Хорошо известно, что иногда существует заметная разница в эффектах энантиомеров одного и того же химического вещества в биологических системах, таких как организмы. В лекарственных средствах, например, часто только один из энантиомеров лекарственного средства отвечает за желаемые физиологические эффекты, тогда как другой энантиомер менее активен, неактивен или иногда обладает противодействием.
Поэтому существуют разные способы получения только желаемого энантиомера. Одна стратегия известна как хиральное разделение. Этот метод включает приготовление соединения в рацемической форме и разделение его на различные энантиомеры. Другой стратегией является асимметричный синтез, т.е. использование различных методик для приготовления желаемого соединения в большом энантиомерном избытке.
Классом молекул, существующих в виде рацемической смеси, являются альфа-галоалкановые кислоты. R- или S-энантиомер этих альфа-галоалкановых кислот может быть разделен для аналитических целей, например, с помощью капиллярной газовой хроматографии, как описано в US 5154738. В целях синтеза наиболее практичным способом получения хиральных альфа-галоалкановых кислот является рацемическое разделение с использованием солей стрихнина или бруцина или путем стереоселективного бромирования R-2-аминомасляных кислот. Однако хиральные предшественники или реагенты для этих реакций, например, [R-2-аминомасляная кислота], не являются экономически выгодными. Кроме того, реакции трудно проводить в более крупном или даже промышленном масштабе. Например, стереоселективное бромирование R-2-аминомасляной кислоты занимает много времени, требует низких температур (от -10 до 5°C), а развитие реакции трудно контролировать. Кроме того, во время реакции образуются азотистые газы, что представляет собой проблему, связанную безопасностью труда.
Таким образом, задача настоящего изобретения состояла в разработке альтернативного, более экономически выгодного, экономичного по времени и безопасного способа, обеспечивающего и/или облегчающего разделение R- и S-энантиомеров альфа-галоалкановых кислот Формулы I.
Настоящее изобретение решает данную задачу путем обеспечения способа селективного гидролиза S-энантиомера альфа-галоалкановой кислоты Формулы I,
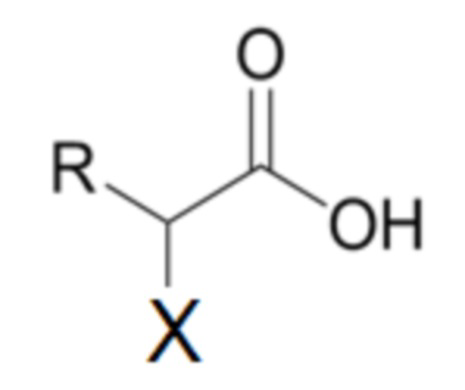
где X представляет собой галоген, и
R представляет собой алкильную цепь, содержащую от 1 до 6 атомов углерода, где эта указанная алкильная цепь может быть неразветвленной или разветвлена на атомах углерода γ или δ,
включающего
- обеспечение рацемата R-энантиомера и S-энантиомера указанной альфа-галоалкановой кислоты,
- обеспечение полипептида, имеющего дегалогеназную активность, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность на по меньшей мере 80% идентичную любой из указанных последовательностей,
- реакцию рацемата в течение 1-8 часов, где
значение pH находится в интервале 9 - 10, и температура находится в интервале 15 - 35°C для полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1, или последовательность, на по меньшей мере 80% идентичную указанной последовательности, или
значение pH находится в интервале 9 - 10, и температура находится в интервале 55 - 65°C для полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную указанной последовательности.
- и где энантиомерный избыток R-энантиомера от 90,0 до 99,9% достигается через 1-8 часов.
Преимущество этого способа состоит в том, что он является альтернативным, более экономически выгодным и экономичным по времени способом, основанным на различном протекании реакции R и S-энантиомеров альфа-галоалкановых кислот Формулы I, позволяющем и/или облегчающем разделение двух энантиомеров, так как полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO.1 или SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, селективно гидролизует S-энантиомер альфа-галоалкановых кислот Формулы I очень эффективно при данных условиях реакции, т.е. достигает высокого пространственно-временного выхода. Селективность реакции гидролиза показана посредством степени энантиомерного избытка оставшегося R-энантиомера от 90 до 99%. Другими словами, было неожиданно обнаружено, что указанные условия способствуют более экономически выгодному, экономичному по времени и более безопасному способу получения R-энантиомера.
Как указано выше полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO.1 или SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, селективно гидролизует S-энантиомер альфа-галоалкановых кислот Формулы I очень эффективно при данных условиях реакции. Во время этой реакции R-энантиомер остается неизменным, тогда как S-энантиомер гидролизуется при превращении его стереоизомерии.
Согласно предпочтительным вариантам осуществления указанного способа полипептид, имеющий дегалогеназную активность, имеет аминокислотную последовательность, как показано в SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную указанной последовательности, и X представляет собой фтор в альфа-галоалкановой кислоте Формулы I. Впервые неожиданно было обнаружено, что при этих условиях реакции и применяя полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO 4, или также последовательность, на по меньшей мере 80% идентичную указанной последовательности, S-энантиомер фторсодержащей альфа-галоалкановой кислоты Формулы I селективно гидролизуется, где энантиомерный избыток R-энантиомера от 90,0 до 99,9% достигается через 1-8 часов.
Термин «содержащий» следует интерпретировать как указывающий наличие указанных частей, стадий или компонентов, но не исключающий наличия одной или более дополнительных частей, стадий или компонентов.
Понятно, что при обращении к слову в единственном числе множественное число также включается в данный документ. Таким образом, ссылка на элемент посредством формы единственного числа не исключает возможности присутствия более одного элемента, если только контекст явно не требует наличия одного и только одного из элементов. Таким образом, форма единственного числа обычно означает «по меньшей мере один».
Используемый в настоящей заявке термин «энантиомер» относится к одному из двух стереоизомеров данной молекулы, которые являются зеркальными изображениями друг друга и не являются накладываемыми друг на друга (не идентичны). Один хиральный атом или подобный структурный признак в соединении приводит к тому, что соединение имеет две возможные структуры, которые не накладываются друг на друга, причем каждая является зеркальным отражением другой.
Используемый в настоящей заявке термин «рацемат» или «рацемический субстрат» относится к смеси, которая имеет равные количества двух стереоизомеров хиральной молекулы.
Как упомянуто выше, в случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1, или последовательность с по меньшей мере 80% идентичностью последовательности с SEQ ID NO. 1, X в Формуле I выбирают из галогена, отличного от тора, и описанный выше способ проводят в течение 1-8 часов, где значение pH находится в интервале 9 - 10, и температура находится в интервале 15 - 35°C.
При использовании этих условий неожиданно было обнаружено, что полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1, или последовательность с по меньшей мере 80% идентичностью последовательности, достоверно достигал значений степени энантиомерного избытка выше 99% (см. Фиг. 3) и пространственно-временного выхода, равного 16,6 гпродукта л-1 ч-1 (при концентрации субстрата, равной 100 г/л)
Следует отметить, что из-за рацемического субстрата максимальный выход R-энантиомера составляет 50%, поэтому максимальный выход при использовании субстрата с концентрацией 100 г/л будет 50 г/л. Более того, пространственно-временной выход был бы еще выше, если бы более низкие концентрации субстрата - например, 50 г/л - применялись. Однако более высокие концентрации субстрата выгодны для производственного процесса в промышленном масштабе.
Согласно предпочтительным вариантам осуществления описанный выше способ в случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1 или последовательность с по меньшей мере 80% идентичностью последовательности с ней, проводят в течение 4-6 часов, где значение pH составляет 9,5, и температура находится в интервале 20 - 30°C. Применяя эти условия, полное превращение, т.е. 90% ee, может быть достигнуто через 3 часа.
Согласно особенно предпочтительному варианту осуществления этой реакции температура составляет 25°C.
Значение pH предпочтительно находится в интервале 9-10, и наиболее предпочтительно при 9,5.
Температура реакции, равная 25°C, и значение pH, равное 9,5, обеспечивали достижение полного превращения, т.е. 90% ee через 1час. Таким образом, эти условия приводили к особенно высокому пространственно-временному выходу. В случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1 или последовательность с по меньшей мере 80% идентичностью последовательности с ней, обеспечиваемого в виде свежей ферментированной биомассы (т.е. полипептид содержится в целых клетках), достигаемый пространственно-временной выход достигал 50 гпродукта л-1 ч-1
Как упомянуто выше, в случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 4 или последовательность с по меньшей мере 80% идентичностью последовательности с ней, описанный выше способ проводят в течение 1-8 часов, где значение pH находится в интервале 9 - 10, и температура находится в интервале 55 - 65°C, и энантиомерный избыток R-энантиомера от 90,0 до 99,9% достигается через 1-8 часов. Согласно предпочтительному варианту осуществления настоящего изобретения описанный выше способ в случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 4 или последовательность с по меньшей мере 80% идентичностью последовательности с ней, проводят в течение 4-6 часов, где значение pH составляет 9,5, и температура находится в интервале 59-61°C.
Согласно особенно предпочтительному варианту осуществления этой реакции температура составляет 60°C.
Значение pH предпочтительно находится в интервале 9-10 и наиболее предпочтительно при 9,5.
Согласно предпочтительному варианту осуществления полипептида с дегалогеназной активностью, описанного в настоящей заявке, указанный полипептид имеет по меньшей мере 80% идентичность последовательности, более предпочтительно по меньшей мере 85% идентичность последовательности, более предпочтительно по меньшей мере 90% идентичность последовательности, более предпочтительно по меньшей мере 95% идентичность последовательности, более предпочтительно по меньшей мере 95%, 96%, 97%, 98%, 99%, и наиболее предпочтительно 100% идентичность последовательности с SEQ ID NO 1, или указанный полипептид имеет по меньшей мере 80% идентичность последовательности, более предпочтительно по меньшей мере 85% идентичность последовательности, более предпочтительно по меньшей мере 90% идентичность последовательности, более предпочтительно по меньшей мере 95% идентичность последовательности, более предпочтительно по меньшей мере 95%, 96%, 97%, 98%, 99%, и наиболее предпочтительно 100% идентичность последовательности с SEQ ID NO 4.
Полипептиды с дегалогеназной активностью, имеющие аминокислотную последовательность, как показано в SEQ ID NO 1 или SEQ ID NO 4, также могут быть охарактеризованы их последовательностями нуклеиновых кислот.
Таким образом, в случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1, обеспеченный полипептид выбран из группы:
a) полипептида, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1, или последовательность с по меньшей мере 80% идентичностью последовательности с указанной последовательностью,
b) выделенного кодон-оптимизированного фрагмента нуклеиновой кислоты, имеющего нуклеотидную последовательность с по меньшей мере 80% идентичностью последовательности с SEQ ID NO. 2
c) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, комплементарную SEQ ID NO. 2,
d) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, которая специфически гибридизуется с указанным выделенным фрагментом нуклеиновой кислоты b) или c)
e) выделенного фрагмента нуклеиновой кислоты, имеющего нуклеотидную последовательность с по меньшей мере 80% идентичностью последовательности с SEQ ID NO. 3
f) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, комплементарную SEQ ID NO. 3,
g) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, которая специфически гибридизуется с указанным выделенным фрагментом нуклеиновой кислоты e) или f).
Таким образом, в случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1, полипептид может быть кодирован различными последовательностями нуклеиновых кислот. Это является результатом вырожденности генетического кода.
В результате этого вырождения генетического кода аминокислоты могут кодироваться одним или более кодонами. В разных организмах кодоны, кодирующие аминокислоту, используются с разными частотами. Адаптация кодонов кодирующей последовательности нуклеиновой кислоты к частоте их использования в организме, в который должна быть интегрирована экспрессируемая последовательность, может способствовать увеличению количества транслируемого белка и/или стабильности рассматриваемой мРНК в конкретных клетках. Частота использования кодонов в рассматриваемых клетках-хозяевах или хозяевах может быть определена специалистом в данной области техники путем изучения как можно большего количества кодирующих последовательностей нуклеиновых кислот рассматриваемого организма с точки зрения частоты, с которой определенные кодоны используются для кодирования определенной аминокислоты. Частота использования кодонов определенных организмов известна специалисту в данной области техники (см. http://www.kazusa.or.jp/codon/) и может быть определена простым и быстрым способом с использованием специально разработанных алгоритмов, реализованных в компьютерных программах (например, Grote et al., 2005, Nucleic Acids Research 33, W526-W531; doi: 10.1093/nar/gki376). Инструменты, использующие такие алгоритмы, являются общедоступными и предоставляются бесплатно в качестве веб-интерфейсов, в том числе во Всемирной паутине, от различных учреждений, таких как Европейский институт биоинформатики (EMBL-EBI) и других (например, http://www.jcat.de; http://gcua.schoedl.de/; http://www.kazusa.or.jp/codon/; http://www.entelechon.com/eng/cutanalysis.html; http://www.ebi.ac.uk/Tools/st/emboss_backtranseq/). Адаптация кодонов кодирующей последовательности нуклеиновой кислоты к частоте их использования в организме, в котором предполагается экспрессия последовательности, может быть осуществлена путем мутагенеза in vitro или, предпочтительно, путем синтеза de novo последовательности гена. Способы синтеза de novo последовательностей нуклеиновых кислот известны специалисту в данной области техники. Синтез de novo может быть осуществлен, например, путем первоначального синтеза отдельных олигонуклеотидов нуклеиновых кислот, их гибридизации с олигонуклеотидами, комплементарными им, так что они образуют двухцепочечную ДНК, а затем лигирования отдельных двухцепочечных олигонуклеотидов так, чтобы получить желаемую последовательность нуклеиновой кислоты. Синтез de novo последовательностей нуклеиновых кислот, включая адаптацию к частоте, с которой кодоны используются для определенного организма-мишени, также может быть предоставлен компаниям, предлагающим этот сервис (например, Eurofins MWG).
Выделенные фрагменты нуклеиновой кислоты, содержащие нуклеотидную последовательность на по меньшей мере 80% идентичную SEQ ID NO. 2 или SEQ ID NO 3, предпочтительно имеют по меньшей мере 85% идентичность последовательности, более предпочтительно по меньшей мере 90% идентичность последовательности, более предпочтительно по меньшей мере 95% идентичность последовательности, более предпочтительно по меньшей мере 95%, 96%, 97%, 98%, 99%, и наиболее предпочтительно 100% идентичность последовательности с SEQ ID NO. 2 или SEQ ID NO 3, соответственно.
В случае полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 4, обеспеченный полипептид выбран из группы:
a) полипептида, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 4, или последовательность с по меньшей мере 80% идентичностью последовательности с указанной последовательностью,
b) выделенного кодон-оптимизированного фрагмента нуклеиновой кислоты, имеющего нуклеотидную последовательность с по меньшей мере 80% идентичностью последовательности с SEQ ID NO. 5
c) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, комплементарную SEQ ID NO. 5,
d) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, которая специфически гибридизуется с указанным выделенным фрагментом нуклеиновой кислоты b) или c)
e) выделенного фрагмента нуклеиновой кислоты, имеющего нуклеотидную последовательность с по меньшей мере 80% идентичностью последовательности с SEQ ID NO. 6
f) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, комплементарную SEQ ID NO. 6,
g) выделенного фрагмента нуклеиновой кислоты, имеющего последовательность, которая специфически гибридизуется с указанным выделенным фрагментом нуклеиновой кислоты e) или f).
Также полипептид с дегалогеназной активностью - в настоящей заявке полипептид, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 4 - может быть кодирован различными последовательностями нуклеиновых кислот. Опять же, это результат вырожденности генетического кода, как описано выше.
Снова, выделенные фрагменты нуклеиновой кислоты, имеющие нуклеотидную последовательность с по меньшей мере 80% идентичностью последовательности с SEQ ID NO. 5 или SEQ ID NO 6, предпочтительно имеют по меньшей мере 85% идентичность последовательности, более предпочтительно по меньшей мере 90% идентичность последовательности, более предпочтительно по меньшей мере 95% идентичность последовательности, более предпочтительно по меньшей мере 95%, 96%, 97%, 98%, 99%, и наиболее предпочтительно 100% идентичность последовательности с SEQ ID NO. 5 или SEQ ID NO 6 , соответственно.
Используемый в настоящей заявке термин «гидролиз» представляет собой ферментативно катализируемое гидролитическое расщепление углерод-галогеновой связи, в которой галоген замещен гидроксильной группой. Некоторые ферменты катализируют гидролитическое дегалогенирование 2-галоалкановых кислот с образованием соответствующих 2-гидроксиалкановых кислот.
Используемый в настоящей заявке термин «селективно гидролизует» относится к тому факту, что некоторые ферменты (т.е. полипептиды), которые катализируют гидролиз, проявляют стереоселективную активность. Другими словами, посредством селективного гидролиза создается энантиомерный избыток либо R-, либо S-энантиомера, в зависимости от предпочтения фермента.
Используемый в настоящей заявке термин «энантиомерный избыток» или «ее значение» относится к измерению чистоты хиральных веществ. Он отражает степень, в которой образец содержит один энантиомер в больших количествах, чем другой. Рацемическая смесь имеет значение ее, равное 0%, в то время как один полностью чистый энантиомер имеет значение, равное ее 100%. Образец с 70% одного энантиомера и 30% другого имеет значение ее, равное 40% (70% - 30%).
Способы определения значения ее известны в данной области техники. Это может, например, определяется с помощью устройств газовой хроматографии, оснащенных хиральной колонкой.
Таким образом, полипептиды, описанные в настоящей заявке, показывают стереоселективную активность при катализе реакции гидролиза Эта стереоселективная активность может быть выражена как значение ee. Как указано выше, полипептиды, имеющие дегалогеназную активность, имеющие аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность на по меньшей мере 80% идентичную любой из указанных последовательностей, показывают стереоселективную активность при катализе реакции гидролиза Эта стереоселективная активность может быть выражена, достигающую очень высоких значений ee.
Надежное утверждение о пригодности данного фермента для данной реакции может быть сделано с использованием параметров: превращение субстрата, образование продукта, энантиомерный избыток (ее) и пространственно-временного выхода (STY). Используемый здесь термин «пространственно-временной выход» относится к количеству продукта, образованного из данной концентрации субстрата в определенном объеме в течение определенного времени.
В некоторых случаях приведенные здесь значения пространственно-временного выхода также установлены в отношении конкретного количества клеточного экстракта или целых клеток, используемых в данной реакции (см. Примеры 3 и 5).
Используемый в настоящей заявке термин «ген» означает последовательность ДНК, состоящую из нуклеотидов, содержащих область (транскрибируемая область), которая транскрибируется в молекулу РНК (например, непосредственно в мРНК без интронных последовательностей) в клетке, функционально связанную с регуляторными областями, способными регулировать экспрессию полипептида. Таким образом, ген может содержать несколько функционально связанных последовательностей, таких как нетранслируемые регуляторные области (например, промотор, энхансер, репрессор), 5'-лидерная последовательность, содержащая, например, последовательности, участвующие в инициации трансляции, кодирующую (белок) область (кДНК или геномную ДНК) и 3'-нетранслируемую последовательность, содержащую, например, сайты терминации транскрипции.
Используемый в настоящей заявке термин «экспрессия гена» или «генная экспрессия» относится к процессу, в котором область ДНК (кодирующая область), которая функционально связана с соответствующими регуляторными областями, в частности с промотором, транскрибируется в молекулу мРНК. Затем молекула мРНК дополнительно обрабатывается (посттранскрипционными процессами) внутри клетки, например, путем инициации трансляции и трансляции в аминокислотную цепь (полипептид) и терминации трансляции стоп-кодонами трансляции.
Используемый в настоящей заявке термин «дикий тип» (также пишется «дикого типа» или «WP») относится к типичной форме фермента или гена, в которой он чаще всего встречается в природе.
Используемый в настоящей заявке термин «полипептид» относится к любому пептиду, полипептиду, олигопептиду или белку. Полипептид состоит из последовательных аминокислот, которые связаны пептидными связями. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, и он может прерываться не аминокислотами. Полипептид может быть человеческим, не человеческим, и искусственным или химическим миметиком соответствующей встречающейся в природе аминокислоты, а также встречающимися в природе аминокислотными полимерами и не встречающимися в природе аминокислотными полимерами. Термин также охватывает аминокислотный полимер, который был модифицирован либо естественными процессами, либо химическими модификациями; например, путем образования дисульфидной связи, гликозилирования, липидирования, ацетилирования, ацилирования, фосфорилирования или любых других манипуляций, таких как конъюгирование с вносящим метку компонентом, таким как, но не ограничиваясь ими, флуоресцентные маркеры, частицы, биотин, шарики, белки, радиоактивные метки, хемилюминесцентные метки, биолюминесцентные метки и тому подобное.
Используемый в настоящей заявке термин «идентичность последовательности» двух родственных нуклеотидных или аминокислотных последовательностей, выраженный в процентах, относится к числу положений в двух оптимально выровненных последовательностях, которые имеют идентичные остатки (х 100), поделенному на число сравниваемых положений. Гэп, то есть позиция в выравнивании, где остаток присутствует в одной последовательности, но не присутствует в другой, рассматривается как позиция с неидентичными остатками. «Оптимальное выравнивание» двух последовательностей достигается путем выравнивания двух последовательностей по всей длине. Другими словами, если две одинаковые последовательности выровнены, значение идентичности последовательности будет 100%.
Выровненные последовательности нуклеотидных или аминокислотных остатков обычно представлены в виде строк в матрице. Гэпы вставляются между остатками так, чтобы идентичные или похожие характеристики были выровнены в последовательных столбцах.
Чтобы определить идентичность последовательности может применяться глобальный алгоритм выравнивания Нидлмана-Вунша (Needleman and Wunsch, 1970, J Mol Biol 48(3):443-53) от The European Molecular Biology Open Software Suite (EMBOSS, Rice et al., 2000, Trends in Genetics 16(6): 276-277; см., например, http://www.ebi.ac.uk/emboss/align/index.html) с использованием параметров по умолчанию (gap opening penalty = 10 (для нуклеотидов) / 10 (для белков) и gap extension penalty = 0,5 (для нуклеотидов) / 0,5 (для белков)). Для нуклеотидов применяемой матрицей замен по умолчанию является EDNAFULL, и для белков матрицей замен по умолчанию является EBLOSUM62.
Термин «молекула нуклеиновой кислоты» предназначен для обозначения любой одноцепочечной или двухцепочечной молекулы нуклеиновой кислоты, содержащей ДНК (кДНК и/или геномную ДНК), РНК (мРНК), PNA, LNA и/или морфолино.
Используемый в настоящей заявке термин «вектор» относится к молекулярному носителю, используемому для переноса чужеродного генетического материала в другую клетку. Сам вектор, как правило, представляет собой последовательность ДНК, которая состоит из вставки (представляющей интерес последовательности) и большей последовательности, которая служит «основой» вектора. Целью вектора для передачи генетической информации в другую клетку, как правило, является выделение, размножение или экспрессия вставки в клетке-мишени.
Используемый в настоящей заявке термин «плазмида» относится к плазмидным векторам, т.е. кольцевой последовательности ДНК, которая способна к автономной репликации в подходящем хозяине благодаря источнику репликации («ORI»). Кроме того, плазмида может содержать селектируемый маркер, указывающий на успех трансформации или других процедур, предназначенных для введения чужеродной ДНК в клетку, и сайт множественного клонирования, который включает множество консенсусных сайтов ферментов рестрикции для обеспечения возможности включения вставки. Плазмидные векторы, называемые клонирующими или донорными векторами, используются для облегчения клонирования и для амплификации представляющей интерес последовательности. Плазмидные векторы, называемые экспрессионными или акцепторными векторами, специально предназначены для экспрессии представляющего интерес гена в определенной клетке-мишени. Эти плазмидные векторы обычно представляют собой экспрессионную кассету, состоящую из промотора, сайта связывания с рибосомой, трансгена и терминаторной последовательности. Для контроля экспрессии эти плазмиды содержат репрессор, который локализован на остове плазмиды. Экспрессионные плазмиды могут представлять собой челночные плазмиды, содержащие элементы, которые обеспечивают размножение и селекцию в разных клетках-хозяевах.
Используемый в настоящей заявке термин «специфически гибридизуется» или «селективно гибридизуется» относится к реакции рассматриваемой последовательности нуклеиновой кислоты в растворе для гибридизации, содержащем 0,5 М натрий-фосфатный буфер, pH 7,2, содержащий 7% SDS, 1 мМ EDTA и 100 мг/мл ДНК из молок лососевых, при 65°С в течение 16 часов, с промывками дважды при 65°С в течение двадцати минут в промывочном растворе, содержащем 9,5 × SSC и 0,1% SDS.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность на по меньшей мере 80% идентичную любой из указанных последовательностей, обеспечивают в виде клеточного лизата, и/или он содержится в целых клетках.
Используемый в настоящей заявке термин «целые клетки» относится к тому факту, что полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность на по меньшей мере 80% идентичную любой из указанных последовательностей, обеспечивается в клетках, в которых он экспрессируется, т.е. что клеточные стенки указанных клеток не были целенаправленно разрушены.
Указанные целые клетки могут представлять собой, например, свежую биомассу, полученную в процессе ферментации. Альтернативно, целые клетки могут быть заморожены перед использованием.
Другими словами, неожиданно было обнаружено, что даже хотя фермент, т.е. полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, который экспрессируется (рекомбинантными) целыми клетками, присутствует только внутри клетки, биодоступность субстрата является достаточной для того, чтобы фермент мог реагировать с субстратом внутри клетки.
Было обнаружено, что клетки E.coli, используемые для синтеза ферментов, продуцируют желаемый фермент при постоянной пропорции общего содержания белка. Другими словами, данное количество общего клеточного белка (например, 5 г) всегда содержит одинаковое количество концентрации растворимого фермента. Также по этой причине в экспериментах можно использовать целые клетки вместо клеточного лизата.
Преимущество использования целых клеток заключается в том, что нет необходимости использовать дорогостоящие и трудоемкие способы для получения клеточного экстракта.
Согласно одному варианту осуществления настоящего изобретения указанные целые клетки представляют собой клетки E.coli, выбранные из группы MG1655, W3110, JM101, BL21DE3, DH5альфа.
Более того, выбранные клетки E.coli предпочтительно несут плазмиду с SEQ ID NO. 7 и экспрессируют фермент, имеющий SEQ ID NO. 1, из плазмиды.
Альтернативно, выбранные клетки E.coli несут плазмиду SEQ ID NO. 8 и экспрессируют фермент, имеющий SEQ ID NO. 4, из плазмиды.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, добавляют в начале реакции с рацематом указанной альфа-галоалкановой кислоты Формулы I, или указанный полипептид с дегалогеназной активностью добавляют в начале и в различные моменты времени в ходе реакции с рацематом указанной альфа-галоалкановой кислоты.
Удивительно, но различные профили дозирования не влияют на производительность процесса. Таким образом, специалист в данной области техники может выбрать профиль дозирования, наиболее подходящий для рассматриваемого процесса. Это свойство заявленных ферментов обеспечивает максимальную гибкость при использовании фермента в данном производственном процессе.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, полипептид с дегалогеназной активностью полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, добавляют в начале и в различные моменты времени в ходе реакции с рацематом указанной альфа-галоалкановой кислоты, где добавляемая концентрация полипептида является одинаковой в каждый момент времени.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, добавляют в начале и в различные моменты времени в ходе реакции с рацематом указанной альфа-галоалкановой кислоты, где добавляемая концентрация полипептида различается в разные моменты времени.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, рацемат добавляют к полипептиду с дегалогеназной активностью полипептид, имеющему дегалогеназную активность, имеющему аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, или указанный полипептид с дегалогеназной активностью добавляют к рацемату.
Если рацемат указанной альфа-галоалкановой кислоты Формулы I добавляют к полипептиду с дегалогеназной активностью, имеющему аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, предпочтительно весь рацемат указанной альфа-галоалкановой кислоты Формулы I добавляют в начале реакции, где значение pH рацемата оптимизировано для условий реакции перед добавлением.
Неожиданно, на производительность процесса не влияет добавляют ли рацемат указанной альфа-галоалкановой кислоты Формулы I, т.е. субстрат, к полипептиду с дегалогеназной активностью, имеющему аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, или указанный полипептид с дегалогеназной активностью добавляют к рацемату указанной альфа-галоалкановой кислоты Формулы I.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, любой из двух полипептидов с дегалогеназной активностью, как описано в настоящей заявке, обеспечивают в виде целых клеток при значении рН, равном 9,5, и рацемат указанной альфа-галоалкановой кислоты Формулы I добавляют к полипептиду, где рацемат указанной альфа-галоалкановой кислоты Формулы I также обеспечивают при значении рН, равном 9,5, и значение pH, равное 9,5, поддерживают постоянным в ходе реакции посредством титрования подходящим основанием.
Подходящее основание может быть выбрано, например, из группы, состоящей из водного КОН или NaOH.
Согласно некоторым вариантам осуществления способа, описанного в настоящей заявке, концентрация рацемата указанной альфа-галоалкановой кислоты Формулы I составляет от 80 до 200 г/л, предпочтительно от 90 до 150 г/л, наиболее предпочтительно она составляет 100 г/л. Другими словами, концентрацию рацемата альфа-галоалкановой кислоты предпочтительно выбирают таким образом, чтобы достигалось полное превращение.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность на по меньшей мере 80% идентичную любой из указанных последовательностей, обеспечивают в виде активных целых клеток, и биомасса указанных активных целых клеток имеет концентрацию между 15 - 200 г/л, предпочтительно между 25 - 100 г/л.
Согласно некоторым вариантам осуществления способа, описанного в настоящей заявке, соотношение рацемата альфа-галоалкановой кислоты Формулы I и биомассы целых клеток, содержащих полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, составляет от 2:1 до 15:1, предпочтительно от 3:1 до 10:1, наиболее предпочтительно 4:1.
Так как полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, ингибируется более высокой концентрацией субстрата, соотношение субстрата, т.е. альфа-галоалкановой кислоты Формулы I, и фермента, т.е. полипептида, имеющего дегалогеназную активность, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, который содержится в целых клетках и следовательно обеспечивается в виде биомассы, увеличивается при более низких концентрациях субстрата.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, галоген X указанной альфа-галоалкановой кислоты Формулы I выбирают из группы, состоящей из бромида и хлорида.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, фрагмент R указанной альфа-галоалкановой кислоты Формулы I представляет собой алкильную цепь, содержащую от 1 до 6 атомов углерода, где эта указанная алкильная цепь представляет собой атомы углерода гамма или дельта, и где атомы углерода, следующие за разветвлением у атомов углерода γ или δ, являются циклическими.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, фрагмент R указанной альфа-галоалкановой кислоты Формулы I выбирают из группы, состоящей из этила, бутила, 2-метил-пропила и метил-циклопропила.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, значение pH поддерживают постоянным посредством титрования подходящим основанием, таким как гидроксид калия или гидроксид натрия.
Поскольку было обнаружено, что условия реакции могут быть легко увеличены, реакционные объемы могут составлять от 1 мл до нескольких тысяч литров.
Согласно одному варианту осуществления способа, описанного в настоящей заявке, указанный полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, представляет собой дегалогеназу галокислоты.
Согласно предпочтительному варианту осуществления настоящего изобретения способа, описанного в настоящей заявке, полипептид с дегалогеназной активностью состоит из последовательности SEQ ID NO 1 и реагирует с рацемическим субстратом, выбранным из группы, состоящей из 2-хлормасляной кислоты, 2-бромгексановой кислоты, 2-бром-4-метилпентановой кислоты и 2-бром-3-циклопропилпропановой кислоты.
Согласно альтернативному предпочтительному варианту осуществления способа, описанного в настоящей заявке, полипептид с дегалогеназной активностью состоит из последовательности SEQ ID NO 4 и реагирует с рацемическим субстратом, выбранным из группы, состоящей из 2-хлормасляной кислоты, 2-фтормасляной кислоты, 2-бромгексановой кислоты и 2-бром-4-метилпентановой кислоты.
После селективного гидролиза S-энантиомера выбранной альфа-галоалкановой кислоты, R-энантиомер и гидролизованный энантиомер присутствуют в виде смеси. Для некоторых применений предпочтительно получать только один, т.е. очищать R-энантиомер. Следовательно, смесь R-энантиомера и гидролизованного энантиомера выбранной альфа-галоалкановой кислоты Формулы I далее обрабатывают.
Другим объектом изобретения, описанного в настоящей заявке, является применение способа, описанного в настоящей заявке, для селективного гидролиза S-энантиомера указанной альфа-галоалкановой кислоты, где энантиомерный избыток R-энантиомера составляет от 90,0 до 99,9.
Неожиданно было обнаружено, что применение полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO 1 или SEQ ID NO. 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей, для контакта с рацематом, содержащим R-энантиомер и S-энантиомер альфа-галоалкановой кислоты Формулы I, где X представляет собой галоген, в случае последовательности SEQ ID NO 1 - галоген, отличный от фтора, и R представляет собой алкильную цепь, содержащую до 6 атомов углерода, где эта указанная алкильная цепь может быть неразветвленной или разветвленной на атомах углерода γ или δ, в течение 1-8 часов, где значение pH находится в интервале 9 - 10, и температура находится в интервале 15 - 35°C для полипептида, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1, или значение pH находится в интервале 9 - 10, и температура находится в интервале 55 - 65°C для полипептида, имеющего аминокислотную последовательность, как показано в SEQ ID No. 4, указанный полипептид будет осуществлять селективный гидролиз S-энантиомера указанной альфа-галоалкановой кислоты, где энантиомерный избыток R-энантиомера составляет от 90,0 до 99,9.
Этот результат был неожиданным, поскольку такие высокие показатели энантиомерного избытка очень необычны, особенно поскольку полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO 4, достигает значений энантиомерного избытка от 90 до 99 за очень короткое время реакции от 1 до 8 часов.
Следовательно, неожиданно было обнаружено, что посредством применения полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную указанным последовательностям, при описанных условиях, обеспечивает альтернативный, более безопасный и более экономически выгодный и экономичный по времени способ, обеспечивающий и/или облегчающий разделение R- и S-энантиомеров альфа-галоалкановых кислот Формулы I на основе различного протекания реакции двух энантиомеров с полипептидом, имеющим аминокислотную последовательность, как показано в SEQ ID NO. 1 или SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную любой из указанных последовательностей.
Другим объектом изобретения, описанного в настоящей заявке, является способ селективного гидролиза энантиомера альфа-галоалкановой кислоты Формулы II,
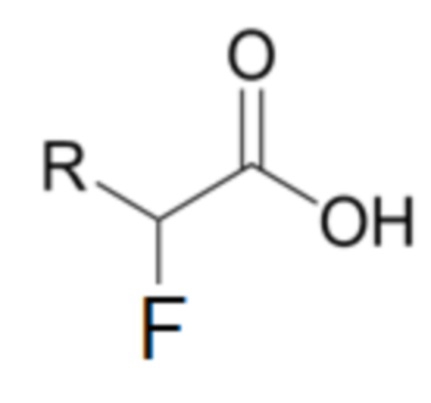
где
R представляет собой алкильную цепь, содержащую от 1 до 6 атомов углерода, где эта указанная алкильная цепь может быть неразветвленной или разветвленной на атомах углерода γ или δ,
применяя полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную указанной последовательности.
Неожиданно было обнаружено, что полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную указанной последовательности селективно гидролизует S-энантиомер альфа-галоалкановых кислот Формулы II, т.е. содержащих атом фтора, очень эффективно и с высоким энантиомерным избытком.
Предпочтительно, описанный выше способ, в котором применяют полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO 4, проводят при 55-65°C и при значении pH, равном 9,5.
Таким образом, в случае полипептида, имеющего дегалогеназную активность, имеющего аминокислотную последовательность, как показано в SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную указанной последовательности, в дополнение к названным выше альфа-галоалкановым кислотам, 2-фтормасляная кислота также является особенно предпочтительной.
Другим объектом описанного в настоящей заявке изобретения является применение полипептида, имеющего дегалогеназную активность, имеющего аминокислотную последовательность, как показано в SEQ ID NO 4, или последовательность, на по меньшей мере 80% идентичную указанной последовательности, для селективного гидролиза S-энантиомера альфа-галоалкановой кислоты Формулы II, где энантиомерный избыток R-энантиомера составляет от 90,0 до 99,9.
Согласно предпочтительному варианту осуществления настоящего изобретения способ проводят следующим образом: полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1, добавляют к 2-бромбутановой кислоте при комнатной температуре при значении рН, равном 9,5. Реакции позволяют протекать в течение 4-6 ч, при этом значение pH поддерживают постоянным посредством титрования 3 M гидроксидом калия.
Согласно особенно предпочтительному варианту осуществления настоящего изобретения способ проводят следующим образом: 10 мг/мл общего клеточного белка в виде клеточного экстракта, содержащего полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1, добавляют к 100 г/л 2-бромбутановой кислоте при комнатной температуре, значении pH, равном 9,5, и в реакционном объеме, равном 6л. Реакции позволяют протекать в течение 4-6 ч, при этом значение pH поддерживают постоянным посредством титрования 3 M гидроксидом калия.
Согласно другому предпочтительному варианту осуществления настоящего изобретения способ проводят следующим образом: полипептид, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 4, добавляют к 2-бромбутановой кислоте при температуре 55°C, значении pH, равном 9,5. Реакции позволяют протекать в течение 4-6 ч, при этом значение pH поддерживают постоянным посредством титрования 3 M гидроксидом калия.
Согласно другому особенно предпочтительному варианту осуществления настоящего изобретения способ проводят следующим образом: 10 мг/мл общего клеточного белка в виде клеточного экстракта, содержащего полипептид с дегалогеназной активностью, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 4, добавляют к 2-фтормасляной кислоте при температуре 55°C, значении pH, равном 9,5.
Краткое описание чертежей
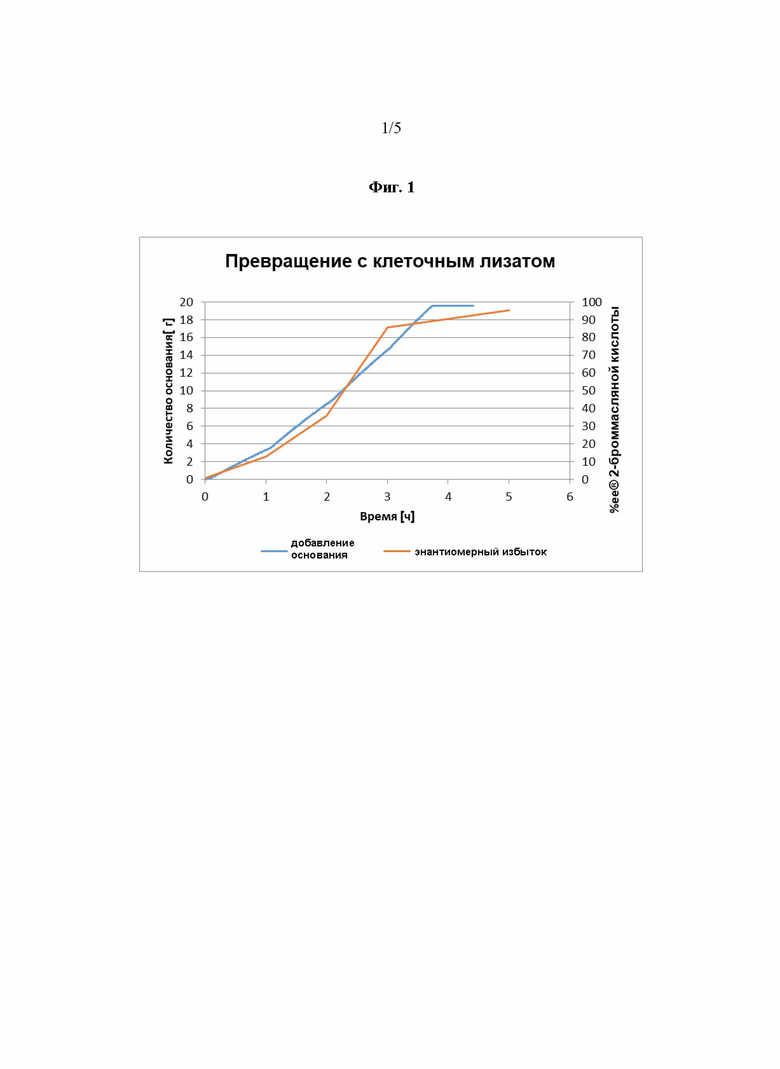
Фиг. 1 показывает реакцию, проводимую в оптимизированных условиях относительно значения рН, т.е. значения 9,5, и температуры, т.е. 25°C. В этом случае в качестве субстрата использовали 100 г/л 2-броммасляной кислоты, присутствовал 20 мМ глицин, и 2 г/л клеточного лизата, т.е. E. coli MG1655, несущий плазмиду pKA81a-HADH-PP-AJ, использовали для экспрессии фермента дегалогеназы. Клеточный лизат добавляли 3 раза, каждый час.
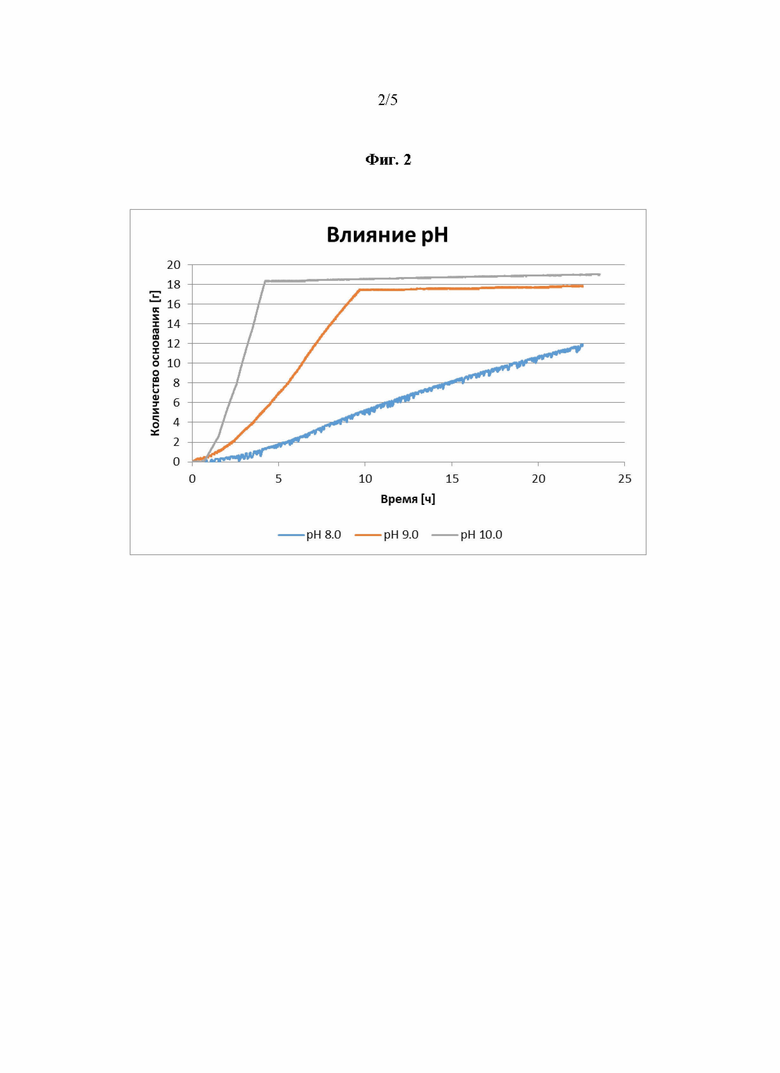
Фиг. 2 демонстрирует, что оптимальное значение pH для реакции согласно Фиг.1 составляет от pH 8 до pH 10.
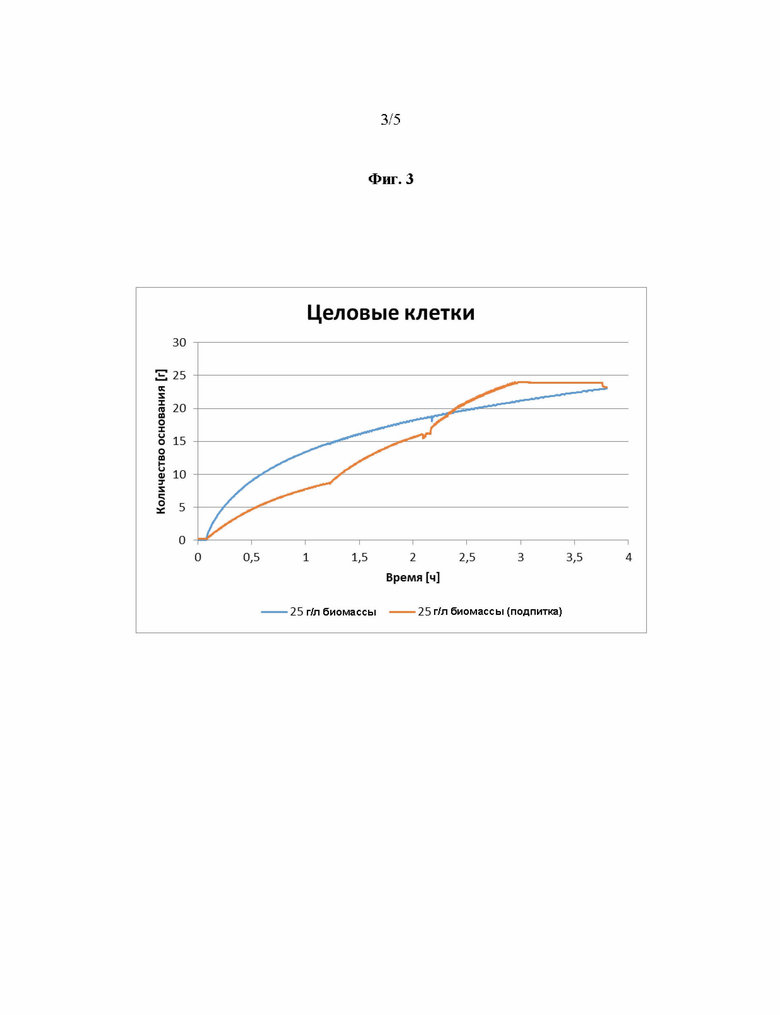
Фиг. 3 демонстрирует, что вместо использования (стерильного) клеточного лизата также для реакции превращения могут быть использованы целые клетки, экспрессирующие заявленный фермент.
Фиг. 4 демонстрирует, что эффективность реакции превращения сохраняется, если субстрат обеспечивают, и фермент добавляют к субстрату.
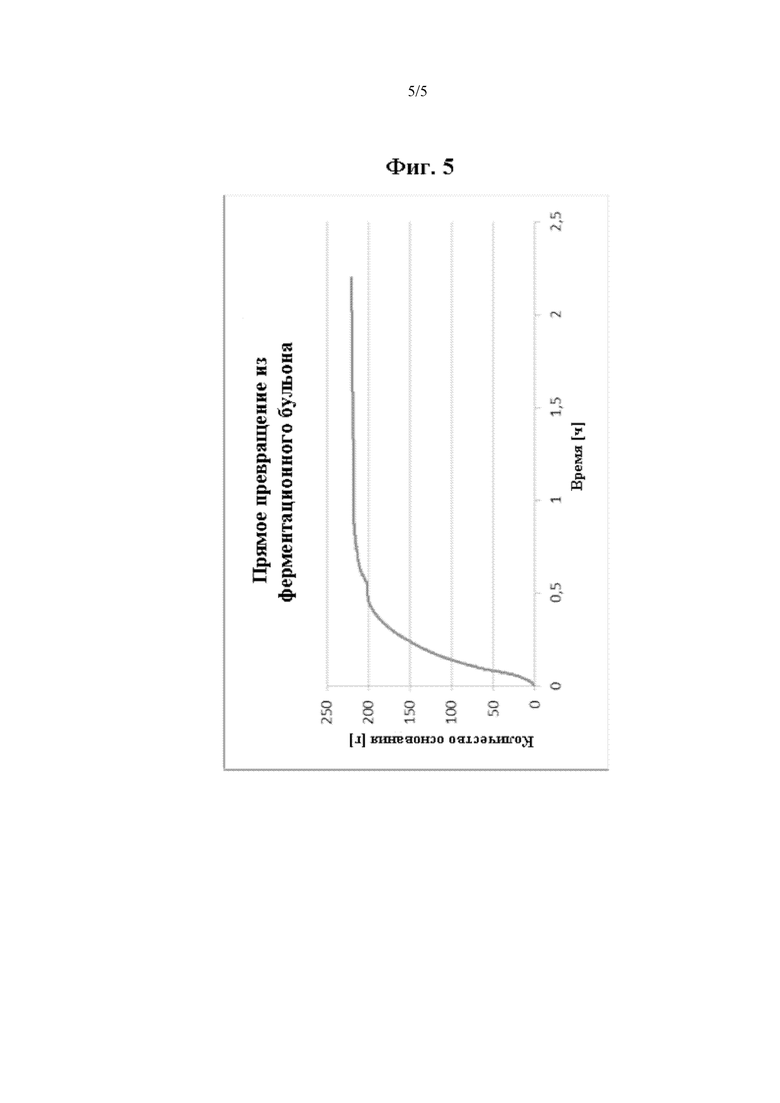
Фиг. 5 демонстрирует, что эффективность реакции превращения сохраняется, если фермент обеспечен в виде ферментационного бульона, и субстрат добавляют.
Примеры
Пример 1. Скрининг активности выбранных дегалогеназ галокислоты
Различные дегалогеназы галокислоты от P. putida AJ, P. putida 109 и S. tokodaii 7 (Jones et al., 1992 (J Gen Microbiol. 1992 Apr;138(4):675-83, Kawasaki et al., 1994 Biosci Biotechnol Biochem. 1994 Jan;58(1):160-3 and Bachas-Daunert et al., 2009 Appl Biochem Biotechnol. 2009 Nov;159(2):382-93) были клонированы в E. coli MG1655 и сверхэкспрессированы посредством IPTG обработки. 50 г/л 2-броммасляной кислоты применяли в качестве субстрата, и 2 x 1,6 г/л клеточного лизата добавляли в начале реакции и через 3,5 часа, причем значение pH составляло 9, и температура была установлена на 37°C.
Активность и стереоселективность, т.е. предпочтительный гидролиз S-энантиомера, были продемонстрированы для всех трех ферментов с помощью аналитики с использованием газовой хроматографии с ферментом от P. putida AJ (SEQ ID NO 1), демонстрирующим наивысшую селективность, достигающую значения ее выше 90%.
Пример 2. Влияние значения pH на исход реакции
Чтобы протестировать оптимальный диапазон pH, устанавливали следующие реакционные условия:
- 100 г/л 2-бромoмасляной кислоты (субстрат)
- 1,5 г/л клеточного лизата клеток E.coli, экспрессирующих дегалогеназу галокислоты (SEQ ID NO 1) от P. putida AJ (6 раз, каждый час)
- 20мМ глицин
- Температура: 25°C
- pH: поддерживали постоянным при 8,0, 9,0, 10,0
Как показано на Фиг. 2, наилучшие результаты были достигнуты при значении pH 9-10.
Пример 3: Дальнейшая оптимизация селективного дегалогенирования для получения высокого энантиомерного избытка с применением дегалогеназы галокислоты от P. putida AJ т.е. полипептида, имеющего дегалогеназную активность, состоящего из SEQ ID NO. 1
В первом случае в качестве субстрата использовали 100 г/л rac-2-броммасляной кислоты в 10 мМ глицине. Реакцию проводили при 25°С, хотя оптимальная температура для активности фермента была при 37°С, поскольку субстрат был более стабильным при 25°С, чем при 37°С, и, следовательно, был предотвращен автогидролиз (данные не показаны).
Фермент обеспечивали в виде клеточного лизата и добавляли либо весь в начале реакции, либо каждый час при концентрации 2 мг/мл. Значение рН поддерживали постоянным на уровне 9,5 путем титрования с помощью 3М КОН до завершения реакции примерно через 4 часа. Затем значение pH доводили до 1,5 с помощью концентрированной H2SO4, и клеточный дебрис фильтровали через Celite. Экстракцию проводили с помощью MTBE, и промывки для удаления оставшейся 2-гидроксимасляной кислоты проводили с помощью водн. CuSO4. Наконец, концентрирование проводили в вакууме.
Неожиданно было обнаружено, что реакция достоверно достигала значения энантиомерного избытка выше 99% (данные не показаны).
Эти высокие степени энантиомерного избытка, т.е. тот факт, что после реакции присутствует только R-энантиомер альфа-галоалкановой кислоты и гидроксилированный продукт предшествующего S-энантиомера альфа-галоалкановой кислоты, делает процесс привлекательным для использования в крупных промышленных масштабах.
Пример 4: Превращение с целыми клетками
Неожиданно было обнаружено, что вместо получения клеточного лизата для превращения можно использовать целые клетки. Это имеет то преимущество, что трудоемкая стадия получения (стерильного) клеточного лизата может быть пропущена.
В первом случае в качестве субстрата использовалась рацемическую 2-броммасляную кислоту при концентрации 100 г/л. Параметры реакции были установлены как: 150 мл, 25°С, 500 оборотов в минуту, 20 мМ глициновый буфер и рН 9,5. Значение рН поддерживали постоянным, используя 3 М КОН. Реакционный сосуд готовили с субстратной смесью, и реакцию начинали добавлением фермента, т.е. добавление целых клеток E.coli MG1655, экспрессирующих фермент дегалогеназу от P. putida AJ, т.е. полипептид, имеющий дегалогеназную активность, состоящий из SEQ ID NO. 1. Клетки добавляли либо в начале реакции, либо поэтапно через 1, 2 и 3 часа, при концентрации 8,3 г/л, соответственно (см. Фиг. 3). Конечная концентрация целых клеток, содержащих фермент, составляла либо 25 г/л, либо 50 г/л. Реакция была завершена через 4 часа. Результаты представлены на Фиг. 3, и приведенная ниже таблица демонстрирует, что в идеале реакция проводится в течение более 1 часа, например, в течение 4 часов.
Затем реакцию повторяли, но биомасса, состоящая из клеток E coli MG1655, экспрессирующих фермент дегалогеназу от P. putida AJ, т.е. полипептид, имеющий дегалогеназную активность, состоящий из SEQ ID NO. 1, добавляли в реакционный сосуд перед добавлением субстрата. Результаты показаны на Фиг. 4 и в таблице ниже. Эти результаты демонстрируют, что можно сначала обеспечить фермент в виде биомассы, содержащей целые клетки, и добавлять субстрат к указанной биомассе.
Более того, превращение проводили с применением ферментационного бульона. Снова клетки E. coli MG1655, несущие плазмиду pKA81a-HADH-PP-AJ, применяли для экспрессии фермента дегалогеназы, т.е. полипептида, имеющего дегалогеназную активность, состоящего из SEQ ID NO. 1. Получение фермента осуществляли путем ферментации E. coli в минимальной среде с использованием стандартного протокола. После генной экспрессии была достигнута концентрация клеток 100 г/л. Затем проводили биотрансформацию с необработанным ферментационным бульоном. Ферментационный бульон охлаждали до 25°С, и значение рН доводили до 9,5 путем добавления 3 М КОН. Реакцию дегалогеназы начинали добавлением субстратной смеси, содержащей 100 г/л 2-броммасляной кислоты, 0,5 об. (об./об.) глицинового буфера при значении рН 9,5 и 2 об. (об./об.) 5 М КОН. Реакция достигла полного превращения, т.е. 90 ее % через 1 ч (см. Фиг. 5).
Из полученной смеси гидроксимасляной кислоты и R-2-броммасляной кислоты около 30% чистой R-2-броммасляной кислоты может быть получено с использованием стандартных методов, таких как подкисление и экстракция.
Пример 5. Тестирование других субстратов
В дополнение к 2-броммасляной кислоте, протестировали другие субстраты, и результаты представлены ниже
SEQ ID NO. 1
SEQ ID NO. 4
Пример 6: Увеличение масштаба
Необходимым условием для увеличения масштаба реакции до 2000 литров было обнаружение возможности использования ферментов, содержащихся в целых клетках, которые были обеспечены в виде биомассы, без необходимости приготовления клеточного лизата. Следовательно, могут быть опущены трудоемкие и дорогостоящие стадии приготовления, такие как фильтрация клеточного лизата, которые являются экономически невыгодными в больших масштабах. Более того, обнаружение того, что реакция может быть начата либо добавлением субстрата (рацемата), либо биомассы, содержащей фермент в целых клетках, означало, что оборудование можно использовать с максимальной гибкостью. Чтобы дополнительно адаптировать процесс, KOH, используемый для титрования pH (см. выше), заменяли на 50% NaOH, растворитель для экстракции продуктов заменяли от MTBE на MIBK, а CuSO4, используемый для удаления побочных продуктов, заменяли на CaCl2, позволяя более простую и дешевую утилизацию отходов.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Bayer Aktiengesellschaft
<120> СПОСОБ ПОЛУЧЕНИЯ ХИРАЛЬНЫХ АЛЬФА-ГАЛОАЛКАНОВЫХ КИСЛОТ
<130> BTS173006
<160> 8
<170> PatentIn version 3.5
<210> 1
<211> 227
<212> PRT
<213> Pseudomonas Putida AJ1
<400> 1
Met Lys Asn Ile Gln Gly Ile Val Phe Asp Leu Tyr Gly Thr Leu Tyr
1 5 10 15
Asp Val His Ser Val Val Gln Ala Cys Glu Glu Val Tyr Pro Gly Gln
20 25 30
Gly Asp Ala Ile Ser Arg Leu Trp Arg Gln Lys Gln Leu Glu Tyr Thr
35 40 45
Trp Leu Arg Ser Leu Met Gly Arg Tyr Val Asn Phe Glu Lys Ala Thr
50 55 60
Glu Asp Ala Leu Arg Phe Thr Cys Thr His Leu Gly Leu Ser Leu Asp
65 70 75 80
Asp Glu Thr His Gln Arg Leu Ser Asp Ala Tyr Leu His Leu Thr Pro
85 90 95
Tyr Ala Asp Thr Ala Asp Ala Val Arg Arg Leu Lys Ala Ala Gly Leu
100 105 110
Pro Leu Gly Ile Ile Ser Asn Gly Ser His Cys Ser Ile Glu Gln Val
115 120 125
Val Thr Asn Ser Glu Met Asn Trp Ala Phe Asp Gln Leu Ile Ser Val
130 135 140
Glu Asp Val Gln Val Phe Lys Pro Asp Ser Arg Val Tyr Ser Leu Ala
145 150 155 160
Glu Lys Arg Met Gly Phe Pro Lys Glu Asn Ile Leu Phe Val Ser Ser
165 170 175
Asn Ala Trp Asp Ala Ser Ala Ala Ser Asn Phe Gly Phe Pro Val Cys
180 185 190
Trp Ile Asn Arg Gln Asn Gly Ala Phe Asp Glu Leu Asp Ala Lys Pro
195 200 205
Thr His Val Val Arg Asn Leu Ala Glu Met Ser Asn Trp Leu Val Asn
210 215 220
Ser Leu Asp
225
<210> 2
<211> 684
<212> ДНК
<213> Pseudomonas Putida AJ1
<400> 2
atgaagaaca ttcagggcat tgtgtttgac ctctatggta cgctgtatga cgtacacagc 60
gtcgttcaag cgtgtgaaga agtttaccct ggtcaaggcg atgcgatttc ccgtttgtgg 120
cgtcagaaac agctggaata cacgtggttg cgttccttga tgggacgcta tgtcaacttc 180
gagaaagcga ctgaggatgc gttacgcttt acctgtacgc acctgggtct gtcccttgac 240
gacgaaaccc atcaacgtct gagcgatgcc tatctccacc tgactccgta cgcagataca 300
gccgatgcag ttcgtcggtt aaaagccgca ggcttaccac tggggatcat cagcaatggc 360
agtcattgca gcattgaaca ggtggtaacc aactcggaaa tgaactgggc tttcgatcag 420
ctgatttcgg tcgaagatgt ccaggtgttc aaacccgatt ctcgcgtgta ttcactggcg 480
gagaaacgca tgggctttcc gaaggagaac atcctcttcg tgagttcgaa tgcttgggat 540
gcgtcagctg cctctaactt tgggtttccg gtttgctgga tcaatcgcca gaatggtgcg 600
tttgacgaac tggatgccaa accgacccat gtggtacgca atctggcaga aatgagcaat 660
tggcttgtga acagtctgga ctaa 684
<210> 3
<211> 684
<212> ДНК
<213> Pseudomonas Putida AJ1
<400> 3
atgaaaaaca tccaaggtat cgttttcgat ttgtatggca cgctctacga cgtgcattcc 60
gtggtgcaag cctgtgaaga ggtctatccg ggccaaggcg acgctatttc tcgcctctgg 120
cggcaaaagc aattggaata cacctggctc aggagcctca tgggccgtta cgtgaacttt 180
gagaaagcaa cagaggatgc cttgcgcttt acctgcacgc atctgggctt gtcgctcgat 240
gatgaaaccc accagcgcct cagtgatgct tatttgcacc tcacccctta tgccgataca 300
gctgacgccg ttcgccgttt gaaagctgcg ggcctaccgc taggcatcat ttcaaatggt 360
tctcattgct cgatcgagca agtcgtgact aactctgaaa tgaattgggc gttcgatcag 420
ctgatcagcg tcgaggatgt gcaagtgttc aaacctgata gtcgcgtcta tagccttgcc 480
gagaagcgca tgggttttcc aaaggaaaac atcctcttcg tttcgtcaaa cgcgtgggat 540
gcgagtgcag ccagtaactt tggtttcccg gtttgctgga tcaatcggca gaacggcgcg 600
tttgatgagc tggatgcaaa gccgacacac gtcgtgcgta atctcgccga aatgtcgaac 660
tggctggtta attcgctcga ttaa 684
<210> 4
<211> 201
<212> PRT
<213> Sulfolobus tokodaii 7
<400> 4
Met Ile Ile Leu Ala Phe Asp Ile Phe Gly Thr Val Leu Asp Thr Ser
1 5 10 15
Thr Val Ile Gln Glu Phe Arg Asn Lys Gln Leu Glu Tyr Thr Trp Leu
20 25 30
Leu Thr Ile Met Gly Lys Tyr Val Glu Phe Glu Glu Ile Thr Lys Ile
35 40 45
Thr Leu Arg Tyr Ile Leu Lys Val Arg Gly Glu Glu Ser Lys Phe Asp
50 55 60
Glu Glu Leu Asn Lys Trp Lys Asn Leu Lys Ala Tyr Glu Asp Thr Lys
65 70 75 80
Tyr Leu Lys Glu Ile Ser Glu Ile Ala Glu Val Tyr Ala Leu Ser Asn
85 90 95
Gly Ser Ile Asn Glu Val Lys Gln His Leu Glu Arg Asn Gly Leu Leu
100 105 110
Arg Tyr Phe Lys Gly Ile Phe Ser Ala Glu Ser Val Lys Glu Tyr Lys
115 120 125
Pro Ser Pro Lys Val Tyr Lys Tyr Phe Leu Asp Ser Ile Gly Ala Lys
130 135 140
Glu Ala Phe Leu Val Ser Ser Asn Ala Phe Asp Val Ile Gly Ala Lys
145 150 155 160
Asn Ala Gly Met Arg Ser Ile Phe Val Asn Arg Lys Asn Thr Ile Val
165 170 175
Asp Pro Ile Gly Gly Lys Pro Asp Val Ile Val Asn Asp Phe Lys Glu
180 185 190
Leu Tyr Glu Trp Ile Leu Arg Tyr Lys
195 200
<210> 5
<211> 606
<212> ДНК
<213> Sulfolobus tokodaii 7
<400> 5
atgatcattc tggcctttga tatctttggt actgtactcg atacatctac cgttatccag 60
gaatttcgca acaaacaact ggagtatact tggctgctga cgattatggg caaatacgtg 120
gaatttgagg aaatcaccaa aattacgtta cgctatatcc tgaaagttcg tggtgaagaa 180
tcgaagtttg acgaagaact gaacaaatgg aagaacctga aagcgtatga agataccaaa 240
taccttaaag agatttcgga aattgccgaa gtttatgcgc tgtcaaatgg gagtattaac 300
gaagtgaaac agcatttgga acgtaatggg ttacttcggt acttcaaagg cattttctcc 360
gcagaaagcg ttaaagagta caaaccgagt ccgaaagtgt ataagtactt tctggatagc 420
attggtgcga aagaagcctt cttggtatct agcaacgcat tcgatgtgat tggcgctaag 480
aatgctggta tgcgttccat ctttgtcaat cgcaagaaca ccattgtcga tcctatcggc 540
ggaaaaccag acgtgatcgt caatgacttc aaagagctgt atgagtggat tctccgctat 600
aaataa 606
<210> 6
<211> 606
<212> ДНК
<213> Sulfolobus tokodaii 7
<400> 6
atgatcattc tagcatttga tatcttcgga acagttcttg atacatctac ggtaattcaa 60
gagtttagga ataagcaatt agagtataca tggttactta caataatggg gaaatatgtg 120
gaatttgagg aaataacaaa gattacttta agatacatct taaaggtaag aggcgaagag 180
agcaaatttg atgaggagtt aaataagtgg aagaatctta aagcttatga agatactaaa 240
tatttaaagg aaatatctga gatagccgag gtctacgcgt tatctaacgg gtctataaat 300
gaggttaaac aacatttaga gcgcaatggt ttgttaagat attttaaggg catatttagt 360
gcagaaagtg ttaaagaata taaaccttca cctaaagtat acaaatattt cctagactcg 420
ataggagcta aagaagcatt cttagtttca tcaaatgcat ttgacgtcat aggagctaaa 480
aacgcgggta tgaggagtat attcgtaaat aggaagaata caatagtcga tcctataggt 540
ggcaaacctg atgttatagt aaatgacttc aaagagttat atgaatggat tttgcgatat 600
aagtga 606
<210> 7
<211> 4515
<212> ДНК
<213> pKA81a-HADH-PP-AJ плазмида
<400> 7
ttagaaaaac tcatcgagca tcaaatgaaa ctgcaattta ttcatatcag gattatcaat 60
accatatttt tgaaaaagcc gtttctgtaa tgaaggagaa aactcaccga ggcagttcca 120
taggatggca agatcctggt atcggtctgc gattccgact cgtccaacat caatacaacc 180
tattaatttc ccctcgtcaa aaataaggtt atcaagtgag aaatcaccat gagtgacgac 240
tgaatccggt gagaatggca aaagtttatg catttctttc cagacttgtt caacaggcca 300
gccattacgc tcgtcatcaa aatcactcgc atcaaccaaa ccgttattca ttcgtgattg 360
cgcctgagcg agacgaaata cgcgatcgct gttaaaagga caattacaaa caggaatcga 420
atgcaaccgg cgcaggaaca ctgccagcgc atcaacaata ttttcacctg aatcaggata 480
ttcttctaat acctggaatg ctgttttccc ggggatcgca gtggtgagta accatgcatc 540
atcaggagta cggataaaat gcttgatggt cggaagaggc ataaattccg tcagccagtt 600
tagtctgacc atctcatctg taacatcatt ggcaacgcta cctttgccat gtttcagaaa 660
caactctggc gcatcgggct tcccatacaa tcgatagatt gtcgcacctg attgcccgac 720
attatcgcga gcccatttat acccatataa atcagcatcc atgttggaat ttaatcgcgg 780
cctagagcaa gacgtttccc gttgaatatg gctcataaca ccccttgtat tactgtttat 840
gtaagcagac agttttattg ttcatgacca aaatccctta acgtgagttt tcgttccact 900
gagcgtcaga ccccgtagaa aagatcaaag gatcttcttg agatcctttt tttctgcgcg 960
taatctgctg cttgcaaaca aaaaaaccac cgctaccagc ggtggtttgt ttgccggatc 1020
aagagctacc aactcttttt ccgaaggtaa ctggcttcag cagagcgcag ataccaaata 1080
ctgtccttct agtgtagccg tagttaggcc accacttcaa gaactctgta gcaccgccta 1140
catacctcgc tctgctaatc ctgttaccag tggctgctgc cagtggcgat aagtcgtgtc 1200
ttaccgggtt ggactcaaga cgatagttac cggataaggc gcagcggtcg ggctgaacgg 1260
ggggttcgtg cacacagccc agcttggagc gaacgaccta caccgaactg agatacctac 1320
agcgtgagct atgagaaagc gccacgcttc ccgaagggag aaaggcggac aggtatccgg 1380
taagcggcag ggtcggaaca ggagagcgca cgagggagct tccaggggga aacgcctggt 1440
atctttatag tcctgtcggg tttcgccacc tctgacttga gcgtcgattt ttgtgatgct 1500
cgtcaggggg gcggagccta tggaaaaacg ccagcaacgc ggccttttta cggttcctgg 1560
ccttttgctg gccttttgct cacatgttct ttcctgcgtt atcccctgat tctgtggata 1620
accgtattac cgcctttgag tgagctgata ccgctcgccg cagccgaacg accgagcgca 1680
gcgagtcagt gagcgaggaa gcggaagagc gcctgatgcg gtattttctc cttacgcatc 1740
tgtgcggtat ttcacaccgc aatggtgcac tctcagtaca atctgctctg atgccgcata 1800
gttaagccag tatacactcc gctatcgcta cgtgactggg tcatggctgc gccccgacac 1860
ccgccaacac ccgctgacgc gccctgacgg gcttgtctgc tcccggcatc cgcttacaga 1920
caagctgtga ccgtctccgg gagctgcatg tgtcagaggt tttcaccgtc atcaccgaaa 1980
cgcgcgaggc agctgcggta aagctcatca gcgtggtcgt gaagcgattc acagatgtct 2040
gcctgttcat ccgcgtccag ctcgttgagt ttctccagaa gcgttaatgt ctggcttctg 2100
ataaagcggg ccatgttaag ggcggttttt tcctgtttgg tcactgatgc ctccgtgtaa 2160
gggggatttc tgttcatggg ggtaatgata ccgatgaaac gagagaggat gctcacgata 2220
cgggttactg atgatgaaca tgcccggtta ctggaacgtt gtgagggtaa acaactggcg 2280
gtatggatgc ggcgggacca gagaaaaatc actcagggtc aatgccagcg ctcatgagcc 2340
cgaagtggcg agcccgatct tccccatcgg tgatgtcggc gatataggcg ccagcaaccg 2400
cacctgtggc gccggtgatg ccggccacga tgcgtccggc gtagaggatc gagatccatt 2460
tacgttgaca ccatcgaatg gtgcaaaacc tttcgcggta tggcatgata gcgcccggaa 2520
gagagtcaat tcagggtggt gaatgtgaaa ccagtaacgt tatacgatgt cgcagagtat 2580
gccggtgtct cttatcagac cgtttcccgc gtggtgaacc aggccagcca cgtttctgcg 2640
aaaacgcggg aaaaagtgga agcggcgatg gcggagctga attacattcc caaccgcgtg 2700
gcacaacaac tggcgggcaa acagtcgttg ctgattggcg ttgccacctc cagtctggcc 2760
ctgcacgcgc cgtcgcaaat tgtcgcggcg attaaatctc gcgccgatca actgggtgcc 2820
agcgtggtgg tgtcgatggt agaacgaagc ggcgtcgaag cctgtaaagc ggcggtgcac 2880
aatcttctcg cgcaacgcgt cagtgggctg atcattaact atccgctgga tgaccaggat 2940
gccattgctg tggaagctgc ctgcactaat gttccggcgt tatttcttga tgtctctgac 3000
cagacaccca tcaacagtat tattttctcc catgaagacg gtacgcgact gggcgtggag 3060
catctggtcg cattgggtca ccagcaaatc gcgctgttag cgggcccatt aagttctgtc 3120
tcggcgcgtc tgcgtctggc tggctggcat aaatatctca ctcgcaatca aattcagccg 3180
atagcggaac gggaaggcga ctggagtgcc atgtccggtt ttcaacaaac catgcaaatg 3240
ctgaatgagg gcatcgttcc cactgcgatg ctggttgcca acgatcagat ggcgctgggc 3300
gcaatgcgcg ccattaccga gtccgggctg cgcgttggtg cggatatttc ggtagtggga 3360
tacgacgata ccgaagacag ctcatgttat atcccgccgt taaccaccat caaacaggat 3420
tttcgcctgc tggggcaaac cagcgtggac cgcttgctgc aactctctca gggccaggcg 3480
gtgaagggca atcagctgtt gcccgtctca ctggtgaaaa gaaaaaccac cctggcttga 3540
gaaatcataa aaaatttatt tgctttgtga gcggataaca attataatag attcaattgt 3600
gagcggataa caatttcaca catctagaaa taattttgtt taactttaag aaggagatat 3660
catatgaaga acattcaggg cattgtgttt gacctctatg gtacgctgta tgacgtacac 3720
agcgtcgttc aagcgtgtga agaagtttac cctggtcaag gcgatgcgat ttcccgtttg 3780
tggcgtcaga aacagctgga atacacgtgg ttgcgttcct tgatgggacg ctatgtcaac 3840
ttcgagaaag cgactgagga tgcgttacgc tttacctgta cgcacctggg tctgtccctt 3900
gacgacgaaa cccatcaacg tctgagcgat gcctatctcc acctgactcc gtacgcagat 3960
acagccgatg cagttcgtcg gttaaaagcc gcaggcttac cactggggat catcagcaat 4020
ggcagtcatt gcagcattga acaggtggta accaactcgg aaatgaactg ggctttcgat 4080
cagctgattt cggtcgaaga tgtccaggtg ttcaaacccg attctcgcgt gtattcactg 4140
gcggagaaac gcatgggctt tccgaaggag aacatcctct tcgtgagttc gaatgcttgg 4200
gatgcgtcag ctgcctctaa ctttgggttt ccggtttgct ggatcaatcg ccagaatggt 4260
gcgtttgacg aactggatgc caaaccgacc catgtggtac gcaatctggc agaaatgagc 4320
aattggcttg tgaacagtct ggactaactc gagcaccacc accaccacca ctgagatccg 4380
gctgctaaca aagcccgaaa ggaagctgag ttggctgctg ccaccgctga gcaataacta 4440
gcataacccc ttggggcctc taaacgggtc ttgaggggtt ttttgctgaa aggaggaact 4500
atatccggat aattc 4515
<210> 8
<211> 4437
<212> ДНК
<213> pKA81a-HADH-вектор с дегалогеназой галокислоты из Sulfolobus tokodaii 7
<400> 8
ttagaaaaac tcatcgagca tcaaatgaaa ctgcaattta ttcatatcag gattatcaat 60
accatatttt tgaaaaagcc gtttctgtaa tgaaggagaa aactcaccga ggcagttcca 120
taggatggca agatcctggt atcggtctgc gattccgact cgtccaacat caatacaacc 180
tattaatttc ccctcgtcaa aaataaggtt atcaagtgag aaatcaccat gagtgacgac 240
tgaatccggt gagaatggca aaagtttatg catttctttc cagacttgtt caacaggcca 300
gccattacgc tcgtcatcaa aatcactcgc atcaaccaaa ccgttattca ttcgtgattg 360
cgcctgagcg agacgaaata cgcgatcgct gttaaaagga caattacaaa caggaatcga 420
atgcaaccgg cgcaggaaca ctgccagcgc atcaacaata ttttcacctg aatcaggata 480
ttcttctaat acctggaatg ctgttttccc ggggatcgca gtggtgagta accatgcatc 540
atcaggagta cggataaaat gcttgatggt cggaagaggc ataaattccg tcagccagtt 600
tagtctgacc atctcatctg taacatcatt ggcaacgcta cctttgccat gtttcagaaa 660
caactctggc gcatcgggct tcccatacaa tcgatagatt gtcgcacctg attgcccgac 720
attatcgcga gcccatttat acccatataa atcagcatcc atgttggaat ttaatcgcgg 780
cctagagcaa gacgtttccc gttgaatatg gctcataaca ccccttgtat tactgtttat 840
gtaagcagac agttttattg ttcatgacca aaatccctta acgtgagttt tcgttccact 900
gagcgtcaga ccccgtagaa aagatcaaag gatcttcttg agatcctttt tttctgcgcg 960
taatctgctg cttgcaaaca aaaaaaccac cgctaccagc ggtggtttgt ttgccggatc 1020
aagagctacc aactcttttt ccgaaggtaa ctggcttcag cagagcgcag ataccaaata 1080
ctgtccttct agtgtagccg tagttaggcc accacttcaa gaactctgta gcaccgccta 1140
catacctcgc tctgctaatc ctgttaccag tggctgctgc cagtggcgat aagtcgtgtc 1200
ttaccgggtt ggactcaaga cgatagttac cggataaggc gcagcggtcg ggctgaacgg 1260
ggggttcgtg cacacagccc agcttggagc gaacgaccta caccgaactg agatacctac 1320
agcgtgagct atgagaaagc gccacgcttc ccgaagggag aaaggcggac aggtatccgg 1380
taagcggcag ggtcggaaca ggagagcgca cgagggagct tccaggggga aacgcctggt 1440
atctttatag tcctgtcggg tttcgccacc tctgacttga gcgtcgattt ttgtgatgct 1500
cgtcaggggg gcggagccta tggaaaaacg ccagcaacgc ggccttttta cggttcctgg 1560
ccttttgctg gccttttgct cacatgttct ttcctgcgtt atcccctgat tctgtggata 1620
accgtattac cgcctttgag tgagctgata ccgctcgccg cagccgaacg accgagcgca 1680
gcgagtcagt gagcgaggaa gcggaagagc gcctgatgcg gtattttctc cttacgcatc 1740
tgtgcggtat ttcacaccgc aatggtgcac tctcagtaca atctgctctg atgccgcata 1800
gttaagccag tatacactcc gctatcgcta cgtgactggg tcatggctgc gccccgacac 1860
ccgccaacac ccgctgacgc gccctgacgg gcttgtctgc tcccggcatc cgcttacaga 1920
caagctgtga ccgtctccgg gagctgcatg tgtcagaggt tttcaccgtc atcaccgaaa 1980
cgcgcgaggc agctgcggta aagctcatca gcgtggtcgt gaagcgattc acagatgtct 2040
gcctgttcat ccgcgtccag ctcgttgagt ttctccagaa gcgttaatgt ctggcttctg 2100
ataaagcggg ccatgttaag ggcggttttt tcctgtttgg tcactgatgc ctccgtgtaa 2160
gggggatttc tgttcatggg ggtaatgata ccgatgaaac gagagaggat gctcacgata 2220
cgggttactg atgatgaaca tgcccggtta ctggaacgtt gtgagggtaa acaactggcg 2280
gtatggatgc ggcgggacca gagaaaaatc actcagggtc aatgccagcg ctcatgagcc 2340
cgaagtggcg agcccgatct tccccatcgg tgatgtcggc gatataggcg ccagcaaccg 2400
cacctgtggc gccggtgatg ccggccacga tgcgtccggc gtagaggatc gagatccatt 2460
tacgttgaca ccatcgaatg gtgcaaaacc tttcgcggta tggcatgata gcgcccggaa 2520
gagagtcaat tcagggtggt gaatgtgaaa ccagtaacgt tatacgatgt cgcagagtat 2580
gccggtgtct cttatcagac cgtttcccgc gtggtgaacc aggccagcca cgtttctgcg 2640
aaaacgcggg aaaaagtgga agcggcgatg gcggagctga attacattcc caaccgcgtg 2700
gcacaacaac tggcgggcaa acagtcgttg ctgattggcg ttgccacctc cagtctggcc 2760
ctgcacgcgc cgtcgcaaat tgtcgcggcg attaaatctc gcgccgatca actgggtgcc 2820
agcgtggtgg tgtcgatggt agaacgaagc ggcgtcgaag cctgtaaagc ggcggtgcac 2880
aatcttctcg cgcaacgcgt cagtgggctg atcattaact atccgctgga tgaccaggat 2940
gccattgctg tggaagctgc ctgcactaat gttccggcgt tatttcttga tgtctctgac 3000
cagacaccca tcaacagtat tattttctcc catgaagacg gtacgcgact gggcgtggag 3060
catctggtcg cattgggtca ccagcaaatc gcgctgttag cgggcccatt aagttctgtc 3120
tcggcgcgtc tgcgtctggc tggctggcat aaatatctca ctcgcaatca aattcagccg 3180
atagcggaac gggaaggcga ctggagtgcc atgtccggtt ttcaacaaac catgcaaatg 3240
ctgaatgagg gcatcgttcc cactgcgatg ctggttgcca acgatcagat ggcgctgggc 3300
gcaatgcgcg ccattaccga gtccgggctg cgcgttggtg cggatatttc ggtagtggga 3360
tacgacgata ccgaagacag ctcatgttat atcccgccgt taaccaccat caaacaggat 3420
tttcgcctgc tggggcaaac cagcgtggac cgcttgctgc aactctctca gggccaggcg 3480
gtgaagggca atcagctgtt gcccgtctca ctggtgaaaa gaaaaaccac cctggcttga 3540
gaaatcataa aaaatttatt tgctttgtga gcggataaca attataatag attcaattgt 3600
gagcggataa caatttcaca catctagaaa taattttgtt taactttaag aaggagatat 3660
catatgatca ttctggcctt tgatatcttt ggtactgtac tcgatacatc taccgttatc 3720
caggaatttc gcaacaaaca actggagtat acttggctgc tgacgattat gggcaaatac 3780
gtggaatttg aggaaatcac caaaattacg ttacgctata tcctgaaagt tcgtggtgaa 3840
gaatcgaagt ttgacgaaga actgaacaaa tggaagaacc tgaaagcgta tgaagatacc 3900
aaatacctta aagagatttc ggaaattgcc gaagtttatg cgctgtcaaa tgggagtatt 3960
aacgaagtga aacagcattt ggaacgtaat gggttacttc ggtacttcaa aggcattttc 4020
tccgcagaaa gcgttaaaga gtacaaaccg agtccgaaag tgtataagta ctttctggat 4080
agcattggtg cgaaagaagc cttcttggta tctagcaacg cattcgatgt gattggcgct 4140
aagaatgctg gtatgcgttc catctttgtc aatcgcaaga acaccattgt cgatcctatc 4200
ggcggaaaac cagacgtgat cgtcaatgac ttcaaagagc tgtatgagtg gattctccgc 4260
tataaataac tcgagcacca ccaccaccac cactgagatc cggctgctaa caaagcccga 4320
aaggaagctg agttggctgc tgccaccgct gagcaataac tagcataacc ccttggggcc 4380
tctaaacggg tcttgagggg ttttttgctg aaaggaggaa ctatatccgg ataattc 4437
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНСТРУКТЫ Т-КЛЕТОЧНОГО РЕЦЕПТОРА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2785954C2 |
| Мутант фосфолипазы С с высокой ферментативной активностью | 2019 |
|
RU2818353C2 |
| РЕКОМБИНАНТНЫЕ КЛЕТКИ, ПРОДУЦИРУЮЩИЕ ГИАЛУРОНОВУЮ КИСЛОТУ | 2022 |
|
RU2839983C2 |
| АНТИТЕЛА В7-Н4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2809243C2 |
| СПОСОБЫ ПРОИЗВОДСТВА ПРОТЕОЛИТИЧЕСКИ ПРОЦЕССИРОВАННЫХ ПОЛИПЕПТИДОВ | 2019 |
|
RU2727402C1 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2741103C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2747978C2 |
| СВЯЗЫВАЮЩИЙ TNF-α ПОЛИПЕПТИД И СПОСОБ ЛЕЧЕНИЯ СВЯЗАННЫХ С TNF-α ЗАБОЛЕВАНИЙ | 2017 |
|
RU2794974C2 |
| СПОСОБЫ ПРОИЗВОДСТВА ПРОТЕОЛИТИЧЕСКИ ПРОЦЕССИРОВАННЫХ ПОЛИПЕПТИДОВ | 2018 |
|
RU2719164C1 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |

Изобретение относится к области биотехнологии. Описана группа изобретений, включающая способ селективного гидролиза S-энантиомера альфа-галоалкановой кислоты (варианты). В одном из вариантов способ включает стадии обеспечения рацемата R-энантиомера и S-энантиомера альфа-галоалкановой кислоты, обеспечения полипептида, имеющего дегалогеназную активность, с аминокислотной последовательностью SEQ ID NO. 1 и реакцию селективного гидролиза рацемата в течение 4-6 часов. Способ обеспечивает получение избытка R-энантиомера от 90,0 до 99,9% через 4-6 часов. Изобретение расширяет арсенал способов селективного гидролиза S-энантиомеров альфа-галоалкановых кислот. 2 н. и 3 з.п. ф-лы, 5 ил., 3 табл., 6 пр.
1. Способ селективного гидролиза S-энантиомера альфа-галоалкановой кислоты Формулы I
 ,
,
где X представляет собой бромид или хлорид и
R представляет собой алкильную цепь, содержащую от 1 до 6 атомов углерода, где эта указанная алкильная цепь может быть неразветвленной или разветвленной на атомах углерода γ или δ относительно карбоксильной группы,
включающий
- обеспечение рацемата R-энантиомера и S-энантиомера указанной альфа-галоалкановой кислоты,
- обеспечение полипептида, имеющего дегалогеназную активность, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1,
- реакцию селективного гидролиза рацемата в течение 4-6 часов, где
значение рН находится в интервале 9-10 и температура составляет 25°С для полипептида с дегалогеназной активностью, имеющего аминокислотную последовательность, как показано в SEQ ID NO. 1,
и где энантиомерный избыток R-энантиомера от 90,0 до 99,9% достигается через 4-6 часов.
2. Способ по п. 1, где соотношение рацемата альфа-галоалкановой кислоты Формулы I и биомассы целых клеток, содержащих полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 1, составляет от 2:1 до 15:1, предпочтительно от 3:1 до 10:1, наиболее предпочтительно 4:1.
3. Способ по любому из пп. 1, 2, где фрагмент R указанной альфа-галоалкановой кислоты Формулы I выбирают из группы, состоящей из этила, бутила, 2-метил-пропила и метил-циклопропила.
4. Способ селективного гидролиза S-энантиомера альфа-галоалкановой кислоты Формулы II
,
где R представляет собой алкильную цепь, содержащую от 1 до 6 атомов углерода, где эта указанная алкильная цепь может быть разветвлена на атомах углерода γ или δ относительно карбоксильной группы, применяя полипептид, имеющий дегалогеназную активность, имеющий аминокислотную последовательность, как показано в SEQ ID NO. 4, в течение 4-6 часов, где значение рН находится в интервале 9-10 и температура находится в интервале 59-61°С.
5. Способ по п. 1 или 4, где указанная алкильная цепь разветвлена на атомах углерода γ или δ и где атомы углерода, следующие после разветвления на атомах углерода γ или δ, являются циклическими.
| Jones D | |||
| H | |||
| A., Barth P | |||
| T., Byrom D., Thomas C | |||
| M | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Journal of General Microbiology, 1992, 138(4), 675-683 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Barth P | |||
| T., Bolton L., Thomson J | |||
| C | |||
| Cloning | |||

