амплифицировали целевой ген и лигировали его в указанный вектор экспрессии путем инфузии. Правильность последовательности рекомбинантного вектора экспрессии подтверждали секвенированием, плазмиды экстрагировали, транзиентно переносили в клетки HEK-293 (нокаут fut8), культивировали в течение 7 суток и супернатант собирали центрифугированием. Полученный клеточный супернатант очищали с помощью аффинной хроматографии с белком А для очистки слитого белка.
7.2. Деградация слитых белков антитело Х/укороченный TGFβR2
Чистоту, а также деградацию слитого белка X/TGFβR2 оценивали с помощью SDS-ПААГ в восстанавливающих условиях. Результаты приведены на Фиг. 31. Различные экспрессированные образцы слитых белков X/TGFβR2 с выбранными укороченными формами внеклеточного домена TGFβR2 были существенно более стабильными и имели намного меньше полос деградировавших продуктов, чем контрольные образцы с полноразмерным внеклеточным доменом TGFβR2. Следует признать, что стабильность образцов обусловлена присутствующими укороченными формами внеклеточного домена TGFβR2, независимо от того, какие виды нацеливающей части, т.е. антител, используются.
7.3. Тенденция деградации слитого белка антитело Х/укороченный TGFβR2
В этом Примере, супернатант клеток 293Е использовали для ускоренной обработки слитого белка, и стабильность в условиях деградации слитого белка X/TGFβR2 с выбранным укороченным внеклеточным доменом TGFβR2 дополнительно определяли как в Примере 3.3. Результаты анализа чистоты образца показаны на Фиг. 32, а чистота образца и процент отщепленных частей показаны в Таблице 24. Результаты указывают на то, что слитые белки X/TGFβR2 с предпочтительной укороченной формой внеклеточного домена TGFβR2 являются более устойчивыми к протеазной деградации, чем слитой белок с полноразмерной формой TGFβR2.
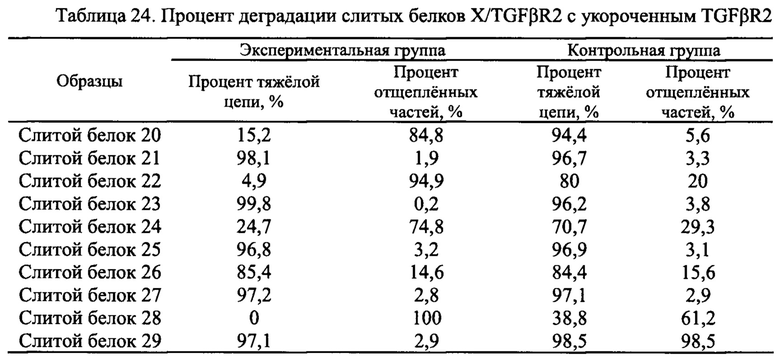
Для оценки стабильности слитых белков X/TGFβR2 с выбранным укороченным внеклеточным доменом TGFβR2 в условиях определенных концентраций, их стабильность при ультрафильтрации анализировали с использованием метода из Примера 6.1. Результаты приведены в Таблице 25. Слитой белок X/TGFβR2 с предпочтительным укороченным внеклеточным доменом TGFβR2 меньше подвержен деградации. После концентрирования процентное содержание отщепленных частей было менее 4,0% (чистота по SDS-ПААГ), что говорит о более высокой стабильности при ультрафильтрации, чем слитой белок X/TGFβR2 с полноразмерным внеклеточным доменом TGFβR2.
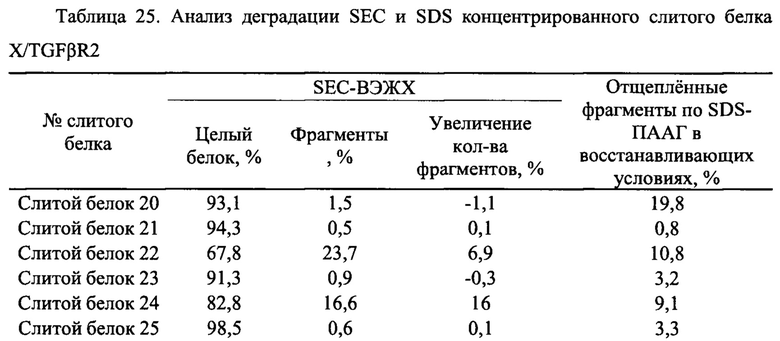

Таким образом, отличная стабильность слитого белка, содержащего предпочтительный укороченный внеклеточный домен TGFβR2, была дополнительно подтверждена с использованием слитых белков с несколькими типами антител против антигенов солидных опухолей в качестве нацеливающей части.
Пример 8: Биологические свойства in vitro слитых белков X/TGFβR2 с нацеливающими на антигены солидных опухолей антителами в качестве нацеливающей части и укороченным TGFβR2 (слитой белок 6)
8.1. Анализ связывания слитого белка X/TGFβR2
Способность к связыванию слитых белков X/TGFβR2 с TGF-β1 измеряли с помощью ИФА в соответствии с методом из Примера 3.4. Как показано на Фиг. 33, способность слитых белков X/TGFβR2 с предпочтительной укороченной формой TGFβR2 блокировать TGF-β1 была немного ниже, чем у слитого белка X/TGFβR2 с полноразмерной формой TGFβR2.
8.2. Анализ слитого белка X/TGFβR2 в нейтрализации TGF-β
Способность слитого белка X/TGFβR2 к нейтрализации TGF-β1 и TGF-β3 проверяли со ссылкой на Пример 3.5. Как показано на Фиг. 34, способность слитого белка X/TGFβR2 с предпочтительной укороченной формой TGFβR2 к нейтрализации как TGF-β1, так и TGF-β3 была выше, чем у слитого белка X/TGFβR2 с полноразмерной формой
8.3. Анализ связывания слитого белка X/TGFβR2 с мишенью части X
Антигены ERBB2-his, VEGF165, VEGFR2-His, CTLA4-his и EGFR-His, которые являются мишенью для нацеливающих частей X, в конечных концентрациях 10 нг/мл, 5 нг/мл, 80 нг/мл, 80 нг/мл, и 40 нг/мл, соответственно, наносили на 96-луночные планшеты в количестве 100 мкл на лунку. Планшеты покрывали ими в течение ночи при 4°С. Планшеты промывали на следующий день, блокировали при комнатной температуре в течение 1 часа и инкубировали с 100 мкл слитого белка X-TGFβR2 в конечной концентрации 13,89 нМ в течение 1 часа. Планшеты промывали для удаления несвязавшихся антител, инкубировали со вторичным антителом козы против hIgG Fc/HRP и повторяли отмывку. Наконец, раствор хромогенного субстрата добавляли для развития окраски, и считывали OD450 с помощью микропланшетного ридера после окончания реакции. Как показано на Фиг. 35, слитой белок X/TGFβR2 с выбранным укороченным TGFβR2 обладал схожей способностью к блокированию соответствующего антигена со стороны части X в слитом белке X/TGFβR2, содержащем полноразмерную форму TGFβR2.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
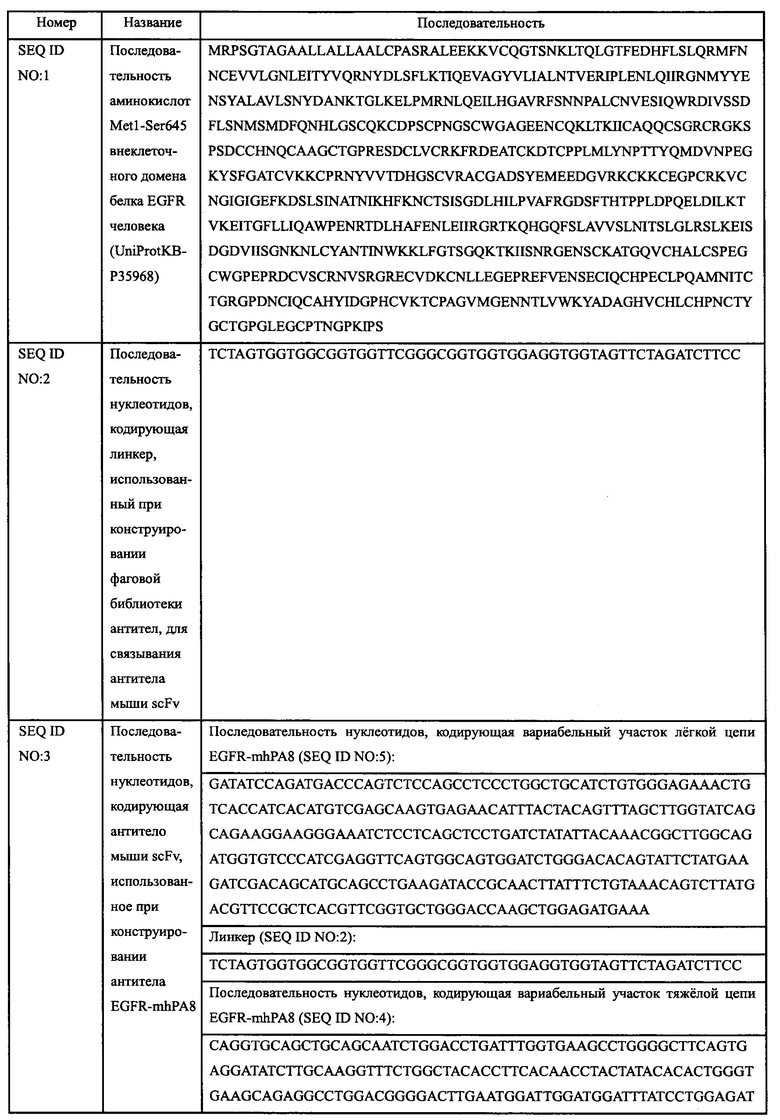















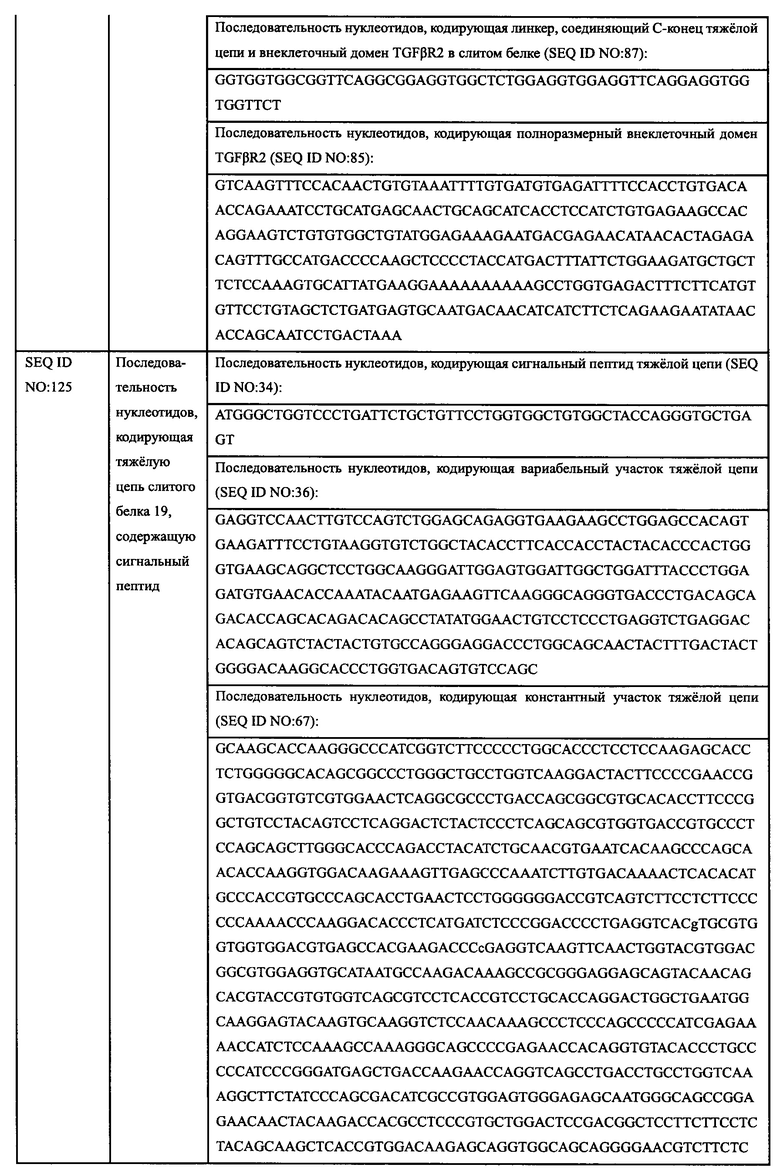
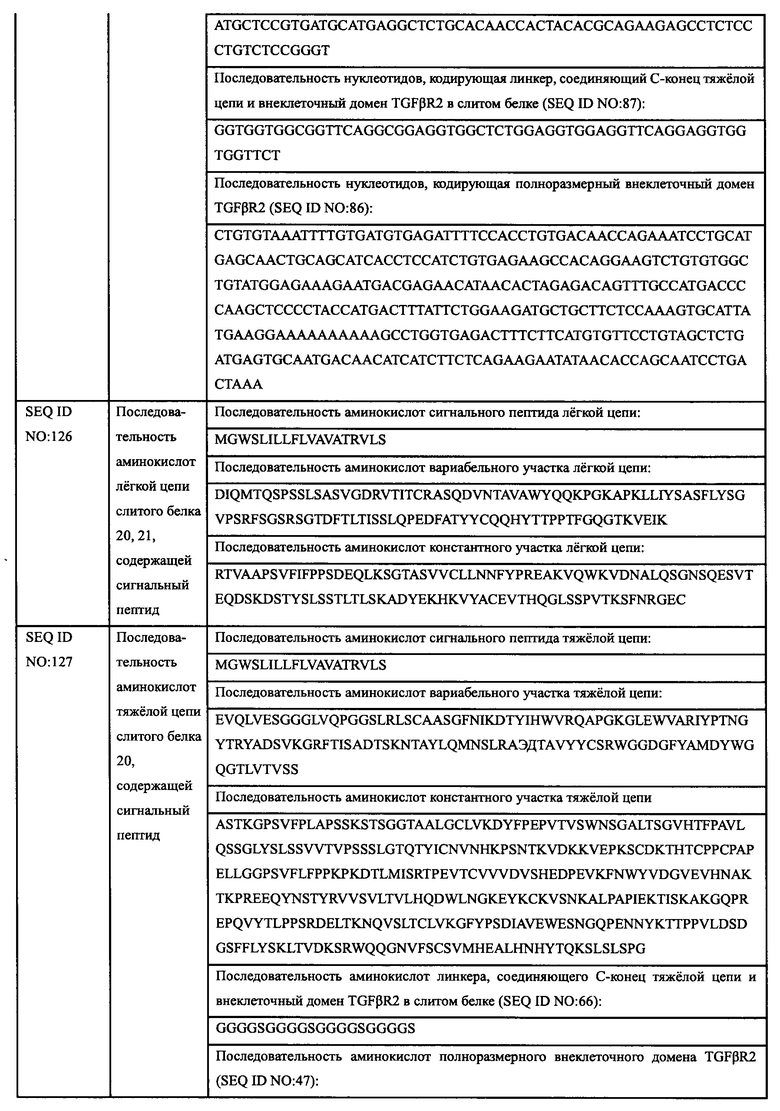
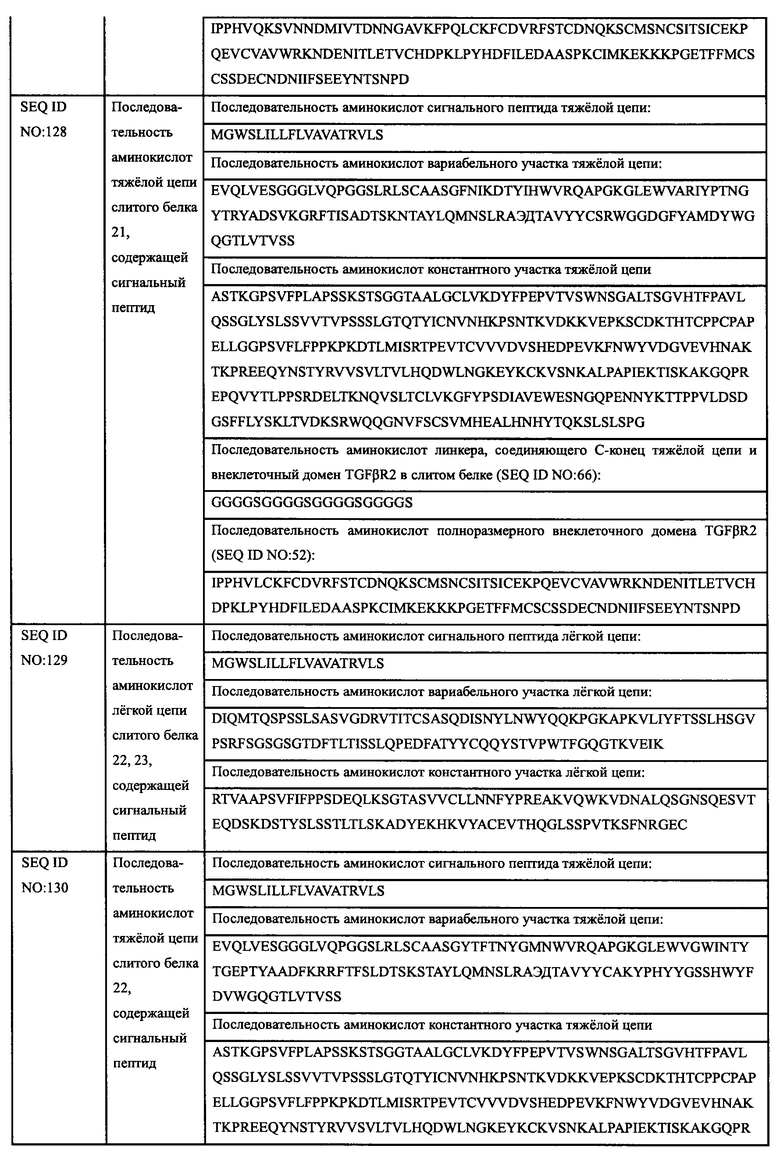

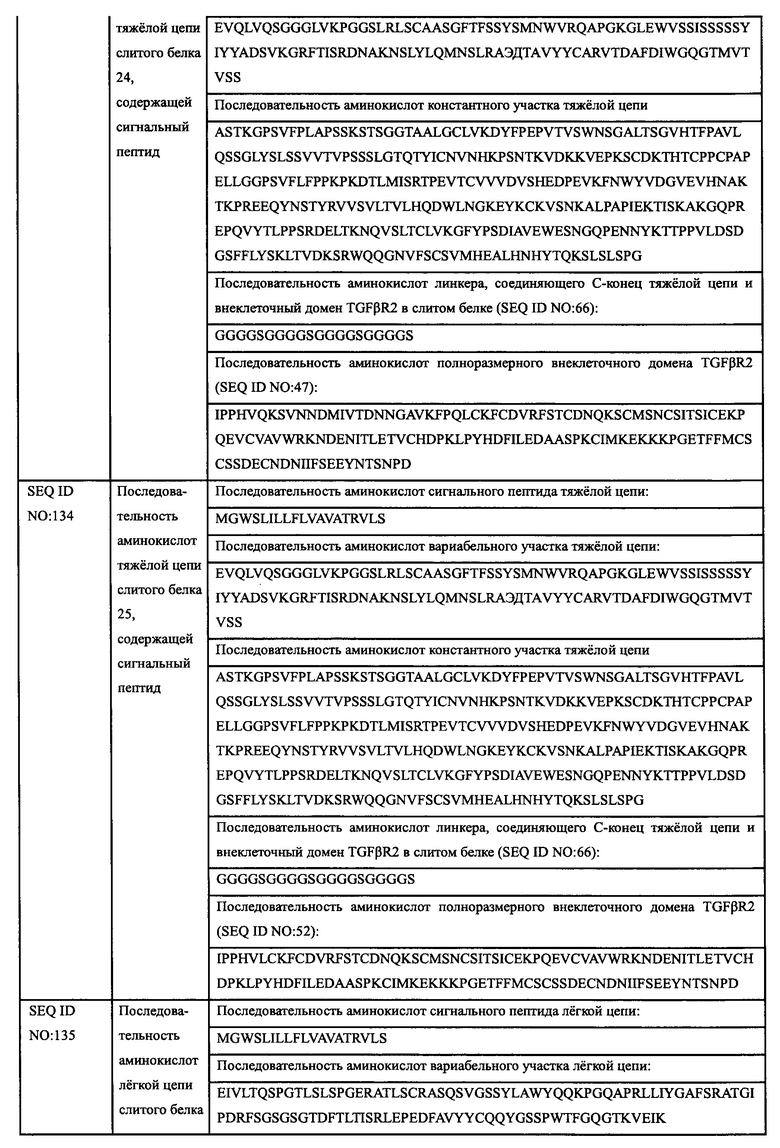


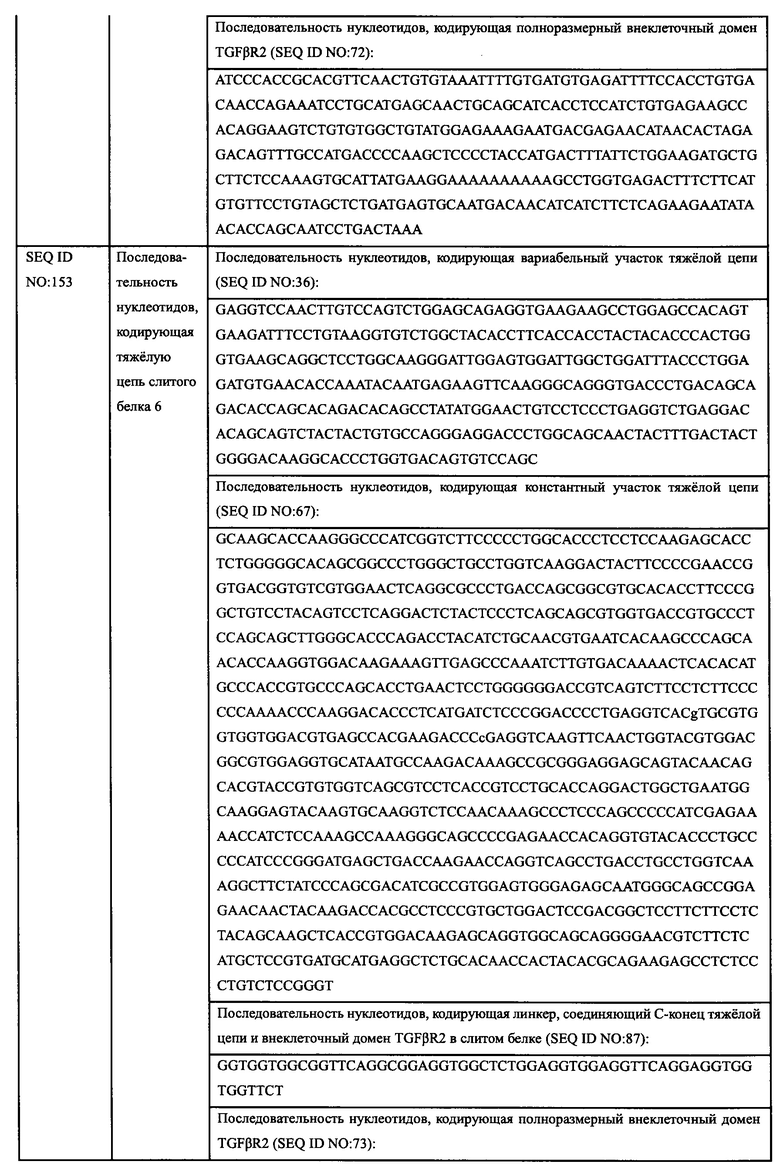








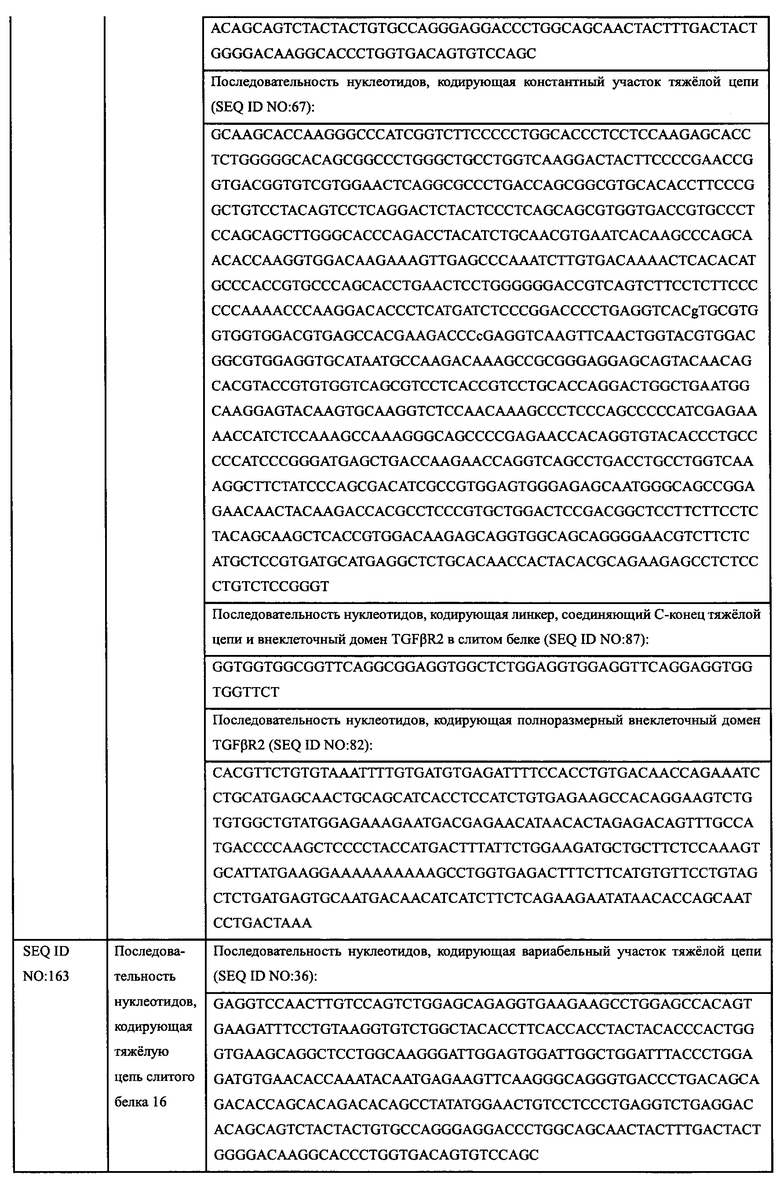
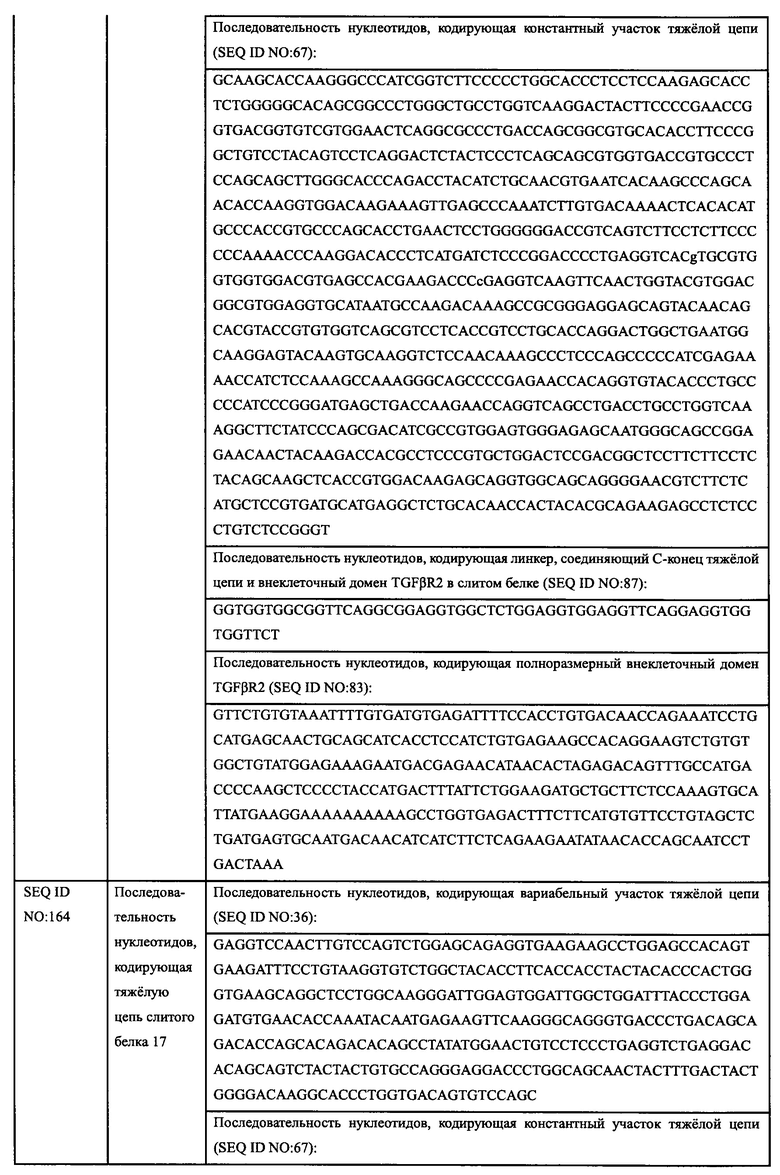
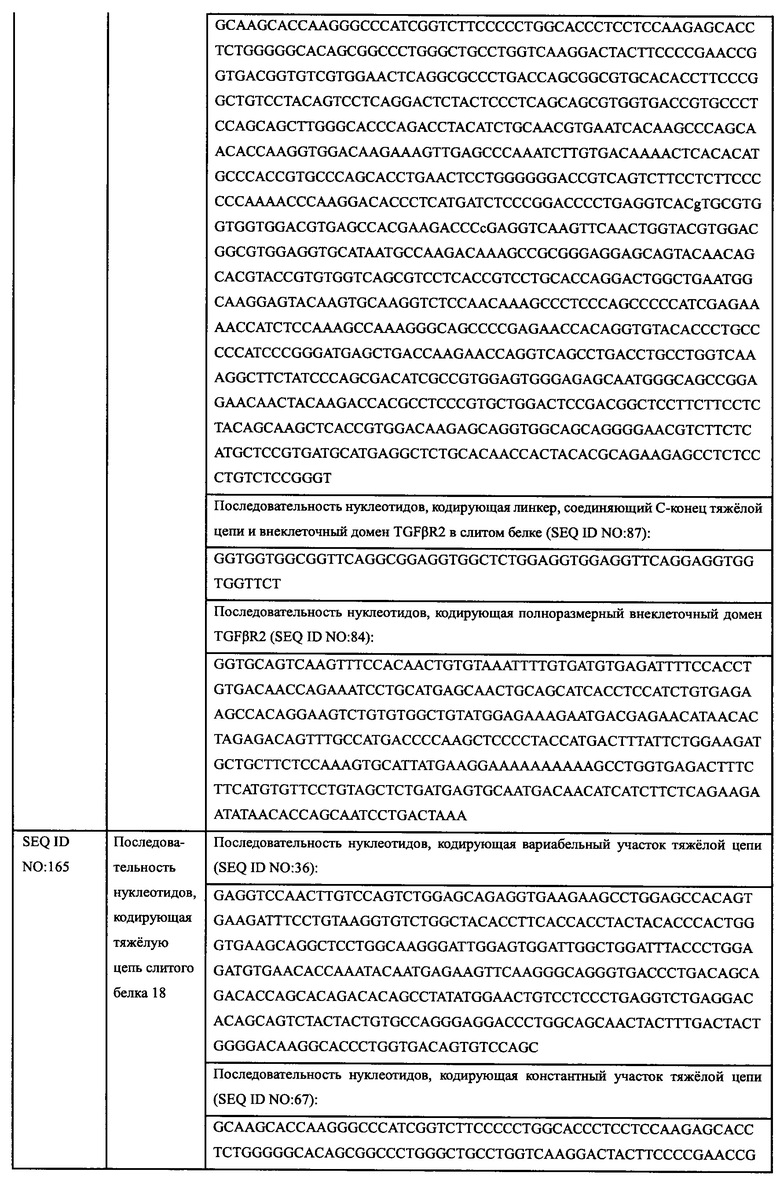


ССЫЛОЧНЫЕ МАТЕРИАЛЫ
1. Xie, E, и др. TGF-beta signaling in cancer metastasis. Acta Biochim Biophys Sin (Shanghai), 2018. 50(1): p. 121-132.
2. Colak, S. и P. Ten Dijke. Targeting TGF-beta Signaling in Cancer. Trends Cancer, 2017. 3(1): p. 56-71.
3. Fabregat, I. и др. TGF-beta signaling in cancer treatment. Curr Pharm Des, 2014. 20(17): p. 2934-47.
4. Batlle, E. и J. Massague, Transforming Growth Factor-beta Signaling in Immunity and Cancer. Immunity, 2019. 50(4): p. 924-940.
5. Laskin, J.J. и A.B. Sandler, Epidermal growth factor receptor: a promising target in solid tumours. Cancer treatment reviews, 2004. 30(1): p. 1-17.
6. Hynes, N. и др. The ErbB receptor tyrosine family as signal integrators. Endocrine-related cancer, 2001. 8(3): p. 151-159.
7. Zandi, R. и др. Mechanisms for oncogenic activation of the epidermal growth factor receptor. Cellular signalling, 2007. 19(10): p. 2013-2023.
8. Seshacharyulu, P. и др. Targeting the EGFR signaling pathway in cancer therapy. Expert opinion on therapeutic targets, 2012. 16(1): p. 15-31.
9. Zhao, Y. и др. TGF -β transactivates EGFR and facilitates mammary cancer migration and invasion through canonical Smad3 and ERK/Sp1 signaling pathways. Molecular oncology, 2018.12(3): p. 305-321.
10. Wendt, M.K., J. A. Smith, и W.P. Schiemann, Transforming growth factors-induced epithelial-mesenchymal transition facilitates epidermal growth factor-dependent mammary cancer progression. Oncogene, 2010. 29(49): p. 6485.
11. Lee, E., и др. Transforming growth factorβ1 transactivates EGFR via an H2O2-dependent mechanism in squamous carcinoma cell line. Cancer letters, 2010. 290(1): p. 43-48.
12. Dunfield, L.D. и M.W. Nachtigal, Inhibition of the antiproliferative effect of TGFβ by EGF in primary human ovarian cancer cells. Oncogene, 2003. 22(30): p. 4745.
13. Kretzschmar, M. и др. A mechanism of repression of TGFβ/Smad signaling by oncogenic Ras. Genes & development, 1999. 13(7): p. 804-816.
14. ten Dijke, P., K. Miyazono, и C.-H. Heldin, Signaling inputs converge on nuclear effectors in TGF-β signaling. Trends in biochemical sciences, 2000. 25(2): p. 64-70.
15. Funaba, M., CM. Zimmerman, и L.S. Mathews, Modulation of Smad2-mediated signaling by extracellular signal-regulated kinase. Journal of Biological Chemistry, 2002. 277(44): p. 41361-41368.
16. Richter, P. и др. EGF/TGFbeta1 co-stimulation of oral squamous cell carcinoma cells causes an epithelial-mesenchymal transition cell phenotype expressing laminin 332. J Oral Pathol Med, 2011. 40(1): p. 46-54.
17. Uttamsingh, S. и др. Synergistic effect between EGF and TGF-beta1 in inducing oncogenic properties of intestinal epithelial cells. Oncogene, 2008. 27(18): p. 2626-34.
18. Xu, Z. и др. TGFbeta and EGF synergistically induce a more invasive phenotype of epithelial ovarian cancer cells. Biochem Biophys Res Commun, 2010. 401(3): p. 376-81.
19. Xiong, J. и др. Epidermal growth factor promotes transforming growth factor-beta 1-induced epithelial-mesenchymal transition in HK-2 cells through a synergistic effect on Snail. Mol Biol Rep, 2014. 41(1): p. 241-50.
20. Buonato, J.M., I.S. Lan, и M.J. Lazzara, EGF augments TGFbeta-induced epithelial-mesenchymal transition by promoting SHP2 binding to GAB1. J Cell Sci, 2015. 128(21): p. 3898-909.
21. Wang, Т. и др. The TGFβ-miR-499a-SHKBP1 pathway induces resistance to EGFR inhibitors in остеосаркома cancer stem cell-like cells. Journal of Experimental & Clinical Cancer Research, 2019. 38(1): p. 226.
22. Jie, H.B. и др. CTLA-4(+) Regulatory T Cells Increased in Cetuximab-Treated Head and Neck Cancer Patients Suppress NK Cell Cytotoxicity and Correlate with Poor Prognois. Cancer Res, 2015. 75(11): p. 2200-10.
23. Bedi, A. и др. Inhibition of TGF-beta enhances the in vivo antitumor efficacy of EGF receptor-targeted therapy. Mol Cancer Ther, 2012. 11(11): p. 2429-39.
24. Zhang, Y. и др. The canonical TGF-beta/Smad signalling pathway is involved in PD-L1-induced primary resistance to EGFR-TKIs in EGFR-mutant non-small-cell lung cancer. Respir Res, 2019. 20(1): p. 164.
25. Jones, S.T. и M.M. Bendig, Rapid PCR-cloning of full-length mouse immunoglobulin variable regions. Biotechnology (N Y), 1991. 9(6): p. 579.
26. Kabat, E.A. и др. Sequences of proteins of immunological interest. 1992: DIANE publishing.
27. Jones, P.T. и др. Replacing the complementarity-determining regions in a human antibody with those from a mouse. Nature, 1986. 321(6069): p. 522.
28. Verhoeyen, M. и L. Riechmann, Engineering of antibodies. BioEssays, 1988. 8(2 - 3): p. 74-78.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SinoCellTech Ltd.
<120> МОЛЕКУЛА TGFбетаR2 С УКОРОЧЕННЫМ ВНЕКЛЕТОЧНЫМ ДОМЕНОМ, СЛИТОЙ БЕЛОК
МОЛЕКУЛЫ TGFбетаR2 С УКОРОЧЕННЫМ ВНЕКЛЕТОЧНЫМ ДОМЕНОМ И АНТИТЕЛА ПРОТИВ EGFR, И
ПРОТИВООПУХОЛЕВОЕ ПРИМЕНЕНИЕ СЛИТОГО БЕЛКА
<130> PCT69149SXB
<160> 190
<170> PatentIn version 3.3
<210> 1
<211> 645
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 1
Met Arg Pro Ser Gly Thr Ala Gly Ala Ala Leu Leu Ala Leu Leu Ala
1 5 10 15
Ala Leu Cys Pro Ala Ser Arg Ala Leu Glu Glu Lys Lys Val Cys Gln
20 25 30
Gly Thr Ser Asn Lys Leu Thr Gln Leu Gly Thr Phe Glu Asp His Phe
35 40 45
Leu Ser Leu Gln Arg Met Phe Asn Asn Cys Glu Val Val Leu Gly Asn
50 55 60
Leu Glu Ile Thr Tyr Val Gln Arg Asn Tyr Asp Leu Ser Phe Leu Lys
65 70 75 80
Thr Ile Gln Glu Val Ala Gly Tyr Val Leu Ile Ala Leu Asn Thr Val
85 90 95
Glu Arg Ile Pro Leu Glu Asn Leu Gln Ile Ile Arg Gly Asn Met Tyr
100 105 110
Tyr Glu Asn Ser Tyr Ala Leu Ala Val Leu Ser Asn Tyr Asp Ala Asn
115 120 125
Lys Thr Gly Leu Lys Glu Leu Pro Met Arg Asn Leu Gln Glu Ile Leu
130 135 140
His Gly Ala Val Arg Phe Ser Asn Asn Pro Ala Leu Cys Asn Val Glu
145 150 155 160
Ser Ile Gln Trp Arg Asp Ile Val Ser Ser Asp Phe Leu Ser Asn Met
165 170 175
Ser Met Asp Phe Gln Asn His Leu Gly Ser Cys Gln Lys Cys Asp Pro
180 185 190
Ser Cys Pro Asn Gly Ser Cys Trp Gly Ala Gly Glu Glu Asn Cys Gln
195 200 205
Lys Leu Thr Lys Ile Ile Cys Ala Gln Gln Cys Ser Gly Arg Cys Arg
210 215 220
Gly Lys Ser Pro Ser Asp Cys Cys His Asn Gln Cys Ala Ala Gly Cys
225 230 235 240
Thr Gly Pro Arg Glu Ser Asp Cys Leu Val Cys Arg Lys Phe Arg Asp
245 250 255
Glu Ala Thr Cys Lys Asp Thr Cys Pro Pro Leu Met Leu Tyr Asn Pro
260 265 270
Thr Thr Tyr Gln Met Asp Val Asn Pro Glu Gly Lys Tyr Ser Phe Gly
275 280 285
Ala Thr Cys Val Lys Lys Cys Pro Arg Asn Tyr Val Val Thr Asp His
290 295 300
Gly Ser Cys Val Arg Ala Cys Gly Ala Asp Ser Tyr Glu Met Glu Glu
305 310 315 320
Asp Gly Val Arg Lys Cys Lys Lys Cys Glu Gly Pro Cys Arg Lys Val
325 330 335
Cys Asn Gly Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn
340 345 350
Ala Thr Asn Ile Lys His Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp
355 360 365
Leu His Ile Leu Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His Thr
370 375 380
Pro Pro Leu Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys Glu
385 390 395 400
Ile Thr Gly Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp
405 410 415
Leu His Ala Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln
420 425 430
His Gly Gln Phe Ser Leu Ala Val Val Ser Leu Asn Ile Thr Ser Leu
435 440 445
Gly Leu Arg Ser Leu Lys Glu Ile Ser Asp Gly Asp Val Ile Ile Ser
450 455 460
Gly Asn Lys Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu
465 470 475 480
Phe Gly Thr Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu
485 490 495
Asn Ser Cys Lys Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser Pro
500 505 510
Glu Gly Cys Trp Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg Asn
515 520 525
Val Ser Arg Gly Arg Glu Cys Val Asp Lys Cys Asn Leu Leu Glu Gly
530 535 540
Glu Pro Arg Glu Phe Val Glu Asn Ser Glu Cys Ile Gln Cys His Pro
545 550 555 560
Glu Cys Leu Pro Gln Ala Met Asn Ile Thr Cys Thr Gly Arg Gly Pro
565 570 575
Asp Asn Cys Ile Gln Cys Ala His Tyr Ile Asp Gly Pro His Cys Val
580 585 590
Lys Thr Cys Pro Ala Gly Val Met Gly Glu Asn Asn Thr Leu Val Trp
595 600 605
Lys Tyr Ala Asp Ala Gly His Val Cys His Leu Cys His Pro Asn Cys
610 615 620
Thr Tyr Gly Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly
625 630 635 640
Pro Lys Ile Pro Ser
645
<210> 2
<211> 54
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 2
tctagtggtg gcggtggttc gggcggtggt ggaggtggta gttctagatc ttcc 54
<210> 3
<211> 732
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 3
gatatccaga tgacccagtc tccagcctcc ctggctgcat ctgtgggaga aactgtcacc 60
atcacatgtc gagcaagtga gaacatttac tacagtttag cttggtatca gcagaaggaa 120
gggaaatctc ctcagctcct gatctatatt acaaacggct tggcagatgg tgtcccatcg 180
aggttcagtg gcagtggatc tgggacacag tattctatga agatcgacag catgcagcct 240
gaagataccg caacttattt ctgtaaacag tcttatgacg ttccgctcac gttcggtgct 300
gggaccaagc tggagatgaa atctagtggt ggcggtggtt cgggcggtgg tggaggtggt 360
agttctagat cttcccaggt gcagctgcag caatctggac ctgatttggt gaagcctggg 420
gcttcagtga ggatatcttg caaggtttct ggctacacct tcacaaccta ctatacacac 480
tgggtgaagc agaggcctgg acggggactt gaatggattg gatggattta tcctggagat 540
gttaatacga agtacaatga gaaattcaag ggcaaggcca cactgactgc agacaaaacc 600
tccagcacag cctacatgca gctcagcagc ctgacctctg aggactctgc ggtctatttc 660
tgtgcaagag aagaccccgg tagtaactac tttgactact ggggccaagg caccactctc 720
acagtctcct ca 732
<210> 4
<211> 357
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 4
caggtgcagc tgcagcaatc tggacctgat ttggtgaagc ctggggcttc agtgaggata 60
tcttgcaagg tttctggcta caccttcaca acctactata cacactgggt gaagcagagg 120
cctggacggg gacttgaatg gattggatgg atttatcctg gagatgttaa tacgaagtac 180
aatgagaaat tcaagggcaa ggccacactg actgcagaca aaacctccag cacagcctac 240
atgcagctca gcagcctgac ctctgaggac tctgcggtct atttctgtgc aagagaagac 300
cccggtagta actactttga ctactggggc caaggcacca ctctcacagt ctcctca 357
<210> 5
<211> 321
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 5
gatatccaga tgacccagtc tccagcctcc ctggctgcat ctgtgggaga aactgtcacc 60
atcacatgtc gagcaagtga gaacatttac tacagtttag cttggtatca gcagaaggaa 120
gggaaatctc ctcagctcct gatctatatt acaaacggct tggcagatgg tgtcccatcg 180
aggttcagtg gcagtggatc tgggacacag tattctatga agatcgacag catgcagcct 240
gaagataccg caacttattt ctgtaaacag tcttatgacg ttccgctcac gttcggtgct 300
gggaccaagc tggagatgaa a 321
<210> 6
<211> 993
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 6
gcaagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacgt gcgtggtggt ggacgtgagc cacgaagacc ccgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 7
<211> 324
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 7
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttag 324
<210> 8
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 8
Gln Val Gln Leu Gln Gln Ser Gly Pro Asp Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Arg Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Thr His Trp Val Lys Gln Arg Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Thr Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 9
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 9
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ala Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Tyr Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Glu Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Ile Thr Asn Gly Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Met Lys Ile Asp Ser Met Gln Pro
65 70 75 80
Glu Asp Thr Ala Thr Tyr Phe Cys Lys Gln Ser Tyr Asp Val Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Met Lys
100 105
<210> 10
<211> 11
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 10
Arg Ala Ser Glu Asn Ile Tyr Tyr Ser Leu Ala
1 5 10
<210> 11
<211> 7
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 11
Ile Thr Asn Gly Leu Ala Asp
1 5
<210> 12
<211> 9
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 12
Lys Gln Ser Tyr Asp Val Pro Leu Thr
1 5
<210> 13
<211> 10
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 13
Gly Tyr Thr Phe Thr Thr Tyr Tyr Thr His
1 5 10
<210> 14
<211> 17
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 14
Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 15
<211> 12
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 15
Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr
1 5 10
<210> 16
<211> 11
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 16
Arg Ala Ser Glu Asn Ile Tyr Tyr Ser Leu Ala
1 5 10
<210> 17
<211> 7
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 17
Ile Thr Asp Gly Leu Ala Asp
1 5
<210> 18
<211> 9
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 18
Lys Gln Ser Tyr Asp Val Pro Leu Thr
1 5
<210> 19
<211> 10
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 19
Gly Tyr Thr Phe Thr Thr Tyr Tyr Thr His
1 5 10
<210> 20
<211> 17
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 20
Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 21
<211> 12
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 21
Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr
1 5 10
<210> 22
<211> 449
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 22
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 23
<211> 214
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 23
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Tyr Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Gln Leu Leu Ile
35 40 45
Tyr Ile Thr Asp Gly Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Lys Ile Asp Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln Ser Tyr Asp Val Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 24
<211> 468
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 24
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Lys
465
<210> 25
<211> 233
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 25
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile
35 40 45
Tyr Tyr Ser Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Gln
50 55 60
Leu Leu Ile Tyr Ile Thr Asp Gly Leu Ala Asp Gly Val Pro Ser Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Lys Ile Asp Ser
85 90 95
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln Ser Tyr Asp
100 105 110
Val Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 26
<211> 19
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 26
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser
<210> 27
<211> 19
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 27
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 28
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 28
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 29
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 29
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Tyr Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Gln Leu Leu Ile
35 40 45
Tyr Ile Thr Asp Gly Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Lys Ile Asp Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln Ser Tyr Asp Val Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 30
<211> 330
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 30
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 31
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 31
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 32
<211> 1407
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 32
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg taaatga 1407
<210> 33
<211> 702
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 33
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagtgac 60
atccagatga cccagagccc atcctccctg tctgcctctg tgggagacag ggtgaccatc 120
acttgtaggg catctgagaa catctactac tccctggctt ggtatcaaca gaagcctggc 180
aaggctccac agctgctgat ttacatcacc gacggactgg ctgatggagt gccaagcagg 240
ttctctggct ctggctctgg cacagactac accctgaaga tcgactccct ccaacctgag 300
gactttgcca cctactactg taagcagtcc tatgatgtgc cactgacctt tggaggaggc 360
accaaggtgg agattaagcg tacggtggct gcaccatctg tcttcatctt cccgccatct 420
gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc 480
agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag 540
agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg 600
agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg 660
agctcgcccg tcacaaagag cttcaacagg ggagagtgtt ga 702
<210> 34
<211> 57
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 34
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagt 57
<210> 35
<211> 57
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 35
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagt 57
<210> 36
<211> 357
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 36
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagc 357
<210> 37
<211> 321
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 37
gacatccaga tgacccagag cccatcctcc ctgtctgcct ctgtgggaga cagggtgacc 60
atcacttgta gggcatctga gaacatctac tactccctgg cttggtatca acagaagcct 120
ggcaaggctc cacagctgct gatttacatc accgacggac tggctgatgg agtgccaagc 180
aggttctctg gctctggctc tggcacagac tacaccctga agatcgactc cctccaacct 240
gaggactttg ccacctacta ctgtaagcag tcctatgatg tgccactgac ctttggagga 300
ggcaccaagg tggagattaa g 321
<210> 38
<211> 993
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 38
gcaagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacgt gcgtggtggt ggacgtgagc cacgaagacc ccgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa tga 993
<210> 39
<211> 324
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 39
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttga 324
<210> 40
<211> 244
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ala Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Tyr Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Glu Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Ile Thr Asn Gly Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Met Lys Ile Asp Ser Met Gln Pro
65 70 75 80
Glu Asp Thr Ala Thr Tyr Phe Cys Lys Gln Ser Tyr Asp Val Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Met Lys Ser Ser Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg Ser Ser Gln Val Gln
115 120 125
Leu Gln Gln Ser Gly Pro Asp Leu Val Lys Pro Gly Ala Ser Val Arg
130 135 140
Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Thr Tyr Tyr Thr His
145 150 155 160
Trp Val Lys Gln Arg Pro Gly Arg Gly Leu Glu Trp Ile Gly Trp Ile
165 170 175
Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn Glu Lys Phe Lys Gly Lys
180 185 190
Ala Thr Leu Thr Ala Asp Lys Thr Ser Ser Thr Ala Tyr Met Gln Leu
195 200 205
Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys Ala Arg Glu
210 215 220
Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu
225 230 235 240
Thr Val Ser Ser
<210> 41
<211> 18
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 41
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg
1 5 10 15
Ser Ser
<210> 42
<211> 1407
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 42
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtcag 60
gtgcagctgc agcaatctgg acctgatttg gtgaagcctg gggcttcagt gaggatatct 120
tgcaaggttt ctggctacac cttcacaacc tactatacac actgggtgaa gcagaggcct 180
ggacggggac ttgaatggat tggatggatt tatcctggag atgttaatac gaagtacaat 240
gagaaattca agggcaaggc cacactgact gcagacaaaa cctccagcac agcctacatg 300
cagctcagca gcctgacctc tgaggactct gcggtctatt tctgtgcaag agaagacccc 360
ggtagtaact actttgacta ctggggccaa ggcaccactc tcacagtctc ctcagcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg taaatga 1407
<210> 43
<211> 702
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 43
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagtgat 60
atccagatga cccagtctcc agcctccctg gctgcatctg tgggagaaac tgtcaccatc 120
acatgtcgag caagtgagaa catttactac agtttagctt ggtatcagca gaaggaaggg 180
aaatctcctc agctcctgat ctatattaca aacggcttgg cagatggtgt cccatcgagg 240
ttcagtggca gtggatctgg gacacagtat tctatgaaga tcgacagcat gcagcctgaa 300
gataccgcaa cttatttctg taaacagtct tatgacgttc cgctcacgtt cggtgctggg 360
accaagctgg agatgaaacg tacggtggct gcaccatctg tcttcatctt cccgccatct 420
gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc 480
agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag 540
agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg 600
agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg 660
agctcgcccg tcacaaagag cttcaacagg ggagagtgtt ag 702
<210> 44
<211> 468
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 44
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Pro Asp Leu Val Lys
20 25 30
Pro Gly Ala Ser Val Arg Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Arg Pro Gly Arg Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Thr Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Lys
465
<210> 45
<211> 233
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 45
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ala Ala
20 25 30
Ser Val Gly Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile
35 40 45
Tyr Tyr Ser Leu Ala Trp Tyr Gln Gln Lys Glu Gly Lys Ser Pro Gln
50 55 60
Leu Leu Ile Tyr Ile Thr Asn Gly Leu Ala Asp Gly Val Pro Ser Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Gln Tyr Ser Met Lys Ile Asp Ser
85 90 95
Met Gln Pro Glu Asp Thr Ala Thr Tyr Phe Cys Lys Gln Ser Tyr Asp
100 105 110
Val Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Met Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 46
<211> 329
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 46
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 47
<211> 136
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 47
Ile Pro Pro His Val Gln Lys Ser Val Asn Asn Asp Met Ile Val Thr
1 5 10 15
Asp Asn Asn Gly Ala Val Lys Phe Pro Gln Leu Cys Lys Phe Cys Asp
20 25 30
Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys
35 40 45
Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val
50 55 60
Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp
65 70 75 80
Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro
85 90 95
Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met
100 105 110
Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu
115 120 125
Glu Tyr Asn Thr Ser Asn Pro Asp
130 135
<210> 48
<211> 124
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 48
Ile Pro Pro His Val Asn Asn Gly Ala Val Lys Phe Pro Gln Leu Cys
1 5 10 15
Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys
20 25 30
Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val
35 40 45
Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr
50 55 60
Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp
65 70 75 80
Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu
85 90 95
Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile
100 105 110
Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
115 120
<210> 49
<211> 122
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 49
Ile Pro Pro His Val Gly Ala Val Lys Phe Pro Gln Leu Cys Lys Phe
1 5 10 15
Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser
20 25 30
Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val
35 40 45
Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys
50 55 60
His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala
65 70 75 80
Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe
85 90 95
Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe
100 105 110
Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
115 120
<210> 50
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 50
Ile Pro Pro His Val Val Lys Phe Pro Gln Leu Cys Lys Phe Cys Asp
1 5 10 15
Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys
20 25 30
Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val
35 40 45
Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp
50 55 60
Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro
65 70 75 80
Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met
85 90 95
Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu
100 105 110
Glu Tyr Asn Thr Ser Asn Pro Asp
115 120
<210> 51
<211> 116
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 51
Ile Pro Pro His Val Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser
1 5 10 15
Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser
20 25 30
Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn
35 40 45
Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro
50 55 60
Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met
65 70 75 80
Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser
85 90 95
Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr
100 105 110
Ser Asn Pro Asp
115
<210> 52
<211> 115
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 52
Ile Pro Pro His Val Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr
1 5 10 15
Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile
20 25 30
Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp
35 40 45
Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr
50 55 60
His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys
65 70 75 80
Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser
85 90 95
Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser
100 105 110
Asn Pro Asp
115
<210> 53
<211> 114
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 53
Ile Pro Pro His Val Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys
1 5 10 15
Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys
20 25 30
Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu
35 40 45
Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His
50 55 60
Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu
65 70 75 80
Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp
85 90 95
Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn
100 105 110
Pro Asp
<210> 54
<211> 116
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 54
Ile Pro Pro His Val Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser
1 5 10 15
Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser
20 25 30
Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn
35 40 45
Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro
50 55 60
Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met
65 70 75 80
Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser
85 90 95
Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr
100 105 110
Ser Asn Pro Asp
115
<210> 55
<211> 114
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 55
Ile Pro Pro His Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys
1 5 10 15
Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys
20 25 30
Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu
35 40 45
Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His
50 55 60
Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu
65 70 75 80
Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp
85 90 95
Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn
100 105 110
Pro Asp
<210> 56
<211> 113
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 56
Ile Pro Pro Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp
1 5 10 15
Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu
20 25 30
Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn
35 40 45
Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp
50 55 60
Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys
65 70 75 80
Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu
85 90 95
Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro
100 105 110
Asp
<210> 57
<211> 112
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 57
Ile Pro Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn
1 5 10 15
Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys
20 25 30
Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile
35 40 45
Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe
50 55 60
Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys
65 70 75 80
Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys
85 90 95
Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
100 105 110
<210> 58
<211> 111
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 58
Ile Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln
1 5 10 15
Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro
20 25 30
Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr
35 40 45
Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile
50 55 60
Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys
65 70 75 80
Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn
85 90 95
Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
100 105 110
<210> 59
<211> 114
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 59
Pro Pro His Val Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys
1 5 10 15
Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys
20 25 30
Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu
35 40 45
Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His
50 55 60
Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu
65 70 75 80
Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp
85 90 95
Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn
100 105 110
Pro Asp
<210> 60
<211> 113
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 60
Pro His Val Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp
1 5 10 15
Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu
20 25 30
Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn
35 40 45
Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp
50 55 60
Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys
65 70 75 80
Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu
85 90 95
Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro
100 105 110
Asp
<210> 61
<211> 112
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 61
His Val Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn
1 5 10 15
Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys
20 25 30
Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile
35 40 45
Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe
50 55 60
Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys
65 70 75 80
Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys
85 90 95
Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
100 105 110
<210> 62
<211> 111
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 62
Val Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln
1 5 10 15
Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro
20 25 30
Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr
35 40 45
Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile
50 55 60
Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys
65 70 75 80
Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn
85 90 95
Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
100 105 110
<210> 63
<211> 117
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 63
Gly Ala Val Lys Phe Pro Gln Leu Cys Lys Phe Cys Asp Val Arg Phe
1 5 10 15
Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr
20 25 30
Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys
35 40 45
Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu
50 55 60
Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile
65 70 75 80
Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys
85 90 95
Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn
100 105 110
Thr Ser Asn Pro Asp
115
<210> 64
<211> 115
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 64
Val Lys Phe Pro Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr
1 5 10 15
Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile
20 25 30
Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp
35 40 45
Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr
50 55 60
His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys
65 70 75 80
Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser
85 90 95
Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser
100 105 110
Asn Pro Asp
115
<210> 65
<211> 110
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 65
Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys
1 5 10 15
Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln
20 25 30
Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu
35 40 45
Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu
50 55 60
Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro
65 70 75 80
Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp
85 90 95
Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
100 105 110
<210> 66
<211> 20
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 66
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 67
<211> 987
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 67
gcaagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacgt gcgtggtggt ggacgtgagc cacgaagacc ccgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggt 987
<210> 68
<211> 412
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 68
atcccaccgc acgttcagaa gtcggttaat aacgacatga tagtcactga caacaacggt 60
gcagtcaagt ttccacaact gtgtaaattt tgtgatgtga gattttccac ctgtgacaac 120
cagaaatcct gcatgagcaa ctgcagcatc acctccatct gtgagaagcc acaggaagtc 180
tgtgtggctg tatggagaaa gaatgacgag aacataacac tagagacagt ttgccatgac 240
cccaagctcc cctaccatga ctttattctg gaagatgctg cttctccaaa gtgcattatg 300
aaggaaaaaa aaaagcctgg tgagactttc ttcatgtgtt cctgtagctc tgatgagtgc 360
aatgacaaca tcatcttctc agaagaatat aacaccagca atcctgacta aa 412
<210> 69
<211> 376
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 69
atcccaccgc acgttaacaa cggtgcagtc aagtttccac aactgtgtaa attttgtgat 60
gtgagatttt ccacctgtga caaccagaaa tcctgcatga gcaactgcag catcacctcc 120
atctgtgaga agccacagga agtctgtgtg gctgtatgga gaaagaatga cgagaacata 180
acactagaga cagtttgcca tgaccccaag ctcccctacc atgactttat tctggaagat 240
gctgcttctc caaagtgcat tatgaaggaa aaaaaaaagc ctggtgagac tttcttcatg 300
tgttcctgta gctctgatga gtgcaatgac aacatcatct tctcagaaga atataacacc 360
agcaatcctg actaaa 376
<210> 70
<211> 370
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 70
atcccaccgc acgttggtgc agtcaagttt ccacaactgt gtaaattttg tgatgtgaga 60
ttttccacct gtgacaacca gaaatcctgc atgagcaact gcagcatcac ctccatctgt 120
gagaagccac aggaagtctg tgtggctgta tggagaaaga atgacgagaa cataacacta 180
gagacagttt gccatgaccc caagctcccc taccatgact ttattctgga agatgctgct 240
tctccaaagt gcattatgaa ggaaaaaaaa aagcctggtg agactttctt catgtgttcc 300
tgtagctctg atgagtgcaa tgacaacatc atcttctcag aagaatataa caccagcaat 360
cctgactaaa 370
<210> 71
<211> 364
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 71
atcccaccgc acgttgtcaa gtttccacaa ctgtgtaaat tttgtgatgt gagattttcc 60
acctgtgaca accagaaatc ctgcatgagc aactgcagca tcacctccat ctgtgagaag 120
ccacaggaag tctgtgtggc tgtatggaga aagaatgacg agaacataac actagagaca 180
gtttgccatg accccaagct cccctaccat gactttattc tggaagatgc tgcttctcca 240
aagtgcatta tgaaggaaaa aaaaaagcct ggtgagactt tcttcatgtg ttcctgtagc 300
tctgatgagt gcaatgacaa catcatcttc tcagaagaat ataacaccag caatcctgac 360
taaa 364
<210> 72
<211> 352
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 72
atcccaccgc acgttcaact gtgtaaattt tgtgatgtga gattttccac ctgtgacaac 60
cagaaatcct gcatgagcaa ctgcagcatc acctccatct gtgagaagcc acaggaagtc 120
tgtgtggctg tatggagaaa gaatgacgag aacataacac tagagacagt ttgccatgac 180
cccaagctcc cctaccatga ctttattctg gaagatgctg cttctccaaa gtgcattatg 240
aaggaaaaaa aaaagcctgg tgagactttc ttcatgtgtt cctgtagctc tgatgagtgc 300
aatgacaaca tcatcttctc agaagaatat aacaccagca atcctgacta aa 352
<210> 73
<211> 349
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 73
atcccaccgc acgttctgtg taaattttgt gatgtgagat tttccacctg tgacaaccag 60
aaatcctgca tgagcaactg cagcatcacc tccatctgtg agaagccaca ggaagtctgt 120
gtggctgtat ggagaaagaa tgacgagaac ataacactag agacagtttg ccatgacccc 180
aagctcccct accatgactt tattctggaa gatgctgctt ctccaaagtg cattatgaag 240
gaaaaaaaaa agcctggtga gactttcttc atgtgttcct gtagctctga tgagtgcaat 300
gacaacatca tcttctcaga agaatataac accagcaatc ctgactaaa 349
<210> 74
<211> 346
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 74
atcccaccgc acgtttgtaa attttgtgat gtgagatttt ccacctgtga caaccagaaa 60
tcctgcatga gcaactgcag catcacctcc atctgtgaga agccacagga agtctgtgtg 120
gctgtatgga gaaagaatga cgagaacata acactagaga cagtttgcca tgaccccaag 180
ctcccctacc atgactttat tctggaagat gctgcttctc caaagtgcat tatgaaggaa 240
aaaaaaaagc ctggtgagac tttcttcatg tgttcctgta gctctgatga gtgcaatgac 300
aacatcatct tctcagaaga atataacacc agcaatcctg actaaa 346
<210> 75
<211> 352
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 75
atcccaccgc acgttcagct gtgtaaattt tgtgatgtga gattttccac ctgtgacaac 60
cagaaatcct gcatgagcaa ctgcagcatc acctccatct gtgagaagcc acaggaagtc 120
tgtgtggctg tatggagaaa gaatgacgag aacataacac tagagacagt ttgccatgac 180
cccaagctcc cctaccatga ctttattctg gaagatgctg cttctccaaa gtgcattatg 240
aaggaaaaaa aaaagcctgg tgagactttc ttcatgtgtt cctgtagctc tgatgagtgc 300
aatgacaaca tcatcttctc agaagaatat aacaccagca atcctgacta aa 352
<210> 76
<211> 346
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 76
atcccaccgc acctgtgtaa attttgtgat gtgagatttt ccacctgtga caaccagaaa 60
tcctgcatga gcaactgcag catcacctcc atctgtgaga agccacagga agtctgtgtg 120
gctgtatgga gaaagaatga cgagaacata acactagaga cagtttgcca tgaccccaag 180
ctcccctacc atgactttat tctggaagat gctgcttctc caaagtgcat tatgaaggaa 240
aaaaaaaagc ctggtgagac tttcttcatg tgttcctgta gctctgatga gtgcaatgac 300
aacatcatct tctcagaaga atataacacc agcaatcctg actaaa 346
<210> 77
<211> 343
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 77
atcccaccgc tgtgtaaatt ttgtgatgtg agattttcca cctgtgacaa ccagaaatcc 60
tgcatgagca actgcagcat cacctccatc tgtgagaagc cacaggaagt ctgtgtggct 120
gtatggagaa agaatgacga gaacataaca ctagagacag tttgccatga ccccaagctc 180
ccctaccatg actttattct ggaagatgct gcttctccaa agtgcattat gaaggaaaaa 240
aaaaagcctg gtgagacttt cttcatgtgt tcctgtagct ctgatgagtg caatgacaac 300
atcatcttct cagaagaata taacaccagc aatcctgact aaa 343
<210> 78
<211> 340
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 78
atcccactgt gtaaattttg tgatgtgaga ttttccacct gtgacaacca gaaatcctgc 60
atgagcaact gcagcatcac ctccatctgt gagaagccac aggaagtctg tgtggctgta 120
tggagaaaga atgacgagaa cataacacta gagacagttt gccatgaccc caagctcccc 180
taccatgact ttattctgga agatgctgct tctccaaagt gcattatgaa ggaaaaaaaa 240
aagcctggtg agactttctt catgtgttcc tgtagctctg atgagtgcaa tgacaacatc 300
atcttctcag aagaatataa caccagcaat cctgactaaa 340
<210> 79
<211> 337
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 79
atcctgtgta aattttgtga tgtgagattt tccacctgtg acaaccagaa atcctgcatg 60
agcaactgca gcatcacctc catctgtgag aagccacagg aagtctgtgt ggctgtatgg 120
agaaagaatg acgagaacat aacactagag acagtttgcc atgaccccaa gctcccctac 180
catgacttta ttctggaaga tgctgcttct ccaaagtgca ttatgaagga aaaaaaaaag 240
cctggtgaga ctttcttcat gtgttcctgt agctctgatg agtgcaatga caacatcatc 300
ttctcagaag aatataacac cagcaatcct gactaaa 337
<210> 80
<211> 346
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 80
ccaccgcacg ttctgtgtaa attttgtgat gtgagatttt ccacctgtga caaccagaaa 60
tcctgcatga gcaactgcag catcacctcc atctgtgaga agccacagga agtctgtgtg 120
gctgtatgga gaaagaatga cgagaacata acactagaga cagtttgcca tgaccccaag 180
ctcccctacc atgactttat tctggaagat gctgcttctc caaagtgcat tatgaaggaa 240
aaaaaaaagc ctggtgagac tttcttcatg tgttcctgta gctctgatga gtgcaatgac 300
aacatcatct tctcagaaga atataacacc agcaatcctg actaaa 346
<210> 81
<211> 343
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 81
ccgcacgttc tgtgtaaatt ttgtgatgtg agattttcca cctgtgacaa ccagaaatcc 60
tgcatgagca actgcagcat cacctccatc tgtgagaagc cacaggaagt ctgtgtggct 120
gtatggagaa agaatgacga gaacataaca ctagagacag tttgccatga ccccaagctc 180
ccctaccatg actttattct ggaagatgct gcttctccaa agtgcattat gaaggaaaaa 240
aaaaagcctg gtgagacttt cttcatgtgt tcctgtagct ctgatgagtg caatgacaac 300
atcatcttct cagaagaata taacaccagc aatcctgact aaa 343
<210> 82
<211> 340
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 82
cacgttctgt gtaaattttg tgatgtgaga ttttccacct gtgacaacca gaaatcctgc 60
atgagcaact gcagcatcac ctccatctgt gagaagccac aggaagtctg tgtggctgta 120
tggagaaaga atgacgagaa cataacacta gagacagttt gccatgaccc caagctcccc 180
taccatgact ttattctgga agatgctgct tctccaaagt gcattatgaa ggaaaaaaaa 240
aagcctggtg agactttctt catgtgttcc tgtagctctg atgagtgcaa tgacaacatc 300
atcttctcag aagaatataa caccagcaat cctgactaaa 340
<210> 83
<211> 337
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 83
gttctgtgta aattttgtga tgtgagattt tccacctgtg acaaccagaa atcctgcatg 60
agcaactgca gcatcacctc catctgtgag aagccacagg aagtctgtgt ggctgtatgg 120
agaaagaatg acgagaacat aacactagag acagtttgcc atgaccccaa gctcccctac 180
catgacttta ttctggaaga tgctgcttct ccaaagtgca ttatgaagga aaaaaaaaag 240
cctggtgaga ctttcttcat gtgttcctgt agctctgatg agtgcaatga caacatcatc 300
ttctcagaag aatataacac cagcaatcct gactaaa 337
<210> 84
<211> 355
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 84
ggtgcagtca agtttccaca actgtgtaaa ttttgtgatg tgagattttc cacctgtgac 60
aaccagaaat cctgcatgag caactgcagc atcacctcca tctgtgagaa gccacaggaa 120
gtctgtgtgg ctgtatggag aaagaatgac gagaacataa cactagagac agtttgccat 180
gaccccaagc tcccctacca tgactttatt ctggaagatg ctgcttctcc aaagtgcatt 240
atgaaggaaa aaaaaaagcc tggtgagact ttcttcatgt gttcctgtag ctctgatgag 300
tgcaatgaca acatcatctt ctcagaagaa tataacacca gcaatcctga ctaaa 355
<210> 85
<211> 349
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 85
gtcaagtttc cacaactgtg taaattttgt gatgtgagat tttccacctg tgacaaccag 60
aaatcctgca tgagcaactg cagcatcacc tccatctgtg agaagccaca ggaagtctgt 120
gtggctgtat ggagaaagaa tgacgagaac ataacactag agacagtttg ccatgacccc 180
aagctcccct accatgactt tattctggaa gatgctgctt ctccaaagtg cattatgaag 240
gaaaaaaaaa agcctggtga gactttcttc atgtgttcct gtagctctga tgagtgcaat 300
gacaacatca tcttctcaga agaatataac accagcaatc ctgactaaa 349
<210> 86
<211> 334
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 86
ctgtgtaaat tttgtgatgt gagattttcc acctgtgaca accagaaatc ctgcatgagc 60
aactgcagca tcacctccat ctgtgagaag ccacaggaag tctgtgtggc tgtatggaga 120
aagaatgacg agaacataac actagagaca gtttgccatg accccaagct cccctaccat 180
gactttattc tggaagatgc tgcttctcca aagtgcatta tgaaggaaaa aaaaaagcct 240
ggtgagactt tcttcatgtg ttcctgtagc tctgatgagt gcaatgacaa catcatcttc 300
tcagaagaat ataacaccag caatcctgac taaa 334
<210> 87
<211> 60
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 87
ggtggtggcg gttcaggcgg aggtggctct ggaggtggag gttcaggagg tggtggttct 60
<210> 88
<211> 623
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 88
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Gln Lys Ser Val
485 490 495
Asn Asn Asp Met Ile Val Thr Asp Asn Asn Gly Ala Val Lys Phe Pro
500 505 510
Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln
515 520 525
Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro
530 535 540
Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr
545 550 555 560
Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile
565 570 575
Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys
580 585 590
Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn
595 600 605
Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
610 615 620
<210> 89
<211> 611
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 89
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Asn Asn Gly Ala
485 490 495
Val Lys Phe Pro Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr
500 505 510
Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile
515 520 525
Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp
530 535 540
Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr
545 550 555 560
His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys
565 570 575
Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser
580 585 590
Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser
595 600 605
Asn Pro Asp
610
<210> 90
<211> 609
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 90
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Gly Ala Val Lys
485 490 495
Phe Pro Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp
500 505 510
Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu
515 520 525
Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn
530 535 540
Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp
545 550 555 560
Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys
565 570 575
Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu
580 585 590
Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro
595 600 605
Asp
<210> 91
<211> 607
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 91
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Val Lys Phe Pro
485 490 495
Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln
500 505 510
Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro
515 520 525
Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr
530 535 540
Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile
545 550 555 560
Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys
565 570 575
Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn
580 585 590
Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600 605
<210> 92
<211> 603
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 92
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Gln Leu Cys Lys
485 490 495
Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met
500 505 510
Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys
515 520 525
Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val
530 535 540
Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala
545 550 555 560
Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr
565 570 575
Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile
580 585 590
Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 93
<211> 602
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 93
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Leu Cys Lys Phe
485 490 495
Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser
500 505 510
Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val
515 520 525
Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys
530 535 540
His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala
545 550 555 560
Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe
565 570 575
Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe
580 585 590
Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 94
<211> 601
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 94
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Cys Lys Phe Cys
485 490 495
Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn
500 505 510
Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala
515 520 525
Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His
530 535 540
Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser
545 550 555 560
Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe
565 570 575
Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser
580 585 590
Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 95
<211> 603
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 95
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Gln Leu Cys Lys
485 490 495
Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met
500 505 510
Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys
515 520 525
Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val
530 535 540
Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala
545 550 555 560
Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr
565 570 575
Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile
580 585 590
Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 96
<211> 601
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 96
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Leu Cys Lys Phe Cys
485 490 495
Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn
500 505 510
Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala
515 520 525
Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His
530 535 540
Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser
545 550 555 560
Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe
565 570 575
Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser
580 585 590
Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 97
<211> 600
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 97
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro Leu Cys Lys Phe Cys Asp
485 490 495
Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys
500 505 510
Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val
515 520 525
Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp
530 535 540
Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro
545 550 555 560
Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met
565 570 575
Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu
580 585 590
Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 98
<211> 599
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 98
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Leu Cys Lys Phe Cys Asp Val
485 490 495
Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser
500 505 510
Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp
515 520 525
Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro
530 535 540
Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys
545 550 555 560
Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys
565 570 575
Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu
580 585 590
Tyr Asn Thr Ser Asn Pro Asp
595
<210> 99
<211> 598
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 99
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Leu Cys Lys Phe Cys Asp Val Arg
485 490 495
Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile
500 505 510
Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg
515 520 525
Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys
530 535 540
Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys
545 550 555 560
Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser
565 570 575
Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr
580 585 590
Asn Thr Ser Asn Pro Asp
595
<210> 100
<211> 601
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 100
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Pro Pro His Val Leu Cys Lys Phe Cys
485 490 495
Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn
500 505 510
Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala
515 520 525
Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His
530 535 540
Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser
545 550 555 560
Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe
565 570 575
Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser
580 585 590
Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 101
<211> 600
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 101
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Pro His Val Leu Cys Lys Phe Cys Asp
485 490 495
Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys
500 505 510
Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val
515 520 525
Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp
530 535 540
Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro
545 550 555 560
Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met
565 570 575
Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu
580 585 590
Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 102
<211> 599
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 102
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser His Val Leu Cys Lys Phe Cys Asp Val
485 490 495
Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser
500 505 510
Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp
515 520 525
Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro
530 535 540
Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys
545 550 555 560
Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys
565 570 575
Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu
580 585 590
Tyr Asn Thr Ser Asn Pro Asp
595
<210> 103
<211> 598
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 103
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Val Leu Cys Lys Phe Cys Asp Val Arg
485 490 495
Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile
500 505 510
Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg
515 520 525
Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys
530 535 540
Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys
545 550 555 560
Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser
565 570 575
Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr
580 585 590
Asn Thr Ser Asn Pro Asp
595
<210> 104
<211> 604
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 104
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Gly Ala Val Lys Phe Pro Gln Leu Cys
485 490 495
Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys
500 505 510
Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val
515 520 525
Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr
530 535 540
Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp
545 550 555 560
Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu
565 570 575
Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile
580 585 590
Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 105
<211> 602
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 105
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Val Lys Phe Pro Gln Leu Cys Lys Phe
485 490 495
Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser
500 505 510
Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val
515 520 525
Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys
530 535 540
His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala
545 550 555 560
Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe
565 570 575
Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe
580 585 590
Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 106
<211> 597
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 106
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Leu Cys Lys Phe Cys Asp Val Arg Phe
485 490 495
Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr
500 505 510
Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys
515 520 525
Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu
530 535 540
Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile
545 550 555 560
Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys
565 570 575
Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn
580 585 590
Thr Ser Asn Pro Asp
595
<210> 107
<211> 1873
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 107
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgttcaga agtcggttaa taacgacatg 1500
atagtcactg acaacaacgg tgcagtcaag tttccacaac tgtgtaaatt ttgtgatgtg 1560
agattttcca cctgtgacaa ccagaaatcc tgcatgagca actgcagcat cacctccatc 1620
tgtgagaagc cacaggaagt ctgtgtggct gtatggagaa agaatgacga gaacataaca 1680
ctagagacag tttgccatga ccccaagctc ccctaccatg actttattct ggaagatgct 1740
gcttctccaa agtgcattat gaaggaaaaa aaaaagcctg gtgagacttt cttcatgtgt 1800
tcctgtagct ctgatgagtg caatgacaac atcatcttct cagaagaata taacaccagc 1860
aatcctgact aaa 1873
<210> 108
<211> 1837
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 108
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgttaaca acggtgcagt caagtttcca 1500
caactgtgta aattttgtga tgtgagattt tccacctgtg acaaccagaa atcctgcatg 1560
agcaactgca gcatcacctc catctgtgag aagccacagg aagtctgtgt ggctgtatgg 1620
agaaagaatg acgagaacat aacactagag acagtttgcc atgaccccaa gctcccctac 1680
catgacttta ttctggaaga tgctgcttct ccaaagtgca ttatgaagga aaaaaaaaag 1740
cctggtgaga ctttcttcat gtgttcctgt agctctgatg agtgcaatga caacatcatc 1800
ttctcagaag aatataacac cagcaatcct gactaaa 1837
<210> 109
<211> 1831
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 109
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgttggtg cagtcaagtt tccacaactg 1500
tgtaaatttt gtgatgtgag attttccacc tgtgacaacc agaaatcctg catgagcaac 1560
tgcagcatca cctccatctg tgagaagcca caggaagtct gtgtggctgt atggagaaag 1620
aatgacgaga acataacact agagacagtt tgccatgacc ccaagctccc ctaccatgac 1680
tttattctgg aagatgctgc ttctccaaag tgcattatga aggaaaaaaa aaagcctggt 1740
gagactttct tcatgtgttc ctgtagctct gatgagtgca atgacaacat catcttctca 1800
gaagaatata acaccagcaa tcctgactaa a 1831
<210> 110
<211> 1825
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 110
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgttgtca agtttccaca actgtgtaaa 1500
ttttgtgatg tgagattttc cacctgtgac aaccagaaat cctgcatgag caactgcagc 1560
atcacctcca tctgtgagaa gccacaggaa gtctgtgtgg ctgtatggag aaagaatgac 1620
gagaacataa cactagagac agtttgccat gaccccaagc tcccctacca tgactttatt 1680
ctggaagatg ctgcttctcc aaagtgcatt atgaaggaaa aaaaaaagcc tggtgagact 1740
ttcttcatgt gttcctgtag ctctgatgag tgcaatgaca acatcatctt ctcagaagaa 1800
tataacacca gcaatcctga ctaaa 1825
<210> 111
<211> 1813
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 111
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgttcaac tgtgtaaatt ttgtgatgtg 1500
agattttcca cctgtgacaa ccagaaatcc tgcatgagca actgcagcat cacctccatc 1560
tgtgagaagc cacaggaagt ctgtgtggct gtatggagaa agaatgacga gaacataaca 1620
ctagagacag tttgccatga ccccaagctc ccctaccatg actttattct ggaagatgct 1680
gcttctccaa agtgcattat gaaggaaaaa aaaaagcctg gtgagacttt cttcatgtgt 1740
tcctgtagct ctgatgagtg caatgacaac atcatcttct cagaagaata taacaccagc 1800
aatcctgact aaa 1813
<210> 112
<211> 1810
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 112
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgttctgt gtaaattttg tgatgtgaga 1500
ttttccacct gtgacaacca gaaatcctgc atgagcaact gcagcatcac ctccatctgt 1560
gagaagccac aggaagtctg tgtggctgta tggagaaaga atgacgagaa cataacacta 1620
gagacagttt gccatgaccc caagctcccc taccatgact ttattctgga agatgctgct 1680
tctccaaagt gcattatgaa ggaaaaaaaa aagcctggtg agactttctt catgtgttcc 1740
tgtagctctg atgagtgcaa tgacaacatc atcttctcag aagaatataa caccagcaat 1800
cctgactaaa 1810
<210> 113
<211> 1807
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 113
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgtttgta aattttgtga tgtgagattt 1500
tccacctgtg acaaccagaa atcctgcatg agcaactgca gcatcacctc catctgtgag 1560
aagccacagg aagtctgtgt ggctgtatgg agaaagaatg acgagaacat aacactagag 1620
acagtttgcc atgaccccaa gctcccctac catgacttta ttctggaaga tgctgcttct 1680
ccaaagtgca ttatgaagga aaaaaaaaag cctggtgaga ctttcttcat gtgttcctgt 1740
agctctgatg agtgcaatga caacatcatc ttctcagaag aatataacac cagcaatcct 1800
gactaaa 1807
<210> 114
<211> 1813
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 114
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacgttcagc tgtgtaaatt ttgtgatgtg 1500
agattttcca cctgtgacaa ccagaaatcc tgcatgagca actgcagcat cacctccatc 1560
tgtgagaagc cacaggaagt ctgtgtggct gtatggagaa agaatgacga gaacataaca 1620
ctagagacag tttgccatga ccccaagctc ccctaccatg actttattct ggaagatgct 1680
gcttctccaa agtgcattat gaaggaaaaa aaaaagcctg gtgagacttt cttcatgtgt 1740
tcctgtagct ctgatgagtg caatgacaac atcatcttct cagaagaata taacaccagc 1800
aatcctgact aaa 1813
<210> 115
<211> 1807
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 115
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg cacctgtgta aattttgtga tgtgagattt 1500
tccacctgtg acaaccagaa atcctgcatg agcaactgca gcatcacctc catctgtgag 1560
aagccacagg aagtctgtgt ggctgtatgg agaaagaatg acgagaacat aacactagag 1620
acagtttgcc atgaccccaa gctcccctac catgacttta ttctggaaga tgctgcttct 1680
ccaaagtgca ttatgaagga aaaaaaaaag cctggtgaga ctttcttcat gtgttcctgt 1740
agctctgatg agtgcaatga caacatcatc ttctcagaag aatataacac cagcaatcct 1800
gactaaa 1807
<210> 116
<211> 1804
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 116
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccaccg ctgtgtaaat tttgtgatgt gagattttcc 1500
acctgtgaca accagaaatc ctgcatgagc aactgcagca tcacctccat ctgtgagaag 1560
ccacaggaag tctgtgtggc tgtatggaga aagaatgacg agaacataac actagagaca 1620
gtttgccatg accccaagct cccctaccat gactttattc tggaagatgc tgcttctcca 1680
aagtgcatta tgaaggaaaa aaaaaagcct ggtgagactt tcttcatgtg ttcctgtagc 1740
tctgatgagt gcaatgacaa catcatcttc tcagaagaat ataacaccag caatcctgac 1800
taaa 1804
<210> 117
<211> 1801
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 117
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcccactg tgtaaatttt gtgatgtgag attttccacc 1500
tgtgacaacc agaaatcctg catgagcaac tgcagcatca cctccatctg tgagaagcca 1560
caggaagtct gtgtggctgt atggagaaag aatgacgaga acataacact agagacagtt 1620
tgccatgacc ccaagctccc ctaccatgac tttattctgg aagatgctgc ttctccaaag 1680
tgcattatga aggaaaaaaa aaagcctggt gagactttct tcatgtgttc ctgtagctct 1740
gatgagtgca atgacaacat catcttctca gaagaatata acaccagcaa tcctgactaa 1800
a 1801
<210> 118
<211> 1798
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 118
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tatcctgtgt aaattttgtg atgtgagatt ttccacctgt 1500
gacaaccaga aatcctgcat gagcaactgc agcatcacct ccatctgtga gaagccacag 1560
gaagtctgtg tggctgtatg gagaaagaat gacgagaaca taacactaga gacagtttgc 1620
catgacccca agctccccta ccatgacttt attctggaag atgctgcttc tccaaagtgc 1680
attatgaagg aaaaaaaaaa gcctggtgag actttcttca tgtgttcctg tagctctgat 1740
gagtgcaatg acaacatcat cttctcagaa gaatataaca ccagcaatcc tgactaaa 1798
<210> 119
<211> 1807
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 119
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tccaccgcac gttctgtgta aattttgtga tgtgagattt 1500
tccacctgtg acaaccagaa atcctgcatg agcaactgca gcatcacctc catctgtgag 1560
aagccacagg aagtctgtgt ggctgtatgg agaaagaatg acgagaacat aacactagag 1620
acagtttgcc atgaccccaa gctcccctac catgacttta ttctggaaga tgctgcttct 1680
ccaaagtgca ttatgaagga aaaaaaaaag cctggtgaga ctttcttcat gtgttcctgt 1740
agctctgatg agtgcaatga caacatcatc ttctcagaag aatataacac cagcaatcct 1800
gactaaa 1807
<210> 120
<211> 1804
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 120
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tccgcacgtt ctgtgtaaat tttgtgatgt gagattttcc 1500
acctgtgaca accagaaatc ctgcatgagc aactgcagca tcacctccat ctgtgagaag 1560
ccacaggaag tctgtgtggc tgtatggaga aagaatgacg agaacataac actagagaca 1620
gtttgccatg accccaagct cccctaccat gactttattc tggaagatgc tgcttctcca 1680
aagtgcatta tgaaggaaaa aaaaaagcct ggtgagactt tcttcatgtg ttcctgtagc 1740
tctgatgagt gcaatgacaa catcatcttc tcagaagaat ataacaccag caatcctgac 1800
taaa 1804
<210> 121
<211> 1801
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 121
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tcacgttctg tgtaaatttt gtgatgtgag attttccacc 1500
tgtgacaacc agaaatcctg catgagcaac tgcagcatca cctccatctg tgagaagcca 1560
caggaagtct gtgtggctgt atggagaaag aatgacgaga acataacact agagacagtt 1620
tgccatgacc ccaagctccc ctaccatgac tttattctgg aagatgctgc ttctccaaag 1680
tgcattatga aggaaaaaaa aaagcctggt gagactttct tcatgtgttc ctgtagctct 1740
gatgagtgca atgacaacat catcttctca gaagaatata acaccagcaa tcctgactaa 1800
a 1801
<210> 122
<211> 1798
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 122
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tgttctgtgt aaattttgtg atgtgagatt ttccacctgt 1500
gacaaccaga aatcctgcat gagcaactgc agcatcacct ccatctgtga gaagccacag 1560
gaagtctgtg tggctgtatg gagaaagaat gacgagaaca taacactaga gacagtttgc 1620
catgacccca agctccccta ccatgacttt attctggaag atgctgcttc tccaaagtgc 1680
attatgaagg aaaaaaaaaa gcctggtgag actttcttca tgtgttcctg tagctctgat 1740
gagtgcaatg acaacatcat cttctcagaa gaatataaca ccagcaatcc tgactaaa 1798
<210> 123
<211> 1816
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 123
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tggtgcagtc aagtttccac aactgtgtaa attttgtgat 1500
gtgagatttt ccacctgtga caaccagaaa tcctgcatga gcaactgcag catcacctcc 1560
atctgtgaga agccacagga agtctgtgtg gctgtatgga gaaagaatga cgagaacata 1620
acactagaga cagtttgcca tgaccccaag ctcccctacc atgactttat tctggaagat 1680
gctgcttctc caaagtgcat tatgaaggaa aaaaaaaagc ctggtgagac tttcttcatg 1740
tgttcctgta gctctgatga gtgcaatgac aacatcatct tctcagaaga atataacacc 1800
agcaatcctg actaaa 1816
<210> 124
<211> 1810
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 124
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tgtcaagttt ccacaactgt gtaaattttg tgatgtgaga 1500
ttttccacct gtgacaacca gaaatcctgc atgagcaact gcagcatcac ctccatctgt 1560
gagaagccac aggaagtctg tgtggctgta tggagaaaga atgacgagaa cataacacta 1620
gagacagttt gccatgaccc caagctcccc taccatgact ttattctgga agatgctgct 1680
tctccaaagt gcattatgaa ggaaaaaaaa aagcctggtg agactttctt catgtgttcc 1740
tgtagctctg atgagtgcaa tgacaacatc atcttctcag aagaatataa caccagcaat 1800
cctgactaaa 1810
<210> 125
<211> 1795
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 125
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtgag 60
gtccaacttg tccagtctgg agcagaggtg aagaagcctg gagccacagt gaagatttcc 120
tgtaaggtgt ctggctacac cttcaccacc tactacaccc actgggtgaa gcaggctcct 180
ggcaagggat tggagtggat tggctggatt taccctggag atgtgaacac caaatacaat 240
gagaagttca agggcagggt gaccctgaca gcagacacca gcacagacac agcctatatg 300
gaactgtcct ccctgaggtc tgaggacaca gcagtctact actgtgccag ggaggaccct 360
ggcagcaact actttgacta ctggggacaa ggcaccctgg tgacagtgtc cagcgcaagc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg tggtggtggc ggttcaggcg gaggtggctc tggaggtgga 1440
ggttcaggag gtggtggttc tctgtgtaaa ttttgtgatg tgagattttc cacctgtgac 1500
aaccagaaat cctgcatgag caactgcagc atcacctcca tctgtgagaa gccacaggaa 1560
gtctgtgtgg ctgtatggag aaagaatgac gagaacataa cactagagac agtttgccat 1620
gaccccaagc tcccctacca tgactttatt ctggaagatg ctgcttctcc aaagtgcatt 1680
atgaaggaaa aaaaaaagcc tggtgagact ttcttcatgt gttcctgtag ctctgatgag 1740
tgcaatgaca acatcatctt ctcagaagaa tataacacca gcaatcctga ctaaa 1795
<210> 126
<211> 233
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 126
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val
35 40 45
Asn Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
50 55 60
Leu Leu Ile Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg
65 70 75 80
Phe Ser Gly Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr
100 105 110
Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 127
<211> 624
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 127
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile
35 40 45
Lys Asp Thr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
225 230 235 240
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
245 250 255
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
275 280 285
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
340 345 350
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
465 470 475 480
Gly Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Gln Lys Ser
485 490 495
Val Asn Asn Asp Met Ile Val Thr Asp Asn Asn Gly Ala Val Lys Phe
500 505 510
Pro Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn
515 520 525
Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys
530 535 540
Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile
545 550 555 560
Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe
565 570 575
Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys
580 585 590
Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys
595 600 605
Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
610 615 620
<210> 128
<211> 603
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 128
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile
35 40 45
Lys Asp Thr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
225 230 235 240
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
245 250 255
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
275 280 285
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
340 345 350
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
465 470 475 480
Gly Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Leu Cys Lys
485 490 495
Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met
500 505 510
Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys
515 520 525
Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val
530 535 540
Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala
545 550 555 560
Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr
565 570 575
Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile
580 585 590
Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 129
<211> 233
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 129
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile
35 40 45
Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
50 55 60
Val Leu Ile Tyr Phe Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr
100 105 110
Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 130
<211> 627
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 130
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Ala Asp Phe Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Lys Tyr Pro His Tyr Tyr Gly Ser Ser His Trp Tyr
115 120 125
Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
130 135 140
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
145 150 155 160
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
165 170 175
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
180 185 190
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
195 200 205
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
210 215 220
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
225 230 235 240
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
245 250 255
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
260 265 270
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
275 280 285
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
290 295 300
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
305 310 315 320
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
325 330 335
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
340 345 350
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
355 360 365
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
370 375 380
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
385 390 395 400
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
405 410 415
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
420 425 430
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
435 440 445
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
450 455 460
Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val
485 490 495
Gln Lys Ser Val Asn Asn Asp Met Ile Val Thr Asp Asn Asn Gly Ala
500 505 510
Val Lys Phe Pro Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr
515 520 525
Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile
530 535 540
Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp
545 550 555 560
Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr
565 570 575
His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys
580 585 590
Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser
595 600 605
Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser
610 615 620
Asn Pro Asp
625
<210> 131
<211> 606
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 131
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Ala Asp Phe Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Lys Tyr Pro His Tyr Tyr Gly Ser Ser His Trp Tyr
115 120 125
Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
130 135 140
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
145 150 155 160
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
165 170 175
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
180 185 190
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
195 200 205
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
210 215 220
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
225 230 235 240
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
245 250 255
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
260 265 270
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
275 280 285
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
290 295 300
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
305 310 315 320
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
325 330 335
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
340 345 350
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
355 360 365
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
370 375 380
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
385 390 395 400
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
405 410 415
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
420 425 430
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
435 440 445
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
450 455 460
Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val
485 490 495
Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys
500 505 510
Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln
515 520 525
Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu
530 535 540
Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu
545 550 555 560
Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro
565 570 575
Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp
580 585 590
Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600 605
<210> 132
<211> 233
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 132
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala
20 25 30
Ser Ile Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile
35 40 45
Asp Asn Trp Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
50 55 60
Leu Leu Ile Tyr Asp Ala Ser Asn Leu Asp Thr Gly Val Pro Ser Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Tyr Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Gln Ala Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ala Lys Ala
100 105 110
Phe Pro Pro Thr Phe Gly Gly Gly Thr Lys Val Asp Ile Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 133
<211> 620
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 133
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Val Thr Asp Ala Phe Asp Ile Trp Gly Gln Gly
115 120 125
Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
130 135 140
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
145 150 155 160
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
165 170 175
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
180 185 190
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
195 200 205
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
210 215 220
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
225 230 235 240
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
465 470 475 480
Gly Gly Gly Ser Ile Pro Pro His Val Gln Lys Ser Val Asn Asn Asp
485 490 495
Met Ile Val Thr Asp Asn Asn Gly Ala Val Lys Phe Pro Gln Leu Cys
500 505 510
Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys
515 520 525
Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val
530 535 540
Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr
545 550 555 560
Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp
565 570 575
Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu
580 585 590
Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile
595 600 605
Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
610 615 620
<210> 134
<211> 599
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 134
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Val Thr Asp Ala Phe Asp Ile Trp Gly Gln Gly
115 120 125
Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
130 135 140
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
145 150 155 160
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
165 170 175
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
180 185 190
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
195 200 205
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
210 215 220
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
225 230 235 240
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
465 470 475 480
Gly Gly Gly Ser Ile Pro Pro His Val Leu Cys Lys Phe Cys Asp Val
485 490 495
Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser
500 505 510
Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp
515 520 525
Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro
530 535 540
Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys
545 550 555 560
Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys
565 570 575
Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu
580 585 590
Tyr Asn Thr Ser Asn Pro Asp
595
<210> 135
<211> 234
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 135
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu
20 25 30
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val
35 40 45
Gly Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
50 55 60
Arg Leu Leu Ile Tyr Gly Ala Phe Ser Arg Ala Thr Gly Ile Pro Asp
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly
100 105 110
Ser Ser Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 136
<211> 622
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 136
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Thr Phe Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile
100 105 110
Tyr Tyr Cys Ala Arg Thr Gly Trp Leu Gly Pro Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
225 230 235 240
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
275 280 285
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Gln Lys Ser Val Asn
485 490 495
Asn Asp Met Ile Val Thr Asp Asn Asn Gly Ala Val Lys Phe Pro Gln
500 505 510
Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys
515 520 525
Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln
530 535 540
Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu
545 550 555 560
Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu
565 570 575
Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro
580 585 590
Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp
595 600 605
Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
610 615 620
<210> 137
<211> 601
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 137
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Thr Phe Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile
100 105 110
Tyr Tyr Cys Ala Arg Thr Gly Trp Leu Gly Pro Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
225 230 235 240
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
275 280 285
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Leu Cys Lys Phe Cys
485 490 495
Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn
500 505 510
Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala
515 520 525
Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His
530 535 540
Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser
545 550 555 560
Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe
565 570 575
Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser
580 585 590
Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 138
<211> 233
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 138
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Asp Ile Gln Met Thr Gln Ser Pro Ala Ala Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile
35 40 45
Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
50 55 60
Leu Leu Ile Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser
85 90 95
Leu Gln Pro Glu Asp Ile Ala Thr Tyr Phe Cys Gln His Phe Asp His
100 105 110
Leu Pro Leu Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 139
<211> 623
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 139
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys
20 25 30
Pro Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val
35 40 45
Ser Ser Gly Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro Gly Lys
50 55 60
Gly Leu Glu Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr Asn Tyr
65 70 75 80
Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr Ser Lys
85 90 95
Thr Gln Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala
100 105 110
Ile Tyr Tyr Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp Ile Trp
115 120 125
Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Gln Lys Ser Val
485 490 495
Asn Asn Asp Met Ile Val Thr Asp Asn Asn Gly Ala Val Lys Phe Pro
500 505 510
Gln Leu Cys Lys Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln
515 520 525
Lys Ser Cys Met Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro
530 535 540
Gln Glu Val Cys Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr
545 550 555 560
Leu Glu Thr Val Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile
565 570 575
Leu Glu Asp Ala Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys
580 585 590
Pro Gly Glu Thr Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn
595 600 605
Asp Asn Ile Ile Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
610 615 620
<210> 140
<211> 602
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 140
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys
20 25 30
Pro Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val
35 40 45
Ser Ser Gly Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro Gly Lys
50 55 60
Gly Leu Glu Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr Asn Tyr
65 70 75 80
Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr Ser Lys
85 90 95
Thr Gln Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala
100 105 110
Ile Tyr Tyr Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp Ile Trp
115 120 125
Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Ile Pro Pro His Val Leu Cys Lys Phe
485 490 495
Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser
500 505 510
Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val
515 520 525
Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys
530 535 540
His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala
545 550 555 560
Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe
565 570 575
Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe
580 585 590
Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<210> 141
<211> 583
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 141
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Thr His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Val Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Asp Pro Gly Ser Asn Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Ile Pro Pro His Val Leu Cys Lys Phe Cys Asp Val
465 470 475 480
Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn Cys Ser
485 490 495
Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala Val Trp
500 505 510
Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His Asp Pro
515 520 525
Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser Pro Lys
530 535 540
Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe Met Cys
545 550 555 560
Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser Glu Glu
565 570 575
Tyr Asn Thr Ser Asn Pro Asp
580
<210> 142
<211> 699
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 142
atgacagtgc tggcgccagc ctggagccca aattcctccc tgttgctgct gttgctgctg 60
ctgagtcctt gcctgcgggg gacacctgac tgttacttca gccacagtcc catctcctcc 120
aacttcaaag tgaagtttag agagttgact gaccacctgc ttaaagatta cccagtcact 180
gtggccgtca atcttcagga cgagaagcac tgcaaggcct tgtggagcct cttcctagcc 240
cagcgctgga tagagcaact gaagactgtg gcagggtcta agatgcaaac gcttctggag 300
gacgtcaaca ccgagataca ttttgtcacc tcatgtacct tccagcccct accagaatgt 360
ctgcgattcg tccagaccaa catctcccac ctcctgaagg acacctgcac acagctgctt 420
gctctgaagc cctgtatcgg gaaggcctgc cagaatttct ctcggtgcct ggaggtgcag 480
tgccagccgg actcctccac cctgctgccc ccaaggagtc ccatagccct agaagccacg 540
gagctcccag agcctcggcc caggcagctg ttgctcctgc tgctgctgct gctgcctctc 600
acactggtgc tgctggcagc cgcctggggc cttcgctggc aaagggcaag aaggaggggg 660
gagctccacc ctggggtgcc cctcccctcc catccctaa 699
<210> 143
<211> 426
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 143
atgtggctgc agaatttact tttcctgggc attgtggtct acagcctctc agcacccacc 60
cgctcaccca tcactgtcac ccggccttgg aagcatgtag aggccatcaa agaagccctg 120
aacctcctgg atgacatgcc tgtcacgttg aatgaagagg tagaagtcgt ctctaacgag 180
ttctccttca agaagctaac atgtgtgcag acccgcctga agatattcga gcagggtcta 240
cggggcaatt tcaccaaact caagggcgcc ttgaacatga cagccagcta ctaccagaca 300
tactgccccc caactccgga aacggactgt gaaacacaag ttaccaccta tgcggatttc 360
atagacagcc ttaaaacctt tctgactgat atcccctttg aatgcaaaaa accaggccaa 420
aaataa 426
<210> 144
<211> 3630
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 144
atgcgaccct ccgggacggc cggggcagcg ctcctggcgc tgctggctgc gctctgcccg 60
gcgagtcggg ctctggagga aaagaaagtt tgccaaggca cgagtaacaa gctcacgcag 120
ttgggcactt ttgaagatca ttttctcagc ctccagagga tgttcaataa ctgtgaggtg 180
gtccttggga atttggaaat tacctatgtg cagaggaatt atgatctttc cttcttaaag 240
accatccagg aggtggctgg ttatgtcctc attgccctca acacagtgga gcgaattcct 300
ttggaaaacc tgcagatcat cagaggaaat atgtactacg aaaattccta tgccttagca 360
gtcttatcta actatgatgc aaataaaacc ggactgaagg agctgcccat gagaaattta 420
caggaaatcc tgcatggcgc cgtgcggttc agcaacaacc ctgccctgtg caacgtggag 480
agcatccagt ggcgggacat agtcagcagt gactttctca gcaacatgtc gatggacttc 540
cagaaccacc tgggcagctg ccaaaagtgt gatccaagct gtcccaatgg gagctgctgg 600
ggtgcaggag aggagaactg ccagaaactg accaaaatca tctgtgccca gcagtgctcc 660
gggcgctgcc gtggcaagtc ccccagtgac tgctgccaca accagtgtgc tgcaggctgc 720
acaggccccc gggagagcga ctgcctggtc tgccgcaaat tccgagacga agccacgtgc 780
aaggacacct gccccccact catgctctac aaccccacca cgtaccagat ggatgtgaac 840
cccgagggca aatacagctt tggtgccacc tgcgtgaaga agtgtccccg taattatgtg 900
gtgacagatc acggctcgtg cgtccgagcc tgtggggccg acagctatga gatggaggaa 960
gacggcgtcc gcaagtgtaa gaagtgcgaa gggccttgcc gcaaagtgtg taacggaata 1020
ggtattggtg aatttaaaga ctcactctcc ataaatgcta cgaatattaa acacttcaaa 1080
aactgcacct ccatcagtgg cgatctccac atcctgccgg tggcatttag gggtgactcc 1140
ttcacacata ctcctcctct ggatccacag gaactggata ttctgaaaac cgtaaaggaa 1200
atcacagggt ttttgctgat tcaggcttgg cctgaaaaca ggacggacct ccatgccttt 1260
gagaacctag aaatcatacg cggcaggacc aagcaacatg gtcagttttc tcttgcagtc 1320
gtcagcctga acataacatc cttgggatta cgctccctca aggagataag tgatggagat 1380
gtgataattt caggaaacaa aaatttgtgc tatgcaaata caataaactg gaaaaaactg 1440
tttgggacct ccggtcagaa aaccaaaatt ataagcaaca gaggtgaaaa cagctgcaag 1500
gccacaggcc aggtctgcca tgccttgtgc tcccccgagg gctgctgggg cccggagccc 1560
agggactgcg tctcttgccg gaatgtcagc cgaggcaggg aatgcgtgga caagtgcaac 1620
cttctggagg gtgagccaag ggagtttgtg gagaactctg agtgcataca gtgccaccca 1680
gagtgcctgc ctcaggccat gaacatcacc tgcacaggac ggggaccaga caactgtatc 1740
cagtgtgccc actacattga cggcccccac tgcgtcaaga cctgcccggc aggagtcatg 1800
ggagaaaaca acaccctggt ctggaagtac gcagacgccg gccatgtgtg ccacctgtgc 1860
catccaaact gcacctacgg atgcactggg ccaggtcttg aaggctgtcc aacgaatggg 1920
cctaagatcc cgtccatcgc cactgggatg gtgggggccc tcctcttgct gctggtggtg 1980
gccctgggga tcggcctctt catgcgaagg cgccacatcg ttcggaagcg cacgctgcgg 2040
aggctgctgc aggagaggga gcttgtggag cctcttacac ccagtggaga agctcccaac 2100
caagctctct tgaggatctt gaaggaaact gaattcaaaa agatcaaagt gctgggctcc 2160
ggtgcgttcg gcacggtgta taagggactc tggatcccag aaggtgagaa agttaaaatt 2220
cccgtcgcta tcaaggaatt aagagaagca acatctccga aagccaacaa ggaaatcctc 2280
gatgaagcct acgtgatggc cagcgtggac aacccccacg tgtgccgcct gctgggcatc 2340
tgcctcacct ccaccgtgca actcatcacg cagctcatgc ccttcggctg cctcctggac 2400
tatgtccggg aacacaaaga caatattggc tcccagtacc tgctcaactg gtgtgtgcag 2460
atcgcaaagg gcatgaacta cttggaggac cgtcgcttgg tgcaccgcga cctggcagcc 2520
aggaacgtac tggtgaaaac accgcagcat gtcaagatca cagattttgg gctggccaaa 2580
ctgctgggtg cggaagagaa agaataccat gcagaaggag gcaaagtgcc tatcaagtgg 2640
atggcattgg aatcaatttt acacagaatc tatacccacc agagtgatgt ctggagctac 2700
ggggtgaccg tttgggagtt gatgaccttt ggatccaagc catatgacgg aatccctgcc 2760
agcgagatct cctccatcct ggagaaagga gaacgcctcc ctcagccacc catatgtacc 2820
atcgatgtct acatgatcat ggtcaagtgc tggatgatag acgcagatag tcgcccaaag 2880
ttccgtgagt tgatcatcga attctccaaa atggcccgag acccccagcg ctaccttgtc 2940
attcaggggg atgaaagaat gcatttgcca agtcctacag actccaactt ctaccgtgcc 3000
ctgatggatg aagaagacat ggacgacgtg gtggatgccg acgagtacct catcccacag 3060
cagggcttct tcagcagccc ctccacgtca cggactcccc tcctgagctc tctgagtgca 3120
accagcaaca attccaccgt ggcttgcatt gatagaaatg ggctgcaaag ctgtcccatc 3180
aaggaagaca gcttcttgca gcgatacagc tcagacccca caggcgcctt gactgaggac 3240
agcatagacg acaccttcct cccagtgcct gaatacataa accagtccgt tcccaaaagg 3300
cccgctggct ctgtgcagaa tcctgtctat cacaatcagc ctctgaaccc cgcgcccagc 3360
agagacccac actaccagga cccccacagc actgcagtgg gcaaccccga gtatctcaac 3420
actgtccagc ccacctgtgt caacagcaca ttcgacagcc ctgcccactg ggcccagaaa 3480
ggcagccacc aaattagcct ggacaaccct gactaccagc aggacttctt tcccaaggaa 3540
gccaagccaa atggcatctt taagggctcc acagctgaaa atgcagaata cctaagggtc 3600
gcgccacaaa gcagtgaatt tattggagca 3630
<210> 145
<211> 232
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 145
Met Thr Val Leu Ala Pro Ala Trp Ser Pro Asn Ser Ser Leu Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Ser Pro Cys Leu Arg Gly Thr Pro Asp Cys Tyr
20 25 30
Phe Ser His Ser Pro Ile Ser Ser Asn Phe Lys Val Lys Phe Arg Glu
35 40 45
Leu Thr Asp His Leu Leu Lys Asp Tyr Pro Val Thr Val Ala Val Asn
50 55 60
Leu Gln Asp Glu Lys His Cys Lys Ala Leu Trp Ser Leu Phe Leu Ala
65 70 75 80
Gln Arg Trp Ile Glu Gln Leu Lys Thr Val Ala Gly Ser Lys Met Gln
85 90 95
Thr Leu Leu Glu Asp Val Asn Thr Glu Ile His Phe Val Thr Ser Cys
100 105 110
Thr Phe Gln Pro Leu Pro Glu Cys Leu Arg Phe Val Gln Thr Asn Ile
115 120 125
Ser His Leu Leu Lys Asp Thr Cys Thr Gln Leu Leu Ala Leu Lys Pro
130 135 140
Cys Ile Gly Lys Ala Cys Gln Asn Phe Ser Arg Cys Leu Glu Val Gln
145 150 155 160
Cys Gln Pro Asp Ser Ser Thr Leu Leu Pro Pro Arg Ser Pro Ile Ala
165 170 175
Leu Glu Ala Thr Glu Leu Pro Glu Pro Arg Pro Arg Gln Leu Leu Leu
180 185 190
Leu Leu Leu Leu Leu Leu Pro Leu Thr Leu Val Leu Leu Ala Ala Ala
195 200 205
Trp Gly Leu Arg Trp Gln Arg Ala Arg Arg Arg Gly Glu Leu His Pro
210 215 220
Gly Val Pro Leu Pro Ser His Pro
225 230
<210> 146
<211> 141
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 146
Met Trp Leu Gln Asn Leu Leu Phe Leu Gly Ile Val Val Tyr Ser Leu
1 5 10 15
Ser Ala Pro Thr Arg Ser Pro Ile Thr Val Thr Arg Pro Trp Lys His
20 25 30
Val Glu Ala Ile Lys Glu Ala Leu Asn Leu Leu Asp Asp Met Pro Val
35 40 45
Thr Leu Asn Glu Glu Val Glu Val Val Ser Asn Glu Phe Ser Phe Lys
50 55 60
Lys Leu Thr Cys Val Gln Thr Arg Leu Lys Ile Phe Glu Gln Gly Leu
65 70 75 80
Arg Gly Asn Phe Thr Lys Leu Lys Gly Ala Leu Asn Met Thr Ala Ser
85 90 95
Tyr Tyr Gln Thr Tyr Cys Pro Pro Thr Pro Glu Thr Asp Cys Glu Thr
100 105 110
Gln Val Thr Thr Tyr Ala Asp Phe Ile Asp Ser Leu Lys Thr Phe Leu
115 120 125
Thr Asp Ile Pro Phe Glu Cys Lys Lys Pro Gly Gln Lys
130 135 140
<210> 147
<211> 1210
<212> PRT
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 147
Met Arg Pro Ser Gly Thr Ala Gly Ala Ala Leu Leu Ala Leu Leu Ala
1 5 10 15
Ala Leu Cys Pro Ala Ser Arg Ala Leu Glu Glu Lys Lys Val Cys Gln
20 25 30
Gly Thr Ser Asn Lys Leu Thr Gln Leu Gly Thr Phe Glu Asp His Phe
35 40 45
Leu Ser Leu Gln Arg Met Phe Asn Asn Cys Glu Val Val Leu Gly Asn
50 55 60
Leu Glu Ile Thr Tyr Val Gln Arg Asn Tyr Asp Leu Ser Phe Leu Lys
65 70 75 80
Thr Ile Gln Glu Val Ala Gly Tyr Val Leu Ile Ala Leu Asn Thr Val
85 90 95
Glu Arg Ile Pro Leu Glu Asn Leu Gln Ile Ile Arg Gly Asn Met Tyr
100 105 110
Tyr Glu Asn Ser Tyr Ala Leu Ala Val Leu Ser Asn Tyr Asp Ala Asn
115 120 125
Lys Thr Gly Leu Lys Glu Leu Pro Met Arg Asn Leu Gln Glu Ile Leu
130 135 140
His Gly Ala Val Arg Phe Ser Asn Asn Pro Ala Leu Cys Asn Val Glu
145 150 155 160
Ser Ile Gln Trp Arg Asp Ile Val Ser Ser Asp Phe Leu Ser Asn Met
165 170 175
Ser Met Asp Phe Gln Asn His Leu Gly Ser Cys Gln Lys Cys Asp Pro
180 185 190
Ser Cys Pro Asn Gly Ser Cys Trp Gly Ala Gly Glu Glu Asn Cys Gln
195 200 205
Lys Leu Thr Lys Ile Ile Cys Ala Gln Gln Cys Ser Gly Arg Cys Arg
210 215 220
Gly Lys Ser Pro Ser Asp Cys Cys His Asn Gln Cys Ala Ala Gly Cys
225 230 235 240
Thr Gly Pro Arg Glu Ser Asp Cys Leu Val Cys Arg Lys Phe Arg Asp
245 250 255
Glu Ala Thr Cys Lys Asp Thr Cys Pro Pro Leu Met Leu Tyr Asn Pro
260 265 270
Thr Thr Tyr Gln Met Asp Val Asn Pro Glu Gly Lys Tyr Ser Phe Gly
275 280 285
Ala Thr Cys Val Lys Lys Cys Pro Arg Asn Tyr Val Val Thr Asp His
290 295 300
Gly Ser Cys Val Arg Ala Cys Gly Ala Asp Ser Tyr Glu Met Glu Glu
305 310 315 320
Asp Gly Val Arg Lys Cys Lys Lys Cys Glu Gly Pro Cys Arg Lys Val
325 330 335
Cys Asn Gly Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn
340 345 350
Ala Thr Asn Ile Lys His Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp
355 360 365
Leu His Ile Leu Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His Thr
370 375 380
Pro Pro Leu Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys Glu
385 390 395 400
Ile Thr Gly Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp
405 410 415
Leu His Ala Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln
420 425 430
His Gly Gln Phe Ser Leu Ala Val Val Ser Leu Asn Ile Thr Ser Leu
435 440 445
Gly Leu Arg Ser Leu Lys Glu Ile Ser Asp Gly Asp Val Ile Ile Ser
450 455 460
Gly Asn Lys Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu
465 470 475 480
Phe Gly Thr Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu
485 490 495
Asn Ser Cys Lys Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser Pro
500 505 510
Glu Gly Cys Trp Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg Asn
515 520 525
Val Ser Arg Gly Arg Glu Cys Val Asp Lys Cys Asn Leu Leu Glu Gly
530 535 540
Glu Pro Arg Glu Phe Val Glu Asn Ser Glu Cys Ile Gln Cys His Pro
545 550 555 560
Glu Cys Leu Pro Gln Ala Met Asn Ile Thr Cys Thr Gly Arg Gly Pro
565 570 575
Asp Asn Cys Ile Gln Cys Ala His Tyr Ile Asp Gly Pro His Cys Val
580 585 590
Lys Thr Cys Pro Ala Gly Val Met Gly Glu Asn Asn Thr Leu Val Trp
595 600 605
Lys Tyr Ala Asp Ala Gly His Val Cys His Leu Cys His Pro Asn Cys
610 615 620
Thr Tyr Gly Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly
625 630 635 640
Pro Lys Ile Pro Ser Ile Ala Thr Gly Met Val Gly Ala Leu Leu Leu
645 650 655
Leu Leu Val Val Ala Leu Gly Ile Gly Leu Phe Met Arg Arg Arg His
660 665 670
Ile Val Arg Lys Arg Thr Leu Arg Arg Leu Leu Gln Glu Arg Glu Leu
675 680 685
Val Glu Pro Leu Thr Pro Ser Gly Glu Ala Pro Asn Gln Ala Leu Leu
690 695 700
Arg Ile Leu Lys Glu Thr Glu Phe Lys Lys Ile Lys Val Leu Gly Ser
705 710 715 720
Gly Ala Phe Gly Thr Val Tyr Lys Gly Leu Trp Ile Pro Glu Gly Glu
725 730 735
Lys Val Lys Ile Pro Val Ala Ile Lys Glu Leu Arg Glu Ala Thr Ser
740 745 750
Pro Lys Ala Asn Lys Glu Ile Leu Asp Glu Ala Tyr Val Met Ala Ser
755 760 765
Val Asp Asn Pro His Val Cys Arg Leu Leu Gly Ile Cys Leu Thr Ser
770 775 780
Thr Val Gln Leu Ile Thr Gln Leu Met Pro Phe Gly Cys Leu Leu Asp
785 790 795 800
Tyr Val Arg Glu His Lys Asp Asn Ile Gly Ser Gln Tyr Leu Leu Asn
805 810 815
Trp Cys Val Gln Ile Ala Lys Gly Met Asn Tyr Leu Glu Asp Arg Arg
820 825 830
Leu Val His Arg Asp Leu Ala Ala Arg Asn Val Leu Val Lys Thr Pro
835 840 845
Gln His Val Lys Ile Thr Asp Phe Gly Leu Ala Lys Leu Leu Gly Ala
850 855 860
Glu Glu Lys Glu Tyr His Ala Glu Gly Gly Lys Val Pro Ile Lys Trp
865 870 875 880
Met Ala Leu Glu Ser Ile Leu His Arg Ile Tyr Thr His Gln Ser Asp
885 890 895
Val Trp Ser Tyr Gly Val Thr Val Trp Glu Leu Met Thr Phe Gly Ser
900 905 910
Lys Pro Tyr Asp Gly Ile Pro Ala Ser Glu Ile Ser Ser Ile Leu Glu
915 920 925
Lys Gly Glu Arg Leu Pro Gln Pro Pro Ile Cys Thr Ile Asp Val Tyr
930 935 940
Met Ile Met Val Lys Cys Trp Met Ile Asp Ala Asp Ser Arg Pro Lys
945 950 955 960
Phe Arg Glu Leu Ile Ile Glu Phe Ser Lys Met Ala Arg Asp Pro Gln
965 970 975
Arg Tyr Leu Val Ile Gln Gly Asp Glu Arg Met His Leu Pro Ser Pro
980 985 990
Thr Asp Ser Asn Phe Tyr Arg Ala Leu Met Asp Glu Glu Asp Met Asp
995 1000 1005
Asp Val Val Asp Ala Asp Glu Tyr Leu Ile Pro Gln Gln Gly Phe
1010 1015 1020
Phe Ser Ser Pro Ser Thr Ser Arg Thr Pro Leu Leu Ser Ser Leu
1025 1030 1035
Ser Ala Thr Ser Asn Asn Ser Thr Val Ala Cys Ile Asp Arg Asn
1040 1045 1050
Gly Leu Gln Ser Cys Pro Ile Lys Glu Asp Ser Phe Leu Gln Arg
1055 1060 1065
Tyr Ser Ser Asp Pro Thr Gly Ala Leu Thr Glu Asp Ser Ile Asp
1070 1075 1080
Asp Thr Phe Leu Pro Val Pro Glu Tyr Ile Asn Gln Ser Val Pro
1085 1090 1095
Lys Arg Pro Ala Gly Ser Val Gln Asn Pro Val Tyr His Asn Gln
1100 1105 1110
Pro Leu Asn Pro Ala Pro Ser Arg Asp Pro His Tyr Gln Asp Pro
1115 1120 1125
His Ser Thr Ala Val Gly Asn Pro Glu Tyr Leu Asn Thr Val Gln
1130 1135 1140
Pro Thr Cys Val Asn Ser Thr Phe Asp Ser Pro Ala His Trp Ala
1145 1150 1155
Gln Lys Gly Ser His Gln Ile Ser Leu Asp Asn Pro Asp Tyr Gln
1160 1165 1170
Gln Asp Phe Phe Pro Lys Glu Ala Lys Pro Asn Gly Ile Phe Lys
1175 1180 1185
Gly Ser Thr Ala Glu Asn Ala Glu Tyr Leu Arg Val Ala Pro Gln
1190 1195 1200
Ser Ser Glu Phe Ile Gly Ala
1205 1210
<210> 148
<211> 1816
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 148
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgttc agaagtcggt taataacgac 1440
atgatagtca ctgacaacaa cggtgcagtc aagtttccac aactgtgtaa attttgtgat 1500
gtgagatttt ccacctgtga caaccagaaa tcctgcatga gcaactgcag catcacctcc 1560
atctgtgaga agccacagga agtctgtgtg gctgtatgga gaaagaatga cgagaacata 1620
acactagaga cagtttgcca tgaccccaag ctcccctacc atgactttat tctggaagat 1680
gctgcttctc caaagtgcat tatgaaggaa aaaaaaaagc ctggtgagac tttcttcatg 1740
tgttcctgta gctctgatga gtgcaatgac aacatcatct tctcagaaga atataacacc 1800
agcaatcctg actaaa 1816
<210> 149
<211> 1780
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 149
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgtta acaacggtgc agtcaagttt 1440
ccacaactgt gtaaattttg tgatgtgaga ttttccacct gtgacaacca gaaatcctgc 1500
atgagcaact gcagcatcac ctccatctgt gagaagccac aggaagtctg tgtggctgta 1560
tggagaaaga atgacgagaa cataacacta gagacagttt gccatgaccc caagctcccc 1620
taccatgact ttattctgga agatgctgct tctccaaagt gcattatgaa ggaaaaaaaa 1680
aagcctggtg agactttctt catgtgttcc tgtagctctg atgagtgcaa tgacaacatc 1740
atcttctcag aagaatataa caccagcaat cctgactaaa 1780
<210> 150
<211> 1774
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 150
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgttg gtgcagtcaa gtttccacaa 1440
ctgtgtaaat tttgtgatgt gagattttcc acctgtgaca accagaaatc ctgcatgagc 1500
aactgcagca tcacctccat ctgtgagaag ccacaggaag tctgtgtggc tgtatggaga 1560
aagaatgacg agaacataac actagagaca gtttgccatg accccaagct cccctaccat 1620
gactttattc tggaagatgc tgcttctcca aagtgcatta tgaaggaaaa aaaaaagcct 1680
ggtgagactt tcttcatgtg ttcctgtagc tctgatgagt gcaatgacaa catcatcttc 1740
tcagaagaat ataacaccag caatcctgac taaa 1774
<210> 151
<211> 1768
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 151
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgttg tcaagtttcc acaactgtgt 1440
aaattttgtg atgtgagatt ttccacctgt gacaaccaga aatcctgcat gagcaactgc 1500
agcatcacct ccatctgtga gaagccacag gaagtctgtg tggctgtatg gagaaagaat 1560
gacgagaaca taacactaga gacagtttgc catgacccca agctccccta ccatgacttt 1620
attctggaag atgctgcttc tccaaagtgc attatgaagg aaaaaaaaaa gcctggtgag 1680
actttcttca tgtgttcctg tagctctgat gagtgcaatg acaacatcat cttctcagaa 1740
gaatataaca ccagcaatcc tgactaaa 1768
<210> 152
<211> 1756
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 152
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgttc aactgtgtaa attttgtgat 1440
gtgagatttt ccacctgtga caaccagaaa tcctgcatga gcaactgcag catcacctcc 1500
atctgtgaga agccacagga agtctgtgtg gctgtatgga gaaagaatga cgagaacata 1560
acactagaga cagtttgcca tgaccccaag ctcccctacc atgactttat tctggaagat 1620
gctgcttctc caaagtgcat tatgaaggaa aaaaaaaagc ctggtgagac tttcttcatg 1680
tgttcctgta gctctgatga gtgcaatgac aacatcatct tctcagaaga atataacacc 1740
agcaatcctg actaaa 1756
<210> 153
<211> 1753
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 153
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgttc tgtgtaaatt ttgtgatgtg 1440
agattttcca cctgtgacaa ccagaaatcc tgcatgagca actgcagcat cacctccatc 1500
tgtgagaagc cacaggaagt ctgtgtggct gtatggagaa agaatgacga gaacataaca 1560
ctagagacag tttgccatga ccccaagctc ccctaccatg actttattct ggaagatgct 1620
gcttctccaa agtgcattat gaaggaaaaa aaaaagcctg gtgagacttt cttcatgtgt 1680
tcctgtagct ctgatgagtg caatgacaac atcatcttct cagaagaata taacaccagc 1740
aatcctgact aaa 1753
<210> 154
<211> 1750
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 154
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgttt gtaaattttg tgatgtgaga 1440
ttttccacct gtgacaacca gaaatcctgc atgagcaact gcagcatcac ctccatctgt 1500
gagaagccac aggaagtctg tgtggctgta tggagaaaga atgacgagaa cataacacta 1560
gagacagttt gccatgaccc caagctcccc taccatgact ttattctgga agatgctgct 1620
tctccaaagt gcattatgaa ggaaaaaaaa aagcctggtg agactttctt catgtgttcc 1680
tgtagctctg atgagtgcaa tgacaacatc atcttctcag aagaatataa caccagcaat 1740
cctgactaaa 1750
<210> 155
<211> 1756
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 155
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacgttc agctgtgtaa attttgtgat 1440
gtgagatttt ccacctgtga caaccagaaa tcctgcatga gcaactgcag catcacctcc 1500
atctgtgaga agccacagga agtctgtgtg gctgtatgga gaaagaatga cgagaacata 1560
acactagaga cagtttgcca tgaccccaag ctcccctacc atgactttat tctggaagat 1620
gctgcttctc caaagtgcat tatgaaggaa aaaaaaaagc ctggtgagac tttcttcatg 1680
tgttcctgta gctctgatga gtgcaatgac aacatcatct tctcagaaga atataacacc 1740
agcaatcctg actaaa 1756
<210> 156
<211> 1750
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 156
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgcacctgt gtaaattttg tgatgtgaga 1440
ttttccacct gtgacaacca gaaatcctgc atgagcaact gcagcatcac ctccatctgt 1500
gagaagccac aggaagtctg tgtggctgta tggagaaaga atgacgagaa cataacacta 1560
gagacagttt gccatgaccc caagctcccc taccatgact ttattctgga agatgctgct 1620
tctccaaagt gcattatgaa ggaaaaaaaa aagcctggtg agactttctt catgtgttcc 1680
tgtagctctg atgagtgcaa tgacaacatc atcttctcag aagaatataa caccagcaat 1740
cctgactaaa 1750
<210> 157
<211> 1747
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 157
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ccgctgtgta aattttgtga tgtgagattt 1440
tccacctgtg acaaccagaa atcctgcatg agcaactgca gcatcacctc catctgtgag 1500
aagccacagg aagtctgtgt ggctgtatgg agaaagaatg acgagaacat aacactagag 1560
acagtttgcc atgaccccaa gctcccctac catgacttta ttctggaaga tgctgcttct 1620
ccaaagtgca ttatgaagga aaaaaaaaag cctggtgaga ctttcttcat gtgttcctgt 1680
agctctgatg agtgcaatga caacatcatc ttctcagaag aatataacac cagcaatcct 1740
gactaaa 1747
<210> 158
<211> 1744
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 158
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatccca ctgtgtaaat tttgtgatgt gagattttcc 1440
acctgtgaca accagaaatc ctgcatgagc aactgcagca tcacctccat ctgtgagaag 1500
ccacaggaag tctgtgtggc tgtatggaga aagaatgacg agaacataac actagagaca 1560
gtttgccatg accccaagct cccctaccat gactttattc tggaagatgc tgcttctcca 1620
aagtgcatta tgaaggaaaa aaaaaagcct ggtgagactt tcttcatgtg ttcctgtagc 1680
tctgatgagt gcaatgacaa catcatcttc tcagaagaat ataacaccag caatcctgac 1740
taaa 1744
<210> 159
<211> 1741
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 159
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctatcctg tgtaaatttt gtgatgtgag attttccacc 1440
tgtgacaacc agaaatcctg catgagcaac tgcagcatca cctccatctg tgagaagcca 1500
caggaagtct gtgtggctgt atggagaaag aatgacgaga acataacact agagacagtt 1560
tgccatgacc ccaagctccc ctaccatgac tttattctgg aagatgctgc ttctccaaag 1620
tgcattatga aggaaaaaaa aaagcctggt gagactttct tcatgtgttc ctgtagctct 1680
gatgagtgca atgacaacat catcttctca gaagaatata acaccagcaa tcctgactaa 1740
a 1741
<210> 160
<211> 1750
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 160
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctccaccg cacgttctgt gtaaattttg tgatgtgaga 1440
ttttccacct gtgacaacca gaaatcctgc atgagcaact gcagcatcac ctccatctgt 1500
gagaagccac aggaagtctg tgtggctgta tggagaaaga atgacgagaa cataacacta 1560
gagacagttt gccatgaccc caagctcccc taccatgact ttattctgga agatgctgct 1620
tctccaaagt gcattatgaa ggaaaaaaaa aagcctggtg agactttctt catgtgttcc 1680
tgtagctctg atgagtgcaa tgacaacatc atcttctcag aagaatataa caccagcaat 1740
cctgactaaa 1750
<210> 161
<211> 1747
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 161
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctccgcac gttctgtgta aattttgtga tgtgagattt 1440
tccacctgtg acaaccagaa atcctgcatg agcaactgca gcatcacctc catctgtgag 1500
aagccacagg aagtctgtgt ggctgtatgg agaaagaatg acgagaacat aacactagag 1560
acagtttgcc atgaccccaa gctcccctac catgacttta ttctggaaga tgctgcttct 1620
ccaaagtgca ttatgaagga aaaaaaaaag cctggtgaga ctttcttcat gtgttcctgt 1680
agctctgatg agtgcaatga caacatcatc ttctcagaag aatataacac cagcaatcct 1740
gactaaa 1747
<210> 162
<211> 1744
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 162
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctcacgtt ctgtgtaaat tttgtgatgt gagattttcc 1440
acctgtgaca accagaaatc ctgcatgagc aactgcagca tcacctccat ctgtgagaag 1500
ccacaggaag tctgtgtggc tgtatggaga aagaatgacg agaacataac actagagaca 1560
gtttgccatg accccaagct cccctaccat gactttattc tggaagatgc tgcttctcca 1620
aagtgcatta tgaaggaaaa aaaaaagcct ggtgagactt tcttcatgtg ttcctgtagc 1680
tctgatgagt gcaatgacaa catcatcttc tcagaagaat ataacaccag caatcctgac 1740
taaa 1744
<210> 163
<211> 1741
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 163
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctgttctg tgtaaatttt gtgatgtgag attttccacc 1440
tgtgacaacc agaaatcctg catgagcaac tgcagcatca cctccatctg tgagaagcca 1500
caggaagtct gtgtggctgt atggagaaag aatgacgaga acataacact agagacagtt 1560
tgccatgacc ccaagctccc ctaccatgac tttattctgg aagatgctgc ttctccaaag 1620
tgcattatga aggaaaaaaa aaagcctggt gagactttct tcatgtgttc ctgtagctct 1680
gatgagtgca atgacaacat catcttctca gaagaatata acaccagcaa tcctgactaa 1740
a 1741
<210> 164
<211> 1759
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 164
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctggtgca gtcaagtttc cacaactgtg taaattttgt 1440
gatgtgagat tttccacctg tgacaaccag aaatcctgca tgagcaactg cagcatcacc 1500
tccatctgtg agaagccaca ggaagtctgt gtggctgtat ggagaaagaa tgacgagaac 1560
ataacactag agacagtttg ccatgacccc aagctcccct accatgactt tattctggaa 1620
gatgctgctt ctccaaagtg cattatgaag gaaaaaaaaa agcctggtga gactttcttc 1680
atgtgttcct gtagctctga tgagtgcaat gacaacatca tcttctcaga agaatataac 1740
accagcaatc ctgactaaa 1759
<210> 165
<211> 1753
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 165
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctgtcaag tttccacaac tgtgtaaatt ttgtgatgtg 1440
agattttcca cctgtgacaa ccagaaatcc tgcatgagca actgcagcat cacctccatc 1500
tgtgagaagc cacaggaagt ctgtgtggct gtatggagaa agaatgacga gaacataaca 1560
ctagagacag tttgccatga ccccaagctc ccctaccatg actttattct ggaagatgct 1620
gcttctccaa agtgcattat gaaggaaaaa aaaaagcctg gtgagacttt cttcatgtgt 1680
tcctgtagct ctgatgagtg caatgacaac atcatcttct cagaagaata taacaccagc 1740
aatcctgact aaa 1753
<210> 166
<211> 1738
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 166
gaggtccaac ttgtccagtc tggagcagag gtgaagaagc ctggagccac agtgaagatt 60
tcctgtaagg tgtctggcta caccttcacc acctactaca cccactgggt gaagcaggct 120
cctggcaagg gattggagtg gattggctgg atttaccctg gagatgtgaa caccaaatac 180
aatgagaagt tcaagggcag ggtgaccctg acagcagaca ccagcacaga cacagcctat 240
atggaactgt cctccctgag gtctgaggac acagcagtct actactgtgc cagggaggac 300
cctggcagca actactttga ctactgggga caaggcaccc tggtgacagt gtccagcgca 360
agcaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacgtgcg tggtggtgga cgtgagccac gaagaccccg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtggtggt ggcggttcag gcggaggtgg ctctggaggt 1380
ggaggttcag gaggtggtgg ttctctgtgt aaattttgtg atgtgagatt ttccacctgt 1440
gacaaccaga aatcctgcat gagcaactgc agcatcacct ccatctgtga gaagccacag 1500
gaagtctgtg tggctgtatg gagaaagaat gacgagaaca taacactaga gacagtttgc 1560
catgacccca agctccccta ccatgacttt attctggaag atgctgcttc tccaaagtgc 1620
attatgaagg aaaaaaaaaa gcctggtgag actttcttca tgtgttcctg tagctctgat 1680
gagtgcaatg acaacatcat cttctcagaa gaatataaca ccagcaatcc tgactaaa 1738
<210> 167
<211> 32
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 167
gtcaccgtcc tgacacgaag cttgccgcca cc 32
<210> 168
<211> 23
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 168
actatagaat agggccctct aga 23
<210> 169
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 169
ggcaaggctc caaagctgct gatttac 27
<210> 170
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 170
gtaaatcagc agctttggag ccttgcc 27
<210> 171
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 171
acctactact gtatgcagtc ctatgat 27
<210> 172
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 172
atcataggac tgcatacagt agtaggt 27
<210> 173
<211> 51
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 173
gctaccaggg tgctgagtga ggtccaactt gtccagtctg gagcagaggt g 51
<210> 174
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 174
cactgtggct ccaggcttct tcacctctgc tcc 33
<210> 175
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 175
cctggagcca cagtgaagat ttcctgtaag gtg 33
<210> 176
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 176
ggtggtgaag gtgtagccag acaccttaca gga 33
<210> 177
<211> 31
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 177
tacaccttca ccacctacta cacccactgg g 31
<210> 178
<211> 32
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 178
cttgccagga gcctgcttca cccagtgggt gt 32
<210> 179
<211> 31
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 179
caggctcctg gcaagggatt ggagtggatt g 31
<210> 180
<211> 32
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 180
atctccaggg taaatccagc caatccactc ca 32
<210> 181
<211> 31
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 181
atttaccctg gagatgtgaa caccaaatac a 31
<210> 182
<211> 32
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 182
cctgcccttg aacttctcat tgtatttggt gt 32
<210> 183
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 183
aagttcaagg gcagggtgac cctgacagca gac 33
<210> 184
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 184
ataggctgtg tctgtgctgg tgtctgctgt cag 33
<210> 185
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 185
acagacacag cctatatgga actgtcctcc ctg 33
<210> 186
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 186
gactgctgtg tcctcagacc tcagggagga cag 33
<210> 187
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 187
gaggacacag cagtctacta ctgtgccagg gag 33
<210> 188
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 188
aaagtagttg ctgccagggt cctccctggc aca 33
<210> 189
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 189
ggcagcaact actttgacta ctggggacaa ggc 33
<210> 190
<211> 51
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность синтезирована искусственно
<400> 190
tgggcccttg gtgcttgcgc tggacactgt caccagggtg ccttgtcccc a 51
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| РЕКОМБИНАНТНЫЕ БЕЛКИ С ДОМЕНАМИ CCN И СЛИТЫЕ БЕЛКИ | 2020 |
|
RU2825102C2 |
| СЛИТЫЕ БЕЛКИ С АЛЬБУМИН-СВЯЗЫВАЮЩИМИ ДОМЕНАМИ | 2018 |
|
RU2786444C2 |
| ВАРИАНТНЫЕ МОЛЕКУЛЫ IL-10 И СПОСОБ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2020 |
|
RU2836849C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800923C2 |
| ПОЛИВАЛЕТНЫЕ И ПОЛИСПЕЦИФИЧНЫЕ GITR-СВЯЗЫВАЮЩИЕ СЛИТЫЕ БЕЛКИ | 2016 |
|
RU2753439C2 |
| ДИСПЛЕЙ ИНТЕГРАЛЬНОГО МЕМБРАННОГО БЕЛКА НА ВНЕКЛЕТОЧНЫХ ОБОЛОЧЕЧНЫХ ВИРИОНАХ ПОКСВИРУСА | 2017 |
|
RU2759846C2 |
| Слитые молекулы, происходящие от Cholix-токсина, для пероральной доставки биологически активных нагрузок | 2015 |
|
RU2723178C2 |
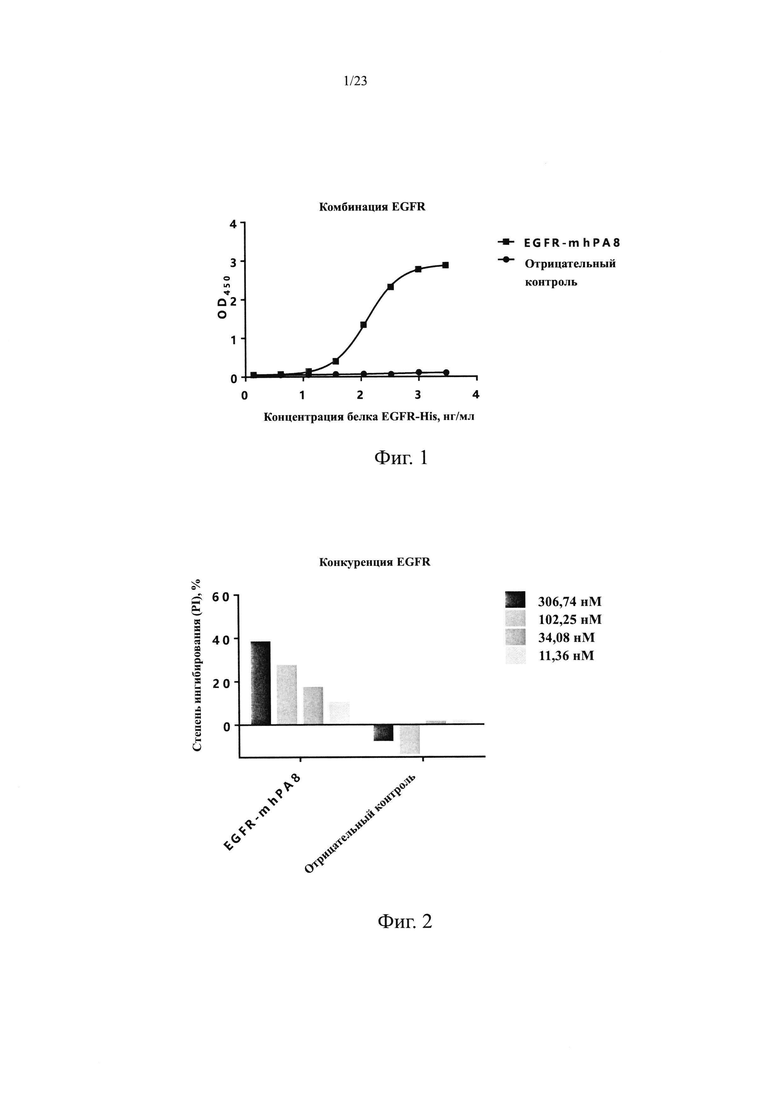
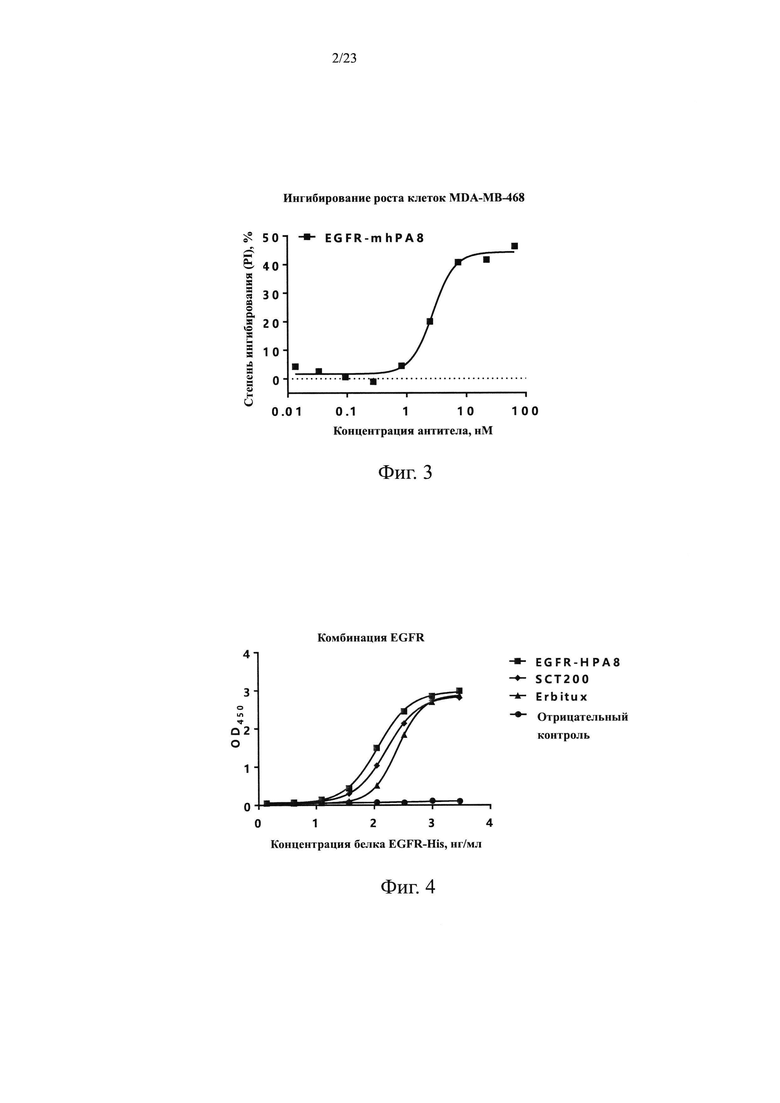
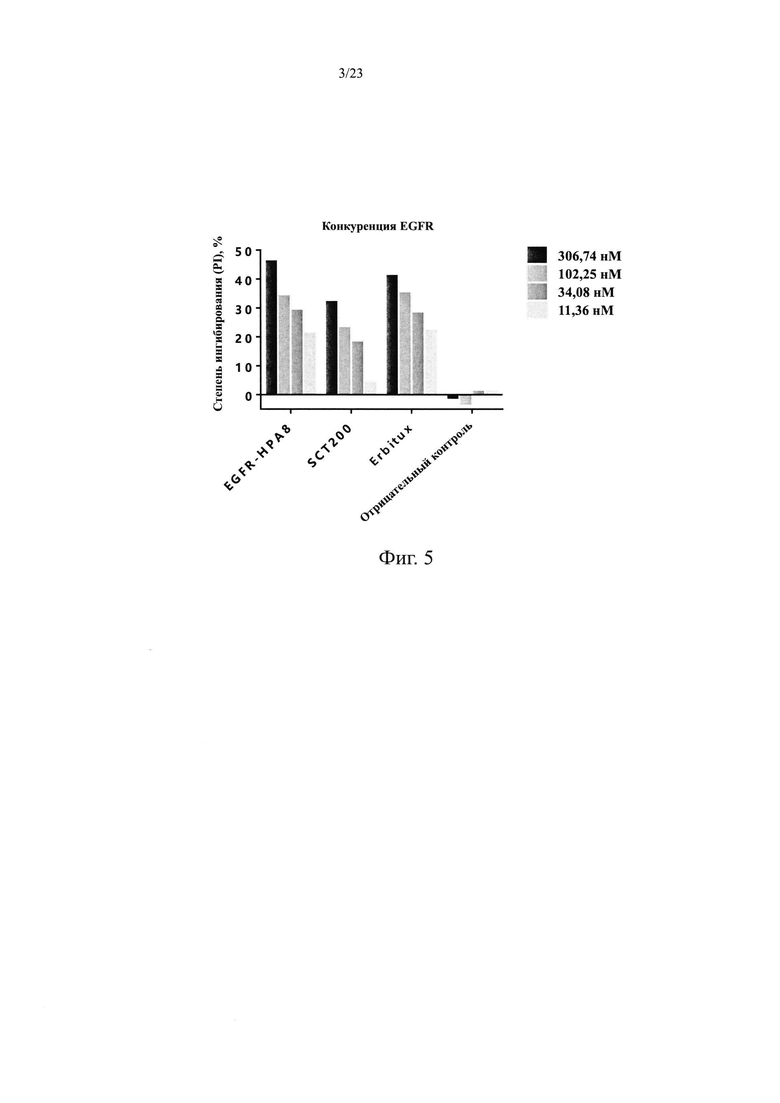

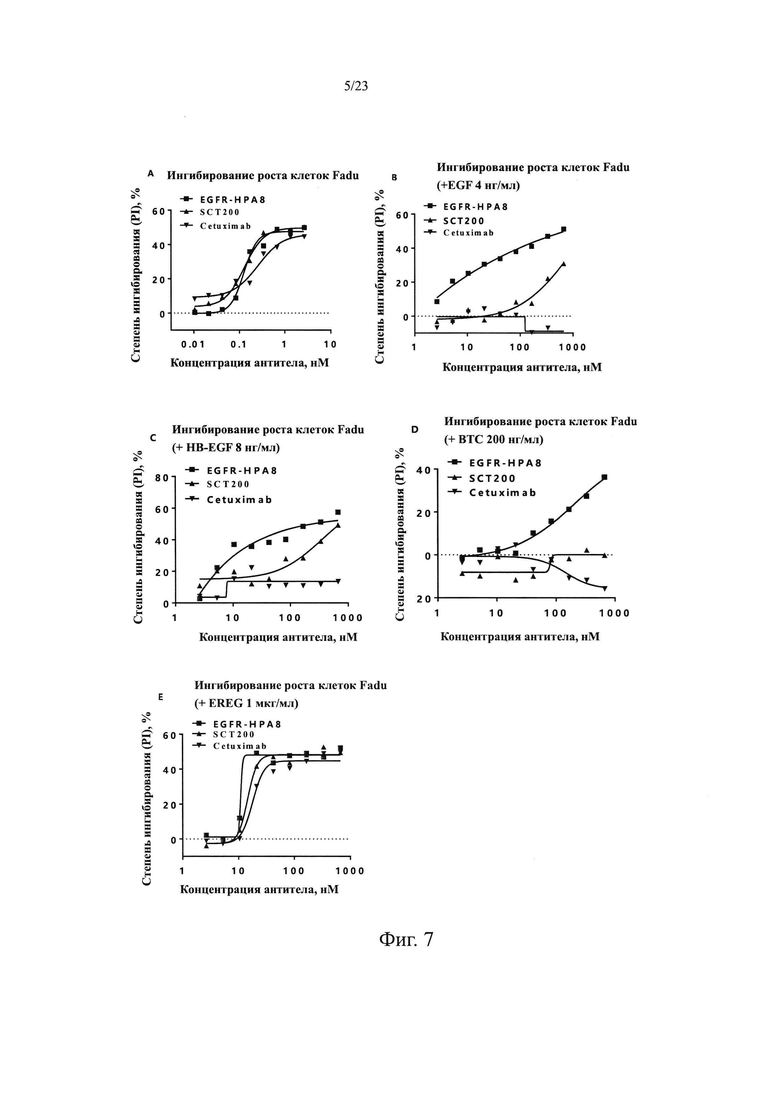
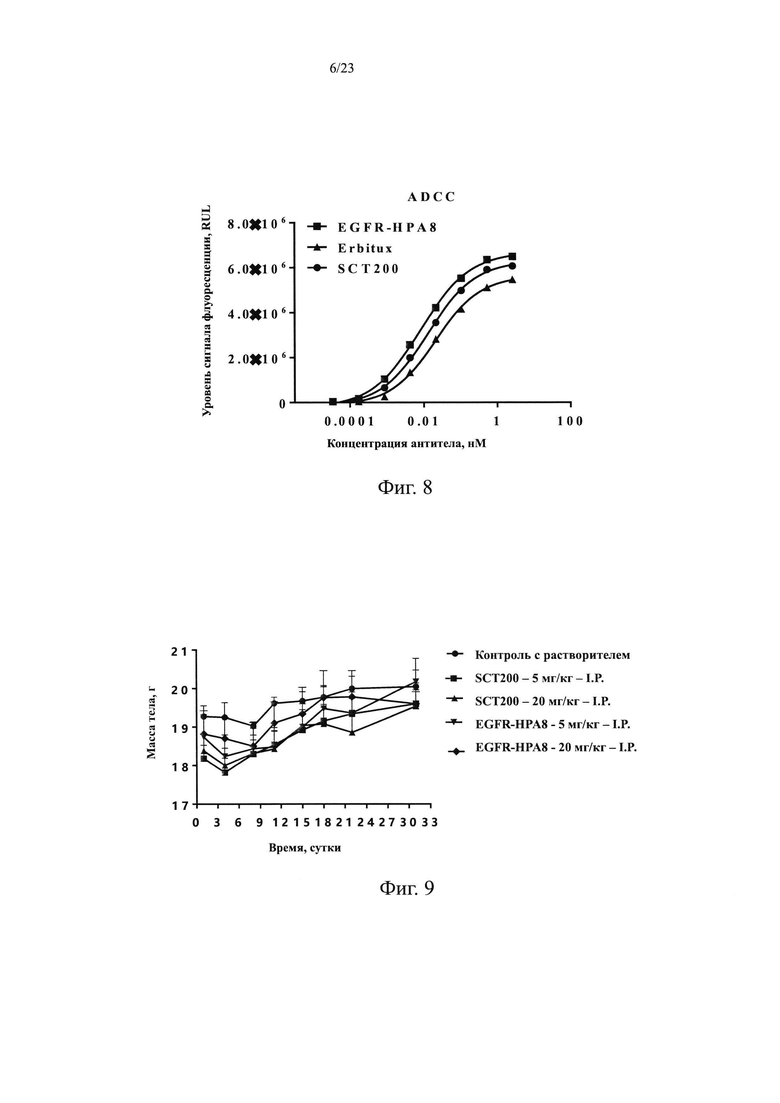
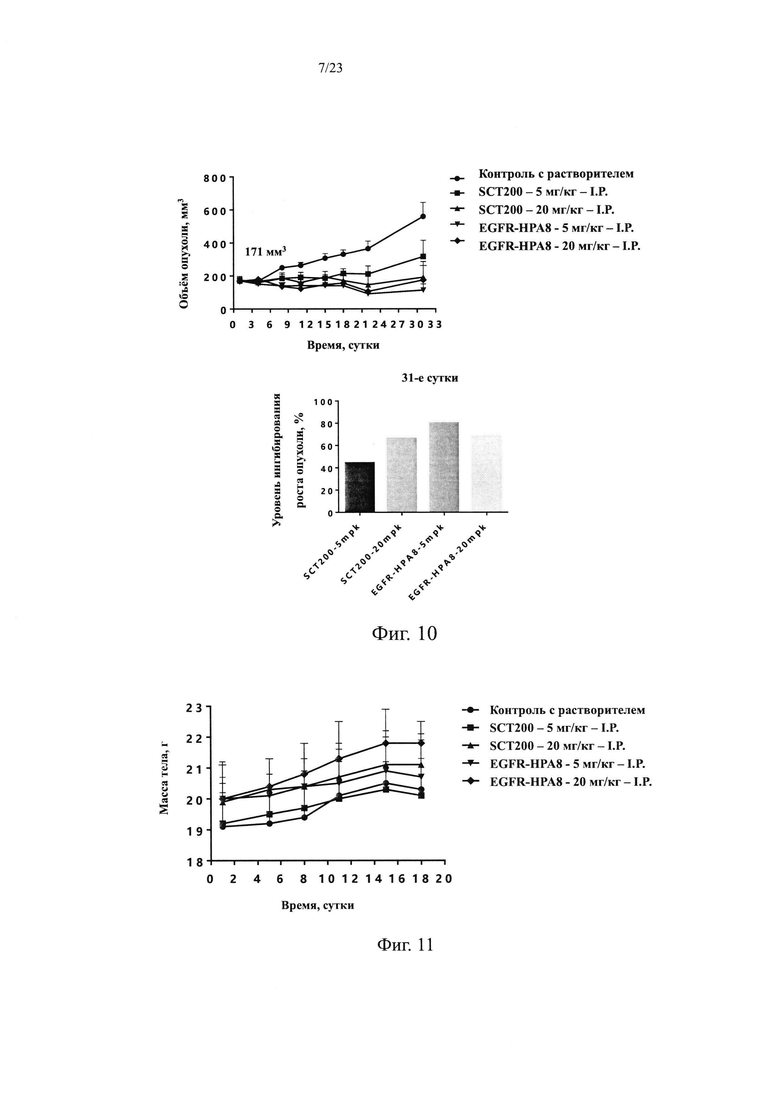
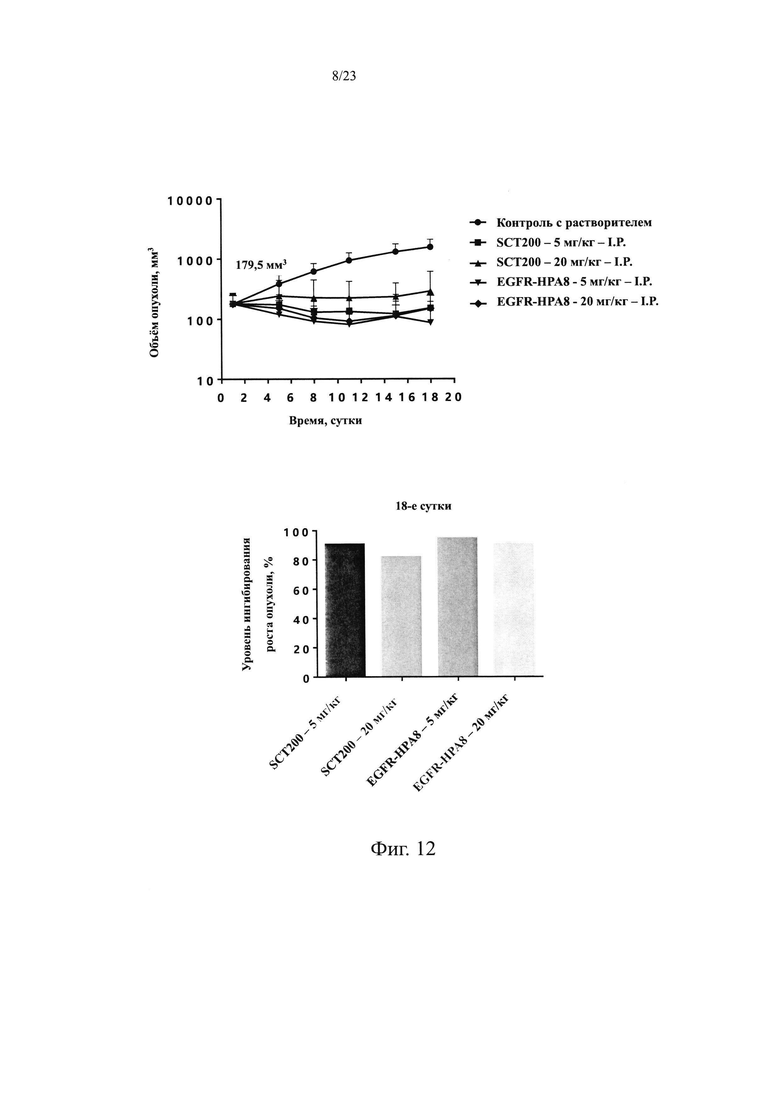
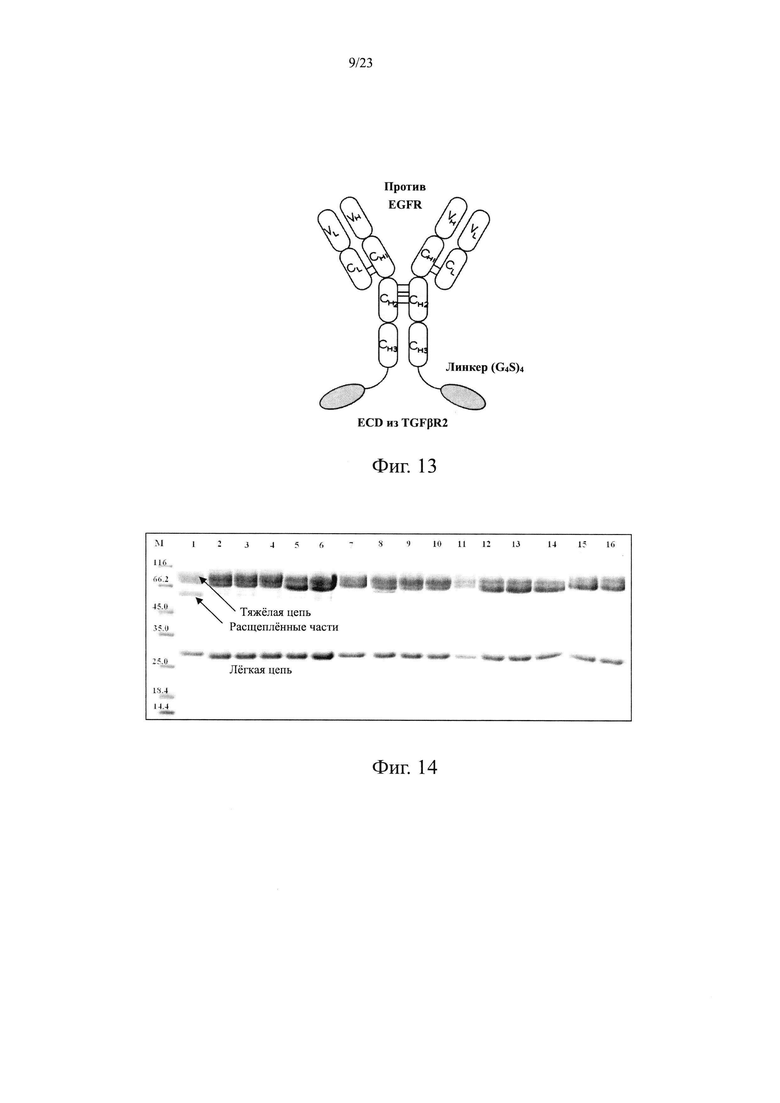
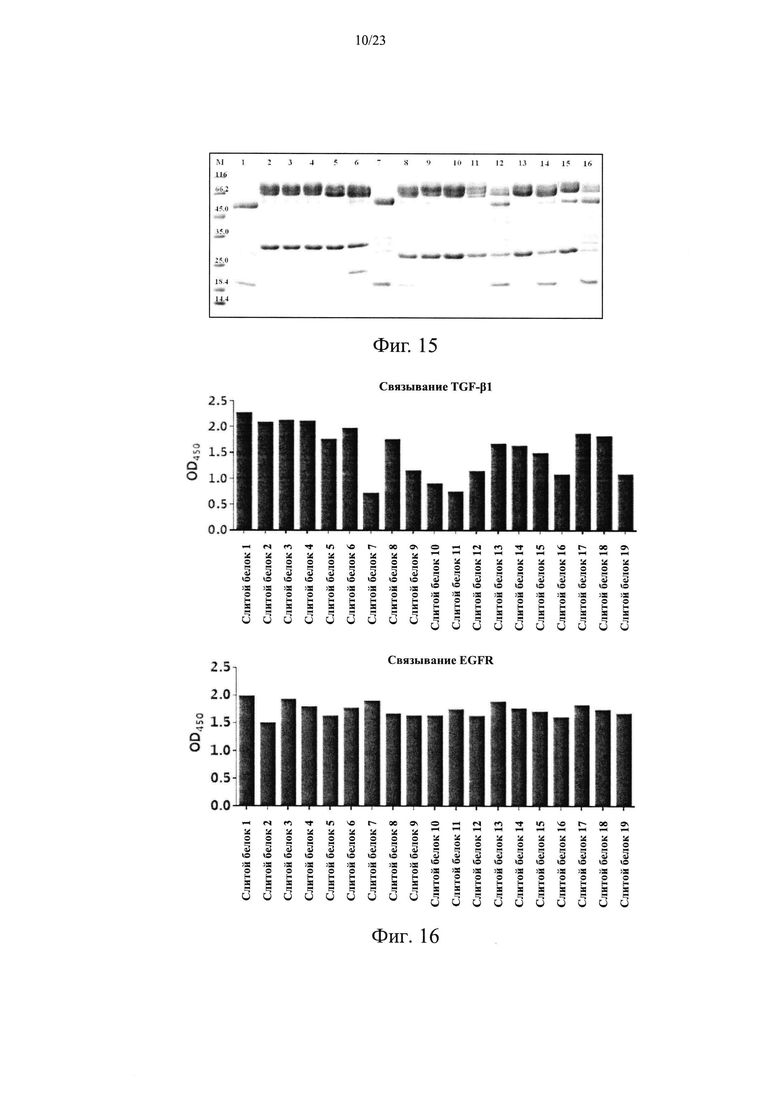
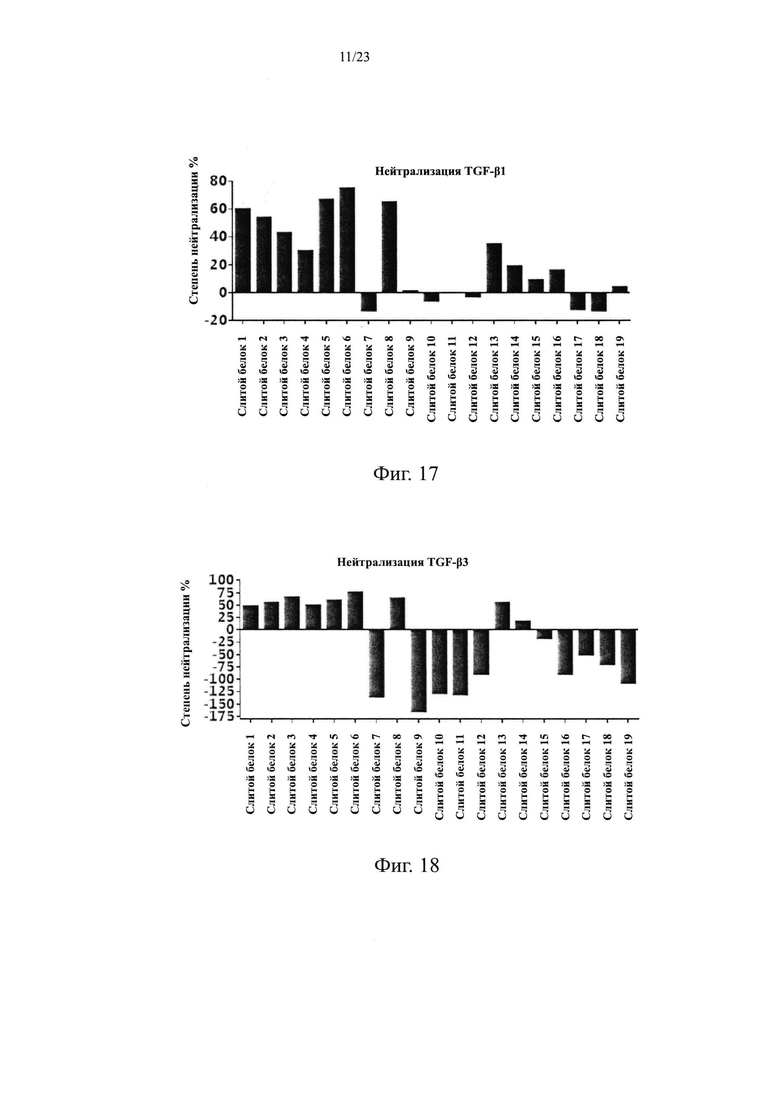
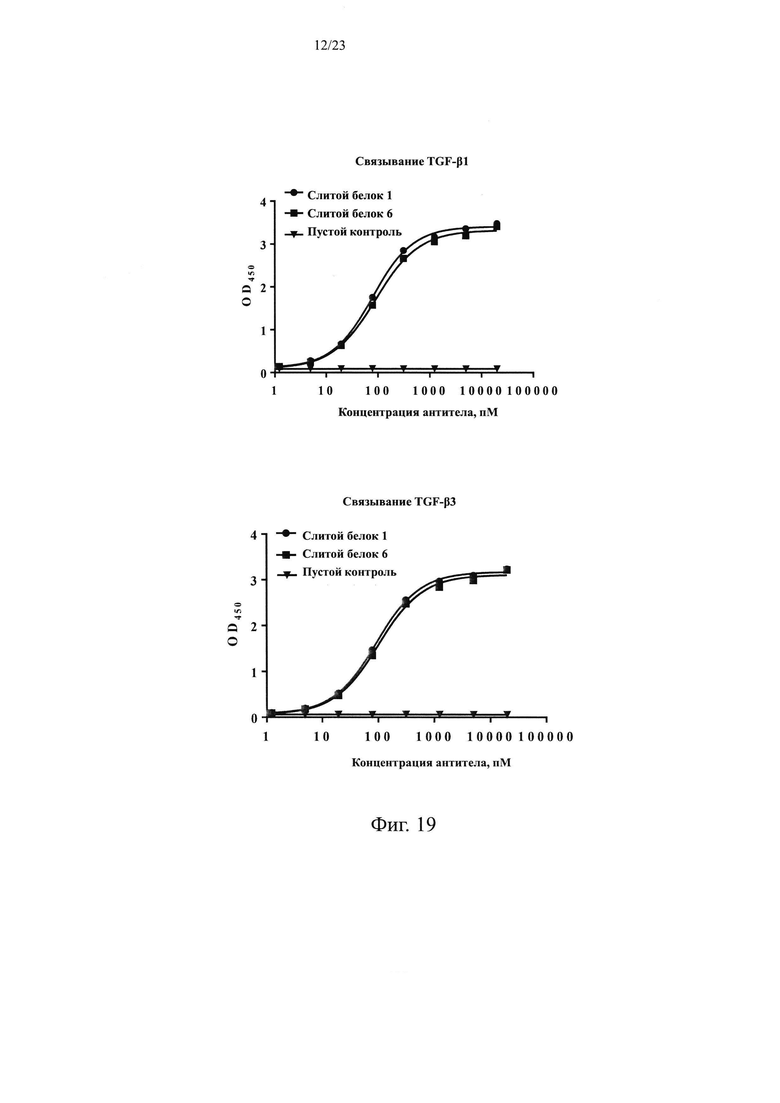
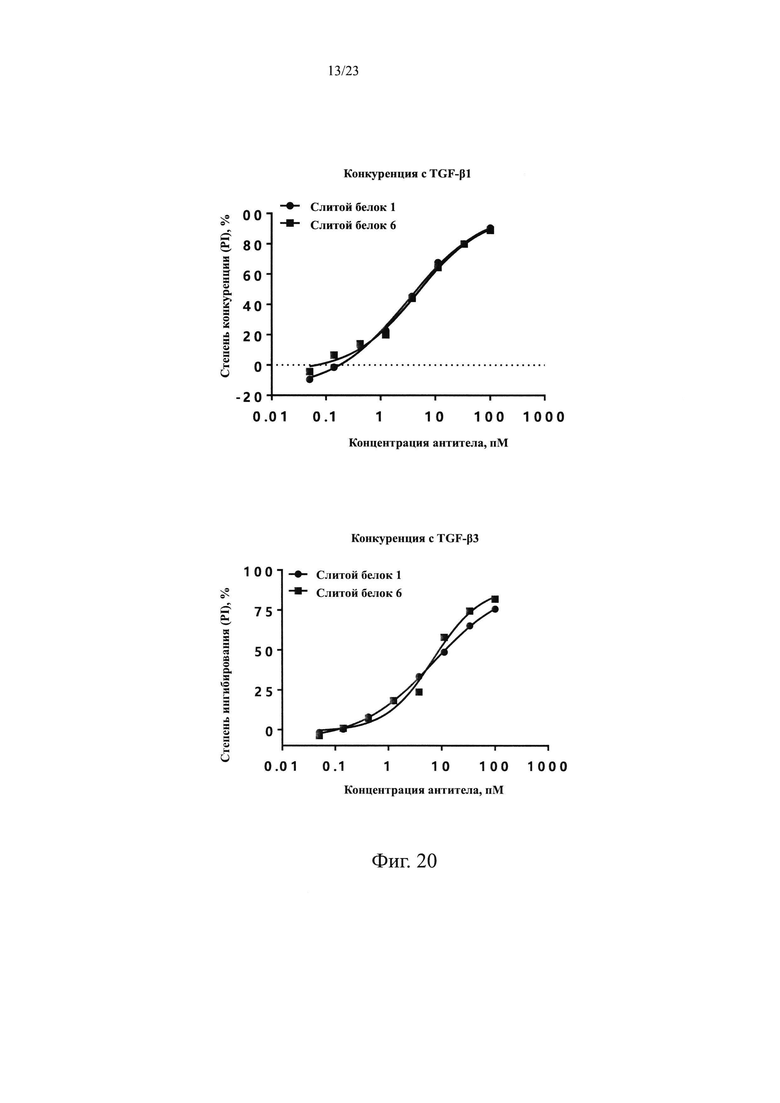
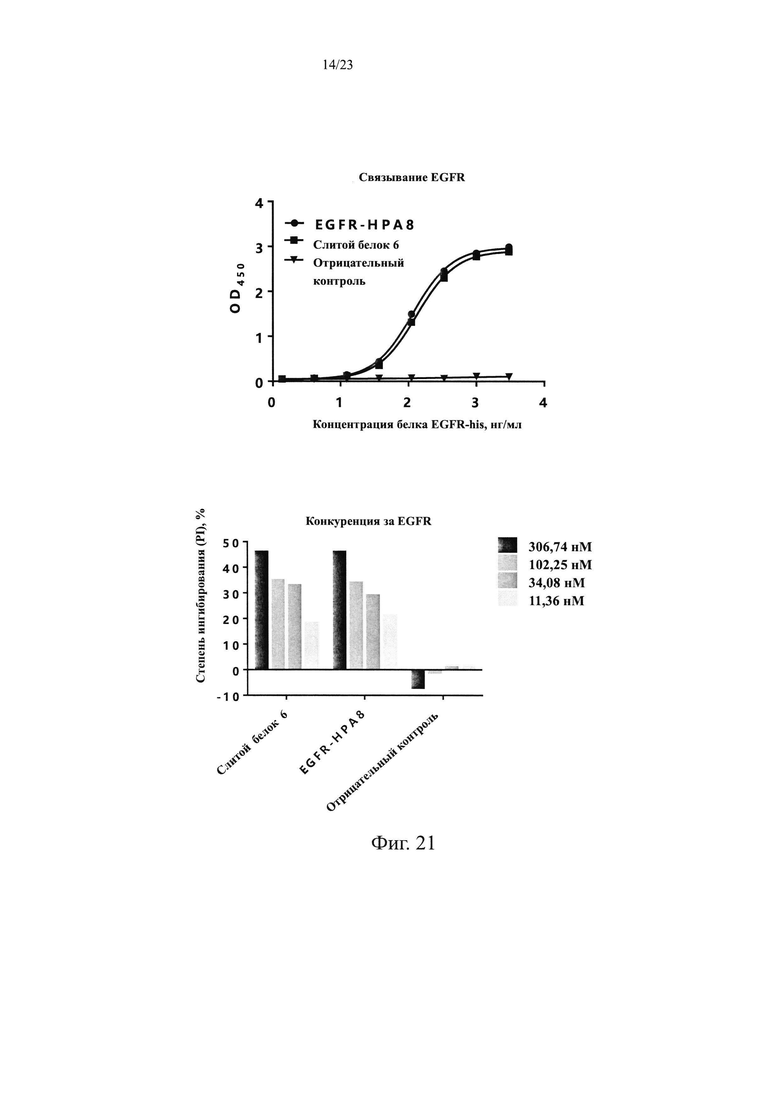
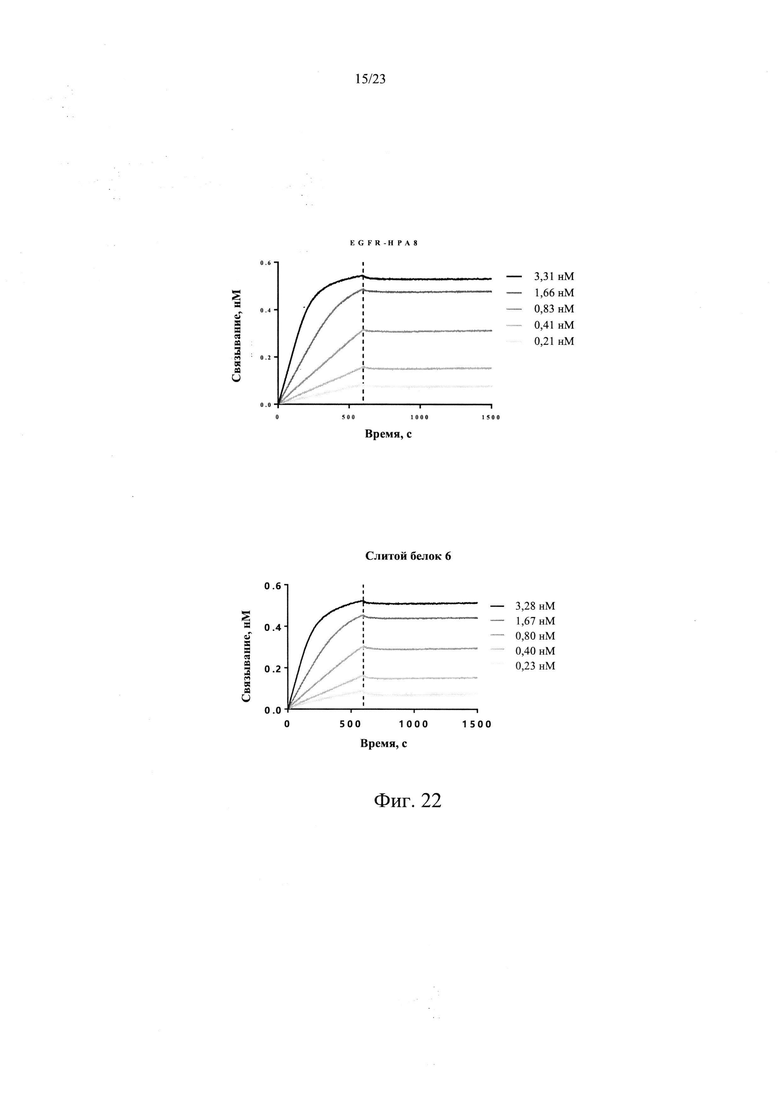
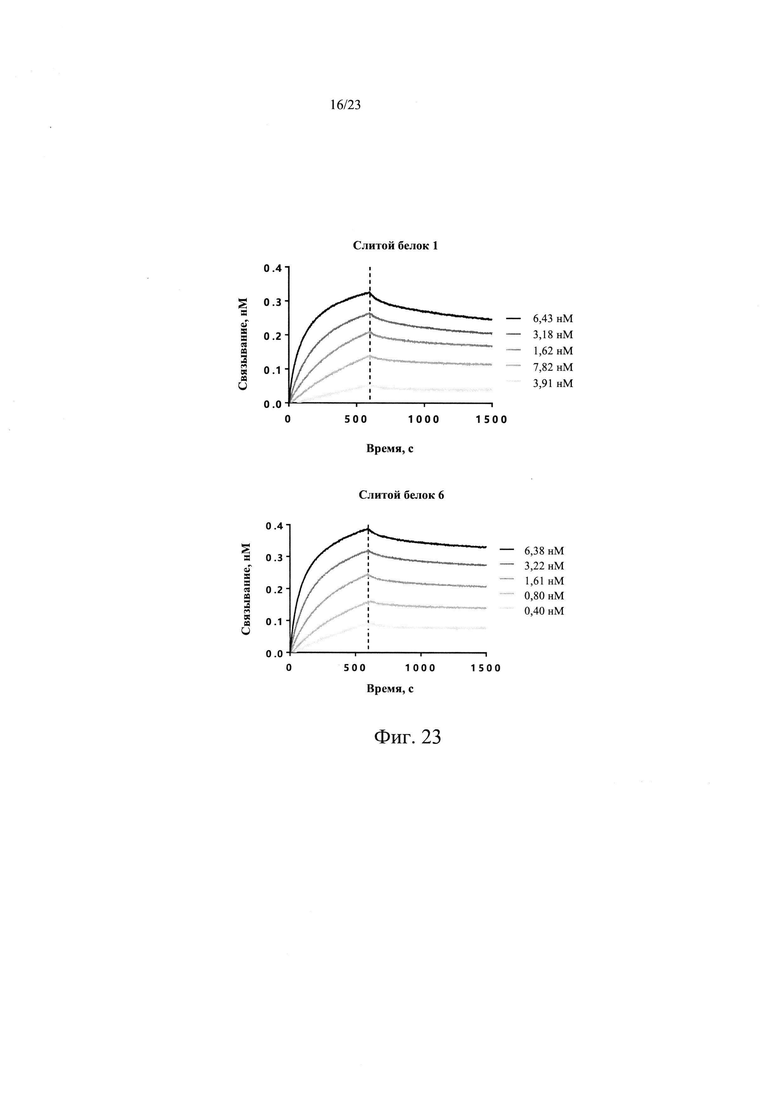
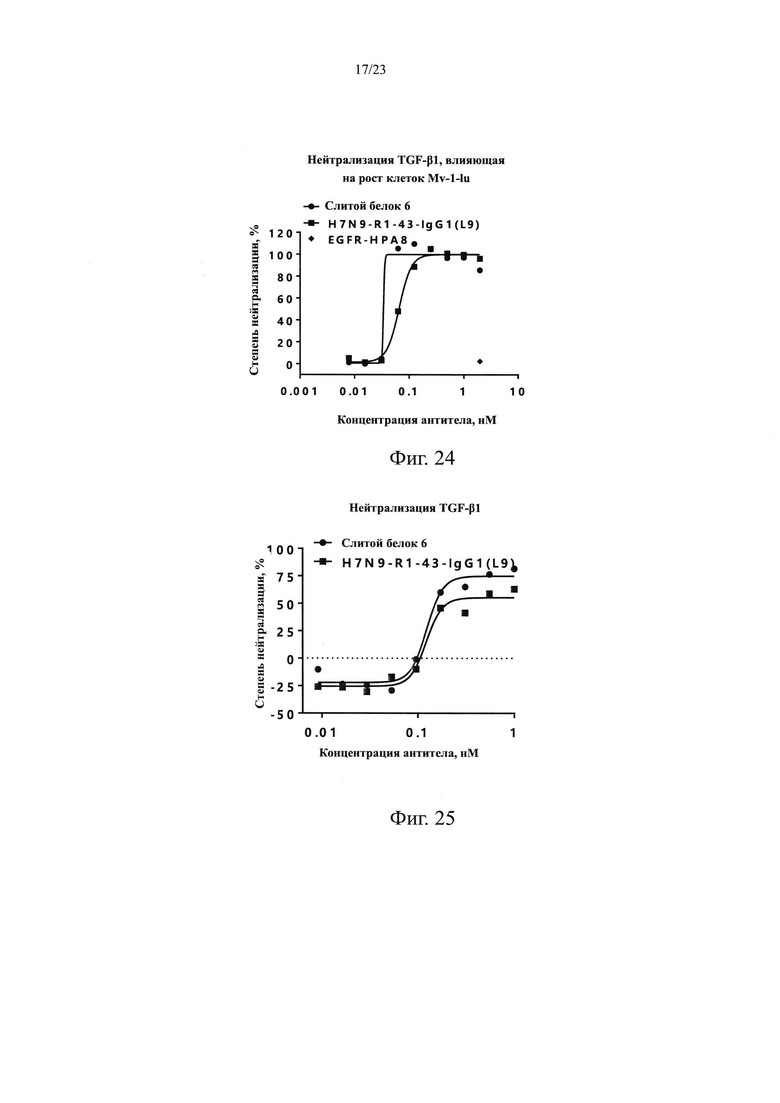
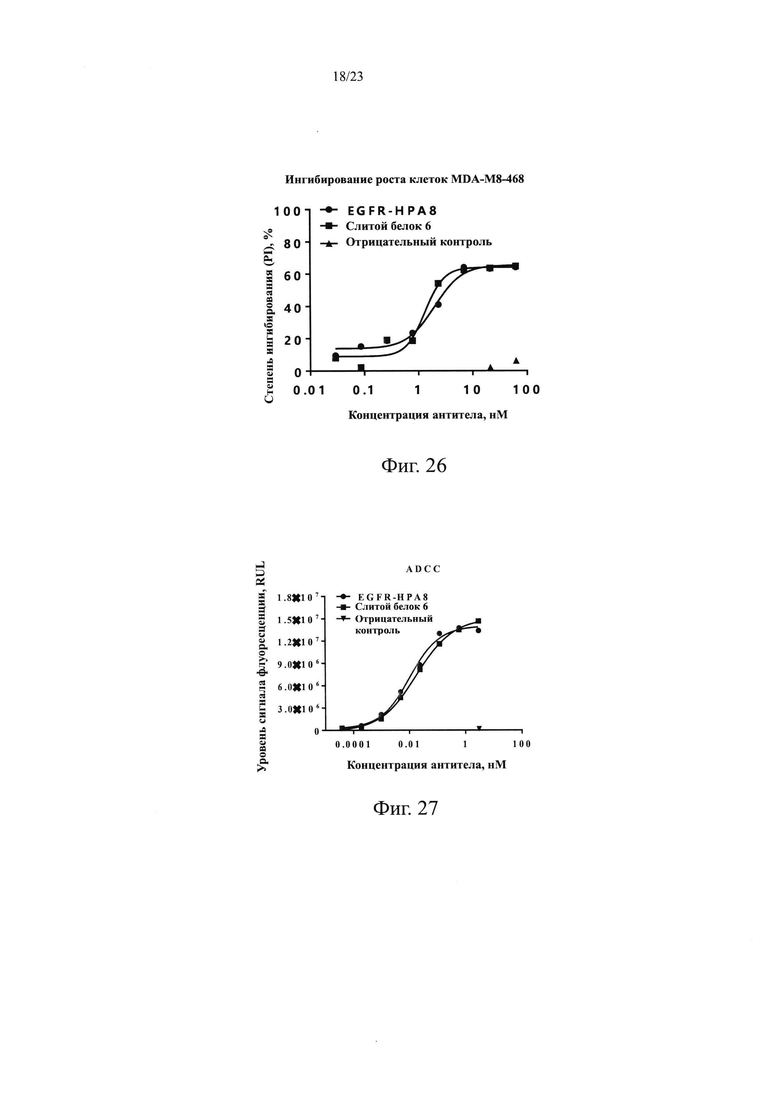
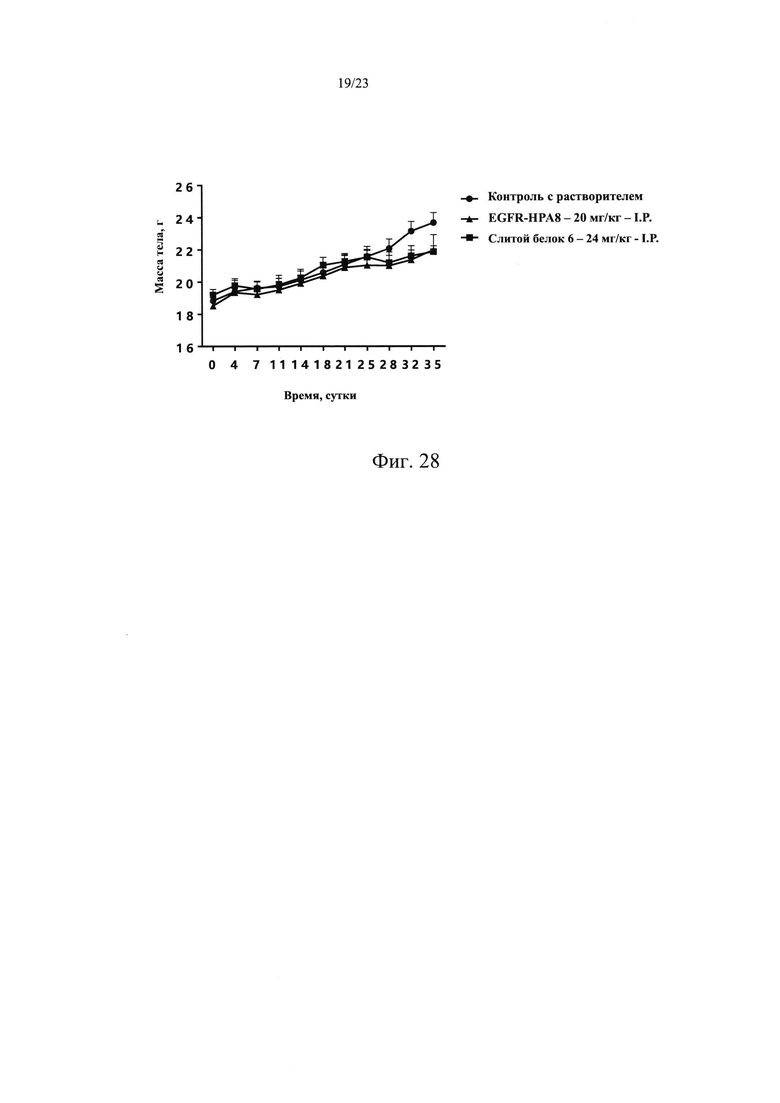
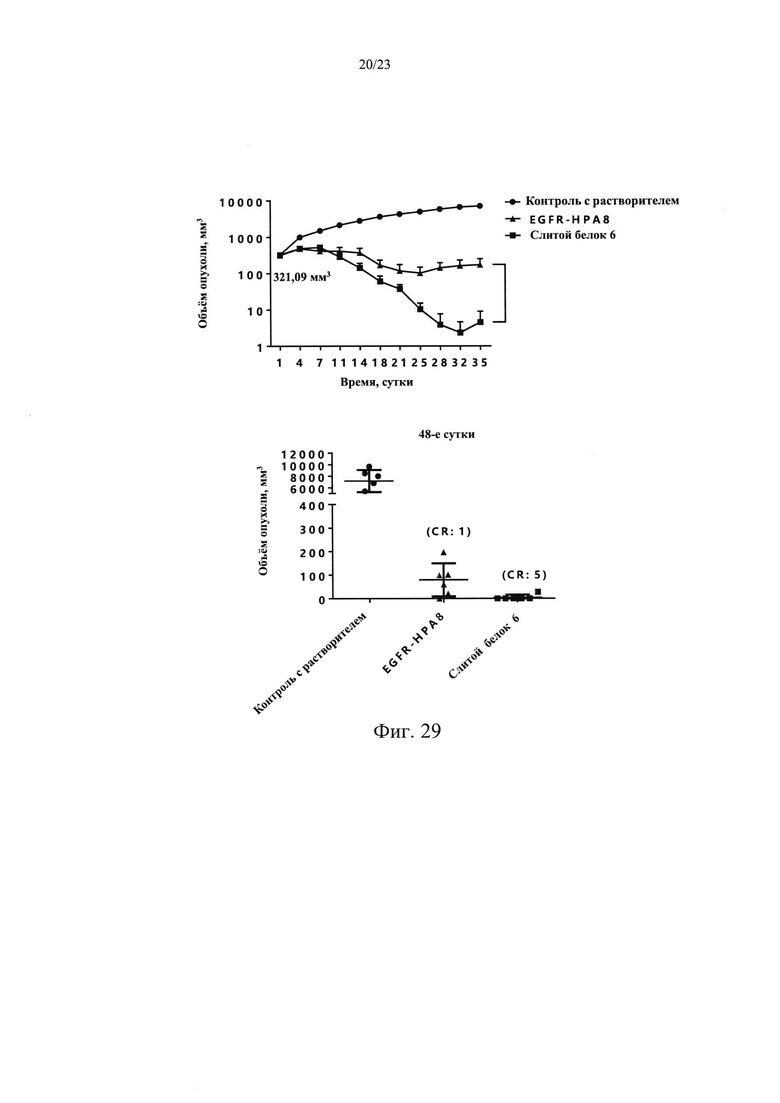
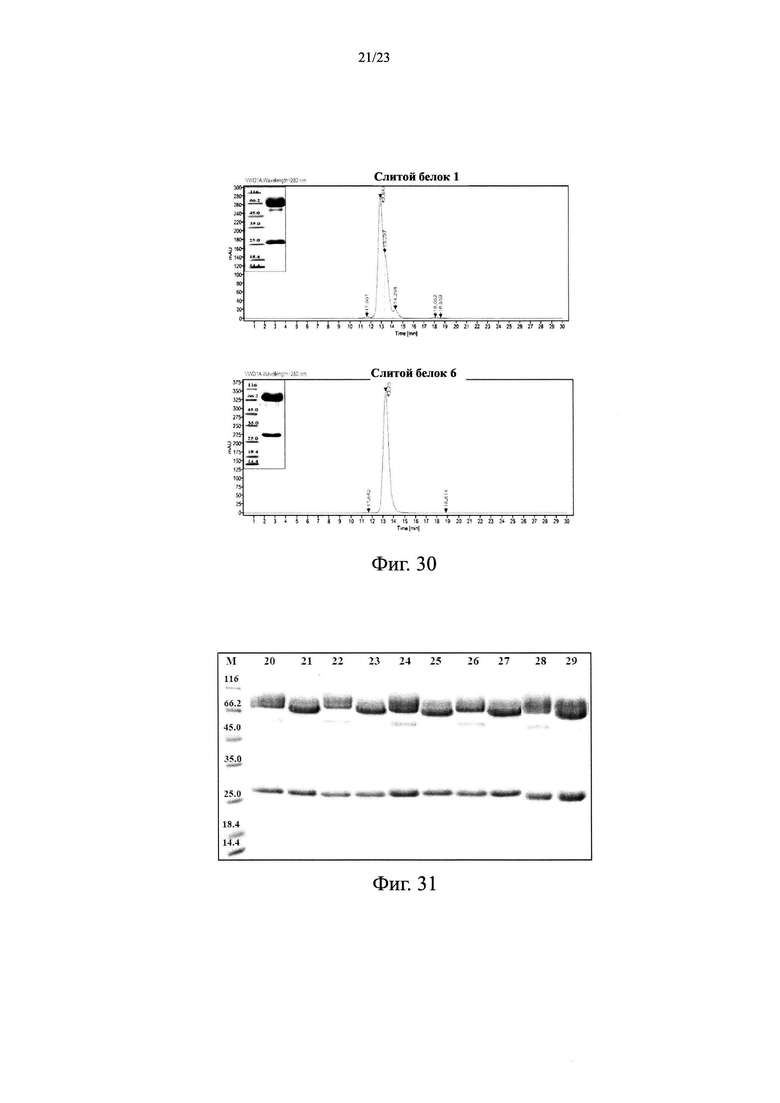
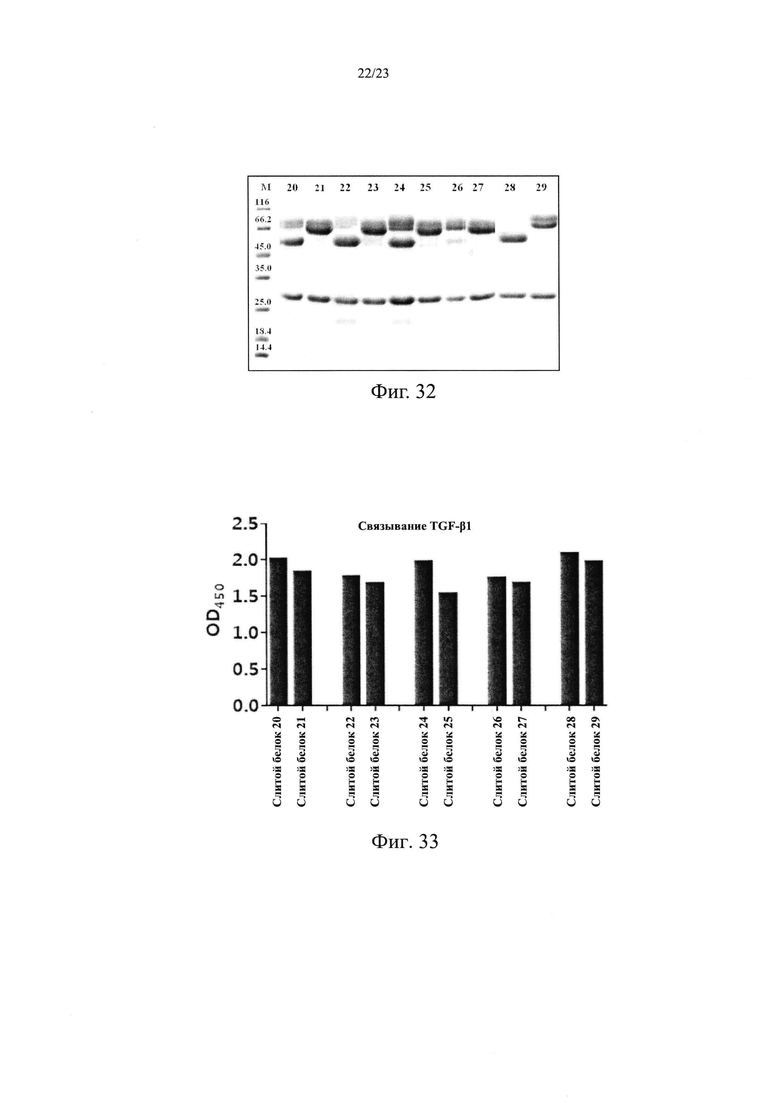
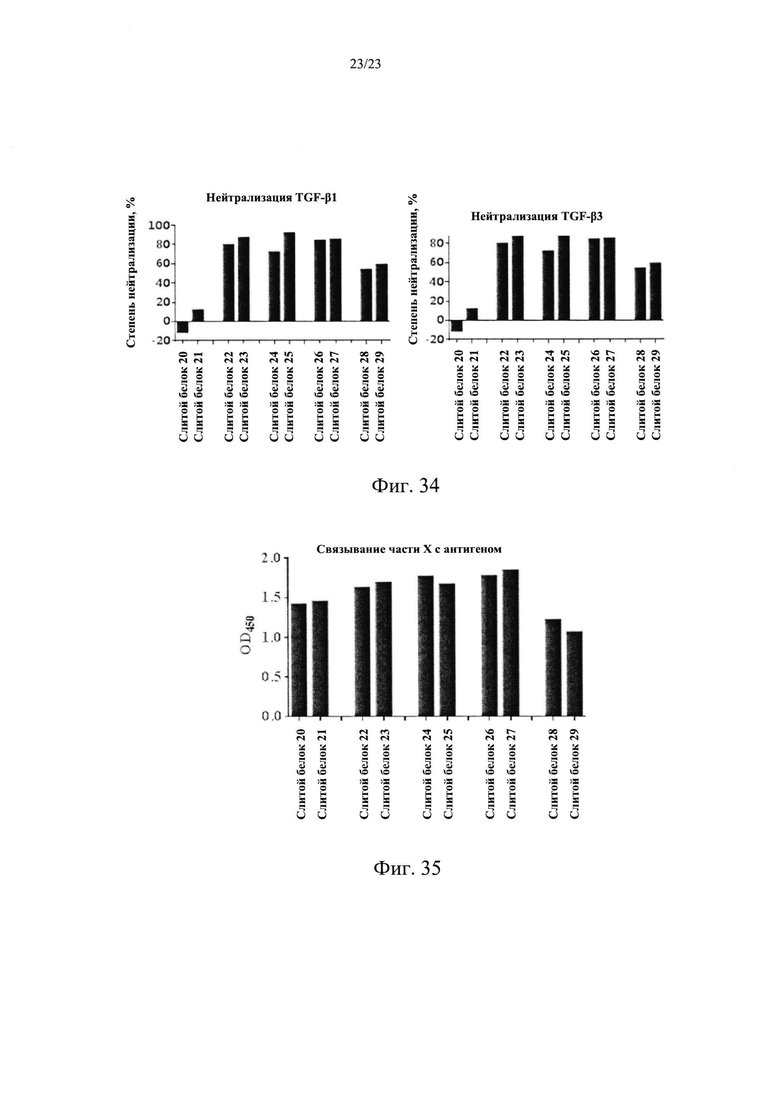
Изобретение относится к области биотехнологии, конкретно к рекомбинантным молекулам рецептора типа II трансформирующего фактора роста бета (TGFβR2), и может быть использовано в медицине для лечения опухоли, ассоциированной с EGFR. Изобретение обеспечивает получение молекулы TGFβR2 с укороченным внеклеточным доменом, которая при соединении с антителом, нацеленным на раковую клетку, является более устойчивой к расщеплению протеазами по сравнению с природной формой молекулы TGFβR2. 5 н. и 7 з.п. ф-лы, 35 ил., 25 табл., 8 пр.
1. Молекула рецептора типа II трансформирующего фактора роста бета (TGFβR2) с укороченным внеклеточным доменом, представленная в любой из SEQ ID NO: 48, 49, 52, 55, 56 и 59, при этом она соединена с антителом, нацеленным на раковую клетку, и указанная молекула является более устойчивой к расщеплению протеазами по сравнению с природной формой молекулы TGFβR2, представленной в SEQ ID NO: 47.
2. Слитый белок для лечения рака, ассоциированного с рецептором эпидермального фактора роста (EGFR), содержащий
а) молекулу TGFβR2 с укороченным внеклеточным доменом по п. 1; и
б) антитело против EGFR в качестве нацеливающей части, при этом N-конец указанной молекулы TGFβR2 с укороченным внеклеточным доменом соединен посредством гибкого пептидного линкера G4S с С-концом указанной тяжелой цепи нацеливающей части.
3. Слитый белок по п. 2, отличающийся тем, что указанный линкер представляет собой пептидный линкер (G4S)4.
4. Слитый белок по п. 2, отличающийся тем, что антитело против EGFR содержит
а) вариабельный участок тяжелой цепи, содержащий участки CDR1 тяжелой цепи, CDR2 тяжелой цепи и CDR3 тяжелой цепи, содержащие SEQ ID NO: 19, 20, и 21, соответственно, и
б) вариабельный участок легкой цепи, содержащий участки CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, содержащие SEQ ID NO: 16, 17, и 18, соответственно.
5. Слитый белок по п. 4, отличающийся тем, что антитело против EGFR содержит
а) вариабельный участок тяжелой цепи, содержащий последовательность SEQ ID NO: 28 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью; и
б) вариабельный участок легкой цепи, содержащий последовательность SEQ ID NO: 29 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью.
6. Слитый белок по п. 4, отличающийся тем, что антитело против EGFR дополнительно включает:
а) константный участок тяжелой цепи, предпочтительно, содержащий последовательность SEQ ID NO: 30 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью; и/или
б) константный участок легкой цепи, предпочтительно, содержащий последовательность, содержащую SEQ ID NO: 31, или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью.
7. Слитый белок по пп. 2, 3, отличающийся тем, что нацеливающая часть выбрана из бевацизумаба, рамуцирумаба, ипилимумаба или панитумумаба.
8. Слитый белок по п. 6, отличающийся тем, что
а) указанная последовательность аминокислот тяжелой цепи содержит SEQ ID NO: 141 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью; и
б) указанная последовательность аминокислот легкой цепи содержит SEQ ID NO: 23 или последовательность, имеющую по меньшей мере 85%, 88%, 90%, 95%, 98%, или 99% идентичность с этой последовательностью;
при этом слитый белок содержит две тяжелых цепи и две легких цепи; дисульфидная связь образована между его первой легкой цепью и первой тяжелой цепью, дисульфидная связь образована между его второй легкой цепью и второй тяжелой цепью, и дисульфидная связь образована между его первой тяжелой цепью и второй тяжелой цепью.
9. Слитый белок по п. 8,
имеющий значение KD 2,92 пМ - 26,3 пМ, предпочтительно 7 пМ - 9 пМ, более предпочтительно 8, 77 пМ по аффинности связывания с белком EGFR человека, и
имеющий значение KD 23 пМ - 288,3 пМ, предпочтительно 64 пМ - 144 пМ, более предпочтительно 96,1 пМ по аффинности связывания с белком TGF-β1 человека.
10. Нуклеиновая кислота, кодирующая молекулу TGFβR2 с укороченным внеклеточным доменом по п. 1, которая представляет собой мРНК или ДНК.
11. Нуклеиновая кислота, кодирующая слитый белок по любому из пп. 2-8, которая представляет собой мРНК или ДНК.
12. Фармацевтическая композиция для лечения опухоли, ассоциированной с EGFR, содержащая
а) слитый белок по любому из пп. 2-8, или нуклеиновую кислоту по п. 11, и
б) фармацевтически приемлемый носитель.
| WO 2018205985 A1, 15.11.2018 | |||
| CN 110785431 A, 11.02.2020 | |||
| СЛИТЫЕ ИММУНОМОДУЛИРУЮЩИЕ БЕЛКИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2014 |
|
RU2662991C2 |
| SU C | |||
| T.-T | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| PAKULA A.A | |||
| et al | |||
| Genetic analysis of protein stability and function | |||

