УРОВЕНЬ ТЕХНИКИ
(1) Область изобретения
Настоящее изобретение относится к способам комбинированного лечения злокачественных новообразований, включающим CTLA-4-блокирующее средство и PD-1-блокирующее средство. В частности, настоящее изобретение относится к способам комбинированного лечения, где CTLA-4-блокирующее средство является антителом против CTLA-4 со сниженной или отсутствующей измеримой эффекторной функцией или фрагментом антитела против CTLA-4, в котором отсутствует Fc-домен, и PD-1-блокирующее средство является антителом против PD-1, фрагментом антитела против PD-1, антителом против PD-L1 или фрагментом антитела против PD-L1.
(2) Описание связанной области
Иммунотерапия опухолей играет все более важную роль в лечении различных онкологических заболеваний. Клинический успех использования блокады антителами иммунных контрольных точек, включающих ингибиторные рецепторы, экспрессирующиеся на T-клетках, такие как антиген-4 цитотоксических T-лимфоцитов (CTLA-4) и рецептор программируемой гибели-1 (PD-1), стимулировал заметное развитие иммунотерапии злокачественных новообразований. Монотерапевтическая блокада CTLA-4 или PD-1 моноклональными антителами (mAb) приводила к повышенным противоопухолевым ответам и благоприятным клиническим исходам в контролируемых рандомизированных клинических испытаниях.
Характерной особенностью блокады иммунных контрольных точек при лечении различных злокачественных новообразований является клинически валидированный благоприятный эффект способов комбинированного лечения, включающих антитела против PD-1 и против CTLA-4. С появлением все большего количества клинических данных становится очевидным, что способы комбинированного лечения с использованием средств против PD-1/CTLA-4 могут обеспечивать превосходную клиническую эффективность по сравнению с таргетингом любого пути контрольных точек в отдельности. Однако иммуноопосредованная токсичность (irAE), ассоциированная с антителами против CTLA-4, является значимой и в условиях монотерапии, и в способах комбинированного лечения с антителами против PD-1. Например, антитело против CTLA-4 ипилимумаб, продаваемое Bristol-Myers Squibb под торговым названием YERVOY и являющееся единственным антителом против CTLA-4, одобренным Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) США, имеет особое предупреждение в черной рамке из-за вероятности индуцированиф тяжелых и фатальных иммуноопосредованных нежелательных явлений, таких как воспаление кишечника, печени, кожи, эндокринных желез и/или глаз. Ипилимумаб также одобрен для способов комбинированного лечения с использованием антитела против PD-1 ниволумаба (продаваемого Bristol-Myers Squibb под торговым названием OPDIVO) для почечноклеточных карцином на поздних стадиях и некоторых типов колоректального рака, но из-за риска значительных irAE, ипилимумаб вводят в низкой или субтерапевтической дозе 1 мг/кг. Субтерапевтическая доза для комбинированного лечения составляет гораздо меньше чем 3 мг/кг монотерапевтической дозы для нерезектабельной или метастатической меланомы или 10 мг/кг монотерапевтической дозы для адъювантной меланомы (см. вкладыш в упаковку и ярлык к YERVOY (июль 2018 года)).
Оба одобренных FDA mAb против PD1, ниволумаб и пембролизумаб, являются гуманизированными антителами против PD1 IgG4 каппа, описанными в патенте США №8008449 и патенте США №8354509, соответственно. Общеизвестно, что Fc-домен изотипа IgG4 имеет низкую детектируемую эффекторную функцию.
Ипилимумаб является антителом человека против CTLA-4 IgG1 каппа, описанным в патенте США №6984720. Константный домен тяжелой цепи (HC) изотипа IgG1 имеет Fc-домен, как известно, имеющий высокую аффинность к Fc-рецепторам (FcR), что придает антителу значительную эффекторную функцию (например, индуцированную антителозависимую клеточную цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP) и/или комплементзависимую цитотоксичность (CDC)). Исследования показали, что эффекторная функция Fc необходима для эффективности антител против CTLA-4. Например, в Ingram et al., Proc. Natl. Acad. Sci. USA 115: 3912-3917 (2018) на модели на мышах показано, что фрагмент антитела альпаки только из тяжелой цепи (VHH) против CTLA-4, в котором отсутствует Fc-домен тяжелой цепи и его сопутствующая эффекторная функция не имели противоопухолевой эффективности; однако, противоопухолевую эффективность молекулы можно восстановить посредством ее слияния с Fc-доменом тяжелой цепи IgG2 мыши, демонстрирующим эффекторную функцию; и в Selby et al., Cancer Immunol. Res. 1: 32-42 (2013) на модели на мышах показано, что антитела против CTLA-4, слитые или связанные с Fc-доменом, подвергнутым мутагенезу для устранения эффекторной функции, не демонстрировали какую-либо противоопухолевую активность. Также см. Simpson et al., J. Exp. Med.; 210: 1695-710 (2013) и международную патентную заявку № WO 2014089113.
Тремелизумаб является антителом IgG2 человека против CTLA-4 человека, описанным в патенте США №8491895. Изотип IgG2 человека выбран для минимизации потенциальной активности эффекторной функции и, таким образом, потенциального снижения irAE. Однако, как показано в Vargas et al., Cancer cells 33: 649-663 (2018), тремелизумаб сохраняет эффекторную функцию; и Bertrand et al., BMC Med. 13: 211-214 (2015) показали, что, хотя тремелизумаб можно вводить в дозе, превышающей дозу ипилимумаба, он все равно может индуцировать irAE, в частности, воспалительную иммуноопосредованную токсичность в кишечнике и коже. Также см. Ribas et al., The Oncologist 12: 873-993 (2007), Schneider-Merck et al., J. Immunol. 184: 512-520 (2010) и  et al., PLoS One. 10: e0145633(2015), где показаны другие антитела IgG2 человека, индуцирующие ADCC и ADCP in vitro схожей величины с изоформой IgG1 человека.
et al., PLoS One. 10: e0145633(2015), где показаны другие антитела IgG2 человека, индуцирующие ADCC и ADCP in vitro схожей величины с изоформой IgG1 человека.
Другие попытки снизить irAE ипилимумаба включают BMS-986249, протело, состоящее из ипилимумаба, связанного с запатентованным маскирующим пептидом, охватывающим активный антигенсвязывающий участок антитела через расщепляемый протеазой линкер. Маскирующий пептид может снижать irAE посредством минимизации способности ипилимумаба связываться с CTLA-4 в нормальных тканях (см. международные патентные заявки №№ WO 2009025846, WO 2010081173, WO 2018222949, WO 2018085555, Pai et al., J. Clin. Invest. 129: 349-363 (2019) и Korman et al., Abstract SY09-01, AACR Annual Meeting Vol. 77, issue 13 (2017)).
В свете исследований, в которых предполагают, что терапевтическая эффективность антител против CTLA-4, подобных ипилимумаб, может включать истощение регуляторных T-клеток (Treg-клеток, предполагают, что антитела против CTLA-4, такие как ипилимумаб, имеющие повышенную активность ADCC, будут обеспечивать более эффективную противоопухолевую активность, чем существующие антитела. В патенте США №10196445 описано несколько вариантов ипилимумаба с повышенной активностью ADCC.
Стандарт лечения для некоторых способов противоопухолевой терапии включает введение антитела против PD-1 в комбинации с химиотерапией. Противоопухолевая активность антитела против CTLA-4 может дополнительно повышать эффективность этих способов терапии; однако, т.к. желудочно-кишечная токсичность является одним из наиболее распространенных побочных эффектов, наблюдаемых во время химиотерапии, добавление антитела против CTLA-4 к терапии, вместо этого, может усиливать желудочно-кишечную токсичность.
Очевидно, что антитела против CTLA-4, делающие возможным введение на более высоких, более оптимальных уровнях без ассоциированных irAE, в частности, воспалительной иммуноопосредованной токсичности в отношении кожи и кишечника, ассоциированной с существующими антителами против CTLA-4, вероятно, сделают возможной более эффективную терапию в комбинации с антагонистами PD-1 и, необязательно, химиотерапевтическими средствами.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения обнаружили, что, хотя некоторые CTLA-4-блокирующие средства, связывающиеся с CTLA-4, имеют сниженную или отсутствующую измеримую противоопухолевую активность при введении в виде монотерапии, они могут демонстрировать клинически значимую противоопухолевую активность при использовании в способах комбинированного лечения с PD-1-блокирующим средством. Авторы настоящего изобретения также обнаружили, что эти некоторые CTLA-4-блокирующие средства могут иметь противоопухолевую активность при комбинированном лечении с блокадой CTLA-4/PD-1 без индуцирования иммуноопосредованных нежелательных явлений (irAE), включая irAE в коже и кишечнике, ассоциированные с одобренными в настоящее время способами комбинированного лечения с блокадой CTLA-4/PD-1. Комбинации с блокадой CTLA-4/PD-1, представленные в настоящем описании, делают возможными способы терапии с повышенным терапевтическим индексом относительно существующих способов комбинированного лечения с блокадой CTLA-4/PD-1, включая способы комбинированного лечения, включающие химиотерапию, что может приводить к более эффективному лечению злокачественных новообразований с улучшенными исходами для пациентов.
Некоторые CTLA-4-блокирующие средства, используемые как часть комбинированного лечения с блокадой CTLA-4/PD-1 по настоящему изобретению, можно выбирать из группы, состоящей из (i) эффекторно-молчащего антитела против CTLA-4 и (ii) эффекторно-молчащего фрагмента антитела против CTLA-4, в котором отсутствует домен кристаллизуемого фрагмента (Fc), или которое имеет Fc-домен, содержащий делеции этих областей в Fc-домене, связывающимся с Fc рецепторами (FcR). Эффекторно-молчащее антитело или фрагмент антитела демонстрирует (i) отсутствие измеримого связывания с одним или более FcR, что можно измерять с помощью анализа Biacore, где константа ассоциации в микромолярном диапазоне указывает на отсутствие измеримого связывания, или (ii) измеримое связывание с одним или более FcR, что можно измерять с помощью анализа Biacore, сниженное по сравнению со связыванием, являющимся типичным для антитела того же изотипа. Эти некоторые CTLA-4-блокирующие средства являются эффекторно-молчащими CTLA-4-блокирующими средствами.
В конкретных вариантах осуществления эти эффекторно-молчащие антитела против CTLA-4 и эффекторно-молчащие фрагменты антител против CTLA-4 могут не демонстрировать измеримую противоопухолевую активность при противоопухолевой монотерапии, но будут демонстрировать измеримую противоопухолевую активность в комбинированной противоопухолевой терапии с использованием PD-1- или PD-L1-блокирующего средства, не демонстрируя irAE, как правило, ассоциированные со способами комбинированного лечения с блокадой CTLA-4/PD-1, в частности, воспалительную иммуноопосредованную токсичность в отношении кожи или кишечника.
Эффекторно-молчащие антитела против CTLA-4 или эффекторно-молчащие фрагменты антител против CTLA-4, представленные в настоящем описании, можно использовать в более высоких дозах и в течение более длительных периодов времени в комбинации с PD-1- или PD-L1-блокирующими средствами без irAE, как правило, ассоциированных со способами комбинированного лечения с блокадой CTLA-4/PD-1, в частности, воспалительной иммуноопосредованной токсичности в отношении кожи или кишечника. В настоящее время доза антитела против CTLA-4 ипилимумаба, одобренного для использования в способах комбинированного лечения с блокадой CTLA-4/PD-1, составляет 1 мг/кг по сравнению с дозой 3 мг/кг или 10 мг/кг, одобренной для использования в монотерапии (см. вкладыш в упаковку и ярлык для YERVOY (июль 2018 года)), или фиксированной дозой 100 мг или менее, предусмотренной для способов комбинированного лечения с блокадой CTLA-4/PD-1 в международной патентной заявке № WO 2018183408. Таким образом, в способах комбинированного лечения с блокадой CTLA-4/PD-1 по настоящему изобретению можно использовать эффекторно-молчащие антитела против CTLA-4 или эффекторно-молчащие фрагменты антител против CTLA-4 в дозах, таких же или более высоких, чем дозы, одобренные в настоящее время для антител против CTLA-4 в монотерапии. Эффекторно-молчащие антитела против CTLA-4 или фрагменты антител против CTLA-4, представленные в настоящем описании, также можно использовать в комбинации с антителами против PD-1 или против PD-L1 в дозах, схожих с используемыми в настоящее время или предусмотренными для способов комбинированного лечения с блокадой CTLA-4/PD-1, но в течение более длительного периода времени, чем для антител против CTLA-4, и без irAE, как правило, ассоциированных со способами комбинированного лечения с блокадой CTLA-4/PD-1, в частности, воспалительной иммуноопосредованной токсичности в отношении кожи или кишечника.
Таким образом, настоящее изобретение относится к комбинированному лечению для лечения злокачественного новообразования у индивидуума, нуждающегося в таком лечении, способу, включающему введение индивидууму со злокачественным новообразованием (i) терапевтической дозы PD-1- или PD-L1-блокирующего средства и (ii) терапевтической дозы эффекторно-молчащего CTLA-4-блокирующего средства для лечения злокачественного новообразования, где эффекторно-молчащее CTLA-4-блокирующее средство демонстрирует противоопухолевую активность при комбинированном лечении, которую оно не демонстрирует при введении индивидууму в виде монотерапии без PD-1- или PD-L1-блокирующего средства. В дополнительных вариантах осуществления комбинированное лечение не индуцирует или имеет сниженный риск индуцирования иммуноопосредованных нежелательных явлений (irAE) в кишечнике или коже в течение курса комбинированного лечения, имеющих степень более 2, что определяют по Общим терминологическим критериям нежелательных явлений (CTCAE) версии 5.0, по сравнению со способами комбинированного лечения, включающими антитело против CTLA-4, демонстрирующее эффекторную функцию. В конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство, используемое как часть комбинированного лечения, не индуцирует irAE в коже или кишечнике, превышающие степень 2, в течение по меньшей мере первых 10 недель комбинированного лечения. В конкретных вариантах осуществления комбинированное лечение не приводит к детектируемым irAE в течение по меньшей мере первых четырех недель комбинированного лечения или irAE степени более 1 в течение по меньшей мере первых четырех недель комбинированного лечения.
В одном из вариантов осуществления эффекторно-молчащее CTLA-4-блокирующее средство, используемое как часть комбинированного лечения, представленного в настоящем описании, является эффекторно-молчащим антителом против CTLA-4 или эффекторно-молчащим фрагментом антитела против CTLA-4.
В другом варианте осуществления PD-1-блокирующее средство, используемое как часть комбинированного лечения, представленного в настоящем описании, является антителом против PD-1 или против PD-L1 или фрагментом антитела против PD-1 или против PD-L1. В конкретных вариантах осуществления антитело против PD-1 или против PD-L1 содержит домен HC, содержащий одну или более мутаций в Fc-домене, делающих антитело эффекторно-молчащим. PD-1-блокирующее средство также может являться фрагментом антитела против PD-1 или против PD-L1, в каждом из которых отсутствует Fc-домен или области Fc-домена, связывающиеся с одним или более FcR, что делает фрагмент антитела эффекторно-молчащим.
Настоящее изобретение дополнительно относится к способам комбинированного лечения злокачественных новообразований, включающим введение индивидууму, нуждающемуся в терапии злокачественных новообразований, (i) первого состава, содержащего PD-1-блокирующее средство, выбранное из группы, состоящей из антитела против PD-1, имеющего Fc-домен IgG4 или IgG2, эффекторно-молчащего антитела против PD-1 и эффекторно-молчащего фрагмента антитела против PD-1; и (ii) второго состава, содержащего эффекторно-молчащее CTLA-4-блокирующее средство, выбранное из группы, состоящей из эффекторно-молчащего антитела против CTLA-4 и эффекторно-молчащего фрагмента антитела против CTLA-4.
Настоящее изобретение дополнительно относится к противоопухолевому комбинированному лечению, включающему введение индивидууму, нуждающемуся в терапии злокачественных новообразований, состава, включающего (i) PD-1-блокирующее средство, выбранное из группы, состоящей из антитела против PD-1, имеющего Fc-домен IgG4 или IgG2, эффекторно-молчащего антитела против PD-1 и эффекторно-молчащего фрагмента антитела против PD-1; и (ii) эффекторно-молчащее CTLA-4-блокирующее средство, выбранное из группы, состоящей из эффекторно-молчащего антитела против CTLA-4 и эффекторно-молчащего фрагмента антитела против CTLA-4.
Настоящее изобретение дополнительно относится к способам комбинированного лечения злокачественных новообразований, включающим введение индивидууму, нуждающемуся в терапии злокачественных новообразований, (i) первого состава, содержащего PD-L1-блокирующее средство, выбранное из группы, состоящей из антитела против PD-L1, имеющего Fc-домен IgG4 или IgG2, эффекторно-молчащего антитела против PD-L1 и эффекторно-молчащего фрагмента антитела против PD-L1; и (ii) второго состава, содержащего эффекторно-молчащее CTLA-4-блокирующее средство, выбранное из группы, состоящей из эффекторно-молчащего антитела против CTLA-4 и эффекторно-молчащего фрагмента антитела против CTLA-4.
Настоящее изобретение дополнительно относится к противоопухолевому комбинированному лечению, включающему введение индивидууму, нуждающемуся в терапии злокачественных новообразований, состава, содержащего (i) PD-L1-блокирующее средство, выбранное из группы, состоящей из антитела против PD-L1 имеющего Fc-домен IgG4 или IgG2, эффекторно-молчащего антитела против PD-L1 и эффекторно-молчащего фрагмента антитела против PD-L1; и (ii) эффекторно-молчащее CTLA-4-блокирующее средство, выбранное из группы, состоящей из эффекторно-молчащего антитела против CTLA-4 и эффекторно-молчащего фрагмента антитела против CTLA-4.
В более конкретных вариантах осуществления комбинированного лечения эффекторно-молчащие антитело против CTLA-4 содержит Fc-домен IgG1, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A, L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, S267E/L328F, S2339D/A330L/I332E, L235G/G236R, N297A/D356E/L358M, L234F/L235E/P331S/D365E/L358M и D265A/N297G, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, S267E/L328F, S2339D/A330L/I332E, L235G/G236R, D356E/L358M, L234F/L235E/P331S/D365E/L358M и D265A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В конкретных вариантах осуществления комбинированного лечения эффекторно-молчащие антитело против CTLA-4 содержит Fc-домен IgG2, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В конкретных вариантах осуществления комбинированного лечения эффекторно-молчащее антитело против CTLA-4 содержит Fc-домен IgG4, содержащий замену аминокислоты S228P и дополнительно содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N267A, P329G и D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из N267A, P329G и D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В дополнительном варианте осуществления комбинированного лечения эффекторно-молчащий фрагмент антитела против CTLA-4, в котором отсутствует Fc-домен, представляет собой или содержит одноцепочечный вариабельный фрагмент (scFv), антигенсвязывающий фрагмент (Fab) или димер антигенсвязывающего фрагмента F(ab')2.
В конкретных вариантах осуществления комбинированного лечения эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит три определяющие комплементарность области (CDR) тяжелой цепи (HC) и три CDR легкой цепи (LC) антитела против CTLA-4, выбранного из группы, состоящей из ипилимумаба, тремелимумаба, REGN4659, AGEN1884w, 8D2/8D2 (RE), 8D2/8D2 (RE)-варианта 1, 8D2H1L1, 8D2H1L1-варианта 1, 8D2H2L2, 8D2H2L2-варианта 1, 8D3H3L3, 8D2H2L15, 8D2H2L15-варианта 1, 8D2H2L17 и 8D2H2L17-варианта 1.
В конкретных вариантах осуществления комбинированного лечения эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит VH и VL ипилимумаба, VH и VL тремелимумаба, VH и VL REGN4659, VH и VL AGEN1884w, VH и VL 8D2/8D2 (RE), VH и VL 8D2/8D2 (RE)-варианта 1, VH и VL 8D2H1L1, VH и VL 8D2H1L1-варианта 1, VH и VL 8D2H2L2, VH и VL 8D2H2L2-варианта 1, VH и VL 8D3H3L3, VH и VL 8D2H2L15, VH и VL 8D2H2L15-варианта 1, VH и VL 8D2H2L17 или VH и VL 8D2H2L17-варианта 1.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит (i) VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 7, и VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 8; (ii) VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 15, и VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 16; (iii) VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 95, и VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 96; или (iv) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 97, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 98.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит (i) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; (ii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; (iii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78; (iv) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80; (v) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 82; (vi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 83, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 84; (vii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 85, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 86; (viii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 87, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 88; (ix) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 89, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 90; (x) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 91, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 92; или (xi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 93, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94.
В дополнительных вариантах осуществления комбинированного лечения эффекторно-молчащее CTLA-4-блокирующее средство является эффекторно-молчащим антителом против CTLA-4, выбранным из эффекторно-молчащих антител против CTLA-4, приведенных в таблицах 4-18.
В дополнительном варианте осуществления комбинированного лечения эффекторно-молчащее CTLA-4-связывающее средство является эффекторно-молчащим фрагментом антитела против CTLA-4, содержащим один или более отдельных вариабельных доменов иммуноглобулинов (ISVD), где каждый ISVD содержит вариабельный домен (VHH) антитела Верблюжьих, содержащего только тяжелую цепь; при условии, что ни один из ISVD не содержит VHH, содержащий CDR1, содержащую аминокислотную последовательность FYGMG (SEQ ID NO: 69), CDR2, содержащую аминокислотную последовательность DIRTSAGRTTYADSVKG (SEQ ID NO: 70), и CDR3, содержащую аминокислотную последовательность EMSGISGWDY (SEQ ID NO: 71) или EPSGISGWDY (SEQ ID NO: 72), эти ISVD описаны в международных патентных заявках №№ WO 2008071447, WO 2017087587 и WO 2017087588, или VHH, содержащий 1, 2 или 3 мутации в CDR3, как описано в WO 2008071447, за исключением ISVD, содержащих указанные CDR в вариантах осуществления, где это условие не исключает один или более ISVD, слитых или связанных с эффекторно-молчащим гетерологичным доменом HC или Fc-доменом, включая, например, любой из доменов HC или Fc-доменов эффекторно-молчащего антитела, представленных в настоящем описании.
В конкретных вариантах осуществления комбинированного лечения антитело против PD-1 или фрагмент антитела против PD-1 содержит три определяющие комплементарность области (CDR) тяжелой цепи и три CDR легкой цепи пембролизумаба, ниволумаба или цемиплимаба-rwlc. В конкретных вариантах осуществления комбинированного лечения антитело против PD-1 содержит (i) VH и VL пембролизумаба; (ii) VH и VL ниволумаба; или (iii) VH и VL цемиплимаба-rwlc.
В дополнительных вариантах осуществления антитело против PD-1 или фрагмент антитела против PD-1 содержит (i) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 29, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 30; (ii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 23, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 24; или (iii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 99, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 100. В дополнительном варианте осуществления антитело против PD1 содержит (i) HC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 27, и LC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 28; (ii) HC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 25, и LC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 26; или (iii) HC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 101, и LC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 102.
В конкретных вариантах осуществления комбинированного лечения антитело против PD-L1 или фрагмент антитела против PD-L1 содержит (i) домены VH и VL атезолизумаба; (ii) домены VH и VL авелумаба; или (iii) домены VH и VL дурвалумаба.
В дополнительных вариантах осуществления антитело против PD-L1 или фрагмент антитела против PD-L1 содержат (i) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 103, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 104; (ii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 105, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 106; или (iii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 107, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 108.
В конкретных вариантах осуществления комбинированного лечения антитело против PD-1 или против PD-L1 может содержать Fc-домен IgG1, IgG2 или IgG4, как представлено в настоящем описании, который может содержать C-концевой лизин или не содержать C-концевой лизин или C-концевой дипептид глицин-лизин.
В конкретных вариантах осуществления комбинированного лечения антитело против PD-1 или против PD-L1 содержит (i) Fc-домен IgG2 или IgG4; (ii) Fc-домен IgG1, IgG2 или IgG4, содержащий мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (iii) Fc-домен IgG1, содержащий замены аминокислот N297A, L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, L235G/G236R, S267E/L328F, S2339D/A330L/I332E или D265A/N297G, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (iv) Fc-домен IgG2, содержащий замены аминокислот N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (v) Fc-домен IgG4, содержащий замену аминокислоты S228P и замену аминокислоты N267A, P329G или D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот определяют в соответствии с нумерацией Eu.
В конкретных вариантах осуществления комбинированного лечения PD-1-блокирующее средство является антителом против PD-1, выбранным из антител против PD-1, приведенных в таблицах 19-27, или антителом против PD-L1, выбранным из антител против PD-L1, приведенных в таблицах 28-36.
В дополнительном варианте осуществления комбинированного лечения фрагмент антитела против PD-1 или фрагмент антитела против PD-L1, в каждом из которых отсутствует Fc-домен, является одноцепочечным вариабельным фрагментом (scFv), антигенсвязывающим фрагментом (Fab) или димером антигенсвязывающего фрагмента F(ab')2.
В дополнительном варианте осуществления комбинированного лечения антитело против PD-1 или фрагмент антитела против PD-L1 содержит один или более ISVD, где каждый ISVD содержит VHH антитела Верблюжьих, содержащего только тяжелую цепь.
В конкретных вариантах осуществления комбинированного лечения CTLA-4-блокирующее средство вводят в дозе, содержащей от приблизительно 1 мг/кг до приблизительно 3 мг/кг CTLA-4-блокирующего средства, или фиксированной дозе CTLA-4-блокирующего средства, независящей от массы тела индивидуума и превышающей приблизительно 100 мг.
В конкретных вариантах осуществления комбинированного лечения CTLA-4-блокирующее средство вводят в дозе, содержащей от 1 мг/кг до 3 мг/кг CTLA-4-блокирующего средства.
В конкретных вариантах осуществления комбинированного лечения CTLA-4-блокирующее средство вводят в дозе, содержащей от 3 мг/кг до 10 мг/кг CTLA-4-блокирующего средства.
В конкретных вариантах осуществления комбинированного лечения CTLA-4-блокирующее средство вводят в дозе, содержащей более приблизительно 10 мг/кг CTLA-4-блокирующего средства.
В конкретных вариантах осуществления комбинированного лечения PD-1-блокирующее средство вводят в дозе, содержащей приблизительно 2 или 3 мг/кг или более, или фиксированной дозе, независящей от массы тела индивидуума и составляющей приблизительно 200 мг или более.
В конкретных вариантах осуществления комбинированного лечения PD-1-блокирующее средство вводят в дозе, независящей от массы тела индивидуума, составляющей от 200 мг до 400 мг.
В конкретных вариантах осуществления комбинированного лечения PD-1-блокирующее средство вводят в дозе, независящей от массы тела индивидуума и составляющей 400 мг.
В конкретных вариантах осуществления комбинированного лечения PD-1-блокирующее средство вводят индивидууму первым и CTLA-4-блокирующее средство вводят индивидууму вторым или CTLA-4-блокирующее средство вводят индивидууму первым и PD-1-блокирующее средство вводят индивидууму вторым. В конкретном варианте осуществления PD-1-блокирующее средство и CTLA-4-блокирующее средство вводят одновременно.
В конкретных вариантах осуществления комбинированного лечения индивидууму вводят химиотерапевтическое средство до, одновременно или после комбинированного лечения. В конкретных вариантах осуществления химиотерапевтическое средство выбрано из группы, состоящей из актиномицина, полностью транс-ретиноевой кислоты, алитретиноина, азацитидина, азатиоприна, бексаротена, блеомицина, бортезомиба, кармофура, карбоплатина, капецитабина, цисплатина, хлорамбуцила, циклофосфамида, цитарабина, дакарбазина, даунорубицина, доцетаксела, доксифлуридина, доксорубицина, эпирубицина, эпотилона, этопозида, фторурацила, гемцитабина, гидроксимочевины, идарубицина, иматиниба, иксабепилона, иринотекана, мехлоретамина, мелфалана, меркаптопурина, метотрексата, митоксантрона, нитрозомочевины, оксалиплатина, паклитаксела, пеметрекседа, ромидепсина, тегафура, темозоломида (перорального дакарбазина), тенипозида, тиогуанина, топотекана, утиделона, валрубицина, вемурафениба, винбластина, винкристина, виндезина, винорелбина и вориностата.
В конкретных вариантах осуществления комбинированного лечения злокачественное новообразование является меланомой, немелкоклеточным раком легких, раком головы и шеи, уротелиальным раком, раком молочной железы, желудочно-кишечным злокачественным новообразованием, множественной миеломой, печеночно-клеточным раком, неходжкинской лимфомой, раком почки, лимфомой Ходжкина, мезотелиомой, раком яичников, мелкоклеточным раком легких, раком пищевода, раком анального канала, раком желчных протоков, колоректальным раком, раком шейки матки, раком щитовидной железы или раком слюнных желез.
В конкретных вариантах осуществления комбинированного лечения злокачественное новообразование является раком поджелудочной железы, раком бронхов, раком предстательной железы, раком поджелудочной железы, раком желудка, раком яичников, раком мочевого пузыря, злокачественным новообразованием головного мозга или центральной нервной системы, злокачественным новообразованием периферической нервной системы, раком матки или эндометрия, раком полости рта или глотки, раком печени, раком почки, раком яичка, раком желчных протоков, раком тонкого кишечника или аппендикса, раком надпочечника, остеосаркомой, хондросаркомой или злокачественным новообразованием кроветворных тканей.
В конкретных вариантах осуществления комбинированного лечения индивидуум является человеком, CTLA-4-блокирующее средство связывается с CTLA-4 человека, PD-1-блокирующее средство связывается с PD-1 человека и PD-L1-блокирующее средство связывается с PD-L1 человека.
Антитела и композиции
Настоящее изобретение дополнительно относится к эффекторно-молчащему антителу против CTLA-4 или эффекторно-молчащему фрагменту антитела против CTLA-4, каждое из которых содержит VH и VL, где VH содержит три CDR тяжелой цепи, и VL содержит три CDR легкой цепи, совместно связывающиеся с CTLA-4. В конкретных вариантах осуществления CTLA-4 является CTLA-4 человека.
В более конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит Fc-домен IgG1, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот при условии, что эффекторно-молчащее антитело против CTLA-4 не включает ипилимумаб, состоящий исключительно из замены N297A; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A, L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, S267E/L328F, S2339D/A330L/I332E, L235G/G236R, N297A/D356E/L358M, L234F/L235E/P331S/D365E/L358M и D265A/N297G, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, S267E/L328F, S2339D/A330L/I332E, L235G/G236R, D356E/L358M, L234F/L235E/P331S/D365E/L358M и D265A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит Fc-домен IgG2, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит Fc-домен IgG4, содержащий замену аминокислоты S228P и дополнительно содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N267A, P329G и D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из N267A, P329G и D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит три определяющие комплементарность области (CDR) тяжелой цепи (HC) и три CDR легкой цепи (LC) антитела против CTLA-4, выбранного из группы, состоящей из ипилимумаба, тремелимумаба, REGN4659, AGEN1884w, 8D2/8D2 (RE), 8D2H1L1, 8D2H2L2, 8D3H3L3, 8D2H2L15 и 8D2H2L17.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит VH и VL ипилимумаба, VH и VL тремелимумаба, VH и VL REGN4659, VH и VL AGEN1884w, VH и VL 8D2/8D2 (RE), VH и VL 8D2H1L1, VH и VL 8D2H2L2, VH и VL 8D3H3L3, VH и VL 8D2H2L15 или VH и VL 8D2H2L17.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит VH и VL 8D2/8D2 (RE)-варианта 1, VH и VL 8D2H1L1-варианта 1, VH и VL 8D2H2L2-варианта 1, VH и VL 8D2H2L15-варианта 1 или VH и VL 8D2H2L17-варианта 1. Эти варианты содержат замену изолейцина метионином в положении 18 в аминокислотной последовательности VH.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит (i) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 7, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 8; (ii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 15, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 16; (iii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 95, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 96; или, (iv) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 97, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 98.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA4 или эффекторно-молчащий фрагмент антитела против CTLA-4 содержит (i) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; (ii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; (iii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78; (iv) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80; (v) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 82; (vi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 83, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 84; (vii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 85, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 86; (viii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 87, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 88; (ix) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 89, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 90; (x) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 91, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 92; или (xi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 93, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94.
В конкретных вариантах осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 выбран из группы, состоящей из F(ab), F(ab')2, Fv и scFv.
В дополнительном варианте осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 содержит один или более отдельных вариабельных доменов иммуноглобулинов (ISVD), где каждый ISVD содержит вариабельный домен (VHH) антитела Верблюжьих, содержащего только тяжелую цепь; при условии, что ни один из ISVD не содержит VHH, содержащий CDR1, содержащую аминокислотную последовательность FYGMG (SEQ ID NO: 69), CDR2, содержащую аминокислотную последовательность DIRTSAGRTTYADSVKG (SEQ ID NO: 70), и CDR3, содержащую аминокислотную последовательность EMSGISGWDY (SEQ ID NO: 71) или EPSGISGWDY (SEQ ID NO: 72), как ISVD, описанные в международной патентной заявке № WO 2008071447, WO 2017087587 и WO 2017087588, или VHH, содержащий 1, 2 или 3 мутации в CDR3, как описано в WO 2008071447, за исключением ISVD, содержащих указанные CDR в вариантах осуществления, где это условие не исключает один или более ISVD, слитых или связанных с эффекторно-молчащим гетерологичным доменом HC или Fc-доменом, включающим, например, любой из доменов HC или Fc-доменов эффекторно-молчащего антитела, представленных в настоящем описании.
Настоящее изобретение дополнительно относится к каждому из эффекторно-молчащих антител против CTLA-4, описанных в таблицах 4-18, при условии, что эффекторно-молчащее антитело против CTLA-4 не включает ипилимумаб, состоящий исключительно из замены N297A.
Настоящее изобретение дополнительно относится к композиции, содержащей эффекторно-молчащее антитело против CTLA-4 или эффекторно-молчащий фрагмент антитела против CTLA-4, как представлено в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение дополнительно относится к антителу против PD-1, содержащему
(a) тяжелую цепь (HC), содержащую вариабельный домен HC (VH), и легкую цепь (LC), содержащую вариабельный домен LC (VL), где (i) VH содержит по меньшей мере три определяющие комплементарность области (CDR) HC пембролизумаба, и VL содержит по меньшей мере три LC-CDR пембролизумаба, (ii) VH содержит по меньшей мере три HC-CDR ниволумаба, и VL содержит по меньшей мере три LC-CDR ниволумаба, или (iii) VH содержит по меньшей мере три HC-CDR цемиплимаба-rwlc, и VL содержит по меньшей мере три LC-CDR цемиплимаба-rwlc, и
(b) Fc-домен IgG1, IgG2 или IgG4, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) Fc-домен IgG1, содержащий замены аминокислот N297A, L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, N297A/D356E/L358M, L234F/L235E/P331S/D356E/L358M или D265A/N297G, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (iii) Fc-домен IgG2, содержащий замены аминокислот N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iv) Fc-домен IgG4, содержащий замену аминокислоты S228P и замену аминокислоты N267A, P329G, D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот определяют в соответствии с нумерацией Eu.
В дополнительных вариантах осуществления указанного выше антитела против PD-1 антитело против PD-1 содержит (i) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 29, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 30, (ii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 23, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 24, или (iii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 99, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 100.
В конкретных вариантах осуществления антитела против PD-1 Fc-домен IgG1, IgG2 или IgG4, как представлено в настоящем описании, может дополнительно содержать C-концевой лизин или не содержать C-концевой лизин или C-концевой дипептид глицин-лизин.
Настоящее изобретение дополнительно относится к фрагменту антитела против PD-1, содержащему тяжелую цепь (HC), содержащую вариабельный домен HC (VH), и легкую цепь (LC), содержащую вариабельный домен LC (VL), где (i) VH содержит по меньшей мере три определяющие комплементарность области HC (CDR) пембролизумаба, и VL содержит по меньшей мере три LC-CDR пембролизумаба, (ii) VH содержит по меньшей мере три HC-CDR ниволумаба, и VL содержит по меньшей мере три LC-CDR ниволумаба, или (iii) VH содержит по меньшей мере три HC-CDR цемиплимаба-rwlc, и VL содержит по меньшей мере три LC-CDR цемиплимаба-rwlc.
В дополнительных вариантах осуществления фрагмента антитела против PD-1 фрагмент антитела против PD-1 содержит (i) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 29, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 30, (ii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 23, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 24, или (iii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 99, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 100.
В конкретных вариантах осуществления указанных выше фрагментов антител против PD-1 фрагмент антитела против PD-1 выбран из группы, состоящей из F(ab), F(ab')2, Fv и scFv.
Настоящее изобретение дополнительно относится к каждому из антител против PD-1, приведенных в таблицах 19-27.
Настоящее изобретение дополнительно относится к композиции, содержащей антитело против PD-1 или фрагмент антитела против PD-1, представленное в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение дополнительно относится к антителу против PD-L1, содержащему (a) тяжелую цепь (HC), содержащую вариабельный домен HC (VH), и легкую цепь (LC), содержащую вариабельный домен LC (VL), где (i) VH содержит по меньшей мере три определяющие комплементарность области (CDR) HC дурвалумаба, и VL содержит по меньшей мере три LC-CDR дурвалумаба, (ii) VH содержит по меньшей мере три HC-CDR авелумаба, и VL содержит по меньшей мере три LC-CDR авелумаба, или (iii) VH содержит по меньшей мере три HC-CDR атезолизумаба, и VL содержит по меньшей мере три LC-CDR атезолизумаба, и
(b) Fc-домен IgG1, IgG2 или IgG4, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) Fc-домен IgG1 содержащий замены аминокислот N297A, L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, N297A/D356E/L358M, L234F/L235E/P331S/D356E/L358M или D265A/N297G, или мутантный Fc-домен содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (iii) Fc-домен IgG2 содержащий замены аминокислот N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iv) Fc-домен IgG4, содержащий замену аминокислоты S228P и замену аминокислоты N267A, P329G, D265A/N297A, или мутантный Fc-домен содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот определяют в соответствии с нумерацией Eu, при условии, что если VH и VL имеют аминокислотные последовательности SEQ ID NO: 107 и SEQ ID NO: 108, соответственно, то константный домен тяжелой цепи (HC) не принадлежит изотипу IgG1 с комбинацией замен N297A/D356E/L358M, или, если VH и VL имеют аминокислотные последовательности SEQ ID NO: 103 и SEQ ID NO: 104, соответственно, то константный домен HC не принадлежит изотипу IgG1 с комбинацией замен L234F/L235E/P331S/D356E/L358M.
В дополнительных вариантах осуществления указанного выше антитела против PD-L1 антитело против PD-L1 содержит (i) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 103, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 104, (ii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 105, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 106, или (iii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 107, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 108.
В конкретных вариантах осуществления антитела против PD-L1 Fc-домен IgG1, IgG2 или IgG4, представленный в настоящем описании может дополнительно содержать C-концевой лизин или не содержать C-концевой лизин или C-концевой дипептид глицин-лизин.
Настоящее изобретение дополнительно относится к фрагменту антитела против PD-L1, содержащему тяжелую цепь (HC), содержащую вариабельный домен HC (VH), и легкую цепь (LC), содержащую вариабельный домен LC (VL), где (i) VH содержит по меньшей мере три определяющие комплементарность области (CDR) HC дурвалумаба, и VL содержит по меньшей мере три LC-CDR дурвалумаба, (ii) VH содержит по меньшей мере три HC-CDR авелумаба, и VL содержит по меньшей мере три LC-CDR авелумаба, или (iii) VH содержит по меньшей мере три HC-CDR атезолизумаба, и VL содержит по меньшей мере три LC-CDR атезолизумаба.
В дополнительных вариантах осуществления фрагмент антитела против PD-L1 содержит (i) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 103, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 104, (ii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 105, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 106, или (iii) VH, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 107, и VL, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 108.
В конкретных вариантах осуществления указанных выше фрагментов антител против PD-L1 фрагмент антитела против PD-1 выбран из группы, состоящей из F(ab), F(ab')2, Fv и scFv.
Настоящее изобретение дополнительно относится к каждому из антител против PD-L1, приведенных в таблицах 28-36, при условии, что, если VH и VL имеют аминокислотные последовательности SEQ ID NO: 107 и SEQ ID NO: 108, соответственно, то константный домен тяжелой цепи (HC) не принадлежит изотипу IgG1 с комбинацией замен N297A/D356E/L358M, или, если VH и VL имеют аминокислотные последовательности SEQ ID NO: 103 и SEQ ID NO: 104, соответственно, то константный домен HC не принадлежит изотипу IgG1 с комбинацией замен L234F/L235E/P331S/D356E/L358M.
Настоящее изобретение дополнительно относится к композиции, содержащей антитело против PD-L1 или фрагмент антитела против PD-L1, представленный в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение дополнительно относится к композиции, содержащей (i) антитело против CTLA-4, представленное в настоящем описании, и антитело против PD-1, представленное в настоящем описании, и фармацевтически приемлемый носитель; или (ii) антитело против CTLA-4, представленное в настоящем описании, и антитело против PD-L1, представленное в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение дополнительно относится к композиции, содержащей (i) фрагмент антитела против CTLA-4, представленный в настоящем описании, и антитело против PD-1, представленное в настоящем описании, и фармацевтически приемлемый носитель, или (ii) фрагмент антитела против CTLA-4, представленный в настоящем описании, и антитело против PD-L1, представленное в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение дополнительно относится к композиции, содержащей (i) фрагмент антитела против CTLA-4, представленный в настоящем описании, и фрагмент антитела против PD-1, представленный в настоящем описании, и фармацевтически приемлемый носитель, или (ii) фрагмент антитела против CTLA-4, представленный в настоящем описании, и фрагмент антитела против PD-L1, представленный в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение дополнительно относится к композиции, содержащей (i) антитело против CTLA-4, представленное в настоящем описании, и фрагмент антитела против PD-1, представленный в настоящем описании, и фармацевтически приемлемый носитель или (ii) антитело против CTLA-4, представленное в настоящем описании, и фрагмент антитела против PD-L1, представленный в настоящем описании, и фармацевтически приемлемый носитель.
Настоящее изобретение дополнительно относится к любому из антител против CTLA-4, против PD-1 или против PD-L1 или композициям, представленным в настоящем описании, для лечения злокачественного новообразования у индивидуума или для получения лекарственного средства для лечения злокачественного новообразования у индивидуума.
Настоящее изобретение дополнительно относится к любому из фрагментов антител против CTLA-4, против PD-1 или против PD-L1 или композициям, представленным в настоящем описании, для лечения злокачественного новообразования у индивидуума или для получения лекарственного средства для лечения злокачественного новообразования у индивидуума.
В конкретных вариантах осуществления злокачественное новообразование является меланомой, немелкоклеточным раком легких, раком головы и шеи, уротелиальным раком, раком молочной железы, желудочно-кишечным злокачественным новообразованием, множественной миеломой, печеночно-клеточным раком, неходжкинской лимфомой, раком почки, лимфомой Ходжкина, мезотелиомой, раком яичников, мелкоклеточным раком легких, раком пищевода, раком анального канала, раком желчных протоков, колоректальным раком, раком шейки матки, раком щитовидной железы или раком слюнных желез.
В конкретных вариантах осуществления злокачественное новообразование является раком поджелудочной железы, раком бронхов, раком предстательной железы, раком поджелудочной железы, раком желудка, раком яичников, раком мочевого пузыря, злокачественным новообразованием головного мозга или центральной нервной системы, злокачественным новообразованием периферической нервной системы, раком матки или эндометрия, раком полости рта или глотки, раком печени, раком почки, раком яичка, раком желчных протоков, раком тонкого кишечника или аппендикса, раком надпочечника, остеосаркомой, хондросаркомой или злокачественным новообразованием кроветворных тканей.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1A-1E: Опосредованный блокадой CTLA-4 колит является Fc-зависимым. Мышам Balb/c дважды в неделю вводили антитела, как указано, в течение 55 дней. Фиг. 1A: Определение профиля экспрессии воспалительных генов кишечника после введения Fc-компетентного антитела против CTLA-4 или эффекторно-молчащего антитела против CTLA-4 включает замену D265A (антитело против CTLA4 (D265S)). После семи недель введения дважды в неделю проксимальные отделы тонкого кишечника собирали для оценки воспалительных маркеров кишечника посредством количественной полимеразной цепной реакции (ПЦР) с обратной транскрипцией. Показана тепловая карта кратного изменения экспрессии воспалительных генов кишечника для двух антител по сравнению с введением изотипического контроля. Экспрессию анализировали во множестве панелей и данные порогового цикла нормализовали по убиквитину в каждой панели. Нормализованные данные для генов, проанализированных как часть множественных панелей, усредняли перед определением кратного изменения относительно изотипического контроля. Фиг. 1B: потеря массы за время эксперимента. Фиг. 1C: Кишечную проницаемость оценивали посредством измерения флуоресценции FITC-декстрана в сыворотке в день 49 и 50. Фиг. 1D: гистологические данные об относительном воспалении кишечника и тяжести колита исследовал и оценивал сертифицированный патолог. Фиг. 1E: типичные микрофотографии окрашенных гематоксилином и эозином гистологических срезов толстого кишечника со дня 55.
Фиг. 2A-2E: Характеризация ISVD против CTLA-4 (nAb). Фиг. 2A: сравнение эффекторно-молчащего ISVD CTLA4 (nAb против CTLA4) с эффекторно-компетентным антителом против CTLA4; фиг. 2B: активированные T-клетки селезенки культивировали в течение трех дней в присутствии nAb против CTLA4 или антитела против CTLA4, как указано. Пролиферацию (фиг. 2C), продукцию ИФНγ (фиг. 2D) и ИЛ-2 (фиг. 22E) измеряли и строили график в виде кратного изменения относительно изотипического контроля (IgG2a мыши). Данные являются типичными для двух-трех независимых экспериментов.
Фиг. 3A-3D: nAb против CTLA4 в комбинации с антителом против PD-1 имеет высокую противоопухолевую эффективность. Фиг. 3A: Несущим опухоль CT26 мышам вводили дозу указанного антитела (антитела против CTLA4, антитела против PD1, антитела против CTLA4 (D265A)) 20 мг/кг и/или nAb против CTLA4 в дозе 30 мг/кг каждые четыре дня всего в пяти дозах, когда опухоли достигали среднего размера 100 мм3 (диапазон 78-125 мм3). Данные приведены как средний объем опухоли в течение 32-дневного периода. Результаты типичны для двух независимых экспериментов (n=10 мышей на группу); фиг. 3B: соотношение CD8+ T-клеток/ Foxp3+ Treg-клеток в опухоли в день один после введения, как указано. Результаты типичны для двух независимых экспериментов (n=7 мышей на группу); фиг. 3C и фиг. 3D: Профиль экспрессии генов всей опухоли в день восемь после введения. Результаты типичны для одного или двух независимых экспериментов (n=5 мышей на группу). *p<0,05, **p<0,01, ***p<0,001 (непарный t-критерий Стьюдента). Планка погрешности ± SEM. Фиг. 3E: показаны объемы опухолей отдельных животных для каждой группа лечения по сравнению с изотипическими контролями. Полные ответы (CR) до дня 39 приведены для групп ответивших на лечение. Данные типичны для двух независимых экспериментов, n=10 мышей на группу.
Фиг. 4A-4D: Опосредованный антителом против CTLA-4 колит является Fc-зависимым. Мышам Balb/c дважды в неделю вводили антитела (антитело против CTLA4, антитело против PD1, антитело против CTLA4 (D265A)) в отдельности или в комбинации с nAb против CTLA4, как указано, в течение 55 дней. Фиг. 4A: потеря массы за время эксперимента; фиг. 4B: гистологическая оценка энтерита в проксимальных отделах тощей кишки в день 55; фиг. 4C: микрофотографии окрашенного гематоксилином и эозином гистологического среза толстого кишечника; фиг. 4D: показана тепловая карта кратного изменения экспрессии воспалительных генов кишечника для указанных образцов по сравнению с изотипическим контролем. Экспрессию анализировали во множественных панелях и данные порогового цикла нормализовали по убиквитину в каждой панели. Нормализованные данные для генов, проанализированных как часть множественных панелей, усредняли перед определением кратного изменения относительно изотипического контроля.
Фиг. 5A-5C: Эффекторная функция Fc антитела против CTLA-4 запускает воспаление в коже, но не системное воспаление. Мышам Balb/c дважды в неделю вводили антитело против CTLA4 или антитело против CTLA4 (D265A), как указано, в течение 55 дней. Фиг. 5A: Микрофотографии окрашенного гематоксилином и эозином гистологического среза кожи уха. Фиг. 5B: Абсолютное количество ИЛ-17-продуцирующих T-клеток, Foxp3+ Treg-клеток и нейтрофилов кожи уха измеряли посредством проточной цитометрии. Фиг. 5C: микрофотографии окрашенного гематоксилином и эозином гистологического среза почки (верхняя панель), печени (средняя панель) и легкого (нижняя панель). Результаты типичны для одного из двух независимых экспериментов (n=4-8 мышей на группу). Масштабные линейки соответствуют 100 мкм. Планка погрешности ± SEM.
Фиг. 6A-6D: Fc-достаточное антитело против CTLA-4 не истощает Foxp3+ Treg-клетки толстого кишечника. Фиг. 6A: внутриклеточное окрашивание CTLA-4 у несущих опухоль CT26 мышей в указанных органах. Фиг. 6B: средняя интенсивность флуоресценции (MFI) CTLA-4 на Foxp3+ Treg-клетках. **p<0,01, ***p<0,001 (парный t-критерий Стьюдента). Фиг. 6C и фиг. 6D: Типичный точечный график и статистика для инфильтрирующих собственную пластинку толстого кишечника и опухоль CT26 Foxp3+ Treg-клеток через 24 часа после введения, как указано. Данные типичны для 2-4 независимых экспериментов (n=4-12 мышей на группу) **p<0,01, ***p<0,001 (непарный t-критерий Стьюдента). Планка погрешности ± SEM.
Фиг. 7A-7D: Опосредованный функцией Fc кишечник при нарушенной антителом против CTLA-4, опосредованной Treg-клетками супрессии колита. CD45Rbhigh наивные T-клетки селезенки переносили мышам-реципиентам CB17-SCID и вводили им антитело против CTLA4 или nAb против CTLA4, как указано. Фиг. 7A: потеря массы за время эксперимента. Фиг. 7B: микрофотографии окрашенного гематоксилином и эозином гистологического среза толстого кишечника, и Фиг. 7C: балльная оценка патологии в день 47(n=14-18 мышей на группу). Фиг. 7D: профиль экспрессии генов во всем толстом кишечнике в день 47 после переноса наивных T-клеток (n=6 мышей на группу). Данные типичны для 1 из 2 независимых экспериментов. Ns=не значимо ****p<0,0001 (непарный t-критерий Стьюдента). Планка погрешности ± SEM.
Фиг. 8A-8E: Вовлечение FcγR и блокада CTLA-4 активируют макрофаги толстого кишечника. Фиг. 8A и фиг. 8B: поверхностная экспрессия CD16/CD32 на макрофагах, выделенных из селезенки, собственной пластинки толстого кишечника и опухоли несущих CT26 мышей, оценивали посредством проточной цитометрии. Фиг. 8C: долю макрофагов (CD45+CD11b+F4/80+) в селезенке, собственной пластинке толстого кишечника и опухоли несущих CT26 мышей оценивали посредством проточной цитометрии. Фиг. 8D: экспрессию мРНК ИЛ-1b, ФНОα, ИФНγ и Stat1 оценивали в толстом кишечнике мышей, которым вводили антитело против CTLA4, антитело против CTLA4 (D265A) или nAb против CTLA4 в день 0, 10 и 18 после введения. Данные типичны для двух независимых экспериментов (n=8-10 мышей на группу). ns=не значимо, **p<0,01, ***p<0,001 (парный t-критерий Стьюдента). Планка погрешности ± SEM. Фиг. 8E: долю ИЛ-17-продуцирующих CD4+ T-клеток (CD45+TCRb+CD4+CD8a-ИЛ-17A+), абсолютное количество ИФНγ-продуцирующих CD8a+ T-клеток (CD45+TCRb+CD4-CD8a+ИФНγ+) и нейтрофилов (CD45+CD11b+Ly6Ghigh) собственной пластинки толстого кишечника измеряли посредством проточной цитометрии. Результаты типичны для 1 из 2 независимых экспериментов (n=4-8 мышей на группу). **p<0,01 (непарный t-критерий Стьюдента). Масштабные линейки соответствуют 100 мкм. Планка погрешности ± SEM.
Фиг. 9A-9B: Противоопухолевая эффективность в исследовании на сингенной модели опухоли мочевого пузыря MB49 на мышах. Фиг. 9A: несущим опухоль MB49 мышам вводили дозу указанного антитела (30 мг/кг nAb против CTLA-4, 10 мг/кг антитела против CTLA4, 5 мг/кг антитела против PD1 или комбинацию nAb против CTLA4 и антитела против PD-1) каждые четыре дня всего в четырех дозах, когда опухоли достигали среднего размера 102 мм3 (диапазон 87-117 мм3). Данные приведены как средний объем опухоли за 21-дневный период. Результаты типичны для двух независимых экспериментов (n=10 мышей на группу). Фиг. 9B: показаны объемы опухолей отдельных животных для каждой группы лечения. Полные ответы (CR) до дня 21 приведены для групп ответивших на лечение. Данные соответствуют результатам эксперимента с использованием n=10 мышей на группу.
Фиг. 10A-10B: Противоопухолевая эффективность в исследовании на сингенной модели опухоли толстого кишечника MC38 на мышах. Фиг. 10A: несущим опухоль MC38 мышам вводили дозу указанного антитела (30 мг/кг nAb против CTLA-4, 10 мг/кг антитела против CTLA4, 5 мг/кг антитела против PD1 или комбинацию nAb против CTLA4 и антитела против PD-1) каждые четыре дня всего в четырех дозах, когда опухоли достигали среднего размера 220 мм3 (диапазон 179-261 мм3). Данные приведены как средний объем опухоли за 23-дневный период. Результаты типичны для двух независимых экспериментов (n=10 мышей на группу). Фиг. 10B: показаны объемы опухолей отдельных животных для каждой группы лечения. Полные ответы (CR) до дня 23 приведены для групп ответивших на лечение. Данные соответствуют результатам эксперимента с использованием n=10 мышей на группу.
Фиг. 11: Индуцирование воспаления кишечника эффекторными клетками. На изображении показано, что Fc-опосредованного индуцирования воспаления кишечника можно достигать с помощью эффекторных T-клеток независимо от истощения Treg-клеток.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определение
В рамках изобретения термин "нежелательное явление" или "AE" приведен в Общих терминологических критериях нежелательных явлений (CTCAE) версии 5.0, опубликованных 27 ноября 2017 года Министерством здравоохранения и социального обеспечения США как любой неблагоприятный и незапланированный признак (включая отклонения результатов лабораторных анализов), симптом или заболевание, по времени ассоциированные с использованием медицинского лечения или способом, у человека, которые можно считать или не считать связанными с медицинским лечением или способом. AE является термином, представляющим собой уникальное представление конкретного явления, используемое для медицинской документации и научных анализов. Медицинское лечение может иметь одно или более ассоциированных AE, и каждое AE может иметь тот же или иной уровень тяжести. Тяжести AE приписывают степень. CTCAE отражает степени от 1 до 5 с уникальными клиническими описаниями тяжести каждого AE на основе общих рекомендаций: степень 1, слабая, или бессимптомная или слабые симптомы, только клиническое или диагностическое наблюдение, или вмешательство не показано; степень 2, умеренная, или минимальная, показано локальное или неинвазивное вмешательство, или соответствующее возрасту ограничение по шкале оценки способности к самообслуживанию (ADL); степень 3, тяжелая или медицински значимая, но не непосредственно угрожающая жизни, или показана госпитализация или пролонгирование госпитализации, или потеря дееспособности, или ограничение способности к самообслуживанию (ADL); степень 4, угрожающие жизни последствия или показано неотложное вмешательство; и степень 5, смерть, связанная с AE.
В рамках изобретения термин "антитело" относится к гликопротеину, содержащему (a) по меньшей мере две тяжелые цепи (HC) и две легкие цепи (LC), соединенные между собой дисульфидными связями, или (b), в случае антител биологических видов Верблюжьих, по меньшей мере две тяжелые цепи (HC), соединенные между собой дисульфидными связями. Каждая HC состоит из вариабельной области или домена тяжелой цепи (VH) и константной области или домена тяжелой цепи. В некоторых природных антителах IgG, IgD и IgA, константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. В целом, основной структурной единицей антитела является тетрамер, содержащий две пары HC/LC, за исключением антител биологических видов Верблюжьих, содержащих только две HC, в случае которых структурной единицей является гомодимер. Каждый тетрамер включает две идентичные пары полипептидных цепей, при этом каждая пара содержит одну цепь LC (приблизительно 25 кДа) и HC (приблизительно 50-70 кДа).
В некоторых природных антителах каждая легкая цепь состоит из вариабельной области или домена LC (VL) и константного домена LC. Константный домен LC состоит из одного домена CL. VH человека включает шесть членов семейства: VH1, VH2, VH3, VH4, VH5 и VH6; и VL человека включает 16 членов семейства: Vκ1, Vκ2, Vκ3, Vκ4, Vκ5, Vκ6, Vλ1, Vλ2, Vλ3, Vλ4, Vλ5, Vλ6, Vλ7, Vλ8, Vλ9 и Vλ10. Каждого из этих членов семейства можно дополнительно разделять на конкретные подтипы. Домены VH и VL можно дополнительно разделять на области гипервариабельности, обозначаемые как определяющие комплементарность области (CDR), перемежающиеся с областями, являющимися более консервативными, обозначаемыми как каркасные области (FR). Каждый из VH и VL состоит из трех областей CDR и четырех областей FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4.
Вариабельные области тяжелых и легких цепей содержат связывающий домен, содержащий CDR, взаимодействующий с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами организма-хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента. Приписывание аминокислот к каждому домену, как правило, соответствует определениям из Sequences of Proteins of Immunological Interest, Kabat, et al.; National Institutes of Health, Bethesda, Md.; 5th ed.; NIH Publ. No. 91-3242 (1991); Kabat (1978) Adv. Prot. Chem. 32: 1-75; Kabat, et al., (1977) J. Biol. Chem. 252: 6609-6616; Chothia, et al., (1987) J Mol. Biol. 196: 901-917 или Chothia, et al., (1989) Nature 342: 878-883.
Как правило, нумерация аминокислот в константный домен тяжелой цепи начинается с числа 118, что соответствует схеме нумерации Eu. Схема нумерации Eu зависит от аминокислотной последовательности IgG1 человека (Eu), содержащей константный домен, начинающийся с положения аминокислоты 118 аминокислотной последовательности IgG1, описанной в Edelman et al., Proc. Natl. Acad. Sci. USA. 63: 78-85 (1969) и показанной для константных доменов IgG1, IgG2, IgG3 и IgG4 в  , et al., Ed. Ginetoux, Correspondence between the IMGT unique numbering for C-DOMAIN, the IMGT exon numbering, the Eu and Kabat numberings: Human IGHG, создано: 17/05/2001, версия: 08/06/2016, доступно по адресу www.imgt.org/IMGTScientificChart/Numbering/Hu_IGHGnber.html#r).
, et al., Ed. Ginetoux, Correspondence between the IMGT unique numbering for C-DOMAIN, the IMGT exon numbering, the Eu and Kabat numberings: Human IGHG, создано: 17/05/2001, версия: 08/06/2016, доступно по адресу www.imgt.org/IMGTScientificChart/Numbering/Hu_IGHGnber.html#r).
В основном, хотя пара VH/VL антитела содержит шесть CDR, три CDR на VH и три CDR на VL, на этом уровне техники известно, что в большинстве случаев область CDR3 тяжелой цепи является первичной детерминантой специфичности антитела, и в этой области известны примеры получения конкретных антител на основе CDR3 тяжелой цепи в отдельности (например, Beiboer et al., J. Mol. Biol. 296: 833-849 (2000); Klimka et al., British J. Cancer 83: 252-260 (2000); Rader et al., Proc. Natl. Acad. Sci. USA 95: 8910-8915 (1998); Xu et al., Immunity 13: 37-45 (2000). См. Kabat et al. Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991) (определение областей CDR антитела по последовательности); также см. Chothia & Lesk J. Mol. Biol. 196: 901-917 (1987) (определение областей CDR антитела по структуре).
Следующие общие правила, описанные на www.bioinf.org.uk: Prof. Andrew C.R. Martin's Group и воспроизведенные в таблице ниже можно использовать для идентификации CDR в последовательности антитела, содержащей эти аминокислоты, специфически взаимодействующие с аминокислотами, содержащими эпитоп в антигене, с которым связывается антитело. Существуют редкие примеры, когда эти, как правило, константные признаки не встречаются; однако, остатки Cys являются наиболее консервативным признаком.
В целом, основная структурная единица антитела содержит тетрамер. Каждый тетрамер включает две идентичные пары полипептидных цепей, где каждая пара содержит одну цепь LC (приблизительно 25 кДа) и одну цепь HC (приблизительно 50-70 кДа). Амино-концевая часть каждой цепи включает вариабельную область из приблизительно от 100 до 110 или более аминокислот, главным образом, отвечающих за распознавание антигена. Карбокси-концевая часть HC может определять константную область, главным образом, отвечающую за эффекторную функцию антитела. Как правило, LC человека классифицируют как LC каппа и лямбда. Кроме того, HC человека, как правило, классифицируют как мю, дельта, гамма, альфа или эпсилон, и они определяют изотип антитела как IgM, IgD, IgG, IgA и IgE, соответственно. В LC и HC вариабельные и константные области соединены областью "J" из приблизительно 12 или более аминокислот, при этом HC также включает область "D" еще из приблизительно 10 аминокислот. В целом, см. Fundamental Immunology, Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989)).
Тяжелая цепь антитела может содержать или не содержать концевой остаток лизина (K) или концевые остатки глицина и лизина (GK). Таким образом, в конкретных вариантах осуществления антитела по изобретению содержат аминокислотную последовательность константной области тяжелой цепи, приведенную в настоящем описании, в которой дополнительно отсутствует концевой лизин, и которая заканчивается остатком глицина, или дополнительные варианты осуществления, в которых также отсутствует концевой остаток глицина. Это является результатом того, что концевой лизин и иногда глицин и лизин вместе могут расщепляться во время экспрессии антитела или отщепляться при введении в организм человека без очевидного неблагоприятного воздействия на эффективность, стабильность или иммуногенность антитела. В некоторых случаях в молекулу нуклеиновой кислоты, кодирующей тяжелую цепь, могут быть намерено не включены кодоны, кодирующие концевой лизин, или кодоны для концевого лизина и глицина.
В рамках изобретения термин "фрагмент антитела" или "антигенсвязывающий фрагмент" относится к фрагментам полноразмерных антител, т.е. фрагментам антител, сохраняющим способность специфически связываться с антигеном, связываемым полноразмерным антителом, но меньшим, чем полноразмерное антитело, и в которых полностью отсутствует Fc-домен или отсутствуют части Fc-домена, обеспечивающие связывание антитела с FcγR. Неограничивающие примеры связывающих фрагментов антител включают Fab-, Fab'-, F(ab')2- и Fv-фрагменты; диатела; молекулы scFv; нанотела и полиспецифические антитела, образующиеся из фрагментов антител.
В рамках изобретения "химерное антитело" является антителом, имеющим вариабельный домен из первого антитела и константный домен из второго антитела, где (i) первое и второе антитела получены из разных биологических видов (патент США №4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)) или (ii) первое и второе антитела принадлежат разным изотипам, например, вариабельный домен из антитела IgG1 и константные домены из антитела IgG4. В одном из аспектов вариабельные домены получены из не принадлежащего человеку антитела, такого как антитело мыши ("родительское антитело"), и последовательности константного домена получены из антитела человека. В дополнительном аспекте вариабельные домены являются гуманизированными вариабельными доменами из антитела мыши и константными доменами антитела человека.
В рамках изобретения термин "комбинированное лечение" относится к лечению человека или животного, включающему введение индивидууму первого терапевтического средства и второго терапевтического средства последовательно или одновременно. В основном, первое и второе терапевтические средства вводят индивидууму раздельно, а не в виде смеси; однако, могут быть варианты осуществления, в которых первое и второе терапевтические средства смешивают перед введением.
В рамках изобретения термин "консервативная замена" относится к заменам аминокислот другими аминокислотами, имеющими схожие характеристики (например, заряд, размер боковой цепи, гидрофобность/гидрофильность, конформацию, жесткость остова и т.д.), таким образом, что часто можно вносить изменения без изменений биологической активности белка. Специалистам в этой области известно, что, в основном, единичные замены аминокислот во второстепенных областях полипептида, по существу, не изменяют биологическую активность (см., например, Watson et al. Molecular biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th Ed.) (1987)). Кроме того, замены структурно или функционально схожих аминокислоты, менее вероятно, нарушат биологическую активность. Примеры консервативных замен приведены в таблице 2.
Термины "антиген-4 цитотоксических T-лимфоцитов", "CTLA-4", "CTLA4", "антиген CTLA-4" и "CD152" (см., например, Murata, Am. J. Pathol. 155: 453-460 (1999)) используют взаимозаменяемо, и они включают варианты, изоформы, видовые гомологи CTLA-4 человека и аналоги, имеющие по меньшей мере один общий эпитоп с CTLA-4 (см., например, Balzano, Int. J. Cancer Suppl. 7: 28-32 (1992)). Полную последовательность нуклеиновой кислоты CTLA-4 можно найти под регистрационным номером GenBank L15006.
В рамках изобретения термин "эффекторная функция" относится к видам биологической активности, приписываемым Fc-области антитела и варьирующимся в зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание C1q и обусловленную комплементом цитотоксичность (CDC); связывание Fc-рецептора; антителозависимую клеточную цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов поверхности клетки (например, B-клеточного рецептора); и активацию B-клеток. Антитела действуют посредством ряда механизмов, большинство из которых затрагивают другие части иммунной системы. Антитела могут просто блокировать взаимодействия молекул или могут активировать классический путь комплемента (известный как обусловленная комплементом цитотоксичность или CDC) посредством взаимодействия C1q в комплексе C1 с кластеризованными антителами. Немаловажно, что антитела также действуют как связующее звено между антитело-опосредованными и клеточно-опосредованными иммунными ответами посредством вовлечения Fc-рецепторов.
В рамках изобретения "эффекторно-молчащие" относится к антителу или фрагменту антитела, демонстрирующему (i) отсутствие измеримого связывания с одним или более Fc-рецепторами (FcR), что можно измерять с помощью анализа Biacore, где константа ассоциации в микромолярном диапазоне свидетельствует об отсутствии измеримого связывания, или (ii) измеримое связывание с одним или более FcR, что можно измерять с помощью анализа Biacore, сниженное по сравнению со связыванием, типичным для антитела того же изотипа. В конкретных вариантах осуществления антитело может содержать одну или более мутаций в константном домене HC и, в частности, Fc-домене таким образом, что мутантное антитело имеет сниженное или отсутствующее измеримое связывание с FcγRIIIa, FcγRIIa и FcγRI по сравнению с антителом дикого типа того же изотипа, что и мутантное антитело. В конкретных вариантах осуществления аффинность или константа ассоциации эффекторно-молчащего антитела с одним или более из FcγRIIIa, FcγRIIa и FcγRI снижена по меньшей мере в 1000 раз по сравнению с аффинностью изотипа дикого типа; снижена по меньшей мере в 100-1000 раз по сравнению с аффинностью изотипа дикого типа, снижена по меньшей мере в 50-100 раза по сравнению с аффинностью изотипа дикого типа; или по меньшей мере 10-50 раз по сравнению с аффинностью изотипа дикого типа. В конкретных вариантах осуществления эффекторно-молчащее антитело не имеет детектируемого или измеримого связывания с одним или более из FcγRIIIa, FcγRIIa и FcγRI по сравнению со связыванием изотипа дикого типа. В основном, в эффекторно-молчащих антителах будет отсутствовать измеримая активность антителозависимой клеточной цитотоксичности (ADCC). В эффекторно-молчащем фрагменте антитела отсутствует Fc-домен или части Fc-домена, придающие связывание с FcR, и, в связи с этим, они не будут демонстрировать детектируемого или измеримого связывания с одним или более из FcγRIIIa, FcγRIIa или FcγRI. В случае эффекторно-молчащего антитела или фрагментов антител связывание измеряют относительно FcR человека.
В рамках изобретения "Fab-фрагмент" состоит из одной LC и VH и CH1 одной HC и исключает остальную часть константного домена HC. CH1 молекулы Fab не могут образовывать дисульфидную связь с другим Fab-фрагментом или HC-содержащей молекулой. "Fab-фрагмент" может являться продуктом расщепления антитела папаином.
В рамках изобретения "Fab'-фрагмент" содержит одну LC и фрагмент одной HC, содержащий домен VH и константный домен HC вплоть до области между доменами CH1 и CH2, и исключает остальную часть константного домена HC, таким образом, что может образовываться межцепочечная дисульфидная связь между двумя HC двух Fab'-фрагментов с образованием молекулы F(ab')2.
В рамках изобретения "F(ab')2-фрагмент" содержит две LC и два фрагмента HC, где каждый фрагмент HC содержит домен VH и константный домен HC вплоть до области между доменами CH1 и CH2 и исключает остальную часть константного домена HC, таким образом, что межцепочечная дисульфидная связь образуется между двумя HC. Таким образом, F(ab')2-фрагмент состоит из двух Fab'-фрагментов, удерживаемых вместе дисульфидной связью между двумя тяжелыми цепями. F(ab')2-фрагмент можно получать посредством расщепления антитела пепсином, расщепляющим антитело в участке между доменами CH1 и CH2.
В рамках изобретения "Fc-домен" или "Fc" является доменом или областью кристаллизуемого фрагмента, полученным из антитела и содержащим домены CH2 и CH3 антитела. В антителе два Fc-домена удерживаются вместе двумя или более дисульфидными связями и гидрофобными взаимодействиями доменов CH3. Fc-домен можно получать посредством расщепления антитела протеазой папаином.
В рамках изобретения "Fc-рецепторы" или "FcR" являются ключевыми иммунными регуляторными рецепторами, связывающими гуморальный иммунный ответ с клеточными эффекторными функциями. Идентифицированы рецепторы для всех классов иммуноглобулинов, включая FcγR (IgG), FcεRI (IgE), FcαRI (IgA), FcμR (IgM) и FcδR (IgD). Существует три класса рецепторов для IgG человека, обнаруживаемые в лейкоцитах: CD64 (FcγRI), CD32 (FcγRIIa, FcγRIIb и FcγRIIc) и CD16 (FcγRIIIa и FcγRIIIb). FcγRI классифицируют как высокоаффинный рецептор (KD наномолярного диапазона), в то время как FcγRII и FcγRIII имеют аффинность от низкой до промежуточной (KD микромолярного диапазона). При антителозависимой клеточной цитотоксичности (ADCC) FcR на поверхности эффекторных клеток (естественных киллеров, макрофагов, моноцитов и эозинофилов) связываются с Fc-областью IgG, которая сама связана с клеткой-мишенью. После связывания путь передачи сигнала активируется, что приводит к секреции различных веществ, таких как литические ферменты, перфорин, гранзимы и фактор некроза опухоли, опосредующие разрушение клетки-мишени. Уровень эффекторной функции ADCC отличается среди подтипов IgG человека. Хотя это зависит от аллотипа и конкретного FcR, в простых терминах ADCC эффекторная функция является высокой для IgG1 и IgG3 человека и низкой для IgG2 и IgG4.
В рамках изобретения "Fv-область" содержит одну VH и пару VL, где полипептид VH и полипептид VL удерживаются вместе дисульфидными связями.
В рамках изобретения "гуманизация" (также называемая реконструированием или пересадкой CDR) представляет собой хорошо известный способ снижения иммуногенности моноклональных антител (mAb) из экзогенных источников (как правило, грызуна) и улучшения эффекторных функций (ADCC, активации комплемента, связывания C1q). Сконструированное mAb конструируют с использованием способов молекулярной биологии, однако, простая пересадка определяющих комплементарность областей (CDR) грызуна на каркасы человека часто приводит к утрате аффинности связывания и/или специфичности исходного mAb. В случае гуманизации антитела дизайн гуманизированного антитела включает такие варианты, как консервативные аминокислотные замены в остатках CDR и обратная замена остатков из mAb грызуна в каркасные области человека (обратные мутации). Положения можно различать или идентифицировать посредством сравнения последовательностей для структурного анализа или посредством анализа модели гомологии трехмерной структуры вариабельных областей. В способе созревания аффинности в последнее время используют фаговые библиотеки для варьирования аминокислот в выбранных положениях. Аналогичным образом используют множество подходов для выбора наиболее подходящих каркасов человека, на которые пересаживают CDR грызуна. С ростом наборов данных известных параметров для структур антител растет сложность и детализация этих способов. Можно использовать консенсусные последовательности или последовательности зародышевой линии из одного антитела или фрагментов каркасных последовательностей в каждой вариабельной области легкой или тяжелой цепи из нескольких разных mAb человека. Другим подходом гуманизации является модификация только поверхностных остатков последовательности грызуна, при этом наиболее распространенные остатки обнаруживают в mAb человека, и его обозначают как "перекладку" или "венирование". Зачастую антитело человека или гуманизированное антитело является, по существу, неиммуногенным для людей.
В рамках изобретения термин "гуманизированное антитело" относится к формам антител или фрагментов антител, содержащим последовательности из антител человека и не принадлежащих человеку антител (например, мыши, крысы). В основном, гуманизированное антитело будет содержать все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых гипервариабельные петли соответствуют петлям не принадлежащего человеку иммуноглобулина, и все или, по существу, все из каркасных (FR) областей являются каркасными областями последовательности иммуноглобулина человека. Гуманизированное антитело, необязательно, может содержать по меньшей мере часть константной области иммуноглобулина человека (например, Fc-домен).
В рамках изобретения термин "гиперпролиферативное заболевание" относится к состояниям, при которых рост клеток повышен относительно нормальных уровней. Например, гиперпролиферативные заболевания или нарушения включают злокачественные заболевания (например, рак пищевода, рак толстого кишечника, рак желчных протоков) и незлокачественные заболевания (например, атеросклероз, доброкачественную гиперплазию, доброкачественную гипертрофию предстательной железы).
В рамках изобретения термин "иммуноопосредованные нежелательные явления" или irAE" относится к AE, являющимся аутоиммунными проявлениями из-за дисбаланса иммунной системы, которые можно приписать использованию одного или более ингибиторов иммунных контрольных точек, таких как антитела против PD-1, против PD-L1 и против CTLA-4.
В рамках изобретения термин "иммунный ответ" относится к действию, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитирующих клеток, гранулоцитов и растворимых макромолекул, продуцируемых указанными выше клетками или печенью (включая антитела, цитокины и комплемент), приводящему к селективному повреждению, нарушению или элиминации из организма человека инвазивных патогенов, клеток или тканей, инфицированных патогенами, злокачественных клеток, или, в случае аутоиммунности или патологического воспаления, нормальных клеток или тканей человека.
В рамках изобретения термин "отдельный вариабельный домен иммуноглобулина" (также обозначаемый как "ISV" или "ISVD"), как правило, используют в отношении вариабельных доменов иммуноглобулина (которые могут являться доменами тяжелой цепи или легкой цепи, включая домены VH, VHH или VL), которые могут образовывать функциональный антигенсвязывающий участок без взаимодействия с другим вариабельным доменом (например, без взаимодействия VH/VL, необходимого между доменами VH и VL общепринятого 4-цепочечного моноклонального антитела). Примеры ISVD включают нанотела (включая VHH, гуманизированный VHH и/или камелизированные VH, такие как камелизированные VH человека), IgNAR, домены, (однодоменные) антитела (такие как dAb™), представляющие собой домены VH или полученные из домена VH, и (однодоменные) антитела (такие как dAb™), представляющие собой домены VL или полученные из домена VL. ISVD, основанные и/или полученные из вариабельных доменов тяжелой цепи (таких как домены VH или VHH), как правило, являются предпочтительными.
В рамках изобретения термин "моноклональное антитело" относится к популяции, по существу, гомогенных антител, т.е. молекулы антител, составляющие популяцию, идентичны по своей аминокислотной последовательности, за исключением возможных природных мутаций, которые могут присутствовать в незначительных количествах. В отличие от этого, общепринятые (поликлональные) препараты антител, как правило, включают множество разных антител, имеющих разные аминокислотные последовательности в своих вариабельных доменах, которые зачастую специфичны для разных эпитопов. Определение "моноклональное" означает природу антитела как получаемого из, по существу, гомогенной популяции антител, и его не следует истолковывать как требующее получения антитела любым конкретным способом. Например, моноклональные антитела, подлежащие использованию по изобретению, можно получать гибридомным способом, впервые описанным в Kohler et al., Nature 256: 495 (1975), или способами рекомбинантной ДНК (см., например, патент США №4816567). "Моноклональные антитела" также можно выделять из фаговых библиотек антител способами, описанными в Clackson et al., Nature 352: 624-628 (1991) и Marks et al., J. Mol. Biol. 222: 581-597 (1991), например. Также см. Presta J. Allergy Clin. Immunol. 116: 731 (2005).
В рамках изобретения термины "нанотело" и "нанотела" являются зарегистрированными торговыми марками Ablynx N.V.
В рамках изобретения термин "не принадлежащие человеку аминокислотные последовательности" в отношении антител или иммуноглобулинов относится к аминокислотной последовательности, являющейся характерной аминокислотной последовательностью не являющегося человеком млекопитающего. Термин не включает аминокислотные последовательности антител или иммуноглобулинов, полученных из библиотеки полностью человеческих антител, где разнообразия библиотеки достигают in silico (см., например, патент США №8877688 или 8691730).
Термин "PD-1" относится к белку программируемой гибели 1 (PD-1), ингибиторному члену расширенного семейства T-клеточных регуляторов CD28/CTLA-4 (Okazaki et al., Curr. Opin. Immunol. 14: 391779-82 (2002); Bennett et al., J. Immunol. 170: 711-8 (2003)). Другие члены семейства CD28 включают CD28, CTLA-4, ICOS и BTLA. Ген PD-1 кодирует трансмембранный белок I типа массой 55 кДа (Agata et al., Intl. Immunol. 8: 765-72 (1996)). Идентифицировано два лиганда для PD-1, PD-L1 (B7-H1) и PD-L2 (B7-DC), которые, как показано, отрицательно регулируют активацию T-клеток после связывания с PD-1 (Freeman et al. (2000) J. Exp. Med. 192: 1027-34; Carter et al. (2002) Eur. J. Immunol. 32: 634-43). PD-1 известен как иммуноингибиторный белок, отрицательно регулирующий сигналы TCR (Ishida, Y. et al., EMBO J. 11: 3887-3895 (1992); Blank, C. et al., Immunol. Immunother. 56(5): 739-745 (Epub 2006 Dec. 29)). Взаимодействие между PD-1 и PD-L1 может действовать как иммунная контрольная точка, приводя, например, к снижению инфильтрирующих опухоль лимфоцитов, снижению опосредованной T-клеточным рецептором пролиферации и/или избеганию злокачественными клетками иммунологического надзора (Dong et al., J. Mol. Med. 81: 281-7 (2003); Blank et al., Cancer Immunol. Immunother. 54: 307-314 (2005); Konishi et al., Clin. Cancer Res. 10: 5094-100 (2004)). Иммунную супрессию можно реверсировать посредством ингибирования локального взаимодействия PD-1 с PD-L1 или PD-L2; эффект является аддитивным, когда также блокируют взаимодействие PD-1 с PD-L2 (Iwai et al., Proc. Nat'l. Acad. Sci. USA 99: 12293-12297 (2002); Brown et al., J. Immunol. 170: 1257-66 (2003)).
Термины "белок программируемой гибели 1", "белок программируемой гибели клеток 1", "белок PD-1", "PD-1", "PD1," "PDCD1", "hPD-1" и "hPD-1" используют взаимозаменяемо, и они включают варианты, изоформы, видовые гомологи PD-1 человека и аналоги, имеющие по меньшей мере один общий эпитоп с PD-1. Полную последовательность PD-1 можно найти под регистрационным номером GenBank U64863.
В рамках изобретения "scFv" или "одноцепочечный вариабельный фрагмент" является слитым белком, содержащим VH и VL, слитые или связанные коротким линкерным пептидом от десяти до приблизительно 25 аминокислот. Линкер, как правило, богат глицином, придающим гибкость, а также серином или треонином, обеспечивающими растворимость, и с помощью него можно соединять N-конец VH с C-концом VL или наоборот. Этот белок сохраняет специфичность исходного иммуноглобулина, несмотря на удаление константных областей и встраивание линкера.
В рамках изобретения термин "субтерапевтическая доза" означает дозу терапевтического соединения (например, антитела), более низкую, чем обычная или типичная доза терапевтического соединения при введении в отдельности для лечения гиперпролиферативного заболевания (например, злокачественного новообразования). Доза терапевтического соединения может варьироваться в зависимости от заболевания, подвергаемого лечению. Например, субтерапевтическая доза антитела против CTLA-4 является однократной дозой антитела менее приблизительно 3 мг/кг, т.е. известной монотерапевтической дозы антитела против CTLA-4 YERVOY для лечения нерезектабельной или метастатической меланомы или, однократной дозой YERVOY менее приблизительно 10 мг/кг, известной монотерапевтической дозы для адъювантной меланомы.
В рамках изобретения термины "лечить" или "лечение" означают внешнее или внутреннее введение терапевтического средства, такого как композиция, содержащая любые из антител или их антигенсвязывающих фрагментов по настоящему изобретению, индивидууму или пациенту, имеющему один или более симптомов заболевания или, как предполагают, имеющему заболевание, в отношении которого средство имеет терапевтическую активность или профилактическую активность. Как правило, средство вводят в количестве, эффективном для облегчения одного или более симптомов заболевания у подвергнутого лечению индивидуума или популяции, посредством индуцирования регрессирования или ингибирования прогрессирования таких симптомов на любую клинически измеримую степень. Количество терапевтического средства, эффективное для облегчения любого конкретного симптома заболевания, может варьироваться в зависимости от факторов, таких как состояние заболевания, возраст и масса тела пациента, и способность лекарственного средства вызывать желаемый ответ у индивидуума. Облегчается ли симптом заболевания, можно оценивать посредством любого клинического измерения, как правило, используемого лечащими врачами или другими медицинскими работниками для оценки тяжести или статуса прогрессирования этих симптомов. Термин дополнительно включает задержку развития симптомов, ассоциированных с нарушением, и/или снижение тяжести симптомов такого нарушения. Термины дополнительно включают улучшение существующих неконтролируемых или нежелательных симптомов, профилактику дополнительных симптомов и улучшение или профилактику основополагающих причин таких симптомов. Таким образом, термины означают, что обеспечен благоприятный результат для человека или животного с нарушением, заболеванием или симптомом или потенциалом развития такого нарушения, заболевания или симптома.
В рамках изобретения термин "терапевтически эффективное количество" относится к количеству конкретного вещества, достаточному для достижения желаемого эффекта у индивидуума, подвергнутого лечению. Например, оно может являться количеством CTLA-4-блокирующего средства, необходимым для ингибирования активации CTLA-4 и индуцирования противоопухолевого ответа, или количеством, необходимым для повышенной отвечаемости против PD-1 или PD-L1 при совместном введении с PD-1 или PD-L1-блокирующим средством, соответственно.
В рамках изобретения "терапевтический индекс", также известный как "терапевтическое окно", "окно безопасности" или "терапевтическое отношение" представляет собой сравнение количества терапевтического средства, вызывающего терапевтический эффект, с количеством терапевтического средства, вызывающим токсичность.
В рамках изобретения термин "лечение", как его используют в отношении человека или животного, относится к терапевтическому лечению, предусматривающему контакт антител или антигенсвязывающих фрагментов с человеком или животным, нуждающимся в лечении антителами или фрагментами антител.
В рамках изобретения термин "VHH" означает, что домен VH получен из антител из HC. Антитела из тяжелых цепей являются функциональными антителами, содержащими две HC и несодержащими LC. Антитела из тяжелых цепей существуют в Верблюжьих, членов биологического семейства Camelidae, из которых их можно получать.
Введение
Антагонисты PD-1, такие как коммерчески доступные антитела против PD-1 KEYTRUDA и OPDIVO, содержат остов IgG4 человека, имеющий сниженную функцию FcγR, т.к. доклинические исследования с использованием антител против PD-1 с функцией связывания FcγR показали плохую противоопухолевую эффективность из-за истощения CD8+ цитотоксических T-клеток (CTL), необходимых для иммунотерапии опухоли (см., например, международную патентную заявку № WO 2014/089113). В отличие от этого, в доклинических экспериментах, в которых сравнивали антитела IgG2a мыши против CTLA-4, имеющие высокую аффинность связывания FcγR, с мутантными антителами IgG1 мыши против CTLA-4, у которых отсутствует измеримая аффинность связывания FcγR, было показано, что в случае монотерапии с использованием антител против CTLA-4 необходима функция FcγR для осуществления сильных противоопухолевых ответов (см. например, Selby et al., Cancer Immunol Res. 1: 32-42 (2013)). Необходимость функции FcγR при монотерапии антагонистом CTLA-4 коррелировала с истощением регуляторных T-клеток (Treg) в моделях опухолей на мышах из-за более высокой экспрессии CTLA-4 на TIL (Simpson et al.,J. Exp. Med. 210: 1695-710 (2013)) по сравнению с популяциями Treg-клеток в селезенке или лимфоузлах.
Авторы настоящего изобретения предполагали, что необходимость функции FcγR для эффективности антитела против CTLA-4 можно преодолевать посредством комбинирования антитела против CTLA-4 с антителом против PD-1. Эта гипотеза подтверждается новыми данными, иллюстрирующими критическую роль CD28-опосредованной костимуляции в опосредованной антагонистом PD-1 активации истощенных CD8+ цитотоксических T-клеток. Антитела против CTLA-4 и против PD-1 осуществляют свою противоопухолевую активность с помощью разных механизмов. Важно, что комбинированные эффекты антител против CTLA-4 и против PD-1 являются не только аддитивными, т.к. комбинированная блокада, вызванная двумя антителами, приводит к активации большого количества генов, включая гены, ассоциированные с пролиферацией, и гены хемокинов, не активируемые любым из антител в отдельности (см., например, фиг. 3C и 4D). Эти данные позволяют предполагать, что механизм действия в случае блокады CTLA-4 при монотерапии отличается от механизма действия в случае комбинации с блокадой PD-1.
Новые данные свидетельствуют о важности CD28-опосредованной костимуляции в активации эффекторных T-клеток (Teff) после блокады PD-1. Передача сигнала PD-1 приводит к дефосфорилированию CD28, а не TCR, как считали ранее, и передача сигнала CD28 необходима для повышенного противоопухолевого ответа, наблюдаемого после блокады PD-1. Таким образом, в то время как монотерапевтическая блокада CTLA-4, главным образом, может быть нацелена на события примирования T-клеток, можно ожидать, что комбинирование блокады CTLA-4 с блокадой PD-1 будет облегчать активацию истощенных T-клеток за пределами ожидаемого при блокаде PD-1 в отдельности. Авторы настоящего изобретения предполагали, что это механизм усиливается антагонистами CTLA-4, делающими возможной повышенную CD28-опосредованную активацию, независимо от функции Fc-рецепторов, и истощение Treg-клеток, что может являться важным фактором для опосредованной irAE токсичности.
Потенциальной проблемой в случае антител против CTLA-4, связывающихся с FcR (Fc-функциональных антител), является то, что истощение Treg-клеток или образование клеточных мостиков между миелоидными клетками и T-клетками может индуцировать нежелательное иммуноопосредованное воспаление. Авторы настоящего изобретения предполагали, что это функция Fc может вносить вклад в наблюдаемые irAE, ассоциированные с иммунотерапией злокачественных новообразований с блокадой CTLA-4. Одним из критических моментов, используемых для аргументации против потенциальной роли функции Fc в индуцировании irAE, является то, что лечение и ипилимумабом (на остове IgG1 человека), и тремелимумабом (на остове IgG2 человека) ассоциировано с воспалением кишечника. Хотя Fc-домен IgG2 человека имеет значимо более низкую аффинность к FcγR человека по сравнению с IgG1 человека, прямое сравнение антител с остовами IgG2 и IgG1 человека показало, что оба вызывали схожие уровни функции Fc с использованием биологических анализов ADCC и ADCP in vitro (например, Vargas et al., Cancer Cell. 33: 649-663 (2018)). Кроме того, истощение Treg-клеток in vivo и противоопухолевая активность химерных антител против CTLA-4 мыши с остовом изотипа IgG1 человека или остовом изотипа IgG2 человека были эквивалентными у мышей с нокином FcγR человека (Vargas et al., там же).
Ключевым препятствием для оценки потенциальной роли функции Fc в индуцировании воспаления кишечника в сингенных моделях опухолей является отсутствие измеримого воспаления и колита при использовании суррогатных антител мыши против CTLA-4. Для преодоления этого препятствия авторы настоящего изобретения использовали панель на основе ПЦР, ранее разработанную Cayatte et al., Clin. Transl. Gastroenterol. 3: e10 (2012) для измерения положительной регуляции воспалительных генов кишечника, ассоциированных с воспалительным заболеванием кишечника (IBD), на модели IBD мыши. Как показано в примерах в настоящем описании, эта панель на основе ПЦР позволила авторам настоящего изобретения определять повышенную экспрессию генов биомаркеров, свидетельствующих о воспалении кишечника у мышей, которым вводили Fc-функциональное антитело против CTLA-4 мыши (антитело против CTLA4), даже в отсутствие явного колита или гистологических признаков повреждения ткани (см. фиг. 1A, 3C и 4D). Это наблюдение субклинической стимуляции путей экспрессии генов воспаления кишечника позволило авторам настоящего изобретения расширить схему лечения для определения того, будет ли лежащее в основе воспаление прогрессировать с развитием клинического колита. Эта модель irAE колита на мышах позволила авторам настоящего изобретения осуществить эмпирические эксперименты для оценки необходимости функции Fc для индуцирования воспаления кишечника в контексте или в отсутствие сопутствующих противоопухолевых ответов в сингенных моделях опухолей, используемых в иммуноонкологической доклинической разработке.
Результаты, описанные в примерах, четко свидетельствуют о том, что в условиях монотерапии ни эффекторно-молчащее антитело против CTLA-4, ни эффекторно-молчащий фрагмент антитела против CTLA-4 не имели измеримую противоопухолевая активность. Однако введение эффекторно-молчащего антитела или эффекторно-молчащего фрагмента антитела в комбинации с антителом против PD1 приводит к противоопухолевой активности, сравнимой с противоопухолевой активностью, вызываемой эффекторно-функциональным антителом против CTLA-4 в отдельности или в комбинации с антителом против PD-1 (см. фиг. 3A) и без irAE кишечника или кожи, наблюдаемых в случае эффекторно-функционального антитела против CTLA-4 в отдельности или в комбинации с антителом против PD-1 (фиг. 4B и 5A), или потери массы (фиг. 4A). В свете этих результатов и обнаружения авторами настоящего изобретения потенциального Treg-независимого механизма, ассоциированного с Fc-опосредованной противоопухолевой активностью и воспалением кишечника, настоящее изобретение делает возможной комбинированную иммунотерапию злокачественных новообразований с блокадой CTLA-4/PD-1 с улучшенным терапевтическим индексом и более обширной пригодностью.
Способы комбинированного лечения
Настоящее изобретение относится к способам комбинированного лечения злокачественных новообразований, включающим введение индивидууму, нуждающемуся в терапии злокачественных новообразований, (i) PD-1-блокирующего средства, выбранного из группы, состоящей из антитела против PD-1, антитела против PD-L1, эффекторно-молчащего антитела против PD-1, эффекторно-молчащего антитела против PD-L1, эффекторно-молчащего фрагмента антитела против PD-1 и эффекторно-молчащего фрагмента антитела против PD-L1; и (ii) эффекторно-молчащего CTLA-4-блокирующего средства, выбранного из группы, состоящей из эффекторно-молчащего антитела против CTLA-4 и эффекторно-молчащего фрагмента антитела против CTLA-4.
Эффекторно-молчащее CTLA-4-блокирующее средство можно вводить в рамках комбинированного лечения с PD-1-блокирующим средством в дозах, превышающих субтерапевтическую дозу 1 мг/кг ипилимумаба, одобренную FDA США для способов комбинированного лечения ипилимумабом/ниволумабом, предназначенным для почечноклеточной карциномы на поздней стадии, или метастатического колоректального рака с высокой микросателлитной нестабильностью или недостаточностью репарации ошибочно спаренных оснований, и с более низким риском индуцирования irAE кожи или кишечника степени более 1-2 в соответствии с критериями, приведенными в Общих терминологических критериях нежелательных явлений (CTCAE) версии 5.0, в течение комбинированного лечения или по меньшей мере части периода времени, когда индивидуума подвергают комбинированному лечению, по сравнению с риском, наблюдаемым в случае способов комбинированного лечения ипилимумабом/ниволумабом. В конкретных вариантах осуществления дозы не индуцируют irAE степени более 1 в течение комбинированного лечения или по меньшей мере части периода времени, когда индивидуума подвергают комбинированному лечению.
Таким образом, в конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство можно вводить индивидууму в дозе более 1 мг/кг. В конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство можно вводить индивидууму в дозе по меньшей мере 3 мг/кг. В конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство можно вводить индивидууму в дозе по меньшей мере 10 мг/кг. В конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство можно вводить индивидууму в дозе по меньшей мере 15 мг/кг. В конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство можно вводить индивидууму в дозе по меньшей мере 20 мг/кг. В конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство можно вводить индивидууму в дозе от 3 мг/кг до 20 мг/кг. В конкретных вариантах осуществления эффекторно-молчащее CTLA-4-блокирующее средство можно вводить индивидууму в фиксированной дозе, независящей от массы тела индивидуума, например, дозе более 100 мг.
В конкретных вариантах осуществления комбинированного лечения эффекторно-молчащее CTLA-4-блокирующее средство является эффекторно-молчащим антителом против CTLA-4 или (b) эффекторно-молчащим фрагментом антитела против CTLA-4. Т.к. активность эффекторной функции нежелательна в случае антитела против CTLA-4, антитело против CTLA-4 имеет домен HC, сконструированный так, чтобы быть "эффекторно-молчащим", т.е. посредством модификации Fc-домена так, чтобы он имел сниженное или отсутствующее измеримое связывание FcR по сравнению с Fc-доменом антитела дикого типа того же изотипа, что и эффекторно-молчащее антитело (например, Fc-доменом немутантного IgG1, IgG2, IgG3 или IgG4), что определяют посредством анализа Biacore. В эффекторно-молчащем фрагменте антитела против CTLA-4 отсутствует Fc-домен или области Fc-домена, связывающиеся с одним или более FcR.
В конкретных вариантах осуществления комбинированному лечению по настоящему изобретению подвергают индивидуума до или после хирургического вмешательства для удаления опухоли, и ее можно использовать до, во время или после лучевой терапии.
В конкретных вариантах осуществления комбинированному лечению по настоящему изобретению подвергают индивидуума, которого ранее не лечили биотерапевтическим или химиотерапевтическим средством, т.е. индивидуум ранее не подвергался лечению. В других вариантах осуществления комбинированному лечению подвергают индивидуума, не достигшего устойчивого ответа после предшествующей терапии биотерапевтическим или химиотерапевтическим средством, т.е. индивидуум ранее подвергался лечению.
В конкретных вариантах осуществления комбинированное лечение по настоящему изобретению используют для лечения опухоли, являющейся достаточно крупной, чтобы ее обнаружить при пальпации или способами визуализации, хорошо известными в этой области, такими как МРТ, ультразвуковое исследование или КТ. В некоторых вариантах осуществления комбинированное лечение по изобретению используют для лечения опухоли на поздней стадии, имеющей размеры по меньшей мере приблизительно 200 мм3, 300 мм3, 400 мм3, 500 мм3, 750 мм3 или до 1000 мм3.
В конкретных вариантах осуществления комбинированному лечению по настоящему изобретению подвергают индивидуума, имеющему злокачественное новообразование, по результатам анализа положительное по экспрессии PD-L1. В некоторых вариантах осуществления экспрессию PD-L1 определяют с использованием диагностического антитела против PD-L1 человека или его антигенсвязывающего фрагмента в иммуногистохимическом (IHC) анализе с помощью фиксированного формалином, погруженного в парафин (FFPE) или замороженного тканевого среза образца опухоли, полученного из индивидуума. Лечащий врач индивидуума может назначать диагностический тест для определения экспрессии PD-L1 в образце опухолевой ткани, полученном из индивидуума перед началом комбинированного лечения по настоящему изобретению, но предполагают, что лечащий врач может назначать первый или последующие диагностические тесты в любое время после начала лечения, например, после завершения цикла лечения.
Комбинированное лечение может включать любой из примеров эффекторно-молчащих антител против CTLA-4 или эффекторно-молчащих фрагментов антител против CTLA-4, представленных в настоящем описании, в комбинации с любым из примеров антител против PD-1 или фрагментов антител против PD-1, представленных в настоящем описании, или любым из примеров антител против PD-L1 или фрагментов антител против PD-L1, представленных в настоящем описании.
(a) Эффекторно-молчащие антитела
Эффекторно-молчащие антитело по настоящему изобретению содержит константный домен HC или его Fc-домен, модифицированный таким образом, что антитело демонстрирует отсутствие измеримого связывания с одним или более FcR или сниженное связывание с одним или более FcR по сравнению с немодифицированным антителом того же изотипа IgG. В дополнительных вариантах осуществления эффекторно-молчащие антитела могут демонстрировать отсутствие измеримого связывания с каждым из FcγRIIIa, FcγRIIa и FcγRI или сниженное связывание с каждым из FcγRIIIa, FcγRIIa и FcγRI по сравнению с немодифицированным антителом того же изотипа IgG. В конкретных вариантах осуществления константный домен HC или Fc-домен является константным доменом HC или Fc-доменом человека.
В конкретных вариантах осуществления эффекторно-молчащее антитело содержит Fc-домен изотипа IgG1 или IgG2, IgG3 или IgG4, модифицированный так, что в нем отсутствует N-гликозилирование остатка аспарагина (Asn) в положении 297 (система нумерации Eu) константного домена HC. Консенсусная последовательность для N-гликозилирования представляет собой Asn-Xaa-Ser/Thr (где Xaa в положении 298 является любой аминокислотой, за исключением Pro); во всех четырех изотипах консенсусная последовательность N-гликозилирования представляет собой Asn-Ser-Thr. Модификации можно достигать посредством замены кодона, кодирующего Asn в положении 297 в молекуле нуклеиновой кислоты, кодирующей константный домен HC, кодоном, кодирующим другую аминокислоту, например Ala, Asp, Gln, Gly или Glu, например N297A, N297Q, N297G, N297E или N297D. Альтернативно, кодон для Ser в положении 298 можно заменять кодоном для Pro или кодон для Thr в положении 299 можно заменять любым кодоном, за исключением кодона для Ser. В дополнительном альтернативном варианте каждую из аминокислот, составляющую консенсусную последовательность N-гликозилирования, заменяют другой аминокислотой. Такие модифицированные молекулы IgG не имеют измеримой эффекторной функции. В конкретных вариантах осуществления эти мутантные молекулы HC могут дополнительно содержать 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями. В дополнительных вариантах осуществления такие IgG, модифицированные так, что в них отсутствует N-гликозилирование в положении 297, могут дополнительно включать одну или более дополнительных мутаций, представленных в настоящем описании, для устранения измеримой эффекторной функции.
Пример константного домена HC IgG1, мутантного в положении 297, что приводит к устранению N-гликозилирования константного домена HC, приведен в SEQ ID NO: 44, пример константного домена HC IgG2, мутантного в положении 297, что приводит к устранению N-гликозилирования константного домена HC, приведен в SEQ ID NO: 50, и пример IgG4 константного домена HC, мутантного в положении 297, что приводит к устранению N-гликозилирования константного домена HC, приведен в SEQ ID NO: 56. В конкретных вариантах осуществления эти мутантные молекулы HC может дополнительно содержать 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления Fc-домен константного домена HC IgG1 IgG2, IgG3 или IgG4, составляющего эффекторно-молчащее антитело, модифицирован так, чтобы включать одну или более замен аминокислот, выбранных из E233P, L234A, L235A, L235E, N297A, N297D, D265S и P331S (где положения определяют в соответствии с нумерацией Eu), и где указанный константный домен HC является эффекторно-молчащим. В конкретных вариантах осуществления модифицированный IgG1 дополнительно содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления константный домен HC содержит замены L234A, L235A и D265S (где положения определяют в соответствии с нумерацией Eu). В конкретных вариантах осуществления константный домен HC содержит замену аминокислоты в положении Pro329 и по меньшей мере одну дополнительную замену аминокислоты, выбранную из E233P, L234A, L235A, L235E, N297A, N297D, D265S и P331S (где положения определяют в соответствии с нумерацией Eu). Эти и другие замены описаны в WO 9428027; WO 2004099249; WO 20121300831, патентах США №№9708406; 8969526; 9296815; Sondermann et al. Nature 406, 267-273 (20 Jul. 2000)).
В указанных выше конкретных вариантах осуществления константный домен HC содержит замены L234A/L235A/D265A; L234A/L235A/P329G; L235E; D265A; D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, где положения определяют в соответствии с нумерацией Eu. В конкретных вариантах осуществления молекулы HC дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления эффекторно-молчащее антитело имеет изотип IgG1, в котором Fc-домен константного домена HC модифицирован так, чтобы быть эффекторно-молчащим, посредством замены аминокислот от положения 233 до положения 236 IgG1 соответствующими аминокислотами HC IgG2 человека и замены аминокислот в положениях 327, 330 и 331 соответствующими аминокислотами HC IgG4 человека, где положения определяют в соответствии с нумерацией Eu (Armour et al., Eur. J. Immunol. 29(8): 2613-24 (1999); Shields et al., J. Biol. Chem. 276(9): 6591-604(2001)). В конкретных вариантах осуществления модифицированный IgG1 дополнительно содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления эффекторно-молчащее антитело содержит домен VH, слитый или связанный с гибридным константным доменом HC иммуноглобулина человека, включающим шарнирную область, домен CH2 и домен CH3 в направлении от N-конца к C-концу, где шарнирная область содержит по меньшей мере частичную аминокислотную последовательность шарнирной области IgD человека или шарнирной области IgG1 человека; и домен CH2 является доменом CH2 IgG4 человека, часть которого в его N-концевой области заменяют 4-37 аминокислотными остатками N-концевой области CH2 IgG2 человека или доменом CH2 IgD человека. Такой гибридный константный домен HC человека описан в патенте США №7867491, включенном в настоящее описание в качестве ссылки в полном объеме.
В конкретных вариантах осуществления эффекторно-молчащее антитело содержит константный домен HC IgG4, в котором серин в положении 228 по системе Eu заменяют пролином, см., например, SEQ ID NO: 52. Эта модификация предотвращает образование возможной межцепочечной дисульфидной связи между цистеинами в положениях Cys226 и Cys229 по системе EU и может мешать образованию правильной внутрицепочечной дисульфидной связи. См. Angal et al. Mol. Imunol. 30: 105 (1993); также см. (Schuurman et al., Mol. Immunol. 38: 1-8, (2001); SEQ ID NO: 14 и 41). В дополнительных вариантах осуществления константный домен IgG4 включает, в дополнение к замене S228P, замену аминокислоты P239G, D265A или D265A/N297G, где положения определяют в соответствии с нумерацией Eu. В указанных выше конкретных вариантах осуществления константный домен HC IgG4 является константным доменом HC человека. В конкретных вариантах осуществления молекулы HC дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
Примеры константных доменов HC IgG1 включают константные домены HC, содержащие аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, приведенных в SEQ ID NO: 38, SEQ ID NO: 39, SEQ ID NO: 40, SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43 и SEQ ID NO: 44. Примеры константных доменов HC IgG2 имеют аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, приведенных в SEQ ID NO: 46, SEQ ID NO: 47, SEQ ID NO: 48 и SEQ ID NO: 49. Примеры константных доменов HC IgG4 имеют аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, приведенных в SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55 и SEQ ID NO: 56.
Более конкретные примеры эффекторно-молчащих антител описаны ниже в комбинации с конкретными примерами эффекторно-молчащих антител против CTLA-4, антител против PD-1 и антител против PD-1.
(b) Примеры эффекторно-молчащих антител против CTLA-4
Примеры эффекторно-молчащих антител против CTLA-4, которые можно использовать в комбинированном лечении по настоящему изобретению и композициях, содержащих эти антитела, включают любое эффекторно-молчащее антитело против CTLA-4, связывающееся с CTLA-4 и ингибирующее связывание CTLA-4 с B7. Конкретные эффекторно-молчащие антитела против CTLA-4 включают следующие эффекторно-молчащие антитела против CTLA-4 и композиции, содержащие любое из этих антител и фармацевтически приемлемый носитель.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит (i) VH, содержащий, три HC-CDR ипилимумаба, слитый или связанный с константным доменом HC, демонстрирующим отсутствие измеримого связывания с FcγRIIIA, FcγRIIA и FcγRI или сниженное связывание по сравнению с полипептидом, содержащим область константного домена IgG дикого типа, что определяют посредством анализа Biacore, и (ii) VL, содержащий три LC-CDR ипилимумаба, слитый или связанный с константным доменом каппа или лямбда LC. Три HC-CDR содержат SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6, соответственно, и три LC-CDR содержат SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, соответственно.
В дополнительных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит (i) VH, содержащий три HC-CDR тремелимумаба, слитый или связанный с константным доменом HC, демонстрирующим отсутствие измеримого связывания с FcγRIIIA, FcγRIIA и FcγRI или сниженное связывание по сравнению с полипептидом, содержащим область константного домена IgG дикого типа, что определяют посредством анализа Biacore, и (ii) VL, содержащий три LC-CDR тремелимумаба, слитый или связанный с константным доменом каппа или лямбда LC. Три HC-CDR содержат SEQ ID NO: 12, SEQ ID NO: 13 и SEQ ID NO: 14, соответственно, и три LC-CDR содержат SEQ ID NO: 9, SEQ ID NO: 10 и SEQ ID NO: 11, соответственно.
В дополнительных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит любые из (i) доменов VH и VL ипилимумаба, (ii) доменов VH и VL тремелимумаба, (iii) доменов VH и VL REGN4659, (iv) доменов VH и VL AGEN1884w или (v) доменов VH и VL клона 2C8 антитела против CTLA-4, описанного в международной патентной заявке № WO 2017194265. Домен VH ипилимумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 7, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 8. Домен VH тремелимумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 15, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 16. Домен VH REGN4659 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 95, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 96. Домен VH AGEN1884w содержит аминокислотную последовательность, приведенную в SEQ ID NO: 97, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 98. В конкретных вариантах осуществления домены VH и VL дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В дополнительных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит домены VH и VL 8D2/8D2 (RE) (см. публикацию патентной заявки США №20170216433 и международную патентную заявку № WO 2018183408), 8D2H1L1, 8D2H2L2, 8D3H3L3, 8D2H2L15 или 8D2H2L17, где домен VH слит или связан с константным доменом HC, демонстрирующим отсутствие измеримого связывания с FcγRIIIA, FcγRIIA и FcγRI или сниженное связывание по сравнению с полипептидом, содержащим область константного домена IgG дикого типа, что определяют посредством анализа Biacore, и домен VL слит или связан с константным доменом каппа или лямбда LC.
В конкретных вариантах осуществления эффекторно-молчащее антитело против CTLA-4 содержит вариант 8D2/8D2 (RE), 8D2H1L1, 8D2H2L2, 8D2H2L15 или 8D2H2L17, где метионин в положении 18 в аминокислотной последовательности VH варианта заменяют изолейцином. Таким образом, эффекторно-молчащее антитело против CTLA-4 может содержать VH и VL 8D2/8D2 (RE)-варианта 1, VH и VL 8D2H1L1-варианта 1, VH и VL 8D2H2L2-варианта 1, VH и VL 8D2H2L15-варианта 1 или VH и VL 8D2H2L17-варианта 1.
В дополнительных вариантах осуществления эффекторно-молчащее антитело против CTLA4 имеет (i) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; (ii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; (iii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78; (iv) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80; (v) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 82; (vi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 83, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 84; (vii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 85, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 86; (viii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 87, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 88; (ix) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 89, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 90; (x) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 91, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 92; или (xi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 93, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94.В конкретных вариантах осуществления домены VH и VL дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В дополнительных вариантах осуществления эффекторно-молчащего антитела против CTLA-4 домен VH слит или связан с константным доменом HC IgG4 или константным доменом HC IgG1, IgG2 или IgG4, модифицированным так, чтобы включать одну или более мутаций, делающих полученное антитело против CTLA4 эффекторно-молчащим.
В одном из вариантов осуществления эффекторно-молчащее антитело против CTLA-4 содержит Fc-домен IgG1, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A, L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, S267E/L328F, S2339D/A330L/I332E, L235G/G236R, N297A/D356E/L358M, L234F/L235E/P331S/D365E/L358M и D265A/N297G, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из L234A/L235A/D265A, L234A/L235A/P329G, L235E, D265A, E233A/L235A, S267E/L328F, S2339D/A330L/I332E, L235G/G236R, D356E/L358M, L234F/L235E/P331S/D365E/L358M и D265A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В другом варианте осуществления эффекторно-молчащее антитело против CTLA-4 содержит Fc-домен IgG2, содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A/D265S, D265A, P329G/D265A/N297G, или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования и мутацию с заменой аминокислоты, выбранной из группы, состоящей из N297A/D265S, D265A, P329G/D265A/N297G или V234A/G237A/P238S/H268A/V309L/A330S/P331S, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В дополнительном варианте осуществления эффекторно-молчащее антитело против CTLA-4 содержит Fc-домен IgG4, содержащий замену аминокислоты S228P и дополнительно содержащий (i) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; (ii) мутацию с заменой аминокислоты, выбранной из группы, состоящей из N267A, P329G и D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот; или (iii) мутацию в участке N-гликозилирования Asn-Xaa-Ser/Thr, начинающуюся в положении аминокислоты 297, устраняющую N-гликозилирование в указанном участке N-гликозилирования, и мутацию с заменой аминокислоты, выбранной из группы, состоящей из N267A, P329G и D265A/N297A, или мутантный Fc-домен, дополнительно содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где положения аминокислот в (i), (ii) и (iii) определяют в соответствии с нумерацией Eu.
В таблицах 4-18 приведены конкретные примеры антител против CTLA-4, которые можно использовать в комбинации с антителом против PD-1 или против PD-L1 в терапии для лечения индивидуума, имеющего злокачественное новообразование. Настоящее изобретение также относится к антителам, приведенным в таблицах, за исключением ипилимумаба, состоящего исключительно из замены N297A, и композициям, где каждая композиция содержит антитело, приведенное в таблицах, и фармацевтически приемлемый носитель, за исключением композиции, содержащей ипилимумаб, состоящий исключительно из замены N297A. Все положения замен аминокислоты HC в таблицах 4-18 соответствуют системе нумерации Eu.
b=75+76,
c=77+78,
d=79+80,
e=81+82,
f=83+84,
g=85+86,
h=87+88,
i=89+90,
j=91+92, или
k=93+94
b=75+76,
c=77+78,
d=79+80,
e=81+82,
f=83+84,
g=85+86,
h=87+88,
i=89+90,
j=91+92, или
k=93+94
b=75+76,
c=77+78,
d=79+80,
e=81+82,
f=83+84,
g=85+86,
h=87+88,
i=89+90,
j=91+92, или
k=93+94
(c) Примеры эффекторно-молчащих фрагментов антител против CTLA-4
Примеры эффекторно-молчащих фрагментов антител против CTLA-4, которые можно использовать в комбинированном лечении по настоящему изобретению и содержащих их композициях включают любой фрагмент антитела, связывающийся с CTLA-4 и ингибирующий связывание CTLA-4 с B7. Конкретные примеры этих фрагментов антител против CTLA-4 включают следующие фрагменты антител против CTLA-4 и композиции, где каждая композиция содержит эффекторно-молчащий фрагмент антитела против CTLA-4 и фармацевтически приемлемый носитель.
В конкретных вариантах осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 является Fv, scFv, F(ab) или F(ab')2, содержащим (i) VH, содержащий три HC-CDR ипилимумаба, и (ii) VL, содержащий три LC-CDR ипилимумаба. Три HC-CDR содержат SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, соответственно, и три LC-CDR содержат SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 7, соответственно.
В конкретных вариантах осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 содержит (i) VH, содержащий три HC-CDR тремелимумаба, и (ii) VL, содержащий три LC-CDR тремелимумаба. Три HC-CDR содержат SEQ ID NO: 9, SEQ ID NO: 10 и SEQ ID NO: 11, соответственно, и три LC-CDR содержат SEQ ID NO: 12, SEQ ID NO: 13 и SEQ ID NO: 14, соответственно.
В конкретных вариантах осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 содержит любые из (i) доменов VH и VL ипилимумаба, (ii) доменов VH и VL тремелимумаба, (iii) доменов VH и VL REGN4659, (iv) доменов VH и VL AGEN1884w или (v) доменов VH и VL клона 2C8 антитела против CTLA-4, описанного в международной патентной заявке № WO 2017194265. Домен VH ипилимумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 7, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 8. Домен VH тремелимумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 15, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 16. Домен VH REGN4659 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 95, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 96. Домен VH AGEN1884w содержит аминокислотную последовательность, приведенную в SEQ ID NO: 97, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 98.
В конкретных вариантах осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 содержит VH и VL ипилимумаба, VH и VL тремелимумаба, VH и VL REGN4659, VH и VL AGEN1884w, VH и VL 8D2/8D2 (RE), VH и VL 8D2H1L1, VH и VL 8D2H2L2, VH и VL 8D3H3L3, VH и VL 8D2H2L15 или VH и VL 8D2H2L17.
В конкретных вариантах осуществления антитело против CTLA-4 или фрагмент антитела против CTLA-4 содержит VH и VL 8D2/8D2 (RE)-вариант 1, VH и VL 8D2H1L1-варианта 1, VH и VL 8D2H2L2-варианта 1, VH и VL 8D2H2L15-варианта 1, или VH и VL 8D2H2L17-варианта 1.
В конкретных вариантах осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 содержит любой (i) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 73, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 74; (ii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 75, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 76; (iii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 77, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 78; (iv) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 79, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 80; (v) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 81, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 82; (vi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 83, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 84; (vii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 85, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 86; (viii) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 87, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 88; (ix) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 89, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 90; (x) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 91, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 92; или (xi) домен VH, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 93, и домен VL, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 94.
В конкретных вариантах осуществления эффекторно-молчащий фрагмент антитела против CTLA-4 содержит один или более отдельных вариабельных доменов иммуноглобулинов (ISVD), где каждый ISVD содержит вариабельный домен (VHH) антитела Верблюжьих, содержащий только тяжелую цепь при условии, что ISVD не содержит CDR1, содержащую аминокислотную последовательность FYGMG (SEQ ID NO: 69, CDR2, содержащую аминокислотную последовательность DIRTSAGRTTYADSVKG (SEQ ID NO: 70), и CDR3, содержащую аминокислотную последовательность EMSGISGWDY (SEQ ID NO: 71) или EPSGISGWDY (SEQ ID NO: 72), как ISVD, описанные в международной патентной заявке № WO 2008071447, WO 2017087587 и WO 2017087588, и варианты ISVD, содержащие 1, 2 или 3 мутаций в CDR3, как описано в WO 2008071447, с таким исключением, что не исключаемыми в силу него являются ISVD, содержащие указанные CDR в вариантах осуществления, где один или более ISVD слиты или связаны с константным доменом или Fc-доменом эффекторно-молчащего антитела, например, любым из константных доменов или Fc-доменов эффекторно-молчащего антитела, представленных в настоящем описании.
(d) Примеры антител против PD-1
Примеры антител против PD-1, которые можно использовать в комбинированном лечении по настоящему изобретению, включают любое антитело, связывающееся с PD-1 и ингибирующее связывание PD-1 с PD-L1. В дополнительном варианте осуществления пример антитела против PD-1 выбран из группы, состоящей из ниволумаба, пембролизумаба и цемиплимаба-rwlc. Примеры антител включают следующие антитела против PD-1 и композиции, содержащие антитело против PD1 и его фармацевтически приемлемую соль.
Пембролизумаб, также известный как KEYTRUDA, ламбролизумаб, MK-3475 или SCH-900475, является гуманизированным антителом против PD-1, описанным в патенте США №8354509 и WO 2009/114335 и, например, в Hamid, et al., New England J. Med. 369 (2): 134-144 (2013). Тяжелые и легкие цепи пембролизумаба представлены аминокислотными последовательностями, приведенными в SEQ ID NO: 27 и 28, соответственно.
Ниволумаб, также известный как OPDIVO, MDX-1106-04, ONO-4538 или BMS-936558, является полностью человеческим антителом IgG4 против PD-1, описанным в WO 2006/121168 и патенте США №8008449. Тяжелые и легкие цепи ниволумаба представлены аминокислотными последовательностями, приведенными в SEQ ID NO: 25 и 26, соответственно.
Цемиплимаб-rwlc, также известный как цемиплимаб, LIBTAYO или REGN2810, является рекомбинантным моноклональным антителом IgG4 человека, описанным в WO 2015112800 и патенте США №9987500. Тяжелые и легкие цепи цемиплимаба-rwlc представлены аминокислотными последовательностями, приведенными в SEQ ID NO: 101 и 102, соответственно.
В конкретных вариантах осуществления антитело против PD-1 содержит (i) VH, содержащий три HC-CDR пембролизумаба, слитый или связанный с эффекторно-молчащим константным доменом HC, и (ii) VL, содержащий три LC-CDR пембролизумаба, слитый или связанный с константным доменом каппа или лямбда LC. Три HC-CDR содержат SEQ ID NO: 31, SEQ ID NO: 32 и SEQ ID NO: 33, соответственно, и три LC-CDR содержат SEQ ID NO: 34, SEQ ID NO: 35 и SEQ ID NO: 36, соответственно.
В конкретных вариантах осуществления антитело против PD-1 содержит (i) VH, содержащий три HC-CDR ниволумаба, слитый или связанный с эффекторно-молчащим константным доменом HC, и (ii) VL, содержащий три LC-CDR ниволумаба, слитый или связанный с константным доменом каппа или лямбда LC. Три HC-CDR содержат SEQ ID NO: 17, SEQ ID NO: 18 и SEQ ID NO: 19, соответственно, и три LC-CDR содержат SEQ ID NO: 20, SEQ ID NO: 21 и SEQ ID NO: 2, соответственно.
В конкретных вариантах осуществления антитело против PD-1 содержит (i) VH, содержащий три HC-CDR цемиплимаба-rwlc, слитый или связанный с эффекторно-молчащим константным доменом HC, и (ii) VL, содержащий три LC-CDR ниволумаба, слитый или связанный с константным доменом каппа или лямбда LC.
В конкретных вариантах осуществления антитело против PD-1 содержит (i) домены VH и VL пембролизумаба, где домен VH слит или связан с эффекторно-молчащим константным доменом HC, и домен VL слит или связан с константным доменом каппа или лямбда LC; (ii) домены VH и VL ниволумаба, где домен VH слит или связан с эффекторно-молчащим константным доменом HC, и домен VL слит или связан с константным доменом каппа или лямбда LC; или (iii) домены VH и VL цемиплимаба-rwlc, где домен VH слит или связан с эффекторно-молчащим константным доменом HC, и домен VL слит или связан с константным доменом каппа или лямбда LC. Домен VH пембролизумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 29, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 30. Домен VH ниволумаб содержит аминокислотную последовательность, приведенную в SEQ ID NO: 23, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 24. Домен VH цемиплимаба-rwlc содержит аминокислотную последовательность, приведенную в SEQ ID NO: 99, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 100. В конкретных вариантах осуществления домены VH и VL могут дополнительно содержать 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления антитело против PD-1 домен VH может быть слитым или связанным с константным доменом HC IgG1, IgG2, IgG3 или IgG4, в настоящее время не связанным с конкретным VH или связанным с константным доменом HC IgG1, IgG2, IgG3 или IgG4, модифицированным так, чтобы включать одну или более мутаций в Fc-домене делающих полученное антитело против PD-1 эффекторно-молчащим.
В определенных вариантах осуществления константный домен HC принадлежит изотипу IgG1, IgG2, IgG3 или IgG4, модифицированному так, что в нем отсутствует N-гликозилирование остатка аспарагина (Asn) в положении 297 константного домена HC посредством замены кодона для Asn в положении 297 в молекуле нуклеиновой кислоты, кодирующей константный домен HC, кодоном для другой аминокислоты, например, Gln. В дополнительных вариантах осуществления такие IgG, модифицированные так, что в них отсутствует N-гликозилирование в положении 297, дополнительно включают одну или более дополнительных мутаций, представленных в настоящем описании, для устранения детектируемой эффекторной функции. В конкретных вариантах осуществления константный домен HC является константным доменом HC человека. В конкретных вариантах осуществления молекулы дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления настоящее изобретение относится к антителу против PD-1, содержащему константный домен HC IgG4, модифицированный так, чтобы содержать замену S228P, и включающему, в дополнение к замене S228P, замены аминокислот P239G, D265A или D265A/N297G, где положения определяют в соответствии с нумерацией Eu. В указанных выше конкретных вариантах осуществления константный домен HC IgG4 является константным доменом HC человека. В конкретных вариантах осуществления молекулы дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В другом варианте осуществления антитело против PD-1 может иметь изотип IgG1 человека, в котором Fc-домен константного домена HC модифицирован так, чтобы быть эффекторно-молчащим, посредством замены аминокислот от положения 233 до положения 236 IgG1 соответствующими аминокислотами HC IgG2 человека и замены аминокислот в положениях 327, 330 и 331 соответствующими аминокислотами HC IgG4 человека, где положения определяют в соответствии с нумерацией Eu. В конкретных вариантах осуществления молекулы HC дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В другом варианте осуществления Fc-домен константного домена HC IgG1 IgG2, IgG3 или IgG4 модифицирован так, чтобы включать одну или более замен аминокислот, выбранных из E233P, L234A, L235A, L235E, N297A, N297D, D265S и P331S, и где указанный полипептид демонстрирует отсутствие измеримого связывания с FcγRIIIA, FcγRIIA и FcγRI или сниженное связывание по сравнению с полипептидом, содержащим область константного домена IgG дикого типа, что определяют посредством анализа Biacore. Эти и другие замены описаны в WO 9428027; WO 2004099249; WO 20121300831, патентах США №№9708406; 8969526; 9296815; Sondermann et al. Nature 406, 267-273 (20 Jul. 2000)). В конкретных вариантах осуществления молекулы HC дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В таблицах 19-27 приведены конкретные примеры антител против PD-1, которые можно использовать в комбинации с антителом против CTLA-4, как представлено в настоящем описании, в терапии для лечения индивидуума, имеющего злокачественное новообразование. Настоящее изобретение также относится к антителам, приведенным в таблицах, и композициям, где каждая композиция содержит антитело, приведенное в таблицах, и фармацевтически приемлемый носитель. Все положения замен аминокислоты HC в таблицах 19-27 соответствуют системе нумерации Eu.
(e) Примеры фрагментов антител против PD-1
Примеры фрагментов антител против PD-1, которые можно использовать в комбинированном лечении по настоящему изобретению, включают любой фрагмент антитела против PD-1, связывающийся с PD-1 и ингибирующий связывание PD-1 с PD-L1, и дополнительно включают следующие фрагменты антител против PD-1, связывающиеся с PD-1, и композиции, содержащие следующие фрагменты антител против PD-1 и фармацевтически приемлемый носитель.
В конкретных вариантах осуществления фрагмент антитела является Fv или scFv, содержащим VH пембролизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 29, и VL пембролизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 30.
В конкретных вариантах осуществления фрагмент антитела против PD-1 является F(ab), содержащим VH пембролизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 29, и VH пембролизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 30.
В конкретных вариантах осуществления фрагмент антитела против PD-1 является F(ab')2, содержащим VH пембролизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 29, и VH пембролизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 30.
В конкретных вариантах осуществления фрагмент антитела против PD-1 является Fv или scFv, содержащим VH ниволумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 23, и VH ниволумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 24.
В конкретных вариантах осуществления фрагмент антитела против PD-1 является F(ab), содержащим VH ниволумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 23, и VH ниволумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 24.
В конкретных вариантах осуществления фрагмент антитела против PD-1 является F(ab')2, содержащим VH ниволумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 23, и VH ниволумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 24.
В конкретных вариантах осуществления фрагмент антитела против PD-1 содержит один или более отдельных вариабельных доменов иммуноглобулинов (ISVD), где каждый ISVD содержит вариабельный домен (VHH) антитела Верблюжьих, содержащего только тяжелую цепь при условии, что ISVD не содержит CDR1, содержащую аминокислотную последовательность THAMG (SEQ ID NO: 73), CDR2, содержащую аминокислотную последовательность VITWSGGITTYADSVKG (SEQ ID NO: 74) или VITVSGGITYYADSVKG (SEQ ID NO: 75), и CDR3, содержащую аминокислотную последовательность DKHQSSWYDY (SEQ ID NO: 76) или DKHQSSFYDY (SEQ ID NO: 77), как ISVD, описанные в международной патентной заявке № WO 2008071447, WO 2017087587 и WO 2017087589, и варианты, содержащие 1, 2 или 3 мутаций в CDR3, как описано в WO 2008071447, с таким исключением, что не исключаемыми в силу него являются ISVD, содержащие указанные CDR в вариантах осуществления, где один или более ISVD слиты или связаны с константным доменом или Fc-доменом эффекторно-молчащего антитела, например, любым из константных доменов или Fc-доменов эффекторно-молчащего антитела, представленных в настоящем описании.
(f) Примеры антител против PD-L1
Примеры антител против PD-L1, которые можно использовать в комбинированном лечении по настоящему изобретению, включают любое антитело против PD-L1, ингибирующее связывание PD-1 с PD-L1, и дополнительно включают следующие антитела против PD-L1 и композиции, содержащие следующие антитела против PD-L1 и фармацевтически приемлемый носитель. В конкретных вариантах осуществления антитело против PD-L1 выбрано из группы, состоящей из атезолизумаба, авелумаба и дурвалумаба.
В конкретных вариантах осуществления антитело против PD-L1 содержит (i) домены VH и VL атезолизумаба, где домен VH слит или связан с константным доменом HC или эффекторно-молчащим константным доменом HC, и домен VL слит или связан с константным доменом каппа или лямбда LC, (ii) домены VH и VL авелумаба, где домен VH слит или связан с константным доменом HC или эффекторно-молчащим константным доменом HC, и домен VL слит или связан с константным доменом каппа или лямбда LC, или (iii) домены VH и VL дурвалумаба, где домен VH слит или связан с константным доменом HC или эффекторно-молчащим константным доменом HC, и домен VL слит или связан с константным доменом каппа или лямбда LC. Домен VH дурвалумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 103, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 104. Домен VH авелумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 105, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 106. Домен VH атезолизумаба содержит аминокислотную последовательность, приведенную в SEQ ID NO: 107, и домен VL содержит аминокислотную последовательность, приведенную в SEQ ID NO: 108. В конкретных вариантах осуществления домены VH и VL дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления домен VH антитела против PD-L1 может быть слитым или связанным с константным доменом HC IgG1, IgG2, IgG3 или IgG4, в настоящее время не связанным с конкретным VH или связанным с константным доменом HC IgG1, IgG2, IgG3 или IgG4, модифицированным так, чтобы включать одну или более мутаций в Fc-домене делающих полученное антитело против PD-L1 эффекторно-молчащим.
В определенных вариантах осуществления константный домен HC принадлежит изотипу IgG1, IgG2, IgG3 или IgG4, модифицированному так, что в нем отсутствует N-гликозилирование остатка аспарагина (Asn) в положении 297 константного домена HC, посредством замены кодона для Asn в положении 297 в молекуле нуклеиновой кислоты, кодирующей константный домен HC, кодоном для другой аминокислоты, например, Gln. Альтернативно, кодон для Ser можно заменять кодоном для Pro или кодон для Thr можно заменять любым кодоном, за исключением кодона для Ser, например, N297A, N297G или N297D. Альтернативно, все три кодона модифицированы. В дополнительных вариантах осуществления такие IgG, модифицированные так, что в них отсутствует N-гликозилирование в положении 297, дополнительно включают одну или более дополнительных мутаций, представленных в настоящем описании, для устранения детектируемой эффекторной функции. В конкретных вариантах осуществления константный домен HC является константным доменом HC человека. В конкретных вариантах осуществления молекулы дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В конкретных вариантах осуществления настоящее изобретение относится к антителу против PD-L1, содержащему константный домен HC IgG4, модифицированный так, чтобы содержать замену S228P, и включающему, в дополнение к замене S228P, замены аминокислот P239G, D265A или D265A/N297G, где положения определяют в соответствии с нумерацией Eu. В указанных выше конкретных вариантах осуществления константный домен HC IgG4 является константным доменом HC человека. В конкретных вариантах осуществления молекулы дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В другом варианте осуществления антитело против PD-L1 может иметь изотип IgG1 человека, в котором Fc-домен константного домена HC модифицирован так, чтобы быть эффекторно-молчащим, посредством замены аминокислот от положения 233 до положения 236 IgG1 соответствующими аминокислотами HC IgG2 человека и замены аминокислот в положениях 327, 330 и 331 соответствующими аминокислотами HC IgG4 человека, где положения определяют в соответствии с нумерацией Eu. В конкретных вариантах осуществления молекулы HC дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В другом варианте осуществления Fc-домен константного домена HC IgG1 IgG2, IgG3 или IgG4 модифицирован так, чтобы включать одну или более замен аминокислот, выбранных из E233P, L234A, L235A, L235E, N297A, N297D, D265S и P331S, и где указанный полипептид демонстрирует отсутствие измеримого связывания с FcγRIIIA, FcγRIIA и FcγRI или сниженное связывание по сравнению с полипептидом, содержащим область константного домена IgG дикого типа, что определяют посредством анализа Biacore. Эти и другие замены описаны в WO 9428027; WO 2004099249; WO 20121300831, патенты США №№9708406; 8969526; 9296815; Sondermann et al. Nature 406, 267-273 (20 Jul. 2000)). В конкретных вариантах осуществления молекулы HC дополнительно содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных замен, инсерций и/или делеций аминокислот, где указанные замены могут являться консервативными мутациями или неконсервативными мутациями.
В таблицах 28-36 приведены примеры антител против PD-L1, которые можно использовать в комбинации с антителом против CTLA-4, как представлено в настоящем описании, в терапии для лечения индивидуума, имеющего злокачественное новообразование. Настоящее изобретение также относится к антителам, приведенным в таблицах, за исключением антител 25-9 и 31-8, и композиции, где каждая композиция содержит антитело, приведенное в таблице, за исключением антител 25-9 и 31-8, и фармацевтически приемлемый носитель. Все положения замен аминокислоты HC в таблицах 28-36 соответствуют системе нумерации Eu.
(g) Примеры фрагментов антител против PD-L1
Примеры фрагментов антител против PD-L1, которые можно использовать в комбинированном лечении по настоящему изобретению, относятся к любому фрагменту антитела против PD-L1, связывающемуся с PD-L1 и ингибирующему связывание PD-L1 с PD-1, и дополнительно включают следующие фрагменты антител против PD-L1 и композиции, где каждая композиция содержит следующий фрагмент антитела против PD-L1 и фармацевтически приемлемый носитель.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является Fv или scFv, содержащим VH дурвалумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 103, и VL дурвалумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 104.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является F(ab), содержащим VH дурвалумаба имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 103, и VH дурвалумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 104.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является F(ab')2, содержащим VH дурвалумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 103, и VH дурвалумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 104.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является Fv или scFv, содержащим VH авелумаб, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 105, и VH авелумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 106.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является F(ab), содержащим VH авелумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 105, и VH авелумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 106.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является F(ab')2, содержащим VH авелумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 105, и VH авелумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 106.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является Fv или scFv, содержащим VH атезолизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 107, и VH атезолизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 108.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является F(ab), содержащим VH атезолизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 107, и VH атезолизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 108.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 является F(ab')2, содержащим VH атезолизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 107, и VH атезолизумаба, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 108.
В конкретных вариантах осуществления фрагмент антитела против PD-L1 содержит один или более отдельных вариабельных доменов иммуноглобулинов (ISVD), где каждый ISVD содержит вариабельный домен (VHH) антитела Верблюжьих, содержащего только тяжелую цепь при условии, что ISVD не содержит ISVD против PD-L1, описанный в международной патентной заявке № WO 2008071447 и содержащий SEQ ID NO: 394-399 в ней или описанный в WO 2009030285, оба из которых включены в настоящее описание в качестве ссылки, с таким исключением, что не исключаемыми условием являются ISVD, где один или более ISVD слиты или связаны с константным доменом или Fc-доменом эффекторно-молчащего антитела, например, любым из константных доменов или Fc-доменов эффекторно-молчащего антитела, представленных в настоящем описании.
(h) Примеры схем комбинированного лечения
Настоящее изобретение относится к противоопухолевым терапевтическим средствам, в которых комбинируют иммуностимулирующие эффекты PD-1-блокирующего средства с противоопухолевыми эффектами CTLA-4-блокирующего средства, но без дерматологических или кишечных irAE, как правило, наблюдаемых в случае CTLA-4-блокирующих средств, вводимых в комбинации с PD-1-блокирующими средствами. Признаком настоящего изобретения является то, что у CTLA-4-блокирующего средства отсутствует измеримое связывание с одним или более FcR, что определяют посредством анализа Biacore, или есть сниженное связывание с одним или более FcR по сравнению с антителом дикого типа того же изотипа, что измеряют посредством анализа Biacore. Таким образом, CTLA-4-блокирующие средства демонстрируют отсутствие измеримой эффекторной функции или сниженную эффекторную функцию, что позволяет использовать эффекторно-молчащие CTLA-4-блокирующие средства в способах комбинированного лечения с PD-1-блокирующими средствами в дозах и в течение периодов времени, недоступных в случае CTLA-4-блокирующих средств, демонстрирующих эффекторную функцию. Этот признак отличает CTLA-4-блокирующие средства по настоящему изобретению от доступных в настоящее время CTLA-4-блокирующих средств.
В типичной схеме введения по настоящему изобретению CTLA-4-блокирующее средство и PD-1-блокирующее средство можно вводить индивидууму одновременно в отдельных дозах и в разных форматах. В основном, CTLA-4-блокирующее средство по настоящему изобретению можно вводить в рамках комбинированного лечения с PD-1-блокирующим средством по меньшей мере в одинаковой дозе, с одинаковой частотой и длительностью лечениями, одобренными в настоящее время FDA США для комбинированного лечения ипилимумабом/ниволумабом по конкретным показаниям. Однако комбинированное лечение не ограничено конкретными показаниями, одобренными FDA США, но может включать любое показание, при котором можно получить пользу от комбинированного лечения по настоящему изобретению. Одобренной в настоящее время дозой является 1 мг/кг ипилимумаба после введения ниволумаба в дозе 3 мг/кг. Затем эту комбинацию доз можно повторять каждые три недели всего в четырех дозах, затем продолжая вводить дозы ниволумаба каждые две недели по мере необходимости. Однако в дополнительных вариантах осуществления CTLA-4-блокирующее средство по настоящему изобретению можно вводить в рамках комбинированного лечения в дозе, составляющей более 1 мг/кг, например, по меньшей мере 3 мг/кг. В дополнительном варианте осуществления доза может составлять по меньшей мере 10 мг/кг, и в дополнительных вариантах осуществления доза может составлять приблизительно от 1 мг/кг до 10 мг/кг. В конкретных вариантах осуществления CTLA-4-блокирующее средство по настоящему изобретению можно вводить с той же частотой и длительностью лечения, что и в одобренном комбинированном лечении ипилимумабом/ниволумабом. В конкретных вариантах осуществления CTLA-4-блокирующие средства по настоящему изобретению можно вводить с той же частотой и длительностью лечения, что и для ниволумаба в одобренном комбинированном лечении ипилимумабом/ниволумабом.
В конкретных вариантах осуществления комбинированного лечения CTLA-4-блокирующее средство вводят в дозе, не основанной на массе тела индивидуума. Таким образом, в конкретных вариантах осуществления CTLA-4-связывающее средство можно вводить в дозе приблизительно от 10 мг до 300 мг. В дополнительном варианте осуществления доза выбрана из группы, состоящей из 10 мг, 25 мг, 50 мг, 75 мг, 100 мг, 150 мг, 200 мг, 250 мг и 300 мг.
В комбинированном лечении по настоящему изобретению PD-1-блокирующее средство можно вводить в той же дозе, с частотой и длительностью лечения, одобренными для PD-1-блокирующего средства в монотерапии для конкретных показаний. Доза CTLA-4-блокирующего средства может являться такой, как указано выше, и CTLA-4-блокирующее средство можно вводить с той же частотой и длительностью лечения, что и указанные выше, или с той же частотой и длительностью лечения, что и для конкретного PD-1-блокирующего средства, используемого в комбинации с CTLA-4-блокирующим средством.
Конкретная доза находящихся в настоящее время на рынке PD-1-блокирующих средств варьируется среди PD-1-блокирующих средств, таким образом, в конкретных вариантах осуществления комбинированного лечения по настоящему изобретению, доза, частота введения и/или длительность лечения могут являться по меньшей мере такими же, что и одобренные FDA США в случае конкретного PD-1-блокирующего средства для конкретных показаний. Например, пембролизумаб одобрен в дозе 200 мг каждые три недели по мере необходимости (детям (от двух до 18 лет) в дозе 2 мг/кг до 200 мг каждые три недели по мере необходимости); ниволумаб одобрен в дозе 3 мг/кг каждые 2 недели; цемиплимаб-rwlc одобрен в дозе 350 мг каждые три недели по мере необходимости; атезолизумаб одобрен в дозе 1200 мг каждые три недели по мере необходимости; авелумаб одобрен в дозе 10 мг/кг или 800 мг каждые две недели по мере необходимости; и дурвалумаб одобрен в дозе 10 мг/кг каждые две недели по мере необходимости.
В конкретных вариантах осуществления комбинированного лечения PD-1-блокирующее средство является антителом против PD-1 или фрагментом антитела против PD-1, которое можно вводить в дозе от приблизительно 150 мг до приблизительно 250 мг, от приблизительно 175 мг до приблизительно 250 мг, от приблизительно 200 мг до приблизительно 250 мг, от приблизительно 150 мг до приблизительно 240 мг, от приблизительно 175 мг до приблизительно 240 мг или от приблизительно 200 мг до приблизительно 240 мг. В некоторых вариантах осуществления доза антитела против PD-1 или его антигенсвязывающего фрагмента составляет 150 мг, 175 мг, 200 мг, 225 мг, 240 мг или 250 мг. В дополнительных вариантах осуществления антитело против PD-1 или фрагмент антитела против PD-1 можно вводить с частотой каждые три недели по мере необходимости. В другом варианте осуществления комбинированного лечения по настоящему изобретению антитело против PD-1 или фрагмент антитела против PD-1 можно вводить в дозе более 250 мг, например, дозе приблизительно 400 мг с частотой каждые шесть недель по мере необходимости.
В конкретных вариантах осуществления комбинированного лечения PD-1-блокирующее средство является антителом против PD-1 или фрагментом антитела против PD-1, которое можно вводить в дозе от приблизительно 10 мг/кг до приблизительно 1200 мг. В дополнительных вариантах осуществления антитело против PD-1 или фрагмент антитела против PD-1 можно вводить с частотой каждые две-три недели по мере необходимости.
Хотя PD-1-блокирующее средство можно вводить по меньшей мере в дозах, с частотой и длительностью лечения, одобренными для находящихся в настоящее время на рынке PD-1-блокирующих средств в монотерапии, точные дозы, частоты и длительность лечения для любой конкретной комбинации по настоящему изобретению могут отличаться от одобренных для монотерапии PD-1-блокирующими средствами. Таким образом, в конкретных вариантах осуществления комбинированного лечения по настоящему изобретению, доза, частота и длительность лечения любым конкретным PD-1-блокирующим средством в комбинированном лечении будут определять с помощью клинических испытаний, проведенных в отношении комбинированного лечения.
В конкретном варианте осуществления комбинированного лечения PD-1-блокирующее средство является ниволумабом или эффекторно-молчащим вариантом ниволумаба, который вводят индивидууму внутривенно в дозе 3 мг/кг в течение от 30 до 60 минут каждые две-три недели по мере необходимости, и где каждую дозу CTLA-4-блокирующего средства вводят внутривенно после введения PD-1-блокирующего средства с той же длительностью лечения, что и PD-1-блокирующее средство или длительностью, меньшей или большей, чем длительность в случае PD-1-блокирующего средства. В конкретном варианте осуществления ниволумаб или эффекторно-молчащий вариант ниволумаба вводят внутривенно индивидууму в начальной дозе 3 мг/кг в течение 30 минут с последующим внутривенным введением CTLA-4-блокирующего средства в течение 30 минут в тот же день, каждые три недели всего в четырех дозах, затем ниволумаб вводят внутривенно в фиксированной дозе 240 мг каждые две недели в течение 30 минут или 480 мг каждые четыре недели в течение 30 минут.
В конкретных вариантах осуществления PD-1-блокирующее средство является пембролизумабом или эффекторно-молчащим вариантом пембролизумаба, который вводят взрослому индивидууму внутривенно в дозе 200 мг в течение 30 минут каждые три недели по мере необходимости или ребенку внутривенно в дозе 2 мг/кг максимально до приблизительно 200 мг в течение 30 минут каждые три недели, где после каждого введения вводят дозу CTLA-4-блокирующего средства, где каждую дозу CTLA-4-блокирующего средства вводят внутривенно после введения PD-1-блокирующего средства с той же длительностью лечения, что и PD-1-блокирующее средство, или длительностью, меньшей или большей, чем длительность в случае PD-1-блокирующего средства.
В конкретных вариантах осуществления PD-1-блокирующее средство является пембролизумабом или эффекторно-молчащим вариантом пембролизумаба, который вводят взрослому индивидууму внутривенно в дозе 400 мг в течение 30 минут каждые шесть недель по мере необходимости, где после каждого введения вводят дозу CTLA-4-блокирующего средства, где каждую дозу CTLA-4-блокирующего средства вводят внутривенно после введения PD-1-блокирующего средства с той же длительностью лечения, что и PD-1-блокирующее средство, или длительностью, меньшей или большей, чем длительность в случае PD-1-блокирующего средства.
В конкретном варианте осуществления комбинированного лечения PD-1-блокирующее средство является цемиплимабом-rwlc или эффекторно-молчащим вариантом цемиплимаба-rwlc, который вводят индивидууму внутривенно в дозе 350 мг в течение 30 минут каждые три недели по мере необходимости, где каждую дозу CTLA-4-блокирующего средства вводят внутривенно после введения PD-1-блокирующего средства с той же длительностью лечения, что и PD-1-блокирующее средство, или длительностью, меньшей или большей, чем длительность в случае PD-1-блокирующего средства. В конкретном варианте осуществления цемиплимаб-rwlc или эффекторно-молчащий вариант цемиплимаба-rwlc вводят внутривенно индивидууму в начальной дозе 350 мг в течение 30 минут с последующим введением CTLA-4-блокирующего средства в течение 30 минут в тот же день каждые три недели по мере необходимости.
В конкретном варианте осуществления комбинированного лечения PD-1-блокирующее средство является атезолизумабом или эффекторно-молчащим вариантом атезолизумаба, который вводят индивидууму внутривенно в дозе 1200 мг в течение 60 минут каждые три недели по мере необходимости, где каждую дозу CTLA-4-блокирующего средства вводят внутривенно после введения PD-1-блокирующего средства с той же длительностью лечения, что и PD-1-блокирующее средство, или длительностью, меньшей или большей, чем длительность в случае PD-1-блокирующего средства. В конкретном варианте осуществления атезолизумаб или эффекторно-молчащий вариант атезолизумаба вводят внутривенно индивидууму в начальной дозе 1200 мг в течение 60 минут с последующим введением CTLA-4-блокирующего средства в течение 30 минут в тот же день каждые три недели по мере необходимости.
В конкретном варианте осуществления комбинированного лечения PD-1-блокирующее средство является авелумабом или эффекторно-молчащим вариантом авелумаба, который вводят индивидууму внутривенно в дозе 10 мг/кг или 800 мг в течение 60 минут каждые две недели по мере необходимости, и где каждую дозу CTLA-4-блокирующего средства вводят внутривенно после введения PD-1-блокирующего средства с той же длительностью лечения, что и PD-1-блокирующее средство, или длительностью, меньшей или большей, чем длительность в случае PD-1-блокирующего средства. В конкретном варианте осуществления авелумаб или эффекторно-молчащий вариант авелумаба вводят внутривенно индивидууму в начальной дозе 10 мг/кг или 800 мг в течение 60 минут с последующим введением CTLA-4-блокирующего средства в течение 30 минут в тот же день каждые две недели по мере необходимости.
В конкретном варианте осуществления комбинированного лечения PD-1-блокирующее средство является дурвалумабом или эффекторно-молчащим вариантом дурвалумаба, который вводят индивидууму внутривенно в дозе 10 мг/кг в течение 60 минут каждые две недели по мере необходимости, и где каждую дозу CTLA-4-блокирующего средства вводят внутривенно после введения PD-1-блокирующего средства с той же длительностью лечения, что и PD-1-блокирующее средство, или длительностью, меньшей или большей, чем длительность в случае PD-1-блокирующего средства. В конкретном варианте осуществления дурвалумаб или эффекторно-молчащий вариант дурвалумаба вводят внутривенно индивидууму в начальной дозе 10 мг/кг в течение 60 минут с последующим введением CTLA-4-блокирующего средства в течение 30 минут в тот же день каждые две недели по мере необходимости.
Хотя одобренные в настоящее время CTLA-4-блокирующие средства и PD-1-блокирующие средства предоставляют в составах с концентрацией, делающей возможным внутривенное введение индивидууму в течение от 30 до 60 минут, в способах комбинированного лечения по настоящему изобретению предусмотрены варианты осуществления, в которых каждое из CTLA-4-блокирующего средства и/или PD-1-блокирующего средства предоставляют в составах в концентрации, позволяющей вводить каждый из них раздельно индивидууму в одной инъекции. Возможность введения по меньшей мере одного из двух блокирующих средств в одной инъекции будет значительно снижать время введения индивидууму обоих блокирующих средств.
В дополнительном варианте осуществления настоящее изобретение относится к комбинированному лечению, в котором CTL-4-блокирующее средство и PD-1-блокирующее средство вводят совместно одновременно. Совместное введение можно осуществлять, предоставляя CTLA-4- и PD-1-блокирующие средства в отдельных составах и одновременно вводя каждый состав индивидууму посредством отдельных IV введений, или смешивания смеси перед введением IV индивидууму, или отдельной инъекции каждого состава индивидууму. Совместное введение также можно осуществлять посредством предоставления CTLA-4- и PD-1-блокирующих средств в одном составе, который затем вводят индивидууму посредством IV введения или одной инъекции.
(i) Комбинированное лечение
Комбинированное лечение по настоящему изобретению можно использовать для лечения любого пролиферативного заболевания, в частности, лечения злокачественного новообразования. В конкретных вариантах осуществления комбинированное лечение по настоящему изобретению можно использовать для лечения меланомы, немелкоклеточного рака легких, рака головы и шеи, уротелиального рака, рака молочной железы, желудочно-кишечного злокачественного новообразования, множественной миеломы, печеночно-клеточного рака, неходжкинской лимфомы, рака почки, лимфомы Ходжкина, мезотелиомы, рака яичников, мелкоклеточного рака легких, рака пищевода, рака анального канала, рака желчных протоков, колоректального рака, рака шейки матки, рака щитовидной железы или рака слюнных желез.
В другом варианте осуществления комбинированное лечение по настоящему изобретению можно использовать для лечения рака поджелудочной железы, рака бронхов, рака предстательной железы, рака поджелудочной железы, рака желудка, рака яичников, рака мочевого пузыря, злокачественного новообразования головного мозга или центральной нервной системы, злокачественного новообразования периферической нервной системы, рака матки или эндометрия, рака полости рта или глотки, рака печени, рака почки, рака яичка, рака желчных протоков, рака тонкого кишечника или аппендикса, рака надпочечника, остеосаркомы, хондросаркомы или злокачественного новообразования кроветворных тканей.
Находящиеся в настоящее время на рынке PD-1-блокирующие средства одобрены FDA США для лечения по меньшей мере одного или более злокачественных новообразований, выбранных из меланомы (метастатической или нерезектабельной), первичной медиастинальной крупноклеточной B-клеточной лимфомы (PMBCL), уротелиальной карциномы, MSIHC, рака желудка, рака шейки матки, печеночноклеточной карциномы (HCC), карциномы из клеток Меркеля (MCC), почечноклеточной карциномы (включая поздние стадии) и плоскоклеточной карциномы кожи. Таким образом, комбинированное лечение по настоящему изобретению можно использовать для лечения по меньшей мере одного или более злокачественных новообразований, выбранных из меланомы (метастатической или нерезектабельной), первичной медиастинальной крупноклеточной B-клеточной лимфомы (PMBCL), уротелиальной карциномы, MSIHC, рака желудка, рака шейки матки, печеночноклеточной карциномы (HCC), карциномы из клеток Меркеля (MCC), почечноклеточной карциномы (включая поздние стадии) и плоскоклеточной карциномы кожи.
(j) Комбинированное лечение в комбинации с химиотерапией
Комбинированное лечение по настоящему изобретению можно вводить индивидууму, имеющему злокачественное новообразование, в комбинации с химиотерапией. Индивидуума можно подвергать химиотерапии одновременно с комбинированным лечением по настоящему изобретению. Индивидуума можно подвергать комбинированному лечению по настоящему изобретению после завершения химиотерапии. Индивидуума можно подвергать химиотерапии после завершения комбинированного лечения. Комбинированное лечение по настоящему изобретению также можно вводить индивидууму, имеющему рецидивирующее или метастатическое злокачественное новообразование с прогрессированием заболевания или рецидивирующее злокачественное новообразование, которого подвергают химиотерапии или завершившего химиотерапии.
Химиотерапия может включать химиотерапевтическое средство, выбранное из группы, состоящей из
(i) алкилирующих средств, включающих, в качестве неограничивающих примеров, бифункциональные алкиляторы, циклофосфамид, мехлоретамин, хлорамбуцил и мелфалан;
(ii) монофункциональных алкиляторов, включающих, в качестве неограничивающих примеров, дакарбазин, нитрозомочевину и темозоломид (пероральный дакарбазин);
(iii) антрациклинов, включающих, в качестве неограничивающих примеров, даунорубицин, доксорубицин, эпирубицин, идарубицин, митоксантрон и валрубицин;
(iv) разрушителей цитоскелета (таксаны), включающих, в качестве неограничивающих примеров, паклитаксел, доцетаксел, абраксан и таксотер;
(v) эпотилонов, включающих, в качестве неограничивающих примеров, иксабепилон и утиделон;
(vi) ингибиторы гистоновых деацетилаз, включающих, в качестве неограничивающих примеров, вориностат и ромидепсин;
(vii) ингибиторов топоизомеразы I, включающих, в качестве неограничивающих примеров, иринотекан и топотекан;
(viii) ингибиторов топоизомеразы II, включающих, в качестве неограничивающих примеров, этопозид, тенипозид и тафлупозид;
(ix) ингибиторов киназ, включающих, в качестве неограничивающих примеров, бортезомиб, эрлотиниб, гефитиниб, иматиниб, вемурафениб и висмодегиб;
(x) аналогов нуклеотидов и их предшественников, включающих, в качестве неограничивающих примеров, азацитидин, азатиоприн, фторпиримидины (например, такие как капецитабин, кармофур, доксифлуридин, фторурацил, и тегафур), цитарабин, гемцитабин, гидроксимочевину, меркаптопурин, метотрексат и тиогуанин;
(xi) пептидных антибиотиков, включающих, в качестве неограничивающих примеров, блеомицин и актиномицин; средства на основе платины, включая, в качестве неограничивающих примеров, карбоплатин, цисплатин и оксалиплатин;
(xii) ретиноидов, включающих, в качестве неограничивающих примеров, третиноин, алитретиноин и бексаротен; и
(xiii) алкалоидов барвинка и их производных, включающих, в качестве неограничивающих примеров, винбластин, винкристин, виндезин и винорелбин.
Выбор дозы химиотерапевтического средства для химиотерапии зависит от нескольких факторов, включая скорость оборота вещества в сыворотке или ткани, уровень симптомов, иммуногенность вещества и доступность целевых клеток, ткани или органа в организме индивидуума, подвергаемого лечению. Доза дополнительного терапевтического средства должна представлять собой количество, обеспечивающее приемлемый уровень побочных эффектов. Таким образом, доза и частота введения каждого дополнительного терапевтического средства будут зависеть частично от конкретного терапевтического средства, тяжести злокачественного новообразования, подвергаемого лечению, и характеристик пациента. Доступны руководства по выбору подходящих доз антител, цитокинов и низкомолекулярных соединений. См., например, Wawrzynczak (1996) Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK; Kresina (ed.) (1991) Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, NY; Bach (ed.) (1993) Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, NY; Baert et al. (2003) New Engl. J. Med. 348: 601-608; Milgrom et al. (1999) New Engl. J. Med. 341: 1966-1973; Slamon et al. (2001) New Engl. J. Med. 344: 783-792; Beniaminovitz et al. (2000) New Engl. J. Med. 342: 613-619; Ghosh et al. (2003) New Engl. J. Med. 348: 24-32; Lipsky et al. (2000) New Engl. J. Med. 343: 1594-1602; Physicians' Desk Reference 2003 (Physicians' Desk Reference, 57th Ed); Medical Economics Company; ISBN: 1563634457; 57th edition (november 2002). Определение подходящей схемы введения может проводить клиницист, например, с использованием параметров или факторов, которые, как известно или как предполагают в этой области, влияют на лечение или, как предполагают, влияют на лечение, и будут зависеть, например, от анамнеза пациента (например, предшествующей терапии), типа и стадии злокачественного новообразования, подлежащего лечению, и биомаркеров ответа на одно или более терапевтических средств в комбинированном лечении.
Например, пембролизумаб одобрен в настоящее время FDA США для комбинированного лечения (i) немелкоклеточного рака легких (NSCLC), включающего пембролизумаб с пеметрекседом и химиотерапии соединениями платины или карбоплатином и паклитакселом или наб-паклитакселом; и (ii) плоскоклеточного рака головы и шеи (HNSCC), включающего пембролизумаб и химиотерапию соединениями платины, и атезолизумаб одобрен в настоящее время для комбинированного лечения NSCLC, включающего бевацизумаб (антитело против VEGF-A, продаваемое под торговым названием AVASTIN), паклитаксел и карбоплатин.
Таким образом, в настоящем изобретении предусмотрены варианты осуществления комбинированного лечения по настоящему изобретению, дополнительно включающие стадию химиотерапии, включающей химиотерапию соединениями платины, пеметрексед и химиотерапию соединениями платины или карбоплатин и паклитаксел или наб-паклитаксел. В конкретных вариантах осуществления комбинированное лечение со стадией химиотерапии можно использовать для лечения, по меньшей мере, NSCLC и HNSCC.
Комбинированное лечение, дополнительно в комбинации со стадией химиотерапии, можно использовать для лечения любого пролиферативного заболевания, в частности, лечения злокачественного новообразования. В конкретных вариантах осуществления комбинированное лечение по настоящему изобретению можно использовать для лечения меланомы, немелкоклеточного рака легких, рака головы и шеи, уротелиального рака, рака молочной железы, желудочно-кишечного злокачественного новообразования, множественной миеломы, печеночно-клеточного рака, неходжкинской лимфомы, рака почки, лимфомы Ходжкина, мезотелиомы, рака яичников, мелкоклеточного рака легких, рака пищевода, рака анального канала, рака желчных протоков, колоректального рака, рака шейки матки, рака щитовидной железы или рака слюнных желез.
В другом варианте осуществления комбинированное лечение, дополнительно в комбинации со стадией химиотерапии, можно использовать для лечения рака поджелудочной железы, рака бронхов, рака предстательной железы, рака поджелудочной железы, рака желудка, рака яичников, рака мочевого пузыря, злокачественного новообразования головного мозга или центральной нервной системы, злокачественного новообразования периферической нервной системы, рака матки или эндометрия, рака полости рта или глотки, рака печени, рака почки, рака яичка, рака желчных протоков, рака тонкого кишечника или аппендикса, рака надпочечника, остеосаркомы, хондросаркомы или злокачественного новообразования кроветворных тканей.
В конкретных вариантах осуществления комбинированное лечение со стадией химиотерапии можно использовать для лечения одного или более злокачественных новообразований, выбранных из меланомы (метастатической или нерезектабельной), первичной медиастинальной крупноклеточной B-клеточной лимфомы (PMBCL), уротелиальной карциномы, MSIHC, рака желудка, рака шейки матки, печеночноклеточной карциномы (HCC), карциномы из клеток Меркеля (MCC), почечноклеточной карциномы (включая поздние стадии) и плоскоклеточной карциномы кожи.
Следующие примеры приведены для лучшего понимания настоящего изобретения.
ПРИМЕР 1
Функция Fc необходима для индуцирования irAE с воспалением кишечника
irAE с воспалением кишечника наблюдают у пациентов со злокачественными новообразованиями во время иммунотерапии с использованием монотерапии антителом против CTLA-4 или его комбинации с антителом против PD-1. Доклинические исследования мышей с дефицитом CTLA-4 в моделях конститутивного и условного нокаута показали развитие выраженного иммуноопосредованного воспалительного заболевания во множестве органов. Однако не показано, что лечение в сингенных моделях опухолей с использованием суррогатных антител против CTLA-4 индуцирует явные irAE, являющиеся прогностическими для токсичности, наблюдаемой у пациентов со злокачественными новообразованиями. Аналогично, гистопатологическую оценку воспаления кишечника осуществляли в сингенной или ксенотрансплантационной модели CT26 (мыши, инокулированные линией клеток карциномы толстого кишечника CT26). Мышам, несущим опухоли CT26, вводили Fc-компетентное mAb против CTLA-4 мыши 9D9-mIgG2a (антитело против CTLA4) (q4×4), что приводило к минимальному гранулоцитарному инфильтрату (степень 1 из 5), наблюдаемому в тканях собственной пластинки кишечника. Не наблюдали гранулоцитарных инфильтратов в когортах, которым вводили Fc-мутантное mAb против mCTLA-4 9D9-mIgG1-D265A (антитело против CTLA-4 (D265A)), ISVD против mCTLA-4 с меньшим Fc F894 (nAb против CTLA-4) или изотипические контроли. Однако не наблюдали сопутствующего изъязвления или другого повреждения ткани.
У Fc-мутантного антитела против CTLA-4 (D265A) отсутствует измеримая аффинность к рецепторам Fcγ (Nimmerjahn et al., Immunity, 23: 41-51 (2005)) и, таким образом, отсутствует Fc-эффекторная функция. У мышей противоопухолевая эффективность монотерапии mAb против CTLA4 мыши зависит от способности антитела опосредовать истощение внутриопухолевых регуляторных T-клеток с помощью эффекторной функции Fc (Selby et al., в цитируемом источнике). В связи с этим, не ожидали, что nAb против CTLA-4 и антитело против CTLA-4 (D265A) будут иметь благоприятный противоопухолевый эффект при монотерапии.
Минимальные результаты гистопатологического анализа побудили авторов настоящего изобретения оценить профили экспрессии генов в качестве потенциально более чувствительных средств для детекции маркеров активации воспалительных клеток в кишечнике. Авторы настоящего изобретения использовали панель экспрессии генов, которую они ранее разработали для протеомного анализа и определения профиля экспрессии генов, ассоциированных с воспаление кишечника, в образцах кала и биоптатах при воспалительном заболевании кишечника (IBD) в доклинических моделях и у пациентов (Cayatte, C, et. al., Clinical and Translational Gastroenterology, 3: e10 (2012)).
Профили экспрессии генов измеряли в образцах ткани тонкого кишечника и толстого кишечника в различные моменты времени после начала лечения антителом против CTLA-4 и сравнивали с антителом против CTLA-4 (D265A) для специфической оценки роли функции Fc в индуцировании воспаления кишечника. Как показано на фиг. 1A, экспрессия многочисленных воспалительных генов кишечника подвергалась положительной регуляции в образцах проксимальных отделов тонкого кишечника мышей, которым вводили антитело против CTLA-4, но не мышей, которым вводили антитело против CTLA-4 (D265A). Результаты свидетельствовали о том, что манифестации воспалительных путей кишечника можно определять по экспрессии генов в тканях тонкого кишечника и толстого кишечника в субклинических условиях.
Положительная регуляция генов, ассоциированных с воспалением кишечника, позволила оценить эффект длительного введения в отношении прогрессирования до клинического энтероколита, наблюдаемого у пациентов, которым вводят ипилимумаб, после лечения в течение 6-7 недель или более (Samaan et al., Nat. Rev. Gastroenterol. Hepatol. 15: 222-234 (2018)). Для оценки относительных эффектов блокады CTLA-4 и функции Fc в отношении воспаления кишечника и прогрессирования до энтероколита мышам BALB/c дважды в неделю вводили антитело против CTLA-4, антитело против CTLA-4 (D265A). Двум группам мышей вводили Fc-компетентное антитело против CTLA-4, одной группе мышей с опухолями CT26 и одной группе наивных мышей BALB/c без опухолей, для оценки потенциального влияния роста опухоли и индуцирования опухолевого иммунитета на индуцирование воспаления кишечника. Массу тела и баллы состояния организма оценивали дважды в неделю на всем протяжении длительного лечения для мониторинга прогрессирования до энтероколита.
Масса тела продолжала повышаться до дня 50 у мышей, которым вводили изотипические антитела и антитело против CTLA-4 (D265A). В отличие от этого, у мышей, которым вводили антитело против CTLA-4, наблюдали снижение средней массы тела через приблизительно 35-40 дней (фиг. 1B). Кишечная проницаемость повышалась в обеих группах, которым вводили антитело против CTLA-4, но не у мышей, которым вводили антитело против CTLA-4 (D265A), что оценивали на мышах, которым через желудочный зонд вводили FITC-декстран в день 49 и 50 (фиг. 1C). Гистологические признаки воспаления в проксимальных отделах тонкого кишечника и толстом кишечнике, оцениваемые патологом (L.A), свидетельствовали о прогрессировании до умеренного и тяжелого энтероколита в группах, которым вводили антитело против CTLA-4 (фиг. 1D и Фиг. 1E), при этом наблюдали обширную инфильтрацию иммунными клетками, утолщение слизистой оболочки и утрату бокаловидных клеток клетки. В отличие от этого, не наблюдали энтероколит у мышей, которым вводили антитело против CTLA-4 (D265A), что является доказательством того, что функция Fc необходима для индуцируемого блокадой CTLA-4 энтероколита. Примечательно, что рост опухоли и противоопухолевые ответы не были необходимыми для индуцируемого антителом против CTLA-4 воспаления кишечника.
Ранее было показано, что антитела против CTLA-4 с сильной функцией FcγR необходимы для сильных противоопухолевых ответов при монотерапии в экспериментах, в которых сравнивали химерные антитела IgG2a мыши с высокой аффинностью к FcγR с мутантными химерами IgG1 без детектируемого связывания FcγR (Selby et al., Cancer Immunol. Res. 1: 32-42 (2013)). Очевидно, специфическое истощение Treg-клеток в опухоли играет ключевую роль при монотерапии антителами против CTLA-4 на остове IgG2a мыши (mIgG2a) (Simpson et al., J. Exp. Med. 210: 1695-710 (2013). Недавние статьи, в которых показана важность CD28 для эффективности против PD-1 (см. ссылки), позволили авторам настоящего изобретения предположить, что повышение функции CD28 посредством блокирования более сильного взаимодействия CTLA-4 с лигандами CD80 и CD86 с использованием антагониста CTLA-4 может обеспечивать высокую комбинированную противоопухолевую эффективность без необходимости истощения Treg-TIL. ISVD, специфические для CTLA-4 мыши (mCTLA-4), способные ингибировать связывание с CD80 и CD86, разрабатывали с использованием однодоменного фрагмента антитела VHH, полученного их антител Верблюжьих, состоящих только из тяжелых цепей. У этих небольших белков массой 15 кДа отсутствуют Fc-области, и, таким образом, они не связываются с FcγR. Для сравнения с полностью Fc-функциональным антителом авторы настоящего изобретения использовали антитело против CTLA-4, как показано ранее, имеющее сильную противоопухолевую эффективность при монотерапии (см. ссылки). Для рассмотрения потенциальных эффектов связывания разных эпитопов CTLA-4 авторы настоящего изобретения использовали антитело против CTLA-4 (D265A), у которого отсутствует детектируемая аффинность к FcγR, для изучения необходимости сильной функции FcγR для противоопухолевой эффективности и переносимости.
nAb против CTLA-4 состояло из двух доменов VHH против CTLA-4, связанных с 35GS-линкером и доменом VHH против альбумина человека в качестве субъединицы для увеличения времени полужизни (HLE) (см. SEQ ID NO: 61). VHH против CTLA-4 имел 96% ингибирования для CD80 и CD86 по сравнению с антителом против CTLA-4, которое использовали в качестве референсного антагониста (100%) (фиг. 2A и фиг. 2B). nAb против CTLA-4 дополнительно сравнивали с антителом против CTLA-4 и эффекторно-молчащим антителом против CTLA-4 (D265A) в биологическом анализе in vitro на основе MLR на способность повышать ответы с пролиферацией (фиг. 2C), продукцией ИФНγ (фиг. 2D) и ИЛ-2 (фиг. 2E).
Функция Fc в случае блокады CTLA-4 не является необходимой для комбинированной иммунотерапии PD-1
Относительную противоопухолевую эффективность, индуцируемую посредством монотерапии тремя антагонистами CTLA-4 и комбинированного лечения с использованием антитела против PD-1 мыши mDX400 (антитела против PD-1) оценивали посредством измерения изменения объема опухоли с течением времени в сингенной модели карциномы толстого кишечника CT26 (фиг. 3A-3E), лишь умеренно отвечающей на монотерапию против PD-1. Не наблюдали противоопухолевой активности в когортах монотерапии, которым вводили nAb против CTLA-4 или антитело против CTLA-4 (D265A) и которые сравнивали с изотипическими контролями (группа 1 на фиг. 3A). Высокую противоопухолевую активность монотерапии наблюдали у мышей, которым вводили антитело против CTLA-4, что соответствует предшествующим данным, свидетельствовавшим о необходимости функции Fc в антителах против CTLA-4 для противоопухолевой эффективности монотерапии в сингенных моделях опухолей на мышах (Simpson et al., J. Exp. Med. 210: 1695-710 (2013)) (группа 2 на фиг. 3A), и лечение антителом против PD-1 в отдельности обеспечивало низкое и умеренное ингибирование роста опухоли по сравнению с антителом против CTLA-4. Однако, комбинированное лечение с использованием nAb против CTLA-4 или антителом против CTLA-4 (D265A) с антителом против PD-1 обеспечивало сильный противоопухолевый благоприятный эффект, сравнимый с наблюдаемым в случае антитела против CTLA-4 в отдельности или антитела против CTLA-4 в комбинации с антителом против PD-1 (группа 2 на фиг. 3A). nAb против CTLA-4 и антитело против CTLA-4 (D265A) связывают отдельные эпитопы на CTLA-4, таким образом, эффект не является специфическим для эпитопа. Дополнительные доказательства устойчивого комбинированного благоприятного эффекта, независящего от функции Fc, были очевидны, т.к. схожие противоопухолевые ответы наблюдали при комбинированном лечении антителом против PD-1 в случае nAb против CTLA-4, антитела против CTLA-4 (D265A) и антитела против CTLA-4. На фиг. 3D показаны отдельные результаты для каждой из 10 мышей, обобщенные на фиг. 3A. Экспансию CD8+ T-клеток и повышенные соотношения CD8+ T-клеток/Treg-клеток наблюдали у мышей, которым вводили антитело против CTLA-4, и мышей, которым вводили комбинацию антитела против CTLA-4 (D265S), и мышей, которым вводили антитело против PD-1, по сравнению с мышами, которым вводили антитело против CTLA-4 (D265S) в отдельности (фиг. 3B). Эти результаты свидетельствуют о том, что антагонисты CTLA-4, у которых отсутствует функция Fc, в комбинации с антагонистом PD-1 обеспечивали противоопухолевую эффективность, превосходящую эффективность, достигаемую с использованием монотерапии антагонистом PD-1, и обеспечивали противоопухолевую эффективность в комбинации, схожую по эффекту с противоопухолевой эффективностью антител против CTLA-4 с функцией Fc при монотерапии.
Авторы настоящего изобретения исследовали регуляцию генов иммунного ответа, ассоциированных с эффективной иммунотерапией злокачественных новообразований, в опухолях на девятый день после начала лечения для выявления потенциальных комплементарных механизмов, ассоциированных с высокой комбинированной активностью. ПЦР-профилирование экспрессии опухолей у мышей, которым вводили антитело против CTLA-4, показало сильную положительную регуляцию многочисленных генов, ассоциированных с эффективной иммунотерапией (фиг. 3C), включая ИФНγ, ИФН-чувствительные гены, хемокины, провоспалительные цитокины и MHC. Наблюдали лишь умеренную положительную регуляцию в опухолях у мышей, которым вводили антитело против CTLA-4 (D265A), что свидетельствует о том, что сильная положительная регуляция, наблюдаемая в опухолях мышей, которым вводили антитело против CTLA-4, по меньшей мере частично зависела от функции Fc. Также наблюдали умеренные ответы в опухолях у мышей, которым вводили nAb против CTLA-4 и антитело против PD-1. В отличие от этого, наблюдали устойчивую положительную регуляцию генов иммунного ответа в опухолях в когортах комбинированного лечения, которым вводили nAb против CTLA-4 или антитело против CTLA-4 (D265A) с антителом против PD-1. На фиг. 3D показано, что ни nAb против CTLA-4, ни антитело против CTLA-4 (D265A) не имели противоопухолевой активности в отсутствие блокады PD-1. Эти данные подтверждают гипотезу о том, что комплементарные механизмы абсолютной блокады CTLA-4 и блокады PD-1 могут обеспечивать сильный комбинированный положительный эффект, независящий от Fc.
Блокада CTLA-4 без функции Fc в комбинации с антителом против PD-1 обеспечивает достижение лучшего терапевтического индекса
Характерной особенностью блокады иммунных контрольных точек является клинически валидированный комбинированный благоприятный эффект антитела против PD-1 и антитела против CTLA-4, приводящий к лучшей клинической эффективности по сравнению с направленным воздействием на пути контрольных точек по отдельности. Однако иммуноопосредованная токсичность (irAE), ассоциированная с комбинированным лечением с блокадой CTLA-4 с антителом против PD-1, ассоциирована с повышенным индуцированием воспаления кишечника у пациентов (Ribas & WO lchok, Science 359: 1350-1355 (2018)). Кроме того, для антитела против CTLA-4 (D265A) и nAb против CTLA-4 с меньшим Fc необходима комбинация с антителом против PD-1, чтобы индуцировать сильный противоопухолевый иммунитет. Для рассмотрения потенциальных эффектов сильного противоопухолевого иммунитета в отношении индуцирования воспаления кишечника авторы настоящего изобретения исследовали профили экспрессии при воспалении кишечника у мышей, подвергаемых комбинированному лечению с использованием антитела против PD-1 с антителом против CTLA-4, антителом против CTLA-4 (D265A) или nAb против CTLA-4, после пяти введений, в день 18 после начала лечения (фиг. 4D).
Для оценки относительных эффектов блокады CTLA-4 и функции Fc в отношении воспаления кишечника наивным мышам BALB/c дважды в неделю вводили антитело против CTLA-4, антитело против CTLA-4 (D265A), nAb против CTLA4 с меньшим Fc, антитело против PD-1 или комбинации антитела против PD-1 с различными антагонистами mCTLA4. Массу тела и баллы состояния организма оценивали дважды в неделю на всем протяжении исследования. Масса тела мышей во всех группах повышалась до приблизительно дня 20 (фиг. 4A). Масса тела продолжала расти до дня 50 у мышей, которым вводили изотипическое антитело, антитело против CTLA-4 (D265A), nAb против CTLA4 или антитело против PD-1 или их комбинации. У мышей, которым вводили антитело против CTLA-4, наблюдали снижение средней массы тела после приблизительно дня 30 приблизительно до уровней до лечения ко дню 50. Введение антитела против CTLA-4 в комбинации с антителом против PD-1 приводило к более быстрому снижению средней массы тела ниже уровней до лечения со дня 20 до дня 50. Примечательно, что у мышей, которым вводили антитело против CTLA-4, баллы состояния организма у 2 из 8 мышей снижались до 2 (неудовлетворительно), начиная со дня 42 и начиная со дня 28 у мышей, которым вводили антитело против CTLA-4 в комбинации с антителом против PD-1. В этих когортах у животных наблюдали лоснящуюся, неряшливую шерсть и опухший живот.
Анализ воспаления был запланирован через семь недель введения, когда у мышей, которым вводили антитело против CTLA-4, наблюдали потерю массы тела за период времени, усиливающуюся при введении в комбинации с антителом против PD-1 (группа B на фиг. 4A). В отличие от этого, ни одно из эффекторно-молчащих CTLA-4-блокирующих средств или антитела против PD-1 не демонстрировало какую-либо значимую потерю массы по сравнению с изотипическими контролями в течение периода времени (группа A на фиг. 4B).
Всех мышей из групп комбинированного лечения и четырех мышей из групп изотипического контроля и введения антитела против PD-1 умерщвляли в день 50 для сбора тканей. Четырех оставшихся мышей из групп изотипического контроля и введения антитела против PD-1 и всех мышей из групп монотерапии умерщвляли в день 54 для сбора тканей. Во время некропсии проксимальные отделы тонкого кишечника и толстый кишечник удаляли для RT-qPCR для определения экспрессии воспалительных генов и оценки воспаления посредством гистопатологического анализа.
Тепловая карта экспрессии генов в проксимальных отделах тонкого кишечника из каждой группы лечения относительно изотипического контроля приведена на фиг. 4D. Введения антитела против CTLA-4 было достаточно для индуцирования положительной регуляции воспалительных генов в тощей кишки (фиг. 4D) и толстом кишечнике (фиг. 4E). Комбинация антитела против CTLA-4 с антителом против PD-1 индуцировала даже более сильную положительную регуляцию воспалительных генов, чем монотерапия антителом против CTLA-4. В отличие от этого, введение nAb против CTLA-4 индуцировало небольшую экспрессию воспалительных генов в кишечнике или не индуцировало ее, а также индуцировало лишь умеренную положительную регуляцию при комбинировании с антителом против PD-1. Аналогично, введение антитела против CTLA-4 (D265A) в отдельности или в комбинации с антителом против PD-1 приводило к минимальному и низкому индуцированию воспалительных генов. Проницаемость кишечника, оцениваемая в сыворотке после введения FITC-декстрана с помощью желудочного зонда, значимо повышалась у мышей, которым вводили антитело против CTLA-4, и мышей, которых подвергали комбинированному лечению антителом против CTLA-4 и антителом против PD-1.
Тяжесть воспаления в проксимальных отделах тонкого кишечника подвергали балльной оценке посредством гистологического анализа энтерита в проксимальных отделах тощей кишки в день 50. По результатам гистопатологической оценки введение антитела против CTLA-4 приводило к воспалению от слабого до тяжелого у большинства мышей. В когортах, которым вводили комбинацию антитела против CTLA-4 и антитела против PD-1, длительное лечение индуцировало воспаление от умеренного до очень тяжелого у всех мышей (фиг. 4B). У мышей с очень тяжелым энтеритом наблюдали еюнит, диффузные нейтрофильные очаги с умеренным количеством тучных клеток и дегенерацией нейронов мейсснерова сплетения. В отличие от этого, введение nAb против CTLA-4 или антитела против CTLA-4 (D265A) не индуцировало воспаление при гистопатологической оценке. Введение nAb против CTLA-4 в комбинации с антителом против PD-1 приводило к отсутствию воспаления или минимальному или слабому воспалению у нескольких мышей. Введение антитела против CTLA-4 (D265A) в комбинации с антителом против PD-1 приводило к слабому воспалению только у одной из восьми мышей. На типичных микрофотографиях показан относительный уровень воспаления в каждой группе лечения (фиг. 4C).
Как показано на фиг. 5A-5C, эффекторная функция Fc антитела против CTLA-4 запускает воспаление кожи (фиг. 5A), но не системное воспаление, т.к. не наблюдали детектируемого воспаления в почках, печени или легких (фиг. 5C). Абсолютное количество ИЛ-17-продуцирующих T-клеток, Foxp3+ Treg-клеток и нейтрофилов кожи уха измеряли посредством проточной цитометрии. Как показано на фиг. 5B, повышенные уровни ИЛ-17-продуцирующих T-клеток, Foxp3+ Treg-клеток и нейтрофилов наблюдали в коже уха мышей, которым вводили антитело против CTLA-4, но не антитело против CTLA-4 (D265A). В совокупности, эти данные подтверждают ключевую роль эффекторной функции Fc в индуцировании воспаления кишечника антителами против CTLA-4 мыши, имеющими эффекторную функцию. Эффекторная функция Fc вносила вклад в индуцируемое антителом против CTLA-4 мыши воспаление кишечника в модели энтероколита на мышах BALB/c, в то время как воспаление кишечника является слабым или отсутствует у мышей, которым вводили nAb против CTLA-4, или мышей, которым вводили антитело против CTLA-4 (D265A).
В совокупности, два признака были ассоциированы с индуцированием воспаления кишечника в модели опухоли CT26 под действием антитела против CTLA-4. Во-первых, необходима специфичность к CTLA-4, т.к. Fc-функциональные изотипические контроли не вызывали экспрессию генов, ассоциированных с воспаление. Однако блокирования связывания CTLA-4 с лигандами CD80/CD86 было недостаточно для индуцирования положительной регуляции воспалительных генов в кишечнике у мышей, которым вводили nAb против CTLA-4 и антитело против CTLA-4 (D265A). Таким образом, высокая активность функции Fc, имеющаяся у изоформы IgG2a в антителе против CTLA-4, необходима для индуцирования воспаления кишечника.
Активация воспаления кишечника инициирована активацией Teff-клеток независимо от истощения Treg-клеток
Механизм действия (MOA) опосредованного антителами против CTLA-4 противоопухолевого иммунитета теоретически опосредован фармакодинамическими (PD) эффектами в отношении популяций регуляторных T-клеток (Treg), а также эффекторных T-клеток (Teff) (CTL, TH1-клеток и т.д.). Истощение Treg-клеток в микроокружении опухоли (TIL) является характерным MOA антител против CTLA-4 в сингенных моделях опухолей на мышах (Simpson et al., в цитируемом источнике). Кроме того, совместное вовлечение Fc-FcγR под действием mAb против CTLA-4 модулирует передачу сигнала T-клеточного рецептора (TCR) и CD28, что приводит к повышенной активации T-клеток, независимо от истощения Treg-клеток (Waight et al., Cancer cell, 33: 1033-1047 (2018)).
Для характеризации дифференциальных эффектов антитела против CTLA-4 и nAb против CTLA-4 в отношении Treg-клеток и эффекторных T-клеток осуществляли проточную цитометрию популяций T-клеток из опухоли (TIL), собственной пластинки толстого кишечника, крови и селезенки через 20 часов после подкожного введения. Авторы настоящего изобретения могли измерять уровни экспрессии CTLA-4 в Treg-клетках мышей, которым вводили антитело против CTLA-4 или антитело против CTLA-4 (D265A), используя клон UC10-4B9 mAb против CTLA-4 (ThermoFisher), т.к. оно не приводит к перекрестному блокированию, что позволяет окрашивать связанный с лекарственным средством CTLA-4. Как и описано в литературе (Selby et al. в цитируемом источнике, Simpson et al. в цитируемом источнике), авторы настоящего изобретения наблюдали дифференциальную экспрессию CTLA-4 в Treg-клетках в селезенке (CTLA-4lo) и микроокружении опухоли (Treghi) у мышей, несущих опухоль CT26. Treg-клетки в PBMC экспрессировали бимодальные уровни CTLA-4lo-mid. Примечательно, что Treg-клетки из собственной пластинки толстого кишечника экспрессировали бимодальные уровни CTLA-4mid-hi. CTLA-4hi Treg-клетки в толстом кишечнике экспрессировали уровни, схожие с популяциями Treg-TIL. Дифференциальные уровни экспрессии CTLA-4 на различных популяциях T-клеток влияют на активность ADCC-опосредованного истощения из-за зависящих от плотности рецепторов механизмов уничтожения. Хотя популяции Treg-клеток в норме экспрессируют более высокие уровни CTLA-4, Treg-клетки в окружении опухоли экспрессируют гораздо более высокие уровни (в 3,3 раза более высокие, MFI=8100 в Treg-TIL в случае изотипических контролей), чем обнаруживаемые в селезенке (MFI=2400). Treg-клетки собственной пластинки толстого кишечника, экспрессирующие более высокие уровни CTLA-4 (MFI CTLA-4hi=10000), напоминали Treg-TIL по относительным уровням экспрессии при использовании проточной цитометрии (фиг. 6A). Как показано на фиг. 6B-6D, значительное истощение Treg-клеток было ограничено TIL из микроокружения опухоли мышей, которым вводили антитело против CTLA-4, имевших наибольшую плотность экспрессии CTLA-4. Введение Fc-мутантного антитела против CTLA-4 (D265A), у которого отсутствует функция Fc, не приводило к истощению Treg-клеток.
Основываясь только на предположении, что клетки, экспрессирующие более высокие уровни CTLA-4, были предрасположены к истощению под действием антитела против CTLA-4, авторы настоящего изобретения прогнозировали, что CTLA-4hi Treg-клетки в собственной пластинке будут истощены, аналогично популяциям Treg-TIL. Неожиданно, только Treg-клетки TIL, выделенные из опухолей мышей, которым вводили антитело против CTLA-4, по-видимому, были истощены. Полученные из собственной пластинки Treg-клетки толстого кишечника мышей, которым вводили антитело против CTLA-4, не выглядели истощенными. Не наблюдали детектируемого истощения полученных из собственной пластинки Treg-клеток толстого кишечника мышей, которым вводили антитело против CTLA-4, что позволяет предполагать, что индуцирование воспаления кишечника не инициировалось утратой Treg-клеток в слизистой оболочке кишечника.
Авторы настоящего изобретения исследовали возможные фенотипические изменения в Treg-клетках, выполняющих супрессорную функцию, которые могут вносить вклад в индуцируемое антителом против CTLA-4 воспаление кишечника. Treg-клетки собственная пластинка (LP) толстого кишечника FoxP3 GLD-репортерных мышей с имплантированной опухолью MC38 подвергали сортировке для ПЦР-профилирования экспрессии генов, ассоциированных с функцией Treg-клеток (фиг. 7A). Однако не наблюдали значимых различий экспрессии генов в Treg-клетках LP мышей, которым вводили антитело против CTLA-4, и мышей, которым вводили антитело против CTLA-4 (D265A). Потенциальный эффект функции Fc в отношении Treg-клеток дополнительно исследовали с использованием модели колита с переносом CD45RBhi T-клеток. Пассивный перенос Treg-клеток с CD45RBhi T-клетками защищал мышей от развития колита (фиг. 7B-7C). Лечение мышей, которым вводили Treg-клетки совместно с CD45RBhi T-клетками, антителом против CTLA-4 приводило к утрате защиты Treg-клетками и развитию колита. В отличие от этого, утрату защиты не наблюдали у мышей, которым вводили nAb против CTLA-4 (фиг. 7B-7C). Профиль экспрессии генов в сортированных посредством проточной цитометрии Foxp3+ Treg-клетках толстого кишечника мышей через 24 часа после введения свидетельствовал о значимой положительной регуляции экспрессии генов у мышей, которым вводили антитело против CTLA-4, что схоже с наблюдаемым в случае CD45RBhigh, по сравнению с экспрессией генов, наблюдаемой в случае клеток, полученных из мышей, которым вводили nAb против CTLA-4 или изотипические контроли. В совокупности, эти результаты позволяют предполагать, что Fc-опосредованное истощение Treg-клеток не является необходимым для индуцирования воспаления кишечника под действием антитела против CTLA-4, но регуляторную функцию Treg-клеток в ответ на воспаление кишечника можно модулировать.
Ограниченная экспрессия CTLA-4 и CD28 на T-клетках и CD80, CD86 и FcγR на антигенпрезентирующих клетках может играть ключевую роль в опосредованной Fc-функциональным nAb против CTLA-4 активации T-клеток, независящей от истощения Treg-клеток (Waight et. al., Cancer cell, 33: 1033-1047, 2018). Fc-усиленная активация в опухолях является предпочтительной, но может вносить вклад в незапланированные irAE в тканях кишечника. Авторы настоящего изобретения исследовали потенциальный вклад функции Fc в активацию эффекторных иммунных клеток в тканях кишечника. Проточный цитометрический анализ экспрессии CD16/32 на макрофагах из опухоли и толстого кишечника показал значимо более высокий уровень FcR на антигенпрезентирующих клетках по сравнению с макрофагами селезенки (фиг. 8A и 8B). Кроме того, доля CD45+CD11b+F4/80+ макрофагов в опухоли и собственной пластинке толстого кишечника была значимо выше, чем в селезенке (фиг. 8C). Функция Fc была необходима для активации цитокиновых ответов ИЛ-1β, ФНОα и ИФНγ в тканях кишечника, что было очевидным уже через 10 дней после начала лечения, задолго до появления явных признаков воспаления кишечника, что позволяет предполагать ее ключевую роль в индуцировании irAE (фиг. 8C).
При цитокиновых ответах в CD4+ T-клетках, выделенных из собственной пластинки толстого кишечника мышей, которым вводили антитело против CTLA-4, после одного месяца лечения наблюдали более высокую долю ИЛ-17-, ФНОα- и ИФНγ-продуцирующих клеток, ассоциированных с Fc-опосредованным воспалением кишечника (фиг. 8E). Кроме того, Fc-зависимое повышение нейтрофилов также наблюдали у мышей, которым вводили антитело против CTLA-4 (фиг. 8E). В совокупности, результаты свидетельствуют о том, что ассоциированное с блокадой CTLA-4 воспаление кишечника индуцировалось Fc-опосредованной активацией эффекторных клеток, усиленной повышением под действием FcγR-антитела образования клеточных мостиков между APC и T-клетками, приводящим к стимуляции воспалительных цитокиновых ответов, как показано на фиг. 11. Fc-опосредованное индуцирование воспаления кишечника может быть вызвано эффекторными T-клетками независимо от истощения Treg-клеток.
Обсуждение
В нескольких недавних статьях подтверждена ключевая роль функции Fc в монотерапии с блокадой CTLA-4 в сингенных моделях иммунотерапии опухолей на мышах (международная патентная заявка № WO 2014/089113; Selby et al., Cancer Immunol. Res. 32-42 (2013); Vargas et al., Cancer cell, 33: 649-663 (2018)). В Ingram et al. Proc. Natl. Acad. Sci. USA 115: 3912-3917, (2018) описано, что для обеспечения ISVD против CTLA-4 противоопухолевой эффективностью необходимо слияние Fc-домена с ISVD. Фактически, авторы настоящего изобретения наблюдали схожее отсутствие эффективности при таргетинге опухолей с помощью nAb против CTLA-4 или антитела против CTLA-4 (D265A) в виде монотерапии. Показано, что специфическое истощение инфильтрирующих опухоль Treg-клеток, экспрессирующих более высокие уровни CTLA-4 на своей поверхности, вносит вклад в противоопухолевую эффективность в моделях опухолей (Simpson et al., J. Exp. Med., 210: 1695-1710 (2013); Selby et al., Cancer Immunol. Res1: 32-42 (2013).
Индуцирование энтероколита антителом против CTLA-4 не было ассоциировано с детектируемым истощением Treg-клеток-резидентов в собственной пластинке. Авторы настоящего изобретения исследовали потенциал блокады CTLA-4 в противоопухолевом лечении посредством демонстрации высокой противоопухолевой активности при комбинировании с антителами против PD-1 без индуцирования сильного воспаления кишечника. Функция Fc не требовалась для достижения комбинированного благоприятного эффекта в случае блокад CTLA-4 и PD-1. Комбинированное лечение, включающее nAb против CTLA-4 или антитело против CTLA-4 (D265A) с антителом против PD-1, индуцировало схожую активацию ИФНγ-ассоциированных генов иммунного ответа в опухолях, с индуцируемой антителом против CTLA-4, имеющим выраженную функцию Fc. В отличие от этого, сильное воспаление кишечника, прогрессирующее до энтероколита, главным образом, наблюдали у мышей, которым вводили антитело против CTLA-4, и оно повышалось при комбинировании с антителом против PD-1 mDX400. Эти результаты свидетельствуют о том, что простой блокады CTLA-4, облегчающей активацию CD28, достаточно для повышения противоопухолевых ответов истощенных T-клеток при комбинировании с блокадой PD-1.
В статье Kamphorst et al. in Science 355: 1423-1427 (2017) показано, что для спасения истощенных CD8+ T-клеток при блокаде PD-1 необходима костимуляция CD28 при активации TCR. Кроме того, в сопутствующей статье Hui et.al. in Science 355: 1428-1433 (2017) показано, что корецептор CD28 является более предпочтительным, чем TCR, в качестве мишени дефосфорилирования PD-1-рекрутированной фосфатазой Shp2, и что CD28 преимущественно дефосфорилирован. Результаты, полученные авторами настоящего изобретения, свидетельствуют о том, что комплементарной активации корецептора TCR CD28, опосредованной простым блокированием CTLA-4 антителами против CTLA-4 без функции Fc, и блокады PD-1-опосредованного дефосфорилирования CD28 может быть достаточно для достижения комбинированного благоприятного эффекта при иммунотерапии злокачественных новообразований. Преимущество простой комбинированной блокады без Fc-опосредованной повышенной активации посредством образования мостиков Fc-FcγR может заключаться в том, что она делает возможным больший терапевтический индекс, что делает возможными большие диапазоны доз и большее время лечения. Это преимущество также может способствовать созданию дополнительных комбинаций с химиотерапевтическими стандартами лечения из-за более низкого риска irAE с воспалением кишечника.
МЕТОДИКА ЭКСПЕРИМЕНТА
Мыши
Мышей C57BL/6J дикого типа получали в Jackson Laboratories. Мышей Balb/c и CB17-SCID дикого типа получали в Taconic. Мышей B6.Foxp3GDL (GFP-DTR-люцифераза) получали и держали в специальных непатогенных условиях в микроизоляторах с фильтруемым воздухом в виварии Merck Research Laboratories (MRL) в Palo Alto, California. Все эксперименты на животных были одобрены Институциональным комитетом по содержанию и использованию животных MRL в соответствии с руководствами Международной ассоциации оценки и аккредитации лабораторных исследований на животных.
Инокуляция опухолевого материала и лечение
Для экспериментов на сингенных опухолях использовали модели опухолей CT26, MC38 и MB49. 8 Мышам Balb/c или C57BL/6J возрастом 8-12 недель подкожно (s.c.) в бок инъецировали 3×105 клеток CT26. Диаметр опухоли измеряли с помощью электронного калипера и вычисляли объем опухоли с помощью уравнения длина × ширина × ширина × 1/2. Лечение начинали, когда опухоли достигали приблизительно 100 мм3. Мышам дважды в неделю подкожно (s.c.) вводили антитело против CTLA-4, антитело против CTLA-4 (D265A), антитело мыши против IgG1-D265A изотипического контроля, антитело мыши против IgG2a изотипического контроля, антитела против PD-1 в дозе 10 мг/кг. Мышам дважды в неделю вводили s.c. nAb против CTLA-4 или контрольный ISVD в дозе 30 мг/кг.
Антитело против CTLA-4 содержит HC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 58, и LC, имеющую аминокислотную последовательность SEQ ID NO: 59.
Антитело против CTLA-4 (D265A) содержит HC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 60, и LC, имеющую аминокислотную последовательность SEQ ID NO: 60.
Антитело против PD-1 содержит HC, имеющую аминокислотную последовательность, приведенную в SEQ ID NO: 63, и LC, имеющую аминокислотную последовательность SEQ ID NO: 64.
nAb против CTLA-4 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 61.
ISVD против PD-1 F037 (nAb против PD-1) содержит аминокислотную последовательность, приведенную в SEQ ID NO: 62.
Индуцирование колита и воспаления кожи
Наивным мышам Balb/c возрастом 8-12 недель дважды в неделю s.c. в течение восьми недель вводили антитело против CTLA-4, антитело против CTLA-4 (D265A), антитело мыши против IgG1-D265A изотипический контроль, антитело мыши против IgG2a изотипического контроля в дозе 20 мг/кг. Мышам дважды в неделю s.c. вводили nAb против CTLA-4 или контрольный ISVD в дозе 30 мг/кг. В день 55 собирали плазму для анализов ELISA и Luminex. Органы собирали и обрабатывали следующим образом: 1) фиксировали 10% нейтральным забуференным формалином и окрашивали погруженные в парафин тканевые срезы гематоксилином и эозином для оценки патологии тканей; 2) быстро замораживали в жидком азоте для дальнейшего выделения РНК; или 3) помещали в HBSS для выделения клеток.
Вызываемый T-клетками колит
Клетки селезенки мышей Balb/c обрабатывали и очищали по CD4 посредством разделения с помощью магнитных частиц (Stem Cell Technologies). TCRb+CD4+CD25-CD45RBhigh T-клетки (CD45RBhigh T-клетки) и TCRb+CD4+CD25+CD45RBlow (Treg-клетки) сортировали с помощью FACS Aria (BD). 3×105 CD45RBhigh T-клеток и 1×105 Treg-клеток инъецировали внутривенно. Мышам i.p. дважды в неделю вводили 350 мкг антитела против CTLA-4 или изотипического контроля или 600 мкг nAb против CTLA-4 или контрольного ISVD. Мышей подвергали мониторингу и взвешивали в течение семи недель после инъекции.
Кишечная проницаемость
Мышам с помощью желудочного зонда вводили FITC-декстран (4 кДа, Sigma-Aldrich) за четыре часа до измерения флуоресценции FITC в сыворотке.
Выделение собственной пластинки толстого кишечника, кожи и опухолевых клеток
Клетки собственной пластинки толстого кишечника выделяли, сначала удаляя эпителиальные клетки посредством инкубации фрагментов ткани кишечника размером 0,5 см в забуференном растворе Хенкса, содержащем 5 мМ ЭДТА и 10 мМ HEPES, в течение 20 минут при 37°C, а затем повторяя эту инкубацию еще раз. Оставшуюся ткань нарезали на небольшие фрагменты, а затем расщепляли с помощью 1-кратной среды HBSS, содержащей 0,250 мг/мл либеразы (Roche), 30 ед./мл ДНКазы I (Sigma-Aldrich) и диспазы (Corning), в тех же условиях. Полученную суспензию клеток наслаивали на градиент 40%/80% PERCOLL и центрифугировали в течение 10 минут при 600×g; клетки LP выделяли с поверхности раздела.
Кожу уха измельчали и расщепляли 1-кратной средой HBSS, содержащей 0,250 мг/мл либеразы (Roche), 30 ед./мл ДНКазы I (Sigma-Aldrich) и диспазы (Corning) в течение 90 минут при 37°C. Суспензию клеток фильтровали и дважды промывали 1-кратным буфером HBSS. Опухоли измельчали и расщепляли 1-кратной средой HBSS, содержащей 0,250 мг/мл либеразы (Roche), 30 ед./мл ДНКазы I (Sigma-Aldrich) и диспазы (Corning) в течение 30 минут при 37°C. Суспензию клеток фильтровали и дважды промывали 1-кратным буфером HBSS.
Гистологические препараты толстого кишечника, кожи уха, печени, легких, почек фиксировали в 10% нейтральном забуференном формалине в течение ночи, переносили в 70% этанол, обрабатывали общепринятым образом, заключали в парафин, получали срезы толщиной 4-5 мкм, затем окрашивали гематоксилином и эозином. Патолог оценивал образцы толстого кишечника в слепом режиме по тяжести заболевания в соответствии с тремя критериями: воспаление: при наличии, оно отличается инфильтрацией большим количеством (60-70%) мононуклеарных клеток (макрофагов и лимфоцитов) и 30-40% нейтрофилов и палочкоядерных гранулоцитов. Балльная оценка воспаления включает тяжесть инфильтрации, утрату желез, эрозию, дилатацию просвета желез, наличие абсцессов крипт и дегенерацию эпителиальных клеток. Воспаление оценивали по шкале 0-4, 0=отрицательно; 1=минимальное, 2=слабое; 3=умеренное; 4=тяжелое. Апоптоз: распространение апоптотических телец оценивали по шкале 0-3, 0=отрицательно; 1=низкое, 2=умеренное; 3=высокое. Регенерация: Оцениваемые регенеративные изменения включают балльную оценку распространения митотических фигур в верхней трети слизистой оболочки, ядерную плотность (скученность ядер) в отдельных железистых структурах, регулярность поверхностного эпителия. Апоптоз оценивали по шкале 0-3, 0=отрицательно; 1=низкий, 2=умеренный; 3=высокий.
Проточная цитометрия и антитела
Клетки ресуспендировали в PBS и окрашивали на льду в течение 30 минут в темноте с помощью красителя Fixable Viability Stain (BD Bioscience). Затем клетки ресуспендировали в буфере для окрашивания (FBS, BD Bioscience) и окрашивали на льду в течение 30 минут с использованием различных комбинаций напрямую конъюгированных с флуорохромом антител. В случае внутриклеточных антигенов поверхностно окрашенные клетки пермеабилизовали, фиксировали с помощью набора буферов для окрашивания Foxp3 (eBiosciences) в течение 30 минут на льду, а затем окрашивали с помощью специфических антител. Антитела мыши: CD45 (30-F11), CD8a (53-6,7), CTLA-4 (UC10-4B9), CD11c (HL3), CD11b (M1/70), TCRβ (H57-597), TCRγδ (GL3), CD4 (RM4-5 или GK1,5), CD25 (PC61), CD45RB (16A), Ly6G (1A8), F4/80 (T45-2342), CD16/32 (2,4G2), ИФНγ (XMG1.2), ИЛ-17A (TC11-18H10), ФНОα (MP6-XT22), Foxp3 (FJK-16s). Все антитела приобретали в BD Biosciences, Biolegend или eBioscience. Для всех образцов данные получали с помощью проточного цитометра LSR II (BD). Данные анализировали с использованием программного обеспечения FLOWJO (Tree Star).
Если продукцию цитокинов измеряли посредством проточной цитометрии, клетки стимулировали 500 нг/мл иономицина, 50 нг/мл PMA (Sigma-Aldrich). Через час добавляли брефелдин A (BD Bioscience) еще на два часа перед окрашиванием.
Анализ аллогенной смешанной лейкоцитарной реакции (MLR) мыши
2×105 T-клеток селезенки мыши C57B6/J (возрастом 8-12 недель, самки) выделяли с использованием набора для выделения T-клеток EASYSEPMouse (STEMCELL) и сокультивировали с 1×105 облученных (2000 рад) спленоцитов мыши Balb/c в присутствии указанной концентрации антитела против CTLA-4, антитела против CTLA-4 (D265A), nAb против CTLA-4 или изотипических контролей. В день три супернатант собирали и измеряли продукцию ИЛ-2 и ИФН-гамма посредством ELISA по инструкциям производителя (Meso Scale Discovery). Затем в клетки вводили [3H]-тимидин (1 мкКи на лунку) в течение шести часов или 16-18 часов. Клетки собирали на фильтры из стекловолокна с использованием харвестера. Фильтры подсчитывали с помощью счетчика MicroBeta (PerkinElmer Microbeta 2450) по инструкциям производителя.
Выделение тотальной РНК из тканей и клеток и последующий анализ экспрессии генов с использованием платформы Fluidigm BIOMARK.
Для анализа посредством ПЦР в реальном времени тотальную РНК выделяли любым из двух способов. Органы гомогенизировали в RNA STAT-60 (Tel-Test Inc., FriendsWO od, TX) с помощью гомогенизатора Polytron, а затем осуществляли выделение РНК с помощью MagMAX-96 для набора Microarrays (ThermoFisher Scientific, Waltham, MA) по инструкциям производителя. В случае образцов клеток РНК выделяли с использованием набора для выделения РНК ARCTURUS PICOPURE по инструкциям производителя (ThermoFisher Scientific, Waltham, MA).
Обработанную ДНКазой тотальную РНК подвергали обратной транскрипции с использованием набора QUANTITECT Reverse Transcription (Qiagen, Valencia, CA) по инструкциям производителя. Праймеры приобретали в ThermoFisher Scientific (Foster City, CA). Гено-специфическую предварительную амплификацию осуществляли с помощью по меньшей мере 2 нг кДНК по инструкциям производителя к Fluidigm BIOMARK (Fluidigm, Foster City). Затем осуществляли количественную ПЦР в реальном времени с помощью Fluidigm BIOMARK с использованием двух немеченых праймеров по 900 нМ каждого, 250 нМ меченого FAM зонда (ThermoFisher Scientific, Foster City, CA) и универсального мастер-микса для ПЦР TAQMAN, содержащего UNG. Образцы и праймеры использовали в формате 48×48 или 96×96 по инструкциям производителя (Fluidigm, Foster City). Уровни убиквитина измеряли в отдельной реакции и использовали их для нормализации данных способом ΔCt (с использованием среднего значения порогового цикла для убиквитина и интересующего гена для каждого образца, для получения нормализованных значений использовали уравнение 1,8^(Ct убиквитина минус Ct интересующего гена) × 104). Референсные последовательности праймеров доступны по запросу.
Статистический анализ
Для вычисления статистической значимости в остальной части настоящего исследования использовали двухсторонний парный и непарный t-критерий Стьюдента. *P<0,05, **P<0,01, ***P<0,001. Статистический анализ осуществляли с использованием программного обеспечения GraphPad PRISM 7.
ПРИМЕР 2
Противоопухолевую эффективность nAb против CTLA-4 оценивали в сингенной модели опухоли MB49 на мышах. Клетки MB49 представляют собой линию клеток уротелиальной карциномы, полученную из взрослой мыши C57BL/6 посредством подвергания первичных эксплантатов клеток эпителия мочевого пузыря воздействию 7,12-диметилбенз[a]антрацена (DMBA) в течение 24 часов с последующим длительным культивированием. Сингенную модель рака мочевого пузыря на мышах широко используют в течение более чем 35 лет.
Клетки рака мочевого пузыря мыши MB49 имплантировали подкожно (s.c.) 80 мышам и приписывали животных к пяти группам лечения по 10 мышей в каждой. Когда медиана начального объема опухоли достигала 103 мм3, мышам делали инъекцию s.c. один раз каждые четыре дня всего в четырех дозах. Неродственный контрольный ISVD (30 мг/кг, лот №01AQL) и 5 мг/кг mAb mIgG1 изотипического контроля (лот №64AIS) вводили в качестве контроля лечения. Лечение включало 30 мг/кг nAb против CTLA-4, 10 мг/кг Fc-компетентного антитела против CTLA-4 (D265A), 5 мг/кг антитела против PD-1 или комбинаций нацеленных на CTLA4 средств и антитела против PD-1. Рост опухоли подвергали мониторингу в течение 21 дня после начала лечения.
На фиг. 9A показаны объемы опухолей отдельных животных в каждой группе лечения. Полные ответы (CR) до дня 21 представлены для групп отвечающих на лечение. На фиг. 9B показан средний объем опухоли и стандартная ошибка среднего для каждой группы лечения (начальное количество n=10/группу). Объемы опухолей у животных, которых исключали из исследования из-за больших объемов опухолей, переносили в среднее значение до проведения последнего измерения в этой группе лечения. На фиг. 9A-9B показано, что, как и в модели опухоли толстого кишечника CT26 (см. фиг. 3A), модели опухоли мочевого пузыря MB49 и моделях опухоли толстого кишечника MC38, комбинированное лечение с использованием nAb против CTLA-4 с меньшим Fc с антителом против PD-1 обеспечивало сильный противоопухолевый благоприятный эффект, независящий от функции Fc.
ПРИМЕР 3
Противоопухолевую эффективность nAb против CTLA-4 оценивали в сингенной модели опухоли MC38 на мышах. Клетки рака толстого кишечника мышь MC38 имплантировали SC 80 мышам и приписывали животных к пяти группам лечения по 10 мышей в каждом. Когда медиана начального объема опухоли достигала 246 мм3, мышам делали инъекцию SC один раз каждые четыре дня всего в четырех дозах. Неродственный контрольный ISVD (30 мг/кг) и 5 мг/кг mAb mIgG1 изотипического контроля вводили в качестве контроля лечения. Лечение включало 30 мг/кг nAb против CTLA-4, 10 мг/кг Fc-компетентного антитела против CTLA-4 (D265A), 5 мг/кг антитела против PD-1 или комбинаций нацеленных на CTLA4 средств и антитела против PD-1. Рост опухоли подвергали мониторингу в течение 23 дней после начала лечения.
На фиг. 10A показаны объемы опухолей отдельных животных для каждой группы лечения. Полные ответы (CR) до дня 23 представлены для групп отвечающих на лечение. На фиг. 10B показан средний объем опухоли и стандартная ошибка среднего для каждой группы лечения (начальное количество n=10/группу). Объемы опухолей у животных, которых исключали из исследования из-за больших объемов опухолей, переносили в среднее значение до проведения последнего измерения в этой группе лечения. На фиг. 10A-10B показано, что, как и в модели опухоли толстого кишечника CT26 (см. фиг. 3A), модели опухоли мочевого пузыря MB49 и моделях опухоли толстого кишечника MC38, комбинированное лечение с использованием nAb против CTLA-4 с меньшим Fc с антителом против PD-1 обеспечивало сильный противоопухолевый благоприятный эффект, независящий от функции Fc.
Последовательности
(все положения аминокислот идентифицированы с использованием нумерации Eu)
PF023700037
X является любой аминокислотой
X является любой аминокислотой
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
ВАРИАНТ 1 M18I
W02019023482
LTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Константные домены HC и LC указаны курсивом.
Ссылки
Azuma et al., B70 antigen is a second ligand for CTLA-4 and CD28. Nature. 1993 Nov 4; 366(6450): 76-9.
Collins et al., The interaction properties of costimulatory molecules revisited. Immunity. 2002 Aug; 17(2): 201-10.
Hui et al., T cell costimulatory receptor CD28 is a primary target for PD-1-mediated inhibition. Science. 2017 Mar 31; 355(6332): 1428-1433.
Ingram et al., Anti-CTLA-4 therapy requires an Fc domain for efficacy. Proc Natl Acad Sci U S A. 2018 Apr 10; 115(15): 3912-3917.
Kamphorst et al., Rescue of exhausted CD8 T cells by PD-1-targeted therapies is CD28-dependent. Science. 2017 Mar 31; 355(6332): 1423-1427.
 et al., Reformatting Rituximab into Human IgG2 and IgG4 Isotypes Dramatically Improves Apoptosis Induction In Vitro. PLoS One. 2015 Dec 29; 10(12): e0145633.
et al., Reformatting Rituximab into Human IgG2 and IgG4 Isotypes Dramatically Improves Apoptosis Induction In Vitro. PLoS One. 2015 Dec 29; 10(12): e0145633.
Lanier et al., CD80 (B7) and CD86 (B70) provide similar costimulatory signals for T cell proliferation, cytokine production, and generation of CTL. J Immunol. 1995 Jan 1; 154(1): 97-105.
Leach et al., Enhancement of antitumor immunity by CTLA-4 blockade. Science. 1996 Mar 22; 271(5256): 1734-6.
Pai et al., Tumor-conditional anti-CTLA4 uncouples antitumor efficacy from immunotherapy-relatedtoxicity. J Clin Invest. 2019 Jan 2; 129(1): 349-363.
Ribas & WO lchok, Cancer immunotherapy using checkpoint blockade. Science. 2018 Mar 23; 359(6382): 1350-1355.
Schneider-Merck et al., Human IgG2 antibodies against epidermal growth factor receptor effectively trigger antibody-dependent cellular cytotoxicity but, in contrast to IgG1, only by cells of myeloid lineage. J Immunol. 2010 Jan 1; 184(1): 512-20.
Selby et al., Anti-CTLA-4 antibodies of IgG2a isotype enhance antitumor activity through reduction of intratumoral regulatory T cells. Cancer Immunol Res. 2013 Jul; 1(1): 32-42.
Sharma et al., Anti-CTLA-4 Immunotherapy Does Not Deplete FOXP3+ Regulatory T Cells (Tregs) in Human Cancers. Clin Cancer Res. 2018 Jul 27.
Simpson et al., Fc-dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti-CTLA-4 therapy against melanoma. J Exp Med. 2013 Aug 26; 210(9): 1695-710.
 & Foltz, Body condition scoring: a rapid and accurate method for assessing health status in mice. Lab Anim Sci. 1999 Jun; 49(3): 319-23.
& Foltz, Body condition scoring: a rapid and accurate method for assessing health status in mice. Lab Anim Sci. 1999 Jun; 49(3): 319-23.
Vargas et al., Fc Effector Function Contributes to the Activity of Human Anti-CTLA-4 Antibodies. Cancer Cell. 2018 Apr 9; 33(4): 649-663.e4.
Waight et al., Selective FcγR Co-engagement on APCs Modulates the Activity of Therapeutic Antibodies Targeting T Cell Antigens. Cancer Cell. 2018 Jun 11; 33(6): 1033-1047.e5.
Хотя настоящее изобретение представлено в настоящем описании со ссылкой на иллюстрированные варианты осуществления, следует понимать, что изобретение ими не ограничено. Специалистам в этой области с учетом идей, изложенных в настоящем описании, будут понятны дополнительные модификации и варианты осуществления в объеме настоящего изобретения. Таким образом, настоящее изобретение ограничено только формулой изобретения, прилагаемой к настоящему описанию.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Merck Sharp & Dohme Corp
LaFace, Drake
Punnonen, Juha
Bowman, Edward
Bauche, David
Chackerian, Alissa
Grein, Jeffery
Mauze, Smita
Sawant, Anandi
Annamalai, Laksmanan
<120> СПОСОБЫ КОМБИНИРОВАННОГО ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ,
ВКЛЮЧАЮЩИЕ СРЕДСТВА, БЛОКИРУЮЩИЕ CTLA-4 И PD-1
<130> 24702
<150> 62/817,749
<151> 2019-03-13
<160> 117
<170> PatentIn version 3.5
<210> 1
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR1 ипилимумаба
<400> 1
Arg Ala Ser Gln Ser Val Gly Ser Ser Tyr Leu Ala
1 5 10
<210> 2
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR2 ипилимумаба
<400> 2
Gly Ala Phe Ser Arg Ala Thr
1 5
<210> 3
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR3 ипилимумаба
<400> 3
Gln Gln Tyr Gly Ser Ser Pro Trp Thr
1 5
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR1 ипилимумаба
<400> 4
Ser Tyr Thr Met His
1 5
<210> 5
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR2 ипилимумаба
<400> 5
Phe Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 6
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR3 ипилимумаба
<400> 6
Thr Gly Trp Leu Gly Pro Phe Asp Tyr
1 5
<210> 7
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH ипилимумаба
<400> 7
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Thr Phe Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Trp Leu Gly Pro Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 8
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL ипилимумаба
<400> 8
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Phe Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 9
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR1 тремелимумаба
<400> 9
Arg Ala Ser Gln Ser Val Gly Ser Ser Tyr Leu Ala
1 5 10
<210> 10
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR2 тремелимумаба
<400> 10
Gly Ala Phe Ser Arg Ala Thr
1 5
<210> 11
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR3 тремелимумаба
<400> 11
Gln Gln Tyr Gly Ser Ser Pro Trp Thr
1 5
<210> 12
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR1 тремелимумаба
<400> 12
Ser Tyr Gly Met His
1 5
<210> 13
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR2 тремелимумаба
<400> 13
Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 14
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR3 тремелимумаба
<400> 14
Tyr Gly Ser Ser Pro
1 5
<210> 15
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH тремелимумаба
<400> 15
Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser
1 5 10 15
Gly Phe Thr Phe Ser Ser Tyr Gly Met His Trp Val Arg Gln Ala Pro
20 25 30
Gly Lys Gly Leu Glu Trp Val Ala Val Ile Trp Tyr Asp Gly Ser Asn
35 40 45
Lys Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
50 55 60
Asn Ser Lys Asn Thr Leu Leu Gln Met Asn Ser Leu Arg Ala Glu Thr
65 70 75 80
Ala Val Tyr Tyr Cys Ala Arg Asp Pro Arg Gly Ala Thr Leu Tyr Tyr
85 90 95
Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 16
<211> 100
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL тремелимумаба
<400> 16
Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
1 5 10 15
Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asp Trp Tyr Gln Gln Lys
20 25 30
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser Leu Gln
35 40 45
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe
50 55 60
Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr
65 70 75 80
Cys Gln Gln Tyr Tyr Ser Thr Pro Phe Thr Phe Gly Pro Gly Thr Lys
85 90 95
Val Glu Ile Lys
100
<210> 17
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR1 ниволумаба
<400> 17
Asn Ser Gly Met His
1 5
<210> 18
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR2 ниволумаба
<400> 18
Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 19
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR3 ниволумаба
<400> 19
Asn Asp Asp Tyr
1
<210> 20
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR1 ниволумаба
<400> 20
Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala
1 5 10
<210> 21
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR2 ниволумаба
<400> 21
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 22
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR3 ниволумаба
<400> 22
Gln Gln Ser Ser Asn Trp Pro Arg Thr
1 5
<210> 23
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH ниволумаба
<400> 23
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Asp Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser
<210> 24
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL ниволумаба
<400> 24
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 25
<211> 440
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC ниволумаба (IgG4 S228P)
<400> 25
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Asp Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser
115 120 125
Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys
180 185 190
Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala
210 215 220
Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
225 230 235 240
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
245 250 255
Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
260 265 270
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
275 280 285
Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
290 295 300
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
305 310 315 320
Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
325 330 335
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr
340 345 350
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
355 360 365
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
370 375 380
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
385 390 395 400
Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe
405 410 415
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
420 425 430
Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 26
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC ниволумаба
<400> 26
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 27
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC пембролизумаба (IgG4 S228P)
<400> 27
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Asn Pro Ser Asn Gly Gly Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Phe Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 28
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC пембролизумаба
<400> 28
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Gly Val Ser Thr Ser
20 25 30
Gly Tyr Ser Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Leu Ala Ser Tyr Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Ser Arg
85 90 95
Asp Leu Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 29
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH пембролизумаба
<400> 29
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Asn Pro Ser Asn Gly Gly Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Phe Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 30
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL пембролизумаба
<400> 30
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Gly Val Ser Thr Ser
20 25 30
Gly Tyr Ser Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Leu Ala Ser Tyr Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Ser Arg
85 90 95
Asp Leu Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 31
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR1 пембролизумаба
<400> 31
Gly Tyr Thr Phe Thr Asn Tyr Tyr Met Tyr
1 5 10
<210> 32
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR2 пембролизумаба
<400> 32
Asn Pro Ser Asn Gly Gly Thr Asn Phe Asn Glu Lys Phe Lys Asn
1 5 10 15
<210> 33
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC-CDR3 пембролизумаба
<400> 33
Arg Asp Tyr Arg Phe Asp Met Gly Phe Asp Tyr
1 5 10
<210> 34
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR1 пембролизумаба
<400> 34
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Ser Tyr Leu His
1 5 10 15
<210> 35
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR2 пембролизумаба
<400> 35
Leu Ala Ser Tyr Leu Glu Ser
1 5
<210> 36
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC-CDR3 пембролизумаба
<400> 36
Cys Gln His Ser Arg Asp Leu Pro Leu Thr
1 5 10
<210> 37
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека
<400> 37
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 38
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (L234A L235A D265S)
<400> 38
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Ser Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 39
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (L234A L235A P329G)
<400> 39
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Ser Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 40
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (L235E)
<400> 40
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 41
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (D265A)
<400> 41
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 42
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (D265A N297G)
<400> 42
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 43
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (E233A/L235A)
<400> 43
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Ala Leu Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 44
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (N297X, где X является любой аминокислотой, иной, чем N)
<220>
<221> прочий_признак
<222> (180)..(180)
<223> Xaa может являться любой природной аминокислотой
<400> 44
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Xaa Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 45
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG2 человека
<400> 45
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 46
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG2 человека (D265S)
<400> 46
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ser
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 47
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG2 человека (P329G)
<400> 47
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Gly
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 48
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG2 человека (D265A)
<400> 48
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 49
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG2 человека (D265A N297G)
<400> 49
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Gly
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 50
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG2 человека (N297X, где X является любой аминокислотой, иной, чем N)
<220>
<221> прочий_признак
<222> (176)..(176)
<223> Xaa может являться любой природной аминокислотой
<400> 50
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Xaa
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 51
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG2 человека (V234A G237A P238S H268A V309L A330S P331S X378S/A)
<220>
<221> прочий_признак
<222> (257)..(257)
<223> Xaa может являться любой природной аминокислотой
<400> 51
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Ala Ala Ala Ser Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser Ala Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ser Ser Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Xaa Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 52
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG4 человека (S228P)
<400> 52
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 53
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG4 человека (S228P P329G)
<400> 53
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Gly Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 54
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG4 человека (S228P D265A)
<400> 54
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Ala Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 55
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG4 человека (S228P D265A N297G)
<400> 55
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Ala Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 56
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG4 человека (S228P N297X, где X является любой аминокислотой, иной, чем N)
<220>
<221> прочий_признак
<222> (177)..(177)
<223> Xaa может являться любой природной аминокислотой
<400> 56
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Xaa Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 57
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен LC каппа человека
<400> 57
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 58
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC IgG2 мыши против CTLA-4 (модифицированное x [CTLA-4_M] mAb мыши (9D9 Balb/c (Igh-1a) гаплотип - основной аллель) IgG2a/каппа—тяжелая цепь))
<400> 58
Glu Ala Lys Leu Gln Glu Ser Gly Pro Val Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met Asn Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Val Ile Asn Pro Tyr Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Ser Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Ile Thr Val Ser Ser Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu
115 120 125
Ala Pro Val Cys Gly Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys
130 135 140
Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser
145 150 155 160
Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp
180 185 190
Pro Ser Gln Ser Ile Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr
195 200 205
Lys Val Asp Lys Lys Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys
210 215 220
Pro Pro Cys Lys Cys Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Ile Phe Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser
245 250 255
Pro Ile Val Thr Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp
260 265 270
Val Gln Ile Ser Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln
275 280 285
Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser
290 295 300
Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys
305 310 315 320
Cys Lys Val Asn Asn Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile
325 330 335
Ser Lys Pro Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro
340 345 350
Pro Pro Glu Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met
355 360 365
Val Thr Asp Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn
370 375 380
Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn
405 410 415
Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu
420 425 430
His Asn His His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
435 440 445
<210> 59
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC каппа антитела мыши против CTLA-4 (12AIL_9D9_LC)
<400> 59
Asp Ile Val Met Thr Gln Thr Thr Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 60
<211> 441
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC антитела мыши против CTLA-4 (D265A) (мышь x [CTLA-4_M] mAb (9D9 мутация D265A) IgG1/каппа—тяжелая цепь)
<400> 60
Glu Ala Lys Leu Gln Glu Ser Gly Pro Val Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met Asn Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Val Ile Asn Pro Tyr Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Ser Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Ile Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu
115 120 125
Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys
130 135 140
Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser
145 150 155 160
Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp
180 185 190
Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr
195 200 205
Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys
210 215 220
Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys
225 230 235 240
Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val
245 250 255
Val Val Ala Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe
260 265 270
Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu
275 280 285
Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His
290 295 300
Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala
305 310 315 320
Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg
325 330 335
Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met
340 345 350
Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro
355 360 365
Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn
370 375 380
Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val
385 390 395 400
Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr
405 410 415
Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu
420 425 430
Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 61
<211> 415
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ISVD мыши против CTLA-4 (54BBI_Llama x [CTLA4_M] [ALB_H] VHH (F023700894) (PI))
<400> 61
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Thr Pro Ser Ile Asn
20 25 30
Tyr Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Phe Val
35 40 45
Ala Thr Ile Arg Ser Gly Gly Ala Thr Asn Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Thr Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Asp Cys Tyr
85 90 95
Thr Gly Gly Gly Gly Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly
145 150 155 160
Leu Val Gln Ala Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
165 170 175
Ser Thr Pro Ser Ile Asn Tyr Met Gly Trp Tyr Arg Gln Ala Pro Gly
180 185 190
Lys Gln Arg Glu Phe Val Ala Thr Ile Arg Ser Gly Gly Ala Thr Asn
195 200 205
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Thr
210 215 220
Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr
225 230 235 240
Ala Val Tyr Asp Cys Tyr Thr Gly Gly Gly Gly Tyr Glu Tyr Trp Gly
245 250 255
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
260 265 270
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
275 280 285
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
290 295 300
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser Leu Arg Leu
305 310 315 320
Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly Met Ser Trp
325 330 335
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser
340 345 350
Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys Gly Arg Phe
355 360 365
Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu Gln Met Asn
370 375 380
Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr Ile Gly Gly
385 390 395 400
Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val Ser Ser
405 410 415
<210> 62
<211> 436
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ISVD мыши против PD-1 PF023700037
<400> 62
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Phe Ser Thr His
20 25 30
Thr Met Gly Trp Phe Arg Gln Gly Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Thr Ile Asn Arg Leu Asp Tyr Thr Tyr Tyr Ala Asn Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Asp Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Ala Asp Ser Glu Arg Arg Leu Gly Val Ile Pro Gly Leu Tyr Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
145 150 155 160
Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
165 170 175
Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Phe Ser Arg Leu Ala Met
180 185 190
Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala Ser
195 200 205
Ile Ser Trp Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
210 215 220
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr Leu Gln
225 230 235 240
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Ala
245 250 255
Ser Arg Glu Tyr Ser Gly Ser Tyr Tyr Tyr Gly Leu Thr Leu Tyr Glu
260 265 270
Tyr Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly
275 280 285
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
290 295 300
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
305 310 315 320
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
325 330 335
Asn Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser
340 345 350
Phe Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
355 360 365
Val Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser
370 375 380
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu
385 390 395 400
Tyr Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr
405 410 415
Cys Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val
420 425 430
Thr Val Ser Ser
435
<210> 63
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC клона DX400 антитела мыши против PD-1
<400> 63
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ser
20 25 30
Gly Leu Ala Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Thr Tyr Asn Gly Thr Ser Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Trp Val Pro Gly Ser Gly Asn Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr
245 250 255
Cys Val Val Val Ala Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 64
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC клона DX400 антитела мыши против PD-1
<400> 64
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Thr Ile Ser
20 25 30
Arg Tyr Thr Leu Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Arg
85 90 95
Glu Ser Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
115 120 125
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
130 135 140
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
145 150 155 160
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
180 185 190
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
195 200 205
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 65
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Остатки после LC-CDR3
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> Xaa = любая аминокислота
<400> 65
Phe Gly Xaa Gly
1
<210> 66
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Остатки перед HC-CDR1
<220>
<221> ВАРИАНТ
<222> (2)..(4)
<223> Xaa = любая аминокислота
<400> 66
Cys Xaa Xaa Xaa
1
<210> 67
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Остатки перед HC-CDR2
<400> 67
Leu Glu Trp Ile Gly
1 5
<210> 68
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Остатки после HC-CDR3
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> Xaa = любая аминокислота
<400> 68
Trp Gly Xaa Gly
1
<210> 69
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR-1 ISVD против CTLA-4
<400> 69
Phe Tyr Gly Met Gly
1 5
<210> 70
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR-2 ISVD против CTLA-4
<400> 70
Asp Ile Arg Thr Ser Ala Gly Arg Thr Thr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 71
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR-3 ISVD против CTLA-4
<400> 71
Glu Met Ser Gly Ile Ser Gly Trp Asp Tyr
1 5 10
<210> 72
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR-3 варианта ISVD против CTLA-4
<400> 72
Glu Pro Ser Gly Ile Ser Gly Trp Asp Tyr
1 5 10
<210> 73
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2/8D2 (RE)
<400> 73
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Gly Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ala
115
<210> 74
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2/8D2 (RE)
<400> 74
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Gly Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Gln Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Phe Gly Ala Thr Asn Leu Ala Asp Gly Met Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Gln Tyr Ser Leu Lys Ile Ser Ser Leu His Pro
65 70 75 80
Asp Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Arg Ser Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105
<210> 75
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2/8D2 RE ВАРИАНТ 1 M18I
<400> 75
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Ile Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Gly Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ala
115
<210> 76
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2/8D2 RE ВАРИАНТ 1 M18I
<400> 76
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Gly Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Gln Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Phe Gly Ala Thr Asn Leu Ala Asp Gly Met Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Gln Tyr Ser Leu Lys Ile Ser Ser Leu His Pro
65 70 75 80
Asp Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Arg Ser Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105
<210> 77
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H1L1
<400> 77
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Met Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 78
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H1L1
<400> 78
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Gln Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Met Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Lys Ile Ser Ser Leu His Pro
65 70 75 80
Asp Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Arg Ser Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 79
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H1L1 ВАРИАНТ 1 M18I
<400> 79
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Ile Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 80
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H1L1 ВАРИАНТ 1 M18I
<400> 80
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Gln Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Met Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Lys Ile Ser Ser Leu His Pro
65 70 75 80
Asp Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Arg Ser Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 81
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H2L2
<400> 81
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Met Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 82
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H2L2
<400> 82
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Val Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Arg Ser Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 83
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H2L2 ВАРИАНТ 1 M18I
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Ile Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 84
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H2L2 ВАРИАНТ 1 M18I
<400> 84
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Val Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Arg Ser Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 85
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D3H3L3
<400> 85
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Glu Tyr Ala Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 86
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D3H3L3
<400> 86
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Leu Arg Ser Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 87
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H2L15
<400> 87
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Met Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 88
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H2L15
<400> 88
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Val Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Arg His Pro
85 90 95
Gly Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 89
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H2L15 ВАРИАНТ 1 M18I
<400> 89
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Ile Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 90
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H2L15 ВАРИАНТ 1 M18I
<400> 90
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Val Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Arg His Pro
85 90 95
Gly Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 91
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H2L17
<400> 91
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Met Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 92
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H2L17
<400> 92
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Val Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Ser Arg Pro
85 90 95
Gly Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 93
<211> 115
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH 8D2H2L17 ВАРИАНТ 1 M18I
<400> 93
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Ile Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Gly Val Tyr
85 90 95
Tyr Cys Thr Ala Gln Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 94
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL 8D2H2L17 ВАРИАНТ 1 M18I
<400> 94
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Glu Asn Ile Tyr Gly Gly
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Ser Gly Val Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Ser Arg Pro
85 90 95
Gly Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 95
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH REGN4659 W02019023482
<400> 95
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Glu Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Arg Thr Ser Gly Thr Thr Lys Tyr Tyr Ala Asp Ser Met
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Gly Thr Phe Leu His Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 96
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL REGN4659
<400> 96
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ala Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Lys Ser Phe Pro Met
85 90 95
Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 97
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH AGEN1884w
<400> 97
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Gly Leu Met Gly Pro Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 98
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL AGEN1884w
<400> 98
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Tyr
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Thr Arg Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 99
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH цемиплимаба-rwlc
<400> 99
Glu Val Gln Leu Leu Glu Ser Gly Gly Val Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Gly Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Gly Gly Arg Asp Thr Tyr Phe Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Lys Trp Gly Asn Ile Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 100
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL цемиплимаба-rwlc
<400> 100
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Ile Thr Ile Thr Cys Arg Ala Ser Leu Ser Ile Asn Thr Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu His Gly Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Thr Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Ser Asn Thr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Val Val Asp Phe Arg
100 105
<210> 101
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC цемиплимаба-rwlc (S228P)
<400> 101
Glu Val Gln Leu Leu Glu Ser Gly Gly Val Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Gly Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Gly Gly Arg Asp Thr Tyr Phe Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Lys Trp Gly Asn Ile Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 102
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC цемиплимаба-rwlc
<400> 102
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Ile Thr Ile Thr Cys Arg Ala Ser Leu Ser Ile Asn Thr Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu His Gly Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Thr Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Ser Asn Thr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Val Val Asp Phe Arg Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 103
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH дурвалумаба
<400> 103
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 104
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL дурвалумаба
<400> 104
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 105
<211> 80
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH авелумаба
<400> 105
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Tyr Pro Ser
1 5 10 15
Gly Gly Ile Thr Phe Tyr Ala Asp Lys Gly Arg Phe Thr Ile Ser Arg
20 25 30
Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
35 40 45
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ile Lys Leu Gly Thr Val
50 55 60
Thr Thr Val Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
65 70 75 80
<210> 106
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL авелумаба
<400> 106
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Arg Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 107
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH атезолизумаба
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 108
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL атезолизумаба
<400> 108
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 109
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC дурвалумаба (IgG1 L234F/L235E/P331S/D356E/L358M)
<400> 109
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 110
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC дурвалумаба
<400> 110
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 111
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC авелумаба (IgG1)
<400> 111
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ile Met Met Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Pro Ser Gly Gly Ile Thr Phe Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Lys Leu Gly Thr Val Thr Thr Val Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 112
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC авелумаба
<400> 112
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Arg Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 113
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HC атезолизумаба (IgG1 N297A/D235E/L358M)
<400> 113
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 114
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LC атезолизумаба
<400> 114
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 115
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (N297A/D356E/L358M)
<400> 115
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 116
<211> 317
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен HC IgG1 человека (L234F/L235E/P331S/D356E/L358M)
<400> 116
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Leu Thr Cys Leu Val Lys
225 230 235 240
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
245 250 255
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
260 265 270
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
275 280 285
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
290 295 300
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
305 310 315
<210> 117
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Константный домен лямбда LC человека
<400> 117
Gly Gln Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2766199C2 |
| КОМБИНИРОВАННЫЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА, НАЦЕЛЕННЫЕ НА PD-1, TIM-3 И LAG-3 | 2018 |
|
RU2795232C2 |
| ИММУНОГЛОБУЛИН С ТАНДЕМНЫМИ FAB-ФРАГМЕНТАМИ И ВАРИАНТЫ ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2767329C2 |
| МУЛЬТСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, МУЛЬТСПЕЦИФИЧЕСКИЕ АКТИВИРУЕМЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2815683C2 |
| СВЯЗЫВАЮЩИЕСЯ С PD-1 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2812280C1 |
| АНТИТЕЛА ПРОТИВ PD-1 И КОМПОЗИЦИИ | 2016 |
|
RU2750675C1 |
| СВЯЗЫВАЮЩИЕСЯ С PD-1 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2773758C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| АНТИТЕЛА ПРОТИВ PD-L1 И ИХ ВАРИАНТЫ | 2017 |
|
RU2770590C2 |
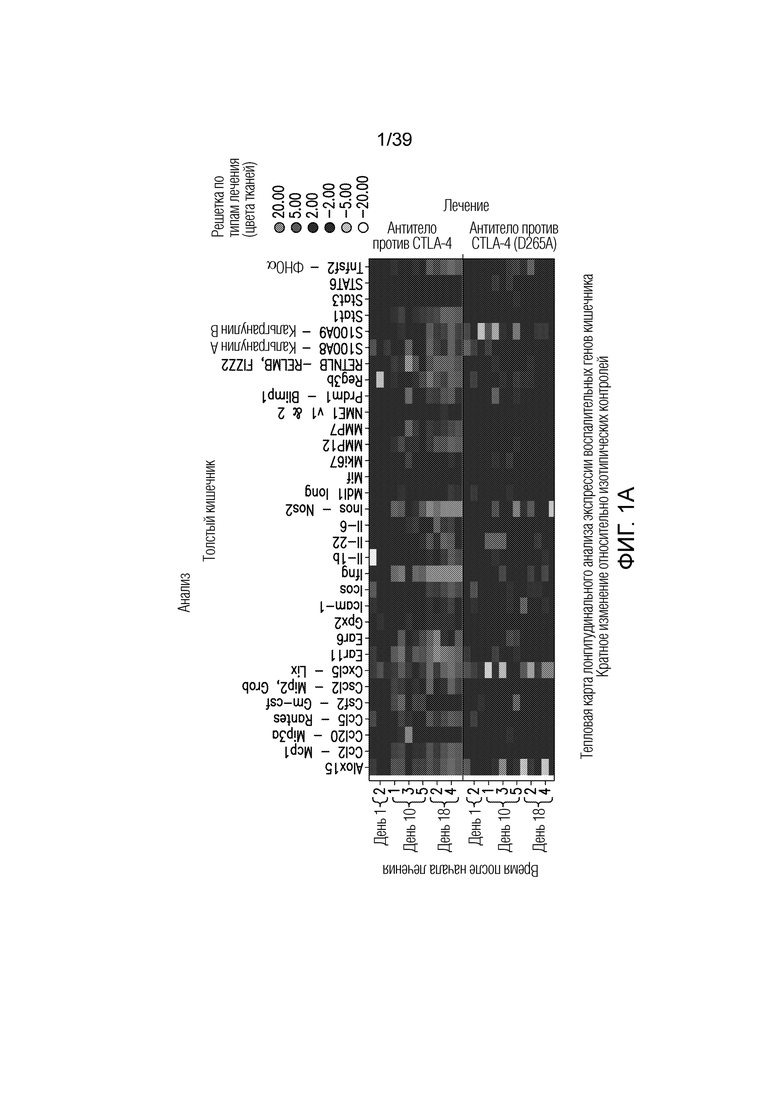
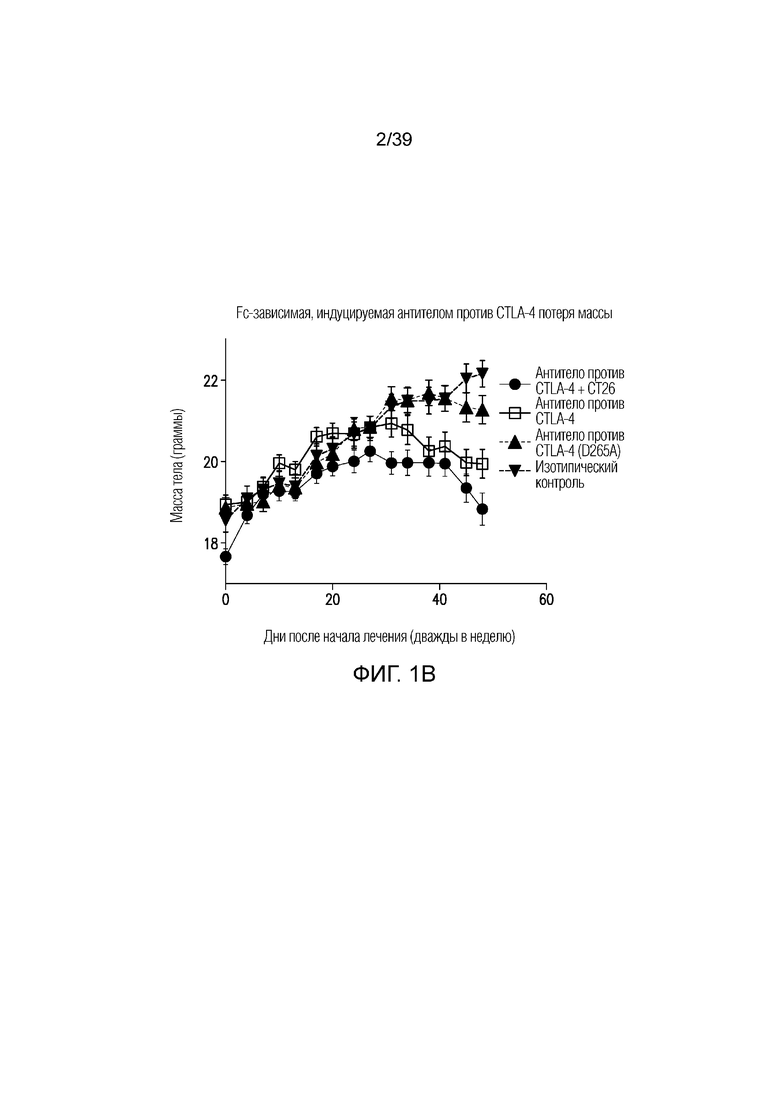
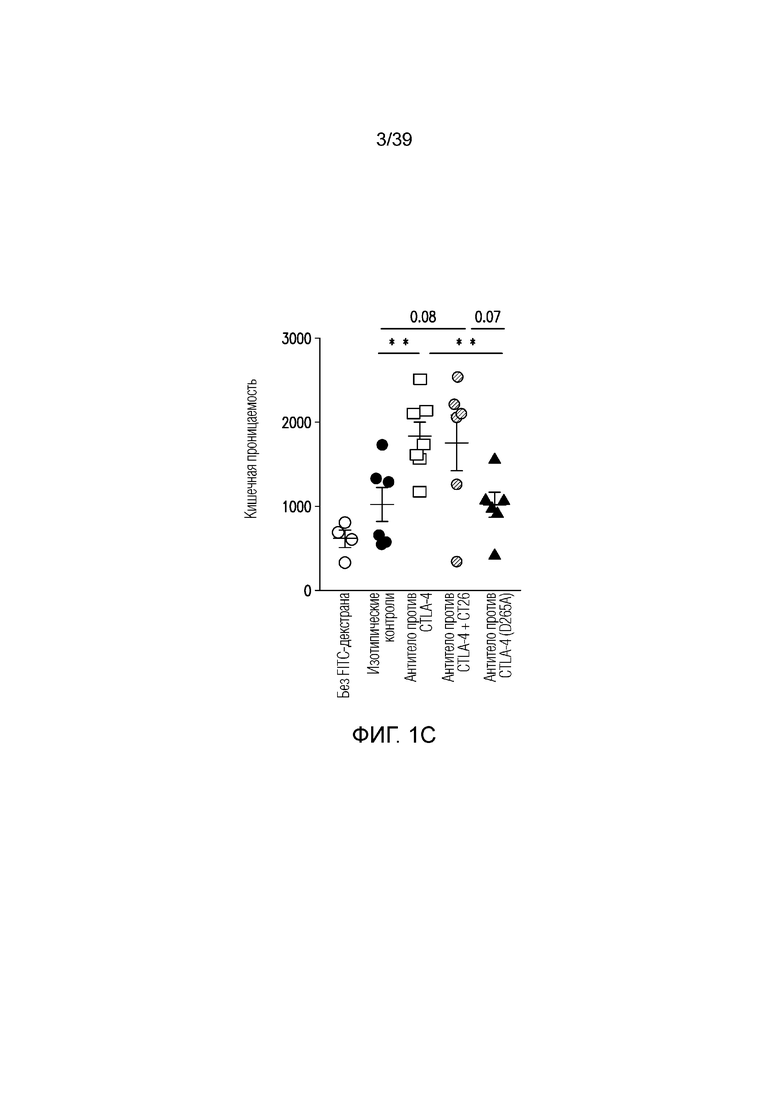
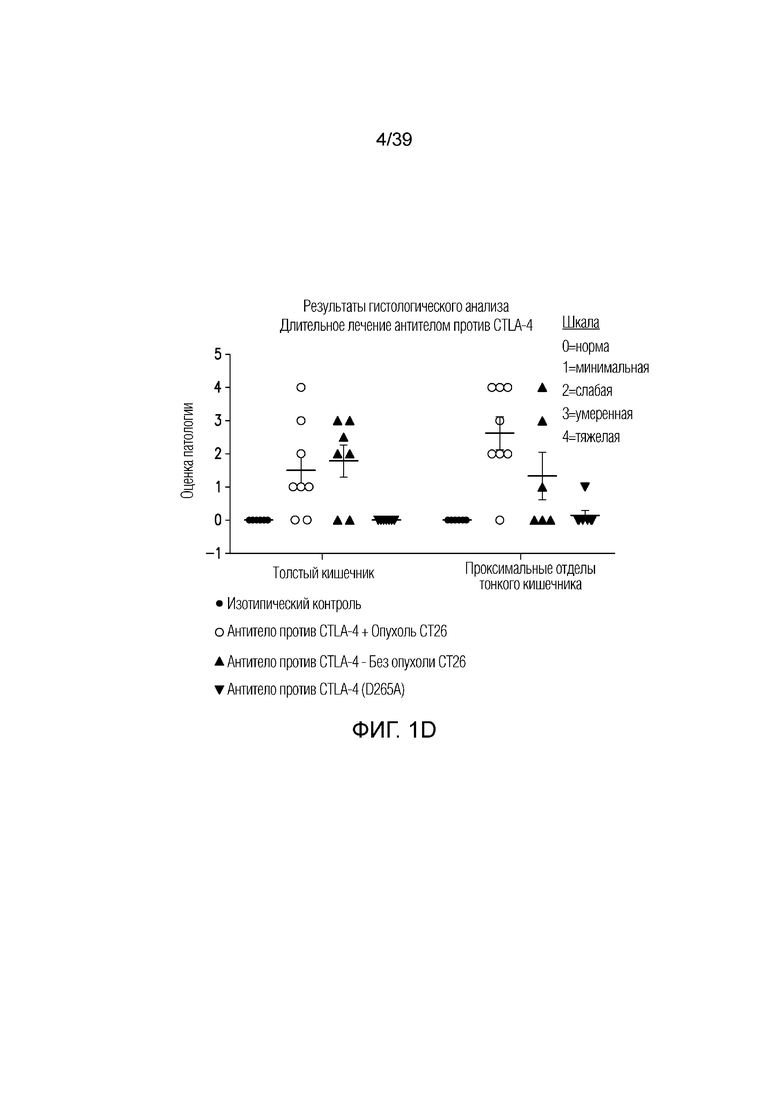
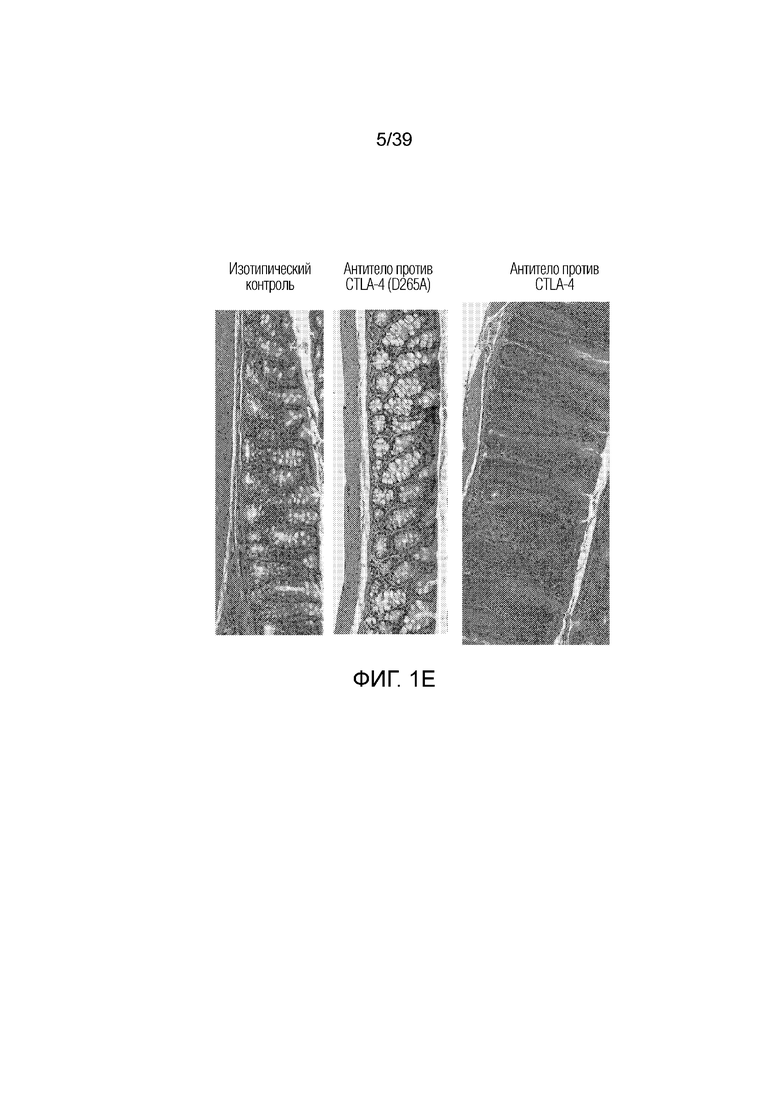
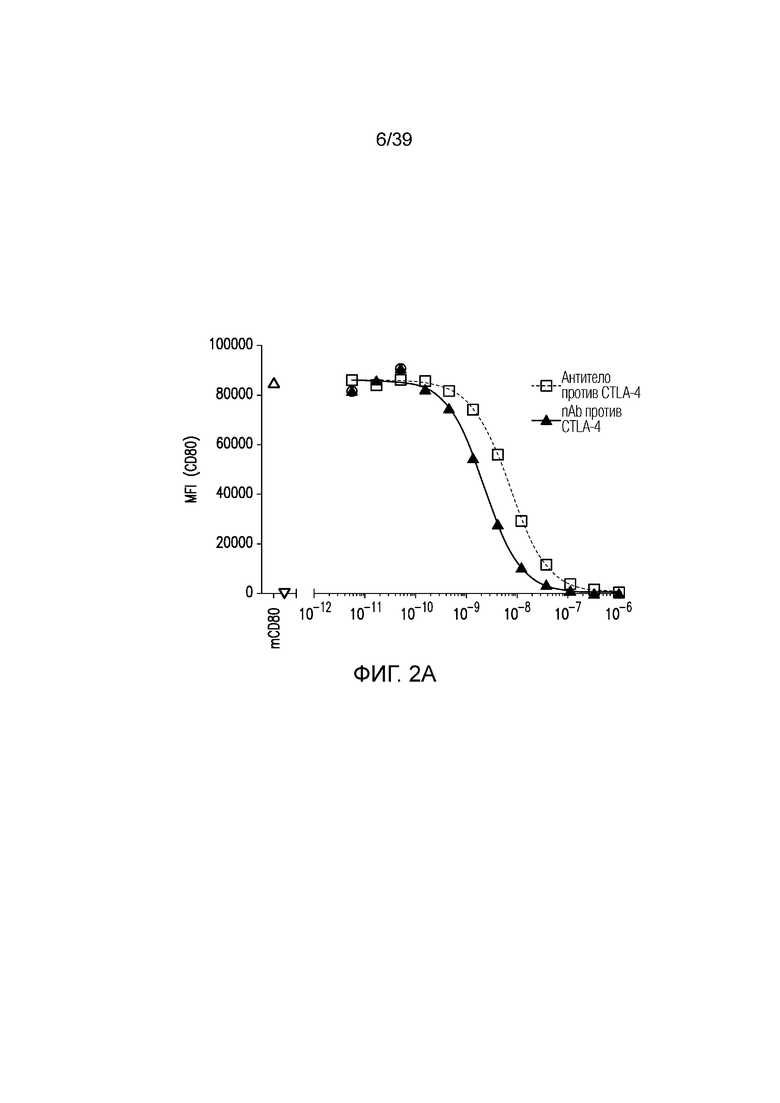
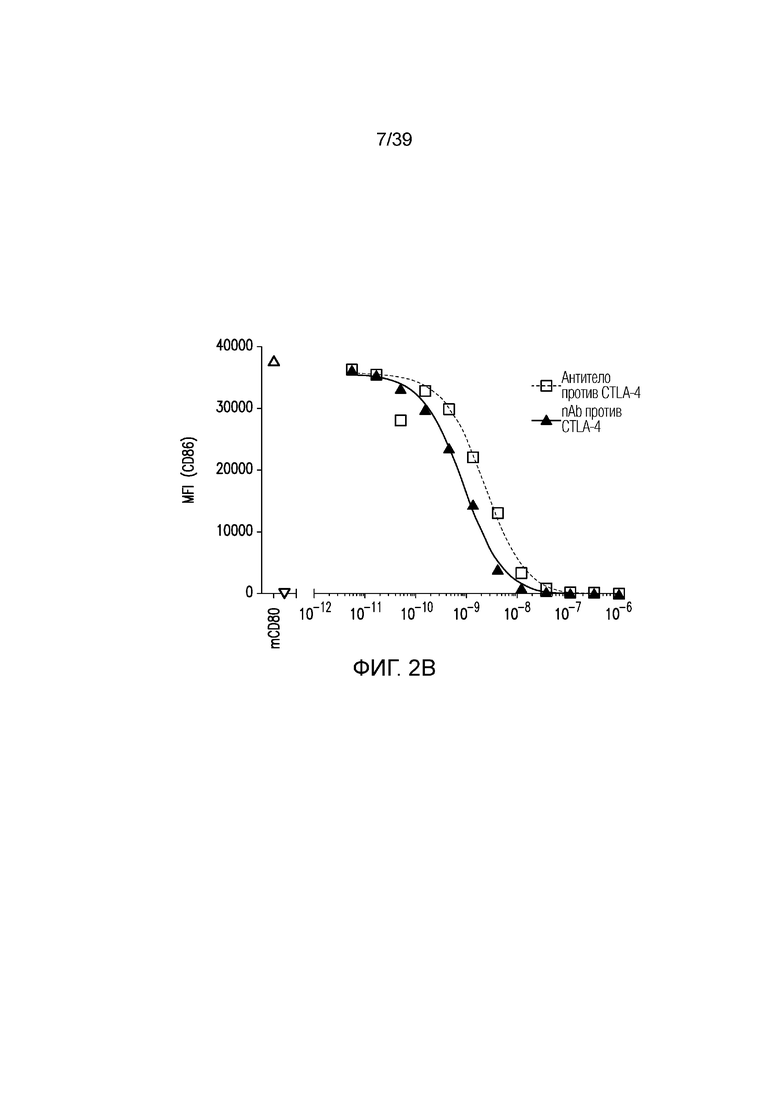
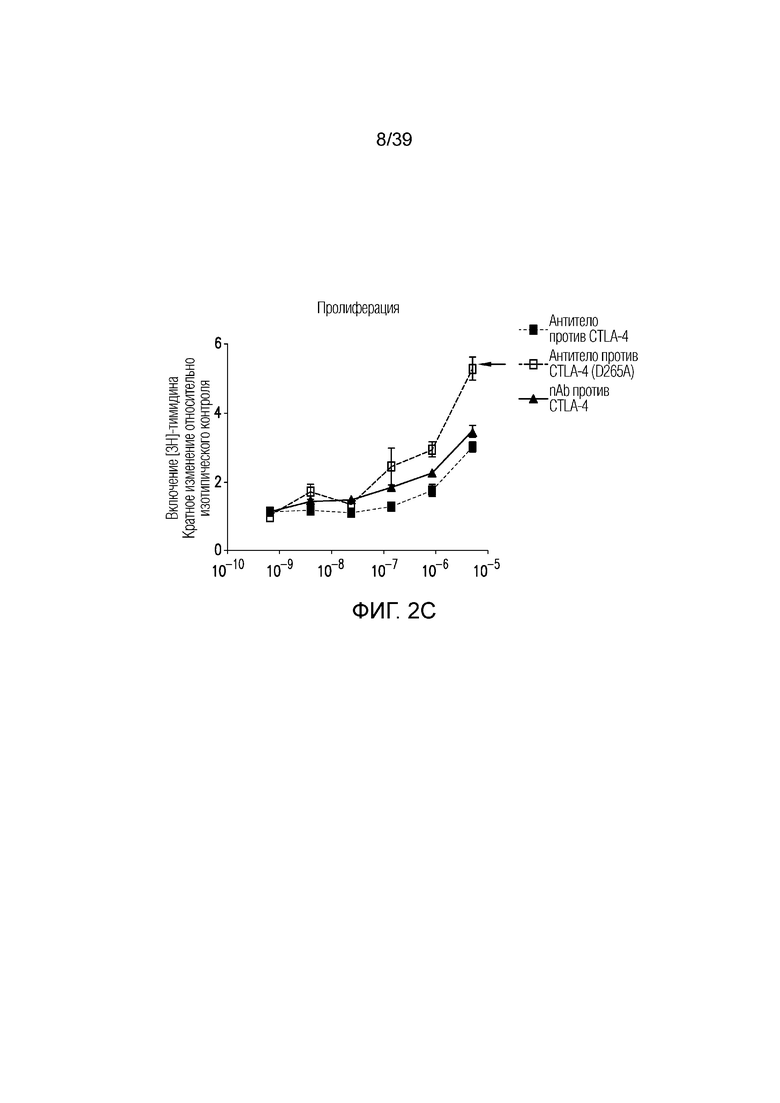
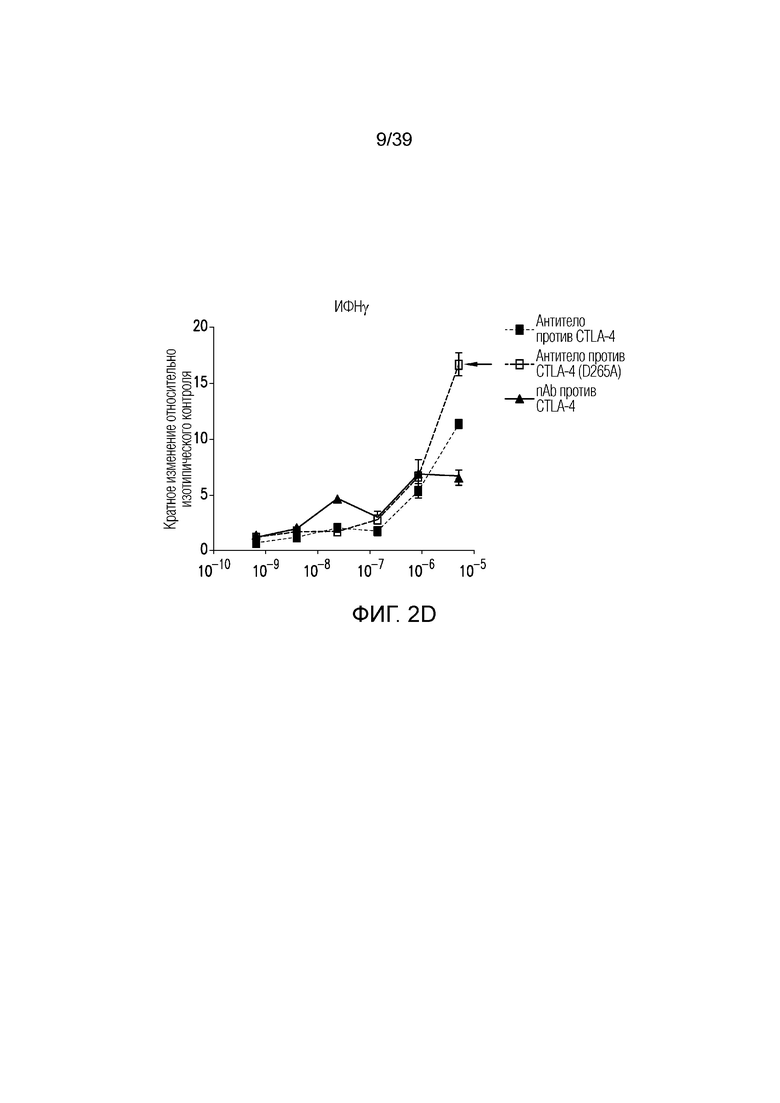
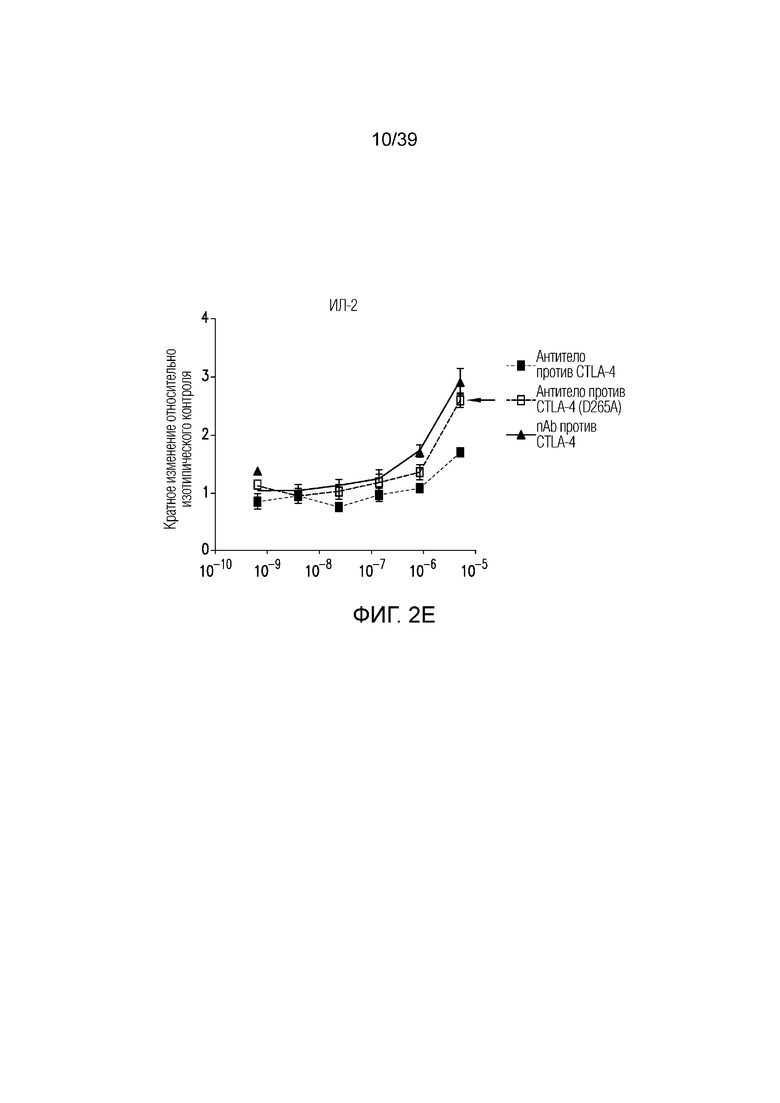
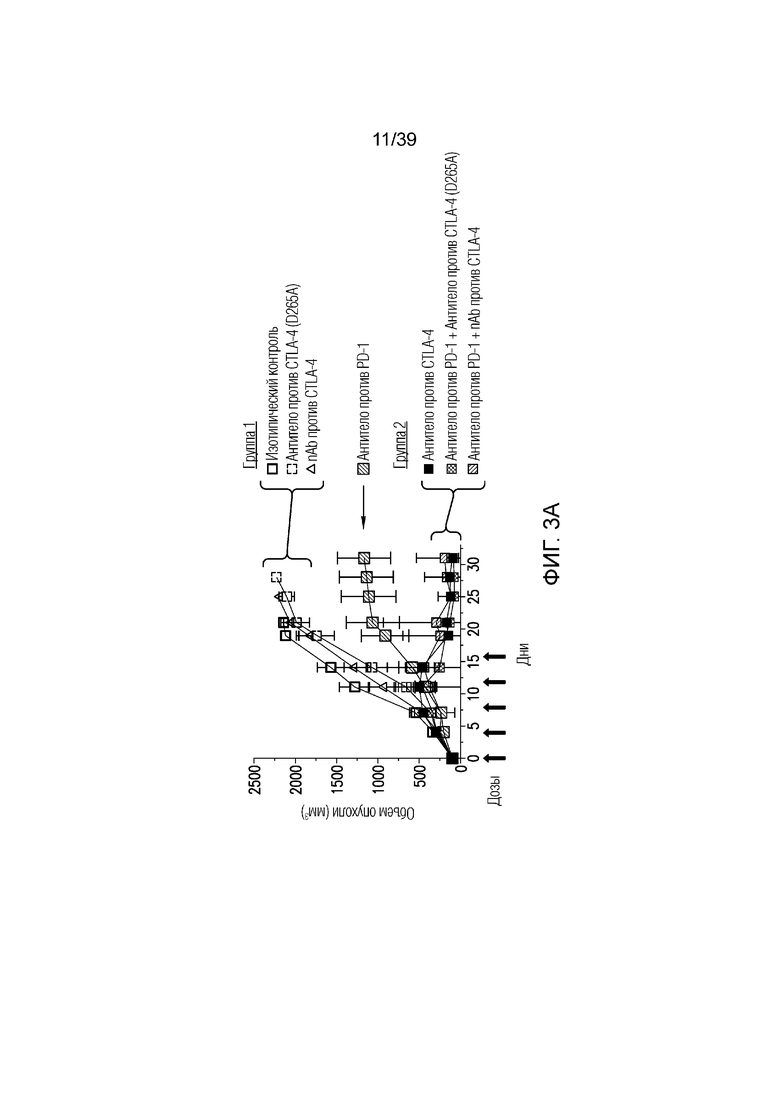
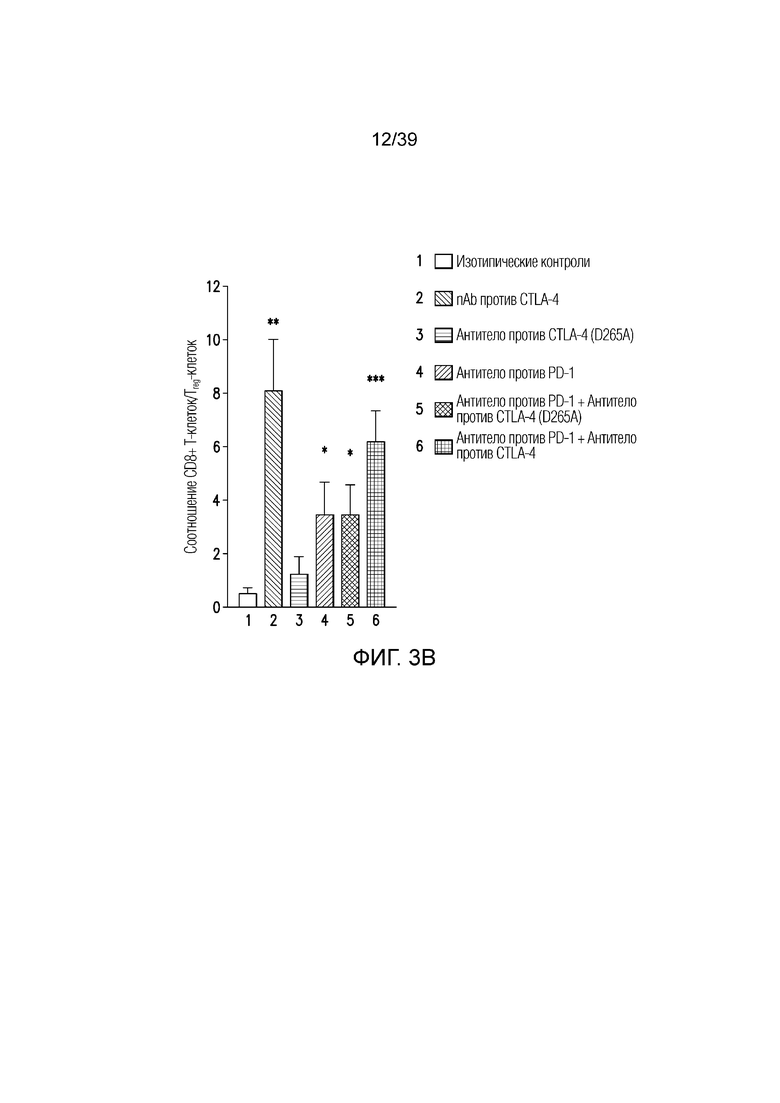

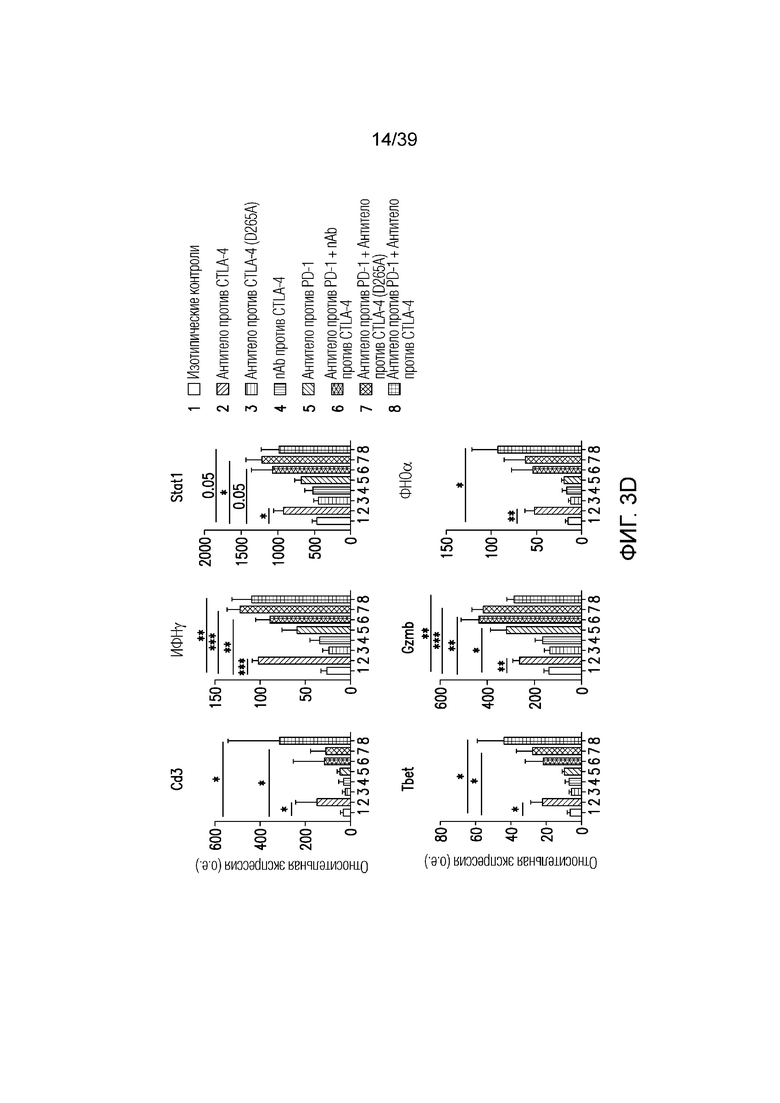
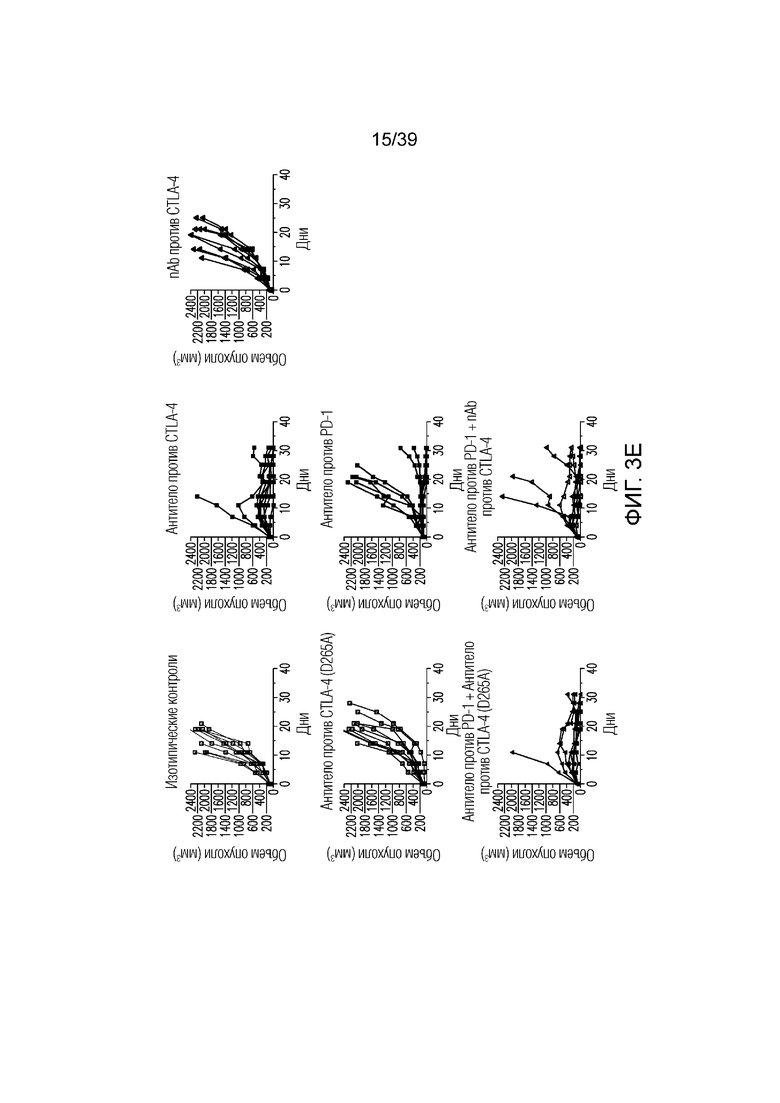
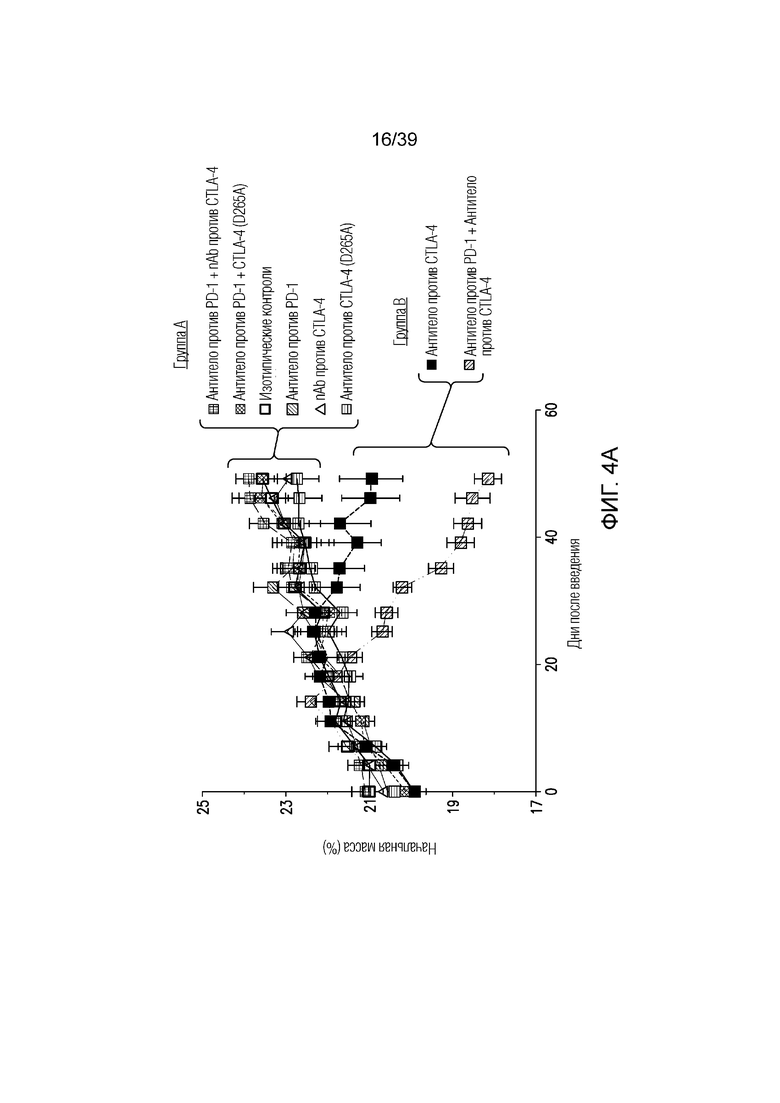
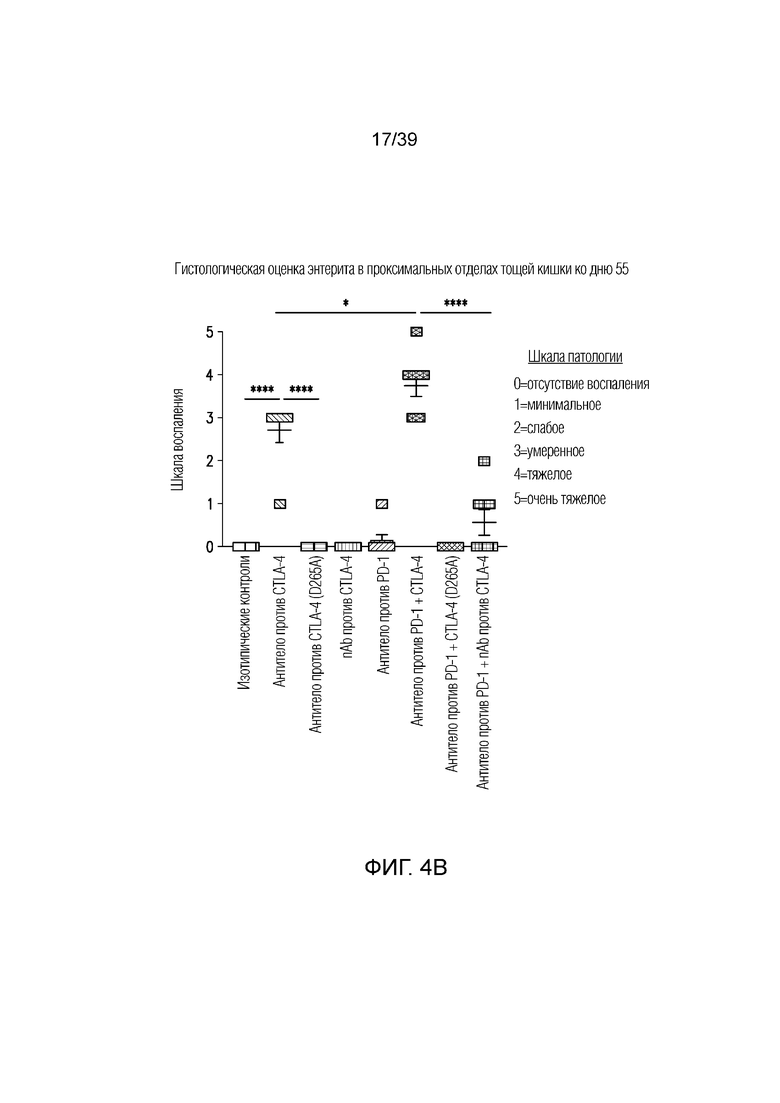
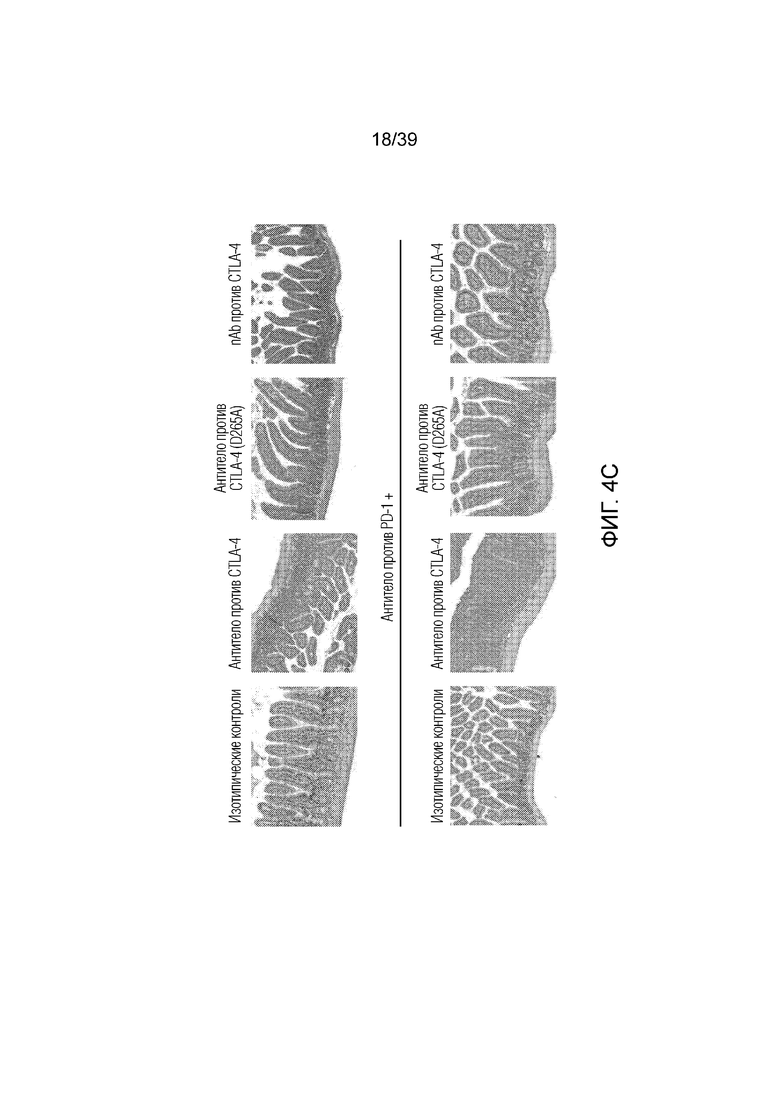


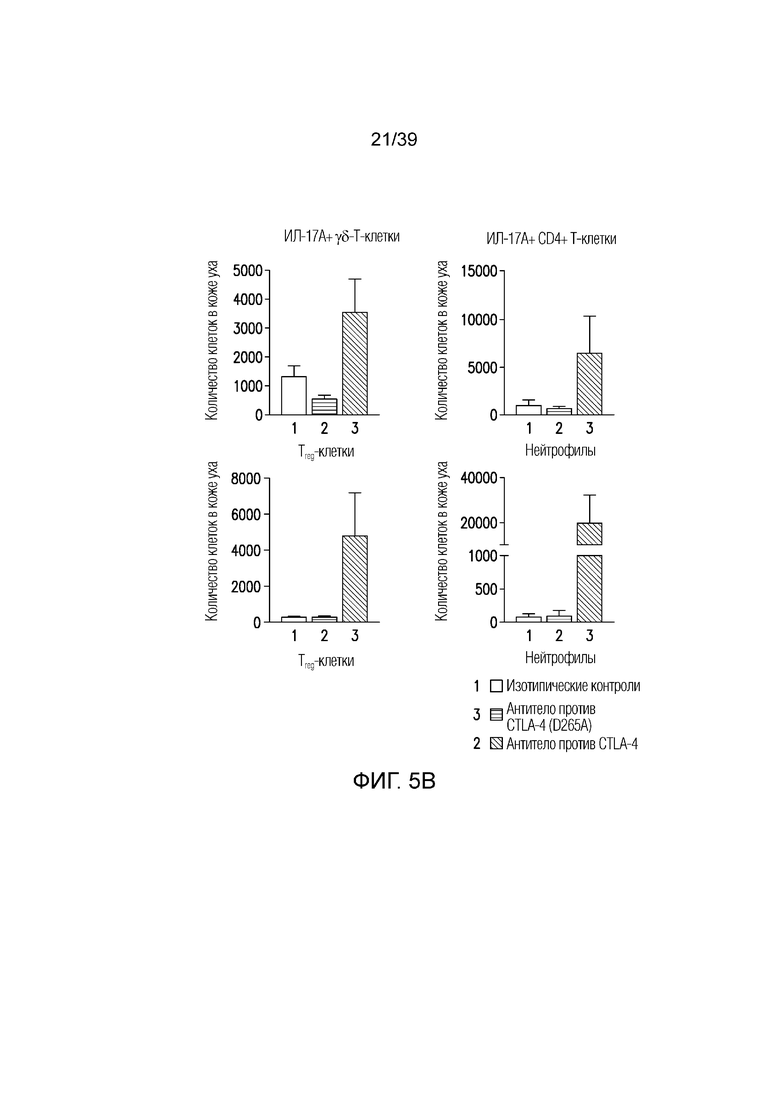
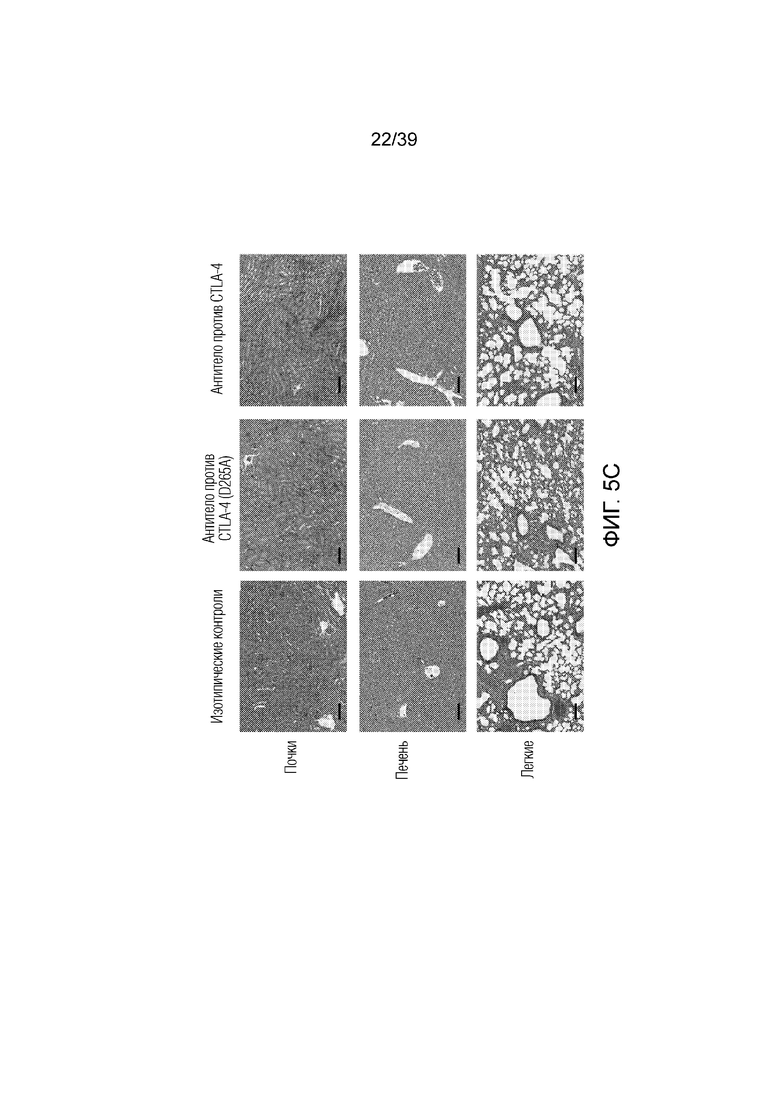
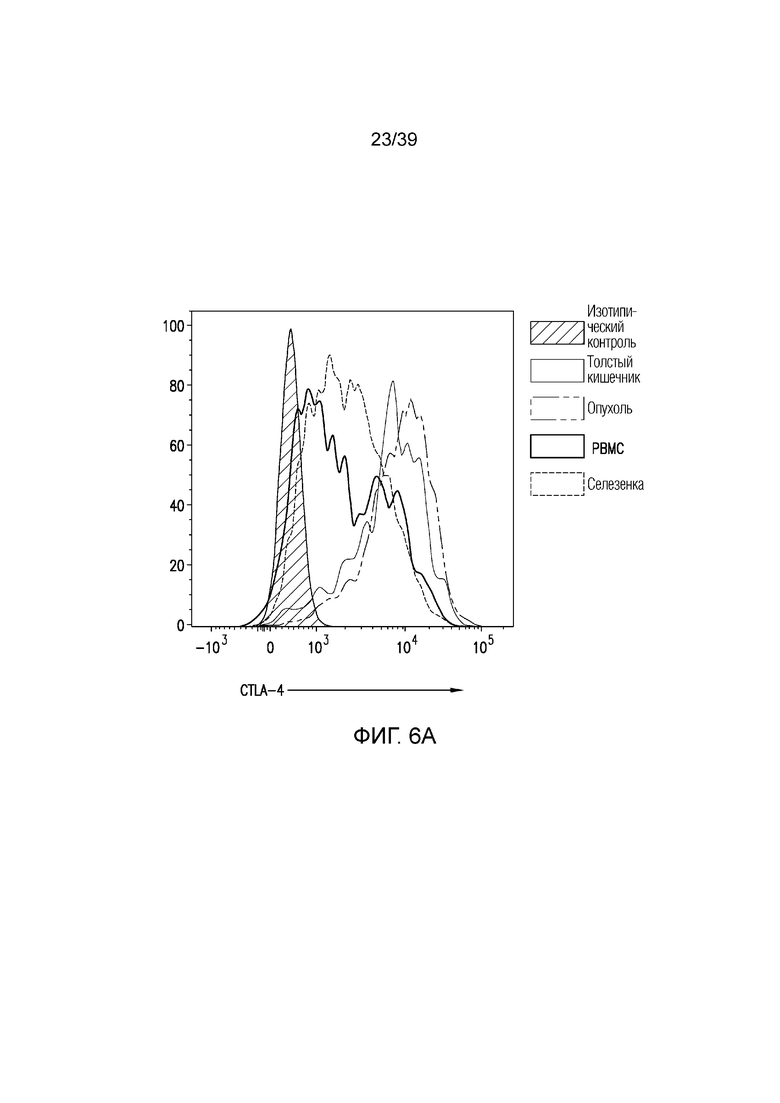
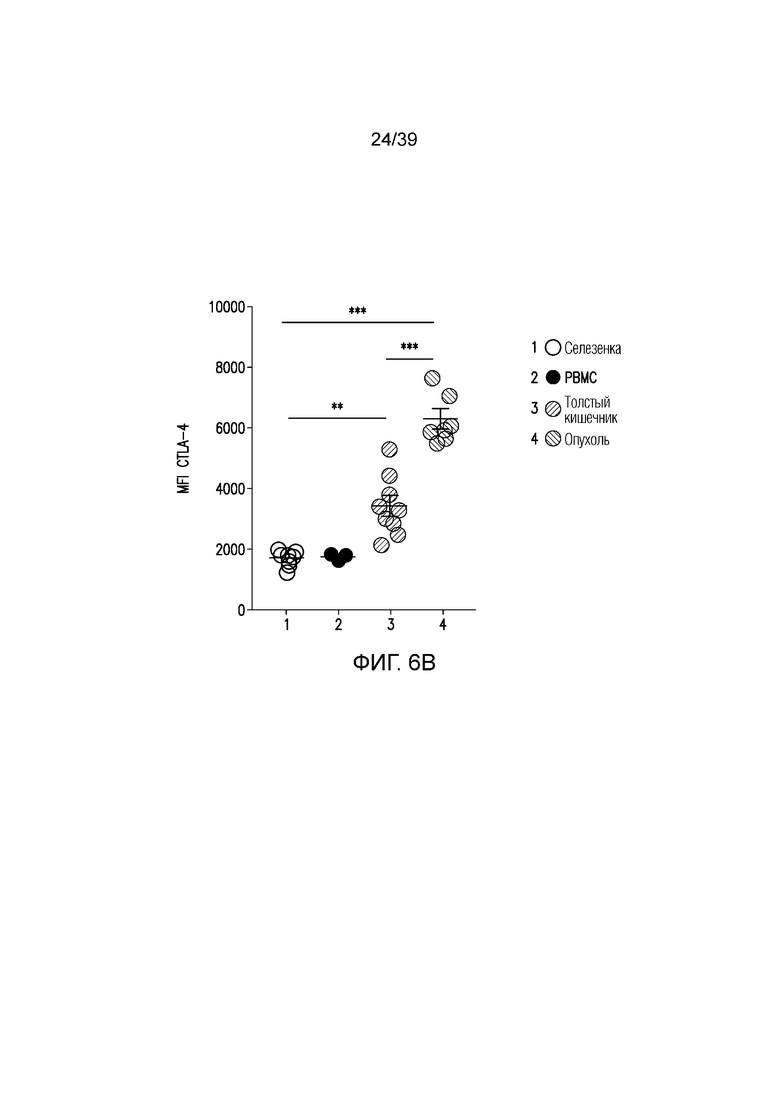
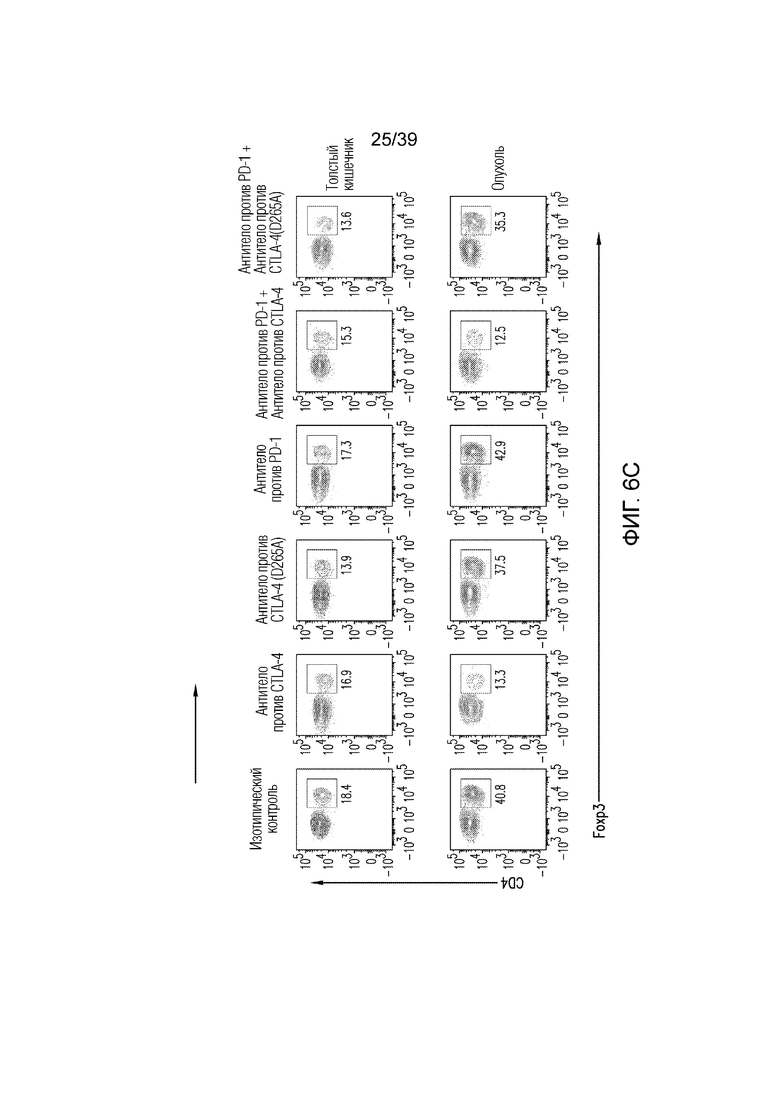
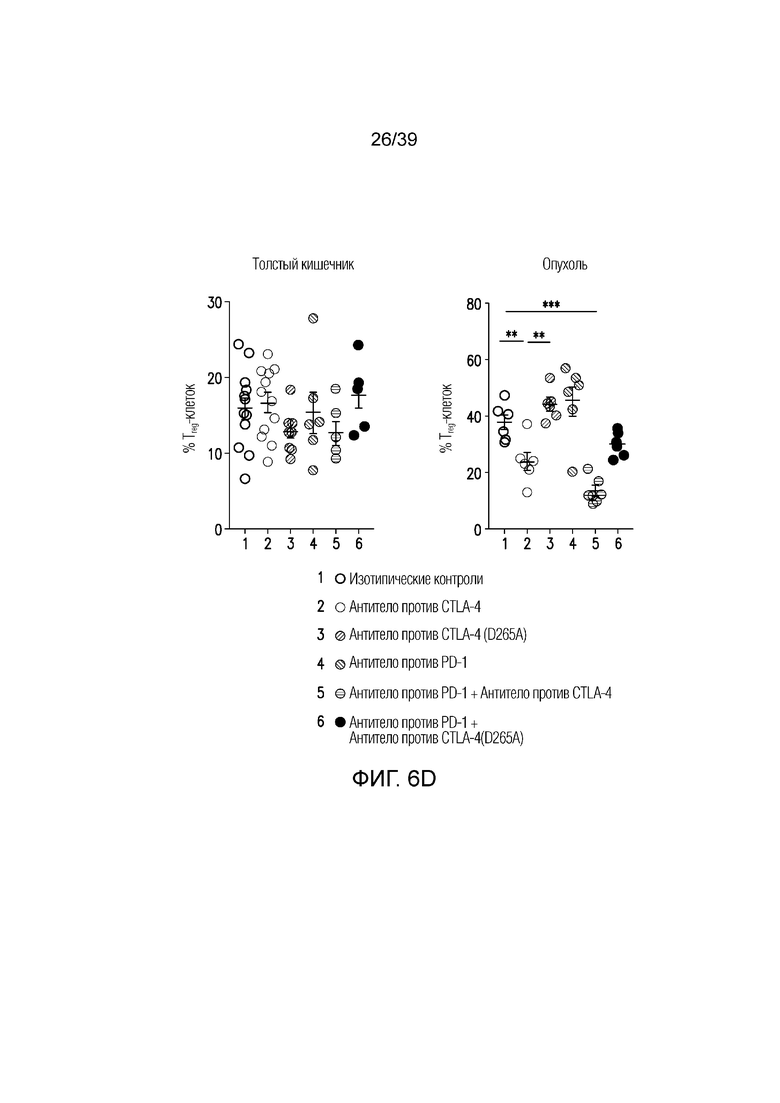
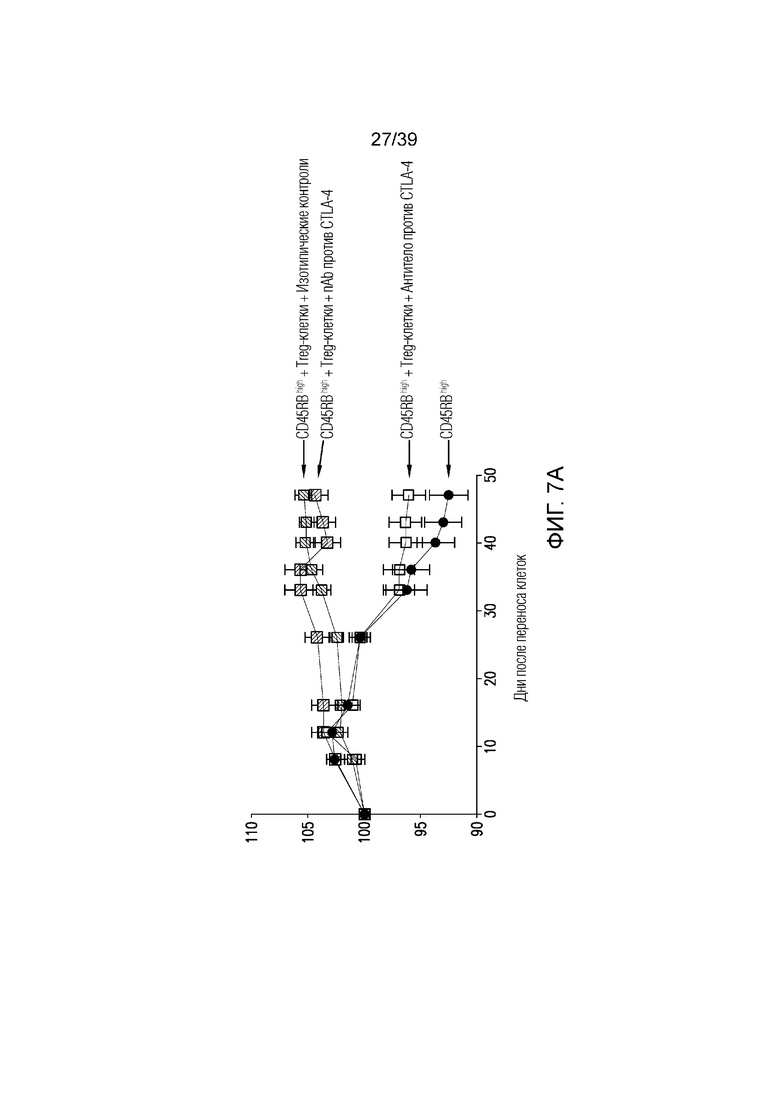

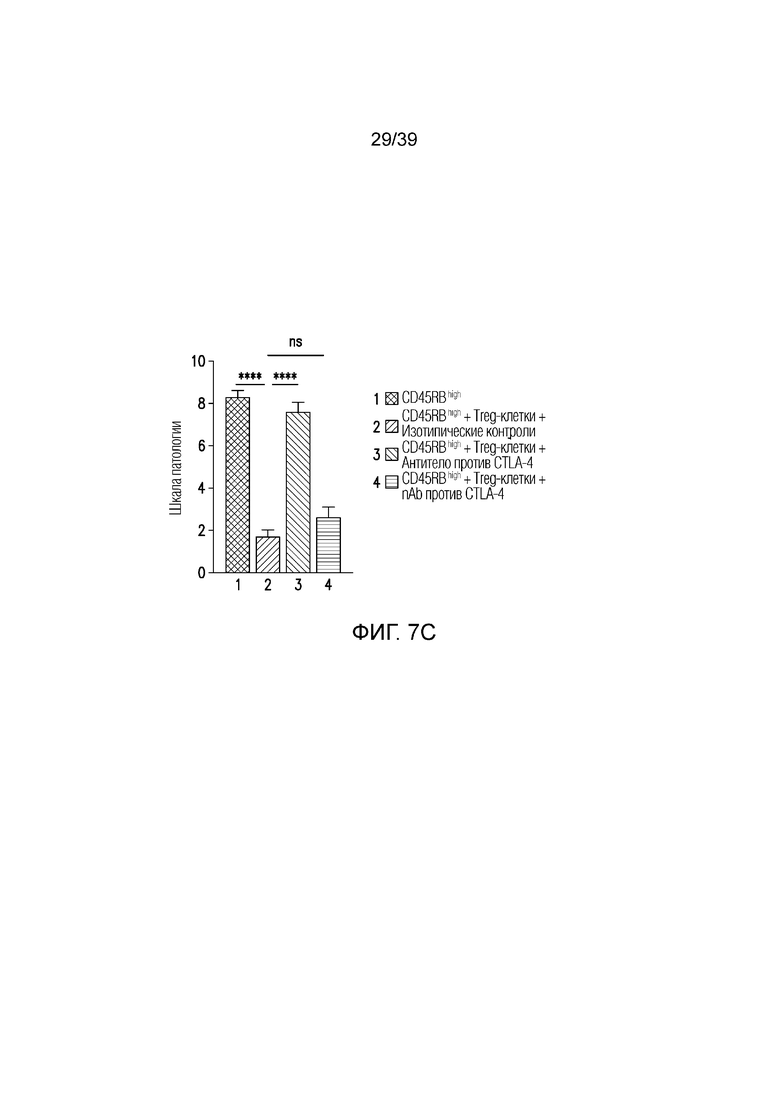
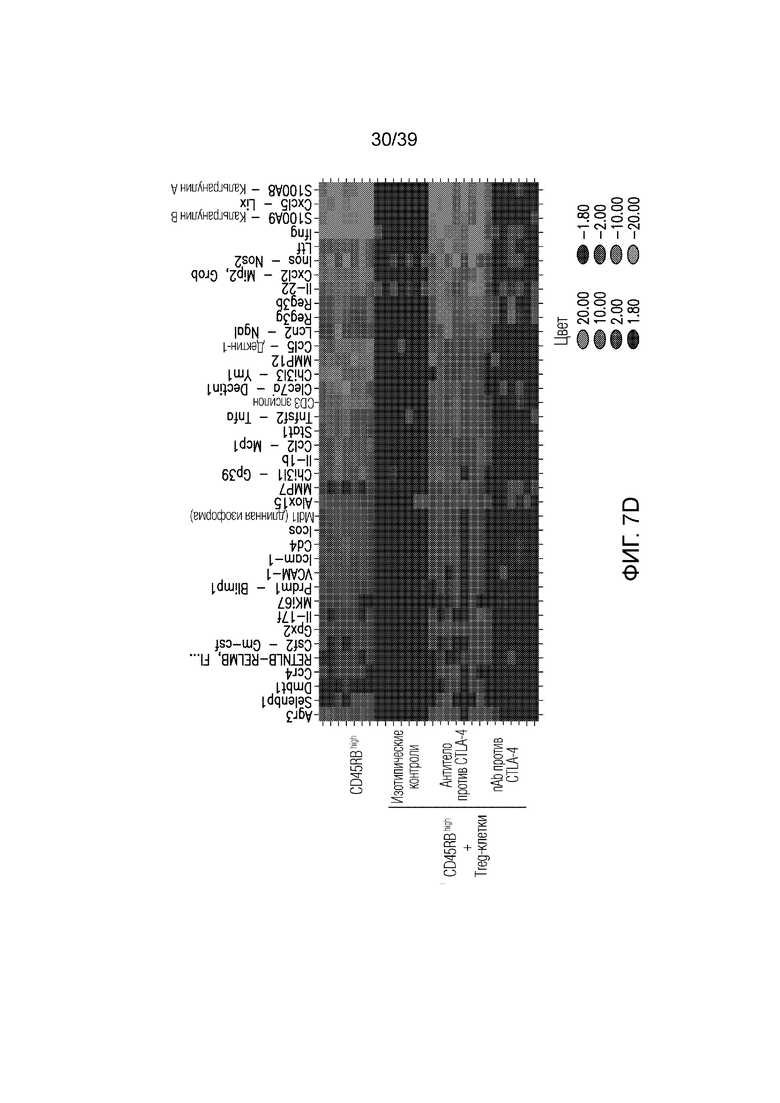
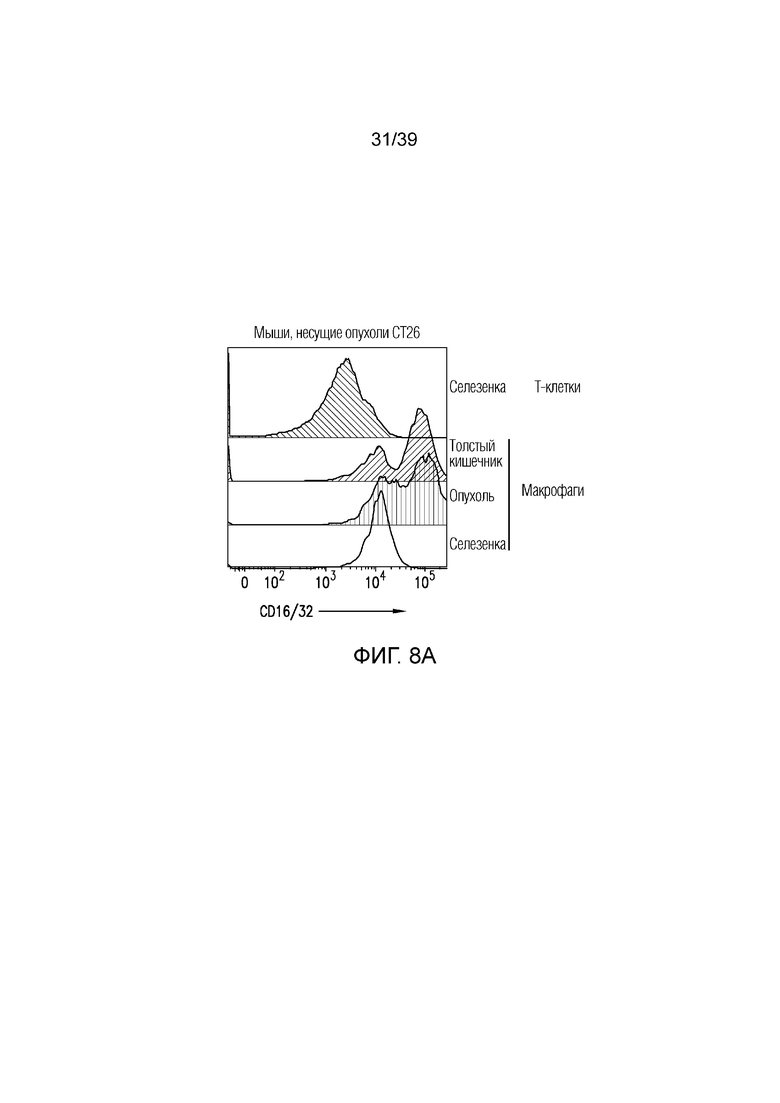
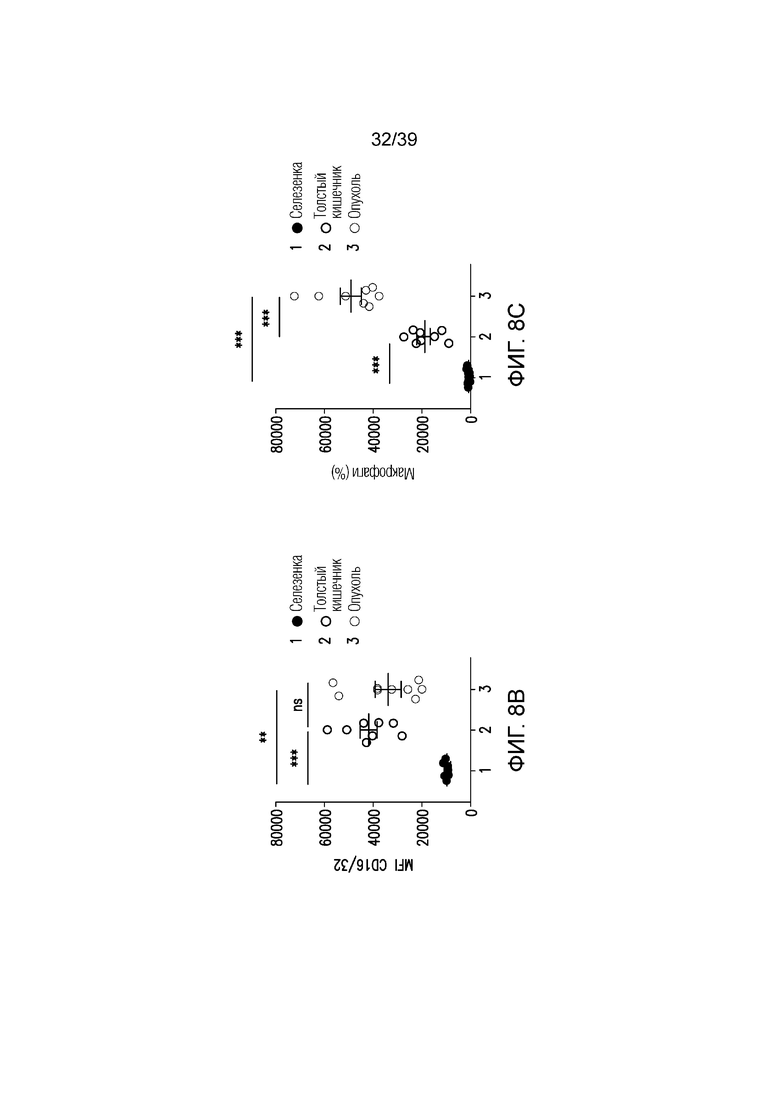
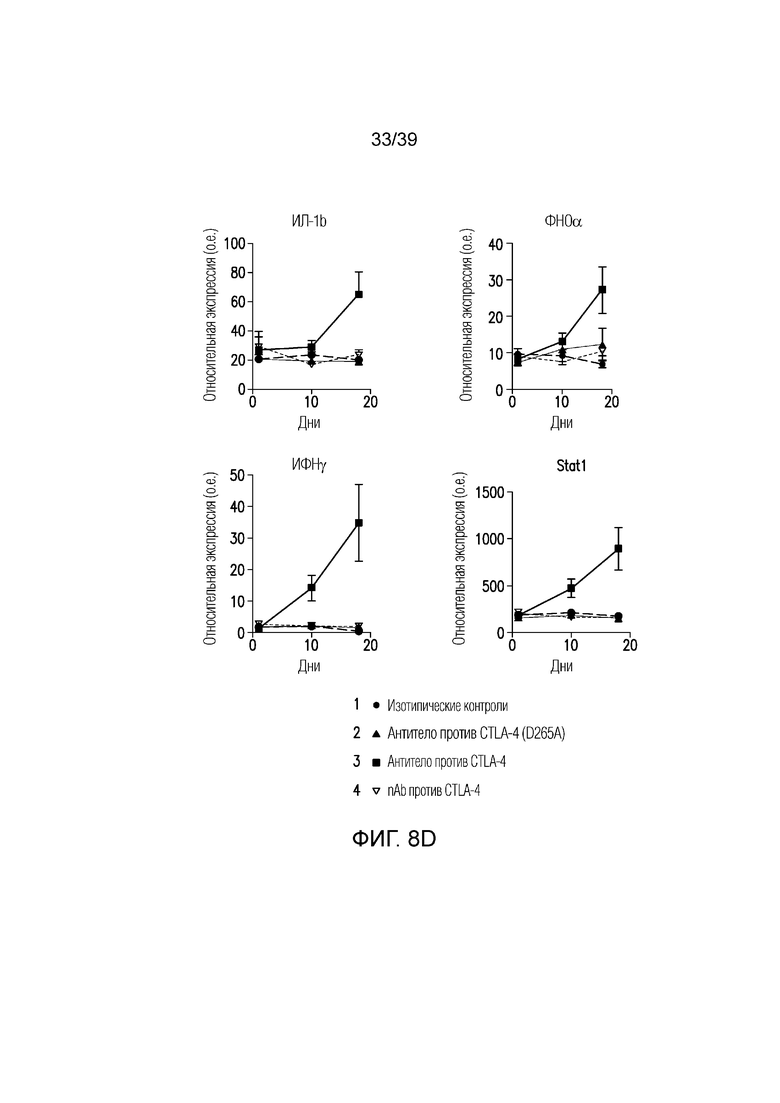

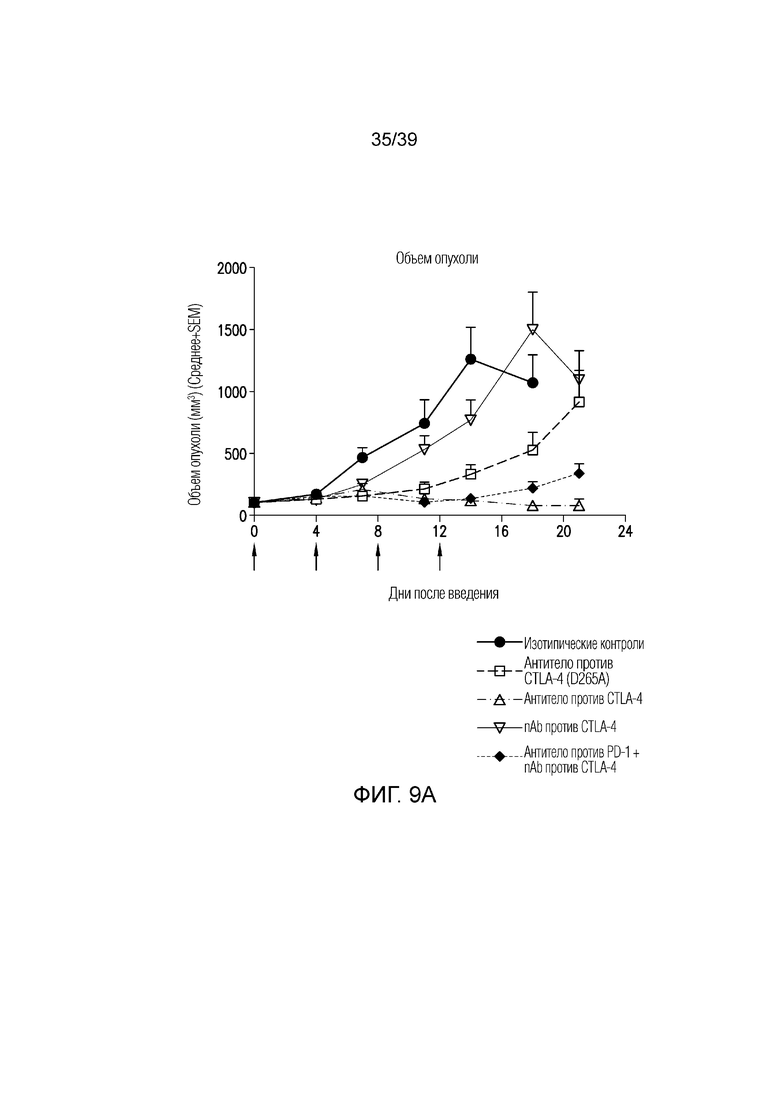
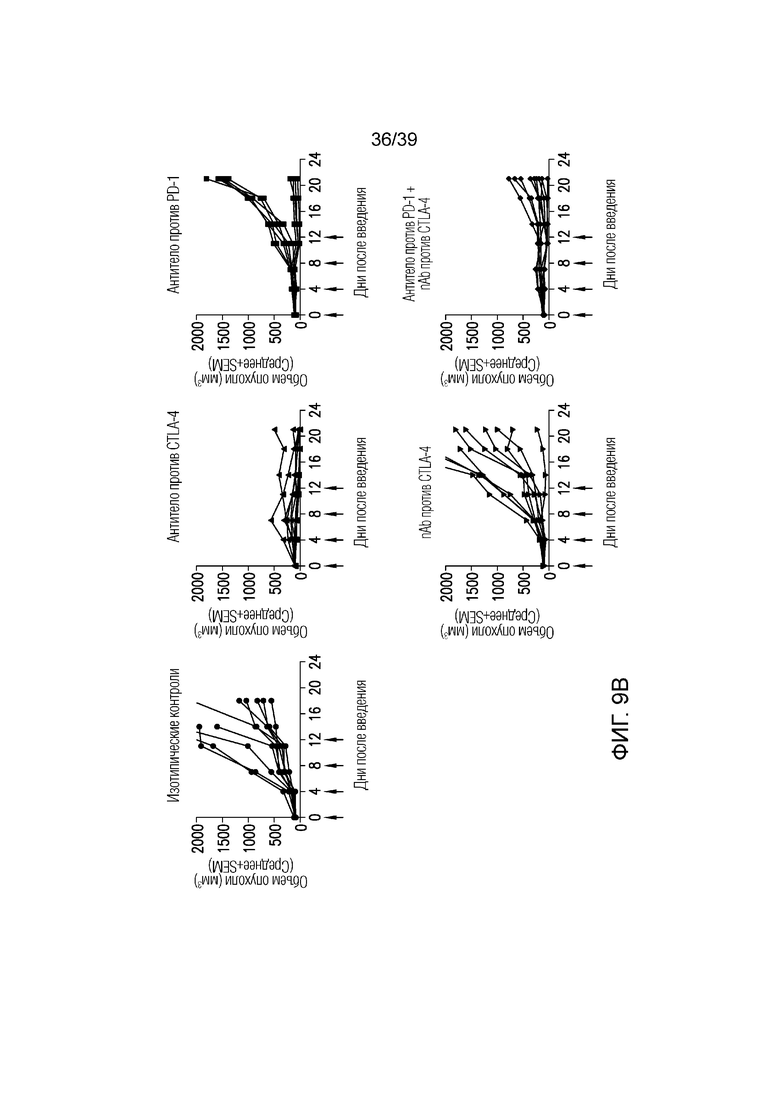
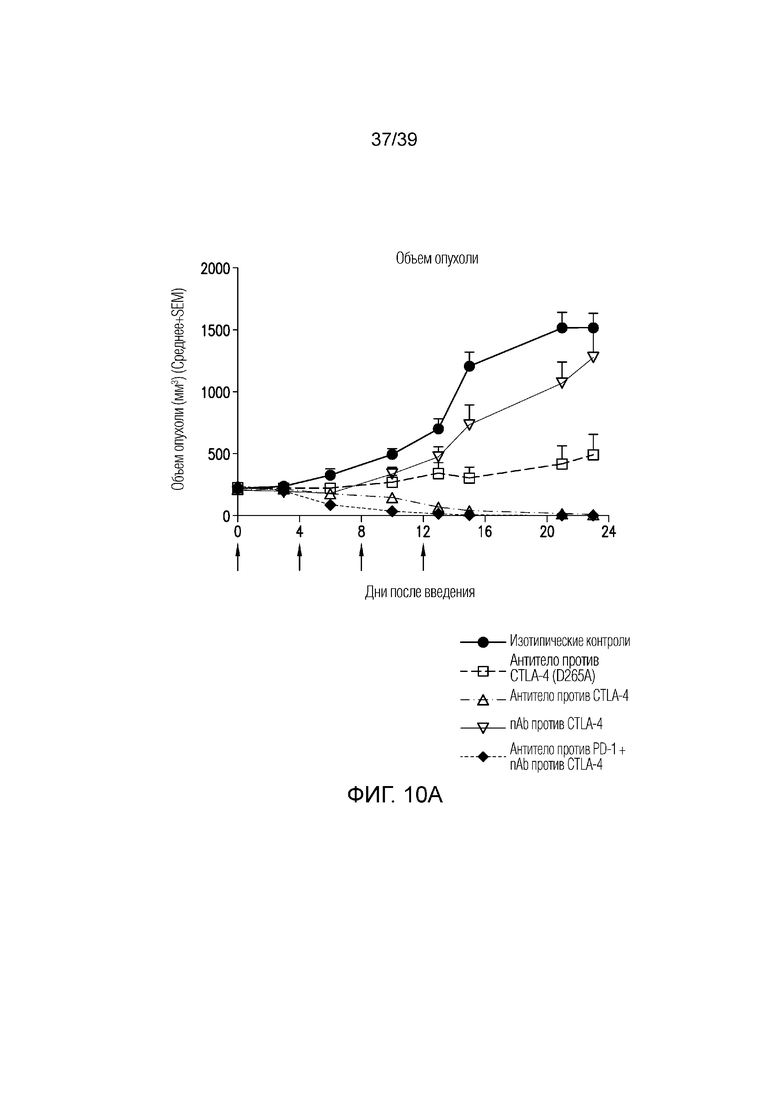
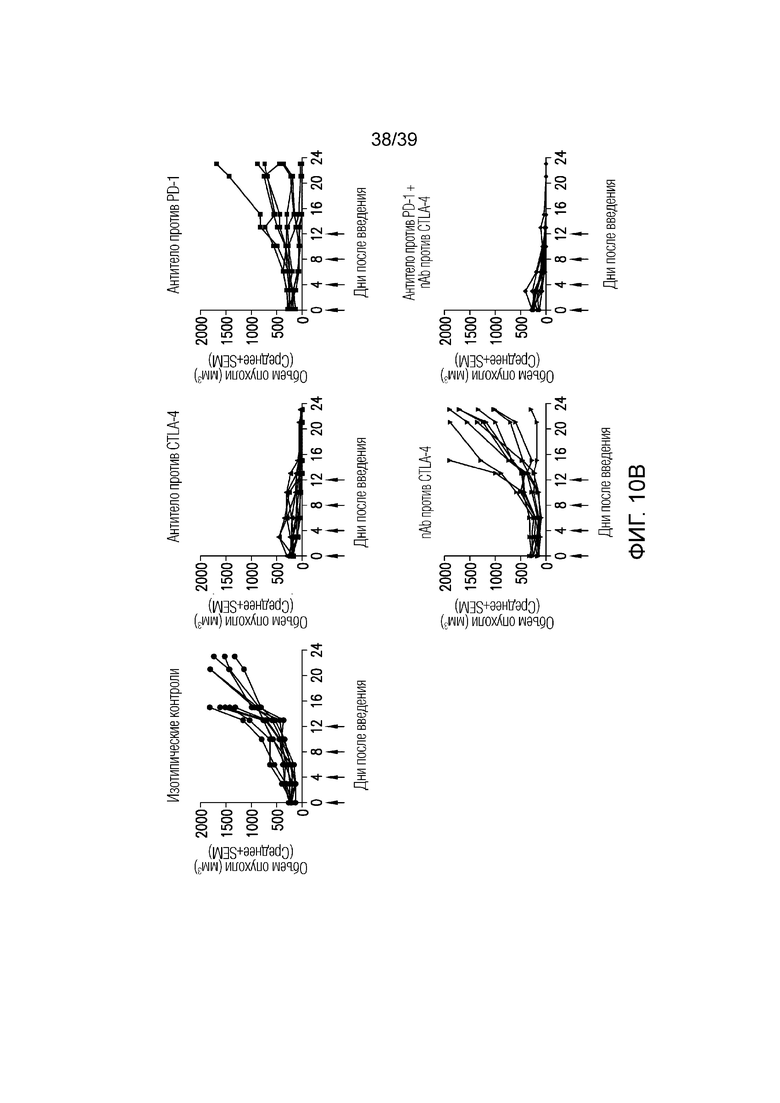
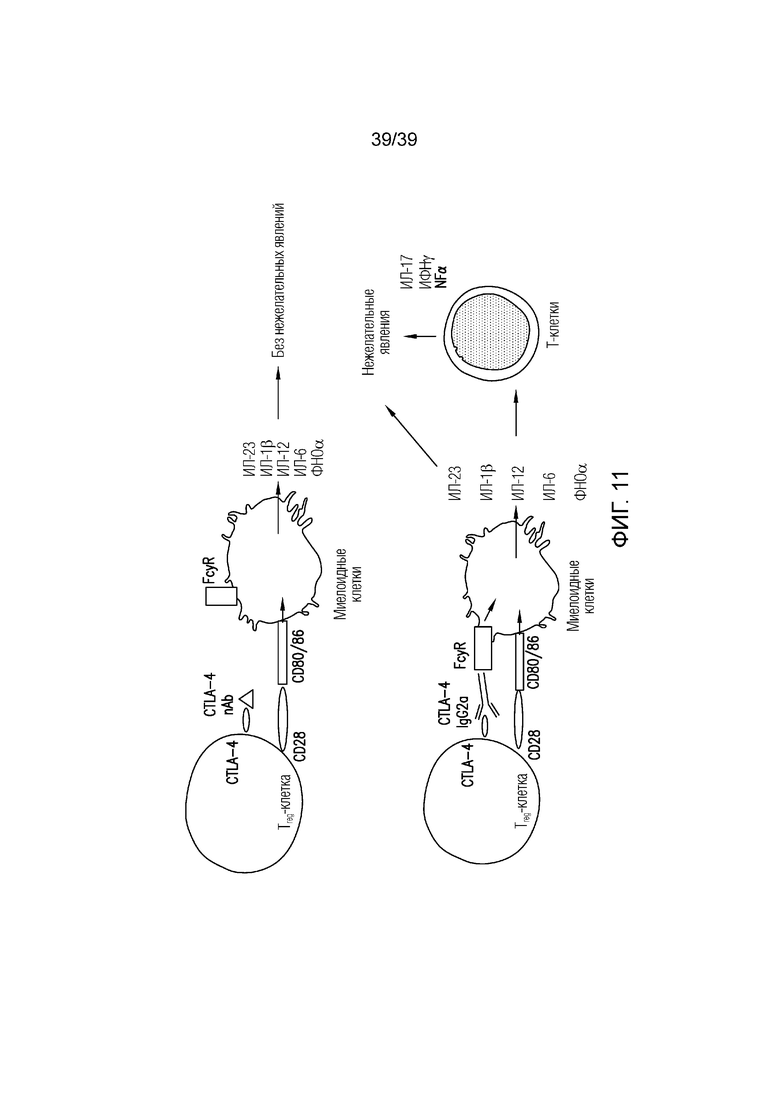
Изобретение относится к области медицины и фармацевтики, а именно к способу лечения злокачественного новообразования у индивидуума, нуждающегося в таком лечении, с использованием комбинации антитела против PD-1 и антитела против CTLA-4, которая не вызывает от умеренной до тяжелой воспалительной иммуноопосредованной токсичности в отношении кожи или кишечника, где указанный способ включает введение комбинации антитела против PD-1, содержащего тяжелую цепь (HC), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 27, и легкую цепь (LC), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 28, и антитела против CTLA-4, содержащего (i) HC, имеющую вариабельный домен HC (VH), содержащий аминокислотную последовательность, представленную в SEQ ID NO: 7, и константный домен HC, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 38, 39, 40, 41, 42, 43 или 44, и (ii) LC, имеющую вариабельный домен LC (VL), содержащий аминокислотную последовательность, представленную в SEQ ID NO: 8, и константный домен LC, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 57. Технический результат заключается в противоопухолевой активности антитела против CTLA-4 при совместном введении с антителом против PD-1 без иммуноопосредованной токсичности в отношении кожи и кишечника. 5 з.п. ф-лы, 11 ил., 36 табл., 3 пр.
1. Способ лечения злокачественного новообразования у индивидуума, нуждающегося в таком лечении, с использованием комбинации антитела против PD-1 и антитела против CTLA-4, которая не вызывает от умеренной до тяжелой воспалительной иммуноопосредованной токсичности в отношении кожи или кишечника, где указанный способ включает введение комбинации антитела против PD-1, содержащего тяжелую цепь (HC), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 27, и легкую цепь (LC), содержащую аминокислотную последовательность, представленную в SEQ ID NO: 28, и антитела против CTLA-4, содержащего (i) HC, имеющую вариабельный домен HC (VH), содержащий аминокислотную последовательность, представленную в SEQ ID NO: 7, и константный домен HC, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 38, 39, 40, 41, 42, 43 или 44, и (ii) LC, имеющую вариабельный домен LC (VL), содержащий аминокислотную последовательность, представленную в SEQ ID NO: 8, и константный домен LC, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 57.
2. Способ по п.1, где антитело против CTLA-4 вводят в дозе, содержащей более 1 мг/кг, или фиксированной дозе, не зависящей от массы тела индивидуума, составляющей 100 мг или более.
3. Способ по п.1, где антитело против PD-1 вводят в дозе, содержащей (i) 2 или 3 мг/кг, или фиксированную дозу, не зависящую от массы тела индивидуума, составляющую 200 мг, (ii) более 3 мг/кг или фиксированную дозу, не зависящую от массы тела индивидуума, составляющую более 200 мг, или (iii) фиксированную дозу, не зависящую от массы тела индивидуума, составляющую от приблизительно 200 мг до приблизительно 400 мг.
4. Способ по п.1, где антитело против PD-1 вводят индивидууму первым и антитело против CTLA-4 вводят индивидууму вторым; антитело против CTLA-4 вводят индивидууму первым и антитело против PD-1 вводят индивидууму вторым; или антитело против PD-1 и антитело против CTLA-4 вводят индивидууму одновременно.
5. Способ по п.1, где злокачественное новообразование является меланомой, немелкоклеточным раком легких, раком головы и шеи, уротелиальным раком, раком молочной железы, желудочно-кишечным злокачественным новообразованием, множественной миеломой, печеночно-клеточным раком, неходжкинской лимфомой, раком почки, лимфомой Ходжкина, мезотелиомой, раком яичников, мелкоклеточным раком легких, раком пищевода, раком анального канала, раком желчных протоков, колоректальным раком, раком шейки матки, раком щитовидной железы или раком слюнных желез.
6. Способ по п.1, где злокачественное новообразование является раком поджелудочной железы, раком бронхов, раком предстательной железы, раком поджелудочной железы, раком желудка, раком яичников, раком мочевого пузыря, злокачественным новообразованием головного мозга или центральной нервной системы, злокачественным новообразованием периферической нервной системы, раком матки или эндометрия, раком полости рта или глотки, раком печени, раком почки, раком яичка, раком желчных протоков, раком тонкого кишечника или аппендикса, раком надпочечника, остеосаркомой, хондросаркомой или злокачественным новообразованием кроветворных тканей.
| US 2018333502 A1, 22.11.2018 | |||
| US 2018319892 A1, 08.11.2018 | |||
| MARKHAM A., DUGGAN S | |||
| Cemiplimab: first global approval // Drugs | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| - Vol | |||
| Парный автоматический сцепной прибор для железнодорожных вагонов | 0 |
|
SU78A1 |
| - P | |||
| Способ укрепления лопаток цепного движителя | 1922 |
|
SU1841A1 |
| WO 2019023482 A1, 31.01.2019 | |||
| WO 2016196237 A1, 08.12.2016 | |||
| WO 2018183408 A1, 04.10.2018 | |||
| US 2006235208 A1, 19.10.2006 | |||
| WO 2017118675 A1, 13.07.2017 | |||
| BENGSCH B., THIMME R | |||
| Success of | |||

