ПЕРЕКРЕСТНАЯ ССЫЛКА
Настоящая заявка заявляет приоритет по предварительной заявке на патент США № 61/937295, поданной 7 февраля 2014 года, и предварительной заявке на патент США № 62/051720, поданной 17 сентября 2014 года, которые включены в данный документ во всей своей полноте посредством ссылки.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПРЕДСТАВЛЕННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
Официальная копия данного перечня последовательностей отправлена в электронном виде с помощью EFS-Web как отформатированный под ASCII перечень последовательностей в файле с названием "6054PCT_Sequence_Listing", созданном 26 января 2015 года и имеющем размер 3 705 килобайтов, и подана одновременно с описанием. Перечень последовательностей, содержащийся в данном отформатированном под ASCII документе, является частью описания и включен в данный документ посредством ссылки во всей своей полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее раскрытие относится к области молекулярной биологии. Представлены новые гены, которые кодируют пестицидные белки. Эти пестицидные белки и последовательности нуклеиновой кислоты, которые их кодируют, применимы в приготовлении пестицидных составов и в получении трансгенных растений, устойчивых к вредителям.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Биологический контроль насекомых-вредителей, имеющих сельскохозяйственное значение, с применением микробного агента, такого как грибы, бактерии или другие виды насекомых, представляет не оказывающую негативного влияния на окружающую среду и коммерчески привлекательную альтернативу синтетическим химическим пестицидам. В целом можно сказать, что применение биопестицидов приводит к меньшему риску загрязнения и неблагоприятных воздействий на окружающую среду, и биопестициды обеспечивают большую специфичность по отношению к мишени, по сравнению со специфичностью, характерной для традиционных химических инсектицидов широкого спектра действия. Кроме того, зачастую производство биопестицидов стоит дешевле и, вследствие этого, улучшается экономически эффективный выход продукции для широкого спектра сельскохозяйственных культур.
Определенные виды микроорганизмов из рода Bacillus, как известно, обладают пестицидной активностью против ряда насекомых-вредителей, в том числе Lepidoptera, Diptera, Coleoptera, Hemiptera и других. Bacillus thuringiensis (Bt) и Bacillus popilliae входят в число наиболее успешных средств биологического контроля, обнаруженных на сегодняшний день. Патогенность в отношении насекомых также приписывалась штаммам B. larvae, B. lentimorbus, B. sphaericus и B. cereus. Микробные инсектициды, в частности полученные из штаммов Bacillus, сыграли важную роль в сельском хозяйстве как альтернатива химическому контролю вредителей.
Были разработаны культурные растения с улучшенной устойчивостью к насекомым при помощи генной инженерии культурных растений с тем, чтобы они вырабатывали пестицидные белки, происходящие из Bacillus. Например, с помощью генной инженерии были созданы растения кукурузы и хлопчатника для выработки пестицидных белков, выделенных из штаммов Bt. Эти сельскохозяйственные культуры, созданные при помощи генной инженерии, на сегодняшний день широко применяются в сельском хозяйстве и предоставляют фермеру не оказывающую негативного влияния на окружающую среду альтернативу традиционным способам контроля насекомых. Несмотря на то, что они были признаны очень успешными с коммерческой точки зрения, эти созданные при помощи генной инженерии устойчивые к насекомым культурные растения предусматривают устойчивость только к узкому диапазону важных в экономическом отношении насекомых-вредителей. В некоторых случаях, насекомые могут развивать устойчивость к различным инсектицидным соединениям, что повышает необходимость в идентификации альтернативных средств биологического контроля для контроля вредителей.
Соответственно, остается потребность в новых пестицидных белках с различными диапазонами инсектицидной активности против насекомых-вредителей, например, инсектицидных белках, которые активны против ряда насекомых из отряда Lepidoptera и/или отряда Coleoptera, в том числе без ограничения насекомых-вредителей, которые развили устойчивость к существующим инсектицидам.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Представлены композиции и способы обеспечения пестицидной активности у бактерий, растений, растительных клеток, тканей и семян. Композиции включают молекулы нуклеиновой кислоты, кодирующие последовательности пестицидных и инсектицидных полипептидов, векторы, содержащие такие молекулы нуклеиновой кислоты, и клетки-хозяева, содержащие векторы. Композиции также включают последовательности пестицидных полипептидов и антитела к таким полипептидам. Последовательности нуклеиновой кислоты можно применять в ДНК-конструкциях или кассетах экспрессии для трансформации и экспрессии в организмах, в том числе микроорганизмах и растениях. Нуклеотидные или аминокислотные последовательности могут представлять собой синтетические последовательности, которые были сконструированы для экспрессии в организме, в том числе без ограничения микроорганизме или растении. Композиции также включают трансформированные бактерии, растения, растительные клетки, ткани и семена.
В частности, предусмотрены выделенные или рекомбинантные молекулы нуклеиновой кислоты, кодирующие полипептиды инсектицидного белка-83 Pteridophyta (PtIP-83), включающие аминокислотные замены, делеции, вставки, его фрагменты. Дополнительно охватываются аминокислотные последовательности, соответствующие полипептидам PtIP-83. Представлены выделенные или рекомбинантные молекулы нуклеиновой кислоты, способные кодировать полипептиды PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, и SEQ ID NO: 769, а также аминокислотные замены, делеции, вставки, его фрагменты, и их комбинации. Также охватываются последовательности нуклеиновой кислоты, которые комплементарны последовательности нуклеиновой кислоты согласно вариантам осуществления или которые гибридизуются с последовательностью согласно вариантам осуществления. Также представлены выделенные или рекомбинантные полипептиды PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, и SEQ ID NO: 769, а также аминокислотные замены, делеции, вставки, его фрагменты, и их комбинации.
Представлены способы получения полипептидов и применения данных полипептидов для контроля или уничтожения вредителей из группы чешуекрылых, жесткокрылых, нематод, грибков и/или двукрылых. Трансгенные растения согласно вариантам осуществления экспрессируют одну или несколько пестицидных последовательностей, раскрытых в данном документе. В различных вариантах осуществления трансгенное растение дополнительно содержит один или несколько дополнительных генов устойчивости к насекомым, например, один или несколько дополнительных генов для контроля вредителей из группы жесткокрылых, чешуекрылых, полужесткокрылых или нематод. Специалисту в данной области будет понятно, что трансгенное растение может содержать какой-либо ген, обеспечивающий агрономический признак, представляющий интерес.
Также включены способы выявления нуклеиновых кислот и полипептидов согласно вариантам осуществления в образце. Представлен набор для выявления присутствия полипептида PtIP-83 или выявления присутствия полинуклеотида, кодирующего полипептид PtIP-83, в образце. Набор может быть представлен вместе со всеми реагентами и контрольными образцами, необходимыми для осуществления способа выявления предполагаемого средства, а также с инструкциями по применению.
Композиции и способы согласно вариантам осуществления применимы для получения организмов с улучшенной устойчивостью к вредителям или переносимостью вредителей. Эти организмы и композиции, содержащие организмы, подходят для сельскохозяйственных целей. Композиции согласно вариантам осуществления также применимы для получения измененных или улучшенных белков, которые обладают пестицидной активностью, или для выявления присутствия полипептидов PtIP-83.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фигуре 1 показана филогения папоротников, основанная на классификации современных папоротников по A. R. Smith et al, TAXON, 55:705-731 (2006).
На фигурах 2a-2j показано выравнивание аминокислотных последовательностей PtIP-83Aa (SEQ ID NO: 1), PtIP-83Ca (SEQ ID NO: 5), PtIP-83Cb (SEQ ID NO: 7), PtIP-83Cc (SEQ ID NO: 9), PtIP-83Cd (SEQ ID NO: 11), PtIP-83Ce (SEQ ID NO: 13), PtIP-83Cf (SEQ ID NO: 15) и PtIP-83Fa (SEQ ID NO: 3); выравнивание прогнозируемой вторичной структуры с помощью PSIPRED, ведущего способа прогнозирования вторичной структуры, для PtIP-83Aa (SEQ ID NO: 1) и PtIP-83Fa (SEQ ID NO: 3); и расположение МОТИВОВ аминокислотной последовательности, прогнозируемых с помощью анализа мотивов MEME, относительно PtIP-83Aa (SEQ ID NO: 1). "H" указывает на прогнозируемую структуру спирали, "E" указывает на структуру бета-тяжа PtIP, а "C" указывает на прогнозируемую структуру клубка.
На фигурах 3a-3b показано выравнивание последовательности PtIP-83Aa (SEQ ID NO: 1) и PtIP-50Aa (SEQ ID NO: 34). Точки кроссинговера в химерах PtIP-83 Aa/PtIP-50Aa, указанные в таблице 13, указаны с помощью стрелки (↓) над аминокислотой.
На фигурах 4a-4d показано выравнивание аминокислотных последовательностей PtIP-83Aa (SEQ ID NO: 1), PtIP-83Fa (SEQ ID NO: 3), PtIP-50Aa (SEQ ID NO: 34), PtIP-50Ba (SEQ ID NO: 35) и PtIP-50Bb (SEQ ID NO: 36). Указаны идентифицированные консервативные мотивы последовательности и аминокислотная последовательность мотивов в PtIP-83Aa (SEQ ID NO: 1) подчеркнута.
На фигурах 5a-5e показано выравнивание аминокислотных последовательностей PtIP-83Aa (SEQ ID NO: 1), PtIP-83Ca (SEQ ID NO: 5), PtIP-83Cb (SEQ ID NO: 7), PtIP-83Cc (SEQ ID NO: 9), PtIP-83Cd (SEQ ID NO: 11), PtIP-83Ce (SEQ ID NO: 13), PtIP-83Cf (SEQ ID NO: 15), PtIP-83Cg (SEQ ID NO: 17) и PtIP-83Da (SEQ ID NO: 19). Различия в последовательностях выделены.
ПОДРОБНОЕ ОПИСАНИЕ
Следует понимать, что настоящее раскрытие не ограничивается конкретными описанными методиками, протоколами, клеточными линиями, родами и реагентами, в связи с этим они могут варьировать. Также следует понимать, что терминология, используемая в данном документе, предназначена лишь для описания конкретных вариантов осуществления и не подразумевается как ограничивающая объем настоящего раскрытия.
Используемые в данном документе формы единственного числа включают ссылки на множественное число, если в контексте явно не указано иное. Так, например, ссылка на "клетку" включает множество таких клеток, а ссылка на "белок" включает ссылку на один или несколько белков и их эквивалентов, известных специалистам в данной области, и т. д. Все технические и научные термины, используемые в данном документе, имеют то же самое значение, как обычно понимается специалистом в области техники, к которой принадлежит настоящее раскрытие, если явно не указано иное.
Настоящее раскрытие относится к композициям и способам контроля вредителей. Способы включают трансформацию организмов последовательностями нуклеиновой кислоты, кодирующими полипептиды PtIP-83. В частности, последовательности нуклеиновой кислоты согласно вариантам осуществления применимы для получения растений и микроорганизмов, которые обладают пестицидной активностью. Таким образом, представлены трансформированные бактерии, растения, растительные клетки, растительные ткани и семена. Композиции представляют собой пестицидные нуклеиновые кислоты и белки из видов бактерий. Последовательности нуклеиновой кислоты находят применение в конструировании векторов экспрессии для последующей трансформации организмов, представляющих интерес, в качестве зондов для выделения других гомологичных (или частично гомологичных) генов и для получения измененных полипептидов PtIP-83 при помощи способов, известных из уровня техники, таких как сайт-направленный мутагенез, замена доменов или ДНК-шаффлинг. PtIP-83 находят применение при контроле или уничтожении популяций вредителей из группы чешуекрылых, жесткокрылых, двукрылых, грибков, полужесткокрылых и нематод и для получения композиций с пестицидной активностью. Насекомые-вредители, представляющие интерес, включают без ограничения виды из отряда Lepidoptera, в том числе без ограничения: совку кукурузную (CEW) (Helicoverpa zea), огневку кукурузную (ECB) (Ostrinia nubilalis), моль капустную, например Helicoverpa zea Boddie; совку соевую, например Pseudoplusia includens Walker; и совку бархатных бобов например Anticarsia gemmatalis Hübner, а также виды из отряда Coleoptera, в том числе без ограничения западного кукурузного жука (Diabrotica virgifera) - WCRW, южного кукурузного жука (Diabrotica undecimpunctata howardi) -SCRW и северного кукурузного жука (Diabrotica barberi) - NCRW.
Под "пестицидным токсином" или "пестицидным белком", как используется в настоящем документе, подразумевают токсин, который обладает токсической активностью против одного или нескольких вредителей, в том числе без ограничения представителей отряда Lepidoptera, Diptera, Hemiptera и Coleoptera или типа Nematoda, или белок, который характеризуется гомологией с таким белком. Пестицидные белки были очищены из организмов, в том числе, например, Bacillus sp., Pseudomonas sp., Photorhabdus sp., Xenorhabdus sp., Clostridium bifermentans и Paenibacillus popilliae. Пестицидные белки включают без ограничения инсектицидные белки из Pseudomonas sp., такие как PSEEN3174 (Monalysin; (2011) PLoS Pathogens 7:1-13); из штамма CHA0 и Pf-5 Pseudomonas protegens (ранее fluorescens) (Pechy-Tarr, (2008) Environmental Microbiology 10:2368-2386; № доступа в GenBank EU400157); из Pseudomonas Taiwanensis (Liu, et al., (2010) J. Agric. Food Chem., 58:12343-12349) и из Pseudomonas pseudoalcligenes (Zhang, et al., (2009) Annals of Microbiology 59:45-50 и Li, et al., (2007) Plant Cell Tiss. Organ Cult. 89:159-168); инсектицидные белки из Photorhabdus sp. и Xenorhabdus sp. (Hinchliffe, et al., (2010) The Open Toxicology Journal, 3:101-118 и Morgan, et al., (2001) Applied and Envir. Micro. 67:2062-2069); патент США № 6048838 и патент США № 6379946; полипептид PIP-1 из публикации заявки на патент США US20140007292; полипептид AfIP-1A и/или AfIP-1B из публикации заявки на патент США US20140033361; полипептид PHI-4 из публикации заявки на патент США US20140274885 и патентной публикации согласно PCT WO2014/150914; полипептид PIP-47 из патентной публикации согласно PCT с серийным номером PCT/US14/51063, полипептид PIP-72 из патентной публикации согласно PCT с серийным номером PCT/US14/55128 и δ-эндотоксины, в том числе без ограничения классы Cry1, Cry2, Cry3, Cry4, Cry5, Cry6, Cry7, Cry8, Cry9, Cry10, Cry11, Cry12, Cry13, Cry14, Cry15, Cry16, Cry17, Cry18, Cry19, Cry20, Cry21, Cry22, Cry23, Cry24, Cry25, Cry26, Cry27, Cry 28, Cry 29, Cry 30, Cry31, Cry32, Cry33, Cry34, Cry35,Cry36, Cry37, Cry38, Cry39, Cry40, Cry41, Cry42, Cry43, Cry44, Cry45, Cry 46, Cry47, Cry49, Cry50, Cry51, Cry52, Cry53, Cry 54, Cry55, Cry56, Cry57, Cry58, Cry59, Cry60, Cry61, Cry62, Cry63, Cry64, Cry65, Cry66, Cry67, Cry68, Cry69, Cry70, Cry71, и Cry 72 генов δ-эндотоксинов и гены цитолитических токсинов cyt1 и cyt2 B. thuringiensis. Представители этих классов инсектицидных белков B. thuringiensis включают без ограничения Cry1Aa1 (№ доступа AAA22353); Cry1Aa2 (№ доступа № доступа AAA22552); Cry1Aa3 (№ доступа BAA00257); Cry1Aa4 (№ доступа CAA31886); Cry1Aa5 (№ доступа BAA04468); Cry1Aa6 (№ доступа AAA86265); Cry1Aa7 (№ доступа AAD46139); Cry1Aa8 (№ доступа I26149); Cry1Aa9 (№ доступа BAA77213); Cry1Aa10 (№ доступа AAD55382); Cry1Aa11 (№ доступа CAA70856); Cry1Aa12 (№ доступа AAP80146); Cry1Aa13 (№ доступа AAM44305); Cry1Aa14 (№ доступа AAP40639); Cry1Aa15 (№ доступа AAY66993); Cry1Aa16 (№ доступа HQ439776); Cry1Aa17 (№ доступа HQ439788); Cry1Aa18 (№ доступа HQ439790); Cry1Aa19 (№ доступа HQ685121); Cry1Aa20 (№ доступа JF340156); Cry1Aa21 (№ доступа JN651496); Cry1Aa22 (№ доступа KC158223); Cry1Ab1 (№ доступа AAA22330); Cry1Ab2 (№ доступа AAA22613); Cry1Ab3 (№ доступа AAA22561); Cry1Ab4 (№ доступа BAA00071); Cry1Ab5 (№ доступа CAA28405); Cry1Ab6 (№ доступа AAA22420); Cry1Ab7 (№ доступа CAA31620); Cry1Ab8 (№ доступа AAA22551); Cry1Ab9 (№ доступа CAA38701); Cry1Ab10 (№ доступа A29125); Cry1Ab11 (№ доступа I12419); Cry1Ab12 (№ доступа AAC64003); Cry1Ab13 (№ доступа AAN76494); Cry1Ab14 (№ доступа AAG16877); Cry1Ab15 (№ доступа AAO13302); Cry1Ab16 (№ доступа AAK55546); Cry1Ab17 (№ доступа AAT46415); Cry1Ab18 (№ доступа AAQ88259); Cry1Ab19 (№ доступа AAW31761); Cry1Ab20 (№ доступа ABB72460); Cry1Ab21 (№ доступа ABS18384); Cry1Ab22 (№ доступа ABW87320); Cry1Ab23 (№ доступа HQ439777); Cry1Ab24 (№ доступа HQ439778); Cry1Ab25 (№ доступа HQ685122); Cry1Ab26 (№ доступа HQ847729); Cry1Ab27 (№ доступа JN135249); Cry1Ab28 (№ доступа JN135250); Cry1Ab29 (№ доступа JN135251); Cry1Ab30 (№ доступа JN135252); Cry1Ab31 (№ доступа JN135253); Cry1Ab32 (№ доступа JN135254); Cry1Ab33 (№ доступа AAS93798); Cry1Ab34 (№ доступа KC156668); Cry1Ab-подобный (№ доступа AAK14336); Cry1Ab-подобный (№ доступа AAK14337); Cry1Ab-подобный (№ доступа AAK14338); Cry1Ab-подобный (№ доступа ABG88858); Cry1Ac1 (№ доступа AAA22331); Cry1Ac2 (№ доступа AAA22338); Cry1Ac3 (№ доступа CAA38098); Cry1Ac4 (№ доступа AAA73077); Cry1Ac5 (№ доступа AAA22339); Cry1Ac6 (№ доступа AAA86266); Cry1Ac7 (№ доступа AAB46989); Cry1Ac8 (№ доступа AAC44841); Cry1Ac9 (№ доступа AAB49768); Cry1Ac10 (№ доступа CAA05505 ); Cry1Ac11 (№ доступа CAA10270); Cry1Ac12 (№ доступа I12418); Cry1Ac13 (№ доступа AAD38701); Cry1Ac14 (№ доступа AAQ06607); Cry1Ac15 (№ доступа AAN07788); Cry1Ac16 (№ доступа AAU87037); Cry1Ac17 (№ доступа AAX18704); Cry1Ac18 (№ доступа AAY88347); Cry1Ac19 (№ доступа ABD37053); Cry1Ac20 (№ доступа ABB89046 ); Cry1Ac21 (№ доступа AAY66992 ); Cry1Ac22 (№ доступа ABZ01836); Cry1Ac23 (№ доступа CAQ30431); Cry1Ac24 (№ доступа ABL01535); Cry1Ac25 (№ доступа FJ513324); Cry1Ac26 (№ доступа FJ617446); Cry1Ac27 (№ доступа FJ617447); Cry1Ac28 (№ доступа ACM90319); Cry1Ac29 (№ доступа DQ438941); Cry1Ac30 (№ доступа GQ227507); Cry1Ac31 (№ доступа GU446674); Cry1Ac32 (№ доступа HM061081); Cry1Ac33 (№ доступа GQ866913); Cry1Ac34 (№ доступа HQ230364); Cry1Ac35 (№ доступа JF340157); Cry1Ac36 (№ доступа JN387137); Cry1Ac37 (№ доступа JQ317685); Cry1Ad1 (№ доступа AAA22340); Cry1Ad2 (№ доступа CAA01880); Cry1Ae1 (№ доступа AAA22410); Cry1Af1 (№ доступа AAB82749); Cry1Ag1 (№ доступа AAD46137); Cry1Ah1 (№ доступа AAQ14326); Cry1Ah2 (№ доступа ABB76664); Cry1Ah3 (№ доступа HQ439779); Cry1Ai1 (№ доступа AAO39719); Cry1Ai2 (№ доступа HQ439780); Cry1A-подобный (№ доступа AAK14339); Cry1Ba1 (№ доступа CAA29898); Cry1Ba2 (№ доступа CAA65003); Cry1Ba3 (№ доступа AAK63251); Cry1Ba4 (№ доступа AAK51084); Cry1Ba5 (№ доступа ABO20894); Cry1Ba6 (№ доступа ABL60921); Cry1Ba7 (№ доступа HQ439781); Cry1Bb1 (№ доступа AAA22344); Cry1Bb2 (№ доступа HQ439782); Cry1Bc1 (№ доступа CAA86568); Cry1Bd1 (№ доступа AAD10292); Cry1Bd2 (№ доступа AAM93496); Cry1Be1 (№ доступа AAC32850); Cry1Be2 (№ доступа AAQ52387); Cry1Be3 (№ доступа ACV96720); Cry1Be4 (№ доступа HM070026); Cry1Bf1 (№ доступа CAC50778); Cry1Bf2 (№ доступа AAQ52380); Cry1Bg1 (№ доступа AAO39720); Cry1Bh1 (№ доступа HQ589331); Cry1Bi1 (№ доступа KC156700); Cry1Ca1 (№ доступа CAA30396); Cry1Ca2 (№ доступа CAA31951); Cry1Ca3 (№ доступа AAA22343); Cry1Ca4 (№ доступа CAA01886); Cry1Ca5 (№ доступа CAA65457); Cry1Ca6 [1] (№ доступа AAF37224 ); Cry1Ca7 (№ доступа AAG50438); Cry1Ca8 (№ доступа AAM00264); Cry1Ca9 (№ доступа AAL79362); Cry1Ca10 (№ доступа AAN16462); Cry1Ca11 (№ доступа AAX53094); Cry1Ca12 (№ доступа HM070027); Cry1Ca13 (№ доступа HQ412621); Cry1Ca14 (№ доступа JN651493); Cry1Cb1 (№ доступа M97880); Cry1Cb2 (№ доступа AAG35409); Cry1Cb3 (№ доступа ACD50894 ); Cry1Cb-подобный (№ доступа AAX63901); Cry1Da1 (№ доступа CAA38099); Cry1Da2 (№ доступа I76415); Cry1Da3 (№ доступа HQ439784); Cry1Db1 (№ доступа CAA80234 ); Cry1Db2 (№ доступа AAK48937 ); Cry1Dc1 (№ доступа ABK35074); Cry1Ea1 (№ доступа CAA37933); Cry1Ea2 (№ доступа CAA39609); Cry1Ea3 (№ доступа AAA22345); Cry1Ea4 (№ доступа AAD04732); Cry1Ea5 (№ доступа A15535); Cry1Ea6 (№ доступа AAL50330); Cry1Ea7 (№ доступа AAW72936); Cry1Ea8 (№ доступа ABX11258); Cry1Ea9 (№ доступа HQ439785); Cry1Ea10 (№ доступа ADR00398); Cry1Ea11 (№ доступа JQ652456); Cry1Eb1 (№ доступа AAA22346); Cry1Fa1 (№ доступа AAA22348); Cry1Fa2 (№ доступа AAA22347); Cry1Fa3 (№ доступа HM070028); Cry1Fa4 (№ доступа HM439638); Cry1Fb1 (№ доступа CAA80235); Cry1Fb2 (№ доступа BAA25298); Cry1Fb3 (№ доступа AAF21767); Cry1Fb4 (№ доступа AAC10641); Cry1Fb5 (№ доступа AAO13295); Cry1Fb6 (№ доступа ACD50892); Cry1Fb7 (№ доступа ACD50893); Cry1Ga1 (№ доступа CAA80233); Cry1Ga2 (№ доступа CAA70506); Cry1Gb1 (№ доступа AAD10291); Cry1Gb2 (№ доступа AAO13756); Cry1Gc1 (№ доступа AAQ52381); Cry1Ha1 (№ доступа CAA80236); Cry1Hb1 (№ доступа AAA79694); Cry1Hb2 (№ доступа HQ439786); Cry1H-подобный (№ доступа AAF01213); Cry1Ia1 (№ доступа CAA44633); Cry1Ia2 (№ доступа AAA22354); Cry1Ia3 (№ доступа AAC36999); Cry1Ia4 (№ доступа AAB00958); Cry1Ia5 (№ доступа CAA70124); Cry1Ia6 (№ доступа AAC26910); Cry1Ia7 (№ доступа AAM73516); Cry1Ia8 (№ доступа AAK66742); Cry1Ia9 (№ доступа AAQ08616); Cry1Ia10 (№ доступа AAP86782); Cry1Ia11 (№ доступа CAC85964 ); Cry1Ia12 (№ доступа AAV53390); Cry1Ia13 (№ доступа ABF83202); Cry1Ia14 (№ доступа ACG63871); Cry1Ia15 (№ доступа FJ617445); Cry1Ia16 (№ доступа FJ617448); Cry1Ia17 (№ доступа GU989199); Cry1Ia18 (№ доступа ADK23801); Cry1Ia19 (№ доступа HQ439787); Cry1Ia20 (№ доступа JQ228426); Cry1Ia21 (№ доступа JQ228424); Cry1Ia22 (№ доступа JQ228427); Cry1Ia23 (№ доступа JQ228428); Cry1Ia24 (№ доступа JQ228429); Cry1Ia25 (№ доступа JQ228430); Cry1Ia26 (№ доступа JQ228431); Cry1Ia27 (№ доступа JQ228432); Cry1Ia28 (№ доступа JQ228433); Cry1Ia29 (№ доступа JQ228434); Cry1Ia30 (№ доступа JQ317686); Cry1Ia31 (№ доступа JX944038); Cry1Ia32 (№ доступа JX944039); Cry1Ia33 (№ доступа JX944040); Cry1Ib1 (№ доступа AAA82114); Cry1Ib2 (№ доступа ABW88019); Cry1Ib3 (№ доступа ACD75515); Cry1Ib4 (№ доступа HM051227); Cry1Ib5 (№ доступа HM070028); Cry1Ib6 (№ доступа ADK38579); Cry1Ib7 (№ доступа JN571740); Cry1Ib8 (№ доступа JN675714); Cry1Ib9 (№ доступа JN675715); Cry1Ib10 (№ доступа JN675716); Cry1Ib11 (№ доступа JQ228423); Cry1Ic1 (№ доступа AAC62933); Cry1Ic2 (№ доступа AAE71691); Cry1Id1 (№ доступа AAD44366); Cry1Id2 (№ доступа JQ228422); Cry1Ie1 (№ доступа AAG43526); Cry1Ie2 (№ доступа HM439636); Cry1Ie3 (№ доступа KC156647); Cry1Ie4 (№ доступа KC156681); Cry1If1 (№ доступа AAQ52382); Cry1Ig1 (№ доступа KC156701); Cry1I-подобный (№ доступа AAC31094); Cry1I-подобный (№ доступа ABG88859); Cry1Ja1 (№ доступа AAA22341); Cry1Ja2 (№ доступа HM070030); Cry1Ja3 (№ доступа JQ228425); Cry1Jb1 (№ доступа AAA98959); Cry1Jc1 (№ доступа AAC31092); Cry1Jc2 (№ доступа AAQ52372); Cry1Jd1 (№ доступа CAC50779); Cry1Ka1 (№ доступа AAB00376); Cry1Ka2 (№ доступа HQ439783); Cry1La1 (№ доступа AAS60191); Cry1La2 (№ доступа HM070031); Cry1Ma1 (№ доступа FJ884067); Cry1Ma2 (№ доступа KC156659); Cry1Na1 (№ доступа KC156648); Cry1Nb1 (№ доступа KC156678); Cry1-подобный (№ доступа AAC31091); Cry2Aa1 (№ доступа AAA22335); Cry2Aa2 (№ доступа AAA83516); Cry2Aa3 (№ доступа D86064); Cry2Aa4 (№ доступа AAC04867); Cry2Aa5 (№ доступа CAA10671); Cry2Aa6 (№ доступа CAA10672); Cry2Aa7 (№ доступа CAA10670); Cry2Aa8 (№ доступа AAO13734); Cry2Aa9 (№ доступа AAO13750 ); Cry2Aa10 (№ доступа AAQ04263); Cry2Aa11 (№ доступа AAQ52384); Cry2Aa12 (№ доступа ABI83671); Cry2Aa13 (№ доступа ABL01536); Cry2Aa14 (№ доступа ACF04939); Cry2Aa15 (№ доступа JN426947); Cry2Ab1 (№ доступа AAA22342); Cry2Ab2 (№ доступа CAA39075); Cry2Ab3 (№ доступа AAG36762); Cry2Ab4 (№ доступа AAO13296 ); Cry2Ab5 (№ доступа AAQ04609); Cry2Ab6 (№ доступа AAP59457); Cry2Ab7 (№ доступа AAZ66347); Cry2Ab8 (№ доступа ABC95996); Cry2Ab9 (№ доступа ABC74968); Cry2Ab10 (№ доступа EF157306); Cry2Ab11 (№ доступа CAM84575); Cry2Ab12 (№ доступа ABM21764); Cry2Ab13 (№ доступа ACG76120); Cry2Ab14 (№ доступа ACG76121); Cry2Ab15 (№ доступа HM037126); Cry2Ab16 (№ доступа GQ866914); Cry2Ab17 (№ доступа HQ439789); Cry2Ab18 (№ доступа JN135255); Cry2Ab19 (№ доступа JN135256); Cry2Ab20 (№ доступа JN135257); Cry2Ab21 (№ доступа JN135258); Cry2Ab22 (№ доступа JN135259); Cry2Ab23 (№ доступа JN135260); Cry2Ab24 (№ доступа JN135261); Cry2Ab25 (№ доступа JN415485); Cry2Ab26 (№ доступа JN426946); Cry2Ab27 (№ доступа JN415764); Cry2Ab28 (№ доступа JN651494); Cry2Ac1 (№ доступа CAA40536); Cry2Ac2 (№ доступа AAG35410); Cry2Ac3 (№ доступа AAQ52385); Cry2Ac4 (№ доступа ABC95997); Cry2Ac5 (№ доступа ABC74969); Cry2Ac6 (№ доступа ABC74793); Cry2Ac7 (№ доступа CAL18690); Cry2Ac8 (№ доступа CAM09325); Cry2Ac9 (№ доступа CAM09326); Cry2Ac10 (№ доступа ABN15104); Cry2Ac11 (№ доступа CAM83895); Cry2Ac12 (№ доступа CAM83896); Cry2Ad1 (№ доступа AAF09583); Cry2Ad2 (№ доступа ABC86927); Cry2Ad3 (№ доступа CAK29504); Cry2Ad4 (№ доступа CAM32331); Cry2Ad5 (№ доступа CAO78739 ); Cry2Ae1 (№ доступа AAQ52362); Cry2Af1 (№ доступа ABO30519); Cry2Af2 (№ доступа GQ866915); Cry2Ag1 (№ доступа ACH91610); Cry2Ah1 (№ доступа EU939453); Cry2Ah2 (№ доступа ACL80665); Cry2Ah3 (№ доступа GU073380); Cry2Ah4 (№ доступа KC156702); Cry2Ai1 (№ доступа FJ788388); Cry2Aj (№ доступа ); Cry2Ak1 (№ доступа KC156660); Cry2Ba1 (№ доступа KC156658); Cry3Aa1 (№ доступа AAA22336); Cry3Aa2 (№ доступа AAA22541); Cry3Aa3 (№ доступа CAA68482); Cry3Aa4 (№ доступа AAA22542); Cry3Aa5 (№ доступа AAA50255); Cry3Aa6 (№ доступа AAC43266); Cry3Aa7 (№ доступа CAB41411); Cry3Aa8 (№ доступа AAS79487); Cry3Aa9 (№ доступа AAW05659); Cry3Aa10 (№ доступа AAU29411); Cry3Aa11 (№ доступа AAW82872); Cry3Aa12 (№ доступа ABY49136 ); Cry3Ba1 (№ доступа CAA34983); Cry3Ba2 (№ доступа CAA00645); Cry3Ba3 (№ доступа JQ397327); Cry3Bb1 (№ доступа AAA22334); Cry3Bb2 (№ доступа AAA74198); Cry3Bb3 (№ доступа I15475); Cry3Ca1 (№ доступа CAA42469); Cry4Aa1 (№ доступа CAA68485); Cry4Aa2 (№ доступа BAA00179); Cry4Aa3 (№ доступа CAD30148); Cry4Aa4 (№ доступа AFB18317); Cry4A-подобный (№ доступа AAY96321); Cry4Ba1 (№ доступа CAA30312); Cry4Ba2 (№ доступа CAA30114); Cry4Ba3 (№ доступа AAA22337); Cry4Ba4 (№ доступа BAA00178); Cry4Ba5 (№ доступа CAD30095); Cry4Ba-подобный (№ доступа ABC47686); Cry4Ca1 (№ доступа EU646202); Cry4Cb1 (№ доступа FJ403208); Cry4Cb2 (№ доступа FJ597622); Cry4Cc1 (№ доступа FJ403207); Cry5Aa1 (№ доступа AAA67694); Cry5Ab1 (№ доступа AAA67693); Cry5Ac1 (№ доступа I34543); Cry5Ad1 (№ доступа ABQ82087); Cry5Ba1 (№ доступа AAA68598); Cry5Ba2 (№ доступа ABW88931); Cry5Ba3 (№ доступа AFJ04417); Cry5Ca1 (№ доступа HM461869); Cry5Ca2 (№ доступа ZP_04123426); Cry5Da1 (№ доступа HM461870); Cry5Da2 (№ доступа ZP_04123980); Cry5Ea1 (№ доступа HM485580); Cry5Ea2 (№ доступа ZP_04124038); Cry6Aa1 (№ доступа AAA22357); Cry6Aa2 (№ доступа AAM46849); Cry6Aa3 (№ доступа ABH03377); Cry6Ba1 (№ доступа AAA22358); Cry7Aa1 (№ доступа AAA22351); Cry7Ab1 (№ доступа AAA21120); Cry7Ab2 (№ доступа AAA21121); Cry7Ab3 (№ доступа ABX24522); Cry7Ab4 (№ доступа EU380678); Cry7Ab5 (№ доступа ABX79555); Cry7Ab6 (№ доступа ACI44005); Cry7Ab7 (№ доступа ADB89216); Cry7Ab8 (№ доступа GU145299); Cry7Ab9 (№ доступа ADD92572); Cry7Ba1 (№ доступа ABB70817); Cry7Bb1 (№ доступа KC156653); Cry7Ca1 (№ доступа ABR67863); Cry7Cb1 (№ доступа KC156698); Cry7Da1 (№ доступа ACQ99547); Cry7Da2 (№ доступа HM572236); Cry7Da3 (№ доступа KC156679); Cry7Ea1 (№ доступа HM035086); Cry7Ea2 (№ доступа HM132124); Cry7Ea3 (№ доступа EEM19403); Cry7Fa1 (№ доступа HM035088); Cry7Fa2 (№ доступа EEM19090); Cry7Fb1 (№ доступа HM572235); Cry7Fb2 (№ доступа KC156682); Cry7Ga1 (№ доступа HM572237); Cry7Ga2 (№ доступа KC156669); Cry7Gb1 (№ доступа KC156650); Cry7Gc1 (№ доступа KC156654); Cry7Gd1 (№ доступа KC156697); Cry7Ha1 (№ доступа KC156651); Cry7Ia1 (№ доступа KC156665); Cry7Ja1 (№ доступа KC156671); Cry7Ka1 (№ доступа KC156680); Cry7Kb1 (№ доступа BAM99306); Cry7La1 (№ доступа BAM99307); Cry8Aa1 (№ доступа AAA21117); Cry8Ab1 (№ доступа EU044830); Cry8Ac1 (№ доступа KC156662); Cry8Ad1 (№ доступа KC156684); Cry8Ba1 (№ доступа AAA21118); Cry8Bb1 (№ доступа CAD57542); Cry8Bc1 (№ доступа CAD57543); Cry8Ca1 (№ доступа AAA21119); Cry8Ca2 (№ доступа AAR98783); Cry8Ca3 (№ доступа EU625349); Cry8Ca4 (№ доступа ADB54826); Cry8Da1 (№ доступа BAC07226); Cry8Da2 (№ доступа BD133574); Cry8Da3 (№ доступа BD133575); Cry8Db1 (№ доступа BAF93483); Cry8Ea1 (№ доступа AAQ73470); Cry8Ea2 (№ доступа EU047597); Cry8Ea3 (№ доступа KC855216); Cry8Fa1 (№ доступа AAT48690); Cry8Fa2 (№ доступа HQ174208); Cry8Fa3 (№ доступа AFH78109); Cry8Ga1 (№ доступа AAT46073); Cry8Ga2 (№ доступа ABC42043); Cry8Ga3 (№ доступа FJ198072); Cry8Ha1 (№ доступа AAW81032); Cry8Ia1 (№ доступа EU381044); Cry8Ia2 (№ доступа GU073381); Cry8Ia3 (№ доступа HM044664); Cry8Ia4 (№ доступа KC156674); Cry8Ib1 (№ доступа GU325772); Cry8Ib2 (№ доступа KC156677); Cry8Ja1 (№ доступа EU625348); Cry8Ka1 (№ доступа FJ422558); Cry8Ka2 (№ доступа ACN87262); Cry8Kb1 (№ доступа HM123758); Cry8Kb2 (№ доступа KC156675); Cry8La1 (№ доступа GU325771); Cry8Ma1 (№ доступа HM044665); Cry8Ma2 (№ доступа EEM86551); Cry8Ma3 (№ доступа HM210574); Cry8Na1 (№ доступа HM640939); Cry8Pa1 (№ доступа HQ388415); Cry8Qa1 (№ доступа HQ441166); Cry8Qa2 (№ доступа KC152468); Cry8Ra1 (№ доступа AFP87548); Cry8Sa1 (№ доступа JQ740599); Cry8Ta1 (№ доступа KC156673); Cry8-подобный (№ доступа FJ770571); Cry8-подобный (№ доступа ABS53003); Cry9Aa1 (№ доступа CAA41122); Cry9Aa2 (№ доступа CAA41425); Cry9Aa3 (№ доступа GQ249293); Cry9Aa4 (№ доступа GQ249294); Cry9Aa5 (№ доступа JX174110); Cry9Aa подобный (№ доступа AAQ52376); Cry9Ba1 (№ доступа CAA52927); Cry9Ba2 (№ доступа GU299522); Cry9Bb1 (№ доступа AAV28716); Cry9Ca1 (№ доступа CAA85764); Cry9Ca2 (№ доступа AAQ52375); Cry9Da1 (№ доступа BAA19948); Cry9Da2 (№ доступа AAB97923); Cry9Da3 (№ доступа GQ249293); Cry9Da4 (№ доступа GQ249297); Cry9Db1 (№ доступа AAX78439); Cry9Dc1 (№ доступа KC156683); Cry9Ea1 (№ доступа BAA34908); Cry9Ea2 (№ доступа AAO12908); Cry9Ea3 (№ доступа ABM21765); Cry9Ea4 (№ доступа ACE88267); Cry9Ea5 (№ доступа ACF04743); Cry9Ea6 (№ доступа ACG63872 ); Cry9Ea7 (№ доступа FJ380927); Cry9Ea8 (№ доступа GQ249292); Cry9Ea9 (№ доступа JN651495); Cry9Eb1 (№ доступа CAC50780); Cry9Eb2 (№ доступа GQ249298); Cry9Eb3 (№ доступа KC156646); Cry9Ec1 (№ доступа AAC63366); Cry9Ed1 (№ доступа AAX78440); Cry9Ee1 (№ доступа GQ249296); Cry9Ee2 (№ доступа KC156664); Cry9Fa1 (№ доступа KC156692); Cry9Ga1 (№ доступа KC156699); Cry9-подобный (№ доступа AAC63366); Cry10Aa1 (№ доступа AAA22614); Cry10Aa2 (№ доступа E00614); Cry10Aa3 (№ доступа CAD30098); Cry10Aa4 (№ доступа AFB18318); Cry10A-подобный (№ доступа DQ167578); Cry11Aa1 (№ доступа AAA22352); Cry11Aa2 (№ доступа AAA22611); Cry11Aa3 (№ доступа CAD30081); Cry11Aa4 (№ доступа AFB18319); Cry11Aa-подобный (№ доступа DQ166531); Cry11Ba1 (№ доступа CAA60504); Cry11Bb1 (№ доступа AAC97162); Cry11Bb2 (№ доступа HM068615); Cry12Aa1 (№ доступа AAA22355); Cry13Aa1 (№ доступа AAA22356); Cry14Aa1 (№ доступа AAA21516); Cry14Ab1 (№ доступа KC156652); Cry15Aa1 (№ доступа AAA22333); Cry16Aa1 (№ доступа CAA63860); Cry17Aa1 (№ доступа CAA67841); Cry18Aa1 (№ доступа CAA67506); Cry18Ba1 (№ доступа AAF89667); Cry18Ca1 (№ доступа AAF89668); Cry19Aa1 (№ доступа CAA68875); Cry19Ba1 (№ доступа BAA32397); Cry19Ca1 (№ доступа AFM37572); Cry20Aa1 (№ доступа AAB93476); Cry20Ba1 (№ доступа ACS93601); Cry20Ba2 (№ доступа KC156694); Cry20-подобный (№ доступа GQ144333); Cry21Aa1 (№ доступа I32932); Cry21Aa2 (№ доступа I66477); Cry21Ba1 (№ доступа BAC06484); Cry21Ca1 (№ доступа JF521577); Cry21Ca2 (№ доступа KC156687); Cry21Da1 (№ доступа JF521578); Cry22Aa1 (№ доступа I34547); Cry22Aa2 (№ доступа CAD43579); Cry22Aa3 (№ доступа ACD93211); Cry22Ab1 (№ доступа AAK50456); Cry22Ab2 (№ доступа CAD43577); Cry22Ba1 (№ доступа CAD43578); Cry22Bb1 (№ доступа KC156672); Cry23Aa1 (№ доступа AAF76375); Cry24Aa1 (№ доступа AAC61891); Cry24Ba1 (№ доступа BAD32657); Cry24Ca1 (№ доступа CAJ43600); Cry25Aa1 (№ доступа AAC61892); Cry26Aa1 (№ доступа AAD25075); Cry27Aa1 (№ доступа BAA82796); Cry28Aa1 (№ доступа AAD24189); Cry28Aa2 (№ доступа AAG00235); Cry29Aa1 (№ доступа CAC80985); Cry30Aa1 (№ доступа CAC80986); Cry30Ba1 (№ доступа BAD00052); Cry30Ca1 (№ доступа BAD67157); Cry30Ca2 (№ доступа ACU24781); Cry30Da1 (№ доступа EF095955); Cry30Db1 (№ доступа BAE80088); Cry30Ea1 (№ доступа ACC95445); Cry30Ea2 (№ доступа FJ499389); Cry30Fa1 (№ доступа ACI22625 ); Cry30Ga1 (№ доступа ACG60020); Cry30Ga2 (№ доступа HQ638217); Cry31Aa1 (№ доступа BAB11757); Cry31Aa2 (№ доступа AAL87458); Cry31Aa3 (№ доступа BAE79808); Cry31Aa4 (№ доступа BAF32571); Cry31Aa5 (№ доступа BAF32572); Cry31Aa6 (№ доступа BAI44026); Cry31Ab1 (№ доступа BAE79809); Cry31Ab2 (№ доступа BAF32570); Cry31Ac1 (№ доступа BAF34368); Cry31Ac2 (№ доступа AB731600); Cry31Ad1 (№ доступа BAI44022); Cry32Aa1 (№ доступа AAG36711); Cry32Aa2 (№ доступа GU063849); Cry32Ab1 (№ доступа GU063850); Cry32Ba1 (№ доступа BAB78601); Cry32Ca1 (№ доступа BAB78602); Cry32Cb1 (№ доступа KC156708); Cry32Da1 (№ доступа BAB78603); Cry32Ea1 (№ доступа GU324274); Cry32Ea2 (№ доступа KC156686); Cry32Eb1 (№ доступа KC156663); Cry32Fa1 (№ доступа KC156656); Cry32Ga1 (№ доступа KC156657); Cry32Ha1 (№ доступа KC156661); Cry32Hb1 (№ доступа KC156666); Cry32Ia1 (№ доступа KC156667); Cry32Ja1 (№ доступа KC156685); Cry32Ka1 (№ доступа KC156688); Cry32La1 (№ доступа KC156689); Cry32Ma1 (№ доступа KC156690); Cry32Mb1 (№ доступа KC156704); Cry32Na1 (№ доступа KC156691); Cry32Oa1 (№ доступа KC156703); Cry32Pa1 (№ доступа KC156705); Cry32Qa1 (№ доступа KC156706); Cry32Ra1 (№ доступа KC156707); Cry32Sa1 (№ доступа KC156709); Cry32Ta1 (№ доступа KC156710); Cry32Ua1 (№ доступа KC156655); Cry33Aa1 (№ доступа AAL26871); Cry34Aa1 (№ доступа AAG50341); Cry34Aa2 (№ доступа AAK64560); Cry34Aa3 (№ доступа AAT29032); Cry34Aa4 (№ доступа AAT29030); Cry34Ab1 (№ доступа AAG41671); Cry34Ac1 (№ доступа AAG50118); Cry34Ac2 (№ доступа AAK64562); Cry34Ac3 (№ доступа AAT29029); Cry34Ba1 (№ доступа AAK64565); Cry34Ba2 (№ доступа AAT29033); Cry34Ba3 (№ доступа AAT29031); Cry35Aa1 (№ доступа AAG50342); Cry35Aa2 (№ доступа AAK64561); Cry35Aa3 (№ доступа AAT29028); Cry35Aa4 (№ доступа AAT29025); Cry35Ab1 (№ доступа AAG41672); Cry35Ab2 (№ доступа AAK64563); Cry35Ab3 (№ доступа AY536891); Cry35Ac1 (№ доступа AAG50117); Cry35Ba1 (№ доступа AAK64566); Cry35Ba2 (№ доступа AAT29027); Cry35Ba3 (№ доступа AAT29026); Cry36Aa1 (№ доступа AAK64558); Cry37Aa1 (№ доступа AAF76376 ); Cry38Aa1 (№ доступа AAK64559); Cry39Aa1 (№ доступа BAB72016); Cry40Aa1 (№ доступа BAB72018); Cry40Ba1 (№ доступа BAC77648); Cry40Ca1 (№ доступа EU381045); Cry40Da1 (№ доступа ACF15199); Cry41Aa1 (№ доступа BAD35157); Cry41Ab1 (№ доступа BAD35163); Cry41Ba1 (№ доступа HM461871); Cry41Ba2 (№ доступа ZP_04099652); Cry42Aa1 (№ доступа BAD35166); Cry43Aa1 (№ доступа BAD15301); Cry43Aa2 (№ доступа BAD95474 ); Cry43Ba1 (№ доступа BAD15303); Cry43Ca1 (№ доступа KC156676); Cry43Cb1 (№ доступа KC156695); Cry43Cc1 (№ доступа KC156696); Cry43-подобный (№ доступа BAD15305); Cry44Aa (№ доступа BAD08532); Cry45Aa (№ доступа BAD22577); Cry46Aa (№ доступа BAC79010); Cry46Aa2 (№ доступа BAG68906); Cry46Ab (№ доступа BAD35170); Cry47Aa (№ доступа AAY24695); Cry48Aa (№ доступа CAJ18351); Cry48Aa2 (№ доступа CAJ86545); Cry48Aa3 (№ доступа CAJ86546 ); Cry48Ab (№ доступа CAJ86548); Cry48Ab2 (№ доступа CAJ86549); Cry49Aa (№ доступа CAH56541); Cry49Aa2 (№ доступа CAJ86541); Cry49Aa3 (№ доступа CAJ86543); Cry49Aa4 (№ доступа CAJ86544); Cry49Ab1 (№ доступа CAJ86542); Cry50Aa1 (№ доступа BAE86999); Cry50Ba1 (№ доступа GU446675); Cry50Ba2 (№ доступа GU446676); Cry51Aa1 (№ доступа ABI14444); Cry51Aa2 (№ доступа GU570697); Cry52Aa1 (№ доступа EF613489); Cry52Ba1 (№ доступа FJ361760); Cry53Aa1 (№ доступа EF633476); Cry53Ab1 (№ доступа FJ361759); Cry54Aa1 (№ доступа ACA52194); Cry54Aa2 (№ доступа GQ140349); Cry54Ba1 (№ доступа GU446677); Cry55Aa1 (№ доступа ABW88932); Cry54Ab1 (№ доступа JQ916908); Cry55Aa2 (№ доступа AAE33526); Cry56Aa1 (№ доступа ACU57499); Cry56Aa2 (№ доступа GQ483512); Cry56Aa3 (№ доступа JX025567); Cry57Aa1 (№ доступа ANC87261); Cry58Aa1 (№ доступа ANC87260); Cry59Ba1 (№ доступа JN790647); Cry59Aa1 (№ доступа ACR43758); Cry60Aa1 (№ доступа ACU24782); Cry60Aa2 (№ доступа EAO57254); Cry60Aa3 (№ доступа EEM99278); Cry60Ba1 (№ доступа GU810818); Cry60Ba2 (№ доступа EAO57253); Cry60Ba3 (№ доступа EEM99279); Cry61Aa1 (№ доступа HM035087); Cry61Aa2 (№ доступа HM132125); Cry61Aa3 (№ доступа EEM19308); Cry62Aa1 (№ доступа HM054509); Cry63Aa1 (№ доступа BAI44028); Cry64Aa1 (№ доступа BAJ05397); Cry65Aa1 (№ доступа HM461868); Cry65Aa2 (№ доступа ZP_04123838); Cry66Aa1 (№ доступа HM485581); Cry66Aa2 (№ доступа ZP_04099945); Cry67Aa1 (№ доступа HM485582); Cry67Aa2 (№ доступа ZP_04148882); Cry68Aa1 (№ доступа HQ113114); Cry69Aa1 (№ доступа HQ401006); Cry69Aa2 (№ доступа JQ821388); Cry69Ab1 (№ доступа JN209957); Cry70Aa1 (№ доступа JN646781); Cry70Ba1 (№ доступа ADO51070); Cry70Bb1 (№ доступа EEL67276); Cry71Aa1 (№ доступа JX025568); Cry72Aa1 (№ доступа JX025569); Cyt1Aa (№ доступа GenBank X03182); Cyt1Ab (№ доступа GenBank X98793); Cyt1B (№ доступа GenBank U37196); Cyt2A (№ доступа GenBank Z14147); и Cyt2B (№ доступа GenBank U52043).
Примеры δ-эндотоксинов также включают без ограничения белки Cry1A из патентов США №№ 5880275 и 7858849; токсин DIG-3 или DIG-11 (варианты с N-терминальной делецией α-спирали 1 и/или α-спирали 2 белков cry, таких как Cry1A, Cry3A) из патентов США №№ 8304604, 8304605 и 8476226; Cry1B из публикации заявки на патент США с серийным номером 10/525318; Cry1C из патента США № 6033874; Cry1F из патентов США №№ 5188960 и 6218188; химеры Cry1A/F из патентов США №№ 7070982; 6962705 и 6713063); белок Cry2, такой как белок Cry2Ab из патента США № 7064249); белок Cry3A, в том числе без ограничения созданный с помощью генной инженерии гибридный инсектицидный белок (eHIP), полученный путем слияния уникальных комбинаций вариабельных участков и консервативных блоков по меньшей мере двух различных белков Cry (публикация заявки на патент США № 2010/0017914); белок Cry4; белок Cry5; белок Cry6; белки Cry8 из патентов США №№ 7329736, 7449552, 7803943, 7476781, 7105332, 7378499 и 7462760; белок Cry9, например, представители семейств Cry9A, Cry9B, Cry9C, Cry9D, Cry9E и Cry9F; белок Cry15 из Naimov, et al., (2008) Applied and Environmental Microbiology, 74:7145-7151; Cry22, белок Cry34Ab1 из патентов США №№ 6127180, 6624145 и 6340593; белок CryET33 и cryET34 из патентов США №№ 6248535, 6326351, 6399330, 6949626, 7385107 и 7504229; гомологи CryET33 и CryET34 из публикации заявок на патент США №№ 2006/0191034, 2012/0278954 и публикации согласно PCT№ WO 2012/139004; белок Cry35Ab1 из патентов США №№ 6083499, 6548291 и 6340593; белок Cry46, белок Cry 51, бинарный токсин Cry; TIC901 и родственный токсин; TIC807 из публикации заявки на патент США № 2008/0295207; ET29, ET37, TIC809, TIC810, TIC812, TIC127, TIC128 из заявки согласно PCT US 2006/033867; AXMI-027, AXMI-036 и AXMI-038 из патента США № 8236757; AXMI-031, AXMI-039, AXMI-040, AXMI-049 из патента США № 7923602; AXMI-018, AXMI-020 и AXMI-021 из WO 2006/083891; AXMI-010 из WO 2005/038032; AXMI-003 из WO 2005/021585; AXMI-008 из публикации заявки на патент США № 2004/0250311; AXMI-006 из публикации заявки на патент США №2004/0216186; AXMI-007 из публикации заявки на патент США №2004/0210965; AXMI-009 из публикации заявки на патент США № 2004/0210964; AXMI-014 из публикации заявки на патент США № 2004/0197917; AXMI-004 из публикации заявки на патент США № 2004/0197916; AXMI-028 и AXMI-029 из WO 2006/119457; AXMI-007, AXMI-008, AXMI-0080rf2, AXMI-009, AXMI-014 и AXMI-004 из WO 2004/074462; AXMI-150 из патента США № 8084416; AXMI-205 из публикации заявки на патент США № 2011/0023184; AXMI-011, AXMI-012, AXMI-013, AXMI-015, AXMI-019, AXMI-044, AXMI-037, AXMI-043, AXMI-033, AXMI-034, AXMI-022, AXMI-023, AXMI-041, AXMI-063 и AXMI-064 из публикации заявки на патент США № 2011/0263488; AXMI-R1 и родственные белки из публикации заявки на патент США № 2010/0197592; AXMI221Z, AXMI222z, AXMI223z, AXMI224z и AXMI225z из WO 2011/103248; AXMI218, AXMI219, AXMI220, AXMI226, AXMI227, AXMI228, AXMI229, AXMI230 и AXMI231 из WO 2011/103247; AXMI-115, AXMI-113, AXMI-005, AXMI-163 и AXMI-184 из патента США № 8334431; AXMI-001, AXMI-002, AXMI-030, AXMI-035 и AXMI-045 из публикации заявки на патент США № 2010/0298211; AXMI-066 и AXMI-076 из публикации заявки на патент США № 2009/0144852; AXMI128, AXMI130, AXMI131, AXMI133, AXMI140, AXMI141, AXMI142, AXMI143, AXMI144, AXMI146, AXMI148, AXMI149, AXMI152, AXMI153, AXMI154, AXMI155, AXMI156, AXMI157, AXMI158, AXMI162, AXMI165, AXMI166, AXMI167, AXMI168, AXMI169, AXMI170, AXMI171, AXMI172, AXMI173, AXMI174, AXMI175, AXMI176, AXMI177, AXMI178, AXMI179, AXMI180, AXMI181, AXMI182, AXMI185, AXMI186, AXMI187, AXMI188, AXMI189 из патента США № 8318900; AXMI079, AXMI080, AXMI081, AXMI082, AXMI091, AXMI092, AXMI096, AXMI097, AXMI098, AXMI099, AXMI100, AXMI101, AXMI102, AXMI103, AXMI104, AXMI107, AXMI108, AXMI109, AXMI110, AXMI111, AXMI112, AXMI114, AXMI116, AXMI117, AXMI118, AXMI119, AXMI120, AXMI121, AXMI122, AXMI123, AXMI124, AXMI1257, AXMI1268, AXMI127, AXMI129, AXMI164, AXMI151, AXMI161, AXMI183, AXMI132, AXMI138, AXMI137 из публикации заявки на патент США № 2010/0005543, белки cry, такие как Cry1A и Cry3A с модифицированными сайтами протеолитического расщепления из патента США № 8319019; белок-токсин Cry1Ac, Cry2Aa и Cry1Ca из штамма VBTS 2528 Bacillus thuringiensis из публикации заявки на патент США № 2011/0064710. Другие белки Cry хорошо известны специалисту в данной области (см. Crickmore, et al., "Bacillus thuringiensis toxin nomenclature" (2011), на сайте lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/, доступ к которому можно получить во всемирной сети Интернет с применением префикса "www"). Инсектицидная активность белков Cry хорошо известна специалисту в данной области (обзор см. у van Frannkenhuyzen, (2009) J. Invert. Path. 101:1-16). Применение белков Cry в качестве признаков трансгенного растения хорошо известно специалисту в данной области, и трансгенные растения с Cry, в том числе без ограничения растения, экспрессирующие Cry1Ac, Cry1Ac+Cry2Ab, Cry1Ab, Cry1A.105, Cry1F, Cry1Fa2, Cry1F+Cry1Ac, Cry2Ab, Cry3A, mCry3A, Cry3Bb1, Cry34Ab1, Cry35Ab1, Vip3A, mCry3A, Cry9c и CBI-Bt, были разрешены контролирующими органами (см. Sanahuja, (2011) Plant Biotech Journal 9:283-300 и CERA. (2010) GM Crop Database Center for Environmental Risk Assessment (CERA), ILSI Research Foundation, Washington D.C. на сайте cera-gmc.org/index.php?action=gm_crop_database, доступ к которым можно получить во всемирной сети Интернет с применением префикса ʺwwwʺ). В растениях также может экспрессироваться два и более пестицидных белков, хорошо известных специалисту в данной области, например, Vip3Ab и Cry1Fa (US2012/0317682); Cry1BE и Cry1F (US2012/0311746); Cry1CA и Cry1AB (US2012/0311745); Cry1F и CryCa (US2012/0317681); Cry1DA и Cry1BE (US2012/0331590); Cry1DA и Cry1Fa (US2012/0331589); Cry1AB и Cry1BE (US2012/0324606); Cry1Fa и Cry2Aa, и Cry1I и Cry1E (US2012/0324605); Cry34Ab/35Ab и Cry6Aa (US20130167269); Cry34Ab/VCry35Ab и Cry3Aa (US20130167268), а также Cry3A и Cry1Ab или Vip3Aa (US20130116170). Пестицидные белки также включают инсектицидные липазы, в том числе липидацильные гидролазы из патента США № 7491869 и холестериноксидазы, например из Streptomyces (Purcell et al. (1993) Biochem Biophys Res Commun 15:1406-1413). Пестицидные белки также включают токсины VIP (инсектицидные белки вегетативной фазы) из патентов США №№ 5877012, 6107279, 6137033, 7244820, 7615686 и 8237020 и т. п. Другие белки VIP хорошо известны специалисту в данной области (см. lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/vip.html, доступ к которому можно получить во всемирной сети Интернет с применением префикса "www"). Пестицидные белки также включают белки токсинового комплекса (TC), которые можно получить из организмов, таких как Xenorhabdus, Photorhabdus и Paenibacillus (см. патенты США №№ 7491698 и 8084418). Некоторые TC-белки обладают ʺсамостоятельнойʺ инсектицидной активностью, а другие TC-белки повышают активность самостоятельных токсинов, производимых тем же данным организмом. Токсичность "самостоятельного" TC-белка (от Photorhabdus, Xenorhabdus или Paenibacillus, например) может повышаться при помощи одного или нескольких TC-белков-"усилителей", полученных от организма-источника из другого рода. Существуют три основных типа TC-белков. Как изложено в данном документе, белки класса A ("белок A") представляют собой самостоятельные токсины. Белки класса B ("белок B") и белки класса C ("белок C") усиливают токсичность белков класса A. Примеры белков класса A представляют собой TcbA, TcdA, XptA1 и XptA2. Примеры белков класса B представляют собой TcaC, TcdB, XptB1Xb и XptC1Wi. Примеры белков класса C представляют собой TccC, XptC1Xb и XptB1Wi. Пестицидные белки также включают белки яда пауков, змей и скорпионов. Примеры пептидов яда пауков включают без ограничения пептиды ликотоксина-1 и его мутантные формы (патент США № 8334366).
В некоторых вариантах осуществления полипептид PtIP-83 включает аминокислотные последовательности, выведенные из последовательностей нуклеиновой кислоты полной длины, раскрытых в данном документе, и аминокислотные последовательности, которые короче, чем последовательности полной длины, либо полученные в результате применения альтернативного сайта инициации, расположенного ниже, либо в результате процессинга, дающего более короткий белок с пестицидной активностью. Процессинг может происходить в организме, в котором экспрессируется белок, или во вредителе после поглощения белка.
Таким образом, в данном документе представлены новые выделенные или рекомбинантные последовательности нуклеиновой кислоты, которые обеспечивают пестицидную активность. Также представлены аминокислотные последовательности полипептидов PtIP-83. Белок, полученный в результате трансляции генов этих полипептидов PtIP-83 в клетках, обеспечивает возможность контроля или уничтожения вредителей, которые поглощают их.
Молекулы нуклеиновой кислоты, а также их варианты и фрагменты
Один аспект относится к выделенным или рекомбинантным молекулам нуклеиновой кислоты, содержащим последовательности нуклеиновой кислоты, кодирующие полипептиды PtIP-83 или их биологически активные части, а также к молекулам нуклеиновой кислоты, подходящим для применения в качестве гибридизационных зондов для идентификации молекул нуклеиновой кислоты, кодирующих белки с участками гомологии последовательностей. Применяемый в данном документе термин "молекула нуклеиновой кислоты" относится к молекулам ДНК (например, рекомбинантной ДНК, кДНК, геномной ДНК, пластидной ДНК, митохондриальной ДНК) или молекулам РНК (например, мРНК) и аналогам ДНК или РНК, полученным с применением аналогов нуклеотидов. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но предпочтительно представляет собой двухцепочечную ДНК.
"Выделенная" молекула нуклеиновой кислоты (или ДНК) применяется в данном документе для обозначения последовательности нуклеиновой кислоты (или ДНК), которая больше не находится в своей естественной среде, например, находится in vitro. "Рекомбинантная" молекула нуклеиновой кислоты (или ДНК) применяется в данном документе для обозначения последовательности нуклеиновой кислоты (или ДНК), которая находится в рекомбинантной бактериальной или растительной клетке-хозяине. В некоторых вариантах осуществления "выделенная" или "рекомбинантная" нуклеиновая кислота не содержит последовательности (предпочтительно, последовательности, кодирующие белок), которые в естественных условиях фланкируют нуклеиновую кислоту (т.е. последовательности, расположенные на 5′- и 3′-концах нуклеиновой кислоты) в геномной ДНК организма, из которого получена нуклеиновая кислота. Для целей настоящего раскрытия "выделенные" или "рекомбинантные", при применении для обозначения молекул нуклеиновой кислоты, исключает выделенные хромосомы. Например, в различных вариантах осуществления рекомбинантная молекула нуклеиновой кислоты, кодирующая полипептид PtIP-83, может содержать менее приблизительно 5 т. н., 4 т. н., 3 т. н., 2 т. н., 1 т. н., 0,5 т. н. или 0,1 т. н. последовательностей нуклеиновой кислоты, которые в естественных условиях фланкируют молекулу нуклеиновой кислоты в геномной ДНК клетки, из которой получена нуклеиновая кислота.
В некоторых вариантах осуществления выделенная молекула нуклеиновой кислоты, кодирующая полипептиды PtIP-83, имеет одно или несколько изменений в последовательности нуклеиновой кислоты относительно нативной или геномной последовательности нуклеиновой кислоты. В некоторых вариантах осуществления изменение в нативной или геномной последовательности нуклеиновой кислоты включает без ограничения изменения в последовательности нуклеиновой кислоты вследствие вырожденности генетического кода; изменения в последовательности нуклеиновой кислоты вследствие аминокислотной замены, вставки, делеции и/или добавления относительно нативной или геномной последовательности; удаление одного или нескольких интронов; делецию одного или нескольких регуляторных участков, расположенных выше или ниже; и делецию 5ʹ- и/или 3ʹ-нетранслируемого участка, ассоциированного с геномной последовательностью нуклеиновой кислоты. В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая полипептид PtIP-83, представляет собой последовательность, отличающуюся от геномной.
Предполагается ряд полинуклеотидов, которые кодируют полипептиды PtIP-83 или родственные белки. Такие полинуклеотиды применимы для получения полипептидов PtIP-83 в клетках-хозяевах, если они функционально связаны с подходящим промотором, терминатором транскрипции и/или последовательностями полиаденилирования. Такие полинуклеотиды также применимы в качестве зондов для выделения гомологичных или фактически гомологичных полинуклеотидов, которые кодируют полипептиды PtIP-83 или родственные белки.
Полинуклеотиды, кодирующие полипептиды PtIP-83
Одним источником полинуклеотидов, которые кодируют полипептиды PtIP-83 или родственные белки, является вид папоротников или других примитивных растений, который содержит полинуклеотид PtIP-83 под SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 717, SEQ ID NO: 738, SEQ ID NO: 739, SEQ ID NO: 740, SEQ ID NO: 741, SEQ ID NO: 742, SEQ ID NO: 743, SEQ ID NO: 744, SEQ ID NO: 745, SEQ ID NO: 746, SEQ ID NO: 747, SEQ ID NO: 748, SEQ ID NO: 749, SEQ ID NO: 750, SEQ ID NO: 751, SEQ ID NO: 752 или SEQ ID NO: 753, кодирующий полипептид PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769. Полинуклеотиды под SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 717, SEQ ID NO: 738, SEQ ID NO: 739, SEQ ID NO: 740, SEQ ID NO: 741, SEQ ID NO: 742, SEQ ID NO: 743, SEQ ID NO: 744, SEQ ID NO: 745, SEQ ID NO: 746, SEQ ID NO: 747, SEQ ID NO: 748, SEQ ID NO: 749, SEQ ID NO: 750, SEQ ID NO: 751, SEQ ID NO: 752 и SEQ ID NO: 753 можно применять для экспрессии полипептидов PtIP-83 в бактериях-хозяевах, которые включают без ограничения бактериальные клетки-хозяева Agrobacterium, Bacillus, Escherichia, Salmonella, Pseudomonas и Rhizobium. Полинуклеотиды также применимы в качестве зондов для выделения гомологичных или фактически гомологичных полинуклеотидов, которые кодируют полипептиды PtIP-83 или родственные белки. Такие зонды можно применять для идентификации гомологичных или фактически гомологичных полинуклеотидов, полученных из видов Pteridophyta.
Полинуклеотиды, которые кодируют полипептиды PtIP-83, также можно синтезировать de novo исходя из последовательности полипептида PtIP-83. Полинуклеотидную последовательность гена можно вывести на основании последовательности полипептида PtIP-83 с применением генетического кода. Компьютерные программы, такие как ʺBackTranslateʺ (GCG™ Package, Acclerys, Inc., Сан-Диего, Калифорния), можно применять для перевода пептидной последовательности в соответствующую нуклеотидную последовательность, кодирующую пептид. Примеры последовательностей полипептида PtIP-83, которые можно применять для получения соответствующих нуклеотидных кодирующих последовательностей, включают без ограничения полипептиды PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, и SEQ ID NO: 769. Кроме того, можно сконструировать синтетические последовательности полинуклеотида PtIP-83 согласно настоящему раскрытию таким образом, что они будут экспрессироваться в растениях. В патенте США № 5500365 описан способ синтеза генов растения для повышения уровня экспрессии белка, кодируемого синтезированным геном. Этот способ относится к модификации последовательностей структурных генов экзогенного трансгена, что приводит к их более эффективной транскрипции, процессингу, трансляции и экспрессии в растении. Характерные особенности генов, которые хорошо экспрессируются в растениях, предусматривают удаление последовательностей, которые могут вызывать нежелательный сплайсинг интронов или полиаденилирование в пределах кодирующего участка генного транскрипта, при этом в существенной мере сохраняется аминокислотная последовательность токсичной части инсектицидного белка. Аналогичный способ для получения повышенной экспрессии трансгенов в однодольных растениях раскрыт в патенте США № 5689052.
В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая полипептид PtIP-83, представляет собой полинуклеотид с последовательностью, изложенной под SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 717, SEQ ID NO: 738, SEQ ID NO: 739, SEQ ID NO: 740, SEQ ID NO: 741, SEQ ID NO: 742, SEQ ID NO: 743, SEQ ID NO: 744, SEQ ID NO: 745, SEQ ID NO: 746, SEQ ID NO: 747, SEQ ID NO: 748, SEQ ID NO: 749, SEQ ID NO: 750, SEQ ID NO: 751, SEQ ID NO: 752, SEQ ID NO: 753, и его варианты, фрагменты и комплементарные ему последовательности. "Комплементарная последовательность" применяется в данном документе для обозначения последовательности нуклеиновой кислоты, которая в достаточной степени комплементарна данной последовательности нуклеиновой кислоты, так что она может гибридизоваться с данной последовательностью нуклеиновой кислоты с образованием, тем самым, стабильного дуплекса. "Варианты полинуклеотидной последовательности" применяется в данном документе для обозначения последовательности нуклеиновой кислоты, которая без учета отличий, связанных с вырожденностью генетического кода, кодирует тот же полипептид.
В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая полипептид PtIP-83, представляет собой последовательность нуклеиновой кислоты, отличающуюся от геномной. Применяемые в данном документе "последовательность нуклеиновой кислоты, отличающаяся от геномной", или "молекула нуклеиновой кислоты, отличающаяся от геномной", или "полинуклеотид, отличающийся от геномного" относятся к молекуле нуклеиновой кислоты, которая имеет одно или несколько изменений в последовательности нуклеиновой кислоты по сравнению с нативной или геномной последовательностью нуклеиновой кислоты. В некоторых вариантах осуществления изменение по отношению к нативной или геномной молекуле нуклеиновой кислоты включает без ограничения: изменения в последовательности нуклеиновой кислоты, обусловленные вырожденностью генетического кода; оптимизацию кодонов последовательности нуклеиновой кислоты для экспрессии в растениях; изменения в последовательности нуклеиновой кислоты для введения по меньшей мере одной аминокислотной замены, вставки, делеции и/или добавления по сравнению с нативной или геномной последовательностью; удаление одного или нескольких интронов, ассоциированных с геномной последовательностью нуклеиновой кислоты; вставку одного или нескольких гетерологичных интронов; делецию одного или нескольких регуляторных участков, расположенных выше или ниже, которые ассоциированы с геномной последовательностью нуклеиновой кислоты; вставку одного или нескольких гетерологичных регуляторных участков, расположенных выше или ниже; делецию 5ʹ- и/или 3ʹ-нетранслируемого участка, ассоциированного с геномной последовательностью нуклеиновой кислоты; вставку гетерологичного 5ʹ- и/или 3ʹ-нетранслируемого участка и модификацию сайта полиаденилирования. В некоторых вариантах осуществления молекула нуклеиновой кислоты, отличающаяся от геномной, представляет собой кДНК. В некоторых вариантах осуществления молекула нуклеиновой кислоты, отличающаяся от геномной, представляет собой синтетическую последовательность нуклеиновой кислоты.
В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая полипептид PtIP-83, представляет собой полинуклеотид, отличающийся от геномного, с нуклеотидной последовательностью, которая по меньшей мере на 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична последовательности нуклеиновой кислоты под SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 717, SEQ ID NO: 738, SEQ ID NO: 739, SEQ ID NO: 740, SEQ ID NO: 741, SEQ ID NO: 742, SEQ ID NO: 743, SEQ ID NO: 744, SEQ ID NO: 745, SEQ ID NO: 746, SEQ ID NO: 747, SEQ ID NO: 748, SEQ ID NO: 749, SEQ ID NO: 750, SEQ ID NO: 751, SEQ ID NO: 752 или SEQ ID NO: 753, где полипептид PtIP-83 обладает инсектицидной активностью.
В некоторых вариантах осуществления полинуклеотид, отличающийся от геномного, не представляет собой последовательность нуклеиновой кислоты под SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 717, SEQ ID NO: 738, SEQ ID NO: 739, SEQ ID NO: 740, SEQ ID NO: 741, SEQ ID NO: 742, SEQ ID NO: 743, SEQ ID NO: 744, SEQ ID NO: 745, SEQ ID NO: 746, SEQ ID NO: 747, SEQ ID NO: 748, SEQ ID NO: 749, SEQ ID NO: 750, SEQ ID NO: 751, SEQ ID NO: 752 или SEQ ID NO: 753.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий аминокислотную последовательность, которая по меньшей мере на 40%, 45%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769, где полипептид PtIP-83 обладает инсектицидной активностью.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий аминокислотную последовательность под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769 с 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70 или более аминокислотными заменами по сравнению с нативной аминокислотой в соответствующем положении SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий аминокислотную последовательность под любым из SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607, SEQ ID NO: 640-645, и SEQ ID NO: 728-737.
В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая полипептид PtIP-83, представляет собой последовательность под любым из SEQ ID NO: 172-235, SEQ ID NO: 300-333, SEQ ID NO: 368-397, SEQ ID NO: 428-517, SEQ ID NO: 634-639, и SEQ ID NO: 718-727.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует вариант полипептида PtIP-83 под SEQ ID NO: 1, где аминокислота в положении 53 представляет собой Val, Ala, Cys или Thr; аминокислота в положении 54 представляет собой Lys, Ala, Cys, Asp, Glu, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser или Thr; аминокислота в положении 55 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 56 представляет собой Leu, Glu, Phe, Ile, Met, Thr или Val; аминокислота в положении 57 представляет собой Tyr, Cys, Ile, Leu, Met, Thr или Val; аминокислота в положении 58 представляет собой Val, Cys, Ile или Leu; аминокислота в положении 59 представляет собой Phe, Leu, Met, Val или Tyr; аминокислота в положении 60 представляет собой Ala, Cys, Gly, Ser, Thr или Val; аминокислота в положении 61 представляет собой Asp, Glu, His или Ser; аминокислота в положении 62 представляет собой Val, Ala, Cys, Ile, Leu или Thr; аминокислота в положении 63 представляет собой Val, Ala, Cys, Ile, Leu, Met или Thr; аминокислота в положении 64 представляет собой Glu, Ala, Cys, Phe, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 65 представляет собой Leu, Ala, Cys, Phe, His, Ile, Met, Asn, Gln, Thr, Val или Trp; аминокислота в положении 66 представляет собой Pro, Asp, Gly, Met, Gln или Arg; аминокислота в положении 363 представляет собой Gln, Ala, Cys, Glu, Phe, Gly, His, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 364 представляет собой Ile, Ala, Cys, Glu, Phe, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 365 представляет собой Leu, Ala, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Arg, Val, Trp или Tyr; аминокислота в положении 366 представляет собой Gly, Ala, Cys, Phe, His, Ile, Lys, Leu, Met, Asn, Ser, Thr или Val; аминокислота в положении 367 представляет собой Ser, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val или Trp; аминокислота в положении 368 представляет собой Tyr, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp; аминокислота в положении 369 представляет собой Leu, Ala, Cys, Asp, Phe, Gly, Ile, Met, Thr или Val; аминокислота в положении 370 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 371 представляет собой Gln, Ala, Cys, Asp, Glu, Phe, Gly, Ile, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 372 представляет собой Gln, Ala, Cys, Asp, Phe, Gly, His, Ile, Leu, Asn, Arg, Ser, Val или Tyr; аминокислота в положении 373 представляет собой Asn, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Gln, Ser, Thr, Val или Trp; аминокислота в положении 556 представляет собой Trp, Phe, Thr или Tyr; аминокислота в положении 557 представляет собой Arg, Cys, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 558 представляет собой Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 559 представляет собой Lys, Ala, Cys, Phe, Gly, His, Ile, Leu, Asn, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 560 представляет собой Cys, Ala, Phe, Gly, Ile, Met, Asn, Arg, Ser, Thr или Val; аминокислота в положении 561 представляет собой Lys, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 562 представляет собой Asn, Cys, Asp, Glu, Gly, His, Leu, Met, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 563 представляет собой Val, Ala, Cys, Asp, Phe, His, Ile, Leu, Met, Asn, Gln, Thr или Trp; аминокислота в положении 564 представляет собой Ala, Cys, Gly, Met, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 646 представляет собой Leu, Ala, Cys, Gly, Ile, Met, Asn, Gln, Ser, Thr или Val; аминокислота в положении 647 представляет собой Leu, Asp, Gly, Met, Asn, Gln или Thr; аминокислота в положении 648 представляет собой Met, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 649 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr; аминокислота в положении 650 представляет собой Thr, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Val или Tyr; аминокислота в положении 651 представляет собой Glu, Ala, Cys, Asp, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 652 представляет собой Leu, Cys, Phe, Ile, Lys, Met, Pro, Arg, Ser, Thr или Val; аминокислота в положении 653 представляет собой Thr, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Pro, Arg, Ser, Val или Trp; аминокислота в положении 654 представляет собой Thr, Ala, Cys, Phe, Ile, Lys, Leu, Met, Pro, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 655 представляет собой Trp, Phe или Tyr; аминокислота в положении 771 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Asn, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 772 представляет собой Arg, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 773 представляет собой Asp, Ala, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 774 представляет собой Gln, Ala, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 775 представляет собой Val, Ala, Cys, Asp, Glu, Gly, His, Ile, Asn, Pro, Gln, Arg, Ser, Thr или Tyr; аминокислота в положении 776 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 777 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 778 представляет собой Phe, Ala, His, Ile, Leu, Met, Asn, Gln, Ser, Val, Trp или Tyr; аминокислота в положении 779 представляет собой Gln, Ala, Cys, Asp, Glu, Gly, His, Lys, Leu, Asn, Pro, Arg, Ser, Thr или Val; аминокислота в положении 780 представляет собой Ala, Cys, Asn, Pro, Gln или Ser; аминокислота в положении 781 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 782 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 783 представляет собой Pro, Ala, Cys, Asp, Glu, Gly, His, Asn, Gln, Arg, Ser, Thr или Val; аминокислота в положении 784 представляет собой Leu, Ala, Glu, Phe, His, Ile, Lys, Met, Asn, Pro, Gln, Ser, Thr, Val или Trp; аминокислота в положении 785 представляет собой Asn, Ala, Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; и аминокислота в положении 786 представляет собой Tyr, Phe, Ile, Leu или Trp.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует вариант полипептида PtIP-83 под SEQ ID NO: 1, где аминокислота в положении 1 представляет собой Met или удалена; аминокислота в положении 2 представляет собой Ala или удалена; аминокислота в положении 3 представляет собой Leu, Val или удалена; аминокислота в положении 4 представляет собой Val, Met или Leu; аминокислота в положении 7 представляет собой Gly или Ser; аминокислота в положении 8 представляет собой Lys или Thr; аминокислота в положении 10 представляет собой Phe или Tyr; аминокислота в положении 11 представляет собой Glu или Arg; аминокислота в положении 18 представляет собой Met или Ile; аминокислота в положении 19 представляет собой Gly, Pro или Ala; аминокислота в положении 20 представляет собой Val или удалена; аминокислота в положении 21 представляет собой Leu или Val; аминокислота в положении 23 представляет собой Arg или Gln; аминокислота в положении 37 представляет собой Val или Leu; аминокислота в положении 38 представляет собой Arg или Asn; аминокислота в положении 40 представляет собой Ala или Ser; аминокислота в положении 43 представляет собой Asn или Asp; аминокислота в положении 45 представляет собой Gly или Ala; аминокислота в положении 46 представляет собой Gln или Glu; аминокислота в положении 48 представляет собой Glu, Pro или Val; аминокислота в положении 51 представляет собой Glu или Gly; аминокислота в положении 52 представляет собой Lys, Arg или Thr; аминокислота в положении 56 представляет собой Leu или Val; аминокислота в положении 59 представляет собой Phe или Leu; аминокислота в положении 66 представляет собой Pro или Ala; аминокислота в положении 67 представляет собой Val, Pro или Thr; аминокислота в положении 68 представляет собой Val, Arg, Phe или Gly; аминокислота в положении 69 представляет собой Glu, Ala или Lys; аминокислота в положении 70 представляет собой Trp, Thr, His, Tyr или Arg; аминокислота в положении 71 представляет собой Arg, Pro или удалена; аминокислота в положении 72 представляет собой Trp, Asp, Leu или удалена; аминокислота в положении 73 представляет собой Pro, Gln, Asn, His или удалена; аминокислота в положении 74 представляет собой Pro, Met или Thr; аминокислота в положении 75 представляет собой Gln, His или Arg; аминокислота в положении 76 представляет собой Ile, Met или Leu; аминокислота в положении 84 представляет собой Ile или Val; аминокислота в положении 91 представляет собой Trp или Phe; аминокислота в положении 93 представляет собой Thr или Ile; аминокислота в положении 94 представляет собой Asp или Gly; аминокислота в положении 96 представляет собой Arg или Ser; аминокислота в положении 97 представляет собой Gln, Phe или Arg; аминокислота в положении 98 представляет собой Ser или удалена; аминокислота в положении 99 представляет собой Asp или Ala; аминокислота в положении 100 представляет собой Thr или Ala; аминокислота в положении 101 представляет собой Glu, Thr или Trp аминокислота в положении 103 представляет собой His, Arg, Glu или Gln; аминокислота в положении 105 представляет собой Thr или Pro; аминокислота в положении 108 представляет собой Lys, Gln или Glu; аминокислота в положении 109 представляет собой Leu или Val; аминокислота в положении 111 представляет собой Ala или Thr; аминокислота в положении 112 представляет собой Ile, Arg, Thr или удалена; аминокислота в положении 113 представляет собой Gln, Ala, Gly или удалена; аминокислота в положении 114 представляет собой Arg, Glu или Ile; аминокислота в положении 115 представляет собой Glu или Gln; аминокислота в положении 116 представляет собой Glu, Asn, Gln или Arg; аминокислота в положении 117 представляет собой Asn, Val, Tyr или Phe; аминокислота в положении 118 представляет собой Arg или Lys; аминокислота в положении 119 представляет собой Trp или Ser; аминокислота в положении 122 представляет собой Thr, Lys или Ala; аминокислота в положении 124 представляет собой Ala или Thr; аминокислота в положении 126 представляет собой Gly или Asp; аминокислота в положении 127 представляет собой Met или Ala; аминокислота в положении 128 представляет собой Asn или Lys; аминокислота в положении 131 представляет собой Val, Ile или Thr; аминокислота в положении 133 представляет собой Ile или Val; аминокислота в положении 134 представляет собой His или Tyr; аминокислота в положении 135 представляет собой Ala или Gly; аминокислота в положении 137 представляет собой Glu или Lys; аминокислота в положении 139 представляет собой Gln или Glu; аминокислота в положении 140 представляет собой Val, Arg или Leu; аминокислота в положении 141 представляет собой Gly или Ser; аминокислота в положении 142 представляет собой Val или Pro; аминокислота в положении 144 представляет собой Thr, Leu, Phe или Tyr; аминокислота в положении 145 представляет собой Met, Pro или Asn; аминокислота в положении 146 представляет собой Ser, Gly или Asn; аминокислота в положении 147 представляет собой Trp или Asn; аминокислота в положении 148 представляет собой Ser, Ala или Pro; аминокислота в положении 149 представляет собой Ser или удалена; аминокислота в положении 150 представляет собой Val, Ile или Tyr; аминокислота в положении 152 представляет собой Arg, Ala, Val или Gly; аминокислота в положении 154 представляет собой Ser, Trp или Glu; аминокислота в положении 156 представляет собой Leu, Asp или Gln; аминокислота в положении 158 представляет собой Ser или Cys; аминокислота в положении 159 представляет собой Val, Thr или Ile; аминокислота в положении 162 представляет собой Ser или Ala; аминокислота в положении 163 представляет собой Gly или удалена; аминокислота в положении 164 представляет собой Phe или удалена; аминокислота в положении 165 представляет собой Arg или Ala; аминокислота в положении 166 представляет собой Ala, Arg, Met или Phe; аминокислота в положении 167 представляет собой Val или His; аминокислота в положении 168 представляет собой Ser или Asn; аминокислота в положении 169 представляет собой Val, His или Thr; аминокислота в положении 170 представляет собой Phe или Val; аминокислота в положении 171 представляет собой Glu, Asn или Asp; аминокислота в положении 172 представляет собой Val, Ala, Arg или Glu; аминокислота в положении 175 представляет собой Ser, Arg или Trp; аминокислота в положении 176 представляет собой Val или Ile; аминокислота в положении 177 представляет собой Arg или Ile; аминокислота в положении 179 представляет собой Thr, Ile, Val или Ser; аминокислота в положении 180 представляет собой Leu, Phe или Thr; аминокислота в положении 181 представляет собой Gly, Thr, Gln или Ser; аминокислота в положении 182 представляет собой Ala, Leu, Phe или Ile; аминокислота в положении 183 представляет собой Thr или Gly; аминокислота в положении 184 представляет собой Leu, Thr, Ser или Arg; аминокислота в положении 185 представляет собой Arg, Gly, Asp или Ala; аминокислота в положении 186 представляет собой Pro, Val или Gln; аминокислота в положении 187 представляет собой Asp, Thr или Ser; аминокислота в положении 188 представляет собой His, Gly или Ala; аминокислота в положении 189 представляет собой Ala, Arg, Pro или удалена; аминокислота в положении 190 представляет собой Leu, Asn или удалена; аминокислота в положении 191 представляет собой Tyr или удалена; аминокислота в положении 192 представляет собой Ser, Ile, Val или Asn; аминокислота в положении 193 представляет собой Thr или Asp; аминокислота в положении 194 представляет собой Thr или Ser; аминокислота в положении 195 представляет собой Met или Thr; аминокислота в положении 196 представляет собой Gln, His, Leu или Ser; аминокислота в положении 197 представляет собой Ala, Gly или Leu; аминокислота в положении 198 представляет собой Thr, Glu или Ala; аминокислота в положении 199 представляет собой Pro или Arg; аминокислота в положении 200 представляет собой Asn, Ser, Thr или Gly; аминокислота в положении 201 представляет собой Ala, Leu, Glu или Trp; аминокислота в положении 202 представляет собой Ser, Asp, Phe или Leu; аминокислота в положении 203 представляет собой His, Pro, Gly или Ser; аминокислота в положении 204 представляет собой Ile, Trp, His или Gly; аминокислота в положении 205 представляет собой Ser, Asn или Ile; аминокислота в положении 206 представляет собой Ala, Gly, Asp, Tyr или Arg; аминокислота в положении 207 представляет собой Phe, Val или Leu; аминокислота в положении 208 представляет собой Asn, Ser, Pro или Leu; аминокислота в положении 210 представляет собой Arg, Asp, Glu или Tyr; аминокислота в положении 211 представляет собой Ile, Ser или Thr; аминокислота в положении 212 представляет собой Val, Ala или Asp; аминокислота в положении 214 представляет собой Pro или Arg; аминокислота в положении 215 представляет собой Ser или Thr; аминокислота в положении 217 представляет собой Tyr или Phe; аминокислота в положении 218 представляет собой Arg или Ser; аминокислота в положении 219 представляет собой Val или Ala; аминокислота в положении 220 представляет собой Cys, Leu или Ser; аминокислота в положении 221 представляет собой Pro или His; аминокислота в положении 222 представляет собой Leu, Arg или Ser; аминокислота в положении 224 представляет собой Asn или Ser; аминокислота в положении 225 представляет собой Asp, Arg или Thr; аминокислота в положении 226 представляет собой Thr или Asn; аминокислота в положении 227 представляет собой Asp, Leu или удалена; аминокислота в положении 228 представляет собой Thr или удалена; аминокислота в положении 229 представляет собой Tyr или удалена; аминокислота в положении 230 представляет собой Leu или удалена; аминокислота в положении 231 представляет собой Gly или удалена; аминокислота в положении 232 представляет собой Ile или удалена; аминокислота в положении 233 представляет собой Pro или удалена; аминокислота в положении 234 представляет собой Ala, Pro или удалена; аминокислота в положении 235 представляет собой Asp, Ile или Val; аминокислота в положении 236 представляет собой Val, Ser или Glu; аминокислота в положении 237 представляет собой Ala, Phe или Tyr; аминокислота в положении 238 представляет собой Ala или Thr; аминокислота в положении 239 представляет собой Val, Ser или Gly; аминокислота в положении 240 представляет собой Leu или Ile; аминокислота в положении 243 представляет собой Asp или Glu; аминокислота в положении 249 представляет собой Asn или Ser; аминокислота в положении 252 представляет собой Leu или Met; аминокислота в положении 257 представляет собой Thr или Ser; аминокислота в положении 259 представляет собой His или Leu; аминокислота в положении 266 представляет собой Ala или Val; аминокислота в положении 267 представляет собой Cys или Gly; аминокислота в положении 268 представляет собой His, Arg или Tyr; аминокислота в положении 272 представляет собой Asp или Glu; аминокислота в положении 273 представляет собой Val, Met, Ile или Leu; аминокислота в положении 274 представляет собой Val или Met; аминокислота в положении 278 представляет собой Gly или Ala; аминокислота в положении 279 представляет собой Glu или Val; аминокислота в положении 281 представляет собой Leu или Ala; аминокислота в положении 282 представляет собой Asn, Leu или Ile; аминокислота в положении 285 представляет собой Asn или Ser; аминокислота в положении 286 представляет собой Lys, Asp или Glu; аминокислота в положении 287 представляет собой Leu или Val; аминокислота в положении 290 представляет собой Pro, Gln или Arg; аминокислота в положении 291 представляет собой Leu или Val; аминокислота в положении 292 представляет собой Lys или Val; аминокислота в положении 293 представляет собой Glu или Gln; аминокислота в положении 294 представляет собой Ser, Asn или Lys; аминокислота в положении 295 представляет собой Thr или Ser; аминокислота в положении 296 представляет собой Gln или His; аминокислота в положении 297 представляет собой Leu или Met; аминокислота в положении 300 представляет собой Ser или Thr; аминокислота в положении 301 представляет собой Glu или Ala; аминокислота в положении 302 представляет собой Ser, Pro или Ala; аминокислота в положении 304 представляет собой Lys или Asn; аминокислота в положении 313 представляет собой Val или Ile; аминокислота в положении 314 представляет собой His, Glu или Gln; аминокислота в положении 315 представляет собой Ala, Cys или Ser; аминокислота в положении 316 представляет собой Ala или Val; аминокислота в положении 317 представляет собой Met или Ile; аминокислота в положении 319 представляет собой Met или Ile; аминокислота в положении 320 представляет собой Val или Gly; аминокислота в положении 321 представляет собой Arg или Pro; аминокислота в положении 322 представляет собой Ile или Phe; аминокислота в положении 323 представляет собой Gly или Val; аминокислота в положении 324 представляет собой Leu или Ser; аминокислота в положении 336 представляет собой Ser или Asn; аминокислота в положении 339 представляет собой Asn, Lys или Arg; аминокислота в положении 350 представляет собой Arg или Gln; аминокислота в положении 351 представляет собой Glu или Asp; аминокислота в положении 353 представляет собой Lys или Arg; аминокислота в положении 354 представляет собой Gln или Arg; аминокислота в положении 355 представляет собой Phe или Leu; аминокислота в положении 356 представляет собой Lys или Arg; аминокислота в положении 360 представляет собой Ile, Val или Ala; аминокислота в положении 365 представляет собой Leu или Phe; аминокислота в положении 371 представляет собой или Glu; аминокислота в положении 372 представляет собой или Lys; аминокислота в положении 374 представляет собой Arg или Lys; аминокислота в положении 376 представляет собой Phe или Leu; аминокислота в положении 378 представляет собой Glu или Asp; аминокислота в положении 381 представляет собой Leu или Val; аминокислота в положении 388 представляет собой Ala или Ser; аминокислота в положении 395 представляет собой Arg или Lys; аминокислота в положении 396 представляет собой Glu, Gln или Gly; аминокислота в положении 399 представляет собой Asp или Asn; аминокислота в положении 400 представляет собой Asn, Thr или Asp; аминокислота в положении 401 представляет собой Thr или Ala; аминокислота в положении 402 представляет собой Phe, Ile или Leu; аминокислота в положении 406 представляет собой Asp или Glu; аминокислота в положении 408 представляет собой Leu или Met; аминокислота в положении 410 представляет собой Gly или Leu; аминокислота в положении 414 представляет собой Ala или Glu; аминокислота в положении 416 представляет собой Ser, Asn или Asp; аминокислота в положении 417 представляет собой Ser, Arg или Gly; аминокислота в положении 423 представляет собой Lys или Gln; аминокислота в положении 431 представляет собой Arg или Lys; аминокислота в положении 432 представляет собой Gln или Glu; аминокислота в положении 436 представляет собой Arg или Glu; аминокислота в положении 440 представляет собой Asn или Arg; аминокислота в положении 442 представляет собой Leu или Val; аминокислота в положении 447 представляет собой Ser, Lys или Arg; аминокислота в положении 448 представляет собой Ala или Ser; аминокислота в положении 451 представляет собой Gln или Met; аминокислота в положении 453 представляет собой Gly или Ala; аминокислота в положении 455 представляет собой Ala или Val; аминокислота в положении 457 представляет собой Leu или Val; аминокислота в положении 467 представляет собой Val или Ala; аминокислота в положении 471 представляет собой Gly или Ala; аминокислота в положении 475 представляет собой Ser или Asn; аминокислота в положении 483 представляет собой Gly или Ala; аминокислота в положении 493 представляет собой Gln или Gly; аминокислота в положении 504 представляет собой Val или Ile; аминокислота в положении 506 представляет собой Asp или His; аминокислота в положении 509 представляет собой Asp или Asn; аминокислота в положении 510 представляет собой Ser или Ala; аминокислота в положении 512 представляет собой Glu или Asp; аминокислота в положении 515 представляет собой Gly или Ser; аминокислота в положении 516 представляет собой Gln или His; аминокислота в положении 517 представляет собой Ile или Leu; аминокислота в положении 519 представляет собой Asp, Gly или Gln; аминокислота в положении 522 представляет собой Val, Glu, Pro или Val; аминокислота в положении 525 представляет собой Glu или Asp; аминокислота в положении 526 представляет собой Leu или Met; аминокислота в положении 539 представляет собой Val или Ile; аминокислота в положении 555 представляет собой Val или Ala; аминокислота в положении 557 представляет собой Arg или Lys; аминокислота в положении 563 представляет собой Val или Met; аминокислота в положении 571 представляет собой Ser или Cys; аминокислота в положении 575 представляет собой Val или Glu; аминокислота в положении 577 представляет собой Met или Ile; аминокислота в положении 579 представляет собой Glu или Gln; аминокислота в положении 583 представляет собой Asp или Glu; аминокислота в положении 589 представляет собой Met или Leu; аминокислота в положении 590 представляет собой Met или Leu; аминокислота в положении 593 представляет собой Met или Ile; аминокислота в положении 595 представляет собой Arg или Gln; аминокислота в положении 596 представляет собой Ser или Thr; аминокислота в положении 597 представляет собой Gln или His; аминокислота в положении 607 представляет собой Ala или Val; аминокислота в положении 608 представляет собой Asp или Asn; аминокислота в положении 612 представляет собой Tyr, His или Phe; аминокислота в положении 617 представляет собой Thr или Ile; аминокислота в положении 618 представляет собой Gln или His; аминокислота в положении 625 представляет собой Arg или Ser; аминокислота в положении 626 представляет собой Met или Ile; аминокислота в положении 628 представляет собой Leu или Ile; аминокислота в положении 633 представляет собой Ile или Met; аминокислота в положении 634 представляет собой Leu или Met; аминокислота в положении 642 представляет собой Arg или Met; аминокислота в положении 648 представляет собой Met или Thr; аминокислота в положении 651 представляет собой Glu или Gln; аминокислота в положении 654 представляет собой Thr, Val или Ala; аминокислота в положении 658 представляет собой Gly или Arg; аминокислота в положении 663 представляет собой Gly или Ala; аминокислота в положении 664 представляет собой Asp или Asn; аминокислота в положении 668 представляет собой Ala или Thr; аминокислота в положении 669 представляет собой Gln или His; аминокислота в положении 671 представляет собой Asn или Ser аминокислота в положении 675 представляет собой Ile, Val или Ser; аминокислота в положении 678 представляет собой Met, Ile, Ala или Thr; аминокислота в положении 682 представляет собой Pro или Gln; аминокислота в положении 683 представляет собой Ser или Pro; аминокислота в положении 685 представляет собой Asp или Asn; аминокислота в положении 694 представляет собой Asp или Gly; аминокислота в положении 697 представляет собой Asn или Ser; аминокислота в положении 704 представляет собой Glu или Gly; аминокислота в положении 714 представляет собой Ala или Gly; аминокислота в положении 721 представляет собой Ser или Phe; аминокислота в положении 722 представляет собой Ser или Asn; аминокислота в положении 724 представляет собой Ser или Thr; аминокислота в положении 734 представляет собой His или Gln; аминокислота в положении 736 представляет собой Val или Ala; аминокислота в положении 737 представляет собой Lys или Gln; аминокислота в положении 739 представляет собой Ala или Ser; аминокислота в положении 740 представляет собой Ser или Met; аминокислота в положении 741 представляет собой Gly или Asn; аминокислота в положении 742 представляет собой Ile или Gly; аминокислота в положении 743 представляет собой Gly или удалена; аминокислота в положении 745 представляет собой Gly или Asp; аминокислота в положении 751 представляет собой Thr, Ser или Ala; аминокислота в положении 753 представляет собой Gln или Arg; аминокислота в положении 754 представляет собой Thr или Ser; аминокислота в положении 756 представляет собой Thr или Ile; аминокислота в положении 757 представляет собой Val или Ile; аминокислота в положении 766 представляет собой Ile или Val; аминокислота в положении 773 представляет собой Asp или Glu; аминокислота в положении 774 представляет собой Gln или Glu; аминокислота в положении 776 представляет собой Leu или Met; аминокислота в положении 777 представляет собой Pro или Thr; аминокислота в положении 782 представляет собой Ala, Asp или Val; аминокислота в положении 786 представляет собой Tyr или Phe; аминокислота в положении 787 представляет собой His или Gln; аминокислота в положении 788 представляет собой Tyr или Met; аминокислота в положении 789 представляет собой Ala или Arg; аминокислота в положении 790 представляет собой Tyr или Thr; аминокислота в положении 791 представляет собой Arg или Ala; аминокислота в положении 792 представляет собой Leu или Ser; аминокислота в положении 796 представляет собой Asp или Glu; аминокислота в положении 797 представляет собой Ser, Thr или Ala аминокислота в положении 802 представляет собой Glu или Gln; аминокислота в положении 806 представляет собой Gln, Asp, Glu или His; аминокислота в положении 810 представляет собой Lys или Thr; аминокислота в положении 819 представляет собой Arg или His; аминокислота в положении 829 представляет собой Lys, Ser, Ala или Pro; аминокислота в положении 832 представляет собой Ala, Lys или Glu; аминокислота в положении 833 представляет собой Gly или Glu; аминокислота в положении 842 представляет собой Leu или Pro; аминокислота в положении 847 представляет собой Gln или Glu; аминокислота в положении 848 представляет собой Ile или Val; аминокислота в положении 849 представляет собой Val или Ala; аминокислота в положении 855 представляет собой Thr или Met; аминокислота в положении 860 представляет собой Ile или Val; и аминокислота в положении 864 представляет собой His или Gln.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует вариант полипептида PtIP-83 под SEQ ID NO: 1, где аминокислота в положении 1 представляет собой Met или удалена; аминокислота в положении 2 представляет собой Ala или удалена; аминокислота в положении 3 представляет собой Leu, Val, Ile или удалена; аминокислота в положении 4 представляет собой Val, Met, Ile или Leu; аминокислота в положении 7 представляет собой Gly, Thr или Ser; аминокислота в положении 8 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 10 представляет собой Phe, Trp или Tyr; аминокислота в положении 11 представляет собой Glu, Asp, Lys или Arg; аминокислота в положении 18 представляет собой Met, Val, Leu или Ile; аминокислота в положении 19 представляет собой Gly, Pro или Ala; аминокислота в положении 20 представляет собой Val, Ile, Leu или удалена; аминокислота в положении 21 представляет собой Leu, Ile или Val; аминокислота в положении 23 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 37 представляет собой Val, Ile или Leu; аминокислота в положении 38 представляет собой Arg, Lys, Gln или Asn; аминокислота в положении 40 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 43 представляет собой Asn, Gln, Glu или Asp; аминокислота в положении 45 представляет собой Gly или Ala; аминокислота в положении 46 представляет собой Gln, Asp, Asn или Glu; аминокислота в положении 48 представляет собой Glu, Asp, Pro, Ile, Leu или Val; аминокислота в положении 51 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 52 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 56 представляет собой Leu, Ile или Val; аминокислота в положении 59 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 66 представляет собой Pro, Gly или Ala; аминокислота в положении 67 представляет собой Val, Pro, Ile, Leu, Ser или Thr; аминокислота в положении 68 представляет собой Val, Arg, Phe, Ile, Leu, Lys или Gly; аминокислота в положении 69 представляет собой Glu, Ala, Asp, Gly, Arg или Lys; аминокислота в положении 70 представляет собой Trp, Thr, His, Tyr, Lys или Arg; аминокислота в положении 71 представляет собой Arg, Pro, Lys или удалена; аминокислота в положении 72 представляет собой Trp, Asp, Leu, Ile, Val, Glu или удалена; аминокислота в положении 73 представляет собой Pro, Gln, Asn, His или удалена; аминокислота в положении 74 представляет собой Pro, Met, Ser или Thr; аминокислота в положении 75 представляет собой Gln, His, Asn, Lys или Arg; аминокислота в положении 76 представляет собой Ile, Met, Val или Leu; аминокислота в положении 84 представляет собой Ile, Leu или Val; аминокислота в положении 91 представляет собой Trp или Phe; аминокислота в положении 93 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 94 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 96 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 97 представляет собой Gln, Phe, Asn, Lys или Arg; аминокислота в положении 98 представляет собой Ser, Thr или удалена; аминокислота в положении 99 представляет собой Asp, Glu, Gly или Ala; аминокислота в положении 100 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 101 представляет собой Glu, Thr, Asp, Ser или Trp аминокислота в положении 103 представляет собой His, Arg, Lys, Glu или Gln; аминокислота в положении 105 представляет собой Thr, Ser или Pro; аминокислота в положении 108 представляет собой Lys, Arg, Asn, Asp, Gln или Glu; аминокислота в положении 109 представляет собой Leu, Ile или Val; аминокислота в положении 111 представляет собой Ala, Ser или Thr; аминокислота в положении 112 представляет собой Ile, Arg, Thr, Leu, Val, Lys, Ser или удалена; аминокислота в положении 113 представляет собой Gln, Ala, Gly, Asn или удалена; аминокислота в положении 114 представляет собой Arg, Glu, Lys, Asp или Ile; аминокислота в положении 115 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 116 представляет собой Glu, Asn, Gln, Asp, Lys или Arg; аминокислота в положении 117 представляет собой Asn, Val, Tyr, Ile, Leu, Gln, Trp или Phe; аминокислота в положении 118 представляет собой Arg или Lys; аминокислота в положении 119 представляет собой Trp, Thr или Ser; аминокислота в положении 122 представляет собой Thr, Lys, Ser, Arg или Ala; аминокислота в положении 124 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 126 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 127 представляет собой Met, Gly или Ala; аминокислота в положении 128 представляет собой Asn, Gln, Arg или Lys; аминокислота в положении 131 представляет собой Val, Ile, Leu, Ser или Thr; аминокислота в положении 133 представляет собой Ile, Leu или Val; аминокислота в положении 134 представляет собой His или Tyr; аминокислота в положении 135 представляет собой Ala или Gly; аминокислота в положении 137 представляет собой Glu, Asp, Arg или Lys; аминокислота в положении 139 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 140 представляет собой Val, Arg, Ile, Lys или Leu; аминокислота в положении 141 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 142 представляет собой Val, Ile, Leu или Pro; аминокислота в положении 144 представляет собой Thr, Leu, Phe, Ile, Val или Tyr; аминокислота в положении 145 представляет собой Met, Pro, Gln или Asn; аминокислота в положении 146 представляет собой Ser, Gly, Thr, Ala, Gln или Asn; аминокислота в положении 147 представляет собой Trp, Gln, Tyr или Asn; аминокислота в положении 148 представляет собой Ser, Ala, Thr, Gly или Pro; аминокислота в положении 149 представляет собой Ser, Thr или удалена; аминокислота в положении 150 представляет собой Val, Ile, Leu или Tyr; аминокислота в положении 152 представляет собой Arg, Ala, Val, Ile, Leu, Lys или Gly; аминокислота в положении 154 представляет собой Ser, Trp, Thr, Asp или Glu; аминокислота в положении 156 представляет собой Leu, Asp, Ile, Val, Asn, Glu или Gln; аминокислота в положении 158 представляет собой Ser, Thr или Cys; аминокислота в положении 159 представляет собой Val, Thr, Leu или Ile; аминокислота в положении 162 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 163 представляет собой Gly, Ala или удалена; аминокислота в положении 164 представляет собой Phe или удалена; аминокислота в положении 165 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 166 представляет собой Ala, Arg, Met, Lys или Phe; аминокислота в положении 167 представляет собой Val, Ile, Leu или His; аминокислота в положении 168 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 169 представляет собой Val, His, Ile, Leu, Ser или Thr; аминокислота в положении 170 представляет собой Phe, Ile, Leu или Val; аминокислота в положении 171 представляет собой Glu, Asn, Gln или Asp; аминокислота в положении 172 представляет собой Val, Ala, Arg, Ile, Leu, Gly, Lys, Asp или Glu; аминокислота в положении 175 представляет собой Ser, Arg, Thr, Lys или Trp; аминокислота в положении 176 представляет собой Val, Leu или Ile; аминокислота в положении 177 представляет собой Arg, Lys, Leu, Val или Ile; аминокислота в положении 179 представляет собой Thr, Ile, Val, Leu или Ser; аминокислота в положении 180 представляет собой Leu, Phe, Ile, Val, Ser или Thr; аминокислота в положении 181 представляет собой Gly, Thr, Gln, Asn или Ser; аминокислота в положении 182 представляет собой Ala, Leu, Phe, Val или Ile; аминокислота в положении 183 представляет собой Thr, Ser, Ala или Gly; аминокислота в положении 184 представляет собой Leu, Thr, Ser, Ile, Val, Lys или Arg; аминокислота в положении 185 представляет собой Arg, Gly, Asp, Lys, Glu или Ala; аминокислота в положении 186 представляет собой Pro, Val, Ile, Leu, Asn или Gln; аминокислота в положении 187 представляет собой Asp, Thr, Glu или Ser; аминокислота в положении 188 представляет собой His, Gly или Ala; аминокислота в положении 189 представляет собой Ala, Arg, Pro, Lys, Gly или удалена; аминокислота в положении 190 представляет собой Leu, Asn, Ile, Val, Gln или удалена; аминокислота в положении 191 представляет собой Tyr или удалена; аминокислота в положении 192 представляет собой Ser, Ile, Val, Leu, Thr или Asn; аминокислота в положении 193 представляет собой Thr, Ser, Glu или Asp; аминокислота в положении 194 представляет собой Thr или Ser; аминокислота в положении 195 представляет собой Met или Thr; аминокислота в положении 196 представляет собой Gln, His, Leu, Asn, Ile, Val, Thr или Ser; аминокислота в положении 197 представляет собой Ala, Gly, Ile, Val или Leu; аминокислота в положении 198 представляет собой Thr, Glu, Ser, Asp, Gly или Ala; аминокислота в положении 199 представляет собой Pro, Lys или Arg; аминокислота в положении 200 представляет собой Asn, Ser, Thr, Gln, Ala или Gly; аминокислота в положении 201 представляет собой Ala, Leu, Glu, Ile, Asp или Trp; аминокислота в положении 202 представляет собой Ser, Asp, Phe, Ile, Val, Thr, Glu или Leu; аминокислота в положении 203 представляет собой His, Pro, Gly, Ala, Thr или Ser; аминокислота в положении 204 представляет собой Ile, Trp, His, Leu, Val, Ala или Gly; аминокислота в положении 205 представляет собой Ser, Asn, Leu, Val, Thr, Gln или Ile; аминокислота в положении 206 представляет собой Ala, Gly, Asp, Tyr, Glu, Lys или Arg; аминокислота в положении 207 представляет собой Phe, Val, Ile или Leu; аминокислота в положении 208 представляет собой Asn, Ser, Pro, Gln, Thr, Val, Ile или Leu; аминокислота в положении 210 представляет собой Arg, Asp, Glu, Lys, Ser или Tyr; аминокислота в положении 211 представляет собой Ile, Ser, Leu, Val или Thr; аминокислота в положении 212 представляет собой Val, Ala, Ile, Leu, Glu, Gly или Asp; аминокислота в положении 214 представляет собой Pro, Lys или Arg; аминокислота в положении 215 представляет собой Ser или Thr; аминокислота в положении 217 представляет собой Tyr или Phe; аминокислота в положении 218 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 219 представляет собой Val, Ile, Leu или Ala; аминокислота в положении 220 представляет собой Cys, Leu, Ile, Val, Thr или Ser; аминокислота в положении 221 представляет собой Pro или His; аминокислота в положении 222 представляет собой Leu, Arg, Lys, Ile, Val, Thr или Ser; аминокислота в положении 224 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 225 представляет собой Asp, Arg, Glu, Lys, Ser или Thr; аминокислота в положении 226 представляет собой Thr, Ser, Gln или Asn; аминокислота в положении 227 представляет собой Asp, Leu, Glu, Ile, Val или удалена; аминокислота в положении 228 представляет собой Thr, Ser или удалена; аминокислота в положении 229 представляет собой Tyr или удалена; аминокислота в положении 230 представляет собой Leu, Ile, Val или удалена; аминокислота в положении 231 представляет собой Gly, Ala или удалена; аминокислота в положении 232 представляет собой Ile, Leu, Val или удалена; аминокислота в положении 233 представляет собой Pro или удалена; аминокислота в положении 234 представляет собой Ala, Pro, Gly или удалена; аминокислота в положении 235 представляет собой Asp, Ile, Leu, Glu или Val; аминокислота в положении 236 представляет собой Val, Ser, Ile, Leu, Thr, Asp или Glu; аминокислота в положении 237 представляет собой Ala, Phe или Tyr; аминокислота в положении 238 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 239 представляет собой Val, Ser, Ile, Leu, Thr, Ala или Gly; аминокислота в положении 240 представляет собой Leu, Val или Ile; аминокислота в положении 243 представляет собой Asp или Glu; аминокислота в положении 249 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 252 представляет собой Leu, Ile, Val или Met; аминокислота в положении 257 представляет собой Thr или Ser; аминокислота в положении 259 представляет собой His, Ile, Val или Leu; аминокислота в положении 266 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 267 представляет собой Cys, Ala или Gly; аминокислота в положении 268 представляет собой His, Arg, Lys или Tyr; аминокислота в положении 272 представляет собой Asp или Glu; аминокислота в положении 273 представляет собой Val, Met, Ile или Leu; аминокислота в положении 274 представляет собой Val, Ile, Leu или Met; аминокислота в положении 278 представляет собой Gly или Ala; аминокислота в положении 279 представляет собой Glu, Asp, Gly или Val; аминокислота в положении 281 представляет собой Leu, Ile, Val, Gly или Ala; аминокислота в положении 282 представляет собой Asn, Leu или Ile; аминокислота в положении 285 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 286 представляет собой Lys, Asp, Arg или Glu; аминокислота в положении 287 представляет собой Leu, Ile или Val; аминокислота в положении 290 представляет собой Pro, Gln, Asn, Lys или Arg; аминокислота в положении 291 представляет собой Leu, Ile или Val; аминокислота в положении 292 представляет собой Lys, Arg, Ile, Leu или Val; аминокислота в положении 293 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 294 представляет собой Ser, Asn, Thr, Gln, Arg или Lys; аминокислота в положении 295 представляет собой Thr или Ser; аминокислота в положении 296 представляет собой Gln, Asn или His; аминокислота в положении 297 представляет собой Leu, Ile, Val или Met; аминокислота в положении 300 представляет собой Ser или Thr; аминокислота в положении 301 представляет собой Glu, Asp, Gly или Ala; аминокислота в положении 302 представляет собой Ser, Pro, Thr, Gly или Ala; аминокислота в положении 304 представляет собой Lys, Arg, Gln или Asn; аминокислота в положении 313 представляет собой Val, Leu или Ile; аминокислота в положении 314 представляет собой His, Glu, Asn, Asp или Gln; аминокислота в положении 315 представляет собой Ala, Cys, Gly, Thr или Ser; аминокислота в положении 316 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 317 представляет собой Met, Leu, Val или Ile; аминокислота в положении 319 представляет собой Met, Leu, Val или Ile; аминокислота в положении 320 представляет собой Val, Ile, Leu, Ala или Gly; аминокислота в положении 321 представляет собой Arg, Lys или Pro; аминокислота в положении 322 представляет собой Ile, Leu, Val или Phe; аминокислота в положении 323 представляет собой Gly, Ile, Leu или Val; аминокислота в положении 324 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 336 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 339 представляет собой Asn, Lys, Gln или Arg; аминокислота в положении 350 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 351 представляет собой Glu или Asp; аминокислота в положении 353 представляет собой Lys или Arg; аминокислота в положении 354 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 355 представляет собой Phe, Ile, Leu или Leu; аминокислота в положении 356 представляет собой Lys или Arg; аминокислота в положении 360 представляет собой Ile, Val, Leu, Gly или Ala; аминокислота в положении 365 представляет собой Leu, Ile, Val или Phe; аминокислота в положении 371 представляет собой или Glu или Asp; аминокислота в положении 372 представляет собой или Lys или Arg; аминокислота в положении 374 представляет собой Arg или Lys; аминокислота в положении 376 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 378 представляет собой Glu или Asp; аминокислота в положении 381 представляет собой Leu, Ile или Val; аминокислота в положении 388 представляет собой Ala, Thr, Gly или Ser; аминокислота в положении 395 представляет собой Arg или Lys; аминокислота в положении 396 представляет собой Glu, Gln, Asp, Asn, Ala или Gly; аминокислота в положении 399 представляет собой Asp, Gln, Glu или Asn; аминокислота в положении 400 представляет собой Asn, Thr, Ser, Glu, Gln или Asp; аминокислота в положении 401 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 402 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 406 представляет собой Asp или Glu; аминокислота в положении 408 представляет собой Leu, Ile, Val или Met; аминокислота в положении 410 представляет собой Gly, Ile, Val, Ala или Leu; аминокислота в положении 414 представляет собой Ala, Gly, Asp или Glu; аминокислота в положении 416 представляет собой Ser, Asn, Thr, Gln, Glu или Asp; аминокислота в положении 417 представляет собой Ser, Arg, Lys, Thr, Ala или Gly; аминокислота в положении 423 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 431 представляет собой Arg или Lys; аминокислота в положении 432 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 436 представляет собой Arg, Lys, Asp или Glu; аминокислота в положении 440 представляет собой Asn, Gln, Lys или Arg; аминокислота в положении 442 представляет собой Leu, Ile или Val; аминокислота в положении 447 представляет собой Ser, Lys, Thr или Arg; аминокислота в положении 448 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 451 представляет собой Gln, Asn или Met; аминокислота в положении 453 представляет собой Gly или Ala; аминокислота в положении 455 представляет собой Ala, Leu, Ile или Val; аминокислота в положении 457 представляет собой Leu, Ile или Val; аминокислота в положении 467 представляет собой Val, Ile, Leu, Gly или Ala; аминокислота в положении 471 представляет собой Gly или Ala; аминокислота в положении 475 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 483 представляет собой Gly или Ala; аминокислота в положении 493 представляет собой Gln, Asn или Gly; аминокислота в положении 504 представляет собой Val, Leu или Ile; аминокислота в положении 506 представляет собой Asp, Glu или His; аминокислота в положении 509 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 510 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 512 представляет собой Glu или Asp; аминокислота в положении 515 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 516 представляет собой Gln, Asn или His; аминокислота в положении 517 представляет собой Ile, Val или Leu; аминокислота в положении 519 представляет собой Asp, Asn, Glu, Gly или Gln; аминокислота в положении 522 представляет собой Val, Glu, Pro, Ile, Leu или Asp; аминокислота в положении 525 представляет собой Glu или Asp; аминокислота в положении 526 представляет собой Leu, Ile, Val или Met; аминокислота в положении 539 представляет собой Val, Leu или Ile; аминокислота в положении 555 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 557 представляет собой Arg или Lys; аминокислота в положении 563 представляет собой Val, Leu, Ile или Met; аминокислота в положении 571 представляет собой Ser, Thr или Cys; аминокислота в положении 575 представляет собой Val, Leu, Ile, Asp или Glu; аминокислота в положении 577 представляет собой Met, Leu, Val или Ile; аминокислота в положении 579 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 583 представляет собой Asp или Glu; аминокислота в положении 589 представляет собой Met, Ile, Val или Leu; аминокислота в положении 590 представляет собой Met, Ile, Val или Leu; аминокислота в положении 593 представляет собой Met, Leu, Val или Ile; аминокислота в положении 595 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 596 представляет собой Ser или Thr; аминокислота в положении 597 представляет собой Gln, Asn или His; аминокислота в положении 607 представляет собой Ala, Gly, Ile, Leu или Val; аминокислота в положении 608 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 612 представляет собой Tyr, His или Phe; аминокислота в положении 617 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 618 представляет собой Gln, Asn или His; аминокислота в положении 625 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 626 представляет собой Met, Leu, Val или Ile; аминокислота в положении 628 представляет собой Leu, Val или Ile; аминокислота в положении 633 представляет собой Ile, Leu, Val или Met; аминокислота в положении 634 представляет собой Leu, Ile, Val или Met; аминокислота в положении 642 представляет собой Arg, Lys или Met; аминокислота в положении 648 представляет собой Met, Ser или Thr; аминокислота в положении 651 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 654 представляет собой Thr, Val, Ser, Ile, Leu, Gly или Ala; аминокислота в положении 658 представляет собой Gly, Lys, Ala или Arg; аминокислота в положении 663 представляет собой Gly или Ala; аминокислота в положении 664 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 668 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 669 представляет собой Gln, Asn или His; аминокислота в положении 671 представляет собой Asn, Gln, Thr или Ser аминокислота в положении 675 представляет собой Ile, Val, Ile, Thr или Ser; аминокислота в положении 678 представляет собой Met, Ile, Ala, Leu, Ser или Thr; аминокислота в положении 682 представляет собой Pro, Asn или Gln; аминокислота в положении 683 представляет собой Ser, Thr или Pro; аминокислота в положении 685 представляет собой Asp, Glu, Asp или Asn; аминокислота в положении 694 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 697 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 704 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 714 представляет собой Ala или Gly; аминокислота в положении 721 представляет собой Ser, Thr или Phe; аминокислота в положении 722 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 724 представляет собой Ser или Thr; аминокислота в положении 734 представляет собой His, Asn или Gln; аминокислота в положении 736 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 737 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 739 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 740 представляет собой Ser, Thr или Met; аминокислота в положении 741 представляет собой Gly, Ala, Gln или Asn; аминокислота в положении 742 представляет собой Ile, Leu, Val, Ala или Gly; аминокислота в положении 743 представляет собой Gly или удалена; аминокислота в положении 745 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 751 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 753 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 754 представляет собой Thr или Ser; аминокислота в положении 756 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 757 представляет собой Val, Leu или Ile; аминокислота в положении 766 представляет собой Ile, Leu или Val; аминокислота в положении 773 представляет собой Asp или Glu; аминокислота в положении 774 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 776 представляет собой Leu, Ile, Val или Met; аминокислота в положении 777 представляет собой Pro, Ser или Thr; аминокислота в положении 782 представляет собой Ala, Asp, Glu, Ile, Leu или Val; аминокислота в положении 786 представляет собой Tyr или Phe; аминокислота в положении 787 представляет собой His, Asn или Gln; аминокислота в положении 788 представляет собой Tyr или Met; аминокислота в положении 789 представляет собой Ala, Lys или Arg; аминокислота в положении 790 представляет собой Tyr или Thr; аминокислота в положении 791 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 792 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 796 представляет собой Asp или Glu; аминокислота в положении 797 представляет собой Ser, Thr или Ala аминокислота в положении 802 представляет собой Glu, Lys, Asp, Asn или Gln; аминокислота в положении 806 представляет собой Gln, Asp, Glu, Asn или His; аминокислота в положении 810 представляет собой Lys, Arg или Thr; аминокислота в положении 819 представляет собой Arg, Lys или His; аминокислота в положении 829 представляет собой Lys, Ser, Ala или Pro; аминокислота в положении 832 представляет собой Ala, Lys, Arg, Asp или Glu; аминокислота в положении 833 представляет собой Gly, Ala, Asp или Glu; аминокислота в положении 842 представляет собой Leu, Ile, Val или Pro; аминокислота в положении 847 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 848 представляет собой Ile, Leu или Val; аминокислота в положении 849 представляет собой Val, Leu, Ile, Gly или Ala; аминокислота в положении 855 представляет собой Thr, Ser или Met; аминокислота в положении 860 представляет собой Ile, Leu или Val; аминокислота в положении 864 представляет собой His, Asn или Gln.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует вариант полипептида PtIP-83 под SEQ ID NO: 1, где аминокислота в положении 1 представляет собой Met или удалена; аминокислота в положении 2 представляет собой Ala или удалена; аминокислота в положении 3 представляет собой Leu, Val, Ile или удалена; аминокислота в положении 4 представляет собой Val, Met, Ile или Leu; аминокислота в положении 7 представляет собой Gly, Thr или Ser; аминокислота в положении 8 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 10 представляет собой Phe, Trp или Tyr; аминокислота в положении 11 представляет собой Glu, Asp, Lys или Arg; аминокислота в положении 18 представляет собой Met, Val, Leu или Ile; аминокислота в положении 19 представляет собой Gly, Pro или Ala; аминокислота в положении 20 представляет собой Val, Ile, Leu или удалена; аминокислота в положении 21 представляет собой Leu, Ile или Val; аминокислота в положении 23 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 37 представляет собой Val, Ile или Leu; аминокислота в положении 38 представляет собой Arg, Lys, Gln или Asn; аминокислота в положении 40 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 43 представляет собой Asn, Gln, Glu или Asp; аминокислота в положении 45 представляет собой Gly или Ala; аминокислота в положении 46 представляет собой Gln, Asp, Asn или Glu; аминокислота в положении 48 представляет собой Glu, Asp, Pro, Ile, Leu или Val; аминокислота в положении 51 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 52 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 53 представляет собой Val, Ala, Cys или Thr; аминокислота в положении 54 представляет собой Lys, Ala, Cys, Asp, Glu, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser или Thr; аминокислота в положении 55 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 56 представляет собой Leu, Glu, Phe, Ile, Met, Thr или Val; аминокислота в положении 57 представляет собой Tyr, Cys, Ile, Leu, Met, Thr или Val; аминокислота в положении 58 представляет собой Val, Cys, Ile или Leu; аминокислота в положении 59 представляет собой Phe, Leu, Met, Val или Tyr; аминокислота в положении 60 представляет собой Ala, Cys, Gly, Ser, Thr или Val; аминокислота в положении 61 представляет собой Asp, Glu, His или Ser; аминокислота в положении 62 представляет собой Val, Ala, Cys, Ile, Leu или Thr; аминокислота в положении 63 представляет собой Val, Ala, Cys, Ile, Leu, Met или Thr; аминокислота в положении 64 представляет собой Glu, Ala, Cys, Phe, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 65 представляет собой Leu, Ala, Cys, Phe, His, Ile, Met, Asn, Gln, Thr, Val или Trp; аминокислота в положении 66 представляет собой Pro, Asp, Gly, Met, Gln или Arg; аминокислота в положении 67 представляет собой Val, Pro, Ile, Leu, Ser или Thr; аминокислота в положении 68 представляет собой Val, Arg, Phe, Ile, Leu, Lys или Gly; аминокислота в положении 69 представляет собой Glu, Ala, Asp, Gly, Arg или Lys; аминокислота в положении 70 представляет собой Trp, Thr, His, Tyr, Lys или Arg; аминокислота в положении 71 представляет собой Arg, Pro, Lys или удалена; аминокислота в положении 72 представляет собой Trp, Asp, Leu, Ile, Val, Glu или удалена; аминокислота в положении 73 представляет собой Pro, Gln, Asn, His или удалена; аминокислота в положении 74 представляет собой Pro, Met, Ser или Thr; аминокислота в положении 75 представляет собой Gln, His, Asn, Lys или Arg; аминокислота в положении 76 представляет собой Ile, Met, Val или Leu; аминокислота в положении 84 представляет собой Ile, Leu или Val; аминокислота в положении 91 представляет собой Trp или Phe; аминокислота в положении 93 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 94 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 96 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 97 представляет собой Gln, Phe, Asn, Lys или Arg; аминокислота в положении 98 представляет собой Ser, Thr или удалена; аминокислота в положении 99 представляет собой Asp, Glu, Gly или Ala; аминокислота в положении 100 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 101 представляет собой Glu, Thr, Asp, Ser или Trp аминокислота в положении 103 представляет собой His, Arg, Lys, Glu или Gln; аминокислота в положении 105 представляет собой Thr, Ser или Pro; аминокислота в положении 108 представляет собой Lys, Arg, Asn, Asp, Gln или Glu; аминокислота в положении 109 представляет собой Leu, Ile или Val; аминокислота в положении 111 представляет собой Ala, Ser или Thr; аминокислота в положении 112 представляет собой Ile, Arg, Thr, Leu, Val, Lys, Ser или удалена; аминокислота в положении 113 представляет собой Gln, Ala, Gly, Asn или удалена; аминокислота в положении 114 представляет собой Arg, Glu, Lys, Asp или Ile; аминокислота в положении 115 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 116 представляет собой Glu, Asn, Gln, Asp, Lys или Arg; аминокислота в положении 117 представляет собой Asn, Val, Tyr, Ile, Leu, Gln, Trp или Phe; аминокислота в положении 118 представляет собой Arg или Lys; аминокислота в положении 119 представляет собой Trp, Thr или Ser; аминокислота в положении 122 представляет собой Thr, Lys, Ser, Arg или Ala; аминокислота в положении 124 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 126 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 127 представляет собой Met, Gly или Ala; аминокислота в положении 128 представляет собой Asn, Gln, Arg или Lys; аминокислота в положении 131 представляет собой Val, Ile, Leu, Ser или Thr; аминокислота в положении 133 представляет собой Ile, Leu или Val; аминокислота в положении 134 представляет собой His или Tyr; аминокислота в положении 135 представляет собой Ala или Gly; аминокислота в положении 137 представляет собой Glu, Asp, Arg или Lys; аминокислота в положении 139 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 140 представляет собой Val, Arg, Ile, Lys или Leu; аминокислота в положении 141 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 142 представляет собой Val, Ile, Leu или Pro; аминокислота в положении 144 представляет собой Thr, Leu, Phe, Ile, Val или Tyr; аминокислота в положении 145 представляет собой Met, Pro, Gln или Asn; аминокислота в положении 146 представляет собой Ser, Gly, Thr, Ala, Gln или Asn; аминокислота в положении 147 представляет собой Trp, Gln, Tyr или Asn; аминокислота в положении 148 представляет собой Ser, Ala, Thr, Gly или Pro; аминокислота в положении 149 представляет собой Ser, Thr или удалена; аминокислота в положении 150 представляет собой Val, Ile, Leu или Tyr; аминокислота в положении 152 представляет собой Arg, Ala, Val, Ile, Leu, Lys или Gly; аминокислота в положении 154 представляет собой Ser, Trp, Thr, Asp или Glu; аминокислота в положении 156 представляет собой Leu, Asp, Ile, Val, Asn, Glu или Gln; аминокислота в положении 158 представляет собой Ser, Thr или Cys; аминокислота в положении 159 представляет собой Val, Thr, Leu или Ile; аминокислота в положении 162 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 163 представляет собой Gly, Ala или удалена; аминокислота в положении 164 представляет собой Phe или удалена; аминокислота в положении 165 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 166 представляет собой Ala, Arg, Met, Lys или Phe; аминокислота в положении 167 представляет собой Val, Ile, Leu или His; аминокислота в положении 168 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 169 представляет собой Val, His, Ile, Leu, Ser или Thr; аминокислота в положении 170 представляет собой Phe, Ile, Leu или Val; аминокислота в положении 171 представляет собой Glu, Asn, Gln или Asp; аминокислота в положении 172 представляет собой Val, Ala, Arg, Ile, Leu, Gly, Lys, Asp или Glu; аминокислота в положении 175 представляет собой Ser, Arg, Thr, Lys или Trp; аминокислота в положении 176 представляет собой Val, Leu или Ile; аминокислота в положении 177 представляет собой Arg, Lys, Leu, Val или Ile; аминокислота в положении 179 представляет собой Thr, Ile, Val, Leu или Ser; аминокислота в положении 180 представляет собой Leu, Phe, Ile, Val, Ser или Thr; аминокислота в положении 181 представляет собой Gly, Thr, Gln, Asn или Ser; аминокислота в положении 182 представляет собой Ala, Leu, Phe, Val или Ile; аминокислота в положении 183 представляет собой Thr, Ser, Ala или Gly; аминокислота в положении 184 представляет собой Leu, Thr, Ser, Ile, Val, Lys или Arg; аминокислота в положении 185 представляет собой Arg, Gly, Asp, Lys, Glu или Ala; аминокислота в положении 186 представляет собой Pro, Val, Ile, Leu, Asn или Gln; аминокислота в положении 187 представляет собой Asp, Thr, Glu или Ser; аминокислота в положении 188 представляет собой His, Gly или Ala; аминокислота в положении 189 представляет собой Ala, Arg, Pro, Lys, Gly или удалена; аминокислота в положении 190 представляет собой Leu, Asn, Ile, Val, Gln или удалена; аминокислота в положении 191 представляет собой Tyr или удалена; аминокислота в положении 192 представляет собой Ser, Ile, Val, Leu, Thr или Asn; аминокислота в положении 193 представляет собой Thr, Ser, Glu или Asp; аминокислота в положении 194 представляет собой Thr или Ser; аминокислота в положении 195 представляет собой Met или Thr; аминокислота в положении 196 представляет собой Gln, His, Leu, Asn, Ile, Val, Thr или Ser; аминокислота в положении 197 представляет собой Ala, Gly, Ile, Val или Leu; аминокислота в положении 198 представляет собой Thr, Glu, Ser, Asp, Gly или Ala; аминокислота в положении 199 представляет собой Pro, Lys или Arg; аминокислота в положении 200 представляет собой Asn, Ser, Thr, Gln, Ala или Gly; аминокислота в положении 201 представляет собой Ala, Leu, Glu, Ile, Asp или Trp; аминокислота в положении 202 представляет собой Ser, Asp, Phe, Ile, Val, Thr, Glu или Leu; аминокислота в положении 203 представляет собой His, Pro, Gly, Ala, Thr или Ser; аминокислота в положении 204 представляет собой Ile, Trp, His, Leu, Val, Ala или Gly; аминокислота в положении 205 представляет собой Ser, Asn, Leu, Val, Thr, Gln или Ile; аминокислота в положении 206 представляет собой Ala, Gly, Asp, Tyr, Glu, Lys или Arg; аминокислота в положении 207 представляет собой Phe, Val, Ile или Leu; аминокислота в положении 208 представляет собой Asn, Ser, Pro, Gln, Thr, Val, Ile или Leu; аминокислота в положении 210 представляет собой Arg, Asp, Glu, Lys, Ser или Tyr; аминокислота в положении 211 представляет собой Ile, Ser, Leu, Val или Thr; аминокислота в положении 212 представляет собой Val, Ala, Ile, Leu, Glu, Gly или Asp; аминокислота в положении 214 представляет собой Pro, Lys или Arg; аминокислота в положении 215 представляет собой Ser или Thr; аминокислота в положении 217 представляет собой Tyr или Phe; аминокислота в положении 218 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 219 представляет собой Val, Ile, Leu или Ala; аминокислота в положении 220 представляет собой Cys, Leu, Ile, Val, Thr или Ser; аминокислота в положении 221 представляет собой Pro или His; аминокислота в положении 222 представляет собой Leu, Arg, Lys, Ile, Val, Thr или Ser; аминокислота в положении 224 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 225 представляет собой Asp, Arg, Glu, Lys, Ser или Thr; аминокислота в положении 226 представляет собой Thr, Ser, Gln или Asn; аминокислота в положении 227 представляет собой Asp, Leu, Glu, Ile, Val или удалена; аминокислота в положении 228 представляет собой Thr, Ser или удалена; аминокислота в положении 229 представляет собой Tyr или удалена; аминокислота в положении 230 представляет собой Leu, Ile, Val или удалена; аминокислота в положении 231 представляет собой Gly, Ala или удалена; аминокислота в положении 232 представляет собой Ile, Leu, Val или удалена; аминокислота в положении 233 представляет собой Pro или удалена; аминокислота в положении 234 представляет собой Ala, Pro, Gly или удалена; аминокислота в положении 235 представляет собой Asp, Ile, Leu, Glu или Val; аминокислота в положении 236 представляет собой Val, Ser, Ile, Leu, Thr, Asp или Glu; аминокислота в положении 237 представляет собой Ala, Phe или Tyr; аминокислота в положении 238 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 239 представляет собой Val, Ser, Ile, Leu, Thr, Ala или Gly; аминокислота в положении 240 представляет собой Leu, Val или Ile; аминокислота в положении 243 представляет собой Asp или Glu; аминокислота в положении 249 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 252 представляет собой Leu, Ile, Val или Met; аминокислота в положении 257 представляет собой Thr или Ser; аминокислота в положении 259 представляет собой His, Ile, Val или Leu; аминокислота в положении 266 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 267 представляет собой Cys, Ala или Gly; аминокислота в положении 268 представляет собой His, Arg, Lys или Tyr; аминокислота в положении 272 представляет собой Asp или Glu; аминокислота в положении 273 представляет собой Val, Met, Ile или Leu; аминокислота в положении 274 представляет собой Val, Ile, Leu или Met; аминокислота в положении 278 представляет собой Gly или Ala; аминокислота в положении 279 представляет собой Glu, Asp, Gly или Val; аминокислота в положении 281 представляет собой Leu, Ile, Val, Gly или Ala; аминокислота в положении 282 представляет собой Asn, Leu или Ile; аминокислота в положении 285 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 286 представляет собой Lys, Asp, Arg или Glu; аминокислота в положении 287 представляет собой Leu, Ile или Val; аминокислота в положении 290 представляет собой Pro, Gln, Asn, Lys или Arg; аминокислота в положении 291 представляет собой Leu, Ile или Val; аминокислота в положении 292 представляет собой Lys, Arg, Ile, Leu или Val; аминокислота в положении 293 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 294 представляет собой Ser, Asn, Thr, Gln, Arg или Lys; аминокислота в положении 295 представляет собой Thr или Ser; аминокислота в положении 296 представляет собой Gln, Asn или His; аминокислота в положении 297 представляет собой Leu, Ile, Val или Met; аминокислота в положении 300 представляет собой Ser или Thr; аминокислота в положении 301 представляет собой Glu, Asp, Gly или Ala; аминокислота в положении 302 представляет собой Ser, Pro, Thr, Gly или Ala; аминокислота в положении 304 представляет собой Lys, Arg, Gln или Asn; аминокислота в положении 313 представляет собой Val, Leu или Ile; аминокислота в положении 314 представляет собой His, Glu, Asn, Asp или Gln; аминокислота в положении 315 представляет собой Ala, Cys, Gly, Thr или Ser; аминокислота в положении 316 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 317 представляет собой Met, Leu, Val или Ile; аминокислота в положении 319 представляет собой Met, Leu, Val или Ile; аминокислота в положении 320 представляет собой Val, Ile, Leu, Ala или Gly; аминокислота в положении 321 представляет собой Arg, Lys или Pro; аминокислота в положении 322 представляет собой Ile, Leu, Val или Phe; аминокислота в положении 323 представляет собой Gly, Ile, Leu или Val; аминокислота в положении 324 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 336 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 339 представляет собой Asn, Lys, Gln или Arg; аминокислота в положении 350 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 351 представляет собой Glu или Asp; аминокислота в положении 353 представляет собой Lys или Arg; аминокислота в положении 354 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 355 представляет собой Phe, Ile, Leu или Leu; аминокислота в положении 356 представляет собой Lys или Arg; аминокислота в положении 360 представляет собой Ile, Val, Leu, Gly или Ala; аминокислота в положении 363 представляет собой Gln, Ala, Cys, Glu, Phe, Gly, His, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 364 представляет собой Ile, Ala, Cys, Glu, Phe, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 365 представляет собой Leu, Ala, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Arg, Val, Trp или Tyr; аминокислота в положении 366 представляет собой Gly, Ala, Cys, Phe, His, Ile, Lys, Leu, Met, Asn, Ser, Thr или Val; аминокислота в положении 367 представляет собой Ser, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val или Trp; аминокислота в положении 368 представляет собой Tyr, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp; аминокислота в положении 369 представляет собой Leu, Ala, Cys, Asp, Phe, Gly, Ile, Met, Thr или Val; аминокислота в положении 370 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 371 представляет собой Gln, Ala, Cys, Asp, Glu, Phe, Gly, Ile, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 372 представляет собой Gln, Ala, Cys, Asp, Phe, Gly, His, Ile, Leu, Asn, Arg, Ser, Val или Tyr; аминокислота в положении 373 представляет собой Asn, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Gln, Ser, Thr, Val или Trp; аминокислота в положении 374 представляет собой Arg или Lys; аминокислота в положении 376 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 378 представляет собой Glu или Asp; аминокислота в положении 381 представляет собой Leu, Ile или Val; аминокислота в положении 388 представляет собой Ala, Thr, Gly или Ser; аминокислота в положении 395 представляет собой Arg или Lys; аминокислота в положении 396 представляет собой Glu, Gln, Asp, Asn, Ala или Gly; аминокислота в положении 399 представляет собой Asp, Gln, Glu или Asn; аминокислота в положении 400 представляет собой Asn, Thr, Ser, Glu, Gln или Asp; аминокислота в положении 401 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 402 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 406 представляет собой Asp или Glu; аминокислота в положении 408 представляет собой Leu, Ile, Val или Met; аминокислота в положении 410 представляет собой Gly, Ile, Val, Ala или Leu; аминокислота в положении 414 представляет собой Ala, Gly, Asp или Glu; аминокислота в положении 416 представляет собой Ser, Asn, Thr, Gln, Glu или Asp; аминокислота в положении 417 представляет собой Ser, Arg, Lys, Thr, Ala или Gly; аминокислота в положении 423 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 431 представляет собой Arg или Lys; аминокислота в положении 432 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 436 представляет собой Arg, Lys, Asp или Glu; аминокислота в положении 440 представляет собой Asn, Gln, Lys или Arg; аминокислота в положении 442 представляет собой Leu, Ile или Val; аминокислота в положении 447 представляет собой Ser, Lys, Thr или Arg; аминокислота в положении 448 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 451 представляет собой Gln, Asn или Met; аминокислота в положении 453 представляет собой Gly или Ala; аминокислота в положении 455 представляет собой Ala, Leu, Ile или Val; аминокислота в положении 457 представляет собой Leu, Ile или Val; аминокислота в положении 467 представляет собой Val, Ile, Leu, Gly или Ala; аминокислота в положении 471 представляет собой Gly или Ala; аминокислота в положении 475 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 483 представляет собой Gly или Ala; аминокислота в положении 493 представляет собой Gln, Asn или Gly; аминокислота в положении 504 представляет собой Val, Leu или Ile; аминокислота в положении 506 представляет собой Asp, Glu или His; аминокислота в положении 509 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 510 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 512 представляет собой Glu или Asp; аминокислота в положении 515 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 516 представляет собой Gln, Asn или His; аминокислота в положении 517 представляет собой Ile, Val или Leu; аминокислота в положении 519 представляет собой Asp, Asn, Glu, Gly или Gln; аминокислота в положении 522 представляет собой Val, Glu, Pro, Ile, Leu или Asp; аминокислота в положении 525 представляет собой Glu или Asp; аминокислота в положении 526 представляет собой Leu, Ile, Val или Met; аминокислота в положении 539 представляет собой Val, Leu или Ile; аминокислота в положении 555 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 556 представляет собой Trp, Phe, Thr или Tyr; аминокислота в положении 557 представляет собой Arg, Cys, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 558 представляет собой Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 559 представляет собой Lys, Ala, Cys, Phe, Gly, His, Ile, Leu, Asn, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 560 представляет собой Cys, Ala, Phe, Gly, Ile, Met, Asn, Arg, Ser, Thr или Val; аминокислота в положении 561 представляет собой Lys, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 562 представляет собой Asn, Cys, Asp, Glu, Gly, His, Leu, Met, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 563 представляет собой Val, Ala, Cys, Asp, Phe, His, Ile, Leu, Met, Asn, Gln, Thr или Trp; аминокислота в положении 564 представляет собой Ala, Cys, Gly, Met, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 571 представляет собой Ser, Thr или Cys; аминокислота в положении 575 представляет собой Val, Leu, Ile, Asp или Glu; аминокислота в положении 577 представляет собой Met, Leu, Val или Ile; аминокислота в положении 579 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 583 представляет собой Asp или Glu; аминокислота в положении 589 представляет собой Met, Ile, Val или Leu; аминокислота в положении 590 представляет собой Met, Ile, Val или Leu; аминокислота в положении 593 представляет собой Met, Leu, Val или Ile; аминокислота в положении 595 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 596 представляет собой Ser или Thr; аминокислота в положении 597 представляет собой Gln, Asn или His; аминокислота в положении 607 представляет собой Ala, Gly, Ile, Leu или Val; аминокислота в положении 608 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 612 представляет собой Tyr, His или Phe; аминокислота в положении 617 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 618 представляет собой Gln, Asn или His; аминокислота в положении 625 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 626 представляет собой Met, Leu, Val или Ile; аминокислота в положении 628 представляет собой Leu, Val или Ile; аминокислота в положении 633 представляет собой Ile, Leu, Val или Met; аминокислота в положении 634 представляет собой Leu, Ile, Val или Met; аминокислота в положении 642 представляет собой Arg, Lys или Met; аминокислота в положении 646 представляет собой Leu, Ala, Cys, Gly, Ile, Met, Asn, Gln, Ser, Thr или Val; аминокислота в положении 647 представляет собой Leu, Asp, Gly, Met, Asn, Gln или Thr; аминокислота в положении 648 представляет собой Met, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 649 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr; аминокислота в положении 650 представляет собой Thr, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Val или Tyr; аминокислота в положении 651 представляет собой Glu, Ala, Cys, Asp, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 652 представляет собой Leu, Cys, Phe, Ile, Lys, Met, Pro, Arg, Ser, Thr или Val; аминокислота в положении 653 представляет собой Thr, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Pro, Arg, Ser, Val или Trp; аминокислота в положении 654 представляет собой Thr, Ala, Cys, Phe, Ile, Lys, Leu, Met, Pro, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 655 представляет собой Trp, Phe или Tyr; аминокислота в положении 658 представляет собой Gly, Lys, Ala или Arg; аминокислота в положении 663 представляет собой Gly или Ala; аминокислота в положении 664 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 668 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 669 представляет собой Gln, Asn или His; аминокислота в положении 671 представляет собой Asn, Gln, Thr или Ser аминокислота в положении 675 представляет собой Ile, Val, Ile, Thr или Ser; аминокислота в положении 678 представляет собой Met, Ile, Ala, Leu, Ser или Thr; аминокислота в положении 682 представляет собой Pro, Asn или Gln; аминокислота в положении 683 представляет собой Ser, Thr или Pro; аминокислота в положении 685 представляет собой Asp, Glu, Asp или Asn; аминокислота в положении 694 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 697 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 704 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 714 представляет собой Ala или Gly; аминокислота в положении 721 представляет собой Ser, Thr или Phe; аминокислота в положении 722 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 724 представляет собой Ser или Thr; аминокислота в положении 734 представляет собой His, Asn или Gln; аминокислота в положении 736 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 737 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 739 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 740 представляет собой Ser, Thr или Met; аминокислота в положении 741 представляет собой Gly, Ala, Gln или Asn; аминокислота в положении 742 представляет собой Ile, Leu, Val, Ala или Gly; аминокислота в положении 743 представляет собой Gly или удалена; аминокислота в положении 745 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 751 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 753 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 754 представляет собой Thr или Ser; аминокислота в положении 756 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 757 представляет собой Val, Leu или Ile; аминокислота в положении 766 представляет собой Ile, Leu или Val; аминокислота в положении 771 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Asn, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 772 представляет собой Arg, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 773 представляет собой Asp, Ala, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 774 представляет собой Gln, Ala, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 775 представляет собой Val, Ala, Cys, Asp, Glu, Gly, His, Ile, Asn, Pro, Gln, Arg, Ser, Thr или Tyr; аминокислота в положении 776 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 777 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 778 представляет собой Phe, Ala, His, Ile, Leu, Met, Asn, Gln, Ser, Val, Trp или Tyr; аминокислота в положении 779 представляет собой Gln, Ala, Cys, Asp, Glu, Gly, His, Lys, Leu, Asn, Pro, Arg, Ser, Thr или Val; аминокислота в положении 780 представляет собой Ala, Cys, Asn, Pro, Gln или Ser; аминокислота в положении 781 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 782 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 783 представляет собой Pro, Ala, Cys, Asp, Glu, Gly, His, Asn, Gln, Arg, Ser, Thr или Val; аминокислота в положении 784 представляет собой Leu, Ala, Glu, Phe, His, Ile, Lys, Met, Asn, Pro, Gln, Ser, Thr, Val или Trp; аминокислота в положении 785 представляет собой Asn, Ala, Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 786 представляет собой Tyr, Phe, Ile, Leu или Trp; аминокислота в положении 787 представляет собой His, Asn или Gln; аминокислота в положении 788 представляет собой Tyr или Met; аминокислота в положении 789 представляет собой Ala, Lys или Arg; аминокислота в положении 790 представляет собой Tyr или Thr; аминокислота в положении 791 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 792 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 796 представляет собой Asp или Glu; аминокислота в положении 797 представляет собой Ser, Thr или Ala аминокислота в положении 802 представляет собой Glu, Lys, Asp, Asn или Gln; аминокислота в положении 806 представляет собой Gln, Asp, Glu, Asn или His; аминокислота в положении 810 представляет собой Lys, Arg или Thr; аминокислота в положении 819 представляет собой Arg, Lys или His; аминокислота в положении 829 представляет собой Lys, Ser, Ala или Pro; аминокислота в положении 832 представляет собой Ala, Lys, Arg, Asp или Glu; аминокислота в положении 833 представляет собой Gly, Ala, Asp или Glu; аминокислота в положении 842 представляет собой Leu, Ile, Val или Pro; аминокислота в положении 847 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 848 представляет собой Ile, Leu или Val; аминокислота в положении 849 представляет собой Val, Leu, Ile, Gly или Ala; аминокислота в положении 855 представляет собой Thr, Ser или Met; аминокислота в положении 860 представляет собой Ile, Leu или Val; и аминокислота в положении 864 представляет собой His, Asn или Gln.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников отдела Pteridophyta. Филогения папоротников, применяемая в данном документе, основана на классификации современных папоротников по A. R. Smith et al, TAXON, 55:705-731 (2006). Консенсусная филогения, основанная на классификации по A. R. Smith, показана на фигуре 1. Другие филогенетические классификации современных папоротников известны специалисту в данной области. Дополнительную информацию, касающуюся филогении папоротников, можно найти на сайте mobot.org/MOBOT/research/APweb/ (доступ к которому можно получить с применением префикса "www") и в работе Schuettpelz E. and Pryer K. M., TAXON 56: 1037-1050 (2007), и она основана на трех пластидных генах. Дополнительные виды папоротников и других примитивных растений можно найти на сайте homepages.caverock.net.nz/~bj/fern/list.htm (доступ к которому можно получить с применением префикса http://).
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников класса Psilotopsida. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников класса Psilotopsida, порядка Psilotales. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников класса Psilotopsida, порядка Ophioglossales. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников класса Psilotopsida, порядка Ophioglossales, семейства Psilotaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников класса Psilotopsida, порядка Ophioglossales семейства Ophioglossaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников рода Ophioglossum L., Botrychium, Botrypus, Helminthostachys, Ophioderma, Cheiroglossa, Sceptridium или Mankyua.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из видов класса Polypodiopsida/Pteridopsida. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Osmundales (королевские папоротники); семейства Osmundaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Hymenophyllales; семейства Hymenophyllaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Gleicheniales; семейства Gleicheniaceae, семейства Dipteridaceae или семейства Matoniaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Schizaeales; семейства Lygodiaceae, семейства Anemiaceae или семейства Schizaeaceae. В некоторых вариантах осуществления нуклеиновую кислоту, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Schizaeales; семейства Schizaeaceae, рода Lygodium, выбранного без ограничения из Lygodium articulatum, Lygodium circinatum, Lygodium conforme, Lygodium cubense, Lygodium digitatum, Lygodium flexuosum, Lygodium heterodoxum, Lygodium japonicum, Lygodium kerstenii, Lygodium lanceolatum, Lygodium longifolium, Lygodium merrilii, Lygodium micans, Lygodium microphyllum, Lygodium microstachyum, Lygodium oligostachyum, Lygodium palmatum, Lygodium polystachyum, Lygodium radiatum, Lygodium reticulatum, Lygodium salicifolium, Lygodium scandens, Lygodium smithianum, Lygodium subareolatum, Lygodium trifurcatum, Lygodium venustum, Lygodium versteeghii, Lygodium volubile и Lygodium yunnanense. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Salviniales; семейства Marsileaceae или семейства Salviniaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Cyatheales; семейства Thyrsopteridaceae, семейства Loxsomataceae, семейства Culcitaceae, семейства Plagiogyriaceae, семейства Cibotiaceae, семейства Cyatheaceae, семейства Dicksoniaceae или семейства Metaxyaceae.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales; семейства Lindsaeaceae, семейства Saccolomataceae, семейства Cystodiaceae, семейства Dennstaedtiaceae, семейства Pteridaceae, семейства Aspleniaceae, семейства Thelypteridaceae, семейства Woodsiaceae, семейства Onocleaceae, семейства Blechnaceae, семейства Dryopteridaceae, семейства Lomariopsidaceae, семейства Tectariaceae, семейства Oleandraceae, семейства Davalliaceae или семейства Polypodiaceae.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Pteridaceae, рода Adiantaceae, выбранного без ограничения из Adiantum aethiopicum, Adiantum aleuticum, Adiantum bonatianum, Adiantum cajennense, Adiantum capillus-junonis, Adiantum capillus-veneris, Adiantum caudatum, Adiantum chienii, Adiantum chilense, Adiantum cuneatum, Adiantum cunninghamii, Adiantum davidii, Adiantum diaphanum, Adiantum edentulum, Adiantum edgeworthii, Adiantum excisum, Adiantum fengianum, Adiantum fimbriatum, Adiantum flabellulatum, Adiantum formosanum, Adiantum formosum, Adiantum fulvum, Adiantum gravesii, Adiantum hispidulum, Adiantum induratum, Adiantum jordanii, Adiantum juxtapositum, Adiantum latifolium, Adiantum leveillei, Adiantum lianxianense, Adiantum malesianum, Adiantum mariesii, Adiantum monochlamys, Adiantum myriosorum, Adiantum obliquum, Adiantum ogasawarense, Adiantum pedatum, Adiantum pentadactylon, Adiantum peruvianum, Adiantum philippense, Adiantum princeps, Adiantum pubescens, Adiantum raddianum, Adiantum reniforme, Adiantum roborowskii, Adiantum serratodentatum, Adiantum sinicum, Adiantum soboliferum, Adiantum subcordatum, Adiantum tenerum, Adiantum terminatum, Adiantum tetraphyllum, Adiantum trapeziforme, Adiantum venustum, Adiantum viridescens и Adiantum viridimontanum.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Aspleniaceae, рода Asplenium.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Aspleniaceae, рода Asplenium, выбранного без ограничения из Asplenium adiantum, Asplenium adulterinum, Asplenium aequibasis, Asplenium aethiopicum, Asplenium africanum, Asplenium × alternifolium, Asplenium angustum, Asplenium antiquum, Asplenium ascensionis, Asplenium attenuatum, Asplenium aureum, Asplenium auritum, Asplenium australasicum, Asplenium azoricum, Asplenium bifrons, Asplenium billottii, Asplenium bipinnatifidum, Asplenium brachycarpum, Asplenium bradleyi, Asplenium bulbiferum, Asplenium caudatum, Asplenium ceterach, Asplenium compressum, Asplenium congestum, Asplenium corderoanum, Asplenium crinicaule, Asplenium cristatum, Asplenium cuneifolium, Asplenium cymbifolium, Asplenium daghestanicum, Asplenium dalhousiae, Asplenium dareoides, Asplenium daucifolium, Asplenium difforme, Asplenium fissum, Asplenium dimorphum, Asplenium divaricatum, Asplenium dregeanum, Asplenium × ebenoides, Asplenium ecuadorense, Asplenium feei Kunze, Asplenium fissum, Asplenium flabellifolium, Asplenium flaccidum, Asplenium fontanum, Asplenium forisiense, Asplenium formosum, Asplenium gemmiferum, Asplenium × germanicum, Asplenium gueinzii, Asplenium goudeyi, Asplenium hemionitis, Asplenium hermannii-christii, Asplenium hookerianum, Asplenium hybridum, Asplenium incisum, Asplenium × jacksonii, Asplenium × kenzoi, Asplenium laciniatum, Asplenium lamprophyllum, Asplenium laserpitiifolium, Asplenium lepidum, Asplenium listeri, Asplenium longissimum, Asplenium lucidum, Asplenium lunulatum, Asplenium lyallii, Asplenium macedonicum, Asplenium majoricum, Asplenium marinum, Asplenium × microdon, Asplenium milnei, Asplenium montanum, Asplenium musifolium, Asplenium nidus, Asplenium normale, Asplenium obliquum, Asplenium oblongifolium, Asplenium obovatum, Asplenium obtusatum, Asplenium oligolepidum, Asplenium oligophlebium, Asplenium onopteris, Asplenium pacificum, Asplenium paleaceum, Asplenium palmeri, Asplenium petrarchae, Asplenium pinnatifidum, Asplenium planicaule, Asplenium platybasis, Asplenium platyneuron, Asplenium polyodon, Asplenium praemorsum, Asplenium prolongatum, Asplenium pteridoides, Asplenium resiliens, Asplenium rhizophyllum, Asplenium richardii, Asplenium ruprechtii, Asplenium ruta-muraria, Asplenium rustifolium, Asplenium sagittatum, Asplenium sandersonii, Asplenium × sarniense, Asplenium schizotrichum, Asplenium schweinfurthii, Asplenium scleroprium, Asplenium scolopendrium (syn. Phyllitis scolopendrium), Asplenium seelosii, Asplenium septentrionale, Asplenium septentrionale × trichomanes, Asplenium serra, Asplenium serratum, Asplenium sessilifolium, Asplenium shuttleworthianum, Asplenium simplicifrons, Asplenium splendens, Asplenium surrogatum, Asplenium tenerum, Asplenium terrestre, Asplenium theciferum, Asplenium thunbergii, Asplenium trichomanes, Asplenium tutwilerae, Asplenium vespertinum, Asplenium vieillardii, Asplenium virens, Asplenium viride, Asplenium vittiforme и Asplenium viviparum.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Blechnaceae, рода Blecnum.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae рода Acrophorus, рода Acrorumohra, рода Anapausia, рода Arachniodes, рода Bolbitis, рода Ctenitis, рода Cyclodium, рода Cyrtogonellum, рода Cyrtomidictyum, рода Cyrtomium, рода Diacalpe, рода Didymochlaena, рода Dryopsis, рода Dryopteris, рода Elaphoglossum, рода Hypodematium, рода Lastreopsis, рода Leptorumohra, рода Leucostegia, рода Lithostegia, рода Lomagramma, рода Maxonia, рода Megalastrum, рода Olfersia, рода Peranema, рода Phanerophlebia, рода Phanerophlebiopsis, рода Polybotrya, рода Polystichopsis, рода Polystichum, рода Rumohra, рода Sorolepidium, рода Stigmatopteris или рода Teratophyllum.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Polystichum. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Polystichum, выбранного без ограничения из Polystichum acanthophyllum, Polystichum acrostichoides, Polystichum aculeatum, Polystichum acutidens, Polystichum acutipinnulum, Polystichum alcicorne, Polystichum aleuticum, Polystichum andersonii, Polystichum atkinsonii, Polystichum australiense, Polystichum bakerianum, Polystichum biaristatum, Polystichum bomiense, Polystichum bonseyi, Polystichum brachypterum, Polystichum braunii, Polystichum brachypterum, Polystichum calderonense, Polystichum californicum, Polystichum capillipes, Polystichum castaneum, Polystichum chilense, Polystichum christii Ching, Polystichum chunii Ching, Polystichum craspedosorum, Polystichum cyclolobum, Polystichum cystostegia, Polystichum deltodon, Polystichum dielsii, Polystichum discretum, Polystichum drepanum, Polystichum dudleyi, Polystichum duthiei, Polystichum echinatum, Polystichum erosum, Polystichum excellens, Polystichum eximium, Polystichum falcatipinnum, Polystichum falcinellum, Polystichum fallax, Polystichum formosanum, Polystichum gongboense, Polystichum grandifrons, Polystichum gymnocarpium, Polystichum haleakalense, Polystichum hancockii, Polystichum hecatopteron, Polystichum herbaceum, Polystichum imbricans, Polystichum incongruum, Polystichum kruckebergii, Polystichum kwakiutlii, Polystichum lachenense, Polystichum lanceolatum, Polystichum lemmonii, Polystichum lentum, Polystichum lonchitis, Polystichum longidens, Polystichum longipaleatum, Polystichum longipes, Polystichum luctuosum, Polystichum macleae, Polystichum macrochlaenum, Polystichum makinoi, Polystichum martini, Polystichum mayebarae, Polystichum mediocre, Polystichum medogense, Polystichum microchlamys, Polystichum mohrioides, Polystichum mollissimum, Polystichum monticola, Polystichum moorei, Polystichum morii, Polystichum moupinense, Polystichum muricatum, Polystichum nakenense, Polystichum neolobatum, Polystichum nepalense, Polystichum ningshenense, Polystichum obliquum, Polystichum omeiense, Polystichum ordinatum, Polystichum orientalitibeticum, Polystichum paramoupinense, Polystichum parvipinnulum, Polystichum piceopaleaceum, Polystichum polyblepharum, Polystichum prescottianum, Polystichum prionolepis, Polystichum proliferum, Polystichum pseudocastaneum, Polystichum pseudomakinoi, Polystichum punctiferum, Polystichum pungens, Polystichum qamdoense, Polystichum retrosopaleaceum, Polystichum rhombiforme, Polystichum rhomboidea, Polystichum richardii, Polystichum rigens, Polystichum rotundilobum, Polystichum scopulinum, Polystichum semifertile, Polystichum setiferum, Polystichum setigerum, Polystichum shensiense, Polystichum silvaticum, Polystichum simplicipinnum, Polystichum sinense, Polystichum squarrosum, Polystichum stenophyllum, Polystichum stimulans, Polystichum submite, Polystichum tacticopterum, Polystichum thomsoni, Polystichum tibeticum, Polystichum transvaalense, Polystichum tripteron, Polystichum tsus-simense, Polystichum vestitum, Polystichum wattii, Polystichum whiteleggei, Polystichum xiphophyllum, Polystichum yadongense и Polystichum yunnanense.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Rumohra. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Rumohra, выбранного без ограничения из Rumohra adiantiformis, Rumohra aristata, Rumohra bartonae, Rumohra berteroana, Rumohra capuronii, Rumohra glandulosa, Rumohra humbertii, Rumohra linearisquamosa, Rumohra lokohensis, Rumohra madagascarica и Rumohra quadrangularis.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae рода Campyloneurum, рода Drynaria, рода Lepisorus, рода Microgramma, рода Microsorum, рода Neurodium, рода Niphidium, рода Pecluma M.G., рода Phlebodium, рода Phymatosorus, рода Platycerium, рода Pleopeltis, рода Polypodium.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Microsorum.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Microsorum, выбранного без ограничения из Microsorum alatum, Microsorum angustifolium, Microsorum aurantiacum, Microsorum australiense, Microsorum baithoense, Microsorum basicordatum, Microsorum biseriatum, Microsorum brassii, Microsorum buergerianum, Microsorum chapaense, Microsorum cinctum, Microsorum commutatum, Microsorum congregatifolium, Microsorum cuneatum, Microsorum cuspidatum, Microsorum dengii, Microsorum egregium, Microsorum emeiensis, Microsorum ensatum, Microsorum ensiforme, Microsorum excelsum, Microsorum fortunei, Microsorum griseorhizoma, Microsorum grossum, Microsorum hemionitideum, Microsorum henryi, Microsorum heterocarpum, Microsorum heterolobum, Microsorum howense, Microsorum insigne, Microsorum intermedium, Microsorum kongtingense, Microsorum krayanense, Microsorum lanceolatum, Microsorum lancifolium, Microsorum lastii, Microsorum latilobatum, Microsorum leandrianum, Microsorum lineare, Microsorum linguiforme, Microsorum longissimum, Microsorum longshengense, Microsorum maculosum, Microsorum maximum, Microsorum membranaceum, Microsorum membranifolium, Microsorum microsorioides, Microsorum minor, Microsorum monstrosum, Microsorum muliense, Microsorum mutense, Microsorum nanchuanense, Microsorum ningpoense, Microsorum normale, Microsorum novae-zealandiae, Microsorum ovalifolium, Microsorum ovatum, Microsorum palmatopedatum, Microsorum pappei, Microsorum papuanum, Microsorum parksii, Microsorum pentaphyllum, Microsorum piliferum, Microsorum pitcairnense, Microsorum powellii, Microsorum pteropodum, Microsorum pteropus, Microsorum punctatum, Microsorum pustulatum, Microsorum rampans, Microsorum revolutum, Microsorum rubidum, Microsorum samarense, Microsorum sapaense, Microsorum sarawakense, Microsorum scandens, Microsorum scolopendria, Microsorum sibomense, Microsorum sinense, Microsorum sopuense, Microsorum спектр, Microsorum steerei, Microsorum subhemionitideum, Microsorum submarginale, Microsorum subnudum, Microsorum superficiale, Microsorum takhtajanii, Microsorum tenuipes, Microsorum tibeticum, Microsorum triglossum, Microsorum truncatum, Microsorum tsaii, Microsorum varians, Microsorum venosum, Microsorum vieillardii, Microsorum x inaequibasis, Microsorum yiliangensis и Microsorum zippelii.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Polypodium L. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Polypodium L., выбранного без ограничения из Polypodium absidatum, Polypodium acutifolium, Polypodium adiantiforme, Polypodium aequale, Polypodium affine, Polypodium albidopaleatum, Polypodium alcicorne, Polypodium alfarii, Polypodium alfredii, Polypodium alfredii var. curtii, Polypodium allosuroides, Polypodium alsophilicola, Polypodium amamianum, Polypodium amoenum, Polypodium amorphum, Polypodium anetioides, Polypodium anfractuosum, Polypodium anguinum, Polypodium angustifolium f. remotifolia, Polypodium angustifolium var. amphostenon, Polypodium angustifolium var. heterolepis, Polypodium angustifolium var. monstrosa, Polypodium angustipaleatum, Polypodium angustissimum, Polypodium anisomeron var. pectinatum, Polypodium antioquianum, Polypodium aoristisorum, Polypodium apagolepis, Polypodium apicidens, Polypodium apiculatum, Polypodium apoense, Polypodium appalachianum, Polypodium appressum, Polypodium arenarium, Polypodium argentinum, Polypodium argutum, Polypodium armatum, Polypodium aromaticum, Polypodium aspersum, Polypodium assurgens, Polypodium atrum, Polypodium auriculatum, Polypodium balaonense, Polypodium balliviani, Polypodium bamleri, Polypodium bangii, Polypodium bartlettii, Polypodium basale, Polypodium bernoullii, Polypodium biauritum, Polypodium bifrons, Polypodium blepharodes, Polypodium bolivari, Polypodium bolivianum, Polypodium bolobense, Polypodium bombycinum, Polypodium bombycinum var. insularum, Polypodium bradeorum, Polypodium bryophilum, Polypodium bryopodum, Polypodium buchtienii, Polypodium buesii, Polypodium bulbotrichum, Polypodium caceresii, Polypodium californicum f. brauscombii, Polypodium californicum f. parsonsiae, Polypodium californicum, Polypodium calophlebium, Polypodium calvum, Polypodium camptophyllarium var. abbreviatum, Polypodium capitellatum, Polypodium carpinterae, Polypodium chachapoyense, Polypodium chartaceum, Polypodium chimantense, Polypodium chiricanum, Polypodium choquetangense, Polypodium christensenii, Polypodium christii, Polypodium chrysotrichum, Polypodium ciliolepis, Polypodium cinerascens, Polypodium collinsii, Polypodium colysoides, Polypodium confluens, Polypodium conforme, Polypodium confusum, Polypodium congregatifolium, Polypodium connellii, Polypodium consimile var. bourgaeanum, Polypodium consimile var. minor, Polypodium conterminans, Polypodium contiguum, Polypodium cookii, Polypodium coriaceum, Polypodium coronans, Polypodium costaricense, Polypodium costatum, Polypodium crassifolium f. angustissimum, Polypodium crassifolium var. longipes, Polypodium crassulum, Polypodium craterisorum, Polypodium cryptum, Polypodium crystalloneuron, Polypodium cucullatum var. planum, Polypodium cuencanum, Polypodium cumingianum, Polypodium cupreolepis, Polypodium curranii, Polypodium curvans, Polypodium cyathicola, Polypodium cyathisorum, Polypodium cyclocolpon, Polypodium daguense, Polypodium damunense, Polypodium dareiformioides, Polypodium dasypleura, Polypodium decipiens, Polypodium decorum, Polypodium delicatulum, Polypodium deltoideum, Polypodium demeraranum, Polypodium denticulatum, Polypodium diaphanum, Polypodium dilatatum, Polypodium dispersum, Polypodium dissectum, Polypodium dissimulans, Polypodium dolichosorum, Polypodium dolorense, Polypodium donnell-smithii, Polypodium drymoglossoides, Polypodium ebeninum, Polypodium eggersii, Polypodium elmeri, Polypodium elongatum, Polypodium enterosoroides, Polypodium erubescens, Polypodium erythrolepis, Polypodium erythrotrichum, Polypodium eurybasis, Polypodium eurybasis var. villosum, Polypodium exornans, Polypodium falcoideum, Polypodium fallacissimum, Polypodium farinosum, Polypodium faucium, Polypodium feei, Polypodium ferrugineum, Polypodium feuillei, Polypodium firmulum, Polypodium firmum, Polypodium flaccidum, Polypodium flagellare, Polypodium flexuosum, Polypodium flexuosum var. ekmanii, Polypodium forbesii, Polypodium formosanum, Polypodium fraxinifolium subsp. articulatum, Polypodium fraxinifolium subsp. luridum, Polypodium fructuosum, Polypodium fucoides, Polypodium fulvescens, Polypodium galeottii, Polypodium glaucum, Polypodium glycyrrhiza, Polypodium gracillimum, Polypodium gramineum, Polypodium grandifolium, Polypodium gratum, Polypodium graveolens, Polypodium griseo-nigrum, Polypodium griseum, Polypodium guttatum, Polypodium haalilioanum, Polypodium hammatisorum, Polypodium hancockii, Polypodium haplophlebicum, Polypodium harrisii, Polypodium hastatum var. simplex, Polypodium hawaiiense, Polypodium heanophyllum, Polypodium helleri, Polypodium hemionitidium, Polypodium henryi, Polypodium herzogii, Polypodium hesperium, Polypodium hessii, Polypodium hombersleyi, Polypodium hostmannii, Polypodium humile, Polypodium hyalinum, Polypodium iboense, Polypodium induens var. subdentatum, Polypodium insidiosum, Polypodium insigne, Polypodium intermedium subsp. masafueranum var. obtuseserratum, Polypodium intramarginale, Polypodium involutum, Polypodium itatiayense, Polypodium javanicum, Polypodium juglandifolium, Polypodium kaniense, Polypodium knowltoniorum, Polypodium kyimbilense, Polypodium l'herminieri var. costaricense, Polypodium lachniferum f. incurvata, Polypodium lachniferum var. glabrescens, Polypodium lachnopus, Polypodium lanceolatum var. complanatum, Polypodium lanceolatum var. trichophorum, Polypodium latevagans, Polypodium laxifrons, Polypodium laxifrons var. lividum, Polypodium lehmannianum, Polypodium leiorhizum, Polypodium leptopodon, Polypodium leuconeuron var. angustifolia, Polypodium leuconeuron var. latifolium, Polypodium leucosticta, Polypodium limulum, Polypodium lindigii, Polypodium lineatum, Polypodium lomarioides, Polypodium longifrons, Polypodium loretense, Polypodium loriceum var. umbraticum, Polypodium loriforme, Polypodium loxogramme f. gigas, Polypodium ludens, Polypodium luzonicum, Polypodium lycopodioides f. obtusum, Polypodium lycopodioides L., Polypodium macrolepis, Polypodium macrophyllum, Polypodium macrosorum, Polypodium macrosphaerum, Polypodium maculosum, Polypodium madrense, Polypodium manmeiense, Polypodium margaritiferum, Polypodium maritimum, Polypodium martensii, Polypodium mayoris, Polypodium megalolepis, Polypodium melanotrichum, Polypodium menisciifolium var. pubescens, Polypodium meniscioides, Polypodium merrillii, Polypodium mettenii, Polypodium mexiae, Polypodium microsorum, Polypodium militare, Polypodium minimum, Polypodium minusculum, Polypodium mixtum, Polypodium mollendense, Polypodium mollissimum, Polypodium moniliforme var. minus, Polypodium monoides, Polypodium monticola, Polypodium montigenum, Polypodium moritzianum, Polypodium moultonii, Polypodium multicaudatum, Polypodium multilineatum, Polypodium multisorum, Polypodium munchii, Polypodium muscoides, Polypodium myriolepis, Polypodium myriophyllum, Polypodium myriotrichum, Polypodium nematorhizon, Polypodium nemorale, Polypodium nesioticum, Polypodium nigrescentium, Polypodium nigripes, Polypodium nigrocinctum, Polypodium nimbatum, Polypodium nitidissimum, Polypodium nitidissimum var. latior, Polypodium nubrigenum, Polypodium oligolepis, Polypodium oligosorum, Polypodium oligosorum, Polypodium olivaceum, Polypodium olivaceum var. elatum, Polypodium oodes, Polypodium oosphaerum, Polypodium oreophilum, Polypodium ornatissimum, Polypodium ornatum, Polypodium ovatum, Polypodium oxylobum, Polypodium oxypholis, Polypodium pakkaense, Polypodium pallidum, Polypodium palmatopedatum, Polypodium palmeri, Polypodium panamense, Polypodium parvum, Polypodium patagonicum, Polypodium paucisorum, Polypodium pavonianum, Polypodium pectinatum var. caliense, Polypodium pectinatum var. hispidum, Polypodium pellucidum, Polypodium pendulum var. boliviense, Polypodium percrassum, Polypodium perpusillum, Polypodium peruvianum var. subgibbosum, Polypodium phyllitidis var. elongatum, Polypodium pichinchense, Polypodium pilosissimum, Polypodium pilosissimum var. glabriusculum, Polypodium pilossimum var. tunguraquensis, Polypodium pityrolepis, Polypodium platyphyllum, Polypodium playfairii, Polypodium plebeium var. cooperi, Polypodium plectolepidioides, Polypodium pleolepis, Polypodium plesiosorum var.i, Polypodium podobasis, Polypodium podocarpum, Polypodium poloense, Polypodium polydatylon, Polypodium polypodioides var. aciculare, Polypodium polypodioides var. michauxianum, Polypodium praetermissum, Polypodium preslianum var. immersum, Polypodium procerum, Polypodium procerum, Polypodium productum, Polypodium productum, Polypodium prolongilobum, Polypodium propinguum, Polypodium proteus, Polypodium pruinatum, Polypodium pseudocapillare, Polypodium pseudofraternum, Polypodium pseudonutans, Polypodium pseudoserratum, Polypodium pulcherrimum, Polypodium pulogense, Polypodium pungens, Polypodium purpusii, Polypodium radicale, Polypodium randallii, Polypodium ratiborii, Polypodium reclinatum, Polypodium recreense, Polypodium repens var. abruptum, Polypodium revolvens, Polypodium rhachipterygium, Polypodium rhomboideum, Polypodium rigens, Polypodium robustum, Polypodium roraimense, Polypodium roraimense, Polypodium rosei, Polypodium rosenstockii, Polypodium rubidum, Polypodium rudimentum, Polypodium rusbyi, Polypodium sablanianum, Polypodium sarmentosum, Polypodium saxicola, Polypodium schenckii, Polypodium schlechteri, Polypodium scolopendria, Polypodium scolopendria, Polypodium scolopendrium, Polypodium scouleri, Polypodium scutulatum, Polypodium segregatum, Polypodium semihirsutum, Polypodium semihirsutum var. fuscosetosum, Polypodium senile var. minor, Polypodium sericeolanatum, Polypodium serraeforme, Polypodium serricula, Polypodium sesquipedala, Polypodium sessilifolium, Polypodium setosum var. calvum, Polypodium setulosum, Polypodium shaferi, Polypodium sibomense, Polypodium siccum, Polypodium simacense, Polypodium simulans, Polypodium singeri, Polypodium sinicum, Polypodium sintenisii, Polypodium skutchii, Polypodium sloanei, Polypodium sodiroi, Polypodium sordidulum, Polypodium sordidum, Polypodium sphaeropteroides, Polypodium sphenodes, Polypodium sprucei, Polypodium sprucei var. furcativenosa, Polypodium steirolepis, Polypodium stenobasis, Polypodium stenolepis, Polypodium stenopterum, Polypodium subcapillare, Polypodium subflabelliforme, Polypodium subhemionitidium, Polypodium subinaequale, Polypodium subintegrum, Polypodium subspathulatum, Polypodium subtile, Polypodium subvestitum, Polypodium subviride, Polypodium superficiale var. attenuatum, Polypodium superficiale var. chinensis, Polypodium sursumcurrens, Polypodium tablazianum, Polypodium taenifolium, Polypodium tamandarei, Polypodium tatei, Polypodium tenuiculum var. acrosora, Polypodium tenuiculum var. brasiliense, Polypodium tenuilore, Polypodium tenuinerve, Polypodium tepuiense, Polypodium teresae, Polypodium tetragonum var. incompletum, Polypodium thysanolepis var. bipinnatifidum, Polypodium thyssanolepis, var. thyssanolepis, Polypodium thyssanolepsi, Polypodium tobagense, Polypodium trichophyllum, Polypodium tridactylum, Polypodium tridentatum, Polypodium trifurcatum var. brevipes, Polypodium triglossum, Polypodium truncatulum, Polypodium truncicola var. major, Polypodium truncicola var. minor, Polypodium tuberosum, Polypodium tunguraguae, Polypodium turquinum, Polypodium turrialbae, Polypodium ursipes, Polypodium vagans, Polypodium valdealatum, Polypodium versteegii, Polypodium villagranii, Polypodium virginianum f. cambroideum, Polypodium virginianum f. peraferens, Polypodium vittarioides, Polypodium vulgare, Polypodium vulgare L., Polypodium vulgare subsp. oreophilum, Polypodium vulgare var. acuminatum, Polypodium vulpinum, Polypodium williamsii, Polypodium wobbense, Polypodium x fallacissimum-guttatum, Polypodium xantholepis, Polypodium xiphopteris, Polypodium yarumalense, Polypodium yungense, и Polypodium zosteriforme.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Platycerium.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из видов отдела Lycophyta. Филогения современных плаунов, как применяется в данном документе, основана на классификации по N. Wikstrom, American Fern Journal, 91:150-156 (2001). Другие филогенетические классификации современных плаунов известны специалисту в данной области. Дополнительную информацию, касающуюся филогении папоротников, можно найти на сайте mobot.org/MOBOT/research/APweb/ (доступ к которому можно получить с применением префикса "www") и в работе Schuettpelz E. and Pryer K. M., TAXON 56: 1037-1050 (2007), и она основана на трех пластидных генах. Дополнительные виды плаунов можно найти на сайте homepages.caverock.net.nz/~bj/fern/list.htm (доступ к которому можно получить с применением префикса http://).
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида класса Isoetopsida или класса Lycopodiopsida.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из видов класса Isoetopsida, порядка Selaginales. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников класса Isoetopsida, порядка Selaginales, семейства Selaginellaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из видов рода Selaginella.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из видов класса Lycopodiopsida, порядка Lycopodiales. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников класса Lycopodiopsida, порядка Lycopodiales семейства Lycopodiaceae или семейства Huperziaceae. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из видов рода Austrolycopodium, Dendrolycopodium, Diphasiastrum, Diphasium, Huperzia, Lateristachys, Lycopodiastrum, Lycopodiella, Lycopodium, Palhinhaea, Pseudodiphasium, Pseudolycopodiella, Pseudolycopodium или Spinulum. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из видов рода Lycopodium.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MP[DE]MPSEADWSIFVNE[IV]EAVAEGMPTEVSEVP[AV]WKAKCKN[MV]AALGREM[SC]I (SEQ ID NO: 646); МОТИВА 2 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PQLQYRMYG[NS]LI[KN]QMAQVAQNYDQ[ED]FKQ[FL]KLFI[IA]QNQI[LF]GSYLLQQN[KR]AF (SEQ ID NO: 647); МОТИВА 3 аминокислотной последовательности, представленного аминокислотной последовательностью формулы NTFMQMTPFTRWRLRLSASASENA[EG]LAFPTATA[PL]DSTT[EQ][IV]VITFHVTAIR (SEQ ID NO: 648); МОТИВА 4 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [DN]FTSRHVVK[GD]IPVSLLLDGEDWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 649); МОТИВА 5 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [ST]G[IT]VYILLQGSTIFHDRRR[DE]EVMTFQAA[DA]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 650); МОТИВА 6 аминокислотной последовательности, представленного аминокислотной последовательностью формулы S[HQ]ADRLAAIQP[AV]DLTN[HY]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI]LRIQNAALMYEY (SEQ ID NO: 651); МОТИВА 7 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [VL]DRVEFSEVMVIHRMYVRL[SA]DL[ND]VGEL[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 652); МОТИВА 8 аминокислотной последовательности, представленного аминокислотной последовательностью формулы A[DE]RELQMESFHSAVISQRRQEL[ND]TA[IF]AKM[DE]R[LM]SLQMEEE[NS]RAMEQAQKEM (SEQ ID NO: 653); МОТИВА 9 аминокислотной последовательности, представленного аминокислотной последовательностью формулы FVTAGATAPGA[AV]ASAGQAVSIAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 654); МОТИВА 10 аминокислотной последовательности, представленного аминокислотной последовательностью формулы DGMNWG[IT]YI[YH]GE[KE]V[EQ]RSPLLPSNAILAVWADRC[TI]ITSARHNH[VF]NAPGR[IV]I (SEQ ID NO: 655); МОТИВА 11 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [KV][VK][CA]RPPSPDM[MV]SAVAEHALWLNDVLLQVVQ[KN]ESQ[LM]QGT[AE]PYNECLALLGR (SEQ ID NO: 656); МОТИВА 12 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PTELT[VA]WPLGMDTV[AG]NLLIAQENAAL[VL]GLIQLGPSS (SEQ ID NO: 657); МОТИВА 13 аминокислотной последовательности, представленного аминокислотной последовательностью формулы RDQ[MT][HQ]MPGSVTVI[IV]LCRLLQFP[IT]DGSQA[TA]T (SEQ ID NO: 658); МОТИВА 14 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TSIPVEVVTDP[SN]ILLGMQTTV[LH]IAEL (SEQ ID NO: 659); МОТИВА 15 аминокислотной последовательности, представленного аминокислотной последовательностью формулы EGLR[EQ]FQNRQVARA[VL]FAVLKAVA[MQ]I[AG] (SEQ ID NO: 660); МОТИВА 16 аминокислотной последовательности, представленного аминокислотной последовательностью формулы W[TS]RVRIRHLEM[QH]F[AV]QEASG (SEQ ID NO: 661); МОТИВА 17 аминокислотной последовательности, представленного аминокислотной последовательностью формулы QISELQY[ED]IWVQG[LM][ML]RDIA (SEQ ID NO: 662); МОТИВА 18 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MDYSTLYRDLNQIS (SEQ ID NO: 664); МОТИВА 20 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LRLPFM[QK]LHARVIEQN[VR]K[SE] (SEQ ID NO: 665); МОТИВА 21 аминокислотной последовательности, представленного аминокислотной последовательностью формулы VDSLEQVG[QH][IL]V[GD]AP (SEQ ID NO: 666); МОТИВА 22 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [IV][EQ][CA]VMK[IM]GRF[VG][SL]VV (SEQ ID NO: 667); МОТИВА 23 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TLTNEPSE[EQ]F (SEQ ID NO: 668); и МОТИВА 24 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LPRQSRNISF (SEQ ID NO: 669).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MP[DE]MPSEADWSIFVNE[IV]EAVAEGMPTEVSEVP[AV]WKAKCKN[MV]AALGREM[SC]I (SEQ ID NO: 646); МОТИВА 2 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PQLQYRMYG[NS]LI[KN]QMAQVAQNYDQ[ED]FKQ[FL]KLFI[IA]QNQI[LF]GSYLLQQN[KR]AF (SEQ ID NO: 647); МОТИВА 3 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой NTFMQMTPFTRWRLRLSASASENA[EG]LAFPTATA[PL]DSTT[EQ][IV]VITFHVTAIR (SEQ ID NO: 648); МОТИВА 4 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [DN]FTSRHVVK[GD]IPVSLLLDGEDWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 649); МОТИВА 5 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой IIHQP[SA]T[RQ][ST]G[IT]VYILLQGSTIFHDRRR[DE]EVMTFQAA[DA]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 650); МОТИВА 6 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой S[HQ]ADRLAAIQP[AV]DLTN[HY]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI]LRIQNAALMYEY (SEQ ID NO: 651); МОТИВА 7 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [VL]DRVEFSEVMVIHRMYVRL[SA]DL[ND]VGEL[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 652); МОТИВА 8 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой A[DE]RELQMESFHSAVISQRRQEL[ND]TA[IF]AKM[DE]R[LM]SLQMEEE[NS]RAMEQAQKEM (SEQ ID NO: 653); МОТИВА 9 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой FVTAGATAPGA[AV]ASAGQAVSIAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 654); МОТИВА 10 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой DGMNWG[IT]YI[YH]GE[KE]V[EQ]RSPLLPSNAILAVWADRC[TI]ITSARHNH[VF]NAPGR[IV]I (SEQ ID NO: 655); МОТИВА 11 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [KV][VK][CA]RPPSPDM[MV]SAVAEHALWLNDVLLQVVQ[KN]ESQ[LM]QGT[AE]PYNECLALLGR (SEQ ID NO: 656); МОТИВА 12 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PTELT[VA]WPLGMDTV[AG]NLLIAQENAAL[VL]GLIQLGPSS (SEQ ID NO: 657); МОТИВА 13 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой RDQ[MT][HQ]MPGSVTVI[IV]LCRLLQFP[IT]DGSQA[TA]T (SEQ ID NO: 658); МОТИВА 14 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TSIPVEVVTDP[SN]ILLGMQTTV[LH]IAEL (SEQ ID NO: 659); МОТИВА 15 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой EGLR[EQ]FQNRQVARA[VL]FAVLKAVA[MQ]I[AG] (SEQ ID NO: 660); МОТИВА 16 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой W[TS]RVRIRHLEM[QH]F[AV]QEASG (SEQ ID NO: 661); МОТИВА 17 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой QISELQY[ED]IWVQG[LM][ML]RDIA (SEQ ID NO: 662); МОТИВА 18 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MDYSTLYRDLNQIS (SEQ ID NO: 664); МОТИВА 20 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LRLPFM[QK]LHARVIEQN[VR]K[SE] (SEQ ID NO: 665); МОТИВА 21 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой VDSLEQVG[QH][IL]V[GD]AP (SEQ ID NO: 666); МОТИВА 22 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [IV][EQ][CA]VMK[IM]GRF[VG][SL]VV (SEQ ID NO: 667); МОТИВА 23 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TLTNEPSE[EQ]F (SEQ ID NO: 668); и МОТИВА 24 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LPRQSRNISF (SEQ ID NO: 669).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AV]W[KR]AKCKN[MV]AALGREM[SC]I (SEQ ID NO: 670); МОТИВА 2 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PQLQYRMYG[NS]LI[KRN]QMAQVAQNYD[QR][ED]FK[QR][FL][KR]LFI[IAVL]QNQI[LF]GSYLL[QE]QN[KR]AF (SEQ ID NO: 671); МОТИВА 3 аминокислотной последовательности, представленного аминокислотной последовательностью формулы N[TK]FMQMTPFT[RH]WRLRLSASA[SPKA]EN[AK][EG]LAFPTATA[PL]DSTT[EQ][IV][VA]ITFHVTAIR (SEQ ID NO: 672); МОТИВА 4 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [DN]FTSRHVVK[GD]IPV[SN]LLLDG[EG]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 673); МОТИВА 5 аминокислотной последовательности, представленного аминокислотной последовательностью формулы IIHQP[SA]T[RQ][ST]G[IT][VI]YILLQGST[IV]FHDRRR[DE][EQ]V[ML]T[FP]QAA[DAV]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 674); МОТИВА 6 аминокислотной последовательности, представленного аминокислотной последовательностью формулы S[HQ]ADRLAAIQP[AV][DN]LTN[HYF]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI][LM]RIQNAAL[MR]YEY (SEQ ID NO: 675); МОТИВА 7 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [VL]D[RQ]VEFSEVMVIHRMYV[N]RL[SA]DL[ND]V[GA][EQ]L[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 676); МОТИВА 8 аминокислотной последовательности, представленного аминокислотной последовательностью формулы A[DE]RELQMESFH[SA]AVISQ[RK]R[QGE]EL[ND][TD][AT][IF]AKM[DE]R[LM]SLQMEEE[NSD][RG]AMEQA[QR]KEM (SEQ ID NO: 677); МОТИВА 9 аминокислотной последовательности, представленного аминокислотной последовательностью формулы F[VL]TAGATAPGA[AV]ASAGQAV[SN]IAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 678); МОТИВА 10 аминокислотной последовательности, представленного аминокислотной последовательностью формулы D[GD][MA][NK]WG[IT]Y[IV][YH][GA]E[KE]V[EQ][RVL]SPL[LYF][PN][SNG][NW][ASP][IY]L[AGV]V[WE]A[DQ]R[CS][TI]IT[SA]A[RFM]HN[HVT][VF][ND][AER]PG[RW][IV][IR] (SEQ ID NO: 679); МОТИВА 11 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [KV][VK][CA][RGC][PHY]PSP[DE][MIL][MV]SAV[AG][EV]HA[LIN]WL[NS][DK]VLL[QR]VVQ[KN]ES[QH][LM]QGT[AE][PSA]YNECLALLGR (SEQ ID NO: 680); МОТИВА 12 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [PN]T[EQ]LT[VAT]WPL[GR]MDTV[AG][ND]LLI[AT][QH]E[NS]AAL[VLS]GL[ITMA]QLG[PQ][SP]S (SEQ ID NO: 681); МОТИВА 13 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [RLC][DLWK][QNPR][MTP][HQR][MIL]PGSVTVI[IV]LCRLLQFP[IT][DG]G[SR][QFR][AS][TAD][TW] (SEQ ID NO: 682); МОТИВА 14 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [TA][SGV][IL]PV[ED]VVTDP[SN]IL[LM]GMQT[TS]V[LH]IAEL (SEQ ID NO: 683); МОТИВА 15 аминокислотной последовательности, представленного аминокислотной последовательностью формулы EGLR[EQ]FQN[RE]QVA[RN]A[VL]FAVL[KS][AS]VA[MQ]I[AG] (SEQ ID NO: 684); МОТИВА 16 аминокислотной последовательности, представленного аминокислотной последовательностью формулы W[TS]RVRIRHLEM[QH]F[AV][QK]E[AS][SM][GN] (SEQ ID NO: 685); МОТИВА 17 аминокислотной последовательности, представленного аминокислотной последовательностью формулы Q[IM]S[EQ]LQY[ED]IWVQG[LM][ML]RD[IM]A (SEQ ID NO: 686); МОТИВА 18 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [MLV]DY[SK][TSK]L[YF][RE]DLNQIS (SEQ ID NO: 687); МОТИВА 20 аминокислотной последовательности, представленного аминокислотной последовательностью формулы L[RHQ]L[PT]FM[QK]LHA[RIT][VQL][IR]E[QER][NF][VR][KWS][SE] (SEQ ID NO: 688); МОТИВА 21 аминокислотной последовательности, представленного аминокислотной последовательностью формулы VV[DN][SA]L[ED]QV[GS][QH][IL]V[GD]AP (SEQ ID NO: 689); МОТИВА 22 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [IV][EQH][CAS][VA][MI]K[IM][GV][RP][FI][VG][SL]VV (SEQ ID NO: 690); МОТИВА 23 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TLTN[EQ]PSE[EQDH]F (SEQ ID NO: 691); и МОТИВА 24 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LP[RS]QS[RT]N[IV]SF (SEQ ID NO: 692).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AV]W[KR]AKCKN[MV]AALGREM[SC]I (SEQ ID NO: 670); МОТИВА 2 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PQLQYRMYG[NS]LI[KRN]QMAQVAQNYD[QR][ED]FK[QR][FL][KR]LFI[IAVL]QNQI[LF]GSYLL[QE]QN[KR]AF (SEQ ID NO: 671); МОТИВА 3 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой N[TK]FMQMTPFT[RH]WRLRLSASA[SPKA]EN[AK][EG]LAFPTATA[PL]DSTT[EQ][IV][VA]ITFHVTAIR (SEQ ID NO: 672); МОТИВА 4 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [DN]FTSRHVVK[GD]IPV[SN]LLLDG[EG]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 673); МОТИВА 5 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой IIHQP[SA]T[RQ][ST]G[IT][VI]YILLQGST[IV]FHDRRR[DE][EQ]V[ML]T[FP]QAA[DAV]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 674); МОТИВА 6 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой S[HQ]ADRLAAIQP[AV][DN]LTN[HYF]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI][LM]RIQNAAL[MR]YEY (SEQ ID NO: 675); МОТИВА 7 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [VL]D[RQ]VEFSEVMVIHRMYV[N]RL[SA]DL[ND]V[GA][EQ]L[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 676); МОТИВА 8 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой A[DE]RELQMESFH[SA]AVISQ[RK]R[QGE]EL[ND][TD][AT][IF]AKM[DE]R[LM]SLQMEEE[NSD][RG]AMEQA[QR]KEM (SEQ ID NO: 677); МОТИВА 9 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой F[VL]TAGATAPGA[AV]ASAGQAV[SN]IAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 678); МОТИВА 10 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой D[GD][MA][NK]WG[IT]Y[IV][YH][GA]E[KE]V[EQ][RVL]SPL[LYF][PN][SNG][NW][ASP][IY]L[AGV]V[WE]A[DQ]R[CS][TI]IT[SA]A[RFM]HN[HVT][VF][ND][AER]PG[RW][IV][IR] (SEQ ID NO: 679); МОТИВА 11 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [KV][VK][CA][RGC][PHY]PSP[DE][MIL][MV]SAV[AG][EV]HA[LIN]WL[NS][DK]VLL[QR]VVQ[KN]ES[QH][LM]QGT[AE][PSA]YNECLALLGR (SEQ ID NO: 680); МОТИВА 12 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [PN]T[EQ]LT[VAT]WPL[GR]MDTV[AG][ND]LLI[AT][QH]E[NS]AAL[VLS]GL[ITMA]QLG[PQ][SP]S (SEQ ID NO: 681); МОТИВА 13 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [RLC][DLWK][QNPR][MTP][HQR][MIL]PGSVTVI[IV]LCRLLQFP[IT][DG]G[SR][QFR][AS][TAD][TW] (SEQ ID NO: 682); МОТИВА 14 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [TA][SGV][IL]PV[ED]VVTDP[SN]IL[LM]GMQT[TS]V[LH]IAEL (SEQ ID NO: 683); МОТИВА 15 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой EGLR[EQ]FQN[RE]QVA[RN]A[VL]FAVL[KS][AS]VA[MQ]I[AG] (SEQ ID NO: 684); МОТИВА 16 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой W[TS]RVRIRHLEM[QH]F[AV][QK]E[AS][SM][GN] (SEQ ID NO: 685); МОТИВА 17 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой Q[IM]S[EQ]LQY[ED]IWVQG[LM][ML]RD[IM]A (SEQ ID NO: 686); МОТИВА 18 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [MLV]DY[SK][TSK]L[YF][RE]DLNQIS (SEQ ID NO: 687); МОТИВА 20 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой L[RHQ]L[PT]FM[QK]LHA[RIT][VQL][IR]E[QER][NF][VR][KWS][SE] (SEQ ID NO: 688); МОТИВА 21 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой V[DN][SA]L[ED]QV[GS][QH][IL]V[GD]AP (SEQ ID NO: 689); МОТИВА 22 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [IV][EQH][CAS][VA][MI]K[IM][GV][RP][FI][VG][SL]VV (SEQ ID NO: 690); МОТИВА 23 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TLTN[EQ]PSE[EQDH]F (SEQ ID NO: 691); и МОТИВА 24 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LP[RS]QS[RT]N[IV]SF (SEQ ID NO: 692).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AVIL]W[KR]AKCKN[MVIL]AALGREM[SCT]I (SEQ ID NO: 693); МОТИВА 2 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PQLQYRMYG[NS]LI[KRNQ]QMAQVAQNYD[QRNK][ED]FK[QRNK][FL][KR]LFI[IAVL]QNQI[LFIV]GSYLL[QEND]QN[KR]AF (SEQ ID NO: 694); МОТИВА 3 аминокислотной последовательности, представленного аминокислотной последовательностью формулы N[TKSR]FMQMTPFT[RHK]WRLRLSASA[SPKATR]EN[AKR][EG]LAFPTATA[PLIV]DSTT[EQND][IVL][VAIL]ITFHVTAIR (SEQ ID NO: 695); МОТИВА 4 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [DNQE]FTSRHVVK[GDE]IPV[SNTQ]LLLDG[EGD]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 696); МОТИВА 5 аминокислотной последовательности, представленного аминокислотной последовательностью формулы IIHQP[SAT]T[RQKN][ST]G[ITLVS][VIL]YILLQGST[IVL]FHDRRR[DE][EQDN]V[MLIV]T[FP]QAA[DAVEIL]PLN[FY][QHN]YAYRLDTG (SEQ ID NO: 697); МОТИВА 6 аминокислотной последовательности, представленного аминокислотной последовательностью формулы S[HQN]ADRLAAIQP[AVIL][DN]LTN[HYF]LEMAT[HQN]MDMRTT[RSKT][MILV]L[ILV]GLLN[MILV][LMIV]RIQNAAL[MRILVK]YEY (SEQ ID NO: 698); МОТИВА 7 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [VLI]D[RQKN]VEFSEVMVIHRMYV[N]RL[SAT]DL[NDQE]V[GA][EQND]L[PED]GA[EGD][RK]VKR[VLI]YV[FLIV]ADVVE (SEQ ID NO: 699); МОТИВА 8 аминокислотной последовательности, представленного аминокислотной последовательностью формулы A[DE]RELQMESFH[SAT]AVISQ[RK]R[QGEND]EL[NDQE][TDSE][ATS][IFLV]AKM[DE]R[LMIV]SLQMEEE[NSDQET][RGK]AMEQA[QRNK]KEM (SEQ ID NO: 700); МОТИВА 9 аминокислотной последовательности, представленного аминокислотной последовательностью формулы F[VLI]TAGATAPGA[AVIL]ASAGQAV[SNTQ]IAGQAAQ[AG]LRRVVEILE[GQN]LEAVMEVVAA[VIL]K (SEQ ID NO: 701); МОТИВА 10 аминокислотной последовательности, представленного аминокислотной последовательностью формулы D[GDE][MA][NKQK]WG[ITLVS]Y[IVL][YH][GA]E[KERD]V[EQND][RVLKI]SPL[LYFIV][PNQ][SNGTQ][NWQ][ASPT][IYLV]L[AGVIL]V[WED]A[DQNE]R[CST][TISLV]IT[SAT]A[RFMK]HN[HVTILS][VFIL][NDQE][AERDK]PG[RWK][IVL][IRLVK] (SEQ ID NO: 702); МОТИВА 11 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [KVRIL][VKRIL][CA][RGCK][PHY]PSP[DE][MILV][MVIL]SAV[AG][EVDIL]HA[LINVQ]WL[NSQT][DKER]VLL[QRNK]VVQ[KNRQ]ES[QHN][LMIV]QGT[AED][PSAT]YNECLALLGR (SEQ ID NO: 703); МОТИВА 12 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [PNQ]T[EQDN]LT[VATILS]WPL[GRK]MDTV[AG][NDQE]LLI[ATS][QHN]E[NSQT]AAL[VLSIT]GL[ITMALVS]QLG[PQN][SPT]S (SEQ ID NO: 704); МОТИВА 13 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [RLCKIV][DLWKEIVR][QNPRK][MTP][HQR][MILV]PGSVTVI[IVL]LCRLLQFP[ITLVS][DGE]G[SRTK][QFRNK][AST][TADES][TWS] (SEQ ID NO: 705); МОТИВА 14 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [TA][SGVTIL][ILV]PV[ED]VVTDP[SNTQ]IL[LMIV]GMQT[TS]V[LHIV]IAEL (SEQ ID NO: 706); МОТИВА 15 аминокислотной последовательности, представленного аминокислотной последовательностью формулы EGLR[EQND]FQN[REKD]QVA[RNKQ]A[VLI]FAVL[KSRT][AST]VA[MQN]I[AG] (SEQ ID NO: 707); МОТИВА 16 аминокислотной последовательности, представленного аминокислотной последовательностью формулы W[TS]RVRIRHLEM[QHN]F[AVIL][QKNR]E[AST][SMT][GNQ] (SEQ ID NO: 708); МОТИВА 17 аминокислотной последовательности, представленного аминокислотной последовательностью формулы Q[IMLV]S[EQND]LQY[ED]IWVQG[LMIV][MLIV]RD[IMLV]A (SEQ ID NO: 709); МОТИВА 18 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [MLVI]DY[SKTR][TSKR]L[YF][REKD]DLNQIS (SEQ ID NO: 710); МОТИВА 20 аминокислотной последовательности, представленного аминокислотной последовательностью формулы L[RHQKN]L[PTS]FM[QKNR]LHA[RITKLVS][VQLIN][IRLVK]E[QERNDK][NFQ][VRILK][KWSRT][SETD] (SEQ ID NO: 711); МОТИВА 21 аминокислотной последовательности, представленного аминокислотной последовательностью формулы V[DNQE][SAT]L[ED]QV[GST][QHN][ILV]V[GDE]AP (SEQ ID NO: 712); МОТИВА 22 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [IVL][EQHND][CAST][VAIL][MILV]K[IMLV][GVIL][RPK][FILV][VGIL][SLTIV]VV (SEQ ID NO: 713); МОТИВА 23 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TLTN[EQDN]PSE[EQDHN]F (SEQ ID NO: 714); и МОТИВА 24 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LP[RSKT]QS[RTKS]N[IVL]SF (SEQ ID NO: 715).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AVIL]W[KR]AKCKN[MVIL]AALGREM[SCT]I (SEQ ID NO: 693); МОТИВА 2 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PQLQYRMYG[NS]LI[KRNQ]QMAQVAQNYD[QRNK][ED]FK[QRNK][FL][KR]LFI[IAVL]QNQI[LFIV]GSYLL[QEND]QN[KR]AF (SEQ ID NO: 694); МОТИВА 3 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой N[TKSR]FMQMTPFT[RHK]WRLRLSASA[SPKATR]EN[AKR][EG]LAFPTATA[PLIV]DSTT[EQND][IVL][VAIL]ITFHVTAIR (SEQ ID NO: 695); МОТИВА 4 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [DNQE]FTSRHVVK[GDE]IPV[SNTQ]LLLDG[EGD]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 696); МОТИВА 5 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой IIHQP[SAT]T[RQKN][ST]G[ITLVS][VIL]YILLQGST[IVL]FHDRRR[DE][EQDN]V[MLIV]T[FP]QAA[DAVEIL]PLN[FY][QHN]YAYRLDTG (SEQ ID NO: 697); МОТИВА 6 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой S[HQN]ADRLAAIQP[AVIL][DN]LTN[HYF]LEMAT[HQN]MDMRTT[RSKT][MILV]L[ILV]GLLN[MILV][LMIV]RIQNAAL[MRILVK]YEY (SEQ ID NO: 698); МОТИВА 7 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [VLI]D[RQKN]VEFSEVMVIHRMYV[N]RL[SAT]DL[NDQE]V[GA][EQND]L[PED]GA[EGD][RK]VKR[VLI]YV[FLIV]ADVVE (SEQ ID NO: 699); МОТИВА 8 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой A[DE]RELQMESFH[SAT]AVISQ[RK]R[QGEND]EL[NDQE][TDSE][ATS][IFLV]AKM[DE]R[LMIV]SLQMEEE[NSDQET][RGK]AMEQA[QRNK]KEM (SEQ ID NO: 700); МОТИВА 9 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой F[VLI]TAGATAPGA[AVIL]ASAGQAV[SNTQ]IAGQAAQ[AG]LRRVVEILE[GQN]LEAVMEVVAA[VIL]K (SEQ ID NO: 701); МОТИВА 10 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой D[GDE][MA][NKQK]WG[ITLVS]Y[IVL][YH][GA]E[KERD]V[EQND][RVLKI]SPL[LYFIV][PNQ][SNGTQ][NWQ][ASPT][IYLV]L[AGVIL]V[WED]A[DQNE]R[CST][TISLV]IT[SAT]A[RFMK]HN[HVTILS][VFIL][NDQE][AERDK]PG[RWK][IVL][IRLVK] (SEQ ID NO: 702); МОТИВА 11 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [KVRIL][VKRIL][CA][RGCK][PHY]PSP[DE][MILV][MVIL]SAV[AG][EVDIL]HA[LINVQ]WL[NSQT][DKER]VLL[QRNK]VVQ[KNRQ]ES[QHN][LMIV]QGT[AED][PSAT]YNECLALLGR (SEQ ID NO: 703); МОТИВА 12 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [PNQ]T[EQDN]LT[VATILS]WPL[GRK]MDTV[AG][NDQE]LLI[ATS][QHN]E[NSQT]AAL[VLSIT]GL[ITMALVS]QLG[PQN][SPT]S (SEQ ID NO: 704); МОТИВА 13 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [RLCKIV][DLWKEIVR][QNPRK][MTP][HQR][MILV]PGSVTVI[IVL]LCRLLQFP[ITLVS][DGE]G[SRTK][QFRNK][AST][TADES][TWS] (SEQ ID NO: 705); МОТИВА 14 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [TA][SGVTIL][ILV]PV[ED]VVTDP[SNTQ]IL[LMIV]GMQT[TS]V[LHIV]IAEL (SEQ ID NO: 706); МОТИВА 15 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой EGLR[EQND]FQN[REKD]QVA[RNKQ]A[VLI]FAVL[KSRT][AST]VA[MQN]I[AG] (SEQ ID NO: 707); МОТИВА 16 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой W[TS]RVRIRHLEM[QHN]F[AVIL][QKNR]E[AST][SMT][GNQ] (SEQ ID NO: 708); МОТИВА 17 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой Q[IMLV]S[EQND]LQY[ED]IWVQG[LMIV][MLIV]RD[IMLV]A (SEQ ID NO: 709); МОТИВА 18 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [MLVI]DY[SKTR][TSKR]L[YF][REKD]DLNQIS (SEQ ID NO: 710); МОТИВА 20 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой L[RHQKN]L[PTS]FM[QKNR]LHA[RITKLVS][VQLIN][IRLVK]E[QERNDK][NFQ][VRILK][KWSRT][SETD] (SEQ ID NO: 711); МОТИВА 21 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой V[DNQE][SAT]L[ED]QV[GST][QHN][ILV]V[GDE]AP (SEQ ID NO: 712); МОТИВА 22 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [IVL][EQHND][CAST][VAIL][MILV]K[IMLV][GVIL][RPK][FILV][VGIL][SLTIV]VV (SEQ ID NO: 713); МОТИВА 23 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TLTN[EQDN]PSE[EQDHN]F (SEQ ID NO: 714); и МОТИВА 24 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LP[RSKT]QS[RTKS]N[IVL]SF (SEQ ID NO: 715).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца, МОТИВ аминокислотной последовательности, выбранный из: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 10 (SEQ ID NO: 655, SEQ ID NO: 679 или SEQ ID NO: 702), MOTIF 18 (SEQ ID NO: 663), MOTIF 24 (SEQ ID NO: 669, SEQ ID NO: 692 или SEQ ID NO: 715), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 11 (SEQ ID NO: 656, SEQ ID NO: 680 или SEQ ID NO: 703), MOTIF 22 (SEQ ID NO: 667, SEQ ID NO: 690 или SEQ ID NO: 713), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца, МОТИВ аминокислотной последовательности, выбранный из: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца, мотивы аминокислотной последовательности: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 10 (SEQ ID NO: 655, SEQ ID NO: 679 или SEQ ID NO: 702), MOTIF 18 (SEQ ID NO: 663), MOTIF 24 (SEQ ID NO: 669, SEQ ID NO: 692 или SEQ ID NO: 715), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 11 (SEQ ID NO: 656, SEQ ID NO: 680 или SEQ ID NO: 703), MOTIF 22 (SEQ ID NO: 667, SEQ ID NO: 690 или SEQ ID NO: 713), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца, мотивы аминокислотной последовательности: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, содержащий МОТИВ аминокислотной последовательности из: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 10 (SEQ ID NO: 655, SEQ ID NO: 679 или SEQ ID NO: 702), MOTIF 18 (SEQ ID NO: 663), MOTIF 24 (SEQ ID NO: 669, SEQ ID NO: 692 или SEQ ID NO: 715), и/или MOTIF 14 с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот, содержащий МОТИВ аминокислотной последовательности из MOTIF 22 (SEQ ID NO: 667, SEQ ID NO: 690 или SEQ ID NO: 713), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), и/или MOTIF 12, со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот, содержащий МОТИВ аминокислотной последовательности из MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и/или MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695), с консенсусной вторичной структурой, содержащий структуру преимущественно бета-тяжа.
В некоторых вариантах осуществления полипептид PtIP-83 содержит МОТИВ аминокислотной последовательности в положениях, показанных в таблице 2.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий 8-10 сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий 6-8 сегментов со структурой преимущественно бета-тяжа. Применяемое в данном документе "главным образом неконсервативная вторичная структура" означает, что участки вторичной структуры не выравниваются единообразно в пределах семейства полипептидов PtIP. Применяемое в данном документе "структура преимущественно альфа-спирали" означает, что прогноз вторичной структуры может иметь один или несколько гэпов размером 1-6 аминокислот со структурой клубка и/или бета-тяжа, вставленных в структуру альфа-спирали. Применяемое в данном документе "структура преимущественно бета-тяжа" означает, что прогноз вторичной структуры может иметь один или несколько гэпов размером 1-6 аминокислот со структурой клубка и/или альфа-спирали, вставленных в структуру бета-тяжа. В некоторых вариантах осуществления вторичную структуру получают с помощью PSIPRED, ведущего способа прогнозирования вторичной структуры (Jones DT. (1999) J. Mol. Biol. 292: 195-202).
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий девять сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов со структурой преимущественно бета-тяжа.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой, где Участок A содержит консервативный бета-тяж 1 (β1a) длиной от приблизительно 4 до приблизительно 12 аминокислот в пределах от приблизительно аминокислотного остатка 30 до приблизительно аминокислотного остатка 130 от N-конца полипептида PtIP-83; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий девять сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов со структурой преимущественно бета-тяжа.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот с гибкой консенсусной вторичной структурой, где Участок A содержит консервативный бета-тяж 1 (β1a) длиной от приблизительно 4 до приблизительно 12 аминокислот, клубок длиной от приблизительно 3 до приблизительно 18 аминокислот и бета-тяж 2 (β1b) длиной от приблизительно 4 до приблизительно 32 аминокислот в пределах от приблизительно аминокислотного остатка 50 до приблизительно аминокислотного остатка 165 от N-конца полипептида PtIP-83; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий девять сегментов преимущественно с структурой альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов преимущественно со структурой бета-тяжа.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) альфа-спираль-1 длиной от приблизительно 10 до приблизительно 26 аминокислот; ii) клубок-1 длиной от приблизительно 2 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-1 и альфа-спиралью-2; iii) альфа-спираль-2 длиной от приблизительно 15 до приблизительно 24 аминокислот; iv) клубок-2 длиной от приблизительно 4 до приблизительно 14 аминокислот, фланкированный альфа-спиралью-2 и альфа-спиралью-3; v) альфа-спираль 3 длиной от приблизительно 15 до приблизительно 27 аминокислот; vi) клубок-3 длиной от приблизительно 11 до приблизительно 13 аминокислот, фланкированный альфа-спиралью-3 и альфа-спиралью-4; vii) альфа-спираль-4 длиной приблизительно 24-180 аминокислот; viii) клубок-4 длиной от приблизительно 4 до приблизительно 5 аминокислот, фланкированный альфа-спиралью-4 и альфа-спиралью-5; ix) альфа-спираль-5 длиной от приблизительно 50 до приблизительно 54 аминокислот ; x) клубок-5 длиной от приблизительно 11 до приблизительно 17 аминокислот, фланкированный альфа-спиралью-5 и альфа-спиралью-6; xi) альфа-спираль-6 длиной от приблизительно 15 до приблизительно 16 аминокислот; xii) клубок-6 длиной от приблизительно 6 до приблизительно 9 аминокислот, фланкированный альфа-спиралью-6 и альфа-спиралью-7; xiii) альфа-спираль-7 длиной от приблизительно 49 до приблизительно 55 аминокислот; xiv) клубок-7 длиной от приблизительно 3 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-7 и альфа-спиралью-8; xv) альфа-спираль-8 длиной от приблизительно 33 до приблизительно 36 аминокислот; xvi) клубок-8 длиной от приблизительно 14 до приблизительно 16 аминокислот, фланкированный альфа-спиралью-8 и альфа-спиралью-9; xvii) альфа-спираль-9 длиной от приблизительно 16 до приблизительно 23 аминокислот; xviii) клубок-9 длиной от приблизительно 21 до приблизительно 28 аминокислот, фланкированный альфа-спиралью-9 и Участком C; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов со структурой преимущественно бета-тяжа.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий десять сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) бета-тяж-1 (β1) длиной от приблизительно 3 аминокислот до приблизительно 5 аминокислот; ii) клубок длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот; iii) бета-тяж-2 (β2) длиной от приблизительно 7 аминокислот до приблизительно 11 аминокислот; iv) клубок длиной от приблизительно 17 аминокислот до приблизительно 23 аминокислот; v) бета-тяж-3 (β3) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; vi) клубок длиной от приблизительно 12 аминокислот до приблизительно 14 аминокислот; vii) бета-тяж-4 (β4) длиной от приблизительно 5 аминокислот до приблизительно 6 аминокислот; viii) клубок длиной от приблизительно 2 аминокислот до приблизительно 7 аминокислот; ix) бета-тяж-5 (β5) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; x) клубок длиной от приблизительно 26 аминокислот до приблизительно 28 аминокислот; xi) бета-тяж-6 (β6) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; xii) клубок длиной от приблизительно 16 аминокислот до приблизительно 20 аминокислот; и xiii) бета-тяж-1 (β7) длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) альфа-спираль-1 длиной от приблизительно 10 до приблизительно 26 аминокислот; ii) клубок-1 длиной от приблизительно 2 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-1 и альфа-спиралью-2; iii) альфа-спираль-2 длиной от приблизительно 15 до приблизительно 24 аминокислот; iv) клубок-2 длиной от приблизительно 4 до приблизительно 14 аминокислот, фланкированный альфа-спиралью-2 и альфа-спиралью-3; v) альфа-спираль 3 длиной от приблизительно 15 до приблизительно 27 аминокислот; vi) клубок-3 длиной от приблизительно 11 до приблизительно 13 аминокислот, фланкированный альфа-спиралью-3 и альфа-спиралью-4; vii) альфа-спираль-4 длиной приблизительно 24-180 аминокислот; viii) клубок-4 длиной от приблизительно 4 до приблизительно 5 аминокислот, фланкированный альфа-спиралью-4 и альфа-спиралью-5; ix) альфа-спираль-5 длиной от приблизительно 50 до приблизительно 54 аминокислот; x) клубок-5 длиной от приблизительно 11 до приблизительно 17 аминокислот, фланкированный альфа-спиралью-5 и альфа-спиралью-6; xi) альфа-спираль-6 длиной от приблизительно 15 до приблизительно 16 аминокислот; xii) клубок-6 длиной от приблизительно 6 до приблизительно 9 аминокислот, фланкированный альфа-спиралью-6 и альфа-спиралью-7; xiii) альфа-спираль-7 длиной от приблизительно 49 до приблизительно 55 аминокислот; xiv) клубок-7 длиной от приблизительно 3 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-7 и альфа-спиралью-8; xv) альфа-спираль-8 длиной от приблизительно 33 до приблизительно 36 аминокислот; xvi) клубок-8 длиной от приблизительно 14 до приблизительно 16 аминокислот, фланкированный альфа-спиралью-8 и альфа-спиралью-9; xvii) альфа-спираль-9 длиной от приблизительно 16 до приблизительно 23 аминокислот; xviii) клубок-9 длиной от приблизительно 21 до приблизительно 28 аминокислот, фланкированный альфа-спиралью-9 и Участком C; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) бета-тяж-1 (β1) длиной от приблизительно 3 аминокислот до приблизительно 5 аминокислот; ii) клубок длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот; iii) бета-тяж-2 (β2) длиной от приблизительно 7 аминокислот до приблизительно 11 аминокислот; iv) клубок длиной от приблизительно 17 аминокислот до приблизительно 23 аминокислот; v) бета-тяж-3 (β3) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; vi) клубок длиной от приблизительно 12 аминокислот до приблизительно 14 аминокислот; vii) бета-тяж-4 (β4) длиной от приблизительно 5 аминокислот до приблизительно 6 аминокислот; viii) клубок длиной от приблизительно 2 аминокислот до приблизительно 7 аминокислот; ix) бета-тяж-5 (β5) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; x) клубок длиной от приблизительно 26 аминокислот до приблизительно 28 аминокислот; xi) бета-тяж-6 (β6) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; xii) клубок длиной от приблизительно 16 аминокислот до приблизительно 20 аминокислот; и xiii) бета-тяж-1 (β7) длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот.
В некоторых вариантах осуществления молекула нуклеиновой кислоты кодирует полипептид PtIP-83, содержащий последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот с гибкой консенсусной вторичной структурой, где Участок A содержит консервативный бета-тяж 1 (β1a) длиной от приблизительно 4 до приблизительно 12 аминокислот в пределах от приблизительно аминокислотного остатка 30 до приблизительно аминокислотного остатка 130 от N-конца полипептида PtIP-83; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) альфа-спираль-1 длиной от приблизительно 10 до приблизительно 26 аминокислот; ii) клубок-1 длиной от приблизительно 2 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-1 и альфа-спиралью-2; iii) альфа-спираль-2 длиной от приблизительно 15 до приблизительно 24 аминокислот; iv) клубок-2 длиной от приблизительно 4 до приблизительно 14 аминокислот, фланкированный альфа-спиралью-2 и альфа-спиралью-3; v) альфа-спираль 3 длиной от приблизительно 15 до приблизительно 27 аминокислот; vi) клубок-3 длиной от приблизительно 11 до приблизительно 13 аминокислот, фланкированный альфа-спиралью-3 и альфа-спиралью-4; vii) альфа-спираль-4 длиной приблизительно 24-180 аминокислот; viii) клубок-4 длиной от приблизительно 4 до приблизительно 5 аминокислот, фланкированный альфа-спиралью-4 и альфа-спиралью-5; ix) альфа-спираль-5 длиной от приблизительно 50 до приблизительно 54 аминокислот; x) клубок-5 длиной от приблизительно 11 до приблизительно 17 аминокислот, фланкированный альфа-спиралью-5 и альфа-спиралью-6; xi) альфа-спираль-6 длиной от приблизительно 15 до приблизительно 16 аминокислот; xii) клубок-6 длиной от приблизительно 6 до приблизительно 9 аминокислот, фланкированный альфа-спиралью-6 и альфа-спиралью-7; xiii) альфа-спираль-7 длиной от приблизительно 49 до приблизительно 55 аминокислот; xiv) клубок-7 длиной от приблизительно 3 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-7 и альфа-спиралью-8; xv) альфа-спираль-8 длиной от приблизительно 33 до приблизительно 36 аминокислот; xvi) клубок-8 длиной от приблизительно 14 до приблизительно 16 аминокислот, фланкированный альфа-спиралью-8 и альфа-спиралью-9; xvii) альфа-спираль-9 длиной от приблизительно 16 до приблизительно 23 аминокислот; xviii) клубок-9 длиной от приблизительно 21 до приблизительно 28 аминокислот, фланкированный альфа-спиралью-9 и Участком C; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной последовательностью, содержащий последовательно: i) бета-тяж-1 (β1) длиной от приблизительно 3 аминокислот до приблизительно 5 аминокислот; ii) клубок длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот; iii) бета-тяж-2 (β2) длиной от приблизительно 7 аминокислот до приблизительно 11 аминокислот; iv) клубок длиной от приблизительно 17 аминокислот до приблизительно 23 аминокислот; v) бета-тяж-3 (β3) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; vi) клубок длиной от приблизительно 12 аминокислот до приблизительно 14 аминокислот; vii) бета-тяж-4 (β4) длиной от приблизительно 5 аминокислот до приблизительно 6 аминокислот; viii) клубок длиной от приблизительно 2 аминокислот до приблизительно 7 аминокислот; ix) бета-тяж-5 (β5) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; x) клубок длиной от приблизительно 26 аминокислот до приблизительно 28 аминокислот; xi) бета-тяж-6 (β6) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; xii) клубок длиной от приблизительно 16 аминокислот до приблизительно 20 аминокислот; и xiii) бета-тяж-1 (β7) длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот.
Также представлены молекулы нуклеиновой кислоты, которые кодируют продукты транскрипции и/или трансляции, которые впоследствии подвергаются сплайсингу с образованием, в конечном итоге, функциональных полипептидов PtIP-83. Сплайсинг может осуществляться in vitro или in vivo, и он может включать цис- или транс-сплайсинг. Субстратом для сплайсинга могут быть полинуклеотиды (например, РНК-транскрипты) или полипептиды. Примером цис-сплайсинга полинуклеотида является ситуация, когда интрон, вставленный в кодирующую последовательность, удаляется и два фланкирующих экзонных участка соединяются с образованием последовательности, кодирующей полипептид PtIP-83. Примером транс-сплайсинга будет ситуация, когда полинуклеотид кодируется с разделением кодирующей последовательности на два или более фрагментов, которые могут транскрибироваться раздельно, а затем соединяться с образованием пестицидной кодирующей последовательности полной длины. Применение последовательности энхансера сплайсинга, которую можно вводить в конструкцию, может облегчать сплайсинг, как при цис-, так и при транс-сплайсинге полипептидов (патенты США №№ 6365377 и 6531316). Таким образом, в некоторых вариантах осуществления полинуклеотиды напрямую не кодируют полипептид PtIP-83 полной длины, а кодируют фрагмент или фрагменты полипептида PtIP-83. Эти полинуклеотиды можно применять для экспрессии функционального полипептида PtIP-83 посредством механизма, включающего сплайсинг, при этом сплайсинг может происходить на уровне полинуклеотида (например, интрон/экзон) и/или полипептида (например, интеин/экстеин). Это может быть применимо, например, при регуляции экспрессии пестицидной активности, поскольку функциональный пестицидный полипептид будет экспрессироваться только в том случае, если все требуемые фрагменты экспрессируются в среде, которая обеспечивает возможность процессов сплайсинга с образованием функционального продукта. В другом примере введение одной или нескольких последовательностей вставок в полинуклеотид может облегчать рекомбинацию с полинуклеотидом с низкой гомологией; применение интрона или интеина для последовательности вставки облегчает удаление встроенной последовательности, что тем самым восстанавливает функцию кодируемого варианта.
Молекулы нуклеиновой кислоты, которые являются фрагментами этих последовательностей нуклеиновой кислоты, кодирующих полипептиды PIP-83, также охватываются вариантами осуществления. Используемый в данном документе ʺфрагментʺ относится к части последовательности нуклеиновой кислоты, кодирующей полипептид PtIP-83. Фрагмент последовательности нуклеиновой кислоты может кодировать биологически активную часть полипептида PtIP-83, или он может представлять собой фрагмент, который можно применять в качестве гибридизационного зонда или ПЦР-праймера с применением раскрытых ниже способов. Молекулы нуклеиновой кислоты, которые являются фрагментами последовательности нуклеиновой кислоты, кодирующей полипептид PtIP-83, содержат по меньшей мере приблизительно 150, 180, 210, 240, 270, 300, 330 или 360 смежных нуклеотидов или вплоть до количества нуклеотидов, присутствующих в последовательности нуклеиновой кислоты полной длины, кодирующей полипептид PtIP-83, раскрытый в данном документе, в зависимости от предполагаемого применения. "Смежные нуклеотиды" применяется в данном документе для обозначения нуклеотидных остатков, которые непосредственно прилегают друг к другу. Фрагменты последовательностей нуклеиновой кислоты согласно вариантам осуществления будут кодировать фрагменты белка, которые сохраняют биологическую активность полипептида PtIP-83 и, вследствие этого, сохраняют инсектицидную активность. "Сохраняет инсектицидную активность" применяют в данном документе для обозначения полипептида, обладающего по меньшей мере приблизительно 10%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 70%, 80%, 90%, 95% или большей инсектицидной активностью полипептида PtIP-83Aa полной длины (SEQ ID NO: 1). В некоторых вариантах осуществления инсектицидная активность представляет собой активность в отношении Lepidoptera. В одном варианте осуществления инсектицидная активность представляет собой активность против видов жесткокрылых. В некоторых вариантах осуществления инсектицидная активность представляет собой активность против одного или нескольких насекомых вредителей из группы кукурузного жука: западного кукурузного жука, Diabrotica virgifera; северного кукурузного жука, D. barberi: блошки одиннадцатиточечной Говарда или жука-блошки одиннадцатиточечной; Diabrotica undecimpunctata howardi, и мексиканского кукурузного жука, D. virgifera zeae. В одном варианте осуществления инсектицидная активность представляет собой активность против вида Diabrotica.
В некоторых вариантах осуществления фрагмент последовательности нуклеиновой кислоты, кодирующей полипептид PtIP-83, кодирующий биологически активную часть белка, будет кодировать по меньшей мере приблизительно 15, 20, 30, 50, 75, 100, 125 смежных аминокислот или вплоть до общего количества аминокислот, присутствующих в полипептиде PtIP-83 полной длины согласно вариантам осуществления. В некоторых вариантах осуществления фрагмент представляет собой N-терминальное и/или C-терминальное усечение по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 или более аминокислот с N-конца и/или C-конца относительно SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23 или SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769 или их вариантов, например, путем протеолиза, вставки старт-кодона, делеции кодонов, кодирующих удаляемые аминокислоты с одновременной вставкой стоп-кодона или посредством вставки стоп-кодона в кодирующую последовательность. В некоторых вариантах осуществления фрагменты, охватываемые данным документом, получены в результате удаления N-терминальных 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более аминокислот с N-конца относительно SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или их вариантов, например, путем протеолиза или путем вставки старт-кодона в кодирующую последовательность.
В некоторых вариантах осуществления фрагменты, охватываемые данным документом, получены в результате удаления N-терминальных 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 аминокислот относительно SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или их вариантов, например, путем протеолиза или путем вставки старт-кодона в кодирующую последовательность.
В некоторых вариантах осуществления полипептид PtIP-83 кодируется последовательностью нуклеиновой кислоты, достаточно гомологичной последовательности нуклеиновой кислоты под SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 717, SEQ ID NO: 738, SEQ ID NO: 739, SEQ ID NO: 740, SEQ ID NO: 741, SEQ ID NO: 742, SEQ ID NO: 743, SEQ ID NO: 744, SEQ ID NO: 745, SEQ ID NO: 746, SEQ ID NO: 747, SEQ ID NO: 748, SEQ ID NO: 749, SEQ ID NO: 750, SEQ ID NO: 751, SEQ ID NO: 752 или SEQ ID NO: 753. "Достаточно гомологичный" применяется в данном документе для обозначения аминокислотной последовательности или последовательности нуклеиновой кислоты, последовательность которой по меньшей мере приблизительно на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более гомологична эталонной последовательности при применении одной из программ выравнивания, описанных в данном документе, с применением стандартных параметров. Специалисту в данной области техники будет понятно, что эти значения можно соответствующим образом скорректировать для определения соответствующей гомологии белков, кодируемых двумя последовательностями нуклеиновой кислоты, принимая во внимание вырожденность кодонов, аминокислотное сходство, расположение рамки считывания и т. п.
В некоторых вариантах осуществления гомология последовательностей определяется в отношении последовательности полной длины полинуклеотида, кодирующего полипептид PtIP-83, или в отношении последовательности полной длины полипептида PtIP-83.
В некоторых вариантах осуществления нуклеиновая кислота, кодирующая полипептид PtIP-83, выбрана из последовательности под SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 28, SEQ ID NO: 29, под любым из SEQ ID NO: 172-235, под любым из SEQ ID NO: 300-333, под любым из SEQ ID NO: 368-397, под любым из SEQ ID NO: 428-517, SEQ ID NO: 717, под любым из SEQ ID NO: 718-727 и под любым из SEQ ID NO: 738-753.
В некоторых вариантах осуществления нуклеиновая кислота кодирует полипептид PtIP-83, последовательность которого по меньшей мере на приблизительно 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична последовательности под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769. В некоторых вариантах осуществления идентичность последовательностей рассчитывают с применением алгоритма ClustalW в модуле ALIGNX® пакета программ Vector NTI® Program Suite (Invitrogen Corporation, Карлсбад, Калифорния) со всеми параметрами по умолчанию. В некоторых вариантах осуществления идентичность последовательностей рассчитывают по всей длине полипептида с применением алгоритма ClustalW в модуле ALIGNX пакета программ Vector NTI Program Suite (Invitrogen Corporation, Карлсбад, Калифорния) со всеми параметрами по умолчанию.
Для определения процентной идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновой кислоты осуществляют выравнивание последовательностей для целей оптимального сравнения. Процентная идентичность двух последовательностей является функцией количества идентичных положений, имеющихся в последовательностях (т. е. процентная идентичность=количество идентичных положений/общее количество положений (например, перекрывающихся положений)×100). В одном варианте осуществления две последовательности имеют одинаковую длину. В другом варианте осуществления сравнение проводят по всей протяженности эталонной последовательности (например, по всей протяженности SEQ ID NO: 1). Процентную идентичность двух последовательностей можно определить с применением методик, аналогичных описанным ниже, которые допускают присутствие гэпов или не допускают присутствия гэпов. При расчете процентной идентичности, как правило, подсчитывают точные совпадения.
Определение процентной идентичности двух последовательностей можно выполнять с применением математического алгоритма. Неограничивающим примером математического алгоритма, используемого для сравнения двух последовательностей, является алгоритм Karlin и Altschul, (1990) Proc. Natl. Acad. Sci. USA 87:2264, в модификации согласно Karlin и Altschul, (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877. Такой алгоритм внедрен в программы BLASTN и BLASTX от Altschul, et al., (1990) J. Mol. Biol. 215:403. Поиски нуклеотидных последовательностей в BLAST можно выполнять с помощью программы BLASTN, балл=100, длина слова=12, с получением последовательностей нуклеиновой кислоты, гомологичных пестицидным молекулам нуклеиновой кислоты согласно вариантам осуществления. Поиски белковых последовательностей в BLAST можно выполнять с помощью программы BLASTX, балл=50, длина слова=3, с получением аминокислотных последовательностей, гомологичных молекулам белка согласно вариантам осуществления. Для получения выравниваний с введением гэпов для целей сравнения можно использовать Gapped BLAST (в BLAST 2.0), как описано в Altschul, et al., (1997) Nucleic Acids Res. 25:3389. В качестве альтернативы, можно применять PSI-Blast для осуществления итерационного поиска, при помощи которого выявляют отдаленные связи между молекулами. См. Altschul, et al., (1997) выше. При использовании программ BLAST, Gapped BLAST и PSI-Blast можно применять параметры по умолчанию соответствующих программ (например, BLASTX и BLASTN). Выравнивание можно проводить вручную путем просмотра.
Другим неограничивающим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм ClustalW (Higgins, et al., (1994) Nucleic Acids Res. 22:4673-4680). ClustalW сравнивает последовательности и выравнивает всю протяженность аминокислотной последовательности или последовательности ДНК, и, таким образом, он может предоставлять данные о консервативности последовательностей для полной аминокислотной последовательности. Алгоритм ClustalW применяется в нескольких коммерчески доступных пакетах программного обеспечения для анализа ДНК/аминокислотных последовательностей, таких как модуль ALIGNX® пакета программ Vector NTI® Program Suite (Invitrogen Corporation, Карлсбад, Калифорния). После выравнивания аминокислотных последовательностей с помощью ClustalW можно оценивать процентную идентичность аминокислотных последовательностей. Неограничивающим примером программного обеспечения, применимого для анализа выравниваний ClustalW, является GENEDOC™. GENEDOC™ (Karl Nicholas) обеспечивает возможность оценки сходства и идентичности аминокислотных (или ДНК) последовательностей нескольких белков. Другим неограничивающим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Myers и Miller, (1988) CABIOS 4:11-17. Такой алгоритм внедрен в программу ALIGN (версия 2.0), которая является частью программного пакета GCG Wisconsin Genetics Software Package, версии 10 (доступного от Accelrys, Inc., 9685 Scranton Rd., Сан-Диего, Калифорния, США). При использовании программы ALIGN для сравнения аминокислотных последовательностей можно применять таблицу весов замен остатков PAM120, штраф за продолжение гэпа 12 и штраф за открытие гэпа 4.
Другим неограничивающим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Needleman и Wunsch, (1970) J. Mol. Biol. 48(3):443-453, использующий программное обеспечение GAP версии 10 для определения идентичности и сходства последовательностей с применением следующих параметров по умолчанию: % идентичности и % сходства для последовательности нуклеиновой кислоты с применением штрафа за открытие гэпа 50, и штрафа за продолжение гэпа 3, и матрицы замен nwsgapdna.cmpii; % идентичности или % сходства для аминокислотной последовательности с применением штрафа за открытие гэпа 8, и штрафа за продолжение гэпа 2, и матрицы замен BLOSUM62. Также можно применять эквивалентные программы. ʺЭквивалентная программаʺ применяется в данном документе для обозначения любой программы для сравнения последовательностей, которая для любых двух исследуемых последовательностей генерирует выравнивание, характеризующееся идентичными совпадениями нуклеотидов и идентичной процентной идентичностью последовательности при сравнении с соответствующим выравниванием, сгенерированным с помощью GAP версии 10.
Варианты осуществления также охватывают молекулы нуклеиновой кислоты, кодирующие варианты полипептида PtIP-83. "Варианты" последовательностей нуклеиновой кислоты, кодирующих полипептид PtIP-83, включают последовательности, которые кодируют полипептиды PtIP-83, раскрытые в данном документе, но которые отличаются консервативными заменами, обусловленными вырожденностью генетического кода, а также последовательности, которые являются достаточно идентичными, как обсуждалось выше. Встречающиеся в природе аллельные варианты можно идентифицировать с применением хорошо известных методик молекулярной биологии, таких как полимеразная цепная реакция (ПЦР) и методики гибридизации, изложенные ниже. Вариантные последовательности нуклеиновой кислоты также включают синтетически полученные последовательности нуклеиновой кислоты, которые были получены, например, с применением сайт-направленного мутагенеза, но которые все еще кодируют раскрытые полипептиды PtIP-83, обсуждаемые ниже.
Настоящее раскрытие предусматривает выделенные или рекомбинантные полинуклеотиды, которые кодируют любые из полипептидов PtIP-83, раскрытых в данном документе. Специалисты в данной области техники легко поймут, что вследствие вырожденности генетического кода существует множество нуклеотидных последовательностей, кодирующих полипептиды PtIP-83 согласно настоящему раскрытию. Таблица 1 представляет собой таблицу кодонов, в которой представлены синонимичные кодоны для каждой аминокислоты. Например, все кодоны AGA, AGG, CGA, CGC, CGG и CGU кодируют аминокислоту аргинин. Таким образом, в каждом положении в нуклеиновых кислотах согласно настоящему раскрытию, в котором аргинин обозначен кодоном, кодон может быть изменен на любой из соответствующих кодонов, описанных выше, без изменения кодируемого полипептида. Понятно, что U в последовательности РНК соответствует T в последовательности ДНК.
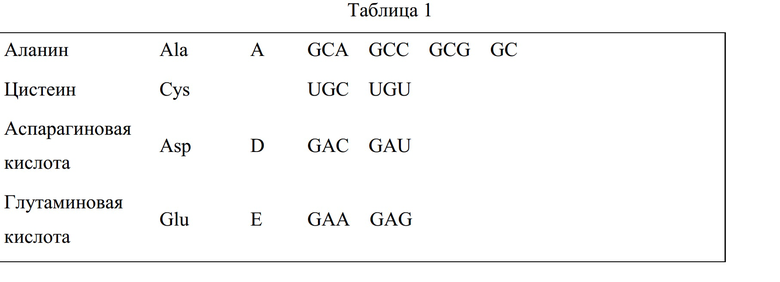
Также специалисту в данной области будет понятно, что изменения можно вводить путем внесения мутаций в последовательности нуклеиновой кислоты, что ведет к изменениям в аминокислотной последовательности кодируемых полипептидов PtIP-83 без изменения биологической активности белка. Таким образом, вариантные молекулы нуклеиновой кислоты можно создавать путем введения одной или нескольких нуклеотидных замен, добавлений и/или делеций в соответствующую последовательность нуклеиновой кислоты, раскрытую в данном документе, так что одна или несколько аминокислотных замен, добавлений или делеций вводятся в кодируемый белок. Мутации можно вводить при помощи стандартных методик, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Такие вариантные последовательности нуклеиновой кислоты также охватываются настоящим раскрытием.
В качестве альтернативы, вариантные последовательности нуклеиновой кислоты можно получать путем введения мутаций случайным образом по всей или части кодирующей последовательности, как, например, путем сайт-насыщающего мутагенеза, и полученных мутантов можно подвергать скринингу в отношении способности обеспечивать пестицидную активность для идентификации мутантов, которые сохраняют активность. После мутагенеза кодируемый белок можно экспрессировать рекомбинантным способом, и активность белка можно определять с применением стандартных методик анализа.
Полинуклеотиды согласно настоящему раскрытию и их фрагменты необязательно применяют в качестве субстратов для ряда реакций рекомбинации и рекуррентной рекомбинации, в дополнение к стандартным способам клонирования, изложенным, например, в Ausubel, Berger и Sambrook, т. е. для получения дополнительных гомологов пестицидных полипептидов и их фрагментов с требуемыми свойствами. Известен ряд таких реакций, в том числе разработанные авторами настоящего изобретения и их сотрудниками. Способы получения варианта любой нуклеиновой кислоты, приведенной в данном документе, включают рекуррентную рекомбинацию такого полинуклеотида со вторым (или большим количеством) полинуклеотидом, таким образом, получение библиотеки вариантных полинуклеотидов также представляет собой варианты осуществления согласно настоящему раскрытию, также как и полученные библиотеки, клетки, содержащие библиотеки, и любой рекомбинантный полинуклеотид, полученный такими способами. Дополнительно, такие способы необязательно включают отбор вариантного полинуклеотида из таких библиотек на основе пестицидной активности, как есть, где такую рекуррентную рекомбинацию осуществляют in vitro или in vivo.
Ряд протоколов создания разнообразия, в том числе протоколы рекуррентной рекомбинации нуклеиновых кислот, доступны и полностью описаны в уровне техники. Методики можно применять отдельно и/или в комбинации для получения одного или нескольких вариантов нуклеиновой кислоты или набора нуклеиновых кислот, а также вариантов кодируемых белков. По отдельности и вместе эти методики обеспечивают надежные, широко используемые способы создания диверсифицированных нуклеиновых кислот и наборов нуклеиновых кислот (в том числе, например, библиотек нуклеиновых кислот), применимых, например, для конструирования или быстрой эволюции нуклеиновых кислот, белков, метаболических путей, клеток и/или организмов с новыми и/или улучшенными характеристиками.
Хотя в ходе следующего обсуждения для ясности проводят разграничение и классификацию, будет принято во внимание, что методики часто не являются взаимоисключающими. Более того, различные способы можно применять по отдельности или в комбинации, одновременно или последовательно, для достижения различных вариантов последовательностей.
Результатом осуществления любой из методик создания разнообразия, описанных в данном документе, может быть создание одной или нескольких нуклеиновых кислот, которые можно подвергнуть отбору или скринингу в отношении нуклеиновых кислот, имеющих требуемые свойства или обеспечивающих их, или нуклеиновых кислот, которые кодируют белки, имеющие требуемые свойства или обеспечивающие их. После диверсификации с помощью одного или нескольких способов, описанных в данном документе или иным образом доступных специалисту в данной области, любые получаемые нуклеиновые кислоты можно подвергать отбору в отношении требуемой активности или свойства, например, пестицидной активности или такой активности при требуемом pH и т. д. Это может включать идентификацию любой активности, которую можно выявить, например, в автоматизированном или автоматизируемом формате, посредством любого из анализов, известных из уровня техники, см., например, обсуждение скрининга в отношении инсектицидной активности, ниже. Можно оценивать ряд связанных (или даже несвязанных) свойств последовательно или одновременно, на усмотрение специалиста-практика.
Описания ряда методик создания разнообразия для создания модифицированных последовательностей нуклеиновой кислоты, например, последовательностей, кодирующих полипептиды с пестицидной активностью или их фрагменты, можно найти в следующих публикациях и литературе, процитированной в них: Soong, et al., (2000) Nat Genet 25(4):436-439; Stemmer, et al., (1999) Tumor Targeting 4:1-4; Ness, et al., (1999) Nat Biotechnol 17:893-896; Chang, et al., (1999) Nat Biotechnol 17:793-797; Minshull and Stemmer, (1999) Curr Opin Chem Biol 3:284-290; Christians, et al., (1999) Nat Biotechnol 17:259-264; Crameri, et al., (1998) Nature 391:288-291; Crameri, et al., (1997) Nat Biotechnol 15:436-438; Zhang, et al., (1997) PNAS USA 94:4504-4509; Patten, et al., (1997) Curr Opin Biotechnol 8:724-733; Crameri, et al., (1996) Nat Med 2:100-103; Crameri, et al., (1996) Nat Biotechnol 14:315-319; Gates, et al., (1996) J Mol Biol 255:373-386; Stemmer, (1996) "Sexual PCR and Assembly PCR" в: The Encyclopedia of Molecular Biology. VCH Publishers, New York. pp. 447-457; Crameri and Stemmer, (1995) BioTechniques 18:194-195; Stemmer, et al., (1995) Gene, 164:49-53; Stemmer, (1995) Science 270: 1510; Stemmer, (1995) Bio/Technology 13:549-553; Stemmer, (1994) Nature 370:389-391 и Stemmer, (1994) PNAS USA 91:10747-10751.
Мутационные способы создания разнообразия включают, например, сайт-направленный мутагенез (Ling, et al., (1997) Anal Biochem 254(2):157-178; Dale, et al., (1996) Methods Mol Biol 57:369-374; Smith, (1985) Ann Rev Genet 19:423-462; Botstein and Shortle, (1985) Science 229:1193-1201; Carter, (1986) Biochem J 237:1-7 и Kunkel, (1987) "The efficiency of oligonucleotide directed mutagenesis" в Nucleic Acids & Molecular Biology (Eckstein and Lilley, eds., Springer Verlag, Berlin)); мутагенез с применением урацил-содержащих матриц (Kunkel, (1985) PNAS USA 82:488-492; Kunkel, et al., (1987) Methods Enzymol 154:367-382 и Bass, et al., (1988) Science 242:240-245); олигонуклеотид-направленный мутагенез (Zoller and Smith, (1983) Methods Enzymol 100:468-500; Zoller and Smith, (1987) Methods Enzymol 154:329-350 (1987); Zoller and Smith, (1982) Nucleic Acids Res 10:6487-6500), мутагенез фосфоротиоат-модифицированной ДНК (Taylor, et al., (1985) Nucl Acids Res 13:8749-8764; Taylor, et al., (1985) Nucl Acids Res 13:8765-8787 (1985); Nakamaye and Eckstein, (1986) Nucl Acids Res 14:9679-9698; Sayers, et al., (1988) Nucl Acids Res 16:791-802 и Sayers, et al., (1988) Nucl Acids Res 16:803-814); мутагенез с применением дуплексной ДНК с брешью (Kramer, et al., (1984) Nucl Acids Res 12:9441-9456; Kramer and Fritz, (1987) Methods Enzymol 154:350-367; Kramer, et al., (1988) Nucl Acids Res 16:7207 и Fritz, et al., (1988) Nucl Acids Res 16:6987-6999).
Дополнительные подходящие способы включают точечную репарацию ошибочно спаренных оснований (Kramer, et al., (1984) Cell 38:879-887), мутагенез с применением штаммов-хозяев с недостаточностью репарации (Carter, et al., (1985) Nucl Acids Res 13:4431-4443 и Carter, (1987) Methods in Enzymol 154:382-403), делеционный мутагенез (Eghtedarzadeh and Henikoff, (1986) Nucl Acids Res 14:5115), рестрикцию-отбор и рестрикцию-очистку (Wells, et al., (1986) Phil Trans R Soc Lond A 317:415-423), мутагенез посредством синтеза полного гена (Nambiar, et al., (1984) Science 223:1299-1301; Sakamar and Khorana, (1988) Nucl Acids Res 14:6361-6372; Wells, et al., (1985) Gene 34:315-323 и Grundström, et al., (1985) Nucl Acids Res 13:3305-3316), репарацию двухцепочечных разрывов (Mandecki, (1986) PNAS USA, 83:7177-7181 и Arnold, (1993) Curr Opin Biotech 4:450-455). Дополнительные сведения по многим из вышеуказанных способов можно найти в Methods Enzymol, том 154, в котором также описаны полезные руководства по поиску и устранению проблем в случае различных способов мутагенеза.
Дополнительные сведения, касающиеся различных способов создания разнообразия, можно найти в следующих патентах США, публикациях и заявках согласно PCT и публикациях EPO: Патент США № 5,723,323, Патент США № 5,763,192, Патент США № 5,814,476, Патент США № 5,817,483, Патент США № 5,824,514, Патент США № 5,976,862, Патент США № 5,605,793, Патент США № 5,811,238, Патент США № 5,830,721, Патент США № 5,834,252, Патент США № 5,837,458, WO 1995/22625, WO 1996/33207, WO 1997/20078, WO 1997/35966, WO 1999/41402, WO 1999/41383, WO 1999/41369, WO 1999/41368, EP 752008, EP 0932670, WO 1999/23107, WO 1999/21979, WO 1998/31837, WO 1998/27230, WO 1998/27230, WO 2000/00632, WO 2000/09679, WO 1998/42832, WO 1999/29902, WO 1998/41653, WO 1998/41622, WO 1998/42727, WO 2000/18906, WO 2000/04190, WO 2000/42561, WO 2000/42559, WO 2000/42560, WO 2001/23401 и PCT/US01/06775.
Нуклеотидные последовательности согласно вариантам осуществления также можно применять для выделения соответствующих последовательностей из папоротников или другого примитивного растения, в частности видов Asplenium, Polypodium, Adiantum, Platycerium, Nephrolepis, Ophioglossum, Colysis, Bolbitis, Blechnum, Selaginella, Lycopodium и Huperzia. Таким образом, такие способы как ПЦР, гибридизация и т.п., можно применять для идентификации таких последовательностей исходя из их гомологии с последовательностями, изложенными в данном документе. Вариантами осуществления охватываются последовательности, выбранные на основе идентичности последовательности с полными последовательностями, изложенными в данном документе, или их фрагментами. Такие последовательности включают последовательности, которые являются ортологами раскрытых последовательностей. Термин "ортологи" относится к генам, происходящим от общего предкового гена и выявляемым у различных видов вследствие видообразования. Гены, обнаруживаемые у различных видов, считаются ортологами, если их нуклеотидные последовательности и/или кодируемые ими белковые последовательности имеют существенную степень идентичности, как определено в других разделах в данном документе. Функции ортологов часто являются высококонсервативными у разных видов.
В случае подхода, основанного на ПЦР, можно сконструировать олигонуклеотидные праймеры для применения в ПЦР-реакциях, чтобы амплифицировать соответствующие последовательностей ДНК на основании кДНК или геномной ДНК, извлеченных из какого-либо организма, представляющего интерес. Способы конструирования ПЦР-праймеров и ПЦР-клонирования в целом известны из уровня техники и раскрыты в Sambrook, et al., (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Plainview, New York), в дальнейшем "Sambrook". См. также Innis, et al., eds. (1990) PCR Protocols: A Guide to Methods and Applications (Academic Press, New York); Innis and Gelfand, eds. (1995) PCR Strategies (Academic Press, New York); и Innis and Gelfand, eds. (1999) PCR Methods Manual (Academic Press, New York). Известные способы ПЦР включают без ограничения способы с применением парных праймеров, гнездовых праймеров, одиночных специфичных праймеров, вырожденных праймеров, ген-специфических праймеров, вектор-специфических праймеров, праймеров, в которых часть нуклеотидов являются ошибочно спаренными и т.п.
Для идентификации потенциальных полипептидов PtIP-83 из коллекций папоротников или мхов, лизаты клеток папоротников или мхов можно подвергнуть скринингу с помощью антител, вырабатываемых против полипептидов PtIP-83 и/или полипептидов PtIP-83, с применением способов вестерн-блоттинга и/или ELISA. Этот тип анализов можно проводить высокопроизводительным образом. Положительные образцы можно дополнительно анализировать при помощи различных методик, таких как очистка и идентификация белков при помощи антител. Способы получения антител хорошо известны из уровня техники, как обсуждается ниже.
В качестве альтернативы, для идентификации гомологов полипептидов PtIP-83 можно применять способ идентификации белков на основе масс-спектрометрии с применением протоколов из литературных источников (Scott Patterson, (1998), 10.22, 1-24, Current Protocol in Molecular Biology, опубликованный John Wiley & Son Inc). Точнее говоря, способ идентификации белков на основе LC-MS/MS применяют для установления связи MS-данных указанных клеточных лизатов или образцов, обогащенных молекулами с требуемым молекулярным весом (вырезанных из геля SDS-PAGE полосок с молекулярным весом, соответствующим полипептидам PtIP-83), с информацией о последовательности полипептидов PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769 и их гомологов. Любое совпадение в пептидных последовательностях указывает на возможность наличия гомологичных белков в образцах. Дополнительные методики (способы очистки белка и методики молекулярной биологии) можно применять для выделения белка и идентификации последовательностей гомологов.
В способах гибридизации для скрининга кДНК или геномных библиотек можно применять всю или часть последовательности пестицидной нуклеиновой кислоты. Способы для конструирования таких кДНК и геномных библиотек в целом известны из уровня техники и раскрыты в Sambrook и Russell, (2001), выше. Так называемые гибридизационные зонды могут представлять собой фрагменты геномной ДНК, фрагменты кДНК, фрагменты РНК или другие олигонуклеотиды, и они могут быть помечены детектируемой группой, такой как 32P, или любым другим детектируемым маркером, таким как другие радиоактивные изотопы, флуоресцентным соединением, ферментом или кофактором фермента. Зонды для гибридизации можно создавать путем мечения синтетических олигонуклеотидов, основанных на известной последовательности нуклеиновой кислоты, кодирующей полипептид PtIP-83, раскрытой в данном документе. Дополнительно можно применять вырожденные праймеры, сконструированные на основе консервативных нуклеотидов или аминокислотных остатков в последовательности нуклеиновой кислоты или кодируемой аминокислотной последовательности. Зонд, как правило, содержит участок последовательности нуклеиновой кислоты, который гибридизуется при жестких условиях по меньшей мере с приблизительно 12, по меньшей мере с приблизительно 25, по меньшей мере с приблизительно 50, 75, 100, 125, 150, 175 или 200 смежными нуклеотидами последовательности нуклеиновой кислоты, кодирующей полипептид PtIP-83 согласно настоящему раскрытию или его фрагмент или вариант. Способы получения зондов для гибридизации, в целом, известны из уровня техники и раскрыты в Sambrook и Russell (2001), выше, включенном в данный документ посредством ссылки.
Например, полную последовательность нуклеиновой кислоты, кодирующей полипептид PtIP-83, раскрытую в данном документе, или одну или несколько ее частей можно применять в качестве зонда, способного специфично гибридизоваться с соответствующими последовательностями нуклеиновой кислоты, кодирующими последовательности, подобные полипептиду PtIP-83, и матричными РНК. Для достижения специфической гибридизации при различных условиях такие зонды включают последовательности, которые являются уникальными, и предпочтительно имеют длину по меньшей мере приблизительно 10 нуклеотидов или по меньшей мере приблизительно 20 нуклеотидов. Такие зонды можно применять для амплификации соответствующих пестицидных последовательностей из выбранного организма с помощью ПЦР. Эту методику можно применять для выделения дополнительных кодирующих последовательностей из требуемого организма или в качестве диагностического анализа для определения присутствия кодирующих последовательностей в организме. Методики гибридизации включают гибридизационный скрининг высеянных ДНК-библиотек (либо бляшек, либо колоний; см., например, Sambrook, et al., (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.). Гибридизацию таких последовательностей можно проводить в жестких условиях. "Жесткие условия" или "жесткие условия гибридизации" применяются в данном документе для обозначения условий, при которых зонд будет гибридизоваться со своей целевой последовательностью в явно большей степени, чем с другими последовательностями (например, по меньшей мере в 2 раза больше по сравнению с фоном). Жесткие условия являются зависимыми от последовательности и будут отличаться при различных обстоятельствах. Путем регулирования жесткости условий гибридизации и/или отмывки можно идентифицировать целевые последовательности, которые на 100% комплементарны зонду (гибридизация с гомологичным зондом). В качестве альтернативы, условия жесткости можно скорректировать для обеспечения некоторого ошибочного спаривания в последовательностях с тем, чтобы выявлять более низкие степени сходства (гибридизация с гетерологичным зондом). В целом, длина зонда составляет менее приблизительно 1000 нуклеотидов, предпочтительно он имеет менее 500 нуклеотидов в длину.
Как правило, жесткие условия будут такими, при которых концентрация соли составляет менее приблизительно 1,5 M ионов Na, как правило, концентрация ионов Na (или других солей) составляет приблизительно 0,01-1,0 M при pH 7,0-8,3, а температура составляет по меньшей мере приблизительно 30°C для коротких зондов (например, 10-50 нуклеотидов) и по меньшей мере приблизительно 60°C для длинных зондов (например, более 50 нуклеотидов). Жесткие условия также могут быть достигнуты с помощью добавления дестабилизирующих средств, таких как формамид. Иллюстративные условия низкой жесткости включают гибридизацию в буферном растворе с 30-35% формамида, 1M NaCl, 1% SDS (додецилсульфата натрия) при 37°C и отмывку в 1× - 2×SSC (20×SSC=3,0 M NaCl/0,3 M цитрата тринатрия) при 50-55°C. Иллюстративные условия умеренной жесткости включают гибридизацию в 40-45% формамиде, 1,0 M NaCl, 1% SDS при 37°C и отмывку в 0,5× - 1×SSC при 55-60°C. Иллюстративные условия высокой жесткости включают гибридизацию в 50% формамиде, 1 M NaCl, 1% SDS при 37°C и отмывку в 0,1×SSC при 60-65°C. Необязательно, отмывочные буферы могут содержать от приблизительно 0,1% до приблизительно 1% SDS. Длительность гибридизации, в целом, составляет менее приблизительно 24 часов, обычно от приблизительно 4 до приблизительно 12 часов.
Специфичность, как правило, зависит от отмывок после гибридизации, причем ключевыми факторами являются ионная сила и температура конечного отмывочного раствора. Для гибридов ДНК-ДНК Tm можно приблизительно рассчитать из уравнения Meinkoth and Wahl, (1984) Anal. Biochem. 138:267-284: Tm=81,5°C+16,6 (log M)+0,41 (% GC) - 0,61 (% формамида) - 500/л; где M представляет собой молярность раствора одновалентных катионов, % GC представляет собой процентную долю гуанозиновых и цитозиновых нуклеотидов в ДНК, % формамида представляет собой процентную долю формамида в растворе для гибридизации, а L представляет собой длину гибрида в парах оснований. Tm представляет собой температуру (при определенной ионной силе и pH), при которой 50% комплементарной целевой последовательности гибридизируется с идеально совпадающим зондом. Tm снижают приблизительно на 1°C для каждого 1% ошибочного спаривания; таким образом, Tm, условия гибридизации и/или отмывки можно скорректировать для гибридизации с последовательностями с требуемой идентичностью. Например, если необходимы последовательности, идентичные на ≧90%, Tm можно снизить на 10°C. Обычно, жесткие условия выбирают так, чтобы они были приблизительно на 5°C ниже температуры плавления (Tm) конкретной последовательности и комплементарной ей последовательности при определенной ионной силе и pH. Однако, при условиях сильной жесткости можно применять гибридизацию и/или отмывку при температуре на 1, 2, 3 или 4°C ниже температуры плавления (Tm); при условиях умеренной жесткости можно применять гибридизацию и/или отмывку при температуре на 6, 7, 8, 9 или 10°C ниже температуры плавления (Tm); при условиях низкой жесткости можно применять гибридизацию и/или отмывку при температуре на 11, 12, 13, 14, 15 или 20°C ниже температуры плавления (Tm). Специалистам будет понятно, что изменения жесткости растворов для гибридизации и/или отмывки по сути описаны с использованием уравнения, композиций для гибридизации и отмывки и требуемой Tm. Если требуемая степень ошибочного спаривания приводит к Tm меньше 45°C (водный раствор) или 32°C (раствор формамида), предпочтительно увеличивать концентрацию SSC так, чтобы можно было применять более высокую температуру. Обширное руководство по гибридизации нуклеиновых кислот приведено в Tijssen, (1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2 (Elsevier, N.Y.) и Ausubel, et al., eds. (1995) Current Protocols in Molecular Biology, Chapter 2 (Greene Publishing and Wiley-Interscience, New York). См. Sambrook, et al., (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.).
В некоторых вариантах осуществления представлены полинуклеотиды, кодирующие полипептид PtIP-83, содержащий аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична последовательности под любым из SEQ ID NO: 786-888.
В некоторых вариантах осуществления представлены полинуклеотиды, кодирующие полипептид PtIP-83, содержащий аминокислотную последовательность под любым из SEQ ID NO: 786-888.
В некоторых вариантах осуществления полинуклеотид, кодирующий полипептид PtIP-83, содержащий аминокислотную последовательность под любым из SEQ ID NO: 786-888, представляет собой последовательность, отличающуюся от геномной.
В некоторых вариантах осуществления полинуклеотид, кодирующий полипептид PtIP-83, содержащий аминокислотную последовательность под любым из SEQ ID NO: 786-888, представляет собой кДНК.
Белки, а также их варианты и фрагменты
Полипептиды PtIP-83 также охвачены настоящим раскрытием. Термины "инсектицидный белок-83 Pteridophyta", "полипептид PtIP-83" и "белок PtIP-83" применяются в данном документе взаимозаменяемо для обозначения полипептида с инсектицидной активностью, в том числе без ограничения инсектицидной активностью против одного или более насекомых-вредителей из отрядов Lepidoptera и/или Coleoptera, и достаточно гомологичен белку под SEQ ID NO: 1. Подразумевается целый ряд полипептидов PtIP-83. Источниками полипептидов PtIP-83 или родственных белков являются виды папоротников, выбранные без ограничения из Polypodium punctatum, Lygodium flexuosum, Microsorum musifolium, Adiantum peruvianum, Adiantum trapeziforme и Adiantum pedatum.
"Достаточно гомологична" применяется в данном документе для обозначения аминокислотной последовательности, последовательность которой по меньшей мере на приблизительно 40%, 45%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более гомологична эталонной последовательности с применением одной из программ выравнивания, описанных в данном документе, с применением стандартных параметров. В некоторых вариантах осуществления гомологию последовательности определяют относительно последовательности полной длины полипептида PtIP-83. В некоторых вариантах осуществления последовательность полипептида PtIP-83 по меньшей мере на приблизительно 40%, 45%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична последовательности под SEQ ID NO: 1. Специалисту в данной области будет понятно, что эти значения можно соответствующим образом скорректировать для определения соответствующей гомологии белков, принимая во внимание аминокислотное сходство и т. п. В некоторых вариантах осуществления идентичность последовательностей рассчитывают с применением алгоритма ClustalW в модуле ALIGNX® пакета программ Vector NTI® Program Suite (Invitrogen Corporation, Карлсбад, Калифорния) со всеми параметрами по умолчанию. В некоторых вариантах осуществления идентичность последовательностей рассчитывают по всей длине полипептида с применением алгоритма ClustalW в модуле ALIGNX® пакета программ Vector NTI® Program Suite (Invitrogen Corporation, Карлсбад, Калифорния) со всеми параметрами по умолчанию.
Применяемые в данном документе термины "белок", "пептидная молекула" или "полипептид" включают любую молекулу, которая содержит пять или более аминокислот. Из уровня техники хорошо известно, что белок, пептид или полипептидные молекулы могут подвергаться модификации, в том числе посттрансляционным модификациям, таким как без ограничения, образование дисульфидных связей, гликозилирование, фосфорилирование или олигомеризация. Таким образом, применяемые в данном документе термины "белок", "пептидная молекула" или "полипептид" включают любой белок, который модифицирован посредством какого-либо биологического или небиологического процесса. Термины "аминокислота" и "аминокислоты" относятся к встречающимся в природе L-аминокислотам.
Термины "рекомбинантный белок" или "рекомбинантный полипептид" применяются в данном документе для обозначения белка, который более не находится в своей естественной среде, а например, in vitro или в рекомбинантной бактериальной или растительной клетке-хозяине. Термины "очищенный белок" или "очищенный полипептид" применяются в данном документе для обозначения белка, который фактически не содержит клеточный материал. Полипептид PtIP-83, который фактически не содержит клеточный материал, включает препараты белка, имеющие менее приблизительно 30%, 20%, 10% или 5% (по сухому весу) белка, не относящегося к пестицидному (также называемого в данном документе ʺзагрязняющий белокʺ).
"Фрагменты" или "биологически активные части" включают фрагменты полипептида, содержащие аминокислотные последовательности достаточно идентичные полипептиду PtIP-83 и проявляющие инсектицидную активность. "Фрагменты" или "биологически активные части" полипептидов PtIP-83включают фрагменты, содержащие аминокислотные последовательности достаточно идентичные аминокислотной последовательности, изложенной под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769, где полипептид PtIP-83 обладает инсектицидной активностью. Такие биологически активные части можно получать при помощи рекомбинантных методик и оценивать в отношении инсектицидной активности. В некоторых вариантах осуществления фрагмент полипептида PtIP-83 представляет собой N-терминальное и/или C-терминальное усечение по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 или более аминокислот с N-конца и/или C-конца относительно SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769, например, путем протеолиза, путем вставки старт-кодона, путем делеции кодонов, кодирующих удаляемые аминокислоты, и одновременной вставки старт-кодона и/или вставки стоп-кодона.
В некоторых вариантах осуществления фрагменты полипептида PtP-83, охваченные в данном документе, получены в результате удаления 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более N-терминальных аминокислот относительно SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или их вариантов, например, путем протеолиза или путем вставки старт-кодона, путем делеции кодонов, кодирующих удаляемые аминокислоты, и одновременной вставки старт-кодона.
Применяемые в данном документе "варианты" обозначают белки или полипептиды с аминокислотной последовательностью, которая по меньшей мере на приблизительно 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична исходной аминокислотной последовательности.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность, которая по меньшей мере на приблизительно 40%, 45%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769, где полипептид PtIP-83 обладает инсектицидной активностью.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность, которая по меньшей мере на приблизительно 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична по всей длине аминокислотной последовательности под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность, которая по меньшей мере на приблизительно 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична по всей длине аминокислотной последовательности под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769, и имеет по меньшей мере одну аминокислотную замену, делецию, вставку и/или добавление на N-конце или C-конце по сравнению с нативной последовательностью под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769 с 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70 или более аминокислотными заменами по сравнению в нативной аминокислотой в соответствующем положении SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления идентичность последовательностей рассчитывают по всей длине полипептида с применением алгоритма ClustalW в модуле ALIGNX® пакета программ Vector NTI® Program Suite (Invitrogen Corporation, Карлсбад, Калифорния) со всеми параметрами по умолчанию.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность с 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59 или 60 аминокислотными заменами, в любой комбинации, по сравнению с нативной аминокислотой в соответствующем положении SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность с 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28 или 29 аминокислотными заменами, в любой комбинации, по сравнению с нативной аминокислотой в соответствующем положении SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 содержит не встречающуюся в природе аминокислотную последовательность. Применяемый в данном документе термин "не встречающаяся в природе аминокислотная последовательность" означает, что аминокислотная последовательность не найдена в природе.
В некоторых вариантах осуществления полипептид PtIP-83 не является полипептидом под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 представляет собой вариант полипептида под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769, где вариант полипептида PtIP-83 имеет по меньшей мере одну аминокислотную замену, делецию, вставку и/или добавление на N-конце или C-конце по сравнению с SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 содержит аминокислотную последовательность под любым из SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607, SEQ ID NO: 640-645, и SEQ ID NO: 728-737.
В некоторых вариантах осуществления полипептид PtIP-83 представляет собой вариант последовательности под SEQ ID NO: 1, где аминокислота в положении 53 представляет собой Val, Ala, Cys или Thr; аминокислота в положении 54 представляет собой Lys, Ala, Cys, Asp, Glu, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser или Thr; аминокислота в положении 55 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 56 представляет собой Leu, Glu, Phe, Ile, Met, Thr или Val; аминокислота в положении 57 представляет собой Tyr, Cys, Ile, Leu, Met, Thr или Val; аминокислота в положении 58 представляет собой Val, Cys, Ile или Leu; аминокислота в положении 59 представляет собой Phe, Leu, Met, Val или Tyr; аминокислота в положении 60 представляет собой Ala, Cys, Gly, Ser, Thr или Val; аминокислота в положении 61 представляет собой Asp, Glu, His или Ser; аминокислота в положении 62 представляет собой Val, Ala, Cys, Ile, Leu или Thr; аминокислота в положении 63 представляет собой Val, Ala, Cys, Ile, Leu, Met или Thr; аминокислота в положении 64 представляет собой Glu, Ala, Cys, Phe, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 65 представляет собой Leu, Ala, Cys, Phe, His, Ile, Met, Asn, Gln, Thr, Val или Trp; аминокислота в положении 66 представляет собой Pro, Asp, Gly, Met, Gln или Arg; аминокислота в положении 363 представляет собой Gln, Ala, Cys, Glu, Phe, Gly, His, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 364 представляет собой Ile, Ala, Cys, Glu, Phe, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 365 представляет собой Leu, Ala, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Arg, Val, Trp или Tyr; аминокислота в положении 366 представляет собой Gly, Ala, Cys, Phe, His, Ile, Lys, Leu, Met, Asn, Ser, Thr или Val; аминокислота в положении 367 представляет собой Ser, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val или Trp; аминокислота в положении 368 представляет собой Tyr, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp; аминокислота в положении 369 представляет собой Leu, Ala, Cys, Asp, Phe, Gly, Ile, Met, Thr или Val; аминокислота в положении 370 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 371 представляет собой Gln, Ala, Cys, Asp, Glu, Phe, Gly, Ile, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 372 представляет собой Gln, Ala, Cys, Asp, Phe, Gly, His, Ile, Leu, Asn, Arg, Ser, Val или Tyr; аминокислота в положении 373 представляет собой Asn, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Gln, Ser, Thr, Val или Trp; аминокислота в положении 556 представляет собой Trp, Phe, Thr или Tyr; аминокислота в положении 557 представляет собой Arg, Cys, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 558 представляет собой Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 559 представляет собой Lys, Ala, Cys, Phe, Gly, His, Ile, Leu, Asn, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 560 представляет собой Cys, Ala, Phe, Gly, Ile, Met, Asn, Arg, Ser, Thr или Val; аминокислота в положении 561 представляет собой Lys, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 562 представляет собой Asn, Cys, Asp, Glu, Gly, His, Leu, Met, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 563 представляет собой Val, Ala, Cys, Asp, Phe, His, Ile, Leu, Met, Asn, Gln, Thr или Trp; аминокислота в положении 564 представляет собой Ala, Cys, Gly, Met, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 646 представляет собой Leu, Ala, Cys, Gly, Ile, Met, Asn, Gln, Ser, Thr или Val; аминокислота в положении 647 представляет собой Leu, Asp, Gly, Met, Asn, Gln или Thr; аминокислота в положении 648 представляет собой Met, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 649 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr; аминокислота в положении 650 представляет собой Thr, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Val или Tyr; аминокислота в положении 651 представляет собой Glu, Ala, Cys, Asp, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 652 представляет собой Leu, Cys, Phe, Ile, Lys, Met, Pro, Arg, Ser, Thr или Val; аминокислота в положении 653 представляет собой Thr, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Pro, Arg, Ser, Val или Trp; аминокислота в положении 654 представляет собой Thr, Ala, Cys, Phe, Ile, Lys, Leu, Met, Pro, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 655 представляет собой Trp, Phe или Tyr; аминокислота в положении 771 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Asn, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 772 представляет собой Arg, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 773 представляет собой Asp, Ala, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 774 представляет собой Gln, Ala, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 775 представляет собой Val, Ala, Cys, Asp, Glu, Gly, His, Ile, Asn, Pro, Gln, Arg, Ser, Thr или Tyr; аминокислота в положении 776 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 777 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 778 представляет собой Phe, Ala, His, Ile, Leu, Met, Asn, Gln, Ser, Val, Trp или Tyr; аминокислота в положении 779 представляет собой Gln, Ala, Cys, Asp, Glu, Gly, His, Lys, Leu, Asn, Pro, Arg, Ser, Thr или Val; аминокислота в положении 780 представляет собой Ala, Cys, Asn, Pro, Gln или Ser; аминокислота в положении 781 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 782 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 783 представляет собой Pro, Ala, Cys, Asp, Glu, Gly, His, Asn, Gln, Arg, Ser, Thr или Val; аминокислота в положении 784 представляет собой Leu, Ala, Glu, Phe, His, Ile, Lys, Met, Asn, Pro, Gln, Ser, Thr, Val или Trp; аминокислота в положении 785 представляет собой Asn, Ala, Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; и аминокислота в положении 786 представляет собой Tyr, Phe, Ile, Leu или Trp.
В некоторых вариантах осуществления полипептид PtIP-83 представляет собой вариант последовательности под SEQ ID NO: 1, где аминокислота в положении 1 представляет собой Met или удалена; аминокислота в положении 2 представляет собой Ala или удалена; аминокислота в положении 3 представляет собой Leu, Val или удалена; аминокислота в положении 4 представляет собой Val, Met или Leu; аминокислота в положении 7 представляет собой Gly или Ser; аминокислота в положении 8 представляет собой Lys или Thr; аминокислота в положении 10 представляет собой Phe или Tyr; аминокислота в положении 11 представляет собой Glu или Arg; аминокислота в положении 18 представляет собой Met или Ile; аминокислота в положении 19 представляет собой Gly, Pro или Ala; аминокислота в положении 20 представляет собой Val или удалена; аминокислота в положении 21 представляет собой Leu или Val; аминокислота в положении 23 представляет собой Arg или Gln; аминокислота в положении 37 представляет собой Val или Leu; аминокислота в положении 38 представляет собой Arg или Asn; аминокислота в положении 40 представляет собой Ala или Ser; аминокислота в положении 43 представляет собой Asn или Asp; аминокислота в положении 45 представляет собой Gly или Ala; аминокислота в положении 46 представляет собой Gln или Glu; аминокислота в положении 48 представляет собой Glu, Pro или Val; аминокислота в положении 51 представляет собой Glu или Gly; аминокислота в положении 52 представляет собой Lys, Arg или Thr; аминокислота в положении 56 представляет собой Leu или Val; аминокислота в положении 59 представляет собой Phe или Leu; аминокислота в положении 66 представляет собой Pro или Ala; аминокислота в положении 67 представляет собой Val, Pro или Thr; аминокислота в положении 68 представляет собой Val, Arg, Phe или Gly; аминокислота в положении 69 представляет собой Glu, Ala или Lys; аминокислота в положении 70 представляет собой Trp, Thr, His, Tyr или Arg; аминокислота в положении 71 представляет собой Arg, Pro или удалена; аминокислота в положении 72 представляет собой Trp, Asp, Leu или удалена; аминокислота в положении 73 представляет собой Pro, Gln, Asn, His или удалена; аминокислота в положении 74 представляет собой Pro, Met или Thr; аминокислота в положении 75 представляет собой Gln, His или Arg; аминокислота в положении 76 представляет собой Ile, Met или Leu; аминокислота в положении 84 представляет собой Ile или Val; аминокислота в положении 91 представляет собой Trp или Phe; аминокислота в положении 93 представляет собой Thr или Ile; аминокислота в положении 94 представляет собой Asp или Gly; аминокислота в положении 96 представляет собой Arg или Ser; аминокислота в положении 97 представляет собой Gln, Phe или Arg; аминокислота в положении 98 представляет собой Ser или удалена; аминокислота в положении 99 представляет собой Asp или Ala; аминокислота в положении 100 представляет собой Thr или Ala; аминокислота в положении 101 представляет собой Glu, Thr или Trp аминокислота в положении 103 представляет собой His, Arg, Glu или Gln; аминокислота в положении 105 представляет собой Thr или Pro; аминокислота в положении 108 представляет собой Lys, Gln или Glu; аминокислота в положении 109 представляет собой Leu или Val; аминокислота в положении 111 представляет собой Ala или Thr; аминокислота в положении 112 представляет собой Ile, Arg, Thr или удалена; аминокислота в положении 113 представляет собой Gln, Ala, Gly или удалена; аминокислота в положении 114 представляет собой Arg, Glu или Ile; аминокислота в положении 115 представляет собой Glu или Gln; аминокислота в положении 116 представляет собой Glu, Asn, Gln или Arg; аминокислота в положении 117 представляет собой Asn, Val, Tyr или Phe; аминокислота в положении 118 представляет собой Arg или Lys; аминокислота в положении 119 представляет собой Trp или Ser; аминокислота в положении 122 представляет собой Thr, Lys или Ala; аминокислота в положении 124 представляет собой Ala или Thr; аминокислота в положении 126 представляет собой Gly или Asp; аминокислота в положении 127 представляет собой Met или Ala; аминокислота в положении 128 представляет собой Asn или Lys; аминокислота в положении 131 представляет собой Val, Ile или Thr; аминокислота в положении 133 представляет собой Ile или Val; аминокислота в положении 134 представляет собой His или Tyr; аминокислота в положении 135 представляет собой Ala или Gly; аминокислота в положении 137 представляет собой Glu или Lys; аминокислота в положении 139 представляет собой Gln или Glu; аминокислота в положении 140 представляет собой Val, Arg или Leu; аминокислота в положении 141 представляет собой Gly или Ser; аминокислота в положении 142 представляет собой Val или Pro; аминокислота в положении 144 представляет собой Thr, Leu, Phe или Tyr; аминокислота в положении 145 представляет собой Met, Pro или Asn; аминокислота в положении 146 представляет собой Ser, Gly или Asn; аминокислота в положении 147 представляет собой Trp или Asn; аминокислота в положении 148 представляет собой Ser, Ala или Pro; аминокислота в положении 149 представляет собой Ser или удалена; аминокислота в положении 150 представляет собой Val, Ile или Tyr; аминокислота в положении 152 представляет собой Arg, Ala, Val или Gly; аминокислота в положении 154 представляет собой Ser, Trp или Glu; аминокислота в положении 156 представляет собой Leu, Asp или Gln; аминокислота в положении 158 представляет собой Ser или Cys; аминокислота в положении 159 представляет собой Val, Thr или Ile; аминокислота в положении 162 представляет собой Ser или Ala; аминокислота в положении 163 представляет собой Gly или удалена; аминокислота в положении 164 представляет собой Phe или удалена; аминокислота в положении 165 представляет собой Arg или Ala; аминокислота в положении 166 представляет собой Ala, Arg, Met или Phe; аминокислота в положении 167 представляет собой Val или His; аминокислота в положении 168 представляет собой Ser или Asn; аминокислота в положении 169 представляет собой Val, His или Thr; аминокислота в положении 170 представляет собой Phe или Val; аминокислота в положении 171 представляет собой Glu, Asn или Asp; аминокислота в положении 172 представляет собой Val, Ala, Arg или Glu; аминокислота в положении 175 представляет собой Ser, Arg или Trp; аминокислота в положении 176 представляет собой Val или Ile; аминокислота в положении 177 представляет собой Arg или Ile; аминокислота в положении 179 представляет собой Thr, Ile, Val или Ser; аминокислота в положении 180 представляет собой Leu, Phe или Thr; аминокислота в положении 181 представляет собой Gly, Thr, Gln или Ser; аминокислота в положении 182 представляет собой Ala, Leu, Phe или Ile; аминокислота в положении 183 представляет собой Thr или Gly; аминокислота в положении 184 представляет собой Leu, Thr, Ser или Arg; аминокислота в положении 185 представляет собой Arg, Gly, Asp или Ala; аминокислота в положении 186 представляет собой Pro, Val или Gln; аминокислота в положении 187 представляет собой Asp, Thr или Ser; аминокислота в положении 188 представляет собой His, Gly или Ala; аминокислота в положении 189 представляет собой Ala, Arg, Pro или удалена; аминокислота в положении 190 представляет собой Leu, Asn или удалена; аминокислота в положении 191 представляет собой Tyr или удалена; аминокислота в положении 192 представляет собой Ser, Ile, Val или Asn; аминокислота в положении 193 представляет собой Thr или Asp; аминокислота в положении 194 представляет собой Thr или Ser; аминокислота в положении 195 представляет собой Met или Thr; аминокислота в положении 196 представляет собой Gln, His, Leu или Ser; аминокислота в положении 197 представляет собой Ala, Gly или Leu; аминокислота в положении 198 представляет собой Thr, Glu или Ala; аминокислота в положении 199 представляет собой Pro или Arg; аминокислота в положении 200 представляет собой Asn, Ser, Thr или Gly; аминокислота в положении 201 представляет собой Ala, Leu, Glu или Trp; аминокислота в положении 202 представляет собой Ser, Asp, Phe или Leu; аминокислота в положении 203 представляет собой His, Pro, Gly или Ser; аминокислота в положении 204 представляет собой Ile, Trp, His или Gly; аминокислота в положении 205 представляет собой Ser, Asn или Ile; аминокислота в положении 206 представляет собой Ala, Gly, Asp, Tyr или Arg; аминокислота в положении 207 представляет собой Phe, Val или Leu; аминокислота в положении 208 представляет собой Asn, Ser, Pro или Leu; аминокислота в положении 210 представляет собой Arg, Asp, Glu или Tyr; аминокислота в положении 211 представляет собой Ile, Ser или Thr; аминокислота в положении 212 представляет собой Val, Ala или Asp; аминокислота в положении 214 представляет собой Pro или Arg; аминокислота в положении 215 представляет собой Ser или Thr; аминокислота в положении 217 представляет собой Tyr или Phe; аминокислота в положении 218 представляет собой Arg или Ser; аминокислота в положении 219 представляет собой Val или Ala; аминокислота в положении 220 представляет собой Cys, Leu или Ser; аминокислота в положении 221 представляет собой Pro или His; аминокислота в положении 222 представляет собой Leu, Arg или Ser; аминокислота в положении 224 представляет собой Asn или Ser; аминокислота в положении 225 представляет собой Asp, Arg или Thr; аминокислота в положении 226 представляет собой Thr или Asn; аминокислота в положении 227 представляет собой Asp, Leu или удалена; аминокислота в положении 228 представляет собой Thr или удалена; аминокислота в положении 229 представляет собой Tyr или удалена; аминокислота в положении 230 представляет собой Leu или удалена; аминокислота в положении 231 представляет собой Gly или удалена; аминокислота в положении 232 представляет собой Ile или удалена; аминокислота в положении 233 представляет собой Pro или удалена; аминокислота в положении 234 представляет собой Ala, Pro или удалена; аминокислота в положении 235 представляет собой Asp, Ile или Val; аминокислота в положении 236 представляет собой Val, Ser или Glu; аминокислота в положении 237 представляет собой Ala, Phe или Tyr; аминокислота в положении 238 представляет собой Ala или Thr; аминокислота в положении 239 представляет собой Val, Ser или Gly; аминокислота в положении 240 представляет собой Leu или Ile; аминокислота в положении 243 представляет собой Asp или Glu; аминокислота в положении 249 представляет собой Asn или Ser; аминокислота в положении 252 представляет собой Leu или Met; аминокислота в положении 257 представляет собой Thr или Ser; аминокислота в положении 259 представляет собой His или Leu; аминокислота в положении 266 представляет собой Ala или Val; аминокислота в положении 267 представляет собой Cys или Gly; аминокислота в положении 268 представляет собой His, Arg или Tyr; аминокислота в положении 272 представляет собой Asp или Glu; аминокислота в положении 273 представляет собой Val, Met, Ile или Leu; аминокислота в положении 274 представляет собой Val или Met; аминокислота в положении 278 представляет собой Gly или Ala; аминокислота в положении 279 представляет собой Glu или Val; аминокислота в положении 281 представляет собой Leu или Ala; аминокислота в положении 282 представляет собой Asn, Leu или Ile; аминокислота в положении 285 представляет собой Asn или Ser; аминокислота в положении 286 представляет собой Lys, Asp или Glu; аминокислота в положении 287 представляет собой Leu или Val; аминокислота в положении 290 представляет собой Pro, Gln или Arg; аминокислота в положении 291 представляет собой Leu или Val; аминокислота в положении 292 представляет собой Lys или Val; аминокислота в положении 293 представляет собой Glu или Gln; аминокислота в положении 294 представляет собой Ser, Asn или Lys; аминокислота в положении 295 представляет собой Thr или Ser; аминокислота в положении 296 представляет собой Gln или His; аминокислота в положении 297 представляет собой Leu или Met; аминокислота в положении 300 представляет собой Ser или Thr; аминокислота в положении 301 представляет собой Glu или Ala; аминокислота в положении 302 представляет собой Ser, Pro или Ala; аминокислота в положении 304 представляет собой Lys или Asn; аминокислота в положении 313 представляет собой Val или Ile; аминокислота в положении 314 представляет собой His, Glu или Gln; аминокислота в положении 315 представляет собой Ala, Cys или Ser; аминокислота в положении 316 представляет собой Ala или Val; аминокислота в положении 317 представляет собой Met или Ile; аминокислота в положении 319 представляет собой Met или Ile; аминокислота в положении 320 представляет собой Val или Gly; аминокислота в положении 321 представляет собой Arg или Pro; аминокислота в положении 322 представляет собой Ile или Phe; аминокислота в положении 323 представляет собой Gly или Val; аминокислота в положении 324 представляет собой Leu или Ser; аминокислота в положении 336 представляет собой Ser или Asn; аминокислота в положении 339 представляет собой Asn, Lys или Arg; аминокислота в положении 350 представляет собой Arg или Gln; аминокислота в положении 351 представляет собой Glu или Asp; аминокислота в положении 353 представляет собой Lys или Arg; аминокислота в положении 354 представляет собой Gln или Arg; аминокислота в положении 355 представляет собой Phe или Leu; аминокислота в положении 356 представляет собой Lys или Arg; аминокислота в положении 360 представляет собой Ile, Val или Ala; аминокислота в положении 365 представляет собой Leu или Phe; аминокислота в положении 371 представляет собой или Glu; аминокислота в положении 372 представляет собой или Lys; аминокислота в положении 374 представляет собой Arg или Lys; аминокислота в положении 376 представляет собой Phe или Leu; аминокислота в положении 378 представляет собой Glu или Asp; аминокислота в положении 381 представляет собой Leu или Val; аминокислота в положении 388 представляет собой Ala или Ser; аминокислота в положении 395 представляет собой Arg или Lys; аминокислота в положении 396 представляет собой Glu, Gln или Gly; аминокислота в положении 399 представляет собой Asp или Asn; аминокислота в положении 400 представляет собой Asn, Thr или Asp; аминокислота в положении 401 представляет собой Thr или Ala; аминокислота в положении 402 представляет собой Phe, Ile или Leu; аминокислота в положении 406 представляет собой Asp или Glu; аминокислота в положении 408 представляет собой Leu или Met; аминокислота в положении 410 представляет собой Gly или Leu; аминокислота в положении 414 представляет собой Ala или Glu; аминокислота в положении 416 представляет собой Ser, Asn или Asp; аминокислота в положении 417 представляет собой Ser, Arg или Gly; аминокислота в положении 423 представляет собой Lys или Gln; аминокислота в положении 431 представляет собой Arg или Lys; аминокислота в положении 432 представляет собой Gln или Glu; аминокислота в положении 436 представляет собой Arg или Glu; аминокислота в положении 440 представляет собой Asn или Arg; аминокислота в положении 442 представляет собой Leu или Val; аминокислота в положении 447 представляет собой Ser, Lys или Arg; аминокислота в положении 448 представляет собой Ala или Ser; аминокислота в положении 451 представляет собой Gln или Met; аминокислота в положении 453 представляет собой Gly или Ala; аминокислота в положении 455 представляет собой Ala или Val; аминокислота в положении 457 представляет собой Leu или Val; аминокислота в положении 467 представляет собой Val или Ala; аминокислота в положении 471 представляет собой Gly или Ala; аминокислота в положении 475 представляет собой Ser или Asn; аминокислота в положении 483 представляет собой Gly или Ala; аминокислота в положении 493 представляет собой Gln или Gly; аминокислота в положении 504 представляет собой Val или Ile; аминокислота в положении 506 представляет собой Asp или His; аминокислота в положении 509 представляет собой Asp или Asn; аминокислота в положении 510 представляет собой Ser или Ala; аминокислота в положении 512 представляет собой Glu или Asp; аминокислота в положении 515 представляет собой Gly или Ser; аминокислота в положении 516 представляет собой Gln или His; аминокислота в положении 517 представляет собой Ile или Leu; аминокислота в положении 519 представляет собой Asp, Gly или Gln; аминокислота в положении 522 представляет собой Val, Glu, Pro или Val; аминокислота в положении 525 представляет собой Glu или Asp; аминокислота в положении 526 представляет собой Leu или Met; аминокислота в положении 539 представляет собой Val или Ile; аминокислота в положении 555 представляет собой Val или Ala; аминокислота в положении 557 представляет собой Arg или Lys; аминокислота в положении 563 представляет собой Val или Met; аминокислота в положении 571 представляет собой Ser или Cys; аминокислота в положении 575 представляет собой Val или Glu; аминокислота в положении 577 представляет собой Met или Ile; аминокислота в положении 579 представляет собой Glu или Gln; аминокислота в положении 583 представляет собой Asp или Glu; аминокислота в положении 589 представляет собой Met или Leu; аминокислота в положении 590 представляет собой Met или Leu; аминокислота в положении 593 представляет собой Met или Ile; аминокислота в положении 595 представляет собой Arg или Gln; аминокислота в положении 596 представляет собой Ser или Thr; аминокислота в положении 597 представляет собой Gln или His; аминокислота в положении 607 представляет собой Ala или Val; аминокислота в положении 608 представляет собой Asp или Asn; аминокислота в положении 612 представляет собой Tyr, His или Phe; аминокислота в положении 617 представляет собой Thr или Ile; аминокислота в положении 618 представляет собой Gln или His; аминокислота в положении 625 представляет собой Arg или Ser; аминокислота в положении 626 представляет собой Met или Ile; аминокислота в положении 628 представляет собой Leu или Ile; аминокислота в положении 633 представляет собой Ile или Met; аминокислота в положении 634 представляет собой Leu или Met; аминокислота в положении 642 представляет собой Arg или Met; аминокислота в положении 648 представляет собой Met или Thr; аминокислота в положении 651 представляет собой Glu или Gln; аминокислота в положении 654 представляет собой Thr, Val или Ala; аминокислота в положении 658 представляет собой Gly или Arg; аминокислота в положении 663 представляет собой Gly или Ala; аминокислота в положении 664 представляет собой Asp или Asn; аминокислота в положении 668 представляет собой Ala или Thr; аминокислота в положении 669 представляет собой Gln или His; аминокислота в положении 671 представляет собой Asn или Ser аминокислота в положении 675 представляет собой Ile, Val или Ser; аминокислота в положении 678 представляет собой Met, Ile, Ala или Thr; аминокислота в положении 682 представляет собой Pro или Gln; аминокислота в положении 683 представляет собой Ser или Pro; аминокислота в положении 685 представляет собой Asp или Asn; аминокислота в положении 694 представляет собой Asp или Gly; аминокислота в положении 697 представляет собой Asn или Ser; аминокислота в положении 704 представляет собой Glu или Gly; аминокислота в положении 714 представляет собой Ala или Gly; аминокислота в положении 721 представляет собой Ser или Phe; аминокислота в положении 722 представляет собой Ser или Asn; аминокислота в положении 724 представляет собой Ser или Thr; аминокислота в положении 734 представляет собой His или Gln; аминокислота в положении 736 представляет собой Val или Ala; аминокислота в положении 737 представляет собой Lys или Gln; аминокислота в положении 739 представляет собой Ala или Ser; аминокислота в положении 740 представляет собой Ser или Met; аминокислота в положении 741 представляет собой Gly или Asn; аминокислота в положении 742 представляет собой Ile или Gly; аминокислота в положении 743 представляет собой Gly или удалена; аминокислота в положении 745 представляет собой Gly или Asp; аминокислота в положении 751 представляет собой Thr, Ser или Ala; аминокислота в положении 753 представляет собой Gln или Arg; аминокислота в положении 754 представляет собой Thr или Ser; аминокислота в положении 756 представляет собой Thr или Ile; аминокислота в положении 757 представляет собой Val или Ile; аминокислота в положении 766 представляет собой Ile или Val; аминокислота в положении 773 представляет собой Asp или Glu; аминокислота в положении 774 представляет собой Gln или Glu; аминокислота в положении 776 представляет собой Leu или Met; аминокислота в положении 777 представляет собой Pro или Thr; аминокислота в положении 782 представляет собой Ala, Asp или Val; аминокислота в положении 786 представляет собой Tyr или Phe; аминокислота в положении 787 представляет собой His или Gln; аминокислота в положении 788 представляет собой Tyr или Met; аминокислота в положении 789 представляет собой Ala или Arg; аминокислота в положении 790 представляет собой Tyr или Thr; аминокислота в положении 791 представляет собой Arg или Ala; аминокислота в положении 792 представляет собой Leu или Ser; аминокислота в положении 796 представляет собой Asp или Glu; аминокислота в положении 797 представляет собой Ser, Thr или Ala аминокислота в положении 802 представляет собой Glu или Gln; аминокислота в положении 806 представляет собой Gln, Asp, Glu или His; аминокислота в положении 810 представляет собой Lys или Thr; аминокислота в положении 819 представляет собой Arg или His; аминокислота в положении 829 представляет собой Lys, Ser, Ala или Pro; аминокислота в положении 832 представляет собой Ala, Lys или Glu; аминокислота в положении 833 представляет собой Gly или Glu; аминокислота в положении 842 представляет собой Leu или Pro; аминокислота в положении 847 представляет собой Gln или Glu; аминокислота в положении 848 представляет собой Ile или Val; аминокислота в положении 849 представляет собой Val или Ala; аминокислота в положении 855 представляет собой Thr или Met; аминокислота в положении 860 представляет собой Ile или Val; и аминокислота в положении 864 представляет собой His или Gln.
В некоторых вариантах осуществления полипептид PtIP-83 представляет собой вариант последовательности под SEQ ID NO: 1, где аминокислота в положении 1 представляет собой Met или удалена; аминокислота в положении 2 представляет собой Ala или удалена; аминокислота в положении 3 представляет собой Leu, Val, Ile или удалена; аминокислота в положении 4 представляет собой Val, Met, Ile или Leu; аминокислота в положении 7 представляет собой Gly, Thr или Ser; аминокислота в положении 8 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 10 представляет собой Phe, Trp или Tyr; аминокислота в положении 11 представляет собой Glu, Asp, Lys или Arg; аминокислота в положении 18 представляет собой Met, Val, Leu или Ile; аминокислота в положении 19 представляет собой Gly, Pro или Ala; аминокислота в положении 20 представляет собой Val, Ile, Leu или удалена; аминокислота в положении 21 представляет собой Leu, Ile или Val; аминокислота в положении 23 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 37 представляет собой Val, Ile или Leu; аминокислота в положении 38 представляет собой Arg, Lys, Gln или Asn; аминокислота в положении 40 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 43 представляет собой Asn, Gln, Glu или Asp; аминокислота в положении 45 представляет собой Gly или Ala; аминокислота в положении 46 представляет собой Gln, Asp, Asn или Glu; аминокислота в положении 48 представляет собой Glu, Asp, Pro, Ile, Leu или Val; аминокислота в положении 51 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 52 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 56 представляет собой Leu, Ile или Val; аминокислота в положении 59 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 66 представляет собой Pro, Gly или Ala; аминокислота в положении 67 представляет собой Val, Pro, Ile, Leu, Ser или Thr; аминокислота в положении 68 представляет собой Val, Arg, Phe, Ile, Leu, Lys или Gly; аминокислота в положении 69 представляет собой Glu, Ala, Asp, Gly, Arg или Lys; аминокислота в положении 70 представляет собой Trp, Thr, His, Tyr, Lys или Arg; аминокислота в положении 71 представляет собой Arg, Pro, Lys или удалена; аминокислота в положении 72 представляет собой Trp, Asp, Leu, Ile, Val, Glu или удалена; аминокислота в положении 73 представляет собой Pro, Gln, Asn, His или удалена; аминокислота в положении 74 представляет собой Pro, Met, Ser или Thr; аминокислота в положении 75 представляет собой Gln, His, Asn, Lys или Arg; аминокислота в положении 76 представляет собой Ile, Met, Val или Leu; аминокислота в положении 84 представляет собой Ile, Leu или Val; аминокислота в положении 91 представляет собой Trp или Phe; аминокислота в положении 93 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 94 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 96 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 97 представляет собой Gln, Phe, Asn, Lys или Arg; аминокислота в положении 98 представляет собой Ser, Thr или удалена; аминокислота в положении 99 представляет собой Asp, Glu, Gly или Ala; аминокислота в положении 100 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 101 представляет собой Glu, Thr, Asp, Ser или Trp аминокислота в положении 103 представляет собой His, Arg, Lys, Glu или Gln; аминокислота в положении 105 представляет собой Thr, Ser или Pro; аминокислота в положении 108 представляет собой Lys, Arg, Asn, Asp, Gln или Glu; аминокислота в положении 109 представляет собой Leu, Ile или Val; аминокислота в положении 111 представляет собой Ala, Ser или Thr; аминокислота в положении 112 представляет собой Ile, Arg, Thr, Leu, Val, Lys, Ser или удалена; аминокислота в положении 113 представляет собой Gln, Ala, Gly, Asn или удалена; аминокислота в положении 114 представляет собой Arg, Glu, Lys, Asp или Ile; аминокислота в положении 115 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 116 представляет собой Glu, Asn, Gln, Asp, Lys или Arg; аминокислота в положении 117 представляет собой Asn, Val, Tyr, Ile, Leu, Gln, Trp или Phe; аминокислота в положении 118 представляет собой Arg или Lys; аминокислота в положении 119 представляет собой Trp, Thr или Ser; аминокислота в положении 122 представляет собой Thr, Lys, Ser, Arg или Ala; аминокислота в положении 124 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 126 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 127 представляет собой Met, Gly или Ala; аминокислота в положении 128 представляет собой Asn, Gln, Arg или Lys; аминокислота в положении 131 представляет собой Val, Ile, Leu, Ser или Thr; аминокислота в положении 133 представляет собой Ile, Leu или Val; аминокислота в положении 134 представляет собой His или Tyr; аминокислота в положении 135 представляет собой Ala или Gly; аминокислота в положении 137 представляет собой Glu, Asp, Arg или Lys; аминокислота в положении 139 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 140 представляет собой Val, Arg, Ile, Lys или Leu; аминокислота в положении 141 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 142 представляет собой Val, Ile, Leu или Pro; аминокислота в положении 144 представляет собой Thr, Leu, Phe, Ile, Val или Tyr; аминокислота в положении 145 представляет собой Met, Pro, Gln или Asn; аминокислота в положении 146 представляет собой Ser, Gly, Thr, Ala, Gln или Asn; аминокислота в положении 147 представляет собой Trp, Gln, Tyr или Asn; аминокислота в положении 148 представляет собой Ser, Ala, Thr, Gly или Pro; аминокислота в положении 149 представляет собой Ser, Thr или удалена; аминокислота в положении 150 представляет собой Val, Ile, Leu или Tyr; аминокислота в положении 152 представляет собой Arg, Ala, Val, Ile, Leu, Lys или Gly; аминокислота в положении 154 представляет собой Ser, Trp, Thr, Asp или Glu; аминокислота в положении 156 представляет собой Leu, Asp, Ile, Val, Asn, Glu или Gln; аминокислота в положении 158 представляет собой Ser, Thr или Cys; аминокислота в положении 159 представляет собой Val, Thr, Leu или Ile; аминокислота в положении 162 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 163 представляет собой Gly, Ala или удалена; аминокислота в положении 164 представляет собой Phe или удалена; аминокислота в положении 165 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 166 представляет собой Ala, Arg, Met, Lys или Phe; аминокислота в положении 167 представляет собой Val, Ile, Leu или His; аминокислота в положении 168 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 169 представляет собой Val, His, Ile, Leu, Ser или Thr; аминокислота в положении 170 представляет собой Phe, Ile, Leu или Val; аминокислота в положении 171 представляет собой Glu, Asn, Gln или Asp; аминокислота в положении 172 представляет собой Val, Ala, Arg, Ile, Leu, Gly, Lys, Asp или Glu; аминокислота в положении 175 представляет собой Ser, Arg, Thr, Lys или Trp; аминокислота в положении 176 представляет собой Val, Leu или Ile; аминокислота в положении 177 представляет собой Arg, Lys, Leu, Val или Ile; аминокислота в положении 179 представляет собой Thr, Ile, Val, Leu или Ser; аминокислота в положении 180 представляет собой Leu, Phe, Ile, Val, Ser или Thr; аминокислота в положении 181 представляет собой Gly, Thr, Gln, Asn или Ser; аминокислота в положении 182 представляет собой Ala, Leu, Phe, Val или Ile; аминокислота в положении 183 представляет собой Thr, Ser, Ala или Gly; аминокислота в положении 184 представляет собой Leu, Thr, Ser, Ile, Val, Lys или Arg; аминокислота в положении 185 представляет собой Arg, Gly, Asp, Lys, Glu или Ala; аминокислота в положении 186 представляет собой Pro, Val, Ile, Leu, Asn или Gln; аминокислота в положении 187 представляет собой Asp, Thr, Glu или Ser; аминокислота в положении 188 представляет собой His, Gly или Ala; аминокислота в положении 189 представляет собой Ala, Arg, Pro, Lys, Gly или удалена; аминокислота в положении 190 представляет собой Leu, Asn, Ile, Val, Gln или удалена; аминокислота в положении 191 представляет собой Tyr или удалена; аминокислота в положении 192 представляет собой Ser, Ile, Val, Leu, Thr или Asn; аминокислота в положении 193 представляет собой Thr, Ser, Glu или Asp; аминокислота в положении 194 представляет собой Thr или Ser; аминокислота в положении 195 представляет собой Met или Thr; аминокислота в положении 196 представляет собой Gln, His, Leu, Asn, Ile, Val, Thr или Ser; аминокислота в положении 197 представляет собой Ala, Gly, Ile, Val или Leu; аминокислота в положении 198 представляет собой Thr, Glu, Ser, Asp, Gly или Ala; аминокислота в положении 199 представляет собой Pro, Lys или Arg; аминокислота в положении 200 представляет собой Asn, Ser, Thr, Gln, Ala или Gly; аминокислота в положении 201 представляет собой Ala, Leu, Glu, Ile, Asp или Trp; аминокислота в положении 202 представляет собой Ser, Asp, Phe, Ile, Val, Thr, Glu или Leu; аминокислота в положении 203 представляет собой His, Pro, Gly, Ala, Thr или Ser; аминокислота в положении 204 представляет собой Ile, Trp, His, Leu, Val, Ala или Gly; аминокислота в положении 205 представляет собой Ser, Asn, Leu, Val, Thr, Gln или Ile; аминокислота в положении 206 представляет собой Ala, Gly, Asp, Tyr, Glu, Lys или Arg; аминокислота в положении 207 представляет собой Phe, Val, Ile или Leu; аминокислота в положении 208 представляет собой Asn, Ser, Pro, Gln, Thr, Val, Ile или Leu; аминокислота в положении 210 представляет собой Arg, Asp, Glu, Lys, Ser или Tyr; аминокислота в положении 211 представляет собой Ile, Ser, Leu, Val или Thr; аминокислота в положении 212 представляет собой Val, Ala, Ile, Leu, Glu, Gly или Asp; аминокислота в положении 214 представляет собой Pro, Lys или Arg; аминокислота в положении 215 представляет собой Ser или Thr; аминокислота в положении 217 представляет собой Tyr или Phe; аминокислота в положении 218 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 219 представляет собой Val, Ile, Leu или Ala; аминокислота в положении 220 представляет собой Cys, Leu, Ile, Val, Thr или Ser; аминокислота в положении 221 представляет собой Pro или His; аминокислота в положении 222 представляет собой Leu, Arg, Lys, Ile, Val, Thr или Ser; аминокислота в положении 224 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 225 представляет собой Asp, Arg, Glu, Lys, Ser или Thr; аминокислота в положении 226 представляет собой Thr, Ser, Gln или Asn; аминокислота в положении 227 представляет собой Asp, Leu, Glu, Ile, Val или удалена; аминокислота в положении 228 представляет собой Thr, Ser или удалена; аминокислота в положении 229 представляет собой Tyr или удалена; аминокислота в положении 230 представляет собой Leu, Ile, Val или удалена; аминокислота в положении 231 представляет собой Gly, Ala или удалена; аминокислота в положении 232 представляет собой Ile, Leu, Val или удалена; аминокислота в положении 233 представляет собой Pro или удалена; аминокислота в положении 234 представляет собой Ala, Pro, Gly или удалена; аминокислота в положении 235 представляет собой Asp, Ile, Leu, Glu или Val; аминокислота в положении 236 представляет собой Val, Ser, Ile, Leu, Thr, Asp или Glu; аминокислота в положении 237 представляет собой Ala, Phe или Tyr; аминокислота в положении 238 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 239 представляет собой Val, Ser, Ile, Leu, Thr, Ala или Gly; аминокислота в положении 240 представляет собой Leu, Val или Ile; аминокислота в положении 243 представляет собой Asp или Glu; аминокислота в положении 249 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 252 представляет собой Leu, Ile, Val или Met; аминокислота в положении 257 представляет собой Thr или Ser; аминокислота в положении 259 представляет собой His, Ile, Val или Leu; аминокислота в положении 266 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 267 представляет собой Cys, Ala или Gly; аминокислота в положении 268 представляет собой His, Arg, Lys или Tyr; аминокислота в положении 272 представляет собой Asp или Glu; аминокислота в положении 273 представляет собой Val, Met, Ile или Leu; аминокислота в положении 274 представляет собой Val, Ile, Leu или Met; аминокислота в положении 278 представляет собой Gly или Ala; аминокислота в положении 279 представляет собой Glu, Asp, Gly или Val; аминокислота в положении 281 представляет собой Leu, Ile, Val, Gly или Ala; аминокислота в положении 282 представляет собой Asn, Leu или Ile; аминокислота в положении 285 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 286 представляет собой Lys, Asp, Arg или Glu; аминокислота в положении 287 представляет собой Leu, Ile или Val; аминокислота в положении 290 представляет собой Pro, Gln, Asn, Lys или Arg; аминокислота в положении 291 представляет собой Leu, Ile или Val; аминокислота в положении 292 представляет собой Lys, Arg, Ile, Leu или Val; аминокислота в положении 293 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 294 представляет собой Ser, Asn, Thr, Gln, Arg или Lys; аминокислота в положении 295 представляет собой Thr или Ser; аминокислота в положении 296 представляет собой Gln, Asn или His; аминокислота в положении 297 представляет собой Leu, Ile, Val или Met; аминокислота в положении 300 представляет собой Ser или Thr; аминокислота в положении 301 представляет собой Glu, Asp, Gly или Ala; аминокислота в положении 302 представляет собой Ser, Pro, Thr, Gly или Ala; аминокислота в положении 304 представляет собой Lys, Arg, Gln или Asn; аминокислота в положении 313 представляет собой Val, Leu или Ile; аминокислота в положении 314 представляет собой His, Glu, Asn, Asp или Gln; аминокислота в положении 315 представляет собой Ala, Cys, Gly, Thr или Ser; аминокислота в положении 316 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 317 представляет собой Met, Leu, Val или Ile; аминокислота в положении 319 представляет собой Met, Leu, Val или Ile; аминокислота в положении 320 представляет собой Val, Ile, Leu, Ala или Gly; аминокислота в положении 321 представляет собой Arg, Lys или Pro; аминокислота в положении 322 представляет собой Ile, Leu, Val или Phe; аминокислота в положении 323 представляет собой Gly, Ile, Leu или Val; аминокислота в положении 324 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 336 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 339 представляет собой Asn, Lys, Gln или Arg; аминокислота в положении 350 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 351 представляет собой Glu или Asp; аминокислота в положении 353 представляет собой Lys или Arg; аминокислота в положении 354 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 355 представляет собой Phe, Ile, Leu или Leu; аминокислота в положении 356 представляет собой Lys или Arg; аминокислота в положении 360 представляет собой Ile, Val, Leu, Gly или Ala; аминокислота в положении 365 представляет собой Leu, Ile, Val или Phe; аминокислота в положении 371 представляет собой или Glu или Asp; аминокислота в положении 372 представляет собой или Lys или Arg; аминокислота в положении 374 представляет собой Arg или Lys; аминокислота в положении 376 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 378 представляет собой Glu или Asp; аминокислота в положении 381 представляет собой Leu, Ile или Val; аминокислота в положении 388 представляет собой Ala, Thr, Gly или Ser; аминокислота в положении 395 представляет собой Arg или Lys; аминокислота в положении 396 представляет собой Glu, Gln, Asp, Asn, Ala или Gly; аминокислота в положении 399 представляет собой Asp, Gln, Glu или Asn; аминокислота в положении 400 представляет собой Asn, Thr, Ser, Glu, Gln или Asp; аминокислота в положении 401 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 402 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 406 представляет собой Asp или Glu; аминокислота в положении 408 представляет собой Leu, Ile, Val или Met; аминокислота в положении 410 представляет собой Gly, Ile, Val, Ala или Leu; аминокислота в положении 414 представляет собой Ala, Gly, Asp или Glu; аминокислота в положении 416 представляет собой Ser, Asn, Thr, Gln, Glu или Asp; аминокислота в положении 417 представляет собой Ser, Arg, Lys, Thr, Ala или Gly; аминокислота в положении 423 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 431 представляет собой Arg или Lys; аминокислота в положении 432 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 436 представляет собой Arg, Lys, Asp или Glu; аминокислота в положении 440 представляет собой Asn, Gln, Lys или Arg; аминокислота в положении 442 представляет собой Leu, Ile или Val; аминокислота в положении 447 представляет собой Ser, Lys, Thr или Arg; аминокислота в положении 448 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 451 представляет собой Gln, Asn или Met; аминокислота в положении 453 представляет собой Gly или Ala; аминокислота в положении 455 представляет собой Ala, Leu, Ile или Val; аминокислота в положении 457 представляет собой Leu, Ile или Val; аминокислота в положении 467 представляет собой Val, Ile, Leu, Gly или Ala; аминокислота в положении 471 представляет собой Gly или Ala; аминокислота в положении 475 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 483 представляет собой Gly или Ala; аминокислота в положении 493 представляет собой Gln, Asn или Gly; аминокислота в положении 504 представляет собой Val, Leu или Ile; аминокислота в положении 506 представляет собой Asp, Glu или His; аминокислота в положении 509 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 510 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 512 представляет собой Glu или Asp; аминокислота в положении 515 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 516 представляет собой Gln, Asn или His; аминокислота в положении 517 представляет собой Ile, Val или Leu; аминокислота в положении 519 представляет собой Asp, Asn, Glu, Gly или Gln; аминокислота в положении 522 представляет собой Val, Glu, Pro, Ile, Leu или Asp; аминокислота в положении 525 представляет собой Glu или Asp; аминокислота в положении 526 представляет собой Leu, Ile, Val или Met; аминокислота в положении 539 представляет собой Val, Leu или Ile; аминокислота в положении 555 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 557 представляет собой Arg или Lys; аминокислота в положении 563 представляет собой Val, Leu, Ile или Met; аминокислота в положении 571 представляет собой Ser, Thr или Cys; аминокислота в положении 575 представляет собой Val, Leu, Ile, Asp или Glu; аминокислота в положении 577 представляет собой Met, Leu, Val или Ile; аминокислота в положении 579 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 583 представляет собой Asp или Glu; аминокислота в положении 589 представляет собой Met, Ile, Val или Leu; аминокислота в положении 590 представляет собой Met, Ile, Val или Leu; аминокислота в положении 593 представляет собой Met, Leu, Val или Ile; аминокислота в положении 595 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 596 представляет собой Ser или Thr; аминокислота в положении 597 представляет собой Gln, Asn или His; аминокислота в положении 607 представляет собой Ala, Gly, Ile, Leu или Val; аминокислота в положении 608 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 612 представляет собой Tyr, His или Phe; аминокислота в положении 617 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 618 представляет собой Gln, Asn или His; аминокислота в положении 625 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 626 представляет собой Met, Leu, Val или Ile; аминокислота в положении 628 представляет собой Leu, Val или Ile; аминокислота в положении 633 представляет собой Ile, Leu, Val или Met; аминокислота в положении 634 представляет собой Leu, Ile, Val или Met; аминокислота в положении 642 представляет собой Arg, Lys или Met; аминокислота в положении 648 представляет собой Met, Ser или Thr; аминокислота в положении 651 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 654 представляет собой Thr, Val, Ser, Ile, Leu, Gly или Ala; аминокислота в положении 658 представляет собой Gly, Lys, Ala или Arg; аминокислота в положении 663 представляет собой Gly или Ala; аминокислота в положении 664 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 668 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 669 представляет собой Gln, Asn или His; аминокислота в положении 671 представляет собой Asn, Gln, Thr или Ser аминокислота в положении 675 представляет собой Ile, Val, Ile, Thr или Ser; аминокислота в положении 678 представляет собой Met, Ile, Ala, Leu, Ser или Thr; аминокислота в положении 682 представляет собой Pro, Asn или Gln; аминокислота в положении 683 представляет собой Ser, Thr или Pro; аминокислота в положении 685 представляет собой Asp, Glu, Asp или Asn; аминокислота в положении 694 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 697 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 704 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 714 представляет собой Ala или Gly; аминокислота в положении 721 представляет собой Ser, Thr или Phe; аминокислота в положении 722 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 724 представляет собой Ser или Thr; аминокислота в положении 734 представляет собой His, Asn или Gln; аминокислота в положении 736 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 737 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 739 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 740 представляет собой Ser, Thr или Met; аминокислота в положении 741 представляет собой Gly, Ala, Gln или Asn; аминокислота в положении 742 представляет собой Ile, Leu, Val, Ala или Gly; аминокислота в положении 743 представляет собой Gly или удалена; аминокислота в положении 745 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 751 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 753 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 754 представляет собой Thr или Ser; аминокислота в положении 756 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 757 представляет собой Val, Leu или Ile; аминокислота в положении 766 представляет собой Ile, Leu или Val; аминокислота в положении 773 представляет собой Asp или Glu; аминокислота в положении 774 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 776 представляет собой Leu, Ile, Val или Met; аминокислота в положении 777 представляет собой Pro, Ser или Thr; аминокислота в положении 782 представляет собой Ala, Asp, Glu, Ile, Leu или Val; аминокислота в положении 786 представляет собой Tyr или Phe; аминокислота в положении 787 представляет собой His, Asn или Gln; аминокислота в положении 788 представляет собой Tyr или Met; аминокислота в положении 789 представляет собой Ala, Lys или Arg; аминокислота в положении 790 представляет собой Tyr или Thr; аминокислота в положении 791 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 792 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 796 представляет собой Asp или Glu; аминокислота в положении 797 представляет собой Ser, Thr или Ala аминокислота в положении 802 представляет собой Glu, Lys, Asp, Asn или Gln; аминокислота в положении 806 представляет собой Gln, Asp, Glu, Asn или His; аминокислота в положении 810 представляет собой Lys, Arg или Thr; аминокислота в положении 819 представляет собой Arg, Lys или His; аминокислота в положении 829 представляет собой Lys, Ser, Ala или Pro; аминокислота в положении 832 представляет собой Ala, Lys, Arg, Asp или Glu; аминокислота в положении 833 представляет собой Gly, Ala, Asp или Glu; аминокислота в положении 842 представляет собой Leu, Ile, Val или Pro; аминокислота в положении 847 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 848 представляет собой Ile, Leu или Val; аминокислота в положении 849 представляет собой Val, Leu, Ile, Gly или Ala; аминокислота в положении 855 представляет собой Thr, Ser или Met; аминокислота в положении 860 представляет собой Ile, Leu или Val; аминокислота в положении 864 представляет собой His, Asn или Gln;
В некоторых вариантах осуществления полипептид PtIP-83 представляет собой вариант последовательности под SEQ ID NO: 1, где аминокислота в положении 1 представляет собой Met или удалена; аминокислота в положении 2 представляет собой Ala или удалена; аминокислота в положении 3 представляет собой Leu, Val, Ile или удалена; аминокислота в положении 4 представляет собой Val, Met, Ile или Leu; аминокислота в положении 7 представляет собой Gly, Thr или Ser; аминокислота в положении 8 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 10 представляет собой Phe, Trp или Tyr; аминокислота в положении 11 представляет собой Glu, Asp, Lys или Arg; аминокислота в положении 18 представляет собой Met, Val, Leu или Ile; аминокислота в положении 19 представляет собой Gly, Pro или Ala; аминокислота в положении 20 представляет собой Val, Ile, Leu или удалена; аминокислота в положении 21 представляет собой Leu, Ile или Val; аминокислота в положении 23 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 37 представляет собой Val, Ile или Leu; аминокислота в положении 38 представляет собой Arg, Lys, Gln или Asn; аминокислота в положении 40 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 43 представляет собой Asn, Gln, Glu или Asp; аминокислота в положении 45 представляет собой Gly или Ala; аминокислота в положении 46 представляет собой Gln, Asp, Asn или Glu; аминокислота в положении 48 представляет собой Glu, Asp, Pro, Ile, Leu или Val; аминокислота в положении 51 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 52 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 53 представляет собой Val, Ala, Cys или Thr; аминокислота в положении 54 представляет собой Lys, Ala, Cys, Asp, Glu, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser или Thr; аминокислота в положении 55 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 56 представляет собой Leu, Glu, Phe, Ile, Met, Thr или Val; аминокислота в положении 57 представляет собой Tyr, Cys, Ile, Leu, Met, Thr или Val; аминокислота в положении 58 представляет собой Val, Cys, Ile или Leu; аминокислота в положении 59 представляет собой Phe, Leu, Met, Val или Tyr; аминокислота в положении 60 представляет собой Ala, Cys, Gly, Ser, Thr или Val; аминокислота в положении 61 представляет собой Asp, Glu, His или Ser; аминокислота в положении 62 представляет собой Val, Ala, Cys, Ile, Leu или Thr; аминокислота в положении 63 представляет собой Val, Ala, Cys, Ile, Leu, Met или Thr; аминокислота в положении 64 представляет собой Glu, Ala, Cys, Phe, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 65 представляет собой Leu, Ala, Cys, Phe, His, Ile, Met, Asn, Gln, Thr, Val или Trp; аминокислота в положении 66 представляет собой Pro, Asp, Gly, Met, Gln или Arg; аминокислота в положении 67 представляет собой Val, Pro, Ile, Leu, Ser или Thr; аминокислота в положении 68 представляет собой Val, Arg, Phe, Ile, Leu, Lys или Gly; аминокислота в положении 69 представляет собой Glu, Ala, Asp, Gly, Arg или Lys; аминокислота в положении 70 представляет собой Trp, Thr, His, Tyr, Lys или Arg; аминокислота в положении 71 представляет собой Arg, Pro, Lys или удалена; аминокислота в положении 72 представляет собой Trp, Asp, Leu, Ile, Val, Glu или удалена; аминокислота в положении 73 представляет собой Pro, Gln, Asn, His или удалена; аминокислота в положении 74 представляет собой Pro, Met, Ser или Thr; аминокислота в положении 75 представляет собой Gln, His, Asn, Lys или Arg; аминокислота в положении 76 представляет собой Ile, Met, Val или Leu; аминокислота в положении 84 представляет собой Ile, Leu или Val; аминокислота в положении 91 представляет собой Trp или Phe; аминокислота в положении 93 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 94 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 96 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 97 представляет собой Gln, Phe, Asn, Lys или Arg; аминокислота в положении 98 представляет собой Ser, Thr или удалена; аминокислота в положении 99 представляет собой Asp, Glu, Gly или Ala; аминокислота в положении 100 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 101 представляет собой Glu, Thr, Asp, Ser или Trp аминокислота в положении 103 представляет собой His, Arg, Lys, Glu или Gln; аминокислота в положении 105 представляет собой Thr, Ser или Pro; аминокислота в положении 108 представляет собой Lys, Arg, Asn, Asp, Gln или Glu; аминокислота в положении 109 представляет собой Leu, Ile или Val; аминокислота в положении 111 представляет собой Ala, Ser или Thr; аминокислота в положении 112 представляет собой Ile, Arg, Thr, Leu, Val, Lys, Ser или удалена; аминокислота в положении 113 представляет собой Gln, Ala, Gly, Asn или удалена; аминокислота в положении 114 представляет собой Arg, Glu, Lys, Asp или Ile; аминокислота в положении 115 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 116 представляет собой Glu, Asn, Gln, Asp, Lys или Arg; аминокислота в положении 117 представляет собой Asn, Val, Tyr, Ile, Leu, Gln, Trp или Phe; аминокислота в положении 118 представляет собой Arg или Lys; аминокислота в положении 119 представляет собой Trp, Thr или Ser; аминокислота в положении 122 представляет собой Thr, Lys, Ser, Arg или Ala; аминокислота в положении 124 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 126 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 127 представляет собой Met, Gly или Ala; аминокислота в положении 128 представляет собой Asn, Gln, Arg или Lys; аминокислота в положении 131 представляет собой Val, Ile, Leu, Ser или Thr; аминокислота в положении 133 представляет собой Ile, Leu или Val; аминокислота в положении 134 представляет собой His или Tyr; аминокислота в положении 135 представляет собой Ala или Gly; аминокислота в положении 137 представляет собой Glu, Asp, Arg или Lys; аминокислота в положении 139 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 140 представляет собой Val, Arg, Ile, Lys или Leu; аминокислота в положении 141 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 142 представляет собой Val, Ile, Leu или Pro; аминокислота в положении 144 представляет собой Thr, Leu, Phe, Ile, Val или Tyr; аминокислота в положении 145 представляет собой Met, Pro, Gln или Asn; аминокислота в положении 146 представляет собой Ser, Gly, Thr, Ala, Gln или Asn; аминокислота в положении 147 представляет собой Trp, Gln, Tyr или Asn; аминокислота в положении 148 представляет собой Ser, Ala, Thr, Gly или Pro; аминокислота в положении 149 представляет собой Ser, Thr или удалена; аминокислота в положении 150 представляет собой Val, Ile, Leu или Tyr; аминокислота в положении 152 представляет собой Arg, Ala, Val, Ile, Leu, Lys или Gly; аминокислота в положении 154 представляет собой Ser, Trp, Thr, Asp или Glu; аминокислота в положении 156 представляет собой Leu, Asp, Ile, Val, Asn, Glu или Gln; аминокислота в положении 158 представляет собой Ser, Thr или Cys; аминокислота в положении 159 представляет собой Val, Thr, Leu или Ile; аминокислота в положении 162 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 163 представляет собой Gly, Ala или удалена; аминокислота в положении 164 представляет собой Phe или удалена; аминокислота в положении 165 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 166 представляет собой Ala, Arg, Met, Lys или Phe; аминокислота в положении 167 представляет собой Val, Ile, Leu или His; аминокислота в положении 168 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 169 представляет собой Val, His, Ile, Leu, Ser или Thr; аминокислота в положении 170 представляет собой Phe, Ile, Leu или Val; аминокислота в положении 171 представляет собой Glu, Asn, Gln или Asp; аминокислота в положении 172 представляет собой Val, Ala, Arg, Ile, Leu, Gly, Lys, Asp или Glu; аминокислота в положении 175 представляет собой Ser, Arg, Thr, Lys или Trp; аминокислота в положении 176 представляет собой Val, Leu или Ile; аминокислота в положении 177 представляет собой Arg, Lys, Leu, Val или Ile; аминокислота в положении 179 представляет собой Thr, Ile, Val, Leu или Ser; аминокислота в положении 180 представляет собой Leu, Phe, Ile, Val, Ser или Thr; аминокислота в положении 181 представляет собой Gly, Thr, Gln, Asn или Ser; аминокислота в положении 182 представляет собой Ala, Leu, Phe, Val или Ile; аминокислота в положении 183 представляет собой Thr, Ser, Ala или Gly; аминокислота в положении 184 представляет собой Leu, Thr, Ser, Ile, Val, Lys или Arg; аминокислота в положении 185 представляет собой Arg, Gly, Asp, Lys, Glu или Ala; аминокислота в положении 186 представляет собой Pro, Val, Ile, Leu, Asn или Gln; аминокислота в положении 187 представляет собой Asp, Thr, Glu или Ser; аминокислота в положении 188 представляет собой His, Gly или Ala; аминокислота в положении 189 представляет собой Ala, Arg, Pro, Lys, Gly или удалена; аминокислота в положении 190 представляет собой Leu, Asn, Ile, Val, Gln или удалена; аминокислота в положении 191 представляет собой Tyr или удалена; аминокислота в положении 192 представляет собой Ser, Ile, Val, Leu, Thr или Asn; аминокислота в положении 193 представляет собой Thr, Ser, Glu или Asp; аминокислота в положении 194 представляет собой Thr или Ser; аминокислота в положении 195 представляет собой Met или Thr; аминокислота в положении 196 представляет собой Gln, His, Leu, Asn, Ile, Val, Thr или Ser; аминокислота в положении 197 представляет собой Ala, Gly, Ile, Val или Leu; аминокислота в положении 198 представляет собой Thr, Glu, Ser, Asp, Gly или Ala; аминокислота в положении 199 представляет собой Pro, Lys или Arg; аминокислота в положении 200 представляет собой Asn, Ser, Thr, Gln, Ala или Gly; аминокислота в положении 201 представляет собой Ala, Leu, Glu, Ile, Asp или Trp; аминокислота в положении 202 представляет собой Ser, Asp, Phe, Ile, Val, Thr, Glu или Leu; аминокислота в положении 203 представляет собой His, Pro, Gly, Ala, Thr или Ser; аминокислота в положении 204 представляет собой Ile, Trp, His, Leu, Val, Ala или Gly; аминокислота в положении 205 представляет собой Ser, Asn, Leu, Val, Thr, Gln или Ile; аминокислота в положении 206 представляет собой Ala, Gly, Asp, Tyr, Glu, Lys или Arg; аминокислота в положении 207 представляет собой Phe, Val, Ile или Leu; аминокислота в положении 208 представляет собой Asn, Ser, Pro, Gln, Thr, Val, Ile или Leu; аминокислота в положении 210 представляет собой Arg, Asp, Glu, Lys, Ser или Tyr; аминокислота в положении 211 представляет собой Ile, Ser, Leu, Val или Thr; аминокислота в положении 212 представляет собой Val, Ala, Ile, Leu, Glu, Gly или Asp; аминокислота в положении 214 представляет собой Pro, Lys или Arg; аминокислота в положении 215 представляет собой Ser или Thr; аминокислота в положении 217 представляет собой Tyr или Phe; аминокислота в положении 218 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 219 представляет собой Val, Ile, Leu или Ala; аминокислота в положении 220 представляет собой Cys, Leu, Ile, Val, Thr или Ser; аминокислота в положении 221 представляет собой Pro или His; аминокислота в положении 222 представляет собой Leu, Arg, Lys, Ile, Val, Thr или Ser; аминокислота в положении 224 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 225 представляет собой Asp, Arg, Glu, Lys, Ser или Thr; аминокислота в положении 226 представляет собой Thr, Ser, Gln или Asn; аминокислота в положении 227 представляет собой Asp, Leu, Glu, Ile, Val или удалена; аминокислота в положении 228 представляет собой Thr, Ser или удалена; аминокислота в положении 229 представляет собой Tyr или удалена; аминокислота в положении 230 представляет собой Leu, Ile, Val или удалена; аминокислота в положении 231 представляет собой Gly, Ala или удалена; аминокислота в положении 232 представляет собой Ile, Leu, Val или удалена; аминокислота в положении 233 представляет собой Pro или удалена; аминокислота в положении 234 представляет собой Ala, Pro, Gly или удалена; аминокислота в положении 235 представляет собой Asp, Ile, Leu, Glu или Val; аминокислота в положении 236 представляет собой Val, Ser, Ile, Leu, Thr, Asp или Glu; аминокислота в положении 237 представляет собой Ala, Phe или Tyr; аминокислота в положении 238 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 239 представляет собой Val, Ser, Ile, Leu, Thr, Ala или Gly; аминокислота в положении 240 представляет собой Leu, Val или Ile; аминокислота в положении 243 представляет собой Asp или Glu; аминокислота в положении 249 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 252 представляет собой Leu, Ile, Val или Met; аминокислота в положении 257 представляет собой Thr или Ser; аминокислота в положении 259 представляет собой His, Ile, Val или Leu; аминокислота в положении 266 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 267 представляет собой Cys, Ala или Gly; аминокислота в положении 268 представляет собой His, Arg, Lys или Tyr; аминокислота в положении 272 представляет собой Asp или Glu; аминокислота в положении 273 представляет собой Val, Met, Ile или Leu; аминокислота в положении 274 представляет собой Val, Ile, Leu или Met; аминокислота в положении 278 представляет собой Gly или Ala; аминокислота в положении 279 представляет собой Glu, Asp, Gly или Val; аминокислота в положении 281 представляет собой Leu, Ile, Val, Gly или Ala; аминокислота в положении 282 представляет собой Asn, Leu или Ile; аминокислота в положении 285 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 286 представляет собой Lys, Asp, Arg или Glu; аминокислота в положении 287 представляет собой Leu, Ile или Val; аминокислота в положении 290 представляет собой Pro, Gln, Asn, Lys или Arg; аминокислота в положении 291 представляет собой Leu, Ile или Val; аминокислота в положении 292 представляет собой Lys, Arg, Ile, Leu или Val; аминокислота в положении 293 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 294 представляет собой Ser, Asn, Thr, Gln, Arg или Lys; аминокислота в положении 295 представляет собой Thr или Ser; аминокислота в положении 296 представляет собой Gln, Asn или His; аминокислота в положении 297 представляет собой Leu, Ile, Val или Met; аминокислота в положении 300 представляет собой Ser или Thr; аминокислота в положении 301 представляет собой Glu, Asp, Gly или Ala; аминокислота в положении 302 представляет собой Ser, Pro, Thr, Gly или Ala; аминокислота в положении 304 представляет собой Lys, Arg, Gln или Asn; аминокислота в положении 313 представляет собой Val, Leu или Ile; аминокислота в положении 314 представляет собой His, Glu, Asn, Asp или Gln; аминокислота в положении 315 представляет собой Ala, Cys, Gly, Thr или Ser; аминокислота в положении 316 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 317 представляет собой Met, Leu, Val или Ile; аминокислота в положении 319 представляет собой Met, Leu, Val или Ile; аминокислота в положении 320 представляет собой Val, Ile, Leu, Ala или Gly; аминокислота в положении 321 представляет собой Arg, Lys или Pro; аминокислота в положении 322 представляет собой Ile, Leu, Val или Phe; аминокислота в положении 323 представляет собой Gly, Ile, Leu или Val; аминокислота в положении 324 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 336 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 339 представляет собой Asn, Lys, Gln или Arg; аминокислота в положении 350 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 351 представляет собой Glu или Asp; аминокислота в положении 353 представляет собой Lys или Arg; аминокислота в положении 354 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 355 представляет собой Phe, Ile, Leu или Leu; аминокислота в положении 356 представляет собой Lys или Arg; аминокислота в положении 360 представляет собой Ile, Val, Leu, Gly или Ala; аминокислота в положении 363 представляет собой Gln, Ala, Cys, Glu, Phe, Gly, His, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 364 представляет собой Ile, Ala, Cys, Glu, Phe, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 365 представляет собой Leu, Ala, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Arg, Val, Trp или Tyr; аминокислота в положении 366 представляет собой Gly, Ala, Cys, Phe, His, Ile, Lys, Leu, Met, Asn, Ser, Thr или Val; аминокислота в положении 367 представляет собой Ser, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val или Trp; аминокислота в положении 368 представляет собой Tyr, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp; аминокислота в положении 369 представляет собой Leu, Ala, Cys, Asp, Phe, Gly, Ile, Met, Thr или Val; аминокислота в положении 370 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 371 представляет собой Gln, Ala, Cys, Asp, Glu, Phe, Gly, Ile, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 372 представляет собой Gln, Ala, Cys, Asp, Phe, Gly, His, Ile, Leu, Asn, Arg, Ser, Val или Tyr; аминокислота в положении 373 представляет собой Asn, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Gln, Ser, Thr, Val или Trp; аминокислота в положении 374 представляет собой Arg или Lys; аминокислота в положении 376 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 378 представляет собой Glu или Asp; аминокислота в положении 381 представляет собой Leu, Ile или Val; аминокислота в положении 388 представляет собой Ala, Thr, Gly или Ser; аминокислота в положении 395 представляет собой Arg или Lys; аминокислота в положении 396 представляет собой Glu, Gln, Asp, Asn, Ala или Gly; аминокислота в положении 399 представляет собой Asp, Gln, Glu или Asn; аминокислота в положении 400 представляет собой Asn, Thr, Ser, Glu, Gln или Asp; аминокислота в положении 401 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 402 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 406 представляет собой Asp или Glu; аминокислота в положении 408 представляет собой Leu, Ile, Val или Met; аминокислота в положении 410 представляет собой Gly, Ile, Val, Ala или Leu; аминокислота в положении 414 представляет собой Ala, Gly, Asp или Glu; аминокислота в положении 416 представляет собой Ser, Asn, Thr, Gln, Glu или Asp; аминокислота в положении 417 представляет собой Ser, Arg, Lys, Thr, Ala или Gly; аминокислота в положении 423 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 431 представляет собой Arg или Lys; аминокислота в положении 432 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 436 представляет собой Arg, Lys, Asp или Glu; аминокислота в положении 440 представляет собой Asn, Gln, Lys или Arg; аминокислота в положении 442 представляет собой Leu, Ile или Val; аминокислота в положении 447 представляет собой Ser, Lys, Thr или Arg; аминокислота в положении 448 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 451 представляет собой Gln, Asn или Met; аминокислота в положении 453 представляет собой Gly или Ala; аминокислота в положении 455 представляет собой Ala, Leu, Ile или Val; аминокислота в положении 457 представляет собой Leu, Ile или Val; аминокислота в положении 467 представляет собой Val, Ile, Leu, Gly или Ala; аминокислота в положении 471 представляет собой Gly или Ala; аминокислота в положении 475 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 483 представляет собой Gly или Ala; аминокислота в положении 493 представляет собой Gln, Asn или Gly; аминокислота в положении 504 представляет собой Val, Leu или Ile; аминокислота в положении 506 представляет собой Asp, Glu или His; аминокислота в положении 509 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 510 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 512 представляет собой Glu или Asp; аминокислота в положении 515 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 516 представляет собой Gln, Asn или His; аминокислота в положении 517 представляет собой Ile, Val или Leu; аминокислота в положении 519 представляет собой Asp, Asn, Glu, Gly или Gln; аминокислота в положении 522 представляет собой Val, Glu, Pro, Ile, Leu или Asp; аминокислота в положении 525 представляет собой Glu или Asp; аминокислота в положении 526 представляет собой Leu, Ile, Val или Met; аминокислота в положении 539 представляет собой Val, Leu или Ile; аминокислота в положении 555 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 556 представляет собой Trp, Phe, Thr или Tyr; аминокислота в положении 557 представляет собой Arg, Cys, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 558 представляет собой Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 559 представляет собой Lys, Ala, Cys, Phe, Gly, His, Ile, Leu, Asn, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 560 представляет собой Cys, Ala, Phe, Gly, Ile, Met, Asn, Arg, Ser, Thr или Val; аминокислота в положении 561 представляет собой Lys, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 562 представляет собой Asn, Cys, Asp, Glu, Gly, His, Leu, Met, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 563 представляет собой Val, Ala, Cys, Asp, Phe, His, Ile, Leu, Met, Asn, Gln, Thr или Trp; аминокислота в положении 564 представляет собой Ala, Cys, Gly, Met, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 571 представляет собой Ser, Thr или Cys; аминокислота в положении 575 представляет собой Val, Leu, Ile, Asp или Glu; аминокислота в положении 577 представляет собой Met, Leu, Val или Ile; аминокислота в положении 579 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 583 представляет собой Asp или Glu; аминокислота в положении 589 представляет собой Met, Ile, Val или Leu; аминокислота в положении 590 представляет собой Met, Ile, Val или Leu; аминокислота в положении 593 представляет собой Met, Leu, Val или Ile; аминокислота в положении 595 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 596 представляет собой Ser или Thr; аминокислота в положении 597 представляет собой Gln, Asn или His; аминокислота в положении 607 представляет собой Ala, Gly, Ile, Leu или Val; аминокислота в положении 608 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 612 представляет собой Tyr, His или Phe; аминокислота в положении 617 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 618 представляет собой Gln, Asn или His; аминокислота в положении 625 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 626 представляет собой Met, Leu, Val или Ile; аминокислота в положении 628 представляет собой Leu, Val или Ile; аминокислота в положении 633 представляет собой Ile, Leu, Val или Met; аминокислота в положении 634 представляет собой Leu, Ile, Val или Met; аминокислота в положении 642 представляет собой Arg, Lys или Met; аминокислота в положении 646 представляет собой Leu, Ala, Cys, Gly, Ile, Met, Asn, Gln, Ser, Thr или Val; аминокислота в положении 647 представляет собой Leu, Asp, Gly, Met, Asn, Gln или Thr; аминокислота в положении 648 представляет собой Met, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 649 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr; аминокислота в положении 650 представляет собой Thr, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Val или Tyr; аминокислота в положении 651 представляет собой Glu, Ala, Cys, Asp, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 652 представляет собой Leu, Cys, Phe, Ile, Lys, Met, Pro, Arg, Ser, Thr или Val; аминокислота в положении 653 представляет собой Thr, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Pro, Arg, Ser, Val или Trp; аминокислота в положении 654 представляет собой Thr, Ala, Cys, Phe, Ile, Lys, Leu, Met, Pro, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 655 представляет собой Trp, Phe или Tyr; аминокислота в положении 658 представляет собой Gly, Lys, Ala или Arg; аминокислота в положении 663 представляет собой Gly или Ala; аминокислота в положении 664 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 668 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 669 представляет собой Gln, Asn или His; аминокислота в положении 671 представляет собой Asn, Gln, Thr или Ser аминокислота в положении 675 представляет собой Ile, Val, Ile, Thr или Ser; аминокислота в положении 678 представляет собой Met, Ile, Ala, Leu, Ser или Thr; аминокислота в положении 682 представляет собой Pro, Asn или Gln; аминокислота в положении 683 представляет собой Ser, Thr или Pro; аминокислота в положении 685 представляет собой Asp, Glu, Asp или Asn; аминокислота в положении 694 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 697 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 704 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 714 представляет собой Ala или Gly; аминокислота в положении 721 представляет собой Ser, Thr или Phe; аминокислота в положении 722 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 724 представляет собой Ser или Thr; аминокислота в положении 734 представляет собой His, Asn или Gln; аминокислота в положении 736 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 737 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 739 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 740 представляет собой Ser, Thr или Met; аминокислота в положении 741 представляет собой Gly, Ala, Gln или Asn; аминокислота в положении 742 представляет собой Ile, Leu, Val, Ala или Gly; аминокислота в положении 743 представляет собой Gly или удалена; аминокислота в положении 745 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 751 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 753 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 754 представляет собой Thr или Ser; аминокислота в положении 756 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 757 представляет собой Val, Leu или Ile; аминокислота в положении 766 представляет собой Ile, Leu или Val; аминокислота в положении 771 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Asn, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 772 представляет собой Arg, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 773 представляет собой Asp, Ala, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 774 представляет собой Gln, Ala, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 775 представляет собой Val, Ala, Cys, Asp, Glu, Gly, His, Ile, Asn, Pro, Gln, Arg, Ser, Thr или Tyr; аминокислота в положении 776 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 777 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 778 представляет собой Phe, Ala, His, Ile, Leu, Met, Asn, Gln, Ser, Val, Trp или Tyr; аминокислота в положении 779 представляет собой Gln, Ala, Cys, Asp, Glu, Gly, His, Lys, Leu, Asn, Pro, Arg, Ser, Thr или Val; аминокислота в положении 780 представляет собой Ala, Cys, Asn, Pro, Gln или Ser; аминокислота в положении 781 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 782 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 783 представляет собой Pro, Ala, Cys, Asp, Glu, Gly, His, Asn, Gln, Arg, Ser, Thr или Val; аминокислота в положении 784 представляет собой Leu, Ala, Glu, Phe, His, Ile, Lys, Met, Asn, Pro, Gln, Ser, Thr, Val или Trp; аминокислота в положении 785 представляет собой Asn, Ala, Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 786 представляет собой Tyr, Phe, Ile, Leu или Trp; аминокислота в положении 787 представляет собой His, Asn или Gln; аминокислота в положении 788 представляет собой Tyr или Met; аминокислота в положении 789 представляет собой Ala, Lys или Arg; аминокислота в положении 790 представляет собой Tyr или Thr; аминокислота в положении 791 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 792 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 796 представляет собой Asp или Glu; аминокислота в положении 797 представляет собой Ser, Thr или Ala аминокислота в положении 802 представляет собой Glu, Lys, Asp, Asn или Gln; аминокислота в положении 806 представляет собой Gln, Asp, Glu, Asn или His; аминокислота в положении 810 представляет собой Lys, Arg или Thr; аминокислота в положении 819 представляет собой Arg, Lys или His; аминокислота в положении 829 представляет собой Lys, Ser, Ala или Pro; аминокислота в положении 832 представляет собой Ala, Lys, Arg, Asp или Glu; аминокислота в положении 833 представляет собой Gly, Ala, Asp или Glu; аминокислота в положении 842 представляет собой Leu, Ile, Val или Pro; аминокислота в положении 847 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 848 представляет собой Ile, Leu или Val; аминокислота в положении 849 представляет собой Val, Leu, Ile, Gly или Ala; аминокислота в положении 855 представляет собой Thr, Ser или Met; аминокислота в положении 860 представляет собой Ile, Leu или Val; и аминокислота в положении 864 представляет собой His, Asn или Gln.
В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников отдела Pteridophyta. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников класса Psilotopsida. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников класса Psilotopsida, порядка Psilotales. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников класса Psilotopsida, порядка Ophioglossales. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников класса Psilotopsida, порядка Ophioglossales, семейства Psilotaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников класса Psilotopsida, порядка Ophioglossales семейства Ophioglossaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников рода Ophioglossum L., Botrychium, Botrypus, Helminthostachys, Ophioderma, Cheiroglossa, Sceptridium или Mankyua. В некоторых вариантах осуществления полипептид PtIP-83 получают из видов класса Polypodiopsida/Pteridopsida. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Osmundales (королевские папоротники); семейства Osmundaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Hymenophyllales; семейства Hymenophyllaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Gleicheniales; семейства Gleicheniaceae, семейства Dipteridaceae или семейства Matoniaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Schizaeales; семейства Lygodiaceae, семейства Anemiaceae или семейства Schizaeaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Schizaeales; семейства Schizaeaceae, рода Lygodium, выбранного без ограничения из Lygodium articulatum, Lygodium circinatum, Lygodium conforme, Lygodium cubense, Lygodium digitatum, Lygodium flexuosum, Lygodium heterodoxum, Lygodium japonicum, Lygodium kerstenii, Lygodium lanceolatum, Lygodium longifolium, Lygodium merrilii, Lygodium micans, Lygodium microphyllum, Lygodium microstachyum, Lygodium oligostachyum, Lygodium palmatum, Lygodium polystachyum, Lygodium radiatum, Lygodium reticulatum, Lygodium salicifolium, Lygodium scandens, Lygodium smithianum, Lygodium subareolatum, Lygodium trifurcatum,, Lygodium venustum, Lygodium versteeghii, Lygodium volubile и Lygodium yunnanense.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Salviniales; семейства Marsileaceae или семейства Salviniaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Cyatheales; семейства Thyrsopteridaceae, семейства Loxsomataceae, семейства Culcitaceae, семейства Plagiogyriaceae, семейства Cibotiaceae, семейства Cyatheaceae, семейства Dicksoniaceae или семейства Metaxyaceae.
Внекоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales; семейства Lindsaeaceae, семейства Saccolomataceae, семейства Cystodiaceae, семейства Dennstaedtiaceae, семейства Pteridaceae, семейства Aspleniaceae, семейства Thelypteridaceae, семейства Woodsiaceae, семейства Onocleaceae, семейства Blechnaceae, семейства Dryopteridaceae, семейства Lomariopsidaceae, семейства Tectariaceae, семейства Oleandraceae, семейства Davalliaceae или семейства Polypodiaceae.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Pteridaceae, рода Adiantaceae, выбранного без ограничения из Adiantum aethiopicum, Adiantum aleuticum, Adiantum bonatianum, Adiantum cajennense, Adiantum capillus-junonis, Adiantum capillus-veneris, Adiantum caudatum, Adiantum chienii, Adiantum chilense, Adiantum cuneatum, Adiantum cunninghamii, Adiantum davidii, Adiantum diaphanum, Adiantum edentulum, Adiantum edgeworthii, Adiantum excisum, Adiantum fengianum, Adiantum fimbriatum, Adiantum flabellulatum, Adiantum formosanum, Adiantum formosum, Adiantum fulvum, Adiantum gravesii, Adiantum hispidulum, Adiantum induratum, Adiantum jordanii, Adiantum juxtapositum, Adiantum latifolium, Adiantum leveillei, Adiantum lianxianense, Adiantum malesianum, Adiantum mariesii, Adiantum monochlamys, Adiantum myriosorum, Adiantum obliquum, Adiantum ogasawarense, Adiantum pedatum, Adiantum pentadactylon, Adiantum peruvianum, Adiantum philippense, Adiantum princeps, Adiantum pubescens, Adiantum raddianum, Adiantum reniforme, Adiantum roborowskii, Adiantum serratodentatum, Adiantum sinicum, Adiantum soboliferum, Adiantum subcordatum, Adiantum tenerum, Adiantum terminatum, Adiantum tetraphyllum, Adiantum trapeziforme, Adiantum venustum, Adiantum viridescens и Adiantum viridimontanum.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Aspleniaceae, рода Asplenium.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Aspleniaceae, рода Asplenium, выбранного без ограничения из Asplenium adiantum, Asplenium adulterinum, Asplenium aequibasis, Asplenium aethiopicum, Asplenium africanum, Asplenium × alternifolium, Asplenium angustum, Asplenium antiquum, Asplenium ascensionis, Asplenium attenuatum, Asplenium aureum, Asplenium auritum, Asplenium australasicum, Asplenium azoricum, Asplenium bifrons, Asplenium billottii, Asplenium bipinnatifidum, Asplenium brachycarpum, Asplenium bradleyi, Asplenium bulbiferum, Asplenium caudatum, Asplenium ceterach, Asplenium compressum, Asplenium congestum, Asplenium corderoanum, Asplenium crinicaule, Asplenium cristatum, Asplenium cuneifolium, Asplenium cymbifolium, Asplenium daghestanicum, Asplenium dalhousiae, Asplenium dareoides, Asplenium daucifolium, Asplenium difforme, Asplenium fissum, Asplenium dimorphum, Asplenium divaricatum, Asplenium dregeanum, Asplenium × ebenoides, Asplenium ecuadorense, Asplenium feei Kunze, Asplenium fissum, Asplenium flabellifolium, Asplenium flaccidum, Asplenium fontanum, Asplenium forisiense, Asplenium formosum, Asplenium gemmiferum, Asplenium × germanicum, Asplenium gueinzii, Asplenium goudeyi, Asplenium hemionitis, Asplenium hermannii-christii, Asplenium hookerianum, Asplenium hybridum, Asplenium incisum, Asplenium × jacksonii, Asplenium × kenzoi, Asplenium laciniatum, Asplenium lamprophyllum, Asplenium laserpitiifolium, Asplenium lepidum, Asplenium listeri, Asplenium longissimum, Asplenium lucidum, Asplenium lunulatum, Asplenium lyallii, Asplenium macedonicum, Asplenium majoricum, Asplenium marinum, Asplenium × microdon, Asplenium milnei Carruth, Asplenium montanum, Asplenium musifolium, Asplenium nidus, Asplenium normale, Asplenium obliquum, Asplenium oblongifolium, Asplenium obovatum, Asplenium obtusatum, Asplenium oligolepidum, Asplenium oligophlebium, Asplenium onopteris, Asplenium pacificum, Asplenium paleaceum, Asplenium palmeri, Asplenium petrarchae, Asplenium pinnatifidum, Asplenium planicaule, Asplenium platybasis, Asplenium platyneuron, Asplenium polyodon, Asplenium praemorsum, Asplenium prolongatum, Asplenium pteridoides, Asplenium resiliens, Asplenium rhizophyllum, Asplenium richardii, Asplenium ruprechtii, Asplenium ruta-muraria, Asplenium rustifolium, Asplenium sagittatum, Asplenium sandersonii, Asplenium × sarniense, Asplenium schizotrichum, Asplenium schweinfurthii, Asplenium scleroprium, Asplenium scolopendrium (syn. Phyllitis scolopendrium), Asplenium seelosii, Asplenium septentrionale, Asplenium septentrionale × trichomanes, Asplenium serra, Asplenium serratum, Asplenium sessilifolium, Asplenium shuttleworthianum, Asplenium simplicifrons, Asplenium splendens, Asplenium surrogatum, Asplenium tenerum, Asplenium terrestre, Asplenium theciferum, Asplenium thunbergii, Asplenium trichomanes, Asplenium tutwilerae, Asplenium vespertinum, Asplenium vieillardii, Asplenium virens, Asplenium viride, Asplenium vittiforme и Asplenium viviparum.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Blechnaceae, рода Blecnum.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae рода Acrophorus, рода Acrorumohra, рода Anapausia, рода Arachniodes, рода Bolbitis, рода Ctenitis, рода Cyclodium, рода Cyrtogonellum, рода Cyrtomidictyum, рода Cyrtomium, рода Diacalpe, рода Didymochlaena, рода Dryopsis, рода Dryopteris, рода Elaphoglossum, рода Hypodematium, рода Lastreopsis, рода Leptorumohra, рода Leucostegia, рода Lithostegia, рода Lomagramma, рода Maxonia, рода Megalastrum, рода Olfersia, рода Peranema, рода Phanerophlebia, рода Phanerophlebiopsis, рода Polybotrya, рода Polystichopsis, рода Polystichum, рода Rumohra, рода Sorolepidium, рода Stigmatopteris или рода Teratophyllum.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Polystichum. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Polystichum, выбранного без ограничения из Polystichum acanthophyllum, Polystichum acrostichoides, Polystichum aculeatum, Polystichum acutidens, Polystichum acutipinnulum, Polystichum alcicorne, Polystichum aleuticum, Polystichum andersonii, Polystichum atkinsonii, Polystichum australiense, Polystichum bakerianum, Polystichum biaristatum, Polystichum bomiense, Polystichum bonseyi, Polystichum brachypterum, Polystichum braunii, Polystichum brachypterum, Polystichum calderonense, Polystichum californicum, Polystichum capillipes, Polystichum castaneum, Polystichum chilense, Polystichum christii Ching, Polystichum chunii Ching, Polystichum craspedosorum, Polystichum cyclolobum, Polystichum cystostegia, Polystichum deltodon, Polystichum dielsii, Polystichum discretum, Polystichum drepanum, Polystichum dudleyi, Polystichum duthiei, Polystichum echinatum, Polystichum erosum, Polystichum excellens, Polystichum eximium, Polystichum falcatipinnum, Polystichum falcinellum, Polystichum fallax, Polystichum formosanum, Polystichum gongboense, Polystichum grandifrons, Polystichum gymnocarpium, Polystichum haleakalense, Polystichum hancockii, Polystichum hecatopteron, Polystichum herbaceum, Polystichum imbricans, Polystichum incongruum, Polystichum kruckebergii, Polystichum kwakiutlii, Polystichum lachenense, Polystichum lanceolatum, Polystichum lemmonii, Polystichum lentum, Polystichum lonchitis, Polystichum longidens, Polystichum longipaleatum, Polystichum longipes, Polystichum luctuosum, Polystichum macleae, Polystichum macrochlaenum, Polystichum makinoi, Polystichum martini, Polystichum mayebarae, Polystichum mediocre, Polystichum medogense, Polystichum microchlamys, Polystichum mohrioides, Polystichum mollissimum, Polystichum monticola, Polystichum moorei, Polystichum morii, Polystichum moupinense, Polystichum muricatum, Polystichum nakenense, Polystichum neolobatum, Polystichum nepalense, Polystichum ningshenense, Polystichum obliquum, Polystichum omeiense, Polystichum ordinatum, Polystichum orientalitibeticum, Polystichum paramoupinense, Polystichum parvipinnulum, Polystichum piceopaleaceum, Polystichum polyblepharum, Polystichum prescottianum, Polystichum prionolepis, Polystichum proliferum, Polystichum pseudocastaneum], Polystichum pseudomakinoi, Polystichum punctiferum, Polystichum pungens, Polystichum qamdoense, Polystichum retrosopaleaceum, Polystichum rhombiforme, Polystichum rhomboidea, Polystichum richardii, Polystichum rigens, Polystichum rotundilobum, Polystichum scopulinum, Polystichum semifertile, Polystichum setiferum, Polystichum setigerum, Polystichum shensiense, Polystichum silvaticum, Polystichum simplicipinnum, Polystichum sinense, Polystichum squarrosum, Polystichum stenophyllum, Polystichum stimulans, Polystichum submite, Polystichum tacticopterum, Polystichum thomsoni, Polystichum tibeticum, Polystichum transvaalense, Polystichum tripteron, Polystichum tsus-simense, Polystichum vestitum, Polystichum wattii, Polystichum whiteleggei, Polystichum xiphophyllum, Polystichum yadongense и Polystichum yunnanense.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Rumohra. В некоторых вариантах осуществления молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, получают из вида папоротников порядка Polypodiales, семейства Dryopteridaceae, рода Rumohra, выбранного без ограничения из Rumohra adiantiformis, Rumohra aristata, Rumohra bartonae, Rumohra berteroana, Rumohra capuronii, Rumohra glandulosa, Rumohra humbertii, Rumohra linearisquamosa, Rumohra lokohensis, Rumohra madagascarica и Rumohra quadrangularis.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Lomariopsidaceae, рода Nephrolepis.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Campyloneurum, рода Drynaria, рода Lepisorus, рода Microgramma, рода Microsorum, рода Neurodium, рода Niphidium, рода Pecluma M.G., рода Phlebodium, рода Phymatosorus, рода Platycerium, рода Pleopeltis, рода Polypodium.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Microsorum.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Microsorum, выбранного без ограничения из Microsorum alatum, Microsorum angustifolium, Microsorum aurantiacum, Microsorum australiense, Microsorum baithoense, Microsorum basicordatum, Microsorum biseriatum, Microsorum brassii, Microsorum buergerianum, Microsorum chapaense, Microsorum cinctum, Microsorum commutatum, Microsorum congregatifolium, Microsorum cuneatum, Microsorum cuspidatum, Microsorum dengii, Microsorum egregium, Microsorum emeiensis, Microsorum ensatum, Microsorum ensiforme, Microsorum excelsum, Microsorum fortunei, Microsorum griseorhizoma, Microsorum grossum, Microsorum hemionitideum, Microsorum henryi, Microsorum heterocarpum, Microsorum heterolobum, Microsorum howense, Microsorum insigne, Microsorum intermedium, Microsorum kongtingense, Microsorum krayanense, Microsorum lanceolatum, Microsorum lancifolium, Microsorum lastii, Microsorum latilobatum, Microsorum leandrianum, Microsorum lineare, Microsorum linguiforme, Microsorum longissimum, Microsorum longshengense, Microsorum maculosum, Microsorum maximum, Microsorum membranaceum, Microsorum membranifolium, Microsorum microsorioides, Microsorum minor, Microsorum monstrosum, Microsorum muliense, Microsorum mutense, Microsorum nanchuanense, Microsorum ningpoense, Microsorum normale, Microsorum novae-zealandiae, Microsorum ovalifolium, Microsorum ovatum, Microsorum palmatopedatum, Microsorum pappei, Microsorum papuanum, Microsorum parksii, Microsorum pentaphyllum, Microsorum piliferum, Microsorum pitcairnense, Microsorum powellii, Microsorum pteropodum, Microsorum pteropus, Microsorum punctatum, Microsorum pustulatum, Microsorum rampans, Microsorum revolutum, Microsorum rubidum, Microsorum samarense, Microsorum sapaense, Microsorum sarawakense, Microsorum scandens, Microsorum scolopendria, Microsorum sibomense, Microsorum sinense, Microsorum sopuense, Microsorum спектр, Microsorum steerei, Microsorum subhemionitideum, Microsorum submarginale, Microsorum subnudum, Microsorum superficiale, Microsorum takhtajanii, Microsorum tenuipes, Microsorum tibeticum, Microsorum triglossum, Microsorum truncatum, Microsorum tsaii, Microsorum varians, Microsorum venosum, Microsorum vieillardii, Microsorum x inaequibasis, Microsorum yiliangensis и Microsorum zippelii.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Polypodium L.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Polypodium L., выбранного без ограничения из Polypodium absidatum, Polypodium acutifolium, Polypodium adiantiforme, Polypodium aequale, Polypodium affine, Polypodium albidopaleatum, Polypodium alcicorne, Polypodium alfarii, Polypodium alfredii, Polypodium alfredii var. curtii, Polypodium allosuroides, Polypodium alsophilicola, Polypodium amamianum, Polypodium amoenum, Polypodium amorphum, Polypodium anetioides, Polypodium anfractuosum, Polypodium anguinum, Polypodium angustifolium f. remotifolia, Polypodium angustifolium var. amphostenon, Polypodium angustifolium var. heterolepis, Polypodium angustifolium var. monstrosa, Polypodium angustipaleatum, Polypodium angustissimum, Polypodium anisomeron var. pectinatum, Polypodium antioquianum, Polypodium aoristisorum, Polypodium apagolepis, Polypodium apicidens, Polypodium apiculatum, Polypodium apoense, Polypodium appalachianum, Polypodium appressum, Polypodium arenarium, Polypodium argentinum, Polypodium argutum, Polypodium armatum, Polypodium aromaticum, Polypodium aspersum, Polypodium assurgens, Polypodium atrum, Polypodium auriculatum, Polypodium balaonense, Polypodium balliviani, Polypodium bamleri, Polypodium bangii, Polypodium bartlettii, Polypodium basale, Polypodium bernoullii, Polypodium biauritum, Polypodium bifrons, Polypodium blepharodes, Polypodium bolivari, Polypodium bolivianum, Polypodium bolobense, Polypodium bombycinum, Polypodium bombycinum var. insularum, Polypodium bradeorum, Polypodium bryophilum, Polypodium bryopodum, Polypodium buchtienii, Polypodium buesii, Polypodium bulbotrichum, Polypodium caceresii, Polypodium californicum f. brauscombii, Polypodium californicum f. parsonsiae, Polypodium californicum, Polypodium calophlebium, Polypodium calvum, Polypodium camptophyllarium var. abbreviatum, Polypodium capitellatum, Polypodium carpinterae, Polypodium chachapoyense, Polypodium chartaceum, Polypodium chimantense, Polypodium chiricanum, Polypodium choquetangense, Polypodium christensenii, Polypodium christii, Polypodium chrysotrichum, Polypodium ciliolepis, Polypodium cinerascens, Polypodium collinsii, Polypodium colysoides, Polypodium confluens, Polypodium conforme, Polypodium confusum, Polypodium congregatifolium, Polypodium connellii, Polypodium consimile var. bourgaeanum, Polypodium consimile var. minor, Polypodium conterminans, Polypodium contiguum, Polypodium cookii, Polypodium coriaceum, Polypodium coronans, Polypodium costaricense, Polypodium costatum, Polypodium crassifolium f. angustissimum, Polypodium crassifolium var. longipes, Polypodium crassulum, Polypodium craterisorum, Polypodium cryptum, Polypodium crystalloneuron, Polypodium cucullatum var. planum, Polypodium cuencanum, Polypodium cumingianum, Polypodium cupreolepis, Polypodium curranii, Polypodium curvans, Polypodium cyathicola, Polypodium cyathisorum, Polypodium cyclocolpon, Polypodium daguense, Polypodium damunense, Polypodium dareiformioides, Polypodium dasypleura, Polypodium decipiens, Polypodium decorum, Polypodium delicatulum, Polypodium deltoideum, Polypodium demeraranum, Polypodium denticulatum, Polypodium diaphanum, Polypodium dilatatum, Polypodium dispersum, Polypodium dissectum, Polypodium dissimulans, Polypodium dolichosorum, Polypodium dolorense, Polypodium donnell-smithii, Polypodium drymoglossoides, Polypodium ebeninum, Polypodium eggersii, Polypodium elmeri, Polypodium elongatum, Polypodium enterosoroides, Polypodium erubescens, Polypodium erythrolepis, Polypodium erythrotrichum, Polypodium eurybasis, Polypodium eurybasis var. villosum, Polypodium exornans, Polypodium falcoideum, Polypodium fallacissimum, Polypodium farinosum, Polypodium faucium, Polypodium feei, Polypodium ferrugineum, Polypodium feuillei, Polypodium firmulum, Polypodium firmum, Polypodium flaccidum, Polypodium flagellare, Polypodium flexuosum, Polypodium flexuosum var. ekmanii, Polypodium forbesii, Polypodium formosanum, Polypodium fraxinifolium subsp. articulatum, Polypodium fraxinifolium subsp. luridum, Polypodium fructuosum, Polypodium fucoides, Polypodium fulvescens, Polypodium galeottii, Polypodium glaucum, Polypodium glycyrrhiza, Polypodium gracillimum, Polypodium gramineum, Polypodium grandifolium, Polypodium gratum, Polypodium graveolens, Polypodium griseo-nigrum, Polypodium griseum, Polypodium guttatum, Polypodium haalilioanum, Polypodium hammatisorum, Polypodium hancockii, Polypodium haplophlebicum, Polypodium harrisii, Polypodium hastatum var. simplex, Polypodium hawaiiense, Polypodium heanophyllum, Polypodium helleri, Polypodium hemionitidium, Polypodium henryi, Polypodium herzogii, Polypodium hesperium, Polypodium hessii, Polypodium hombersleyi, Polypodium hostmannii, Polypodium humile, Polypodium hyalinum, Polypodium iboense, Polypodium induens var. subdentatum, Polypodium insidiosum, Polypodium insigne, Polypodium intermedium subsp. masafueranum var. obtuseserratum, Polypodium intramarginale, Polypodium involutum, Polypodium itatiayense, Polypodium javanicum, Polypodium juglandifolium, Polypodium kaniense, Polypodium knowltoniorum, Polypodium kyimbilense, Polypodium l'herminieri var. costaricense, Polypodium lachniferum f. incurvata, Polypodium lachniferum var. glabrescens, Polypodium lachnopus, Polypodium lanceolatum var. complanatum, Polypodium lanceolatum var. trichophorum, Polypodium latevagans, Polypodium laxifrons, Polypodium laxifrons var. lividum, Polypodium lehmannianum, Polypodium leiorhizum, Polypodium leptopodon, Polypodium leuconeuron var. angustifolia, Polypodium leuconeuron var. latifolium, Polypodium leucosticta, Polypodium limulum, Polypodium lindigii, Polypodium lineatum, Polypodium lomarioides, Polypodium longifrons, Polypodium loretense, Polypodium loriceum var. umbraticum, Polypodium loriforme, Polypodium loxogramme f. gigas, Polypodium ludens, Polypodium luzonicum, Polypodium lycopodioides f. obtusum, Polypodium lycopodioides L., Polypodium macrolepis, Polypodium macrophyllum, Polypodium macrosorum, Polypodium macrosphaerum, Polypodium maculosum, Polypodium madrense, Polypodium manmeiense, Polypodium margaritiferum, Polypodium maritimum, Polypodium martensii, Polypodium mayoris, Polypodium megalolepis, Polypodium melanotrichum, Polypodium menisciifolium var. pubescens, Polypodium meniscioides, Polypodium merrillii, Polypodium mettenii, Polypodium mexiae, Polypodium microsorum, Polypodium militare, Polypodium minimum, Polypodium minusculum, Polypodium mixtum, Polypodium mollendense, Polypodium mollissimum, Polypodium moniliforme var. minus, Polypodium monoides, Polypodium monticola, Polypodium montigenum, Polypodium moritzianum, Polypodium moultonii, Polypodium multicaudatum, Polypodium multilineatum, Polypodium multisorum, Polypodium munchii, Polypodium muscoides, Polypodium myriolepis, Polypodium myriophyllum, Polypodium myriotrichum, Polypodium nematorhizon, Polypodium nemorale, Polypodium nesioticum, Polypodium nigrescentium, Polypodium nigripes, Polypodium nigrocinctum, Polypodium nimbatum, Polypodium nitidissimum, Polypodium nitidissimum var. latior, Polypodium nubrigenum, Polypodium oligolepis, Polypodium oligosorum, Polypodium oligosorum, Polypodium olivaceum, Polypodium olivaceum var. elatum, Polypodium oodes, Polypodium oosphaerum, Polypodium oreophilum, Polypodium ornatissimum, Polypodium ornatum, Polypodium ovatum, Polypodium oxylobum, Polypodium oxypholis, Polypodium pakkaense, Polypodium pallidum, Polypodium palmatopedatum, Polypodium palmeri, Polypodium panamense, Polypodium parvum, Polypodium patagonicum, Polypodium paucisorum, Polypodium pavonianum, Polypodium pectinatum var. caliense, Polypodium pectinatum var. hispidum, Polypodium pellucidum, Polypodium pendulum var. boliviense, Polypodium percrassum, Polypodium perpusillum, Polypodium peruvianum var. subgibbosum, Polypodium phyllitidis var. elongatum, Polypodium pichinchense, Polypodium pilosissimum, Polypodium pilosissimum var. glabriusculum, Polypodium pilossimum var. tunguraquensis, Polypodium pityrolepis, Polypodium platyphyllum, Polypodium playfairii, Polypodium plebeium var. cooperi, Polypodium plectolepidioides, Polypodium pleolepis, Polypodium plesiosorum var.i, Polypodium podobasis, Polypodium podocarpum, Polypodium poloense, Polypodium polydatylon, Polypodium polypodioides var. aciculare, Polypodium polypodioides var. michauxianum, Polypodium praetermissum, Polypodium preslianum var. immersum, Polypodium procerum, Polypodium procerum, Polypodium productum, Polypodium productum, Polypodium prolongilobum, Polypodium propinguum, Polypodium proteus, Polypodium pruinatum, Polypodium pseudocapillare, Polypodium pseudofraternum, Polypodium pseudonutans, Polypodium pseudoserratum, Polypodium pulcherrimum, Polypodium pulogense, Polypodium pungens, Polypodium purpusii, Polypodium radicale, Polypodium randallii, Polypodium ratiborii, Polypodium reclinatum, Polypodium recreense, Polypodium repens var. abruptum, Polypodium revolvens, Polypodium rhachipterygium, Polypodium rhomboideum, Polypodium rigens, Polypodium robustum, Polypodium roraimense, Polypodium roraimense, Polypodium rosei, Polypodium rosenstockii, Polypodium rubidum, Polypodium rudimentum, Polypodium rusbyi, Polypodium sablanianum, Polypodium sarmentosum, Polypodium saxicola, Polypodium schenckii, Polypodium schlechteri, Polypodium scolopendria, Polypodium scolopendria, Polypodium scolopendrium, Polypodium scouleri, Polypodium scutulatum, Polypodium segregatum, Polypodium semihirsutum, Polypodium semihirsutum var. fuscosetosum, Polypodium senile var. minor, Polypodium sericeolanatum, Polypodium serraeforme, Polypodium serricula, Polypodium sesquipedala, Polypodium sessilifolium, Polypodium setosum var. calvum, Polypodium setulosum, Polypodium shaferi, Polypodium sibomense, Polypodium siccum, Polypodium simacense, Polypodium simulans, Polypodium singeri, Polypodium sinicum, Polypodium sintenisii, Polypodium skutchii, Polypodium sloanei, Polypodium sodiroi, Polypodium sordidulum, Polypodium sordidum, Polypodium sphaeropteroides, Polypodium sphenodes, Polypodium sprucei, Polypodium sprucei var. furcativenosa, Polypodium steirolepis, Polypodium stenobasis, Polypodium stenolepis, Polypodium stenopterum, Polypodium subcapillare, Polypodium subflabelliforme, Polypodium subhemionitidium, Polypodium subinaequale, Polypodium subintegrum, Polypodium subspathulatum, Polypodium subtile, Polypodium subvestitum, Polypodium subviride, Polypodium superficiale var. attenuatum, Polypodium superficiale var. chinensis, Polypodium sursumcurrens, Polypodium tablazianum, Polypodium taenifolium, Polypodium tamandarei, Polypodium tatei, Polypodium tenuiculum var. acrosora, Polypodium tenuiculum var. brasiliense, Polypodium tenuilore, Polypodium tenuinerve, Polypodium tepuiense, Polypodium teresae, Polypodium tetragonum var. incompletum, Polypodium thysanolepis var. bipinnatifidum, Polypodium thyssanolepis, var. thyssanolepis, Polypodium thyssanolepsi, Polypodium tobagense, Polypodium trichophyllum, Polypodium tridactylum, Polypodium tridentatum, Polypodium trifurcatum var. brevipes, Polypodium triglossum, Polypodium truncatulum, Polypodium truncicola var. major, Polypodium truncicola var. minor, Polypodium tuberosum, Polypodium tunguraguae, Polypodium turquinum, Polypodium turrialbae, Polypodium ursipes, Polypodium vagans, Polypodium valdealatum, Polypodium versteegii, Polypodium villagranii, Polypodium virginianum f. cambroideum, Polypodium virginianum f. peraferens, Polypodium vittarioides, Polypodium vulgare, Polypodium vulgare L., Polypodium vulgare subsp. oreophilum, Polypodium vulgare var. acuminatum, Polypodium vulpinum, Polypodium williamsii, Polypodium wobbense, Polypodium x fallacissimum-guttatum, Polypodium xantholepis, Polypodium xiphopteris, Polypodium yarumalense, Polypodium yungense, и Polypodium zosteriforme.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида папоротников порядка Polypodiales, семейства Polypodiaceae, рода Platycerium.
Внекоторых вариантах осуществления полипептид PtIP-83 получают из вида отдела Lycophyta.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида класса Isoetopsida или класса Lycopodiopsida.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида класса Isoetopsida порядка Selaginales. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида класса Isoetopsida, порядка Selaginales, семейства Selaginellaceae. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида рода Selaginella. В некоторых вариантах осуществления полипептид PtIP-83 получают из вида класса Lycopodiopsida, порядка Lycopodiales.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида класса Lycopodiopsida, порядка Lycopodiales семейства Lycopodiaceae или семейства Huperziaceae.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида рода Austrolycopodium, Dendrolycopodium, Diphasiastrum, Diphasium, Huperzia, Lateristachys, Lycopodiastrum, Lycopodiella, Lycopodium, Palhinhaea, Pseudodiphasium, Pseudolycopodiella, Pseudolycopodium или Spinulum.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида рода Lycopodium.
В некоторых вариантах осуществления полипептид PtIP-83 получают из вида рода Huperzia.
Филогенетический анализ, анализ мотивов последовательности и структурный анализ семейств инсектицидных белков
Используемый способ анализа последовательности и структуры состоит из четырех компонентов: построение филогенетического дерева, обнаружение мотивов белковой последовательности, прогнозирование вторичной структуры и выравнивание белковых последовательностей и вторичных структур. Детали о каждом компоненте проиллюстрированы ниже.
Построение филогенетического дерева
Филогенетический анализ осуществляли с применением программного обеспечения MEGA5. Белковые последовательности подвергали анализу ClustalW версия 2 (Larkin M.A et al. (2007) Bioinformatics 23(21): 2947-2948) для множественного выравнивания последовательностей. Затем определяли эволюционную историю с помощью способа максимального правдоподобия, исходя из модели на основе JTT-матрицы. Получали дерево с самым высоким логарифмическим правдоподобием, экспортировали его в формат Newick, и дополнительно обрабатывали для извлечения ID последовательностей в том же порядке, как они появляются на дереве. Некоторые клады, представляющие подсемейства идентифицировали вручную для каждого семейства инсектицидных белков.
Обнаружение мотивов белковой последовательности
Белковые последовательности перегруппировывали согласно ранее построенному филогенетическому дереву и передавали в инструмент для анализа МОТИВОВ MEME (Multiple EM for MOTIF Elicitation) (Bailey T.L., and Elkan C., Proceedings of the Second International Conference on Intelligent Systems for Molecular Biology, pp. 28-36, AAAI Press, Menlo Park, California, 1994.) для идентификации ключевых мотивов последовательности. Начальные установки MEME были следующими: минимальное количество сайтов 2, минимальный размер мотива 5 и максимальное количество мотивов 30. Мотивы последовательности, уникальные для каждого подсемейства, идентифицировали с помощью визуального осмотра. Распределение МОТИВОВ по всему семейству генов можно было визуализировать на HTML-вебстранице. МОТИВЫ пронумеровали согласно расположению E-значения для каждого МОТИВА. МОТИВЫ аминокислотной последовательности, идентифицированные для каждого полипептида PtIP-83, и диапазоны номеров остатков, определяющие МОТИВЫ, касающиеся каждого из соответствующих идентификаторов последовательности (SEQ ID NO:), показаны в таблице 2. На фигуре 2 показано выравнивание полипептидов PtIP-83, PtIP-83Aa (SEQ ID NO: 1), PtIP-83Ca (SEQ ID NO: 5), PtIP-83Cb (SEQ ID NO: 7), PtIP-83Cc (SEQ ID NO: 9), PtIP-83Cd (SEQ ID NO: 11), PtIP-83Ce (SEQ ID NO: 13), PtIP-83Cf (SEQ ID NO: 15) и PtIP-83Fa (SEQ ID NO: 3), и местоположение относительно PtIP-83Aa (SEQ ID NO: 1) МОТИВОВ аминокислотной последовательности, присутствующих в PtIP-83Aa (SEQ ID NO: 1).
Прогнозирование вторичной структуры
PSIPRED, ведущий способ прогнозирования вторичной структуры (Jones DT. (1999) J. Mol. Biol. 292: 195-202), инсталлировали на локальном Linux-сервере, и применяли для прогнозирования вторичной структуры белка. Инструмент обеспечивает точное прогнозирование структуры с применением двух нейронных цепей прямой связи, исходя из результата PSI-BLAST. Базу данных для PSI-BLAST создавали путем удаления участков низкой сложности, трансмембранного участка и участков суперспирали в Uniref100. Результаты PSIPRED содержат вторичную структуру PtIP (альфа-спираль: H, бета-тяж: E и клубок: C) и соответствующую степень уверенности для каждой аминокислоты в данной белковой последовательности. На фигуре 2 показано выравнивание аминокислотных последовательностей полипептида PtIP-83 и участки консервативной вторичной структуры.
Таблица 2
n. p.=не присутствует
Выравнивание белковых последовательностей и вторичных структур
Разрабатывали индивидуальный скрипт для создания выравнивания вторичных структур с гэпами согласно множественному выравниванию белковых последовательностей из этапа 1 для всех белков. Все подвергшиеся выравниванию белковые последовательности и структуры комбинировали в единый FASTA-файл, и затем импортировали в MEGA для визуализации и идентификации консервативных структур.
В некоторых вариантах осуществления полипептид PtIP-83 содержит МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MP[DE]MPSEADWSIFVNE[IV]EAVAEGMPTEVSEVP[AV]WKAKCKN[MV]AALGREM[SC]I (SEQ ID NO: 646); МОТИВА 2 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PQLQYRMYG[NS]LI[KN]QMAQVAQNYDQ[ED]FKQ[FL]KLFI[IA]QNQI[LF]GSYLLQQN[KR]AF (SEQ ID NO: 647); МОТИВА 3 аминокислотной последовательности, представленного аминокислотной последовательностью формулы NTFMQMTPFTRWRLRLSASASENA[EG]LAFPTATA[PL]DSTT[EQ][IV]VITFHVTAIR (SEQ ID NO: 648); МОТИВА 4 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [DN]FTSRHVVK[GD]IPVSLLLDGEDWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 649); МОТИВА 5 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [ST]G[IT]VYILLQGSTIFHDRRR[DE]EVMTFQAA[DA]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 650); МОТИВА 6 аминокислотной последовательности, представленного аминокислотной последовательностью формулы S[HQ]ADRLAAIQP[AV]DLTN[HY]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI]LRIQNAALMYEY (SEQ ID NO: 651); МОТИВА 7 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [VL]DRVEFSEVMVIHRMYVRL[SA]DL[ND]VGEL[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 652); МОТИВА 8 аминокислотной последовательности, представленного аминокислотной последовательностью формулы A[DE]RELQMESFHSAVISQRRQEL[ND]TA[IF]AKM[DE]R[LM]SLQMEEE[NS]RAMEQAQKEM (SEQ ID NO: 653); МОТИВА 9 аминокислотной последовательности, представленного аминокислотной последовательностью формулы FVTAGATAPGA[AV]ASAGQAVSIAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 654); МОТИВА 10 аминокислотной последовательности, представленного аминокислотной последовательностью формулы DGMNWG[IT]YI[YH]GE[KE]V[EQ]RSPLLPSNAILAVWADRC[TI]ITSARHNH[VF]NAPGR[IV]I (SEQ ID NO: 655); МОТИВА 11 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [KV][VK][CA]RPPSPDM[MV]SAVAEHALWLNDVLLQVVQ[KN]ESQ[LM]QGT[AE]PYNECLALLGR (SEQ ID NO: 656); МОТИВА 12 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PTELT[VA]WPLGMDTV[AG]NLLIAQENAAL[VL]GLIQLGPSS (SEQ ID NO: 657); МОТИВА 13 аминокислотной последовательности, представленного аминокислотной последовательностью формулы RDQ[MT][HQ]MPGSVTVI[IV]LCRLLQFP[IT]DGSQA[TA]T (SEQ ID NO: 658); МОТИВА 14 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TSIPVEVVTDP[SN]ILLGMQTTV[LH]IAEL (SEQ ID NO: 659); МОТИВА 15 аминокислотной последовательности, представленного аминокислотной последовательностью формулы EGLR[EQ]FQNRQVARA[VL]FAVLKAVA[MQ]I[AG] (SEQ ID NO: 660); МОТИВА 16 аминокислотной последовательности, представленного аминокислотной последовательностью формулы W[TS]RVRIRHLEM[QH]F[AV]QEASG (SEQ ID NO: 661); МОТИВА 17 аминокислотной последовательности, представленного аминокислотной последовательностью формулы QISELQY[ED]IWVQG[LM][ML]RDIA (SEQ ID NO: 662); МОТИВА 18 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MDYSTLYRDLNQIS (SEQ ID NO: 664); МОТИВА 20 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LRLPFM[QK]LHARVIEQN[VR]K[SE] (SEQ ID NO: 665); МОТИВА 21 аминокислотной последовательности, представленного аминокислотной последовательностью формулы VDSLEQVG[QH][IL]V[GD]AP (SEQ ID NO: 666); МОТИВА 22 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [IV][EQ][CA]VMK[IM]GRF[VG][SL]VV (SEQ ID NO: 667); МОТИВА 23 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TLTNEPSE[EQ]F (SEQ ID NO: 668); и МОТИВА 24 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LPRQSRNISF (SEQ ID NO: 669).
В некоторых вариантах осуществления полипептид PtIP-83 содержит МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MP[DE]MPSEADWSIFVNE[IV]EAVAEGMPTEVSEVP[AV]WKAKCKN[MV]AALGREM[SC]I (SEQ ID NO: 646); МОТИВА 2 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PQLQYRMYG[NS]LI[KN]QMAQVAQNYDQ[ED]FKQ[FL]KLFI[IA]QNQI[LF]GSYLLQQN[KR]AF (SEQ ID NO: 647); МОТИВА 3 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой NTFMQMTPFTRWRLRLSASASENA[EG]LAFPTATA[PL]DSTT[EQ][IV]VITFHVTAIR (SEQ ID NO: 648); МОТИВА 4 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [DN]FTSRHVVK[GD]IPVSLLLDGEDWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 649); МОТИВА 5 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой IIHQP[SA]T[RQ][ST]G[IT]VYILLQGSTIFHDRRR[DE]EVMTFQAA[DA]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 650); МОТИВА 6 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой S[HQ]ADRLAAIQP[AV]DLTN[HY]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI]LRIQNAALMYEY (SEQ ID NO: 651); МОТИВА 7 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [VL]DRVEFSEVMVIHRMYVRL[SA]DL[ND]VGEL[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 652); МОТИВА 8 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой A[DE]RELQMESFHSAVISQRRQEL[ND]TA[IF]AKM[DE]R[LM]SLQMEEE[NS]RAMEQAQKEM (SEQ ID NO: 653); МОТИВА 9 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой FVTAGATAPGA[AV]ASAGQAVSIAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 654); МОТИВА 10 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой DGMNWG[IT]YI[YH]GE[KE]V[EQ]RSPLLPSNAILAVWADRC[TI]ITSARHNH[VF]NAPGR[IV]I (SEQ ID NO: 655); МОТИВА 11 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [KV][VK][CA]RPPSPDM[MV]SAVAEHALWLNDVLLQVVQ[KN]ESQ[LM]QGT[AE]PYNECLALLGR (SEQ ID NO: 656); МОТИВА 12 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PTELT[VA]WPLGMDTV[AG]NLLIAQENAAL[VL]GLIQLGPSS (SEQ ID NO: 657); МОТИВА 13 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой RDQ[MT][HQ]MPGSVTVI[IV]LCRLLQFP[IT]DGSQA[TA]T (SEQ ID NO: 658); МОТИВА 14 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TSIPVEVVTDP[SN]ILLGMQTTV[LH]IAEL (SEQ ID NO: 659); МОТИВА 15 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой EGLR[EQ]FQNRQVARA[VL]FAVLKAVA[MQ]I[AG] (SEQ ID NO: 660); МОТИВА 16 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой W[TS]RVRIRHLEM[QH]F[AV]QEASG (SEQ ID NO: 661); МОТИВА 17 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой QISELQY[ED]IWVQG[LM][ML]RDIA (SEQ ID NO: 662); МОТИВА 18 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MDYSTLYRDLNQIS (SEQ ID NO: 664); МОТИВА 20 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LRLPFM[QK]LHARVIEQN[VR]K[SE] (SEQ ID NO: 665); МОТИВА 21 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой VDSLEQVG[QH][IL]V[GD]AP (SEQ ID NO: 666); МОТИВА 22 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [IV][EQ][CA]VMK[IM]GRF[VG][SL]VV (SEQ ID NO: 667); МОТИВА 23 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TLTNEPSE[EQ]F (SEQ ID NO: 668); и МОТИВА 24 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LPRQSRNISF (SEQ ID NO: 669).
В некоторых вариантах осуществления полипептид PtIP-83 содержит МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AV]W[KR]AKCKN[MV]AALGREM[SC]I (SEQ ID NO: 670); МОТИВА 2 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PQLQYRMYG[NS]LI[KRN]QMAQVAQNYD[QR][ED]FK[QR][FL][KR]LFI[IAVL]QNQI[LF]GSYLL[QE]QN[KR]AF (SEQ ID NO: 671); МОТИВА 3 аминокислотной последовательности, представленного аминокислотной последовательностью формулы N[TK]FMQMTPFT[RH]WRLRLSASA[SPKA]EN[AK][EG]LAFPTATA[PL]DSTT[EQ][IV][VA]ITFHVTAIR (SEQ ID NO: 672); МОТИВА 4 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [DN]FTSRHVVK[GD]IPV[SN]LLLDG[EG]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 673); МОТИВА 5 аминокислотной последовательности, представленного аминокислотной последовательностью формулы IIHQP[SA]T[RQ][ST]G[IT][VI]YILLQGST[IV]FHDRRR[DE][EQ]V[ML]T[FP]QAA[DAV]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 674); МОТИВА 6 аминокислотной последовательности, представленного аминокислотной последовательностью формулы S[HQ]ADRLAAIQP[AV][DN]LTN[HYF]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI][LM]RIQNAAL[MR]YEY (SEQ ID NO: 675); МОТИВА 7 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [VL]D[RQ]VEFSEVMVIHRMYV[N]RL[SA]DL[ND]V[GA][EQ]L[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 676); МОТИВА 8 аминокислотной последовательности, представленного аминокислотной последовательностью формулы A[DE]RELQMESFH[SA]AVISQ[RK]R[QGE]EL[ND][TD][AT][IF]AKM[DE]R[LM]SLQMEEE[NSD][RG]AMEQA[QR]KEM (SEQ ID NO: 677); МОТИВА 9 аминокислотной последовательности, представленного аминокислотной последовательностью формулы F[VL]TAGATAPGA[AV]ASAGQAV[SN]IAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 678); МОТИВА 10 аминокислотной последовательности, представленного аминокислотной последовательностью формулы D[GD][MA][NK]WG[IT]Y[IV][YH][GA]E[KE]V[EQ][RVL]SPL[LYF][PN][SNG][NW][ASP][IY]L[AGV]V[WE]A[DQ]R[CS][TI]IT[SA]A[RFM]HN[HVT][VF][ND][AER]PG[RW][IV][IR] (SEQ ID NO: 679); МОТИВА 11 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [KV][VK][CA][RGC][PHY]PSP[DE][MIL][MV]SAV[AG][EV]HA[LIN]WL[NS][DK]VLL[QR]VVQ[KN]ES[QH][LM]QGT[AE][PSA]YNECLALLGR (SEQ ID NO: 680); МОТИВА 12 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [PN]T[EQ]LT[VAT]WPL[GR]MDTV[AG][ND]LLI[AT][QH]E[NS]AAL[VLS]GL[ITMA]QLG[PQ][SP]S (SEQ ID NO: 681); МОТИВА 13 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [RLC][DLWK][QNPR][MTP][HQR][MIL]PGSVTVI[IV]LCRLLQFP[IT][DG]G[SR][QFR][AS][TAD][TW] (SEQ ID NO: 682); МОТИВА 14 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [TA][SGV][IL]PV[ED]VVTDP[SN]IL[LM]GMQT[TS]V[LH]IAEL (SEQ ID NO: 683); МОТИВА 15 аминокислотной последовательности, представленного аминокислотной последовательностью формулы EGLR[EQ]FQN[RE]QVA[RN]A[VL]FAVL[KS][AS]VA[MQ]I[AG] (SEQ ID NO: 684); МОТИВА 16 аминокислотной последовательности, представленного аминокислотной последовательностью формулы W[TS]RVRIRHLEM[QH]F[AV][QK]E[AS][SM][GN] (SEQ ID NO: 685); МОТИВА 17 аминокислотной последовательности, представленного аминокислотной последовательностью формулы Q[IM]S[EQ]LQY[ED]IWVQG[LM][ML]RD[IM]A (SEQ ID NO: 686); МОТИВА 18 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [MLV]DY[SK][TSK]L[YF][RE]DLNQIS (SEQ ID NO: 687); МОТИВА 20 аминокислотной последовательности, представленного аминокислотной последовательностью формулы L[RHQ]L[PT]FM[QK]LHA[RIT][VQL][IR]E[QER][NF][VR][KWS][SE] (SEQ ID NO: 688); МОТИВА 21 аминокислотной последовательности, представленного аминокислотной последовательностью формулы VV[DN][SA]L[ED]QV[GS][QH][IL]V[GD]AP (SEQ ID NO: 689); МОТИВА 22 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [IV][EQH][CAS][VA][MI]K[IM][GV][RP][FI][VG][SL]VV (SEQ ID NO: 690); МОТИВА 23 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TLTN[EQ]PSE[EQDH]F (SEQ ID NO: 691); и МОТИВА 24 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LP[RS]QS[RT]N[IV]SF (SEQ ID NO: 692).
В некоторых вариантах осуществления полипептид PtIP-83 содержит МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AV]W[KR]AKCKN[MV]AALGREM[SC]I (SEQ ID NO: 670); МОТИВА 2 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PQLQYRMYG[NS]LI[KRN]QMAQVAQNYD[QR][ED]FK[QR][FL][KR]LFI[IAVL]QNQI[LF]GSYLL[QE]QN[KR]AF (SEQ ID NO: 671); МОТИВА 3 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой N[TK]FMQMTPFT[RH]WRLRLSASA[SPKA]EN[AK][EG]LAFPTATA[PL]DSTT[EQ][IV][VA]ITFHVTAIR (SEQ ID NO: 672); МОТИВА 4 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [DN]FTSRHVVK[GD]IPV[SN]LLLDG[EG]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 673); МОТИВА 5 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой IIHQP[SA]T[RQ][ST]G[IT][VI]YILLQGST[IV]FHDRRR[DE][EQ]V[ML]T[FP]QAA[DAV]PLN[FY][QH]YAYRLDTG (SEQ ID NO: 674); МОТИВА 6 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой S[HQ]ADRLAAIQP[AV][DN]LTN[HYF]LEMAT[HQ]MDMRTT[RS][MI]L[IL]GLLN[MI][LM]RIQNAAL[MR]YEY (SEQ ID NO: 675); МОТИВА 7 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [VL]D[RQ]VEFSEVMVIHRMYV[N]RL[SA]DL[ND]V[GA][EQ]L[PE]GA[EG][RK]VKR[VL]YV[FL]ADVVE (SEQ ID NO: 676); МОТИВА 8 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой A[DE]RELQMESFH[SA]AVISQ[RK]R[QGE]EL[ND][TD][AT][IF]AKM[DE]R[LM]SLQMEEE[NSD][RG]AMEQA[QR]KEM (SEQ ID NO: 677); МОТИВА 9 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой F[VL]TAGATAPGA[AV]ASAGQAV[SN]IAGQAAQ[AG]LRRVVEILE[GQ]LEAVMEVVAA[VI]K (SEQ ID NO: 678); МОТИВА 10 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой D[GD][MA][NK]WG[IT]Y[IV][YH][GA]E[KE]V[EQ][RVL]SPL[LYF][PN][SNG][NW][ASP][IY]L[AGV]V[WE]A[DQ]R[CS][TI]IT[SA]A[RFM]HN[HVT][VF][ND][AER]PG[RW][IV][IR] (SEQ ID NO: 679); МОТИВА 11 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [KV][VK][CA][RGC][PHY]PSP[DE][MIL][MV]SAV[AG][EV]HA[LIN]WL[NS][DK]VLL[QR]VVQ[KN]ES[QH][LM]QGT[AE][PSA]YNECLALLGR (SEQ ID NO: 680); МОТИВА 12 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [PN]T[EQ]LT[VAT]WPL[GR]MDTV[AG][ND]LLI[AT][QH]E[NS]AAL[VLS]GL[ITMA]QLG[PQ][SP]S (SEQ ID NO: 681); МОТИВА 13 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [RLC][DLWK][QNPR][MTP][HQR][MIL]PGSVTVI[IV]LCRLLQFP[IT][DG]G[SR][QFR][AS][TAD][TW] (SEQ ID NO: 682); МОТИВА 14 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [TA][SGV][IL]PV[ED]VVTDP[SN]IL[LM]GMQT[TS]V[LH]IAEL (SEQ ID NO: 683); МОТИВА 15 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой EGLR[EQ]FQN[RE]QVA[RN]A[VL]FAVL[KS][AS]VA[MQ]I[AG] (SEQ ID NO: 684); МОТИВА 16 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой W[TS]RVRIRHLEM[QH]F[AV][QK]E[AS][SM][GN] (SEQ ID NO: 685); МОТИВА 17 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой Q[IM]S[EQ]LQY[ED]IWVQG[LM][ML]RD[IM]A (SEQ ID NO: 686); МОТИВА 18 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [MLV]DY[SK][TSK]L[YF][RE]DLNQIS (SEQ ID NO: 687); МОТИВА 20 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой L[RHQ]L[PT]FM[QK]LHA[RIT][VQL][IR]E[QER][NF][VR][KWS][SE] (SEQ ID NO: 688); МОТИВА 21 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой V[DN][SA]L[ED]QV[GS][QH][IL]V[GD]AP (SEQ ID NO: 689); МОТИВА 22 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [IV][EQH][CAS][VA][MI]K[IM][GV][RP][FI][VG][SL]VV (SEQ ID NO: 690); МОТИВА 23 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TLTN[EQ]PSE[EQDH]F (SEQ ID NO: 691); и МОТИВА 24 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LP[RS]QS[RT]N[IV]SF (SEQ ID NO: 692).
В некоторых вариантах осуществления полипептид PtIP-83 содержит МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, представленного аминокислотной последовательностью формулы MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AVIL]W[KR]AKCKN[MVIL]AALGREM[SCT]I (SEQ ID NO: 693); МОТИВА 2 аминокислотной последовательности, представленного аминокислотной последовательностью формулы PQLQYRMYG[NS]LI[KRNQ]QMAQVAQNYD[QRNK][ED]FK[QRNK][FL][KR]LFI[IAVL]QNQI[LFIV]GSYLL[QEND]QN[KR]AF (SEQ ID NO: 694); МОТИВА 3 аминокислотной последовательности, представленного аминокислотной последовательностью формулы N[TKSR]FMQMTPFT[RHK]WRLRLSASA[SPKATR]EN[AKR][EG]LAFPTATA[PLIV]DSTT[EQND][IVL][VAIL]ITFHVTAIR (SEQ ID NO: 695); МОТИВА 4 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [DNQE]FTSRHVVK[GDE]IPV[SNTQ]LLLDG[EGD]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 696); МОТИВА 5 аминокислотной последовательности, представленного аминокислотной последовательностью формулы IIHQP[SAT]T[RQKN][ST]G[ITLVS][VIL]YILLQGST[IVL]FHDRRR[DE][EQDN]V[MLIV]T[FP]QAA[DAVEIL]PLN[FY][QHN]YAYRLDTG (SEQ ID NO: 697); МОТИВА 6 аминокислотной последовательности, представленного аминокислотной последовательностью формулы S[HQN]ADRLAAIQP[AVIL][DN]LTN[HYF]LEMAT[HQN]MDMRTT[RSKT][MILV]L[ILV]GLLN[MILV][LMIV]RIQNAAL[MRILVK]YEY (SEQ ID NO: 698); МОТИВА 7 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [VLI]D[RQKN]VEFSEVMVIHRMYV[N]RL[SAT]DL[NDQE]V[GA][EQND]L[PED]GA[EGD][RK]VKR[VLI]YV[FLIV]ADVVE (SEQ ID NO: 699); МОТИВА 8 аминокислотной последовательности, представленного аминокислотной последовательностью формулы A[DE]RELQMESFH[SAT]AVISQ[RK]R[QGEND]EL[NDQE][TDSE][ATS][IFLV]AKM[DE]R[LMIV]SLQMEEE[NSDQET][RGK]AMEQA[QRNK]KEM (SEQ ID NO: 700); МОТИВА 9 аминокислотной последовательности, представленного аминокислотной последовательностью формулы F[VLI]TAGATAPGA[AVIL]ASAGQAV[SNTQ]IAGQAAQ[AG]LRRVVEILE[GQN]LEAVMEVVAA[VIL]K (SEQ ID NO: 701); МОТИВА 10 аминокислотной последовательности, представленного аминокислотной последовательностью формулы D[GDE][MA][NKQK]WG[ITLVS]Y[IVL][YH][GA]E[KERD]V[EQND][RVLKI]SPL[LYFIV][PNQ][SNGTQ][NWQ][ASPT][IYLV]L[AGVIL]V[WED]A[DQNE]R[CST][TISLV]IT[SAT]A[RFMK]HN[HVTILS][VFIL][NDQE][AERDK]PG[RWK][IVL][IRLVK] (SEQ ID NO: 702); МОТИВА 11 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [KVRIL][VKRIL][CA][RGCK][PHY]PSP[DE][MILV][MVIL]SAV[AG][EVDIL]HA[LINVQ]WL[NSQT][DKER]VLL[QRNK]VVQ[KNRQ]ES[QHN][LMIV]QGT[AED][PSAT]YNECLALLGR (SEQ ID NO: 703); МОТИВА 12 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [PNQ]T[EQDN]LT[VATILS]WPL[GRK]MDTV[AG][NDQE]LLI[ATS][QHN]E[NSQT]AAL[VLSIT]GL[ITMALVS]QLG[PQN][SPT]S (SEQ ID NO: 704); МОТИВА 13 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [RLCKIV][DLWKEIVR][QNPRK][MTP][HQR][MILV]PGSVTVI[IVL]LCRLLQFP[ITLVS][DGE]G[SRTK][QFRNK][AST][TADES][TWS] (SEQ ID NO: 705); МОТИВА 14 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [TA][SGVTIL][ILV]PV[ED]VVTDP[SNTQ]IL[LMIV]GMQT[TS]V[LHIV]IAEL (SEQ ID NO: 706); МОТИВА 15 аминокислотной последовательности, представленного аминокислотной последовательностью формулы EGLR[EQND]FQN[REKD]QVA[RNKQ]A[VLI]FAVL[KSRT][AST]VA[MQN]I[AG] (SEQ ID NO: 707); МОТИВА 16 аминокислотной последовательности, представленного аминокислотной последовательностью формулы W[TS]RVRIRHLEM[QHN]F[AVIL][QKNR]E[AST][SMT][GNQ] (SEQ ID NO: 708); МОТИВА 17 аминокислотной последовательности, представленного аминокислотной последовательностью формулы Q[IMLV]S[EQND]LQY[ED]IWVQG[LMIV][MLIV]RD[IMLV]A (SEQ ID NO: 709); МОТИВА 18 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [MLVI]DY[SKTR][TSKR]L[YF][REKD]DLNQIS (SEQ ID NO: 710); МОТИВА 20 аминокислотной последовательности, представленного аминокислотной последовательностью формулы L[RHQKN]L[PTS]FM[QKNR]LHA[RITKLVS][VQLIN][IRLVK]E[QERNDK][NFQ][VRILK][KWSRT][SETD] (SEQ ID NO: 711); МОТИВА 21 аминокислотной последовательности, представленного аминокислотной последовательностью формулы V[DNQE][SAT]L[ED]QV[GST][QHN][ILV]V[GDE]AP (SEQ ID NO: 712); МОТИВА 22 аминокислотной последовательности, представленного аминокислотной последовательностью формулы [IVL][EQHND][CAST][VAIL][MILV]K[IMLV][GVIL][RPK][FILV][VGIL][SLTIV]VV (SEQ ID NO: 713); МОТИВА 23 аминокислотной последовательности, представленного аминокислотной последовательностью формулы TLTN[EQDN]PSE[EQDHN]F (SEQ ID NO: 714); и МОТИВА 24 аминокислотной последовательности, представленного аминокислотной последовательностью формулы LP[RSKT]QS[RTKS]N[IVL]SF (SEQ ID NO: 715).
В некоторых вариантах осуществления полипептид PtIP-83 содержит МОТИВ аминокислотной последовательности, выбранный из: МОТИВА 1 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой MP[DE]MP[ST][ED]ADWSIFVNE[IVL]EAVAEGMPTEVSEVP[AVIL]W[KR]AKCKN[MVIL]AALGREM[SCT]I (SEQ ID NO: 693); МОТИВА 2 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой PQLQYRMYG[NS]LI[KRNQ]QMAQVAQNYD[QRNK][ED]FK[QRNK][FL][KR]LFI[IAVL]QNQI[LFIV]GSYLL[QEND]QN[KR]AF (SEQ ID NO: 694); МОТИВА 3 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой N[TKSR]FMQMTPFT[RHK]WRLRLSASA[SPKATR]EN[AKR][EG]LAFPTATA[PLIV]DSTT[EQND][IVL][VAIL]ITFHVTAIR (SEQ ID NO: 695); МОТИВА 4 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [DNQE]FTSRHVVK[GDE]IPV[SNTQ]LLLDG[EGD]DWEFEIPVQ[AG]GMSSFP (SEQ ID NO: 696); МОТИВА 5 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой IIHQP[SAT]T[RQKN][ST]G[ITLVS][VIL]YILLQGST[IVL]FHDRRR[DE][EQDN]V[MLIV]T[FP]QAA[DAVEIL]PLN[FY][QHN]YAYRLDTG (SEQ ID NO: 697); МОТИВА 6 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой S[HQN]ADRLAAIQP[AVIL][DN]LTN[HYF]LEMAT[HQN]MDMRTT[RSKT][MILV]L[ILV]GLLN[MILV][LMIV]RIQNAAL[MRILVK]YEY (SEQ ID NO: 698); МОТИВА 7 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [VLI]D[RQKN]VEFSEVMVIHRMYV[N]RL[SAT]DL[NDQE]V[GA][EQND]L[PED]GA[EGD][RK]VKR[VLI]YV[FLIV]ADVVE (SEQ ID NO: 699); МОТИВА 8 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой A[DE]RELQMESFH[SAT]AVISQ[RK]R[QGEND]EL[NDQE][TDSE][ATS][IFLV]AKM[DE]R[LMIV]SLQMEEE[NSDQET][RGK]AMEQA[QRNK]KEM (SEQ ID NO: 700); МОТИВА 9 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой F[VLI]TAGATAPGA[AVIL]ASAGQAV[SNTQ]IAGQAAQ[AG]LRRVVEILE[GQN]LEAVMEVVAA[VIL]K (SEQ ID NO: 701); МОТИВА 10 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой D[GDE][MA][NKQK]WG[ITLVS]Y[IVL][YH][GA]E[KERD]V[EQND][RVLKI]SPL[LYFIV][PNQ][SNGTQ][NWQ][ASPT][IYLV]L[AGVIL]V[WED]A[DQNE]R[CST][TISLV]IT[SAT]A[RFMK]HN[HVTILS][VFIL][NDQE][AERDK]PG[RWK][IVL][IRLVK] (SEQ ID NO: 702); МОТИВА 11 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [KVRIL][VKRIL][CA][RGCK][PHY]PSP[DE][MILV][MVIL]SAV[AG][EVDIL]HA[LINVQ]WL[NSQT][DKER]VLL[QRNK]VVQ[KNRQ]ES[QHN][LMIV]QGT[AED][PSAT]YNECLALLGR (SEQ ID NO: 703); МОТИВА 12 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [PNQ]T[EQDN]LT[VATILS]WPL[GRK]MDTV[AG][NDQE]LLI[ATS][QHN]E[NSQT]AAL[VLSIT]GL[ITMALVS]QLG[PQN][SPT]S (SEQ ID NO: 704); МОТИВА 13 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [RLCKIV][DLWKEIVR][QNPRK][MTP][HQR][MILV]PGSVTVI[IVL]LCRLLQFP[ITLVS][DGE]G[SRTK][QFRNK][AST][TADES][TWS] (SEQ ID NO: 705); МОТИВА 14 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [TA][SGVTIL][ILV]PV[ED]VVTDP[SNTQ]IL[LMIV]GMQT[TS]V[LHIV]IAEL (SEQ ID NO: 706); МОТИВА 15 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой EGLR[EQND]FQN[REKD]QVA[RNKQ]A[VLI]FAVL[KSRT][AST]VA[MQN]I[AG] (SEQ ID NO: 707); МОТИВА 16 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой W[TS]RVRIRHLEM[QHN]F[AVIL][QKNR]E[AST][SMT][GNQ] (SEQ ID NO: 708); МОТИВА 17 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой Q[IMLV]S[EQND]LQY[ED]IWVQG[LMIV][MLIV]RD[IMLV]A (SEQ ID NO: 709); МОТИВА 18 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TFTLGSGVTGITSMHGEPSLDPWNGVSLDSASPTAF (SEQ ID NO: 663); МОТИВА 19 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [MLVI]DY[SKTR][TSKR]L[YF][REKD]DLNQIS (SEQ ID NO: 710); МОТИВА 20 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой L[RHQKN]L[PTS]FM[QKNR]LHA[RITKLVS][VQLIN][IRLVK]E[QERNDK][NFQ][VRILK][KWSRT][SETD] (SEQ ID NO: 711); МОТИВА 21 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой V[DNQE][SAT]L[ED]QV[GST][QHN][ILV]V[GDE]AP (SEQ ID NO: 712); МОТИВА 22 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой [IVL][EQHND][CAST][VAIL][MILV]K[IMLV][GVIL][RPK][FILV][VGIL][SLTIV]VV (SEQ ID NO: 713); МОТИВА 23 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой TLTN[EQDN]PSE[EQDHN]F (SEQ ID NO: 714); и МОТИВА 24 аминокислотной последовательности, последовательность которого по меньшей мере на 90% идентична аминокислотной последовательности, представленной формулой LP[RSKT]QS[RTKS]N[IVL]SF (SEQ ID NO: 715).
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца МОТИВ аминокислотной последовательности, выбранный из: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 10 (SEQ ID NO: 655, SEQ ID NO: 679 или SEQ ID NO: 702), MOTIF 18 (SEQ ID NO: 663), MOTIF 24 (SEQ ID NO: 669, SEQ ID NO: 692 или SEQ ID NO: 715), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 11 (SEQ ID NO: 656, SEQ ID NO: 680 или SEQ ID NO: 703), MOTIF 22 (SEQ ID NO: 667, SEQ ID NO: 690 или SEQ ID NO: 713), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца МОТИВ аминокислотной последовательности, выбранный из: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца МОТИВЫ аминокислотной последовательности: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 10 (SEQ ID NO: 655, SEQ ID NO: 679 или SEQ ID NO: 702), MOTIF 18 (SEQ ID NO: 663), MOTIF 24 (SEQ ID NO: 669, SEQ ID NO: 692 или SEQ ID NO: 715), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 11 (SEQ ID NO: 656, SEQ ID NO: 680 или SEQ ID NO: 703), MOTIF 22 (SEQ ID NO: 667, SEQ ID NO: 690 или SEQ ID NO: 713), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца МОТИВЫ аминокислотной последовательности: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 14 (SEQ ID NO: 659, SEQ ID NO: 683 или SEQ ID NO: 706), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), MOTIF 12 (SEQ ID NO: 657, SEQ ID NO: 681 или SEQ ID NO: 704), MOTIF 4 (SEQ ID NO: 649, SEQ ID NO: 673 или SEQ ID NO: 696), MOTIF 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и MOTIF 3 (SEQ ID NO: 648, SEQ ID NO: 672 или SEQ ID NO: 695).
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, содержащий МОТИВ аминокислотной последовательности из: MOTIF 19 (SEQ ID NO: 664, SEQ ID NO: 687 или SEQ ID NO: 710), MOTIF 7 (SEQ ID NO: 652, SEQ ID NO: 676 или SEQ ID NO: 699), MOTIF 13 (SEQ ID NO: 658, SEQ ID NO: 682 или SEQ ID NO: 705), MOTIF 20 (SEQ ID NO: 665, SEQ ID NO: 688 или SEQ ID NO: 711), MOTIF 10 (SEQ ID NO: 655, SEQ ID NO: 679 или SEQ ID NO: 702), MOTIF 18 (SEQ ID NO: 663), MOTIF 24 (SEQ ID NO: 669, SEQ ID NO: 692 или SEQ ID NO: 715), и/или МОТИВА 14 с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот, содержащий МОТИВ аминокислотной последовательности из МОТИВА 22 (SEQ ID NO: 667, SEQ ID NO: 690 или SEQ ID NO: 713), MOTIF 2 (SEQ ID NO: 647, SEQ ID NO: 671 или SEQ ID NO: 694), MOTIF 8 (SEQ ID NO: 653, SEQ ID NO: 677 или SEQ ID NO: 700), MOTIF 15 (SEQ ID NO: 660, SEQ ID NO: 684 или SEQ ID NO: 707), MOTIF 9 (SEQ ID NO: 654, SEQ ID NO: 678 или SEQ ID NO: 701), MOTIF 21 (SEQ ID NO: 666, SEQ ID NO: 689 или SEQ ID NO: 712), MOTIF 1 (SEQ ID NO: 646, SEQ ID NO: 670 или SEQ ID NO: 693), MOTIF 17 (SEQ ID NO: 662, SEQ ID NO: 686 или SEQ ID NO: 709), MOTIF 6 (SEQ ID NO: 651, SEQ ID NO: 675 или SEQ ID NO: 698), и/или МОТИВА 12 со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот, содержащий МОТИВ аминокислотной последовательности из МОТИВА 16 (SEQ ID NO: 661, SEQ ID NO: 685 или SEQ ID NO: 708), MOTIF 5 (SEQ ID NO: 650, SEQ ID NO: 674 или SEQ ID NO: 697), MOTIF 23 (SEQ ID NO: 668, SEQ ID NO: 691 или SEQ ID NO: 714), и/или МОТИВА 3 с консенсусной вторичной структурой, содержащий структуру преимущественно бета-тяжа.
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащей 8-10 сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий 6-8 сегментов со структурой преимущественно бета-тяжа. Применяемое в данном документе "главным образом неконсервативная вторичная структура" означает, что участки вторичной структуры не выравниваются единообразно в пределах семейства полипептидов PtIP. Применяемое в данном документе "структура преимущественно альфа-спирали" означает, что прогноз вторичной структуры может иметь один или несколько гэпов размером 1-4 аминокислот со структурой клубка и/или бета-тяжа, вставленных в структуру альфа-спирали. Применяемое в данном документе "структура преимущественно бета-тяжа" означает, что прогноз вторичной структуры может иметь гэп размером 1-4 аминокислот со структурой клубка и/или альфа-спирали, вставленных в структуру бета-тяжа. В некоторых вариантах осуществления вторичную структуру получают с помощью PSIPRED, ведущего способа прогнозирования вторичной структуры (Jones DT. (1999) J. Mol. Biol. 292: 195-202).
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий девять сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов со структурой преимущественно бета-тяжа.
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом гибкой консенсусной вторичной структурой, где Участок A содержит консервативный бета-тяж 1 (β1a) длиной от приблизительно 4 до приблизительно 12 аминокислот в пределах от приблизительно аминокислотного остатка 30 до приблизительно аминокислотного остатка 130 от N-конца полипептида PtIP-83; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий девять сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов со структурой преимущественно бета-тяжа. Применяемый в данном документе термин "приблизительно", при применении в контексте нижнего/верхнего предела длины элемента вторичной структуры означает наибольшее из -/+ целого числа, составляющего -/+ 20% длины элемента вторичной структуры или -/+ 1 аминокислота. В качестве примера, элемент вторичной структуры длиной от приблизительно 3 аминокислот до приблизительно 23 аминокислот означает элемент вторичной структуры длиной от 2 до 27 аминокислот.
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот с гибкой консенсусной вторичной структурой, где Участок A содержит консервативный бета-тяж 1 (β1a) длиной от приблизительно 4 до приблизительно 12 аминокислот, клубок длиной от приблизительно 10 до приблизительно 20 аминокислот и бета-тяж 2 (β1b) длиной от приблизительно 4 до приблизительно 12 аминокислот в пределах от приблизительно аминокислотного остатка 30 до приблизительно аминокислотного остатка 165 от N-конца полипептида PtIP-83; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий девять сегментов преимущественно с структурой альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов преимущественно со структурой бета-тяжа.
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) альфа-спираль-1 длиной от приблизительно 10 до приблизительно 26 аминокислот; ii) клубок-1 длиной от приблизительно 2 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-1 и альфа-спиралью-2; iii) альфа-спираль-2 длиной от приблизительно 15 до приблизительно 24 аминокислот; iv) клубок-2 длиной от приблизительно 4 до приблизительно 14 аминокислот, фланкированный альфа-спиралью-2 и альфа-спиралью-3; v) альфа-спираль 3 длиной от приблизительно 15 до приблизительно 27 аминокислот; vi) клубок-3 длиной от приблизительно 11 до приблизительно 13 аминокислот, фланкированный альфа-спиралью-3 и альфа-спиралью-4; vii) альфа-спираль-4 длиной приблизительно 180 аминокислот; viii) клубок-4 длиной от приблизительно 4 до приблизительно 5 аминокислот, фланкированный альфа-спиралью-4 и альфа-спиралью-5; ix) альфа-спираль-5 длиной от приблизительно 50 до приблизительно 54 аминокислот ; x) клубок-5 длиной от приблизительно 11 до приблизительно 17 аминокислот, фланкированный альфа-спиралью-5 и альфа-спиралью-6; xi) альфа-спираль-6 длиной от приблизительно 15 до приблизительно 16 аминокислот; xii) клубок-6 длиной от приблизительно 6 до приблизительно 9 аминокислот, фланкированный альфа-спиралью-6 и альфа-спиралью-7; xiii) альфа-спираль-7 длиной от приблизительно 49 до приблизительно 55 аминокислот; xiv) клубок-7 длиной от приблизительно 3 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-7 и альфа-спиралью-8; xv) альфа-спираль-8 длиной от приблизительно 33 до приблизительно 36 аминокислот; xvi) клубок-8 длиной от приблизительно 14 до приблизительно 16 аминокислот, фланкированный альфа-спиралью-8 и альфа-спиралью-9; xvii) альфа-спираль-9 длиной от приблизительно 16 до приблизительно 23 аминокислот; xviii) клубок-9 длиной от приблизительно 21 до приблизительно 28 аминокислот, фланкированный альфа-спиралью-9 и Участком C; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий семь сегментов со структурой преимущественно бета-тяжа.
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий десять сегментов со структурой преимущественно альфа-спирали; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) бета-тяж-1 (β1) длиной от приблизительно 3 аминокислот до приблизительно 5 аминокислот; ii) клубок длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот; iii) бета-тяж-2 (β2) длиной от приблизительно 7 аминокислот до приблизительно 11 аминокислот; iv) клубок длиной от приблизительно 17 аминокислот до приблизительно 23 аминокислот; v) бета-тяж-3 (β3) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; vi) клубок длиной от приблизительно 12 аминокислот до приблизительно 14 аминокислот; vii) бета-тяж-4 (β4) длиной от приблизительно 5 аминокислот до приблизительно 6 аминокислот; viii) клубок длиной от приблизительно 2 аминокислот до приблизительно 7 аминокислот; ix) бета-тяж-5 (β5) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; x) клубок длиной от приблизительно 26 аминокислот до приблизительно 28 аминокислот; xi) бета-тяж-6 (β6) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; xii) клубок длиной от приблизительно 16 аминокислот до приблизительно 20 аминокислот; и xiii) бета-тяж-1 (β7) длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот.
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот, с главным образом неконсервативной вторичной структурой; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) альфа-спираль-1 длиной от приблизительно 10 до приблизительно 26 аминокислот; ii) клубок-1 длиной от приблизительно 2 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-1 и альфа-спиралью-2; iii) альфа-спираль-2 длиной от приблизительно 15 до приблизительно 24 аминокислот; iv) клубок-2 длиной от приблизительно 4 до приблизительно 14 аминокислот, фланкированный альфа-спиралью-2 и альфа-спиралью-3; v) альфа-спираль 3 длиной от приблизительно 15 до приблизительно 27 аминокислот; vi) клубок-3 длиной от приблизительно 11 до приблизительно 13 аминокислот, фланкированный альфа-спиралью-3 и альфа-спиралью-4; vii) альфа-спираль-4 длиной приблизительно 24-180 аминокислот; viii) клубок-4 длиной от приблизительно 4 до приблизительно 5 аминокислот, фланкированный альфа-спиралью-4 и альфа-спиралью-5; ix) альфа-спираль-5 длиной от приблизительно 50 до приблизительно 54 аминокислот; x) клубок-5 длиной от приблизительно 11 до приблизительно 17 аминокислот, фланкированный альфа-спиралью-5 и альфа-спиралью-6; xi) альфа-спираль-6 длиной от приблизительно 15 до приблизительно 16 аминокислот; xii) клубок-6 длиной от приблизительно 6 до приблизительно 9 аминокислот, фланкированный альфа-спиралью-6 и альфа-спиралью-7; xiii) альфа-спираль-7 длиной от приблизительно 49 до приблизительно 55 аминокислот; xiv) клубок-7 длиной от приблизительно 3 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-7 и альфа-спиралью-8; xv) альфа-спираль-8 длиной от приблизительно 33 до приблизительно 36 аминокислот; xvi) клубок-8 длиной от приблизительно 14 до приблизительно 16 аминокислот, фланкированный альфа-спиралью-8 и альфа-спиралью-9; xvii) альфа-спираль-9 длиной от приблизительно 16 до приблизительно 23 аминокислот; xviii) клубок-9 длиной от приблизительно 21 до приблизительно 28 аминокислот, фланкированный альфа-спиралью-9 и Участком C; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) бета-тяж-1 (β1) длиной от приблизительно 3 аминокислот до приблизительно 5 аминокислот; ii) клубок длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот; iii) бета-тяж-2 (β2) длиной от приблизительно 7 аминокислот до приблизительно 11 аминокислот; iv) клубок длиной от приблизительно 17 аминокислот до приблизительно 23 аминокислот; v) бета-тяж-3 (β3) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; vi) клубок длиной от приблизительно 12 аминокислот до приблизительно 14 аминокислот; vii) бета-тяж-4 (β4) длиной от приблизительно 5 аминокислот до приблизительно 6 аминокислот; viii) клубок длиной от приблизительно 2 аминокислот до приблизительно 7 аминокислот; ix) бета-тяж-5 (β5) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; x) клубок длиной от приблизительно 26 аминокислот до приблизительно 28 аминокислот; xi) бета-тяж-6 (β6) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; xii) клубок длиной от приблизительно 16 аминокислот до приблизительно 20 аминокислот; и xiii) бета-тяж-1 (β7) длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот.
В некоторых вариантах осуществления полипептид PtIP-83 содержит последовательно от N-конца до C-конца: Участок A длиной от приблизительно 200 до приблизительно 300 аминокислот с гибкой консенсусной вторичной структурой, где Участок A содержит консервативный бета-тяж 1 (β1a) длиной от приблизительно 4 до приблизительно 12 аминокислот в пределах от приблизительно аминокислотного остатка 30 до приблизительно аминокислотного остатка 130 от N-конца полипептида PtIP-83; Участок B длиной от приблизительно 380 до приблизительно 465 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) альфа-спираль-1 длиной от приблизительно 10 до приблизительно 26 аминокислот; ii) клубок-1 длиной от приблизительно 2 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-1 и альфа-спиралью-2; iii) альфа-спираль-2 длиной от приблизительно 15 до приблизительно 24 аминокислот; iv) клубок-2 длиной от приблизительно 4 до приблизительно 14 аминокислот, фланкированный альфа-спиралью-2 и альфа-спиралью-3; v) альфа-спираль 3 длиной от приблизительно 15 до приблизительно 27 аминокислот; vi) клубок-3 длиной от приблизительно 11 до приблизительно 13 аминокислот, фланкированный альфа-спиралью-3 и альфа-спиралью-4; vii) альфа-спираль-4 длиной приблизительно 24-180 аминокислот; viii) клубок-4 длиной от приблизительно 4 до приблизительно 5 аминокислот, фланкированный альфа-спиралью-4 и альфа-спиралью-5; ix) альфа-спираль-5 длиной от приблизительно 50 до приблизительно 54 аминокислот; x) клубок-5 длиной от приблизительно 11 до приблизительно 17 аминокислот, фланкированный альфа-спиралью-5 и альфа-спиралью-6; xi) альфа-спираль-6 длиной от приблизительно 15 до приблизительно 16 аминокислот; xii) клубок-6 длиной от приблизительно 6 до приблизительно 9 аминокислот, фланкированный альфа-спиралью-6 и альфа-спиралью-7; xiii) альфа-спираль-7 длиной от приблизительно 49 до приблизительно 55 аминокислот; xiv) клубок-7 длиной от приблизительно 3 до приблизительно 8 аминокислот, фланкированный альфа-спиралью-7 и альфа-спиралью-8; xv) альфа-спираль-8 длиной от приблизительно 33 до приблизительно 36 аминокислот; xvi) клубок-8 длиной от приблизительно 14 до приблизительно 16 аминокислот, фланкированный альфа-спиралью-8 и альфа-спиралью-9; xvii) альфа-спираль-9 длиной от приблизительно 16 до приблизительно 23 аминокислот; xviii) клубок-9 длиной от приблизительно 21 до приблизительно 28 аминокислот, фланкированный альфа-спиралью-9 и Участком C; и Участок C длиной от приблизительно 150 до приблизительно 180 аминокислот с консенсусной вторичной структурой, содержащий последовательно: i) бета-тяж-1 (β1) длиной от приблизительно 3 аминокислот до приблизительно 5 аминокислот; ii) клубок длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот; iii) бета-тяж-2 (β2) длиной от приблизительно 7 аминокислот до приблизительно 11 аминокислот; iv) клубок длиной от приблизительно 17 аминокислот до приблизительно 23 аминокислот; v) бета-тяж-3 (β3) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; vi) клубок длиной от приблизительно 12 аминокислот до приблизительно 14 аминокислот; vii) бета-тяж-4 (β4) длиной от приблизительно 5 аминокислот до приблизительно 6 аминокислот; viii) клубок длиной от приблизительно 2 аминокислот до приблизительно 7 аминокислот; ix) бета-тяж-5 (β5) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; x) клубок длиной от приблизительно 26 аминокислот до приблизительно 28 аминокислот; xi) бета-тяж-6 (β6) длиной от приблизительно 5 аминокислот до приблизительно 7 аминокислот; xii) клубок длиной от приблизительно 16 аминокислот до приблизительно 20 аминокислот; и xiii) бета-тяж-1 (β7) длиной от приблизительно 13 аминокислот до приблизительно 17 аминокислот.
В некоторых вариантах осуществления полипептид PtIP-83 имеет расчетный молекулярный вес от приблизительно 70 кДа до приблизительно 120 кДа, от приблизительно 75 кДа до приблизительно 110 кДа, и от приблизительно 80 кДа до приблизительно 105 кДа, и от приблизительно 85 кДа до приблизительно 105 кДа.
В некоторых вариантах осуществления полипептид PtIP-83 характеризуется модифицированным физическим свойством. Применяемый в данном документе термин "физическое свойство" относится к любому параметру, который подходит для описания физико-химических характеристик белка. Применяемые данном документе термины "физическое свойство, представляющее интерес" и "свойство, представляющее интерес" используются взаимозаменяемо для обозначения физических свойств белков, подлежащих исследованию и/или модификации. Примеры физических свойств включают без ограничения суммарный поверхностный заряд и распределение зарядов на поверхности белка, суммарную гидрофобность и распределение гидрофобных остатков на поверхности белка, плотность поверхностного заряда, плотность гидрофобности поверхности, общее число поверхностных ионизируемых групп, поверхностное натяжение, размер белка и его распределение в растворе, температуру плавления, теплоемкость и второй вириальный коэффициент. Примеры физических свойств также включают без ограничения растворимость, фолдинг, стабильность и усвояемость. В некоторых вариантах осуществления полипептид PtIP-83 характеризуется повышенной перевариваемостью фрагментов, полученных в результате протеолитического расщепления, в кишечнике насекомого. Модели для переваривания при помощи искусственного желудочного сока известны специалисту в данной области (Fuchs, R.L. and J.D. Astwood. Food Technology 50: 83-88, 1996; Astwood, J.D., et al Nature Biotechnology 14: 1269-1273, 1996; Fu TJ et al J. Agric Food Chem. 50: 7154-7160, 2002).
В некоторых вариантах осуществления варианты включают полипептиды, которые отличаются аминокислотной последовательностью вследствие мутагенеза. Вариантные белки, охваченные настоящим раскрытием, являются биологически активными, то есть они по-прежнему обладают требуемой биологической активностью (т. е. пестицидной активностью) нативного белка. В некоторых вариантах осуществления вариант будет обладать по меньшей мере приблизительно 10%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 80% или более инсектицидной активности нативного белка. В некоторых вариантах осуществления варианты могут обладать усиленной активностью по сравнению с нативным белком.
Бактериальные гены довольно часто имеют несколько метиониновых кодонов инициации трансляции вблизи стартового сайта открытой рамки считывания. Зачастую, инициация трансляции по одному или нескольким из этих старт-кодонов будет приводить к образованию функционального белка. Эти старт-кодоны могут включать кодоны ATG. Однако, бактерии, такие как Bacillus sp., также распознают кодон GTG в качестве старт-кодона, и белки, трансляция которых инициируется по кодонам GTG, в качестве первой аминокислоты содержат метионин. В редких случаях, трансляция в бактериальных системах может инициироваться по кодону TTG, хотя в этом случае TTG кодирует метионин. Кроме того, зачастую a priori не определено, какой из этих кодонов естественным образом используется в бактерии. Таким образом, понятно, что применение одного из альтернативных метиониновых кодонов может также приводить к образованию пестицидных белков. Эти пестицидные белки охватываются настоящим раскрытием и могут применяться в способах согласно настоящему раскрытию. Будет понятно, что при экспрессии в растениях будет необходимо изменить альтернативный старт-кодон на ATG для полноценной трансляции.
В другом аспекте полипептид PtIP-83 может экспрессироваться в виде белка-предшественника с вставочной последовательностью, которая катализирует многостадийный, посттрансляционный сплайсинг белка. Сплайсинг белка включает вырезание вставочной последовательности из полипептида с одновременным соединением фланкирующих последовательностей с получением нового полипептида (Chong, et al., (1996) J. Biol. Chem., 271:22159-22168). Эта вставочная последовательность или элемент сплайсинга белка, называемые интеинами, которые катализируют свое собственное вырезание посредством трех согласованных реакций на N-терминальной и C-ерминальной границах сплайсинга: ацильной перестройки N-терминального цистеина или серина; реакции переэтерификации между двумя концами с образованием разветвленного сложноэфирного или тиоэфирного промежуточного соединения и расщепления пептидной связи, сопряженного с образованием кольца с участием C-терминального аспарагина интеина с высвобождением интеина (Evans, et al., (2000) J. Biol. Chem., 275:9091-9094. Выяснение механизма сплайсинга белка привело к возникновению целого ряда применений, связанных с интеинами (Comb, et al., Патент США № 5,496,714; Comb, et al., Патент США № 5,834,247; Camarero and Muir, (1999) J. Amer. Chem. Soc. 121:5597-5598; Chong, et al., (1997) Gene 192:271-281, Chong, et al., (1998) Nucleic Acids Res. 26:5109-5115; Chong, et al., (1998) J. Biol. Chem. 273:10567-10577; Cotton, et al., (1999) J. Am. Chem. Soc. 121:1100-1101; Evans, et al., (1999) J. Biol. Chem. 274:18359-18363; Evans, et al., (1999) J. Biol. Chem. 274:3923-3926; Evans, et al., (1998) Protein Sci. 7:2256-2264; Evans, et al., (2000) J. Biol. Chem. 275:9091-9094; Iwai and Pluckthun, (1999) FEBS Lett. 459:166-172; Mathys, et al., (1999) Gene 231:1-13; Mills, et al., (1998) Proc. Natl. Acad. Sci. USA 95:3543-3548; Muir, et al., (1998) Proc. Natl. Acad. Sci. USA 95:6705-6710; Otomo, et al., (1999) Biochemistry 38:16040-16044; Otomo, et al., (1999) J. Biolmol. NMR 14:105-114; Scott, et al., (1999) Proc. Natl. Acad. Sci. USA 96:13638-13643; Severinov and Muir, (1998) J. Biol. Chem. 273:16205-16209; Shingledecker, et al., (1998) Gene 207:187-195; Southworth, et al., (1998) EMBO J. 17:918-926; Southworth, et al., (1999) Biotechniques 27:110-120; Wood, et al., (1999) Nat. Biotechnol. 17:889-892; Wu, et al., (1998a) Proc. Natl. Acad. Sci. USA 95:9226-9231; Wu, et al., (1998b) Biochim Biophys Acta 1387:422-432; Xu, et al., (1999) Proc. Natl. Acad. Sci. USA 96:388-393; Yamazaki, et al., (1998) J. Am. Chem. Soc., 120:5591-5592). Относительно применения интеиноов в растительных трансгенах, см., Yang, et al., (Transgene Res 15:583-593 (2006)) и Evans, et al., (Annu. Rev. Plant Biol. 56:375-392 (2005)).
В другом аспекте полипептид PtIP-83 может кодироваться двумя отдельными генами, при этом интеин белка-предшественника берет начало от двух генов, он называется сплит-интеин, и две части предшественника соединяются при образовании пептидной связи. Это образование пептидной связи осуществляется при помощи транс-сплайсинга, опосредованного интеином. Для этой цели, первая и вторая кассета экспрессии, содержащие два отдельных гена, дополнительно кодируют интеины, способные опосредовать транс-сплайсинг белков. При помощи транс-сплайсинга белки и полипептиды, кодируемые первым и вторым фрагментами, могут быть соединены посредством образования пептидной связи. Интеины для транс-сплайсинга можно выбирать из ядерного генома или генома органелл различных организмов, в том числе эукариот, архебактерий и эубактерий. Интеины, которые можно применять, перечислены на сайте neb.com/neb/inteins.html, доступ к которому можно получить во всемирной сети Интернет с применением префикса "www"). Нуклеотидную последовательность, кодирующую интеин, можно расщеплять на 5′- и 3′-часть, которые кодируют 5′- и 3′-часть интеина соответственно. Части последовательности, которые не являются необходимыми для интеин-сплайсинга (например, домен хоминг-эндонуклеазы), могут быть удалены. Интеин-кодирующая последовательность расщепляют, так что 5′- и 3′-части способны к транс-сплайсингу. Для выбора подходящего сайта расщепления интеин-кодирующей последовательности можно следовать рекомендациям, опубликованным Southworth, et al., (1998) EMBO J. 17:918-926. При конструировании первой и второй кассеты экспрессии 5′ интеин-кодирующую последовательность соединяют с 3′-концом первого фрагмента, кодирующего N-концевую часть полипептида PtIP-83, а 3′ интеин-кодирующую последовательность соединяют с 5′-концом второго фрагмента, кодирующего C-концевую часть полипептида PtIP-83.
В целом, партнеров для транс-сплайсинга можно конструировать с применением любого сплит-интеина, в том числе любых встречающихся в природе или искусственно расщепленных сплит-интеинов. Известны несколько встречающихся в природе сплит-интеинов, например: сплит-интеин гена DnaE PCC6803 Synechocystis sp. (см. Wu, et al., (1998) Proc Natl Acad Sci USA. 95(16):9226-31 и Evans, et al., (2000) J Biol Chem. 275(13):9091-4 и гена DnaE из Nostoc punctiforme (см. Iwai, et al., (2006) FEBS Lett. 580(7):1853-8). Интеины, не относящиеся к сплит-интеинам, были искусственно расщеплены в лаборатории с созданием новых сплит-интеинов, например, искусственно расщепленный интеин Ssp DnaB (см. Wu, et al., (1998) Biochim Biophys Acta. 1387:422-32), и расщепленный интеин Sce VMA (см. Brenzel, et al., (2006) Biochemistry. 45(6):1571-8), и искусственно расщепленный мини-интеин грибкового происхождения (см. Elleuche, et al., (2007) Biochem Biophys Res Commun. 355(3):830-4). Также доступны базы данных по интеинам, в которых перечислены известные интеины (см., например, базу данных, доступную в режиме реального времени на сайте bioinformatics.weizmann.ac.il/˜pietro/inteins/Inteinstable.html, доступ к которому можно получить во всемирной сети Интернет с применением префикса "www").
Встречающиеся в природе интеины, не относящиеся к сплит-интеинам, могут обладать эндонуклеазной или другой ферментативной активностью, которую, как правило, можно устранять при конструировании искусственно расщепленного сплит-интеина. Такие мини-интеины или минимизированные сплит-интеины хорошо известны из уровня техники и, как правило, они состоят из менее 200 аминокислотных остатков (см. Wu, et al., (1998) Biochim Biophys Acta. 1387:422-32). Подходящие сплит-интеины могут иметь другие обеспечивающие очистку элементы полипептида, добавляемые к их структуре, при условии, что такие элементы не ингибируют сплайсинг сплит-интеина, или они добавлены так, что обеспечивается возможность их удаления до сплайсинга. Сообщалось о сплайсинге белка с применением белков, которые включают бактериальные интеин-подобные (BIL) домены (см. Amitai, et al., (2003) Mol Microbiol. 47:61-73) и самопроцессирующиеся домены hedgehog (Hog) (последние при объединении с интеинами называют суперсемейство Hog/интеин или семейство HINT (см. Dassa, et al., (2004) J Biol Chem. 279:32001-7), и такие домены, которые можно также применять для получения искусственно расщепленных интеинов. В частности, не подвергающихся сплайсингу представителей таких семейств можно модифицировать при помощи методик молекулярной биологии для введения или восстановления активности сплайсинга в таких родственных разновидностях. Последние исследования показывают, что сплайсинг можно наблюдать при обеспечении реакции N-терминального компонента сплит-интеина с C-терминальным компонентом сплит-интеина, при этом в естественных условиях он не является его "партнером"; например, сплайсинг наблюдали при использовании партнеров, которые всего на 30-50% гомологичны "естественному" сплайсинг-партнеру (см. Dassa, et al., (2007) Biochemistry. 46(1):322-30). Было показано, что другие такие смеси несовместимых партнеров сплит-интеинов не реагируют друг с другом (см. Brenzel, et al., (2006) Biochemistry. 45(6):1571-8). Однако, специалист в соответствующей области техники может определить, сможет ли конкретная пара полипептидов связываться друг с другом с обеспечением функционального интеина, с применением стандартных способов и без использования изобретательских навыков.
В другом аспекте полипептид PtIP-83 представляет собой вариант с круговыми перестановками. В определенных вариантах осуществления полипептид PtIP-83 представляет собой вариант с круговыми перестановками полипептида под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
Разработка способов с применением рекомбинантной ДНК обеспечила возможность исследовать эффекты транспозиции последовательностей на фолдинг, структуру и функцию белка. Подход, применяемый при создании новых последовательностей, напоминает то, что происходит со встречающимися в природе парами белков, которые связываются посредством линейной реорганизации их аминокислотных последовательностей (Cunningham, et al.,(1979) Proc. Natl. Acad. Sci. U.S.A. 76:3218-3222; Teather and Erfle, (1990) J. Bacteriol. 172:3837-3841; Schimming, et al., (1992) Eur. J. Biochem. 204:13-19; Yamiuchi and Minamikawa, (1991) FEBS Lett. 260:127-130; MacGregor, et al., (1996) FEBS Lett. 378:263-266). Первое in vitro применение данного типа перестановки у белков описали Goldenberg и Creighton (J. Mol. Biol. 165:407-413, 1983). При создании варианта с круговыми перестановками новый N-конец выбирают во внутреннем сайте (точечный разрыв) оригинальной последовательности, при этом новая последовательность имеет такой же порядок аминокислот, что и оригинальная, от точечного разрыва до тех пор, пока она не достигает аминокислоты, которая находится на оригинальном C-конце или вблизи него. В этой точке новая последовательность соединяется, либо напрямую, либо через дополнительную часть последовательности (линкер), с аминокислотой, которая находится на оригинальном N-конце или вблизи него, и новая последовательность продолжается с такой же последовательностью, что и оригинальная до тех пор, пока она не достигнет точки, которая находится в месте аминокислоты, которая была N-терминальной по отношению к сайту точечного разрыва оригинальной последовательности или вблизи него, причем этот остаток образует новый C-конец цепи. Длину аминокислотной последовательности линкера можно выбирать эмпирически, или исходя из информации о структуре, или путем применения комбинации двух этих подходов. Если информация о структуре является недоступной, можно получить небольшие серии линкеров для тестирования с применением конструкции, длина которой варьирует, чтобы охватить диапазон от 0 до 50 Å, и последовательность которой выбрана таким образом, чтобы соответствовать доступности поверхностных групп (гидрофильность, Hopp and Woods, (1983) Mol. Immunol. 20:483-489; Kyte and Doolittle, (1982) J. Mol. Biol. 157:105-132; площади поверхности, доступной воздействию растворителя, Lee and Richards, (1971) J. Mol. Biol. 55:379-400) и способности принимать необходимую конформацию без нарушения конфигурации пестицидного полипептида (конформационно подвижный; Karplus and Schulz, (1985) Naturwissenschaften 72:212-213). При условии, что при трансляции средняя длина остатка составляет 2,0-3,8 Å, это будет означать, что длина, подлежащая тестированию, будет составлять от 0 до 30 остатков, при этом предпочтительным диапазоном является 0-15 остатков. Примером такой эмпирической серии будет конструирование линкеров с применением кассетной последовательности, такой как Gly-Gly-Gly-Ser, повторяемой n раз, где n равно 1, 2, 3 или 4. Специалисты в данной области поймут, что существует множество таких последовательностей, варьирующих в длину или по составу, которые могут служить в качестве линкеров, исходя из таких первичных соображений, что они не являются ни чрезмерно длинными, ни чрезмерно короткими (см. также, Sandhu, (1992) Critical Rev. Biotech. 12:437-462); причем, если они являются слишком длинными, энтропийные эффекты, по-видимому, будут дестабилизировать трехмерную укладку и также могут делать фолдинг кинетически невыполнимым, а если они являются слишком короткими, они, вероятно, будут дестабилизировать молекулу вследствие скручивающей или стерической деформации. Специалистам, разбирающимся в анализе информации о структуре белка, будет понятно, что расстояние между концами цепей, определяемое как расстояние между c-альфа атомами углерода, можно применять для определения длины применяемой последовательности или по меньшей мере для ограничения числа возможностей, которые требуется протестировать, при эмпирическом отборе линкеров. Они также будут понимать, что иногда встречается случай, когда положения концов полипептидной цепи являются нечеткими в структурных моделях, полученных с помощью данных рентгеноструктурного анализа или ядерной магнитно-резонансной спектроскопии, и при такой ситуации, следовательно, ее необходимо принимать во внимание для правильной оценки длины требуемого линкера. На основании остатков, положение которых четко определено, выбирают два остатка, которые близки по последовательности к концам цепи, и расстояние между их c-альфа атомами углерода применяют для расчета приблизительной длины линкера между ними. С применением расчетной длины в качестве предварительных данных, далее отбирают линкеры в пределах диапазона количества остатков (из расчета длины остатка 2-3,8 Å). Эти линкеры можно составлять из оригинальной последовательности, укороченной или удлиненной, в случае необходимости, и, в случае удлинения, можно выбирать дополнительные остатки, которые являются гибкими и гидрофильными, как описано выше; или, необязательно, оригинальная последовательность может быть замещена с применением серии линкеров, причем одним примером является упомянутый выше подход с применением кассеты Gly-Gly-Gly-Ser; или, необязательно, можно применять комбинацию оригинальной последовательности и новой последовательности, имеющих подходящую общую длину. Последовательности пестицидных полипептидов, способных к фолдингу с образованием биологически активных состояний, можно получать путем соответствующего отбора начальных (амино-конец) и концевых (карбоксильный конец) положений внутри оригинальной полипептидной цепи, при этом с применением линкерной последовательности, как описано выше. Амино- и карбоксильные концы выбирают из общего отрезка последовательности, называемого участком точечного разрыва, с применением рекомендаций, описанных ниже. Новую аминокислотную последовательность, таким образом, получают путем отбора амино- и карбоксильных концов из такого же участка точечного разрыва. Во многих случаях выбор новых концов будет таким, что оригинальное положение карбоксильного конца непосредственно предшествует положению амино-конца. Однако, специалистам в данной области техники будет понятно, что выбор концов в каком-либо месте в пределах участка может оказывать действие, и что он, фактически, будет приводить либо к делециям, либо к добавлениям к амино- или карбоксильным частям новой последовательности. Основным положением молекулярной биологии является то, что первичная аминокислотная последовательность белка обуславливает фолдинг в трехмерную структуру, необходимую для проявления его биологической функции. Специалистам в данной области известны способы получения и интерпретации информации о трехмерной структуре с применением рентгеноструктурного анализа одиночных кристаллов белка или ядерной магнитно-резонансной спектроскопии растворов белка. Примеры информации о структуре, которая подходит для идентификации участков точечного разрыва, включают расположение и тип вторичной структуры белка (альфа- и 3-10 спирали, параллельные и антипараллельные бета-складчатые слои, обращения или повороты цепи и петли; Kabsch and Sander, (1983) Biopolymers 22:2577-2637; степень доступности аминокислотных остатков для растворителя, масштаб и тип взаимодействий остатков друг с другом (Chothia, (1984) Ann. Rev. Biochem. 53:537-572) и статическое и динамическое распределение конформаций на протяжении полипептидной цепи (Alber and Mathews, (1987) Methods Enzymol. 154:511-533). В некоторых случаях известна дополнительная информация о доступности остатков для растворителя; причем одним примером является сайт посттрансляционного присоединения углеводов, который обязательно находится на поверхности белка. Если экспериментальная информация о структуре не доступна или ее невозможно получить, также доступны способы для анализа первичной аминокислотной последовательности для того, чтобы делать прогнозы о третичной и вторичной структуре белка, доступности для растворителя и наличия поворотов и петель. Если прямые способы определения структуры не применимы, для эмпирического определения доступности поверхностных групп также иногда применяют биохимические способы; например, с использованием идентификации сайтов деполимеризации после ограниченного протеолиза, чтобы делать заключение о доступности поверхностных групп (Gentile and Salvatore, (1993) Eur. J. Biochem. 218:603-621). Таким образом, путем применения либо информации о структуре, полученной экспериментальным путем, либо прогностических способов (например, Srinivisan and Rose, (1995) Proteins: Struct., Funct. & Genetics 22:81-99) проводят исследование исходной аминокислотной последовательности для классификации участков в отношении того, важны ли они для поддержания вторичной и третичной структуры. Наличия последовательностей в участках, которые, как известно, вовлечены в периодическую вторичную структуру (альфа- и 3-10 спирали, параллельные и антипараллельные бета-складчатые слои), следует избегать. Аналогично, участки аминокислотной последовательности PtIP, которые, как наблюдается, обладают низкой степенью доступности для действия растворителя, наиболее вероятно, являются частью так называемого гидрофобного ядра белка, и их следует также избегать при выборе амино- или карбоксильных концов. В отличие от этого, участки PtIP, которые, как известно, находятся в поверхностных поворотах или петлях, и, в частности, участки, о которых известно, что они не требуются для биологической активности, являются предпочтительными сайтами для расположения противоположных концов полипептидной цепи. Непрерывные нити аминокислотной последовательности, которые являются предпочтительными на основании вышеприведенных критериев, называют участком точечного разрыва. Полинуклеотиды, кодирующие полипептиды PtIP-83 с круговыми перестановками с новым N-концом/C-концом, которые содержат линкерный участок, отделяющий оригинальный C-конец и N-конец, фактически, можно получать по способу, описанному в Mullins, et al., (1994) J. Am. Chem. Soc. 116:5529-5533. Несколько стадий амплификации посредством полимеразной цепной реакции (ПЦР) применяют для перестройки последовательности ДНК, кодирующей первичную аминокислотную последовательность белка. Полинуклеотиды, кодирующие полипептиды PtIP-83 с круговыми перестановками с новым N-концом/C-концом, которые содержат линкерный участок, отделяющий оригинальный C-конец и N-конец, можно создавать на основе способа тандемных повторов, описанного в Horlick, et al., (1992) Protein Eng. 5:427-431. Амплификацию посредством полимеразной цепной реакции (ПЦР) генов с новыми N-концом/C-концом проводят с применением ДНК-матрицы с тандемными повторами.
В другом аспекте представлены белки слияния, в аминокислотную последовательность которых включена аминокислотная последовательность, содержащая полипептид PtIP-83, в том числе без ограничения полипептид под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769, и его активные фрагменты.
Способы разработки и конструирования белков слияния (и кодирующих их полинуклеотидов) известны специалистам в данной области. Полинуклеотиды, кодирующие полипептид PtIP-83, могут быть слиты с сигнальными последовательностями, которые будут управлять локализацией полипептида PtIP-83 в конкретных компартментах прокариотической или эукариотической клетки и/или управлять секрецией полипептида PtIP-83 согласно вариантам осуществления из прокариотической или эукариотической клетки. Например, в E. coli, может потребоваться направить экспрессию белка в периплазматическое пространство. Примеры сигнальных последовательностей или белков (или их фрагментов), с которыми можно сливать полипептид PtIP-83 с тем, чтобы направлять экспрессию полипептида в периплазматическое пространство бактерий, включают без ограничения сигнальную последовательность pelB, сигнальную последовательность белка, связывающего мальтозу (MBP), MBP, сигнальную последовательность ompA, сигнальную последовательность B-субъединицы периплазматического неустойчивого к нагреванию энтеротоксина E. coli и сигнальную последовательность щелочной фосфатазы. Для конструирования белков слияния коммерчески доступны несколько векторов, которые будут управлять локализацией белка, такие как серия векторов pMAL (в частности, серия pMAL-p), доступная от New England Biolabs. В конкретном варианте осуществления полипептид PtIP-83 можно сливать с сигнальной последовательностью пектатлиазы pelB для повышения эффективности экспрессии и очистки таких полипептидов у грамотрицательных бактерий (см. патенты США №№ 5576195 и 5846818). Слияния пластидный транзитный пептид растения/полипептид хорошо известны из уровня техники (см. патент США № 7193133). Апопластные транзитные пептиды, такие как сигнальная последовательность для секреции альфа-амилазы риса или ячменя, также хорошо известны из уровня техники. Пластидный транзитный пептид, как правило, сливают со стороны N-конца с полипептидом, подлежащим нацеливанию (например, партнером слияния). В одном варианте осуществления белок слияния фактически состоит из пластидного транзитного пептида и полипептида PtIP-83, подлежащего нацеливанию. В другом варианте осуществления белок слияния состоит из пластидного транзитного пептида и полипептида, подлежащего нацеливанию. В таких вариантах осуществления пластидный транзитный пептид, предпочтительно, находится на N-конце белка слияния. Однако, дополнительные аминокислотные остатки могут находиться на N-конце относительно пластидного транзитного пептида при условии, что белок слияния по меньшей мере частично нацеливается на пластиду. В конкретном варианте осуществления пластидный транзитный пептид находится на N-терминальной половине, N-терминальной трети или N-терминальной четверти белка слияния. Большая часть или весь пластидный транзитный пептид, как правило, отщепляется от белка слияния после попадания в пластиду. Положение сайта отщепления может незначительно варьировать у разных видов растений, на различных стадиях развития растения, в результате специфических внутриклеточных условий или конкретной комбинации применяемого транзитного пептида/партнера слияния. В одном варианте осуществления сайт отщепления пластидного транзитного пептида является гомогенным, так что сайт отщепления является идентичным в группе белков слияния. В другом варианте осуществления сайт отщепления пластидного транзитного пептида не является гомогенным, так что сайт отщепления варьирует в пределах 1-10 аминокислот в группе белков слияния. Пластидный транзитный пептид можно рекомбинантно сливать со вторым белком с применением одного из нескольких путей. Например, сайт распознавания рестрикционной эндонуклеазы можно вводить в нуклеотидную последовательность транзитного пептида в положении, соответствующем его C-терминальному концу, и такой же или совместимый сайт можно вводить с помощью методик генной инженерии в нуклеотидную последовательность белка, подлежащего нацеливанию, по его N-терминальному концу. При конструировании этих сайтов нужно заботиться о том, чтобы кодирующие последовательности транзитного пептида и второго белка содержались "в рамке" для обеспечения синтеза требуемого белка слияния. В некоторых случаях предпочтительным может быть удаление инициаторного метионинового кодона второго белка при введении нового сайта рестрикции. Введение сайтов распознавания рестрикционной эндонуклеазы в обе исходные молекулы и их последующее связывание с помощью методик с применением рекомбинантной ДНК может приводить к добавлению одной или нескольких дополнительных аминокислот между транзитным пептидом и вторым белком. Это, как правило, не влияет на активность в отношении нацеливания, поскольку сайт отщепления транзитного пептида остается доступным и функционирование второго белка не изменится при добавлении этих дополнительных аминокислот по его N-концу. В качестве альтернативы, специалист в данной области техники может создать точный сайт отщепления между транзитным пептидом и вторым белком (с наличием инициаторного метионина или без него) с применением синтеза генов (Stemmer, et al., (1995) Gene 164:49-53) или аналогичных способов. В дополнение, слияние транзитного пептида может целенаправленно включать аминокислоты ниже сайта отщепления. Аминокислоты на N-конце зрелого белка могут воздействовать на способность транзитного пептида нацеливать белки в пластиды и/или на эффективность отщепления после импорта белков. Это может зависеть от белка, подлежащего нацеливанию. См., например, Comai, et al., (1988) J. Biol. Chem. 263(29):15104-9.
В некоторых вариантах осуществления представлены белки слияния, содержащие полипептид PtIP-83 и инсектицидный полипептид, соединенные аминокислотным линкером. В некоторых вариантах осуществления представлены белки слияния, представленные формулой, выбранной из группы, состоящей из:
R1-L-R2, R2-L-R1, R1-R2 или R2-R1,
где R1 представляет собой полипептид PtIP-83, R2 представляет собой белок, представляющий интерес. Полипептид R1 слит либо непосредственно, либо через линкерный (L) сегмент с полипептидом R2. Термин "непосредственно" обозначает слияния, в которых полипептиды соединены без пептидного линкера. Таким образом, "L" представляет собой химическую связь или полипептидный сегмент, с которым оба R1 и R2 слиты в рамке, при этом наиболее часто L представляет собой линейный пептид, с которым R1 и R2 связаны посредством амидных связей, связывающих карбоксильный конец R1 с амино-концом L и карбоксильный конец L с амино-концом R2. Под "слиты в рамке" подразумевается, что между рамками считывания R1 и R2 отсутствуют сайты терминации трансляции или разрыв. Связывающая группа (L) обычно представляет собой полипептид от 1 до 500 аминокислот в длину. Линкеры, соединяющие две молекулы, предпочтительно, конструируют так, (1) чтобы они давали возможность двум молекулам сворачиваться и действовать независимо друг от друга, (2) чтобы они не характеризовались склонностью к развитию упорядоченной вторичной структуры, которая может негативно воздействовать на функциональные домены двух белков, (3) чтобы они имели минимальную гидрофобную или характеристику в отношении зарядов, которые влияют на функциональные домены белка, и (4) чтобы они обеспечивали стерическое разделение R1 и R2, так что R1 и R2 могли одновременно взаимодействовать со своими соответствующими рецепторами на одной клетке. Как правило поверхностные аминокислоты в гибких участках белка включают Gly, Asn и Ser. По сути, любое сочетание аминокислотных последовательностей, содержащих Gly, Asn и Ser, как ожидается, будет удовлетворять вышеуказанным критериям для линкерной последовательности. Другие нейтральные аминокислоты, такие как Thr и Ala, также можно применять в линкерной последовательности. Дополнительные аминокислоты также можно включать в линкеры, поскольку добавление уникальных сайтов рестрикции в линкерную последовательность облегчает конструирование слияний.
В некоторых вариантах осуществления линкеры содержат последовательности, выбранные из группы формул: (Gly3Ser)n, (Gly4Ser)n, (Gly5Ser)n, (GlynSer)n или (AlaGlySer)n, где n представляет собой целое число. Одним примером очень гибкого линкера является спейсерный участок, богатый (GlySer), присутствующий в белке pIII нитевидных бактериофагов, например, бактериофагов M13 или fd (Schaller, et al., 1975). Этот участок обеспечивает длинный гибкий спейсерный участок между двумя доменами поверхностного белка pIII. Также включены линкеры, в состав которых входит последовательность распознавания эндопептидазы. Такой сайт расщепления может быть необходимым для разделения индивидуальных компонентов слияния для определения того, являются ли они правильным образом свернутыми и активными in vitro. Примеры различных эндопептидаз включают без ограничения плазмин, энтерокиназу, калликреин, урокиназу, тканевой активатор плазминогена, клострипаин, химозин, коллагеназу, протеазу яда гадюки Рассела, фермент расщепления постпролина, протеазу V8, тромбин и фактор Xa. В некоторых вариантах осуществления линкер содержит аминокислоты EEKKN (SEQ ID NO: 37) из мультигенного экспрессионного "проводника" (MGEV), который расщепляется протеазами вакуолей, как раскрыто в публикации заявки на патент США № 2007/0277263. В других вариантах осуществления пептидные линкерные сегменты из шарнирного участка тяжелой цепи иммуноглобулинов IgG, IgA, IgM, IgD или IgE обеспечивают угловое взаимодействие между прикрепленными полипептидами. В частности, применимыми являются такие шарнирные участки, в которых цистеины замещены на серины. Линкеры согласно настоящему раскрытию включают последовательности, полученные из шарнирного участка IgG гамма 2b мыши, в котором цистеины были заменены на серины. Белки слияния не ограничиваются формой, размером или количеством применяемых линкерных последовательностей, а единственным требованием для линкера является то, что функционально он не влияет негативным образом на фолдинг и функционирование отдельных молекул слияния.
В другом аспекте представлены химерные полипептиды PtIP-83, которые создают путем соединения двух или более частей генов PtIP-83, изначально кодирующих отдельные белки PtIP-83, с созданием химерного гена. Трансляция химерного гена приводит к единому химерному полипептиду PtIP-83 с участками, мотивами или доменами, полученными из каждого из исходных полипептидов. В определенных вариантах осуществления химерный белок содержит части, мотивы или домены полипептидов PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769, в любой комбинации.
Понятно, что последовательности ДНК можно изменять различными способами, и что эти изменения могут приводить к образованию последовательностей ДНК, кодирующих белки с аминокислотными последовательностями, отличающимися от тех, которые кодируют пестицидный белок дикого типа (или нативный). В некоторых вариантах осуществления полипептид PtIP-83 можно изменять различными способами, в том числе с помощью аминокислотных замен, делеций, усечений и вставок одной или нескольких аминокислот, включая до 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145 или более аминокислотных замен, делеций и/или вставок или их комбинаций по сравнению с последовательностью под любым из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления вариант полипептида PtIP-83 содержит одну или несколько аминокислотных замен из таблицы 13, таблицы 14, таблицы 15, таблицы 16, таблицы 17, таблицы 18, таблицы 20, таблицы 21, таблицы 23, таблицы 24 или их комбинации по сравнению с нативной аминокислотой из PtIP-83Aa (SEQ ID NO: 1) в соответствующем остатке.
В некоторых вариантах осуществления вариант полипептида PtIP-83 выбран без ограничения из последовательности под любым из SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607 и SEQ ID NO: 728-737.
В некоторых вариантах осуществления полипептид PtIP-83 содержит делецию 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или более аминокислот с N-конца полипептида PtIP-83 относительно аминокислотного положения в последовательности под любым из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
В некоторых вариантах осуществления полипептид PtIP-83 содержит делецию 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или более аминокислот с C-конца полипептида PtIP-83 относительно аминокислотного положения в последовательности под любым из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769.
Способы осуществления таких манипуляций, как правило, известны из уровня техники. Например, варианты аминокислотной последовательности полипептида PtIP-83 можно получать при помощи мутаций в ДНК. Это также можно осуществлять при помощи одной из нескольких форм мутагенеза и/или путем направленной эволюции. В некоторых аспектах изменения, закодированные в аминокислотной последовательности, не будут существенно влиять на функцию белка. Такие варианты будут обладать требуемой пестицидной активностью. Однако, понятно, что способность полипептида PtIP-83 обеспечивать пестицидную активность можно повышать путем применения таких методик в композициях согласно настоящему раскрытию.
Например, можно делать консервативные аминокислотные замены по одному или нескольким несущественным аминокислотным остаткам PtIP. "Несущественный" аминокислотный остаток представляет собой остаток, который можно изменять относительно последовательности дикого типа полипептида PtIP-83 без изменения биологической активности. "Консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток замещен на аминокислотный остаток, имеющий аналогичную боковую цепь. Семейства аминокислотных остатков, имеющие аналогичные боковые цепи, были определены в уровне техники. Эти семейства включают: аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин); кислотными боковыми цепями (например, аспарагиновую кислоту, глутаминовую кислоту); полярными отрицательно заряженными остатками и их амидами (например, аспарагиновую кислоту, аспарагин, глутаминовую кислоту, глутамин); незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин); небольшими алифатическими неполярными или слабополярными остатками (например, аланин, серин, треонин, пролин, глицин); неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан); крупными алифатическими неполярными остатками (например, метионин, лейцин, изолейцин, валин, цистин); бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин); ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин); крупными ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан).
Аминокислотные замены можно проводить в неконсервативных участках, которые сохраняют функцию. В целом, такие замены не следует проводить для консервативных аминокислотных остатков или для аминокислотных остатков, находящихся в консервативном мотиве, где такие остатки являются существенными для активности белка. Примеры остатков, которые являются консервативными и которые могут быть существенными для активности белка, включают, например, остатки, которые идентичны у всех белков, содержащихся в выравнивании аналогичных или родственных токсинов с последовательностями согласно вариантам осуществления (например, остатки, которые идентичны при выравнивании гомологичных белков). Примеры остатков, которые являются консервативными, но которые могут позволять консервативные аминокислотные замены и все еще сохранять активность, включают, например, остатки, которые характеризуются только консервативными заменами у всех белков, содержащихся в выравнивании аналогичных или родственных токсинов с последовательностями согласно вариантам осуществления (например, остатки, которые характеризуются только консервативными заменами у всех белков, содержащихся в выравнивании гомологичных белков). Однако, специалисту в данной области будет понятно, что функциональные варианты могут иметь незначительные консервативные или неконсервативные изменения по консервативным остаткам. Указания в отношении подходящих аминокислотных замен, которые не влияют на биологическую активность белка, представляющего интерес, можно найти в модели Dayhoff, et al., (1978) Atlas of Protein Sequence and Structure (Natl. Biomed. Res. Found., Washington, D.C.), включенной в данный документ посредством ссылки. При осуществлении таких изменений можно учитывать индекс гидропатичности аминокислот. Важность индекса гидропатичности аминокислот для обеспечения согласованной биологической функции белка, в целом, понятна из уровня техники (Kyte and Doolittle, (1982) J Mol Biol. 157(1):105-32). Принято считать, что относительная гидропатичность аминокислоты вносит вклад во вторичную структуру полученного в результате белка, что, в свою очередь, определяет взаимодействие белка с другими молекулами, например, ферментами, субстратами, рецепторами, ДНК, антителами, антигенами и т. п.
Из уровня техники известно, что определенные аминокислоты можно замещать другими аминокислотами, имеющими аналогичный индекс или показатель гидропатичности, и в результате это все еще приводит к белку с аналогичной биологической активностью, т. е. по-прежнему получают белок с эквивалентной биологической функцией. Каждой аминокислоте был присвоен индекс гидропатичности на основании ее гидрофобности и характеристик в отношении заряда (Kyte and Doolittle, там же). Они являются следующими: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серин (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9) и аргинин (-4,5). При осуществлении таких изменений предпочтительной является замена аминокислот, индексы гидропатичности которых находятся в пределах до +2, особенно предпочтительной с индексами в пределах до +1 и наиболее предпочтительной с индексами в пределах до +0,5.
Из уровня техники также понятно, что замену подобных аминокислот можно эффективно проводить на основании гидрофильности. В патенте США № 4554101 заявляется, что наибольшая локальная средняя гидрофильность белка, определяемая гидрофильностью смежных аминокислот, коррелирует с биологическим свойством белка.
Как подробно описано в патенте США № 4554101, аминокислотным остаткам были присвоены следующие значения гидрофильности: аргинин (+3,0); лизин (+3,0); аспартат (+3,0.+0,1); глутамат (+3,0.+0,1); серин (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5.+0,1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5); триптофан (-3,4).
В качестве альтернативы, изменения можно производить в белковой последовательности многих белков на амино- или карбокси-конце без существенного воздействия на активность. Они могут включать вставки, делеции или изменения, введенные при помощи современных молекулярных способов, таких как ПЦР, в том числе амплификации посредством ПЦР, которые изменяют или удлиняют последовательность, кодирующую белок, посредством включения последовательностей, кодирующих аминокислоты, в олигонуклеотиды, используемые при амплификации посредством ПЦР. В качестве альтернативы, добавленные белковые последовательности могут включать последовательности, кодирующие весь белок, например, такие, которые обычно применяются в уровне техники для получения белковых слияний. Такие белки слияния часто применяют для (1) усиления экспрессии белка, представляющего интерес, (2) введения связывающего домена, ферментативной активности или эпитопа для облегчения либо очистки белка, либо выявления белка или других экспериментальных применений, известных из уровня техники, (3) нацеливания секреции или трансляции белка во внутриклеточную органеллу, такую как периплазматическое пространство грамотрицательных бактерий, митохондрии или хлоропласты растений или эндоплазматический ретикулум эукариотических клеток, причем последнее зачастую приводит к гликозилированию белка.
Вариантные нуклеотидные и аминокислотные последовательности согласно настоящему раскрытию также охватывают последовательности, полученные с помощью процедур, приводящих к мутациям и рекомбинациям, таких как ДНК-шаффлинг. С помощью такой процедуры один или несколько различных кодирующих участков полипептида PtIP-83 можно применять для создания нового полипептида PtIP-83, обладающего требуемыми свойствами. Таким образом, библиотеки рекомбинантных полинуклеотидов создают из группы родственных по последовательностям полинуклеотидов, содержащих участки последовательностей, которые характеризуются значительной идентичностью последовательности и могут подвергаться гомологичной рекомбинации in vitro или in vivo. Например, с применением этого подхода, мотивы с последовательностями, кодирующими домен, представляющий интерес, можно подвергать шаффлингу между пестицидным геном и другими известными пестицидными генами с получением нового гена, кодирующего белок с улучшенным свойством, представляющим интерес, таким как повышенная инсектицидная активность. Стратегии для такого ДНК-шаффлинга известны из уровня техники. См., например, Stemmer, (1994) Proc. Natl. Acad. Sci. USA 91:10747-10751; Stemmer, (1994) Nature 370:389-391; Crameri, et al., (1997) Nature Biotech. 15:436-438; Moore, et al., (1997) J. Mol. Biol. 272:336-347; Zhang, et al., (1997) Proc. Natl. Acad. Sci. USA 94:4504-4509; Crameri, et al., (1998) Nature 391:288-291; и патенты США №№ 5605793 и 5837458.
Замена доменов или шаффлинг представляет собой другой механизм получения измененных полипептидов PtIP-83. Можно проводить замену доменов между полипептидами PtIP-83, что дает гибридные или химерные токсины с повышенной пестицидной активностью или расширенным спектром мишеней. Способы получения рекомбинантных белков и тестирования их в отношении пестицидной активности хорошо известны из уровня техники (см., например, Naimov, et al., (2001) Appl. Environ. Microbiol. 67:5328-5330; de Maagd, et al., (1996) Appl. Environ. Microbiol. 62:1537-1543; Ge, et al., (1991) J. Biol. Chem. 266:17954-17958; Schnepf, et al., (1990) J. Biol. Chem. 265:20923-20930; Rang, et al., 91999) Appl. Environ. Microbiol. 65:2918-2925).
Выравнивание гомологов PtIP-83 (фигура 2) обеспечивает возможность идентификации остатков, которые являются высококонсервативными среди естественных гомологов в данном семействе.
В некоторых вариантах осуществления представлены полипептиды PtIP-83, содержащие аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична последовательности под любым из SEQ ID NO: 786-888.
В некоторых вариантах осуществления представлены полипептиды PtIP-83, содержащие аминокислотную последовательность под любым из SEQ ID NO: 786-888.
В некоторых вариантах осуществления полипептид PtIP-83 не является аминокислотной последовательностью под любым из SEQ ID NO: 786-888.
Композиции
Также охвачены композиции, содержащие полипептид PtIP-83 согласно настоящему раскрытию. В некоторых вариантах осуществления композиция содержит полипептид PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769 или его вариант. В некоторых вариантах осуществления композиция содержит белок слияния PtIP-83.
В некоторых вариантах осуществления представлены композиции, содержащие полипептид PtIP-83, содержащий аминокислотную последовательность под любым из SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607, SEQ ID NO: 640-645 и SEQ ID NO: 728-737, или его вариант.
В некоторых вариантах осуществления представлены композиции, содержащие полипептид PtIP-83, содержащий аминокислотную последовательность под любым из SEQ ID NO: 786-888 или его варианта.
В некоторых вариантах осуществления раскрыты композиции полипептидов PtIP-83 для применения в сельском хозяйстве. В вариантах осуществления трансформированный микроорганизм (который включает целые организмы, клетки, спору(ы), полипептид(ы) PtIP-83, пестицидный компонент(ы), оказывающий воздействие на вредителя компонент(ы), вариант(ы), живые или мертвые клетки и клеточные компоненты, в том числе смеси живых и мертвых клеток и клеточных компонентов, и в том числе разрушенные клетки и клеточные компоненты) или выделенный полипептид(ы) PtIP-83 можно составить с приемлемым носителем в пестицидную композицию(и), которая представляет собой, например, суспензию, раствор, эмульсию, распыляемый порошок, диспергируемые гранулы или пеллеты, смачиваемый порошок и эмульгируемый концентрат, аэрозоль или распыляемый раствор, пропитанные гранулы, вспомогательное вещество, наносимую в качестве покрытия пасту, коллоид, а также инкапсулированные формы, например, в полимерных веществах. Такие составленные композиции можно получать с помощью таких общепринятых способов, как высушивание, лиофилизация, гомогенизация, экстракция, фильтрование, центрифугирование, седиментация или концентрирование культуры клеток, содержащих полипептид.
Такие композиции, раскрытые выше, можно получать с помощью добавления поверхностно-активного средства, инертного носителя, консерванта, увлажнителя, стимулятора поедания, аттрактанта, средства для инкапсуляции, связующего, эмульгатора, красителя, защитного средства от УФ, буфера, средства, повышающего текучесть, или удобрения, донора питательных микроэлементов или других препаратов, которые влияют на рост растения. Один или несколько агрохимикатов, в том числе без ограничения гербициды, инсектициды, фунгициды, бактерициды, нематоциды, моллюскоциды, акарициды, регуляторы роста растений, вспомогательные средства для сбора урожая и удобрения, можно комбинировать с носителями, поверхностно-активными веществами или вспомогательными средствами, традиционно используемыми в области составления, или другими компонентами для облегчения обращения с продуктом и его применения по отношению к конкретным целевым вредителям. Подходящие носители и вспомогательные средства могут быть твердыми или жидкими и соответствуют веществам, обычно используемым в технологии составления, например, природным или регенерированным минеральным веществам, растворителям, диспергирующим веществам, смачивающим средствам, веществам, придающим клейкость, связующим или удобрениям. Активные ингредиенты согласно вариантам осуществления обычно применяют в виде композиций и их можно применять по отношению к возделываемой площади, растению или семени, подлежащим обработке. Например, композиции согласно вариантам осуществления можно применять по отношению к зерну при подготовке к хранению или во время хранения в зерновом бункере или элеваторе и т.д. Композиции согласно вариантам осуществления можно применять одновременно или последовательно с другими соединениями. Способы применения активного ингредиента согласно вариантам осуществления или агрохимической композиции согласно вариантам осуществления, которая содержит по меньшей мере один из вариантных полипептидов Cyt1A, вырабатываемых штаммами бактерий согласно вариантам осуществления, включают без ограничения внекорневое применение, нанесение покрытия на семена и применение по отношению к почве. Количество применений и норма применения зависят от интенсивности поражения соответствующим вредителем.
Подходящие поверхностно-активные средства включают без ограничений анионные соединения, такие как карбоксилат, например, металла; карбоксилат длинноцепочечной жирной кислоты; N-ацилсаркозинат; сложные моно- или диэфиры фосфорной кислоты с этоксилатами жирных спиртов или соли таких сложных эфиров; сульфаты жирных спиртов, такие как додецилсульфат натрия, октадецилсульфат натрия или цетилсульфат натрия; сульфаты этоксилированных жирных спиртов; этоксилированные алкилфенолсульфаты; лигнинсульфонаты; нефтяные сульфонаты; алкиларилсульфонаты, такие как алкил-бензолсульфонаты или низшие алкилнафталинсульфонаты, например, бутилнафталинсульфонат; соли сульфированных нафталин-формальдегидных конденсатов; соли сульфированных фенол-формальдегидных конденсатов; более сложные сульфонаты, такие как сульфонаты амидов, например, сульфированный продукт конденсации олеиновой кислоты и N-метилтаурина; или диалкилсульфосукцинаты, например, сульфонат натрия или диоктилсукцинат. Неионогенные средства включают продукты конденсации сложных эфиров жирных кислот, жирных спиртов, амидов жирных кислот или жирных алкил- или алкенилзамещенных фенолов с этиленоксидом, сложные эфиры жирной кислоты и эфиров многоатомных спиртов, например, сложные эфиры сорбита и жирных кислот, продукты конденсации таких сложных эфиров с этиленоксидом, например, сложные эфиры полиоксиэтиленсорбитана и жирных кислот, блок-сополимеры этиленоксида и пропиленоксида, ацетиленовые гликоли, такие как 2,4,7,9-тетраэтил-5-децин-4,7-диол, или этоксилированные ацетиленовые гликоли. Примеры катионного поверхностно-активного средства включают, к примеру, алифатический моно-, ди- или полиамин, такой как ацетат, нафтенат или олеат; или кислородсодержащий амин, такой как аминоксид полиоксиэтиленалкиламина; амины с амидными связями, полученные путем конденсации карбоновой кислоты с ди- или полиамином; или соль четвертичного аммония.
Примеры инертных материалов включают без ограничений неорганические минеральные вещества, такие как каолин, филлосиликаты, карбонаты, сульфаты, фосфаты, слюду, аморфный силикагель, тальк, глину, вулканический пепел, или растительные материалы, такие как пробка, измельченные в порошок стержни кукурузных початков, шелуха арахиса, шелуха риса и скорлупа грецких орехов. В качестве материалов-носителей применимы каолины, такие как каолинит, диккит, накрит, анауксит, галлуазит и энделлит. В качестве материалов-носителей применимы монтмориллониты, такие как бейделлит, нонтронит, монтмориллонит, гекторит, сапонит, сауконит и бентонит. В качестве материалов-носителей применимы вермикулиты, такие как биотит.
Композиции согласно вариантам осуществления могут находиться в форме, подходящей для непосредственного применения, или в виде концентрата первичной композиции, для которого требуется разведение подходящим количеством воды или другого разбавителя перед применением. Концентрация пестицида будет варьировать в зависимости от природы конкретного состава, в частности от того, является ли он концентратом или должен применяться непосредственно. Композиция содержит 1-98% твердого или жидкого инертного носителя и 0-50% или 0,1-50% поверхностно-активного вещества. Эти композиции будут вводить из расчета обозначенной нормы для коммерческого продукта, например, приблизительно 0,01 фунта - 5,0 фунтов на акр при применении в сухой форме и расчета приблизительно 0,01 пинты - 10 пинт на акр при применении в жидкой форме.
Антитела
Также охвачены антитела к полипептиду PtIP-83 согласно вариантам осуществления или к его вариантам или фрагментам. Антитела согласно настоящему раскрытию включают поликлональные и моноклональные антитела, а также их фрагменты, которые сохраняют свою способность связываться с полипептидом PtIP-83, обнаруженным в кишечнике насекомого. Считается, что антитело, моноклональное антитело или его фрагмент способны связывать молекулу, если они способны специфично взаимодействовать с молекулой, тем самым, связывать молекулу с антителом, моноклональным антителом или его фрагментом. Как подразумевается, термин "антитело" (Ab) или "моноклональное антитело" (Mab) включает интактные молекулы, а также их фрагменты, или связывающие участки, или домены (такие как, например, фрагменты Fab и F(ab)2), которые способны связывать гаптен. Такие фрагменты, как правило, получают при помощи протеолитического расщепления, например, с помощью папаина или пепсина. В качестве альтернативы, гаптен-связывающие фрагменты можно получать посредством применения технологии рекомбинантной ДНК или при помощи синтетической химии. Способы получения антител согласно настоящему раскрытию, как правило, известны из уровня техники. Например, см. Antibodies, A Laboratory Manual, Ed Harlow and David Lane (eds.) Cold Spring Harbor Laboratory, N.Y. (1988), а также процитированные в нем литературные источники. Стандартные справочные издания, излагающие общие принципы иммунологии, включают: Klein, J. Immunology: The Science of Cell-Noncell Discrimination, John Wiley & Sons, N.Y. (1982); Dennett, et al., Monoclonal Antibodies, Hybridoma: A New Dimension in Biological Analyses, Plenum Press, N.Y. (1980) и Campbell, "Monoclonal Antibody Technology," в Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 13, Burdon, et al., (eds.), Elsevier, Amsterdam (1984). См. также патенты США №№ 4196265; 4609893; 4713325; 4714681; 4716111; 4716117 и 4720459. Антитела к полипептиду PtIP-83 или их антиген-связывающие части можно получать при помощи ряда методик, в том числе традиционных методик получения моноклональных антител, например, стандартной методики гибридизации соматических клеток согласно Kohler and Milstein, (1975) Nature 256:495. Также можно использовать другие методики получения моноклонального антитела, такие как вирусная или онкогенная трансформация B-лимфоцитов. Система для получения гибридом с применением животных представляет собой систему с применением мышей. Из уровня техники известны протоколы и методики выделения спленоцитов иммунизированных животных для слияния. Также известны партнеры для слияния (например, клетки миеломы мыши) и процедуры слияния. Антитело и моноклональные антитела согласно настоящему раскрытию можно получить путем использования полипептида PtIP-83 в качестве антигенов.
Представлен набор для выявления присутствия полипептида PtIP-83 или выявления присутствия нуклеотидной последовательности, кодирующей полипептид PtIP-83, в образце. В одном варианте осуществления в наборе имеются реагенты на основе антитела для выявления присутствия полипептида PtIP-83 в образце ткани. В другом варианте осуществления в наборе имеются меченые зонды из нуклеиновой кислоты, применяемые для выявления присутствия одного или нескольких полинуклеотидов, кодирующих полипептид PtIP-83. В наборе также имеются соответствующие реагенты и контрольные образцы для проведения способа выявления, а также инструкции для применения набора.
Идентификация и выделение рецепторов
Также охвачены рецепторы к полипептиду PtIP-83 согласно вариантам осуществления или к его вариантам или фрагментам. Способы идентификации рецепторов, хорошо известные из уровня техники (см. Hofmann, et. al., (1988) Eur. J. Biochem. 173:85-91; Gill, et al., (1995) J. Biol. Chem. 27277-27282), можно использовать для идентификации и выделения рецептора, который распознает полипептид PtIP-83, с применением мембранных везикул щеточной каемки от восприимчивых насекомых. В дополнение к способу радиоактивного мечения, приведенному в процитированных литературных источниках, полипептид PtIP-83 можно метить с помощью флуоресцентного красителя и других общепринятых меток, таких как стрептавидин. Мембранные везикулы щеточной каемки (BBMV) восприимчивых насекомых, таких как соевая совка и щитники, можно получать в соответствии с протоколами, приведенными в ссылочных документах, и разделять в геле SDS-PAGE, и переносить на подходящую мембрану. Меченые полипептиды PtIP-83 можно инкубировать с мембраной, на которой находятся подвергнутые блоттингу BBMV, и при этом меченые полипептиды PtIP-83 можно идентифицировать при помощи меченых репортеров. Идентификацию белковой полоски(полосок), которая взаимодействуют с полипептидами PtIP-83, можно проводить при помощи секвенирования N-терминальных аминокислотных остатков в газовой фазе или способа идентификации белков на основе масс-спектрометрии (Patterson, (1998) 10.22, 1-24, Current Protocol in Molecular Biology published by John Wiley & Son Inc). После идентификации белка соответствующий ген можно клонировать из библиотеки геномной ДНК или кДНК восприимчивых насекомых, и сродство связывания можно оценивать непосредственно при помощи полипептидов PtIP-83. Рецепторную функцию в осуществлении инсектицидной активности полипептидов PtIP-83 можно подтвердить путем осуществления способа нокаута гена при помощи RNAi (Rajagopal, et al., (2002) J. Biol. Chem. 277:46849-46851).
Нуклеотидные конструкции, кассеты экспрессии и векторы
Применение термина "нуклеотидные конструкции" в данном документе не предназначено ограничивать варианты осуществления нуклеотидными конструкциями, содержащими ДНК. Специалистам в данной области будет понятно, что нуклеотидные конструкции, в частности полинуклеотиды и олигонуклеотиды, состоящие из рибонуклеотидов и комбинаций рибонуклеотидов и дезоксирибонуклеотидов, также можно использовать в способах, раскрытых в данном описании. Нуклеотидные конструкции, нуклеиновые кислоты и нуклеотидные последовательности согласно вариантам осуществления дополнительно охватывают все комплементарные формы таких конструкций, молекул и последовательностей. Кроме того, нуклеотидные конструкции, нуклеотидные молекулы и нуклеотидные последовательности согласно вариантам осуществления охватывают все нуклеотидные конструкции, молекулы и последовательности, которые можно использовать в способах согласно вариантам осуществления для трансформации растений, в том числе без ограничения состоящие из дезоксирибонуклеотидов, рибонуклеотидов и их комбинаций. Такие дезоксирибонуклеотиды и рибонуклеотиды предусматривают как встречающиеся в природе молекулы, так и синтетические аналоги. Нуклеотидные конструкции, нуклеиновые кислоты и нуклеотидные последовательности согласно вариантам осуществления также охватывают все формы нуклеотидных конструкций, в том числе без ограничения одноцепочечные формы, двухцепочечные формы, шпильки, структуры "стебель-и-петля" и т. п.
Дополнительный вариант осуществления относится к трансформированному организму, такому как организм, выбранный из растительных клеток или клеток насекомых, бактерий, дрожжей, бакуловируса, простейших, нематод и водорослей. Трансформированный организм содержит молекулу ДНК согласно вариантам осуществления, кассету экспрессии, содержащую молекулу ДНК, или вектор, содержащий кассету экспрессии, которые могут быть стабильно встроенными в геном трансформированного организма.
В некоторых вариантах осуществления представлены трансгенные клетки-хозяева, трансформированные полинуклеотидом, кодирующим полипептид PtIP-83 согласно настоящему раскрытию. В некоторых вариантах осуществления клетка-хозяин представляет собой растительную клетку. В некоторых вариантах осуществления клетка-хозяин представляет собой бактерию.
Последовательности согласно вариантам осуществления представлены в составе ДНК-конструкций для экспрессии в организме, представляющем интерес. Конструкции будут включать 5' и 3' регуляторные последовательности, функционально связанные с последовательностью согласно вариантам осуществления. Применяемый в данном документе термин "функционально связанные" относится к функциональной связи между промотором и второй последовательностью, где промоторная последовательность инициирует и опосредует транскрипцию последовательности ДНК, соответствующей второй последовательности. Как правило, функционально связанные означает, что последовательности нуклеиновой кислоты, подлежащие связыванию, являются смежными и, при необходимости, два кодирующих белок участка соединяют в одной рамке считывания. Конструкция может дополнительно содержать по меньшей мере один дополнительный ген, подлежащий введению в организм путем котрансформации. В качестве альтернативы, дополнительный ген(ы) может обеспечиваться на множественных ДНК-конструкциях.
Такая ДНК-конструкция снабжена несколькими сайтами рестрикции для вставки последовательности гена полипептида PtIP-83, транскрипция которой будет регулироваться регуляторными участками. ДНК-конструкция может дополнительно содержать гены селектируемых маркеров.
В направлении транскрипции 5' - 3' ДНК-конструкция, как правило, будет включать: участок инициации транскрипции и трансляции (т. е. промотор), последовательность ДНК согласно вариантам осуществления и участок терминации транскрипции и трансляции (т. е. участок терминации), функционирующие в организме, выступающем в качестве хозяина. Участок инициации транскрипции (т.е. промотор) может быть нативным, аналогичным, чужеродным или гетерологичным относительно организма-хозяина и/или последовательности согласно вариантам осуществления. Кроме того, промотор может быть природной последовательностью или, в качестве альтернативы, синтетической последовательностью. Применяемый в данном документе термин "чужеродный" указывает на то, что промотор не найден в нативном организме, в который введен промотор. Если промотор является "чужеродным" или "гетерологичным" относительно последовательности согласно вариантам осуществления, предполагается, что промотор не является нативным или встречающимся в природе промотором для функционально связанной последовательности согласно вариантам осуществления. Применяемый в данном документе химерный ген содержит кодирующую последовательность, функционально связанную с участком инициации транскрипции, который является гетерологичным для кодирующей последовательности. Если промотор является нативной или природной последовательностью, экспрессия функционально связанной последовательности изменена по сравнению с экспрессией дикого типа, что приводит к изменению фенотипа.
В некоторых вариантах осуществления ДНК-конструкция может также включать последовательность транскрипционного энхансера. Применяемый в данном документе термин "энхансер" относится к последовательности ДНК, которая может стимулировать активность промотора и которая может представлять собой характерный элемент промотора или гетерологичный элемент, вставленный для повышения уровня активности или тканевой специфичности промотора. Из уровня техники известны различные энхансеры, например, также могут применяться интроны со свойствами улучшения генной экспрессии в растениях (публикация заявки на патент США № 2009/0144863, интрон убиквитина (т.е. интрон 1 убиквитина маиса (см., например, последовательность NCBI S94464)), омега-энхансер или омега-штрих-энхансер (Gallie, et al., (1989) Molecular Biology of RNA ed. Cech (Liss, New York) 237-256 и Gallie, et al., (1987) Gene 60:217-25), энхансер CaMV 35S (см., например, Benfey, et al., (1990) EMBO J. 9:1685-96) и энхансеры из патента США № 7803992, каждый из которых включен посредством ссылки. Приведенный выше перечень транскрипционных энхансеров не предназначен для ограничения. В вариантах осуществления можно применять любой подходящий транскрипционный энхансер.
Участок терминации может быть нативным относительно участка инициации транскрипции, может быть нативным относительно функционально связанной последовательности ДНК, представляющей интерес, может быть нативным относительно растения-хозяина или может быть получен из другого источника (т. e. чужеродный или гетерологичный для промотора, последовательности, представляющей интерес, растения-хозяина или какой-либо их комбинации).
Подходящие участки терминации доступны из Ti-плазмиды A. tumefaciens, такие как участки терминации генов октопинсинтазы и нопалинсинтазы. См. также, Guerineau, et al., (1991) Mol. Gen. Genet. 262:141-144; Proudfoot, (1991) Cell 64:671-674; Sanfacon, et al., (1991) Genes Dev. 5:141-149; Mogen, et al., (1990) Plant Cell 2:1261-1272; Munroe, et al., (1990) Gene 91:151-158; Ballas, et al., (1989) Nucleic Acids Res. 17:7891-7903 и Joshi, et al., (1987) Nucleic Acid Res. 15:9627-9639.
При необходимости нуклеиновую кислоту можно оптимизировать для усиления экспрессии в организме-хозяине. Таким образом, если организм-хозяин является растением, для усиления экспрессии можно синтезировать синтетические нуклеиновые кислоты с применением кодонов, предпочтительных для растений. См., например, Campbell и Gowri, (1990) Plant Physiol. 92:1-11, где обсуждается применение кодонов, предпочтительных для растений. Например, хотя последовательности нуклеиновой кислоты согласно вариантам осуществления могут экспрессироваться у видов как однодольных, так и двудольных растений, последовательности можно модифицировать с учетом специфических предпочтений кодонов и предпочтений по содержанию GC у однодольных или двудольных, если было показано, что предпочтения отличаются (Murray et al. (1989) Nucleic Acids Res. 17:477-498). Таким образом, кодон конкретной аминокислоты, предпочтительный для маиса, можно установить из известных генных последовательностей маиса. Данные о частоте использования кодонов в маисе для 28 генов из растений маиса приведены в таблице 4 из Murray, et al., выше. Из уровня техники доступны способы синтеза генов, предпочтительных для растений. См., например, патенты США №№ 5380831 и 5436391, и Murray, et al., (1989) Nucleic Acids Res. 17:477-498, и Liu H et al. Mol Bio Rep 37:677-684, 2010, включенные в данный документ посредством ссылки. Таблицу частоты использования кодонов Zea maize также можно найти на сайте kazusa.or.jp/codon/cgi-bin/showcodon.cgi?species=4577, доступ к которому можно получить с применением префикса www. В таблице 3 показан анализ оптимальных кодонов маиса (адаптировано из Liu H et al. Mol Bio Rep 37:677-684, 2010).
Таблица 3
Частоту использования кодонов сравнивали с применением критерия сопряженности хи-квадрат для идентификации оптимальных кодонов. Кодоны, которые встречаются статистически более часто (P\0,01), обозначены звездочкой.
Таблица 4
Таблица частоты использования кодонов Glycine max показана в виде таблицы 4, и ее также можно найти на сайте kazusa.or.jp/codon/cgi-bin/showcodon.cgi?species=3847&aa=1&style=N, доступ к которому можно получить с применением префикса www.
В некоторых вариантах осуществления рекомбинантная молекула нуклеиновой кислоты, кодирующая полипептид PtIP-83, имеет кодоны, оптимизированные для маиса.
Известны дополнительные модификации последовательности для усиления экспрессии гена у клеточного хозяина. Они включают устранение последовательностей, кодирующих ложные сигналы полиаденилирования, сигналы сайта сплайсинга экзонов и интронов, транспозон-подобные повторы и другие хорошо изученные последовательности, которые могут быть вредны для экспрессии гена. Содержание GC в последовательности можно скорректировать до уровней, средних для данного клеточного хозяина, рассчитываемых с учетом известных генов, экспрессируемых в клетке-хозяине. Применяемый в данном документе термин "клетка-хозяин" относится к клетке, которая содержит вектор и поддерживает репликацию и/или экспрессию предполагаемых векторов экспрессии. Клетки-хозяева могут быть прокариотическими клетками, такими как E. coli, или эукариотическими клетками, такими как клетки дрожжей, насекомых, амфибий или млекопитающих, или клетками однодольных или двудольных растений. Примером клетки-хозяина, относящейся к однодольному растению, является клетка-хозяин маиса. Если возможно, последовательность PtIP модифицируют для того, чтобы избежать образования шпилечных вторичных структур мРНК.
Кассеты экспрессии могут дополнительно содержать 5'-лидерные последовательности. Такие лидерные последовательности могут способствовать усилению трансляции. Лидерные последовательности трансляции известны из уровня техники и включают: лидерные последовательности пикорнавирусов, например, лидерную последовательность EMCV (5'-некодирующий участок вируса энцефаломиокардита) (Elroy-Stein, et al., (1989) Proc. Natl. Acad. Sci. USA 86:6126-6130); лидерные последовательности потивирусов, например, лидерную последовательность TEV (вирус гравировки табака) (Gallie, et al., (1995) Gene 165(2):233-238), лидерную последовательность MDMV (вирус карликовой мозаики кукурузы), белок, связывающий тяжелую цепь иммуноглобулина человека (BiP) (Macejak, et al., (1991) Nature 353:90-94); нетранслируемую лидерную последовательность из мРНК белка оболочки вируса мозаики люцерны (AMV RNA 4) (Jobling, et al., (1987) Nature 325:622-625); лидерную последовательность вируса табачной мозаики (TMV) (Gallie, et al., (1989) в Molecular Biology of RNA, ed. Cech (Liss, New York), pp. 237-256) и лидерную последовательность вируса хлорозной мозаичности маиса (MCMV) (Lommel, et al., (1991) Virology 81:382-385). См. также Della Cioppa et al. (1987) Plant Physiology 84:965 968. Такие конструкции могут также содержать "сигнальную последовательность" или "лидерную последовательность" для облегчения сопряженного с трансляцией или посттрансляционного транспорта пептида в определенные внутриклеточные структуры, такие как хлоропласт (или другая пластида), эндоплазматический ретикулум или аппарат Гольджи.
Применяемая в данном документе "сигнальная последовательность" относится к последовательности, которая, как известно или как ожидается, приводит к сопряженному с трансляцией или посттрансляционному транспорту пептида через клеточную мембрану. У эукариот это, как правило, подразумевает секрецию в пузырьках аппарата Гольджи, при этом происходит определенное гликолизирование. Инсектицидные токсины бактерий зачастую синтезируются в виде протоксинов, которые активируются под действием протеолиза в кишечнике целевого вредителя (Chang, (1987) Methods Enzymol. 153:507-516). В некоторых вариантах осуществления сигнальная последовательность расположена в нативной последовательности или может быть получена из последовательности согласно вариантам осуществления. Применяемая в данном документе "лидерная последовательность" относится к любой последовательности, которая при трансляции приводит к аминокислотной последовательности, способной запускать сопряженный с трансляцией транспорт пептидной цепи во внутриклеточную органеллу. Таким образом, этот термин включает лидерные последовательности, нацеливающие транспорт и/или гликозилирование путем перехода в эндоплазматический ретикулум, перехода в вакуоли, пластиды, в том числе хлоропласты, митохондрии и т. п. Кодируемые в ядре белки, нацеленные в компартмент полости тилакоида хлоропластов, имеют характерный двойной транзитный пептид, состоящий из сигнального пептида, нацеливающего на строму, и сигнального пептида, нацеливающего на полость. Информация для нацеливания на строму находится в ближайшей к амино-концу части транзитного пептида. Сигнальный пептид, нацеливающий на полость, находится в ближайшей к карбоксильному концу части транзитного пептида и содержит всю информацию для нацеливания на полость. Последние исследования по протеомике хлоропластов высших растений добились успеха в идентификации многочисленных кодируемых в ядре белков полости (Kieselbach et al. FEBS LETT 480:271-276, 2000; Peltier et al. Plant Cell 12:319-341, 2000; Bricker et al. Biochim. Biophys Acta 1503:350-356, 2001), при этом их сигнальный пептид, нацеливающий на полость, можно потенциально применять в соответствии с настоящим раскрытием. О приблизительно 80 белках из Arabidopsis, а также гомологичных белках из шпината и гороха огородного сообщается в Kieselbach et al., Photosynthesis Research, 78:249-264, 2003. В частности, в таблице 2 этой публикации, которая включена в настоящее описание посредством ссылки, раскрыто 85 белков из полости хлоропласта, идентифицированных по их № доступа (см. также публикацию заявки на патент США 2009/09044298). В дополнение, опубликованная недавно предварительная версия генома риса (Goff et al, Science 296:92-100, 2002) является подходящим источником для информации о сигнальном пептиде, нацеливающим на полость, который можно применять в соответствии с настоящим раскрытием.
Подходящие транзитные пептиды хлоропластов (CTP), хорошо известные специалисту в данной области, также включают химерные CTP, содержащие без ограничения N-терминальный домен, центральный домен или C-терминальный домен CTP из 1-дезокси-D ксилоза-5-фосфатсинтазы Oryza sativa, супероксиддисмутазы Oryza sativa, синтазы растворимого крахмала Oryza sativa, NADP-зависимого фермента, действующего на яблочную кислоту, Oryza sativa, фосфо-2-дегидро-3-дезоксигептонатальдолазы 2 Oryza sativa, L-аскорбатпероксидазы 5 Oryza sativa, фосфоглюкан-вода-дикиназы Oryza sativa, ssRUBISCO Zea Mays, бета-глюкозидазы Zea Mays, малатдегидрогеназы Zea Mays, тиоредоксина M-типа Zea Mays из патентной публикации США 2012/0304336).
Ген полипептида PtIP-83, подлежащего нацеливанию в хлоропласт, можно оптимизировать для экспрессии в хлоропласте с учетом отличий по использованию кодонов между растительным ядром и этой органеллой. Таким образом, нуклеиновые кислоты, представляющие интерес, можно синтезировать с применением кодонов, предпочтительных для хлоропласта. См., например, патент США № 5380831, включенный в данный документ посредством ссылки.
При получении кассеты экспрессии с различными фрагментами ДНК можно проводить манипуляции так, чтобы получить последовательности ДНК в надлежащей ориентации и, при необходимости, в надлежащей рамке считывания. С этой целью, для соединения фрагментов ДНК можно использовать адаптеры или линкеры, или можно задействовать другие манипуляции для обеспечения подходящих сайтов рестрикции, удаления избыточной ДНК, удаления сайтов рестрикции и т. п. С этой целью можно задействовать мутагенез in vitro, репарацию с помощью праймеров, рестрикцию, гибридизацию, повторные замены, например, транзиции и трансверсии.
При практическом осуществлении вариантов осуществления можно применять ряд промоторов. Промоторы можно выбирать, исходя из требуемого результата. Нуклеиновые кислоты можно комбинировать с конститутивными, предпочтительными для ткани, индуцируемыми или другими промоторами для экспрессии в организме-хозяине. Подходящие конститутивные промоторы для применения в растительной клетке-хозяине включают, например, коровый промотор промотора Rsyn7 и другие конститутивные промоторы, раскрытые в WO 1999/43838 и патенте США № 6072050; коровый промотор CaMV 35S (Odell, et al., (1985) Nature 313:810-812); промотор актина риса (McElroy, et al., (1990) Plant Cell 2:163-171); убиквитина (Christensen, et al., (1989) Plant Mol. Biol. 12:619-632 и Christensen, et al., (1992) Plant Mol. Biol. 18:675-689); pEMU (Last, et al., (1991) Theor. Appl. Genet. 81:581-588); MAS (Velten, et al., (1984) EMBO J. 3:2723-2730); промотор гена ALS (патент США № 5659026) и т.п. Другие конститутивные промоторы включают, например, раскрытые в патентах США №№ 5608149; 5608144; 5604121; 5569597; 5466785; 5399680; 5268463; 5608142 и 6177611.
В зависимости от требуемого результата, может быть полезно экспрессировать ген при помощи индуцируемого промотора. Особый интерес для регуляции экспрессии нуклеотидных последовательностей согласно вариантам осуществления у растений представляют индуцируемые ранением промоторы. Такие индуцируемые ранением промоторы могут реагировать на повреждение, вызванное питанием насекомого, и они включают ген ингибитора протеиназы картофеля (pin II) (Ryan, (1990) Ann. Rev. Phytopath. 28:425-449; Duan, et al., (1996) Nature Biotechnology 14:494-498); wun1 и wun2, патент США № 5428148; win1 и win2 (Stanford, et al., (1989) Mol. Gen. Genet. 215:200-208); системин (McGurl, et al., (1992) Science 225:1570-1573); WIP1 (Rohmeier, et al., (1993) Plant Mol. Biol. 22:783-792; Eckelkamp, et al., (1993) FEBS Letters 323:73-76); ген MPI (Corderok, et al., (1994) Plant J. 6(2):141-150) и т.п., включенные в данный документ посредством ссылки.
Кроме того, в способах и нуклеотидных конструкциях согласно вариантам осуществления можно использовать индуцируемые патогеном промоторы. Такие индуцируемые патогеном промоторы включают промоторы из белков, связанных с патогенезом (PR-белки), которые индуцируются после заражения патогеном; например, PR-белки, SAR-белки, бета-1,3-глюканаза, хитиназа и т. д. См., например, Redolfi, et al., (1983) Neth. J. Plant Pathol. 89:245-254; Uknes, et al., (1992) Plant Cell 4: 645-656 и Van Loon, (1985) Plant Mol. Virol. 4:111-116. См. также WO 1999/43819, включенную в данный документ посредством ссылки.
Представляют интерес промоторы, которые экспрессируются локально в месте заражения патогеном или рядом с ним. См., например, Marineau, et al., (1987) Plant Mol. Biol. 9:335-342; Matton, et al., (1989) Molecular Plant-Microbe Interactions 2:325-331; Somsisch, et al., (1986) Proc. Natl. Acad. Sci. USA 83:2427-2430; Somsisch, et al., (1988) Mol. Gen. Genet. 2:93-98 и Yang, (1996) Proc. Natl. Acad. Sci. USA 93:14972-14977. См. также, Chen, et al., (1996) Plant J. 10:955-966; Zhang, et al., (1994) Proc. Natl. Acad. Sci. USA 91:2507-2511; Warner, et al., (1993) Plant J. 3:191-201; Siebertz, et al., (1989) Plant Cell 1:961-968; патент США № 5750386 (индуцируемый нематодами), а также источники, цитируемые в этих документах. Особый интерес представляет индуцируемый промотор для гена маиса PRms, экспрессия которого индуцируется патогеном Fusarium moniliforme (см, например, Cordero, et al., (1992) Physiol. Mol. Plant Path. 41:189-200).
Регулируемые химическими веществами промоторы можно применять для модуляции экспрессии гена в растении посредством применения экзогенного химического регулятора. В зависимости от цели промотор может представлять собой индуцируемый химическим веществом промотор, при этом применение химического вещества индуцирует экспрессию гена, или репрессируемый химическим веществом промотор, при этом применение химического вещества подавляет экспрессию гена. Индуцируемые химическими веществами промоторы известны из уровня техники и включают без ограничения промотор In2-2 маиса, который активируется антидотами к бензолсульфонамидным гербицидам, промотор GST маиса, который активируется гидрофобными электрофильными соединениями, которые применяются в качестве предвсходовых гербицидов, и промотор PR-1a табака, который активируется салициловой кислотой. Другие регулируемые химическими веществами промоторы, представляющие интерес, включают чувствительные к стероидам промоторы (см., например, индуцируемый глюкокортикоидом промотор в Schena, et al., (1991) Proc. Natl. Acad. Sci. USA 88:10421-10425 и McNellis, et al., (1998) Plant J. 14(2):247-257), а также индуцируемые тетрациклинами и репрессируемые тетрациклинами промоторы (см., например, Gatz, et al., (1991) Mol. Gen. Genet. 227:229-237 и патенты США №№ 5814618 и 5789156), включенные в данный документ посредством ссылки.
Для целенаправленного воздействия на усиленную экспрессию полипептида PtIP-83 в конкретной ткани растения можно использовать предпочтительные для ткани промоторы. Предпочтительные для ткани промоторы включают промоторы, обсуждаемые в Yamamoto, et al., (1997) Plant J. 12(2)255-265; Kawamata, et al., (1997) Plant Cell Physiol. 38(7):792-803; Hansen, et al., (1997) Mol. Gen Genet. 254(3):337-343; Russell, et al., (1997) Transgenic Res. 6(2):157-168; Rinehart, et al., (1996) Plant Physiol. 112(3):1331-1341; Van Camp, et al., (1996) Plant Physiol. 112(2):525-535; Canevascini, et al., (1996) Plant Physiol. 112(2):513-524; Yamamoto, et al., (1994) Plant Cell Physiol. 35(5):773-778; Lam, (1994) Results Probl. Cell Differ. 20:181-196; Orozco, et al., (1993) Plant Mol Biol. 23(6):1129-1138; Matsuoka, et al., (1993) Proc Natl. Acad. Sci. USA 90(20):9586-9590 и Guevara-Garcia, et al., (1993) Plant J. 4(3):495-505. Такие промоторы можно модифицировать, при необходимости, для слабой экспрессии.
Из уровня техники известны предпочтительные для листьев промоторы. См., например, Yamamoto, et al., (1997) Plant J. 12(2):255-265; Kwon, et al., (1994) Plant Physiol. 105:357-67; Yamamoto, et al., (1994) Plant Cell Physiol. 35(5):773-778; Gotor, et al., (1993) Plant J. 3:509-18; Orozco, et al., (1993) Plant Mol. Biol. 23(6):1129-1138 и Matsuoka, et al., (1993) Proc. Natl. Acad. Sci. USA 90(20):9586-9590.
Известны промоторы, предпочтительные для корней или специфические в отношении корней, и их можно выбирать из многих доступных из литературы или выделенных de novo из различных совместимых видов. См., например, Hire, et al., (1992) Plant Mol. Biol. 20(2):207-218 (soybean root-specific glutamine synthetase gene); Keller и Baumgartner, (1991) Plant Cell 3(10):1051-1061 (root-specific control element in the GRP 1,8 gene of French bean); Sanger, et al., (1990) Plant Mol. Biol. 14(3):433-443 (root-specific promoter of the mannopine synthase (MAS) gene of Agrobacterium tumefaciens) and Miao, et al., (1991) Plant Cell 3(1):11-22 (full-length cDNA clone encoding cytosolic glutamine synthetase (GS), which is expressed in roots and root nodules of soybean). См. также Bogusz et al. (1990) Plant Cell 2(7):633-641, где описаны два специфические в отношении корней промотора, выделенные из генов гемоглобина из азотфиксирующего растения Parasponia andersonii, не относящегося к бобовым, и родственного растения Trema tomentosa, не относящегося к бобовым и не являющегося азотфиксирующим. Промоторы этих генов были связаны с репортерным геном β-глюкуронидазы и введены как в Nicotiana tabacum, не относящееся к бобовым, так и в бобовое Lotus corniculatus, и при этом в обоих случаях специфическая в отношении корней промоторная активность сохранялась. Leach and Aoyagi (1991) описывают свой анализ промоторов, обеспечивающих высокий уровень экспрессии генов roIC и roID Agrobacterium rhizogenes, индуцирующих разрастание корней (см. Plant Science (Limerick) 79(1):69-76). Авторы пришли к выводу, что в этих промоторах энхансер и предпочтительные для ткани ДНК-детерминанты разделены. Teeri et al. (1989) применяли слияние гена с lacZ для того, чтобы показать, что ген из T-ДНК Agrobacterium, кодирующий октопинсинтазу, является особенно активным в эпидермисе кончика корня, и что ген TR2' является специфическим в отношении корней в интактном растении и стимулируется при ранении в ткани листа, что является особенно целесообразной комбинацией характеристик для применения с инсектицидным или ларвицидным геном (смотрите EMBO J. 8(2):343-350). Ген TR1', слитый с nptII (неомицинфосфотрансфераза II), продемонстрировал аналогичные характеристики. Дополнительные предпочтительные для корней промоторы включают промотор гена VfENOD-GRP3 (Kuster, et al., (1995) Plant Mol. Biol. 29(4):759-772) и промотор rolB (Capana, et al., (1994) Plant Mol. Biol. 25(4):681-691. См. также патенты США №№ 5837876; 5750386; 5633363; 5459252; 5401836; 5110732 и 5023179. Предпочтительные для корней регуляторные последовательности Arabidopsis thaliana раскрыты в US20130117883.
"Предпочтительные для семени" промоторы включают как промоторы, "специфические в отношении семени" (промоторы, которые активны во время развития семени, такие как промоторы запасных белков семени), так и промоторы "прорастания семени" (промоторы, которые активны во время прорастания семени). См. Thompson, et al., (1989) BioEssays 10:108, включенную в настоящий документ посредством ссылки. Такие предпочтительные для семени промоторы включают без ограничения Cim1 (индуцируемый цитокинином транскрипт); cZ19B1 (19 кДа зеин маиса) и milps (мио-инозитол-1-фосфатсинтаза); (см. патент США № 6225529, включенный в настоящий документ посредством ссылки). Гамма-зеин и Glb-1 представляют собой промоторы, специфические в отношении эндосперма. В случае двудольных растений, промоторы, специфические в отношении семени, включают без ограничения промотор гена ингибитора трипсина Кунитца 3 (KTi3) (Jofuku and Goldberg, (1989) Plant Cell 1:1079-1093), β-фазеолина бобов, напина, β-конглицинина, глицинина 1, лектина сои, круциферина и т. п. В случае однодольных растений, промоторы, специфические в отношении семени, включают без ограничения промоторы из генов 15 кДа зеина маиса, 22 кДа зеина, 27 кДа зеина, g-зеина, waxy, shrunken 1, shrunken 2, глобулина 1 и т. д. См. также WO 2000/12733, где раскрыты предпочтительные для семени промоторы из генов end1 и end2, включенную в данный документ посредством ссылки. У двудольных растений, промоторы, специфические в отношении семени, включают без ограничения промотор гена белков семенной оболочки из Arabidopsis, pBAN; а также промоторы генов раннего развития семян из Arabidopsis, p26, p63 и p63tr (патенты США №№ 7294760 и 7847153). Промотор, который характеризуется "предпочтительной" экспрессией в конкретной ткани, экспрессируется в такой ткани в большей степени, чем по меньшей мере в одной другой растительной ткани. Для некоторых предпочтительных для ткани промоторов показана экспрессия почти исключительно в конкретной ткани.
Если требуется низкий уровень экспрессии, будут применяться слабые промоторы. Как правило, применяемый в данном документе термин "слабый промотор" относится к промотору, который управляет экспрессией кодирующей последовательности на низком уровне. Под низким уровнем экспрессии подразумевают уровни от приблизительно 1/1000 транскриптов до приблизительно 1/100000 транскриптов, до приблизительно 1/500000 транскриптов. В качестве альтернативы, понятно, что термин "слабые промоторы" также охватывает промоторы, которые управляют экспрессией только в небольшом количестве клеток и не управляют в других, при этом обеспечивается общий низкий уровень экспрессии. Если промотор управляет экспрессией с неприемлемо высокими уровнями, части промоторной последовательности можно удалить или модифицировать для снижения уровней экспрессии.
Такие слабые конститутивные промоторы включают, например, коровый промотор промотора Rsyn7 (WO 1999/43838 и патент США № 6072050), коровый промотор 35S CaMV и т. п. Другие конститутивные промоторы включают, например, раскрытые в патентах США №№ 5608149; 5608144; 5604121; 5569597; 5466785; 5399680; 5268463; 5608142 и 6177611, включенных в данный документ посредством ссылки.
Приведенный выше перечень промоторов не предназначен для ограничения. В вариантах осуществления можно применять любой подходящий промотор.
Как правило, кассета экспрессии будет содержать ген селектируемого маркера для отбора трансформированных клеток. Гены селектируемых маркеров используют для отбора трансформированных клеток или тканей. Гены маркеров включают гены, кодирующие устойчивость к антибиотику, такие как гены, кодирующие неомицинфосфотрансферазу II (NEO) и гигромицинфосфотрансферазу (HPT), а также гены, обеспечивающие устойчивость к гербицидным соединениям, таким как глуфосинат аммония, бромоксинил, имидазолиноны и 2,4-дихлорфеноксиацетат (2,4-D). Дополнительные примеры генов подходящих селектируемых маркеров включают без ограничения гены, кодирующие устойчивость к хлорамфениколу (Herrera Estrella, et al., (1983) EMBO J. 2:987-992); метотрексату (Herrera Estrella, et al., (1983) Nature 303:209-213 и Meijer, et al., (1991) Plant Mol. Biol. 16:807-820); стрептомицину (Jones, et al., (1987) Mol. Gen. Genet. 210:86-91); спектиномицину (Bretagne-Sagnard, et al., (1996) Transgenic Res. 5:131-137); блеомицину (Hille, et al., (1990) Plant Mol. Biol. 7:171-176); сульфонамиду (Guerineau, et al., (1990) Plant Mol. Biol. 15:127-136); бромоксинилу (Stalker, et al., (1988) Science 242:419-423); глифосату (Shaw, et al., (1986) Science 233:478-481 и заявки на патент США с серийными №№ 10/004357 и 10/427692); фосфинотрицин (DeBlock, et al., (1987) EMBO J. 6:2513-2518). Cм., в целом, Yarranton, (1992) Curr. Opin. Biotech. 3:506-511; Christopherson, et al., (1992) Proc. Natl. Acad. Sci. USA 89:6314-6318; Yao, et al., (1992) Cell 71:63-72; Reznikoff, (1992) Mol. Microbiol. 6:2419-2422; Barkley, et al., (1980) в The Operon, pp. 177-220; Hu, et al., (1987) Cell 48:555-566; Brown, et al., (1987) Cell 49:603-612; Figge, et al., (1988) Cell 52:713-722; Deuschle, et al., (1989) Proc. Natl. Acad. Sci. USA 86:5400-5404; Fuerst, et al., (1989) Proc. Natl. Acad. Sci. USA 86:2549-2553; Deuschle, et al., (1990) Science 248:480-483; Gossen, (1993) Ph.D. Thesis, University of Heidelberg; Reines, et al., (1993) Proc. Natl. Acad. Sci. USA 90:1917-1921; Labow, et al., (1990) Mol. Cell. Biol. 10:3343-3356; Zambretti, et al., (1992) Proc. Natl. Acad. Sci. USA 89:3952-3956; Baim, et al., (1991) Proc. Natl. Acad. Sci. USA 88:5072-5076; Wyborski, et al., (1991) Nucleic Acids Res. 19:4647-4653; Hillenand-Wissman, (1989) Topics Mol. Struc. Biol. 10:143-162; Degenkolb, et al., (1991) Antimicrob. Agents Chemother. 35:1591-1595; Kleinschnidt, et al., (1988) Biochemistry 27:1094-1104; Bonin, (1993) Ph.D. Thesis, University of Heidelberg; Gossen, et al., (1992) Proc. Natl. Acad. Sci. USA 89:5547-5551; Oliva, et al., (1992) Antimicrob. Agents Chemother. 36:913-919; Hlavka, et al., (1985) Handbook of Experimental Pharmacology, Vol. 78 (Springer-Verlag, Berlin) и Gill, et al., (1988) Nature 334:721-724. Такие раскрытия включены в данный документ посредством ссылки.
Приведенный выше перечень генов селектируемых маркеров не предназначен для ограничения. Любой ген селектируемого маркера можно применять в вариантах осуществления.
Трансформация растений
Способы согласно вариантам осуществления включают введение полипептида или полинуклеотида в растение. Применяемый в данном документе термин "введение" означает представление растению полинуклеотида или полипептида таким образом, что последовательность попадает внутрь клетки растения. Способы согласно вариантам осуществления не зависят от конкретного способа введения полинуклеотида или полипептида в растение, необходимо только, чтобы полинуклеотид или полипептиды получили доступ внутрь по меньшей мере одной клетки растения. Из уровня техники известны способы введения полинуклеотида или полипептидов в растения, в том числе без ограничений способы стабильной трансформации, способы временной трансформации и способы трансформации, опосредованной вирусами.
Применяемый в данном документе термин "стабильная трансформация" означает, что нуклеотидная конструкция, введенная в растение, интегрируется в геном растения и может наследоваться его потомством. Применяемый в данном документе термин "временная трансформация" означает, что полинуклеотид вводится в растение и не интегрируется в геном растения, или в растение вводится полипептид. Применяемый в данном документе термин "растение" относится к целым растениям, органам растения (например, листьям, стеблям, корням и т. д.), семенам, растительным клеткам, частям растения для вегетативного размножения, зародышам и их потомству. Растительные клетки могут быть дифференцированными или недифференцированными (например, клетками каллюса, клетками суспензионных культур, протопластами, клетками листьев, клетками корней, клетками флоэмы и пыльцой).
Протоколы трансформации, а также протоколы для введения нуклеотидных последовательностей в растения могут изменяться в зависимости от типа растения или растительной клетки, т.е. однодольного или двудольного растения, которые планируется подвергнуть трансформации. Подходящие способы введения нуклеотидных последовательностей в клетки растений и последующей вставки в геном растения включают микроинъекцию (Crossway, et al., (1986) Biotechniques 4:320-334), электропорацию (Riggs, et al., (1986) Proc. Natl. Acad. Sci. USA 83:5602-5606), трансформацию, опосредованную Agrobacterium (патенты США №№ 5563055 и 5981840), прямой перенос гена (Paszkowski, et al., (1984) EMBO J. 3:2717-2722) и баллистическое ускорение частиц (см., например, патенты США №№ 4945050; 5879918; 5886244 и 5932782; Tomes, et al., (1995) в Plant Cell, Tissue, and Organ Culture: Fundamental Methods, ed. Gamborg and Phillips, (Springer-Verlag, Berlin) и McCabe, et al., (1988) Biotechnology 6:923-926) и трансформацию гена Lecl (WO 00/28058). Что касается трансформации картофеля, см. Tu, et al., (1998) Plant Molecular Biology 37:829-838 и Chong, et al., (2000) Transgenic Research 9:71-78. Дополнительные процедуры трансформации можно найти у Weissinger, et al., (1988) Ann. Rev. Genet. 22:421-477; Sanford, et al., (1987) Particulate Science and Technology 5:27-37 (лук); Christou, et al., (1988) Plant Physiol. 87:671-674 (соя); McCabe, et al., (1988) Bio/Technology 6:923-926 (соя); Finer and McMullen, (1991) In Vitro Cell Dev. Biol. 27P:175-182 (соя); Singh, et al., (1998) Theor. Appl. Genet. 96:319-324 (соя); Datta, et al., (1990) Biotechnology 8:736-740 (рис); Klein, et al., (1988) Proc. Natl. Acad. Sci. USA 85:4305-4309 (маис); Klein, et al., (1988) Biotechnology 6:559-563 (маис); патенты США №№ 5240855; 5322783 и 5324646; Klein, et al., (1988) Plant Physiol. 91:440-444 (маис); Fromm, et al., (1990) Biotechnology 8:833-839 (маис); Hooykaas-Van Slogteren, et al., (1984) Nature (London) 311:763-764; патент США № 5,736,369 (злаки); Bytebier, et al., (1987) Proc. Natl. Acad. Sci. USA 84:5345-5349 (Liliaceae); De Wet, et al., (1985) в The Experimental Manipulation of Ovule Tissues, ed. Chapman, et al., (Longman, New York), pp. 197-209 (пыльца); Kaeppler, et al., (1990) Plant Cell Reports 9:415-418 и Kaeppler, et al., (1992) Theor. Appl. Genet. 84:560-566 (трансформация, опосредованная вискерами); D'Halluin, et al., (1992) Plant Cell 4:1495-1505 (электропорация); Li, et al., (1993) Plant Cell Reports 12:250-255 и Christou and Ford, (1995) Annals of Botany 75:407-413 (рис); Osjoda, et al., (1996) Nature Biotechnology 14:745-750 (трансформация маиса с помощью Agrobacterium tumefaciens); все из которых включены в данный документ посредством ссылки.
В конкретных вариантах осуществления последовательности согласно вариантам осуществления можно вводить в растение с применением ряда способов временной трансформации. Такие способы временной трансформации включают без ограничения введение полинуклеотида PtIP-83 или его вариантов и фрагментов непосредственно в растение или введение транскрипта полипептида PtIP-83 в растение. Такие способы включают, например, микроинъекцию или бомбардировку частицами. См, например, Crossway, et al., (1986) Mol Gen. Genet. 202:179-185; Nomura, et al., (1986) Plant Sci. 44:53-58; Hepler, et al., (1994) Proc. Natl. Acad. Sci. 91:2176-2180 и Hush, et al., (1994) The Journal of Cell Science 107:775-784, все из которых включены посредством ссылки. В качестве альтернативы, растение можно временно трансформировать с помощью полинуклеотида для полипептида PtIP-83 с применением методик, известных из уровня техники. Такие методики включают применение вирусной векторной системы и осаждение полинуклеотида таким способом, который исключает последующее высвобождение ДНК. Таким образом, может происходить транскрипция ДНК, связанной с частицами, но частота, с которой она высвобождается для интеграции в геном, в значительной степени снижена. Такие способы включают применение частиц, покрытых полиэтиленимином (PEI; Sigma, кат. № P3143).
Из уровня техники известны способы нацеленной вставки полинуклеотида в конкретное местоположение в геноме растения. В одном варианте осуществления вставку полинуклеотида в требуемое местоположение в геноме выполняют с использованием системы сайт-специфической рекомбинации. См., например, WO 1999/25821, WO 1999/25854, WO 1999/25840, WO 1999/25855 и WO 1999/25853, все из которых включены в данный документ посредством ссылки. Вкратце, полинуклеотид согласно вариантам осуществления может содержаться в кассете для переноса, фланкированной двумя неидентичными сайтами рекомбинации. Кассету для переноса вводят в растение, имеющее в своем геноме стабильно встроенный целевой сайт, который фланкирован двумя неидентичными сайтами рекомбинации, которые соответствуют сайтам кассеты для переноса. Обеспечивают подходящую рекомбиназу, и кассета для переноса интегрируется в целевой сайт. Полинуклеотид, представляющий интерес, таким образом интегрируется в определенное хромосомное положение в геноме растения.
Векторы для трансформации растений могут состоять из одного или нескольких ДНК-векторов, необходимых для достижения трансформации растения. Например, общепринятой практикой в уровне техники является использование векторов для трансформации растений, которые состоят из двух и более смежных сегментов ДНК. В уровне техники эти векторы зачастую называют "бинарными векторами". Бинарные векторы, а также векторы с плазмидами-помощниками наиболее часто применяются при трансформации, опосредованной Agrobacterium, где размер и сложность сегментов ДНК, необходимых для достижения эффективной трансформации, являются очень большими, и предпочтительным является разделение функций на отдельных молекулах ДНК. Бинарные векторы, как правило, содержат плазмидный вектор, который содержит действующие в цис-положении последовательности, требуемые для переноса Т-ДНК (как, например, левой границы и правой границы), селектируемый маркер, который разрабатывают таким образом, что он может экспрессироваться в растительной клетке, и "ген, представляющий интерес" (ген, разработанный таким образом, что он может экспрессироваться в растительной клетке, из которой требуется получить трансгенные растения). Также в этом плазмидном векторе присутствуют последовательности, требуемые для репликации в бактериях. Действующие в цис-положении последовательности расположены так, чтобы обеспечить возможность эффективного переноса в растительные клетки и экспрессии в них. Например, ген селектируемого маркера и пестицидный ген расположены между левой и правой границами. Часто второй плазмидный вектор содержит действующие в транс-положении факторы, которые опосредуют перенос T-ДНК из Agrobacterium в растительные клетки. Эта плазмида часто содержит функции вирулентности (гены Vir), что обеспечивает возможность заражения растительных клеток Agrobacterium и перенос ДНК путем расщепления по граничным последовательностям и переноса ДНК, опосредованного vir, как понятно из уровня техники (Hellens and Mullineaux, (2000) Trends in Plant Science 5:446-451). Несколько типов штаммов Agrobacterium (например, LBA4404, GV3101, EHA101, EHA105 и т.п.) можно применять для трансформации растений. Второй плазмидный вектор не является необходимым для трансформации растений другими способами, такими как бомбардировка микрочастицами, микроинъекция, электропорация, с помощью полиэтиленгликоля и т.д.
В целом, способы трансформации растений включают перенос гетерологичной ДНК в целевые растительные клетки (например, незрелые или зрелые зародыши, суспензионные культуры, недифференцированный каллюс, протопласты и т.д.) с последующим применением соответствующего отбора с максимальным пороговым значением (в зависимости от гена селектируемого маркера) для выделения трансформированных растительных клеток из группы нетрансформированной клеточной массы. После интеграции гетерологичной чужеродной ДНК в растительные клетки затем применяют соответствующий отбор с максимальным пороговым значением в среде для уничтожения нетрансформированных клеток и отделения и размножения предположительно трансформированных клеток, которые переживают эту обработку с отбором, посредством регулярного переноса на свежую среду. При помощи продолжающегося пассирования и проверки при помощи соответствующего отбора идентифицируют и размножают клетки, которые трансформированы плазмидным вектором. Затем можно применять молекулярные и биохимические способы для подтверждения присутствия интегрированного гетерологичного гена, представляющего интерес, в геноме трансгенного растения.
Эксплантаты, как правило, переносят на свежую порцию той же среды и культивируют обычным способом. Впоследствии трансформированные клетки дифференцируются в побеги после помещения в среду для регенерации, дополненную средством отбора с максимальным пороговым значением. Побеги затем переносят на селективную среду для выращивания растений с получением укоренившихся побегов или саженцев. Затем трансгенный саженец выращивают до зрелого растения и получают фертильные семена (например, Hiei, et al., (1994) Plant Journal 6:271-282; Ishida, et al., (1996) Nature Biotechnology 14:745-750). Эксплантаты, как правило, переносят на свежую порцию той же среды и культивируют обычным способом. Общее описание методик и способов получения трансгенных растений приведены в Ayres and Park, (1994) Critical Reviews in Plant Science 13:219-239 и Bommineni and Jauhar, (1997) Maydica 42:107-120. Поскольку трансформированный материал содержит множество клеток, как трансформированные, так и нетрансформированные клетки присутствуют в любой части подвергнутого воздействию целевого каллюса, или ткани, или группы клеток. Возможность уничтожать нетрансформированные клетки и обеспечивать размножение трансформированных клеток приводит в результате к трансформированным растительным культурам. Зачастую, возможность удаления нетрансформированных клеток является ограничивающим фактором для быстрого выделения трансформированных растительных клеток и успешного выращивания трансгенных растений.
Из клеток, которые были трансформированы, можно вырастить растения в соответствии с традиционными способами. См., например, McCormick, et al., (1986) Plant Cell Reports 5:81-84. Эти растения затем можно выращивать и опылять либо с использованием такого же трансформированного штамма, либо другого штамма и идентифицировать полученный гибрид с конститутивной или индуцируемой экспрессией требуемой фенотипической характеристики. Можно вырастить два или более поколений для того, чтобы убедиться в том, что экспрессия требуемой фенотипической характеристики стабильно поддерживается и наследуется, а затем собрать семена, чтобы убедиться в том, что экспрессия требуемой фенотипической характеристики была достигнута.
Нуклеотидные последовательности согласно вариантам осуществления можно вводить в растение путем приведения растения в контакт с вирусом или вирусными нуклеиновыми кислотами. Как правило, такие способы включают встраивание нуклеотидной конструкции, представляющей интерес, в молекулу вирусной ДНК или РНК. Понятно, что рекомбинантные белки согласно вариантам осуществления можно первоначально синтезировать как часть вирусного полипротеина, который позже может быть процессирован путем протеолиза in vivo или in vitro с получением требуемого полипептида PtIP-83. Также понятно, что такой вирусный полипротеин, содержащий по меньшей мере часть аминокислотной последовательности PtIP-83 согласно вариантам осуществления, может обладать требуемой пестицидной активностью. Такие вирусные полипротеины и нуклеотидные последовательности, которые их кодируют, охватываются вариантами осуществления. Способы введения в растения нуклеотидных конструкций и получения кодируемых белков в растениях, которые включают молекулы вирусной ДНК или РНК, известны из уровня техники. См., например, патенты США №№ 5889191; 5889190; 5866785; 5589367 и 5316931; включенные в данный документ посредством ссылки.
Способы трансформации хлоропластов известны из уровня техники. См., например, Svab, et al., (1990) Proc. Natl. Acad. Sci. USA 87:8526-8530; Svab and Maliga, (1993) Proc. Natl. Acad. Sci. USA 90:913-917; Svab and Maliga, (1993) EMBO J. 12:601-606. Способ основан на доставке генной пушкой ДНК, содержащей селектируемый маркер, и нацеливании ДНК на геном пластид с помощью гомологичной рекомбинации. Кроме того, трансформацию пластид можно осуществлять трансактивацией молчащего трансгена, находящегося в пластидах, путем осуществляемой преимущественно в определенной ткани экспрессии РНК-полимеразы, кодируемой ядерным геномом и функционирующей в пластиде. О такой системе сообщали в McBride, et al., (1994) Proc. Natl. Acad. Sci. USA 91:7301-7305.
Варианты осуществления дополнительно относятся к материалу для размножения трансформированного растения согласно вариантам осуществления, в том числе без ограничения семенам, клубням, клубнелуковицам, луковицам, листьям и черенкам корней и побегов.
Варианты осуществления можно применять для трансформации любых видов растений, в том числе без ограничения однодольных и двудольных. Примеры растений, представляющих интерес, включают без ограничений кукурузу (Zea mays), Brassica sp. (например, B. napus, B. rapa, B. juncea), в частности виды Brassica, пригодные в качестве источников масла из семян растений, люцерну (Medicago sativa), рис (Oryza sativa), рожь (Secale cereale), сорго (Sorghum bicolor, Sorghum vulgare), просо (например, пеннисетум рогозовидный (Pennisetum glaucum), просо культурное (Panicum miliaceum), щетинник итальянский (Setaria italica), просо пальчатое (Eleusine coracana)), подсолнечник (Helianthus annuus), сафлор (Carthamus tinctorius), пшеницу (Triticum aestivum), сою (Glycine max), табак (Nicotiana tabacum), картофель (Solanum tuberosum), разновидности арахиса (Arachis hypogaea), хлопчатник (Gossypium barbadense, Gossypium hirsutum), сладкий картофель (Ipomoea batatus), маниок (Manihot esculenta), кофейное дерево (Coffea spp.), кокосовую пальму (Cocos nucifera), ананас (Ananas comosus), цитрусовые деревья (Citrus spp.), шоколадное дерево (Theobroma cacao), чайный куст (Camellia sinensis), банан (Musa spp.), авокадо (Persea americana), инжир (Ficus casica), гуаву (Psidium guajava), манго (Mangifera indica), маслину (Olea europaea), папайю (Carica papaya), кешью (Anacardium occidentale), макадамию (Macadamia integrifolia), миндаль (Prunus amygdalus), разновидности сахарной свеклы (Beta vulgaris), сахарный тростник (Saccharum spp.), разновидности овса, ячмень, овощи, декоративные растения и хвойные деревья.
Овощи включают томаты (Lycopersicon esculentum), латук (например, Lactuca sativa), зеленую фасоль (Phaseolus vulgaris), лимскую фасоль (Phaseolus limensis), горох (Lathyrus spp.) и представителей рода Cucumis, таких как огурец (C. sativus), канталупа (C. cantalupensis) и дыня мускусная (C. melo). Декоративные растения включают азалию (Rhododendron spp.), гортензию (Macrophylla hydrangea), гибискус (Hibiscus rosasanensis), розы (Rosa spp.), тюльпаны (Tulipa spp.), нарциссы (Narcissus spp.), петунии (Petunia hybrida), гвоздику (Dianthus caryophyllus), пуансеттию (Euphorbia pulcherrima) и хризантему. Хвойные, которые можно использовать при практическом осуществлении вариантов осуществления, включают, например, виды сосны, такие как сосна ладанная (Pinus taeda), сосна Эллиота (Pinus elliotii), сосна желтая (Pinus ponderosa), сосна скрученная широкохвойная (Pinus contorta) и сосна лучистая (Pinus radiata); псевдотсугу Мензиса (Pseudotsuga menziesii); тсугу канадскую (Tsuga canadensis); ель сизую (Picea glauca); секвойю (Sequoia sempervirens); виды пихты, такие как пихта миловидная (Abies amabilis) и пихта бальзамическая (Abies balsamea), и виды туйи, такие как туя гигантская (Thuja plicata) и каллитропсис нутканский (Chamaecyparis nootkatensis). Растения согласно вариантам осуществления включают культурные растения (например, кукурузу, люцерну, подсолнечник, Brassica, сою, хлопчатник, сафлор, арахис, сорго, пшеницу, просо, табак и т.д.), такие как растения кукурузы и сои.
Разновидности газонной травы включают без ограничения: мятлик однолетний (Poa annua); райграс однолетний (Lolium multiflorum); мятлик сплюснутый (Poa compressa); овсяницу красную Чуинга (Festuca rubra); полевицу тонкую (Agrostis tenuis); полевицу болотную (Agrostis palustris); житняк пустынный (Agropyron desertorum); житняк гребенчатый (Agropyron cristatum); овсяницу длиннолистную (Festuca longifolia); мятлик луговой (Poa pratensis); ежу сборную (Dactylis glomerata); плевел многолетний (Lolium perenne); овсяницу красную (Festuca rubra); полевицу белую (Agrostis alba); мятлик обыкновенный (Poa trivialis); овсяницу овечью (Festuca ovina); костер безостый (Bromus inermis); овсяницу тростниковую (Festuca arundinacea); тимофеевку луговую (Phleum pratense); полевицу собачью (Agrostis canina); бескильницу расставленную (Puccinellia distans); пырей Смита (Agropyron smithii); свинорой пальчатый (Cynodon spp.); узкобороздник однобокий (Stenotaphrum secundatum); зойсию (Zoysia spp.); гречку заметную (Paspalum notatum); аксонопус афинский (Axonopus affinis); эремохлою змеехвостую (Eremochloa ophiuroides); кикуйю (Pennisetum clandesinum); паспалум влагалищный (Paspalum vaginatum); москитную траву (Bouteloua gracilis); бизонову траву (Buchloe dactyloids); боковой овес (Bouteloua curtipendula).
Растения, представляющие интерес, включают зерновые растения, которые дают семена, представляющие интерес, масличные растения и бобовые растения. Семена, представляющие интерес, включают семена зерновых культур, таких как кукуруза, пшеница, ячмень, рис, сорго, рожь, просо и т.д. Масличные растения включают хлопчатник, сою, сафлор, подсолнечник, Brassica, маис, люцерну, пальму, кокосовую пальму, лен, клещевину, маслину и т.д. Бобовые растения включают разновидности бобов и разновидности гороха. Бобы включают гуар, рожковое дерево, пажитник, сою, разновидности обыкновенной фасоли, вигну китайскую, золотистую фасоль, лимскую фасоль, стручковую фасоль, разновидности чечевицы, турецкий горох и т.д.
Оценка трансформации растений
После введения гетерологичной чужеродной ДНК в растительные клетки трансформацию или интеграцию гетерологичного гена в геном растения подтверждают при помощи различных способов, таких как анализ нуклеиновых кислот, белков и метаболитов, связанных с интегрированным геном.
ПЦР-анализ представляет собой быстрый способ скрининга трансформированных клеток, ткани или побегов в отношении присутствия встроенного гена на ранней стадии перед высаживанием в почву (Sambrook and Russell, (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). ПЦР проводят с применением олигонуклеотидных праймеров, специфических в отношении гена, представляющего интерес, или в отношении исходных последовательностей вектора на основе Agrobacterium и т.д.
Трансформацию растений можно подтвердить с помощью Саузерн-блот анализа геномной ДНК (Sambrook и Russell, (2001) выше). В целом, общую ДНК экстрагируют из трансформанта, разрезают соответствующими ферментами рестрикции, фракционируют в агарозном геле и переносят на нитроцеллюлозную или найлоновую мембрану. Затем мембрану или "блот" анализируют с помощью зонда, например, целевого фрагмента ДНК с радиоактивной меткой 32P для подтверждения интеграции внедренного гена в геном растения в соответствии со стандартными методиками (Sambrook и Russell, (2001) выше).
При нозерн-блот анализе РНК выделяют из специфических тканей трансформанта, фракционируют в агарозном геле, содержащем формальдегид, и переносят на найлоновый фильтр в соответствии со стандартными методиками, которые обычно применяются в уровне техники (Sambrook и Russell, (2001) выше). Экспрессию РНК, кодируемой пестицидным геном, затем тестируют при помощи гибридизации фильтра с радиоактивным зондом, полученным из пестицидного гена, посредством способов, известных из уровня техники (Sambrook и Russell, (2001) выше).
Вестерн-блот, биохимические анализы и подобные можно проводить в отношении трансгенных растений для подтверждения присутствия белка, кодируемого пестицидным геном, при помощи стандартных процедур (Sambrook и Russell, 2001, выше) с применением антител, которые связываются с одним или несколькими эпитопами, присутствующими на полипептиде PtIP-83.
Пакетирование признаков в трансгенном растении
Трансгенные растения могут содержать пакет из одного или нескольких инсектицидных полинуклеотидов, раскрытых в данном документе, с одним или несколькими дополнительными полинуклеотидами, приводящими к продукции или подавлению нескольких полипептидных последовательностей. Трансгенные растения, содержащие пакеты из полинуклеотидных последовательностей, можно получить либо при помощи традиционных способов скрещивания, либо посредством способов генной инженерии, либо применяя и то, и другое. Эти способы включают без ограничения скрещивание индивидуальных линий, при этом каждая содержит полинуклеотид, представляющий интерес, трансформацию трансгенного растения, содержащего ген, раскрытый в данном документе, дополнительным геном и котрансформацию генами одной растительной клетки. Применяемый в данном документе термин "пакетированный" включает состояние, при котором в одном растении присутствуют несколько признаков (т.е. оба признака внедрены в ядерный геном, один признак внедрен в ядерный геном и один признак внедрен в геном пластиды, или оба признака внедрены в геном пластиды). В одном неограничивающем примере "пакетированные признаки" включают в себя молекулярный пакет, в котором последовательности физически граничат друг с другом. Применяемый в данном документе термин "признак" относится к фенотипу, обусловленному конкретной последовательностью или группами последовательностей. Котрансформацию генов можно проводить с применением единичных векторов для трансформации, содержащих несколько генов, или нескольких векторов, которые несут отдельные гены. Если последовательности пакетированы с помощью генетической трансформации растений, то представляющие интерес полинуклеотидные последовательности можно комбинировать в любое время и в любом порядке. Признаки можно вводить одновременно в протоколе котрансформации с представляющими интерес полинуклеотидами, представленными любой комбинацией кассет для трансформации. Например, если будут вводиться две последовательности, то эти две последовательности могут содержаться в отдельных кассетах для трансформации (транс) или содержаться в одной кассете для трансформации (цис). Экспрессия этих последовательностей может управляться одним и тем же промотором или различными промоторами. В определенных случаях может потребоваться введение кассеты для трансформации, которая будет подавлять экспрессию полинуклеотида, представляющего интерес. Ее можно комбинировать с любой комбинацией других кассет для супрессии или кассет для сверхэкспрессии с целью получения требуемой комбинации признаков в растении. Кроме того, считается, что полинуклеотидные последовательности можно пакетировать в требуемом местоположении в геноме с применением системы для сайт-специфической рекомбинации. См., например, WO 1999/25821, WO 1999/25854, WO 1999/25840, WO 1999/25855 и WO 1999/25853, все из которых включены в данный документ посредством ссылки.
В некоторых вариантах осуществления полинуклеотиды, кодирующие полипептид PtIP-83, раскрытые в данном документе, отдельно или пакетированные с одним или несколькими дополнительными признаками устойчивости к насекомым, можно пакетировать с одним или несколькими дополнительными вводимыми признаками, влияющими на затраты (например, устойчивость к гербициду, устойчивость к грибам, устойчивость к вирусам, переносимость стрессов, устойчивость к заболеваниям, мужская стерильность, сила стебля и т. п.), или признаками, влияющими на выход (например, повышенная урожайность, модифицированные разновидности крахмала, улучшенный профиль масел, сбалансированные аминокислоты, высокое содержание лизина или метионина, повышенная усвояемость, улучшенное качество волокон, устойчивость к засухе и т. п.). Таким образом, варианты осуществления полинуклеотида можно применять для обеспечения полного комплекса агротехнических признаков, обеспечивающих улучшенное качество сельскохозяйственной культуры, с возможностью гибкого и экономически эффективного контроля любого числа сельскохозяйственных вредителей.
Трансгены, пригодные для пакетирования, без ограничения включают следующие.
1. Трансгены, которые обеспечивают устойчивость к насекомым или заболеваниям и которые кодируют следующее.
(A) Гены устойчивости растения к заболеваниям. Защитные механизмы растений зачастую активируются посредством специфического взаимодействия между продуктом гена устойчивости к заболеваниям (R) в растении и продуктом соответствующего гена авирулентности (Avr) в патогене. Разновидность растений можно трансформировать с помощью клонированного гена устойчивости для разработки растений, которые устойчивы к специфическим штаммам патогена. См., например, Jones, et al., (1994) Science 266:789 (клонирование гена Cf-9 томата, обеспечивающего устойчивость к Cladosporium fulvum); Martin, et al., (1993) Science 262:1432 (ген Pto томата, обеспечивающий устойчивость к Pseudomonas syringae pv. томата, кодирует протеинкиназу); Mindrinos, et al., (1994) Cell 78:1089 (ген RSP2 Arabidopsis, обеспечивающий устойчивость к Pseudomonas syringae), McDowell and Woffenden, (2003) Trends Biotechnol. 21(4):178-83 и Toyoda, et al., (2002) Transgenic Res. 11(6):567-82. Растение, устойчивое к заболеванию, представляет собой растение, которое более устойчиво к патогену в сравнении с растением дикого типа.
(B) Гены, кодирующие белок Bacillus thuringiensis, его производное или синтетический полипептид, смоделированный на его основе. См., например, Geiser, et al., (1986) Gene 48:109, которые раскрывают клонирование и нуклеотидную последовательность гена дельта-эндотоксина Bt. Кроме того, молекулы ДНК, кодирующие гены дельта-эндотоксина, можно приобрести в Американской коллекции типовых культур (Роквилл, Мэриленд), например, под номерами доступа ATCC® 40098, 67136, 31995 и 31998. Другие неограничивающие примеры трансгенов Bacillus thuringiensis, разработанных c помощью методик генной инженерии, приведены в следующих патентах и заявках на патент и, тем самым, включены посредством ссылки для этой цели: Патенты США №№ 5,188,960; 5,689,052; 5,880,275; 5,986,177; 6,023,013, 6,060,594, 6,063,597, 6,077,824, 6,620,988, 6,642,030, 6,713,259, 6,893,826, 7,105,332; 7,179,965, 7,208,474; 7,227,056, 7,288,643, 7,323,556, 7,329,736, 7,449,552, 7,468,278, 7,510,878, 7,521,235, 7,544,862, 7,605,304, 7,696,412, 7,629,504, 7,705,216, 7,772,465, 7,790,846, 7,858,849 и WO 1991/14778; WO 1999/31248; WO 2001/12731; WO 1999/24581 и WO 1997/40162.
Гены, кодирующие пестицидные белки, можно также пакетировать в том числе без ограничения: с инсектицидными белками из Pseudomonas sp., такими как PSEEN3174 (Monalysin, (2011) PLoS Pathogens, 7:1-13), из штамма CHA0 и Pf-5 Pseudomonas protegens (ранее fluorescens) (Pechy-Tarr, (2008) Environmental Microbiology 10:2368-2386: № доступа в GenBank EU400157); из Pseudomonas Taiwanensis (Liu, et al., (2010) J. Agric. Food Chem. 58:12343-12349) и из Pseudomonas pseudoalcligenes (Zhang, et al., (2009) Annals of Microbiology 59:45-50 и Li, et al., (2007) Plant Cell Tiss. Organ Cult. 89:159-168); инсектицидными белками из Photorhabdus sp. и Xenorhabdus sp. (Hinchliffe, et al., (2010) The Open Toxinology Journal 3:101-118 и Morgan, et al., (2001) Applied and Envir. Micro. 67:2062-2069); патент США № 6048838 и патент США № 6379946; полипептидом PIP-1 из публикации заявки на патент США US20140007292; полипептидом AfIP-1A и/или AfIP-1B из публикации заявки на патент США US20140033361; полипептидом PHI-4 из публикации заявки на патент США US20140274885 или патентной публикации согласно PCT WO2014/150914; полипептидом PIP-47 из патентной публикации согласно PCT с серийным номером PCT/US14/51063, полипептидом PIP-72 из патентной публикации согласно PCT с серийным номером PCT/US14/55128 и δ-эндотоксинами, в том числе без ограничения из классов Cry1, Cry2, Cry3, Cry4, Cry5, Cry6, Cry7, Cry8, Cry9, Cry10, Cry11, Cry12, Cry13, Cry14, Cry15, Cry16, Cry17, Cry18, Cry19, Cry20, Cry21, Cry22, Cry23, Cry24, Cry25, Cry26, Cry27, Cry 28, Cry 29, Cry 30, Cry31, Cry32, Cry33, Cry34, Cry35,Cry36, Cry37, Cry38, Cry39, Cry40, Cry41, Cry42, Cry43, Cry44, Cry45, Cry 46, Cry47, Cry49, Cry50, Cry51, Cry52, Cry53, Cry 54, Cry55, Cry56, Cry57, Cry58, Cry59, Cry60, Cry61, Cry62, Cry63, Cry64, Cry65, Cry66, Cry67, Cry68, Cry69, Cry70, Cry71, и Cry 72 генов δ-эндотоксинов и генов Cyt1 и Cyt2 цитолитических токсинов B. thuringiensis. Представители этих классов инсектицидных белков B. thuringiensis включают без ограничения Cry1Aa1 (№ доступа AAA22353); Cry1Aa2 (№ доступа № доступа AAA22552); Cry1Aa3 (№ доступа BAA00257); Cry1Aa4 (№ доступа CAA31886); Cry1Aa5 (№ доступа BAA04468); Cry1Aa6 (№ доступа AAA86265); Cry1Aa7 (№ доступа AAD46139); Cry1Aa8 (№ доступа I26149); Cry1Aa9 (№ доступа BAA77213); Cry1Aa10 (№ доступа AAD55382); Cry1Aa11 (№ доступа CAA70856); Cry1Aa12 (№ доступа AAP80146); Cry1Aa13 (№ доступа AAM44305); Cry1Aa14 (№ доступа AAP40639); Cry1Aa15 (№ доступа AAY66993); Cry1Aa16 (№ доступа HQ439776); Cry1Aa17 (№ доступа HQ439788); Cry1Aa18 (№ доступа HQ439790); Cry1Aa19 (№ доступа HQ685121); Cry1Aa20 (№ доступа JF340156); Cry1Aa21 (№ доступа JN651496); Cry1Aa22 (№ доступа KC158223); Cry1Ab1 (№ доступа AAA22330); Cry1Ab2 (№ доступа AAA22613); Cry1Ab3 (№ доступа AAA22561); Cry1Ab4 (№ доступа BAA00071 ); Cry1Ab5 (№ доступа CAA28405); Cry1Ab6 (№ доступа AAA22420); Cry1Ab7 (№ доступа CAA31620); Cry1Ab8 (№ доступа AAA22551); Cry1Ab9 (№ доступа CAA38701); Cry1Ab10 (№ доступа A29125); Cry1Ab11 (№ доступа I12419); Cry1Ab12 (№ доступа AAC64003); Cry1Ab13 (№ доступа AAN76494); Cry1Ab14 (№ доступа AAG16877); Cry1Ab15 (№ доступа AAO13302); Cry1Ab16 (№ доступа AAK55546); Cry1Ab17 (№ доступа AAT46415); Cry1Ab18 (№ доступа AAQ88259); Cry1Ab19 (№ доступа AAW31761); Cry1Ab20 (№ доступа ABB72460); Cry1Ab21 (№ доступа ABS18384); Cry1Ab22 (№ доступа ABW87320); Cry1Ab23 (№ доступа HQ439777); Cry1Ab24 (№ доступа HQ439778); Cry1Ab25 (№ доступа HQ685122); Cry1Ab26 (№ доступа HQ847729); Cry1Ab27 (№ доступа JN135249); Cry1Ab28 (№ доступа JN135250); Cry1Ab29 (№ доступа JN135251); Cry1Ab30 (№ доступа JN135252); Cry1Ab31 (№ доступа JN135253); Cry1Ab32 (№ доступа JN135254); Cry1Ab33 (№ доступа AAS93798); Cry1Ab34 (№ доступа KC156668); Cry1Ab-подобный (№ доступа AAK14336); Cry1Ab-подобный (№ доступа AAK14337); Cry1Ab-подобный (№ доступа AAK14338); Cry1Ab-подобный (№ доступа ABG88858); Cry1Ac1 (№ доступа AAA22331); Cry1Ac2 (№ доступа AAA22338); Cry1Ac3 (№ доступа CAA38098); Cry1Ac4 (№ доступа AAA73077); Cry1Ac5 (№ доступа AAA22339); Cry1Ac6 (№ доступа AAA86266); Cry1Ac7 (№ доступа AAB46989); Cry1Ac8 (№ доступа AAC44841); Cry1Ac9 (№ доступа AAB49768); Cry1Ac10 (№ доступа CAA05505 ); Cry1Ac11 (№ доступа CAA10270); Cry1Ac12 (№ доступа I12418); Cry1Ac13 (№ доступа AAD38701); Cry1Ac14 (№ доступа AAQ06607); Cry1Ac15 (№ доступа AAN07788); Cry1Ac16 (№ доступа AAU87037); Cry1Ac17 (№ доступа AAX18704); Cry1Ac18 (№ доступа AAY88347); Cry1Ac19 (№ доступа ABD37053); Cry1Ac20 (№ доступа ABB89046 ); Cry1Ac21 (№ доступа AAY66992 ); Cry1Ac22 (№ доступа ABZ01836); Cry1Ac23 (№ доступа CAQ30431); Cry1Ac24 (№ доступа ABL01535); Cry1Ac25 (№ доступа FJ513324); Cry1Ac26 (№ доступа FJ617446); Cry1Ac27 (№ доступа FJ617447); Cry1Ac28 (№ доступа ACM90319); Cry1Ac29 (№ доступа DQ438941); Cry1Ac30 (№ доступа GQ227507); Cry1Ac31 (№ доступа GU446674); Cry1Ac32 (№ доступа HM061081); Cry1Ac33 (№ доступа GQ866913); Cry1Ac34 (№ доступа HQ230364); Cry1Ac35 (№ доступа JF340157); Cry1Ac36 (№ доступа JN387137); Cry1Ac37 (№ доступа JQ317685); Cry1Ad1 (№ доступа AAA22340); Cry1Ad2 (№ доступа CAA01880); Cry1Ae1 (№ доступа AAA22410); Cry1Af1 (№ доступа AAB82749); Cry1Ag1 (№ доступа AAD46137); Cry1Ah1 (№ доступа AAQ14326); Cry1Ah2 (№ доступа ABB76664); Cry1Ah3 (№ доступа HQ439779); Cry1Ai1 (№ доступа AAO39719); Cry1Ai2 (№ доступа HQ439780); Cry1A-подобный (№ доступа AAK14339); Cry1Ba1 (№ доступа CAA29898); Cry1Ba2 (№ доступа CAA65003); Cry1Ba3 (№ доступа AAK63251); Cry1Ba4 (№ доступа AAK51084); Cry1Ba5 (№ доступа ABO20894); Cry1Ba6 (№ доступа ABL60921); Cry1Ba7 (№ доступа HQ439781); Cry1Bb1 (№ доступа AAA22344); Cry1Bb2 (№ доступа HQ439782); Cry1Bc1 (№ доступа CAA86568); Cry1Bd1 (№ доступа AAD10292); Cry1Bd2 (№ доступа AAM93496); Cry1Be1 (№ доступа AAC32850); Cry1Be2 (№ доступа AAQ52387); Cry1Be3 (№ доступа ACV96720); Cry1Be4 (№ доступа HM070026); Cry1Bf1 (№ доступа CAC50778); Cry1Bf2 (№ доступа AAQ52380); Cry1Bg1 (№ доступа AAO39720); Cry1Bh1 (№ доступа HQ589331); Cry1Bi1 (№ доступа KC156700); Cry1Ca1 (№ доступа CAA30396); Cry1Ca2 (№ доступа CAA31951); Cry1Ca3 (№ доступа AAA22343); Cry1Ca4 (№ доступа CAA01886); Cry1Ca5 (№ доступа CAA65457); Cry1Ca6 [1] (№ доступа AAF37224 ); Cry1Ca7 (№ доступа AAG50438); Cry1Ca8 (№ доступа AAM00264); Cry1Ca9 (№ доступа AAL79362); Cry1Ca10 (№ доступа AAN16462); Cry1Ca11 (№ доступа AAX53094); Cry1Ca12 (№ доступа HM070027); Cry1Ca13 (№ доступа HQ412621); Cry1Ca14 (№ доступа JN651493); Cry1Cb1 (№ доступа M97880); Cry1Cb2 (№ доступа AAG35409); Cry1Cb3 (№ доступа ACD50894 ); Cry1Cb-подобный (№ доступа AAX63901); Cry1Da1 (№ доступа CAA38099); Cry1Da2 (№ доступа I76415); Cry1Da3 (№ доступа HQ439784); Cry1Db1 (№ доступа CAA80234 ); Cry1Db2 (№ доступа AAK48937 ); Cry1Dc1 (№ доступа ABK35074); Cry1Ea1 (№ доступа CAA37933); Cry1Ea2 (№ доступа CAA39609); Cry1Ea3 (№ доступа AAA22345); Cry1Ea4 (№ доступа AAD04732); Cry1Ea5 (№ доступа A15535); Cry1Ea6 (№ доступа AAL50330); Cry1Ea7 (№ доступа AAW72936); Cry1Ea8 (№ доступа ABX11258); Cry1Ea9 (№ доступа HQ439785); Cry1Ea10 (№ доступа ADR00398); Cry1Ea11 (№ доступа JQ652456); Cry1Eb1 (№ доступа AAA22346); Cry1Fa1 (№ доступа AAA22348); Cry1Fa2 (№ доступа AAA22347); Cry1Fa3 (№ доступа HM070028); Cry1Fa4 (№ доступа HM439638); Cry1Fb1 (№ доступа CAA80235); Cry1Fb2 (№ доступа BAA25298); Cry1Fb3 (№ доступа AAF21767); Cry1Fb4 (№ доступа AAC10641); Cry1Fb5 (№ доступа AAO13295); Cry1Fb6 (№ доступа ACD50892); Cry1Fb7 (№ доступа ACD50893); Cry1Ga1 (№ доступа CAA80233); Cry1Ga2 (№ доступа CAA70506); Cry1Gb1 (№ доступа AAD10291); Cry1Gb2 (№ доступа AAO13756); Cry1Gc1 (№ доступа AAQ52381); Cry1Ha1 (№ доступа CAA80236); Cry1Hb1 (№ доступа AAA79694); Cry1Hb2 (№ доступа HQ439786); Cry1H-подобный (№ доступа AAF01213); Cry1Ia1 (№ доступа CAA44633); Cry1Ia2 (№ доступа AAA22354); Cry1Ia3 (№ доступа AAC36999); Cry1Ia4 (№ доступа AAB00958); Cry1Ia5 (№ доступа CAA70124); Cry1Ia6 (№ доступа AAC26910); Cry1Ia7 (№ доступа AAM73516); Cry1Ia8 (№ доступа AAK66742); Cry1Ia9 (№ доступа AAQ08616); Cry1Ia10 (№ доступа AAP86782); Cry1Ia11 (№ доступа CAC85964 ); Cry1Ia12 (№ доступа AAV53390); Cry1Ia13 (№ доступа ABF83202); Cry1Ia14 (№ доступа ACG63871); Cry1Ia15 (№ доступа FJ617445); Cry1Ia16 (№ доступа FJ617448); Cry1Ia17 (№ доступа GU989199); Cry1Ia18 (№ доступа ADK23801); Cry1Ia19 (№ доступа HQ439787); Cry1Ia20 (№ доступа JQ228426); Cry1Ia21 (№ доступа JQ228424); Cry1Ia22 (№ доступа JQ228427); Cry1Ia23 (№ доступа JQ228428); Cry1Ia24 (№ доступа JQ228429); Cry1Ia25 (№ доступа JQ228430); Cry1Ia26 (№ доступа JQ228431); Cry1Ia27 (№ доступа JQ228432); Cry1Ia28 (№ доступа JQ228433); Cry1Ia29 (№ доступа JQ228434); Cry1Ia30 (№ доступа JQ317686); Cry1Ia31 (№ доступа JX944038); Cry1Ia32 (№ доступа JX944039); Cry1Ia33 (№ доступа JX944040); Cry1Ib1 (№ доступа AAA82114); Cry1Ib2 (№ доступа ABW88019); Cry1Ib3 (№ доступа ACD75515); Cry1Ib4 (№ доступа HM051227); Cry1Ib5 (№ доступа HM070028); Cry1Ib6 (№ доступа ADK38579); Cry1Ib7 (№ доступа JN571740); Cry1Ib8 (№ доступа JN675714); Cry1Ib9 (№ доступа JN675715); Cry1Ib10 (№ доступа JN675716); Cry1Ib11 (№ доступа JQ228423); Cry1Ic1 (№ доступа AAC62933); Cry1Ic2 (№ доступа AAE71691); Cry1Id1 (№ доступа AAD44366); Cry1Id2 (№ доступа JQ228422); Cry1Ie1 (№ доступа AAG43526); Cry1Ie2 (№ доступа HM439636); Cry1Ie3 (№ доступа KC156647); Cry1Ie4 (№ доступа KC156681); Cry1If1 (№ доступа AAQ52382); Cry1Ig1 (№ доступа KC156701); Cry1I-подобный (№ доступа AAC31094); Cry1I-подобный (№ доступа ABG88859); Cry1Ja1 (№ доступа AAA22341); Cry1Ja2 (№ доступа HM070030); Cry1Ja3 (№ доступа JQ228425); Cry1Jb1 (№ доступа AAA98959); Cry1Jc1 (№ доступа AAC31092); Cry1Jc2 (№ доступа AAQ52372); Cry1Jd1 (№ доступа CAC50779); Cry1Ka1 (№ доступа AAB00376); Cry1Ka2 (№ доступа HQ439783); Cry1La1 (№ доступа AAS60191); Cry1La2 (№ доступа HM070031); Cry1Ma1 (№ доступа FJ884067); Cry1Ma2 (№ доступа KC156659); Cry1Na1 (№ доступа KC156648); Cry1Nb1 (№ доступа KC156678); Cry1-подобный (№ доступа AAC31091); Cry2Aa1 (№ доступа AAA22335); Cry2Aa2 (№ доступа AAA83516); Cry2Aa3 (№ доступа D86064); Cry2Aa4 (№ доступа AAC04867); Cry2Aa5 (№ доступа CAA10671); Cry2Aa6 (№ доступа CAA10672); Cry2Aa7 (№ доступа CAA10670); Cry2Aa8 (№ доступа AAO13734); Cry2Aa9 (№ доступа AAO13750 ); Cry2Aa10 (№ доступа AAQ04263); Cry2Aa11 (№ доступа AAQ52384); Cry2Aa12 (№ доступа ABI83671); Cry2Aa13 (№ доступа ABL01536); Cry2Aa14 (№ доступа ACF04939); Cry2Aa15 (№ доступа JN426947); Cry2Ab1 (№ доступа AAA22342); Cry2Ab2 (№ доступа CAA39075); Cry2Ab3 (№ доступа AAG36762); Cry2Ab4 (№ доступа AAO13296 ); Cry2Ab5 (№ доступа AAQ04609); Cry2Ab6 (№ доступа AAP59457); Cry2Ab7 (№ доступа AAZ66347); Cry2Ab8 (№ доступа ABC95996); Cry2Ab9 (№ доступа ABC74968); Cry2Ab10 (№ доступа EF157306); Cry2Ab11 (№ доступа CAM84575); Cry2Ab12 (№ доступа ABM21764); Cry2Ab13 (№ доступа ACG76120); Cry2Ab14 (№ доступа ACG76121); Cry2Ab15 (№ доступа HM037126); Cry2Ab16 (№ доступа GQ866914); Cry2Ab17 (№ доступа HQ439789); Cry2Ab18 (№ доступа JN135255); Cry2Ab19 (№ доступа JN135256); Cry2Ab20 (№ доступа JN135257); Cry2Ab21 (№ доступа JN135258); Cry2Ab22 (№ доступа JN135259); Cry2Ab23 (№ доступа JN135260); Cry2Ab24 (№ доступа JN135261); Cry2Ab25 (№ доступа JN415485); Cry2Ab26 (№ доступа JN426946); Cry2Ab27 (№ доступа JN415764); Cry2Ab28 (№ доступа JN651494); Cry2Ac1 (№ доступа CAA40536); Cry2Ac2 (№ доступа AAG35410); Cry2Ac3 (№ доступа AAQ52385); Cry2Ac4 (№ доступа ABC95997); Cry2Ac5 (№ доступа ABC74969); Cry2Ac6 (№ доступа ABC74793); Cry2Ac7 (№ доступа CAL18690); Cry2Ac8 (№ доступа CAM09325); Cry2Ac9 (№ доступа CAM09326); Cry2Ac10 (№ доступа ABN15104); Cry2Ac11 (№ доступа CAM83895); Cry2Ac12 (№ доступа CAM83896); Cry2Ad1 (№ доступа AAF09583); Cry2Ad2 (№ доступа ABC86927); Cry2Ad3 (№ доступа CAK29504); Cry2Ad4 (№ доступа CAM32331); Cry2Ad5 (№ доступа CAO78739 ); Cry2Ae1 (№ доступа AAQ52362); Cry2Af1 (№ доступа ABO30519); Cry2Af2 (№ доступа GQ866915); Cry2Ag1 (№ доступа ACH91610); Cry2Ah1 (№ доступа EU939453); Cry2Ah2 (№ доступа ACL80665); Cry2Ah3 (№ доступа GU073380); Cry2Ah4 (№ доступа KC156702); Cry2Ai1 (№ доступа FJ788388); Cry2Aj (№ доступа ); Cry2Ak1 (№ доступа KC156660); Cry2Ba1 (№ доступа KC156658); Cry3Aa1 (№ доступа AAA22336); Cry3Aa2 (№ доступа AAA22541); Cry3Aa3 (№ доступа CAA68482); Cry3Aa4 (№ доступа AAA22542); Cry3Aa5 (№ доступа AAA50255); Cry3Aa6 (№ доступа AAC43266); Cry3Aa7 (№ доступа CAB41411); Cry3Aa8 (№ доступа AAS79487); Cry3Aa9 (№ доступа AAW05659); Cry3Aa10 (№ доступа AAU29411); Cry3Aa11 (№ доступа AAW82872); Cry3Aa12 (№ доступа ABY49136 ); Cry3Ba1 (№ доступа CAA34983); Cry3Ba2 (№ доступа CAA00645); Cry3Ba3 (№ доступа JQ397327); Cry3Bb1 (№ доступа AAA22334); Cry3Bb2 (№ доступа AAA74198); Cry3Bb3 (№ доступа I15475); Cry3Ca1 (№ доступа CAA42469); Cry4Aa1 (№ доступа CAA68485); Cry4Aa2 (№ доступа BAA00179); Cry4Aa3 (№ доступа CAD30148); Cry4Aa4 (№ доступа AFB18317); Cry4A-подобный (№ доступа AAY96321); Cry4Ba1 (№ доступа CAA30312); Cry4Ba2 (№ доступа CAA30114); Cry4Ba3 (№ доступа AAA22337); Cry4Ba4 (№ доступа BAA00178); Cry4Ba5 (№ доступа CAD30095); Cry4Ba-подобный (№ доступа ABC47686); Cry4Ca1 (№ доступа EU646202); Cry4Cb1 (№ доступа FJ403208); Cry4Cb2 (№ доступа FJ597622); Cry4Cc1 (№ доступа FJ403207); Cry5Aa1 (№ доступа AAA67694); Cry5Ab1 (№ доступа AAA67693); Cry5Ac1 (№ доступа I34543); Cry5Ad1 (№ доступа ABQ82087); Cry5Ba1 (№ доступа AAA68598); Cry5Ba2 (№ доступа ABW88931); Cry5Ba3 (№ доступа AFJ04417); Cry5Ca1 (№ доступа HM461869); Cry5Ca2 (№ доступа ZP_04123426); Cry5Da1 (№ доступа HM461870); Cry5Da2 (№ доступа ZP_04123980); Cry5Ea1 (№ доступа HM485580); Cry5Ea2 (№ доступа ZP_04124038); Cry6Aa1 (№ доступа AAA22357); Cry6Aa2 (№ доступа AAM46849); Cry6Aa3 (№ доступа ABH03377); Cry6Ba1 (№ доступа AAA22358); Cry7Aa1 (№ доступа AAA22351); Cry7Ab1 (№ доступа AAA21120); Cry7Ab2 (№ доступа AAA21121); Cry7Ab3 (№ доступа ABX24522); Cry7Ab4 (№ доступа EU380678); Cry7Ab5 (№ доступа ABX79555); Cry7Ab6 (№ доступа ACI44005); Cry7Ab7 (№ доступа ADB89216); Cry7Ab8 (№ доступа GU145299); Cry7Ab9 (№ доступа ADD92572); Cry7Ba1 (№ доступа ABB70817); Cry7Bb1 (№ доступа KC156653); Cry7Ca1 (№ доступа ABR67863); Cry7Cb1 (№ доступа KC156698); Cry7Da1 (№ доступа ACQ99547); Cry7Da2 (№ доступа HM572236); Cry7Da3 (№ доступа KC156679); Cry7Ea1 (№ доступа HM035086); Cry7Ea2 (№ доступа HM132124); Cry7Ea3 (№ доступа EEM19403); Cry7Fa1 (№ доступа HM035088); Cry7Fa2 (№ доступа EEM19090); Cry7Fb1 (№ доступа HM572235); Cry7Fb2 (№ доступа KC156682); Cry7Ga1 (№ доступа HM572237); Cry7Ga2 (№ доступа KC156669); Cry7Gb1 (№ доступа KC156650); Cry7Gc1 (№ доступа KC156654); Cry7Gd1 (№ доступа KC156697); Cry7Ha1 (№ доступа KC156651); Cry7Ia1 (№ доступа KC156665); Cry7Ja1 (№ доступа KC156671); Cry7Ka1 (№ доступа KC156680); Cry7Kb1 (№ доступа BAM99306); Cry7La1 (№ доступа BAM99307); Cry8Aa1 (№ доступа AAA21117); Cry8Ab1 (№ доступа EU044830); Cry8Ac1 (№ доступа KC156662); Cry8Ad1 (№ доступа KC156684); Cry8Ba1 (№ доступа AAA21118); Cry8Bb1 (№ доступа CAD57542); Cry8Bc1 (№ доступа CAD57543); Cry8Ca1 (№ доступа AAA21119); Cry8Ca2 (№ доступа AAR98783); Cry8Ca3 (№ доступа EU625349); Cry8Ca4 (№ доступа ADB54826); Cry8Da1 (№ доступа BAC07226); Cry8Da2 (№ доступа BD133574); Cry8Da3 (№ доступа BD133575); Cry8Db1 (№ доступа BAF93483); Cry8Ea1 (№ доступа AAQ73470); Cry8Ea2 (№ доступа EU047597); Cry8Ea3 (№ доступа KC855216); Cry8Fa1 (№ доступа AAT48690); Cry8Fa2 (№ доступа HQ174208); Cry8Fa3 (№ доступа AFH78109); Cry8Ga1 (№ доступа AAT46073); Cry8Ga2 (№ доступа ABC42043); Cry8Ga3 (№ доступа FJ198072); Cry8Ha1 (№ доступа AAW81032); Cry8Ia1 (№ доступа EU381044); Cry8Ia2 (№ доступа GU073381); Cry8Ia3 (№ доступа HM044664); Cry8Ia4 (№ доступа KC156674); Cry8Ib1 (№ доступа GU325772); Cry8Ib2 (№ доступа KC156677); Cry8Ja1 (№ доступа EU625348); Cry8Ka1 (№ доступа FJ422558); Cry8Ka2 (№ доступа ACN87262); Cry8Kb1 (№ доступа HM123758); Cry8Kb2 (№ доступа KC156675); Cry8La1 (№ доступа GU325771); Cry8Ma1 (№ доступа HM044665); Cry8Ma2 (№ доступа EEM86551); Cry8Ma3 (№ доступа HM210574); Cry8Na1 (№ доступа HM640939); Cry8Pa1 (№ доступа HQ388415); Cry8Qa1 (№ доступа HQ441166); Cry8Qa2 (№ доступа KC152468); Cry8Ra1 (№ доступа AFP87548); Cry8Sa1 (№ доступа JQ740599); Cry8Ta1 (№ доступа KC156673); Cry8-подобный (№ доступа FJ770571); Cry8-подобный (№ доступа ABS53003); Cry9Aa1 (№ доступа CAA41122); Cry9Aa2 (№ доступа CAA41425); Cry9Aa3 (№ доступа GQ249293); Cry9Aa4 (№ доступа GQ249294); Cry9Aa5 (№ доступа JX174110); Cry9Aa подобный (№ доступа AAQ52376); Cry9Ba1 (№ доступа CAA52927); Cry9Ba2 (№ доступа GU299522); Cry9Bb1 (№ доступа AAV28716); Cry9Ca1 (№ доступа CAA85764); Cry9Ca2 (№ доступа AAQ52375); Cry9Da1 (№ доступа BAA19948); Cry9Da2 (№ доступа AAB97923); Cry9Da3 (№ доступа GQ249293); Cry9Da4 (№ доступа GQ249297); Cry9Db1 (№ доступа AAX78439); Cry9Dc1 (№ доступа KC156683); Cry9Ea1 (№ доступа BAA34908); Cry9Ea2 (№ доступа AAO12908); Cry9Ea3 (№ доступа ABM21765); Cry9Ea4 (№ доступа ACE88267); Cry9Ea5 (№ доступа ACF04743); Cry9Ea6 (№ доступа ACG63872 ); Cry9Ea7 (№ доступа FJ380927); Cry9Ea8 (№ доступа GQ249292); Cry9Ea9 (№ доступа JN651495); Cry9Eb1 (№ доступа CAC50780); Cry9Eb2 (№ доступа GQ249298); Cry9Eb3 (№ доступа KC156646); Cry9Ec1 (№ доступа AAC63366); Cry9Ed1 (№ доступа AAX78440); Cry9Ee1 (№ доступа GQ249296); Cry9Ee2 (№ доступа KC156664); Cry9Fa1 (№ доступа KC156692); Cry9Ga1 (№ доступа KC156699); Cry9-подобный (№ доступа AAC63366); Cry10Aa1 (№ доступа AAA22614); Cry10Aa2 (№ доступа E00614); Cry10Aa3 (№ доступа CAD30098); Cry10Aa4 (№ доступа AFB18318); Cry10A-подобный (№ доступа DQ167578); Cry11Aa1 (№ доступа AAA22352); Cry11Aa2 (№ доступа AAA22611); Cry11Aa3 (№ доступа CAD30081); Cry11Aa4 (№ доступа AFB18319); Cry11Aa-подобный (№ доступа DQ166531); Cry11Ba1 (№ доступа CAA60504); Cry11Bb1 (№ доступа AAC97162); Cry11Bb2 (№ доступа HM068615); Cry12Aa1 (№ доступа AAA22355); Cry13Aa1 (№ доступа AAA22356); Cry14Aa1 (№ доступа AAA21516); Cry14Ab1 (№ доступа KC156652); Cry15Aa1 (№ доступа AAA22333); Cry16Aa1 (№ доступа CAA63860); Cry17Aa1 (№ доступа CAA67841); Cry18Aa1 (№ доступа CAA67506); Cry18Ba1 (№ доступа AAF89667); Cry18Ca1 (№ доступа AAF89668); Cry19Aa1 (№ доступа CAA68875); Cry19Ba1 (№ доступа BAA32397); Cry19Ca1 (№ доступа AFM37572); Cry20Aa1 (№ доступа AAB93476); Cry20Ba1 (№ доступа ACS93601); Cry20Ba2 (№ доступа KC156694); Cry20-подобный (№ доступа GQ144333); Cry21Aa1 (№ доступа I32932); Cry21Aa2 (№ доступа I66477); Cry21Ba1 (№ доступа BAC06484); Cry21Ca1 (№ доступа JF521577); Cry21Ca2 (№ доступа KC156687); Cry21Da1 (№ доступа JF521578); Cry22Aa1 (№ доступа I34547); Cry22Aa2 (№ доступа CAD43579); Cry22Aa3 (№ доступа ACD93211); Cry22Ab1 (№ доступа AAK50456); Cry22Ab2 (№ доступа CAD43577); Cry22Ba1 (№ доступа CAD43578); Cry22Bb1 (№ доступа KC156672); Cry23Aa1 (№ доступа AAF76375); Cry24Aa1 (№ доступа AAC61891); Cry24Ba1 (№ доступа BAD32657); Cry24Ca1 (№ доступа CAJ43600); Cry25Aa1 (№ доступа AAC61892); Cry26Aa1 (№ доступа AAD25075); Cry27Aa1 (№ доступа BAA82796); Cry28Aa1 (№ доступа AAD24189); Cry28Aa2 (№ доступа AAG00235); Cry29Aa1 (№ доступа CAC80985); Cry30Aa1 (№ доступа CAC80986); Cry30Ba1 (№ доступа BAD00052); Cry30Ca1 (№ доступа BAD67157); Cry30Ca2 (№ доступа ACU24781); Cry30Da1 (№ доступа EF095955); Cry30Db1 (№ доступа BAE80088); Cry30Ea1 (№ доступа ACC95445); Cry30Ea2 (№ доступа FJ499389); Cry30Fa1 (№ доступа ACI22625 ); Cry30Ga1 (№ доступа ACG60020); Cry30Ga2 (№ доступа HQ638217); Cry31Aa1 (№ доступа BAB11757); Cry31Aa2 (№ доступа AAL87458); Cry31Aa3 (№ доступа BAE79808); Cry31Aa4 (№ доступа BAF32571); Cry31Aa5 (№ доступа BAF32572); Cry31Aa6 (№ доступа BAI44026); Cry31Ab1 (№ доступа BAE79809); Cry31Ab2 (№ доступа BAF32570); Cry31Ac1 (№ доступа BAF34368); Cry31Ac2 (№ доступа AB731600); Cry31Ad1 (№ доступа BAI44022); Cry32Aa1 (№ доступа AAG36711); Cry32Aa2 (№ доступа GU063849); Cry32Ab1 (№ доступа GU063850); Cry32Ba1 (№ доступа BAB78601); Cry32Ca1 (№ доступа BAB78602); Cry32Cb1 (№ доступа KC156708); Cry32Da1 (№ доступа BAB78603); Cry32Ea1 (№ доступа GU324274); Cry32Ea2 (№ доступа KC156686); Cry32Eb1 (№ доступа KC156663); Cry32Fa1 (№ доступа KC156656); Cry32Ga1 (№ доступа KC156657); Cry32Ha1 (№ доступа KC156661); Cry32Hb1 (№ доступа KC156666); Cry32Ia1 (№ доступа KC156667); Cry32Ja1 (№ доступа KC156685); Cry32Ka1 (№ доступа KC156688); Cry32La1 (№ доступа KC156689); Cry32Ma1 (№ доступа KC156690); Cry32Mb1 (№ доступа KC156704); Cry32Na1 (№ доступа KC156691); Cry32Oa1 (№ доступа KC156703); Cry32Pa1 (№ доступа KC156705); Cry32Qa1 (№ доступа KC156706); Cry32Ra1 (№ доступа KC156707); Cry32Sa1 (№ доступа KC156709); Cry32Ta1 (№ доступа KC156710); Cry32Ua1 (№ доступа KC156655); Cry33Aa1 (№ доступа AAL26871); Cry34Aa1 (№ доступа AAG50341); Cry34Aa2 (№ доступа AAK64560); Cry34Aa3 (№ доступа AAT29032); Cry34Aa4 (№ доступа AAT29030); Cry34Ab1 (№ доступа AAG41671); Cry34Ac1 (№ доступа AAG50118); Cry34Ac2 (№ доступа AAK64562); Cry34Ac3 (№ доступа AAT29029); Cry34Ba1 (№ доступа AAK64565); Cry34Ba2 (№ доступа AAT29033); Cry34Ba3 (№ доступа AAT29031); Cry35Aa1 (№ доступа AAG50342); Cry35Aa2 (№ доступа AAK64561); Cry35Aa3 (№ доступа AAT29028); Cry35Aa4 (№ доступа AAT29025); Cry35Ab1 (№ доступа AAG41672); Cry35Ab2 (№ доступа AAK64563); Cry35Ab3 (№ доступа AY536891); Cry35Ac1 (№ доступа AAG50117); Cry35Ba1 (№ доступа AAK64566); Cry35Ba2 (№ доступа AAT29027); Cry35Ba3 (№ доступа AAT29026); Cry36Aa1 (№ доступа AAK64558); Cry37Aa1 (№ доступа AAF76376 ); Cry38Aa1 (№ доступа AAK64559); Cry39Aa1 (№ доступа BAB72016); Cry40Aa1 (№ доступа BAB72018); Cry40Ba1 (№ доступа BAC77648); Cry40Ca1 (№ доступа EU381045); Cry40Da1 (№ доступа ACF15199); Cry41Aa1 (№ доступа BAD35157); Cry41Ab1 (№ доступа BAD35163); Cry41Ba1 (№ доступа HM461871); Cry41Ba2 (№ доступа ZP_04099652); Cry42Aa1 (№ доступа BAD35166); Cry43Aa1 (№ доступа BAD15301); Cry43Aa2 (№ доступа BAD95474 ); Cry43Ba1 (№ доступа BAD15303); Cry43Ca1 (№ доступа KC156676); Cry43Cb1 (№ доступа KC156695); Cry43Cc1 (№ доступа KC156696); Cry43-подобный (№ доступа BAD15305); Cry44Aa (№ доступа BAD08532); Cry45Aa (№ доступа BAD22577); Cry46Aa (№ доступа BAC79010); Cry46Aa2 (№ доступа BAG68906); Cry46Ab (№ доступа BAD35170); Cry47Aa (№ доступа AAY24695); Cry48Aa (№ доступа CAJ18351); Cry48Aa2 (№ доступа CAJ86545); Cry48Aa3 (№ доступа CAJ86546 ); Cry48Ab (№ доступа CAJ86548); Cry48Ab2 (№ доступа CAJ86549); Cry49Aa (№ доступа CAH56541); Cry49Aa2 (№ доступа CAJ86541); Cry49Aa3 (№ доступа CAJ86543); Cry49Aa4 (№ доступа CAJ86544); Cry49Ab1 (№ доступа CAJ86542); Cry50Aa1 (№ доступа BAE86999); Cry50Ba1 (№ доступа GU446675); Cry50Ba2 (№ доступа GU446676); Cry51Aa1 (№ доступа ABI14444); Cry51Aa2 (№ доступа GU570697); Cry52Aa1 (№ доступа EF613489); Cry52Ba1 (№ доступа FJ361760); Cry53Aa1 (№ доступа EF633476); Cry53Ab1 (№ доступа FJ361759); Cry54Aa1 (№ доступа ACA52194); Cry54Aa2 (№ доступа GQ140349); Cry54Ba1 (№ доступа GU446677); Cry55Aa1 (№ доступа ABW88932); Cry54Ab1 (№ доступа JQ916908); Cry55Aa2 (№ доступа AAE33526); Cry56Aa1 (№ доступа ACU57499); Cry56Aa2 (№ доступа GQ483512); Cry56Aa3 (№ доступа JX025567); Cry57Aa1 (№ доступа ANC87261); Cry58Aa1 (№ доступа ANC87260); Cry59Ba1 (№ доступа JN790647); Cry59Aa1 (№ доступа ACR43758); Cry60Aa1 (№ доступа ACU24782); Cry60Aa2 (№ доступа EAO57254); Cry60Aa3 (№ доступа EEM99278); Cry60Ba1 (№ доступа GU810818); Cry60Ba2 (№ доступа EAO57253); Cry60Ba3 (№ доступа EEM99279); Cry61Aa1 (№ доступа HM035087); Cry61Aa2 (№ доступа HM132125); Cry61Aa3 (№ доступа EEM19308); Cry62Aa1 (№ доступа HM054509); Cry63Aa1 (№ доступа BAI44028); Cry64Aa1 (№ доступа BAJ05397); Cry65Aa1 (№ доступа HM461868); Cry65Aa2 (№ доступа ZP_04123838); Cry66Aa1 (№ доступа HM485581); Cry66Aa2 (№ доступа ZP_04099945); Cry67Aa1 (№ доступа HM485582); Cry67Aa2 (№ доступа ZP_04148882); Cry68Aa1 (№ доступа HQ113114); Cry69Aa1 (№ доступа HQ401006); Cry69Aa2 (№ доступа JQ821388); Cry69Ab1 (№ доступа JN209957); Cry70Aa1 (№ доступа JN646781); Cry70Ba1 (№ доступа ADO51070); Cry70Bb1 (№ доступа EEL67276); Cry71Aa1 (№ доступа JX025568); Cry72Aa1 (№ доступа JX025569).
Примеры δ-эндотоксинов также включают без ограничения белки Cry1A из патентов США №№ 5880275 и 7858849; токсин DIG-3 или DIG-11 (варианты с N-терминальной делецией α-спирали 1 и/или α-спирали 2 белков Cry, таких как Cry1A) из патентов США №№ 8304604 и 8304605; Cry1B из заявки на патент США с серийным номером 10/525318; Cry1C из патента США № 6033874; Cry1F из патентов США №№ 5188960 и 6218188; химеры Cry1A/F из патентов США №№ 7070982; 6962705 и 6713063); белок Cry2, такой как белок Cry2Ab из патента США № 7064249); белок Cry3A, в том числе без ограничения созданный с помощью генной инженерии гибридный инсектицидный белок (eHIP), полученный путем слияния уникальных комбинаций вариабельных участков и консервативных блоков по меньшей мере двух различных белков Cry (публикация заявки на патент США № 2010/0017914); белок Cry4; белок Cry5; белок Cry6; белки Cry8 из патентов США №№ 7329736, 7449552, 7803943, 7476781, 7105332, 7378499 и 7462760; белок Cry9, например, представители семейств Cry9A, Cry9B, Cry9C, Cry9D, Cry9E и Cry9F; белок Cry15 из Naimov, et al., (2008) Applied and Environmental Microbiology 74:7145-7151; Cry22, белок Cry34Ab1 из патентов США №№ 6127180, 6624145 и 6340593; белок CryET33 и CryET34 из патентов США №№ 6248535, 6326351, 6399330, 6949626, 7385107 и 7504229; гомологи CryET33 и CryET34 из публикации заявок на патент США №№ 2006/0191034, 2012/0278954 и публикации согласно PCT№ WO 2012/139004; белок Cry35Ab1 из патентов США №№ 6083499, 6548291 и 6340593; белок Cry46, белок Cry 51, бинарный токсин Cry; TIC901 или родственный токсин; TIC807 из US 2008/0295207; ET29, ET37, TIC809, TIC810, TIC812, TIC127, TIC128 из PCT US 2006/033867; AXMI-027, AXMI-036 и AXMI-038 из патента США № 8236757; AXMI-031, AXMI-039, AXMI-040, AXMI-049 из US7923602; AXMI-018, AXMI-020 и AXMI-021 из WO 2006/083891; AXMI-010 из WO 2005/038032; AXMI-003 из WO 2005/021585; AXMI-008 из US 2004/0250311; AXMI-006 из US 2004/0216186; AXMI-007 из US 2004/0210965; AXMI-009 из US 2004/0210964; AXMI-014 из US 2004/0197917; AXMI-004 из US 2004/0197916; AXMI-028 и AXMI-029 из WO 2006/119457; AXMI-007, AXMI-008, AXMI-0080rf2, AXMI-009, AXMI-014 и AXMI-004 из WO 2004/074462; AXMI-150 из патента США № 8084416; AXMI-205 из US20110023184; AXMI-011, AXMI-012, AXMI-013, AXMI-015, AXMI-019, AXMI-044, AXMI-037, AXMI-043, AXMI-033, AXMI-034, AXMI-022, AXMI-023, AXMI-041, AXMI-063, и AXMI-064 из US 2011/0263488; AXMI-R1 и родственные белки из US 2010/0197592; AXMI221Z, AXMI222z, AXMI223z, AXMI224z и AXMI225z of WO 2011/103248; AXMI218, AXMI219, AXMI220, AXMI226, AXMI227, AXMI228, AXMI229, AXMI230, и AXMI231 из WO11/103247; AXMI-115, AXMI-113, AXMI-005, AXMI-163 и AXMI-184 из патента США № 8334431; AXMI-001, AXMI-002, AXMI-030, AXMI-035 и AXMI-045 из US 2010/0298211; AXMI-066 и AXMI-076 из US20090144852; AXMI128, AXMI130, AXMI131, AXMI133, AXMI140, AXMI141, AXMI142, AXMI143, AXMI144, AXMI146, AXMI148, AXMI149, AXMI152, AXMI153, AXMI154, AXMI155, AXMI156, AXMI157, AXMI158, AXMI162, AXMI165, AXMI166, AXMI167, AXMI168, AXMI169, AXMI170, AXMI171, AXMI172, AXMI173, AXMI174, AXMI175, AXMI176, AXMI177, AXMI178, AXMI179, AXMI180, AXMI181, AXMI182, AXMI185, AXMI186, AXMI187, AXMI188, AXMI189 из патента США № 8318900; AXMI079, AXMI080, AXMI081, AXMI082, AXMI091, AXMI092, AXMI096, AXMI097, AXMI098, AXMI099, AXMI100, AXMI101, AXMI102, AXMI103, AXMI104, AXMI107, AXMI108, AXMI109, AXMI110, AXMI111, AXMI112, AXMI114, AXMI116, AXMI117, AXMI118, AXMI119, AXMI120, AXMI121, AXMI122, AXMI123, AXMI124, AXMI1257, AXMI1268, AXMI127, AXMI129, AXMI164, AXMI151, AXMI161, AXMI183, AXMI132, AXMI138, AXMI137 из US 2010/0005543; и белки Cry, такие как Cry1A и Cry3A с модифицированными сайтами протеолитического расщепления из патента США № 8319019; и белок-токсин Cry1Ac, Cry2Aa и Cry1Ca из штамма VBTS 2528 Bacillus thuringiensis из публикации заявки на патент США № 2011/0064710. Другие белки Cry хорошо известны специалисту в данной области (см. Crickmore, et al., "Bacillus thuringiensis toxin nomenclature" (2011), на сайте lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/, доступ к которому можно получить во всемирной сети Интернет с применением префикса "www"). Инсектицидная активность белков Cry хорошо известна специалисту в данной области (обзор см. у van Frannkenhuyzen, (2009) J. Invert. Path. 101:1-16). Применение белков Cry в качестве признаков трансгенного растения хорошо известно специалисту в данной области техники, и трансгенные растения с Cry, в том числе без ограничения с Cry1Ac, Cry1Ac+Cry2Ab, Cry1Ab, Cry1A.105, Cry1F, Cry1Fa2, Cry1F+Cry1Ac, Cry2Ab, Cry3A, mCry3A, Cry3Bb1, Cry34Ab1, Cry35Ab1, Vip3A, mCry3A, Cry9c и CBI-Bt были разрешены контролирующими органами (см., Sanahuja, (2011) Plant Biotech Journal 9:283-300 и CERA (2010) GM Crop Database Center for Environmental Risk Assessment (CERA), ILSI Research Foundation, Washington D.C. на сайте cera-gmc.org/index.php?action=gm_crop_database, доступ к которому можно получить во всемирной сети Интернет с применением префикса "www"). В растениях также может экспрессироваться два и более пестицидных белков, хорошо известных специалисту в данной области, например, Vip3Ab и Cry1Fa (US2012/0317682), Cry1BE и Cry1F (US2012/0311746), Cry1CA и Cry1AB (US2012/0311745), Cry1F и CryCa (US2012/0317681), Cry1DA и Cry1BE (US2012/0331590), Cry1DA и Cry1Fa (US2012/0331589), Cry1AB и Cry1BE (US2012/0324606) и Cry1Fa и Cry2Aa, Cry1I или Cry1E (US2012/0324605). Пестицидные белки также включают инсектицидные липазы, в том числе липидацильные гидролазы из патента США № 7491869 и холестериноксидазы, например из Streptomyces (Purcell et al. (1993) Biochem Biophys Res Commun 15:1406-1413). Пестицидные белки также включают токсины VIP (инсектицидные белки вегетативной фазы) из патентов США №№ 5877012, 6107279, 6137033, 7244820, 7615686 и 8237020 и т. п. Другие белки VIP хорошо известны специалисту в данной области (см. lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/vip.html, доступ к которому можно получить во всемирной сети Интернет с применением префикса "www"). Пестицидные белки также включают белки токсинового комплекса (TC), которые можно получить из организмов, таких как Xenorhabdus, Photorhabdus и Paenibacillus (см. патенты США №№ 7491698 и 8084418). Некоторые TC-белки обладают ʺсамостоятельнойʺ инсектицидной активностью, а другие TC-белки повышают активность самостоятельных токсинов, производимых тем же данным организмом. Токсичность "самостоятельного" TC-белка (от Photorhabdus, Xenorhabdus или Paenibacillus, например) может повышаться при помощи одного или нескольких TC-белков-"усилителей", полученных от организма-источника из другого рода. Существуют три основных типа TC-белков. Как изложено в данном документе, белки класса A ("белок A") представляют собой самостоятельные токсины. Белки класса B ("белок B") и белки класса C ("белок C") усиливают токсичность белков класса A. Примеры белков класса A представляют собой TcbA, TcdA, XptA1 и XptA2. Примеры белков класса B представляют собой TcaC, TcdB, XptB1Xb и XptC1Wi. Примеры белков класса C представляют собой TccC, XptC1Xb и XptB1Wi. Пестицидные белки также включают белки яда пауков, змей и скорпионов. Примеры пептидов яда пауков включают без ограничения пептиды ликотоксина-1 и его мутантные формы (патент США № 8334366).
(C) Полинуклеотид, кодирующий специфический для насекомых гормон или феромон, такой как экдистероид и ювенильный гормон, его вариант, миметик на его основе, или его антагонист или агонист. См., например, раскрытие Hammock, et al., (1990) Nature 344:458, качающееся бакуловирусной системы экспрессии клонированной эстеразы ювенильного гормона, деактиватора ювенильного гормона.
(D) Полинуклеотид, кодирующий специфический для насекомых пептид, который при экспрессии нарушает физиологию насекомого, на которого оказывают воздействие. Например, см. раскрытия Regan, (1994) J. Biol. Chem. 269:9 (экспрессионное клонирование приводит к ДНК, кодирующей рецептор диуретического гормона насекомых); Pratt, et al., (1989) Biochem. Biophys. Res. Comm. 163:1243 (аллостатин, обнаруженный у Diploptera puntata); Chattopadhyay, et al., (2004) Critical Reviews in Microbiology 30(1):33-54; Zjawiony, (2004) J Nat Prod 67(2):300-310; Carlini and Grossi-de-Sa, (2002) Toxicon 40(11):1515-1539; Ussuf, et al., (2001) Curr Sci. 80(7):847-853 и Vasconcelos and Oliveira, (2004) Toxicon 44(4):385-403. См. также патент США № 5266317, Tomalski, et al., в котором раскрываются гены, кодирующие специфические для насекомых токсины.
(E) Полинуклеотид, кодирующий фермент, ответственный за гипернакопление монотерпена, сесквитерпена, стероида, гидроксамовой кислоты, производного фенилпропаноида или другой небелковой молекулы с инсектицидной активностью.
(F) Полинуклеотид, кодирующий фермент, вовлеченный в модификацию, в том числе посттрансляционную модификацию, биологически активной молекулы; например, гликолитический фермент, протеолитический фермент, липолитический фермент, нуклеаза, циклаза, трансаминаза, эстераза, гидролаза, фосфатаза, киназа, фосфорилаза, полимераза, эластаза, хитиназа и глюканаза, либо натуральные, либо синтетические. См. заявку согласно PCT WO 1993/02197 от имени Scott, et al., в которой раскрыта нуклеотидная последовательность гена каллазы. Молекулы ДНК, которые содержат последовательности, кодирующие хитиназу, можно получить, например, из ATCC® под номерами доступа 39637 и 67152. См. также Kramer, et al., (1993) Insect Biochem. Molec. Biol. 23:691, в которой содержатся сведения о нуклеотидной последовательности кДНК, кодирующей хитиназу табачного бражника, и Kawalleck, et al., (1993) Plant Molec. Biol. 21:673, в которой предложена нуклеотидная последовательность гена полиубиквитина ubi4-2 петрушки, и патенты США №№ 6563020; 7145060 и 7087810.
(G) Полинуклеотид, кодирующий молекулу, которая стимулирует сигнальную трансдукцию. Например, см. раскрытие в Botella, et al., (1994) Plant Molec. Biol. 24:757, каcающееся нуклеотидных последовательностей клонов кДНК кальмодулина из маша, и Griess, et al., (1994) Plant Physiol. 104:1467, в которой представлена нуклеотидная последовательность клона кДНК кальмодулина маиса.
(H) Полинуклеотид, кодирующий пептид с гидрофобным моментом. См. заявку согласно PCT WO 1995/16776 и патент США № 5580852, раскрывающие пептидные производные тахиплезина, которые ингибируют грибные патогены растений, и заявку согласно PCT WO 1995/18855, а также патент США № 5607914 (в котором раскрыты синтетические противомикробные пептиды, которые придают устойчивость к заболеваниям).
(I) Полинуклеотид, кодирующий мембранную пермеазу, каналообразователь или блокатор каналов. Например, см. раскрытие в Jaynes, et al., (1993) Plant Sci. 89:43, касающееся гетерологичной экспрессии аналога цекропин-бета литического пептида для придания трансгенным растениям табака устойчивости к Pseudomonas solanacearum.
(J) Ген, кодирующий вирусный инвазивный белок или сложный токсин, полученный из него. Например, накопление белков вирусной оболочки в трансформированных растительных клетках придает устойчивость к вирусной инфекции и/или развитию заболевания, вызванного вирусом, из которого получен ген белка оболочки, а также родственными вирусами. См. Beachy, et al., (1990) Ann. Rev. Phytopathol. 28:451. Устойчивость, опосредованная белком оболочки, была обеспечена у трансформированных растений в отношении вируса мозаики люцерны, вируса мозаики огурца, вируса полосчатости табака, вируса X картофеля, вируса Y картофеля, вируса гравировки табака, вируса погремковости табака и вируса табачной мозаики. Там же.
(K) Ген, кодирующий антитело, специфическое в отношении насекомого, или иммунотоксин, полученный из него. Таким образом, антитело, нацеленное на критическую метаболическую функцию в кишке насекомого, будет инактивировать фермент, на который оказывается воздействие, с уничтожением насекомого. Ср. Taylor, et al., Abstract #497, SEVENTH INT′L SYMPOSIUM ON MOLECULAR PLANT-MICROBE INTERACTIONS (Edinburgh, Scotland, 1994) (ферментативная инактивация в трансгенном табаке посредством выработки одноцепочечных фрагментов антител).
(L) Ген, кодирующий антитело, специфическое в отношении вируса. См., например, Tavladoraki, et al., (1993) Nature 366:469, где показано, что трансгенные растения, экспрессирующие гены рекомбинантного антитела, защищены от нападения вируса.
(M) Полинуклеотид, кодирующий останавливающий развитие белок, вырабатываемый в природе патогеном или паразитом. Таким образом, грибные эндо-альфа-1,4-D-полигалактуроназы облегчают грибную колонизацию и высвобождение питательных веществ растения путем солюбилизации гомо-альфа-1,4-D-галактуроназы клеточной стенки растения. См. Lamb, et al., (1992) Bio/Technology 10:1436. Клонирование и определение характеристик гена, который кодирует белок, ингибирующий эндополигалактуроназу бобов, описан в Toubart, et al., (1992) Plant J. 2:367.
(N) Полинуклеотид, кодирующий останавливающий развитие белок, вырабатываемый в природе растением. Например, Logemann, et al., (1992) Bio/Technology 10:305 показали, что трансгенные растения, экспрессирующие ген ячменя, инактивирующий рибосомы, обладали повышенной устойчивостью к грибковым заболеваниям.
(O) Гены, вовлеченные в реакцию системной приобретенной устойчивости (SAR), и/или гены, связанные с патогенезом. Briggs, (1995) Current Biology 5(2), Pieterse and Van Loon, (2004) Curr. Opin. Plant Bio. 7(4):456-64 и Somssich, (2003) Cell 113(7):815-6.
(P) Противогрибковые гены (Cornelissen and Melchers, (1993) Pl. Physiol. 101:709-712, и Parijs, et al., (1991) Planta 183:258-264, и Bushnell, et al., (1998) Can. J. of Plant Path. 20(2):137-149. Также см. заявки на патент США №№ 09/950933; 11/619645; 11/657710; 11/748994; 11/774121 и патенты США №№ 6891085 и 7306946. Киназы, подобные рецептору LysM, для распознавания фрагментов хитина, как первый этап в защитной реакции растения против грибов-патогенов (US 2012/0110696).
(Q) Гены системы детоксикации, такие как кодирующие фумонизин, беауверицин, монилиформин и зеараленон и их структурно родственные производные. Например, см. патенты США №№ 5716820; 5792931; 5798255; 5846812; 6083736; 6538177; 6388171 и 6812380.
(R) Полинуклеотид, кодирующий цистатин и ингибиторы цистеиновой протеиназы. См. патент США № 7205453.
(S) Гены дефензинов. См. WO 2003/000863 и патенты США №№ 6911577; 6855865; 6777592 и 7238781.
(T) Гены, обеспечивающие устойчивость к нематодам. См., например, заявку согласно PCT WO 1996/30517; заявку согласно PCT WO 1993/19181, WO 2003/033651 и Urwin, et al., (1998) Planta 204:472-479, Williamson, (1999) Curr Opin Plant Bio. 2(4):327-31; патенты США №№ 6284948 и 7301069 и гены miR164 (WO 2012/058266).
(U) Гены, которые обеспечивают устойчивость к корневой гнили, вызываемой Phytophthora, такие как Rps 1, Rps 1-a, Rps 1-b, Rps 1-c, Rps 1-d, Rps 1-e, Rps 1-k, Rps 2, Rps 3-a, Rps 3-b, Rps 3-c, Rps 4, Rps 5, Rps 6, Rps 7 и другие гены Rps. См., например, Shoemaker, et al., Phytophthora Root Rot Resistance Gene Mapping in Soybean, Plant Genome IV Conference, San Diego, Calif. (1995).
(V) Гены, которые обеспечивают устойчивость к бурой стеблевой гнили, такие как описаны в патенте США № 5689035, включенном посредством ссылки для этой цели.
(W) Гены, которые обеспечивают устойчивость к Colletotrichum, такие как описанные в публикации заявки на патент США US 2009/0035765, включенной посредством ссылки для этой цели. Они включают локус Rcg, который можно использовать как конверсию одного локуса.
2. Трансгены, которые обеспечивают устойчивость к гербициду, например.
(A) Полинуклеотид, кодирующий устойчивость к гербициду, который ингибирует конус нарастания или меристему, такому как имидазолинон или сульфонилмочевина. Иллюстративные гены в этой категории кодируют мутантный фермент ALS и AHAS, как описано, например, у Lee, et al., (1988) EMBO J. 7:1241 и Miki, et al., (1990) Theor. Appl. Genet. 80:449, соответственно. См. также патенты США №№ 5605011; 5013659; 5141870; 5767361; 5731180; 5304732; 4761373; 5331107; 5928937 и 5378824; заявку на патент США № 11/683737 и публикацию международной заявки WO 1996/33270.
(B) Полинуклеотид, кодирующий белок для устойчивости к глифосату (устойчивость придают мутантные гены 5-енолприрувил-3-фосфошикиматсинтазы (EPSP) и aroA, соответственно) и другим соединениям с фосфоновыми группами, таким как глуфосинат (гены фосфинотрицин-ацетилтрансферазы (PAT) и фосфинотрицин-ацетилтрансферазы (bar) Streptomyces hygroscopicus), и пиридинокси- или феноксипропионовым кислотам и циклогексонам (гены, кодирующие ингибитор ACCазы). См., например, патент США № 4940835, Shah, et al., в котором раскрыта нуклеотидная последовательность формы EPSPS, которая может придавать устойчивость к глифосату. В патенте США № 5627061, Barry, et al., также описаны гены, кодирующие ферменты EPSPS. См. также патенты США №№ 6566587; 6338961; 6248876 B1; 6040497; 5804425; 5633435; 5145783; 4971908; 5312910; 5188642; 5094945, 4940835; 5866775; 6225114 B1; 6130366; 5310667; 4535060; 4769061; 5633448; 5510471; Re. 36449; RE 37287 E и 5491288, а также публикации международных заявок EP 1173580; WO 2001/66704; EP 1173581 и EP 1173582, которые включены в данный документ посредством ссылки для этой цели. Устойчивость к глифосату также придается растениям, которые экспрессируют ген, кодирующий фермент глифосат-оксидоредуктазу, как более подробно описано в патентах США №№ 5776760 и 5463175, которые включены в данный документ посредством ссылки для этой цели. Кроме того, устойчивость к глифосату может придаваться растениям посредством сверхэкспрессии генов, кодирующих глифосат-N-ацетилтрансферазу. См., например, патенты США №№ 7462481; 7405074 и публикацию заявки на патент США № US 2008/0234130. Молекулу ДНК, кодирующую мутантный ген aroA, можно получить под номером доступа ATCC® 39256, а нуклеотидная последовательность мутантного гена раскрыта в патенте США № 4769061, выданном Comai. В заявке EP № 0333033, Kumada, et al., и патенте США № 4975374, выданном Goodman, et al., раскрыты нуклеотидные последовательности генов глутаминсинтетазы, придающие устойчивость к гербицидам, таким как L-фосфинотрицин. Нуклеотидная последовательность гена фосфинотрицин-ацетилтрансферазы представлена в заявках EP №№ 0242246 и 0242236, Leemans, et al.; De Greef, et al., (1989) Bio/Technology 7:61, где описано получение трансгенных растений, которые экспрессируют химерные гены bar, кодирующие фосфинотрицин-ацетилтрансферазную активность. См. также патенты США №№ 5969213; 5489520; 5550318; 5874265; 5919675; 5561236; 5648477; 5646024; 6177616 B1 и 5879903, которые включены в данный документ посредством ссылки с этой целью. Иллюстративные гены, придающие устойчивость к феноксипропионовым кислотам и циклогексонам, таким как сетоксидим и галоксифоп, представляют собой гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные у Marshall, et al., (1992) Theor. Appl. Genet. 83:435.
(C) Полинуклеотид, кодирующий белок, обеспечивающий устойчивость к гербициду, который ингибирует фотосинтез, такому как триазин (гены psbA и gs+) и бензонитрил (ген нитрилазы). Przibilla, et al., (1991) Plant Cell 3:169 описывают трансформацию Chlamydomonas с помощью плазмид, кодирующих мутантные гены psbA. Нуклеотидные последовательности генов нитрилазы раскрыты в патенте США № 4810648, выданном Stalker, и молекулы ДНК, содержащие эти гены, доступны под номерами доступа ATCC® 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, описаны у Hayes, et al., (1992) Biochem. J. 285:173.
(D) В ряд растений был введен полинуклеотид, кодирующий белок, обеспечивающий устойчивость к синтазе ацетогидроксикислот, которая, как было обнаружено, делает растения, которые экспрессируют этот фермент, устойчивыми к нескольким типам гербицидов (см., например, Hattori, et al., (1995) Mol Gen Genet. 246:419). Другие гены, которые придают устойчивость к гербицидам, включают ген, кодирующий химерный белок цитохрома P4507A1 крысы и NADPH-цитохром P450-оксидоредуктазу дрожжей (Shiota, et al., (1994) Plant Physiol 106:17), гены, кодирующие глутатионредуктазу и супероксиддисмутазу (Aono, et al., (1995) Plant Cell Physiol 36:1687), и гены, кодирующие различные фосфотрансферазы (Datta, et al., (1992) Plant Mol Biol 20:619).
(E) Полинуклеотид, кодирующий устойчивость к гербициду, целенаправленно воздействующему на протопорфириноген-оксидазу (protox), которая необходима для выработки хлорофилла. Фермент protox служит в качестве мишени для целого ряда гербицидных соединений. Эти гербициды также ингибируют рост всех различных присутствующих видов растений, вызывая их полное разрушение. Разработка растений, характеризующихся измененной protox-активностью, которые устойчивы к этим гербицидам, описана в патентах США №№ 6288306 B1; 6282837 B1 и 5767373 и публикации международной заявки WO 2001/12825.
(F) Ген aad-1 (изначально из Sphingobium herbicidovorans) кодирует белок арилоксиалканоат-диоксигеназу (AAD-1). Признак обеспечивает переносимость гербицидов на основе 2,4-дихлорфеноксиуксусной кислоты и арилоксифеноксипропионата (обычно называемых "фоп"-гербициды, таких как квизалофоп). Ген aad-1, как таковой, обеспечивающий переносимость гербицида у растений, впервые был раскрыт в WO 2005/107437 (см. также US 2009/0093366). Ген aad-12, полученный из Delftia acidovorans, который кодирует белок арилоксиалканоат-диоксигеназу (AAD-12), обеспечивающий переносимость гербицидов на основе 2,4-дихлорфеноксиуксусной кислоты и пиридилоксиацетата посредством инактивации некоторых гербицидов с арилоксиалканоатным фрагментом, в том числе фенокси-ауксина (например, 2,4-D, MCPA), а также видов пиридилокси-ауксина (например, флуроксипира, триклопира).
(G) Полинуклеотид, кодирующий монооксигеназу, устойчивую к гербициду дикамба, раскрытый в публикации заявки на патент США 2003/0135879, для придания переносимости дикамбы.
(H) Полинуклеотидная молекула, кодирующая бромоксинилнитрилазу (Bxn), раскрытая в патенте США № 4810648, для придания переносимости бромоксинила.
(I) Полинуклеотидная молекула, кодирующая фитоен (crtl), описанный у Misawa, et al., (1993) Plant J. 4:833-840 и у Misawa, et al., (1994) Plant J. 6:481-489, для переносимости норфлуразона.
3. Трансгены, которые обеспечивают измененные характеристики зерна или вносят в них вклад
Такие как представленные ниже.
(A) Измененные жирные кислоты, например, посредством следующего.
(1) Подавления стеароил-ACP для повышения содержания стеариновой кислоты в растении. См. Knultzon, et al., (1992) Proc. Natl. Acad. Sci. USA 89:2624 и WO 1999/64579 (Genes to Alter Lipid Profiles in Corn).
(2) Повышения содержания олеиновой кислоты посредством модификации гена FAD-2 и/или снижения содержания линоленовой кислоты посредством модификации гена FAD-3 (см. патенты США №№ 6063947; 6323392; 6372965 и WO 1993/11245).
(3) Изменения содержания конъюгированных линоленовой или линолевой кислоты, например, как в WO 2001/12800.
(4) Изменения LEC1, AGP, Dek1, Superal1, mi1 ps, различных генов Ipa, таких как Ipa1, Ipa3, hpt или hggt. Например, см. WO 2002/42424, WO 1998/22604, WO 2003/011015, WO 2002/057439, WO 2003/011015, патенты США №№ 6423886, 6197561, 6825397 и публикации заявки на патент США №№ US 2003/0079247, US 2003/0204870 и Rivera-Madrid, et al., (1995) Proc. Natl. Acad. Sci. 92:5620-5624.
(5) Генов, кодирующих дельта-8-десатуразу для получения длинноцепочечных полиненасыщенных жирных кислот (патенты США №№ 8058571 и 8338152), дельта-9 десатуразу для снижения содержания насыщенных жиров (патент США № 8063269), Δ6-десатуразу примулы для улучшения профилей омега-3-жирных кислот.
(6) Выделенных нуклеиновых кислот и белков, ассоциированных с регуляцией метаболизма липидов и сахаров, в частности, белка липидного метаболизма (LMP), применяемых в способах получения трансгенных растений и модуляции уровней запасных веществ семени, в том числе липидов, жирных кислот, видов крахмала или запасных белков семени, и их применения в способах модуляции размера семени, количества семян, веса семян, длины корней и размера листьев растений (EP 2404499).
(7) Изменения экспрессии белка, индуцируемого сахарами 2 (HSI2), с высоким уровнем экспрессии в растении для повышения или снижения экспрессии HSI2 в растении. Повышение экспрессии HSI2 повышает содержание масла, тогда как понижение экспрессии HSI2 снижает восприимчивость к абсцизовой кислоте и/или повышает устойчивость к засухе (публикация заявки на патент США № 2012/0066794).
(8) Экспрессии цитохрома b5 (Cb5) отдельно или вместе с FAD2 для модуляции содержания масла в семени растения, в частности, для повышения уровней омега-3-жирных кислот и улучшения соотношения омега-6- к омега-3-жирным кислотам (публикация заявки на патент США № 2011/0191904).
(9) Молекул нуклеиновой кислоты, кодирующих wrinkled1-подобные полипептиды для модуляции метаболизма сахаров (патент США № 8217223).
(B) Измененное содержание фосфора, например, посредством следующего.
(1) Введения гена, кодирующего фитазу, при этом будет улучшаться распад фитата, что приводит к большему количеству свободного фосфата в трансформированном растении. Например, см. Van Hartingsveldt, et al., (1993) Gene 127:87, касательно раскрытия нуклеотидной последовательности гена фитазы Aspergillus niger.
(2) Модуляции гена, который снижает содержание фитата. Например, в случае маиса это можно осуществлять при помощи клонирования, а затем повторного введения ДНК, ассоциированной с одним или несколькими аллелями, такими как аллели LPA, идентифицированные у мутантов маиса, характеризующихся низкими уровнями фитиновой кислоты, как, например, в WO 2005/113778, и/или посредством изменения активности инозитолкиназы, как в WO 2002/059324, публикации заявки на патент США № 2003/0009011, WO 2003/027243, публикации заявки на патент США № 2003/0079247, WO 1999/05298, патенте США № 6197561, патенте США № 6291224, патенте США № 6391348, WO 2002/059324, публикации заявки на патент США № 2003/0079247, WO 1998/45448, WO 1999/55882, WO 2001/04147.
(C) Измененные углеводы, на которые воздействовали, например, посредством изменения гена, кодирующего фермент, который воздействует на паттерн ветвления крахмала, или гена, изменяющего тиоредоксин, такого как NTR и/или TRX (см. патент США № 6531648, который включен посредством ссылки с этой целью), и/или нокаута гамма-зеина, или использования мутанта, такого как cs27, или TUSC27, или en27 (см. патент США № 6858778 и публикацию заявки на патент США № 2005/0160488, публикацию заявки на патент США № 2005/0204418, которые включены посредством ссылки для этой цели). См. Shiroza, et al., (1988) J. Bacteriol. 170:810 (нуклеотидная последовательность мутантного гена фруктозилтрансферазы Streptococcus), Steinmetz, et al., (1985) Mol. Gen. Genet. 200:220 (нуклеотидная последовательность гена левансахаразы Bacillus subtilis), Pen, et al., (1992) Bio/Technology 10:292 (получение трансгенных растений, которые экспрессируют альфа-амилазу Bacillus licheniformis), Elliot, et al., (1993) Plant Molec. Biol. 21:515 (нуклеотидные последовательности генов инвертазы томата), Søgaard, et al., (1993) J. Biol. Chem. 268:22480 (сайт-направленный мутагенез гена альфа-амилазы ячменя) и Fisher, et al., (1993) Plant Physiol. 102:1045 (фермент ветвления крахмала II эндосперма маиса), WO 1999/10498 (улучшенная усвояемость и/или извлечение крахмала путем модификации UDP-D-ксилоза 4-эпимеразы, Fragile 1 и 2, Ref1, HCHL, C4H), патент США № 6,232,529 (способ получения семян с высоким содержанием масла путем модификации уровней крахмала (AGP)). Гены модификации жирных кислот, упомянутые в данном документе, также можно применять для воздействия на содержание и/или состав крахмала благодаря взаимосвязи путей метаболизма крахмала и масла.
(D) Измененное содержание или состав антиоксидантов, как, например, изменение токоферола или токотриенолов. Например, см. патент США № 6787683, публикацию заявки на патент США № 2004/0034886 и WO 2000/68393, предусматривающие манипуляцию с уровнями антиоксидантов, и WO 2003/082899, благодаря изменению гомогентизатгеранил-геранилтрансферазы (hggt).
(E) Измененные незаменимые аминокислоты семян. Например, см. патент США № 6127600 (способ повышения накопления незаменимых аминокислот в семенах), патент США № 6080913 (бинарные способы повышения накопления незаменимых аминокислот в семенах), патент США № 5990389 (высокое содержание лизина), WO 1999/40209 (изменение аминокислотного состава семян), WO 1999/29882 (способы изменения аминокислотного состава белков), патент США № 5850016 (изменение аминокислотного состава семян), WO 1998/20133 (белки с повышенными уровнями незаменимых аминокислот), патент США № 5885802 (высокое содержание метионина), патент США № 5885801 (высокое содержание треонина), патент США № 6664445 (растительные ферменты биосинтеза аминокислот), патент США № 6459019 (повышенное содержание лизина и треонина), патент США № 6441274 (бета-субъединица растительной триптофансинтазы), патент США № 6346403 (ферменты метаболизма метионина), патент США № 5939599 (высокое содержание серы), патент США № 5912414 (повышенное содержание метионина), WO 1998/56935 (растительные ферменты биосинтеза аминокислот), WO 1998/45458 (разработанный с помощью генной инженерии белок семени, имеющий более высокую процентную долю незаменимых аминокислот), WO 1998/42831 (повышенное содержание лизина), патент США № 5633436 (повышение содержания серосодержащих аминокислот), патент США № 5559223 (синтетические запасные белки с определенной структурой, содержащие программируемые уровни незаменимых аминокислот для улучшения питательной ценности растений), WO 1996/01905 (повышенное содержание треонина), WO 1995/15392 (повышенное содержание лизина), публикацию заявки на патент США № 2003/0163838, публикацию заявки на патент США № 2003/0150014, публикацию заявки на патент США № 2004/0068767, патент США № 6803498, WO 2001/79516.
4. Гены, контролирующие мужскую стерильность
Доступно несколько способов обеспечения генетической мужской стерильности, как, например, несколько мутантных генов в отдельных положениях в пределах генома, которые придают мужскую стерильность, как раскрыто в патентах США №№ 4654465 и 4727219, Brar, et al., и хромосомные транслокации, как описано Patterson в патентах США №№ 3861709 и 3710511. В дополнение к этим способам, Albertsen, et al. в патенте США № 5432068 описывают систему ядерной мужской стерильности, которая включает: идентификацию гена, который крайне важен для мужской фертильности; сайленсинг этого нативного гена, который крайне важен для мужской фертильности; удаление нативного промотора из гена, важного для мужской фертильности, и замещение его на индуцируемый промотор; вставку этого полученного с помощью генной инженерии гена обратно в растение и, таким образом, создание растения, которое характеризуется мужской стерильностью, поскольку индуцируемый промотор не "включен", в результате чего ген мужской фертильности не транскрибируется. Фертильность восстанавливают посредством индуцирования или "включения" промотора, который, в свою очередь, осуществляет транскрипцию гена, который обеспечивает мужскую фертильность.
(A) Введение гена деацетилазы под управлением промотора, специфического в отношении тапетума, и с применением химического N-Ac-PPT (WO 2001/29237).
(B) Введение различных промоторов, специфических в отношении тычинок (WO 1992/13956, WO 1992/13957).
(C) Введение барназы и гена барстара (Paul, et al., (1992) Plant Mol. Biol. 19:611-622).
Дополнительные примеры систем и генов ядерной мужской и женской стерильности см. также в патентах США №№ 5859341; 6297426; 5478369; 5824524; 5850014 и 6265640, все из которых, тем самым, включены посредством ссылки.
5. Гены, которые создают сайт для сайт-специфической интеграции ДНК
Подразумевается введение сайтов FRT, которые можно применять в системе FLP/FRT, и/или сайтов Lox, которые можно применять в системе Cre/Loxp. Например, см. Lyznik, et al., (2003) Plant Cell Rep 21:925-932 и WO 1999/25821, которые включены в данный документ посредством ссылки. Другие системы, которые можно применять, включают рекомбиназу Gin из фага Mu (Maeser, et al., (1991) Vicki Chandler, The Maize Handbook ch. 118 (Springer-Verlag 1994), рекомбиназу Pin из E. coli (Enomoto, et al., 1983) и систему R/RS из плазмиды pSRi (Araki, et al., 1992).
6. Гены, которые воздействуют на устойчивость к абиотическому стрессу
В том числе без ограничений на цветение, развитие початка и семени, улучшение эффективности использования азота, измененную реактивность в отношении азота, устойчивость к засухе или ее переносимость, устойчивость к холоду или его переносимость, а также устойчивость к засолению или его переносимость и повышенную урожайность при стрессе.
(A) Например, см. WO 2000/73475, в котором эффективность использования воды изменена благодаря изменению малата; Патенты США №№ 5,892,009, 5,965,705, 5,929,305, 5,891,859, 6,417,428, 6,664,446, 6,706,866, 6,717,034, 6,801,104, WO 2000/060089, WO 2001/026459, WO 2001/035725, WO 2001/034726, WO 2001/035727, WO 2001/036444, WO 2001/036597, WO 2001/036598, WO 2002/015675, WO 2002/017430, WO 2002/077185, WO 2002/079403, WO 2003/013227, WO 2003/013228, WO 2003/014327, WO 2004/031349, WO 2004/076638, WO 199809521.
(B) WO 199938977, в которой описаны гены, в том числе гены CBF, и факторы транскрипции, эффективные в ослаблении негативных эффектов замораживания, высокого содержания солей и засухи на растения, а также обеспечивающие другие положительные эффекты в отношении фенотипа растения.
(C) Публикация заявки на патент США № 2004/0148654 и WO 2001/36596, где в растениях изменяется содержание абсцизовой кислоты, что приводит к улучшенному фенотипу растения, такому как повышенная урожайность и/или повышенная переносимость абиотического стресса.
(D) WO 2000/006341, WO 2004/090143, патенты США №№ 7531723 и 6992237, где экспрессия цитокинина модифицируется, что приводит к растениям с повышенной переносимостью стрессов, как, например, переносимостью засухи, и/или повышенной урожайностью. Также см. WO 2002/02776, WO 2003/052063, JP 2002/281975, патент США № 6084153, WO 2001/64898, патент США № 6177275 и патент США № 6107547 (улучшение использования азота и измененная реактивность в отношении азота).
(E) Что касается изменения содержания этилена, см. публикацию заявки на патент США № 2004/0128719, публикацию заявки на патент США № 2003/0166197 и WO 2000/32761.
(F) Что касается растительных факторов транскрипции или транскрипционных регуляторов, связанных с реакцией на абиотический стресс, см., например, публикацию заявки на патент США № 2004/0098764 или публикацию заявки на патент США № 2004/0078852.
(G) Гены, которые повышают экспрессию вакуолярной пирофосфатазы, такие как AVP1 (патент США № 8058515), для повышенного урожая; нуклеиновая кислота, кодирующая полипептиды HSFA4 или HSFA5 (фактор теплового шока класса A4 или A5), полипептид, подобный белку транспортера олигопептидов, (OPT4-подобный); пластохрон-2-подобный (PLA2-подобный) полипептид или Wuschel-родственный гомеобокс 1-подобный (WOX1-подобный) полипептид (публикация заявки на патент США № US 2011/0283420).
(H) Подавление полинуклеотидов, кодирующих белки поли-(АДФ-рибоза)-полимеразы (PARP), с целью модуляции запрограммированной гибели клеток (патент США № 8058510) для повышения мощности.
(I) Полинуклеотид, кодирующий полипептиды DTP21, для обеспечения устойчивости к засухе (публикация заявки на патент США № US 2011/0277181).
(J) Нуклеотидные последовательности, кодирующие белки ACC синтазы 3 (ACS3) для модуляции развития, модуляции реакции на стресс и модуляции переносимости стрессов (публикация заявки на патент США № US 2010/0287669).
(K) Полинуклеотиды, которые кодируют белки, которые обеспечивают фенотип переносимости засухи (DTP), для обеспечения устойчивости к засухе (WO 2012/058528).
(L) Гены токоферолциклазы (TC) для обеспечения переносимости засухи и засоления (публикация заявки на патент США № 2012/0272352).
(M) Белки семейства протеаз, нацеленные на CAAX-концевые аминокислоты, для придания переносимости стрессов (патент США № 8338661).
(N) Мутации в гене, кодирующем SAL1, характеризовались повышенной переносимостью стрессов, в том числе характеризовались повышенной устойчивостью к засухе (публикация заявки на патент США № 2010/0257633).
(O) Экспрессия последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептида GRF, RAA1-подобного полипептида, полипептида SYR, полипептида ARKL и полипептида YTP, усиливающих признаки, связанные с урожайностью (публикация заявки на патент США № 2011/0061133).
(P) Модуляция экспрессии в растении нуклеиновой кислоты, кодирующей полипептид трегалозо-фосфатфосфатазу (TPP) класса III для улучшения признаков, связанных с урожайностью, у растений, в частности, повышения урожайности семян (публикация заявки на патент США № 2010/0024067).
Другие гены и факторы транскрипции, которые воздействуют на рост и агротехнические признаки растений, такие как урожайность, цветение, рост растения и/или структура растения, можно вводить или интрогрессировать в растения, см., например, WO 1997/49811 (LHY), WO 1998/56918 (ESD4), WO 1997/10339 и патент США № 6573430 (TFL), патент США № 6713663 (FT), WO 1996/14414 (CON), WO 1996/38560, WO 2001/21822 (VRN1), WO 2000/44918 (VRN2), WO 1999/49064 (GI), WO 2000/46358 (FR1), WO 1997/29123, патент США № 6794560, патент США № 6307126 (GAI), WO 1999/09174 (D8 и Rht), и WO 2004/076638, и WO 2004/031349 (факторы транскрипции).
7. Гены, которые обеспечивают повышенную урожайность
(A) Трансгенное культурное растение, трансформированное при помощи нуклеиновой кислоты, кодирующей 1-аминоциклопропан-1-карбоксилатдезаминаза-подобный полипептид (ACCDP), где экспрессия последовательности нуклеиновой кислоты в культурном растении приводит к повышенному росту корней, и/или повышенной урожайности, и/или повышенной переносимости стрессов под влиянием факторов окружающей среды у растения по сравнению с разновидностью дикого типа растения (патент США № 8097769).
(B) Сверхэкспрессия гена белков "цинковых пальцев" маиса (Zm-ZFP1) с применением промотора, предпочтительного для семян, как было показано, усиливает рост растения, увеличивает количество зерен и общий вес зерен на растение (публикация заявки на патент США № 2012/0079623).
(C) Конститутивная сверхэкспрессия белка с доменом границ латеральных органов (LOB) (Zm-LOBDP1) маиса, как было показано, увеличивает количество зерен и общий вес зерен на растение (публикация заявки на патент США № 2012/0079622).
(D) Улучшение признаков, связанных с урожайностью, у растений посредством модуляции экспрессии у растения нуклеиновой кислоты, кодирующей VIM1- (вариант с метилированием 1) подобный полипептид или VTC2-подобный (GDP-L-галактоза-фосфорилаза) полипептид, или полипептид DUF1685, или ARF6-подобный (восприимчивый к ауксину фактор) полипептид (WO 2012/038893).
(E) Модуляция экспрессии в растении нуклеиновой кислоты, кодирующей Ste20-подобный полипептид или его гомолог, позволяет растениям производить повышенный урожай относительно контрольных растений (EP 2431472).
(F) Гены, кодирующие полипептиды нуклеозиддифосфаткиназы (NDK) и их гомологи для модификации строения корня растения (публикация заявки на патент США № 2009/0064373).
8. Гены, которые обеспечивают усвояемость растения
(A) Изменение уровня ксилана, присутствующего в клеточной стенке растения, посредством модуляции экспрессии ксилансинтазы (патент США № 8173866).
В некоторых вариантах осуществления пакетированный признак может представлять собой признак или трансгенный объект, который получил разрешение контролирующих органов, в том числе без ограничения является трансгенным объектом из таблицы 5A - 5F.
Таблица 5A. Люцерна Medicago sativa
Таблица 5B. Подсолнечник Helianthus annuus
Таблица 5C. Рис Oryza sativa
Таблица 5D. Пшеница Triticum aestivum
Таблица 5E. Соя Glycine max L.
Таблица 5F. Маис Zea mays L.
Другие трансгенные объекты, разрешенные контролирующими органами, хорошо известны специалисту в данной области, и их можно найти на сайте Центра оценки риска для окружающей среды (cera-gmc.org/?action=gm_crop_database, доступ к которому можно получить с применением префикса www) и на сайте Международной службы по приобретению агробиотехнологических приложений (isaaa.org/gmapprovaldatabase/default.asp, доступ к которому можно получить с применением префикса www).
Сайленсинг генов
В некоторых вариантах осуществления пакетированный признак может представлять собой форму, применяемую для сайленсинга одного или нескольких полинуклеотидов, представляющих интерес, что приводит к супрессии одного или нескольких целевых полипептидов вредителя. В некоторых вариантах осуществления сайленсинг достигается благодаря применению супрессионной ДНК-конструкции.
В некоторых вариантах осуществления один или несколько полинуклеотидов, кодирующих полипептиды из полипептида PtIP-83 или их фрагментов или вариантов, могут быть пакетированы с одним или несколькими полинуклеотидами, кодирующими один или несколько полипептидов с инсектицидной активностью или агрономическими признаками, изложенными выше, и необязательно может дополнительно предусматривать один или несколько полинуклеотидов, предусмотренных для сайленсинга генов одного или нескольких целевых полинуклеотидов, как обсуждается ниже.
"Супрессионная ДНК-конструкция" представляет собой рекомбинантную ДНК-конструкцию, которая при трансформации или стабильной интеграции в геном растения приводит к "сайленсингу" целевого гена в растении. Целевой ген может быть эндогенным или трансгенным по отношению к растению. "Сайленсинг", применяемый в данном документе по отношению к целевому гену, обычно относится к супрессии уровней мРНК или белка/фермента, экспрессируемых целевым геном, и/или уровня активности фермента или функционирования белка. Термин "супрессия" предусматривает понижение, снижение, ухудшение, уменьшение, ингибирование, устранение и предотвращение. "Сайленсинг" или "сайленсинг генов" не определяет механизм и включает, без ограничения, супрессию, опосредованную антисмысловыми олигонуклеотидами, косупрессию, супрессию, опосредованную вирусами, супрессию, опосредованную шпильками, супрессию, опосредованную структурами типа "стебель-петля", подходы на основе RNAi и подходы на основе низкомолекулярных РНК.
Супрессионная ДНК-конструкция может содержать участок, полученный из целевого гена, представляющего интерес, и может содержать всю последовательность нуклеиновой кислоты смысловой нити (или антисмысловой нити) целевого гена, представляющего интерес, или ее часть. В зависимости от подхода, который будет использоваться, участок может быть на 100% идентичным или менее чем на 100% идентичным (например, по меньшей мере на 50% или на любое целое число от 51% до 100% идентичным) всей смысловой нити (или антисмысловой нити) гена, представляющего интерес, или ее части.
Супрессионные ДНК-конструкции хорошо известны из уровня техники, легко конструируются сразу после выбора целевого гена, представляющего интерес, и включают без ограничения косупрессионные конструкции, антисмысловые конструкции, вирусные супрессионные конструкции, шпильковые супрессионные конструкции, супрессионные конструкции типа "стебель-петля", конструкции, вырабатывающие двухнитевые РНК, и, в более общем смысле, конструкции для RNAi (РНК-интерференции) и конструкции на основе малых РНК, такие как конструкции на основе siRNA (коротких интерферирующих РНК) и miRNA (микроРНК).
"Антисмысловое ингибирование" относится к продуцированию антисмысловых РНК-транскриптов, способных супрессировать экспрессию целевого белка.
"Антисмысловая РНК" относится к РНК-транскрипту, который комплементарен всему или части целевого первичного транскрипта или мРНК и который блокирует экспрессию фрагмента целевой выделенной нуклеиновой кислоты (патент США № 5107065). Комплементарность антисмысловой РНК может быть с любой частью конкретного генного транскрипта, т.е. с 5'-некодирующей последовательностью, 3'-некодирующей последовательностью, интронами или кодирующей последовательностью.
"Косупрессия" относится к выработке смысловых РНК-транскриптов, способных супрессировать экспрессию целевого белка. "Смысловая" РНК относится к РНК-транскрипту, который включает мРНК, и она может быть транслирована с образованием белка в клетке или in vitro. Конструкции для косупрессии у растений ранее разрабатывались с основным вниманием к сверхэкспрессии последовательностей нуклеиновой кислоты, имеющих гомологию с нативной мРНК, в смысловой ориентации, что приводит к снижению уровней всех РНК, имеющих гомологию со сверхэкспрессированной последовательностью (см., Vaucheret, et al., (1998) Plant J. 16:651-659 и Gura, (2000) Nature 404:804-808).
В другом варианте описывается применение последовательностей растительных вирусов для управления супрессией проксимальных элементов мРНК-кодирующих последовательностей (публикация согласно PCT WO 1998/36083).
В недавней работе было описано применение "шпильковых" структур, которые включают всю или часть мРНК-кодирующей последовательности в комплементарной ориентации, что приводит к потенциальной структуре "стебель-петля" у экспрессированной РНК (публикация согласно PCT WO 1999/53050). В данном случае стебель формируется полинуклеотидами, соответствующими гену, представляющему интерес, в смысловой или в антисмысловой ориентации по отношению к промотору, а петля формируется несколькими полинуклеотидами гена, представляющего интерес, которые не имеют комплементарной последовательности в конструкции. Это повышает частоту косупрессии или сайленсинга в регенерированных трансгенных растениях. Обзор шпильковой супрессии см. в Wesley, et al., (2003) Methods in Molecular Biology, Plant Functional Genomics: Methods and Protocols 236:273-286.
Конструкция, в которой стебель образован по меньшей мере 30 нуклеотидами из гена, который подлежит супрессии, и петля образована случайной нуклеотидной последовательностью, также эффективно применялась для супрессии (публикация согласно PCT WO 1999/61632).
Также было описано применение последовательностей поли-T и поли-A для получения стебля в структуре "стебель-петля" (публикация согласно PCT WO 2002/00894).
Еще один вариант включает применение синтетических повторов, способствующих образованию стебля в структуре "стебель-петля". Было показано, что трансгенные организмы, полученные с такими фрагментами рекомбинантной ДНК, характеризуются сниженными уровнями белка, кодируемого нуклеотидным фрагментом, который образует петлю, как описано в публикации согласно РСТ WO 2002/00904.
РНК-интерференция относится к процессу специфичного в отношении последовательности посттранскрипционного сайленсинга генов у животных, который опосредован короткими интерферирующими РНК (siRNA) (Fire, et al., (1998) Nature 391:806). Соответствующий процесс у растений обычно называют посттранскрипционным сайленсингом генов (PTGS) или РНК-опосредованным сайленсингом, и также называют подавлением в случае грибов. Процесс посттранскрипционного сайленсинга генов считается эволюционно-консервативным механизмом клеточной защиты, применяющимся для предотвращения экспрессии чужеродных генов, и он широко распространен у различных представителей растительного мира и таксономических групп растений (Fire, et al., (1999) Trends Genet. 15:358). Такая защита от экспрессии чужеродных генов могла развиться в ответ на выработку двухцепочечных РНК (dsRNA) в результате вирусной инфекции или в результате случайной интеграции транспозонных элементов в геном хозяина путем клеточного ответа, который приводит к специфическому разрушению гомологичной одноцепочечной РНК вирусной геномной РНК. Присутствие dsRNA в клетках вызывает RNAi-ответ через механизм, который полностью еще не охарактеризован.
Присутствие длинных dsRNA в клетках стимулирует активность фермента рибонуклеазы III, называемого дайсер. Дайсер вовлечен в процессинг dsRNA с образованием коротких фрагментов dsRNA, называемых короткими интерферирующими РНК (siRNA) (Berstein et al., Nature 409:363, 2001). Короткие интерферирующие РНК, полученные в результате активности дайсера, как правило, имеют длину от приблизительно 21 до приблизительно 23 нуклеотидов и содержат дуплексы приблизительно из 19 пар оснований (Elbashir et al., (2001) Genes Dev. 15:188). Кроме того, дайсер вовлечен в вырезание 21- и 22-нуклеотидных малых временных РНК (stRNA) из РНК-предшественника консервативной структуры, которые вовлечены в контроль на уровне трансляции (Hutvagner, et al., (2001) Science 293:834). Для RNAi-ответа также характерен эндонуклеазный комплекс, обычно называемый комплексом РНК-индуцированного сайленсинга (RISC), который опосредует расщепление одноцепочечной РНК, последовательность которой комплементарна антисмысловой цепи дуплекса siRNA. Расщепление целевой РНК происходит в середине участка, комплементарного антисмысловой цепи дуплекса siRNA (Elbashir et al., (2001) Genes Dev. 15:188). Кроме того, РНК-интерференция также может включать сайленсинг генов, опосредованный малыми РНК (например, miRNA), предположительно посредством клеточных механизмов, которые регулируют структуру хроматина, и, тем самым, предотвращают транскрипцию последовательностей целевых генов (см., например, Allshire, (2002) Science 297:1818-1819; Volpe, et al., (2002) Science 297:1833-1837; Jenuwein, (2002) Science 297:2215-2218 и Hall, et al., (2002) Science 297:2232-2237). Таким образом, молекулы miRNA согласно настоящему раскрытию можно применять для опосредования сайленсинга генов путем взаимодействия с РНК-транскриптами или, в качестве альтернативы, взаимодействия с конкретными генными последовательностями, где такое взаимодействие приводит к сайленсингу генов либо на транскрипционном, либо на посттранскрипционном уровне.
Дополнительно представлены способы и композиции, которые обеспечивают возможность увеличения количества RNAi, полученной с помощью элемента сайленсинга. В таких вариантах осуществления в способах и композициях используется первый полинуклеотид, содержащий элемент сайленсинга для целевой последовательности вредителя, функционально связанный с промотором, активным в растительной клетке; и второй полинуклеотид, содержащий усиливающий супрессию элемент, содержащий целевую последовательность вредителя или ее активный вариант или фрагмент, функционально связанные с промотором, активным в растительной клетке. Комбинированная экспрессия элемента сайленсинга с усиливающим супрессию элементом ведет к увеличению амплификации ингибиторных РНК, вырабатываемых на основе элемента сайленсинга сверх того уровня, который достигается при экспрессии только элемента сайленсинга самого по себе. В дополнение к увеличенной амплификации специфических видов RNAi самих по себе, способы и композиции дополнительно обеспечивают возможность выработки отличной группы видов RNAi, которые могут повышать эффективность нарушения экспрессии целевого гена. Таким образом, когда усиливающий супрессию элемент экспрессируется в растительной клетке в комбинации с элементом сайленсинга, способы и композиции могут обеспечивать возможность системной выработки RNAi в растении; выработку больших количеств RNAi, чем можно было бы наблюдать в случае только конструкции с элементом сайленсинга отдельно; и повышенную нагрузку RNAi во флоэме растения, таким образом, обеспечивается лучший контроль насекомых, питающихся флоэмой, при помощи подхода с RNAi. Таким образом, различные способы и композиции обеспечивают улучшенные способы доставки ингибиторных РНК в целевой организм. См., например, публикацию заявки на патент США 2009/0188008.
Применяемый в данном документе "усиливающий супрессию элемент" включает полинуклеотид, содержащий целевую последовательность, подлежащую супрессии, или ее активный фрагмент или вариант. Понятно, что усиливающий супрессию элемент не должен быть идентичным целевой последовательности, а скорее, усиливающий супрессию элемент может содержать вариант целевой последовательности, при условии, что последовательность усиливающего супрессию элемента в достаточной степени идентична целевой последовательности, чтобы обеспечить возможность увеличенного уровня RNAi, вырабатываемых на основе элемента сайленсинга, сверх того уровня, который достигается при экспрессии только элемента сайленсинга. Аналогично, усиливающий супрессию элемент может содержать фрагмент целевой последовательности, где фрагмент имеет достаточную длину, чтобы обеспечить возможность увеличенного уровня RNAi, вырабатываемых на основе элемента сайленсинга, сверх того уровня, который достигается при экспрессии только элемента сайленсинга.
Понятно, что можно использовать несколько усиливающих супрессию элементов из одной целевой последовательности, или из различных целевых последовательностей, или из различных участков одной целевой последовательности. Например, используемые усиливающие супрессию элементы могут содержать фрагменты целевой последовательности, полученные из другого участка целевой последовательности (т. е., из 3′UTR, кодирующей последовательности, интрона и/или 5′UTR). Кроме того, усиливающий супрессию элемент может содержаться в кассете экспрессии, как описано в другой части данного документа, и в определенных вариантах осуществления усиливающий супрессию элемент находится в том же самом или в другом ДНК-векторе или конструкции по отношению к элементу сайленсинга. Усиливающий супрессию элемент может быть функционально связан с промотором, раскрытым в данном документе. Понятно, что усиливающий супрессию элемент может экспрессироваться конститутивно, или, в качестве альтернативы, он может вырабатываться специфичным в отношении стадии образом, с использованием различных индуцируемых, или предпочтительных для определенной ткани, или регулируемых в процессе развития промоторов, которые рассмотрены в другой части данного документа.
В конкретных вариантах осуществления при использовании как элемента сайленсинга, так и усиливающего супрессию элемента системная выработка RNAi происходит по всему растению. В дополнительных вариантах осуществления растение или части растения согласно настоящему раскрытию имеют повышенную нагрузку RNAi во флоэме растения, по сравнению с той, которую можно было бы наблюдать при экспрессии конструкции элемента сайленсинга отдельно, и, таким образом, обеспечивается лучший контроль насекомых, питающихся за счет флоэмы, при помощи подхода с RNAi. В определенных вариантах осуществления растения, части растения и растительные клетки согласно настоящему раскрытию можно дополнительно охарактеризовать, как обеспечивающие возможность выработки разнообразных видов RNAi, которые могут повышать эффективность нарушения экспрессии целевого гена.
В конкретных вариантах осуществления комбинированная экспрессия элемента сайленсинга и усиливающего супрессию элемента повышает концентрацию ингибиторных РНК в растительной клетке, растении, части растения, растительной ткани или флоэме сверх того уровня, который достигается при экспрессии элемента сайленсинга отдельно.
Применяемый в данном документе "повышенный уровень ингибиторных РНК" включает какое-либо статистически значимое повышение уровня RNAi, вырабатываемых в растении, обладающем комбинированной экспрессией при сравнении с подходящим контрольным растением. Например, повышение уровня RNAi в растении, части растения или растительной клетке может включать по меньшей мере приблизительно 1%, приблизительно 1%-5%, приблизительно 5%-10%, приблизительно 10%-20%, приблизительно 20%-30%, приблизительно 30%-40%, приблизительно 40%-50%, приблизительно 50%-60%, приблизительно 60-70%, приблизительно 70%-80%, приблизительно 80%-90%, приблизительно 90%-100% или большее повышение уровня RNAi в растении, части растения, растительной клетке или флоэме при сравнении с подходящим контролем. В других вариантах осуществления повышение уровня RNAi в растении, части растения, растительной клетке или флоэме может включать по меньшей мере приблизительно 1-кратное, приблизительно 1-кратное - 5-кратное, приблизительно 5-кратное - 10-кратное, приблизительно 10-кратное - 20-кратное, приблизительно 20-кратное - 30-кратное, приблизительно 30-кратное - 40-кратное, приблизительно 40-кратное - 50-кратное, приблизительно 50-кратное - 60-кратное, приблизительно 60-кратное - 70-кратное, приблизительно 70-кратное - 80-кратное, приблизительно 80-кратное - 90-кратное, приблизительно 90-кратное - 100-кратное или большее повышение уровня RNAi в растении, части растения, растительной клетке или флоэме при сравнении с подходящим контролем. Примеры комбинированной экспрессии элемента сайленсинга с усиливающим супрессию элементом для контроля щитников и Lygus можно найти в публикации заявки на патент США 2011/0301223 и публикации заявки на патент США 2009/0192117.
Некоторые варианты осуществления относятся к подавлению экспрессии целевых генов у видов насекомых-вредителей при помощи молекул интерферирующей рибонуклеиновой кислоты (РНК). В публикации согласно PCT WO 2007/074405 описаны способы ингибирования экспрессии целевых генов у беспозвоночных вредителей, в том числе колорадского жука. В публикации согласно PCT WO 2005/110068 описаны способы ингибирования экспрессии целевых генов у беспозвоночных вредителей, в том числе, в частности, у западного кукурузного жука, в качестве средства контроля заражения насекомыми. Кроме того, в публикации согласно PCT WO 2009/091864 описаны композиции и способы супрессии целевых генов из видов насекомых-вредителей, в том числе вредителей из рода Lygus. Молекулы нуклеиновой кислоты, в том числе RNAi для нацеливания на H-субъединицу вакуолярной АТФазы, применимы для контроля популяции вредителей из группы жесткокрылых и заражения ими, как описано в публикации заявки на патент США 2012/0198586. В публикации согласно PCT WO 2012/055982 описана рибонуклеиновая кислота (РНК или двухцепочечная РНК), которая ингибирует или обеспечивает подавление экспрессии целевого гена, который кодирует: рибосомальный белок насекомого, такой как рибосомальный белок L19, рибосомальный белок L40 или рибосомальный белок S27A; субъединицу протеасомы насекомого, такую как белок Rpn6, Pros 25, белок Rpn2, белок бета 1 субъединицы протеасомы или белок бета 2 Pros; β-коатомер COPI-везикулы насекомого, γ-коатомер COPI-везикулы, β'-коатомерный белок или ζ-коатомер COPI-везикулы; белок тетраспанин 2 A насекомого, который представляет собой предполагаемый белок трансмембранного домена; белок насекомого, принадлежащий к семейству актина, такой как актин 5C; белок убиквитин-5E насекомого; белок Sec23 насекомого, который представляет собой активатор ГТФазы, вовлеченной во внутриклеточный транспорт белков; белок "crinkled" насекомого, который представляет собой нестандартный миозин, который вовлечен в двигательную активность; белок "crooked neck" насекомого, который вовлечен в регуляцию ядерного альтернативного сплайсинга мРНК; белок G-субъединицы вакуолярной H+-АТФазы насекомого и Tbp-1 насекомого, такой как Tat-связывающий белок. В публикациях заявок на патент США 2012/029750, US 20120297501 и 2012/0322660 описаны интерферирующие рибонуклеиновые кислоты (РНК или двухцепочечные РНК), которые функционируют при поглощении видами насекомых-вредителей с подавлением экспрессии целевого гена в указанном насекомом-вредителе, где РНК содержит по меньшей мере один элемент сайленсинга, где элемент сайленсинга представляет собой участок двухцепочечной РНК, содержащий гибридизованные комплементарные цепи, из которых одна цепь содержит или состоит из последовательности нуклеотидов, которая по меньшей мере частично комплементарна целевой нуклеотидной последовательности в пределах целевого гена. В публикации заявки на патент США 2012/0164205 описаны потенциальные мишени для интерферирующих двухцепочечных рибонуклеиновых кислот для подавления беспозвоночных вредителей, в том числе: гомологичная последовательность Chd3, гомологичная последовательность бета-тубулина, a 40 kDa гомологичная последовательность 40 кДа V-АТФазы, гомологичная последовательность EF1α, гомологичная последовательность субъединицы p28 протеосомы 26S, гомологичная последовательность эпоксидгидролазы ювенильного гормона, гомологичная последовательность белка хлоридных каналов, зависимых от набухания, гомологичная последовательность белка глюкоза-6-фосфат-1-дегидрогеназы, гомологичная последовательность белка Act42A, гомологичная последовательность фактора 1 АДФ-рибозилирования, гомологичная последовательность белка фактора транскрипции IIB, гомологичные последовательности хитиназы, гомологичная последовательность фермента, конъюгирующего убиквитин, гомологичная последовательность глицеральдегид-3-фосфатдегидрогеназы, гомологичная последовательность убиквитина B, гомолог эстеразы ювенильного гормона и гомологичная последовательность альфа-тубулина.
Применение в пестицидном контроле
Из уровня техники известны общие способы использования штаммов, содержащих последовательность нуклеиновой кислоты согласно вариантам осуществления или ее вариант, в пестицидном контроле или при конструировании других организмов в качестве пестицидных средств. См., например, патент США № 5039523 и EP 0480762A2.
Могут быть выбраны микроорганизмы-хозяева, которые, как известно, заселяют "фитосферу" (филлоплан, филлосферу, ризосферу и/или ризоплан) одной или нескольких сельскохозяйственных культур, представляющих интерес. Эти микроорганизмы выбирают таким образом, чтобы они могли успешно конкурировать в конкретной окружающей среде с микроорганизмами дикого типа, обеспечивать стабильное поддержание и экспрессию гена, экспрессирующего полипептид PtIP-83, и, желательно, обеспечивать улучшенную защиту пестицида от разрушения и инактивации в окружающей среде.
Такие микроорганизмы включают бактерии, водоросли и грибы. Особый интерес представляют микроорганизмы, такие как бактерии, например, Pseudomonas, Erwinia, Serratia, Klebsiella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylius, Agrobacterium, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc и Alcaligenes, грибы, в частности дрожжи, например, Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotorula и Aureobasidium. Особый интерес представляют такие виды бактерий фитосферы как Pseudomonas syringae, Pseudomonas fluorescens, Pseudomonas chlororaphis, Serratia marcescens, Acetobacter xylinum, Agrobacteria, Rhodopseudomonas spheroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes entrophus, Clavibacter xyli и Azotobacter vinelandii и виды дрожжей фитосферы, такие как Rhodotorula rubra, R. glutinis, R. marina, R. aurantiaca, Cryptococcus albidus, C. diffluens, C. laurentii, Saccharomyces rosei, S. pretoriensis, S. cerevisiae, Sporobolomyces roseus, S. odorus, Kluyveromyces veronae и Aureobasidium pollulans. Особый интерес представляют пигментированные микроорганизмы. Организмы-хозяева, представляющие особый интерес, включают дрожжи, такие как Rhodotorula spp., Aureobasidium spp., Saccharomyces spp. (такие как S. cerevisiae), Sporobolomyces spp., организмы филлоплана, такие как Pseudomonas spp. (такие как P. aeruginosa, P. fluorescens, P. chlororaphis), Erwinia spp., и Flavobacterium spp., и другие такие организмы, в том числе Agrobacterium tumefaciens, E. coli, Bacillus subtilis, Bacillus cereus и т.п.
Гены, кодирующие полипептид PtIP-83 согласно вариантам осуществления можно вводить в микроорганизмы, которые размножаются на растениях (эпифиты) для доставки полипептида PtIP-83 в организм потенциальных целевых вредителей. Эпифиты, например, могут представлять собой грамположительные или грамотрицательные бактерии.
Бактерии, колонизирующие корни, например, можно выделять из растений, представляющих интерес, при помощи способов, известных из уровня техники. А именно, штамм Bacillus cereus, который колонизирует корни, можно выделять из корней растения (см., например, Handelsman et al. (1991) Appl. Environ. Microbiol. 56:713-718). Гены, кодирующие полипептид PtIP-83 согласно вариантам осуществления можно вводить в Bacillus cereus, колонизирующие корни, при помощи стандартных способов, известных из уровня техники.
Гены, кодирующие полипептиды PtIP-83, можно вводить, например, в Bacillus, колонизирущие корни, посредством электротрансформации. А именно, гены, кодирующие полипептиды PtIP-83, можно клонировать в челночный вектор, например, pHT3101 (Lerecius, et al., (1989) FEMS Microbiol. Letts. 60:211-218. При помощи челночного вектора pHT3101, содержащего кодирующую последовательность конкретного гена полипептида PtIP-83, можно, например, трансформировать Bacillus, колонизирующие корни, посредством электропорации (Lerecius, et al., (1989) FEMS Microbiol. Letts. 60:211-218).
Системы экспрессии можно конструировать таким образом, что полипептиды PtIP-83 секретируются вне цитоплазмы грамотрицательных бактерий, таких как, например, E. coli. Преимущества секреции полипептидов PtIP-83 являются следующими: (1) предупреждение потенциальных цитотоксических эффектов экспрессированного полипептида PtIP-83; и (2) повышение эффективности очистки полипептида PtIP-83, в том числе без ограничения, повышение эффективности выделения и очистки белка на объем клеточного бульона и снижение времени и/или затрат на выделение и очистку на индивидуальный белок.
Для целей секреции у E. coli, полипептиды PtIP-83 можно создавать, например, путем слияния соответствующего сигнального пептида E. coli с амино-терминальным концом полипептида PtIP-83. Сигнальные пептиды, распознаваемые E. coli, можно найти в белках, которые, как уже известно, секретируются у E. coli, например, белок OmpA (Ghrayeb, et al., (1984) EMBO J, 3:2437-2442). OmpA представляет собой главный белок внешней мембраны E. coli, и, таким образом, полагают, что его сигнальный пептид является эффективным в процессе транслокации. Также, сигнальный пептид OmpA не нужно модифицировать перед процессингом, как может быть в случае других сигнальных пептидов, например, сигнального пептида липопротеинов (Duffaud, et al., (1987) Meth. Enzymol. 153:492).
Полипептиды PtIP-83 согласно вариантам осуществления можно ферментировать в бактерии-хозяине, а полученные бактерии обрабатывать и использовать в качестве микробных распыляемых растворов таким же образом, что и штаммы Bt, которые применялись в качестве инсектицидных распыляемых растворов. В случае полипептида(ов) PtIP-83, который секретируется Bacillus, сигнал секреции удаляют или подвергают мутации с применением методик, известных из уровня техники. Такие мутации и/или делеции предотвращают секрецию полипептида(ов) PtIP-83 в ростовую среду во время процесса ферментации. Полипептид PtIP-83 остается в клетке, и клетки затем обрабатывают с получением инкапсулированного полипептида PtIP-83. Для этой цели можно применять любой подходящий микроорганизм. Pseudomonas применяли для экспрессии токсинов Bt в качестве инкапсулированных белков и полученные клетки обрабатывали и распыляли в качестве инсектицида (Gaertner, et al., (1993), в: Advanced Engineered Pesticides, ed. Kim).
В качестве альтернативы, полипептид PtIP-83 получают путем введения гетерологичного гена в клеточного хозяина. Экспрессия гетерологичного гена приводит, прямо или опосредованно, к выработке и сохранению пестицида внутри клетки. Эти клетки затем обрабатывают при условиях, которые продлевают активность токсина, вырабатываемого в клетке, когда осуществляют применение клетки по отношению к среде целевого вредителя(ей). Полученный продукт сохраняет токсичность токсина. Эти инкапсулированные естественным образом полипептиды PtIP-83 затем можно составлять в соответствии с традиционными методиками для внесения в среду пребывания целевого вредителя, например, в почву, воду и на листву растений. См., например, EPA 0192319 и ссылочные документы, процитированные в нем.
Пестицидные композиции
В некоторых вариантах осуществления активные ингредиенты можно наносить в форме композиции и можно наносить на возделываемую площадь или растение, подлежащие обработке, одновременно или последовательно с другими соединениями. Эти соединения могут представлять собой удобрения, средства борьбы с сорняками, криопротекторы, поверхностно-активные вещества, детергенты, пестицидные мыла, масла, применяемые во время состояния покоя, полимеры и/или составы с носителем с замедленным высвобождением или биоразлагаемым носителем, который обеспечивает длительное дозированное внесение на целевой площади после однократного внесения состава. Они также могут представлять собой селективные гербициды, химические инсектициды, вируциды, микробоциды, амебоциды, пестициды, фунгициды, бактерициды, нематоциды, моллюскоциды или смеси из нескольких этих препаратов, при необходимости, вместе с дополнительными приемлемыми с точки зрения сельского хозяйства носителями, поверхностно-активными веществами или вспомогательными средствами, способствующими нанесению, традиционно используемыми в области техники, связанной с получением составов. Подходящие носители и вспомогательные средства могут быть твердыми или жидкими и соответствуют веществам, обычно используемым в технологии составления, например, природным или регенерированным минеральным веществам, растворителям, диспергирующим веществам, смачивающим средствам, веществам, придающим клейкость, связующим или удобрениям. Аналогично, составы можно готовить в виде съедобных "приманок" или формировать в "ловушки" для вредителей, что обеспечивает поедание или заглатывание пестицидного состава целевым вредителем.
Способы нанесения активного ингредиента или агрохимической композиции, которая содержит по меньшей мере один из полипептидов PtIP-83, вырабатываемых штаммами бактерий, включают нанесение на листья, покрытие семян и внесение в почву. Количество применений и норма применения зависят от интенсивности поражения соответствующим вредителем.
Композицию можно составлять в виде порошка, дуста, пеллеты, гранулы, распыляемого раствора, эмульсии, коллоидного вещества, раствора и т.п., и ее можно получать при помощи таких традиционных способов как сушка, лиофилизация, гомогенизация, экстракция, фильтрация, центрифугирование, осаждение или концентрирование культуры клеток, содержащих полипептид. Во всех таких композициях, которые содержат по меньшей мере один такой пестицидный полипептид, полипептид может присутствовать в концентрации от приблизительно 1% до приблизительно 99% по весу.
При помощи способов согласно настоящему раскрытию можно уничтожать вредителей из группы чешуекрылых, двукрылых, настоящих клопов, нематод, полужесткокрылых или жесткокрылых или уменьшать их количество на указанной площади, или такие композиции можно вносить профилактически в окружающую среду для предотвращения заражения восприимчивым вредителем. Предпочтительно, вредитель заглатывает пестицидно эффективное количество полипептида или контактирует с ним. Применяемый в данном документе термин "пестицидно эффективное количество" относится к количеству пестицида, которое может приводить к гибели по меньшей мере одного вредителя или к заметно сниженному росту, питанию или нормальному физиологическому развитию вредителя. Это количество будет варьировать в зависимости от таких факторов, как, например, специфические целевые вредители, подлежащие контролю, специфическая среда, местоположение, растение, культура или сельскохозяйственный участок, подлежащие обработке, условия окружающей среды и способ, норма, концентрация, стабильность и количество внесений пестицидно эффективной полипептидной композиции. Составы также могут варьировать в зависимости от климатических условий, экологических соображений, и/или частоты внесения, и/или тяжести заражения вредителями.
Описанные пестицидные композиции можно создавать путем составления либо суспензии бактериальных клеток, кристаллов и/или спор, либо выделенного белкового компонента с требуемым носителем, приемлемым с точки зрения сельского хозяйства. Композиции можно составлять перед введением с помощью надлежащих способов, таких как лиофилизация, сублимационная сушка, высушивание, или в водном носителе, среде или подходящем растворителе, таком как солевой раствор или другой буфер. Составленные композиции могут находиться в форме дуста, или гранулированного материала, или суспензии в масле (растительном или минеральном), или водных эмульсий или эмульсий масло-в-воде, или в виде смачиваемого порошка, или в комбинации с любым другим материалом носителя, подходящим для сельскохозяйственного применения. Подходящие носители, приемлемые с точки зрения сельского хозяйства, могут быть твердыми или жидкими и хорошо известны из уровня техники. Термин "приемлемый с точки зрения сельского хозяйства носитель" охватывает все адъюванты, инертные компоненты, диспергирующие вещества, поверхностно-активные вещества, вещества, придающие клейкость, связующие и т.д., которые обычно применяются в технологии составления пестицидов; причем они хорошо известны специалистам по составлению пестицидов. Составы можно смешивать с одним или несколькими твердыми или жидкими вспомогательными средствами и получать при помощи различных способов, например, путем равномерного перемешивания, смешивания и/или размалывания пестицидной композиции с подходящими вспомогательными средствами с применением традиционных методик составления. Подходящие составы и способы нанесения описаны в патенте США № 6468523, включенном в данный документ посредством ссылки. Растения можно также обрабатывать одной или несколькими химическими композициями, в том числе одним или несколькими гербицидами, инсектицидами или фунгицидами. Иллюстративные химические композиции включают следующее. Гербициды для фруктов/овощей: атразин, бромацил, диурон, глифосат, линурон, метрибузин, симазин, трифлуралин, флуазифоп, глуфосинат, галосульфурон от Gowan, паракват, пропизамид, сетоксидим, бутафенацил, галосульфурон, индазифлам; инсектициды для фруктов/овощей: альдикарб, Bacillus thuriengiensis, карбарил, карбофуран, хлорпирифос, циперметрин, дельтаметрин, диазинон, малатион, абамектин, цифлутрин/бета-цифлутрин, эсфенвалерат, лямбда-цигалотрин, ацеквиноцил, бифеназат, метоксифенозид, новалурон, кромафенозид, тиаклоприд, динотефуран, флуакрипирим, толфенпирад, клотианидин, спиродиклофен, гамма-цигалотрин, спиромезифен, спиносад, ринаксипир, циазипир, спинотерам, трифлумурон, спиротетрамат, имидаклоприд, флубендиамид, тиодикарб, метафлумизон, сульфоксафлор, цифлуметофен, цианопирафен, имидаклоприд, клотианидин, тиаметоксам, спиноторам, тиодикарб, флоникамид, метиокарб, эмамектин-бензоат, индоксакарб, фортиазат, фенамифос, кадусафос, пирипроксифен, фенбутатин-оксид, гекстиазокс, метомил, 4-[[(6-хлорпиридин-3-ил)метил](2,2-дифторэтил)амино]фуран-2(5H)-он; фунгициды для фруктов/овощей: карбендазим, хлороталонил, EBDC, сера, тиофанат-метил, азоксистробин, цимоксанил, флуазинам, фосетил, ипродион, крезоксим-метил, металаксил/мефеноксам, трифлоксистробин, этабоксам, ипроваликарб, трифлоксистробин, фенгексамид, окспоконазола фумарат, циазофамид, фенамидон, зоксамид, пикоксистробин, пираклостробин, цифлуфенамид, боскалид; гербициды для злаков: изопротурон, бромоксинил, иоксинил, фенокси-соединения, хлорсульфурон, клодинафоп, диклофоп, дифлуфеникан, феноксапроп, флорасулам, флуроксипир, метсульфурон, триасульфурон, флукарбазон, йодосульфурон, пропоксикарбазон, пиколинафен, мезосульфурон, бефлубутамид, пиноксаден, амидосульфурон, тифенсульфурон-метил, трибенурон, флупирсульфурон, сульфосульфурон, пирасульфотол, пироксулам, флуфенацет, тралкоксидим, пироксасульфон; фунгициды для злаков: карбендазим, хлороталонил, азоксистробин, ципроконазол, ципродинил, фенпропиморф, эпоксиконазол, крезоксим-метил, квиноксифен, тебуконазол, трифлоксистробин, симеконазол, пикоксистробин, пираклостробин, димоксистробин, протиоконазол, флуоксастробин; инсектициды для злаков: диметоат, лямбда-цигалотрин, дельтаметрин, альфа-циперметрин, β-цифлутрин, бифентрин, имидаклоприд, клотианидин, тиаметоксам, тиаклоприд, ацетамиприд, динетофуран, хлорпирифос, метамидофос, оксидеметон-метил, пиримикарб, метиокарб; гербициды для маиса: атразин, алахлор, бромоксинил, ацетохлор, дикамба, клопиралид, (S-)диметенамид, глуфосинат, глифосат, изоксафлутол, (S-)метолахлор, мезотрион, никосульфурон, примисульфурон, римсульфурон, сулькотрион, форамсульфурон, топрамезон, темботрион, сафлуфенацил, тиенкарбазон, флуфенацет, пироксасульфон; инсектициды для маиса: карбофуран, хлорпирифос, бифентрин, фипронил, имидаклоприд, лямбда-цигалотрин, тефлутрин, тербуфос, тиаметоксам, клотианидин, спиромезифен, флубендиамид, трифлумурон, ринаксипир, дельтаметрин, тиодикарб, β-цифлутрин, циперметрин, бифентрин, люфенурон, трифлуморон, тефлутрин, тебупиримфос, этипрол, циазипир, тиаклоприд, ацетамиприд, динетофуран, авермектин, метиокарб, спиродиклофен, спиротетрамат; фунгициды для маиса: фенитропан, тирам, протиоконазол, тебуконазол, трифлоксистробин; гербициды для риса: бутахлор, пропанил, азимсульфурон, бенсульфурон, цигалофоп, даимурон, фентразамид, имазосульфурон, мефенацет, оксазикломефон, пиразосульфурон, пирибутикарб, квинклорак, тиобенкарб, инданофан, флуфенацет, фентразамид, галосульфурон, оксазикломефон, бензобициклон, пирифталид, пеноксулам, биспирибак, оксадиаргил, этоксисульфурон, претилахлор, мезотрион, тефурилтрион, оксадиазон, феноксапроп, пиримисульфан; инсектициды для риса: диазинон, фенитротион, фенобукарб, монокротофос, бенфуракарб, бупрофезин, динотефуран, фипронил, имидаклоприд, изопрокарб, тиаклоприд, кромафенозид, тиаклоприд, динотефуран, клотианидин, этипрол, флубендиамид, ринаксипир, дельтаметрин, ацетамиприд, тиаметоксам, циазипир, спиносад, спиноторам, эмамектин-бензоат, циперметрин, хлорпирифос, картап, метамидофос, этофенпрокс, триазофос, 4-[[(6-хлорпиридин-3-ил)метил](2,2-дифторэтил)амино]фуран-2(5H)-он, карбофуран, бенфуракарб; фунгициды для риса: тиофанат-метил, азоксистробин, карпропамид, эдифенфос, феримзон, ипробенфос, изопротиолан, пенцикурон, пробеназол, пироквилон, трициклазол, трифлоксистробин, диклоцимет, феноксанил, симеконазол, тиадинил; гербициды для хлопчатника: диурон, флуометурон, MSMA, оксифлуорфен, прометрин, трифлуралин, карфентразон, клетодим, флуазифоп-бутил, глифосат, норфлуразон, пендиметалин, пиритиобак-натрий, трифлоксисульфурон, тепралоксидим, глуфосинат, флумиоксазин, тидиазурон; инсектициды для хлопчатника: ацефат, альдикарб, хлорпирифос, циперметрин, дельтаметрин, малатион, монокротофос, абамектин, ацетамиприд, эмамектин бензоат, имидаклоприд, индоксакарб, лямбда-цигалотрин, спиносад, тиодикарб, гамма-цигалотрин, спиромезифен, пиридалил, флоникамид, флубендиамид, трифлумурон, ринаксипир, бета-цифлутрин, спиротетрамат, клотианидин, тиаметоксам, тиаклоприд, динетофуран, флубендиамид, циазипир, спиносад, спиноторам, гамма-цигалотрин, 4-[[(6-хлорпиридин-3-ил)метил](2,2-дифторэтил)амино]фуран-2(5H)-он, тиодикарб, авермектин, флоникамид, пиридалил, спиромезифен, сульфоксафлор, профенофос, триазофос, эндосульфан; фунгициды для хлопчатника: этридиазол, металаксил, квинтозен; гербициды для сои: алахлор, бентазон, трифлуралин, хлоримурон-этил, хлорансулам-метил, феноксапроп, фомесафен, флуазифоп, глифосат, имазамокс, имазаквин, имазетапир, (S-)метолахлор, метрибузин, пендиметалин, тепралоксидим, глуфосинат; инсектициды для сои: лямбда-цигалотрин, метомил, паратион, тиокарб, имидаклоприд, клотианидин, тиаметоксам, тиаклоприд, ацетамиприд, динетофуран, флубендиамид, ринаксипир, циазипир, спиносад, спиноторам, эмамектин-бензоат, фипронил, этипрол, дельтаметрин, β-цифлутрин, гамма- и лямбда-цигалотрин, 4-[[(6-хлорпиридин-3-ил)метил](2,2-дифторэтил)амино]фуран-2(5H)-он, спиротетрамат, спинодиклофен, трифлумурон, флоникамид, тиодикарб, бета-цифлутрин; фунгициды для сои: азоксистробин, ципроконазол, эпоксиконазол, флутриафол, пираклостробин, тебуконазол, трифлоксистробин, протиоконазол, тетраконазол; гербициды для сахарной свеклы: хлоридазон, десмедифам, этофумезат, фенмедифам, триаллат, клопиралид, флуазифоп, ленацил, метамитрон, квинмерак, циклоксидим, трифлусульфурон, тепралоксидим, квизалофоп; инсектициды для сахарной свеклы: имидаклоприд, клотианидин, тиаметоксам, тиаклоприд, ацетамиприд, динетофуран, дельтаметрин, β-цифлутрин, гамма/лямбда-цигалотрин, 4-[[(6-хлорпиридин-3-ил)метил](2,2-дифторэтил)амино]фуран-2(5H)-он, тефлутрин, ринаксипир, циаксипир, фипронил, карбофуран; гербициды для канолы: клопиралид, диклофоп, флуазифоп, глуфосинат, глифосат, метазахлор, трифлуралин, этаметсульфурон, квинмерак, квизалофоп, клетодим, тепралоксидим; фунгициды для канолы: азоксистробин, карбендазим, флудиоксонил, ипродион, прохлораз, винклозолин; инсектициды для канолы: карбофурановые фосфороорганические соединения, пиретроиды, тиаклоприд, дельтаметрин, имидаклоприд, клотианидин, тиаметоксам, ацетамиприд, динетофуран, β-цифлутрин, гамма и лямбда-цигалотрин, тау-флувалериат, этипрол, спиносад, спиноторам, флубендиамид, ринаксипир, циазипир, 4-[[(6-хлорпиридин-3-ил)метил](2,2-дифторэтил)амино]фуран-2(5H)-он.
В некоторых вариантах осуществления гербицид представляет собой атразин, бромацил, диурон, хлорсульфурон, метсульфурон, тифенсульфурон-метил, трибенурон, ацетохлор, дикамба, изоксафлутол, никосульфурон, римсульфурон, пиритиобак-натрий, флумиоксазин, хлоримурон-этил, метрибузин, квизалофоп, S-метолахлор, гексазинон или их комбинации.
В некоторых вариантах осуществления инсектицид представляет собой эсфенвалерат, хлорантранилипрол, метомил, индоксакарб, оксамил или их комбинации.
Пестицидная и инсектицидная активность
"Вредитель" включает без ограничений насекомых, грибы, бактерии, нематод, клещей, иксодовых клещей и т. п. Насекомые-вредители включают насекомых, выбранных из отрядов Coleoptera, Diptera, Hymenoptera, Lepidoptera, Mallophaga, Homoptera, Hemiptera, Orthroptera, Thysanoptera, Dermaptera, Isoptera, Anoplura, Siphonaptera, Trichoptera и т.д., в частности Lepidoptera и Coleoptera.
Специалистам в данной области будет понятно, что не все соединения в равной степени эффективны против всех вредителей. Соединения согласно вариантам осуществления проявляют активность против насекомых-вредителей, которые могут включать экономически важных вредителей агрономических, лесных, тепличных продуктов, продуктов питомников декоративных растений, продуктов пищи и волокнистых продуктов, продуктов, связанных со здоровьем людей и животных, продуктов, связанных с домашней и коммерческой структурой, товаров для дома и продуктов для хранения.
Личинки из отряда Lepidoptera включают без ограничения совок, подгрызающих совок, пядениц и гелиотин семейства Noctuidae Spodoptera frugiperda JE Smith (совка травяная); S. exigua Hübner (совка малая); S. litura Fabricius (табачная совка, гусеница, пожирающая соцветия); Mamestra configurata Walker (совка Берта); M. brassicae Linnaeus (совка капустная); Agrotis ipsilon Hufnagel (совка-ипсилон); A. orthogonia Morrison (совка прямоугольная); A. subterranea Fabricius (совка зернистая); Alabama argillacea Hübner (совка хлопковая американская); Trichoplusia ni Hübner (совка ни); Pseudoplusia includens Walker (соевая совка); Anticarsia gemmatalis Hübner (совка бархатных бобов); Hypena scabra Fabricius (совка клеверная); Heliothis virescens Fabricius (табачная листовертка); Pseudaletia unipuncta Haworth (совка луговая); Athetis mindara Barnes и Mcdunnough (шершавая совка); Euxoa messoria Harris (чернобокая гусеница совки); Earias insulana Boisduval (совка хлопковая египетская); E. vittella Fabricius (совка пятнистая); Helicoverpa armigera Hübner (совка щетинконогая резедовая); H. zea Boddie (совка кукурузная или хлопковая совка); Melanchra picta Harris (совка); Egira (Xylomyges) curialis Grote (цитрусовая совка); огневок, чехлоносок, бабочек, строящих паутинные гнезда, конусных бабочек и вредителей, скелетирующих листья, из семейства Pyralidae Ostrinia nubilalis Hübner (мотылек стеблевой кукурузный); Amyelois transitella Walker (гусеницы, повреждающие рубчики цитрусовых); Anagasta kuehniella Zeller (огневка мельничная); Cadra cautella Walker (огневка сухофруктовая); Chilo suppressalis Walker (желтая рисовая огневка); C. partellus (сорговая огневка); Corcyra cephalonica Stainton (огневка рисовая); Crambus caliginosellus Clemens (огневка кукурузная); C. teterrellus Zincken (огневка мятликовая); Cnaphalocrocis medinalis Guenée (листовертка рисовая); Desmia funeralis Hübner (виноградная листовертка); Diaphania hyalinata Linnaeus (дынная огневка); D. nitidalis Stoll (огневка огурцов-пикули); Diatraea grandiosella Dyar (огневка кукурузная юго-западная), D. saccharalis Fabricius (огневка сахарного тростника); Eoreuma loftini Dyar (мексиканская рисовая огневка); Ephestia elutella Hübner (огневка зерновая (какао)); Galleria mellonella Linnaeus (большая восковая моль); Herpetogramma licarsisalis Walker (огневка-травянка); Homoeosoma electellum Hulst (огневка подсолнечниковая); Elasmopalpus lignosellus Zeller (малая кукурузная огневка); Achroia grisella Fabricius (малая восковая моль); Loxostege sticticalis Linnaeus (луговой мотылек); Orthaga thyrisalis Walker (чайная моль); Maruca testulalis Geyer (огневка акациевая); Plodia interpunctella Hübner (моль индийская мучная); Scirpophaga incertulas Walker (стеблевая рисовая огневка); Udea rubigalis Guenée (огневка ржаво-коричневая); и листоверток, листоверток-почкоедов, плодожорок и гусениц-вредителей плодов из семейства Tortricidae Acleris gloverana Walsingham (западная черноголовая листовертка); A. variana Fernald (восточная черноголовая листовертка); Archips argyrospila Walker (листовертка плодовых деревьев); A. rosana Linnaeus (европейская листовертка) и другие виды Archips, Adoxophyes orana Fischer von Rösslerstamm (листовертка сетчатая); Cochylis hospes Walsingham (полосатая подсолнечниковая моль); Cydia latiferreana Walsingham (лещинная плодожорка); C. pomonella Linnaeus (яблонная плодожорка); Platynota flavedana Clemens (листовертка изменчивая); P. stultana Walsingham (листовертка всеядная); Lobesia botrana Denis & Schiffermüller (листовертка европейская виноградная); Spilonota ocellana Denis & Schiffermüller (листовертка почковая); Endopiza viteana Clemens (листовертка виноградная); Eupoecilia ambiguella Hübner (листовертка гроздевая); Bonagota salubricola Meyrick (листовертка бразильская яблочная); Grapholita molesta Busck (плодожорка восточная персиковая); Suleima helianthana Riley (листовертка подсолнечниковая); Argyrotaenia spp.; Choristoneura spp.
Другие выбранные сельскохозяйственные вредители из отряда Lepidoptera включают без ограничения Alsophila pometaria Harris (осенний плодовый червь); Anarsia lineatella Zeller (моль фруктовая полосатая); Anisota senatoria J.E. Smith (сатурния оранжевая дубовая); Antheraea pernyi Guérin-Méneville (китайский дубовый шелкопряд); Bombyx mori Linnaeus (тутовый шелкопряд); Bucculatrix thurberiella Busck (кривоусая хлопковая моль); Colias eurytheme Boisduval (люцерновая желтушка); Datana integerrima Grote & Robinson (хохлатка ореховая); Dendrolimus sibiricus Tschetwerikov (сибирский шелкопряд), Ennomos subsignaria Hübner (пяденица ильмовая); Erannis tiliaria Harris (пяденица липовая); Euproctis chrysorrhoea Linnaeus (шелкопряд золотистый); Harrisina americana Guérin-Méneville (пироморфида американская); Hemileuca oliviae Cockrell (гусеница бабочки-сатурнии); Hyphantria cunea Drury (американская белая бабочка); Keiferia lycopersicella Walsingham (томатная моль); Lambdina fiscellaria fiscellaria Hulst (пяденица гемлоковая восточная); L. fiscellaria lugubrosa Hulst (пяденица гемлоковая западная); Leucoma salicis Linnaeus (волнянка ивовая); Lymantria dispar Linnaeus (непарный шелкопряд); Manduca quinquemaculata Haworth (бражник пятиточечный, томатный бражник); M. sexta Haworth (томатный бражник, табачный бражник); Operophtera brumata Linnaeus (пяденица зимняя); Paleacrita vernata Peck (пяденица весенняя); Papilio cresphontes Cramer (парусник кресфонтес, «апельсиновая собака»); Phryganidia californica Packard (коконопряд кольчатый калифорнийский); Phyllocnistis citrella Stainton (цитрусовая мушка-минер); Phyllonorycter blancardella Fabricius (моль-пестрянка плодовая нижнесторонняя); Pieris brassicae Linnaeus (белянка капустная большая); P. rapae Linnaeus (белянка капустная малая); P. napi Linnaeus (белянка брюквенная); Platyptilia carduidactyla Riley (пальцекрылка артишоковая); Plutella xylostella Linnaeus (моль капустная); Pectinophora gossypiella Saunders (розовый коробочный червь); Pontia protodice Boisduval and Leconte (клетчатая белянка); Sabulodes aegrotata Guenée (всеядная пяденица); Schizura concinna J.E. Smith (хохлатка); Sitotroga cerealella Olivier (моль ячменная ангумуазская); Thaumetopoea pityocampa Schiffermuller (походный шелкопряд сосновый); Tineola bisselliella Hummel (моль комнатная); Tuta absoluta Meyrick (томатная моль); Yponomeuta padella Linnaeus (горностаевая моль плодовая); Heliothis subflexa Guenée; Malacosoma spp. и Orgyia spp.
Представляют интерес личинки и имаго из отряда Coleoptera, в том числе долгоносики из семейств Anthribidae, Bruchidae и Curculionidae (в том числе без ограничения: Anthonomus grandis Boheman (долгоносик хлопковый); Lissorhoptrus oryzophilus Kuschel (долгоносик рисовый водяной); Sitophilus granarius Linnaeus (долгоносик амбарный); S. oryzae Linnaeus (долгоносик рисовый); Hypera punctata Fabricius (долгоносик точечный); Cylindrocopturus adspersus LeConte (долгоносик подсолнечниковый стеблевой); Smicronyx fulvus LeConte (красный долгоносик подсолнечника); S. sordidus LeConte (серый долгоносик подсолнечника); Sphenophorus maidis Chittenden (долгоносик маисовый)); земляные блошки, блошки, корневые черви, листоеды, картофельные жуки и листовые минеры семейства Chrysomelidae (в том числе без ограничения: Leptinotarsa decemlineata Say (колорадский жук); Diabrotica virgifera virgifera LeConte (западный кукурузный жук); D. barberi Smith and Lawrence (северный кукурузный жук); D. undecimpunctata howardi Barber (южный кукурузный жук); Chaetocnema pulicaria Melsheimer (земляная кукурузная блошка); Phyllotreta cruciferae Goeze (блошка крестоцветная); Phyllotreta striolata (полосатая блошка); Colaspis brunnea Fabricius (листоед виноградный); Oulema melanopus Linnaeus (пьявица красногрудая); Zygogramma exclamationis Fabricius (подсолнечниковый листоед)); жуки семейства Coccinellidae (в том числе без ограничения: Epilachna varivestis Mulsant (мексиканская фасолевая коровка)); хрущи и другие жуки из семейства Scarabaeidae (в том числе без ограничения: Popillia japonica Newman (хрущик японский); Cyclocephala borealis Arrow (дупляк северный, хрущ); C. immaculata Olivier (дупляк южный, хрущ); Rhizotrogus majalis Razoumowsky (нехрущ майский); Phyllophaga crinita Burmeister (хрущ); Ligyrus gibbosus De Geer (жук морковный)); кожееды из семейства Dermestidae; проволочники из семейства Elateridae, Eleodes spp., Melanotus spp.; Conoderus spp.; Limonius spp.; Agriotes spp.; Ctenicera spp.; Aeolus spp.; короеды из семейства Scolytidae и жуки из семейства Tenebrionidae.
Представляют интерес имаго и незрелые особи отряда Diptera, в том числе минирующие мушки Agromyza parvicornis Loew (кукурузная минирующая мушка); галлицы (в том числе без ограничения: Contarinia sorghicola Coquillett (галлица сорговая); Mayetiola destructor Say (гессенская муха); Sitodiplosis mosellana Géhin (оранжевая зерновая галлица); Neolasioptera murtfeldtiana Felt, (подсолнечниковая галлица)); плодовые мушки (Tephritidae), Oscinella frit Linnaeus (плодовые мушки); цветочные мухи (в том числе без ограничения: Delia platura Meigen (муха ростковая); D. coarctata Fallen (муха озимая) и другие Delia spp., Meromyza americana Fitch (американская меромиза); Musca domestica Linnaeus (комнатная муха); Fannia canicularis Linnaeus, F. femoralis Stein (малые комнатные мухи); Stomoxys calcitrans Linnaeus (жигалка осенняя)); мухи полевые, жигалки, мухи мясные, Chrysomya spp.; Phormia spp. и другие мухи-вредители надсемейства Muscoidea, слепни Tabanus spp.; носоглоточные оводы Gastrophilus spp.; Oestrus spp.; бычьи оводы Hypoderma spp.; пестряки Chrysops spp.; Melophagus ovinus Linnaeus (рунец овечий) и другие Brachycera, комары Aedes spp.; Anopheles spp.; Culex spp.; мошки Prosimulium spp.; Simulium spp.; мокрецы, москиты, сциариды и другие Nematocera.
В качестве насекомых, представляющих интерес, включены имаго и личинки из отрядов Hemiptera и Homoptera, такие как без ограничения хермесы из семейства Adelgidae, слепняки из семейства Miridae, цикады из семейства Cicadidae, цикадки, Empoasca spp.; из семейства Cicadellidae, насекомые надсемейства Fulgoroidea из семейств Cixiidae, Flatidae, Fulgoroidea, Issidae и Delphacidae, горбатки из семейства Membracidae, листоблошки из семейства Psyllidae, белокрылки из семейства Aleyrodidae, тли из семейства Aphididae, филлоксеры из семейства Phylloxeridae, мучнистые червецы из семейства Pseudococcidae, червецы из семейств Asterolecanidae, Coccidae, Dactylopiidae, Diaspididae, Eriococcidae Ortheziidae, Phoenicococcidae и Margarodidae, кружевницы из семейства Tingidae, щитники из семейства Pentatomidae, клопы-черепашки, Blissus spp.; и другие наземники из семейства Lygaeidae, пенницы из семейства Cercopidae, краевики из семейства Coreidae и красноклопы и красноклопы хлопковые из семейства Pyrrhocoridae.
Важные с точки зрения сельского хозяйства представители отряда Homoptera дополнительно включают без ограничения: Acyrthisiphon pisum Harris (тля гороховая); Aphis craccivora Koch (тля люцерновая); A. fabae Scopoli (тля свекловичная); A. gossypii Glover (тля хлопковая, тля бахчевая); A. maidiradicis Forbes (тля кукурузная корневая); A. pomi De Geer (тля яблонная); A. spiraecola Patch (тля зеленая цитрусовая); Aulacorthum solani Kaltenbach (тля картофельная обыкновенная); Chaetosiphon fragaefolii Cockerell (тля земляничная американская); Diuraphis noxia Kurdjumov/Mordvilko (русская пшеничная тля); Dysaphis plantaginea Paaserini (тля яблоневая розовая); Eriosoma lanigerum Hausmann (тля яблонная кровяная); Brevicoryne brassicae Linnaeus (капустная тля); Hyalopterus pruni Geoffroy (тля сливовая опыленная); Lipaphis erysimi Kaltenbach (тля ложнокапустная); Metopolophium dirrhodum Walker (розанно-злаковая тля); Macrosiphum euphorbiae Thomas (большая картофельная тля); Myzus persicae Sulzer (тля персиковая, тля оранжерейная); Nasonovia ribisnigri Mosley (тля салатная зеленая); Pemphigus spp. (тли корневые и тли галловые); Rhopalosiphum maidis Fitch (тля сорговая); R. padi Linnaeus (тля черемуховая обыкновенная); Schizaphis graminum Rondani (тля злаковая обыкновенная); Sipha flava Forbes (тля желтая сахарного тростника); Sitobion avenae Fabricius (тля листовая); Therioaphis maculata Buckton (пятнистая люцерновая тля); Toxoptera aurantii Boyer de Fonscolombe (тля померанцевая) и T. citricida Kirkaldy (тля цитрусовая); Adelges spp. (хермесы); Phylloxera devastatrix Pergande (филлоксера гикори); Bemisia tabaci Gennadius (белокрылка табачная, белокрылка хлопковая); B. argentifolii Bellows и Perring (белокрылка магнолиевая); Dialeurodes citri Ashmead (белокрылка цитрусовая); Trialeurodes abutiloneus (белокрылка полосатокрылая) и T. vaporariorum Westwood (белокрылка тепличная); Empoasca fabae Harris (цикадка картофельная); Laodelphax striatellus Fallen (темная цикадка); Macrolestes quadrilineatus Forbes (цикадка астровая); Nephotettix cinticeps Uhler (цикадка зеленая); N. nigropictus Stål (рисовая цикадка); Nilaparvata lugens Stål (бурая рисовая цикадка); Peregrinus maidis Ashmead (цикадка кукурузная); Sogatella furcifera Horvath (цикадка белоспинная); Sogatodes orizicola Muir (дельфацид рисовый); Typhlocyba pomaria McAtee (цикадка яблонная); Erythroneoura spp. (цикадки виноградные); Magicicada septendecim Linnaeus (периодическая цикада); Icerya purchasi Maskell (червец австралийский желобчатый); Quadraspidiotus perniciosus Comstock (щитовка калифорнийская); Planococcus citri Risso (мучнистый червец виноградный); Pseudococcus spp. (другие мучнистые червецы); Cacopsylla pyricola Foerster (листоблошка грушевая); Trioza diospyri Ashmead (листоблошка хурмовая).
Важные с точки зрения сельского хозяйства виды из отряда Hemiptera включают без ограничения: Acrosternum hilare Say (щитник зеленый); Anasa tristis De Geer (клоп-ромбовик печальный); Blissus leucopterus leucopterus Say (клоп-черепашка); Corythuca gossypii Fabricius (кружевница хлопковая); Cyrtopeltis modesta Distant (клоп томатный); Dysdercus suturellus Herrich-Schäffer (красноклоп хлопковый); Euschistus servus Say (клоп коричневый вонючий); E. variolarius Palisot de Beauvois (щитник однопятнистый); Graptostethus spp. (комплекс клопов-наземников); Leptoglossus corculus Say (клоп-краевик сосновый); Lygus lineolaris Palisot de Beauvois (клоп луговой); L. Hesperus Knight (слепняк западный матовый); L. pratensis Linnaeus (клопик полевой); L. rugulipennis Poppius (клоп травяной); Lygocoris pabulinus Linnaeus (зеленый слепняк); Nezara viridula Linnaeus (зеленый овощной клоп); Oebalus pugnax Fabricius (клоп-щитник рисовый); Oncopeltus fasciatus Dallas (клоп молочайный большой); Pseudatomoscelis seriatus Reuter (клоп-слепняк хлопковый).
Кроме того, варианты осуществления могут быть эффективными против Hemiptera, например, Calocoris norvegicus Gmelin (клопик картофельный); Orthops campestris Linnaeus; Plesiocoris rugicollis Fallen (клоп яблонный северный); Cyrtopeltis modestus Distant (томатный клоп); Cyrtopeltis notatus Distant (клоп-слепняк); Spanagonicus albofasciatus Reuter (слепняк белоточечный); Diaphnocoris chlorionis Say (клоп-слепняк гледичии); Labopidicola allii Knight (слепняк луковый); Pseudatomoscelis seriatus Reuter (слепняк хлопковый); Adelphocoris rapidus Say (клоп быстрый); Poecilocapsus lineatus Fabricius (слепняк четырехлинейный); Nysius ericae Schilling (низиус вересковый); Nysius raphanus Howard (ложная черепашка); Nezara viridula Linnaeus (зеленый овощной клоп); Eurygaster spp.; Coreidae spp.; Pyrrhocoridae spp.; Tinidae spp.; Blostomatidae spp.; Reduviidae spp. и Cimicidae spp.
Также включены имаго и личинки из отряда Acari (клещи), такие как Aceria tosichella Keifer (галловый клещ пшеничный); Petrobia latens Müller (петробия многоядная); клещики паутинные и клещики красные семейства Tetranychidae, Panonychus ulmi Koch (красный плодовый клещ); Tetranychus urticae Koch (обыкновенный паутинный клещ); (T. mcdanieli McGregor (клещик Макданиела); T. cinnabarinus Boisduval (красный паутинный клещик); T. turkestani Ugarov & Nikolski (туркестанский паутинный клещик); плоские клещи семейства Tenuipalpidae, Brevipalpus lewisi McGregor (оранжевый клещ); ржавчинные и почковые клещи семейства Eriophyidae и другие клещи, питающиеся листьями, и клещи, опасные для здоровья человека и животных, т. е., пылевые клещи семейства Epidermoptidae, железницы семейства Demodicidae, зерновые клещи семейства Glycyphagidae, иксодовые клещи отряда Ixodidae. Ixodes scapularis Say (черноногий клещ); I. holocyclus Neumann (австралийский паралитический клещ); Dermacentor variabilis Say (клещ иксодовый собачий); Amblyomma americanum Linnaeus (иксодовый клещ Amblyomma) и конские и чесоточные клещи семейств Psoroptidae, Pyemotidae и Sarcoptidae.
Представляющими интерес насекомыми-вредителями из отряда Thysanura являются, например, Lepisma saccharina Linnaeus (чешуйница); Thermobia domestica Packard (термобия).
Дополнительные охваченные вредители-артроподы включают: пауков из отряда Araneae, таких как Loxosceles reclusa Gertsch and Mulaik (бурый паук-отшельник) и Latrodectus mactans Fabricius (черная вдова), и многоножек из отряда Scutigeromorpha, таких как Scutigera coleoptrata Linnaeus (обыкновенная мухоловка).
Насекомое-вредитель, представляющее интерес, включает надсемейство щитников и других родственных насекомых, в том числе без исключения виды, принадлежащие к семейству Pentatomidae (Nezara viridula, Halyomorpha halys, Piezodorus guildini, Euschistus servus, Acrosternum hilare, Euschistus heros, Euschistus tristigmus, Acrosternum hilare, Dichelops furcatus, Dichelops melacanthus, и Bagrada hilaris (клоп из рода Bagrada)), семейству Plataspidae (Megacopta cribraria - полушаровидный щитник) и семейству Cydnidae (Scaptocoris castanea - коричневый клоп-землекоп), и виды Lepidoptera, в том числе без ограничения: моль капустную, e.g., Helicoverpa zea Boddie; соевую совку, например, Pseudoplusia includens Walker, и совку бархатных бобов, например, Anticarsia gemmatalis Hübner.
Способы измерения пестицидной активности хорошо известны из уровня техники. См., например, Czapla and Lang, (1990) J. Econ. Entomol. 83:2480-2485; Andrews, et al., (1988) Biochem. J. 252:199-206; Marrone, et al., (1985) J. of Economic Entomology 78:290-293 и патент США № 5743477, все из которых включены в данный документ посредством ссылки во всей своей полноте. Как правило, белок смешивают и применяют в анализах питания. См., например, Marrone, et al., (1985) J. of Economic Entomology 78:290-293. Такие анализы могут включать приведение растений в контакт с одним или несколькими вредителями и определение способности растения выживать и/или вызывать гибель вредителей.
Нематоды включают паразитических нематод, таких как галловые, цистообразующие и ранящие нематоды, в том числе Heterodera spp., Meloidogyne spp. и Globodera spp.; в частности представителей цистообразующих нематод, в том числе без ограничения, Heterodera glycines (соевая цистообразующая нематода); Heterodera schachtii (цистообразующая нематода свеклы); Heterodera avenae (цистообразующая нематода злаков), и Globodera rostochiensis, и Globodera pailida (цистообразующые нематоды картофеля). Ранящие нематоды включают Pratylenchus spp.
Средство для обработки семян
Для защиты и для повышения урожайности и улучшения технологий усовершенствования признаков дополнительные средства для обработки семян могут обеспечивать дополнительную приспособляемость культурных растений и экономически эффективный контроль насекомых, сорняков и заболеваний. Семенной материал можно обрабатывать, как правило, обрабатывать поверхность с помощью композиции, содержащей комбинации химических или биологических гербицидов, антидотов гербицидов, инсектицидов, фунгицидов, ингибиторов и усилителей прорастания, питательных веществ, регуляторов и активаторов роста растений, бактерицидов, нематоцидов, авицидов и/или моллюскоцидов. Эти соединения, как правило, составляют вместе с дополнительными носителями, поверхностно-активными веществами или вспомогательными средствами, способствующими нанесению, традиционно используемыми в области техники, связанной с получением составов. Покрытия можно наносить с помощью пропитки материала для размножения жидким составом или с помощью покрытия комбинированным влажным или сухим составом. Примеры различных типов соединений, которые можно применять в качестве средств для обработки семян, представлены в The Pesticide Manual: A World Compendium, C.D.S. Tomlin Ed., Published by the British Crop Production Council, который включен в данный документ посредством ссылки.
Некоторые средства для обработки семян, которые можно применять на семенах сельскохозяйственных культур, включают без ограничения один или несколько из абсцизовой кислоты, ацибензолар-S-метила, авермектина, амитрола, азаконазола, азоспириллума, азадирахтина, азоксистробина, Bacillus spp. (в том числе один или несколько из видов cereus, firmus, megaterium, pumilis, sphaericus, subtilis и/или thuringiensis), Bradyrhizobium spp. (в том числе один или несколько из betae, canariense, elkanii, iriomotense, japonicum, liaonigense, pachyrhizi и/или yuanmingense), каптана, карбоксина, хитозана, клотианидина, меди, циазипира, дифеноконазола, этидиазола, фипронила, флудиоксонила, флуоксастробина, флуквинконазола, флуразола, флуксофенима, белка гарпина, имазалила, имидаклоприда, ипконазола, изофлавеноидов, липохитоолигосахарида, манкозеба, марганца, манеба, мефеноксама, металаксила, метконазола, миклобутанила, PCNB, пенфлуфена, пинециллума, пентиопирада, перметрина, пикоксистробина, протиоконазола, пираклостробина, ринаксипира, S-метолахлора, сапонина, седаксана, TCMTB, тебуконазола, тиабендазола, тиаметоксама, тиокарба, тирама, толклофос-метила, триадименола, триходермы, трифлоксистробина, тритиконазола и/или цинка. Средство для покрытия семян PCNB, относящееся к номеру регистрации EPA 00293500419, содержит квинтозен и терразол. TCMTB называется 2-(тиоцианометилтио)бензотиазол.
Сорта семян и семена со специфическими трансгенными признаками можно тестировать для определения того, какие дополнительные варианты обработки семян и нормы внесения могут дополнять такие сорта и трансгенные признаки для повышения урожайности. Например, сорт с хорошей потенциальной урожайностью, но восприимчивостью к пыльной головне, может выиграть от применения средства для обработки семян, которое обеспечивает защиту от пыльной головни, сорт с хорошей потенциальной урожайностью, но восприимчивостью к цистообразующим нематодам, может выиграть от применения средства обработки семян, которое обеспечивает защиту от цистообразующей нематоды и т.д. Аналогично, сорт, включающий трансгенный признак, обеспечивающий устойчивость к насекомым, может выиграть от второго механизма действия, придаваемого средством для обработки семян, сорт, включающий трансгенный признак, придающий устойчивость к гербициду, может выиграть от средства обработки семян с антидотом, который повышает устойчивость растений к такому гербициду и т.д. К тому же, хорошее укоренение и ранняя всхожесть, которые являются результатом правильного применения средства для обработки семян, могут приводить к более эффективному использованию азота, лучшей способности переносить засуху и общему повышению потенциальной урожайности сорта или сортов, содержащих определенный признак, в комбинации со средством для обработки семян.
Способы уничтожения насекомого-вредителя и контроля популяции насекомых
В некоторых вариантах осуществления представлены способы уничтожения насекомого-вредителя, включающие приведение в контакт насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного полипептида PtIP-83. В некоторых вариантах осуществления предусмотрены способы уничтожения насекомого-вредителя, включающие приведение в контакт насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного пестицидного белка под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или его варианта.
В некоторых вариантах осуществления представлены способы уничтожения насекомого-вредителя, включающие приведение в контакт насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного полипептида PtIP-83 под SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607, SEQ ID NO: 640-645 и SEQ ID NO: 728-737.
В некоторых вариантах осуществления представлены способы уничтожения насекомого-вредителя, включающие приведение в контакт насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного полипептида PtIP-83, содержащего аминокислотную последовательность под любым из SEQ ID NO: 786-888 или его варианта.
В некоторых вариантах осуществления представлены способы контроля популяции насекомого-вредителя, включающие приведение в контакт популяции насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного полипептида PtIP-83. В некоторых вариантах осуществления представлены способы контроля популяции насекомого-вредителя, включающие приведение в контакт популяции насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного полипептида PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или его варианта. Применяемые в данном документе термины "контроль популяции вредителя" или "контролировать вредителя" относятся к любому эффекту в отношении вредителя, который приводит к ограничению вреда, который наносит вредитель. Контроль вредителя включает без ограничений уничтожение вредителя, подавление развития вредителя, изменение плодовитости или роста вредителя таким образом, что вредитель оказывает меньше вреда в отношении растения, снижение количества производимого потомства, получение менее приспособленных вредителей, получение вредителей, более восприимчивых к нападению хищников или удержание вредителей от поедания растения.
В некоторых вариантах осуществления представлены способы контроля популяции насекомого-вредителя, включающие приведение в контакт популяции насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного полипептида PtIP-83 под любым из SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607, SEQ ID NO: 640-645 и SEQ ID NO: 728-737.
В некоторых вариантах осуществления представлены способы контроля популяции насекомого-вредителя, включающие приведение в контакт популяции насекомого-вредителя с инсектицидно эффективным количеством рекомбинантного полипептида PtIP-83 под любым из SEQ ID NO: 786-888 или его варианта.
В некоторых вариантах осуществления представлены способы защиты растения от насекомого-вредителя, включающие экспрессию в растении или его клетке рекомбинантного полинуклеотида, кодирующего полипептид PtIP-83. В некоторых вариантах осуществления представлены способы защиты растения от насекомого-вредителя, включающие экспрессию в растении или его клетке рекомбинантного полинуклеотида, кодирующего полипептид PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768 или SEQ ID NO: 769 или его варианты.
В некоторых вариантах осуществления представлены способы защиты растения от насекомого-вредителя, включающие экспрессию в растении или его клетке рекомбинантного полинуклеотида, кодирующего полипептид PtIP-83. В некоторых вариантах осуществления представлены способы защиты растения от насекомого-вредителя, включающие экспрессию в растении или его клетке рекомбинантного полинуклеотида, кодирующего полипептид PtIP-83 под любым из SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607, SEQ ID NO: 640-645 и SEQ ID NO: 728-737.
В некоторых вариантах осуществления представлены способы защиты растения от насекомого-вредителя, включающие экспрессию в растении или его клетке рекомбинантного полинуклеотида, кодирующего полипептид PtIP-83. В некоторых вариантах осуществления представлены способы защиты растения от насекомого-вредителя, включающие экспрессию в растении или его клетке рекомбинантного полинуклеотида, кодирующего полипептид PtIP-83 под любым из SEQ ID NO: 786-888 или его варианта.
Стратегии управления устойчивостью насекомых (IRM)
Было доказано, что экспрессия δ-эндотоксинов B. thuringiensis в трансгенных растениях кукурузы является эффективным средством контроля важных с точки зрения сельского хозяйства насекомых-вредителей (Perlak, et al., 1990; 1993). Однако возникли насекомые, которые устойчивы к δ-эндотоксинам B. thuringiensis, экспрессирующимся в трансгенных растениях. Такая устойчивость, если она станет широко распространенной, будет явно ограничивать коммерческое значение идиоплазмы, содержащей гены, кодирующие такие δ-эндотоксины B. thuringiensis.
Одним способом повышения эффективности трансгенных инсектицидов против целевых вредителей и одновременного снижения развития устойчивых к инсектицидам вредителей является создание нетрансгенных (т. е. не содержащих инсектицидные белок) рефугиев (участок земли с сельскохозяйственными культурами/кукурузой, не содержащими инсектицидный белок) для применения вместе с трансгенными сельскохозяйственными культурами, вырабатывающими один инсектицидный белок, активный против целевых вредителей. Управление по охране окружающей среды Соединенных Штатов (epa.gov/oppbppdl/biopesticides/pips/bt_corn_refuge_2006.htm, доступ к которому можно получить с применением префикса www) публикует требования по применению трансгенных сельскохозяйственных культур, вырабатывающих один Bt-белок, активный против целевых вредителей. В дополнение, Национальная ассоциация кукурузоводов на своем веб-сайте: (ncga.com/insect-resistance-management-fact-sheet-bt-corn, доступ к которому можно получить с применением префикса www) также предлагает аналогичные руководства, касающиеся требований к рефугиям. Из-за потерь, обусловленных насекомыми в пределах зоны рефугия, более крупные рефугии могут снижать общую урожайность.
Другим способом повышения эффективности трансгенных инсектицидов против целевых вредителей и одновременного снижения развития устойчивых к инсектицидам вредителям может быть создание хранилища инсектицидных генов, которые эффективны против групп насекомых-вредителей и которые проявляют свои эффекты посредством отличающихся механизмов действия.
Экспрессия в растении двух или большего числа инсектицидных композиций, токсичных для одного вида насекомых, при этом каждый инсектицид экспрессируется на эффективных уровнях, будет представлять собой другой способ для достижения контроля развития устойчивости. Это основано на принципе, что эволюция устойчивости к двум отдельным механизмам действия значительно менее вероятна, чем только к одному. Roush, например, описывает стратегию двух токсинов, также называемую "создание пирамиды" или "пакетирование," для управления инсектицидными трансгенными сельскохозяйственными культурами. (The Royal Society. Phil. Trans. R. Soc. Lond. B. (1998) 353:1777-1786). Пакетирование или создание пирамиды из двух различных белков, каждый из которых эффективен против целевых вредителей и при этом отсутствует перекрестная устойчивость или она невелика, может обеспечивать возможность применения меньшего рефугия. Управление по охране окружающей среды США требует значительно меньший (как правило 5%) структурированный рефугий для высаживания кукурузы, не являющейся Bt, чем для продуктов с одним признаком (как правило 20%). Существуют различные способы обеспечения эффектов IRM рефугия, в том числе различные геометрические паттерны высаживания в полях и смеси семян "в мешке", как дополнительно обсуждается у Roush.
В некоторых вариантах осуществления полипептиды PtIP-83 по настоящему раскрытию применимы ы в качестве стратегии управления устойчивостью насекомых в комбинации (т. е. в составе пирамиды) с другими пестицидными белками, включающими без ограничения Bt-токсины, инсектицидные белки Xenorhabdus sp. или Photorhabdus sp. и т. п.
Предусмотрены способы контроля заражения насекомыми отряда Lepidoptera и/или Coleoptera трансгенного растения, которые обеспечивают управление устойчивостью насекомых, включающие экспрессию в растении по меньшей мере двух различных инсектицидных белков, имеющих отличающиеся механизмы действия.
В некоторых вариантах осуществления способы контроля заражения насекомыми из отряда Lepidoptera и/или Coleoptera трансгенного растения и содействия в управлении устойчивостью насекомых по меньшей мере к одному из инсектицидных белков включают полипептид PIP-83, инсектицидный в отношении насекомых из отряда Lepidoptera и/или Coleoptera.
В некоторых вариантах осуществления способы контроля заражения насекомыми из отряда Lepidoptera и/или Coleoptera трансгенного растения и содействия в управлении устойчивостью насекомых по меньшей мере к одному из инсектицидных белков включают полипептид PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или его варианты, инсектицидные в отношении насекомых из отряда Lepidoptera и/или Coleoptera.
В некоторых вариантах осуществления способы контроля заражения насекомыми из отряда Lepidoptera и/или Coleoptera трансгенного растения и содействия в управлении устойчивостью насекомых по меньшей мере к одному из инсектицидных белков включают полипептид PtIP-83 под любым из SEQ ID NO: 236-299, SEQ ID NO: 334-367, SEQ ID NO: 398-427, SEQ ID NO: 518-607, SEQ ID NO: 640-645 и SEQ ID NO: 728-737.
В некоторых вариантах осуществления способы контроля заражения насекомыми из отряда Lepidoptera и/или Coleoptera трансгенного растения и содействия в управлении устойчивостью насекомых включают экспрессию в трансгенном растении полипептида PtIP-83 и белка Cry, инсектицидных в отношении насекомых из отряда Lepidoptera и/или Coleoptera, причем они имеют отличающиеся механизмы действия.
В некоторых вариантах осуществления способы контроля заражения насекомыми из отряда Lepidoptera и/или Coleoptera трансгенного растения и содействия в управлении устойчивостью насекомых включают экспрессию в трансгенном растении полипептида PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или его вариантов и белка Cry, инсектицидных в отношении насекомых из отряда Lepidoptera и/или Coleoptera, причем они имеют отличающиеся механизмы действия.
Также представлены способы снижения вероятности появления устойчивости у насекомых из отряда Lepidoptera и/или Coleoptera к трансгенным растениям, экспрессирующим в растениях инсектицидные белки для контроля видов насекомых, включающие экспрессию полипептида PtIP-83, инсектицидного в отношении вида насекомых, в комбинации со вторым белком, инсектицидным в отношении вида насекомых, причем они имеют отличающиеся механизмы действия.
Также представлены средства для эффективного управления устойчивостью насекомых из отряда Lepidoptera и/или Coleoptera к трансгенным растениям, включающие совместную экспрессию на высоких уровнях в растениях двух или более инсектицидных белков, токсичных для насекомых из отряда Lepidoptera и/или Coleoptera, но при этом каждый характеризуется отличающимся механизмом осуществления его активности в отношении уничтожения, при этом два или более инсектицидных белка включают полипептид PtIP-83 и белок Cry. Также предусмотрены средства для эффективного управления устойчивостью насекомых из отряда Lepidoptera и/или Coleoptera к трансгенным растениям, включающие совместную экспрессию на высоких уровнях в растениях двух или более инсектицидных белков, токсичных для насекомых из отряда Lepidoptera и/или Coleoptera, но при этом каждый характеризуется отличающимся механизмом осуществления его активности применительно к уничтожению, при этом два или более инсектицидных белка включают полипептид PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или его варианты и белок Cry.
В дополнение, представлены способы получения разрешения контролирующих органов для выращивания или коммерческой реализации растений, экспрессирующих белки, инсектицидные в отношении насекомых из отряда Lepidoptera и/или Coleoptera, включающие стадию обращения, предоставления или ссылки на данные анализов связывания белков насекомых, демонстрирующие, что полипептид PtIP-83 не конкурирует с сайтами связывания белков Cry у таких насекомых. В дополнение, предусмотрены способы получения разрешения контролирующих органов для выращивания или коммерческой реализации растений, экспрессирующих белки, инсектицидные в отношении насекомых из отряда Lepidoptera и/или Coleoptera, включающие стадию обращения, предоставления или ссылки на данные анализов связывания белков насекомых, показывающих, что полипептид PtIP-83 под SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 716, SEQ ID NO: 754, SEQ ID NO: 755, SEQ ID NO: 756, SEQ ID NO: 757, SEQ ID NO: 758, SEQ ID NO: 759, SEQ ID NO: 760, SEQ ID NO: 761, SEQ ID NO: 762, SEQ ID NO: 763, SEQ ID NO: 764, SEQ ID NO: 765, SEQ ID NO: 766, SEQ ID NO: 767, SEQ ID NO: 768, SEQ ID NO: 769 или его варианты не конкурируют с сайтами связывания для белков Cry у таких насекомых.
Способы повышения урожайности растения
Представлены способы повышения урожайности. Способы включают получение растения или растительной клетки, экспрессирующих полинуклеотид, кодирующий пестицидную полипептидную последовательность, раскрытую в данном документе, и выращивание растения или его семени в поле, зараженном вредителем, против которого полипептид обладает пестицидной активностью. В некоторых вариантах осуществления полипептид обладает пестицидной активностью против вредителя из группы чешуекрылых, жесткокрылых, двукрылых, полужесткокрылых или нематод, и поле заражено вредителем из группы чешуекрылых, полужесткокрылых, жесткокрылых, двукрылых или нематод.
Как определяется в данном документе, "урожайностью растения называют качество и/или количество биомассы, продуцируемой растением. Применяемый в данном документе термин "биомасса" относится к любому измеренному продукту растения. Увеличением продукции биомассы является любое улучшение урожайности измеренного продукта растения. Увеличение урожайности растения имеет несколько коммерческих применений. Например, увеличение биомассы листьев растения может приводить к увеличению урожайности листовых овощей для потребления человеком или животными. Кроме того, увеличение биомассы листьев можно применять для увеличения производства фармацевтических или промышленных продуктов растительного происхождения. Увеличение урожайности может предусматривать любое статистически значимое увеличение, в том числе без ограничения, по меньшей мере 1% повышение, по меньшей мере 3% повышение, по меньшей мере 5% повышение, по меньшей мере 10% повышение, по меньшей мере 20% повышение, по меньшей мере 30% повышение, по меньшей мере 50% повышение, по меньшей мере 70% повышение, по меньшей мере 100% или большее повышение урожайности по сравнению с растением, не экспрессирующим пестицидную последовательность.
В конкретных способах урожайность растения повышается в результате улучшенной устойчивости к вредителю растения, экспрессирующего полипептид PtIP-83, раскрытый в данном документе. Экспрессия полипептида PtIP-83 приводит к сниженной способности вредителя к заражению растения или питанию на растении, таким образом, улучшая урожайность растения.
Способы переработки
Дополнительно представлены способы переработки растения, части растения или семени с получением пищевого или кормового продукта из растения, части растения или семени, содержащих полипептид PtIP-83. Растения, части растения или семена, представленные в данном документе, можно перерабатывать с получением масла, белковых продуктов и/или побочных продуктов, которые являются производными, полученными путем переработки, которые имеют коммерческое значение. Неограничивающие примеры включают трансгенные семена, содержащие молекулу нуклеиновой кислоты, кодирующую полипептид PtIP-83, который можно перерабатывать с получением соевого масла, соевых продуктов и/или соевых побочных продуктов.
"Переработка" относится к любым физическим и химическим способам, применяемым для получения какого-либо соевого продукта, и они включают без ограничения кондиционирование нагреванием, вальцевание и измельчение, экструзию, экстракцию растворителем или вымачивание в воде и экстракцию цельных или дробленых семян.
Следующие примеры представлены для иллюстрирования, а не для ограничения.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Пример 1. Идентификация инсектицидного белка, активного против широкого спектра насекомых из группы чешуекрылых, из папоротника, Adiantum pedatum, (PS-7140)
Инсектицидный белок PtIP-83Aa (SEQ ID NO: 1) идентифицировали с помощью очистки белка, масс-спектрометрии (MS) и ПЦР-клонирования из Adiantum pedatum, (PS-7140) следующим образом.
Adiantum pedatum собирал сотрудник, и ему присвоили идентификационный номер PS-7140. PS-7140 собирали, подвергали быстрому замораживанию в жидком N2 и хранили при -800 C. После хранения его измельчали в мелкий порошок при температуре жидкого N2 с помощью шаровой мельницы Geno (SPEX, Метачен, Нью-Джерси). Для экстракции белка к каждым 5 г сырого веса ткани добавляли 20 мл 50 мМ буфера Tris, pH 8,0, 150 мМ KCl, 2,5 мМ EDTA, 1,5% поливинилполипирролидона (PVPP) и смесь ингибиторов протеаз (Roche Diagnostics, Германия). Гомогенат центрифугировали для удаления клеточного дебриса, фильтровали через 0,22 мкм фильтры и обессоливали с применением 10 мл колонки для обессоливания Zeba (Thermo Scientific, Иллинойс)
Биологические анализы в отношении трех видов вредителей, совки соевой (SBL) (Chrysodeixis includens), совки кукурузной (CEW) (Helicoverpa zea) и огневки кукурузной (ECB) (Ostrinia nubilalis), проводили с применением обессоленного экстракта белков, помещенного на питательную среду для Lepidoptera на основе агара (Southland Products Inc., Лейк-Виллидж, Арканзас) в формате 96-луночного планшета. Применяли по шесть повторностей на каждый образец. Образцы оставляли высыхать на поверхности питательной среды и от двух до пяти новорожденных насекомых помещали в каждую лунку обработанного планшета. После четырех дней инкубации при 27°C личинок оценивали в отношении смертности или тяжести остановки роста. Баллы регистрировали количественно как погибшие (3), со значительной остановкой роста (2) (незначительный рост или отсутствие роста, но живые и эквивалентные личинкам 1ого возраста), с остановкой роста (1) (рост до второго возраста, но не эквивалентны контролям) или нормальные (0). Воздействие на образец протеиназы K и тепловая обработка приводили к потере активности, что указывает на то, что действующее начало было белковым по природе. Результаты биологических анализов показаны в таблице 6.
Таблица 6
Для очистки белка вайи PS-7140 измельчали в мелкий порошок при температуре жидкого N2 с помощью шаровой мельницы Geno (SPEX, Метачен, Нью-Джерси). Белок экстрагировали в 100 мМ буфере Tris, pH 8,0, 150 мМ KCl, 2,5 мМ EDTA, 1,5% PVPP и смеси ингибиторов протеаз (Roche Diagnostics, Германия). Экстрагированный материал центрифугировали для удаления клеточного дебриса, фильтровали через Miracloth™ (Calbiochem) и затем добавляли до 35% сульфат аммония и давали отстояться. Суспензию центрифугировали и полученный осадок ресуспендировали в малом объеме 20 мМ буфера Tris, pH 8. После осветления путем центрифугирования его подвергали обессоливанию с применением колонки Sephadex G25 (GE, Пискатавей, Нью-Джерси), уравновешенной в 20 мМ буфере Tris, pH 8. Пул обессоленной белковой фракции загружали на 1 мл колонку Mono Q (GE, Пискатавей, Нью-Джерси) и элюировали с помощью линейного (60 CV (объемов колонки) градиента от 0 M до 0,7 M NaCl в 20 мМ Tris, pH 8,0. Фракции, активные против SBL и ECB, объединяли и подвергали обессоливанию в 25 мМ MOPS, pH 6,7. Затем их загружали на 4 мл колонку Mono P (буфер A: 25 мМ MOPS, pH 6,7; буфер B: Polybuffer 74, pH 4) с применением 4 CV линейного градиента (0% буфера B), за которым следовала отмывка с помощью 15 CV 100% буфера B. Собирали фракции по 1 мл. Фракции 47 и 48 демонстрировали активность против ECB и SBL. Исходя из LDS-PAGE, эти активные фракции содержали белковую полоску на уровне приблизительно 95 кДа. Белок, представляющие полоску 95 кДа, назвали PtIP-83Aa (SEQ ID NO: 1).
Идентификацию белка проводили посредством MS-анализа после расщепления белка с помощью трипсина. Белки для идентификации с помощью MS получали после прогона образца на геле LDS-PAGE, окрашенном с помощью красителя бриллиантовый синий G-250. Полоски, представляющие интерес, вырезали из геля, удаляли краситель, восстанавливали с помощью дитиотреитола и затем алкилировали с помощью йодоацетамида. После расщепления с помощью трипсина в течение ночи, образцы анализировали с помощью наножидкостной хроматографии/тандемной масс-спектрометрии (нано-LC/ES-MSMS) на масс-спектрометре Thermo Q Exactive Orbitrap (Thermo Fisher Scientific), сопряженной с системой для наножидкостной хроматографии Eksigent NanoLC Ultra 1-D Plus и автодозатором nanolc-as2 (AB Sciex). Идентификацию белков проводили путем поиска данных нано-LC/MSMS в сравнении с внутрилабораторной базой данных транскриптомов, содержащей транскрипты из исходных растительных материалов и открытой базы данных белков Swiss-Prot с применением средства поиска Mascot (Matrix Science).
Аминокислотную последовательность под SEQ ID NO: 1 подвергали поиску BLAST (средство поиска основного локального выравнивания; Altschul, et al., (1993) J. Mol. Biol. 215:403-410; см. также ncbi.nlm.nih.gov/BLAST/, доступ к которому можно получить с применением префикса www) в сравнении с открытой и внутренней базой данных DUPONT-PIONEER, которая включала последовательности растительных белков. Аминокислотные последовательности выравнивали с белками в проприетарной базе данных белков от DUPONT-PIONEER.
Пример 2. Транскриптомное секвенирование PtIP-83Aa
Транскриптом для Adiantum pedatum, PS-7140, получали следующим образом. Общие РНК выделяли из замороженных тканей путем применения набора Qiagen® RNeasy® для выделения общей РНК. Библиотеки для секвенирования из полученных общих РНК получали с применением набора для секвенирования мРНК TruSeq™ и протокола от Illumina®, Inc. (Сан-Диего, Калифорния). Вкратце, мРНК выделяли путем присоединения к гранулам с олиго(dT), фрагментировали до среднего размера 180 нуклеотидов, подвергали обратной транскрипции в кДНК с помощью случайных гексамерных праймеров, подвергали репарации концов, добавляли 3' A-хвост и лигировали с помощью пронумерованных адаптеров TruSeq™ от Illumina®. Лигированные фрагменты кДНК подвергали ПЦР-амплификации с применением праймеров TruSeq™ от Illumina® и очищенные продукты ПЦР проверяли в отношении качества и количества на чипе Agilent Bioanalyzer® DNA 7500. После оценки качества и количества по 100 нг библиотек транскриптов нормализовали путем обработки с помощью дуплекс-специфичной нуклеазы (DSN) (Evrogen®, Москва, Россия). Нормализацию завершали путем добавления 200 мМ буфера Hepes, за которым следовала тепловая денатурация и пять часов гибридизации при 68°C. Гибридизированную библиотеку обрабатывали с помощью 2 мкл фермента DSN в течение 25 минут, очищали с помощью колонок Qiagen® MinElute® в соответствии с протоколами производителя, и амплифицировали двенадцать циклов с применением специфичных в отношении адаптера праймеров от Illumina®. Конечные продукты очищали с помощью гранул Ampure® XP (Beckman Genomics, Данверс, Массачуссетс) и проверяли в отношении качества и количества на чипе Agilent Bioanalyzer® DNA 7500.
Нормализованные библиотеки транскриптов секвенировали в соответствии с протоколами производителя на Illumina® Genome Analyzer IIx. Каждую библиотеку гибридизировали в проточных ячейках с двумя дорожками и амплифицировали, блокировали, линеализировали и гибридизировали с праймером с применением процесса получения клональных кластеров на cBot® от Illumina. Секвенирование завершали на Genome Analyzer IIx, который давал шестьдесят миллионов ридов со спаренными концами из 75 п.о. на нормализованную библиотеку.
В отношении пептидной последовательности, идентифицированной для PtIP-83Aa (SEQ ID NO: 1) с помощью LC-MS/MS/MS-секвенирования (описано в примере 1), проводили поиск в сравнении с последовательностями белков, прогнозируемых по открытым рамкам считывания (ORF) из внутренних транскриптомных сборок для PS-7140CF. Пептиды давали точное совпадение с транскриптом, соответствующим PtIP-83Aa (SEQ ID NO: 2). Кодирующие последовательности применяли для разработки следующих праймеров для клонирования кодирующей последовательности PtIP-83Aa:
CCATGGCTCTCGTGGATTACGGCAAG (95KD N-T Nco I for) (SEQ ID NO: 30) и GTTAACCTACTCTTCGTCGTGCCGCCAGTC (95KD C-T Hpa I REV) (SEQ ID NO: 31). Данный клон получали с помощью полимеразной цепной реакции с применением набора для ПЦР KOD Hot Start DNA polymerase® (Novagen, Merck KGaA, Дармштадт, Германия) и общей РНК из Adiantum pedatum в качестве шаблона. Клонированный продукт ПЦР подтверждали путем секвенирования
На основе результатов секвенирования ДНК и белка полинуклеотидная последовательность PtIP-83Aa показана как SEQ ID NO: 2, а полипептидная последовательность как SEQ ID NO: 1.
Пример 3. Идентификация гомологов PtIP-83
Генную идентичность можно определить посредством проведения поисков BLAST® (средство поиска основного локального выравнивания; Altschul, et al., (1993) J. Mol. Biol. 215:403-410; см. также ncbi.nlm.nih.gov/BLAST/, доступ к которому можно получить с применением префикса www) с параметрами по умолчанию в отношении сходства с последовательностями. Анализировали полинуклеотидную последовательность PtIP-83Aa (SEQ ID NO: 2). В открытых базах данных не идентифицировали близких гомологов PtIP-83Aa (SEQ ID NO: 1).
Определение генной идентичности, проводимое с помощью BLAST® в внутренней базе данных транскриптомов папоротников и других примитивных растений DUPONT PIONEER, идентифицировало гомологи PtIP-83Aa (SEQ ID NO: 1). Гомологи PtIP-83Aa и организм, в котором они были идентифицированы, показаны в таблице 7.
Таблица 7
'Angustifolia'
'Angustifolia'
'Angustifolia'
'Angustifolia'
'Angustifolia'
'Angustifolia'
кДНК получали из организмов-источников с идентифицированными гомологами с помощью обратной транскрипции из общей РНК. Гомологи подвергали ПЦР-амплификации за счет их соответствующих кДНК с применением праймеров, разработанных для кодирующих последовательностей каждого гомолога, и субклонировали в вектор для временной экспрессии в растении, содержащий промотор DMMV. Клонированные продукты ПЦР подтверждали путем секвенирования. Праймеры клонирования показаны в таблице 8. кДНК для гомологов PtIP-83Fb синтезировали (SEQ ID NO: 717) на основании транскриптомной сборки PS-9319 и субклонировали в вектор для временной экспрессии в растении.
Таблица 8
(SEQ ID NO: 32)
(SEQ ID NO: 33)
Дополнительное разнообразие восстанавливали посредством ПЦР-амплификации с праймерами, разработанными для некодирующих участков, фланкирующих идентифицированные гомологи. Праймеры разрабатывали для консервативных последовательностей в 5' и 3' нетранслируемых участках BLAST-хитов для PtIP-83Ca в пределах внутренней базы данных транскриптомов DUPONT PIONEER. Пары праймеров (1) GCCTTTATCGACTCCTAATTCACACC (SEQ ID NO: 626) и CCACATTGTGCATTACGACCAC (SEQ ID NO: 627), и (2) CCAGTGATTTGAGTTCCTTCATTATG (SEQ ID NO: 628) и GAACAGTACATTGACTGCATGTGC (SEQ ID NO: 629) применяли для получения продуктов ПЦР за счет кДНК PS-11481 и PS-12345. Полученные продукты ПЦР подвергали клонированию по тупым концам с применением набора для ПЦР-клонирования Zero Blunt® TOPO® (Invitrogen) и анализу последовательности. На основании этого анализа идентифицировали гомологи PtIP-83Cg, Da, Ea и Eb, и их субклонировали в вектор для временной экспрессии в растении. Праймеры клонирования показаны в таблице 8.
Значения идентичности аминокислотных последовательностей гомологов PtIP-83Aa, рассчитанная с применением алгоритма Нидлмана-Вунша, реализованного в программе Needle (набор инструментов EMBOSS), показаны в таблицах 9a - 9c.
Таблица 9a
SEQ ID NO: 5
SEQ ID NO: 7
SEQ ID NO: 9
SEQ ID NO: 11
SEQ ID NO: 13
SEQ ID NO: 15
SEQ ID NO: 17
SEQ ID NO: 755
SEQ ID NO: 754
SEQ ID NO: 759
SEQ ID NO: 1
SEQ ID NO: 5
SEQ ID NO: 7
SEQ ID NO: 9
SEQ ID NO: 11
SEQ ID NO: 13
SEQ ID NO: 15
SEQ ID NO: 17
SEQ ID NO: 755
SEQ ID NO: 754
SEQ ID NO: 759
SEQ ID NO: 758
SEQ ID NO: 763
SEQ ID NO: 762
SEQ ID NO: 19
SEQ ID NO: 21
SEQ ID NO: 23
SEQ ID NO: 3
SEQ ID NO: 716
SEQ ID NO: 756
SEQ ID NO: 757
SEQ ID NO: 761
SEQ ID NO: 760
SEQ ID NO: 766
SEQ ID NO: 767
SEQ ID NO: 769
SEQ ID NO: 768
SEQ ID NO: 765
Таблица 9b
SEQ ID NO: 758
SEQ ID NO: 763
SEQ ID NO: 762
SEQ ID NO: 19
SEQ ID NO: 21
SEQ ID NO: 23
SEQ ID NO: 3
SEQ ID NO: 716
SEQ ID NO: 756
SEQ ID NO: 1
SEQ ID NO: 5
SEQ ID NO: 7
SEQ ID NO: 9
SEQ ID NO: 11
SEQ ID NO: 13
SEQ ID NO: 15
SEQ ID NO: 17
SEQ ID NO: 755
SEQ ID NO: 754
SEQ ID NO: 759
SEQ ID NO: 758
SEQ ID NO: 763
SEQ ID NO: 762
SEQ ID NO: 19
SEQ ID NO: 21
SEQ ID NO: 23
SEQ ID NO: 3
SEQ ID NO: 716
SEQ ID NO: 756
SEQ ID NO: 757
SEQ ID NO: 761
SEQ ID NO: 760
SEQ ID NO: 766
SEQ ID NO: 767
SEQ ID NO: 769
SEQ ID NO: 768
SEQ ID NO: 765
Таблица 9c
SEQ ID NO: 757
SEQ ID NO: 761
SEQ ID NO: 760
SEQ ID NO: 766
SEQ ID NO: 767
SEQ ID NO: 769
SEQ ID NO: 768
SEQ ID NO: 765
SEQ ID NO: 764
SEQ ID NO: 1
SEQ ID NO: 5
SEQ ID NO: 7
SEQ ID NO: 9
SEQ ID NO: 11
SEQ ID NO: 13
SEQ ID NO: 15
SEQ ID NO: 17
SEQ ID NO: 755
SEQ ID NO: 754
SEQ ID NO: 759
SEQ ID NO: 758
SEQ ID NO: 763
SEQ ID NO: 762
SEQ ID NO: 19
SEQ ID NO: 21
SEQ ID NO: 23
SEQ ID NO: 3
SEQ ID NO: 716
SEQ ID NO: 756
SEQ ID NO: 757
SEQ ID NO: 761
SEQ ID NO: 760
SEQ ID NO: 766
SEQ ID NO: 767
SEQ ID NO: 769
SEQ ID NO: 768
SEQ ID NO: 765
Пример 4. Временная экспрессия в листьях и биологический анализ на насекомых
Для подтверждения активности PtIP-83Aa (SEQ ID NO: 1) использовали систему для временной экспрессии под контролем вирусного промотора pDMMV и/или AtUBQ10 (Dav, et. al., (1999) Plant Mol. Biol. 40:771-782; Norris SR et al (1993) Plant Mol Biol. 21(5):895-906). Из уровня техники хорошо известен способ инфильтрации агробактериями путем введения клеточной суспензии Agrobacterium в растительные клетки интактных тканей для того, чтобы можно было измерить или изучить воспроизводимые инфицирование и дальнейшую экспрессию трансгена, полученного из растения (Kapila, et. al., (1997) Plant Science 122:101-108). Вкратце, фасоль кустовую (фасоль обыкновенная, Phaseolus vulgaris) или сою(Glycine max) на стадии одного листа подвергали инфильтрации агробактериями с помощью нормализованных культур бактериальных клеток тестируемых и контрольных штаммов. Через 4-7 дней из каждого проростка вырезали листовые диски и заражали их по отдельности 2 новорожденными особями совки соевой (SBL) (Chrysodeixis includens), 2 новорожденными особями совки кукурузной (CEW) (Helicoverpa zea), 2 новорожденными особями совки травяной (Spodoptera frugiperda) или 4 новорожденными особями огневки кукурузной (ECB) (Ostrinia nubilalis). Контрольные листовые диски получали с Agrobacterium, содержащими только флуоресцентный маркер DsRed2 (Clontech™, 1290 Terra Bella Ave. Маунтин-Вью, Калифорния 94043). Листовые диски от не подвергнутых инфильтрации растений включали в качестве второго контроля. Потребление зеленых тканей листа оценивали через два (CEW, FAW) или три (ECB, SBL, FAW) дня после заражения и присваивали баллы от 0 до 9. Временно экспрессируемый PtIP-83Aa (SEQ ID NO: 1) защищал листовые диски от потребления насекомыми, которыми их заражали, в то же время полное потребление зеленой ткани наблюдали для ткани отрицательного контроля и необработанной ткани (таблица 10). Временную экспрессию белка PtIP-83Aa (SEQ ID NO: 1) подтверждали с помощью идентификации белка на основе масс-спектрометрии с применением трипсинизированных белковых экстрактов из подвергнутых инфильтрации тканей листа (Patterson, (1998) 10(22):1-24, Current Protocol in Molecular Biology published by John Wiley & Son Inc). Временную экспрессию можно было проверить путем применения меченых HA или EPEA полипептидов PtIP-83 с помощью вестерн-блот анализа в отношении меток HA или EPEA.
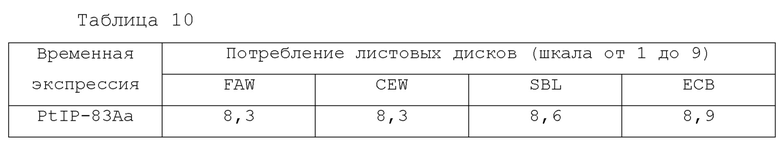

Активность PtIP-83Fa (SEQ ID NO: 4) подтверждали с применением системы временной экспрессии на фасоли кустовой и обнаружили, что она сравнима с PtIP-83Aa (SEQ ID NO: 2). PtIP-83Fa (SEQ ID NO: 4) демонстрировал аналогичный спектр инсектицидной активности, за исключением отсутствия активности против FAW (таблица 11).
Таблица 11
Активность PtIP-83Ca (SEQ ID NO: 5), PtIP-83Cb (SEQ ID NO: 7), PtIP-83Cc (SEQ ID NO: 9), PtIP-83Cd (SEQ ID NO: 11), PtIP-83Ce (SEQ ID NO: 13), PtIP-83Cf (SEQ ID NO: 15), PtIP-83Cg (SEQ ID NO: 17), PtIP-83Da (SEQ ID NO: 19), PtIP-83Ea (SEQ ID NO: 21), PtIP-83Eb (SEQ ID NO: 23), PtIP-83Fb (SEQ ID NO: 716), PtIP-83Ch (SEQ ID NO: 754), PtIP-83Ch-like (SEQ ID NO: 755), PtIP-83Fd (SEQ ID NO: 756), PtIP-83Fe (SEQ ID NO: 757), PtIP-83Ci (SEQ ID NO: 758), PtIP-83Ci-подобного (SEQ ID NO: 759), PtIP-83Ff (SEQ ID NO: 760) и PtIP-83Ff-подобного (SEQ ID NO: 761) также подтверждали с применением системы временной экспрессии на фасоли кустовой. Спектр активности для всех гомологов PtIP-83 подытожен в таблице 12, где "+" указывает на средний балл активности, составляющий <=60% потребленных листовых дисков, а "-" указывает на средний балл активности, составляющий >=60% потребленных листовых дисков.
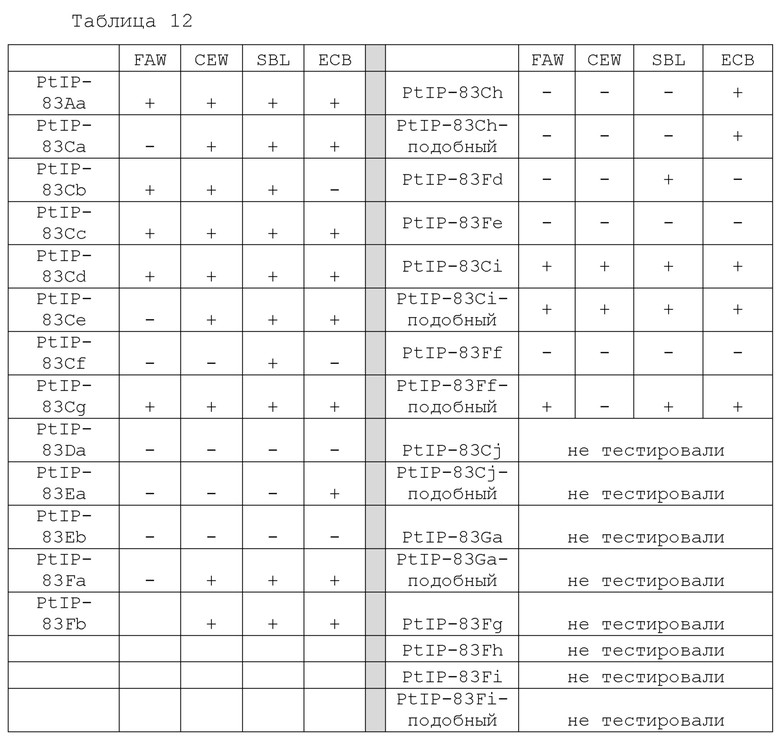
Пример 5. Бакуловирусная экспрессия полипептидов PtIP-83Aa
Ген, кодирующий PtIP-83Aa под SEQ ID NO: 2, субклонировали в двойной вектор pFastBac™ (Invitrogen®) с удаленным стоп-кодоном для C-терминальной трансляции добавления метки из 10x-гистидина (SEQ ID NO: 27), и последовательность полипептида PtIP-83Aa с гистидиновой меткой изложена как SEQ ID NO: 2. Этот вектор трансформировали в клетки DH10Bac с получением бакуловирусов. Эти бакуловирусы применяли для инфицирования клеток насекомых sf9 и инкубировали в течение 72 часов при 27°C. Инфицированные клетки насекомых собирали путем центрифугирования. Осадок клеточной культуры суспендировали в 100 мл буфера для лизиса (1XPBS, 10% глицерина, с ингибитором протеаз и бензоназой) и инкубировали при 4°C в течение 5 мин при перемешивании, затем дважды гомогенизировали. Лизат центрифугировали при 16000 об./мин. в течение 20 мин. Супернатант сохраняли и загружали на 2 мл колонки со смолой Ni-NTA Hi-Bind (Novagen, № по кат. 70666), предварительно уравновешенные с помощью буфера для элюирования (1XPBS, 10% глицерина). Затем колонки последовательно элюировали с помощью 10 мл буфера для элюирования, содержащего 10, 20, 50 и 250 мМ имидазола. образцы анализировали с помощью SDS-PAGE. Очищенные фракции (E250) концентрировали с применением фильтров для ультрацентрифугирования 100K Amicon® (Millipore) до ~0,5 мг/мл, и они демонстрировали инсектицидную активность против CEW, ECB, FAW и SBL, аналогичную спектру активности, полученному с применением листовых дисков с временной экспрессией.
Пример 6. Опосредованная Agrobacterium трансформация маиса и регенерация трансгенных растений
Для опосредованной Agrobacterium трансформации маиса с помощью нуклеотидных последовательностей PtIP-83, таких как PtIP-83Aa (SEQ ID NO: 2), PtIP-83Aa ModA (кодон-оптимизированная) (SEQ ID NO: 28) и PtIP-83Aa ModB (кодон-оптимизированная (SEQ ID NO: 29) можно применять способ Zhao (патент США № 5981840 и патентная публикация согласно PCT № WO 1998/32326; содержание которых включено в данный документ посредством ссылки). Вкратце, незрелые зародыши выделяли из маиса и зародыши приводили в контакт с суспензией Agrobacterium в условиях, при которых бактерии способны перенести нуклеотидную последовательность (PtIP-83Aa SEQ ID NO: 2, PtIP-83Aa ModA (кодон-оптимизированная) -SEQ ID NO: 28 и PtIP-83Aa ModB (кодон-оптимизированная - SEQ ID NO: 29) по меньшей мере в одну клетку по меньшей мере одного из незрелых зародышей (стадия 1: стадия инфицирования). На этой стадии незрелые зародыши можно погружать в суспензию Agrobacterium для инициации инокуляции. Зародыши в течение определенного времени культивировали совместно с Agrobacterium (стадия 2: стадия совместного культивирования). Незрелые зародыши можно культивировать на твердой среде после стадии инфицирования. После периода совместного культивирования предполагалась необязательная стадия "покоя". На этой стадии покоя зародыши инкубировали в присутствии по меньшей мере одного антибиотика, который, как известно, ингибирует рост Agrobacterium, без добавления селективного средства для трансформации растений (стадия 3: стадия покоя). Незрелые зародыши культивировали на твердой среде с антибиотиком, но без селективного средства, для элиминации Agrobacterium и в течение фазы покоя для инфицированных клеток. Затем, инокулированные зародыши культивировали на среде, содержащей селективное средство, и выделяли растущий трансформированный каллюс (стадия 4: стадия отбора). Незрелые зародыши культивировали на твердой среде с селективным средством, что приводило к селективному росту трансформированных клеток. Затем каллюс регенерировали с получением растений (стадия 5: стадия регенерации), и каллюсы, выращенные на селективной среде, культивировали на твердой среде для регенерации растений.
Пример 7. Трансформация и регенерация сои (Glycine max)
Трансгенные линии сои получали с помощью способа бомбардировки генной пушкой (Klein et al., Nature (London) 327:70-73 (1987); патент США № 4945050) с применением устройства BIORAD Biolistic PDS1000/He и либо плазмиды, либо фрагмента ДНК. Для трансформации и регенерации растений сои применяют следующие исходные растворы и среды:
Исходные растворы:
Исходный 100 X раствор сульфатов:
37 г MgSO4.7 H2O, 1,69 г MnSO4.H2O, 0,86 г ZnSO4.7 H2O, 0,0025 г CuSO4.5 H2O
Исходный 100 X раствор галогенидов:
30,0 г CaCl2.2 H2O, 0,083 г KI, 0,0025 г CoCl2.6 H2O
Исходный 100X раствор P, B, Mo:
18,5 г KH2PO4, 0,62 г H3BO3, 0,025 г Na2MoO4.2 H2O
Исходный 100X раствор Fe EDTA:
3,724 г Na2EDTA, 2,784 г FeSO4.7 H2O
Исходный раствор 2,4-D:
10 мг/мл витамина
Исходный 1000X раствор витаминов B5:
100,0 г мио-инозитола, 1,0 г никотиновой кислоты, 1,0 г пиридоксин-HCl, 10 г тиамин.HCL.
Среда (на литр):
Твердая среда SB199:
1 упаковка солей MS (Gibco/ BRL - № по кат. 11117-066), 1 мл исходного 1000X раствора витаминов B5, 30 г сахарозы, 4 мл 2,4-D (конечная концентрация 40 мг/л), pH 7,0, 2 г Gelrite
Твердая среда SB1:
1 упаковка солей MS (Gibco/ BRL - № по кат. 11117-066), 1 мл исходного 1000X раствора витаминов B5, 31,5 г глюкозы, 2 мл 2,4-D (конечная концентрация 20 мг/л), pH 5,7, 8 г агара TC
SB196:
по 10 мл каждого вышеуказанных исходных растворов 1-4, 1 мл исходного раствора витаминов B5, 0,463 г (NH4)2 SO4, 2,83 г KNO3, 1 мл исходного раствора 2,4 D, 1 г аспарагина, 10 г сахарозы, pH 5,7
SB71-4:
соли Гамборга B5, 20 г сахарозы, 5 г агара TC, рН 5,7.
SB103:
1 упаковка смеси солей Murashige & Skoog, 1 мл исходного раствора витаминов B5, 750 мг MgCl2 гексагидрата, 60 г мальтозы, 2 g Gelrite™, pH 5,7.
SB166:
SB103, дополненная активированных углем из расчета 5 г на литр.
Инициация эмбриогенных суспензионных культур сои
Бобы с незрелыми семенами из доступных растений сои собирали через 45-55 дней после высаживания, очищали от оболочек и помещали в стерильный контейнер Magenta. Семена сои стерилизовали путем встряхивания в течение 15 мин. в 5% растворе Clorox® с 1 каплей мыла Ivory™ (т.е. 95 мл автоклавированной дистиллированной воды плюс 5 мл Clorox® и 1 капля мыла, тщательно перемешанных). Семена промывали с использованием 2 л стерильной дистиллированной воды и семена размером менее 3 мм помещали на отдельные предметные стекла. Срезали верхний конец семени и семядоли выдавливали из семенной оболочки. Семядоли переносили на чашки, содержащие среду SB199 (25-30 семядолей на чашку), на две недели, затем переносили на SB1 на 2-4 недели. Чашки обертывали волокнистой лентой. По прошествии этого времени отрезали вторичные зародыши и помещали в жидкую среду SB196 на 7 дней.
Условия культивирования:
Эмбриогенные суспензионные культуры сои (сорт 93Y21) поддерживали в 50 мл жидкой среды SB196 на ротационном шейкере, 100-150 об/мин, 26°C с фотопериодом 16:8 часов день/ночь при интенсивности света 80-100 мкЭрг/м2/с. Культуры пассировали через каждые 7-14 дней путем инокуляции количества ткани размером до ½ десятицентовой монеты (скопления, растущие вместе) в 50 мл свежей жидкой среды SB196.
Получение ДНК для бомбардировки
При процедурах бомбардировки генной пушкой возможно применение очищенных 1) плазмидной ДНК целиком; или 2) фрагментов ДНК, содержащих только кассету(ы) экспрессии рекомбинантной ДНК, представляющую интерес. Для каждых семнадцати трансформаций путем бомбардировки готовили 85 мкл суспензии, содержащей 1-90 пикограммов (пг) плазмидной ДНК на пару оснований каждой ДНК-плазмиды. ДНК-плазмиды или фрагменты совместно осаждали на частицах золота следующим образом. ДНК в суспензии добавляли к 50 мкл суспензии с 10-60 мг/мл 0,6 мкм частиц золота и затем объединяли с 50 мкл CaCl2 (2,5 M) и 20 мкл спермидина (0,1 M). Смесь перемешивали на вихревой мешалке в течение 5 секунд, центрифугировали в микроцентрифуге в течение 5 секунд, и супернатант удаляли. Затем частицы, покрытые ДНК, однократно промывали с помощью 150 мкл 100% этанола, перемешивали на вихревой мешалке и снова центрифугировали в микроцентрифуге, затем ресуспендировали в 85 мкл безводного этанола. Затем по пять мкл частиц золота, покрытых ДНК, загружали на каждый диск макроносителя.
Получение тканей и бомбардировка ДНК
Приблизительно 100 мг двухнедельной суспензионной культуры помещали в пустую 60 мм X 15 мм чашку Петри и остаточную жидкость удаляли из ткани с применением пипетки. Ткань располагали на расстоянии приблизительно 3,5 дюйма от задерживающего экрана и каждую чашку с тканью бомбардировали один раз. Давление разрыва мембраны устанавливали на 650 фунтов на квадратный дюйм и из камеры откачивали воздух до давления -28 дюймов ртутного столба. После бомбардировки ткань из каждой чашки разделяли между двумя колбами, помещали обратно в жидкую среду и культивировали как описано выше.
Отбор трансформированных зародышей и регенерация растений:
После бомбардировки ткань из каждой подвергнутой бомбардировке чашки разделяли и помещали в две колбы с жидкой средой для поддержания культуры SB196 на чашку с подвергнутой бомбардировке ткани. Через семь дней после бомбардировки жидкую среду в каждой колбе заменяли свежей средой SB196 для поддержания культуры с добавлением 100 нг/мл селективного средства (селективной среды). Для отбора трансформированных клеток сои применяемое селективное средство может представлять собой соединение сульфонилмочевины (SU) с химическим названием 2-хлор-N-((4-метокси-6 метил-1,3,5-триазин-2-ил)аминокарбонил) бензолсульфонамид (обычные названия: DPX-W4189 и хлорсульфурон). Хлорсульфурон представляет собой активный ингредиент гербицида на основе сульфонил мочевины GLEAN® от DuPont. Селективную среду, содержащую SU, заменяли через каждые две недели в течение 8 недель. После 8-недельного периода отбора наблюдали, что островки зеленой, трансформированной ткани растут из нетрансформированных, некротических эмбриогенных кластеров. Эти предполагаемые трансгенные объекты выделяли и поддерживали в жидкой среде SB196 с SU из расчета 100 нг/мл в течение еще 5 недель, при этом среду меняли каждые 1-2 недели, с получением новых, клонально размножающихся, трансформированных эмбриогенных суспензионных культур. Зародыши проводили суммарно приблизительно 13 недель в контакте с SU. Суспензионные культуры пассировали и поддерживали в виде скоплений незрелых зародышей, и также регенерировали в целые растения путем созревания и проращивания отдельных соматических зародышей.
Соматические зародыши становились пригодными для проращивания после четырех недель в среде для созревания (1 неделя в SB166, затем 3 недели в SB103). Затем их извлекали из среды для созревания и подсушивали в пустых чашках Петри до семи дней. Затем подсушенные зародыши высаживали в среду SB71-4, где их оставляли прорастать в тех же световых и температурных условиях, описываемых выше. Проросшие зародыши переносили в горшечную среду и выращивали до зрелости с получением семян.
Пример 8. Трансформация посредством бомбардировки частицами и регенерация трансгенных растений
Незрелые зародыши маиса от тепличных растений-доноров бомбардировали плазмидой, содержащей нуклеотидную последовательность, кодирующую инсектицидный белок. Початки очищали от листовой обертки и подвергали поверхностной стерилизации в 30% отбеливателе Clorox® вместе с 0,5% моющего средства Micro в течение 20 минут, и ополаскивали два раза стерильной водой. Незрелые зародыши вырезали и помещали рубчиком вниз (щитком вверх), 25 зародышей на чашку, на среду 560Y на 4 часа и затем выравнивали в пределах целевой зоны 2,5 см при подготовке к бомбардировке. ДНК плазмидного вектора, содержащую нуклеотидную последовательность, кодирующую инсектицидный белок, функционально связанный с промотором, осаждали на 1,1 мкм (средний диаметр) вольфрамовые гранулы с применением процедуры преципитации с CaCl2 следующим образом: 100 мкл подготовленных частиц вольфрама в воде; 10 мкл (1 мкг) ДНК в буфере Tris EDTA (1 мкг общей ДНК); 100 мкл 2,5 M CaCl2 и 10 мкл 0,1 M спермидина.
Каждый реагент добавляли последовательно к суспензии частиц вольфрама, при этом она находилась на вихревой мешалке для множества пробирок. Конечную смесь подвергали непродолжительной ультразвуковой обработке и обеспечивали инкубацию при постоянном перемешивании на вихревой мешалке в течение 10 минут. После периода осаждения пробирки непродолжительно центрифугировали, жидкость удаляли, промывали с помощью 500 мл 100% этанола и центрифугировали в течение 30 секунд. Жидкость вновь удаляли и к конечному осадку частиц вольфрама добавляли 105 мкл 100% этанола. Для бомбардировки генной пушкой частицы вольфрама/ДНК подвергали непродолжительной ультразвуковой обработке, и вносили 10 мкл на центр каждого макроносителя, и давали высохнуть приблизительно 2 минуты перед бомбардировкой. Чашки с образцами бомбардировали с помощью генной пушки на уровне № 4. Все образцы получали однократный выстрел из расчета 650 фунтов/кв. дюйм, при этом общее количество аликвот, взятых из каждой пробирки с полученными частицами/ДНК, составляло десять.
После бомбардировки зародыши поддерживали на среде 560Y в течение 2 дней, затем переносили на среду для отбора 560R, содержащую 3 мг/л биалафоса, и пассировали каждые 2 недели. Спустя приблизительно 10 недель отбора каллюсные клоны, устойчивость которых была выявлена в результате отбора, переносили на среду 288J для инициирования регенерации растений. После созревания соматических зародышей (2-4 недели) хорошо развитые соматические зародыши переносили в среду для проращивания и переносили в освещаемую комнату для культивирования. Спустя приблизительно 7-10 дней развивающиеся проростки переносили на безгормональную среду 272V в пробирках на 7-10 дней до хорошего укоренения проростков. Затем растения переносили во вставки в лотки (эквивалентны горшку на 2,5 дюйма), содержащие почвенную горшечную смесь, и выращивали в течение 1 недели в вегетационной камере, впоследствии выращивали в течение дополнительных 1-2 недель в теплице, затем переносили в горшки Classic 600 (1,6 галлона) и выращивали до зрелости. За растениями вели наблюдение и проводили оценку в отношении экспрессии полипептида PtIP-83 с помощью анализов, известных из уровня техники, таких как, например, иммунологические анализы и вестерн-блоттинг.
Трансгенные растения маиса, положительные в отношении экспрессии инсектицидных белков, тестировали на пестицидную активность с применением стандартных биологических анализов, известных из уровня техники. Такие способы включают, например, биологические анализы с иссечением корня и биологические анализы целого растения. См., например, публикацию заявки на патент США № US 2003/0120054 и международную публикацию № WO 2003/018810.
Среда для бомбардировки (560Y) содержала 4,0 г/л основных солей N6 (SIGMA C-1416), 1,0 мл/л витаминной смеси Эрикссона (1000X SIGMA-1511), 0,5 мг/л тиамина-HCl, 120,0 г/л сахарозы, 1,0 мг/л 2,4-D и 2,88 г/л L-пролина (доведенных до объема с помощью D-I H2O после доведения pH до 5,8 с помощью KOH); 2,0 г/л Gelrite (добавленного после доведения до объема с помощью D-I H2O) и 8,5 мг/л нитрата серебра (добавленного после стерилизации среды и охлаждения до комнатной температуры). Среда для отбора (560R) содержала 4,0 г/л основных солей N6 (SIGMA C-1416), 1,0 мл/л витаминной смеси Эрикссона (1000X SIGMA-1511), 0,5 мг/л тиамина-HCl, 30,0 г/л сахарозы и 2,0 мг/л 2,4-D (доведенных до объема с помощью D-I H2O после доведения pH до 5,8 с помощью KOH); 3,0 г/л Gelrite (добавленного после доведения до объема с помощью D-I H2O) и 0,85 мг/л нитрата серебра и 3,0 мг/л биалафоса (оба из которых добавлены после стерилизации среды и охлаждения до комнатной температуры).
Среда для регенерации растений (288J) содержала 4,3 г/л солей по MS (GIBCO 11117-074), 5,0 мл/л исходного раствора витаминов по MS (0,100 г никотиновой кислоты, 0,02 г/л тиамина-HCL, 0,10 г/л пиридоксина-HCL и 0,40 г/л глицина, доведенных до объема с помощью очищенной D-I H2O) (Murashige and Skoog (1962) Physiol. 15:473), 100 мг/л миоинозитола, 0,5 мг/л зеатина, 60 г/л сахарозы и 1,0 мл/л 0,1 мМ абсцизовой кислоты (доведенных до объема с помощью очищенной D-I H2O после доведения pH до 5,6); 3,0 г/л Gelrite (добавленного после доведения до объема с помощью D-I H2O), и 1,0 мг/л индолилуксусной кислоты, и 3,0 мг/л биалафоса (добавленных после стерилизации среды и охлаждения до 60°C). Безгормональная среда (272V) содержала 4,3 г/л солей по MS (GIBCO 11117-074), 5,0 мл/л исходного раствора витаминов по MS (0,100 г/л никотиновой кислоты, 0,02 г/л тиамина-HCL, 0,10 г/л пиридоксина-HCL и 0,40 г/л глицина, доведенных до объема с помощью очищенной D-I H2O), 0,1 г/л миоинозитола и 40,0 г/л сахарозы (доведенных до объема с помощью очищенной D-I H2O после доведения pH до 5,6); и 6 г/л бактоагара (добавленного после доведения до объема с помощью очищенной D-I H2O), ее стерилизовали и охлаждали до 60°C.
Пример 9. Эффективность контроля насекомых у стабильно трансформированных растений сои и кукурузы против широкого спектра насекомых из группы чешуекрылых
Из трансформированных растений вырезали листовые диски и тестировали их в отношении инсектицидной активности полипептидов PtIP-83 против совки кукурузной (SBL) (Chrysodeixis includens), совки кукурузной, (CEW) (Helicoverpa zea), огневки кукурузной (ECB) (Ostrinia nubilalis), совки бархатных бобов (VBC) (Anticarsia gemmatalis) и совки травяной (Spodoptera frugiperda).
Пример 10. Химеры между PtIP-83Aa (SEQ ID NO: 1) и PtIP-50Aa (SEQ ID NO: 34)
Для получения однокомпонентных активных вариантов с диверсифицированными последовательностями получали химеры между PtIP-83Aa (SEQ ID NO: 1) и PtIP50Aa (SEQ ID NO: 34) с помощью сборки множественных перекрывающихся ПЦР-фрагментов (набор для клонирования Gibson Assembly, New England Biolabs Inc.). Всего сконструировали 6 химер в таблице 13 показаны точки кроссинговера и % идентичности последовательности с PtIP-83Aa (SEQ ID NO: 1).
Таблица 13
SEQ ID NO: 640
SEQ ID NO: 641
SEQ ID NO: 642
SEQ ID NO: 643
SEQ ID NO: 644
SEQ ID NO: 645
Номера положений кроссинговера основаны на выравнивании, показанном на фигуре 3
Гены химер клонировали в векторы для временной экспрессии в растении и проводили анализы в отношении активности против SBL, как описано в примере 4.
Пример 11. Идентификация мотивов, влияющих на функционирование белка PtIP-83Aa
Для идентификации промежутка последовательности, влияющего на структурную стабильность и инсектицидную функцию PtIP-83Aa, с помощью аминокислотного выравнивания идентифицировали пять уникальных мотивов среди PtIP-83Aa, PtIP-50Aa (SEQ ID NO: 34), PtIP-50Ba (SEQ ID NO: 35), PtIP-50Bb (SEQ ID NO: 36) и PtIP-83Fa (SEQ ID NO: 3) (фигуры 4a-4d). Три консервативных мотива: аминокислоты V53-P66 из SEQ ID NO: 1 определили как Мотив A, аминокислоты Q363-N373 из SEQ ID NO: 1 как Мотив B и аминокислоты W556-A564 из SEQ ID NO: 1 как Мотив C. Также отобрали другие два вариабельные мотива: аминокислоты L646-W655 из SEQ ID NO: 1 как Мотив D и аминокислоты R771-Y786 из SEQ ID NO: 1 как Мотив E. Для этих пяти мотивов разрабатывали праймеры для насыщающего мутагенеза, показанные в таблице 14, таблице 15, таблице 16, таблице 17 и таблице 18. Насыщающий мутагенез проводили с применением набора для молниеносного сайт-направленного мутагенеза QuikChange® от Agilent. Мутации вводили путем амплификации вектора для экспрессии в растении, содержащего полинуклеотид под SEQ ID NO: 2, кодирующий полипептид PtIP-83Aa (SEQ ID NO: 1), с применением комплементирующих прямого и обратного праймеров, содержащих вырожденный кодон NNK в целевом положении. Аминокислотные замены в каждом целевом положении идентифицировали с помощью секвенирования ДНК. Временную экспрессию в растении и анализы активности в отношении SBL проводили, как описано в примере 4. Аминокислотные замены, идентифицированные в Мотиве A, Мотиве B, Мотиве C, Мотиве D и Мотиве E PtIP-83Aa (SEQ ID NO: 1) и аминокислотные замены, обусловливающие инсектицидную активность, показаны в таблице 14, таблице 15, таблице 16, таблице 17 и таблице 18, соответственно.
Таблица 14
† Активные замены: средний балл активности <=60% съеденного листового диска
Таблица 15
Таблица 16
Таблица 17
Таблица 18
Пример 12. Варианты PtIP-83Aa с множественными аминокислотными заменами в Мотиве A или Мотиве C
Варианты PtIP-83Aa с множественными аминокислотными заменами в пределах мотива конструировали для Мотива A и Мотива C с применением либо набора для молниеносного сайт-направленного мутагенеза QuikChange® (Agilent) (как описано в примере 11), либо с помощью сборки множественных перекрывающихся ПЦР-фрагментов (набор для клонирования Gibson Assembly, New England Biolabs Inc.) в векторе для временной экспрессии в растении, содержащем вирусный промотор DMMV. Праймеры, применяемые в конструировании комбинаций мутаций в пределах отдельного мотива, обобщены в таблице 19. Полученные комбинаторные библиотеки трансформировали в Agrobacterium для временной экспрессии в растении и последующих анализов активности в отношении SBL, как описано в примере 4. Активные варианты, определяемые как имеющие средний балл активности <=60% съеденного листового диска, посылали на идентификацию последовательности.
Таблица 19
Две библиотеки с отличающимися частотами мутаций подвергали скринингу в отношении Мотива A. Мотив A Combi-1 содержит потенциальные мутации в положениях K54, R55, L56, Y57, V58, F59, A60, V62, V63, E64 и L65 в PtIP-83Aa (SEQ ID NO: 1): Мотив A Combi-2 содержит потенциальные мутации в положениях K54, R55, V58, A60, V62, V63 и E64 в PtIP-83Aa (SEQ ID NO: 1): Из последовательностей Мотив A Combi -1 восстановили 5 активных уникальных вариантов (всего подвергли скринингу 96). Из последовательностей Мотив A Combi -2 восстановили 59 активных уникальных вариантов (всего подвергли скринингу 192). В таблице 20 обобщены аминокислотные замены полученных активных вариантов Мотива A PtIP-83Aa по сравнению с PtIP-83Aa.
Таблица 20
SEQ ID NO: 1
SEQ ID NO: 236
Одну библиотеку комбинаций для Мотива C, содержащую потенциальные мутации в положениях R557, A558, K559, K561 и N562 PtIP-83Aa (SEQ ID NO: 1) подвергали скринингу. Из суммарно 233 вариантов, подвергнутых скринингу из Мотив C Combi, идентифицировали 34 активных уникальных вариантов последовательности. В таблице 21 обобщены аминокислотные замены полученных активных вариантов Мотива C PtIP-83Aa по сравнению с PtIP-83Aa.
Таблица 21
SEQ ID NO: 1
Пример 13. Варианты PtIP-83Aa с множественными аминокислотными заменами в Мотиве A и Мотиве C
Дополнительное разнообразие последовательностей создавали путем комбинирования активных комбинаций для Мотива A с активными комбинациями для Мотива C. Двадцать четыре варианта уникальных активных комбинаций для Мотива A и 11 варианта уникальных активных комбинаций для Мотива C отбирали для конструирования библиотеки Мотив A x Мотив C (таблица 22).
Таблица 22. Варианты последовательности, применяемые для конструирования библиотеки комбинаций Мотив A x Мотив C

Плазмидную ДНК выделяли из этих вариантов и объединяли в пул по мотиву. Комбинации для Мотива A подвергали ПЦР-амплификации из пула Мотива A с применением следующих праймеров: AATCTCTCATCTAAGAGGCTGGATCCTAGGATGGCTCTCGTGGATTACGGC (SEQ ID NO: 630) и GCAGCCACAACCTCCATCACAGC (SEQ ID NO: 631). Комбинации для Мотива C подвергали ПЦР-амплификации из пула Мотива C с применением следующих праймеров: TGGCCAATCCAGAAGATGGACAAGTCTAGACTACTCTTCGTCGTGCCGCCAG (SEQ ID NO: 632) и GCTGTGATGGAGGTTGTGGCTGC (SEQ ID NO: 633). Затем эти два продукта ПЦР собирали с помощью множественных перекрывающихся ПЦР-фрагментов (набор для клонирования Gibson Assembly, New England Biolabs Inc) в вектор для временной экспрессии в растении, содержащий вирусный промотор DMMV. 94 варианта из этой библиотеки подвергали скринингу для определения временной экспрессии в растении и анализов активности в отношении SBL, как описано в примере 4. 30 уникальных активных вариантов, определяемых как имеющие средний балл активности <=60% съеденного листового диска, идентифицировали с помощью секвенирования. В таблице 23 обобщены аминокислотные замены полученных активных вариантов Мотив A x Мотив C PtIP-83Aa по сравнению с PtIP-83Aa.
Таблица 23
SEQ ID NO: 1
SEQ ID NO: 399
SEQ ID NO: 400
SEQ ID NO: 401
SEQ ID NO: 402
SEQ ID NO: 403
SEQ ID NO: 404
SEQ ID NO: 405
SEQ ID NO: 406
Пример 14. Варианты PtIP-83Aa с множественными аминокислотными заменами
Для создания вариантов PtIP-83Aa (SEQ ID NO: 1) с множественными изменениями аминокислот получали библиотеки вариантов с помощью шаффлинга семейства (Chia-Chun J. Chang et al, 1999, Nature Biotechnology 17, 793-797) полинуклеотида (SEQ ID NO: 2), кодирующего PtIP-83Aa (SEQ ID NO: 1), и полинуклеотида (SEQ ID NO: 2), кодирующего PtIP-83Cb (SEQ ID NO: 7). Частоты мутаций в библиотеках контролировали путем варьирования участков PtIP-83Aa (SEQ ID NO: 1) и PtIP-83Cb (SEQ ID NO: 7), которые включали в реакцию шаффлинга библиотеки. Между этими двумя вариантами получили пять библиотек. В первой библиотеке (83FS-1), генную последовательность полной длины от обоих PtIP-83Aa (SEQ ID NO: 2) и PtIP-83Cb (SEQ ID NO: 8) включали в реакцию шаффлинга. Во второй библиотеке (83FS-2) фрагменты, содержащие нуклеотиды 1114-2604 из SEQ ID NO: 2 (кодирующим PtIP-83Aa (SEQ ID NO: 1)) и нуклеотиды 1072-2559 из SEQ ID NO: 8 (кодирующим PtIP-83Cb (SEQ ID NO: 7)) подвергали шаффлингу. Затем подвергнутые шаффлингу фрагменты комбинировали с нуклеотидами: 1-1113 из SEQ ID NO: 2 (кодирующим PtIP-83Aa (SEQ ID NO: 1)) с помощью Gibson Assembly (New England Biolabs). В третьей библиотеке (83FS-3) 5'- и 3'-участки двух вариантов подвергали шаффлингу в отдельных реакциях. Реакцию с 5'-участками, содержащую подвергнутые шаффлингу фрагменты с нуклеотидами: 1-1113 из SEQ ID NO: 2 (кодирующим PtIP-83Aa (SEQ ID NO: 1)) и нуклеотиды 1-1071 из SEQ ID NO: 8 (кодирующим PtIP-83Cb (SEQ ID NO: 7)), и реакцию с 3'-участками, содержащую подвергнутые шаффлингу фрагменты с нуклеотидами: 1114-2604 из SEQ ID NO: 2 (кодирующим PtIP-83Aa (SEQ ID NO: 1)) и нуклеотидами 1072-2559 из SEQ ID NO: 8 (кодирующим PtIP-83Cb (SEQ ID NO: 7)), затем комбинировали с помощью Gibson Assembly. В четвертой библиотеке (83FS-4) фрагменты из 5'-участка каждого варианта (нуклеотиды 1-1113 из SEQ ID NO: 2 (кодирующим PtIP-83Aa (SEQ ID NO: 1)) и нуклеотиды 1-1071 из SEQ ID NO: 8 (кодирующим PtIP-83Cb (SEQ ID NO: 7)) подвергали шаффлингу с дополнительными праймерами, добавляемыми в реакцию сборки для содействия кроссинговерам между двумя вариантами. Подвергнутый шаффлингу 5'-участок затем комбинировали с помощью Gibson assembly с 3'-участком либо SEQ ID NO: 2 (кодирующим PtIP-83Aa (SEQ ID NO: 1)), либо SEQ ID NO: 8 (кодирующим PtIP-83Cb (SEQ ID NO: 7)). В пятой библиотеке (83FS-5) варианты с одиночным кроссинговером получали вручную между SEQ ID NO: 2 (кодирующим PtIP-83Aa (SEQ ID NO: 1)) и SEQ ID NO: 8 (кодирующим PtIP-83Cb (SEQ ID NO: 7)) с применением Gibson assembly.
Все подвергнутые шаффлингу библиотеки клонировали в вектор для временной экспрессии в растении, содержащий вирусный промотор dMMV для последующей временной экспрессии в фасоли кустовой и анализов активности в отношении SBL, как описано в примере 4. Активные варианты, определяемые как имеющие средний балл активности ≤ 60% съеденного листового диска, посылали на идентификацию последовательности. В библиотеке 83FS-1 подвергли скринингу 197 и идентифицировали последовательность 45 активных уникальных вариантов. В библиотеке 83FS-2 подвергли скринингу 96 и идентифицировали последовательность 32 активных уникальных вариантов. В библиотеке 83FS-3 подвергли скринингу 192 и идентифицировали последовательность 13 активных уникальных вариантов. В библиотеке 83FS-4 подвергли скринингу 96 вариантов и идентифицировали последовательность 6 уникальных активных вариантов. В библиотеке 83FS-5 обнаружили, что четыре из одиннадцати сконструированных вариантов с одним кроссинговером являются активными.
Идентичность последовательностей активных вариантов с PtIP-83Aa рассчитывали с применением алгоритма Нидлмана-Вунша, реализованного в программе Needle (набор инструментов EMBOSS). Процентная идентичность по сравнению с PtIP-83Aa (SEQ ID NO: 1), обозначение варианта, нуклеотидные последовательности и аминокислотные последовательности полученных активных вариантов полипептида PtIP-83Aa из всех библиотек обобщены в таблице 24. В таблице 25 обобщен % идентичности активных вариантов по сравнению с PtIP-83Aa (SEQ ID NO: 1), число вариантов с каждой % идентичностью и идентификация варианта.
Таблица 24
Таблица 25
Приведенное выше описание различных проиллюстрированных вариантов осуществления согласно настоящему раскрытию не подразумевается как исчерпывающее или служащее для ограничения объема точной раскрытой формой. Хотя конкретные варианты осуществления и примеры описаны в данном документе для иллюстративных целей, в пределах объема настоящего раскрытия возможны различные эквивалентные модификации, как будет понятно специалистам в данной области. Представленные в данном документе идеи можно применять для других целей, отличающихся от примеров, описанных выше. В свете вышеизложенных идей возможны многочисленные модификации и вариации, и, следовательно, они находятся в пределах объема прилагаемой формулы изобретения.
Эти и другие изменения можно выполнять в свете вышеизложенного подробного описания. В целом, в следующей формуле изобретения используемые выражения не должны рассматриваться как ограничивающие объем настоящего изобретения конкретными вариантами осуществления, раскрытыми в описании и формуле изобретения.
Полное раскрытие каждого процитированного документа (в том числе патенты, патентные заявки, статьи из научных журналов, рефераты, руководства, книги или другие раскрытия) в Предпосылках изобретения, Подробном описании и Примерах включены в данный документ посредством ссылки во всей их полноте.
Были сделаны попытки обеспечения точности в отношении применяемых чисел (например, количества, температуры, концентрации и т.д.), но могут предусматриваться некоторые экспериментальные ошибки и отклонения. Если не указано иное, части представляют собой части по весу, молекулярный вес представляет собой средний молекулярный вес; температура приведена в градусах по Цельсию и давление является атмосферным или практически атмосферным.
| название | год | авторы | номер документа |
|---|---|---|---|
| Белки, являющиеся модификаторами вкуса и вкусоароматических свойств | 2019 |
|
RU2823578C2 |
| НОВЫЙ ПОЛИПЕПТИД И СПОСОБ ПОЛУЧЕНИЯ L-ЛЕЙЦИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2021 |
|
RU2811433C1 |
| ВАРИАНТЫ СКВАЛЕНГОПЕНЦИКЛАЗЫ (SHC) | 2020 |
|
RU2829857C1 |
| Слитые молекулы, происходящие от Cholix-токсина, для пероральной доставки биологически активных нагрузок | 2015 |
|
RU2723178C2 |
| Мутантная нитрилгидратаза, нуклеиновая кислота, кодирующая указанную мутантную нитрилгидратазу, вектор экспрессии и трансформант, содержащие указанную нуклеиновую кислоту, способ получения указанной мутантной нитрилгидратазы и способ получения амидного соединения | 2017 |
|
RU2736086C1 |
| МУТАНТНЫЙ БЕЛОК F RSV И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2807742C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
| НОВЫЕ ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2666914C2 |
| ВАРИАНТЫ IgG-FC ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2814952C2 |





















Изобретение относится к области биохимии, в частности к очищенному полипептиду PtIP-83, обладающему инсектицидной активностью против совки кукурузной (Helicoverpa zea). Также раскрыты рекомбинантный полинуклеотид, кодирующий указанный белок; трансгенное растение и трансгенная клетка, содержащие указанный полинуклеотид; ДНК-конструкция, содержащая указанный полинуклеотид; композиция, содержащая указанный полипептид; белок слияния, содержащий указанный полипептид. Раскрыты способы снижения популяции совки кукурузной (Helicoverpa zea), подавления роста или уничтожения совки кукурузной (Helicoverpa zea) с помощью указанного полипептида; применение полипептида PtIP-83. Изобретение обладает способностью эффективно бороться с совкой кукурузной (Helicoverpa zea). 14 н. и 16 з.п. ф-лы, 14 пр., 25 табл., 5 ил.
1. Очищенный полипептид PtIP-83 с последовательностью, по меньшей мере на 90% идентичной по всей длине аминокислотной последовательности PtIP-83Aa, представленной на Фигурах 2a-2j, где полипептид PtIP-83 обладает инсектицидной активностью против совки кукурузной (Helicoverpa zea).
2. Очищенный полипептид PtIP-83 по п. 1, где последовательность полипептида PtIP-83 по меньшей мере на 95% идентична по всей длине аминокислотной последовательности PtIP-83Aa, представленной на Фигурах 2a-2j.
3. Очищенный полипептид PtIP-83 по п. 1, где полипептид PtIP-83 имеет по меньшей мере одну аминокислотную замену, делецию, вставку и/или добавление на N-конце или C-конце по сравнению с PtIP-83Aa, представленной на Фигурах 2a-2j.
4. Очищенный полипептид PtIP-83 по п. 1, где полипептид PtIP-83 представляет собой не встречающуюся в природе аминокислотную последовательность.
5. Очищенный полипептид PtIP-83 по любому из пп. 1-4, где полипептид PtIP-83 специфически связывается с мембранной везикулой щеточной каемки, выделенной от Lepidoptera.
6. Очищенный полипептид PtIP-83 по любому из пп. 1-4, где полипептид PtIP-83 нарушает ионный баланс по сторонам мембранной везикулы щеточной каемки, выделенной от Lepidoptera.
7. Очищенный полипептид PtIP-83 по п. 1 или 2, где полипептид PtIP-83 получен из вида папоротника или примитивного растения отдела Lycophyta.
8. Очищенный полипептид PtIP-83 по п. 7, где папоротник относится к роду Lygodium.
9. Очищенный полипептид PtIP-83 по п. 7, где папоротник относится к роду Adiantaceae.
10. Очищенный полипептид PtIP-83 по п. 7, где папоротник относится к роду Polypodium.
11. Очищенный полипептид PtIP-83 по любому из пп. 1-4, где полипептид PtIP-83 представляет собой вариант полипептида PtIP-83, имеющий по меньшей мере одну аминокислотную замену по сравнению с PtIP-83Aa, представленной на Фигурах 2a-2j, где аминокислота в положении 1 представляет собой Met или удалена; аминокислота в положении 2 представляет собой Ala или удалена; аминокислота в положении 3 представляет собой Leu, Val, Ile или удалена; аминокислота в положении 4 представляет собой Val, Met, Ile или Leu; аминокислота в положении 7 представляет собой Gly, Thr или Ser; аминокислота в положении 8 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 10 представляет собой Phe, Trp или Tyr; аминокислота в положении 11 представляет собой Glu, Asp, Lys или Arg; аминокислота в положении 18 представляет собой Met, Val, Leu или Ile; аминокислота в положении 19 представляет собой Gly, Pro или Ala; аминокислота в положении 20 представляет собой Val, Ile, Leu или удалена; аминокислота в положении 21 представляет собой Leu, Ile или Val; аминокислота в положении 23 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 37 представляет собой Val, Ile или Leu; аминокислота в положении 38 представляет собой Arg, Lys, Gln или Asn; аминокислота в положении 40 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 43 представляет собой Asn, Gln, Glu или Asp; аминокислота в положении 45 представляет собой Gly или Ala; аминокислота в положении 46 представляет собой Gln, Asp, Asn или Glu; аминокислота в положении 48 представляет собой Glu, Asp, Pro, Ile, Leu или Val; аминокислота в положении 51 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 52 представляет собой Lys, Arg, Ser или Thr; аминокислота в положении 53 представляет собой Val, Ala, Cys или Thr; аминокислота в положении 54 представляет собой Lys, Ala, Cys, Asp, Glu, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser или Thr; аминокислота в положении 55 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 56 представляет собой Leu, Glu, Phe, Ile, Met, Thr или Val; аминокислота в положении 57 представляет собой Tyr, Cys, Ile, Leu, Met, Thr или Val; аминокислота в положении 58 представляет собой Val, Cys, Ile или Leu; аминокислота в положении 59 представляет собой Phe, Leu, Met, Val или Tyr; аминокислота в положении 60 представляет собой Ala, Cys, Gly, Ser, Thr или Val; аминокислота в положении 61 представляет собой Asp, Glu, His или Ser; аминокислота в положении 62 представляет собой Val, Ala, Cys, Ile, Leu или Thr; аминокислота в положении 63 представляет собой Val, Ala, Cys, Ile, Leu, Met или Thr; аминокислота в положении 64 представляет собой Glu, Ala, Cys, Phe, Gly, His, Ile, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 65 представляет собой Leu, Ala, Cys, Phe, His, Ile, Met, Asn, Gln, Thr, Val или Trp; аминокислота в положении 66 представляет собой Pro, Asp, Gly, Met, Gln или Arg; аминокислота в положении 67 представляет собой Val, Pro, Ile, Leu, Ser или Thr; аминокислота в положении 68 представляет собой Val, Arg, Phe, Ile, Leu, Lys или Gly; аминокислота в положении 69 представляет собой Glu, Ala, Asp, Gly, Arg или Lys; аминокислота в положении 70 представляет собой Trp, Thr, His, Tyr, Lys или Arg; аминокислота в положении 71 представляет собой Arg, Pro, Lys или удалена; аминокислота в положении 72 представляет собой Trp, Asp, Leu, Ile, Val, Glu или удалена; аминокислота в положении 73 представляет собой Pro, Gln, Asn, His или удалена; аминокислота в положении 74 представляет собой Pro, Met, Ser или Thr; аминокислота в положении 75 представляет собой Gln, His, Asn, Lys или Arg; аминокислота в положении 76 представляет собой Ile, Met, Val или Leu; аминокислота в положении 84 представляет собой Ile, Leu или Val; аминокислота в положении 91 представляет собой Trp или Phe; аминокислота в положении 93 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 94 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 96 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 97 представляет собой Gln, Phe, Asn, Lys или Arg; аминокислота в положении 98 представляет собой Ser, Thr или удалена; аминокислота в положении 99 представляет собой Asp, Glu, Gly или Ala; аминокислота в положении 100 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 101 представляет собой Glu, Thr, Asp, Ser или Trp; аминокислота в положении 103 представляет собой His, Arg, Lys, Glu или Gln; аминокислота в положении 105 представляет собой Thr, Ser или Pro; аминокислота в положении 108 представляет собой Lys, Arg, Asn, Asp, Gln или Glu; аминокислота в положении 109 представляет собой Leu, Ile или Val; аминокислота в положении 111 представляет собой Ala, Ser или Thr; аминокислота в положении 112 представляет собой Ile, Arg, Thr, Leu, Val, Lys, Ser или удалена; аминокислота в положении 113 представляет собой Gln, Ala, Gly, Asn или удалена; аминокислота в положении 114 представляет собой Arg, Glu, Lys, Asp или Ile; аминокислота в положении 115 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 116 представляет собой Glu, Asn, Gln, Asp, Lys или Arg; аминокислота в положении 117 представляет собой Asn, Val, Tyr, Ile, Leu, Gln, Trp или Phe; аминокислота в положении 118 представляет собой Arg или Lys; аминокислота в положении 119 представляет собой Trp, Thr или Ser; аминокислота в положении 122 представляет собой Thr, Lys, Ser, Arg или Ala; аминокислота в положении 124 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 126 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 127 представляет собой Met, Gly или Ala; аминокислота в положении 128 представляет собой Asn, Gln, Arg или Lys; аминокислота в положении 131 представляет собой Val, Ile, Leu, Ser или Thr; аминокислота в положении 133 представляет собой Ile, Leu или Val; аминокислота в положении 134 представляет собой His или Tyr; аминокислота в положении 135 представляет собой Ala или Gly; аминокислота в положении 137 представляет собой Glu, Asp, Arg или Lys; аминокислота в положении 139 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 140 представляет собой Val, Arg, Ile, Lys или Leu; аминокислота в положении 141 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 142 представляет собой Val, Ile, Leu или Pro; аминокислота в положении 144 представляет собой Thr, Leu, Phe, Ile, Val или Tyr; аминокислота в положении 145 представляет собой Met, Pro, Gln или Asn; аминокислота в положении 146 представляет собой Ser, Gly, Thr, Ala, Gln или Asn; аминокислота в положении 147 представляет собой Trp, Gln, Tyr или Asn; аминокислота в положении 148 представляет собой Ser, Ala, Thr, Gly или Pro; аминокислота в положении 149 представляет собой Ser, Thr или удалена; аминокислота в положении 150 представляет собой Val, Ile, Leu или Tyr; аминокислота в положении 152 представляет собой Arg, Ala, Val, Ile, Leu, Lys или Gly; аминокислота в положении 154 представляет собой Ser, Trp, Thr, Asp или Glu; аминокислота в положении 156 представляет собой Leu, Asp, Ile, Val, Asn, Glu или Gln; аминокислота в положении 158 представляет собой Ser, Thr или Cys; аминокислота в положении 159 представляет собой Val, Thr, Leu или Ile; аминокислота в положении 162 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 163 представляет собой Gly, Ala или удалена; аминокислота в положении 164 представляет собой Phe или удалена; аминокислота в положении 165 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 166 представляет собой Ala, Arg, Met, Lys или Phe; аминокислота в положении 167 представляет собой Val, Ile, Leu или His; аминокислота в положении 168 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 169 представляет собой Val, His, Ile, Leu, Ser или Thr; аминокислота в положении 170 представляет собой Phe, Ile, Leu или Val; аминокислота в положении 171 представляет собой Glu, Asn, Gln или Asp; аминокислота в положении 172 представляет собой Val, Ala, Arg, Ile, Leu, Gly, Lys, Asp или Glu; аминокислота в положении 175 представляет собой Ser, Arg, Thr, Lys или Trp; аминокислота в положении 176 представляет собой Val, Leu или Ile; аминокислота в положении 177 представляет собой Arg, Lys, Leu, Val или Ile; аминокислота в положении 179 представляет собой Thr, Ile, Val, Leu или Ser; аминокислота в положении 180 представляет собой Leu, Phe, Ile, Val, Ser или Thr; аминокислота в положении 181 представляет собой Gly, Thr, Gln, Asn или Ser; аминокислота в положении 182 представляет собой Ala, Leu, Phe, Val или Ile; аминокислота в положении 183 представляет собой Thr, Ser, Ala или Gly; аминокислота в положении 184 представляет собой Leu, Thr, Ser, Ile, Val, Lys или Arg; аминокислота в положении 185 представляет собой Arg, Gly, Asp, Lys, Glu или Ala; аминокислота в положении 186 представляет собой Pro, Val, Ile, Leu, Asn или Gln; аминокислота в положении 187 представляет собой Asp, Thr, Glu или Ser; аминокислота в положении 188 представляет собой His, Gly или Ala; аминокислота в положении 189 представляет собой Ala, Arg, Pro, Lys, Gly или удалена; аминокислота в положении 190 представляет собой Leu, Asn, Ile, Val, Gln или удалена; аминокислота в положении 191 представляет собой Tyr или удалена; аминокислота в положении 192 представляет собой Ser, Ile, Val, Leu, Thr или Asn; аминокислота в положении 193 представляет собой Thr, Ser, Glu или Asp; аминокислота в положении 194 представляет собой Thr или Ser; аминокислота в положении 195 представляет собой Met или Thr; аминокислота в положении 196 представляет собой Gln, His, Leu, Asn, Ile, Val, Thr или Ser; аминокислота в положении 197 представляет собой Ala, Gly, Ile, Val или Leu; аминокислота в положении 198 представляет собой Thr, Glu, Ser, Asp, Gly или Ala; аминокислота в положении 199 представляет собой Pro, Lys или Arg; аминокислота в положении 200 представляет собой Asn, Ser, Thr, Gln, Ala или Gly; аминокислота в положении 201 представляет собой Ala, Leu, Glu, Ile, Asp или Trp; аминокислота в положении 202 представляет собой Ser, Asp, Phe, Ile, Val, Thr, Glu или Leu; аминокислота в положении 203 представляет собой His, Pro, Gly, Ala, Thr или Ser; аминокислота в положении 204 представляет собой Ile, Trp, His, Leu, Val, Ala или Gly; аминокислота в положении 205 представляет собой Ser, Asn, Leu, Val, Thr, Gln или Ile; аминокислота в положении 206 представляет собой Ala, Gly, Asp, Tyr, Glu, Lys или Arg; аминокислота в положении 207 представляет собой Phe, Val, Ile или Leu; аминокислота в положении 208 представляет собой Asn, Ser, Pro, Gln, Thr, Val, Ile или Leu; аминокислота в положении 210 представляет собой Arg, Asp, Glu, Lys, Ser или Tyr; аминокислота в положении 211 представляет собой Ile, Ser, Leu, Val или Thr; аминокислота в положении 212 представляет собой Val, Ala, Ile, Leu, Glu, Gly или Asp; аминокислота в положении 214 представляет собой Pro, Lys или Arg; аминокислота в положении 215 представляет собой Ser или Thr; аминокислота в положении 217 представляет собой Tyr или Phe; аминокислота в положении 218 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 219 представляет собой Val, Ile, Leu или Ala; аминокислота в положении 220 представляет собой Cys, Leu, Ile, Val, Thr или Ser; аминокислота в положении 221 представляет собой Pro или His; аминокислота в положении 222 представляет собой Leu, Arg, Lys, Ile, Val, Thr или Ser; аминокислота в положении 224 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 225 представляет собой Asp, Arg, Glu, Lys, Ser или Thr; аминокислота в положении 226 представляет собой Thr, Ser, Gln или Asn; аминокислота в положении 227 представляет собой Asp, Leu, Glu, Ile, Val или удалена; аминокислота в положении 228 представляет собой Thr, Ser или удалена; аминокислота в положении 229 представляет собой Tyr или удалена; аминокислота в положении 230 представляет собой Leu, Ile, Val или удалена; аминокислота в положении 231 представляет собой Gly, Ala или удалена; аминокислота в положении 232 представляет собой Ile, Leu, Val или удалена; аминокислота в положении 233 представляет собой Pro или удалена; аминокислота в положении 234 представляет собой Ala, Pro, Gly или удалена; аминокислота в положении 235 представляет собой Asp, Ile, Leu, Glu или Val; аминокислота в положении 236 представляет собой Val, Ser, Ile, Leu, Thr, Asp или Glu; аминокислота в положении 237 представляет собой Ala, Phe или Tyr; аминокислота в положении 238 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 239 представляет собой Val, Ser, Ile, Leu, Thr, Ala или Gly; аминокислота в положении 240 представляет собой Leu, Val или Ile; аминокислота в положении 243 представляет собой Asp или Glu; аминокислота в положении 249 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 252 представляет собой Leu, Ile, Val или Met; аминокислота в положении 257 представляет собой Thr или Ser; аминокислота в положении 259 представляет собой His, Ile, Val или Leu; аминокислота в положении 266 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 267 представляет собой Cys, Ala или Gly; аминокислота в положении 268 представляет собой His, Arg, Lys или Tyr; аминокислота в положении 272 представляет собой Asp или Glu; аминокислота в положении 273 представляет собой Val, Met, Ile или Leu; аминокислота в положении 274 представляет собой Val, Ile, Leu или Met; аминокислота в положении 278 представляет собой Gly или Ala; аминокислота в положении 279 представляет собой Glu, Asp, Gly или Val; аминокислота в положении 281 представляет собой Leu, Ile, Val, Gly или Ala; аминокислота в положении 282 представляет собой Asn, Leu или Ile; аминокислота в положении 285 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 286 представляет собой Lys, Asp, Arg или Glu; аминокислота в положении 287 представляет собой Leu, Ile или Val; аминокислота в положении 290 представляет собой Pro, Gln, Asn, Lys или Arg; аминокислота в положении 291 представляет собой Leu, Ile или Val; аминокислота в положении 292 представляет собой Lys, Arg, Ile, Leu или Val; аминокислота в положении 293 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 294 представляет собой Ser, Asn, Thr, Gln, Arg или Lys; аминокислота в положении 295 представляет собой Thr или Ser; аминокислота в положении 296 представляет собой Gln, Asn или His; аминокислота в положении 297 представляет собой Leu, Ile, Val или Met; аминокислота в положении 300 представляет собой Ser или Thr; аминокислота в положении 301 представляет собой Glu, Asp, Gly или Ala; аминокислота в положении 302 представляет собой Ser, Pro, Thr, Gly или Ala; аминокислота в положении 304 представляет собой Lys, Arg, Gln или Asn; аминокислота в положении 313 представляет собой Val, Leu или Ile; аминокислота в положении 314 представляет собой His, Glu, Asn, Asp или Gln; аминокислота в положении 315 представляет собой Ala, Cys, Gly, Thr или Ser; аминокислота в положении 316 представляет собой Ala, Ile, Leu или Val; аминокислота в положении 317 представляет собой Met, Leu, Val или Ile; аминокислота в положении 319 представляет собой Met, Leu, Val или Ile; аминокислота в положении 320 представляет собой Val, Ile, Leu, Ala или Gly; аминокислота в положении 321 представляет собой Arg, Lys или Pro; аминокислота в положении 322 представляет собой Ile, Leu, Val или Phe; аминокислота в положении 323 представляет собой Gly, Ile, Leu или Val; аминокислота в положении 324 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 336 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 339 представляет собой Asn, Lys, Gln или Arg; аминокислота в положении 350 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 351 представляет собой Glu или Asp; аминокислота в положении 353 представляет собой Lys или Arg; аминокислота в положении 354 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 355 представляет собой Phe, Ile, Leu или Leu; аминокислота в положении 356 представляет собой Lys или Arg; аминокислота в положении 360 представляет собой Ile, Val, Leu, Gly или Ala; аминокислота в положении 363 представляет собой Gln, Ala, Cys, Glu, Phe, Gly, His, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 364 представляет собой Ile, Ala, Cys, Glu, Phe, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 365 представляет собой Leu, Ala, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Arg, Val, Trp или Tyr; аминокислота в положении 366 представляет собой Gly, Ala, Cys, Phe, His, Ile, Lys, Leu, Met, Asn, Ser, Thr или Val; аминокислота в положении 367 представляет собой Ser, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val или Trp; аминокислота в положении 368 представляет собой Tyr, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp; аминокислота в положении 369 представляет собой Leu, Ala, Cys, Asp, Phe, Gly, Ile, Met, Thr или Val; аминокислота в положении 370 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 371 представляет собой Gln, Ala, Cys, Asp, Glu, Phe, Gly, Ile, Lys, Leu, Asn, Arg, Ser, Thr, Val или Trp; аминокислота в положении 372 представляет собой Gln, Ala, Cys, Asp, Phe, Gly, His, Ile, Leu, Asn, Arg, Ser, Val или Tyr; аминокислота в положении 373 представляет собой Asn, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Gln, Ser, Thr, Val или Trp; аминокислота в положении 374 представляет собой Arg или Lys; аминокислота в положении 376 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 378 представляет собой Glu или Asp; аминокислота в положении 381 представляет собой Leu, Ile или Val; аминокислота в положении 388 представляет собой Ala, Thr, Gly или Ser; аминокислота в положении 395 представляет собой Arg или Lys; аминокислота в положении 396 представляет собой Glu, Gln, Asp, Asn, Ala или Gly; аминокислота в положении 399 представляет собой Asp, Gln, Glu или Asn; аминокислота в положении 400 представляет собой Asn, Thr, Ser, Glu, Gln или Asp; аминокислота в положении 401 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 402 представляет собой Phe, Ile, Val или Leu; аминокислота в положении 406 представляет собой Asp или Glu; аминокислота в положении 408 представляет собой Leu, Ile, Val или Met; аминокислота в положении 410 представляет собой Gly, Ile, Val, Ala или Leu; аминокислота в положении 414 представляет собой Ala, Gly, Asp или Glu; аминокислота в положении 416 представляет собой Ser, Asn, Thr, Gln, Glu или Asp; аминокислота в положении 417 представляет собой Ser, Arg, Lys, Thr, Ala или Gly; аминокислота в положении 423 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 431 представляет собой Arg или Lys; аминокислота в положении 432 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 436 представляет собой Arg, Lys, Asp или Glu; аминокислота в положении 440 представляет собой Asn, Gln, Lys или Arg; аминокислота в положении 442 представляет собой Leu, Ile или Val; аминокислота в положении 447 представляет собой Ser, Lys, Thr или Arg; аминокислота в положении 448 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 451 представляет собой Gln, Asn или Met; аминокислота в положении 453 представляет собой Gly или Ala; аминокислота в положении 455 представляет собой Ala, Leu, Ile или Val; аминокислота в положении 457 представляет собой Leu, Ile или Val; аминокислота в положении 467 представляет собой Val, Ile, Leu, Gly или Ala; аминокислота в положении 471 представляет собой Gly или Ala; аминокислота в положении 475 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 483 представляет собой Gly или Ala; аминокислота в положении 493 представляет собой Gln, Asn или Gly; аминокислота в положении 504 представляет собой Val, Leu или Ile; аминокислота в положении 506 представляет собой Asp, Glu или His; аминокислота в положении 509 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 510 представляет собой Ser, Thr, Gly или Ala; аминокислота в положении 512 представляет собой Glu или Asp; аминокислота в положении 515 представляет собой Gly, Ala, Thr или Ser; аминокислота в положении 516 представляет собой Gln, Asn или His; аминокислота в положении 517 представляет собой Ile, Val или Leu; аминокислота в положении 519 представляет собой Asp, Asn, Glu, Gly или Gln; аминокислота в положении 522 представляет собой Val, Glu, Pro, Ile, Leu или Asp; аминокислота в положении 525 представляет собой Glu или Asp; аминокислота в положении 526 представляет собой Leu, Ile, Val или Met; аминокислота в положении 539 представляет собой Val, Leu или Ile; аминокислота в положении 555 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 556 представляет собой Trp, Phe, Thr или Tyr; аминокислота в положении 557 представляет собой Arg, Cys, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 558 представляет собой Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 559 представляет собой Lys, Ala, Cys, Phe, Gly, His, Ile, Leu, Asn, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 560 представляет собой Cys, Ala, Phe, Gly, Ile, Met, Asn, Arg, Ser, Thr или Val; аминокислота в положении 561 представляет собой Lys, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 562 представляет собой Asn, Cys, Asp, Glu, Gly, His, Leu, Met, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 563 представляет собой Val, Ala, Cys, Asp, Phe, His, Ile, Leu, Met, Asn, Gln, Thr или Trp; аминокислота в положении 564 представляет собой Ala, Cys, Gly, Met, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 571 представляет собой Ser, Thr или Cys; аминокислота в положении 575 представляет собой Val, Leu, Ile, Asp или Glu; аминокислота в положении 577 представляет собой Met, Leu, Val или Ile; аминокислота в положении 579 представляет собой Glu, Asp, Asn или Gln; аминокислота в положении 583 представляет собой Asp или Glu; аминокислота в положении 589 представляет собой Met, Ile, Val или Leu; аминокислота в положении 590 представляет собой Met, Ile, Val или Leu; аминокислота в положении 593 представляет собой Met, Leu, Val или Ile; аминокислота в положении 595 представляет собой Arg, Lys, Asn или Gln; аминокислота в положении 596 представляет собой Ser или Thr; аминокислота в положении 597 представляет собой Gln, Asn или His; аминокислота в положении 607 представляет собой Ala, Gly, Ile, Leu или Val; аминокислота в положении 608 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 612 представляет собой Tyr, His или Phe; аминокислота в положении 617 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 618 представляет собой Gln, Asn или His; аминокислота в положении 625 представляет собой Arg, Lys, Thr или Ser; аминокислота в положении 626 представляет собой Met, Leu, Val или Ile; аминокислота в положении 628 представляет собой Leu, Val или Ile; аминокислота в положении 633 представляет собой Ile, Leu, Val или Met; аминокислота в положении 634 представляет собой Leu, Ile, Val или Met; аминокислота в положении 642 представляет собой Arg, Lys или Met; аминокислота в положении 646 представляет собой Leu, Ala, Cys, Gly, Ile, Met, Asn, Gln, Ser, Thr или Val; аминокислота в положении 647 представляет собой Leu, Asp, Gly, Met, Asn, Gln или Thr; аминокислота в положении 648 представляет собой Met, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 649 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr; аминокислота в положении 650 представляет собой Thr, Ala, Cys, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Val или Tyr; аминокислота в положении 651 представляет собой Glu, Ala, Cys, Asp, Gly, His, Ile, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 652 представляет собой Leu, Cys, Phe, Ile, Lys, Met, Pro, Arg, Ser, Thr или Val; аминокислота в положении 653 представляет собой Thr, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Pro, Arg, Ser, Val или Trp; аминокислота в положении 654 представляет собой Thr, Ala, Cys, Phe, Ile, Lys, Leu, Met, Pro, Arg, Ser, Val, Trp или Tyr; аминокислота в положении 655 представляет собой Trp, Phe или Tyr; аминокислота в положении 658 представляет собой Gly, Lys, Ala или Arg; аминокислота в положении 663 представляет собой Gly или Ala; аминокислота в положении 664 представляет собой Asp, Glu, Gln или Asn; аминокислота в положении 668 представляет собой Ala, Gly, Ser или Thr; аминокислота в положении 669 представляет собой Gln, Asn или His; аминокислота в положении 671 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 675 представляет собой Ile, Val, Ile, Thr или Ser; аминокислота в положении 678 представляет собой Met, Ile, Ala, Leu, Ser или Thr; аминокислота в положении 682 представляет собой Pro, Asn или Gln; аминокислота в положении 683 представляет собой Ser, Thr или Pro; аминокислота в положении 685 представляет собой Asp, Glu, Asp или Asn; аминокислота в положении 694 представляет собой Asp, Glu, Ala или Gly; аминокислота в положении 697 представляет собой Asn, Gln, Thr или Ser; аминокислота в положении 704 представляет собой Glu, Asp, Ala или Gly; аминокислота в положении 714 представляет собой Ala или Gly; аминокислота в положении 721 представляет собой Ser, Thr или Phe; аминокислота в положении 722 представляет собой Ser, Thr, Gln или Asn; аминокислота в положении 724 представляет собой Ser или Thr; аминокислота в положении 734 представляет собой His, Asn или Gln; аминокислота в положении 736 представляет собой Val, Leu, Ile или Ala; аминокислота в положении 737 представляет собой Lys, Arg, Asn или Gln; аминокислота в положении 739 представляет собой Ala, Gly, Thr или Ser; аминокислота в положении 740 представляет собой Ser, Thr или Met; аминокислота в положении 741 представляет собой Gly, Ala, Gln или Asn; аминокислота в положении 742 представляет собой Ile, Leu, Val, Ala или Gly; аминокислота в положении 743 представляет собой Gly или удалена; аминокислота в положении 745 представляет собой Gly, Ala, Glu или Asp; аминокислота в положении 751 представляет собой Thr, Ser, Gly или Ala; аминокислота в положении 753 представляет собой Gln, Asn, Lys или Arg; аминокислота в положении 754 представляет собой Thr или Ser; аминокислота в положении 756 представляет собой Thr, Ser, Leu, Val или Ile; аминокислота в положении 757 представляет собой Val, Leu или Ile; аминокислота в положении 766 представляет собой Ile, Leu или Val; аминокислота в положении 771 представляет собой Arg, Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Asn, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 772 представляет собой Arg, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 773 представляет собой Asp, Ala, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 774 представляет собой Gln, Ala, Asp, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 775 представляет собой Val, Ala, Cys, Asp, Glu, Gly, His, Ile, Asn, Pro, Gln, Arg, Ser, Thr или Tyr; аминокислота в положении 776 представляет собой Leu, Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Asn, Pro, Gln, Arg, Ser, Thr, Val или Tyr; аминокислота в положении 777 представляет собой Pro, Ala, Cys, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Gln, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 778 представляет собой Phe, Ala, His, Ile, Leu, Met, Asn, Gln, Ser, Val, Trp или Tyr; аминокислота в положении 779 представляет собой Gln, Ala, Cys, Asp, Glu, Gly, His, Lys, Leu, Asn, Pro, Arg, Ser, Thr или Val; аминокислота в положении 780 представляет собой Ala, Cys, Asn, Pro, Gln или Ser; аминокислота в положении 781 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 782 представляет собой Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 783 представляет собой Pro, Ala, Cys, Asp, Glu, Gly, His, Asn, Gln, Arg, Ser, Thr или Val; аминокислота в положении 784 представляет собой Leu, Ala, Glu, Phe, His, Ile, Lys, Met, Asn, Pro, Gln, Ser, Thr, Val или Trp; аминокислота в положении 785 представляет собой Asn, Ala, Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Gln, Arg, Ser, Thr, Val, Trp или Tyr; аминокислота в положении 786 представляет собой Tyr, Phe, Ile, Leu или Trp; аминокислота в положении 787 представляет собой His, Asn или Gln; аминокислота в положении 788 представляет собой Tyr или Met; аминокислота в положении 789 представляет собой Ala, Lys или Arg; аминокислота в положении 790 представляет собой Tyr или Thr; аминокислота в положении 791 представляет собой Arg, Lys, Gly или Ala; аминокислота в положении 792 представляет собой Leu, Ile, Val, Thr или Ser; аминокислота в положении 796 представляет собой Asp или Glu; аминокислота в положении 797 представляет собой Ser, Thr или Ala; аминокислота в положении 802 представляет собой Glu, Lys, Asp, Asn или Gln; аминокислота в положении 806 представляет собой Gln, Asp, Glu, Asn или His; аминокислота в положении 810 представляет собой Lys, Arg или Thr; аминокислота в положении 819 представляет собой Arg, Lys или His; аминокислота в положении 829 представляет собой Lys, Ser, Ala или Pro; аминокислота в положении 832 представляет собой Ala, Lys, Arg, Asp или Glu; аминокислота в положении 833 представляет собой Gly, Ala, Asp или Glu; аминокислота в положении 842 представляет собой Leu, Ile, Val или Pro; аминокислота в положении 847 представляет собой Gln, Asn, Asp или Glu; аминокислота в положении 848 представляет собой Ile, Leu или Val; аминокислота в положении 849 представляет собой Val, Leu, Ile, Gly или Ala; аминокислота в положении 855 представляет собой Thr, Ser или Met; аминокислота в положении 860 представляет собой Ile, Leu или Val; и аминокислота в положении 864 представляет собой His, Asn или Gln, по сравнению с PtIP-83Aa, представленной на Фигурах 2a-2j.
12. Рекомбинантный полинуклеотид, кодирующий полипептид PtIP-83 по любому из пп. 1-11.
13. Рекомбинантный полинуклеотид по п. 12, где полинуклеотид представляет собой полинуклеотид, отличающийся от геномного.
14. Рекомбинантный полинуклеотид по п. 13, где полинуклеотид представляет собой кДНК.
15. Рекомбинантный полинуклеотид по п. 13, где полинуклеотид представляет собой синтетический полинуклеотид.
16. Рекомбинантный полинуклеотид по п. 13, где полинуклеотид имеет кодоны, оптимизированные для экспрессии в культуре, важной с точки зрения сельского хозяйства.
17. Трансгенное растение, содержащее полинуклеотид по любому из пп. 12-16, где трансгенное растение обладает инсектицидной активностью против совки кукурузной (Helicoverpa zea).
18. ДНК-конструкция, содержащая полинуклеотид по любому из пп. 12-16, функционально связанный с гетерологичным регуляторным элементом, где ДНК-конструкция придает инсектицидную активность против совки кукурузной (Helicoverpa zea).
19. Трансгенное растение, содержащая ДНК-конструкцию по п. 18, где трансгенное растение обладает инсектицидной активностью против совки кукурузной (Helicoverpa zea).
20. Трансгенная растительная клетка, содержащая ДНК-конструкцию по п. 18, где трансгенная растительная клетка обладает инсектицидной активностью против совки кукурузной (Helicoverpa zea).
21. Композиция для применения в сельском хозяйстве, содержащая инсектицидно эффективную дозу полипептида PtIP-83 по любому из пп. 1-11, где композиция для применения в сельском хозяйстве обладает инсектицидной активностью против совки кукурузной (Helicoverpa zea).
22. Композиция для применения в сельском хозяйстве по п. 21, дополнительно содержащая по меньшей мере один приемлемый с точки зрения сельского хозяйства носитель.
23. Белок слияния, содержащий полипептид PtIP-83 по любому из пп. 1-11, где белок слияния обладает инсектицидной активностью против совки кукурузной (Helicoverpa zea).
24. Способ снижения популяции совки кукурузной (Helicoverpa zea), включающий приведение в контакт популяции совки кукурузной (Helicoverpa zea) с полипептидом PtIP-83 по любому из пп. 1-11.
25. Способ подавления роста или уничтожения совки кукурузной (Helicoverpa zea), включающий приведение в контакт совки кукурузной (Helicoverpa zea) с композицией, содержащей полипептид PtIP-83 по любому из пп. 1-11.
26. Способ снижения заражения совкой кукурузной (Helicoverpa zea) трансгенного растения и обеспечения управления устойчивостью совки кукурузной (Helicoverpa zea), включающий экспрессию в растении полипептида PtIP-83 по любому из пп. 1-11.
27. Способ снижения популяции совки кукурузной (Helicoverpa zea), включающий приведение в контакт популяции совки кукурузной (Helicoverpa zea) с трансгенным растением по п. 19 или трансгенной растительной клеткой по п. 20.
28. Способ подавления роста или уничтожения совки кукурузной (Helicoverpa zea), включающий приведение в контакт совки кукурузной (Helicoverpa zea) с трансгенным растением по п. 19 или трансгенной растительной клеткой по п. 20.
29. Способ по любому из пп. 24-28, где совка кукурузная (Helicoverpa zea) или популяция совки кукурузной (Helicoverpa zea) устойчивы по меньшей мере к одному Bt-токсину.
30. Применение полипептида PtIP-83 по любому из пп. 1-11 для ингибирования роста или уничтожения совки кукурузной (Helicoverpa zea) или популяции совки кукурузной (Helicoverpa zea).
| US 8581047 B2, 12.11.2013 | |||
| US 20130025006 A1, 24.01.2013 | |||
| US 20120302495 A1, 29.11.2012 | |||
| US 20120148524 A1, 14.06.2012 | |||
| НОВЫЕ ПЕСТИЦИДНЫЕ ПРОТЕИНЫ И ШТАММЫ | 1995 |
|
RU2196824C2 |

