ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области секвенирования нуклеиновых кислот. В частности, настоящее изобретение относится к основанному на самолюминесценции способу одноканального секвенирования, в котором сигнал самолюминесценции используют для установления различий последовательного включения различных нуклеотидов, таким образом, реализуя определение последовательности полинуклеотида.
УРОВЕНЬ ТЕХНИКИ
Технология секвенирования ДНК включает технологию секвенирования ДНК первого поколения, представленную способом секвенирования по Сенгеру, и технологию секвенирования ДНК второго поколения, представленную Illumina Hiseq2500, Roche 454, ABI Solid, BGISEQ-500 и т.д. В 1977 г., Сенгер изобрел способ секвенирования с дидезокси-концевой терминацией и стал представителем первого поколения технологии секвенирования. В 2001 г., на основе технологии секвенирования первого поколения, завершен проект генома человека. Секвенирование по Сенгеру имеет характеристики простой экспериментальной операции, интуитивных и точных результатов, и короткого экспериментального периода. Оно имеет широкий ряд применений в клинической детекции мутаций генов и генотипировании, которые требуют высокой своевременности результатов детекции. Однако недостатками способа секвенирования по Сенгеру являются низкая производительность и высокая стоимость, которые ограничивают его применение в крупномасштабном секвенировании генов. Для преодоления недостатков способа секвенирования по Сенгеру, возникла технология секвенирования второго поколения. По сравнению с технологией секвенирования ДНК первого поколения, технология секвенирования ДНК второго поколения имеет характеристики высокой производительности секвенирования, низкой стоимости, высокой степени автоматизации и секвенирования отдельной молекулы. Принимая технологию секвенирования Hiseq2500V2 в качестве примера, в одном экспериментальном процессе можно получать данные для 10-200 миллиардов оснований, и средняя стоимость секвенирования на основание составляет менее 1/1000 от стоимости секвенирования для способа секвенирования по Сенгеру, и полученные результаты секвенирования можно обрабатывать и анализировать напрямую посредством компьютера. Таким образом, технология секвенирования ДНК второго поколения является очень подходящей для крупномасштабного секвенирования.
Технологии секвенирования ДНК второго поколения, разработанные в настоящее время, в основном включают технологию секвенирования посредством лигирования (SBL) и технологию секвенирования посредством синтеза (SBS). Типичные примеры этих технологий секвенирования включают способ секвенирования SOLiD, разработанный в Applied Biosystems, способ комбинаторного лигирования зонда и якоря (cPAL), независимо разработанный в Complete Genomics, и способ комбинаторного синтеза зонда и якоря (cPAS), разработанный в Beijing Genomics Institute (BGI), способ секвенирования Illumina, разработанный совместно компанией Illumina и компанией Solexa Technology, и т.д. В этих способах секвенирования, Illumina и Complate Genomics принимают способ детекции световых сигналов, и чтобы реализовать идентификацию и дифференцировку 4 видов оснований (A, T/U, C и G), обычно необходимо использовать 4 вида флуоресцентных красителей для мечения, соответственно, этих 4 оснований. В этом случае, для считывания флуоресцентного сигнала, который несет каждое основание, устройство для секвенирования должно быть оборудовано по меньшей мере двумя источниками монохроматического света возбуждения и по меньшей мере двумя камерами, что приводит к дорогой стоимости изготовления и большому объему устройства для секвенирования.
В последние 10 лет технологии секвенирования генов второго поколения постепенно выросли от появляющихся технологий до основного направления способов секвенирования, и постепенно стали важными инструментами тестирования в клинической области, таким образом, играя увеличивающуюся роль в предотвращении и контроле инфекционных заболеваний, диагностике генетических заболеваний, и неинвазивном пренатальном скрининге. Для дальнейшего расширения рынка секвенирования и популяризации секвенаторов, разработка дешевых и миниатюризованных секвенаторов постепенно стала тенденцией развития в области секвенирования. В качестве классического способа технологии секвенирования второго поколения, три способа секвенирования на четырехканальной, двухканальной и одноканальной основе имеют свои собственные преимущества; при их сравнении, одноканальное секвенирование имеет преимущества меньшего потребления расходных материалов, более низкой стоимости, более простого получения миниатюризации и портативных устройств, и постепенно становится тенденцией развития в области секвенирования. В настоящее время, продукты, основанные на монохромном канале, на рынке в основном включают секвенаторы серий с ионным потоком, секвенаторы 454 и последний Iseq100 от Illumina.
Среди современных технологий секвенирования на одноканальной основе, использование серий устройств с ионным потоком ограничено из-за высокой частоты ошибок секвенирования структуры полимера. Сходным образом, устройство 454 от Roche также постепенно убирают с рынка секвенирования из-за недостаточной точности секвенирования и высокой стоимости секвенирования. Illumina's Iseq100 основан на технологии монохроматической флуоресценции и полупроводниковой технологии для реализации миниатюризации секвенатора и сохранения высокого качества секвенирования. Однако, поскольку оптический сигнал излучается лазером, устройство оборудовано дополнительным лазером, который увеличивает объем устройства. Кроме того, чтобы удалить фоновое значение, образованное светом возбуждения, специальную обработку необходимо проводить на полупроводниковом чипе, чтобы отфильтровывать фон, образованный светом возбуждения, и эта обработка может приводить к высоким затратам и увеличенной стоимости секвенирования.
Таким образом, в данной области существует необходимость в дешевом способе секвенирования, который не требует внешнего источника света возбуждения и не нуждается во внедрении дополнительного дизайна, чтобы отфильтровывать фон, образованный лазерным источником света.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Для преодоления вышеупомянутых технических проблем, авторы настоящего изобретения разработали новый способ секвенирования, в котором используют сигнал системы самолюминесценции, чтобы различать четыре основания A, (T/U), C и G. Таким образом, сигнал люминесценции, используемый для осуществления способа секвенирования по настоящему изобретению, происходит из биолюминесценции или хемилюминесценции, таким образом, нет необходимости в конфигурации дополнительного лазера, и нет необходимости внедрять дополнительный дизайн, чтобы отфильтровывать фон, образованный лазерным источником света, что уменьшает стоимость секвенирования. Устройство для секвенирования, используемое в способе секвенирования по настоящему изобретению, можно даже удобно переносить для немедленной детекции/детекции на месте. Кроме того, 3'-концевой гидроксил из дезоксирибонуклеотида, используемого в способе секвенирования по настоящему изобретению, является модифицированным и блокированным. Во время процесса секвенирования, только один дезоксирибонуклеотид можно синтезировать в каждой реакции, гарантируя связывание только одного дезоксирибонуклеотида в каждой реакции, таким образом, улучшая точность секвенирования.
В одном аспекте, настоящее изобретение относится к способу секвенирования молекулы нуклеиновой кислоты, включающему следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3'-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3'-конец растущей цепи нуклеиновой кислоты;
(5) обеспечение возможности контакта дуплекса из предыдущей стадии с двумя различными люциферазами и проведение реакции лигирования, где две люциферазы можно специфически лигировать с первой молекулярной меткой и второй молекулярной меткой, соответственно; затем обеспечение возможности подвергания люцифераз флуоресцентной реакции в присутствии субстрата, и детекция излучаемого сигнала флуоресценции;
(6) удаление молекулярной метки каждого нуклеотида;
(7) необязательно, повторение стадий (3) - (7) для получения информации о последовательности молекулы нуклеиновой кислоты.
В одном варианте осуществления настоящего изобретения, детекцию самолюминесценции нуклеотида, подлежащего секвенированию, осуществляют посредством лигирования люциферазы и производного нуклеотида, так что нет необходимости в дополнительном источнике света возбуждения. В конкретном варианте осуществления, лигирование люциферазы и нуклеотида осуществляют посредством специфического связывания метки на люциферазе и соответствующей метки на производном нуклеотида. В конкретном варианте осуществления, первый нуклеотид лигируют с первой люциферазой, второй нуклеотид лигируют с второй люциферазой, третий нуклеотид лигируют с первой люциферазой и второй люциферазой, и четвертый нуклеотид не лигируют ни с какой люциферазой. Затем сигналы люминесценции четырех оснований детектируют посредством пропускания соответствующих субстратов двух люцифераз; когда субстрат первой люциферазы пропускают, первый и третий нуклеотиды излучают свет, когда субстрат второй люциферазы пропускают, второй и третий нуклеотиды излучают свет, так что основания можно идентифицировать в соответствии с люминесценцией четырех нуклеотидов.
Таким образом, в иллюстративном варианте осуществления, способ секвенирования молекулы нуклеиновой кислоты по настоящему изобретению включает следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов, и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3’-конец растущей цепи нуклеиновой кислоты;
(5) удаление фазы раствора из реакционной системы на предыдущей стадии, сохранение дуплекса связанным с подложкой, и добавление двух различных люцифераз для проведения реакции лигирования, где две люциферазы можно специфически лигировать с первой молекулярной меткой и второй молекулярной меткой, соответственно;
(6) удаление несвязанной люциферазы посредством использования буфера для элюции;
(7) добавление субстрата первой люциферазы и детекция флуоресцентного сигнала в то же самое время;
(8) удаление раствора из реакции предыдущей стадии;
(9) добавление субстрата второй люциферазы и детекция флуоресцентного сигнала в то же самое время;
(10) удаление раствора из реакции предыдущей стадии;
(11) удаление молекулярной метки и 3'-защитной группы каждого нуклеотида;
(12) необязательно, удаление раствора из реакции предыдущей стадии;
(13) необязательно, повторение стадий (3) - (12) для получения информации о последовательности молекулы нуклеиновой кислоты.
В конкретном варианте осуществления, лигирование двух люцифераз с нуклеотидами можно также проводить отдельно. Например, первую люциферазу, меченную первым способом, можно добавлять первой, чтобы позволять ей подвергаться реакции лигирования с нуклеотидом, меченным первой молекулой, затем несвязанную первую люциферазу удаляют посредством использования буфера для элюции, добавляют субстрат первой люциферазы, и флуоресцентный сигнал детектируют в то же самое время; затем, вторую люциферазу, меченную вторым способом, можно добавлять, чтобы позволять ей подвергаться реакции лигирования с нуклеотидом, меченным второй молекулой, затем несвязанную вторую люциферазу удаляют посредством использования буфера для элюции, добавляют субстрат второй люциферазы, и флуоресцентный сигнал детектируют в то же самое время. Конкретно, настоящее изобретение относится к способу секвенирования молекулы нуклеиновой кислоты, включающему следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов, и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3’-конец растущей цепи нуклеиновой кислоты;
(5) удаление фазы раствора из реакционной системы на предыдущей стадии, сохранение дуплекса связанным с подложкой, и добавление первой люциферазы для проведения реакции лигирования, где первая люцифераза может специфически связываться с первой молекулярной меткой;
(6) удаление несвязанной первой люциферазы посредством использования буфера для элюции;
(7) добавление субстрата первой люциферазы и детекция флуоресцентного сигнала в то же самое время;
(8) удаление раствора из реакции предыдущей стадии;
(9) добавление второй люциферазы для проведения реакции лигирования, где вторая люцифераза может специфически связываться с второй молекулярной меткой;
(10) удаление несвязанной второй люциферазы посредством использования буфера для элюции;
(11) добавление субстрата второй люциферазы и детекция флуоресцентного сигнала в то же самое время;
(12) удаление раствора из реакции предыдущей стадии;
(13) необязательно, удаление молекулярной метки и 3’-защитной группы каждого нуклеотида;
(14) необязательно, повторение стадий (3) - (13) или (3) - (11) один или несколько раз для получения информации о последовательности молекулы нуклеиновой кислоты.
В конкретном варианте осуществления, две люциферазы могут являться одинаковыми, которые могут специфически связываться с первой молекулярной меткой и второй молекулярной меткой, соответственно, то есть используют только одну люциферазу и субстрат, и первую люциферазу, меченную первым способом, и вторую люциферазу, меченную вторым способом, отдельно лигируют с нуклеотидом. Например, первую люциферазу, меченную первым способом, добавляют первой, позволяют ей подвергаться реакции лигирования с нуклеотидом, меченным первой молекулой, затем несвязанную первую люциферазу удаляют с использованием буфера для элюции, добавляют субстрат люциферазы, и флуоресцентный сигнал детектируют в то же самое время; затем вторую люциферазу, меченную вторым способом, добавляют, позволяют ей подвергаться реакции лигирования с нуклеотидом, меченным второй молекулой, затем несвязанную вторую люциферазу удаляли с использованием буфера для элюции, добавляют субстрат люциферазы, и флуоресцентный сигнал детектируют в то же самое время. Конкретно, настоящее изобретение относится к способу секвенирования молекулы нуклеиновой кислоты, включающему следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов, и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3’-конец растущей цепи нуклеиновой кислоты;
(5) удаление фазы раствора из реакционной системы на предыдущей стадии, сохранение дуплекса связанным с подложкой, и добавление первой люциферазы для проведения реакции лигирования, где первая люцифераза может специфически связываться с первой молекулярной меткой;
(6) удаление несвязанной первой люциферазы посредством использования буфера для элюции;
(7) добавление субстрата люциферазы и детекция флуоресцентного сигнала в то же самое время;
(8) удаление раствора из реакции предыдущей стадии;
(9) добавление реагента для денатурации люциферазы;
(10) удаление раствора из реакции предыдущей стадии;
(11) добавление второй люциферазы для проведения реакции лигирования, где вторая люцифераза может специфически связываться с второй молекулярной меткой;
(12) удаление несвязанной второй люциферазы посредством использования буфера для элюции;
(13) добавление субстрата люциферазы и детекция флуоресцентного сигнала в то же самое время;
(14) необязательно, удаление раствора из реакции предыдущей стадии;
(15) необязательно, удаление молекулярной метки и 3’-защитной группы каждого нуклеотида;
(16) необязательно, повторение стадий (3) - (15) или (3) - (13) один или несколько раз для получения информации о последовательности молекулы нуклеиновой кислоты.
В другом аспекте настоящее изобретение также относится к набору для секвенирования полинуклеотида, содержащему:
(a) четыре соединения, которые представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой; и
(b) две люциферазы, которые могут специфически связываться с первой молекулярной меткой и второй молекулярной меткой, соответственно, и две люциферазы могут быть одинаковыми или различными.
В некоторых предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит: реагент и/или устройство для выделения молекулы нуклеиновой кислоты из образца; реагент для предварительной обработки молекулы нуклеиновой кислоты; подложку для связывания молекулы нуклеиновой кислоты, подлежащей секвенированию; реагент для связывания (например, ковалентного или нековалентного связывания) молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой; праймер для инициации реакции полимеризации нуклеотидов; полимеразу для проведения реакции полимеризации нуклеотидов; один или несколько буферных растворов; один или несколько растворов для промывки; или любую их комбинацию.
ПОДРОБНОЕ ОПИСАНИЕ
Если не определено иное, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, которое является общепринятым для специалиста в области, к которой относится настоящее изобретение. Полное содержание всех патентов, заявок и других публикаций, упомянутых в настоящем описании, приведено в качестве ссылки. Если определения, приведенные в настоящем описании, имеют противоречие или несоответствие с определениями, описанными в патентах, заявках и других публикациях, содержание которых приведено в настоящем описании в качестве ссылки, определения, описанные в настоящем описании, должны иметь преимущество.
Как используют в настоящем описании, термин «полинуклеотид» относится к дезоксирибонуклеиновой кислоте (ДНК), рибонуклеиновой кислоте (РНК) или их аналогам. Полинуклеотиды могут являться одноцепочечными, двухцепочечными или содержать как одноцепочечные, так и двухцепочечные последовательности. Молекулы полинуклеотидов могут происходить из двухцепочечной формы ДНК (дцДНК) (например, геномной ДНК, продуктов ПЦР и амплификации и т.д.), или могут происходить из одноцепочечной формы ДНК (оцДНК) или РНК, и их можно переводить в форму дцДНК, и наоборот. Точная последовательность молекулы полинуклеотида может являться известной или неизвестной. Следующее представляет собой иллюстративные примеры полинуклеотидов: ген или фрагмент гена (например, зонд, праймер, метка EST или SAGE), геномная ДНК, фрагмент геномной ДНК, экзон, интрон, матричная РНК (мРНК), транспортная РНК, рибосомальная РНК, рибозим, кДНК, рекомбинантный полинуклеотид, синтетический полинуклеотид, разветвленный полинуклеотид, плазмида, вектор, выделенная ДНК любой последовательности, выделенная РНК любой последовательности и нуклеиновая кислота-зонд, праймер или амплифицированная копия любой из вышеуказанных последовательностей.
Полинуклеотиды могут включать нуклеотиды или аналоги нуклеотидов. Нуклеотид обычно содержит сахарид (т.е. рибозу или дезоксирибозу), основание, и по меньшей мере одну фосфатную группу. Нуклеотид может являться лишенным азотистого основания (т.е. свободным от основания). Нуклеотид включает дезоксирибонуклеотид, модифицированный дезоксирибонуклеотид, рибонуклеотид, модифицированный рибонуклеотид, пептидный нуклеотид, модифицированный пептидный нуклеотид, нуклеозид с модифицированным фосфатно-сахаридным остовом и их смеси. Примеры нуклеотида включают, например, аденозинмонофосфат (AMP), аденозиндифосфат (ADP), аденозинтрифосфат (ATP), тимидинмонофосфат (TMP), тимидиндифосфат (TDP), тимидинтрифосфат (TTP), цитидинмонофосфат (CMP), цитидиндифосфат (CDP), цитидинтрифосфат (CTP), гуанозинмонофосфат (GMP), гуанозиндифосфат (GDP), гуанозинтрифосфат (GTP), уридинмонофосфат (UMP), уридиндифосфат (UDP), уридинтрифосфат (UTP), дезоксиаденозинмонофосфат (dAMP), дезоксиаденозиндифосфат (dADP), дезоксиаденозинтрифосфат (dATP), дезокситимидинмонофосфат (dTMP), дезокситимидиндифосфат (dTDP), дезокситимидинтрифосфат (dTTP), дезоксицитидиндифосфат (dCDP), дезоксицитидинтрифосфат (dCTP), дезоксигуанозинмонофосфат (dGMP), дезоксигуанозиндифосфат (dGDP), дезоксигуанозинтрифосфат (dGTP), дезоксиуридинмонофосфат (dUMP), дезоксиуридиндифосфат (dUDP) и дезоксиуридинтрифосфат (dUTP). Аналоги нуклеотидов, содержащие модифицированные основания, можно также использовать в способах, описанных в настоящем описании. Независимо от того, имеет ли он природный остов или сходную структуру, иллюстративное модифицированное основание, которое может содержаться в полинуклеотиде, включает, например, инозин, ксатанин, гипоксатанин, изоцитозин, изогуанин, 2-аминопурин, 5-метилцитозин, 5-гидроксиметилцитозин, 2-аминоаденин, 6-метиладенин, 6-метилгуанин, 2-пропилгуанин, 2-пропиладенин, 2-тиоурацил, 2-тиотимин, 2-тиоцитозин, 15-галогенированный урацил, 15-галогенированный цитозин, 5-пропинилурацил, 5-пропинилцитозин, 6-азоурацил, 6-азоцитозин, 6-азотимин, 5-урацил, 4-тиоурацил, 8-галогенированный аденин или гуанин, 8-аминоаденин или 8-аминогуанин, 8-тиоаденин или 8-тиогуанин, 8-тиоалкиладенин или 8-тиоалкилгуанин, 8-гидроксиаденин или 8-гидроксигуанин, 5-галогенированный урацил или цитозин, 7-метилгуанин, 7-метиладенин, 8-азагуанин, 8-азааденин, 7-деазагуанин, 7-деазааденин, 3-деазагуанин, 3-деазааденин и т.д. Как известно в данной области, конкретные аналоги нуклеотидов, например, аналоги нуклеотидов, такие как аденозин-5'-фосфорилсульфат, нельзя включать в полинуклеотид.
Говоря в общем, нуклеотиды включают нуклеотиды A, C, G, T или U. Как используют в настоящем описании, термин «нуклеотид A» относится к нуклеотиду, содержащему аденин (A) или его модифицированную форму или аналог, такому как ATP, dATP. «Нуклеотид G» относится к нуклеотиду, содержащему гуанин (G) или его модифицированную форму или аналог, такому как GTP, dGTP. «Нуклеотид C» относится к нуклеотиду, содержащему цитозин (C) или его модифицированную форму или аналог, такому как CTP, dCTP. «Нуклеотид T» относится к нуклеотиду, содержащему тимин (T) или его модифицированную форму или аналог, такому как TTP, dTTP. «Нуклеотид U» относится к нуклеотиду, содержащему урацил (U) или его модифицированную форму или аналог, такому как UTP, dUTP.
Мечение нуклеотида
Настоящее изобретение относится к мечению нуклеотида с использованием различных меток, индивидуально или в комбинации, так что различные люциферазы можно лигировать с нуклеотидом. Как используют в настоящем описании, молекулярная метка для мечения нуклеотида и метка, специфически связывающаяся с ней, могут представлять собой любые образующие пары молекулы, которые могут специфически связываться друг с другом. Специфическое связывание между образующими пару членами реализует лигирование нуклеотида с люциферазой. Иллюстративные образующие пару члены включают, но без ограничения: (a) гаптен или антигенное соединение в комбинации с соответствующим антителом или его связывающей частью или фрагментом, такие как дигоксин-антитело против дигоксина, N3G-антитело против N3G, FITC-антитело против FITC; (b) нуклеиновая кислота-аптамер и белок; (c) неиммунную связывающуюся пару (например, биотин-авидин, биотин-стрептавидин, биотин-нейтравидин); (d) гормон-связывающий гормон белок; (e) рецептор-агонист или антагонист рецептора; (f) лектин-углевод; (g) фермент-кофактор фермента; (h) фермент-ингибитор фермента; и (i) пару комплементарных олигонуклеотидов или полинуклеотидов, способных формировать дуплекс нуклеиновой кислоты.
В конкретном варианте осуществления, первая молекулярная метка и вторая молекулярная метка представляют собой низкомолекулярные метки, которые выбраны из биотина, дигоксина, N3G или FITC. Две люциферазы могут специфически связываться с первой молекулярной меткой и второй молекулярной меткой, соответственно. Например, в конкретном варианте осуществления, первая молекулярная метка представляет собой биотин, тогда первая люцифераза может представлять собой меченную стрептавидином люциферазу; вторая молекулярная метка представляет собой дигоксин, вторая люцифераза может представлять собой меченную антителом против дигоксина люциферазу, которая является отличной от первой люциферазы, и вторая люцифераза может также представлять собой меченную антителом против дигоксина люциферазу, которая является такой же, как первая люцифераза. Источник люциферазы включает, но без ограничения, светляка, gaussia, Renilla и другие организмы. Например, меченная стрептавидином люцифераза может представлять собой SA-Gluc: стрептавидин-люцифераза Gaussia princeps от Adivity Company. Меченная антителом против дигоксина люцифераза может представлять собой антитело против дигоксина-Gluc или антитело против дигоксина-Nluc.
Как используют в настоящем описании, выражение «первое соединение связано с первой молекулярной меткой» означает, что все первое соединение связано с первой молекулярной меткой, или часть первого соединения связана с первой молекулярной меткой, в то время как остальное первое соединение не является связанным с молекулярной меткой. Таким же образом, выражение «второе соединение связано с второй молекулярной меткой» означает, что все второе соединение связано с второй молекулярной меткой, или часть второго соединения связана с второй молекулярной меткой, в то время как остальное второе соединение не является связанным с молекулярной меткой. Выражение «третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой» означает, что все третье соединение связано с первой молекулярной меткой и второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой и второй молекулярной меткой, в то время как остальное третье соединение не является связанным с молекулярными метками.
Секвенирование полинуклеотида
Предпочтительно, нуклеотиды, лигированные с различными люциферазами по настоящему изобретению, являются пригодными для секвенирования посредством синтеза. Способы секвенирования посредством синтеза, как используют в настоящем описании, представляют собой различные способы секвенирования посредством синтеза, хорошо известные в данной области. В основном, секвенирование посредством синтеза включает: сначала гибридизацию молекулы нуклеиновой кислоты, подлежащей секвенированию, с праймером для секвенирования, и затем, в присутствии полимеразы, полимеризацию нуклеотидов, лигированных с различными люциферазами, как описано в настоящем описании, на 3’-конце праймера для секвенирования, с использованием молекулы нуклеиновой кислоты, подлежащей секвенированию, в качестве матрицы. После полимеризации, нуклеотид идентифицируют посредством детекции флуоресцентного сигнала, излучаемого люциферазой. После удаления люциферазы из меченого нуклеотида, проводят следующий цикл секвенирования полимера.
Способ определения последовательности полинуклеотида-мишени можно осуществлять следующим образом: денатурация последовательности полинуклеотида-мишени, приведение полинуклеотида-мишени в контакт с различными нуклеотидами, соответственно, таким образом, чтобы формировать нуклеотид, комплементарный нуклеотиду-мишени, и детекция включенного нуклеотида. В способе используют полимеризацию, позволяющую полимеразе продлевать комплементарную цепь посредством включения корректных нуклеотидов, комплементарных мишени. Реакция полимеризации требует также специального праймера для инициации полимеризации.
Для каждого цикла реакции, включение нуклеотида осуществляют посредством полимеразы, и затем измеряют событие включения. Существует много различных полимераз, и специалисту в данной области легко определять наиболее подходящую полимеразу. Предпочтительные ферменты включают ДНК-полимеразу I, фрагмент Кленова, ДНК-полимеразу III, ДНК-полимеразу T4 или T7, Taq-полимеразу или полимеразу vent. Можно также использовать полимеразы, сконструированные для наличия специфических свойств.
Способ секвенирования предпочтительно осуществляют для полинуклеотида-мишени, аранжированного на твердой подложке. Посредством линкерных молекул, множество полинуклеотидов-мишеней можно иммобилизовывать на твердой подложке, или можно присоединять к частицам, таким как микросферы, и частицы можно также присоединять к материалу твердой подложки.
Полинуклеотид можно присоединять к твердой подложке множеством способов, включая использование взаимодействия биотин-стрептавидин. Способы иммобилизации полинуклеотидов на твердой подложке хорошо известны в данной области, и включают способы литографии и нанесение пятна каждого полинуклеотида в конкретном положении на твердой подложке. Пригодные твердые подложки известны в данной области и включают стеклянные пластины и бусины, керамические и силиконовые поверхности, и пластиковые материалы. Подложка обычно является плоской, хотя микробусины (микросферы) также можно использовать, и последние можно также присоединять к другим твердым подложкам известными способами. Микросферы могут иметь любой пригодный размер, и их диаметр обычно составляет 10-100 нанометров. В предпочтительном варианте осуществления, полинуклеотид напрямую прикреплен на плоской поверхности, предпочтительно, на плоской стеклянной поверхности. Связывание предпочтительно осуществляют в форме ковалентной связи. Используемый массив, предпочтительно, представляет собой массив отдельных молекул, который включает полинуклеотиды, локализованные в уникальных оптически различимых областях, например, как описано в Международной патентной заявке No. WO00/06770.
Необходимые условия для полимеризации хорошо известны специалисту в данной области. Для проведения полимеразной реакции, обычно последовательность праймера должна сначала гибридизоваться с полинуклеотидом-мишенью. Последовательность праймера поддается узнаванию полимеразой и служит в качестве участка инициации для последующего удлинения комплементарной цепи. Последовательность праймера можно добавлять в качестве независимого компонента, относительно полинуклеотида-мишени. Кроме того, каждый из праймера и полинуклеотида-мишени может составлять часть одноцепочечной молекулы, и часть праймера и часть мишени формируют внутримолекулярный дуплекс, то есть, структуру стебля-петли. Структура может быть иммобилизована на твердой подложке посредством любого положения молекулы. Другие условия, необходимые для полимеразной реакции, хорошо известны специалисту в данной области, и эти условия включают температуру, pH и состав буфера.
Впоследствии, меченые нуклеотиды по настоящему изобретению приводят в контакт с полинуклеотидом-мишенью, чтобы позволить полимеризацию. Нуклеотиды можно добавлять последовательно, то есть, каждый тип нуклеотида (A, C, G или T/U) добавляют отдельно или добавляют в то же самое время.
Стадии полимеризации позволяют протекать в течение периода времени, достаточного для включения одного нуклеотида.
Затем, невключенные нуклеотиды удаляют, например, посредством удаления фазы раствора из реакционной системы на предыдущей стадии, оставляя дуплексы прикрепленными к подложке.
Впоследствии, две люциферазы, содержащие различные люциферазы, можно добавлять для проведения реакции лигирования. Две люциферазы могут специфически связываться с молекулярными метками для мечения нуклеотидов, соответственно, таким образом, реализуя лигирование люциферазы с включенным нуклеотидом. Затем, посредством добавления соответствующих субстратов люцифераз и детекции флуоресцентного сигнала, осуществляют идентификацию включенного нуклеотида. Можно также добавлять два вида люцифераз, содержащих одинаковую люциферазу, для проведения реакции лигирования. Люциферазы могут специфически связываться с молекулярными метками для мечения нуклеотидов, соответственно, таким образом, реализуя лигирование люциферазы с включенным нуклеотидом. Затем, посредством добавления соответствующего субстрата люциферазы и детекции флуоресцентного сигнала, реализуют идентификацию включенного нуклеотида.
В конкретном варианте осуществления, четыре аналога дезоксирибонуклеотидов являются меченными с использованием различных низкомолекулярных меток, т.е., биотина (сокращенно обозначенного как B) и дигоксина (сокращенно обозначенного как D), например, нуклеотид A является меченным с использованием B, нуклеотид C является меченным с использованием B и D, нуклеотид T является меченным с использованием D, и нуклеотид G не является меченным. 3'-концевые гидроксильные группы четырех аналогов дезоксирибонуклеотидов, меченных с использованием различных малых молекул, все блокируют, чтобы гарантировать, что только один дезоксирибонуклеотид связывается в ходе каждой реакции секвенирования. В ходе реакции секвенирования, сначала вводят смесь четырех меченых аналогов дезоксирибонуклеотидов и полимеразы для секвенирования, под действием полимеразы, один аналог дезоксирибонуклеотида включают в 3’-конец растущей цепи нуклеиновой кислоты, в соответствии с принципом комплементарного спаривания оснований. Посредством удаления фазы раствора из реакционной системы на предыдущей стадии, оставляя дуплекс связанным с подложкой, несвязанные аналоги дезоксирибонуклеотидов можно удалять. Затем добавляют две люциферазы, содержащие различные люциферазы, где первая люцифераза является меченной с использованием стрептавидина, который связывается с нуклеотидом A или нуклеотидом C, меченным с использованием малой молекулы B, и вторая люцифераза является меченной с использованием антитела против дигоксина, которое связывается с нуклеотидом C или нуклеотидом T, меченным с использованием малой молекулы D. После удаления несвязанных люцифераз с использованием буфера для элюции, добавляют субстрат первой люциферазы, нуклеотид, лигированный с первой люциферазой, излучает свет, и сигнал детектируют посредством детектора; добавляют субстрат второй люциферазы, нуклеотид, лигированный с второй люциферазой, излучает свет, и сигнал детектируют посредством детектора, так что получают люминесценцию, показанную в следующей таблице, и основание можно идентифицировать.
В конкретном варианте осуществления, лигирование двух люцифераз, содержащих различные люциферазы, с мечеными нуклеотидами и детекцию сигнала можно осуществлять отдельно. Сначала, добавляют первую люциферазу, которая является меченной с использованием стрептавидина и связывается с нуклеотидом A или нуклеотидом C, меченным с использованием малой молекулы B. После удаления несвязанной первой люциферазы с использованием буфера для элюции, добавляют субстрат первой люциферазы, нуклеотид, лигированный с первой люциферазой, излучает свет, и сигнал детектируют посредством детектора. После удаления реакционного раствора, добавляют вторую люциферазу, меченную с использованием антитела против дигоксина, которая связывается с нуклеотидом C или нуклеотидом T, меченным с использованием малой молекулы D, и затем несвязанную вторую люциферазу удаляют с использованием буфера для элюции, добавляют субстрат второй люциферазы, нуклеотид, лигированный с второй люциферазой, излучает свет, и сигнал детектируют посредством детектора. Таким образом, получают люминесценцию, показанную в таблице выше, и можно осуществлять идентификацию оснований.
В другом конкретном варианте осуществления, лигирование двух люцифераз, содержащих одинаковую люциферазу, с мечеными нуклеотидами и детекцию сигнала осуществляют отдельно. Сначала, добавляют первую люциферазу, которая является меченной с использованием стрептавидина и связывается с нуклеотидом A или нуклеотидом C, меченным с использованием малой молекулы B. После удаления несвязанной первой люциферазы с использованием буфера для элюции, добавляют субстрат люциферазы, нуклеотид, лигированный с первой люциферазой, излучает свет, и сигнал детектируют посредством детектора. Затем, добавляют реагент, который денатурирует первую люциферазу; после удаления реакционного раствора, добавляют вторую люциферазу, которая является меченной с использованием антитела против дигоксина и связывается с нуклеотидом C или нуклеотидом T, меченным с использованием малой молекулы D, затем несвязанную вторую люциферазу удаляют посредством использования буфера для элюции, добавляют субстрат люциферазы, основание, лигированное с второй люциферазой, излучает свет, и сигнал детектируют посредством детектора. Таким образом, получают люминесценцию, показанную в таблице выше, и можно осуществлять идентификацию оснований.
Детекция сигнала флуоресценции
Способ детекции флуоресцентного сигнала хорошо известен в данной области. Например, он может быть реализован посредством устройства, детектирующего длину волны флуоресценции. Такие устройства хорошо известны в данной области. Например, такое устройство может представлять собой конфокальный сканирующий микроскоп, который сканирует поверхность твердой подложки с использованием лазера, чтобы визуализировать флуорофор, напрямую связанный с молекулой нуклеиновой кислоты, подлежащей секвенированию. Кроме того, чувствительный 2-D детектор, такой как детектор с зарядовой связью (CCD), можно использовать, например, для наблюдения каждого полученного сигнала. Можно использовать также, например, другие способы, такие как ближнепольная сканирующая оптическая микроскопия (SNOM).
Удаление метки
После детекции, подходящие условия можно использовать для удаления метки, присоединенной к нуклеотидам.
В конкретном варианте осуществления, меченые нуклеотиды по настоящему изобретению также имеют 3’-защитную группу. В некоторых вариантах осуществления настоящего изобретения, защитная группа и метка обычно представляют собой две различные группы на 3'-блокированном меченом нуклеотиде, но в других вариантах осуществления, защитная группа и метка могут также представлять собой одинаковую группу.
Как используют в настоящем описании, термин «защитная группа» относится к группе, которая препятствует полимеразе (которая включает нуклеотид, содержащий группу, в синтезируемую полинуклеотидную цепь) безостановочно катализировать включение другого нуклеотида после включения нуклеотида, содержащего группу, в синтезируемую полинуклеотидную цепь. Такая защитная группа также обозначена в настоящем описании как 3'-OH защитная группа. Нуклеотид, содержащий такую защитную группу, также обозначен в настоящем описании как 3’-блокированный нуклеотид. Защитная группа может представлять собой любую пригодную группу, которую можно добавлять к нуклеотиду, при условии, что защитная группа может препятствовать включению дополнительной молекулы нуклеотида в полинуклеотидную цепь и может быть легко удалена из сахаридной части нуклеотида без разрушения полинуклеотидной цепи. Кроме того, нуклеотид, модифицированный с использованием защитной группы, должен является устойчивым к полимеразам или другим подходящим ферментам в отношении включения модифицированного нуклеотида в полинуклеотидную цепь. Таким образом, идеальная защитная группа имеет длительную стабильность, может быть эффективно включена посредством полимеразы, предотвращает вторичное или последующее включение нуклеотидов, и может быть удалена в мягких условиях, предпочтительно, в водных условиях, которые не разрушают структуру полинуклеотида.
На предшествующем уровне техники описано множество защитных групп, соответствующих приведенному выше описанию. Например, в WO 91/06678 описано, что 3'-OH-защитные группы включают сложные эфиры и эфиры, -F, -NH2, -OCH3, -N3, -OPO3, -NHCOCH3, 2-нитрофенилкарбонат, 2,4-гипосульфонилдинитро и тетрагидрофурановый эфир. В Metzker et al. (Nucleic Acids Research, 22(20): 4259-4267, 1994) описаны синтез и применение восьми 3'-модифицированных 2-дезоксирибонуклеозид-5'-трифосфатов (3'-модифицированных dNTP). В WO2002/029003 описано применение аллильной защитной группы для кэпирования 3'-OH-группы растущей цепи ДНК в полимеразной реакции. Предпочтительно, можно использовать различные защитные группы, опубликованные в Публикациях международных заявок WO2014139596 и WO2004/018497, включая, например, защитные группы, проиллюстрированные на фигуре 1A, и защитные группы 3'-гидроксила (т.е., защитные группы), определенные в формуле изобретения WO2014139596, и например, защитные группы, проиллюстрированные на фигурах 3 и 4, и группы, определенные в формуле изобретения WO2004/018497. Полное содержание вышеуказанных ссылок приведено в настоящем описании в качестве ссылки.
Специалисту в данной области понятно, как присоединять подходящую защитную группу к рибозному кольцу, чтобы блокировать взаимодействие с 3'-OH. Защитную группу можно напрямую присоединять в 3'-положении, или можно присоединять в 2'-положении (защитная группа имеет достаточный размер или заряд для блокирования взаимодействия в 3'-положении). Кроме того, защитную группу можно присоединять в 3'- и 2'-положениях, и можно отщеплять для экспонирования 3'-OH-группы.
После успешного включения 3'-блокированного нуклеотида в растущую цепь нуклеиновой кислоты, способ секвенирования требует удаления защитной группы для получения пригодного к использованию 3'-OH-участка для непрерванного синтеза цепи. Реагенты, которые могут удалять защитные группы из модифицированных нуклеотидов, как используют в настоящем описании, зависят, в большой степени, от используемых защитных групп. Например, удаление защитной группы сложного эфира с 3'-гидроксильной группы обычно осуществляют посредством щелочного гидролиза. Простота удаления защитных групп сильно меняется; как правило, чем больше электроотрицательность заместителя на карбонильном атоме углерода, тем больше простота удаления. Например, высоко электроотрицательную группу трифторуксусной кислоты можно быстро отщеплять от 3'-гидроксильной группы при pH 7 в метаноле (Cramer et al., 1963), поскольку она является нестабильной во время полимеризации при этом pH. Группу феноксиацетата отщепляют в пределах менее чем 1 минуты, но необходим значительно более высокий pH, например, с использованием NH-/метанола (Reese and Steward, 1968). Различные защитные группы гидрокси можно избирательно отщеплять с использованием химических способов, отличных от щелочного гидролиза. 2,4-динитрофенилтио-группу можно быстро отщеплять посредством обработки с использованием нуклеофилов, таких как тиофенол и тиосульфат (Letsinger et al., 1964). Аллильный эфир можно отщеплять посредством обработки с использованием Hg(II) в ацетоне/воде (Gigg and Warren, 1968). Тетрагидротианиловый эфир можно удалять в нейтральных условиях с использованием Ag(I) или Hg (II) (Cohen and Steele, 1966; Cruse et al., 1978). Фотохимическое снятие блокировки можно использовать с фотохимически отщепляемыми защитными группами. Существует несколько защитных групп, которые можно использовать в этом способе. Использование o-нитробензильного эфира в качестве защитной группы для функциональной группы 2'-гидроксила рибонуклеозида известно и подтверждено (Ohtsuka et al., 1978); и ее удаляют посредством облучения при 260 нм. Защитную группу алкилкарбоната o-нитробензилкарбоната также удаляют посредством облучения при pH 7 (Cama and Christensen, 1978). Ферментное снятие блокировки защитной группы 3'-OH также является возможным. Показано, что полинуклеотидкиназа T4 может переводить 3'-фосфатный конец в 3'-гидроксильный конец, который затем можно использовать в качестве праймера для ДНК-полимеразы I (Henner et al., 1983). Эту активность 3'-фосфатазы используют для удаления 3'-защитной группы этих аналогов dNTP, содержащих фосфат в качестве защитной группе.
Другие реагенты, которые могут удалять защитные группы с 3'-блокированных нуклеотидов включают, например, фосфины (например, трис(гидроксиметил)фосфин (THP)), которые могут, например, удалять защитную группу 3'-OH, содержащую азид, с нуклеотида (это применение фосфина см., например, в описании WO2014139596, полное содержание которого приведено в настоящем описании в качестве ссылки). Другие реагенты, которые могут удалять защитные группы с 3'-блокированных нуклеотидов, также включают, например, соответствующие реагенты, описанные на страницах 114-116 в описании WO2004/018497, для удаления 3'-аллила, 3,4-диметоксибензилоксиметила или фторметоксиметила, используемых в качестве защитной группы 3'-OH.
В одном варианте осуществления настоящего изобретения, метку нуклеотида предпочтительно удаляют вместе с защитной группой после детекции.
В конкретных вариантах осуществления, метку можно включать в защитную группу, таким образом, позволяя ее удаление вместе с защитной группой после включения 3'-блокированного нуклеотида в цепь нуклеиновой кислоты.
В других вариантах осуществления, метку можно присоединять к нуклеотиду с использованием связывающей группы и защитную группу отдельно. Такую метку можно, например, присоединять к пуриновому или пиримидиновому основанию нуклеотида. В конкретных вариантах осуществления, используемая связывающая группа является отщепляемой. Использование отщепляемой связывающей группы обеспечивает то, что метку можно удалять после детекции, что исключает любую интерференцию сигналов с любыми мечеными нуклеотидами, включенными впоследствии. В других вариантах осуществления, можно использовать неотщепляемую связывающую группу, поскольку после включения меченого нуклеотида в цепь нуклеиновой кислоты, нет необходимости последующего включения нуклеотида, таким образом, нет необходимости удаления метки с нуклеотида.
В других вариантах осуществления, метка и/или связывающая группа могут иметь размер или структуру, достаточную для блокирования включения других нуклеотидов в полинуклеотидную цепь (то есть, метка сама может служить в качестве защитной группы). Блокирование может быть обусловлено стерическими затруднениями, или оно может быть обусловлено комбинацией размера, заряда и структуры.
Отщепляемые связывающие группы хорошо известны в данной области, и общепринятые химические способы можно использовать для присоединения связывающей группы к нуклеотидному основанию и метки. Связывающую группу можно присоединять к любому положению нуклеотидного основания, при условии, что все еще можно осуществлять уотсон-криковское спаривание оснований. Для пуриновых оснований, является предпочтительным, чтобы связывающая группа была присоединена через положение 7 пуринов или предпочтительных аналогов деазапурина, через 8-модифицированные пурины, через N-6-модифицированные аденины или N-2-модифицированные гуанины. Для пиримидинов, является предпочтительным связывание через положение 5 цитозина, тимина и урацила, и положение N-4 цитидина.
Использование термина «отщепляемая связывающая группа» не означает, что всю связывающую группу необходимо удалять (например, с нуклеотидного основания). Когда метка присоединена к основанию, участок отщепления нуклеозида может быть локализован в некотором положении на связывающей группе, где положение может обеспечивать то, что часть связывающей группы остается присоединенной к нуклеотидному основанию после отщепления.
Пригодные связывающие группы включают, но без ограничения, дисульфидные связывающие группы, кислотолабильные связывающие группы (включая диалкоксибензильные связывающие группы, связывающие группы Зибера, индольные связывающие группы, трет-бутиловые связывающие группы Зибера), электрофильно отщепляемые связывающие группы, нуклеофильно отщепляемые связывающие группы, фотоотщепляемые связывающие группы, связывающие группы, которые можно отщеплять в восстанавливающих и окисляющих условиях, связывающие группы «с предохранителем», и связывающие группы, которые можно отщеплять посредством механизмов элиминирования. Пригодные связывающие группы можно модифицировать с использованием стандартных химических защитных групп, как описано в следующих документах: Greene & Wuts, protecting groups in Organic Synthesis, John Wiley & Sons. В Guillier et al. описаны другие пригодные отщепляемые связывающие группы для твердофазного синтеза (Chem. Rev. 100:2092-2157, 2000).
Связывающую группу можно отщеплять любым пригодным способом, включая подвергание воздействию кислот, оснований, нуклеофилов, электрофилов, свободных радикалов, металлов, восстанавливающих или окисляющих реагентов, света, температуры, ферментов и т.д., и пригодные способы отщепления различных отщепляемых связывающих групп иллюстративно описаны ниже. Как правило, отщепляемую связывающую группу можно отщеплять в таких же условиях, что и условия для защитной группы, так что только одна обработка является необходимой для удаления метки и защитной группы.
Связывающую группу для электрофильного отщепления, как правило, отщепляют посредством протона, что включает чувствительное к кислотам отщепление. Пригодные электрофильно отщепляемые связывающие группы включают модифицированные бензильные системы, такие как тритил, п-гидрокарбонилоксибензильный сложный эфир и п-гидрокарбонилоксибензиламид. Другие пригодные связывающие группы включают группы трет-бутоксикарбонила (Boc) и ацетальные системы. Для получения пригодных связывающих молекул, можно также рассматривать использование тиофильных металлов, таких как никель, серебро или ртуть, в отщеплении тиоацеталей или других содержащих серу защитных групп. Связывающая группа для нуклеофильного отщепления включает группу, которая является нестабильной в воде (т.е., может быть легко отщеплена при щелочном pH), такую как сложный эфир, и группу, которая является нестабильной по отношению к неводному нуклеофилу. Ионы фторида можно использовать для расщепления связей кремний-кислород в таких группах, как триизопропилсилан (TIPS) или трет-бутилдиметилсилан (TBDMS). Фотолизируемые связывающие группы широко используют в химии сахаридов. Предпочтительно, свет, необходимый для активации отщепления, не влияет на другие компоненты в модифицированном нуклеотиде. Например, если флуорофор используют в качестве метки, является предпочтительным, чтобы флуорофор поглощал свет длины волны, отличной от света, необходимого для отщепления связывающей молекулы. Пригодные связывающие группы включают группы, основанные на O-нитробензильных соединениях и нитровератроловых соединениях. Связывающие группы, основанные на химических реакциях бензоина, также можно использовать (Lee et al., J. Org. Chem. 64:3454-3460, 1999). Известно множество связывающих групп, чувствительных к восстановительному отщеплению. Каталитическую гидрогенизацию с использованием катализаторов на основе палладия использовали для отщепления бензильных и бензилоксикарбонильных групп. Восстановление дисульфидной связи также известно в данной области. Способы, основанные на окислении, хорошо известны в данной области. Эти способы включают окисление гидрокарбонилоксибензильной группы и окисление связывающих групп, основанных на сере и селене. В объем настоящего изобретения включено также использование водного иода для отщепления дисульфидных и других основанных на сере или селене связывающих групп. Линкеры «с предохранителем» представляют собой линкеры, которые отщепляются в две стадии. В предпочтительной системе, первая стадия представляет собой получение реакционноспособного нуклеофильного центра, и последующая вторая стадия включает внутримолекулярную циклизацию, которая приводит к отщеплению. Например, левулинатную связь можно обрабатывать с использованием гидразина или фотохимических способов для высвобождения активного амина, и амин затем можно циклизовать для расщепления сложного эфира в другом месте в молекуле (Burgess et al., J. Org. Chem. 62: 5165 -5168, 1997). Реакции элиминирования также можно использовать для отщепления связывающих групп. Можно использовать катализируемое основанием элиминирование групп, таких как флуоренилметоксикарбонил и цианоэтил, и катализируемое палладием восстановительное элиминирование аллильных систем.
В конкретных вариантах осуществления, связывающая группа может содержать спейсерное звено. Длина связывающей группы не является важной, при условии, что метку и нуклеотид поддерживают на достаточном расстоянии, чтобы не создавать помех для взаимодействия между нуклеотидом и ферментом.
В конкретных вариантах осуществления, связывающая группа может состоять из функциональной группы, сходной с защитной группой 3’-OH. Это может позволять то, что только одна обработка является необходимой для удаления метки и защитной группы. Особенно предпочтительной связывающей группой является содержащая азид связывающая группа, которую можно отщеплять посредством фосфина.
Реагенты, которые могут удалять метку с модифицированного нуклеотида, как используют в настоящем описании, в большой степени зависят от используемой метки. Например, в случае, когда защитная группа включена в метку, реагент для удаления защитной группы, описанный выше, используют для удаления метки. Альтернативно, когда метка присоединена к основанию нуклеотида посредством отщепляемой связывающей группы, метку удаляют с использованием реагента, который отщепляет связывающую группу, как описано выше. В предпочтительном варианте осуществления, одинаковый реагент используют для удаления метки и защитной группы с модифицированного нуклеотида, например, в случае, когда связывающая группа состоит из функциональной группы, сходной с защитной группой 3’-OH.
Набор
Настоящее изобретение также относится к набору для секвенирования полинуклеотида, который содержит:
(a) четыре соединения, которые представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксильная группа (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищена посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой; и
(b) две различные люциферазы, которые могут специфически связываться с первой молекулярной меткой и второй молекулярной меткой, соответственно.
В конкретном варианте осуществления, молекулярные метки, используемые для мечения четырех соединений, и метки, используемые для мечения двух люцифераз, являются такими, как определено выше.
В некоторых предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит: реагент и/или устройство для выделения молекулы нуклеиновой кислоты из образца; реагент для предварительной обработки молекулы нуклеиновой кислоты, подлежащей секвенированию; подложку для связывания молекулы нуклеиновой кислоты, подлежащей секвенированию; реагент для связывания (например, ковалентного или нековалентного связывания) молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой; праймер для инициации реакции полимеризации нуклеотидов; полимеразу для проведения реакции полимеризации нуклеотидов; один или несколько буферных растворов; один или несколько растворов для промывки; или любую их комбинацию.
В некоторых предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит реагент и/или устройство для выделения молекулы нуклеиновой кислоты из образца. Способы выделения молекул нуклеиновой кислоты из образцов хорошо известны в данной области. Таким образом, различным реагентам и/или устройствам для выделения молекул нуклеиновой кислоты, таким как реагенты для разрушения клеток, реагенты для преципитации ДНК, реагенты для промывки ДНК, реагенты для растворения ДНК, реагенты для преципитации РНК, реагенты для промывки РНК, реагенты для растворения РНК, реагенты для удаления белков, реагенты для удаления ДНК (например, когда молекула нуклеиновой кислоты - мишень представляет собой РНК), реагенты для удаления РНК (например, когда молекула нуклеиновой кислоты - мишень представляет собой ДНК), и любой их комбинации, можно придавать конфигурацию набора по настоящему изобретению, по необходимости.
В некоторых предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит реагент для предварительной обработки молекулы нуклеиновой кислоты. В наборе по настоящему изобретению, реагенты, используемые для предварительной обработки молекул нуклеиновой кислоты, не подвергаются дополнительным ограничениям, и могут быть выбраны в соответствии с фактическими нуждами. Реагенты, используемые для предварительной обработки молекул нуклеиновой кислоты, включают, например, реагенты для фрагментации молекул нуклеиновой кислоты (например, ДНКазу I), реагенты для дополнения концов молекул нуклеиновой кислоты (например, ДНК-полимеразу, такую как ДНК-полимераза T4, ДНК-полимераза Pfu, ДНК-полимераза Кленова), линкерные молекулы, молекулы метки, реагенты для связывания линкерных молекул с молекулами нуклеиновой кислоты - мишенями (например, лигазу, такую как ДНК-лигаза T4), реагенты для репарации разрывов нуклеиновой кислоты (например, ДНК-полимеразу, которая лишена 3'-5'-экзонуклеазной активности, но имеет 5'-3'-экзонуклеазную активность), реагенты для амплификации молекул нуклеиновой кислоты (например, ДНК-полимеразу, праймер, dNTP), реагенты (например, хроматографические колонки) для разделения и очистки молекул нуклеиновой кислоты, и любую их комбинацию.
В некоторых предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит подложку для связывания молекулы нуклеиновой кислоты, подлежащей секвенированию. Подложка может иметь любые технические признаки и любую их комбинацию, подробно описанные выше для подложки.
Например, по настоящему изобретению, подложка может быть изготовлена из различных пригодных материалов. Такие материалы включают, например, неорганические материалы, природные полимеры, синтетические полимеры и любую их комбинацию. Конкретные примеры включают, но без ограничения: целлюлозу, производные целлюлозы (например, нитроцеллюлозу), акриловую смолу, стекло, силикагель, полистирол, желатин, поливинилпирролидон, сополимер винила и акриламида, и полистирол, перекрестно сшитый, например, с использованием дивинилбензола (см., например, Merrifield Biochemistry 1964, 3, 1385-1390), полиакриламид, латекс, декстран, резину, силикон, пластик, природную губку, металлопластик, перекрестно сшитый декстран (например, Sephadex™), агарозный гель (Sepharose™) и другие подложки, известные специалисту в данной области.
В некоторых предпочтительных вариантах осуществления, подложка, используемая для связывания молекулы нуклеиновой кислоты, подлежащей секвенированию, может представлять собой твердую подложку, включая инертный субстрат или матрикс (например, стеклянную пластину, полимерные бусины и т.д.). Инертный субстрат или матрикс функционализируют, например, с использованием промежуточных материалов, содержащих реакционноспособные группы, позволяющие ковалентное прикрепление биомолекул, таких как полинуклеотиды. Примеры такой подложки включают, но без ограничения, полиакриламидные гидрогели, поддерживаемые на инертном субстрате, таком как стекло, в частности полиакриламидные гидрогели, описанные в WO2005/065814 и US2008/0280773, где полное содержание патентных заявок приведено в настоящем описании в качестве ссылки. В таких вариантах осуществления, биомолекулы (например, полинуклеотиды) могут быть напрямую ковалентно прикреплены к промежуточному материалу (например, гидрогелю), и сам промежуточный материал может быть нековалентно прикреплен к субстрату или матриксу (например, стеклянному субстрату). В некоторых предпочтительных вариантах осуществления, подложка представляет собой стеклянную пластину или кремниевую вафлю, поверхность которой является модифицированной с использованием слоя химически групп на основе авидина, амино, акриламидсилана или альдегида.
По настоящему изобретению, подложка или твердая подложка не ограничена по своему размеру, форме и конфигурации. В некоторых вариантах осуществления, подложка или твердая подложка представляет собой плоскую структуру, такую как пластина, чип, микрочип и/или массив. Поверхность такой подложки может находиться в форме плоского слоя. В некоторых вариантах осуществления, подложка или ее поверхность является неплоской, например, внутренняя или внешняя поверхность пробирки или контейнера. В некоторых вариантах осуществления, подложка или твердая подложка включает микросферы или бусины. В конкретных предпочтительных вариантах осуществления, подложка, используемая для связывания молекулы нуклеиновой кислоты, подлежащей секвенированию, представляет собой массив бусин или лунок.
В некоторых предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит реагент для связывания (например, ковалентного или нековалентного связывания) молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой. Такой реагент включает, например, реагенты, которые активируют или модифицируют молекулу нуклеиновой кислоты (например, ее 5'-конец), такие как фосфорная кислота, тиол, амин, карбоновая кислота или альдегид; реагенты, которые активируют или модифицируют поверхность подложки, такие как амино-алкоксисилан (например, аминопропилтриметоксисилан, аминопропилтриэтоксисилан, 4-аминобутилтриэтоксисилан и т.д.); перекрестно сшивающее средство, такое как янтарный ангидрид, фенилдиизотиоцианат (Guo et al., 1994), малеиновый ангидрид (Yang et al., 1998), гидрохлорид 1-этил-3-(3-диметиламинопропил)-карбодиимида (EDC), эфир мета-малеимидобензойной кислоты-N-гидроксисукцинимида (MBS), N-сукцинимидил[4-иодацетил]аминобензойная кислота (SIAB), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), N-γ-малеимидобутирилокси-сукцинимидный сложный эфир (GMBS), сукцинимидил-4-(п-малеимидофенил)бутират (SMPB); и любую их комбинацию.
В конкретных предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит праймер для инициации реакции полимеризации нуклеотидов. По настоящему изобретению, праймер не подвергается дополнительным ограничениям при условии, что он может специфически гибридизоваться с областью молекулы нуклеиновой кислоты - мишени. В некоторых иллюстративных вариантах осуществления, длина праймера может составлять 5-50 п.о., например, 5-10, 10-15, 15-20, 20-25, 25-30, 30-35, 35-40, 40-45, 45-50 п.о. В некоторых иллюстративных вариантах осуществления, праймер может содержать природные или неприродные нуклеотиды. В некоторых иллюстративных вариантах осуществления, праймер содержит или состоит из природных нуклеотидов. В некоторых иллюстративных вариантах осуществления, праймер содержит модифицированный нуклеотид, например, запертую нуклеиновую кислоту (ЗНК). В конкретных предпочтительных вариантах осуществления, праймер содержит универсальную последовательность праймера.
В конкретных предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит полимеразу для проведения реакции полимеризации нуклеотидов. По настоящему изобретению, можно использовать различные пригодные полимеразы. В некоторых иллюстративных вариантах осуществления, полимераза может использовать ДНК в качестве матрицы для синтеза новой цепи ДНК (например, ДНК-полимераза). В некоторых иллюстративных вариантах осуществления, полимераза может использовать РНК в качестве матрицы для синтеза новой цепи ДНК (например, обратная транскриптаза). В некоторых иллюстративных вариантах осуществления, полимераза может использовать ДНК или РНК в качестве матрицы для синтеза новой цепи РНК (например, РНК-полимераза). Таким образом, в конкретных предпочтительных вариантах осуществления, полимераза выбрана из группы, состоящей из ДНК-полимеразы, РНК-полимеразы и обратной транскриптазы.
В конкретных предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит один или несколько буферных растворов. Такие буферы включают, но без ограничения, буферный раствор для ДНКазы I, буферный раствор для ДНК-полимеразы, буферный раствор для лигазы, буферный раствор для элюции молекулы нуклеиновой кислоты, буферный раствор для растворения молекулы нуклеиновой кислоты, буферный раствор для проведения реакции полимеризации нуклеотидов (например, ПЦР) и буферный раствор для реакции лигирования. Набор по настоящему изобретению может содержать любой один или несколько из вышеупомянутых буферных растворов.
В конкретных предпочтительных вариантах осуществления, набор по настоящему изобретению дополнительно содержит один или несколько растворов для промывки. Примеры таких растворов для промывки включают, но без ограничения, фосфатный буфер, цитратный буфер, буфер Трис-HCl, ацетатный буфер, карбонатный буфер и т.п. Набор по настоящему изобретению может содержать любой один или несколько из вышеупомянутых растворов для промывки.
Благоприятные эффекты настоящего изобретения
По сравнению с предшествующим уровнем техники, техническое решение по настоящему изобретению имеет следующие благоприятные эффекты:
(1) В способе по настоящему изобретению используют только две молекулярные метки для реализации мечения четырех нуклеотидов, и реализуют самолюминесценцию с использованием люциферазы. Таким образом, для устройства для секвенирования, используемого в способе секвенирования по настоящему изобретению, как нет необходимости в оборудовании источником света возбуждения, так и нет необходимости во внедрении дополнительного дизайна, чтобы отфильтровывать фон, образованный лазерным источником света. С одной стороны, стоимость изготовления устройства для секвенирования сильно уменьшается, что является полезным для продвижения и применения устройства для секвенирования и способа секвенирования; с другой стороны, объем устройства для секвенирования значительно уменьшается, что делает устройство для секвенирования более легким и более простым для переноски.
(2) 3'-гидроксильная группа дезоксирибонуклеотида, используемого в способе секвенирования по настоящему изобретению, является модифицированной и блокированной. В ходе процесса секвенирования, только один дезоксирибонуклеотид можно синтезировать на реакцию, что гарантирует, что только один дезоксирибонуклеотид можно включать на реакцию, таким образом, улучшая точность секвенирования.
Несмотря на то, что конкретные варианты осуществления настоящего изобретения описаны подробно, специалисту в данной области понятно, что различные модификации и изменения можно вносить в детали, в соответствии со всеми объяснениями, которые были описаны, и эти изменения включены в объем защиты настоящего изобретения. Полный объем настоящего изобретения приведен посредством пунктов прилагаемой формулы изобретения и любых их эквивалентов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ:
Фигура 1: способ конъюгации антитела и Nluc, где фермент на фигуре обозначает Nluc, и антитело на фигуре обозначает антитело против дигоксина.
ПРИМЕРЫ
Пример 1
1. Конструирование библиотеки для секвенирования

Для удобства конструирования библиотеки, последовательности олигонуклеотидов (жирный шрифт) добавлены на двух концах последовательности, и линкерная последовательность (закрашенная часть) из BGISEQ-500 вставлена в среднюю часть, выделенная жирным курсивом часть показывает первые 10 п.о. оснований последовательности, подлежащей секвенированию. Вышеуказанную последовательность синтезировали в GenScript Biotechnology Company, и для неограниченного использования последовательности, синтезированную последовательность вставляли в вектор pUC57 и трансформировали в клетки E. coli.
(2) Подходящее количество E.coli, содержащей известную библиотеку, культивировали, и плазмиды выделяли; сконструировали следующие пары праймеров: GATATCTGCAGGCAT (SEQ ID NO: 2, праймер 1), GATATCACAGGCTGA (SEQ ID NO: 3, праймер 2), известную последовательность амплифицировали в соответствии со следующими системой (Таблица 1) и способом (Таблица 2), и продукт ПЦР очищали с использованием магнитных бусин. К очищенному продукту ПЦР добавляли составной олигонуклеотид (ATGCCTGCAGATATCGATATCACAGGCTGA, SEQ ID NO: 4) и подвергали циркуляризации и конструированию библиотеки в соответствии с инструкцией и способом из набора для конструирования библиотек с использованием циркуляризации BGISEQ-500 SE50 (MGI) для последующего использования;
Таблица 1: (использовали ферменты собственного производства BGI)
Таблица 2:
2. Амплификация последовательностей библиотеки
96-луночный планшет, покрытый стрептавидином, закупали из Thermo Fisher Company, и 100 мкл 1 мкМ модифицированного на 5’-конце с использованием биотина праймера GCCATGTCGTTCTGTGAGCCAAGG (SEQ ID NO: 5) инкубировали в одной из лунок при комнатной температуре в течение 30 минут, реакционную жидкость отбрасывали, добавляли 6 нг библиотеки, сконструированной в разделе 1 выше, и 20 мкл буфера для получения DNB I в наборе BGISEQ-500 (изготовленном в MGI), гибридизацию праймера с использованием вышеуказанного модифицированного с использованием биотина праймера проводили при 60°C в течение 5 минут, добавляли 40 мкл полимеразы I DNB в наборе для секвенирования BGISEQ-500 (изготовленном в MGI) и 4 мкл полимеразы II DNB, реакцию проводили при 30°C в течение 60 минут, после нагревания до 65°C, реакцию останавливали, и реакционный раствор осторожно отбрасывали. Добавляли 100 мкл 5 мкМ праймера для секвенирования GCTCACAGAACGACATGGCTACGATCCGACTT (SEQ ID NO: 6), гибридизацию проводили при комнатной температуре в течение 30 минут, и реакционный раствор осторожно отбрасывали;
3. Секвенирование
(1) Четыре dNTP, как показано ниже, синтезировали в Acme Bioscience в качестве внешней компании:
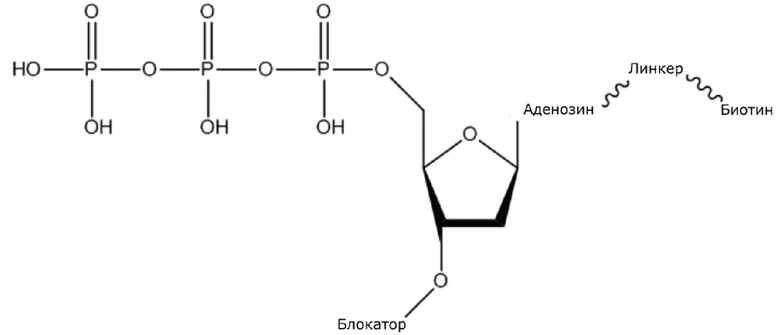
dATP-линкер-биотин
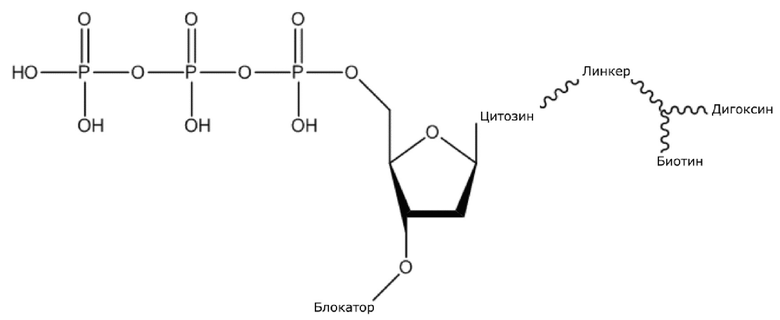
dCTP-линкер-биотин-дигоксин
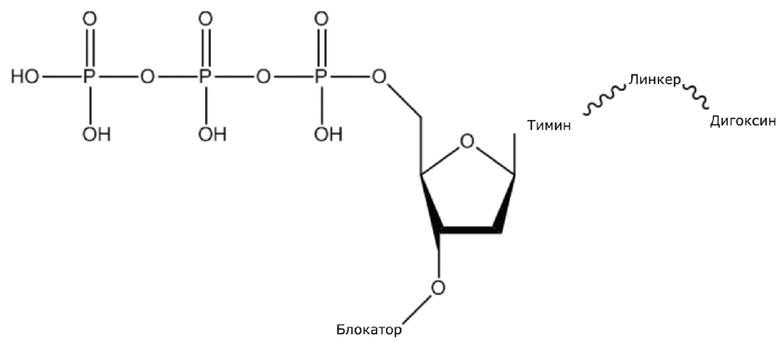
dTTP-линкер-дигоксин
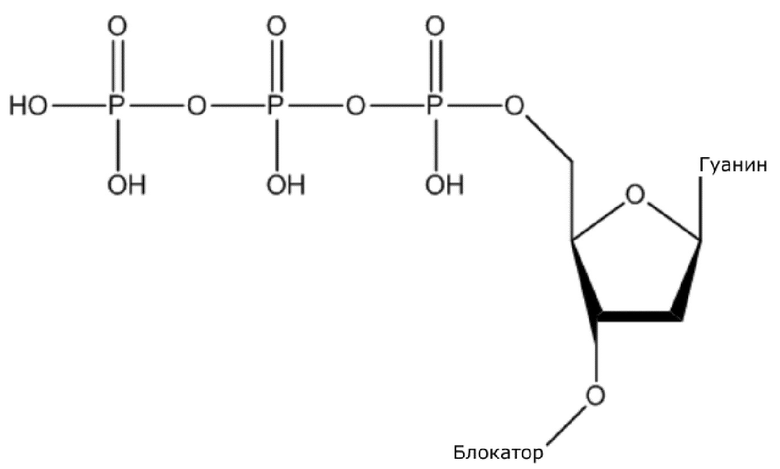
dGTP
(2) Получение двух люцифераз:
a. SA-Gluc (закуплена из компании adivity)
b. Антитело (закуплено из Abcam), конъюгировано с Nluc (закуплена из adivity) для получения Ab-Nluc
Набор для связывания белков закупали из Thermo, и конъюгацию антитела и Nluc проводили в соответствии с инструкциями и способом, как показано на фигуре 1.
(3) Получение реагентов:
Получение других реагентов, необходимых в реакции секвенирования
Реакционный раствор для полимеризации: 50 мМ Трис-Hcl, 50 мМ NaCl, 10 мМ (NH4)2SO4, 0,02 мг/мл полимеразы BG9 (BGI), 3 мМ MgSO4, 1 мМ ЭДТА, 1 мкМ каждый из вышеуказанных четырех dNTP
Буфер для элюции: 5XSSC, 0,05% Tween-20;
Реакционный раствор для связывания ферментов: вышеуказанные два фермента разводили в буфере TBST, и конечная концентрация каждого из двух ферментов составляла 2 мкг/мл;
Реакционный раствор субстрата 1: получали буфер 50 мМ трис-Hcl 0,5 мМ Nacl, и 50X коэлентеразин (nanolight) разводили до 1X;
Реакционный раствор субстрата 2: получали буфер 50 мМ трис-Hcl 0,5 мМ Nacl, и 50X субстрат NLuc FLASH (nanolight) разводили до 1X;
Буфер для отрезания: 20 мМ THPP, 0,5M NaCl, 50 мМ Трис-HCl, pH 9,0, 0,05% tween-20;
(4) Реакция секвенирования:
Способ секвенирования:
a. Полимеризация: 100 мкл раствора для полимеразной реакции добавляли в каждую лунку амплифицированной библиотеки, температуру считывателя для микропланшетов повышали до 55°C, и реакцию проводили в течение 3 минут, так что четыре dNTP полимеризовались на амплифицированной библиотеке. После осторожного отбрасывания реакционного раствора, добавляли 100 мкл реакционного раствора для элюции, осторожно пипетировали несколько раз для удаления реакционного раствора для элюции;
b. Лигирование люциферазы: добавляли 100 мкл реакционного раствора для связывания фермента, инкубировали при 35°C в течение 30 минут, так что SA-gluc подвергалась лигированию с меченными биотином производными dCTP и dATP, и Ab-Nluc подвергалась лигированию с меченными дигоксином производными dCTP и dTTP, реакционный раствор отбрасывали, раствор для элюции добавляли и осторожно пипетировали несколько раз для удаления раствора для элюции;
c. Детекция сигнала фермента 1: устанавливали подходящие параметры считывателя для микропланшетов, добавляли реакционный раствор субстрата 1, проводили детекцию сигнала фермента 1, и регистрировали наивысшее значение сигнала;
d. Детекция сигнала фермента 2: реакционный раствор субстрата 1 удаляли, устанавливали подходящие параметры считывателя для микропланшетов, реакционный раствор субстрата 2 добавляли, проводили детекцию сигнала фермента 2, и регистрировали наивысшее значение сигнала;
e. Отрезание; реакционный раствор субстрата 2 удаляли, добавляли 200 мкл буфера для элюции, после осторожного пипетирования несколько раз, буфер для элюции отбрасывали, добавляли 100 мкл реакционного раствора для отрезания, реакцию проводили при 55°C в течение 3 минут, реакционный раствор для отрезания отбрасывали; 200 мкл буфера для элюции добавляли для промывки, и промывку повторяли три раза;
f. Стадии a-e повторяли для следующего цикла секвенирования; и проводили секвенирование всего 10 п.о.
(5) Результаты секвенирования
a. Значения сигналов для двух видов люминесценции являлись следующими:
b. Анализ результатов секвенирования:
В соответствии с значениями сигнала люминесценции, цикл 1 и цикл 7 имели сигналы, только когда вводили субстрат 2, так что можно было заключить, что биотин-dTTP подвергался полимеризации в этих двух циклах, и первое и седьмое основание библиотеки, подлежащей тестированию, представляли собой основание T;
Для цикла 2, цикла 4, цикла 5 и цикла 8, сигналы наблюдали, только когда вводили субстрат 1, так что можно было заключить, что дигоксин-dATP подвергался полимеризации в этих четырех циклах, и второе, четвертое, пятое и восьмое основания библиотеки, подлежащей тестированию, представляли собой основание A;
Для цикла 3, цикла 6 и цикла 9, сигналы наблюдали, когда вводили субстрат 1 и субстрат 2, так что можно было заключить, что биотин-дигоксин-dCTP подвергался полимеризации в этих трех циклах, и третье, шестое и девятое основания библиотеки, подлежащей тестированию, представляли собой основание C;
Для цикла 10, люминесценции не наблюдали, когда вводили два субстрата, так что можно было заключить, что холодный G подвергался полимеризации в этом цикле, и десятое основание библиотеки представляло собой основание G.
В общем, первые 10 оснований последовательности, подлежащей тестированию, представляли собой: TACAACTACG (SEQ ID NO: 7), что на 100% совпадало с последовательностью оснований TACAACTACG (SEQ ID NO: 7) первых 10 п.о. библиотеки, подлежащей тестированию.
Пример 2
Конструирование библиотеки для секвенирования и амплификация последовательностей библиотеки являлись такими же, как показано в примере 1.
Секвенирование:
(1) Четыре dNTP: как показано в примере 1
(2) Получение люцифераз и родственных белков
a. SA-Gluc (закуплена из компании adivity)
b. Антитело (закуплено из Abcam), и в соответствии с инструкциями для NHS-s-s-биотина от thermo fisher, антитело метили с использованием биотиновой метки для получения антитела-s-s-биотина.
(3) Получение реагентов
Получение других реагентов, необходимых в реакции секвенирования
Реакционный раствор для полимеризации: 50 мМ Трис-Hcl, 50 мМ NaCl, 10 мМ (NH4)2SO4, 0,02 мг/мл полимеразы BG9 (BGI), 3 мМ MgSO4, 1 мМ ЭДТА, 1 мкМ каждый из вышеуказанных четырех dNTP
Буфер для элюции: 5XSSC, 0,05% Tween-20;
Реакционный раствор для связывания фермента: вышеуказанные два белка SA-Gluc и антитело-s-s-биотин разводили в буфере TBST, соответственно, и конечная концентрация каждого из двух белков составляла 2 мкг/мл;
Реакционный раствор субстрата: получали буфер 50 мМ Трис-HCl 0,5 мМ NaCl, и 50X коэлентеразин (nanolight) разводили до 1X;
Буфер для инактивации фермента: 5 мМ DTT, 50 мМ Трис-HCl, pH 9,0;
Буфер для отрезания: 20 мМ THPP, 50 мМ Трис-HCl, pH 9,0, 0,5M NaCl, 0,05% Tween-20;
(4) Реакция секвенирования
a. Полимеризация: 100 мкл раствора для полимеразной реакции добавляли в каждую лунку амплифицированной библиотеки, температуру считывателя для микропланшетов повышали до 55°C, реакцию проводили в течение 3 минут для полимеризации четырех dNTP на амплифицированной библиотеке, реакционный раствор осторожно удаляли, добавляли 100 мкл реакционного раствора для элюции и осторожно пипетировали несколько раз, и реакционный раствор для элюции удаляли;
b. Лигирование люциферазы: добавляли 100 мкл реакционного раствора для связывания люциферазы SA-Gluc, инкубировали в течение 30 мин при 35°C, так что SA-gluc подвергалась лигированию с меченными биотином производными dCTP и dATP, реакционный раствор удаляли, раствор для элюции добавляли и осторожно пипетировали несколько раз, и раствор для элюции удаляли;
c. Первая детекция сигнала: устанавливали подходящие параметры считывателя для микропланшетов, реакционный раствор субстрата добавляли, проводили детекцию сигнала и регистрировали наивысшее значение сигнала;
d. Инактивация фермента: реакционный раствор субстрата удаляли, добавляли буфер для инактивации фермента и инкубировали при 35°C в течение 10 минут, реакционный раствор удаляли, и затем буфер для элюции добавляли для элюции;
e. Второе лигирование люциферазы: реакционный раствор антитела-S-S-биотина добавляли и инкубировали при 35°C в течение 30 минут, так что антитело-s-s-биотин подвергалось лигированию с меченными дигоксином производными dCTP и dTTP, реакционный раствор удаляли, буфер для элюции добавляли и осторожно пипетировали несколько раз, раствор для элюции удаляли, добавляли 100 мкл реакционного раствора для связывания SA-Gluc и инкубировали при 35°C в течение 30 минут, затем реакционный раствор удаляли, добавляли буфер для элюции, после завершения элюции, буфер для элюции удаляли;
f. Вторая детекция сигнала: устанавливали подходящие параметры считывателя для микропланшетов, реакционный раствор субстрата добавляли, проводили детекцию сигнала и регистрировали наивысшее значение сигнала;
g. Отрезание: реакционный раствор субстрата 2 удаляли, добавляли 200 мкл буфера для элюции и осторожно пипетировали несколько раз, буфер для элюции удаляли, добавляли 100 мкл реакционного раствора для отрезания, реакцию проводили при 55°C в течение 3 минут, реакционный раствор для отрезания удаляли; 200 мкл буфера для элюции добавляли для промывки, промывку повторяли три раза;
h. Стадии a-g повторяли для следующего цикла секвенирования; и проводили секвенирование всего 10 п.о.
(5) Результаты секвенирования
a. Результаты детекции сигнала 10 п.о. являлись следующими:
b. Анализ результатов секвенирования:
Для цикла 1 и цикла 7, значения сигналов получены только при второй детекции, так что можно было заключить, что биотин-dTTP подвергался полимеризации в этих двух циклах, и таким образом, первое и седьмое основания библиотеки, подлежащей тестированию, представляли собой T;
Для цикла 2, цикла 4, цикла 5 и цикла 8, сигналы наблюдали только при первой детекции, так что можно было заключить, что дигоксин-dATP подвергался полимеризации в этих четырех циклах, и таким образом, второе, четвертое, пятое и восьмое основания библиотеки, подлежащей тестированию, представляли собой основание A;
Для цикла 3, цикла 6 и цикла 9, сигналы получали как при первой детекции, так и при второй детекции, так что можно было заключить, что биотин-дигоксин-dCTP подвергался полимеризации в этих двух циклах, и таким образом, третье, шестое и девятое основания библиотеки, подлежащей тестированию, представляли собой основание C;
Для цикла 10, свет не излучался при двух детекциях сигналов, так что можно было заключить, что холодный G подвергался полимеризации в этом цикле, и таким образом, десятое основание этой библиотеки представляло собой основание G.
В общем, первые 10 оснований последовательности, подлежащей тестированию, представляли собой: TACAACTACG (SEQ ID NO: 7), что на 100% совпадало с последовательностью оснований TACAACTACG (SEQ ID NO: 7) первых 10 п.о. библиотеки, подлежащей тестированию.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> EGI TECH (SHEN ZHEN) CO., LIMITED
<120> Способ одноканального секвенирования на основе самолюминесценции
<130> PCT/CN2019/086974
<160> 7
<170> PatentIn версии 3.5
<210> 1
<211> 233
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Сконструированная последовательность ДНК
<400> 1
gatatctgca ggcatagaat gaatattatt gaatcaataa ttaaagtcgg aggccaagcg 60
gtcttaggaa gacaacaact ccttggctca cagaacgaca tggctacgat ccgactttac 120
aactacgata atgggctgga tacatggaat gattatagat atattaagga ataatgttaa 180
ttaatgccta aattaattaa tctaaggggg ttaatacttc agcctgtgat atc 233
<210> 2
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 2
gatatctgca ggcat 15
<210> 3
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 3
gatatcacag gctga 15
<210> 4
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> составной олигонуклеотид
<400> 4
atgcctgcag atatcgatat cacaggctga 30
<210> 5
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 5
gccatgtcgt tctgtgagcc aagg 24
<210> 6
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 6
gctcacagaa cgacatggct acgatccgac tt 32
<210> 7
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность оснований первых 10 п.о.
<400> 7
tacaactacg 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СЕКВЕНИРОВАНИЯ НА ОСНОВЕ ОДНОГО ФЛУОРЕСЦЕНТНОГО КРАСИТЕЛЯ | 2017 |
|
RU2760737C2 |
| АНАЛИЗ МНОЖЕСТВА АНАЛИТОВ С ИСПОЛЬЗОВАНИЕМ ОДНОГО АНАЛИЗА | 2019 |
|
RU2824049C2 |
| СПОСОБ И ПРОДУКТ ДЛЯ ЛОКАЛИЗОВАННОЙ ИЛИ ПРОСТРАНСТВЕННОЙ ДЕТЕКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ В ОБРАЗЦЕ ТКАНИ | 2012 |
|
RU2603074C2 |
| ПАНЕЛЬ И СПОСОБ ПОЛУЧЕНИЯ ПРОСТРАНСТВЕННОЙ ИНФОРМАЦИИ О НУКЛЕИНОВЫХ КИСЛОТАХ | 2020 |
|
RU2803202C2 |
| СПОСОБЫ ИНКАПСУЛИРОВАНИЯ ОДИНОЧНЫХ КЛЕТОК, ИНКАПСУЛИРОВАННЫЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2793717C2 |
| СПОСОБЫ И СРЕДСТВА ПОЛУЧЕНИЯ БИБЛИОТЕКИ ДЛЯ СЕКВЕНИРОВАНИЯ | 2019 |
|
RU2815513C2 |
| СПОСОБЫ ИНКАПСУЛИРОВАНИЯ ОДИНОЧНЫХ КЛЕТОК, ИНКАПСУЛИРОВАННЫЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2750567C2 |
| СПОСОБЫ И ПРИБОРЫ ДЛЯ ОБНАРУЖЕНИЯ И ИДЕНТИФИКАЦИИ ЗАКОДИРОВАННЫХ ГРАНУЛ И БИОЛОГИЧЕСКИХ МОЛЕКУЛ | 2007 |
|
RU2487169C2 |
| ГЕНОМНЫЙ ОТБОР И СЕКВЕНИРОВАНИЕ С ПОМОЩЬЮ КОДИРОВАННЫХ МИКРОНОСИТЕЛЕЙ | 2010 |
|
RU2609630C2 |
| БИБЛИОТЕКИ ДЛЯ СЕКВЕНИРОВАНИЯ НОВОГО ПОКОЛЕНИЯ | 2014 |
|
RU2698125C2 |
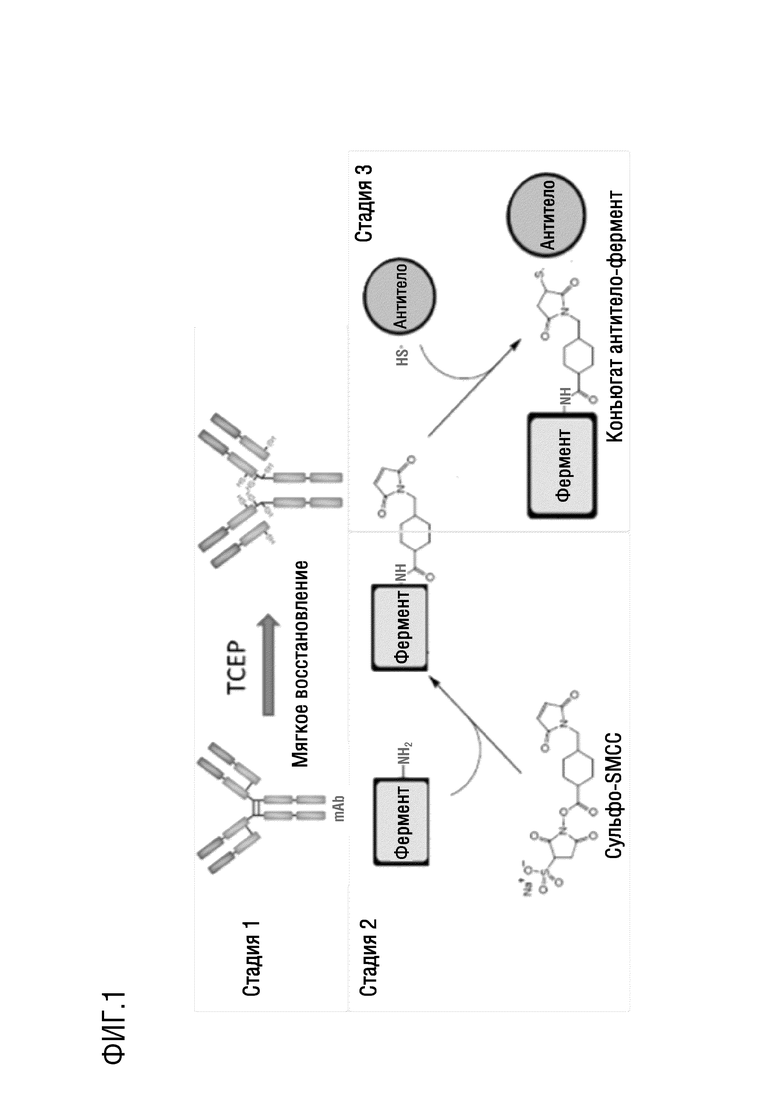
Изобретение относится к биотехнологии, в частности к способу секвенирования, основанному на одном флуоресцентном красителе, в котором сигнал самолюминесценции используют для установления различий последовательного включения различных нуклеотидов, таким образом, реализуя определение последовательности полинуклеотида. 2 н. и 12 з.п. ф-лы, 1 ил., 2 табл., 2 пр.
1. Способ секвенирования молекулы нуклеиновой кислоты, включающий следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3’-конец растущей цепи нуклеиновой кислоты;
(5) обеспечение возможности контакта дуплекса из предыдущей стадии с двумя различными люциферазами и проведение реакции лигирования, где две люциферазы можно специфически лигировать с первой молекулярной меткой и второй молекулярной меткой, соответственно; затем обеспечение возможности подвергания люциферазы флуоресцентной реакции в присутствии субстрата, и детекция излучаемого сигнала флуоресценции;
(6) удаление молекулярной метки каждого нуклеотида;
(7) необязательно, повторение стадий (3) - (6) или (3) - (5) один или более раз для получения информации о последовательности молекулы нуклеиновой кислоты.
2. Способ по п. 1, где на стадии (5), дуплекс приводят в контакт с двумя различными люциферазами в одностадийной реакции и подвергают реакции лигирования; или на стадии (5) дуплекс последовательно приводят в контакт с двумя люциферазами и подвергают реакции лигирования.
3. Способ по п. 1 или 2, включающий следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов, и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3’-конец растущей цепи нуклеиновой кислоты;
(5) удаление фазы раствора из реакционной системы на предыдущей стадии, сохранение дуплекса связанным с подложкой, и добавление двух различных люцифераз для проведения реакции лигирования, где две люциферазы можно специфически лигировать с первой молекулярной меткой и второй молекулярной меткой, соответственно;
(6) удаление несвязанной люциферазы посредством использования буфера для элюции;
(7) добавление субстрата первой люциферазы и детекция флуоресцентного сигнала в то же самое время;
(8) удаление раствора из реакции предыдущей стадии;
(9) добавление субстрата второй люциферазы и детекция флуоресцентного сигнала в то же самое время;
(10) удаление раствора из реакции предыдущей стадии;
(11) удаление молекулярной метки и 3’-защитной группы каждого нуклеотида;
(12) необязательно, удаление раствора из реакции предыдущей стадии;
(13) необязательно, повторение стадий (3) - (12) или (3) - (9) один или более раз для получения информации о последовательности молекулы нуклеиновой кислоты.
4. Способ по п. 1 или 2, включающий следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов, и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3’-конец растущей цепи нуклеиновой кислоты;
(5) удаление фазы раствора из реакционной системы на предыдущей стадии, сохранение дуплекса связанным с подложкой и добавление первой люциферазы для проведения реакции лигирования, где первая люцифераза может специфически связываться с первой молекулярной меткой;
(6) удаление несвязанной первой люциферазы посредством использования буфера для элюции;
(7) добавление субстрата первой люциферазы и детекция флуоресцентного сигнала в то же самое время;
(8) удаление раствора из реакции предыдущей стадии;
(9) добавление второй люциферазы для проведения реакции лигирования, где вторая люцифераза может специфически связываться с второй молекулярной меткой;
(10) удаление несвязанной второй люциферазы посредством использования буфера для элюции;
(11) добавление субстрата второй люциферазы и детекция флуоресцентного сигнала в то же самое время;
(12) удаление раствора из реакции предыдущей стадии;
(13) необязательно, удаление молекулярной метки каждого нуклеотида;
(14) необязательно, повторение стадий (3) - (13) или (3) - (11) один или более раз для получения информации о последовательности молекулы нуклеиновой кислоты.
5. Способ по п. 3 или 4, где первая люцифераза и вторая люцифераза содержат различные люциферазы.
6. Способ по п. 1 или 2, включающий следующие стадии:
(1) предоставление молекулы нуклеиновой кислоты, подлежащей секвенированию, которая является связанной с подложкой, или связывание молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой;
(2) добавление праймера для инициации реакции полимеризации нуклеотидов, полимеразы для проведения реакции полимеризации нуклеотидов, и четырех соединений для формирования реакционной системы, содержащей фазу раствора и твердую фазу; где четыре соединения представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой;
(3) гибридизация праймера с молекулой нуклеиновой кислоты, подлежащей секвенированию, и формирование дуплекса, связанного с подложкой, посредством использования праймера в качестве исходной растущей цепи нуклеиновой кислоты, вместе с молекулой нуклеиновой кислоты, подлежащей секвенированию;
(4) использование полимеразы для осуществления реакции полимеризации нуклеотидов в условиях, позволяющих полимеразе осуществлять реакцию полимеризации нуклеотидов, таким образом, включение одного из четырех соединений в 3’-конец растущей цепи нуклеиновой кислоты;
(5) удаление фазы раствора из реакционной системы на предыдущей стадии, сохранение дуплекса связанным с подложкой и добавление первой люциферазы для проведения реакции лигирования, где первая люцифераза может специфически связываться с первой молекулярной меткой;
(6) удаление несвязанной первой люциферазы посредством использования буфера для элюции;
(7) добавление субстрата люциферазы и детекция флуоресцентного сигнала в то же самое время;
(8) удаление раствора из реакции предыдущей стадии;
(9) добавление реагента для денатурации люциферазы;
(10) удаление раствора из реакции предыдущей стадии;
(11) добавление второй люциферазы для проведения реакции лигирования, где вторая люцифераза может специфически связываться с второй молекулярной меткой;
(12) удаление несвязанной второй люциферазы посредством использования буфера для элюции;
(13) добавление субстрата люциферазы и детекция флуоресцентного сигнала в то же самое время;
(14) необязательно, удаление раствора из реакции предыдущей стадии;
(15) необязательно, удаление молекулярной метки и 3’-защитной группы каждого нуклеотида;
(16) необязательно, повторение стадий (3) - (15) или (3) - (13) один или более раз для получения информации о последовательности молекулы нуклеиновой кислоты.
7. Способ по п. 6, где первая люцифераза и вторая люцифераза могут содержать одинаковую люциферазу.
8. Способ по любому из пп. 1-7, где первая молекулярная метка и вторая молекулярная метка выбраны из группы, состоящей из биотина, дигоксина, N3G или FITC, и первая люцифераза и вторая люцифераза являются, соответственно, меченными стрептавидином, антителом против дигоксина, антителом против N3G или антителом против FITC.
9. Способ по любому из пп. 1-8, где указанные четыре соединения показаны в формуле I, формуле II, формуле III или формуле IV:
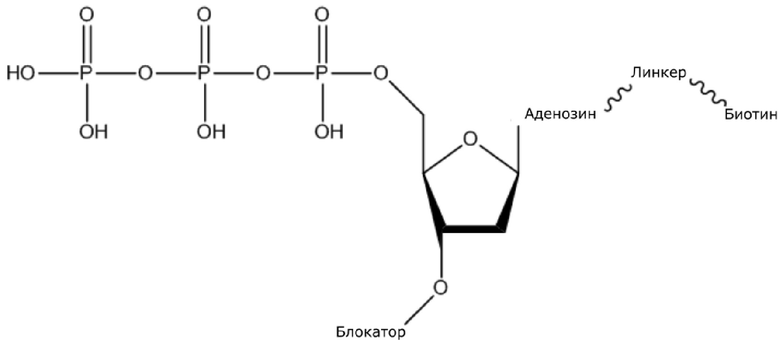
Формула I
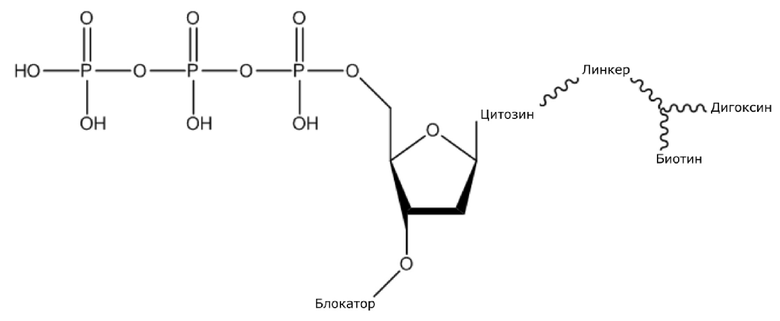
Формула II
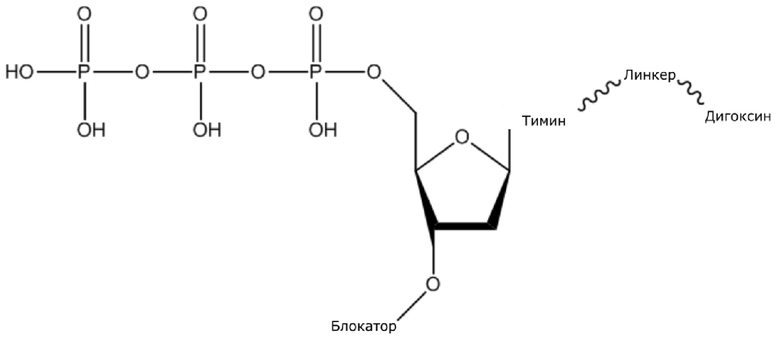
Формула III
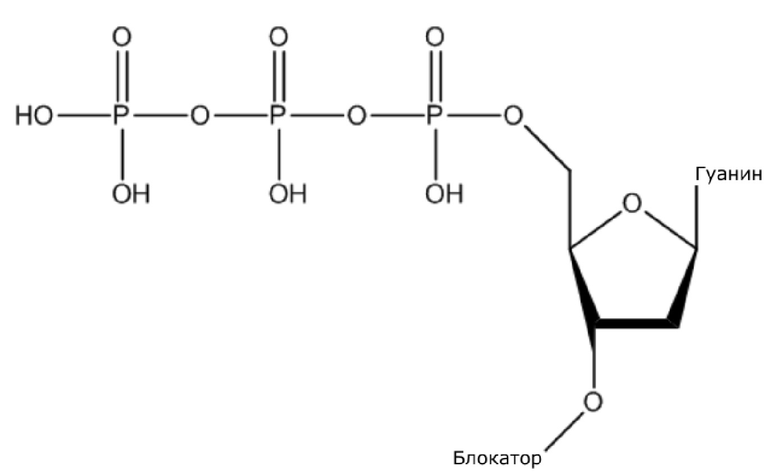
Формула IV
10. Набор для секвенирования полинуклеотида, содержащий:
(a) четыре соединения, которые представляют собой, соответственно, производные нуклеотидов A, (T/U), C и G, и имеют способность комплементарного спаривания оснований; и гидроксил (-OH) в 3’-положении рибозы или дезоксирибозы четырех соединений защищен посредством защитной группы; и
первое соединение связано с первой молекулярной меткой,
второе соединение связано с второй молекулярной меткой,
третье соединение связано с первой молекулярной меткой и с второй молекулярной меткой, или часть третьего соединения связана с первой молекулярной меткой, и другая часть третьего соединения связана с второй молекулярной меткой,
четвертое соединение не связано ни с какой молекулярной меткой; и
(b) две различные люциферазы, которые могут специфически связываться с первой молекулярной меткой и второй молекулярной меткой, соответственно.
11. Набор по п. 10, где две различные люциферазы могут содержать одинаковую люциферазу или различные люциферазы.
12. Набор по п. 10 или 11, где первая молекулярная метка и вторая молекулярная метка выбраны из группы, состоящей из биотина, дигоксина, N3G или FITC, и первая люцифераза и вторая люцифераза являются, соответственно, меченными стрептавидином, антителом против дигоксина, антителом против N3G или антителом против FITC.
13. Набор по любому из пп. 10-12, дополнительно содержащий: реагент и/или устройство для выделения молекулы нуклеиновой кислоты из образца; реагент для предварительной обработки молекулы нуклеиновой кислоты; подложку для связывания молекулы нуклеиновой кислоты, подлежащей секвенированию; реагент для связывания (например, ковалентного или нековалентного связывания) молекулы нуклеиновой кислоты, подлежащей секвенированию, с подложкой; праймер для инициации реакции полимеризации нуклеотидов; полимеразу для проведения реакции полимеризации нуклеотидов; один или более буферных растворов; один или более растворов для промывки; или любую их комбинацию.
14. Набор по любому из пп. 10-13, где указанные четыре соединения показаны в формуле I, формуле II, формуле III или формуле IV:
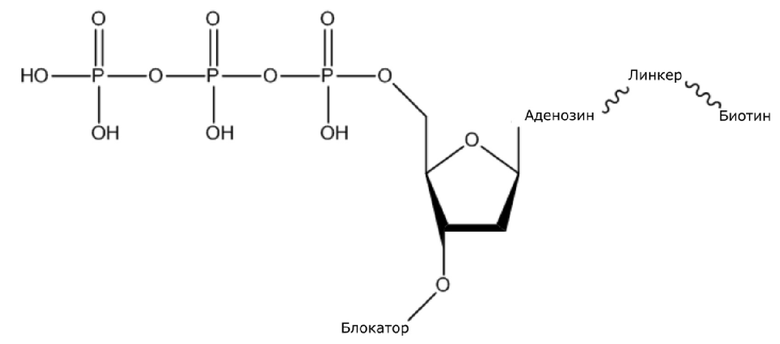
Формула I
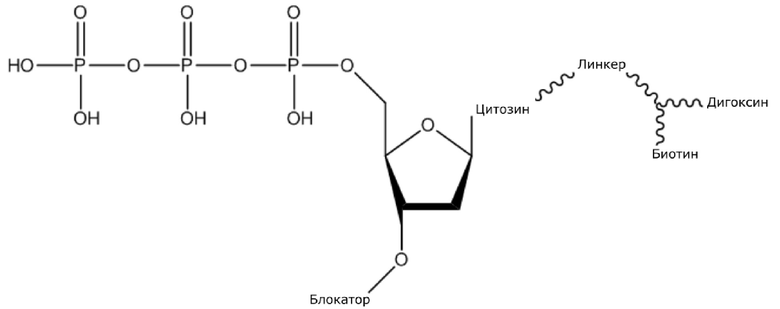
Формула II
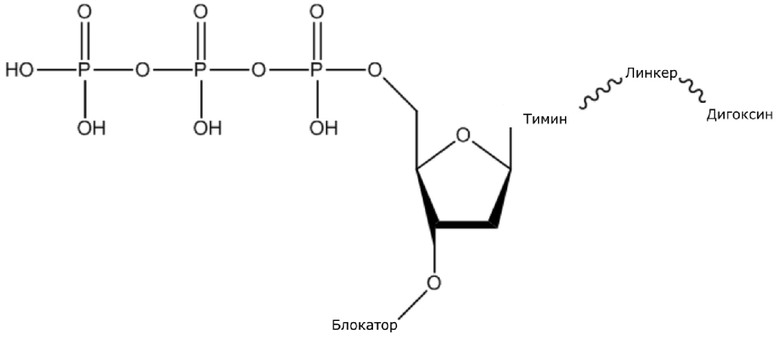
Формула III
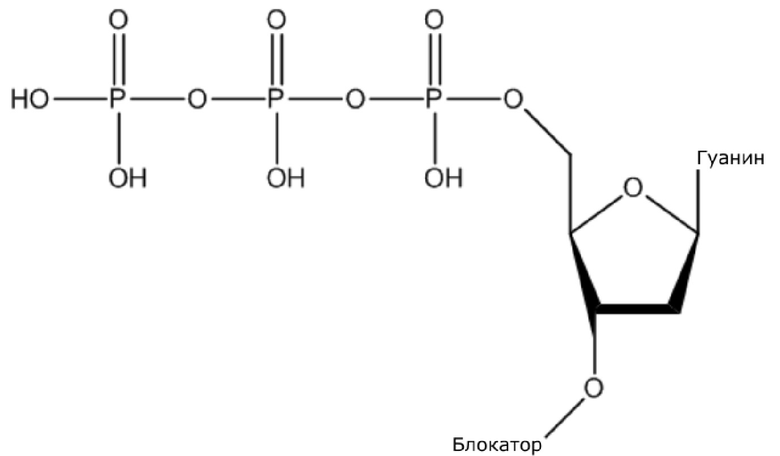
Формула IV
| CN1854308 A 1.11.2006 (формула, описание стр | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| CN108697499 A 23.10.2018 (формула, реферат) | |||
| СПОСОБ ВЫЯВЛЕНИЯ МУТАЦИЙ В СЛОЖНЫХ СМЕСЯХ ДНК | 2014 |
|
RU2613489C2 |
