Область техники
Изобретение относится к областям молекулярных биотехнологии и медицины и касается двух линий животных, гомозиготных по нуль-аллелю гена P2rx3, с полным отсутствием экспрессии пуринергического рецептора P2X3, а также способа их получения. Изобретение может быть использовано для тестирования возможных веществ-кандидатов на роль блокатора P2X3.
Изобретение создано при финансовой поддержке Министерства Науки и Высшего образования Российской Федерации в рамках Соглашения № 075-15-2019-1661 от 31.10.2019 на базе Центра высокоточного редактирования и генетических технологий для биомедицины (ЦВРГТБ) ИБГ РАН.
Уровень техники
Одним из основных пуринергических рецепторов, вовлеченных в процесс патогенеза хронических болевых ощущений, является P2X3 (Bernier L.-P., Ase A. R., Séguéla P. P2X receptor channels in chronic pain pathways. British Journal of Pharmacology (2018) 175 2219-2230). Гомомеры P2X3 и гетеромеры P2X2/3 преимущественно локализованы на периферических первичных сенсорных афферентах, в немиелинизированных С-волокнах малого диаметра. Особое значение при хронических болевых заболеваниях имеет экспрессия рецепторов Р2Х3 в центральных окончаниях задних корешковых ганглиев (ДРГ) (или тройничных ганглиев), оканчивающихся во внутренней пластинке II (желатинозной субстанции) задних рогов спинного мозга. Поскольку рецепторы P2X3 могут усиливать ноцицепцию за счет прямой сенсибилизации болевых волокон, нарушение регуляции этого пуринергического пути связано с патологической болью. Таким образом, разработка обезболивающих препаратов, воздействующих непосредственно на P2X3, является одной из важных задач современной фармакологии (Krajewski J. L. P2X3-Containing Receptors as Targets for the Treatment of Chronic Pain. Neurotherapeutics (2020) 17:826-838). В исследованиях возможных веществ-кандидатов на роль блокатора P2X3 незаменимы лабораторные животные с инактивированным геном P2rx3, поскольку эксперименты с их использованием позволяют однозначно установить, является ли молекула P2X3 мишенью исследуемого вещества.
Мыши с инактивированным геном P2rx3 описаны в литературе, однако они получены с помощью инъекции генетически модифицированных эмбриональных стволовых клеток в полость бластоцисты (Cockayne DA, Dunn PM, Zhong Y, et al. P2X2 knockout mice and P2X2/P2X3 double knockout mice reveal a role for the P2X2 receptor subunit in mediating multiple sensory effects of ATP. J Physiol. 2005; 567(Pt 2):621-639) и недоступны для заказа в России. Таким образом, создание мышей, несущих в геноме нуль-аллель (нефункциональную копию) гена P2rx3, является актуальной задачей для развития современной фармакологии в России.
Краткое описание изобретения
Для получения нокаута по гену P2rx3 мы использовали систему CRISPR/Cas9, состоящую из белка spCas9 и гидовой РНК, ПАМ-сайт которой расположен в первом экзоне гена P2rx3. Нокаут образуется благодаря микроинъекциям в пронуклеус эмбрионов системы CRISPR/Cas9 в форме плазмид, кодирующих белок spCas9 и две гидовые РНК.
Задачей заявляемого изобретения является создание линии генетически модифицированных мышей, гомозиготных по нуль-аллелю гена P2rx3.
Задача решается тем, что с использованием системы CRISPR/Cas9 получена новая линия генетически модифицированных мышей с нуль-аллелем гена P2rx3, для чего:
1. На основе вектора px330 (SEQ ID NO: 1) были получены плазмидные конструкции px330+P2rx3-sg1 и px330+P2rx3-sg2, содержащая гидовые РНК sg1 и sg2 (SEQ ID NO: 2 и SEQ ID NO: 3 соответственно) против соответствующего участка гена P2rx3 под контролем U6 промотора и ОРС гена spCas9 под контролем CAG промотора.
2. Был разработан способ получения линии мышей с нуль-аллелем гена P2rx3, характеризующийся следующими стадиями:
а) микроинъекция конструкции px330+P2rx3-sg1 и px330+P2rx3-sg2, содержащая гидовые РНК sg1 и sg2 из п. 1 в пронуклеус оплодотворенной яйцеклетки мыши гибрида F1(CBA×C57BL/6);
б) выявление жизнеспособных зигот;
в) пересадка выживших зигот из пункта (б) псевдобеременным самкам-реципиентам, имеющим копулятивную пробку после ссаживания с вазэктомированными самцами;
г) получение новорожденных мышей на 19 день после пересадки зигот из пункта (в);
д) проведение анализа наличия сдвига рамки считывания в первом экзоне гена P2rx3 через 8-14 дней после рождения мышей (г), путем амплификации целевого участка гена P2rx3 с праймерами SEQ ID NO: 4 и SEQ ID NO: 5 с последующим секвенированием фрагмента с использованием праймера SEQ ID NO: 6;
е) отбор мышей, несущих целевую мутацию;
ж) проведение скрещивания мышей из пункта (е) с мышами дикого типа для получения самок-гетерозигот или мутантных самцов, несущих целевую мутацию.
з) проведение скрещивания мышей из пункта (ж) для получения итоговой линии мышей, гомозиготных по нуль-аллелю гена P2rx3.
Технический результат изобретения: разработан способ получения генетически модифицированных мышей с нуль-аллелем гена P2rx3 и выведены соответствующие линии мышей.
Краткое описание чертежей
Фигура 1. Схема расположения промотора (5’utr) и первого экзона p2rx3, а также подобранных сгРНК и праймеров для генотипирования. Ген расположен на (-) цепи. Указаны места посадки гидовых РНК, ПАМ-сайт, праймеры для генотипирования, 1-й экзон и его кодирующая белок часть.
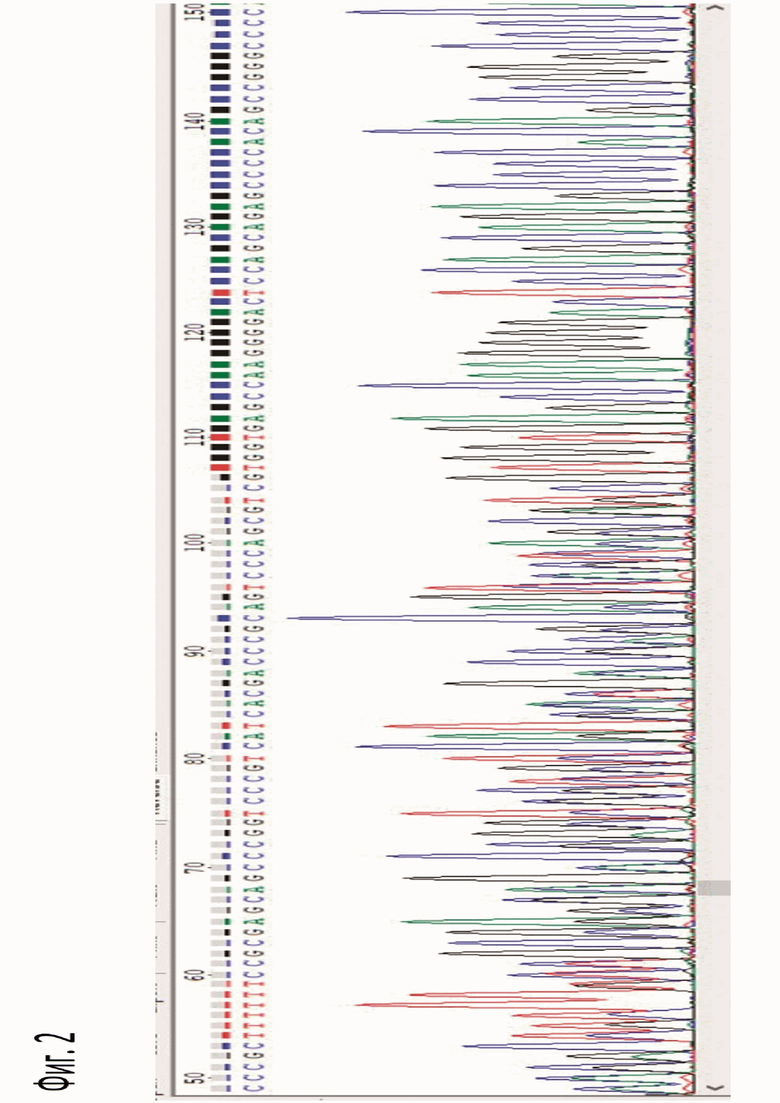
Фигура 2. Хроматограмма, полученная в результате секвенирования образцов тканей мозаичной мыши поколения F0.
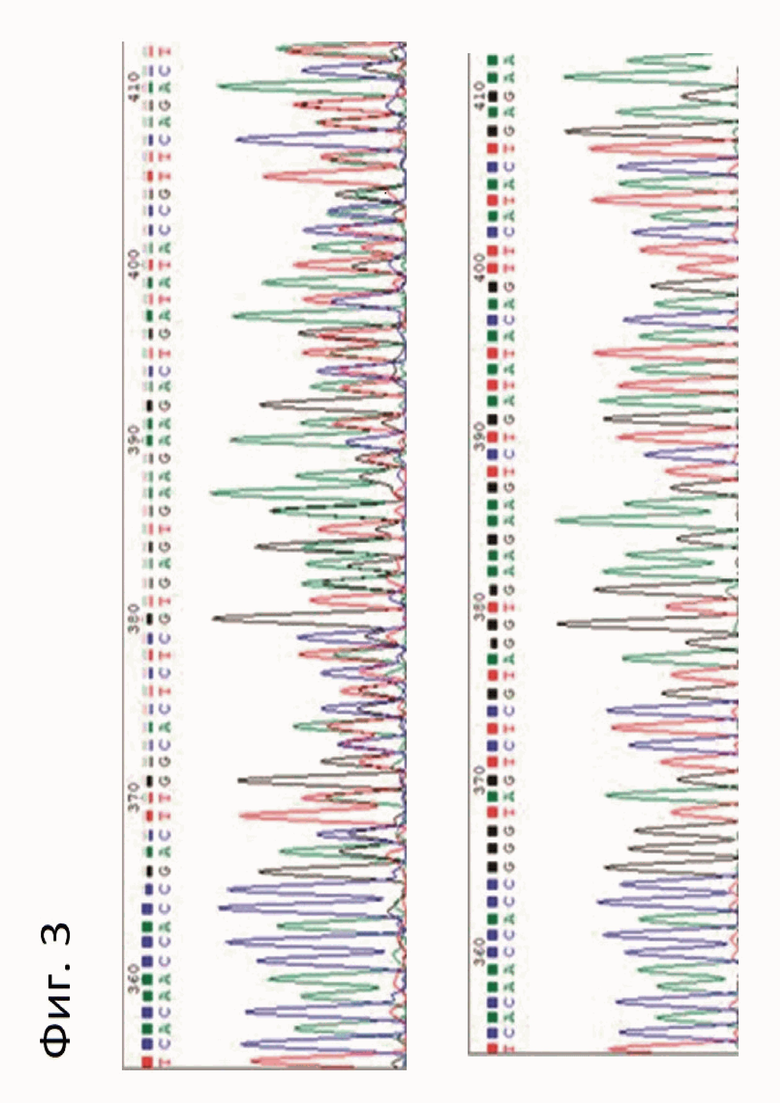
Фигура 3. Хроматограммы, полученные в результате секвенирования образцов тканей мышей F1 (верхняя панель) и F2 (нижняя панель) линии delACTT.
Фигура 4. Хроматограмма, полученные в результате секвенирования образцов тканей мышей F1 (верхняя панель) и F2 (нижняя панель) линии insT.
Фигура 5. Суммарные амплификационные кривые с образцов эмбрионов линии delACTT и эмбрионов дикого типа, а также эмбриональных фибробластов дикого типа. Синий - дикий вариант, оранжевый - пробы эмбрионов линии delACTT.
Осуществление изобретения
Способ осуществляется следующим образом:
1. Подбор гидовых РНК для внесения разреза в первый экзон гена P2rx3 с целью внесения стоп-кодона осуществлялся с использованием онлайн инструмента CRISPOR TEFOR (http://crispor.tefor.net/) с учетом последующего использования нуклеазы SpCas9. Были подобраны две разнонаправленных гидовых РНК (sg1 и sg2, последовательности SEQ ID NO: 2 и SEQ ID NO: 3, соответственно) к первому экзону гена P2rx3. В результате NHEJ-репарации после их разрезания должен образовываться сдвиг рамки считывания в первом экзоне, приводящий к изменению аминокислотной последовательности белка и преждевременному образованию стоп-кодона (фиг. 1). Выбранную сгРНК мы заклонировали в плазмидный вектор px330, предназначенный для одновременной экспрессии белка spCas9 и сгРНК. Последовательности каждой гидовой РНК были синтезированы с использованием двух олигонуклеотидов SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10.
Разрезание вектора проводили с использованием рестриктазы BbsI-HF (New England Biolabs), дефосфорилирование вектора с помощью фермента FastAP (Thermofisher Scientific), затем очистка в агарозном геле и последующее выделение с помощью набора Monarch DNA Gel Extraction Kit.
Олигонуклеотиды фосфорилировались с использованием фермента PNK kinase (Thermofisher Scientific), отжигались друг с другом и лигировались с приготовленным ранее вектором и использованием фермента T4 DNA ligase (Thermofisher Scientific). Лигазная смесь трансформировалась в компетентные клетки штамма XL1blue (Евроген). Скрининг колоний осуществлялся с использованием пцр-наборов компании Изоген и праймеров U6-forv (SEQ ID NO: 11) и реверсного праймера, использовавшегося для клонирования. По две колонии, содержащие сгРНК, наращивались в ночной культуре и выделялись с помощью набора Monarch Plasmid Miniprep Kit. Анализ полученных плазмид проводили методом секвенирования по Сэнгеру. Сиквенс полученных плазмид осуществлялся с использованием праймера U6-forv (SEQ ID NO: 11) компанией Евроген. Анализ сиквенсов проводился с использованием программного обеспечения Chromas.
Проверенную конструкцию разводили до концентрации, используемой при микроинъекциях - 1нг/мкл.
2. Получение яйцеклеток мышей. Яйцеклетки для микроинъекции получали методом индукции суперовуляции. Для этого неполовозрелым самкам гибридам F1(CBA×C57BL/6) весом 12-13 г внутрибрюшинно вводили 8 ед. гонадотропина сыворотки жеребых кобыл (ГСЖК) и через 48 час - 8 ед. хорионического гонадотропина человека (ХгЧ). После такой обработки самок ссаживали с самцами-производителями F1(CBA×C57BL/6). Факт спаривания констатировали на следующее утро по наличию копулятивной пробки.
Схема индукции суперовуляции: 12:00 - ГСЖК, через 48 часов - ХгЧ. В 17:00 этого же дня - подсадка к самцам-производителям. Отбор доноров производили на следующий день в 9:00. Световой режим в виварии был установлен с 7:00 до 19:00.
Отобранных самок-доноров умерщвляли, извлекали яйцеводы, затем вымывали яйцеклетки в среде HEPES-KSOM с добавлением гиалуронидазы. Процедуру проводили под бинокуляром (Zeiss Stemi DV4) с увеличением в 32 раза. Для вымывания яйцеклеток использовали стеклянные капилляры с внутренним диаметром примерно 100 мкм, изготовленные на пуллере Narishige PC-10 (Япония) и микрокузнице Narishige MF-900 (Япония).
3. Микроинъекции. Полученные зиготы культивировали в течение двух часов при t=37°C и 5% СО2 в капле среды KSOM под минеральным маслом (Sigma, США), затем помещали в микроинъекционную камеру. Микроинъекции проводили в среде HEPES-KSOM под микроскопом Zeiss Axiovert 200М при увеличении в 400-600 раз, используя микроманипуляторы Narishige. Для изготовления игл для микроинъекций использовали пуллер Sutter instrument Со Р-97 (США), для изготовления удерживающей пипетки использовали пуллер Narishige PC-10 и микрокузницу Narishige MF-900.
После окончания микроинъекций выжившие зиготы переносили в каплю среды KSOM под минеральное масло (Sigma) и культивировали в течение 1 часа для выявления жизнеспособных эмбрионов.
4. Получение самок-реципиентов. Самок-реципиентов яйцеклеток получали следующим образом: половозрелых самок F1(CBA×C57BL/6) весом не менее 24 г ссаживали с вазэктомированными самцами той же линии. Через 18 часов псевдобеременных реципиентов отбирали по наличию копулятивных пробок. Выжившие после микроинъекции зиготы трансплантировали в яйцеводы псевдобеременной самки. Одной псевдобеременной самке пересаживали 6-10 эмбрионов. Операцию проводили под наркозом (смесь золетила и рометара, вводился внутрибрюшинно).
5. Операция вазэктомирования. Операцию вазэктомирования проводили заранее под наркозом (смесь золетила и рометара, вводился внутрибрюшинно). Через надрез в коже и брюшной стенке вытягивали из брюшной полости семенник, придатки семенника и семявыносящий проток, после чего раскаленным пинцетом разрушали семявыносящий проток. Органы возвращали в брюшную полость, и повторяли всю процедуру на другом семявыносящем протоке. На завершающем этапе на брюшную стенку и на кожу накладывали швы с последующей антисептической обработкой операционного поля.
6. Получение новорожденных мышей. На 19 день после пересадки микроинъецированных эмбрионов реципиента умерщвляли путем цервикальной дислокации и проводили кесарево сечение, после чего выживших детенышей помещали к заранее подготовленной кормилице. Через 8-14 дней после рождения у мышат брали образец ткани, выделяли ДНК и анализировали наличие сдвига рамки считывания в целевой области генома методом секвенирования.
7. Для получения животных, гомозиготных по нокауту гена P2rx3, сначала скрещивали мышей F0 со сдвигом рамки считывания в целевой области генома с мышами дикого типа, а полученное F1 скрещивали между собой для получения животных поколения F2.
Анализ наличия сдвига рамки считывания в целевой области генома мышей производили методом секвенирования ПЦР-фрагмента. ДНК для ПЦР выделяли из тканей по стандартному протоколу. Амплификацию фрагментов ДНК проводили с помощью набора GenPak™ PCR Core (Isogene) в присутствии 10 пМ праймеров SEQ ID NO: 4 и SEQ ID NO: 5 в следующих режимах: денатурация 95°С - 3 мин; далее следовало 35 циклов: 95°С - 40 сек, 58°С - 40 сек; 72°С - 40 сек; и последний синтез 72°С - 2 мин. После прохождения реакции ПЦР-смесь очищали в 1,5% агарозном геле, полученный продукт выделяли набором Monarch DNA Gel Extraction Kit и секвенировали в компании Евроген и использованием праймера SEQ ID NO: 5. Полученные сиквенсы можно подразделить на три категории: сиквенсы дикого типа, последовательность в которых соответствует дикому типу, двоящиеся сиквенсы (фиг. 2), соответствующие мозаичным животным F0 и гетерозиготным трансгенным животным последующих поколений, и сиквенсы, на которых присутствует только мутантный вариант ДНК, соответствующие гомозиготным трансгенным животным последующих поколений.
Пример 1. Получение линии мышей с нуль-аллелем гена P2rx3, получившей название delACTT. Среди животных F0 по результатам секвенирования по Сэнгеру был выбран вариант с делецией 4х нуклеотидов в позициях 40-43, затем его скрестили с диким типом для получения генетически модифированных животных F1, полученных животных F1 скрещивали между собой для получения гомозиготных по нуль-аллелю гена P2rx3 животных (согласно п.7). Результаты секвенирования по Сэнгеру мышей поколений F1 и F2 линии delACTT представлены на фигуре 3.
Пример 2. Получение линии мышей с нуль-аллелем гена P2rx3, получившей название insT. Среди животных F0 по результатам секвенирования по Сэнгеру был выбран вариант с вставкой одного нуклеотида в позиции 40, его скрестили с диким типом для получения генетически модифированных животных F1, полученных животных F1 скрещивали между собой для получения гомозиготных по нуль-аллелю гена P2rx3 животных (согласно п.7). Результаты секвенирования по Сэнгеру мышей поколений F1 и F2 линии insT представлены на фигуре 4.
Пример 3. Тест-система для детекции отсутствия экспрессии гена P2rx3 у мышей линии delACTT на основе праймеров и флуоресцентных зондов для дискриминирования вариантов кДНК.
Исследование экспрессии P2rx3 проводилось с помощью количественного ПЦР с обратной транскрипцией с флуоресцентными зондами. Тотальная РНК была выделена из головной части эмбрионов мышей на E10-E14 линии delACTT, а также мышей дикого типа (wt) набором ExtractRNA (Евроген). Кроме того, РНК была выделена из эмбриональных (E17.5) фибробластов мыши дикого типа. Была синтезирована кДНК с использованием праймеров oligo(dT) ((SEQ ID NO: 12) ревертазой MMLV (Евроген) с 1 мкг РНК.
С кДНК была поставлена рутинная ПЦР с праймерами к началу первого экзона и перекрытию первого и второго экзонов кДНК P2rx3 (P2rx3 expr f (SEQ ID NO: 13), P2rx3 expr r (SEQ ID NO: 14). для предамплификации целевого фрагмента, поскольку проведение ПЦР в реальном времени без предамплификации давало неудовлетворительные результаты: подъем фрагмента на поздних циклах, неспецифические продукты, нестабильность фоновых линий флуоресценции. Ампликоны были использованы как матрица для дальнейшего qPCR после разведения в 200000 раз.
Для ПЦР в реальном времени использовались те же праймеры зонды ROX-delACTT и FAM-wt (SEQ ID NO: 15; SEQ ID NO: 16), комплементарные соответствующим аллелям. Реакция проводилась при температуре отжига 60.5° длительностью 20 секунд и элонгации 71° длительностью 25 секунд на полимеразе qPCR-HS (Евроген) в течение 40 циклов. Выбор температуры обусловлен повышением специфичности без снижения чувствительности метода.
Была подтверждена экспрессия мутантного P2rx3 одновременно с отсутствием экспрессии гена дикого типа в эмбрионах линии delACTT (фиг. 5).
--->
Информация о перечнях последовательностей:
Версия DTD: V1_3
Название файла: Способ получения генно-модифицированных лабораторных
животных с нуль-аллелем гена P2rx3.xml
Название программного обеспечения: WIPO Sequence
Версия программного обеспечения: 2.1.2
Дата производства: 2022-09-08
Общая информация:
Текущая заявка / Номер дела заявителя: non
Имя заявителя: Федеральное государственное бюджетное учреждение
науки Институт биологии гена Российской академии наук
Имя заявителя / Язык: ru
Имя заявителя / Имя латиницей: Institute of Gene Biology Russian
Academy of Sciences
Название изобретения: Способ получения генно-модифицированных
лабораторных животных с нуль-аллелем гена P2rx3 ( ru )
Общее количество последовательностей: 16
Последовательности:
Номер последовательности (ID): 1
Длина: 8484
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..8484
> mol_type, other DNA
> organism, synthetic construct
Остатки:
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
cgaaacaccg ggtcttcgag aagacctgtt ttagagctag aaatagcaag ttaaaataag 300
gctagtccgt tatcaacttg aaaaagtggc accgagtcgg tgcttttttg ttttagagct 360
agaaatagca agttaaaata aggctagtcc gtttttagcg cgtgcgccaa ttctgcagac 420
aaatggctct agaggtaccc gttacataac ttacggtaaa tggcccgcct ggctgaccgc 480
ccaacgaccc ccgcccattg acgtcaatag taacgccaat agggactttc cattgacgtc 540
aatgggtgga gtatttacgg taaactgccc acttggcagt acatcaagtg tatcatatgc 600
caagtacgcc ccctattgac gtcaatgacg gtaaatggcc cgcctggcat tgtgcccagt 660
acatgacctt atgggacttt cctacttggc agtacatcta cgtattagtc atcgctatta 720
ccatggtcga ggtgagcccc acgttctgct tcactctccc catctccccc ccctccccac 780
ccccaatttt gtatttattt attttttaat tattttgtgc agcgatgggg gcgggggggg 840
ggggggggcg gggcgagggg cggggcgggg cgaggcggag aggtgcggcg gcagccaatc 900
agagcggcgc gctccgaaag tttcctttta tggcgaggcg gcggcggcgg cggccctata 960
aaaagcgaag cgcgcggcgg gcgggagtcg ctgcgcgctg ccttcgcccc gtgccccgct 1020
ccgccgccgc ctcgcgccgc ccgccccggc tctgactgac cgcgttactc ccacaggtga 1080
gcgggcggga cggcccttct cctccgggct gtaattagct gagcaagagg taagggttta 1140
agggatggtt ggttggtggg gtattaatgt ttaattacct ggagcacctg cctgaaatca 1200
ctttttttca ggttggaccg gtgccaccat ggactataag gaccacgacg gagactacaa 1260
ggatcatgat attgattaca aagacgatga cgataagatg gccccaaaga agaagcggaa 1320
ggtcggtatc cacggagtcc cagcagccga caagaagtac agcatcggcc tggacatcgg 1380
caccaactct gtgggctggg ccgtgatcac cgacgagtac aaggtgccca gcaagaaatt 1440
caaggtgctg ggcaacaccg accggcacag catcaagaag aacctgatcg gagccctgct 1500
gttcgacagc ggcgaaacag ccgaggccac ccggctgaag agaaccgcca gaagaagata 1560
caccagacgg aagaaccgga tctgctatct gcaagagatc ttcagcaacg agatggccaa 1620
ggtggacgac agcttcttcc acagactgga agagtccttc ctggtggaag aggataagaa 1680
gcacgagcgg caccccatct tcggcaacat cgtggacgag gtggcctacc acgagaagta 1740
ccccaccatc taccacctga gaaagaaact ggtggacagc accgacaagg ccgacctgcg 1800
gctgatctat ctggccctgg cccacatgat caagttccgg ggccacttcc tgatcgaggg 1860
cgacctgaac cccgacaaca gcgacgtgga caagctgttc atccagctgg tgcagaccta 1920
caaccagctg ttcgaggaaa accccatcaa cgccagcggc gtggacgcca aggccatcct 1980
gtctgccaga ctgagcaaga gcagacggct ggaaaatctg atcgcccagc tgcccggcga 2040
gaagaagaat ggcctgttcg gaaacctgat tgccctgagc ctgggcctga cccccaactt 2100
caagagcaac ttcgacctgg ccgaggatgc caaactgcag ctgagcaagg acacctacga 2160
cgacgacctg gacaacctgc tggcccagat cggcgaccag tacgccgacc tgtttctggc 2220
cgccaagaac ctgtccgacg ccatcctgct gagcgacatc ctgagagtga acaccgagat 2280
caccaaggcc cccctgagcg cctctatgat caagagatac gacgagcacc accaggacct 2340
gaccctgctg aaagctctcg tgcggcagca gctgcctgag aagtacaaag agattttctt 2400
cgaccagagc aagaacggct acgccggcta cattgacggc ggagccagcc aggaagagtt 2460
ctacaagttc atcaagccca tcctggaaaa gatggacggc accgaggaac tgctcgtgaa 2520
gctgaacaga gaggacctgc tgcggaagca gcggaccttc gacaacggca gcatccccca 2580
ccagatccac ctgggagagc tgcacgccat tctgcggcgg caggaagatt tttacccatt 2640
cctgaaggac aaccgggaaa agatcgagaa gatcctgacc ttccgcatcc cctactacgt 2700
gggccctctg gccaggggaa acagcagatt cgcctggatg accagaaaga gcgaggaaac 2760
catcaccccc tggaacttcg aggaagtggt ggacaagggc gcttccgccc agagcttcat 2820
cgagcggatg accaacttcg ataagaacct gcccaacgag aaggtgctgc ccaagcacag 2880
cctgctgtac gagtacttca ccgtgtataa cgagctgacc aaagtgaaat acgtgaccga 2940
gggaatgaga aagcccgcct tcctgagcgg cgagcagaaa aaggccatcg tggacctgct 3000
gttcaagacc aaccggaaag tgaccgtgaa gcagctgaaa gaggactact tcaagaaaat 3060
cgagtgcttc gactccgtgg aaatctccgg cgtggaagat cggttcaacg cctccctggg 3120
cacataccac gatctgctga aaattatcaa ggacaaggac ttcctggaca atgaggaaaa 3180
cgaggacatt ctggaagata tcgtgctgac cctgacactg tttgaggaca gagagatgat 3240
cgaggaacgg ctgaaaacct atgcccacct gttcgacgac aaagtgatga agcagctgaa 3300
gcggcggaga tacaccggct ggggcaggct gagccggaag ctgatcaacg gcatccggga 3360
caagcagtcc ggcaagacaa tcctggattt cctgaagtcc gacggcttcg ccaacagaaa 3420
cttcatgcag ctgatccacg acgacagcct gacctttaaa gaggacatcc agaaagccca 3480
ggtgtccggc cagggcgata gcctgcacga gcacattgcc aatctggccg gcagccccgc 3540
cattaagaag ggcatcctgc agacagtgaa ggtggtggac gagctcgtga aagtgatggg 3600
ccggcacaag cccgagaaca tcgtgatcga aatggccaga gagaaccaga ccacccagaa 3660
gggacagaag aacagccgcg agagaatgaa gcggatcgaa gagggcatca aagagctggg 3720
cagccagatc ctgaaagaac accccgtgga aaacacccag ctgcagaacg agaagctgta 3780
cctgtactac ctgcagaatg ggcgggatat gtacgtggac caggaactgg acatcaaccg 3840
gctgtccgac tacgatgtgg accatatcgt gcctcagagc tttctgaagg acgactccat 3900
cgacaacaag gtgctgacca gaagcgacaa gaaccggggc aagagcgaca acgtgccctc 3960
cgaagaggtc gtgaagaaga tgaagaacta ctggcggcag ctgctgaacg ccaagctgat 4020
tacccagaga aagttcgaca atctgaccaa ggccgagaga ggcggcctga gcgaactgga 4080
taaggccggc ttcatcaaga gacagctggt ggaaacccgg cagatcacaa agcacgtggc 4140
acagatcctg gactcccgga tgaacactaa gtacgacgag aatgacaagc tgatccggga 4200
agtgaaagtg atcaccctga agtccaagct ggtgtccgat ttccggaagg atttccagtt 4260
ttacaaagtg cgcgagatca acaactacca ccacgcccac gacgcctacc tgaacgccgt 4320
cgtgggaacc gccctgatca aaaagtaccc taagctggaa agcgagttcg tgtacggcga 4380
ctacaaggtg tacgacgtgc ggaagatgat cgccaagagc gagcaggaaa tcggcaaggc 4440
taccgccaag tacttcttct acagcaacat catgaacttt ttcaagaccg agattaccct 4500
ggccaacggc gagatccgga agcggcctct gatcgagaca aacggcgaaa ccggggagat 4560
cgtgtgggat aagggccggg attttgccac cgtgcggaaa gtgctgagca tgccccaagt 4620
gaatatcgtg aaaaagaccg aggtgcagac aggcggcttc agcaaagagt ctatcctgcc 4680
caagaggaac agcgataagc tgatcgccag aaagaaggac tgggacccta agaagtacgg 4740
cggcttcgac agccccaccg tggcctattc tgtgctggtg gtggccaaag tggaaaaggg 4800
caagtccaag aaactgaaga gtgtgaaaga gctgctgggg atcaccatca tggaaagaag 4860
cagcttcgag aagaatccca tcgactttct ggaagccaag ggctacaaag aagtgaaaaa 4920
ggacctgatc atcaagctgc ctaagtactc cctgttcgag ctggaaaacg gccggaagag 4980
aatgctggcc tctgccggcg aactgcagaa gggaaacgaa ctggccctgc cctccaaata 5040
tgtgaacttc ctgtacctgg ccagccacta tgagaagctg aagggctccc ccgaggataa 5100
tgagcagaaa cagctgtttg tggaacagca caagcactac ctggacgaga tcatcgagca 5160
gatcagcgag ttctccaaga gagtgatcct ggccgacgct aatctggaca aagtgctgtc 5220
cgcctacaac aagcaccggg ataagcccat cagagagcag gccgagaata tcatccacct 5280
gtttaccctg accaatctgg gagcccctgc cgccttcaag tactttgaca ccaccatcga 5340
ccggaagagg tacaccagca ccaaagaggt gctggacgcc accctgatcc accagagcat 5400
caccggcctg tacgagacac ggatcgacct gtctcagctg ggaggcgaca aaaggccggc 5460
ggccacgaaa aaggccggcc aggcaaaaaa gaaaaagtaa gaattcctag agctcgctga 5520
tcagcctcga ctgtgccttc tagttgccag ccatctgttg tttgcccctc ccccgtgcct 5580
tccttgaccc tggaaggtgc cactcccact gtcctttcct aataaaatga ggaaattgca 5640
tcgcattgtc tgagtaggtg tcattctatt ctggggggtg gggtggggca ggacagcaag 5700
ggggaggatt gggaagagaa tagcaggcat gctggggagc ggccgcagga acccctagtg 5760
atggagttgg ccactccctc tctgcgcgct cgctcgctca ctgaggccgg gcgaccaaag 5820
gtcgcccgac gcccgggctt tgcccgggcg gcctcagtga gcgagcgagc gcgcagctgc 5880
ctgcaggggc gcctgatgcg gtattttctc cttacgcatc tgtgcggtat ttcacaccgc 5940
atacgtcaaa gcaaccatag tacgcgccct gtagcggcgc attaagcgcg gcgggtgtgg 6000
tggttacgcg cagcgtgacc gctacacttg ccagcgcctt agcgcccgct cctttcgctt 6060
tcttcccttc ctttctcgcc acgttcgccg gctttccccg tcaagctcta aatcgggggc 6120
tccctttagg gttccgattt agtgctttac ggcacctcga ccccaaaaaa cttgatttgg 6180
gtgatggttc acgtagtggg ccatcgccct gatagacggt ttttcgccct ttgacgttgg 6240
agtccacgtt ctttaatagt ggactcttgt tccaaactgg aacaacactc aactctatct 6300
cgggctattc ttttgattta taagggattt tgccgatttc ggtctattgg ttaaaaaatg 6360
agctgattta acaaaaattt aacgcgaatt ttaacaaaat attaacgttt acaattttat 6420
ggtgcactct cagtacaatc tgctctgatg ccgcatagtt aagccagccc cgacacccgc 6480
caacacccgc tgacgcgccc tgacgggctt gtctgctccc ggcatccgct tacagacaag 6540
ctgtgaccgt ctccgggagc tgcatgtgtc agaggttttc accgtcatca ccgaaacgcg 6600
cgagacgaaa gggcctcgtg atacgcctat ttttataggt taatgtcatg ataataatgg 6660
tttcttagac gtcaggtggc acttttcggg gaaatgtgcg cggaacccct atttgtttat 6720
ttttctaaat acattcaaat atgtatccgc tcatgagaca ataaccctga taaatgcttc 6780
aataatattg aaaaaggaag agtatgagta ttcaacattt ccgtgtcgcc cttattccct 6840
tttttgcggc attttgcctt cctgtttttg ctcacccaga aacgctggtg aaagtaaaag 6900
atgctgaaga tcagttgggt gcacgagtgg gttacatcga actggatctc aacagcggta 6960
agatccttga gagttttcgc cccgaagaac gttttccaat gatgagcact tttaaagttc 7020
tgctatgtgg cgcggtatta tcccgtattg acgccgggca agagcaactc ggtcgccgca 7080
tacactattc tcagaatgac ttggttgagt actcaccagt cacagaaaag catcttacgg 7140
atggcatgac agtaagagaa ttatgcagtg ctgccataac catgagtgat aacactgcgg 7200
ccaacttact tctgacaacg atcggaggac cgaaggagct aaccgctttt ttgcacaaca 7260
tgggggatca tgtaactcgc cttgatcgtt gggaaccgga gctgaatgaa gccataccaa 7320
acgacgagcg tgacaccacg atgcctgtag caatggcaac aacgttgcgc aaactattaa 7380
ctggcgaact acttactcta gcttcccggc aacaattaat agactggatg gaggcggata 7440
aagttgcagg accacttctg cgctcggccc ttccggctgg ctggtttatt gctgataaat 7500
ctggagccgg tgagcgtgga agccgcggta tcattgcagc actggggcca gatggtaagc 7560
cctcccgtat cgtagttatc tacacgacgg ggagtcaggc aactatggat gaacgaaata 7620
gacagatcgc tgagataggt gcctcactga ttaagcattg gtaactgtca gaccaagttt 7680
actcatatat actttagatt gatttaaaac ttcattttta atttaaaagg atctaggtga 7740
agatcctttt tgataatctc atgaccaaaa tcccttaacg tgagttttcg ttccactgag 7800
cgtcagaccc cgtagaaaag atcaaaggat cttcttgaga tccttttttt ctgcgcgtaa 7860
tctgctgctt gcaaacaaaa aaaccaccgc taccagcggt ggtttgtttg ccggatcaag 7920
agctaccaac tctttttccg aaggtaactg gcttcagcag agcgcagata ccaaatactg 7980
ttcttctagt gtagccgtag ttaggccacc acttcaagaa ctctgtagca ccgcctacat 8040
acctcgctct gctaatcctg ttaccagtgg ctgctgccag tggcgataag tcgtgtctta 8100
ccgggttgga ctcaagacga tagttaccgg ataaggcgca gcggtcgggc tgaacggggg 8160
gttcgtgcac acagcccagc ttggagcgaa cgacctacac cgaactgaga tacctacagc 8220
gtgagctatg agaaagcgcc acgcttcccg aagggagaaa ggcggacagg tatccggtaa 8280
gcggcagggt cggaacagga gagcgcacga gggagcttcc agggggaaac gcctggtatc 8340
tttatagtcc tgtcgggttt cgccacctct gacttgagcg tcgatttttg tgatgctcgt 8400
caggggggcg gagcctatgg aaaaacgcca gcaacgcggc ctttttacgg ttcctggcct 8460
tttgctggcc ttttgctcac atgt 8484
Номер последовательности (ID): 2
Длина: 23
Тип молекулы: RNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other RNA
> organism, Mus musculus
Остатки:
caccgacttg gtagtctcgt agg 23
Номер последовательности (ID): 3
Длина: 23
Тип молекулы: RNA
Характеристики Местоположение/Квалификаторы:
- source, 1..23
> mol_type, other RNA
> organism, Mus musculus
Остатки:
ctacgagact accaagtcgg tgg 23
Номер последовательности (ID): 4
Длина: 20
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..20
> mol_type, other DNA
> organism, synthetic construct
Остатки:
gaaatgtggg gaggggtctg 20
Номер последовательности (ID): 5
Длина: 20
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..20
> mol_type, other DNA
> organism, synthetic construct
Остатки:
agctggtgag ctgggataga 20
Номер последовательности (ID): 6
Длина: 20
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..20
> mol_type, other DNA
> organism, synthetic construct
Остатки:
cggtgcttgt ggctttaacc 20
Номер последовательности (ID): 7
Длина: 25
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..25
> mol_type, other DNA
> organism, synthetic construct
Остатки:
caccgcaccg acttggtagt ctcgt 25
Номер последовательности (ID): 8
Длина: 25
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..25
> mol_type, other DNA
> organism, synthetic construct
Остатки:
aaacacgaga ctaccaagtc ggtgc 25
Номер последовательности (ID): 9
Длина: 25
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..25
> mol_type, other DNA
> organism, synthetic construct
Остатки:
caccgctacg agactaccaa gtcgg 25
Номер последовательности (ID): 10
Длина: 24
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..24
> mol_type, other DNA
> organism, synthetic construct
Остатки:
aaacccgact tggtagtctc gtag 24
Номер последовательности (ID): 11
Длина: 21
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..21
> mol_type, other DNA
> organism, synthetic construct
Остатки:
gagggcctat ttcccatgat t 21
Номер последовательности (ID): 12
Длина: 15
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..15
> mol_type, other DNA
> organism, synthetic construct
Остатки:
tttttttttt ttttt 15
Номер последовательности (ID): 13
Длина: 21
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..21
> mol_type, other DNA
> organism, synthetic construct
Остатки:
ccattaagca gcccactcca g 21
Номер последовательности (ID): 14
Длина: 21
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..21
> mol_type, other DNA
> organism, synthetic construct
Остатки:
gtgcaagaaa acccacccca c 21
Номер последовательности (ID): 15
Длина: 21
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..21
> mol_type, other DNA
> organism, synthetic construct
Остатки:
ctacgagact acccggtggt t 21
Номер последовательности (ID): 16
Длина: 25
Тип молекулы: DNA
Характеристики Местоположение/Квалификаторы:
- source, 1..25
> mol_type, other DNA
> organism, synthetic construct
Остатки:
ctacgagact accaagtcgg tggtt 25
END
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения мышиной модели для изучения миодистрофии Дюшенна и вариантов ее терапии | 2023 |
|
RU2815936C1 |
| Способ получения генно-модифицированных мышей, экспрессирующих миниген антитромбина III человека, с помощью микроинъекций TelN-линеаризованного фрагмента ДНК | 2022 |
|
RU2806568C1 |
| Способ получения генно-модифицированных лабораторных животных с индуцируемой системной и тканеспецифической экспрессией циклофилин А человека | 2022 |
|
RU2800264C1 |
| Способ получения генно-модифицированных кроликов с нокаутом гена LEPR с помощью системы CRISPR/Cas9 | 2023 |
|
RU2836438C1 |
| ЛИНИЯ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ МЫШЕЙ ДЛЯ МОДЕЛИРОВАНИЯ БОЛЕЗНИ АЛЬЦГЕЙМЕРА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ЕЕ ПРИМЕНЕНИЕ, sgPHK И КОМБИНАЦИЯ crRNA/tracrRNA, ИСПОЛЬЗУЕМЫЕ В СПОСОБЕ, И НАБОР ПРАЙМЕРОВ | 2024 |
|
RU2835497C1 |
| Способ получения нокаута гена CD209 в эмбрионах Bos taurus путем трансдукции зигот адено-ассоциированными вирусами, кодирующими saCas9 и соответствующую гидовую РНК | 2022 |
|
RU2800917C1 |
| Способ анализа соматических мутаций в генах IDH1 и IDH2 с использованием LNA-блокирующей ПЦР и гибридизации с биологическим микрочипом | 2024 |
|
RU2839291C1 |
| Нуклеотидная последовательность, кодирующая фермент литиказу, и панель олигонуклеотидов для получения синтетической нуклеотидной последовательности гена литиказы | 2023 |
|
RU2826150C1 |
| Способ мультиплексной идентификации 32 генетических маркеров льна | 2022 |
|
RU2804939C1 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ, НАЦЕЛЕННЫЕ НА ГЕН CAV3.1, И ИХ ПРИМЕНЕНИЯ | 2022 |
|
RU2839721C2 |



Изобретение относится к области биотехнологии, в частности к плазмидной конструкции px330+P2rx3-sg1 для получения трансгенных мышей, гомозиготных по нуль-аллелю гена P2rx3. Также раскрыта линия мышей, гомозиготных по нуль-аллелю гена P2rx3, а также способ ее получения. Изобретение эффективно для тестирования веществ на роль блокатора P2X3. 5 н.п. ф-лы, 5 ил., 3 пр.
1. Плазмидная конструкция px330+P2rx3-sg1 для получения трансгенных мышей, гомозиготных по нуль-аллелю гена P2rx3, полученная на основе вектора px330 последовательностью SEQ ID NO: 1, кодирующая ОРС гена spCas9 под контролем CAG промотора и экспрессирующая гидовую РНК sg1 последовательностью SEQ ID NO: 2 против соответствующего участка гена P2rx3, под контролем U6 промотора.
2. Плазмидная конструкция px330+P2rx3-sg2 для получения трансгенных мышей, гомозиготных по нуль-аллелю гена P2rx3, полученная на основе вектора px330 последовательностью SEQ ID NO: 1, кодирующая ОРС гена spCas9 под контролем CAG промотора и экспрессирующая гидовую РНК sg2 последовательностью SEQ ID NO: 3 против соответствующего участка гена P2rx3, под контролем U6 промотора.
3. Линия мышей, гомозиготных по нуль-аллелю гена P2rx3, с полным отсутствием экспрессии пуринергического рецептора P2X3 для тестирования веществ на роль блокатора P2X3, трансформированная плазмидной конструкцией по пп.1, 2 и несущая нуль-аллель гена P2rx3, отличающаяся тем, что ген P2rx3 несет делецию четырех нуклеотидов в позициях 40-43.
4. Линия мышей, гомозиготных по нуль-аллелю гена P2rx3, с полным отсутствием экспрессии пуринергического рецептора P2X3 для тестирования веществ на роль блокатора P2X3, трансформированная плазмидной конструкцией по пп.1, 2 и несущая нуль-аллель гена P2rx3, отличающаяся тем, что ген P2rx3 несет вставку одного нуклеотида в позиции 40.
5. Способ получения двух линий мышей по пп.3 и 4, гомозиготных по нуль-аллелю гена P2rx3, с полным отсутствием экспрессии пуринергического рецептора P2X3 для тестирования веществ на роль блокатора P2X3, характеризующийся следующими стадиями:
а) микроинъекция одновременно двух плазмидных конструкций по пп.1, 2 в пронуклеус оплодотворенной яйцеклетки мыши гибрида первого поколения CBA×C57BL/6;
б) выявление жизнеспособных зигот;
в) пересадка выживших зигот из пункта (б) псевдобеременным самкам-реципиентам, имеющим копулятивную пробку после ссаживания с вазэктомированными самцами;
г) получение новорожденных мышей на 19 день после пересадки зигот из пункта (в);
д) проведение анализа наличия сдвига рамки считывания в первом экзоне гена P2rx3 через 8-14 дней после рождения мышей (г) путем амплификации целевого участка гена P2rx3 с праймерами, имеющими последовательности SEQ ID NO: 4 и SEQ ID NO: 5, с последующим секвенированием фрагмента с использованием праймера последовательностью SEQ ID NO: 6;
е) отбор мышей, несущих целевую мутацию;
ж) проведение скрещивания мышей из пункта (е) с мышами дикого типа для получения самок-гетерозигот или мутантных самцов, несущих целевую мутацию.
з) проведение скрещивания мышей из пункта (ж) для получения итоговой линии мышей, гомозиготных по нуль-аллелю гена P2rx3.
| ZHIRONG WANG et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| COCKAYNE DA et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

