Ссылка на родственные заявки
Согласно настоящей заявке испрашивается приоритет и преимущество в соответствии с предварительной заявкой на выдачу патента США №62/738687, поданной 28 сентября 2018 г., и предварительной заявкой на выдачу патента США №62/599299, поданной 15 декабря 2017 г., которые полностью включены в настоящий документ посредством ссылки.
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к композициям Т-клеток и способам их применения в контексте терапии и лечения. В частности, настоящее изобретение относится к Т-клеткам, которые являются модифицированными (например, генетически и/или функционально) для поддержания функциональности в условиях, при которых немодифицированные Т-клетки демонстрируют истощение. Композиции и способы, раскрытые в настоящем документе, находят применение для предотвращения истощения сконструированных (например, Т-клеток с химерным антигенным рецептором (CAR)), а также не сконструированных Т-клеток, тем самым усиливая функцию Т-клеток (например, активность против рака или инфекционного заболевания). Композиции и способы согласно настоящему изобретению находят применение как в клинических, так и в исследовательских условиях, например, в областях биологии, иммунологии, медицины и онкологии.
Предшествующий уровень техники настоящего изобретения
Т-клетки представляют собой иммунные клетки, которые активируются посредством передачи сигналов и костимуляции рецептора Т-клеток (TCR) после взаимодействия с антигеном. Физиологическая активация через рецептор Т-клеток делает Т-клетки способными оказывать сильное противоопухолевое и/или противоинфекционное действие. Во время разрешения острого воспалительного ответа подгруппа активированных эффекторных Т-клеток дифференцируется в долгоживущие клетки памяти. В отличие от этого, у пациентов с хроническими инфекциями или раком Т-клетки нередко подвергаются патологической дифференцировке в сторону состояния дисфункции, которое называется истощением Т-клеток. Истощение Т-клеток характеризуется выраженными изменениями в метаболической функции, транскрипционном программировании, потере эффекторной функции (например, секреции цитокинов, способности к уничтожению) и коэкспрессии множественных поверхностных ингибирующих рецепторов. Основной причиной истощения Т-клеток является постоянное воздействие антигена, ведущее к непрерывной передаче сигналов TCR. Профилактика или устранение истощения Т-клеток долгое время являлись искомыми в качестве средства для повышения эффективности Т-клеток (например, у пациентов с раком или хроническими инфекциями).
Т-клетки с химерным антигенным рецептором (CAR) демонстрируют впечатляющие показатели ответа при злокачественных опухолях В-клеток, но длительный контроль над заболеваниями наблюдается только приблизительно у 50% пациентов с В-ALL1 и крупноклеточной В-клеточной лимфомой (ссылка 2; полностью включена в настоящее описание посредством ссылки), и еще реже наблюдается при CLL (ссылка 3; полностью включена в настоящий документ посредством ссылки). Более того, несмотря на многочисленные испытания, CAR Т-клетки не опосредовали устойчивых противоопухолевых эффектов в солидных опухолях (ссылка 4; полностью включена в настоящий документ посредством ссылки). Многочисленные факторы ограничивают эффективность CAR Т-клеток, включая в себя гетерогенную экспрессию антигена и потребность в высокой плотности антигена для оптимальной функции CAR, что позволяет обеспечить быстрый выбор вариантов устранения антигена (ссылки 5-7; полностью включенные в настоящий документ посредством ссылки), супрессивное микроокружение опухоли (ссылка 8; полностью включена в настоящий документ посредством ссылки) и истинную дисфункцию Т-клеток в результате истощения Т-клеток (ссылки 3, 9, 10; полностью включенные в настоящий документ посредством ссылки). Истощение Т-клеток все чаще рассматривается как причина дисфункции Т-клеток в CAR Т-клетках. Тоническая антиген-независимая передача сигналов, благодаря агрегации scFv, обычно происходит в Т-клетках, экспрессирующих CAR, и может вызывать быстрое истощение (ссылка 9; полностью включена в настоящий документ посредством ссылки). Интеграция эндодомена CD28 в рецепторы CAR Т-клеток второго поколения усиливает экспансию, но также предрасполагает CAR Т-клетки к истощению как в условиях тонически передающих сигнал рецепторов, так и в CD19-28z CAR Т-клетках, подвергающихся воздействию высоких опухолевых нагрузок (ссылка 9; полностью включена в настоящий документ посредством ссылки). Недавно было продемонстрировано, что повышенная частота встречаемости Т-клеток, обладающих характеристиками истощения, содержащихся в трансплантатах CD19-BBz CAR, позволяет отличить не отвечающих на лечение пациентов от отвечающих пациентов, получавших лечение в отношении CLL3. Широкая база данных из разнообразных исследований указывает на истинную дисфункцию Т-клеток вследствие истощения Т-клеток как основного фактора, ограничивающего эффективность терапии CAR Т-клетками, и повышает вероятность того, что разработка резистентных к истощению CAR Т-клеток может значительно улучшить клинические результаты.
Краткое раскрытие настоящего изобретения
Настоящее изобретение относится к композициям и способам применения для предотвращения истощения сконструированных (например, Т-клеток, сконструированных для экспрессии синтетического рецептора, такого как сконструированный рецептор Т-клеток или химерный антигенный рецептор (CAR)), а также не сконструированных (например, нативных) Т-клеток. Т-клетки, модифицированные (например, для предотвращения истощения Т-клеток) согласно настоящему изобретению, содержащие их композиции и способы их применения усиливают функциональность Т-клеток (например, активность против рака или инфекционного заболевания).
CAR Т-клетки опосредуют противоопухолевые эффекты у небольшой группы пациентов с раком, но дисфункция из-за истощения Т-клеток является важным барьером для прогресса. Чтобы исследовать биологию истощения в Т-клетках человека, экспрессирующих рецепторы CAR, проведены эксперименты при разработке вариантов осуществления настоящего изобретения с использованием модельной системы, использующей тонически передающий сигналы CAR, который вызывает признаки истощения, описанные в других условиях. Результаты демонстрируют, что истощение было связано с выраженным дефектом в продукции IL-2 наряду с повышенной доступностью хроматина мотивов фактора транскрипции АР-1 и избыточной экспрессией многочисленных факторов транскрипции bZIP и IRF, которые были вовлечены в ингибирующую активность. Инженерия CAR Т-клеток для избыточной экспрессии фактора АР-1 (например, c-Jun) увеличивала потенциал экспансии, увеличивала функциональную способность, уменьшала терминальную дифференцировку и улучшала противоопухолевую активность в многочисленных моделях опухолей in vivo.
Эксперименты, проведенные во время разработки вариантов осуществления в настоящем документе, дополнительно демонстрируют, что функциональный дефицит фактора АР-1 (например, c-Jun) опосредует дисфункцию в истощенных Т-клетках человека, и инженерия CAR Т-клеток для избыточной экспрессии фактора АР-1 (например, с-Jun) делает их устойчивыми к истощению, устраняя тем самым серьезный барьер на пути прогресса для этого нового класса терапевтических средств.
Эксперименты, проведенные во время разработки вариантов осуществления в настоящем документе, дополнительно демонстрируют, что нокдаун IRF4 резко увеличивает функциональную активность истощенных HA-28z CAR Т-клеток, что усиленная in vivo функция c-Jun-модифицированных HA-28z CAR Т-клеток не может быть воспроизведена ex vivo обеспечением IL-2, который c-Jun усиливал активность Her2-BBz CAR Т-клеток в супрессивном микроокружении солидной опухоли, что эта избыточная экспрессия c-Jun повышает устойчивость к опосредованной TGFβ супрессии истощенных HA-28z CAR Т-клеток, и что транскрипционные изменения в клетках, модифицированных c-Jun, согласуются с уменьшением истощения и усилением формирования памяти.
Как описано в настоящем документе, предусмотрены сконструированные Т-клетки (например, Т-клетки, сконструированные для экспрессии синтетического рецептора, такого как сконструированный рецептор Т-клеток или CAR), а также не сконструированные (например, нативные, природные) Т-клетки, которые модифицированы, чтобы избыточно экспрессировать и/или содержать повышенные содержания (например, созданные, чтобы характеризоваться физиологически повышенными содержаниями) одного или нескольких факторов транскрипции белка-активатор а 1 (АР-1) (например, семейства c-Fos, c-Jun, активирующий фактор транскрипции (ATF) и белок димеризации Jun (JDP)) и/или модифицированные (например, генетически) для сниженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса (например, JunB и BATF3 и другие представители семейства BATF, IRF4, и представители семейства ATF).
Соответственно, согласно одному аспекту настоящее изобретение относится к Т-клеткам, модифицированным, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1. Например, согласно одному варианту осуществления c-Jun экспрессируется в Т-клетках, которые сконструированы для экспрессии синтетического рецептора, такого как сконструированный рецептор Т-клеток или химерный антигенный рецептор (CAR). Согласно другому варианту осуществления c-Jun экспрессируется в Т-клетках с нативным, природным Т-клеточным рецептором. Настоящее изобретение не ограничено средствами экспрессии одного или нескольких факторов транскрипции АР-1. Согласно одному варианту осуществления, при коэкспрессии со сконструированным TCR или CAR, c-Jun (и/или другой фактор транскрипции АР-1) и сконструированный рецептор коэкспрессируются из различных вирусных векторов. Согласно другому варианту осуществления они экспрессируются из одного векторного конструкта с использованием бицистронного вектора. c-Jun (и/или другой фактор транскрипции АР-1) может быть экспрессирован конститутивно или регулируемым образом (например, с использованием системы для удаленной регуляции экспрессии с помощью малой молекулы или с использованием эндогенно регулируемой системы). Согласно другому варианту осуществления гены c-Jun и/или других факторов транскрипции АР-1 могут быть генетически интегрированы в клеточную ДНК с использованием ретровирусного, лентивирусного или другого вирусного вектора или посредством системы на основе CRISPR/Cas9. Согласно еще одному варианту осуществления c-Jun и/или другие транскрипционные факторы АР-1 экспрессируется/экспрессируются посредством РНК или онколитического вируса или другой системы транзиентной экспрессии, известной в настоящей области техники. c-Jun и/или другие транскрипционные факторы АР-1 можно доставлять ex vivo в Т-клетки для адоптивного переноса или доставлять посредством генетического переноса in vivo.
Аналогичным образом, настоящее изобретение не ограничено типом Т-клеток, модифицированных, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или модифицированных (например, генетически) для снижения экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса (например, JunB и BATF3 и другие представители семейства BATF, представители семейства IRF4 и ATF). Согласно некоторым вариантам осуществления Т-клетки представляют собой CD3+ Т-клетки (например, комбинацию CD4+ и CD8+ Т-клеток). Согласно определенным вариантам осуществления Т-клетки представляют собой CD8+ Т-клетки. Согласно другим вариантам осуществления Т-клетки представляют собой CD4+ Т-клетки. Согласно некоторым вариантам осуществления Т-клетки представляют собой Т-клетки -естественные киллеры (NK). Согласно некоторым вариантам осуществления Т-клетки представляют собой альфа-бета Т-клетки. Согласно некоторым вариантам осуществления Т-клетки представляют собой гамма-дельта Т-клетки. Согласно некоторым вариантам осуществления Т-клетки представляют собой комбинацию CD4+ и CD8+ Т-клеток (например, которые представляют собой CD3+). Согласно определенным вариантам осуществления Т-клетки представляют собой Т-клетки памяти. Согласно определенным вариантам осуществления Т-клетки памяти представляют собой центральные Т-клетки памяти. Согласно определенным вариантам осуществления Т-клетки памяти представляют собой эффекторные Т-клетки памяти. Согласно некоторым вариантам осуществления Т-клетки представляют собой инфильтрирующие опухоль лимфоциты. Согласно определенным вариантам осуществления Т-клетки представляют собой комбинацию CD8+ Т-клеток, CD4+ Т-клеток, Т-клеток - NK, Т-клеток памяти и/или гамма-дельта Т-клеток. Согласно некоторым вариантам осуществления Т-клетки представляют собой индуцированные цитокинами клетки-киллеры.
Согласно некоторым вариантам осуществления Т-клетка представляет собой противоопухолевую Т-клетку (например, Т-клетка с активностью против опухоли (например, аутологичной опухоли), которая становится активированной и подвергается экспансии в ответ на антиген). Согласно одному варианту осуществления противоопухолевые Т-клетки (например, применимые для адоптивного переноса Т-клеток) в себя Т-клетки, полученные из периферической крови, генетически модифицированные рецепторами, которые распознают и реагируют на опухолевые антигены. Такие рецепторы, как правило, состоят из внеклеточных доменов, содержащих одноцепочечное антитело (scFv), специфическое в отношении опухолевого антигена, связанное с мотивами внутриклеточной передачи сигналов Т-клеток (см., например, Westwood, J. A. et al, 2005, Proc. Natl. Acad. Sci., USA, 102(52): 19051-19056). Другие противоопухолевые Т-клетки включают в себя Т-клетки, полученные из резецированных опухолей или биоптатов опухолей (например, инфильтрирующие опухоль лимфоциты (TIL)). Согласно другому варианту осуществления Т-клетка представляет собой поликлональную или моноклональную реакцонноспособную в отношении опухоли Т-клетку (например, полученную посредством афереза, подвергнутую экспансии ex vivo против опухолевых антигенов, презентированных аутологичными или искусственными
антигенпрезентирующими клетками). Согласно другому варианту осуществления Т-клетка сконструирована для экспрессии Т-клеточного рецептора человеческого или мышиного происхождения, который распознает опухолевый антиген. Настоящее изобретение не ограничено типом опухолевого антигена, распознанного таким образом. Действительно, любая Т-клетка, содержащая рецептор, который распознает опухолевый антиген, находит применение в композициях и способах согласно настоящему изобретению. Примеры включают в себя без ограничения следующее: Т-клетки, экспрессирующие рецептор (например, нативный или встречающийся в природе рецептор, или рецептор, сконструированный для экспрессии синтетического рецептора, такого как сконструированный рецептор Т-клетки или CAR), который распознает антиген, выбранный из следующего: CD19, CD20, CD22, орфанный рецептор 1, подобный рецепторной тирозинкиназе (ROR1), дисиалоганглиозид 2 (GD2), белок или антиген вируса Эпштейна-Барр (EBV), рецептор фолата, мезотелин, карциноэмбриональный антиген человека (СБА), CD33/IL3Rα, тирозин-протеинкиназа Met (c-Met) или рецептор фактора роста гепатоцитов (HGFR), простатический специфический мембранный антиген (PSMA), гликолипид F77, вариант III рецептора эпидермального фактора роста (EGFRvIII), NY-ESO-1, представитель A3 семейства генов антигена меланомы (MAGE) (MAGE-А3), антиген 1 меланомы, распознаваемый Т-клетками (MART-1), GP1000, р53 или другие опухолевые антигены, описанные в настоящем документе.
Согласно некоторым вариантам осуществления Т-клетка сконструирована для экспрессии CAR. Настоящее изобретение не ограничено типом CAR. Действительно, любой CAR, который специфически связывается с требуемым антигеном (например, опухолевым антигеном), может быть модифицирован, как раскрыто и описано в настоящем документе, чтобы избыточно экспрессировать и/или содержать повышенные содержания (например, характеризоваться физиологически повышенными содержаниями) одного или нескольких факторов транскрипции АР-1 (например, c-Jun). Согласно определенным вариантам осуществления CAR содержит антигенсвязывающий домен. Согласно определенным вариантам осуществления антигенсвязывающий домен представляет собой одноцепочечный вариабельный фрагмент (scFv), содержащий вариабельные области тяжелой и легкой цепей, которые специфически связываются с требуемым антигеном. Согласно некоторым вариантам осуществления CAR дополнительно содержит транс мембранный домен (например, трансмембранный домен Т-клеток (например, транс мембранный домен CD28)) и сигнальный домен, содержащий один или несколько иммунорецепторных мотивов активации на основе тирозина (ITAM) (например, сигнальный домен Т-клеточного рецептора (например, дзета-цепь TCR). Согласно некоторым вариантам осуществления CAR содержит один или несколько костимулирующих доменов (например, доменов, которые обеспечивают второй сигнал для стимуляции активации Т-клеток). Настоящее изобретение не ограничено типом костимулирующего домена. Действительно, можно использовать любой костимулирующий домен, известный в настоящей области техники, включая в себя без ограничения следующее: CD28, OX40/CD134, 4-1 ВВ/CD137/TNFRSF9, гамма-субъединица высокоаффинного рецептора иммуноглобулина Ε (FcERIγ, ICOS/CD278, бета-субъединица интерлейкина 2 (ILRβ) или CD122, общая гамма-субъединица цитокинового рецептора (IL-2Rγ) или CD132 и CD40. Согласно одному варианту осуществления костимулирующий домен представляет собой 4-1 ВВ.
Согласно одному аспекту настоящее изобретение относится к способу лечения заболевания или состояния у субъекта, предусматривающему введение субъекту (например, пациенту), характеризующемуся наличием заболевания или состояния, эффективного количества Т-клеток, модифицированных чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или модифицированных (например, генетически) для снижения экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса (например, JunB и BATF3 и другие представители семейства BATF, IRF4 и представители семейства ATF). Настоящее изобретение не ограничено типом заболевания или состояния, которое лечат. Действительно, любое заболевание или состояние, которое поддается лечению (например, для которого при лечении уменьшается интенсивность признаков или симптомов заболевания) посредством введения Т-клеток, можно лечить улучшенным и более эффективным способом с использованием композиций и способов согласно настоящему изобретению (например, содержащие и/или использующие Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1). Согласно одному варианту осуществления заболевание или состояние представляет собой рак. Согласно другому варианту осуществления заболевание или состояние представляет собой инфекционное заболевание. Настоящее изобретение не ограничено типом рака или типом инфекционного заболевания. Действительно, любой рак, известный в настоящей области техники, в отношении которого для лечения используют Т-клеточную терапию, можно лечить композициями и способами согласно настоящему изобретению. Подобным образом, любое инфекционное заболевание, известное в настоящей области техники, в отношении которого для лечения используют Т-клеточную терапию, можно лечить с помощью композиций и способов согласно настоящему изобретению. Согласно одному варианту осуществления введение субъекту (например, пациенту), характеризующемуся наличием заболевания или состояния, эффективного количества Т-клеток, модифицированных чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса, ингибирует истощение Т-клеток (например, по сравнению с субъектом, получающим такое же количество сконструированных Т-клеток (например, CAR Т-клеток или Т-клеток, содержащих рекомбинантный TCR), не модифицированных, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 или чтобы характеризоваться пониженной экспрессией и/или активностью одного или нескольких представителей ингибирующего АР-1 комплекса).
Таким образом, согласно одному варианту осуществления настоящее изобретение относится к способу ингибирования истощения Т-клеток (например, поддержание функциональности Т-клеток, подвергающихся воздействию избыточного антигена (например, в контексте лечения заболевания или состояния)) посредством модификации Т-клеток, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса (например, по сравнению с контрольными Т-клетками, так не модифицированными). Согласно одному варианту осуществления Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 (например, c-Jun), демонстрируют повышенную функциональность и/или активность (например, повышенную антиген-индуцированную продукцию цитокинов, усиленную способность к уничтожению (например, повышенное распознавание опухолевых мишеней с низким поверхностным антигеном), повышенное образование клеток памяти и/или усиленная пролиферация в ответ на антиген) и/или снижение признаков истощения (например, более низкие содержания маркеров, указывающих на истощение (например, PD-1, TIM-3, LAG-3) и/или более низкие уровни запрограммированной клеточной смерти). Согласно некоторым вариантам осуществления Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса, описанных в настоящем документе, значительно усиливают клиническую эффективность (например, сконструированных Т-клеток (например, CAR Т-клеток) и/или не сконструированных природных Т-клеток).
Согласно определенным вариантам осуществления настоящее изобретение демонстрирует, что лечение субъекта, характеризующегося наличием рака, терапевтически эффективным количеством композиции, содержащей Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, превосходит лечение у субъекта, характеризующегося наличием рака, с помощью Т-клеток, экспрессирующих нормальные количества одного или нескольких факторов транскрипции АР-1. Согласно некоторым вариантам осуществления лечение животных (например, людей), страдающих от рака, терапевтически эффективными количествами иммунотерапевтических композиций, содержащих Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, ингибирует развитие или рост раковых клеток или делает раковые клетки популяцией, более восприимчивой к другим видам лечения (например, к вызывающей клеточную смерть активности противораковых лекарственных средств или лучевой терапии). Соответственно, композиции и способы согласно настоящему изобретению можно использовать в качестве монотерапии (например, для уничтожения раковых клеток и/или уменьшения или ингибирования роста раковых клеток, индукции апоптоза и/или остановки клеточного цикла в раковых клетках) или при введении в комбинации с одним или несколькими дополнительными средствами, такими как другие противораковые средства (например, противораковые терапевтические лекарственные средства, индуцирующие клеточную смерть или разрушающие клеточный цикл, или лучевая терапия), чтобы сделать большую долю раковых клеток восприимчивой к уничтожению, ингибированному росту раковых клеток, индуцированному апоптозу и/или остановке клеточного цикла по сравнению с соответствующей долей клеток у животного, получавшего только противораковое лекарственное средство или только лучевую терапию.
Соответственно, согласно определенным вариантам осуществления настоящее изобретение относится к способам лечения или отсрочки прогрессирования рака у пациента, предусматривающим введение пациенту терапевтически эффективного количества композиции, содержащей Т-клетки, модифицированные (например, генетически), чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 (например, c-Jun) и/или модифицированные (например, генетически) для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса (например, JunB и BATF3 и другие представители семейства BATF, IRF4 и представители семейства ATF). Согласно определенным вариантам осуществления терапевтически эффективное количество композиции модифицированных Т-клеток снижает количество раковых клеток у пациента после такого лечения. Согласно определенным вариантам осуществления терапевтически эффективное количество композиции модифицированных Т-клеток снижает и/или устраняет опухолевую нагрузку у пациента после такого лечения. Согласно определенным вариантам осуществления способ дополнительно предусматривает введение лучевой терапии пациенту. Согласно определенным вариантам осуществления лучевую терапию вводят до, одновременно и/или после получения пациентом терапевтически эффективного количества композиции модифицированных Т-клеток. Согласно определенным вариантам осуществления способ дополнительно предусматривает введение пациенту одного или нескольких противораковых средств и/или одного или нескольких химиотерапевтических средств. Согласно определенным вариантам осуществления одно или несколько противораковых средств и/или одно или несколько химиотерапевтических средств вводят до, одновременно и/или после получения пациентом терапевтически эффективного количества композиции модифицированных Т-клеток. Согласно определенным вариантам осуществления комбинированное лечение пациента с помощью терапевтически эффективного количества модифицированных Т-клеток и курс противоракового средства дает больший ответ опухоли и клиническую эффективность у такого пациента по сравнению с теми пациентами, которые получают модифицированные Т-клетки или противораковые лекарственные средства/лучевую терапию отдельно. Поскольку дозы для всех одобренных противораковых лекарственных средств и лучевой терапии известны, настоящее изобретение предусматривает их различные комбинации с модифицированными Т-клетками.
Согласно определенным вариантам осуществления настоящее изобретение относится к терапевтически эффективному количеству композиции (например, иммунотерапевтической композиции), содержащей Т-клетки, модифицированные согласно настоящему раскрытию (например, для применения в лечении или отсрочке прогрессирования рака у субъекта). Как описано в настоящем документе, композиция может дополнительно содержать одно или несколько противораковых средств, например, одно или несколько химиотерапевтических средств. Настоящее изобретение также относится к применению композиции для индукции остановки клеточного цикла и/или апоптоза. Настоящее изобретение также относится к применению композиций для сенсибилизации клеток к дополнительному(ым) средству(ам), такому(им) как индукторы апоптоза и/или остановки клеточного цикла и химиопротекции нормальных клеток путем индукции остановки клеточного цикла перед лечением с помощью химиотерапевтических средств. Композиции согласно настоящему изобретению применимы для лечения, уменьшения интенсивности или профилактики нарушений, таких как любой тип рака или инфекционное заболевание, и дополнительно любых клеток, чувствительных к индукции апоптотической клеточной смерти (например, нарушений, характеризующихся нарушением регуляции апоптоза, включая в себя гиперпролиферативные заболевания, такие как рак). Согласно определенным вариантам осуществления композиции можно использовать для лечения, уменьшения интенсивности или профилактики рака, который дополнительно характеризуется устойчивостью к противораковой терапии (например, таких раковых клеток, которые являются устойчивыми в химиотерапии, лучевой терапии, гормональной терапии и т.п.). Настоящее изобретение также относится к фармацевтическим композициям, содержащим композицию (например, иммунотерапевтические композиции), содержащую модифицированные Т-клетки согласно настоящему изобретению в фармацевтически приемлемом носителе.
Согласно другому варианту осуществления настоящее изобретение относится к композиции, в которой Т-клетки модифицированы введением нуклеиновой кислоты, кодирующей c-Jun, и где c-Jun и рекомбинантный рецептор экспрессируются из отдельных экспрессионных векторных конструктов.
Согласно другому варианту осуществления настоящее изобретение относится к композиции, в которой Т-клетки модифицированы введением нуклеиновой кислоты, кодирующей c-Jun, и где c-Jun и рекомбинантный рецептор коэкспрессируются из одного экспрессионного векторного конструкта.
Согласно другому варианту осуществления настоящее изобретение относится к способу лечения или отсрочки прогрессирования рака у пациента, предусматривающему введение пациенту терапевтически эффективного количества композиции, содержащей Т-клетки, модифицированные (например, генетически), чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 (например, c-Jun) в комбинации с терапевтически эффективным количеством ингибитора передачи сигналов TCR (например, для предотвращения истощения Т-клеток). Согласно определенным вариантам осуществления ингибитор передачи сигналов TCR представляет собой ингибитор тирозинкиназы. Согласно другому варианту осуществления ингибитор тирозинкиназы ингибирует киназу Lck. Ингибитор передачи сигналов TCR можно вводить с помощью любого подходящего способа введения, но, как правило, его вводят перорально. Субъекту можно вводить многократные циклы лечения. Согласно определенным вариантам осуществления ингибитор передачи сигналов TCR вводят в соответствии со стандартной схемой введения доз (например, ежедневно или периодически). Согласно другому варианту осуществления ингибитор передачи сигналов TCR вводят в течение периода времени, достаточного для восстановления, по меньшей мере частичной функции Т-клеток, затем прекращают.
Эти и другие варианты осуществления предмета изобретения будут легко понятны специалистам в настоящей области техники с учетом раскрытия в настоящем документе.
Описание графических материалов
На фиг. 1А-В показано, что транскрипционные факторы АР-1 c-Fos и c-Jun отрицательно регулируются в истощенных GD2-28Z CAR Т-клетках.
На фиг. 2 А-Е показано, что усиленная экспрессия АР-1 снижает признаки истощения в CAR Т-клетках.
На фиг. 3A-F показано, что функциональный благоприятный эффект АР-1 главным образом связан с экспрессией c-Jun.
На фиг. 4А-Е показано, что бицистронная экспрессия c-Jun с CAR усиливает функциональную активность CAR Т-клеток.
На фиг. 5А-С показано, что бицистронная экспрессия c-Jun с CAR усиливает функциональную активность CAR Т-клеток и фенотип центральных Т-клеток памяти.
ФИГ. 6A-F показано, что бицистронная экспрессия c-Jun с CAR усиливает продукцию CAR Т-клетками провоспалительных цитокинов и уменьшает IL-10.
ФИГ. 7А-Е показано, что бицистронная экспрессия c-Jun усиливает CD19 и CD22 CAR Т-клеточную активность в ответ на опухолевые клетки с низкими уровнями антигена.
ФИГ. 8A-D показано, что нокдаун ингибирующих представителей семейства АР-1 JunB и BATF3 увеличивает продукцию IL2 в истощенных CAR Т-клетках. (A) CRISPR генный нокаут (KO) JunB в истощенных HA-28Z CAR Т-клетках значительно увеличивает продукцию IL2 (вверху) и IFNg (внизу) после воздействия GD2+ клеточных линий Nalm6-GD2 (слева), остеосаркомы 143 В (в центре) и нейробластомы Келли (справа). Это увеличение было даже больше, чем для избыточной экспрессии c-Jun (ОЕ) отдельно. Двойные JUNB-KO и c-JUN-OE Т-клетки не показали никакого благоприятного эффекта по сравнению с JUNB-KO отдельно. (В) CRISPR генный нокаут (KO) JunB в GD2-BBZ CAR Т-клетка значительно увеличивает продукцию IL2 (вверху) и IFNg (внизу) после воздействия GD2+ клеточных линий Nalm6-GD2 (слева), остеосаркомы 143 В (в центре) и нейробластомы Келли (справа), однако, избыточно экспрессирующие c-Jun (ОЕ) GD2-BBZ CAR Т-клетки показали наибольший функциональный благоприятный эффект.Двойные JUNB-KO и cJUN-OE Т-клетки не показали никакого благоприятного эффекта по сравнению с cJUN-OE отдельно. (С) CRISPR генный нокаут (KO) JunB в CD19-28Z (слева) или CD19-BBZ (справа) CAR Т-клетках не влиял на продукцию IL2 (вверху) после воздействия клеток лейкоза Nalm6-GD2, позволяя предположить, что JunB является мощным ингибитором только в тонически сигнализирующих/истощенных GD2 CAR Т-клетках. (D) CRISPR генный нокаут (KO) BATF3 в истощенных HA-28Z CAR Т-клетках повышает продукцию IL2 (вверху) после воздействия Nalm6-GD2 (слева) и нейробластомы Келли (справа), в то время как продукция IFNγ не изменяется. Истощенные HA-28Z CAR Т-клетки, отредактированные с использованием трех независимых гРНК, нацеленных на BATF3, показали повышенную продукцию IL2 по сравнению с контрольными или отредактированными ZB2 контролями.
На фиг. 9А-В показано, что экспрессирующие c-Jun HA-GD2 CAR Т-клетки проявляют превосходную лечебную активность in vivo по сравнению с ^модифицированными HA-GD2 CAR Т-клетками. Рост клеток лейкоза Nalm6-GD2, стабильно экспрессирующих люциферазу светлячка, отслеживали in vivo с использованием биолюминесцентной визуализации после адоптивного переноса 2xl06CAR+Т-клеток или Т-клеток-имитации (нетрансдуцированных). (А) Количественная биолюминесценция с течением времени. (В) Изображения, показывающие отдельных мышей, п=5 мышей на группу, (все шкалы 1×104-1×106, за исключением того, что шкалы имитации d32 регулировали).
На фиг. 10 показано, что модифицированные c-Jun GD2-BBZ CAR Т-клетки проявляют превосходную активность in vivo на модели агрессивной солидной опухоли 143 В остеосаркомы. Рост внутримышечно имплантированных опухолевых клеток остеосаркомы 143 В отслеживали in vivo, используя измерения штангенциркулем после адоптивного переноса 1×107 CAR+ Т-клеток или Т-клеток-имитации (нетрансдуцированных). (А) Количественный рост опухоли с течением времени для n=5 мышей на группу.
На фиг. 11A-D показано, что модифицированные c-Jun CD19 CAR Т-клетки проявляют повышенную активность in vivo против лейкоза CD19low Nalm6. ЗхЮ6 CAR+Т-клеток доставляли в/в мышам с опухолью - лейкозом Nalm6, CD19-low клон F. (А-В) Рост опухоли (А) и выживаемость (В) мышей, получивших лечение с помощью CD19-BBZ CAR Т-клеток +/- c-Jun. Модифицированные c-Jun CD19-BBZ CAR Т-клетки демонстрируют сниженный рост опухоли и значительно повышенную выживаемость. (C-D) Рост опухоли (С) и выживаемость (D) мышей, получивших лечение с помощью CD19-28Z CAR Т-клеток +/- c-Jun. Модифицированные c-Jun CD19-28Z CAR Т-клетки рано показывают снижение роста опухоли, но С019-отрицательное заболевание в конечном итоге вырастает в обеих группах и не дает никаких благоприятных эффектов в отношении выживаемости (р>0,05).
На фиг. 12A-J показано, что HA-28z CAR Т-клетки проявляют фенотипические, функциональные, транскрипционные и эпигенетические характерные признаки истощения Т-клеток. а) Снижение экспансии HA-28z по сравнению с CD19-28z CAR Т-клетками во время первичной экспансии культуры. D0 = активация микрочастицы, D2 = трансдукция. Планки погрешностей представляют собой среднее ± SEM от n=10 доноров. Ь) Поверхностная экспрессия маркеров, связанных с истощением (D10). с) CD19-28z в основном содержат стволовые Т-клетки памяти (CD45RA+CD62L+) и центральные клетки памяти (CD45RA- CD62L+), тогда как HA-28z в основном содержат эффекторные клетки памяти CD45RA-CD62L- (D10). d) Высвобождение IL-2 (слева) и IFNg (справа) после 24-часового совместного культивирования с клетками лейкоза CD19+GD2+Nalm6- GD2. Планки погрешностей представляют собой среднее значение ± стандартное отклонение от трех лунок. Один репрезентативный донор показан для каждого анализа, е) Принципиальный компонентный анализ (РСА) глобальных транскрипционных профилей полученных из наивных и CM CD19 или НА CAR Т-клеток, на 7, 10 и 14 дни в культуре. РС1 (39,3% дисперсии) отделяет CD19 от НА CAR Т-клеток. е) Экспрессия генов 200 верхних генов, управляющих РС1. Представляющие интерес гены в каждом кластере перечислены выше, g) Дифференциально доступные области хроматина в CD8+CD19 и HA-28z CAR Т-клетках (D10). Как N, так и СМ подмножества включены для каждого CAR. h) РСА доступности хроматина ATAC-seq в CD19 или HA-28z CAR Т-клетках (D10). PCI (76,9% дисперсия) отделяет CD 19 от образцов НА CAR. i) Глобальный профиль доступности хроматина CD19 и HA-28z CAR Т-клеток, полученных из подмножества (D10). Наиболее результативные 5000 дифференциально доступных областей (пиков), j) Дифференциально доступные энхансерные области в CD 19 и НА CAR Т-клетках в локусах CTLA4 (вверху) или IL7R (внизу). N - наивные, СМ - центральная память. * р<,05, ** р<,01, ***p<,001.ns p>,05.
На фиг. 13А-Е показана сигнатура семейства АР-1 в истощенных CAR Т-клетках. а) Наиболее результативные 25 баллов отклонения мотива транскрипционного фактора в день 10 по сравнению с Т-клетками, экспрессирующими CD19-CAR, с помощью анализа chromVAR показывают многочисленные представители семейства АР-1 (bZIP) в CD4+ и CD8+ Т-клетках, полученных из подмножеств N или CM (D10). Ь) Анализ обогащения мотивов TF в полученных из наивных CD8+HA-28z CAR Т-клетках демонстрирует мотивы семейства АР-1 (bZIP) как наиболее значительно обогащенные, с) Экспрессия массового РНК-секвенирования (FPKM) указанных представителей семейства АР-1 (bZIP) и IRF в CD 19 и HA-28z CAR Т-клетках. Планки погрешностей представляют собой среднее ± SEM от n=6 образцов у 3 доноров, показывающих парную экспрессию CD 19 по сравнению с НА для каждого гена, р-значения получали с использованием критерия ранговых знаков согласованных пар Уилкоксона. d) CD19-28z и HA-28z CAR Т-клетки лизировали и экспрессию указанных белков семейства АР-1 оценивали вестерн-блоттингом. Повышенная экспрессия белка JunB, BATF3 и IRF4 в HA-28z CAR Т-клетках по сравнению с CD 19 CAR Т-клетками была подтверждена на 7, 10 и 14 дни культивирования, е) Корреляционная сеть транскрипционных факторов, связанных с истощением, в полученных из нативных CD8+ (слева) и CD4+ (справа) GD2-28z CAR Т-клетки с использованием анализа РНК-секвенирования одиночных клеток. Гены фактора транскрипции, идентифицированные как дифференциально экспрессированные (р<0,05) с помощью DESeq2, образуют узлы сети. Цвета представляют log2 кратное изменение (FC) (GD2 по сравнению с CD 19 CAR). Толщина края представляет величину корреляции в экспрессии между соответствующей парой генов в клетках. Балл корреляции>0,1 использовали для построения сетей. * р<,05, **р<,01, ***р<,001. ns р>,05.
На фиг. 14А-К показано, что избыточная экспрессия c-Jun усиливает функцию истощенных CAR Т-клеток. а) Схема вектора экспрессии JUN-P2A-CAR. Ь) Внутриклеточная проточная цитометрия, демонстрирующая общую экспрессию c-Jun в контроле и JUN-модифицированных CAR Т-клетках (D10). с) Вестерн-блоттинг для общего c-Jun и фосфо-с-тип8ег73 в контроле и JUN-модифицированных CD 19 и НА CAR Т-клетках (D10). (d) IL-2 и (е) Продукция IFNg после 24-часового совместного культивирования контрольных (синий) или JUN-модифицированных (красный) CD 19 и НА CAR Т-клеток в ответ на антиген+опухолевые клетки. Планки погрешностей представляют собой среднее значение ± стандартное отклонение для лунок в трех повторностях. Показаны данные от одного репрезентативного донора. Кратность увеличения продукции IL-2 или IFNg в JUN по сравнению с контрольными CAR Т-клетками у нескольких доноров можно найти на фигуре 24. f) Слева: графики проточной цитометрии, показывающие репрезентативную экспрессию CD45RA/CD62L в контрольных по сравнению с JUN-CAR Т-клетками (D10), Справа: относительная частота эффекторных клеток (Е, RA+62L-), стволовых клеток памяти (SCM, RA+62L+), центральных клеток памяти (RA-62L+) и эффекторных клеток памяти (RA-62L-) в CD4+(вверху) или CD8+(ниже) контрольных или JUN-HA-28z CAR Т-клетках. п=6 доноров из независимых экспериментов. Линии обозначают парные образцы от одного и того же донора. Проводили парные двусторонние t-критерии, g) На 39-й день культивирования 1хЮ6 жизнеспособных Т-клеток из фиг. 24 с заново высевали и культивировали в течение 7 дней с IL-2 или без него, h-j) Фенотип клеточной поверхности контрольных или JUN-CD19-28z CAR Т-клеток из (g) на 46 день, h) Экспрессия CD4 по сравнению с CD8. i) CD8+JUN-модифицированные CD19-28z CAR Т-клетки с поздней экспансией характеризуются фенотипом стволовых клеток памяти (CD45RA+CD62L+). j) CD8+JUN-модифицированные CD19-28z CAR Т-клетки с поздней экспансией характеризуются пониженной экспрессией маркера истощения по сравнению с контролем, к) Т-клетки из g криоконсервировали на D10, оттаивали и оставляли на ночь в IL-2. Здоровым мышам NSG вводили 5×106 контрольных или JUN-модифицированных CD 19-28z или CD 19- BBz CAR Т-клеток путем внутривенной инъекции. На 25 день после инфузии количество Т-клеток периферической крови определяли методом проточной цитометрии. Планки погрешностей представляют собой среднее ± SEM для п=5 мышей на группу. * р<,05, ** р<,01, *** р<,001. ns р>,05. НТМ-шарнирный/трансмембранный. ICD -внутриклеточный домен.
На фиг. 15A-J показано, что для функционального восстановления истощенных НА-28z CAR Т-клеток требуется присутствие c-Jun во время как хронической, так и острой стимуляции Т-клеток, и оно не зависит от JNP. а) Предлагаемые механизмы c-Jun-опосредованного восстановления истощения Т-клеток. АР-1-i указывает на ингибирующий комплекс АР-1. б) Схема регулируемого DD экспрессирующего вектора JUN. в) Схема лекарственной стабилизации экспрессии JUN-DD. Желтый ромб - молекула, стабилизирующая ТМР. d) Общая экспрессия c-Jun в контрольных, JUN-WT и JUN-DD НА-28z CAR Т-клетках (D10) с помощью внутриклеточной проточной цитометрии (слева) и вестерн-блоттинга (справа), е) Продукция IL-2 (слева) и IFNg (справа) в контрольных, JUN-WT, или JUN-DD (OFF, ON) модифицированных HA-28z CAR Т-клетках через 24 часа после стимуляции клетками-мишенями Nalm6-GD2 или 143 В или средой отдельно (исходный уровень) (D10). В d-e) OFF указывает на отсутствие ТМР, ON указывает на Т-клетки, культивируемые в присутствии 10 мкМ ТМР из D4 и во время совместного культивирования. В f-g) ТМР добавляли либо во время экспансии Т-клеток (начиная с D4), либо только во время совместного культивирования с опухолевыми клетками, как указано в f. Для условий ON-OFF и OFF-ON ТМР удаляли/добавляли за 18 ч до совместного культивирования, чтобы обеспечить полную деградацию/стабилизацию c-Jun, соответственно, до воздействия антигена, g) Экспрессия IL-2 у одного репрезентативного донора (слева, SD для лунок в трех повторностях) и кратное увеличение IL-2 (SEM n=6 независимых экспериментов, представляющих 3 разных донора, относительно условия OFF-OFF). h-j) Повышенная функциональная активность JUN-CAR Т-клеток не зависит от N-концевого фосфорилирования Jun (JNP). h) Схема белка c-Jun, показывающая N-концевой трансактивирующий домен (TAD). Звездочки обозначают сайты JNP в положениях Ser63 и Ser73, которые мутированы на аланин в мутанте JUN-AA. i) Вестерн-блот всего c-Jun и c-Jun-PSer73 в контрольных, JUN-WT и JUN-AA HA-28z CAR Т-клетках. j) Высвобождение IL-2 (слева) и IFNg (справа) в контрольных, JUN-WT и JUN-AA HA-28z CAR Т-клетках после 24-часовой стимуляции клетками-мишенями Nalm6-GD2 или 143 В или только средой (исходный уровень). Планки погрешностей представляют собой среднее значение ± стандартное отклонение для лунок в трех повторностях. Репрезентативный результат 3 независимых экспериментов. * р<,05, ** р<,01, *** р<,001. ns р>,05. НТМ - шарнирный/трансмембранный, ICD - внутриклеточный домен, DD - дестабилизирующий домен из DHFR Е. coli, ТМР - триметоприм, WT - дикий тип.
На фиг. 16А-Н показано, что JUN-модифицированные CAR Т-клетки увеличивают активность in vivo против лейкоза и солидных опухолей. В а-с мышей NSG инокулировали lx Nalm6-GD2 клетками лейкоза посредством внутривенной инъекции. 3×106 имитации, HA-28z или JUN-HA-28z CAR+Т-клеток вводили внутривенно на день 3. Прогрессирование опухоли контролировали с использованием биолюминесцентного изображения (а-b). Шкалы нормализованы для всех временных точек, с) JUN-HA-28z CAR Т-клетки индуцировали долгосрочную выживаемость без опухолей. Планки погрешностей представляют собой среднее ±SEM для n=5 мышей/группа. Это открытие было воспроизведено в >3 независимых экспериментах, однако в некоторых экспериментах длительная выживаемость уменьшалась из-за роста клонов GD2(-) Nalm6. г) Схема ретровирусного векторного конструкта JUN-Her2-BBz. е) Лизис Her2-BBz CAR Т-клетками клеток-мишеней GFP+Nalm6-Her2B соотношении Е:Т, составляющем 1:8. Планки погрешностей представляют собой среднее значение ± стандартное отклонение для лунок в трех повторностях. Репрезентативный результат 2 независимых экспериментов. В f-h мышей NSG инокулировали с помощью 1×106 клеток остеосаркомы 143b-19 посредством внутримышечной инъекции. 1х107 клеток-имитаций, Her2-BBz, или JUN-Her2-BBz CAR Т-клеток вводили в/в на день 7 (d7). е) Рост опухоли контролировали измерениями штангенциркулем, g) Мыши, получившие лечение с помощью JUN-Her2-BBz CAR Т-клеток, сохраняли длительную выживаемость без опухолей, h) На d20 после имплантации опухолевых клеток Т-клетки периферической крови определяли количественно у мышей, получавших лечение, как в f. Планки погрешностей представляют собой среднее ± SEM для n=5 мышей/группа. Репрезентативный результат 2 независимых экспериментов с аналогичными результатами. * р<,05, ** р<,01, *** р<,001. НТМ - шарнирный/ транс мембранный. ICD - внутриклеточный домен.
На фиг. 17А-1 показано, что JUN-CAR Т-клетки усиливают функцию Т-клеток при субоптимальной стимуляции, а) Продукция IL-2 и b) IFNg после 24-часовой стимуляции контрольных или JUN-модифицированных HA-28z CAR Т-клеток иммобилизованным идиотипическим антителом 1А7 к CAR. Каждая кривая соответствовала нелинейной кинетике доза-ответ для определения ЕС50. Меньшие графики справа визуализируют кривую, где концентрация антител составляет 0-1 мкг/мл. в) Векторная схема ретровирусного векторного конструкта JUN-CD22-BBz. d) Поверхностная экспрессия CD22 на исходных Nalm6, Nalm6-22KO и Nalm6-22KO+CD221ow. е) Высвобождение IL-2 (слева) и IFNg (справа) после совместного культивирования контрольных или JUN CD22-BBz CAR Т-клеток, подвергнутых воздействию Nalm6 и Nalm6-221ow. Планки погрешностей представляют собой среднее значение ± стандартное отклонение для лунок в трех повторностях. Репрезентативный результат 3 независимых экспериментов. В f-i) мышей NSG инокулировали с помощью 1×106 клеток лейкоза Nalm6-221ow в день 0. В день 4 3×106 контрольных клеток или JUN-CD22-BBz CAR+ Т-клеток или 3×106 ложно трансдуцированных Т-клеток переносили в/в. Рост опухоли контролировали с помощью биолюминесцентного изображения (f) с изображениями (i). g) У мышей, получающих JUN-22BBz CAR Т-клетки, на 23-й день наблюдается увеличение Т-клеток периферической крови, h) Экспрессия JUN значительно улучшает долгосрочную выживаемость мышей, получивших лечение с помощью CAR. На f-g планки погрешностей представляют собой среднее ± SEM для n=5 мышей на группу. Репрезентативный результат 2 независимых экспериментов с аналогичными результатами. * р<,05, ** р<,01, *** р<,001. НТМ - шарнирный/трансмембранный. ICD - внутриклеточный домен.
На фиг. 18А-С показано, что 14g2a-GD2E101K CAR Т-клетки с высокой аффинностью (НА) проявляют избыточную сигнатуру истощения по сравнению с исходным 14g2a-GD2 CAR. а) Экспрессия ингибирующего поверхностно рецептора в CD 19, GD2, и HA-GD2E101K CAR Т-клетках на 10-й день культивирования. Мутация с высокой аффинностью Е101К приводит к увеличению экспрессии ингибирующего рецептора в CD4+и CD8+CAR Т-клетках по сравнению с исходным GD2 CAR. b) Секреция IL-2 после 24-часового совместного культивирования HA-GD2E101K или исходных GD2-28z CAR Т-клеток с GD2+ клетками-мишенями. Профиль повышенного истощения НА-GD2E101K CAR Т-клеток соответствует сниженной функциональной активности, что измеряется по способности продуцировать IL-2 при стимуляции. Планки погрешностей представляют собой среднее значение ± стандартное отклонение для лунок в трех повторностях. Репрезентативный результат по меньшей мере 4 независимых экспериментов с аналогичными результатами, с) РСА массового РНК-секвенирования демонстрирует большую дисперсию между НА- GD2E101K и CD 19 CAR Т-клетками, тогда как GD2-28z(sh) CAR Т-клетки являются промежуточными. Слева - CD4+ Т-клетки. Справа - CD8+ CAR Т-клетки, полученные из наивных, d- е) Экспрессия HA-GD2E101K CAR вызывает усиление экспрессии ингибирующего рецептора (d) и снижение формирования памяти (е) в CD4+ CAR Т-клетках. (Данные CD8+ на фигуре 12). f) РСА РНК-секвенирования на фиг. 12е, показывающий, что разделение РС2 обусловлено разделением СМ по сравнению с N и РС3, обусловленным CD4 по сравнению с CD8. g) GSEA: наборы генов, положительно регулируемых на 10-й день культивирования HA-28z CAR Т-клетки по сравнению с CD19-28z CAR Т-клетками, демонстрировали значительное перекрытие с генами, положительно регулируемыми в истощенных по сравнению с клетками памяти CD8+ (слева), истощенных по сравнению с эффекторными CD8+ (справа) и истощенных по сравнению с наивными клетками CD8+ (справа) в модели хронической вирусной инфекции на мышах (Wherry et al. Immunity, 2007). * p<,05, ** p<,01, *** p<,001. РСА - принципиальный компонентный анализ, NES - нормализованный показатель обогащения.
На фиг. 19А-С показано, что GD2-28z CAR Т-клетки показывают сигнатуру истощения на уровне отдельных клеток, а) Диаграмма Венна, показывающая перекрывающиеся гены в анализе дифференциальной экспрессии данных отдельных клеток (красный) и 200 наиболее результативных генов, определяющих разделение CD 19 и НА-28z CAR Т-клеток в массовом РНК-секвенировании. 79 из 200 наиболее результативных генов из массового РНК-секвенирования дифференциально экспрессируются с помощью анализа DESeq2 в GD2-28z по сравнению с CD19-28z одиночными клетками. Выделенные гены из пересечения включают в себя ингибирующие рецепторы (CTLA4, LAG3, GITR, эффекторные молекулы CD25, IFNG, GZMB и цитокины IL13 и ILIA и транскрипционные факторы BATF3 и IRF4 семейства АР-1/bZIP. В) Тепловая карта, кластеризующая наиболее результативные 50 дифференциально экспрессируемых генов в транскриптомном анализе одиночных клеток GD2-28z по сравнению с CD19-28z. Каждый ряд представляет одну клетку, с) Скрипичные графики, изображающие экспрессию индивидуальных генов в CD8+ GD2-28z и CD19-28z одиночных CAR Т-клетках. Гены, положительно регулируемые в GD2 CAR Т-клетках, включают в себя ингибирующие рецепторы, эффекторные молекулы и факторы транскрипции семейства АР-1, в то время как CD19 CAR Т-клетки обладают повышенной экспрессией генов, связанных с памятью. Р-значения, которые отображены для каждого гена выше отдельных графиков, рассчитывали с использованием непарного двустороннего U-критерия Уилкоксона-Манна-Уитни.
На фиг. 20A-D показан контроль качества данных ATAC-seq. а) Дины вставки и Ь) расстояние вставки от сайта начала транскрипции (TSS) для комбинированных (вверху) и отдельных образцов (внизу), с) Корреляция между образцами в повторностях. d) Расположение сопоставленных пиков в каждом образце по общему количеству пиков (вверху) и частоте встречаемости от общего (ниже).
На фиг. 21A-D показано, что семейство АР-1 содержит наиболее значительно обогащенные мотивы транскрипционных факторов в истощенных HA-28z CAR Т-клетках. а) Дифференциально доступные области хроматина в CD4+ CD 19 и НА CAR Т-клетках (D10). Как N, так и СМ подмножества включены для каждого CAR. b) РСА из фигуры 1h, показывающий, что разделение РС2 обусловлено разделением СМ по сравнению с N и разделение РС3 обусловлено CD4 по сравнению с CD8. с) Наиболее результативные мотивы факторов транскрипции, обогащенные областями хроматина, дифференциально доступными в HA-28z CAR Т-клетках, содержат факторы семейства АР-1/bZIP во всех исходных подмножествах Т-клеток. Наивное подмножество CD8+ клеток показано на фигуре 2. d) Кластеризация пиков по общему регуляторному мотиву (слева) и тепловая карта обогащения мотивов факторов транскрипции (справа) в каждом кластере. 10 различных кластеров, включая в себя кластеры, связанные с истощенными (ЕХ1-ЕХ4) или здоровыми (HLT1-HLT2) CAR Т-клетками, СМ (СМ) или N (наивным) исходным подмножеством и подмножеством CD4 или CD8 Т-клеток. Представляющие интерес гены в каждом кластере выделены справа. (N наивные клетки, СМ - центральные клетки памяти).
На фиг. 22А-С показано, что факторы транскрипции семейства АР-1/bZIP активируются в HA-28z CAR Т-клетках и образуют иммунорегуляторные комплексы, а) Кратное изменение экспрессии генов (HA/CD19) для указанных генов семейства АР-1/bZIP и IRF по данным РНК-секвенирования на фигуре 2. Планки погрешностей представляют собой среднее ± SEM для n=6 образцов у 3 независимых доноров. b) Кратность изменения экспрессии белка (HA/CD19) для указанных белков семейства АР-1/bZIP и IRF определяли денситометрическим анализом вестерн-блоттинга. Планки погрешностей представляют собой среднее ± SEM из n=4 экспериментов на 3 независимых донорах. * р<,05, ** р<,01, *** р<,001. с) Вестерн-блоттинг для указанных белков - представителей семейства АР-1/bZIP и IRF на входе (левые столбцы) или после иммунопреципитации для c-Jun (средние столбцы) или JunB (правые столбцы) в CD19 и HA-28z CAR Т-клетках. Числа ниже представляют кратность увеличения экспрессии белка для НА по сравнению CD19 при каждом условии, а цветные формы представляют комплексы, определенные в масштабе. IP-вестерн-блоты демонстрируют повышенное присутствие комплексов c-Jun/JunB, с-Jun/IRF4, c-Jun/BATF и c-Jun/BATF3 в HA-28z CAR Т-клетках. IRF4 также связывается с аналогичным отношением с JunB, в то время как BATF и BATF3 показывают преимущественное комплексообразование с JunB.
На фиг. 23А-Е показано, что усиленная активность API-модифицированных CAR Т-клеток зависит от c-Jun, но не от c-Fos. а-с) CAR Т-клетки совместно трансдуцировали с помощью (API) или без него (контроль) лентивирусным вектором, кодирующим оба фактора транскрипции API, Fos и Jun, и усеченный поверхностный селекционный маркер NGFR (tNGFR). а) Схема лентивирусного конструкта. b) Репрезентативная эффективность трансдукции API-модифицированных CAR Т-клеток, измеренная по поверхностной экспрессии NGFR в указанных CD4+ и CD8+ CAR Т-клетках. с) Продукция IL-2 в контрольных или API-модифицированных CAR Т-клетках после 24-часовой стимуляции клетками-мишенями 143В-19. API-модифицированные HA-28z CAR Т-клетки проявляют повышенную продукцию IL-2 по сравнению с контрольными CAR Т-клетками. Репрезентативный эксперимент 2 независимых экспериментов с аналогичными результатами, d-e) CAR Т-клетки совместно трансдуцировали лентивирусными векторами, кодирующими любой из факторов транскрипции API, Fos или Jun, и усеченный поверхностный селекционный маркер NGFR (tNGFR). d) Схемы лентивирусных конструктов Fos и Jun. е) Продукция IL-2 в контрольных, модифицированных Fos или Jun CAR Т-клетках после 24-часовой стимуляции клетками-мишенями Nalm6-GD2. Планки погрешностей представляют собой среднее значение ± стандартное отклонение для лунок в трех повторностях. Репрезентативный эксперимент 2 независимых экспериментов с аналогичными результатами. B a и d* обозначает стоп-кодон. *р<,05, **р<,01, ***р<,001, ns p>0,05.
На фиг. 24А-С показана расширенная функциональная оценка JUN-модифицированных CAR Т-клеток. а-b) Кратность увеличения высвобождения IL-2 (а) и IFNg (b) после 24-часового совместного культивирования с указанными клетками-мишенями в JUN по сравнению с контрольными CD19 и HA-28z CAR Т-клетками. Каждая точка представляет один независимый эксперимент от разных доноров, с) Расширенная экспансия контрольных или JUN-модифицированных CAR Т-клеток in vitro в 3 независимых экспериментах с 3 различными здоровыми донорами. В указанные моменты времени Т-клетки повторно высевали в свежую среду для Т-клеток + 100 МЕ/мл IL2. Т-клетки подсчитывали и количественно доводили для поддержания клеток в концентрации 0,5×106/мл каждые 2-3 дня. Для донора-1 5×106 жизнеспособных Т-клеток повторно высевали на 14 и 28 дни. Для донор-2 5×106 жизнеспособных Т-клеток повторно высевали на 14, 28, 42 и 56 дней. Для донор-3 5×106 жизнеспособных Т-клетки повторно высевали на 10, 17, 24 и 31 дни.
На фиг. 25А-Е показано, что избыточная экспрессия c-Jun снижает распространенность и комплексообразование ингибирующих представителей семейства АР-1: JunB, BATF и BATF3. а) Кинетика индуцированной лекарственным средством стабильности c-Jun в JUN-DD CAR Т-клетках, оцененная вестерн-блоттингом. В момент времени 0 10 мкМ ТМР либо добавляли к необработанным клеткам (ON), либо отмывали из предварительно обработанных клеток (OFF). Клетки удаляли из каждого условия через 1, 2, 4, 8, 24 и 48 ч и готовили для вестерн-блот-анализа экспрессии c-Jun. Наблюдаемая полоса соответствует размеру JUN-DD. б) Денситометрический анализ проводили на блотах из (а) и нормализовали по контролю нагрузки. Экспрессию наносили на графики в зависимости от времени и подбирали кривые кинетики первого порядка для данных, чтобы определить t1/2 для кинетики OFF и ON. с) Анализ вестерн-блоттинг для указанных белков-представителей семейства АР-1/bZIP и IRF в контрольных и JUN-CAR Т-клетках (D10). Числа ниже представляют кратность изменения экспрессии белка по сравнению с CD19. d) Соответствующее снижение экспрессии мРНК BATF, BATF3 и JUNB в JUN-HA-28z CAR Т-клетках по сравнению с HA-28z. n=3 донора, нормализованных по мРНК CD19. е) Избыточная экспрессия c-Jun снижает ингибирующие комплексы JunB/BATF и JunB/BATF3 с помощью анализа IP-вестерн-блоттинг. Ввод (левые столбцы), иммунопреципитация для c-Jun (средние столбцы) или JunB (правые столбцы) в контрольных или JUN-HA-28z CAR Т-клетках. Белок IRF4, мРНК и комплексообразование с c-Jun не изменяются.
На фиг. 26А-Е показано, что JUN-CAR Т-клетки усиливают функцию GD2-BBz CAR Т-клеток в солидных опухолях, а) Схема вектора - ретровирусного векторного конструкта JUN-GD2-BBz. b) Продукция IL-2 (слева) и IFNg (справа) в JUN-модифицированных (красный) или контрольных (синий) GD2-BBz CAR Т-клетках, после 24-часовой стимуляции клетками-мишенями Nalm6-GD2 или 143В. с) Лизис GD2-BBz CAR Т-клетками клеток-мишеней GFP+Nalm6-GD2 при соотношении Е:Т, составляющем 1:1 (слева) или 1:4 (справа). На а-с планки погрешности представляют среднее значение ± стандартное отклонение для лунок в трех повторностях. Репрезентативный результат как минимум 3 независимых экспериментов. В d-e мышей NSG инокулировали с помощью 0,5×106 клеток остеосаркомы 143В-19 посредством внутримышечной инъекции. 1×107 имитации, GD2-BBz, или JUN-GD2-BBz CAR Т-клеток вводили в/в на 3-й день, d) Рост опухоли контролировали измерениями штангенциркулем, е) Количество CD4+ (выше) или CD8+ (ниже) Т-клеток периферической крови на 14 день после приживления опухоли. Планки погрешности представляют собой среднее ± SEM для n=5 мышей на группу. Репрезентативный результат двух независимых экспериментов, хотя ранние смерти (не связанные с размером опухоли) исключали кривые выживаемости в обеих моделях. *р<,05, **р<,01, ***р<,001. НТМ - шарнирный/трансмембранный. ICD - внутриклеточный домен.
На фиг. 27А-Е показано, что N-концевые мутации c-Jun способны восстанавливать истощенные HA-28z CAR Т-клетки. а) Различные мутации c-Jun, клонированные в вектор HA-28z CAR Т-клетки. b) Секреция IL-2 (вверху) и IFNg (внизу) после 24-часового совместного культивирования с клетками-мишенями остеосаркомы GD2+ 143В. с) Лизис in vitro клеток-мишеней GFP+ Nalm6-GD2 или 143В измеряли в течение 5 дней при соотношении эффектор : мишень, составляющем 1:1, 1:2 или 1:4 (Е:Т). При низких соотношениях Е:Т и поздних временных точках JUN-WT, JUN-AA, JUN-Dd и JUN-DTAD демонстрируют улучшенный контроль роста опухоли по сравнению с JUN-De, JUN-Dbasic, JUN-DLeu и JUN-DbZIP CAR Т-клетками. d) Повышенную продукцию цитокинов подтверждали как в CD4+, так и в CD8+ HA-28z CAR Т-клетках путем внутриклеточного окрашивания цитокинов и проточной цитометрии после 5-часовой стимуляции Nalm6-GD2. е) Количественное определение количества периферических Т-клеток через 12 дней после инфузии Т-клеток мышам NSG с лейкозом Nalm6-GD2.
На фиг. 28 показано, что нокдаун IRF4 резко увеличивает функциональную активность истощенных HA-28z CAR Т-клеток.
На фиг. 29А-С показано, что транскрипционный мутант (JUN-AA) также восстанавливает функциональную активность и пролиферативную способность в CD19 CAR Т-клетках.
На фиг. 30 показано, что усиленная функция in vivo c-Jun-модифицированных НА-28z CAR Т-клеток не может быть воспроизведена предоставлением IL-2 ex vivo.
На фиг. 31А-Е показано, что c-Jun усиливал активность Her2-BBz CAR Т-клеток в супрессивном микроокружении солидной опухоли.
На фиг. 32 показано, что избыточная экспрессия c-Jun повышает устойчивость к TGFβ-опосредованной супрессии истощенных HA-28z CAR Т-клеток.
На фиг. 33A-D показано, что транскрипционные изменения в клетках, модифицированных c-Jun, согласуются с уменьшенным истощением и повышенным формированием памяти.
Определения
Для целей толкования настоящего описания будут применять следующие определения, и при необходимости термины, используемые в единственном числе, будут также включать в себя множественное число и наоборот. В случае, если какое-либо определение, изложенное ниже, вступает в противоречие с любым документом, включенным в настоящий документ посредством ссылки, определение, изложенное ниже, должно иметь преимущественную силу.
Используемые в настоящем документе термины «заболевание» и «патологическое состояние» используют взаимозаменяемо, если в настоящем документе не указано иное, для описания отклонения от состояния, рассматриваемого как нормальное или среднее для представителей вида или группы (например, людей), и которое является вредным для пораженного индивидуума в условиях, которые не являются вредными для большинства индивидуумов этого вида или группы. Такое отклонение может проявляться как состояние, признаки и/или симптомы (например, диарея, тошнота, лихорадка, боль, волдыри, фурункулы, сыпь, подавление иммунитета, воспаление и т.д.), которые связаны с любым нарушением нормального состояния субъекта или любого из его органов или тканей, что прерывает или изменяет выполнение нормальных функций. Заболевание или патологическое состояние может быть вызвано или может являться результатом контакта с микроорганизмом (например, патогеном или другим инфекционным агентом (например, вирусом или бактериями)), может являться реакцией на факторы окружающей среды (например, недоедание, опасные производственные факторы и/или климат), может являться реакцией на врожденный или скрытый дефект в организме (например, генетические аномалии) или на комбинации этих и других факторов.
Термины «хозяин», «субъект» или «пациент» используют в настоящем документе взаимозаменяемо для обозначения индивидуума, которого лечат (например, которому вводят) с помощью композиций и способов согласно настоящему изобретению. Субъекты включают в себя без ограничения следующее: млекопитающие (например, люди, мыши, крысы, обезьяны, лошади, коровы, свиньи, собаки, кошки и тому подобное). В контексте настоящего изобретения термин «субъект», как правило, относится к индивидууму, которому будут вводить или которому ввели одну или несколько композиций согласно настоящему изобретению (например, модифицированные или сконструированные (например, генетически) Т-клетки, описанные в настоящем документе).
«Истощение Т-клеток» относится к потере функции Т-клеток, которая может возникнуть в результате инфекции (например, хронической инфекции) или заболевания.
Истощение Т-клеток связано с повышенной экспрессией PD-1, TIM-3 и LAG-3, апоптозом и сниженной секрецией цитокинов. Соответственно, термины «облегчают истощение Т-клеток», «ингибируют истощение Т-клеток», «снижают истощение Т-клеток» и тому подобное относятся к состоянию восстановленной функциональности Т-клеток, характеризующемуся одним или несколькими из следующего: сниженная экспрессия и/или содержание одного или нескольких из PD-1, TIM-3 и LAG-3; увеличенное образование клеток памяти и/или поддержание маркеров памяти (например, CD62L); профилактика апоптоза; увеличенная антиген-индуцированная продукция и/или секреция цитокинов (например, IL-2); усиленная способность уничтожать; повышенное распознавание опухолевых мишеней с низкой плотностью поверхностного антигена; усиленная пролиферация в ответ на антиген.
Термины «буфер» или «буферные агенты» относятся к материалам, которые при добавлении в раствор вызывают устойчивость раствора к изменениям рН.
Используемые в настоящем документе термины «рак» и «опухоль» относятся к ткани, содержащей клетки, которые утратили способность контролировать рост и пролиферацию или росту таких клеток. Раковые и опухолевые клетки, как правило, характеризуются потерей контактного торможения, могут быть инвазивными и могут проявлять способность метастазировать (например, они утратили способность к адгезии к другим клеткам/тканям). Настоящее изобретение не ограничено типом рака или типом лечения (например, профилактическим и/или терапевтическим лечением). Действительно, различные формы рака можно лечить с помощью композиций и способов, описанных в настоящем документе, включая в себя без ограничения рак головного мозга или другие формы рака центральной нервной системы, меланомы, лимфомы, рак костей, эпителиальный рак, рак молочной железы, рак яичников, рак эндометрия, рак толстой и прямой кишки, рак легкого, рак почки, меланома, рак почки, рак предстательной железы, саркомы, карциномы и/или их комбинации.
Используемый в настоящем документе термин «метастазирование» относится к процессу, посредством которого рак распространяется или переносится из места происхождения в другие области тела с развитием аналогичного ракового поражения в новом месте. «Метастатическая» или «метастазирующая» клетка представляет собой клетку, которая теряет адгезивные контакты с соседними клетками и мигрирует через кровоток или лимфу из первичного участка заболевания, проникая в ткани в других частях тела.
Используемый в настоящем документе термин «противораковое средство» относится к любому терапевтическому средству (например, химиотерапевтическим соединениям и/или молекулярным терапевтическим соединениям), антисмысловой терапии, лучевой терапии и тому подобному, используемому при лечении гиперпролиферативных заболеваний, таких как рак (например, у млекопитающих, например у людей).
«Эффективное количество» относится к количеству фармацевтической композиции, противоракового средства или другого лекарственного средства, эффективному в дозировках и в течение периодов времени, необходимых для достижения требуемого терапевтического или профилактического результата (например, ослабления некоторых или всех симптомов заболевания, подлежащего лечению).
Используемый в настоящем документе термин «терапевтически эффективное количество» относится к такому количеству терапевтического средства, которое является достаточным для того, чтобы привести к уменьшению интенсивности одного или нескольких симптомов нарушения или предотвратить развитие нарушения или вызвать регрессию нарушения. Например, что касается лечения рака, согласно одному варианту осуществления терапевтически эффективное количество будет относиться к количеству терапевтического средства, которое уменьшает скорость роста опухоли (например, снижает и/или устраняет опухолевую нагрузку у пациента), уменьшает массу опухоли, уменьшает количество метастазов, уменьшает прогрессирование опухоли или увеличивает продолжительность выживания по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 100%.
Используемые в настоящем документе термины «сенсибилизировать» и «сенсибилизирующий» относятся к тому, чтобы сделать посредством введения первого средства животное или клетку животного более восприимчивыми или более чувствительными к биологическим эффектам (например, стимулирование или замедление аспекта клеточной функции, включая в себя без ограничения деление клеток, рост клеток, пролиферацию, инвазию, ангиогенез, некроз или апоптоз) второго средства. Сенсибилизирующее действие первого средства на клетку-мишень можно измерить как разницу в предполагаемом биологическом эффекте (например, стимулирование или замедление аспекта клеточной функции, включая в себя без ограничения рост клеток, пролиферацию, инвазию, ангиогенез, или апоптоз), наблюдаемый при введении второго средства с введением и без введения первого средства. Ответ сенсибилизированной клетки может быть увеличен по меньшей мере приблизительно на 10%, по меньшей мере приблизительно на 20%, по меньшей мере приблизительно на 30%, по меньшей мере приблизительно на 40%, по меньшей мере приблизительно на 50%, по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 100%, по меньшей мере приблизительно на 150%, по меньшей мере приблизительно на 200%, по меньшей мере приблизительно на 250%, по меньшей мере на 300%, по меньшей мере приблизительно на 350%, по меньшей мере приблизительно на 400%, по меньшей мере приблизительно на 450%, или по меньшей мере приблизительно на 500% по сравнению с ответом при отсутствии первого средства.
Используемые в настоящем документе термины «очищенный» или «очищать» относятся к удалению примесей или нежелательных соединений из образца или композиции. Используемый в настоящем документе термин «по существу очищенный» относится к удалению от приблизительно 70 до 90%, вплоть до 100%, примесей или нежелательных соединений из образца или композиции.
Используемые в настоящем документе термины «введение» и «осуществление введения» относятся к акту предоставления композиции субъекту. Иллюстративные пути введения в организм человека включают в себя без ограничения введение через глаза (офтальмологическое), рот (оральное), кожу (трансдермальное), нос (назальное), легкие (ингаляционное), слизистую оболочку рта (трансбуккальное), ухо, ректальное, путем инъекции (например, внутривенно, подкожно, внутрибрюшинно, внутриопухолево и т.д.), местно и тому подобное. Согласно одному варианту осуществления введение Т-клеток согласно настоящему изобретению осуществляют посредством внутривенной инфузии.
Используемые в настоящем документе термины «совместное введение» и «осуществление совместного введения» относятся к введению по меньшей мере двух средств (например, генетически модифицированных иммунных клеток и одного или нескольких других средств, например, противораковых средств) или видов терапии субъекту. Согласно некоторым вариантам осуществления совместное введение двух или более средств или видов лечения является одновременным. Согласно другим вариантам осуществления первое средство/терапию вводят перед вторым средством/терапией. Согласно некоторым вариантам осуществления совместное введение можно осуществлять одним и тем же или другим путем введения. Специалистам в настоящей области техники понятно, что составы и/или пути введения различных используемых средств или способов лечения могут варьироваться. Подходящая дозировка для совместного введения может быть легко определена специалистом в настоящей области техники. Согласно некоторым вариантам осуществления, когда средства или способы лечения вводят совместно, соответствующие средства или виды лечения вводят в более низких дозировках, чем подходящие для их введения отдельно. Таким образом, совместное введение является особенно желательным в вариантах осуществления, где совместное введение средств или видов лечения снижает необходимую дозировку потенциально вредного (например, токсического) средства(средств) и/или когда совместное введение двух или более средств приводит к сенсибилизации субъекта к благоприятным эффектам одного из средств посредством совместного введения другого средства.
Используемые в настоящем документе термины «фармацевтически приемлемый» или «фармакологически приемлемый» относятся к композициям, которые по существу не вызывают побочных реакций (например, токсических, аллергических или других иммунологических реакций) при введении субъекту.
Используемый в настоящем документе термин «фармацевтически приемлемый носитель» относится к любому из стандартных фармацевтических носителей, включая в себя без ограничения физиологический раствор с фосфатным буфером, воду и различные типы увлажняющих агентов (например, лаурилсульфат натрия), любой и все растворители, дисперсионные среды, покрытия, лаурилсульфат натрия, изотонические и замедляющие абсорбцию агенты, разрыхлители (например, картофельный крахмал или крахмалгликолят натрия), полиэтиленгликоль и тому подобное. Композиции также могут включать в себя стабилизаторы и консерванты. Примеры носителей, стабилизаторов и адъювантов были описаны и известны в настоящей области техники (см., например, Martin, Remington's Pharmaceutical Sciences, 15-е изд., Mack Publ. Co., Easton, Pa. (1975), включенный в настоящий документ посредством ссылки).
Используемый в настоящем документе термин «фармацевтически приемлемая соль» относится к любой соли (например, полученной в результате реакции с кислотой или основанием) композиции согласно настоящему изобретению, которая физиологически переносится у целевого субъекта. «Соли» композиций согласно настоящему изобретению могут быть получены из неорганических или органических кислот и оснований. Примеры кислот включают в себя без ограничения следующее: соляная, бромистоводородная, серная, азотная, хлорная, фумаровая, малеиновая, фосфорная, гликолевая, молочная, салициловая, янтарная, толуол-п-сульфоновая, винная, уксусная, лимонная, метансульфоновая, этансульфоновая, муравьиная, бензойная, малоновая, сульфоновая, нафталин-2-сульфоновая, бензолсульфоновая кислота и тому подобное. Другие кислоты, такие как щавелевая кислота, хотя сами по себе они не являются фармацевтически приемлемыми, могут быть использованы при получении солей, применимых в качестве промежуточных соединений при получении композиций согласно настоящему изобретению, и их фармацевтически приемлемых солей присоединения кислоты. Примеры оснований включают в себя без ограничения гидроксиды щелочных металлов (например, натрия), гидроксиды щелочноземельных металлов (например, магния), аммиак и соединения формулы NW4+, где W представляет собой С1-4 алкил, и тому подобное.
Примеры солей включают в себя без ограничения: ацетат, адипат, альгинат, аспартат, бензоат, бензолсульфонат, бисульфат, бутират, цитрат, камфорат, камфорсульфонат, циклопентанпропионат, диглюконат, додецилсульфат, этансульфонат, фумарат, флюкогептаноат, глицерофосфат, гемисульфат, гептаноат, гексаноат, хлорид, бромид, йодид, 2-гидроксиэтансульфонат, лактат, малеат, метансульфонат, 2-нафталинсульфонат, никотинат, оксалат, пальмоат, пектинат, персульфат, фенилпропионат, пикрат, пивалат, пропионат, сукцинат, тартрат, тиоцианат, тозилат, ундеканоат и тому подобное. Другие примеры солей включают в себя анионы соединений согласно настоящему изобретению, соединенные с подходящим катионом, таким как Na+, NH4+ и NW4+ (где W представляет собой С1-4 алкильную группу), и тому подобное. Для терапевтического применения соли соединений согласно настоящему изобретению рассматриваются как фармацевтически приемлемые. Однако соли кислот и оснований, которые не являются фармацевтически приемлемыми, также могут найти применение, например, при получении или очистке фармацевтически приемлемого соединения.
Для терапевтического применения соли композиций согласно настоящему изобретению рассматриваются как фармацевтически приемлемые. Однако соли кислот и оснований, которые не являются фармацевтически приемлемыми, также могут найти применение, например, при получении или очистке фармацевтически приемлемой композиции.
Используемый в настоящем документе термин «подверженный риску заболевания» относится к субъекту, который предрасположен к тому, чтобы испытывать конкретное заболевание (например, инфекционное заболевание). Эта предрасположенность может быть генетической (например, особая генетическая тенденция испытывать заболевание, такое как наследственные нарушения), или вследствие других факторов (например, условий окружающей среды, воздействия вредных соединений, присутствующих в окружающей среде, и т.д.). Таким образом, не предполагается, что настоящее изобретение ограничено каким-либо конкретным риском (например, субъект может быть «подвержен риску заболевания», просто подвергаясь воздействию других людей и взаимодействуя с ними), а также не предполагается, что настоящее изобретение будет ограничивается каким-либо конкретным заболеванием (например, раком).
Используемый в настоящем документе термин «набор» относится к любой системе доставки для доставки материалов. В контексте иммунотерапевтических средств такие системы доставки включают в себя системы, которые позволяют хранить, транспортировать или доставлять иммуногенные средства и/или вспомогательные материалы (например, письменные инструкции по использованию материалов и т.д.) из одного места в другое. Например, наборы включают в себя одно или несколько емкостей-вкладок (например, коробок), содержащих соответствующие иммунотерапевтические средства (например, модифицированные Т-клетки и/или вспомогательные материалы). Используемый в настоящем документе термин «фрагментированный набор» относится к системам доставки, содержащим два или более отдельных контейнера, каждый из которых содержит часть всех компонентов набора. Контейнеры могут быть доставлены предполагаемому получателю вместе или по отдельности. Например, первый контейнер может содержать композицию, содержащую иммунотерапевтическую композицию для конкретного применения, тогда как второй контейнер содержит второе средство (например, химиотерапевтическое средство). Действительно, любая система доставки, содержащая два или более отдельных контейнера, каждый из которых содержит часть компонентов всего набора, включена в термин «фрагментированный набор». Напротив, «комбинированный набор» относится к системе доставки, содержащей все компоненты, необходимые для конкретного использования, в одном контейнере (например, в одной коробке, в которой находится каждый из требуемых компонентов). Термин «набор» включает в себя как фрагментированные, так и комбинированные наборы.
Используемый в настоящем документе термин «иммуноглобулин» или «антитело» относится к белкам, которые связывают один или несколько эпитопов на конкретном антигене. Иммуноглобулины включают в себя без ограничения поликлональные, моноклональные, химерные и гуманизированные антитела, а также фрагменты Fab и фрагменты F(ab')2 следующих классов: IgG, IgA, IgM, IgD, IgE и секретируемые иммуноглобулины (sIg). Иммуноглобулины, как правило, содержат две идентичные тяжелые цепи и две легкие цепи. Однако термины «антитело» и «иммуноглобулин» также охватывают одноцепочечные антитела и двухцепочечные антитела.
«Вариабельная область» или «вариабельный домен» антитела относится к аминоконцевым доменам тяжелой или легкой цепи антитела. Вариабельный домен тяжелой цепи может обозначаться как «VH». Вариабельный домен легкой цепи может обозначаться как «VL». Эти домены, как правило, являются наиболее вариабельными частями антитела и содержат антигенсвязывающие сайты.
«Одноцепочечные Fv» или «scFv» фрагменты антитела содержат домены VH и VL антитела, причем эти домены присутствуют в одной полипептидной цепи. Как правило, полипептид scFv дополнительно содержит полипептидный линкер между доменами VH и VL, который позволяет scFv формировать структуру для связывания антигена. Для обзора scFv см., например, Pluckthun, в The Pharmacology of Monoclonal Antibodies, том. 113, Rosenburg and Moore eds., (Springer-Verlag, New York, 1994), c. 269-315.
Используемый в настоящем документе термин «антигенсвязывающий белок» относится к белкам, которые связываются с конкретным антигеном. «Антигенсвязывающие белки» включают в себя без ограничения иммуноглобулины, включая в себя поликлональные, моноклональные, химерные и гуманизированные антитела; фрагменты Fab, фрагменты F(ab')2 и библиотеки экспрессии Fab; и одноцепочечные антитела.
Используемый в настоящем документе термин «эпитоп» относится к той части антигена, которая вступает в контакт с конкретным иммуноглобулином.
Термины «специфическое связывание» или «специфически связывающийся» при использовании в отношении взаимодействия антитела (или его части (например, scFv)) и белка или пептида означает, что взаимодействие зависит от присутствия конкретной последовательности или структуры (например, антигенной детерминанты или эпитопа) на белке, иными словами, антитело (или его часть (например, scFv)) распознает и связывается с определенной последовательностью или структурой белка, а не с белками в целом. Например, если антитело является специфическим в отношении эпитопа «А», присутствие белка, содержащего эпитоп А (или свободного, не меченного А), в реакции, содержащей меченый «А», и антитело, будет уменьшать количество меченого А, связанного с антителом.
Используемые в настоящем документе термины «неспецифическое связывание» и «фоновое связывание» при использовании в отношении взаимодействия антитела и белка или пептида относятся к взаимодействию, которое не зависит от наличия конкретной структуры (т.е. антитело связывается с белками в целом, а не с конкретной структурой, такой как эпитоп).
Используемый в настоящем документе термин «субъект, подозреваемый в наличии рака» относится к субъекту, у которого присутствует один или несколько симптомов, указывающих на рак (например, заметное уплотнение или масса), или который проходит скрининг в отношении рака (например, во время медосмотра). Субъект, подозреваемый на наличие рака, также может характеризоваться наличием одного или нескольких факторов риска развития рака. Субъект, подозреваемый на наличие рака, как правило не тестировался на рак. Тем не менее, «субъект, подозреваемый на наличие рака» включает в себя индивидуума, который получил предварительный диагноз (например, компьютерная томография, показывающая массу), но для которого подтверждающий тест (например, биопсия и/или гистология) не был проведен или для которого тип и/или стадия рака неизвестны. Термин также включает в себя людей, у которых ранее был рак (например, индивидуум с ремиссией). «Субъекту, подозреваемому на наличие рака» иногда диагностируют рак, а иногда обнаруживают, что у него нет рака.
Используемый в настоящем документе термин «субъект, у которого диагностирован рак», относится к субъекту, который был протестирован и у которого обнаружены раковые клетки. Рак может быть диагностирован с использованием любого подходящего способа, включая в себя без ограничения биопсию, рентген, анализ крови и т.д.
Используемый в настоящем документе термин «послеоперационная опухолевая ткань» относится к раковой ткани (например, ткани органа), которая была удалена у субъекта (например, во время операции).
Используемый в настоящем документе термин «субъект с риском развития рака» относится к субъекту с одним или несколькими факторами риска развития конкретного рака. Факторы риска включают в себя без ограничения пол, возраст, генетическую предрасположенность, воздействие окружающей среды и предыдущие случаи рака, ранее существовавшие незлокачественные заболевания и образ жизни.
Используемый в настоящем документе термин «определение характеристик рака у субъекта» относится к идентификации одного или нескольких свойств образца рака у субъекта, включая в себя без ограничения наличие доброкачественной, предраковой или раковой ткани и стадии рака.
Используемый в настоящем документе термин «определение характеристик ткани у субъекта» относится к идентификации одного или нескольких свойств образца ткани (например, включая в себя без ограничения наличие раковой ткани, наличие предраковой ткани, которая может стать раковой, и наличие раковой ткани, которая может метастазировать).
Используемый в настоящем документе термин «стадия рака» относится к качественной или количественной оценке уровня развития рака. Критерии, используемые для определения стадии рака, включают в себя без ограничения размер опухоли, распространилась ли опухоль на другие части тела и где распространился рак (например, в пределах того же органа или области тела или на другой орган).
Используемый в настоящем документе термин «первичная опухолевая клетка» относится к раковой клетке, которая выделена из опухоли у млекопитающего и не была интенсивно культивирована in vitro.
Используемые в настоящем документе термины «лечение», «терапевтическое применение» или «лекарственное применение» относятся к любому и всем применениям композиций и способов согласно настоящему изобретению, которые устраняют болезненное состояние или симптомы или иным образом предотвращают, препятствуют, замедляют или устраняют прогрессирование заболевания или других нежелательных симптомов каким-либо образом. Например, термины «лечение рака» или «лечение опухоли» или грамматические эквиваленты в настоящем документе означают подавление, регрессию или частичное или полное исчезновение ранее существовавшего рака или опухоли. Подразумевается, что определение включает в себя любое уменьшение размера, агрессивности или скорости роста ранее существовавшего рака или опухоли.
Используемые в настоящем документе термины «улучшенный терапевтический эффект» и «повышенная терапевтическая эффективность» по отношению к раку относятся к замедлению или уменьшению роста раковых клеток или солидной опухоли или уменьшению общего количества раковых клеток или общей опухолевой нагрузки. «Улучшенный терапевтический эффект» или «повышенная терапевтическая эффективность» означает, что состояние индивидуума улучшается в соответствии с любыми клинически приемлемыми критериями, включая в себя купирование развившейся опухоли, увеличение продолжительности жизни или улучшение качества жизни.
Используемый в настоящем документе термин «система переноса генов» относится к любым средствам доставки композиции, содержащей последовательность нуклеиновой кислоты, в клетку или ткань. Например, системы переноса генов включают в себя без ограничения следующее: векторы (например, системы доставки на основе ретровирусов, аденовирусов, лентивирусов, аденоассоциированных вирусов и другие системы доставки на основе нуклеиновых кислот), микроинъекция голой нуклеиновой кислоты, системы доставки на основе полимеров (например, системы на основе липосом и металлических частиц), биолистическая инъекция, трансдукция с помощью систем на основе транспозазы для интеграции генов, интеграция генов, опосредованная Crispr/Cas9, неинтегрирующие векторы, такие как РНК или аденоассоциированные вирусы и тому подобное.
Используемый в настоящем документе термин «вирусная система переноса генов» относится к системам переноса генов, содержащим вирусные элементы (например, интактные вирусы, модифицированные вирусы и вирусные компоненты, такие как нуклеиновые кислоты или белки) для облегчения доставки образца в требуемую клетку или ткань. Неограничивающими примерами вирусных систем переноса генов, применяемых в композициях и способах согласно настоящему изобретению, являются лентивирусные и ретровирусные системы переноса генов.
Используемый в настоящем документе термин «сайт-специфические последовательности-мишени рекомбинации» относится к последовательностям нуклеиновых кислот, которые обеспечивают последовательности распознавания для факторов рекомбинации, и локализацию, где происходит рекомбинация.
Используемый в настоящем документе термин «молекула нуклеиновой кислоты» относится к любой молекуле, содержащей нуклеиновую кислоту, включая в себя без ограничения ДНК или РНК. Термин охватывает последовательности, которые включают в себя любой из известных аналогов оснований ДНК и РНК, включая в себя без ограничения следующее: 4-ацетил цитозин, 8-гидрокси-N6-метиладенозин, азиридинилцитозин, псевдоизоцитозин, 5-(карбоксигидроксилметил)-урацил, 5-фторурацил, 5-бромурацил, 5-карбоксиметиламинометил-2-тиоурацил, 5-карбоксиметиламинометилурацил, дигидроурацил, инозин, N6-изопентениладенин, 1-метиладенин, 1-метилпсевдоурацил, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метил цитозин, 5-метилцитозин, N6-метиладенин, 7-метил гуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеуозин, 5'-метоксикарбонилметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусная кислота, оксибутоксозин, псевдоурацил, квеуозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир N-урацил-5-оксиуксусной кислоты и 2,6-диаминопурин.
Используемый в настоящем документе термин «гетерологичный ген» относится к гену, который отсутствует в естественной для него среде. Например, гетерологичный ген включает в себя ген одного вида, введенный другому виду. Гетерологичный ген также включает в себя ген, нативный для организма, который каким-то образом изменен (например, мутирован, добавлен в нескольких копиях, связан с ненативными регуляторными последовательностями и т.д.). Гетерологичные гены отличаются от эндогенных генов тем, что последовательности гетерологичных генов, как правило, присоединяются к последовательностям ДНК, которые не обнаруживаются в природе ассоциированными с последовательностями генов в хромосоме или связаны с частями хромосомы, которых нет в природе (например, гены, экспрессируемые в локусы, где ген обычно не экспрессируется). Клетки, которые содержат гетерологичный ген, описаны в настоящем документе как «модифицированные» или «сконструированные» клетки. Например, Т-клетки, которые содержат гетерологичный ген фактора транскрипции АР-1 (например, конструкт экспрессии гетерологичного гена фактора транскрипции АР-1 (например, используется для избыточной экспрессии фактора транскрипции АР-1 в Т-клетках)) и/или гетерологичный ген рецептора (например, конструкт экспрессии гетерологичного гена рецептора Т-клетки или конструкт экспрессии гетерологичного гена химерный антигенного рецептора) описаны в настоящем документе как модифицированные и/или сконструированные Т-клетки.
Используемый в настоящем документе термин «экспрессия гена» относится к процессу преобразования генетической информации, закодированной в гене, в РНК (например, мРНК, рРНК, тРНК или мяРНК) посредством «транскрипции» гена (т.е. посредством ферментативного действия РНК-полимеразы) и для генов, кодирующих белок, в белок посредством «трансляции» мРНК. Экспрессия генов может регулироваться на многих стадиях процесса. «Положительная регуляция» или «активация» относится к регуляции, которая увеличивает продукцию продуктов экспрессии генов (т.е. РНК или белка), тогда как «отрицательная регуляция» или «репрессия» относится к регуляции, которая уменьшает продукцию. Молекулы (например, факторы транскрипции), которые участвуют в положительной или отрицательной регуляции, часто называют «активаторами» и «репрессорами», соответственно.
Помимо содержания интронов, геномные формы гена могут также включать в себя последовательности, расположенные как на 5', так и на 3' концах последовательностей, которые присутствуют в транскрипте РНК. Эти последовательности называются «фланкирующими» последовательностями или областями (эти фланкирующие последовательности расположены 5' или 3' от нетранслируемых последовательностей, присутствующих в транскрипте мРНК). 5'-фланкирующая область может содержать регуляторные последовательности, такие как промоторы и энхансеры, которые контролируют или влияют на транскрипцию гена. 3'-фланкирующая область может содержать последовательности, которые управляют терминацией транскрипции, посттранскрипционным расщеплением и полиаденилированием.
Используемые в настоящем документе термины «кодирование молекулы нуклеиновой кислоты», «кодирование последовательности ДНК» и «кодирование ДНК» относятся к порядку или последовательности дезоксирибонуклеотидов вдоль цепи дезоксирибонуклеиновой кислоты. Порядок этих дезоксирибонуклеотидов определяет порядок аминокислот вдоль полипептидной (белковой) цепи. Таким образом, последовательность ДНК кодирует аминокислотную последовательность.
Используемые в настоящем документе термины «олигонуклеотид, характеризующийся нуклеотидной последовательностью, кодирующей ген» и «полинуклеотид, характеризующийся нуклеотидной последовательностью, кодирующей ген», означают последовательность нуклеиновой кислоты, содержащую кодирующую область гена, или, другими словами, последовательность нуклеиновой кислоты, которая кодирует продукт гена. Кодирующая область может присутствовать в форме кДНК, геномной ДНК или РНК. Когда она присутствует в форме ДНК, олигонуклеотид или полинуклеотид может быть одноцепочечным (т.е. смысловая цепь) или двухцепочечным. Подходящие контролирующие элементы, такие как энхансеры/промоторы, границы сплайсинга, сигналы полиаденилирования и т.д., могут размещаться в непосредственной близости от кодирующей области гена, если это необходимо, для обеспечения надлежащей инициации транскрипции и/или правильного процессинга первичной транскрипции РНК. Альтернативно, кодирующая область, используемая в экспрессионных векторах согласно настоящему изобретению, может содержать эндогенные энхансеры/промоторы, границы сплайсинга, вставочные последовательности, сигналы полиаденилирования и т.д. или комбинацию как эндогенных, так и экзогенных контролирующих элементов.
Используемые в настоящем документе термины «в функциональной комбинации», «в функциональном порядке» и «функционально связанные» относятся к связыванию последовательностей нуклеиновых кислот таким образом, что молекула нуклеиновой кислоты способна направлять транскрипцию данного гена и/или происходит синтез требуемой молекулы белка. Термин также относится к связыванию аминокислотных последовательностей таким образом, что производится функциональный белок.
Термин «выделенный» при использовании по отношению к нуклеиновой кислоте, как во фразе «выделенный олигонуклеотид» или «выделенный полинуклеотид», относится к последовательности нуклеиновой кислоты, которая идентифицирована и отделена по меньшей мере от одного компонента или примеси, с которыми она обычно связана в своем естественном источнике. Выделенная нуклеиновая кислота - это такая нуклеиновая кислота, которая находится в форме или условиях, отличающихся от тех, в которых она встречается в природе. Напротив, невыделенные нуклеиновые кислоты представляют собой такие нуклеиновые кислоты, такие как ДНК и РНК, которые находятся в состоянии, в котором они существуют в природе. Например, данная последовательность ДНК (например, ген) обнаружена на хромосоме клетки-хозяина в непосредственной близости от соседних генов; последовательности РНК, такие как конкретная последовательность мРНК, кодирующая конкретный белок, обнаруживаются в клетке в виде смеси с множеством других мРНК, которые кодируют множество белков. Однако выделенная нуклеиновая кислота, кодирующая данный белок, включает в себя, в качестве примера, такую нуклеиновую кислоту в клетках, которые обычно экспрессируют данный белок, где нуклеиновая кислота находится в хромосомной локализации, отличной от таковой в естественных клетках, или иным образом фланкирована другой последовательностью нуклеиновой кислоты, чем та, которая встречается в природе. Выделенная нуклеиновая кислота, олигонуклеотид или полинуклеотид могут присутствовать в одноцепочечной или двухцепочечной форме. Когда для экспрессии белка следует использовать выделенную нуклеиновую кислоту, олигонуклеотид или полинуклеотид, олигонуклеотид или полинуклеотид будет содержать как минимум смысловую или кодирующую цепь (т.е. олигонуклеотид или полинуклеотид может являться одноцепочечным), но может содержать как смысловую, так и антисмысловую цепи (т.е. олигонуклеотид или полинуклеотид могут являться двухцепочечными).
Используемый в настоящем документе термин «нативный белок» указывает, что белок не содержит аминокислотных остатков, кодируемых векторными последовательностями; то есть нативный белок содержит только те аминокислоты, которые содержатся в белке, как это происходит в природе. Нативный белок можно получить рекомбинантными способами или можно выделить из встречающегося в природе источника.
Используемый в настоящем документе термин «часть» по отношению к белку (как во фразе «часть данного белка») относится к фрагментам этого белка. Фрагменты могут характеризоваться размером от четырех аминокислотных остатков до всей аминокислотной последовательности минус одна аминокислота.
Используемый в настоящем документе термин «вектор» предназначен для обозначения молекулы нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. Один тип вектора представляет собой «плазмиду», которая относится к кольцевой двухцепочечной ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, характеризующиеся бактериальной точкой начала репликации, и эписомные векторы млекопитающих). Можно использовать лентивирусные векторы или ретровирусные векторы (например, для введения ДНК, кодирующей один или несколько факторов транскрипции АР-1 и/или конструкт CAR, в клетки (например, Т-клетки)). Другие векторы (например, неэписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, реплицируются вместе с геномом-хозяином. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы в настоящем документе называются «рекомбинантные экспрессионные векторы» или просто «экспрессионные векторы». В целом, экспрессионные векторы, используемые в техниках рекомбинантной ДНК, часто находятся в форме плазмид. В настоящем описании термины «плазмида» и «вектор» могут использовать взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора.
Используемый в настоящем документе термин «экспрессионный вектор» относится к молекуле рекомбинантной ДНК, содержащей требуемую кодирующую последовательность и соответствующие последовательности нуклеиновой кислоты, необходимые для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Последовательности нуклеиновых кислот, необходимые для экспрессии в прокариотах, как правило, включают в себя промотор, оператор (необязательно) и сайт связывания рибосомы, часто вместе с другими последовательностями. Известно, что эукариотические клетки используют промоторы, энхансеры и сигналы терминации и полиаденилирования.
Используемый в настоящем документе термин «трансфекция» относится к введению чужеродной ДНК в эукариотические клетки. Трансфекцию можно осуществить различными способами, известными в настоящей области техники, включая в себя совместное осаждение фосфата кальция и ДНК, опосредованную DEAE-декстраном трансфекцию, опосредованную полибреном трансфекцию, электропорацию, микроинъекцию, слияние липосом, липофекцию, слияние протопластов, ретровирусную инфекцию и биобаллистику.
Термин «стабильная трансфекция» или «стабильно трансфицированный» относится к введению и интеграции чужеродной ДНК в геном трансфицированной клетки. Термин «стабильный трансфектант» относится к клетке, которая стабильно интегрировала чужеродную ДНК в геномную ДНК.
Термин «транзиентная трансфекция» или «транзиентно трансфицированный» относится к введению чужеродной ДНК в клетку, где чужеродная ДНК не может интегрироваться в геном трансфицированной клетки. Инородная ДНК сохраняется в ядре трансфицированной клетки (например, в течение нескольких дней). В течение этого времени чужеродная ДНК подвергается действию регуляторных контролирующих элементов, которые регулируют экспрессию эндогенных генов в хромосомах. Термин «транзиентный трансфектант» относится к клеткам, которые захватили чужеродную ДНК, но не смогли интегрировать эту ДНК.
Используемый в настоящем документе термин «селектируемый маркер» относится к применению гена, который кодирует ферментативную активность, которая придает способность расти в среде, в которой отсутствует то, что в противном случае было бы необходимым питательным веществом; кроме того, селектируемый маркер может придавать устойчивость к антибиотику или лекарственному средству клетке, в которой экспрессируется селектируемый маркер. Селектируемые маркеры могут являться «доминантными»; доминантный селектируемый маркер кодирует ферментативную активность, которая может быть обнаружена в любой эукариотической клеточной линии. Примеры доминантных селектируемых маркеров включают в себя ген бактериальной аминогликозид-3'-фосфотрансферазы (также называемый геном neo), который придает устойчивость к лекарственному средству G418 в клетках млекопитающих, ген бактериальной фосфотрансферазы гигромицина G (hyg), который придает устойчивость к антибиотику гигромицину, и ген бактериальной ксантин-гуанин-фосфорибозилтрансферазы (также называемый геном gpt), который придает способность расти в присутствии микофеноловой кислоты. Другие селектируемые маркеры не являются доминантными в том смысле, что их использование должно быть связано с клеточной линией, у которой отсутствует активность соответствующего фермента. Примеры недоминантных селектируемых маркеров включают в себя ген тимидинкиназы (tk), который используют в сочетании с tk-отрицательными (tk-) клеточными линиями, ген CAD, который используют в сочетании с CAD-дефицитными клетками, и ген гипоксантин-гуанинфосфорибозилтрансферазы млекопитающих (hprt), который используют в сочетании с hprt-отрицательными (hprt-) клеточными линиями. Обзор применения селектируемых маркеров в клеточных линиях млекопитающих представлен в Sambrook, J. et al., Molecular Cloning: A Laboratory Manual, 2-е изд., Cold Spring Harbor Laboratory Press, New York (1989), c. 16.9-16.15.
Используемый в настоящем документе термин «in vitro» относится к искусственной среде и процессам или реакциям, которые происходят в искусственной среде. Среды in vitro могут без ограничения состоять из пробирок и клеточных культур. Термин «in vivo» относится к природной среде (например, животному или клетке) и процессам или реакциям, которые происходят в естественной среде.
Используемый в настоящем документе термин «клеточная культура» относится к любой культуре клеток in vitro. В этот термин включены непрерывные клеточные линии (например, с бессмертным фенотипом), первичные клеточные культуры, трансформированные клеточные линии, перевиваемые клеточные линии (например, нетрансформированные клетки) и любая другая клеточная популяция, поддерживаемая in vitro.
Используемый в настоящем документе термин «образец» используют в самом широком смысле. В одном смысле подразумевается, что он включает в себя образец или культуру, полученные из любого источника, а также биологические образцы и образцы окружающей среды. Биологические образцы можно получить от животных (включая в себя людей) и включают в себя жидкости, твердые вещества, ткани и газы. Биологические образцы включают в себя продукты крови, такие как плазма, сыворотка и тому подобное. Такие примеры, однако, не должны рассматриваться как ограничивающие типы образцов, применимые к настоящему изобретению.
Подробное раскрытие вариантов осуществления
Настоящее изобретение основано на открытии того факта, что Т-клетки, модифицированные (например, генетически), чтобы избыточно экспрессировать и/или содержать повышенные (например, супрафизиологические) содержания одного или нескольких факторов транскрипции АР-1 (например, c-Jun), демонстрируют пониженные уровни истощения Т-клеток (например, по сравнению с немодифицированными Т-клетками, экспрессирующими нормальные содержания факторов транскрипции АР-1). Как подробно описано в настоящем документе, экспрессия транскрипционных факторов АР-1 Fos и c-Jun снижается/отрицательно регулируется в истощенных Т-клетках, а Т-клетки, модифицированные, чтобы избыточно экспрессировать и/или содержать повышенные содержания c-Jun или другого фактора транскрипции АР-1, показывают пониженные уровни истощения Т-клеток. Например, избыточная экспрессия транскрипционных факторов АР-1 (в частности, c-Jun) предотвращала развитие истощения Т-клеток и поддерживала функциональность Т-клеток (например, даже после воздействия высоких уровней антигена) (см. примеры 1-6). GD2 CAR Т-клетки, экспрессирующие повышенные содержания факторов транскрипции АР-1 (в частности, c-Jun), демонстрируют пониженные признаки истощения (включая в себя более низкие содержания маркеров истощения, повышенное формирование памяти и увеличенную продукцию цитокинов), что указывает на то, что Т-клетки, модифицированные, чтобы избыточно экспрессировать и/или содержать повышенные (например, супрафизиологические) содержания одного или нескольких факторов транскрипции АР-1, демонстрируют повышенную клиническую эффективность при множественных злокачественных новообразованиях (см., например, примеры 1-6). Кроме того, как CD19 CAR Т-клетки, модифицированные, чтобы избыточно экспрессировать c-Jun, так и CD22 CAR Т-клетки, модифицированные, чтобы избыточно экспрессировать c-Jun, показали повышенное распознавание CAR Т-клетками целевых клеток лейкоза с низкими уровнями поверхностного антигена (см., например, пример 6). CAR Т-клетки, модифицированные, чтобы избыточно экспрессировать факторы транскрипции АР-1, демонстрировали снижение истощения Т-клеток и усиленную функциональность в трех отдельных моделях опухолей in vivo с использованием cJUN-модифицированных CAR Т-клеток (см. пример 8). Таким образом, избыточная экспрессия факторов транскрипции АР-1 (в частности, c-Jun) предотвращала развитие истощения Т-клеток и поддерживала функциональность Т-клеток. В совокупности эти результаты показывают, что композиции и способы согласно настоящему изобретению являются широко применимыми и устраняют многие из существующих барьеров для успешной CAR Т-клеточной терапии.
Соответственно, настоящее изобретение относится к модифицированным Т-клеткам (например, генетически и/или функционально модифицированным (например, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса)), которые поддерживают функциональность в условиях, при которых немодифицированные Т-клетки демонстрируют истощение Т-клеток. Таким образом, композиции и способы согласно настоящему изобретению можно использовать для предотвращения истощения сконструированных Т-клеток (например, сконструированных для экспрессии специфического Т-клеточного рецептора или сконструированных для экспрессии химерного антигенного рецептора (CAR)) и для предотвращения истощения не сконструированных Т-клеток (например, нативных или природных Т-клеток (например, выделенных из организма субъекта), тем самым усиливая функциональность (например, активность против рака или инфекционного заболевания) сконструированных, а также не сконструированных Т-клеток.
Модификация Т-клеток для избыточной экспрессии и/или содержания повышенных содержаний одного или нескольких факторов транскрипции АР-1 может предотвратить истощение факторов транскрипции АР-1 в Т-клетке (например, это происходит, когда Т-клетки истощаются (см., например, пример 1) и/или приводят к повышенным (например, супрафизиологическим) содержаниям факторов транскрипции АР-1). Факторы транскрипции АР-1 (например, c-Jun) являются факторами, которые индуцируются после активации Т-клеток и участвуют в продукции и секреция цитокинов (например, интерлейкина-2) Т-клетками. Т-клетки, которые экспрессируют CAR, подвергаются тонической, антиген-независимой передаче сигналов из-за кластеризации рецепторов и воспроизводят фундаментальный биологический механизм истощения Т-клеток, о чем свидетельствуют высокие уровни экспрессии PD-1, TIM-3 и LAG-3, уменьшенная антиген-индуцированная продукция цитокинов и избыточная запрограммированная клеточная смерть. Факторы транскрипции АР-1 были идентифицированы и охарактеризованы как сниженные в истощенных Т-клетках (см., например, пример 1). Модификация Т-клеток для избыточной экспрессии и/или содержания повышенных содержаний одного или нескольких факторов транскрипции АР-1 значительно увеличивала функциональность Т-клеток в условиях, которые вызывают истощение Т-клеток (см., например, примеры 2-5). Хотя понимание механизма не требуется для практического применения настоящего изобретения, и хотя настоящее изобретение не ограничено каким-либо конкретным механизмом действия, поддержание и/или повышение содержаний фактора транскрипции АР-1 (например, содержания c-Jun) внутри Т-клеток предотвращает дисфункцию Т-клеток, связанную с истощением Т-клеток (например, минимальный эффект избыточной экспрессии фактора транскрипции АР-1 наблюдался в неистощенных Т-клетках, что указывает на то, что относительный или абсолютный дефицит c-Jun представляет собой важный фактор дисфункции, связанной с истощением Т-клеток).
Согласно некоторым вариантам осуществления Т-клетки являются модифицированными, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких мутированных и/или усеченных факторов транскрипции АР-1 для предотвращения истощения факторов транскрипции АР-1 в Т-клетке. Эксперименты, проведенные во время разработки вариантов осуществления в настоящем документе, демонстрируют, что части белков семейства АР-1 (например, c-Jun) могут быть мутированы и/или усечены без влияния на способность мутированных/усеченных факторов АР-1 опосредовать восстановление дисфункциональных Т-клеток. Например, полипептиды c-Jun с N-концевыми делециями и мутациями (например, в домене трансактивации) сохраняют свою способность восстанавливать функцию истощенных HA-28z CAR Т-клеток и сохраняют эквивалентное увеличение продукции цитокинов по сравнению с c-Jun. Согласно некоторым вариантам осуществления полипептид АР-1 (например, c-Jun) содержит мутации или делеции в доменах транс активации и/или дельта. Согласно некоторым вариантам осуществления полипептид АР-1 (например, c-Jun) содержит мутации или делеции, которые делают домены трансактивации и/или дельта неактивными или не присутствующими. Согласно некоторым вариантам осуществления мутированный/усеченный полипептид АР-1 (например, c-Jun) содержит 70% (например, 70%, 75%, 80%, 85%, 90%, 95%, 99%, 100% или диапазоны между ними) или большую идентичность последовательности с С-концевыми аминокислотными остатками (например, 50 остатков, 75 остатков, 100 остатков, 150 остатков, 200 остатков 250 остатков или диапазоны между ними), С-концевой частью (например, с четвертой, третьей частью, половиной) или С-концевыми доменами (например, эпсилон, bZIP и их С-концевые аминокислоты) фактора транскрипции АР-1 дикого типа (например, c-Jun). Согласно некоторым вариантам осуществления N-концевые аминокислотные остатки (например, 50 остатков, 75 остатков, 100 остатков, 150 остатков или интервалы между ними), N-концевая часть (например, четвертая, третья часть, половина) или N-концевые домены (например, дельта, трансактивирующий домен и его N-конец аминокислоты) фактора транскрипции АР-1 дикого типа (например, c-Jun) подвергают делеции, мутации или инактивации иным образом. Любые варианты осуществления, описанные в настоящем документе со ссылкой на фактор транскрипции АР-1, могут содержать мутированный/усеченный полипептид АР-1 (например, c-Jun), что согласуется, например, с вышеуказанным.
Хотя усиленная экспрессия факторов транскрипции АР-1, таких как c-Fos и c-Jun, увеличивала функциональность модифицированных Т-клеток, существуют другие ингибирующие представители семейства АР-1, экспрессируемые в истощенных активированных Т-клетках. Ингибирование/нокдаун представителей ингибирующего АР-1 комплекса (например, для увеличения доступности канонических факторов АР-1) также уменьшало истощение Т-клеток (например, см. пример 7). В частности, ингибирование/нокдаун представителей ингибирующего АР-1 комплекса приводило к значительному улучшению функции Т-клеток (например, усилению продукции и/или экспрессии цитокинов) истощенных Т-клеток, но не в здоровых Т-клетках (например, см. фиг. 8A-D). Таким образом, согласно некоторым вариантам осуществления настоящее изобретение относится к композициям и способам ингибирования истощения Т-клеток посредством ингибирования экспрессии и/или активности представителей ингибирующего АР-1 комплекса (например, BATF3, и другие представители семейства BATF (BATF 1 и 2), IRF4, IRF8, и другие представители семейства IRF (IRF 1, 2, 3, 5,6, 7 или 9), и представители семейства ATF (ATF 1, 2, 3, 4, 5, 6, или 7)). Настоящее изобретение не ограничено средствами ингибирования экспрессии и/или активности представителей ингибирующего АР-1 комплекса (например, генов). Например, согласно некоторым вариантам осуществления экспрессия и/или активность представителей ингибирующего АР-1 комплекса, происходит на геномном уровне (например, посредством нарушения экспрессии гена). Согласно другим вариантам осуществления ингибирование экспрессии и/или активности представителей ингибирующего АР-1 комплекса происходит на уровне экспрессии и/или активности белка (например, посредством нарушения экспрессии и/или активности белка). Иллюстративные композиции и способы ингибирования экспрессии и/или активности представителей ингибирующего АР-1 комплекса включают в себя без ограничения следующее: нуклеазное разрушение, системы CRISPR-Cas9, нацеливание на цинк-пальцевые нуклеазы, редактирование генома TALEN, кшРНК, миРНК, нацеливание микроРНК, дегроны, регулируемые промоторы, белковые ингибиторы (например, химические ингибиторы, низкомолекулярные ингибиторы, антитела и т.д.), экспрессия доминантных негативов и тому подобное. Иллюстративные композиции для ингибирования экспрессии и/или активности представителей ингибирующего АР-1 комплекса представлены в таблице 1 ниже. CAS расшифровывается как Chemical Abstracts Service и является уникальным числовым идентификатором, присваиваемым каждому химическому веществу.
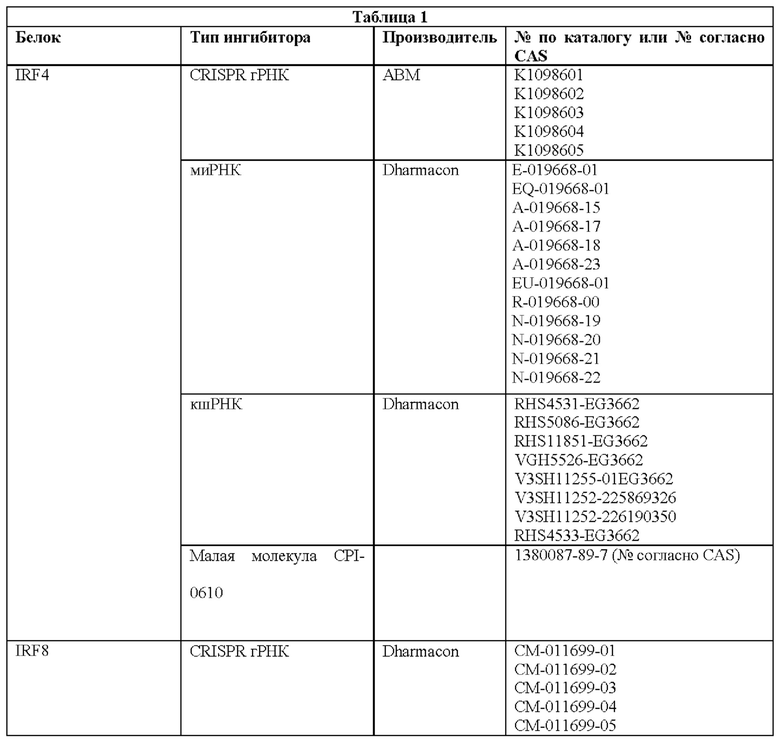
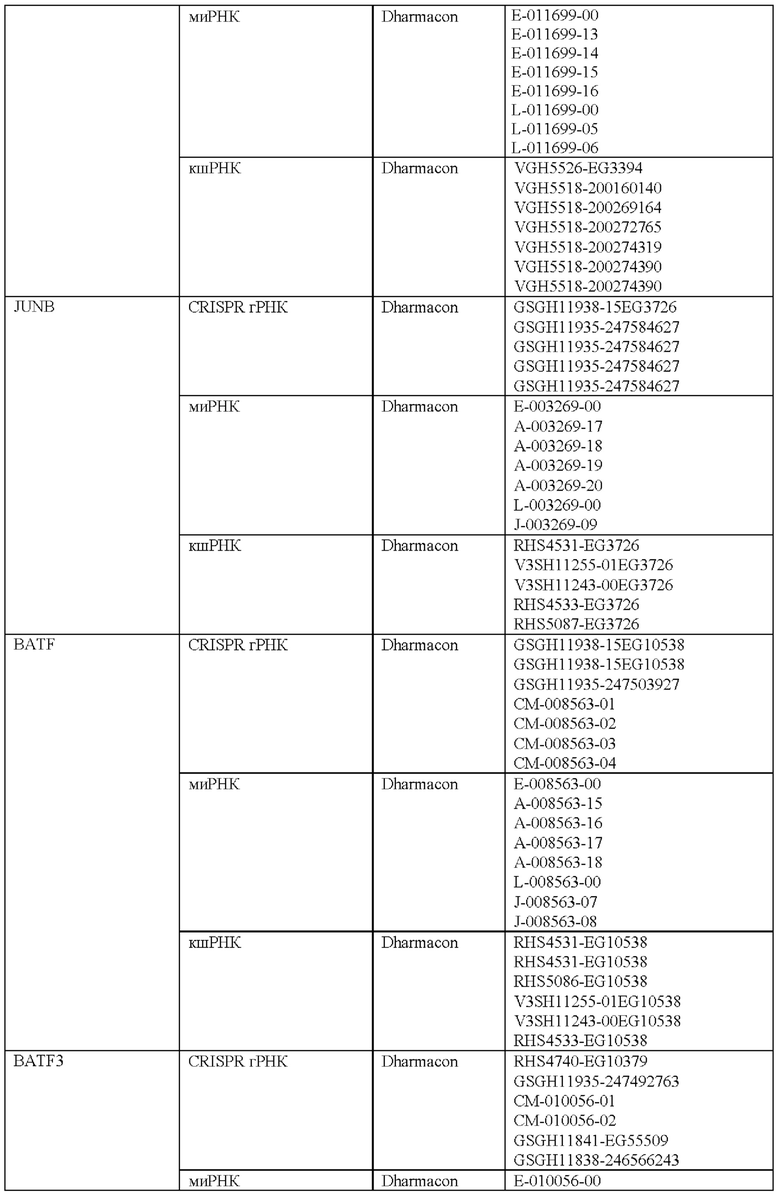
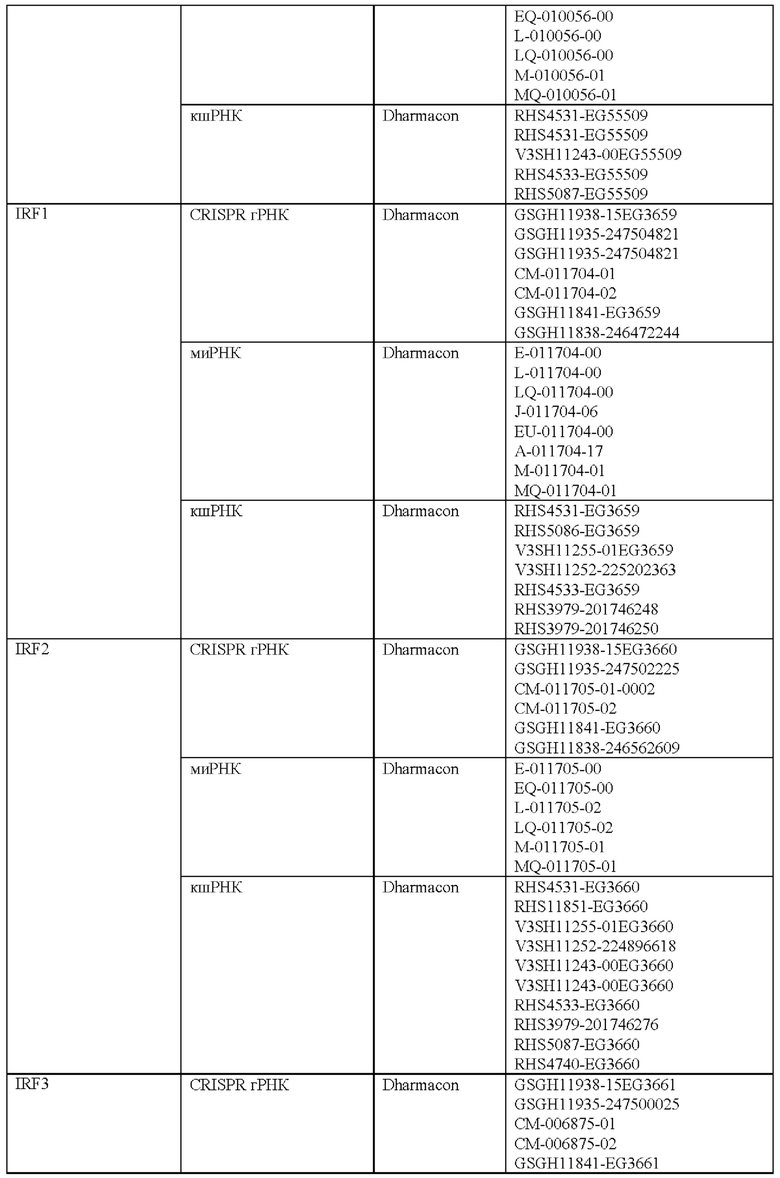
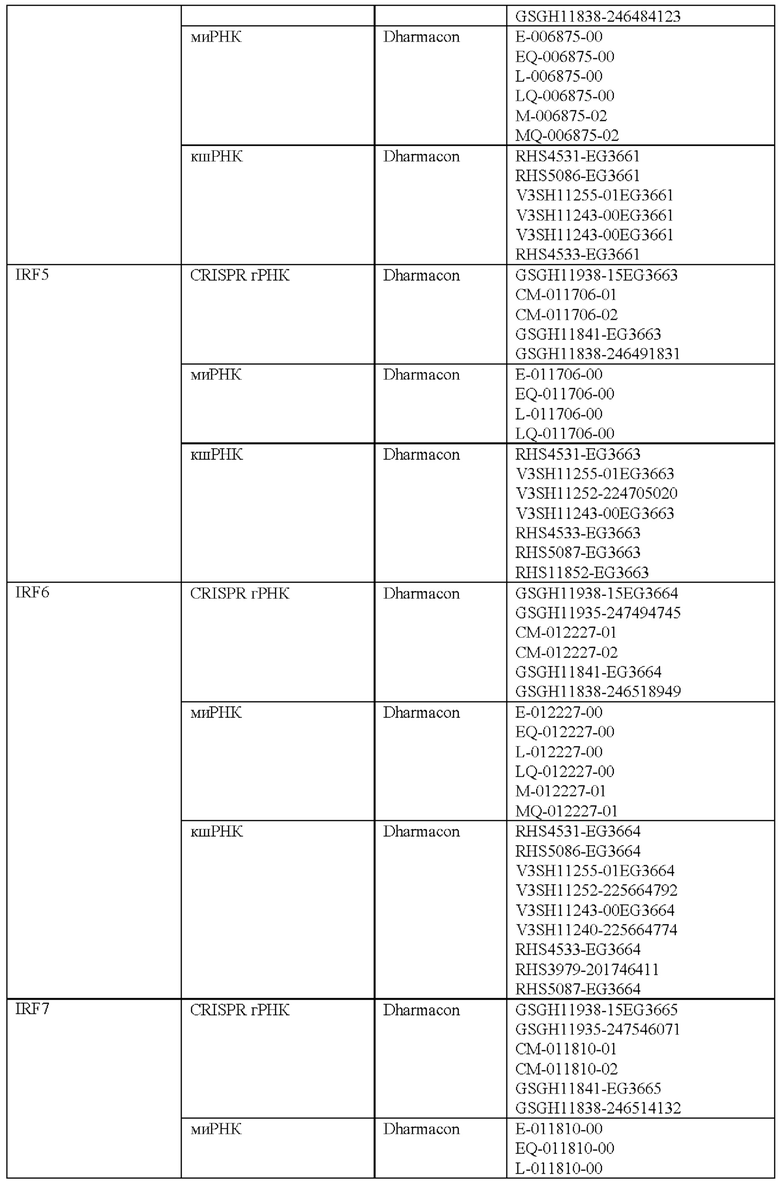
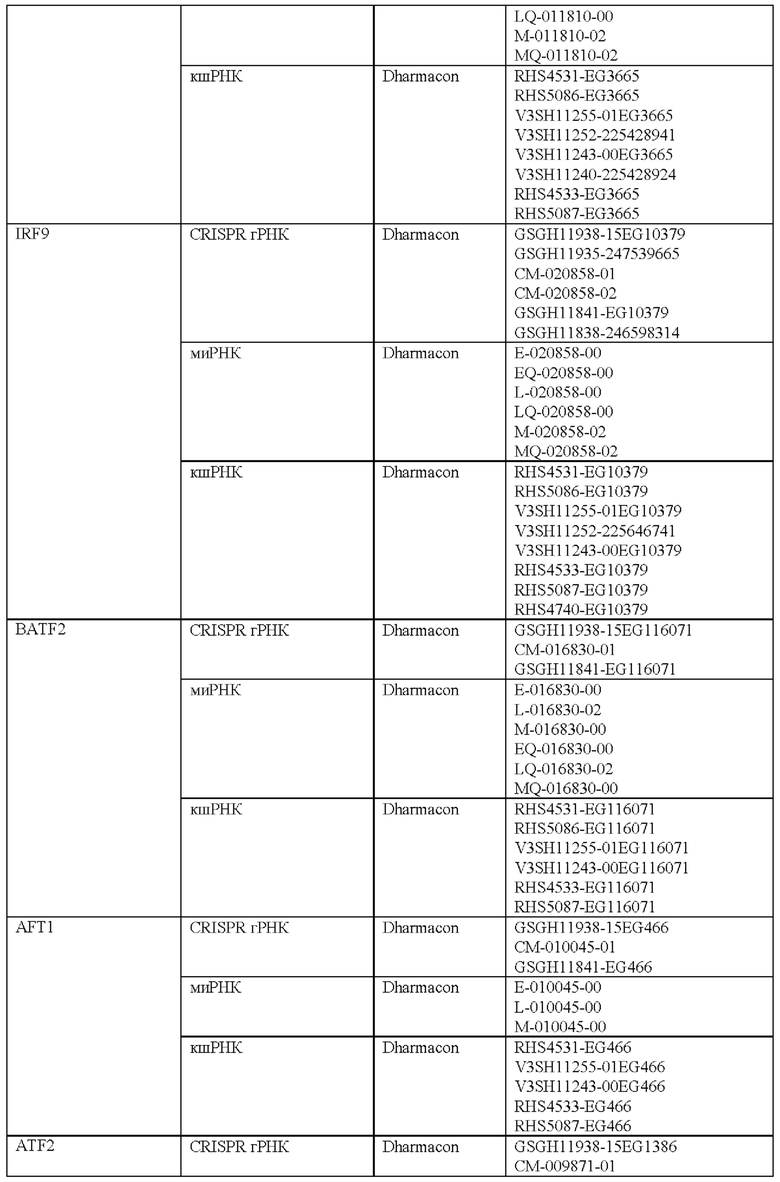
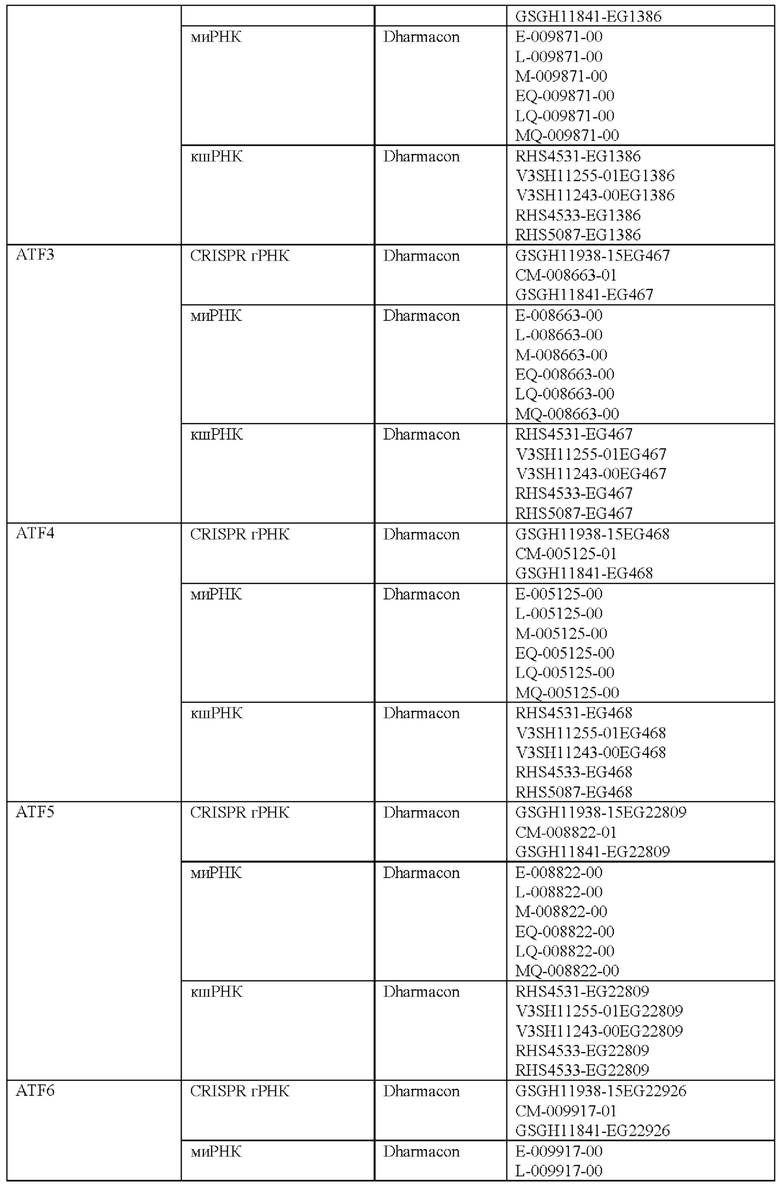
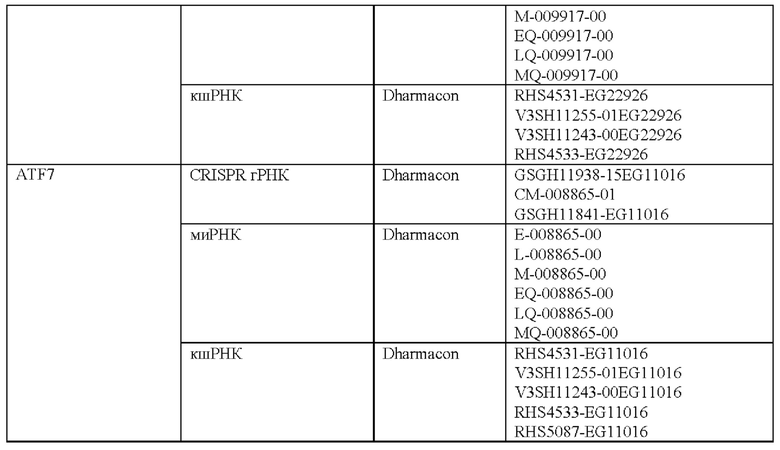
Таким образом, настоящее изобретение относится композициям и способам снижения истощения Т-клеток, содержащим Т-клетки, модифицированные, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или модифицированные для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса. Настоящее изобретение не ограничено заболеванием или состоянием, которое можно лечить с использованием модифицированных Т-клеток согласно настоящему изобретению. Действительно, композиции и способы, представленные в настоящем документе, могут являться применимыми при лечении любого заболевания, для которого повышенная активность Т-клеток может обеспечить терапевтический эффект.
Соответственно, настоящее изобретение относится к композиции, содержащей Т-клетки, модифицированные, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или модифицированные (например, генетически) для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса (например, JunB, BATF3 и другие представители семейства BATF, IRF4, IRF8 и другие представители семейства IRF, и представители семейства ATF). Как описано подробно в настоящем документе, Т-клетки могут представлять собой сконструированные или не сконструированные Т-клетки (например, инфильтрирующие опухоль лимфоциты (TIL)). Более того, настоящее изобретение не ограничено типом Т-клетки, модифицированной, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1. Согласно некоторым вариантам осуществления Т-клетки представляют собой CD3+ Т-клетки (например, комбинация CD4+ и CD8+ Т-клеток). Согласно определенным вариантам осуществления Т-клетки представляют собой CD8+Т-клетки. Согласно другим вариантам осуществления Т-клетки представляют собой CD4+ Т-клетки. Согласно некоторым вариантам осуществления Т-клетки представляют собой Т-клетки - естественные киллеры. Согласно некоторым вариантам осуществления Т-клетки представляют собой гамма-дельта Т-клетки. Согласно некоторым вариантам осуществления Т-клетки представляют собой комбинацию CD4+ и CD8+ Т-клеток (например, которые представляют собой CD3+). Согласно определенным вариантам осуществления Т-клетки представляют собой Т-клетки памяти. Согласно определенным вариантам осуществления Т-клетки представляют собой комбинацию CD8+ Т-клеток, CD4+ Т-клеток, Т-клеток - NK, Т-клеток памяти и/или гамма-дельта Т-клеток. Согласно некоторым вариантам осуществления Т-клетки представляют собой индуцированные цитокинами клетки-киллеры. Согласно некоторым вариантам осуществления Т-клетки являются сконструированными для экспрессии химерного антигенного рецептора. Согласно другому варианту осуществления Т-клетки являются сконструированными для экспрессии специфического Т-клеточного рецептора (например, со специфичностью в отношении опухолевого антигена или антигена инфекционного заболевания). Согласно некоторым вариантам осуществления Т-клетки представляют собой противоопухолевые Т-клетки. Композиция может содержать фармацевтически приемлемый носитель (например, буфер). Композиция может дополнительно содержать одно или несколько других средств (например, химиотерапевтическое средство (например, химиотерапевтическое средство, описанные в настоящем документе) и/или антимикробное средство. Иллюстративные антимикробные средства включают в себя без ограничения антитела, хлорид бензалкония, хлорид бензетония, бензиловый спирт, хлорид цетилпиридиния, хлорбутанол, фенол, фенилэтиловый спирт, нитрат фенилртути, тиомерсал и/или их комбинации. Композиция может необязательно включать в себя одно или несколько дополнительных средств, таких как другие лекарственные средства для лечения истощения Т-клеток (например, ингибитор контрольной точки - антитело к PD-1, такое как ниволумаб), или другие лекарственные средства, используемые для лечения субъекта от инфекции или заболевания, связанного с истощением Т-клеток (например, противовирусные средства, антибиотики, противомикробные или противораковые средства).
Антимикробные терапевтические средства можно использовать в качестве терапевтических средств в композиции согласно настоящему изобретению. Можно использовать любое средство, которое может уничтожать, ингибировать или иным образом ослаблять функцию микроорганизмов, а также любое средство, которое, как предполагается, обладает такими активностями. Антимикробные средства включают в себя без ограничения природные и синтетические антибиотики, антитела, ингибирующие белки (например, дефензины), антисмысловые нуклеиновые кислоты, разрушающие мембрану средства и тому подобное, используемые отдельно или в комбинации. Действительно, можно использовать любой тип антибиотика, включая в себя без ограничения антибактериальные средства, противовирусные средства, противогрибковые средства и тому подобное.
В настоящей области техники известно несколько стратегий нацеливания на опухолевые антигены. Например, иммунотерапия, нацеленная на GD2, в настоящее время проходит клинические и доклинические исследования по нескольким заболеваниям, включая в себя нейробластому, остеосаркому и меланому (см., например, Thomas et al., PLoS One, 2016. 11(3): p.e0152196; Long et al., Nature Medicine, 2015. 21(6): p. 581-590; Long et al., Cancer Immunology Research, 2016. 4(10): p. 869-880; Yu et al., N Engl J Med, 2010. 363(14): p. 1324-34; Perez Horta et al., Immunotherapy, 2016. 8(9): p. 1097-117; Heczey et al, Molecular Therapy). В отличие от mAb, которые не успешно проникают через гематоэнцефалический барьер, активированные CAR Т-клетки эффективно проникают в ЦНС после адоптивного переноса. Соответственно, согласно одному варианту осуществления любую CAR Т-клетку можно модифицировать в соответствии с композициями и способами согласно настоящему изобретению (например, модифицированные, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 (например, тем самым усиливая функциональность CAR Т-клеток и/или ингибирование истощения CAR Т-клеток)).
Например, как описано в настоящем документе, модифицированная Т-клетка согласно настоящему изобретению (например, модифицированная, чтобы избыточно экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и/или модифицированная (например, генетически) для пониженной экспрессии и/или активности одного или нескольких представителей ингибирующего АР-1 комплекса (например, JunB и BATF3 и другие представители семейства BATF, IRF4 и представители семейства ATF)) может также являться сконструированной, чтобы содержать CAR, который содержит специфический в отношении мишени связывающий элемент, иначе называемый антигенсвязывающим фрагментом. Выбор фрагмента зависит от типа и количества лигандов, которые определяют поверхность клетки-мишени. Например, антигенсвязывающий домен можно выбрать для распознавания лиганда, который действует как маркер клеточной поверхности на клетках-мишенях, связанных с конкретным болезненным состоянием. Примеры маркеров клеточной поверхности, которые могут действовать в качестве лигандов для домена антигенного фрагмента в CAR согласно настоящему изобретению, включают в себя маркеры, связанные с вирусными, бактериальными и паразитарными инфекциями, аутоиммунным заболеванием и раковыми клетками.
Например, CAR может быть сконструирован для нацеливания на представляющий интерес опухолевый антиген путем конструирования требуемого антигенсвязывающего фрагмента, который специфически связывается с антигеном на опухолевой клетке. Используемый в настоящем документе термин «опухолевый антиген» или «антиген гиперпролиферативного нарушения» или «антиген, ассоциированный с гиперпролиферативным нарушением» или «раковый антиген» относится к антигенам, которые являются общими для конкретных гиперпролиферативных нарушений, таких как рак. Иллюстративные антигены, упомянутые в настоящем документе, включены в качестве примера. Этот перечень не предназначен для того, чтобы быть исключительным, и дополнительные примеры будут очевидны для специалистов в настоящей области техники.
Опухолевые антигены представляют собой белки, которые вырабатываются опухолевыми клетками, которые вызывают иммунный ответ, в частности, опосредованные Т-клетками иммунные ответы. Таким образом, антигенсвязывающий фрагмент можно выбрать на основании конкретного типа рака, подлежащего лечению. Опухолевые антигены хорошо известны в настоящей области техники и включают в себя, например, глиома-ассоциированный антиген, карциноэмбриональный антиген (СЕА), бета-субъединица хорионического гонадотропина человека, альфафетопротеин (AFP), лектин-реактивный AFP, тиреоглобулин, RAGE-1, MN-CA IX, обратная транскриптаза теломеразы человеческая, RU1, RU2 (AS), кишечная карбоксилэстераза, mut hsp70-2, M-CSF, простатический специфический антиген (PSA), PAP, NY-ESO-1, LAGE-1a, р53, простеин, PSMA, Her2/neu, сурвивин и теломераза, опухолевый антиген 1 карциномы предстательной железы, MAGE, ELF2M, нейтрофильная эластаза, эфрин-В2, CD22, инсулиновый фактор роста (IGF)-I, IGF-II, рецептор IGF-I и мезотелин.
Опухолевый антиген может содержать один или несколько антигенных раковых антигенов/эпитопов, ассоциированных со злокачественной опухолью. Злокачественные опухоли экспрессируют ряд белков, которые могут служить антигенами-мишенями для иммунной атаки. Эти молекулы включают в себя без ограничения следующее: тканеспецифические антигены, такие как MART-1, тирозиназа и GP100 при меланоме и простатическая кислая фосфатаза (РАР) и простатический специфический антиген (PSA) при раке предстательной железы. Другие молекулы-мишени относятся к группе связанных с трансформацией молекул, таких как онкоген HER-2/Neu/ErbB-2. Еще одной группой антигенов-мишеней являются онкофетальные антигены, такие как карциноэмбриональный антиген (СЕА). При В-клеточной лимфоме иммуноглобулин специфического для опухоли идиотипа представляет собой действительно опухолеспецифический антиген иммуноглобулина, который является уникальным для отдельной опухоли. Антигены дифференцировки В-клеток, такие как CD 19, CD20 и CD37, являются другими кандидатами на антигены-мишени при В-клеточной лимфоме.
Опухолевый антиген также может являться опухолеспецифическим антигеном (TSA) или опухолевым антигеном (ТАА). TSA является уникальным для опухолевых клеток и не встречается в других клетках организма. ТАА не является уникальным для опухолевой клетки и вместо этого также экспрессируется на некоторых нормальных клетках в условиях, которые не могут вызвать состояние иммунологической толерантности к антигену. Экспрессия антигена в опухоли может происходить в условиях, которые позволяют иммунной системе реагировать на антиген. ТАА могут являться антигенами, которые экспрессируются на нормальных клетках во время развития плода, когда иммунная система является незрелой и неспособной реагировать, или они могут являться антигенами, которые, как правило, присутствуют на чрезвычайно низких уровнях в нормальных клетках, но которые экспрессируются на гораздо более высоких уровнях на опухолевых клетках.
Примеры TSA или ТАА включают в себя без ограничения антигены дифференцировки, такие как MART-1/MelanA (MART-1), gp100 (Pmel 17), тирозиназа, TRP-1, TRP-2 и опухолеспецифичные мультилинеиные антигены, такие как как MAGE-1, MAGE-3, BAGE, GAGE-1, GAGE-2, р 15; избыточно экспрессированные эмбриональные антигены, такие как СЕА; избыточно экспрессированные онкогены и мутированные гены-супрессоры опухолей, такие как р53, Ras, HER-2/neu; уникальные опухолевые антигены, возникающие в результате хромосомных транслокаций; такие как BCR-ABL, E2A-PRL, Н4-RET, IGH-IGK, MYL-RAR; и вирусные антигены, такие как антигены вируса Эпштейна-Барр EBVA и антигены вируса папилломы человека (HPV) Е6 и Е7. Другие крупные антигены на основе белка включают в себя TSP-180, MAGE-4, MAGE-5, MAGE-6, RAGE, NY-ESO-1, pl85erbB2, pl80erbB-3, c-met, nm-23Hl, PSA, TAG-72, CA 19-9, CA 72-4, CAM 17.1, NuMa, K-ras, бета-катенин, CDK4, Mum-1, p 15, p 16, 43-9F, 5T4, 791Tgp72, альфа-фетопротеин, бета-HCG, BCA225, BTAA, CA 125, CA 15-3\CA 27.291\BCAA, CA 195, CA 242, CA-50, CAM43, CD68\P1, CO-029, FGF-5, G250, Ga733\EpCAM, HTgp-175, M344, MA-50, MG7-Ag, MOV 18, NB/70K, NY-CO-1, RCAS1, SDCCAG16, TA-90\Mac-2 связывающий белок\циклофилин С-ассоциированный белок, TAAL6, TAG72, TLP и TPS.
В зависимости от требуемого антигена, на который необходимо нацеленно воздействовать, CAR может быть сконструирован так, чтобы включать соответствующий антигенсвязывающий фрагмент, специфический для требуемого антигена-мишени. Например, если CD19 является требуемым антигеном, который должен быть нацелен, антитело к CD19 можно использовать в качестве антигенсвязывающего фрагмента для включения в CAR согласно настоящему изобретению.
Способы лечения
Согласно другому аспекту в настоящем документе предусмотрены способы лечения заболевания или состояния у субъекта, предусматривающие введение субъекту (например, пациенту), характеризующемуся наличием заболевания или состояния, эффективного количества Т-клеток, модифицированных, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1. Настоящее изобретение не ограничено типом заболевания или состояния, которое лечат.Действительно, любое заболевание или состояние, которое поддается лечению (например, для которого признаки или симптомы заболевания уменьшается в интенсивности при лечении) посредством введения Т-клеток, можно лечить улучшенным и более эффективным способом с использованием композиций и способов согласно настоящему изобретению (например, содержащих и/или использующих Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1). Согласно одному варианту осуществления заболевание или состояние представляет собой рак. Согласно другому варианту осуществления заболевание или состояние представляет собой инфекционное заболевание. Настоящее изобретение не ограничено типом рака или типом инфекционного заболевания. Действительно, любой рак, известный в настоящей области техники, для лечения которого используют Т-клеточную терапию, можно лечить с помощью композиций и способов согласно настоящему изобретению. Аналогично, любое инфекционное заболевание, известное в настоящей области техники, для лечения которого используют Т-клеточную терапию, можно лечить с помощью композиций и способов согласно настоящему изобретению. Согласно одному варианту осуществления введение эффективного количества Т-клеток, модифицированных, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, субъекту (например, пациенту), характеризующемуся наличием заболевания или состояния, ингибирует истощение Т-клеток у субъекта (например, что могло в противном случае произойти, когда такое же количество немодифицированных Т-клеток вводят субъекту). Например, согласно определенным вариантам осуществления способы предусматривают способы лечения субъекта, характеризующегося наличием заболевания или состояния, реагирующего на лечение с помощью адоптивной клеточной терапии. Согласно определенным вариантам осуществления способы лечения субъекта, характеризующегося наличием заболевания или состояния, реагирующего на лечение с помощью адоптивной клеточной терапии, предусматривают стадию введения композиции, содержащей эффективное количество Т-клеток, генетически модифицированных, чтобы экспрессировать повышенные содержания одного или нескольких факторов транскрипции АР-1, причем Т-клетки, генетически модифицированные, чтобы экспрессировать повышенные содержания одного или нескольких факторов транскрипции АР-1, демонстрируют пониженные уровни истощения Т-клеток по сравнению с Т-клетками, экспрессирующими нормальные уровни одного или нескольких факторы транскрипции АР-1. Согласно определенным вариантам осуществления заболевание или состояние включает в себя рак.
Согласно одному аспекту настоящее изобретение относится к способам лечения форм рака/опухолей с использованием Т-клеток, модифицированных/сконструированных для экспрессии одного или нескольких факторов транскрипции АР-1. Согласно различным аспектам настоящего изобретения предусмотрены способы лечения рака/опухолей, причем способы предусматривают введение пациенту, характеризующемуся наличием такого рака или опухоли, эффективного количества Т-клеток, модифицированных/сконструированных для экспрессии одного или нескольких факторов транскрипции АР-1. Настоящее изобретение не ограничено типом рака и/или опухоли, подлежащих лечению. Используемый в настоящем документе термин «рак» относится к различным типам злокачественных новообразований, большинство из которых могут проникать в окружающие ткани и могут метастазировать в разные области (см., например, PDR Medical Dictionary, 1-е издание (1995), включенный в настоящий документ посредством ссылки в полном объеме для любых целей). Термины «новообразование» и «опухоль» относятся к аномальной ткани, которая растет за счет клеточной пролиферации быстрее, чем обычно, и продолжает расти после удаления стимулов, инициирующих пролиферацию. Такая аномальная ткань демонстрирует частичное или полное отсутствие структурной организации и функциональной координации с нормальной тканью, которая может быть либо доброкачественной (т.е. доброкачественная опухоль), либо злокачественной (т.е. злокачественная опухоль). Примеры общих категорий рака включают в себя без ограничения карциномы (например, злокачественные опухоли, происходящие из эпителиальных клеток, такие как, например, распространенные формы рака молочной железы, предстательной железы, легких и толстой кишки), саркомы (например, злокачественные опухоли, происходящие из соединительной ткани или мезенхимальных клеток), лимфомы (например, злокачественные новообразования, происходящие из гемопоэтических клеток), лейкозы (например, злокачественные новообразования, происходящие из гемопоэтических клеток), герминогенные опухоли (например, опухоли, происходящие из тотипотентных клеток). У взрослых чаще всего встречаются в яичке или яичнике; у плодов, младенцев и детей младшего возраста чаще всего обнаруживаются по средней линии тела, особенно на кончике копчика), бластные опухоли (например, типично злокачественная опухоль, которая напоминает незрелую или эмбриональную ткань) и тому подобное. Дополнительные примеры опухолей и/или новообразований, которые можно лечить с использованием композиций и способов согласно настоящему изобретению, включают в себя без ограничения новообразования, связанные с раком нервной ткани, кроветворной ткани, молочной железы, кожи, кости, предстательной железы, яичников, матки, шейки матки, печени, легких, головного мозга, гортани, желчного пузыря, поджелудочной железы, прямой кишки, паращитовидной железы, щитовидной железы, надпочечников, иммунной системы, головы и шеи, толстой кишки, желудка, бронхов и/или почек. Согласно некоторым вариантам осуществления рак представляет собой скрытый рак, ранее диагностированный первичный рак или метастатический рак.
Согласно определенным вариантам осуществления присутствие повышенных содержаний одного или нескольких факторов транскрипции АР-1 в модифицированных Т-клетках (например, CAR Т-клетках (например, по сравнению с Т-клетками, которые не являются модифицированными)) приводит к более эффективному лечению (например, уничтожению и/или ингибированию прогрессирования) форм рака/опухолей по сравнению с лечением с использованием Т-клеток (например, CAR Т-клеток), которые не являются модифицированными, чтобы содержать повышенные содержания одного или нескольких факторов транскрипции АР-1.
Согласно определенным вариантам осуществления настоящее изобретение относится к способам лечения (например, ингибирования роста и/или уничтожению) форм рака/опухолей с использованием Т-клеток (например, CD3 + Т-клеток)), генетически сконструированных, чтобы избыточно экспрессировать и/или содержать повышенные (например, супрафизиологические) содержания одного или нескольких факторов транскрипции АР-1 и которые сконструированы для экспрессии рецептора, который распознает поверхностный опухолевый антиген (например, CD19, CD20, CD22, ROR1, GD2, белок или антиген EBV, рецептор фолата, мезотелин, канцероэмбриональный антиген человека, CD33/IL3Ra, c-Met, PSMA, гликолипид F77, EGFRvIII, NY-ESO-1, MAGE-A3, MART-1, GP1000 и/или p53) и передают сигнал, который активирует Т-клетку, чтобы вызвать экспансию Т-клетки и/или уничтожение опухоли. Неограничивающим примером рецептора является химерный антигенный рецептор (CAR), который включает в себя scFv, полученный из mAb, который распознает GD2, а также трансмембранный домен и один или несколько внутриклеточных сигнальных доменов. CAR может быть дополнительно сконструирован для включения других сигнальных элементов, которые облегчают экспансию сконструированных клеток после столкновения с антигеном опухолевых клеток (например, антигеном GD2), а также с элементов, которые обеспечивают долговременную персистенцию сконструированных клеток. Настоящее изобретение, кроме того, предусматривает композиции, содержащие генетически сконструированный клетки (например, иммунотерапевтические композиции, производимые и вводимые в количестве, достаточном для достижения форм рака и/или опухолей).
Конструирование модифицированных Т-клеток.
Как описано в настоящем документе, настоящее изобретение относится к композициям, содержащим Т-клетки, которые модифицированы (например, генетически (например, трансдуцированы)) для экспрессии (например, гетерологически) одного или нескольких факторов транскрипции АР-1. Например, настоящее изобретение относится к трансдуцированным Т-клеткам. «Трансдуцированная клетка» представляет собой клетку, в которую была введена молекула нуклеиновой кислоты с использованием техник молекулярной биологии. Т-клетки, которые являются
модифицированными/трансдуцированными для экспрессии одного или нескольких гетерологичных факторов транскрипции АР-1, могут быть трансдуцированы с помощью любой техники, за счет которой молекула нуклеиновой кислоты может быть введена в такую клетку, включая в себя без ограничения трансфекцию вирусными векторами (например, ретровирусный, лентивирусный или другой вирусный вектор) через систему на основе CRISPR/Cas9, трансформацию плазмидными векторами и/или введение голой ДНК путем электропорации, липофекции и ускорения с помощью генной пушки. Вирусные и/или плазмидные векторы можно использовать для экспрессии in vitro, in vivo и/или ex vivo. Фактор транскрипции АР-1 можно коэкспрессировать со сконструированным TCR или CAR, причем как фактор транскрипции, так и TCR и/или CAR коэкспрессируются из различных вирусных векторов. Согласно другому варианту осуществления они экспрессируются из одной векторного конструкта с использованием бицистронного вектора. c-Jun (и/или другой фактор транскрипции АР-1) можно экспрессировать конститутивно или регулируемым образом (например, с использованием системы для удаленного регулирования экспрессии с помощью малой молекулы или с использованием эндогенно регулируемой системы). Согласно другому варианту осуществления гены c-Jun и/или других факторов транскрипции АР-1 могут быть генетически интегрированы в клеточную ДНК с использованием ретровирусного, лентивирусного или другого вирусного вектора или посредством системы на основе CRTSPR/Cas9. Согласно еще одному варианту осуществления c-Jun и/или другой фактор транскрипции АР-1 экспрессируется с помощью РНК или онколитического вируса или другой системы транзиентной экспрессии, известной в настоящей области техники. c-Jun и/или другой факт транскрипции АР 1 может быть доставлен ex vivo в Т-клетки (например, где Т-клетки используются для адоптивного переноса) или доставлен посредством генетического переноса in vivo.
Настоящее изобретение не ограничено химерным антигенным рецептором (CAR), экспрессируемым в Т-клетках (например, конструктом CAR, используемым в способах согласно настоящему изобретению). Согласно одному варианту осуществления CAR содержит белок слияния вариабельных областей тяжелой (VH) и легкой цепей (VL) (например, одноцепочечный вариабельный фрагмент (scFv)) иммуноглобулина, который специфично связывается с опухолевым антигеном (например, GD2). Специалистам в настоящей области техники известно, что scFv представляет собой белок слияния вариабельных областей тяжелых (VH) и легких цепей (VL) иммуноглобулинов, связанных с линкерным пептидом (например, приблизительно 10 - приблизительно 25 аминокислот в длину). Настоящее изобретение не ограничено типом линкера. Согласно некоторым вариантам осуществления линкер обогащен глицином (например, для гибкости). Согласно некоторым вариантам осуществления линкер содержит серии и/или треонин (например, для растворимости). Согласно некоторым вариантам осуществления линкер содержит часть, богатую глицином, и часть, содержащую серии и/или треонин.
Любое антитело/иммуноглобулин, который специфически связывается с опухолевым антигеном (например, GD2), можно использовать для конструирования CAR (например, с использованием областей VH и VL для конструирования белка слияния scFv) для экспрессии в иммунных клетках, используемых в терапевтических способах согласно настоящему изобретению. Примеры таких антител/иммуноглобулинов включают в себя без ограничения для GD2: 14G2a, ch14.18, hu14.18K322A, m3F8, hu3F8-IgG1, hu3F8-IgG4, HM3F8, UNITUXIN, DMAb-20 или любое другое антитело, которое связывается со специфичностью с GD2 (например, известный или описанный в настоящей области техники, или еще не идентифицированный). CAR опухолевого антигена (например, GD2) может содержать рецептор, включающий в себя варианты в пределах scFv антитела к опухолевому антигену (например, GD2), генерируемого для усиления аффинности и/или уменьшения тонической передачи сигналов. CAR опухолевого антигена (например, GD2) может включать в себя переменные длины шарнирных областей (например, между scFv и сигнальными доменами) и/или различные трансмембранные домены. Настоящее изобретение не ограничено используемым трансмембранным доменом. Действительно, можно использовать любой трансмембранный домен, включая в себя без ограничения весь или часть трансмембранного домена дзета-цепи TCR (CD3ζ, CD28, OX40/CD134, 4-1BB/CD137/TNFRSF9, FcERIy, ICOS/CD278, ILRB/CD122, IL-2RG/CD132 или CD40.
Конструкт CAR согласно настоящему изобретению может включать в себя внутриклеточный сигнальный домен (например, дзета CD3 комплекса нативного Т-клеточного рецептора и/или другой сигнальный домен (например, сигнальный домен MyD88)), который трансдуцирует событие связывания лиганда с внутриклеточным сигналом (например, который активирует (например, частично) Т-клетку. При отсутствии костимулирующих сигналов, связывание рецептор-лиганд часто является недостаточным для полной активации и пролиферации Т-клетки. Таким образом, конструкт CAR может включать в себя один или несколько костимулирующих доменов (например, которые обеспечивают второй сигнал для стимуляции полной активации Т-клеток). Согласно одному варианту осуществления используют костимулирующий домен, который увеличивает продукцию цитокинов иммунных CAR Т-клеток. Согласно другому варианту осуществления используют костимулирующий домен, который облегчает репликацию Т-клеток. Согласно еще одному варианту осуществления используют костимулирующий домен, который предотвращает истощение Т-клеток CAR. Согласно другому варианту осуществления используют костимулирующий домен, который повышает противоопухолевую активность Т-клеток. Согласно еще одному варианту осуществления используют костимулирующий домен, который повышает выживаемость CAR Т-клеток (например, после инфузии пациентам). Примеры белков или доменов или их частей, которые можно использовать для обеспечения костимулирующих сигналов, включают в себя без ограничения: B7-1/CD80; CD28; B7-2/CD86; CTLA-4; B7-H1/PD-L1; ICOS/CD278; ILRB/CD122; IL-2RG/CD132; В7-Н2; PD-1; В7-Н3; PD-L2; В7-Н4; PDCD6; BTLA; 4-1BB/TNFRSF9/CD137; FcERIγ; CD40 лиганд/TNFSFS; 4-1 ВВ лиганд/TNFSF9; GITR/TNFRSF18; BAFF/BLyS/TNFSF13B; GITR лиганд/TNFSF18; BAFF R/TNFRSF13C; Н VEM/TNFRS F14; CD27/TNFRSF7; LIGHT/TNFSF14; CD27 лиганд/ТНР8Р7; OX40/TNFRSF4; CD30/TNFRSF8; ОХ40 лиганд/ТКР8Р4; CD30 лиганд/TNFSFS; TAC1/TNFRSF13B; CD40/TNFRSF5; 2B4/CD244/SLAMF4; CD84/SLAMF5; BLAME/SLAMF8; CD229/SLAMF3; CD2 CRACC/SLAMF7; CD2F-10/SLAMF9; NTB-A/SLAMF6; CD48/SLAMF2; SLAM/CD150; CD58/LFA-3; CD2; Ikaros; CD53; интегрин альфа 4/CD49d; CD82/Kai-1; интегрин альфа 4 бета 1; CD90/Thyl; интегрин альфа 4 бета 7/LPAM-1; CD96; LAG-3; CD160; LMIR1/CD300A; CRTAM; TCL1A; DAP 12; TIM-1/KIM-1/HAVCR; дектин-1/CLEC7A; TIM-4; DPPIV/CD26; TSLP; EphB6; TSLP R и HLA-DR. Согласно одному варианту осуществления конструкт CAR, экспрессируемый в Т-клетках, используемых в композициях и способах согласно настоящему изобретению, включает в себя эндодомен CD28, эндодомен 4-1ВВ и/или эндодомен ОХ40. Согласно определенным вариантам осуществления конструкт CAR, специфический в отношении опухолевого антигена (например, GD2) согласно настоящему изобретению, содержит scFv антитела, которое специфично связывается с опухолевым антигеном (например, GD2), транс мембранный домен (например, CD8), внутриклеточный сигнальный домен Т-клеточного рецептора (например, дзета-цепь TCR (CD3-дзета)) и по меньшей мере один костимулирующий домен (например, 4-1ВВ).
Настоящее изобретение не ограничено средствами генетической экспрессии TCR, CAR и/или одного или нескольких факторов транскрипции АР-1 в Т-клетках. Действительно, можно использовать любые средства, известные в настоящей области техники и/или описанные в настоящем документе. Неограничивающие примеры способов генной инженерии Т-клеток включают в себя без ограничения опосредованную ретровирусами или лентивирусами трансдукцию, трансдукцию с использованием систем на основе транспозазы для интеграции генов, Crispr/Cas9-опосредованную интеграцию генов, неинтегрирующие векторы, такие как РНК или аденоассоциированные вирусы, или другие способы, описанные в настоящем документе. Композиции, содержащие сконструированные Т-клетки, могут включать в себя не сконструированные Т-клетки или другие иммунные клетки или подклассы Т-клеток, выбранные для большей способности к экспансии или персистенции. Для уменьшения токсичности может быть предусмотрено включение элементов, которые позволяют уничтожать сконструированные клетки. Для уменьшения токсичности и/или усиления эффективности может быть предусмотрено включение элементов, которые позволяют регулировать экспрессию белка в сконструированных клетках.
Противораковые терапевтические средства, композиции и комбинированная терапия.
Согласно определенным вариантам осуществления настоящее изобретение относится к способам лечения или отсрочки прогрессирования рака или лечения или отсрочки прогрессирования инфекционного заболевания у индивидуума, предусматривающим введение индивидууму эффективного количества модифицированных Т-клеток согласно настоящему изобретению. Согласно некоторым вариантам осуществления лечение приводит к устойчивому ответу у индивидуума после прекращения лечения. Способы, описанные в настоящем документе, могут найти применение при лечении состояний, при которых желательна повышенная иммуногенность, например, повышение иммуногенности опухоли для лечения рака. В настоящем документе также предусмотрены способы усиления иммунной функции у индивидуума, характеризующегося наличием рака, предусматривающие введение индивидууму эффективного количества модифицированных Т-клеток согласно настоящему изобретению. В этих способах можно использовать любой тип Т-клеток, генетически модифицированных для экспрессии CAR и/или TCR, известных в настоящей области техники или описанных в настоящем документе.
Согласно некоторым вариантам осуществления индивидуум характеризуется наличием рака, который является устойчивым (например, было продемонстрировано, что он является устойчивым) к одной или нескольким другим формам противоракового лечения (например, химиотерапия, иммунотерапия и т.д.). Согласно некоторым вариантам осуществления резистентность включает в себя рецидив рака или рефрактерный рак. Рецидив может относиться к повторному появлению рака на исходном или новом месте после лечения. Согласно некоторым вариантам осуществления резистентность включает в себя прогрессирование рака во время лечения химиотерапией. Согласно некоторым вариантам осуществления резистентность включает в себя рак, который не реагирует на традиционное или общепринятое лечение химиотерапевтическим средством. Рак может являться резистентным в начале лечения или он может стать резистентным во время лечения. Согласно некоторым вариантам осуществления рак находится на ранней стадии или на поздней стадии.
Согласно определенным вариантам осуществления настоящее изобретение, что воздействие на животных (например, людей), страдающих от рака/опухолей, терапевтически эффективных количеств иммунотерапевтических композиций, содержащих Т-клетки, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, ингибирует рост таких раковых клеток прямо и/или делает такие клетки популяцией, более восприимчивой к противораковым лекарственным средствам или лучевой терапии (например, к их вызывающей клеточную смерть активности). Иммунотерапевтические композиции и способы согласно настоящему изобретению можно использовать для лечения, уменьшения интенсивности или профилактики нарушений, таких как любой тип рака.
Согласно определенным вариантам осуществления иммунотерапевтические композиции, содержащие Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, используют для лечения, уменьшения интенсивности или профилактики рака, который характеризуется устойчивостью к одному или нескольким традиционным методам лечения рака (например, те раковые клетки, которые являются химиорезистентными, резистентными к лучевой терапии, гормонорезистентными и т.п.). Как описано в настоящем документе, любые Т-клетки, генетически модифицированные для экспрессии опухолеспецифического CAR, можно использовать в иммунотерапевтических композициях и способах согласно настоящему изобретению.
Иммунотерапевтические композиции (например, содержащие Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1) и согласно настоящему изобретению можно использовать для индукции цитотоксических активностей против опухолевых клеток и/или для стимулирования выживания и функционирования клеток (например, выживание и функционирование модифицированных иммунных клеток). Например, иммунотерапевтические композиции и способы согласно настоящему изобретению можно использовать для индукции интерлейкина-2 (IL-2) для стимулирования выживания Т-клеток; для индукции Fas-лиганда (FasL) и/или индуцирующего апоптоз лиганда, связанного с фактором некроза опухоли (TRAIL) (например, для индукции апоптоза опухолевых клеток); и/или для индукции интерферона (IFN)-гамма (например, для активации врожденного иммунного ответа (например, против рака)). Согласно некоторым вариантам осуществления композиции и способы согласно настоящему изобретению используют для индукции остановки клеточного цикла и/или апоптоза, а также для усиления индукции остановки клеточного цикла и/или апоптоза либо отдельно, либо в ответ на дополнительные сигналы индукции апоптоза. Согласно некоторым вариантам осуществления Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, повышают чувствительность раковых клеток к индукции остановки клеточного цикла и/или апоптоза, включая в себя клетки, которые обычно являются устойчивыми к таким индуцирующим стимулам.
Согласно некоторым вариантам осуществления композиции и способы согласно настоящему изобретению используют для лечения патологических клеток, тканей, органов или патологических состояний и/или болезненных состояний у животного (например, пациента - млекопитающего, включая в себя без ограничения людей и животных-компаньонов). В связи с этим различные заболевания и патологии поддаются лечению или профилактике с использованием настоящих способов и композиций. Согласно некоторым вариантам осуществления раковые клетки, подвергаемые лечению, являются метастатическими. Согласно другим вариантам осуществления раковые клетки, подвергаемые лечению, являются устойчивыми к противораковым средствам.
Согласно некоторым вариантам осуществления настоящего изобретения предусмотрены способы введения эффективного количества Т-клеток, модифицированных, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1 и по меньшей мере одного дополнительного терапевтического средства (включая в себя без ограничения химиотерапевтические противоопухолевые средства, модулирующие апоптоз средства, противомикробные средства, противовирусные средства, противогрибковые средства и противовоспалительные средства) и/или терапевтической техники (например, хирургическое вмешательство и/или лучевая терапия). Согласно конкретному варианту осуществления дополнительное терапевтическое средство(а) представляет(ют) собой противораковое средство.
Ряд подходящих противораковых средств предусмотрен для применения в способах согласно настоящему изобретению. Действительно, настоящее изобретение предусматривает без ограничения введение многочисленных противоопухолевых средств, таких как следующее: средства, которые вызывают апоптоз; полинуклеотиды (например, антисмысловые, рибозимы, миРНК); полипептиды (например, ферменты и антитела); биологические миметики; алкалоиды; алкилирующие средства; противоопухолевые антибиотики; антиметаболиты; гормоны; соединения платины; моноклональные или поликлональные антитела (например, антитела, конъюгированные с противораковыми лекарственными средствами, токсинами, дефензинами), токсины; радионуклиды; модификаторы биологического ответа (например, интерфероны (например, IFN-α) и интерлейкины (например, IL-2)); средства адоптивной иммунотерапии; гемопоэтические факторы роста; средства, которые вызывают дифференцировку опухолевых клеток (например, полностью транс-ретиноевая кислота); реагенты для генной терапии (например, реагенты для антисмысловой терапии и нуклеотиды); противоопухолевые вакцины; ингибиторы ангиогенеза; ингибиторы протеасомы; модуляторы NF-KB; соединения против CDK; ингибиторы HDAC; и тому подобное. Специалистам в настоящей области техники известны многочисленные другие примеры химиотерапевтических соединений и противораковой терапии, подходящие для совместного введения с раскрытыми соединениями.
Согласно определенным вариантам осуществления противораковые средства включают в себя средства, которые индуцируют или стимулируют апоптоз. Средства, которые индуцируют апоптоз, включают в себя без ограничения облучение (например, рентгеновские лучи, гамма-лучи, УФ-излучение); факторы, связанные с фактором некроза опухоли (TNF) (например, рецепторные белки семейства TNF, лиганды семейства TNF, TRAIL, антитела к TRAIL-R1 или TRAIL-R2); ингибиторы киназы (например, ингибитор киназы рецептора эпидермального фактора роста (EGFR), ингибитор киназы рецептора фактора роста эндотелия сосудов (VGFR), ингибитор киназы рецептора фактора роста фибробластов (FGFR), ингибитор киназы рецептора тромбоцитарного фактора роста (PDGFR) и ингибиторы киназы Bcr-Abl (такие как гливек (GLEEVEC)); антисмысловые молекулы; антитела (например, герцептин, ритуксан, зевалин и авастин); антиэстрогены (например, ралоксифен и тамоксифен); антиандрогены (например, флутамид, бикалутамид, финастерид, аминоглутетамид, кетоконазол и кортикостероиды); ингибиторы циклооксигеназы 2 (СОХ-2) (например, целекоксиб, мелоксикам, NS-398 и нестероидные противовоспалительные препараты (НПВП)); противовоспалительные лекарственные средства (например, бутазолидин, декадрон, дельтазон, дексаметазон, дексаметазон интенсол, дексон, гексадрол, гидроксихлорохин, метикортен, орадексон, оразон, оксифенбутазон, педиапред, фенилбутазон, плаквенил, преднизолон, преднизон, прелон и тандеарил); и противоопухолевые химиотерапевтические лекарственные средства (например, иринотекан (камптосар), СРТ-11, флударабин (флудара), дакарбазин (DTIC), дексаметазон, митоксантрон, милотарг, VP-16, цисплатин, карбоплатин, оксалиплатин, 5-FU, доксорубицин, гемцитабин, бортезомиб, гефитиниб, бевацизумаб, таксотер или таксол); клеточные сигнальные молекулы; керамиды и цитокины; стауроспорин и тому подобное.
Согласно другим вариантам осуществления композиции и способы согласно настоящему изобретению используют вместе по меньшей мере с одним антигиперпролиферативным или противоопухолевым средством, выбранным из алкилирующих средств, антиметаболитов и натуральных продуктов (например, трав и других соединений растительного и/или животного происхождения).
Алкилирующие средства, подходящие для применения в настоящих композициях и способах, включают в себя без ограничения следующее: 1) азотистые иприты (например, мехлоретамин, циклофосфамид, ифосфамид, мелфалан (L-сарколизин) и хлорамбуцил); 2) этиленимины и метилмеламины (например, гексаметилмеламин и тиотепа); 3) алкилсульфонаты (например, бусульфан); 4) нитрозомочевины (например, кармустин (BCNU); ломустин (CCNU); семустин (метил-CCNU) и стрептозоцин (стрептозотоцин)); и 5) триазены (например, дакарбазин (DTIC; диметилтриазеноимид-азолкарбоксамид).
Согласно некоторым вариантам осуществления антиметаболиты, подходящие для применения в настоящих композициях и способах, включают в себя без ограничения следующее: 1) аналоги фолиевой кислоты (например, метотрексат (аметоптерин)); 2) аналоги пиримидина (например, фторурацил (5-фторурацил; 5-FU), флоксуридин (фтор-оксиуридин; FudR) и цитарабин (цитозинарабинозид)); и 3) аналоги пурина (например, меркаптопурин (6-меркаптопурин; 6-МР), тиогуанин (6-тиогуанин; TG) и пентостатин (2'-дезоксикоформицин)).
Согласно другим вариантам осуществления химиотерапевтические средства, подходящие для применения в композициях и способах согласно настоящему изобретению, включают в себя без ограничения следующее: 1) алкалоиды барвинка (например, винбластин (VBL), винкристин); 2) эпиподофиллотоксины (например, этопозид и тенипозид); 3) антибиотики (например, дактиномицин (актиномицин D), даунорубицин (дауномицин; рубидомицин), доксорубицин, блеомицин, пликамицин (митрамицин) и митомицин (митомицин С)); 4) ферменты (например, L-аспарагиназа); 5) модификаторы биологического ответа (например, интерферон-альфа); 6) координационные комплексы платины (например, цисплатин (цис-DDP) и карбоплатин); 7) антрацендионы (например, митоксантрон); 8) замещенные мочевины (например, гидроксимочевина); 9) производные метилгидразина (например, прокарбазин (N-метилгидразин; MIH)); 10) адренокортикальные супрессивные средства (например, митотан (o,p'-DDD) и аминоглютетимид); 11) адренокортикостероиды (например, преднизон); 12) прогестины (например, капроат гидроксипрогестерона, ацетат медроксипрогестерона и ацетат мегестрола); 13) эстрогены (например, диэтилстилбестрол и этинилэстрадиол); 14) антиэстрогены (например, тамоксифен); 15) андрогены (например, пропионат тестостерона и флуоксиместерон); 16) антиандрогены (например, флутамид) и 17) аналоги гонадотропин-рилизинг-гормона (например, лейпролид).
Любое онколитическое средство, которое обычно используют в контексте терапии рака, также можно использовать в композициях и способах согласно настоящему изобретению. Например, Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США поддерживает фармакологический справочник онколитических средств, одобренный для использования в Соединенных Штатах. Международные агентства-партнеры FDA США поддерживают подобные фармакологические справочники.
Противораковые средства дополнительно включают в себя соединения, которые, как было установлено, обладают противораковой активностью. Примеры включают в себя без ограничения следующее: 3-АР, 12-O-тетрадеканоилфорбол-13-ацетат, 17AAG, 852А, ABI-007, ABR-217620, АВТ-751, ADI-PEG 20, АЕ-941, AG-013736, AGRO100, аланозин, AMG 706, антитело G250, антинеопластоны, АР23573, апазиквон, АРС8015, атипримод (atiprimod), ATN-161, атрасентен, азацитидин, ВВ-10901, ВСХ-1777, бевацизумаб, BG00001, бикалутамид, BMS 247550, бортезомиб, бриостатин-1, бусерелин, кальцитриол, CCI-779, CDB-2914, цефиксим, цетуксимабат CG0070, циленгитид, клофарабин, комбретастатин А4-фосфат, СР-675,206, СР-724,714, CpG 7909, куркумин, децитабин, DENSPM, доксеркальциферол, Е7070, Е7389, эктеинасцидин 743, эфапроксирал, эфлорнитин, EKB-569, энзастаурин, эрлотиниб, экзисулинд, фенретинид, флавопиридол, флударабин, флутамид, фотемустин, FR901228, G17DT, галиксимаб, гефитиниб, генистеин, глуфосфамид, GTI-2040, гистрелин, HKI-272, гомогаррингтонин, HSPPC-96, белок слияния hu14.18-интерлейкин-2, HuMax-CD4, илопрост, имиквимод, инфликсимаб, интерлейкин-12, IPI-504, ирофульвен, иксабепилон, лапатиниб, леналидомид, лестауртиниб, лейпролид, иммунотоксин LMB-9, лонафарниб, луниликсимаб, мафосфамид, МВ07133, MDX-010, MLN2704, моноклональное антитело 3F8, моноклональное антитело J591, мотексафин, MS-275, MVA-MUC1-IL2, нилутамид, нитрокамптотецин, нолатрексед дигидрохлорид, нолвадекс, NS-9, Об-бензилгуанин, облимерсен натрия, ONYX-015, ореговомаб, OSI-774, панитумумаб, параплатин, PD-0325901, пеметрексед, PHY906, пиоглитазон, пирфенидон, пиксантрон, PS-341, PSC 833, PXD101, пиразол о акридин, R115777, RAD001, ранпирназа, аналог ребеккамицина, белок rhu-ангиостатин, rhuMab 2С4, росиглитазон, рубитекан, S-1, S-8184, сатраплатин, SB-, 15992, SGN-0010, SGN-40, сорафениб, SR31747A, ST1571, SU011248, субероиланилидгидроксамовая кислота, сурамин, талабостат, талампанель, тариквидар, темсиролимус, иммунотоксин TGFa-PE38, талидомид, тимальфазин, типифарниб, тирапазамин, TLK286, трабектедин, триметрексат глюкуронат, TroVax, UCN-1, вальпроевая кислота, винфлунин, VNP40101M, волоциксимаб, вориностат, VX-680, ZD1839, ZD6474, зилеутон и зосуквидар тригидрохлорид.
Настоящее изобретение относится к способам введения композиций и способы согласно настоящему изобретению вместе с (например, до, во время или после) лучевой терапии. Настоящее изобретение не ограничено типами, количествами или системами доставки и введения, используемыми для доставки терапевтической дозы облучения животному. Например, животное может получать фотонную лучевую терапию, лучевую терапию пучком частиц, другие виды лучевой терапии и их комбинации. Согласно некоторым вариантам осуществления облучение доставляется животному с использованием линейного ускорителя. Согласно другим вариантах осуществления облучение доставляется с использованием гамма-ножа.
Источник излучения может быть внешним или внутренним по отношению к животному. Внешняя лучевая терапия является наиболее распространенной и включает в себя направление пучка высокоэнергетического излучения в участок опухоли через кожу с использованием, например, линейного ускорителя. Хотя пучок излучения локализуется в месте опухоли, почти невозможно избежать воздействия на нормальные, здоровые ткани. Однако внешнее излучение, как правило, хорошо переносится животными. Внутренняя лучевая терапия включает в себя имплантацию источника излучения, такого как шарики, стержни, пеллеты, капсулы, частицы и тому подобное, внутри тела в или около места опухоли, включая в себя использование систем доставки, которые специально нацелены на раковые клетки (например, с использованием частиц, прикрепленных к лигандам, связывающим раковые клетки). Такие имплантаты могут быть удалены после лечения или оставлены в организме неактивными. Типы внутренней лучевой терапии включают в себя без ограничения брахитерапию, внутритканевое облучение, внутриполостное облучение, радиоиммунотерапию и тому подобное.
Животное может необязательно получать радиосенсибилизаторы (например, метронидазол, мизонидазол, внутриартериальный 5-бромдезоксиуридин (Budr), внутривенный йоддезоксиуридин (IudR), нитроимидазол, 5-замещенные-4-нитроимидазолы, 2Н-изоиндолдионы, [[(2-бромэтил)-амино]метил]-нитро-1Н-имидазол-1-этанол, производные нитроанилина, селективные цитотоксины ДНК-аффинной гипоксии, галогенированный ДНК-лиганд, оксиды 1,2,4-бензотриазина, производные 2-нитроимидазола, фторсодержащие производные нитроазола, бензамид, никотинамид, акридин-интеркалятор, производное 5-тиотретразола, 3-нитро-1,2,4-триазол, производное 4,5-динитроимидазола, гидроксилированные тексафрины, цисплатин, митомицин, тирипазамин, нитрозомочевина, меркаптопурин, метотрексат, фторурацил, блеомицин, винкристин, карбоплатин, эпирубицин, доксорубицин, циклофосфамид, виндезин, этопозид, паклитаксел, тепло (гипертермия) и тому подобное), радиопротекторы (например, цистеамин, аминоалкилдигидрофосфоротиоаты, амифостин (WR2721), IL-1, IL-6 и тому подобное). Радиосенсибилизаторы усиливают уничтожение опухолевых клеток. Радиопротекторы защищают здоровые ткани от вредного воздействия облучения.
Любой тип облучения может быть введен животному, при условии, что доза облучения переносится животным без неприемлемых отрицательных побочных эффектов. Подходящие типы лучевой терапии включают в себя, например, ионизирующую (электромагнитную) лучевую терапию (например, рентгеновское излучение или гамма-излучение) или лучевую терапию пучком частиц (например, излучение с высокой линейной энергией). Ионизирующее излучение определяется как излучение, содержащее частицы или фотоны, которые имеют достаточную энергию для создания ионизации, т.е. приобретения или потери электронов (как описано, например, в патенте США №5770581, полностью включенном в настоящий документ посредством ссылки). Влияние облучения может по меньшей мере частично контролироваться врачом. Согласно одному варианту осуществления дозу облучения фракционируют для максимального воздействия на клетки-мишени и снижения токсичности.
Согласно одному варианту осуществления общая доза облучения, вводимая животному, составляет от приблизительно 0,01 Грэй (Гр) до приблизительно 100 Гр. Согласно другому варианту осуществления от приблизительно 10 Гр до приблизительно 65 Гр (например, приблизительно 15 Гр, 20 Гр, 25 Гр, 30 Гр, 35 Гр, 40 Гр, 45 Гр, 50 Гр, 55 Гр или 60 Гр) вводят в течение курса лечения. Хотя согласно некоторым вариантам осуществления полную дозу облучения можно вводить в течение одного дня, оптимально общую дозу фракционируют и вводят в течение нескольких дней. Желательно, чтобы лучевая терапия проводилась в течение по меньшей мере приблизительно 3 дней, например, по меньшей мере 5, 7,10, 14, 17, 21, 25, 28, 32, 35, 38, 42, 46, 52 или 56 дней (приблизительно 1-8 недель). Соответственно, суточная доза облучения будет составлять приблизительно 1-5 Гр (например, приблизительно 1 Гр, 1,5 Гр, 1,8 Гр, 2 Гр, 2,5 Гр, 2,8 Гр, 3 Гр, 3,2 Гр, 3,5 Гр, 3,8 Гр, 4 Гр, 4,2 Гр, или 4,5 Гр), или 1-2 Гр (например, 1,5-2 Гр). Суточная доза облучения должна быть достаточной, чтобы вызвать разрушение клеток-мишеней. При удлинении периода согласно одному варианту осуществления облучение вводят не каждый день, что позволяет животному отдыхать и реализовываться эффектам терапии. Например, облучение желательно вводить в течение 5 последовательных дней, и не вводить в течение 2 дней, в течение каждой недели лечения, тем самым обеспечивая 2 дня отдыха в неделю. Однако облучение можно вводить 1 день в неделю, 2 дня в неделю, 3 дня в неделю, 4 дня в неделю, 5 дней в неделю, 6 дней в неделю или все 7 дней в неделю, в зависимости от реакции животного и любых потенциальных побочных эффектов. Лучевая терапия может быть начата в любое время в терапевтическом периоде. Согласно одному варианту осуществления облучение начинают на неделе 1 или неделе 2 и вводят в течение оставшейся продолжительности терапевтического периода. Например, облучение вводят на 1-6 неделях или на 2-6 неделях терапевтического периода, включающего в себя 6 недель, для лечения, например, солидной опухоли. Альтернативно, облучение вводят на 1-5 неделях или 2-5 неделях терапевтического периода, включающего в себя 5 недель. Эти иллюстративные схемы введения лучевой терапии не предназначены, однако, для ограничения настоящего изобретения.
Согласно некоторым вариантам осуществления настоящего изобретения Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, и одно или несколько терапевтических средств или противораковых средств вводят животному при одном или нескольких из следующих условий: с различной периодичностью, с различной продолжительностью, с различными концентрациями, разными путями введения и т.д. Согласно некоторым вариантам осуществления Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, вводят перед терапевтическим или противораковым средством, например, за 0,5, 1, 2, 3, 4, 5, 10, 12, 18 часов или более, за 1, 2, 3, 4, 5, 6 или более дней или за 1, 2, 3, 4, 5, 6 или более недель до введения терапевтического или противоракового средства. Согласно некоторым вариантам осуществления Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, вводят после терапевтического или противоопухолевого средства, например, через 0,5, 1, 2, 3, 4, 5, 10, 12, 18 или более часов, через 1, 2, 3, 4, 5, 6 или более дней или через 1, 2, 3, 4, 5, 6 или более недель после введения противоракового средства. Согласно некоторым вариантам осуществления Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, и терапевтическое или противораковое средство вводят одновременно, но по разным схемам, например, модифицированные иммунные клетки вводят ежедневно, в то время как терапевтическое или противораковое средство вводят один раз в неделю, один раз каждые две недели, один раз каждые три недели, один раз каждые четыре недели или больше. Согласно другим вариантам осуществления Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, вводят один раз в неделю, тогда как терапевтическое или противораковое средство вводят ежедневно, один раз в неделю, один раз каждые две недели, один раз каждые три недели, раз в четыре недели или больше.
Композиции в пределах объема настоящего изобретения включают в себя все композиции, в которых Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, содержатся в количестве, которое эффективно для достижения предполагаемой цели. Хотя индивидуальные потребности варьируются, определение оптимальных диапазонов эффективных количеств каждого компонента находится в пределах квалификации в настоящей области техники. В одном неограничивающем примере Т-клетки, модифицированные, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, можно вводить млекопитающим, например, людям, чтобы обеспечить человека от 1000 до 1010 Т-клеток в день (например, для лечения рака). Согласно другому варианту осуществления от 1000 до 1010 модифицированных Т-клеток вводят для лечения, уменьшения интенсивности или профилактики рака (например, для предотвращения метастазирования, рецидива и/или прогрессирования рака). Единичную дозу можно вводить за одно или несколько введений один или несколько раз в день (например, в течение 1, 2, 3, 4, 5, 6 или больше дней или недель).
Т-клетки можно вводить как часть фармацевтического препарата, содержащего подходящие фармацевтически приемлемые носители, содержащие вспомогательные вещества и вспомогательные средства, которые облегчают обработку и/или введение модифицированных клеток в препараты, которые можно использовать фармацевтически. Иммунные Т-клетки и/или фармацевтические препараты, содержащие их, можно вводить внутривенно, внутримышечно, подкожно, внутриопухолево, внутрибрюшинно, интратекально или интравентрикулярно. Эффективное количество Т-клеток и/или фармацевтических препаратов, содержащих их, можно вводить для профилактики или лечения заболевания. Подходящую дозировку можно определить на основе типа заболевания, подлежащего лечению, типа модифицированной Т-клетки, тяжести и течения заболевания, клинического состояния индивидуума, истории болезни индивидуума и ответа на лечение, а также по усмотрению лечащего врача.
Эффективность любого из способов, описанных в настоящем документе (например, лечение Т-клетками, модифицированными, чтобы экспрессировать и/или содержать повышенные содержания одного или нескольких факторов транскрипции АР-1, отдельно или в комбинации с одним или несколькими химиотерапевтическими средствами, описанными в настоящем документе), можно испытывать в различных моделях, известных в настоящей области техники, таких как клинические или доклинические модели. Подходящие доклинические модели приведены в настоящем документе в качестве примера. Для любой иллюстративной модели после развития опухолей мышей случайным образом набирают в группы лечения, получающие лечение или контрольное лечение. Размер опухоли (например, объем опухоли) измеряют в течение курса лечения, а также отслеживают общую выживаемость.
Согласно некоторым вариантам осуществления образец получают до лечения с помощью Т-клеток (например, отдельно или в комбинации с другой терапией, описанной в настоящем документе) в качестве исходного значения для измерения ответа на лечение. Согласно некоторым вариантам осуществления образец представляет собой образец ткани (например, фиксированный формалином и залитый парафином (FFPE), архивный, свежий или замороженный). Согласно некоторым вариантам осуществления образец представляет собой цельную кровь. Согласно некоторым вариантам осуществления цельная кровь содержит иммунные клетки, циркулирующие опухолевые клетки и любые их комбинации.
Восприимчивость к лечению может относиться к любому одному или нескольким из следующего: увеличение выживаемости (включая в себя общую выживаемость и выживаемость без прогрессирования заболевания); обеспечение объективного ответа (включая в себя полный ответ или частичный ответ); или улучшение признаков или симптомов рака. Согласно некоторым вариантам осуществления восприимчивость может относиться к улучшению одного или нескольких факторов в соответствии с опубликованным набором рекомендаций RECIST для определения состояния опухоли у пациента с раком, т.е. отвечаемость, стабилизации или прогрессирования. Для более подробного обсуждения этих рекомендаций см. Eisenhauer et al., Eur J Cancer 2009; 45: 228-47; Topalian et al., N Engl J Med 2012; 366: 2443-54; Wolchok et al., Clin Can Res 2009; 15: 7412-20; and Therasse, P., et al. J. Natl. Cancer Inst. 92: 205-16 (2000). Восприимчивый субъект может относиться к субъекту, у которого рак показывает улучшение, например, согласно одному или нескольким факторам, основанным на критериях RECIST. Не восприимчивый субъект может относиться к субъекту, у которого рак не показывает улучшения, например, согласно одному или нескольким факторам, основанным на критериях RECIST.
Общепринятые критерии ответа могут являться недостаточными для характеристики противоопухолевой активности иммунотерапевтических средств, которые могут вызывать отсроченные ответы, которым может предшествовать первоначальное явное радиологическое прогрессирование, включая в себя появление новых очагов поражения. Таким образом, были разработаны модифицированные критерии ответа, которые учитывают возможное появление новых очагов поражения и позволяют подтвердить радиологическое прогрессирование при последующей оценке. Соответственно, согласно некоторым вариантам осуществления восприимчивость может относиться к улучшению одного из нескольких факторов в соответствии с иммунозаисимыми критериями ответа 2 (irRC). См., например, Wolchok et al., Clin Can Res 2009; 15: 7412-20. Согласно некоторым вариантам осуществления новые очаги поражения добавляются в уже определенную опухолевую нагрузку и отслеживаются, например, в отношении радиологического прогрессирования при последующей оценке. Согласно некоторым вариантам осуществления наличие нецелевых очагов поражения включено в оценку полного ответа и не включено в оценку радиологического прогрессирования. Согласно некоторым вариантам осуществления радиологическое прогрессирование можно определить только на основе измеримого заболевания и/или можно подтвердить с помощью последовательной оценки через >4 недель от впервые задокументированной даты.
Настоящее описание изобретения считается достаточным для того, чтобы позволить специалисту в настоящей области техники реализовать настоящее изобретение. Различные модификации согласно настоящему изобретению в дополнение к тем, которые показаны и описаны в настоящем документе, станут очевидными для специалистов в настоящей области техники из предшествующего описания, и они подпадают под объем прилагаемой формулы изобретения. Все публикации, патенты и заявки на выдачу патентов, процитированные настоящем документе, тем самым полностью включены посредством ссылки для всех целей.
ПРИМЕРЫ
Следующие примеры иллюстрируют, но не ограничивают соединения, композиции и способы согласно настоящему изобретению. Другие подходящие модификации и адаптации множества состояний и параметров, обычно встречающихся в клинической терапии и которые очевидны для специалистов в настоящей области техники, находятся в пределах сущности и объема настоящего изобретения.
Материалы и методы
Конструирование вирусного вектора
Ранее были описаны ретровирусные векторы MSGV, кодирующие следующие CAR: CD19-28z, CD19-BBz, GD2-28z, GD2-BBz, Her2-BBz и CD22-BBz. Чтобы создать HA-28z CAR, в 14G2a scFv плазмиды CAR GD2-28z вводили точечную мутацию для создания мутации E101K. Мутации «4/2NQ» 46 вводили в домены СН2СН3 спейсерной области IgG1, чтобы уменьшить распознавание рецептора Fc для использования in vivo HA-28z CAR Т-клеток. Оптимизированные по кодонам кДНК, кодирующие c-Jun (JUN), c-Fos (FOS) и усеченный NGFR (tNGFR), синтезировали с помощью IDT и клонировали в лентивирусные векторы экспрессии для создания JUN-P2A-FOS, а также отдельных векторов экспрессии JUN и FOS, коэкспрессирующих tNGFR под контролем отдельного промотора PGK. Затем JUN-P2A субклонировали в сайт XhoI векторов MSGV CAR с использованием набора для клонирования In-Fusion HD (Takara) выше лидерной последовательности CAR для создания ретровирусных векторов JUN-P2A-CAR. Для JUN-AA вводили точечные мутации для преобразования Ser63 и Ser73 в Ala. Последовательность Е. coli DHFR-DD вставляли выше Jun для создания конструктов JUN-DD. В некоторых случаях кДНК GFP также субклонировали выше от CAR для создания контролен вектора GFP-P2A-CAR.
Продукция вирусного вектора
Ретровирусный супернатант получали в пакующей клеточной линии 293GP, как описано ранее. Вкратце, 20 см планшеты 293GP с 70% конфлюентностью котрансфицировали 20 мкг векторной плазмиды MSGV и 10 мкг оболочечной плазмидной ДНК RD114 с использованием липофектамина 2000. Среду заменяли через 24 и 48 часов после трансфекции. Вирусные супернатанты 48 ч и 72 ч собирали, центрифугировали для удаления клеточного дебриса и замораживали при -80°С для дальнейшего использования. Самоинактивирующийся лентивирусный супернатант третьего поколения получали в пакующей клеточной линии 293Т, как описано ранее. Вкратце, 20 см планшеты 293Т с 70% конфлюентностью котрансфицировали векторной плазмидой 18 мкг ДНК векторной плазмиды pELNS и 18 мкг pRSV-Rev, 18 мкг pMDLg/pRRE (Gag/Pol) и 7 мкг pMD2.G ДНК (оболочка VSVG) пакующей плазмиды с использованием липофектамина 2000. Среду заменяли через 24 часа после трансфекции. 24 ч и 48 ч вирусные супернатанты собирали, объединяли и концентрировали ультрацентрифугированием при 28000 об/мин в течение 2,5 часов. Концентрированные лентивирусные исходные материалы замораживали при -80°С для дальнейшего использования.
Выделение Т-клеток
Первичные Т-клетки человека выделяли у здоровых доноров с использованием набора для обогащения Т-клеток человека RosetteSep (Stem Cell Technologies). Лейкоцитарные пленки приобретали в Стэнфордском центре крови и обрабатывали в соответствии с протоколом производителя с использованием среды с градиентом плотности Lymphoprep и пробирок SepMate-50. Выделенные Т-клетки криоконсервировали при 2×107 Т-клеток на флакон в криоконсервирующей среде CryoStor CS10 (Stem Cell Technologies).
Продукция CAR Т-клеток
Криоконсервированные Т-клетки оттаивали и активировали в тот же день с помощью парамагнитных микрочастиц Human T-Expander CD3/CD28 Dynabeads (Gibco) в соотношении микрочастиц к клеткам, составляющем 3:1, в среде Т-клеток (AIMV с добавлением 5% FBS, 10 мМ HEPES, 2 мМ GlutaMAX, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина (Gibco)). Рекомбинантный IL-2 человека (Peprotech) вводили в концентрации 100 ед/мл. Т-клетки трансдуцировали ретровирусным вектором на 2 и 3 дни после активации и поддерживали на уровне 0,5-1×106 клеток на мл в Т-клеточной среде с IL-2. Если не указано иное, CAR Т-клетки использовали для анализов in vitro или переносили мышам на 10-11 день после активации.
Ретровирусная трансдукция
12-луночные планшеты, не обработанные тканевой культурой, покрывали в течение ночи при 4°С 1 мл ретронектина (Takara) в концентрации 25 мкг/мл в PBS. Планшеты промывали PBS и блокировали 2% BSA в течение 15 минут. Оттаявший ретровирусный супернатант добавляли при ~1 мл на лунку и центрифугировали в течение 2 часов при 32°С и 3200 об/мин перед добавлением клеток.
Клеточные линии
Клеточные линии нейробластомы Келли, саркомы Юинга EW8, остеосаркомы 143b и ТС32 первоначально получали от АТСС. Некоторые клеточные линии стабильно трансдуцировали с помощью GFP и люциферазы светлячка (GL). Клеточная линия CD19+CD22+Nalm6-GL В-ALL предоставлена Дэвидом Барреттом (David Barrett). Nalm6-GD2 создавали путем совместной трансдукции Nalm6-GL с кДНК для синтазы GD2 и синтазы GD3. Затем выбирали клон из одной клетки для экспрессии GD2 на высоком уровне. Nalm6-22KO и 221ow были ранее описаны и любезно предоставлены Терри Фраем (Terry Fry). Все клеточные линии культивировали в полной среде (RPMI, дополненный 10% FBS, 10 мМ HEPES, 2 мМ GlutaMAX, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина (Gibco)).
Проточная цитометрия
CAR CD22 и Her2 обнаруживали с использованием рекомбинантных белков CD22-Fc и Her2-Fc человека (R&D). Идиотипические антитела и Fc-слитые белки конъюгировали с наборами для маркировки антител Dylight488 и/или 650 (Thermo Fisher). Фенотип поверхности Т-клеток оценивали с использованием следующих антител:
От BioLegend: CD4-APC-Cy7 (клон OKT4), CD8-PerCp-Cy5.5 (клон SKI), TIM3-BV510 (клон F38-2E2), CD39-FITC или АРС-Су7 (клон A1), CD95-PE (клон DX2), CD3-PacBlue (клон HIT3a), от eBioscience: PD1-PE-Cy7 (клон eBio J105), LAG3-PE (клон 3DS223H), CD45RO-PE-Cy7 (клон UCHL1), CD45-PerCp-Cy5.5 (клон HI30), от BD: CD45RA-FITC или BV711 (клон HI100), CCR7-BV421 (клон 150503), CD122-BV510 (клон Mik-b3), CD62L-BV605 (клон DREG-56), CD4-BUV395 (клон SK3), CD8-BUV805 (клон SKI).
Продукция цитокинов
1×105 CAR+Т-клеток и 1×105 опухолевых клеток культивировали в 200 мкл СМ в 96-луночных планшетах с плоским дном в течение 24 часов. Для стимуляции идиотипа серийные разведения 1А7 поперечно сшивали в 1-кратном покрывающем буфере (BioLegend) в течение ночи при 4°С на 96-луночных планшетах для ELISA Nunc Maxisorp (Thermo Scientific). Лунки промывали один раз PBS и 1×105 CAR+ Т-клеток высевали в 200 мкл СМ и культивировали в течение 24 часов. Лунки в трех повторностях высевали для каждого условия. Супернатанты культур собирали и анализировали в отношении IFNg и IL-2 с помощью ELISA (BioLegend).
Анализ лизиса
5×104 GFP + клетки лейкоза или 2,5×104 GFP + адгезивные опухолевые клетки совместно культивировали с CAR Т-клетками в 200 мкл СМ в 96-луночных планшетах с плоским дном в течение до 96 часов. Лунки в трех повторностях высевали для каждого условия. Планшеты визуализировали каждые 2-3 часа с использованием системы анализа живых клеток IncuCyte ZOOM (Essen Bioscience). 4 изображения на лунку при 10-кратном увеличении собирали в каждый момент времени. Общую интегрированную интенсивность GFP на лунку оценивали как количественную меру живых GFP + опухолевых клеток. Значения нормализовали к исходному измерению и наносили на график в зависимости от времени. Отношения Е:Т указаны на легендах фигуры.
Вестерн-блоттинг и иммунопреципитация
Белковые лизаты цельных клеток получали в неденатурирующем буфере (150 ммоль/л NaCl, 50 ммоль/л Трис-рН8, 1% NP-10, 0,25% дезоксихолата натрия). Концентрации белка оценивали с помощью колориметрического анализа Bio-Rad. Иммуноблоттинг осуществляли путем загрузки 20 мкг белка в гели с 11% PAGE с последующим переносом на мембраны PVF. Сигналы обнаруживали с помощью усиленной хемилюминесценции (Pierce) или системы визуализации Odyssey. Показаны репрезентативные блоты. Следующие используемые первичные антитела приобретали у Cell Signaling: c-Jun (60А8), P-c-JunSer73 (D47G9), JunB (C37F9), BATF (D7C5) и IRF4 (4964). Антитело BATF3 (AF7437) приобретали у R&D. Иммунопреципитацию проводили в 100 мг лизатов цельных клеток в 150 мкл неденатурирующего буфера и 7,5 мг агар конъюгированных антител c-Jun (G4) или JunB (С11) (Sant Craz Biotechnology). После инкубации в течение ночи при 4°С микрочастицы промывали 3 раза неденатурирующим буфером, а белки элюировали в буфере для образцов Laemmli, кипятили и наносили на гели PAGE. Детекцию иммунопреципитированных белков проводили с помощью вышеупомянутых реагентов и антител.
Мыши
Мышей с ослабленным иммунитетом NOD/SCID/IL2Rg-/- (NSG) приобретали у JAX и выращивали самостоятельно. Всех мышей выращивали, содержали и лечили в соответствии с протоколами, утвержденными IACUC Стэнфордского университета (APLAC). Мышам в возрасте 6-8 недель инокулировали либо 1×106 клеток лейкоза Nalm6-GL посредством внутривенной (в/в), либо 0,5-1×106 клеток остеосаркомы 143В посредством внутримышечной (IM) инъекции. Все CAR Т-клетки были вводили с помощью внутривенной инъекции. Время и доза лечения указаны на легендах фигур. Прогрессирование лейкоза измеряли с помощью биолюминесцентной визуализации с использованием системы визуализации IVIS. Значения проанализировали с использованием программного обеспечения Living Image. Прогрессирование солидной опухоли отслеживали с помощью штангенциркуля для измерения площади задней лапы, в которую ввели инъекцию. 5 мышей на группу получали лечение в каждом эксперименте, и каждый эксперимент повторяли 2 или 3 раза, как указано. Мышей рандомизировали для обеспечения равного опухолевой нагрузки до лечения перед лечением CAR Т-клетками.
Анализ крови и ткани
Забор периферической крови проводили путем ретроорбитального забора крови под анестезией изофлураном в указанные моменты времени. 50 мкл крови метили с помощью CD45, CD3, CD4 и CD8, лизировали с использованием лизирующего раствора BD FACS и количественно определяли с использованием гранул CountBright Absolute Counting (Thermo Fisher) на проточном цитометре BD Fortessa.
Анализ ATAC-seq
Подготовку библиотеки ATAC-seq проводили, как описано ранее48. Вкратце, 100000 клеток из каждого образца сортировали с помощью FACS в СМ, центрифугировали при 500 g при 4°С, затем ресуспендировали в буфере для ресуспендирования ATAC-seq (RSB) (10 мМ трис-HCl, 10 мМ NaCl, 3 мМ MgCl2) с добавлением 0,1% NP-40,0,1% Tween-20 и 0,01% дигитонина. Образцы разделяли на две повторности каждый перед всеми последующими стадиями. Образцы инкубировали на льду в течение 3 минут, затем промывали 1 мл RSB с добавлением 0,1% Tween -20. Ядра осаждали при 500g в течение 10 минут при 4°С. Осадок ядер ресуспендировали в 50 мкл смеси для транспозиции (25 мкл 2× буфера TD, 2,5 мкл транспозазы (Illumina), 16,5 мкл PBS, 0,5 мкл 1% дигитонина, 0,5 мкл 10% Tween-20, 5 мкл Н2О) и инкубировали при 37°С в течение 30 минут в термомиксере при встряхивании 1000 об/мин. Реакционную смесь очищали с использованием набора для очистки ПЦР Qiagen MinElute. Библиотеки амплифицировали с помощью ПЦР, используя мастер-микс высокоточной ПЦР NEBNext и специальные праймеры (IDT), как описано ранее20. Библиотеки являлись достаточно амплифицированными после 5 циклов ПЦР, как показано с помощью кривых флуоресценции кПЦР20. Библиотеки очищали с помощью набора для очистки ПЦР Qiagen MinElute и количественно определяли с помощью набора для количественного определения библиотек KAPA. Библиотеки секвенировали на Illumina NextSeq в Стэнфордском центре функциональной геномики с парными прочтениями 75 п.н. Последовательности адаптера обрезали с использованием SeqPurge и выравнивали по геному hg19 с использованием bowtie2. Эти прочтения затем фильтровали по митохондриальным прочтениям, низкому качеству картирования (Q>=20) и дубликатам ПЦР с использованием инструментов Picard. Затем авторы настоящего изобретения преобразовали значения ВАМ в BED и получили скорректированные по Tn5 сайты вставки («+» цепочечный + 4 п. н., «-» цепочечный - 5 п. н.). Чтобы определить пики, авторы настоящего изобретения назвали пики для каждого образца с использованием MACS2 "--shift -75 --extsize 150 --nomodel --call-summits --nolambda -- keep-dup all - p 0,00001", используя значения BED вставки. Чтобы получить набор пиков объединения, авторы настоящего изобретения (1) расширили все вершины до 500 п.н., (2) объединили все файлы вершин BED, а затем (3) использовали кластер bedtools и выбрали вершину с наибольшим баллом MACS2. Затем он был отфильтрован с помощью черного списка ENCODE hg19 (https://www.encodeproject.org/annotations/ENCSR636HFF/), чтобы удалить пики, выходящие за пределы концов хромосом. Затем авторы настоящего изобретения аннотировали эти пики, используя HOMER, и вычислили возникновение мотива TF, используя motifmatchr в R, с набором chromVARMotifs HOMER. Чтобы создать треки секвенирования, авторы настоящего изобретения считывали скорректированные по Tn5 сайты вставки в R и создавали скопление перекрытия, сгруппированное каждые 100 п.н., используя rtracklayer. Затем авторы настоящего изобретения подсчитали все вставки, которые попали в каждый пик, чтобы получить матрицу подсчетов (пик × образцы). Для определения дифференциальных пиков авторы настоящего изобретения сначала использовали пики, которые были обозначены как «TSS» в качестве контрольных генов или «служебных пиков» для DESeq2, а затем вычислили дифференциальные пики с помощью этой нормализации. Всю кластеризацию выполняли с использованием регуляризованных значений логарифмического преобразования из DESeq2. Анализ отклонения мотива транскрипционного фактора проводили с использованием chromVAR, как описано ранее21. Обогащение мотивов TF рассчитывали с использованием гипергеометрического теста в R, проверяющего представление мотива (из motifmatchr выше) в подмножестве пиков по сравнению со всеми пиками.
РНК-секвенирование подмножеств
Для специфического для подмножеств Т-клеток РНК-секвенирования Т-клетки выделяли из лейкоцитарных оболочек здоровых доноров, как описано выше. Перед активацией выделяли подмножества наивных клеток и центральных клеток памяти CD4 + или CD8 + с использованием сортировщика клеток BD FACSAria (Stem Cell FACS Core, Медицинский факультет Стэнфордского университета) с использованием следующих маркеров: наивные (CD45RA+CD45RO-, CD62L+, CCR7+, CD95- и CD122-), центральные клетки памяти (CD45RA-CD45RO+, CD62L+, CCR7+). Отсортированные исходные популяции активировали, трансдуцировали и культивировали, как описано выше. На 7, 10 и 14 дни культивирования клетки CAR+CD4+ и CD8+ клетки сортировали и выделяли РНК с использованием набора Qiagen mRNEasy. Образцы предварительно подготовили в библиотеке и секвенировали с помощью платформы для секвенирования парных прочтений Illumina NextSeq с помощью Stanford Functional Genomics Core.
Массовое РНК-секвенирование
Для выделения общей РНК здоровые донорные Т-клетки получали, как описано. На 10 или 11 день культивирования общую мРНК выделяли из 2×106 общих CAR T-клеток с использованием мини-набора для выделения Qiagen RNEasy Plus. Массовое секвенирование РНК выполняли с помощью BGI America (Кембридж, Массачусетс) с использованием платформы BGISEQ-500, длина прочтения одного конца 50 п. н., 30×106 прочтений на образец. Анализ основных компонентов выполняли с использованием пакета stats и графиков с пакетом ggplot2 в R (версия 3.5) 49. Анализ обогащения генного набора проводили с использованием программного обеспечения GSEA (Broad Institute), как описано 50, 51.
РНК-секвенирование одиночных клеток
Чтобы сравнить экспрессию генов в отдельных CD19-CAR и GD2-CAR Т-клетках, авторы настоящего изобретения отсортировали исходное подмножество Т-клеток в день 0 для последующего анализа одиночных клеток в день 10 с использованием платформы Chromium (10× Genomics) и реагентов набора Chromium Single Cell 3' v2 в соответствии с инструкциями производителя. Библиотеки кДНК готовили отдельно для клеток CD19-CAR и GD2-CAR, и клетки CD4+ и клетки CD8+ объединяли в каждом опыте для того, чтобы биоинформативно разделить далее. Секвенирование выполняли в системе Illumina NextSeq (парные прочтения, 26 п.н. в прочтении 1 и 98 п.н. в прочтении 2) до глубины >100000 прочтений на клетку. Прочтения секвенирования РНК одиночных клеток выравнивали в соответствии с версией 38 консорциума референсного генома человека (GRCh38), нормализовали в отношении групповых эффектов и отфильтровывали в отношении событий в клетках с использованием программного обеспечения Cell Ranger (10Х Genomics). В общей сложности 804 CD19-CAR и 726 GD2-CAR Т-клеток секвенировали в среднем до 350587 прочтений после нормализации на клетку. Матрицу клеточных генов дополнительно обрабатывали с использованием программного обеспечения Cell Ranger R Kit (10X Genomics), как описано52. Вкратце, авторы настоящего изобретения сначала отобрали гены с числом уникальных молекулярных идентификаторов (UMI)≥1 в любой данной клетке. Затем показатели UMI нормализовывали к суммам UMI для каждой клетки и умножали на медианное число UMI по клеткам. Затем данные преобразовывали путем взятия натурального логарифма полученной матрицы данных.
Статистический анализ
Если не указано иное, статистический анализ значимых различий между группами проводили с использованием непарных двусторонних t-критериев без учета согласованности SD с использованием GraphPad Prism7. Для массового РНК-секвенирования на фигуре 2С использовали непараметрический критерий ранговых знаков согласованных пар Уилкоксона. Кривые выживаемости сравнивали с использованием логрангового критерия Кокса-Мантеля. Таблицу с полным статистическим анализом, включая в себя точные значения р, отношение t и dof, можно найти в дополнительных материалах.
Пример 1
Анализ экспрессии генов истощения Т-клеток
Антиген-независимая тоническая передача сигналов химерными антигенными рецепторами (CAR) может усиливать дифференцировку и истощение Т-клеток, ограничивая их активность. Например, было описано, что специфические для GD2 CAR самоагрегируются при отсутствии антигена, что приводит к активации хронического нижележащего каскада передачи сигналов активации Т-клеток. В то время как GD2-CAR, включающие в себя костимулирующий домен CD28, быстро развивают характерные признаки истощения Т-клеток, GD2-CAR, включающие в себя костимулирующий домен 4-1ВВ, показывают меньшее количество признаков истощения Т-клеток и сохраняют большую функциональность, несмотря на сходную агрегацию и передачу сигналов. Наиболее характерные признаки истощения Т-клеток у экспрессирующих GD2-28z Т-клеток включают в себя повышенную поверхностную экспрессию ингибирующих рецепторов (например, PD1, TIM3, LAG3, CD39), сниженную экспрессию маркеров памяти (например, CD62L и CCR7) и сниженную продукцию цитокинов (особенно IL2) при стимуляции антигеном.
Эксперименты проводили во время разработки вариантов осуществления настоящего изобретения для оценки анализа транскрипции генов в истощенных и не истощенных Т-клетках. Сниженная экспрессия представителей семейства АР-1 была обнаружена в GD2- 28Z CAR Т-клетках по сравнению с GD2-BBZ (не истощенный CAR) (см. фиг. 1а). Классические партнеры АР-1 FOS и JUN также были среди наиболее отрицательно регулируемых генов в GD2-28Z CAR Т-клетках по сравнению со здоровыми CD19-28Z CAR Т-клетками с помощью анализа секвенирования РНК (см. фиг. 1b). Семейство транскрипционных факторов АР-1 активируется после передачи сигналов TCR и регулирует широкий и разнообразный набор ключевых функций Т-клеток, включая в себя рост, апоптоз, продукцию цитокинов и эффекторные функции. Дополнительные эксперименты проводили во время разработки вариантов осуществления настоящего изобретения, чтобы оценить и охарактеризовать функциональные роли представителей семейства АР-1 в CAR Т-клетках (например, вносит ли вклад ли отсутствие представителей семейства АР-1 в CAR Т-клетках в их CAR Т-клеточный фенотип).
Пример 2
Конструирование CAR Т-клеток с усиленной экспрессией c-Jun и c-Fos
Чтобы определить, может ли замена АР-1 ослабить симптомы истощения в GD2-28Z CAR Т-клетках, сконструировали лентивирусный экспрессионный конструкт с усиленной экспрессией c-Jun и c-Fos под контролем конститутивного промотора (см. фиг. 2а). Этот конструкт также кодировал экспрессионную кассету усеченного рецептора фактора роста нервов (NGFR (tNGFR)) в качестве поверхностного маркера трансдукции Т-клеток. Впоследствии активированные первичные Т-клетки человека трансдуцировали с помощью CD 19, GD2-BBZ или высокоаффинного (НА) GD2-28Z CAR с (АР-1) или без (wo) экспрессионного вектора АР-1.
Пример 3
Экспрессия c-Jun и c-Fos в CAR Т-клетках
На 8 день культивирования Т-клеток трансдуцированные АР-1 CD4 или CD8 CAR Т-клетки сортировали с использованием NGFR. Конститутивная экспрессия АР-1 снижала частоту связанных с истощением ингибирующих рецепторов PD1, TIM3, LAG3 и CD39 и увеличивала маркер памяти CD62L как в CD4 (см. фиг.2b), так и в CD8 (см. фиг. 2с) CAR Т-клетках. CAR Т-клетки совместно культивировали с (АР-1) или без (wo) АР-1 совместно с опухолевыми клетками, экспрессирующими антиген CD19 и GD2, для оценки функциональных изменений в трансдуцированных АР-1 CAR Т-клетках. В большинстве случаев трансдуцированные АР-1 CAR Т-клетки высвобождали больше IL2 (см. фиг. 2d) и IFNγ (см. фиг. 2е) по сравнению с клетками без него.
Пример 4
CAR Т-клетки с усиленной экспрессией c-Jun или c-Fos
Чтобы оценить, требует ли функциональный благоприятный результат экспрессии АР-1 как экспрессии c-Fos, так и c-Jun, создавали отдельные лентивирусные векторы, кодирующие только c-Jun или c-Fos (см. фиг. 3а). CAR Т-клетки трансдуцировали с помощью кодирующих c-Fos или c-Jun векторов.
После стимуляции CAR Т-клеток антиген-положительными опухолевыми клетками только CAR Т-клетки, экспрессирующие c-Jun, приводили к увеличению IL2, тогда как экспрессии только c-Fos было недостаточно (см. фиг. 3b). Избыточная экспрессия c-Jun влияла на секрецию IFNγ аналогично, но менее драматично (см. фиг. 3с). Чтобы подтвердить повышенную экспрессию цитокинов в экспрессирующих c-Jun CAR Т-клетках на уровне отдельных клеток, проводили внутриклеточное цитокиновое окрашивание (ICS) методом проточной цитометрии после 6-часовой стимуляции Т-клеток антиген-положительными опухолевыми клетками. В большинстве случаев увеличение частоты и/или средней интенсивности флуоресценции (MFI) IL2 (см. фиг. 3d), IFNγ (см. фиг. 3е) и TNFα (см. фиг. 3f) в трансдуцированных c-Jun CAR Т-клетках обнаруживали по сравнению с клетками без (wo), в то время как трансдукция c-Fos, как правило, приводила к снижению продукции цитокинов по сравнению с контролями. Эти данные указывают на то, что избыточная экспрессия c-Jun может увеличивать как частоту CAR Т-клеток, отвечающих на стимуляцию антигена, так и содержание цитокинов, продуцируемых отдельной CAR Т-клеткой при столкновении с антигеном.
Пример 5
Конструирование бицистронных векторов, коэкспрессирующих c-Jun и CAR из одного и того же вектора
Поскольку было показано, что c-Jun в первую очередь ответственен за повышенную активность трансдуцированных АР-1 CAR Т-клеток, создали бицистронные ретровирусные векторы, которые коэкспрессируют c-Jun и CAR из одного и того же вектора, разделенного последовательностью проскока рибосомы для вырезания вирусных пептидов 2А (см. фиг. 4а). Эти конструкты гарантировали, что любая CAR+Т-клетка также будет коэкспрессировать c-Jun, и устраняли необходимость в совместной трансдукции и сортировке для достижения чистых дважды положительных популяций. Восемь различных векторов CAR отдельно клонировали в основную цепь c-Jun: CD19-28Z, CD19-BBZ, CD22-28Z, CD22-BBZ, GD2-28Zshort, GD2-28ZLong, HA(GD2)-28ZLong и GD2-BBZ, для того, чтобы проверить функциональное влияние повторной экспрессии c-Jun на множественные антигенные специфичности и показания к применению для различных опухолей.
Экспрессия c-Jun в сочетании с большинством CAR приводила к снижению экспрессии поверхностных маркеров истощения (см. фиг. 4b и 5а). Повышенная секреция цитокинов наблюдалась в GD2 CAR Т-клетках, коэкспрессирующих c-Jun, по сравнению с контрольной CAR Т-клеткой, экспрессирующей GFP (см. фиг. 4d-e, 5с и 6а-b).
c-Jun опосредует повышенную секрецию цитокинов в ответ как на лейкоз (Nalm6-GD2), так и на GD2+ солидные опухоли у детей, саркому Юинга (EW8), остеосаркому (143В) и нейробластому (Келли), что подчеркивает широкую клиническую применимость усиленных c-Jun GD2 CAR Т-клеток. Кроме того, избыточная экспрессия c-Jun способствует увеличению частоты центральных CAR Т-клеток памяти (см. фиг. 5b). Используя внутриклеточное цитокиновое окрашивание (ICS), обнаружили, что повышенная продукция провоспалительных цитокинов также наблюдалась на уровне отдельных клеток (фиг.4 с и 6с-е) как в CD4+, так и в CD8+ c-Jun-CAR Т-клетках. Авторы настоящего изобретения также отметили снижение выработки противовоспалительного цитокина IL10 в c-Jun CAR Т-клетках по сравнению с контролями (см. фиг. 6f), что также указывает на то, что усиленная экспрессия c-Jun может вносить вклад в усиленный профиль цитокинов Thl.
Пример 6
Замена c-Jun и функциональная активность в различных CAR Т-клетках
Как подробно описано в настоящем документе, замена c-Jun в истощенных CAR Т-клетках (например, GD2 CAR Т-клетках) может ослаблять истощение в CAR Т-клетках. Тем не менее, тенденции к снижению маркеров истощения и повышенной функциональной активности также наблюдались в здоровых CD19 CAR Т-клетках. В то время как CD19 CAR Т-клетки опосредовали значительные клинические ответы у пациентов с В-ALL, растущее число рецидивов происходит у 30% пациентов с заболеванием с низкой экспрессией CD19 или отрицательным по CD19 заболеванием. CD22 CAR, альтернативная стратегия для нацеливания на В-клеточные злокачественные новообразования, также может быть ограничена низкой плотностью антигена CD22 в некоторых клетках лейкоза пациента. Таким образом, во время разработки вариантов осуществления настоящего изобретения проводили эксперименты для оценки и характеристики активности c-Jun-CD19 и c-Jun-CD22 CAR Т-клеток в отношении опухолевых клеток с нормальной (Nalm6) или низкой экспрессией антигена (Nalm6-F и Z для CD19 или Nalm6-221ow) (см. фиг. 7).
Хотя c-Jun не усиливал активность CD19 CAR против высоких уровней антигена на Nalm6, наблюдали значительные улучшения в IL2 (см. фиг. 7а) и IFNγ (см. фиг. 7b) в ответ на клоны Z и F Nalm6 с низкой экспрессией CD19. В анализе уничтожения иммунных клеток INCUCYTE (см. фиг. 7с) 3 клеточные линии GFP+Nalm6 совместно культивировали с CAR Т-клетками в течение 92 часов. Все 4 CD19 CAR уничтожали исходную опухоль Nalm6 (что измеряли по потере интенсивности GFP с течением времени). Как CD19-28Z, так и c-Jun-CD19-28Z были способны уничтожать опухолевые клетки с низким уровнем CD19, клоны F и Z, однако CD19-BBZ CAR Т-клетки оказывали только умеренный эффект. Добавление c-Jun-CD19-BBZ CAR Т-клеток показало значительное увеличение уничтожения клонов с низким уровнем CD19 по сравнению с 19-BBZ отдельно. Повышенная секреция цитокинов CD22 CAR Т-клетками (особенно CD22-BBZ), коэкспрессирующими c-Jun, также наблюдалась в ответ как на исходные Nalm6, так и на Nalm6-22low (см. фиг. 7d-е).
Ингибирование представителей ингибирующего АР-1 комплекса снижает истощение Т-клеток
Хотя усиление экспрессии c-Fos и c-Jun может увеличить функциональность модифицированных Т-клеток, существуют другие ингибирующих представители семейства АР-1, экспрессируемые в истощенных активированных Т-клетках. Соответственно, эксперименты проводили во время разработки вариантов осуществления настоящего изобретения, чтобы определить и охарактеризовать, может ли ингибирование/нокдаун представителей ингибирующего АР-1 комплекса (например, для увеличения доступности канонических факторов АР-1) снизить истощение Т-клеток (например, увеличить функцию Т-клеток). Системы гРНК CRISPR-Cas9 разработали для нацеливания на потенциальных ингибирующих АР-1 представителей. Продукцию цитокинов оценивали из CAR Т-клеток с отредактированным геном JUNB и BATF3 (нокаут, КО). Нокдаун JUNB значительно увеличивал продукцию IL2 и IFNγ из истощенных HA-GD2-28Z и GD2-BBZ CAR Т-клеток, в то время как он не воздействовал на CD19 CAR Т-клетки (см. фиг. 8А-С). Это указывало на то, что нокдаун JUNB положительно влиял на истощенные, но не здоровые Т-клетки. Аналогично, нокаут BATF3 также увеличивает продукцию IL2 (но не IFNγ) из истощенных HA-28Z CAR Т-клеток (см. фиг. 80) по сравнению с контрольными отредактированными Т-клетками.
Пример 8
In vivo эффективность c-Jun-модифицированных CAR Т-клеток
Эффективность in vivo c-Jun-модифицированных CAR Т-клеток оценивали на нескольких различных моделях опухолей. Экспрессирующие c-Jun HA-GD2 истощенные CAR Т-клетки проявляли превосходную лечебную активность in vivo по сравнению с модифицированными HA-GD2 CAR Т-клетками в модели лейкоза Nalm6, сконструированной для экспрессии GD2 (N6-GD2) (см. фиг. 9). c-Jun-модифицированные GD2-BBZ CAR Т-клетки проявляют превосходную активность in vivo на модели солидной опухоли агрессивной остеосаркомы (см. фиг. 10). Наконец, CD19 CAR Т-клетки с экспрессией c-Jun демонстрируют усиленную активность in vivo против клона F Nalm6 с низкой плотностью антигена (см. фиг. 11).
Экспрессия HA-28z CAR в Т-клетках человека быстро индуцирует характерные признаки истощения Т-клеток
CAR GD2-28z представляет собой истощенный фенотип в Т-клетках человека после экспрессии CAR, включающего в себя GD2-специфический 14g2a scFv, TCR дзета и эндодомен CD28, в результате тонической передачи сигналов, опосредованной через антиген-независимую агрегацию (ссылка 9; в настоящий документ полностью включенная посредством ссылки). Эксперименты, проведенные во время разработки вариантов осуществления в настоящем документе, демонстрируют, что экспрессия CAR, содержащего 14g2a scFv, несущего точечную мутацию E101K, которая обеспечивает взаимодействие с более высокой аффинностью (НА) с GD219 (HA-28z CAR), аналогичным образом индуцирует истощение в Т-клетках человека, хотя и с более тяжелым фенотипом (фиг. 12 и 18а-с). В отличие от CD19-28z CAR Т-клеток, HA-28z CAR Т-клетки обладали значительными фенотипическими и функциональными признаками истощения, включая в себя снижение экспансии в культуре (фиг. 12а), повышенную поверхностную экспрессию ингибирующих рецепторов PD-1, TIM-3, LAG-3 и CD39 (фиг. 12b и 18d), избыточную дифференцировку эффекторов и плохое формирование памяти (фиг. 12с и 18е), а также снижение IFN-g и значительное снижение продукции IL-2 при стимуляции с помощью клеток лейкоза CD19+GD2+Nalm6 (фиг. 12d).
Чтобы лучше объяснить молекулярные основы истощения Т-клеток в этой системе, транскриптом HA-28z сравнивали с CD19-28z-CAR Т-клетками. Очищенные наивные (N) и центральные Т-клетки памяти (СМ) трансдуцировали с помощью НА или CD19-28z CAR, а затем выделяли РНК на 7, 10 и 14 дни культивирования. Сортировка предварительно выбранных подгрупп позволила оценить влияние состояния дифференцировки Т-клеток и различия между истощением CD4 и CD8 в развитии истощения Т-клеток в этой модели. Принципиальный компонентный анализ (РСА) во всех 24 образцах показал, что самым сильным фактором дисперсии было наличие НА по сравнению с CD19-28z CAR (PC1, 39,3% дисперсии, фиг. 12е), что согласуется с моделью, в которой тоническая передача сигналов в HA-28z CAR Т-клетках вызывают истощение во всех изученных подгруппах Т-клеток. Различия, однако, наблюдали на основании исходного состояния дифференцировки, поскольку N по сравнению с СМ было отражено в РС2 (дисперсия 22,88%) (фиг. 18f) и между популяциями CD4 и CD8, которые приводили к РС3 (дисперсия 11,9%) (фиг. 12е и 18f).
Среди 200 наиболее результативных генов, управляющих РС1 (наиболее дифференцированно экспрессируемых в НА- по сравнению с CD19-28z CAR Т-клетках во всех подгруппах) (фиг. 12f), были гены, связанные с активацией (IFNG, GZMB, IL2RA), ингибирующими рецепторами (LAG3, CTLA4) и некоторыми воспалительными хемокинами/цитокинами (CXCL8, IL13, ILIA), в то время как гены, отрицательно регулируемые в HA-28z CAR Т-клетках, включают в себя гены, связанные с наивными Т-клетками и Т-клетками памяти (IL7R, TCF7, LEF1 и KLF2). Используя GSEA, продемонстрировали, что гены, положительно регулируемые в 10-дневных HA-28z CAR Т-клетках по сравнению с CD19-28z CAR Т-клетками, перекрываются со связанными с истощением наборами генов, ранее описанными на моделях хронического LCMV с использованием мышей13 (фиг. 18g). Хотя степень истощения в GD2-28z CAR Т-клетках менее выражена, анализ дифференциальной экспрессии генов на уровне одиночных клеток GD2-28z по сравнению с CD19-28z CAR Т-клетками выявил сходный профиль экспрессии генов (фиг. 19). Вместе эти данные подтверждают основание рекомендовать экспрессирующие HA-28z и GD2-28z Т-клетки в качестве моделей для исследования истощения Т-клеток человека.
Истощение Т-клеток связано с изменениями доступности хроматина на мышиной модели и у пациентов-людей с хроническими вирусными инфекциями и раком (ссылки 12, 17; полностью включенные в настоящий документ посредством ссылки). Анализ доступности хроматина с использованием ATAC-seq20 (фиг. 20) CD4+ и CD8+HA-28z, полученных из N или CM CD4+ и CD8+HA-28z по сравнению с CD19-28z CAR Т-клетками продемонстрировал значительные изменения в эпигенетической сигнатуре на 10-й день культивирования (фиг. 12g), при этом CD8+HA-28z CAR Т-клетки проявляли >20000 уникальных доступных областей хроматина (пики), по сравнению с <3000 уникальных пиков в CD8+CD19-28z CAR Т-клетках (FDR<0,1 и log2FC>1). Эти закономерности изменений доступности хроматина, вызванных истощением, были сходными в CD4+ Т-клетках (фиг. 21а). Подобно транскриптомному анализу, РСА выявил НА- по сравнению с CD19-CAR как наиболее сильный стимулирующий фактор дифференциальных состояний хроматина (дисперсия РС1 79,6%, фиг. 12h), с более слабыми, но существенными различиями, наблюдаемыми между клетками N и СМ (дисперсия РС2 7,4%), а также между подгруппами CD4 и CD8 (РС3 дисперсия 6,5%) (фиг. 21b). Кластеризация наиболее результативных 5000 дифференциально доступных областей (пиков) выявила глобально сходную доступность хроматина в HA-28z CAR Т-клетках независимо от исходной подгруппы (фиг. 12i). HA-28z CAR Т-клетки продемонстрировали повышенную доступность хроматина в регуляторных сайтах вблизи генов, связанных с истощением, таких как CTLA4, и пониженную доступность в регуляторных сайтах вблизи генов, связанных с памятью, таких как IL7R (фиг. 12j).
Эпигенетический и транскрипционный анализы показывают сильную сигнатуру АР-1 в истощенных CAR Т-клетках
Чтобы идентифицировать транскрипционные программы, которые, по прогнозам, не регулируются эпигенетическими изменениями, вызванными в истощенных Т-клетках, сравнивали отклонение мотива транскрипционного фактора (TF) между открытым хроматином истощенных и здоровых CAR Т-клеток. Используя анализ ChromVAR (ссылка 21; полностью включена в настоящий документ посредством ссылки), идентифицировали 25 наиболее отличительных мотивов во всех 8 образцах, и обнаружили, что многие из них принадлежат к семейству АР-1 (bZIP) (фиг. 13а). Аналогичным образом, анализ обогащения мотивов TF выявил мотивы связывания АР-1/bZIP и bZIP/IRF как одни из наиболее обогащенных в истощенных CAR Т-клетках (фиг. 13b и фиг. 21с).
Кластеризация дифференциально доступных пиков за счет совместного обогащения мотива TF позволила выявить 4 кластера, связанных с истощенными HA-28z CAR Т-клетками (фиг. 21d, ЕХ1-ЕХ4). Связанные с истощением кластеры содержали пики в непосредственной близости от генов, таких как BTLA, CD39, IFNG и CTLA4, что указывает на общую регуляцию TF генов, связанных с истощением. Все 4 кластера, связанных с истощением, показали сильное обогащение в отношении TF семейства АР-1 и связанного с АР-1 семейства, что указывает на широко распространенную модуляцию TF АР-1 регуляции генов, связанных с истощением. Сильное обогащение для мотивов TF семейств NFkB, NFAT и RUNX также наблюдалось в некоторых кластерах истощения, что указывает на то, что подгруппа генов, связанных с истощением, может регулироваться этими транскрипционными программами и воспроизводить эпигенетические сигнатуры истощения, наблюдаемые в других моделях (ссылки 12, 17, 22; полностью включены в настоящее описание посредством ссылки). Кластеры, ассоциированные со здоровыми CD19-28z CAR Т-клетками (HLT1-2), показали профиль, сходный с кластером, в значительной мере связанным с исходной наивной подгруппой. Это наблюдение согласуется с идеей о том, что здоровые CAR Т-клетки поддерживают эпигенетическую сигнатуру, более похожую на полученные из наивных Т-клетки, подгруппу, связанную с повышенной устойчивостью и эффективностью адоптивной Т-клеточной терапии (ссылка 23; полностью включена в настоящее описание посредством ссылки), тогда как хроническая антигенная стимуляция приводит к широкому расхождению в эпигенетическом перепрограммировании.
Связанные с АР-1 TF координируются с образованием разнообразного набора гомо- и гетеродимеров посредством взаимодействий в общем домене bZIP и могут димеризоваться с факторами транскрипции IRF (ссылки 14, 24; полностью включены в настоящее описание посредством ссылки). Комплексы фактора АР-1 конкурируют за связывание с элементами ДНК, содержащими согласованные ядерные консенсусные мотивы TGA-G/C-TCA. Активирующие комплексы, такие как комплексы, содержащие классически описанный гетеродимер c-Fos и c-Jun АР-1, приводят в действие транскрипцию IL-2. И наоборот, другие представители семейства АР-1 и IRF могут непосредственно противодействовать активности c-Jun и/или управлять экспрессией иммунорегуляторных генов в Т-клетках (ссылки 14, 24-29; полностью включены в настоящее описание посредством ссылки). Чтобы оценить, были ли изменения в доступности связывающего АР-1 хроматина связаны с повышенной доступностью активирующих и ингибирующих TF bZIP и IRF, во время разработки вариантов осуществления настоящего изобретения провели эксперименты для сравнения уровней транскриптов представителей семейств bZIP и IRF с использованием РНК-секвенирования в истощенных HA-28z по сравнению со здоровыми CD19-28z CAR Т-клетками. Парный анализ РНК-секвенирования у 3 разных доноров выявил единую закономерность избыточной экспрессии представителей семейств bZIP и IRF, которая была наиболее значимой для JUNB, FOSL1, BATF, BATF3, ATF3, ATF4 и IRF4 (фиг. 13 с и фиг. 22а). Анализ вестерн-блоттинг подтвердил устойчивую избыточную экспрессию белка JunB, IRF4 и BATF3 в клетках НА- по сравнению с CD 19 CAR Т-клетками (фиг. 13d и фиг. 22b), при этом иммунорегуляторные TF BATF/IRF демонстрируют более высокие уровни экспрессии по сравнению с c-Jun. Биологическую значимость повышенных содержаний ингибирующих представителей семейства bZIP/IRF дополнительно продемонстрировали демонстрацией того, что вестерн-блоттинг иммунопреципитатов Jun (IP) показал, что несколько ингибирующих представителей семейства находятся в прямом комплексе с c-Jun и JunB в истощенных HA-28z CAR Т-клетках (фиг. 22в). Сравнение профилей TF с помощью анализа РНК-секвенирования в формате одиночной клетки CD8+ Т-клеток, экспрессирующих CD19-28z по сравнению с GD2-28z CAR, подтвердило, что представители семейства bZIP: JUN, JUNB, JUND и ATF4, были одними из наиболее дифференцированно экспрессированных и широко связанных в сетях истощенных GD2-28z CAR Т-клеток (фиг. 13е и фиг. 19).
Избыточная экспрессия (ОЕ) c-Jun снижает функциональное истощение в CAR Т-клетках
На основании данных о том, что истощенные CAR Т-клетки проявляют очень низкую продукцию IL-2 (фиг. 12d) и преимущественно избыточную экспрессию транскрипционных факторов bZIP и IRF, которые управляют иммунорегуляторными и ассоциированными с истощением программами, можно предположить, что дисфункция Т-клеток в истощенных клетках может быть обусловлена относительным дефицитом гетеродимеров c-Jun/c-Fos, необходимых для запуска транскрипции IL-2. HA-28z и CD19-28z CAR Т-клетки совместно трансдуцировали с помощью бицистронного лентивирусного вектора для избыточной экспрессии c-Jun и c-Fos. HA-28z CAR Т-клетки, сконструированные для избыточной экспрессии АР-1, продемонстрировали повышенную продукцию IL-2 при стимуляции антигеном (фиг. 23а-с). Однако при использовании отдельных векторов экспрессии усиленная функциональность наблюдалась только при ОЕ c-Jun в клетках HA-28z CAR Т-клетках, тогда как трансдукция с помощью отдельного вектора экспрессии c-Fos не давала стабильного улучшения функции (фиг. 23d-e).
Для дальнейшего изучения возможности c-Jun усиливать функцию истощенных CAR Т-клеток и обеспечивать конститутивную экспрессию c-Jun во всех Т-клетках, экспрессирующих CAR, создавали бицистронные векторы, которые коэкспрессируют трансгены c-Jun и CAR, разделенные вирусным пропускающим пептидом Р2А (JUN-CAR, фиг. 14а). Эти экспрессионные векторы увеличивали экспрессию c-Jun как в CD19, так и в HA-28z CAR Т-клетки (фиг. 14b), хотя c-Jun был преимущественно активирован (фосфорилирован) в JUN-HA CAR Т-клетках (HA-28z), что согласуется с N-концевым фосфорилированием c-Jun (JNP) белками JNK, активированными ниже по ходу транскрипции от тонического сигнального каскада TCR, распространяемого через HA-28z CAR30. После стимуляции GD2+ линиями опухолевых клеток JUN-HA-28z CAR Т-клетки продемонстрировали заметное увеличение продукции IL-2 и IFNg по сравнению с контрольными HA-28z CAR Т-клетками (фиг. 14d-e). Кратное увеличение продукции цитокинов в условиях ОЕ c-Jun было по существу больше для JUN-HA CAR по сравнению с JUN-CD19 CAR Т-клетками (фиг. 24а-b). Аналогично, JUN-HA CAR Т-клетки продемонстрировали повышенную частоту подгрупп SCM/CM в отличие от Е/ЕМ по сравнению с HA-28z CAR Т-клетками (фиг. 14f), в то время как между CD19 и JUN-CD19 CAR Т-клетками не наблюдалось значительного различия в составе подгрупп на 10 день культивирования. В совокупности данные согласуются с моделью, в которой ОЕ c-Jun является функционально более значимой в истощенных Т-клетках, которые избыточно экспрессируют ингибирующие TF bZIP и IRF.
Чтобы оценить, увеличивает ли ОЕ c-Jun долгосрочную пролиферативную способность; которая связана с противоопухолевыми эффектами при солидных опухолях (ссылка 31; полностью включена в настоящее описание посредством ссылки), и проверить, может ли ОЕ c-Jun усиливать функцию в CAR Т-клетках без тонической передачи сигналов (CD19-28z, CD19-BBz) или в клетках с более низкими уровнями тонической передачи сигналов (GD2-BBz), экспансию in vitro JUN-CAR Т-клеток измеряли у 3 разных здоровых доноров в течение длительного периода (фиг. 24с). В присутствии ОЕ c-Jun наблюдалась закономерность усиления длительной пролиферативного потенциала, которая оставалась зависимой от IL-2, так как эти клетки немедленно прекращали экспансию при отсутствии IL-2 (фиг. 14g). В соответствии со способностью c-Jun вызывать устойчивость к истощению, CD8+ JUN- CD19-28z CAR Т-клетки с поздней экспансией демонстрировали пониженную экспрессию маркеров истощения и поддерживали устойчивую подгруппу клеток, обладающих фенотипом стволовых клеток памяти (SCM) (CD45RA+CD62L+) по сравнению с контрольными CD19-28z CAR Т-клетками, протестированными в тот же момент времени (фиг. 14h-j). Оценивали гомеостатическую экспансию JUN-CAR Т-клеток, адоптивно перенесенных мышам NSG без опухолей. Количество Т-клеток периферической крови было увеличено у мышей, получивших лечение с помощью как JUN-CD19-28z, так и JUN-CD19-BBz CAR Т-клеток, по сравнению с контрольной группой через 25 дней после инфузии (фиг. 14k), что привело к ускоренному GVHD у мышей, получивших лечение с помощью JUN-CD19-BBz CAR Т-клеток. Совокупно данные показывают, что ОЕ c-Jun уменьшает истощение Т-клеток в многочисленных протестированных CAR, включая в себя те, которые включают в себя ко стимулирующие домены CD28 или 4-1ВВ, и независимо от того, вызвано ли истощение принудительной долгосрочной экспансией или тонической передачей сигналов.
Пример 12
Молекулярные требования для c-Jun-опосредованного восстановления истощения CAR Т-клеток
Эксперименты проводили во время разработки вариантов осуществления в настоящем документе, чтобы определить, может ли ОЕ c-Jun восстанавливать истощенные Т-клетки двумя различными механизмами, которые не являются взаимоисключающими. ОЕ c-Jun может напрямую усиливать опосредованную АР-1 транскрипцию генов путем увеличения доступности димеров Fos/Jun или Jun/Jun или может работать опосредовано, нарушая ингибирующие транскрипционные комплексы AP-1/IRF (AP-1-i)14,24, которые управляют экспрессией генов, связанных с истощением (фиг. 15а). Чтобы лучше понять механизмы, с помощью которых ОЕ c-Jun смягчает истощение Т-клеток, домен дестабилизации (DD), полученный из дигидрофолатредуктазы (DHFR) Е. coli, был слит с N-концом c-Jun, чтобы временно регулировать экспрессию c-Jun (JUN-DD, фиг. 15b). DD стабилизируется в присутствии проницаемого для клеток низкомолекулярного триметоприма (ТМР) и приводит к стабильной экспрессии c-Jun, но при отсутствии ТМР DD дестабилизируется, вызывая протеасомную деградацию всего слитого белка (фиг. 15с). JUN-DD CAR Т-клетки быстро увеличивали экспрессию c-Jun в присутствии ТМР (1/2max через 6,76 часа после воздействия лекарственного средства), тогда как JUN-DD быстро становился не обнаруживаемым при отсутствии ТМР (t1/2 из 1,84 часа) (фиг. 15d и фиг. 25а-b). JUN-DD-CAR Т-клетки опосредовали увеличение продукции IL-2 и IFNg только в присутствии ТМР, подтверждая критически важную роль содержаний c-Jun в модулировании функциональности CAR Т-клеток (фиг. 15е). Было высказано мнение, что прямое влияние c-Jun на транскрипцию можно было бы осуществить быстро, и поэтому было проверили, будет ли функциональное восстановление происходить, когда ОЕ c-Jun в HA-28z CAR Т-клетках ограничена периодом острой антигенной стимуляции (OFF-ON). Напротив, если ОЕ c-Jun была необходима для конкуренции с ингибирующими комплексами АР-1 (АР-1i) во время индукции истощения, может потребоваться более длительное воздействие во время экспансии Т-клеток (ON-OFF) (фиг. 15f). По сравнению с HA-28z CAR Т-клетками, которые никогда не испытывали ОЕ c-Jun (OFF-OFF), оба условия опосредовали частичное восстановление, однако полное восстановление функции IL-2 требовало ОЕ c-Jun как во время экспансии Т-клеток, так и при стимуляции антигеном (ON-ON) (фиг. 15g). Это открытие согласуется с моделью, в которой ОЕ c-Jun напрямую усиливает транскрипцию генов во время острой стимуляции после столкновения с антигеном, а также опосредованно модулирует молекулярное перепрограммирование во время развития истощения. Кроме того, наблюдалось снижение экспрессии как белка, так и мРНК представителей семейства JUNB и BATF/BATF3 при избыточной экспрессии c-Jun (фиг. 25c-d), а также снижение комплексов JunB/BATF посредством IP (фиг. 25е).
Косвенная модель c-Jun-опосредованного разрушения ингибирующих комплексов АР-1 не зависит от прямой транскрипциональной активации c-Jun. Чтобы проверить, необходима ли прямая c-Jun-опосредованная активация гена для функционального восстановления истощения Т-клеток, создали JNP-дефицитный c-Jun с заменами аланина Ser63 и Ser73 в домене трансактивации c-Jun для предотвращения фосфорилирования в этих сайтах (c-JunAA), которые, как было показано, важны для опосредованной c-Jun транскрипции гена (ссылки 33, 34; полностью включены в настоящее описание посредством ссылки) (фиг. 15h-i). JUNAA-HA-28z CAR Т-клетки продемонстрировали эквивалентное увеличение продукции IL-2 и IFNg по сравнению с JUN-CAR Т-клетками дикого типа (фиг. 25j). Вместе эти данные согласуются с моделью, в которой c-Jun-опосредованное восстановление истощенных не требует прямой активации гена.
Пример 13
JUN-CAR Т-клетки опосредуют повышенную противоопухолевую активность in vivo
Эксперименты проводили во время разработки вариантов осуществления в настоящем документе, чтобы определить, будут ли JUN-CAR Т-клетки демонстрировать усиленную активность in vivo. Клетки лейкоза Nalm6-GD2+ прививали мышам и обрабатывали контрольными HA-28z или JUN-HA-28z CAR Т-клетками на 3-й день. В то время как HA-28z CAR Т-клетки проявляли некоторую противоопухолевую активность, лечение в конечном итоге оказалось неудачным, поскольку все мыши скончался от болезни (медиана выживаемости d59). Напротив, JUN-HA-28z CAR Т-клетки опосредовали полную регрессию опухоли к 24 дню и обеспечили долгосрочную выживаемость без опухолей (фиг. 16а-с). Чтобы определить, может ли ОЕ c-Jun усиливать функциональность CAR, нацеленных на солидные опухоли, эффект JUN-CAR оценивали с использованием нацеленных на Нег2 и GD2 CAR, включающих в себя костимулирующий домен 4-1 ВВ, который стал предпочтительным сигнальным доменом для придания долгосрочной персистенции (ссылки 35-37; полностью включены в настоящий документ посредством ссылки). В длительном анализе уничтожения ех vivo остеосаркомы 143b JUN-Her2-BBz CAR Т-клетки продемонстрировали значительно более сильную активность уничтожения при соотношении эффектор : мишень, составляющем 1:8 (Е : Т), что согласуется с повышенной активностью в расчете на клетку (фиг. 16d-e). Аналогично, JUN-Her2-BBz CAR Т-клетки предотвращали рост опухоли in vivo и приводили к значительному улучшению долгосрочной выживаемости, что было связано с увеличением экспансии Т-клеток in vivo (фиг. 16f-h). Аналогичные результаты наблюдались при сравнении GD2-BBz и JUN-GD2-BBz CAR Т-клеток против остеосаркомы 143b (фиг. 26), подтверждая преимущество ОЕ c-Jun в CAR Т-клетках, отвечающих на солидные опухоли, и в CAR Т-клетках, включающих в себя сигнальные домены 4-1 ВВ.
Пример 14
Избыточная экспрессия c-Jun снижает порог активации CAR Т-клеток и позволяет распознавать опухолевые клетки с более низкой плотностью антигена
Повышенные содержания ингибирующих представителей семейства API в истощенных CAR Т-клетках повысили вероятность того, что дисфункция в истощенных CAR Т-клетках связана, по меньшей мере частично, с более высоким порогом активации, который можно нормализовать путем восстановления баланса активации по сравнению с ингибирующими представителями семейства API. Чтобы проверить это, сравнивали продукцию цитокинов HA-CAR по сравнению с JUN-HA-CAR Т-клетками в ответ на серийные разведения связанного с планшетом 1А7, антиидиотипического антитела, которое связывает scFv 14g2a, что позволяет контролировать силу стимула. ОЕ c-Jun существенно увеличивала максимальную продукцию IL-2 и IFNg, а также существенно снижала количество 1А7, необходимое для индукции секреции IL-2, что согласуется со сниженным порогом активации в JUN-HA-CAR Т-клетках (фиг. 17а-b).
Ограничение уровней экспрессии антигена-мишени на опухолевых клетках все чаще признается фактором, ограничивающим функциональность CAR (ссылки 5, 6, 38; включены в настоящий документ посредством ссылки). Недавно сообщалось о рецидивах CD22dim у пациентов с лейкозом после первоначальных ответов на терапию CD22 CAR. Поскольку ОЕ c-Jun снижает порог активации в тонически сигнализирующих HA-28z CAR Т-клетках, оценивали, будут ли JUN-CAR распознавать и уничтожать опухолевые клетки с более низкой плотностью антигена, которые могут избежать распознавания контрольными CAR Т-клетками. Когда JUN-CD22-BBz CAR (фиг. 17) Т-клетки подвергали сенсибилизации клетками лейкоза CD22low (фиг. 17d), JUN-CAR Т-клетки демонстрировали повышенную продукцию цитокинов in vitro (фиг. 17е) и резко увеличивали противоопухолевую активность in vivo. (фиг. 17f-i). Контрольные CD22-BBz CAR Т-клетки демонстрировали начальную активность при введении в дозе, составляющей 3×106 CAR Т-клеток, но это лечение в конечном итоге не позволило контролировать рост опухоли (средняя выживаемость d45). Напротив, JUN-CD22-BBz CAR Т-клетки опосредовали значительные противоопухолевые эффекты и являлись полностью излечивающими. Таким образом, ОЕ c-Jun демонстрирует значительно улучшенный противоопухолевый контроль на 4 моделях опухолей и связана с улучшенной экспансией, устойчивостью к истощению и улучшенной способностью распознавать мишени с низким содержанием антигена.
Пример 15
Усеченные белки JUN
Эксперименты проводили во время разработки вариантов осуществления в настоящем документе для определения необходимых доменов JUN, которые отвечают за опосредование восстановления дисфункциональных Т-клеток. Создали серию мутантов JUN с удаленными различными доменами (фиг. 27а). Мутации JUN с N-концевыми делециями и усечениями (JUN-AA, JUN-Dd и JUN-DTAD) сохраняют способность восстанавливать функцию истощенных HA-28z CAR Т-клеток, тогда как С-концевые мутации в доменах эпсилон и bZIP не восстанавливают функциональность. Конструкты JUN-AA, JUN-Dd и JUN-DTAD сохраняют эквивалентное увеличение продукции цитокинов по сравнению с JUN-WT (фиг. 27b, d) и улучшают долговременное уничтожение при низких отношениях Е:Т (фиг. 27с) по сравнению с мутациями JUN-De, JUN-Dbasic, JUN-DLeu и JUN-DbZIP. В соответствии с улучшенной функциональной активностью in vitro HA-28z CAR Т-клетки, экспрессирующие JUN-WT и JUN-DTAD, демонстрируют повышенную пролиферацию in vivo по сравнению с JUN-DLeu и JUN-DbZIP при введении мышам с лейкозом N6-GD2.
Пример 16
Нокдаун IRF4 резко увеличивает функциональную активность истощенных HA-28z CAR Т-клеток
Проводили эксперименты, которые оптимизировали гРНК CRISPR для 9 различных компонентов семейства bZIP/IRF и протестировали способность нокдауна восстанавливать функциональную активность (секрецию цитокинов) истощенных HA-28z CAR Т-клеток. Сводные данные, показывающие результаты 3-6 независимых экспериментов со здоровыми донорами, представлены на фиг. 28. Как показано на фиг. 28, в то время как нокаут JunB и BATF3 улучшал функциональную активность у некоторых доноров, нокаут IRF4 значительно улучшал секрецию IL-2 в стимулированных HA-28z CAR Т-клетках (слева и посередине) у всех протестированных доноров. IRF4-KO CAR Т-клетки даже показали улучшенную исходную секрецию IL-2 при тонической передаче сигналов.
Пример 17
Транскрипционный мутант (JUN-AA) также восстанавливает функциональную активность и пролиферативную способность в CD19 CAR Т-клетках
Проводили эксперименты, предусматривающие создание мутаций JUN-AA в CD19 CAR Т-клетках. JUN-AA сохранил повышенную реактивность против антигена с низкой плотностью (фиг. 29А) и увеличил длительную пролиферацию в культуре (фиг. 29В). In vivo c-Jun продемонстрировал улучшение выживаемости мышей с лейкозом Nalm6, получавших низкую дозу «стресс-теста» дозы CD19 CAR Т-клеток (фиг. 29С).
Пример 18
Улучшенная функция in vivo c-Jun-модифицированных HA-28z CAR Т-клеток не может быть воспроизведена предоставлением IL-2 ех vivo
Хотя продукция IL-2 является отличным биомаркером для устойчивой к истощению клетки, усиленная функция JUN-CAR Т-клеток не может быть воспроизведена с помощью IL-2 отдельно (фиг. 30). В день 7, 9, 11, 13 и 15 после приживления опухоли IP получали 250000 МЕ/мышь. 1×106 CAR+ Т-клеток вводили внутривенно на 7-й день. Репрезентативный результат от 3 независимых экспериментов с аналогичными результатами показан на фиг. 30
Пример 19
c-Jun усиливает активность Her2-BBz CAR Т-клеток в супрессивном микроокружении солидной опухоли
JUN-CAR Т-клетки демонстрируют пониженное истощение и повышенную функциональную активность ех vivo. Ксенотрансплантаты остеосаркомы 143В имплантировали посредством внутримышечной инъекции мышам NSG. После измерения массы солидной опухоли (на 14-й день после инокуляции опухоли) внутривенно вводили 1×107 контрольных клеток Her2-BBz (синий) CAR Т-клеток или JUN-Her2-BBz (красный) CAR Т-клеток. На фиг. 31А показано, что JUN-Her2-BBz CAR Т-клетки опосредовали регрессию крупных, развившихся солидных опухолей 143В, тогда как Her2-BBz CAR Т-клетки не продемонстрировали никакого изменения по сравнению с контрольными нетрансдуцированными Т-клетками - имитацией. Через две недели после инъекции Т-клеток (до полного уничтожения опухолей у мышей JUN) мышей умерщвляли и ткань солидной опухоли извлекали и механически диссоциировали (n = 6-8 мышей на группу). На фиг. 31В показано, что гидролизаты солидных опухолей от мышей, обработанных JUN, содержали значительно более высокий процент CD8+ Т-клеток (частота встречаемости всех жизнеспособных клеток) и сохраняли более высокую частоту встречаемости CAR+ CD8+ Т-клеток. На фиг. 31С показано, что локализованные в опухоли CD8+Т-клетки демонстрировали пониженную экспрессию (и коэкспрессию) связанных с истощением ингибирующих рецепторов PD-1 и CD39 в JUN-Her2-BBz (n = 6 мышей на группу, слева) (репрезентативные диаграммы проточной цитометрии справа).
Затем провели эксперименты, в которых оценивали функциональную способность локализованных в опухоли CAR Т-клеток путем повторной стимуляции ex vivo клетками-мишенями Nalm6-Her2+ (фиг. 31D и Е). Как показано на фиг. 31D, 5×104 отсортированные с помощью FACS CD45+ Т-клеток повторно стимулировали с помощью 5×104 клеток-мишеней, и секрецию IL-2 измеряли после 24 часов с помощью ELISA. Как показано на фиг. 31Е, 3×105 гидролизаты одиночных клеток опухоли повторно стимулировали 3×105 выделенными Nalm6-Her2+ Т-клетками в течение 6 часов в присутствии монензина и антитела к CD107a. После 6 часов продукцию одиночными клетками CD107a, IL-2, IFNγ и TNFα оценивали путем внутриклеточного цитокинового окрашивания. Стимулированные ех vivo JUN-Her2-BBz CAR Т-клетки демонстрировали значительно большую частоту клеток, продуцирующих CD107a, IL-2, IFNγ, TNFα, а также клеток, продуцирующих двойные цитокины, по сравнению с контрольными Her2-BBz.
Пример 20
Избыточная экспрессия с-Jun повышает устойчивость к TGFβ-опосредованной супрессии истощенных HA-28z CAR Т-клеток
Контрольные и JUN-WT или JUN-AA модифицированные HA-28z CAR Т-клетки стимулировали клетками-мишенями Nalm6-GD2 в присутствии или при отсутствии 5 нМ TGFβ. Как показано на фиг. 32 слева, секрецию IL-2 измеряли с помощью ELISA, показан один репрезентативный донор. Как показано на фиг. 32 справа, кратное снижение секреции IL-2 в TGFβ+ условиях по сравнению с условиями без TGFβ (n = 3 независимых донора из 3 независимых экспериментов).
Пример 21
Транскрипционные изменения в клетках, модифицированных c-Jun, согласуются с уменьшением истощения и усилением формирования памяти
На фиг. 33А показана тепловая карта генов, дифференциально регулируемых в истощенных HA-28z CAR Т-клетках при добавлении избыточной экспрессии c-Jun (193 гена отрицательно регулируются c-Jun, 176 генов положительно регулируются c-Jun р(скорректированное) < 0,05. n = 3 независимых донора). Представляющие интерес гены выделены справа. Гены, отрицательно регулируемые в Jun-клетках, включают в себя ингибирующие рецепторы (LAG3, ENTPD1) и другие потенциально ингибирующие представители семейства АР-1 (BATF3, JUNB), а также другие гены, связанные с истощением (IL13, GZMB, LTA, TNFRSF9). Гены, положительно регулируемые c-Jun, связаны с наивными и Т-клетками памяти (IL7R, SELL (CD62L), CD44 и транскрипционные факторы LEF1 и foxp1).
На фиг. 33В показаны диаграммы Венна, показывающие перекрытие генов, отрицательно регулируемых в c-Jun-HA-28z CAR Т-клетках, и генов, стимулирующих истощение в РС1, тогда как на фиг. 33С показано, что гены, положительно регулируемые в c-Jun, показывают перекрытие с генами, связанными со здоровыми (CD 19) CAR Т-клетками (ниже). Приблизительно 25% наиболее результативных генов, определяющих различия между здоровыми и истощенными Т-клетками, можно модулировать с помощью избыточной экспрессии c-Jun, что позволяет предположить, что семейство АР-1 может составлять ~ 25% истощенного фенотипа в этой модели.
На фиг. 33D показана тепловая карта, показывающая экспрессию генов 50 генов в областях перекрытия от b-c во всех 12 образцах (n = 3 донора на условие).
В таблице 2 представлен полный перечень генов, измененных с помощью c-Jun
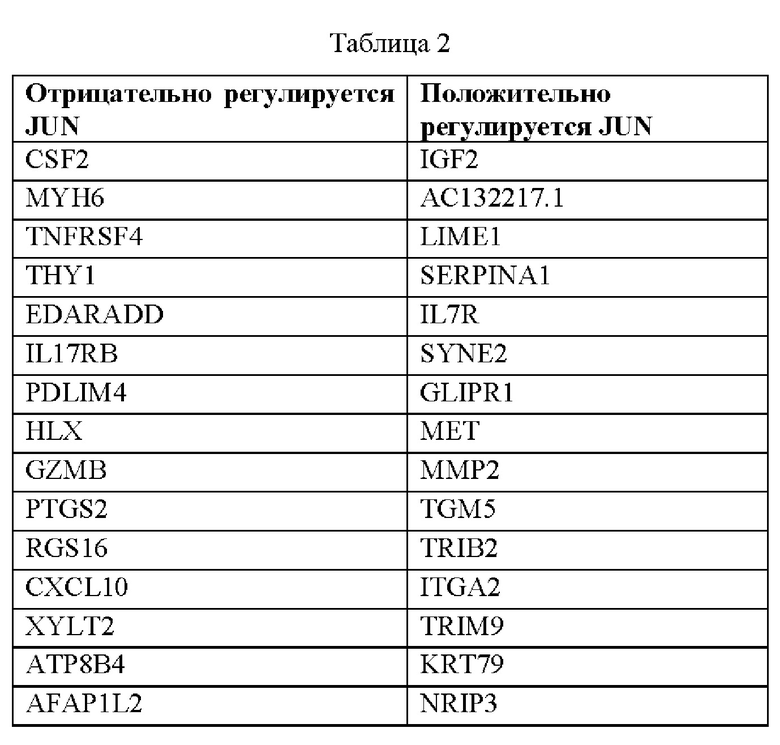
Пример 22
Ингибирование IRF4 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF4, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF4, по сравнению с контрольными Т-клетками.
Пример 23
Ингибирование IRF8 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF8, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF8, по сравнению с контрольными Т-клетками.
Пример 24
Ингибирование BATF снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов BATF, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами BATF, по сравнению с контрольными Т-клетками.
Пример 25
Ингибирование BATF3 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов BATF3, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами BATF3, по сравнению с контрольными Т-клетками.
Пример 26
Ингибирование JUNB снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов JUNB, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами JUNB, по сравнению с контрольными Т-клетками.
Пример 27
Ингибирование IRF1 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF1, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF1, по сравнению с контрольными Т-клетками.
Пример 28
Ингибирование IRF2 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF2, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF2, по сравнению с контрольными Т-клетками.
Пример 29
Ингибирование IRF3 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF3, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF3, по сравнению с контрольными Т-клетками.
Пример 30
Ингибирование IRF5 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF5, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF5, по сравнению с контрольными Т-клетками.
Пример 31
Ингибирование IRF6 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF6, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF6, по сравнению с контрольными Т-клетками.
Пример 32
Ингибирование IRF7 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF7, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF7, по сравнению с контрольными Т-клетками.
Пример 33
Ингибирование IRF9 снижает истощение Т-клеток
Контрольные и истощенные Т-клетки приводят в контакт с эффективными количествами (0,1-1000 мкМ) ингибиторов IRF9, показанных в таблице 1. Образцы собирают и обрабатывают для анализов продукции цитокина и интерферона. Увеличение продукции цитокина IL2 и/или интерферона IFNγ наблюдают в истощенных Т-клетках, обработанных ингибиторами IRF9, по сравнению с контрольными Т-клетками.
Все публикации и патенты, упомянутые в приведенном выше описании, включены в настоящее описание посредством ссылки. Различные модификации и вариации описанного способа и системы согласно настоящему изобретению будут очевидны для специалистов в настоящей области техники без отступления от объема и сущности настоящего изобретения. Хотя настоящее изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что заявляемое изобретение не должно чрезмерно ограничиваться такими конкретными вариантами осуществления. Действительно, различные модификации описанных способов осуществления настоящего изобретения, которые очевидны для специалистов в соответствующих областях, должны находиться в пределах объема следующей формулы изобретения.
Ссылки
Следующие ссылки, некоторые из которых процитированы выше с номером (например, ссылка X), полностью включены в настоящий документ посредством ссылки.
1. Maude, S. L. et al. Tisagenlecleucel in Children and Young Adults with B-Cell Lymphoblastic Leukemia. N Engl J Med 378, 439-448, doi: 10.1056/NEJMoal709866 (2018).
2. Neelapu, S. S. et al. Axicabtagene Ciloleucel CAR T-Cell Therapy in Refractory Large B- Cell Lymphoma. N Engl J Med 377, 2531-2544, doi: 10.1056/NEJMoa1707447 (2017).
3. Fraietta, J. A. et al. Determinants of response and resistance to CD 19 chimeric antigen receptor (CAR) T cell therapy of chronic lymphocytic leukemia. Nat Med 24, 563-571, doi:10.1038/s41591-018-0010-l (2018).
4. June, С.H., O'Connor, R. S., Kawalekar, O. U., Ghassemi, S. & Milone, M. C. CAR T cell immunotherapy for human cancer. Science 359, 1361-1365 (2018).
5. Walker, A. J. et al. Tumor Antigen and Receptor Densities Regulate Efficacy of a Chimeric Antigen Receptor Targeting Anaplastic Lymphoma Kinase. Mol Ther 25, 2189-2201, doi:10.1016/j.ymthe.2017.06.008(2017).
6. Fry, T. J. et al. CD22-targeted CAR T cells induce remission in B-ALL that is naive or resistant to CD 19-targeted CAR immunotherapy. Nat Med 24, 20-28, (2018).
7. Hegde, M., Moll, A. J., Byrd, Т. Т., Louis, C. U. & Ahmed, N. Cellular immunotherapy for pediatric solid tumors. Cytotherapy 17, 3-17, (2015).
8. Long, A.H. et al. Reduction of MDSCs with All-trans Retinoic Acid Improves CAR Therapy
Efficacy for Sarcomas. Cancer Immunol Res 4, 869-880, doi:10.1158/2326-6066.CIR-15-0230 (2016).
9. Long, A. H. et al. 4-IBB costimulation ameliorates T cell exhaustion induced by tonic signaling of chimeric antigen receptors. Nat Med 21, 581-590, doi:10.1038/nm.3838 (2015).
10. Eyquem, J. et al. Targeting a CAR to the TRAC locus with CRISPR/Cas9 enhances tumour rejection. Nature 543, 113-117, doi: 10.1038/nature21405 (2017).
11. Fraietta, J. A. et al. Disruption of TET2 promotes the therapeutic efficacy of CD 19-targeted T cells. Nature 558, 307-312, doi:10.1038/s41586-018-0178-z (2018).
12. Sen, D.R. et al. The epigenetic landscape of T cell exhaustion. Science 354, 1165-1169, doi:10.1126/science.aae0491 (2016).
13. Wherry, E.J. et al. Molecular signature of CD8+T cell exhaustion during chronic viral infection. Immunity 27, 670-684, doi:10.1016/j.immuni.2007.09.006 (2007).
14. Man, K. et al. Transcription Factor IRF4 Promotes CD8(+) T Cell Exhaustion and Limits the Development of Memory-like T Cells during Chronic Infection. Immunity 47, 1129-1141 ell25, doi:10.1016/j.immuni.2017.11.021 (2017).
15. Wherry, E. J. & Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol 15, 486-499, doi: 10.1038/nri3862 (2015).
16. Bengsch, B. et al. Epigenomic-Guided Mass Cytometry Profiling Reveals Disease-Specific Features of Exhausted CD8 T Cells. Immunity 48, 1029-1045 el 025, doi:10.1016/j.immuni.2018.04.026 (2018).
17. Pauken, К. E. et al. Epigenetic stability of exhausted T cells limits durability of reinvigoration by PD-1 blockade. Science 354, 1160-1165, doi:10.1126/science.aaf2807 (2016).
18. Heczey, A. et al. CAR T Cells Administered in Combination with Lymphodepletion and PD-1 Inhibition to Patients with Neuroblastoma. Mol Ther 25, 2214-2224, doi:10.1016/j.ymthe.2017.05.012(2017).
19. Horwacik, I. et al. Structural Basis of GD2 Ganglioside and Mimetic Peptide Recognition by 14G2a Antibody. Mol Cell Proteomics 14, 2577-2590, doi: 10.1074/mcp.Ml 15.052720 (2015).
20. Buenrostro, J. D., Giresi, P. G., Zaba, L. C, Chang, H. Y. & Greenleaf, W. J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open
chromatin, DNA- binding proteins and nucleosome position. Nat Methods 10, 1213-1218, doi:10.1038/nmeth.2688 (2013).
21. Schep, A. N., Wu, В., Buenrostro, J. D. & Greenleaf, W. J. chromVAR: inferring transcription-factor-associated accessibility from single-cell epigenomic data. Nat Methods 14, 975-978, doi: 10.1038/nmeth.4401 (2017).
22. Philip, M. et al. Chromatin states define tumour-specific T cell dysfunction and reprogramming. Nature 545, 452-456, doi: 10.1038/nature22367 (2017).
23. Singh, N., Perazzelli, J., Grupp, S. A. & Barrett, D. M. Early memory phenotypes drive T cell proliferation in patients with pediatric malignancies. Sci Transl Med 8, 320ra323, doi: 10.1126/scitranslmed. aad5222 (2016).
24. Murphy, T. L., Tussiwand, R. & Murphy, К. M. Specificity through cooperation: BATF-IRF interactions control immune-regulatory networks. Nat Rev Immunol 13, 499-509, doi:10.1038/nri3470 (2013).
25. Meixner, A., Karreth, F., Kenner, L. & Wagner, E. F. JunD regulates lymphocyte proliferation and T helper cell cytokine expression. EMBO J 23, 1325-1335, doi:10.1038/sj.emboj.7600133 (2004).
26. Chiu, R., Angel, P. & Karin, M. Jun-B differs in its biological properties from, and is a negative regulator of, c-Jun. Cell 59, 979-986 (1989).
27. Echlin, D. R., Tae, H. J., Mitin, N. & Taparowsky, E. J. B-ATF functions as a negative regulator of AP-1 mediated transcription and blocks cellular transformation by Ras and Fos. Oncogene 19, 1752-1763, doi: 10.1038/sj.one. 1203491 (2000).
28. Li, P. et al. BATF-JUN is critical for IRF4-mediated transcription in T cells. Nature 490, 543-546, doi:10.1038/naturell530 (2012).
29. Quigley, M. et al. Transcriptional analysis of HIV-specific CD8+T cells shows that PD-1 inhibits T cell function by upregulating BATF. Nat Med 16, 1147-1151, doi: 10.1038/nm.2232 (2010).
30. Derijard, B. et al. JNK1: a protein kinase stimulated by UV light and Ha-Ras that binds and phosphorylates the c-Jun activation domain. Cell 76, 1025-1037 (1994).
31. S, P. D. A. et al. Antitumor Activity Associated with Prolonged Persistence of Adoptively Transferred NY-ESO-l(c259)T Cells in Synovial Sarcoma. Cancer Discov, doi: 10.1158/2159-8290.CD-17-1417 (2018).
32. Iwamoto, M., Bjorklund, Т., Lundberg, C, Kirik, D. & Wandless, T. J. A general chemical method to regulate protein stability in the mammalian central nervous system. Chem Biol 17, 981-988, doi:10.1016/j.chembiol.2010.07.009 (2010).
33. Bannister, A. J., Oehler, Т., Wilhelm, D., Angel, P. & Kouzarides, T. Stimulation of c-Jun activity by СВР: c-Jun residues Ser63/73 are required for СВР induced stimulation in vivo and СВР binding in vitro. Oncogene 11, 2509-2514 (1995).
34. Weiss, C. et al. JNK phosphorylation relieves HDAC3-dependent suppression of the transcriptional activity of c-Jun. EMBO J 22, 3686-3695, doi: 10.1093/emboj/cdg364 (2003).
35. Milone, M. C. et al. Chimeric receptors containing CD137 signal transduction domains mediate enhanced survival of T cells and increased antileukemic efficacy in vivo. Mol Ther 17, 1453-1464, doi:10.1038/mt.2009.83 (2009).
36. Porter, D. L. et al. Chimeric antigen receptor T cells persist and induce sustained remissions in relapsed refractory chronic lymphocytic leukemia. Sci Transl Med 7, 303ra139, doi:10.1126/scitranslmed.aac5415 (2015).
37. Davila, M. L. et al. Efficacy and toxicity management of 19-28z CAR T cell therapy in В cell acute lymphoblastic leukemia. Sci Transl Med 6, 224ra225, doi: 10.1126/scitranslmed.3008226 (2014).
38. Majzner, R. G. & Mackall, C. L. Tumor Antigen Escape from CAR T-cell Therapy. Cancer Discov, doi:10.1158/2159-8290.CD-18-0442 (2018).
39. Martinez, G. J. et al. The transcription factor NFAT promotes exhaustion of activated
CD8(+) T cells. Immunity 42, 265-278, doi:10.1016/j.immuni.2015.01.006 (2015).
40. Roychoudhuri, R. et al. BACH2 regulates CD8(+) T cell differentiation by controlling access of AP-1 factors to enhancers. Nat Immunol 17, 851-860, doi:10.1038/ni.3441 (2016).
41. Bohmann, D. et al. Human proto-oncogene c-jun encodes a DNA binding protein with structural and functional properties of transcription factor AP-1. Science 238, 1386-1392 (1987).
42. Mariani, O. et al. JUN oncogene amplification and overexpression block adipocytic differentiation in highly aggressive sarcomas. Cancer Cell 11, 361-374, doi:10.1016/j.ccr.2007.02.007 (2007).
43. Shaulian, E. AP-l~The Jun proteins: Oncogenes or tumor suppressors in disguise? Cell Signal 22, 894-899, doi:10.1016/j.cellsig.2009.12.008 (2010).
44. Behrens, A., Jochum, W., Sibilia, M. & Wagner, E. F. Oncogenic transformation by ras and fos is mediated by c-Jun N-terminal phosphorylation. Oncogene 19, 2657-2663, doi: 10.1038/sj.onc. 1203603 (2000).
45. Di Stasi, A. et al. Inducible apoptosis as a safety switch for adoptive cell therapy. N Engl J Med 365, 1673-1683, doi: 10.1056/NEJMoal 106152 (2011).
46. Hudecek, M. et al. The nonsignaling extracellular spacer domain of chimeric antigen receptors is decisive for in vivo antitumor activity. Cancer Immunol Res 3, 125-135, doi: 10.1158/2326-6066.CIR-14-0127 (2015).
47. Jena, B. et al. Chimeric antigen receptor (CAR)-specific monoclonal antibody to detect CD19-specific T cells in clinical trials. PLoS One 8, e57838, doi:10.1371/journal.pone.0057838 (2013).
48. Corces, M. R. et al. An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. Nat Methods 14, 959-962, doi: 10.1038/nmeth.4396 (2017).
49. Wickham, H. Ggplot2: elegant graphics for data analysis. (Springer, 2009).
50. Mootha, V. K. et al. PGC-1 alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat Genet 34, 267-273, doi:10.1038/ng1180 (2003).
51. Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci USA 102, 15545-
15550, doi:10.1073/pnas.0506580102 (2005).
52. Zheng, G. X. et al. Massively parallel digital transcriptional profiling of single cells. NatCommun 8, 14049, doi: 10.1038/ncomms 14049 (2017).
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ К CD22 | 2012 |
|
RU2644243C2 |
| АНТИГЕННЫЙ РЕЦЕПТОР | 2020 |
|
RU2832189C2 |
| МОДИФИЦИРОВАННЫЕ МОНОЦИТЫ/МАКРОФАГИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2766690C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ М971 | 2013 |
|
RU2658485C2 |
| УЛУЧШЕННЫЕ ИММУННЫЕ КЛЕТКИ С ДВОЙНОЙ кшРНК И КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ИХ | 2019 |
|
RU2793922C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| МАТЕРИАЛЫ И СПОСОБЫ ЛЕЧЕНИЯ РАКА | 2019 |
|
RU2787828C2 |
| СПОСОБЫ EX VIVO ЭКСПАНСИИ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2814083C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ГУМАНИЗИРОВАННОГО АНТИ-CD19 ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА | 2014 |
|
RU2711975C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА К CD33 | 2015 |
|
RU2747384C2 |
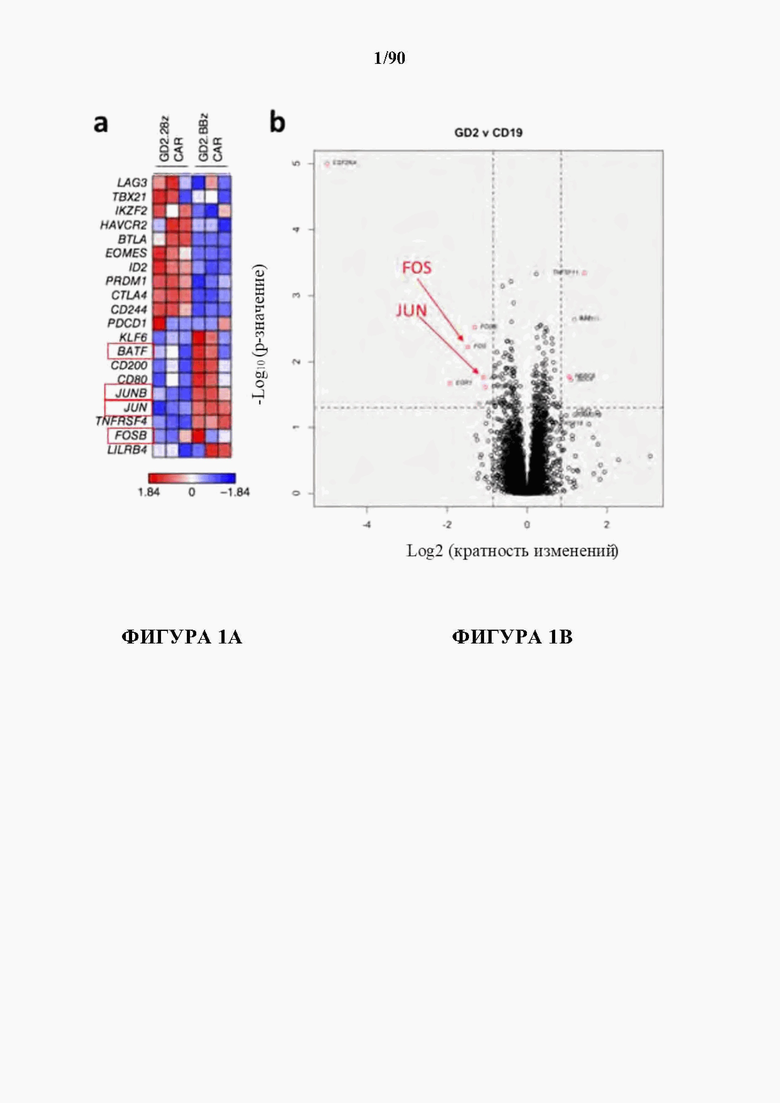
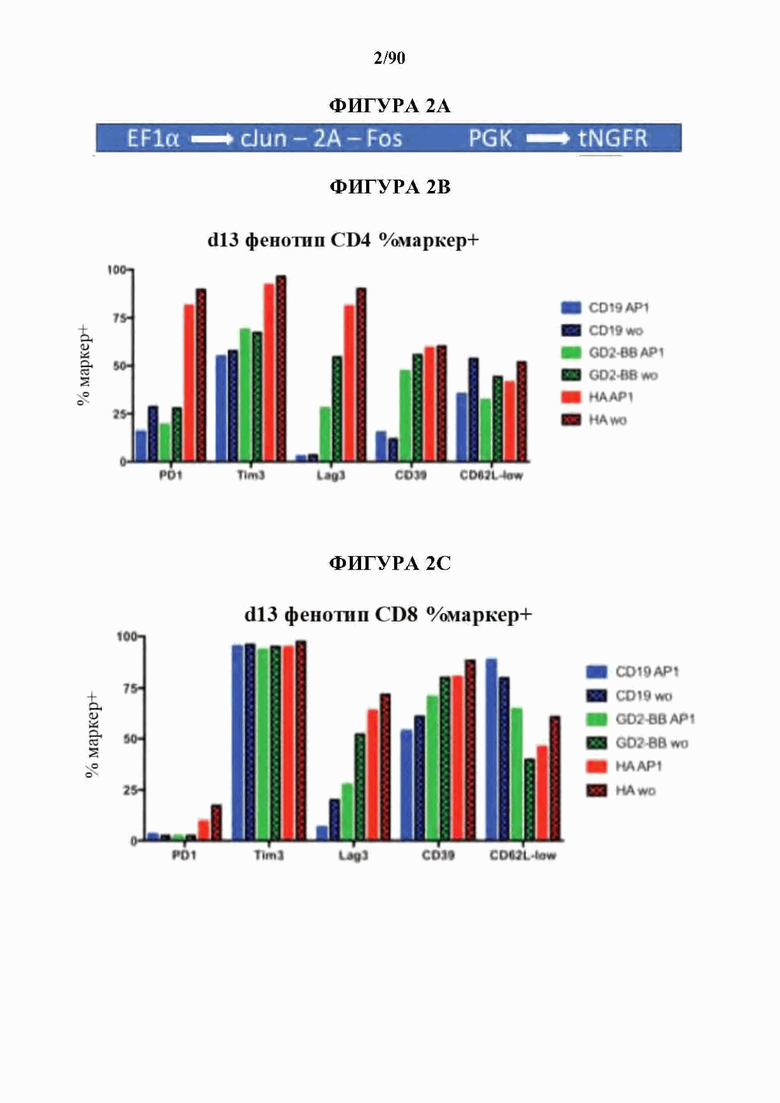
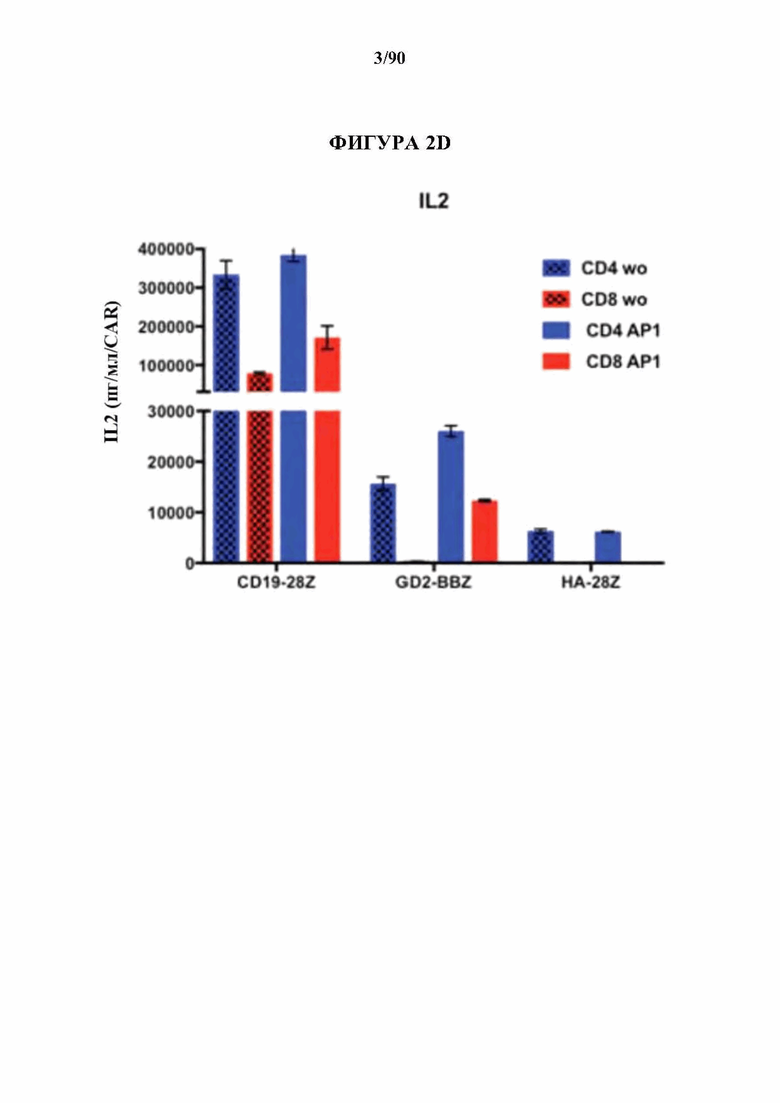
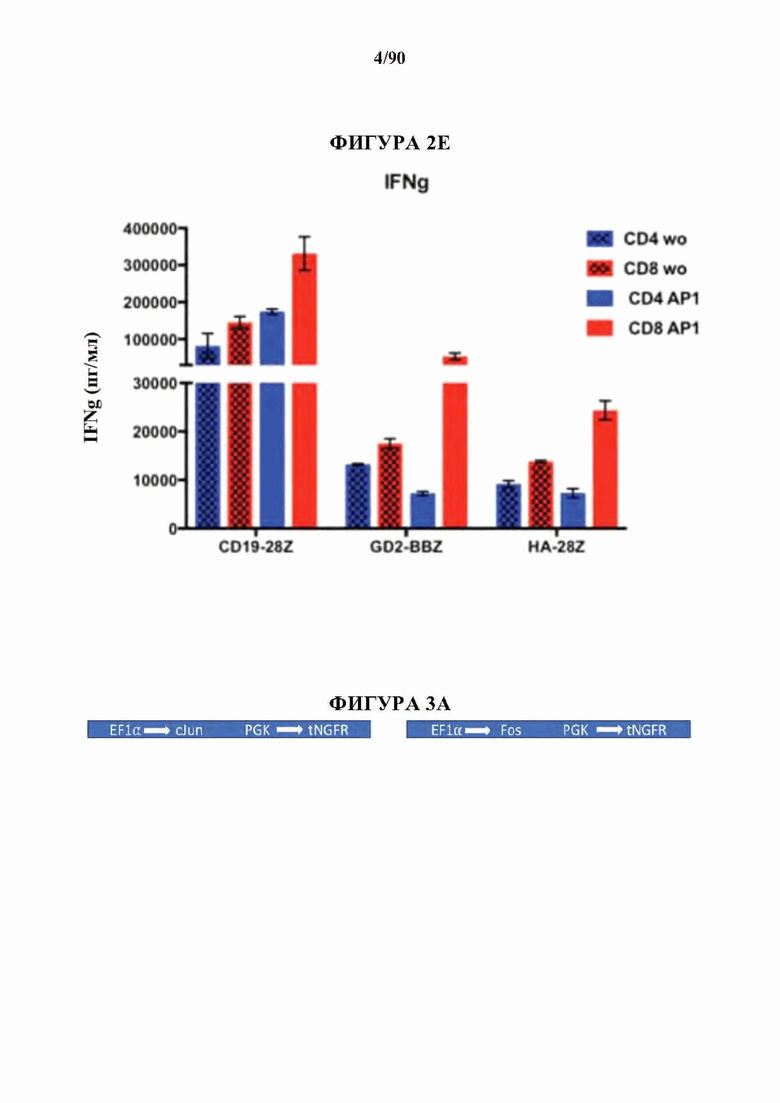
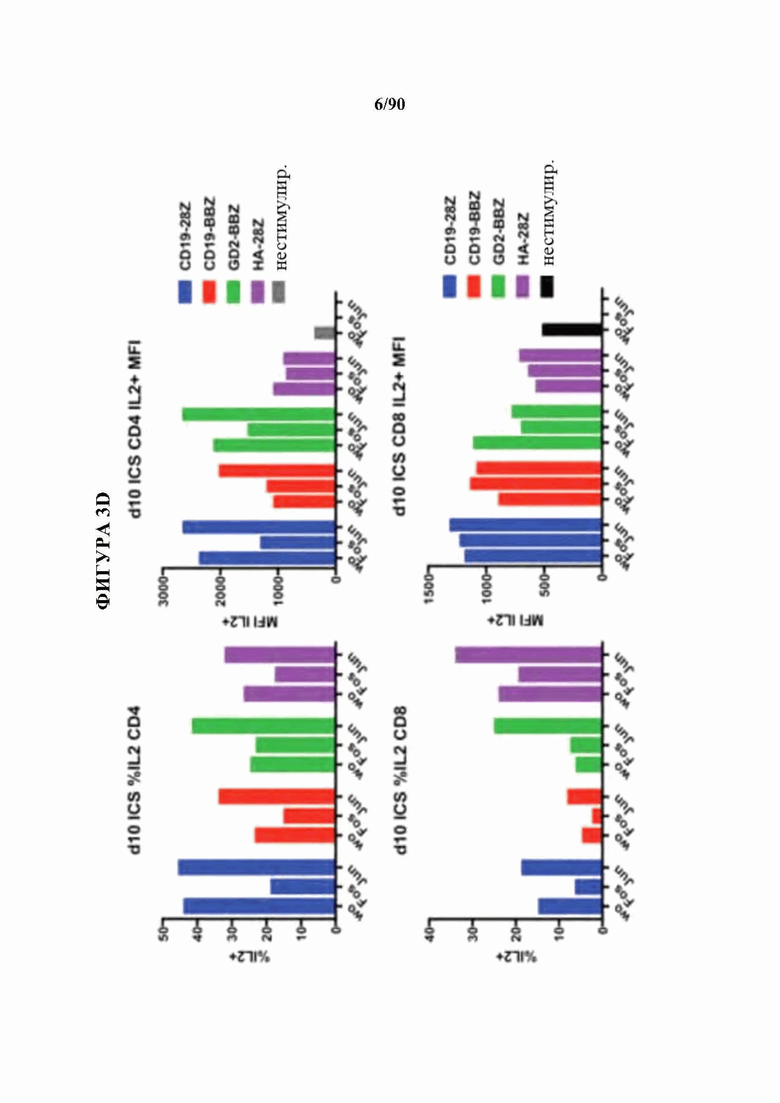
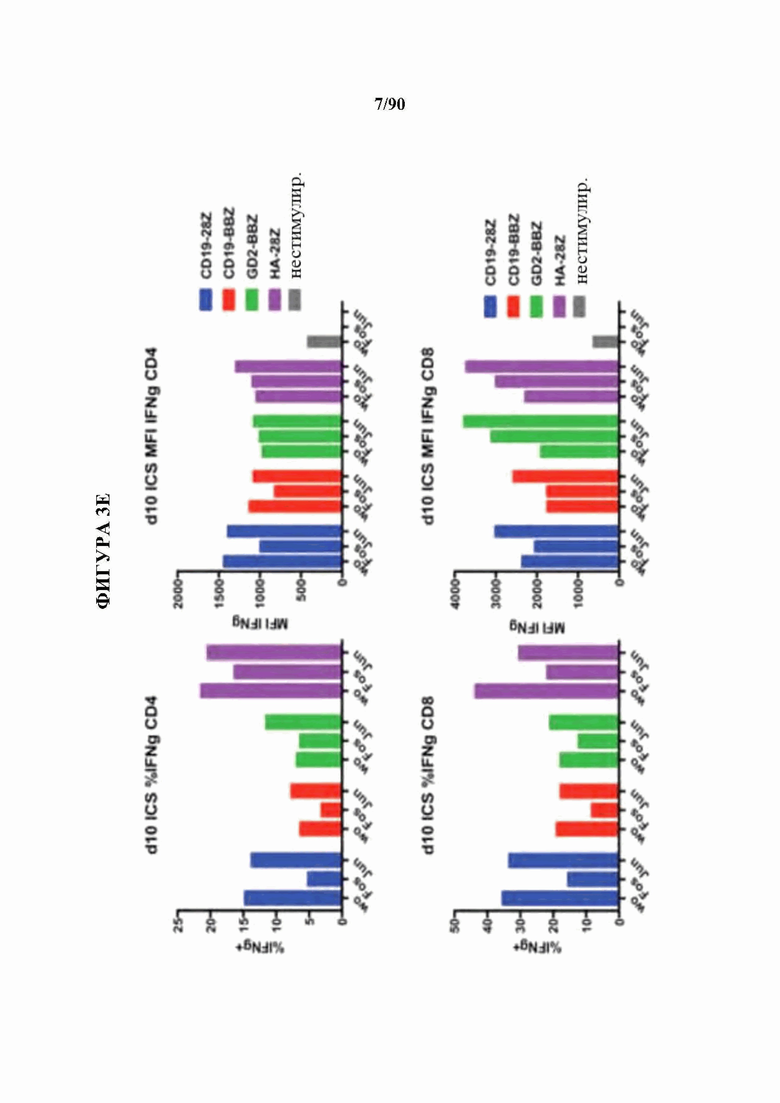
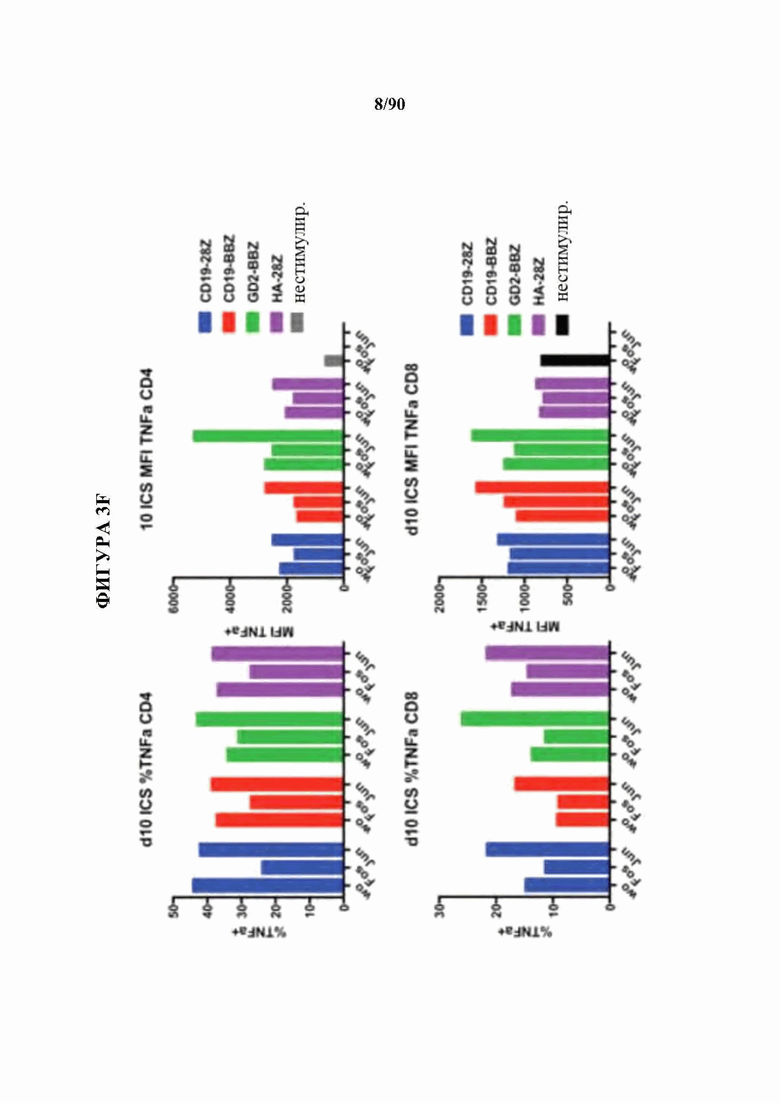
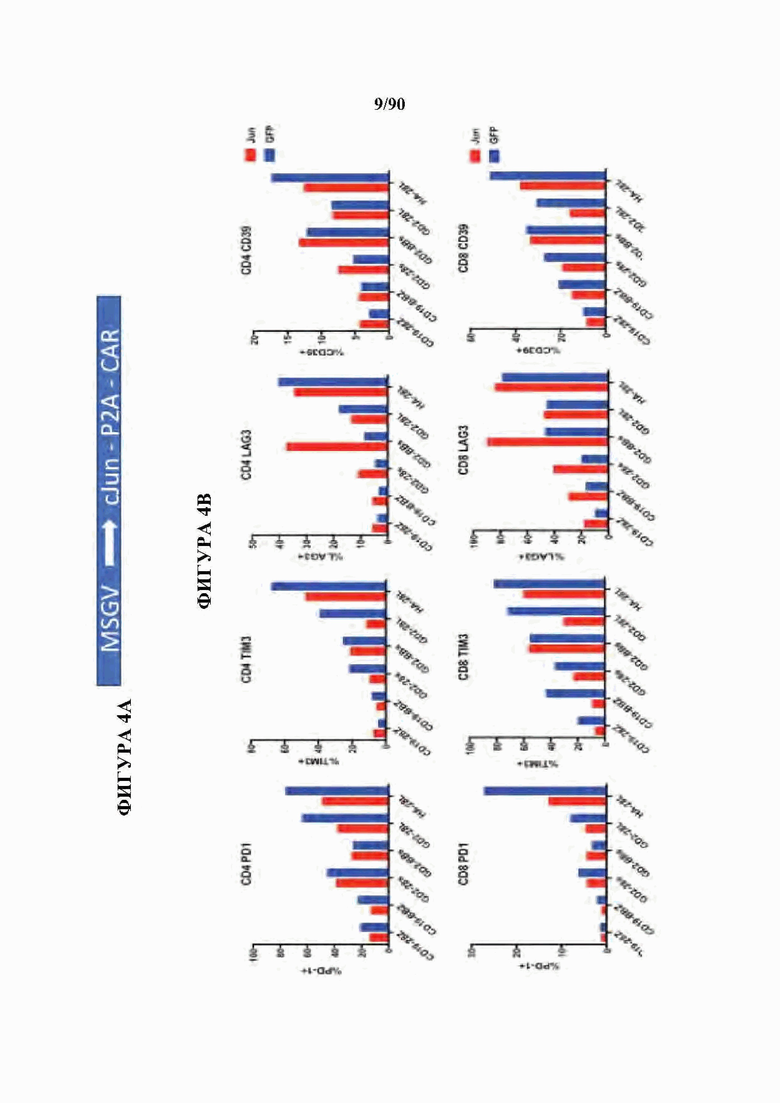
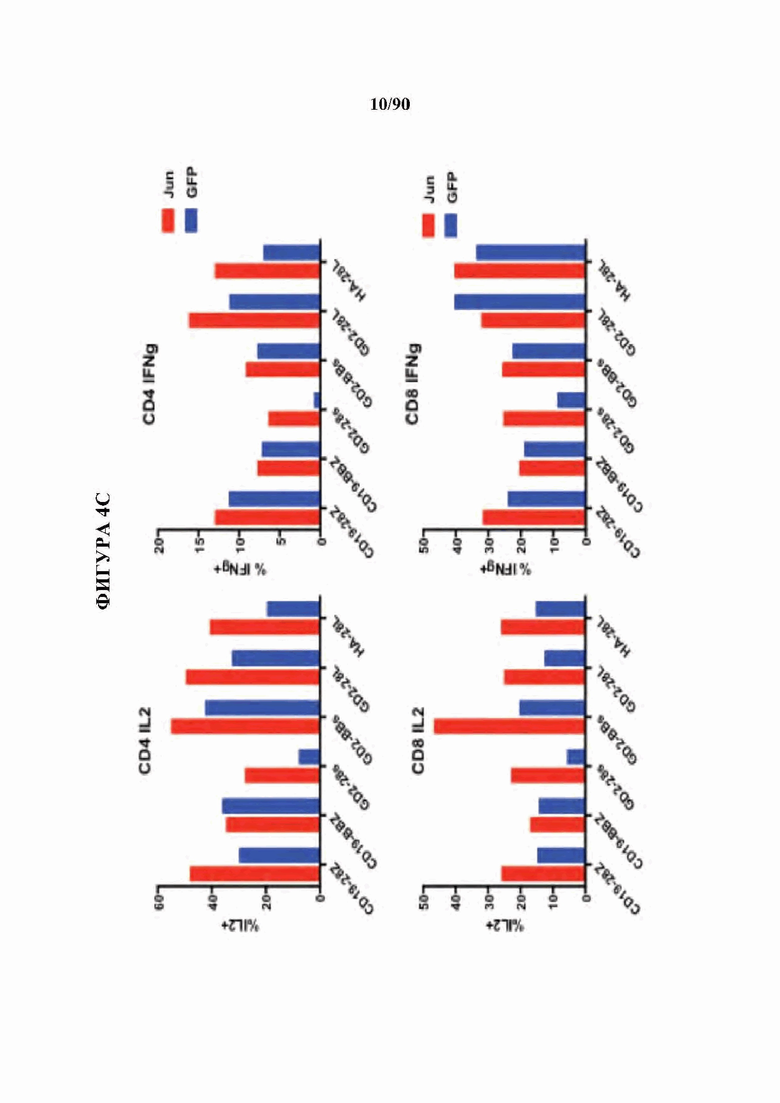
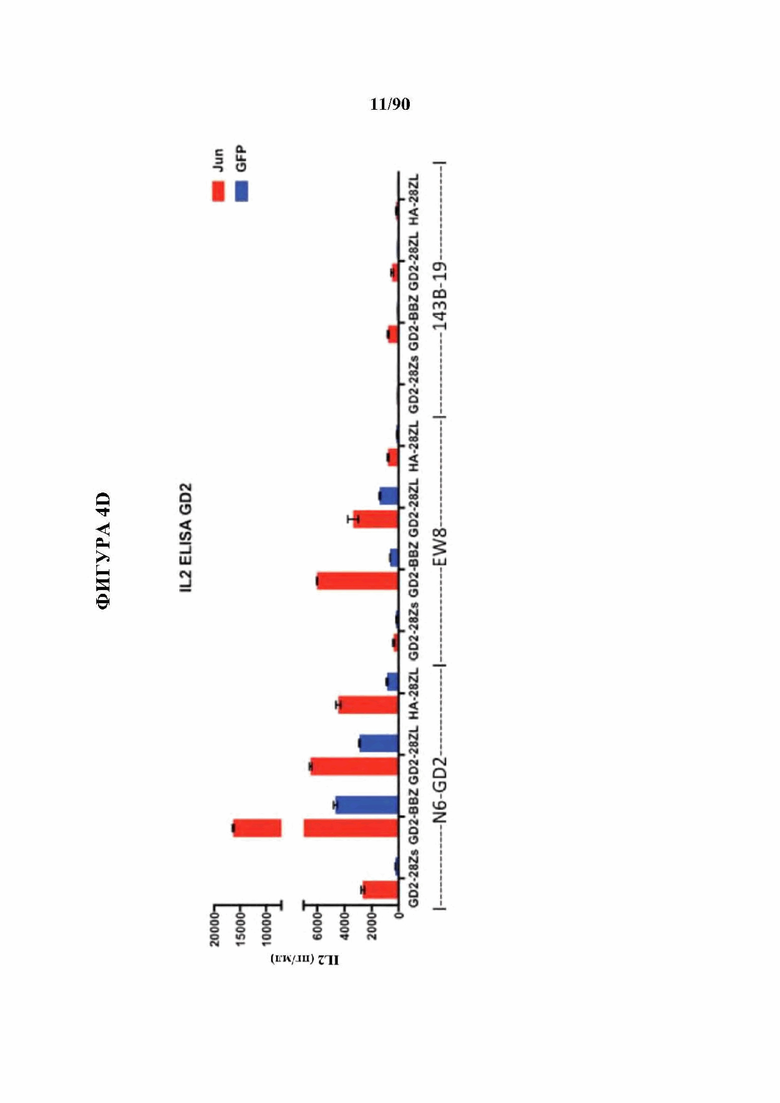
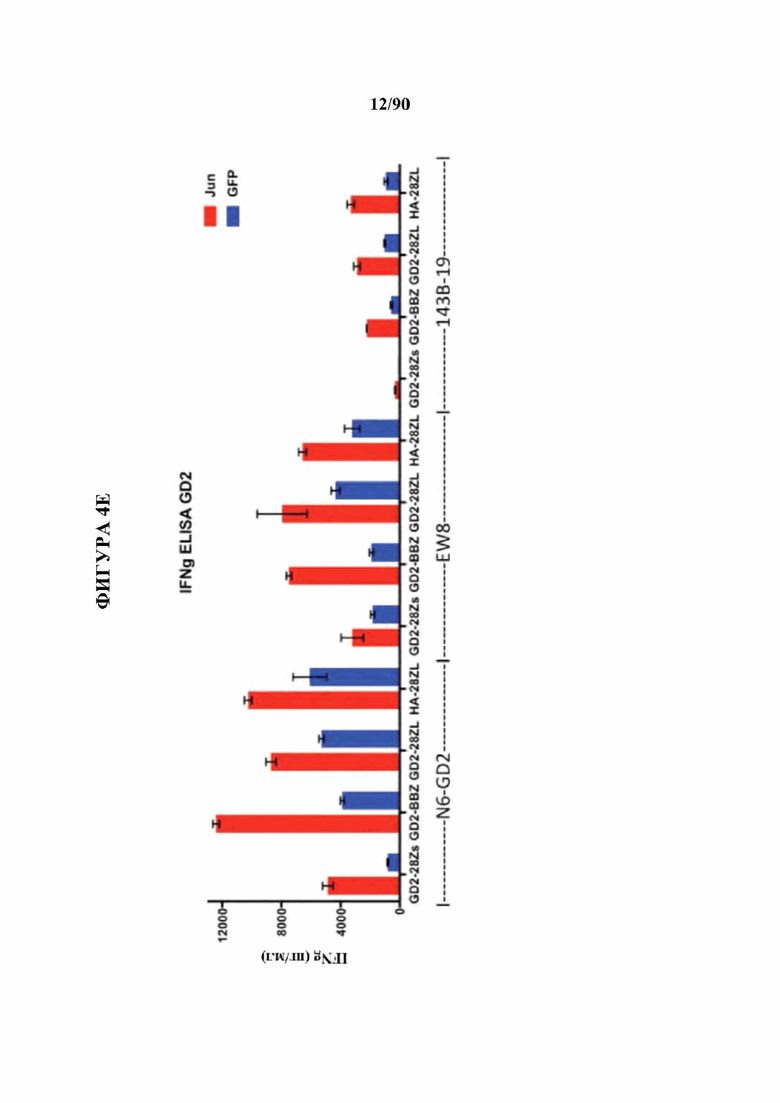
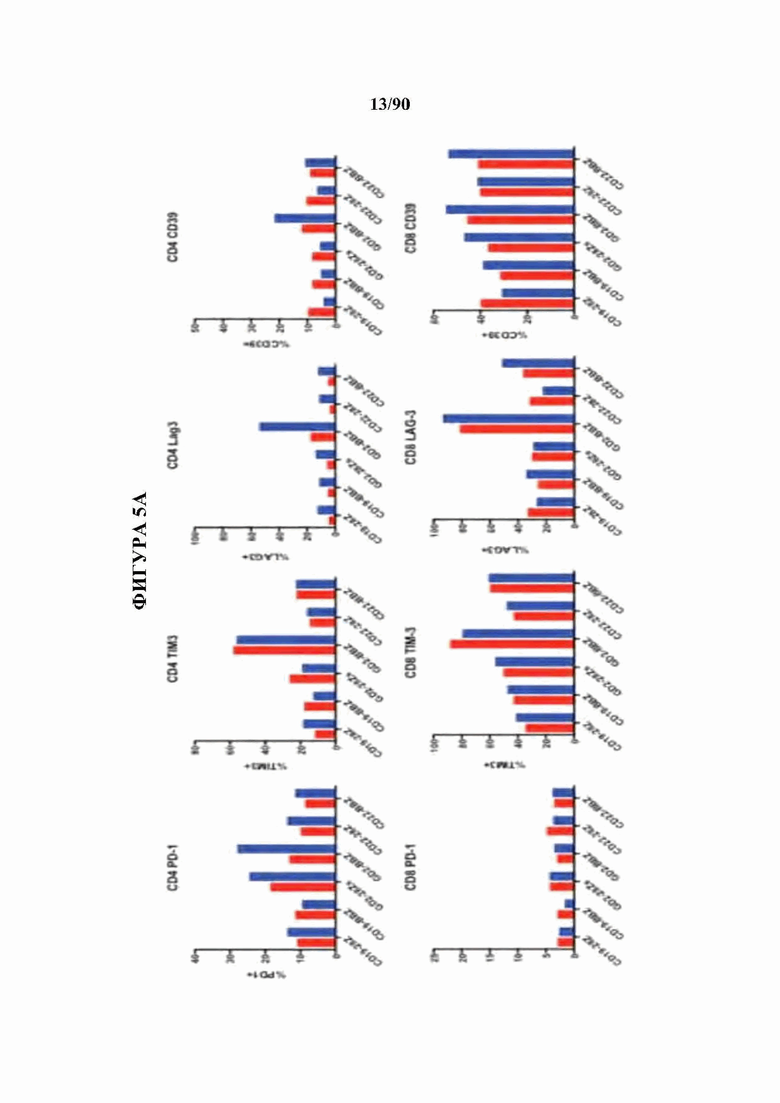
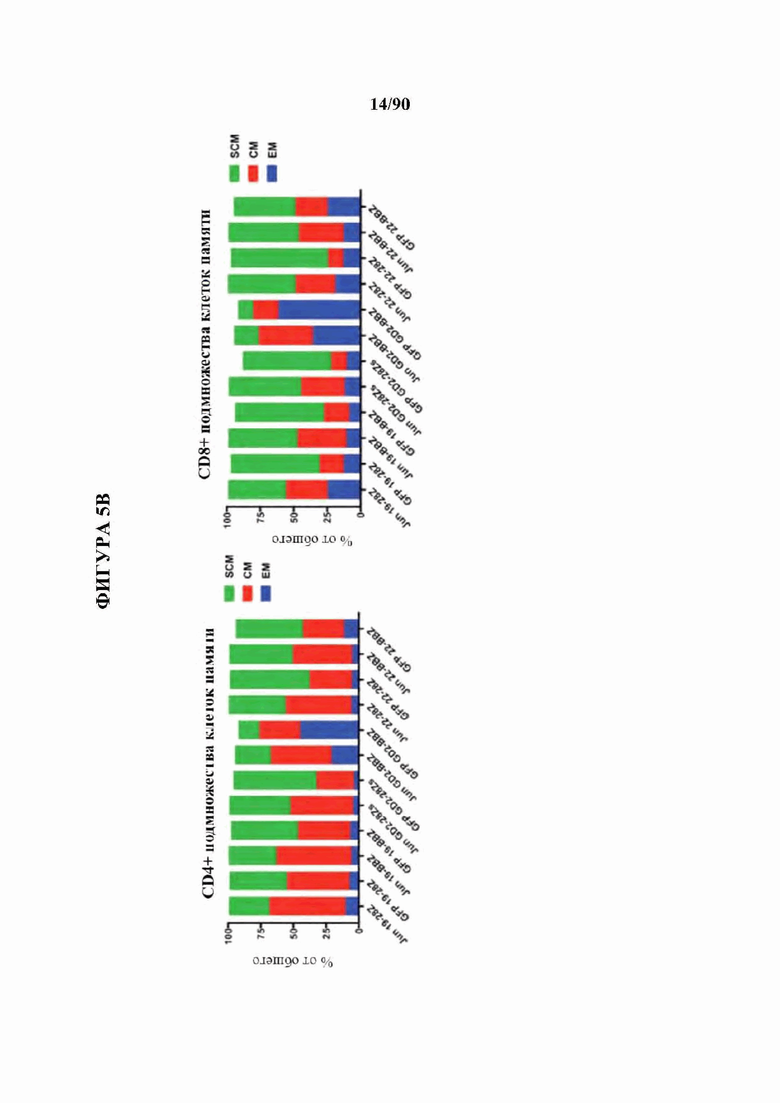
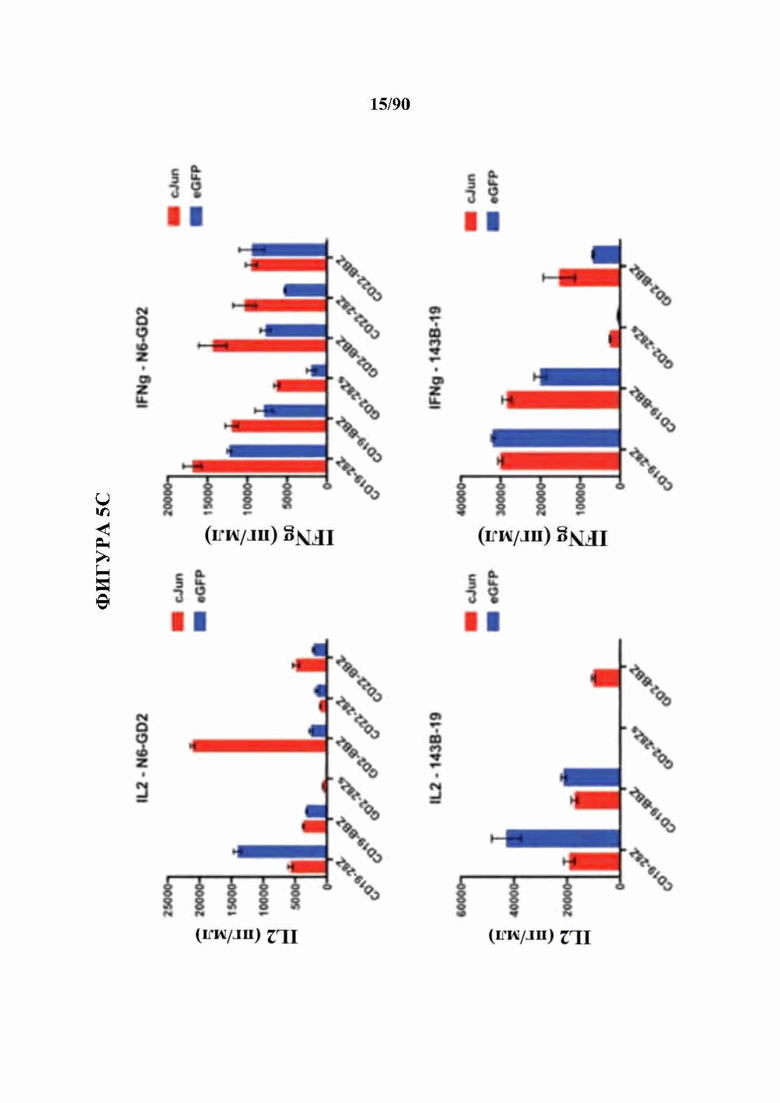
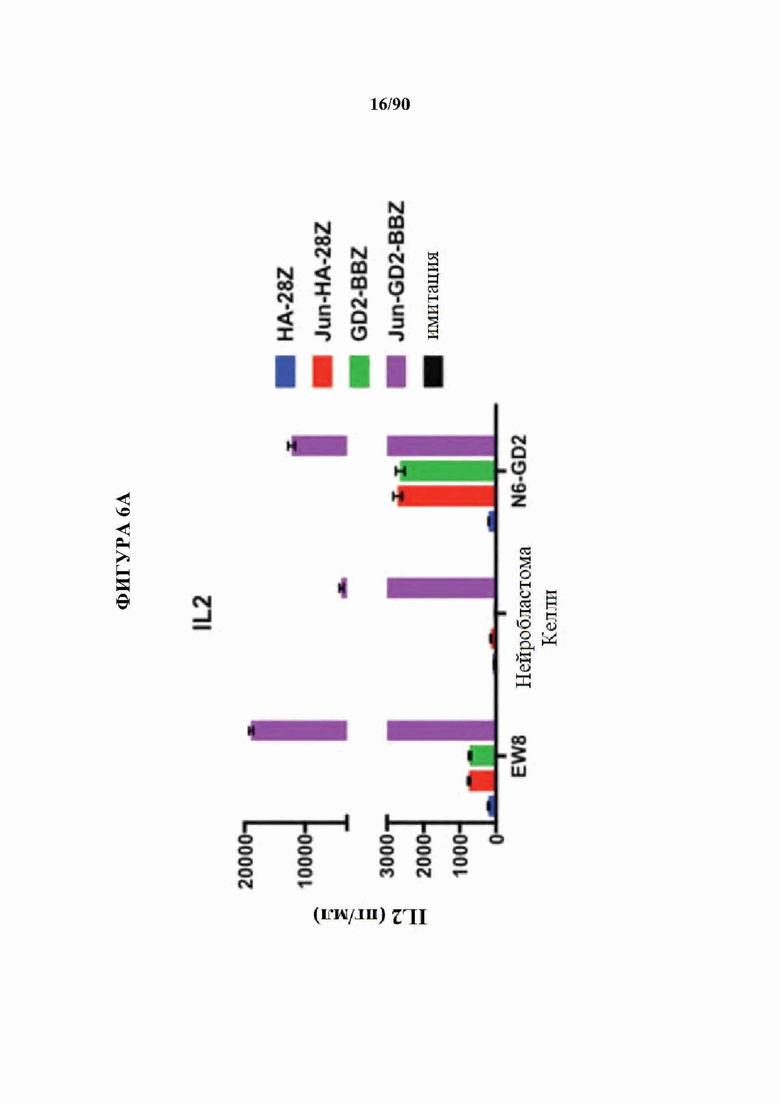
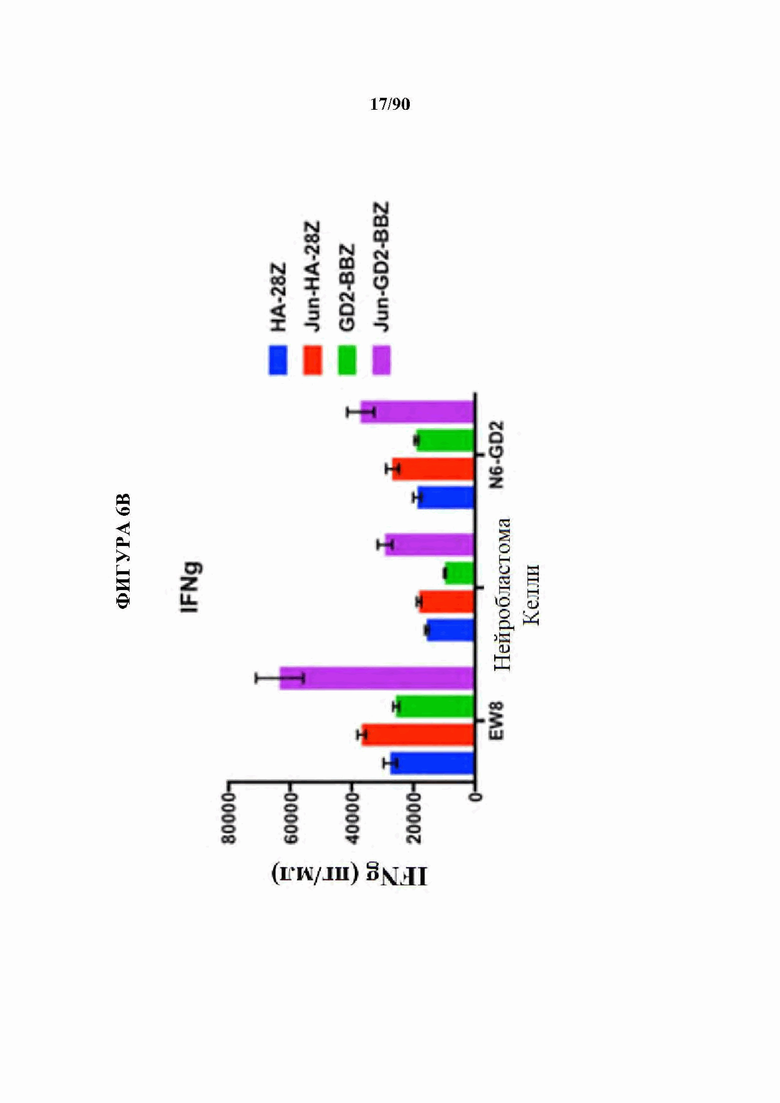
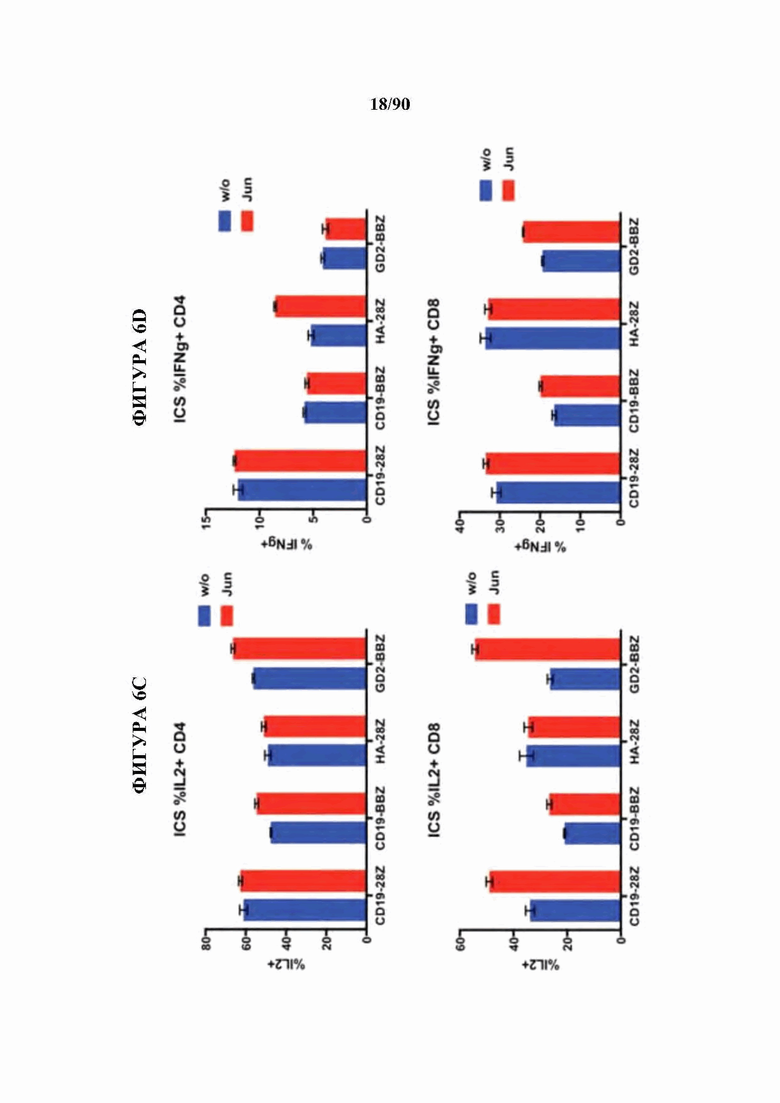
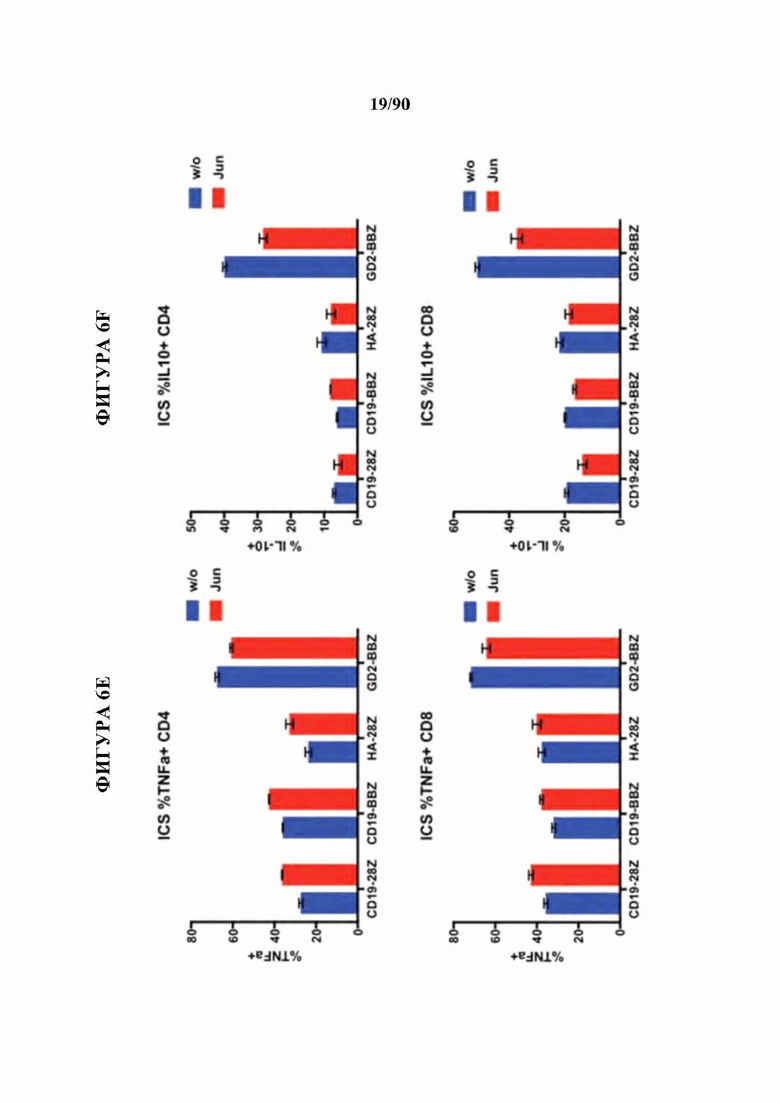
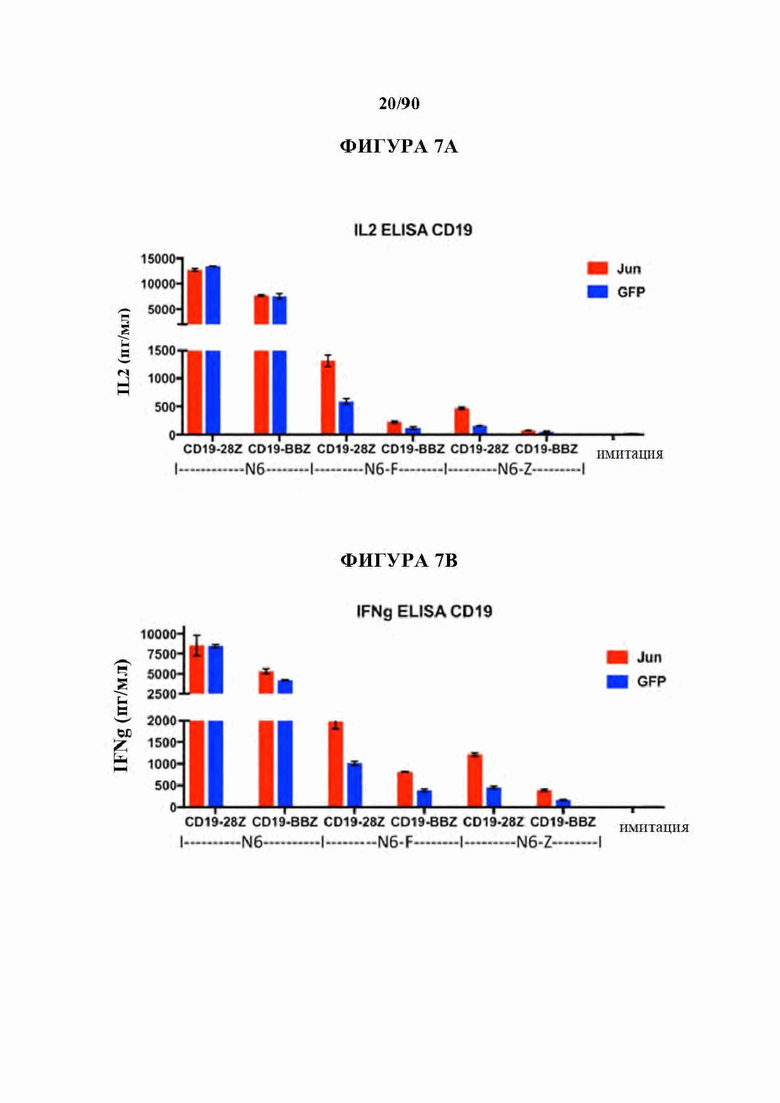
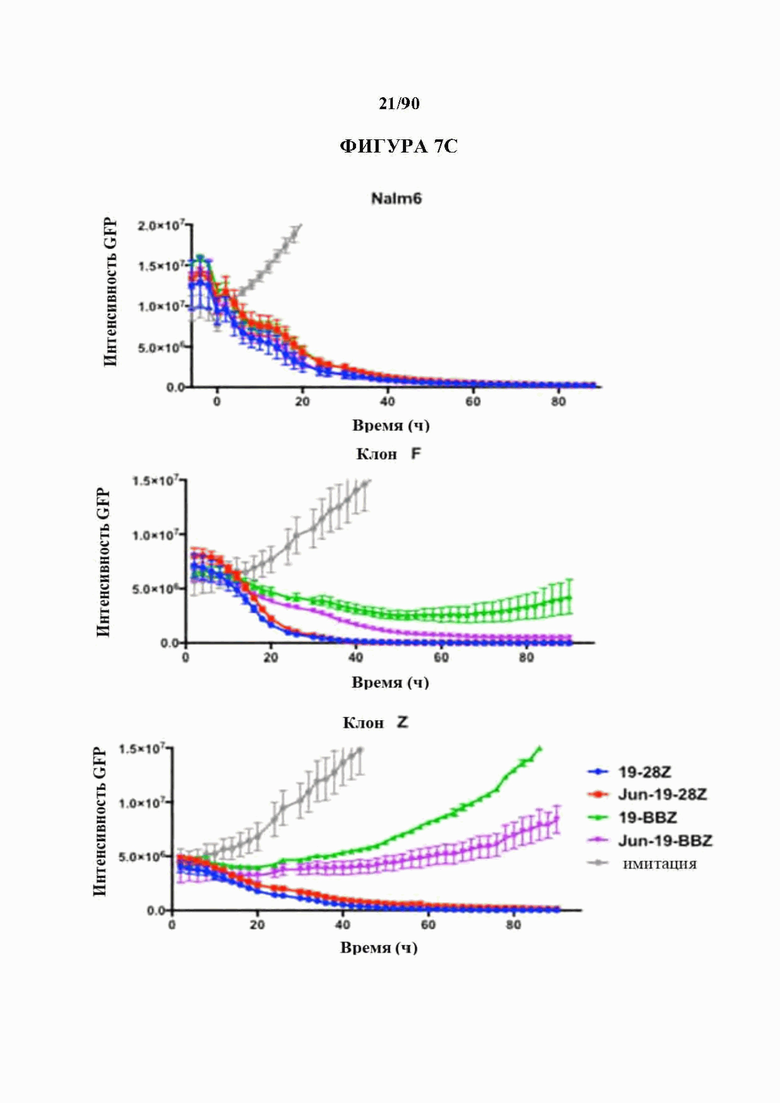
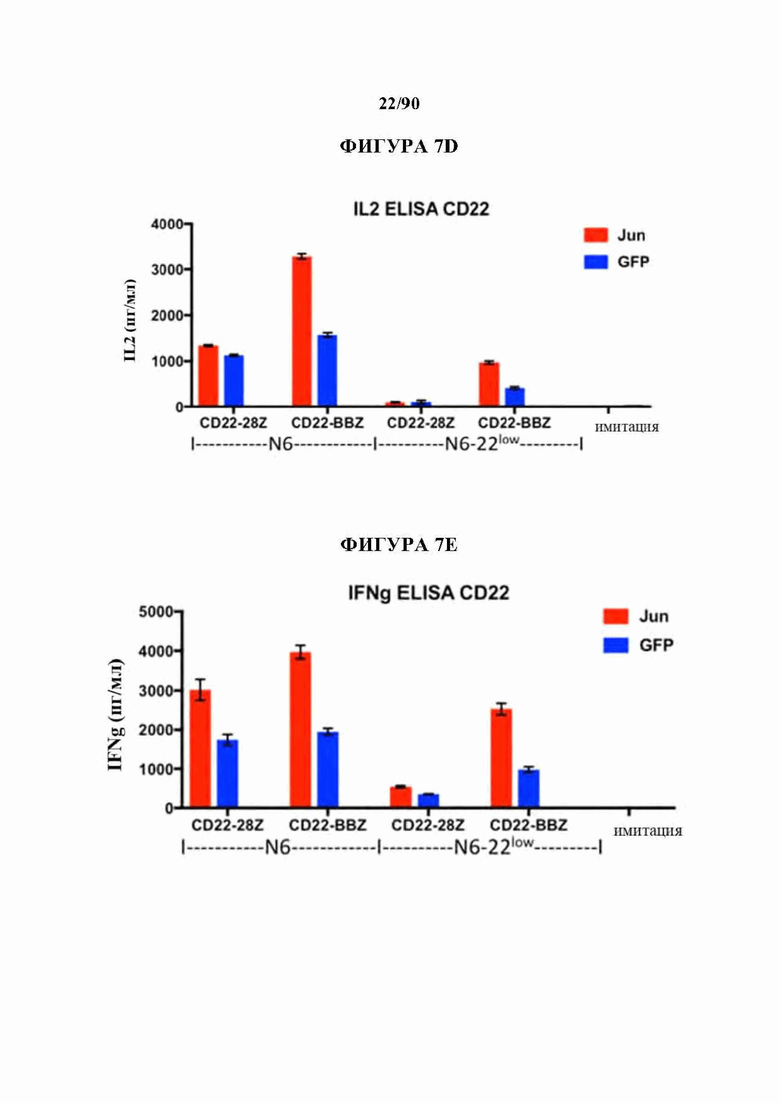
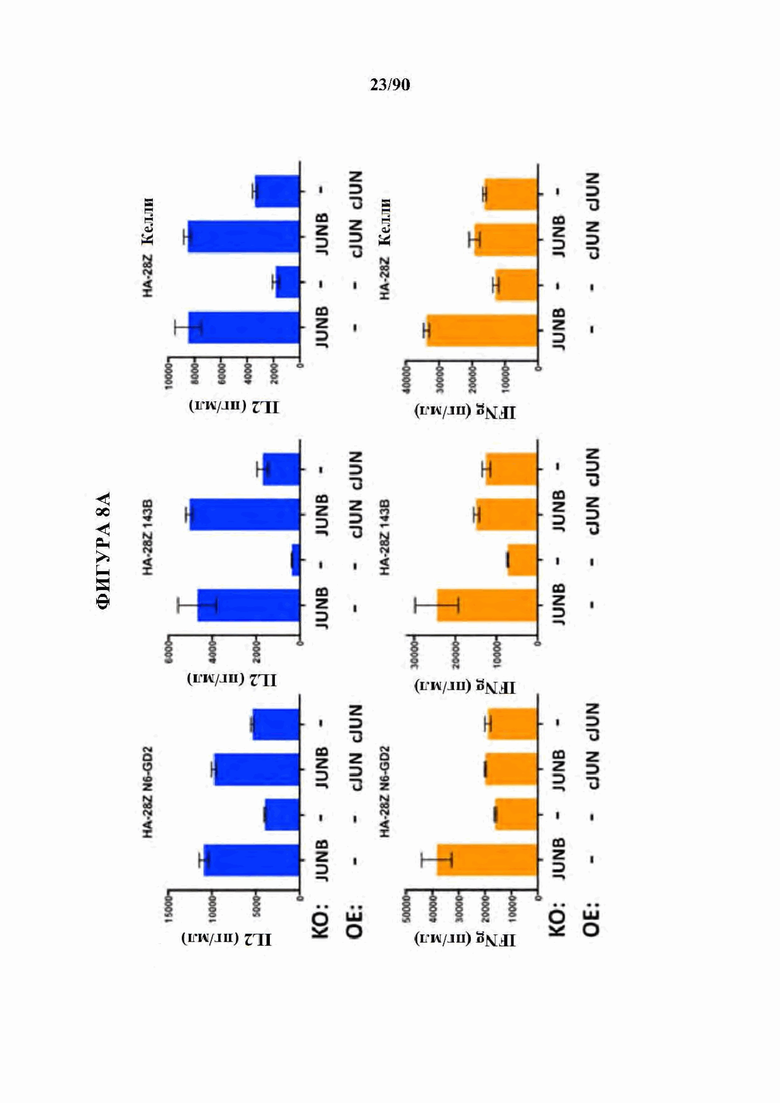
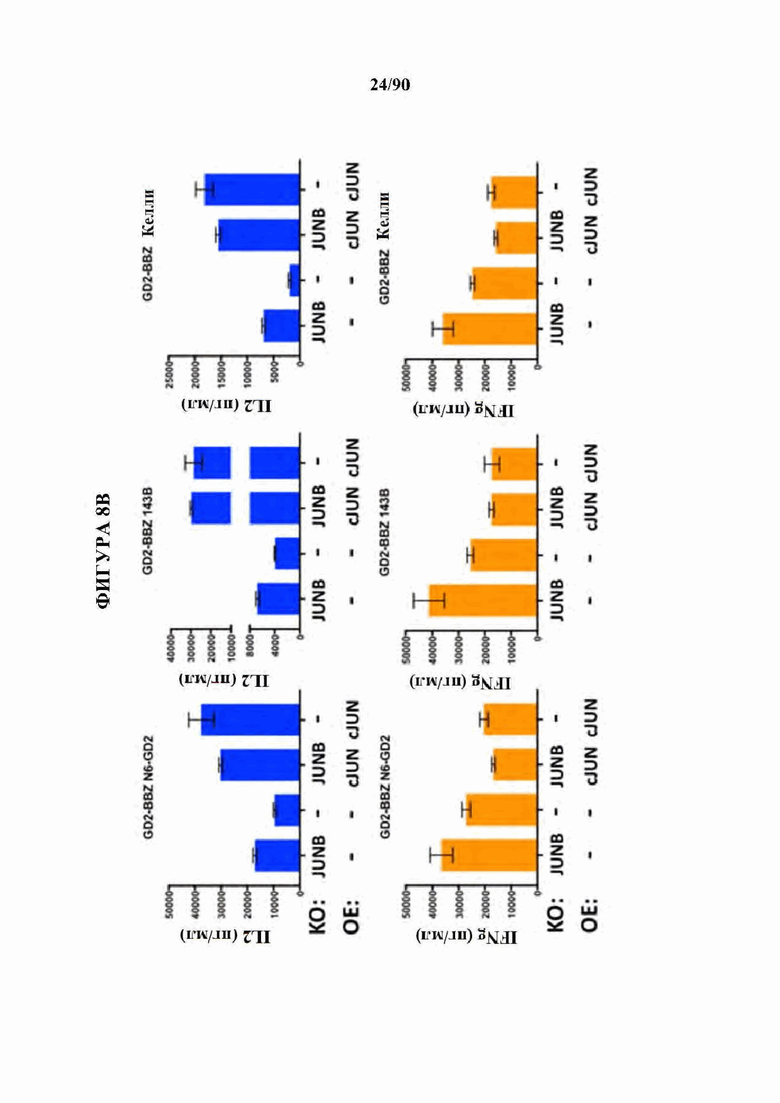
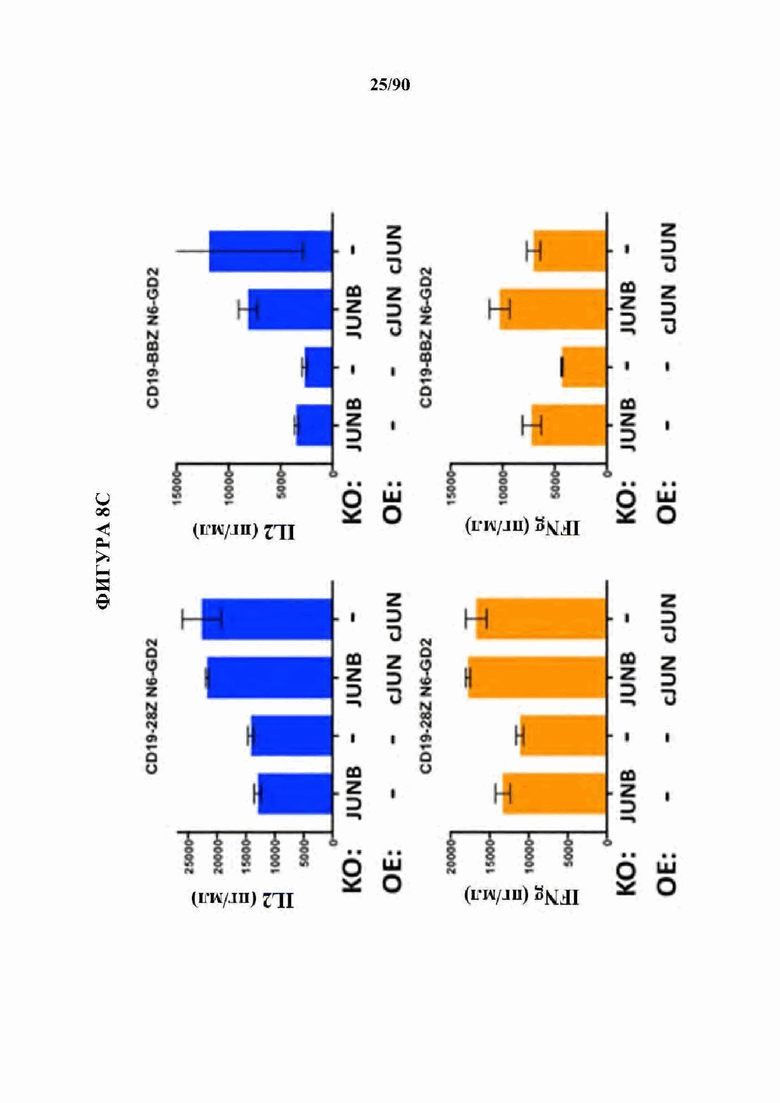
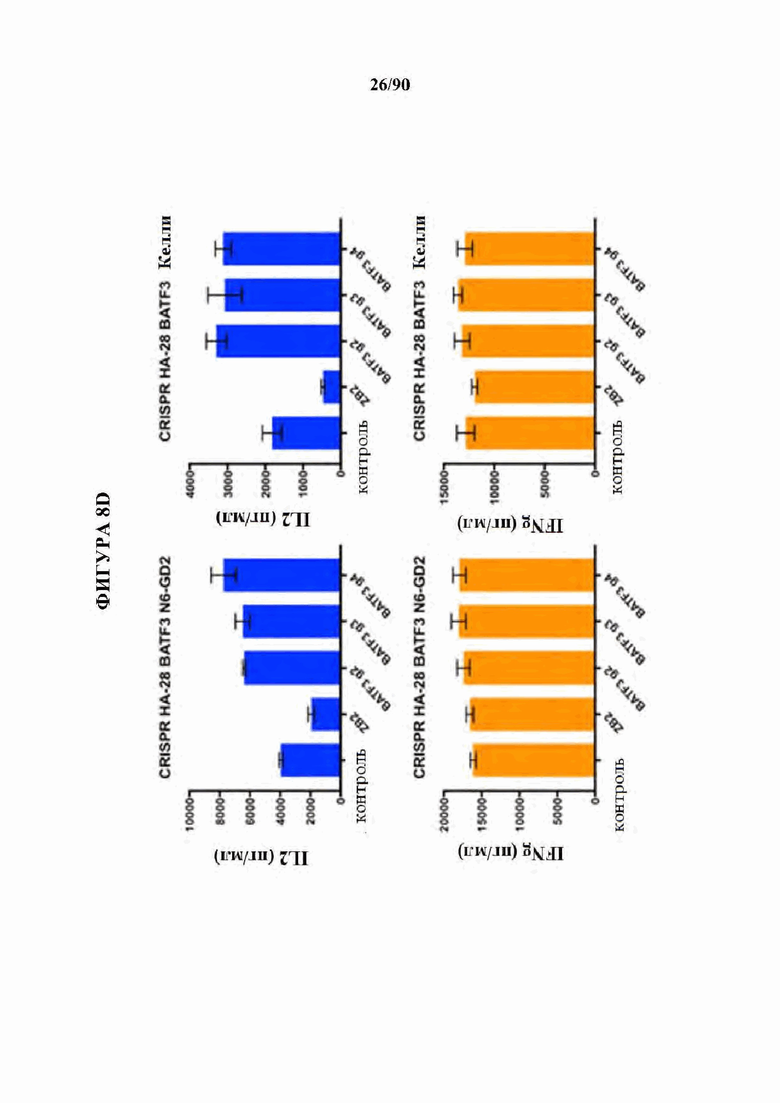
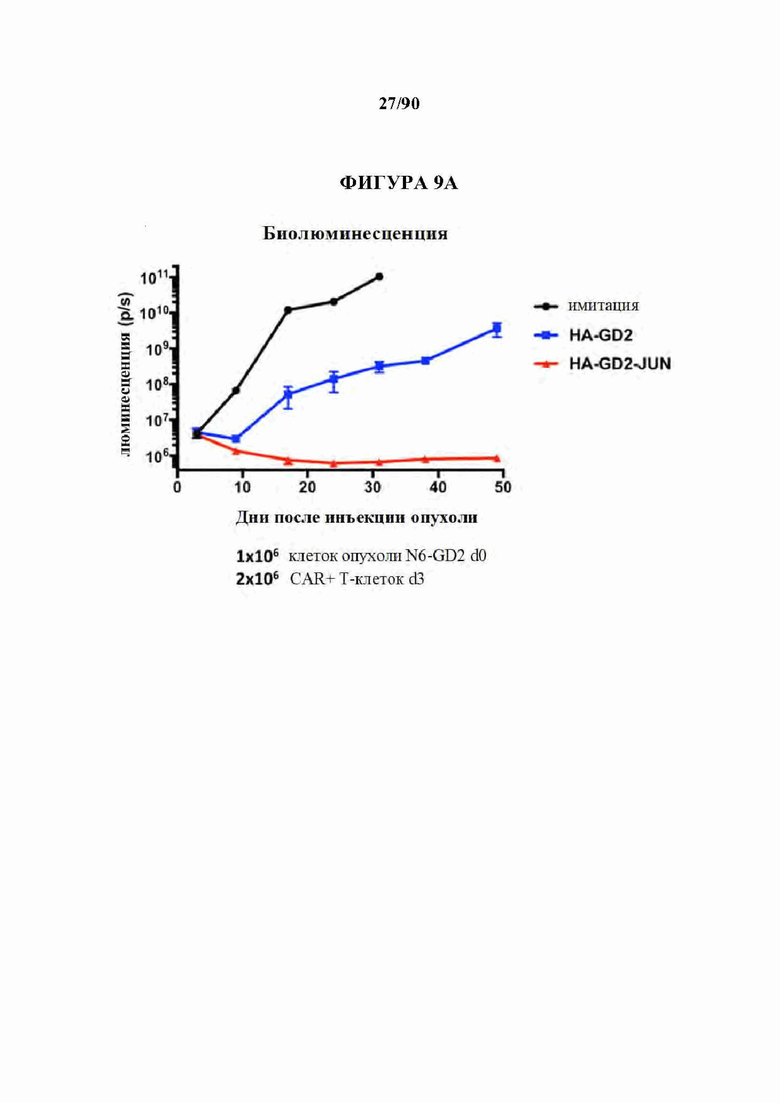
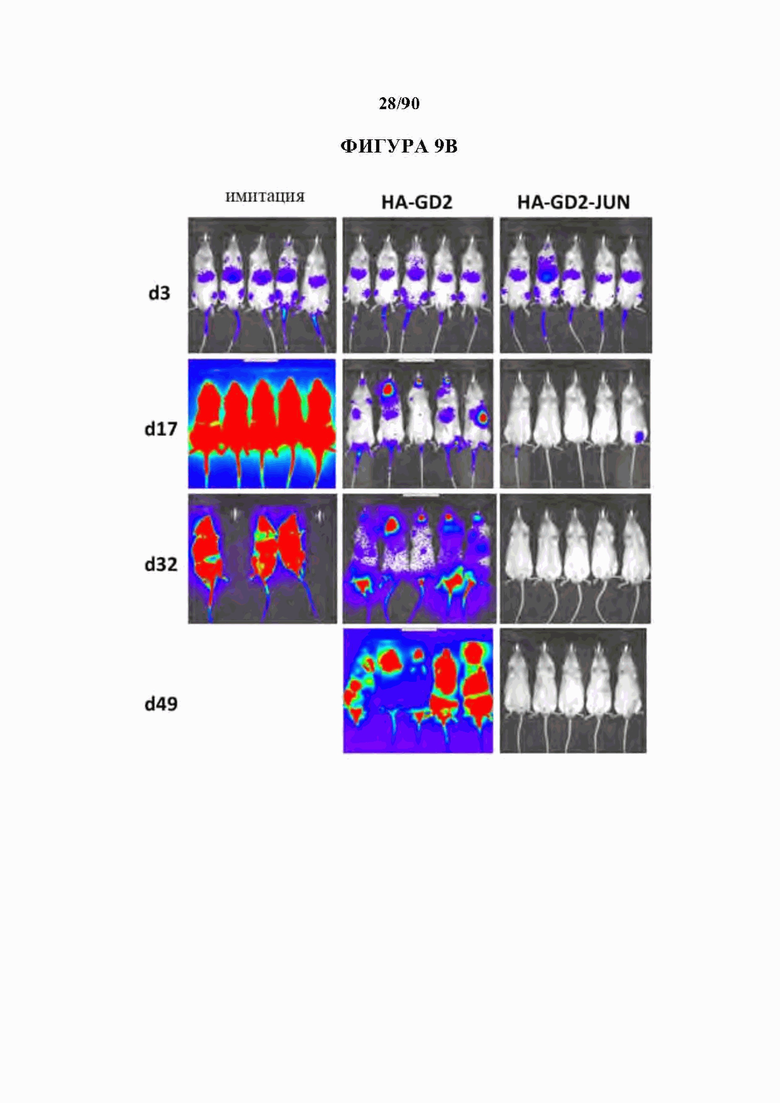
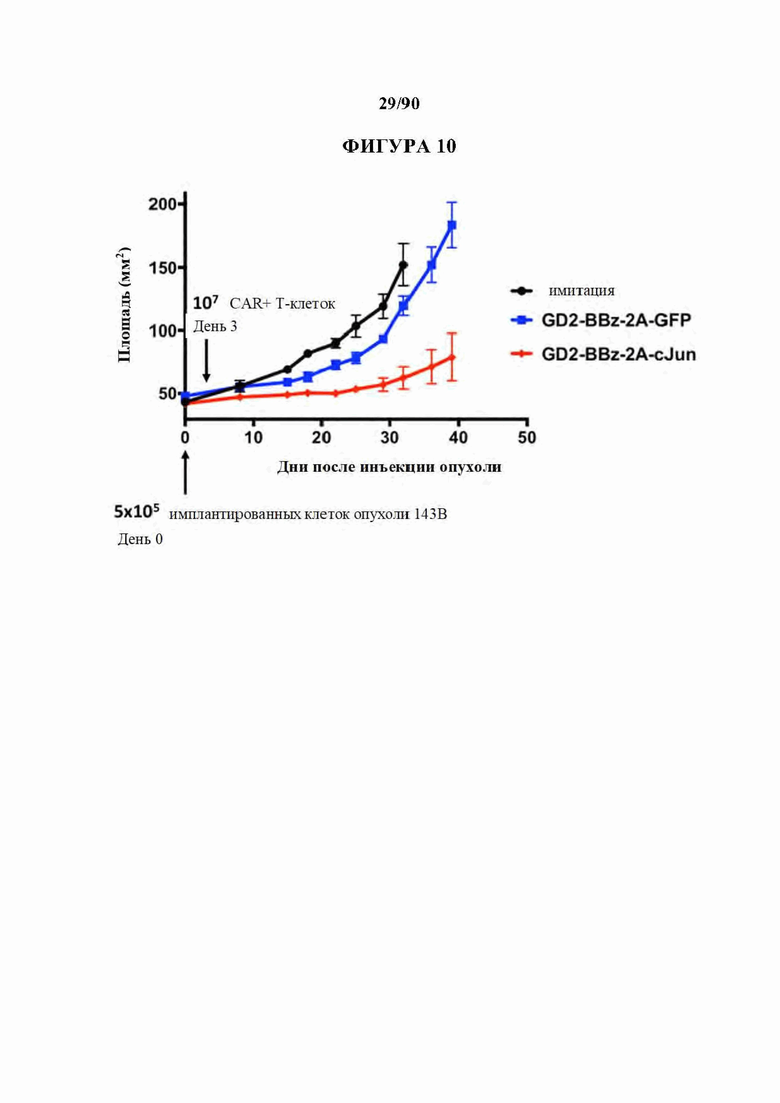
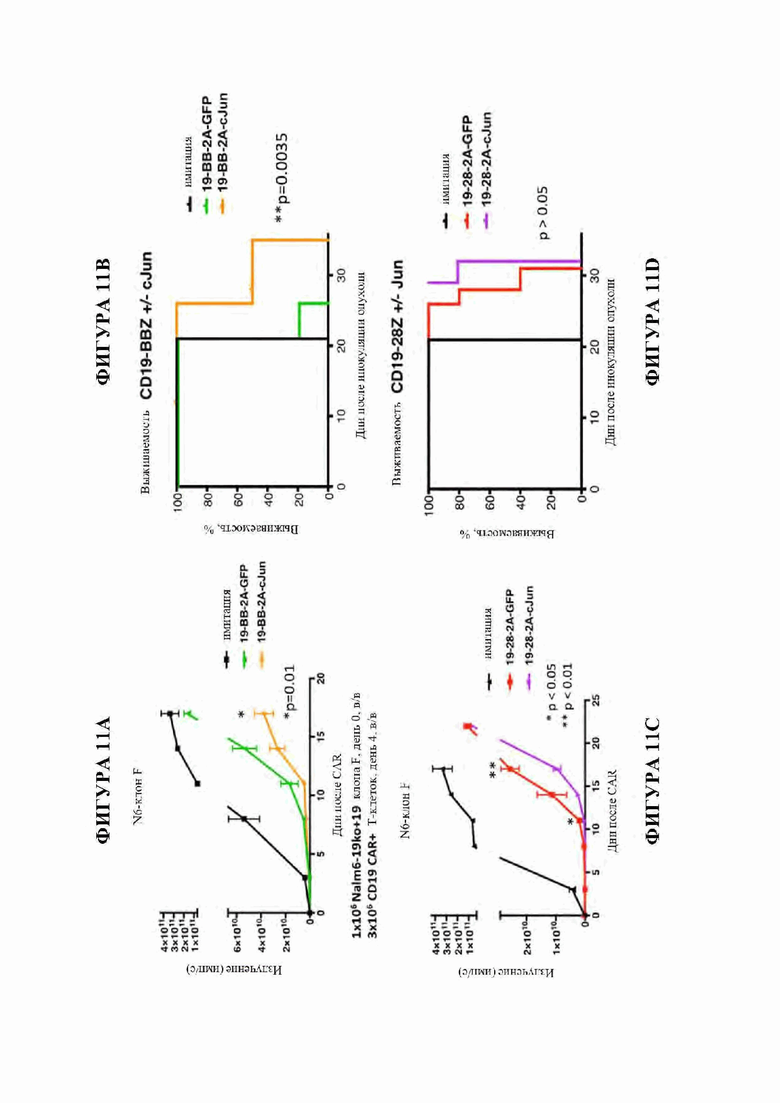
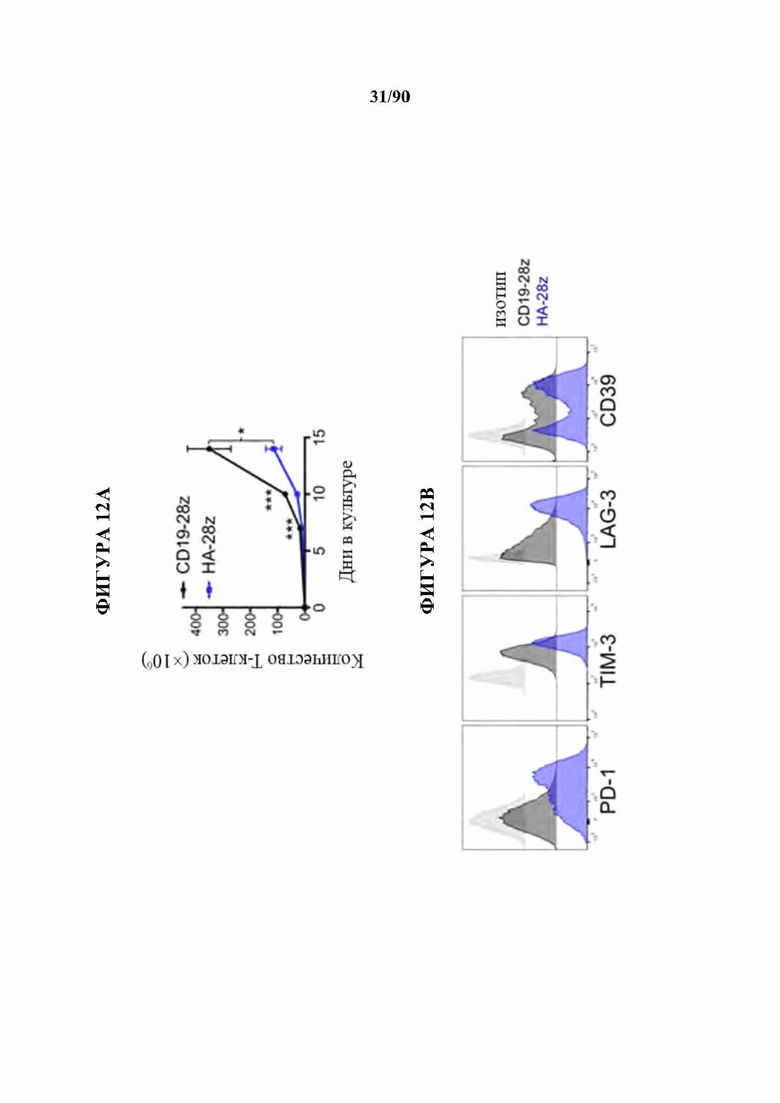
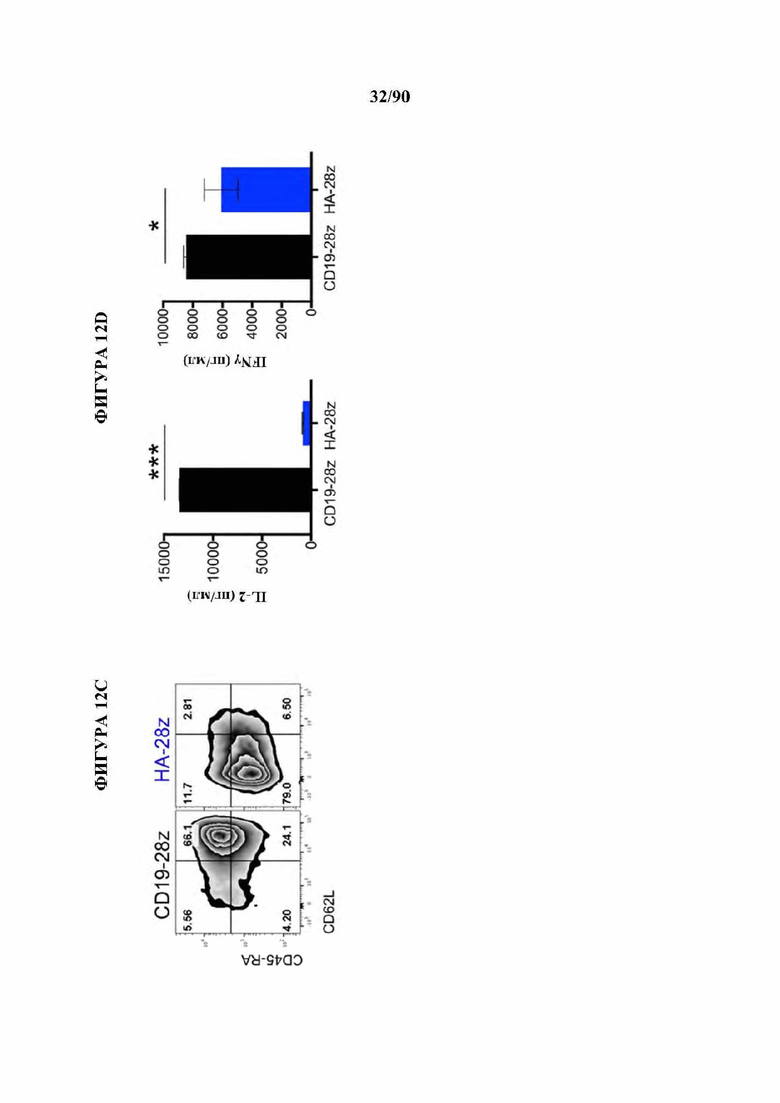
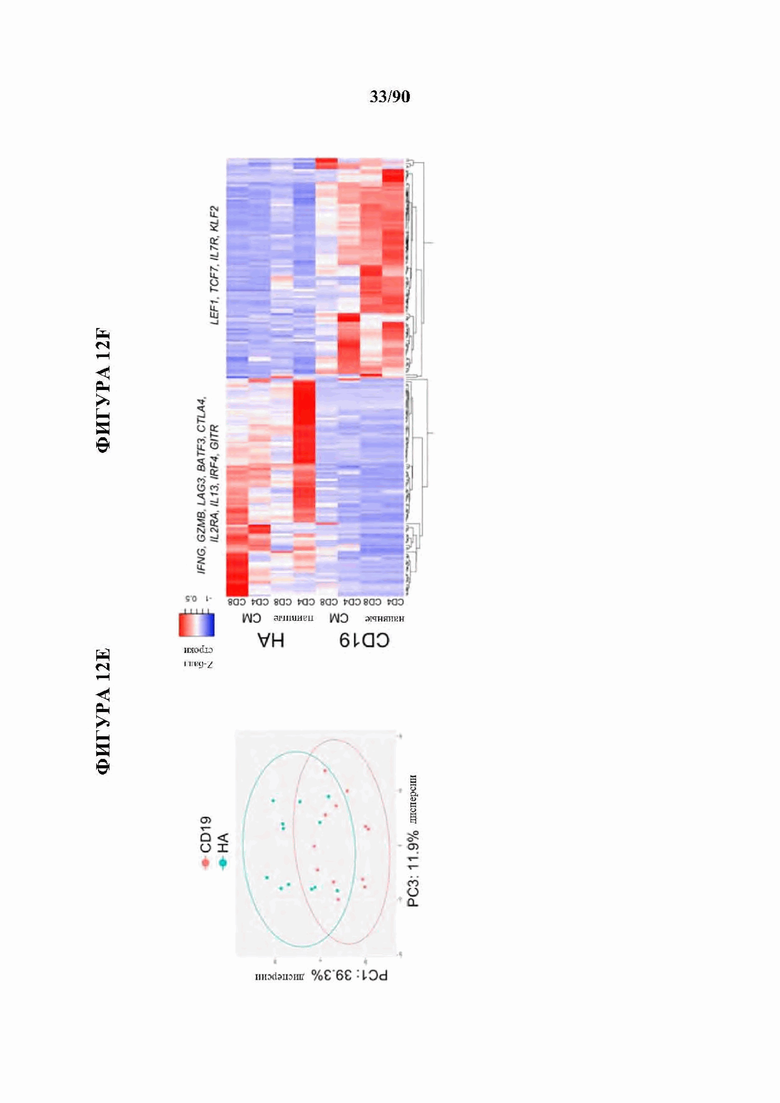
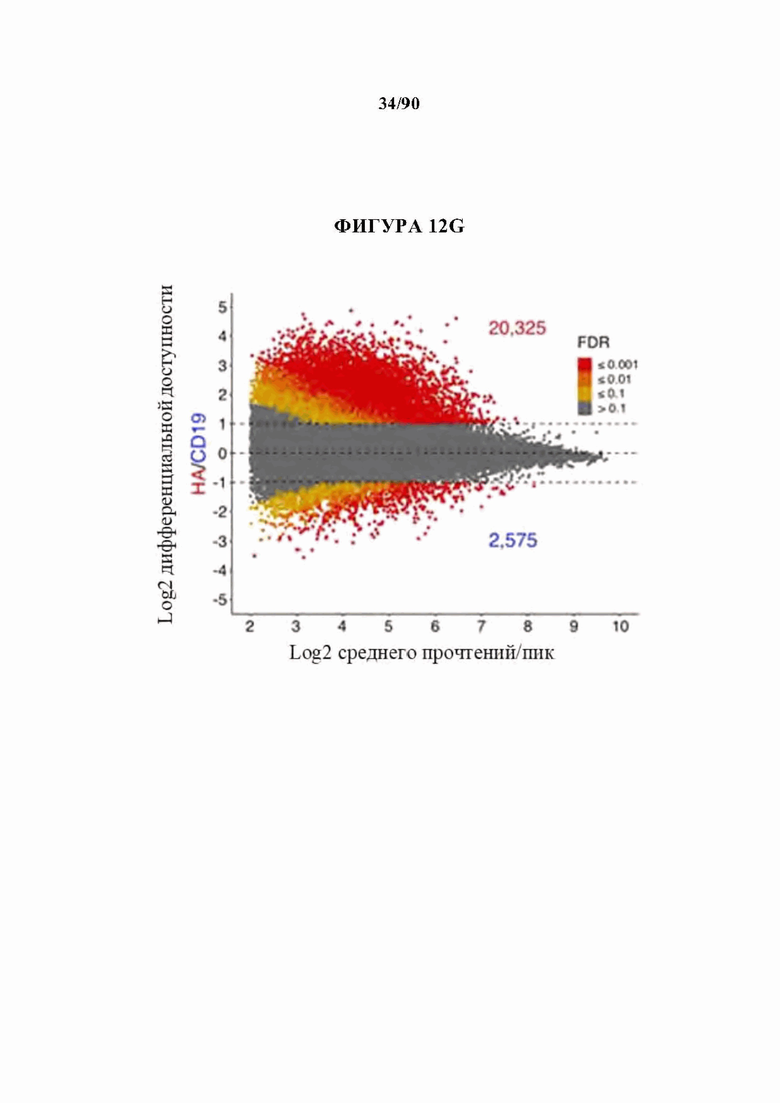
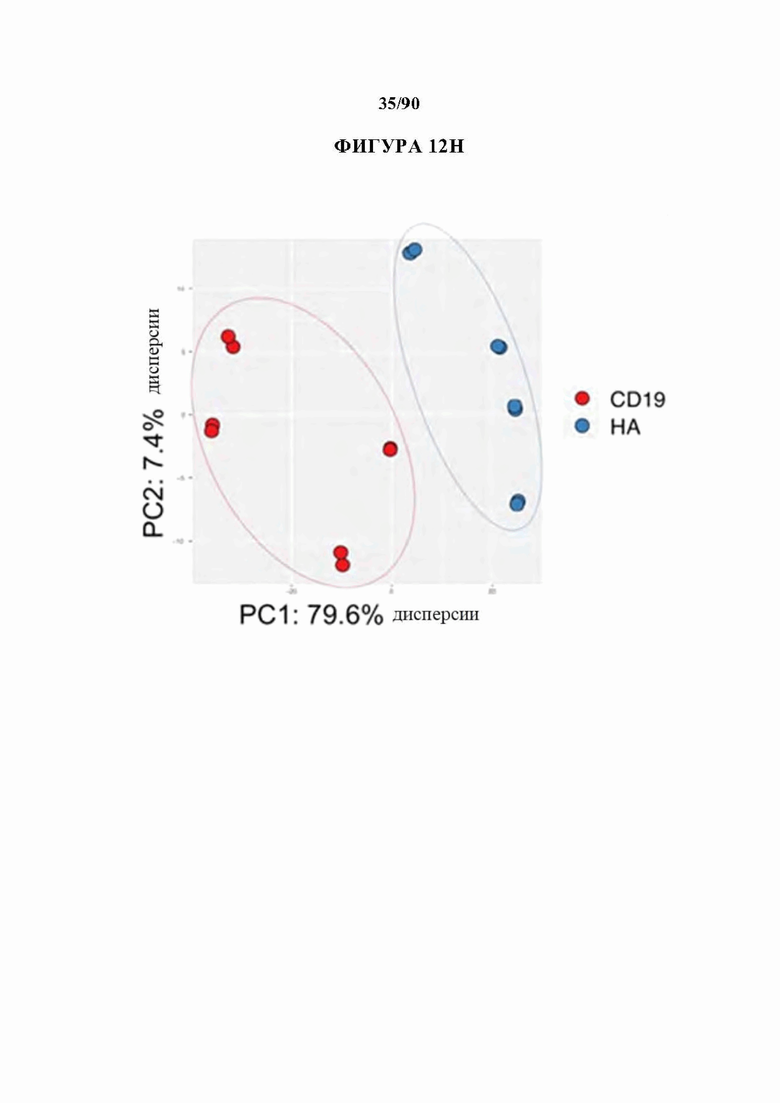
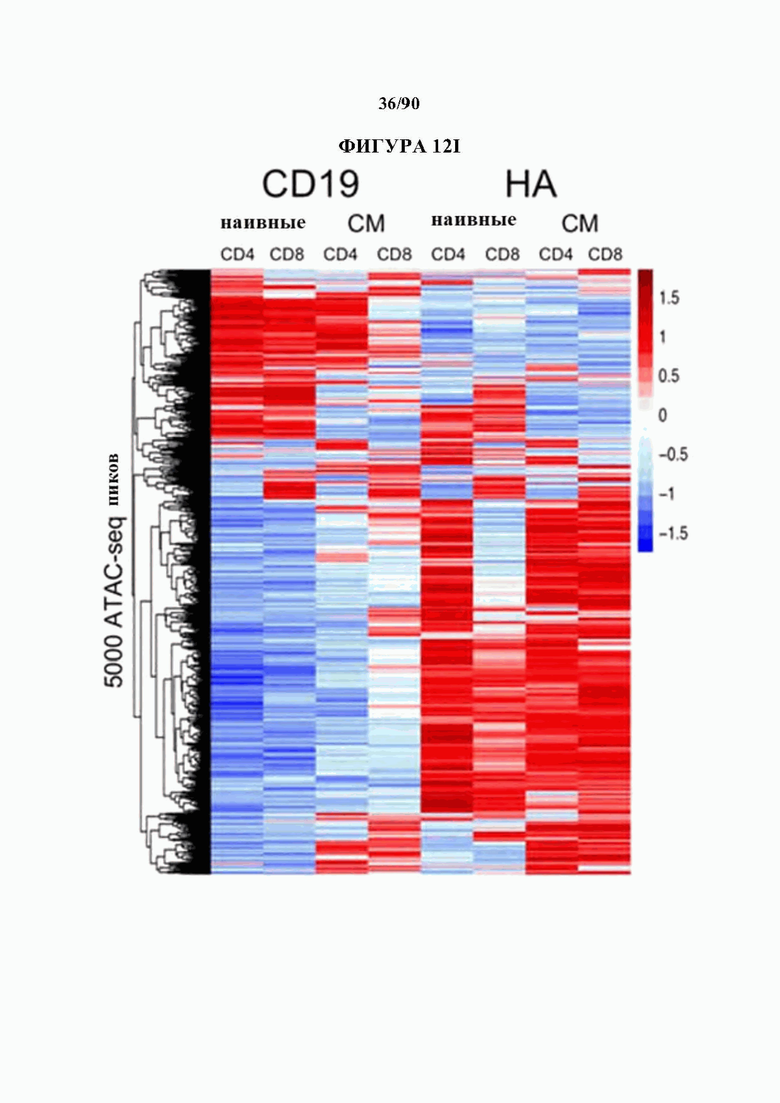

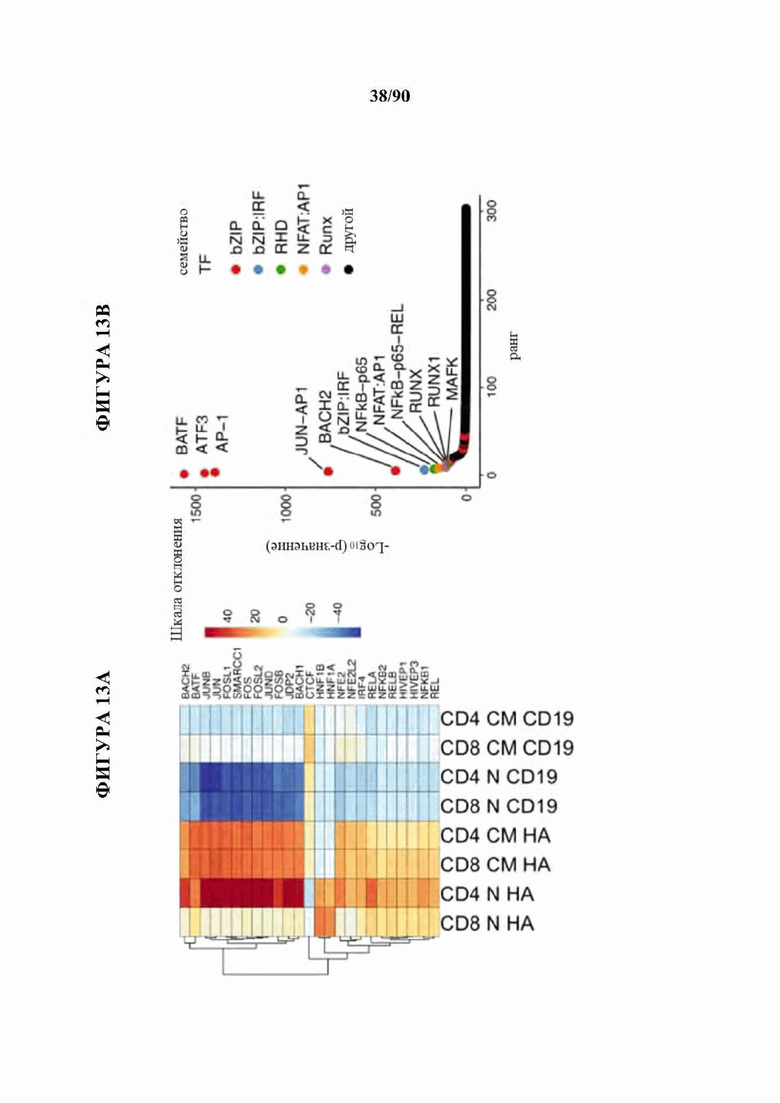
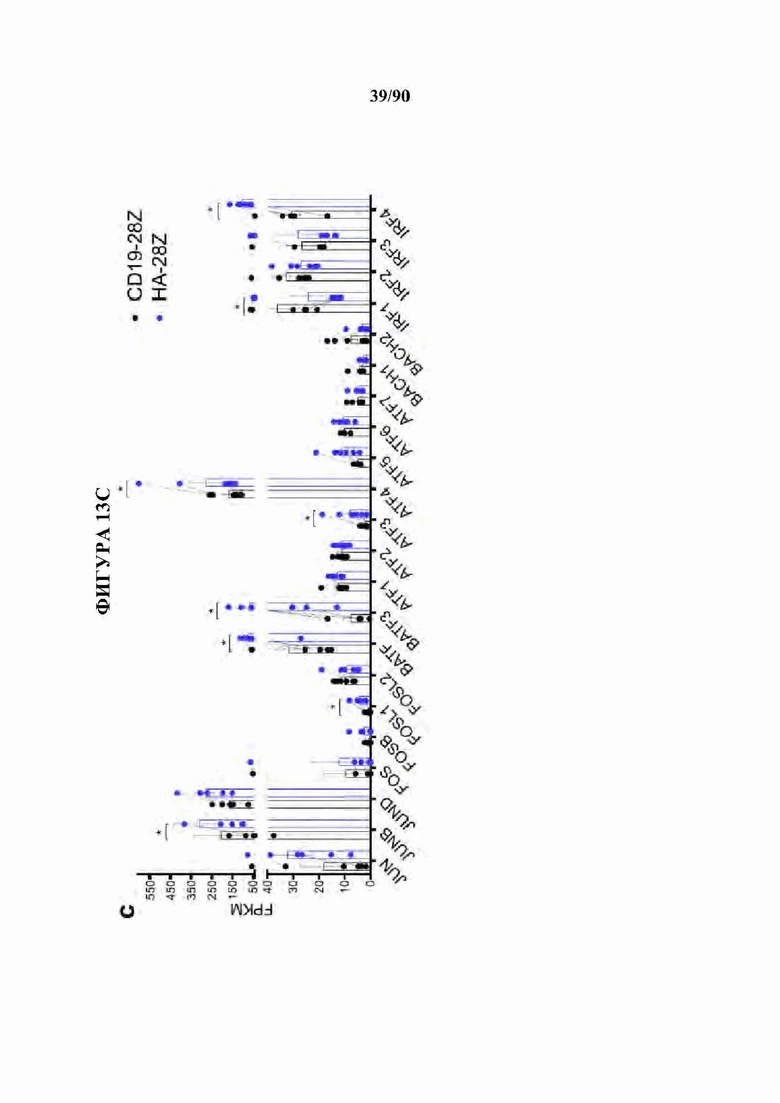
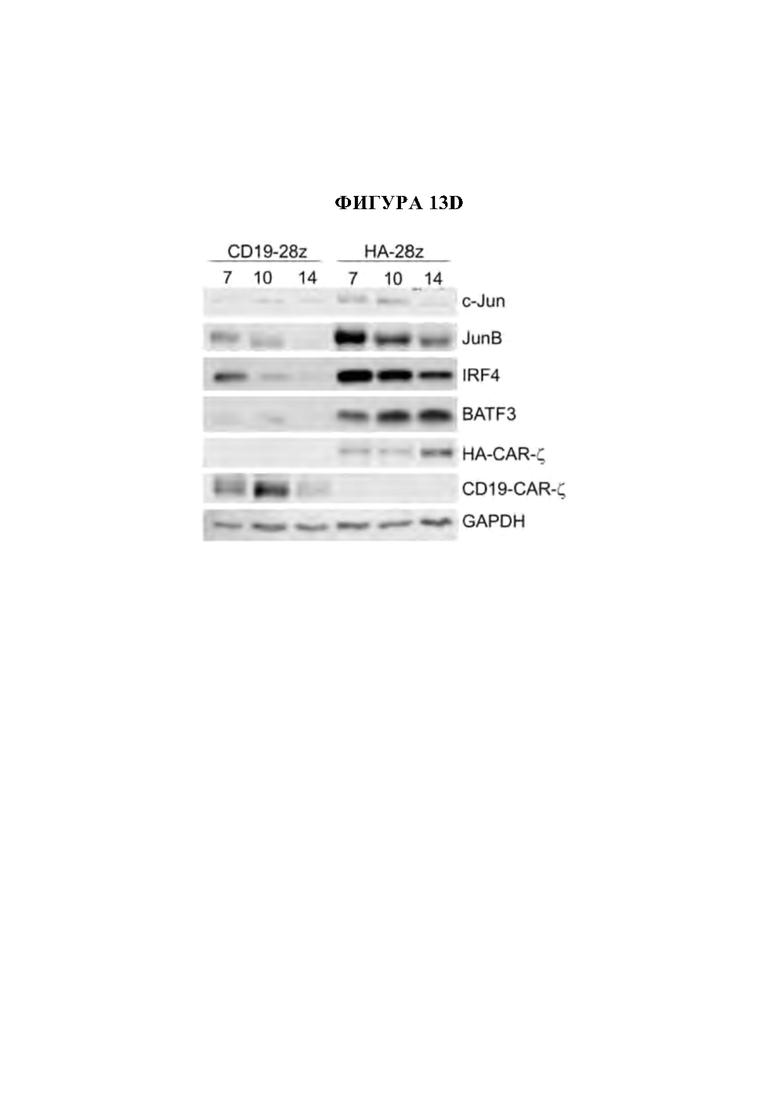
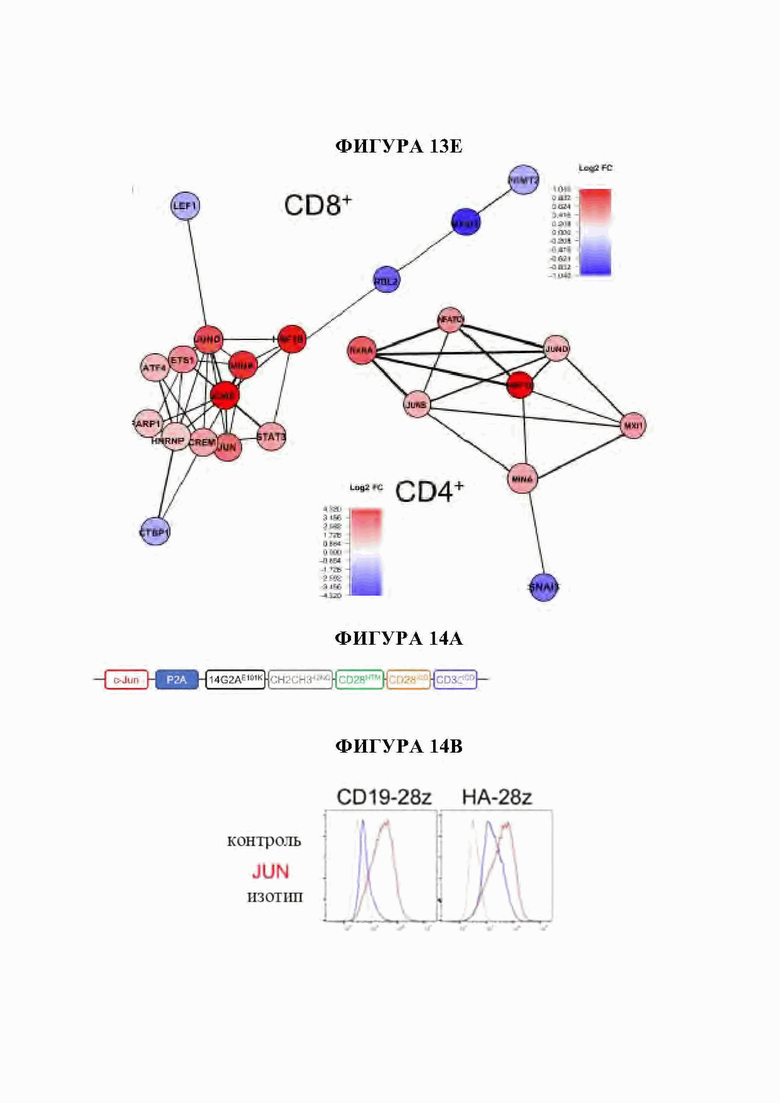
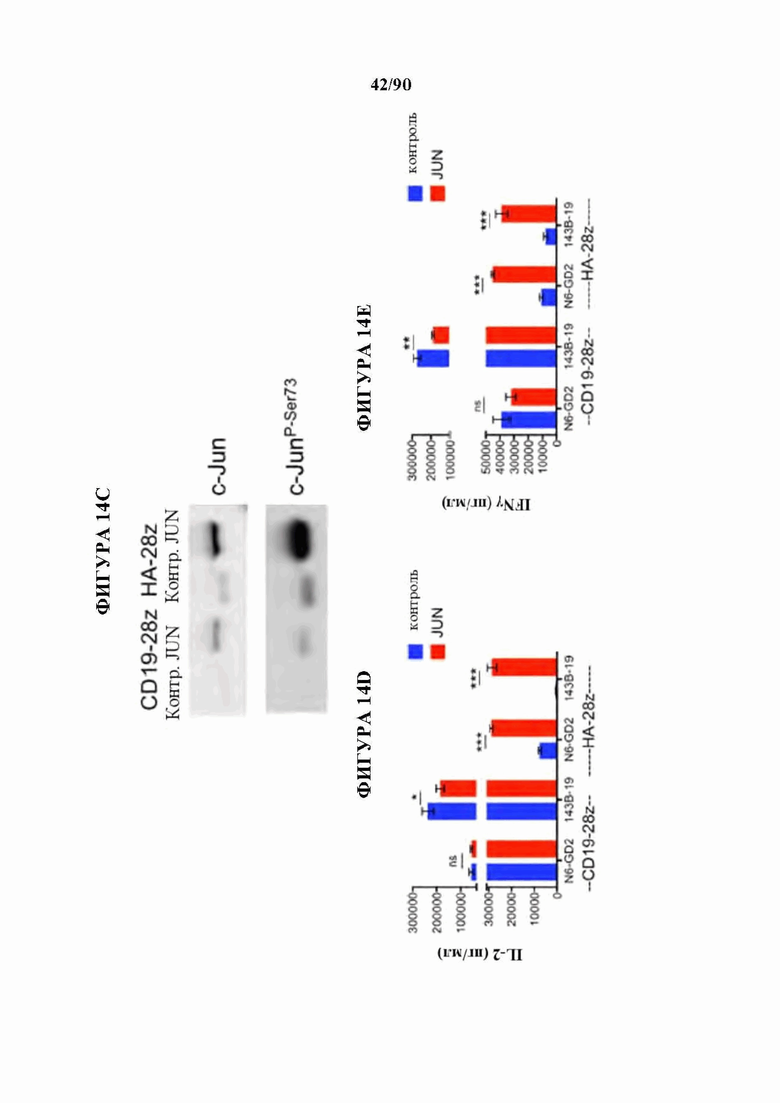
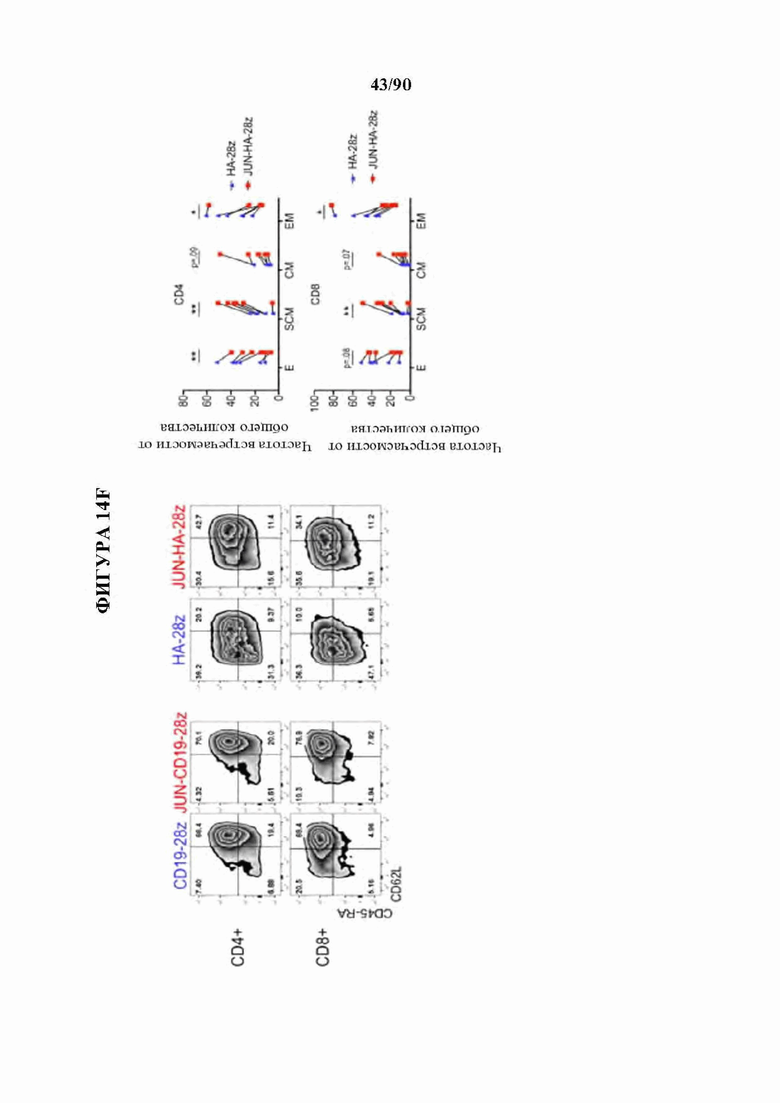
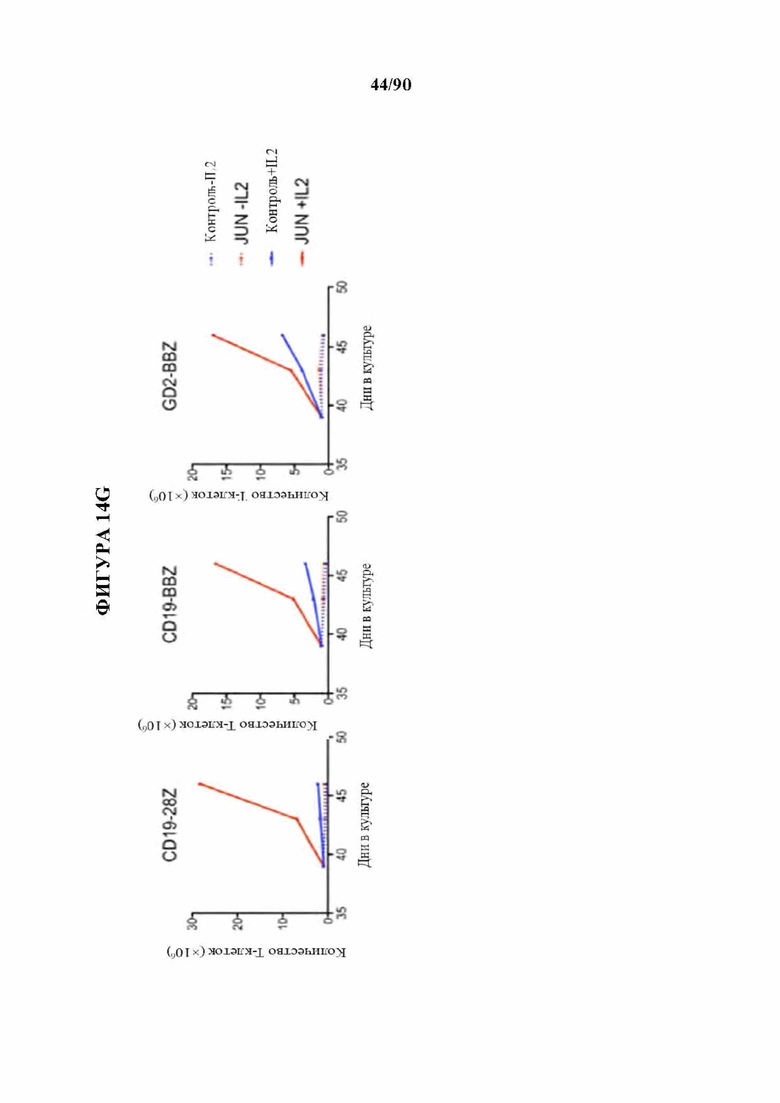
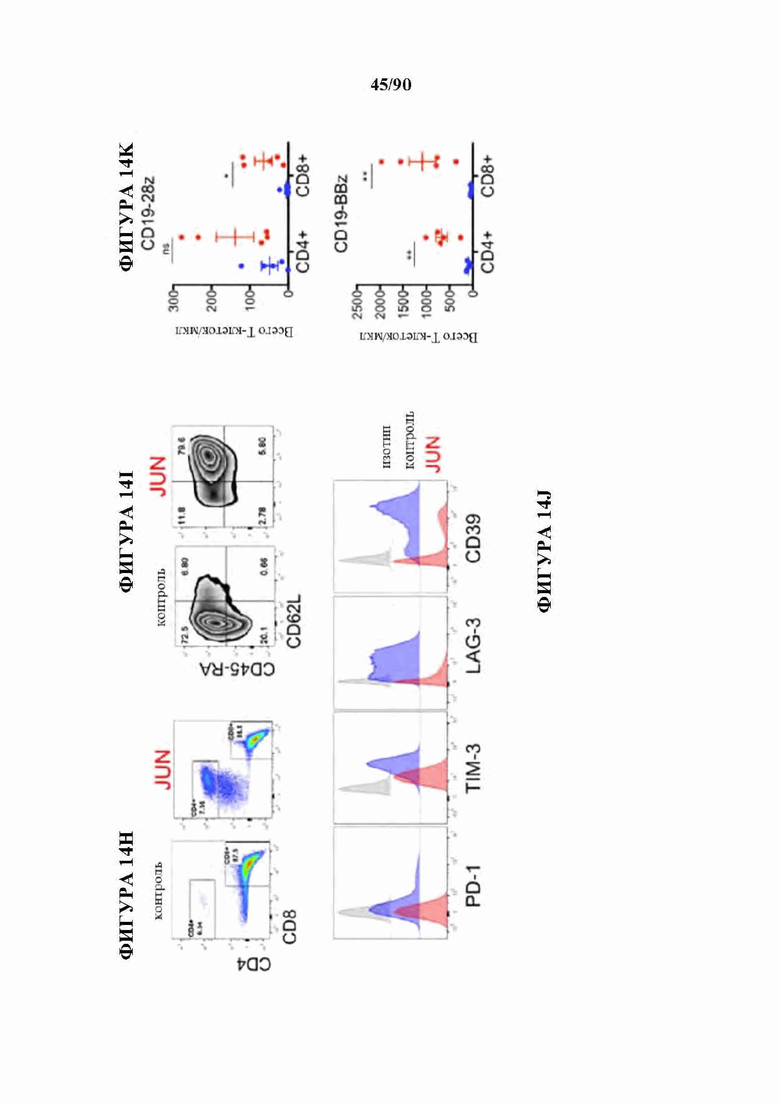
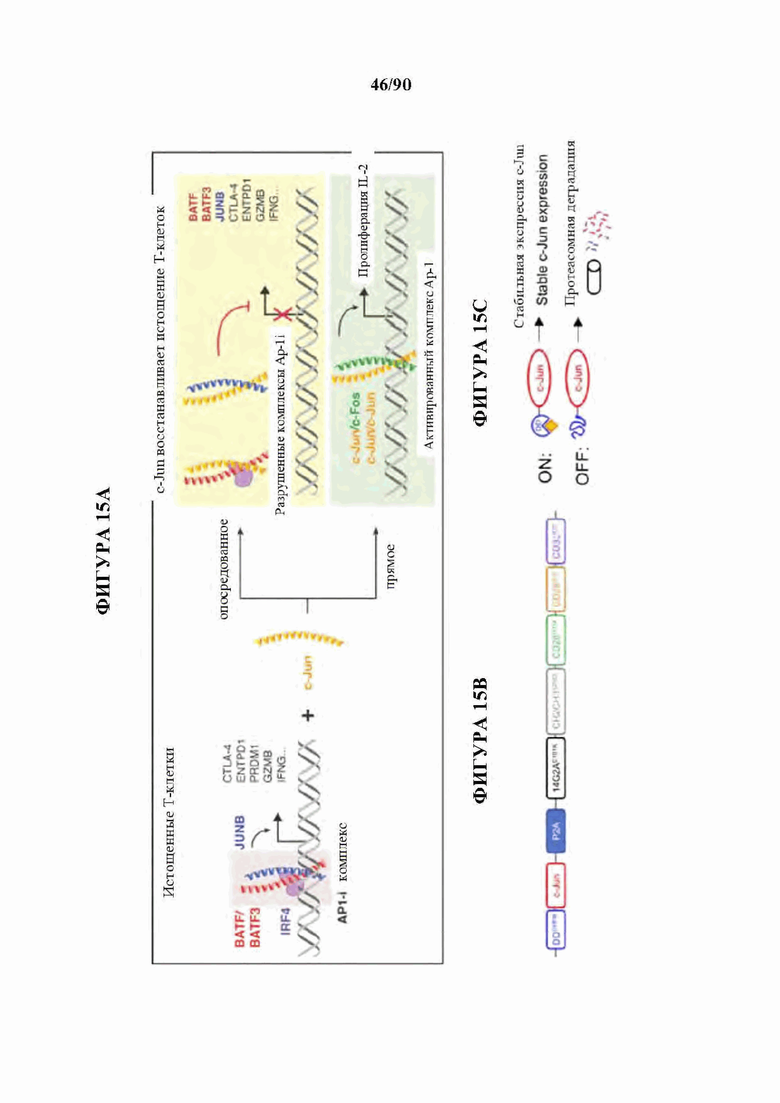
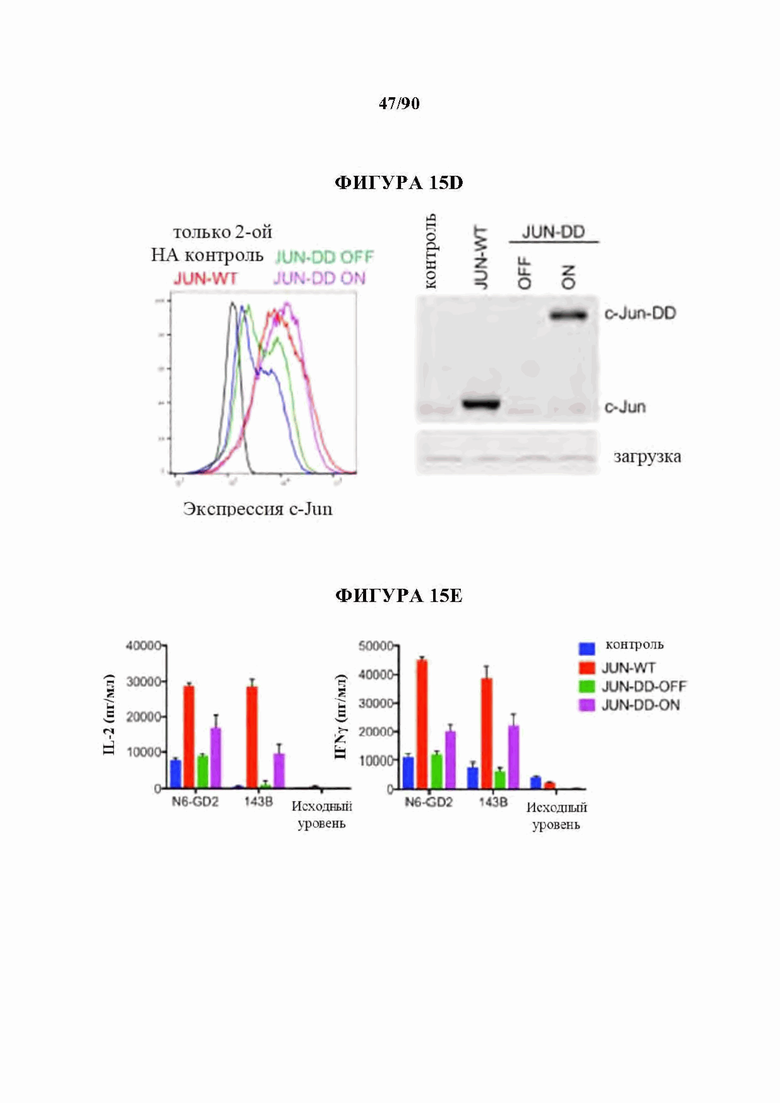
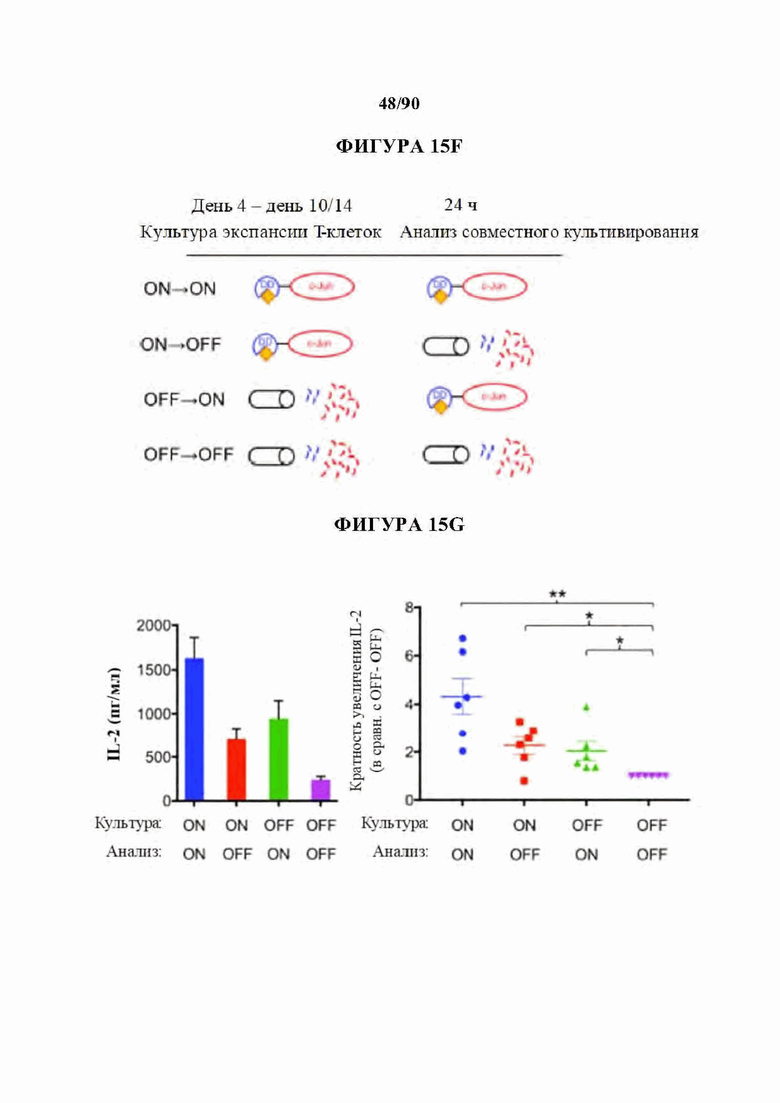
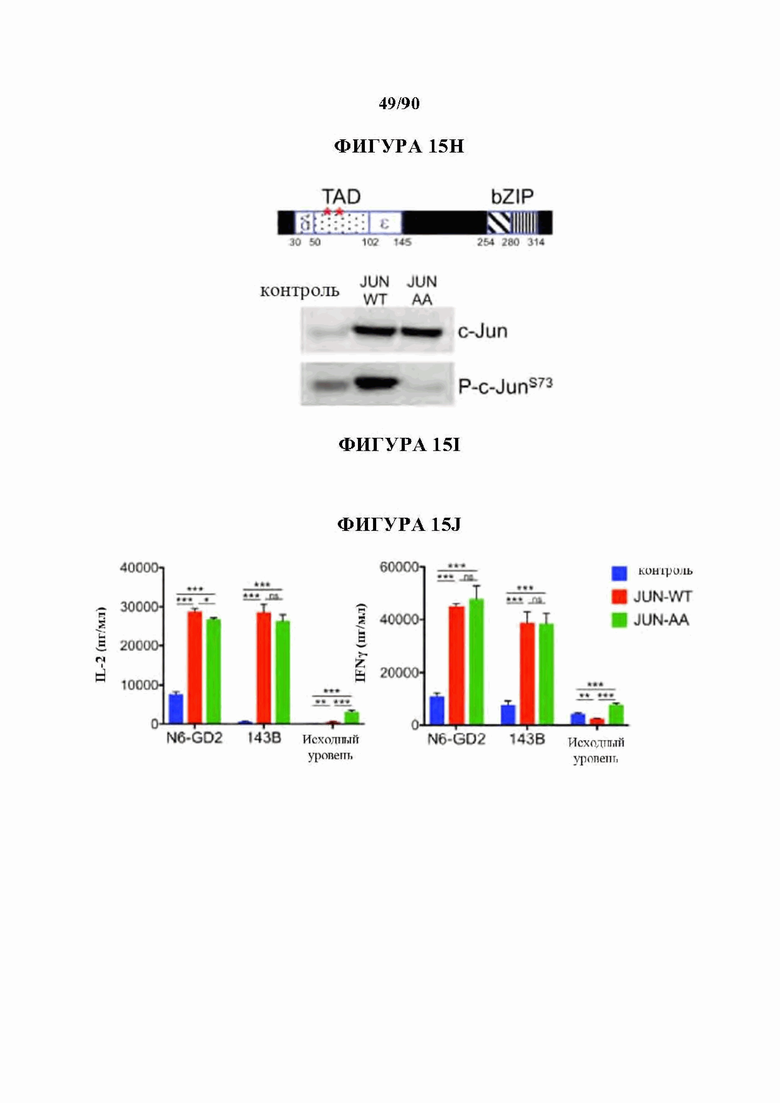
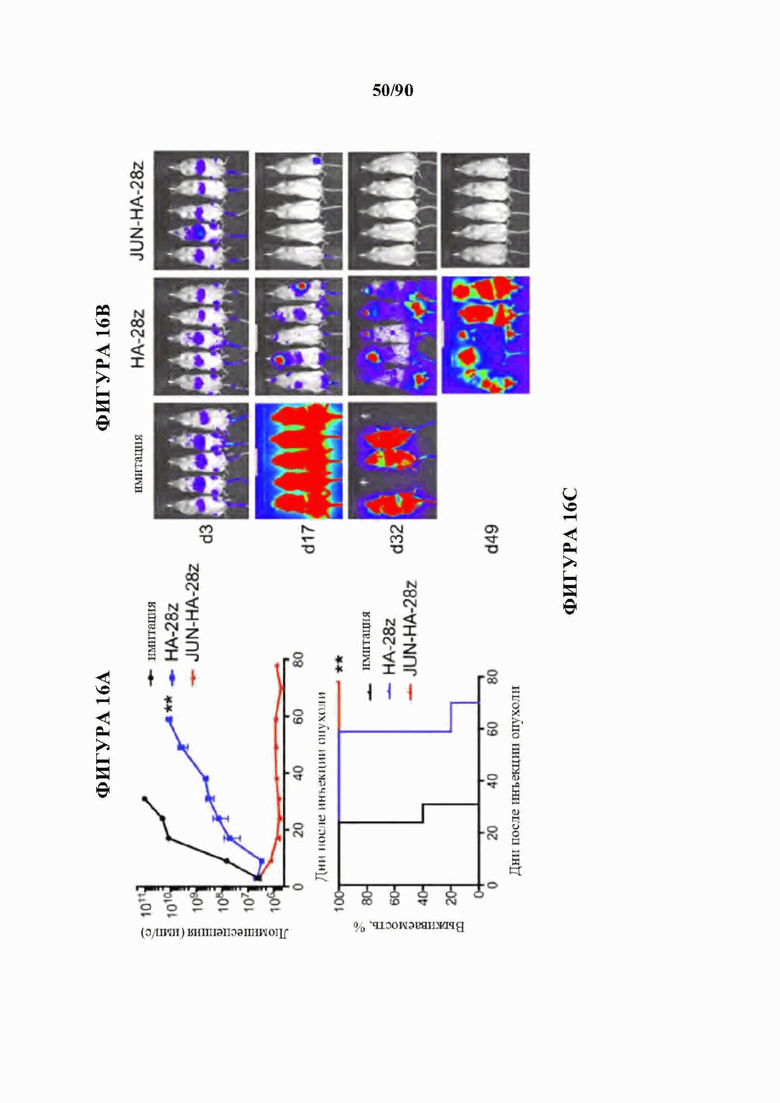
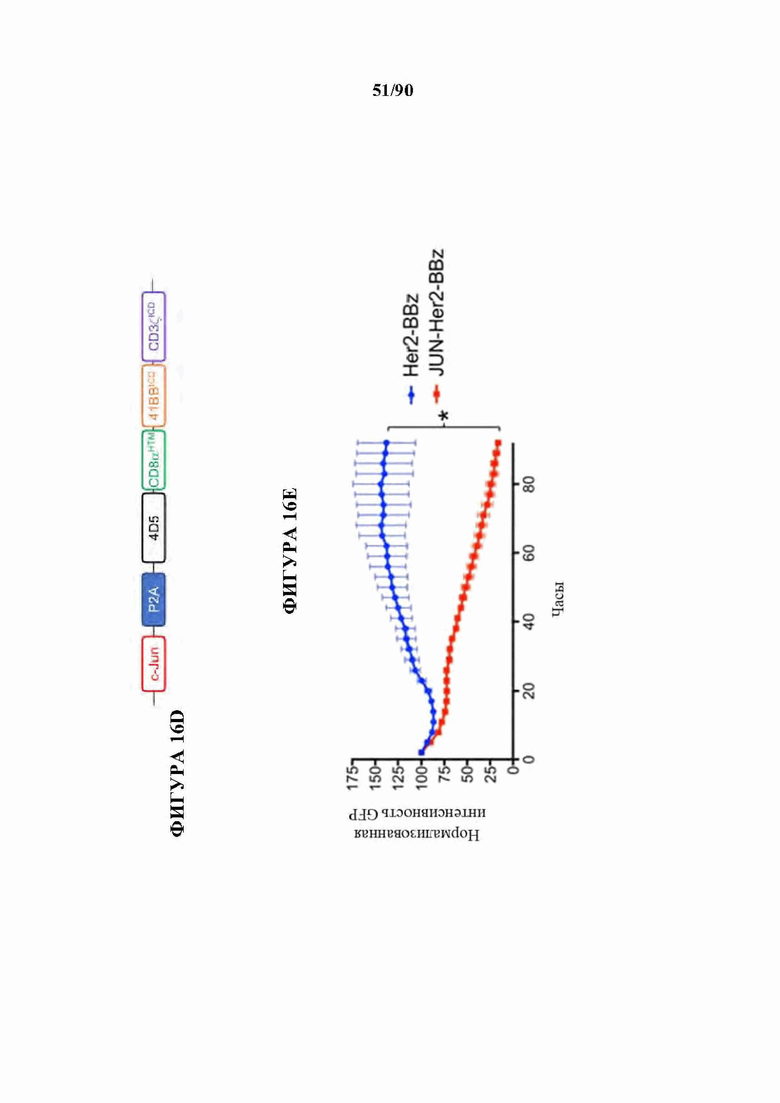
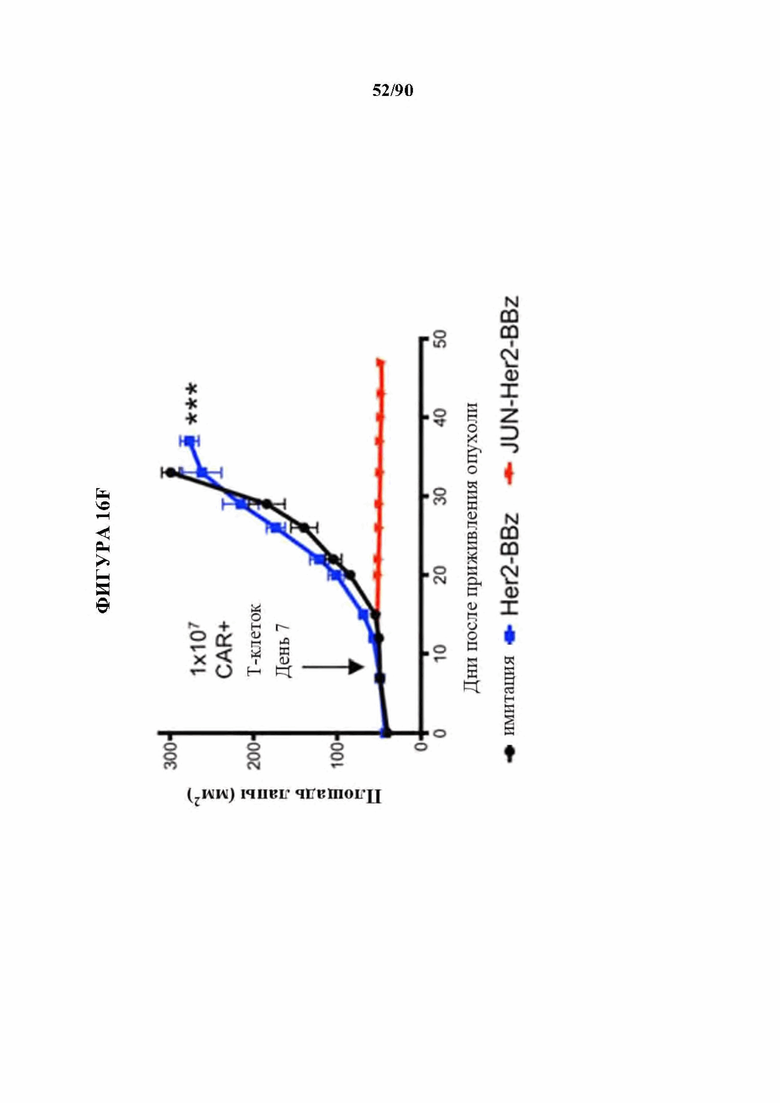
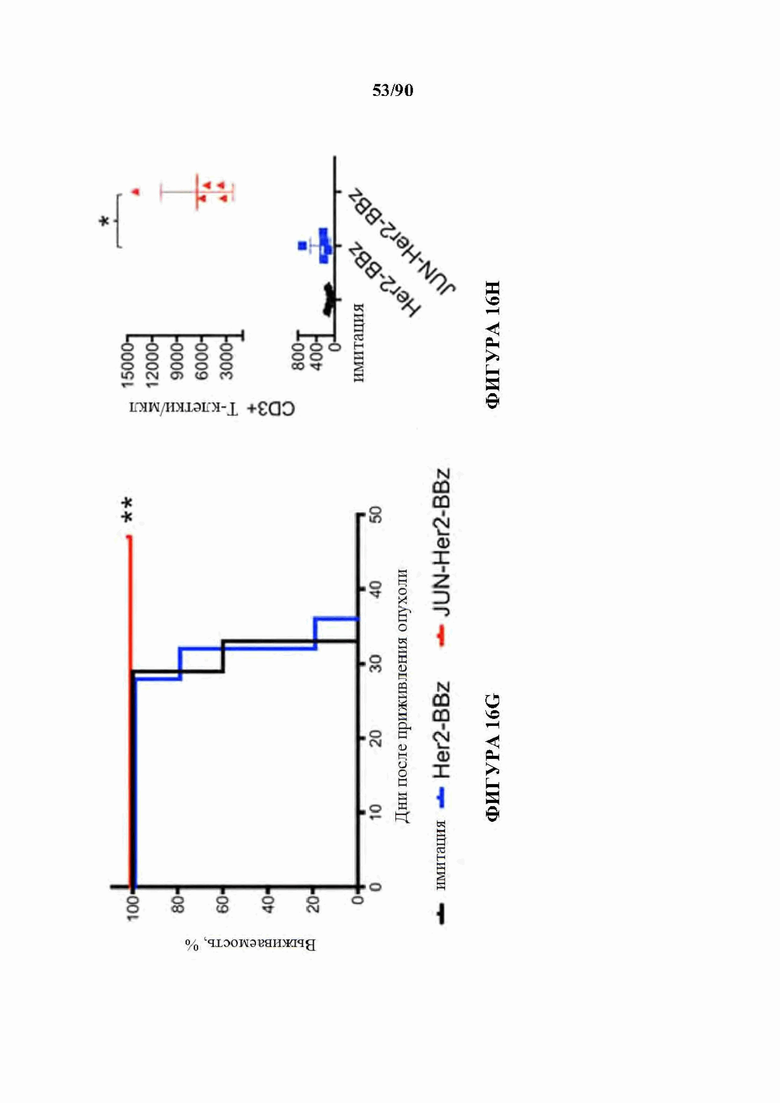

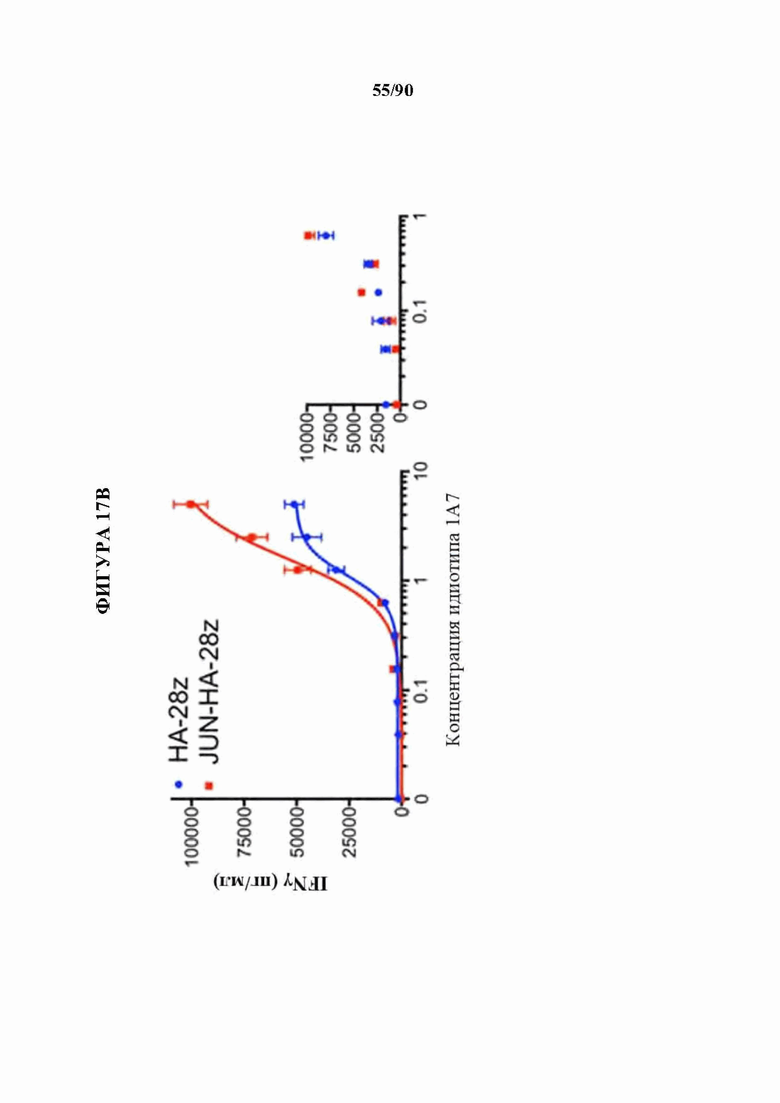
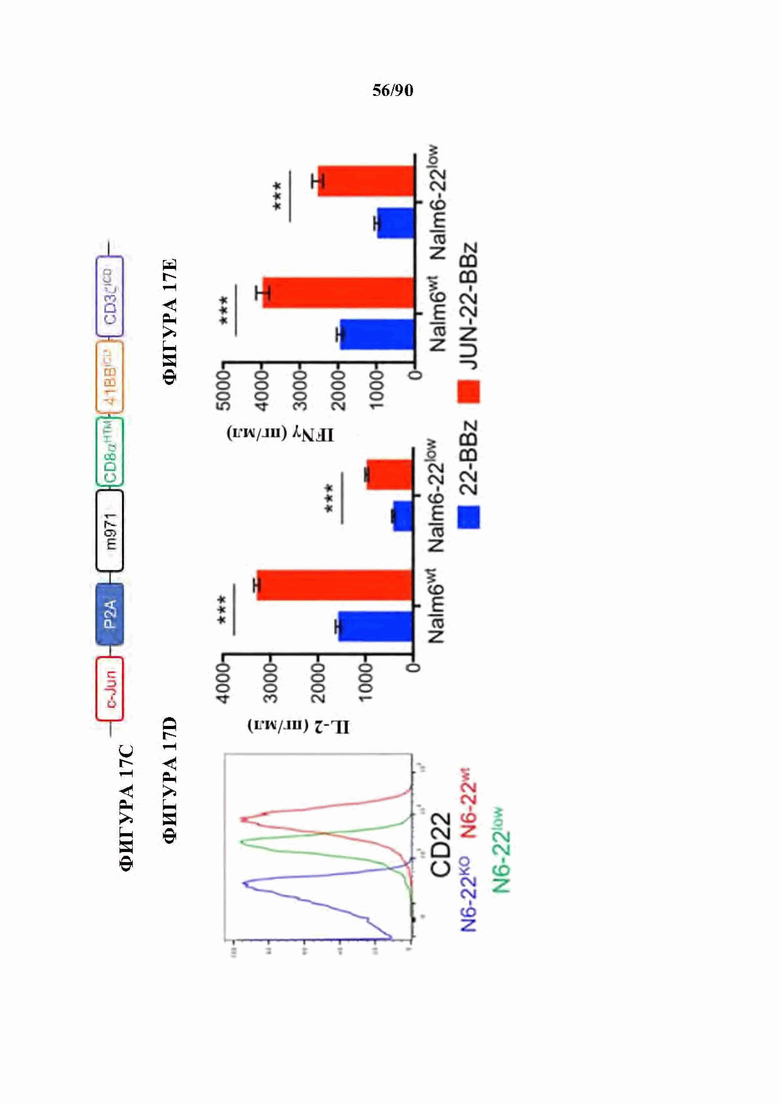
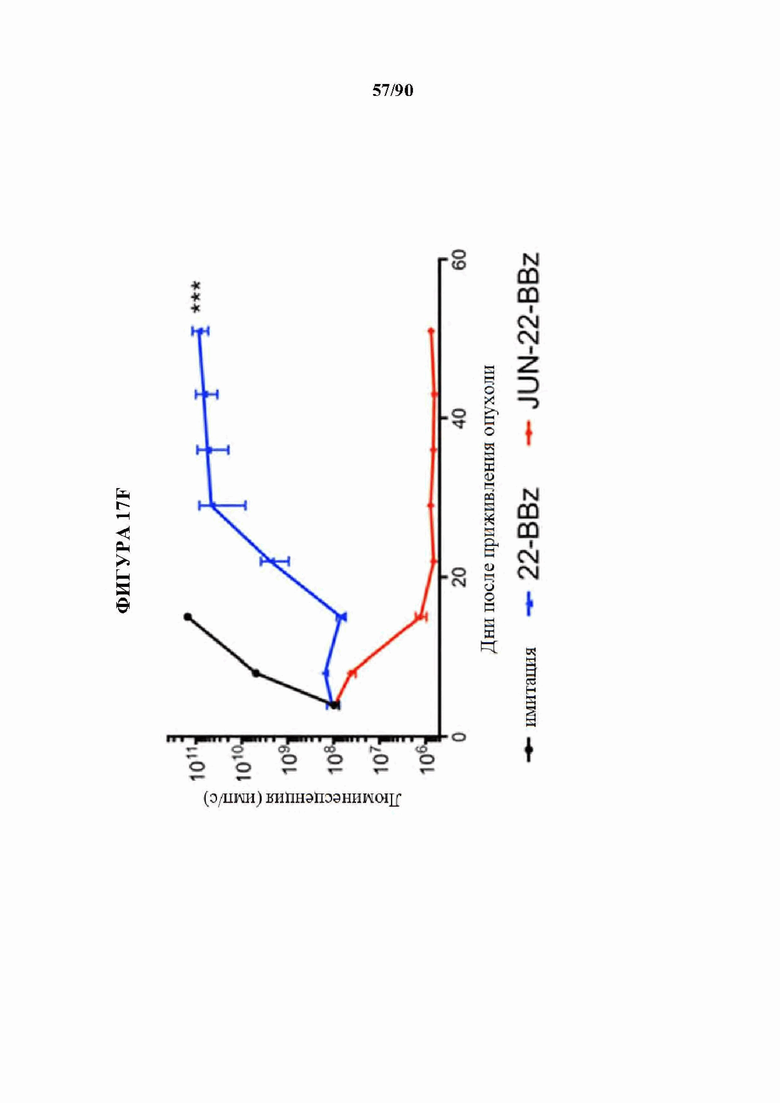
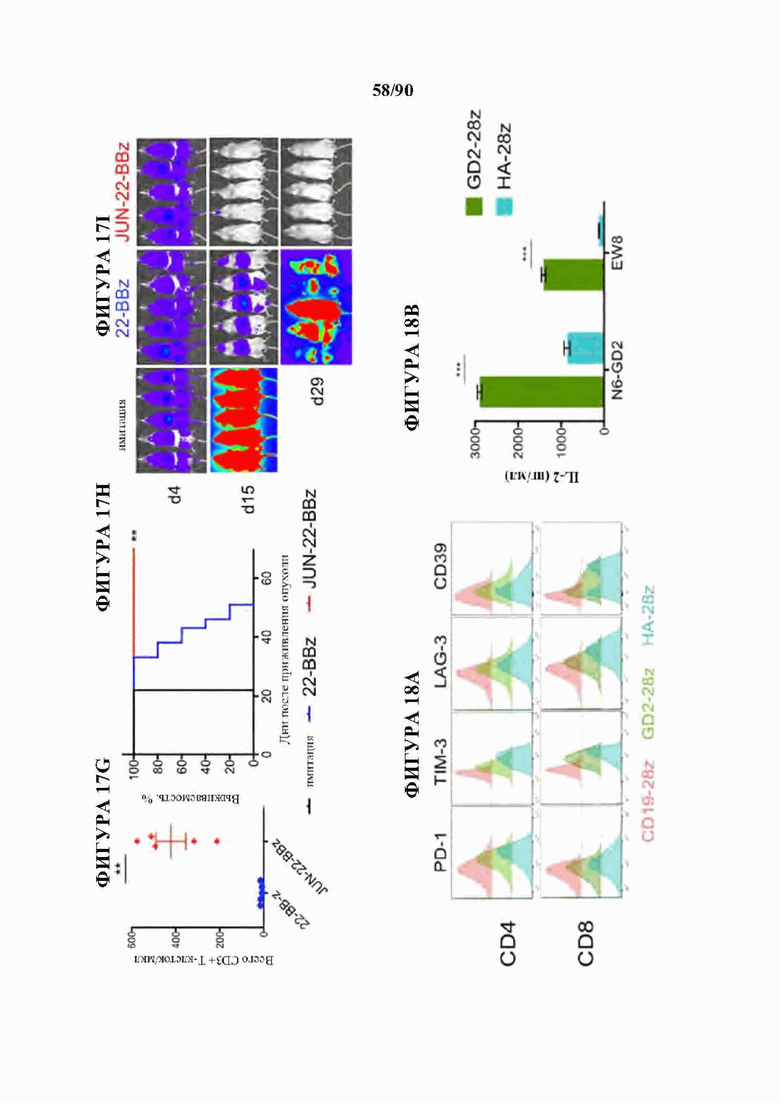
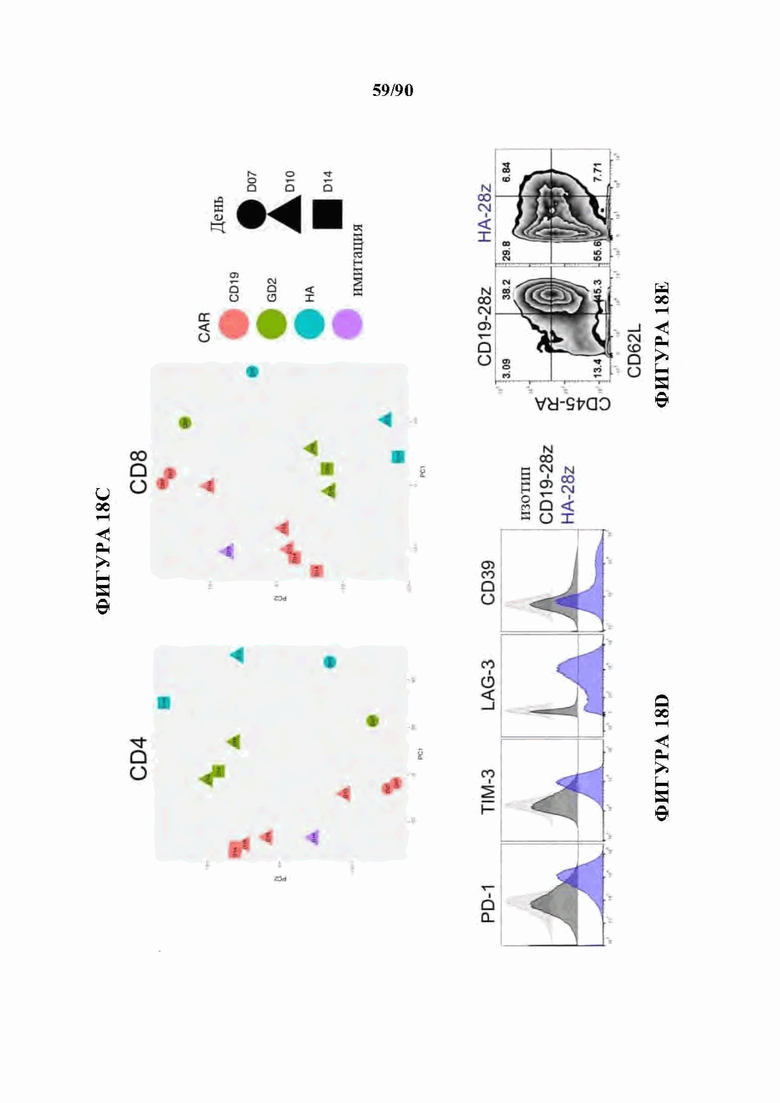
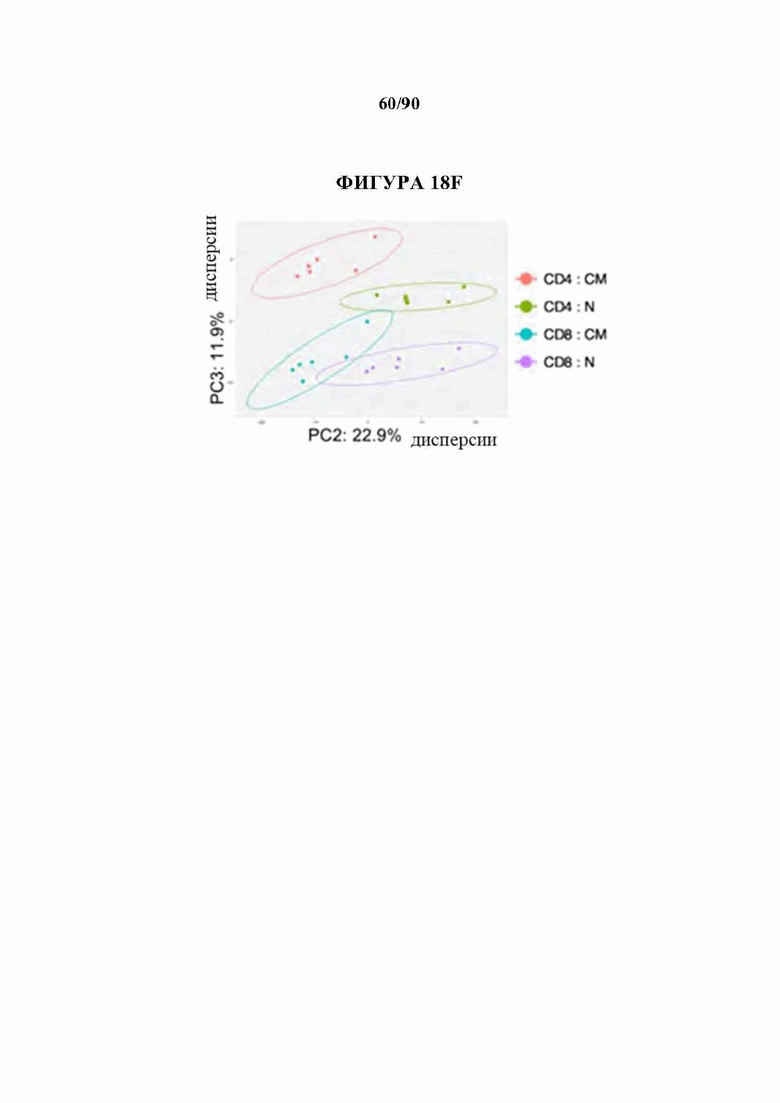
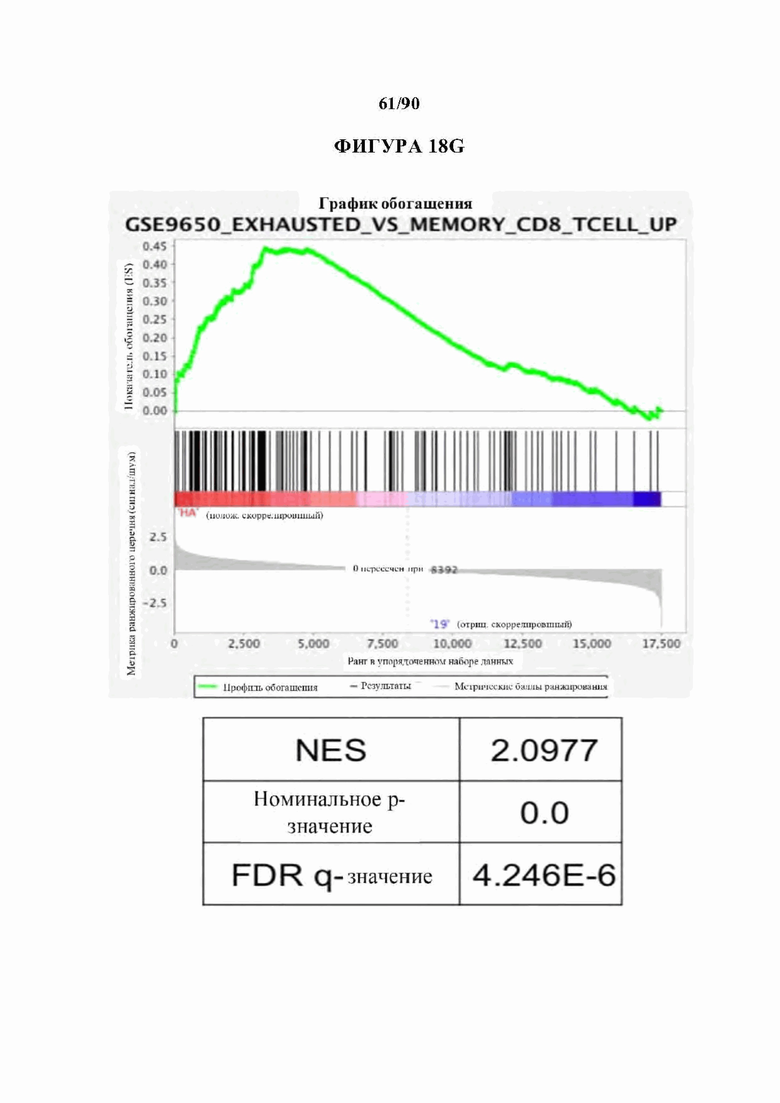
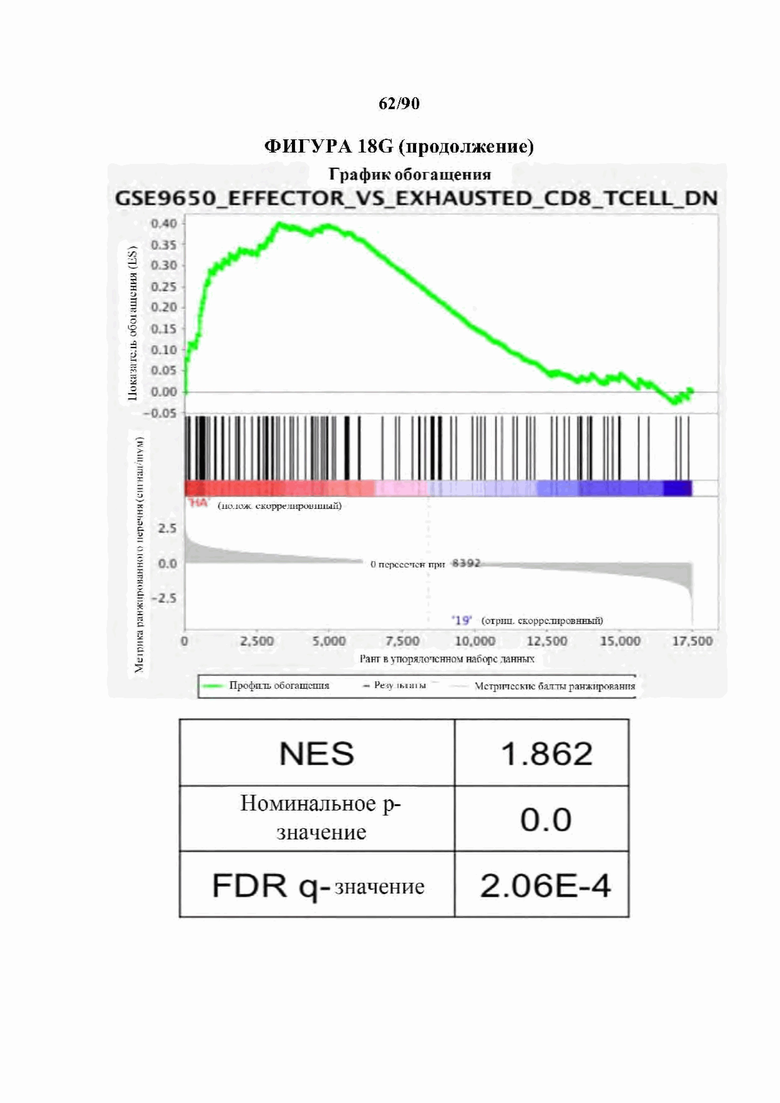
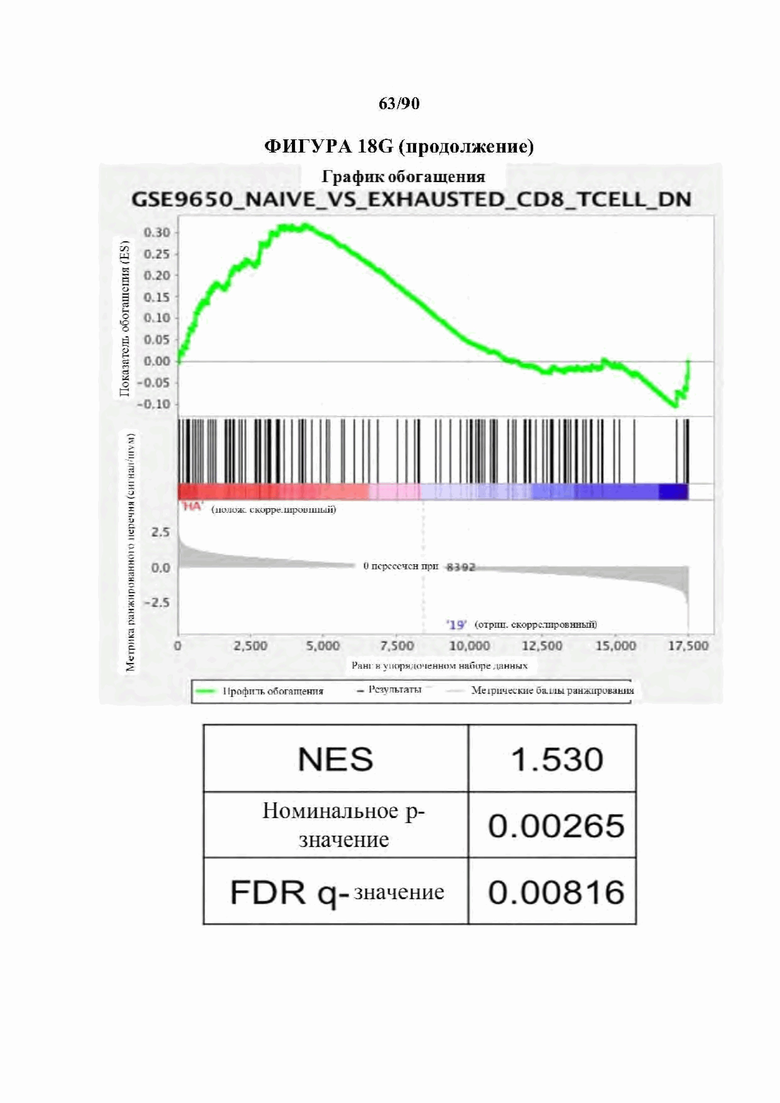
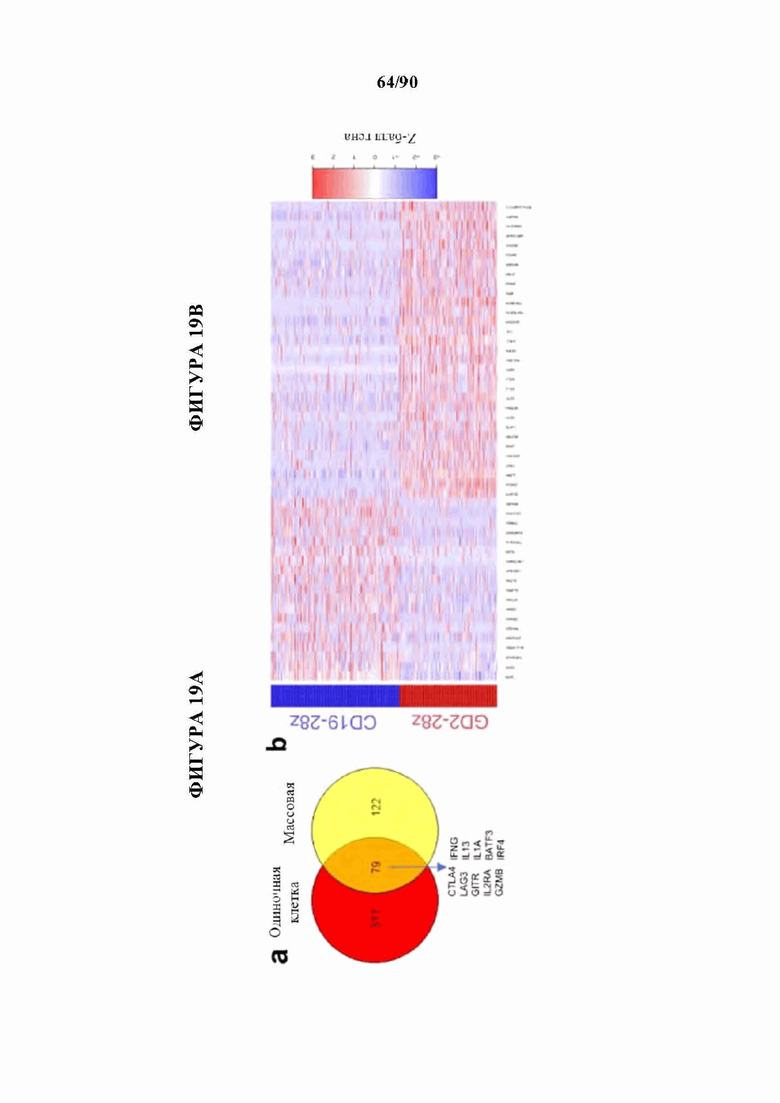
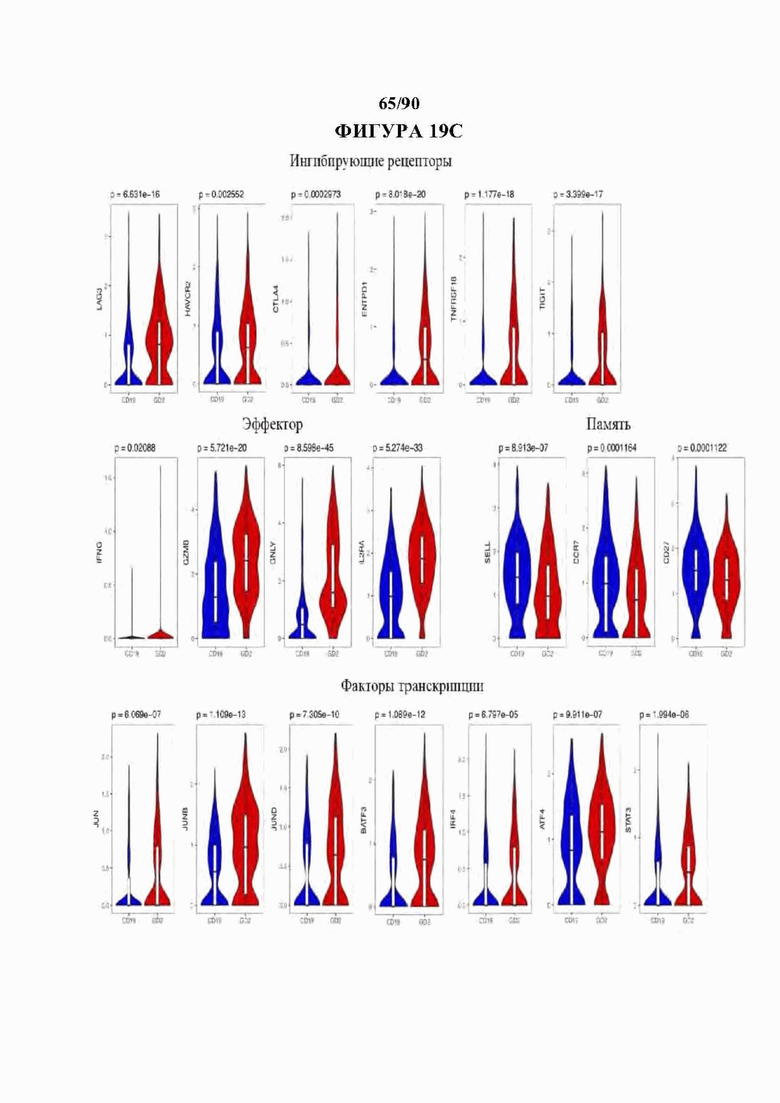
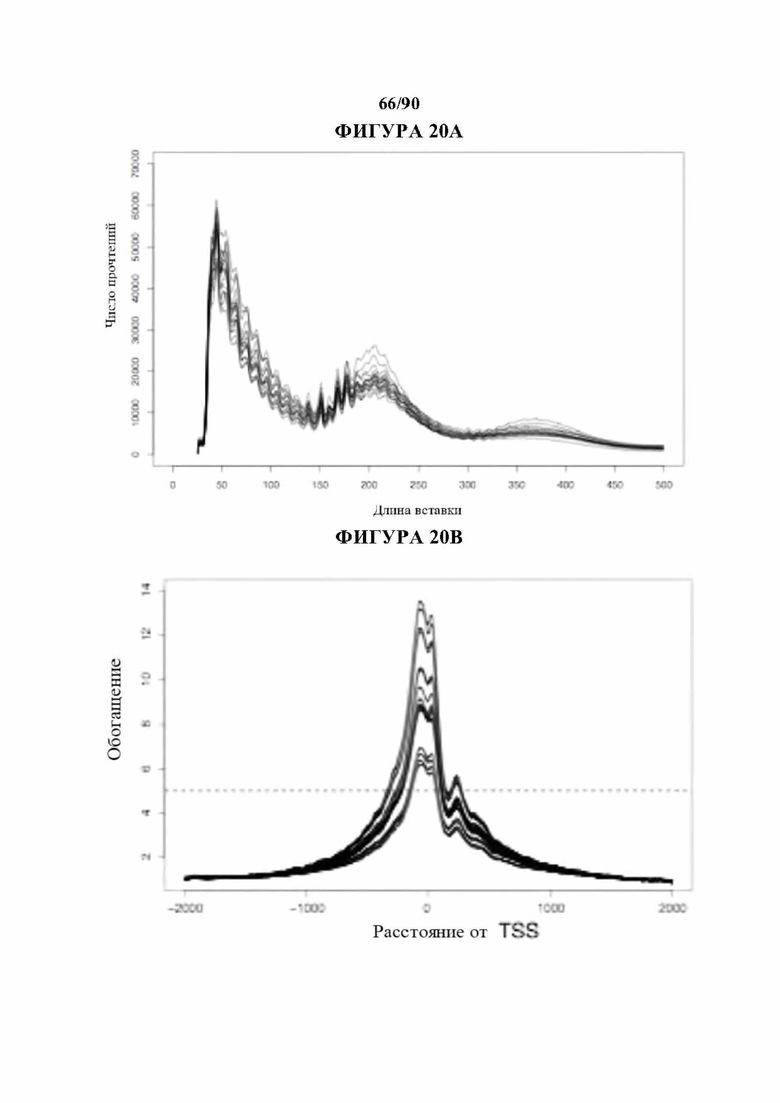
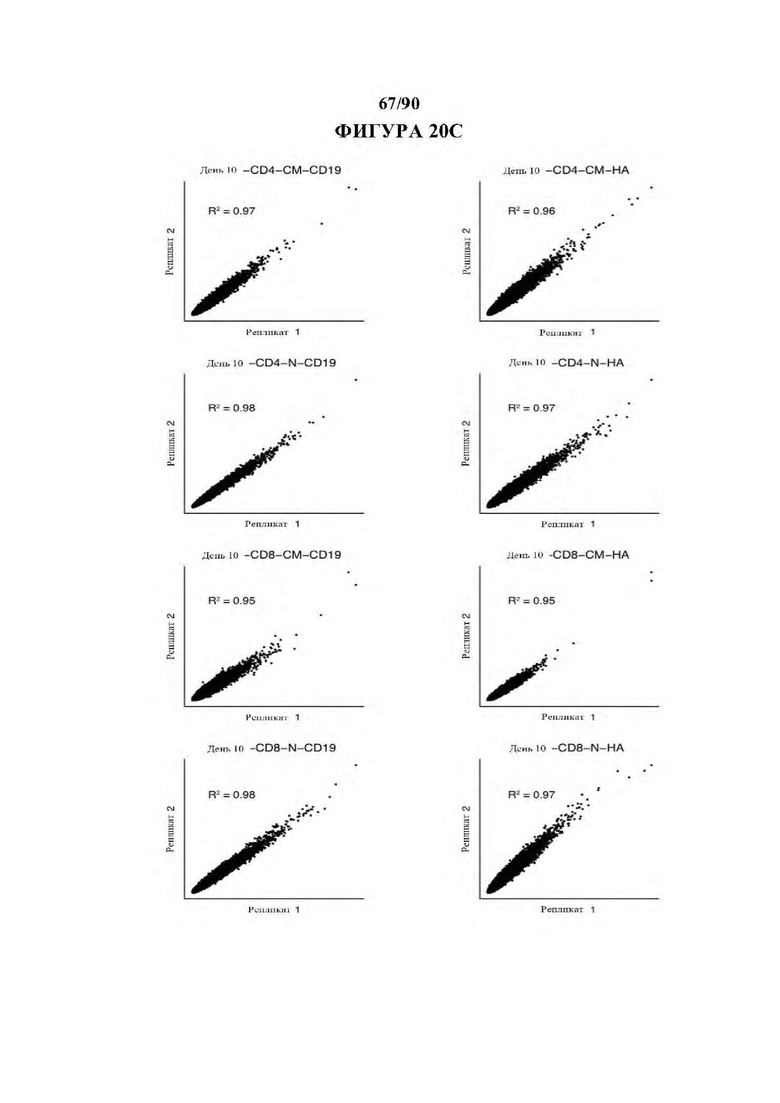
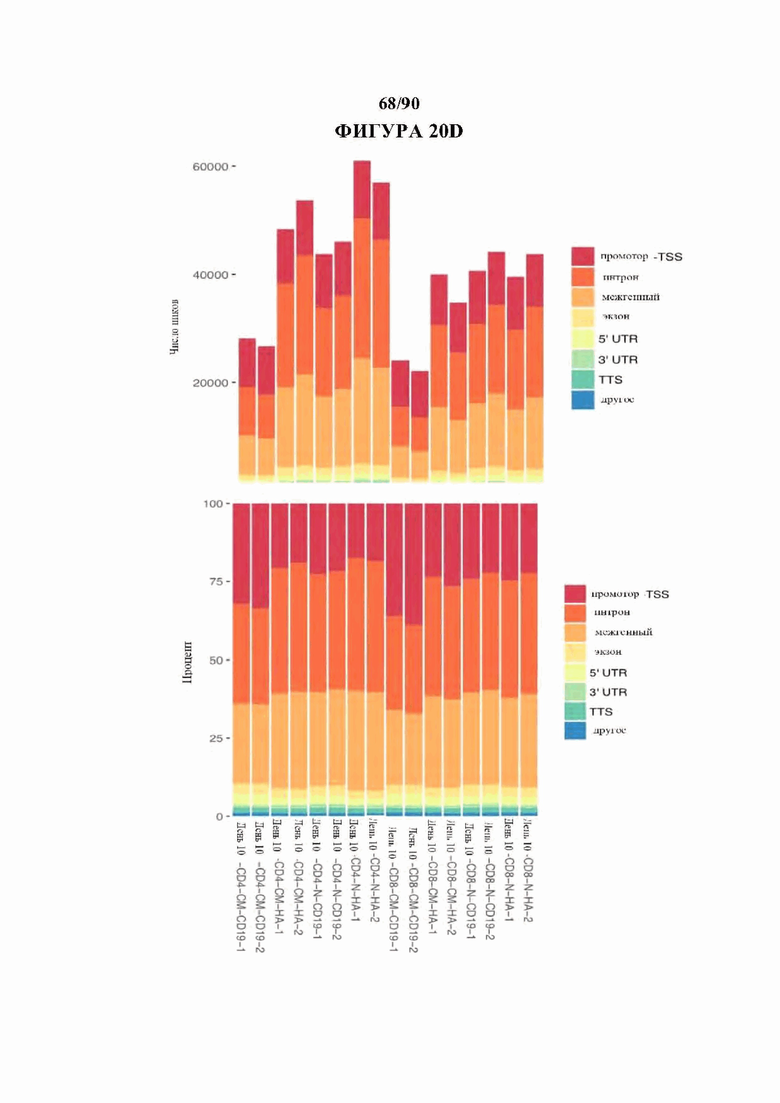
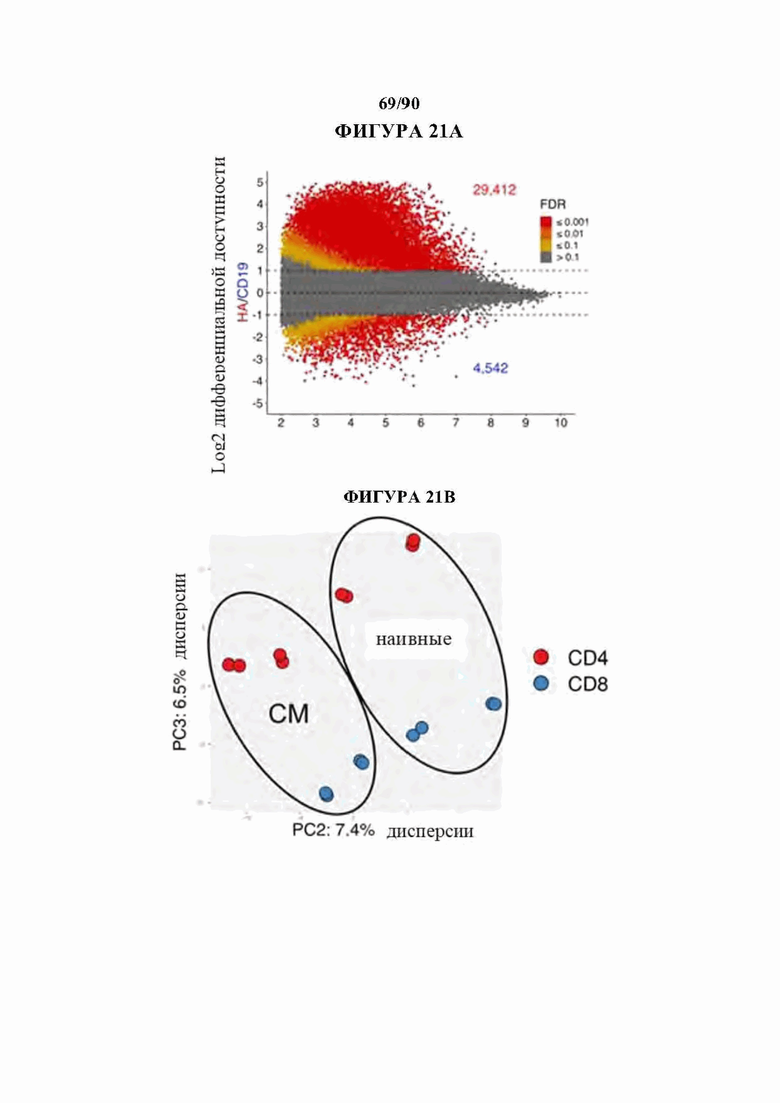
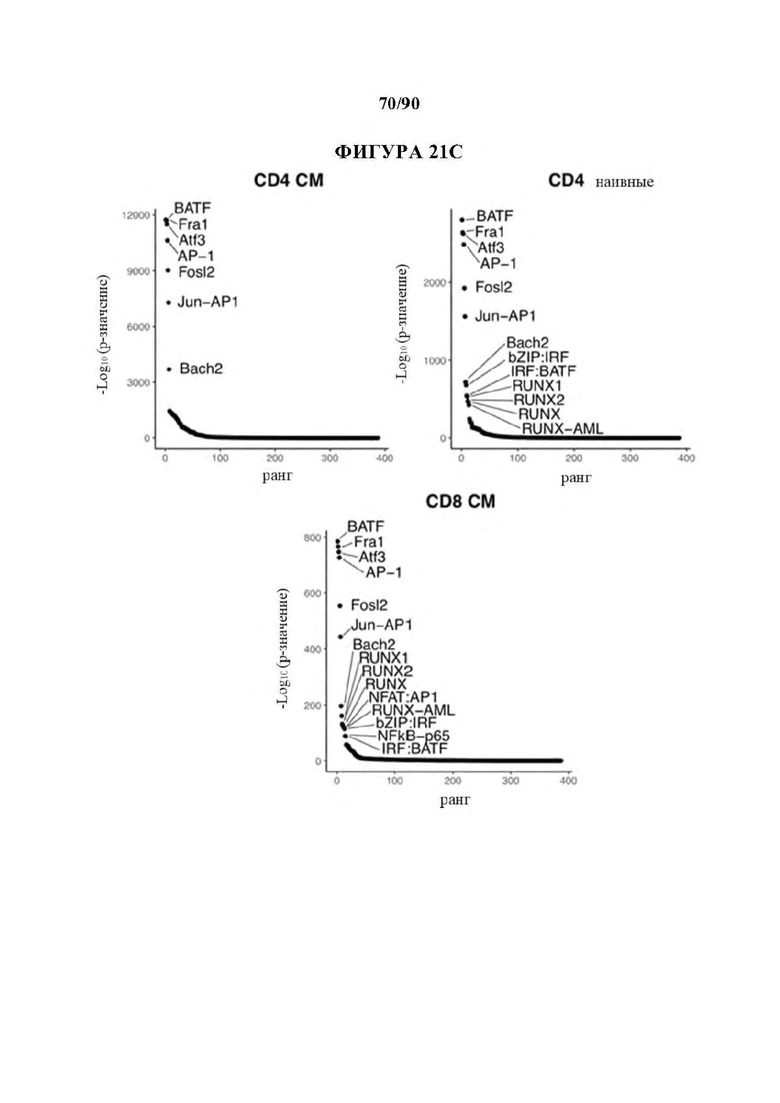

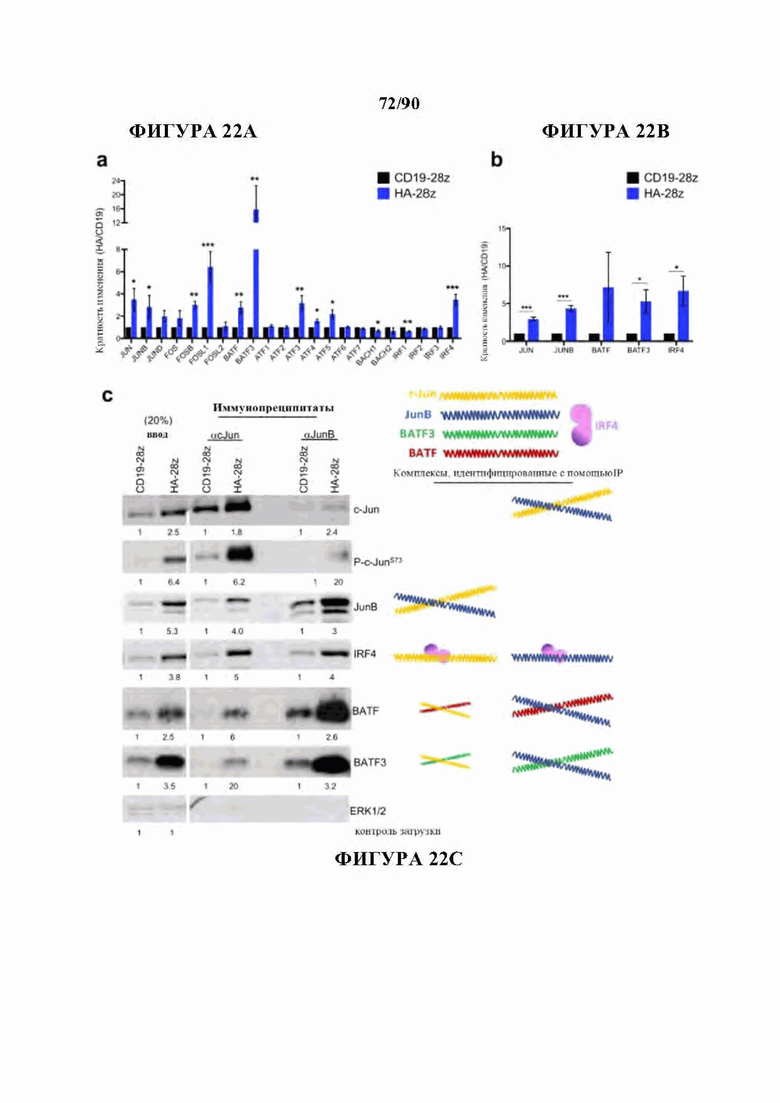
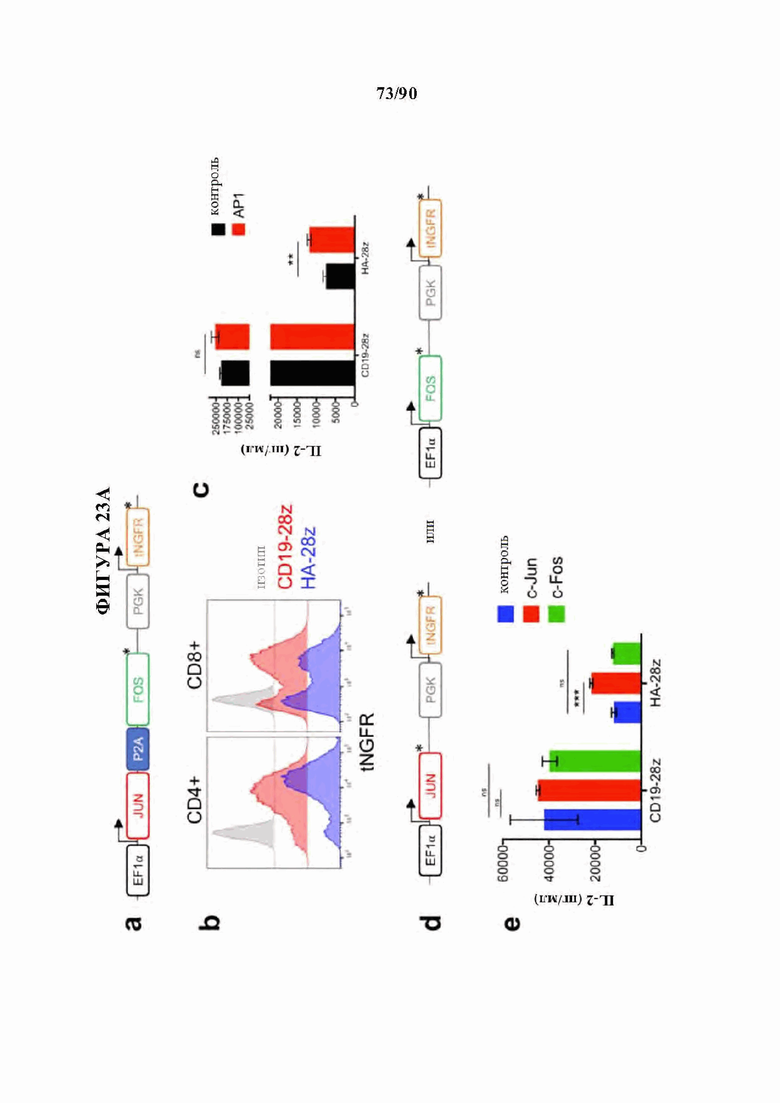
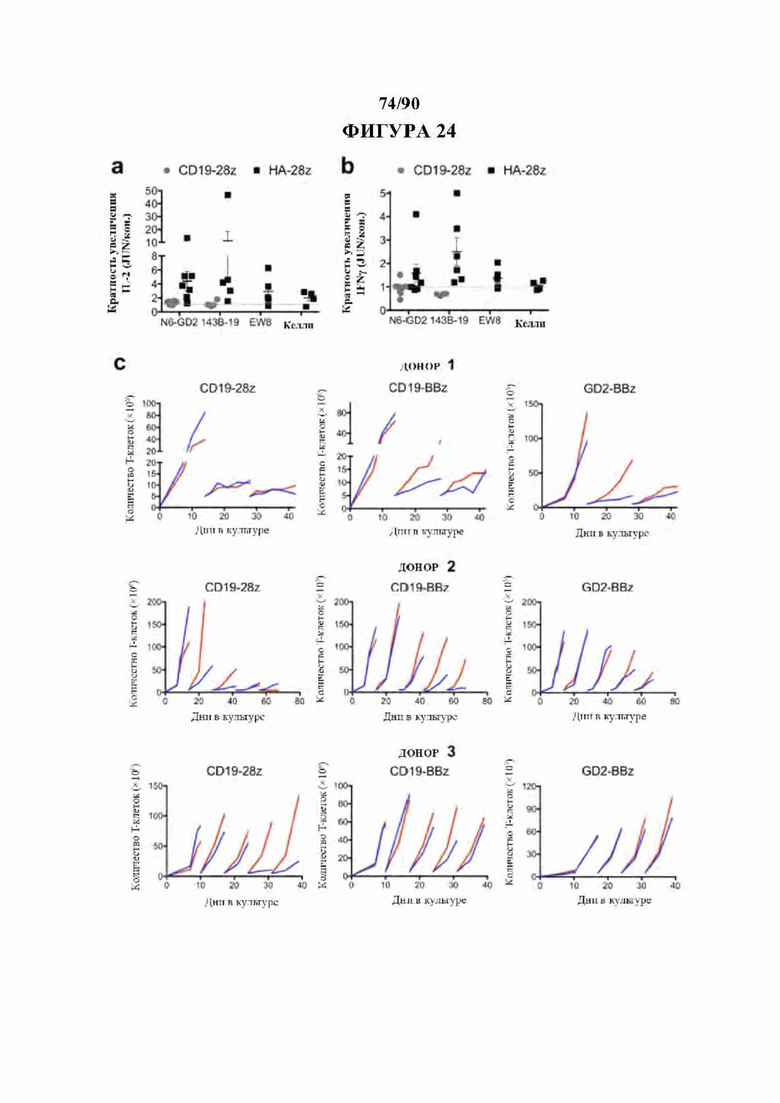
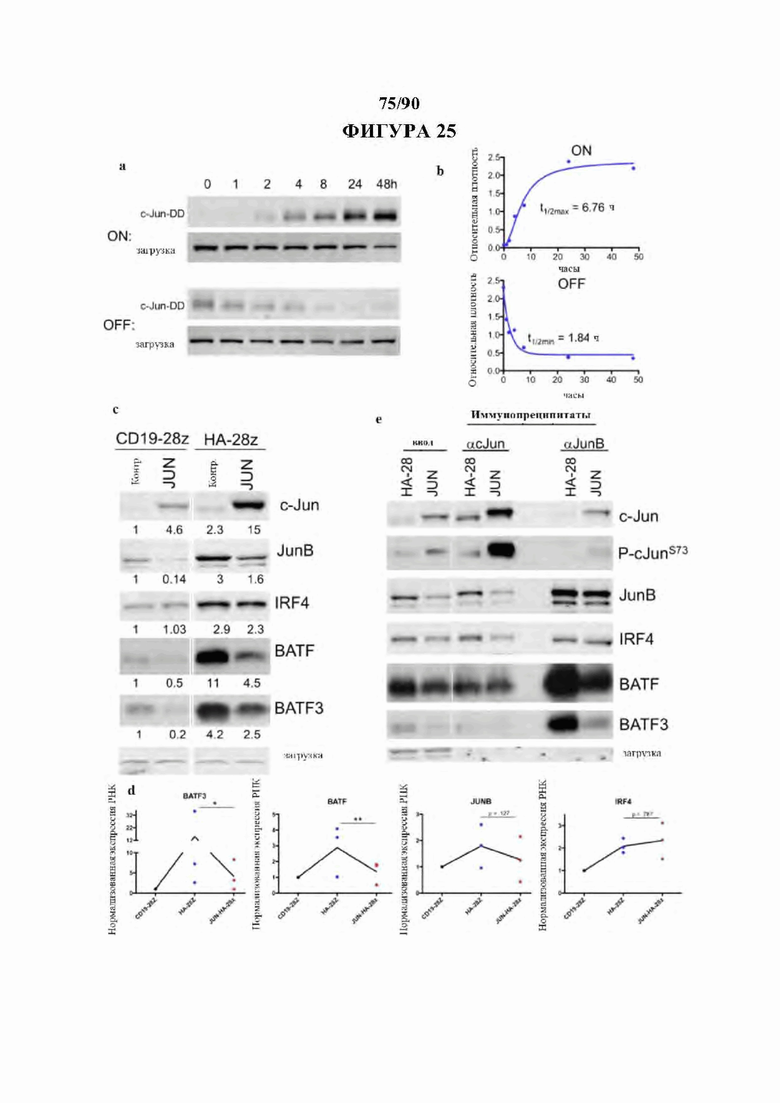
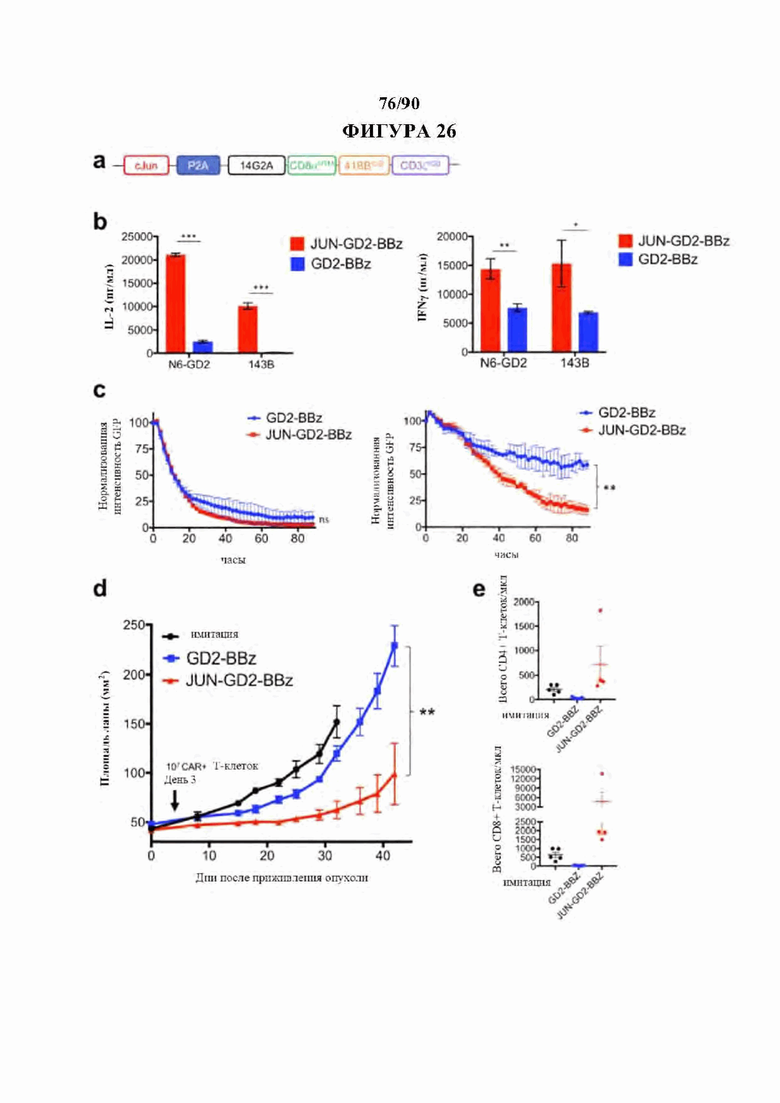
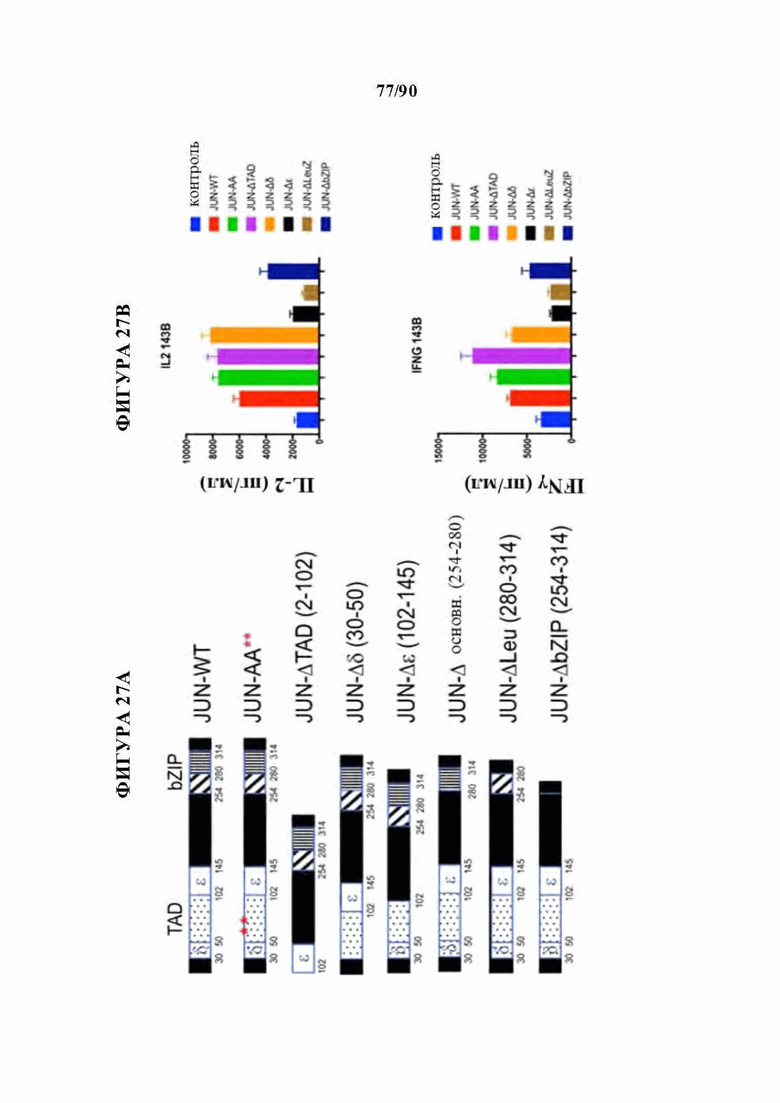
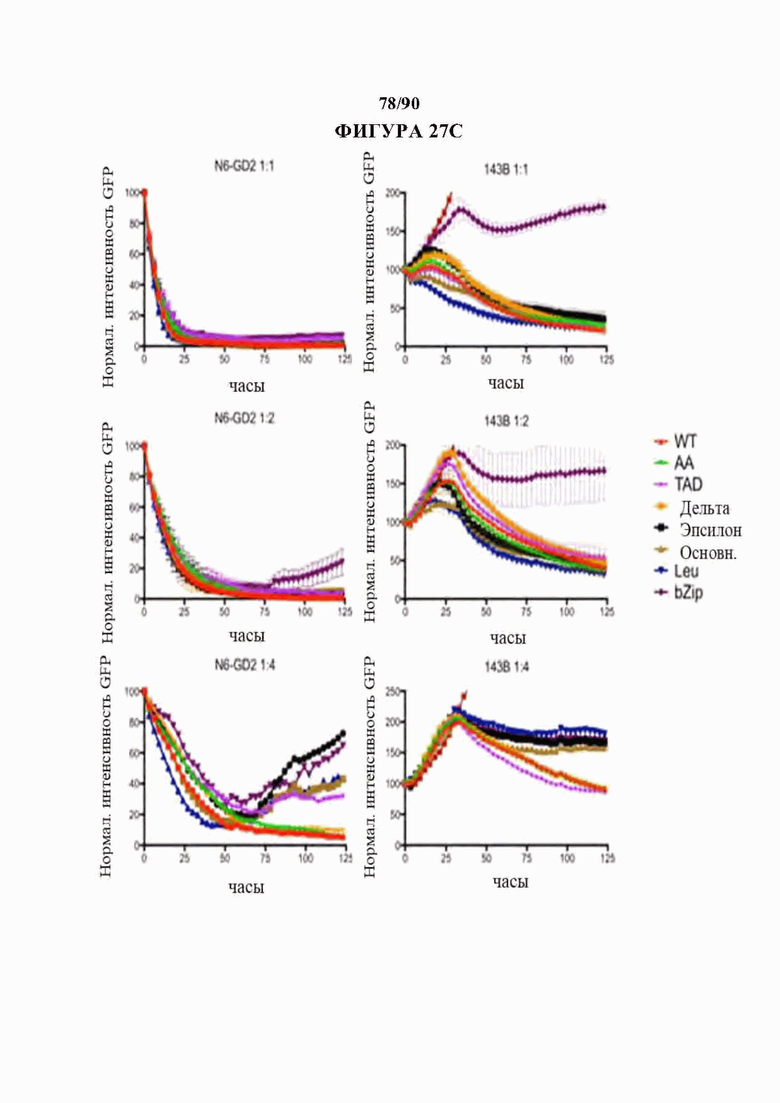
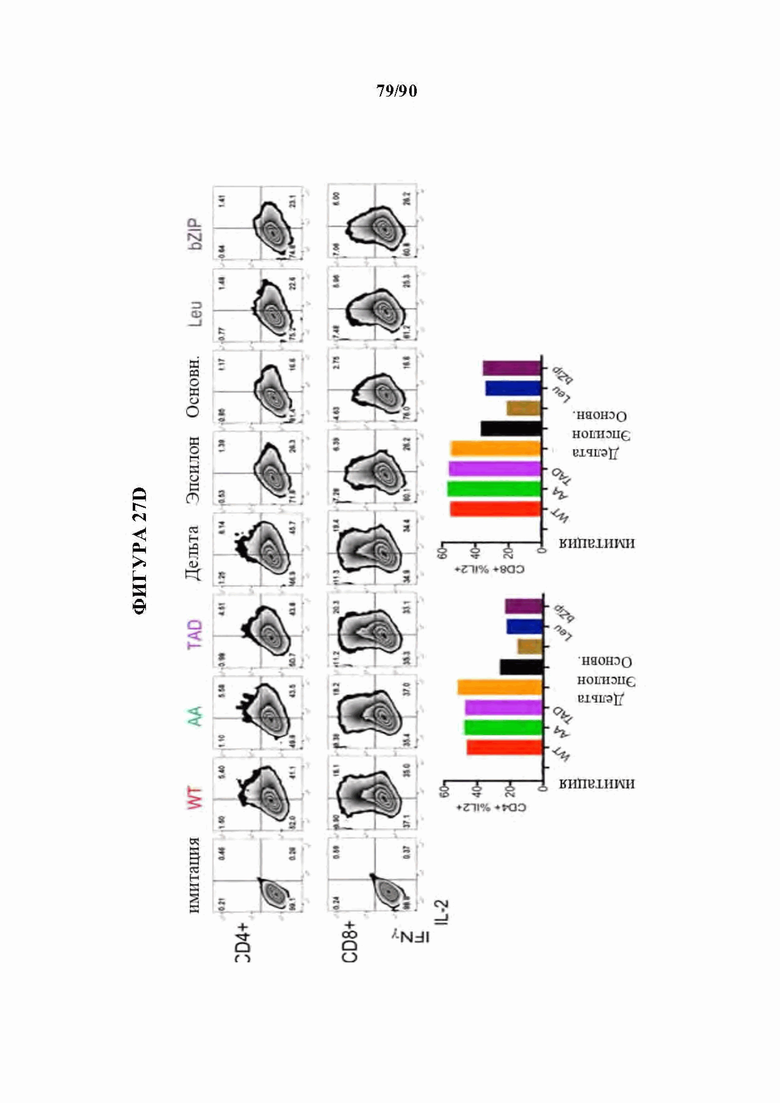
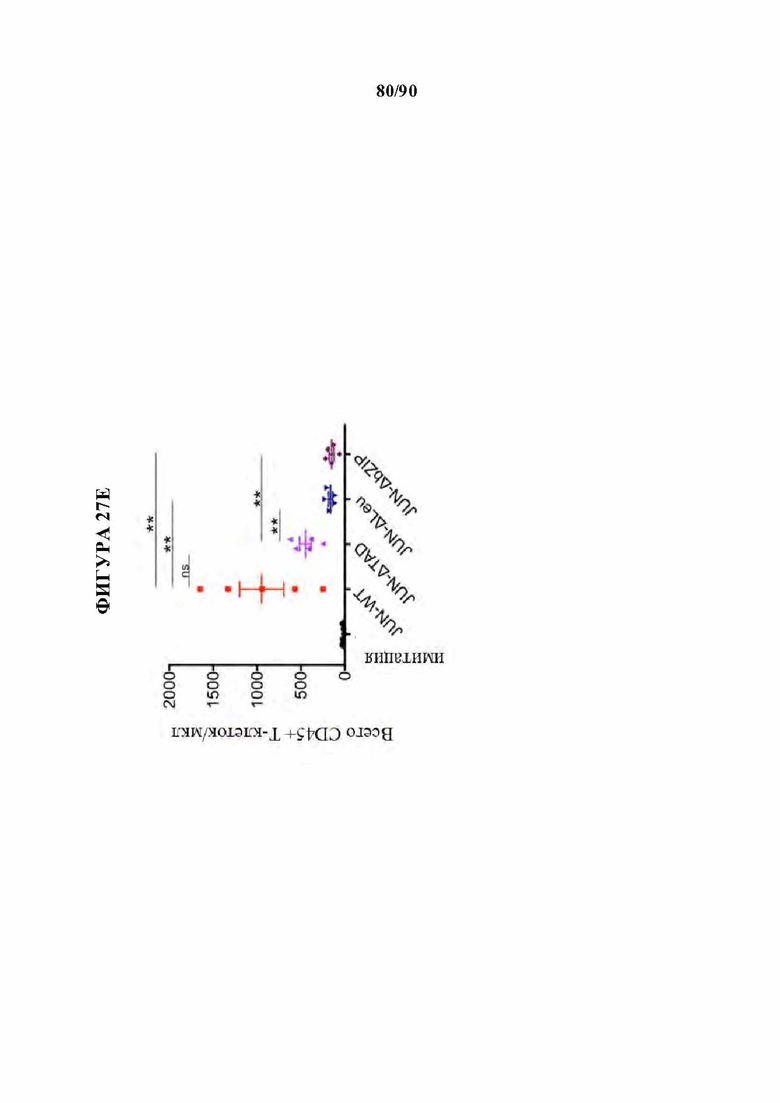
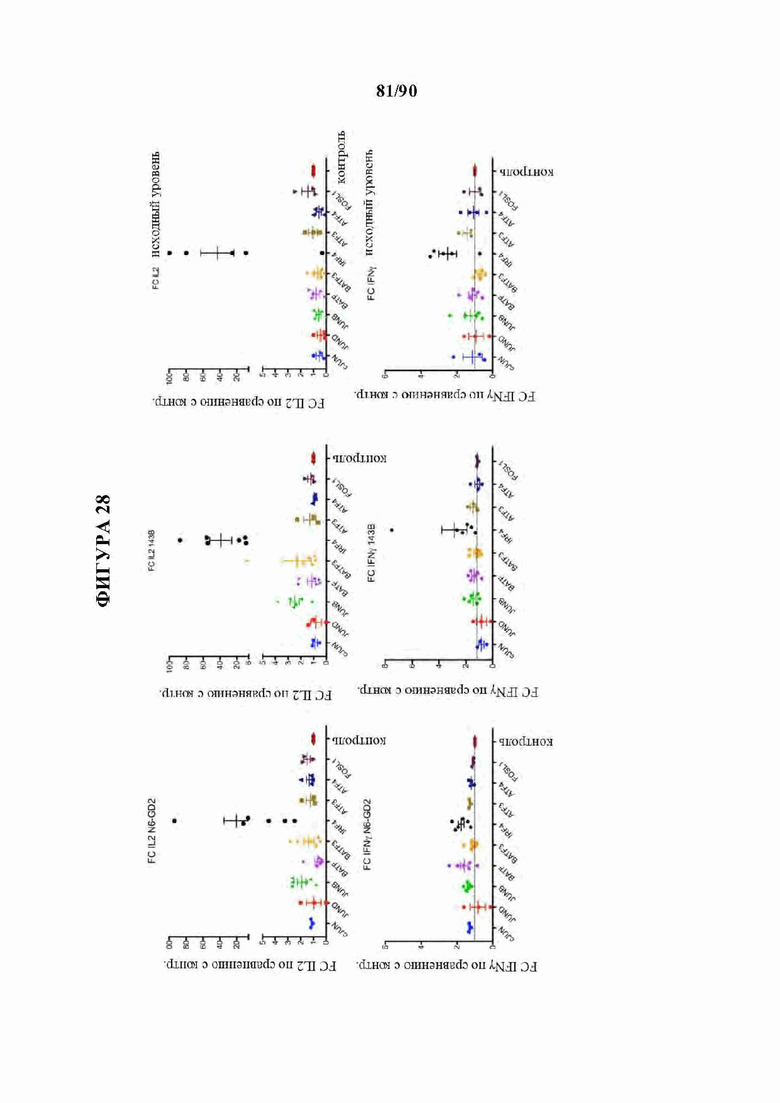
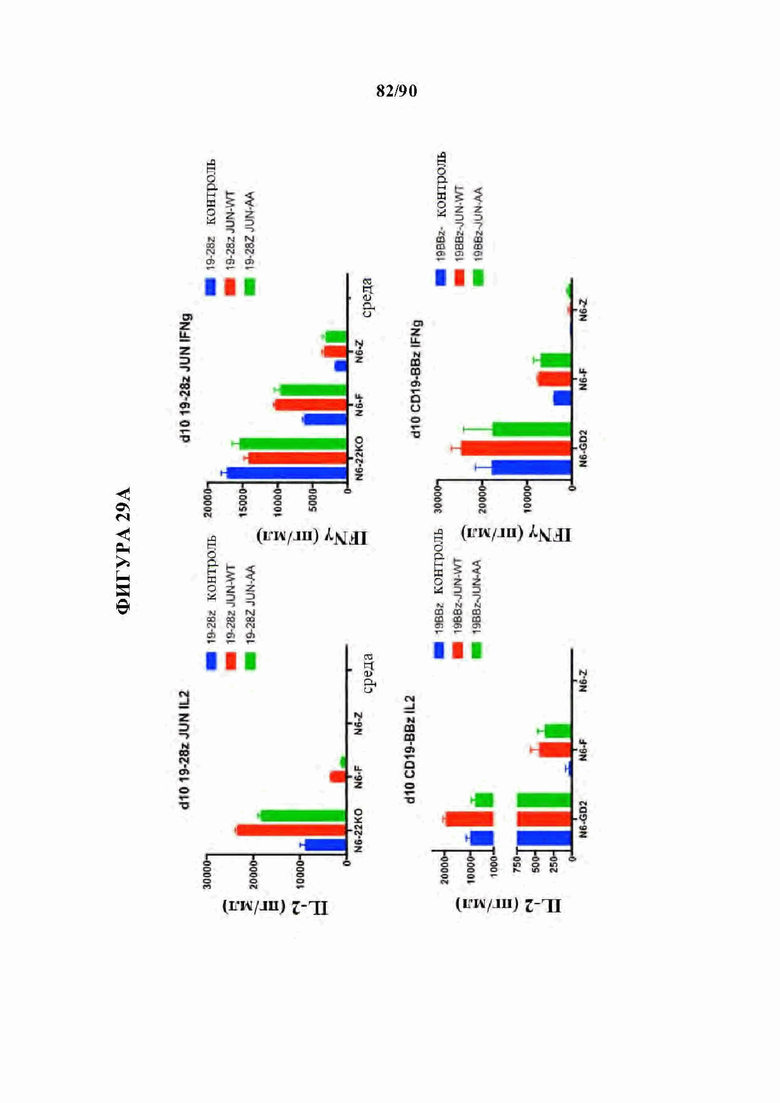
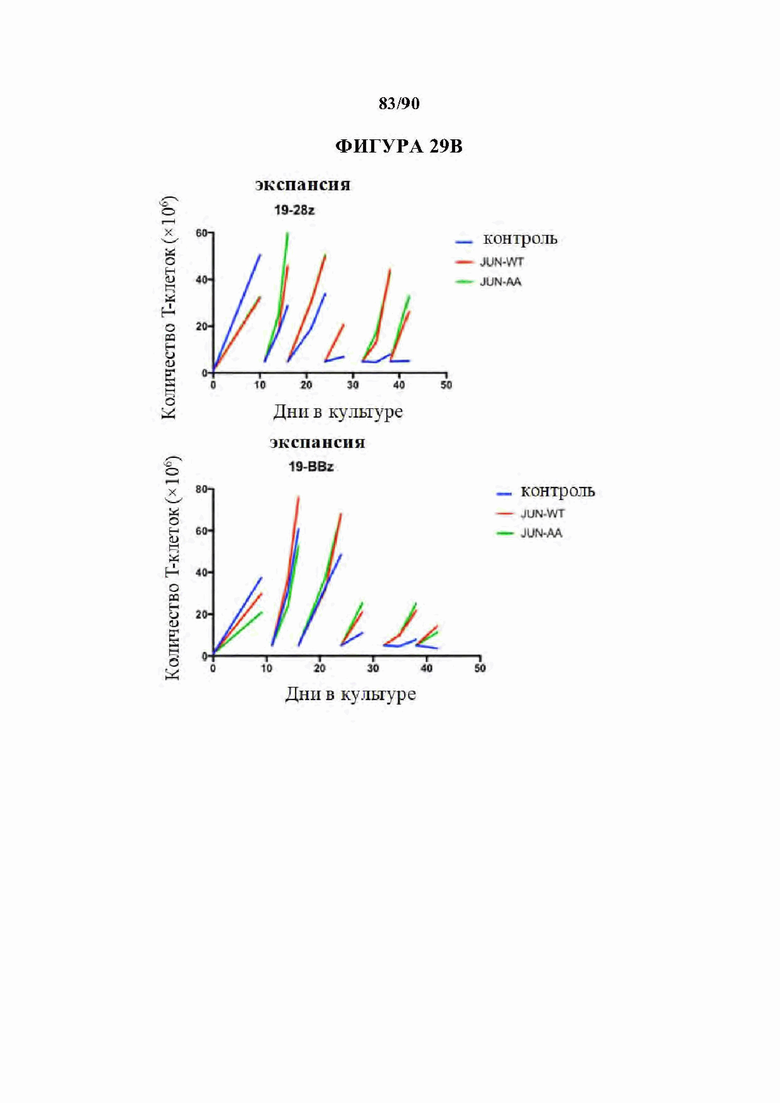
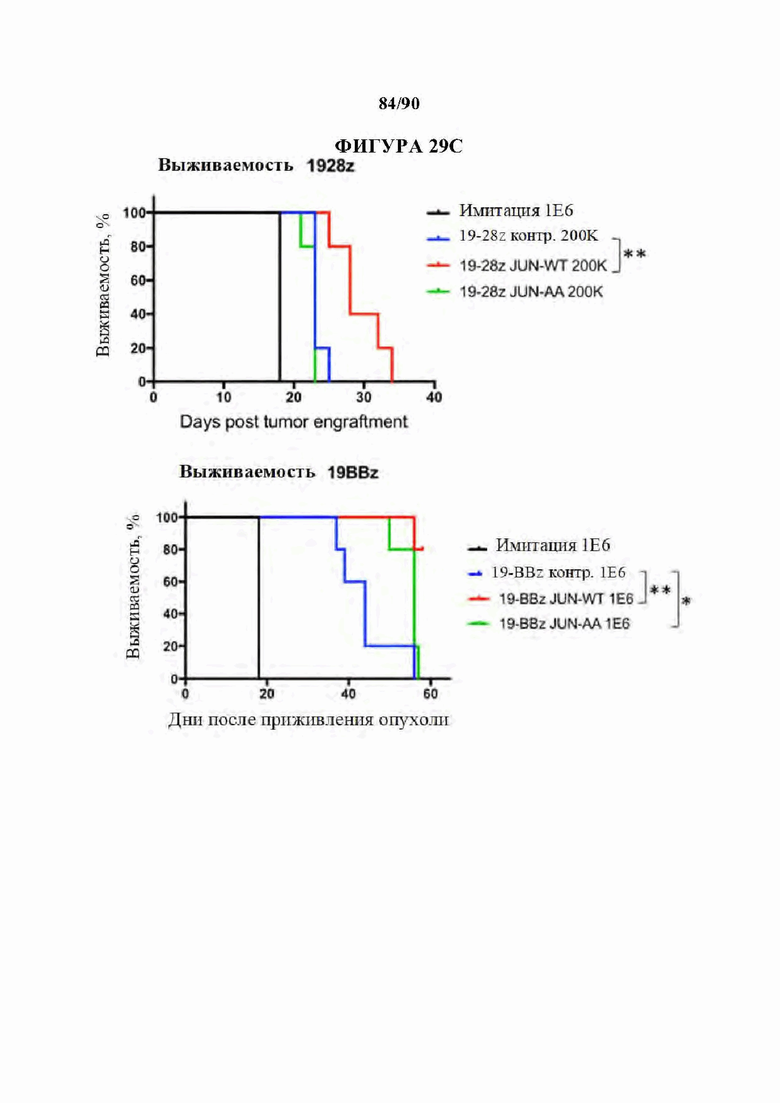
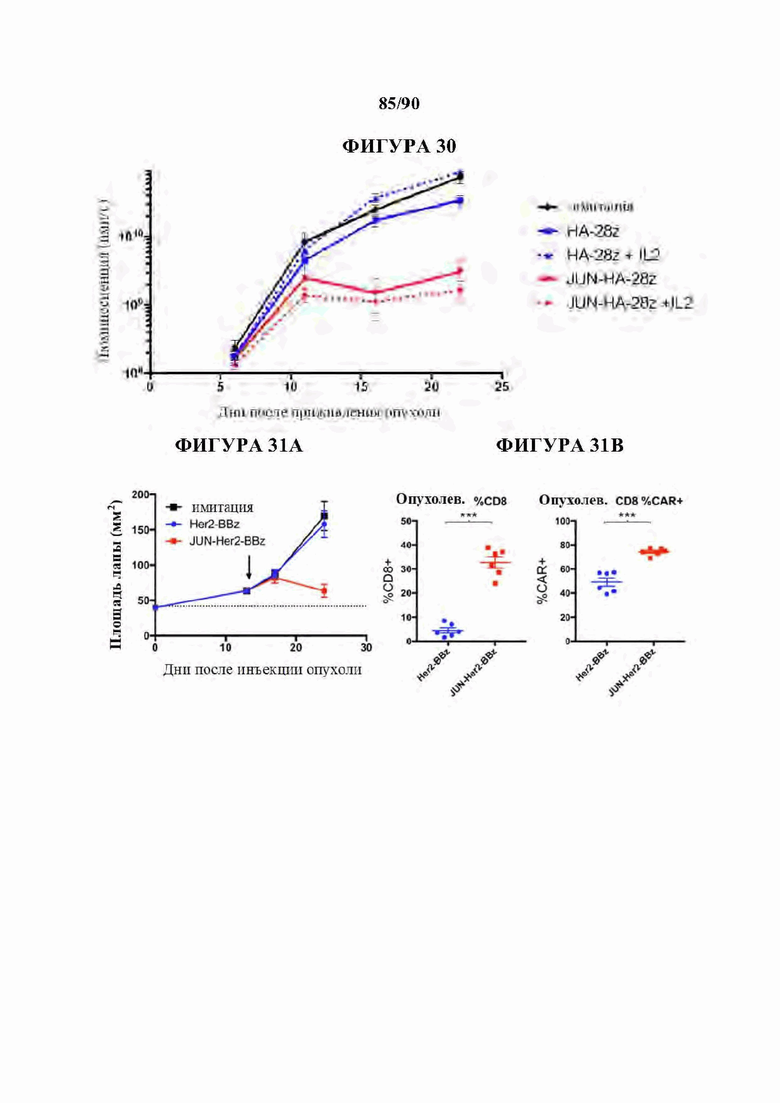
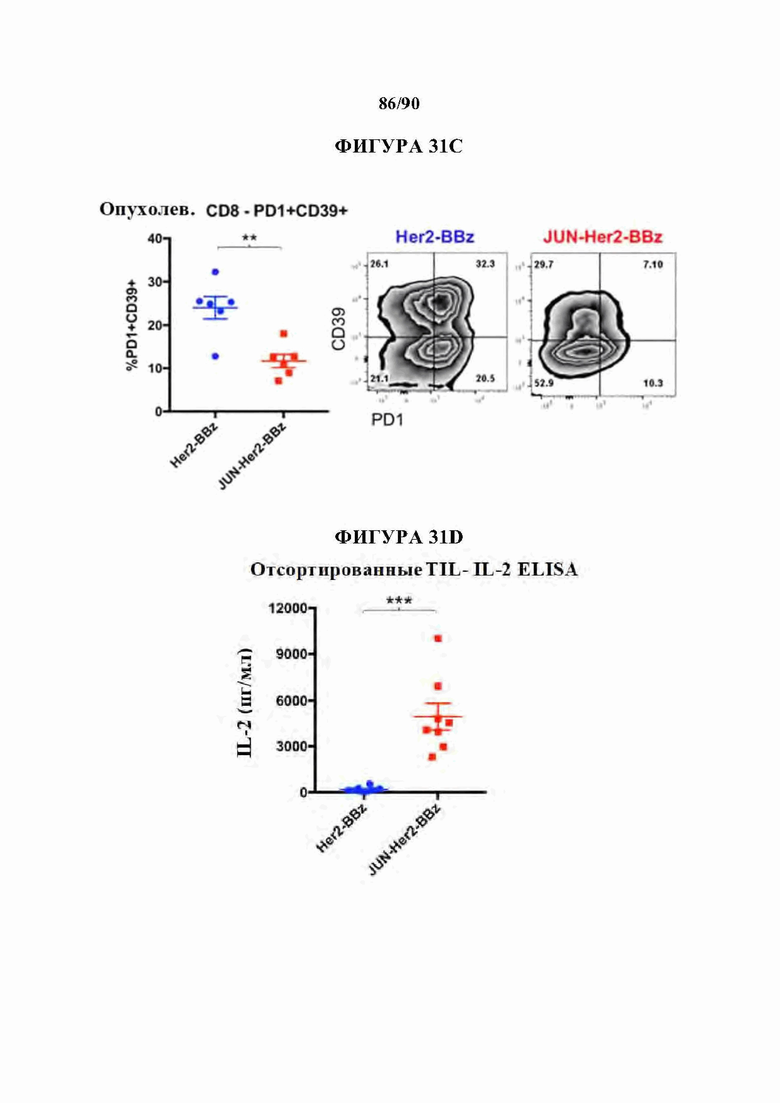
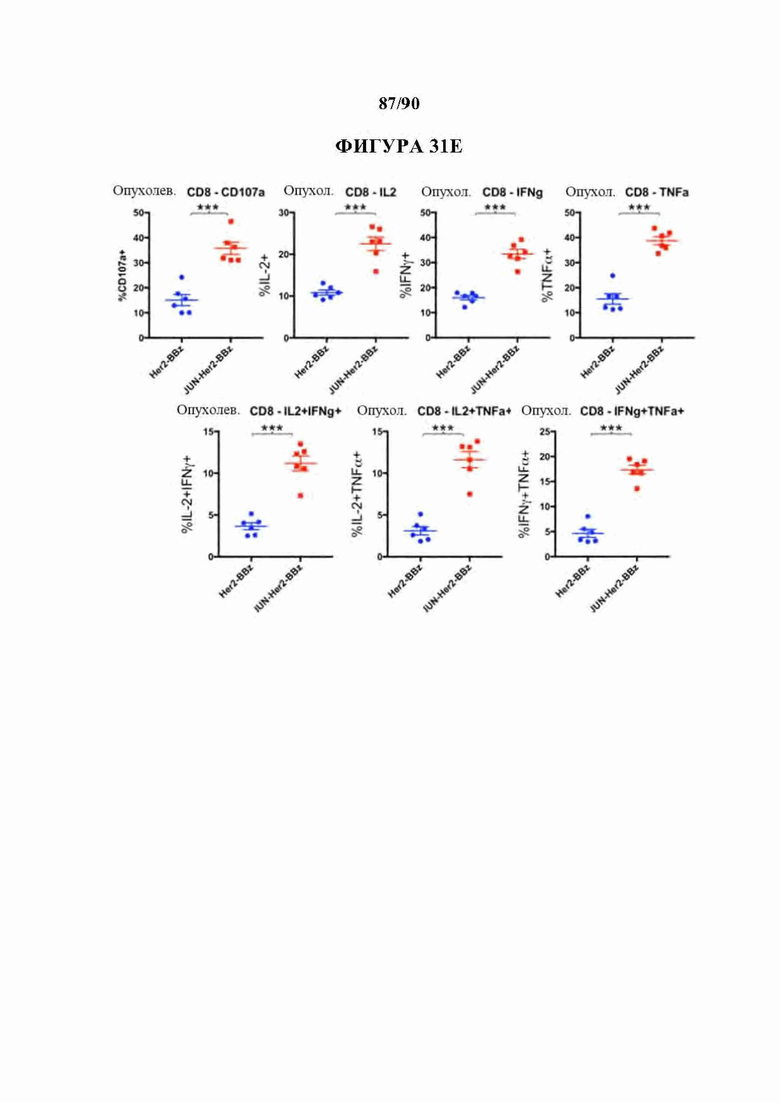

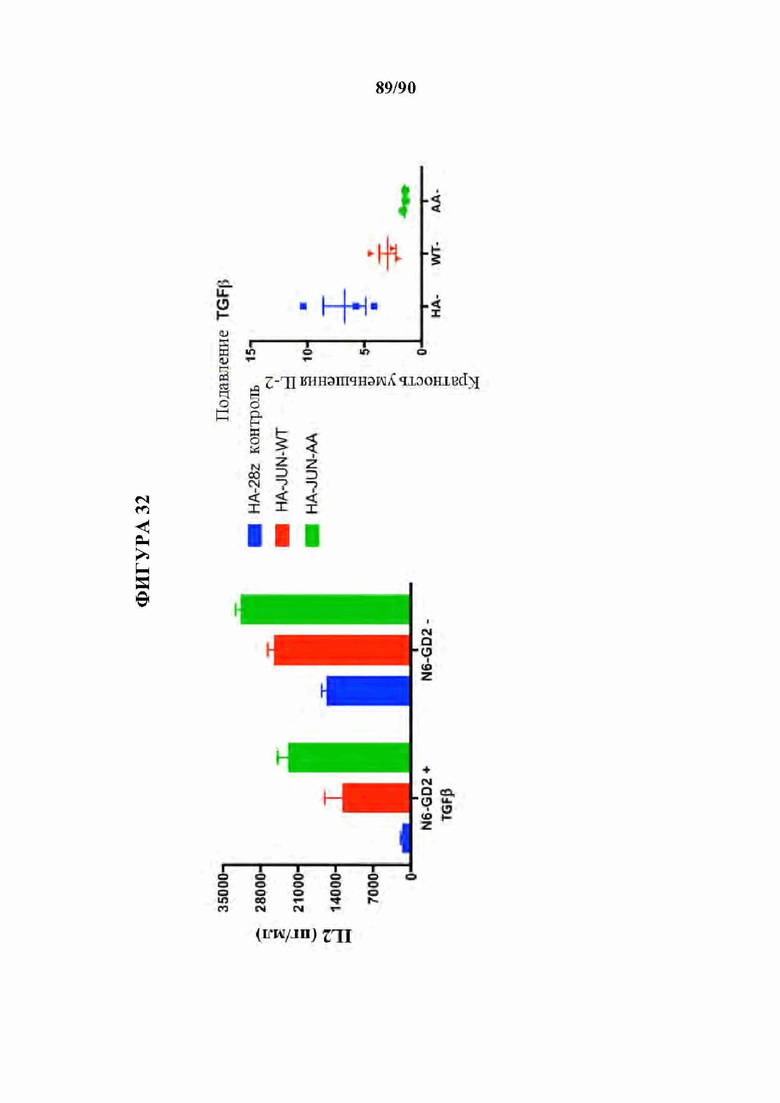
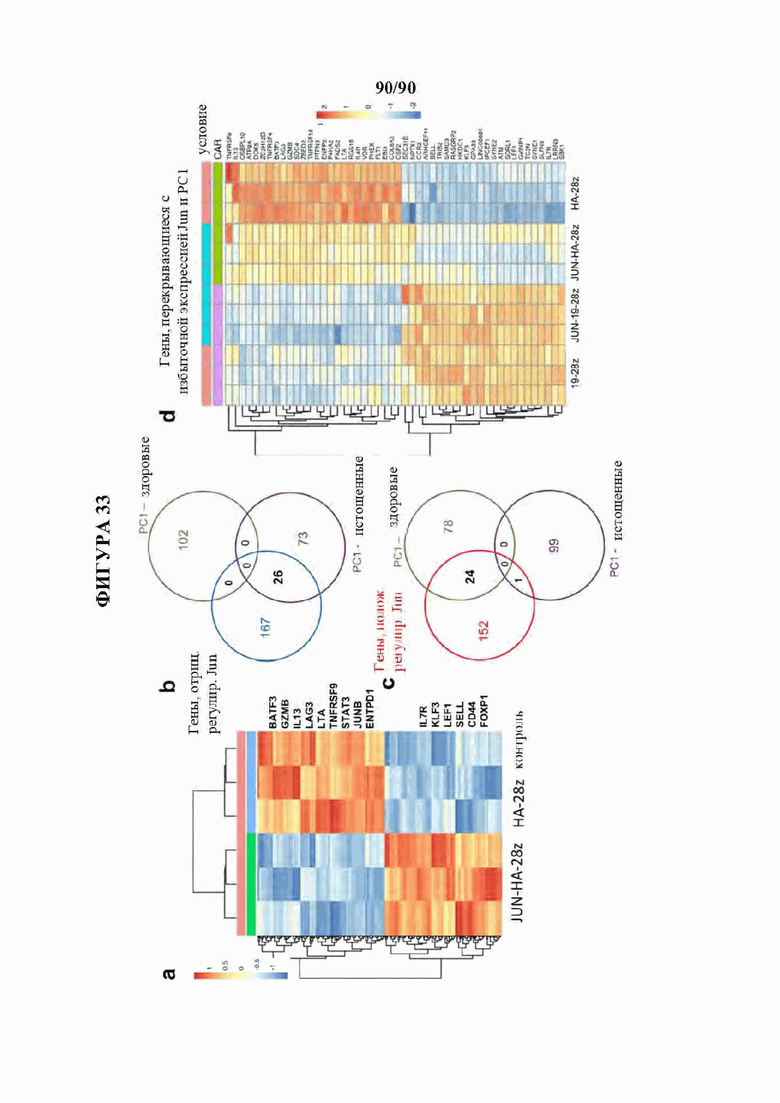
Группа изобретений относится к области медицины и раскрывает композицию для лечения заболевания или патологического состояния у субъекта, поддающегося лечению введением T-клеток, где композиция содержит выделенные T-клетки и где выделенные T-клетки содержат введенную нуклеиновую кислоту для избыточной экспрессии c-Jun. Также раскрыт способ лечения заболевания или патологического состояния у субъекта, предусматривающий введение субъекту, характеризующемуся наличием заболевания или патологического состояния, эффективного количества композиции, где заболевание или патологическое состояние поддается лечению введением T-клеток. Техническим результатом группы изобретений является уменьшение истощения выделенных Т-клеток. 2 н. и 29 з.п. ф-лы, 2 табл., 33 ил., 33 пр.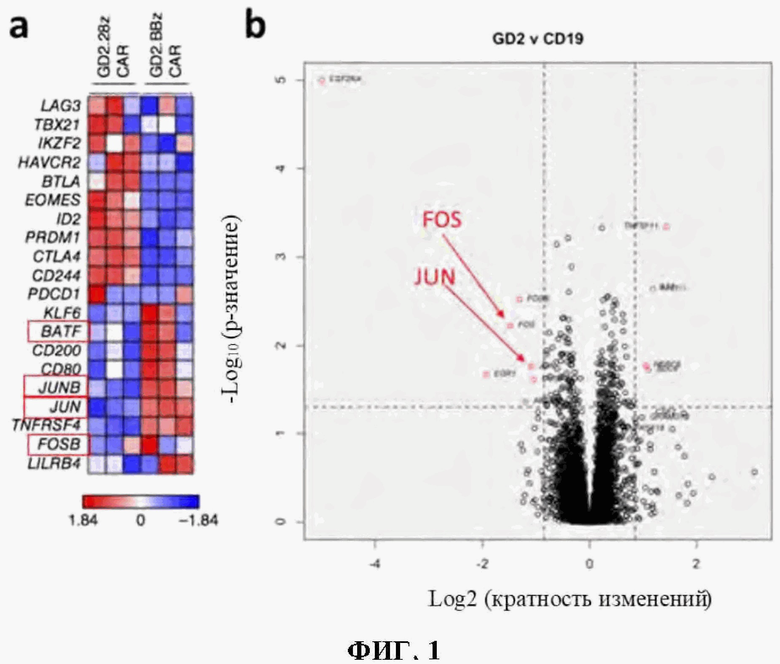
1. Композиция для лечения заболевания или патологического состояния у субъекта, поддающегося лечению введением T-клеток, где композиция содержит выделенные T-клетки и где выделенные T-клетки содержат введенную нуклеиновую кислоту для избыточной экспрессии c-Jun.
2. Композиция по п. 1, в которой выделенные T-клетки экспрессируют рекомбинантный рецептор.
3. Композиция по п. 2, в которой рекомбинантный рецептор представляет собой T-клеточный рецептор (TCR).
4. Композиция по п. 2, в которой рекомбинантный рецептор представляет собой химерный антигенный рецептор (CAR).
5. Композиция по любому из пп. 2-4, в которой рекомбинантный рецептор является специфическим для опухолевого антигена.
6. Композиция по п. 5, в которой опухолевый антиген выбран из группы, состоящей из следующего: CD19, CD20, CD22, ROR1, GD2, белок или антиген EBV, рецептор фолата, мезотелин, карциноэмбриональный антиген человека, CD33/IL3Ra, c-Met, PSMA, гликолипид F77, EGFRvIII, NY-ESO-1, MAGE-A3, MART-1, GP1000 и p53.
7. Композиция по любому из пп. 1-6, в которой выделенные T-клетки представляют собой нативные, встречающиеся в природе T-клетки.
8. Композиция по п. 7, в которой нативные, встречающиеся в природе T-клетки, представляют собой инфильтрирующие опухоль лимфоциты (TILs).
9. Композиция по п. 7, в которой нативные, встречающиеся в природе T-клетки получают с помощью лейкафереза образца крови.
10. Композиция по п. 9, в которой экспансию T-клеток осуществляют ex vivo.
11. Композиция по любому из пп. 1-10, в которой T-клетки выбраны из группы, состоящей из следующего: CD3+ T-клетки, CD8+ T-клетки, CD4+ T-клетки, Т-клетки - естественные киллеры, гамма-дельта T-клетки, комбинация CD4+ и CD8+ Т-клеток, T-клетки памяти, индуцированные цитокинами клетки-киллеры и их комбинации.
12. Композиция по любому из пп. 1-11, в которой c-Jun представляет собой мутированный/усеченный c-Jun.
13. Композиция по п. 12, в которой мутированный/усеченный c-Jun содержит N-концевую делецию.
14. Композиция по п. 12, в которой мутированный/усеченный c-Jun (i) не содержит трансактивирующий домен или (ii) содержит неактивный трансактивирующий домен.
15. Композиция по любому из пп. 1-11, в которой c-Jun представляет c-Jun дикого типа.
16. Композиция по любому из пп. 2-15, в которой Т-клетки модифицированы введением нуклеиновой кислоты, кодирующей c-Jun, и где c-Jun и рекомбинантный рецептор экспрессируются из отдельных экспрессионных векторных конструктов.
17. Композиция по любому из пп. 2-15, в которой Т-клетки модифицированы введением нуклеиновой кислоты, кодирующей c-Jun, и где c-Jun и рекомбинантный рецептор коэкспрессируются из одного экспрессионного векторного конструкта.
18. Композиция по любому из пп. 1-17, в которой выделенные T-клетки представляют собой T-клетки со специфичностью в отношении опухоли и активностью против нее.
19. Композиция по п. 18, в которой T-клетки представляют собой T-клетки периферической крови, генетически модифицированные для экспрессии рецептора, который распознает и отвечает на опухоль.
20. Композиция по любому из пп. 1-19, в которой T-клетки являются модифицированными для снижения и/или устранения экспрессии и/или активности одного или нескольких представителей ингибирующего AP-1 комплекса.
21. Композиция по п. 20, в которой представитель ингибирующего AP-1 комплекса выбран из группы, состоящей из JunB, представителя семейства BATF, IRF4, представителя семейства ATF или их комбинации.
22. Композиция по п. 20, в которой представитель ингибирующего AP-1 комплекса представляет собой JunB и/или BATF3.
23. Способ лечения заболевания или патологического состояния у субъекта, предусматривающий введение субъекту, характеризующемуся наличием заболевания или патологического состояния, эффективного количества композиции по любому из пп. 1-22, где заболевание или патологическое состояние поддается лечению введением T-клеток.
24. Способ по п. 23, при котором T-клетки в композиции являются менее подверженными истощению Т-клеток.
25. Способ по п. 23 или 24, при котором T-клетки в композиции характеризуются повышенной реактивностью в отношении антигена с низкой плотностью на клетках-мишенях Т-клеток.
26. Способ по любому из пп. 23-25, при котором T-клетки в композиции являются менее подверженными снижению формирования памяти.
27. Способ по любому из пп. 23-26, при котором T-клетки в композиции являются менее подверженными снижению пролиферативного потенциала.
28. Способ по любому из пп. 23-27, при котором заболевание или состояние представляет собой опухоль или рак.
29. Способ по п. 28, дополнительно предусматривающий введение пациенту одного или нескольких противораковых средств и/или одного или нескольких химиотерапевтических средств.
30. Способ по п. 29, при котором введение происходит до, одновременно и/или после получения пациентом лучевой терапии.
31. Способ по любому из пп. 23-27, при котором заболевание или состояние представляет собой вирусную, бактериальную и/или паразитарную инфекцию.
| WO 2017165412 A2, 28.09.2017 | |||
| Long AH et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Nat Med., 2015, 21(6), pp.581-590 | |||
| WO 2013063019 A1, 02.05.2013 | |||
| WO 2016179283 A1, 10.11.2016 | |||
| BEHRENS, A et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
