ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0001] CD73 или экто-5'-нуклеотидаза (5'-NT) повсеместно экспрессируется в ряде тканей.
Данный белок заякорен на клеточной мембране посредством гликозилфосфатидилинозитольной (GPI) связи, обладает эктоферментной активностью и играет роль в передаче сигналов. Основной функцией CD73 является преобразование внеклеточных нуклеотидов (например, 5'-АМР), для которых клетки в основном непроницаемы, в их соответствующие нуклеозиды (например, аденозин), которые могут легко проникать в большинство клеток. Было показано, что CD73-выработка аденозина посредством дефосфорилирования AMP регулирует связывание аденозиновых рецепторов во многих тканях, указывая на то, что аденозин выполняет свои функции при цитопротекции, клеточном росте, ангиогенезе и иммуносупрессии, а также играет роль в онкогенезе.
[0002] Сообщалось, что экспрессия CD73 в опухолевых клетках присутствует при нескольких типах рака, в том числе колоректальном раке, раке поджелудочной железы, раке мочевого пузыря, лейкемии, лимфоме, глиоме, глиобластоме, меланоме, раке яичников, раке щитовидной железы, раке пищевода, раке предстательной железы и раке молочной железы. Повышенная экспрессия CD73 была связана также с инвазивностью опухоли, метастазами и сокращением времени выживания пациентов. CD73 образует иммуносупрессивную среду, характеризующуюся повышенными уровнями аденозина, которые способствуют развитию и прогрессированию рака. Примечательно, что экспрессия CD73 была связана с прометастатическим фенотипом при меланоме и раке молочной железы.
[0003] Ингибиторы иммунных контрольных точек обладают большим потенциалом в качестве средств для терапии рака. Тем не менее, клинические преимущества ингибирования иммунных контрольных точек были незначительными. Одно из возможных объяснений заключается в том, что в опухолях задействованы неперекрывающиеся механизмы иммуносупрессии для облегчения ускользания от иммунного ответа. Соответственно, существует острая необходимость в улучшенных композициях и способах ослабления иммуносупрессии, опосредованной опухолью.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0004] В настоящем изобретении предусматриваются выделенные связывающие молекулы или их антигенсвязывающие фрагменты, которые специфически связываются с CD73. Согласно некоторым аспектам такие CD73-связывающие молекулы представляют собой, например, антитела или их антигенсвязывающие фрагменты. Согласно конкретным вариантам осуществления антитела к CD73 по настоящему изобретению (например, MEDI9447) являются пригодными для ослабления иммуносупрессии, опосредованной опухолью. Соответственно, в настоящем изобретении также предусматриваются терапевтические комбинации, содержащие антитела к CD73 (например, MEDI9447) и другие средства, оказывающие целенаправленное воздействие на дополнительные компоненты противоракового иммунного цикла (т.е. антитела к PD-1 или PD-L1, антитела к CTLA4, антагонисты A2aR, ингибиторы STAT-3), при этом способы применения таких комбинаций являются пригодными для ослабления иммуносупрессии, опосредованной опухолью.
[0005] Согласно одному аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с эпитопом CD73, при этом связывающая молекула специфически связывается с тем же эпитопом CD73, что и антитело или его антигенсвязывающий фрагмент, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL) антитела, выбранного из CD730002, CD730003, CD730004, CD730008, CD730010, CD730011, CD730021, CD730042, CD730046, CD730047 или CD730058.
[0006] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73 и конкурентно ингибируют связывание CD73 антителом или его антигенсвязывающим фрагментом, содержащим VH и VL CD730002, CD730003, CD730004, CD730008, CD730010, CD730011, CD730021, CD730042, CD730046, CD730047 или CD730058.
[0007] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они содержат VL антитела, где VL имеет аминокислотную последовательность:
[FW1]SGSLSNIGRNX1VN[FW2]LX2NX3RX4X5[FW3]ATWDDSX6X7GWX8[FW4],
где [FW1], [FW2], [FW3] и [FW4] представляют собой каркасные области VL, и где X1 представляет собой аминокислотные остатки пролина (Р), глутаминовой кислоты (Е) или аспарагиновой кислоты (D); Х2 представляет собой аминокислотные остатки аспарагина (N) или аспарагиновой кислоты (D);
Х3 представляет собой аминокислотные остатки глутамина (Q) или лейцина (L);
Х4 представляет собой аминокислотные остатки лейцина (L) или пролина (Р);
Х5 представляет собой аминокислотные остатки глицина (G) или серина (S);
Х6 представляет собой аминокислотные остатки лейцина (L) или гистидина (Н);
Х7 представляет собой аминокислотные остатки лизина (K), пролина (Р), изолейцина (I) или аспарагина (N); и
Х8 представляет собой аминокислотные остатки лейцина (L) или треонина (Т).
Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенная связывающая молекула или ее антигенсвязывающий фрагмент по пункту 6, где FW1 содержит SEQ ID NO: 25 или 26, FW2 содержит SEQ ID NO: 27 или 28, FW3 содержит SEQ ID NO: 29, и FW4 содержит SEQ ID NO: 30.
[0008] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они содержат VH антитела, где VH имеет аминокислотную последовательность:
[FW5]SYAX9S[FW6]X10IX11GSX12GX13TYYADSVKG[FW7]LGYX14X15X16DX17[FW8],
где [FW5], [FW6], [FW7] и [FW8] представляют собой каркасные области VH, и где
Х9 представляет собой аминокислотные остатки метионина (М) или тирозина (Y);
Х10 представляет собой аминокислотные остатки лейцина (L) или аланина (А);
Х11 представляет собой аминокислотные остатки триптофана (W) или серина (S);
Х12 представляет собой аминокислотные остатки триптофана (W) или глицина (G);
Х13 представляет собой аминокислотные остатки серина (S) или аргинина (R);
X14 представляет собой аминокислотные остатки глицина (G) или серина (S);
X15 представляет собой аминокислотные остатки аргинина (R) или треонина (Т);
Х16 представляет собой аминокислотные остатки валина (V) или изолейцина (I); и
Х17 представляет собой аминокислотные остатки тирозина (Y), лизина (K), метионина (М), лейцина (L) или глутаминовой кислоты (Е).
[0009] Согласно другому аспекту в настоящем изобретении предусматриваются связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они содержат VL антитела и VH антитела, где VL содержит аминокислотную последовательность:
[FW1]SGSLSNIGRNX1VN[FW2]LX2NX3RX4X5[FW3]ATWDDSX6X7GWX8[FW4],
где [FW1], [FW2], [FW3] и [FW4] представляют собой каркасные области VL, и где
X1 представляет собой аминокислотные остатки пролина (Р), глутаминовой кислоты (Е) или аспарагиновой кислоты (D);
Х2 представляет собой аминокислотные остатки аспарагина (N) или аспарагиновой кислоты (D);
Х3 представляет собой аминокислотные остатки глутамина (Q) или лейцина (L);
X4 представляет собой аминокислотные остатки лейцина (L) или пролина (Р);
Х5 представляет собой аминокислотные остатки глицина (G) или серина (S);
Х6 представляет собой аминокислотные остатки лейцина (L) или гистидина (Н);
Х7 представляет собой аминокислотные остатки лизина (K), пролина (Р), изолейцина (I) или аспарагина (N); и
X8 представляет собой аминокислотные остатки лейцина (L) или треонина (Т);
и где VH содержит аминокислотную последовательность:
[FW5]SYAX9S[FW6]X10IX11GSX12GX13TYYADSVKG[FW7]LGYX14X15X16DX17[FW8],
где [FW5], [FW6], [FW7] и [FW8] представляют собой каркасные области VH, и где
Х9 представляет собой аминокислотные остатки метионина (М) или тирозина (Y);
Х10 представляет собой аминокислотные остатки лейцина (L) или аланина (А);
Х11 представляет собой аминокислотные остатки триптофана (W) или серина (S);
Х12 представляет собой аминокислотные остатки триптофана (W) или глицина (G);
Х13 представляет собой аминокислотные остатки серина (S) или аргинина (R);
Х14 представляет собой аминокислотные остатки глицина (G) или серина (S);
X15 представляет собой аминокислотные остатки аргинина (R) или треонина (Т);
X16 представляет собой аминокислотные остатки валина (V) или изолейцина (I); и
X17 представляет собой аминокислотные остатки тирозина (Y), лизина (K), метионина (М), лейцина (L) или глутаминовой кислоты (Е).
[0010] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VL антитела, где VL имеет аминокислотную последовательность определяющей комплементарность области-2 VL (VL-CDR2), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены по отношению к SEQ ID NO: 49, SEQ ID NO: 50, SEQ ID NO: 51 или SEQ ID NO: 52.
[0011] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются CD73, при этом они имеют VL антитела, где VL имеет аминокислотную последовательность определяющей комплементарность области-3 (VL-CDR3), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены по отношению к SEQ ID NO: 53, SEQ ID NO: 54, SEQ ID NO: 55, или SEQ ID NO: 56.
[0012] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VH антитела, где VH имеет аминокислотную последовательность определяющей комплементарность области-1 (VH-CDR1), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены по отношению к SEQ ID NO: 35 или SEQ ID NO: 36.
[0013] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VH антитела, где VH имеет аминокислотную последовательность определяющей комплементарность области-2 (VH-CDR2), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены по отношению к SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39, или SEQ ID NO: 40.
[0014] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VH антитела, где VH имеет аминокислотную последовательность определяющей комплементарность области-3 (VH-CDR3), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены по отношению к SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43, SEQ ID NO: 44, или SEQ ID NO: 45.
[0015] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VL антитела, где VL имеет аминокислотные последовательности VL-CDR1, VL-CDR2 и VL-CDR3, идентичные или идентичные за исключением четырех, трех, двух или одной аминокислотной замены в одном или нескольких VL-CDR по отношению к SEQ ID NOs: 46, 49 и 53; SEQ ID NOs: 47, 49, и 53; SEQ ID NOs: 47, 49, и 54; SEQ ID NOs: 46, 50, и 54; SEQ ID NOs: 46, 51, и 55; SEQ ID NOs: 48, 52, и 54; SEQ ID NOs: 46, 49, и 56; SEQ ID NOs: 47, 49, и 56; SEQ ID NOs: 46, 50, и 56; SEQ ID NOs: 46, 51, и 56; или SEQ ID NOs: 48, 52, и 56, соответственно.
[0016] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VH антитела, где VH имеет аминокислотные последовательности VH-CDR1, VH-CDR2 и VH-CDR3, идентичные или идентичные за исключением четырех, трех, двух или одной аминокислотной замены в одном или нескольких VH-CDR по отношению к SEQ ID NOs: 35, 37 и 41; SEQ ID NOs: 36, 37, и 42; SEQ ID NOs: 36, 38, и 43; SEQ ID NOs: 36, 39, и 44; SEQ ID NOs: 36, 40, и 44; SEQ ID NOs: 35, 37, и 45; SEQ ID NOs: 36, 37, и 45; SEQ ID NOs: 36, 38, и 45; SEQ ID NOs: 36, 39, и 45; или SEQ ID NOs: 36, 40, и 45 соответственно.
[0017] Согласно другому аспекту в настоящем изобретении предусматриваются выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VL и VH, имеющие аминокислотные последовательности VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3, идентичные или идентичные за исключением четырех, трех, двух или одной аминокислотной замены по отношению к SEQ ID NOs: 46, 49, 53, 35, 37, и 41; SEQ ID NOs: 47, 49, 53, 35, 37, и 41; SEQ ID NOs: 47, 49, 54, 36, 37, и 42; SEQ ID NOs: 46, 50, 54, 36, 38, и 43; SEQ ID NOs: 46, 51, 55, 36, 39, и 44; SEQ ID NOs: 48, 52, 54, 36, 40, и 44; SEQ ID NOs: 46, 49, 56, 35, 37, и 41; SEQ ID NOs: 46, 49, 53, 35, 37, и 45; SEQ ID NOs: 47, 49, 56, 36, 37, и 45; SEQ ID NOs: 46, 50, 56, 36, 38, и 45; SEQ ID NOs: 46, 51, 56, 36, 39, и 45; SEQ ID NOs: 48, 52, 56, 36, 40, и 45; или SEQ ID NOs: 46,49, 56, 35, 37, и 45.
[0018] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VL и VH антитела, где VL имеет аминокислотную последовательность, по меньшей мере от приблизительно 90% до приблизительно 100% идентичную эталонной аминокислотной последовательности, выбранной из SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, и SEQ ID NO: 70..
[0019] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они имеют VL и VH антитела, где VH имеет аминокислотную последовательность, по меньшей мере от приблизительно 90% до приблизительно 100% идентичную эталонной аминокислотной последовательности, выбранной из SEQ ID NO: 71, SEQ ID NO: 72, SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83 и SEQ ID NO: 84..
[0020] Согласно другому аспекту в настоящем изобретении предусматриваются выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом антитело или антигенсвязывающий фрагмент имеют VL, имеющую последовательность, по меньшей мере от приблизительно 90% до приблизительно 100% идентичную эталонной аминокислотной последовательности, выбранной из SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, и SEQ ID NO: 70, и при этом антитело или антигенсвязывающий фрагмент имеют VH, имеющую последовательность, по меньшей мере от приблизительно 90% до приблизительно 100% идентичную эталонной аминокислотной последовательности, выбранной из SEQ ID NO: 71, SEQ ID NO: 72, SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83 и SEQ ID NO: 84..
[0021] Согласно другому аспекту в настоящем изобретении предусматриваются выделенное антитело или его антигенсвязывающий фрагмент, которые содержат VL, состоящую по сути из SEQ ID NO: 57, и VH, состоящую по сути из SEQ ID NO: 71.
[0022] Согласно другому аспекту в настоящем изобретении предусматриваются выделенное антитело или его антигенсвязывающий фрагмент, которые содержат VL, состоящую по сути из SEQ ID NO: 68, и VH, состоящую по сути из SEQ ID NO: 82.
[0023] Согласно другому аспекту в настоящем изобретении предусматриваются выделенное антитело или его антигенсвязывающий фрагмент, которые содержат VL, состоящий из SEQ ID NO: 57, и VH, состоящий из SEQ ID NO: 71.
[0024] Согласно другому аспекту в настоящем изобретении предусматриваются выделенное антитело или его антигенсвязывающий фрагмент, которые содержат VL, состоящий из SEQ ID NO: 68, и VH, состоящую из SEQ ID NO: 82.
[0025] Согласно другому аспекту в настоящем изобретении предусматривается композиция, содержащая выделенное антитело или его антигенсвязывающий фрагмент в соответствии с настоящим изобретением и носитель.
[0026] Согласно другому аспекту в настоящем изобретении предусматривается нуклеиновая кислота, содержащая последовательность, кодирующую выделенное антитело или его антигенсвязывающий фрагмент в соответствии с настоящим изобретением.
[0027] Согласно другому аспекту в настоящем изобретении предусматривается композиция, включающая нуклеиновую кислоту в соответствии с настоящим изобретением.
[0028] Согласно другому аспекту в настоящем изобретении предусматривается вектор, содержащий нуклеиновую кислоту в соответствии с настоящим изобретением.
[0029] Согласно другому аспекту в настоящем изобретении предусматривается клетка-хозяин, содержащая последовательность нуклеиновой кислоты, композицию или вектор в соответствии с настоящим изобретением.
[0030] Согласно другому аспекту в настоящем изобретении предусматривается способ получения антитела или его антигенсвязывающего фрагмента в соответствии с настоящим изобретением, включающий культивирование клетки, содержащей последовательность нуклеиновой кислоты, композицию или вектор в соответствии с настоящим изобретением, и выделение антитела или его антигенсвязывающего фрагмента.
[0031] Согласно другому аспекту в настоящем изобретении предусматривается диагностический реагент, содержащий выделенное антитело или антигенсвязывающий фрагмент в соответствии с настоящим изобретением, которые являются мечеными.
[0032] Согласно другому аспекту в настоящем изобретении предусматривается набор, содержащий выделенное антитело или его антигенсвязывающий фрагмент, композицию или диагностический реагент в соответствии с настоящим изобретением.
[0033] Согласно другому аспекту в настоящем изобретении предусматривается способ подавления роста клетки, экспрессирующей CD73, включающий приведение клетки в контакт с антителом или его антигенсвязывающим фрагментом в соответствии с настоящим изобретением.
[0034] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения рака у нуждающегося в этом субъекта, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента в соответствии с настоящим изобретением.
[0035] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения рака у субъекта, включающий введение субъекту терапевтически эффективного количества первого средства, которое представляет собой антитело или антигенсвязывающий фрагмент в соответствии с настоящим изобретением, в комбинации с терапевтически эффективным количеством второго средства, которое представляет собой противораковое средство, отличное от первого средства.
[0036] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения, включающий введение антитела к CD73 или его антигенсвязывающего фрагмента субъекту, идентифицированному как имеющий опухоль, которая характеризуется повышенной экспрессией CD73 по сравнению с эталоном.
[0037] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения, включающий введение антитела к CD73 или его антигенсвязывающего фрагмента и антитела к PD-1, антитела к PD-L1 или антитела к CTLA-4 или их антигенсвязывающих фрагментов субъекту, идентифицированному как имеющий опухоль, которая характеризуется повышенной экспрессией CD73 по сравнению с эталоном.
[0038] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения, включающий введение hIgG1 MEDI9447 или Phen0203 или их антигенсвязывающих фрагментов и пембролизумаба (Keytruda®) или ниволумаба (Opdiva®) или их антигенсвязывающих фрагментов субъекту, идентифицированному как имеющий опухоль, которая характеризуется повышенной экспрессией CD73 по сравнению с эталоном.
[0039] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения, включающий введение hIgG1 MEDI9447 или Phen0203 или их антигенсвязывающих фрагментов и MEDI4736 или его антигенсвязывающего фрагмента субъекту, идентифицированному как имеющий опухоль, которая характеризуется повышенной экспрессией CD73 по сравнению с эталоном.
[0040] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения, включающий введение hIgG1 MEDI9447 или Phen0203 или их антигенсвязывающих фрагментов и тремелимумаба или его антигенсвязывающего фрагмента субъекту, идентифицированному как имеющий опухоль, которая характеризуется повышенной экспрессией CD73 по сравнению с эталоном.
[0041] Согласно другому аспекту в настоящем изобретении предусматривается способ идентификации субъекта, имеющего рак, реагирующий на терапию антителом к CD73, при этом способ включает выявление повышенного уровня экспрессии или активности CD73 в опухолевой клетке или клетке крови субъекта по сравнению с эталоном, идентифицируя таким образом указанный рак как реагирующий на терапию антителом к CD73.
[0042] Согласно другому аспекту в настоящем изобретении предусматривается способ идентификации субъекта, имеющего рак, реагирующий на терапию антителом к CD73 в комбинации с одной или несколькими из видов терапии антителоми к PD-1, PD-L1 или CTLA4, при этом способ включает выявление повышенного уровня экспрессии или активности CD73 в опухолевой клетке или клетке крови субъекта по сравнению с эталоном, идентифицируя таким образом указанный рак как реагирующий на терапию антителом к CD73 в комбинации с одним или несколькими из видов терапии антителами к PD-1, PD-L1 или CTLA4.
[0043] Согласно другому аспекту в настоящем изобретении предусматривается способ идентификации субъекта, имеющего рак, реагирующий на терапию антителами к PD-1, PD-L1 или CTLA4, при этом способ включает выявление сниженного уровня экспрессии или активности CD73 в опухолевой клетке или клетке крови субъекта по сравнению с эталоном, идентифицируя таким образом указанный рак как реагирующий на терапию антителами к PD-1, PD-L1 или CTLA4.
[0044] Согласно другому аспекту в настоящем изобретении предусматривается способ подавления роста опухоли у субъекта, при этом способ включает введение нуждающемуся в этом субъекту антитела к CD73 или его антигенсвязывающего фрагмента и одного или нескольких из антитела к PD-1, антитела к PD-L1, антитела к CTLA4 или их антигенсвязывающих фрагментов.
[0045] Согласно другому аспекту в настоящем изобретении предусматривается способ усиления противоопухолевого иммунного ответа у субъекта, при этом способ включает введение нуждающемуся в этом субъекту антитела к CD73 или его антигенсвязывающего фрагмента и одного или нескольких из антитела к PD-1, антитела к PD-L1, антитела к CTLA4 или их антигенсвязывающих фрагментов.
[0046] Согласно другому аспекту в настоящем изобретении предусматривается способ лечения опухоли у субъекта, при этом способ включает введение нуждающемуся в этом субъекту антитела к CD73 или его антигенсвязывающего фрагмента и одного или нескольких из антитела к PD-1, антитела к PD-L1, антитела к CTLA4 или их антигенсвязывающих фрагментов.
[0047] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество антитела к CD73 или его антигенсвязывающего фрагмента и антитела к PD-1 или его антигенсвязывающего фрагмента.
[0048] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество MEDI9447 или его антигенсвязывающего фрагмента и пембролизумаба (Keytruda®) или его антигенсвязывающего фрагмента.
[0049] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество MEDI9447 или его антигенсвязывающего фрагмента и ниволумаба (Opdiva®) или его антигенсвязывающего фрагмента.
[0050] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество hIgG1 Phen0203 или его антигенсвязывающего фрагмента и пембролизумаба (Keytruda®) или его антигенсвязывающего фрагмента.
[0051] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество hIgG1 Phen0203 или его антигенсвязывающего фрагмента и ниволумаба (Opdiva®) или его антигенсвязывающего фрагмента.
[0052] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество антитела к CD73 или его антигенсвязывающего фрагмента и антитела к PD-L1 или его антигенсвязывающего фрагмента.
[0053] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество MEDI9447 или его антигенсвязывающего фрагмента и MEDI4736 или его антигенсвязывающего фрагмента.
[0054] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество hIgG1 Phen0203 или его антигенсвязывающего фрагмента и MEDI4736 или его антигенсвязывающего фрагмента.
[0055] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество антитела к CD73 или его антигенсвязывающего фрагмента и антитела к CTLA4 или его антигенсвязывающего фрагмента.
[0056] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество MEDI9447 или его антигенсвязывающего фрагмента и тремелимумаба или его антигенсвязывающего фрагмента.
[0057] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество MEDI9447 или его антигенсвязывающего фрагмента и ипилимумаба или его антигенсвязывающего фрагмента.
[0058] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество hIgG1 Phen0203 или его антигенсвязывающего фрагмента и тремелимумаба или его антигенсвязывающего фрагмента.
[0059] Согласно другому аспекту в настоящем изобретении предусматривается фармацевтический состав, содержащий эффективное количество hIgG1 Phen0203 или его антигенсвязывающего фрагмента и ипилимумаба или его антигенсвязывающего фрагмента.
[0060] Согласно другому аспекту в настоящем изобретении предусматривается набор для повышения противоопухолевой активности, при этом набор содержит антитело к CD73 или его антигенсвязывающий фрагмент и антитело к PD-1 или его антигенсвязывающий фрагмент.
[0061] Согласно другому аспекту в настоящем изобретении предусматривается набор для повышения противоопухолевой активности, при этом набор содержит антитело к CD73 или его антигенсвязывающий фрагмент и антитело к PD-L1 или его антигенсвязывающий фрагмент.
[0062] Согласно другому аспекту в настоящем изобретении предусматривается набор для повышения противоопухолевой активности, при этом набор содержит антитело к CD73 или его антигенсвязывающий фрагмент и антитело к CTLA4 или его антигенсвязывающий фрагмент.
[0063] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, VL и VH CD730002 представляют собой или включают в себя SEQ ID NO: 1 и 2 соответственно, a VL и VH CD730010 представляют собой или включают в себя SEQ ID NO: 3 и 4 соответственно.
[0064] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенная связывающая молекула или ее антигенсвязывающий фрагмент предусматривает антитело или его антигенсвязывающий фрагмент.
[0065] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, связывающая молекула является молекулой с созревшей аффинностью.
[0066] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, FW5 представляют собой или включает в себя SEQ ID NO: 31, FW6 представляет собой или включает в себя SEQ ID NO: 32, FW7 представляет собой или включает в себя SEQ ID NO: 33 и FW8 представляет собой или включает в себя SEQ ID NO: 34.
[0067] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, FW1 представляет собой или включает в себя SEQ ID NO: 25 или 26, FW2 представляет собой или включает в себя SEQ ID NO: 27 или 28, FW3 представляет собой или включает в себя SEQ ID NO: 29, FW4 представляет собой или включает в себя SEQ ID NO: 30, FW5 представляет собой или включает в себя SEQ ID NO: 31, FW6 представляет собой или включает в себя SEQ ID NO: 32, FW7 представляет собой или включает в себя SEQ ID NO: 33 и FW8 представляет собой или включает в себя SEQ ID NO: 34.
[0068] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, VL включает в себя аминокислотную последовательность определяющей комплементарность области-1 VL (VL-CDR1), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены по отношению к SEQ ID NO: 46, SEQ ID NO: 47 или SEQ ID NO: 48.
[0069] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенное антитело или его антигенсвязывающий фрагмент содержат VL, содержащий SEQ ID NO: 57, и VH, содержащий SEQ ID NO: 71.
[0070] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенное антитело или его антигенсвязывающий фрагмент содержат VL, содержащий SEQ ID NO: 68, и VH, содержащий SEQ ID NO: 82.
[0071] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенное антитело или его антигенсвязывающий фрагмент включают в себя константную область тяжелой цепи или ее фрагмент.
[0072] Согласно различным вариантам осуществления константная область тяжелой цепи или ее фрагмент представляют собой константную область IgG, в том числе, например, константную область IgG1, константную область IgG2, константную область IgG3 или константную область IgG4.
[0073] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенное антитело или его антигенсвязывающий фрагмент включаю в себя константную область легкой цепи, выбранную из константной области каппа-цепи человека и константной области лямбда-цепи человека.
[0074] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, константная область IgG имеет одну или несколько аминокислотных замен относительно константной области IgG дикого типа, при этом модифицированный IgG характеризуется увеличенным временем полужизни по сравнению с временем полужизни IgG с константной областью IgG дикого типа.
[0075] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, константная область IgG содержит одну или несколько аминокислотных замен в положениях аминокислотных остатков 251-257, 285-290, 308-314, 385-389 и 428-436, при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat.
[0076] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, по меньшей мере одна аминокислотная замена в константной области IgG выбрана из:
(a) замены аминокислоты по положению 252 на тирозин (Y), фенилаланин (F), триптофан (W) или треонин (Т);
(b) замены аминокислоты по положению 254 на треонин (Т);
(c) замены аминокислоты по положению 256 на серии (S), аргинин (R), глутамин (Q), глутаминовую кислоту (Е), аспарагиновую кислоту (D) или треонин (Т);
(d) замены аминокислоты по положению 257 на лейцин (L);
(e) замены аминокислоты по положению 309 на пролин (Р);
(f) замены аминокислоты по положению 311 на серии (S);
(g) замены аминокислоты по положению 428 на треонин (Т), лейцин (L), фенилаланин (F) или серии (S);
(h) замены аминокислоты по положению 433 на аргинин (R), серии (S), изолейцин (I), пролин (Р) или глутамин (Q);
(i) замены аминокислоты по положению 434 на триптофан (W), метионин (М), серии (S), гистидин (Н), фенилаланин (F) или тирозин и
(j) комбинации двух или более указанных замен,
при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat.
[0077] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, константная область IgG человека содержит аминокислотные замены относительно константной области человеческого IgG дикого типа по положениям 252, 254 и 256, где
(a) аминокислота по положению 252 заменена на тирозин (Y),
(b) аминокислота по положению 254 заменена на треонин (Т) и
(c) аминокислота по положению 256 заменена на глутаминовую кислоту (Е), при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat.
[0078] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, аминокислота по положению 434 заменена на аминокислоту, выбранную из триптофана (W), метионина (М), тирозина (Y) и серина (S), и при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat.
[0079] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, аминокислота по положению 428 заменена на аминокислоту, выбранную из треонина (Т), лейцина (L), фенилаланина (F) и серина (S), и при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat.
[0080] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, аминокислота по положению 257 заменена на лейцин (L) и аминокислота по положению 434 согласно Kabat заменена на тирозин (Y), и при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat.
[0081] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, аминокислота по положению 428 согласно Kabat заменена на лейцин (L) и аминокислота по положению 434 согласно Kabat заменена на серии (S).
[0082] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, константная область человеческого IgG содержит аминокислотные замены относительно константной области человеческого IgG дикого типа по положениям 252, 254 и 256, при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat, и при этом
(a) аминокислота по положению 252 заменена на тирозин (Y),
(b) аминокислота по положению 254 заменена на треонин (Т) и
(c) аминокислота по положению 256 заменена на глутаминовую кислоту (Е).
[0083] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, антитело представляет собой полностью человеческое антитело, гуманизированное антитело, химерное антитело, моноклональное антитело, поликлональное антитело, рекомбинантное антитело, мультиспецифичное антитело или их антигенсвязывающий фрагмент.
[0084] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, антигенсвязывающий фрагмент представляет собой Fv, Fab, F(ab')2, Fab', dsFv, scFv или sc(Fv)2.
[0085] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенное антитело или его антигенсвязывающий фрагмент конъюгированы по меньшей мере с одним гетерологичным средством, в том числе, например, противораковым средством.
[0086] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, композиция в соответствии с настоящим изобретением дополнительно содержит противораковое средство.
[0087] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенное антитело или его антигенсвязывающий фрагмент не индуцируют антителозависимую клеточно-опосредованную цитотоксичность (ADCC).
[0088] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенное антитело или его антигенсвязывающий фрагмент являются антагонистами CD73.
[0089] Согласно различным вариантам осуществления выделенное антитело или его антигенсвязывающий фрагмент являются антагонистами CD73 в клетках, выбранных из MB-MDA-231, 4Т1, MK1 или комбинации двух или более перечисленных клеток.
[0090] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, CD73 представляет собой CD73 человека.
[0091] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, связывание антитела или антигенсвязывающего фрагмента с CD73 может снижать пролиферацию клеток.
[0092] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, антитело или антигенсвязывающий фрагмент к CD73 могут связываться с CD73 человека, CD73 яванского макака и CD73 мыши.
[0093] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, рак выбран из колоректального рака, рака поджелудочной железы, рака мочевого пузыря, лейкемии, лимфомы, глиомы, глиобластомы, меланомы, рака яичников, рака щитовидной железы, рака пищевода, рака предстательной железы и рака молочной железы.
[0094] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, рак характеризуется прометастатическим фенотипом, в том числе меланома или рак молочной железы.
[0095] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, субъектом является человек.
[0096] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, комбинация первого средства и второго средства характеризуется превосходной противоопухолевой активностью, при этом она может быть аддитивной или синергической.
[0097] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, второе средство представляет собой антитело или его антигенсвязывающий фрагмент.
[0098] Согласно различным вариантам осуществления второе средство специфически связывается с PD-1 (белком-1 запрограммированной смерти клетки), PD-L1 (лигандом-1 белка-1 запрограммированной смерти клетки), PD-L2 (лигандом-2 белка-1 запрограммированной смерти клетки) или CTLA-4 (белком-антигеном-4 цитотоксических Т-лимфоцитов).
[0099] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, второе средство представляет собой антитело к CTLA-4 или его антигенсвязывающий фрагмент, в том числе, например, ипилимумаб, тремелимумаб (тицилимумаб, СР-675,206) или их антигенсвязывающие фрагменты.
[00100] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, второе средство представляет собой антитело к PD-1 или его антигенсвязывающий фрагмент, в том числе, например, пембролизумаб (Keytruda®, ламбролизумаб, MK-3475), ниволумаб (Opdiva®, BMS-936558, MDX-1106, ONO-4538), АМР-224 или их антигенсвязывающие фрагменты.
[00101] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, второе средство представляет собой антитело к PD-L1 или его антигенсвязывающий фрагмент, в том числе, например, MEDI4736, BMS-936559, MPDL3280A или их антигенсвязывающие фрагменты.
[00102] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, антитело к CD73 представляет собой MEDI9447, hIgG1 Phen0203 или их антигенсвязывающие фрагменты.
[00103] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, субъект подвергается, подвергался или будет подвергаться терапии, направленной в отношении PD-1, PD-L1 или CTLA4.
[00104] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, терапия, направленная в отношении PD-1, PD-L1 или CTLA4, предусматривает введение антитела к PD-1, антитела к PD-L1 или антитела к CTLA4 или их антигенсвязывающих фрагментов соответственно.
[00105] Согласно различным вариантам осуществления антитело к PD-1 представляет собой пембролизумаб (Keytruda®, ламбролизумаб, MK-3475), ниволумаб (Opdiva®, BMS-936558, MDX-1106, ONO-4538), АМР-224 или их антигенсвязывающие фрагменты.
[00106] Согласно различным вариантам осуществления антитело к PD-L1 представляет собой MEDI4736, BMS-936559, MPDL3280A или их антигенсвязывающие фрагменты.
[00107] Согласно различным вариантам осуществления антитело к CTLA-4 представляет собой ипилимумаб, тремелимумаб (тицилимумаб, СР-675,206) или их антигенсвязывающие фрагменты.
[00108] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, опухоль представляет собой рак толстой кишки, меланому, рак молочной железы, лимфому, немелкоклеточный рак легкого, лимфому Ходжкина, неходжкинскую лимфому и лимфому Беркитта, рак яичников, рак молочной железы, формы рака головы и шеи или рак поджелудочной железы.
[00109] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, экспрессию или активность CD73 выявляют в образце опухоли, образце крови или образце лимфы.
[00110] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, экспрессию CD73 выявляют в опухолевой клетке или клетке периферической крови, включая субпопуляции лимфоидных или миелоидных клеток (т.е. одна или несколько из В-лимфоцитов, CD4+, FoxP3+ лимфоцитов или супрессорной клетки миелоидного происхождения (MDSC)).
[00111] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, экспрессию CD73 выявляют с помощью проточной цитометрии, иммуногистохимии (IHC) или активности фермента CD73 или уровней содержания растворимого CD73 в образцах.
[00112] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, антитело к CD73 или его антигенсвязывающий фрагмент и антитела к PD-1, PD-L1 или CTLA4 или их антигенсвязывающие фрагменты вводят одновременно.
[00113] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, способ обеспечивает индуцирование или усиление опухолеспецифичного иммунного ответа.
[00114] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, способ обеспечивает уменьшение иммуносупрессивных эффектов пути АМР/СD73/аденозин.
[00115] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, опухоль представляет собой опухоль, сверхэкспрессирующую CD73.
[00116] Согласно другому аспекту в настоящем изобретении предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом антитело содержит VL и VH, которые специфически связываются с эпитопом белка CD73, содержащего одну или несколько аминокислот, соответствующих Val144, Lys180, и Asn185..
[00117] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенная связывающая молекула или ее антигенсвязывающий фрагмент дополнительно содержат одну или несколько аминокислот, соответствующих Tyr135, Lys136, и Asn187..
[00118] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенная связывающая молекула или ее антигенсвязывающий фрагмент содержат аминокислоты, соответствующие Tyr135, Lys136, и Asn187..
[00119] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенная связывающая молекула или ее антигенсвязывающий фрагмент содержат аминокислоты, соответствующие Tyr135, Lys136, Asn187, Tyr135, Lys136, и Asn187..
[00120] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенная связывающая молекула или ее антигенсвязывающий фрагмент связывают эпитоп в пределах одной или нескольких из следующих областей белка CD73: Tyr132-Val144 и/или Lys180-Asn187..
[00121] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, выделенная связывающая молекула или ее антигенсвязывающий фрагмент содержатся или находятся в пределах аминокислотных последовательностей при Tyr132-Val144 и/или Lys180-Asn187..
[00122] Согласно другому аспекту в настоящем изобретении предусматривается конформационный эпитоп на поверхности белка CD73, содержащий одну или несколько аминокислот, соответствующих Val144, Lys180 и Asn185, при этом белок CD73, содержащий данный эпитоп, может быть специфически связан моноклональным антителом MEDI9447 или его антигенсвязывающим фрагментом, вариантом, аналогом или производным.
[00123] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, конформационный эпитоп дополнительно содержит одну или несколько аминокислот, соответствующих Tyr135, Lys136, и Asn18..
[00124] Согласно различным вариантам осуществления любого аспекта, описанного
в данном документе, конформационный эпитоп содержит аминокислоты, соответствующие Tyr135, Lys136, и Asn187..
[00125] Согласно различным вариантам осуществления любого аспекта, описанного
в данном документе, конформационный эпитоп содержит аминокислоты, соответствующие Tyr135, Lys136, Asn187, Tyr135, Lys136, и Asn187..
[00126] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, конформационный эпитоп находится в пределах одной или нескольких из следующих областей белка CD73: Tyr132-Val144 и/или Lys180-Asn187.
[00127] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, конформационный эпитоп содержится или находится в пределах аминокислотных последовательностей при Tyr132-Val144 и/или Lys180-Asn187..
[00128] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, MEDI9447 связывается с белком CD73 в неактивном или каталитически активном состоянии, либо в открытом или закрытом состоянии.
[00129] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, белок CD73 представляет собой CD73 человека.
[00130] Согласно различным вариантам осуществления любого аспекта, описанного в данном документе, предусматриваются выделенная связывающая молекула или ее антигенсвязывающий фрагмент, где VL и VH представляют собой VL и VH MED 19447.
[00131] Композиции и изделия, определенные настоящим изобретением, были выделены или изготовлены в соответствии с примерами, представленными ниже. Другие признаки и преимущества настоящего изобретения будут очевидны из подробного описания и формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ/ФИГУР
[00132] На фигуре 1А показана нуклеотидная последовательность и трансляция аминокислот VH-домена MEDI9447 с CDR, показанные на основе системы нумерации по Kabat.
[00133] На фигуре 1В показана нуклеотидная последовательность и трансляция аминокислот VL-домена MEDI9447 с CDR, показанные на основе системы нумерации по Kabat.
[00134] На фигуре 1С показано выравнивание VH MEDI9447 с ближайшими последовательностями зародышевой линии VH и JH человека. CDR на основе системы нумерации по Kabat выделены, а остатки, отличающиеся от последовательностей зародышевой линии, заключены в рамку.
[00135] На фигуре 1D показано выравнивание VL MEDI9447 с ближайшими последовательностями зародышевой линии VL и JL человека. CDR на основе системы нумерации по Kabat выделены, а остатки, отличающиеся от последовательностей зародышевой линии, заключены в рамку.
[00136] На фигуре 2 представлены два графика, показывающие антителоопосредованную интернализацию цитотоксического реагента FabZAP в клетки MDA-MB-231 и клетки 4Т1, где антитела представляют собой MEDI9447, а контрольное антитело - R347.
[00137] Фигура 3А представляет собой график, показывающий ингибирование 5'-эктонуклеотидазы с помощью антитела к CD73 MEDI9447.
[00138] Фигура 3В представляет собой график, показывающий ингибирование гидролиза AMP с помощью антитела к CD73 CD370010.
[00139] Фигура 4 представляет собой график, показывающий, что MEDI9447 ингибирует рост опухоли в модели сингенной опухоли СТ26. Самкам мышей Balb/C имплантировали подкожно в правый бок опухолевые клетки СТ26 мыши. Опухолям давали расти в течение 3 дней и обрабатывали с помощью MEDI9447 или изотипическим контролем дважды в неделю в течение двух недель. На 16 день опухоли собирали для анализа посредством проточной цитометрии.
[00140] Фигура 5 представляет собой график, показывающий, что MEDI9447 ингибирует супрессорные клетки миелоидного происхождения, инфильтрирующие опухоль (MDSC). Мышей-опухоленосителей СТ26, обработанных MEDI9447, умерщвляли и опухоли собирали на 16-й день исследования. Опухоли разделяли на отдельные клетки, окрашивали на маркеры CD45 и MDSC, и анализировали с помощью проточной цитометрии.
[00141] На фигуре 6 представлены шесть лепестковых диаграмм, показывающих эффект mIgG1 MEDI9447, антитела к PD-1 или комбинации в отношении объема опухоли. Контрольные антитела включают в себя rIgG2a, которое представляет собой контрольное моноклональное антитело IgG2a крысы, специфичное к β-галактозидазе Е. coli (β-Gal), и изотипический контроль IgG1 мыши. Объемы опухолей из каждой группы животных отображали графически для отдельных животных на 40-й день исследования. К концу 40-дневного периода исследования ни у одной из мышей контрольной группы не отсутствовала опухоль. В результате обработки только антителом к CD73 в конце исследования опухоль отсутствовала у 10% животных. Также в результате обработки только антителом к PD1 в конце исследования опухоль отсутствовала у 10% животных. Примечательно, что в результате обработки комбинацией антитела к CD73 и антитела к PD опухоль отсутствовала у 60% мышей. К концу исследования ни у одной из мышей контрольной группы не отсутствовала опухоль.
[00142] Фигура 7 представляет собой график, показывающий эффект mIgG1 MEDI9447, антитела к PD1 или комбинации в отношении выживания.
[00143] Фигура 8 представляет собой график, показывающий, что комбинация MEDI9447 и антитела к PD-1 значительно усиливает ингибирование роста опухоли (р<0,05) при сравнении с любым отдельным средством в опухолях, характерных для колоректальной карциномы. Мышам подкожно инъецировали сингенные клетки MC38-OVA колоректальной карциномы и дважды в неделю обрабатывали с помощью 10 мг на кг MEDI9447 или 10 мг на кг только антитела к PD-1 или комбинации обоих антител. Объем опухоли измеряли дважды в неделю.
[00144] Фигура 9 представляет собой график, показывающий, что антитело к PD-1 индуцировало богатое на CD73 микроокружение опухоли, что измеряли с помощью экспрессии CD73 на опухолевых клетках, выделенных из мышей-опухоленосителей. Мышам (n=4) подкожно инъецировали сингенные клетки СТ26 колоректальной карциномы и дважды в неделю обрабатывали с помощью 10 мг на кг антитела к PD-1 или нерелевантного антитела изотипического контроля. Через один день после первой обработки опухоли выделяли, клетки диссоциировали и анализировали в отношении поверхностного фенотипа с помощью проточной цитометрии.
[00145] Фигура 10 представляет собой график, показывающий, что антитело к PD-1 индуцировало богатое на CD73 микроокружение опухоли, что измеряли с помощью экспрессии CD73 на супрессорных клетках миелоидного происхождения (MDSC), выделенных из мышей-опухоленосителей. Мышам (n=4) подкожно инъецировали сингенные клетки СТ26 колоректальной карциномы и дважды в неделю обрабатывали с помощью 10 мг на кг антитела к PD-1 или нерелевантного антитела изотипического контроля. Через один день после первой обработки опухоли выделяли, опухолевые клетки выделяли, собирали цельные клетки периферической крови и анализировали в отношении поверхностной экспрессии CD73 с помощью проточной цитометрии.
[00146] Фигура 11 представляет собой график, показывающий, что антитело к PD-1 индуцировало богатое на CD73 микроокружение опухоли, что измеряли с помощью экспрессии CD73 на CD4+, FoxP3+ лимфоцитах, выделенных из мышей-опухоленосителей. Мышам (n=4) подкожно инъецировали сингенные клетки СТ26 колоректальной карциномы и дважды в неделю обрабатывали с помощью 10 мг на кг антитела к PD-1 или нерелевантного антитела изотипического контроля. Через три дня после первой обработки опухоли выделяли, собирали цельные клетки периферической крови и анализировали в отношении поверхностной экспрессии CD73 с помощью проточной цитометрии.
[00147] Фигура 12 представляет собой график, показывающий, что комбинация MEDI9447 и антитела к PD-L1 значительно усиливает ингибирование роста опухоли (р<0,05) при сравнении с любым отдельным средством в опухолях, характерных для меланомы. Мышам подкожно инъецировали сингенные клетки B16F10 меланомы и дважды в неделю обрабатывали с помощью 10 мг на кг MEDI9447 или 10 мг на кг только антитела к PD-L1 или комбинации обоих антител. Объем опухоли измеряли дважды в неделю.
[00148] Фигура 13 представляет собой график, показывающий, что комбинация MEDI9447 и антитела к PD-L1 значительно усиливает ингибирование роста опухоли (р<0,01) при сравнении с любым отдельным средством в опухолях лимфомы. Мышам подкожно инъецировали сингенные клетки EG7-OVA лимфомы и дважды в неделю обрабатывали с помощью 10 мг на кг MEDI9447 или 10 мг на кг только антитела к PD-L1 или комбинации обоих антител. Объем опухоли измеряли дважды в неделю.
[00149] Фигура 14 представляет собой график, показывающий, что антитело к PD-L1 индуцировало богатое на CD73 микроокружение опухоли, что измеряли с помощью поверхностной экспрессии CD73 на В-лимфоцитах дренирующих лимфатических узлов. Мышам (n=4) подкожно инъецировали сингенные клетки СТ26 колоректальной карциномы и дважды в неделю обрабатывали с помощью 10 мг на кг антитела к PD-L1 или нерелевантного антитела изотипического контроля. Через день после первой обработки клетки выделяли из дренирующих лимфатических узлов и анализировали в отношении поверхностного фенотипа с помощью проточной цитометрии.
[00150] Фигура 15 представляет собой график, показывающий, что антитело к PD-L1 индуцировало богатое на CD73 микроокружение опухоли, что измеряли с помощью поверхностной экспрессии CD73 на инфильтрующих опухоль CD4+, FoxP3+ лимфоцитах. Мышам (n=4) подкожно инъецировали сингенные клетки СТ26 колоректальной карциномы и дважды в неделю обрабатывали с помощью 10 мг на кг антитела к PD-L1 или нерелевантного антитела изотипического контроля. Через три дня после первой обработки опухоли выделяли, клетки диссоциировали и анализировали в отношении поверхностного фенотипа с помощью проточной цитометрии.
[00151] Фигуры 16А и В представляют собой графики, показывающие, что MEDI9447 отдельно или в комбинации с антителом к PD-L1 снижало экспрессию CD73 в лимфоидных клетках, инфильтрующих опухоль. Мышей, несущих колоректальные сингенные опухоли СТ26, дважды в неделю (12-й день и 16-й день) обрабатывали с помощью либо 30 мг/кг MEDI9447, либо 30 мг/кг антитела к PD-L1 отдельно, либо комбинации как MEDI9447, так и антитела к PD-L1. На 17-й день опухоли собирали и анализировали в отношении поверхностной экспрессии CD73 с помощью проточной цитометрии. Экспрессия CD73 в опухолевых инфильтратах (A) CD4+ FoxP3+ Treg и (В) CD8+ Т-клетки.
[00152] Фигуры 17А и В представляют собой графики, показывающие, что MEDI9447 отдельно или в комбинации с антителом к PD-L1 снижало активность CD73 в (А) опухолевых клетках и (В) цельных клетках периферической крови. Мышей, несущих колоректальные сингенные опухоли СТ26, дважды в неделю (12-й день и 16-й день) обрабатывали с помощью либо 30 мг/кг MEDI9447, либо 30 мг/кг антитела к PD-L1 отдельно, либо комбинации как MEDI9447, так и антитела к PD-L1. На 17-й день опухоли и цельные клетки периферической крови собирали и анализировали в отношении поверхностной экспрессии CD73, в отношении ферментативной активности путем применения Cell-Titre Glo.
[00153] Фигура 18 представляет собой набор графиков, изображающих цитокиновые профили мононуклеарных клеток периферической крови, обработанных с помощью MEDI9447 и антител или гибридных белков, специфичных в отношении CTLA4, ОХ40, PD-1 и PD-L1. Первичные мононуклеарные клетки периферической крови человека инкубировали в течение 72 часов при смешанной лейкоцитарной реакции с MEDI9447 и/или антителами или гибридными белками, специфичными в отношении указанных мишеней. Цитокины (IFN-γ, IL-1β, TNF-α) в двойных супернатантах количественно определяли с помощью ELISA. Представленные данные представляют собой оптимальные комбинации доз антитела к CD73 с 4 различными соучаствующими средствами. Комбинация антитела к PD-1 и антитела к CD73 продемонстрировала значимую (р<0,05) синергию, которую определяли с помощью способа поверхностного отклика по Блиссу (Zhao et al.). Цитокиновый профиль указывает, что влиянию подвергались как миелоидные, так и лимфоидные линии. Были протестированы более 50 пар доноров.
[00154] На фигурах 19А и 19В показаны результаты анализа CD73 в комплексе с MEDI9447 посредством MS, основанной на водород-дейтериевом обмене (HDX-MS). На фигуре 19А показана теплокарта водород-дейтериевого обмена, показывающая те области CD73 (от N- до С-конца), которые подвергались уменьшенному поглощению дейтерия при связывании с MEDI9447. Относительный обмен между связанным с антителом и несвязанным CD73 изображен в зависимости от времени воздействия, при этом сниженный обмен изображен красным, повышенный обмен изображен синим и отсутствие изменений изображено белым. N-концевые области в положениях 132-143 и 182-187 проявляли наивысшую степень дифференциального обмена. На фигуре 8В показана кристаллическая структура мономера CD73, изображающая расположение HDX-идентифицируемого связывающего поверхностного участка (зеленовато-голубой) в N-концевом домене (желтый). Линкерная область CD73 и С-концевой домен представлены соответственно оранжевым и синим.
[00155] На фигурах 20А-20Е изображены результаты анализа посредством MS, основанного на водород-дейтериевом обмене (HDX-MS), показывающие области CD73 и MEDI9447, которые подвергались дифференциальному водородному обмену в свободном и связанном состояниях. На фигуре 20А изображены графики, отражающие относительное поглощение дейтерия (изменение массы в дальтонах) в зависимости от времени воздействия дейтерия внутри пептидов, охватывающих область 132-143. На фигуре 20В изображены графики, отражающие относительное поглощение дейтерия (изменение массы в дальтонах) в зависимости от времени воздействия дейтерия внутри пептидов, охватывающих область 182-187. На фигурах 20А и 20В поглощение только для CD73 показано в квадратах, а поглощение для CD73, связанного с Fab MEDI9447, показано красным. Пептидная последовательность, положение и масса указаны в поле графика. Чтобы сузить область, которая содержит последовательность, отображающую изменение в водородном обмене, и которая, как предсказано, формирует эпитоп, сравнивалось относительное изменение массы перекрывающихся пептидов. Например, пептиды, охватывающие положения 173-186, отображали дифференциальный обмен, тогда как в пептиде, охватывающем 173-181, различий не было. Таким образом, было установлено, что остатки, расположенные выше от 182, не являются дифференциально меченными. На фигуре 20С изображена разностная диаграмма DynamX для тяжелой цепи Fab MEDI9447. На фигуре 20D изображена разностная диаграмма DynamX для легкой цепи Fab MEDI9447. Что касается фигур 20С и 20D каждая точка на графике показывает разницу в поглощении дейтерия между комплексом CD73+Fab (положительные значения по оси y) и только Fab (отрицательные значения по оси y). Вертикальная черта представляет собой сумму разниц поглощения по моментам времени воздействия. Указаны CDR, демонстрирующие более низкое относительное поглощение при связывании Fab с CD73. На фигуре 9Е изображена разностная диаграмма DynamX для CD73 в отдельности (отрицательные значения по оси у) по сравнению с CD73, связанным с Fab (положительные значения по оси у). Показаны области Е1 (аа 132-143) и Е2 (аа 182-187). Горизонтальная ось соответствует проанализированным пептидам от N- до С-конца (слева направо). На график наложена пунктирная линия, показывающая 1,6 дальтон, 98% доверительный интервал отсечки для статистически значимых изменений.
[00156] На фигурах 21А-21Н изображены данные сенсорного чипа, показывающие, что эпитоп MEDI9447 находится в N-концевом домене CD73. Фигура 21А представляет собой график, отображающий данные сенсорного чипа для белка CD73 дикого типа. Белок CD73 дикого типа иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью поверхностного плазмонного резонанса (SPR). Фигура 21В представляет собой график, отображающий данные сенсорного чипа для белка CD73 с заменой N-концевого домена. Белок CD73 с заменой N-концевого домена иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью SPR. MEDI9447 не связывалось с CD73 при замене N-концевого домена. Фигура 21С представляет собой график, отображающий данные сенсорного чипа для белка CD73 с заменой N-концевого и С-концевого доменов. Белок CD73 с заменой N-концевого и С-концевого доменов иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью SPR. MEDI9447 не связывалось с CD73 при замене как N-концевого, так и С-концевого доменов. Фигура 21D представляет собой график, отображающий данные сенсорного чипа для белка CD73 с заменой линкерной области. Белок CD73 с заменой линкерной области иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью SPR. Замена только линкерной области не влияла на связывание. Фигура 21Е представляет собой график, отображающий данные сенсорного чипа для белка CD73 с заменой С-концевого домена. Белок CD73 с заменой С-концевого домена иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью SPR. Замена только С-концевого домена не влияла на связывание. Фигура 21F представляет собой график, отображающий данные сенсорного чипа для белка CD73 с заменой поверхностного участка Е1 (аа 132-143). Белок CD73 с заменой поверхностного участка E1 (аа 132-143) иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью SPR. Фигура 21G представляет собой график, отображающий данные сенсорного чипа для белка CD73 с заменой поверхностного участка Е2 (аа 182-187). Белок CD73 с заменой поверхностного участка Е2 (аа 182-187) иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью SPR. Фигура 21Н представляет собой график, отображающий данные сенсорного чипа для белка CD73 с заменой поверхностных участков Е1 (аа 132-143) и Е2 (аа 182-187). Белок CD73 с заменой поверхностных участков Е1 (аа 132-143) и Е2 (аа 182-187) иммобилизовали на сенсорном чипе HTG и связывание MEDI9447 в разбавлениях (от 5 нМ до 0,3 нМ) измеряли с помощью SPR. Что касается фигур 21F-21H, замена HDX поверхностного участка Е1 (аа 132-143) (фигура 21F) оказывала незначительное влияние на связывание, в отличие от замены HDX поверхностного участка Е2 (аа 182-187) отдельно (фигура 21G) или в комбинации с Е1 (фигура 21Н). Что касается фигур 21А-21Н, сенсограммы и наложенные подобранные кривые показаны в соответствующих цветах. Показатели кинетики для каждого анализа связывания приведены в таблице 16.
[00157] На фигуре 22 показано выравнивание последовательностей белка CD73 человека и курицы. Показаны только последовательности зрелого белка. В последовательности курицы выделены неконсервативные остатки. Аннотируются области с заменой последовательности курицы на последовательность человека для получения нокаутных вариантов (например, DSla, DSlb и т.д.).
[00158] На фигуре 23 показан результат связывания MEDI9447 с вариантами CD73. На фигуре 23 представлена таблица данных, показывающая связывание MEDI9447 с вариантами CD73. KD для вариантов, выделенных синим, изменяется более чем в 2 раза по сравнению с исходной конструкцией WT или КО. *Показатели кинетики, полученные из подбора 2:1 гетерогенного лиганда. **Нумерация соответствует последовательности курицы (129=133, 140=144 и 181=185 у человека).
[00159] На фигурах 24A-24F показано, что эпитоп MEDI9447 расположен на вершине N-концевого домена. На рисунке 13А показано, что в результате оценки связывания MEDI9447 с панелью вариантов CD73 (см. фигуры 22 и 23) выявили шесть положений, которые составляют сайт взаимодействия. Два из трех наиболее значимых остатков (выделенные блоки) расположены за пределами областей поверхностных участков HDX (выделены синим). Три менее важных остатка (розовые блоки) расположены внутри поверхностного участка HDX. На фигуре 24В приведена таблица, показывающая, что включение N185 и V144 (К180 является консервативным) в конструкцию CD73, содержащую последовательность N- и С-концевых доменов курицы, восстанавливает связывание менее чем в 20 раз от значения KD для CD73 дикого типа (разбавления MEDI9447 от 5 нМ до 0,3 нМ; в сравнение с фигурой 10В). На фигуре 24С в увеличенном масштабе изображены остатки эпитопов, расположенных в N-концевом домене CD73. Наиболее важные остатки для связывания выделены, а менее значимые положения (Y135, K136 и N187) показаны розовым. Поверхностный участок HDX закрашен синим. На фигуре 24D изображено поверхностное представление, показывающее, что эпитоп образует почти непрерывную связывающую поверхность. На фигуре 24Е показана кристаллическая структура открытой конформации CD73, показывающая положение эпитопа на апикальной, боковой поверхности N-концевого домена. На фигуре 24F показано, что расположение эпитопа находится на расстоянии от сайта связывания субстрата (аденозин, изображенный в кружках) и координационного сайта иона цинка (серый кружок) (боковые цепи - зеленовато-голубым цветом). Во всех кристаллических структурах N-концевой домен CD73, линкерная область и С-концевой домен изображены соответственно желтым, оранжевым и синим.
[00160] На фигурах 25А-25С показано, что MEDI9447 является неконкурентным ингибитором CD73-гидролиза AMP. На фигуре 25А представлен график, изображающий кинетику СБ73-фосфогидролиза AMP, измеренную в присутствии MEDI9447 или изотипически сходного контрольного mAb. Фигура 25В представляет собой график, показывающий, что MEDI9447 действует как неконкурентный ингибитор, так как он эквивалентно ингибирует гидролиз, независимо от концентрации субстрата. Напротив, АРСР, известный конкурентный ингибитор CD73, увеличивает Km, но не Vmax. Фигура 25С представляет собой график, изображающий эффект дозы IgG, Fab MEDI9447 или контрольного IgG на ингибирование CD73-гидролиза AMP. IgG MEDI9447 достигал максимального ингибирования при молярной стехиометрии 1:1с димером CD73 (стрелка). При высоких концентрациях, когда IgE MEDI9447 дан в избытке (>10 нМ), наблюдалась утрата ингибирования или "хук-эффект". Fab MEDI9447 и контрольный IgG не ингибировали CD73. Все эксперименты выполняли с использованием анализа CellTiterGlo, как описано в данном документе (RLU, относительные световые единицы).
[00161] На фигурах 26А-26С показано, что связывание mAb к CD73 (клон 0069) зависит от остатков N- и С-концевых доменов CD73. Фигура 26А представляет собой график, показывающий данные сенсорного чипа для CD73, меченного гистидином. CD73, меченный гистидином, иммобилизовали на биосенсоре HIS2 и связывание mAb А измеряли с помощью интерферометрии биослоя (BLI). Связывание mAb А с CD7 WT (синяя сенсограмма), нокаутным CD73 с заменой N-концевого домена (KO_1-291, зеленая сенсограмма) и нокаутным CD7 с заменой С-концевого домена (СО_311-523, зеленовато-голубая сенсограмма) показывает, что на связывание mAb влияют остатки как в N-, так и в С-концевом доменах. На фигуре 26В показана кристаллическая структура открытого и закрытого CD73 с указанием положения связывания mAb А с выделением "горячей точки" (аа 114-134 и 153-170), которая расположена вблизи поверхностного участка N- и С-терминальных доменов (N-концевой домен - желтый, линкер - оранжевый и С-концевой домен - синий). Картирование основывалось на данных связывания, показанных на фигурах 26А и 26С. На фигуре 26С показаны сенсограммы связывания mAb А с различными нокаутными вариантами CD73 с заменой доменов. Замена подобластей DS2c (аа 114-134) или DS3a (аа 153-170) инактивирует связывание. Все анализы связывания выполняли на приборе Octet QK384, как описано в данном документе.
[00162] На фигурах 27А-27С показано, что MEDI9447 ингибировал переход CD73 в конформационно-активную структуру. Фигура 27А представляет собой график, показывающий данные биосенсора для CD73 дикого типа. CD73 дикого типа иммобилизовали на биосенсоре HIS2 и связывание MEDI9447 (синяя сенсограмма) и антитела mAb А к CD73 (коричневая сенсограмма) измеряли с помощью BLI на Octet QK384. Когда CD73 предварительно инкубировали с Zn2+ и АРСР, у MEDI9447 сохранялось связывание (черная сенсограмма), однако у mAb А связывание утрачивалось (оранжевая сенсограмма). Фигура 27В представляет собой график, показывающий, что хотя предварительная инкубация CD73 с Zn2+ и АРСР вызывала утрату связывания mAb А (оранжевая сенсограмма), предварительная инкубация с MEDI9447 перед добавлением Zn2+ и АРСР восстанавливала связывание (фиолетовая сенсограмма). Связывание mAb А только с CD73 и CD73, предварительно инкубированным с MEDI9447 (но не Zn2+ и АРСР), показаны соответственно на синих и коричневых сенсограммах. На фигуре 27С показана предполагаемая модель, изображающая как MEDI9447 препятствует CD73 принять полностью закрытую, активную конформацию, индуцированную Zn2+ и АРСР. MEDI9947 может ограничивать переход в промежуточное состояние с более низкой аффинностью к mAb А.
[00163] На фигурах 28А и 28В показано связывание MEDI9447 или mAb А с CD73 при различных условиях, измеренное с помощью ВLI, как описано в данном документе, если иное не указано ниже. Фигура 28А представляет собой график, изображающий связывание антитела mAb А к CD73 с меченым гистидином CD73 дикого типа, иммобилизованным на биосенсоре HIS2. Через 100 сек. от исходного уровня захваченный CD73 инкубировали с Zn2+, АРСР и/или EDTA в течение 900 сек, а затем биосенсор инкубировали в 30 нМ mAb А в течение 600 сек. с целью измерения связывания. mAb А связывалось с CD73 (синяя сенсограмма), но не с CD73, предварительно инкубированным с Zn2+ и АРСР (фиолетовая сенсограмма). mAb А поддерживало связывание с CD73, предварительно инкубированным с АРСР и EDTA (зеленая сенсограмма) или Zn2+, АРСР и EDTA (золотистая сенсограмма). Хелатирующий эффект EDTA показывает, что для утраты связывания mAb А при инкубации CD73 с Zn2+ и АРСР требовался двухвалентный катион. Фигура 28В представляет собой график, показывающий, что Fab MEDI9447 или контрольный IgG не восстанавливали связывание mAb А с CD73, предварительно инкубированным с Zn2+ и АРСР. Анализ выполняли, как показано на фигуре 27В. Fab MEDI9447 или изотипически сходный контрольный IgG перед добавлением Zn2+ и АРСР предварительно инкубировали с CD73.
Иммобилизовали mAb А на биосенсоре, связанном только с CD73 (синяя сенсограмма), CD73, предварительно инкубированным либо с Fab MEDI9447 (голубая сенсограмма), либо с контрольным IgG (черная сенсограмма), но не с CD73, инкубированным с Zn2+ и АРСР (коричневая сенсограмма), либо с Fab (золотистая сенсограмма), либо контрольным IgG (фиолетовая сенсограмма), предварительно инкубированными с CD73 перед добавлением Zn2+ и АРСР.
[00164] На фигурах 29А и 29В показано, что связывание антитела mAb В к CD73 зависит от остатков в подобластях DS2b (аа 92-134) или DS2c (аа 114-134). Фигура 29А представляет собой таблицу, показывающую данные SEC-MALS, соответствующие фигурам 30А-30С. Для каждой смеси CD73 и либо MEDI9447, либо mAb В показаны соответствующее время удерживания SEC, Mw и полидисперсность образовавшихся комплексов. На фигуре 29В показано определение "горячей точки" связывания mAb В с CD73. Связывание mAb В с вариантами CD73, иммобилизованными на биосенсорах HIS2, измеряли с помощью ВLI, как описано для mAb А (клон 0069) на фигурах 26А-26С в соответствии с описанными в данном документе способами. Сенсограммы связывания показали, что замена либо подобласти DS2b (аа 92-134), либо DS2c (аа 114-134) инактивировала связывание mAb В.
[00165] На фигурах 30А-30С показано, что MEDI9447 образует междимерные мостики между растворимыми молекулами CD73. CD73 инкубировали с различными количествами MEDI9447 или mAb В к CD73 и анализировали с помощью SEC-MALS. Показаны SEC УФ-хроматограммы с временем удерживания белка на оси х и молярной массой, определенной с помощью MALS, на оси у. Фигура 30А представляет собой хроматограмму, показывающую, что при молярном соотношении 1:1 (зеленый след) MEDI9447 образовывал комплексы с CD73 при ~1,7 (∧) и ~6,6 (+) мегадальтон. Сопоставимые по размерам комплексы образовывались при более низких соотношениях MEDI9447 : CD73 (0,5:1 в синем, 0,1:1 в пурпурном). MEDI9447 и CD73 отдельно представлены соответственно в черном и красном УФ-следах. Фигура 30В представляет собой вид сверху вниз кристаллической структуры димера CD73, показывающий "горячую точку" связывания mAb В (фиолетовая) и эпитоп MEDI9447 (пурпурный и розовый). MAb В связывается с сайтом, расположенным близко к центральной борозде между димерами в открытой конформации. Фигура 30С представляет собой хроматограмму, показывающую, что при связывании CD73 с mAb В образовывался один преобладающий комплекс ~270-290 кДа (пик при ~7,2 мин.). Показанные УФ-следы представляют собой 1:1 mAb B:CD73 (красный), 0,5:1 (синий) и 0,1:1 (зеленый). Отдельно mAb А и CD73 имеют соответственно пурпурный и черный цвет.
[00166] На фиг. 31А-31D изображено, что связанный с поверхностью CD73 ингибировался с помощью форматов IgG и Fab MEDI9447. Фигура 31А представляет собой график, изображающий ингибирование гидролиза AMP иммобилизованным CD73 с помощью IgG, Fab MEDI9447 или контрольных антител. CD73 иммобилизовали посредством С-концевой гистидиновой метки на микротитрационном планшете, покрытом никелем, и измеряли ингибирование гидролиза AMP с помощью IgG, Fab MEDI9447 или контрольных антител, применяя набор для анализа с малахитовым зеленым, как описано в данном документе. IgG MEDI9447, но не контрольный IgG, дозозависимым образом ингибировал CD73-гидролиз AMP. Fab MEDI9447 также ингибировал активность CD73, но в значительно меньшей степени. На фигуре 31В изображены комплексы, содержащие Fab MEDI9447 (зеленый), связанный с антителом к Fd (xFd, красный). Когда Fab MEDI9447 (зеленый) связывали с одним плечом антитела к Fd (xFd, красный), а другое плечо связывали с неспецифичным поликлональным Fab (pFab, оранжевый), то ингибирование увеличивалась до уровня, сопоставимого с IgG MEDI9447 (Fab+XFd+pFab против IgG MEDI9447 и IgG MEDI9947+xFd+pFab) (см. фигуру 31 А). Фигура 31С представляет собой график, изображающий ингибирование гидролиза AMP GPI-якорным CD73 с помощью IgG, Fab MEDI9447 или контрольных антител. Ферментативную активность эндогенно экспрессируемого CD73 в клетках MDA-MB-231 измеряли с помощью анализа CellTiterGlo. По аналогии с иммобилизованным рекомбинантным CD73, IgG MEDI9447 ингибирует гидролиз AMP в большей степени, чем Fab, однако увеличение эффективного размера Fab MEDI9447 путем образования комплекса с антителом к Fd усиливает ингибирование. Фигура 31D представляет собой график, изображающий ингибирование гидролиза AMP растворимым CD73 (sCD73) с помощью IgG, Fab MEDI9447 или контрольных антител. Чтобы проверить, может ли xFd+MEDI9447 ингибировать растворимый CD73, гидролиз AMP измеряли, применяя набор для анализа с малахитовым зеленым. Fab MEDI9447 либо отдельно, либо связанный с одним плечом xFd, не ингибировал активность растворимого CD73. Напротив, связывание Fab MEDI9947 с обоими плечами xFd (Fab MEDI9447 + xFd) обеспечивало двухвалентность, приводящую к ингибированию CD73.
[00167] Фигура 32 представляет собой график, показывающий, что IgG MEDI94447 и Fab ингибировали CD73-гидролиз AMP. Активность CD73 измеряли в присутствии возрастающих концентраций антитела, применяя набор для анализа с малахитовым зеленым, как описано в данном документе. IgG MEDI9447 ингибировал гидролитическую активность CD73 дозозависимым образом, и хук-эффект или утрата ингибирования не наблюдались. Fab MEDI9447 также ингибировал функцию CD73, однако до более низкого уровня максимального ингибирования. Эксперимент выполняли дважды с сопоставимыми результатами. Данные показаны только от одного эксперимента.
[00168] На фигуре 33 изображена модель, показывающая, что ингибирование гидролитической активности CD73 с помощью MEDI9447 происходит посредством двойного механизма. IgG MEDI9447 (зеленый) ингибирует растворимый CD73 путем образования междимерных мостиков, которые препятствуют конформационному переходу в закрытое состояние. Моновалентно связанные IgG или Fab не ингибируют растворимый CD73. Когда CD73 связан с поверхностью, ингибирование может происходить посредством образования мостика между соседними димерами CD73 или стерического блокирования благодаря комплексу моновалентно связанного IgG или Fab/xFd (красный).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00169] В настоящем изобретении предусматриваются выделенные связывающие молекулы или их антигенсвязывающие фрагменты, которые специфически связываются с CD73. Согласно некоторым аспектам такие молекулы представляют собой антитела и их антигенсвязывающие фрагменты, которые специфически связываются с CD73. Также предусматриваются соответствующие полинуклеотиды, векторы, фармацевтические композиции, содержащие антитела к CD73 или их антигенсвязывающие фрагменты. Также предусматриваются способы получения, а также способы применения антител к CD73 и антигенсвязывающих фрагментов, раскрытых в данном документе, например, способы диагностики и способы лечения рака у субъекта (в виде непосредственной терапии, адъювантной терапии или комбинированной терапии). В настоящем изобретении также предусматриваются конъюгаты на основе антитело-лекарственное средство, полученные из CD73-связывающих молекул, раскрытых в данном документе. Кроме того, в настоящем изобретении предусматриваются терапевтические комбинации, содержащие антитела к CD73 (например, MEDI9447) и одно или несколько средств, оказывающих целенаправленное воздействие на дополнительные компоненты противоракового иммунного цикла, такие как антитела к PD-1, антитела к PD-L1 (например, MEDI4736), антитела к CTLA4; и способы применения таких комбинаций для снижения иммуносупрессии, опосредованной опухолью.
[00170] Для возможности более легкого понимания настоящего раскрытия сперва приведены определения некоторых терминов. В ходе подробного описания изложены дополнительные определения.
I. Определения
[00171] Прежде чем подробно описывать настоящее изобретение, следует понимать, что настоящее изобретение не ограничивается конкретными композициями или стадиями способов, поскольку таковые могут изменяться. Используемые в настоящем описании и прилагаемой формуле изобретения формы единственного числа включают определяемые объекты и во множественном числе, если из контекста явно не следует иное. Термины "один или несколько" и "по меньшей мере один" можно использовать в данном документе взаимозаменяемо.
[00172] Кроме того, "и/или" при использовании в данном документе следует рассматривать как конкретное раскрытие каждого из двух указанных признаков или компонентов с другим или без другого. Таким образом, подразумевается, что термин "и/или", используемый в данном документе в такой фразе как "А и/или В", включает "А и В", "А или В", "А" (отдельно) и "В" (отдельно). Аналогично, подразумевается, что термин "и/или", используемый в такой фразе, как "А, В и/или С", охватывает каждый из следующих аспектов: А, В и С; А, В или С; А или С; А или В; В или С; А и С; А и В; В и С; А (отдельно); В (отдельно) и С (отдельно).
[00173] Если не определено иное, то все используемые в данном документе технические и научные термины имеют то же значение, которое обычно понимает специалист обычной квалификации в данной области техники, к которой относится данное раскрытие. Например, Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show, 2nd ed., 2002, CRC Press; The Dictionary of Cell and Molecular Biology, 3rd ed., 1999, Academic Press и Oxford Dictionary Of Biochemistry And Molecular Biology, Revised, 2000, Oxford University Press обеспечивают специалиста общим словарем многих терминов, используемых в настоящем раскрытии.
[00174] Единицы измерения, префиксы и символы обозначены в их форме, принятой согласно Международной системе единиц (SI). Числовые диапазоны включают числа, определяющие диапазон. Если не указано иное, то аминокислотные последовательности записаны слева направо в направлении от амино- к карбокси-концу. Приведенные в данном документе заголовки не ограничивают различные аспекты, которые могут обеспечиваться ссылкой на описание в целом. Соответственно, термины, приведенные непосредственно ниже, более полно определены в полном объеме с помощью ссылки на описание.
[00175] Следует понимать, что какие бы аспекты ни описывались в данном документе формулировкой "содержащий", другие аналогичные аспекты, описываемые терминами "состоящий из" и/или "состоящий по сути из", также предусмотрены.
[00176] В данном документе аминокислоты обозначены либо с помощью их общеизвестных трехбуквенных символов, либо с помощью однобуквенных символов, рекомендованных Комиссией по биохимической номенклатуре IUPAC-IUB. Аналогично, нуклеотиды обозначены с помощью их общепринятых однобуквенных кодов.
[00177] Термин "полипептид CD73", используемый в данном документе, относится к белку CD73 (кластер дифференцировки 73), называемому также в литературе 5'-нуклеотидазой (5'-NT) или экто-5'-нуклеотидазой, который кодируется геном NT5E. См., например, Misumi et al. Eur. J. Biochem. 191(3): 563-9 (1990). Соответствующие последовательности форм CD73 человека и мыши доступны в базе данных Uniprot под номерами доступа Р21589 и Q61503 соответственно. При определении любых эпитопов антитела к CD73 используемая нумерация аминокислот представляет собой аминокислотный остаток зрелого белка CD73, который не содержит остатков сигнальной последовательности. Соответственно, антитело, связывающее аминокислоты Val144, Lys180 и Asn185, относится, например, к положениям аминокислот после расщепления сигнальной последовательности, т.е. к аминокислоте в зрелом белке.
[00178] Иллюстративный полипептид CD73 приведен ниже:

[00179] Были идентифицированы растворимые и мембранносвязанные формы CD73. См. Klemens et al., Biochem. Biophys. Res. Commun. 172(3): 1371-7 (1990). Кроме того, было идентифицировано несколько различных изоферментов. См. Rosi et al. Life Sci. 62(25):2257-66 (1998). Полноразмерный белок CD73 содержит 574 аминокислоты. Зрелый белок CD73 получают после удаления сигнальной последовательности (положения 1-26) и С-концевой области пропептида (положения 550-574). Коме того, аминокислоты 404-453 удаляются в изоформе 2 CD73 после альтернативного сплайсинга. Природные варианты также известны, например, вариант C358Y, вариант Т376А и вариант М379Т. См. Misumi et al., Eur. J. Biochem. 191:563-569 (1990); Otsuki et al. DNA Res. 12:117-126 (2005); Mungall et al. Nature 425:805-811 (2003); Hansen et al. Gene 167:307-312 (1995); Klemens et al. Biochem. Biophys. Res. Commun. 172:1371-1377 (1990); Knapp et al. Structure 20:2161-2173 (2012) или St. Hilaire et al. N. Engl. J. Med. 364:432-442 (2011), все из которых в полном объеме включены в данный документ с помощью ссылки.
[00180] Типичными заболеваниями, приводящими к изменению у пациента уровня CD73 в тканевых жидкостях, особенно в сыворотке, являются: травма ткани; реперфузионные повреждения, вызванные инфарктом миокарда или инсультом, трансплантацией органов или другими хирургическими операциями; рак или метастаз рака; или воспалительные состояния, вызванные вышеупомянутыми травмами или реперфузионными повреждениями, или хронические состояния, в том числе аллергические состояния, аутоиммунные заболевания и воспалительные заболевания. В качестве примеров таких хронических состояний можно упомянуть артрит, аллергические состояния, такие как астма, воспалительные состояния, такие как воспалительное заболевание кишечника или воспалительное состояние кожи, псориаз, болезнь Паркинсона, болезнь Алыдгеймера, аутоиммунные заболевания, диабет I типа или II типа, атеросклероз, рассеянный склероз, болезнь Крона или реакции отторжения из-за трансплантации органов. В частности, синдром системной воспалительной реакции на воспалительные заболевания (SIRS), острое повреждение легких (ALI), полиорганная недостаточность (MOF), ишемически-реперфузионное повреждение (IRI) и неблагоприятная лекарственная реакция (ADRS) приводят к изменениям уровня белка CD73 в тканевой жидкости. Соответственно, раскрытые в данном документе CD73-связывающие молекулы можно применять, например, для лечения или диагностики рака (например, рака толстой кишки, меланомы, рака молочной железы, лимфомы, немелкоклеточного рака легкого, лимфомы Ходжкина, неходжкинской лимфомы и лимфомы Беркитта, рака яичников, рака молочной железы, форм рака головы и шеи и рака поджелудочной железы). Кроме того, измерение уровней содержания CD73 в образце от пациента (например, тканевой жидкости) с применением CD73-связывающих молекул, раскрытых в данном документе, может применяться для отслеживания развития вышеописанных заболеваний, для оценки эффективности видов терапии, выбора пациентов для лечения конкретной терапией или принятия медицинских решений, например, о начале, окончании, прерывании или изменении определенного лечения.
[00181] Термины "ингибировать", "блокировать", "подавлять" и их грамматические варианты используются в данном документе взаимозаменяемо и относятся к любому статистически значимому снижению биологической активности, в том числе полному блокированию активности. Например, выражение "ингибирование" может относиться к снижению биологической активности приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100%. Соответственно, если термины "ингибирование" или "подавление" применяются, например, для описания эффекта в отношении ферментативной активности CD73, то данные термины относятся к способности антитела к CD73 или его антигенсвязывающего фрагмента статистически значимо снижать 5'-нуклеотидазную активность CD73 (катаболизирующую гидролиз аденозинмонофосфата, AMP, до аденозина) относительно CD73-опосредованной 5'-нуклеотидазой активности в необработанной (контрольной) клетке. Клетка, которая экспрессирует CD73, может быть встречающейся в природе клеткой или линией клеток (например, раковой клеткой), или может быть получена рекомбинантным путем посредством введения в клетку-хозяина нуклеиновой кислоты, кодирующей CD73. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент могут статистически значимо снижать 5'-нуклеотидазную активность растворимой формы CD73 в биологической жидкости. Согласно одному аспекту молекула, связывающаяся с CD73, например, антитело или его антигенсвязывающий фрагмент, ингибирует CD73-опосредованную 5'-нуклеотидазную активность по меньшей мере на 10%, по меньшей мере на 15% или по меньшей мере на 20%, по меньшей мере на 25% или по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40%, по меньшей мере на 45% или по меньшей мере на 50%, по меньшей мере на 55% или по меньшей мере на 60%, по меньшей мере на 65% или по меньшей мере на 70%, по меньшей мере на 75% или по меньшей мере на 80%, по меньшей мере на 85% или по меньшей мере на 90%, по меньшей мере на 95% или приблизительно на 100%, как определено, например, с помощью способов, описанных в примерах ниже, и/или способов, известных из уровня техники.
[00182] Термин "подавление активности CD73", используемый в данном документе, относится к способности молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента, статистически значимо снижать CD73-зависимую 5'-нуклеотидазную активность в клетке, экспрессирующей CD73, или образце, содержащем CD73. Согласно некоторым аспектам подавление активности CD73 может представлять собой по меньшей мере 10%, или по меньшей мере 15%, или по меньшей мере 20%, или по меньшей мере 25%, или по меньшей мере 30%, или по меньшей мере 35%, или по меньшей мере 40%, или по меньшей мере 45%, или по меньшей мере 50%, или по меньшей мере 55%, или по меньшей мере 60%, или по меньшей мере 65%, или по меньшей мере 70%, или по меньшей мере 75%, или по меньшей мере 80%, или по меньшей мере 85%, или по меньшей мере 90%, или по меньшей мере 95%, или приблизительно 100% снижение при приведении клетки или образца в контакт с молекулой, связывающейся с CD73, например, антителом или его антигенсвязывающим фрагментом по настоящему раскрытию, относительно активности CD73, измеренной при отсутствии молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента (контрольные условия).
[00183] Термины "антитело" или "иммуноглобулин", используемые в данном документе взаимозаменяемо, включают целые антитела и любой их антигенсвязывающий фрагмент или отдельные цепи.
[00184] Типичное антитело содержит по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (в данном документе сокращенно VH или VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов: CH1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (в данном документе сокращенно VL или VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена CL. Области VH и VL можно дополнительно подразделять на области гипервариабельности, называемые определяющими комплементарность областями (CDR), которые чередуются с более консервативными областями, называемыми каркасными областями (FW). Каждая VH и VL состоит из трех CDR и четырех FW, расположенных от амино-конца к карбокси- концу в следующем порядке: FW1, CDR1, FW2, CDR2, FW3, CDR3, FW4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента. Иллюстративные антитела по настоящему раскрытию включают антитела к CD73 (исходные и модифицированные на уровне генов зародышевой линии), клоны, оптимизированные по аффинности, оптимизированные антитела, не обладающие ADCC, конъюгированные антитела (например, ADC) и другие оптимизированные антитела (например, антитела, оптимизированные по времени полужизни в сыворотке крови, в том числе, например, с YTE мутациями, см. Dall'Acqua et al., J. Biol. Chem. 281:23514-24 (2006) и патент США № 7083784, которые включены в данный документ в полном объеме с помощью ссылки).
[00185] Термин "модифицирование на уровне генов зародышевой линии" означает, что аминокислоты в конкретных положениях в антителе подвергнуты обратной мутации с получением свойственных зародышевой линии аминокислот.
[00186] Термин "антитело" означает молекулу иммуноглобулина, которая распознает и специфически связывается с мишенью, такой как белок, полипептид, пептид, углевод, полинуклеотид, липид или комбинации вышеуказанных, посредством по меньшей мере одного антигенраспознающего сайта в пределах вариабельной области молекулы иммуноглобулина. Термин "антитело", используемый в данном документе, охватывает интактные поликлональные антитела, интактные моноклональные антитела, фрагменты антител (такие как Fab, Fab', F(ab')2 и Fv-фрагменты), одноцепочечные мутантные формы Fv (scFv), полиспецифичные антитела, такие как биспецифичные антитела, полученные по меньшей мере из двух интактных антител, химерные антитела, гуманизированные антитела, человеческие антитела, гибридные белки, содержащие антиген-определяющую часть антитела, а также любую другую модифицированную молекулу иммуноглобулина, содержащую антигенраспознающий сайт, при условии, что антитела проявляют требуемую биологическую активность.
[00187] Антитело может быть любым из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM или их подклассов (изотипов) (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2), исходя из идентичности их константных доменов тяжелой цепи, обозначаемых соответственно как альфа, дельта, эпсилон, гамма и мю. Разные классы иммуноглобулинов имеют разные и хорошо известные структуры субъединиц и пространственные конфигурации. Антитела могут быть "голыми" или конъюгированными с другими молекулами, такими как токсины, радиоактивные изотопы и т.д., с образованием ADC.
[00188] "Блокирующее" антитело или "антитело-антагонист" представляет собой антитело, которое ингибирует или снижает биологическую активность антигена, с которым оно связывается, такого как CD73. Согласно определенным аспектам блокирующие антитела или антитела-антагонисты практически или полностью ингибируют биологическую активность антигена. Желательно, чтобы биологическая активность снижалась по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или даже на 100%.
[00189] Термины "антитело CD73", "антитело, которое связывается с CD73" или "антитело к CD73" относится к антителу или его антигенсвязывающему фрагменту, которые способны к связыванию CD73 с достаточной аффинностью, благодаря чему данная молекула является пригодной в качестве терапевтического средства или диагностического реагента при целенаправленном воздействии на CD73. Степень связывания антитела к CD73 с неродственным белком, отличным от CD73, составляет менее чем приблизительно 10% от связывания антитела с CD73, как измерено, например, с помощью радиоиммунологического анализа (RIA), BIACORE™ (с применением рекомбинантного CD73 в качестве аналита и антитела в качестве лиганда или наоборот), либо других анализов связывания, известных из уровня техники. Согласно определенным аспектам антитело, которое связывается с CD73, характеризуется константой диссоциации (KD), составляющей ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤10 пМ, ≤1 пМ или ≤0,1 пМ. Термин "антитело к CD73" широко охватывает также молекулы, включающие в себя, например CDR антител, раскрытых в данном документе, включенные в каркас. Таким образом, фраза "выделенная связывающая молекула или ее антигенсвязывающий фрагмент, которые специфически связываются с CD73" будет относиться не только к антителам и их антигенсвязывающим фрагментам, но также будет относиться к молекуле, содержащей, например, один или несколько каркасов (таких как домен фибронектина III из фибронектина или тенасцина-3), включающих CDR антител, раскрытых в данном документе. См., например, публ. патента США № 20150098955, которая в полном объеме включена в данный документ с помощью ссылки.
[00190] Согласно одному варианту осуществления антитело к CD73 относится к антителу в формате IgG1-TM, так что домен Fc IgG1 содержит мутации L234, L235E и Р331, связывает растворимый и представленный на поверхности клетки CD73 и ингибирует ферментативную активность CD73. На фигурах 1A-1D представлены нуклеотидные и аминокислотные последовательности доменов VH и VL MEDI9447.
[00191] Под "полипептидом CTLA4" подразумевается полипептид, имеющий последовательность, по меньшей мере на 85% идентичную аминокислотной последовательности с № доступа в GenBank AAL07473.1, или его фрагмент, обладающий ингибирующей активностью в отношении Т-клеток. Последовательность AAL07473.1 приведена ниже:
Последовательность полипептида CTLA4 [Homo sapiens]

[00192] Под "молекулой нуклеиновой кислоты CTLA4" подразумевается полинуклеотид, кодирующий полипептид CTLA4. Иллюстративная последовательность молекулы нуклеиновой кислоты CTLA4 приведена в GenBank под № доступа AAL07473.
[00193] Под "антителом к CTLA4" подразумевается антитело, которое селективно связывает полипептид CTLA4. Иллюстративные антитела к CTLA4 описываются, например, в патентах США №№ 6682736, 7109003, 7123281, 7411057, 7824679, 8143379, 7807797 и 8491895 (в них тремелимумаб обозначается 11.2.1), которые включены в данный документ с помощью ссылки. Тремелимумаб является иллюстративным антителом к CTLA4. Последовательности тремелимумаба приводятся в перечне последовательностей, представленном в данном документе ниже (SEQ ID NO: 130-137).
[00194] Под "полипептидом PD-1" подразумевается полипептид или его фрагмент, имеющие последовательность, по меньшей мере на 85% идентичную аминокислотной последовательности из NCBI с № доступа NP_005009, и обладающие активностью связывания с PD-L1 и PD-L2. Последовательность NP_005009 приведена ниже.
Последовательность полипептида PD-1
№ доступа в NCBI NP_005009

[00195] Под "молекулой нуклеиновой кислоты PD-1" подразумевается полинуклеотид, кодирующий полипептид PD-1. Иллюстративная последовательность молекулы нуклеиновой кислоты PD-1 приведена в NCBI под № доступа NM_005018.
[00196] Под "антителом к PD-1" подразумевается антитело или его антигенсвязывающий фрагмент, которые селективно связывают полипептид PD-1. Иллюстративные антитела к PD-1 включают, например, пембролизумаб (KEYTRUDA®, ламбролизумаб, MK-3475), ниволумаб (Opdiva®, BMS-936558, MDX-1106, ONO-4538) или АМР-224.
[00197] Под "полипептидом PD-L1" подразумевается полипептид или его фрагмент, имеющие последовательность, по меньшей мере на 85%, 95% или 100% идентичную аминокислотной последовательности из NCBI с № доступа NP_001254635, и обладающие активностью связывания с PD-1 и CD80. Последовательность NP_001254635 приведена ниже.
Последовательность полипептида PD-L1
№ доступа в NCBI NP_001254 635

[00198] Под "молекулой нуклеиновой кислоты PD-L1" подразумевается полинуклеотид, кодирующий полипептид PD-L1. Иллюстративная последовательность молекулы нуклеиновой кислоты PD-L1 приведена в NCBI под № доступа NM_001267706.
[00199] Под "антителом к PD-L1" подразумевается антитело или его антигенсвязывающий фрагмент, которые селективно связывают полипептид PD-L1. Иллюстративные антитела к PD-L1 описаны, например, в US20130034559/US8779108 и US20140356353, которые включены в данный документ с помощью ссылки. MEDI4736 является иллюстративным антителом к PD-L1. Последовательности MEDI4736 приводятся в перечне последовательностей, представленном в данном документе ниже (SEQ ID NO: 138-145).
[00200] Термин "антигенсвязывающий фрагмент" относится к молекуле, содержащей часть интактного антитела, и, в частности, относится к молекуле, содержащей антиген-определяющие вариабельные области интактного антитела. Из уровня техники известно, что антигенсвязывающая функция антитела может быть выполнена фрагментами полноразмерного антитела. Примеры фрагментов антитела включают без ограничения Fab, Fab', F(ab')2 и Fv-фрагменты, линейные антитела, одноцепочечные антитела и полиспецифические антитела, образованные из фрагментов антитела.
[00201] "Моноклональное антитело" относится к гомогенной популяции антител, вовлеченных в высокоспецифичное распознавание и связывание одной антигенной детерминанты или эпитопа. Этим они отличаются от поликлональных антител, которые, как правило, включают разные антитела, направленные против разных антигенных детерминант.
[00202] Термин "моноклональное антитело" охватывает как интактные, так и полноразмерные моноклональные антитела, а также фрагменты антител (такие как Fab, Fab', F(ab')2, Fv), одноцепочечные вариабельные фрагменты (scFv), гибридные белки, содержащие часть антитела, и любую другую модифицированную молекулу иммуноглобулина, содержащую сайт распознавания антигена. Кроме того, "моноклональное антитело" относится к таким антителам, которые получены любым из ряда способов, в том числе без ограничения с помощью гибридомы, селекции с помощью фагового дисплея, рекомбинантной экспрессии и с использованием трансгенных животных (например, экспрессии человеческого антитела в трансгенной мыши).
[00203] Термин "гуманизированное антитело" относится к антителу, полученному из иммуноглобулина, отличного от иммуноглобулина человека (например, мыши), которое было сконструировано так, чтобы содержание последовательностей не человеческого (например, мышиного) происхождения было минимальным. Как правило, гуманизированные антитела представляют собой иммуноглобулины человека, в которых остатки CDR заменены остатками CDR отличных от человека видов (например, мыши, крысы, кролика или хомяка), которые характеризуются требуемой специфичностью, аффинностью и способностью (Jones et al., 1986, Nature, 321:522-525; Riechmann etal, 1988, Nature, 332:323-327; Verhoeyen etal, 1988, Science, 239:1534-1536). В некоторых случаях аминокислотные остатки каркаса (FW) иммуноглобулина человека заменены соответствующими остатками антитела из отличных от человека видов, которые характеризуются требуемой специфичностью, и/или аффинностью, и/или способностью.
[00204] Гуманизированное антитело можно дополнительно модифицировать с помощью замены дополнительных остатков в каркасной области Fv и/или в замененных остатках нечеловеческого происхождения для усовершенствования и оптимизации специфичности, аффинности и/или способности антитела. В целом, гуманизированное антитело будет содержать практически все из по меньшей мере одного, и, как правило, двух или трех вариабельных доменов, содержащих все или практически все CDR-области, которые соответствуют таковым в иммуноглобулине, отличном от иммуноглобулина человека, тогда как все или практически все из FW-областей являются таковыми из консенсусной последовательности иммуноглобулина человека. Гуманизированное антитело также может содержать по меньшей мере часть константной области или домена (Fc) иммуноглобулина, как правило, иммуноглобулина человека. Примеры способов, используемых для получения гуманизированных антител, описаны в патентах США №№ 5225539 или 5639641.
[00205] "Вариабельная область" антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела либо отдельно, либо в комбинации.
Каждые из вариабельных областей тяжелой и легкой цепей состоят из четырех FW-областей, соединенных с тремя CDR-областями. В каждой цепи CDR удерживаются в непосредственной близости с помощью FW-областей, и вместе с CDR из другой цепи участвуют в образовании антигенсвязывающего участка антител. Существует по меньшей мере две методики определения CDR: (1) подход, основанный на межвидовой вариабельности последовательностей (т.е. Kabat et al. Sequences of Proteins of Immunological Interest, (5th ed., 1991, National Institutes of Health, Bethesda Md.)); и (2) подход, основанный на кристаллографических исследованиях комплексов антиген-антитело (Al-lazikani et al. (1997) J. Molec. Biol. 273:927-948)). Кроме того, для определения CDR в данной области техники иногда используют комбинации этих двух подходов.
[00206] В основном систему нумерации согласно Kabat применяют при обозначении остатка в вариабельном домене (примерно остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., Sequences of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)).
[00207] Фразы "нумерация положений аминокислот согласно Kabat", "положение по Kabat" и их грамматические варианты относятся к системе нумерации, применяемой к вариабельным доменам тяжелой цепи или вариабельным доменам легкой цепи, при компиляции антител согласно Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991). При применении данной системы нумерации фактическая линейная аминокислотная последовательность может содержать меньшее количество аминокислот или дополнительные аминокислоты, соответствующие укорочению FW или CDR вариабельного домена или вставке в них. Например, вариабельный домен тяжелой цепи может содержать вставку одной аминокислоты (остаток 52а согласно Kabat) после остатка 52 в Н2 и вставленные остатки (например, остатки 82а, 82b и 82с и т.д. согласно Kabat) после остатка 82 FW тяжелой цепи.
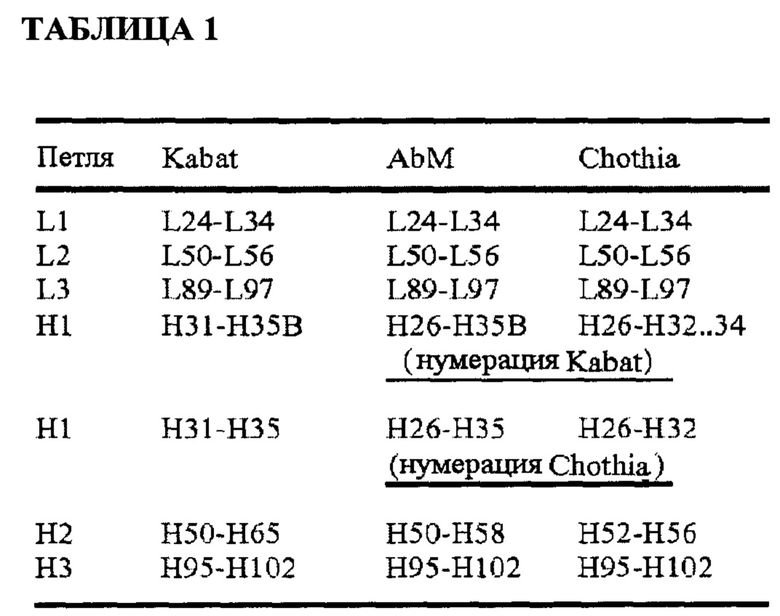
[00208] Нумерацию остатков согласно Kabat можно определить для данного антитела с помощью выравнивания областей антитела с гомологией последовательности антитела со "стандартной" последовательностью, пронумерованной согласно Kabat. В отличие от этого, Chothia обращается к расположению структурных петель (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987)). Конец петли CDR-H1 согласно Chothia при нумерации с использованием правила нумерации согласно Kabat варьирует от Н32 до Н34, в зависимости от длины петли (это обусловлено тем, что согласно схеме нумерации Kabat вставки размещаются в Н35А и Н35В; при этом если как 35А, так и 35В отсутствуют, то петля заканчивается на 32; если присутствует только 35А, то петля заканчивается на 33; если присутствуют как 35А, так и 35В, то петля заканчивается на 34). Определение гипервариабельных областей согласно АbМ представляет собой компромисс между определением CDR согласно Kabat и структурных петель согласно Chothia, и применяется в программном обеспечении для моделирования антител с помощью Oxford Molecular's AbM.
[00209] IMGT (ImMunoGeneTics) предусматривает также систему нумерации для вариабельных областей иммуноглобулинов, в том числе CDR. См., например, Lefranc, М.Р. et al., Dev. Comp. Immunol. 27: 55-77(2003), которая включена в данный документ с помощью ссылки. Система нумерации IMGT была основана на выравнивании более 5000 последовательностей, данных о структуре и определении характеристик гипервариабельных петель, и она обеспечивает возможность легкого сравнения вариабельных и CDR-областей для всех видов. В соответствии со схемой нумерации IMGT VH-CDR1 находится в положениях 26-35, VH-CDR2 находится в положениях 51-57, VH-CDR3 находится в положениях 93-102, VL-CDR1 находится в положениях 27-32, VL-CDR2 находится в положениях 50-52 и VL-CDR3 находится в положениях 89-97.
[00210] EU-индекс или система нумерации EU основаны на последовательной нумерации первого секвенированного IgG человека (антитело EU). Поскольку наиболее частой ссылкой для данной системы является руководство по последовательностям согласно Kabat (Kabat et al., 1991), EU-индекс иногда ошибочно используют синонимично с индексом Kabat. EU-индекс не предусматривает вставки и делеции, и поэтому в некоторых случаях сравнения положений IgG среди подклассов IgG и видов могут быть неясными, в частности в шарнирных областях. Тем не менее, эта система подходит для прямого сравнения между Fc-областями в многочисленных исследованиях структурной функции Fc. Соответственно, схема нумерации, используемая в данном описании для замен и вставок в Fc-областях, представляет собой EU-индекс по Kabat. Напротив, схема нумерации, используемая в данном описании для вариабельных областей (VH и VL), представляет собой обычную нумерацию согласно Kabat.
[00211] Описанные последовательности VH CDR, используемые в настоящем описании, соответствуют местоположениям согласно классической нумерации по Kabat, а именно VH-CDR1 согласно Kabat находится в положениях 31-35, VH-CDR2 находится в положениях 50-65 и VH- CDR3 находится в положениях 95-102. VL-CDR1, VL-CDR2 и VL-CDR3 также соответствуют местоположениям согласно классической нумерации по Kabat, а именно положениям 24-34, 50-56 и 89-97 соответственно.
[00212] Как используется в данном документе, Fc-область включает в себя полипептиды, содержащие константную область антитела за исключением первого домена константной области иммуноглобулина. Таким образом, Fc относится к последним двум доменам константной области иммуноглобулинов IgA, IgD и IgG и последним трем доменам константной области иммуноглобулинов IgE и IgM, а также гибкой шарнирной области на N-конце этих доменов. В случае IgA и IgM, Fc может включать в себя J-цепь. В случае IgG, Fc содержит домены иммуноглобулина С-гамма-2 и С-гамма-3 (Сγ2 и Сγ3) и шарнирный участок между С-гамма-1 (Cγ1) и С-гамма-2 (Сγ1).
[00213] Несмотря на то, что границы Fc-области могут варьировать, Fc-область тяжелой цепи IgG человека обычно определяется как содержащая остатки С226 или Р230 на ее карбоксильном конце, при этом нумерация приведена в соответствии с EU-индексом, как изложено у Kabat (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). Fc может относиться к данной области в отдельности или к данной области в контексте антитела, фрагмента антитела или Fc-гибридного белка. Полиморфизмы наблюдались в ряде различных положений Fc, в том числе без ограничения в положениях 270, 272, 312, 315, 356 и 358, пронумерованных согласно EU-индексу, и следовательно могут существовать незначительные различия между представленной последовательностью и последовательностями из уровня техники.
[00214] Термин "антитело человека" означает антитело, вырабатываемое в организме человека, или антитело с аминокислотной последовательностью, соответствующей антителу, вырабатываемому в организме человека, полученное с использованием любых методик, известных из уровня техники (например, рекомбинантной экспрессии в культивируемых клетках или экспрессии в трансгенных животных). Таким образом, термин "антитело человека" охватывает также антитело с аминокислотной последовательностью, соответствующей антителу, изначально вырабатываемому в организме человека (или его сконструированному варианту или производному), но экспрессированному в системе, не имеющей отношения к человеку (например, полученное с помощью химического синтеза; экспрессированное рекомбинантным путем в клетках микроорганизмов, млекопитающих или насекомых или экспрессированное в организме животного). Соответственно, антитело, полученное от субъекта-человека или из клеток человека (например, гибридомы или линии клеток, экспрессирующих рекомбинантное антитело или его фрагмент) и затем экспрессированное в организме животного, например мыши, считается человеческим антителом. Это определение человеческого антитела включает в себя интактные или полноразмерные антитела, их фрагменты и/или антитела, содержащие по меньшей мере один полипептид тяжелой и/или легкой цепи человеческого происхождения, как, например, антитело, содержащее полипептиды легкой цепи мышиного происхождения и тяжелой цепи человеческого происхождения.
[00215] Термин "химерные антитела" относится к антителам, в которых аминокислотная последовательность молекулы иммуноглобулина происходит от двух или более видов животных. Как правило, вариабельная область как легкой, так и тяжелой цепей соответствует вариабельной области антител, происходящих от одного вида млекопитающих (например, мыши, крысы, кролика и т.д.) с необходимой специфичностью, и/или аффинностью, и/или способностью, в то время как константные области гомологичны последовательностям антител, происходящих от другого вида (обычно человека), во избежание вызывания иммунного ответа у тех видов.
[00216] Термин "эпитоп", используемый в данном документе, относится к антигенной белковой детерминанте, способной связываться с антителом к CD73 или с CD73-связывающей молекулой, раскрытой в данном документе. Эпитопы обычно состоят из химически активных поверхностных групп молекул, таких как боковые цепи аминокислот или Сахаров, и обычно обладают специфическими характеристиками трехмерной структуры, а также специфическими характеристиками заряда. Часть антитела или связывающей молекулы, которая распознает эпитоп, называется паратопом. Эпитопы белковых антигенов разделяют на две категории, конформационные эпитопы и линейные эпитопы, исходя из их структуры и взаимодействия с паратопом. Конформационный эпитоп состоит из прерывистых участков аминокислотной последовательности антигена. Эти эпитопы взаимодействуют с паратопом за счет 3D-особенностей поверхности и формы или третичной структуры антигена. В отличие от этого, линейные эпитопы взаимодействуют с паратопом за счет их первичной структуры. Линейный эпитоп образован непрерывной последовательностью аминокислот антигена.
[00217] Термин "антителосвязывающий сайт" относится к области в антигене (например, CD73), содержащей непрерывный или прерывистый участок (т.е. эпитоп), с которым специфически связывается комплементарное антитело. Таким образом, антителосвязывающий сайт может содержать дополнительные участки в антигене, которые находятся вне эпитопа и которые могут определять такие свойства, как аффинность связывания и/или стабильность, или влияют на такие свойства, как ферментативная активность антигена или димеризация. Соответственно, даже если два антитела связываются с одним и тем же эпитопом в пределах антигена, если между молекулами антител устанавливаются отличающиеся межмолекулярные контакты с аминокислотами вне эпитопа, то такие антитела рассматриваются как связывающиеся с разными антителосвязывающими сайтами.
[00218] В основном "аффинность связывания" относится к силе суммы всех нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не указано иное, "аффинность связывания", как используется в данном документе, относится к присущей аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающейся пары (например, антителом и антигеном). В основном аффинность молекулы X к ее партнеру Y можно представить константой диссоциации (KD). Аффинность можно измерить с помощью способов, общеизвестных из уровня техники, в том числе описанных в данном документе. В основном антитела с низкой аффинностью связывают антиген медленно и склонны легко диссоциировать, тогда как антитела с высокой аффинностью, как правило, связывают антиген быстрее и склонны оставаться дольше в связанном состоянии. Из уровня техники известен ряд способов измерения аффинности связывания, любой из которых можно применять для целей настоящего раскрытия.
[00219] Как правило, "эффективность" выражают в виде значения IC50 в нМ, если не указано иное. IC50 представляет собой медианную ингибирующую концентрацию антигенсвязывающей молекулы. В функциональных анализах IC50 представляет собой концентрацию, которая снижает биологический ответ на 50% от его максимума. В исследованиях по связыванию лиганда IC50 представляет собой концентрацию, которая снижает уровень связывания рецептора на 50% от максимального уровня специфического связывания. IC50 можно рассчитать любым из ряда способов, известных в данной области техники. Повышение эффективности может быть определено с помощью измерения, например, относительно исходного антитела (например, исходного антитела перед модифицированием на уровне генов зародышевой линии или исходного антитела перед оптимизацией по аффинности).
[00220] Кратность повышения эффективности антитела или полипептидов по настоящему раскрытию по сравнению с исходным антителом может быть по меньшей мере приблизительно 2-кратной, по меньшей мере приблизительно 4-кратной, по меньшей мере приблизительно 6-кратной, по меньшей мере приблизительно 8-кратной, по меньшей мере приблизительно 10-кратной, по меньшей мере приблизительно 15-кратной, по меньшей мере приблизительно 20-кратной, по меньшей мере приблизительно 25-кратной, по меньшей мере приблизительно 30-кратной, по меньшей мере приблизительно 40-кратной, по меньшей мере приблизительно 50-кратной, по меньшей мере приблизительно 60-кратной, по меньшей мере приблизительно 70-кратной, по меньшей мере приблизительно 80-кратной, по меньшей мере приблизительно 90-кратной, по меньшей мере приблизительно 100-кратной, по меньшей мере приблизительно 110-кратной, по меньшей мере приблизительно 120-кратной, по меньшей мере приблизительно 130-кратной, по меньшей мере приблизительно 140-кратной, по меньшей мере приблизительно 150- кратной, по меньшей мере приблизительно 160-кратной, по меньшей мере приблизительно 170-кратной или по меньшей мере приблизительно 180-кратной или больше.
[00221] "Антителозависимая клеточноопосредованная цитотоксичность" или "ADCC" относится к форме цитотоксичности, при которой секретируемые иммуноглобулины, связываемые Fc-рецепторами (FcR), представленными на определенных цитотоксических клетках (например, естественных клетках-киллерах (NK), нейтрофилах и макрофагах), дают возможность этим цитотоксическим эффекторным клеткам специфично связываться с клеткой-мишенью, несущей антиген, а затем уничтожать клетку-мишень с помощью цитотоксинов. Специфические антитела IgG с высокой аффинностью, направленные на поверхность клеток-мишеней, "активизируют" цитотоксические клетки и исключительно необходимы для такого уничтожения. Лизис клетки-мишени является внеклеточным, требует непосредственного межклеточного контакта и не вовлекает систему комплемента. Предполагается, что в дополнение к антителам другие белки, содержащие Fc-области, в частности Fc-гибридные белки, со способностью специфично связываться с клеткой-мишенью, несущей антиген, будут способны влиять на клеточно-опосредованную цитотоксичность. Для краткости клеточноопосредованная цитотоксичность, возникающая в результате активности Fc-гибридного белка, также упоминается в данном документе как ADCC-активность.
[00222] Полипептид, антитело, полинуклеотид, вектор, клетка или композиция, которые являются "выделенными", представляют собой полипептид, антитело, полинуклеотид, вектор, клетку или композицию, которые находятся в форме, не встречающейся в природе. Выделенные полипептиды, антитела, полинуклеотиды, векторы, клетки или композиции включают таковые, которые были очищены до такой степени, что они больше не находятся в такой форме, в которой они встречаются в природе. Согласно некоторым аспектам выделенное антитело, полинуклеотид, вектор, клетка или композиция являются практически чистыми.
[00223] Выражение "субъект" относится к любому животному (например, млекопитающему), в том числе без ограничения к людям, отличным от человека приматам, грызунам и т.п., которое будет реципиентом конкретного лечения. Как правило, термины "субъект" и "пациент" используются в данном документе взаимозаменяемо, когда ссылаются на субъекта-человека.
[00224] Термин "фармацевтическая композиция" относится к препарату, который находится в такой форме, которая обеспечивает эффективность биологической активности активного ингредиента (например, раскрытой в данном документе молекулы, связывающейся с CD73), и который не содержит дополнительных компонентов, которые неприемлемо токсичны для субъекта, которому будут вводить композицию. Такая композиция может быть стерильной.
(00225] "Эффективное количество" раскрытой в данном документе молекулы, связывающейся с CD73, представляет собой количество, достаточное для осуществления конкретно поставленной цели. "Эффективное количество" в отношении поставленной цели можно определить опытным путем и стандартным способом.
[00226] Термин "терапевтически эффективное количество" относится к количеству раскрытой в данном документе молекулы, связывающейся с CD73, или другого лекарственного средства, эффективному для "лечения" заболевания или нарушения у субъекта или млекопитающего.
[00227] Слово "метка" при использовании в данном документе относится к выявляемому соединению или композиции, которые являются слитыми (например, генетически слитыми) или конъюгированными (например, химически конъюгированными) прямо или опосредованно с раскрытой в данном документе молекулой, связывающейся с CD73, так, что образуют "меченую" молекулу, связывающуюся с CD73. Метка может быть выявляемой сама по себе (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментной метки, может катализировать химическое превращение субстратного соединения или композиции, которое является выявляемым.
[00228] Такие термины как "дериватизируемая группа" и "дериватизируемая функциональная группа" используются взаимозаменяемо и относятся к функциональной группе, которая способна вступать в реакцию с возможностью образования ковалентной связи между раскрытой в данном документе молекулой, связывающейся с CD73 (например, антителом к CD73), и другим веществом. Согласно некоторым аспектам такое вещество представляет собой терапевтическое средство (например, цитотоксин), выявляемую метку, полимер (например, PEG) и т.д. Иллюстративные дериватизируемые группы включают тиольную, гидроксильную, амино, карбокси и амидную, а также их модифицированные формы, такие как активированные или защищенные формы.
[00229] Такие термины как "осуществление лечения", или "лечение", или "лечить" или "облегчение", или "облегчать" относятся как к (1) терапевтическим мероприятиям, с помощью которых излечивают, замедляют, ослабляют симптомы диагностированного патологического состояния или нарушения и/или останавливают его прогрессирование, так и к (2) профилактическим или предупреждающим мероприятиям, с помощью которых предупреждают и/или замедляют развитие целевого патологического состояния или нарушения. Таким образом, к нуждающимся в лечении относят тех, у кого уже есть нарушение; тех, кто предрасположен к развитию нарушения, и тех, у кого нужно предупредить развитие нарушения. Согласно определенным аспектам субъекта успешно "лечат" от рака в соответствии со способами по настоящему раскрытию, если пациент характеризуется, например, полной, частичной или временной ремиссией определенного типа рака.
[00230] Термины "рак", "опухоль", "раковый" и "злокачественный" относятся к физиологическому состоянию у млекопитающих, которое, как правило, характеризуется неконтролируемым клеточным ростом, или описывают его. Примеры форм рака включают без ограничения карциному, в том числе формы аденокарциномы, лимфомы, бластомы, меланомы, саркомы и лейкемии. Более конкретно примеры таких форм рака включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, рак желудочно-кишечного тракта, лимфому Ходжкина и неходжкинскую лимфому, рак поджелудочной железы, глиобластому, глиому, рак шейки матки, рак яичников, рак печени, например, печеночную карциному и гепатому, рак мочевого пузыря, рак молочной железы (в том числе гормонально опосредованный рак молочной железы, см., например, Innes et al. (2006) Br. J. Cancer 94:1057-1065), рак толстой кишки, колоректальный рак, карциному эндометрия, миелому (например, множественную миелому), карциному слюнных желез, рак почки, например, почечно-клеточную карциному и формы опухоли Вильмса, базальноклеточную карциному, меланому, рак предстательной железы, рак вульвы, рак щитовидной железы, рак яичка, рак пищевода, различные формы рака головы и шеи, а также формы рака муцинозного происхождения, например, муцинозный рак яичников, холангиокарциному (печени) и папиллярную карциному почки. Согласно некоторым аспектам термин "рак", используемый в данном документе, относится в частности к форме рака, характеризующейся экспрессией CD73. Согласно некоторым конкретным аспектам термин рак относится к формам рака, характеризующимся экспрессией CD73 на низких уровнях. Согласно некоторым аспектам термин рак, используемый в данном документе, конкретно относится к раку, характеризующемуся экспрессией CD73 (например, рак толстой кишки, рак молочной железы, лимфома, немелкоклеточная карцинома).
[00231] "Полинуклеотид" или "нуклеиновая кислота", как используется в данном документе взаимозаменяемо, относятся к полимерам из нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания, и/или их аналоги, или любой субстрат, который можно встроить в полимер с помощью ДНК- или РНК-полимеразы. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Предыдущее описание применимо ко всем указанным в данном документе полинуклеотидам, в том числе РНК и ДНК.
[00232] Термин "вектор" означает конструкцию, способную доставлять в клетку-хозяина, и согласно некоторым аспектам экспрессировать в клетке-хозяине один или несколько представляющих интерес ген(генов) или последовательность(последовательностей). Примеры векторов включают без ограничения вирусные векторы, векторы экспрессии "голой" ДНК или РНК, плазмиды, космиды или фаговые векторы, векторы экспрессии ДНК или РНК, ассоциированные с катионными конденсирующими средствами, векторы экспрессии ДНК или РНК, инкапсулированные в липосомы, а также определенные эукариотические клетки, такие как клетки-продуценты.
[00233] Термины "полипептид", "пептид" и "белок" используются в данном документе взаимозаменяемо для обозначения полимеров из аминокислот любой длины. Полимер может быть линейным или разветвленным, может содержать модифицированные аминокислоты и может быть разделен не аминокислотами. Термины также охватывают полимер из аминокислот, который был модифицирован в природных условиях или посредством вмешательства; например, посредством образования дисульфидной связи, гликозилирования, липидизации, ацетилирования, фосфорилирования или любой другой манипуляции или модификации, такой как конъюгация с метящим компонентом. В данное определение также включены, например, полипептиды, содержащие один или несколько аналогов аминокислоты (в том числе, например, не встречающиеся в природе аминокислоты и т.п.), а также другие модификации, известные из уровня техники. Следует понимать, что поскольку полипептиды по настоящему раскрытию исходят из антител, согласно определенным аспектам данные полипептиды могут встречаться в виде одиночных цепей или связанных цепей.
[00234] "Рекомбинантный" полипептид или белок относятся к полипептиду или белку, полученным посредством технологии рекомбинантных ДНК. Полученные рекомбинантным путем полипептиды и белки, экспрессирующиеся в сконструированных клетках-хозяевах, считаются выделенными, поскольку являются нативными или рекомбинантными полипептидами, разделенными, фракционированными или частично либо в значительной степени очищенными с помощью любой подходящей методики. Раскрытые в данном документе полипептиды можно получить рекомбинантным путем с применением известных в данной области техники способов. В качестве альтернативы, раскрытые в данном документе белки и пептиды можно синтезировать химическим путем.
[00235] Термин "аминокислотная замена" относится к замене остатка аминокислоты, находящегося в исходной последовательности, на остаток другой аминокислоты. Аминокислоту можно заменить в исходной последовательности, например, посредством химического синтеза пептидов или посредством рекомбинантных способов, известных из уровня техники. Следовательно, ссылки на выражения "замещение в положении X" или "замещение в положении X" относятся к замещению аминокислоты, находящейся в положении X, на альтернативный аминокислотный остаток. Согласно некоторым аспектам модели замещения можно описать согласно схеме AXY, где А представляет собой однобуквенный код, соответствующий аминокислоте, находящейся в природных условиях в положении X, и Y представляет собой замещающий аминокислотный остаток. Согласно другим аспектам модели замещения можно описать согласно схеме XY, где Y представляет собой однобуквенный код, соответствующий аминокислотному остатку, замещающему аминокислоту, находящуюся в природных условиях в положении X.
[00236] "Консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток замещается аминокислотным остатком со сходной боковой цепью. Семейства аминокислотных остатков со сходными боковыми цепями определены в данной области техники и включают в себя основные боковые цепи (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагиновую кислоту, глутаминовую кислоту), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, если аминокислота в полипептиде заменена на другую аминокислоту из того же семейства боковых цепей, то замена считается консервативной. Согласно другому аспекту ряд аминокислот можно консервативно заменить на структурно похожий ряд, который отличается по порядку и/или составу представителей семейства боковых цепей.
[00237] Неконсервативные замены включают замены, в которых (i) остаток, имеющий электроположительную боковую цепь (например, Arg, His или Lys), заменяется на электроотрицательный остаток или с помощью него (например, Glu или Asp), (ii) гидрофильный остаток (например, Ser или Thr) заменяется на гидрофобный остаток или с помощью него (например, Ala, Leu, Ile, Phe или Val), (iii) цистеин или пролин заменяются на любой другой остаток или с помощью них или (iv) остаток, имеющий объемную гидрофобную или ароматическую боковую цепь (например, Val, His, Ilе или Тrр), заменяется на остаток, имеющий меньшую боковую цепь (например, Ala, Ser), или без боковой цепи, или с помощью него (например, Gly).
[00238] Другие замены могут быть легко идентифицированы специалистами обычной квалификации в данной области техники. Например, для аминокислоты аланин может быть взята любая замена из D-аланина, глицина, бета-аланина, L-цистеина и D-цистеина. Для лизина заместитель может быть любым из D-лизина, аргинина, D-аргинина, гомо-аргинина, метионина, D-метионина, орнитина или D-орнитина. В основном заменами в функционально важных областях, которые, как можно ожидать, вызывают изменения свойств выделенных полипептидов, являются такие, в которых (i) полярный остаток, например серии или треонин, заменяется на гидрофобный остаток (или с помощью него), например, лейцин, изолейцин, фенилаланин или аланин; (ii) остаток цистеина заменяется на любой другой остаток (или с помощью него); (iii) остаток, имеющий электроположительную боковую цепь, например лизин, аргинин или гистидин, заменяется на остаток, имеющий электроотрицательную боковую цепь (или с помощью него), например глутаминовую кислоту или аспарагиновую кислоту; или (iv) остаток, имеющий объемную боковую цепь, например фенилаланин, заменяется на остаток, не имеющий такой боковой цепи (или с помощью него), например глицин. Вероятность того, что одна из вышеуказанных неконсервативных замен может изменить функциональные свойства белка также коррелирует с положением замены относительно функционально важных областей белка; при этом некоторые неконсервативные замены могут, соответственно, оказывать незначительный эффект в отношении биологических свойств или не оказывать такого эффекта.
[00239] Термин "вставка аминокислоты" относится к введению нового аминокислотного остатка между двумя аминокислотными остатками, находящимися в исходной последовательности. Аминокислоту можно вставить в исходную последовательность, например, посредством химического синтеза пептидов или посредством рекомбинантных способов, известных из уровня техники. Соответственно, используемые в данном документе фразы "вставка между положениями X и Y'' или "вставка между положениями согласно Kabat X и Y'', где X и Y соответствуют аминокислотным положениям (например, вставка аминокислоты цистеина между положениями 239 и 240), относятся к вставке аминокислоты между положениями X и Y, а также к вставке в нуклеотидную последовательность кодона, кодирующего аминокислоту, между кодонами, кодирующими аминокислоты в положениях X и Y. Модели вставок можно описать согласно схеме AXins, где А представляет собой однобуквенный код, соответствующий подлежащей вставке аминокислоте, а X представляет собой положение, предшествующее вставке.
[00240] Термин "процентная идентичность последовательностей" между двумя полипептидными или полинуклеотидными последовательностями относится к числу идентичных совпадающих положений, общих для последовательностей в окне сравнения с учетом добавлений или делеций (т.е. гэпов), которые должны быть введены для оптимального выравнивания двух последовательностей. Совпадающее положение представляет собой любое положение, в котором идентичные нуклеотид или аминокислота представлены как в целевой, так и в эталонной последовательности. Гэпы, представленные в целевой последовательности, не учитываются, поскольку гэпы не являются нуклеотидами или аминокислотами. Аналогично, гэпы, представленные в эталонной последовательности, не учитываются, поскольку учитываются нуклеотиды или аминокислоты целевой последовательности, а не нуклеотиды или аминокислоты эталонной последовательности.
[00241] Процентное значение идентичности последовательностей рассчитывают путем определения числа положений, в которых идентичные аминокислотный остаток или основание нуклеиновой кислоты присутствуют в обеих последовательностях, с получением числа совпадающих положений, деления числа совпадающих положений на общее число положений в окне сравнения и умножения результата на 100 с получением процентного значения идентичности последовательностей. Сравнение последовательностей и определение процентной идентичности последовательностей между двумя последовательностями можно осуществлять с помощью программного обеспечения, легко доступного как для применения в режиме онлайн, так и для загрузки. Подходящие программы из системы программного обеспечения доступны из различных источников и предназначены для выравнивания как белковых, так и нуклеотидных последовательностей. Одной подходящей программой для определения процентной идентичности последовательностей является bl2seq, часть пакета программ BLAST, доступного на веб-сайте BLAST Национального центра биотехнологической информации правительства США (blast.ncbi.nlm.nih.gov). Bl2seq выполняет сравнение между двумя последовательностями с помощью алгоритмов BLASTN или BLASTP. BLASTN применяют для сравнения нуклеотидных последовательностей, тогда как BLASTP применяют для сравнения аминокислотных последовательностей. Другими подходящими программами являются, например, Needle, Stretcher, Water или Matcher, являющиеся частью пакета биоинформатических программ EMBOSS, и доступные также от Европейского института биоинформатики (EBI) на www.ebi.ac.uk/Tools/psa.
[00242] Каждая из различных областей в одной целевой полинуклеотидной или полипептидной последовательности, выравниваемой относительно эталонной полинуклеотидной или полипептидной последовательности, может обладать собственной процентной идентичностью последовательностей. Следует отметить, что значение процентной идентичности последовательностей округляют до ближайших десятых. Например, 80,11, 80,12, 80,13 и 80,14 округляют в меньшую сторону до 80,1, тогда как 80,15, 80,16, 80,17, 80,18 и 80,19 округляют в большую сторону до 80,2. Также следует отметить, что значение длины всегда будет целым числом.
[00243] Согласно определенным аспектам процент идентичности "X" первой аминокислотной последовательности со второй аминокислотной последовательностью рассчитывают как 100 х (Y/Z), где Y представляет собой количество аминокислотных остатков, подсчитанных как идентичные совпадения при выравнивании первой и второй последовательностей (выравненные с помощью визуальной проверки или специальной программы выравнивания последовательностей), и Z представляет собой общее число остатков во второй последовательности. Если длина первой последовательности больше, чем второй последовательности, то процентная идентичность первой последовательности со второй последовательностью будет выше, чем процентная идентичность второй последовательности с первой последовательностью.
[00244] Специалисту в данной области техники будет понятно, что построение выравнивания последовательностей для расчета процентной идентичности последовательностей не ограничивается бинарным сравнением двух последовательностей, основанным исключительно на данных о первичных последовательностях. Выравнивание последовательностей можно получить из множественного выравнивания последовательностей. Одной подходящей программой для построения множественного выравнивания последовательностей является ClustalW2, доступная на www.clustal.org. Другой подходящей программой является MUSCLE, доступная на www.drive5.com/muscle. ClustalW2 и MUSCLE альтернативно доступны, например, от EBI.
[00245] Также следует понимать, что выравнивание последовательностей можно построить с помощью интеграции данных о последовательностях с данными из гетерогенных источников, таких как данные о структуре (например, кристаллическая структура белка), данные о функции (например, локализация мутаций) или данные филогенетики. Подходящей программой, в которой интегрируют гетерогенные данные с построением множественного выравнивания последовательностей, является T-Coffee, доступная на www.tcoffee.org и альтернативно доступная, например, от EBI. Также следует понимать, что конечное выравнивание, применяемое для расчета процентной идентичности последовательностей, можно проверять либо автоматически, либо вручную.
[00246] Термин "консенсусная последовательность", используемый в данном документе применительно к вариабельным областям легкой цепи (VL) и тяжелой цепи (VH), относится к составной или обобщенной VL- или VH-последовательности, определенной с учетом информации о том, какие аминокислотные остатки в VL- или VH-цепи поддаются модификации без ущерба для связывания антигена. Таким образом, в "консенсусной последовательности" для VL- или VH-цепи определенные положения аминокислот заняты одним из нескольких возможных аминокислотных остатков в данном положении. Например, если аргинин (R) или серии (S) находятся в определенном положении, то данное конкретное положение в пределах консенсусной последовательности может быть либо аргинином, либо серином (R или S). Консенсусные последовательности для VH- и VL-цепей могут быть определены, например, с помощью созревания аффинности in vitro (например, рандомизирования каждого положения аминокислоты в определенной CDR с использованием вырожденных кодирующих праймеров), с помощью сканирующего мутагенеза (например, аланин-сканирующего мутагенеза) аминокислотных остатков в CDR антитела или любых других способов, известных из уровня техники, с последующей оценкой связывания мутантых форм с антигеном для определения того, влияет ли положение подверженной мутированию аминокислоты на связывание антигена. Согласно некоторым аспектам мутации вводят в CDR-области. Согласно другим аспектам мутации вводят в каркасные области. Согласно некоторым другим аспектам мутации вводят в CDR и каркасные области.
II. CD73-связывающие молекулы
[00247] В настоящем раскрытии предусматриваются CD73-связывающие молекулы, например, антитела и их антигенсвязывающие фрагменты, которые специфически связывают CD73, например, CD73 человека. Полноразмерная аминокислотная (аа) и нуклеотидная (nt) последовательности CD73 известны из уровня техники (см. например, в UniProt под № доступа Р21589 для CD73 человека или в UniProt под № доступа Q61503 для CD73 мыши). Согласно некоторым аспектам молекулы, связывающиеся с CD73, представляют собой антитела человека (например, клон антитела 10.3, клон антитела 2С5, MEDI9447). Согласно определенным аспектам CD73-связывающие молекулы представляют собой антитела или их антигенсвязывающие фрагменты.
[00248] Согласно некоторым аспектам CD73-связывающие молекулы, например, антитела или их антигенсвязывающие фрагменты, содержат Fab, Fab', F(ab')2, Fd, одноцепочечный Fv или scFv, связанный дисульфидной связью Fv, домен V-NAR, IgNar, интратело, IgG СН2, миниантитело, F(ab')3, тетратело, триатело, диатело, однодоменное антитело, DVD-Ig, Fcab, mAb2, (scFv)2 или scFv-Fc. Согласно некоторым аспектам антитело представлено типом IgG, например типом IgG1.
[00249] Согласно некоторым аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент содержат константную область тяжелой цепи или ее фрагмент. Согласно некоторым конкретным аспектам константная область тяжелой цепи представляет собой константную область IgG. Константная область IgG может содержать константную область легкой цепи, выбранную из группы, состоящей из константной области каппа-цепи и константной области лямбда-цепи.
[00250] Согласно определенным аспектам антитела к CD73 или их антигенсвязывающие фрагменты, раскрытые в данном документе, модифицированы относительно исходного антитела, например, антитела CD730010 или антитела CD730002. Согласно некоторым аспектам исходное антитело представляет собой CD730010. Согласно другим аспектам исходное антитело представляет собой CD730002. Согласно другим аспектам исходное антитело представляет собой CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068 или CD730069. Модификации могут включать в себя мутации в CDR-областях и/или в FW-областях по сравнению с исходным антителом, например, CD730010 или CD730002.
[00251] Фраза "антитело CD730002" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: l, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 2.
[00252] Фраза "антитело CD730004" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 104, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 103.
[00253] Фраза "антитело CD730008" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 106, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 107.
[00254] Фраза "антитело CD730010" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 3, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 4.
[00255] Фраза "антитело CD730011" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 5, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 6.
[00256] Фраза "антитело CD730021" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 7, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 8.
[00257] Фраза "антитело CD730042" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 9, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 10.
[00258] Фраза "антитело CD730046" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 11, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 12.
[00259] Фраза "антитело CD730047" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 13, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 14.
[00260] Фраза "антитело CD730068" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: 108, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 107.
[00261] Фраза "антитело CD730069" относится к IgG1, содержащему два VL-домена, содержащих аминокислотную последовательность SEQ ID NO: l 10, и два VH-домена, содержащих аминокислотную последовательность SEQ ID NO: 109.
(i) Антитела к CD73, полученные из CD730010
[00262] Согласно определенным аспектам антитело к CD73 по настоящему раскрытию содержит модификации в CDR1, и/или CDR2, и/или CDR3 легкой цепи антитела CD730010, в том числе без ограничения:
1) CDR1 легкой цепи, содержащей консенсусную последовательность SGSLSNIGRNX1VN, где X1 представляет собой аминокислотные остатки пролина (Р), глутаминовой кислоты (Е) или аспарагиновой кислоты (D); и/или
2) CDR2 легкой цепи, содержащей консенсусную последовательность LX2NX3RX4X5, где Х2 представляет собой аминокислотные остатки аспарагина (N) или аспарагиновой кислоты (D), Х3 представляет собой аминокислотные остатки глутамина (Q) или лейцина (L), Х4 представляет собой аминокислотные остатки лейцина (L) или пролина (Р) и Х5 представляет собой аминокислотные остатки глицина (G) или серина (S); и/или
3) СDR3 легкой цепи, содержащей консенсусную последовательность ATWDDSX6X7GWX8, где Х6 представляет собой аминокислотные остатки лейцина (L) или гистидина (Н), Х7 представляет собой аминокислотные остатки лизина (K), пролина (Р), изолейцина (I) или аспарагина (N), и X8 представляет собой аминокислотные остатки лейцина (L) или треонина (Т).
[00263] Согласно определенным аспектам антитело к CD73 по настоящему раскрытию содержит модификации в CDR1, и/или CDR2, и/или CDR3 тяжелой цепи антитела CD730010, в том числе без ограничения:
1) CDR1 тяжелой цепи, содержащей консенсусную последовательность SYAX9S, где Х9 представляет собой аминокислотные остатки метионина (М) или тирозина (Y); и/или
2) CDR2 тяжелой цепи, содержащей консенсусную последовательность X10IX11GSX12GX13TYYADSVKG, где Х10 представляет собой аминокислотные остатки лейцина (L) или аланина (А), Х11 представляет собой аминокислотные остатки триптофана (W) или серина (S), Х12 представляет собой аминокислотные остатки триптофана (W) или глицина (G) и Х13 представляет собой аминокислотные остатки серина (S) или аргинина (R); и/или
3) CDR3 тяжелой цепи, содержащей консенсусную последовательность LGYX14X15X16DX17, где Х14 представляет собой аминокислотные остатки глицина (G) или серина (S), X15 представляет собой аминокислотные остатки аргинина (R) или треонина (Т), X16 представляет собой аминокислотные остатки валина (V) или изолейцина (I), и Х17 представляет собой аминокислотные остатки тирозина (Y), лизина (К), метионина (М), лейцина (L) или глутаминовой кислоты (Е).
[00264] Согласно одному аспекту антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-область, содержащую консенсусную аминокислотную последовательность:
[FW1]SGSLSNIGRNX1VN[FW2]LX2NX3RX4X5[FW3]ATWDDSX6X7GWX8[FW4]
где [FW]], [FW2], [FW3] и [FW4] представляют собой аминокислотные остатки каркасной области 1 VL (SEQ ID NO: 25 или 26), каркасной области 2 VL (SEQ ID NO: 27 или 28), каркасной области 3 VL (SEQ ID NO: 29) и каркасной области 4 VL (SEQ ID NO: 30) соответственно, и где
X1 представляет собой аминокислотные остатки пролина (Р), глутаминовой кислоты (Е) или аспарагиновой кислоты (D);
Х2 представляет собой аминокислотные остатки аспарагина (N) или аспарагиновой кислоты (D);
Х3 представляет собой аминокислотные остатки глутамина (Q) или лейцина (L);
Х4 представляет собой аминокислотные остатки лейцина (L) или пролина (Р);
Х5 представляет собой аминокислотные остатки глицина (G) или серина (S);
Х6 представляет собой аминокислотные остатки лейцина (L) или гистидина (Н);
Х7 представляет собой аминокислотные остатки лизина (К), пролина (Р), изолейцина (I) или аспарагина (N); и
X8 представляет собой аминокислотные остатки лейцина (L) или треонина (Т).
[00265] Согласно одному аспекту антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-область, содержащую консенсусную аминокислотную последовательность:
[FW5]SYAX9S[FW6]X10IX11GSX12GX13TYYADSVKG[FW7]LGYX14X15X16DX17[FW8]
где [FW5], [FW6], [FW7] и [FW8] представляют собой аминокислотные остатки каркасной области 1 VH (SEQ ID NO: 31), каркасной области 2 VH (SEQ ID NO: 32), каркасной области 3 VH (SEQ ID NO: 33) и каркасной области 4 VH (SEQ ID NO: 34) соответственно, и где
Х9 представляет собой аминокислотные остатки метионина (М) или тирозина (Y);
Х10 представляет собой аминокислотные остатки лейцина (L) или аланина (А);
X11 представляет собой аминокислотные остатки триптофана (W) или серина (S);
Х12 представляет собой аминокислотные остатки триптофана (W) или глицина (G);
Х13 представляет собой аминокислотные остатки серина (S) или аргинина (R);
Х14 представляет собой аминокислотные остатки глицина (G) или серина (S);
Х15 представляет собой аминокислотные остатки аргинина (R) или треонина (Т);
Х16 представляет собой аминокислотные остатки валина (V) или изолейцина (I);
Х17 представляет собой аминокислотные остатки тирозина (Y), лизина (K), метионина (М),
лейцина (L) или глутаминовой кислоты (Е).
[00266] Согласно одному аспекту антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-область, содержащую консенсусную аминокислотную последовательность:
[FW5]SGSLSNIGRNX1VN[FW2]LX2NX3RX4X5[FW3]ATWDDSX6X7GWX8[FW4]
где [FW1], [FW2], [FW3] и [FW4] представляют собой аминокислотные остатки каркасной области 1 VL (SEQ ID NO: 25 или 26), каркасной области 2 VL (SEQ ID NO: 27 или 28), каркасной области 3 VL (SEQ ID NO: 29) и каркасной области 4 VL (SEQ ID NO: 30) соответственно, и где
X1 представляет собой аминокислотные остатки пролина (Р), глутаминовой кислоты (Е) или аспарагиновой кислоты (D);
Х2 представляет собой аминокислотные остатки аспарагина (N) или аспарагиновой кислоты (D);
Х3 представляет собой аминокислотные остатки глутамина (Q) или лейцина (L);
Х4 представляет собой аминокислотные остатки лейцина (L) или пролина (Р);
Х5 представляет собой аминокислотные остатки глицина (G) или серина (S);
Х6 представляет собой аминокислотные остатки лейцина (L) или гистидина (Н);
Х7 представляет собой аминокислотные остатки лизина (К), пролина (Р), изолейцина (I) или аспарагина (N); и
Х8 представляет собой аминокислотные остатки лейцина (L) или треонина (Т);
и где антитело к CD73 или его антигенсвязывающий фрагмент дополнительно содержат VH-область, содержащую консенсусную аминокислотную последовательность:
[FW5]SYAX9S[FW6]X10IX11GSX12GX13TYYADSVKG[FW7]LGYX14X15X16DX17[FW8]
где [FW5], [FW6], [FW7] и [FW8] представляют собой аминокислотные остатки каркасной области 1 VH (SEQ ID NO: 31), каркасной области 2 VH (SEQ ID NO: 32), каркасной области 3 VH (SEQ ID NO: 33) и каркасной области 4 VH (SEQ ID NO: 34) соответственно, и где
Х9 представляет собой аминокислотные остатки метионина (М) или тирозина (Y);
Х10 представляет собой аминокислотные остатки лейцина (L) или аланина (А);
Х11 представляет собой аминокислотные остатки триптофана (W) или серина (S);
X12 представляет собой аминокислотные остатки триптофана (W) или глицина (G);
Х13 представляет собой аминокислотные остатки серина (S) или аргинина (R);
Х14 представляет собой аминокислотные остатки глицина (G) или серина (S);
Х15 представляет собой аминокислотные остатки аргинина (R) или треонина (Т);
X16 представляет собой аминокислотные остатки валина (V) или изолейцина (I);
Х17 представляет собой аминокислотные остатки тирозина (Y), лизина (K), метионина (М),
лейцина (L) или глутаминовой кислоты (Е).
[00267] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48.
[00268] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52.
[00269] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53, 54, 55 и 56. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53, 54, 55 и 56.
[00270] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36.
[00271] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40.
[00272] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45.
[00273] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48, за исключением одной, двух, трех или четырех аминокислотных замен.
[00274] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52, за исключением одной, двух, трех или четырех аминокислотных замен.
[00275] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53, 54, 55 и 56, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53, 54, 55 и 56, за исключением одной, двух, трех или четырех аминокислотных замен.
[00276] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36, за исключением одной, двух, трех или четырех аминокислотных замен.
[00277] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40, за исключением одной, двух, трех или четырех аминокислотных замен.
[00278] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45, за исключением одной, двух, трех или четырех аминокислотных замен.
[00279] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52, и VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53, 54, 55 и 56. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52; и VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53,54, 55 и 56.
[00280] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40, и VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40; VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45.
[00281] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48, за исключением одной, двух, трех или четырех аминокислотных замен; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52, за исключением одной, двух, трех или четырех аминокислотных замен; и VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53, 54, 55 и 56, за исключением одной, двух, трех или четырех аминокислотных замен.
[00282] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 47 и 48, за исключением одной, двух, трех или четырех аминокислотных замен; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, 50, 51 и 52, за исключением одной, двух, трех или четырех аминокислотных замен; и VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 53, 54, 55 и 56, за исключением одной, двух, трех или четырех аминокислотных замен.
[00283] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40, за исключением одной, двух, трех или четырех аминокислотных замен; и VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45, за исключением одной, двух, трех или четырех аминокислотных замен.
[00284] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35 и 36, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 38, 39 и 40, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 42, 43, 44 и 45, за исключением одной, двух, трех или четырех аминокислотных замен.
[00285] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат модификации в CDR1, и/или CDR2, и/или CDR3 тяжелой и/или легкой цепей и дополнительно содержат модификации FW1, и/или FW2, и/или FW3, и/или FW4 тяжелой и/или легкой цепей.
[00286] Согласно некоторым аспектам FW1 содержит SEQ ID NO: 25 или 26, FW2 содержит SEQ ID NO: 27 или 28, FW3 содержит SEQ ID NO: 29, FW4 содержит SEQ ID NO: 30, FW5 содержит SEQ ID NO: 31, FW6 содержит SEQ ID NO: 32, FW7 содержит SEQ ID NO: 33, и FW8 содержит SEQ ID NO: 34.
[00287] Согласно некоторым аспектам FW1 содержит SEQ ID NO: 25 или 26, за исключением одной, двух, трех или четырех аминокислотных замен; FW2 содержит SEQ ID NO: 27 или 28, за исключением одной, двух, трех или четырех аминокислотных замен; FW3 содержит SEQ ID NO: 29, за исключением одной, двух, трех или четырех аминокислотных замен; FW4 содержит SEQ ID NO: 30, за исключением одной, двух, трех или четырех аминокислотных замен; FW5 содержит SEQ ID NO: 31, за исключением одной, двух, трех или четырех аминокислотных замен; FW6 содержит SEQ ID NO: 32, за исключением одной, двух, трех или четырех аминокислотных замен; FW7 содержит SEQ ID NO: 33, за исключением одной, двух, трех или четырех аминокислотных замен; и FW8 содержит SEQ ID NO: 34.
[00288] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL и VH, содержащие аминокислотные последовательности VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3, идентичные или идентичные за исключением одной, двух, трех или четырех аминокислотных замен в одном или нескольких CDR, где такие VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3 представляют собой:
SEQ ID NOs: 46, 49, 53, 35, 37, и 41; или,
SEQ ID NOs: 47, 49, 53, 35, 37 и 41; или,
SEQ ID NOs: 47, 49, 54, 36, 37 и 42; или,
SEQ ID NOs: 46, 50, 54, 36, 38 и 43; или,
SEQ ID NOs: 46, 51, 55, 36, 39 и 44; или,
SEQ ID NOs: 48, 52, 54, 36, 40 и 44; или,
SEQ ID NOs: 46, 49, 56, 35, 37 и 41; или,
SEQ ID NOs: 46, 49, 53, 35, 37 и 45; или,
SEQ ID NOs: 47, 49, 56, 36, 37 и 45; или,
SEQ ID NOs: 46, 50, 56, 36, 38 и 45; или,
SEQ ID NOs: 46, 51, 56, 36, 39 и 45; или,
SEQ ID NOs: 48, 52, 56, 36, 40 и 45; или
SEQ ID NOs: 46, 49, 56, 35, 37 и 45, соответственно.
[00289] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL антитела и VH антитела, где VL содержит аминокислотную последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, и SEQ ID NO: 70.
[00290] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL антитела и VH антитела, где VH содержит аминокислотную последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 71, SEQ ID NO: 72, SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83 и SEQ ID NO: 84.
[00291] Согласно другим аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL, содержащую последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, и SEQ ID NO: 70,, и дополнительно содержат VH, содержащую последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 71, SEQ ID NO: 72, SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, SEQ ID NO: 83 и SEQ ID NO: 84.
[00292] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL, содержащую последовательность SEQ ID NO: 68, и VH, содержащую последовательность SEQ ID NO: 82. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL, содержащую последовательность SEQ ID NO: 68, и VH, содержащую последовательность SEQ ID NO: 82.
[00293] "Клон антитела 10.3" (также обозначаемый "73combo3" или "MEDI9447") представляет собой IgG1, содержащий две легкие цепи (VL), полученные из CD730010, с SEQ ID NO: 68 (содержащие три CDR - CDR1, CDR2 и CDR3, с последовательностями SEQ ID NO: 46, 51 и 56 соответственно), и две тяжелые цепи (VH), полученные из CD730010, с SEQ ID NO: 82 (содержащие три CDR - CDR1, CDR2 и CDR3, с последовательностями SEQ ID NO: 36, 39 и 45 соответственно).
[00294] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент, раскрытые в данном документе, связывают CD73 с практически такой же или более высокой аффинностью, что и антитело 10.3, содержащее тяжелую цепь VH 10.3 с SEQ ID NO: 82 и легкую цепь VL 10.3 с SEQ ID NO: 68.
(i) Антитела к CD73, полученные из CD730002
[00295] Согласно определенным аспектам антитело к CD73 по настоящему раскрытию содержит модификации в CDR1, и/или CDR2, и/или СDR3 легкой цепи антитела CD730002, в том числе без ограничения:
1) CDR1 легкой цепи, содержащей последовательность SGDKVGDKYAS; и/или
2) CDR2 легкой цепи, содержащей консенсусную последовательность EDX18KX19X20S, где X18 представляет собой аминокислотные остатки серина (S) или треонина (Т), Х19 представляет собой аминокислотные остатки аргинина (R) или тирозина (Y) и Х20 представляет собой аминокислотные остатки гистидина (Н), пролина (Р) или лейцина (L); и/или,
3) СDR3 легкой цепи, содержащей последовательность QAWDTSFWV.
[00296] Согласно определенным аспектам антитело к CD73 по настоящему раскрытию содержит модификации в CDR1, и/или CDR2, и/или CDR3 тяжелой цепи CD730002, в том числе без ограничения:
1) CDR1 тяжелой цепи, содержащей последовательность SX21A X22S, где Х21 представляет собой аминокислотные остатки тирозина (Y) или валина (V) и Х22 представляет собой аминокислотные остатки метионина (М) или аргинина (R); и/или
2) CDR2 тяжелой цепи, содержащей последовательность AISGSGGSX23YY X24DSVKX25, где Х23 представляет собой аминокислотные остатки треонина (Т) или пролина (Р); Х24 представляет собой аминокислотные остатки аланина (А) или G (глицина) и Х25 представляет собой аминокислотные остатки глицина (G) или аргинина (R); и/или
3) CDR3 тяжелой цепи, содержащей последовательность DKGYYWYM.
[00297] Согласно одному аспекту антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-область, содержащую консенсусную аминокислотную последовательность:
[FW9]SGDKVGDKYAS[FW10]EDX18KX19X20S[FW11]QAWDTSFWV[FW12]
где [FW9], [FW10], [FW11] и [FW12] представляют собой аминокислотные остатки каркасной области 1 VL (SEQ ID NO: 90 или 91), каркасной области 2 VL (SEQ ID NO: 92), каркасной области 3 VL (SEQ ID NO: 93, 94 или 122) и каркасной области 4 VL (SEQ ID NO: 30) соответственно; и где Х18 представляет собой аминокислотные остатки пролина (Р) или лейцина (L); Х19 представляет собой аминокислотные остатки аргинина (R) или тирозина (Y) и Х20 представляет собой аминокислотные остатки гистидина (Н), пролина (Р) или лейцина (L).
[00298] Согласно одному аспекту антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-область, содержащую консенсусную аминокислотную последовательность:
[FW13]SX21AX22S[FW14]AISGSGGSX23YYX24DSVKX25[FW15]DKGYYWYM[FW16]
где [FW13], [FW14], [FW15] и [FW16] представляют собой аминокислотные остатки каркасной области 1 VH (SEQ ID NO: 31), каркасной области 2 VH (SEQ ID NO: 32), каркасной области 3 VH (SEQ ID NO: 33) и каркасной области 4 VH (SEQ ID NO: 89) соответственно; и где X21 представляет собой аминокислотные остатки тирозина (Y) или валина (V); Х22 представляет собой аминокислотные остатки метионина (М) или аргинина (R); Х23 представляет собой аминокислотные остатки треонина (Т) или пролина (Р); Х24 представляет собой аминокислотные остатки аланина (А) или G (глицина) и Х25 представляет собой аминокислотные остатки глицина (G) или аргинина (R).
[00299] Согласно одному аспекту антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-область, содержащую консенсусную аминокислотную последовательность:
[FW9]SGDKVGDKYAS[FW10]EDX18KX19X20S[FW11]QAWDTSFWV[FW12]
где [FW9], [FW10], [FW11] и [FW12] представляют собой аминокислотные остатки каркасной области 1 VL (SEQ ID NO: 90 или 91), каркасной области 2 VL (SEQ ID NO: 92), каркасной области 3 VL (SEQ ID NO: 93, 94 или 122) и каркасной области 4 VL (SEQ ID NO: 30) соответственно; и где X18 представляет собой аминокислотные остатки пролина (Р) или лейцина (L); Х19 представляет собой аминокислотные остатки аргинина (R) или тирозина (Y) и Х20 представляет собой аминокислотные остатки гистидина (Н), пролина (Р) или лейцина (L),
и где антитело к CD73 или его антигенсвязывающий фрагмент дополнительно содержат VH-область, содержащую консенсусную аминокислотную последовательность:
[FW13]SX21AX22S[FW14]AISGSGGSX23YYX24DSVKX25[FW15]DKGYYWYM[FW16]
где [FW13], [FW14], [FW15] и [FW16] представляют собой аминокислотные остатки каркасной области 1 VH (SEQ ID NO: 31), каркасной области 2 VH (SEQ ID NO: 32), каркасной области 3 VH (SEQ ID NO: 33) и каркасной области 4 VH (SEQ ID NO: 89) соответственно; и где Х21 представляет собой аминокислотные остатки тирозина (Y) или валина (V); Х22 представляет собой аминокислотные остатки метионина (М) или аргинина (R); Х23 представляет собой аминокислотные остатки треонина (Т) или пролина (Р); Х24 представляет собой аминокислотные остатки аланина (А) или G (глицина) и Х25 представляет собой аминокислотные остатки глицина (G) или аргинина (R).
[00300] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 97. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, состоящую из SEQ ID NO: 97.
[00301] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129.
[00302] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 100. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 100.
[00303] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124.
[00304] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126.
[00305] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 96. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 96.
[00306] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 97, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, состоящую из SEQ ID NO: 97, за исключением одной, двух, трех или четырех аминокислотных замен.
[00307] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129, за исключением одной, двух, трех или четырех аминокислотных замен.
[00308] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 100, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 100, за исключением одной, двух, трех или четырех аминокислотных замен.
[00309] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124, за исключением одной, двух, трех или четырех аминокислотных замен.
[00310] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126, за исключением одной, двух, трех или четырех аминокислотных замен.
[00311] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 96, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 96, за исключением одной, двух, трех или четырех аминокислотных замен.
[00312] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 97; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129; и VL-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 100. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, состоящую из SEQ ID NO: 97; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129; и VL-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 100.
[00313] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126; и VH-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 96. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126; VH-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 96.
[00314] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 97, за исключением одной, двух, трех или четырех аминокислотных замен; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129, за исключением одной, двух, трех или четырех аминокислотных замен; и VL-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 100, за исключением одной, двух, трех или четырех аминокислотных замен.
[00315] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1, содержащую последовательность, состоящую из SEQ ID NO: 97, за исключением одной, двух, трех или четырех аминокислотных замен; VL-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, 99, 127, 128 и 129, за исключением одной, двух, трех или четырех аминокислотных замен; и VL-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 100, за исключением одной, двух, трех или четырех аминокислотных замен.
[00316] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126, за исключением одной, двух, трех или четырех аминокислотных замен; и VH-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 96, за исключением одной, двух, трех или четырех аминокислотных замен.
[00317] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 123 и 124, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 37, 95, 125 и 126, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR3, содержащую последовательность, состоящую из SEQ ID NO: 96, за исключением одной, двух, трех или четырех аминокислотных замен.
[00318] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат модификации в CDR1, и/или CDR2, и/или CDR3 тяжелой и/или легкой цепей и дополнительно содержат модификации FW1, и/или FW2, и/или FW3, и/или FW4 тяжелой и/или легкой цепей.
[00319] Согласно некоторым аспектам FW9 содержит SEQ ID NO: 90 или 91, FW10 содержит SEQ ID NO: 92, FW11 содержит SEQ ID NO: 93, 94 или 122, FW12 содержит SEQ ID NO: 30, FW13 содержит SEQ ID NO: 31, FW14 содержит SEQ ID NO: 32, FW15 содержит SEQ ID NO: 33 и FW16 содержит SEQ ID NO: 89.
[00320] Согласно некоторым аспектам FW9 содержит SEQ ID NO: 90 или 91, за исключением одной, двух, трех или четырех аминокислотных замен; FW10 содержит SEQ ID NO: 92, за исключением одной, двух, трех или четырех аминокислотных замен; FW11 содержит SEQ ID NO: 93, 94 или 122, за исключением одной, двух, трех или четырех аминокислотных замен; FW12 содержит SEQ ID NO: 30, за исключением одной, двух, трех или четырех аминокислотных замен; FW13 содержит SEQ ID NO: 31, за исключением одной, двух, трех или четырех аминокислотных замен; FW14 содержит SEQ ID NO: 32, за исключением одной, двух, трех или четырех аминокислотных замен; FW15 содержит SEQ ID NO: 33, за исключением одной, двух, трех или четырех аминокислотных замен; и FW16 содержит SEQ ID NO: 89.
[00321] Согласно определенным аспектам антитело к CD733 или его антигенсвязывающий фрагмент содержат VL и VH, содержащие аминокислотные последовательности VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3, идентичные или идентичные за исключением одной, двух, трех или четырех аминокислотных замен в одном или нескольких CDR, где такие VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3 представляют собой:
SEQ ID NOs: 97, 98, 100, 35, 37, и 96; или,
SEQ ID NOs: 97, 99, 100, 35, 95 и 96; или,
SEQ ID NOs: 97, 98, 100, 35, 37, и 96; или,
SEQ ID NOs: 97, 99, 100, 123, 37, и 96; или,
SEQ ID NOs: 97, 99, 100, 124, 37, и 96; или,
SEQ ID NOs: 97, 99, 100, 35, 125, и 96; или,
SEQ ID NOs: 97, 99, 100, 35, 126, и 96; или,
SEQ ID NOs: 97, 99, 100, 35, 95, и 96; или,
SEQ ID NOs: 97, 127, 100, 35, 95, и 96; или,
SEQ ID NOs: 97, 128, 100, 35, 95, и 96; или,
SEQ ID NOs: 97, 129, 100, 35, 95, и 96; или,
SEQ ID NOs: 97, 99, 100, 35, 95, и 96; соответственно.
[00322] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL антитела и VH антитела, где VL содержит аминокислотную последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 86, 88, 112, 118, 119, 120, и 121.
[00323] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL антитела и VH антитела, где VH содержит аминокислотную последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 85, 87, 111, 113, 114, 115, 116, и 117.
[00324] Согласно другим аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL, содержащую последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 86, 88, 112, 118, 119, 120, и 121;, и дополнительно содержат VH, содержащую последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из SEQIDNOs: 85, 87, 111, 113, 114, 115, 116, и 117.
[00325] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL, содержащую последовательность SEQ ID NO: 88, и VH, содержащую последовательность SEQ ID NO: 87. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL, содержащую последовательность SEQ ID NO: 87, и VH, содержащую последовательность SEQ ID NO: 87.
[00326] "Клон антитела 2С5" представляет собой IgG1, содержащий две легкие цепи (VL), полученные из CD730002, с SEQ ID NO: 88 (содержащие три CDR - CDR1, CDR2 и CDR3, с последовательностями SEQ ID NO: 97, 99 и 100 соответственно), и тяжелые цепи (VH), полученные из CD7300002, с SEQ ID NO: 87 (содержащие три CDR - CDR1, CDR2 и CDR3, с последовательностями SEQ ID NO: 35, 95 и 96 соответственно).
[00327] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент, раскрытые в данном документе, связывают CD73 с практически такой же или более высокой аффинностью, что и антитело 2С5, содержащее тяжелую цепь VH 2С5 с SEQ ID NO: 87 и легкую цепь VL 2С5 с SEQ ID NO: 88.
(iii) Антитела к CD73 на основе исходных антител, отличных от CD730002 или CD730010
[00328] Согласно другим аспектам исходное антитело для антитела к CD73 или антигенсвязывающего фрагмента, раскрытых в данном документе, представляет собой CD730004 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 104 и VH с SEQ ID NO: 103), CD730008 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 106 и VH с SEQ ID NO: 107), CD7300011 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 5 и VH с SEQ ID NO: 6), CD730021 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 7 и VH с SEQ ID NO: 8), CD730042 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 9 и VH с SEQ ID NO: 10), CD730046 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 11 и VH с SEQ ID NO: 12), CD730047 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 13 и VH с SEQ ID NO: 14), CD730068 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 108 и VH с SEQ ID NO: 107) или CD730069 (т.е. антитело к CD73, содержащее VL с SEQ ID NO: 110 и VH с SEQ ID NO: 109). Модификации в исходных антителах могут включать в себя мутации в CDR-областях и/или в FW-областях по сравнению с исходным антителом, например, CD730004.
[00329] Согласно определенным аспектам антитело к CD73 содержит модификации в CDR1, и/или CDR2, и/или CDR3 легкой цепи CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068 или CD730069.
[00330] Согласно определенным аспектам антитело к CD73 содержит модификации в CDR1, и/или CDR2, и/или CDR3 тяжелой цепи CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00331] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00332] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00333] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR2 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR3 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен.
[00334] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR2 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен. Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR3 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен.
[00335] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069; VL-CDR2 H3CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069; и VL-CDR3 H3CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00336] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069; VH-CDR2 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069; и VH-CDR3 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00337] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен; VL-CDR2 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен; и VL-CDR3 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен.
[00338] Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VH-CDR1 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR2 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен; и VH-CDR3 из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, за исключением одной, двух, трех или четырех аминокислотных замен.
[00339] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат модификации в CDR1, и/или CDR2, и/или CDR3 тяжелой и/или легкой цепей из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, и дополнительно содержат модификации в FW1, и/или FW2, и/или FW3, и/или FW4 тяжелой и/или легкой цепей из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00340] Согласно определенным аспектам антитело к CD733 или его антигенсвязывающий фрагмент содержат VL и VH, содержащие аминокислотные последовательности VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3, идентичные или идентичные за исключением одной, двух, трех или четырех аминокислотных замен в одном или нескольких CDR, где такие VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3 происходят из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00341] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL антитела и VH антитела, где VL содержит аминокислотную последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из последовательностей VL из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00342] Согласно другим аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL антитела и VH антитела, где VH содержит аминокислотную последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из последовательностей VH из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00343] Согласно другим аспектам антитело к CD73 или его антигенсвязывающий фрагмент содержат VL, содержащую последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из последовательностей VL из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069, и дополнительно содержат VH, содержащую последовательность, по меньшей мере приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98%, приблизительно на 99% или приблизительно на 100% идентичную эталонной аминокислотной последовательности, выбранной из последовательностей VH из CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069.
[00344] Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент, раскрытые в данном документе, связывают CD73 с практически такой же или более высокой аффинностью, что и антитела CD730004, CD730008, CD7300011, CD730021, CD730042, CD730046, CD730047, CD730068, или CD730069 .
(iv) Смешанные и подобранные антитела к CD73
[00345] Последовательности VH и VL из раскрытых в данном документе молекул, связывающихся с CD73 (например, CD730002, CD730004, CD730008, CD730010, CD730011, CD73021, CD730042, CD730046, CD73004, CD730068, или CD730069), или VH и VL из вариантов таких последовательностей (например, клон 10 GL9, клон 10 Р32Е, клон 10 С1, клон 10 С2, клон 10 D3, клон 10 G10, клон 10 НРТ, клон 10 GRVE, клон 10 combo1, клон 10 combo2, клон 10 combo3, клон 10 combo5 или клон combo6) могут быть "смешаны и подобраны" с образованием других молекул, связывающихся с CD73.
[00346] Согласно определенным аспектам последовательности VH антитела 10.3 и антитела 2С5 смешивают и подбирают. Согласно другому аспекту последовательности VL антитела 10.03 и антитела 2С5 могут быть смешаны и подобраны. В дополнение или в качестве альтернативы, раскрытые в данном документе последовательности VL и/или VH вариантов клона 10 (CD730010) могут быть смешаны и подобраны. В дополнение или в качестве альтернативы, раскрытые в данном документе последовательности VL и/или VH вариантов клона 2 (CD730002) могут быть смешаны и подобраны. В дополнение или в качестве альтернативы, раскрытые в данном документе последовательности VL и/или VH вариантов клона 10 (CD730010) и клона 2 (CD730002) могут быть смешаны и подобраны.
[00347] Согласно некоторым аспектам смешивание и подбор VL и/или VH может происходить между последовательностями, полученными из антител, сгруппированных в одну эпитопную группу (см. пример 2). Используемый в данном документе термин "эпитопная группа" относится к группировке антител или их антигенсвязывающих фрагментов, которые связываются с одним и тем же эпитопом или перекрывающимся эпитопом, или конкурируют друг с другом за связывание с одним и тем же эпитопом или перекрывающимся эпитопом. Например, последовательности из CD730003, CD730010, CD730021, CD730042, CD730046 и CD730047, все из которых представляют собой антитела, принадлежащие к "эпитопной группе В", могут быть смешаны и подобраны. Согласно другим аспектам смешивание и подбор VL и/или VH может происходить между последовательностями, полученными из антител к CD73, сгруппированных в разные эпитопные группы. Соответственно, последовательности из антител, принадлежащих к "эпитопной группе В", могут быть смешаны и подобраны с последовательностями из антител к CD73 из "эпитопной группы A" (CD730002, CD730004, CD730008 и CD730011) или "эпитопной группы С" (CD730068 и CD730069).
(v) Мутантные антитела к CD73
[00348] Согласно определенным аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, раскрытые в данном документе, содержат мутации, которые улучшают связывание с FcRn человека и увеличивают время полужизни антитела к CD73 или его антигенсвязывающего фрагмента. Согласно некоторым аспектам такие мутации представляют собой мутацию "метионин (М) на тирозин (Y)" в положении 252, мутацию "серии (S) на треонин (Т)" в положении 254 и мутацию "треонин (Т) на глутаминовую кислоту (Е)" в положении 256, пронумерованных в соответствии с EU-индексом по Kabat (Kabat, et al. (1991) Sequences of Proteins of Immunological Interest, U.S. Public Health Service, National Institutes of Health, Washington, D.C.), введенные в константный домен IgGl. См. патент США № 7658921, который включен в данный документ с помощью ссылки. Было показано, что данный тип мутантного IgG, называемый "YTE-мутантом", характеризуется временем полужизни, увеличенным примерно в четыре раза по сравнению с вариантами того же антитела дикого типа (Dall'Acqua et al., J. Biol. Chem. 281:23514-24 (2006)). Согласно некоторым аспектам антитело к CD73 или его антигенсвязывающий фрагмент, содержащие константный домен IgG, содержат одну или несколько аминокислотных замен аминокислотных остатков по положениям 251-257, 285-290, 308-314, 385-389 и 428-436, пронумерованных в соответствии с EU-индексом по Kabat, при этом такие мутации увеличивают время полужизни в сыворотке крови антитела к CD73 или его антигенсвязывающего фрагмента.
[00349] Согласно некоторым аспектам YTE-мутант дополнительно содержит замену по положению 434 константного домена IgG, пронумерованному в соответствии с EU-индексом по Kabat, на аминокислоту, выбранную из группы, состоящей из триптофана (W), метионина (М), тирозина (Y) и серина (S). Согласно другим аспектам YTE-мутант дополнительно содержит замену по положению 434 константного домена IgG, пронумерованному в соответствии с EU-индексом по Kabat, на аминокислоту, выбранную из группы, состоящей из триптофана (W), метионина (М), тирозина (Y) и серина (S), и замену по положению 428 константного домена IgG, пронумерованному в соответствии с EU-индексом по Kabat, на аминокислоту, выбранную из группы, состоящей из треонина (Т), лейцина (L), фенилаланина (F) и серина (S).
[00350] Согласно еще одному аспекту YTE-мутант дополнительно содержит замену по положению 434 константного домена IgG, пронумерованному в соответствии с EU-индексом по Kabat, на тирозин (Y) и замену по положению 257 константного домена IgG, пронумерованному в соответствии с EU-индексом по Kabat, на лейцин (L). Согласно некоторым аспектам YTE-мутант дополнительно содержит замену по положению 434 константного домена IgG, пронумерованному в соответствии с EU-индексом по Kabat, на серии (S) и замену по положению 428 константного домена IgG, пронумерованному в соответствии с EU-индексом по Kabat, на лейцин (L).
[00351] Согласно конкретным аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, раскрытые в данном документе, содержат константный домен IgG1, содержащий мутацию метионина (М) на тирозин (Y) в положении 252, мутацию серина (S) на треонин (Т) в положении 254 и мутацию треонина (Т) на глутаминовую кислоту (Е) в положении 256 константного домена IgG1, пронумерованных в соответствии с EU-индексом по Kabat.
[00352] Согласно определенным аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, раскрытые в данном документе, содержат по меньшей мере одну аминокислотную замену в константном домене IgG, выбранную из группы, состоящей из:
(a) замены аминокислоты по положению 252 на тирозин (Y), фенилаланин (F), триптофан (W) или треонин (Т);
(b) замены аминокислоты по положению 254 на треонин (Т);
(c) замены аминокислоты по положению 256 на серии (S), аргинин (R), глутамин (Q), глутаминовую кислоту (Е), аспарагиновую кислоту (D) или треонин (Т);
(d) замены аминокислоты по положению 257 на лейцин (L);
(e) замены аминокислоты по положению 309 на пролин (Р);
(f) замены аминокислоты по положению 311 на серии (S);
(g) замены аминокислоты по положению 428 на треонин (Т), лейцин (L), фенилаланин (F) или серии (S);
(h) замены аминокислоты по положению 433 на аргинин (R), серии (S), изолейцин (I), пролин (Р) или глутамин (Q);
(i) замены аминокислоты по положению 434 на триптофан (W), метионин (М), серии (S), гистидин (Н), фенилаланин (F) или тирозин и
(j) комбинации двух или более указанных замен,
где положения пронумерованы в соответствии с EU-индексом по Kabat, и где модифицированный IgG характеризуется увеличенным временем полужизни в сыворотке крови по сравнению с временем полужизни в сыворотке крови IgG, имеющего константный домен IgG дикого типа.
[00353] Согласно некоторым аспектам аминокислотная последовательность VH и/или VL антитела к CD73 (например, MEDI9447, клона антитела 10.3 или клона антитела 2С5) или его антигенсвязывающего фрагмента, раскрытых в данном документе, может быть на 85%, 90%, 95%, 96%, 97%, 98% или 99% сходной с вышеизложенными последовательностями VH и VL и содержать 1, 2, 3, 4, 5 или более консервативных замен. Антитело к CD73, содержащее VH- и VL-области, характеризующиеся высокой (т.е. 80% или больше) идентичностью последовательностей или сходством последовательностей соответственно с VH-областью с SEQ ID NOs: 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 87, 111, 113, 114, 115, 116, или 117, и/или VL-областью с SEQ ID NOs: 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 86, 88, 112, 118, 119, 120, или 121, может быть получено посредством мутагенеза (например, сайт-направленного или ПЦР-опосредованного мутагенеза) молекул нуклеиновых кислот, кодирующих SEQ ID NOs: 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, или 121, с последующим тестированием измененного кодируемого антитела в отношении сохранения функции с помощью функциональных анализов, описанных в данном документе.
[00354] Согласно некоторым аспектам Fc-домен антитела к CD73, раскрытого в данном документе, или Fc-домен гибридного белка, содержащего CD73-связывающий фрагмент антитела, раскрытого в данном документе, характеризуются пониженным связыванием с Fc-рецептором с обеспечением снижения цитотоксичности, например, посредством ADCC. Согласно некоторым аспектам Fc-домен антитела или Fc-гибридного белка характеризуются повышенным связыванием с Fc-рецептором с обеспечением повышения цитотоксичности, например, посредством ADCC. Согласно некоторым аспектам Fc-домен антитела или Fc-гибридного белка содержат не встречающийся в природе аминокислотный остаток, снижающий ADCC, в одном или нескольких положениях, выбранных из группы, состоящей из 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245,247, 251, 252, 254, 255, 256, 262, 263, 264, 265, 266, 267, 269, 279, 280, 284, 292, 296, 297, 298, 299, 305, 313, 316, 325, 326, 327, 328, 329, 330, 331 , 332, 333, 334, 339, 341, 343, 370, 373, 378, 392, 416, 419, 421, 440, и 443, пронумерованных согласно EU-индексу, как указано у Kabat. Многочисленные специфические мутации, способные снижать ADCC-активность антитела, известны из уровня техники и включают, например, 234F, 235Е, 235F, 235Q (или 235Y), 239А, 332Q, 331S и их комбинации. Например, см. мутации, описанные в патентах США №№ 5624821, 5648260, 7597889, 8961967, 7371826, 7785791, 7790858, публикациях патентов США №№2014/0378663, 2013/0071390, 2011/0212087, 2015/0118227, 2006/0194290, 2006/0194291, 2008/0274105, 2008/0274506, US 2013/0089541 и US 2013/0108623, которые в полном объеме включены в данный документ с помощью ссылки. Антитела со сниженной эффекторной функцией ADCC также включают антитела с заменой одного или нескольких остатков Fc-области 238, 265, 269, 270, 297, 327 и 329 (см., например, патент США №6737056). Такие Fc-мутанты также включают в себя Fc-мутантов с заменами по двух или более аминокислотным положениям 265, 269, 270, 297 и 327, в том числе Fc-мутанта с заменой остатков 265 и 297 на аланин (см., например, патент США №7332581). Необязательно могут быть введены мутации, которые снижают как ADCC, так и CDC. Согласно некоторым аспектам раскрытые в данном документе антитела к CD73 или их антигенсвязывающий фрагмент, содержащие мутации, которые снижают или предотвращают ADCC и/или CDC, можно применять для получения конъюгатов антитело-лекарственное средство (ADC).
[00355] Согласно одному аспекту в настоящем раскрытии предусматривается антитело к CD73, при этом антитело представляет собой IgG1, IgG2 или IgG3 и содержит по меньшей мере одну модификацию по одному или нескольким положениям, выбранным из группы, состоящей из 234, 235 и 331, пронумерованных согласно EU-индексу, как указано у Kabat. Согласно еще одному конкретному аспекту Fc-область представляет собой Fc-область IgG1, IgG2 или IgG3 и не встречающиеся в природе аминокислоты выбраны из группы, состоящей из 234F, 235Е, 235F, 235Q (или 235Y), 239А, 332Q, 33 IS, 332Q, пронумерованные согласно EU-индексу, как указано у Kabat.
[00356] Согласно другому аспекту в настоящем раскрытии предусматривается антитело к CD73, при этом антитело представляет собой IgG4 и содержит по меньшей мере одну модификацию по одному или нескольким положениям, выбранным из группы, состоящей из 228 и 235, пронумерованных согласно EU-индексу, как указано у Kabat. Согласно еще одному конкретному аспекту Fc-область представляет собой Fc-область IgG4 и не встречающиеся в природе аминокислоты выбраны из группы, состоящей из 228Р, 235Е и 235Y, пронумерованные согласно EU-индексу, как указано у Kabat. Согласно конкретным аспектам в настоящем раскрытии предусматривается антитело к CD73, при этом антитело представляет собой IgG1, IgG2 или IgG3 и содержит модификации по положениям (i) 234F, 235Е и 331S; (ii) 234F, 235F и 33IS; (iii) 234F, 235Q и 322Q. Согласно другому конкретному аспекту в настоящем раскрытии предусматривается антитело к CD73, при этом антитело представляет собой IgG4 и содержит модификации 228Р и 235Е.
III. CD73-связывающие молекулы, конкурирующие за эпитоп
[00357] Согласно другому аспекту в настоящем раскрытии предусматриваются CD73-связывающие молекулы, которые связываются с тем же эпитопом, что и различные описанные в данном документе антитела к CD73, например, молекулы, которые связываются с тем же эпитопом, что и MEDI9447, клон антитела 10.3 или с тем же эпитопом, что и клон антитела 2С5.
[00358] Такие антитела можно идентифицировать исходя из их способности к перекрестному конкурированию (например, к конкурентному ингибированию связывания статистически значимым образом) с раскрытыми в данном документе антителами к CD73, такими как антитело CD730010, антитело CD730002, антитело CD730004 и их антигенсвязывающие фрагменты, при стандартных анализах связывания CD73 (например, анализах с помощью проточной цитометрии, поверхностного плазмонного резонанса или анализах в растворе).
[00359] Соответственно, согласно одному аспекту в настоящем раскрытии предусматриваются антитела к CD73 и их антигенсвязывающие фрагменты, например, моноклональные антитела человека, которые конкурируют за связывание с CD73 с другим антителом к CD73 или его антигенсвязывающим фрагментом, таким как антитело CD730010, антитело CD730002, антитело CD730004, их варианты (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты. Способность тестируемого антитела ингибировать связывание, например, антитела CD730010 (или клона антитела 10.3 или его антигенсвязывающего фрагмента) или антитела CD730002 (или клона антитела 2С5 или его антигенсвязывающего фрагмента), демонстрирует, что тестируемое антитело может конкурировать с данным антителом за связывание с CD73; при этом такое антитело может, в соответствии с теорией, не имеющей ограничительного характера, связываться с тем же или родственным (например, структурно сходным или пространственно близким) эпитопом CD73, что и антитело к CD73 или его антигенсвязывающий фрагмент, с которыми оно конкурирует. Согласно одному аспекту антитело к CD73 или его антигенсвязывающий фрагмент, которые связываются с тем же эпитопом CD73, что и, например, антитело CD730010 (или клон антитела 10.3 или его антигенсвязывающий фрагмент) или антитело CD730002 (или клон антитела 2С5 или его антигенсвязывающий фрагмент), представляет собой моноклональное антитело человека.
[00360] Как описано в данном документе, эпитоп MEDI9447, моноклонального антитела, которое непосредственно ингибирует ферментативную активность CD73, был идентифицирован для изучения механизма действия MEDI9447. Эпитоп находится в пределах апикальной поверхности N-концевого домена CD73, области, более отдаленной от остатков сайта связывания субстрата и активного сайта. В исследованиях по структуре и механизму действия выявили, что MEDI9447 антагонизирует CD73 посредством двойного механизма, который препятствует тому, чтобы CD73 принимал каталитически активную конформацию. Эти результаты обеспечивают первые сведения о точно картируемом эпитопе, на который можно осуществлять целенаправленное воздействие в отношении селективного, потенциального и неконкурентного ингибирования CD73 в качестве способа модуляции аденозиновой передачи сигнала в микроокружении опухоли.
[00361] С помощью масс-спектрометрии (MS), основанной на водород-дейтериевом обмене (HDX), и стратегий мутагенеза, авторы настоящего изобретения определили эпитоп MEDI9447, а также исследовали потенциальные эффекты связывания антитела с глобальной структурой CD73. Антитело связывается с сайтом в N-концевом домене CD73, что обеспечивает неконкурентное ингибирование гидролиза AMP. Согласно различным аспектам эпитоп содержит один или несколько аминокислотных остатков CD73, соответствующих V144, К180 и N185. Согласно различным аспектам эпитоп дополнительно содержит один или несколько аминокислотных остатков CD73, соответствующих Y135, К136 и N187 в CD73. Примечательно, что эпитоп расположен таким образом, что связывание антитела препятствует преобразованию CD73 из открытого конформера в каталитически активный закрытый конформер. Кроме того, исследования авторов настоящего изобретения показывают, что MEDI9447 может ингибировать как заякоренный, так и растворимый CD73 посредством двойного механизма ингибирования, который опосредован валентностью антитела, взаимодействующего с CD73.
IV. Функциональные характеристики антител к CD73
[00362] Аффинность или авидность антитела к антигену можно определить экспериментально с помощью любого подходящего хорошо известного из уровня техники способа, например, проточной цитометрии, твердофазного иммуноферментного анализа (ELISA), или радиоиммунологического анализа (RIA), или кинетических анализов (например, анализа BIACORE™). С легкостью можно применять анализы в формате прямого связывания, а также анализы в формате конкурентного связывания. (См., например, Berzofsky et al., "Antibody-Antigen Interactions," в Fundamental Immunology, Paul, W.E., Ed., Raven Press: New York, N.Y. (1984); Kuby, Immunology, W.H. Freeman and Company: New York, N.Y. (1992) и описанные в данном документе способы. Измеряемая аффинность взаимодействия конкретных антитела-антигена может варьировать при измерении в различных условиях (например, концентрация солей, рН, температура). Таким образом, измерения аффинности и других параметров связывания антигена (например, KD или Kd, Kon, Koff) выполняют со стандартизированными растворами антитела и антигена и стандартизированным буфером, известным из уровня техники, а также с таким буфером, как описан в данном документе.
[00363] Также из уровня техники известно, что показатели аффинности, измеряемые с помощью анализа, основанного на поверхностном плазмонном резонансе (например, BIACORE™), могут варьировать в зависимости от того, какой из реагентов связан с чипом. В связи с этим аффинность можно измерять с использованием формата, при котором целенаправленно воздействующее антитело (например, клон антитела 10.3 или клон антитела 2С5) иммобилизовано на чипе (упоминается как формат "IgG внизу"), или с использованием формата, при котором белок-мишень (например, CD73) иммобилизован на чипе (упоминается как, например, формат "CD73 внизу").
[00364] Согласно одному аспекту настоящего раскрытия антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент специфически связывают CD73 и/или его антигенные фрагменты с константой диссоциации или (kof/kon), составляющей менее 10-6 М, или менее 10-7 М, или менее 10-8 М, или менее 10-9 М, или менее 10-10 М, или менее 10-11 М, или менее 10-12 М, или менее 10-13 М.
[00365] Согласно другому аспекту антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73 и/или его антигенными фрагментами со значением Koff, составляющим менее 1×10-3 с-1 или менее 2×10-3 с-1. Согласно другим аспектам антитело к CD73 или его антигенсвязывающий фрагмент связываются с CD73 и его антигенными фрагментами со значением Koff, составляющим менее 10-3 с-1, менее 5×10-3 с-1, менее 10-4 с-1, менее 5×10-4 с-1, менее 10-5 с-1, менее 5×10-5 с-1, менее 10-6 с-1, менее 5×10-6 с-1, менее 5×10-7 с-1, менее 10-8 с-1, менее 5×10-8 с-1, менее 10-9 с-1, менее 5×10-9 с-1 или менее 10-10 с-1.
[00366] Согласно другому аспекту антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73 и/или его антигенными фрагментами с константой скорости ассоциации или коэффициентом kon, составляющим по меньшей мере 105 М-1 с-1, по меньшей мере 5×105M-1 с-1, по меньшей мере 106М-1 с-1, по меньшей мере 5×106М-1 с-1, по меньшей мере 107М-1 с-1, по меньшей мере 5×107М-1 с-1, или по меньшей мере 108 М-1 с-1, или по меньшей мере 109 М-1 с-1.
[00367] Согласно некоторым аспектам антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73, присутствующим на поверхности клеток MB-MDA-231, со значением KD, составляющим по меньшей мере приблизительно 60 пМ, по меньшей мере приблизительно 70 пМ, по меньшей мере приблизительно 80 пМ, по меньшей мере приблизительно 90 пМ, по меньшей мере приблизительно 100 пМ, по меньшей мере приблизительно 110 пМ, по меньшей мере приблизительно 120 пМ, по меньшей мере прибл изитель но 130 пМ, по меньшей мере приблизительно 140 пМ, по меньшей мере приблизительно 150 пМ, по меньшей мере приблизительно 160 пМ или по меньшей мере приблизительно 170 пМ, измеренным с помощью проточной цитометрии. Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73, присутствующим на поверхности клеток МВ-MDA-231, со значением KD, составляющим приблизительно 150 пМ, измеренным с помощью проточной цитометрии. Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73, присутствующим на поверхности клеток МВ-MDA-231, со значением KD, составляющим приблизительно 80 пМ, измеренным с помощью проточной цитометрии.
[00368] Согласно некоторым аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73, присутствующим на поверхности клеток 3Т1 мыши, со значением KD, составляющим по меньшей мере приблизительно 40 пМ, по меньшей мере приблизительно 50 пМ, по меньшей мере приблизительно 60 пМ, по меньшей мере приблизительно 70 пМ, по меньшей мере приблизительно 80 пМ, по меньшей мере приблизительно 90 пМ, по меньшей мере приблизительно 100 пМ, по меньшей мере приблизительно 120 пМ или по меньшей мере приблизительно 130 пМ, измеренным с помощью проточной цитометрии. Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73, присутствующим на поверхности клеток 3Т1 мыши, со значением KD, составляющим приблизительно 110 пМ, измеренным с помощью проточной цитометрии. Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73, присутствующим на поверхности клеток 3Т1 мыши, со значением KD, составляющим приблизительно 55 пМ, измеренным с помощью проточной цитометрии.
[00369] Согласно некоторым аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73, присутствующим на поверхности клеток MK-1 яванского макака, со значением KD, составляющим по меньшей мере приблизительно 40 пМ, по меньшей мере приблизительно 50 пМ, по меньшей мере приблизительно 60 пМ, по меньшей мере приблизительно 70 пМ, по меньшей мере приблизительно 80 пМ, по меньшей мере приблизительно 90 пМ или по меньшей мере приблизительно 100 пМ, измеренным с помощью проточной цитометрии. Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73, присутствующим на поверхности клеток MK-1 яванского макака, со значением КD, составляющим приблизительно 80 пМ, измеренным с помощью проточной цитометрии. Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73, присутствующим на поверхности клеток MK-1 яванского макака, со значением KD, составляющим приблизительно 60 пМ, измеренным с помощью проточной цитометрии.
[00370] Согласно некоторым аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73 человека со значением KD, составляющим по меньшей мере приблизительно 3 пМ, по меньшей мере приблизительно 4 пМ, по меньшей мере приблизительно 5 пМ, по меньшей мере приблизительно 6 пМ, по меньшей мере приблизительно 7 пМ, по меньшей мере приблизительно 8 пМ, по меньшей мере приблизительно 9 пМ или по меньшей мере приблизительно 10 пМ, измеренным с помощью поверхностного плазменного резонанса (PROTEON®). Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73 человека со значением KD, составляющим приблизительно 4 пМ, измеренным с помощью поверхностного плазмонного резонанса (PROTEON®). Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73 человека со значением KD, составляющим приблизительно 9 пМ, измеренным с помощью поверхностного плазмонного резонанса (PROTEON®).
[00371] Согласно некоторым аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, раскрытые в данном документе, связываются с CD73 мыши со значением KD, составляющим по меньшей мере приблизительно 1 пМ, по меньшей мере приблизительно 2 пМ, по меньшей мере приблизительно 3 пМ, по меньшей мере приблизительно 4 пМ, по меньшей мере приблизительно 5 пМ, по меньшей мере приблизительно 6 пМ, по меньшей мере приблизительно 7 пМ, по меньшей мере приблизительно 8 пМ, по меньшей мере приблизительно 9 пМ, по меньшей мере приблизительно 10 пМ, по меньшей мере приблизительно 11 пМ, по меньшей мере приблизительно 12 пМ, по меньшей мере приблизительно 13 пМ, по меньшей мере приблизительно 14 пМ, по меньшей мере приблизительно 15 пМ, по меньшей мере приблизительно 16 пМ, по меньшей мере приблизительно 17 пМ, по меньшей мере приблизительно 18 пМ, по меньшей мере приблизительно 19 пМ, по меньшей мере приблизительно 20 пМ, по меньшей мере приблизительно 21 пМ, по меньшей мере приблизительно 22 пМ, по меньшей мере приблизительно 23 пМ, по меньшей мере приблизительно 24 пМ или по меньшей мере приблизительно 25 пМ, измеренным с помощью поверхностного плазмонного резонанса (PROTEON®). Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73 мыши со значением KD, составляющим приблизительно 1 пМ, измеренным с помощью поверхностного плазмонного резонанса (PROTEON®). Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73 мыши со значением KD, составляющим приблизительно 22 пМ, измеренным с помощью поверхностного плазмонного резонанса (PROTEON®).
[00372] Согласно некоторым аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73 яванского макака со значением KD, составляющим по меньшей мере приблизительно 3 пМ, по меньшей мере приблизительно 4 пМ, по меньшей мере приблизительно 5 пМ, по меньшей мере приблизительно 6 пМ, по меньшей мере приблизительно 7 пМ, по меньшей мере приблизительно 8 пМ, по меньшей мере приблизительно 9 пМ или по меньшей мере приблизительно 10 пМ, измеренным с помощью поверхностного плазмонного резонанса (PROTEON®). Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73 яванского макака со значением KD, составляющим приблизительно 7 пМ, измеренным с помощью SPR (Proteon). Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73 яванского макака со значением КD, составляющим приблизительно 9 пМ, измеренным с помощью поверхностного плазмонного резонанса (PROTEON®).
[00373] Согласно некоторым аспектам антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73 человека со значением KD, составляющим по меньшей мере приблизительно 40 пМ, по меньшей мере приблизительно 50 пМ, по меньшей мере приблизительно 60 пМ, по меньшей мере приблизительно 70 пМ, по меньшей мере приблизительно 80 пМ, по меньшей мере приблизительно 90 пМ, по меньшей мере приблизительно 100 пМ или по меньшей мере приблизительно 110 пМ, измеренным с помощью связывания в растворе. Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73 человека со значением KD, составляющим приблизительно 80 пМ, измеренным с помощью связывания в растворе. Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73 человека со значением КD, составляющим приблизительно 80 пМ, измеренным с помощью связывания в растворе.
[00374] Согласно некоторым аспектам антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73 мыши со значением КD, составляющим по меньшей мере приблизительно 100 пМ, по меньшей мере приблизительно 200 пМ, по меньшей мере приблизительно 300 пМ, по меньшей мере приблизительно 400 пМ, по меньшей мере приблизительно 500 пМ, по меньшей мере приблизительно 600 пМ, по меньшей мере приблизительно 700 пМ, по меньшей мере приблизительно 800 пМ, по меньшей мере приблизительно 900 пМ, по меньшей мере приблизительно 1000 пМ, по меньшей мере приблизительно 1100 пМ, по меньшей мере приблизительно 1200 пМ, по меньшей мере приблизительно 1300 пМ, по меньшей мере приблизительно 1400 пМ, по меньшей мере приблизительно 1500 пМ, по меньшей мере приблизительно 1600 пМ или по меньшей мере приблизительно 1700 пМ, измеренным с помощью связывания в растворе. Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73 мыши со значением KD, составляющим приблизительно 130 пМ, измеренным с помощью связывания в растворе. Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73 мыши со значением KD, составляющим приблизительно 1500 пМ, измеренным с помощью связывания в растворе.
[00375] Согласно некоторым аспектам антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент связываются с CD73 яванского макака со значением KD, составляющим по меньшей мере приблизительно 60 пМ, по меньшей мере приблизительно 70 пМ, по меньшей мере приблизительно 80 пМ, по меньшей мере приблизительно 90 пМ, по меньшей мере приблизительно 100 пМ, по меньшей мере приблизительно 110 пМ или по меньшей мере приблизительно 120 пМ, измеренным с помощью связывания в растворе. Согласно одному конкретному аспекту антитело к CD73 представляет собой клон антитела 10.3 и оно связывается с CD73 яванского макака со значением Ко, составляющим приблизительно 90 пМ, измеренным с помощью связывания в растворе. Согласно другому конкретному аспекту антитело к CD73 представляет собой клон антитела 2С5 и оно связывается с CD73 яванского макака со значением KD, составляющим приблизительно 100 пМ, измеренным с помощью связывания в растворе. Согласно конкретному аспекту MEDI9447 связывается с CD73 со значением KD, составляющим приблизительно 1×10-12, 5×10-12, 10×10-12, 100×10-12 или 150×10-12.
[00376] Согласно некоторым аспектам CD73-связывающая молекула, раскрытая в данном документе, например, антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, могут облегчать АМР-опосредованное подавление деления Т-клеток. Согласно другим аспектам СВ73-связывающая молекула, раскрытая в данном документе, например, антитело к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, могут восстанавливать АМР-индуцированное подавление Teff с помощью Treg.
[00377] Согласно некоторым аспектам CD73-связывающая молекула, раскрытая в данном документе, например, антитело к CD73 или его антигенсвязывающий фрагмент (например, клон антитела 10.3 или клон антитела 2С5), могут в значительной степени ингибировать рост сингенной опухоли. Согласно одному аспекту опухоль представляет собой опухоль, характерную для немелкоклеточного рака легкого, рака яичников, рака молочной железы, рака головы и шеи, рака поджелудочной железы, колоректального рака, опухоль, характерную для меланомы, опухоль, характерную для лимфомы. Согласно одному аспекту опухоль представляет собой сингенную CRC-опухоль СТ26 мыши, опухоль B16F10, характерную для меланомы, опухоль EG7-OVA, характерную для лимфомы, или опухоль LL2 (характерную для карциномы легких Льюиса). Согласно некоторым аспектам CD73-связывающая молекула, например, антитело к CD73 или его антигенсвязывающий фрагмент, раскрытые в данном документе (например, клон антитела 10.3 или клон антитела 2С5), может в значительной степени ингибировать рост опухоли, при этом опухоль является нечувствительной к терапии антителом к PD-1, и/или антителом к PD-L1, и/или антителом к PD-L2, и/или антителом к CTLA-4. Согласно некоторым аспектам CD73-связывающая молекула, раскрытая в данном документе, например, антитело к CD73 или его антигенсвязывающий фрагмент (например, клон антитела 10.3 или клон антитела 2С5), может в значительной степени ингибировать рост опухоли в случае введения при концентрации PD1, составляющей приблизительно 1 мг/кг, приблизительно 2 мг/кг, приблизительно 3 мг/кг, приблизительно 4 мг/кг, приблизительно 5 мг/кг, приблизительно 6 мг/кг, приблизительно 7 мг/кг, приблизительно 8 мг/кг, приблизительно 9 мг/кг или приблизительно 10 мг/кг.
[00378] Согласно некоторым аспектам CD73-связывающая молекула, раскрытая в данном документе, например, антитело к CD73 или его антигенсвязывающий фрагмент (например, клон антитела 10.3 или клон антитела 2С5), может интернализоваться после связывания с клетками. Согласно некоторым аспектам CD73-связывающая молекула представляет собой конъюгат антитело-лекарственное средство (ADC).
V. Получение антител к CD73 и их антигенсвязывающих фрагментов
[00379] Моноклональные антитела к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) и их антигенсвязывающие фрагменты можно получать с помощью способов с использованием гибридомы, таких как описанные у Kohler и Milstein (1975) Nature 256:495. В способе с использованием гибридомы мышь, хомяка или другого подходящего животного-хозяина иммунизируют, как описано выше, чтобы вызвать выработку лимфоцитами антител, которые будут специфически связываться с иммунизирующим антигеном. Лимфоциты также можно подвергать иммунизации in vitro. После иммунизации лимфоциты выделяют и сливают с подходящей линией клеток миеломы, например с применением полиэтиленгликоля, с образованием клеток гибридомы, которые затем могут быть отобраны из неслитых лимфоцитов и клеток миеломы. Гибридомы, которые вырабатывают моноклональные антитела, специфически направленные против выбранного антигена, как определяется с помощью иммунопреципитации, иммуноблоттинга или с использованием анализа связывания in vitro (например, радиоиммунологического анализа (RIA), твердофазного иммуноферментного анализа (ELISA)), затем можно размножать либо в культуре in vitro с использованием стандартных способов (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, 1986), либо in vivo в виде асцитных опухолей у животных. Затем моноклональные антитела можно выделить из культуральной среды или асцитной жидкости, как описано выше для поликлональных антител.
[00380] В качестве альтернативы, моноклональные антитела к CD73 (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) и их антигенсвязывающие фрагменты также можно получать с применением способов рекомбинантных ДНК, как описано, например, в патенте США №4816567. Полинуклеотиды, кодирующие моноклональное антитело, выделяют из зрелых В-клеток или клеток гибридомы, например, с помощью RT-PCR с использованием олигонуклеотидных праймеров, которые специфически амплифицируют гены, кодирующие тяжелую и легкую цепи антитела, и их последовательность определяют с помощью обычных процедур. Выделенные полинуклеотиды, кодирующие тяжелую и легкую цепи, затем клонируют в подходящие векторы экспрессии, которые при трансфекции в клетки-хозяева, такие как клетки Е. coli, клетки COS обезьяны, клетки яичника китайского хомячка (СНО) или клетки миеломы, которые в других случаях не вырабатывают белок, представляющий собой иммуноглобулин, обеспечивают выработку моноклональных антител с помощью клеток-хозяев. Также рекомбинантные моноклональные антитела к CD73 или их антигенсвязывающие фрагменты требуемых видов можно выделить с использованием библиотек фагового дисплея, экспрессирующих CDR требуемых видов, как описано (McCafferty et al., 1990, Nature, 348:552-554; Clarkson etal, 1991, Nature, 352:624-628 и Marks etal, 1991, J. Mol. Biol., 222:581-597).
[00381] Полинуклеотид (полинуклеотиды), кодирующий(кодирующие) антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, можно дополнительно модифицировать с помощью ряда различных способов с применением технологии рекомбинантных ДНК с получением альтернативных антител. Согласно некоторым аспектам константные домены легкой и тяжелой цепей, например моноклонального антитела мыши, можно заменить (1) на таковые области, например антитела человека, с получением химерного антитела или (2) на отличный от иммуноглобулина полипептид с получением гибридного антитела. Согласно некоторым аспектам константные области усекают или удаляют с получением требуемого фрагмента антитела из моноклонального антитела. Сайт-направленный или высокопроизводительный мутагенез вариабельной области можно применять для оптимизации специфичности, аффинности и т.д. моноклонального антитела.
[00382] Согласно определенным аспектам антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент представляют собой антитело человека или его антигенсвязывающий фрагмент. Антитела человека можно непосредственно получать с помощью различных методик, известных из уровня техники. Можно получать иммортализованные В-лимфоциты человека, иммунизированные in vitro или выделенные у иммунизированного индивидуума, у которого вырабатываются антитела к целевому антигену (см., например. Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boemer et al., 1991, J. Immunol., 147 (1):86-95 и патент США №5750373).
[00383] Также антитело к CD73 человека (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент могут быть отобраны из фаговой библиотеки, при этом в такой фаговой библиотеке экспрессируются антитела человека, как описано, например, в Vaughan et al., 1996, Nat. Biotech., 14:309-314, Sheets et al., 1998, Proc. Nat'l. Acad. Sci., 95:6157-6162, Hoogenboom and Winter, 1991, J. Mol. Biol., 227:381 и Marks et al., 1991, J. Mol. Biol., 222:581). Методики получения и применения фаговых библиотек антител также описаны в патентах США №№5,969,108, 6,172,197, 5,885,793, 6,521,404; 6,544,731; 6,555,313; 6,582,915; 6,593,081; 6,300,064; 6,653,068; 6,706,484; и 7264963; а также Rothe et al., 2007, J. Mol. Bio., doi:10.1016/j.jmb.2007.12.018 (каждый из которых в полном объеме включен в данный документ с помощью ссылки).
[00384] Стратегии созревания аффинности и стратегии перестановки цепей (Marks et al., 1992, Bio/Technology 10:779-783, включенная в полном объеме с помощью ссылки) известны из уровня техники и могут использоваться для получения высокоаффинных антител человека или их антигенсвязывающих фрагментов.
[00385] Согласно некоторым аспектам моноклональное антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) может представлять собой гуманизированное антитело. Также можно применять способы конструирования, гуманизации или изменения поверхности антител, не являющихся антителами человека, или антител человека, и они хорошо известны из уровня техники. Гуманизированное, с измененной поверхностью или подобным образом сконструированное антитело может иметь один или несколько аминокислотных остатков, отличных от таковых человеческого происхождения, например, без ограничения мыши, крысы, кролика, отличного от человека примата или другого млекопитающего. Эти аминокислотные остатки, не являющиеся аминокислотными остатками человека, заменяют остатками, часто называемыми "импортированными" остатками, которые, как правило, взяты из "импортированного" вариабельного, константного или другого домена известной последовательности человека. Такие импортированные последовательности можно применять для снижения иммуногенности или снижения, повышения или модификации связывания, аффинности, скорости ассоциации, скорости диссоциации, авидности, специфичности, времени полужизни или любых других подходящих характеристик, известных из уровня техники. В целом, остатки CDR непосредственно и в наиболее значительной степени участвуют во влиянии на связывание CD73. Соответственно, сохраняются часть или все из последовательностей CDR, не относящихся или относящихся к человеку, тогда как последовательности вариабельных и константных областей, не относящихся к человеку, могут быть заменены на человеческие или другие аминокислотные последовательности.
[00386] Антитела также необязательно можно гуманизировать, изменять поверхность, конструировать, или антитела человека можно конструировать с сохранением высокой аффинности к антигену CD73 и других предпочтительных биологических свойств. Для достижения данной цели гуманизированные (или человеческие) или сконструированные антитела к CD73 и антитела с измененной поверхностью можно необязательно получать посредством способа с анализом исходных последовательностей и гуманизированных и сконструированных различными подходами продуктов с использованием трехмерных моделей исходных, сконструированных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются общедоступными и известны специалистам в данной области техники. Существуют компьютерные программы, в которых иллюстрируются и отображаются возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулина-кандидата. Рассмотрение этих изображений позволяет проанализировать вероятную роль остатков в функционировании последовательности иммуноглобулина-кандидата, т.е. проанализировать остатки, которые влияют на способность иммуноглобулина-кандидата связывать его антиген, такой как CD73. Таким образом, остатки каркасных областей (FW) можно выбирать и комбинировать из консенсусной и импортированной последовательностей так, чтобы обеспечить требуемые характеристики антитела, такие как повышенная аффинность к целевому антигену(антигенам).
[00387] Гуманизацию, изменение поверхности или конструирование антител к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающих фрагментов можно выполнять с применением любого известного способа, например, без ограничения способами, описанными в Jones et al., Nature 321:522 (1986); Riechmann et al., Nature 332:323 (1988); Verhoeyen et al., Science 239:1534 (1988)), Sims et al., J. Immunol. 151: 2296 (1993); Chothia and Lesk, J. Mol. Biol. 196:901 (1987), Carter et al., Proc. Natl. Acad. Sci. U.S.A. 89:4285 (1992); Presta et al., J. Immunol. 151:2623 (1993), патентах США №№5,639,641, 5,723,323; 5,976,862; 5,824,514; 5,817,483; 5,814,476; 5,763,192; 5,723,323; 5,766,886; 5,714,352; 5,9,55,358; 6,204,023; 6,180,370; 6,331,431; 5,693,762; 5,530,101; 5,585,089; 5,225,539; 4,816,567; 5,969,108; 7,635,666; 7,723,270; 7,557,189; 7,538,195; и 7,342,110; Международных заявках №№ PCT/US98/16280; PCT/US91/05939; PCT/US94/01234; PCT/GB92/01755;, публикациях Международных заявках на патент №№ WO90/14443; WO90/14424; WO90/14430; и публикации европейского патента № ЕР 229246, каждый из которых в полном объеме включен в данный документ с помощью ссылки, включая цитируемые в них ссылки.
[00388] Гуманизированные антитела к CD73 и их антигенсвязывающие фрагменты также можно получать с использованием трансгенных мышей, содержащих локусы иммуноглобулинов человека, которые способны после иммунизации вырабатывать полный спектр антител человека при отсутствии выработки эндогенных иммуноглобулинов. Данный подход описан в патентах США №№5,545,807; 5,545,806; 5,569,825; 5,625,126; 5,633,425; и 5,661,016.
[00389] Согласно определенным аспектам предусматривается фрагмент антитела к CD73 (например, фрагмент из клона антитела 10.3 или из клона антитела 2С5). Известны различные методики получения фрагментов антител. Традиционно эти фрагменты получают посредством протеолитического расщепления интактных антител (например, Morimoto et al., 1993, Journal of Biochemical and Biophysical Methods 24:107-117; Brennan et al., 1985, Science, 229:81). Согласно определенным аспектам фрагменты антитела к CD73 получают рекомбинантным путем. Все Fab-, Fv- и scFv-фрагменты антител могут экспрессироваться и секретироваться Е. coli или другими клетками-хозяевами, что обеспечивает таким образом получение больших количеств этих фрагментов. Такие фрагменты антител к CD73 также можно выделять из фаговых библиотек антител, обсуждаемых выше. Фрагменты антител к CD73 также могут представлять собой линейные антитела, описанные в патенте США №5641870. Другие методики получения фрагментов антител, например химический синтез, будут очевидны практикующему специалисту.
[00390] В соответствии с настоящим раскрытием методики можно адаптировать для получения одноцепочечных антител, специфичных в отношении CD73 (см., например, патент США №4946778). Кроме того, способы можно адаптировать для создания экспрессионных библиотек Fab (см., например, Huse et al., Science 246:1275-1281 (1989)), с возможностью быстрой и эффективной идентификации моноклональных Fab-фрагментов с требуемой специфичностью в отношении CD73 или их производных, фрагментов, аналогов или гомологов. Фрагменты антител можно получать с помощью методик, известных из уровня техники, при этом они включают без ограничения: (а) F(аb')2-фрагмент, полученный посредством расщепления молекулы антитела пепсином; (b) Fab-фрагмент, полученный посредством восстановления дисульфидных мостиков F(аb')2-фрагмента, (с) Fab-фрагмент, полученный посредством обработки молекулы антитела папаином и восстанавливающим средством, и (d) Fv-фрагменты.
[00391] Антитело к CD733 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, раскрытые в данном документе, можно модифицировать с целью увеличения его времени полужизни в сыворотке. Этого можно достичь, например, посредством введения эпитопа, связывающегося с рецептором реутилизации, в антитело или фрагмент антитела с помощью мутации соответствующей области в антителе или фрагменте антитела, или посредством введения эпитопа в пептидную метку, которую затем сливают с антителом или фрагментом антитела либо на любом конце или в середине (например, с помощью синтеза ДНК или пептида), либо с помощью мутации YTE. Из уровня техники известны другие способы увеличения времени полужизни в сыворотке крови антитела или его антигенсвязывающего фрагмента, например, конъюгация с гетерологичной молекулой, такой как PEG.
[00392] Гетероконъюгатные антитела к CD73 (например, клон антитела 10.3 или клон антитела 2С5) и их антигенсвязывающие фрагменты также находятся в пределах объема настоящего раскрытия. Гетероконъюгатные антитела состоят из двух ковалентно связанных антител. Например, такие антитела были предложены для направления иммунных клеток к нежелательным клеткам (см., например, патент США №4676980). Предполагается, что гетероконъюгатные антитела к CD73 (например, клон антитела 10.3 или клон антитела 2С5) и их антигенсвязывающие фрагменты можно получать in vitro с применением известных способов химического синтеза белков, в том числе способов, включающих средства для перекрестного связывания. Например, иммунотоксины можно сконструировать с помощью реакции дисульфидного обмена или посредством образования тиоэфирной связи. Примеры подходящих для этой цели реагентов включают иминотиолат и метил-4-меркаптобутиримидат.
[00393] Согласно определенным аспектам раскрытые в данном документе CD73-связывающие молекулы, например, антитела (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты, можно комбинировать с другими терапевтическими средствами (например, в комбинированной терапии), или их можно сливать (например, генетически сливать с образованием гибридного белка) или конъюгировать (например, конъюгировать химическим или ферментативным путем) по меньшей мере с одним гетерологичным фрагментом. Таким образом, раскрытые в данном документе CD73-связывающие молекулы можно сливать или конъюгировать с другими терапевтическими средствами или токсинами с образованием иммуноконъюгатов и/или гибридных белков. В настоящем раскрытии также предусматриваются конъюгаты антитело-лекарственное средство (ADC), содержащие по меньшей мере одну из раскрытых в данном документе CD73-связывающих молекул, которая была дериватизирована или связана (например, химическим или рекомбинантным путем) с другой молекулой (например, пептидом, низкомолекулярным лекарственным средством, выявляемой молекулой и т.д.). В целом, антитела к CD73 или их части дериватизируют так, что дериватизация или введение метки не сказывается отрицательно на их связывании с CD73. Соответственно предполагается, что антитела и части антител к CD73 по настоящему раскрытию включают в себя как интактные, так и модифицированные формы описанных в данном документе молекул, связывающихся с CD73. Например, раскрытую в данном документе молекулу, связывающуюся с CD73, или ее CD73-связывающую часть можно функционально связывать (посредством химического сочетания, генетического слияния, нековалентного соединения или иным образом) с одной или несколькими другими молекулярными структурами, такими как цитотоксическое средство, фармацевтическое средство, выявляемое средство и/или белок или пептид, которые могут опосредовать соединение молекулы, связывающейся с CD73, с другой молекулой (такой как центральная область стрептавидина или полигистидиновая метка).
[00394] Один из типов дериватизированных молекул можно получить посредством перекрестного связывания двух или более молекулярных структур, например, раскрытой в данном документе молекулы, связывающейся с CD73, и терапевтического средства (например, цитотоксина, такого как тубулизин или MEDI 1508). Подходящие средства для перекрестного связывания включают средства, которые являются гетеробифункциональными, т.е. имеющими две разные реакционноспособные группы, разделенные подходящим промежуточным звеном (например, сложный эфир м-малеимидобензоил-N-гидроксисукцинимида), или гомобифункциональными (например, дисукцинимидилсуберат). Такие средства для перекрестного связывания доступны, например, от Pierce Chemical Company, Рокфорд, Иллинойс. Дополнительные бифункциональные средства для сочетания включают N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат, иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCL), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис-(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол).
[00395] Другой тип дериватизированной молекулы можно получить посредством включения выявляемой метки. Пригодные средства для выявления включают флуоресцентные соединения (например, флуоресцеин, флуоресцеина изоцианат, родамин, 5-диметиламин-1-нафталинсульфонилхлорид, фикоэритрин, люминофоры на основе комплексов лантанидов и т.п.), ферменты, пригодные для выявления (например, пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза, глюкозооксидаза и т.п.), эпитопы, распознаваемые вторичным репортером (например, парные последовательности "лейциновых застежек", сайты связывания для вторичных антител, домены связывания металлов, эпитопные метки и т.д.). Согласно некоторым аспектам выявляемые метки можно присоединять с помощью по меньшей мере одной спейсерной группы. Спейсерные группы могут быть различной длины с целью уменьшения потенциальных стерических затруднений.
[00396] Раскрытые в данном документе молекулы, связывающиеся с CD73, также можно метить радиоактивно меченой аминокислотой. Радиоактивную метку можно использовать как в диагностических, так и в терапевтических целях. Например, радиоактивную метку можно использовать для выявления клеток, экспресеирующих CD73, с помощью рентгеновской или других диагностических методик, таких как позитронно-эмиесионная томография (PET).
[00397] Кроме того, радиоактивную метку можно использовать в терапевтических целях в качестве токсина для клеток, экспрессирующих CD73, например таких, которые вызывают нежелательный иммунный ответ. Примеры меток для полипептидов включают без ограничения следующие радиоактивные изотопы или радионуклиды: 3H, 14С, 15N, 35S, 90Y, 99Тc, 111In, 125I и 131I. Согласно некоторым аспектам молекулу, связывающуюся с CD73, можно метить парамагнитным, радиоактивным или флуорогенным ионом, выявляемым при визуализации. Согласно некоторым аспектам парамагнитный ион представляет собой хром (III), марганец (II), железо (III), железо (II), кобальт (II), никель (II), медь (II), неодим (III), самарий (III), иттербий (III), гадолиний (III), ванадий (II), тербий (III), диспрозий (III), гольмий (III) или эрбий (III). Согласно другим аспектам радиоактивный ион представляет собой йод-123, технеций-99, индий-111, рений-188, рений-186, медь-67, йод-131, иттрий-90, йод-125, астат-211 и галлий-67. Согласно другим аспектам молекулу, связывающуюся с CD73, метят средством для рентгеновской визуализации, таким как лантан (III), золото (III), свинец (II) и висмут (III). Раскрытые в данном документе молекулы, связывающиеся с CD73, можно также дериватизировать с помощью химической группы, например полимера, такого как полиэтиленгликоль (PEG), метильной группы, этильной группы или углеводной группы. Эти группы пригодны для улучшения биологических характеристик антитела, например, для увеличения времени полужизни в сыворотке крови или для повышения связывания в тканях.
[00398] Термин "цитотоксическое средство", используемый в данном документе, определяется в широком смысле и относится к веществу, которое ингибирует или препятствует выполнению функции клеток, и/или вызывает разрушение клеток (гибель клеток), и/или оказывает противоопухолевые/антипролиферативные эффекты. Например, цитотоксическое средство может прямо или опосредованно препятствовать развитию, созреванию или распространению неопластических клеток опухоли. Термин включает также средства, вызывающие не только цитотоксический, но и цитостатический эффект. Термин включает химиотерапевтические средства, описанные ниже, а также другие антагонисты CD73, антиангиогенные средства, ингибиторы тирозинкиназы, ингибиторы протеинкиназы А, представителей семейства цитокинов, радиоактивные изотопы и токсины, такие как ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения.
[00399] Термин "химиотерапевтическое средство" является подмножеством термина "цитотоксическое средство", включающим природные или синтетические химические соединения. Примеры химиотерапевтических средств включают алкилирующие средства, например азотистые иприты, этилениминовые соединения, алкилсульфонаты и другие соединения с алкилирующим действием, такие как нитрозомочевины, цисплатин и дакарбазин; антиметаболиты, например антагонисты фолиевой кислоты, пурина или пиримидина; митотические ингибиторы, например алкалоиды представителей рода Vinca и производные подофиллотоксина; цитотоксические антибиотики и производные камптотецина. Другие химиотерапевтические средства представляют собой амифостин (ETHYOL®), цисплатин, дакарбазин (DTIC), дактиномицин, мехлорэтамин (азотистый иприт), стрептозоцин, циклофосфамид, кармустин (BCNU), ломустин (CCNU), доксорубицин (ADRIAMYCIN®), инкапсулированный в липосомы доксорубицин (DOXIL®), гемцитабин (GEMZAR®), даунорубицин, инкапсулированный в липосомы даунорубицин (DAUNOXOME®), прокарбазин, митомицин, цитарабин, этопозид, метотрексат, 5-фторурацил (5-FU), винбластин, винкристин, блеомицин, паклитаксел (TAXOL®), доцетакеел (TAXOTERE®), альдеслейкин, аспарагиназу, бусульфан, карбоплатин, кладрибин, камптотецин, СРТ-11, 10-гидрокси-7-этилкамптотецин (SN38), гефитиниб (IRESSA®), дакарбазин, флоксуридин, флударабин, гидроксимочевину, ифосфамид, идарубицин, месна, интерферон альфа, интерферон бета, иринотекан, митоксантрон, топотекан, лейпролид, мегестрол, мелфалан, меркаптопурин, пликамицин, митотан, пэгаспаргаза, пентостатин, пипоброман, пликамицин, стрептозоцин, тамоксифен, тенипозид, тестолактон, тиогуанин, тиотепу, урациловый иприт, винорелбин, хлорамбуцил, ингибиторы ароматазы и их комбинации.
[00400] Для целей настоящего раскрытия следует понимать, что модифицированные антитела к CD73 или их антигенсвязывающие фрагменты могут содержать вариабельную область любого типа, которая предусматривается для соединения антитела или полипептида с CD73. В связи с этим вариабельная область может содержаться или может происходить от млекопитающего любого типа, у которого может быть вызван гуморальный ответ и выработка иммуноглобулинов против требуемого опухолеассоциированного антигена. Таким образом, вариабельная область модифицированных антител к CD73 или их антигенсвязывающих фрагментов может происходить, например, от человека, мыши, отличного от человека примата (например, яванских макаков, макаков и т.д.) или представителя семейства волчьих. Согласно некоторым аспектам как вариабельные, так и константные области модифицированных антител к CD73 или их антигенсвязывающих фрагментов происходят от человека. Согласно другим аспектам вариабельные области совместимых антител (обычно происходящие от источника, отличного от человека) можно сконструировать или специфически оптимизировать с целью улучшения свойств связывания или снижения иммуногенности молекулы. В связи с этим вариабельные области могут быть гуманизированы или иным образом изменены посредством включения импортируемых аминокислотных последовательностей.
[00401] Согласно определенным аспектам вариабельные домены как тяжелых, так и легких цепей антитела к CD73 (например, клона антитела 10.3 или клона антитела 2С5) или его антигенсвязывающего фрагмента изменены посредством по меньшей мере частичного замещения одной или нескольких CDR и, если необходимо, посредством частичного замещения каркасной области и изменения последовательности. Хотя CDR могут быть получены из антитела того же класса или даже подкласса, что и антитело, из которого получены каркасные области, предусматривается, что CDR будут получены из антитела другого класса и согласно некоторым аспектам из антитела от другого вида. Для передачи антигенсвязывающей способности одного вариабельного домена другому, замещение всех CDR полными CDR из донорной вариабельной области не является необходимым. Наоборот, необходимым является только перенос тех остатков, которые являются необходимыми для сохранения активности антигенсвязывающего сайта. С учетом пояснений, изложенных в патентах США №№5585089, 5693761 и 5693762, получение функционального антитела со сниженной иммуногенностью либо с помощью проведения стандартных экспериментов, либо с помощью исследования методом проб и ошибок будет находиться полностью в пределах компетенции специалиста в данной области техники.
[00402] Безотносительно к изменениям в вариабельной области, специалистам в данной области техники будет понятно, что модифицированные антитела к CD73 (например, модифицированный клон антитела 10.3 или модифицированный клон антитела 2С5) или их антигенсвязывающие фрагменты будут предусматривать антитела (например, полноразмерные антитела или их иммунореактивные фрагменты), в которых по меньшей мере часть одного или нескольких доменов константной области были удалены или иным образом изменены с обеспечением требуемых биохимических характеристик, таких как усиленная локализация в опухоли или сокращенное время полужизни в сыворотке крови, по сравнению с антителом примерно с такой же иммуногенностью, содержащим нативную или неизмененную константную область. Согласно некоторым аспектам константная область модифицированных антител будет содержать константную область человеческого происхождения. Модификации константной области, совместимые с этими молекулами антитела к CD73, раскрытыми в данном документе, включают добавления, делеции или замены одной или нескольких аминокислот в одном или нескольких доменах. То есть раскрытые в данном документе модифицированные антитела могут содержать изменения или модификации одного или нескольких из трех константных доменов тяжелой цепи (СН1, СН2 или СН3) и/или константного домена легкой цепи (CL). Согласно некоторым аспектам предполагаются модифицированные константные области, в которых один или несколько доменов частично или полностью удалены. Согласно некоторым аспектам модифицированные антитела будут содержать конструкции с удаленным доменом или варианты, в которых полный СН2-домен был удален (ΔCH2-конструкции). Согласно некоторым аспектам исключенный домен константной области будет замещен коротким аминокислотным спейсером (например, из 10 остатков), что обеспечивает некоторую гибкость молекулы, которую, как правило, придает отсутствующая константная область.
[00403] Помимо его конфигурации, из уровня техники известно, что константная область опосредует некоторые эффекторные функции. Например, связывание С1-компонента системы комплемента с антителами активирует систему комплемента. Активация комплемента важна для опсонизации и лизиса патогенов в клетке. Активация комплемента также стимулирует воспалительный ответ и также может быть вовлечена в аутоиммунные реакции гиперчувствительности. Кроме того, антитела связываются с клетками посредством Fc-области, при этом сайт связывания Fc-рецептора Fc-области антитела связывается с Fc-рецептором (FcR) клетки. Существует ряд Fc-рецепторов, являющихся специфичными в отношении антител различных классов, в том числе IgG (гамма-рецепторы), IgE (эта-рецепторы), IgA (альфа-рецепторы) и IgM (мю-рецепторы). Связывание антитела с Fc-рецепторами на поверхностях клеток запускает ряд важных и разнообразных биологических ответов, в том числе поглощение и разрушение нагруженных антителами частиц, клиренс иммунных комплексов, лизис нагруженных антителами клеток-мишеней клетками-киллерами (называемый антителозависимой клеточноопосредованной цитотоксичностью или ADCC), высвобождение медиаторов воспаления, плацентарный перенос и контроль выработки иммуноглобулинов.
[00404] Согласно определенным аспектам антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент предусматриваются с измененными эффекторными функциями, которые в свою очередь влияют на биологические параметры введенного антитела или его антигенсвязывающего фрагмента. Например, делеция или инактивация (посредством точечных мутаций или других средств) домена константной области может ослаблять связывание циркулирующего модифицированного антитела с Fc-рецепторами, усиливая таким образом локализацию в опухоли. В других случаях модификации константной области, соответствующие настоящему раскрытию, могут ослаблять связывание комплемента и таким образом сокращать время полужизни в сыворотке крови и неспецифичное соединение конъюгированного цитотоксина. Еще несколько модификаций константной области можно применять для устранения дисульфидных связей или олигосахаридных фрагментов, что обеспечивает возможность усиления локализации в связи с повышенной специфичностью по отношению к антигену или гибкостью антитела. Аналогичным образом модификации константной области в соответствии с настоящим раскрытием с легкостью можно осуществлять с применением хорошо известных методик биохимической или молекулярной инженерии, которые полностью находятся в пределах компетенции специалиста в данной области техники.
[00405] Согласно определенным аспектам раскрытая в данном документе CD73-связывающая молекула, то есть антитело (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, не обладает одной или несколькими эффекторными функциями. К примеру, согласно некоторым аспектам антитело или его антигенсвязывающий фрагмент не обладают активностью в виде антителозависимой клеточной цитотоксичности (ADCC) и/или активностью в виде комплементзависимой цитотоксичности (CDC). Согласно определенным аспектам антитело к CD73 или его антигенсвязывающий фрагмент не связываются с Fc-рецептором и/или факторами комплемента. Согласно определенным аспектам антитело или его антигенсвязывающий фрагмент не обладают эффекторной функцией.
[00406] Следует отметить, что согласно определенным аспектам модифицированные антитела к CD73 или их антигенсвязывающие фрагменты можно конструировать для слияния СН3-домена непосредственно с шарнирной областью соответствующих модифицированных антител или их фрагментов. В других конструкциях может потребоваться обеспечение вставки пептидного спейсера между шарнирной областью и модифицированными СН2 и/или СН3-доменами. Например, можно было бы экспрессировать совместимые конструкции, в которых СН2-домен был удален и оставшийся СН3-домен (модифицированный или немодифицированный) соединен с шарнирной областью с помощью спейсера из 5-20 аминокислот. Такой спейсер можно добавлять, к примеру, чтобы убедиться, что регуляторные элементы константного домена остаются свободными и доступными или что шарнирная область остается гибкой. Однако следует отметить, что аминокислотные спейсеры могут в некоторых случаях оказаться иммуногенными и вызывать нежелательный иммунный ответ в отношении конструкции. Соответственно, согласно определенным аспектам любой спейсер, добавленный в конструкцию, будет относительно неиммуногенным или вовсе не будет включен с целью сохранения требуемых биохимических свойств модифицированных антител.
[00407] Наряду с делецией целых доменов константной области, следует принять во внимание, что антитела к CD73 и их антигенсвязывающие фрагменты по настоящему раскрытию могут быть получены посредством частичной делеции или замены нескольких или даже одной аминокислоты. Например, мутации одной аминокислоты в выбранных областях СН2-домена может быть достаточно, чтобы существенно ослабить связывание Fc и тем самым усилить локализацию в опухоли. Аналогичным образом, может потребоваться просто удалить ту часть одной или нескольких доменов константных областей, которые контролируют эффекторную функцию (например, связывание с C1Q комплемента), подлежащую модулированию. Такие частичные делеции константных областей могут обеспечивать улучшение выбранных характеристик антитела или его антигенсвязывающего фрагмента (например, время полужизни в сыворотке крови), сохраняя при этом другие требуемые функции, ассоциированные с соответствующим доменом константной области, в неизменном виде. Кроме того, как указывалось выше, константные области раскрытых антител к CD73 и их антигенсвязывающих фрагментов можно модифицировать посредством мутации или замены одной или нескольких аминокислот, что улучшает профиль полученной конструкции. В связи с этим можно блокировать активность, обеспечиваемую консервативным сайтом связывания (например, Fc-связывания), при этом существенно сохраняя конфигурацию и иммуногенный профиль модифицированного антитела или его антигенсвязывающего фрагмента. Определенные аспекты могут включать добавление одной или нескольких аминокислот к константной области с улучшением требуемых характеристик, таких как ослабление или усиление эффекторной функции или обеспечение присоединения большего количества цитотоксинов или углеводов. Согласно таким аспектам могут потребоваться вставка или повторение специфических последовательностей, полученных из выбранных доменов константных областей.
[00408] В настоящем раскрытии также предусматриваются варианты и эквиваленты, которые являются практически гомологичными химерным, гуманизированным и человеческим антителам к CD73 или их антигенсвязывающим фрагментам, изложенным в данном документе. Они могут содержать, например, мутации в виде консервативных замен, т.е. замену одной или нескольких аминокислот на сходные аминокислоты. Например, консервативная замена относится к замене аминокислоты на другую из того же общего класса, как, например, одной кислой аминокислоты на другую кислую аминокислоту, одной основной аминокислоты на другую основную аминокислоту или одной нейтральной аминокислоты на другую нейтральную аминокислоту. Из уровня техники хорошо известно, что подразумевается под консервативной заменой аминокислоты.
[00409] Антитело к CD73 или его антигенсвязывающий фрагмент можно дополнительно модифицировать, чтобы содержать дополнительные химические фрагменты, не являющиеся в обычных условиях частью белка. Данные дериватизированные фрагменты могут улучшать растворимость, биологическое время полужизни или абсорбцию белка. Данные фрагменты могут также уменьшать или устранять любые требуемые побочные эффекты белков и т.п. Обзор таких фрагментов можно найти в Remington's Pharmaceutical Sciences, 20th ed.. Mack Publishing Co., Easton, PA (2000).
VI. Полинуклеотиды, кодирующие CD73-связывающие молекулы
[00410] Согласно определенным аспектам настоящее раскрытие охватывает полинуклеотиды, содержащие последовательности нуклеиновой кислоты, кодирующие полипептид, который специфически связывает CD73 или его антигенсвязывающий фрагмент. Например, в настоящем раскрытии предусматривается полинуклеотид, содержащий последовательность нуклеиновой кислоты, которая кодирует антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или кодирует антигенсвязывающий фрагмент такого антитела. Такие полинуклеотиды по настоящему раскрытию могут быть в виде РНК или в виде ДНК. ДНК включает кДНК, геномную ДНК и синтетическую ДНК и может быть двунитевой или однонитевой, при этом если она однонитевая, то может представлять собой кодирующую нить или некодирующую (антисмысловую) нить.
[00411] Согласно определенным аспектам полинуклеотиды являются выделенными. Согласно определенным аспектам полинуклеотиды являются практически чистыми. Согласно определенным аспектам полинуклеотиды содержат последовательность, кодирующую зрелый полипептид, слитую в той же рамке считывания с полинуклеотидом, который способствует, например, экспрессии и секреции полипептида из клетки-хозяина (например, лидерной последовательностью, которая функционирует как последовательность сигнала секреции для регулирования транспорта полипептида из клетки). Полипептид с лидерной последовательностью является белком-предшественником и может иметь лидерную последовательность, расщепляемую клеткой-хозяином, с образованием зрелой формы полипептида. Полинуклеотиды также могут кодировать CD73-связывающий пробелок, который представляет собой зрелый белок с дополнительными аминокислотными остатками на 5'-конце.
[00412] Согласно определенным аспектам полинуклеотиды содержат последовательность, кодирующую зрелый CD73-связывающий полипептид, например, антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, слитую в той же рамке считывания с маркерной последовательностью, что обеспечивает возможность, например, очистки кодируемого полипептида. Например, маркерная последовательность может представлять собой гексагистидиновую метку, доставляемую вектором pQE-9, для обеспечения очистки слитого с маркером зрелого полипептида в случае хозяина-бактерии, или маркерная последовательность может представлять собой гемагглютининовую (НА) метку, полученную из белка гемагглютинина вируса гриппа, если используется хозяин-млекопитающее (например, клетки COS-7).
[00413] В настоящем раскрытии также предусматриваются варианты описанных полинуклеотидов, кодирующих, например, CD73-связывающие фрагменты, аналоги и производные раскрытых в данном документе CD73-связывающих молекул (например, клон антитела 10.3 или клон антитела 2С5).
[00414] Варианты полинуклеотидов могут содержать изменения в кодирующих областях, некодирующих областях, или как в тех, так и в других. Согласно некоторым аспектам варианты полинуклеотидов содержат изменения, которые приводят к "молчащим" заменам, добавлениям или делециям, однако не изменяют свойства или виды активностей кодируемого полипептида. Согласно некоторым аспектам нуклеотидные варианты получены с помощью "молчащих" замен из-за вырожденности генетического кода. Варианты полинуклеотидов можно получать в силу ряда причин, например, для оптимизации экспрессии кодонов для конкретного хозяина (замена кодонов в mRNA человека на кодоны, предпочтительные для хозяина-бактерии, такого как Е. coli). Также предусмотрены векторы и клетки, содержащие полинуклеотиды, описанные в данном документе.
[00415] Согласно некоторым аспектам последовательность ДНК, кодирующую CD73-связывающую молекулу, например, антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, можно конструировать посредством химического синтеза с применением олигонуклеотидного синтезатора. Такие олигонуклеотиды можно конструировать на основе аминокислотной последовательности требуемого полипептида и отбирая те кодоны, которые являются предпочтительными для клеток-хозяев, в которых будет вырабатываться рекомбинантный полипептид, представляющий интерес. Для синтеза выделенной полинуклеотидной последовательности, кодирующей выделенный полипептид, представляющий интерес, можно применять стандартные способы. Например, полную аминокислотную последовательность можно применять для конструирования обратно транслируемого гена. Затем можно синтезировать ДНК-олигомер, содержащий нуклеотидную последовательность, кодирующую конкретный выделенный полипептид. Например, можно синтезировать несколько небольших олигонуклеотидов, кодирующих части требуемого полипептида, а затем лигировать их. Отдельные олигонуклеотиды, как правило, содержат 5' или 3' "липкие" концы для комплементарной сборки.
[00416] Сразу после сборки (посредством синтеза, сайт-направленного мутагенеза или другого способа) полинуклеотидные последовательности, кодирующие конкретный выделенный полипептид, представляющий интерес, будут вставлены в вектор экспрессии и функционально связаны с контролирующей экспрессию последовательностью, соответствующей экспрессии белка в требуемом хозяине. Правильность сборки можно проверить посредством секвенирования нуклеотидов, рестрикционного картирования и экспрессии биологически активного полипептида в подходящем хозяине. Как хорошо известно из уровня техники, для того чтобы получить высокие уровни экспрессии трансфицированного гена в хозяине, ген должен быть функционально связан с последовательностями, контролирующими транскрипцию и трансляцию, которые являются функциональными в выбранном для экспрессии хозяине.
[00417] Согласно определенным аспектам рекомбинантные векторы экспрессии применяют для амплификации и экспрессии ДНК, кодирующей антитела к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты. Рекомбинантные векторы экспрессии представляют собой реплицируемые ДНК-конструкции, которые имеют синтетические или полученные из кДНК фрагменты ДНК, кодирующие полипептидную цепь антитела к CD73 или/и его антигенсвязывающий фрагмент, функционально связанные с подходящими элементами, регулирующими транскрипцию или трансляцию, полученными из генов млекопитающих, микроорганизмов, вирусов или насекомых.
[00418] В основном транскрипционная единица содержит сборку из (1) генетического элемента или элементов, имеющих регуляторную роль в экспрессии генов, например, промоторы или энхансеры транскрипции, (2) структурной или кодирующей последовательности, которая транскрибируется в mRNA и транслируется в белок, и (3) соответствующих последовательностей инициации транскрипции и трансляции, а также последовательностей терминации, подробно описанных ниже. Такие регуляторные элементы могут включать последовательность оператора для контроля транскрипции. Способность к репликации в хозяине обычно обеспечивается точкой начала репликации, при этом можно дополнительно встроить ген селекции для облегчения распознавания трансформантов. Области ДНК являются функционально связанными, когда они функционально зависят друг от друга. Например, ДНК сигнального пептида (секреторная лидерная последовательность) функционально связана с ДНК полипептида, если она экспрессируется в виде предшественника, который участвует в секреции полипептида; промотор функционально связан с кодирующей последовательностью, если он контролирует транскрипцию последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы обеспечивать трансляцию. Структурные элементы, предназначенные для применения в дрожжевых системах экспрессии, включают лидерную последовательность, обеспечивающую внеклеточную секрецию транслируемого белка клеткой-хозяином. В качестве альтернативы, если рекомбинантный белок экспрессируется без лидерной или транспортной последовательности, он может включать остаток метионина на N-конце. Этот остаток может впоследствии быть необязательно отщеплен от экепрессируемого рекомбинантного белка с получением конечного продукта.
[00419] Выбор последовательности, контролирующей экспрессию, и вектора экспрессии будет зависеть от выбора хозяина. Для экспрессии можно использовать большое разнообразие комбинаций хозяин/вектор. Пригодные векторы экспрессии для эукариотических хозяев включают, например, векторы, содержащие последовательности, контролирующие экспрессию, из SV40, вируса папилломы крупного рогатого скота, аденовируса и цитомегаловируса. Пригодные векторы экспрессии для хозяев-бактерий включают известные бактериальные плазмиды, такие как плазмиды из Е. соli, в том числе pCR1, pBR322, pMB9 и их производные, плазмиды для более широкого круга хозяев, такие как М13 и нитевидные фаги, содержащие однонитевую ДНК.
[00420] Подходящие клетки-хозяева для экспрессии CD73-связывающей молекулы, например, антитела к CD73 (например, клона антитела 10.3 или клона антитела 2С5) или его антигенсвязывающего фрагмента, под контролем соответствующих промоторов, включают клетки прокариот, дрожжей, насекомых или высших эукариот. Прокариоты включают грамотрицательные или грамположительные организмы, например, Е. coli или бациллы. Клетки высших эукариот включают устойчивые линии клеток, происходящие от млекопитающих, описанные ниже. Можно также использовать бесклеточные системы трансляции. Соответствующие векторы для клонирования и экспрессии для применения в клетках-хозяевах бактерий, грибков, дрожжей и млекопитающих описаны Pouwels et al. (Cloning Vectors: A Laboratory Manual, Elsevier, N.Y., 1985), соответствующее раскрытие которого включено в данный документ с помощью ссылки. Дополнительную информацию касательно способов получения белков, в том числе получения антител, можно найти, например, в публикации патента США №2008/0187954, патентах США №№6413746, 6660501 и 7932087, каждый из которых в полном объеме включен в данный документ с помощью ссылки.
[00421] Различные системы культивирования клеток млекопитающих или насекомых можно с успехом использовать также для экспрессии рекомбинантных CD73-связывающих молекул, например, антител к CD73 (например, клона антитела 10.3 или клона антитела 2С5) или их антигеневязывающих фрагментов. Экспрессию рекомбинантных белков можно осуществлять в клетках млекопитающих, поскольку такие белки в основном правильно свернуты, модифицированы соответствующим образом и полностью функциональны.
[00422] Примеры подходящих линий клеток-хозяев млекопитающих включают НЕK-293 и НЕK-293Т, линии клеток почки обезьяны COS-7, описанные Gluzman (Cell 23:175, 1981), и другие линии клеток, в том числе, например, L-клетки, линии клеток С127, 3Т3, яичника китайского хомячка (СНО), NSO, HeLa и ВНK. Векторы экспрессии для млекопитающих могут содержать нетранскрибируемые элементы, такие как точка начала репликации, подходящий промотор и энхансер, связанный с геном, подлежащим экспрессии, и другие 5' и 3'-фланкированные нетранскрибируемые последовательности, а также 5' или 3'-нетранслируемые последовательности, такие как необходимые сайты связывания рибосом, сайт полиаденилирования, донорные и акцепторные сайты сплайсинга и последовательности терминации транскрипции. Бакуловирусные системы для выработки гетерологичных белков в клетках насекомых рассматриваются в Luckow & Summers, BioTechnology 6:47 (1988).
[00423] СD73-связывающие молекулы, например, антитела к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты, вырабатываемые трансформированным хозяином, можно очистить в соответствии с любым подходящим способом. Такие стандартные способы включают хроматографию (например, ионообменную, аффинную и эксклюзионную колоночную хроматографию), центрифугирование, очистку на основе различной растворимости или любую другую стандартную методику очистки белков. Аффинные метки, такие как гексагистидин, мальтозосвязывающий домен, последовательность белка оболочки вируса гриппа и глутатион-S-трансфераза, можно присоединять к белку для обеспечения возможности легкой очистки с помощью пропускания через соответствующую аффинную колонку. Выделенные белки можно также физически характеризовать с помощью таких методик, как протеолиз, ядерный магнитный резонанс и рентгеноструктурная кристаллография.
[00424] Например, супернатанты из систем, которые секретируют рекомбинантный белок в культуральную среду, можно сначала сконцентрировать с помощью коммерчески доступного фильтра для концентрирования белка, например, установки для ультрафильтрации amicon® или pellicon® от Millipore. Следом за стадией концентрирования концентрат можно нанести на подходящую матрицу для очистки. В качестве альтернативы можно использовать анионообменную смолу, например, матрицу или субстрат с боковыми диэтиламиноэтильными (DEAE) группами. Матрицы могут быть акриламидными, агарозными, декстрановыми, целлюлозными или других типов, обычно используемых для очистки белков. В качестве альтернативы можно использовать стадию катионного обмена. Подходящие катионообменники включают различные нерастворимые матрицы, содержащие сульфопропильные или карбоксиметильные группы. Наконец, для дополнительной очистки CD73-связывающей молекулы (например, клона антитела 10.3 или клона антитела 2С5) можно использовать одну или несколько стадий обращенно-фазовой высокоэффективной жидкостной хроматографии (RP-HPLC), в которых используют гидрофобные среды для RP-HPLC, например, силикагель с боковыми метальными или другими алифатическими группами. Некоторые или все из вышеизложенных стадий очистки в различных комбинациях можно также использовать для получения гомогенного рекомбинантного белка.
[00425] Рекомбинантный CD73-связывающий белок, например, антитело к CD73 (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, получаемый в бактериальной культуре, можно выделить, например, с помощью изначального выделения из клеточных осадков с последующей одной или несколькими стадиями концентрирования, высаливания, ионообменной или эксклюзионной хроматографии в водной среде. На конечных стадиях очистки можно использовать высокоэффективную жидкостную хроматографию (HPLC). Клетки микроорганизмов, используемые для экспрессии рекомбинантного белка, можно разрушать с помощью любого удобного способа, в том числе проведения циклов замораживания-размораживания, обработки ультразвуком, механического разрушения или применения средств для лизиса клеток.
[00426] Известные из уровня техники способы очистки антител и других белков также включают, например, способы, описанные в публикациях патентов США №№2008/0312425, 2008/0177048 и 2009/0187005, каждый из которых в полном объеме включен в данный документ с помощью ссылки.
[00427] Согласно определенным аспектам CD73-cвязывaющaя молекула представляет собой полипептид, который не является антителом. Ряд способов идентификации и получения полипептидов, не являющихся антителами, которые связываются с высокой аффинностью с белком-мишенью, известны из уровня техники. См., например, Skerra, Curr. Opin. Biotechnol., 18:295-304 (2007), Hosse et al., Protein Science, 15:14-27 (2006), Gill et al., Curr. Opin. Biotechnol., 17:653-658 (2006), Nygren, FEBS J., 275:2668-76 (2008) и Skerra, FEBS J., 275:2677-83 (2008), каждый из которых в полном объеме включен в данный документ с помощью ссылки. Согласно определенным аспектам для идентификации/получения CD73-связывающего полипептида можно применять технологию фагового дисплея. Согласно определенным аспектам полипептид содержит белковый каркас типа, выбранного из группы, состоящей из белка А, липокалина, домена фибронектина (например, домена фибронектина, такого как Fn III-домен тенасцина-3), домена анкирина с консенсусными повторами и тиоредоксина.
VI. Способы лечения с применением терапевтических антител к CD73
[00428] В настоящем раскрытии предусматриваются способы, направленные на применение молекул, связывающихся с CD73, например, антител, в том числе их антигенсвязывающих фрагментов, вариантов и производных (например, клона антитела 10.3 или клона антитела 2С5), для лечения пациентов с заболеванием, ассоциированным с экспрессией CD73, или клетками, экспрессирующими CD73, например, рака. Согласно некоторым конкретным аспектам такой рак представляет собой рак легкого, рак молочной железы, рак яичников, колоректальный рак, рак мочевого пузыря, рак поджелудочной железы, рак почки, рак желудка, рак предстательной железы, рак молочной железы, рак легкого, рак толстой кишки и лимфому.
[00429] "Клетка, экспрессирующая CD73" означает клетку, экспрессирующую CD73. CD73 может быть связанным с мембранной посредством заякоревания с помощью гликозилфосфатидилинозитола, а также присутствовать в виде растворимого белка. Способы выявления экспрессии CD73 в клетках и других подходящих образцах хорошо известны из уровня техники и включают без ограничения иммуногистохимию, проточную цитометрию, вестерн-блоттинг, ELISA и т.п.
[00430] Хотя нижеследующее обсуждение относится к способам диагностики и лечения различных заболеваний и расстройств с помощью CD73-cвязывaющeй молекулы по настоящему раскрытию (например, клона антитела 10.3 или клона антитела 2С5), описанные в данном документе способы применимы также к любым другим антителам к CD73 и антигенсвязывающим фрагментам, вариантам и производным (например, гибридным белкам или конъюгатам) этих антител к CD73, которые сохраняют требуемые свойства антител к CD73, раскрытых в данном документе, например, способность к специфическому связыванию с CD73 и нейтрализации его 5'-нуклеотидазной активности. Согласно некоторым аспектам CD73-связывающие молекулы представляют собой человеческие или гуманизированные антитела, которые не опосредуют ADCC человека, или представляют собой антитела к CD73, которые сконструированы так, что они не опосредуют ADCC.
[00431] Согласно некоторым аспектам CD73-связывающая молекула представляет собой антитело CD730010 или его антигенсвязывающий фрагмент, клон антитела 10.3 или его антигенсвязывающий фрагмент, антитело CD730002 или его антигенсвязывающий фрагмент, клон антитела 2С5 или его антигенсвязывающий фрагмент или антитело CD73004 или его антигенсвязывающий фрагмент. Согласно другим аспектам CD73-связывающая молекула представляет собой клон мутантного антитела 10.3. Согласно некоторым аспектам CD73-связывающая молекула представляет собой клон моноклонального антитела 10.3. Согласно некоторым аспектам CD73-связывающая молекула представляет собой клон моноклонального антитела 10.3, сконструированный для продления времени полужизни в сыворотке крови. Согласно другим аспектам CD73-связывающая молекула представляет собой клон YTE-мутантного антитела 10.3. Согласно другим аспектам CD73-cвязывaющaя молекула представляет собой клон мутантного антитела 2С5. Согласно некоторым аспектам CD73-связывающая молекула представляет собой клон моноклонального антитела 2С5. Согласно некоторым аспектам CD73-связывающая молекула представляет собой клон моноклонального антитела 2С5, сконструированный для продления времени полужизни в сыворотке крови. Согласно другим аспектам CD73-cвязывaющaя молекула представляет собой клон YTE-мутантного антитела 2С5.
[00432] Согласно одному аспекту лечение предусматривает применение или введение субъекту или пациенту молекулы, связывающейся с CD73, например, антитела (например, клона антитела 10.3 или клона антитела 2С5) или его антигенсвязывающего фрагмента, варианта или производного по настоящему раскрытию, или применение или введение молекулы, связывающейся с CD73, в выделенную ткань или линию клеток от субъекта или пациента, при этом субъект или пациент имеют заболевание, симптом заболевания или предрасположенность к заболеванию. Согласно другому аспекту предполагается, что лечение также предусматривает применение или введение субъекту или пациенту фармацевтической композиции, содержащей молекулу, связывающуюся с CD73, например, антитела или его антигенсвязывающего фрагмента, варианта или производного по настоящему раскрытию, или применение или введение фармацевтической композиции, содержащей молекулу, связывающуюся с CD73, в выделенную ткань или линию клеток от субъекта или пациента, которые имеют заболевание, симптом заболевания или предрасположенность к заболеванию.
[00433] Молекулы, связывающиеся с CD73, например, антитела (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты, варианты или производные по настоящему раскрытию, являются пригодными для лечения различных форм рака. Согласно одному аспекту в настоящем раскрытии предусматриваются молекулы, связывающиеся с CD73, например, антитела (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты, варианты или производные, для применения в качестве лекарственного препарата, в частности для применения при лечении или профилактике рака (например, рака толстой кишки, меланомы, рака молочной железы, лимфомы или немелкоклеточного рака легкого, лимфомы Ходжкина, неходжкинской лимфомы и лимфомы Беркитта, рака яичников, рака молочной железы, форм рака головы и шеи и рака поджелудочной железы). Согласно некоторым аспектам рак характеризуется прометастатическим фенотипом. Согласно некоторым аспектам рак, характеризующийся прометастатическим фенотипом, представляет собой меланому или рак молочной железы. Согласно некоторым аспектам рак представляет собой метастатический рак. Согласно некоторым аспектам раскрытые в данном документе молекулы, связывающиеся с CD73, могут запускать адаптивную противоопухолевую активность и/или ингибировать метастаз. Согласно некоторым конкретным аспектам раскрытые в данном документе молекулы, связывающиеся с CD73, могут ингибировать метастаз при раке молочной железы.
[00434] В соответствии со способами по настоящему раскрытию по меньшей мере одну молекулу, связывающуюся с CD73, например, антитело (например, клон антитела 10.3 или клон антитела 2С5) или его антигенсвязывающий фрагмент, вариант или производное, определяемые в другом месте данного документа, применяют для стимуляции положительного терапевтического ответа в отношении рака. Термин "положительный терапевтический ответ" в отношении лечения рака относится к ослаблению заболевания в связи с активностью этих молекул, связывающихся с CD73, например, антител или их антигенсвязывающих фрагментов, вариантов или производных, и/или к ослаблению симптомов, ассоциированных с заболеванием. Таким образом, например, ослабление заболевания можно охарактеризовать как полный ответ. Под "полным ответом" подразумевается отсутствие клинически обнаруживаемого заболевания с нормализацией относительно результатов любых ранее проведенных тестов. В качестве альтернативы, ослабление заболевания может быть классифицировано как частичный ответ. "Положительный терапевтический ответ" охватывает уменьшение или замедление прогрессирования и/или длительности рака, уменьшение или облегчение тяжести рака и/или облегчение одного или нескольких его симптомов в результате введения раскрытой в данном документе молекулы, связывающейся с CD73
[00435] Согласно конкретным аспектам такие термины относятся к одному, двум или трем или более результатам после введения раскрытых в данном документе молекул, связывающихся с CD73: (1) стабилизация, уменьшение или устранение популяции раковых клеток; (2) стабилизация или замедление роста раковой опухоли; (3) нарушение формирования раковой опухоли; (4) ликвидация, устранение или контроль первичного, местнораспространенного рака и/или метастатического рака; (5) снижение уровня смертности; (6) увеличение выживаемости без признаков заболевания, без рецидива, без прогрессирования заболевания и/или общей выживаемости, продолжительности жизни или процента выживаемости; (7) увеличение частоты ответа, длительности ответа или числа пациентов с ответом или на стадии ремиссии; (8) снижение частоты госпитализаций; (9) снижение продолжительности госпитализаций; (10) размер раковой опухоли сохраняется и не увеличивается или увеличивается менее чем на 10%, предпочтительно менее чем на 5%, предпочтительно менее чем на 4%, предпочтительно менее чем на 2% и (12) увеличение числа пациентов с ремиссией.
[00436] Клинический ответ можно оценивать с использованием скрининговых методик, таких как сканирование с помощью магнитно-резонансной томографии (MRI), рентгенография, сканирование с помощью компьютерной томографии (СТ), проточная цитометрия или анализ с применением клеточного сортера с активацией флуоресценции (FACS), гистология, макропатология и биохимический анализ крови, в том числе без ограничения изменения, выявляемые с помощью ELISA, RIA, хроматографии и т.п. В дополнение к этим положительным терапевтическим ответам, субъект, подвергаемый лечению с использованием молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, может испытывать благоприятный эффект в виде ослабления симптомов, ассоциированных с заболеванием.
[00437] Молекулы, связывающиеся с CD73, например, антитела (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты, варианты, или производные, раскрытые в данном документе, можно применять в комбинации с любыми известными видами терапии рака, в том числе с любым средством или комбинацией средств, которые, как известно, пригодны или которые использовалась или используются в настоящее время для лечения рака, например, рака толстой кишки, меланомы, рака молочной железы, лимфомы, немелкоклеточного рака легкого, лимфомы Ходжкина, неходжкинской лимфомы и лимфомы Беркитта, рака яичников, рака молочной железы, форм рака головы и шеи и рака поджелудочной железы). Второе средство или комбинация средств фармацевтического комбинированного состава или режима дозирования предпочтительно характеризуются взаимосвязанными видами активности в отношении антитела или полипептида по настоящему раскрытию, с учетом того, что они не оказывают отрицательного воздействия друг на друга.
[00438] Противораковые средства включают лекарственные средства, применяемые для лечения злокачественных новообразований, таких как раковая опухоль. Медикаментозная терапия может применяться отдельно или в комбинации с другими видами лечения, такими как оперативное вмешательство или радиационная терапия. При лечении рака можно применять лекарственные средства нескольких классов, в зависимости от типа пораженного органа. Например, формы рака молочной железы обычно стимулируются эстрогенами, и их можно лечить с применением лекарственных средств, которые инактивируют половые гормоны. Аналогичным образом, рак предстательной железы можно лечить с применением лекарственных средств, которые инактивируют андрогены - мужские половые гормоны. Противораковые средства для применения в определенных способах по настоящему раскрытию включают, среди прочих, антитела (например, антитела, которые связывают IGF-1R, антитела, которые связывают EGFR, антитела, которые связывают Her2, или антитела, которые связывают сМЕТ), малые молекулы, целенаправленно воздействующие на IGF1R, малые молекулы, целенаправленно воздействующие на EGFR, малые молекулы, целенаправленно воздействующие на Her2, антиметаболиты, алкилирующие средства, ингибиторы топоизомеразы, средства, целенаправленно воздействующие на микротрубочки, ингибиторы киназ, ингибиторы синтеза белка, иммунотерапевтические средства, виды гормональной терапии, глюкокортикоиды, ингибиторы ароматазы, ингибиторы mTOR, химиотерапевтические средства, ингибиторы протеинкиназы В, ингибиторы фосфатидилинозитол-3-киназы (PI3K), ингибиторы циклинзависимой киназы (CDK), RLr9, CD289, ингибиторы ферментов, антитела к TRAIL, ингибиторы МЕK и т.д.
[00439] Согласно конкретным аспектам раскрытые в данном документе CD73-связывающие молекулы, например, антитела (например, клон антитела 10.3 или клон антитела 2С5) или их антигенсвязывающие фрагменты, можно водить в комбинации с антителами или фрагментами антител, целенаправленно воздействующими, например, на PD-1 (белок-1 запрограммированной смерти клетки), его два лиганда PD-L1 (лиганд-1 белка запрограммированной смерти клетки) и/или PD-L2 или CTLA-4 (белок-антиген-4 цитотоксических Т-лимфоцитов). См., например, Stagg et al. PNAS 107:1547-1552 (2010); Jin et al. Cancer Res. 70(6): (2010); Allard et al. Clin. Cancer Res. 19:5626 (2013), которые включены в полном объеме в данный документ с помощью ссылки. Согласно некоторым аспектам антитело к CTLA-4 представляет собой ипилимумаб или его антигенсвязывающий фрагмент. Согласно другим аспектам антитело к CTLA-4 представляет собой тремелимумаб (тицилимумаб, СР-675,206) или его антигенсвязывающий фрагмент. Согласно некоторым аспектам антитело к PD-1 представляет собой пембролизумаб (keytruda®, ранее ламбролизумаб, также известный как МK-3475) или его антигенсвязывающий фрагмент. Согласно некоторым аспектам антитело к PD-1 представляет собой ниволумаб (BMS-936558, MDX-1106, ONO-4538, opdiva®) или его антигенсвязывающий фрагмент. Согласно некоторым аспектам антитело к PD-L1 представляет собой BMS-936559 или его антигенсвязывающий фрагмент. Согласно другим аспектам антитело к PD-L1 представляет собой MPDL3280A. Согласно другим аспектам антитело к PD-1 представляет собой АМР-224 (Fc-гибридный белок антитела к PD-1) или его антигенсвязывающий фрагмент. Согласно различным аспектам антитело к PD-L1 представляет собой MEDI4736 или его антигенсвязывающий фрагмент.
[00440] Согласно некоторым аспектам раскрытые в данном документе CD73-связывающие молекулы (например, клон антитела 10.3 или клон антитела 2С5) можно водить в комбинации с антителом к PD-1 или PD-L1. Согласно различным вариантам осуществления антитело к CD73 вводится при концентрации приблизительно 1 мг/кг, приблизительно 2 мг/кг, приблизительно 3 мг/кг, приблизительно 4 мг/кг, приблизительно 5 мг/кг, приблизительно 6 мг/кг, приблизительно 7 мг/кг, приблизительно 8 мг/кг, приблизительно 9 мг/кг, приблизительно 10 мг/кг, приблизительно 11 мг/кг, приблизительно 12 мг/кг, приблизительно 13 мг/кг, приблизительно 14 мг/кг, приблизительно 15 мг/кг, приблизительно 16 мг/кг, приблизительно 17 мг/кг, приблизительно 18 мг/кг, приблизительно 19 мг/кг или приблизительно 20 мг/кг. Согласно некоторым аспектам раскрытые в данном документе СD73-связывающие молекулы (например, клон антитела 10.3 или клон антитела 2С5) можно водить в комбинации с антителом к PD-1, PD-L1 или CTLA4, при этом антитело к PD-1, PD-L1 или CTLA4 вводится при концентрации приблизительно 1 мг/кг, приблизительно 2 мг/кг, приблизительно 3 мг/кг, приблизительно 4 мг/кг, приблизительно 5 мг/кг, приблизительно 6 мг/кг, приблизительно 7 мг/кг, приблизительно 8 мг/кг, приблизительно 9 мг/кг, приблизительно 10 мг/кг, приблизительно 11 мг/кг, приблизительно 12 мг/кг, приблизительно 13 мг/кг, приблизительно 14 мг/кг, приблизительно 15 мг/кг, приблизительно 16 мг/кг, приблизительно 17 мг/кг, приблизительно 18 мг/кг, приблизительно 19 мг/кг или приблизительно 20 мг/кг. Согласно некоторым аспектам антитело к CD73 и антитело к PD-1, PD-L1 или CTLA4 вводятся при соотношении приблизительно 1:1, 1:2, 1:3 или 1:4. Согласно некоторым аспектам антитело к CD73 и антитело к PD-1, PD-L1 или CTLA4 вводятся при соотношении приблизительно 1:2. Согласно конкретным аспектам концентрация антитела к CD73 (например, клона антитела 10.3 или клона антитела 2С5) составляет приблизительно 10 мг/кг, а концентрация антитела к PD-1 составляет приблизительно 20 мг/кг. Согласно некоторым аспектам раскрытые в данном документе CD73-связывающие молекулы (например, клон антитела 10.3 или клон антитела 2С5) можно водить в комбинации с антителом к PD-1. Согласно некоторым аспектам введение комбинированного средства для лечения, содержащего раскрытую в данном документе CD73-связывающую молекулу (например, MEDI9447, клон антитела 10.3 или клон антитела 2С5) в комбинации с антителом к PD-1, PD-L1 или CTLA4, может увеличивать показатель выживаемости на приблизительно 10%, приблизительно 20%, приблизительно 30%, приблизительно 40%, приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 90% или приблизительно 100% по сравнению с субъектами, которых не подвергали лечению, или субъектами, подвергаемыми лечению с помощью монотерапии (например, антителом к PD-1, PD-L1 или CTLA4 без антитела к CD73). Согласно некоторым аспектам введение комбинированного средства для лечения, содержащего раскрытую в данном документе CD73-связывающую молекулу (например, клон антитела 10.3 или клон антитела 2С5) в комбинации с антителом к PD-1, PD-L1 или CTLA4, может увеличивать показатель выживаемости приблизительно в 2 раза, приблизительно в 3 раза, приблизительно в 4 раза, приблизительно в 5 раз, приблизительно в 6 раз, приблизительно в 7 раз, приблизительно в 8 раз, приблизительно в 9 раз или приблизительно в 10 раз по сравнению с субъектами, которых не подвергали лечению, или субъектами, подвергаемыми лечению с помощью монотерапии (например, антителом к PD-1, PD-L1 или CTLA4 без антитела к CD73).
[00441] Если комбинированные виды терапии включают введение молекулы, связывающейся с CD73, в комбинации с введением другого терапевтического средства (например, антитела к PD-1, PD-L1 или CTLA4), то раскрытые в данном документе способы охватывают совместное введение с применением отдельных составов или одного фармацевтического состава и последовательное введение в любом порядке. Согласно некоторым аспектам описанные в данном документе антитела к CD73 (например, клон антитела 10.3 или клон антитела 2С5) вводят в комбинации с другими лекарственными средствами, при этом антитело или его антигенсвязывающий фрагмент, вариант или производное и терапевтическое средство(средства) можно вводить последовательно в любом порядке или одновременно (т.е. параллельно или в пределах одного периода времени).
[00442] Комбинированная терапия способна обеспечивать "синергизм" и оказывает "синергический" эффект, т.е. когда достигаемый при совместном применении активных компонентов эффект больше, чем сумма эффектов, получаемых при раздельном применении соединений. Синергический эффект можно получить, когда активные компоненты: (1) совместно составляют и вводят или доставляют одновременно в виде комбинированного состава со стандартной дозировкой; (2) доставляют поочередно или параллельно в виде отдельных составов или (3) по какой-либо другой схеме. При доставке в виде чередующейся терапии синергический эффект можно достичь, когда соединения вводят или доставляют последовательно, например, посредством разных инъекций, в отдельных шприцах. В целом, в ходе чередующейся терапии эффективную дозу каждого активного ингредиента вводят последовательно, т.е серийно, в то время как при комбинированной терапии эффективные дозы двух или более активных ингредиентов вводят совместно.
[00443] Согласно другим аспектам раскрытые в данном документе CD73-связывающие молекулы (например, клон антитела 10.3 или клон антитела 2С5) можно водить в комбинации с ингибиторами тирозинкиназы. Согласно некоторым другим конкретным аспектам раскрытые в данном документе CD73-связывающие молекулы можно вводить в комбинации с ингибиторами тирозинкиназной активности, ассоциированной с EGFR и/или HER2/neu, например, лапатинибом. Согласно некоторым аспектам раскрытые в данном документе CD73-связывающие молекулы можно водить в комбинации с антимитотическими средствами. Согласно некоторым конкретным аспектам раскрытые в данном документе CD73-связывающие молекулы можно водить в комбинации со средствами, которые стабилизируют сборку микротрубочек митотического веретена, например, паклитакселом или доцетакселом. Дополнительный аспект предусматривает применение молекул, связывающихся с CD73, например, антител или их антигенсвязывающих фрагментов, вариантов или производных (например, клона антитела 10.3 или клона антитела 2С5), для диагностического мониторинга уровней белка в ткани как части процедуры клинического исследования, например для определения эффективности схемы лечения. Например, выявление можно облегчить с помощью связывания антитела с выявляемым веществом.
[00444] Примеры выявляемых веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы и радиоактивные материалы. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, β-галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных материалов включают умбеллиферон, флуоресцеин, флуоресцеина изотиоцианат, родамин, флуоресцеина дихлортриазиниламин, дансилхлорид или фикоэритрин; пример люминесцентного материала включает люминол; примеры биолюминесцентных материалов включают люциферазу, люциферин и экворин и примеры подходящих радиоактивных материалов включают 125I, 131I, 35S или 3H.
VIII. Терапевтические комбинации антитела к CD73 и совместная терапия
[00445] В настоящем раскрытии предусматриваются способы, направленные на применение терапевтических комбинаций, содержащих молекулы, связывающиеся с CD73, например, антитела, в том числе их антигенсвязывающие фрагменты, варианты и производные (например, клон антитела 10.3 или клон антитела 2С5), для лечения пациентов с раком (в том числе раком толстой кишки, меланомой, раком молочной железы, лимфомой, немелкоклеточным раком легкого, лимфомой Ходжкина, неходжкинской лимфомой, лимфомой Беркитта, раком яичников, раком молочной железы, формами рака головы и шеи и раком поджелудочной железы).
[00446] Хотя нижеследующее обсуждение относится к терапевтическим комбинациям, содержащим CD73-связывающую молекулу по настоящему раскрытию (например, клон антитела 10.3 или клон антитела 2С5), описанные в данном документе способы применимы также к любым другим антителам к CD73 и антигенсвязывающим фрагментам, вариантам и производным (например, гибридным белкам или конъюгатам) этих антител к CD73, которые сохраняют требуемые свойства антител к CD73, раскрытых в данном документе, например, способность к специфическому связыванию с CD73 и нейтрализации его 5'-нуклеотидазной активности. Согласно некоторым аспектам CD73-связывающие молекулы представляют собой человеческие или гуманизированные антитела, которые не опосредуют ADCC человека, или представляют собой антитела к CD73, которые сконструированы так, что они не опосредуют ADCC.
[00447] Лечение пациента с солидной опухолью с применением комбинации по настоящему изобретению, такой как антитело к CD73 или его антигенсвязывающий фрагмент в комбинации с антителом к PD-1, PD-L1 или CTLA4 или их антигенсвязывающими фрагментами, может приводить в результате к аддитивному или синергическому эффекту. Термин "синергический", используемый в данном документе, относится к комбинации терапевтических средств (например, комбинации антитела к CD73 (например, MEDI9447) и антитела к PD-1, PD-L1 или CTLA4), которая является более эффективной, чем аддитивные эффекты отдельных терапевтических средств.
[00448] Синергический эффект комбинации терапевтических средств (например, комбинации антитела к CD73 (например, MEDI9447) и антитела к PD-1, PD-L1, или CTLA4) делает возможным применение более низких дозировок одного или нескольких терапевтических средств и/или введение указанных терапевтических средств с меньшей частотой пациенту с солидной опухолью. Возможность применения более низких доз терапевтических средств и/или введения указанных терапевтических средств с меньшей частотой снижает токсичность, ассоциированную с введением указанных терапевтических средств субъекту без снижения эффективности указанных терапевтических средств при лечении солидной опухоли. Кроме того, синергический эффект может приводить к улучшенной эффективности терапевтических средств для контроля, лечения или облегчения состояния при солидной опухоли. С помощью синергического эффекта комбинации терапевтических средств можно избежать неблагоприятных или нежелательных побочных эффектов, ассоциированных с применением любого одного терапевтического средства, или снизить их.
[00449] При совместной терапии комбинация антитела к CD73 (например, MEDI9447) или его антигенсвязывающего фрагмента и антитела к PD-1, PD-L1 или CTLA4 или их антигеневязывающих фрагментов может быть необязательно включена в одну и ту же фармацевтическую композицию или может быть включена в отдельную фармацевтическую композицию. В последнем случае фармацевтическая композиция, содержащая антитело к CD73 (например, MEDI9447) или его антигенсвязывающий фрагмент, подходит для введения до, одновременно с или после введения фармацевтической композиции, содержащей антитело к PD-1, PD-L1 или CTLA4 или его антигенсвязывающий фрагмент. В определенных случаях антитело к CD73 (например, MEDI9447) или его антигенсвязывающий фрагмент и антитело к PD-1, PD-L1 или CTLA4 вводят в перекрывающиеся периоды времени в отдельной композиции.
[00450] Антитело к CD73 (например, MEDI9447) или его антигенсвязывающий фрагмент и антитело к PD-1, PD-L1 или CTLA4 или его антигенсвязывающий фрагмент можно вводить только один раз или нечасто с обеспечением при этом пользы для пациента. Согласно дополнительным аспектам пациенту вводят дополнительные дозы со ступенчатым возрастанием концентрации. Дозы со ступенчатым возрастанием концентрации можно вводить с различными интервалами времени, в зависимости от возраста, веса пациента, клинических анализов, массы опухоли и/или других факторов, в том числе оценки лечащего врача.
[00451] С помощью способов, представленных в данном документе, можно замедлять или затормаживать рост опухоли. Согласно некоторым аспектам замедление или затормаживание может быть статистически значимым. Замедление роста опухоли можно измерять с помощью сравнения роста опухоли пациента на исходном уровне с ожидаемым ростом опухоли, с ожидаемым ростом опухоли с учетом большой группы пациентов или с ростом опухоли в контрольной группе. Согласно другим вариантам осуществления с помощью способов по настоящему изобретению повышают показатель выживаемости.
IX. Антитела к PD-L1
[00452] Антитела, которые специфически связывают и ингибируют активность PD-L1 (например, связывание с PD-1 и/или CD80), применимы для лечения опухолей. В7-Н1, также известный как PD-L1, представляет собой трансмембранный белок типа I размером примерно 53 кДа. У людей В7-Н1 экспрессируется на нескольких типах иммунных клеток, в том числе активированных и анергических/истощенных Т-клетках, на непримированных и активированных В-клетках, а также на миелоидных дендритных клетках (DC), моноцитах и тучных клетках. Он также экспрессируется на клетках, отличных от иммунных, в том числе панкреатических островках, купферовских клетках печени, эндотелии сосудов и отдельных видах эпителия, например, эпителия дыхательных путей и эпителия почечных канальцев, при этом его экспрессия повышается во время воспалительных эпизодов. Экспрессия В7-Н1 также обнаруживается при повышенных уровнях во многих опухолях, в том числе без ограничения при раке молочной железы, раке толстой кишки, колоректальном раке, раке легкого, раке почки, в том числе почечно-клеточной карциноме, раке желудка, раке мочевого пузыря, немелкоклеточном раке легкого (NSCLC), гепатоцеллюлярном раке (НСС) и раке поджелудочной железы, а также меланоме.
[00453] Известно, что В7-Н1 связывает два альтернативных лиганда, первый из которых, PD-1, представляет собой трансмембранный рецептор типа I размером 50-55 кДа, который первоначально был идентифицирован в линии Т-клеток, подверженной апоптозу, индуцированному активацией. PD-1 экспрессируется на активированных Т-клетках, В-клетках и моноцитах, а также на других клетках иммунной системы, и связывает как В7-Н1 (PD-L1), так и родственный B7-DC (PD-L2). Второй является членом В7-1 семейства В7, который экспрессируется на активированных Т-клетках, В-клетках, моноцитах и антигенпрезентирующих клетках.
[00454] Полагают, что передача сигналов по пути PD-1/B7-H1 обеспечивает важные неизбыточные функции в иммунной системе путем отрицательной регулировки ответов Т-клеток. Полагают, что экспрессия В7-Н1 в опухолевых клетках способствует опухолям в уклонении от обнаружения и уничтожения иммунной системой. В этом отношении В7-Н1 функционирует посредством нескольких альтернативных механизмов, в том числе динамического истощения и анергии Т-лимфоцитов, инфильтрующих опухоль, стимулирования секреции иммунных репрессивных цитокинов в микроокружение опухоли, стимулирования репрессивной регуляторной функции Т-клеток и защиты опухолевых клеток, экспрессирующих В7-Н1, от лизиса цитотоксическими Т-клетками, специфическими в отношении опухолевых клеток.
[00455] MEDI4736 представляет собой иллюстративное антитело к PD-L1, которое является селективным в отношении В7-Н1 и блокирует связывание В7-Н1 с рецепторами PD-1 и CD80. MEDI4736 может облегчать В7-Н1-опосредованную супрессию активации Т-клеток человека in vitro и ингибировать рост опухолей в ксенотрансплантатной модели посредством Т-клеточно-зависимого механизма. Другие средства, которые можно было бы применять, включают средства, которые ингибируют PD-L1 и/или PD-1 (АВ или другой).
[00456] Информацию, касающуюся MEDI4736 (или его фрагментов) для применения в способах, представленных в данном документе, можно найти в US 2013/0034559, US 8779108 и US 2014/0356353, раскрытия которых в полном объеме включены в данный документ с помощью ссылки. Домен кристаллизующегося фрагмента (Fc) MEDI4736 содержит тройную мутацию в константном домене тяжелой цепи IgG1, что снижает степень связывания с компонентом комплемента C1q и Fcγ-рецепторами, ответственными за опосредование антителозависимой клеточноопосредованной цитотоксичности (ADCC).
[00457] MEDI4736 и его антигенсвязывающие фрагменты для применения в способах, представленных в данном документе, содержат тяжелую цепь и легкую цепь или вариабельную область тяжелой цепи и вариабельную область легкой цепи. Согласно конкретному аспекту MEDI4736 или его антигенсвязывающий фрагмент для применения в способах, представленных в данном документе, содержат вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:130, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:131. Согласно конкретному аспекту MEDI4736 или его антигенсвязывающий фрагмент для применения в способах, представленных в данном документе, содержат вариабельную область тяжелой цепи и вариабельную область легкой цепи, при этом вариабельная область тяжелой цепи содержит определенные согласно Kabat последовательности CDR1, CDR2 и CDR3 с SEQ ID NO:132-134, и при этом вариабельная область легкой цепи содержит определенные согласно Kabat последовательности CDR1, CDR2 и CDR3 с SEQ ID NO:135-137. Специалисты обычной квалификации в данной области техники с легкостью смогут идентифицировать CDR, определенные согласно Chothia, определенные согласно Abm или определенные согласно другим системам, известным специалистам обычной квалификации в данной области техники. Согласно конкретному аспекту MEDI4736 или его антигенсвязывающий фрагмент для применения в способах, представленных в данном документе, содержат последовательности CDR вариабельной области тяжелой цепи и вариабельной области легкой цепи антитела 2.14Н9OРТ, раскрытого в US 2013/0034559, US 8779108 и US 2014/0356353, которые в полном объеме включены в данный документ с помощью ссылки.
X. Антитела к CTLA4
[00458] Соответственно, согласно одному варианту осуществления терапевтические комбинации по настоящему изобретению содержат антитело, блокирующее CTLA4 (например, тремелимумаб), и/или антитела, которые снижают взаимодействия PD1/PD-L1. В двух Т-клеточно-модулирующих путях, привлекающих в настоящее время особое внимание, сигнал передается через антиген-4 цитотоксических Т-лимфоцитов (CTLA4, CD 152) и лиганд-1 белка запрограммированной смерти клетки (PD-L1, также известный как В7Н-1 или CD274).
[00459] CTLA4 экспрессируетея на активированных Т-клетках и служит в качестве коингибитора для удерживания Т-клеточных ответов под контролем после CD28-опосредованной активации Т-клеток. Полагают, что CTLA4 регулирует амплитуду ранней активации непримированных Т-клеток и Т-клеток памяти после вовлечения TCR и является частью центрального ингибиторного пути, который влияет как на противоопухолевый иммунитет, так и на аутоиммунитет. CTLA4 экспрессируетея главным образом на Т-клетках, и экспрессия его лигандов CD80 (В71) и CD86 (В7.2) в значительной степени ограничивается антигенпрезентирующими клетками, Т-клетками и другими клетками, опосредующими иммунитет. Сообщалось о том, что антагонистические антитела к CTLA4, которые блокируют сигнальный путь CTLA4, усиливают активацию Т-клеток. Одно такое антитело, ипилимумаб, было одобрено FDA в 2011 году для лечения метастатической меланомы. Другое антитело к CTLA4, тремелимумаб, тестировали в III фазе испытаний для лечения меланомы на поздней стадии, но оно в то же время не существенно повышало общую выживаемость пациентов по сравнению со стандартом лечения (темозоломидом или дакарбазином).
[00460] Информацию, касающуюся тремелимумаба (или его антиген-связывающих фрагментов) для применения в способах, представленных в данном документе, можно найти в US 6682736 (где он обозначен как 11.2.1), раскрытие которого в полном объеме включено в настоящий документ с помощью ссылки. Тремелимумаб (также известный как СР-675,206, СР-675, СР-675206 и тицилимумаб) представляет собой человеческое моноклональное антитело IgG2, которое является высокоселективным в отношении CTLA4 и блокирует связывание CTLA4 с CD80 (В7.1) и CD86 (В7.2). Было показано, что оно приводит к активации иммунитета in vitro, и у некоторых пациентов, которых лечили тремелимумабом, наблюдали регрессию опухоли.
[00461] Тремелимумаб для применения в способах, представленных в данном документе, содержит тяжелую цепь и легкую цепь или вариабельную область тяжелой цепи и вариабельную область легкой цепи. Согласно конкретному аспекту Тремелимумаб или его антигенсвязывающий фрагмент для применения в способах, представленных в данном документе, содержат вариабельную область легкой цепи и вариабельную область тяжелой цепи. Согласно конкретному аспекту Тремелимумаб или его антигенсвязывающий фрагмент для применения в способах, представленных в данном документе, содержат вариабельную область тяжелой цепи и вариабельную область легкой цепи, рассмотренные в данном документе. Специалисты обычной квалификации в данной области техники с легкостью смогут идентифицировать CDR, определенные согласно Chothia, определенные согласно Abm или определенные согласно другим системам, известным специалистам обычной квалификации в данной области техники. Согласно конкретному аспекту Тремелимумаб или его антигенсвязывающий фрагмент для применения в способах, представленных в данном документе, содержат последовательности CDR вариабельной области тяжелой цепи и вариабельной области легкой цепи антитела 11.2.1, раскрытого в US 6682736, который в полном объеме включен в настоящий документ с помощью ссылки.
XI. VII. Фармацевтические композиции и способы введения
[00462] Способы получения и введения молекул, связывающихся с CD73, например, антител или их антигенсвязывающих фрагментов, вариантов или производных (например, клона антитела 10.3 или клона антитела 2С5), нуждающемуся в этом субъекту хорошо известны специалистам в данной области техники или без труда определяются ними. Путь введения молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, может быть, например, пероральным, парентеральным, ингаляционным или местным. Термин "парентеральный", используемый в данном документе, включает, например, внутривенное, внутриартериальное, внутрибрюшинное, внутримышечное, подкожное, ректальное или вагинальное введение. Тем не менее, в других способах, совместимых с идеями данного документа, молекулы, связывающиеся с CD73, например, антитела или их антигенсвязывающие фрагменты, варианты или производные по настоящему раскрытию, можно доставлять непосредственно в место локализации нежелательной популяции клеток, увеличивая тем самым воздействие терапевтического средства на пораженную ткань.
[00463] Как обсуждается в данном документе, молекулы, связывающиеся с CD73, например, антитела или их антигенсвязывающие фрагменты, варианты или производные по настоящему раскрытию (например, клон антитела 10.3 или клон антитела 2С5), можно вводить в фармацевтически эффективном количестве для лечения in vivo заболеваний, опосредованных клетками, экспрессирующими CD73, таких как определенные типы рака.
[00464] Способы получения и введения терапевтических комбинаций, содержащих молекулы, связывающиеся с CD73, например, антитела или их антигенсвязывающие фрагменты, варианты или производные (например, клон антитела 10.3 или клон антитела 2С5), в комбинации с антителом к PD-1, PD-L1 и/или CTLA4 или их антигенсвязывающими фрагментами, нуждающемуся в этом субъекту хорошо известны специалистам в данной области техники или без труда определяются ними. Путь введения их комбинации может быть, например, пероральным, парентеральным, ингаляционным или местным. Термин "парентеральный", используемый в данном документе, включает, например, внутривенное, внутриартериальное, внутрибрюшинное, внутримышечное, подкожное, ректальное или вагинальное введение. Тем не менее, в других способах, совместимых с идеями данного документа, комбинацию по настоящему раскрытию можно доставлять непосредственно в место локализации нежелательной популяции клеток, увеличивая тем самым воздействие терапевтического средства на пораженную ткань. Как обсуждается в данном документе, комбинацию антитела к CD73 (например, MEDI9447) и антитела к PD-1, PD-L1 и/или CTLA4 можно вводить в фармацевтически эффективном количестве для лечения in vivo заболеваний, опосредованных клетками, экспрессирующими CD73, таких как определенные типы рака.
[00465] Фармацевтические композиции, применяемые согласно настоящему раскрытию, могут содержать фармацевтически приемлемые носители, в том числе, например, воду, ионообменные вещества, белки, буферные вещества и соли. Также могут присутствовать консерванты и другие добавки. Носитель может представлять собой растворитель или дисперсионную среду. Подходящие составы для применения в терапевтических способах, раскрытых в данном документе, описаны в Remington's Pharmaceutical Sciences (Mack Publishing Co.) 16th ed. (1980).
[00466] В любом случае стерильные растворы для инъекций можно получать путем включения активного соединения терапевтической комбинации по настоящему изобретению (например, антитела к CD73 или его антигенсвязывающего фрагмента, варианта или производного, например, клона антитела 10.3 или клона антитела 2С5, самих по себе или в комбинации с другими активными средствами) в требуемом количестве с соответствующим растворителем с последующей стерилизацией фильтрованием. Кроме того, препараты можно упаковывать и реализовывать в форме набора. Такие готовые изделия могут иметь этикетки или листки-вкладыши, на которых указано, что соответствующие композиции пригодны для лечения субъекта, страдающего от заболевания или нарушения или предрасположенного к ним.
[00467] Составы для парентерального введения могут представлять собой однократную болюсную дозу, инфузионную или нагрузочную болюсную дозу с последующей поддерживающей дозой. Эти композиции можно вводить с конкретными фиксированными или переменными интервалами, например, раз в день или по мере необходимости.
[00468] Композицию можно вводить в виде однократной дозы, многократных доз или в течение установленного периода времени в виде инфузии. Схемы дозирования можно также корректировать таким образом, чтобы обеспечить оптимальный требуемый ответ (например, терапевтический или профилактический ответ).
[00469] Терапевтически эффективные дозы композиций по настоящему раскрытию для лечения заболеваний, опосредованных клетками, экспреесирующими CD73, таких как определенные типы рака, в том числе, например, рака толстой кишки, меланомы, рака молочной железы, лимфомы, немелкоклеточного рака легкого, лимфомы Ходжкина, неходжкинской лимфомы и лимфомы Беркитта, рака яичников, рака молочной железы, форм рака головы и шеи и рака поджелудочной железы, изменяются в зависимости от множества различных факторов, в том числе способа введения, целевого сайта, физиологического состояния пациента, будь то пациент человеком или животным, других вводимых лекарственных препаратов и того, является ли лечение профилактическим или терапевтическим. Обычно пациентом является человек, однако также можно лечить отличных от человека млекопитающих, в том числе трансгенных млекопитающих. Для оптимизации безопасности и эффективности дозировки для лечения можно подбирать с помощью обычных способов, известных специалистам в данной области техники.
[00470] Специалист обычной квалификации в данной области техники без излишних экспериментов, с учетом настоящего раскрытия с легкостью определит количество по меньшей мере одной молекулы, связывающейся с CD73, например, антитела, или его антигенсвязывающего фрагмента, варианта или производного (например, клона антитела 10.3 или клона антитела 2С5), или терапевтической комбинации по настоящему изобретению, подлежащих введению. Факторы, влияющие на способ введения и соответствующее количество по меньшей мере одной молекулы, связывающейся с CD73, например, антитела, его антигеневязывающего фрагмента, варианта или производного, или терапевтической комбинации по настоящему изобретению, включают без ограничения тяжесть заболевания, историю болезни и возраст, рост, вес, состояние здоровья и физическое состояние индивидуума, подвергаемого терапии. Аналогичным образом, количество молекулы, связывающейся с CD73, например, антитела или его фрагмента, варианта или производного, или терапевтической комбинации по настоящему изобретению, подлежащих введению, будет зависеть от способа введения и от того, будет ли субъект получать одну дозу или несколько доз данного средства.
[00471] В настоящем раскрытии также предусматривается применение молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента, варианта или производного (например, клона антитела 10.3 или клона антитела 2С5), или терапевтической комбинации по настоящему изобретению, в изготовлении лекарственного препарата для лечения определенного типа рака, в том числе, например, рака толстой кишки, меланомы, рака молочной железы, лимфомы, немелкоклеточного рака легкого, лимфомы Ходжкина, неходжкинской лимфомы и лимфомы Беркитта, рака яичников, рака молочной железы, форм рака головы и шеи и рака поджелудочной железы.
[00472] В настоящем раскрытии также предусматривается применение молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента, варианта или производного (например, клона антитела 10.3 или клона антитела 2С5), в изготовлении лекарственного препарата для лечения субъекта с определенным типом рака. Согласно определенным аспектам лекарственный препарат применяют у субъекта, который ранее проходил лечение с помощью по меньшей мере одного другого вида терапии.
[00473] Под "ранее проходивший лечение" или "предшествующее лечение" подразумевается, что субъект получал один или несколько других видов терапии (например, подвергался лечению с помощью по меньшей мере одного другого вида противораковой терапии) перед получением лекарственного препарата, содержащего молекулу, связывающуюся с CD73, например, антитело или его антигенсвязывающий фрагмент, вариант или производное (например, клон антитела 10.3 или клон антитела 2С5). Необязательно, чтобы субъект отвечал на предшествующее лечение с помощью предшествующего вида терапии или видов терапии. Таким образом, субъект, который получает лекарственный препарат, содержащий молекулу, связывающуюся с CD73, например, антитело или его антигенсвязывающий фрагмент, вариант или производное, мог отвечать или мог не отвечать на предшествующее лечение с помощью предшествующей терапии или на один или несколько предшествующих видов терапии, если предшествующее лечение включало несколько видов терапии.
[00474] В настоящем раскрытии также предусматривается совместное введение молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента, варианта или производного (например, клона антитела 10.3 или клона антитела 2С5), и по меньшей мере одного другого терапевтического средства. Антитело к CD73 и по меньшей мере одно другое терапевтическое средство можно совместно вводить друг с другом в одной композиции или можно совместно вводить друг с другом в одно и то же время или в перекрывающиеся периоды времени в отдельных композициях. Согласно некоторым аспектам антитело к CD73 можно совместно вводить, например, с антителом, которое целенаправленно воздействует на PD-1 (белок-1 запрограммированной смерти клетки). В настоящем раскрытии также предусматривается применение молекулы, связывающейся с CD73, например, антитела или его антигенсвязывающего фрагмента, варианта или производного (например, клона антитела 10.3 или клона антитела 2С5), в изготовлении лекарственного препарата для лечения субъекта с раком, при этом молекулу, связывающуюся с CD73, вводят до того, как субъект получает лечение с помощью по меньшей мере одного другого терапевтического средства.
VIII. Диагностика
[00475] В настоящем раскрытии дополнительно предусматриваются диагностические способы, пригодные для диагностики заболеваний, опосредованных клетками, экспрессирующими CD73, таких как определенные типы рака, которые включают измерение уровня экспрессии белка CD73 в ткани или других клетках, или жидкости организма от индивидуума, и сравнение измеренного уровня экспрессии со стандартным уровнем экспрессии CD73 в нормальной ткани или жидкости организма, при этом повышение уровня экспрессии по сравнению со стандартным свидетельствует о нарушении.
[00476] Антитела к CD73, раскрытые в данном документе, и их антигенсвязывающие фрагменты, варианты и производные (например, клон антитела 10.3 или клон антитела 2С5) можно применять для анализа уровней белка CD73 в биологическом образце с применением классических иммуногистологических способов, известных специалистам в данной области техники (например, см. Jalkanen, et al., J. Cell. Biol. 707:976-985 (1985); Jalkanen et al., J. Cell Biol. 105:3087-3096 (1987)). Другие способы на основе применения антител, пригодные для обнаружения экспрессии белка CD73, включают иммунологические анализы, такие как твердофазный иммуноферментный анализ (ELISA), иммунопреципитация или вестерн-блоттинг. Подходящие анализы более подробно описаны в другом месте данного документа.
[00477] Под "анализом уровня экспрессии полипептида CD73" подразумевается качественное или количественное измерение или оценка уровня полипептида CD73 в первом биологическом образце либо непосредственно (например, путем определения или оценки абсолютного уровня белка), либо относительно (например, путем сравнения с уровнем ассоциированного с заболеванием полипептида во втором биологическом образце). Уровень экспрессии полипептида CD73 в первом биологическом образце можно измерить или оценить и сравнить со стандартным уровнем полипептида CD73, при этом стандарт берут из второго биологического образца, полученного от индивидуума, не имеющего нарушения, или определяют путем усреднения уровней от группы индивидуумов, не имеющих нарушения. Как будет понятно в данной области техники, если "стандартный" уровень полипептида CD73 известен, то его можно использовать повторно в качестве стандарта для сравнения.
[00478] Под "биологическим образцом" подразумевается любой биологический образец, полученный от индивидуума, линии клеток, культуры ткани или другого источника клеток, потенциально экспрессирующих CD73. Способы получения биоптатов тканей и жидкостей организма от млекопитающих хорошо известны в данной области техники.
IX. Наборы, содержащие СD73-связывающие молекулы
[00479] В настоящем раскрытии также предусматриваются наборы, содержащие по меньшей мере одну из раскрытых в данном документе CD73-связывающих молекул, например, антитела к CD73 или их антигенсвязывающий фрагмент, варианты или производные раскрытых в данном документе молекул (например, клона антитела 10.3 или клона антитела 2С5), которые можно применять для выполнения способов, описанных в данном документе. Согласно определенным аспектам набор содержит по меньшей мере одно очищенное антитело к CD73 или его антигенсвязывающий фрагмент в одном или нескольких контейнерах. Согласно некоторым аспектам наборы содержат все компоненты, необходимые и/или достаточные для выполнения анализа на обнаружение, в том числе все контрольные образцы, указания для выполнения анализов и любое необходимое программное обеспечение для анализа и представления результатов. Специалист в данной области техники без труда поймет, что раскрытая CD73-связывающая молекула, например, антитело к CD73 или его антигенсвязывающий фрагмент по настоящему раскрытию (например, клон антитела 10.3 или клон антитела 2С5), может быть легко включена в один из установленных форматов набора, которые хорошо известны в данной области техники.
X. Иммунологические анализы
[00480] Раскрытые в данном документе молекулы, связывающиеся с CD73, например, антитела к CD73 или их антигенсвязывающие фрагменты, варианты или производные раскрытых в данном документе молекул (например, клона антитела 10.3 или клона антитела 2С5), можно анализировать в отношении иммуноепецифичного связывания любым способом, известным в данной области техники. Иммунологические анализы, которые можно применять, включают без ограничения системы конкурентного и неконкурентного анализа с использованием таких методик, как вестерн-блоттинг, радиоиммунологические анализы, ELISA (твердофазный иммуноферментный анализ), Иммунологические "сэндвич"-анализы, анализы иммунопреципитации, реакции преципитации, реакции диффузионной преципитации в геле, анализы иммунодиффузии, анализы агглютинации, анализы фиксации комплемента, иммунорадиометрические анализы, флуоресцентные Иммунологические анализы, Иммунологические анализы с белком А и многие другие. Такие анализы являются обычными и хорошо известными в данной области техники (см., например, Ausubel et al., eds, (1994) Current Protocols in Molecular Biology (John Wiley & Sons, Inc., NY) Vol. 1, который в полном объеме включен в данный документ с помощью ссылки).
[00481] CD73-связывающие молекулы, например, антитела к CD73 или их антигенсвязывающие фрагменты, а также их варианты или производные (например, клон антитела 10.3 или клон антитела 2С5), можно использовать в гистологических анализах, как, например, в иммунофлуоресценции, иммуноэлектронной микроскопии или отличных от иммунологических анализах, для обнаружения in situ CD73 или его консервативных вариантов или пептидных фрагментов. Обнаружение in situ можно осуществлять путем отбора гистологического образца у пациента и нанесения на него меченой CD73-связывающей молекулы, например, антитела к CD73 или его антигенсвязывающего фрагмента, варианта или производного, предпочтительно путем покрытия биологического образца меченой CD73-связывающей молекулой (например, антителом или фрагментом). Благодаря применению такой процедуры можно определить не только присутствие CD73 или его консервативных вариантов или пептидных фрагментов, но также и его распределение в исследуемой ткани. Используя настоящее раскрытие, специалисты обычной квалификации в данной области техники без труда поймут, что любой из широкого спектра гистологических способов (таких как методы окрашивания) можно модифицировать для того, чтобы добиться такого обнаружения in situ.
[00482] Активность связывания данной партии CD73-связывающей молекулы, например, антитела к CD73 (например, клона антитела 10.3 или клона антитела 2С5) или его антигенсвязывающего фрагмента, варианта или производного, можно определить в соответствии с широко известными способами. Специалисты в данной области техники смогут определить рабочие и оптимальные условия анализа для каждого определения с использованием стандартных экспериментов.
[00483] Способы и реагенты, подходящие для определения характеристик связывания выделенной CD73-связывающей молекулы, например, антитела к CD73 (например, клона антитела 10.3 или клона антитела 2С5) или его антигенсвязывающего фрагмента, варианта, или его измененного/мутантного производного, известны в данной области техники и/или коммерчески доступны. Оборудование и программное обеспечение, предназначенное для таких кинетических анализов, являются коммерчески доступными (например, BIAcore, программное обеспечение BIAevaluation, GE Healthcare; программное обеспечение KinExa, Sapidyne Instruments).
[00484] В практических аспектах настоящего раскрытия будут использоваться, если не указано иное, традиционные методики клеточной биологии, культивирования клеток, молекулярной биологии, биологии трансгенных организмов, микробиологии, рекомбинантных ДНК и иммунологии, которые находятся в пределах компетентности специалиста в данной области техники. Такие методики в полном объеме объясняются в литературе. См., например, Sambrook et al., ed. (1989) Molecular Cloning A Laboratory Manual (2nd ed.; Cold Spring Harbor Laboratory Press); Sambrook et al., ed. (1992) Molecular Cloning: A Laboratory Manual, (Cold Springs Harbor Laboratory, NY); D.N. Glover ed., (1985) DNA Cloning, Volumes I and II; Gait, ed. (1984) Oligonucleotide Synthesis; Mullis et al., патент США №4683195; Hames and Higgins, eds. (1984) Nucleic Acid Hybridization; Hames and Higgins, eds. (1984) Transcription And Translation; Freshney (1987) Culture Of Animal Cells (Alan R. Liss, Inc.); Immobilized Cells And Enzymes (IRL Press) (1986); Perbal (1984) A Practical Guide To Molecular Cloning; научный трактат, Methods In Enzymology (Academic Press, Inc., NY); Miller and Calos eds. (1987) Gene Transfer Vectors For Mammalian Cells, (Cold Spring Harbor Laboratory); Wu et al., eds., Methods In Enzymology, Vols. 154 and 155; Mayer and Walker, eds. (1987) Immunochemical Methods In Cell And Molecular Biology (Academic Press, London); Weir and Blackwell, eds., (1986) Handbook Of Experimental Immunology, Volumes I-IV; Manipulating the Mouse Embryo, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., (1986); и в Ausubel et al (1989) Current Protocols in Molecular Biology (John Wiley and Sons, Baltimore, Md.).
[00485] Общие принципы конструирования антител изложены в Borrebaeck, ed. (1995) Antibody Engineering (2nd ed.; Oxford Univ. Press). Общие принципы конструирования белков изложены в Riсkwood et al., eds. (1995) Protein Engineering, A Practical Approach (IRL Press at Oxford Univ. Press, Oxford, Eng.). Общие принципы относительно антител и связывания антител с гаптенами изложены в Nisonoff (1984) Molecular Immunology (2nd ed.; Sinauer Associates, Sunderland, Mass.) и Steward (1984) Antibodies, Their Structure and Function (Chapman and Hall, New York, N.Y.). Кроме того, стандартные способы в иммунологии, известные в данной области техники и не описанные конкретно, в основном соответствуют изложенному в Current Protocols in Immunology, John Wiley & Sons, New York; Stites et al., eds. (1994) Basic and Clinical Immunology (8th ed; Appleton & Lange, Norwalk, Conn.) и Mishell and Shiigi (eds) (1980) Selected Methods in Cellular Immunology (W.H. Freeman and Co., NY).
[00486] Авторитетные справочные издания, в которых изложены общие принципы иммунологии, включают Current Protocols in Immunology, John Wiley & Sons, New York; Klein (1982) J., Immunology: The Science of Self-Nonself Discrimination (John Wiley & Sons, NY); Kennett et al., eds. (1980) Monoclonal Antibodies, Hybridoma: A New Dimension in Biological Analyses (Plenum Press, NY; Campbell (1984) "Monoclonal Antibody Technology" в Laboratory Techniques in Biochemistry and Molecular Biology, ed. Burden et al., (Elsevere, Amsterdam); Goldsby et al., eds. (2000) Kuby Immunology (4th ed.; H. Freemand & Co.); Roitt et al. (2001) Immunology (6th ed.; London: Mosby); Abbas et al. (2005) Cellular and Molecular Immunology (5th ed.; Elsevier Health Sciences Division); Kontermann and Dubel (2001) Antibody Engineering (Springer Verlan); Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press); Lewin (2003) Genes VIII (Prentice Hall2003); Harlow and Lane (1988) Antibodies: A Laboratory Manual (Cold Spring Harbor Press); Dieffenbach and Dveksler (2003) PCR Primer (Cold Spring Harbor Press).
[00487] Все из цитируемых выше ссылок, а также все ссылки, цитируемые в данном документе, в полном объеме включены в данный документ с помощью ссылки.
[00488] Следующие примеры предлагаются в качестве иллюстрации, но не в качестве ограничения.
ПРИМЕРЫ
[00489] Аспекты настоящего раскрытия могут быть дополнительно определены с помощью ссылки на следующие неограничивающие примеры, в которых подробно описывается получение определенных антител по настоящему раскрытию и способы применения антител по настоящему раскрытию. Специалистам в данной области техники будет очевидно, что многочисленные модификации как материалов, так и способов можно осуществлять на практике без отступления от объема настоящего раскрытия.
[00490] CD73 (кластер дифференцировки 73), также известный как экто-5'-нуклеотидаза (NT5E), представляет собой трансмембранный рецептор, обнаруженный на опухолевых клетках, а также в нормальных стромальных клетках, таких как эндотелиальные клетки и определенные лейкоциты. CD73 катализирует превращение аденозинмонофосфата до аденозина и органического фосфата. Связывание внеклеточной части аденозиновых рецепторов передает сигнал через циклический AMP для ингибирования активации рецепторов Т-клеток (рассмотрено Linden и Cekic, 2012). CD73, как полагают, выполняет роль в опосредовании ингибирующей функции регуляторных В- и Т-лимфоцитов (Saze et al, 2013), а также в поддержании целостности эндотелия (рассмотрено Jalkanen и Salmi, 2008).
[00491] В дополнение к своей роли в биологии в норме, CD73 и аденозин влияют на биологию опухолей. Наличие внеклеточного аденозина в микроокружении опухоли описано как иммуносупрессивное "гало" (Antonioli et al, 2013). Было показано, что в соответствии с данной ролью для аденозина, нокаутные мыши с отсутствием аденозиновых рецепторов легче отторгают опухоли, чем нормальные мыши (Ohta et al, 2006). Первичным источником внеклеточного аденозина внутри опухолей, как полагают, является CD73 (Augusto et al, 2013). В соответствии с данной гипотезой, а также исследованиями на мышах с дефицитом А2А, у нокаутных мышей с отсутствующим CD73 противоопухолевый иммунитет повышался (Stagg et al, 2011), и они продемонстрировали снижение канцерогенеза (Stagg et al, 2012) по сравнению с нормальными мышами. В частности, внеклеточный аденозин, как полагают, опосредует иммуносупрессивные эффекты, среди прочих, как регуляторных Т-клеток, так и супрессорных клеток миелоидного происхождения (MDSC) (рассмотрено Antonioli et al, 2013). В совокупности с другими исследованиями, показывающими, что молекулярное ингибирование CD73 с помощью небольших молекул или антител способно ингибировать образование, рост и метастазирование опухоли (рассмотрено Young et al, 2014), предполагается, что опухоли используют CD73 для выработки аденозина и, таким образом, подавляют противоопухолевый иммунитет. Соответственно, антитела к CD73, которые еелективно связываются с CD73 и ингибируют его эктонуклеотидазную активность, вероятно являются пригодными для повышения противоопухолевого иммунного ответа.
ПРИМЕР 1. Выделение и идентификация антител к CD73
[00492] Фаг-дисплейные библиотеки scFv человека подвергали пэннингу с биотинилированным внеклеточным доменом CD73 (ECD) с целью выделения антител, связывающихся с CD73 человека, яванского макака и мыши. Было показано, что лидерное антитело CD730010 специфично связывается с CD73-экспрессирующими клетками человека, мыши и яванского макака (с помощью проточной цитометрии) и ингибирует активность рекомбинантного растворимого ECD CD73, а также нативного CD73, представленного на клетках. Аффинное созревание CD730010 инициировали для повышения аффинности связывания CD730010 с CD73 человека.
[00493] Перед оптимизацией аффинности была предпринята попытка восстановить столько же остатков каркаса CD730010 в ближайшие последовательности зародышевой линии человека (основанные на репертуаре IMGT) без ухудшения аффинности. Это было сделано с целью минимизации потенциальной иммуногенности конечного лекарственного средства на основе антител у людей. Все остатки каркаса VL-домена и все кроме одного остатка каркаса VH-домена могут быть восстановлены в соответствии с аминокислотной последовательностью IGLV1-44, IGLJ3, IGHV3-23 и IGHJ2 зародышевых линий человека. Лизин в положении 94 (нумерация согласно Kabat; Kabat, 1991) VH-домена CD730010 не мог быть восстановлен без утраты аффинности.
[00494] Аффинность и эффективность антитела CD730010, модифицированного на уровне генов зародышевой линии, оптимизировали с помощью создания библиотек вариантов CDR и тестирования вариантов в отношении улучшенного связывания с CD73. Несколько мутаций с наилучшим улучшением аффинности объединяли с целью создания кандидатного лекарственного средства MEDI9447. Нуклеотидные и предсказанные аминокислотные последовательности MEDI9447 показаны на фигурах 1A-1D.
[00495] CD73-специфичные scFv антител выделяли из фаг-дисплейной библиотеки scFv человека в сериях повторяющихся альтернативных циклов отбора по биотинилированном внеклеточном домене (ECD) CD73 человека и мыши, продуцируемого внутри клеток млекопитающего, в сущности как ранее описано у Lloyd et al., PEDS 22:159-68 (2009). Гены ScFv из 2-го и 3-го раундов продуктов отбора партией превращали в бактериальные векторы экспрессии scFv-Fc или Fab. Супернатанты бактериальной культуры, несущие растворимые scFv-Fc или Fab, подвергали скринингу на их связывание с ECD CD73 человека, мыши и яванского макака с помощью ELISA или гомогенной флуоресценции с временным разрешением (HTRF). Наилучшие совпадения, демонстрирующие перекрестную реактивность, отбирали, подвергали секвенированию ДНК и превращали в цельный иммуноглобулин G1 формата тройного мутантного антитела ("IgG-TM", Fc-последовательность IgG1, включающая мутации L234F, L235E и P331S). Антитела IgG1 TM экспрессировали в клетках млекопитающих, очищали с помощью аффинной хроматографии и оценивали на основании их характеристик в анализах связывания и функциональности.
ПРИМЕР 2. Эпитоп-специфическая сортировка антител к CD73
[00496] Способность антител к CD73 конкурировать друг с другом за связывание с ECD CD73 человека оценивали на приборе Octet, в сущности как описано (Abdiche YN et al., Anal Biochem 386: 172-80 (2009). Белок ECD CD73 и первое антитело к CD73 предварительно инкубировали и добавляли к биотинилированному второму антителу к CD73, захваченному на стрептавидиновом сенсоре. Если первое антитело к CD73 блокировало связывание ECD CD73 со вторым антителом к CD73, то оба антитела помещали в одни и те же или перекрывающиеся эпитопные группы. Если оба антитела могли одновременно связываться с ECD CD73, то их помещали в неперекрывающиеся эпитопные группы. Попарное тестирование антител к CD73 показало, что они относятся к 3 неперекрывающимся эпитопным группам (таблица 2).
[00497] 
ПРИМЕР 3. Связывание антител к CD73 с CD73
[00498] Аффинность связывания и специфичность антител к CD73 определяли с помощью поверхностного плазменного резонанса (SPR) и проточной цитометрии.
[00499] Прибор ProteOn XPR36 применяли, чтобы охарактеризовать связывание MEDI9447 с ECD CD73 человека, мыши и яванского макака. MEDI9447 захватывали с учетом аффинности с помощью антитела к Fc человека. ECD CD73 находился в мобильной фазе. Ассоциацию и диссоциацию CD73 с MEDI9447 можно было точно описать с помощью модели Ленгмюра 1:1. Результаты, представленные в таблице 3, демонстрируют, что аффинность MEDI9447 к ECD CD73 от трех видов является сопоставимой и находится в низком пикомолярном диапазоне.
[00500] 
ka - константа скорости ассоциации; kd - константа скорости диссоциации; KD - константа диссоциации
[00501] Связывание MEDI9447 с нативным CD73, экспрессируемым на линиях клеток человека, мыши и яванского макака, характеризовали с помощью проточной цитометрии. Клетки инкубировали с различными концентрациями MEDI9447 и отслеживали связывание антитела с антителом к Fc человека, меченым флуорофором. График средней интенсивности флуоресценции в зависимости от концентрации MEDI9447 нелинейно аппроксимировали с применением изотермической модели односайтового связывания для расчета равновесной константы диссоциации. Анализом с помощью проточной цитометрии подтверждали связывание MEDI9447 с CD73 человека, мыши и яванского макака с сопоставимыми значениями аффинности (таблица 4), однако значения KD были 13-126 раз больше, чем определенные с помощью SPR, вероятно из-за конформационных различий между рекомбинантным и нативным CD73.
[00502] 
[00503] Для определения специфичности MEDI9447 в отношении CD73 человека с помощью проточной цитометрии, получали линии клеток. Клетки MDA-MB-231, которые представляют собой клетки рака молочной железы человека, выведенные из плеврального выпота, трансфицировали короткой шпилечной РНК (shRNA) CD73 человека с целью инактивирования экспрессии CD73 на поверхности клеток. Клетки Jurkat, линию Т-клеток, полученных из клеток лимфомы Беркитта, трансфицировали плазмидой, экспрессирующей mRNA CD73 человека, с целью активации экспрессии CD73 на поверхности клеток. Клетки Jurkat экспрессировали небольшое количество эндогенного CD73.
[00504] Специфичность MEDI9447 в отношении CD73 человека определяли по соотношению значений связывания MEDI9447 с линией клеток с высокой экспрессией CD73 (MDA-MB-231) и линией клеток с низкой экспрессией (MDA-MB-231, CD73-shRNA). Специфичность MEDI9447 в отношении CD73 человека определяли по соотношению значений в клеточной линии с высокой экспрессией CD73 (Jurkat - нокин CD73) и клеточной линии Jurkat с низкой экспрессией.
[00505] Специфичность MEDI9447 в отношении CD73 мыши (mCD73) определяли с помощью проточной цитометрии, сравнивая линию клеток 4Т1 мыши (высокая экспрессия mCD73) с линией клеток, подвергнутой инактивированию (4Т1 mCD73 - shRNA). В дополнение, специфичность MEDI9447 в отношении клеток Jurkat с CD73 мыши, подвергнутым нокину, сравнивали с клетками Jurkat дикого типа (без CD73 мыши).
[00506] 
ПРИМЕР 4. Интернализация CD73 с помощью антитела к CD73 MEDI9447
[00507] Антителоопосредованную интернализацию или "слущивание" CD73 оценивали с помощью проточной цитометрии. Клетки MDA-MB-231 инкубировали в присутствии 100 нМ MEDI9447 или антитела R347 отрицательного контроля в среде для выращивания при 37°С в течение 0-4 часов. Клетки промывали и ресуспендировали в ледяном PBS. Наличие CD73 на поверхности клеток выявляли путем добавления 10 нМ детекторного антитела, меченого DyLight488 Клетки инкубировали в течение 15 минут, промывали и анализировали с помощью проточной цитометрии. Детекторное антитело связывается с эпитопом CD73, который отличается от эпитопа MEDI9447, и оба антитела одновременно связываются с CD73 без интерференции. Экспрессия CD73 на поверхности клеток снижалась до 73% от ее исходного значения после 4 часов инкубации с MEDI9447, указывая на то, что 27% CD73 было интернализовано или "слущенно" при связывании MEDI9447 (таблица 3.5-1).
[00508] 

[00509] Интернализацию MEDI9447 в линии клеток MDA-MB-231 (карцинома молочной железы человека) и 4Т1 (карцинома молочной железы мыши) оценивали с применением набора для интернализации антител человека, который коммерчески реализуется как анализ FabZAP (Advanced Targeting Systems, Сан-Диего, Калифорния). Серийные разбавления MEDI9447 или антитела R347 отрицательного контроля предварительно инкубировали с 40 нМ реагента FabZAP (Fab-фрагмент поликлонального антитела к IgG человека, конъюгированный с цитотоксическим белком сапорином), а затем добавляли к линиям клеток. Через 3 дня культивирования пролиферацию клеток измеряли с использованием люминесцентного анализа жизнеспособности клеток, коммерчески реализуемого как анализ CellTiter-Glo (Promega, Мэдисон, Висконсин). Этот анализ использовали для расчета значений EC50 и максимальной токсичности. Реагент FabZAP не мог самостоятельно интернализоваться в клетки. Он связывался с тестируемым антителом (например, MEDI9447) и проявлял цитотоксичность только после интернализации тестируемого антитела. MEDI9447 приводило к интернализации FabZAP и ингибировало пролиферацию клеток дозозавиеимым образом.
[00510]

[00511] Пролиферацию клеток MDA-MB-231 и клеток 4Т1, обработанных серийными разбавлениями тестируемого антитела и реактивом FabZAP, измеряли с помощью анализа CellTiter-Glo (фигура 2). Сигнал от антитела R347 отрицательного контроля при анализе CellTiter-Glo вычитали из сигнала MEDI9447, а значение ЕС50 и максимальную токсичность рассчитывали путем подбора кривой зависимости доза-эффект с помощью анализа методом нелинейной регрессии.
ПРИМЕР 5. Ингибирование 5'-эктонуклеотидазной активности антителом к CD73 MEDI9447
[00512] В данном исследовании функциональную активность MEDI9447 определяли в анализе in vitro, в котором измеряли СD73-катализируемый гидролиз AMP, используя линию клеток немелкоклеточной карциномы человека NCI-H322. Состав MEDI9447 получали путем разбавления его исходного раствора в среде RPMI, не содержащей сыворотку, до конечной концентрации 1 мкМ. Состав R347 получали путем разбавления его исходного раствора в RPMI до конечной концентрации 1 мкМ.
[00513] Клетки NCI-H322 подвергали центрифугированию в течение 5 минут при 1500об./мин. Супернатант удаляли и заменяли средой RPMI, не содержащей сыворотку. Суспензию клеток подсчитывали с применением счетчика клеток ViCell (Beckman, Coulter). Клетки высевали в 96-луночные планшеты при плотности клеток, составляющей 10000 клеток на 100 мкл на лунку. Добавляли 50 мкл 4Х концентрированного AMP (200 мкМ). Затем планшеты инкубировали при 37°С, 5% СО2 в течение 24 часов. Планшеты центрифугировали и 50 мкл супернатанта культуры переносили последовательно в каждую лунку круглодонных 96-луночных непрозрачных планшетов. Затем добавляли 2Х АТР. CellTiterGlo® (Promega) добавляли в соответствии с указаниями производителя. Ингибирование клеточного фермента 5'-эктонуклеотидазы измеряли на многоканальном ридере в установке Envision от Perkin-Elmer. Образцы анализировали с использованием программного обеспечения Prism.
[00514] MEDI9447 специфически ингибировало дефосфорилирование аденозинмонофосфата (AMP) в системе in vitro человека. В клеточном анализе CD73, экспрессируемого на поверхности, преобразование аденозинмонофосфата в аденозин снижалось дозозависимым образом с помощью MEDI9447, но не с помощью нерелевантного антитела изотипического контроля (фигура 3). Результаты, изображенные на фигуре 3, получали с использованием CD73-экспрессирующих клеток NSCLC, которые высевали в 96-луночные планшеты, необработанные тканевой культурой (Falcon 3788), в количестве 10000 клеток на лунку в 100 мкл среды RPMI без добавок. Антитела добавляли в двух повторах вместе с AMP (конечная концентрация 200 мкМ) и планшеты инкубировали при 37°С, 5% СО2 в течение 24 часов. Планшеты центрифугировали в течение 3 минут при 1500 об./мин. Супернатанты собирали в новый 96-луночный планшет (Costar #3605) и АТР добавляли до конечной концентрации 100 мкМ. Реагент CellTiter-Glo® (Promega) добавляли при 1:1 и АМР-фосфорилазную активность клеточного фермента CD73 определяли путем измерения уровней АТР с применением люминесцентного планшетного ридера Envision (Perkin Elmer). В качестве отрицательного контроля использовали буфер, содержащий только АТР и AMP. Анализ повторяли, и результаты, показанные на фигуре 3, являются репрезентативными для двух подобных экспериментов, в которых использовали другие линии раковых клеток человека.
[00515] Эти результаты показывают, что MEDI9447 ингибировало выработку аденозина раковыми клетками. Аденозин, как полагают, опосредует иммуносупрессивные эффекты опухоли в микроокружении опухоли.
ПРИМЕР 6. Сокращение количества супрессорных клеток миелоидного происхождения, инфильтрующих опухоль, с помощью MEDI9447
[00516] Клетки СТ26, полученные из раковой опухоли толстой кишки мыши, вводили посредством подкожной (SC) инъекции в количестве 5×105 клеток, суспендированных в 0,1 мл PBS, в правые бока 4-6 недельных самок мышей. Мышей обрабатывали MEDI9447 или контрольным антителом.
[00517] В данном исследовании использовали 10 мышей на группу. Животных произвольно разделяли на группы. Животные в группе 1 были необработанными, а группе 2 вводили изотипический контроль. MEDI9447 вводили группе 3. Тестируемые образцы вводили внутрибрюшинно дважды в неделю, начиная с 3-го дня. На 16-й день 5 животных из каждой группы подвергали вскрытию и выделяли опухоли.
[00518] Обозначение групп и уровни доз представлены в таблице 8.
[00519] 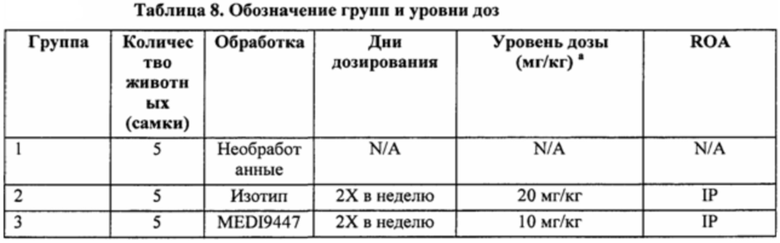
N/A = не применяется; ROA = путь введения.
Опухоли измеряли в дни 1, 7, 9, 12, 14 и 16 с помощью штангенциркуля, и объемы опухолей рассчитывали следующим образом:
(1) (длина опухоли (мм)×(ширина опухоли)2(мм))/2.
Противораковые эффекты MEDI9447 выражали в виде процента ингибирования роста опухоли, который рассчитывали следующим образом:
(2) (средний объем опухоли для MEDI9447/cpeдний объемом опухоли для R347-TM)×100.
[00520] Опухоли выделяли для анализа проточной цитометрией. Опухоли иссекали из мышей-опухоленосителей СТ26 на 16-й день исследования. Опухоли разрезали на мелкие кусочки и расщепляли коллагеназой. После 30-минутной инкубации расщепленный образец пропускали через 70-микронный фильтр. Диссоциированные клетки осаждали при 1000 об./мин в течение 5 минут при 4°С и ресуспендировали в буфере для сортировки флуоресцентно-активированных клеток (FACS). Клетки подсчитывали на Vi-Cell с использованием настроек по умолчанию. Высевали 1×106 клеток на лунку. Клетки метили с помощью антитела к CD45 (для обнаружения всех лейкоцитов), антитела к GR1 (для обнаружения MDSC) и антитела к Ly6g (Gran MDSC). Данные получали на проточном цитометре LSRII. Подтверждающие значимость p-значения, если таковые имеются, полученные в результате анализов MDSC, представлены на фигуре 4 рядом с описательными статистическими показателями (т.е. средним значением и стандартным отклонением).
[00521] MEDI9447 ингибировало рост опухоли в сингенной модели опухоли СТ26 у мышей Balb/C (фигура 4).
[00522] MEDI9447 снижало долю MDSC, инфильтрующих опухоль, в сингенной модели опухоли СТ26 у мышей Balb/C (фигура 5).
[00523] MEDI9447 ингибировало рост сингенных опухолей СТ26 у мышей. Кроме того, количество супрессорных клеток миелоидного происхождения снижалось в сингенных опухолях СТ26, характерных для карциномы толстой кишки, после обработкой MEDI9447. Внутриопухолевые MDSC оказывают иммуносупрессивный эффект на микроокружение опухоли, что обеспечивает усиленный рост опухоли. Наблюдаемое сокращение количества внутриопухолевых MDSC после обработки MEDI9447 подтверждает механизм, посредством которого обработка MEDI9447 ослабляет иммунную супресеию опухоли.
ПРИМЕР 7. Комбинация mIgG1 MEDI9447 и антитела к PD-1 обеспечивала снижение роста опухоли и повышение показателя выживаемости
[00524] Сингенные опухоли формировали путем подкожной (SC) инъекции 0,1 мл 5×106 клеток СТ26/мл, суспендированных в HBSS, в правые бока 8-10-недельных животных. Опухоли измеряли с помощью штангенциркуля, и объемы опухолей (TV) рассчитывали с помощью следующей формулы:
[00525] (1) TV=(L×W2)/2,
[00526] где L представляет собой длину опухоли в миллиметрах и W представляет собой ширину опухоли в миллиметрах.
[00527] Мышей произвольно распределяли в группы с учетом веса тела. Замен животных не производили. В данном исследовании использовали 60 самок мышей Balb/c.
[00528] Животных произвольно разделяли на 6 групп. Животным вводили MEDI9447 (mIgG1). Тестируемые образцы вводили с помощью внутрибрюшинной (IP) инъекции дважды в неделю, начиная с 3-го дня. Обозначение групп и уровни доз представлены в таблице 9.
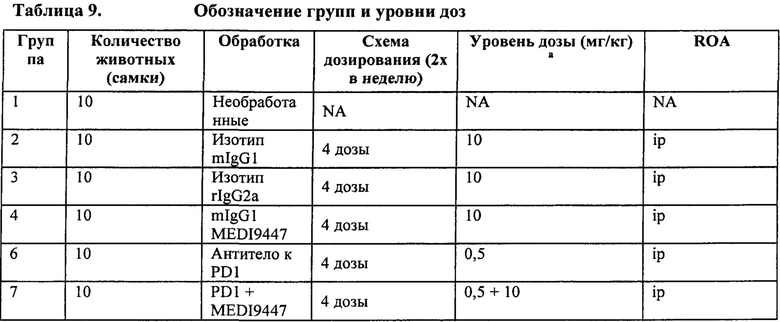
F = самки; IV = внутривенное; М = самцы; ROA = путь введения.
а Объем дозы: 10 мл/кг.
(а) Результаты исследования ингибирования опухолей in vivo
[00529] СТ26-карциному толстой кишки мыши имплантировали сингенным мышам Balb/C и обрабатывали антителом к CD73 (mIgG1 MEDI9447), антителом к PD1 или их комбинацией. Обработка комбинацией значимо ингибировала рост опухоли по сравнению с антителом к CD73 отдельно (p=0,015, ANOVA) (фигура 6). Значения объемов опухолей из каждой группы животных представляли в виде графика для отдельных животных до 40-го дня исследования. К концу 40-дневного периода исследования ни у одной из мышей контрольной группы не отсутствовала опухоль. В результате обработки только антителом к CD73 в конце исследования опухоль отсутствовала у 10% животных. Также в результате обработки только антителом к PD1 в конце исследования опухоль отсутствовала у 10% животных. Примечательно, что в результате обработки комбинацией антитела к CD73 и антитела к PD опухоль отсутствовала у 60% мышей. К концу исследования ни у одной из мышей контрольной группы не отсутствовала опухоль. Опухоли СТ26 измеряли у мышей, обработанных антителом к CD73 (mIgG1 MEDI9447), антителом к PD1 или комбинацией антитела к CD73 и антитела к PD1. У мышей проводили замеры до 40-го дня исследования и гуманно умерщвляли, как только опухоли достигали 2000 мм3. Обработка комбинацией антитела к CD73 и антитела к PD1 в совокупности приводила к статистически значимому увеличению показателя выживаемости по сравнению с обработкой антителом к CD73 или антителом к PD1 отдельно (p-значение=0,005 и p=0,038 соответственно, логарифмический ранговый критерий) (фигура 7). Медиана выживаемости увеличивалась от 25 до 33 дня (антитело к CD73 и антитело к PD1 соответственно) по сравнению с "неопределенной" на 40-й день для комбинации (таблица 10).
[00530]
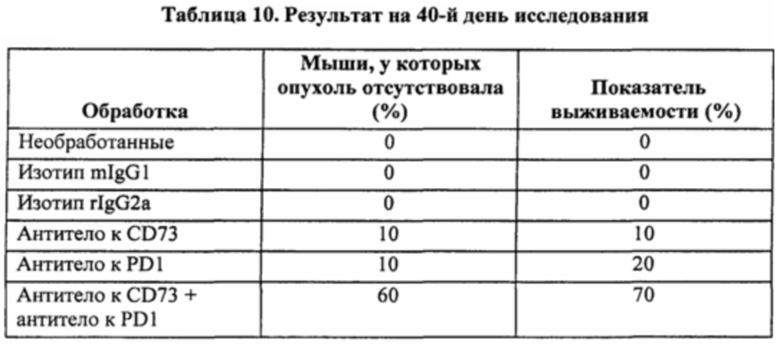
[00531] В итоге антитело к CD73, mIgG1 MEDI9447, продемонстрировало повышенную противоопухолевую активность при комбинировании с антителом к PD-1 в сингенной мышиной модели карциномы толстой кишки СТ26. Кроме того, в результате обработки комбинацией антитела к CD73 и антитела к PD опухоль отсутствовала у 60% мышей. Обработка комбинацией антитела к CD73 и антитела к PD1 в совокупности также приводила к статистически значимому увеличению показателя выживаемости по сравнению с обработкой антителом к CD73 или антителом к PD1 отдельно.
[00532] ПРИМЕР 8 Антитело к PD-1 индуцировало богатое на CD73 микроокружение опухоли, что измеряли с помощью экспрессии CD73 на опухолевых клетках (фигура 9), супрессорных клетках миелоидного происхождения (MDSC) и инфильтрующих опухоль CD4+, FoxP3+ лимфоцитах (фигура 15).
[00533] Сингенные опухоли формировали путем подкожной (SC) инъекции сингенных клеток B16F10 меланомы или сингенных клеток EG7-OVA лимфомы. Мышей дважды в неделю обрабатывали MEDI9447 (10 мг/кг), антителом к PD-L1 (10 мг/кг) или комбинацией MEDI9447 (10 мг/кг) и антитела к PD-L1 (10 мг/кг). Объем опухоли измеряли дважды в неделю. Введение MEDI9447 и антитела к PD-L1 в комбинации значительно усиливало ингибирование роста в опухолях, характерных для меланомы (фигура 12), и опухолях, характерных для лимфомы (фигура 13).
[00534] Чтобы понять эффект антитела к PD-L1 в отношении микроокружения опухоли, исследовали экспрессию CD73 на лимфоцитах. Мышам (n=4) подкожно инъецировали сингенные клетки СТ26 колоректальной карциномы и дважды в неделю обрабатывали с помощью 10 мг на кг антитела к PD-L1 или нерелевантного антитела изотипического контроля. Через день после первой обработки клетки выделяли из дренирующих лимфатических узлов и анализировали в отношении поверхностного фенотипа с помощью проточной цитометрии. Через три дня после первой обработки опухоли выделяли, клетки диссоциировали и анализировали в отношении поверхностного фенотипа с помощью проточной цитометрии. Антитело к PD-1 индуцировало богатое на CD73 микроокружение опухоли, что измеряли с помощью поверхностной экспрессии CD73 на В-лимфоцитах дренирующих лимфатических узлов (фигура 14) и инфильтрующих опухоль CD4+, FoxP3+ лимфоцитах (фигура 15).
[00535] Мышей, несущих колоректальные сингенные опухоли СТ26, дважды в неделю обрабатывали MEDI9447 (30 мг/кг), или антителом к PD-L1 (30 мг/кг), или комбинацией MEDI9447 (30 мг/кг) и антитела к PD-L1 (30 мг/кг). На 16-й день опухоли и цельные клетки периферической крови собирали и анализировали в отношении поверхностной экспрессии CD73 с помощью проточной цитометрии и ферментативной активности. MEDI9447 отдельно или в комбинации с антителом к PD-L1 снижало экспрессию CD73 на цельных клетках периферической крови (фигура 16), инфильтрующих опухоль CD4+, FoxP3+ лимфоцитах (фигура 17) и инфильтрующих опухоль CD8+ лимфоцитах (фигура 18). MEDI9447 отдельно или в комбинации с антителом к PD-L1 также снижало экспрессию CD73 на опухолевых клетках (фигура 19).
[00536] MEDI9447 и антитела или гибридные белки, специфичные в отношении CTLA4, OХ40, PD-1 и PD-L1, инкубировали в течение 72 ч. с первичными мононуклеарными клетками периферической крови человека при смешанной лейкоцитарной реакции. Указанные цитокины в двойных супернатантах количественно определяли с помощью ELISA. Представленные данные представляют собой оптимальные комбинации доз антитела к CD73 с 4 различными соучаствующими средствами. Комбинация антитела к PD-1 и антитела к CD73 продемонстрировала значимую (p<0,05) синергию (фигура 20), которую определяли с помощью способа поверхностного отклика по Блиссу (Zhao et al.). Цитокиновый профиль указывает, что влиянию подвергались как миелоидные, так и лимфоидные линии. Были протестированы более 50 пар доноров.
[00537] В итоге антитела к PD-1 и антитела к PD-L1 создавали богатое на CD73 микроокружение опухоли, обнаруживаемое на периферии и обратимое при обработке антителом к CD73, mIgG1 MEDI9447. В частности, уровни экспрессии CD73 на поверхности клеток и ферментативной активности резко возрастали в мышиных опухолях СТ26, когда мышей, несущих эти опухоли, обрабатывали антителом к PD-1 или антителом к PD-L1. Кроме того, уровни экспрессии и активности снижались в результате обработки антителом к CD73 MEDI9447 отдельно или в комбинации с антителом к PD-L1. Эти изменения наблюдали как в опухолях, так и в циркулирующих цельных клетках периферической крови. Таким образом, экспрессия CD73 и активность могут служить в качестве фармакодинамического маркера как антитела к PDL1, так и антитела к CD73, или в качестве предиктивного биомаркера для сегментации пациентов, получавших антитело к PD-1 или антитело к PD-L1, у которых опухоли "ускользнули" в результате нерегулируемой экспрессии CD73 и активности. Важно отметить, что антитело к CD73, MEDI9447, в комбинации с антителом к PD1 или антителом к PD-L1, MEDI 4736, продемонстрировали повышенную противоопухолевую активность.
[00538] В совокупности, эти результаты подтверждают различными показателями, что антитела к PD-1 и антитела к PD-L1 индуцировали "богатое на CD73" микроокружение опухоли и обеспечивали веское обоснование для комбинирования MEDI9447 с терапевтическими средствами, которые оказывающие целенаправленное воздействие на PD-1 /PD-L1.
Пример 9. Антитело к CD730010 и варианты антитела
[00539] 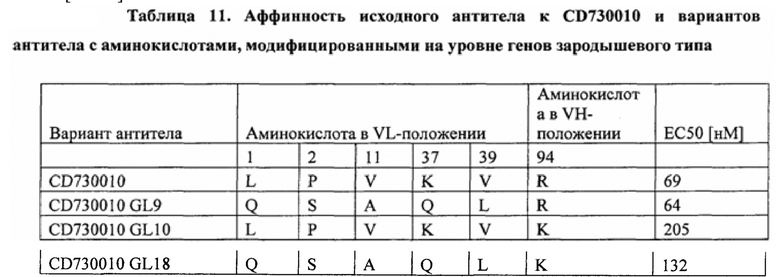
[00540] Для CD730002 ближайшими генами зародышевого типа были IGHV3-23 и IGHJ3 для VH-домена и IGLV3-1 и IGLJ3 для VL-домена. За пределами CDR-областей идентифицировали четыре несвойственных зародышевому типу остатка: R94 в VH и Т20, R57, L81 и F87 в VL (нумерация согласно Кабат) (таблица 11). Нуклеотиды в векторе экспрессии IgG1-TM CD730002 подвергали мутации к первоначальному виду стандартными методиками молекулярной биологии, так что полученные векторы экспрессии кодировали свойственные зародышевому типу аминокислоты в этих положениях (K94 в VH и S20, G57, М81 и Y87 в VL). Варианты белка IgG1-TM CD730002 экспрессировали, очищали и тестировали в отношении связывания с рекомбинантным CD73 человека и мыши с помощью проточной цитометрии. Все 4 несвойственных зародышевому типу аминокислоты в VL могли быть заменены на их остатки зародышевого типа без ухудшения связывания. Однако R94 в VH важен для связывания и его замена на K препятствует связыванию. Вариант SGMY CD730002 (V, немодифицированная на уровне генов зародышевого типа, VL, полностью модифицированная на уровне генов зародышевого типа) использовали в качестве матрицы для получения вариантов антител с оптимизированной аффинностью.
ПРИМЕР 10. Оптимизация по аффинности антител к CD73 CD730010GL9
[00541] IgG1-TM CD730010GL9 оптимизировали посредством скрининга библиотек Fab, содержащих варианты CDR-последовательностей с единичными аминокислотными мутациями. Каждое из 61 положения в шести CDR индивидуально рандомизировали на 19 аминокислот (все природные аминокислоты, за исключением цистеина), создавая библиотеку с теоретическим разнообразием из 1159 уникальных клонов (19 аминокислот на положение умножить на 61 положение). Бактериальные Fab-фрагменты получали из 4224 клонов библиотеки и подвергали скринингу в отношении связывания с белком CD73 человека и мыши с помощью ELISA с захватом (анализ 2). Отбирали 180 клонов с повышенным сигналом связывания по сравнению с исходным IgG1-TM CD730010 GL9 и мутации в VH- или VL-домене идентифицировали путем секвенирования ДНК. Концентрацию Fab в бактериальных супернатантах нормализовали и связывание нормализованных супернатантов с белком CD73 человека и мыши оценивали с помощью прямого ELISA (анализ 1). В таблице 12 перечислены выбранные полезные единичные аминокислотные замены и их эффект в отношении связывания с рекомбинантным белком CD73.
[00542] 
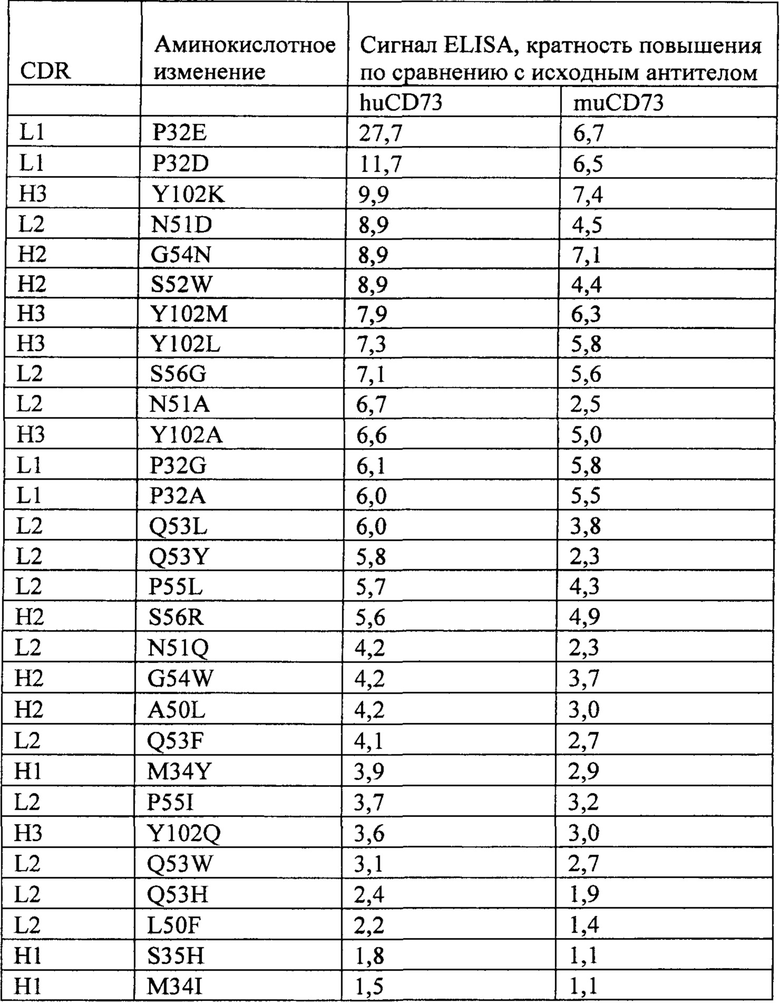
[00543] Для дополнительного улучшения аффинности антитела к CD73 несколько отдельных аминокислотных изменений, которые улучшали связывание по сравнению с исходным CD730010GL9, объединяли для создания комбинаторной библиотеки Fab (анализ 4). Fab-фрагменты 4224 клонов комбинаторной библиотеки получали в E. coli и подвергали скринингу в отношении связывания с белком CD73 человека и мыши с помощью ELISA с захватом. Для дальнейшего определения характеристик отбирали наилучшие 20 клонов из каждого скринингового анализа. Концентрацию Fab в супернатантах нормализовали и серийные разбавления нормализованных супернатантов тестировали в отношении связывания с CD73 человека и мыши с помощью ELISA с захватом и прямого ELISA. Клоны C1, C2, D3 и G10 продемонстрировали сильное связывание с CD73 человека и мыши и были отобраны для дальнейшего определения характеристик.
[00544] Связывание CD730010GL9 с антигеном также оптимизировали с помощью отборов с использованием фагового дисплея на основе аффинности. Большие библиотеки scFv, полученные из лидерной последовательности CD730010 GL9, создавали с помощью олигонуклеотид-направленного мутагенеза определяющей комплементарность области-3 (CDR3), вариабельной области тяжелой цепи (VH) или CDR3 вариабельной области легкой цепи (VL) с применением стандартных методик молекулярной биологии, как описано (Finch et al., JMB 411, 791-807 (2011)). Библиотеки подвергали пэннингу в сериях повторяющихся альтернативных циклов отбора с биотинилированным белком внеклеточного домена CD73 человека и мыши. Гены ScFv из 3-го раунда продукта отбора партиями преобразовывали в бактериальный вектор экспрессии IgG. Супернатанты бактериальной культуры, содержащие растворимый IgG, подвергали скринингу в отношении его связывания с CD73 человека и мыши. Варианты IgG со значительно улучшенным связыванием с CD73 по сравнению с исходным CD730010 GL9 подвергали секвенированию ДНК. Для дальнейшего определения характеристик отбирали два варианта - GRVE и НРТ.
[00545] Для получения дополнительных улучшений аффинности полезные мутации, идентифицированные из комбинаторной библиотеки Fab и из отбора с использованием фагового дисплея на основе аффинности, объединяли, создавая варианты 73combo1 - 73combo6.
ПРИМЕР 11. Оптимизация по аффинности антител к CD73 CD730002SGMY
[00546] IgG1-TM CD730002SGMY оптимизировали посредством скрининга библиотек Fab, содержащих варианты CDR-последовательностей с единичными аминокислотными мутациями, как описано для CD730010GL9. Идентифицировали пять аминокислотных мутаций в VH-домене и четыре аминокислотные мутации в VL-домене, которые приводили к усиленному сигналу связывания с рекомбинантным CD73. В таблице 13 перечислены полезные единичные аминокислотные замены и их эффект в отношении связывания с рекомбинантным CD73.
[00547] 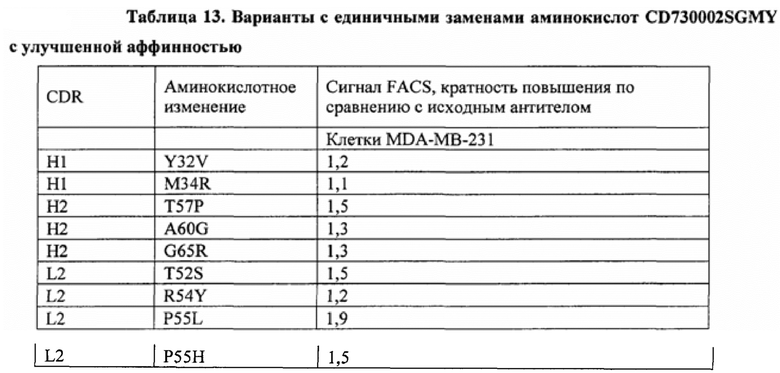
[00548] Для дополнительного улучшения аффинности антитела к CD73 CD730002SGMY получали варианты IgG, которые содержали одно полезное изменение аминокислоты в VH-домене и одно полезное изменение аминокислоты в VL-домене. Варианты антитела получали с помощью транзиентной трансфекции клеток 293F и подвергали скринингу в отношении связывания с клетками MDA-MB-231 с помощью проточной цитометрии. Клон 2С5 имел в 3 раза более низкое значение EC50, чем исходный CD730002SGMY.
ПРИМЕР 12. Аффинность оптимизированных антител к CD73
[00549] Аффинность оптимизированных антител к CD73 (в формате IgG1-TM) в отношении CD73 человека, мыши и яванского макака определяли с помощью проточной цитометрии и поверхностного плазменного резонанса (SPR) (таблица 14). Оптимизированные антитела характеризовались аффинностью в отношении клеточного и рекомбинантного CD73 от трех видов на пМ уровне.
[00550] 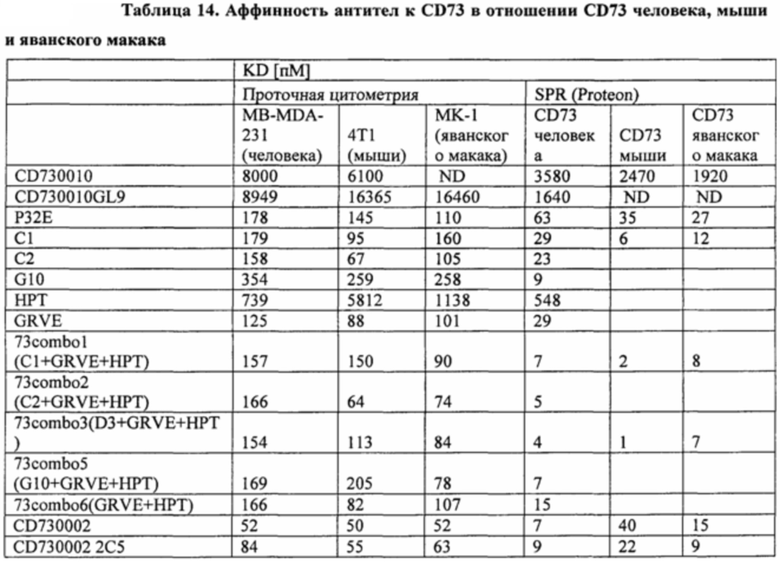
ПРИМЕР 13. Интернализация антител к CD73
[00551] Интернализацию антител к CD73 в линии клеток MDA-MB-231 и 4Т1 оценивали с помощью анализа FabZAP (Advanced Targeting Systems, Сан-Диего, Калифорния). Линии клеток инкубировали в присутствии антител к CD73 и реагента FabZAP. Через 3 дня измеряли пролиферацию клеток для расчета значений EC50 и максимальной токсичности (таблица 15). Данные FACS показывают, что степень интернализации оптимизированных по аффинности антител является низкой.
[00552]
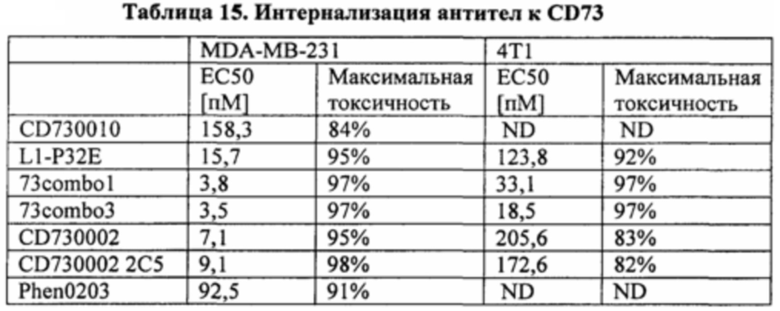
[00553] В другом эксперименте интернализацию антител в линии клеток оценивали с помощью анализа цитотоксического эффекта конъюгатов антитела к CD73/сапорина на CD73-положительные линии клеток (таблица 15). Антитела к CD73 непосредственно конъюгировали с токсином сапорином с применением химических веществ S-HyNic и 4FB (Solulink, Сан-Диего, Калифорния) и определяли концентрацию конъюгата антитело/сапорин, требуемую для ингибирования клеточного роста на 50%.
[00554] 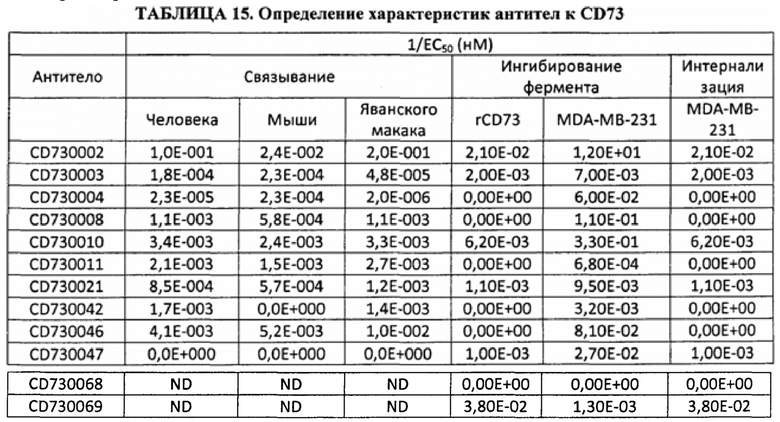
ПРИМЕР 14. Антитело к CD73 человека, hIgG1 Phen0203, ингибировало АМР-опосредованную супрессию пролиферации CD4+CD25- Т-клеток in vitro зависимым от концентрации образом
[00555] Данное исследование проводили с целью определения способности антитела к CD73 (Phen0203) снижать АМР-опосредованную супрессию Т-клеток in vitro. В данном исследовании in vitro изучали способность антитела к CD73 человека (hIgG1 Phen0203) ингибировать катализ аденозинмонофосфата до аденозина и органического фосфата с помощью CD73 и последующее влияние на функцию Т-клеток. hIgG1 Phen0203 имеет схожие функциональные свойства с MEDI9447, в том числе способность ингибировать клеточную и биохимическую ферментативную активность CD73 in vitro.
[00556] Антитело hIgG1 Phen0203 представляет собой mAb IgG1 человека без инженерии в константной области тяжелой цепи. Аналогично MEDI9447 оно также селективно связывается и ингибирует выработку иммуносупрессивного аденозина с помощью эктонуклеотидазной активности CD73 человека. Однако Phen0203 не обладает перекрестной реактивностью в отношении CD73 мыши.
[00557] В анализе на АМР-опосредованную супрессию Т-клеток первичные CD4+ Т-клетки человека, истощенные клетками CD25+, выделяли из содержимого конических лейкоцитарных фильтров и использовали в качестве эффекторных клеток; при этом каждый конус обрабатывали отдельно. Вкратце, содержимое из конического лейкоцитарного фильтра разбавляли в PBS, затем разделяли на слои с помощью Ficoll-Paque Plus (GE Healthcare, Чалфонт Сейнт Джайлс, Великобритания) и центрифугировали при 400×g в течение 40 минут с выключенным торможением. Затем мононуклеарные клетки периферической крови (РВМС) выделяли из поверхности раздела и промывали PBS с помощью центрифугирования при 200×g в течение 10 минут. Супернатант удаляли и клетки суепендировали в PBS. Определяли жизнеспособные клетки, затем осаждали при 350×g в течение 5 минут и суепендировали в буфере Robosep (Stem Cell, Гренобль, Франция) в концентрации 5×107 на мл. CD4+ Т-клетки выделяли из РВМС путем отрицательного отбора с применением набора EasySep для обогащения CD4+ Т-клетками человека (Stem Cell, Гренобль, Франция) и RoboSep (Stem Cell, Гренобль, Франция). Очищенные CD4+ Т клетки осаждали и ресуспендировали при 1,5×107 на мл в буфере Robosep. Частицы Dynabeads CD25 (компонент набора для анализа регуляторных CD4+CD25+ Т-клеток Dynabeads; Life Technologies, Пейсли, Великобритания) добавляли в количестве 200 мкл на 1,5×107 клеток и инкубировали в течение 25 минут при 4°С с непрерывным перемешиванием. Затем клетки помещали на магнит DynaMag-15 (Life Technologies, Пейсли, Великобритания) на 1 минуту и супернатант, содержащий эффекторные клетки CD4+CD25-, переносили в новую пробирку.
[00558] Выделенные эффекторные клетки метили зондом CFSE (3 мкМ) с применением набора для детекции пролиферации клеток CellTrace CFSE (Life Technologies, Пейсли, Великобритания) при плотности клеток 1×106 клеток на мл в PBS, содержащем 0,1% BSA, в течение периода инкубации 15 минут при 37°С. Клетки дважды промывали теплой средой X-Vivo 15 и суспендировали при 5×105 клеток на мл в той же среде. Меченые эффекторные клетки активировали в течение 1 часа при 37°С путем добавления 25 мкл микрогранул, покрытых антителом к CD3 и антителом к CD28 (набор для активации Т-клеток CD3/CD28 человека Dynabeads; Life Technologies, Пейсли, Великобритания) на 1×106 клеток и 60 МЕ/мл rhIL-2. После этого активированные клетки CD4+CD25- (приблизительно 50000 в 100 мкл) добавляли в лунки стерильных круглодонных 96-луночных планшетов. Серийные разбавления следующих реагентов выполняли в среде X-Vivo 15 (Lonza, Слау, Великобритания): тестируемый образец hIgG1 Phen0203 и контрольное антитело R347.
[00559] К клеткам в планшете добавляли 50 мкл разбавленных реагентов, а затем 50 мкл X-Vivo 15 (Lonza, Слау, Великобритания), содержащей 400 мкМ или 800 мкМ AMP (Sigma-Aldrich, Джиллингем, Великобритания). Также включали следующие контрольные лунки: активированные CFSE-меченые клетки CD4+CD25- без AMP (активированный контроль); CFSE-меченые клетки CD4+CD25- с AMP, но без тестируемых/контрольных образцов (необработанный контроль), и неактивированные (контроль в состоянии покоя) CFSE-меченные клетки CD4+CD25- без AMP. Клетки в анализе осторожно осаждали с помощью центрифугирования при 100×g в течение 2 минут и помещали в увлажненный инкубатор для тканевых культур с 5% СО2 при 37°С на 72 часа.
[00560] После 72 часов инкубации клетки осаждали с помощью центрифугирования при 380g в течение 4 минут, промывали один раз с помощью 100 мкл буфера FACS (eBioscienee, Хэтфилд, Великобритания) и наконец суспендировали в 100 мкл PBS, содержащего 3,7% формальдегида для анализа по методу проточной цитометрии на BD FACSCanto II (BD Biosciences, Оксфорд, Великобритания). Покоящиеся клетки CFSE+ CD4+CD25- из лунок без AMP использовали для идентификации клеток, которые подверглись клеточному делению (делящиеся клетки).
[00561] Было обнаружено, что CD73 экспреесировался в субпопуляции CD4+ Т-клеток. В присутствии внеклеточного AMP CD73+ Т-клетки обладали потенциалом для активации паракринных/аутокринных путей, в которые вовлекается биотрансформация AMP в аденозин с помощью CD73, с последующей активацией аденозиновых рецепторов и последующим регулированием функции Т-клеток. Этот CD73/аденозиновый путь моделировали in vitro с применением очищенных первичных CD4+CD25- Т-клеток человека, активированных путем передачи сигналов TCR и rhIL-2. Пролиферация Т-клеток подавлялась в присутствии 100 или 200 мкМ внеклеточного AMP.
[00562] Антитело к CD73 человека hIgG1 Phen0203 было способным ингибировать АМР-опосредованную супрессию пролиферации CD4+CD25- Т-клеток in vitro зависимым от концентрации образом. Эти данные обеспечивают научное обоснование для подхода с использованием антитела к CD73, для которого целью является иммуносупрессивные эффекты АМР/СD73/аденозинового пути.
ПРИМЕР 15. Картирование эпитопов и паратопов MEDI9447
[00563] Чтобы идентифицировать связывание поверхностных участков MEDI9447 и CD73, выполняли анализ MS, основанный на водород-дейтериевом обмене (HDX-MS), с использованием Fab MEDI9447 и рекомбинантного растворимого CD73 (sCD73) либо отдельно, либо в комплексе (фигуры 19А, 19В и 20А-20Е). В последнее время MS, основанная на водород-дейтериевом обмене (HDX-MS), зарекомендовала себя как мощный инструмент для картирования сайтов белок-белкового взаимодействия и характеризации структуры белка и конформационной динамики. С помощью дифференциального мечения областей белка способом, обусловленным доступностью растворителя, эпитопы антител успешно картируют с применением HDX-MS. Путем сравнения кинетики водородного обмена между свободным и комплексным CD73 выявили две области, расположенные в N-концевом домене sCD73 (аминокислоты (аа) 132-143 и 182-187), которые характеризовались сниженным поглощением дейтерия при связывании с Fab (фигуры 19А, 20А и 20В). Область 132-143 (HDX_E1) характеризовалась значительным изменением поглощения только в самые короткие периоды времени воздействия. При более длительном времени воздействия разницы в обмене не было. Без привязки к конкретной теории полагают, что это указывает на то, что сайт только частично защищен от растворителя. Напротив, степень дифференциального поглощения дейтерия в области 182-187 (HDX_E2) увеличивалась с увеличением времени воздействия. Без привязки к конкретной теории полагают, что это согласуется с высокой аффинностью белок-белкового взаимодействия, которое уменьшает доступность растворителя (фигура 19А). Анализ Fab показал, что определяющие комплементарность области 1 и 3 (CDR) тяжелой цепи, а также CDR1 и CDR2 легкой цепи демонстрировали наибольшее снижение обмена, находясь в комплексе с CD73, что указывало на то, что эти области являлись основными составляющими паратопа (фигуры 20С и 20D). Хотя аа 132-143 (HDX_E1) и 182-187 (HDX_E2) CD73 были прерывистыми в пространстве последовательностей, они являются смежными при картировании на свернутую структуру CD73 (фигура 19В). Различия в водородном обмене наблюдали в других областях в пределах CD73. Однако большинство изменений массы, в том числе пептидов, содержащих известные остатки сайта связывания субстрата и активного сайта, не были статистически значимыми (фигуры 19А и 20Е).
[00564] Поскольку при использовании HDX сообщалось о контекстно-зависимых изменениях в воздействии растворителя на полипептидный остов, необходимой являлась валидация, чтобы отличить различия в водородном обмене из-за маскировки остова внутри поверхностного участка эпитоп-паратоп по сравнению с конформационными изменениями опосредованно или аллостерически индуцированными путем связывания антитела. Для определения того, составляют ли HDX_E1 и HDX_E2 эпитоп MEDI9447, оценивали связывание антитела с химерными мутантами CD73 с заменой домена (фигуры 21А-21Н и таблица 16).
[00565] 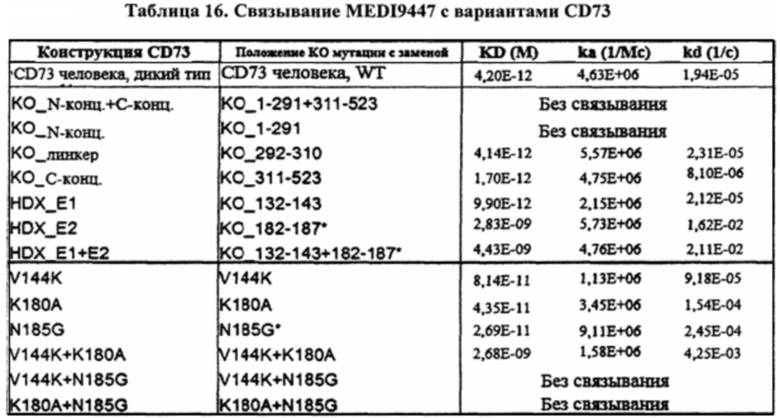
*Значения кинетики, полученные из подбора 2:1
[00566] Замена домена представляет собой методику, при которой гомолог белка-мишени (т.е. CD73 человека), который не связывается с антителом, представляющим интерес, применяют для создания химер, которые выявляют эти остатки, составляющие эпитоп. В отличие от введения делеций, замена последовательности между гомологами сводит к минимуму вероятность глобального разрушения структуры белка или предотвращения экспрессии белка. CD73 Gallus gallus (курицы) обладает ~65% идентичностью последовательности зрелого CD73 человека (фигура 22). При инактивации N-концевого домена CD73 человека (KO_1-291) или как N-, так и C-концевых доменов (KO_1-291+311-523) путем замены на соответствующую последовательность курицы, связывание замененных химер с MEDI9447 утрачивалось (фигуры 21А-21Н и таблица 16). Инактивация только C-концевого домена (аа 311-523) или альфа-спиральной линкерной области (аа 292-310) не влияла на связывание по сравнению с белком дикого типа (WT) (фигуры 21А, 21D, 21Е и таблица 16). Такие результаты согласуются с расположением эпитопа в N-концевом домене CD73. Далее, каждую область поверхностного участка HDX инактивировали отдельно и в комбинации. Связывание MEDI9447 с KO_HDX_E1 было сопоставимо с CD73 WT, хотя наблюдалось незначительное снижение скорости ассоциации ka (фигура 21F и таблица 16). Напротив, аффинность связывания с KO_HDX_E2 была значительно слабее, чем у CD73 WT, что обусловлено более быстрой диссоциацией (фигура 21F и таблица 16). Инактивация обеих областей (KO_HDX_E1+Е2) приводила лишь к небольшому уменьшению связывания по сравнению с областью Е2 отдельно (фигура 21G и таблица 16), Эти данные показывают, что хотя область 182-187 и в меньшей степени 132-143 содержат остатки, важные для связывания антител, дополнительные остатки вне поверхностного участка, идентифицированного с помощью HDX, составляют эпитоп.
[00567] Чтобы полностью определить эпитоп MEDI9447, создавали панель химер с замененными последовательностями, заменяя фрагменты из ~70 аа N-концевого домена CD73 человека на соответствующую последовательность курицы (фигуры 22, 23А и 23В). Инактивация либо области-2 (DS2), либо области-3 (DS3) в отдельности снижала связывание, и одновременная замена обеих областей (DS2_3) инактивировала связывание (фигуры 23А и 23В). Инактивация области 4 (DS4) или части области 1 (DS1a) (замена полной области предотвращала экспрессию) не влияла на связывание (фигуры 23А и 23В). Анализ связывания с химерами, содержащими замененные области из ~20-30 аа, показал, что подобласти DS2d (аа 135-152) и DS3b (аа 171-188) содержат остатки, которые влияют на связывание MEDI9447 (фигуры 23А и 23В). Когда часть подобластей 2d (DS2dmod) заменяли на область 3b (DS2dmod_3b), связывание утрачивалось (фигуры 23А и 23В), Примечательно, что две данные подобласти охватывают поверхностный участок, определенный в исследовании с помощью HDX (фигура 22). Наблюдение того, что одновременная замена обеих HDX-областей не устраняла связывание, указывает, что дополнительные остатки, опосредующие связывание, расположены в подобластях 2d и 3b, но не в HDX-идентифицированных сайгах. Чтобы картировать эти дополнительные остатки эпитопа, оценивали связывание антитела с панелью химер CD73, содержащих либо одноточечные мутации, либо комбинацию точечных мутаций и замены области. Аланиновый мутагенез также выполняли на остатках в пределах и пространственно близких к поверхностному участку HDX, которые являются консервативными среди CD73 курицы и человека (фигуры 22, 23А и 23В). Эти анализы показали, что V144, K180 и N185 являются основными остатками эпитопа, причем наиболее важным является N185 (фигуры 23А, 23В и 24). Комбинирование мутации N185G либо с K180А, либо с V144K устраняло связывание, тогда как мутирование K180 и V144 вместе приводило к уменьшению связывания (фигуры 23А и 23В). В дополнение к K180 было обнаружено, что три других консервативных остатка, Y135, K136 и N187, влияли на связывание, но в меньшей степени (фигуры 23А, 23В и 24А). Влияние этих последних трех остатков выявляли посредством их мутации на аланин в контексте фона КО с заменой домена; как исключительные точечные мутации они оказывали минимальное влияние или вообще не влияли на аффинность (фигуры 23А и 23В). Для проверки V144, K180 и N185 в качестве важных компонентов эпитопа, CD73 курицы заменяли на V144 и N185 (K180 сохранялся). Присутствие этих трех остатков обеспечивало связывание с помощью MEDI9947 при еубнаномолярной аффинности (KD=79 пМ) (фигуры 23А, 23В и 24В). Без привязки к конкретной теории полагают, что это указывает на то, что связывание MEDI9447 с CD73 в основном опосредовано этими тремя положениями аминокислот. Примечательно, что хотя в анализе с помощью HDX идентифицировали общее расположение связывающего поверхностного участка, два из трех важных остатков эпитопа не содержались в пептидах, проявляющих изменение в водородном обмене (фигуры 20А и 20В).
[00568] Наложение идентифицированного эпитопа на структуру CD73 показывает, что сайт связывания расположен на апикальной боковой поверхности открытой конформации CD73 (фигуры 24С-24Е), N185 расположен вблизи вершины N-концевого домена в области петли, простирающейся наружу от спирали G, которая содержит K180 (фигура 24С). Y135 и K136 расположены на β-нити 6 и примыкают к K180. V144 расположен в пределах β-нити 7 в тесном контакте с N187 (фигура 24С). В совокупности, боковые цепи этих остатков образуют почти непрерывную связывающую поверхность (фигура 24D). Если рассматривать CD73 в открытой мономерной структуре, то очевидно, что эпитоп находится как на противоположной стороне, так и пространственно удален от сайта связывания субстрата (фигура 24F). Кроме того, сайт связывания не охватывает любые остатки активного сайта, в том числе те, которые координируют взаимодействие с кофактором Zn2+ (фигура 24F). Без привязки к конкретной теории было предсказано, что MEDI9447 не конкурирует за связывание AMP, а вместо этого ингибирует ферментативную активность CD73 посредством потенциального аллостеричеекого механизма, основанного на положении эпитопа.
ПРИМЕР 14. MEDI9447 ингибировал ферментативную активность CD73
[00569] Чтобы охарактеризовать механизм ингибирования MEDI9447, сначала изучали кинетику гидролиза AMP посредством sCD73 в присутствии либо MEDI9447, либо негидролизируемого ингибитора CD73, АРСР. MEDI9447 неконкурентно ингибировал sCD73, о чем свидетельствовало снижение Vmax (4,59±0,26 в сравнении с 1,21±0,03), независимо от концентрации субстрата (фигура 25А). Напротив, АРСР увеличивал константу Михаэлиса Km (75,85±3,36 в сравнении с 26,03±3,87), но не уменьшал Vmax (3,35±0,04 в сравнении с 3,50±0,11), что согласовалось с предыдущими результатами о том, что АРСР является конкурентным ингибитором CD73 (фигура 25В). Такие результаты показывают, что MEDI9447 блокировал способность sCD73 гидролизовать AMP. Кроме того, они указывают, что MEDI9447 не блокировал связывание субстрата AMP, что согласовалось с положением эпитопа.
[00570] Далее, ингибирование sCD73 тестировали в зависимости от повышения концентрации MEDI9447 в формате либо IgG, либо Fab. IgG MEDI9447 ингибировал активность sCD73 дозозависимым образом, при этом максимальное ингибирование достигалось при молярном соотношении IgG и димера sCD73, составляющем ~1:1 (фигура 25С). Однако, когда IgG был в стехиометрическом избытке относительно sCD73, наблюдалась утрата ингибирования (фигура 25С). Этот так называемый «хук-эффект» наблюдали в других иммунологических анализах, и он может возникать в результате связывания моновалентных антител за счет плечей Fab на той же молекуле IgG, конкурирующих за ограничение сайтов связывания на целевом антигене. В соответствии с этим наблюдением Fab-формат MEDI9447 не ингибировал активность sCD73 (фигура 25С). В совокупности, такие результаты указывают, что для ингибирования функции sCD73 необходимо бивалентное взаимодействие MEDI9447.
ПРИМЕР 15. MEDI9447 препятствовал конформационному переходу CD73 в закрытое состояние
[00571] В предыдущих исследованиях структуры CD73 выявили, что ферментативная активность CD73 зависит от перехода от "открытой" в "закрытую" конформацию. В открытом состоянии фермент неактивен, тогда как в закрытом состоянии образуется активный сайт, обеспечивающий гидролиз субстрата. Определение кристаллической структуры CD73 человека показало, что для перехода между открытым и закрытым конформерами требуется значительный изгиб и вращение альфа-спиральной линкерной области, чтобы позволить N-концевому домену переместиться относительно С-концевого домена с образованием активного сайта. Было предположено, что связывание каждого Fab-плеча MEDI9447 с двумя N-концевыми доменами CD73 могло образовывать мостик, который ограничивал переход CD73 из неактивного открытого состояния в каталитически активное закрытое состояние. Чтобы проверить, ингибирует ли MEDI9447 конформационный переход CD73, использовали антитело mAb А, которое разрабатывали в качестве репортера конформации CD73. Картирование связывающего поверхностного участка mAb А показало, что он взаимодействует как с N-, так и с С-концевым доменом CD73 (фигуры 26А-26С). Принимая во внимание расположение области связывания, было предположено, что эпитоп будет находиться в открытом конформере CD73, но разрушаться в закрытом конформере (фигура 26В). Чтобы проверить это, измеряли связывание mAb А с открытым, несвязанным с субстратом sCD73 и закрытым sCD73 (фигуры 27А-27С). Чтобы индуцировать закрытую конформацию, sCD73 предварительно инкубировали с Zn2+ и АРСР; данный кофактор и негидролизуемый субстрат ранее использовали для получения кристаллической структуры закрытого конформера CD73 человека. Анализ связывания показал, что mAb А связывался с открытым sCD73, но связывание прекращалось, когда CD73 предварительно инкубировали с Zn2+ и АРСР (фигура 27А). Без привязки к конкретной теории полагают, что данный результат согласуется с тем, что эпитоп mAb А присутствует только в открытой структуре CD73. Напротив, связывание MEDI9447 не было чувствительным к состоянию CD73, что указывает на то, что он мог связываться с CD73, несвязанным либо связанным с субстратом (фигура 27А). Утрата связывания mAb А зависела как от Zn2+, так и от АРСР (фигура 28А). Установив, что связывание mAb А свидетельствует о конформационном состоянии CD73, затем тестировали эффект MEDI9447 в отношении структурного перехода CD73, индуцированного Zn2+/APCP. mAb А связывался с sCD73, который предварительно инкубировали с MEDI9447, что указывало на то, что два антитела связывались с разными эпитопами (фигура 27В). Важно отметить, что когда комплекс sCD73-MEDI9447 впоследствии инкубировали с Zn2+ и АРСР, связывание mAb А сохранялось (фигура 27В). Напротив, контрольный IgG и Fab-формат MEDI9447 не восстанавливали связывание mAb А при предварительном образовании комплекса с sCD73 перед добавлением Zn2+ и АРСР (фигура 28В). Такие результаты подтверждают гипотезу о том, что связывание бивалентного MEDI9447 предотвращает переход sCD73 из открытого состояния в полностью закрытую гидролитически активную конформацию. Наблюдение того, что связывание mAb А только частично сохранялось при связывании MEDI9447, указывало на то, что Zn2+ и АРСР могли индуцировать принятие CD73 каталитически неактивной формы, которая связывалась с репортерным антителом с более низкой аффинностью (фигура 27С).
ПРИМЕР 16. sCD73 и MEDI9447 образовывали междимерные мостиковые комплексы
[00572] Наблюдаемая ингибирующая активность MEDI9447 может осуществляться посредством N-концевого домена, образования междимерных мостиков с одной молекулой CD73 или посредством образования междимерных мостиков с отдельными молекулами CD73. Чтобы различать эти процессы, характеризовали размер образованных в растворе комплексов. Исходя из измеренной массы несвязанных MEDI9447 и CD73 (145 и 125 килодальтон (кДа) соответственно), предсказанный размер междимерного мостикового комплекса на основе антитела и CD73 1:1 будет составлять ~270 кДа (фигура 29А). Когда MEDI9447 и sCD73 связывались при молярном соотношении 1:1, образовывались два комплекса. Преобладающие виды имели средневесовую молярную массу (Mw) ~1,74 мегадальтон, а менее распространенные виды имели Mw ~0,66 кДа (фигура 30А). Mw самого большого комплекса соответствовала олигомеру, содержащему семь димеров CD73 и шесть IgG (7×125 кДа +6×150 кДа = 1,745 мегадальтон). Когда MEDI9447 ограничивали (0,5:1, 0,1:1), образовались комплексы с сопоставимой Mw, но относительная разница в численности каждого вида была менее выраженной (фигура 30А). Комплексы с MEDI9447 сравнивали с комплексами, образованными с другим антителом к CD73 mAb В. Анализ связывания mAb В с панелью химер CD73 с заменами доменов показал, что оно связывается с областью в N-концевом домене CD73, которая, в отличие от MEDI9447, находится вблизи борозды, образованной между мономерами CD73 (фигуры 29В и 19В). Было предположено, что связывание на этой расположенной внутри поверхности может препятствовать тому, чтобы Fab-плечи mAb В образовывали мостики через димеры CD73. Действительно, с помощью SEC-MALS продемонстрировали, что mAb В образует комплексы размером ~270-295 кДа, при этом значение Mw было близким к предсказанному для взаимодействия 1:1 (фигура 30С). В совокупности результаты указывают на то, что MEDI9447 образует междимерные мостики между несколькими димерами sCD73, и что образование этих олигомеров обеспечивается эпитопом.
ПРИМЕР 17. MEDI9447 ингибировал заякоренный CD73 посредством моновалентного взаимодействия
[00573] Хотя CD73 "слущивался" из поверхности клеток in vivo и сохранял ферментативную активность в его растворимой форме, большая часть нативного CD73 существовала в GPI-заякоренном формате. В свете этого и того факта, что предыдущие исследования выполняли с растворимой формой CD73, необходимо было охарактеризовать активность MEDI9447 с CD73 в иммобилизованном состоянии. Захват рекомбинантного CD73 посредством С-концевой шесть-гистидиновой метки на микротитрационных планшетах, покрытых никелем, позволил авторам настоящего изобретения проанализировать ферментативную активность иммобилизованного CD73, пространственно ориентированного так, как это было сделано в отношении CD73, заякоренного посредством GPI на поверхности клетки. Аналогично предыдущим результатам авторов настоящего изобретения, IgG MEDI9447 ингибировал гидролиз AMP дозозависимым образом (фигура 31А). Однако когда антитело находилось в молярном избытке относительно димера CD73, утрата ингибирования или хук-эффект не наблюдались (фигуры 31А и 32). Неожиданно Fab MEDI9447 также ингибировал активность CD73, однако при более низком максимальном ингибировании по сравнению с IgG MEDI9447 (фигуры 31А и 32). Такие результаты указывают на то, что в отличие от растворимого CD73, заякоренный CD73 может ингибироватьея посредством взаимодействия с моновалентным антителом. Различие в ингибировании между IgG и Fab дает основание предположить, что размер молекулы антитела может определять эффективность. Для изучения этого. Fab MEDI9447 предварительно инкубировали с антителом к Fd (xFd) в условиях, при которых одно Fab-плечо антитела xFd связывалось с Fab MEDI9447, а другое плечо связывалось с неспецифичным поликлональным Fab (pFab) (фигура 31В). Образование данного комплекса эффективно увеличивало размер Fab MEDI9447 при одновременном сохранении моновалентности к CD73. Этот xFd-связанный Fab ингибировал активность CD73 в равной степени, как IgG MEDI9447 (фигура 31А). Чтобы подтвердить это наблюдение, ингибирование антителом эндогенно экспрессируемого CD73 измеряли в линии эпителиальных клеток MBA-MD-231 рака молочной железы человека. Аналогично иммобилизованному рекомбинантному CD73, GPI-заякоренный CD73 сильно ингибировалея с помощью IgG MEDI9447, умеренно с помощью Fab MEDI9447, и предварительное связывание Fab с антителом xFd увеличивало максимальное ингибирование до уровня, эквивалентного IgG (фигура 31С). Наконец, тестировали ингибирование sCD73 с помощью Fab MEDI9447, связанного либо с одним, либо с двумя плечами антитела xFd. В отличие от связанного с поверхностью CD73, sCD73 не ингибировалея с помощью Fab MEDI9447, связанного с одним плечом xFd (фигура 31D). Однако обеспечение бивалентности посредством связывания Fab MEDI9447 с обоими плечами xFd приводило к ингибированию sCD73, сопоставимому с IgG MEDI9447 (фигуры 31В и 31D). Данные результаты показывают, что заякоренный на поверхности CD73 может ингибироваться в результате связывания с моновалентным антителом, и что активность опосредуетея размером антитела. Этим он отличается от sCD73, который ингибируется только с помощью MEDI9447 посредством бивалентного взаимодействия.
ПРИМЕР 18. MEDI9447 ингибирует конфирмационное изменение CD73 и гидролиз AMP с помощью двойного неконкурентного механизма действия
[00574] Как описано в данном документе, определяли эпитоп и механизм действия терапевтического моноклонального антитела, которое ингибирует ферментативную активность CD73. По результатам данного исследования выявили сайт связывания в N-концевом домене CD73, что обеспечивало возможность осуществления ингибирования посредством двух разных механизмов. Важно отметить, что данный признак придает MEDI9447 возможность блокировать как растворимый, так и заякоренный на поверхности клеток CD73 неконкурентным способом.
[00575] С помощью HDX-MS идентифицировали связывающий поверхностный участок между CD73 и Fab MEDI9447. Из-за относительно крупных пептидов, полученных в результате расщепления пепсином, сайт взаимодействия, идентифицированный с помощью анализа обменов, был относительно широким, охватывая в общей сложности восемнадцать аминокислот в пределах двух прерывистых пептидов. Эксперименты по замене доменов и мутагенезу показали, что только подгруппа этих остатков оказывала влияние на связывание антител, при этом две из трех наиболее влияющих аминокислот (V144 и K180) содержались в пептидах, которые не проявляли дифференциального водородного обмена. Одно из объяснений данного несоответствия состоит в том, что пока V144 и K180 боковых цепей могли быть закрыты из-за контакта с остатками CDR антитела, атомы водорода амида связанного полипептидного остова, которые подвергались обмену на дейтерий, могли относительно подвергаться воздействию растворителя. Результаты HDX-MS также показали, что только область, состоящая из аа 132-143, проявляла значительное изменение кинетики обмена за самые короткие периоды времени воздействия. Без привязки к конкретной теории полагают, что это указывает на то, что связывание в данной области являлось относительно слабым. Действительно, два остатка, которые, как обнаружилось, влияли на связывание в пределах данной области (Y135 и K136), оказывали только незначительный эффект в отношении аффинности MEDI9947 (фигуры 23А и 23В). Напротив, область аа 182-187 с наиболее значительным дифференциальным обменом содержала наиболее важный остаток для связывания (N185). При отсутствии сокристаллической структуры нельзя формально исключить то, что некоторые остатки, которые учитывались как взаимодействующие с MEDI9447, действительно образовывали контакты с паратопом. Однако обширный анализ связывания мутантов указывал на то, что данные потенциальные остатки могут вносить минимальный вклад в термодинамику связывания. Поскольку CDR3 тяжелой цепи играет важную роль в образовании контактирующих с антигеном сайтов, эта CDR проявляла наибольшую степень дифференциального обмена. Результаты HDX также указывают на то, что легкая цепь важна для связывания антигена, в частности CDR1 и 2 (фигуры 20С и 20D). В совокупности эти результаты подчеркивают как полезность HDX-MS при картировании эпитопов, так и важность проверки предсказанного связывающего поверхностного участка с помощью ортогональной методики, такой как мутагенез.
[00576] Расположение эпитопа в положении внутри N-концевого домена, который удален от остатков сайга связывания субстрата CD73 и активного сайта, согласовывалось с результатами, показывающими, что MEDI9447 характеризуется неконкурентным способом ингибирования. Исходя из области узнавания одного эпитопа, можно было предположить, что антитело действует как классический аллостерический ингибитор, вызывая долгосрочное конформационное изменение в CD73, которое искажает остатки активного сайта таким образом, что это предотвращает гидролитическую активность. Тем не менее, по данным HDX-MS не выявили значительных изменений структуры CD73 в областях, находящихся вне связывающего поверхностного участка, которые бы поддерживали такую форму аллостерии. В качестве альтернативы, эпитоп мог свидетельствовать о том, что связывание MEDI9447 ограничивает перемещение петель, β-нитей или α-спиралей внутри N-концевого домена, что необходимо для каталитической активности. В противоположность этому, сообщалось о незначительной разнице или ее отсутствии во вторичной или третичной структуре в N-концевом домене между открытыми и закрытыми структурами CD73. Локальные междоменные конформационные изменения, которые при этом происходят, ограничиваются линкером и С-терминальным доменом.
[00577] В качестве альтернативы к индукции структурных изменений, считалось, что расположение эпитопа находилось бы в правильном положении, чтобы обеспечить возможность образования антителами мостиков или поперечного связывания димеров CD73, что ограничивало бы необходимое изменение конформации. Такой подход, заключающийся в образовании мостиков, подтверждался данными, показывающими, что IgG был необходим для ингибирования растворимой формы CD73, и что MEDI9447 образует комплексы, содержащие несколько димеров CD73. Этот последний результат отличался от mAb В, которое не образовывало междимерные мостики, подчеркивая важность эпитопа MEDI9447 для обеспечения этой активности поперечного связывания. Рационально, если два N-концевых домена CD73 связывались Fab-доменами одного и того же IgG, то это физически ограничивало бы перемещение N-концевого домена и линкерной области, что необходимо для того, чтобы фермент принял свою закрытую активную структуру. Путем применения mAb А, которое действовало как конформационный зонд состояния CD73, было продемонстрировано, что связывание MEDI9447 ингибировало CD73 от принятия полностью закрытого конформера, индуцированного с помощью Zn2+ и АРСР. Промежуточный уровень связывания с помощью репортерного mAb А при предварительном образовании комплекса MEDI9447 с CD73 указывал на то, что конформационный переход в некоторой степени по прежнему индуцировали Zn2+ и АРСР. Сниженное связывание mAb А могло отражать то, что его эпитоп был частично изменен в данном промежуточном состоянии, однако все еще достаточным чтобы обеспечивать связывание. Учитывая высокую степень гибкости шарнирной области, неудивительно, что даже когда MEDI9447 образовывал мостики с димерами CD73, структурные изменения CD73 по-прежнему будут присутствовать вследствие связывания субстрата. Без привязки к конкретной теории трудно представить, что связанный IgG будет захватывать CD73 в полностью жесткой конформации.
[00578] Неожиданный результат данной работы состоит в том, что в дополнение к образованию мостиков посредством бивалентного взаимодействия, MEDI9447 может также ингибировать CD73, когда он связан с поверхностью посредством моновалентного связывания. Будучи изначально сложным, второй пространственно-опосредованный механизм блокирования активности заякоренного CD73 согласовывался с наблюдениями, описанными в данном документе. Для образования каталитически активного GPI-заякоренного CD73 требовалось, чтобы N-концевой домен вращался вниз в положение, ближайшее к поверхности клетки. Нативный димер CD73 составлял ~130 кДа. Сравнивая с размером IgG или Fab (~150 кДа и ~50 кДа соответственно), есть основания полагать, что антитело, связанное с N-концевым доменом, сможет пространственно блокировать CD73 от полного вращения для принятия замкнутой конформации. Данный механизм подтверждался двумя наблюдениями. Во-первых, факт, что Fab демонстрировал более низкое максимальное ингибирование по сравнению с IgG, согласовался с пространственным эффектом, зависимым от размера. Разница между Fab и IgG вряд ли объясняется различиями в связывании, так как аффинность Fab по-прежнему была субнаномолярной (KD=327 пМ, данные не показаны). Кроме того, повышенная эффективность, обеспечиваемая путем увеличения эффективного размера Fab посредством конъюгирования с xFd антитела, также оказывала сильную поддержку в отношении пространственного механизма, зависимого от размера, а не валентности или авидности. Второе наблюдение, которое поддерживало данный механизм ингибирования, заключалось в том, что хук-эффект или утрату ингибирования не наблюдали с IgG MEDI9447, когда CD73 был связан с поверхностью. Предположительно это связано с тем, что антитело могло блокировать гидролиз AMP заякоренным CD73 посредством либо би-, либо моновалентного взаимодействия (фигура 33). Напротив, хук-эффект наблюдали, когда CD73 был растворимым из-за отсутствия твердой фазы, необходимой для моновалентного связывания IgG или Fab, чтобы пространственно заблокировать вращение N-концевого домена.
[00579] Таким образом, предлагается модель, в которой MEDI9447 противодействует функционированию растворимого и GPI-заякоренного CD73 посредством двойного механизма ингибирования, который полностью связан с его эпитопом (фигура 33). Хотя описанные в данном документе исследования показывают, что MEDI9447 может блокировать как растворимый, так и связанный CD73 in vivo, неизвестным является то, будет ли GPI-заякоренный CD73 ингибироваться посредством одного или обоих механизмов. Предположительно плотность, ориентация и расстояние между димерами CD73 на поверхности клеток обусловливают доминирующий способ ингибирования. Учитывая, что большинство раковых клеток сверхэкспрессируют CD73, что увеличивает вероятность непосредственной близости димеров, ожидается, что MEDI9447 будет участвовать как в би-, так и в моновалентном взаимодействиях. На нормальной неопухолевой ткани, где экспрессия CD73 происходила бы при относительно низкой поверхностной плотности, MEDI9447 мог бы ингибировать гидролиз AMP, главным образом, посредством механизма пространственного блокирования.
[00580] С точки зрения механизма действия блокирование CD73 является стратегией иммуноонкологии, отличной от стратегии сопоставимых мишеней, таких как PD-1 и CTLA-4, для которых уже имеются одобренные лекарственные средства. С терапевтической точки зрения активность MEDI9447 является преимущественной. Он неконкурентно ингибирует CD73, и, следовательно, не должен конкурировать с эндогенным нуклеотидным связыванием путем блокирования активного сайта. Это позволяет избежать потенциальной перекрестной реактивности по отношению к другим нуклеотидным/боковым связывающим белкам со структурно консервативными активными сайгами. Кроме того, MEDI9447 может ингибировать как растворимые, так и мембранносвязанные CD73 посредством одно- или бивалентного взаимодействия. Как ожидается, обе эти особенности будут способствовать эффективности in vivo. Данное терапевтическое mAb обладает иммуномодулирующим потенциалом для лечения рака как отдельно, так и в комбинации с существующими химиотерапевтическими средствами, которые целенаправленно воздействуют на комплементарные пути иммуномодуляции.
Вышеописанные результаты выполнялись с применением следующих материалов и способов.
Анализ 1. Прямой ELISA
[00581] 384-луночные планшеты для ELISA покрывали с помощью приблизительно 1,5 нг/лунка рекомбинантного белка CD73, блокировали с помощью 1% BSA/0,1% Tween-20 / PBS и инкубировали с образцами антител в течение 90 минут при комнатной температуре. Затем следовала инкубация с конъюгатом антитела козы к Ig-лямбда с пероксидазой хрена (HRP) в течение 30 мин. при комнатной температуре. Активность HRP выявляли с помощью субстрата тетраметилбензидина (ТМВ) и реакцию останавливали с помощью 1 М HCl. Планшеты считывали при 450 нм.
Анализ 2. ELISA с захватом
[00582] 384-луночные планшеты для ELISA покрывали с помощью приблизительно 3 нг/лунка антитела овцы к Fd человека (для скрининга антител в формате Fab), блокировали с помощью 1% BSA/0,1% Tween-20/PBS и инкубировали с образцами в течение 90 минут при комнатной температуре. Затем добавляли биотинилированный белок CD73 на 1 ч. при комнатной температуре. Затем следовала инкубация с конъюгатом стрептавидина с пероксидазой хрена (HRP) в течение 30 мин. при комнатной температуре. Активность HRP выявляли с помощью субстрата тетраметилбензидина (ТМВ) и реакцию останавливали с помощью 1 М HCl. Планшеты считывали при 450 нм.
[00583] Для скрининга клонов с единичными мутациями аминокислот использовали приблизительно 50 нг/лунка биотинилированного рекомбинантного CD73. Для скрининга клонов из комбинаторной библиотеки использовали 10 нг/лунка биотинилированного рекомбинантного CD73.
Анализ 3. Анализ связывания с помощью проточной цитометрии
[00584] Все эксперименты с помощью проточной цитометрии проводили при 4°С и реагенты получали в буфере PBS/1% FBS. 10000 клеток инкубировали с тестируемым антителом в объеме 50 мкл в течение 4 часов. Клетки промывали дважды и инкубировали с 50 мкл конъюгата антитела козы к IgGFc человека с AlexaFluor647 в течение 15 минут. Клетки промывали, ресуспендировали в буфере, дополненном Dapi, и анализировали на проточном цитометре. Нежизнеспособные клетки, идентифицированные по высокоинтенсивному окрашиванию Dapi, исключали из анализа. Для определения значений KD, график средней интенсивности флуоресценции в зависимости от концентрации тестируемого антитела нелинейно аппроксимировали с применением изотермической модели односайтового связывания.
Анализ 4. Создание библиотеки антител с единичными аминокислотными изменениями
[00585] Сайт-направленный мутагенез кодонов CDR CD730010 или CD730002 выполняли с применением набора для множественного сайт-направленного мутагенеза QuikChange Lightning (Agilent) и праймеров. Каждый кодон подвергали мутагенезу с помощью праймера, который заменял кодон дикого типа на кодон NNS. Подвергнутые мутагенезу гены VH и VL клонировали в вектор Fab для бактериальной экспрессии. Штамм BL21 (DE3) Е. coli трансформировали с помощью библиотеки антител, отдельные колонии отбирали и культивировали в среде Magic Media (Invitrogen) в течение 24 часов при комнатной температуре с получением бактериальных Fab-фрагментов. Получали бактериальный супернатант и использовали его для скрининга библиотеки антител в анализах связывания ELISA.
Анализ 5. Создание библиотеки антител с комбинаторными аминокислотными изменениями
[00586] Гены VH и VL CD730010GL9 клонировали в вектор для экспрессии бактериального Fab. Сайт-направленный мутагенез кодонов CDR CD730010GL9 выполняли либо с применением набора для множественного сайт-направленного мутагенеза QuikChange Lightning (Agilent), либо перекрывающейся ПЦР и вырожденных праймеров. Вырожденные праймеры конструировали так, чтобы они кодировали выбранные аминокислотные изменения, а также исходную аминокислоту в одном и том же положении. Штамм BL21 (DE3) Е. coli трансформировали с помощью библиотеки Fab, отдельные колонии культивировали в среде Magic Media (Invitrogen) в течение 24 часов при комнатной температуре с получением бактериальных Fab-фрагментов. Бактериальные супернатанты использовали для скрининга библиотеки антител в анализах связывания ELISA.
Анализ 6. Анализ FabZAP
[00587] 1000 клеток/лунка культивировали в 96-луночных планшетах в RPMI/10% FBS. Серийные разбавления антител к CD73, начиная с 5 нМ, смешивали с реагентом FabZAP (Advanced Targeting Systems, Сан-Диего, Калифорния) и добавляли к клеткам. После 3 дней инкубации при 37°С пролиферацию клеток измеряли с использованием анализа CellTiter-Glo (Promega, Мэдисон, Висконсин).
[00588] Следующие анализы (анализы 7-11) выполняли с применением следующих реагентов CD73 и антител.
[00589] Конструировали плазмидный вектор экспрессии млекопитающего, кодирующий рекомбинантный зрелый CD73 человека (положения аминокислот 1-526) (MedImmune). Чтобы достичь экспрессии растворимой секретируемой формы CD73, сигнальный пептид GPI-якоря удаляли и заменяли С-концевой 6х-гистидиновой меткой. Нумерация последовательности CD73 основана на зрелом белке без сигнального пептида (NT5E человека, эталонная последовательность в NCBI NP_002517.1). Плазмиды, кодирующие рекомбинантные "нокаутные" (KО) химерные мутанты с заменой доменов человека/курицы, конструировали с применением синтетической ДНК gBlocks (IDT, Inc.), кодирующей кодон-оптимизированную последовательность ДНК CD73 курицы (NT5E курицы, эталонная последовательность в NCBI ХР_004940453.1). На основе выравнивания с последовательностью белка CD73 человека положение 1 кодируемой аминокислоты в конструкциях, содержащих N-концевой домен курицы, соответствовало положению 20 в незрелом CD73 курицы, содержащем предсказанный сигнальный пептид. Полноразмерные KO ДНК-конструкции получали с помощью однократной ПЦР с перекрывающимися праймерами gBlocks и ПЦР-ампликонами CD73 человека. Все конструкции содержали 6х-гистидиновую метку на С-конце. Единичные и множественные точечные мутации в человеческих и химерных конструкциях осуществляли с помощью сайт-направленного мутагенеза с применением набора для множественного сайт-направленного мутагенеза Quick Change Lightning (Stratagene). Все конструкции CD73 экспрессировали в суспензионных клетках НЕK293. Человеческий CD73 дикого типа, меченый гистидином (без сигнальной последовательности GPI-якоря) были очищены с применением никелевой аффинной колонки HisTrap (GE Healthcare Life Sciences) исследовательской группой MedImmune Protein Sciences. Было подтверждено, что белок является димером в растворе с молярной массой ~125 кДа (см. фигуры 18А и 18В). Все конструкции мутантного CD73 экепрессировали с помощью транзиентной трансфекции суспензионных клеток НЕK293 с применением 293Fectin (Life Technologies). Клетки выращивали и трансфицировали в бессывороточной среде 293Freestyle (Life Technologies) в 24-луночных блоках с глубокими лунками. Неочищенные супернатанты клеток собирали через шесть дней после трансфекции и фильтровали через 0,45 мкм фильтр с удалением клеточного дебриса перед применением. Концентрации вариантов CD73 в супернатанте определяли путем измерения связывания белков, меченных гиетидином, с биосенсорами HIS2 ( /Pall Life Sciences) на приборе для интерферометрии биоелоя Octet QK384 (/Pall Life Sciences). Концентрации рассчитывали с применением программного обеспечения для анализа данных Octet, сравнивая сигнал связывания со стандартной кривой, построенной по значениям разбавлений очищенного рекомбинантного CD73 человека, меченого 6х-гистидиновой меткой, с известной концентрацией. MEDI9447 (форматы IgG1 человека и IgG1 мыши), mAb A (IgG1 человека) и mAb В (IgG1 человека) и Fab MEDI9447 (IgG1 человека) были экспрессированы и очищены исследовательскими группами MedImmune Protein Sciences and Expression. IgG экспрессировали в клетках млекопитающего и очищали с помощью белка А и эксклюзионной хроматографии. Для получения Fab MEDI9447 10 мг IgG расщепляли в течение 5 ч. при 37°С с помощью иммобилизированного папаина (Thermo Scientific/Life Technologies), и Fab очищали с применением колонки HiTrap Q (GE Healthcare Life Sciences).
/Pall Life Sciences) на приборе для интерферометрии биоелоя Octet QK384 (/Pall Life Sciences). Концентрации рассчитывали с применением программного обеспечения для анализа данных Octet, сравнивая сигнал связывания со стандартной кривой, построенной по значениям разбавлений очищенного рекомбинантного CD73 человека, меченого 6х-гистидиновой меткой, с известной концентрацией. MEDI9447 (форматы IgG1 человека и IgG1 мыши), mAb A (IgG1 человека) и mAb В (IgG1 человека) и Fab MEDI9447 (IgG1 человека) были экспрессированы и очищены исследовательскими группами MedImmune Protein Sciences and Expression. IgG экспрессировали в клетках млекопитающего и очищали с помощью белка А и эксклюзионной хроматографии. Для получения Fab MEDI9447 10 мг IgG расщепляли в течение 5 ч. при 37°С с помощью иммобилизированного папаина (Thermo Scientific/Life Technologies), и Fab очищали с применением колонки HiTrap Q (GE Healthcare Life Sciences).
Анализ 7. Анализ HDX-MS
[00590] Образцы рекомбинантного CD73 человека и Fab MEDI9447 получали в концентрации 2 мг/мл. Комплекс CD73+Fab образовывали путем предварительной инкубации при соотношении концентраций 1:1. Все эксперименты HDX проводили с применением устройства для HDX от Waters (Waters Corporation), оснащенного модулем автоматизации Leap. Вкратце, 1,25 мкл образцов белка разбавляли двадцать раз либо Н2О, либо буфером D2O (10 мМ фосфата, рН 7,0) при 20°С. После различных периодов времени инкубации (0 секунд для экспериментов без дейтерирования или 0,5, 1, 5, 10, 30, 60 и 120 минут для экспериментов с дейтерированием), меченые образцы гасили путем добавления равного объема ледяного раствора с 4,0 М гуанидин-HCl (Pierce Biotechnology), 500 мМ трис(2-карбоксиэтил)фосфина гидрохлоридом (ТСЕР) (Pierce Biotechnology), pH 2,4. Сразу после этого образцы расщепляли с применением картриджа Poroszyme с иммобилизированным пепсином (Applied Biosystems) при 20°С. Пепсиновые фрагменты собирали и высаливали с применением предколонки ACQUITY ВЕН С18 VanGuard (2,1×5 мм, Waters) и элюировали в колонку ACQUITY ВЕН С18 (1,7 мкм, 1,0×100 мм, Waters) при 0°С. Пептиды, разделенные на колонке, анализировали на масс-спектрометре SYNAPT G2 (Waters). Для анализа данных пептиды идентифицировали с применением программного обеспечения ProteinLynx Global Server (Waters) и уровни включения дейтерия для каждого пепсинового пептида с каждого момента времени мечения рассчитывали с применением DynamX (Waters). С каждым белок выполняли четыре эксперимента без дейтерирования и три полных эксперимента HDX. Значения значимой разницы (±1,6 дальтон) рассчитывали с использованием значения погрешности эксперимента и 98% доверительного интервала, как описано выше, за исключением того, что вместо среднего значения отдельных стандартных отклонений использовали объединенную дисперсию стандартных отклонений. Значения относительного фракционного поглощения между комплексом CD73 + Fab и CD73 получали с помощью программного обеспечения DynamX (Waters) и экспортировали в PyMOL ( , Inc.) для моделирования структуры. Все фигуры, изображающие структуру CD73 человека, получали с использованием PyMol и выявленных кристаллических структур открытой и закрытой конформаций CD73 (ссылочные номера в PDB: 4H2F и 4H2I соответственно).
, Inc.) для моделирования структуры. Все фигуры, изображающие структуру CD73 человека, получали с использованием PyMol и выявленных кристаллических структур открытой и закрытой конформаций CD73 (ссылочные номера в PDB: 4H2F и 4H2I соответственно).
Анализ 8. Анализ связывания с помощью SPR и BLI
[00591] Связывание MEDI9447 с белками CD73 дикого типа и мутантным измеряли с помощью поверхностного плазменного резонанса (SPR) с применением прибора ProteOn (BioRad). Образцы неочищенного супернатанта белков CD73 из клеток разбавляли до 3 мкг/мл в PBS, 0,005% Tween-20, pH 7,4 (BioRad) и иммобилизовали до ~400 RU на сенсорном чипе HTG tris-NTA (BioRad), предварительно активированном с помощью 10 мМ NiSO4, 10 мМ MES, рН 6,0 (BioRad). Эквивалентно разбавленный образец неочищенного супернатанта клеток из нетрансфицированных клеток включали в качестве эталонного контроля канала. Сенсограммы регистрировали с помощью проточных двукратных разбавлений MEDI9447, полученного в PBS, 0,005% Tween-20, рН 7,4, в диапазоне от 5 нМ до 0,31 нМ. В некоторых случаях для вариантов CD73, которые слабо связывались с MEDI9447, разбавления антител варьировали от 20 нМ до 1,25 нМ. Связывание антитела измеряли при скорости потока 100 мкл/мин с 3-минутной фазой ассоциации и 20-минутной фазой диссоциации. Поверхность сенсорного чипа регенерировали между каждым прогоном путем впрыскивания 300 мМ EDTA, рН 8,5 (BioRad) в течение 800 с при скорости потока 30 мкл/мин. Кинетику связывания анализировали с применением программного обеспечения ProteOn для анализа данных. Осуществляли двойное реферирование, и если не указано иное, то для подбора данных использовали модель связывания Ленгмюра 1:1. Некоторые варианты CD73, содержащие мутации в пределах аминокислотной области 171-188, плохо подходили для модели 1:1 (т, е. значения Хи2>10% от Rmax). В случаях, где это отмечено, данные варианты подходили модели гетерогенного антигена (2:1). Из-за высокой аффинности MEDI9447 (~4 пМ) и ограничений чувствительности ProteOn, при сравнении связывания MEDI9447 с вариантами CD73 значимыми считались только >2-кратные изменения в измеренных значениях кинетики. Из-за формата анализа (иммобилизованный антиген) и димерного состояния CD73 значения KD могли быть преувеличены из-за эффектов авидности. Однако не предполагалось, что это влияет на ранжирование связывания MEDI9447 с различными вариантами CD73.
[00592] Картирование "горячих точек" связывания mAb А и mAb В антитела к CD73 осуществляли с помощью BLI, с применением прибора Octet QK384. Все белки получали в 1х буфере для кинетического анализа (/Pall Life Sciences). Меченые по С-концевому гистидину варианты CD73 из неочищенных супернатантов клеток разбавляли до 6 мкг/мл и иммобилизовали на биосенсорах HIS2 до порога реакции на связывание 0,8 нм. После 300-секундной исходной стадии датчики погружали в 30 нМ антитела. Периоды времени ассоциации и диссоциации составлял 600 с. Эталонный контроль в виде супернатанта нетрансфицированных клеток включали для вычитания фонового связывания во время анализа данных. С применением программного обеспечения для анализа данных ForteBio обрабатывали данные и получали графики.
Анализ 9. Анализы ферментативной активности CD73
[00593] CD73-катализируемый гидролиз AMP в аденозин и неорганический фосфат анализировали с помощью либо количественного анализа неорганического фосфата (анализ с использованием малахитового зеленого, R&D Systems), либо измерения АТФ-зависимого окисления люциферина, которое ингибируется AMP (анализ CellTiterGlo, Promega). Графики данных и измерения ферментативной кинетики (нелинейная регрессия Михаэлиса-Ментена) получали с применением программного обеспечения Prism (Graphpad). Эксперименты проводили либо в двух повторах, либо в трех повторах.
[00594] Для измерений растворимого рекомбинантного CD73 с использованием анализа CellTiterGlo, 400 пМ рекомбинантного CD73 и различные концентрации антитела к CD73 инкубировали в аналитическом буфере (25 мМ Трис, рН 7,5, 5 мМ MgCl2, 0,005% Tween-20) в течение 1 часа при 37°С перед добавлением равного объема 200 мкМ AMP/600 мкМ АТФ (в аналитическом буфере). После инкубации в течение 1 часа при 37°С концентрацию AMP в образце определяли с использованием анализа CellTiterGlo в соответствии с инструкциями производителя.
[00595] Для измерений растворимого рекомбинантного CD73, применяя анализ с использованием малахитового зеленого, 1 нМ рекомбинантного CD73 и либо 1 нМ антитела, либо 40 мкМ аденозин 5'-(α, β-метилен)дифосфата (АРСР, Sigma-Aldrich) инкубировали в аналитическом буфере (25 мМ Трис, рН 7,5, 5 мМ MgCl2, 0,005% Tween-20) в течение 1 часа при комнатной температуре. Добавляли равный объем 400 мкМ AMP (для антител к CD73) или 3 мМ AMP (для АРСР) в аналитическом буфере, и образцы инкубировали в течение 15 минут при комнатной температуре. Концентрацию неорганического фосфата определяли, применяя анализ с использованием малахитового зеленого в соответствии с инструкциями производителя.
[00596] Для измерений иммобилизованного рекомбинантного CD73, 50 мкл CD73 при содержании 100 нг/мл в аналитическом буфере (25 мМ Трис, рН 7,5, 5 мМ MgCl2, 0,005% Tween-20), дополненном 100 мкг/мл BSA, иммобилизовали на планшетах, покрытых никелем (Life Technologies). Несвязанный CD73 смывали и добавляли 50 мкл антитела к CD73 (в аналитическом буфере). После инкубирования в течение 1 часа при комнатной температуре, планшеты снова промывали, добавляли 100 мкл АМФ при 500 мкМ (в аналитическом буфере) и образцы инкубировали в течение 15 минут при комнатной температуре. Концентрацию неорганического фосфата определяли, применяя анализ с использованием малахитового зеленого в соответствии с инструкциями производителя. В некоторых экспериментах с иммобилизованным и растворимым CD73, перед добавлением к CD73 антитело к CD73 (IgG и Fab) предварительно инкубировали в течение по меньшей мере 2 часов с 10-кратным молярным избытком поликлонального Fab-фрагмента человека (Bethyl Laboratories) и 100-кратным молярным избытком антитела овцы к IgG человека (Fd) (Meridian Life Sciences).
[00597] Для измерения эндогенной активности CD73 в культивируемых клетках, 20000 клеток MDA-MB-231 на лунку высевали в RPMI/ 10% FBS (Life Technologies) в 96-луночном планшете. После инкубации в течение ночи лунки промывали 3 раза RPMI, не содержащей сыворотку, и добавляли 50 мкл антител (в RPMI, не содержащей сыворотку). После инкубации в течение 30 минут при 37°С, добавляли 25 мкл 1,2 мМ AMP на лунку (в RPMI, не содержащей сыворотку). Планшеты инкубировали в течение 3 часов при 37°С. Смешивали 25 мкл клеточного супернатанта и 25 мкл 100 мМ АТР, и концентрацию AMP в образце определяли с использованием анализа CellTiterGlo в соответствии с инструкциями производителя.
Анализ 10. Анализ конформационного перехода CD73 с помощью репортера mAb A
[00598] Связывание MEDI9447 и mAb А с очищенным рекомбинантным CD73 человека осуществляли на приборе Octet QK384. Для сравнения связывания mAb А и MEDI9447, CD73 разбавляли до 6 мкг/мл в PBS, рН 7,4 (Life Technologies) e 0,5% альбумина бычьей сыворотки (PBSB, Sigma-Aldrich) и загружали на биосенсоры HIS2 до порога различимости сигнала связывания, составляющего 1,0 нм. Затем биосенсоры переносили либо только в PBSB, либо в PBSB с 10 мкМ ZnCl2 (Sigma-Aldrich), APCP и/или 2 мМ этилендиаминтетрауксусной кислоты (EDTA) (Life Technologies) и инкубировали в течение 15 мин. Затем биосенсоры переносили в PBSB, содержащий MEDI9447 или mAb А, разбавленное до 30 нМ в PBSB, и ассоциацию антител измеряли в течение 10 мин.
[00599] Чтобы проверить влияние MEDI9447 на конформационный переход CD73 в присутствии ZnCl2 и APCP, mAb А, разбавленное до 10 мкг/мл в PBS, рН 7,4, иммобилизовали на биосенсоре АНС для антитела к Fc человека ( /Pall Life Sciences). После 400-секундной стадии иммобилизации биосенсоры блокировали в течение 10 мин. в неспецифичном поликлональном IgG человека (Jackson ImmunoResearch Laboratories) при 50 мкг/мл в PBS, рН 7,4. После 4 мин. исходной стадии связывание mAb А измеряли путем инкубирования биосенсоров в течение 600 с в лунках, содержащих CD73, разбавленный до 250 нМ (с учетом молекулярного веса димера) только в PBS или в PBS, содержащем 10 мкМ ZnCl2 и/или 0,5 мМ APCP. В случае образцов, содержащих MEDI9447, связанный с CD73, вариант MEDI9447, представляющий собой IgG1 мыши (используемый во избежание связывания с биосенсором для антитела к Fc человека), разбавленный до 250 нМ в PBS, перед инкубацией с ZnCl2 и APCP предварительно инкубировали с CD73 в течение 15 мин. при комнатной температуре. Fab MEDI9447 и изотипически сходный контрольный IgG1 мыши (полученный в MedImmune), тестировали при концентрации 500 нМ. Скорость встряхивания для всех стадий анализа составляла 1000 об./мин. Анализ связывания и графики данных получали с применением программного обеспечения для анализа данных ForteBio.
/Pall Life Sciences). После 400-секундной стадии иммобилизации биосенсоры блокировали в течение 10 мин. в неспецифичном поликлональном IgG человека (Jackson ImmunoResearch Laboratories) при 50 мкг/мл в PBS, рН 7,4. После 4 мин. исходной стадии связывание mAb А измеряли путем инкубирования биосенсоров в течение 600 с в лунках, содержащих CD73, разбавленный до 250 нМ (с учетом молекулярного веса димера) только в PBS или в PBS, содержащем 10 мкМ ZnCl2 и/или 0,5 мМ APCP. В случае образцов, содержащих MEDI9447, связанный с CD73, вариант MEDI9447, представляющий собой IgG1 мыши (используемый во избежание связывания с биосенсором для антитела к Fc человека), разбавленный до 250 нМ в PBS, перед инкубацией с ZnCl2 и APCP предварительно инкубировали с CD73 в течение 15 мин. при комнатной температуре. Fab MEDI9447 и изотипически сходный контрольный IgG1 мыши (полученный в MedImmune), тестировали при концентрации 500 нМ. Скорость встряхивания для всех стадий анализа составляла 1000 об./мин. Анализ связывания и графики данных получали с применением программного обеспечения для анализа данных ForteBio.
Анализ 11. Анализ SEC-MALS
[00600] Для экспериментов по анализу комплексов, образованных с CD73 и MEDI9447 или mAb В, 900 пмоль CD73 инкубировали с 900, 450, 90 или 0 пмоль антитела, разбавленного в PBS, рН 7,4. Также получали отдельный образец только с 900 пмоль антитела. Образцы инкубировали в течение 30 мин. при комнатной температуре, а затем 100 мкл каждого образца разделяли на HP 1100 HPLC (Agilent), используя колонку TSKgel G3000W×L 5 мкм, 7,88 мм ×30 см (Tosoh Bioscience, LLC), при скорости потока 1 мл/мин в течение 20 мин. Подвижный буфер для образца представлял собой 0,1 М NaPi, 0,1 М NaSO4, pH 6,8. После разделения с помощью HPLC все образцы анализировали с применением MALS-детектора Dawn Heleos II и детектора по показателю преломления Optilab T-rEx (Wyatt). Данные анализировали с применением программного обеспечения Astra (Wyatt).
[00601]
[00602] Предыдущее описание конкретных аспектов настолько полно раскрывает общий характер изобретения, что другие могут, применяя знания в пределах уровня техники, легко модифицировать и/или адаптировать для различных применений эти конкретные аспекты без излишнего экспериментирования, не отступая от общей концепции настоящего изобретения. Следовательно, такие адаптации и модификации предназначены находиться в пределах значения и диапазона эквивалентов раскрытых аспектов, основанных на идее и принципе, представленных в данном документе. Следует понимать, что формулировки или терминология в данном документе предназначены для целей описания, а не ограничения, поэтому терминологию или формулировки в настоящем описании квалифицированному специалисту следует интерпретировать в свете идей и принципов.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
>SEQ ID NO:1 CD730002 VL
QSVLTQPPSVSVSPGQTATITCSGDKVGDKYASWYQQKPGQSPVLVIYEDTKRPSRIPERFSGSNSGNTATLTISGTQALDEADYFCQAWDTSFWVFGGGTKLTVL
>SEQ ID NO:2 CD730002 VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:3 CD730010 VL
LPVLTQPPSVSGTPGQRVTISCSGSLSNIGRNPVNWYKQVPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLNGWLFGGGTKLTVL
>SEQ ID NO:4 CD730010 VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDYWGRGTLVTVSS
>SEQ ID NO:5 CD730011 VL
NFMLTQPHSVSESPGKTVTISCTRSSGSIASKYVQWYQQRPGSSPMAVIYKDNQRSSGVPDRFSGSIDSSSNSASLTISGLKPEDEADYYCQSYDASNYYVFGTGTKVTVL
>SEQ ID NO:6 CD730011 VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARRGHGLYFDLWGQGTTVTVSS
>SEQ ID NO:7 CD730021 VL
QSVLTQPPSASGTPGQRVTISCSGSRPNIGGNTVNWYQQLPGAAPKLLIYSNSQRPSGVPDRFSGSKYGTSASLAISGLQSDDEADYYCGTWDDSLNGPVFGRGTKLTVL
>SEQ ID NO:8 CD730021 VH
QVQLQESGPGLVRPSETLSLTCTVSGGSISSSSYYWAWVRQSPGKGLEWIGNIYYRGSTYYNPSLKSRVTMSVDMSKHQFSLKLSSLNAADTAVYYCASLYSGTYVFDYWGRGTLVTVSS
>SEQ ID NO:9 CD730042 VL
QSVLTQPASVSGSPGQSITISCAGTSSDVGGYNYVSWYQQHPGKAPKLMIYEGSKRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYYCSSYTTRSTRVFGGGTKLTVL
>SEQ ID NO:10 CD730042 VH
GVQLVQSGGGLVQPGGSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVISYDGSIKYYADSVKGRFTISRDDSKNALYLQMNSLRAEDTAVYYCSTLSGSYGYFDYWGRGTLVTVSS
>SEQ ID NO:11 CD730046 VL
QSVLTQPPSASGTPGQRVTISCSGSSSNIGSNTVSWYQHLPGTAPQLLIYTNNHRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCAAWDDSLSGYVFGTGTKVTVL
>SEQ ID NO:12 CD730046 VH
QVQLVQSGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVISYDGSIKYYADSVKGRFTISRDNSKNSLFLQMNSLRDDDTATYYCARGHLLRIGDIFYYSLDVWGQGTLVTVSS
>SEQ ID NO:13 CD730047 VL
QSVLTQPPSASGAPGQRVTISCSGSSSNIGSNTVNWYQRLPGAAPQLLIYNNDQRPSGIPDRFSGSKSGTSGSLVISGLQSEDEADYYCAAWDDSLSGNVFGTGTKVTVL
>SEQ ID NO:14 CD730047 VH
EVQLVQSGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVISYDGSIKYYADSVKGRFAISRDNAKNTLYLQMNSLRREDTAMYYCASGLTGVAGALGVWGRGTLVTVSS
>SEQ ID NO:15 CD730058 VL
QSVLTQPPSVSVSPGQTATITCSGDRLRNEFVSWYQQRPGQSPVVVIYQDIYRPSGIPDRFSGSKSGNTATLTISGPQTVDEADYYCQAWDSNTVVFGGGTKLTVL
>SEQ ID NO:16 CD730058 VH
QLQLQESGSGLVKPSQTLSLICAVSGGSITSGGNAWNWIRQSPGAGLEWIGYIFSNGATYYNPSLESRVTISADTSKNQFSLTLTSVTAADTAVYYCARGDFWTGKGVFDPWGQGTLVTVSS
>SEQ ID NO:17 10,3AA_HC
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSGGRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPASIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGPYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
>SEQ ID NO:18 10,3Nt_HC
GAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCTATAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGAACATACTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTGTATTACTGTGCGAGATTAGGGTATGGGCGGGTGGACGAGTGGGGCAGGGGAACCCTGGTCACCGTCTCGAGTGCGTCGACCAAGGGCCCATCCGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCcTGGAACTCAGGCGCtCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAGAGTTGAGCCCAAATCTTGTGACAAAACTCACACATgcCCacCGTGCCCAGCACCTGAATTCGAGGGGGGAcCGTCAGTCTTCCTCTTCCCCCCAAAACCCaaGgACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCTCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTcTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTATAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA
>SEQ ID NO:19 10,3AA_LC
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLDNLRLSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
>SEQ ID NO:20 10,3Nt-LC
CAGTCTGTGCTGACTCAGCCACCCTCAGCGTCTGGGACCCCCGGGCAGAGGGTCACCATCTCTTGTTCTGGAAGCCTCTCCAACATCGGAAGGAATCCTGTTAACTGGTATCAGCAGCTCCCAGGGACGGCCCCCAAACTCCTCATCTATCTTGATAATCTACGGCTAAGTGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGAACCTCAGCCTCCCTGGCCATCAGTGGGCTCCAGTCTGAGGATGAGGCTGATTATTACTGTGCAACATGGGATGACAGCCACCCCGGGTGGACGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCGGCGCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCCTGGAAGGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTACCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCA
>SEQ ID NO:21 10,3AA_VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSGGRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSS
>SEQ ID NO:22 10,3Nt_VH
GAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCTATAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGAACATACTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTGTATTACTGTGCGAGATTAGGGTATGGGCGGGTGGACGAGTGGGGCAGGGGAACCCTGGTCACCGTCTCGAGT
>SEQ ID NO:23 10,3AA_VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLDNLRLSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVL
>SEQ ID NO:24 10,3Nt-VL
CAGTCTGTGCTGACTCAGCCACCCTCAGCGTCTGGGACCCCCGGGCAGAGGGTCACCATCTCTTGTTCTGGAAGCCTCTCCAACATCGGAAGGAATCCTGTTAACTGGTATCAGCAGCTCCCAGGGACGGCCCCCAAACTCCTCATCTATCTTGATAATCTACGGCTAAGTGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGAACCTCAGCCTCCCTGGCCATCAGTGGGCTCCAGTCTGAGGATGAGGCTGATTATTACTGTGCAACATGGGATGACAGCCACCCCGGGTGGACGTTCGGCGGAGGGACCAAGCTGACCGTCCTA
>SEQ ID NO:25 CD7300010 исходный FW1-VL
LPVLTQPPSVSGTPGQRVTISC
>SEQ ID NO:26 CD7300010 GL FW1-VL
QSVLTQPPSASGTPGQRVTISC
>SEQ ID NO:27 CD7300010 исходный FW2-VL
WYKQVPGTAPKLLIY
>SEQ ID NO:28 CD7300010 GL FW2-VL
WYQQLPGTAPKLLIY
>SEQ ID NO:29 CD7300010 исходный/GL FW3-VL
GVPDRFSGSKSGTSASLAISGLQSEDEADYYC
>SEQ ID NO:30 CD7300010 исходный/GL FW4-VL (также CD370002 и 2С5)
FGGGTKLTVL
>SEQ ID NO:31 CD7300010 исходный/GL FW1-VH (также CD370002 и 2С5)
EVQLLESGGGLVQPGGSLRLSCAASGFTFS
>SEQ ID NO:32 CD7300010 исходный/GL FW2-VH (также CD370002 и 2С5)
WVRQAPGKGLEWVS
>SEQ ID NO:33 CD7300010 исходный/GL FW3-VH (также CD370002 и 2С5)
RFTISRDNSKNTLYLQMNSLRAEDTAVYYCAR
>SEQ ID NO:34 CD7300010 исходный/GL FW4-VH
WGRGTLVTVSS
>SEQ ID NO:35 CD7300010 CDR1-VH (также CD370002 и 2С5)
SYAMS
>SEQ ID NO:36 CD7300010 CDR1-VH
SYAYS
>SEQ ID NO:37 CDR2-VH (также CD370002)
AISGSGGSTYYADSVKG
>SEQ ID NO:38 CDR2-VH
LIWGSWGSTYYADSVKG
>SEQ ID NO:39 CDR2-VH
AISGSGGRTYYADSVKG
>SEQ ID NO:40 CDR2-VH
AISGSWGRTYYADSVKG
>SEQ ID NO:41 CDR3-VH
LGYSTIDY
>SEQ ID NO:42 CDR3-VH
LGYSTIDK
>SEQ ID NO:43 CDR3-VH
LGYSTIDM
>SEQ ID NO:44 CDR3-VH
LGYSTIDL
>SEQ ID NO:45 CDR3-VH
LGYGRVDE
>SEQ ID NO:46 CDR1-VL
SGSLSNIGRNPVN
>SEQ ID NO:47 CDR1-VL
SGSLSNIGRNEVN
>SEQ ID NO:48 CDR1-VL
SGSLSNIGRNDVN
>SEQ ID NO:49 CDR2-VL
LNNQRPS
>SEQ ID NO:50 CDR2-VL
LDNLRLG
>SEQ ID NO:51 CDR2-VL
LDNLRLS
>SEQ ID NO:52 CDR2-VL
LNNQRLG
>SEQ ID NO:53 CDR3-VL
ATWDDSLNGWL
>SEQ ID NO:54 CDR3-VL
ATWDDSLKGWL
>SEQ ID NO:55 CDR3-VL
ATWDDSLIGWL
>SEQ ID NO:56 CDR3-VL
ATWDDSHPGWT
>SEQ ID NO:57 CD730010-VL
LPVLTQPPSVSGTPGQRVTISCSGSLSNIGRNPVNWYKQVPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLNGWLFGGGTKLTVL
>SEQ ID NO:58 CD730010GL9-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLNGWLFGGGTKLTVL
>SEQ ID NO:59 P32E-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNEVNWYQQLPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLNGWLFGGGTKLTVL
>SEQ ID NO:60 C1-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNEVNWYQQLPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLKGWLFGGGTKLTVL
>SEQ ID NO:61 C2-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLDNLRLGGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLKGWLFGGGTKLTVL
>SEQ ID NO:62 D3-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLDNLRLSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLIGWLFGGGTKLTVL
>SEQ ID NO:63 G10-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNDVNWYQQLPGTAPKLLIYLNNQRLGGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLKGWLFGGGTKLTVL
>SEQ ID NO:64 HPT-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVL
>SEQ ID NO:65 GRVE-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSLNGWLFGGGTKLTVL
>SEQ ID NO:66 73combol (C1+GRVE+HPT)-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNEVNWYQQLPGTAPPCLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVL
>SEQ ID NO:67 73combo2 (C2+GRVE+HPT)-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLDNLRLGGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVL
>SEQ ID NO:68 73combo3(D3+GRVE+HPT)-VL [10.3]
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLDNLRLSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVL
>SEQ ID NO:69 73combo5 (G10+GRVE+HPT)-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNDVNWYQQLPGTAPKLLIYLNNQRLGGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVL
>SEQ ID NO:70 73combo6(GRVE+HPT)-VL
QSVLTQPPSASGTPGQRVTISCSGSLSNIGRNPVNWYQQLPGTAPKLLIYLNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCATWDDSHPGWTFGGGTKLTVL
>SEQ ID NO:71 CD730010-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDYWGRGTLVTVSS
>SEQ ID NO:72 CD730010GL9-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDYWGRGTLVTVSS
>SEQ ID NO:73 P32E-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDYWGRGTLVTVSS
>SEQ ID NO:74 C1-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDKWGRGTLVTVSS
>SEQ ID NO:75 C2-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSLIWGSWGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDMWGRGTLVTVSS
>SEQ ID NO:76 D3-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSGGRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTmLWGRGTLVTVSS
>SEQ ID NO:77 G10-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSWGRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDLWGRGTLVTVSS
>SEQ ID NO:78 HPT-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYSTIDYWGRGTLVTVSS
>SEQ ID NO:79 GRVE-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSS
>SEQ ID NO:80 73combo1 (C1+GRVE+HPT)-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSS
>SEQ ID NO:81 73combo2 (C2+GRVE+HPT)-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSLIWGSWGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSS
>SEQ ID NO:82 73combo3 (D3+GRVE+HPT)-VH [10.3]
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSGGRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSS
>SEQ ID NO:83 73combo5 (G10+GRVE+HPT)-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAYSWVRQAPGKGLEWVSAISGSWGRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSS
>SEQ ID NO:84 73combo6 (GRVE+HPT)-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLGYGRVDEWGRGTLVTVSS
>SEQ ID NO:85 CD730002VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:86 CD730002VL
QSVLTQPPSVSVSPGQTATITCSGDKVGDKYASWYQQKPGQSPVLVIYEDTKRPSRIPERFSGSNSGNTATLTISGTQALDEADYFCQAWDTSFWVFGGGTKLTVL
>SEQ ID NO:87 2C5VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKRRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:88 2C5VL
QSVLTQPPSVSVSPGQTASITCSGDKVGDKYASWYQQKPGQSPVLVIYEDTKRLSGIPERFSGSNSGNTATLTISGTQAMDEADYYCQAWDTSFWVFGGGTKLTVL
>SEQ ID NO:89 CD730002 исходный/2С5 FW4-VH
WGQGTMVTVSS
>SEQ ID NO:90 CD730002 исходный FW1-VL
QSVLTQPPSVSVSPGQTATITC
>SEQ ID NO:91 CD7300002 2C5 FW1-VL
QSVLTQPPSVSVSPGQTASITC
>SEQ ID NO:92 CD730002 исходный/2С5 FW2-VL
WYQQKPGQSPVLVIY
>SEQ ID NO:93 CD730002 исходный FW3-VL
RIPERFSGSNSGNTATLTISGTQALDEADYFC
>SEQ ID NO:94 CD730002/2C5 FW3-VL
GIPERFSGSNSGNTATLTISGTQAMDEADYYC
>SEQ ID NO:95 CD730002/2C5 CDR2-VH
AISGSGGSTYYADSVKR
>SEQ ID NO:96 CD730002 исходный/2С5 CDR3-VH
DKGYYWYMDV
>SEQ ID NO:97 CD730002 исходный/2С5 CDR1-VL
SGDKVGDKYAS
>SEQ ID NO:98 CD730002 исходный CDR2-VL
EDTKRPS
>SEQ ID NO:99 CD730002 2C5 CDR2-VL
EDTKRLS
>SEQ ID NO:100 CD730002 исходный/2С5 CDR3-VL
QAWDTSFWV
>SEQ ID NO:101
Phen0203-VH
EVQLVQSGGGVVQPGRSLRLSCAASGFRFSDFAMHWVRQAPGKGLEWVAGISYDGGNKYYADSVKGRFTISRDNSNNTLYLQMNSLRAEDTAVYYCAKDHGYSGYYGHLDYWGRGTLVTVSS
>SEQ ID NO:102 Phen0203-VL
QSVVTQPPSASGTPGQRVTISCSGSSSNIGSNTVNWYQQLPGTAPKLLIYSNNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCAAWDDSLNGRVFGTGTKLTVL
>SEQ ID NO:103 CD730004-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDRPNYYGASGSYYKQGGDHWGQGTMVTVss
>SEQ ID NO:104 CD730004-VL
NFMLTQPHSVSESPGQTVTISCTRSSGSIASKYVQWYQKRPGSSPTTVIYEDTQRPSGVPDRFSGSIDISSNSASLTISGLRTEDEADYYCQSYDSTNWVFGGGTKVTVL
>SEQ ID NO:105 CD730008-VH
EVQLLETGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGNYGNLDHWGKGTLVTVSS
>SEQ ID NO:106 CD730008-VL
QSVLTQPPSVSVSPGQTASITCSGDKVGDKYASWYQQKPGQSPVLVIYQDRKRPSGIPERFSGSNSGNTATLTISGTQPMDEADYYCQAWDSSHWVFGGGTKLTVL
>SEQ ID NO:107 CD730068-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSNYIMGWVRQAPGKGLEWVSSISSSGGATIYADSVKGRFTISRDNSKNTLYLQMNSLRAEDMAVYYCAKDHLGGHGMDVWGQGTTVTVSS
>SEQ ID NO:108 CD730068-VL
DIQMTQSPSSLSASVGDRVTITCRASQDISNYLAWFQQKPGKAPKSLIFAASSLESGVPSKFSGSGSGTDFTLTISSLQPEDSATYYCQQYNSYPLTFGGGTKVEIK
>SEQ ID NO:109 CD730069-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSEYQMGWVRQAPGKGLEWVSYIRSSGGQTIYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARRTYSSGWHIDYWGQGTLVTVSS
>SEQ ID NO:110 CD730069-VL
DIQMTQSPDSLSASVGDRVTITCRASQSISRYLNWYQQKPGKAPKLLIYGASSLQSGVPSRFSGSGSGTDFSLTISSLQLEDFATYYCQQSYRTPLTFGGGTKVEIQ
>SEQ ID NO:111 клон 2 SGMY-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:112 клон 2 SGMY-VL
QSVLTQPPSVSVSPGQTASITCSGDKVGDKYASWYQQKPGQSPVLVIYEDTKRPSGIPERFSGSNSGNTATLTISGTQAMDEADYYCQAWDTSFWVFGGGTKLTVL
>SEQ ID NO:113 CDRH1 Y32V-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSVAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:114 CDRH1 M34R-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYARSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:115 CDRH2 T57P-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSPYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:116 CDRH2 A60G-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYGDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:117 CDRH2 G65R-VH
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKRRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDKGYYWYMDVWGQGTMVTVSS
>SEQ ID NO:118 CDRL2 T52S-VL
QSVLTQPPSVSVSPGQTASITCSGDKVGDKYASWYQQKPGQSPVLVIYEDSKRPSGIPERFSGSNSGNTATLTISGTQAMDEADYYCQAWDTSFWVFGGGTKLTVLL
>SEQ ID NO:119 CDRL2 R54Y-VL
QSVLTQPPSVSVSPGQTASITCSGDKVGDKYASWYQQKPGQSPVLVIYEDTKYPSGIPERFSGSNSGNTATLTISGTQAMDEADYYCQAWDTSFWVFGGGTKLTVLVL
>SEQ ID NO:120 CDRL2 P55H-VL
QSVLTQPPSVSVSPGQTASITCSGDKVGDKYASWYQQKPGQSVLVIYEDTKRHSGIPERFSGSNSGNTATLTISGTQAMDEADYYCQAWDTSFWVFGGGTKLTVLVL
>SEQ ID NO:121 CDRL2 P55L-VL
QSVLTQPPSVSVSPGQTASITCSGDKVGDKYASWYQQKPGQSPVLVIYEDTKRLSGIPERFSGSNRGNTATLTISGTQAMDEADYYCQAWDTSFWVFGGGTKLTVLVL
>SEQ ID NO:122 2SGMY FW3-VL
GIPERFSGSNRGNTATLTISGTQAMDEADYYC
>SEQ ID NO:123 CDRH1 Y32V CDR1-VH
SVAMS
>SEQ ID NO:124 CDRH1 M34R CDR1-VH
SYARS
>SEQ ID NO:125 CDRH2 T57P CDR2-VH
AISGSGGSPYYADSVKG
>SEQ ID NO:126 CDRH2 A60G CDR2-VH
AISGSGGSTYYGDSVKG
>SEQ ID NO:127 CDRL2 T52S CDR2-VL
EDSKRPS
>SEQ ID NO:128 CDRL2 R54H CDR2-VL
EDTKYPS
>SEQ ID NO:129 CDRL2 P55H CDR2-VL
EDTKRLS
>SEQ ID NO:130 VL тремелимумаба из U.S. 6682736
PSSLSASVGDRVTITCRASQSINSYLDWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYSTPFTFGPGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYP REAKV
>SEQ ID NO:131 VH тремелимумаба из U.S. 6682736
GVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVIWYDGSNKYYADSVKGRFTISR DNSKNTLYLQMNSLRAEDTAVYYCARDPRGATLYYYYYGMDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVH
>SEQ ID NO:132 CDR1 VH тремелимумаба из U.S. 6682736
GFTFSSYGMH
>SEQ ID NO:133 CDR2 VH тремелимумаба из U.S. 6682736
VIWYDGSNKYYADSV
>SEQ ID NO:134 CDR3 VH тремелимумаба из U.S. 6682736
DPRGATLYYYYYGMDV
>SEQ ID NO:135 CDR1 VL тремелимумаба из U.S. 6682736
RASQSINSYLD
>SEQ ID NO:136 CDR2 VL тремелимумаба из U.S. 6682736
AASSLQS
>SEQ ID NO:137 CDR3 VL тремелимумаба из U.S. 6682736
QQYYSTPFT
>SEQ ID NO:138
US 2013/0034559_77, последовательность 77 из US 2013/0034559, организм: Homo sapiens
EIVLTQSPGTLSLSPGERATLSCRASQRVSSSYLAWYQQKPGQAPRLLIYDASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSLPWTFGQGTKVEIK
>SEQ ID NO:139
US 2013/0034559_72, последовательность 72 из US 2013/0034559, организм: Homo sapiens
EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWMSWVRQAPGKGLEWVANIKQDGSEKYYVDSVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCAREGGWFGELAFDYWGQGTLVTVSS
>SEQ ID NO:140 - VHCDR1
US 2013/0034559_73, последовательность 73 из US 2013/0034559, организм: Homo sapiens
RYWMS
>SEQ ID NO:141 - VH CDR2
US 2013/0034559_74, последовательность 74 из US 2013/0034559, организм: Homo sapiens
NIKQDGSEKYYVDSVKG
>SEQ ID NO:142 - VH CDR3
US 2013/0034559J75, последовательность 75 из US 2013/0034559, организм: Homo sapiens
EGGWFGELAFDY
>SEQ ID NO:143 - VL CDR1
US 2013/0034559_78, последовательность 78 из US 2013/0034559, организм: Homo sapiens
RASQRVSSSYLA
>SEQ ID NO:144 - VL CDR2
US 2013/0034559_79, последовательность 79 из US 2013/0034559, организм: Homo sapiens
DASSRAT
>SEQ ID NO:145 - VL CDR3
US 2013/0034559_80: последовательность 80 из US 2013/0034559, организм: Homo sapiens
QQYGSLPWT
<---
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MEDIMMUNE LIMITED
<120> СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ CD73, И ПУТИ ИХ ПРИМЕНЕНИЯ
<130> CD73-100WO1
<140>
<141>
<150> 62/188,999
<151> 2015-07-06
<150> 62/147,329
<151> 2015-04-14
<150> 62/077,486
<151> 2014-11-10
<160> 178
<170> PatentIn версия 3.5
<210> 1
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 1
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Thr Lys Arg Pro Ser Arg Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Leu
65 70 75 80
Asp Glu Ala Asp Tyr Phe Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 2
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 2
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 3
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 3
Leu Pro Val Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Lys Gln Val Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 4
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 4
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Tyr Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 5
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 5
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Ile Ala Ser Lys
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ser Pro Met Ala Val
35 40 45
Ile Tyr Lys Asp Asn Gln Arg Ser Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ala
85 90 95
Ser Asn Tyr Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 6
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 6
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly His Gly Leu Tyr Phe Asp Leu Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 7
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 7
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Arg Pro Asn Ile Gly Gly Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Ala Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Ser Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Tyr Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Asp Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Arg Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 8
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 8
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Ala Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Asn Ile Tyr Tyr Arg Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Met Ser Val Asp Met Ser Lys His Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Leu Asn Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Ser Leu Tyr Ser Gly Thr Tyr Val Phe Asp Tyr Trp Gly Arg
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 9
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 9
Gln Ser Val Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Ala Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Gly Ser Lys Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Thr Arg
85 90 95
Ser Thr Arg Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 10
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 10
Gly Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ala Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Thr Leu Ser Gly Ser Tyr Gly Tyr Phe Asp Tyr Trp Gly Arg Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 11
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 11
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Ser Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Gln Leu Leu
35 40 45
Ile Tyr Thr Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Ser Gly Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 12
<211> 125
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 12
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ser Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Asp Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Gly His Leu Leu Arg Ile Gly Asp Ile Phe Tyr Tyr Ser Leu
100 105 110
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 13
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 13
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Arg Leu Pro Gly Ala Ala Pro Gln Leu Leu
35 40 45
Ile Tyr Asn Asn Asp Gln Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Gly Ser Leu Val Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Ser Gly Asn Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 14
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 14
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ile Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Arg Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Ser Gly Leu Thr Gly Val Ala Gly Ala Leu Gly Val Trp Gly Arg
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 15
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 15
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Ser Gly Asp Arg Leu Arg Asn Glu Phe Val
20 25 30
Ser Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Val Val Ile Tyr
35 40 45
Gln Asp Ile Tyr Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Lys Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Pro Gln Thr Val
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Asn Thr Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 16
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 16
Gln Leu Gln Leu Gln Glu Ser Gly Ser Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Ile Cys Ala Val Ser Gly Gly Ser Ile Thr Ser Gly
20 25 30
Gly Asn Ala Trp Asn Trp Ile Arg Gln Ser Pro Gly Ala Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Phe Ser Asn Gly Ala Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Glu Ser Arg Val Thr Ile Ser Ala Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Thr Leu Thr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Gly Asp Phe Trp Thr Gly Lys Gly Val Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 17
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 17
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 18
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 18
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcct atagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag aacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagattaggg 300
tatgggcggg tggacgagtg gggcagggga accctggtca ccgtctcgag tgcgtcgacc 360
aagggcccat ccgtcttccc cctggcaccc tcctccaaga gcacctctgg gggcacagcg 420
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc ctggaactca 480
ggcgctctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac 540
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc 600
aacgtgaatc acaagcccag caacaccaag gtggacaaga gagttgagcc caaatcttgt 660
gacaaaactc acacatgccc accgtgccca gcacctgaat tcgagggggg accgtcagtc 720
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca 780
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac 840
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac 900
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag 960
tgcaaggtct ccaacaaagc cctcccagcc tccatcgaga aaaccatctc caaagccaaa 1020
gggcagcccc gagaaccaca ggtctacacc ctgcccccat cccgggagga gatgaccaag 1080
aaccaggtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 1140
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 1200
gacggctcct tcttcctcta tagcaagctc accgtggaca agagcaggtg gcagcagggg 1260
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc 1320
ctctccctgt ctccgggtaa a 1341
<210> 19
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 19
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asp Asn Leu Arg Leu Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 20
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<400> 20
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcctctc caacatcgga aggaatcctg ttaactggta tcagcagctc 120
ccagggacgg cccccaaact cctcatctat cttgataatc tacggctaag tggggtccct 180
gaccgattct ctggctccaa gtctggaacc tcagcctccc tggccatcag tgggctccag 240
tctgaggatg aggctgatta ttactgtgca acatgggatg acagccaccc cgggtggacg 300
ttcggcggag ggaccaagct gaccgtccta ggtcagccca aggcggcgcc ctcggtcact 360
ctgttcccgc cctcctctga ggagcttcaa gccaacaagg ccacactggt gtgtctcata 420
agtgacttct acccgggagc cgtgacagtg gcctggaagg cagatagcag ccccgtcaag 480
gcgggagtgg agaccaccac accctccaaa caaagcaaca acaagtacgc ggccagcagc 540
tacctgagcc tgacgcctga gcagtggaag tcccacagaa gctacagctg ccaggtcacg 600
catgaaggga gcaccgtgga gaagacagtg gcccctacag aatgttca 648
<210> 21
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 21
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 22
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<220>
<221> CDS
<222> (1)..(351)
<400> 22
gag gtg cag ctg ttg gag tct ggg gga ggc ttg gta cag cct ggg ggg 48
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
tcc ctg aga ctc tcc tgt gca gcc tct gga ttc acc ttt agc agc tat 96
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
gcc tat agc tgg gtc cgc cag gct cca ggg aag ggg ctg gag tgg gtc 144
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
tca gct att agt ggt agt ggt ggt aga aca tac tac gca gac tcc gtg 192
Ser Ala Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
aag ggc cgg ttc acc atc tcc aga gac aat tcc aag aac acg ctg tat 240
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
ctg caa atg aac agc ctg aga gcc gag gac acg gcc gtg tat tac tgt 288
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
gcg aga tta ggg tat ggg cgg gtg gac gag tgg ggc agg gga acc ctg 336
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
gtc acc gtc tcg agt 351
Val Thr Val Ser Ser
115
<210> 23
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 23
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asp Asn Leu Arg Leu Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 24
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полинуклеотид"
<220>
<221> CDS
<222> (1)..(330)
<400> 24
cag tct gtg ctg act cag cca ccc tca gcg tct ggg acc ccc ggg cag 48
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
agg gtc acc atc tct tgt tct gga agc ctc tcc aac atc gga agg aat 96
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
cct gtt aac tgg tat cag cag ctc cca ggg acg gcc ccc aaa ctc ctc 144
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
atc tat ctt gat aat cta cgg cta agt ggg gtc cct gac cga ttc tct 192
Ile Tyr Leu Asp Asn Leu Arg Leu Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
ggc tcc aag tct gga acc tca gcc tcc ctg gcc atc agt ggg ctc cag 240
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
tct gag gat gag gct gat tat tac tgt gca aca tgg gat gac agc cac 288
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
ccc ggg tgg acg ttc ggc gga ggg acc aag ctg acc gtc cta 330
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 25
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 25
Leu Pro Val Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys
20
<210> 26
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 26
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys
20
<210> 27
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 27
Trp Tyr Lys Gln Val Pro Gly Thr Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 28
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 28
Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 29
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 29
Gly Val Pro Asp Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser
1 5 10 15
Leu Ala Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 30
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 30
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 31
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 31
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 32
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 32
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 33
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 33
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 34
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 34
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 35
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 35
Ser Tyr Ala Met Ser
1 5
<210> 36
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 36
Ser Tyr Ala Tyr Ser
1 5
<210> 37
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 37
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 38
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 38
Leu Ile Trp Gly Ser Trp Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 39
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 39
Ala Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 40
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 40
Ala Ile Ser Gly Ser Trp Gly Arg Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 41
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 41
Leu Gly Tyr Ser Thr Ile Asp Tyr
1 5
<210> 42
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 42
Leu Gly Tyr Ser Thr Ile Asp Lys
1 5
<210> 43
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 43
Leu Gly Tyr Ser Thr Ile Asp Met
1 5
<210> 44
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 44
Leu Gly Tyr Ser Thr Ile Asp Leu
1 5
<210> 45
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 45
Leu Gly Tyr Gly Arg Val Asp Glu
1 5
<210> 46
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 46
Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn Pro Val Asn
1 5 10
<210> 47
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 47
Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn Glu Val Asn
1 5 10
<210> 48
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 48
Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn Asp Val Asn
1 5 10
<210> 49
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 49
Leu Asn Asn Gln Arg Pro Ser
1 5
<210> 50
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 50
Leu Asp Asn Leu Arg Leu Gly
1 5
<210> 51
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 51
Leu Asp Asn Leu Arg Leu Ser
1 5
<210> 52
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 52
Leu Asn Asn Gln Arg Leu Gly
1 5
<210> 53
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 53
Ala Thr Trp Asp Asp Ser Leu Asn Gly Trp Leu
1 5 10
<210> 54
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 54
Ala Thr Trp Asp Asp Ser Leu Lys Gly Trp Leu
1 5 10
<210> 55
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 55
Ala Thr Trp Asp Asp Ser Leu Ile Gly Trp Leu
1 5 10
<210> 56
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 56
Ala Thr Trp Asp Asp Ser His Pro Gly Trp Thr
1 5 10
<210> 57
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 57
Leu Pro Val Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Lys Gln Val Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 58
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 58
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 59
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 59
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Glu Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 60
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 60
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Glu Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Lys Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 61
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 61
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asp Asn Leu Arg Leu Gly Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Lys Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 62
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 62
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asp Asn Leu Arg Leu Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Ile Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 63
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 63
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Asp Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Leu Gly Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Lys Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 64
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 64
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 65
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 65
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 66
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 66
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Glu Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 67
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 67
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asp Asn Leu Arg Leu Gly Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 68
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 68
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asp Asn Leu Arg Leu Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 69
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 69
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Asp Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Leu Gly Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 70
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 70
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser His
85 90 95
Pro Gly Trp Thr Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 71
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 71
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Tyr Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 72
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 72
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Tyr Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 73
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 73
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Tyr Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 74
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 74
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Lys Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 75
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Leu Ile Trp Gly Ser Trp Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Met Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 76
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 76
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Leu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 77
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 77
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Trp Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Leu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 78
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 78
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Ser Thr Ile Asp Tyr Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 79
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 79
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 80
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 80
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 81
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 81
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Leu Ile Trp Gly Ser Trp Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 82
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 82
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 83
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 83
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Tyr Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Trp Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 84
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 84
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Glu Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 85
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 85
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 86
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 86
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Thr Lys Arg Pro Ser Arg Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Leu
65 70 75 80
Asp Glu Ala Asp Tyr Phe Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 87
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 87
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Arg Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 88
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 88
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Thr Lys Arg Leu Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 89
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 89
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
1 5 10
<210> 90
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 90
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys
20
<210> 91
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 91
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys
20
<210> 92
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 92
Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 93
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 93
Arg Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Thr Gln Ala Leu Asp Glu Ala Asp Tyr Phe Cys
20 25 30
<210> 94
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 94
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 95
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 95
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Arg
<210> 96
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 96
Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val
1 5 10
<210> 97
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 97
Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala Ser
1 5 10
<210> 98
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 98
Glu Asp Thr Lys Arg Pro Ser
1 5
<210> 99
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 99
Glu Asp Thr Lys Arg Leu Ser
1 5
<210> 100
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 100
Gln Ala Trp Asp Thr Ser Phe Trp Val
1 5
<210> 101
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 101
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Phe Ser Asp Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gly Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Asn Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp His Gly Tyr Ser Gly Tyr Tyr Gly His Leu Asp Tyr Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 102
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 102
Gln Ser Val Val Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Arg Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 103
<211> 128
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 103
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Pro Asn Tyr Tyr Gly Ala Ser Gly Ser Tyr Tyr Lys
100 105 110
Gln Gly Gly Asp His Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120 125
<210> 104
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 104
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Ile Ala Ser Lys
20 25 30
Tyr Val Gln Trp Tyr Gln Lys Arg Pro Gly Ser Ser Pro Thr Thr Val
35 40 45
Ile Tyr Glu Asp Thr Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ile Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Arg Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser
85 90 95
Thr Asn Trp Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 105
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 105
Glu Val Gln Leu Leu Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asn Tyr Gly Asn Leu Asp His Trp Gly Lys Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 106
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 106
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Arg Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Pro Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Ser His Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 107
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 107
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ile Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Gly Gly Ala Thr Ile Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp His Leu Gly Gly His Gly Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 108
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 108
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Ser Leu Ile
35 40 45
Phe Ala Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Lys Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ser Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 109
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 109
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Glu Tyr
20 25 30
Gln Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Arg Ser Ser Gly Gly Gln Thr Ile Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Ser Ser Gly Trp His Ile Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 110
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 110
Asp Ile Gln Met Thr Gln Ser Pro Asp Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Ser Leu Thr Ile Ser Ser Leu Gln Leu
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Arg Thr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Gln
100 105
<210> 111
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 111
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 112
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 112
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Thr Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 113
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 113
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Val
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 114
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 114
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Arg Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 115
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 115
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Pro Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 116
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 116
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 117
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 117
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Arg Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 118
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 118
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Leu
100 105
<210> 119
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 119
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Thr Lys Tyr Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Val Leu
100 105
<210> 120
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 120
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Val Leu Val Ile Tyr Glu
35 40 45
Asp Thr Lys Arg His Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn
50 55 60
Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp
65 70 75 80
Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Thr Ser Phe Trp Val Phe
85 90 95
Gly Gly Gly Thr Lys Leu Thr Val Leu Val Leu
100 105
<210> 121
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 121
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Thr Lys Arg Leu Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Arg Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Val Leu
100 105
<210> 122
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 122
Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Arg Gly Asn Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 123
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 123
Ser Val Ala Met Ser
1 5
<210> 124
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 124
Ser Tyr Ala Arg Ser
1 5
<210> 125
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 125
Ala Ile Ser Gly Ser Gly Gly Ser Pro Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 126
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 126
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Gly Asp Ser Val Lys
1 5 10 15
Gly
<210> 127
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 127
Glu Asp Ser Lys Arg Pro Ser
1 5
<210> 128
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 128
Glu Asp Thr Lys Tyr Pro Ser
1 5
<210> 129
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 129
Glu Asp Thr Lys Arg Leu Ser
1 5
<210> 130
<211> 139
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 130
Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
1 5 10 15
Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asp Trp Tyr Gln Gln Lys
20 25 30
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser Leu Gln
35 40 45
Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe
50 55 60
Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr
65 70 75 80
Cys Gln Gln Tyr Tyr Ser Thr Pro Phe Thr Phe Gly Pro Gly Thr Lys
85 90 95
Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
100 105 110
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
115 120 125
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
130 135
<210> 131
<211> 167
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 131
Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser
1 5 10 15
Gly Phe Thr Phe Ser Ser Tyr Gly Met His Trp Val Arg Gln Ala Pro
20 25 30
Gly Lys Gly Leu Glu Trp Val Ala Val Ile Trp Tyr Asp Gly Ser Asn
35 40 45
Lys Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
50 55 60
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
65 70 75 80
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Pro Arg Gly Ala Thr Leu
85 90 95
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His
165
<210> 132
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 132
Gly Phe Thr Phe Ser Ser Tyr Gly Met His
1 5 10
<210> 133
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 133
Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
1 5 10 15
<210> 134
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 134
Asp Pro Arg Gly Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val
1 5 10 15
<210> 135
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 135
Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asp
1 5 10
<210> 136
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 136
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 137
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 137
Gln Gln Tyr Tyr Ser Thr Pro Phe Thr
1 5
<210> 138
<211> 108
<212> БЕЛОК
<213> Homo sapiens
<400> 138
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 139
<211> 121
<212> БЕЛОК
<213> Homo sapiens
<400> 139
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 140
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 140
Arg Tyr Trp Met Ser
1 5
<210> 141
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 141
Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val Lys
1 5 10 15
Gly
<210> 142
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 142
Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr
1 5 10
<210> 143
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 143
Arg Ala Ser Gln Arg Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 144
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 144
Asp Ala Ser Ser Arg Ala Thr
1 5
<210> 145
<211> 9
<212> БЕЛОК
<213> Homo sapiens
<400> 145
Gln Gln Tyr Gly Ser Leu Pro Trp Thr
1 5
<210> 146
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(22)
<223> /примечание="Данная область может охватывать "LPVLTQPPSVSGTPGQRVTISC" или "QSVLTQPPSASGTPGQRVTISC"
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> /замена="Gln"
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> /замена="Ser"
<220>
<221> ВАРИАНТ
<222> (10)..(10)
<223> /замена="Ala"
<220>
<221> ВАРИАНТ
<222> (33)..(33)
<223> /замена="Glu" или "Asp"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (36)..(50)
<223> /примечание="Данная область может охватывать "WYKQVPGTAPKLLIY" или "WYQQLPGTAPKLLIY"
<220>
<221> ВАРИАНТ
<222> (38)..(38)
<223> /замена="Gln"
<220>
<221> ВАРИАНТ
<222> (40)..(40)
<223> /замена="Leu"
<220>
<221> ВАРИАНТ
<222> (52)..(52)
<223> /замена="Asp"
<220>
<221> ВАРИАНТ
<222> (54)..(54)
<223> /замена="Leu"
<220>
<221> ВАРИАНТ
<222> (56)..(56)
<223> /замена="Pro"
<220>
<221> ВАРИАНТ
<222> (57)..(57)
<223> /замена="Ser"
<220>
<221> ВАРИАНТ
<222> (96)..(96)
<223> /замена="His"
<220>
<221> ВАРИАНТ
<222> (97)..(97)
<223> /замена="Pro", или "Ile", или "Asn"
<220>
<221> ВАРИАНТ
<222> (100)..(100)
<223> /замена="Thr"
<220>
<221> другой_признак
<222> (1)..(110)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 146
Leu Pro Val Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn
20 25 30
Pro Val Asn Trp Tyr Lys Gln Val Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Leu Asn Asn Gln Arg Leu Gly Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Lys Gly Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 147
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<220>
<221> ВАРИАНТ
<222> (34)..(34)
<223> /замена="Tyr"
<220>
<221> ВАРИАНТ
<222> (50)..(50)
<223> /замена="Ala"
<220>
<221> ВАРИАНТ
<222> (52)..(52)
<223> /замена="Ser"
<220>
<221> ВАРИАНТ
<222> (55)..(55)
<223> /замена="Gly"
<220>
<221> ВАРИАНТ
<222> (57)..(57)
<223> /замена="Arg"
<220>
<221> ВАРИАНТ
<222> (102)..(102)
<223> /замена="Ser"
<220>
<221> ВАРИАНТ
<222> (103)..(103)
<223> /замена="Thr"
<220>
<221> ВАРИАНТ
<222> (104)..(104)
<223> /замена="Ile"
<220>
<221> ВАРИАНТ
<222> (106)..(106)
<223> /замена="Lys", или "Met", или "Leu", или "Glu"
<220>
<221> другой_признак
<222> (1)..(117)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 147
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Leu Ile Trp Gly Ser Trp Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Tyr Gly Arg Val Asp Tyr Trp Gly Arg Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 148
<211> 574
<212> БЕЛОК
<213> Homo sapiens
<400> 148
Met Cys Pro Arg Ala Ala Arg Ala Pro Ala Thr Leu Leu Leu Ala Leu
1 5 10 15
Gly Ala Val Leu Trp Pro Ala Ala Gly Ala Trp Glu Leu Thr Ile Leu
20 25 30
His Thr Asn Asp Val His Ser Arg Leu Glu Gln Thr Ser Glu Asp Ser
35 40 45
Ser Lys Cys Val Asn Ala Ser Arg Cys Met Gly Gly Val Ala Arg Leu
50 55 60
Phe Thr Lys Val Gln Gln Ile Arg Arg Ala Glu Pro Asn Val Leu Leu
65 70 75 80
Leu Asp Ala Gly Asp Gln Tyr Gln Gly Thr Ile Trp Phe Thr Val Tyr
85 90 95
Lys Gly Ala Glu Val Ala His Phe Met Asn Ala Leu Arg Tyr Asp Ala
100 105 110
Met Ala Leu Gly Asn His Glu Phe Asp Asn Gly Val Glu Gly Leu Ile
115 120 125
Glu Pro Leu Leu Lys Glu Ala Lys Phe Pro Ile Leu Ser Ala Asn Ile
130 135 140
Lys Ala Lys Gly Pro Leu Ala Ser Gln Ile Ser Gly Leu Tyr Leu Pro
145 150 155 160
Tyr Lys Val Leu Pro Val Gly Asp Glu Val Val Gly Ile Val Gly Tyr
165 170 175
Thr Ser Lys Glu Thr Pro Phe Leu Ser Asn Pro Gly Thr Asn Leu Val
180 185 190
Phe Glu Asp Glu Ile Thr Ala Leu Gln Pro Glu Val Asp Lys Leu Lys
195 200 205
Thr Leu Asn Val Asn Lys Ile Ile Ala Leu Gly His Ser Gly Phe Glu
210 215 220
Met Asp Lys Leu Ile Ala Gln Lys Val Arg Gly Val Asp Val Val Val
225 230 235 240
Gly Gly His Ser Asn Thr Phe Leu Tyr Thr Gly Asn Pro Pro Ser Lys
245 250 255
Glu Val Pro Ala Gly Lys Tyr Pro Phe Ile Val Thr Ser Asp Asp Gly
260 265 270
Arg Lys Val Pro Val Val Gln Ala Tyr Ala Phe Gly Lys Tyr Leu Gly
275 280 285
Tyr Leu Lys Ile Glu Phe Asp Glu Arg Gly Asn Val Ile Ser Ser His
290 295 300
Gly Asn Pro Ile Leu Leu Asn Ser Ser Ile Pro Glu Asp Pro Ser Ile
305 310 315 320
Lys Ala Asp Ile Asn Lys Trp Arg Ile Lys Leu Asp Asn Tyr Ser Thr
325 330 335
Gln Glu Leu Gly Lys Thr Ile Val Tyr Leu Asp Gly Ser Ser Gln Ser
340 345 350
Cys Arg Phe Arg Glu Cys Asn Met Gly Asn Leu Ile Cys Asp Ala Met
355 360 365
Ile Asn Asn Asn Leu Arg His Thr Asp Glu Met Phe Trp Asn His Val
370 375 380
Ser Met Cys Ile Leu Asn Gly Gly Gly Ile Arg Ser Pro Ile Asp Glu
385 390 395 400
Arg Asn Asn Gly Thr Ile Thr Trp Glu Asn Leu Ala Ala Val Leu Pro
405 410 415
Phe Gly Gly Thr Phe Asp Leu Val Gln Leu Lys Gly Ser Thr Leu Lys
420 425 430
Lys Ala Phe Glu His Ser Val His Arg Tyr Gly Gln Ser Thr Gly Glu
435 440 445
Phe Leu Gln Val Gly Gly Ile His Val Val Tyr Asp Leu Ser Arg Lys
450 455 460
Pro Gly Asp Arg Val Val Lys Leu Asp Val Leu Cys Thr Lys Cys Arg
465 470 475 480
Val Pro Ser Tyr Asp Pro Leu Lys Met Asp Glu Val Tyr Lys Val Ile
485 490 495
Leu Pro Asn Phe Leu Ala Asn Gly Gly Asp Gly Phe Gln Met Ile Lys
500 505 510
Asp Glu Leu Leu Arg His Asp Ser Gly Asp Gln Asp Ile Asn Val Val
515 520 525
Ser Thr Tyr Ile Ser Lys Met Lys Val Ile Tyr Pro Ala Val Glu Gly
530 535 540
Arg Ile Lys Phe Ser Thr Gly Ser His Cys His Gly Ser Phe Ser Leu
545 550 555 560
Ile Phe Leu Ser Leu Trp Ala Val Ile Phe Val Leu Tyr Gln
565 570
<210> 149
<211> 223
<212> БЕЛОК
<213> Homo sapiens
<400> 149
Met Ala Cys Leu Gly Phe Gln Arg His Lys Ala Gln Leu Asn Leu Ala
1 5 10 15
Thr Arg Thr Trp Pro Cys Thr Leu Leu Phe Phe Leu Leu Phe Ile Pro
20 25 30
Val Phe Cys Lys Ala Met His Val Ala Gln Pro Ala Val Val Leu Ala
35 40 45
Ser Ser Arg Gly Ile Ala Ser Phe Val Cys Glu Tyr Ala Ser Pro Gly
50 55 60
Lys Ala Thr Glu Val Arg Val Thr Val Leu Arg Gln Ala Asp Ser Gln
65 70 75 80
Val Thr Glu Val Cys Ala Ala Thr Tyr Met Met Gly Asn Glu Leu Thr
85 90 95
Phe Leu Asp Asp Ser Ile Cys Thr Gly Thr Ser Ser Gly Asn Gln Val
100 105 110
Asn Leu Thr Ile Gln Gly Leu Arg Ala Met Asp Thr Gly Leu Tyr Ile
115 120 125
Cys Lys Val Glu Leu Met Tyr Pro Pro Pro Tyr Tyr Leu Gly Ile Gly
130 135 140
Asn Gly Thr Gln Ile Tyr Val Ile Asp Pro Glu Pro Cys Pro Asp Ser
145 150 155 160
Asp Phe Leu Leu Trp Ile Leu Ala Ala Val Ser Ser Gly Leu Phe Phe
165 170 175
Tyr Ser Phe Leu Leu Thr Ala Val Ser Leu Ser Lys Met Leu Lys Lys
180 185 190
Arg Ser Pro Leu Thr Thr Gly Val Tyr Val Lys Met Pro Pro Thr Glu
195 200 205
Pro Glu Cys Glu Lys Gln Phe Gln Pro Tyr Phe Ile Pro Ile Asn
210 215 220
<210> 150
<211> 288
<212> БЕЛОК
<213> Homo sapiens
<400> 150
Met Gln Ile Pro Gln Ala Pro Trp Pro Val Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Arg Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp
35 40 45
Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg
85 90 95
Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg
100 105 110
Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val
130 135 140
Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro
145 150 155 160
Arg Pro Ala Gly Gln Phe Gln Thr Leu Val Val Gly Val Val Gly Gly
165 170 175
Leu Leu Gly Ser Leu Val Leu Leu Val Trp Val Leu Ala Val Ile Cys
180 185 190
Ser Arg Ala Ala Arg Gly Thr Ile Gly Ala Arg Arg Thr Gly Gln Pro
195 200 205
Leu Lys Glu Asp Pro Ser Ala Val Pro Val Phe Ser Val Asp Tyr Gly
210 215 220
Glu Leu Asp Phe Gln Trp Arg Glu Lys Thr Pro Glu Pro Pro Val Pro
225 230 235 240
Cys Val Pro Glu Gln Thr Glu Tyr Ala Thr Ile Val Phe Pro Ser Gly
245 250 255
Met Gly Thr Ser Ser Pro Ala Arg Arg Gly Ser Ala Asp Gly Pro Arg
260 265 270
Ser Ala Gln Pro Leu Arg Pro Glu Asp Gly His Cys Ser Trp Pro Leu
275 280 285
<210> 151
<211> 176
<212> БЕЛОК
<213> Homo sapiens
<400> 151
Met Arg Ile Phe Ala Val Phe Ile Phe Met Thr Tyr Trp His Leu Leu
1 5 10 15
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
20 25 30
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
35 40 45
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
50 55 60
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
65 70 75 80
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
85 90 95
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
100 105 110
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Thr His Leu Val
115 120 125
Ile Leu Gly Ala Ile Leu Leu Cys Leu Gly Val Ala Leu Thr Phe Ile
130 135 140
Phe Arg Leu Arg Lys Gly Arg Met Met Asp Val Lys Lys Cys Gly Ile
145 150 155 160
Gln Asp Thr Asn Ser Lys Lys Gln Ser Asp Thr His Leu Glu Glu Thr
165 170 175
<210> 152
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (11)..(11)
<223> /замена="Glu" или "Asp"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(13)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 152
Ser Gly Ser Leu Ser Asn Ile Gly Arg Asn Pro Val Asn
1 5 10
<210> 153
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> /замена="Asp"
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> /замена="Leu"
<220>
<221> ВАРИАНТ
<222> (6)..(6)
<223> /замена="Pro"
<220>
<221> ВАРИАНТ
<222> (7)..(7)
<223> /замена="Ser"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(7)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 153
Leu Asn Asn Gln Arg Leu Gly
1 5
<210> 154
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (7)..(7)
<223> /замена="His"
<220>
<221> ВАРИАНТ
<222> (8)..(8)
<223> /замена="Pro", или "Ile", или "Asn"
<220>
<221> ВАРИАНТ
<222> (11)..(11)
<223> /замена="Thr"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(11)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 154
Ala Thr Trp Asp Asp Ser Leu Lys Gly Trp Leu
1 5 10
<210> 155
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> /замена="Tyr"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(5)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 155
Ser Tyr Ala Met Ser
1 5
<210> 156
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> /замена="Ala"
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> /замена="Ser"
<220>
<221> ВАРИАНТ
<222> (6)..(6)
<223> /замена="Gly"
<220>
<221> ВАРИАНТ
<222> (8)..(8)
<223> /замена="Arg"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(17)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 156
Leu Ile Trp Gly Ser Trp Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 157
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> /замена="Ser"
<220>
<221> ВАРИАНТ
<222> (5)..(5)
<223> /замена="Thr"
<220>
<221> ВАРИАНТ
<222> (6)..(6)
<223> /замена="Ile"
<220>
<221> ВАРИАНТ
<222> (8)..(8)
<223> /замена="Lys", или "Met", или "Leu", или "Glu"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(8)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 157
Leu Gly Tyr Gly Arg Val Asp Tyr
1 5
<210> 158
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (3)..(3)
<223> /замена="Thr"
<220>
<221> ВАРИАНТ
<222> (5)..(5)
<223> /замена="Tyr"
<220>
<221> ВАРИАНТ
<222> (6)..(6)
<223> /замена="Pro" или "Leu"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(7)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 158
Glu Asp Ser Lys Arg His Ser
1 5
<210> 159
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> /замена="Val"
<220>
<221> ВАРИАНТ
<222> (4)..(4)
<223> /замена="Arg"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(5)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 159
Ser Tyr Ala Met Ser
1 5
<210> 160
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (9)..(9)
<223> /замена="Pro"
<220>
<221> ВАРИАНТ
<222> (12)..(12)
<223> /замена="Gly"
<220>
<221> ВАРИАНТ
<222> (17)..(17)
<223> /замена="Arg"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(17)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 160
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 161
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 161
Asp Lys Gly Tyr Tyr Trp Tyr Met
1 5
<210> 162
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(22)
<223> /примечание="Данная область может охватывать "QSVLTQPPSVSVSPGQTATITC" или "QSVLTQPPSVSVSPGQTASITC"
<220>
<221> ВАРИАНТ
<222> (19)..(19)
<223> /замена="Ser"
<220>
<221> ВАРИАНТ
<222> (51)..(51)
<223> /замена="Leu"
<220>
<221> ВАРИАНТ
<222> (53)..(53)
<223> /замена="Tyr"
<220>
<221> ВАРИАНТ
<222> (54)..(54)
<223> /замена="Pro" или "Leu"
<220>
<221> ВАРИАНТ
<222> (56)..(56)
<223> /замена="Gly"
<220>
<221> ВАРИАНТ
<222> (66)..(66)
<223> /замена="Arg"
<220>
<221> ВАРИАНТ
<222> (80)..(80)
<223> /замена="Met"
<220>
<221> ВАРИАНТ
<222> (86)..(86)
<223> /замена="Tyr"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (56)..(87)
<223> /примечание="Данная область может охватывать "RIPERFSGSNSGNTATLTISGTQALDEADYFC", "GIPERFSGSNSGNTATLTISGTQAMDEADYYC" или "GIPERFSGSNRGNTATLTISGTQAMDEADYYC"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(106)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 162
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Ser Gly Asp Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Pro Lys Arg His Ser Arg Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Leu
65 70 75 80
Asp Glu Ala Asp Tyr Phe Cys Gln Ala Trp Asp Thr Ser Phe Trp Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 163
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<220>
<221> ВАРИАНТ
<222> (32)..(32)
<223> /замена="Val"
<220>
<221> ВАРИАНТ
<222> (34)..(34)
<223> /замена="Arg"
<220>
<221> ВАРИАНТ
<222> (58)..(58)
<223> /замена="Pro"
<220>
<221> ВАРИАНТ
<222> (61)..(61)
<223> /замена="Gly"
<220>
<221> ВАРИАНТ
<222> (66)..(66)
<223> /замена="Arg"
<220>
<221> ДРУГОЙ_ПРИЗНАК
<222> (1)..(117)
<223> /примечание="Остатки вариантов, приведенные в последовательности, не имеют предпочтения по отношению к тем, которые указаны в аннотациях для положений вариантов"
<400> 163
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Tyr Tyr Trp Tyr Met Trp Gly Gln Gly Thr Met
100 105 110
Val Thr Val Ser Ser
115
<210> 164
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетическая метка 6xHis"
<400> 164
His His His His His His
1 5
<210> 165
<211> 115
<212> БЕЛОК
<213> Homo sapiens
<400> 165
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Tyr Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 166
<211> 108
<212> БЕЛОК
<213> Homo sapiens
<400> 166
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 167
<211> 526
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 167
Trp Glu Leu Thr Ile Leu His Thr Asn Asp Val His Ser Arg Leu Glu
1 5 10 15
Gln Thr Ser Glu Asp Ser Ser Lys Cys Val Asn Ala Ser Arg Cys Met
20 25 30
Gly Gly Val Ala Arg Leu Phe Thr Lys Val Gln Gln Ile Arg Arg Ala
35 40 45
Glu Pro Asn Val Leu Leu Leu Asp Ala Gly Asp Gln Tyr Gln Gly Thr
50 55 60
Ile Trp Phe Thr Val Tyr Lys Gly Ala Glu Val Ala His Phe Met Asn
65 70 75 80
Ala Leu Arg Tyr Asp Ala Met Ala Leu Gly Asn His Glu Phe Asp Asn
85 90 95
Gly Val Glu Gly Leu Ile Glu Pro Leu Leu Lys Glu Ala Lys Phe Pro
100 105 110
Ile Leu Ser Ala Asn Ile Lys Ala Lys Gly Pro Leu Ala Ser Gln Ile
115 120 125
Ser Gly Leu Tyr Leu Pro Tyr Lys Val Leu Pro Val Gly Asp Glu Val
130 135 140
Val Gly Ile Val Gly Tyr Thr Ser Lys Glu Thr Pro Phe Leu Ser Asn
145 150 155 160
Pro Gly Thr Asn Leu Val Phe Glu Asp Glu Ile Thr Ala Leu Gln Pro
165 170 175
Glu Val Asp Lys Leu Lys Thr Leu Asn Val Asn Lys Ile Ile Ala Leu
180 185 190
Gly His Ser Gly Phe Glu Met Asp Lys Leu Ile Ala Gln Lys Val Arg
195 200 205
Gly Val Asp Val Val Val Gly Gly His Ser Asn Thr Phe Leu Tyr Thr
210 215 220
Gly Asn Pro Pro Ser Lys Glu Val Pro Ala Gly Lys Tyr Pro Phe Ile
225 230 235 240
Val Thr Ser Asp Asp Gly Arg Lys Val Pro Val Val Gln Ala Tyr Ala
245 250 255
Phe Gly Lys Tyr Leu Gly Tyr Leu Lys Ile Glu Phe Asp Glu Arg Gly
260 265 270
Asn Val Ile Ser Ser His Gly Asn Pro Ile Leu Leu Asn Ser Ser Ile
275 280 285
Pro Glu Asp Pro Ser Ile Lys Ala Asp Ile Asn Lys Trp Arg Ile Lys
290 295 300
Leu Asp Asn Tyr Ser Thr Gln Glu Leu Gly Lys Thr Ile Val Tyr Leu
305 310 315 320
Asp Gly Ser Ser Gln Ser Cys Arg Phe Arg Glu Cys Asn Met Gly Asn
325 330 335
Leu Ile Cys Asp Ala Met Ile Asn Asn Asn Leu Arg His Ala Asp Glu
340 345 350
Met Phe Trp Asn His Val Ser Met Cys Ile Leu Asn Gly Gly Gly Ile
355 360 365
Arg Ser Pro Ile Asp Glu Arg Asn Asn Gly Thr Ile Thr Trp Glu Asn
370 375 380
Leu Ala Ala Val Leu Pro Phe Gly Gly Thr Phe Asp Leu Val Gln Leu
385 390 395 400
Lys Gly Ser Thr Leu Lys Lys Ala Phe Glu His Ser Val His Arg Tyr
405 410 415
Gly Gln Ser Thr Gly Glu Phe Leu Gln Val Gly Gly Ile His Val Val
420 425 430
Tyr Asp Leu Ser Arg Lys Pro Gly Asp Arg Val Val Lys Leu Asp Val
435 440 445
Leu Cys Thr Lys Cys Arg Val Pro Ser Tyr Asp Pro Leu Lys Met Asp
450 455 460
Glu Val Tyr Lys Val Ile Leu Pro Asn Phe Leu Ala Asn Gly Gly Asp
465 470 475 480
Gly Phe Gln Met Ile Lys Asp Glu Leu Leu Arg His Asp Ser Gly Asp
485 490 495
Gln Gly Ile Asn Val Val Ser Thr Tyr Ile Ser Lys Met Lys Val Ile
500 505 510
Tyr Pro Ala Val Glu Gly Arg Ile Lys Phe Ser Thr Gly Ser
515 520 525
<210> 168
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 168
Ser Ala Asn Ile Lys Ala Lys Gly Pro Leu Ala Ser Gln Ile Ser Gly
1 5 10 15
Leu
<210> 169
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 169
Val Leu Pro Val Lys Val Leu Pro Val Gly Asp Glu
1 5 10
<210> 170
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 170
Leu Pro Tyr Lys Val Leu Pro Val Gly Asp
1 5 10
<210> 171
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 171
Glu Val Val Gly Ile Val Gly
1 5
<210> 172
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 172
Ala Leu Gln Pro Glu Val Asp Lys Leu
1 5
<210> 173
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 173
Ala Leu Gln Pro Glu Val Asp Lys Leu Lys Thr Leu Asn Val
1 5 10
<210> 174
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 174
Lys Leu Lys Thr Leu Asn Val Asn Lys Ile Ile Ala
1 5 10
<210> 175
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 175
Asn Lys Ile Ile Ala Leu Gly His Ser Gly Phe
1 5 10
<210> 176
<211> 523
<212> БЕЛОК
<213> Homo sapiens
<400> 176
Trp Glu Leu Thr Ile Leu His Thr Asn Asp Val His Ser Arg Leu Glu
1 5 10 15
Gln Thr Ser Glu Asp Ser Ser Lys Cys Val Asn Ala Ser Arg Cys Met
20 25 30
Gly Gly Val Ala Arg Leu Phe Thr Lys Val Gln Gln Ile Arg Arg Ala
35 40 45
Glu Pro Asn Val Leu Leu Leu Asp Ala Gly Asp Gln Tyr Gln Gly Thr
50 55 60
Ile Trp Phe Thr Val Tyr Lys Gly Ala Glu Val Ala His Phe Met Asn
65 70 75 80
Ala Leu Arg Tyr Asp Ala Met Ala Leu Gly Asn His Glu Phe Asp Asn
85 90 95
Gly Val Glu Gly Leu Ile Glu Pro Leu Leu Lys Glu Ala Lys Phe Pro
100 105 110
Ile Leu Ser Ala Asn Ile Lys Ala Lys Gly Pro Leu Ala Ser Gln Ile
115 120 125
Ser Gly Leu Tyr Leu Pro Tyr Lys Val Leu Pro Val Gly Asp Glu Val
130 135 140
Val Gly Ile Val Gly Tyr Thr Ser Lys Glu Thr Pro Phe Leu Ser Asn
145 150 155 160
Pro Gly Thr Asn Leu Val Phe Glu Asp Glu Ile Thr Ala Leu Gln Pro
165 170 175
Glu Val Asp Lys Leu Lys Thr Leu Asn Val Asn Lys Ile Ile Ala Leu
180 185 190
Gly His Ser Gly Phe Glu Met Asp Lys Leu Ile Ala Gln Lys Val Arg
195 200 205
Gly Val Asp Val Val Val Gly Gly His Ser Asn Thr Phe Leu Tyr Thr
210 215 220
Gly Asn Pro Pro Ser Lys Glu Val Pro Ala Gly Lys Tyr Pro Phe Ile
225 230 235 240
Val Thr Ser Asp Asp Gly Arg Lys Val Pro Val Val Gln Ala Tyr Ala
245 250 255
Phe Gly Lys Tyr Leu Gly Tyr Leu Lys Ile Glu Phe Asp Glu Arg Gly
260 265 270
Asn Val Ile Ser Ser His Gly Asn Pro Ile Leu Leu Asn Ser Ser Ile
275 280 285
Pro Glu Asp Pro Ser Ile Lys Ala Asp Ile Asn Lys Trp Arg Ile Lys
290 295 300
Leu Asp Asn Tyr Ser Thr Gln Glu Leu Gly Lys Thr Ile Val Tyr Leu
305 310 315 320
Asp Gly Ser Ser Gln Ser Cys Arg Phe Arg Glu Cys Asn Met Gly Asn
325 330 335
Leu Ile Cys Asp Ala Met Ile Asn Asn Asn Leu Arg His Ala Asp Glu
340 345 350
Met Phe Trp Asn His Val Ser Met Cys Ile Leu Asn Gly Gly Gly Ile
355 360 365
Arg Ser Pro Ile Asp Glu Arg Asn Asn Gly Thr Ile Thr Trp Glu Asn
370 375 380
Leu Ala Ala Val Leu Pro Phe Gly Gly Thr Phe Asp Leu Val Gln Leu
385 390 395 400
Lys Gly Ser Thr Leu Lys Lys Ala Phe Glu His Ser Val His Arg Tyr
405 410 415
Gly Gln Ser Thr Gly Glu Phe Leu Gln Val Gly Gly Ile His Val Val
420 425 430
Tyr Asp Leu Ser Arg Lys Pro Gly Asp Arg Val Val Lys Leu Asp Val
435 440 445
Leu Cys Thr Lys Cys Arg Val Pro Ser Tyr Asp Pro Leu Lys Met Asp
450 455 460
Glu Val Tyr Lys Val Ile Leu Pro Asn Phe Leu Ala Asn Gly Gly Asp
465 470 475 480
Gly Phe Gln Met Ile Lys Asp Glu Leu Leu Arg His Asp Ser Gly Asp
485 490 495
Gln Asp Ile Asn Val Val Ser Thr Tyr Ile Ser Lys Met Lys Val Ile
500 505 510
Tyr Pro Ala Val Glu Gly Arg Ile Lys Phe Ser
515 520
<210> 177
<211> 519
<212> БЕЛОК
<213> Gallus gallus
<400> 177
Leu Arg Leu Arg Leu Leu His Thr Asn Asp Val His Ala His Val Glu
1 5 10 15
Ala Arg Gly Cys Ala Glu Gly Pro Arg Gly Cys Phe Gly Gly Val Ala
20 25 30
Arg Arg Ala Ala Arg Val Ala Ala Glu Arg Ala Ala Gln Arg Asn Val
35 40 45
Leu Leu Leu Asp Ala Gly Asp Gln Tyr Gln Gly Ser Val Trp Phe Ser
50 55 60
Arg Phe Lys Gly Gln Glu Ala Val His Phe Met Asn Leu Leu Arg Tyr
65 70 75 80
Asp Ala Met Ala Leu Gly Asn His Glu Phe Asp Glu Gly Val Arg Gly
85 90 95
Leu Leu Asn Pro Leu Leu Arg Asn Ala Ser Phe Ala Ile Leu Ser Ala
100 105 110
Asn Ile Lys Gly Lys Thr Pro Leu Gly Asn Gln Met Met Lys Tyr Val
115 120 125
His Pro Tyr Lys Ile Leu His Ile Asp Ser Glu Lys Ile Gly Ile Val
130 135 140
Gly Tyr Thr Thr Gln Glu Thr Ser Phe Leu Ser Gln Pro Gly Asn Asp
145 150 155 160
Val Ile Phe Glu Asp Glu Ile Glu Ala Leu Gln Val Gln Val Asn Lys
165 170 175
Leu Thr Ala Met Gly Val Asn Lys Ile Ile Ala Leu Gly His Ser Gly
180 185 190
Phe Thr Val Asp Ile Asn Ile Ala Gln Lys Val Lys Gly Val Asp Val
195 200 205
Val Ile Gly Gly His Thr Asn Thr Phe Leu Tyr Thr Gly Thr Pro Pro
210 215 220
Ser Thr Glu Gln Pro Ala Gly Pro Tyr Pro Phe Met Val Asp Ser Asp
225 230 235 240
Asp Gly Arg Lys Val Pro Val Val Gln Ala Tyr Ala Tyr Gly Lys Tyr
245 250 255
Leu Gly Tyr Leu Asn Val Thr Phe Asp Glu Lys Gly Asn Val Val Glu
260 265 270
Ala Val Gly Asn Pro Ile Leu Leu Asp Ser Ser Val Pro Glu Asp Glu
275 280 285
Gln Ile Lys Glu Glu Val Glu Lys Trp Arg Lys Asn Leu Gly Asn Tyr
290 295 300
Ser Lys Glu Ile Gly Thr Thr Ser Val Tyr Leu Asn Gly Thr Ser Glu
305 310 315 320
Ala Cys Arg Phe Gln Glu Cys Asn Met Gly Asn Leu Leu Cys Asp Ala
325 330 335
Met Leu Tyr Glu Asn Val Arg Arg Pro Asp Arg Lys Ser Trp Asn His
340 345 350
Val Ser Leu Cys Ile Leu Asn Gly Gly Gly Ile Arg Ala Ser Ile Asp
355 360 365
Glu Arg Asn Ala Asn Gly Ser Ile Thr Met Glu Asp Leu Leu Ser Val
370 375 380
Leu Pro Phe Gly Gly Arg Phe Asp Leu Val Thr Leu Lys Gly Ser Thr
385 390 395 400
Leu Lys Glu Ala Phe Glu His Ser Val Arg Arg Tyr Gly Arg Gly Thr
405 410 415
Gly Glu Leu Leu Gln Val Gly Gly Ile His Val Val Tyr Asp Leu Ser
420 425 430
Arg Ala Pro Gly His Arg Ala Val Ser Ile Glu Val Leu Cys Thr Ala
435 440 445
Cys Arg Val Pro Ala Tyr Val Pro Leu Glu Met Asp Glu Val Tyr Asn
450 455 460
Val Thr Leu Pro Ser Tyr Met Leu Phe Gly Gly Asp Gly Tyr Tyr Met
465 470 475 480
Leu Arg Asp Asn His Ile Thr Tyr Ser Lys Gly Glu Pro Asp Ile Glu
485 490 495
Val Val Ser Arg Tyr Leu Asp Arg Met Lys Arg Val Tyr Pro Ala Val
500 505 510
Glu Gly Arg Ile Lys Phe Ser
515
<210> 178
<211> 60
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 178
Ser Gly Leu Tyr Leu Pro Tyr Lys Val Leu Pro Val Gly Asp Glu Val
1 5 10 15
Val Gly Ile Val Gly Tyr Thr Ser Lys Glu Thr Pro Phe Leu Ser Asn
20 25 30
Pro Gly Thr Asn Leu Val Phe Glu Asp Glu Ile Thr Ala Leu Gln Pro
35 40 45
Glu Val Asp Lys Leu Lys Thr Leu Asn Val Asn Lys
50 55 60
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО К CD73 ЧЕЛОВЕКА | 2017 |
|
RU2754058C2 |
| АНТИТЕЛА, БЛОКИРУЮЩИЕ CD73 | 2020 |
|
RU2819204C2 |
| АНТИТЕЛА К ENTPD2, ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ И СПОСОБЫ ПРИМЕНЕНИЯ АНТИТЕЛ И ВИДОВ КОМБИНИРОВАННОЙ ТЕРАПИИ | 2019 |
|
RU2790991C2 |
| СПОСОБЫ И КОМПОЗИЦИИ НА ОСНОВЕ АНТИТЕЛ К CD73 И ИХ ВАРИАНТОВ | 2020 |
|
RU2841372C2 |
| МОЛЕКУЛЫ АНТИТЕЛ К CD73 И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2791192C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2811477C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| БИФУНКЦИОНАЛЬНЫЕ БЕЛКИ, КОМБИНИРУЮЩИЕ БЛОКАДУ КОНТРОЛЬНОЙ ТОЧКИ, ДЛЯ ТАРГЕТНОЙ ТЕРАПИИ | 2019 |
|
RU2756899C1 |


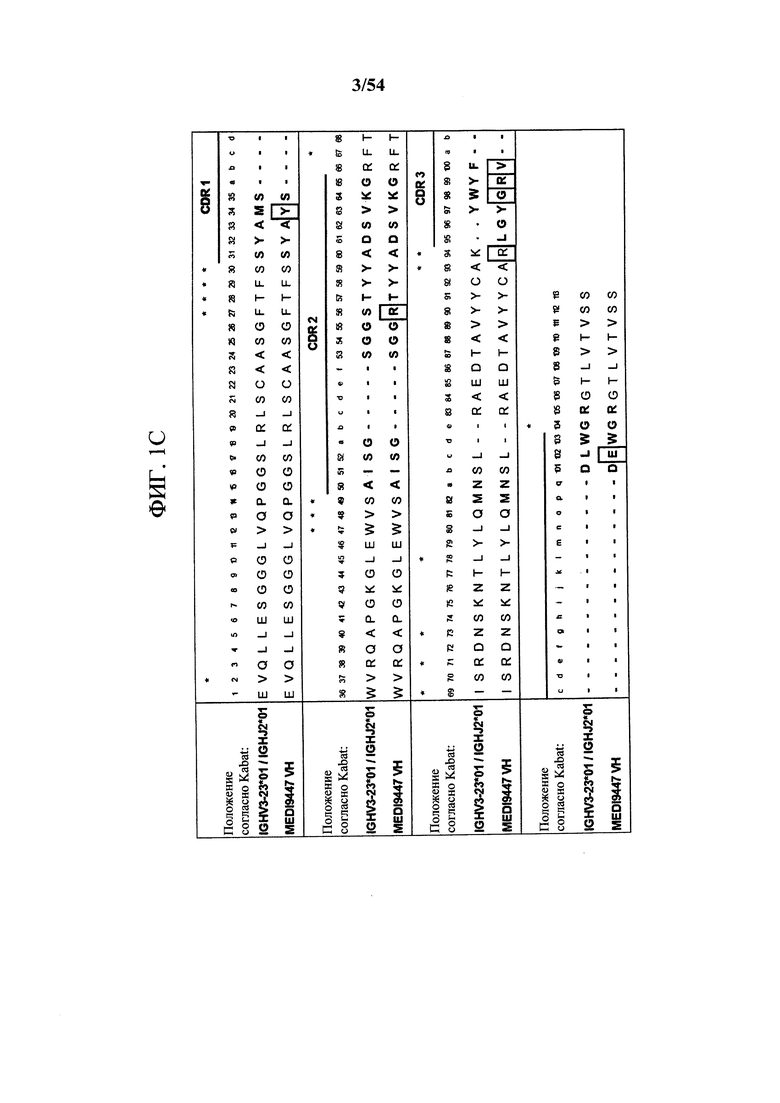
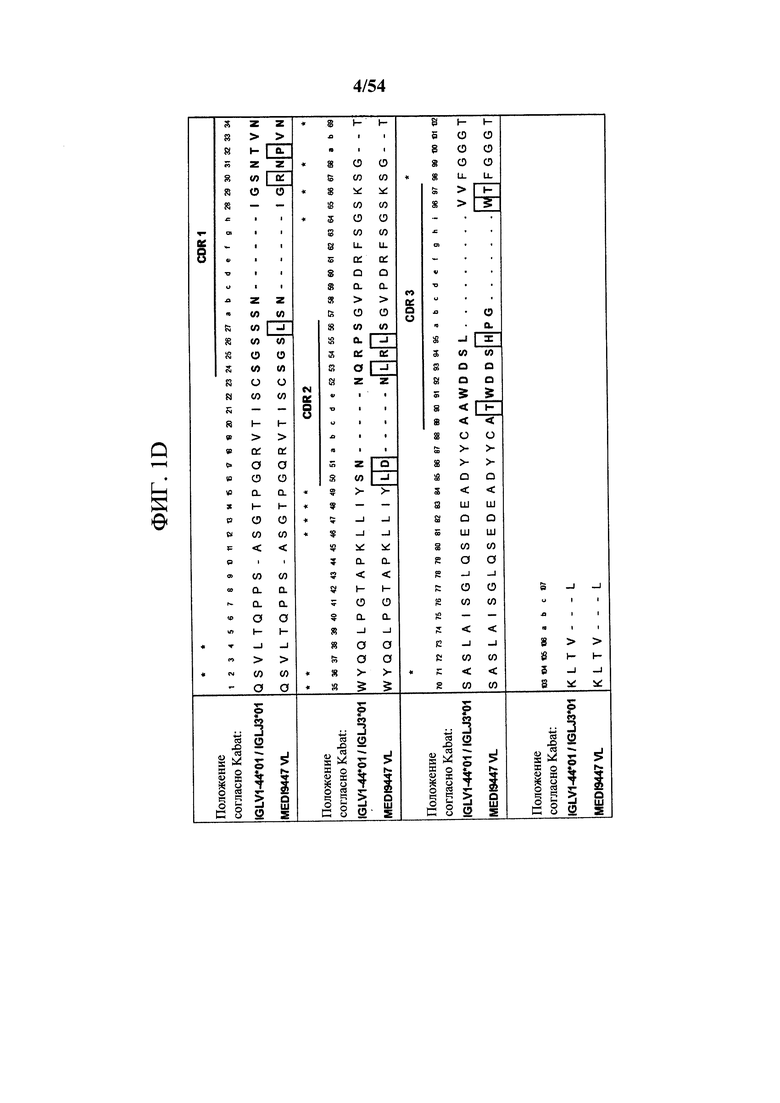
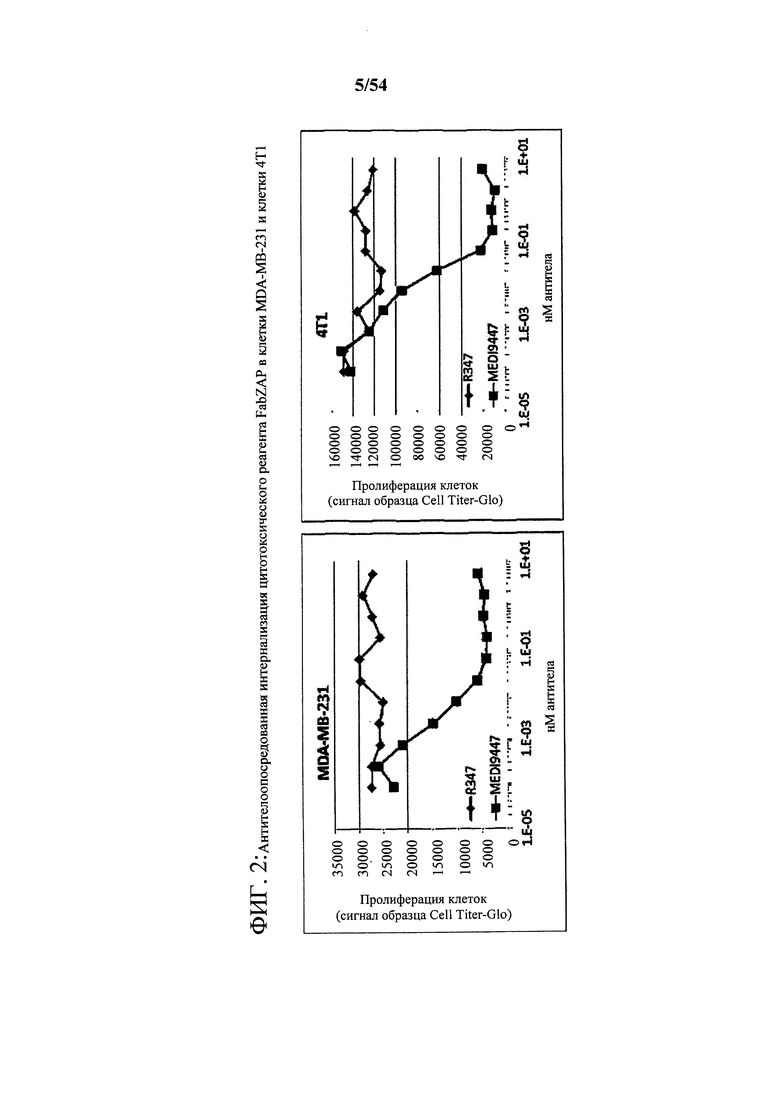
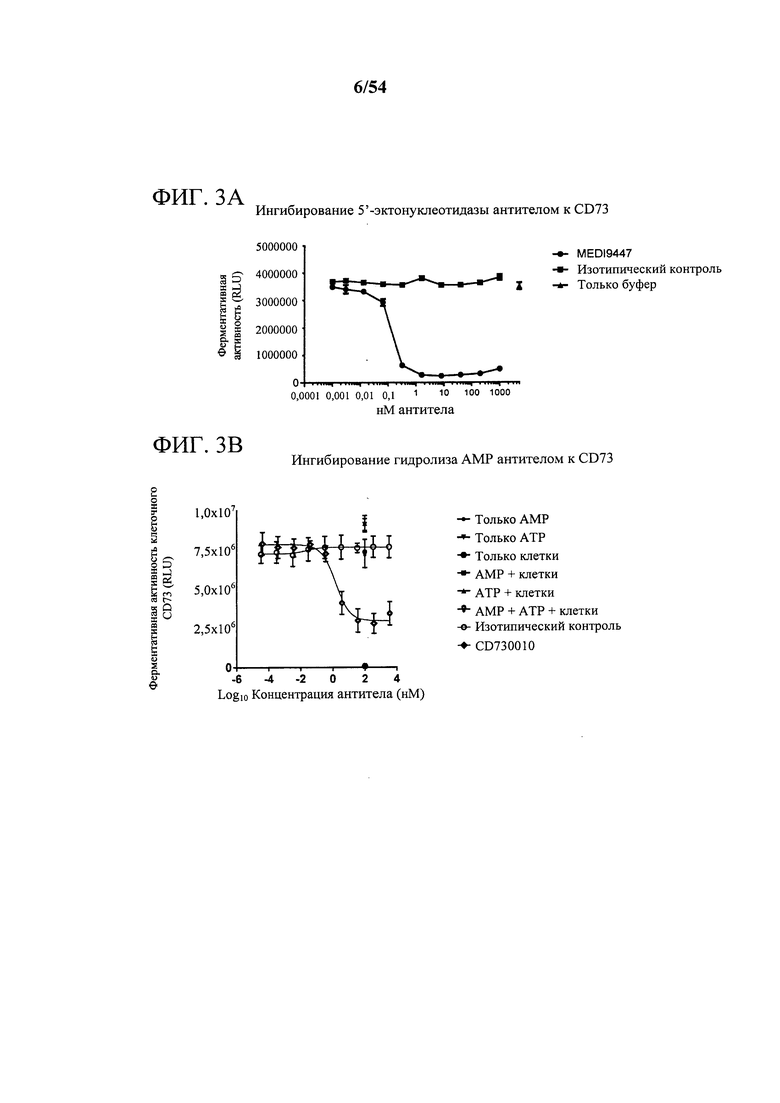
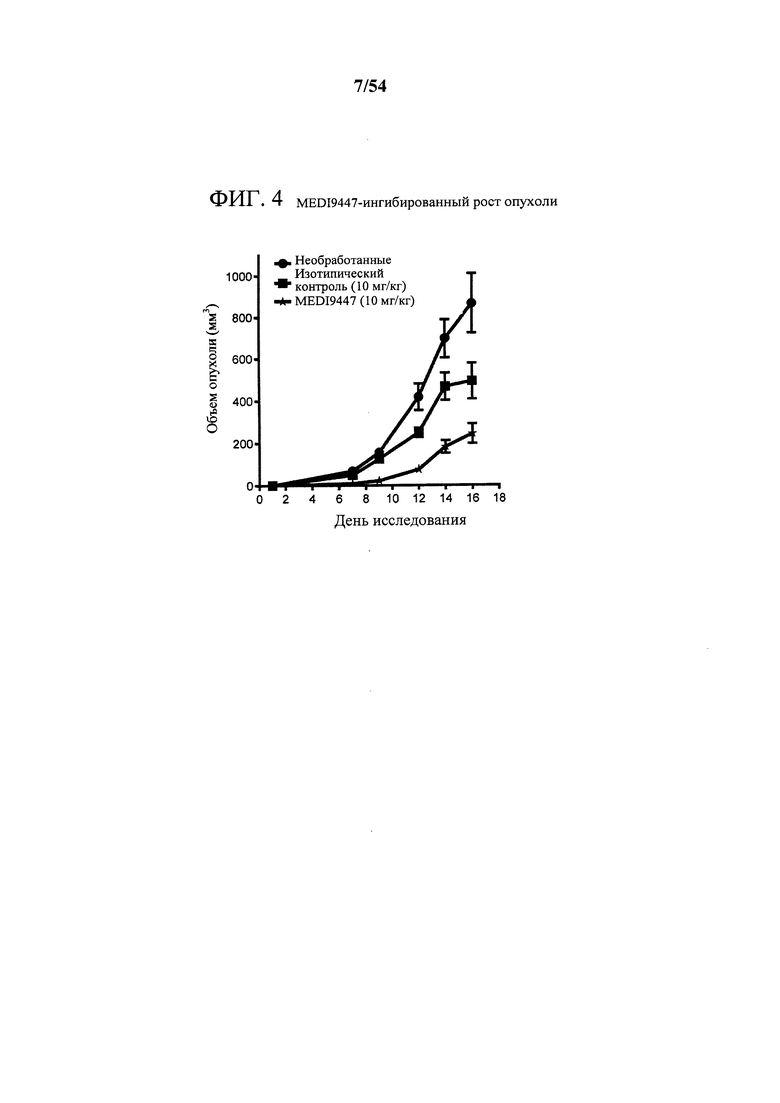
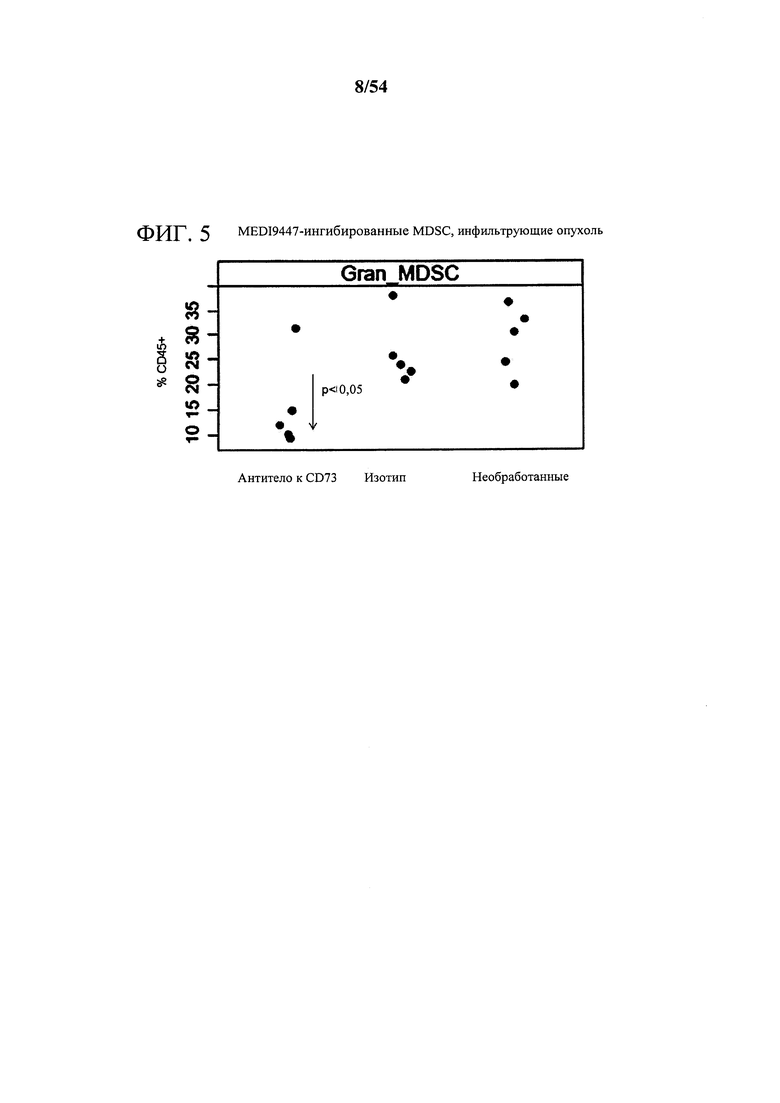
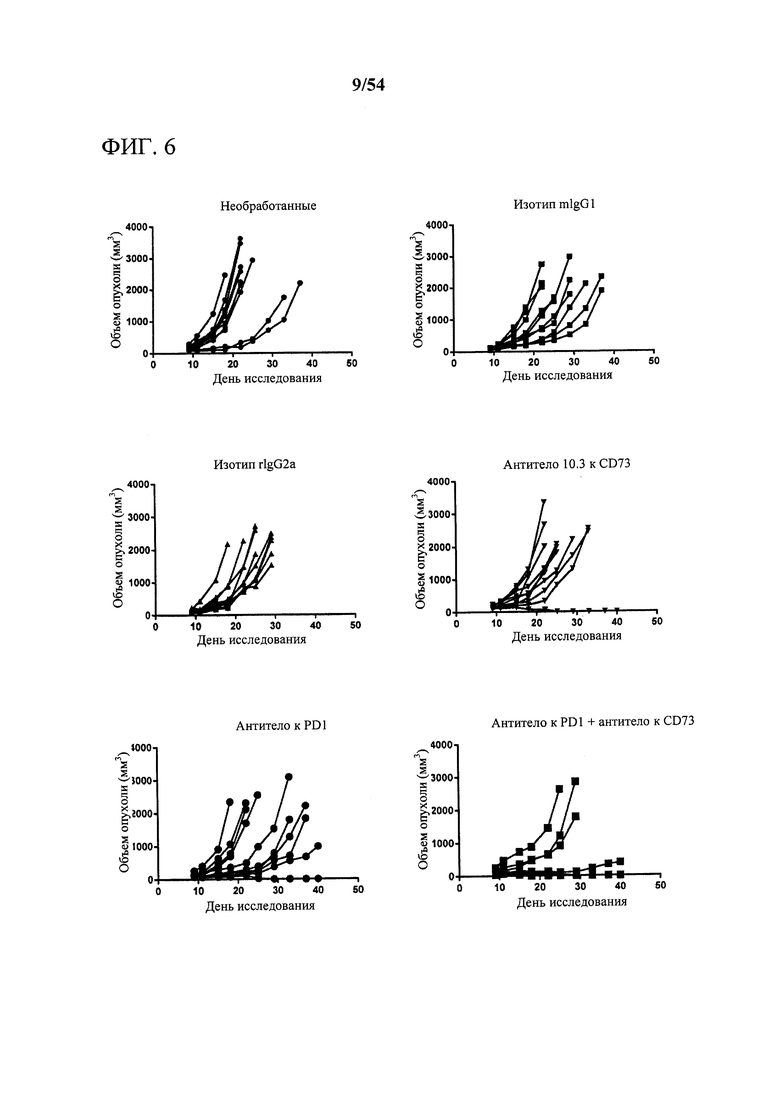
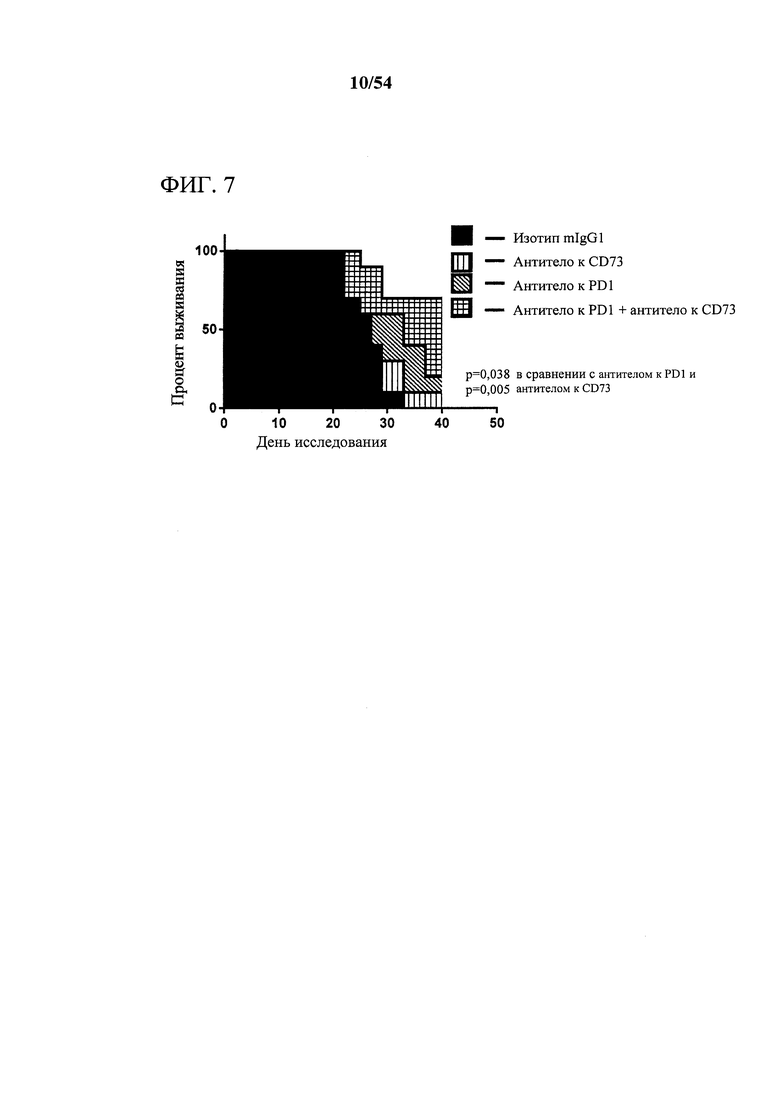
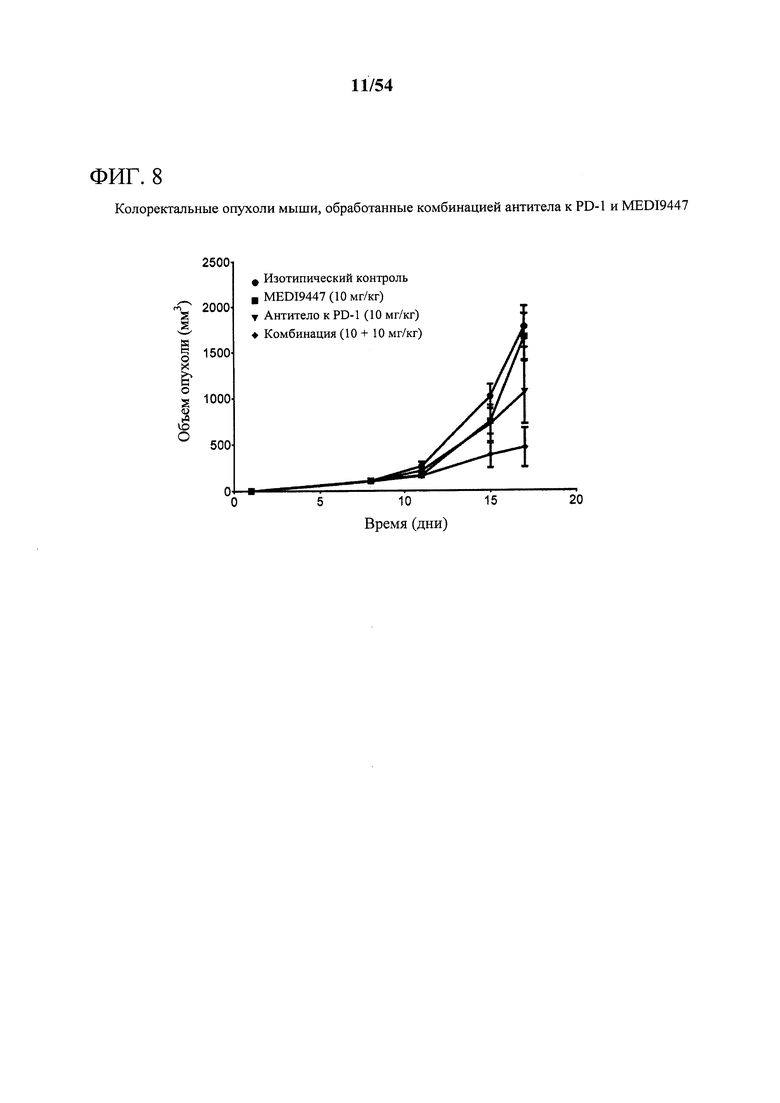
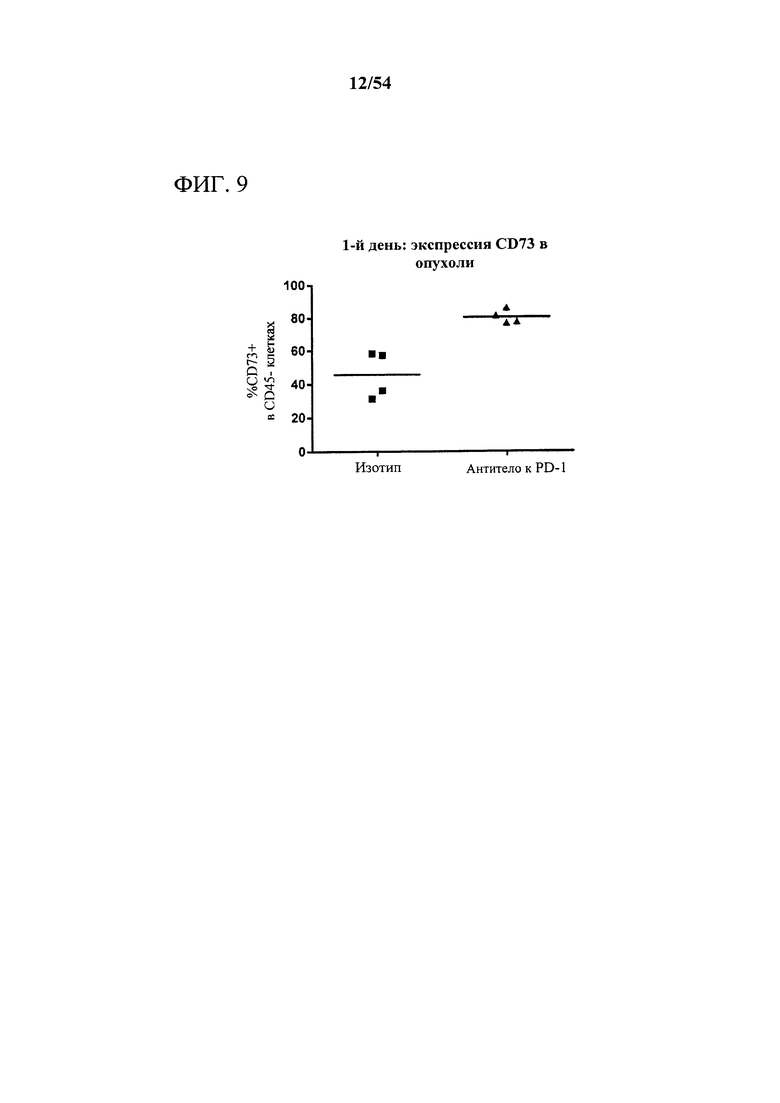
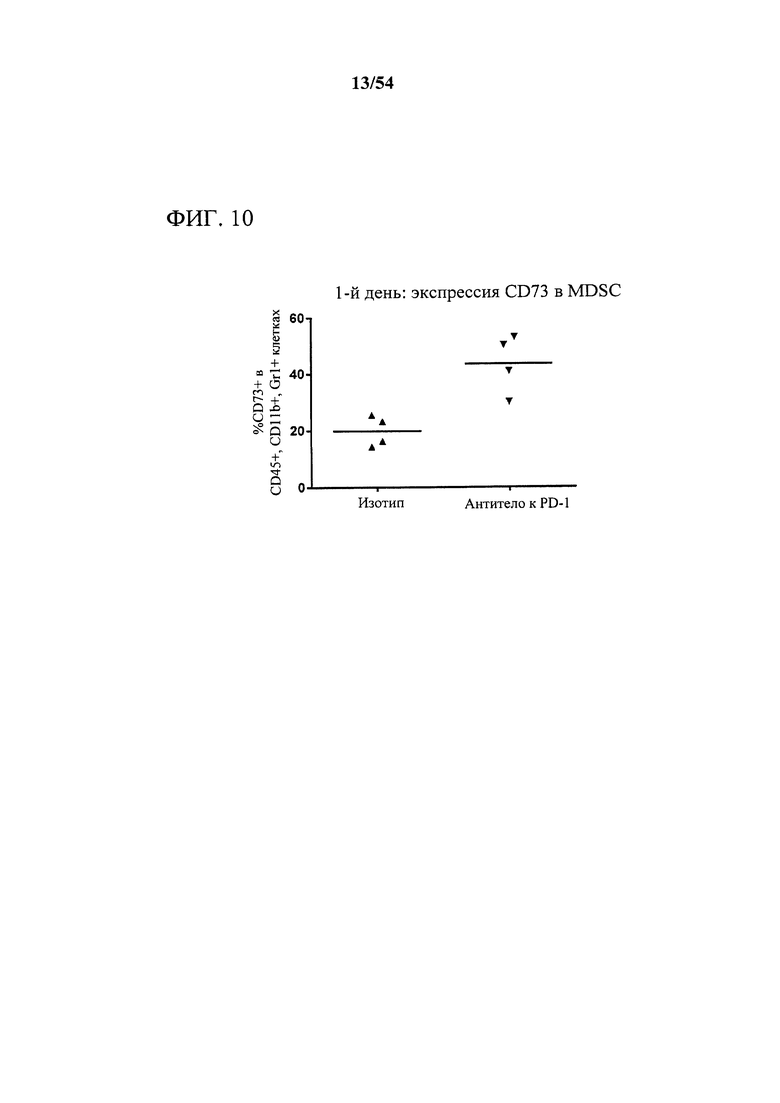
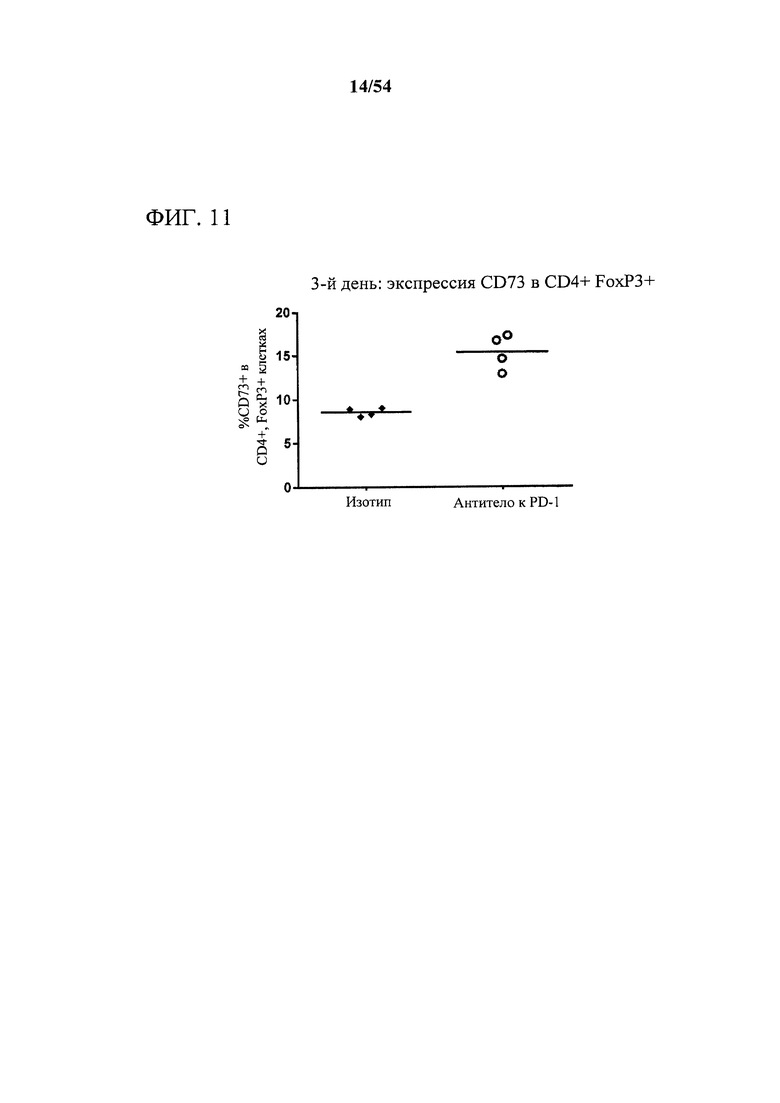
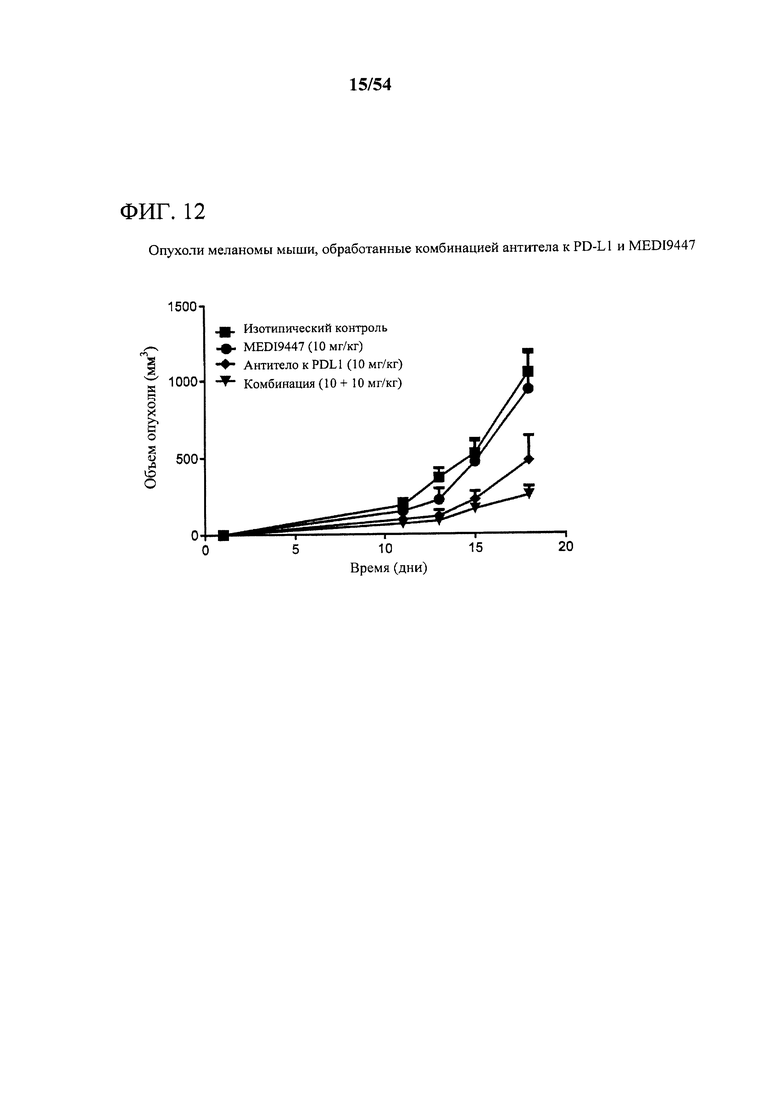
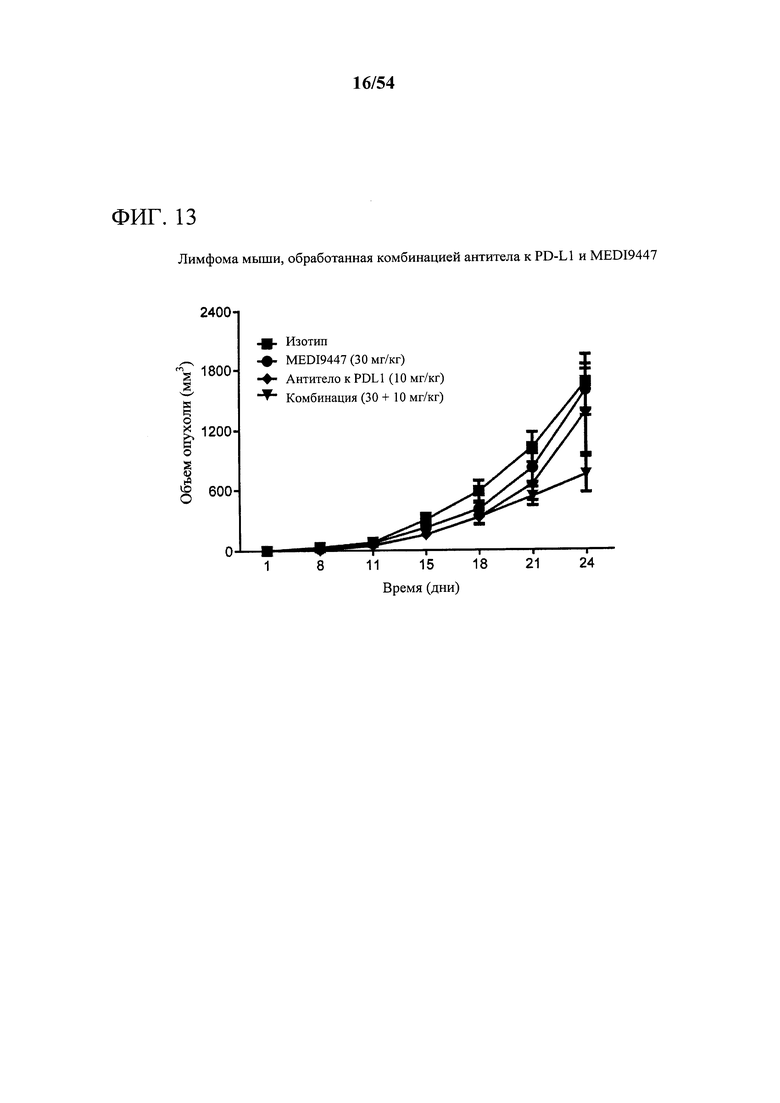
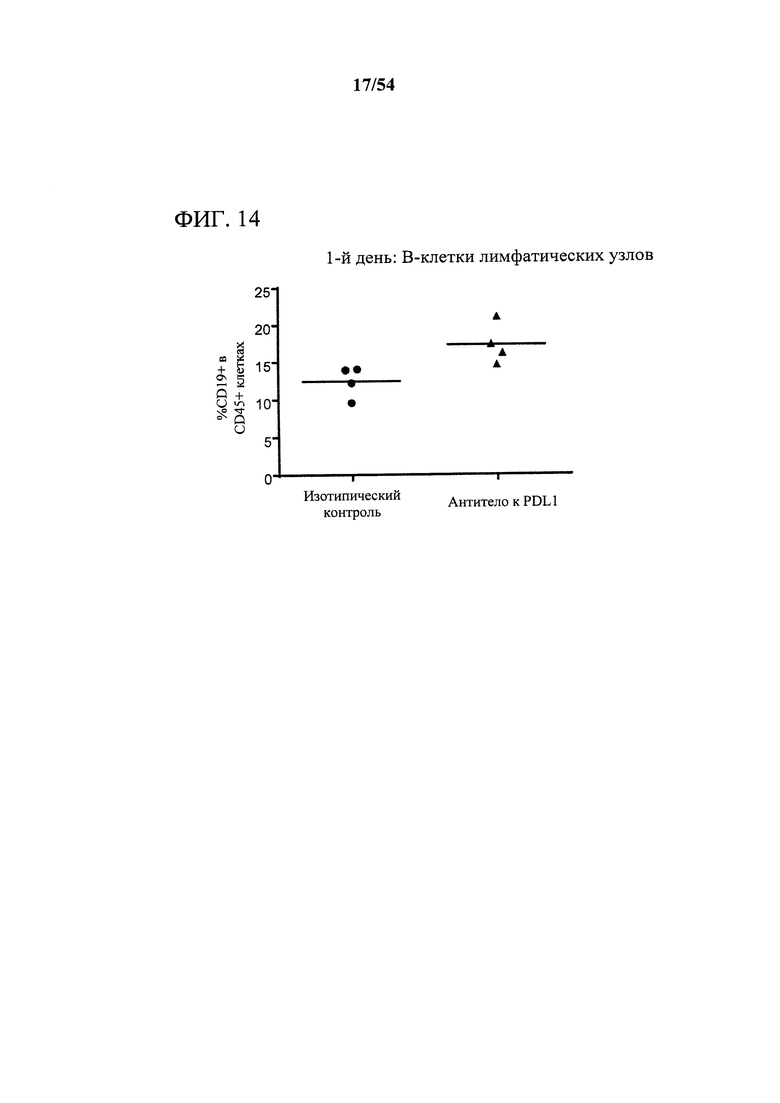
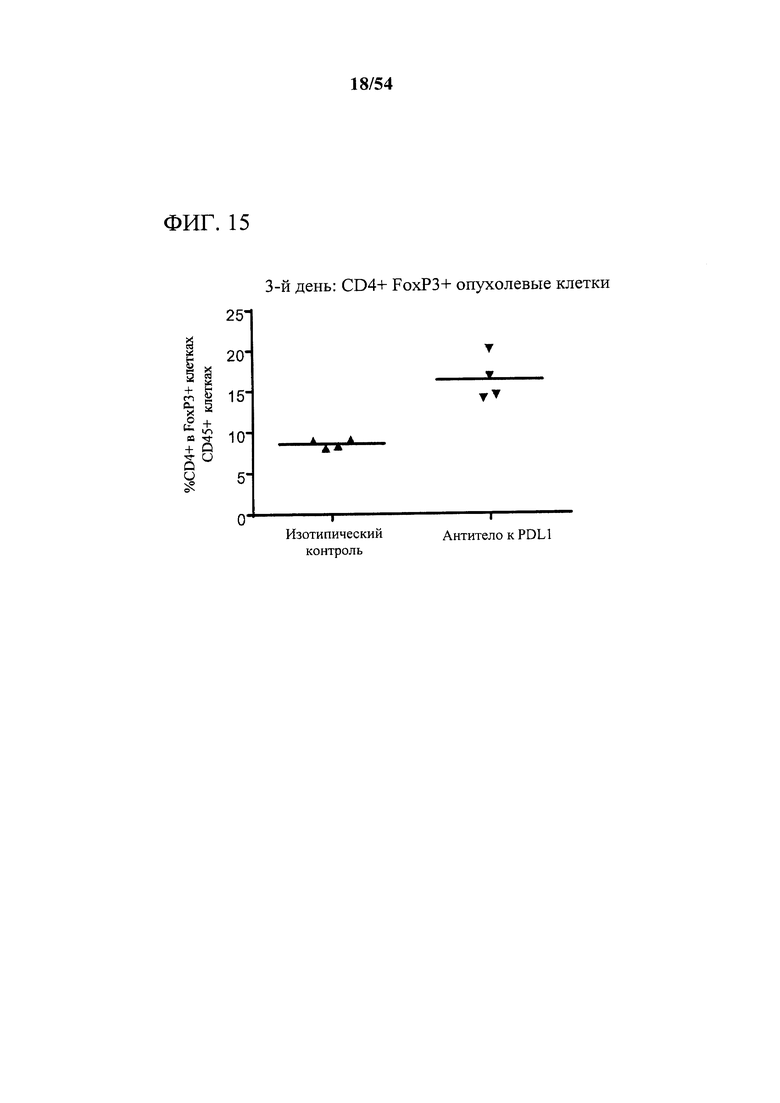
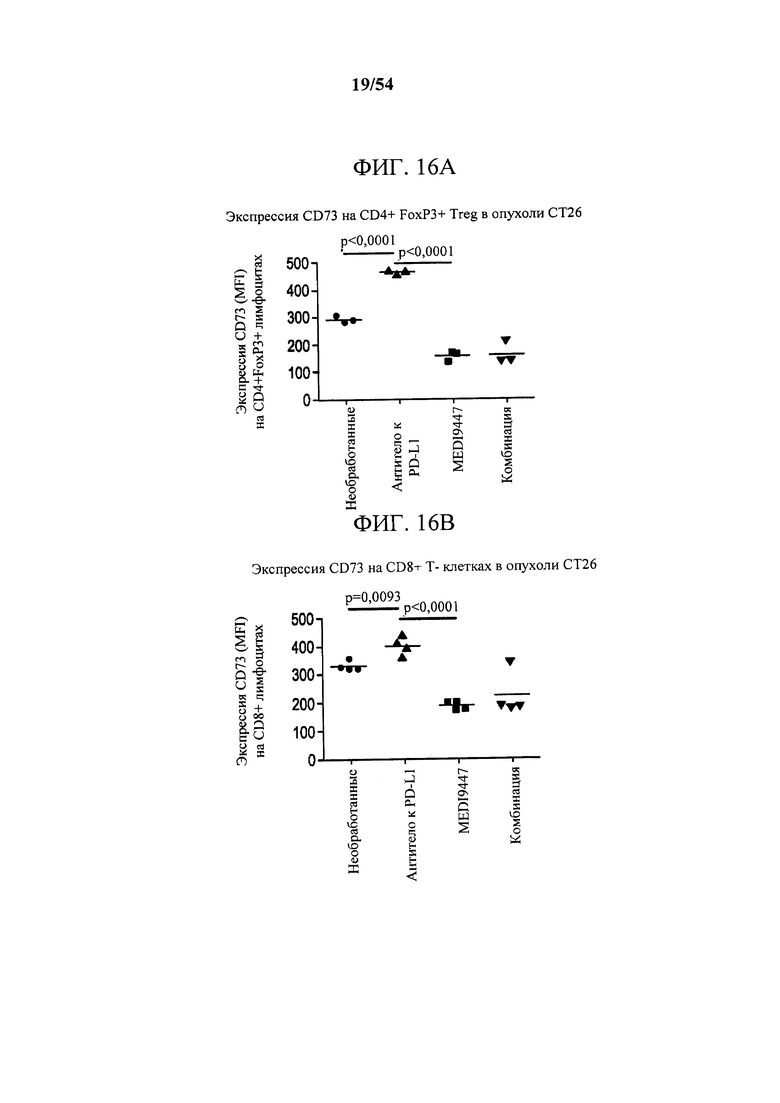
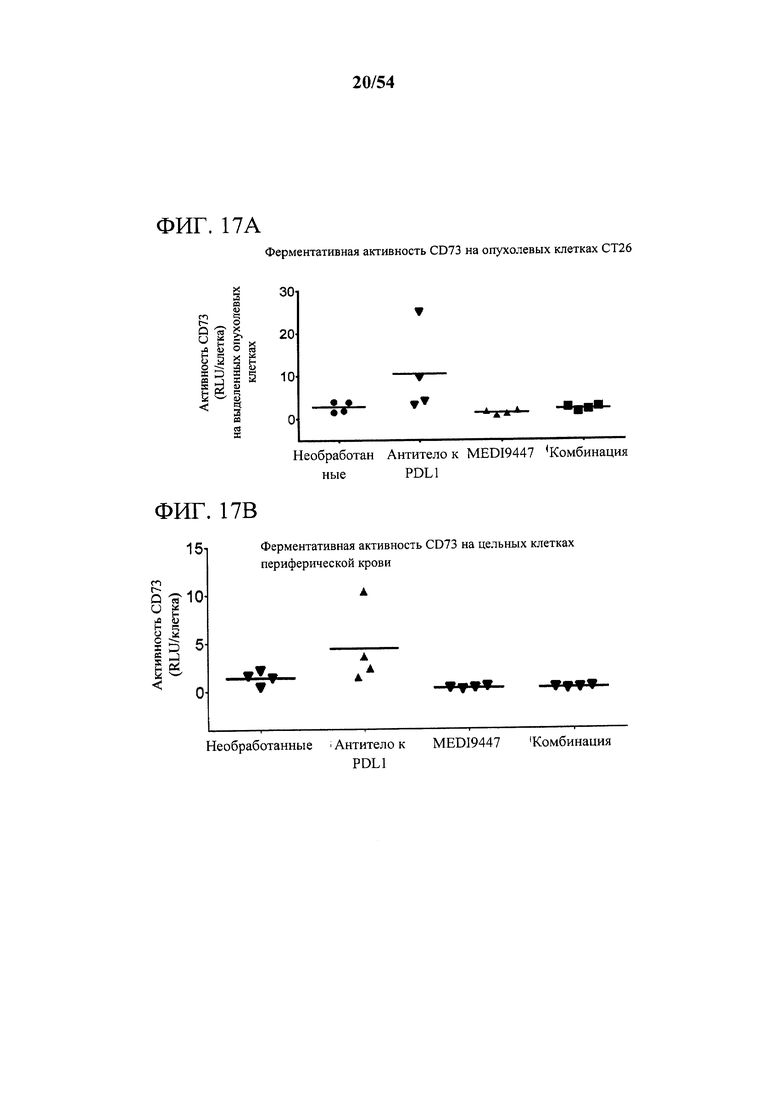



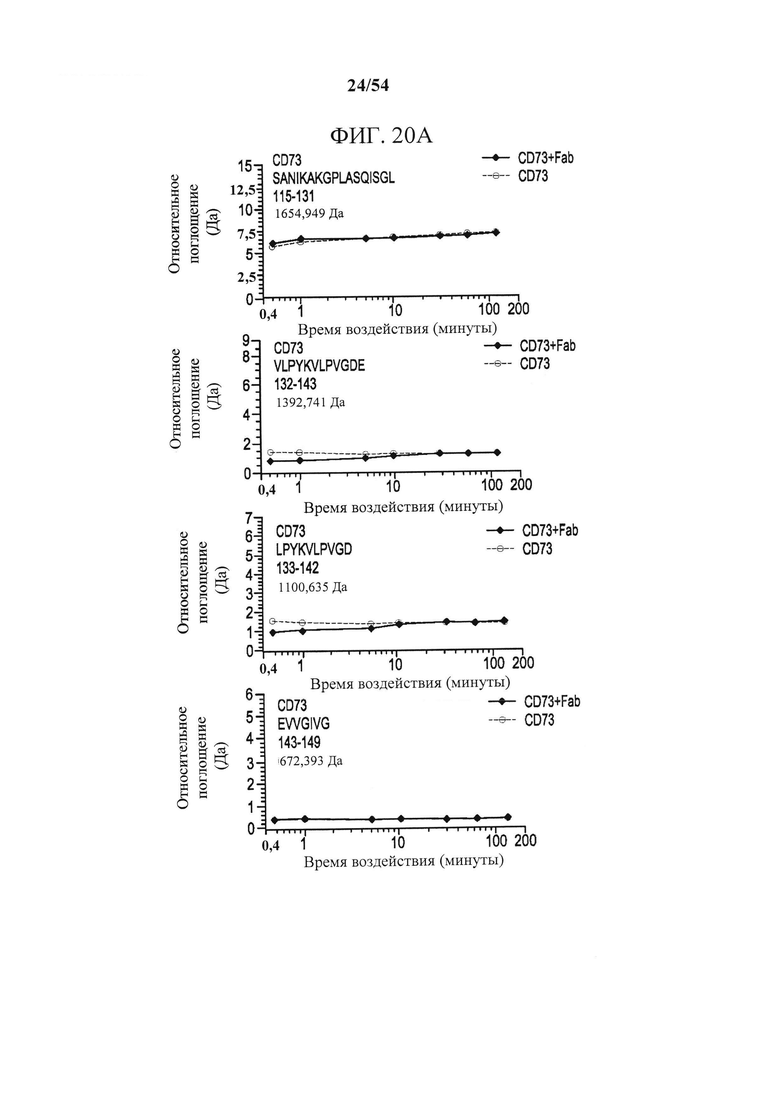

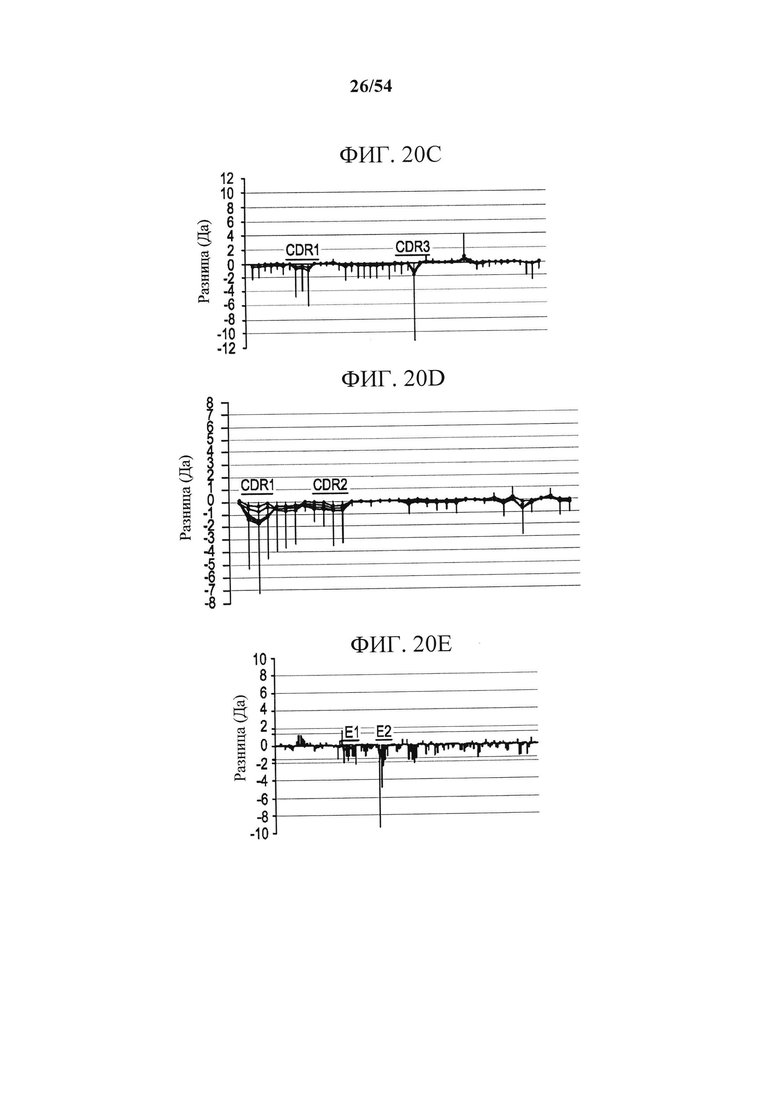
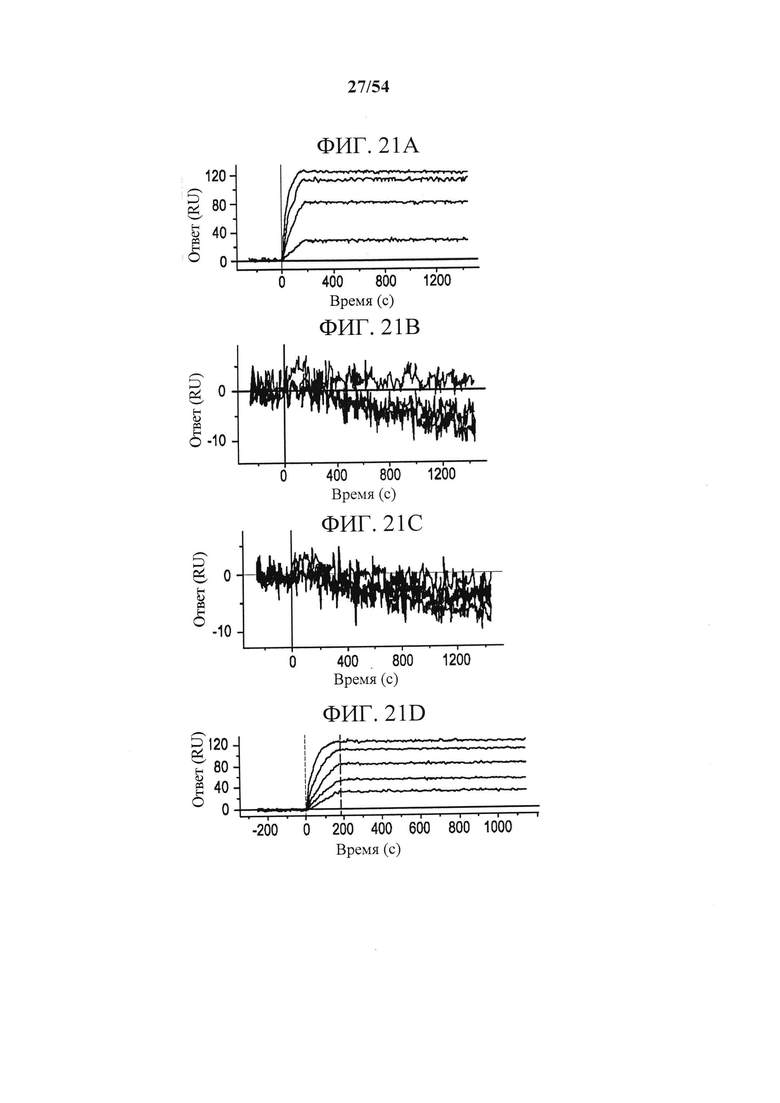

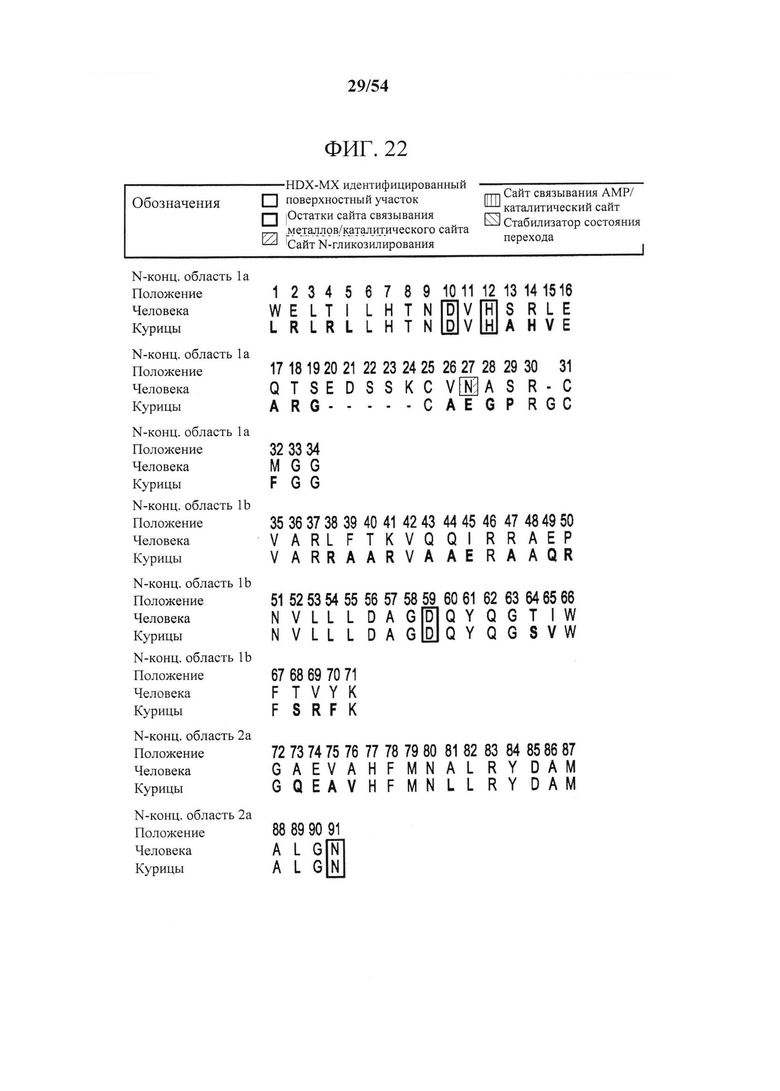




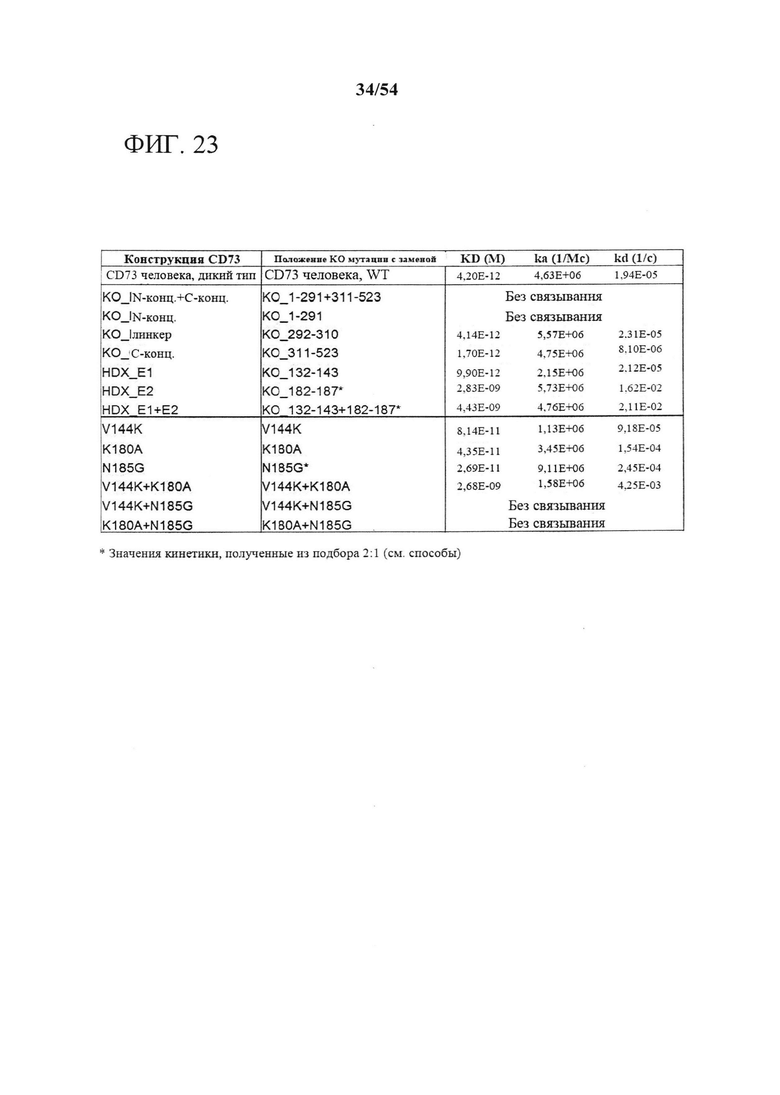
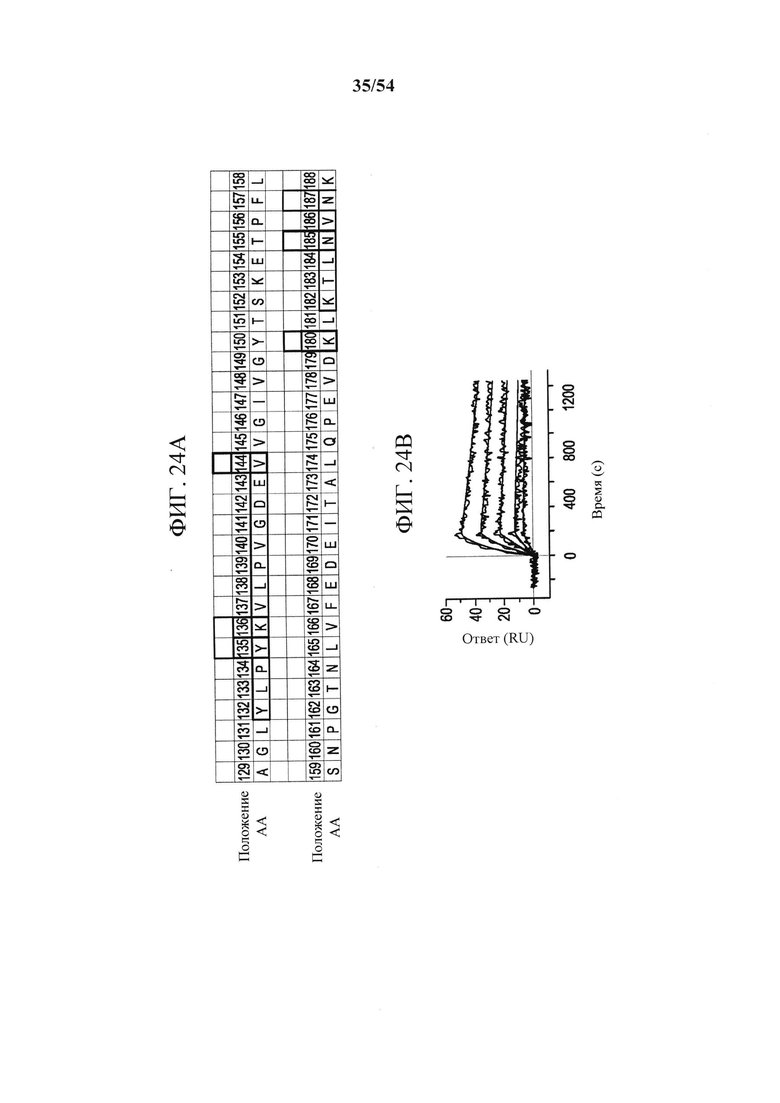
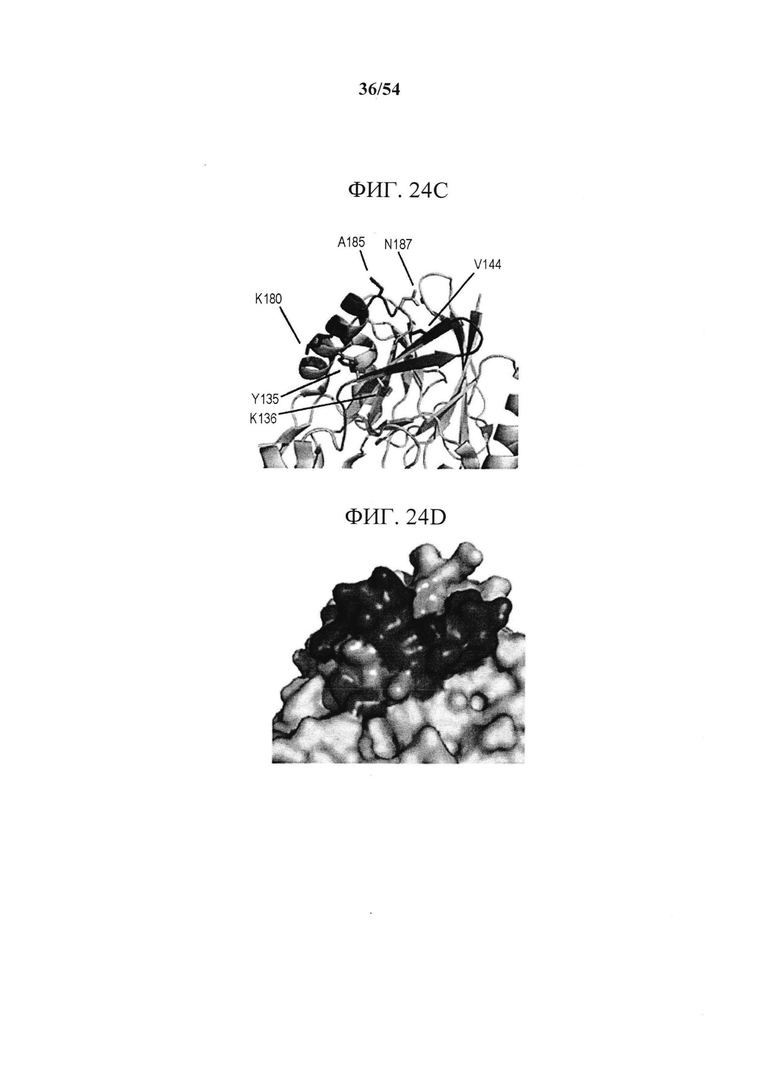
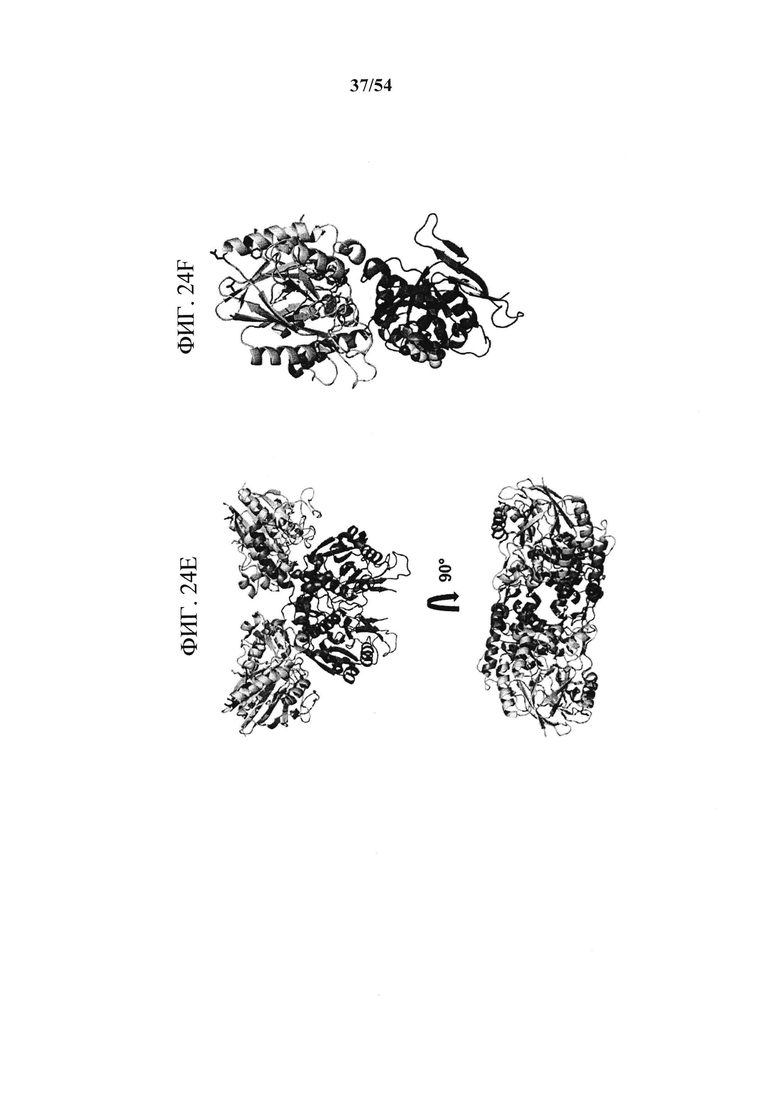
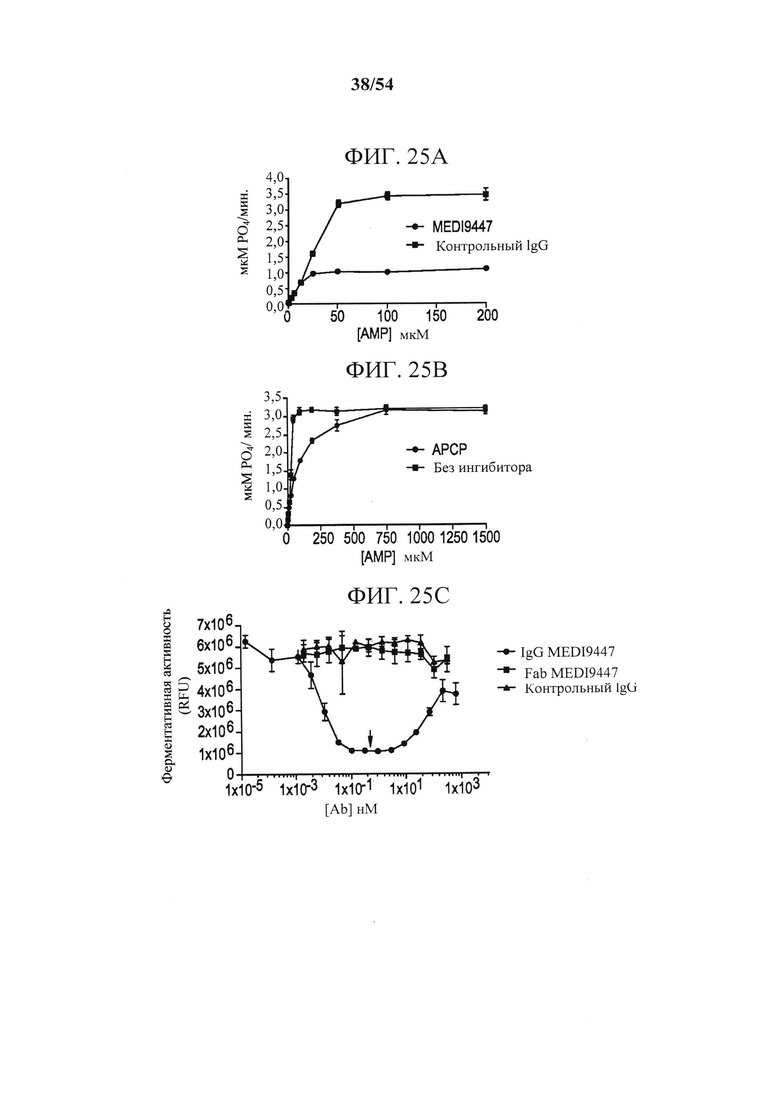
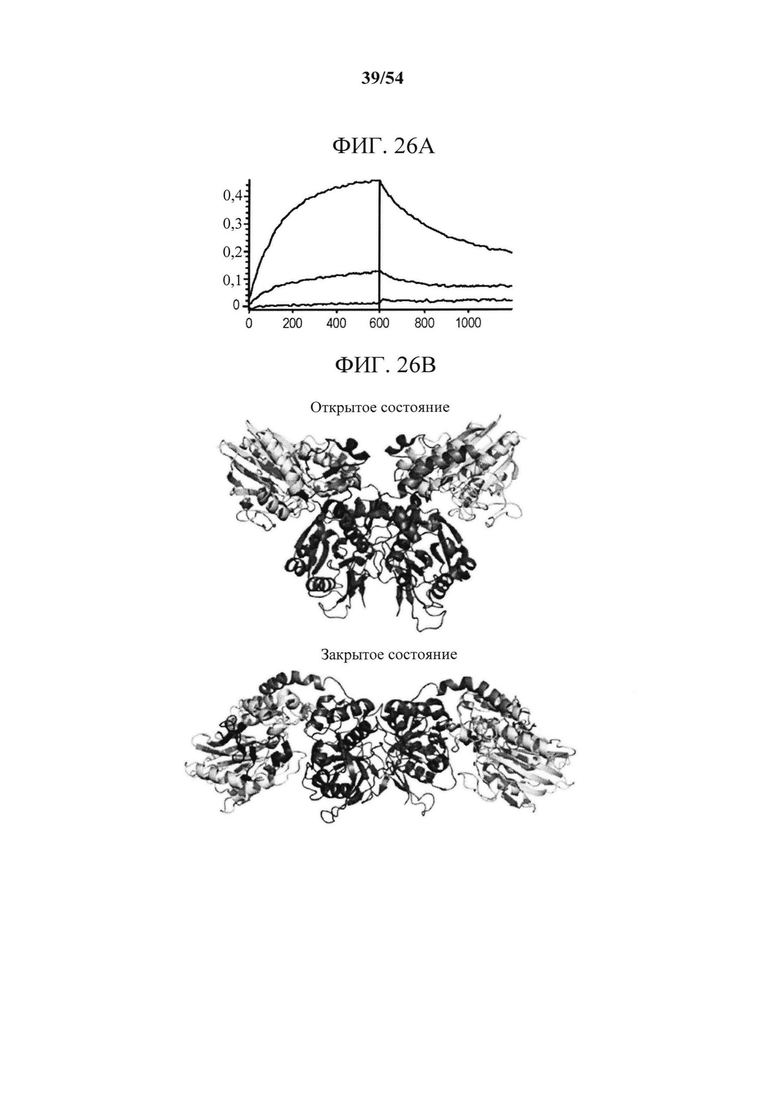

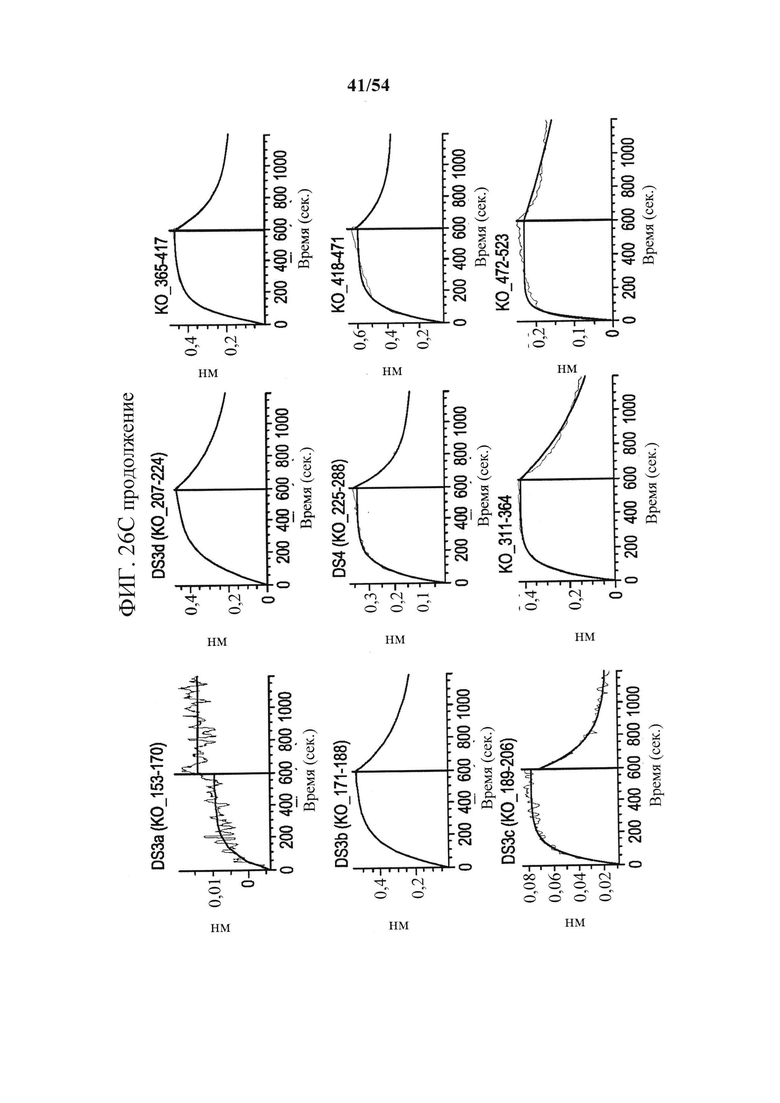
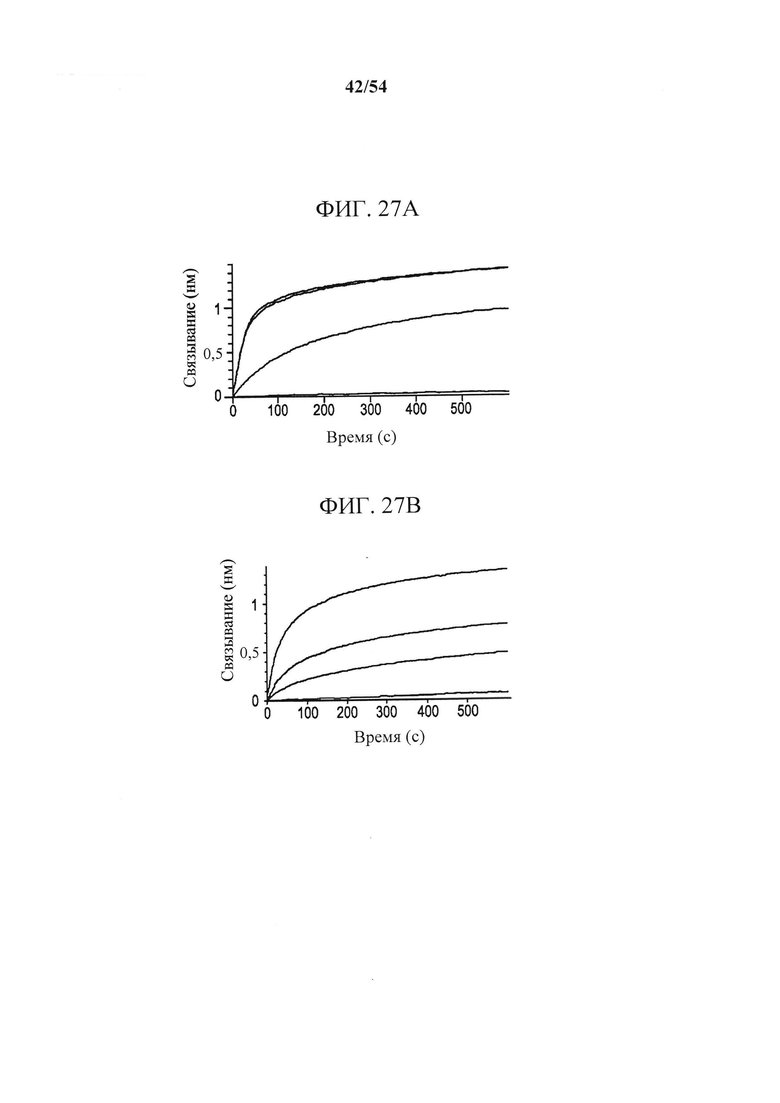


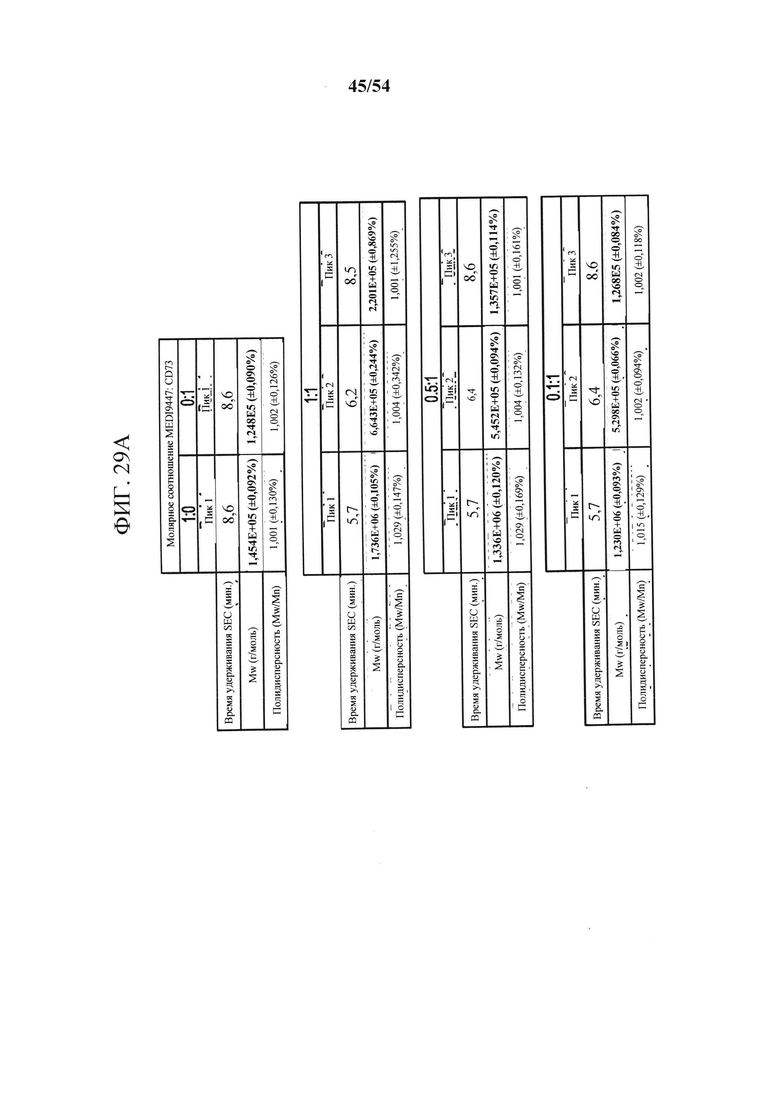
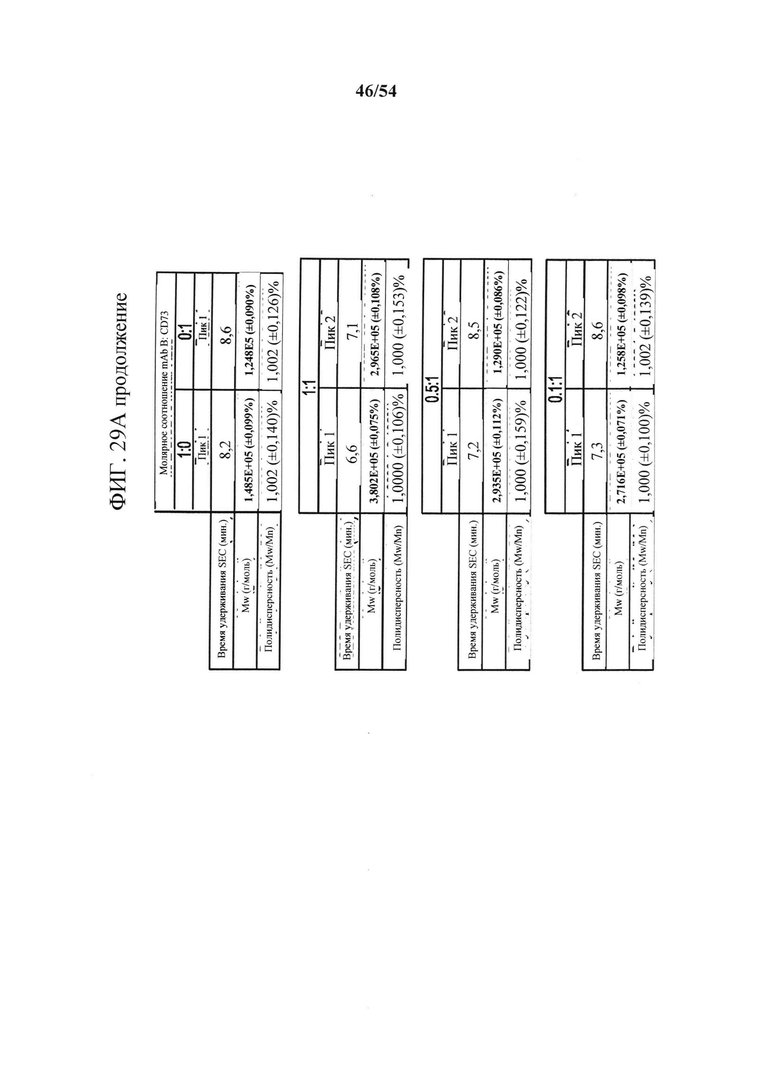
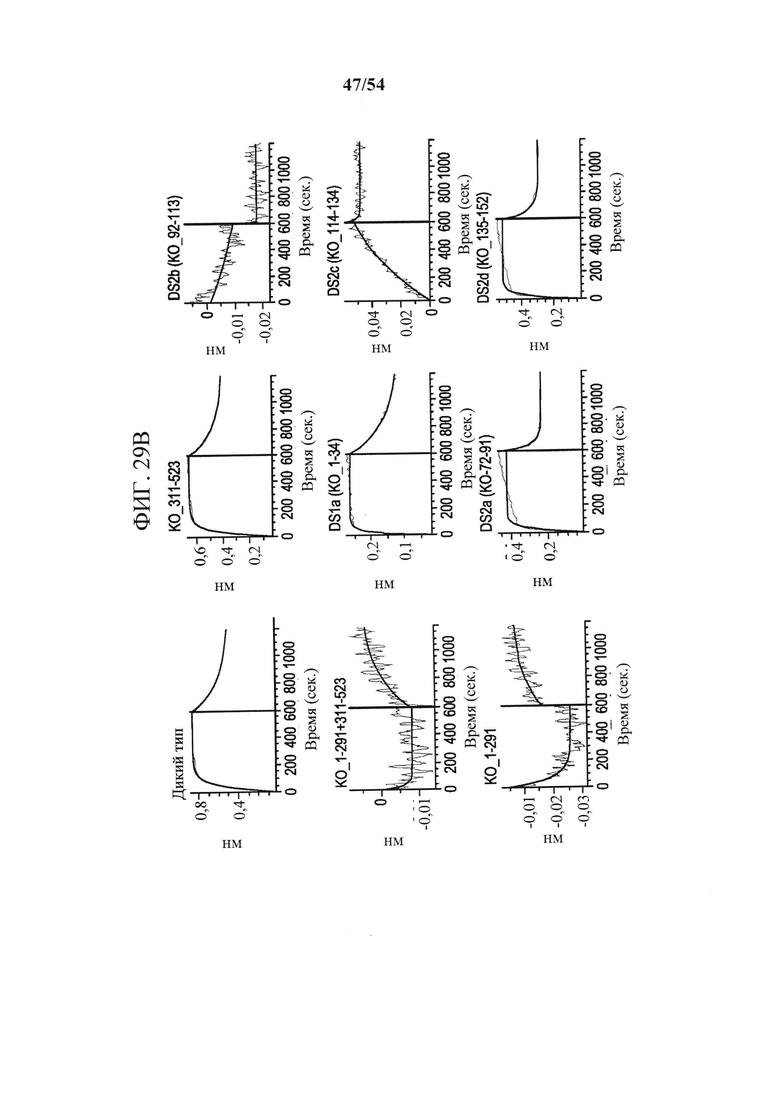
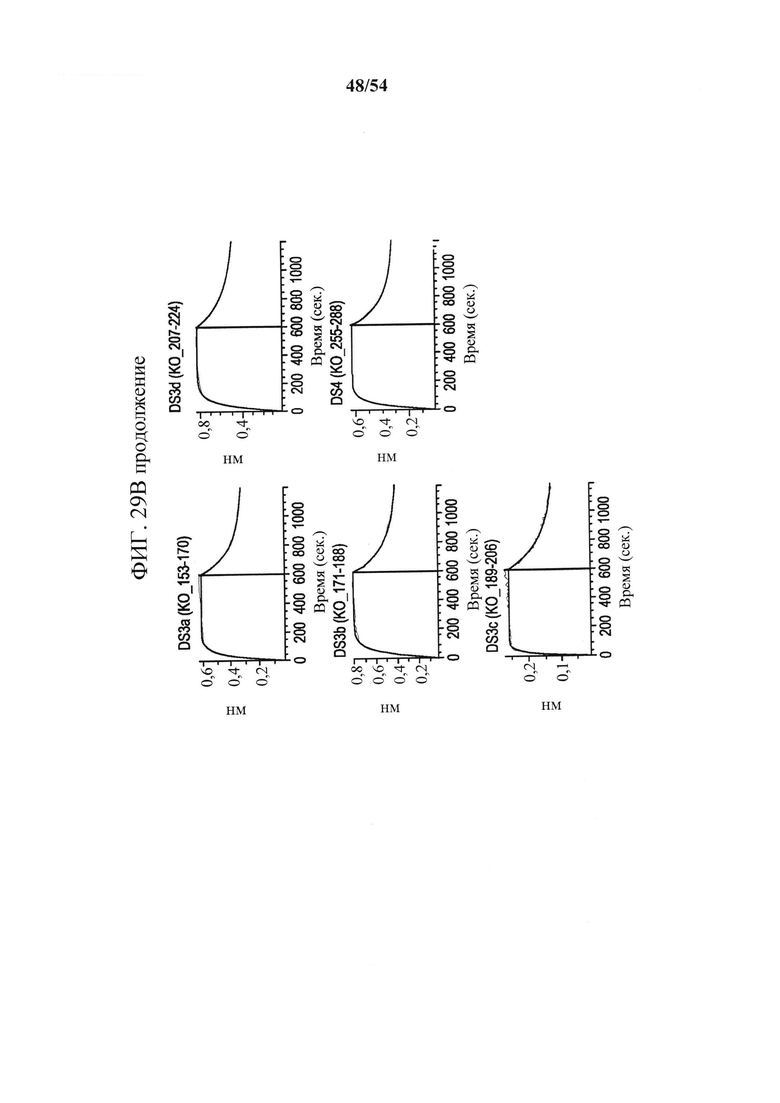
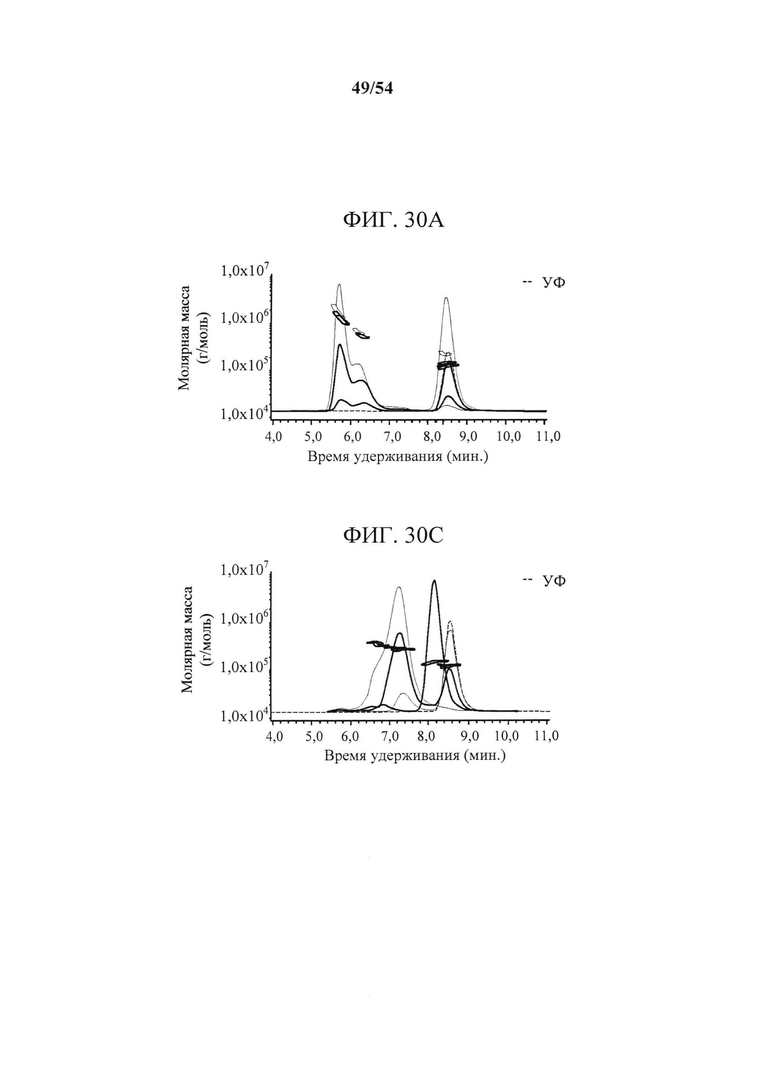

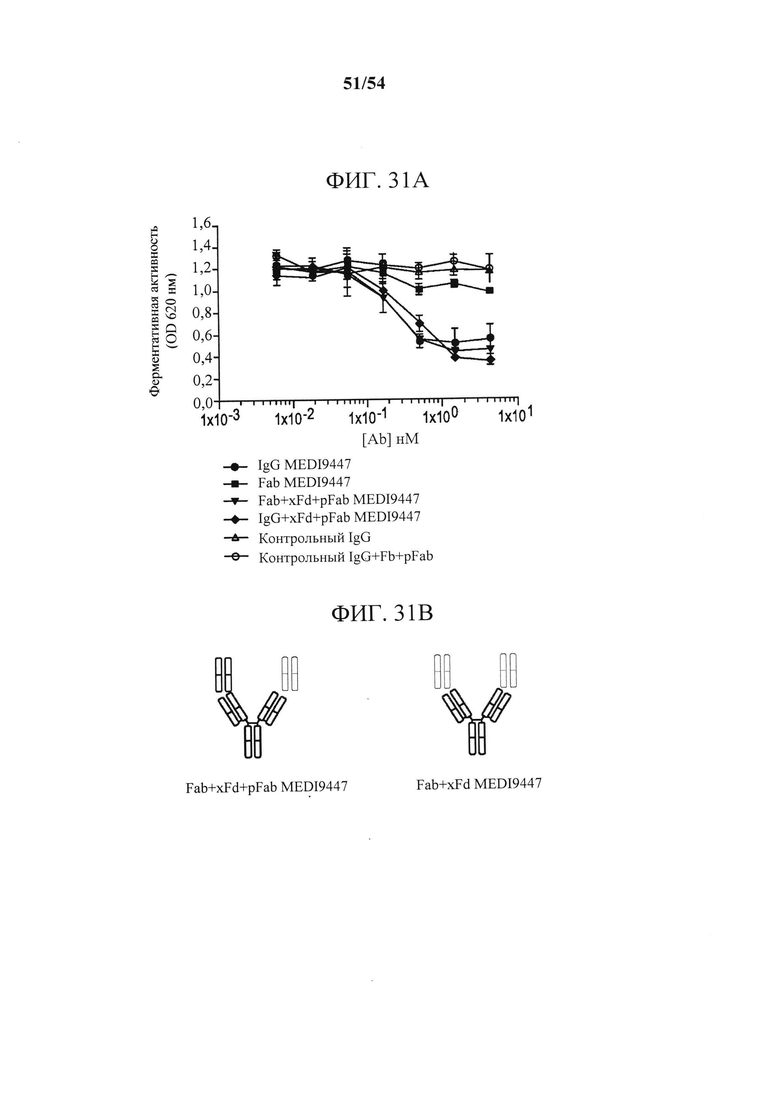
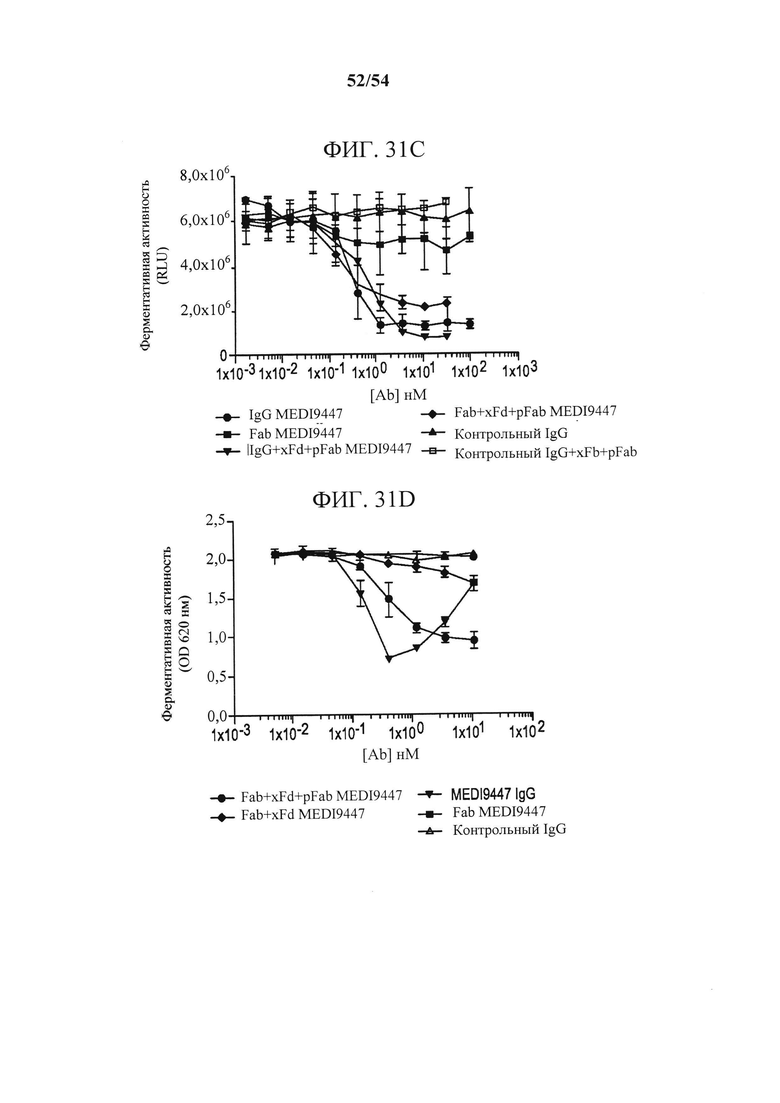
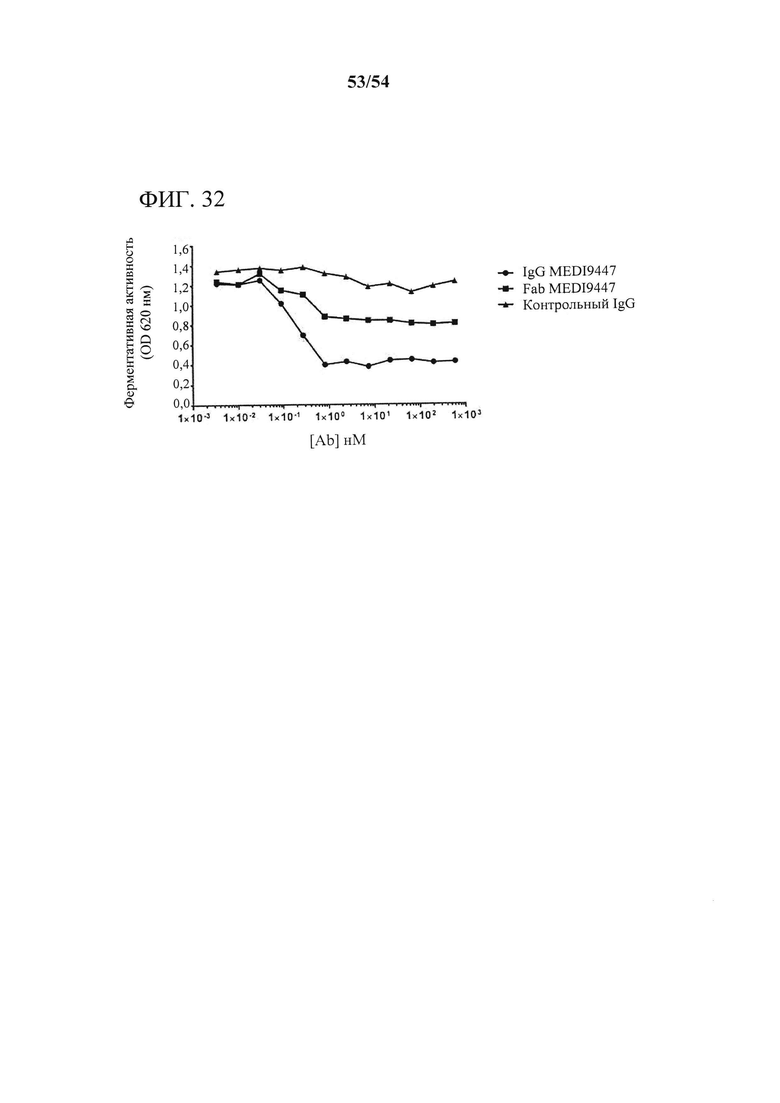
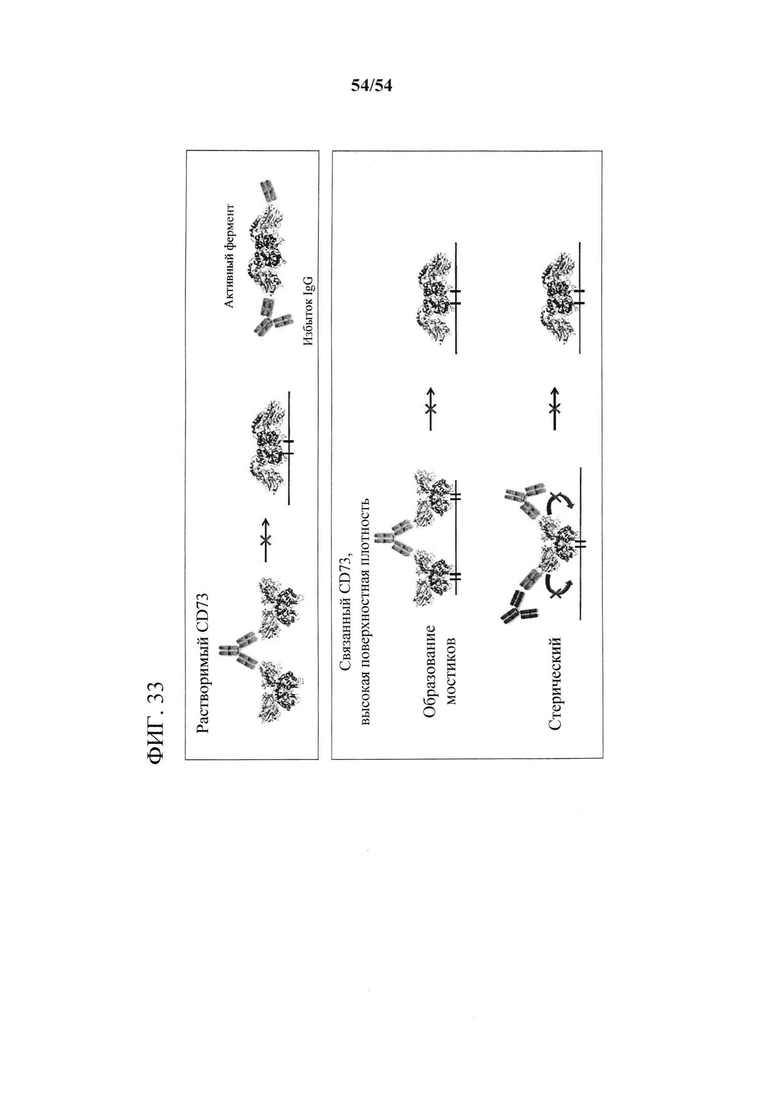
Изобретение относится к области биохимии, в частности к выделенному антителу или его антигенсвязывающему фрагменту, которые специфически связываются с CD73, а также к фармацевтическому составу, его содержащему. Также раскрыт способ лечения, предусматривающий использование вышеуказанного антитела или его фрагмента. Изобретение позволяет эффективно осуществлять лечение субъекта, идентифицированного как имеющий опухоль, которая характеризуется повышенной экспрессией CD73. 7 н. и 18 з.п. ф-лы, 33 ил., 17 табл., 18 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они содержат VL антитела и VH антитела, где VL содержит аминокислотную последовательность SEQ ID NO: 68, и где VH содержит аминокислотную последовательность SEQ ID NO: 82.
2. Выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они содержат VL и VH, содержащие VL-CDR1, VL-CDR2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3, где VL-CDR1 содержит SEQ ID NO: 46, VL-CDR2 содержит SEQ ID NO: 51, VL-CDR3 содержит SEQ ID NO: 56, VH-CDR1 содержит SEQ ID NO: 36, VH-CDR2 содержит SEQ ID NO: 39 и VH-CDR3 содержит SEQ ID NO: 45.
3. Выделенное антитело или его антигенсвязывающий фрагмент по п. 1 или 2, которые содержат константную область тяжелой цепи или ее фрагмент.
4. Выделенное антитело или его антигенсвязывающий фрагмент по п. 1 или 2, которые представляют собой антагонист CD73.
5. Выделенное антитело или его антигенсвязывающий фрагмент по п. 4, где CD73 представляет собой CD73 человека.
6. Фармацевтический состав для лечения субъекта, идентифицированного как имеющий опухоль, которая характеризуется повышенной экспрессией CD73, содержащий эффективное количество антитела к CD73 или его антигенсвязывающего фрагмента и антитела к PD-L1 или его антигенсвязывающего фрагмента, где антитело к CD73 представляет собой антитело по любому из пп. 1 или 2 или его антигенсвязывающий фрагмент.
7. Фармацевтический состав по п. 6, где антитело к PD-L1 представляет собой MEDI4736, BMS-936559 или MPDL3280A или их антигенсвязывающий фрагмент.
8. Фармацевтический состав для лечения субъекта, идентифицированного как имеющий опухоль, которая характеризуется повышенной экспрессией CD73, содержащий эффективное количество антитела к CD73 или его антигенсвязывающего фрагмента и антитела к CTLA-4 или его антигенсвязывающего фрагмента, где антитело к CD73 представляет собой антитело по любому из пп. 1 или 2 или его их антигенсвязывающий фрагмент.
9. Фармацевтический состав по п. 8, где антитело к CTLA-4 представляет собой ипилимумаб или тремелимумаб (тицилимумаб, CP-675,206) или их антигенсвязывающий фрагмент.
10. Способ лечения рака, ассоциированного с экспрессией CD73, у нуждающегося в этом субъекта, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента, которые специфически связываются с CD73, при этом они содержат VH-домен тяжелой цепи и VL-домен легкой цепи, содержащие:
VH-CDR1, имеющий аминокислотную последовательность SEQ ID NO: 36;
VH-CDR2, имеющий аминокислотную последовательность SEQ ID NO: 39;
VH-CDR3, имеющий аминокислотную последовательность SEQ ID NO: 45;
VL-CDR1, имеющий аминокислотную последовательность SEQ ID NO: 46;
VL-CDR2, имеющий аминокислотную последовательность SEQ ID NO: 51; и
VL-CDR3, имеющий аминокислотную последовательность SEQ ID NO: 56.
11. Способ по п. 10, где VL-домен антитела содержит аминокислотную последовательность SEQ ID NO: 68, и где VH-домен антитела содержит аминокислотную последовательность SEQ ID NO: 82.
12. Способ по п. 10, где рак выбран из группы, состоящей из колоректального рака, рака поджелудочной железы, рака мочевого пузыря, лейкемии, лимфомы, глиомы, глиобластомы, меланомы, рака яичников, рака щитовидной железы, рака пищевода, рака предстательной железы и рака молочной железы.
13. Способ лечения рака, ассоциированного с экспрессией CD73, у субъекта, включающий введение субъекту терапевтически эффективного количества первого средства, которое представляет собой антитело или его антигенсвязывающий фрагмент, которые специфически связываются с CD73, при этом они содержат VH-домен тяжелой цепи и VL домен легкой цепи, содержащие:
VH-CDR1, имеющий аминокислотную последовательность SEQ ID NO: 36;
VH-CDR2, имеющий аминокислотную последовательность SEQ ID NO: 39;
VH-CDR3, имеющий аминокислотную последовательность SEQ ID NO: 45;
VL-CDR1, имеющий аминокислотную последовательность SEQ ID NO: 46;
VL-CDR2, имеющий аминокислотную последовательность SEQ ID NO: 51; и
VL-CDR3, имеющий аминокислотную последовательность SEQ ID NO: 56, в комбинации с терапевтически эффективным количеством второго средства, которое
представляет собой противораковое средство, отличное от первого средства.
14. Способ по п. 13, где VL-домен антитела содержит аминокислотную последовательность SEQ ID NO: 68, и где VH-домен антитела содержит аминокислотную последовательность SEQ ID NO: 82.
15. Способ по п. 13, где комбинация первого средства и второго средства является аддитивно или синергически эффективной.
16. Способ по п. 13, где второе средство специфически связывается с PD-1 (белком-1 запрограммированной смерти клетки), PD-L1 (лигандом-1 белка-1 запрограммированной смерти клетки), PD-L2 (лигандом-2 белка-1 запрограммированной смерти клетки) или CTLA-4 (белком-антигеном-4 цитотоксических Т-лимфоцитов).
17. Способ по п. 13, где второе средство представляет собой антитело к CTLA-4 или его антигенсвязывающий фрагмент.
18. Способ по п. 17, где антитело к CTLA-4 представляет собой тремелимумаб (тицилимумаб, CP-675,206).
19. Способ по п. 13, где второе средство представляет собой антитело к PD-1 или его антигенсвязывающий фрагмент.
20. Способ по п. 19, где антитело к PD-L1 представляет собой MEDI4736.
21. Способ лечения рака, ассоциированного с экспрессией CD73, включающий введение антитела к CD73 или его антигенсвязывающего фрагмента субъекту, идентифицированному как имеющий опухоль, которая характеризуется повышенной экспрессией CD73 по сравнению с эталоном, и где антитело к CD73 представляет собой антитело по любому из пп. 1 или 2.
22. Способ по п. 21, где субъект подвергается, подвергался или будет подвергаться терапии, направленной в отношении PD-1, PD-L1 или CTLA-4.
23. Способ по п. 22, где терапия, направленная в отношении PD-1, PD-L1 или CTLA-4, предусматривает введение антитела к PD-1, антитела к PD-L1 или антитела к CTLA-4 или их антигенсвязывающих фрагментов.
24. Способ по п. 23, где антитело к PD-1 представляет собой MEDI4736, BMS936559 или MPDL3280A или их антигенсвязывающие фрагменты.
25. Способ по п. 23, где антитело к CTLA-4 представляет собой ипилимумаб, тремелимумаб (тицилимумаб, CP-675,206) или их антигенсвязывающие фрагменты.
| BERTRAND ALLARD et al | |||
| Способ подготовки рафинадного сахара к высушиванию | 0 |
|
SU73A1 |
| PAOLO A | |||
| ASCIERTO et al | |||
| Biomarkers for Immunostimulatory Monoclonal Antibodies in Combination Strategies for Melanoma and Other Tumor Types, Clin Cancer Res, 2013, Vol.19, N.5, pp.1009-1020 | |||
| US 2008233116 | |||

