ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к получению белка и/или других макромолекул с использованием микроорганизмов. В частности, настоящее изобретение относится к новым бактериальным штаммам и способам непрерывного культивирования для получения белка или биомассы с использованием бактерий, при которых в клетки поступают газы и минеральные вещества. Настоящее изобретение также относится к продуктам этих способов и применению этих продуктов, например, в пищевых продуктах или кормах.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Растущее население мира, изменение климата и нехватка воды все чаще представляют угрозу для традиционного сельского хозяйства и, следовательно, для достаточного снабжения продовольствием и кормами. Поэтому исследуются альтернативные источники органических молекул, таких как белки. Потенциальной альтернативой является получение единичных клеток, то есть получение белка и/или других макромолекул с использованием микроорганизмов.
Описаны хемоавтотрофные микроорганизмы, способные расти на минимальной минеральной среде с газообразным водородом в качестве источника энергии и углекислым газом в качестве единственного источника углерода. Для обзора этих микроорганизмов см., например, Shively et al. (1998) Annu Rev Microbiol 52:191. В Международной патентной заявке WO2018144965 описаны различные микроорганизмы и биопроцессы для преобразования газообразных субстратов в высокобелковую биомассу. В работе Andersen et al. (1979) Biochim Biophys Acta 585:1-11 описаны мутантные штаммы Alcaligenes eutrophus, водородной бактерии, которая легко растет в гетеротрофных и автотрофных условиях. Были охарактеризованы мутанты с измененной активностью рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (rubisco). В работе Ohmiya et al. (2003) J. Biosci. Bioeng. 95:549-561 описано применение микробных генов для утилизации неразлагаемой биомассы. В работе Yu Jian et al. (2013) Int J Hydrogen Ener 38:8683-8690 описана фиксация диоксида углерода бактериальным изолятом, окисляющим водород. Высокая энергетическая эффективность 50% была измерена при умеренной концентрации кислорода (10 мол.%).
Однако различные хемоавтотрофные микроорганизмы обладают различными свойствами с точки зрения скорости роста, выхода, состава биомассы, а также свойств, связанных с использованием в качестве пищевого ингредиента, таких как безопасность для потребления человеком, вкус, запах, ощущение во рту, технические и функциональные свойства в приготовление пищи и т.д. Не каждый хемоавтотрофный микроорганизм имеет достаточную скорость роста и обеспечивает достаточный выход, и не каждый способ можно реально масштабировать до экономически жизнеспособного крупномасштабного процесса. Чтобы иметь достаточный выход функционального белка, т.е. для пищевых продуктов или кормов важно найти подходящий продуцирующий организм и подходящий способ, который можно было бы осуществлять в больших масштабах. Эта потребность решается настоящим изобретением.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В первом основном аспекте настоящее изобретение относится к выделенному бактериальному штамму VTT-E-193585 или его производному.
В дополнительных аспектах настоящее изобретение относится к культуре, содержащей бактериальный штамм настоящего изобретения или его производное. Кроме того, настоящее изобретение относится к способу получения биомассы и/или белка, где указанный способ включает в себя культивирование бактериального штамма настоящего изобретения или его производного.
В ещё одном аспекте настоящее изобретение относится к способу получения биомассы и/или белка, где указанный способ включает в себя культивирование бактериального штамма рода Xanthobacter в непрерывной культуре с водородом в качестве источника энергии и источником неорганического углерода, где источник неорганического углерода включает в себя двуокись углерода.
В дополнительных основных аспектах настоящее изобретение относится к белку в виде большого количества, биомассе или небелковым клеточным или химическим компонентам, полученным или получаемым способом настоящего изобретению, и к пищевому продукту или кормовому продукту, полученному или получаемому способом настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1. Оптическая плотность, измеренная при длине волны 600 нм (черные кружки), и показания датчика оптической плотности во время хемоавтотрофного культивирования в объёме 200 литров выделенного бактериального штамма, депонированного как VTT-E-193585.
Фиг. 2. Оптическая плотность, измеренная при длине волны 600 нм во время параллельного хемоавтотрофного культивирования в объёме 200 мл выделенного бактериального штамма, депонированного как VTT-E-193585, на различных источниках азота.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Используемый здесь термин «выделенный», например, в контексте штамма означает выделенный из его естественной среды. Предпочтительно выделенный штамм является чистым, т.е. свободным от других штаммов.
Термин «производный», используемый здесь в контексте штамма, относится к штамму, который получен из эталонного штамма, т.е. создан с использованием эталонного штамма в качестве отправной точки. Например, генетически модифицированный или иным образом мутировавший или генетически модифицированный штамм представляет собой осуществление такого производного. Генетические модификации включают в себя точечные мутации, а также вставки или делеции, включая вставки или делеции целых локусов или их фрагментов. Производное предпочтительно имеет менее 10 генетических модификаций, т.е. менее 5, например 4, 3, 2 или 1 генетические модификации по сравнению с эталонным штаммом.
Используемый здесь термин «культура» относится к суспензии жизнеспособных клеток в жидкой среде.
Термин «биомасса» имеет свое обычное значение в области бактериальной ферментации и относится к клеточному материалу.
Используемый здесь термин «непрерывное культивирование» относится к процессу культивирования, при котором свежая среда непрерывно добавляется к культуре, а среда с бактериальной культурой непрерывно удаляется практически с одинаковой скоростью.
Аспекты и варианты осуществления настоящего изобретения
В первом основном аспекте настоящее изобретение относится к выделенному бактериальному штамму VTT-E-193585 или его производному.
Штамм VTT-E-193585 был выделен с побережья Балтийского моря в Наантали, Финляндия. Этот организм способен расти в подходящих условиях биореактора с минимальной минеральной средой с водородом в качестве источника энергии и углекислым газом в качестве источника углерода в условиях ограниченного содержания кислорода. Секвенирование 16S и метагеномное секвенирование Illumina показали, что этот штамм, скорее всего, относится к роду Xanthobacter, но не является известным видом. Бактериальный штамм очень подходит для пищевых продуктов и кормов, поскольку порошок высушенных клеток имеет высокое содержание белка и содержит все незаменимые аминокислоты. Он также содержит больше ненасыщенных жирных кислот, чем насыщенных, и высокий уровень витаминов группы В. Он имеет низкие уровни пептидогликанов и липополисахаридов, которые могут вызывать аллергию или токсичность. Был проведен анализ токсичности, и для штамма не наблюдалось генотоксичности или цитотоксичности. Кроме того, штамм в целом чувствителен к антибиотикам.
Штамм VTT-E-193585 (SoF1) был депонирован 11 июня 2019 г. в Коллекции культур VTT в Центре технических исследований VTT в Финляндии, P.O. Box 1000, FI-02044 VTT, Финляндия, Международный депозитарный орган в соответствии с Будапештским договором. Дополнительная информация о характеристиках штамма и способах культивирования штамма представлена в приведенных здесь Примерах.
В предпочтительном варианте осуществления изобретения, если штамм является производным штамма VTT-E-193585, то это производное сохраняет способность расти с использованием газообразного водорода в качестве источника энергии и диоксида углерода в качестве единственного источника углерода.
В одном варианте осуществления изобретения, если штамм является производным штамма VTT-E-193585, то это производное содержит 16S рибосомальную РНК, указанную в SEQ ID NO:1, или 16S рибосомальную РНК, имеющую до 20 нуклеотидных различий с SEQ ID NO:1, например от 1 до 10, например от 1 до 5, например одно, два или три нуклеотидных отличия с SEQ ID NO:1.
SEQ ID NO:1. Последовательность 16S рибосомальной РНК штамма VTT-E-193585:
CTTGAGAGTTTGATCCTGGCTCAGAGCGAACGCTGGCGGCAGGCCTAACACATGCAAGTCGAGCGCCCAGCAATGGGAGCGGCAGACGGGTGAGTAACGCGTGGGGATGTGCCCAATGGTACGGAATAACCCAGGGAAACTTGGACTAATACCGTATGAGCCCTTCGGGGGAAAGATTTATCGCCATTGGATCAACCCGCGTCTGATTAGCTAGTTGGTGGGGTAACGGCCCACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGGACAATGGGCGCAAGCCTGATCCAGCCATGCCGCGTGTGTGATGAAGGCCTTAGGGTTGTAAAGCACTTTCGCCGGTGAAGATAATGACGGTAACCGGAGAAGAAGCCCCGGCTAACTTCGTGCCAGCAGCCGCGGTAATACGAAGGGGGCTAGCGTTGCTCGGAATCACTGGGCGTAAAGCGCACGTAGGCGGATCGTTAAGTCAGGGGTGAAATCCTGGAGCTCAACTCCAGAACTGCCCTTGATACTGGCGACCTTGAGTTCGAGAGAGGTTGGTGGAACTGCGAGTGTAGAGGTGAAATTCGTAGATATTCGCAAGAACACCAGTGGCGAAGGCGGCCAACTGGCTCGATACTGACGCTGAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGGATGCTAGCCGTTGGGCAGCTTGCTGTTCAGTGGCGCAGCTAACGCATTAAGCATCCCGCCTGGGGAGTACGGTCGCAAGATTAAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGAAGCAACGCGCAGAACCTTACCAGCCTTTGACATGGCAGGACGATTTCCAGAGATGGATCTCTTCCAGCAATGGACCTGCACACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGCCTCTAGTTGCCAGCATTCAGTTGGGCACTCTAGAGGGACTGCCGGTGATAAGCCGAGAGGAAGGTGGGGATGACGTCAAGTCCTCATGGCCCTTACGGGCTGGGCTACACACGTGCTACAATGGTGGTGACAGTGGGATGCGAAAGGGCGACCTCTAGCAAATCTCCAAAAGCCATCTCAGTTCGGATTGTACTCTGCAACTCGAGTGCATGAAGTTGGAATCGCTAGTAATCGTGGATCAGCATGCCACGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTTGGCTTTACCCGAAGGCGCTGCGCTAACCCGCAAGGGAGGCAGGCGACCACGGTAGGGTCAGCGACTGGGGTGAAGTCGTAACAAGGTAGCCGTAGGGGAACCTGCGGCTGGATCACCTCCTTT
В ещё одном аспекте настоящее изобретение относится к культуре, содержащей бактериальный штамм настоящего изобретения или его производное. В предпочтительном варианте осуществления изобретения объём культуры составляет 100 мл или более, например 1 л или более, например 10 л или более, например 1000 л или более, например 10 000 л или более, например 50 000 л или более, например 100 000 л или более, например 200 000 л или более.
В ещё одном аспекте настоящее изобретение относится к способу получения биомассы и/или белка, где указанный способ включает в себя культивирование бактериального штамма настоящего изобретения или его производного. В одном варианте осуществления изобретения способ предназначен для производства биомассы. В другом варианте осуществления изобретения способ предназначен для получения белка. В одном варианте осуществления изобретения способ включает в себя культивирование штамма в непрерывной культуре с водородом в качестве источника энергии и источником неорганического углерода, где источник неорганического углерода включает в себя диоксид углерода. В дополнительном варианте осуществления изобретения способ предназначен для производства биомассы и включает в себя культивирование штамма в непрерывной культуре с водородом в качестве источника энергии и источником неорганического углерода, где источник неорганического углерода включает в себя диоксид углерода. Ниже описаны различные дополнительные варианты осуществления способа.
В ещё одном основном аспекте настоящее изобретение относится к способу получения биомассы и/или белка, где указанный способ включает в себя культивирование бактериального штамма рода Xanthobacter в непрерывной культуре с водородом в качестве источника энергии и источником неорганического углерода, где источник неорганического углерода включает в себя двуокись углерода. В одном варианте осуществления изобретения способ предназначен для производства биомассы. В другом варианте осуществления изобретения способ предназначен для получения белка. Ниже описаны различные дополнительные варианты осуществления способа.
Судя по последовательности генома, штамм, депонированный под номером VTT-E-193585, скорее всего, использует цикл Кальвина-Бенсона-Бэсшема для фиксации углерода, в котором молекула углекислого газа соединяется с 5-углеродной цепью рибулозо-1,5-бисфосфата, образуя две молекулы глицерат-3-фосфата. Это позволяет штамму синтезировать все другие органические молекулы, необходимые для роста. Энергия водорода поступает в клетку, скорее всего, через NAD+-восстанавливающие гидрогеназы и/или NiFeSe-гидрогеназы. По сути, это окислительно-восстановительная реакция, в которой водород (H2) окисляется до H+, а NAD+ восстанавливается до NADH. Помимо АТФ, NADH является одним из основных переносчиков энергии внутри живых организмов. В качестве альтернативы, некоторый другой энергетический эквивалент восстанавливается другим ферментом гидрогеназы с использованием H2. Цикл Кальвина-Бенсона-Бэсшема требует энергии в виде АТФ и NADH/NADPH для связывания СО2. Штамм, скорее всего, генерирует АТФ посредством окислительного фосфорилирования, которое состоит из четырех белковых комплексов, создающих протонный градиент через мембрану. Протонный градиент генерируется в основном за счет энергии NADH. Протонный градиент управляет комплексом АТФ-синтазы, генерирующим АТФ. Судя по последовательности генома, штамм имеет бактериальную АТФ-синтазу F-типа.
Следует понимать, что когда указано, что способ включает в себя культивирование штамма с источником неорганического углерода, то источник неорганического углерода является основным источником углерода в культуре. Таким образом, в культуре могут присутствовать незначительные количества источников органического углерода, но основной метаболизм и рост культуры основаны на использовании источника неорганического углерода, предпочтительно двуокиси углерода, в качестве источника углерода. Предпочтительно доля углерода, подаваемого в культуру, которая является органической, составляет менее 5%, например менее 1%, например менее 0,1% всего углерода, поступающего в культуру в процессе реализации способа. Предпочтительно в процессе реализации способа не подаются никакие источники органического углерода.
Точно так же следует понимать, что когда указано, что способ включает в себя культивирование штамма с водородом (H2) в качестве источника энергии, то водород является основным источником энергии в культуре. Таким образом, в культуре могут присутствовать другие второстепенные источники энергии, такие как аммиак, который может являться источником азота, или небольшие количества органических соединений, но основной метаболизм и рост культуры основаны на использовании водорода в качестве источник энергии. В общем процессе водород предпочтительно получают электролизом воды; т.е. путем разделения воды электричеством на газообразный водород и кислород. Таким образом, газообразные водород и кислород подаются в биореактор из расположенного поблизости электролизёра. Альтернативно, электроды могут быть помещены внутрь биореактора для производства водорода и кислорода в биореакторе, а не в отдельном электролизёре.
Источник неорганического углерода, содержащий диоксид углерода, может включать в себя другие источники неорганического углерода, такие как, например, монооксид углерода. В одном варианте осуществления изобретения в культуру подают только источники углерода в газообразной форме. В предпочтительном варианте осуществления изобретения диоксид углерода является единственным источником неорганического углерода и, действительно, единственным источником углерода, подаваемым в культуру. В одном варианте осуществления изобретения в культуру подаются только газы и минералы, а уровень диоксида углерода в подаваемом газе составляет от 10% до 50%, т.е. от 15% до 45%, например от 20% до 40%, например от 25% до 35%, например, от 26% до 30%.
В другом варианте осуществления изобретения в культуру подают газы и минералы, а уровень водорода (H2) в подаваемом газе составляет от 30% до 80%, например от 35% до 75%, например от 40% до 70%, например от 45% до 65%, например от 50% до 60%.
В другом варианте осуществления изобретения в культуру подают газы и минералы, а уровень кислорода (O2) в подаваемом газе составляет от 10% до 25%, например от 15% до 20%, например, от 16% до 18%. В другом варианте осуществления изобретения уровень подаваемого кислорода таков, что уровень растворенного кислорода в культуре поддерживается на уровне от 5% до 10%.
В предпочтительном варианте осуществления изобретения в культуру подают только газы и минералы, и подаваемый газ содержит H2, CO2 и O2, где процентное содержание H2 составляет от 40% до 70%, процентное содержание CO2 составляет от 18% до 28% и процентное содержание O2 составляет от 12% до 22%.
Как правило, способ настоящего изобретения включает в себя добавление источника азота. Источник азота может быть, например, представлен в виде гидроксида аммония, соли аммония, такой как сульфат аммония или хлорид аммония, аммиака, мочевины или нитрата, например нитрата калия. В других вариантах осуществления изобретения в качестве источника азота используют газообразный азот (N2). В предпочтительном варианте осуществления изобретения источник азота представляет собой гидроксид аммония или соль аммония, такую как сульфат аммония.
В одном варианте осуществления изобретения подаваемый источник азота представляет собой гидроксид аммония в концентрации от 100 мг/л до 10 г/л, например, от 250 мг/л до 4 г/л, например от 0,5 г/л до 2 г/л, например от 0,75 г/л до 1,5 г/л.
Как правило, способ настоящего изобретения включает в себя добавление минералов, таких как минералы, содержащие аммоний, фосфат, калий, натрий, ванадий, железо, сульфат, магний, кальций, молибден, марганец, бор, цинк, кобальт, селен, йод, медь и/или никель. Подходящие минеральные среды хорошо известны в данной области техники и, например, имеют, были описаны в Thermophilic Bacteria, CRC Press, Boca Raton, FL, Jacob K. Kristjansson, ed., 1992, например, на странице 87, в таблице 4.
В одном варианте осуществления изобретения добавленные минералы включают в себя один или несколько из следующих минералов: аммиак, аммоний (например, хлорид аммония (NH4Cl), сульфат аммония ((NH4)2SO4)), нитрат (например, нитрат калия (KNO3)), мочевина или источник органического азота; фосфат (например, динатрийфосфат (Na2HPO4), фосфат калия (KH2PO4), фосфорная кислота (H3PO4), дитиофосфат калия (K3PS2O2), ортофосфат калия (K3PO4), динатрийфосфат (Na2HPO4•2H2O), дикалийфосфат (K2HPO4) или монокалийфосфат (KH2PO4); сульфат; дрожжевой экстракт; хелатное железо (например, с ЭДТА или лимонной кислотой); калий (например, фосфат калия (KH2PO4), нитрат калия (KNO3), йодид калия (KI), бромид калия (KBr)); и другие неорганические соли, минералы и микроэлементы (например, хлорид натрия (NaCl), сульфат магния (MgSO4•7H2O) или хлорид магния (MgCl2), хлорид кальция (CaCl2), сульфат кальция (CaSO4) или карбонат кальция (CaCO3), сульфат марганца (MnSO4•7H2O) или хлорид марганца (MnCl2), хлорид железа (FeCl2), сульфат железа (FeSO4•7H2O) или хлорид железа (FeCl2 •4H2O), гидрокарбонат натрия (NaHCO3) или карбонат натрия (Na2CO3), сульфат цинка (ZnSO4) или хлорид цинка (ZnCl2), молибдат аммония (NH4MoO4) или молибдат натрия (Na2MoO4•2H2O), сульфат меди (CuSO4) или хлорид меди (CuCl2•2H2O), хлорид кобальта (CoCl2•6H2O) или сульфат кобальта (CoSO4), хлорид алюминия (AlCl3•6H2O), хлорид лития (LiCl), борная кислота (H3BO3), хлорид никеля (NiCl2•6H2O) или сульфат никеля (NiSO4), хлорид олова (SnCl2•H2O), хлорид бария (BaCl2•2H2O), селенат меди (CuSeO4·5H2O), селенат натрия (Na2SeO4) или селенит натрия (Na2SeO3), метаванадат натрия (NaVO3), соли хрома).
В предпочтительном варианте осуществления изобретения способ настоящего изобретения включает в себя добавление одного, нескольких или всех из: NH4OH, KH2PO4, Na2HPO4•2H2O, NaVO3•H2O, FeSO4×7H2O, MgSO4•7H2O, CaSO4, Na2MoO4•2H2O, MnSO4•7H2O, ZnSO4. •7H2O, H3BO3, CoSO4, CuSO4, NiSO4.
В одном варианте осуществления изобретения среда, подаваемая клеткам, содержит менее 1 г/л хлоридных солей, например менее 0,25 г/л хлоридных солей, например менее 0,1 г/л хлоридных солей, например менее 0,025 г/л хлоридных солей, например менее 0,01 г/л хлорида. В одном варианте осуществления изобретения в культуру не добавляют хлоридные соли.
В другом варианте осуществления изобретения во время осуществления способа не добавляют витамины, т.е. среда, подаваемая в культуру, не содержит витаминов.
В другом варианте осуществления изобретения во время осуществления способа не добавляют аминокислоты, т.е. среда, подаваемая в культуру, не содержит аминокислот.
В другом варианте осуществления изобретения во время осуществления способа не добавляют никаких органических соединений, т.е. среда, подаваемая в культуру, не содержит никаких органических соединений.
В некоторых вариантах осуществления изобретения значение pH бактериальной культуры регулируется на определенном уровне. В некоторых вариантах осуществления изобретения значение рН регулируют в пределах оптимального диапазона для поддержания и/или роста бактерий, и/или образования органических соединений. В одном варианте осуществления изобретения значение pH в культуре поддерживается в диапазоне от 5,5 до 8,0, например, от 6,5 до 7,0, например, 6,8.
В некоторых вариантах осуществления изобретения контролируют температуру бактериальной культуры. В некоторых вариантах осуществления изобретения температуру регулируют в пределах оптимального диапазона для поддержания и/или роста бактерий, и/или образования органических соединений. В одном варианте осуществления изобретения культуру выращивают при температуре от 25°С до 40°С, например, от 28°С до 32°С, например, при 30°С.
Как правило, способ изобретения осуществляют в биореакторе. Биореактор используется для культивирования клеток, которые могут поддерживаться на определенных фазах кривой их роста. Использование биореакторов выгодно во многих отношениях для культивирования в условиях хемоавтотрофного роста. Как правило, в биореакторе облегчается контроль условий роста, включая контроль растворенного диоксида углерода, кислорода и других газов, таких как водород, а также других растворенных питательных веществ, микроэлементов, температуры и рН. Питательные среды, а также газы можно добавлять в биореактор или в виде разового добавления, или периодически, или в ответ на обнаруженное истощение или в заданный запрограммированный момент времени, либо непрерывно в течение периода выращивания и/или поддержания культуры. В непрерывном процессе культивирования в биореактор непрерывно добавляют питательные среды, а также газы. Кроме того, из биореактора постоянно удаляют среду, содержащую бактерии.
В предпочтительном варианте осуществления изобретения объём бактериальной культуры составляет 100 мл или более, например 1 л или более, например 10 л или более, например 100 л или более, например 1000 л или более, например 10000 л или более, например 50000 л или более, например, 100000 л или более, например 200000 л или более.
В одном варианте осуществления изобретения продуктивность культуры составляет более 0,1 г сухой массы клеток на литр в час, например более 0,2, например более 0,3, например более 0,4, например более 0,5, например более 0,6, например более 0,7, например более 0,8, например более 0,9, например более 1 г на литр в час.
Бактерии можно инокулировать непосредственно из банка клеток или с помощью посевной культуры в меньшем масштабе. Желательно, чтобы подача свежей среды к культуре и удаление израсходованной среды с бактериями происходили с одинаковой скоростью, чтобы объём в биореакторе оставался неизменным.
В одном варианте осуществления изобретения после начальной фазы достижения подходящей плотности клеток бактерии растут в стационарном или псевдостационарном состоянии, постоянно оставаясь в своей логарифмической фазе при ОП600 выше 5, например выше 10, например выше 20, например от 50 до 200, например от 50 до 100.
В одном варианте осуществления способа настоящего изобретения бактериальный штамм имеет скорость роста 0,04-0,12 ч-1.
В другом варианте осуществления способа настоящего изобретения скорость подачи жидкости в непрерывной фазе составляет 50-80% от скорости роста.
Xanthobacter представляет собой род грамотрицательных бактерий из семейства Xanthobacteraceae.В одном варианте осуществления изобретения штамм Xanthobacter, используемый в способе по изобретению, представляет собой штамм, который использует путь Кальвина-Бенсона-Бэсшема для превращения диоксида углерода в органические соединения, например, глюкозу, необходимые для живых организмов.
В одном варианте осуществления изобретения штамм Xanthobacter, используемый в способе настоящего изобретения, представляет собой штамм, который использует гидрогеназы NiFeSe для превращения водорода (H2) в эквиваленты клеточной энергии.
В одном варианте осуществления изобретения штамм Xanthobacter, используемый в способе настоящего изобретения, представляет собой штамм, который использует NAD+-восстанавливающие гидрогеназы для превращения водорода (H2) в эквиваленты клеточной энергии.
В одном варианте осуществления изобретения штамм Xanthobacter, используемый в способе настоящего изобретения, способен к фиксации азота.
В одном варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, выбран из группы, состоящей из: X.agilis, X.aminoxidans, X.autotrophicus, X.flavus, X.tagetidis, X.viscosus, Xanthobacter sp. 126, Xanthobacter sp. 91 и штамма VTT-E-193585.
В предпочтительном варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, представляет собой VTT-E-193585 или X. tagetidis. Наиболее предпочтительно штамм, используемый в способе настоящего изобретения, представляет собой штамм VTT-E-193585.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит 16S рибосомальную РНК, указанную в SEQ ID NO:1, или 16S рибосомальную РНК, имеющую до 20 нуклеотидных различий с SEQ ID NO:1, например от 1 до 10, например от 1 до 5, например одно, два или три нуклеотидных отличия с SEQ ID NO:1.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий большую цепь рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (rubisco), имеющую последовательность, указанную в SEQ ID NO:3, или последовательность, имеющую более 93% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98% идентичности, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:3.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий малую цепь рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (rubisco), имеющую последовательность, указанную в SEQ ID NO:5, или последовательность, имеющую более 83% идентичность по последовательности, например более 86% идентичности, например более 90% идентичности, например более 95% идентичности, например, более 96% идентичности, например более 97% идентичности, например, более 98% идентичности, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:5.
SEQ ID NO:2:
Нуклеотидная последовательность большой цепи рибулозобисфосфаткарбоксилазы:
ATGGGTGCCGAAGCAACCGTCGGGCAGATCACGGACGCCAAGAAGAGATACGCCGCCGGCGTGCTGAAGTACGCCCAGATGGGCTACTGGAACGGCGACTACGTTCCCAAGGACACCGACCTCCTGGCGGTGTTCCGCATCACCCCCCAGGCGGGCGTGGACCCGGTGGAAGCCGCCGCGGCGGTCGCCGGCGAAAGCTCCACCGCTACCTGGACCGTGGTGTGGACCGACCGGCTCACCGCCGCCGACGTCTACCGCGCCAAGGCCTACAAGGTGGAGCCGGTGCCGGGCCAGGAAGGCCAGTATTTCTGCTACATCGCCTATGATCTCGATTTGTTCGAGGAAGGCTCCATCGCCAACCTCACGGCGTCGATCATCGGCAACGTCTTCTCCTTCAAGCCGCTGAAGGCGGCGCGGCTGGAGGACATGCGGCTTCCCGTCGCCTATGTGAAGACCTTCCGCGGCCCGCCCACCGGCATCGTGGTCGAGCGCGAGCGCCTGGACAAGTTCGGCCGCCCCCTTCTGGGCGCCACCACCAAGCCGAAGCTTGGCCTCTCGGGCAAGAATTACGGCCGCGTGGTCTATGAGGCCCTCAAGGGCGGCCTCGACTTCGTGAAGGACGACGAGAACATCAACTCGCAGCCCTTCATGCACTGGCGCGATCGCTTCCTCTATTGCATGGAGGCCGTCAACAAGGCCCAGGCCGAGACCGGCGAGGTGAAGGGGCACTATCTCAACATCACCGCCGGGACCATGGAGGAGATGTACCGCCGCGCCGAGTTCGCCAAGGAACTGGGCTCCGTGGTGGTGATGGTGGATCTCATCATCGGCTGGACCGCCATCCAGTCCATGTCCAACTGGTGCCGCGAGAACGACATGATCCTGCACATGCACCGTGCGGGCCATGGCACCTACACGCGCCAGAAGAGCCACGGCGTCTCCTTCCGCGTCATCGCCAAGTGGCTGCGGCTCGCCGGCGTCGACCACCTGCACACCGGCACCGCCGTGGGCAAGCTGGAAGGCGACCCCATGACCGTGCAGGGCTTCTACAATGTCTGCCGCGAGACGACGACGCAGCAGGACCTCACCCGCGGCCTGTTCTTCGAGCAGGACTGGGGCGGCATCCGCAAGGTGATGCCGGTGGCCTCCGGCGGCATCCATGCGGGCCAGATGCACCAGCTCATCGACCTGTTCGGCGAGGACGTGGTGCTCCAGTTCGGCGGCGGCACCATCGGCCACCCGGACGGCATCCAGGCCGGCGCCACCGCCAACCGCGTGGCGCTGGAAACCATGATCCTCGCCCGCAACGAGGGCCGCGACATCAGGAACGAGGGCCCGGAAATCCTGGTGGAAGCCGCCAAATGGTGCCGTCCGCTGCGCGCGGCGCTCGATACCTGGGGCGAGGTGACCTTCAACTACGCCTCCACCGACACGTCCGATTACGTGCCCACCGCGTCCGTCGCCTGA
SEQ ID NO: 3:
Аминокислотная последовательность большой цепи рибулозобисфосфаткарбоксилазы
MGAEATVGQITDAKKRYAAGVLKYAQMGYWNGDYVPKDTDLLAVFRITPQAGVDPVEAAAAVAGESSTATWTVVWTDRLTAADVYRAKAYKVEPVPGQEGQYFCYIAYDLDLFEEGSIANLTASIIGNVFSFKPLKAARLEDMRLPVAYVKTFRGPPTGIVVERERLDKFGRPLLGATTKPKLGLSGKNYGRVVYEALKGGLDFVKDDENINSQPFMHWRDRFLYCMEAVNKAQAETGEVKGHYLNITAGTMEEMYRRAEFAKELGSVVVMVDLIIGWTAIQSMSNWCRENDMILHMHRAGHGTYTRQKSHGVSFRVIAKWLRLAGVDHLHTGTAVGKLEGDPMTVQGFYNVCRETTTQQDLTRGLFFEQDWGGIRKVMPVASGGIHAGQMHQLIDLFGEDVVLQFGGGTIGHPDGIQAGATANRVALETMILARNEGRDIRNEGPEILVEAAKWCRPLRAALDTWGEVTFNYASTDTSDYVPTASVA
SEQ ID NO: 4:
Нуклеотидная последовательность малой цепи рибулозобисфосфаткарбоксилазы:
ATGCGCATCACCCAAGGCTCCTTCTCCTTCCTGCCGGACCTCACCGACACGCAGATCAAGGCCCAGGTGCAATATTGCCTGGACCAGGGCTGGGCGGTCTCGGTGGAGCACACCGACGATCCCCACCCGCGCAACACCTATTGGGAGATGTGGGGCCCGCCCATGTTCGATCTGCGCGACGCGGCCGGCGTCTTCGGCGAGATCGAAGCCTGCCGGGCCGCCAATCCCGAGCATTATGTGCGGGTGAACGCCTTCGATTCCAGCCGCGGATGGGAGACGATCCGCCTGTCCTTCATCGTTCAGCGGCCCACCGTGGAAGAGGGCTTCCGCCTCGACCGCACCGAAGGCAAGGGCCGCAACCAGAGCTACGCCATGCGCTACCGGGCGCAGTTCGCGCCGCGCTGA
SEQ ID NO: 5:
Аминокислотная последовательность малой цепи рибулозобисфосфаткарбоксилазы:
MRITQGSFSFLPDLTDTQIKAQVQYCLDQGWAVSVEHTDDPHPRNTYWEMWGPPMFDLRDAAGVFGEIEACRAANPEHYVRVNAFDSSRGWETIRLSFIVQRPTVEEGFRLDRTEGKGRNQSYAMRYRAQFAPR
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий альфа-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:7, или последовательность, имеющую более 70% идентичности по последовательности, например более 80% идентичности, например более 90% идентичности, например более 95% идентичности, например более 96% идентичности, например, более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:7.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий бета-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:9, или последовательность, имеющую более 77% идентичности по последовательности, например более 80% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например, более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:9.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий гамма-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:11, или последовательность, имеющую более 70% идентичности по последовательности, например более 80% идентичности, например более 90% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:11.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий дельта-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:13, или последовательность, имеющую более 79% идентичности по последовательности, например более 80% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например, более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:13.
SEQ ID NO:6:
Нуклеотидная последовательность альфа-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
ATGATGCCATCTGAGCCGCACGGCGCGGGCATGCCGCCCCCACGGGAAGCGGCCGCGGTTCCCACCCCCCAGGAGGTGAGCGCGGTGGTGGCCGAGGTGGTCGCGGATGCCGTGGCATCGGTGGGCGGCGCACGCACCCGGCTCATGGACATCGTCCAGCTGGCCCAGCAGCGTCTCGGCCATCTCTCCGAAGAGACCATGGCGGCCATTGCCGCGCGGCTCGCCATTCCGCCGGTGGAAGTGGCGGACATGGTGTCCTTCTACGCCTTCCTGAACCGCGCGCCCAAGGGCCGCTACCACATCCGCCTGTCGCGCAGCCCCATCTCGCTGATGAAGGGCGCCGAGGCGGTGGCTGCCGCCTTCTGCCAGATCCTCGGCATCGCCATGGGCGAGACCTCGCAGGATGGCGACTTCACCCTGGAATGGACCAACGACATCGGCATGGCCGACCAGGAGCCGGCCGCCCTCGTCAACGGCACGGTGATGACGCAGCTCGCGCCCGGCGATGCGGCCATCATCGTCGGCCGGCTGCGGGCCCATCACGCGCCCAATGCCCTGCCGCTGTTCCCTGGAGCCGGCGTGGCCGGCTCCGGCCTGCCCCATGCCCGGATCCGCCCCAGCCTGGTGATGCCGGGACAGCTTCTGTTCCGCGAGGACCACACGACGCCGGGCGCCGGCATCAAGGCGGCACTCGCCCTCACCCCGGACGAAGTGGTGCAGAAGGTCTCCGCCGCGCGCCTGCGCGGGCGGGGTGGCGCCGGCTTTCCCACCGGTCTCAAATGGAAGCTCTGCCGCCAGTCGCCCGCCACCACCCGCCATGTGATCTGCAATGCGGACGAGGGCGAGCCCGGCACCTTCAAGGATCGCGTGCTGCTCACGCAGGCGCCGCACCTCATGTTCGACGGCATGACCATCGCCGGCTACGCCTTGGGGGCGCGGGAGGGCGTGGTCTATCTGCGCGGCGAGTACGCCTATCTGTGGGAGCCTCTGCATGCGGTCCTGCGCGAGCGCTATGGGCTCGGGCTCGCCGGCGCGAACATCCTGGGACACGCGGGCTTCGACTTCGACATCCGCATCCAGCTGGGCGCCGGCGCCTATATCTGCGGCGAGGAATCCGCGCTGGTGGAATCGCTGGAAGGCAAGCGCGGCTCGCCCCGCGACCGCCCCCCCTTCCCCACCGTGCGCGGCCATCTCCAGCAGCCCACCGCCGTGGACAATGTGGAGACCTTCGCCTGCGCCGCCCGCATCCTGGAGGATGGCGTGGAGGCGTTCGCGGGCATCGGCACGCCCGAATCCGCCGGCACGAAGCTCCTCTCGGTGTCGGGCGATTGCCCGCGCCCCGGCGTGTATGAGGTGCCCTTCGGCCTCACGGTGAACGCGCTGCTCGACCTTGTCGGCGCGCCGGACGCCGCCTTCGTGCAGATGGGTGGGCCGTCCGGCCAATGCGTGGCGCCGAAGGATTACGGCCGCCGCATCGCCTTCGAGGACCTGCCCACCGGCGGCTCGGTGATGGTGTTCGGCCCGGGGCGCGACGTGCTCGCCATGGTGCGCGAGTTCGCGGATTTCTTCGCCGGCGAATCCTGCGGCTGGTGCACGCCCTGCCGGGTGGGCACCACCTTGCTCAAGGAAGAGCTGGACAAGCTCCTCGCCAACCGCGCCACCCTCGCCGACATCCGCGCGCTGGAGACCCTGGCCACGACCGTCTCCCGCACCAGCCGCTGCGGCCTCGGCCAGACGGCGCCCAACCCCATCCTTTCCACCATGCGCAACCTGCCGGAAGCCTATGAGGCGAGGCTGAGGCCCGAAGACTTCCTGCCCTGGGCCTCGCTCGACGAGGCGCTGAAGCCCGCCATCGTCATCCAGGGCCGCGCGCCCGTGCCGGAGGAAGAGGCATGA
SEQ ID NO: 7:
Аминокислотная последовательность альфа-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
MMPSEPHGAGMPPPREAAAVPTPQEVSAVVAEVVADAVASVGGARTRLMDIVQLAQQRLGHLSEETMAAIAARLAIPPVEVADMVSFYAFLNRAPKGRYHIRLSRSPISLMKGAEAVAAAFCQILGIAMGETSQDGDFTLEWTNDIGMADQEPAALVNGTVMTQLAPGDAAIIVGRLRAHHAPNALPLFPGAGVAGSGLPHARIRPSLVMPGQLLFREDHTTPGAGIKAALALTPDEVVQKVSAARLRGRGGAGFPTGLKWKLCRQSPATTRHVICNADEGEPGTFKDRVLLTQAPHLMFDGMTIAGYALGAREGVVYLRGEYAYLWEPLHAVLRERYGLGLAGANILGHAGFDFDIRIQLGAGAYICGEESALVESLEGKRGSPRDRPPFPTVRGHLQQPTAVDNVETFACAARILEDGVEAFAGIGTPESAGTKLLSVSGDCPRPGVYEVPFGLTVNALLDLVGAPDAAFVQMGGPSGQCVAPKDYGRRIAFEDLPTGGSVMVFGPGRDVLAMVREFADFFAGESCGWCTPCRVGTTLLKEELDKLLANRATLADIRALETLATTVSRTSRCGLGQTAPNPILSTMRNLPEAYEARLRPEDFLPWASLDEALKPAIVIQGRAPVPEEEA
SEQ ID NO: 8:
Нуклеотидная последовательность бета-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
ATGAGCCGGGGATCCCCCGATGCCGGGAAAGACCGCACCATGAGCGCCACCGACGGCACCACCGCCCCCCGCAAGATCGTCATCGATCCGGTGACCCGCGTGGAGGGCCACGGCAAGGTCACCATCCGCCTGGATGAAGCCGGCGCGGTGGAGGATGCGCGTTTCCACATCGTGGAGTTCCGCGGCTTCGAGCGGTTCATCCAGGGCCGGATGTACTGGGAAGTGCCCCTTATCATCCAGCGGCTGTGCGGCATCTGCCCGGTGAGCCACCATCTGGCGGCGGCGAAAGCCATGGACCAGGTGGCGGGCGTGGACCGCGTACCGCCCACCGCCGAGAAACTGCGCCGGCTGATGCATTATGGGCAGGTGCTGCAATCCAACGCTTTGCACATCTTCCACCTCGCCTCGCCCGACCTCCTGTTCGGCTTCGACGCGCCGGCCGAGCAGCGCAACATCATCGCCGTGCTCCAGCGTTATCCGGAGATCGGCAAATGGGCGATCTTCATCAGGAAGTTCGGCCAGGAGGTCATCAAGGCCACCGGCGGGCGCAAGATCCATCCCACCAGCGCCATTCCCGGCGGGGTCAACCAGAACCTCGCCGTGGAGGACCGCGACGCCCTGCGCGCCAAGGTGGGCGAGATCATCAGCTGGTGCATGGCGGCGCTGGACCATCACAAGGCCTATGTGGCGGAAAACCGGGCGCTGCATGACAGCTTCGCCGCCTTCCCCTCCGCCTTCATGAGCCTCGTGGGGCCGGATGGCGGCATGGACCTTTATGACGGCACCCTGCGGGTGATCGATGCCGAGGGCGCCCCCCTCATCGAAGGCGCGCCGCCCGCCTCCTACCGCGACCACCTCATCGAGGAGGTGCGGCCCTGGAGCTATCTGAAATTCCCCCATCTGCGCGCCTTCGGCCGCGACGATGGCTGGTATCGGGTCGGCCCCCTCGCCCAGGTCAATTGCGCCGCGTCCATCGACACGCCCCGCGCCGAGGCGGCCCGGCGGGACTTCATGGCCGAGGGCGGCGGCAAGCCGGTGCATGCCACCCTCGCTTATCACTGGGCGCGGCTCATCGTGCTGGTCCATTGCGCGGAGAAGATCGAACAGCTGCTGTTCGACGACGACCTGCAAGGCTGCGATCTGCGTGCGGAGGGCACCCGGCGCGGGGAAGGCGTCGCCTGGATCGAGGCGCCGCGCGGCACCCTCATCCACCATTACGAGGTGGACGAGAACGACCAGGTGCGCCGCGCCAACCTCATCGTCTCCACCACCCACAATAACGAGGCCATGAACCGCGCCGTGCGGCAGGTGGCGAAGACGGACCTTTCCGGTCGCGAGATCACCGAAGGGCTGCTGAACCATATCGAGGTGGCCATCCGCGCCTTCGACCCCTGCCTGTCCTGCGCCACCCATGCGCTGGGCCAGATGCCGCTGATCGTGACGCTTGAAGATGCCTCCGGCGCAGAGATCGCCCGCGGAGTGAAGGAATGA
SEQ ID NO: 9:
Аминокислотная последовательность бета-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
MSRGSPDAGKDRTMSATDGTTAPRKIVIDPVTRVEGHGKVTIRLDEAGAVEDARFHIVEFRGFERFIQGRMYWEVPLIIQRLCGICPVSHHLAAAKAMDQVAGVDRVPPTAEKLRRLMHYGQVLQSNALHIFHLASPDLLFGFDAPAEQRNIIAVLQRYPEIGKWAIFIRKFGQEVIKATGGRKIHPTSAIPGGVNQNLAVEDRDALRAKVGEIISWCMAALDHHKAYVAENRALHDSFAAFPSAFMSLVGPDGGMDLYDGTLRVIDAEGAPLIEGAPPASYRDHLIEEVRPWSYLKFPHLRAFGRDDGWYRVGPLAQVNCAASIDTPRAEAARRDFMAEGGGKPVHATLAYHWARLIVLVHCAEKIEQLLFDDDLQGCDLRAEGTRRGEGVAWIEAPRGTLIHHYEVDENDQVRRANLIVSTTHNNEAMNRAVRQVAKTDLSGREITEGLLNHIEVAIRAFDPCLSCATHALGQMPLIVTLEDASGAEIARGVKE
SEQ ID NO: 10:
Нуклеотидная последовательность гамма-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
ATGAGCGAGACCCCCTTCACCTTTACCGTGGACGGCATCGCGGTCCCGGCCACCCCCGGCCAGAGCGTCATCGAGGCGTGCGATGCGGCGGGCATCTATATCCCGCGCCTGTGCCACCACCCGGACCTGCCGCCGGCGGGCCATTGCCGGGTGTGCACCTGCATCATCGACGGGCGGCCGGCCAGCGCCTGCACCATGCCCGCCGCCAGGGGCATGGTGGTGGAGAACGAGACGCCCGCTTTGCTGGCGGAGCGGCGCACGCTGATCGAGATGCTGTTCGCGGAAGGCAACCATTTCTGCCAGTTCTGCGAGGCGAGCGGCGATTGCGAATTGCAGGCGCTGGGCTACCTGTTCGGCATGGTGGCCCCGCCCTTCCCCCATCTGTGGCCGAAGCGGCCGGTGGATGCCAGCCATCCGGATATCTATATCGACCACAATCGCTGCATCCTGTGCTCGCGCTGCGTGCGCGCCTCGCGCACCCTGGACGGCAAGTCCGTGTTCGGCTTCGAGGGGCGCGGCATCGAGATGCATCTGGCGGTGACCGGCGGGCACCTGGACGACAGCGCCATCGCCGCCGCCGACAGGGCGGTTGAGATGTGCCCGGTGGGCTGCATCGTCCTCAAGCGCACCGGCTACCGCACGCCCTATGGCCGGCGGCGCTACGACGCCGCGCCCATCGGCTCCGACATCACCGCCCGGCGCGGCGGCGCGAAGGACTGA
SEQ ID NO: 11:
Аминокислотная последовательность гамма-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
MSETPFTFTVDGIAVPATPGQSVIEACDAAGIYIPRLCHHPDLPPAGHCRVCTCIIDGRPASACTMPAARGMVVENETPALLAERRTLIEMLFAEGNHFCQFCEASGDCELQALGYLFGMVAPPFPHLWPKRPVDASHPDIYIDHNRCILCSRCVRASRTLDGKSVFGFEGRGIEMHLAVTGGHLDDSAIAAADRAVEMCPVGCIVLKRTGYRTPYGRRRYDAAPIGSDITARRGGAKD
SEQ ID NO: 12:
Нуклеотидная последовательность дельта-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
ATGGCCAAGCCCAAACTCGCCACCTGCGCGCTGGCCGGCTGCTTCGGCTGCCACATGTCCTTCCTGGACATGGACGAGCGCATCGTCGAGCTCATCGACCTGGTGGACCTCGACGTCTCGCCCCTCGACGACAAGAAAAACTTCACCGGCATGGTGGAAATCGGCCTGGTGGAAGGCGGCTGCGCCGACGAGCGCCATGTGAAGGTGCTGCGCGAGTTCCGCGAGAAATCCCGCATCCTGGTGGCGGTGGGCGCCTGCGCCATCACCGGCGGCATCCCGGCATTGCGCAACCTCGCCGGCCTCGACGAATGCCTGAGGGAAGCCTACCTCACCGGCCCCACGGTGGAAGGCGGCGGGCTCATTCCCAACGACCCGGAGCTGCCGCTGCTGCTGGACAAGGTCTATCCGGTGCAGGACTTCGTGAAGATCGACCATTTCCTGCCCGGCTGCCCGCCCTCGGCCGACGCCATCTGGGCGGCTCTGAAGGCGCTGCTGACCGGCACCGAGCCGCATCTGCCCTACCCGCTTTTCAAGTACGAATGA
SEQ ID NO: 13:
Аминокислотная последовательность дельта-субъединицы NAD+-восстанавливающей гидрогеназы HoxS:
MAKPKLATCALAGCFGCHMSFLDMDERIVELIDLVDLDVSPLDDKKNFTGMVEIGLVEGGCADERHVKVLREFREKSRILVAVGACAITGGIPALRNLAGLDECLREAYLTGPTVEGGGLIPNDPELPLLLDKVYPVQDFVKIDHFLPGCPPSADAIWAALKALLTGTEPHLPYPLFKYE
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий большую субъединицу гидрогеназы NiFeSe, имеющую последовательность, указанную в SEQ ID NO:15, или последовательность, имеющую более 84% идентичности по последовательности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например, более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:15.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий малую субъединицу гидрогеназы NiFeSe, имеющую последовательность, указанную в SEQ ID NO:17, или последовательность, имеющую более 90% идентичности, например более 95% идентичности, например более 96% идентичности, например, более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:17.
SEQ ID NO: 14:
Нуклеотидная последовательность большой субъединицы периплазматической гидрогеназы [NiFeSe]:
TCCAGACCCGGGCAACATTGCTCCATGTGCTGGGCACCCTGGCCGGCCGCTGGCCCCATACCCTCGCGCTCCAGCCCGGCGGGGTGACCCGAAGCGCCGACCAGCACGACCGCATGCGCCTGCTCGCGACGCTGAAGGCGGTGCGGGCGGCGCTGGAAGAGACCTTGTTCGGCGCGCCTTTGGAAGAGGTGGCGGCCCTGGACGGCGCCGCCGCCGTGGAGGCCTGGCGCGCCAACGGCCCGGAAGGGGATTTCCGCCTGTTCCTGGAGATCGCCGCCGACCTGGAGCTGGACCGGCTCGGCCGCGCGCACGACCGCTTTCTCTCCTTCGGCGCCTACGCCCAGGACGAGGGGCGCCTTTATGGCGCCGGCACCTTCGAGGCCGGGACGGCGGGAGGGCTCGATCCCAACGCCATCACCGAGGACCACGCCTTCGCCCGCATGGAGGACCGCGCGGCGCCCCATGCGCCCTTTGACGGCTCCACCTTCCCCGATGCCGACGACACCGAGGGCTACACCTGGTGCAAGGCGCCGCGCCTTGCCGGCCTGCCCTTCGAGACCGGCGCCTTCGCCCGGCAGGTGGTGGCGGGCCATCCGCTCGCCCGGGACCTCGTGACGCGGGAAGGCGGCACTGTGCGCAGCCGCGTGGTCGGCCGGCTGCTGGAAACCGCGCGCACCCTGATCGCCATGGAGGGCTGGGTGAAGGAACTGCGGCCCGAAGGGCCCTGGTGCGCCCAGGGCCACCTGCCCCAGGAAGGCCGCGCCTTCGGCCTCACCGAGGCGGCGCGCGGGGCGCTCGGCCACTGGATGGTGGTGGAGAAGGGCCGCATTGCCCGCTACCAGATCATCGCCCCCACCACCTGGAACTTCTCCCCCCGCGACGGCGCGGGCCTGCCCGGCCCGCTGGAGACGGCCCTGGTGGGCGCGCCCGTGCGGCAGGGAGAGACGACGCCCGTGAGCGTGCAGCACATCGTGCGCTCCTTCGACCCGTGCATGGTCTGCACTGTGCATTGA
SEQ ID NO: 15:
Аминокислотная последовательность большой субъединицы периплазматической гидрогеназы [NiFeSe]:
MSAETRRLVVGPFNRVEGDLEVRLDVQDGRVQQAFVSSPLFRGFERILEGRDPRDALVIAPRICGICSVSQSHAAALALAGLQGIAPTHDGRIATNLIVAAENVADHLTHFHVFFMPDFARAVYEDRPWFAQAARRFKANQGVSVRRALQTRATLLHVLGTLAGRWPHTLALQPGGVTRSADQHDRMRLLATLKAVRAALEETLFGAPLEEVAALDGAAAVEAWRANGPEGDFRLFLEIAADLELDRLGRAHDRFLSFGAYAQDEGRLYGAGTFEAGTAGGLDPNAITEDHAFARMEDRAAPHAPFDGSTFPDADDTEGYTWCKAPRLAGLPFETGAFARQVVAGHPLARDLVTREGGTVRSRVVGRLLETARTLIAMEGWVKELRPEGPWCAQGHLPQEGRAFGLTEAARGALGHWMVVEKGRIARYQIIAPTTWNFSPRDGAGLPGPLETALVGAPVRQGETTPVSVQHIVRSFDPCMVCTVH
SEQ ID NO: 16:
Нуклеотидная последовательность малой субъединицы периплазматической гидрогеназы [NiFeSe]
ACGGGGGAGGAAGCCCGCGCCATCTTCGACGCCATCCTTGCCGGCGTTATCGTCCTCGACGCCCTGTGCGTGGAAGGCGCGCTGCTGCGCGGGCCGAACGGCACCGGGCGCTTCCATGTGCTGGCGGGCACGGACACCCCCACCATCGACTGGGCGCGGCAGCTCGCCGGCATGGCGCGCCACGTGGTGGCGGTGGGCACCTGCGCCGCCTATGGGGGCGTGACGGCGGCGGGCATCAACCCCACCGATGCCTGCGGCCTCCAGTTCGACGGACGCCGGAAGGGTGGGGCGCTGGGGGCGGACTTCCGCTCCCGCTCGGGGCTTCCGGTCATCAATGTGGCCGGCTGCCCCACCCATCCCAACTGGGTGACGGAAACCCTGATGCTGCTCGCCTGCGGCCTGCTGGGCGAGGCCGACCTCGACGTCTATGGCCGCCCGCGCTTCTATGCGGACCTGCTGGTGCATCACGGCTGCCCGCGCAACGAATACTATGAATACAAGGCGAGCGCCGAGAAGATGAGCGACCTCGGCTGCATGATGGAGCATCTGGGCTGCCTCGGCACCCAGGCCCACGCCGACTGCAACACGCGCCTTTGGAATGGCGAGGGCTCGTGCACCCGCGGCGGCTATGCCTGCATCAACTGCACGGCGCCGGAATTCGAGGAGCCGGGCCACGCCTTCCTGGAGACGCCCAAGATCGGCGGCATCCCCATCGGCCTGCCCACCGACATGCCCAAGGCCTGGTTCATCGCCTTGTCCTCCCTCGCCAAGGCGGCGACGCCGGAGCGGCTGCGCAAGAACGCGGTGTCCGACCATGTGGTCACGCCGCCCGCCGTCAAGGACATCAAGCGGCGATGA
SEQ ID NO: 17:
Аминокислотная последовательность малой субъединицы периплазматической гидрогеназы [NiFeSe]
MSTPFSVLWLQSGGCGGCTMSLLCAEAPDLATTLDAAGIGFLWHPALSEETGEEARAIFDAILAGVIVLDALCVEGALLRGPNGTGRFHVLAGTDTPTIDWARQLAGMARHVVAVGTCAAYGGVTAAGINPTDACGLQFDGRRKGGALGADFRSRSGLPVINVAGCPTHPNWVTETLMLLACGLLGEADLDVYGRPRFYADLLVHHGCPRNEYYEYKASAEKMSDLGCMMEHLGCLGTQAHADCNTRLWNGEGSCTRGGYACINCTAPEFEEPGHAFLETPKIGGIPIGLPTDMPKAWFIALSSLAKAATPERLRKNAVSDHVVTPPAVKDIKRR
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий гамма-цепь АТФ-синтазы atpG_1, имеющую последовательность, указанную в SEQ ID NO:19, или последовательность, имеющую более 70% идентичности, например более 80% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:19.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий альфа-субъединицу АТФ-синтазы atpA_1, имеющую последовательность, указанную в SEQ ID NO:21, или последовательность, имеющую более 78% идентичности, например более 80% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:21.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретению, содержит ген, кодирующий субъединицу b АТФ-синтазы atpF_1, имеющую последовательность, указанную в SEQ ID NO:23, или последовательность, имеющую более 62% идентичности, например более 70% идентичности, например, более 80% идентичности, например более 90% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:23.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий субъединицу c специфичной к ионам натрия АТФ-синтазы atpE_1, имеющую последовательность, указанную в SEQ ID NO:25, или последовательность, имеющую 90% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:25.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий субъединицу a АТФ-синтазы atpB_1, имеющую последовательность, указанную в SEQ ID NO:27, или последовательность, имеющую более 80% идентичности, например более 90% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:27.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий эпсилон-цепь АТФ-синтазы atpC_1, имеющую последовательность, указанную в SEQ ID NO: 29, или последовательность, имеющую более 71% идентичности, например более 80% идентичности, например более 90% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:29.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий бета-субъединицу АТФ-синтазы atpD_1, имеющую последовательность, указанную в SEQ ID NO:31, или последовательность, имеющую более 84% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:31.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий бета-субъединицу АТФ-синтазы atpD_2, имеющую последовательность, указанную в SEQ ID NO:33, или последовательность, имеющую более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:33.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретению, содержит ген, кодирующий гамма-цепь АТФ-синтазы atpG_2, имеющую последовательность, указанную в SEQ ID NO:35, или последовательность, имеющую более 86% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:35.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий субъединицу альфа АТФ-синтазы atpA_2, имеющую последовательность, указанную в SEQ ID NO: 37, или последовательность, имеющую более 98%, например более 99% идентичности по последовательности с последовательностью, указанной в SEQ ID NO:37.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий дельта-субъединицу АТФ-синтазы atpH, имеющую последовательность, указанную в SEQ ID NO:39, или последовательность, имеющую более 85% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:39.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий субъединицу b АТФ-синтазы atpF_2, имеющую последовательность, указанную в SEQ ID NO:41, или последовательность, имеющую более 87% идентичности, например более 90% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:41.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий субъединицу b' АТФ-синтазы atpG_3, имеющую последовательность, указанную в SEQ ID NO:43, или последовательность, имеющую более 81% идентичности, например более 90% идентичности, например более 95% идентичностиь, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:43.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий субъединицу c АТФ-синтазы atpE_2, имеющую последовательность, указанную в SEQ ID NO:45, или последовательность, имеющую более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:45.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий субъединицу a АТФ-синтазы atpB_2, имеющую последовательность, указанную в SEQ ID NO:47, или последовательность, имеющую более 92% идентичности, например более 95% идентичности, например более 96% идентичности, например, более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:47.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий белок I АТФ-синтазы atpI, имеющий последовательность, указанную в SEQ ID NO: 49, или последовательность, имеющую более 60% идентичности, например более 70% идентичности, например, более 80% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например, более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:49.
SEQ ID NO: 18:
Нуклеотидная последовательность гамма-цепи АТФ-синтазы atpG_1
GTGACCGAGCGCCTGTCCGACGTCAACGCCCGCATCGCCTCGGTGCGGCAGCTCTCATCGGTCATCACGGCCATGCGGGGCATTGCGGCGGCGCGGGCGCGGGAGGCGCGGGGTCGGCTCGACGGCATCCGCGCCTATGCGCAGACCATCGCCGAGGCCATCGGCCATGTGCTCGCCGTGCTGCCCGAGGAGGCCCGCGCCCGGTCCTCCGGGCACCGGCATCGGGGCCATGCGGTCATCGCCCTGTGCGCGGAGCAGGGCTTTGCCGGCGTCTTCAACGAGCGGGTGCTGGACGAGGCCGCCCGGCTGCTGACCGGCGGGGCGGGGCCGGCCGAGCTGCTGCTGGTGGGCGACCGGGGCCTGATGGTGGCCCGCGAGCGGGGGCTCGATGTCTCCTGGTCGGTGCCCATGGTGGCCCATGCGGGCCAGGCCTCGGCGCTGGCGGACCGCATCAGCGAGGAGCTCTACCGGCGGATCGATGCGGGACGGGTGACGCGGGTGTCGGTGGTGCACGCCGAGCCCGCCGCGTCCGCCGCCATCGAGACGGTGGTGAAAGTGCTGGTGCCGTTCGACTTCGCCCGCTTCCCCCTGGCGCGGGTGGCATCCGCCCCGCTCATGACCATGCCGCCGCCGCGGCTGCTGGCCCAGCTGTCGGAGGAATATGTGTTCGCCGAGCTGTGCGAGGCGCTCACCTTGTCCTTCGCGGCGGAGAACGAGGCCCGCATGCGGGCCATGATCGCCGCCCGCGCCAATGTGGCCGATACCCTGGAGGGCCTCGTCGGCCGCGCCCGGCAGATGCGCCAGGAGGAGATCACCAACGAGATCATCGAGCTGGAAGGCGGCGCCGGCAGCGCCCGGCATGCGGATTGA
SEQ ID NO: 19:
Аминокислотная последовательность гамма-цепи АТФ-синтазы atpG_1
MTERLSDVNARIASVRQLSSVITAMRGIAAARAREARGRLDGIRAYAQTIAEAIGHVLAVLPEEARARSSGHRHRGHAVIALCAEQGFAGVFNERVLDEAARLLTGGAGPAELLLVGDRGLMVARERGLDVSWSVPMVAHAGQASALADRISEELYRRIDAGRVTRVSVVHAEPAASAAIETVVKVLVPFDFARFPLARVASAPLMTMPPPRLLAQLSEEYVFAELCEALTLSFAAENEARMRAMIAARANVADTLEGLVGRARQMRQEEITNEIIELEGGAGSARHAD
SEQ ID NO: 20:
Нуклеотидная последовательность альфа-субъединицы АТФ-синтазы atpA_1
ATGAGCACGGGCGCGCAAGCGAGCGAGGATTGGCTCACCCGGAGCCGGGCGGCCCTGGCCGGGACGCGCCTTTCCCAGCAATCCCAATCGGTGGGCCGGGTGGAGGAGATGGCCGACGGCATCGCCCGCGTCTCCGGCCTGCCGGATGTGCGGCTCGACGAGCTTCTCACCTTCGAGGGCGGCCAGACCGGCTATGCCCTCACCCTCGATCGCACCGAGATCGCCGTGGTGCTGCTGGATGACGCCTCCGGCGTGGAGGCGGGCGCCCGGGTGTTCGGCACCGGCGAGGTGGTGAAGGTGCCGGTGGGGCCGGGGCTGCTGGGCCGCATCGTCGACCCCCTCGGCCGGCCCATGGACCGCTCCGAGCCGGTGGTGGCGCAGGCGCACCATCCCATCGAGCGGCCGGCGCCGGCCATCATCGCCCGCGACCTGGTCTCGCAGCCGGTTCAGACCGGCACGCTGGTGGTGGATGCGCTGTTCTCCCTCGGCCGGGGCCAGCGCGAGCTCATCATCGGCGACCGGGCTACCGGCAAGACCGCCATCGCGGTGGACACCATCATCAGCCAGAAGCATTCGGACATCGTGTGCATCTACGTGGCGGTGGGCCAGCGCGCCGCCGCCGTGGAGCGGGTGGTGGAGGCGGTGCGCGCCCACGGGGCGATCGAGCGCTGCATCTTCGTGGTCGCCTCGGCCGCCGCCTCGCCAGGGCTGCAATGGATCGCGCCGTTCGCCGGCATGACCATGGCGGAATATTTCCGCGACAACGGCCAGCATGCGCTCATCATCATCGATGATCTCACCAAGCATGCGGCCACCCATCGCGAGCTGGCGCTGCTCACCCACGAGCCGCCGGGCCGCGAGGCCTATCCCGGCGACATCTTCTATGTGCACGCCCGCCTTCTGGAGCGGGCCGCCAAGCTCTCCGCCGAGCTGGGCGGTGGCTCGCTCACGGCCCTGCCCATCGCGGAGACGGACGCGGGAAACCTCTCCGCCTATATCCCCACCAACCTCATCTCCATCACCGATGGGCAGATCGTGCTGGATTCGCGGCTGTTCGCGGCCAACCAGCGCCCGGCGGTGGATGTGGGCCTCTCCGTGAGCCGGGTGGGCGGCAAGGCGCAGCATCCCGCGCTTCGGGCCGTGTCCGGGCGCATCCGGCTCGATTATTCCCAGTTCCTGGAGCTGGAAATGTTCACCCGCTTCGGCGGCATCACCGATACCCGCGTGAAGGCGCAGATCACCCGGGGCGAGCGCATCCGCGCGCTGCTCACCCAGCCGCGCTTTTCCACCCTGCGCCTTCAGGACGAGGTGGCGCTGCTGGCCGCGCTGGCGGAGGGGGTGTTCGACACTTTGGCCCCGGGGCTGATGGGCGCCGTGCGTGCCCGCATTCCGGCCCAGCTGGATGCGCAGGTGAAGGACGTGGCCTCGGCCCTCGCCGAGGGCAAGGTGCTGGAGGAGGGCTTGCACGCCCGTCTCGTGGCGGCCGTGCGGGCCGTCGCGGCGGACGTGGCCGCGACCGCGAAGGCCGGGCCGTGA
SEQ ID NO: 21:
Аминокислотная последовательность альфа-субъединицы АТФ-синтазы atpA_1
MSTGAQASEDWLTRSRAALAGTRLSQQSQSVGRVEEMADGIARVSGLPDVRLDELLTFEGGQTGYALTLDRTEIAVVLLDDASGVEAGARVFGTGEVVKVPVGPGLLGRIVDPLGRPMDRSEPVVAQAHHPIERPAPAIIARDLVSQPVQTGTLVVDALFSLGRGQRELIIGDRATGKTAIAVDTIISQKHSDIVCIYVAVGQRAAAVERVVEAVRAHGAIERCIFVVASAAASPGLQWIAPFAGMTMAEYFRDNGQHALIIIDDLTKHAATHRELALLTHEPPGREAYPGDIFYVHARLLERAAKLSAELGGGSLTALPIAETDAGNLSAYIPTNLISITDGQIVLDSRLFAANQRPAVDVGLSVSRVGGKAQHPALRAVSGRIRLDYSQFLELEMFTRFGGITDTRVKAQITRGERIRALLTQPRFSTLRLQDEVALLAALAEGVFDTLAPGLMGAVRARIPAQLDAQVKDVASALAEGKVLEEGLHARLVAAVRAVAADVAATAKAGP
SEQ ID NO: 22:
Нуклеотидная последовательность субъединицы b АТФ-синтазы atpF_1
ATGCAGATCGACTGGTGGACGCTGGGCCTGCAGACGGTCAACGTCCTCGTTCTCATCTGGCTCCTGAGCCGCTTCCTGTTCAAGCCGGTGGCGCAGGTCATCGCGCAGCGCCGTGCCGAGATCGAGAAGCTGGTGGAGGATGCGCGCGCCGCCAAGGCCGCCGCCGAGGCCGAGCGGGACACGGCGAAGGCGGAGGAGGCGCGCCTTGCCGCCGAGCGCGGCGCCCGCATGGCGGCGGTCGCCAAGGAGGCGGAGGCGCAGAAGGCGGCATTGCTGGCCGCCGCCAAGACCGAGGCCGAGGCCCTGCACGCGGCCGCGGAAGCGGCCATCGTCCGGGCGCGGGCGAGCGAGGAGGAAGCCGCCGCCGACCGCGCCAGCCGCCTTGCCGTGGACATCGCCGCCAAGCTGCTGGACCGGCTGCCCGACGACGCCCGGGTCGCGGGCTTCATCGATGGCCTCGCCGAGGGGCTTGAAGCCCTGCCCGAGGCGAGCCGGGCGGTGATCGGCGTCGACGGCGCGCCAGTGCGCGTGACGGCCGCGCGCGCCCTTATGCCGGCGGAGGAGGAGGCCTGCCGCACGCGGCTCTCCCAGGCGCTGGGCCGTCCGGTGACGCTGGCCGTGACCATCGACCCCGCCCTCATCGCCGGCCTGGAGATGGAGACGCCCCACGCGGTGGTGCGCAATTCCTTCAAGGCCGATCTCGACCGCGTCACCGCGGCGCTCACCCATCATGGGACCTGA
SEQ ID NO: 23:
Аминокислотная последовательность субъединицы b АТФ-синтазы atpF_1
MQIDWWTLGLQTVNVLVLIWLLSRFLFKPVAQVIAQRRAEIEKLVEDARAAKAAAEAERDTAKAEEARLAAERGARMAAVAKEAEAQKAALLAAAKTEAEALHAAAEAAIVRARASEEEAAADRASRLAVDIAAKLLDRLPDDARVAGFIDGLAEGLEALPEASRAVIGVDGAPVRVTAARALMPAEEEACRTRLSQALGRPVTLAVTIDPALIAGLEMETPHAVVRNSFKADLDRVTAALTHHGT
SEQ ID NO: 24:
Нуклеотидная последовательность субъединицы c специфичной к ионам натрия АТФ-синтазы atpE_1
ATGACTGTCGAGATGGTCAGCATCTTCGCGGCGGCGCTCGCCGTCTCCTTCGGCGCCATCGGGCCGGCCCTGGGCGAGGGCCGGGCGGTGGCCGCGGCCATGGACGCCATCGCCCGCCAGCCGGAGGCGGCCGGAACCTTGTCGCGCACGCTCTTCGTCGGCCTCGCCATGATCGAGACCATGGCGATCTACTGCCTGGTGATCGCGCTCCTGGTGCTCTTCGCCAATCCGTTCGTGAAGTGA
SEQ ID NO: 25:
Аминокислотная последовательность субъединицы c специфичной к ионам натрия АТФ-синтазы atpE_1
MTVEMVSIFAAALAVSFGAIGPALGEGRAVAAAMDAIARQPEAAGTLSRTLFVGLAMIETMAIYCLVIALLVLFANPFVK
SEQ ID NO: 26:
Нуклеотидная последовательность субъединицы a АТФ-синтазы atpB_1
ATGGGCTCGCCGCTGATCCTCGAACCCCTGTTCCATATCGGGCCCGTGCCCATCACCGCGCCGGTGGTGGTCACCTGGCTCATCATGGCCGCCTTCATTGGGCTGGCGCGGCTCATCACCCGGAAGCTTTCCACCGATCCCACCCGGACCCAGGCGGCGGTGGAAACGGTGCTGACCGCCATCGATTCCCAGATCGCCGACACCATGCAGGCCGATCCCGCGCCTTATCGCGCGCTCATCGGCACCATCTTCCTTTATGTGCTGGTGGCCAACTGGTCCTCGCTCATCCCGGGCATCGAGCCGCCCACGGCGCATATCGAGACCGATGCGGCGCTCGCTTTCATCGTGTTCGCCGCCACCATCGGGTTCGGGTTGAAGACAAGGGGTGTGAAGGGCTATCTCGCCACCTTCGCCGAACCCTCCTGGGTGATGATCCCGCTCAATGTGGTGGAGCAGATCACCCGGACCTTCTCGCTCATCGTGCGCCTGTTCGGCAACATCATGAGCGGGGTGTTCGTGGTCGGCATCATCCTGTCCCTCGCCGGGCTGCTGGTGCCCATCCCCCTCATGGCGCTCGATCTCCTGACCGGCGCCGTGCAGGCCTACATCTTCGCGGTGCTGGCCTGCGTGTTCATCGGCGCGGCCATTGGCGAGGCGCCGGCAAAGCCCCAATCGAAGGAGCCAGGGAAAACATCATGA
SEQ ID NO: 27:
Аминокислотная последовательность субъединицы a АТФ-синтазы atpB_1
MGSPLILEPLFHIGPVPITAPVVVTWLIMAAFIGLARLITRKLSTDPTRTQAAVETVLTAIDSQIADTMQADPAPYRALIGTIFLYVLVANWSSLIPGIEPPTAHIETDAALAFIVFAATIGFGLKTRGVKGYLATFAEPSWVMIPLNVVEQITRTFSLIVRLFGNIMSGVFVVGIILSLAGLLVPIPLMALDLLTGAVQAYIFAVLACVFIGAAIGEAPAKPQSKEPGKTS
SEQ ID NO: 28:
Нуклеотидная последовательность эпсилон-цепи АТФ-синтазы atpC_1
GTGAGCGCGCCGCTGCACCTCACCATCACCACGCCGGCCGCCGTTCTGGTGGACCGTGCCGACATCGTGGCCCTGCGTGCCGAGGACGAGAGCGGCAGCTTCGGCATCCTGCCCGGCCATGCGGATTTCCTGACCGTTCTGGAGGCCTGCGTGGTGCGCTTCAAGGATGGGGCCGACGGCGTGCATTATTGTGCTCTCAGTGGTGGCGTGCTGTCGGTCGAGGAGGGCCGGCGCATCGCCATCGCCTGCCGTCAGGGCACGGTGAGCGACGACCTGGTCGCCCTGGAAGGGGCGGTGGACGCCATGCGTTCGGCGGAGAGCGATGCCGACAAGCGGGCCCGGGTGGAGCAGATGCGCCTTCATGCCCACGCCGTGCGCCAGCTCCTGCACTATCTGCGGCCCGGCCGGGCCGGCGGCGTGGCGCCGGCCGCCGCGCCGGAGGAGGGGCCGTCATGA
SEQ ID NO: 29:
Аминокислотная последовательность эпсилон-цепи АТФ-синтазы atpC_1
MSAPLHLTITTPAAVLVDRADIVALRAEDESGSFGILPGHADFLTVLEACVVRFKDGADGVHYCALSGGVLSVEEGRRIAIACRQGTVSDDLVALEGAVDAMRSAESDADKRARVEQMRLHAHAVRQLLHYLRPGRAGGVAPAAAPEEGPS
SEQ ID NO: 30:
Нуклеотидная последовательность бета-субъединицы АТФ-синтазы atpD_1
ATGGCAGCGGCAGATGAGGAGGCGCAATCGGCCGCCGGCCCCGCCTCGGGCCGGGTGGTGGCCGTGCGCGGCGCGGTGATCGACATCGCCTTTGCCCAGCCTCCGCTGCCGCCGCTGGACGACGCCCTTCTCATCACCGACGGCCGGGGCGGCACGGTGCTGGTGGAGGTGCAGAGCCATATGGATCGGCACACGGTGCGCGCCATCGCCCTTCAGGCCACCACCGGCCTCAGCCGGGGGCTGGAGGCGGCGCGGGTGGGCGGGCCGGTGAAGGTGCCGGTGGGAGACCATGTGCTCGGCCGCCTCCTGGATGTCACCGGCGCCATCGGCGACAAGGGCGGGCCGCTGCCGGCCGACGTGCCCACGCGGCCGATCCACCACGCGCCGCCATCCTTCGCCGCGCAGGGCGGCACGTCCGATCTGTTTCGCACCGGCATCAAGGTCATCGACCTCCTGGCGCCCCTCGCCCAGGGCGGCAAGGCGGCCATGTTCGGCGGGGCCGGCGTGGGCAAGACCGTGCTGGTGATGGAGCTGATCCACGCCATGGTGGCGAGCTACAAGGGCATCTCGGTGTTTGCCGGCGTGGGGGAGCGCTCCCGCGAGGGCCACGAGATGCTGCTGGACATGACCGATTCCGGCGTGCTCGACCGCACCGTTCTGGTCTATGGCCAGATGAACGAGCCCCCCGGGGCCCGCTGGCGGGTGCCCATGACGGCGCTGACCATCGCCGAATATTTCCGCGACGAGAAGCACCAGAACGTCCTGCTGCTGATGGACAACATCTTCCGCTTCGTCCAGGCGGGGGCGGAGGTCTCCGGCCTTTTGGGCCGTCCGCCCTCCCGGGTGGGATACCAGCCGACGCTGGCGAGCGAGGTGGCGGCGCTCCAGGAACGCATCACCTCCGTGGGCGAGGCCTCGGTGACCGCCATCGAGGCGGTCTACGTGCCGGCGGATGACTTCACCGATCCCGCCGTGACCACCATCGCCGCCCACGTGGATTCCATGGTGGTGCTCTCCCGCGCCATGGCGGCGGAGGGCATGTATCCGGCGGTGGACCCCATCTCCTCCTCGTCGGTGCTGCTCGACCCGCTCATCGTGGGGGACGAGCATGCGCGCGTCGCCAACGAGGTGCGCCGGACCATCGAGCATTATCGCGAGCTTCAGGATGTGATCTCGCTGCTGGGCATGGAGGAATTGGGCACCGAGGATCGCCGCATCGTGGAGCGGGCGCGCCGGCTCCAGCGCTTCCTCACCCAGCCCTTCACGGTCACCGAGGCCTTCACCGGCGTGCCCGGCCGCTCGGTGGCCATCGCCGACACCATCGCCGGCTGCAGGATGATCCTGTCCGGCGCCTGCGACGACTGGCAGGAAAGCGCCCTCTACATGGTGGGCACCATCGACGAGGCCCGCCAGAAGGAGGAGGCCGCTCGCGCCAAGGCGGGGCAGGGCGCCCCGGCCGGGACGGCAGCCGAGACGGCGGAGGCCGCCCCGTGA
SEQ ID NO: 31:
Аминокислотная последовательность бета-субъединицы АТФ-синтазы atpD_1
MAAADEEAQSAAGPASGRVVAVRGAVIDIAFAQPPLPPLDDALLITDGRGGTVLVEVQSHMDRHTVRAIALQATTGLSRGLEAARVGGPVKVPVGDHVLGRLLDVTGAIGDKGGPLPADVPTRPIHHAPPSFAAQGGTSDLFRTGIKVIDLLAPLAQGGKAAMFGGAGVGKTVLVMELIHAMVASYKGISVFAGVGERSREGHEMLLDMTDSGVLDRTVLVYGQMNEPPGARWRVPMTALTIAEYFRDEKHQNVLLLMDNIFRFVQAGAEVSGLLGRPPSRVGYQPTLASEVAALQERITSVGEASVTAIEAVYVPADDFTDPAVTTIAAHVDSMVVLSRAMAAEGMYPAVDPISSSSVLLDPLIVGDEHARVANEVRRTIEHYRELQDVISLLGMEELGTEDRRIVERARRLQRFLTQPFTVTEAFTGVPGRSVAIADTIAGCRMILSGACDDWQESALYMVGTIDEARQKEEAARAKAGQGAPAGTAAETAEAAP
SEQ ID NO: 32:
Нуклеотидная последовательность бета-субъединицы АТФ-синтазы atpD_2
ATGGCGAACAAGGTCGGACGCATCACCCAGATCATCGGCGCCGTCGTCGACGTGCAGTTCGACGGGCATCTGCCGGCGATTCTCAACGCGATCGAGACCACCAACCAGGGCAACCGGCTGGTGCTCGAAGTGGCTCAGCATCTCGGCGAGAACACCGTGCGCTGCATCGCCATGGATGCCACTGAAGGCCTGGTGCGTGGCCAGGAGGTGGCCGACACCGATGCGCCCATCCAGGTGCCCGTGGGCGCCGCCACCCTCGGCCGCATCATGAACGTGATCGGCGAGCCGGTGGACGAGCTGGGCCCCATCGAGGGCGAAGCGCTGCGCGGCATCCATCAGCCGGCCCCCTCCTATGCGGAGCAGGCCACGGAAGCTGAGATCCTCGTCACCGGCATCAAGGTGGTGGATCTGCTGGCGCCCTATTCCAAGGGCGGCAAGGTGGGCCTGTTCGGCGGCGCCGGCGTGGGCAAGACCGTGCTCATCATGGAGCTGATCAACAACGTGGCCAAGGCGCACGGCGGCTATTCCGTGTTCGCCGGCGTGGGTGAGCGCACCCGCGAGGGCAACGACCTCTACCACGAGATGATCGAGTCCAACGTGAACAAGGACCCGCACGAGAACAATGGCTCGGCGGCCGGTTCCAAGTGCGCCCTGGTCTATGGCCAGATGAACGAGCCGCCCGGCGCCCGCGCCCGCGTGGCCCTCACCGGCCTCACCGTCGCCGAGCATTTCCGCGACCAGGGCCAGGACGTGCTGTTCTTCGTGGACAACATCTTCCGCTTCACCCAGGCGGGCTCCGAGGTGTCGGCGCTTCTCGGCCGCATCCCCTCGGCGGTGGGCTACCAGCCGACGCTGGCCACCGACATGGGCCAGCTGCAGGAGCGCATCACCACCACCACCAAGGGCTCCATCACCTCGGTGCAGGCCATCTACGTGCCGGCGGACGATCTGACCGATCCGGCGCCGGCCGCCTCCTTCGCCCATCTGGACGCCACCACGGTGCTGTCGCGCTCCATCGCGGAGAAGGGCATCTACCCGGCGGTGGATCCGCTGGACTCCACCTCGCGCATGCTGTCTCCCGCCATCCTCGGCGACGAGCACTACAACACCGCGCGCCAGGTGCAGCAGACCCTGCAGCGCTACAAGGCGCTCCAGGACATCATCGCCATCCTGGGCATGGACGAACTCTCCGAAGAGGACAAGCTCACCGTGGCCCGCGCCCGCAAGATCGAGCGCTTCCTCTCCCAGCCCTTCCACGTGGCCGAGGTGTTCACCGGTTCGCCCGGCAAGCTGGTCGACCTCGCCGACACCATCAAGGGCTTCAAGGGCCTGGTGGACGGCAAGTACGACTACCTGCCCGAGCAGGCCTTCTACATGGTGGGCACCATCGAAGAAGCCATCGAGAAGGGCAAGAAGCTGGCGGCCGAGGCGGCCTGA
SEQ ID NO: 33:
Аминокислотная последовательность бета-субъединицы АТФ-синтазы atpD_2
MANKVGRITQIIGAVVDVQFDGHLPAILNAIETTNQGNRLVLEVAQHLGENTVRCIAMDATEGLVRGQEVADTDAPIQVPVGAATLGRIMNVIGEPVDELGPIEGEALRGIHQPAPSYAEQATEAEILVTGIKVVDLLAPYSKGGKVGLFGGAGVGKTVLIMELINNVAKAHGGYSVFAGVGERTREGNDLYHEMIESNVNKDPHENNGSAAGSKCALVYGQMNEPPGARARVALTGLTVAEHFRDQGQDVLFFVDNIFRFTQAGSEVSALLGRIPSAVGYQPTLATDMGQLQERITTTTKGSITSVQAIYVPADDLTDPAPAASFAHLDATTVLSRSIAEKGIYPAVDPLDSTSRMLSPAILGDEHYNTARQVQQTLQRYKALQDIIAILGMDELSEEDKLTVARARKIERFLSQPFHVAEVFTGSPGKLVDLADTIKGFKGLVDGKYDYLPEQAFYMVGTIEEAIEKGKKLAAEAA
SEQ ID NO: 34:
Нуклеотидная последовательность гамма-цепи АТФ-синтазы atpG_2
ATGGCGAGTCTGAAGGACCTGAGAAACCGCATTGCCTCGGTGAAGGCGACGCAGAAGATCACCAAGGCGATGCAGATGGTCGCCGCGGCGAAGCTGCGTCGCGCCCAGGCGGCGGCTGAAGCGGCCCGTCCCTATGCGGAACGCATGGAGACGGTGCTCGGAAATCTTGCCTCCGGCATGGTGGTGGGCGCGCAGGCGCCTGTTCTCATGACCGGGACGGGCAAGAGCGACACCCACCTGCTGCTGGTGTGCACCGGCGAGCGCGGCCTGTGCGGCGCCTTCAACTCGTCCATCGTGCGCTTCGCCCGCGAGCGGGCGCAGCTGCTGCTGGCCGAGGGCAAGAAGGTGAAAATCCTGTGCGTGGGCCGCAAGGGCCACGAGCAGCTGCGCCGCATCTACCCGGACAACATCATCGACGTGGTGGACCTGCGCGCGGTGCGCAACATCGGCTTCAAGGAGGCCGACGCCATCGCCCGCAAGGTGCTGGCCCTGCTCGATGAAGGCGCATTCGACGTCTGCACGCTCTTCTACTCCCACTTCAGGAGCGTGATCGCCCAGGTGCCGACGGCCCAGCAGCTCATTCCGGCCACCTTCGACGAGCGGCCGGCCGTCGCCGATGCGCCGGTCTATGAATATGAGCCGGAGGAGGAGGAGATCCTCGCCGAGCTGCTGCCGCGCAACGTGGCGGTGCAGATCTTCAAGGCCCTCCTCGAGAACCAGGCTTCTTTCTATGGCTCCCAGATGAGCGCCATGGACAACGCCACGCGCAATGCGGGCGAGATGATCAAGAAGCAGACGCTCACCTACAACCGTACCCGCCAGGCCATGATCACGAAGGAACTCATCGAGATCATCTCCGGCGCCGAGGCCGTCTGA
SEQ ID NO: 35:
Аминокислотная последовательность гамма-цепи АТФ-синтазы atpG_2
MASLKDLRNRIASVKATQKITKAMQMVAAAKLRRAQAAAEAARPYAERMETVLGNLASGMVVGAQAPVLMTGTGKSDTHLLLVCTGERGLCGAFNSSIVRFARERAQLLLAEGKKVKILCVGRKGHEQLRRIYPDNIIDVVDLRAVRNIGFKEADAIARKVLALLDEGAFDVCTLFYSHFRSVIAQVPTAQQLIPATFDERPAVADAPVYEYEPEEEEILAELLPRNVAVQIFKALLENQASFYGSQMSAMDNATRNAGEMIKKQTLTYNRTRQAMITKELIEIISGAEAV
SEQ ID NO: 36:
Нуклеотидная последовательность альфа-субъединицы АТФ-синтазы atpA_2
ATGGACATTCGAGCCGCTGAAATCTCTGCCATCCTGAAAGAGCAGATCCAGAATTTCGGCCAGGAGGCGGAAGTCTCCGAGGTGGGTCAGGTTCTGTCCGTGGGTGACGGCATCGCGCGCGTCTACGGCCTCGACAACGTCCAGGCGGGCGAGATGGTCGAGTTCGAGAACGGCACGCGCGGCATGGCGCTGAACCTCGAGCTCGACAATGTCGGCATCGTGATCTTCGGTTCCGACCGCGAGATCAAGGAAGGCCAGACCGTCAAGCGGACCGGCGCCATCGTGGACGCCCCCGTCGGCAAGGGCCTGCTCGGCCGCGTCGTGGACGCTCTCGGCAACCCGATCGACGGCAAGGGCCCGATCATGTTCACCGAGCGTCGCCGGGTCGACGTGAAGGCGCCGGGCATCATCCCGCGCAAGTCGGTGCACGAGCCCATGCAGACCGGCCTGAAGGCCATCGATGCGCTCATCCCCATCGGCCGCGGCCAGCGCGAGCTCATCATCGGCGACCGCCAGACCGGCAAGACCGCCGTGGCGCTCGACTCGATCCTGAACCAGAAGCCCATCAACCAGGGCGACGACGAGAAGGCCAAGCTCTACTGCGTCTATGTCGCGGTGGGCCAGAAGCGTTCCACTGTCGCGCAGTTCGTGAAGGTGCTCGAGGAGCACGGCGCGCTGGAATATTCCATCGTCGTCGCCGCCACCGCCTCGGACGCGGCCCCCATGCAGTTCCTGGCGCCGTTCACCGGCACCGCCATGGGCGAGTATTTCCGCGACAACGGCATGCACGCCCTCATCATCCATGATGACCTGTCCAAGCAGGCCGTGGCCTACCGCCAGATGTCGCTGCTGCTGCGCCGCCCGCCGGGCCGCGAGGCCTATCCCGGCGATGTGTTCTACCTGCACTCCCGCCTCTTGGAGCGCGCCGCCAAGCTCAATGACGAGCACGGCGCCGGCTCGCTGACCGCCCTGCCGGTGATCGAGACCCAGGCCAACGACGTGTCGGCCTACATCCCGACCAACGTGATCTCCATCACCGACGGTCAGATCTTCCTTGAATCCGATCTGTTCTACCAGGGCATCCGCCCGGCGGTGAACGTGGGCCTGTCGGTGTCGCGCGTGGGCTCTTCGGCCCAGATCAAGGCGATGAAGCAGGTGGCCGGCAAGATCAAGGGCGAGCTCGCCCAGTATCGCGAGCTGGCGGCCTTCGCCCAGTTCGGTTCGGACCTGGACGCGGCCACCCAGAAGCTGCTGAACCGCGGCGCCCGCCTCACCGAGCTGCTGAAGCAGAGCCAGTTCTCGCCCCTCAAGGTGGAGGAGCAGGTGGCGGTGATCTATGCCGGCACCAATGGCTATCTCGATCCGCTGCCGGTCTCCAAGGTGCGCGAGTTCGAGCAGGGTCTGCTCCTGTCGCTGCGCTCGCAGCATCCGGAGATCCTGGACGCCATCCGCACGTCCAAGGAGCTTTCCAAGGACACCGCCGAGAAGCTGACGAAGGCCATCGACGCCTTCGCCAAGAGCTTCTCCTGA
SEQ ID NO: 37:
Аминокислотная последовательность альфа-субъединицы АТФ-синтазы atpA_2
MDIRAAEISAILKEQIQNFGQEAEVSEVGQVLSVGDGIARVYGLDNVQAGEMVEFENGTRGMALNLELDNVGIVIFGSDREIKEGQTVKRTGAIVDAPVGKGLLGRVVDALGNPIDGKGPIMFTERRRVDVKAPGIIPRKSVHEPMQTGLKAIDALIPIGRGQRELIIGDRQTGKTAVALDSILNQKPINQGDDEKAKLYCVYVAVGQKRSTVAQFVKVLEEHGALEYSIVVAATASDAAPMQFLAPFTGTAMGEYFRDNGMHALIIHDDLSKQAVAYRQMSLLLRRPPGREAYPGDVFYLHSRLLERAAKLNDEHGAGSLTALPVIETQANDVSAYIPTNVISITDGQIFLESDLFYQGIRPAVNVGLSVSRVGSSAQIKAMKQVAGKIKGELAQYRELAAFAQFGSDLDAATQKLLNRGARLTELLKQSQFSPLKVEEQVAVIYAGTNGYLDPLPVSKVREFEQGLLLSLRSQHPEILDAIRTSKELSKDTAEKLTKAIDAFAKSFS
SEQ ID NO: 38:
Нуклеотидная последовательность дельта-субъединицы АТФ-синтазы atpH
GTGGCGGAAACGATCGTGTCAGGCATGGCGGGACGCTATGCGACCGCGCTGTTCGAGCTGGCGGACGAAGCCGGTGCCATCGATTCCGTCCAGGCGGATCTTGATCGCCTGTCCGGCCTTCTGGCCGAGAGCGCGGATCTGGCGCGGCTGGTCAAGAGCCCGGTCTTCACCGCCGAGCAGCAGCTCGGCGCGATGGCGGCCATTCTCGATCAAGCAGGCATTTCCGGCCTTGCGGGCAAATTCGTGAAGCTGGTGGCGCAGAACCGCCGCCTGTTCGCACTGCCGCGCATGATTGCCGAATACGCCGTCCTGGTGGCCCGGAAGAAGGGCGAGACCTCGGCGAGCGTGACCGTTGCCACCCCCCTGAGCGATGAGCATCTGGCCACGCTCAAGGCGGCCCTGGCTGAAAAGACCGGCAAGGACGTGAAGCTCGACGTCACCGTCGATCCGTCCATCCTCGGTGGTCTCATCGTGAAGCTCGGCTCGCGCATGGTCGATGCTTCCCTGAAGACCAAACTCAATTCTATCCGGCATGCGATGAAAGAGGTCCGCTGA
SEQ ID NO: 39:
Аминокислотная последовательность дельта-субъединицы АТФ-синтазы atpH
MAETIVSGMAGRYATALFELADEAGAIDSVQADLDRLSGLLAESADLARLVKSPVFTAEQQLGAMAAILDQAGISGLAGKFVKLVAQNRRLFALPRMIAEYAVLVARKKGETSASVTVATPLSDEHLATLKAALAEKTGKDVKLDVTVDPSILGGLIVKLGSRMVDASLKTKLNSIRHAMKEVR
SEQ ID NO: 40:
Нуклеотидная последовательность субъединицы b АТФ-синтазы atpF_2
ATGACCGAAATGGAACTGGCTGAGCTCTGGGTCGCCATCGCCTTCCTGGTTTTCGTAGGCCTCCTGATCTATGCGGGCGCCCACCGCGCCATCGTCTCCGCCCTGGATTCCCGCGGCTCGCGCATCGCCTCGGAACTGGAGGAGGCCCGTCGGCTCAAGGAAGAGGCCCAGAAGCTGGTGGCCGAATTCAAGCGCAAGCAGCGCGAGGCCGAGGCCGAGGCCGAATCCATCGTCACCGGCGCCAAGGCCGAGGCCGAGCGCCTCGCCGCCGAGGCCAAGGCGAAGATCGAGGATTTCGTCACCCGCCGCACCAAGATGGCCGAGGACAAGATCGCCCAGGCCGAGCATCAGGCTCTGGCGGACGTGAAGTCCATCGCCGCCGAGGCGGCGGCCAAGGCGGCCGAGGTGATCCTCGGCGCCCAGGCCACCGGCGCGGTGGCGGAGCGTCTGCTGTCGGGCGCCATCTCCGAGGTCAAGACCAAGCTCAACTGA
SEQ ID NO: 41:
Аминокислотная последовательность субъединицы b АТФ-синтазы atpF_2
MTEMELAELWVAIAFLVFVGLLIYAGAHRAIVSALDSRGSRIASELEEARRLKEEAQKLVAEFKRKQREAEAEAESIVTGAKAEAERLAAEAKAKIEDFVTRRTKMAEDKIAQAEHQALADVKSIAAEAAAKAAEVILGAQATGAVAERLLSGAISEVKTKLN
SEQ ID NO: 42:
Нуклеотидная последовательность субъединицы b' АТФ-синтазы atpG_3
ATGATGATTGCATGGAAGCGGACCTTCGCAGTCGTGACCTTCGGGGCCGCCCTGATGGCCATGCCCGTCGCGGGCGTGGTCGCAGCTGAGACTTCTCCCGCTCCGGCGGCAGTGGCGCAGGCCGATCATGCGGTGCCCACCGAGGCGGCCGGCCAGGGCACCGCCGATGCGGCCCATGCCGCCGCGCCGGGCGAGGCCGCCCATGGTGGCGCGGCCAAGCACGAAACCCATTTCCCGCCCTTCGACGGCACCACCTTCGCCTCCCAGTTGCTGTGGCTCGCCGTCACCTTCGGCCTGCTTTACTACCTCATGAGCAAGGTCACGCTGCCGCGCATCGGCCGCATCCTGGAAGAGCGCCACGACCGCATCGCCGATGATCTGGAGGAAGCCTCCAAGCATCGCGCCGAGAGCGAGGCCGCCCAGCGGGCCTATGAGAAGGCGCTGAGCGAGGCCCGCGCGAAGGCCCATTCCATCGCCGCGGAAACCCGCGACCGCCTTGCCGCCCACGCCGACACCAACCGCAAGGCGCTGGAGAGCGAGCTCACCGCCAAGCTGCAGGCGGCCGAGGAGCGCATCGCCACCACCAAGAGCGAAGCCCTCACCCATGTGCGCGGCATCGCGGTGGACGCCACCCAATCCATCGTCTCCACCCTCATCGGTGTCGCGCCCGCGGCGGCCGACGTGGAAAAAGCGGTGGACGGCGCCCTGTCCCAGCACGGCCAGGCCTGA
SEQ ID NO: 43:
Аминокислотная последовательность субъединицы b' АТФ-синтазы atpG_3
MMIAWKRTFAVVTFGAALMAMPVAGVVAAETSPAPAAVAQADHAVPTEAAGQGTADAAHAAAPGEAAHGGAAKHETHFPPFDGTTFASQLLWLAVTFGLLYYLMSKVTLPRIGRILEERHDRIADDLEEASKHRAESEAAQRAYEKALSEARAKAHSIAAETRDRLAAHADTNRKALESELTAKLQAAEERIATTKSEALTHVRGIAVDATQSIVSTLIGVAPAAADVEKAVDGALSQHGQA
SEQ ID NO: 44:
Нуклеотидная последовательность субъединицы c АТФ-синтазы atpE_2
ATGGAAGCGGAAGCTGGAAAGTTCATCGGTGCCGGCCTCGCCTGCCTCGGCATGGGTCTCGCTGGCGTCGGCGTCGGTAACATCTTCGGTAACTTCCTCTCCGGCGCCCTGCGCAACCCGTCCGCTGCCGACGGCCAGTTCGCCCGCGCCTTCATCGGCGCCGCCCTCGCGGAAGGTCTCGGCATCTTCTCGCTGGTCGTTGCGCTCGTCCTGCTGTTCGTGGCCTGA
SEQ ID NO: 45:
Аминокислотная последовательность субъединицы c АТФ-синтазы c atpE_2
MEAEAGKFIGAGLACLGMGLAGVGVGNIFGNFLSGALRNPSAADGQFARAFIGAALAEGLGIFSLVVALVLLFVA
SEQ ID NO: 46:
Нуклеотидная последовательность субъединицы a АТФ-синтазы atpB_2
ATGACCGTCGATCCGATCCACCAGTTCGAGATCAAGCGCTACGTGGATCTGCTGAACGTCGGCGGTGTCCAGTTCTCCTTCACCAACGCAACGGTGTTCATGATTGGCATCGTCCTGGTGATTTTCTTCTTCCTGACTTTCGCGACACGCGGTCGCACCCTTGTGCCGGGCCGGATGCAGTCGGCGGCGGAGCTGAGCTACGAGTTCATCGCCAAGATGGTGCGCGACGCGGCCGGCAGCGAGGGAATGGTGTTCTTTCCCTTCGTCTTCTCGCTCTTCATGTTCGTGCTGGTGGCGAACGTATTGGGGCTCATCCCCTACACCTTCACGGTGACCGCCCACCTCATCGTCACCGCCGCCCTGGCGGCGACGGTGATCCTCACCGTCATCATCTACGGCTTCGTGCGGCACGGCACCCACTTCCTGCACCTGTTCGTGCCGTCGGGCGTGCCGGGCTTCCTCCTGCCCTTCCTCGTGGTGATCGAGGTGGTGTCGTTCCTGTCGCGGCCCATCAGCCTCTCGCTGCGTCTGTTCGCCAACATGCTGGCGGGCCACATCGCCCTCAAGGTGTTCGCCTTCTTCGTCGTGGGACTGGCCTCGGCCGGCGCGATCGGCTGGTTCGGCGCCACCCTGCCCTTCTTCATGATCGTGGCGCTCACCGCGCTGGAGCTGCTGGTGGCGGTGCTGCAGGCCTACGTGTTCGCGGTGCTGACCTCGATCTACCTCAACGACGCCATCCATCCCGGCCACTGA
SEQ ID NO: 47:
Аминокислотная последовательность субъединицы a АТФ-синтазы atpB_2
MTVDPIHQFEIKRYVDLLNVGGVQFSFTNATVFMIGIVLVIFFFLTFATRGRTLVPGRMQSAAELSYEFIAKMVRDAAGSEGMVFFPFVFSLFMFVLVANVLGLIPYTFTVTAHLIVTAALAATVILTVIIYGFVRHGTHFLHLFVPSGVPGFLLPFLVVIEVVSFLSRPISLSLRLFANMLAGHIALKVFAFFVVGLASAGAIGWFGATLPFFMIVALTALELLVAVLQAYVFAVLTSIYLNDAIHPGH
SEQ ID NO: 48:
Нуклеотидная последовательность белка I АТФ-синтазы atpI
ATGTCCGAGCCGAATGATCCATCCCGCAGGGACGGTGCGAAGGCGAAAGACGAGACGCAGGACTCCCGGCCCGGTGAGGCGGATCTTGCTCGGCGCCTCGATGCGCTCGGCACCTCCATCGGTCAGGTCAAGTCCAGAAGCGGGGAGCCCGCGGCGACGCCGCGCAAGGACACCTCCTCGGCCTCCGGCGCGGCCCTGGCGTTTCGGCTGGGCGCCGAGTTTGTTTCAGGCGTGCTGGTGGGCTCGCTCATCGGCTACGGGTTGGATTATGCGTTTGCGATTTCGCCCTGGGGGCTGATCGCCTTCACGCTGATCGGCTTTGCCGCCGGCGTCCTGAACATGCTGCGCGTGGCGAACAGCGATGCCAAGCGCCACAGCGCGGACAGGTGA
SEQ ID NO: 49:
Аминокислотная последовательность белка I АТФ-синтазы atpI
MSEPNDPSRRDGAKAKDETQDSRPGEADLARRLDALGTSIGQVKSRSGEPAATPRKDTSSASGAALAFRLGAEFVSGVLVGSLIGYGLDYAFAISPWGLIAFTLIGFAAGVLNMLRVANSDAKRHSADR
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий альфа-цепь молибден-железо белка нитрогеназы nifD_1, имеющую последовательность, указанную в SEQ ID NO:51, или последовательность, имеющую более 60% идентичности, например более 70% идентичности, например более 92% идентичности, например более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:51.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий альфа-цепь молибден-железо белка нитрогеназы nifD_2, имеющую последовательность, указанную в SEQ ID NO:53, или последовательность, имеющую более 60% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:53.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий бета-цепь молибден-железо белка нитрогеназы nifK_1, имеющую последовательность, указанную в SEQ ID NO:55, или последовательность, имеющую более 87% идентичности, например более 90% идентичности, например, более 95% идентичности, например более 96% идентичности, например, более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:55.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий бета-цепь молибден-железо белка нитрогеназы nifK_2, имеющую последовательность, указанную в SEQ ID NO: 57, или последовательность, имеющую более 95% идентичности, например более 96% идентичности, например более 97% идентичности, например более 98%, например более 99% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:57.
В другом варианте осуществления изобретения бактериальный штамм, используемый в способе настоящего изобретения, содержит ген, кодирующий железо белок нитрогеназы nifH, имеющий последовательность, указанную в SEQ ID NO: 59, или последовательность, имеющую более 98,5% идентичности по последовательности с последовательностью, указанной в SEQ ID NO: 59.
SEQ ID NO: 50:
Нуклеотидная последовательность альфа-цепи молибден-железо белка нитрогеназы nifD_1
ATGAGTTCGCTCTCCGCCACTATTCAACAGGTCTTCAACGAGCCGGGCTGCGCGAAGAACCAGAATAAGTCCGAGGCGGAGAAGAAGAAGGGCTGCACCAAGCAGCTGCAACCCGGCGGAGCGGCCGGCGGCTGCGCGTTCGACGGCGCGAAGATCGCGCTCCAGCCCTTGACCGACGTCGCCCACCTGGTGCACGGCCCCATCGCCTGCGAAGGCAATTCCTGGGACAATCGTGGCGCCAAGTCCTCCGGCTCGAACATCTGGCGCACCGGCTTCACCACGGACATCAACGAAACCGACGTGGTGTTCGGCGGCGAGAAGCGTCTGTTCAAGTCCATCAAGGAAATCATCGAGAAGTACGACCCGCCGGCCGTCTTCGTCTATCAGACCTGCGTCCCCGCCATGATCGGCGACGACATCGACGCGGTGTGCAAGGCGGCCAGGGAGAAGTTCGGAAAGCCGGTGATCCCGATCAATTCCCCCGGCTTCGTGGGGCCGAAGAATCTCGGCAACAAGCTCGCCGGCGAGGCGCTCCTCGACCATGTGATCGGCACCGAGGAGCCCGATTACACGACGGCCTACGACATCAACATCATCGGCGAATACAATCTCTCCGGCGAGTTGTGGCAGGTGAAGCCGCTGCTGGACGAGCTGGGCATCCGCATCCTCGCCTGCATCTCCGGCGACGGGAAGTACAAGGATGTGGCGTCCTCCCACCGCGCCAAGGCGGCGATGATGGTGTGCTCCAAGGCCATGATCAACGTGGCCCGCAAGATGGAGGAGCGCTACGACATCCCCTTCTTCGAAGGCTCCTTCTACGGCATCGAGGATAGCTCCGATTCCCTGCGCGAGATTGCGCGCATGCTCATCGAGAAGGGCGCCGATCCGGAGCTGATGGACCGCACCGAGGCGCTGATTGAGCGGGAAGAGAAGAAGGCGTGGGACGCCATCGCCGCCTACAAGCCCCGCTTCAAGGACAAGAAGGTGCTGCTCATCACCGGCGGCGTGAAATCCTGGTCGGTGGTGGCAGCGCTCCAGGAAGCCGGCCTCGAACTGGTGGGCACCTCGGTGAAGAAGTCCACCAAGGAGGACAAGGAGCGCATCAAGGAACTGATGGGCCAGGACGCCCACATGATCGACGACATGACGCCCCGCGAAATGTACAAGATGCTGAAGGACGCCAAGGCGGACATCATGCTCTCGGGCGGGCGCTCGCAATTCATCGCGCTCAAGGCCGCCATGCCCTGGCTCGACATCAACCAGGAGCGCCACCACGCCTATATGGGCTATGTGGGCATGGTGAAGCTGGTCGAGGAGATCGACAAGGCGCTCTACAATCCCGTGTGGGAACAGGTGCGCAAGCCCGCCCCGTGGGAAAATCCGGAAGACACCTGGCAGGCCCGTGCGCTCGCCGAAATGGAGGCGGAGGCCGCCGCGCTCGCCGCCGATCCGGTGCGCGCGGAAGAGGTGCGCCGGTCCAAGAAGATCTGCAATTGCAAGAGCGTCGACCTCGGAACCATTGAGGACGCCATCAAGGCTCACGCGCTGACCACCGTGGAGGGTGTGCGAGAGCACACCAATGCCTCGGGAGGCTGCGGAGCCTGCAGCGGGCGGATCGAGGAGATCTTCGAGGCCGTGGGCGTTGTCGCCGCCCCGCCTCCCGCGGAGGCCGCCCCGTCTCCGCAGGAGATCGCGCCCGATCCGCTCGCTGCGGAGGAAAAGCGCCGCGCCAAGAAGGCCTGCGGCTGCAAGGAGGTAGCGGTCGGCACCATTGAGGATGCCATCCGCGCCAAGGGTCTGCGAAACATCGCGGAGGTGCGTGCGGCCACCGATGCCAACACCGGCTGCGGCAATTGCCAGGAGCGGGTGGAGGGCATCCTCGACCGGGTTCTCGCCGAGGCGGCCTCAGAACTCCAGGCGGCGGAATAG
SEQ ID NO: 51:
Аминокислотная последовательность альфа-цепи молибден-железо белка нитрогеназы nifD_1
MSSLSATIQQVFNEPGCAKNQNKSEAEKKKGCTKQLQPGGAAGGCAFDGAKIALQPLTDVAHLVHGPIACEGNSWDNRGAKSSGSNIWRTGFTTDINETDVVFGGEKRLFKSIKEIIEKYDPPAVFVYQTCVPAMIGDDIDAVCKAAREKFGKPVIPINSPGFVGPKNLGNKLAGEALLDHVIGTEEPDYTTAYDINIIGEYNLSGELWQVKPLLDELGIRILACISGDGKYKDVASSHRAKAAMMVCSKAMINVARKMEERYDIPFFEGSFYGIEDSSDSLREIARMLIEKGADPELMDRTEALIEREEKKAWDAIAAYKPRFKDKKVLLITGGVKSWSVVAALQEAGLELVGTSVKKSTKEDKERIKELMGQDAHMIDDMTPREMYKMLKDAKADIMLSGGRSQFIALKAAMPWLDINQERHHAYMGYVGMVKLVEEIDKALYNPVWEQVRKPAPWENPEDTWQARALAEMEAEAAALAADPVRAEEVRRSKKICNCKSVDLGTIEDAIKAHALTTVEGVREHTNASGGCGACSGRIEEIFEAVGVVAAPPPAEAAPSPQEIAPDPLAAEEKRRAKKACGCKEVAVGTIEDAIRAKGLRNIAEVRAATDANTGCGNCQERVEGILDRVLAEAASELQAAE
SEQ ID NO: 52:
Нуклеотидная последовательность альфа-цепи молибден-железо белка нитрогеназы nifD_2
ATGAGTGTCGCACAGTCCCAGAGCGTCGCCGAGATCAAGGCGCGCAACAAGGAACTCATCGAAGAGGTCCTCAAGGTCTATCCCGAGAAGACCGCCAAGCGCCGCGCCAAGCACCTGAACGTCCACGAAGCCGGCAAGTCCGACTGCGGCGTGAAGTCCAACATCAAGTCCATCCCGGGCGTGATGACCATCCGCGGTTGCGCTTATGCCGGCTCCAAGGGTGTGGTGTGGGGTCCCATCAAGGACATGATCCACATCTCCCACGGCCCGGTGGGCTGCGGCCAGTATAGCTGGGCCGCCCGCCGCAACTACTATATCGGCACGACCGGCATCGACACCTTCGTGACGATGCAGTTCACCTCCGACTTCCAGGAGAAGGACATCGTCTTCGGCGGCGACAAGAAGCTCGCCAAGATCATGGACGAGATCCAGGAGCTGTTCCCGCTGAACAACGGCATCACCGTTCAGTCCGAGTGCCCCATCGGCCTCATCGGCGACGACATCGAGGCCGTCTCCAAGCAGAAGTCCAAGGAGTATGAGGGCAAGACCATCGTGCCGGTGCGCTGCGAGGGCTTCCGCGGCGTGTCCCAGTCCCTGGGCCACCACATCGCCAACGACGCCATCCGCGATTGGGTGTTCGACAAGATCGCGCCCGACGCCGAGCCGCGCTTTGAGCCGACCCCGTACGACGTCGCCATCATCGGCGACTACAATATCGGTGGTGACGCCTGGTCGTCCCGTATCCTCCTGGAGGAGATGGGCCTGCGCGTGATCGCCCAGTGGTCCGGCGACGGTTCGCTCGCTGAGCTGGAGGCCACCCCGAAGGCCAAGCTCAACGTGCTGCACTGCTACCGCTCCATGAACTACATCTCGCGCCACATGGAAGAGAAGTACGGTATCCCGTGGTGCGAGTACAACTTCTTCGGTCCTTCCAAGATCGCCGAGTCCCTGCGCAAGATCGCCAGCTACTTCGACGACAAGATCAAGGAAGGCGCGGAGCGCGTCATCGCCAAGTATCAGCCGCTCATGGATGCGGTGATCGCGAAGTATCGTCCCCGCCTCGAGGGCAAGACCGTGATGCTGTACGTGGGCGGCCTGCGTCCCCGTCACGTCATCGGCGCCTACGAGGACCTGGGCATGGAAGTGGTCGGCACGGGCTACGAGTTCGCCCATAACGACGACTACCAGCGCACCGCCCAGCACTACGTCAAGGATGGCACCATCATCTATGACGACGTGACCGGCTACGAGTTCGAGAAGTTCGTCGAGAAGATCCAGCCGGACCTGGTCGGTTCGGGCATCAAGGAAAAGTACGTCTTCCAGAAGATGGGCGTGCCGTTCCGCCAGATGCACTCCTGGGACTACTCGGGCCCGTACCACGGCTATGACGGCTTCGCGATCTTCGCGCGCGACATGGACATGGCCATCAACAGCCCCGTGTGGAAGATGACCCAGGCTCCGTGGAAGAGCGTCCCCAAGCCGACGATGCTCGCGGCTGAATGA
SEQ ID NO: 53:
Аминокислотная последовательность альфа-цепи молибден-железо белка нитрогеназы nifD_2
MSVAQSQSVAEIKARNKELIEEVLKVYPEKTAKRRAKHLNVHEAGKSDCGVKSNIKSIPGVMTIRGCAYAGSKGVVWGPIKDMIHISHGPVGCGQYSWAARRNYYIGTTGIDTFVTMQFTSDFQEKDIVFGGDKKLAKIMDEIQELFPLNNGITVQSECPIGLIGDDIEAVSKQKSKEYEGKTIVPVRCEGFRGVSQSLGHHIANDAIRDWVFDKIAPDAEPRFEPTPYDVAIIGDYNIGGDAWSSRILLEEMGLRVIAQWSGDGSLAELEATPKAKLNVLHCYRSMNYISRHMEEKYGIPWCEYNFFGPSKIAESLRKIASYFDDKIKEGAERVIAKYQPLMDAVIAKYRPRLEGKTVMLYVGGLRPRHVIGAYEDLGMEVVGTGYEFAHNDDYQRTAQHYVKDGTIIYDDVTGYEFEKFVEKIQPDLVGSGIKEKYVFQKMGVPFRQMHSWDYSGPYHGYDGFAIFARDMDMAINSPVWKMTQAPWKSVPKPTMLAAE
SEQ ID NO: 54:
Нуклеотидная последовательность бета-цепи молибден-железо белка нитрогеназы nifK_1
ATGGCCACCGTTTCCGTCTCCAAGAAGGCCTGCGCGGTCAACCCCCTCAAGATGAGCCAGCCGGTGGGCGGCGCGCTCGCCTTCATGGGCGTGCGCAAGGCCATGCCGCTGCTGCACGGCTCGCAGGGCTGCACCTCCTTCGGCCTGGTGCTGTTCGTGCGCCACTTCAAGGAAGCCATCCCCATGCAGACCACCGCCATGAGCGAGGTGGCGACGGTTCTGGGCGGCCTTGAGAATGTGGAGCAGGCCATTCTCAACATCTACAATCGCACCAAGCCGGAGATCATCGGCATCTGCTCCACCGGCGTCACCGAGACCAAGGGCGATGATGTCGACGGCTACATCAAGCTGATCCGGGACAAGTATCCCCAGCTGGCCGACTTCCCGCTGGTCTATGTCTCCACCCCCGATTTCAAGGACGCCTTCCAGGACGGTTGGGAGAAGACCGTGGCGAAGATGGTGGAGGCGCTGGTGAAGCCCGCCGCCGACAAGCAGAAGGACAAGACCCGCGTCAACGTCCTGCCCGGCTGCCACCTCACGCCCGGCGATCTGGATGAGATGCGGACCATCTTCGAGGATTTCGGGCTCACACCCTATTTCCTGCCGGATCTGGCCGGCTCGCTGGATGGGCATATCCCCGAGGACTTCTCGCCCACCACCATCGGCGGCATCGGCATCGATGAGATCGCCACCATGGGCGAGGCGGCCCACACCATCTGCATCGGCGCGCAGATGCGCCGGGCGGGCGAGGCCATGGAGAAGAAGACCGGCATTCCCTTCAAGCTGTTCGAGCGCCTGTGCGGCCTGGAGGCGAACGACGCCTTCATCATGCACCTGTCGCAGATCTCCGGCCGGCCGGTGCCGGTGAAGTATCGCCGGCAGCGGGGCCAGCTGGTGGATGCCATGCTGGACGGCCACTTCCATCTGGGCGGTCGCAAGGTGGCCATGGGGGCGGAGCCGGACCTGCTCTACGACGTGGGCTCCTTCCTGCACGAGATGGGCGCCCACATCCTTTCCGCGGTCACCACCACCCAGTCGCCGGTGCTGGCGCGCCTGCCTGCCGAGGAGGTGCTTATCGGCGACCTGGAGGATCTGGAGACCCAGGCGAAGGCGCGCGGATGCGATCTCCTGCTCACCCATTCCCATGGGCGCCAGGCGGCGGAGCGCCTCCACATCCCCTTCTACCGGATCGGCATTCCCATGTTTGACCGGCTGGGGGCGGGGCATCTGTTGTCGGTGGGCTATCGCGGCACCCGCGACCTCATCTTCCATCTCGCCAACCTTGTGATCGCCGACCACGAGGAAAATCACGAGCCGACGCCCGACACCTGGGCCACCGGCCATGGCGAGCATGCCGCCGCCCCCACTTCCCATTGA
SEQ ID NO: 55:
Аминокислотная последовательность бета-цепи молибден-железо белка нитрогеназы nifK_1
MATVSVSKKACAVNPLKMSQPVGGALAFMGVRKAMPLLHGSQGCTSFGLVLFVRHFKEAIPMQTTAMSEVATVLGGLENVEQAILNIYNRTKPEIIGICSTGVTETKGDDVDGYIKLIRDKYPQLADFPLVYVSTPDFKDAFQDGWEKTVAKMVEALVKPAADKQKDKTRVNVLPGCHLTPGDLDEMRTIFEDFGLTPYFLPDLAGSLDGHIPEDFSPTTIGGIGIDEIATMGEAAHTICIGAQMRRAGEAMEKKTGIPFKLFERLCGLEANDAFIMHLSQISGRPVPVKYRRQRGQLVDAMLDGHFHLGGRKVAMGAEPDLLYDVGSFLHEMGAHILSAVTTTQSPVLARLPAEEVLIGDLEDLETQAKARGCDLLLTHSHGRQAAERLHIPFYRIGIPMFDRLGAGHLLSVGYRGTRDLIFHLANLVIADHEENHEPTPDTWATGHGEHAAAPTSH
SEQ ID NO: 56:
Нуклеотидная последовательность бета-цепи молибден-железо белка нитрогеназы nifK_2
ATGCCACAAAATGCTGACAATGTGCTCGATCACTTCGAGCTCTTCCGTGGTCCCGAATACCAGCAGATGCTGGCCAATAAGAAAAAGATGTTCGAGAACCCCCGCGATCCGGCCGAAGTCGAGCGCGTGCGGGAATGGGCGAAGACTCCTGAATACAAGGAGCTGAACTTCGCCCGCGAGGCGCTCACCGTGAATCCGGCCAAGGCTTGTCAGCCGCTGGGCGCGGTGTTCGTCGCCGTCGGCTTCGAGAGCACGATCCCCTTCGTGCACGGCTCGCAGGGTTGCGTCGCGTATTACCGCTCGCACCTCTCCCGCCACTTCAAGGAGCCGTCCTCCTGCGTCTCCTCGTCCATGACCGAGGATGCGGCGGTGTTCGGCGGCCTCAACAACATGATTGACGGCCTCGCCAACACCTACAACATGTACAAGCCGAAGATGATCGCCGTCTCCACCACCTGCATGGCGGAAGTCATCGGCGACGATCTGAACGCCTTCATCAAGACCGCGAAGGAAAAGGGCTCGGTTCCGGCCGAATACGACGTGCCCTTCGCCCACACCCCGGCGTTCGTCGGCAGCCATGTCACCGGCTACGACAATGCGCTCAAGGGCATCCTCGAGCACTTCTGGGACGGCAAGGCCGGCACCGCGCCGAAGCTGGAGCGCGTTCCCAACGAGAAGATCAACTTCATCGGCGGCTTCGACGGCTACACCGTCGGCAACACTCGCGAAGTGAAGCGCATCTTCGAGGCGTTCGGCGCCGATTACACCATCCTCGCCGACAATTCCGAAGTGTTCGACACCCCGACCGACGGCGAGTTCCGCATGTATGACGGCGGCACGACCCTGGAGGACGCGGCGAACGCGGTGCACGCCAAGGCCACCATCTCCATGCAGGAATACTGCACGGAGAAGACCCTGCCCATGATCGCCGGTCATGGCCAGGACGTGGTCGCCCTCAACCACCCCGTGGGCGTGGGCGGCACCGACAAGTTCCTCATGGAGATCGCCCGCCTCACCGGCAAGGAGATCCCCGAGGAGCTGACCCGCGAGCGCGGCCGTCTCGTGGACGCTATCGCGGACTCTTCCGCGCACATCCACGGCAAGAAGTTCGCCATCTACGGCGATCCGGATCTGTGCCTGGGCCTCGCCGCGTTCCTGCTGGAGCTGGGCGCCGAGCCGACCCATGTGCTGGCCACCAACGGCACCAAGAAGTGGGCCGAGAAGGTTCAGGAACTGTTCGACTCTTCGCCGTTCGGCGCCAACTGCAAGGTCTATCCCGGCAAGGACCTGTGGCACATGCGCTCGCTCCTGTTCGTGGAGCCGGTGGATTTCATCATCGGCAACACCTACGGCAAGTATCTCGAGCGCGACACGGGCACCCCGCTGATCCGTATCGGCTTCCCGGTGTTCGACCGTCACCACCACCACCGCCGTCCGGTGTGGGGCTATCAGGGCGGCATGAACGTCCTGATCACGATCCTCGACAAGATCTTTGACGAGATCGACCGCAACACCAACGTGCCGGCCAAGACCGACTACTCGTTCGACATCATTCGTTGA
SEQ ID NO: 57:
Аминокислотная последовательность бета-цепи молибден-железо белка нитрогеназы nifK_2
MPQNADNVLDHFELFRGPEYQQMLANKKKMFENPRDPAEVERVREWAKTPEYKELNFAREALTVNPAKACQPLGAVFVAVGFESTIPFVHGSQGCVAYYRSHLSRHFKEPSSCVSSSMTEDAAVFGGLNNMIDGLANTYNMYKPKMIAVSTTCMAEVIGDDLNAFIKTAKEKGSVPAEYDVPFAHTPAFVGSHVTGYDNALKGILEHFWDGKAGTAPKLERVPNEKINFIGGFDGYTVGNTREVKRIFEAFGADYTILADNSEVFDTPTDGEFRMYDGGTTLEDAANAVHAKATISMQEYCTEKTLPMIAGHGQDVVALNHPVGVGGTDKFLMEIARLTGKEIPEELTRERGRLVDAIADSSAHIHGKKFAIYGDPDLCLGLAAFLLELGAEPTHVLATNGTKKWAEKVQELFDSSPFGANCKVYPGKDLWHMRSLLFVEPVDFIIGNTYGKYLERDTGTPLIRIGFPVFDRHHHHRRPVWGYQGGMNVLITILDKIFDEIDRNTNVPAKTDYSFDIIR
SEQ ID NO: 58:
Нуклеотидная последовательность железо белка нитрогеназы nifH
GTGGAGTCCGGTGGTCCTGAGCCGGGCGTGGGCTGCGCCGGCCGCGGCGTGATCACCTCCATCAACTTCCTGGAGGAGAACGGCGCCTACGAGGACATCGACTATGTGTCCTACGACGTGCTGGGCGACGTGGTGTGCGGCGGCTTCGCCATGCCCATCCGCGAGAACAAGGCGCAGGAAATCTACATCGTGATGTCCGGCGAGATGATGGCCATGTATGCGGCCAACAACATCTCCAAGGGCATCCTGAAGTATGCCAATTCCGGCGGCGTGCGCCTGGGCGGGCTGGTCTGCAACGAGCGCCAGACCGACAAGGAGCTGGAGCTGGCGGAGGCTCTGGCGAAGAAGCTCGGCACCGAGCTGATCTACTTCGTGCCGCGCGACAACATCGTGCAGCATGCCGAGCTGCGCCGCATGACAGTGATCGAGTATGCGCCCGATTCCGCCCAGGCCCAGCACTACCGGAACCTGGCCGAGAAGGTGCACGCCAACAAGGGCAACGGCATCATCCCGACCCCGATCACCATGGACGAGCTGGAAGACATGCTCATGGAGCACGGCATCATGAAGGCCGTGGACGAGAGCCAGATCGGCAAGACCGCCGCCGAGCTCGCCGTCTGA
SEQ ID NO: 59:
Аминокислотная последовательность железо белка нитрогеназы nifH
MESGGPEPGVGCAGRGVITSINFLEENGAYEDIDYVSYDVLGDVVCGGFAMPIRENKAQEIYIVMSGEMMAMYAANNISKGILKYANSGGVRLGGLVCNERQTDKELELAEALAKKLGTELIYFVPRDNIVQHAELRRMTVIEYAPDSAQAQHYRNLAEKVHANKGNGIIPTPITMDELEDMLMEHGIMKAVDESQIGKTAAELAV
Дальнейшая обработка
В одном варианте осуществления изобретения способ настоящего изобретения включает в себя дополнительный этап сбора биомассы, полученной во время культивирования. Биомасса может, например, быть собрана путем осаждения (осаждения под действием силы тяжести), фильтрации, центрифугирования или флокуляции. Для флокуляции может потребоваться добавление флокулянта. Центрифугирование можно проводить, например, с помощью центрифуги непрерывного действия.
В одном варианте осуществления изобретения собранную биомассу затем сушат. Сушка может быть выполнена, например, с использованием хорошо известных методов, включая центрифугирование, барабанную сушку, выпаривание, сушку вымораживанием, нагревание, распылительную сушку, вакуумную сушку и/или вакуумную фильтрацию. Высушенная биомасса впоследствии может быть использована в продукте, например, в виде пищевого или кормового продукта или кормового или пищевого ингредиента.
В другом варианте осуществления изобретения клетки собранной биомассы лизируют. В некоторых вариантах осуществления изобретения лизат может быть разделен на нерастворимую и растворимую фракции, каждая из которых или обе могут быть впоследствии сконцентрированы или высушены, а затем использованы в продукте, например, в пищевом или кормовом продукте.
В одном варианте осуществления изобретения биомассу собирают и из указанной биомассы выделяют белки, в результате чего получают белковую фракцию и фракцию, содержащую небелковые компоненты. Таким образом, в одном варианте осуществления изобретения способ предназначен для получения белка и включает в себя этап культивирования штамма VTT-E-193585 или его производного с последующим этапом сбора биомассы и дополнительным этапом выделения белков из указанной биомассы. В другом варианте осуществления изобретения способ предназначен для получения белка и включает в себя культивирование бактериального штамма рода Xanthobacter в непрерывной культуре с водородом в качестве источника энергии и источником неорганического углерода, где источник неорганического углерода включает в себя диоксид углерода, с последующим этапом сбора биомассы, и дальнейший этап выделения белков из указанной биомассы. В зависимости от способа выделения белков, полученные фракции могут быть более или менее чистыми. Таким образом, термин «белковая фракция» означает фракцию, обогащенную белками. Белковая фракция может по-прежнему содержать значительные количества других компонентов, а также значительные количества белка могут оказаться во «фракции, содержащей небелковые компоненты».
Выделение белков можно проводить с использованием любого подходящего способа. Например, в одном варианте осуществления изобретения белки выделяют путем механического разрушения клеток и отделения белка от клеточного дебриса с помощью одного или нескольких этапов фильтрации, например путем последовательной фильтрации через несколько фильтров с уменьшающимся размером пор. Механическое разрушение может быть осуществлено с использованием любого подходящего способа, например при помощи шаровой мельницы, обработки ультразвуком, гомогенизации, гомогенизации под высоким давлением, механической резки и т. д. Полученная отфильтрованная белковая фракция будет обогащена белками, но также будет содержать другие более мелкие компоненты. Белок может быть необязательно дополнительно очищен от этой фракции с использованием любого подходящего способа.
В другом варианте осуществления изобретения белковую фракцию выделяют, проводя экстракцию этанолом с последующей одной или несколькими этапами фильтрации. Такие способы известно, например, для получения соевых белков (см., например, главу 5 «Soybean Protein Concentrates» в «Technology of production of edible flours and protein products from soybeans» by Berk FAO Agricultural Services Bulletin No. 97 (1992)). Полученная белковая фракция будет обогащена белками, но также содержать другие компоненты. Белок может быть необязательно дополнительно очищен от этой фракции с использованием любого подходящего способа.
В одном варианте осуществления изобретения способ настоящего изобретения включает в себя дополнительный этап гидролиза белковой фракции, полученной способом настоящего изобретения, с получением аминокислот и малых пептидов.
В одном варианте осуществления способа настоящего изобретения способ включает в себя дополнительный этап получения пищевого продукта или кормового продукта из указанной биомассы, из указанной белковой фракции или из указанной фракции, содержащей небелковые компоненты. Указанный дополнительный этап может просто включать в себя введение указанной биомассы, белковой фракции или фракции, содержащей небелковые компоненты, в пищевой продукт или кормовой продукт путем их добавления во время производства пищевого продукта или кормового продукта. В других вариантах осуществления изобретения дальнейшую очистку или модификацию биомассы или ее фракции осуществляют в процессе ее включения в пищевой или кормовой продукт.
В ещё одном аспекте настоящее изобретение относится к продукту, такому как биомасса, белку или небелковым компонентам, полученным или получаемым способом настоящего изобретения.
В одном варианте осуществления изобретения продукт, полученный способом настоящего изобретения, содержит более 40% белка, например, от 40% до 99% белка, например от 40% до 90% белка, например, от 40% до 60% белка. В конкретном варианте осуществления изобретения продукт содержит от 25% до 75% белка, от 0% до 20% липидов и от 5% до 40% углеводов. В другом варианте осуществления изобретения продукт содержит от 40 до 60% белка, от 0 до 15% липидов и от 10 до 25% углеводов. В ещё одном варианте осуществления изобретения продукт, полученный способом настоящего изобретения, содержит от 45% до 55% белка, от 5% до 10% липидов и от 10% до 20% углеводов.
Как описано выше, в дополнительном аспекте настоящее изобретение относится к пищевому продукту или кормовому продукту, полученному или получаемому способом настоящего изобретения. Подразумевается, что используемые здесь термины «пищевой продукт» и «корм» включают в себя не только обычные пищевые продукты и кормовые продукты, такие как переработанные пищевые продукты, но и сопутствующие продукты, такие как пищевые и кормовые добавки, например, протеиновые батончики, порошки или коктейли, заменители мяса, пищевые ингредиенты, пробиотики, пребиотики, нутрицевтики и тому подобное. В некоторых вариантах осуществления изобретения указанную биомассу, указанную белковую фракцию или указанную фракцию, содержащую небелковые компоненты, используют при производстве вегетарианского или веганского пищевого продукта.
Изобретение дополнительно проиллюстрировано следующими неограничивающими примерами:
ПРИМЕРЫ
Пример 1. Выделение бактериального штамма, способного к хемоавтотрофному росту
Образец объёмом 50 мл, содержащий почву и морскую воду, был собран в стерильную пробирку Falcon с берега Балтийского моря в Наантали в Финляндии. Часть образца почвы смешивали с 10 мл минеральной среды в стерильной колбе Эрленмейера. Среда содержала 1 г/л NH4OH, 0,23 г/л KH2PO4, 0,29 г/л Na2HPO4 • 2H2O, 0,005 г/л NaVO3 • H2O, 0,2 г/л FeSO4 • 7H2O, 0,5 г/л MgSO4 • 7 H2O, 0,01 г/л CaSO4, 0,00015 г/л Na2MoO4 • 2H2O, 0,005 г/л MnSO4, 0,0005 г/л ZnSO4 • 7H2O, 0,0015 г/л H3BO3, 0,001 г/л CoSO4, 0,00005 г/л CuSO4 и 0,0001 г/л NiSO4, и была приготовлена на водопроводной воде. Суспензию почвы и среды инкубировали в инкубаторе-качалке при температуре +30°С в герметичном стальном боксе, непрерывно продуваемом газовой смесью: 150 мл/мин N2, 18 мл/мин H2, 3 мл/мин O2 и 6 мл/мин CO2. Культивирование возобновляли с семидневными интервалами путем взятия 1 мл суспензии, которую в стерильных условиях добавляли к 9 мл среды в колбе Эрленмейера, а затем помещали обратно в инкубационный бокс. После четвертого разведения в суспензии не осталось заметного загрязнения почвой. Объём клеточной суспензии увеличивали до 100 мл с целью наращивания биомассы для культивирования в биореакторе. Оптическая плотность (ОП600) суспензии составила 1,53 при её инокуляции в 190 мл минеральной среды в системе параллельного биореактора объёмом 200 мл с 15 сосудами (Medicel Explorer, Medicel Oy, Финляндия). Условиями культивирования были перемешивание со скоростью 800 об/мин, температура +30°C, и значение рН доводили до 6,8, регулируя его с помощью 1 М NaOH. Газ подавали через барботер в виде газовой смеси, состоящей из 14 мл/мин Н2, 3 мл/мин О2 и 6 мл/мин СО2. Головное пространство реактора продували воздухом со скоростью 300 мл/мин. Непрерывное культивирование подпитывали минеральной средой со скоростью 6 мл/ч, клеточную суспензию отбирали из реактора через капилляр, поддерживая постоянный объём на уровне 200 мл. Суспензию клеток, извлеченную из реактора, хранили при температуре +4°С. Образец отбирали из биореактора автоматически каждый день и измеряли оптическую плотность при длине волны 600 нм для отслеживания роста. После 498 часов культивирования в биореакторе в асептических условиях отбирали образцы, суспензию разбавляли и высевали на чашки с агаризованной минеральной средой, содержащей вышеуказанные минералы и 2% бактериологического агара. Чашки инкубировали в тех же условиях, что были описаны выше для колб Эрленмейера. Затем колонии собирали с чашек с агаром и высевали штрихами на новые чашки с агаром, чтобы выделить один микроорганизм в одной колонии. Эту операцию повторяли дважды. Отдельные колонии собирали и суспендировали в 200 мкл среды в 96-луночном титрационном микропланшете. Суспензию инкубировали при температуре +30°C и встряхивали со скоростью 625 об/мин в газонепроницаемом боксе EnzyScreen, который непрерывно продували 150 мл/мин N2, 18 мл/мин H2, 3 мл/мин O2 и 6 мл/мин СО2. Суспензию из одной лунки переносили в колбу Эрленмейера и дополняли свежей средой. Объём увеличивали до тех пор, пока не было достаточно биомассы для проведения культивирования в биореакторе. Организм депонировали в коллекции культур VTT как VTT-E-193585.
Секвенирование 16S рРНК образца показало, что образец содержал только один организм. Тот же образец был использован для секвенирования Illumina NextSeq, что позволило получить метагеномные последовательности методом дробовика размером 1x150 п.н. С помощью программы Unicycler (Wick et al., 2017 PLoS computer biology 13:e1005595) была создана сборка de novo для метагеномных последовательностей, состоящих из 101 контига. Общая длина генома составила 4846739 п.н., а содержание GC — 67,9 %. Предсказания генов и функциональные аннотации были выполнены с использованием программы Prokka (Seemann, 2014 Bioinformatics 30:2068). Аннотация генома выявила 4429 генов. Пангеномное выравнивание Roary (Page et al., 2015 Bioinformatics 31:3691) сгруппировало VTT-E-193585 среди видов Xanthobacter. Таким образом, штамм был идентифицирован как Xanthobacter sp., ближайшим геномом для которого является Xanthobacter tagetidis. Основанный на выравнивании расчёт средней идентичности нуклеотидов, учитывающий только ортологичные фрагменты (OrthoANI) (Lee et al., 2016 Int J Syst Evol Microbiol 66:1100), дал наилучшее совпадение 80,4% с Xanthobacter tagetidis (ATCC 700314; GCF_003667445.1), тогда как предлагаемая граница видов составляет 95-96% (см., например, Chun et al., 2018 Int J Syst Evol Microbiol, 68: 461-466). Xanthobacter autotrophicus Py2 дал совпадение на 79,6%, в то время как совпадение для Xanthobacter sp. 91 было 79,0%. Таким образом, можно сделать вывод, что выделенный бактериальный штамм, депонированный как VTT-E-193585, принадлежит к филуму: Proteobacteria; к классу: Альфа-протеобактерии; и к порядку: Rhizobiales. Наиболее вероятным семейством является Xanthobacteraceae и род Xanthobacter. Бактериальный штамм VTT-E-193585 не может быть однозначно отнесен к какому-либо известному виду.
Проводили поиск предполагаемых генов устойчивости к противомикробным препаратам. Инструмент ABRicate (https://github.com/tseemann/abricate) использовали для геномного поиска в базах данных Arg-Annot, NCBI, ResFinder, ecOH, Megares и VFDB с использованием blastn или blastp. Порог 50% был установлен как для идентичности, так и для охвата как на уровне нуклеотидов, так и на уровне белка. Были идентифицированы только два предполагаемых гена устойчивости к противомикробным препаратам. Эти два гена не содержат аминокислотных изменений, связанных с устойчивостью к антибиотикам, и, таким образом, резистентный фенотип не ожидается.
Пример 2. Пилотное культивирование и анализ выделенного бактериального штамма
Выделенный бактериальный штамм, депонированный как VTT-E-193585, культивировали в обычном 200-литровом биореакторе с мешалкой (MPF-U, Marubishi Ltd, Япония). Перемешивание осуществляли крыльчатками Раштона, вращающимися со скоростью 400 об/мин. Температуру при культивировании поддерживали на уровне +30°С. Значение рН поддерживали на уровне 6,8±0,2 путем добавления 8 М NaOH или 3,6 М H3PO4 с программным управлением. Среда культивирования содержала 1 г/л NH4OH, 0,23 г/л KH2PO4, 0,29 г/л Na2HPO4 • 2H2O, 0,005 г/л NaVO3 • H2O, 0,2 г/л FeSO4 • 7H2O, 0,5 г/л MgSO4 • 7H2O, 0,01 г/л CaSO4, 0,00015 г/л Na2MoO4 • 2H2O, 0,005 г/л MnSO4, 0,0005 г/л ZnSO4 • 7H2O, 0,0015 г/л H3BO3, 0,001 г/л CoSO4, 0,00005 г/л CuSO4 и 0,0001 г/л NiSO4, и была приготовлена на водопроводной воде. В качестве основного источника энергии и углерода постоянно подавали смесь, содержащую 1,8-10,5 л/мин газообразного водорода, 0,6-2,5 л/мин газообразного кислорода и 1,8-5 л/мин газообразного диоксида углерода. Уровень растворенного кислорода поддерживали на уровне 7,2±0,5% путем регулирования состава газовой смеси. Инокулят для культивирования готовили, как описано в Примере 1. Рост контролировали, отбирая образцы вручную и анализируя плотность клеток как оптическую плотность путем измерения поглощения при длине волны 600 нм (УФ/видимый свет спектрофотометр Ultrospec 2100 pro, Biochrom Ltd., Англия) и путем измерения сухой массы клеток (CDW) путем сушки в печи в течение ночи при температуре 105°C. Оптическую плотность также контролировали с помощью датчика поглощения in situ (Trucell 2, Finesse Ltd, США). Кривая роста культивирования представлена на Фигуре 1. Максимальная скорость роста в периодической фазе составила 0,06 ч-1. Максимальная плотность клеток составила 4,5 г_CDW/л через 92 часа. После 92 ч культивирования начинали подачу свежей культуральной среды, как описано выше, со скоростью разбавления 0,01 ч-1. Во время непрерывной подпитки плотность клеток составляла в среднем 2,9 г_CDW/л. Культуральную жидкость постоянно собирали в охлаждаемый (+10°С) резервуар, откуда порциями по 300 л подавали в центробежный сепаратор непрерывного действия (BTPX-205, Alfa-Laval AB, Швеция). Собранную из сепаратора суспензию, содержащую клетки, подавали в атмосферную двухбарабанную сушилку (Buflovak 6x8 ADDD, Hebeler Process Solutions Llc., США), нагреваемую паром под давлением 4 бар и барабанами, вращающимися со скоростью 3,5 об/мин. Это привело к получению высушенного клеточного порошка с содержанием сухого вещества приблизительно 96%. Результаты анализа высушенного клеточного порошка представлены в Таблице 1 для приблизительного состава, в Таблице 2 для аминокислотного состава, в Таблице 3 для жирнокислотного состава и в Таблице 4 для содержания витаминов. Анализы показали, что высушенный клеточный порошок имеет высокое содержание белка со всеми незаменимыми аминокислотами. Он также содержит больше ненасыщенных жирных кислот, чем насыщенных, и много витаминов группы В. Содержание пептидогликана составляет всего 0,002 мг/г_CDW, а содержание липополисахарида составляет 0,01 мг/г_CDW. Желательно, чтобы эти концентрации были как можно меньше. Для сравнения, в коммерческом препарате молочнокислых бактерий, проанализированном в то же время, содержание пептидогликана составило 0,244 мг/г_DW, а содержание липополисахарида составило 0,015 мг/г_DW. Анализы цитотоксичности и генотоксичности проводили с использованием надосадочных образцов культивирования. Никакой цитотоксичности в отношении клеточных линий человека HepG2 или HeLa229 не наблюдалось. Генотоксичности в отношении штаммов Escherichia coli WP2 trp- или CM871 uvrA recA lexA не наблюдалось.
Таблица 1. Результаты анализа порошка высушенных клеток выделенного бактериального штамма, депонированного как VTT-E-193585
Таблица 2. Аминокислотный состав высушенного клеточного порошка выделенного бактериального штамма, депонированного как VTT-E-193585
Таблица 3. Жирнокислотный состав высушенного клеточного порошка выделенного бактериального штамма, депонированного как VTT-E-193585
Таблица 4. Содержание витаминов в высушенном клеточном порошке выделенного бактериального штамма, депонированного как VTT-E-193585
Пример 3. Культивирование выделенного бактериального штамма на различных источниках азота
Выделенный бактериальный штамм, депонированный как VTT-E-193585, культивировали в системе параллельных биореакторов с 15 сосудами объёмом 200 мл (Medicel Explorer, Medicel Oy, Финляндия). Перемешивание осуществляли крыльчатками Раштона, вращающимися со скоростью 800 об/мин. Температуру при культивировании поддерживали на уровне +30°С. Значение pH поддерживали на уровне 6,8 добавлением 1 М NaOH. Среда культивирования содержала 0,23 г/л KH2PO4, 0,29 г/л Na2HPO4 • 2 H2O, 0,005 г/л NaVO3 • H2O, 0,2 г/л FeSO4 • 7H2O, 0,5 г/л MgSO4 • 7H2O, 0,01 г/л CaSO4, 0,00015 г/л Na2MoO4 • 2H2O, 0,005 г/л MnSO4, 0,0005 г/л ZnSO4 • 7H2O, 0,0015 г/л H3BO3, 0,001 г/л CoSO4, 0,00005 г/л CuSO4 и 0,0001 г/л NiSO4, и была приготовлена на водопроводной воде. Кроме того, источник азота при культивировании варьировался так, что четыре культивирования содержали 18,7 мМ NH4OH, четыре культивирования содержали 9,34 мМ мочевины (OC(NH2)2), четыре культивирования содержали 18,7 мМ нитрата калия (KNO3) и три культивирования оставались без источника азота в среде. В качестве основного источника энергии и углерода постоянно подавали смесь, содержащую 22 мл/мин газообразного водорода, 3,2 мл/мин воздуха и 6,4 мл/мин газообразного диоксида углерода. Таким образом, вместе с воздухом все варианты культивирования получали и газообразный азот. Рост контролировали, автоматически отбирая образцы и анализируя плотность клеток как оптическую плотность путем измерения поглощения при длине волны 600 нм (УФ/видимый свет спектрофотометр Ultrospec 2100 pro, Biochrom Ltd., Англия). Кривые роста культивирования представлены на фиг. 2. Рост на аммиаке и мочевине был сопоставим. Рост на нитратах или газообразном азоте был явно медленнее, чем на аммиаке или мочевине. К концу культивирования рост на нитратах был лучше, чем на газообразном азоте как единственном источнике азота. Тем не менее, также наблюдался рост в культурах, в которых газообразный азот был единственным источником азота, демонстрируя, что выделенный бактериальный штамм, депонированный как VTT-E-193585, способен фиксировать азот.
Пример 4. Характеристика чувствительности к антибиотикам
Чувствительность к антибиотикам гентамицин, канамицин, стрептомицин, тетрациклин, ампициллин, ципрофлоксацин, колистин и фосфомицин для выделенного бактериального штамма, депонированного как VTT-E-193585, анализировали в соответствии со стандартом CLSI M07-A111 (Институт клинических и лабораторных стандартов. Методы разбавления в тестах на чувствительность к противомикробным препаратам для бактерий, которые растут в аэробных условиях, 11-е изд., стандарт CLSI M07, 2018 г.) с приготовленной авторами настоящего изобретения чашкой для микроразведений для ампициллина, ципрофлоксацина и колистина, с чашкой VetMIC Lact-1 (Национальный ветеринарный институт SVA, Уппсала, Швеция) для гентамицина, канамицина, стрептомицина и тетрациклина с использованием метода микроразведения в бульоне и фосфомицина с использованием метода разведения в агаре в аэробных условиях при температуре +35 ± 2°C в течение 48 ± 1 часов с использованием бульона Мюллера-Хинтона с отрегулированным содержанием катионов (LabM, LAB114, катионы Mg2+ и Ca2+ добавляются отдельно). В качестве штамма контроля качества использовали штамм Escherichia coli ATCC 25922, который инкубировали в аэробных условиях при температуре +35 ± 2°C в течение 18 ± 2 часов. Результаты определения чувствительности штаммов к антибиотикам представлены в Таблице 5. Установлено, что выделенный бактериальный штамм в целом чувствителен к антибиотикам. Для гентамицина, канамицина, стрептомицина и тетрациклина значения минимальной ингибирующей концентрации (МИК) для VTT-E-193585 были ниже или сопоставимы с E. coli ATCC 25922, в то время как для ампициллина, ципрофлоксацина, колистина и фосфомицина значения МИК были выше для VTT-E-193585.
Таблица 5. Минимальная ингибирующая концентрация (МИК, мкг/мл) – значения антибиотиков для штамма VTT-E-193585 и Escherichia coli ATCC 25922
48 ч ± 1 ч
18 ч ± 2 ч
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> СОЛАР ФУДС ОЙ
<120> ШТАММЫ И СПОСОБЫ ДЛЯ ПОЛУЧЕНИЯ БЕЛКА ОДНОКЛЕТОЧНЫХ ОРГАНИЗМОВ ИЛИ БИОМАССЫ
<130> Solar002
<160> 59
<170> Патентная версия 3.5
<210> 1
<211> 1482
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 1
cttgagagtt tgatcctggc tcagagcgaa cgctggcggc aggcctaaca catgcaagtc 60
gagcgcccag caatgggagc ggcagacggg tgagtaacgc gtggggatgt gcccaatggt 120
acggaataac ccagggaaac ttggactaat accgtatgag cccttcgggg gaaagattta 180
tcgccattgg atcaacccgc gtctgattag ctagttggtg gggtaacggc ccaccaaggc 240
gacgatcagt agctggtctg agaggatgat cagccacact gggactgaga cacggcccag 300
actcctacgg gaggcagcag tggggaatat tggacaatgg gcgcaagcct gatccagcca 360
tgccgcgtgt gtgatgaagg ccttagggtt gtaaagcact ttcgccggtg aagataatga 420
cggtaaccgg agaagaagcc ccggctaact tcgtgccagc agccgcggta atacgaaggg 480
ggctagcgtt gctcggaatc actgggcgta aagcgcacgt aggcggatcg ttaagtcagg 540
ggtgaaatcc tggagctcaa ctccagaact gcccttgata ctggcgacct tgagttcgag 600
agaggttggt ggaactgcga gtgtagaggt gaaattcgta gatattcgca agaacaccag 660
tggcgaaggc ggccaactgg ctcgatactg acgctgaggt gcgaaagcgt ggggagcaaa 720
caggattaga taccctggta gtccacgccg taaacgatgg atgctagccg ttgggcagct 780
tgctgttcag tggcgcagct aacgcattaa gcatcccgcc tggggagtac ggtcgcaaga 840
ttaaaactca aaggaattga cgggggcccg cacaagcggt ggagcatgtg gtttaattcg 900
aagcaacgcg cagaacctta ccagcctttg acatggcagg acgatttcca gagatggatc 960
tcttccagca atggacctgc acacaggtgc tgcatggctg tcgtcagctc gtgtcgtgag 1020
atgttgggtt aagtcccgca acgagcgcaa ccctcgcctc tagttgccag cattcagttg 1080
ggcactctag agggactgcc ggtgataagc cgagaggaag gtggggatga cgtcaagtcc 1140
tcatggccct tacgggctgg gctacacacg tgctacaatg gtggtgacag tgggatgcga 1200
aagggcgacc tctagcaaat ctccaaaagc catctcagtt cggattgtac tctgcaactc 1260
gagtgcatga agttggaatc gctagtaatc gtggatcagc atgccacggt gaatacgttc 1320
ccgggccttg tacacaccgc ccgtcacacc atgggagttg gctttacccg aaggcgctgc 1380
gctaacccgc aagggaggca ggcgaccacg gtagggtcag cgactggggt gaagtcgtaa 1440
caaggtagcc gtaggggaac ctgcggctgg atcacctcct tt 1482
<210> 2
<211> 1467
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 2
atgggtgccg aagcaaccgt cgggcagatc acggacgcca agaagagata cgccgccggc 60
gtgctgaagt acgcccagat gggctactgg aacggcgact acgttcccaa ggacaccgac 120
ctcctggcgg tgttccgcat caccccccag gcgggcgtgg acccggtgga agccgccgcg 180
gcggtcgccg gcgaaagctc caccgctacc tggaccgtgg tgtggaccga ccggctcacc 240
gccgccgacg tctaccgcgc caaggcctac aaggtggagc cggtgccggg ccaggaaggc 300
cagtatttct gctacatcgc ctatgatctc gatttgttcg aggaaggctc catcgccaac 360
ctcacggcgt cgatcatcgg caacgtcttc tccttcaagc cgctgaaggc ggcgcggctg 420
gaggacatgc ggcttcccgt cgcctatgtg aagaccttcc gcggcccgcc caccggcatc 480
gtggtcgagc gcgagcgcct ggacaagttc ggccgccccc ttctgggcgc caccaccaag 540
ccgaagcttg gcctctcggg caagaattac ggccgcgtgg tctatgaggc cctcaagggc 600
ggcctcgact tcgtgaagga cgacgagaac atcaactcgc agcccttcat gcactggcgc 660
gatcgcttcc tctattgcat ggaggccgtc aacaaggccc aggccgagac cggcgaggtg 720
aaggggcact atctcaacat caccgccggg accatggagg agatgtaccg ccgcgccgag 780
ttcgccaagg aactgggctc cgtggtggtg atggtggatc tcatcatcgg ctggaccgcc 840
atccagtcca tgtccaactg gtgccgcgag aacgacatga tcctgcacat gcaccgtgcg 900
ggccatggca cctacacgcg ccagaagagc cacggcgtct ccttccgcgt catcgccaag 960
tggctgcggc tcgccggcgt cgaccacctg cacaccggca ccgccgtggg caagctggaa 1020
ggcgacccca tgaccgtgca gggcttctac aatgtctgcc gcgagacgac gacgcagcag 1080
gacctcaccc gcggcctgtt cttcgagcag gactggggcg gcatccgcaa ggtgatgccg 1140
gtggcctccg gcggcatcca tgcgggccag atgcaccagc tcatcgacct gttcggcgag 1200
gacgtggtgc tccagttcgg cggcggcacc atcggccacc cggacggcat ccaggccggc 1260
gccaccgcca accgcgtggc gctggaaacc atgatcctcg cccgcaacga gggccgcgac 1320
atcaggaacg agggcccgga aatcctggtg gaagccgcca aatggtgccg tccgctgcgc 1380
gcggcgctcg atacctgggg cgaggtgacc ttcaactacg cctccaccga cacgtccgat 1440
tacgtgccca ccgcgtccgt cgcctga 1467
<210> 3
<211> 488
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 3
Met Gly Ala Glu Ala Thr Val Gly Gln Ile Thr Asp Ala Lys Lys Arg
1 5 10 15
Tyr Ala Ala Gly Val Leu Lys Tyr Ala Gln Met Gly Tyr Trp Asn Gly
20 25 30
Asp Tyr Val Pro Lys Asp Thr Asp Leu Leu Ala Val Phe Arg Ile Thr
35 40 45
Pro Gln Ala Gly Val Asp Pro Val Glu Ala Ala Ala Ala Val Ala Gly
50 55 60
Glu Ser Ser Thr Ala Thr Trp Thr Val Val Trp Thr Asp Arg Leu Thr
65 70 75 80
Ala Ala Asp Val Tyr Arg Ala Lys Ala Tyr Lys Val Glu Pro Val Pro
85 90 95
Gly Gln Glu Gly Gln Tyr Phe Cys Tyr Ile Ala Tyr Asp Leu Asp Leu
100 105 110
Phe Glu Glu Gly Ser Ile Ala Asn Leu Thr Ala Ser Ile Ile Gly Asn
115 120 125
Val Phe Ser Phe Lys Pro Leu Lys Ala Ala Arg Leu Glu Asp Met Arg
130 135 140
Leu Pro Val Ala Tyr Val Lys Thr Phe Arg Gly Pro Pro Thr Gly Ile
145 150 155 160
Val Val Glu Arg Glu Arg Leu Asp Lys Phe Gly Arg Pro Leu Leu Gly
165 170 175
Ala Thr Thr Lys Pro Lys Leu Gly Leu Ser Gly Lys Asn Tyr Gly Arg
180 185 190
Val Val Tyr Glu Ala Leu Lys Gly Gly Leu Asp Phe Val Lys Asp Asp
195 200 205
Glu Asn Ile Asn Ser Gln Pro Phe Met His Trp Arg Asp Arg Phe Leu
210 215 220
Tyr Cys Met Glu Ala Val Asn Lys Ala Gln Ala Glu Thr Gly Glu Val
225 230 235 240
Lys Gly His Tyr Leu Asn Ile Thr Ala Gly Thr Met Glu Glu Met Tyr
245 250 255
Arg Arg Ala Glu Phe Ala Lys Glu Leu Gly Ser Val Val Val Met Val
260 265 270
Asp Leu Ile Ile Gly Trp Thr Ala Ile Gln Ser Met Ser Asn Trp Cys
275 280 285
Arg Glu Asn Asp Met Ile Leu His Met His Arg Ala Gly His Gly Thr
290 295 300
Tyr Thr Arg Gln Lys Ser His Gly Val Ser Phe Arg Val Ile Ala Lys
305 310 315 320
Trp Leu Arg Leu Ala Gly Val Asp His Leu His Thr Gly Thr Ala Val
325 330 335
Gly Lys Leu Glu Gly Asp Pro Met Thr Val Gln Gly Phe Tyr Asn Val
340 345 350
Cys Arg Glu Thr Thr Thr Gln Gln Asp Leu Thr Arg Gly Leu Phe Phe
355 360 365
Glu Gln Asp Trp Gly Gly Ile Arg Lys Val Met Pro Val Ala Ser Gly
370 375 380
Gly Ile His Ala Gly Gln Met His Gln Leu Ile Asp Leu Phe Gly Glu
385 390 395 400
Asp Val Val Leu Gln Phe Gly Gly Gly Thr Ile Gly His Pro Asp Gly
405 410 415
Ile Gln Ala Gly Ala Thr Ala Asn Arg Val Ala Leu Glu Thr Met Ile
420 425 430
Leu Ala Arg Asn Glu Gly Arg Asp Ile Arg Asn Glu Gly Pro Glu Ile
435 440 445
Leu Val Glu Ala Ala Lys Trp Cys Arg Pro Leu Arg Ala Ala Leu Asp
450 455 460
Thr Trp Gly Glu Val Thr Phe Asn Tyr Ala Ser Thr Asp Thr Ser Asp
465 470 475 480
Tyr Val Pro Thr Ala Ser Val Ala
485
<210> 4
<211> 405
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 4
atgcgcatca cccaaggctc cttctccttc ctgccggacc tcaccgacac gcagatcaag 60
gcccaggtgc aatattgcct ggaccagggc tgggcggtct cggtggagca caccgacgat 120
ccccacccgc gcaacaccta ttgggagatg tggggcccgc ccatgttcga tctgcgcgac 180
gcggccggcg tcttcggcga gatcgaagcc tgccgggccg ccaatcccga gcattatgtg 240
cgggtgaacg ccttcgattc cagccgcgga tgggagacga tccgcctgtc cttcatcgtt 300
cagcggccca ccgtggaaga gggcttccgc ctcgaccgca ccgaaggcaa gggccgcaac 360
cagagctacg ccatgcgcta ccgggcgcag ttcgcgccgc gctga 405
<210> 5
<211> 134
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 5
Met Arg Ile Thr Gln Gly Ser Phe Ser Phe Leu Pro Asp Leu Thr Asp
1 5 10 15
Thr Gln Ile Lys Ala Gln Val Gln Tyr Cys Leu Asp Gln Gly Trp Ala
20 25 30
Val Ser Val Glu His Thr Asp Asp Pro His Pro Arg Asn Thr Tyr Trp
35 40 45
Glu Met Trp Gly Pro Pro Met Phe Asp Leu Arg Asp Ala Ala Gly Val
50 55 60
Phe Gly Glu Ile Glu Ala Cys Arg Ala Ala Asn Pro Glu His Tyr Val
65 70 75 80
Arg Val Asn Ala Phe Asp Ser Ser Arg Gly Trp Glu Thr Ile Arg Leu
85 90 95
Ser Phe Ile Val Gln Arg Pro Thr Val Glu Glu Gly Phe Arg Leu Asp
100 105 110
Arg Thr Glu Gly Lys Gly Arg Asn Gln Ser Tyr Ala Met Arg Tyr Arg
115 120 125
Ala Gln Phe Ala Pro Arg
130
<210> 6
<211> 1896
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 6
atgatgccat ctgagccgca cggcgcgggc atgccgcccc cacgggaagc ggccgcggtt 60
cccacccccc aggaggtgag cgcggtggtg gccgaggtgg tcgcggatgc cgtggcatcg 120
gtgggcggcg cacgcacccg gctcatggac atcgtccagc tggcccagca gcgtctcggc 180
catctctccg aagagaccat ggcggccatt gccgcgcggc tcgccattcc gccggtggaa 240
gtggcggaca tggtgtcctt ctacgccttc ctgaaccgcg cgcccaaggg ccgctaccac 300
atccgcctgt cgcgcagccc catctcgctg atgaagggcg ccgaggcggt ggctgccgcc 360
ttctgccaga tcctcggcat cgccatgggc gagacctcgc aggatggcga cttcaccctg 420
gaatggacca acgacatcgg catggccgac caggagccgg ccgccctcgt caacggcacg 480
gtgatgacgc agctcgcgcc cggcgatgcg gccatcatcg tcggccggct gcgggcccat 540
cacgcgccca atgccctgcc gctgttccct ggagccggcg tggccggctc cggcctgccc 600
catgcccgga tccgccccag cctggtgatg ccgggacagc ttctgttccg cgaggaccac 660
acgacgccgg gcgccggcat caaggcggca ctcgccctca ccccggacga agtggtgcag 720
aaggtctccg ccgcgcgcct gcgcgggcgg ggtggcgccg gctttcccac cggtctcaaa 780
tggaagctct gccgccagtc gcccgccacc acccgccatg tgatctgcaa tgcggacgag 840
ggcgagcccg gcaccttcaa ggatcgcgtg ctgctcacgc aggcgccgca cctcatgttc 900
gacggcatga ccatcgccgg ctacgccttg ggggcgcggg agggcgtggt ctatctgcgc 960
ggcgagtacg cctatctgtg ggagcctctg catgcggtcc tgcgcgagcg ctatgggctc 1020
gggctcgccg gcgcgaacat cctgggacac gcgggcttcg acttcgacat ccgcatccag 1080
ctgggcgccg gcgcctatat ctgcggcgag gaatccgcgc tggtggaatc gctggaaggc 1140
aagcgcggct cgccccgcga ccgccccccc ttccccaccg tgcgcggcca tctccagcag 1200
cccaccgccg tggacaatgt ggagaccttc gcctgcgccg cccgcatcct ggaggatggc 1260
gtggaggcgt tcgcgggcat cggcacgccc gaatccgccg gcacgaagct cctctcggtg 1320
tcgggcgatt gcccgcgccc cggcgtgtat gaggtgccct tcggcctcac ggtgaacgcg 1380
ctgctcgacc ttgtcggcgc gccggacgcc gccttcgtgc agatgggtgg gccgtccggc 1440
caatgcgtgg cgccgaagga ttacggccgc cgcatcgcct tcgaggacct gcccaccggc 1500
ggctcggtga tggtgttcgg cccggggcgc gacgtgctcg ccatggtgcg cgagttcgcg 1560
gatttcttcg ccggcgaatc ctgcggctgg tgcacgccct gccgggtggg caccaccttg 1620
ctcaaggaag agctggacaa gctcctcgcc aaccgcgcca ccctcgccga catccgcgcg 1680
ctggagaccc tggccacgac cgtctcccgc accagccgct gcggcctcgg ccagacggcg 1740
cccaacccca tcctttccac catgcgcaac ctgccggaag cctatgaggc gaggctgagg 1800
cccgaagact tcctgccctg ggcctcgctc gacgaggcgc tgaagcccgc catcgtcatc 1860
cagggccgcg cgcccgtgcc ggaggaagag gcatga 1896
<210> 7
<211> 631
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 7
Met Met Pro Ser Glu Pro His Gly Ala Gly Met Pro Pro Pro Arg Glu
1 5 10 15
Ala Ala Ala Val Pro Thr Pro Gln Glu Val Ser Ala Val Val Ala Glu
20 25 30
Val Val Ala Asp Ala Val Ala Ser Val Gly Gly Ala Arg Thr Arg Leu
35 40 45
Met Asp Ile Val Gln Leu Ala Gln Gln Arg Leu Gly His Leu Ser Glu
50 55 60
Glu Thr Met Ala Ala Ile Ala Ala Arg Leu Ala Ile Pro Pro Val Glu
65 70 75 80
Val Ala Asp Met Val Ser Phe Tyr Ala Phe Leu Asn Arg Ala Pro Lys
85 90 95
Gly Arg Tyr His Ile Arg Leu Ser Arg Ser Pro Ile Ser Leu Met Lys
100 105 110
Gly Ala Glu Ala Val Ala Ala Ala Phe Cys Gln Ile Leu Gly Ile Ala
115 120 125
Met Gly Glu Thr Ser Gln Asp Gly Asp Phe Thr Leu Glu Trp Thr Asn
130 135 140
Asp Ile Gly Met Ala Asp Gln Glu Pro Ala Ala Leu Val Asn Gly Thr
145 150 155 160
Val Met Thr Gln Leu Ala Pro Gly Asp Ala Ala Ile Ile Val Gly Arg
165 170 175
Leu Arg Ala His His Ala Pro Asn Ala Leu Pro Leu Phe Pro Gly Ala
180 185 190
Gly Val Ala Gly Ser Gly Leu Pro His Ala Arg Ile Arg Pro Ser Leu
195 200 205
Val Met Pro Gly Gln Leu Leu Phe Arg Glu Asp His Thr Thr Pro Gly
210 215 220
Ala Gly Ile Lys Ala Ala Leu Ala Leu Thr Pro Asp Glu Val Val Gln
225 230 235 240
Lys Val Ser Ala Ala Arg Leu Arg Gly Arg Gly Gly Ala Gly Phe Pro
245 250 255
Thr Gly Leu Lys Trp Lys Leu Cys Arg Gln Ser Pro Ala Thr Thr Arg
260 265 270
His Val Ile Cys Asn Ala Asp Glu Gly Glu Pro Gly Thr Phe Lys Asp
275 280 285
Arg Val Leu Leu Thr Gln Ala Pro His Leu Met Phe Asp Gly Met Thr
290 295 300
Ile Ala Gly Tyr Ala Leu Gly Ala Arg Glu Gly Val Val Tyr Leu Arg
305 310 315 320
Gly Glu Tyr Ala Tyr Leu Trp Glu Pro Leu His Ala Val Leu Arg Glu
325 330 335
Arg Tyr Gly Leu Gly Leu Ala Gly Ala Asn Ile Leu Gly His Ala Gly
340 345 350
Phe Asp Phe Asp Ile Arg Ile Gln Leu Gly Ala Gly Ala Tyr Ile Cys
355 360 365
Gly Glu Glu Ser Ala Leu Val Glu Ser Leu Glu Gly Lys Arg Gly Ser
370 375 380
Pro Arg Asp Arg Pro Pro Phe Pro Thr Val Arg Gly His Leu Gln Gln
385 390 395 400
Pro Thr Ala Val Asp Asn Val Glu Thr Phe Ala Cys Ala Ala Arg Ile
405 410 415
Leu Glu Asp Gly Val Glu Ala Phe Ala Gly Ile Gly Thr Pro Glu Ser
420 425 430
Ala Gly Thr Lys Leu Leu Ser Val Ser Gly Asp Cys Pro Arg Pro Gly
435 440 445
Val Tyr Glu Val Pro Phe Gly Leu Thr Val Asn Ala Leu Leu Asp Leu
450 455 460
Val Gly Ala Pro Asp Ala Ala Phe Val Gln Met Gly Gly Pro Ser Gly
465 470 475 480
Gln Cys Val Ala Pro Lys Asp Tyr Gly Arg Arg Ile Ala Phe Glu Asp
485 490 495
Leu Pro Thr Gly Gly Ser Val Met Val Phe Gly Pro Gly Arg Asp Val
500 505 510
Leu Ala Met Val Arg Glu Phe Ala Asp Phe Phe Ala Gly Glu Ser Cys
515 520 525
Gly Trp Cys Thr Pro Cys Arg Val Gly Thr Thr Leu Leu Lys Glu Glu
530 535 540
Leu Asp Lys Leu Leu Ala Asn Arg Ala Thr Leu Ala Asp Ile Arg Ala
545 550 555 560
Leu Glu Thr Leu Ala Thr Thr Val Ser Arg Thr Ser Arg Cys Gly Leu
565 570 575
Gly Gln Thr Ala Pro Asn Pro Ile Leu Ser Thr Met Arg Asn Leu Pro
580 585 590
Glu Ala Tyr Glu Ala Arg Leu Arg Pro Glu Asp Phe Leu Pro Trp Ala
595 600 605
Ser Leu Asp Glu Ala Leu Lys Pro Ala Ile Val Ile Gln Gly Arg Ala
610 615 620
Pro Val Pro Glu Glu Glu Ala
625 630
<210> 8
<211> 1491
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 8
atgagccggg gatcccccga tgccgggaaa gaccgcacca tgagcgccac cgacggcacc 60
accgcccccc gcaagatcgt catcgatccg gtgacccgcg tggagggcca cggcaaggtc 120
accatccgcc tggatgaagc cggcgcggtg gaggatgcgc gtttccacat cgtggagttc 180
cgcggcttcg agcggttcat ccagggccgg atgtactggg aagtgcccct tatcatccag 240
cggctgtgcg gcatctgccc ggtgagccac catctggcgg cggcgaaagc catggaccag 300
gtggcgggcg tggaccgcgt accgcccacc gccgagaaac tgcgccggct gatgcattat 360
gggcaggtgc tgcaatccaa cgctttgcac atcttccacc tcgcctcgcc cgacctcctg 420
ttcggcttcg acgcgccggc cgagcagcgc aacatcatcg ccgtgctcca gcgttatccg 480
gagatcggca aatgggcgat cttcatcagg aagttcggcc aggaggtcat caaggccacc 540
ggcgggcgca agatccatcc caccagcgcc attcccggcg gggtcaacca gaacctcgcc 600
gtggaggacc gcgacgccct gcgcgccaag gtgggcgaga tcatcagctg gtgcatggcg 660
gcgctggacc atcacaaggc ctatgtggcg gaaaaccggg cgctgcatga cagcttcgcc 720
gccttcccct ccgccttcat gagcctcgtg gggccggatg gcggcatgga cctttatgac 780
ggcaccctgc gggtgatcga tgccgagggc gcccccctca tcgaaggcgc gccgcccgcc 840
tcctaccgcg accacctcat cgaggaggtg cggccctgga gctatctgaa attcccccat 900
ctgcgcgcct tcggccgcga cgatggctgg tatcgggtcg gccccctcgc ccaggtcaat 960
tgcgccgcgt ccatcgacac gccccgcgcc gaggcggccc ggcgggactt catggccgag 1020
ggcggcggca agccggtgca tgccaccctc gcttatcact gggcgcggct catcgtgctg 1080
gtccattgcg cggagaagat cgaacagctg ctgttcgacg acgacctgca aggctgcgat 1140
ctgcgtgcgg agggcacccg gcgcggggaa ggcgtcgcct ggatcgaggc gccgcgcggc 1200
accctcatcc accattacga ggtggacgag aacgaccagg tgcgccgcgc caacctcatc 1260
gtctccacca cccacaataa cgaggccatg aaccgcgccg tgcggcaggt ggcgaagacg 1320
gacctttccg gtcgcgagat caccgaaggg ctgctgaacc atatcgaggt ggccatccgc 1380
gccttcgacc cctgcctgtc ctgcgccacc catgcgctgg gccagatgcc gctgatcgtg 1440
acgcttgaag atgcctccgg cgcagagatc gcccgcggag tgaaggaatg a 1491
<210> 9
<211> 496
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 9
Met Ser Arg Gly Ser Pro Asp Ala Gly Lys Asp Arg Thr Met Ser Ala
1 5 10 15
Thr Asp Gly Thr Thr Ala Pro Arg Lys Ile Val Ile Asp Pro Val Thr
20 25 30
Arg Val Glu Gly His Gly Lys Val Thr Ile Arg Leu Asp Glu Ala Gly
35 40 45
Ala Val Glu Asp Ala Arg Phe His Ile Val Glu Phe Arg Gly Phe Glu
50 55 60
Arg Phe Ile Gln Gly Arg Met Tyr Trp Glu Val Pro Leu Ile Ile Gln
65 70 75 80
Arg Leu Cys Gly Ile Cys Pro Val Ser His His Leu Ala Ala Ala Lys
85 90 95
Ala Met Asp Gln Val Ala Gly Val Asp Arg Val Pro Pro Thr Ala Glu
100 105 110
Lys Leu Arg Arg Leu Met His Tyr Gly Gln Val Leu Gln Ser Asn Ala
115 120 125
Leu His Ile Phe His Leu Ala Ser Pro Asp Leu Leu Phe Gly Phe Asp
130 135 140
Ala Pro Ala Glu Gln Arg Asn Ile Ile Ala Val Leu Gln Arg Tyr Pro
145 150 155 160
Glu Ile Gly Lys Trp Ala Ile Phe Ile Arg Lys Phe Gly Gln Glu Val
165 170 175
Ile Lys Ala Thr Gly Gly Arg Lys Ile His Pro Thr Ser Ala Ile Pro
180 185 190
Gly Gly Val Asn Gln Asn Leu Ala Val Glu Asp Arg Asp Ala Leu Arg
195 200 205
Ala Lys Val Gly Glu Ile Ile Ser Trp Cys Met Ala Ala Leu Asp His
210 215 220
His Lys Ala Tyr Val Ala Glu Asn Arg Ala Leu His Asp Ser Phe Ala
225 230 235 240
Ala Phe Pro Ser Ala Phe Met Ser Leu Val Gly Pro Asp Gly Gly Met
245 250 255
Asp Leu Tyr Asp Gly Thr Leu Arg Val Ile Asp Ala Glu Gly Ala Pro
260 265 270
Leu Ile Glu Gly Ala Pro Pro Ala Ser Tyr Arg Asp His Leu Ile Glu
275 280 285
Glu Val Arg Pro Trp Ser Tyr Leu Lys Phe Pro His Leu Arg Ala Phe
290 295 300
Gly Arg Asp Asp Gly Trp Tyr Arg Val Gly Pro Leu Ala Gln Val Asn
305 310 315 320
Cys Ala Ala Ser Ile Asp Thr Pro Arg Ala Glu Ala Ala Arg Arg Asp
325 330 335
Phe Met Ala Glu Gly Gly Gly Lys Pro Val His Ala Thr Leu Ala Tyr
340 345 350
His Trp Ala Arg Leu Ile Val Leu Val His Cys Ala Glu Lys Ile Glu
355 360 365
Gln Leu Leu Phe Asp Asp Asp Leu Gln Gly Cys Asp Leu Arg Ala Glu
370 375 380
Gly Thr Arg Arg Gly Glu Gly Val Ala Trp Ile Glu Ala Pro Arg Gly
385 390 395 400
Thr Leu Ile His His Tyr Glu Val Asp Glu Asn Asp Gln Val Arg Arg
405 410 415
Ala Asn Leu Ile Val Ser Thr Thr His Asn Asn Glu Ala Met Asn Arg
420 425 430
Ala Val Arg Gln Val Ala Lys Thr Asp Leu Ser Gly Arg Glu Ile Thr
435 440 445
Glu Gly Leu Leu Asn His Ile Glu Val Ala Ile Arg Ala Phe Asp Pro
450 455 460
Cys Leu Ser Cys Ala Thr His Ala Leu Gly Gln Met Pro Leu Ile Val
465 470 475 480
Thr Leu Glu Asp Ala Ser Gly Ala Glu Ile Ala Arg Gly Val Lys Glu
485 490 495
<210> 10
<211> 720
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 10
atgagcgaga cccccttcac ctttaccgtg gacggcatcg cggtcccggc cacccccggc 60
cagagcgtca tcgaggcgtg cgatgcggcg ggcatctata tcccgcgcct gtgccaccac 120
ccggacctgc cgccggcggg ccattgccgg gtgtgcacct gcatcatcga cgggcggccg 180
gccagcgcct gcaccatgcc cgccgccagg ggcatggtgg tggagaacga gacgcccgct 240
ttgctggcgg agcggcgcac gctgatcgag atgctgttcg cggaaggcaa ccatttctgc 300
cagttctgcg aggcgagcgg cgattgcgaa ttgcaggcgc tgggctacct gttcggcatg 360
gtggccccgc ccttccccca tctgtggccg aagcggccgg tggatgccag ccatccggat 420
atctatatcg accacaatcg ctgcatcctg tgctcgcgct gcgtgcgcgc ctcgcgcacc 480
ctggacggca agtccgtgtt cggcttcgag gggcgcggca tcgagatgca tctggcggtg 540
accggcgggc acctggacga cagcgccatc gccgccgccg acagggcggt tgagatgtgc 600
ccggtgggct gcatcgtcct caagcgcacc ggctaccgca cgccctatgg ccggcggcgc 660
tacgacgccg cgcccatcgg ctccgacatc accgcccggc gcggcggcgc gaaggactga 720
<210> 11
<211> 239
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 11
Met Ser Glu Thr Pro Phe Thr Phe Thr Val Asp Gly Ile Ala Val Pro
1 5 10 15
Ala Thr Pro Gly Gln Ser Val Ile Glu Ala Cys Asp Ala Ala Gly Ile
20 25 30
Tyr Ile Pro Arg Leu Cys His His Pro Asp Leu Pro Pro Ala Gly His
35 40 45
Cys Arg Val Cys Thr Cys Ile Ile Asp Gly Arg Pro Ala Ser Ala Cys
50 55 60
Thr Met Pro Ala Ala Arg Gly Met Val Val Glu Asn Glu Thr Pro Ala
65 70 75 80
Leu Leu Ala Glu Arg Arg Thr Leu Ile Glu Met Leu Phe Ala Glu Gly
85 90 95
Asn His Phe Cys Gln Phe Cys Glu Ala Ser Gly Asp Cys Glu Leu Gln
100 105 110
Ala Leu Gly Tyr Leu Phe Gly Met Val Ala Pro Pro Phe Pro His Leu
115 120 125
Trp Pro Lys Arg Pro Val Asp Ala Ser His Pro Asp Ile Tyr Ile Asp
130 135 140
His Asn Arg Cys Ile Leu Cys Ser Arg Cys Val Arg Ala Ser Arg Thr
145 150 155 160
Leu Asp Gly Lys Ser Val Phe Gly Phe Glu Gly Arg Gly Ile Glu Met
165 170 175
His Leu Ala Val Thr Gly Gly His Leu Asp Asp Ser Ala Ile Ala Ala
180 185 190
Ala Asp Arg Ala Val Glu Met Cys Pro Val Gly Cys Ile Val Leu Lys
195 200 205
Arg Thr Gly Tyr Arg Thr Pro Tyr Gly Arg Arg Arg Tyr Asp Ala Ala
210 215 220
Pro Ile Gly Ser Asp Ile Thr Ala Arg Arg Gly Gly Ala Lys Asp
225 230 235
<210> 12
<211> 543
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 12
atggccaagc ccaaactcgc cacctgcgcg ctggccggct gcttcggctg ccacatgtcc 60
ttcctggaca tggacgagcg catcgtcgag ctcatcgacc tggtggacct cgacgtctcg 120
cccctcgacg acaagaaaaa cttcaccggc atggtggaaa tcggcctggt ggaaggcggc 180
tgcgccgacg agcgccatgt gaaggtgctg cgcgagttcc gcgagaaatc ccgcatcctg 240
gtggcggtgg gcgcctgcgc catcaccggc ggcatcccgg cattgcgcaa cctcgccggc 300
ctcgacgaat gcctgaggga agcctacctc accggcccca cggtggaagg cggcgggctc 360
attcccaacg acccggagct gccgctgctg ctggacaagg tctatccggt gcaggacttc 420
gtgaagatcg accatttcct gcccggctgc ccgccctcgg ccgacgccat ctgggcggct 480
ctgaaggcgc tgctgaccgg caccgagccg catctgccct acccgctttt caagtacgaa 540
tga 543
<210> 13
<211> 180
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 13
Met Ala Lys Pro Lys Leu Ala Thr Cys Ala Leu Ala Gly Cys Phe Gly
1 5 10 15
Cys His Met Ser Phe Leu Asp Met Asp Glu Arg Ile Val Glu Leu Ile
20 25 30
Asp Leu Val Asp Leu Asp Val Ser Pro Leu Asp Asp Lys Lys Asn Phe
35 40 45
Thr Gly Met Val Glu Ile Gly Leu Val Glu Gly Gly Cys Ala Asp Glu
50 55 60
Arg His Val Lys Val Leu Arg Glu Phe Arg Glu Lys Ser Arg Ile Leu
65 70 75 80
Val Ala Val Gly Ala Cys Ala Ile Thr Gly Gly Ile Pro Ala Leu Arg
85 90 95
Asn Leu Ala Gly Leu Asp Glu Cys Leu Arg Glu Ala Tyr Leu Thr Gly
100 105 110
Pro Thr Val Glu Gly Gly Gly Leu Ile Pro Asn Asp Pro Glu Leu Pro
115 120 125
Leu Leu Leu Asp Lys Val Tyr Pro Val Gln Asp Phe Val Lys Ile Asp
130 135 140
His Phe Leu Pro Gly Cys Pro Pro Ser Ala Asp Ala Ile Trp Ala Ala
145 150 155 160
Leu Lys Ala Leu Leu Thr Gly Thr Glu Pro His Leu Pro Tyr Pro Leu
165 170 175
Phe Lys Tyr Glu
180
<210> 14
<211> 1013
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 14
tccagacccg ggcaacattg ctccatgtgc tgggcaccct ggccggccgc tggccccata 60
ccctcgcgct ccagcccggc ggggtgaccc gaagcgccga ccagcacgac cgcatgcgcc 120
tgctcgcgac gctgaaggcg gtgcgggcgg cgctggaaga gaccttgttc ggcgcgcctt 180
tggaagaggt ggcggccctg gacggcgccg ccgccgtgga ggcctggcgc gccaacggcc 240
cggaagggga tttccgcctg ttcctggaga tcgccgccga cctggagctg gaccggctcg 300
gccgcgcgca cgaccgcttt ctctccttcg gcgcctacgc ccaggacgag gggcgccttt 360
atggcgccgg caccttcgag gccgggacgg cgggagggct cgatcccaac gccatcaccg 420
aggaccacgc cttcgcccgc atggaggacc gcgcggcgcc ccatgcgccc tttgacggct 480
ccaccttccc cgatgccgac gacaccgagg gctacacctg gtgcaaggcg ccgcgccttg 540
ccggcctgcc cttcgagacc ggcgccttcg cccggcaggt ggtggcgggc catccgctcg 600
cccgggacct cgtgacgcgg gaaggcggca ctgtgcgcag ccgcgtggtc ggccggctgc 660
tggaaaccgc gcgcaccctg atcgccatgg agggctgggt gaaggaactg cggcccgaag 720
ggccctggtg cgcccagggc cacctgcccc aggaaggccg cgccttcggc ctcaccgagg 780
cggcgcgcgg ggcgctcggc cactggatgg tggtggagaa gggccgcatt gcccgctacc 840
agatcatcgc ccccaccacc tggaacttct ccccccgcga cggcgcgggc ctgcccggcc 900
cgctggagac ggccctggtg ggcgcgcccg tgcggcaggg agagacgacg cccgtgagcg 960
tgcagcacat cgtgcgctcc ttcgacccgt gcatggtctg cactgtgcat tga 1013
<210> 15
<211> 485
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 15
Met Ser Ala Glu Thr Arg Arg Leu Val Val Gly Pro Phe Asn Arg Val
1 5 10 15
Glu Gly Asp Leu Glu Val Arg Leu Asp Val Gln Asp Gly Arg Val Gln
20 25 30
Gln Ala Phe Val Ser Ser Pro Leu Phe Arg Gly Phe Glu Arg Ile Leu
35 40 45
Glu Gly Arg Asp Pro Arg Asp Ala Leu Val Ile Ala Pro Arg Ile Cys
50 55 60
Gly Ile Cys Ser Val Ser Gln Ser His Ala Ala Ala Leu Ala Leu Ala
65 70 75 80
Gly Leu Gln Gly Ile Ala Pro Thr His Asp Gly Arg Ile Ala Thr Asn
85 90 95
Leu Ile Val Ala Ala Glu Asn Val Ala Asp His Leu Thr His Phe His
100 105 110
Val Phe Phe Met Pro Asp Phe Ala Arg Ala Val Tyr Glu Asp Arg Pro
115 120 125
Trp Phe Ala Gln Ala Ala Arg Arg Phe Lys Ala Asn Gln Gly Val Ser
130 135 140
Val Arg Arg Ala Leu Gln Thr Arg Ala Thr Leu Leu His Val Leu Gly
145 150 155 160
Thr Leu Ala Gly Arg Trp Pro His Thr Leu Ala Leu Gln Pro Gly Gly
165 170 175
Val Thr Arg Ser Ala Asp Gln His Asp Arg Met Arg Leu Leu Ala Thr
180 185 190
Leu Lys Ala Val Arg Ala Ala Leu Glu Glu Thr Leu Phe Gly Ala Pro
195 200 205
Leu Glu Glu Val Ala Ala Leu Asp Gly Ala Ala Ala Val Glu Ala Trp
210 215 220
Arg Ala Asn Gly Pro Glu Gly Asp Phe Arg Leu Phe Leu Glu Ile Ala
225 230 235 240
Ala Asp Leu Glu Leu Asp Arg Leu Gly Arg Ala His Asp Arg Phe Leu
245 250 255
Ser Phe Gly Ala Tyr Ala Gln Asp Glu Gly Arg Leu Tyr Gly Ala Gly
260 265 270
Thr Phe Glu Ala Gly Thr Ala Gly Gly Leu Asp Pro Asn Ala Ile Thr
275 280 285
Glu Asp His Ala Phe Ala Arg Met Glu Asp Arg Ala Ala Pro His Ala
290 295 300
Pro Phe Asp Gly Ser Thr Phe Pro Asp Ala Asp Asp Thr Glu Gly Tyr
305 310 315 320
Thr Trp Cys Lys Ala Pro Arg Leu Ala Gly Leu Pro Phe Glu Thr Gly
325 330 335
Ala Phe Ala Arg Gln Val Val Ala Gly His Pro Leu Ala Arg Asp Leu
340 345 350
Val Thr Arg Glu Gly Gly Thr Val Arg Ser Arg Val Val Gly Arg Leu
355 360 365
Leu Glu Thr Ala Arg Thr Leu Ile Ala Met Glu Gly Trp Val Lys Glu
370 375 380
Leu Arg Pro Glu Gly Pro Trp Cys Ala Gln Gly His Leu Pro Gln Glu
385 390 395 400
Gly Arg Ala Phe Gly Leu Thr Glu Ala Ala Arg Gly Ala Leu Gly His
405 410 415
Trp Met Val Val Glu Lys Gly Arg Ile Ala Arg Tyr Gln Ile Ile Ala
420 425 430
Pro Thr Thr Trp Asn Phe Ser Pro Arg Asp Gly Ala Gly Leu Pro Gly
435 440 445
Pro Leu Glu Thr Ala Leu Val Gly Ala Pro Val Arg Gln Gly Glu Thr
450 455 460
Thr Pro Val Ser Val Gln His Ile Val Arg Ser Phe Asp Pro Cys Met
465 470 475 480
Val Cys Thr Val His
485
<210> 16
<211> 858
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 16
acgggggagg aagcccgcgc catcttcgac gccatccttg ccggcgttat cgtcctcgac 60
gccctgtgcg tggaaggcgc gctgctgcgc gggccgaacg gcaccgggcg cttccatgtg 120
ctggcgggca cggacacccc caccatcgac tgggcgcggc agctcgccgg catggcgcgc 180
cacgtggtgg cggtgggcac ctgcgccgcc tatgggggcg tgacggcggc gggcatcaac 240
cccaccgatg cctgcggcct ccagttcgac ggacgccgga agggtggggc gctgggggcg 300
gacttccgct cccgctcggg gcttccggtc atcaatgtgg ccggctgccc cacccatccc 360
aactgggtga cggaaaccct gatgctgctc gcctgcggcc tgctgggcga ggccgacctc 420
gacgtctatg gccgcccgcg cttctatgcg gacctgctgg tgcatcacgg ctgcccgcgc 480
aacgaatact atgaatacaa ggcgagcgcc gagaagatga gcgacctcgg ctgcatgatg 540
gagcatctgg gctgcctcgg cacccaggcc cacgccgact gcaacacgcg cctttggaat 600
ggcgagggct cgtgcacccg cggcggctat gcctgcatca actgcacggc gccggaattc 660
gaggagccgg gccacgcctt cctggagacg cccaagatcg gcggcatccc catcggcctg 720
cccaccgaca tgcccaaggc ctggttcatc gccttgtcct ccctcgccaa ggcggcgacg 780
ccggagcggc tgcgcaagaa cgcggtgtcc gaccatgtgg tcacgccgcc cgccgtcaag 840
gacatcaagc ggcgatga 858
<210> 17
<211> 335
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 17
Met Ser Thr Pro Phe Ser Val Leu Trp Leu Gln Ser Gly Gly Cys Gly
1 5 10 15
Gly Cys Thr Met Ser Leu Leu Cys Ala Glu Ala Pro Asp Leu Ala Thr
20 25 30
Thr Leu Asp Ala Ala Gly Ile Gly Phe Leu Trp His Pro Ala Leu Ser
35 40 45
Glu Glu Thr Gly Glu Glu Ala Arg Ala Ile Phe Asp Ala Ile Leu Ala
50 55 60
Gly Val Ile Val Leu Asp Ala Leu Cys Val Glu Gly Ala Leu Leu Arg
65 70 75 80
Gly Pro Asn Gly Thr Gly Arg Phe His Val Leu Ala Gly Thr Asp Thr
85 90 95
Pro Thr Ile Asp Trp Ala Arg Gln Leu Ala Gly Met Ala Arg His Val
100 105 110
Val Ala Val Gly Thr Cys Ala Ala Tyr Gly Gly Val Thr Ala Ala Gly
115 120 125
Ile Asn Pro Thr Asp Ala Cys Gly Leu Gln Phe Asp Gly Arg Arg Lys
130 135 140
Gly Gly Ala Leu Gly Ala Asp Phe Arg Ser Arg Ser Gly Leu Pro Val
145 150 155 160
Ile Asn Val Ala Gly Cys Pro Thr His Pro Asn Trp Val Thr Glu Thr
165 170 175
Leu Met Leu Leu Ala Cys Gly Leu Leu Gly Glu Ala Asp Leu Asp Val
180 185 190
Tyr Gly Arg Pro Arg Phe Tyr Ala Asp Leu Leu Val His His Gly Cys
195 200 205
Pro Arg Asn Glu Tyr Tyr Glu Tyr Lys Ala Ser Ala Glu Lys Met Ser
210 215 220
Asp Leu Gly Cys Met Met Glu His Leu Gly Cys Leu Gly Thr Gln Ala
225 230 235 240
His Ala Asp Cys Asn Thr Arg Leu Trp Asn Gly Glu Gly Ser Cys Thr
245 250 255
Arg Gly Gly Tyr Ala Cys Ile Asn Cys Thr Ala Pro Glu Phe Glu Glu
260 265 270
Pro Gly His Ala Phe Leu Glu Thr Pro Lys Ile Gly Gly Ile Pro Ile
275 280 285
Gly Leu Pro Thr Asp Met Pro Lys Ala Trp Phe Ile Ala Leu Ser Ser
290 295 300
Leu Ala Lys Ala Ala Thr Pro Glu Arg Leu Arg Lys Asn Ala Val Ser
305 310 315 320
Asp His Val Val Thr Pro Pro Ala Val Lys Asp Ile Lys Arg Arg
325 330 335
<210> 18
<211> 870
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 18
gtgaccgagc gcctgtccga cgtcaacgcc cgcatcgcct cggtgcggca gctctcatcg 60
gtcatcacgg ccatgcgggg cattgcggcg gcgcgggcgc gggaggcgcg gggtcggctc 120
gacggcatcc gcgcctatgc gcagaccatc gccgaggcca tcggccatgt gctcgccgtg 180
ctgcccgagg aggcccgcgc ccggtcctcc gggcaccggc atcggggcca tgcggtcatc 240
gccctgtgcg cggagcaggg ctttgccggc gtcttcaacg agcgggtgct ggacgaggcc 300
gcccggctgc tgaccggcgg ggcggggccg gccgagctgc tgctggtggg cgaccggggc 360
ctgatggtgg cccgcgagcg ggggctcgat gtctcctggt cggtgcccat ggtggcccat 420
gcgggccagg cctcggcgct ggcggaccgc atcagcgagg agctctaccg gcggatcgat 480
gcgggacggg tgacgcgggt gtcggtggtg cacgccgagc ccgccgcgtc cgccgccatc 540
gagacggtgg tgaaagtgct ggtgccgttc gacttcgccc gcttccccct ggcgcgggtg 600
gcatccgccc cgctcatgac catgccgccg ccgcggctgc tggcccagct gtcggaggaa 660
tatgtgttcg ccgagctgtg cgaggcgctc accttgtcct tcgcggcgga gaacgaggcc 720
cgcatgcggg ccatgatcgc cgcccgcgcc aatgtggccg ataccctgga gggcctcgtc 780
ggccgcgccc ggcagatgcg ccaggaggag atcaccaacg agatcatcga gctggaaggc 840
ggcgccggca gcgcccggca tgcggattga 870
<210> 19
<211> 289
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 19
Met Thr Glu Arg Leu Ser Asp Val Asn Ala Arg Ile Ala Ser Val Arg
1 5 10 15
Gln Leu Ser Ser Val Ile Thr Ala Met Arg Gly Ile Ala Ala Ala Arg
20 25 30
Ala Arg Glu Ala Arg Gly Arg Leu Asp Gly Ile Arg Ala Tyr Ala Gln
35 40 45
Thr Ile Ala Glu Ala Ile Gly His Val Leu Ala Val Leu Pro Glu Glu
50 55 60
Ala Arg Ala Arg Ser Ser Gly His Arg His Arg Gly His Ala Val Ile
65 70 75 80
Ala Leu Cys Ala Glu Gln Gly Phe Ala Gly Val Phe Asn Glu Arg Val
85 90 95
Leu Asp Glu Ala Ala Arg Leu Leu Thr Gly Gly Ala Gly Pro Ala Glu
100 105 110
Leu Leu Leu Val Gly Asp Arg Gly Leu Met Val Ala Arg Glu Arg Gly
115 120 125
Leu Asp Val Ser Trp Ser Val Pro Met Val Ala His Ala Gly Gln Ala
130 135 140
Ser Ala Leu Ala Asp Arg Ile Ser Glu Glu Leu Tyr Arg Arg Ile Asp
145 150 155 160
Ala Gly Arg Val Thr Arg Val Ser Val Val His Ala Glu Pro Ala Ala
165 170 175
Ser Ala Ala Ile Glu Thr Val Val Lys Val Leu Val Pro Phe Asp Phe
180 185 190
Ala Arg Phe Pro Leu Ala Arg Val Ala Ser Ala Pro Leu Met Thr Met
195 200 205
Pro Pro Pro Arg Leu Leu Ala Gln Leu Ser Glu Glu Tyr Val Phe Ala
210 215 220
Glu Leu Cys Glu Ala Leu Thr Leu Ser Phe Ala Ala Glu Asn Glu Ala
225 230 235 240
Arg Met Arg Ala Met Ile Ala Ala Arg Ala Asn Val Ala Asp Thr Leu
245 250 255
Glu Gly Leu Val Gly Arg Ala Arg Gln Met Arg Gln Glu Glu Ile Thr
260 265 270
Asn Glu Ile Ile Glu Leu Glu Gly Gly Ala Gly Ser Ala Arg His Ala
275 280 285
Asp
<210> 20
<211> 1536
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 20
atgagcacgg gcgcgcaagc gagcgaggat tggctcaccc ggagccgggc ggccctggcc 60
gggacgcgcc tttcccagca atcccaatcg gtgggccggg tggaggagat ggccgacggc 120
atcgcccgcg tctccggcct gccggatgtg cggctcgacg agcttctcac cttcgagggc 180
ggccagaccg gctatgccct caccctcgat cgcaccgaga tcgccgtggt gctgctggat 240
gacgcctccg gcgtggaggc gggcgcccgg gtgttcggca ccggcgaggt ggtgaaggtg 300
ccggtggggc cggggctgct gggccgcatc gtcgaccccc tcggccggcc catggaccgc 360
tccgagccgg tggtggcgca ggcgcaccat cccatcgagc ggccggcgcc ggccatcatc 420
gcccgcgacc tggtctcgca gccggttcag accggcacgc tggtggtgga tgcgctgttc 480
tccctcggcc ggggccagcg cgagctcatc atcggcgacc gggctaccgg caagaccgcc 540
atcgcggtgg acaccatcat cagccagaag cattcggaca tcgtgtgcat ctacgtggcg 600
gtgggccagc gcgccgccgc cgtggagcgg gtggtggagg cggtgcgcgc ccacggggcg 660
atcgagcgct gcatcttcgt ggtcgcctcg gccgccgcct cgccagggct gcaatggatc 720
gcgccgttcg ccggcatgac catggcggaa tatttccgcg acaacggcca gcatgcgctc 780
atcatcatcg atgatctcac caagcatgcg gccacccatc gcgagctggc gctgctcacc 840
cacgagccgc cgggccgcga ggcctatccc ggcgacatct tctatgtgca cgcccgcctt 900
ctggagcggg ccgccaagct ctccgccgag ctgggcggtg gctcgctcac ggccctgccc 960
atcgcggaga cggacgcggg aaacctctcc gcctatatcc ccaccaacct catctccatc 1020
accgatgggc agatcgtgct ggattcgcgg ctgttcgcgg ccaaccagcg cccggcggtg 1080
gatgtgggcc tctccgtgag ccgggtgggc ggcaaggcgc agcatcccgc gcttcgggcc 1140
gtgtccgggc gcatccggct cgattattcc cagttcctgg agctggaaat gttcacccgc 1200
ttcggcggca tcaccgatac ccgcgtgaag gcgcagatca cccggggcga gcgcatccgc 1260
gcgctgctca cccagccgcg cttttccacc ctgcgccttc aggacgaggt ggcgctgctg 1320
gccgcgctgg cggagggggt gttcgacact ttggccccgg ggctgatggg cgccgtgcgt 1380
gcccgcattc cggcccagct ggatgcgcag gtgaaggacg tggcctcggc cctcgccgag 1440
ggcaaggtgc tggaggaggg cttgcacgcc cgtctcgtgg cggccgtgcg ggccgtcgcg 1500
gcggacgtgg ccgcgaccgc gaaggccggg ccgtga 1536
<210> 21
<211> 511
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 21
Met Ser Thr Gly Ala Gln Ala Ser Glu Asp Trp Leu Thr Arg Ser Arg
1 5 10 15
Ala Ala Leu Ala Gly Thr Arg Leu Ser Gln Gln Ser Gln Ser Val Gly
20 25 30
Arg Val Glu Glu Met Ala Asp Gly Ile Ala Arg Val Ser Gly Leu Pro
35 40 45
Asp Val Arg Leu Asp Glu Leu Leu Thr Phe Glu Gly Gly Gln Thr Gly
50 55 60
Tyr Ala Leu Thr Leu Asp Arg Thr Glu Ile Ala Val Val Leu Leu Asp
65 70 75 80
Asp Ala Ser Gly Val Glu Ala Gly Ala Arg Val Phe Gly Thr Gly Glu
85 90 95
Val Val Lys Val Pro Val Gly Pro Gly Leu Leu Gly Arg Ile Val Asp
100 105 110
Pro Leu Gly Arg Pro Met Asp Arg Ser Glu Pro Val Val Ala Gln Ala
115 120 125
His His Pro Ile Glu Arg Pro Ala Pro Ala Ile Ile Ala Arg Asp Leu
130 135 140
Val Ser Gln Pro Val Gln Thr Gly Thr Leu Val Val Asp Ala Leu Phe
145 150 155 160
Ser Leu Gly Arg Gly Gln Arg Glu Leu Ile Ile Gly Asp Arg Ala Thr
165 170 175
Gly Lys Thr Ala Ile Ala Val Asp Thr Ile Ile Ser Gln Lys His Ser
180 185 190
Asp Ile Val Cys Ile Tyr Val Ala Val Gly Gln Arg Ala Ala Ala Val
195 200 205
Glu Arg Val Val Glu Ala Val Arg Ala His Gly Ala Ile Glu Arg Cys
210 215 220
Ile Phe Val Val Ala Ser Ala Ala Ala Ser Pro Gly Leu Gln Trp Ile
225 230 235 240
Ala Pro Phe Ala Gly Met Thr Met Ala Glu Tyr Phe Arg Asp Asn Gly
245 250 255
Gln His Ala Leu Ile Ile Ile Asp Asp Leu Thr Lys His Ala Ala Thr
260 265 270
His Arg Glu Leu Ala Leu Leu Thr His Glu Pro Pro Gly Arg Glu Ala
275 280 285
Tyr Pro Gly Asp Ile Phe Tyr Val His Ala Arg Leu Leu Glu Arg Ala
290 295 300
Ala Lys Leu Ser Ala Glu Leu Gly Gly Gly Ser Leu Thr Ala Leu Pro
305 310 315 320
Ile Ala Glu Thr Asp Ala Gly Asn Leu Ser Ala Tyr Ile Pro Thr Asn
325 330 335
Leu Ile Ser Ile Thr Asp Gly Gln Ile Val Leu Asp Ser Arg Leu Phe
340 345 350
Ala Ala Asn Gln Arg Pro Ala Val Asp Val Gly Leu Ser Val Ser Arg
355 360 365
Val Gly Gly Lys Ala Gln His Pro Ala Leu Arg Ala Val Ser Gly Arg
370 375 380
Ile Arg Leu Asp Tyr Ser Gln Phe Leu Glu Leu Glu Met Phe Thr Arg
385 390 395 400
Phe Gly Gly Ile Thr Asp Thr Arg Val Lys Ala Gln Ile Thr Arg Gly
405 410 415
Glu Arg Ile Arg Ala Leu Leu Thr Gln Pro Arg Phe Ser Thr Leu Arg
420 425 430
Leu Gln Asp Glu Val Ala Leu Leu Ala Ala Leu Ala Glu Gly Val Phe
435 440 445
Asp Thr Leu Ala Pro Gly Leu Met Gly Ala Val Arg Ala Arg Ile Pro
450 455 460
Ala Gln Leu Asp Ala Gln Val Lys Asp Val Ala Ser Ala Leu Ala Glu
465 470 475 480
Gly Lys Val Leu Glu Glu Gly Leu His Ala Arg Leu Val Ala Ala Val
485 490 495
Arg Ala Val Ala Ala Asp Val Ala Ala Thr Ala Lys Ala Gly Pro
500 505 510
<210> 22
<211> 741
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 22
atgcagatcg actggtggac gctgggcctg cagacggtca acgtcctcgt tctcatctgg 60
ctcctgagcc gcttcctgtt caagccggtg gcgcaggtca tcgcgcagcg ccgtgccgag 120
atcgagaagc tggtggagga tgcgcgcgcc gccaaggccg ccgccgaggc cgagcgggac 180
acggcgaagg cggaggaggc gcgccttgcc gccgagcgcg gcgcccgcat ggcggcggtc 240
gccaaggagg cggaggcgca gaaggcggca ttgctggccg ccgccaagac cgaggccgag 300
gccctgcacg cggccgcgga agcggccatc gtccgggcgc gggcgagcga ggaggaagcc 360
gccgccgacc gcgccagccg ccttgccgtg gacatcgccg ccaagctgct ggaccggctg 420
cccgacgacg cccgggtcgc gggcttcatc gatggcctcg ccgaggggct tgaagccctg 480
cccgaggcga gccgggcggt gatcggcgtc gacggcgcgc cagtgcgcgt gacggccgcg 540
cgcgccctta tgccggcgga ggaggaggcc tgccgcacgc ggctctccca ggcgctgggc 600
cgtccggtga cgctggccgt gaccatcgac cccgccctca tcgccggcct ggagatggag 660
acgccccacg cggtggtgcg caattccttc aaggccgatc tcgaccgcgt caccgcggcg 720
ctcacccatc atgggacctg a 741
<210> 23
<211> 246
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 23
Met Gln Ile Asp Trp Trp Thr Leu Gly Leu Gln Thr Val Asn Val Leu
1 5 10 15
Val Leu Ile Trp Leu Leu Ser Arg Phe Leu Phe Lys Pro Val Ala Gln
20 25 30
Val Ile Ala Gln Arg Arg Ala Glu Ile Glu Lys Leu Val Glu Asp Ala
35 40 45
Arg Ala Ala Lys Ala Ala Ala Glu Ala Glu Arg Asp Thr Ala Lys Ala
50 55 60
Glu Glu Ala Arg Leu Ala Ala Glu Arg Gly Ala Arg Met Ala Ala Val
65 70 75 80
Ala Lys Glu Ala Glu Ala Gln Lys Ala Ala Leu Leu Ala Ala Ala Lys
85 90 95
Thr Glu Ala Glu Ala Leu His Ala Ala Ala Glu Ala Ala Ile Val Arg
100 105 110
Ala Arg Ala Ser Glu Glu Glu Ala Ala Ala Asp Arg Ala Ser Arg Leu
115 120 125
Ala Val Asp Ile Ala Ala Lys Leu Leu Asp Arg Leu Pro Asp Asp Ala
130 135 140
Arg Val Ala Gly Phe Ile Asp Gly Leu Ala Glu Gly Leu Glu Ala Leu
145 150 155 160
Pro Glu Ala Ser Arg Ala Val Ile Gly Val Asp Gly Ala Pro Val Arg
165 170 175
Val Thr Ala Ala Arg Ala Leu Met Pro Ala Glu Glu Glu Ala Cys Arg
180 185 190
Thr Arg Leu Ser Gln Ala Leu Gly Arg Pro Val Thr Leu Ala Val Thr
195 200 205
Ile Asp Pro Ala Leu Ile Ala Gly Leu Glu Met Glu Thr Pro His Ala
210 215 220
Val Val Arg Asn Ser Phe Lys Ala Asp Leu Asp Arg Val Thr Ala Ala
225 230 235 240
Leu Thr His His Gly Thr
245
<210> 24
<211> 243
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 24
atgactgtcg agatggtcag catcttcgcg gcggcgctcg ccgtctcctt cggcgccatc 60
gggccggccc tgggcgaggg ccgggcggtg gccgcggcca tggacgccat cgcccgccag 120
ccggaggcgg ccggaacctt gtcgcgcacg ctcttcgtcg gcctcgccat gatcgagacc 180
atggcgatct actgcctggt gatcgcgctc ctggtgctct tcgccaatcc gttcgtgaag 240
tga 243
<210> 25
<211> 80
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 25
Met Thr Val Glu Met Val Ser Ile Phe Ala Ala Ala Leu Ala Val Ser
1 5 10 15
Phe Gly Ala Ile Gly Pro Ala Leu Gly Glu Gly Arg Ala Val Ala Ala
20 25 30
Ala Met Asp Ala Ile Ala Arg Gln Pro Glu Ala Ala Gly Thr Leu Ser
35 40 45
Arg Thr Leu Phe Val Gly Leu Ala Met Ile Glu Thr Met Ala Ile Tyr
50 55 60
Cys Leu Val Ile Ala Leu Leu Val Leu Phe Ala Asn Pro Phe Val Lys
65 70 75 80
<210> 26
<211> 699
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 26
atgggctcgc cgctgatcct cgaacccctg ttccatatcg ggcccgtgcc catcaccgcg 60
ccggtggtgg tcacctggct catcatggcc gccttcattg ggctggcgcg gctcatcacc 120
cggaagcttt ccaccgatcc cacccggacc caggcggcgg tggaaacggt gctgaccgcc 180
atcgattccc agatcgccga caccatgcag gccgatcccg cgccttatcg cgcgctcatc 240
ggcaccatct tcctttatgt gctggtggcc aactggtcct cgctcatccc gggcatcgag 300
ccgcccacgg cgcatatcga gaccgatgcg gcgctcgctt tcatcgtgtt cgccgccacc 360
atcgggttcg ggttgaagac aaggggtgtg aagggctatc tcgccacctt cgccgaaccc 420
tcctgggtga tgatcccgct caatgtggtg gagcagatca cccggacctt ctcgctcatc 480
gtgcgcctgt tcggcaacat catgagcggg gtgttcgtgg tcggcatcat cctgtccctc 540
gccgggctgc tggtgcccat ccccctcatg gcgctcgatc tcctgaccgg cgccgtgcag 600
gcctacatct tcgcggtgct ggcctgcgtg ttcatcggcg cggccattgg cgaggcgccg 660
gcaaagcccc aatcgaagga gccagggaaa acatcatga 699
<210> 27
<211> 232
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 27
Met Gly Ser Pro Leu Ile Leu Glu Pro Leu Phe His Ile Gly Pro Val
1 5 10 15
Pro Ile Thr Ala Pro Val Val Val Thr Trp Leu Ile Met Ala Ala Phe
20 25 30
Ile Gly Leu Ala Arg Leu Ile Thr Arg Lys Leu Ser Thr Asp Pro Thr
35 40 45
Arg Thr Gln Ala Ala Val Glu Thr Val Leu Thr Ala Ile Asp Ser Gln
50 55 60
Ile Ala Asp Thr Met Gln Ala Asp Pro Ala Pro Tyr Arg Ala Leu Ile
65 70 75 80
Gly Thr Ile Phe Leu Tyr Val Leu Val Ala Asn Trp Ser Ser Leu Ile
85 90 95
Pro Gly Ile Glu Pro Pro Thr Ala His Ile Glu Thr Asp Ala Ala Leu
100 105 110
Ala Phe Ile Val Phe Ala Ala Thr Ile Gly Phe Gly Leu Lys Thr Arg
115 120 125
Gly Val Lys Gly Tyr Leu Ala Thr Phe Ala Glu Pro Ser Trp Val Met
130 135 140
Ile Pro Leu Asn Val Val Glu Gln Ile Thr Arg Thr Phe Ser Leu Ile
145 150 155 160
Val Arg Leu Phe Gly Asn Ile Met Ser Gly Val Phe Val Val Gly Ile
165 170 175
Ile Leu Ser Leu Ala Gly Leu Leu Val Pro Ile Pro Leu Met Ala Leu
180 185 190
Asp Leu Leu Thr Gly Ala Val Gln Ala Tyr Ile Phe Ala Val Leu Ala
195 200 205
Cys Val Phe Ile Gly Ala Ala Ile Gly Glu Ala Pro Ala Lys Pro Gln
210 215 220
Ser Lys Glu Pro Gly Lys Thr Ser
225 230
<210> 28
<211> 456
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 28
gtgagcgcgc cgctgcacct caccatcacc acgccggccg ccgttctggt ggaccgtgcc 60
gacatcgtgg ccctgcgtgc cgaggacgag agcggcagct tcggcatcct gcccggccat 120
gcggatttcc tgaccgttct ggaggcctgc gtggtgcgct tcaaggatgg ggccgacggc 180
gtgcattatt gtgctctcag tggtggcgtg ctgtcggtcg aggagggccg gcgcatcgcc 240
atcgcctgcc gtcagggcac ggtgagcgac gacctggtcg ccctggaagg ggcggtggac 300
gccatgcgtt cggcggagag cgatgccgac aagcgggccc gggtggagca gatgcgcctt 360
catgcccacg ccgtgcgcca gctcctgcac tatctgcggc ccggccgggc cggcggcgtg 420
gcgccggccg ccgcgccgga ggaggggccg tcatga 456
<210> 29
<211> 151
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 29
Met Ser Ala Pro Leu His Leu Thr Ile Thr Thr Pro Ala Ala Val Leu
1 5 10 15
Val Asp Arg Ala Asp Ile Val Ala Leu Arg Ala Glu Asp Glu Ser Gly
20 25 30
Ser Phe Gly Ile Leu Pro Gly His Ala Asp Phe Leu Thr Val Leu Glu
35 40 45
Ala Cys Val Val Arg Phe Lys Asp Gly Ala Asp Gly Val His Tyr Cys
50 55 60
Ala Leu Ser Gly Gly Val Leu Ser Val Glu Glu Gly Arg Arg Ile Ala
65 70 75 80
Ile Ala Cys Arg Gln Gly Thr Val Ser Asp Asp Leu Val Ala Leu Glu
85 90 95
Gly Ala Val Asp Ala Met Arg Ser Ala Glu Ser Asp Ala Asp Lys Arg
100 105 110
Ala Arg Val Glu Gln Met Arg Leu His Ala His Ala Val Arg Gln Leu
115 120 125
Leu His Tyr Leu Arg Pro Gly Arg Ala Gly Gly Val Ala Pro Ala Ala
130 135 140
Ala Pro Glu Glu Gly Pro Ser
145 150
<210> 30
<211> 1494
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 30
atggcagcgg cagatgagga ggcgcaatcg gccgccggcc ccgcctcggg ccgggtggtg 60
gccgtgcgcg gcgcggtgat cgacatcgcc tttgcccagc ctccgctgcc gccgctggac 120
gacgcccttc tcatcaccga cggccggggc ggcacggtgc tggtggaggt gcagagccat 180
atggatcggc acacggtgcg cgccatcgcc cttcaggcca ccaccggcct cagccggggg 240
ctggaggcgg cgcgggtggg cgggccggtg aaggtgccgg tgggagacca tgtgctcggc 300
cgcctcctgg atgtcaccgg cgccatcggc gacaagggcg ggccgctgcc ggccgacgtg 360
cccacgcggc cgatccacca cgcgccgcca tccttcgccg cgcagggcgg cacgtccgat 420
ctgtttcgca ccggcatcaa ggtcatcgac ctcctggcgc ccctcgccca gggcggcaag 480
gcggccatgt tcggcggggc cggcgtgggc aagaccgtgc tggtgatgga gctgatccac 540
gccatggtgg cgagctacaa gggcatctcg gtgtttgccg gcgtggggga gcgctcccgc 600
gagggccacg agatgctgct ggacatgacc gattccggcg tgctcgaccg caccgttctg 660
gtctatggcc agatgaacga gccccccggg gcccgctggc gggtgcccat gacggcgctg 720
accatcgccg aatatttccg cgacgagaag caccagaacg tcctgctgct gatggacaac 780
atcttccgct tcgtccaggc gggggcggag gtctccggcc ttttgggccg tccgccctcc 840
cgggtgggat accagccgac gctggcgagc gaggtggcgg cgctccagga acgcatcacc 900
tccgtgggcg aggcctcggt gaccgccatc gaggcggtct acgtgccggc ggatgacttc 960
accgatcccg ccgtgaccac catcgccgcc cacgtggatt ccatggtggt gctctcccgc 1020
gccatggcgg cggagggcat gtatccggcg gtggacccca tctcctcctc gtcggtgctg 1080
ctcgacccgc tcatcgtggg ggacgagcat gcgcgcgtcg ccaacgaggt gcgccggacc 1140
atcgagcatt atcgcgagct tcaggatgtg atctcgctgc tgggcatgga ggaattgggc 1200
accgaggatc gccgcatcgt ggagcgggcg cgccggctcc agcgcttcct cacccagccc 1260
ttcacggtca ccgaggcctt caccggcgtg cccggccgct cggtggccat cgccgacacc 1320
atcgccggct gcaggatgat cctgtccggc gcctgcgacg actggcagga aagcgccctc 1380
tacatggtgg gcaccatcga cgaggcccgc cagaaggagg aggccgctcg cgccaaggcg 1440
gggcagggcg ccccggccgg gacggcagcc gagacggcgg aggccgcccc gtga 1494
<210> 31
<211> 497
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 31
Met Ala Ala Ala Asp Glu Glu Ala Gln Ser Ala Ala Gly Pro Ala Ser
1 5 10 15
Gly Arg Val Val Ala Val Arg Gly Ala Val Ile Asp Ile Ala Phe Ala
20 25 30
Gln Pro Pro Leu Pro Pro Leu Asp Asp Ala Leu Leu Ile Thr Asp Gly
35 40 45
Arg Gly Gly Thr Val Leu Val Glu Val Gln Ser His Met Asp Arg His
50 55 60
Thr Val Arg Ala Ile Ala Leu Gln Ala Thr Thr Gly Leu Ser Arg Gly
65 70 75 80
Leu Glu Ala Ala Arg Val Gly Gly Pro Val Lys Val Pro Val Gly Asp
85 90 95
His Val Leu Gly Arg Leu Leu Asp Val Thr Gly Ala Ile Gly Asp Lys
100 105 110
Gly Gly Pro Leu Pro Ala Asp Val Pro Thr Arg Pro Ile His His Ala
115 120 125
Pro Pro Ser Phe Ala Ala Gln Gly Gly Thr Ser Asp Leu Phe Arg Thr
130 135 140
Gly Ile Lys Val Ile Asp Leu Leu Ala Pro Leu Ala Gln Gly Gly Lys
145 150 155 160
Ala Ala Met Phe Gly Gly Ala Gly Val Gly Lys Thr Val Leu Val Met
165 170 175
Glu Leu Ile His Ala Met Val Ala Ser Tyr Lys Gly Ile Ser Val Phe
180 185 190
Ala Gly Val Gly Glu Arg Ser Arg Glu Gly His Glu Met Leu Leu Asp
195 200 205
Met Thr Asp Ser Gly Val Leu Asp Arg Thr Val Leu Val Tyr Gly Gln
210 215 220
Met Asn Glu Pro Pro Gly Ala Arg Trp Arg Val Pro Met Thr Ala Leu
225 230 235 240
Thr Ile Ala Glu Tyr Phe Arg Asp Glu Lys His Gln Asn Val Leu Leu
245 250 255
Leu Met Asp Asn Ile Phe Arg Phe Val Gln Ala Gly Ala Glu Val Ser
260 265 270
Gly Leu Leu Gly Arg Pro Pro Ser Arg Val Gly Tyr Gln Pro Thr Leu
275 280 285
Ala Ser Glu Val Ala Ala Leu Gln Glu Arg Ile Thr Ser Val Gly Glu
290 295 300
Ala Ser Val Thr Ala Ile Glu Ala Val Tyr Val Pro Ala Asp Asp Phe
305 310 315 320
Thr Asp Pro Ala Val Thr Thr Ile Ala Ala His Val Asp Ser Met Val
325 330 335
Val Leu Ser Arg Ala Met Ala Ala Glu Gly Met Tyr Pro Ala Val Asp
340 345 350
Pro Ile Ser Ser Ser Ser Val Leu Leu Asp Pro Leu Ile Val Gly Asp
355 360 365
Glu His Ala Arg Val Ala Asn Glu Val Arg Arg Thr Ile Glu His Tyr
370 375 380
Arg Glu Leu Gln Asp Val Ile Ser Leu Leu Gly Met Glu Glu Leu Gly
385 390 395 400
Thr Glu Asp Arg Arg Ile Val Glu Arg Ala Arg Arg Leu Gln Arg Phe
405 410 415
Leu Thr Gln Pro Phe Thr Val Thr Glu Ala Phe Thr Gly Val Pro Gly
420 425 430
Arg Ser Val Ala Ile Ala Asp Thr Ile Ala Gly Cys Arg Met Ile Leu
435 440 445
Ser Gly Ala Cys Asp Asp Trp Gln Glu Ser Ala Leu Tyr Met Val Gly
450 455 460
Thr Ile Asp Glu Ala Arg Gln Lys Glu Glu Ala Ala Arg Ala Lys Ala
465 470 475 480
Gly Gln Gly Ala Pro Ala Gly Thr Ala Ala Glu Thr Ala Glu Ala Ala
485 490 495
Pro
<210> 32
<211> 1437
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 32
atggcgaaca aggtcggacg catcacccag atcatcggcg ccgtcgtcga cgtgcagttc 60
gacgggcatc tgccggcgat tctcaacgcg atcgagacca ccaaccaggg caaccggctg 120
gtgctcgaag tggctcagca tctcggcgag aacaccgtgc gctgcatcgc catggatgcc 180
actgaaggcc tggtgcgtgg ccaggaggtg gccgacaccg atgcgcccat ccaggtgccc 240
gtgggcgccg ccaccctcgg ccgcatcatg aacgtgatcg gcgagccggt ggacgagctg 300
ggccccatcg agggcgaagc gctgcgcggc atccatcagc cggccccctc ctatgcggag 360
caggccacgg aagctgagat cctcgtcacc ggcatcaagg tggtggatct gctggcgccc 420
tattccaagg gcggcaaggt gggcctgttc ggcggcgccg gcgtgggcaa gaccgtgctc 480
atcatggagc tgatcaacaa cgtggccaag gcgcacggcg gctattccgt gttcgccggc 540
gtgggtgagc gcacccgcga gggcaacgac ctctaccacg agatgatcga gtccaacgtg 600
aacaaggacc cgcacgagaa caatggctcg gcggccggtt ccaagtgcgc cctggtctat 660
ggccagatga acgagccgcc cggcgcccgc gcccgcgtgg ccctcaccgg cctcaccgtc 720
gccgagcatt tccgcgacca gggccaggac gtgctgttct tcgtggacaa catcttccgc 780
ttcacccagg cgggctccga ggtgtcggcg cttctcggcc gcatcccctc ggcggtgggc 840
taccagccga cgctggccac cgacatgggc cagctgcagg agcgcatcac caccaccacc 900
aagggctcca tcacctcggt gcaggccatc tacgtgccgg cggacgatct gaccgatccg 960
gcgccggccg cctccttcgc ccatctggac gccaccacgg tgctgtcgcg ctccatcgcg 1020
gagaagggca tctacccggc ggtggatccg ctggactcca cctcgcgcat gctgtctccc 1080
gccatcctcg gcgacgagca ctacaacacc gcgcgccagg tgcagcagac cctgcagcgc 1140
tacaaggcgc tccaggacat catcgccatc ctgggcatgg acgaactctc cgaagaggac 1200
aagctcaccg tggcccgcgc ccgcaagatc gagcgcttcc tctcccagcc cttccacgtg 1260
gccgaggtgt tcaccggttc gcccggcaag ctggtcgacc tcgccgacac catcaagggc 1320
ttcaagggcc tggtggacgg caagtacgac tacctgcccg agcaggcctt ctacatggtg 1380
ggcaccatcg aagaagccat cgagaagggc aagaagctgg cggccgaggc ggcctga 1437
<210> 33
<211> 478
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 33
Met Ala Asn Lys Val Gly Arg Ile Thr Gln Ile Ile Gly Ala Val Val
1 5 10 15
Asp Val Gln Phe Asp Gly His Leu Pro Ala Ile Leu Asn Ala Ile Glu
20 25 30
Thr Thr Asn Gln Gly Asn Arg Leu Val Leu Glu Val Ala Gln His Leu
35 40 45
Gly Glu Asn Thr Val Arg Cys Ile Ala Met Asp Ala Thr Glu Gly Leu
50 55 60
Val Arg Gly Gln Glu Val Ala Asp Thr Asp Ala Pro Ile Gln Val Pro
65 70 75 80
Val Gly Ala Ala Thr Leu Gly Arg Ile Met Asn Val Ile Gly Glu Pro
85 90 95
Val Asp Glu Leu Gly Pro Ile Glu Gly Glu Ala Leu Arg Gly Ile His
100 105 110
Gln Pro Ala Pro Ser Tyr Ala Glu Gln Ala Thr Glu Ala Glu Ile Leu
115 120 125
Val Thr Gly Ile Lys Val Val Asp Leu Leu Ala Pro Tyr Ser Lys Gly
130 135 140
Gly Lys Val Gly Leu Phe Gly Gly Ala Gly Val Gly Lys Thr Val Leu
145 150 155 160
Ile Met Glu Leu Ile Asn Asn Val Ala Lys Ala His Gly Gly Tyr Ser
165 170 175
Val Phe Ala Gly Val Gly Glu Arg Thr Arg Glu Gly Asn Asp Leu Tyr
180 185 190
His Glu Met Ile Glu Ser Asn Val Asn Lys Asp Pro His Glu Asn Asn
195 200 205
Gly Ser Ala Ala Gly Ser Lys Cys Ala Leu Val Tyr Gly Gln Met Asn
210 215 220
Glu Pro Pro Gly Ala Arg Ala Arg Val Ala Leu Thr Gly Leu Thr Val
225 230 235 240
Ala Glu His Phe Arg Asp Gln Gly Gln Asp Val Leu Phe Phe Val Asp
245 250 255
Asn Ile Phe Arg Phe Thr Gln Ala Gly Ser Glu Val Ser Ala Leu Leu
260 265 270
Gly Arg Ile Pro Ser Ala Val Gly Tyr Gln Pro Thr Leu Ala Thr Asp
275 280 285
Met Gly Gln Leu Gln Glu Arg Ile Thr Thr Thr Thr Lys Gly Ser Ile
290 295 300
Thr Ser Val Gln Ala Ile Tyr Val Pro Ala Asp Asp Leu Thr Asp Pro
305 310 315 320
Ala Pro Ala Ala Ser Phe Ala His Leu Asp Ala Thr Thr Val Leu Ser
325 330 335
Arg Ser Ile Ala Glu Lys Gly Ile Tyr Pro Ala Val Asp Pro Leu Asp
340 345 350
Ser Thr Ser Arg Met Leu Ser Pro Ala Ile Leu Gly Asp Glu His Tyr
355 360 365
Asn Thr Ala Arg Gln Val Gln Gln Thr Leu Gln Arg Tyr Lys Ala Leu
370 375 380
Gln Asp Ile Ile Ala Ile Leu Gly Met Asp Glu Leu Ser Glu Glu Asp
385 390 395 400
Lys Leu Thr Val Ala Arg Ala Arg Lys Ile Glu Arg Phe Leu Ser Gln
405 410 415
Pro Phe His Val Ala Glu Val Phe Thr Gly Ser Pro Gly Lys Leu Val
420 425 430
Asp Leu Ala Asp Thr Ile Lys Gly Phe Lys Gly Leu Val Asp Gly Lys
435 440 445
Tyr Asp Tyr Leu Pro Glu Gln Ala Phe Tyr Met Val Gly Thr Ile Glu
450 455 460
Glu Ala Ile Glu Lys Gly Lys Lys Leu Ala Ala Glu Ala Ala
465 470 475
<210> 34
<211> 876
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 34
atggcgagtc tgaaggacct gagaaaccgc attgcctcgg tgaaggcgac gcagaagatc 60
accaaggcga tgcagatggt cgccgcggcg aagctgcgtc gcgcccaggc ggcggctgaa 120
gcggcccgtc cctatgcgga acgcatggag acggtgctcg gaaatcttgc ctccggcatg 180
gtggtgggcg cgcaggcgcc tgttctcatg accgggacgg gcaagagcga cacccacctg 240
ctgctggtgt gcaccggcga gcgcggcctg tgcggcgcct tcaactcgtc catcgtgcgc 300
ttcgcccgcg agcgggcgca gctgctgctg gccgagggca agaaggtgaa aatcctgtgc 360
gtgggccgca agggccacga gcagctgcgc cgcatctacc cggacaacat catcgacgtg 420
gtggacctgc gcgcggtgcg caacatcggc ttcaaggagg ccgacgccat cgcccgcaag 480
gtgctggccc tgctcgatga aggcgcattc gacgtctgca cgctcttcta ctcccacttc 540
aggagcgtga tcgcccaggt gccgacggcc cagcagctca ttccggccac cttcgacgag 600
cggccggccg tcgccgatgc gccggtctat gaatatgagc cggaggagga ggagatcctc 660
gccgagctgc tgccgcgcaa cgtggcggtg cagatcttca aggccctcct cgagaaccag 720
gcttctttct atggctccca gatgagcgcc atggacaacg ccacgcgcaa tgcgggcgag 780
atgatcaaga agcagacgct cacctacaac cgtacccgcc aggccatgat cacgaaggaa 840
ctcatcgaga tcatctccgg cgccgaggcc gtctga 876
<210> 35
<211> 291
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 35
Met Ala Ser Leu Lys Asp Leu Arg Asn Arg Ile Ala Ser Val Lys Ala
1 5 10 15
Thr Gln Lys Ile Thr Lys Ala Met Gln Met Val Ala Ala Ala Lys Leu
20 25 30
Arg Arg Ala Gln Ala Ala Ala Glu Ala Ala Arg Pro Tyr Ala Glu Arg
35 40 45
Met Glu Thr Val Leu Gly Asn Leu Ala Ser Gly Met Val Val Gly Ala
50 55 60
Gln Ala Pro Val Leu Met Thr Gly Thr Gly Lys Ser Asp Thr His Leu
65 70 75 80
Leu Leu Val Cys Thr Gly Glu Arg Gly Leu Cys Gly Ala Phe Asn Ser
85 90 95
Ser Ile Val Arg Phe Ala Arg Glu Arg Ala Gln Leu Leu Leu Ala Glu
100 105 110
Gly Lys Lys Val Lys Ile Leu Cys Val Gly Arg Lys Gly His Glu Gln
115 120 125
Leu Arg Arg Ile Tyr Pro Asp Asn Ile Ile Asp Val Val Asp Leu Arg
130 135 140
Ala Val Arg Asn Ile Gly Phe Lys Glu Ala Asp Ala Ile Ala Arg Lys
145 150 155 160
Val Leu Ala Leu Leu Asp Glu Gly Ala Phe Asp Val Cys Thr Leu Phe
165 170 175
Tyr Ser His Phe Arg Ser Val Ile Ala Gln Val Pro Thr Ala Gln Gln
180 185 190
Leu Ile Pro Ala Thr Phe Asp Glu Arg Pro Ala Val Ala Asp Ala Pro
195 200 205
Val Tyr Glu Tyr Glu Pro Glu Glu Glu Glu Ile Leu Ala Glu Leu Leu
210 215 220
Pro Arg Asn Val Ala Val Gln Ile Phe Lys Ala Leu Leu Glu Asn Gln
225 230 235 240
Ala Ser Phe Tyr Gly Ser Gln Met Ser Ala Met Asp Asn Ala Thr Arg
245 250 255
Asn Ala Gly Glu Met Ile Lys Lys Gln Thr Leu Thr Tyr Asn Arg Thr
260 265 270
Arg Gln Ala Met Ile Thr Lys Glu Leu Ile Glu Ile Ile Ser Gly Ala
275 280 285
Glu Ala Val
290
<210> 36
<211> 1530
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 36
atggacattc gagccgctga aatctctgcc atcctgaaag agcagatcca gaatttcggc 60
caggaggcgg aagtctccga ggtgggtcag gttctgtccg tgggtgacgg catcgcgcgc 120
gtctacggcc tcgacaacgt ccaggcgggc gagatggtcg agttcgagaa cggcacgcgc 180
ggcatggcgc tgaacctcga gctcgacaat gtcggcatcg tgatcttcgg ttccgaccgc 240
gagatcaagg aaggccagac cgtcaagcgg accggcgcca tcgtggacgc ccccgtcggc 300
aagggcctgc tcggccgcgt cgtggacgct ctcggcaacc cgatcgacgg caagggcccg 360
atcatgttca ccgagcgtcg ccgggtcgac gtgaaggcgc cgggcatcat cccgcgcaag 420
tcggtgcacg agcccatgca gaccggcctg aaggccatcg atgcgctcat ccccatcggc 480
cgcggccagc gcgagctcat catcggcgac cgccagaccg gcaagaccgc cgtggcgctc 540
gactcgatcc tgaaccagaa gcccatcaac cagggcgacg acgagaaggc caagctctac 600
tgcgtctatg tcgcggtggg ccagaagcgt tccactgtcg cgcagttcgt gaaggtgctc 660
gaggagcacg gcgcgctgga atattccatc gtcgtcgccg ccaccgcctc ggacgcggcc 720
cccatgcagt tcctggcgcc gttcaccggc accgccatgg gcgagtattt ccgcgacaac 780
ggcatgcacg ccctcatcat ccatgatgac ctgtccaagc aggccgtggc ctaccgccag 840
atgtcgctgc tgctgcgccg cccgccgggc cgcgaggcct atcccggcga tgtgttctac 900
ctgcactccc gcctcttgga gcgcgccgcc aagctcaatg acgagcacgg cgccggctcg 960
ctgaccgccc tgccggtgat cgagacccag gccaacgacg tgtcggccta catcccgacc 1020
aacgtgatct ccatcaccga cggtcagatc ttccttgaat ccgatctgtt ctaccagggc 1080
atccgcccgg cggtgaacgt gggcctgtcg gtgtcgcgcg tgggctcttc ggcccagatc 1140
aaggcgatga agcaggtggc cggcaagatc aagggcgagc tcgcccagta tcgcgagctg 1200
gcggccttcg cccagttcgg ttcggacctg gacgcggcca cccagaagct gctgaaccgc 1260
ggcgcccgcc tcaccgagct gctgaagcag agccagttct cgcccctcaa ggtggaggag 1320
caggtggcgg tgatctatgc cggcaccaat ggctatctcg atccgctgcc ggtctccaag 1380
gtgcgcgagt tcgagcaggg tctgctcctg tcgctgcgct cgcagcatcc ggagatcctg 1440
gacgccatcc gcacgtccaa ggagctttcc aaggacaccg ccgagaagct gacgaaggcc 1500
atcgacgcct tcgccaagag cttctcctga 1530
<210> 37
<211> 509
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 37
Met Asp Ile Arg Ala Ala Glu Ile Ser Ala Ile Leu Lys Glu Gln Ile
1 5 10 15
Gln Asn Phe Gly Gln Glu Ala Glu Val Ser Glu Val Gly Gln Val Leu
20 25 30
Ser Val Gly Asp Gly Ile Ala Arg Val Tyr Gly Leu Asp Asn Val Gln
35 40 45
Ala Gly Glu Met Val Glu Phe Glu Asn Gly Thr Arg Gly Met Ala Leu
50 55 60
Asn Leu Glu Leu Asp Asn Val Gly Ile Val Ile Phe Gly Ser Asp Arg
65 70 75 80
Glu Ile Lys Glu Gly Gln Thr Val Lys Arg Thr Gly Ala Ile Val Asp
85 90 95
Ala Pro Val Gly Lys Gly Leu Leu Gly Arg Val Val Asp Ala Leu Gly
100 105 110
Asn Pro Ile Asp Gly Lys Gly Pro Ile Met Phe Thr Glu Arg Arg Arg
115 120 125
Val Asp Val Lys Ala Pro Gly Ile Ile Pro Arg Lys Ser Val His Glu
130 135 140
Pro Met Gln Thr Gly Leu Lys Ala Ile Asp Ala Leu Ile Pro Ile Gly
145 150 155 160
Arg Gly Gln Arg Glu Leu Ile Ile Gly Asp Arg Gln Thr Gly Lys Thr
165 170 175
Ala Val Ala Leu Asp Ser Ile Leu Asn Gln Lys Pro Ile Asn Gln Gly
180 185 190
Asp Asp Glu Lys Ala Lys Leu Tyr Cys Val Tyr Val Ala Val Gly Gln
195 200 205
Lys Arg Ser Thr Val Ala Gln Phe Val Lys Val Leu Glu Glu His Gly
210 215 220
Ala Leu Glu Tyr Ser Ile Val Val Ala Ala Thr Ala Ser Asp Ala Ala
225 230 235 240
Pro Met Gln Phe Leu Ala Pro Phe Thr Gly Thr Ala Met Gly Glu Tyr
245 250 255
Phe Arg Asp Asn Gly Met His Ala Leu Ile Ile His Asp Asp Leu Ser
260 265 270
Lys Gln Ala Val Ala Tyr Arg Gln Met Ser Leu Leu Leu Arg Arg Pro
275 280 285
Pro Gly Arg Glu Ala Tyr Pro Gly Asp Val Phe Tyr Leu His Ser Arg
290 295 300
Leu Leu Glu Arg Ala Ala Lys Leu Asn Asp Glu His Gly Ala Gly Ser
305 310 315 320
Leu Thr Ala Leu Pro Val Ile Glu Thr Gln Ala Asn Asp Val Ser Ala
325 330 335
Tyr Ile Pro Thr Asn Val Ile Ser Ile Thr Asp Gly Gln Ile Phe Leu
340 345 350
Glu Ser Asp Leu Phe Tyr Gln Gly Ile Arg Pro Ala Val Asn Val Gly
355 360 365
Leu Ser Val Ser Arg Val Gly Ser Ser Ala Gln Ile Lys Ala Met Lys
370 375 380
Gln Val Ala Gly Lys Ile Lys Gly Glu Leu Ala Gln Tyr Arg Glu Leu
385 390 395 400
Ala Ala Phe Ala Gln Phe Gly Ser Asp Leu Asp Ala Ala Thr Gln Lys
405 410 415
Leu Leu Asn Arg Gly Ala Arg Leu Thr Glu Leu Leu Lys Gln Ser Gln
420 425 430
Phe Ser Pro Leu Lys Val Glu Glu Gln Val Ala Val Ile Tyr Ala Gly
435 440 445
Thr Asn Gly Tyr Leu Asp Pro Leu Pro Val Ser Lys Val Arg Glu Phe
450 455 460
Glu Gln Gly Leu Leu Leu Ser Leu Arg Ser Gln His Pro Glu Ile Leu
465 470 475 480
Asp Ala Ile Arg Thr Ser Lys Glu Leu Ser Lys Asp Thr Ala Glu Lys
485 490 495
Leu Thr Lys Ala Ile Asp Ala Phe Ala Lys Ser Phe Ser
500 505
<210> 38
<211> 555
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 38
gtggcggaaa cgatcgtgtc aggcatggcg ggacgctatg cgaccgcgct gttcgagctg 60
gcggacgaag ccggtgccat cgattccgtc caggcggatc ttgatcgcct gtccggcctt 120
ctggccgaga gcgcggatct ggcgcggctg gtcaagagcc cggtcttcac cgccgagcag 180
cagctcggcg cgatggcggc cattctcgat caagcaggca tttccggcct tgcgggcaaa 240
ttcgtgaagc tggtggcgca gaaccgccgc ctgttcgcac tgccgcgcat gattgccgaa 300
tacgccgtcc tggtggcccg gaagaagggc gagacctcgg cgagcgtgac cgttgccacc 360
cccctgagcg atgagcatct ggccacgctc aaggcggccc tggctgaaaa gaccggcaag 420
gacgtgaagc tcgacgtcac cgtcgatccg tccatcctcg gtggtctcat cgtgaagctc 480
ggctcgcgca tggtcgatgc ttccctgaag accaaactca attctatccg gcatgcgatg 540
aaagaggtcc gctga 555
<210> 39
<211> 184
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 39
Met Ala Glu Thr Ile Val Ser Gly Met Ala Gly Arg Tyr Ala Thr Ala
1 5 10 15
Leu Phe Glu Leu Ala Asp Glu Ala Gly Ala Ile Asp Ser Val Gln Ala
20 25 30
Asp Leu Asp Arg Leu Ser Gly Leu Leu Ala Glu Ser Ala Asp Leu Ala
35 40 45
Arg Leu Val Lys Ser Pro Val Phe Thr Ala Glu Gln Gln Leu Gly Ala
50 55 60
Met Ala Ala Ile Leu Asp Gln Ala Gly Ile Ser Gly Leu Ala Gly Lys
65 70 75 80
Phe Val Lys Leu Val Ala Gln Asn Arg Arg Leu Phe Ala Leu Pro Arg
85 90 95
Met Ile Ala Glu Tyr Ala Val Leu Val Ala Arg Lys Lys Gly Glu Thr
100 105 110
Ser Ala Ser Val Thr Val Ala Thr Pro Leu Ser Asp Glu His Leu Ala
115 120 125
Thr Leu Lys Ala Ala Leu Ala Glu Lys Thr Gly Lys Asp Val Lys Leu
130 135 140
Asp Val Thr Val Asp Pro Ser Ile Leu Gly Gly Leu Ile Val Lys Leu
145 150 155 160
Gly Ser Arg Met Val Asp Ala Ser Leu Lys Thr Lys Leu Asn Ser Ile
165 170 175
Arg His Ala Met Lys Glu Val Arg
180
<210> 40
<211> 492
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 40
atgaccgaaa tggaactggc tgagctctgg gtcgccatcg ccttcctggt tttcgtaggc 60
ctcctgatct atgcgggcgc ccaccgcgcc atcgtctccg ccctggattc ccgcggctcg 120
cgcatcgcct cggaactgga ggaggcccgt cggctcaagg aagaggccca gaagctggtg 180
gccgaattca agcgcaagca gcgcgaggcc gaggccgagg ccgaatccat cgtcaccggc 240
gccaaggccg aggccgagcg cctcgccgcc gaggccaagg cgaagatcga ggatttcgtc 300
acccgccgca ccaagatggc cgaggacaag atcgcccagg ccgagcatca ggctctggcg 360
gacgtgaagt ccatcgccgc cgaggcggcg gccaaggcgg ccgaggtgat cctcggcgcc 420
caggccaccg gcgcggtggc ggagcgtctg ctgtcgggcg ccatctccga ggtcaagacc 480
aagctcaact ga 492
<210> 41
<211> 163
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 41
Met Thr Glu Met Glu Leu Ala Glu Leu Trp Val Ala Ile Ala Phe Leu
1 5 10 15
Val Phe Val Gly Leu Leu Ile Tyr Ala Gly Ala His Arg Ala Ile Val
20 25 30
Ser Ala Leu Asp Ser Arg Gly Ser Arg Ile Ala Ser Glu Leu Glu Glu
35 40 45
Ala Arg Arg Leu Lys Glu Glu Ala Gln Lys Leu Val Ala Glu Phe Lys
50 55 60
Arg Lys Gln Arg Glu Ala Glu Ala Glu Ala Glu Ser Ile Val Thr Gly
65 70 75 80
Ala Lys Ala Glu Ala Glu Arg Leu Ala Ala Glu Ala Lys Ala Lys Ile
85 90 95
Glu Asp Phe Val Thr Arg Arg Thr Lys Met Ala Glu Asp Lys Ile Ala
100 105 110
Gln Ala Glu His Gln Ala Leu Ala Asp Val Lys Ser Ile Ala Ala Glu
115 120 125
Ala Ala Ala Lys Ala Ala Glu Val Ile Leu Gly Ala Gln Ala Thr Gly
130 135 140
Ala Val Ala Glu Arg Leu Leu Ser Gly Ala Ile Ser Glu Val Lys Thr
145 150 155 160
Lys Leu Asn
<210> 42
<211> 729
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 42
atgatgattg catggaagcg gaccttcgca gtcgtgacct tcggggccgc cctgatggcc 60
atgcccgtcg cgggcgtggt cgcagctgag acttctcccg ctccggcggc agtggcgcag 120
gccgatcatg cggtgcccac cgaggcggcc ggccagggca ccgccgatgc ggcccatgcc 180
gccgcgccgg gcgaggccgc ccatggtggc gcggccaagc acgaaaccca tttcccgccc 240
ttcgacggca ccaccttcgc ctcccagttg ctgtggctcg ccgtcacctt cggcctgctt 300
tactacctca tgagcaaggt cacgctgccg cgcatcggcc gcatcctgga agagcgccac 360
gaccgcatcg ccgatgatct ggaggaagcc tccaagcatc gcgccgagag cgaggccgcc 420
cagcgggcct atgagaaggc gctgagcgag gcccgcgcga aggcccattc catcgccgcg 480
gaaacccgcg accgccttgc cgcccacgcc gacaccaacc gcaaggcgct ggagagcgag 540
ctcaccgcca agctgcaggc ggccgaggag cgcatcgcca ccaccaagag cgaagccctc 600
acccatgtgc gcggcatcgc ggtggacgcc acccaatcca tcgtctccac cctcatcggt 660
gtcgcgcccg cggcggccga cgtggaaaaa gcggtggacg gcgccctgtc ccagcacggc 720
caggcctga 729
<210> 43
<211> 242
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 43
Met Met Ile Ala Trp Lys Arg Thr Phe Ala Val Val Thr Phe Gly Ala
1 5 10 15
Ala Leu Met Ala Met Pro Val Ala Gly Val Val Ala Ala Glu Thr Ser
20 25 30
Pro Ala Pro Ala Ala Val Ala Gln Ala Asp His Ala Val Pro Thr Glu
35 40 45
Ala Ala Gly Gln Gly Thr Ala Asp Ala Ala His Ala Ala Ala Pro Gly
50 55 60
Glu Ala Ala His Gly Gly Ala Ala Lys His Glu Thr His Phe Pro Pro
65 70 75 80
Phe Asp Gly Thr Thr Phe Ala Ser Gln Leu Leu Trp Leu Ala Val Thr
85 90 95
Phe Gly Leu Leu Tyr Tyr Leu Met Ser Lys Val Thr Leu Pro Arg Ile
100 105 110
Gly Arg Ile Leu Glu Glu Arg His Asp Arg Ile Ala Asp Asp Leu Glu
115 120 125
Glu Ala Ser Lys His Arg Ala Glu Ser Glu Ala Ala Gln Arg Ala Tyr
130 135 140
Glu Lys Ala Leu Ser Glu Ala Arg Ala Lys Ala His Ser Ile Ala Ala
145 150 155 160
Glu Thr Arg Asp Arg Leu Ala Ala His Ala Asp Thr Asn Arg Lys Ala
165 170 175
Leu Glu Ser Glu Leu Thr Ala Lys Leu Gln Ala Ala Glu Glu Arg Ile
180 185 190
Ala Thr Thr Lys Ser Glu Ala Leu Thr His Val Arg Gly Ile Ala Val
195 200 205
Asp Ala Thr Gln Ser Ile Val Ser Thr Leu Ile Gly Val Ala Pro Ala
210 215 220
Ala Ala Asp Val Glu Lys Ala Val Asp Gly Ala Leu Ser Gln His Gly
225 230 235 240
Gln Ala
<210> 44
<211> 228
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 44
atggaagcgg aagctggaaa gttcatcggt gccggcctcg cctgcctcgg catgggtctc 60
gctggcgtcg gcgtcggtaa catcttcggt aacttcctct ccggcgccct gcgcaacccg 120
tccgctgccg acggccagtt cgcccgcgcc ttcatcggcg ccgccctcgc ggaaggtctc 180
ggcatcttct cgctggtcgt tgcgctcgtc ctgctgttcg tggcctga 228
<210> 45
<211> 75
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 45
Met Glu Ala Glu Ala Gly Lys Phe Ile Gly Ala Gly Leu Ala Cys Leu
1 5 10 15
Gly Met Gly Leu Ala Gly Val Gly Val Gly Asn Ile Phe Gly Asn Phe
20 25 30
Leu Ser Gly Ala Leu Arg Asn Pro Ser Ala Ala Asp Gly Gln Phe Ala
35 40 45
Arg Ala Phe Ile Gly Ala Ala Leu Ala Glu Gly Leu Gly Ile Phe Ser
50 55 60
Leu Val Val Ala Leu Val Leu Leu Phe Val Ala
65 70 75
<210> 46
<211> 753
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 46
atgaccgtcg atccgatcca ccagttcgag atcaagcgct acgtggatct gctgaacgtc 60
ggcggtgtcc agttctcctt caccaacgca acggtgttca tgattggcat cgtcctggtg 120
attttcttct tcctgacttt cgcgacacgc ggtcgcaccc ttgtgccggg ccggatgcag 180
tcggcggcgg agctgagcta cgagttcatc gccaagatgg tgcgcgacgc ggccggcagc 240
gagggaatgg tgttctttcc cttcgtcttc tcgctcttca tgttcgtgct ggtggcgaac 300
gtattggggc tcatccccta caccttcacg gtgaccgccc acctcatcgt caccgccgcc 360
ctggcggcga cggtgatcct caccgtcatc atctacggct tcgtgcggca cggcacccac 420
ttcctgcacc tgttcgtgcc gtcgggcgtg ccgggcttcc tcctgccctt cctcgtggtg 480
atcgaggtgg tgtcgttcct gtcgcggccc atcagcctct cgctgcgtct gttcgccaac 540
atgctggcgg gccacatcgc cctcaaggtg ttcgccttct tcgtcgtggg actggcctcg 600
gccggcgcga tcggctggtt cggcgccacc ctgcccttct tcatgatcgt ggcgctcacc 660
gcgctggagc tgctggtggc ggtgctgcag gcctacgtgt tcgcggtgct gacctcgatc 720
tacctcaacg acgccatcca tcccggccac tga 753
<210> 47
<211> 250
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 47
Met Thr Val Asp Pro Ile His Gln Phe Glu Ile Lys Arg Tyr Val Asp
1 5 10 15
Leu Leu Asn Val Gly Gly Val Gln Phe Ser Phe Thr Asn Ala Thr Val
20 25 30
Phe Met Ile Gly Ile Val Leu Val Ile Phe Phe Phe Leu Thr Phe Ala
35 40 45
Thr Arg Gly Arg Thr Leu Val Pro Gly Arg Met Gln Ser Ala Ala Glu
50 55 60
Leu Ser Tyr Glu Phe Ile Ala Lys Met Val Arg Asp Ala Ala Gly Ser
65 70 75 80
Glu Gly Met Val Phe Phe Pro Phe Val Phe Ser Leu Phe Met Phe Val
85 90 95
Leu Val Ala Asn Val Leu Gly Leu Ile Pro Tyr Thr Phe Thr Val Thr
100 105 110
Ala His Leu Ile Val Thr Ala Ala Leu Ala Ala Thr Val Ile Leu Thr
115 120 125
Val Ile Ile Tyr Gly Phe Val Arg His Gly Thr His Phe Leu His Leu
130 135 140
Phe Val Pro Ser Gly Val Pro Gly Phe Leu Leu Pro Phe Leu Val Val
145 150 155 160
Ile Glu Val Val Ser Phe Leu Ser Arg Pro Ile Ser Leu Ser Leu Arg
165 170 175
Leu Phe Ala Asn Met Leu Ala Gly His Ile Ala Leu Lys Val Phe Ala
180 185 190
Phe Phe Val Val Gly Leu Ala Ser Ala Gly Ala Ile Gly Trp Phe Gly
195 200 205
Ala Thr Leu Pro Phe Phe Met Ile Val Ala Leu Thr Ala Leu Glu Leu
210 215 220
Leu Val Ala Val Leu Gln Ala Tyr Val Phe Ala Val Leu Thr Ser Ile
225 230 235 240
Tyr Leu Asn Asp Ala Ile His Pro Gly His
245 250
<210> 48
<211> 390
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 48
atgtccgagc cgaatgatcc atcccgcagg gacggtgcga aggcgaaaga cgagacgcag 60
gactcccggc ccggtgaggc ggatcttgct cggcgcctcg atgcgctcgg cacctccatc 120
ggtcaggtca agtccagaag cggggagccc gcggcgacgc cgcgcaagga cacctcctcg 180
gcctccggcg cggccctggc gtttcggctg ggcgccgagt ttgtttcagg cgtgctggtg 240
ggctcgctca tcggctacgg gttggattat gcgtttgcga tttcgccctg ggggctgatc 300
gccttcacgc tgatcggctt tgccgccggc gtcctgaaca tgctgcgcgt ggcgaacagc 360
gatgccaagc gccacagcgc ggacaggtga 390
<210> 49
<211> 129
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 49
Met Ser Glu Pro Asn Asp Pro Ser Arg Arg Asp Gly Ala Lys Ala Lys
1 5 10 15
Asp Glu Thr Gln Asp Ser Arg Pro Gly Glu Ala Asp Leu Ala Arg Arg
20 25 30
Leu Asp Ala Leu Gly Thr Ser Ile Gly Gln Val Lys Ser Arg Ser Gly
35 40 45
Glu Pro Ala Ala Thr Pro Arg Lys Asp Thr Ser Ser Ala Ser Gly Ala
50 55 60
Ala Leu Ala Phe Arg Leu Gly Ala Glu Phe Val Ser Gly Val Leu Val
65 70 75 80
Gly Ser Leu Ile Gly Tyr Gly Leu Asp Tyr Ala Phe Ala Ile Ser Pro
85 90 95
Trp Gly Leu Ile Ala Phe Thr Leu Ile Gly Phe Ala Ala Gly Val Leu
100 105 110
Asn Met Leu Arg Val Ala Asn Ser Asp Ala Lys Arg His Ser Ala Asp
115 120 125
Arg
<210> 50
<211> 1929
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 50
atgagttcgc tctccgccac tattcaacag gtcttcaacg agccgggctg cgcgaagaac 60
cagaataagt ccgaggcgga gaagaagaag ggctgcacca agcagctgca acccggcgga 120
gcggccggcg gctgcgcgtt cgacggcgcg aagatcgcgc tccagccctt gaccgacgtc 180
gcccacctgg tgcacggccc catcgcctgc gaaggcaatt cctgggacaa tcgtggcgcc 240
aagtcctccg gctcgaacat ctggcgcacc ggcttcacca cggacatcaa cgaaaccgac 300
gtggtgttcg gcggcgagaa gcgtctgttc aagtccatca aggaaatcat cgagaagtac 360
gacccgccgg ccgtcttcgt ctatcagacc tgcgtccccg ccatgatcgg cgacgacatc 420
gacgcggtgt gcaaggcggc cagggagaag ttcggaaagc cggtgatccc gatcaattcc 480
cccggcttcg tggggccgaa gaatctcggc aacaagctcg ccggcgaggc gctcctcgac 540
catgtgatcg gcaccgagga gcccgattac acgacggcct acgacatcaa catcatcggc 600
gaatacaatc tctccggcga gttgtggcag gtgaagccgc tgctggacga gctgggcatc 660
cgcatcctcg cctgcatctc cggcgacggg aagtacaagg atgtggcgtc ctcccaccgc 720
gccaaggcgg cgatgatggt gtgctccaag gccatgatca acgtggcccg caagatggag 780
gagcgctacg acatcccctt cttcgaaggc tccttctacg gcatcgagga tagctccgat 840
tccctgcgcg agattgcgcg catgctcatc gagaagggcg ccgatccgga gctgatggac 900
cgcaccgagg cgctgattga gcgggaagag aagaaggcgt gggacgccat cgccgcctac 960
aagccccgct tcaaggacaa gaaggtgctg ctcatcaccg gcggcgtgaa atcctggtcg 1020
gtggtggcag cgctccagga agccggcctc gaactggtgg gcacctcggt gaagaagtcc 1080
accaaggagg acaaggagcg catcaaggaa ctgatgggcc aggacgccca catgatcgac 1140
gacatgacgc cccgcgaaat gtacaagatg ctgaaggacg ccaaggcgga catcatgctc 1200
tcgggcgggc gctcgcaatt catcgcgctc aaggccgcca tgccctggct cgacatcaac 1260
caggagcgcc accacgccta tatgggctat gtgggcatgg tgaagctggt cgaggagatc 1320
gacaaggcgc tctacaatcc cgtgtgggaa caggtgcgca agcccgcccc gtgggaaaat 1380
ccggaagaca cctggcaggc ccgtgcgctc gccgaaatgg aggcggaggc cgccgcgctc 1440
gccgccgatc cggtgcgcgc ggaagaggtg cgccggtcca agaagatctg caattgcaag 1500
agcgtcgacc tcggaaccat tgaggacgcc atcaaggctc acgcgctgac caccgtggag 1560
ggtgtgcgag agcacaccaa tgcctcggga ggctgcggag cctgcagcgg gcggatcgag 1620
gagatcttcg aggccgtggg cgttgtcgcc gccccgcctc ccgcggaggc cgccccgtct 1680
ccgcaggaga tcgcgcccga tccgctcgct gcggaggaaa agcgccgcgc caagaaggcc 1740
tgcggctgca aggaggtagc ggtcggcacc attgaggatg ccatccgcgc caagggtctg 1800
cgaaacatcg cggaggtgcg tgcggccacc gatgccaaca ccggctgcgg caattgccag 1860
gagcgggtgg agggcatcct cgaccgggtt ctcgccgagg cggcctcaga actccaggcg 1920
gcggaatag 1929
<210> 51
<211> 642
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 51
Met Ser Ser Leu Ser Ala Thr Ile Gln Gln Val Phe Asn Glu Pro Gly
1 5 10 15
Cys Ala Lys Asn Gln Asn Lys Ser Glu Ala Glu Lys Lys Lys Gly Cys
20 25 30
Thr Lys Gln Leu Gln Pro Gly Gly Ala Ala Gly Gly Cys Ala Phe Asp
35 40 45
Gly Ala Lys Ile Ala Leu Gln Pro Leu Thr Asp Val Ala His Leu Val
50 55 60
His Gly Pro Ile Ala Cys Glu Gly Asn Ser Trp Asp Asn Arg Gly Ala
65 70 75 80
Lys Ser Ser Gly Ser Asn Ile Trp Arg Thr Gly Phe Thr Thr Asp Ile
85 90 95
Asn Glu Thr Asp Val Val Phe Gly Gly Glu Lys Arg Leu Phe Lys Ser
100 105 110
Ile Lys Glu Ile Ile Glu Lys Tyr Asp Pro Pro Ala Val Phe Val Tyr
115 120 125
Gln Thr Cys Val Pro Ala Met Ile Gly Asp Asp Ile Asp Ala Val Cys
130 135 140
Lys Ala Ala Arg Glu Lys Phe Gly Lys Pro Val Ile Pro Ile Asn Ser
145 150 155 160
Pro Gly Phe Val Gly Pro Lys Asn Leu Gly Asn Lys Leu Ala Gly Glu
165 170 175
Ala Leu Leu Asp His Val Ile Gly Thr Glu Glu Pro Asp Tyr Thr Thr
180 185 190
Ala Tyr Asp Ile Asn Ile Ile Gly Glu Tyr Asn Leu Ser Gly Glu Leu
195 200 205
Trp Gln Val Lys Pro Leu Leu Asp Glu Leu Gly Ile Arg Ile Leu Ala
210 215 220
Cys Ile Ser Gly Asp Gly Lys Tyr Lys Asp Val Ala Ser Ser His Arg
225 230 235 240
Ala Lys Ala Ala Met Met Val Cys Ser Lys Ala Met Ile Asn Val Ala
245 250 255
Arg Lys Met Glu Glu Arg Tyr Asp Ile Pro Phe Phe Glu Gly Ser Phe
260 265 270
Tyr Gly Ile Glu Asp Ser Ser Asp Ser Leu Arg Glu Ile Ala Arg Met
275 280 285
Leu Ile Glu Lys Gly Ala Asp Pro Glu Leu Met Asp Arg Thr Glu Ala
290 295 300
Leu Ile Glu Arg Glu Glu Lys Lys Ala Trp Asp Ala Ile Ala Ala Tyr
305 310 315 320
Lys Pro Arg Phe Lys Asp Lys Lys Val Leu Leu Ile Thr Gly Gly Val
325 330 335
Lys Ser Trp Ser Val Val Ala Ala Leu Gln Glu Ala Gly Leu Glu Leu
340 345 350
Val Gly Thr Ser Val Lys Lys Ser Thr Lys Glu Asp Lys Glu Arg Ile
355 360 365
Lys Glu Leu Met Gly Gln Asp Ala His Met Ile Asp Asp Met Thr Pro
370 375 380
Arg Glu Met Tyr Lys Met Leu Lys Asp Ala Lys Ala Asp Ile Met Leu
385 390 395 400
Ser Gly Gly Arg Ser Gln Phe Ile Ala Leu Lys Ala Ala Met Pro Trp
405 410 415
Leu Asp Ile Asn Gln Glu Arg His His Ala Tyr Met Gly Tyr Val Gly
420 425 430
Met Val Lys Leu Val Glu Glu Ile Asp Lys Ala Leu Tyr Asn Pro Val
435 440 445
Trp Glu Gln Val Arg Lys Pro Ala Pro Trp Glu Asn Pro Glu Asp Thr
450 455 460
Trp Gln Ala Arg Ala Leu Ala Glu Met Glu Ala Glu Ala Ala Ala Leu
465 470 475 480
Ala Ala Asp Pro Val Arg Ala Glu Glu Val Arg Arg Ser Lys Lys Ile
485 490 495
Cys Asn Cys Lys Ser Val Asp Leu Gly Thr Ile Glu Asp Ala Ile Lys
500 505 510
Ala His Ala Leu Thr Thr Val Glu Gly Val Arg Glu His Thr Asn Ala
515 520 525
Ser Gly Gly Cys Gly Ala Cys Ser Gly Arg Ile Glu Glu Ile Phe Glu
530 535 540
Ala Val Gly Val Val Ala Ala Pro Pro Pro Ala Glu Ala Ala Pro Ser
545 550 555 560
Pro Gln Glu Ile Ala Pro Asp Pro Leu Ala Ala Glu Glu Lys Arg Arg
565 570 575
Ala Lys Lys Ala Cys Gly Cys Lys Glu Val Ala Val Gly Thr Ile Glu
580 585 590
Asp Ala Ile Arg Ala Lys Gly Leu Arg Asn Ile Ala Glu Val Arg Ala
595 600 605
Ala Thr Asp Ala Asn Thr Gly Cys Gly Asn Cys Gln Glu Arg Val Glu
610 615 620
Gly Ile Leu Asp Arg Val Leu Ala Glu Ala Ala Ser Glu Leu Gln Ala
625 630 635 640
Ala Glu
<210> 52
<211> 1503
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 52
atgagtgtcg cacagtccca gagcgtcgcc gagatcaagg cgcgcaacaa ggaactcatc 60
gaagaggtcc tcaaggtcta tcccgagaag accgccaagc gccgcgccaa gcacctgaac 120
gtccacgaag ccggcaagtc cgactgcggc gtgaagtcca acatcaagtc catcccgggc 180
gtgatgacca tccgcggttg cgcttatgcc ggctccaagg gtgtggtgtg gggtcccatc 240
aaggacatga tccacatctc ccacggcccg gtgggctgcg gccagtatag ctgggccgcc 300
cgccgcaact actatatcgg cacgaccggc atcgacacct tcgtgacgat gcagttcacc 360
tccgacttcc aggagaagga catcgtcttc ggcggcgaca agaagctcgc caagatcatg 420
gacgagatcc aggagctgtt cccgctgaac aacggcatca ccgttcagtc cgagtgcccc 480
atcggcctca tcggcgacga catcgaggcc gtctccaagc agaagtccaa ggagtatgag 540
ggcaagacca tcgtgccggt gcgctgcgag ggcttccgcg gcgtgtccca gtccctgggc 600
caccacatcg ccaacgacgc catccgcgat tgggtgttcg acaagatcgc gcccgacgcc 660
gagccgcgct ttgagccgac cccgtacgac gtcgccatca tcggcgacta caatatcggt 720
ggtgacgcct ggtcgtcccg tatcctcctg gaggagatgg gcctgcgcgt gatcgcccag 780
tggtccggcg acggttcgct cgctgagctg gaggccaccc cgaaggccaa gctcaacgtg 840
ctgcactgct accgctccat gaactacatc tcgcgccaca tggaagagaa gtacggtatc 900
ccgtggtgcg agtacaactt cttcggtcct tccaagatcg ccgagtccct gcgcaagatc 960
gccagctact tcgacgacaa gatcaaggaa ggcgcggagc gcgtcatcgc caagtatcag 1020
ccgctcatgg atgcggtgat cgcgaagtat cgtccccgcc tcgagggcaa gaccgtgatg 1080
ctgtacgtgg gcggcctgcg tccccgtcac gtcatcggcg cctacgagga cctgggcatg 1140
gaagtggtcg gcacgggcta cgagttcgcc cataacgacg actaccagcg caccgcccag 1200
cactacgtca aggatggcac catcatctat gacgacgtga ccggctacga gttcgagaag 1260
ttcgtcgaga agatccagcc ggacctggtc ggttcgggca tcaaggaaaa gtacgtcttc 1320
cagaagatgg gcgtgccgtt ccgccagatg cactcctggg actactcggg cccgtaccac 1380
ggctatgacg gcttcgcgat cttcgcgcgc gacatggaca tggccatcaa cagccccgtg 1440
tggaagatga cccaggctcc gtggaagagc gtccccaagc cgacgatgct cgcggctgaa 1500
tga 1503
<210> 53
<211> 500
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 53
Met Ser Val Ala Gln Ser Gln Ser Val Ala Glu Ile Lys Ala Arg Asn
1 5 10 15
Lys Glu Leu Ile Glu Glu Val Leu Lys Val Tyr Pro Glu Lys Thr Ala
20 25 30
Lys Arg Arg Ala Lys His Leu Asn Val His Glu Ala Gly Lys Ser Asp
35 40 45
Cys Gly Val Lys Ser Asn Ile Lys Ser Ile Pro Gly Val Met Thr Ile
50 55 60
Arg Gly Cys Ala Tyr Ala Gly Ser Lys Gly Val Val Trp Gly Pro Ile
65 70 75 80
Lys Asp Met Ile His Ile Ser His Gly Pro Val Gly Cys Gly Gln Tyr
85 90 95
Ser Trp Ala Ala Arg Arg Asn Tyr Tyr Ile Gly Thr Thr Gly Ile Asp
100 105 110
Thr Phe Val Thr Met Gln Phe Thr Ser Asp Phe Gln Glu Lys Asp Ile
115 120 125
Val Phe Gly Gly Asp Lys Lys Leu Ala Lys Ile Met Asp Glu Ile Gln
130 135 140
Glu Leu Phe Pro Leu Asn Asn Gly Ile Thr Val Gln Ser Glu Cys Pro
145 150 155 160
Ile Gly Leu Ile Gly Asp Asp Ile Glu Ala Val Ser Lys Gln Lys Ser
165 170 175
Lys Glu Tyr Glu Gly Lys Thr Ile Val Pro Val Arg Cys Glu Gly Phe
180 185 190
Arg Gly Val Ser Gln Ser Leu Gly His His Ile Ala Asn Asp Ala Ile
195 200 205
Arg Asp Trp Val Phe Asp Lys Ile Ala Pro Asp Ala Glu Pro Arg Phe
210 215 220
Glu Pro Thr Pro Tyr Asp Val Ala Ile Ile Gly Asp Tyr Asn Ile Gly
225 230 235 240
Gly Asp Ala Trp Ser Ser Arg Ile Leu Leu Glu Glu Met Gly Leu Arg
245 250 255
Val Ile Ala Gln Trp Ser Gly Asp Gly Ser Leu Ala Glu Leu Glu Ala
260 265 270
Thr Pro Lys Ala Lys Leu Asn Val Leu His Cys Tyr Arg Ser Met Asn
275 280 285
Tyr Ile Ser Arg His Met Glu Glu Lys Tyr Gly Ile Pro Trp Cys Glu
290 295 300
Tyr Asn Phe Phe Gly Pro Ser Lys Ile Ala Glu Ser Leu Arg Lys Ile
305 310 315 320
Ala Ser Tyr Phe Asp Asp Lys Ile Lys Glu Gly Ala Glu Arg Val Ile
325 330 335
Ala Lys Tyr Gln Pro Leu Met Asp Ala Val Ile Ala Lys Tyr Arg Pro
340 345 350
Arg Leu Glu Gly Lys Thr Val Met Leu Tyr Val Gly Gly Leu Arg Pro
355 360 365
Arg His Val Ile Gly Ala Tyr Glu Asp Leu Gly Met Glu Val Val Gly
370 375 380
Thr Gly Tyr Glu Phe Ala His Asn Asp Asp Tyr Gln Arg Thr Ala Gln
385 390 395 400
His Tyr Val Lys Asp Gly Thr Ile Ile Tyr Asp Asp Val Thr Gly Tyr
405 410 415
Glu Phe Glu Lys Phe Val Glu Lys Ile Gln Pro Asp Leu Val Gly Ser
420 425 430
Gly Ile Lys Glu Lys Tyr Val Phe Gln Lys Met Gly Val Pro Phe Arg
435 440 445
Gln Met His Ser Trp Asp Tyr Ser Gly Pro Tyr His Gly Tyr Asp Gly
450 455 460
Phe Ala Ile Phe Ala Arg Asp Met Asp Met Ala Ile Asn Ser Pro Val
465 470 475 480
Trp Lys Met Thr Gln Ala Pro Trp Lys Ser Val Pro Lys Pro Thr Met
485 490 495
Leu Ala Ala Glu
500
<210> 54
<211> 1377
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 54
atggccaccg tttccgtctc caagaaggcc tgcgcggtca accccctcaa gatgagccag 60
ccggtgggcg gcgcgctcgc cttcatgggc gtgcgcaagg ccatgccgct gctgcacggc 120
tcgcagggct gcacctcctt cggcctggtg ctgttcgtgc gccacttcaa ggaagccatc 180
cccatgcaga ccaccgccat gagcgaggtg gcgacggttc tgggcggcct tgagaatgtg 240
gagcaggcca ttctcaacat ctacaatcgc accaagccgg agatcatcgg catctgctcc 300
accggcgtca ccgagaccaa gggcgatgat gtcgacggct acatcaagct gatccgggac 360
aagtatcccc agctggccga cttcccgctg gtctatgtct ccacccccga tttcaaggac 420
gccttccagg acggttggga gaagaccgtg gcgaagatgg tggaggcgct ggtgaagccc 480
gccgccgaca agcagaagga caagacccgc gtcaacgtcc tgcccggctg ccacctcacg 540
cccggcgatc tggatgagat gcggaccatc ttcgaggatt tcgggctcac accctatttc 600
ctgccggatc tggccggctc gctggatggg catatccccg aggacttctc gcccaccacc 660
atcggcggca tcggcatcga tgagatcgcc accatgggcg aggcggccca caccatctgc 720
atcggcgcgc agatgcgccg ggcgggcgag gccatggaga agaagaccgg cattcccttc 780
aagctgttcg agcgcctgtg cggcctggag gcgaacgacg ccttcatcat gcacctgtcg 840
cagatctccg gccggccggt gccggtgaag tatcgccggc agcggggcca gctggtggat 900
gccatgctgg acggccactt ccatctgggc ggtcgcaagg tggccatggg ggcggagccg 960
gacctgctct acgacgtggg ctccttcctg cacgagatgg gcgcccacat cctttccgcg 1020
gtcaccacca cccagtcgcc ggtgctggcg cgcctgcctg ccgaggaggt gcttatcggc 1080
gacctggagg atctggagac ccaggcgaag gcgcgcggat gcgatctcct gctcacccat 1140
tcccatgggc gccaggcggc ggagcgcctc cacatcccct tctaccggat cggcattccc 1200
atgtttgacc ggctgggggc ggggcatctg ttgtcggtgg gctatcgcgg cacccgcgac 1260
ctcatcttcc atctcgccaa ccttgtgatc gccgaccacg aggaaaatca cgagccgacg 1320
cccgacacct gggccaccgg ccatggcgag catgccgccg cccccacttc ccattga 1377
<210> 55
<211> 458
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 55
Met Ala Thr Val Ser Val Ser Lys Lys Ala Cys Ala Val Asn Pro Leu
1 5 10 15
Lys Met Ser Gln Pro Val Gly Gly Ala Leu Ala Phe Met Gly Val Arg
20 25 30
Lys Ala Met Pro Leu Leu His Gly Ser Gln Gly Cys Thr Ser Phe Gly
35 40 45
Leu Val Leu Phe Val Arg His Phe Lys Glu Ala Ile Pro Met Gln Thr
50 55 60
Thr Ala Met Ser Glu Val Ala Thr Val Leu Gly Gly Leu Glu Asn Val
65 70 75 80
Glu Gln Ala Ile Leu Asn Ile Tyr Asn Arg Thr Lys Pro Glu Ile Ile
85 90 95
Gly Ile Cys Ser Thr Gly Val Thr Glu Thr Lys Gly Asp Asp Val Asp
100 105 110
Gly Tyr Ile Lys Leu Ile Arg Asp Lys Tyr Pro Gln Leu Ala Asp Phe
115 120 125
Pro Leu Val Tyr Val Ser Thr Pro Asp Phe Lys Asp Ala Phe Gln Asp
130 135 140
Gly Trp Glu Lys Thr Val Ala Lys Met Val Glu Ala Leu Val Lys Pro
145 150 155 160
Ala Ala Asp Lys Gln Lys Asp Lys Thr Arg Val Asn Val Leu Pro Gly
165 170 175
Cys His Leu Thr Pro Gly Asp Leu Asp Glu Met Arg Thr Ile Phe Glu
180 185 190
Asp Phe Gly Leu Thr Pro Tyr Phe Leu Pro Asp Leu Ala Gly Ser Leu
195 200 205
Asp Gly His Ile Pro Glu Asp Phe Ser Pro Thr Thr Ile Gly Gly Ile
210 215 220
Gly Ile Asp Glu Ile Ala Thr Met Gly Glu Ala Ala His Thr Ile Cys
225 230 235 240
Ile Gly Ala Gln Met Arg Arg Ala Gly Glu Ala Met Glu Lys Lys Thr
245 250 255
Gly Ile Pro Phe Lys Leu Phe Glu Arg Leu Cys Gly Leu Glu Ala Asn
260 265 270
Asp Ala Phe Ile Met His Leu Ser Gln Ile Ser Gly Arg Pro Val Pro
275 280 285
Val Lys Tyr Arg Arg Gln Arg Gly Gln Leu Val Asp Ala Met Leu Asp
290 295 300
Gly His Phe His Leu Gly Gly Arg Lys Val Ala Met Gly Ala Glu Pro
305 310 315 320
Asp Leu Leu Tyr Asp Val Gly Ser Phe Leu His Glu Met Gly Ala His
325 330 335
Ile Leu Ser Ala Val Thr Thr Thr Gln Ser Pro Val Leu Ala Arg Leu
340 345 350
Pro Ala Glu Glu Val Leu Ile Gly Asp Leu Glu Asp Leu Glu Thr Gln
355 360 365
Ala Lys Ala Arg Gly Cys Asp Leu Leu Leu Thr His Ser His Gly Arg
370 375 380
Gln Ala Ala Glu Arg Leu His Ile Pro Phe Tyr Arg Ile Gly Ile Pro
385 390 395 400
Met Phe Asp Arg Leu Gly Ala Gly His Leu Leu Ser Val Gly Tyr Arg
405 410 415
Gly Thr Arg Asp Leu Ile Phe His Leu Ala Asn Leu Val Ile Ala Asp
420 425 430
His Glu Glu Asn His Glu Pro Thr Pro Asp Thr Trp Ala Thr Gly His
435 440 445
Gly Glu His Ala Ala Ala Pro Thr Ser His
450 455
<210> 56
<211> 1560
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 56
atgccacaaa atgctgacaa tgtgctcgat cacttcgagc tcttccgtgg tcccgaatac 60
cagcagatgc tggccaataa gaaaaagatg ttcgagaacc cccgcgatcc ggccgaagtc 120
gagcgcgtgc gggaatgggc gaagactcct gaatacaagg agctgaactt cgcccgcgag 180
gcgctcaccg tgaatccggc caaggcttgt cagccgctgg gcgcggtgtt cgtcgccgtc 240
ggcttcgaga gcacgatccc cttcgtgcac ggctcgcagg gttgcgtcgc gtattaccgc 300
tcgcacctct cccgccactt caaggagccg tcctcctgcg tctcctcgtc catgaccgag 360
gatgcggcgg tgttcggcgg cctcaacaac atgattgacg gcctcgccaa cacctacaac 420
atgtacaagc cgaagatgat cgccgtctcc accacctgca tggcggaagt catcggcgac 480
gatctgaacg ccttcatcaa gaccgcgaag gaaaagggct cggttccggc cgaatacgac 540
gtgcccttcg cccacacccc ggcgttcgtc ggcagccatg tcaccggcta cgacaatgcg 600
ctcaagggca tcctcgagca cttctgggac ggcaaggccg gcaccgcgcc gaagctggag 660
cgcgttccca acgagaagat caacttcatc ggcggcttcg acggctacac cgtcggcaac 720
actcgcgaag tgaagcgcat cttcgaggcg ttcggcgccg attacaccat cctcgccgac 780
aattccgaag tgttcgacac cccgaccgac ggcgagttcc gcatgtatga cggcggcacg 840
accctggagg acgcggcgaa cgcggtgcac gccaaggcca ccatctccat gcaggaatac 900
tgcacggaga agaccctgcc catgatcgcc ggtcatggcc aggacgtggt cgccctcaac 960
caccccgtgg gcgtgggcgg caccgacaag ttcctcatgg agatcgcccg cctcaccggc 1020
aaggagatcc ccgaggagct gacccgcgag cgcggccgtc tcgtggacgc tatcgcggac 1080
tcttccgcgc acatccacgg caagaagttc gccatctacg gcgatccgga tctgtgcctg 1140
ggcctcgccg cgttcctgct ggagctgggc gccgagccga cccatgtgct ggccaccaac 1200
ggcaccaaga agtgggccga gaaggttcag gaactgttcg actcttcgcc gttcggcgcc 1260
aactgcaagg tctatcccgg caaggacctg tggcacatgc gctcgctcct gttcgtggag 1320
ccggtggatt tcatcatcgg caacacctac ggcaagtatc tcgagcgcga cacgggcacc 1380
ccgctgatcc gtatcggctt cccggtgttc gaccgtcacc accaccaccg ccgtccggtg 1440
tggggctatc agggcggcat gaacgtcctg atcacgatcc tcgacaagat ctttgacgag 1500
atcgaccgca acaccaacgt gccggccaag accgactact cgttcgacat cattcgttga 1560
<210> 57
<211> 519
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 57
Met Pro Gln Asn Ala Asp Asn Val Leu Asp His Phe Glu Leu Phe Arg
1 5 10 15
Gly Pro Glu Tyr Gln Gln Met Leu Ala Asn Lys Lys Lys Met Phe Glu
20 25 30
Asn Pro Arg Asp Pro Ala Glu Val Glu Arg Val Arg Glu Trp Ala Lys
35 40 45
Thr Pro Glu Tyr Lys Glu Leu Asn Phe Ala Arg Glu Ala Leu Thr Val
50 55 60
Asn Pro Ala Lys Ala Cys Gln Pro Leu Gly Ala Val Phe Val Ala Val
65 70 75 80
Gly Phe Glu Ser Thr Ile Pro Phe Val His Gly Ser Gln Gly Cys Val
85 90 95
Ala Tyr Tyr Arg Ser His Leu Ser Arg His Phe Lys Glu Pro Ser Ser
100 105 110
Cys Val Ser Ser Ser Met Thr Glu Asp Ala Ala Val Phe Gly Gly Leu
115 120 125
Asn Asn Met Ile Asp Gly Leu Ala Asn Thr Tyr Asn Met Tyr Lys Pro
130 135 140
Lys Met Ile Ala Val Ser Thr Thr Cys Met Ala Glu Val Ile Gly Asp
145 150 155 160
Asp Leu Asn Ala Phe Ile Lys Thr Ala Lys Glu Lys Gly Ser Val Pro
165 170 175
Ala Glu Tyr Asp Val Pro Phe Ala His Thr Pro Ala Phe Val Gly Ser
180 185 190
His Val Thr Gly Tyr Asp Asn Ala Leu Lys Gly Ile Leu Glu His Phe
195 200 205
Trp Asp Gly Lys Ala Gly Thr Ala Pro Lys Leu Glu Arg Val Pro Asn
210 215 220
Glu Lys Ile Asn Phe Ile Gly Gly Phe Asp Gly Tyr Thr Val Gly Asn
225 230 235 240
Thr Arg Glu Val Lys Arg Ile Phe Glu Ala Phe Gly Ala Asp Tyr Thr
245 250 255
Ile Leu Ala Asp Asn Ser Glu Val Phe Asp Thr Pro Thr Asp Gly Glu
260 265 270
Phe Arg Met Tyr Asp Gly Gly Thr Thr Leu Glu Asp Ala Ala Asn Ala
275 280 285
Val His Ala Lys Ala Thr Ile Ser Met Gln Glu Tyr Cys Thr Glu Lys
290 295 300
Thr Leu Pro Met Ile Ala Gly His Gly Gln Asp Val Val Ala Leu Asn
305 310 315 320
His Pro Val Gly Val Gly Gly Thr Asp Lys Phe Leu Met Glu Ile Ala
325 330 335
Arg Leu Thr Gly Lys Glu Ile Pro Glu Glu Leu Thr Arg Glu Arg Gly
340 345 350
Arg Leu Val Asp Ala Ile Ala Asp Ser Ser Ala His Ile His Gly Lys
355 360 365
Lys Phe Ala Ile Tyr Gly Asp Pro Asp Leu Cys Leu Gly Leu Ala Ala
370 375 380
Phe Leu Leu Glu Leu Gly Ala Glu Pro Thr His Val Leu Ala Thr Asn
385 390 395 400
Gly Thr Lys Lys Trp Ala Glu Lys Val Gln Glu Leu Phe Asp Ser Ser
405 410 415
Pro Phe Gly Ala Asn Cys Lys Val Tyr Pro Gly Lys Asp Leu Trp His
420 425 430
Met Arg Ser Leu Leu Phe Val Glu Pro Val Asp Phe Ile Ile Gly Asn
435 440 445
Thr Tyr Gly Lys Tyr Leu Glu Arg Asp Thr Gly Thr Pro Leu Ile Arg
450 455 460
Ile Gly Phe Pro Val Phe Asp Arg His His His His Arg Arg Pro Val
465 470 475 480
Trp Gly Tyr Gln Gly Gly Met Asn Val Leu Ile Thr Ile Leu Asp Lys
485 490 495
Ile Phe Asp Glu Ile Asp Arg Asn Thr Asn Val Pro Ala Lys Thr Asp
500 505 510
Tyr Ser Phe Asp Ile Ile Arg
515
<210> 58
<211> 621
<212> ДНК
<213> Неизвестный
<220>
<223> Новый штамм
<400> 58
gtggagtccg gtggtcctga gccgggcgtg ggctgcgccg gccgcggcgt gatcacctcc 60
atcaacttcc tggaggagaa cggcgcctac gaggacatcg actatgtgtc ctacgacgtg 120
ctgggcgacg tggtgtgcgg cggcttcgcc atgcccatcc gcgagaacaa ggcgcaggaa 180
atctacatcg tgatgtccgg cgagatgatg gccatgtatg cggccaacaa catctccaag 240
ggcatcctga agtatgccaa ttccggcggc gtgcgcctgg gcgggctggt ctgcaacgag 300
cgccagaccg acaaggagct ggagctggcg gaggctctgg cgaagaagct cggcaccgag 360
ctgatctact tcgtgccgcg cgacaacatc gtgcagcatg ccgagctgcg ccgcatgaca 420
gtgatcgagt atgcgcccga ttccgcccag gcccagcact accggaacct ggccgagaag 480
gtgcacgcca acaagggcaa cggcatcatc ccgaccccga tcaccatgga cgagctggaa 540
gacatgctca tggagcacgg catcatgaag gccgtggacg agagccagat cggcaagacc 600
gccgccgagc tcgccgtctg a 621
<210> 59
<211> 206
<212> Белок
<213> Неизвестный
<220>
<223> Новый штамм
<400> 59
Met Glu Ser Gly Gly Pro Glu Pro Gly Val Gly Cys Ala Gly Arg Gly
1 5 10 15
Val Ile Thr Ser Ile Asn Phe Leu Glu Glu Asn Gly Ala Tyr Glu Asp
20 25 30
Ile Asp Tyr Val Ser Tyr Asp Val Leu Gly Asp Val Val Cys Gly Gly
35 40 45
Phe Ala Met Pro Ile Arg Glu Asn Lys Ala Gln Glu Ile Tyr Ile Val
50 55 60
Met Ser Gly Glu Met Met Ala Met Tyr Ala Ala Asn Asn Ile Ser Lys
65 70 75 80
Gly Ile Leu Lys Tyr Ala Asn Ser Gly Gly Val Arg Leu Gly Gly Leu
85 90 95
Val Cys Asn Glu Arg Gln Thr Asp Lys Glu Leu Glu Leu Ala Glu Ala
100 105 110
Leu Ala Lys Lys Leu Gly Thr Glu Leu Ile Tyr Phe Val Pro Arg Asp
115 120 125
Asn Ile Val Gln His Ala Glu Leu Arg Arg Met Thr Val Ile Glu Tyr
130 135 140
Ala Pro Asp Ser Ala Gln Ala Gln His Tyr Arg Asn Leu Ala Glu Lys
145 150 155 160
Val His Ala Asn Lys Gly Asn Gly Ile Ile Pro Thr Pro Ile Thr Met
165 170 175
Asp Glu Leu Glu Asp Met Leu Met Glu His Gly Ile Met Lys Ala Val
180 185 190
Asp Glu Ser Gln Ile Gly Lys Thr Ala Ala Glu Leu Ala Val
195 200 205
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕНЫ И АНТИГЕННЫЕ КОМПОЗИЦИИ | 2013 |
|
RU2727476C2 |
| ВЫДЕЛЕНИЕ НОВОГО ПЕСТИВИРУСА, ВЫЗЫВАЮЩЕГО ВРОЖДЕННЫЙ ТРЕМОР А | 2017 |
|
RU2765658C2 |
| ГОМОЛОГ ГЛИЦЕРОЛ-3-ФОСФАТ-АЦИЛТРАНСФЕРАЗЫ И ЕГО ИСПОЛЬЗОВАНИЕ | 2011 |
|
RU2545375C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ АРЕНАВИРУСА, А ТАКЖЕ МУТАНТОВ АРЕНАВИРУСА С ПРОТИВООПУХОЛЕВЫМИ СВОЙСТВАМИ | 2019 |
|
RU2812046C2 |
| АТТЕНУИРОВАННЫЙ ШТАММ ВИРУСА РЕПРОДУКТИВНО-РЕСПИРАТОРНОГО СИНДРОМА СВИНЕЙ (РРСС) И ЕГО ВОЗМОЖНОЕ ПРИМЕНЕНИЕ В СРЕДСТВАХ ДЛЯ ИММУНИЗАЦИИ | 2015 |
|
RU2723040C2 |
| АBC-ТРАНСПОРТЕРЫ ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОИЗВОДСТВА РЕБАУДИОЗИДОВ | 2020 |
|
RU2795855C2 |
| ПРОМОТОРЫ ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ В ПОКСВИРУСАХ | 2016 |
|
RU2753884C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| ОПТИМИЗИРОВАННЫЙ ПОЛИПЕПТИД ДЛЯ СУБЪЕДИНИЧНОЙ ВАКЦИНЫ ПРОТИВ РЕОВИРУСА ПТИЦ | 2017 |
|
RU2742250C2 |
| Новый вариант О-сукцинилгомосеринтрансферазы и способ получения О-сукцинилгомосерина с использованием этого варианта | 2018 |
|
RU2747493C1 |
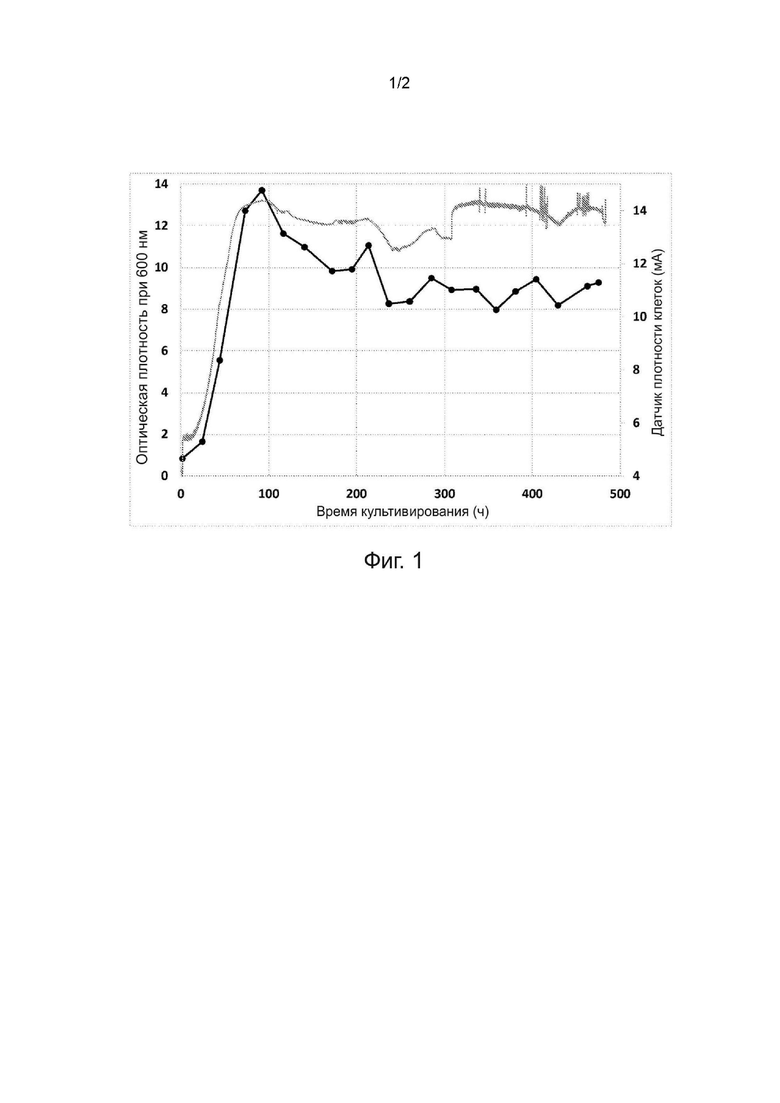
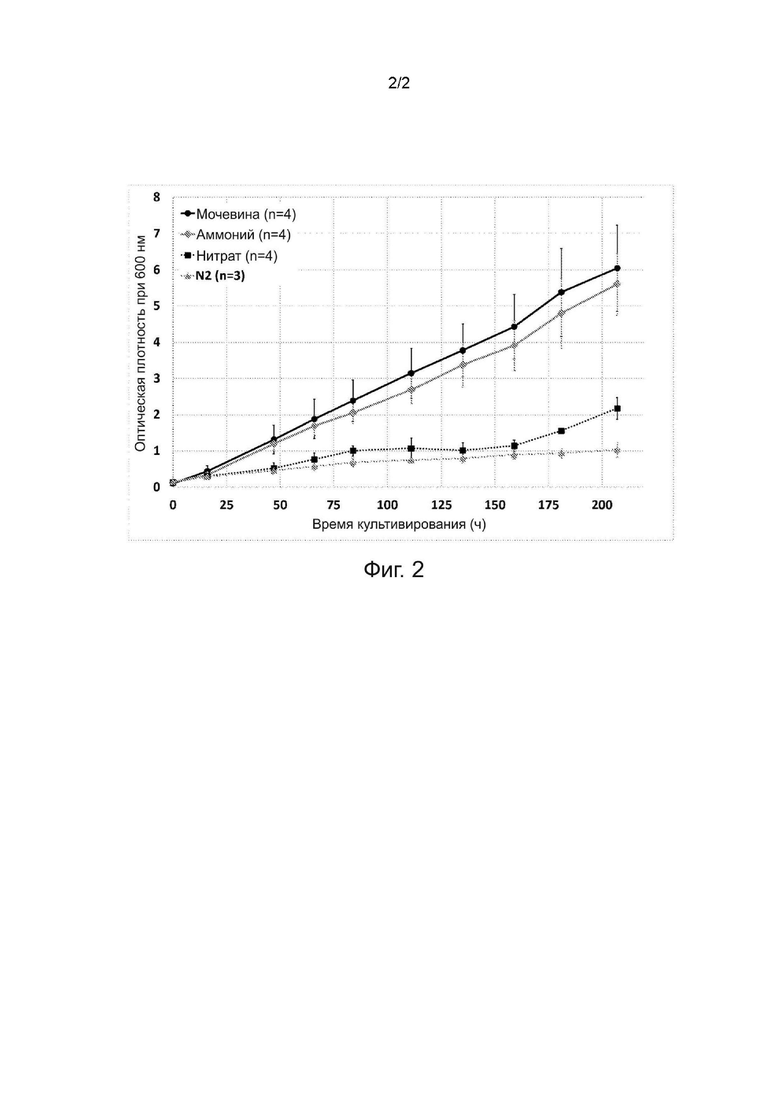
Изобретение относится к биотехнологии. Предложен выделенный бактериальный штамм рода Xanthobacter, депонированный под номером VTT-E-193585 и пригодный для применения в кормах и пищевых продуктах. Также предложена культура, содержащая указанный штамм и пригодная для применения в кормах и пищевых продуктах. Также предложены способ получения биомассы указанного штамма и способ получения белка, предусматривающий получение биомассы, включающее в себя культивирование указанного штамма. Изобретение обеспечивает высокое содержания белка в клетках указанного штамма. 4 н. и 12 з.п. ф-лы, 2 ил., 5 табл., 4 пр.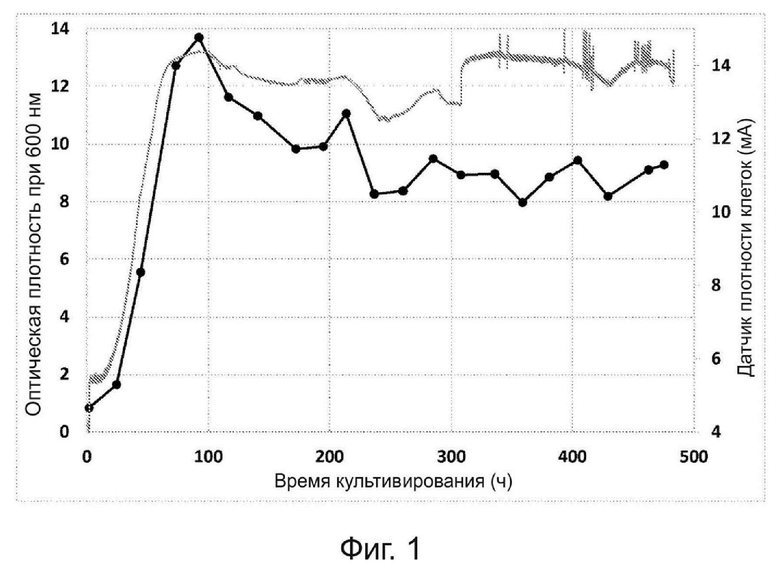
1. Выделенный бактериальный штамм рода Xanthobacter, депонированный под номером VTT-E-193585, пригодный для применения в кормах и пищевых продуктах.
2. Культура, пригодная для применения в кормах и пищевых продуктах, содержащая бактериальный штамм по п. 1 и жидкую среду, содержащую соли аммония, фосфаты, сульфаты, соли калия, натрия, ванадия, железа, магния, кальция, молибдена, марганца, бора, цинка, кобальта, селена, йода, меди или никеля.
3. Способ получения биомассы, включающий в себя культивирование бактериального штамма по п. 1.
4. Способ по п. 3, включающий в себя культивирование бактериального штамма в непрерывной культуре с водородом в качестве источника энергии и источником неорганического углерода, где источник неорганического углерода включает в себя диоксид углерода.
5. Способ по п. 4, отличающийся тем, что содержание растворенного кислорода в культуре поддерживают на уровне от 5 до 10%.
6. Способ по п. 4 или 5, отличающийся тем, что в качестве источника азота используют аммиак, мочевину, нитрат и/или газообразный азот.
7. Способ по п. 3, отличающийся тем, что минералы содержат менее 1 г/л хлоридов, например, менее 0,25 г/л хлористых солей, или менее 0,1 г/л хлоридов, предпочтительно не содержат хлоридов.
8. Способ по п. 7, отличающийся тем, что культуральная среда не содержит витаминов.
9. Способ по п. 3, отличающийся тем, что значение рН в культуре поддерживают в пределах от 5,5 до 8,0, или от 6,5 до 7,0, такое как 6,8.
10. Способ по п. 3, отличающийся тем, что указанную культуру выращивают при температуре от 25°С до 40°С, или от 28°С до 32°С, такой как 30°С.
11. Способ по п. 3, отличающийся тем, что бактериальный штамм имеет скорость роста 0,04-0,12 ч-1.
12. Способ по п. 3, отличающийся тем, что бактериальный штамм содержит 16S рибосомальную РНК, указанную в SEQ ID NO:1.
13. Способ по п. 3, отличающийся тем, что бактериальный штамм содержит:
i) ген, кодирующий большую цепь рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (rubisco), имеющую последовательность, указанную в SEQ ID NO:3, и
ii) ген, кодирующий малую цепь рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (rubisco), имеющую последовательность, указанную в SEQ ID NO:5,
и
iii) ген, кодирующий альфа-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:7,
и
iv) ген, кодирующий бета-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:9,
и
v) ген, кодирующий гамма-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:11, и
vi) ген, кодирующий дельта-субъединицу NAD+-восстанавливающей гидрогеназы HoxS, имеющую последовательность, указанную в SEQ ID NO:13
и
vii) ген, кодирующий большую субъединицу гидрогеназы NiFeSe, имеющую последовательность, указанную в SEQ ID NO:15, и
viii) ген, кодирующий малую субъединицу гидрогеназы NiFeSe, имеющую последовательность, указанную в SEQ ID NO:17,
и
ix) ген, кодирующий гамма-цепь АТФ-синтазы atpG_1, имеющую последовательность, указанную в SEQ ID NO:19,
и
x) ген, кодирующий альфа-субъединицу АТФ-синтазы atpA_1, имеющую последовательность, указанную в SEQ ID NO:21,
и
xi) ген, кодирующий субъединицу b АТФ-синтазы atpF_1, имеющую последовательность, указанную в SEQ ID NO:23,
и
xii) ген, кодирующий субъединицу c специфичной к ионам натрия АТФ-синтазы atpE_1, имеющую последовательность, указанную в SEQ ID NO:25, и
xiii) ген, кодирующий субъединицу aАТФ-синтазыatpB_1, имеющую последовательность, указанную в SEQ ID NO:27,
и
xiv) ген, кодирующий эпсилон-цепь АТФ-синтазы atpC_1, имеющую последовательность, указанную в SEQ ID NO: 29,
и
xv) ген, кодирующий бета-субъединицу АТФ-синтазыatpD_1, имеющую последовательность, указанную в SEQ ID NO:31,
или последовательность, имеющую более 84% идентичности по последовательности с последовательностью, представленной в SEQ ID NO:31, и
xvi) ген, кодирующий бета-субъединицу АТФ-синтазы atpD_2, имеющую последовательность, указанную в SEQ ID NO:33,
и
xvii) ген, кодирующий гамма-цепь АТФ-синтазы atpG_2, имеющую последовательность, указанную в SEQ ID NO:35,
и
xviii) ген, кодирующий субъединицу альфа АТФ-синтазы atpA_2, имеющую последовательность, указанную в SEQ ID NO: 37,
и
xix) ген, кодирующий дельта-субъединицу АТФ-синтазы atpH, имеющую последовательность, указанную в SEQ ID NO:39,
и
xx) ген, кодирующий субъединицу b АТФ-синтазы atpF_2, имеющую последовательность, указанную в SEQ ID NO:41,
и
xxi) ген, кодирующий субъединицу b' АТФ-синтазы atpG_3, имеющую последовательность, указанную в SEQ ID NO:43,
и
xxii) ген, кодирующий субъединицу cАТФ-синтазы atpE_2, имеющую последовательность, указанную в SEQ ID NO:45,
и
xxiii) ген, кодирующий субъединицуaАТФ-синтазы atpB_2, имеющую последовательность, указанную в SEQ ID NO:47,
и
xxiv) ген, кодирующий белок I АТФ-синтазы atpI, имеющий последовательность, указанную в SEQ ID NO: 49,
и
xxv) ген, кодирующий альфа-цепь молибден-железо белка нитрогеназыnifD_1, имеющую последовательность, указанную в SEQ ID NO:51,
и
xxvi) ген, кодирующий альфа-цепь молибден-железо белка нитрогеназы nifD_2, имеющую последовательность, указанную в SEQ ID NO:53,
и
xxvii) ген, кодирующий бета-цепь молибден-железо белка нитрогеназы nifK_1, имеющую последовательность, указанную в SEQ ID NO:55,
и
xxviii) ген, кодирующий бета-цепь молибден-железо белка нитрогеназы nifK_2, имеющую последовательность, указанную в SEQ ID NO: 57,
и
xxix) ген, кодирующий железо белок нитрогеназы nifH, имеющий последовательность, указанную в SEQ ID NO: 59.
14. Способ по п. 3, включающий в себя дополнительный этап сбора биомассы, полученной во время культивирования, необязательно включающий в себя дополнительный этап сушки биомассы.
15. Способ получения белка, предусматривающий получение биомассы, включающее в себя культивирование бактериального штамма по п. 1, сбор биомассы, полученной во время культивирования, и дополнительный этап выделения белка из указанной биомассы.
16. Способ по п. 14 или 15, включающий в себя дополнительный этап получения пищевого продукта или кормового продукта из указанной биомассы.
| OHMIYA K | |||
| ET AL | |||
| Application of microbial genes to recalcitrant biomass utilization and environmental conservation | |||
| J Biosci Bioeng | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| JIANYU ET AL | |||
| The energy efficiency of carbon dioxide fixation by a | |||
