По настоящей заявке испрашивается преимущество приоритета предварительной заявки США с серийным номером 61/870137, поданной 26 августа 2013 г., полное содержание которой включено в данное описание в качестве ссылки.
Это изобретение сделано при поддержке правительства с номером гранта CA-128362, присужденного Национальным институтом рака, NIH. Правительство имеет определенные права на данное изобретение.
УРОВЕНЬ ТЕХНИКИ, К КОТОРОМУ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение в целом относится к антителам, направленным против сиалированного антигена Льюисаа (sLea), и более конкретно к полинуклеотидам, кодирующим антитела против sLea, и соответствующим кодируемым антителам или их фрагментам.
Пассивное введение антител, направленных против опухолевых антигенов, может уничтожать опухолевые клетки и ранние метастазы в процессе развития рака. Это лечение может также иметь значительное влияние на рецидивы рака. Антитела, направленные против специфических опухолевых углеводов, могут быть полезными кандидатами при таком лечении рака. Например, многие моноклональные антитела, направленность которых ограничена опухолью, вырабатываемые в результате иммунизации мышей раковыми клетками человека, как показано, направлены против углеводных антигенов, экспрессируемых на клеточной поверхности в виде гликолипидов или гликопротеинов. Углевод sLea, как показано, экспрессируется в опухолях желудочно-кишечного тракта. Экспрессия sLea, как также обнаружено, вносит вклад в метастатический потенциал и коррелирует с повышенным метастатическим потенциалом при развитии рака толстой кишки человека и аденокарциномы поджелудочной железы. Тем не менее, химия углеводов является довольно сложной, и клиническая разработка антител, которые распознают такие специфические опухолевые углеводы, развивается медленно.
Карцинома поджелудочной железы является одной из самых агрессивных аденокарцином и часто связана с плохим прогнозом. Карцинома поджелудочной железы рассматривается как четвертая ведущая причина смертности от рака. Несмотря на успехи в скрининге различных карцином, надежность обнаружения злокачественных поражений, возникающих в поджелудочной железе, остается на низком уровне. Позитронно-эмиссионная томография с использованием фтордезоксиглюкозы (FDG-PET) требуется для выявления и определения стадии рака поджелудочной железы. Тем не менее, FDG-PET не чувствительна к дифференциальной диагностике панкреатита и злокачественных новообразований, и проблематичным остается определение стадии при небольших первичных поражениях (<7 мм) и печеночных метастазах (<1 см). Один диагностический метод скрининга, используемый для мониторинга статуса протоковой аденокарциномы поджелудочной железы (PDAC) у больных, включает обнаружение повышенных уровней циркулирующего антигена sLea в сыворотке крови. Циркулирующий антиген sLea>37 Ед/мл у больных указывает на рецидив рака. Однако развитие альтернативных диагностических подходов, которые используют такие специфические опухолевые углеводы, происходит медленно.
Таким образом, существует необходимость в идентификации и выработке антител, которые специфически распознают специфические опухолевые углеводы, такие как sLea, для лечения рецидивирующих форм рака и для обнаружения злокачественных поражений и метастазов. Настоящее изобретение удовлетворяет эту потребность и обеспечивает связанные с этим преимущества.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии с настоящим изобретением в настоящем документе предлагаются композиции для получения антител или их функциональных фрагментов, которые связываются с sLea. Композиции включают выделенный полинуклеотид, кодирующий антитело или его функциональный фрагмент, где антитело включает вариабельный домен тяжелой цепи (VH), который имеет аминокислотную последовательность, представленную в настоящем документе. Выделенный полинуклеотид по изобретению может также включать последовательность нуклеиновой кислоты, представленную в данном описании, где последовательность нуклеиновой кислоты кодирует домен VH антитела или его функционального фрагмента.
В другом варианте осуществления настоящего изобретения выделенный полинуклеотид может кодировать антитело или его функциональный фрагмент, где антитело включает вариабельный домен легкой цепи (VL), который имеет аминокислотную последовательность, представленную в настоящем документе. Выделенный полинуклеотид по изобретению может также включать последовательность нуклеиновой кислоты, представленную в данном описании, где последовательность нуклеиновой кислоты кодирует домен VL антитела или его функционального фрагмента.
Композиции по настоящему изобретению также включают выделенное антитело или его функциональный фрагмент, где антитело связывается с sLea. В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, который связывается с sLea, где антитело или его функциональный фрагмент включает домен VH, имеющий аминокислотную последовательность, представленную в настоящем документе.
В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, который связывается с sLea, где антитело или его функциональный фрагмент включает домен VL, имеющий аминокислотную последовательность, представленную в настоящем документе.
В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, который связывается с sLea, где антитело или его функциональный фрагмент включает как домен VH, так и домен VL, где домен VH и домен VL соответственно включают аминокислотную последовательность соответствующих доменов VH и VL изолятов клонов, представленных в настоящем описании.
В некоторых вариантах осуществления в настоящем изобретении предлагается конъюгат, включающий антитело или его функциональный фрагмент, предлагаемые в данном документе, конъюгированные или соединенные рекомбинантным способом с диагностическим агентом, детектируемым агентом или терапевтическим агентом. В некоторых аспектах настоящего изобретения целью является конъюгат по настоящему изобретению, включающий детектируемый агент, который может быть использован в способе обнаружения и/или диагностики образования опухоли. Такие способы могут включать введение эффективного количества конъюгата нуждающемуся в этом индивидууму.
В некоторых вариантах осуществления настоящее изобретение относится к фармацевтическим композициям, включающим одно или более антитело или его функциональный фрагмент по изобретению и фармацевтически приемлемый носитель. В некоторых аспектах настоящее изобретение относится также к способу лечения или профилактики заболевания у нуждающегося в этом индивидуума путем введения терапевтически эффективного количества фармацевтической композиции по настоящему изобретению. В еще одном аспекте настоящее изобретение относится к введению второго терапевтического агента одновременно или последовательно с антителом или его функциональным фрагментом по изобретению.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена тяжелой (VH) цепи клона 5B1 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 1 и аминокислотной последовательностью SEQ ID NO: 2. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 2 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена легкой (VL) цепи клона 5B1 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 3 и аминокислотной последовательностью SEQ ID NO: 4. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 3 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена тяжелой (VH) цепи клона 9H3 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 5 и аминокислотной последовательностью SEQ ID NO: 6. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 4 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена легкой (VL) цепи клона 9H3 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 7 и аминокислотной последовательностью SEQ ID NO: 8. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 5 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена тяжелой (VH) цепи клона 5H11 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 9 и аминокислотной последовательностью SEQ ID NO: 10. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 6 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена легкой (VL) цепи клона 5H11 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 11 и аминокислотной последовательностью SEQ ID NO: 12. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 7 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена тяжелой (VH) цепи клона 7E3 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 13 и аминокислотной последовательностью SEQ ID NO: 14. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 8 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность вариабельного домена легкой (VL) цепи клона 7E3 и лидирующая последовательность, которая может быть использована для рекомбинантной экспрессии. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 15 и аминокислотной последовательностью SEQ ID NO: 16. Идентифицированы также три области, определяющие комплементарность (CDR1, CDR2 и CDR3).
На фиг. 9 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность диатела, обозначаемого как 5B1CysDb, имеющего CDR1, CDR2 и CDR2 как вариабельного домена тяжелой (VH) цепи, так и вариабельного домена легкой (VL) цепи клона 5B1. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 17 и аминокислотной последовательностью SEQ ID NO: 18. Идентифицированы три области, определяющие комплементарность (CDR1, CDR2 и CDR3), обоих доменов VH и VL, обозначенные жирным шрифтом и подчеркиванием текста. Последовательность линкера и полигистидиновой метки (Poly His-Tag) с добавленными аминокислотами также обозначена курсивом и подчеркиванием текста.
На фиг. 10 показана нуклеотидная последовательность и кодируемая аминокислотная последовательность диатела, обозначаемого как 7E3CysDb, имеющего CDR1, CDR2 и CDR2 как вариабельного домена тяжелой (VH) цепи, так и вариабельного домена легкой (VL) цепи клона 7E3. В верхней части фигуры показано выравнивание между нуклеотидной последовательностью SEQ ID NO: 19 и аминокислотной последовательностью SEQ ID NO: 20. Идентифицированы три области, определяющие комплементарность (CDR1, CDR2 и CDR3), обоих доменов VH и VL, обозначенные жирным шрифтом и подчеркиванием текста. Последовательность линкера и полигистидиновой метки (Poly His-Tag) с добавленными аминокислотами также обозначена курсивом и подчеркиванием текста.
На фиг. 11, панели A-E, показано связывание антител человека против sLea с опухолевыми клетками, проанализированное с помощью проточной цитометрии. На панели A показаны клетки DMS-79, окрашенные рекомбинантными (r) антителами 5B1, 9H3, 5H11 и 7Е3. На панелях B-F, соответственно, показаны клетки HT29, BXPC3, SW626, SK-MEL28 и Colo205-Luc, окрашенные 1-2 мкг/мл r5Bl или r7E3 плюс IgG- или IgM-специфичными вторичными антителами, как описано в примере I.
На фиг. 12, панели А и В, показана активность CDC антител r5Bl и r7E3 по сравнению с мышиными 121SLE (IgM) в присутствии комплемента человека (Hu C’) при измерении относительно клеток DMS-79. Контрольные антитела изотипа человека, Hu IgG  и Hu IgM
и Hu IgM  , показали цитотоксичность <4%. Дозозависимый ответ антител r5Bl IgG
, показали цитотоксичность <4%. Дозозависимый ответ антител r5Bl IgG  , r7E3 IgM
, r7E3 IgM  и 121SLE mIgM
и 121SLE mIgM  показан на панели А. Рассчитанная ЕС50 (мкг/мл) для антител r5Bl (IgG), r7E3 (IgM) и 121SLE (mIgM) показана на панели B.
показан на панели А. Рассчитанная ЕС50 (мкг/мл) для антител r5Bl (IgG), r7E3 (IgM) и 121SLE (mIgM) показана на панели B.
На фиг. 13, панели A-C, показана зависимая от антител опосредуемая клетками цитотоксичность (ADCC) антител r5Bl. На панели А показана опосредуемая r5Bl ADCC РВМС человека против клеток DMS-79. РВМС тестировали в отношениях E:T от 100:1 до 12,5:1 с опухолевыми клетками DMS-79 в присутствии или в отсутствие 2 мкг/мл r5Bl. На панели B показана опосредуемая r5Bl ADCC первичных NK-клеток человека против клеток DMS-79. NK-клетки тестировали при более низких отношениях E:T от 5:1 до 0,6:1 с опухолевыми клетками DMS-79 в присутствии или в отсутствие 2 мкг/мл r5Bl. На панели С показана ADCC антитела r5Bl при различных концентрациях PBMCs от 2-доноров при отношении E:T=1:100 против опухолевых клеток DMS-79 в присутствии указанных концентраций r5Bl.
На фиг. 14 показана интернализация sLea в клетки BxPC3. Опухолевые клетки BxPC3 поджелудочной железы выращивали в присутствии антител r5Bl (против sLea) или rlB7 (против GD2), связанных с Hum-ZAP, конъюгированным с сапорином IgG против антигенов человека. Через 3 дня измеряли жизнеспособность клеток методом с использованием бромида 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия (МТТ), и значения образцов нормализовывали на значения необработанных культур.
На фиг. 15 показана активность антитела r5Bl в модели ксенотрансплантата с использованием клеток Colo205-luc. Мыши с тяжелым сочетанным иммунодефицитом (SCID) (5 на группу) получали 0,5 миллиона клеток Colo205-luc путем инъекции в хвостовую вену на день 0. Мыши получали 100 мкг r5Bl путем внутрибрюшинной инъекции в дни 1, 7, 14 и 21 (эксперимент 1, Exp1) или в дни 1, 4, 7, 10, 14 и 21 (эксперимент 2, Exp2) в суммарной дозе 600 мкг. Контрольные (Ctrl) животные получали ложные инъекции PBS.
На фиг. 16 показано влияние r5Bl на опухоли Colo205-luc у мышей SCID. Мыши получали 100 мкг  , 300 мкг
, 300 мкг  или 1 мг
или 1 мг  антитела r5Bl на одну инъекцию, как описано в примере I. Контрольные животные
антитела r5Bl на одну инъекцию, как описано в примере I. Контрольные животные  получали ложные инъекции PBS.
получали ложные инъекции PBS.
На фиг. 17 показана флуоресцентная визуализация пяти мышей на группу для мышей, получавших r5Bl, имеющих опухоли Colo205-luc, в день 0 и через 5 недель. Схема получения препаратов мышами представлена на фиг. 16 и описана в примере I.
На фиг. 18, панели А и В, показана противоопухолевая активность в терапевтической модели подкожного ксенотрансплантата с использованием клеток DMS-79. На панели А показано подавление или регрессия опухоли у мышей, получавших 5B1 (только 5B1 или 5B1+cRGD ), по сравнению с получением IgG человека (только IgG или IgG+RGD ) и с контролем с введением PBS . Стрелки указывают на дни введения антитела или PBS. На панели В показаны репрезентативные изображения обработанных мышей. Стрелки указывают на отсутствие какой-либо видимой опухоли.
На фиг. 19, панели A-F, показано связывание 5B1 с различными типами опухолей. Панель А представляет собой поджелудочную железу, протоковую аденокарциному, стадию III опухоли. Панель B представляет собой сигмовидную кишку, карциному стадии IIIB опухоли. Панель С представляет собой легкое, аденокарциному, стадию IB опухоли. Панель D представляет собой мочевой пузырь, мукозную аденокарциному, стадию IV опухоли. Панель Е представляет собой яичник, метастатическую карциному опухоли толстой кишки. Панель F представляет собой лимфатический узел, метастатическую карциному, стадию IIIA опухоли.
На фиг. 20 показаны серийные изображения проекции максимальной интенсивности (MIP) PET, полученные через 2-120 час после внутривенного введения антитела 5Bl, радиоактивно меченного 89Zr (89Zr-5Bl), самкам мышей SCID с подкожно имплантированными опухолями поджелудочной железы BxPC3. Изображение PET-MIP демонстрирует высокий захват опухолью с клиренсом неспецифически связанной метки уже через 24 часа после инъекции (h p.i.)
На фиг. 21 приведены результаты биологического распределения, которые согласуются с данными PET фиг. 20, с наблюдаемым захватом опухолью 84,73±2,28% ID/г. Из-за малой массы опухоли гистограмма захвата опухолью, выраженная в % ID, в зависимости от времени, изображена на гистограмме во вставке. % ID опухоли показывает значительный захват опухолью 89Zr-5Bl во всех временных точках, и, по меньшей мере, семикратное превышение неспецифического захвата 89Zr-IgG. Конкурентное ингибирование немеченым 5B1 (200 мкг) показывает уменьшение аккумуляции в опухоли.
На фиг. 22, панели A-C, показаны изображения PET-MIP мышей, несущих DMS79 (панель А) и ксенотрансплантатов Colo205-luc (панель B). Указана визуализация PET-MIP очертаний опухоли (T), сердца (H) и печени (L) с помощью 89Zr-5Bl. В модели ксенотрансплантатов Colo205-luc колоректального рака продемонстрирована аккумуляция 89Zr-5Bl, достигающая максимума через 24 час, с последующим снижением, в то время как выявлен рост неспецифического связывания в печени (панель С).
На фиг. 23 показано зависимое от дозы ингибирование и регрессия роста опухоли в модели ксенотрансплантата мелкоклеточной легочной карциномы DMS-79, при лечении последовательным совместным введением антитела 5B1 и таксола (паклитаксела). Большие стрелки на оси Х указывают на лечение 5B1. Совместное введение антитела 5B1 и таксола существенно ограничивало рост опухоли и приводило к регрессии опухоли по сравнению с контрольным IgG человека (HuIgG) или антителом 5B1 и таксолом, вводимыми по отдельности. Показаны значимые отличия от контроля с помощью 2-факторного дисперсионного анализа ANOVA с р<0,01 (**) и р<0,001 (***). N=5.
На фиг. 24 показано ингибирование роста опухоли в модели ксенотрансплантата карциномы поджелудочной железы BxPC3 при лечении последовательным совместным введением антитела 5B1 и таксола (паклитаксела). Большие стрелки на оси Х указывают на лечение таксолом плюс 5B1, в то время как маленькие стрелки указывают на лечение только 5B1. Совместное введение антитела 5B1 и таксола существенно ограничивало рост опухоли по сравнению с контролями (PBS - Ctrl; IgG человека - HuIgG) или антителом 5B1 и таксолом, вводимыми по отдельности.
На фиг. 25, панели А и В, показаны репрезентативные изображения мышей, которым ортотопически трансплантировали ксенотрансплантаты опухолей поджелудочной железы BxPC3-luc. Панель A: совместная регистрация с помощью FDG-PET и компьютерной томографии (КТ) (слева) планарных секций FDG-PET с изображением только (справа) минимально обнаруживаемой метки в опухоли с высоким захватом в тканях с высоким метаболизмом (т.е. в сердце, Н, и мочевом пузыре, В). Панель B: PET изображение той же мыши, выявляемое с помощью радиоактивно меченного 89Zr антитела 5Bl (89Zr-5Bl), регистрируемое совместно с КТ, показало детекцию исключительно опухоли в случае ксенотрансплантатов опухоли BxPC3-luc.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Углеводы, экспрессируемые на поверхности опухолевых клеток, могут представлять собой мишень для пассивной иммунотерапии. Композиции, представленные в настоящем описании, основаны, по меньшей мере, частично, на идентификации и характеристике антител человека, которые вырабатываются лимфоцитами крови индивидуумов, иммунизированных вакциной, представляющей собой конъюгат сиалированного антигена Льюисаа и гемоцианина фиссуреллии (sLea-KLH). Идентифицированы, по меньшей мере, четыре антитела с высоким сродством к sLea (5B1, 9H3, 5H11 и 7E3). Два из этих антител экспрессировали в виде рекомбинантных антител (r5Bl и r7E3) и дополнительно характеризовали в моделях in vitro и in vivo. Оба антитела при анализе характеризовались сильной, зависимой от комплемента цитотоксичностью (CDC), и антитело 5B1 также было высокоактивным в зависимой от антител цитотоксичности. Эффективность антител in vivo тестировали в двух моделях ксенотрансплантатов с использованием либо опухолевых клеток Colo205, либо опухолевых клеток DMS-79, пересаживаемых мышам с тяжелым сочетанным иммунодефицитом (SCID). Междисциплинарная актуальность осуществления изобретения, предлагаемого в настоящем документе, рассматривается в 2 аспектах: во-первых, подход, предлагаемый в настоящем описании, показывает, что ответ в виде антител, вызванный вакциной sLea-KLH, пригоден в качестве вакцины как таковой. Во-вторых, самые мощные антитела, которые вырабатываются при клиническом испытании, могут быть сохранены и, в конечном счете, использованы в качестве терапевтических агентов или для создания терапевтических агентов для таргетной терапии рака у населения. Высокое сродство антител, представленных в настоящем описании, и их высокие эффекторные функции поддерживают этот междисциплинарный потенциал.
Используемый в данном описании термин «антитело» предназначен для обозначения полипептидного продукта В-клеток в пределах иммуноглобулинового класса полипептидов, который способен связываться с конкретной антигенной молекулой, и состоит из двух идентичных пар полипептидных цепей, где каждая пара имеет одну тяжелую цепь (приблизительно 50-70 кДа) и одну легкую цепь (приблизительно 25 кДа), и каждый аминоконцевой участок каждой цепи включает вариабельную область от приблизительно 100 до приблизительно 130 или более аминокислот и каждый карбоксиконцевой участок каждой цепи включает константную область (смотри Borrebaeck (ed.) (1995) Antibody Engineering, Second Edition, Oxford University Press.; Kuby (1997) Immunology, Third Edition, W.H. Freeman and Company, New York). В контексте настоящего изобретения конкретная антигенная молекула, которая может связаться с антителом по настоящему изобретению, включает таргетный углевод sLea.
Термин «от человека» при использовании в отношении антитела или его функционального фрагмента обозначает антитело или его функциональный фрагмент, которые имеют вариабельную область от человека и/или константную область от человека или их часть, соответствующую последовательностям иммуноглобулинов зародышевой линии человека. Такие последовательности иммуноглобулинов зародышевой линии человека описаны Kabat et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242. Антитело человека в контексте настоящего изобретения может включать антитело, которое связывается с sLea и кодируется последовательностью нуклеиновой кислоты, которая представляет собой существующий в природе соматический вариант последовательности нуклеиновой кислоты иммуноглобулина зародышевой линии человека. Иллюстративные способы получения антител человека приведены в примере I, но может быть использован любой способ, хорошо известный специалистам в данной области техники.
Термин «моноклональное антитело» обозначает антитело, которое является продуктом одного клона клеток или гибридомы или популяции клеток, происходящих от одной клетки. Моноклональные антитела также предназначены для обозначения антитела, продуцируемого рекомбинантными методами из генов тяжелых и легких цепей, кодирующих иммуноглобулин, с получением одного варианта молекулы иммуноглобулина. Аминокислотные последовательности антител в пределах препарата моноклонального антитела по существу однородны, и связывающая активность антител в таком препарате демонстрирует, по существу, одну и ту же антигенсвязывающую активность. В противоположность этому поликлональные антитела получают из различных В-клеток в популяции, они представляют собой сочетание молекул иммуноглобулинов, которые связывают специфический антиген. Каждый иммуноглобулин поликлональных антител может связываться с отличным эпитопом того же самого антигена. Способы получения как моноклональных антител, так и поликлональных антител хорошо известны в данной области техники (Harlow and Lane., Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989) and Borrebaeck (ed.), Antibody Engineering: A Practical Guide, W.H. Freeman and Co., Publishers, New York, pp. 103-120 (1991)).
Используемый в данном описании термин «функциональный фрагмент» при применении в отношении антитела предназначен для обозначения части антитела, включая полипептиды тяжелой или легкой цепи, которые сохраняют некоторую или всю связывающую активность антитела, из которого этот фрагмент произошел. Такие функциональные фрагменты могут включать, например, Fd, Fv, Fab, F(аb'), F(аb)2, F(аb')2, одноцепочечный Fv (scFv), диатело, триатело, тетратело и минитело. Другие функциональные фрагменты могут включать, например, полипептиды тяжелой или легкой цепи, полипептиды вариабельной области или полипептиды CDR, или их части, до тех пор, пока такие функциональные фрагменты сохраняют связывающую активность. Такие связывающие фрагменты антител могут быть найдены как описанные, например, в Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York (1989); Myers (ed.), Molec. Biology and Biotechnology: A Comprehensive Desk Reference, New York: VCH Publisher, Inc.; Huston et al., Cell Biophysics, 22:189-224 (1993); Plückthun and Skerra, Meth. Enzymol., 178:497-515 (1989) и в Day, E.D., Advanced Immunochemistry, Second Ed., Wiley-Liss, Inc., New York, NY (1990).
Термин «тяжелая цепь» при использовании в отношении антитела относится к полипептидной цепи приблизительно 50-70 кДа, в которой аминоконцевой участок включает вариабельную область от приблизительно 120 до 130 или более аминокислот и карбоксиконцевой участок, который включает константную область. Константная область может представлять собой один из пяти различных типов, обозначаемых как альфа (α), дельта (δ), эпсилон (ε), гамма (γ) и мю (μ), на основании аминокислотной последовательности константной области тяжелой цепи. Различные тяжелые цепи отличаются по размеру: α, δ и γ содержат приблизительно 450 аминокислот, а μ и ε содержат приблизительно 550 аминокислот. В сочетании с легкой цепью эти различные типы тяжелых цепей приводят к пяти хорошо известным классам антител, IgA, IgD, IgE, IgG и IgM, соответственно, включая четыре подкласса IgG, а именно: IgG1, IgG2, IgG3 и IgG4. Тяжелая цепь может представлять собой тяжелую цепь от человека.
Термин «легкая цепь» при использовании в отношении антитела относится к полипептидной цепи приблизительно 25 кДа, в которой аминоконцевой участок включает вариабельную область от приблизительно 100 до приблизительно 110 или более аминокислот и карбоксиконцевой участок включает константную область. Приблизительная длина легкой цепи составляет от 211 до 217 аминокислот. Существует два различных типа, называемых каппа (κ) или лямбда (λ), на основе аминокислотной последовательности константных доменов. Аминокислотные последовательности легкой цепи хорошо известны в данной области техники. Легкая цепь может представлять собой легкую цепь от человека.
Термин «вариабельный домен» или «вариабельная область» относится к части легких или тяжелых цепей антитела, которая, как правило, расположена на аминоконце легкой или тяжелой цепи и имеет длину от приблизительно 120 до 130 аминокислот в тяжелой цепи и приблизительно от 100 до 110 аминокислот в легкой цепи, и используется для связывания и специфичности каждого конкретного антитела в отношении его конкретного антигена. Вариабельные домены в значительной степени различаются по последовательности между различными антителами. Вариабельность последовательности сконцентрирована в CDR, в то время как менее вариабельные участки в вариабельной области называются каркасными областями (FR). CDRs легкой и тяжелой цепей ответственны в первую очередь за взаимодействие антитела с антигеном. Нумерация положений аминокислот, используемая в данном документе, находится в соответствии с индексом EU, как у Kabat et al. (1991) Sequences of proteins of immunological interest. (U.S. Department of Health and Human Services, Washington, D.C.) 5th ed. Вариабельная область может представлять собой вариабельную область от человека.
CDR относится к одной из трех гипервариабельных областей (H1, H2 или H3) в пределах некаркасной области β-складок VH каркаса иммуноглобулина (Ig или антитела) или к одной из трех гипервариабельных областей (L1, L2 или L3) в пределах некаркасной области β-складок VL каркаса антитела. Соответственно, CDRs представляют собой последовательности вариабельных областей, рассеянные в пределах последовательностей каркасных областей. Области CDR хорошо известны специалистам в данной области техники и были определены, например, Kabat как области наибольшей гипервариабельности в пределах вариабельных (V) доменов антител (Kabat et al., J. Biol. Chem. 252:6609-6616 (1977); Kabat, Adv. Prot. Chem. 32:1-75 (1978)). Последовательности области CDR также были идентифицированы структурно по Chothia как те остатки, которые не являются частью консервативного каркаса β-складок, и, таким образом, способны адаптироваться к различным конформациям (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987)). Оба варианта терминологии хорошо известны специалистам в данной области техники. Положения CDRs в пределах канонического вариабельного домена антитела определены путем сравнения многочисленных структур (Al-Lazikani et al., J. Mol. Biol. 273:927-948 (1997); Morea et al., Methods 20:267-279 (2000)). Поскольку количество остатков в пределах гипервариабельной области варьируется в различных антителах, дополнительные остатки по отношению к каноническим положениям обычно нумеруются как a, b, c и так далее после номера остатка в канонической схеме нумерации вариабельных доменов (Al-Lazikani et al., (1997) выше). Такая номенклатура также хорошо известна специалистам в данной области техники.
Например, обозначения CDRs, определяемые в соответствии либо с Kabat (гипервариабельные), либо с Chothia (структурные), приведены в представленной ниже таблице 1.
Обозначения CDR
2Нумерация остатков по номенклатуре Chothia et al., выше
Одна или более CDR также может быть включена в молекулу либо ковалентно, либо нековалентно, чтобы сделать ее иммуноадгезином. Иммуноадгезин может включать CDR(s) в качестве части более крупной полипептидной цепи, может ковалентно связывать CDR(s) с другой полипептидной цепью или может включать CDR(s) нековалентно. CDRs позволяют иммуноадгезину связываться с определенным антигеном, представляющим интерес.
Используемый в данном описании термин «выделенный» при применении по отношению к антителу, функциональному фрагменту антитела или полинуклеотиду предназначен для обозначения того, что референсная молекула свободна, по меньшей мере, от одного компонента, встречаемого с ней в природе. Термин включает антитело, функциональный фрагмент антитела или полинуклеотид, которые отделяются от некоторых или всех других компонентов, встречаемых с ними в естественном окружении. Компоненты природной среды для антитела включают, например, эритроциты, лейкоциты, тромбоциты, плазму, белки, нуклеиновые кислоты, соли и питательные вещества. Компоненты природной среды для функционального фрагмента антитела или полинуклеотида включают, например, липидные мембраны, клеточные органеллы, белки, нуклеиновые кислоты, соли и питательные вещества. Антитело, функциональный фрагмент антитела или полинуклеотид по изобретению также могут быть свободны или практически по существу свободны от всех этих компонентов или любого другого компонента клеток, из которых они выделены или получены рекомбинантным способом.
Используемый в данном описании термин «изотип» относится к классу антител, который кодируется генами константных областей тяжелых цепей. Тяжелые цепи данного антитела или функционального фрагмента определяют класс этого антитела или функционального фрагмента: IgM, IgG, IgA, IgD или IgE. Каждый класс может иметь либо κ, либо λ легкие цепи. Термин «подкласс» относится к минорным различиям в аминокислотных последовательностях тяжелых цепей, которые определяют деление на подклассы. У человека существует два подкласса IgA (подклассы IgA1 и IgA2) и четыре подкласса IgG (подклассы IgG1, IgG2, IgG3 и IgG4). Такие классы и подклассы хорошо известны специалистам в данной области техники.
Термины «связывается» или «связывание», используемые в данном описании, относятся к взаимодействию между молекулами с образованием комплекса. Взаимодействие может быть, например, нековалентным взаимодействием, включающим водородные связи, ионные связи, гидрофобные взаимодействия и/или взаимодействия Ван-дер-Ваальса. Комплекс может также включать связывание двух или более молекул, удерживаемых вместе с помощью ковалентных или нековалентных связей, взаимодействий или сил. Связывание антитела или его функционального фрагмента может быть определено с помощью, например, твердофазного иммуноферментного анализа, метода, представленного в примере I, или любого другого из ряда методов, которые хорошо известны специалистам в данной области техники.
Сила суммарных нековалентных взаимодействий между одним антигенсвязывающим сайтом на антителе или его функциональном фрагменте и одним эпитопом молекулы-мишени, такой как sLea, представляет собой сродство антитела или его функционального фрагмента к этому эпитопу. Отношение ассоциации антитела или его функционального фрагмента c одновалентным антигеном (k1) к их диссоциации (k-1) (k1/k-1) представляет собой константу ассоциации К, которая является мерой сродства. Величина К варьируется для различных комплексов антитела или его функционального фрагмента с антигеном и зависит как от k1, так и от и k-1. Константа ассоциации К для антитела или его функционального фрагмента по изобретению может быть определена с использованием любого из методов, предлагаемых в данном документе, или с помощью любого другого метода, хорошо известного специалистам в этой области техники.
Сродство к одному сайту связывания не всегда отражает истинную силу взаимодействия между антителом или его функциональным фрагментом и антигеном. Когда сложные антигены, содержащие множественные, повторяющиеся антигенные детерминанты, такие как поливалентный sLea, вступают в контакт с антителами, содержащими множественные сайты связывания, взаимодействие антитела или его функционального фрагмента с антигеном на одном сайте увеличивает вероятность взаимодействия на втором сайте. Прочность таких множественных взаимодействий между поливалентным антителом и антигеном называется авидностью. Авидность антитела или его функционального фрагмента может быть лучшим показателем его связывающей способности, чем сродство его отдельных участков связывания. Например, высокая авидность может компенсировать низкую аффинность, как иногда обнаруживается для пентамерных антител IgM, которые могут иметь более низкое сродство, чем IgG, но высокая авидность IgM, происходящая в результате их мультивалентности, позволяет им эффективно связывать антиген.
Специфичность антитела или его функционального фрагмента относится к способности отдельного антитела или его функционального фрагмента к взаимодействию только с одним антигеном. Антитело или его функциональный фрагмент можно рассматривать как специфические, когда они могут отличать различия в первичной, вторичной или третичной структуре антигена или изомерных форм антигена.
Термин «полинуклеотид» относится к полимерной форме нуклеотидов любой длины, или дезоксирибонуклеотидов, или рибонуклеотидов, или их аналогов. Последовательность полинуклеотида состоит из четырех нуклеотидных оснований: аденина (А); цитозина (С); гуанина (G); тимина (Т); и урацила (U) вместо тимина, когда полинуклеотид представляет собой РНК. Таким образом, термины «нуклеотидная последовательность» или «последовательность нуклеиновой кислоты» представляют собой буквенное представление полинуклеотида. Полинуклеотид может включать ген или фрагмент гена (например, зонд, праймер, EST или тег SAGE), экзоны, интроны, матричную РНК (мРНК), транспортную РНК, рибосомную РНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенную ДНК любой последовательности, выделенную РНК любой последовательности, зонды нуклеиновых кислот и праймеры. Полинуклеотид также относится как к двухцепочечным, так и к одноцепочечным молекулам. Если не указано или не требуется иное, любой вариант осуществления настоящего изобретения, в котором предлагается полинуклеотид, охватывает как двухцепочечную форму, так и каждую из двух комплементарных одноцепочечных форм, известных или предсказанных как составляющие двухцепочечную форму. Понятно, что выделенные полинуклеотиды и нуклеиновые кислоты, описанные в настоящем документе, направлены на не встречающиеся в природе полинуклеотиды и нуклеиновые кислоты. Не встречающиеся в природе полинуклеотиды и нуклеиновые кислоты могут включать, но не ограничиваются этим, кДНК и химически синтезированные молекулы.
Термин «кодировать» или его грамматические эквиваленты при использовании в отношении полинуклеотидов относится к полинуклеотиду в его нативном состоянии или к полинуклеотиду, который может быть транскрибирован с получением мРНК, которая затем транслируется в полипептид и/или его фрагмент, при использовании методов, хорошо известных специалистам в этой области техники. Антисмысловая цепь комплементарна такому полинуклеотиду, и кодирующая последовательность может быть выведена из нее.
Выражение «терапевтический агент» относится к любому агенту, который может использоваться для лечения, управления или облегчения заболевания, связанного с экспрессией sLea, и/или симптома, связанного с ним. В некоторых вариантах осуществления терапевтический агент относится к антителу или его функциональному фрагменту по изобретению. В других вариантах осуществления терапевтический агент относится к агенту, отличному от антитела или его функционального фрагмента по изобретению. Терапевтический агент может представлять собой агент, который хорошо известен как пригодный для лечения, управления или облегчения заболевания, связанного с экспрессией sLea, и/или симптома, связанного с ним, или применялся, или в настоящее время применяется для этих целей.
Выражение «диагностический агент» относится к веществу, вводимому индивидууму с целью диагностики заболевания. Такие вещества могут быть использованы для выявления, точного определения и/или определения локализации процесса, вызывающего заболевание. В некоторых вариантах осуществления диагностический агент включает вещество, конъюгируемое с антителом или с его функциональным фрагментом согласно изобретению, которое при введении индивидууму или при контакте с образцом от индивидуума помогает в диагностике рака или образования опухоли.
Выражение «детектируемый агент» относится к веществу, которое может быть использовано для установления наличия или присутствия желаемой молекулы, такой как антитело или его функциональный фрагмент согласно изобретению, в образце или у индивидуума. Детектируемый агент может представлять собой вещество, которое можно визуализировать, или вещество, которое можно определить и/или измерить другим способом (например, путем количественной оценки).
«Эффективное количество» обозначает количество, достаточное для индукции благоприятных или желаемых результатов. Эффективное количество можно вводить в один или несколько приемов, употреблений или дозировок. Такая доставка зависит от числа переменных, включая период времени, в течение которого должна использоваться индивидуальная единица лекарственной формы, биодоступность агента, путь введения и т.п.
Выражение «терапевтически эффективное количество», используемое в данном описании, относится к количеству терапевтического агента (например, антитела или его функционального фрагмента, предлагаемого в настоящем описании, или любого другого терапевтического агента, предлагаемого в настоящем описании), которое является достаточным для уменьшения и/или ослабления тяжести и/или продолжительности данного заболевания и/или симптома, связанного с ним. Терапевтически эффективное количество терапевтического агента может быть количеством, необходимым для уменьшения или облегчения распространения или прогрессирования данного заболевания, снижения или уменьшения рецидива, развития или возникновения данного заболевания, и/или для улучшения или усиления профилактического или терапевтического действия другого лечения (например, лечения, отличного от введения антитела или его функционального фрагмента, представленного в настоящем описании).
Соединение «сиалированный антиген Льюисаа» (sLea), также известное как сиалил-sLea, сиалил-Льюиса A, сиалированный антиген Льюиса a и СА 19.9, представляет собой тетрасахарид с молекулярной формулой C31H52N2O23 и молярной массой 820,74 г/моль. Структура sLea может включать Neu5Aca2-3Galβ1-3(Fucαl-4)GlcNAcβ и Neu5Gcα2-3Galβ1-3(Fucαl-4) GlcNAcβ. sLea широко экспрессируется на опухолях желудочно-кишечного тракта и используется в качестве опухолевого маркера при раке поджелудочной железы и толстой кишки. sLea также представляет собой известный лиганд для E-селектина, также известного как молекула эндотелиальной адгезии лейкоцитов (ELAM).
В некоторых вариантах осуществления в настоящем изобретении предлагается выделенный полинуклеотид, кодирующий тяжелую или легкую цепь антитела, или его функциональный фрагмент, где антитело или его функциональный фрагмент получены с использованием тяжелой или легкой цепи антитела, связывающейся с sLea. Соответственно, в некоторых вариантах осуществления настоящее изобретение относится к выделенному полинуклеотиду, кодирующему антитело или его функциональный фрагмент, где антитело включает домен VH, который имеет аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-142 SEQ ID NO: 2, остатков 20-142 SEQ ID NO: 6, остатков 20-142 SEQ ID NO: 10, и остатков 20-145 SEQ ID NO: 14. Выделенный полинуклеотид по изобретению также может включать последовательность нуклеиновой кислоты из остатков 58-426 последовательности SEQ ID NO: 1, остатков 58-426 SEQ ID NO: 5, остатков 58-426 SEQ ID NO: 9 или остатков 58-435 из SEQ ID NO: 13, где последовательность нуклеиновой кислоты кодирует домен VH антитела или его функционального фрагмента.
В другом варианте осуществления настоящего изобретения выделенный полинуклеотид может кодировать антитело или его функциональный фрагмент, где антитело включает домен VL, который имеет аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-130 SEQ ID NO: 4, остатков 20-129 SEQ ID NO: 8, остатков 20-130 SEQ ID NO: 12 и остатков 23-130 SEQ ID NO: 16. Выделенный полинуклеотид по изобретению может также включать последовательность нуклеиновой кислоты из остатков 58-390 SEQ ID NO: 3, остатков 58-387 SEQ ID NO: 7, остатков 58-390 SEQ ID NO: 11 или остатков 67-390 SEQ ID NO: 15, где последовательность нуклеиновой кислоты кодирует домен VL антитела или его функционального фрагмента.
В другом варианте осуществления в настоящем изобретении предлагается выделенный полинуклеотид, кодирующий тяжелую или легкую цепь антитела или его функциональный фрагмент, где тяжелая или легкая цепь антитела или его функциональный фрагмент, кодируемые полинуклеотидом по изобретению, имеют одну или более областей, определяющих комплементарность (CDRs), изображенных на фиг. 1-8 или приведенных в таблице 2. Антитело или его функциональный фрагмент, которые включают одну или более CDRs, могут специфически связываться с sLea, как описано в настоящем документе. Специфическое связывание с sLea может включать специфичность, аффинность и/или авидность, как представлено в примере I, для любого из антител, представленных в настоящем описании. В другом аспекте антитело или его функциональный фрагмент, кодируемые полинуклеотидами по изобретению, могут включать активность, связанную с зависимой от комплемента цитотоксичностью (CDC), и/или активность, связанную с зависимой от антител клеточно-опосредуемой цитотоксичностью (ADCC) любого из изолятов клонов 5B1, 9H3, 5H11 или 7Е3, описанных в настоящем документе. Методы оценки специфичности, аффинности и/или авидности антитела или его функционального фрагмента хорошо известны в данной области техники, и иллюстративные методы приведены в данном описании.
CDRs изолятов клонов
(SEQ ID NO:)
(SEQ ID NO:)
(NO: 1)
(NO: 1)
(NO: 1)
(NO: 2)
(NO: 2)
(NO: 3)
(NO: 3)
(NO: 3)
(NO: 4)
(NO: 4)
(NO: 5)
(NO: 5)
(NO: 5)
(NO: 6)
(NO: 6)
(NO: 7)
(NO: 7)
(NO: 7)
(NO: 8)
(NO: 8)
(NO: 9)
(NO: 9)
(NO: 9)
(NO: 10)
(NO: 10)
(NO: 11)
(NO: 11)
(NO: 11)
(NO: 12)
(NO: 12)
(NO: 13)
(NO: 13)
(NO: 13)
(NO: 13)
(NO: 13)
(NO: 15)
(NO: 15)
(NO: 15)
(NO: 16)
(NO: 16)
В некоторых вариантах осуществления антитело или его функциональный фрагмент по изобретению включает менее шести CDRs. В некоторых вариантах осуществления антитело или его функциональный фрагмент включает одну, две, три, четыре или пять CDRs, выбранных из группы, состоящей из VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и/или VL CDR3. В конкретных вариантах осуществления антитело или его функциональный фрагмент включает одну, две, три, четыре или пять CDRs, выбранных из группы, состоящей из VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и/или VL CDR3 изолятов клонов 5B1, 9H3, 5H11 или 7Е3, описанных в настоящем документе.
В некоторых вариантах осуществления в настоящем изобретении предлагается выделенный полинуклеотид, который кодирует антитело или его функциональный фрагмент, где антитело или его функциональный фрагмент включает вариабельный домен тяжелой (VH) цепи, имеющий аминокислотные последовательности CDR1, CDR2 и CDR3, изолятов клонов 5B1, 9H3, 5H11 или 7Е3. Такие домены VH могут включать аминокислотные остатки 55-62, 70-77 и 116-131 SEQ ID NO: 2 или, альтернативно, аминокислотные остатки 45-52, 70-77 и 116-131 SEQ ID NO: 6, или, альтернативно, аминокислотные остатки 45-52, 70-77 и 116-131 SEQ ID NO: 10, или, альтернативно, аминокислотные остатки 45-52, 70-77 и 116-134 SEQ ID NO: 14. В другом аспекте нуклеотидная последовательность, кодирующая CDR1, CDR2 и CDR3 домена VH, может соответственно включать нуклеотидную последовательность остатков 133-156, 208-231 и 346-393 SEQ ID NO: 1, или, альтернативно, нуклеотидную последовательность остатков 133-156, 208-231 и 346-393 SEQ ID NO: 5, или, альтернативно, нуклеотидную последовательность остатков 133-156, 208-231 и 346-393 SEQ ID NO: 9, или, альтернативно, нуклеотидную последовательность остатков 133-156, 208-231, 346-402 SEQ ID NO: 13.
В другом варианте осуществления в настоящем изобретении предлагается выделенный полинуклеотид, кодирующий антитело или его функциональный фрагмент, где антитело включает вариабельный домен легкой (VL) цепи, имеющий аминокислотную последовательность CDR1, CDR2 и CDR3 изолятов клонов 5B1, 9H3, 5H11 или 7Е3. Такой домен VL может включать аминокислотные остатки 45-52, 70-72 и 109-120 SEQ ID NO: 4, или, альтернативно, аминокислотные остатки 45-52, 70-72 и 109-119 SEQ ID NO: 8, или, альтернативно, аминокислотные остатки 45-52, 70-72 и 109-120 SEQ ID NO: 12, или, альтернативно, аминокислотные остатки 49-53, 72-74 и 111-120 SEQ ID NO: 16. В другом аспекте нуклеотидная последовательность, кодирующая CDR1, CDR2 и CDR3 домена VH, может соответственно включать нуклеотидную последовательность остатков 133-156, 208-216 и 325-360 SEQ ID NO: 3, или, альтернативно, нуклеотидную последовательность остатков 133-156, 208-216 и 325-357 SEQ ID NO: 7, или, альтернативно, нуклеотидную последовательность остатков 134-156, 208-216 и 325-360 SEQ ID NO: 11, или, альтернативно, нуклеотидную последовательность остатков 145-162, 214-222 и 331-360 SEQ ID NO: 15.
В другом варианте осуществления в настоящем изобретении предлагается вариант полинуклеотидов, представленных в настоящем описании. Вариант при использовании в отношении полинуклеотида включает полинуклеотид, имеющий один или более модифицированных нуклеотидов, таких как, но не ограничиваясь этим, метилированный нуклеотид или нуклеотидный аналог. Кроме того, вариант полинуклеотида может включать полинуклеотид, который прерывается ненуклеотидными компонентами. Модификации полинуклеотида могут быть сделаны до или после сборки полинуклеотида с использованием способов, хорошо известных специалистам в данной области техники. Например, полинуклеотид может быть модифицирован после полимеризации путем конъюгации с меченым компонентом с использованием либо ферментативных, либо химических методов (например, как описано Gottfried and Weinhold, 2011, Biochem. Soc. Trans., 39(2):523-628; Paredes et al., 2011, Methods, 54(2):251-259).
Полинуклеотиды могут быть получены, и нуклеотидная последовательность полинуклеотидов определена любым способом, хорошо известным в данной области техники. Так как аминокислотные последовательности вариабельных доменов тяжелой и легкой цепи 5B1, 9H3, 5H11 и 7Е3 известны (смотри, например, SEQ ID NO: 2, 4, 6, 8, 10, 12, 14 и 16), нуклеотидные последовательности, кодирующие антитела и модифицированные варианты этих антител, могут быть определены с использованием способов, хорошо известных в данной области техники, т.е. кодоны нуклеотидов, известные как кодирующие конкретные аминокислоты, составляют таким образом, чтобы создать нуклеиновую кислоту, которая кодирует антитело. Такой полинуклеотид, кодирующий антитело, может быть составлен из химически синтезированных олигонуклеотидов (например, как описано Kutmeier et al., 1994, BioTechniques 17:242), что вкратце включает синтез перекрывающихся олигонуклеотидов, содержащих части последовательности, кодирующей антитело, его фрагменты или варианты, отжиг и лигирование этих олигонуклеотидов, а затем амплификацию лигированных олигонуклеотидов с помощью ПЦР.
Полинуклеотид, кодирующий антитело или его функциональный фрагмент по изобретению, может быть получен с использованием последовательности нуклеиновой кислоты вариабельных доменов тяжелой и/или легкой цепи изолятов 5B1, 9H3, 5Н11 или 7Е3 (например, SEQ ID NO: 1, 3, 5, 7, 9, 11, 13 и 15). Нуклеиновая кислота, кодирующая антитело или его функциональный фрагмент, может быть химически синтезирована или получена из подходящего источника (например, кДНК, выделенной из клеток, экспрессирующих антитело или его функциональный фрагмент, таких как клетки гибридомы, выбранные как экспрессирующие антитело или его функциональный фрагмент) с помощью ПЦР-амплификации с использованием синтетических праймеров, гибридизуемых с 3'- и 5'-концами последовательности, или с помощью клонирования с использованием олигонуклеотидного зонда, специфичного для конкретной последовательности нуклеиновой кислоты. Амплифицированные нуклеиновые кислоты, полученные с помощью ПЦР, могут быть затем клонированы в реплицируемые клонирующие векторы, с использованием любого способа, хорошо известного в данной области техники.
В некоторых вариантах осуществления в настоящем изобретении предлагается выделенное антитело или его функциональный фрагмент, где антитело связывается с sLea. Соответственно, в некоторых аспектах настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, которые связываются с sLea, где антитело или его функциональный фрагмент включает домен VH, имеющий аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-142 SEQ ID NO: 2, остатков 20-142 SEQ ID NO: 6, остатков 20-142 SEQ ID NO: 10 и остатков 20-145 SEQ ID NO: 14.
В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, которые связываются с sLea, где антитело или его функциональный фрагмент включает домен VL, имеющий аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-130 SEQ ID NO: 4, остатков 20-129 SEQ ID NO: 8, остатков 20-130 SEQ ID NO: 12 и остатков 23-130 SEQ ID NO: 16.
В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, которые связываются с sLea, где антитело или его функциональный фрагмент включает как домен VH, так и домен VL, где домен VH и домен VL соответственно включают аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-142 SEQ ID NO: 2 и остатков 20-130 SEQ ID NO: 4; остатков 20-142 SEQ ID NO: 6 и остатков 20-129 SEQ ID NO: 8; остатков 20-142 SEQ ID NO: 10 и остатков 20-130 SEQ ID NO: 12; и остатков 20-145 SEQ ID NO: 14 и остатков 23-130 SEQ ID NO: 16.
В некоторых вариантах осуществления для того, чтобы связать sLea, антитело или его функциональный фрагмент по изобретению имеет одну или более CDRs, изображенных на фиг. 1-8 или приведенных в таблице 2. Антитело или его функциональный фрагмент, которые включают одну или более CDRs, в частности CDR3, могут специфически связываться с sLea, как описано в настоящем документе. Специфическое связывание с sLea может включать специфичность и сродство, как это представлено в примере I, для любого из антител, предлагаемых в настоящем описании. В некоторых аспектах антитело или его функциональный фрагмент по настоящему изобретению может включать активность CDC и/или активностью ADCC любого из изолятов клонов 5B1, 9H3, 5H11 или 7Е3, описанных в настоящем документе.
В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, где антитело включает домен цепи VH, имеющий аминокислотную последовательность CDR1, CDR2 и CDR3 изолята клонов 5B1, 9H3, 5H11 или 7E3. Такие домены VH могут включать аминокислотные остатки 55-62, 70-77 и 116-131 SEQ ID NO: 2 или, альтернативно, аминокислотные остатки 45-52, 70-77 и 116-131 SEQ ID NO: 6, или, альтернативно, аминокислотные остатки 45-52, 70-77 и 116-131 SEQ ID NO: 10, или, альтернативно, аминокислотные остатки 45-52, 70-77 и 116-134 SEQ ID NO: 14.
В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его функциональному фрагменту, где антитело включает домен цепи VL, имеющий аминокислотную последовательность CDR1, CDR2 и CDR3 изолята клонов 5B1, 9H3, 5H11 или 7E3. Такой домен VL может включать аминокислотные остатки 45-52, 70-72 и 109-120 SEQ ID NO: 4, или, альтернативно, аминокислотные остатки 45-52, 70-72 и 109-119 SEQ ID NO: 8, или, альтернативно, аминокислотные остатки 45-52, 70-72 и 109-120 SEQ ID NO: 12, или, альтернативно, аминокислотные остатки 49-53, 72-74 и 111-120 SEQ ID NO: 16.
В некоторых аспектах изобретения выделенное антитело или его функциональный фрагмент представляет собой моноклональное антитело. В некоторых аспектах изобретения выделенное антитело или его функциональный фрагмент, предлагаемые в настоящем описании, представляют собой изотип IgG или IgM. В еще одном аспекте настоящего изобретения антитело или его функциональный фрагмент представляет собой антитело подкласса IgG1.
В некоторых вариантах осуществления функциональный фрагмент антитела согласно изобретению может представлять собой, но не ограничивается этим, Fab, Fab', F(аb')2, Fabc, scFv, диатело, триатело, минитело или однодоменное антитело (sdAB). В некоторых аспектах в настоящем изобретении предлагается диатело, которое включает аминокислотную последовательность SEQ ID NO: 18 или 20. Такие диатела по настоящему изобретению в некоторых аспектах могут кодироваться полинуклеотидом, имеющим последовательность нуклеиновой кислоты SEQ ID NO: 17 или 19. Что касается антител и их функциональных фрагментов, различных форм, изменений и модификаций, они хорошо известны в данной области техники. Специфичные для sLea фрагменты антитела по настоящему изобретению могут включать любую из таких различных форм, изменений и модификаций антитела. Примеры таких различных форм и терминов, в том виде как они известны в данной области техники, приведены ниже.
В некоторых вариантах осуществления настоящее изобретение относится к способу получения антитела или его функционального фрагмента по изобретению. Способ по настоящему изобретению может включать введение полинуклеотида по изобретению в клетку-хозяина, культивирование клетки-хозяина в условиях и в течение периода времени, достаточного для продукции кодируемой тяжелой и/или легкой цепи антитела или его функционального фрагмента по изобретению, и очистку тяжелой и/или легкой цепи антитела или его функционального фрагмента.
Рекомбинантная экспрессия антитела или его функционального фрагмента по изобретению, которые связываются с антигеном sLea, может включать конструирование экспрессионного вектора, содержащего полинуклеотид, который кодирует тяжелую и/или легкую цепь антитела или его функционального фрагмента по изобретению. После того, как полинуклеотид, кодирующий антитело или его функциональный фрагмент (предпочтительно, но не обязательно, содержащий вариабельный домен тяжелой и/или легкой цепи) по настоящему изобретению, получен, может быть получен вектор для продукции антитела или его функционального фрагмента методом рекомбинантной ДНК с использованием методов, хорошо известных в данной области техники. Способы получения белка путем экспрессии полинуклеотида, содержащего нуклеотидную последовательность, кодирующую антитело или его функциональный фрагмент, описаны в данном документе.
Методы, которые хорошо известны специалистам в данной области техники, могут быть использованы для конструирования экспрессионных векторов, содержащих кодирующие последовательности антитела или его функциональных фрагментов и подходящие сигналы, контролирующие транскрипцию и трансляцию. Эти методы включают, например, методы рекомбинантных ДНК in vitro, методы синтеза и генетической рекомбинации in vivo. В изобретении, таким образом, предлагаются реплицируемые векторы, включающие нуклеотидную последовательность, кодирующую антитело или его функциональный фрагмент по изобретению, функционально связанную с промотором. Такие векторы могут включать нуклеотидную последовательность, кодирующую константную область молекулы антитела (смотри, например, международные публикации Nos. WO 86/05807 и WO 89/01036; и патент США No. 5122464), и вариабельный домен антитела может быть клонирован в такой вектор для экспрессии всей тяжелой, всей легкой цепи, или как всей тяжелой, так и всей легкой цепей.
Экспрессионный вектор может быть введен в клетку-хозяина с помощью обычных методов, и трансфецированные клетки затем культивируют обычными методами для получения антитела или его функционального фрагмента по настоящему изобретению. Таким образом, изобретение включает клетки-хозяева, содержащие полинуклеотид, кодирующий антитело или его функциональный фрагмент по изобретению, функционально связанный с гетерологичным промотором. В некоторых вариантах осуществления для экспрессии двухцепочечных антител, векторы, кодирующие как тяжелую, так и легкую цепи, могут совместно экспрессироваться в клетке-хозяине для экспрессии всей молекулы иммуноглобулина, как подробно описано ниже.
Разнообразные экспрессирующиеся в хозяине векторные системы могут быть использованы для экспрессии антитела или его функциональных фрагментов по изобретению (смотри, например, патент США No. 5807715). Такие экспрессирующиеся в хозяине системы представляют собой носители, с помощью которых кодирующие последовательности, представляющие интерес, могут быть получены и затем очищены, но также представляют собой клетки, которые могут при трансформации или трансфекции подходящими кодирующими нуклеотидными последовательностями экспрессировать молекулу антитела согласно изобретению in situ. Они включают, но не ограничиваются этим, микроорганизмы, такие как бактерии (например, E. coli и B. subtilis), трансформированные рекомбинантной ДНК бактериофага, экспрессионные векторы с плазмидной ДНК или космидной ДНК, содержащие кодирующие последовательности антитела; дрожжи (например, Saccharomyces Pichia), трансформированные рекомбинантными экспрессионными векторами дрожжей, содержащими кодирующие последовательности антитела; системы клеток насекомых, инфицированных рекомбинантными вирусными (например, бакуловирусными) экспрессионными векторами, содержащими кодирующие последовательности антитела; системы растительных клеток, инфицированных рекомбинантными вирусными (например, вирусом мозаики цветной капусты, CaMV; вирусом табачной мозаики, ВТМ) экспрессионными векторами или трансформированные рекомбинантными плазмидными экспрессионными векторами (например, плазмидой Ti), содержащие кодирующие последовательности антитела; или системы клеток млекопитающих (например, клеток COS, CHO, BHK, 293, NS0 и 3Т3), несущих рекомбинантные экспрессионные конструкции, содержащие промоторы, происходящие из генома клеток млекопитающих (например, промотор металлотионеина) или из вирусов млекопитающих (например, поздний промотор аденовируса, промотор вируса коровьей оспы 7.5K). В некоторых аспектах для экспрессии рекомбинантного антитела или его функционального фрагмента, особенно для экспрессии полного рекомбинантного антитела, используются бактериальные клетки, такие как Escherichia coli, или эукариотные клетки. Например, клетки млекопитающих, такие как клетки яичника китайского хомячка (СНО), в сочетании с вектором, таким как главный промоторный элемент промежуточного раннего гена цитомегаловируса человека, являются эффективной системой экспрессии антител (Foecking et al., 1986, Gene 45:101; и Cockett et al., 1990, Bio/Technology 8:2). В некоторых вариантах осуществления антитела или их фрагменты по изобретению получают в клетках СНО. В одном варианте осуществления экспрессия нуклеотидных последовательностей, кодирующих антитела или их функциональные фрагменты по изобретению, которые связываются с sLea, регулируется конститутивным промотором, индуцируемым промотором или тканеспецифическим промотором.
В бактериальных системах ряд экспрессионных векторов может быть преимущественно выбран в зависимости от предполагаемого использования молекулы антитела, предназначенной для экспрессии. Например, когда планируется продукция большого количества такого антитела для создания фармацевтических композиций молекулы антитела, могут быть желательны векторы, которые управляют высокими уровнями экспрессии гибридных белковых продуктов, которые легко очищать. Такие векторы включают, но не ограничиваются этим, экспрессионный вектор pUR278 E. coli (Ruther et al., 1983, EMBO 12:1791), в котором кодирующая последовательность антитела может быть лигирована индивидуально в вектор в рамке с кодирующей областью lac Z таким образом, что образуется гибридный белок; векторы pIN (Inouye & Inouye, 1985, Nucleic Acids Res. 13:3101-3109; Van Heeke & Schuster, 1989, J. Biol. Chem. 24:5503-5509); и тому подобное. Векторы pGEX также могут быть использованы для экспрессии чужеродных полипептидов в виде гибридных белков с глутатион-S-трансферазой (GST). В целом такие гибридные белки являются растворимыми и могут быть легко очищены от лизированных клеток путем адсорбции и связывания с матриксом глутатион-агарозных шариков с последующим элюированием в присутствии свободного глутатиона. Векторы pGEX созданы для включения тромбина или сайтов расщепления протеазой фактора Xa таким образом, чтобы клонированный продукт гена-мишени мог высвобождаться от части GST.
В системе насекомых вирус ядерного полиэдроза Autographa californica (AcNPV) используют в качестве вектора для экспрессии чужеродных генов. Вирус растет в клетках Spodoptera frugiperda. Кодирующая последовательность антитела или его функционального фрагмента может быть клонирована индивидуально в несущественных областях (например, гена полиэдрина) вируса и помещена под контроль промотора AcNPV (например, промотора полиэдрина).
В клетках-хозяевах млекопитающих может быть использован ряд экспрессионных систем на основе вирусов. В тех случаях, когда в качестве экспрессионного вектора используют аденовирус, кодирующую последовательность представляющего интерес антитела можно лигировать с аденовирусным комплексом, контролирующим транскрипцию/трансляцию, например, поздним промотором и трехраздельной лидирующей последовательностью. Этот гибридный ген затем можно вставить в геном аденовируса путем рекомбинации in vitro или in vivo. Вставка в несущественную область вирусного генома (например, в область E1 или Е3) должна привести к получению рекомбинантного вируса, который является жизнеспособным и способен экспрессировать молекулу антитела в инфицированных хозяевах (например, смотри Logan & Shenk, 1984, Proc. Natl. Acad. Sci. USA 8 1:355-359). Специфические сигналы инициации также могут быть использованы для эффективной трансляции вставленных кодирующих последовательностей антител. Эти сигналы включают инициирующий кодон ATG и смежные последовательности. Кроме того, инициирующий кодон должен совпадать по фазе с рамкой считывания желаемой кодирующей последовательности для обеспечения трансляции всей вставки. Эти экзогенные сигналы контроля трансляции и кодоны инициации могут иметь различное происхождение, как природное, так и синтетическое. Эффективность экспрессии может быть повышена за счет включения соответствующих элементов повышения транскрипции, терминации транскрипции и т.п. (смотри, например, Bittner et al., 1987, Methods in Enzymol. 153:51-544).
Кроме того, может быть выбран штамм клеток-хозяев, который модулирует экспрессию вставленных последовательностей или модифицирует и осуществляет процессинг генного продукта конкретным желаемым способом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важны для функционирования антитела или его функционального фрагмента. Различные клетки-хозяева имеют характерные и специфические механизмы посттрансляционного процессинга и модификации белков и генных продуктов. Подходящие клеточные линии или системы хозяев могут быть выбраны для обеспечения корректной модификации и процессинга экспрессируемого чужеродного белка. С этой целью могут быть использованы эукариотные клетки-хозяева, которые обладают клеточным механизмом для правильного процессинга первичного транскрипта, гликозилирования и фосфорилирования генного продукта. Такие клетки-хозяева млекопитающих включают, но не ограничиваются этим, клетки CHO, VERY, ВНК, HeLa, COS, MDCK, 293, 3Т3, W138, BT483, HS578T, HTB2, BT2O и T47D, NS0 (клеточная линия миеломы мыши, которая эндогенно не продуцирует какие-либо цепи иммуноглобулинов), CRL7030 и клетки HsS78Bst.
Для длительной продукции с высоким выходом рекомбинантных белков предпочтительной является стабильная экспрессия. Например, могут быть сконструированы клеточные линии, которые стабильно экспрессируют антитело или его функциональный фрагмент по изобретению. Вместо использования экспрессионных векторов, которые содержат вирусные сайты инициации репликации, клетки-хозяева могут быть трансформированы ДНК под контролем соответствующих контролирующих экспрессию элементов (например, промотора, энхансера, последовательностей, терминаторов транскрипции, сайтов полиаденилирования и т.д.), а также маркера селекции. После введения чужеродной ДНК сконструированным клеткам можно давать расти в течение 1-2 дней в обогащенной среде, а затем пересевать на селективную среду. Маркер селекции в рекомбинантной плазмиде придает устойчивость к отбору и позволяет клеткам стабильно интегрировать плазмиду в свои хромосомы и расти с образованием фокусов, которые, в свою очередь, могут быть клонированы и размножены в клеточные линии. Этот метод с успехом может быть использован для конструирования клеточных линий, которые экспрессируют молекулу антитела.
Может быть использован ряд систем селекции, включая, но, не ограничиваясь этим, гены тимидинкиназы вируса простого герпеса (Wigler et al., 1977, Cell 11:223), гипоксантингуанинфосфорибозилтрансферазы (Szybalska & Szybalski, 1992, Proc. Natl. Acad. Sci. USA 48:202) и аденинфосфорибозилтрансферазы (Lowy et al., 1980, Cell 22:8-17), которые могут быть использованы в tk-, hgprt- или Aprt-клетках, соответственно. Кроме того, может быть использована устойчивость к антиметаболитам в качестве основы селекции по следующим генам: dhfr, который придает устойчивость к метотрексату (Wigler et al., 1980, Proc. Natl. Acad. Sci. U S A. 77(6):3567-70; O’Hare et al., 1981, Proc. Natl. Acad. Sci. USA 78:1527); глутаминсинтетазе (GS), которая представляет собой фермент, ответственный за биосинтез глутамина с использованием глутамата и аммиака (Bebbington et al., 1992, Biotechnology 10:169); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan & Berg, 1981, Proc. Natl. Acad. Sci. USA 78:2072); neo, который придает устойчивость к аминогликозиду G-418 (Wu and Wu, 1991, Biotherapy 3:87-95; Tolstoshev, 1993, Ann. Rev. Pharmacol. Toxicol. 32:573-596; Mulligan, 1993, Science 260:926-932; и Morgan and Anderson, 1993, Ann. Rev. Biochem. 62:191-217; May, 1993, TIB TECH 11(5):155-215); и hygro, который придает устойчивость к гигромицину (Santerre et al., 1984, Gene 30:147). Методы, хорошо известные в области технологии рекомбинантной ДНК, могут обычно применяться для выбора желаемого рекомбинантного клона, и такие методы описаны, например, в Ausubel et al. (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, NY (1993); Kriegler, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY (1990); и в главах 12 и 13, Dracopoli et al. (eds.), Current Protocols in Human Genetics, John Wiley & Sons, NY (1994); Colberre-Garapin et al., 1981, J. Mol. Biol. 150:1, которые включены в настоящее описание в качестве ссылки в полном объеме.
Уровни экспрессии молекулы антитела могут быть увеличены с помощью амплифицирующего вектора (для обзора смотри Bebbington and Hentschel, The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells in DNA cloning, Vol. 3 (Academic Press, New York, 1987)). Когда маркер в векторной системе экспрессии антитела или его функционального фрагмента является амплифицируемым, увеличение уровня ингибитора, присутствующего в культуре клетки-хозяина, будет увеличивать количество копий маркерного гена. Поскольку амплифицируемая область связана с геном антитела, продукция антитела также будет увеличиваться (Crouse et al., 1983, Mol. Cell. Biol. 3:257).
Клетка-хозяин может быть котрансфецирована двумя экспрессионными векторами по изобретению, первым вектором, кодирующим полипептид, происходящий от тяжелой цепи, и вторым вектором, кодирующим полипептид, происходящий от легкой цепи. Эти два вектора могут содержать идентичные маркеры селекции, которые создают возможность равной экспрессии полипептидов тяжелой и легкой цепей. Альтернативно, может быть использован один вектор, который кодирует и способен экспрессировать полипептиды как тяжелой, так и легкой цепей. В таких ситуациях легкая цепь может быть помещена перед тяжелой цепью, чтобы избежать избыточной токсичности свободной тяжелой цепи (Proudfoot, 1986, Nature 322:52; и Kohler, 1980, Proc. Natl. Acad. Sci. USA 77:2197-2199). Кодирующие последовательности тяжелой и легкой цепей могут включать кДНК или геномную ДНК.
Кроме того, полинуклеотиды, кодирующие тяжелую и/или легкую цепь антитела или его функционального фрагмента по изобретению, могут быть подвергнуты оптимизации кодонов с использованием методов, хорошо известных в данной области техники, для достижения оптимизированной экспрессии антитела или его функционального фрагмента по изобретению в желаемой клетке-хозяине. Например, в одном из способов оптимизации кодонов нативный кодон заменяют наиболее часто встречающимся кодоном из эталонного набора генов, где создается высокая скорость трансляции кодонов для каждой аминокислоты. Дополнительные иллюстративные способы создания кодон-оптимизированных полинуклеотидов для экспрессии желаемого белка, которые могут быть применены к тяжелой и/или легкой цепи антитела или его функционального фрагмента по изобретению, описаны в статьях Kanaya et al., Gene, 238:143-155 (1999), Wang et al., Mol. Biol. Evol., 18(5):792-800 (2001), патенте США 5795737, публикации США 2008/0076161 и WO 2008/000632.
После того, как молекула антитела по настоящему изобретению получена путем рекомбинантной экспрессии, она может быть очищена любым способом, известным в данной области техники для очистки молекулы иммуноглобулина, например, с помощью хроматографии (например, ионообменной, аффинной, особенно с аффинностью для специфического антигена после протеина А, и с помощью колоночной хроматографии с разделением по размеру), с помощью центрифугирования, дифференциальной растворимости или с помощью любого другого стандартного метода очистки белков. Кроме того, для облегчения очистки антитела или их функциональные фрагменты по настоящему изобретению могут быть соединены с гетерологичными полипептидными последовательностями, представленными в настоящем описании, или очищены иным способом, известным в данной области техники. Например, антитело или его функциональный фрагмент по изобретению может быть очищен с помощью рекомбинантного добавления среди прочего полигистидиновой метки (His-тега), FLAG-тега, тега гемагглютинина (HA-тега) или Myc-тега, которые являются коммерчески доступными, и использования методов очистки, хорошо известных специалистам в данной области техники.
Фрагмент Fab относится к одновалентному фрагменту, состоящему из доменов VL, VH, CL и CH1; F(аb')2-фрагмент представляет собой двухвалентный фрагмент, включающий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; фрагмент Fd состоит из доменов VH и CH1; фрагмент Fv состоит из доменов VL и VH одного плеча антитела; и фрагмент dAb (Ward et al., Nature 341:544-546, (1989)), состоит из домена VH.
Антитело может иметь один или несколько сайтов связывания. Если есть более чем один сайт связывания, связывающие сайты могут быть идентичны друг другу или могут отличаться. Например, природный иммуноглобулин имеет два идентичных сайта связывания, одноцепочечное антитело или Fab-фрагмент имеет один сайт связывания, в то время как «биспецифическое» или «бифункциональное» антитело имеет два различных сайта связывания.
Одноцепочечное антитело (scFv), относится к антителу, в котором области VL и VH соединены через линкер (например, синтетическую последовательность аминокислотных остатков) с образованием непрерывной полипептидной цепи, где линкер является достаточно длинным для возможности обратной укладки белковой цепи относительно себя с образованием одновалентного сайта связывания антигена (смотри, например, Bird et al., Science 242:423-26 (1988) и Huston et al., Proc. Natl. Acad. Sci. USA 85:5879-83 (1988)). Диатела относятся к двухвалентным антителам, включающим две полипептидные цепи, где каждая полипептидная цепь включает домены VH и VL, соединенные с помощью линкера, который является слишком коротким для того, чтобы позволить образовывать пары между двумя доменами на одной и той же цепи, что позволяет каждому домену образовывать пару с комплементарным доменом на другой полипептидной цепи (см, например, Holliger et al., Proc. Natl. Acad. Sci. USA 90:6444-48 (1993), и Poljak et al., Structure 2:1121-23 (1994)). Если две полипептидные цепи диатела идентичны, то диатело в результате образования их пар будет иметь два идентичных антигенсвязывающих участка. Полипептидные цепи, имеющие различные последовательности, могут быть использованы для получения диатела с двумя различными антигенсвязывающими сайтами. Сходно триатела и тетратела представляют собой антитела, включающие три и четыре полипептидные цепи, соответственно, и образующие три и четыре антигенсвязывающих сайта, соответственно, которые могут быть одинаковыми или разными.
Настоящее изобретение также относится к антителу или его функциональному фрагменту, происходящим от 5B1, 9H3, 5H11 и/или 7E3, где антитело или его функциональный фрагмент связывается с sLea. Стандартные методы, хорошо известные специалистам в этой области техники, могут быть использованы для введения мутаций в нуклеотидную последовательность, кодирующую антитело или его функциональный фрагмент по изобретению, включая, например, сайт-направленный мутагенез и ПЦР-опосредованный мутагенез, что приводит к аминокислотным заменам. В некоторых аспектах производное включает менее 25 аминокислотных замен, менее 20 аминокислотных замен, менее 15 аминокислотных замен, менее 10 аминокислотных замен, менее 5 аминокислотных замен, менее 4 аминокислотных замен, менее 3 аминокислотных замен или менее 2 аминокислотных замен относительно исходной молекулы.
В некоторых вариантах осуществления настоящее изобретение относится к антителу или его функциональному фрагменту, имеющим модифицированные формы природных аминокислот, консервативные замены, не встречающиеся в природе аминокислоты, аминокислотные аналоги и миметики, пока это антитело или его функциональный фрагмент сохраняет функциональную активность как определено в настоящем описании. В одном варианте осуществления производное имеет консервативные аминокислотные замены, которые сделаны в одном или более предсказанных аминокислотных остатках, не являющихся незаменимыми. Консервативная аминокислотная замена представляет собой замену, в которой аминокислотный остаток заменен на аминокислотный остаток, имеющий боковую цепь со сходным зарядом. Семейства аминокислотных остатков, имеющих боковые цепи со сходными зарядами, были определены в данной области техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Альтернативно, мутации могут быть введены случайным образом вдоль всей или части кодирующей последовательности, например, путем насыщающего мутагенеза, и полученные мутанты могут быть подвергнуты скринингу на биологическую активность с целью идентификации мутантов, которые сохраняют активность. После мутагенеза кодируемое антитело или его функциональный фрагмент может быть экспрессирован, и может быть определена активность антитела или его функционального фрагмента.
В некоторых вариантах осуществления настоящее изобретение относится к антителу или его функциональному фрагменту, имеющим модифицированный фукозилированием, галактозилированием и/или сиалированием фрагмент Fc, содержащийся в антителе или его функциональном фрагменте по изобретению. Такие модификации фрагмента Fc могут влиять на активность, опосредуемую рецептором Fc, как описано Peipp et al., Blood, 112(6):2390-2399 (2008). Например, терапевтические антитела со сконструированным гликозилированием, лишенные коровых остатков фукозы в N-гликанах Fc, обнаруживают сильную ADCC при более низких концентрациях с гораздо более высокой эффективностью по сравнению с фукозилированными аналогами. Shields et al., J. Biol. Chem., 277(30):26733-40 (2002); Okazaki et al., J Mol Biol., 336:1239-1249 (2004); Natsume et al., J. Immunol. Methods., 306:93-103 (2005). Способы модификации фукозилирования, галактозилирования и/или сиалирования антитела или его функционального фрагмента хорошо известны в данной области техники. Например, подходы к дефукозилированию могут быть сгруппированы в три метода (1) переход от пути N-гликозилирования клеток немлекопитающих к «гуманизированному» пути отсутствия фукозилирования; (2) инактивация пути фукозилирования N-гликанов в клетках млекопитающих и (3) химический синтез in vitro нефукозилированного N-гликопротеида или ферментативная модификация N-гликанов до нефукозилированных форм, как это описано Yamane-Ohnuki et al., MAbs., 1(3):230-236 (2009). Понятно, что любой из этих способов или любой другой способ, который хорошо известен в данной области техники, может быть использован для получения антитела или его функционального фрагмента с модифицированным фукозилированием, галактозилированием и/или сиалированием.
Антитела или их функциональные фрагменты по изобретению, которые связываются с sLea, могут быть получены любым способом, известным в данной области техники для синтеза антител, в частности, путем химического синтеза или с помощью методов рекомбинантной экспрессии. В практике настоящего изобретения используются, если не указано иное, обычные методы молекулярной биологии, микробиологии, генетического анализа, рекомбинантной ДНК, органической химии, биохимии, ПЦР, синтеза и модификации олигонуклеотидов, гибридизации нуклеиновых кислот и подобные методы в пределах данной области техники. Эти методы описаны в ссылках, приведенных в данном документе, и полностью описаны в литературе. Смотри, например, руководства Maniatis et al. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press; Sambrook et al. (1989), Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press; Sambrook et al. (2001) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons (1987 и ежегодные обновленные издания); Current Protocols in Immunology, John Wiley & Sons (1987 и ежегодные обновленные издания) Gait (ed.) (1984) Oligonucleotide Synthesis: A Practical Approach, IRL Press; Eckstein (ed.) (1991) Oligonucleotides and Analogues: A Practical Approach, IRL Press; Birren et al. (eds.) (1999) Genome Analysis: A Laboratory Manual, Cold Spring Harbor Laboratory Press; Borrebaeck (ed.) (1995) Antibody Engineering, Second Edition, Oxford University Press; Lo (ed.) (2006) Antibody Engineering: Methods and Protocols (Methods in Molecular Biology); Vol. 248, Humana Press, Inc; каждое из которых включено в настоящее описание в качестве ссылки в полном объеме.
Моноклональные антитела могут быть получены с использованием широкого спектра методов, известных в данной области техники, включая использование гибридомы и рекомбинантных технологий, или их сочетания. Например, моноклональные антитела могут быть получены с использованием гибридомных методов, включая методы, известные в данной области техники и описанные, например, в руководствах Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563 681 (Elsevier, N.Y., 1981), каждое из которых включено в настоящее описание в качестве ссылки в полном объеме. Моноклональное антитело не ограничивается антителами, полученными с помощью гибридомного метода. Другие иллюстративные способы получения моноклональных антител известны в данной области техники. Дополнительные иллюстративные способы получения моноклональных антител приведены в настоящем документе в примере I.
Функциональные фрагменты антител, которые связываются с sLea, могут быть получены любым способом, хорошо известным специалистам в данной области техники. Например, Fab и F(аb')2-фрагменты по изобретению могут быть получены путем протеолитического расщепления молекул иммуноглобулина с использованием ферментов, таких как папаин (для получения Fab-фрагментов) или пепсин (для получения F(аb')2-фрагментов). F(аb')2-фрагменты содержат вариабельную область, константную область легкой цепи и домен CH1 тяжелой цепи.
Функциональные фрагменты антител по изобретению также могут быть получены с использованием различных способов фагового дисплея, известных в данной области техники. Например, в методах фагового дисплея функциональные домены антител, такие как вариабельные области тяжелой и/или легкой цепи, имеющие одну, две, три, четыре, пять или шесть CDRs, предлагаемые в настоящем описании, экспонируются на поверхности фаговых частиц, которые несут кодирующие их полинуклеотидные последовательности. ДНК, кодирующую домены VH и VL, подвергают рекомбинации вместе с линкером scFv с помощью ПЦР и клонируют в фагемидный вектор. Вектор электропорируют в E. coli, и E. coli инфицируют хелперным фагом. Фаги, используемые в этих методах, обычно представляют собой нитчатые фаги, включая fd и M13, и домены VH и VL обычно рекомбинантно соединяют либо с геном III, либо с геном VIII фага. Фаг, экспрессирующий антигенсвязывающий домен, который связывается с конкретным антигеном, таким как sLea, может быть выбран или идентифицирован с помощью антигена, например, с использованием меченого антигена или антигена, связанного или захваченного твердой поверхностью или шариком. Примеры способов фагового дисплея, которые могут быть использованы для получения функциональных фрагментов антитела по настоящему изобретению, включают способы, которые раскрыты в Brinkman et al., 1995, J. Immunol. Methods 182:41-50; Ames et al., 1995, J. Immunol. Methods 184:177-186; Kettleborough et al., 1994, Eur. J. Immunol. 24:952-958; Persic et al., 1997, Gene 187:9-18; Burton et al., 1994, Advances in Immunology 57:191-280; заявке РСТ PCT/GB91/01134; международных публикациях Nos. WO 90/02809, WO 91/10737, WO 92/01047, WO 92/18619, WO 93/1 1236, WO 95/15982, WO 95/20401 и W097/13844; и патентах США Nos. 5698426, 5223409, 5403484, 5580717, 5427908, 5750753, 5821047, 5571698, 5427908, 5516637, 5780225, 5658727, 5733743 и 5969108, каждый из которых включен в настоящее описание в качестве ссылки в полном объеме.
Как описано в приведенных выше ссылках, после фаговой селекции кодирующие области антитела из фага могут быть выделены и использованы для создания полных антител, включая антитела человека, или любого другого желаемого антигенсвязывающего фрагмента, и экспрессированы в любом желаемом хозяине, включая клетки млекопитающих, клетки насекомых, клетки растений, дрожжи и бактерии, например, как описано в настоящем описании.
Могут также применяться методы рекомбинантного получения Fab, Fab' и F(аb')2-фрагментов с использованием способов, известных в данной области техники, таких как раскрытые в публикации РСТ WO 92/22324; Mullinax et al., 1992, BioTechniques 12(6):864-869; Sawai et al., 1995, AJRI 34:26-34; and Better et al., 1988, Science 240:1041-1043, каждая из которых включена в качестве ссылки в полном объеме.
Для создания полных антител ПЦР-праймеры, включающие нуклеотидные последовательности VH или VL, сайт рестрикции, а также фланкирующие последовательности для защиты сайта рестрикции, могут быть использованы для амплификации последовательностей VH или VL в клонах scFv. При использовании методов клонирования, хорошо известных специалистам в данной области техники, ПЦР-амплифицированные домены VH могут быть клонированы в векторы, экспрессирующие константную область VH, например, константную область гамма 1 человека, и ПЦР-амплифицированные домены VL могут быть клонированы в векторы, экспрессирующие константную область VL, например, константные области каппа или лямбда человека. Домены VH и VL могут быть также клонированы в один вектор, экспрессирующий необходимые константные области. Векторы трансформации тяжелой цепи и векторы трансформации легкой цепи затем котрансфецируют в клеточные линии, чтобы создать стабильные или транзиторные клеточные линии, которые экспрессируют полноразмерные антитела, например, IgG, с использованием методов, хорошо известных специалистам в данной области техники.
В некоторых вариантах осуществления антитело или его функциональный фрагмент по изобретению конъюгируют (с помощью ковалентной или нековалентной конъюгации) или рекомбинантным способом соединяют с одним или более диагностическим агентом, детектируемым агентом или терапевтическим агентом или любой другой желаемой молекулой. Конъюгированное или соединенное рекомбинантно антитело или его функциональный фрагмент могут быть полезны для мониторинга или диагностики начала, развития, прогрессии и/или тяжести заболевания, связанного с экспрессией sLea, такого как развитие рака или опухоли, в качестве части клинической процедуры тестирования, такой как определение эффективности конкретного вида терапии.
Обнаружение и диагностика может быть выполнена, например, путем связывания антитела или его функционального фрагмента по изобретению с детектируемыми веществами, включая, но, не ограничиваясь этим, радиоактивные вещества, такие как, но, не ограничиваясь этим, цирконий (89Zr), йод (131I, 125I, 124I, 123I и 121I), углерод (14C, 11C), сера (35S), тритий (3H), индий (115In, 113In, 112In и 111In), технеций (99Tc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (133Xe), фтор (18F), 15O, 13N, 64Cu, 94mТс, 153Sm, 177Lu, 159Gd, 149Pm, 140La, 175Yb, 166Хо, 86Y, 90Y, 47Sc, 186Re, 188Re, 142Pr, 105Rh, 97Ru, 68Ge, 57Co, 65Zn, 85Sr, 32P, 153Gd, 169Yb, 51Cr, 54Mn, 75Se, 113Sn и 117Sn; и излучающие позитроны металлы при использовании различных вариантов позитронно-эмиссионной томографии, различные ферменты, такие как, но, не ограничиваясь этим, пероксидаза хрена, щелочная фосфатаза, бета-галактозидаза или ацетилхолинэстераза; простетические группы, такие как, но, не ограничиваясь этим, стрептавидин/биотин и авидин/биотин; флуоресцентные вещества, такие как, но, не ограничиваясь этим, умбеллиферон, флуоресцеин, флуоресцеина изотиоцианат, родамин, флуоресцеина дихлортриазиниламин, дансилхлорид или фикоэритрин; люминесцентные вещества, такие как, но, не ограничиваясь этим, люминол; биолюминесцентные вещества, такие как, но, не ограничиваясь этим, люцифераза, люциферин и экворин, и нерадиоактивные ионы парамагнитных металлов.
Настоящее изобретение дополнительно охватывает терапевтическое применение антитела или его функционального фрагмента по изобретению, конъюгированного (с помощью ковалентного или нековалентного конъюгирования) или соединенного рекомбинантным способом, с одним или более терапевтическим агентом. В этом контексте, например, антитело может быть конъюгировано или рекомбинантным способом соединено с терапевтическим агентом, таким как цитотоксин, например, цитостатический или цитотоксический агент, или с ионом радиоактивного металла, например, с альфа-излучателем. Цитотоксин или цитотоксический агент включает любой агент, который является вредным для клеток. Терапевтический агент может представлять собой химиотерапевтический агент, такой как, но не ограничиваясь этим, антрациклин (например, доксорубицин и даунорубицин (ранее дауномицин)); таксан (например, паклитаксел (таксол) и доцетаксел (таксотер), антиметаболит (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил и декарбазин), или алкилирующий агент (например, мехлорэтамин, тиоэпа-хлорамбуцил, мелфалан, кармустин (BCNU), ломустин (CCNU), циклофосфамид, бусульфан, дибромоманнит, стрептозотоцин, митомицин С, цисдихлордиамин платины (II), (DDP) и цисплатин); антибиотик (например, актиномицин D, блеомицин, митрамицин, и антрамицин (AMC)); молекулу ауристатин (например, ауристатин РНЕ, бриостатин 1, соластатин 10, монометилауристатин E (ММАЕ) и монометилауристатин F (MMAF)); гормон (например, глюкокортикоиды, прогестины, андрогены и эстрогены), нуклеозидный аналог (например, гемцитабин), ингибитор фермента репарации ДНК (например, этопозид и топотекан), ингибитор киназы (например, соединение ST1571, также известное как гливек или иматиниба мезилат); цитотоксический агент (например, майтансин, паклитаксел, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин и их аналоги или гомологи, и те соединения, которые описаны в патентах США Nos. 6245759, 6399633, 6383790, 6335156, 6271242, 6242196, 6218410, 6218372, 6057300, 6034053, 5985877, 5958769, 5925376, 5922844, 5911995, 5872223, 5863904, 5840745, 5728868, 5648239, 5587459); ингибитор фарнезилтрансферазы (например, R115777, BMS-214662 и ингибиторы, которые раскрыты, например, в патентах США Nos: 6458935, 6451812, 6440974, 6436960, 6432959, 6420387, 6414145, 6410541, 6410539, 6403581, 6399615, 6387905, 6372747, 6369034, 6362188, 6342765, 6342487, 6300501, 6268363, 6265422, 6248756, 6239140, 6232338, 6228865, 6228856, 6225322, 6218406, 6211193, 6187786, 6169096, 6159984, 6143766, 6133303, 6127366, 6124465, 6124295, 6103723, 6093737, 6090948, 6080870, 6077853, 6071935, 6066738, 6063930, 6054466, 6051582, 6051574 и 6040305); ингибитор топоизомеразы (например, камптотецин, иринотекан, SN-38, топотекан, 9-аминокамптотецин, GG-211 (GI 147211), DX-8951f, IST-622, рубитекан, пиразолакридин, XR-5000, саинтопин, UCE6, UCE1022, TAN-1518A, 1518B TAN, KT6006, KT6528, ED-110, NB-506, ED-110, NB-506, фагаронин, коралин, бета-лапакон и ребеккамицин); агент, связывающийся с малой бороздкой ДНК (например, краситель Hoescht 33342 и краситель Hoechst 33258); ингибиторы аденозиндезаминазы (например, флударабина фосфат и 2-хлордезоксиаденозин); или их фармацевтически приемлемые соли, сольваты, клатраты, или их пролекарства. Терапевтический агент может представлять собой иммунотерапевтический агент, такой как, но, не ограничиваясь этим, цетуксимаб, бевацизумаб, герцептин, ритуксимаб).
Кроме того, антитело или его функциональный фрагмент согласно изобретению могут быть конъюгированы с терапевтическим агентом, таким как ион радиоактивного металла, такой как альфа-излучатель, такой как 213Bi или макроциклические хелаторы, пригодные для конъюгации ионов радиоактивного металла, включая, но не ограничиваясь этим, 131В, 131LU, 131Y, 131Ho, 131Sm; или макроциклический хелатор, такой как 1,4,7,10- тетраазациклододекан-N,N',N',N''’-тетрауксусная кислота (DOTA), которая может быть присоединена к антителу или его функциональному фрагменту через линкерную молекулу. Такие линкерные молекулы, как правило, известны в данной области техники и описаны в Denardo et al., 1998, Clin Cancer Res. 4(10):2483-90; Peterson et al., 1999, Bioconjug. Chem. 10(4):553-7; и Zimmerman et al., 1999, Nucl. Med. Biol. 26(8):943-50.
Кроме того, антитело или его функциональный фрагмент по изобретению могут быть конъюгированы (с помощью ковалентного или нековалентного конъюгирования) или соединены рекомбинантным способом с терапевтическим агентом, который модифицирует данный биологический ответ. Таким образом, терапевтические агенты не следует рассматривать как ограничивающиеся классическими химическими терапевтическими агентами. Например, терапевтический агент может представлять собой белок, пептид или полипептид, обладающий желаемой биологической активностью. Такие белки могут включать, например, токсин (например, абрин, рицин A, экзотоксин Pseudomonas, холерный токсин и дифтерийный токсин); такой белок, как фактор некроза опухоли, γ-интерферон, α-интерферон, фактор роста нервов, тромбоцитарный фактор роста, тканевый активатор плазминогена, апоптотический агент (например, TNF-γ, AIM I, AIM II, Fas-лиганд и VEGF), антиангиогенный агент (например, ангиостатин, эндостатин и компонент пути свертывания, такой как тканевый фактор); модификатор биологической реакции (например, цитокин, такой как гамма-интерферон, интерлейкин-1, интерлейкин-2, интерлейкин-5, интерлейкин-6, интерлейкин-7, интерлейкин-9, интерлейкин-10, интерлейкин-12, интерлейкин-15, интерлейкин-23, гранулоцит-макрофаг-колониестимулирующий фактор и гранулоцит-колониестимулирующий фактор); фактор роста (например, гормон роста) или коагуляторный агент (например, кальций, витамин К, тканевые факторы, такие как, но не ограничиваясь этим, фактор Хагемана (фактор XII), высокомолекулярный кининоген (HMWK), прекалликреин (PK), фактор II коагуляции белков (протромбин), фактор V, XIIa, VIII, XIIIa, XI, XIa, IX, IXa, X, фосфолипид и мономер фибрина).
Настоящее изобретение охватывает антитела или их функциональные фрагменты по изобретению, рекомбинантно соединенные или химически конъюгированные (с помощью ковалентной или нековалентной конъюгации) с гетерологичным белком или полипептидом, для создания гибридных белков. В некоторых аспектах такой полипептид может составлять приблизительно 10, приблизительно 20, приблизительно 30, приблизительно 40, приблизительно 50, приблизительно 60, приблизительно 70, приблизительно 80, приблизительно 90 или приблизительно 100 аминокислот в длину. В некоторых аспектах в настоящем изобретении предлагаются гибридные белки, имеющие функциональный фрагмент антитела по изобретению (например, Fab-фрагмент, Fd-фрагмент, Fv-фрагмент, F(аb)2-фрагмент, домен VH, CDR VH, домен VL или CDR VL) и гетерологичный белок или полипептид. В одном варианте осуществления гетерологичный белок или полипептид, с которым соединено антитело или его функциональный фрагмент, служит для направленной доставки антитела или функционального фрагмента к клеткам конкретного типа, таким как клетки, которые экспрессируют sLea.
Конъюгированный или гибридный белок по настоящему изобретению включает любое антитело или его функциональный фрагмент по изобретению, предлагаемый в настоящем описании, конъюгированный (с помощью ковалентной или нековалентной конъюгации) или соединенный рекомбинантным способом с диагностическим агентом, детектируемым агентом или терапевтическим агентом. В одном из вариантов осуществления конъюгированный или гибридный белок по настоящему изобретению включает антитело 5B1, 9H3, 5H11 или 7Е3 и диагностический агент, детектируемый агент или терапевтический агент. В другом варианте осуществления конъюгированный или гибридный белок по настоящему изобретению включает функциональный фрагмент антител 5В1, 9H3, 5H11 или 7Е3 и диагностический агент, детектируемый агент или терапевтический агент. В другом варианте осуществления конъюгированный или гибридный белок по настоящему изобретению включает домен VH, имеющий аминокислотную последовательность любого из доменов VH, представленную остатками 20-142 SEQ ID NO: 2, остатками 20-142 SEQ ID NO: 6, остатками 20-142 SEQ ID NO: 10 или остатками 20-145 SEQ ID NO: 14, и/или домен VL, имеющий аминокислотную последовательность любого из доменов VL, представленную остатками 20-130 SEQ ID NO: 4, остатками 20-129 SEQ ID NO: 8, остатками 20-130 SEQ ID NO: 12 или остатками 23-130 SEQ ID NO: 16, и диагностический агент, детектируемый агент или терапевтический агент. В другом варианте осуществления конъюгированный или гибридный белок по настоящему изобретению включает одну или более CDRs VH, имеющих аминокислотную последовательность любой из CDRs VH, представленных в SEQ ID NO: 2, 6, 10 или 14, и диагностический агент, детектируемый агент или терапевтический агент. В другом варианте осуществления конъюгированный или гибридный белок включает одну или более CDRs VL, имеющих аминокислотную последовательность любой из CDRs VL, представленных в SEQ ID NO: 4, 8, 12 или 16, а также диагностический агент, детектируемый агент или терапевтический агент. В другом варианте осуществления конъюгированный или гибридный белок по настоящему изобретению включает, по меньшей мере, один домен VH и, по меньшей мере, один домен VL, представленные остатками 20-142 SEQ ID NO: 2 и остатками 20-130 SEQ ID NO: 4; остатками 20-142 SEQ ID NO: 6 и остатками 20-129 SEQ ID NO: 8; остатками 20-142 SEQ ID NO: 10 и остатками 20-130 SEQ ID NO: 12; или остатками 20-145 SEQ ID NO: 14 и остатками 23-130 SEQ ID NO: 16, соответственно, и диагностический агент, детектируемый агент или терапевтический агент.
Способы соединения или конъюгации диагностических агентов, детектируемых агентов или терапевтических агентов (включая полипептиды) с антителами хорошо известны, смотри, например Arnon et al., «Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy», in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., «Antibodies For Drug Delivery», in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review», in Monoclonal Antibodies 84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); «Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy», in Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), Thorpe et al., 1982, Immunol. Rev. 62:119-58; патенты США Nos. 5336603, 5622929, 5359046, 5349053, 5447851, 5723125, 5783181, 5908626, 5844095, 5112946, 7981695, 8039273, 8142784; публикации США 2009/0202536, 2010/0034837, 2011/0137017, 2011/0280891, 2012/0003247; EP 307434; EP 367166; EP 394827; публикации РСТ WO 91/06570, WO 96/04388, WO 96/22024, WO 97/34631 и WO 99/04813; Ashkenazi et al., Proc. Natl. Acad. Sci. USA, 88: 10535-10539, 1991; Traunecker et al., Nature, 331:84-86, 1988; Zheng et al., J. Immunol., 154:5590-5600, 1995; Vil et al., Proc. Natl. Acad. Sci. USA, 89:11337-11341, 1992; и Senter, Current Opinion in Chemical Biology, 13:235-244 (2009), которые включены в данное описание в качестве ссылки в полном объеме.
В другом аспекте диагностический агент, детектируемый агент или терапевтический агент может быть присоединен к шарнирной области восстановленного компонента антитела путем образования дисульфидных связей. Альтернативно, такие агенты могут быть присоединены к компоненту антител с использованием гетеробифункционального кросс-линкера, такого как N-сукцинил-3-(2-пиридилдитио)пропионат (SPDP). Yu et al., Int. J. Cancer 56: 244 (1994). Общие методы такой конъюгации хорошо известны в данной области техники. Смотри, например, Wong, CHEMISTRY OF PROTEIN CONJUGATION AND CROSS-LINKING (CRC Press 1991); Upeslacis et al., «Modification of Antibodies by Chemical Methods,» in MONOCLONAL ANTIBODIES: PRINCIPLES AND APPLICATIONS, Birch et al. (eds.), pages 187-230 (Wiley-Liss, Inc. 1995); Price, «Production and Characterization of Synthetic Peptide-Derived Antibodies,» in MONOCLONAL ANTIBODIES: PRODUCTION, ENGINEERING AND CLINICAL APPLICATION, Ritter et al. (eds.), pages 60-84 (Cambridge University Press 1995).
Альтернативно диагностический агент, детектируемый агент или терапевтический агент может быть конъюгирован через углеводную часть в Fc-области антитела. Способы конъюгации пептидов с компонентами антител через углеводную часть антитела хорошо известны специалистам в данной области техники. Смотри, например, Shih et al., Int. J. Cancer. 41:832-839 (1988); Shih et al., Int. J. Cancer. 46:1101-1106 (1990); и Shih et al., патент США No. 5057313, все они включены в качестве ссылки в полном объеме. Общий способ включает взаимодействие компонента антитела, имеющего окисленную часть углевода, с полимером-носителем, который имеет, по меньшей мере, одну свободную аминогруппу и который нагружен множеством молекул пептида. Эта реакция приводит к первоначальному образованию связи основания Шиффа (имина), которая может быть стабилизирована путем восстановления до вторичного амина с образованием конечного конъюгата.
Однако, если Fc-область отсутствует, например, если желателен функциональный фрагмент антитела, как представлено в настоящем описании, все еще возможно присоединить диагностический агент, детектируемый агент или терапевтический агент. Углеводная часть может быть введена в вариабельную область легкой цепи полноразмерного антитела или фрагмента антитела. Смотри, например, Leung et al., J. Immunol., 154: 5919 (1995); патенты США Nos. 5443953 и 6254868, все они включены в качестве ссылки в полном объеме. Сконструированная углеводная часть используется для присоединения диагностического агента, детектируемого агента или терапевтического агента.
Терапевтический агент, конъюгированный или соединенный рекомбинантным способом с функциональным фрагментом антитела согласно изобретению, который связывается с sLea, может быть выбран для достижения желаемого профилактического или терапевтического эффекта(ов). Понятно, что при решении того, какой терапевтический агент конъюгировать или соединять рекомбинантным способом с антителом или его функциональным фрагментом согласно изобретению, в пределах уровня квалификации врача или другого медицинского персонала находится рассмотрение следующего: характера заболевания, тяжести заболевания и состояния индивидуума.
Конъюгат или гибридное антитело или его функциональный фрагмент по изобретению, которые помечены детектируемой меткой, как это предлагается в настоящем документе, и которые связываются с sLea, могут быть использованы в диагностических целях для обнаружения, диагностики или мониторинга заболевания, где клетки, которые вызывают заболевание или связаны с ним, экспрессируют sLea. Например, как представлено в настоящем документе, экспрессируют sLea, как показано, такие раковые клетки и опухоли, как, но, не ограничиваясь этим, опухоли желудочно-кишечного тракта, рак молочной железы, рак яичников, рак толстой кишки, колоректальная аденокарцинома, рак поджелудочной железы, аденокарцинома поджелудочной железы, немелкоклеточный рак легкого, аденокарцинома мочевого пузыря, метастатический рак толстой кишки, колоректальный рак, перстневидноклеточный рак яичников и метастатический рак. Соответственно, настоящее изобретение относится к способам обнаружения злокачественной опухоли или образования опухоли у индивидуума, путем введения эффективного количества конъюгата или гибридного антитела или его функционального фрагмента по изобретению нуждающемуся в этом индивидууму. В некоторых аспектах способ обнаружения может дополнительно включать определение экспрессии sLea на клетках или в образце ткани индивидуума с использованием одного или более конъюгатов или гибридных антител или их функциональных фрагментов согласно изобретению, которые связываются с sLea; и сравнение уровня sLea с контрольным уровнем, например, уровнями в образцах нормальной ткани (например, от индивидуума, не страдающего заболеванием, или от того же индивидуума до начала заболевания), в результате чего повышение определяемого уровня sLea по сравнению с контрольным уровнем sLea свидетельствует о наличии заболевания. Такие диагностические методы могут позволить работникам здравоохранения использовать профилактические меры или агрессивное лечение раньше, чем возможно в противном случае, тем самым предотвращая развитие или дальнейшее прогрессирование заболевания.
Антитело или его функциональный фрагмент по данному изобретению также могут быть использованы для анализа уровней антигена sLea в биологическом образце с использованием классических иммуногистологических методов, как предлагается в настоящем документе, или как хорошо известно специалистам в данной области техники (смотри, например, Jalkanen et al., 1985, J. Cell. Biol. 101:976-985; and Jalkanen et al., 1987, J. Cell. Biol. 105:3087-3096). Другие методы на основе антител, используемые для определения sLea, включают иммунологические методы, такие как твердофазный иммуноферментный анализ (ELISA) и радиоиммуноанализ (RIA). Подходящие метки для анализа антител известны в данной области техники и включают ферментные метки, такие как глюкозооксидаза; радиоизотопы, такие как йод (125I, 121I), углерод (14С), сера (35S), тритий (3Н), индий (121В) и технеций (99Тс); люминесцентные метки, такие как люминол; и флуоресцентные метки, такие как флуоресцеин и родамин, и биотин.
В одном аспекте настоящее изобретение относится к обнаружению и диагностике заболеваний у человека. В одном варианте осуществления диагностика включает: a) введение (например, парентерально, подкожно или внутрибрюшинно) индивидууму эффективного количества конъюгата или гибридного белка по настоящему изобретению, которые связываются с sLea; b) ожидание в течение временного интервала, следующего за введением, для предоставления возможности конъюгату или гибридному белку предпочтительно сконцентрироваться в тех местах индивидуума, где экспрессируется sLea (и, в некоторых аспектах, для того, чтобы несвязанный конъюгат или гибридный белок вымывались до уровня фона); с) определение уровня фона; и d) определение конъюгата или гибридного белка у индивидуума таким образом, что определение конъюгата или гибридного белка выше уровня фона указывает на то, что индивидуум страдает заболеванием. Уровень фона может быть определен различными методами, включая сравнение определяемого количества конъюгата или гибридного белка со стандартным значением, определенным ранее для конкретной системы.
Понятно, что размер индивидуума и используемая система визуализации будут определять величину фрагмента изображения, необходимого для получения диагностических изображений, и это может быть легко определено специалистом в данной области техники. Например, в случае радиоактивного изотопа, конъюгированного с антителом или его функциональным фрагментом по изобретению, для индивидуума, являющегося человеком, количество вводимой радиоактивности обычно находится в пределах от приблизительно 18,5×1010 до 74×1010 мБк 99Тс. Конъюгат должен затем аккумулироваться в месте локализации клеток, которые экспрессируют sLea. Визуализация опухолей in vivo описана в S.W. Burchiel et al., «Immunopharmacokinetics of Radiolabeled Antibodies and Their Fragments.» (Chapter 13 in Tumor Imaging: The Radiochemical Detection of Cancer, S.W. Burchiel and B.A. Rhodes, eds., Masson Publishing Inc. (1982).
В зависимости от нескольких переменных, включая используемый тип детектируемого агента и способ введения, интервал времени после введения для представления возможности конъюгату преимущественно концентрироваться в сайтах индивидуума и для представления возможности несвязанному конъюгату вымыться до фонового уровня составляет от 6 до 48 часов или от 6 до 24 часов или от 6 до 12 часов. В другом варианте осуществления интервал времени после введения составляет от 5 до 20 дней или от 5 до 10 дней. В одном варианте осуществления мониторинг заболевания осуществляется путем повторения метода диагностики, как представлено в данном документе, например, через один месяц после первоначального диагноза, через шесть месяцев после первоначального диагноза, через один год после первоначального диагноза или дольше.
Присутствие конъюгата или гибридного белка может быть обнаружено у индивидуума с использованием способов, известных в данной области техники для сканирования in vivo. Эти способы зависят от типа используемого детектируемого агента. Опытный специалист сможет определить соответствующий способ для обнаружения конкретного детектируемого агента. Способы и устройства, которые могут быть использованы в диагностических способах по изобретению, включают, но не ограничиваются этим, компьютерную томографию (КТ), сканирование всего тела, например, с помощью позитронно-эмиссионной томографии (PET), магнитно-резонансную томографию (МРТ) и эхографию. В одном варианте осуществления антитело или его функциональный фрагмент согласно изобретению конъюгируют с радиоактивным изотопом и детектируют у индивидуума с использованием чувствительного к излучению хирургического инструмента. В другом варианте осуществления антитело или его функциональный фрагмент согласно изобретению конъюгируют с флуоресцентным соединением и детектируют у индивидуума с помощью реагирующего на флуоресценцию сканирующего прибора. В другом варианте осуществления антитело или его функциональный фрагмент согласно изобретению конъюгируют с позитронно-активным металлом, таким как цирконий (99Zr), или с любым другим позитронно-активным металлом, представленным в настоящем описании или хорошо известным в данной области техники как детектируемый с помощью позитронно-эмиссионной томографии, и детектируют у индивидуума с использованием позитронно-эмиссионной томографии. В еще одном варианте осуществления антитело или его функциональный фрагмент согласно изобретению конъюгируют с парамагнитной меткой и детектируют у индивидуума с использованием магнитно-резонансной томографии (МРТ).
В одном варианте осуществления настоящее изобретение относится к фармацевтической композиции, включающей антитело или его функциональный фрагмент по изобретению и фармацевтически приемлемый носитель. Фармацевтически приемлемый носитель, который может быть использован в фармацевтических композициях по настоящему изобретению, включает любой из стандартных фармацевтических носителей, известных в данной области техники, такой как забуференный фосфатом физиологический раствор, вода и эмульсии, такие как эмульсия масла и воды, а также различные типы смачивающих агентов. Эти фармацевтические композиции могут быть получены в жидких единицах лекарственной формы или в любых других единицах лекарственной формы, которые достаточны для доставки антитела или его функционального фрагмента по изобретению к целевой области нуждающегося в лечении индивидуума. Например, фармацевтические композиции могут быть получены любым способом, подходящим для выбранного пути введения, например, внутрисосудистого, внутримышечного, подкожного, внутрибрюшинного и т.д. Другие необязательные компоненты, например, стабилизаторы фармацевтической степени чистоты, буферы, консерванты, наполнители и тому подобное могут быть легко выбраны специалистом в данной области техники. Получение фармацевтической композиции, имеющей должные рН, изотоничность, стабильность и тому подобное, находится в пределах уровня компетентности в данной области техники.
Фармацевтические композиции, содержащие одно или более антител или их функциональных фрагментов по изобретению, предлагаемые в настоящем описании, могут быть получены для хранения путем смешивания антитела, имеющего требуемую степень чистоты, с необязательными физиологически приемлемыми носителями, наполнителями или стабилизаторами (Remington’s Pharmaceutical Sciences (1990) Mack Publishing Co., Easton, PA) в виде лиофилизированных композиций или водных растворов. Приемлемые носители, наполнители или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях и включают буферы, такие как фосфатный, цитратный, и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол и м-крезол); полипептиды низкой молекулярной массы (приблизительно менее 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы с металлами (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как твин™, Pluronics™ или полиэтиленгликоль (ПЭГ).
Таким образом, в некоторых вариантах осуществления настоящее изобретение относится к способу лечения или профилактики заболевания у нуждающегося в этом индивидуума. Способы по настоящему изобретению могут включать введение терапевтически эффективного количества фармацевтической композиции, представленной в данном описании, индивидууму. Например, фармацевтическая композиция может включать одно или более антитело или его функциональный фрагмент, представленные в данном документе. Заболевания, которые можно лечить или предотвращать с использованием способов по изобретению, включают рак, образование опухолей и/или метастазирование. В частности, способы по настоящему изобретению могут быть использованы для лечения рака или образования опухолей, где раковые клетки или опухоли экспрессируют углевод sLea. Неограничивающие примеры видов рака или опухолей, которые можно лечить или предотвращать с использованием способов по изобретению, включают опухоли желудочно-кишечного тракта, например, рак толстой кишки, колоректальную аденокарциному, метастатический рак толстой кишки, колоректальный рак, рак поджелудочной железы или аденокарциному поджелудочной железы; мелкоклеточный рак легкого; аденокарциному мочевого пузыря; перстневидноклеточный рак яичников; рак яичников, метастатическую карциному; и аденокарциному желудка, пищевода, гортани, мочеполового тракта или молочной железы.
Соответственно, в некоторых аспектах настоящее изобретение относится к способу лечения рака или профилактики метастазов опухоли у нуждающегося в этом индивидуума путем введения терапевтически эффективного количества фармацевтической композиции, включающей антитело или его функциональный фрагмент, где антитело или его функциональный фрагмент связывается с sLea и включает домен VH, имеющий аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-142 SEQ ID NO: 2, остатков 20-142 SEQ ID NO: 6, остатков 20-142 SEQ ID NO: 10 и остатков 20-145 SEQ ID NO: 14. В другом аспекте настоящее изобретение относится к способу лечения рака или профилактики метастазов опухоли у нуждающегося в этом индивидуума путем введения терапевтически эффективного количества фармацевтической композиции, включающей антитело или его функциональный фрагмент, где антитело или его функциональный фрагмент связывается с sLea и включает домен VL, имеющий аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-130 SEQ ID NO: 4, остатков 20-129 SEQ ID NO: 8, остатков 20-130 SEQ ID NO: 12 и остатков 23-130 SEQ ID NO: 16. В еще одном аспекте настоящее изобретение относится к способу лечения рака или профилактики метастазов опухоли у нуждающегося в этом индивидуума путем введения терапевтически эффективного количества фармацевтической композиции, включающей антитело или его функциональный фрагмент, где антитело или его функциональный фрагмент связывается с sLea и включает как домен VH, так и домен VL, где домен VH и домен VL соответственно включают аминокислотную последовательность, выбранную из группы, состоящей из остатков 20-142 SEQ ID NO: 2 и остатков 20-130 SEQ ID NO: 4; остатков 20-142 SEQ ID NO: 6 и остатков 20-129 SEQ ID NO: 8; остатков 20-142 SEQ ID NO: 10 и остатков 20-130 SEQ ID NO: 12; и остатков 20-145 SEQ ID NO: 14 и остатков 23-130 SEQ ID NO: 16.
Составы, такие как описанные в настоящем документе, могут также содержать более одного активного соединения, если необходимо для конкретного заболевания, подлежащего лечению. В некоторых вариантах осуществления составы включают антитело или его функциональный фрагмент по изобретению и один или более активных соединений с дополнительной активностью, которые не оказывают отрицательного влияния друг на друга. Такие молекулы могут подходящим образом присутствовать в сочетании и в количествах, которые эффективны для предназначенной цели. Например, антитело или его функциональный фрагмент по изобретению могут быть объединены с одним или более другими терапевтическими агентами. Такие сочетанные терапевтические агенты можно вводить индивидууму одновременно или последовательно.
Таким образом, в некоторых аспектах изобретение относится к способу лечения или профилактики заболевания путем введения терапевтически эффективного количества фармацевтической композиции, представленной в данном документе, нуждающемуся в этом индивидууму, где фармацевтическая композиция включает антитело или его функциональный фрагмент по изобретению и второй терапевтический агент. Подходящий второй терапевтический агент может быть легко определен обычным специалистом в данной области техники, как описано в настоящем документе. Как указано в настоящем описании в примере IV, в некоторых аспектах настоящего изобретения второй терапевтический агент может представлять собой таксол.
Фармацевтические композиции, предлагаемые в данном документе, содержат терапевтически эффективные количества одного или более антител по изобретению, предлагаемых в настоящем описании, и необязательно один или более дополнительных терапевтических агентов в фармацевтически приемлемом носителе. Такие фармацевтические композиции пригодны для профилактики, лечения, управления или облегчения заболевания, такого как рак или образование опухоли, или одного или более из его симптомов.
Фармацевтические композиции могут содержать одно или более антител или его функциональных фрагментов по данному изобретению. В одном варианте осуществления антитела или их функциональные фрагменты составляют в виде подходящих фармацевтических препаратов, таких как стерильные растворы или суспензии для парентерального введения. В одном варианте осуществления антитела или их функциональные фрагменты, представленные в настоящем описании, составляют в виде фармацевтических композиций с использованием методов и процедур, хорошо известных в данной области техники (смотри, например, Ansel (1985) Introduction to Pharmaceutical Dosage Forms, 4th Ed., p. 126).
Антитело или его функциональный фрагмент по изобретению могут быть включены в фармацевтическую композицию в терапевтически эффективном количестве, достаточном для проявления терапевтически полезного эффекта в отсутствии нежелательных побочных эффектов у подвергаемого лечению индивидуума. Терапевтически эффективную концентрацию можно определить эмпирически путем тестирования соединений в системах in vitro и in vivo с использованием стандартных способов и последующей экстраполяции на основе этого доз для человека. Концентрация антитела или его функционального фрагмента в фармацевтической композиции будет зависеть, например, от физико-химических характеристик антитела или его функционального фрагмента, схемы дозирования и количества вводимого препарата, а также от других факторов, хорошо известных специалистам в данной области техники.
В одном варианте осуществления терапевтически эффективная доза обеспечивает концентрацию в сыворотке антитела или его функционального фрагмента от приблизительно 0,1 нг/мл до приблизительно 50-100 мкг/мл. Фармацевтические композиции в другом варианте осуществления обеспечивают дозу от приблизительно 0,001 мг до приблизительно 500 мг антитела на кг массы тела в день. Фармацевтические единицы лекарственных форм могут быть получены для обеспечения от приблизительно 0,01 мг, 0,1 мг или 1 мг до приблизительно 30 мг, 100 мг или 500 мг, и в одном варианте осуществления от приблизительно 10 мг до приблизительно 500 мг антитела или его функционального фрагмента и/или сочетания с другими дополнительными существенными ингредиентами в виде стандартной единицы лекарственной формы.
Антитело или его функциональный фрагмент по изобретению можно вводить однократно, или доза может быть разделена на несколько меньших доз, подлежащих введению через интервалы времени. Понятно, что точная дозировка и продолжительность лечения зависит от заболевания, подлежащего лечению, и может быть определена эмпирически с использованием известных протоколов тестирования или с помощью экстраполяции из данных испытаний in vivo или in vitro. Следует отметить, что концентрации и величины доз также могут варьироваться в зависимости от тяжести состояния, подлежащего облегчению. Следует также понимать, что для любого конкретного индивидуума конкретные режимы дозирования могут быть скорректированы с течением времени в соответствии с индивидуальной потребностью и профессиональной оценкой лица, назначающего или наблюдающего за введением композиций, и что диапазоны концентраций изложены в настоящем описании только в качестве примера и не предназначены для ограничения объема или применения заявленных композиций.
При смешивании или добавлении антитела или его функционального фрагмента по изобретению полученная в результате смесь может представлять собой раствор, суспензию или тому подобное. Форма полученной смеси зависит от ряда факторов, включая предполагаемый способ введения и растворимость соединения в выбранном носителе или наполнителе. Эффективная концентрация представляет собой концентрацию, достаточную для ослабления симптомов заболевания, расстройства или состояния, подлежащего лечению, и может быть определена эмпирически.
Фармацевтические композиции предлагаются для введения человеку и животным в единицах дозированных лекарственных форм, таких как стерильные парентеральные растворы или суспензии, содержащие подходящие количества соединений или их фармацевтически приемлемых производных. Антитело или его функциональный фрагмент в одном варианте осуществления могут быть составлены и введены в виде единицы лекарственной формы или множественных единиц лекарственной формы. Единица лекарственной формы относится к физически дискретным единицам, подходящим для людей или животных и упакованным по отдельности, как известно в данной области техники. Каждая единица лекарственной формы содержит предварительно определенное количество антитела или его функционального фрагмента по изобретению, достаточное для получения желаемого терапевтического эффекта, в сочетании с требуемым фармацевтическим носителем, наполнителем или разбавителем. Примеры единиц лекарственных форм включают ампулы и шприцы. Единицы лекарственной формы можно вводить в виде долей или их кратных величин. Множественные единицы лекарственной формы представляют собой множество идентичных единиц лекарственной формы, упакованных в один контейнер для введения в виде отдельных единиц лекарственной формы. Примеры множественных единиц лекарственной формы включают флаконы или бутыли пинт или галлонов. Следовательно, множественная единица лекарственной формы представляет собой множество единиц лекарственной формы, которые не отделены в упаковке.
В одном варианте осуществления одно или более антител или их функциональных фрагментов по изобретению находятся в жидкой фармацевтической композиции. Вводимые жидкие фармацевтические композиции, например, могут быть получены путем растворения, диспергирования или смешивания иным способом антитела или его функционального фрагмента, предлагаемых в настоящем описании, и необязательных фармацевтических адъювантов в носителе, таком как, например, вода, физиологический раствор, водный раствор декстрозы, глицерин, гликоли, этанол и тому подобное, чтобы в результате получился раствор. При желании фармацевтическая композиция, предназначенная для введения, может также содержать незначительные количества нетоксичных вспомогательных веществ, таких как смачивающие агенты, эмульгаторы, солюбилизирующие агенты, буферные рН агенты и тому подобное, например, ацетат, цитрат натрия, производные циклодекстрина, сорбитанмонолаурат, триэтаноламин ацетат натрия, олеат триэтаноламина и другие подобные агенты. Современные способы получения таких лекарственных форм известны или будут очевидны для специалистов в данной области техники; например, смотри Remington’s Pharmaceutical Sciences (1990) Mack Publishing Co., Easton, PA.
Способы введения фармацевтической композиции по настоящему изобретению хорошо известны в данной области техники. Понятно, что подходящий способ введения фармацевтической композиции может быть легко определен квалифицированным врачом. Типичные пути введения включают внутривенное введение, внутримышечное введение, внутрикожное введение или подкожное введение. Кроме того, следует понимать, что состав фармацевтической композиции можно легко скорректировать для адаптации к пути введения. Изобретение также предусматривает, что после введения фармацевтической композиции по настоящему изобретению, отставленные, последовательные и/или повторные дозы одной или более фармацевтических композиций могут быть введены индивидууму, как представлено в настоящем описании.
Способы по настоящему изобретению для лечения заболевания предназначены для включения (1) предотвращения заболевания, т.е. индукции предотвращения развития клинических симптомов заболевания у индивидуума, который может быть предрасположен к заболеванию, но еще не испытывает или не проявляет симптомов заболевания; (2) ингибирования заболевания, т.е. прекращения или ослабления развития заболевания или его клинических симптомов; или (3) ослабления заболевания, т.е. стимуляции регрессии заболевания или его клинических симптомов. Способы профилактики заболевания по настоящему изобретению предназначены для включения предупреждения клинической симптоматики, указывающей на рак или образование опухоли. Такое предупреждение включает, например, поддержание нормальных физиологических показателей у индивидуума. Таким образом, предотвращение может включать профилактическое лечение индивидуума для защиты его от появления метастазов опухоли.
Терапевтически эффективное количество фармацевтической композиции, используемое в способах по настоящему изобретению, будет изменяться в зависимости от используемой фармацевтической композиции, заболевания и его тяжести и возраста, веса и т.д. индивидуума, подлежащего лечению, все это находится в пределах компетенции лечащего врача. Индивидуум, которого можно лечить с помощью способов по изобретению, включает позвоночное животное, предпочтительно млекопитающее, более предпочтительно человека.
Понятно, что модификации, которые не оказывают существенного влияния на действие различных вариантов осуществления настоящего изобретения, также предлагаются в рамках определения настоящего изобретения, представленного в настоящем описании. Соответственно, следующие примеры предназначены для иллюстрации, но не ограничивают настоящее изобретение.
ПРИМЕР I
Моноклональные антитела человека против sLea SLE обладают сильной противоопухолевой активностью
Углеводный антиген sLea широко экспрессируется на эпителиальных опухолях желудочно-кишечного тракта, молочной железы, поджелудочной железы и на клетках мелкоклеточного рака легких. Так как гиперэкспрессия sLea, как представляется, является ключевым событием в инвазии и метастазировании многих опухолей и приводит к восприимчивости к опосредованному антителами лизису, sLea представляет собой привлекательную мишень для молекулярной терапии опухолей. Соответственно, как описано в настоящем документе, были получены и охарактеризованы полностью человеческие моноклональные антитела (mAb) из лимфоцитов крови индивидуумов, иммунизированных вакциной sLea-KLH. Несколько mAbs было отобрано на основе ELISA и FACS, включая два моноклональных антитела с высоким сродством к sLea (5B1 и 7E3, связывающая аффинность 0,14 и 0,04 нмоль/л, соответственно) и дополнительно охарактеризовано. Оба антитела были специфичными для Neu5Acα2-3Galβ1-3(Fucα1-4) GlcNAcβ и Neu5Gcα2-3Galβ1-3(Fucα1-4)GlcNAcβ при определении матричным анализом гликанов. Зависимая от комплемента цитотоксичность против клеток DMS-79 была выше (ЕС50 0,1 мкг/мл по сравнению с 1,7 мкг/мл) у r7E3 (IgM), чем у r5Bl (IgG1). Кроме того, антитела r5Bl показали высокий уровень активности в отношении зависимой от антител опосредуемой клетками цитотоксичности на клетках DMS-79 с NK-клетками человека или мононуклеарными клетками периферической крови. Для оценки эффективности in vivo антитела были протестированы в модели ксенотрансплантата опухолевых клеток Colo205 или опухолевых клеток DMS-79, трансплантированных мышам с тяжелым комбинированным иммунодефицитом (SCID). В модели ксенотрансплантата Colo205 лечение в течение первых 21 дней четырьмя дозами r5Bl (100 мкг на дозу) удваивало медиану времени выживаемости до 207 дней, и три из пяти животных выживали в течение получения шести доз. В модели ксенотрансплантата DSM-79 рост выявленных опухолей DSM-79 подавлялся или регрессировал у животных, которых лечили антителом r5Bl. На основе потенциала sLea в качестве мишени для иммунной атаки и сродства, специфичности и эффекторных функций 5Bl и 7Е3 они имеют клиническое применение при лечении рака.
Материалы, клетки и антитела
Клеточные линии DMS-79 (Pettengill et al., Cancer, 45:906-18 (1980)), SW626, EL4, HT29, BxPC3, SK-MEL28 и P3× 63Ag8.653 приобретали у американской коллекции типовых культур (АТСС), клетки Colo205-Luc (Bioware ultra) получали от Caliper Life Sciences. Контрольные мышиные mAb 121SLE (IgM) приобретали у GeneTex. Тетрасахарид sLea (Cat # S2279) приобретали у Sigma-Aldrich. Конъюгат sLea-HSA (сывороточный альбумин человека) (Cat # 07-011), одновалентный биотинилированный sLea (sLea-sp-биотин; Cat # 02-044), поливалентный биотинилированный sLea-PAA (Cat # 01-044), меченный биотином Lea-PAA (Cat # 01-035), и sLex-PAA-биотин (Cat # 01-045) приобретали у GlycoTech. В поливалентной презентации тетрасахарид включается в матрицу полиакриламида (PAA), таким образом создавая 30-кДа поливалентный полимер с приблизительно каждой пятой амидной группой полимерной цепи N-замещенного биотина в соотношении 4:1 и приблизительно 20% содержанием углеводов. Другие гликоконъюгаты HSA или BSA, используемые в данном исследовании, получали собственноручно с использованием sLea-пентенилгликозида, как описано. Ragupathi et al., Cancer Immunol Immunother, 58:1397-405 (2009). GD3, фукозил-GM1, GM2 и GM3 приобретали у Matreya и GD2 приобретали у Advanced ImmunoChemical.
Создание гибридом, продуцирующих mAb против sLea
Образцы крови получали от 3 больных в текущем испытании вакцины конъюгированного sLea-KLH у больных раком молочной железы, инициированном в MSKCC по утвержденному MSKCC и FDA IRB протоколу и IND. Образцы крови отбирали от 2 больных после 3 или 4 вакцинации, которые показали титры антител против sLea 1/160 и 1/320, соответственно. Эти сыворотки (и мышиное mAb 19,9) хорошо реагируют с sLea-позитивными клеточными линиями в анализах FACS и опосредуют сильную CDC. Ragupathi et al., Cancer Immunol Immunother, 58:1397-405 (2009). Мононуклеарные клетки периферической крови (РВМС) выделяли из приблизительно от 80 до 90 мл крови путем градиентного центрифугирования на Histopaque-1077 (Sigma-Aldrich).
PBMCs культивировали в среде RPMI-1640, дополненной L-глутамином, заменимыми аминокислотами, пируватом натрия, витамином, пенициллин/стрептомицином, 10% FBS (Omega Scientific), 10 г/мл IL-21 (Biosource) и 1 мкг/мл mAb против CD40 (супернатанта гибридомы G28-5; АТСС). Клетки сливали с помощью электропорации с клетками миеломы P3×63Ag8.653.
ELISA sLea
Для ELISA sLea планшеты покрывали либо конъюгатом 1 мкг/мл sLea-HSA, одновалентным биотинилированным sLea, либо поливалентным биотинилированным sLea-PAA, захватываемыми на планшетах с покрытием Neutr-Avidin. Непокрытые лунки (PBS) и лунки, покрытые HSA, использовали в качестве контролей. Связанные антитела первоначально детектировали козьими антителами против IgA+G+M человека, меченными пероксидазой хрена (HRP) (Jackson ImmunoResearch), и положительные лунки затем зондировали с помощью IgG-Fc- или IgM-специфических вторичных антител для определения изотипов.
Анализ специфичности углеводов
Перекрестную реактивность против близкородственных антигенов Lea и sLex оценивали с помощью поверхностного плазмонного резонанса (SPR) и подтверждали с помощью ELISA с использованием меченного биотином Lea-PAA и биотин-sLex-РАА. Связывание с ганглиозидами GD2, GD3, фукозил-GM1, GM2 и GM3 тестировали с помощью ELISA. Конкурентный ELISA использовали для оценки специфичности mAbs против нескольких других родственных углеводных частей. Вкратце, планшеты покрывали 2 мкг/мл конъюгата sLea-HSA с последующим блокированием 3% BSA в PBS. Затем 30 мкл различных углеводных остатков (40 мкг/мл в PBS, полученных из 1 мг/мл маточного раствора), либо неконъюгированных, либо конъюгированных с HSA или BSA, смешивали отдельно с 30 мкл тестируемого антитела и инкубировали при комнатной температуре в планшете с образцами. Через 30 минут 50 мкл смеси переносили в аналитический планшет с покрытием и инкубировали в течение 1 часа с последующей инкубацией с козьими антителами против IgA+G+М человека, меченными HRP, промывкой и колориметрическим определением связанного антитела с использованием спектрофлуориметра VersaMax (все стадии осуществляли при комнатной температуре). Тестируемые углеводные фрагменты включали globo H, антиген Льюиса Y, антиген Льюиса X, сиалил-Thomson-Nouveaux (STN), кластерный sTn, Thomson Friedenreich (TF), муцин Tighe Leb/Ley, муцин подчелюстной железы свиньи (PSM) и тетрасахарид sLea и конъюгат sLea-HSA. Для определения высокой специфичности антител осуществляли матричный анализ гликанов группой Consortium for Functional Glycomics Core H. Антитела 5B1 и 7E3 тестировали в концентрации 10 мкг/мл с использованием версии 4.1 печатной матрицы, состоящей из 465 гликанов в 6 повторах.
Клонирование кДНК иммуноглобулинов и экспрессия рекомбинантных антител
кДНК вариабельной области тяжелой и легкой цепи mAb человека получали путем ОТ-ПЦР из отдельной гибридомной клеточной линии и субклонировали в экспрессионный вектор тяжелой цепи IgG1 или IgM или легкой цепи IgK или IgL, как описано ранее. Sawada-Hirai et al., J. Immune Based Ther. Vaccines, 2:5 (2004). Экспрессионный вектор тяжелой цепи или легкой цепи Ig дважды расщепляли Not I и Sal I, и затем оба фрагмента лигировали с образованием экспрессионного вектора двойного гена. Клетки СНО в 6-луночных планшетах трансфецировали экспрессионным вектором двойного гена с использованием липофектамина 2000 (Invitrogen). Через 24 часа трансфецированные клетки переносили в 10-см чашку с селективной средой [DMEM, дополненной 10% диализованной FBS (Invitrogen), 50 ммоль/л L-метионинсульфоксимином (MSX), добавкой GS (Sigma-Aldrich) и пенициллин/стрептомицином (Omega Scientific)]. Через две недели устойчивых к MSX трансфектантов выделяли и наращивали. Клоны с высокой продукцией антител против sLea отбирали путем измерения уровней антител в супернатантах с помощью теста ELISA, специфичного для sLea, и их наращивали для крупномасштабной продукции mAb.
Очистка mAb человека
Антитела очищали с использованием пропускания через систему Akta Explorer (GE Healthcare) с программным обеспечением Unicorn 5.0. Вкратце стабильные клоны 5B1 или 7E3 выращивали в бессывороточной культуральной среде в биореакторе Wave, и собранный супернатант осветляли путем центрифугирования и фильтрации и хранили в замороженном виде до использования. Антитела IgG человека очищали на колонках с протеином А соответствующих размеров с использованием 10 ммоль/л PBS и 150 ммоль/л NaCl подвижного буфера. Антитела IgM человека очищали на колонке гидроксиапатита, и IgM элюировали с градиентом от 500 ммоль/л фосфата. Концентрации антител определяли при ОП280 с использованием E1% 1,4 и 1,18 для IgG и IgM, соответственно, для расчета концентрации. Чистоту каждого препарата оценивали с помощью анализа SDS-PAGE (1-5 мкг на полосу) в восстанавливающих условиях, и чистота составляла более 90% в пересчете на сумму тяжелых и легких цепей.
Проточная цитометрия
sLea-положительные или sLea-отрицательные линии опухолевых клеток (0,5×106 клеток на режим) промывали в PBS/2% FBS (PBSF). Затем добавляли тестируемые или контрольные mAb человека (1-2 мкг/мл в полной среде), и инкубировали на льду в течение 30 минут. Gilewski et al., Clin Cancer Res, 6:1693-701 (2000); Gilewski et al. Proc. Natl. Acad. Sci. U.S.A., 98:3270-5 (2001). После промывки в PBSF клетки инкубировали с Alexa-488 против IgM-Fcγ человека или против IgMμ человека (Invitrogen) в течение 30 минут на льду. Клетки дважды промывали в PBSF и анализировали с помощью проточной цитометрии с использованием Guava Personal Cell Analysis-96 (PCA-96) System (Millipore). Клетки Colo205-Luc инкубировали с 2 мкг/мл первичного антитела с последующим окрашиванием вторичными антителами от SouthernBiotech, и анализировали на приборе Becton Dickinson FACS Advantage IV с использованием программного обеспечения FlowJo 7.2.4.
Определение аффинности
Константы сродства определяли с использованием принципа SPR с Biacore 3000 (GE Healthcare). Меченный биотином одновалентный sLea (Cat # 02-044) или поливалентный sLea-PAA-биотин (Cat # 01-044) соединяли с отдельными проточными ячейками чипа SPA биосенсора в соответствии с инструкциями изготовителя. Проточную ячейку блокировали HSA, и использовали культуральную среду, содержащую свободный биотин, в качестве эталонной ячейки. Кинетические параметры связывания определяли для нескольких известных концентраций антитела, разведенного в HBS-EP буфере (10 ммоль/л HEPES, рН 7,4, 150 ммоль/л NaCl, 3,4 ммоль/л ЭДТА, 0,005% поверхностно-активного вещества Р20), с использованием проточной ячейки, покрытой sLea-PAA-биотином. Программное обеспечение для подгонки кривой, предоставленное Biacore instrument, использовали для получения оценок скоростей ассоциации и диссоциации, из которых рассчитывали сродство.
Анализ CDC
sLea-положительные или sLea-отрицательные линии опухолевых клеток использовали для 90-минутного теста на цитотоксичность (Guava PCA-96 Cell-Toxicity kit; Millipore; Cat # 4500-0200) с использованием комплемента человека (Quidel; Cat # Al13) и очищенных mAbs человека в различных разведениях (0,1-25 мкг/мл) или mAbs положительного контроля, как описано ранее (Ragupathi et al. Clin Cancer Res 2003, 9:5214; Ragupathi et al. Int J Cancer 2000, 85:659; Dickler et al. Cancer Res 1999, 5:2773). Вкратце 2,5×106 клеток-мишеней окрашивали сукцинимиловым эфиром карбоксифлуоресцеиндиацетата (CSFE) с получением зеленых/желтых флуоресцентных клеток-мишеней. Окрашенные клетки (1×105/50 мкл образца) инкубировали в течение 40 минут с 100 мкл антител на льду. Затем 50 мкл комплемента человека разводили 1:2 в полной среде (RPMI-1640, 10% FCS) или добавляли только среду к трем параллельным образцам и инкубировали в течение 90 минут при 37°С. Таким образом, конечное разведение комплемента в тесте составляло 1:8. Клетки, которые погибали во время этого периода инкубации, метили путем добавления не проницаемого для мембраны красителя 7-амино-актиномицина D (7-AAD), и образцы анализировали с помощью двойной цветовой иммунофлуоресценции, используя программный модуль Guava CellToxicity. Контрольные образцы, которые инкубировали с NP40, использовали для определения максимальной гибели, и образцы только с комплементом служили в качестве показателя фонового уровня. Процент погибших клеток определяли с помощью соответствующего гейтинга и вычисляли по следующей формуле: % погибших=[(%выборки-%комплемента в покое)/(%NP40-%комплемента в покое)]×100.
Анализ зависимой от антител клеточно-опосредованной цитотоксичности
Эффекторные клетки РВМС выделяли с помощью центрифугирования в градиенте плотности Ficoll-Hypaque из образцов крови, полученных по протоколу, одобренному MSKCC IRB. Клетки-мишени 5×106 клеток/мл инкубировали в полной среде роста с 15 мкл раствора 0,1% кальцеин-АМ (Sigma-Aldrich) в течение 30 минут при температуре 37°С в присутствии 5% СO2. Клетки дважды промывали 15 мл PBS-0,02% ЭДТА и ресуспендировали в 1 мл полной среды роста. Пятьдесят микролитров (10000 клеток) меченых клеток-мишеней высевали в 96-луночный планшет в присутствии или в отсутствие антител в концентрациях, описанных на фиг. 13, и инкубировали с 50 мкл свежевыделенных мононуклеарных клеток периферической крови (эффекторные клетки, при отношении R/T 100:1) соответственно. После 2 часовой инкубации планшет центрифугировали при 300×g в течение 10 минут, и 75 мкл супернатанта переносили в новый 96-луночный планшет с плоским дном. Флуоресценцию в надосадочной жидкости измеряли при 485 нм возбуждения и 535 нм излучения в Fluoroskan Ascent (Thermo Scientific). Спонтанное высвобождение из клеток-мишеней определяли в среде RPMI-1640 с 30% FBS без эффекторных клеток, и максимальное высвобождение из клеток-мишеней определяли в среде RPMI-1640 с 30% FBS и 6% тритоном Х-100 без эффекторных клеток. Процент цитотоксичности рассчитывали как [(уровень флуоресценции в образце - спонтанное высвобождение)/(максимальная флуоресценция - спонтанное высвобождение)] × 100.
Анализ интернализации mAb
Интернализацию антитела 5B1 оценивали путем измерения цитотоксической активности r5Bl и комплекса вторичного конъюгата Hum-Zap (Advanced Targeting Systems) против клеток BxPC3, экспрессирующих sLea, которые высевали в 96-луночный планшет (2000 клеток/90 мкл/лунка) и инкубировали в течение ночи в двух параллельных пробах. Различные концентрации антитела 5B1 инкубировали с вторичными конъюгатами Hum-Zap при RT в соответствии с инструкцией изготовителя. Затем к клеткам добавляли 10 мкл/лунка r5Bl и комплекса Hum-Zap и инкубировали в течение 3-х дней. Двадцать пять микролитров раствора тиазолила синего тетразолия бромида (Sigma-Aldrich) (5 мг/мл в PBS) добавляли в каждую лунку и инкубировали при 37°С. После инкубации в течение 2 часов добавляли 100 мкл/лунка раствора для солюбилизации (20% SDS/50% N,N-диметилформамид) в каждую лунку и инкубировали в течение еще 16 часов при 37°C. ОП измеряли при 570/690 нм, и полученные величины с одной средой использовали для вычитания фона планшета. Восемь параллельных культур без антитела использовали для нормализации величин образцов (образец/среднее необработанных × 100).
Модель трансплантации ксенотрансплантата
Самок мышей СВ17 SCID (5-8 недель) приобретали у Taconic. Для модели ксенотрансплантата Colo205 клетки Colo205-luc (0,5×106) в 0,1 мл полной среды роста вводили через хвостовую вену на день 0 с помощью инсулинового шприца BD с иглой 28G (Becton Dickinson & Co). Для первого исследования сто микрограммов mAb 5B1 вводили внутрибрюшинно в дни 1, 7, 14 и 21 (эксперимент 1) или в дни 1, 4, 7, 10, 14 и 21 (эксперимент 2). Для второго исследования 100 мкг, 300 мкг или 1 мг mAb 5B1 вводили внутрибрюшинно на 4-й день после инъекции опухолевых клеток, а затем два раза в неделю в течение первых двух недель и один раз в неделю в течение следующих 7 недель. У мышей отслеживали развитие опухоли. Для модели ксенотрансплантата DMS-79, клетки DMS-79 (1×106) вводили подкожно самкам мышей СВ17 SCID, и начинали лечение мышей на 19 день после того, как длина опухоли достигала 5 мм (~20 мм2). Животные затем получали IgG человека или антитела 5B1, вводимые путем внутрибрюшинной инъекции 200 мкг на дозу, плюс cRGD путем внутривенной инъекции для увеличения проницаемости сосудов первоначально 80 мкг, затем 5 дней в неделю 40 мкг на дозу до 37 дня.
Все процедуры осуществляли в соответствии с протоколом, утвержденным онкологическим центром института по уходу за животными и комитетом по их использованию Memorial Sloan Kettering. Кривые выживаемости Каплана-Мейера получали с использованием GraphPad Prism 5.1 (GraphPad Software) и анализировали с помощью лог-рангового критерия Mantel-Haenszel.
Результаты
Идентификация моноклональных антител человека с помощью ELISA и создание рекомбинантных антител
Образцы крови от 3 вакцинированных больных использовали для достижения создания гибридом, и выявили много положительных лунок в тестах ELISA, специфичных для антигена (таблица 3). Обширный скрининг использовали для устранения антител, которые проявляли незначительное или неспецифическое связывание. Восемь гибридомных клеток, экспрессирующих антитело человека (1 IgM и 7 IgG), с сильной реактивностью против sLea первоначально выбрали, нарастили и субклонировали для дальнейшей характеристики. Два антитела (9H1 и 9H3) показали сильное связывание с конъюгатами sLea-HSA, но не с планшетами, покрытыми sLea-PAA. Три антитела (5B1, 5H11 и 7Е3) показали сильное связывание с одновалентным и поливалентным sLea и конъюгатами sLea-HSA при измерении с помощью анализов ELISA (таблица 4).
Связывание кандидатных супернатантов гибридом, содержащих моноклональные антитела IgG или IgM, с конъюгатом sLea-ацетилфенилендиамин(APD)-сывороточный альбумин человека (HSA) (sLea-HuSA).
Связывание выбранных антител против sLea, представленных в виде одновалентных (моно-)sLea, поливалентных (поли)sLea или в форме sLea-HSA.
HuSa представляет собой контроль сывороточного альбумина человека. PBS представляет собой контроль физиологического раствора, забуференного фосфатом. NAV представляет собой контроль нейтрального авидина.
Вариабельные области тяжелой и легкой цепи от 4 выбранных антител выделяли с помощью ОТ-ПЦР и клонировали в экспрессионные векторы изобретателей полноразмерных IgG1 или IgM. Молекулярный анализ последовательности с использованием IMGT/V-Quest (Brochet et al., Nucleic Acids Res., 36:W503-8 (2008)) показал, что 3 выбранных антитела IgG 5B1 (IgG/λ), 9H3 (IgG/λ), и 5H11 (IgG/λ) происходят от одного и того же семейства VH и все используют легкие цепи лямбда. Эти антитела IgG1 показали различные последовательности CDR с 16, 5 или 3 мутациями, отклоняющимися от зародышевой линии, соответственно (фиг 1-6; таблица 5). Антитело IgM (7Е3) использует легкую цепь каппа и имеет 6 мутаций тяжелой цепи (фиг 7-8; таблица 5). Повышенные мутации в 5B1 свидетельствуют о созревании аффинности. Рекомбинантные антитела продуцировали в клеточных линиях CHO в системе волнового биореактора и очищали с использованием протеина A или хроматографии на гидроксиапатите для IgG и IgM, соответственно. Очищенные рекомбинантные антитела сохраняли свойства исходных антител, происходящих из гибридомы, в отношении связывания в ELISA и специфичности.
Классификация кДНК выбранных антител человека против sLea, полученных от вакцинированных доноров крови
Анализ связывания опухолевых клеток
Связывание на клеточной поверхности имеет решающее значение для цитотоксической активности и, следовательно, тестировалось в последующем. Проточная цитометрия показала сильное связывание рекомбинантных антител 5В1, 9H3, 5H11 и 7E3 с клетками DMS-79, с суспензией клеточной линии мелкоклеточного рака легких (фиг. 11А). Связывание r5Bl и r7E3 также было подтверждено на клетках НТ29 рака толстой кишки (фиг. 11B), клетках BxPC3 рака поджелудочной железы (фиг. 11C), клетках SW626 рака яичников (фиг. 11D) и клетках Colo205-luc рака толстой кишки (фиг. 11F). Эти антитела не смогли связаться с sLea-негативными (SLE121-негативными) клетками СК-MEL28 меланомы (фиг. 11E) или клетками EL4 лимфомы мыши (данные не показаны).
Измерения аффинности
Относительное сродство/авидность связывания с sLea зондировали с помощью SPR с использованием биосенсорного чипа, покрытого стрептавидином, для захвата биотинилированного sLea-PPA. Как показано в таблице 6, r5Bl и r7E3 быстро связываются с sLea-PPA и показывают значительно более медленную скорость диссоциации по сравнению с 121SLE, коммерчески доступным мышиным антителом IgM против sLea, которое использовали для сравнения. Сродство 5B1 измеряли при 0,14 нмоль/л, и кажущаяся аффинность/авидность 7E3 была приблизительно в 4 раза выше (таблица 6). Определение сродства 9H3 было затруднено, так как антитела 9H3 (нативные и рекомбинантные) не способны связываться с биосенсорным чипом, покрытым sLea-PAA.
Определение кинетических параметров связывания антител против sLea с помощью SPR
моль/л
1/моль/л
kd, 1/сек
Анализ специфичности
Предварительные анализы для исследования углеводной специфичности показали, что 5B1, 9H3 и 7E3 не связывались с близко родственными антигенами sLeX, Lea или LeY, или ганглиозидами GD2, GD3, фукозил-GM1, GM2 и GM3 при измерении с помощью ELISA или SPR. Дополнительный анализ связывания 7E3, 5B1 и 121SLE с sLea-PAA-биотином или с sLea-sp-биотином, захваченным авидиновым чипом Biacore, показал, что все три антитела связываются с поливалентной формой sLea, в то время как 7E3 и 5B1, как показано, связываются с одновалентной формой. Связывание 5B1 с sLea-PAA ингибировалось также тетрасахаридом sLea дозозависимым образом при анализе Biacore с сериями концентраций (данные не показаны). Эти результаты согласуются с предыдущими наблюдениями, что сыворотки с высокими тирами антител против sLea, как обнаружено с помощью ELISA, являются специфичными для sLea, то есть не взаимодействуют с ганглиозидами GM2, GD2, GD3, фукозил-GM1 или с нейтральными гликолипидами Globo H и Ley. Ragupathi et al., Cancer Immunol Immunother 58:1397-405 (2009). В конкурентном анализе с 9 различными родственными углеводными компонентами в различных представлениях (например, в виде церамида или конъюгатов с BSA или HSA), только тетрасахарид sLea и конъюгат sLea-HSA были способны ингибировать связывание с конъюгатом sLea-HSA (таблица 7).
Связывание со sLeA-PAA-HSA в присутствии различных родственных гликоконъюгатов.
Для изучения углеводной специфичности более подробно антитела 5B1 и 7E3 также тестировали с помощью матричного анализа гликанов, проведенного группой Consortium for Functional Glycomics Core H. Оба антитела тестировали в концентрации 10 мкг/мл на печатных матрицах, состоящих из 465 гликанов в 6 повторах. Результаты подтвердили высокую специфичность обоих антител с селективным узнаванием тетрасахарида sLea, Neu5Acα2-3Galβ1-3(Fucαl-4)GlcNAcβ и Neu5Gcα2-3Galβ1-3(Fucα1-4)GlcNAcβ и фактическое отсутствие связывания близкородственных антигенов, которые присутствовали в матрице, включая sLex, Lea, Lex и Ley. Результаты суммированы в таблице 8, в которой показано 5 лучших структур 465 гликанов, которые узнавались соответствующими антителами.
Анализ специфичности углеводов путем скрининга матрицы гликанов.
A. 5B1
Активность CDC
Для оценки функциональной активности 5B1 и 7E3 изобретатели тестировали цитотоксическую активность на клетках DMS-79 в присутствии сыворотки крови человека в качестве источника комплемента. Оба антитела показали в некоторых анализах близкую к 100% киллерную активность при 10 мкг/мл, в то время как контрольное антитело с отличной специфичностью (1B7, mAb против IgG1 GD2) не оказывало влияния при тех же концентрациях (данные не показаны). Активность CDC зависит от концентрации, и 7Е3 было значительно более активно по сравнению с 5B1 в этом анализе (фиг. 12), что ожидалось, так как антитела IgM, как известно, более эффективны в опосредуемой комплементом цитотоксичности. ЕС50 (50% цитотоксичность) составляла 1,7 мкг/мл для 5B1 и 0,1 мкг/мл для 7E3, что переводится в молях в приблизительно в 85 раз более высокую эффективность для 7E3 (фиг. 12).
Активность ADCC
В то время как 7Е3 является значительно более эффективным в анализе CDC, IgG антитела, как известно, обладают активностью в отношении зависимой от антител клеточно-опосредованной цитотоксичности (ADCC), которая считается важной для уничтожения опухолей in vivo. Высокие уровни цитотоксичности были измерены при использовании антитела 5B1 в отношении РВМС человека и клетками-мишенями DMS-79 при различных соотношениях Е:Т (фиг. 13А). Сходные уровни цитотоксичности наблюдались при более низких соотношениях Е:Т в отношении первичных NK-клеток (фиг. 13В). Эксперимент по дозозависимости с РВМС от 2 доноров при измерении при соотношении Е/Т 100:1 показал аналогичную эффективность, и цитотоксичность более 85% была достигнута при концентрациях 5B1 0,5 мкг/мл или более (фиг 13C). Цитотоксичность, опосредуемая 5B1, требует рецепторов FcyRIII, так как она может быть заблокирована антителами 3G8 против CD16. Высокие уровни цитотоксичности также были измерены с использованием антитела 5B1 с РВМС человека против клеток Colo205-luc при соотношении E:Т 100:1. ADCC-активность, достигаемая при 1 мкг/мл антител 5B1, превосходила активность, наблюдавшуюся с антителами против GM2, фукозил-GM1, Globo H или полисиаловой кислоты. Как и следовало ожидать, 7Е3 и мышиный 121SLE (оба IgM) были неактивны в этом анализе.
Анализ интернализации 5B1
Конъюгаты антитела, направленного на антиген «близкородственный» антигену Льюиса Y, как показано ранее, быстро интернализуются и очень эффективны в моделях на животных. Hellstrom et al., Cancer Res 50:2183-90 (1990); Trail et al., Science 261:212-5 (1993). Для проверки интернализации sLea изобретатели инкубировали клеточную линию BxPC3 поджелудочной железы с 5B1 и затем добавляли Hum-Zap против IgG человека, конъюгированный с сапорином, белком инактивации рибосом. Kohls et al., Biotechniques 28:162-5 (2000). Клетки, которые интернализуют содержащий сапорин комплекс, гибнут, в то время как неинтернализованный сапорин покидает клетки без повреждения. Как показано на фиг. 14, клетки BxPC3 эффективно уничтожались в присутствии возрастающих доз 5B1, в то время как присутствие сходного по изотипу антитела IgG1, направленного против GD2, который не экспрессируется на этих клетках, не уничтожает клетки.
Активность в животной модели ксенотрансплантата для метастазирования
Для того чтобы оценить активность 5B1 in vivo антитела тестировали в двух моделях ксенотрансплантата с использованием либо опухолевых клеток Colo205-luc, либо опухолевых клеток DMS-79 у мышей SCID. Для модели ксенотрансплантата с использованием опухолевых клеток Colo205-luc пяти мышам на группу вводили 0,5×106 клеток в хвостовую вену на день 0, и успешность введения клеток подтверждали путем получения изображения животных с помощью системы визуализации in vivo IVIS 200 (Caliper Life Sciences). Через день животные получали антитела 5B1, вводимые внутрибрюшинно, или пустую инъекцию PBS. В эксперименте 1 100 мкг 5B1 давали на 1, 7, 14 и 21 день (400 мкг общая доза), и в эксперименте 2 животные получали 100 мкг 5B1 на 1, 4, 7, 10, 14 и 21 день (600 мкг общая доза). Средняя медиана выживаемости животных без лечения составила 102 дня в 2-х экспериментах, и все животные без лечения умерли в течение 155 дней (фиг. 15). Лечение животных значительно повышало выживаемость: медиана выживаемости удваивалась к 207 дню в группе, которая получала 4 дозы 5B1, и 2 из 5 животных дожили до окончания эксперимента после 301 дня (лог-ранговый критерий, P=0,0499; HR=3,46). Доля выживших дополнительно увеличивалась до 3-х из 5 мышей при введении 6 доз (лог-ранговый критерий, P=0,0064; HR =6,375). Второй эксперимент был прекращен через 308 дней, и у выживших животных не возможно было выявить опухоли Colo205-luc при самой высокой чувствительности системы визуализации (данные не показаны).
Во втором исследовании мыши, которым аналогичным образом вводили опухолевые клетки Colo205-luc, как описано выше, получали увеличивающиеся дозы антител 5B1 или 7Е3 (100 мкг, 300 мкг или 1 мг). Все животные получали первоначально внутрибрюшинно антитела 5B1 или 7E3 или пустую инъекцию PBS (контроль) на 4-й день после инъекции опухолевых клеток, а затем два раза в неделю в течение первых двух недель, и один раз в неделю в течение последующих 7 недель. Отсроченное лечение различными дозами 5B1 показало зависимую от дозы защиту до полного излечения у мышей SCID с трансплантированными опухолевыми клетками Colo205-luc (фиг. 16 и 17). Лечение антителами 7Е3 не показало более высокую степень защиты, несмотря на повышенное кажущееся сродство (данные не показаны).
В модели ксенотрансплантата с использованием клеток DMS-79 пяти мышам на группу вводили подкожно 1×106 клеток на день 0, и начинали лечение на 19-й день после того, как длина опухоли достигала 5 мм (~20 мм2). Животные затем получали IgG человека или антитела 5B1 путем внутрибрюшинной инъекции 200 мкг на дозу, плюс cRGD путем внутривенной инъекции первоначально 80 мкг, затем 5 дней в неделю 40 мкг на дозу до 37 дня. Рост выявленных опухолей DMS-79 подавлялся или регрессировал у животных, получавших 5B1 или сочетание 5B1 плюс cRGD (фиг. 18А и 18В). Лечение животных 5B1 в день трансплантации клеток DMS-79 в модели подкожной трансплантации полностью предотвращало рост опухоли (данные не показаны).
Приведенные выше данные демонстрируют значительную способность вызывать подавление или регрессию выяленных опухолей и обеспечивать преимущество в выживаемости при использовании лечения антителом 5B1.
ПРИМЕР II
Иммуно-PET обнаружение и диагностика рака поджелудочной железы и других sLea-позитивных аденокарцином при использовании радиоактивно меченного моноклонального антитела 5B1
Аденокарциномы являются ведущей причиной смерти от рака. Обнаружение рака поджелудочной железы остается особенно трудным с диагнозом, часто сделанным на поздней стадии. Подходы к более раннему выявлению первичного и метастатического рака поджелудочной железы могут иметь значительный клинический выход. В клинической практике повышенные уровни антигена sLea контролируются для выявления предполагаемого неидентифицированного злокачественного новообразования у больных раком поджелудочной железы. Как описано в настоящем документе, исследовался потенциал нового зонда immunoPET визуализации, направленного на sLea, в доклинических моделях рака поджелудочной железы и других sLea-позитивных аденокарциномах. Положительное окрашивание моноклональным антителом 5B1 человека против sLea выявлено в аденокарциномах человека, известных как sLea-позитивные, но не в sLea-негативных злокачественных опухолях или большинстве нормальных тканей. 89Zr-радиоактивно меченное 5B1 (89Zr-5Bl) продемонстрировало высокую степень метки (>80%) и выходы очистки (>95%). Визуализация с помощью 89Zr-5Bl была исследована при подкожных, ортотопических и метастатических ксенотрансплантатах рака поджелудочной железы самкам мышей SCID. Полученные изображения PET и исследования биораспределения продемонстрировали исключительную специфичность и локализацию 89Zr-5Bl в sLea-гиперэкспрессирующих ксенотрансплантатах BxPC3 с минимальным неспецифическим связыванием со здоровыми тканями. Дальнейший анализ в моделях подкожных ксенотрансплантатов рака толстой кишки и немелкоклеточного рака легкого привел также к отличному очерчиванию опухоли 89Zr-5Bl. Соответственно, эти результаты показывают, что 89Zr-5Bl может быть использован в качестве молекулярного зонда для раннего обнаружения экспрессирующих sLea злокачественных новообразований в клинике.
Клеточные линии и культура ткани
Все манипуляции с культурами ткани выполняли, следуя приемам стерилизации. Клетки DMS79 мелкоклеточного рака легких и клетки BxPC3 рака поджелудочной железы получали от американской коллекции типовых культур (АТСС, Manassas, VA). Клетки Colo205-luc колоректального рака (Bioware Ultra) приобретали у Caliper Life Sciences (CLS, Hopkinton, MA). Все клетки выращивали в соответствии с рекомендациями АТСС и CLS при 37°С с 5% СO2 в увлажненной атмосфере.
Оценка уровней экспрессии sLea с помощью FACS
Проточную цитометрию указанных культивируемых линий раковых клеток проводили, как описано в настоящем документе в примере I. Вкратце, клонированные клеточные суспензии 1×106 культивируемых опухолевых клеток на пробирку отмывали в PBS с 3% фетальной бычьей сывороткой (FBS). Затем добавляли моноклональные антитела человека r5Bl (IgG против sLea) 20 мкг/мл на пробирку, и инкубировали на льду в течение 30 мин. После промывки в PBS с 3% FBS добавляли 20 мкл разбавленного 1:25 козьего антитела против IgG человека, меченного изотиоцианатом флуоресцеина (FITC, Southern Biotechnology, Birmingham, AL), и смесь инкубировали в течение еще 30 минут на льду. После окончательной промывки позитивную популяцию и медиану интенсивности флуоресценции окрашенных клеток дифференцировали с помощью сканирования FACS (Becton & Dickinson, San Jose, CA). Клетки, окрашенные только козьим антителом против IgG, меченным изотиоцианатом флуоресцеина, использовали для настройки FACScan, приводившей в результате к 1% как к фону для сравнения процента позитивных клеток, окрашенных первичным mAb.
Получение антител, меченных 89Zr
Рекомбинантные антитела 5B1 получали и очищали, как описано в настоящем документе. Антитела 5B1 и неспецифические IgG человека функционализировали п-изотиоцианатобензил-дезферриоксамином (DFO-Bz-NCS, Macrocyclics, Inc., Dallas, TX) в отношении 1:4 mAb:DFO-BZ-NCS. Например, к 300 мкл 5B1 (1,23 мг в PBS, рН~9), добавляли объем 7,2 мкл DFO-BZ-NCS (4,25 мМ в ДМСО). Реакционную смесь инкубировали при 37°С в течение 1-1,5 час. Функционализированные антитела очищали либо с помощью обессоливающей колонки PD10 (GE Healthcare), либо с помощью 10 кДа центробежного фильтра (Amicon).
Zr-89 получали с помощью бомбардировки протонным пучком фольги иттрия и выделяли с высокой степенью чистоты в виде оксалата Zr-89 в MSKCC в соответствии с ранее разработанным методом. Holland et al., Nuclear Medicine and Biology 36:729-39 (2009). Мечение антител происходило с помощью методов, описанных Holland et al., Journal of Nuclear Medicine official publication, Society of Nuclear Medicine 51:1293-300 (2010). В общем, оксалат Zr-89 нейтрализовали до рН 7,0-7,2 с помощью 1 М Na2CO3. Затем добавляли DFO-антитела. Реакционную смесь инкубировали при комнатной температуре в течение 1-2 часов. Последующую очистку проводили с использованием любой обессоливающей колонки PD10 с 0,9% физиологическим раствором.
Эксперименты in vitro
89Zr-5Bl исследовали на стабильность in vitro в 0,9% физиологическом растворе и в 1% бычьем сывороточном альбумине в течение 5 дней при температуре 37°C. Изменения радиохимической чистоты контролировали при t=0-5 дней с помощью излучающей iTLC с 50 мМ DTPA в качестве подвижной фазы. Тесты на иммунореактивность in vitro проводили в соответствии с протоколом, описанным Lindmo et al., Journal of Immunological Methods 72:77-89 (1984), для демонстрации целостности антител, меченных радиоактивным Zr-89.
Животные модели
Все исследования на животных были проведены в соответствии с руководящими принципами, установленными комитетом по уходу и использованию животных института. У самок мышей CB17SC-F SCID (Jackson Laboratories, 6-8 недель, 20-22 г) или голых бестимусных (nu/nu) мышей индуцировали развитие опухолей на задних лапах. Все клеточные линии инокулировали подкожно в 200 мкл 1:1 среда:раствор матригеля (BD Biosciences), и выращивали до максимального объема опухоли 250 мм3 перед использованием.
Исследования биораспределения
Исследования биораспределения проводили на нескольких когортах мышей-носителей отдельных ксенотрансплантатов Colo205-luc колоректального рака, BxPC3 рака поджелудочной железы и DMS79 мелкоклеточного рака легких (n=3-5). Zr-89 mAbs (370-740×109 мкБк, 1-2 мкг) в 100 мкл 0,9% физиологического раствора вводили внутривенно в латеральную вену. Дополнительное немеченое mAb (10-50 мкг) вводили совместно с меченым. Исследование блокирования с помощью 250 мкг избытка немеченого mAb проводили с целью изучения специфичности антитела против sLea в когорте мышей. После каждой временной точки (t= 24, 48, 120 час после введения) мышей усыпляли асфиксией СO2. Кровь собирали немедленно посредством пункции сердца, наряду с этим собирали опухоли вместе с выбранными органами. Измеряли сырую массу каждой ткани, и радиоактивность, связанную с каждым органом, подсчитывали с помощью гамма-счетчика Wizard2 2480 (Perkin Elmer). Процент захвата метки, выраженный как % вводимой дозы на грамм (%ID/г), рассчитывали как активность, связанную с тканью, на массу органа, на фактически вводимую дозу с коррекцией на распад к времени подсчета.
Иммуно-PET для мелких животных
Эксперименты по визуализации выполняли с помощью microPET Focus 120 или сканера R4 (Concorde Microsystems). Мышам (n=3-5) вводили антитела, меченные Zr-89 (7400-11100 мкБк, 15-25 мкг), в 100-200 мкл составов 0,9% физиологического раствора путем инъекций в латеральную хвостовую вену. Полученные PET-изображения всего тела мышей записывали через 24-96 час после инъекций при анестезии 1,5-2,0% изофлураном (Baxter Healthcare) в кислороде. Изображения анализировали с помощью программного обеспечения ASIPro VM™ (Concorde) Microsystems. Получали изображения областей, представляющих интерес (ROI), и представляли в зависимости от времени.
Иммуногистохимия
Биотинилированные 5B1 получали путем инкубации с 20× молярным избытком сульфо-NHS-LC-биотина (Thermo Scientific/Pierce Cat # 21327) в течение 30 минут при комнатной температуре. Свободный биотин удаляли с помощью обессоливающих спин-колонок Zebra™ (Thermo Scientific/Pierce, Cat # 89889) в соответствии с инструкциями изготовителя. Проводили замену буфера антител на PBS, содержащий 0,01% азид натрия в концентрации 1,1 мг/мл. Связывание на клетках DMS79 подтверждали FACS, и оно было сравнимо с родительским антителом 5B1.
Предварительные условия иммуногистохимического окрашивания определяли с использованием клеток Colo205 в качестве положительного контроля и клеток SK-MEL28 в качестве отрицательного контроля. Получали клеточную массу, фиксировали формалином и заключали в парафин. Срезы инкубировали с биотинилированными 5B1, разведенными в 10% (об/об) нормальной сыворотке человека в PBS (Jackson ImmunoResearch Labs; cat# 009-000-121). Окрашивание проводили с помощью автоматизированной системы Ventana (Discovery XT platform-Ventana Medical Systems, Inc, Tucson, AZ) стандартным стрептавидин-биотин-иммунопероксидазным методом и с помощью DAB-системы детектирования в качестве метода окрашивания. Открытие антигена проводили с использованием нагревания и кондиционирующего раствора Ventana’s CC1. Мышиное моноклональное антитело CA 19,9 (клон 116-NS-19-9) от Signet (Covance) давало сопоставимые результаты в пилотном исследовании. Клетки Colo205 имели интенсивную положительную окраску биотинилированными 5B1 при использовании в концентрации 10 мкг/мл, в то время как клетки SKMEL28 были негативными по окраске. Тканевые микроматрицы Histo-Array™ приобретали у Imgenex (San Diego, CA). Использовали следующие срезы, содержащие центральную массу биопсии опухолей, а также центральную массу некоторых нормальных тканей: IMH-327 (наиболее распространенные виды рака, 59 образцов), IMH-359 (колоректальная: рак-метастаз-нормальная ткань; 59 образцов) и IMH-324 (метастатический рак яичника). Центральные массы опухолевой ткани поджелудочной железы присутствовали в IMH-327.
Концентрации sLea в сыворотке in vivo
У мышей, несущих ксенотрансплантаты Colo205, BxPC3 и DMS79, брали кровь для анализа антигена sLea. Группа мышей, не имеющих опухолей, служила в качестве контроля. Уровни sLea в сыворотке мышей измеряли с использованием набора ST AIA-PACK CA19.9 (Cat # 025271, TOSOH Bioscience Inc, South San Francisco, CA). Принцип анализа основан на двухслойном количественном иммуноферментном анализе. Анализ проводили, как описано в инструкции изготовителя. Оптическую плотность иммуноаналитических планшетов измеряли на автоматическом иммуноаналитическом анализаторе TOSOH AIA2000 (Tosoh Bioscience, Inc., San Francisco, CA).
Статистический анализ
Значения данных выражали как среднее ± стандартное отклонение, если не указано иное. Статистический анализ проводили с использованием программного обеспечения GraphPad Prism версии 5.03 с использованием однофакторного ANOVA с последующим критерием Dunnett. Значения р<0,05 рассматривались как статистически значимые.
Результаты
Специфичность связывания 5B1 проверяли путем окрашивания выбранных микрочипов злокачественных и нормальных тканей. Реактивность 5B1 была ограничена злокачественными опухолями и случайными нормальными тканями, ранее известными как гиперэкспрессирующие sLea (Фиг. 19; таблица 9). В большинстве нормальные ткани были полностью негативными (таблица 9). В противоположность этому, сильное положительное окрашивание было обнаружено в 21/34 аденокарциномах толстой кишки (62%), 33/57 метастазах аденокарциномы в яичники (58%) и 7/9 протокового рака поджелудочной железы (66%) на различных стадиях (таблица 10). Как показано на фиг. 19, типичная реактивность представляла собой диффузное окрашивание цитоплазмы с некоторыми опухолевыми клетками, четко демонстрирующими окрашивание клеточной мембраны. Кроме того, некоторые варианты перстневидноклеточного рака яичников и некоторые виды рака легких и молочной железы также оказались сильно положительными. В противоположность этому, только 4/43 образцов рака простаты и 0/51 случаев GIST были позитивными (данные не показаны).
Обзор связывания 5B1 с нормальными тканями
Окрашивание 5B1 протоковых аденокарцином поджелудочной железы
Высокая специфичность иммуноокрашивания 5B1 раковых тканей, экспрессирующих sLea, послужила основанием для использования этого mAb в качестве PET-зонда. Создавали модификацию 5B1 бензилизотиоцианатным аналогом дезфероксамина (DFO-BZ-NCS) в соотношении 4:1 (хелат:mAb) с последующей очисткой с помощью центробежной фильтрации с использованием физиологического раствора в качестве промывочного буфера. Легко достижимое введение радиоактивной метки Zr-89 проходило при комнатной температуре после доведения рН до 7,0-7,2. Более узкий диапазон рН, близкий к нейтральному, необходим для достижения оптимального >80% выхода при введении радиоактивной метки. Свободный, несвязанный Zr-89 удаляли с помощью обессоливающей колонки PD10. Концентрирование продукта осуществляли с использованием центробежной фильтрации (MWCO: 10 кДа). Установлена относительно высокая удельная активность 447,7×109±40,7×109 мБк/мг. Перед использованием обеспечивали радиохимическую чистоту более чем на 95%. Анализы иммунореактивности показали сохранение активности sLea (72,4±1,1%, n=3). Стабильность в бычьем сывороточном альбумине при 37°С поддерживалась на уровне >95% в течение 5 дней (данные не показаны). В физиологическом растворе деметаллирование наблюдалось уже через 24 час (>85% в целом) с приблизительно >75% связанного радиоактивного металла через 120 час при 37°С.
Исследования PET-визуализации мелких животных и биораспределения проводили с использованием самок мышей SCID с подкожно имплантированными ксенотрансплантатами BxPC3 рака поджелудочной железы на левой задней ноге. Полученные PET-изображения подтвердили с помощью 89Zr-5Bl существенное ограничение sLea, связанное с опухолью. Из проекций максимальной интенсивности (MIP) на фиг. 20 ксенотрансплантаты BxPC3 (n=3) показали чрезвычайную аккумуляцию радиоактивной метки, вводимой внутривенно. Области, представляющие интерес (ROI), обрисовывающие опухоль в изображениях PET, показали захват 5,0±0,4% ID/г (2 час), 16,2±2,5% ID/г (24 час), 23,8±4,7% ID/г (48 час), 36,8±6,1% ID/г (96 час) и 49,5±7,7% ID/г (120 час). Связывающая активность пула крови и нормальных тканей очевидно исчезала через 24 час после введения. Результаты экспериментов по биораспределению согласуются с данными PET. Наблюдался высокий уровень локализации в опухоли 89Zr-5Bl через 24 час (84,7±12,3% ID/г, n=4); повышенный захват наблюдался далее через 120 час после введения (114,1±23,1% ID/г, n=4) (фиг. 21). Захват опухолью превышает 100% из-за малой массы (62,4±0,03 мг). % ID через 24 час после инъекции, как установлено, был в десять раз выше, чем у неспецифически связывающего IgG в сходных временных точках (фиг. 21, вставка). Конкурентное ингибирование с помощью 250 мкг немеченого 5B1 через 24 час после инъекции блокировало аккумуляцию метки, указывая на специфичность захвата. Наблюдалось минимальное связывание 89Zr-5Bl с нормальной поджелудочной железой и остальными собранными нормальными тканями, что обеспечивает высокий контраст между опухолью и тканями во всех временных точках.
Следуя приведенным выше результатам, 89Zr-5Bl анализировали в ортотопической модели опухоли поджелудочной железы BxPC3. Ортотопические модели являются клинически значимыми и предоставляют клинически приемлемое тестирование эффективности зонда PET. После инокуляции в поджелудочную железу рост опухоли контролировали еженедельно с помощью биолюминесцентного оптического изображения. Эксперименты по PET-визуализации проводили после того, как опухоли становились пальпируемыми. Сравнение свойств очерчивающего опухоли зонда было сделано между FDG-PET и 89Zr-5Bl (фиг. 25). Компьютерная томография (КТ) в сочетании с PET обеспечивала улучшенную визуализацию анатомической области, представляющей интерес.
Для оценки 89Zr-5Bl в качестве зонда PET для других аденокарцином, экспрессирующих sLea, 89Zr-5Bl анализировали в моделях рака легких и толстой кишки. Эксперименты на мелких животных проводили с использованием клеток DMS79 мелкоклеточного рака легких и клеток Colo205-luc рака толстой кишки, вводимых подкожно в правую заднюю лапу самок мышей SCID. PET-изображения MIP получали через 24-120 час после внутривенного введения 7400-11100 мкБк (16-25 мкг). Гетерогенный захват опухолью DMS79 продемонстрирован при 38,15±2,12% ID/г уже через 24 час после введения с прекрасным сигналом относительно фона (фиг. 22A). Повышение аккумуляции метки в опухоли происходило через 48 час после введения (44,60±6,47% ID/г) с сохранением через 120 час после введения (41,97±12,23% ID/г). Неспецифически связанное 89Zr-5Bl быстро вымывалось из нормальных тканей с от минимального захвата до отсутствия фонового захвата через 48 час после введения. Кроме того, очерчивание опухоли наблюдалось у ксенотрансплантатов Colo205-luc, как показано на фиг. 22B через 24-120 час после введения. ROIs продемонстрировали накопление в опухоли с 10,5±0,76, 23,5±2,7, 24,8±4,0, 18,4±4,7, 16,5±2,3% ID/г через 2, 24, 48, 96 и 120 час, соответственно. Наблюдаемое повышение аккумуляции в печени происходило с течением времени с последующим снижением захвата опухолью, как показано для области, представляющей интерес, из изображений PET (фиг. 22C). Данные, полученные из исследований по биораспределению, хорошо коррелируют с наблюдаемыми результатами PET (данные не показаны). Уровень sLea в сыворотке мышей количественно оценивали по мере прогрессии опухолей. Осуществляли получение крови у мышей SCID, несущих ксенотрансплантаты Colo205, DMS79 и BxPC3, в качестве контроля служила группа, не несущая опухоли. Величины sLea были на высоком уровне у мышей, нагруженных Colo205, по сравнению с мышами с имплантацией BxPC3 поджелудочной железы и DSM79 (таблица 11).
Величины sLea у мышей, несущих ксенотрансплантаты опухолей прямой кишки (Colo205), поджелудочной железы (BxPC3) и мелкоклеточного рака легкого (DMS79), по сравнению с контролем.
Эти результаты показывают, что антитело против Slea (89Zr-5B1) является специфическим для обнаружения и диагностики аденокарциномы поджелудочной железы и других sLea-позитивных аденокарцином. 89Zr-5Bl получали с прекрасным выходом и чистотой наряду с высокой удельной активностью и удерживаемой иммунореактивностью. Оценка 89Zr-5Bl в подкожных, ортотопических и метастатических моделях опухолей поджелудочной железы давала отличное разграничение опухоли и диагностику. Доклиническая оценка этой радиоактивной метки у мелких животных-опухоленосителей, несущих опухоль толстой кишки и мелкоклеточную опухоль легкого, продемонстрировала универсальную пригодность этой метки для злокачественных опухолей, экспрессирующих sLea.
ПРИМЕР III
Диатела против sLea, связывающиеся с различными клеточными линиями рака
Получено два диатела с использованием доменов VH и VL изолятов клонов 5B1 и 7E3, описанных в настоящем документе, обозначенных 5BlCysDb и 7E3CysDb, соответственно (фиг. 9 и 10). Оба диатела содержали пять областей аминокислотных линкеров между доменами VL и VH. Полигистидиновую метку на C-конце, которую использовали для очистки и детекции, также включали в оба диатела.
Связывание 5BlCysDb и 7E3CysDb с тремя линиями раковых клеток: (1) клетками DMS-79, суспензионной клеточной линии мелкоклеточного рака легкого; (2) клетками Capan-2, клетками аденокарциномы поджелудочной железы; и (3) клетками BxPC3, клетками рака поджелудочной железы, анализировали путем инкубации 0,25 миллиона клеток в 0,2 мл с 10 мкг/мл 5BlCysDb или 7E3CysDb, соответственно. Сочетания клеток и диатела инкубировали в течение 40 минут на льду в PBS/2% FBS.
После промывки клетки инкубировали в течение 40 мин с 0,2 мл ALEXA-488-меченным антителом против His, разведенным 1:1000 (Life Technology, Cat # A21215). После второй промывки клетки анализировали на проточном цитометре Guava. Как 5BlCysDb, так и 7E3CysDb продемонстрировали значительное связывание с DMS-79, Capan-2 и клетками BxPC3 (таблица 12).
Связывание 5BlCysDb и 7E3CysDb с клеточными линиями
ПРИМЕР IV
Введение 5B1 и таксола ингибирует рост опухоли
Противоопухолевую активность вводимых совместно антитела против sLea (5B1) и химиотерапевтического агента таксола (паклитаксела) оценивали в моделях ксенотрансплантатов рака поджелудочной железы и мелкоклеточного рака легких. Как описано в настоящем документе выше, 1 миллион клеток BxPC3 (клеток опухоли поджелудочной железы) или 5 миллионов клеток DMS-79 (клеток мелкоклеточного рака легких) вводили в задний пах 6-недельным самкам мышей СВ17 SCID (день 0, N=5), опухолям DMS79 давали расти в течение 21 дня, до тех пор, пока средний размер опухоли составил 193±64 мм3. IgG человека или 5B1 (0,5 или 1 мг) вводили в./б. дважды в неделю (начиная на 21-й день), и таксол (0,2 мг/доза) вводили в./в. на 23, 30, 37 и 44 дни. В модели ксенотрансплантата DMS-79 совместное введение антитела 5B1 и таксола значительно ограничивало рост опухоли и приводило к регрессии опухоли по сравнению с контрольным IgG человека или с индивидуальным введением антитела 5B1 и таксола (фиг. 23).
В модели ксенотрансплантата BxPC3 опухоли росли в течение 14 дней, после чего они достигали в среднем 126±30 мм3. Таксол вводили внутривенно на 14, 21, 28 и 34 дни (еженедельно) и 5B1 давали дважды в неделю, начиная с 14-го дня. Совместное введение антитела 5B1 и таксола значительно ограничивало рост опухоли по сравнению с контролями или с индивидуальным введением антитела 5B1 и таксола (фиг. 24). Эти результаты демонстрируют синергичный эффект антитела против sLea и химиотерапевтического агента в предотвращении роста опухоли и/или уменьшении размера опухолей рака поджелудочной железы и мелкоклеточного рака легкого.
В данной заявке даны ссылки на различные публикации. Раскрытия этих публикаций тем самым включены в данную заявку в полном объеме в качестве ссылки для того, чтобы более полно описать состояние техники, к которой относится данное изобретение. Несмотря на то, что изобретение описано со ссылками на примеры, представленные выше, следует понимать, что различные модификации могут быть сделаны без отхода от сущности настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MabVax Therapeutics, Inc.
<120> Нуклеиновые кислоты, кодирующие антитела против
сиалированного антигена Льюисаа человека
<130> 12967-033-228
<140> TBA
<141> 2014-08-26
<150> US 61/870,137
<151> 2013-08-26
<160> 20
<170> Патент версии 3.5
<210> 1
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 5B1
<400> 1
atggagtttg ggctgagctg gctttttctt gtggctattt taaaaggcgt acagtgccag 60
gtgcagctgg tggagtctgg gggaggctcg gtgcagcctg gcaggtccct gagactctcc 120
tgtgaagcct ctggattcac ctttgaggcc tatgccatgc actgggtccg gcaacctcca 180
gggaagggcc tggagtgggt ctcaagtatt aattggaata gtggtcgcat agcctatgcg 240
gactctgtga agggccgatt caccatctcc agagacaacg ccaggaattc cctgtatctg 300
caaatgaaca gtctgagact tgaggacacg gccttctatt actgtgcaaa agatatacgg 360
aggtttagta ccgggggggc ggagtttgag tactggggcc agggaaccct ggtcaccgtc 420
tcctca 426
<210> 2
<211> 142
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 5B1
<400> 2
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Glu Ala Ser Gly Phe Thr Phe
35 40 45
Glu Ala Tyr Ala Met His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ser Ile Asn Trp Asn Ser Gly Arg Ile Ala Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Leu Glu Asp Thr Ala Phe
100 105 110
Tyr Tyr Cys Ala Lys Asp Ile Arg Arg Phe Ser Thr Gly Gly Ala Glu
115 120 125
Phe Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
130 135 140
<210> 3
<211> 390
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 5B1
<400> 3
atggccggct tccctctcct cctcaccctc ctcactcact gtgcagggtc ttgggcccag 60
tctgtgctga ctcagccgcc ctcagcgtct gggacccccg ggcagagggt caccatctct 120
tgttctggaa gcagctccaa catcggaagt aattttgtat actggtacca gcagctccca 180
ggaacggccc ccaaactcct catatatagg aataatcagc ggccctcagg ggtccctgac 240
cgattctctg gctccaggtc tggcacctca gcctccctgg ccatcagtgg actccggtcc 300
gaggatgagg ctgattatta ctgtgcagca tgggatgaca gcctgggagg ccattatgtc 360
ttcggaactg ggaccaaggt caccgtcctt 390
<210> 4
<211> 130
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 5B1
<400> 4
Met Ala Gly Phe Pro Leu Leu Leu Thr Leu Leu Thr His Cys Ala Gly
1 5 10 15
Ser Trp Ala Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
20 25 30
Pro Gly Gln Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile
35 40 45
Gly Ser Asn Phe Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
50 55 60
Lys Leu Leu Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp
65 70 75 80
Arg Phe Ser Gly Ser Arg Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
85 90 95
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp
100 105 110
Asp Ser Leu Gly Gly His Tyr Val Phe Gly Thr Gly Thr Lys Val Thr
115 120 125
Val Leu
130
<210> 5
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 9H3
<400> 5
atggagtttg ggctgagctg gctttttctt gtggctattt taaaaggcgt acagtgcgaa 60
gtgcagctgt tggagtctgg gggaggcttg gtacagcctg gcaggtccct gagactctcc 120
tgtgcggcct ctggatttac ctttgatgat tatgtcatgc actgggtccg gcaagctcca 180
gggaagggcc tggagtgggt ctcaagtatt agttggaata gtggtagcat aggctatgcg 240
gactctgtga agggccgatt catcatctcc agagacaacg ccaagaactc cctgtatctg 300
caaatgaaca gtctgagagc tgaggacacg gccttgtatt actgtgcaaa agatcgtcgt 360
attaggggtg actcggggtt cgagggtgac tactggggcc agggaaccct ggtcaccgtc 420
tcctca 426
<210> 6
<211> 142
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 9H3
<400> 6
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Asp Asp Tyr Val Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ser Ile Ser Trp Asn Ser Gly Ser Ile Gly Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu
100 105 110
Tyr Tyr Cys Ala Lys Asp Arg Arg Ile Arg Gly Asp Ser Gly Phe Glu
115 120 125
Gly Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
130 135 140
<210> 7
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 9H3
<400> 7
atggccggct tccctctcct cctcaccctc ctcactcact gtgcagggtc ttgggcccag 60
tctgtgttga cgcagccgcc ctcagcgtct gggacccccg ggcagagggt caccatctct 120
tgttctggaa gcagctccaa catcggaagt aattatgtat actggtacca gcagctccca 180
ggaacggccc ccaaactcct catctatagg aataatcagc ggccctcagg ggtccctgac 240
cgattctctg gctccaagtc tggcacctca gcctccctgg ccatcagtgg gctccggtcc 300
gaggatgagg ctgattatta ctgtgcagca tgggatgcca gcctgagtgg tgtggtattc 360
ggcggaggga ccaagctgac cgtccta 387
<210> 8
<211> 129
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 9H3
<400> 8
Met Ala Gly Phe Pro Leu Leu Leu Thr Leu Leu Thr His Cys Ala Gly
1 5 10 15
Ser Trp Ala Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
20 25 30
Pro Gly Gln Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile
35 40 45
Gly Ser Asn Tyr Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
50 55 60
Lys Leu Leu Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp
65 70 75 80
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
85 90 95
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp
100 105 110
Ala Ser Leu Ser Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val
115 120 125
Leu
<210> 9
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 5H11
<400> 9
atggagtttg ggctgagctg gctttttctt gtggctattt taaaaggcgt acagtgccag 60
gtgcagctgt tggagtctgg gggaggcttg gtacagcctg gcaggtccct gagactctcc 120
tgtgcagcct ctggattcac ctttgatgaa tatgccatgc actgggtccg gcaagctcca 180
gggaagggcc tggagtgggt ctcaagtgtt agttggaata gtggtagcat aggctatgcg 240
gactctgtga agggccgatt caccatctcc agagacaacg ccaagaactc cctgtatcta 300
caaatgaaca gtctgagagc tgaggacacg gccttgtatt actgtgcaaa agatatacgg 360
acctatagca ccgggggggc ggagtttgcc tcctggggcc agggaaccct ggtcaccgcc 420
tcctca 426
<210> 10
<211> 142
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 5H11
<400> 10
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Asp Glu Tyr Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ser Val Ser Trp Asn Ser Gly Ser Ile Gly Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu
100 105 110
Tyr Tyr Cys Ala Lys Asp Ile Arg Thr Tyr Ser Thr Gly Gly Ala Glu
115 120 125
Phe Ala Ser Trp Gly Gln Gly Thr Leu Val Thr Ala Ser Ser
130 135 140
<210> 11
<211> 390
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 5H11
<400> 11
atggccggct tccctctcct cctcaccctc ctcactcact gtgcagggtc ttgggcccag 60
tctgtgttga cgcagccgcc ctcagcgtct gggacccccg ggcagagggt caccatctct 120
tgttctggaa gcagctccaa catcggaagt aattatgtat actggtacca gcaggtccca 180
ggaacggccc ccaaactcct catctatagg aataatcagc ggccctcagg ggtccctgac 240
cgattctctg gctccaagtc tggcacctca gcctccctgg ccatcagtgg gctccggtcc 300
gaggatgagg ctgattatta ctgtgcagca tgggatgaca gcctgagtgg ccattatgtc 360
ttcggaactg ggaccaaggt caccgtccta 390
<210> 12
<211> 130
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 5H11
<400> 12
Met Ala Gly Phe Pro Leu Leu Leu Thr Leu Leu Thr His Cys Ala Gly
1 5 10 15
Ser Trp Ala Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
20 25 30
Pro Gly Gln Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile
35 40 45
Gly Ser Asn Tyr Val Tyr Trp Tyr Gln Gln Val Pro Gly Thr Ala Pro
50 55 60
Lys Leu Leu Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp
65 70 75 80
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
85 90 95
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp
100 105 110
Asp Ser Leu Ser Gly His Tyr Val Phe Gly Thr Gly Thr Lys Val Thr
115 120 125
Val Leu
130
<210> 13
<211> 435
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 7E3
<400> 13
atggagtttg ggctgagctg gctttttctt gtggctattt taaaaggcgt acagtgccaa 60
gtgcagctgt tggagtctgg gggaggcgtg gtccagcctg ggaggtccct gagactctcc 120
tgtgcagcct ctggattcac cttcagtttc tatggcatgc actgggtccg ccaggctcca 180
ggcaaggggc tggagtgggt ggcagctata tcatatgatg gaagtaataa atactatgca 240
gactccgtga agggccgatt caccatctcc agagacaatt ccaagaacac gctgtatctg 300
caaatgaaca gcctgagagc tgaggacacg gctgtgtatt actgtgcgaa aaggcccaac 360
caattttatt gtagtgatgg tagatgctac tccattgact actggggcca gggaaccctg 420
gtcaccgtct cctca 435
<210> 14
<211> 145
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VH клона 7E3
<400> 14
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Val Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Phe Tyr Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ala Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Lys Arg Pro Asn Gln Phe Tyr Cys Ser Asp Gly Arg
115 120 125
Cys Tyr Ser Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
130 135 140
Ser
145
<210> 15
<211> 390
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 7E3
<400> 15
atggacatga gggtccccgc tcagctcctg gggctcctgc tactctggct ccgaggtgcc 60
cggtgtgaaa ttgtaatgac gcagtctcca gccaccctgt ctgtgtctcc aggggagaga 120
gccaccctct cctgcagggc cagtcagagt gttagcagca acttagcctg gtaccagcag 180
aaacctggcc aggctcccag gctcctcatc tatggtgcat ccaccagggc cactggtatc 240
ccagccaggt tcagtggcag tgggtctggg acagacttca ctctcaccat cagcagcctg 300
cagtctgtag attctgcagt ttattactgt cagcagtata ataactggcc tccgtacact 360
tttggccagg ggaccaagct ggagatcaaa 390
<210> 16
<211> 130
<212> PRT
<213> Искусственная последовательность
<220>
<223> Домен цепи VL клона 7E3
<400> 16
Met Asp Met Arg Val Pro Ala Gln Leu Leu Gly Leu Leu Leu Leu Trp
1 5 10 15
Leu Arg Gly Ala Arg Cys Glu Ile Val Met Thr Gln Ser Pro Ala Thr
20 25 30
Leu Ser Val Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser
35 40 45
Gln Ser Val Ser Ser Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
50 55 60
Ala Pro Arg Leu Leu Ile Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile
65 70 75 80
Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Ser Val Asp Ser Ala Val Tyr Tyr Cys Gln Gln
100 105 110
Tyr Asn Asn Trp Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
115 120 125
Ile Lys
130
<210> 17
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность диатела 5B1CysDb
<400> 17
cagtctgtgc tgacgcagcc gccctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcagctc caacatcgga agtaattttg tatactggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatatat aggaataatc agcggccctc aggggtccct 180
gaccgattct ctggctccag gtctggcacc tcagcctccc tggccatcag tggactccgg 240
tccgaggatg aggctgatta ttactgtgca gcatgggatg acagcctggg aggccattat 300
gtcttcggaa ctgggaccaa ggtcaccgtc ctttctggtg gtggtggtca ggtgcagctg 360
gtggagtctg ggggaggctc ggtgcagcct ggcaggtccc tgagactctc ctgtgaagcc 420
tctggattca cctttgaggc ctatgccatg cactgggtcc ggcaacctcc agggaagggc 480
ctggagtggg tctcaagtat taattggaat agtggtcgca tagcctatgc ggactctgtg 540
aagggccgat tcaccatctc cagagacaac gccaggaatt ccctgtatct gcaaatgaac 600
agtctgagac ttgaggacac ggccttctat tactgtgcaa aagatatacg gaggtttagt 660
accggggggg cggagtttga gtactggggc cagggaaccc tggtcaccgt ctcctcaggt 720
tctcaccatc accatcacca tggcggttgc 750
<210> 18
<211> 250
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность диатела 5B1CysDb
<400> 18
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Arg Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Gly Gly His Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Ser
100 105 110
Gly Gly Gly Gly Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val
115 120 125
Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Glu Ala Ser Gly Phe Thr
130 135 140
Phe Glu Ala Tyr Ala Met His Trp Val Arg Gln Pro Pro Gly Lys Gly
145 150 155 160
Leu Glu Trp Val Ser Ser Ile Asn Trp Asn Ser Gly Arg Ile Ala Tyr
165 170 175
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg
180 185 190
Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Leu Glu Asp Thr Ala
195 200 205
Phe Tyr Tyr Cys Ala Lys Asp Ile Arg Arg Phe Ser Thr Gly Gly Ala
210 215 220
Glu Phe Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
225 230 235 240
Ser His His His His His His Gly Gly Cys
245 250
<210> 19
<211> 750
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность диатела 7E3CysDb
<400> 19
gatgttgtgc tgacgcagtc tccagccacc ctgtctgtgt ctccagggga gagagccacc 60
ctctcctgca gggccagtca gagtgttagc agcaacttag cctggtacca gcagaaacct 120
ggccaggctc ccaggctcct catctatggt gcatccacca gggccactgg tatcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctgcagtct 240
gaagattctg cagtttatta ctgtcagcag tataataact ggcctccgta cacttttggc 300
caggggacca aggtggatat caaatctggt ggtggtggtg aagtgcagct ggtggagtct 360
gggggaggcg tggtccagcc tgggaggtcc ctgagactct cctgtgcagc ctctggattc 420
accttcagtt tctatggcat gcactgggtc cgccaggctc caggcaaggg gctggagtgg 480
gtggcagcta tatcatatga tggaagtaat aaatactatg cagactccgt gaagggccga 540
ttcaccatct ccagagacaa ttccaagaac acgctgtatc tgcaaatgaa cagcctgaga 600
gctgaggaca cggctgtgta ttactgtgcg aaaaggccca accaatttta ttgtagtgat 660
ggtagatgct actccattga ctactggggc cagggaaccc tggtcaccgt ctcctcaggt 720
tctcaccatc accatcacca tggcggttgc 750
<210> 20
<211> 249
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность диатела 7E3CysDb
<400> 20
Asp Val Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gln Ala Pro Arg Leu Leu Ile Tyr
35 40 45
Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser Glu
65 70 75 80
Asp Ser Ala Val Tyr Tyr Cys Gln Gln Tyr Asn Asn Trp Pro Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Ser Gly Gly Gly Gly
100 105 110
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
115 120 125
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Phe Tyr
130 135 140
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
145 150 155 160
Ala Ala Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
165 170 175
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
180 185 190
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
195 200 205
Ala Lys Arg Pro Asn Gln Phe Tyr Cys Ser Asp Gly Arg Cys Tyr Ser
210 215 220
Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Ser
225 230 235 240
His His His His His His Gly Gly Cys
245
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ АНТИТЕЛА ПРОТИВ СИАЛИРОВАННОГО АНТИГЕНА ЛЬЮИСА ЧЕЛОВЕКА | 2014 |
|
RU2699289C2 |
| АНТИТЕЛА ПРОТИВ CD40 И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2796413C2 |
| АНТИТЕЛА ПРОТИВ CTLA4 И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2779312C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РАКОВЫМИ КЛЕТКАМИ И ОБЕСПЕЧИВАЮТ НАПРАВЛЕННЫЙ ПЕРЕНОС РАДИОНУКЛИДОВ К УКАЗАННЫМ КЛЕТКАМ | 2020 |
|
RU2833191C2 |
| Связывающие молекулы, специфичные в отношении CD73, и пути их применения | 2015 |
|
RU2730665C2 |
| АНТИТЕЛА ПРОТИВ OX40 И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783314C2 |
| АНТИТЕЛО К SIRPα И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2822496C1 |
| СПОСОБЫ КОМБИНИРОВАННОГО ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ, ВКЛЮЧАЮЩИЕ СРЕДСТВА, БЛОКИРУЮЩИЕ CTLA-4 И PD-1 | 2020 |
|
RU2833523C2 |
| АНТИТЕЛА ПРОТИВ PD-1 И КОМПОЗИЦИИ | 2016 |
|
RU2750675C1 |
| БИПАРАТОПНЫЕ FR-АЛЬФА АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ | 2020 |
|
RU2832083C2 |











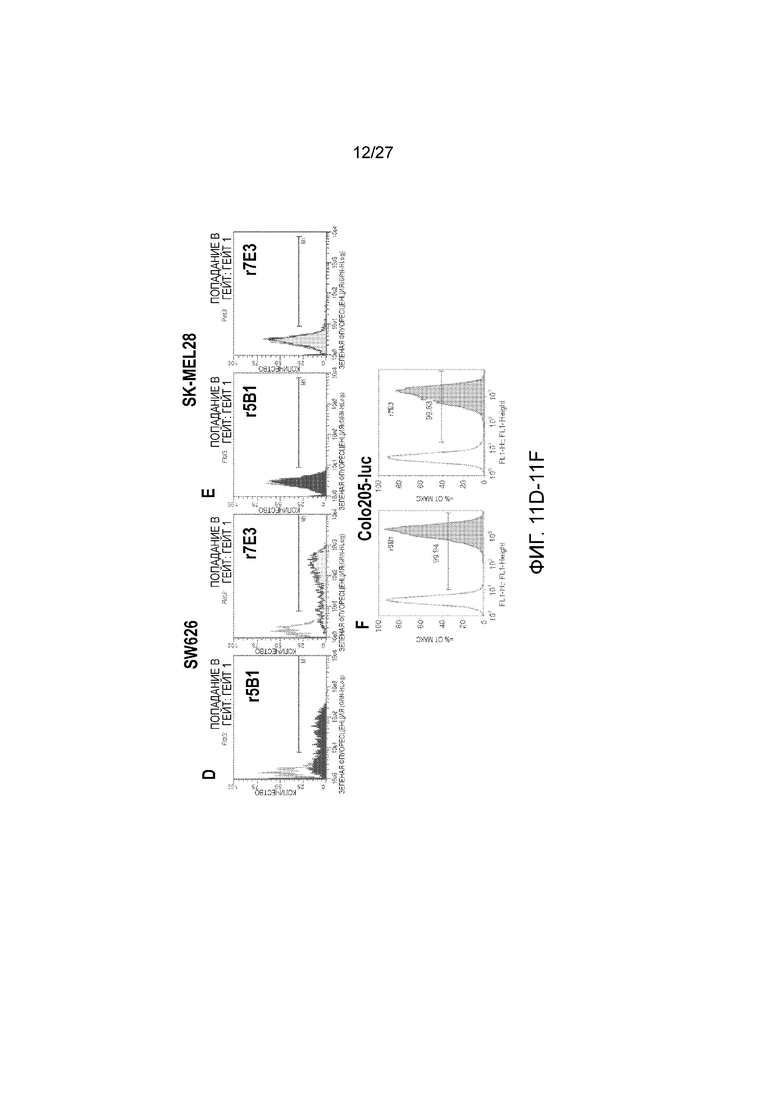
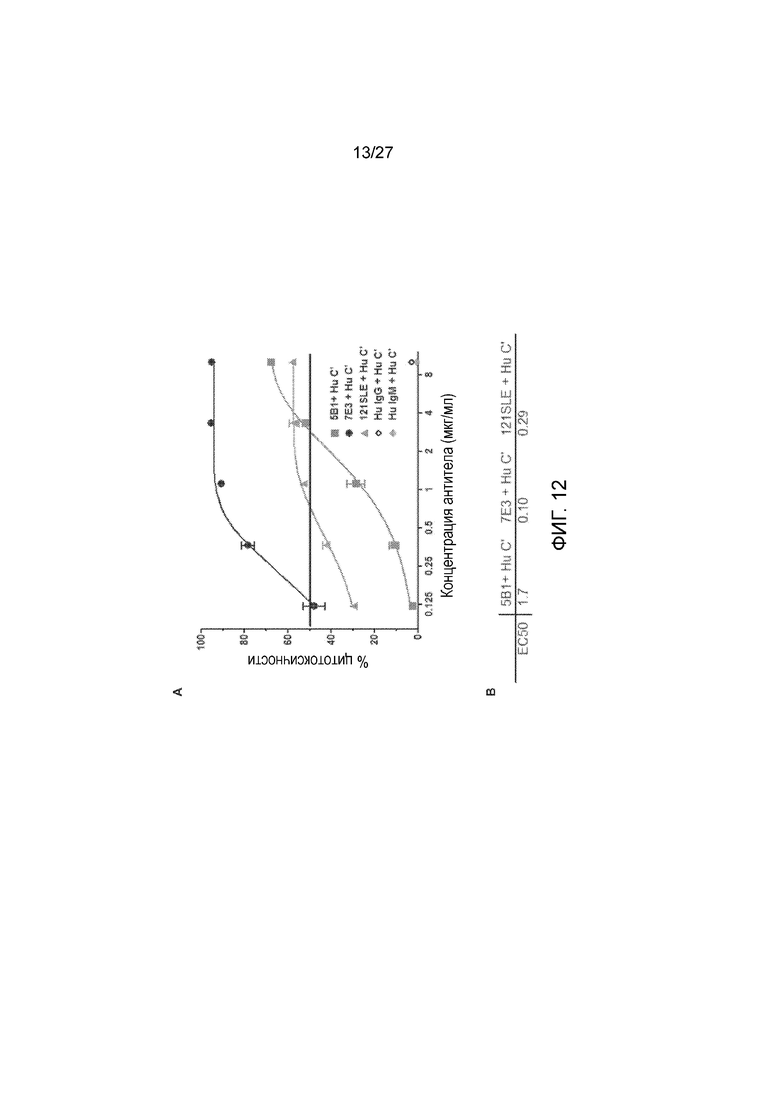
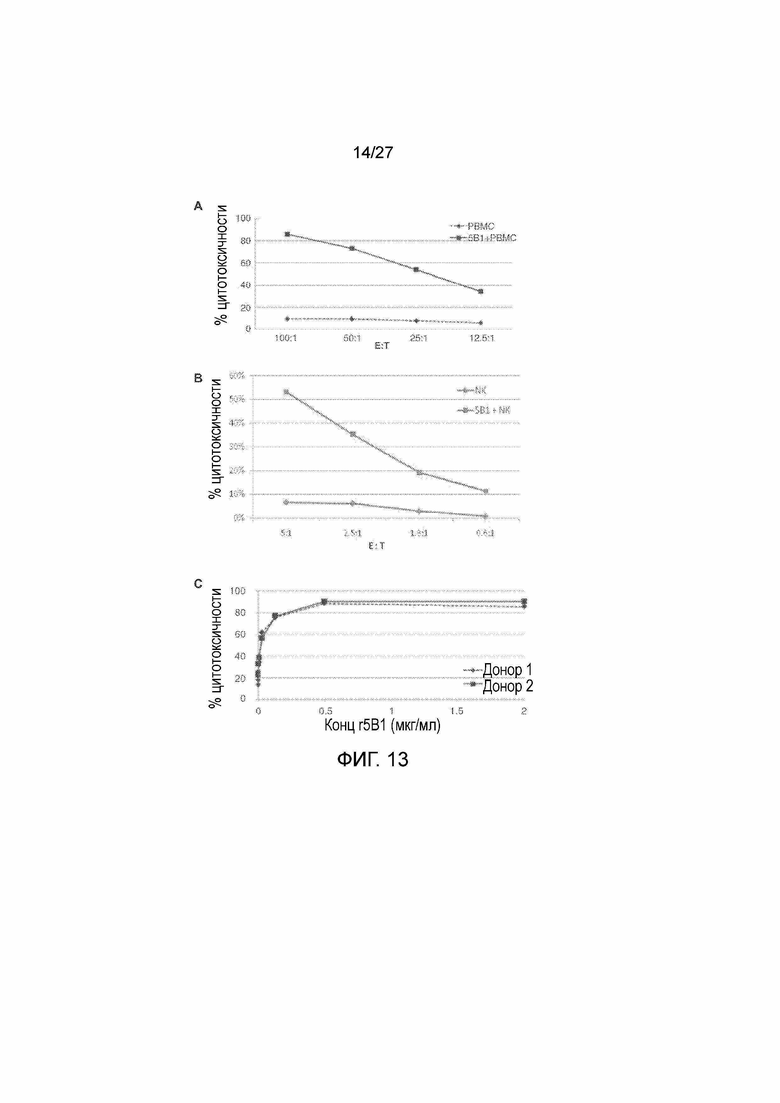
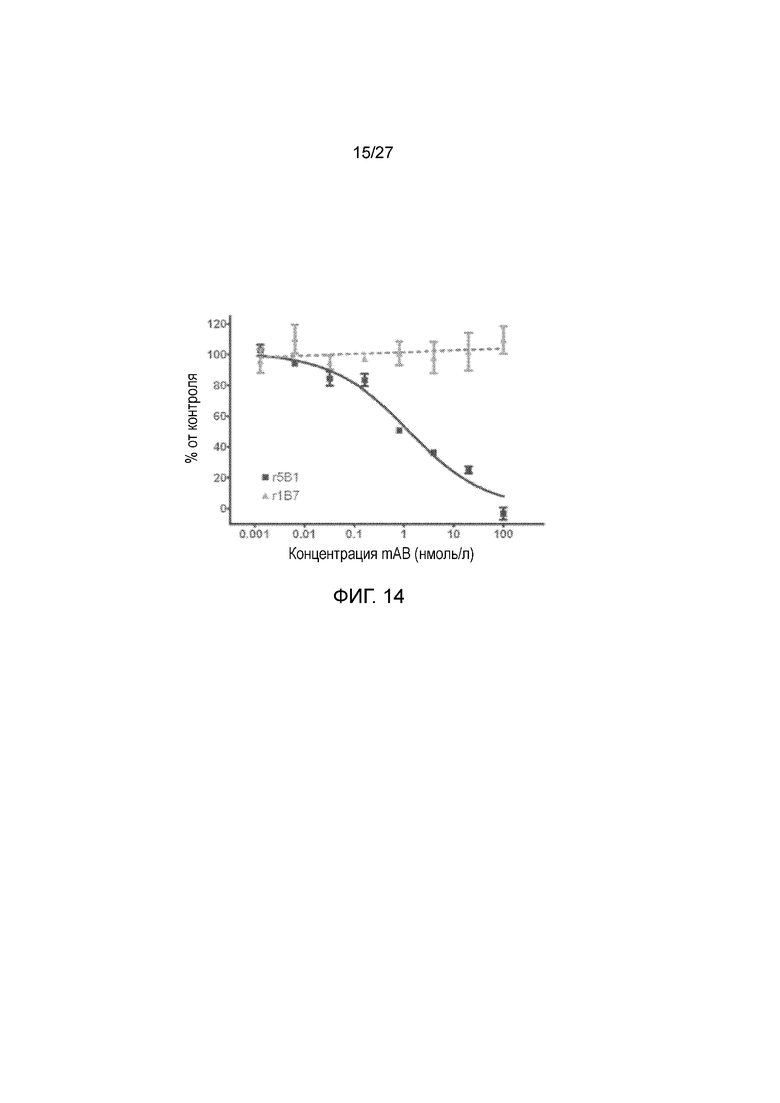
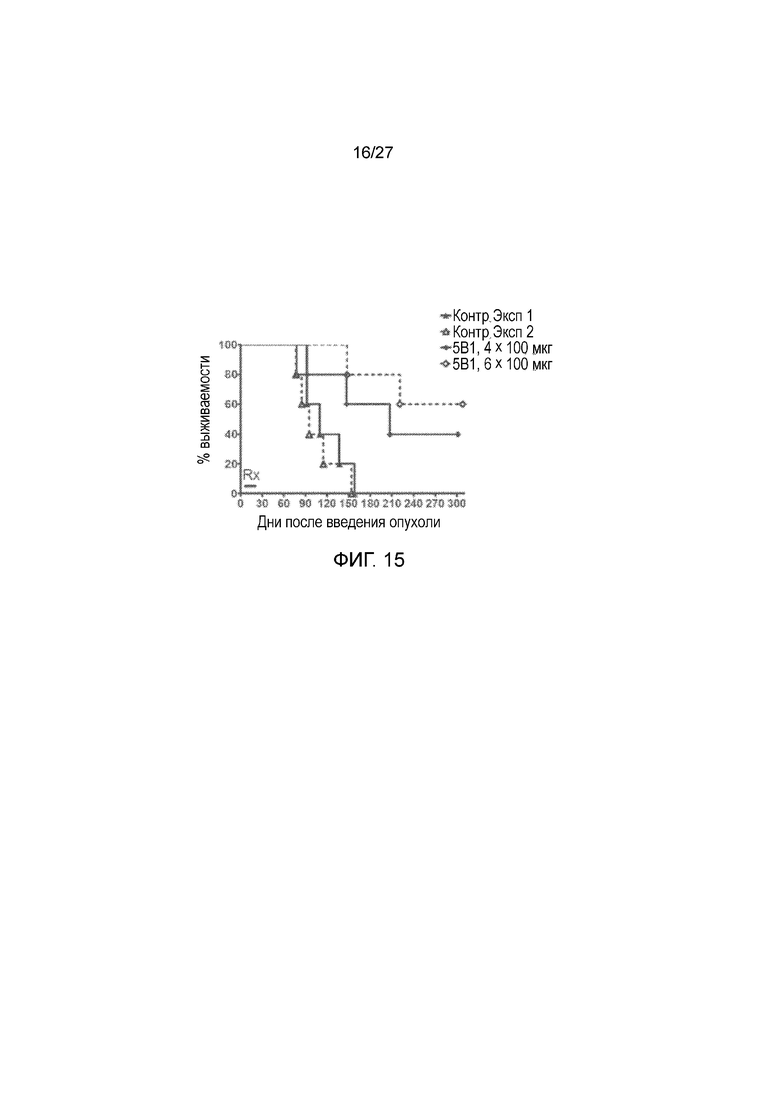
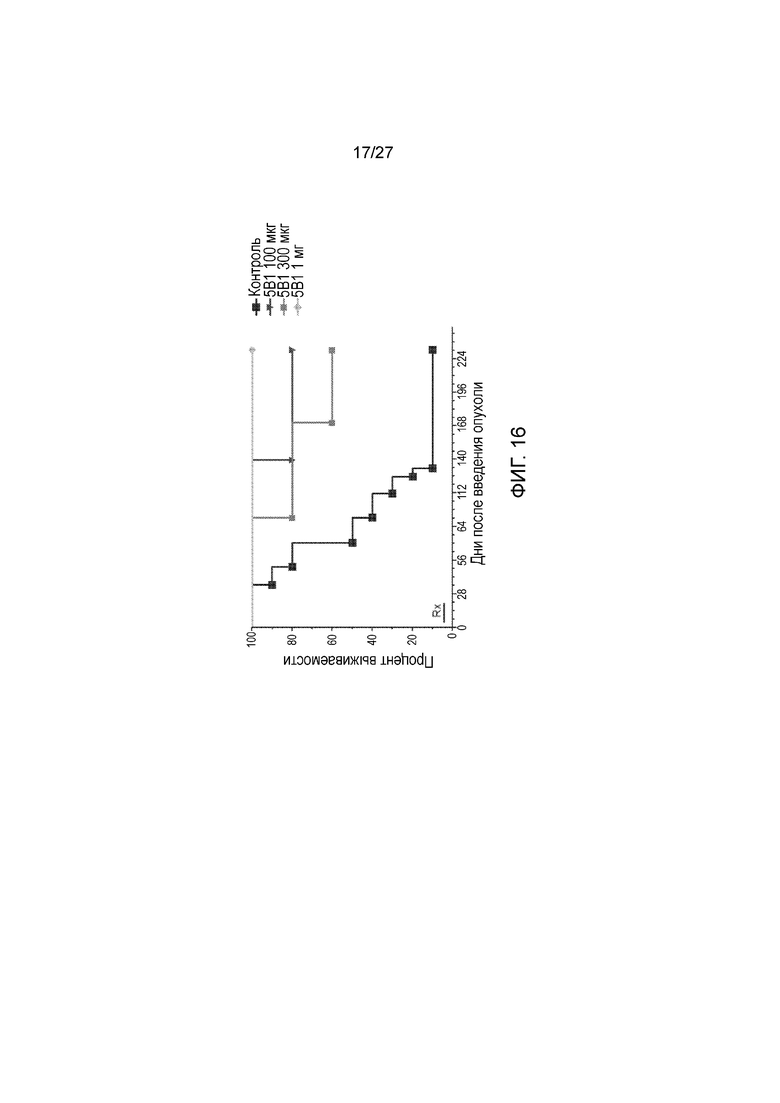
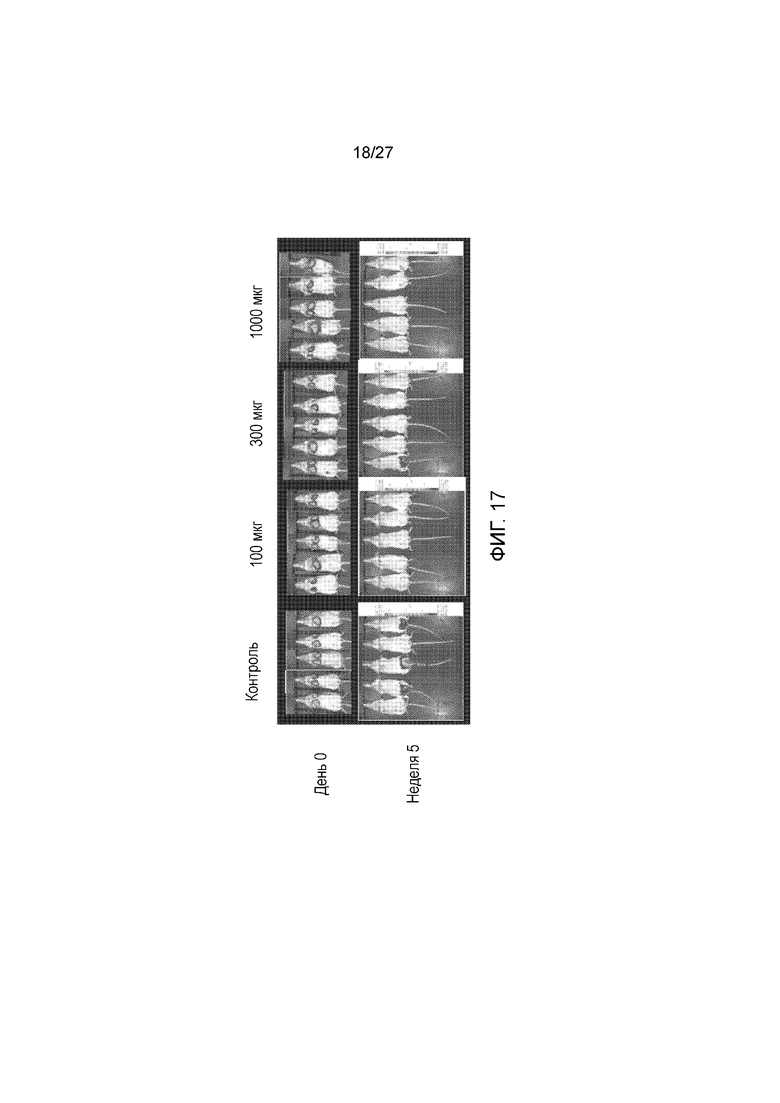

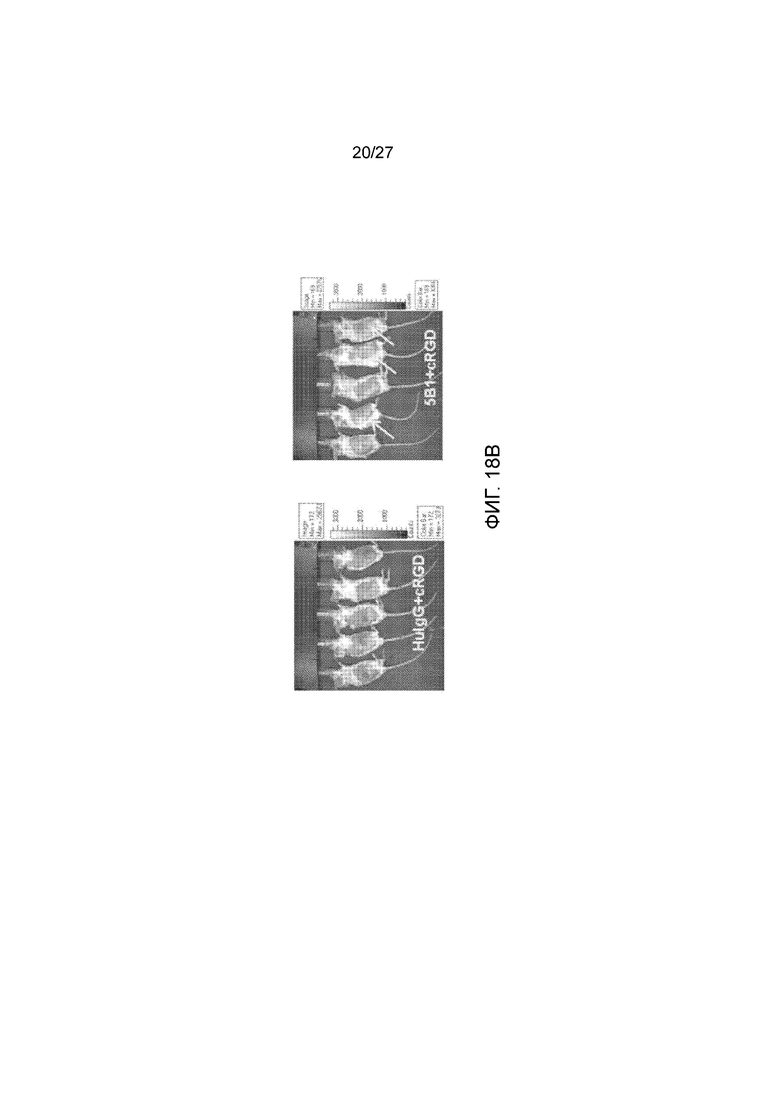
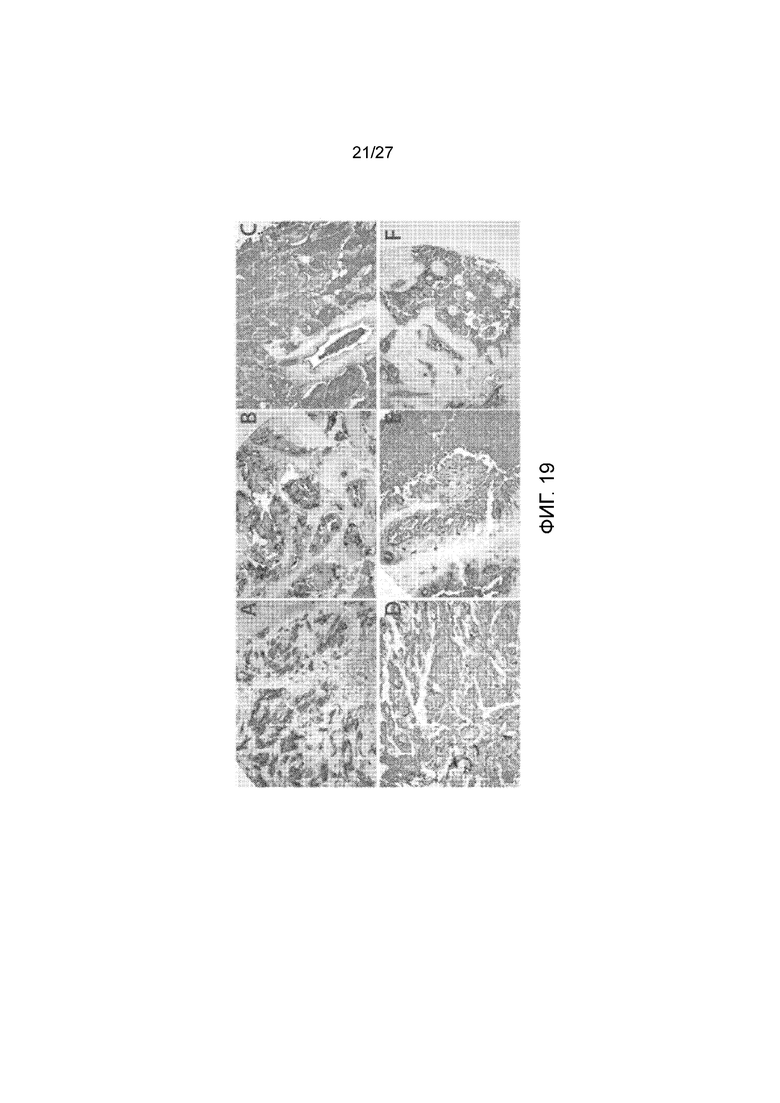

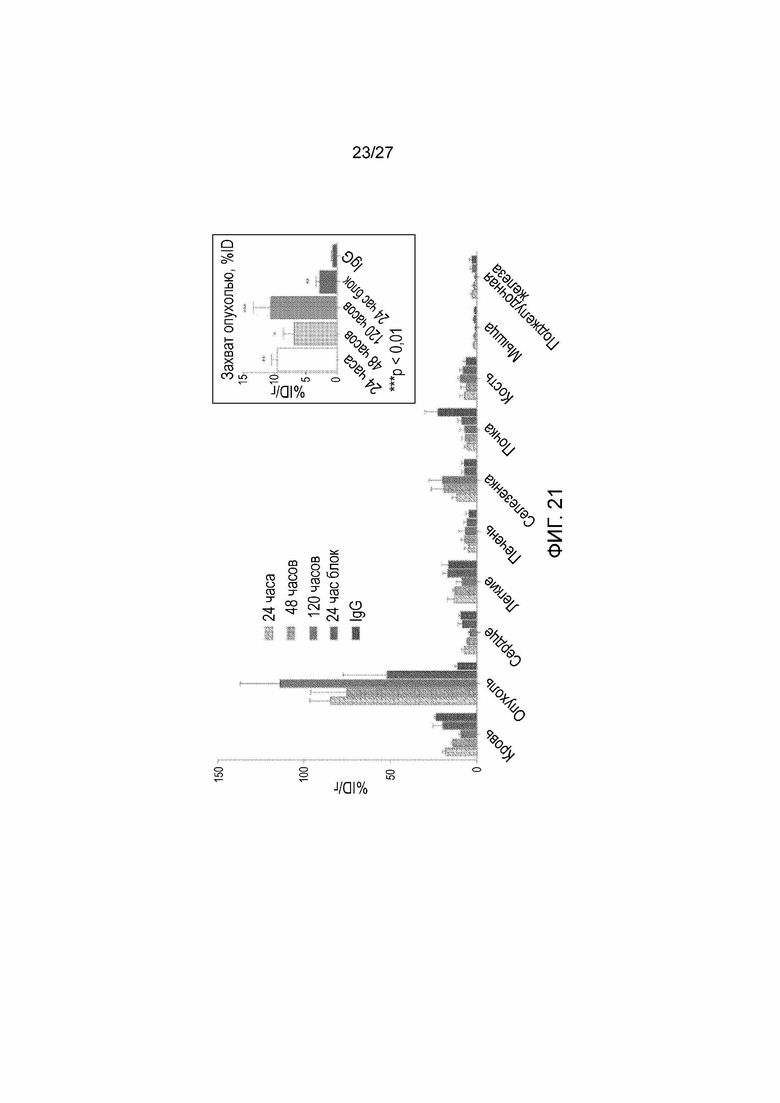
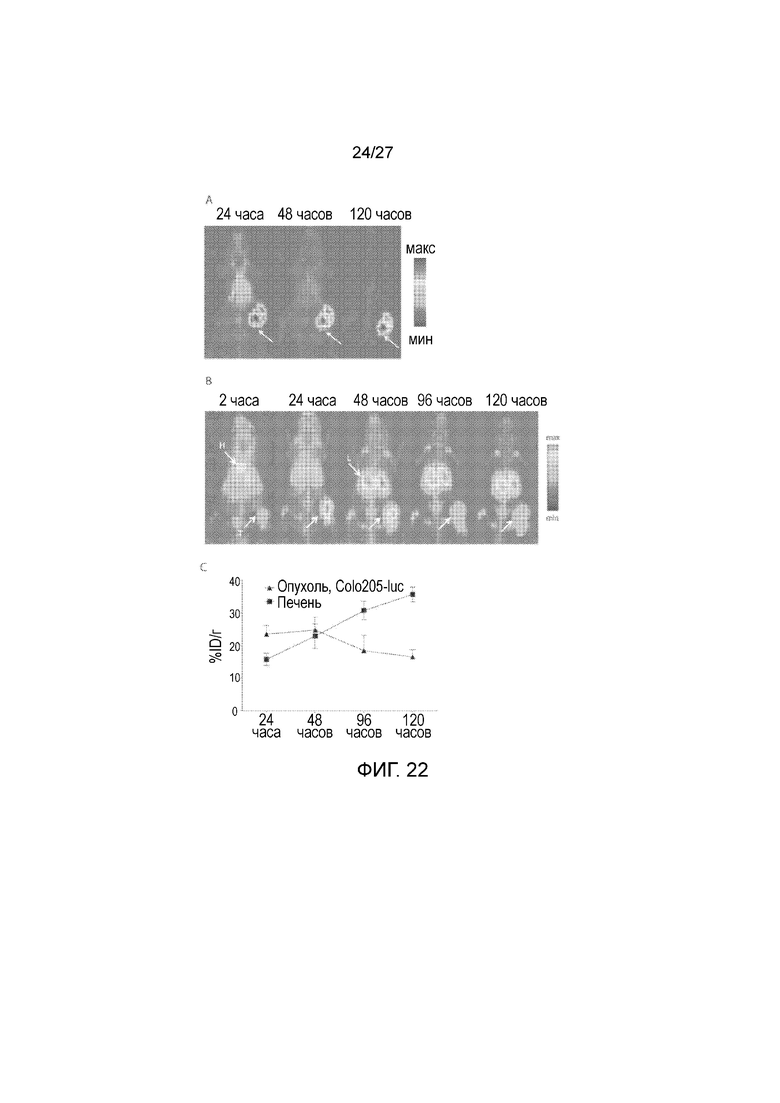
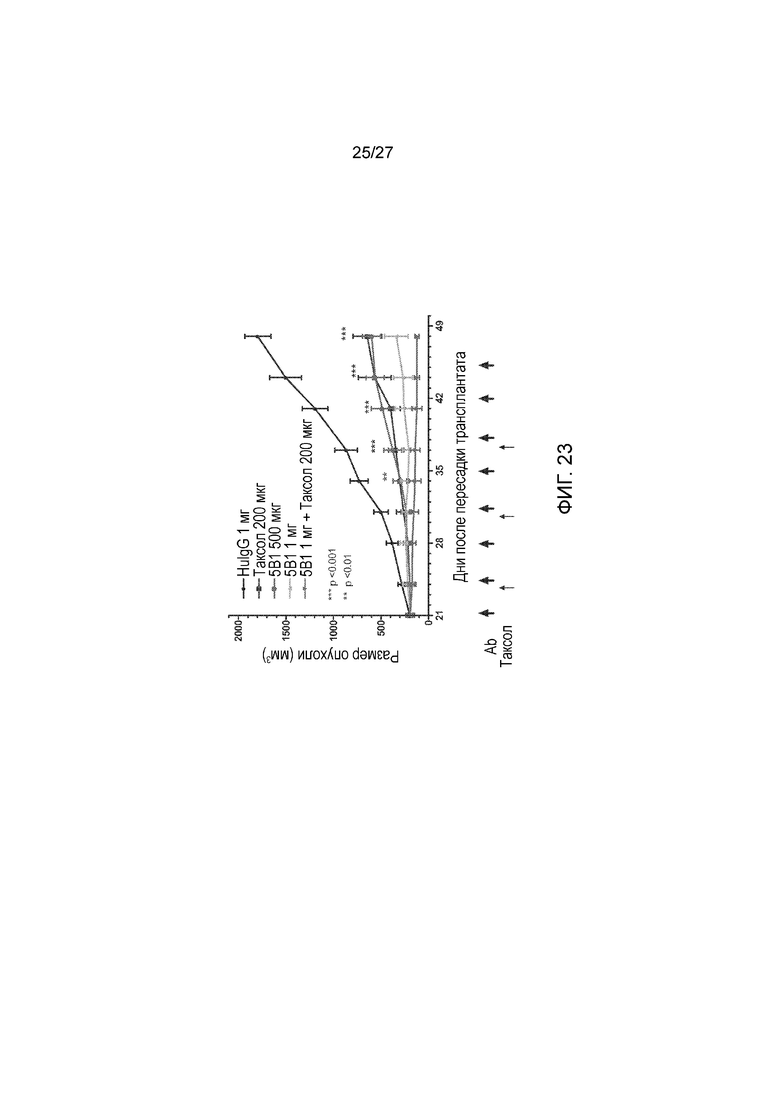
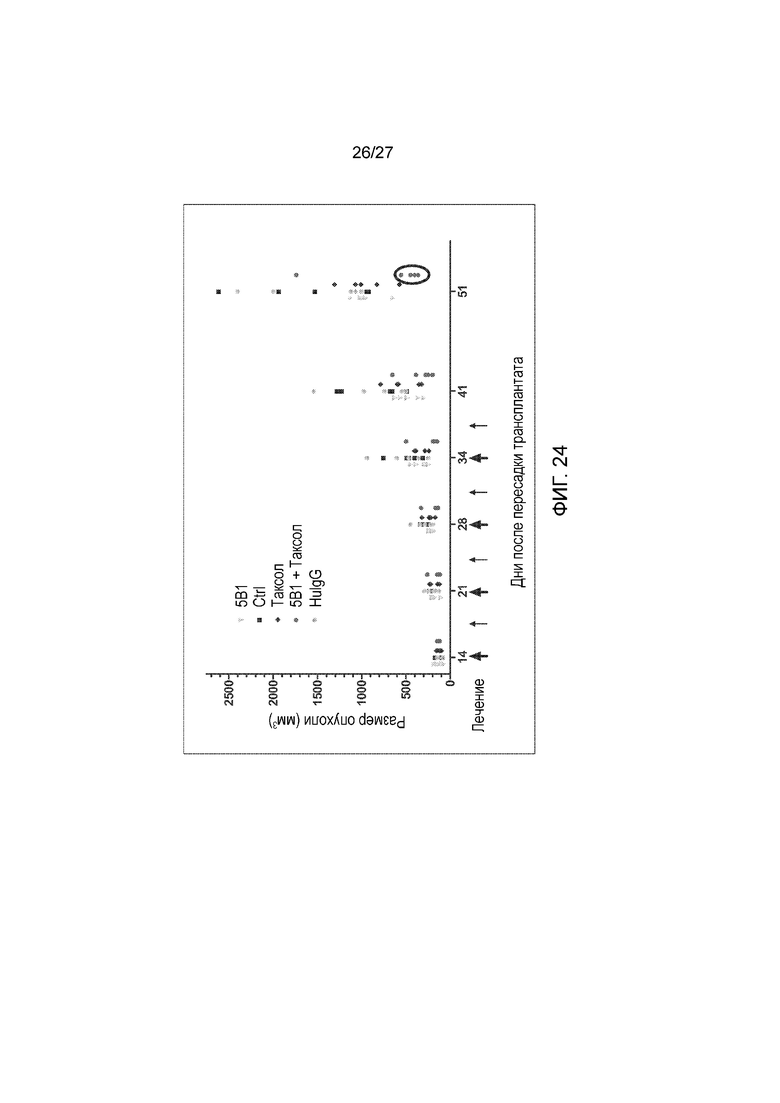

Изобретение относится к области биотехнологии. Описана группа изобретений, включающая в себя выделенный полинуклеотид, кодирующий полипептид, содержащий вариабельной домен тяжелой цепи (VH) антитела, которое связывается с сиалированным антигеном Льюисаа (варианты), выделенный полипептид, содержащий антигенсвязывающий сайт антитела, которое связывается с сиалированным антигеном Льюисаа, фармацевтическую композицию для лечения заболевания, где заболевание представляет собой злокачественное новообразование или опухолевое образование с клетками, экспрессирующими сиалированный антиген Льюисаа и способ лечения или профилактики заболевания, включающий введение фармацевтической композиции. Изобретение расширяет арсенал средств, специфически связывающихся с сиалированным антигеном Льюисаа. 5 н. и 10 з.п. ф-лы, 25 ил., 12 табл., 4 пр.
1. Выделенный полинуклеотид, кодирующий полипептид, содержащий вариабельной домен тяжелой цепи (VH) антитела, которое связывается с сиалированным антигеном Льюисаа,
где указанный вариабельный домен тяжелой цепи (VH) содержит CDR1, CDR2 и CDR3, выбранные из группы, состоящей из:
(а) CDR1, CDR2 и CDR3 с SEQ ID NO: 2, состоящей из аминокислотных остатков 55-62, 70-77 и 116-131,
(b) CDR1, CDR2 и CDR3 с SEQ ID NO: 6, состоящей из аминокислотных остатков 45-52, 70-77 и 116-131,
(c) CDR1, CDR2 и CDR3 с SEQ ID NO: 10, состоящей из аминокислотных остатков 45-52, 70-77 и 116-131,
(d) CDR1, CDR2 и CDR3 с SEQ ID NO: 14, состоящей из аминокислотных остатков 45-52, 70-77 и 116-134.
2. Выделенный полинуклеотид, кодирующий полипептид, содержащий вариабельной домен легкой цепи (VL) антитела, которое связывается с сиалированным антигеном Льюисаа,
где указанный вариабельный домен легкой цепи (VL) содержит CDR1, CDR2 и CDR3, выбранные из группы, состоящей из:
(а) CDR1, CDR2 и CDR3 с SEQ ID NO: 4, состоящей из аминокислотных остатков 45-52, 70-72 и 109-120,
(b) CDR1, CDR2 и CDR3 с SEQ ID NO: 8, состоящей из аминокислотных остатков 45-52, 70-72 и 109-119,
(c) CDR1, CDR2 и CDR3 с SEQ ID NO: 12, состоящей из аминокислотных остатков 45-52, 70-72 и 109-120,
(d) CDR1, CDR2 и CDR3 с SEQ ID NO: 16, состоящей из аминокислотных остатков 49-53, 72-74 и 111-120.
3. Выделенный полипептид, содержащий антигенсвязывающий сайт антитела, которое связывается с сиалированным антигеном Льюисаа,
где указанный антигенсвязывающий сайт содержит вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL),
где указанный домен VH и указанный домен VL соответственно содержит CDR1, CDR2 и CDR3, выбранные из группы, состоящей из:
(а) CDR1, CDR2 и CDR3 с SEQ ID NO: 2, состоящей из аминокислотных остатков 55-62, 70-77 и 116-131,
(b) CDR1, CDR2 и CDR3 с SEQ ID NO: 6, состоящей из аминокислотных остатков 45-52, 70-77 и 116-131,
(c) CDR1, CDR2 и CDR3 с SEQ ID NO: 10, состоящей из аминокислотных остатков 45-52, 70-77 и 116-131,
(d) CDR1, CDR2 и CDR3 с SEQ ID NO: 14, состоящей из аминокислотных остатков 45-52, 70-77 и 116-134,
(e) CDR1, CDR2 и CDR3 с SEQ ID NO: 4, состоящей из аминокислотных остатков 45-52, 70-72 и 109-120,
(f) CDR1, CDR2 и CDR3 с SEQ ID NO: 8, состоящей из аминокислотных остатков 45-52, 70-72 и 109-119,
(g) CDR1, CDR2 и CDR3 с SEQ ID NO: 12, состоящей из аминокислотных остатков 45-52, 70-72 и 109-120,
(h) CDR1, CDR2 и CDR3 с SEQ ID NO: 16, состоящей из аминокислотных остатков 49-53, 72-74 и 111-120.
4. Полипептид по п.3, где указанный полипептид выбран из группы, состоящей из антитела, Fab, Fab', F(ab')2, scFV, диатела, триатела и минитела.
5. Полипептид по п.3 или 4, где антитело является человеческим.
6. Полипептид по п.4 или 5, где полипептид является диателом.
7. Полипептид по п.6, где указанное диатело содержит аминокислотную последовательность SEQ ID NO: 18 или 20.
8. Полипептид по любому из пп.3-5, где указанное антитело представляет собой моноклональное антитело.
9. Полипептид по любому из пп.3-5 и 8, где указанное антитело представляет собой изотип IgG или IgМ.
10. Полипептид по п.9, где указанное IgG антитело относится к подклассу IgG1.
11. Фармацевтическая композиция для лечения заболевания, содержащая эффективное количество полипептида по любому из пп.3-10 и фармацевтически приемлемый носитель, где указанное заболевание представляет собой злокачественное новообразование или опухолевое образование с клетками, экспрессирующими сиалированный антиген Льюисаа.
12. Способ лечения или профилактики заболевания, включающий введение терапевтически эффективного количества фармацевтической композиции по п.11 пациенту, где указанным заболеванием является злокачественное или опухолевое образование с клетками, экспрессирующими сиалированный антиген Льюисаа.
13. Способ по п.12, где указанное злокачественное или опухолевое образование выбрано из группы, состоящей из опухоли желудочно-кишечного тракта, рака толстой кишки, колоректальной аденокарциномы, метастатического рака толстой кишки, колоректального рака, рака поджелудочной железы, аденокарциномы поджелудочной железы, рака протоков поджелудочной железы, мелкоклеточного рака легких, аденокарциномы мочевого пузыря, перстневидноклеточного рака яичников, рака яичников, метастатического рака, аденокарциномы желудка, аденокарциномы пищевода, аденокарциномы горла, аденокарциномы мочеполового тракта и аденокарциномы молочной железы.
14. Способ по п.12 или 13, где указанный способ дополнительно включает одновременное или последовательное введение второго терапевтического средства.
15. Способ по п.14, где указанное второе терапевтическое средство представляет собой химиотерапевтическое средство или иммунотерапевтическое средство.
| US 20110311517 A1 22.12.2011 | |||
| US 20130209481 A1 15.08.2013 | |||
| US 20090203538 A1 13.08.2009 | |||
| РЫБАЛЬСКИЙ Н.Г., СЕРОВА М | |||
| А., ИГНАТЬЕВА Г.А., СТАРЧЕУС А.П | |||
| "Моноклональные антитела и гибридомы", Москва: ВАСХНИЛ, 1989, стр.23-44. |

